ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
В соответствии с § 119 статьи 35 закона США, в настоящей заявке испрашивается приоритет по дате подачи предварительной патентной заявки США № 60/341723, поданной 21 июня 2001 и описание которой включено в настоящую заявку в виде ссылки.
Правительство США может пользоваться правом владения данным изобретением в соответствии с грантом номер 9875939.
ВВЕДЕНИЕ
Область изобретения
Областью настоящего изобретения являются флуоресцирующие белки.
Основа создания изобретения
Мечение является инструментом для маркировки представляющего интерес белка, клетки или организма и играет существенную роль во многих применениях в области биохимии, молекулярной биологии и медицинской диагностики. Создано обширное разнообразие различных меток, включая радиоактивные метки, хромометки, флуоресцентные метки, хемилюминесцентные метки и т.д. Тем не менее ощущается не прекращающийся интерес к созданию новых меток. Особый интерес представляет собой развитие новых белковых меток, включая хромопротеины и/или флуоресцирующие белковые метки.
Новым разрабатываемым в настоящее время важным классом белков являются флуоресцирующие белки рифовых кораллов, как описано у Matz, M.V., et al. (1999) Nature Biotechnol., 17:969-973. Несмотря на то что эти флуоресцирующие белки обладают многими положительными свойствами, особый к ним интерес связан с развитием вариантов флуоресцирующих белков этого очень важного класса, которые обладали бы дополнительными ценными свойствами, например быстрым созреванием. Настоящее изобретение связано с удовлетворением такой потребности.
Релевантная литература
Интерес в этой связи представляют патенты США, включающие в себя патенты под следующими номерами: 6066476; 6020192; 5985577; 5976796; 5968750; 5968738; 5958713; 5919445; 5874304 и 5491084. Представляющие интерес Международные патентные публикации включают в себя публикации WO 00/46233; WO 99/49019 и DE 19718640A. Интересны также публикации Anderluh et al., Biochemical and Biophysical Research Communications (1996) 220:437-442; Dove et al., Biological Bulletin (1995) 189:288-297; Fradkov et al., FEBS Lett. (2000) 479 (3):127-30; Gurskaya et al., FEBS Lett., (2001) 507 (1):16-20; Gurskaya et al., BMC Biochem. (2001) 2:6; Lukyanov, K., et al. (2000) J. Biol. Chemistry 275 (34):25879-25882; Macek et al., Eur. J. Biochem. (1995) 234:329-335; Martynov et al., J. Biol. Chem. (2001) 276:21012-6; Matz, M. V., et al. (1999) Nature Biotechnol., 17:969-973; Terskikh et al., Science (2000) 290:1585-8; Tsien, Annual Rev. of Biochemistry (1998) 67:509-544; Tsien, Nat. Biotech. (1999) 17:956-957; Ward et al., J.Biol. Chem. (1979) 254:781-788; Wiedermann et al., Jarhrestagung der Deutschen Gesellschact fur Tropenokologie-gto. Ulm. 17-19.02. 1999. Poster P-4.20; Yarbrough et al., Proc. Natl. Acad. Sci. U. S. A. (2001) 98:462-7.
КРАТКОЕ СОДЕРЖАНИЕ ИЗОБРЕТЕНИЯ
Композиции нуклеиновых кислот, кодирующие быстросозревающие флуоресцирующие белки, а также их неагрегирующие варианты (и их мутанты) и кодируемые ими белки, составляют существо настоящего изобретения. Белки, о которых идет речь, являются флуоресцирующими белками, причем это их свойство возникает в результате взаимодействия двух или более остатков белковой молекулы. Белки, о которых идет речь, дополнительно характеризуются тем, что, в определенных воплощениях они обнаруживаются в составе белков дикого типа или представляют собой мутанты белков дикого типа, которые получены либо из небиолюминесцентных видов Cnidaria, например, Anthozoa (коралловые полипы), либо получены из видов Anthozoa non-Pennatulacea (коралловые полипы, не являющиеся пеннатулярией). В определенных воплощениях белки, о которых идет речь, представляют собой мутанты флуоресцирующего белка "red" Discosoma sp. дикого типа, продаваемого под коммерческим названием "DsRed". Также интерес представляют белки, которые по существу подобны описанным выше специфическим белкам, или их мутанты. Изобретение связано также с фрагментами нуклеиновых кислот и кодируемыми ими пептидами, а также антителами против белков, о которых идет речь, и трансгенными клетками и организмами. Композиции таких белков и нуклеиновых кислот имеют множество разнообразных применений. Наконец, изобретение связано также и с наборами для таких применений, например, таких, которые включают в себя композиции нуклеиновых кислот согласно изобретению.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА:
Фигура 1. Нормализованное колебание и спектры эмиссии (испускания, излучения) представителей вариантов DsRed. (A) Мутирующий остаток N42 изменяет спектральные свойства DsRed. Спектры показаны для DsRed1 и вариантов N42H и N42Q. Все три белка были полностью зрелыми. (B) Спектры оптимизированных вариантов DsRed.T3 и DsRed.T4.
Фигура 2. Кинетика созревания вариантов DsRed. В логарифмической фазе роста культуры E. coli обрабатывали индуктором изопропил-P-D-тиогалактопиранозидом (IPTG) в течение 30 мин для генерации пульса экспрессии для каждого из вариантов. Затем инициировали чейз (во время 0 на графиках) путем добавления ингибиторов белкового синтеза и продолжали инкубирование при 37°C. В указанных временных точках отбирали аликвоты культур, которые впоследствии анализировали методом проточной цитометрии на предмет определения средней интенсивности красной флуоресценции на клетку. Базовую флуоресценцию (пунктирная линия) измеряли, используя клетки, несущие пустую плазмиду pQE81. На двух этих графиках наносились (A) грубые значения флуоресценции, или (B) значения, полученные путем вычитания флуоресценции, имевшейся во время 0 и нормализованной к максимальному сигналу в 100% для каждого из вариантов DsRed. Слабое отклонение в поздних временных точках средних значений флуоресценции для DsRed.T3 и DsRed.T4, возможно, отражает клеточный лизис. В контрольной культуре ингибиторы белкового синтеза добавлялись к клеткам, несущим плазмиду, экспрессирующую DsRed.T3; как и ожидалось, эти клетки оставались нефлуоресцирующими (данные не показаны). Методом иммуноблоттинга было показано, что в течение чейз-периода количество белка DsRed2, DsRed.T3, и DsRed.T4 в культуре оставалось в основном постоянным, в то время как количество белка DsRed1 прогрессивно уменьшалось приблизительно до половины его первоначального уровня (данные не показаны).
Фигура 3. Одновременная визуализация DsRed.T4 и EGFP у дрожжей. DsRed.T4 был нацелен на митохондриальный матрикс Saccharomyces cerevisiae путем слияния с предпоследовательностью Cox4p. Слитый белок pCox4-DsRed.T4 был продуцирован в штамме, который содержал Sec7p-eGFP, маркер цистерн аппарата Гольджи. Клетки из логарифмической фазы роста культуры были визуализованы либо с помощью набора фильтров Texas Red (красный), либо с помощью набора фильтров EGFP (зеленый). Кроме того, клетки были визуализованы с помощью дифференциальной интерференционно-контрастной (DIC) микроскопии. Как показано на картинке слияния, сигналы DsRed.T4 и EGFP легко разделяются. Масштаб отрезка - 2 мкм.
Фигура 4. Уменьшение результирующего заряда вблизи N-конца DsRed уменьшает агрегацию белка. (A) Неденатурирующий SDS-PAGE очищенного DsRed1 (WT), варианта цикла 1 (R1), варианта цикла 3 (R3), варианта цикла 4 (R4), DsRed.T1 (T1), DsRed.T3 (T3) и DsRed.T4 (T4). 1 мкг каждого очищенного варианта DsRed смешивали на льду с образцом буфера, содержащего SDS, и немедленно подвергали электрофорезу при 4°C в 10% полиакриламидном геле, а затем окрашивали Coomassie Blue. WT* и T4*: Дополнительные аликвоты DsRed1 и DsRed.T4 были денатурированы в результате кипячения перед электрофорезом. МВ: предварительно окрашенные стандартные белки широкого диапазона молекулярных весов (Bio-Rad). (B) Для измерения растворимостей флуоресцирующих белков в E. coli клетки, несущие экспрессирующие векторы на основе pREP4 плюс pQE31, кодирующие DsRed1, DsRed2, вариант цикла 3, вариант цикла 4 или EGFP, выращивали до значения оптической плотности OD600 порядка 0,5, индуцированной IPTG в течение 7 ч, затем лизировали с помощью B-PER II и центрифугировали в течение 20 мин при 27000×g. Эквивалентные количества фракций осадка и супернатанта подвергали SDS-PAGE, а затем - иммуноблоттингу в присутствии анти-гексагистидинового моноклонального антитела (Qiagen). Связанное антитело определяли с помощью набора ECL-Plus (Amersham) и установки Molecular Dynamics Storm 860 phosphorimager. Для каждого флуоресцирующего белка анализировали серийные разведения из бактериального экстракта, а для системы детектирования выбирали образец внутри линейной области. Затем количественно определяли процентное содержание каждого белка во фракции супернатанта. На диаграмме представлены средние значения по результатам двух отдельных экспериментов; значения, полученные для каждого флуоресцирующего белка в двух экспериментах, отличались друг от друга не более чем на 10%.
ОПРЕДЕЛЕНИЯ
В соответствии с настоящим изобретением, могут быть использованы общепринятые для специалистов технологии молекулярной биологии, микробиологии и рекомбинантной ДНК. Такие технологии полностью представлены в литературе. См., например, Maniatis, Fritsch & Sambrook, "Molecular Cloning: A Laboratory Manual (1982); "DNA Cloning: A Practical Approach," Volumes I and II (D. N. Glover ed. 1985); "Oligonucleotide Synthesis" (M. J. Gait ed. 1984); "Nucleic Acid Hybridization" (B.D. Hames & S. J. Higgins eds. (1985)); "Transcription and Translation" (B. D. Hames & S. J. Higgins eds. (1984)); "Animal Cell Culture" (R.I. Freshney, ed. (1986)); "Immobilized Cells and Enzymes" (IRL Press, (1986)); B. Perbal, "A Practical Guide To Molecular Cloning" (1984).
"Вектор" представляет собой репликон, такой как плазмида, фаг или космид, к которому может быть прикреплен другой сегмент ДНК, так чтобы вызвать репликацию прикрепленного сегмента.
Термин "молекула ДНК" относится к полимерной форме дезоксирибонуклеотидов (аденина, гуанина, тимина или цитозина), находящейся либо в одноцепочечной форме, либо в виде двухцепочечной спирали. Этот термин относится только к первичной и вторичной структуре молекулы, не ограничивая ее никакими конкретными формами третичной структуры. Таким образом, этот термин включает в себя двухцепочечную ДНК, обнаруживаемую помимо прочего и в составе линейных молекул ДНК (например, рестрикционные фрагменты), вирусов, плазмид и хромосом.
"Кодирующей последовательностью" ДНК считается последовательность ДНК, которая транскрибируется и транслируется в полипептид in vivo, когда она находится под контролем соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяются инициирующим кодоном на 5'(амино) конце и терминирующим кодоном трансляции на 3'(карбоксильном) конце. Кодирующая последовательность может включать в себя, не ограничиваясь этим, прокариотические последовательности, кДНК из эукариотических мРНК, геномные последовательности ДНК из эукариотических ДНК (например, ДНК млекопитающих) и искусственные последовательности ДНК. Сигнал полиаденилирования и последовательность терминации транскрипции может быть локализована на 3'-конце кодирующей последовательности.
Здесь термин "гибридизация" относится к процессу ассоциации двух цепей нуклеиновых кислот с образованием антипараллельного дуплекса, стабилизированного гидрофобными взаимодействиями между остатками противопоставленных цепей нуклеиновой кислоты.
Термин "олигонуклеотид" относится к короткой (менее 100 оснований в длину) молекуле нуклеиновой кислоты.
Здесь под "регуляторными последовательностями ДНК" подразумеваются последовательности, осуществляющие контроль процессов транскрипции и трансляции, такие как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и тому подобное, которые обеспечивают и/или регулируют экспрессию кодирующей последовательности в хозяйской клетке.
"Промоторной последовательностью" является регуляторная область ДНК, способная к связыванию РНК-полимеразы в клетке и инициирующая транскрипцию в прямом направлении в кодирующей последовательности. В целях, связанных с настоящим изобретением, промоторная последовательность связывается на своем 3'-конце с сайтом инициации транскрипции и простирается в обратном направлении (в направлении к 5'-концу) для включения минимального количества оснований или элементов, необходимых для инициации транскрипции на уровнях, определяемых над фоном. В рамках промоторной последовательности будет находиться сайт инициации транскрипции, так же как и белок-связывающие домены, ответственные за связывание РНК-полимеразы. Эукариотические промоторы часто, но не всегда будут содержать "TATA"-боксы и "CAT"-боксы. Различные промоторы, включая индуцибельные промоторы, могут быть использованы в качестве драйвера для различных векторов согласно изобретению.
Здесь термины "рестрикционные эндонуклеазы" и "ферменты рестрикции" относятся к бактериальным ферментам, каждый из которых разрезает двойную спираль ДНК в строго специфической нуклеотидной последовательности или вблизи нее.
Говорят, что клетка подверглась "трансформации" или была "трансфицирована" экзогенной или гетерологичной ДНК, если такая ДНК была введена внутрь клетки. Трансформирующая ДНК может быть или может не быть интегрирована (ковалентно связана) в клеточный геном. Например, в клетках прокариот, дрожжей и млекопитающих трансформирующая ДНК может быть сохранена на эписомном элементе, таком как плазмида. Что касается эукариотических клеток, стабильно трансформированной клеткой является такая клетка, в которой трансформирующая ДНК интегрировалась в хромосому, так что при хромосомной репликации она передается дочерним клеткам. Такая стабильность демонстрируется возможностью получения из эукариотической клетки клеточных линий или клонов, состоящих из популяции дочерних клеток, содержащих трансформирующую ДНК. "Клон" представляет собой популяцию клеток, полученных из одной клетки или общего предшественника в результате митоза. "Клеточная линия" представляет собой клон первоначальной клетки, который способен к стабильному росту in vitro на протяжении многих поколений.
Термин "гетерологичная" область ДНК-конструкта представляет собой идентифицируемый сегмент ДНК в составе более крупной молекулы ДНК, который в природе в ассоциации с этой более крупной молекулой ДНК не встречается. Таким образом, если гетерологичная область кодирует ген млекопитающего, этот ген, как правило, бывает фланкирован ДНК, которая не фланкирует геномную ДНК млекопитающего в геноме того организма, из которого она получена. В качестве другого примера гетерологичная ДНК включает в себя кодирующую последовательность в конструкте, где части генов из двух различных источников объединяются таким образом, чтобы в результате продуцировался слитый белок. Согласно используемому здесь определению аллельные вариации или природно возникающие мутационные явления не приводят к образованию гетерологичной области ДНК.
Здесь термин "репортерный ген" относится к кодирующей последовательности, которая прикреплена к гетерологичному промотору или энхансерным элементам и продукт которой может быть с легкостью определен и подвергнут количественной оценке, когда этот конструкт вводят в ткани или клетки.
Предпочтительно, чтобы описываемые здесь аминокислоты были представлены в виде "L"-изомера. Аминокислотные последовательности приводятся в однобуквенной кодировке (A: аланин; C: цистеин; D: аспарагиновая кислота; E: глутаминовая кислота; F: фенилаланин; G: глицин; H: гистидин; I: изолейцин; K: лизин; L: лейцин; M: метионин; N: аспарагин; P: пролин; Q: глутамин; R: аргинин; S: серин; T: треонин; V: валин; W: триптофан; Y: тирозин; X: любой остаток). NH2 относится к свободной аминогруппе, присутствующей на амино-конце полипептида. COOH относится к свободной карбокси-группе, присутствующей на карбокси-конце полипептида. Придерживаясь стандартной полипептидной номенклатуре, использовали представления, описанные в публикации J. Biol. Chem., 243 (1969), 3552-59.
Термином "иммунологически активный" определяется способность природного, рекомбинантного или искусственного хромо/флуоресцирующего белка или любого его олигопептида индуцировать специфический иммунный ответ у соответствующих животных или со стороны соответствующих клеток и связываться со специфическими антителами. Здесь под термином "антигенная аминокислотная последовательность" подразумевается аминокислотная последовательность, которая либо сама по себе, либо в ассоциации с молекулой-носителем может вызвать антительный ответ у млекопитающего. Термин "специфическое связывание" в контексте связывания антигена с антителом является термином, хорошо понимаемым в данной области и относящимся к связыванию антитела с антигеном, против которого это антитело было выработано, но не с другими неродственными антигенами.
Под термином "изолированный" здесь подразумевается обозначение полинуклеотида, полипептида, антитела или клетки-хозяина, которые находятся в среде, отличной от той, в которой указанные полинуклеотид, полипептид, антитело или клетка-хозяин встречаются в природе.
Биолюминесценция (BL) определяется как излучение живыми организмами света, который хорошо виден в темноте и действует на визуальное поведение животных (см., например, Harvey, E. N. (1952). Bioluminescence. New York: Academic Press; Hastings, J. W. (1995). Bioluminescence. In: Cell Physiology (ed. by N. Speralakis). pp.651-681. New York: Academic Press.; Wilson, T. and Hastings, J. W. (1998). Bioluminescence. Annu Rev Cell Dev Biol 14, 197-230). Под биолюминесценцией не подразумевается так называемое сверхслабое излучение света, которое с помощью чувствительного люминометрического оборудования может быть обнаружено фактически во всех живых структурах (Murphy, M. E. and Sies, H. (1990). Visible-range low-level chemiluminescence in biological systems. Meth. Enzymol. 186, 595-610; Radotic, K, Radenovic, C, Jeremic, M. (1998.) Spontaneous ultra-weak bioluminescence in plants: origin, mechanisms and properties. Gen Physiol Biophys 17, 289-308), и слабое излучение света, которое, наиболее вероятно, не играет какой-либо экологической роли, такое как свечение растущих бамбуковых побегов (Totsune, H., Nakano, M., Inaba, H. (1993). Chemiluminescence from bamboo shoot cut. Biochem. Biophys. Res Comm. 194, 1025-1029) или излучение света в процессе оплодотворения яиц животных (Klebanoff, S. J., Froeder, C. A., Eddy, E. M., Shapiro, B.M. (1979). Metabolic similarities between fertilization and phagocytosis. Conservation of peroxidatic mechanism. J. Exp. Med. 149, 938-953; Schomer, B. and Epel, D. (1998). Redox changes during fertilization and maturation of marine invertebrate eggs. Dev Biol 203, 1-11).
ОПИСАНИЕ ОСОБЫХ ВОПЛОЩЕНИЙ
В данном изобретении представлены композиции нуклеиновой кислоты, кодирующей быстросозревающие флуоресцирующие белки, а также их неагрегирующие варианты (и их мутанты), и кодируемые ею белки. Белки, о которых идет речь, являются флуоресцирующими белками, причем это их свойство возникает в результате взаимодействия двух или более остатков белковой молекулы. Белки согласно изобретению дополнительно характеризуются тем, что в определенных воплощениях они обнаруживаются в составе белков дикого типа или представляют собой мутанты белков дикого типа, которые получены либо из небиолюминесцирующих видов Cnidaria, например Anthozoa, либо получены из видов Anthozoa non-Pennatulacea. В определенных воплощениях белки согласно изобретению представляют собой мутанты флуоресцирующего белка «red» Discosoma sp. дикого типа. Представляют интерес также белки, которые по существу подобны описанным выше специфическим белкам, или их мутанты. Изобретение связано также с фрагментами нуклеиновых кислот и кодируемыми ими пептидами, а также антителами против белков согласно изобретению и трансгенными клетками и организмами. Композиции белков и нуклеиновых кислот согласно изобретению имеют множество разнообразных применений. Наконец, изобретение связано также и с наборами для использования при таких применениях, которые включают в себя, например, композиции нуклеиновых кислот, о которых идет речь.
Прежде чем приступить к дальнейшему описанию объекта изобретения, следует пояснить, что изобретение не ограничивается частными воплощениями настоящего изобретения, описанными ниже, поскольку могут быть созданы варианты частных воплощений, которые также охватываются объемом, защищаемым прилагаемой формулой изобретения. Следует пояснить также и то, что используемая терминология нужна для описания конкретных воплощений, а не для ограничения объема, охватываемого данным изобретением. Наоборот, объем, охватываемый настоящим изобретением, будет определяться прилагаемой формулой изобретения.
В настоящем описании и прилагаемой формуле изобретения формы единственного числа подразумевают также и множественное число, если только в контексте прямо не оговорено иное.
Если не определено иначе, все используемые здесь технические и научные термины имеют такое же значение, которое обычно вкладывают в него специалисты в данной области, к которым и обращено настоящее изобретение.
Когда речь идет об области значений, подразумевается, если в контексте специально не указано обратное, что каждое входящее в эту область значение между верхним и нижним пределами этой области, вплоть до десятой доли нижнего предельного значения, и любое другое заявленное или входящее в эту заявленную область значение охватывается настоящим изобретением. Верхний и нижний пределы этих более малых областей могут независимо быть включены в эти более малые области и также охватываются настоящим изобретением, становясь предметом любого специфически исключенного предела в заявленной области. Когда заявленная область включает в себя одно или более из предельных значений, области, исключающие какое-либо одно или оба из этих включенных предельных значений, также охватываются настоящим изобретением.
Если не указано иначе, все используемые здесь технические и научные термины имеют такое же значение, которое обычно вкладывают в него специалисты в данной области, к которым и обращено настоящее изобретение. Несмотря на то что в осуществлении данного изобретения или для его проверки могут быть использованы любые методы, устройства и материалы, сходные с таковыми, описываемыми здесь, или эквивалентные им, тем не менее предпочтительные методы, устройства и материалы будут здесь описаны.
Все публикации, указанные здесь, включены в настоящее описание путем цитирования с целью описания и раскрытия клеточных линий, векторов, методологий и других компонентов изобретения, которые описаны в публикациях, которые могут быть использованы в связи с описываемым здесь изобретением.
В дальнейшем, описывая настоящее изобретение, в первую очередь будут описаны композиции нуклеиновых кислот согласно изобретению, затем будут обсуждаться белковые композиции, антительные композиции и трансгенные клетки/организмы. Далее следует обзор репрезентативных методов, в которых нашли применение белки согласно изобретению.
КОМПОЗИЦИИ НУКЛЕИНОВЫХ КИСЛОТ
Как было сказано выше, настоящее изобретение обеспечивает композиции нуклеиновых кислот, кодирующие быстросозревающие хромо/флуоропротеины и их мутанты, а также фрагменты и гомологи этих белков. Под быстросозревающим хромо/флуоропротеином подразумевается белок, который является окрашенным и/или флуоресцирующим, например он может проявлять флуоресценцию в низкой, средней или высокой степени при облучении светом с длиной волны возбуждения. Более того, поскольку белок является быстросозревающим, он приобретает свои конечные свойства хромо/флуоресцирующих белков приблизительно менее чем за 72 часа, иногда менее чем за 48 часов, а иногда и менее чем за 24 часа. В определенных воплощениях белок может достичь зрелого состояния за период менее 20 часов, например 18 часов, 16 часов, 14 часов, 12 часов, 10 часов, 8 часов и т.д.
Во всяком случае, представляющими интерес белками согласно изобретению являются такие белки, свойство окрашивания у которых, например, хромо- и/или флуоресцентное свойство, является таким свойством, которое появляется при взаимодействии двух или более остатков белка, но не возникает на основе какого-то одного остатка белка, более конкретно, одной боковой цепи одного остатка белка. Как таковые флуоресцирующие белки согласно изобретению не включают в себя белки, которые проявляют флуоресцентные свойства только из-за тех остатков, которые сами по себе действуют как внутренние флуоресцирующие агенты, например, триптофан, тирозин и фенилаланин. Как таковые флуоресцирующие белки согласно изобретению являются флуоресцирующими белками, флуоресцентное свойство которых возникает на основе некоторой структуры в белке, которая отличается от структуры указанных выше отдельных остатков, например оно возникает при взаимодействии двух или более остатков.
Под композицией нуклеиновой кислоты подразумевается композиция, содержащая последовательность ДНК, имеющая открытую рамку считывания, которая кодирует хромо/флуорополипептиды согласно изобретению, например хромо/флуоропротеиновый ген, и может при определенных условиях экспрессироваться в виде хромо/флуоропротеина согласно изобретению. Кроме того, настоящим изобретением охватываются также нуклеиновые кислоты, которые гомологичны, по существу подобны или идентичны нуклеиновым кислотам согласно изобретению. Таким образом, настоящее изобретение связано с генами и кодируемыми ими последовательностями, которые кодируют белки согласно изобретению, а также их гомологи. Нуклеиновые кислоты согласно изобретению, когда они возникают естественным путем, находятся в непривычном для них окружении (например, они могут быть изолированы, могут присутствовать в повышенной концентрации и т.д.) по сравнению с их природным окружением, например организмом, из которого они получены.
Дополнительно эти нуклеиновые кислоты характеризуются тем, что когда они кодируют белки, либо происходящие от белков, либо являющиеся мутантами белков, которые происходят от (1) небиолюминесцентных видов, часто небиолюминесцентного вида Cnidaria sp., например небиолюминесцентного вида Anthozoa sp.; или (2) от видов Anthozoa sp., которые не принадлежат виду Pennatulacea sp., например, такие, которые не являются морскими перьями. Как таковые нуклеиновые кислоты могут кодировать белки, которые происходят из белков или являются мутантами белков, которые происходят от биолюминесцентных видов Anthozoan sp., при условии, что эти виды не являются Pennatulacea sp., например, что они не являются видами Renilla или Ptilosarca sp. Особый интерес в определенных воплощения представляют собой быстросозревающие мутанты следующих специфических белков дикого типа (или их мутантов): (1) amFP485, cFP484, zFP506, zFP540, drFP585, dsFP484, asFP600, dgFP512, dmFP592, как описано в заявке с серийным номером № 10/006922, описание которой включено в настоящее описание в виде ссылки; (2) hcFP640, как описано в заявке с серийным номером № 09/976673, описание которой включено в настоящее описание в виде ссылки; (3) CgCP, как описано в заявке с серийным номером № 60/255533, описание которой включено в настоящее описание в виде ссылки; и (4) hcriGFP, zoanRFP, scubGFP1, scubGFP2, rfloRFP, rfloGFP, mcavRFP, mcavGFP, cgigGFP, afraGFP, rfloGFP2, mcavGFP2, mannFP, как описано в заявке с серийным номером № 60/332980, описание которой включено в настоящее описание в виде ссылки.
В определенных воплощениях белки, кодируемые нуклеиновыми кислотами согласно изобретению, представляют собой мутанты (drFP585) флуоресцирующего белка "red" дикого типа Discosoma sp., причем кодирующая последовательность нуклеиновой кислоты и аминокислотная последовательность этого белка описаны в заявке с серийным номером № 10/006922, описание которой включено в настоящее описание в виде ссылки. DsRED дикого типа кодируется нуклеиновой кислотой, имеющей следующую последовательность:
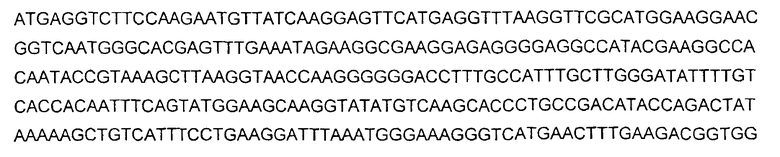
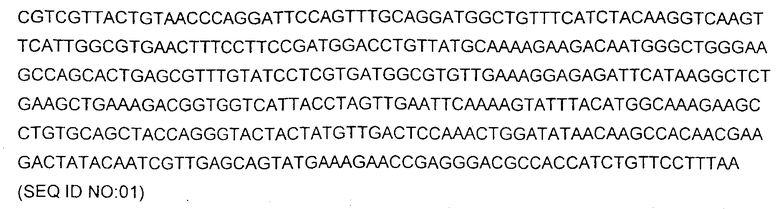
и имеет аминокислотную последовательность:

Представители быстросозревающих мутантов "DsRed" включают в себя, не ограничиваясь ими, точковые мутации в положении 42 относительно стартового остатка, например N42H, N42Q и т.д.; точковые мутации в положении 41 относительно стартового остатка, например, H41L, H41T и т.д.; точковые мутации в положении 44 относительно стартового остатка, например, V44A, и т.д.; точковые мутации в положении 21 относительно стартового остатка, например, T21 S, и т.д.
Одной из последовательностей-представителей последовательности нуклеиновой кислоты согласно изобретению, которая кодирует мутант DsRed.T1, описанный ниже более подробно, включает в себя следующую последовательность:

в которой выделенный/подчеркнутый кодон ATG является стартовым кодоном, а выделенный/подчеркнутый кодон TAG является стоп-кодоном.
Как было сказано ранее, помимо описанных выше быстросозревающих мутантов DsRed, представляют интерес также и быстросозревающие мутанты других видов. Таким мутанты или варианты имеют точковые мутации наподобие тех, которые были описаны выше, в аналогичных или соответствующих положениях в их последовательности относительно специфических положений, идентифицированных в указанных выше представителях мутантов DsRed. Аналогичные или соответствующие положения для образования точковых мутаций в последовательности данного белка легко определяются путем выравнивания прилагаемой специфической последовательности мутантов DsRed и последовательностей белка дикого типа из представляющих интерес видов с зеленым флуоресцирующим белком Aquoria victoria с помощью описанной ниже методики, а также, как показано на Фигуре 1, приведенной у Matz et al., Nature Biotechnology (1999) 969-973. Специфические представители быстросозревающих мутантов других видов включают в себя, не ограничиваясь ими, следующие (где нумерация следующих точковых мутаций соответствует нумерации "GFP", иллюстрируемой на Фигуре 1, приведенной у Matz et al., выше): (1) быстросозревающие мутанты dsFP483, имеющие одну или более точковых мутаций, выбранных из N42, например Q или H, V44, например, A, T21, например быстросозревающие мутанты zFP506, имеющие одну или более точковых мутаций, выбранных из K41, например L или T, I44, например, A, C21, например S; быстросозревающие мутанты aFP538, имеющие одну или более точковых мутаций, выбранных из K41, например Lor T, 144, например A, C21, например S; быстросозревающие мутанты amFP483, имеющие одну или более точковых мутаций, выбранных из C21, например S; и быстросозревающие мутанты cFP484, имеющие одну или более точковых мутаций, выбранных из N21, например S, L44, например A; и т.д.
Помимо описанных выше специфических композиций нуклеиновых кислот, представляют интерес также и гомологи приведенных выше последовательностей. Что касается гомологов нуклеиновых кислот согласно изобретению, источником гомологичных генов могут быть любые виды растений или животных или последовательность может быть полностью или частично синтетической. В определенных воплощениях сходство последовательностей между гомологами приблизительно составляет по меньшей мере 20%, иногда приблизительно составляет по меньшей мере 25%, и может составлять 30%, 35%, 40%, 50%, 60%, 70% или выше, включая 75%, 80%, 85%, 90% и 95% или выше. Сходство последовательностей рассчитывается на основе эталонной (ссылочной) последовательности, которая может представлять собой субпоследовательность более длинной последовательности, такую как консервативный мотив, кодирующая область, фланкирующая область и т.д. Обычно эталонная последовательность составляет в длину по меньшей мере приблизительно 18 нуклеотидов (н.), чаще - по меньшей мере приблизительно 30 н. в длину, а может достигать в длину и полноразмерной последовательности, которая предназначена для сравнения. Алгоритмы, используемые для анализа последовательностей, такие как BLAST, известны в данной области и описаны у Altschul et al. (1990), J. Mol. Biol. 215:403-10 (с использованием параметров по умолчанию, например, параметров w=4 и T=17). Представленные здесь последовательности являются существенными для узнавания родственных и гомологичных нуклеиновых кислот в поисковой базе данных.
Особый интерес в определенных воплощениях представляют нуклеиновые кислоты, имеющие по существу такую же длину, что и нуклеиновые кислоты, идентифицируемые как SEQ ID NO: 01 или 02, где под «по существу такой же длиной» подразумевается то, что какое бы то ни было различие в длине не должно превосходить приблизительно 20 числовых %, обычно не выше, чем приблизительно 10 числовых %, а еще чаще - не выше, чем приблизительно 5 числовых %; и имеют последовательность, идентичную любой из этих последовательностей по меньшей мере приблизительно на 90%, обычно по меньшей мере приблизительно на 95%, а чаще - по меньшей мере приблизительно на 99% относительно полной длины нуклеиновой кислоты. Во многих воплощениях нуклеиновые кислоты имеют последовательность, которая по существу подобна (например, та же самая) или идентична последовательности SEQ ID NO: 01 или 02. Под по существу подобной последовательностью подразумевается то, что идентичность последовательности обычно будет составлять приблизительно по меньшей мере 60%, обычно по меньшей мере приблизительно 75%, а чаще - приблизительно по меньшей мере 80, 85, 90, или даже 95%.
Предусмотрены также нуклеиновые кислоты, которые кодируют белки, кодируемые описанными выше нуклеиновыми кислотами, но отличаются от описанных выше нуклеиновых кислот в связи с вырожденностью генетического кода.
Предусмотрены также нуклеиновые кислоты, которые в жестких условиях гибридизуются с описанной выше нуклеиновой кислотой: примером жестких условий гибридизации является гибридизация при 50°C или выше и 0,1×SSC (15 мМ хлорид натрия/1,5 мМ цитрат натрия). Другим примером жестких условий гибридизации является инкубация в течение ночи при 42°C в растворе, содержащем 50% формамида, 5×SSC (150 мМ хлорид натрия/15 мМ трицитрат натрия), 50 мМ фосфата натрия (pH 7,6), 5 × раствор Денхардта, 10% декстрансульфата и 20 мкг/мл денатурированной, расщепленной ДНК спермы лосося, после чего следует промывание фильтров в 0,1×SSC приблизительно при 65°C. Жесткие условия гибридизации представляют собой такие условия гибридизации, которые являются по меньшей мере столь же жесткими, как описанные выше условия, причем условия можно считать "по меньшей мере такими же жесткими" в том случае, если они по меньшей мере приблизительно на 80% столь же жесткие, обычно по меньшей мере приблизительно на 90% столь же жесткие, насколько жесткими являются описанные выше жесткие условия. Другие жесткие условия гибридизации также известны специалистам в данной области, и эти условия также могут быть использованы для идентификации нуклеиновых кислот конкретно данного воплощения настоящего изобретения.
Предусмотрены также нуклеиновые кислоты, кодирующие мутантные белки согласно изобретению. Мутантные нуклеиновые кислоты могут быть получены в результате неспецифического мутагенеза или направленного мутагенеза с использованием хорошо известных специалистам технологий, которые стали уже рутинными в данной области. В некоторых воплощениях хромопротеины или флуоресцирующие белки, кодируемые нуклеиновыми кислотами, кодирующими гомологи или мутанты, имеют такие же флуоресцентные свойства, что и флуоресцирующие белки дикого типа. В других воплощениях гомологичные или мутантные нуклеиновые кислоты кодируют хромопротеины или флуоресцирующие белки с измененными спектральными свойствами, что описано здесь более подробно.
Одной из категорий мутантов, которые представляют особый интерес, являются неагрегирующие мутанты. Во многих воплощениях неагрегирующие мутанты отличаются от последовательности дикого типа тем, что мутация у них локализована на N-конце, что вызывает модуляцию зарядов, появляющихся на боковых группах N-концевых остатков, например, для реверсии или нейтрализации заряда, таким способом, который можно было бы считать удовлетворительным для продуцирования неагрегирующего мутанта природно возникающего белка или мутанта, причем конкретный белок считается неагрегирующим, если его относят к неагрегирующим белкам на основании анализа, описанного в патентной заявке США под серийным номером № 10/081864, описание которой включено в настоящее описание посредством ссылки, и опубликованного в публикации PCT под номером WO 02/068459.
В некоторых воплощениях нуклеиновые кислоты такого воплощения кодируют неагрегирующие полипептиды, которые in vivo проявляют пониженные агрегационные свойства. "Пониженная агрегация in vivo" относится к пониженной агрегации в клетке. В некоторых воплощениях неагрегирующий полипептид способен к агрегации, которая составляет менее чем приблизительно 90%, менее чем приблизительно 80%, менее чем приблизительно 70%, менее чем приблизительно 60%, менее чем приблизительно 50%, менее чем приблизительно 40%, менее чем приблизительно 30%, менее чем приблизительно 25%, менее чем приблизительно 20%, менее чем приблизительно 15%, менее чем приблизительно 10% или менее чем приблизительно 5% по отношению к степени агрегации, реализуемой его соответствующим аналогом в тех же самых условиях in vivo, например, в другой эукариотической клетке из той же самой линии, в идентичной прокариотической клетке или же в эукариотической клетке или клеточной популяции того же самого типа клеток. Обычно приблизительно менее 60%, приблизительно менее 50%, приблизительно менее 40%, приблизительно менее 30%, приблизительно менее 20%, приблизительно менее 10% или приблизительно менее 5% неагрегирующего полипептида согласно изобретению, присутствующего в клетке или клеточной популяции, является агрегированным.
Способы измерения степени агрегации известны специалистам в данной области; любой из известных способов может быть использован для определения того, обладает ли тот или иной мутант пониженной способностью к агрегации по сравнению с соответствующим агрегирующим аналогом, например, по сравнению с соответствующим агрегирующим полипептидом дикого типа. Такие способы включают в себя, не ограничиваясь ими, гель-электрофорез "псевдонативных" белков; гель-фильтрацию; ультрацентрифугирование; циркулярный дихроизм и светорассеяние. Агрегация может быть измерена по рассеянию света. Для неагрегирующих белков отношение поглощения при более коротких длинах волн к поглощению при более длинной длине волны приближается к нулю. В некоторых воплощениях отношение поглощения неагрегирующего полипептида при 400 нм к поглощению при 566 нм заключено в пределах приблизительно от 0,01 до 0,1, приблизительно от 0,015 до 0,09, приблизительно от 0,02 до 0,08, приблизительно от 0,025 до 0,07 или приблизительно от 0,03 до 0,06.
Во многих воплощениях нуклеиновые кислоты кодируют неагрегирующие быстросозревающие полипептиды, которые имеют аминокислотные последовательности, которые отличаются от соответствующих им последовательностей дикого типа, в результате мутации на N-конце, которая вызывает модуляцию зарядов, появляющихся на боковых группах N-концевых остатков, например, для реверсии или нейтрализации заряда, таким способом, который можно было бы считать удовлетворительным для продуцирования неагрегирующего мутанта природно возникающего белка или его агрегирующего мутанта. Более конкретно, основные остатки, локализованные рядом с N-концевыми остатками белков, замещены, например, остатки лизина или аргинина, примыкающие к N-концу, замещены отрицательно заряженными или нейтральными остатками. Под N-концом подразумевается область, насчитывающая приблизительно 50 остатков от N-конца, часто насчитывающая приблизительно 25 остатков от N-конца, а еще чаще - область, насчитывающая приблизительно 15 остатков от N-конца, причем во многих воплощениях данного изобретения модификации остатков имеют место в области, насчитывающей приблизительно 10 остатков N-конца. Во многих воплощениях изобретения представляющие интерес специфические остатки включают в себя 2, 3, 4, 5, 6, 7, 8, 9 и 10 остатки.
В случае когда белок, кодируемый нуклеиновой кислотой, представляет собой мутант DsRed, как описано выше, представляющие интерес специфические точковые мутации, вызывающие отсутствие агрегации, включают в себя, не ограничиваясь ими, мутации в положении 2, например, R2H, R2L, R2A и т.д.; мутации в положении 5, например, K5E, K5Q, K5M и т.д.; мутации в положении 6, например, N6D и т.д.
Другой категорией мутантов, представляющих особый интерес, являются мутанты с измененной олигомеризацией. Мутант считается относящимся к мутантам с измененной олигомеризацией в том случае, если его способность к олигомеризации отличается от таковой у белка дикого типа. Например, если тот или иной мутант способен к олигомеризации в большей или меньшей степени, чем полипептид дикого типа, он считается олигомеризующимся мутантом. Особый интерес представляют олигомеризующиеся мутанты, которые не олигомеризуются, например являются мономерами при физиологических (например, внутриклеточных) условиях или олигомеризуются в меньшей степени, чем [их аналог] дикого типа, например являются димерами или тримерами во внутриклеточных условиях. Предметом особого интереса являются нуклеиновые кислоты, которые кодируют мономерные варианты быстросозревающих белков согласно изобретению. Одним из представителей мономерного варианта быстросозревающих описанных здесь белков DsRed является мутант под названием mRFP1 (мономерный красный флуоресцирующий белок) который описан у Campbell et al., Proc. Natl. Acad. Sci. USA. 2002 June 11; 99(12):7877-7882. Этот специфический мутант в целом содержит 33 мутации по сравнению с DsRed, 13 из которых являются внутренними по отношению к β-структуре (N42Q, V44A, V71A, K83L, F124L, L150M, K163M, V175A, F177V, S179T, V195T, S1971 и T217A); три из которых являются мутациями, уменьшающими агрегацию, из T1 (R2A, K5E и N6D), три представляют собой интерфейс AB-мутации (1125R, V127T и 1180T), десять представляют собой интерфейс AC-мутации (R153E, H162K, A164R, L174D, Y192A, Y194K, H222S, L223T, F224G и L225A), являются дополнительными полезными мутациями (T21S, H41T, C117E и V156A). Последовательность нуклеиновой кислоты и аминокислотная последовательность этого белка депонированы в Банке Генов под идентификационным номером № AF506027.
Нуклеиновые кислоты согласно изобретению могут представлять собой кДНК или геномную ДНК или ее фрагмент. В определенных воплощениях нуклеиновые кислоты согласно изобретению содержат одну или более открытых рамок считывания, кодирующих специфические флуоресцирующие белки и полипептиды, а также интроны, так же как и смежные 5'- и 3'-некодирующие нуклеотидные последовательности, участвующие в регуляции экспрессии, приблизительно вплоть до более чем на 20 т.н. превышающие кодирующую область, но кроме того, возможно, в том или другом направлении. Нуклеиновые кислоты согласно изобретению могут быть вставлены в соответствующий вектор для внехромосомного пребывания в клетке или же для интеграции в хозяйский геном, как более подробно описано ниже.
Здесь под термином "кДНК" подразумеваются все нуклеиновые кислоты, которые участвуют в аранжировке элементов последовательности, содержащихся в образцах нативной зрелой мРНК, где элементами последовательности являются экзоны и 5'- и 3'-некодирующие области. Обычно образцы мРНК содержат смежные экзоны со включенными в них интронами, которые, если они присутствуют, должны быть удалены путем сплайсинга ядерной РНК для получения открытой рамки считывания, кодирующей белок.
Представляющая интерес геномная последовательность содержит нуклеиновую кислоту, расположенную между инициирующим кодоном и стоп-кодоном, как определено в списке последовательностей, включая все интроны, которые в норме присутствуют в нативной хромосоме. Она может дополнительно содержать 5'- и 3'-нетранслируемые области, обнаруживаемые в зрелой мРНК. Кроме того, она может включать в себя транскрипционную и трансляционную регуляторные последовательности, такие как промоторы, энхансеры и т.д., включая приблизительно 1 т.н., но возможно, и больше, фланкирующей геномной ДНК либо на 5'-, либо на 3'-конце транскрибируемой области. Геномная ДНК может быть изолирована в виде фрагмента длиной 100 т.п.н. или менее; и быть при этом по существу свободной от фланкирующей хромосомной последовательности. Геномная ДНК, фланкирующая кодирующую область, либо 3'-, либо 5'-, или внутренние регуляторные последовательности, обнаруживаемые иногда в интронах, содержат последовательности, необходимые собственно для тканевой и стадиеспецифической экспрессии.
Композиции нуклеиновых кислот согласно изобретению могут полностью или частично кодировать белки согласно изобретению. Двухцепочечные или одноцепочечные фрагменты могут быть получены из последовательности ДНК путем химического синтеза олигонуклеотидов в соответствии с общепринятыми методами путем разрезания рестрикционным ферментом с помощью ПЦР амплификации и т.д. По большей части ДНК-фрагменты будут иметь в длину по меньшей мере приблизительно 15 н., обычно по меньшей мере приблизительно 18 н. или приблизительно 25 н., а могут составлять в длину и по меньшей мере приблизительно 50 н. В некоторых воплощениях молекулы нуклеиновой кислоты согласно изобретению могут составлять в длину по меньшей мере приблизительно 100 н., приблизительно 200 н., приблизительно 300 н., приблизительно 400 н., приблизительно 500 н., приблизительно 600 н., приблизительно 700 н. или приблизительно 720 н. Нуклеиновые кислоты согласно изобретению могут кодировать фрагменты белков согласно изобретению или же полноразмерные белки, например нуклеиновые кислоты согласно изобретению могут кодировать полипептиды, приблизительно содержащие 25 aa, приблизительно 50 aa, приблизительно 75 aa, приблизительно 100 aa, приблизительно 125 aa, приблизительно 150 aa, приблизительно 200 aa, приблизительно 210 aa, приблизительно 220 aa, приблизительно 230 aa или приблизительно 240 aa, вплоть до полноразмерного белка.
Нуклеиновые кислоты согласно изобретению выделены и получены по существу в чистом виде, обычно отличном от того, какими они представлены в интактной хромосоме. Обычно ДНК получают в виде, по существу свободном от других последовательностей нуклеиновой кислоты, которые не содержат нуклеиновую кислоту согласно изобретению или ее фрагмент, обычно составляющем по меньшей мере приблизительно 50%, обычно по меньшей мере приблизительно 90% степень чистоты, и обычно являющейся "рекомбинантной", т. e. фланкированной одним или более нуклеотидами, с которыми в норме она обычно не ассоциирована в составе хромосомы.
Предусмотрены также полинуклеотиды согласно изобретению (например, полинуклеотид, имеющий последовательность SEQ ID NO: 01), соответствующие кДНК, полноразмерный ген и конструкции полинуклеотидов согласно изобретению. Эти молекулы могут быть получены синтетически с помощью целого ряда методик, известных специалистам в данной области. Соответствующие полинуклеотидные конструкции, очищенные с помощью стандартной технологии рекомбинантной ДНК, описаны, например, у Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., (1989) Cold Spring Harbor Press, Cold Spring Harbor, NY, а с помощью развиваемых в настоящее время технологий описаны в разделе HHS по изучению рекомбинантной ДНК в Руководстве Национального Института Здоровья (NIH) США.
Предусмотрены также нуклеиновые кислоты, которые кодируют слитые белки согласно изобретению, или их фрагменты, которые слиты со вторым белком, например деградировавшая последовательность, сигнальный пептид и т.д. Например, представляют интерес слитые белки согласно изобретению с быстро деградирующей последовательностью, такие как описанные в патенте США № 6306600 (описание которого включено сюда в виде ссылки), деградировавший домен мышиной орнитиндекарбоксилазы (MODC), который содержит последовательность PEST. Репрезентативные слитые белки согласно данному воплощению продаются под названием "Destabilized DsRed-Express" фирмой BD Biosciences Clontech (Palo Alto CA). Слитые белки могут содержать полипептид согласно изобретению или его фрагмент, а также неантозойный (non-Anthozoan) полипептид ("партнер слияния"), слитый в пределах рамки считывания на N-конце и/или C-конце полипептида согласно изобретению. Партнеры слияния включают в себя, не ограничиваясь ими, полипептиды, которые могут связывать антитело, специфичное в отношении партнера слияния (например, метки эпитопа); антитела или их связывающие фрагменты; полипептиды, которые обеспечивают каталитическую функцию или индуцируют клеточный ответ; лиганды или рецепторы или их миметики; и тому подобное. В такого рода слитых белках партнер слияния обычно в природе не ассоциирован с частью слитого белка коралловых полипов Anthozoa согласно изобретению, а точнее, чаще не с белком Anthozoa или производным/фрагментом этого белка, т. e. он обычно не обнаруживается в видах Anthozoa.
Предусмотрены также конструкции, содержащие нуклеиновые кислоты согласно изобретению, включенные в вектор, причем такие конструкции могут иметь целый ряд различных применений, включая размножение, продуцирование белка и т.д. Могут быть получены и использованы вирусные и невирусные векторы, включая плазмиды. Выбор вектора будет зависеть от типа клеток, в которых желательно проводить размножение, а также от целей, в которых проводится размножение. Определенные векторы используются для амплификации и наработки нужных количеств требуемой последовательности ДНК. Другие векторы являются подходящими для экспрессии в клетках в культуре. Другие векторы также подходят для переноса и экспрессии в клетках цельного организма животного или человека. Выбор соответствующего вектора специалисту в данной области хорошо известен. Многие из таких векторов являются коммерчески доступными. Для получения конструктов часть полинуклеотида или полноразмерный полинуклеотид вставляется в вектор, обычно путем присоединения с помощью ДНК-лигазы к расщепленному рестрикционным ферментом сайту в векторе. Альтернативно требуемая нуклеотидная последовательность может быть вставлена путем гомологической рекомбинации in vivo. Обычно это выполняется путем присоединения гомологичных областей к вектору на фланкирующих последовательностях требуемого нуклеотида. Гомологичные участки присоединяются путем лигирования олигонуклеотидов или с помощью полимеразной цепной реакции с помощью праймеров, содержащих как гомологичный участок, так и, к примеру, часть требуемой нуклеотидной последовательности. Примеры представляющих интерес специфических векторов включают в себя, не ограничиваясь ими, экспрессирующий вектор pCMV-DsRed; экспрессирующий вектор pDsRED и экспрессирующий вектор pDsRed-Express-1; все они имеются в продаже от компании BD Biosciences Clontech (Palo Alto CA).
Предусмотрены также экспрессирующие кассеты или системы, которые, помимо прочих вариантов использования, находят свое применение в синтезе белков согласно изобретению. Для экспрессии продукт гена, кодируемый полинуклеотидом согласно изобретению, экспрессируется в любой подходящей экспрессирующей системе, включая, к примеру, бактериальную и дрожжевую системы, системы насекомых, амфибий и млекопитающих. Подходящие векторы и хозяйские клетки описаны в патенте США № 5654173. В экспрессирующем векторе полинуклеотид согласно изобретению, например соответствующий последовательностям SEQ ID NO: 01 или 02, соответствующим образом связан с регуляторной последовательностью для достижения необходимых экспрессирующих свойств. Указанные регуляторные последовательности могут включать в себя промоторы (присоединенные либо к 5'-концу смысловой цепи, либо к 3'-концу антисмысловой цепи), энхансеры, терминаторы, операторы, репрессоры и индукторы. Промоторы могут быть регулируемыми или конститутивными. В некоторых ситуациях может оказаться полезным использование промоторов, активных при определенных условиях (кондиционально активных промоторов, таких как тканеспецифические промоторы или промоторы, специфичные в отношении определенной стадии развития. Их соединяют с требуемой нуклеотидной последовательностью с помощью технологий, описанных выше для связывания с векторами. Могут быть использованы любые технологии, известные в данной области. Иными словами, экспрессирующий вектор обеспечит участок инициации транскрипции и трансляции, который может быть индуцибельным или конститутивным, причем кодирующая область под транскрипционным контролем оперативно связана с областью инициации транскрипции и с терминирующей областью транскрипции и трансляции. Указанные контролирующие области могут быть нативными для видов согласно изобретению, из которых получены нуклеиновые кислоты согласно изобретению, или же они могут быть получены из экзогенных источников.
Экспрессирующие векторы, как правило, имеют удобные сайты рестрикции, расположенные рядом с промоторной последовательностью, чтобы обеспечить вставку последовательности нуклеиновой кислоты, кодирующей гетерологичные белки. Может присутствовать селективный маркер, оперативный в экспрессии хозяйской системы. Как сказано выше, экспрессирующие векторы могут быть использованы, помимо всего остального, для продуцирования слитых белков.
Могут быть получены экспрессирующие кассеты, содержащие область инициации транскрипции, ген или его фрагмент, и область терминации транскрипции. Особый интерес представляет применение последовательностей, которые способствуют экспрессии функциональных эпитопов или доменов, обычно насчитывающих в длину по меньшей мере приблизительно 8 аминокислот, чаще по меньшей мере приблизительно 15 аминокислот в длину, приблизительно до 25 аминокислот, и вплоть до полноразмерной открытой рамки считывания гена. После введения ДНК клетки, содержащие конструкцию, могут быть отобраны с помощью селективного маркера, размножены, а затем использованы для экспрессии.
Описанные выше экспрессирующие системы могут быть из соображений удобства использованы как в прокариотах, так и в эукариотах, в зависимости от целей экспрессии. Для широкомасштабного производства белка в качестве экспрессирующей хозяйской клетки может быть использован одноклеточный организм, такой как E.coli, В. subtilis, S. cerevisiae, клетки насекомых в сочетании с бакуловирусными векторами или же клетки высших организмов, таких как позвоночные, например, клетки COS 7, HEK 293, CHO, ооциты Xenopus и т.д. В некоторых ситуациях желательно экспрессировать ген в эукариотических клетках, в которых экспрессированный белок будет приобретать преимущество от нативной пространственной упаковки и посттрансляционных модификаций. Малые пептиды могут быть синтезированы также в лаборатории. Полипептиды, которые имеют частичные последовательности полноразмерной белковой последовательности, могут быть использованы для идентификации и изучения частей функционально важных белков.
Представляющие интерес специфические экспрессирующие системы включают в себя экспрессирующие системы, полученные из бактериальных и дрожжевых клеток, клеток насекомых и клеток млекопитающих. Примеры систем из каждой из указанных категорий приведены ниже:
Бактерии. Экспрессирующие системы в бактериях включают в себя таковые, описанные в публикациях Chang et al., Nature (1978) 275:615; Goeddel et al., Nature (1979) 281:544; Goeddel et al., Nucleic Acids Res. (1980) 8:4057; EP 0036776; патент США № 4551433; De Boer et al., Proc. Natl. Acad. Sci. (USA) (1983) 80:21-25; и Siebenlist et al., Cell (1980) 20:269.
Дрожжи. Экспрессирующие системы в дрожжах включают в себя таковые, описанные в публикациях Hinnen et al., Proc. Natl. Acad.Sci. (USA) (1978) 75:1929; Ito et al., J. Bacteriol. (1983) 153:163; Kurtz et al., Mol. Cell. Biol. (1986) 6:142; Kunze etal., J. Basic Microbiol. (1985) 25:141; Gleeson et al., J. Gen.Microbiol. (1986) 132:3459; Roggenkamp et al., Mol. Gen.Genet. (1986) 202:302; Das et al., J. Bacteriol. (1984) 158:1165; De Louvencourt et al., J. Bacteriol. (1983) 154:737; Van den Berg et al., Bio/Technology (1990) 8:135; Kunze et al., J. Basic Microbiol. (1985) 25:141; Cregg et al., Mol. Cell. Biol. (1985) 5:3376; патенты США №№ 4837148 и 4929555; Beach and Nurse, Nature (1981) 300:706; Davidow et al., Curr. Genet. (1985) 10:380; Gaillardin et al., Curr. Genet. (1985) 10:49; Ballance et al., Biochem. Biophys. Res. Commun. (1983) 112:284-289; Tilburn et al., Gene (1983) 26:205-221; Yelton et al., Proc. Natl. Acad. Sci. (USA) (1984) 81:1470-1474; Kelly and Hynes, EMBO J. (1985) 4:475479; EP 0244234; и международная заявка WO 91/00357.
Клетки насекомых. Экспрессия гетерологичных генов у насекомых производится, как описано в патенте США № 4745051; Friesen et al., "The Regulation of Baculovirus Gene Expression", in: The Molecular Biology Of Baculoviruses (1986) (W. Doerfler, ed.); EP 0127839; EP 0155476; and Vlak et al., J. Gen. Virol. (1988) 69:765-776; Miller et al., Ann. Rev. Microbiol. (1988) 42:177; Carbonell et al., Gene (1988) 73:409; Maeda et al., Nature (1985) 315:592-594; Lebacq-Verheyden et al., Mol. Cell. Biol. (1988) 8:3129; Smith et al., Proc. Natl. Acad. Sci. (USA) (1985) 82:8844; Miyajima et al., Gene (1987) 58:273; and Martin et al., ДНК (1988) 7:99. Множество бакуловирусных штаммов и вариантов и соответствующих пермиссивных хозяйских клеток насекомых из хозяев описано у Luckow et al., Bio/Technology (1988) 6:47-55, Miller et al., Generic Engineering (1986) 8:277-279, и Maeda et al., Nature (1985) 315:592-594.
Клетки млекопитающих. Экспрессия в клетках млекопитающих производится, как описано в публикациях Dijkema et al., EMBO J. (1985) 4:761, Gorman et al., Proc. Natl. Acad. Sci. (USA) (1982) 79:6777, Boshart et al., Cell (1985) 41:521 и патенте США № 4399216. Другие признаки экспрессии в клетках млекопитающих в облегченном варианте представлены в публикациях Ham and Wallace, Meth. Enz. (1979) 58:44, Barnes and Sato, Anal. Biochem. (1980) 102:255, патентах США №№ 4767704, 4657866, 4927762, 4560655, международных заявках WO 90/103430, WO 87/00195 и в переизданном патентном документе США 30985.
При использовании любой из перечисленных выше хозяйских клеток или организмов для репликации и/или экспрессии полинуклеотидов или нуклеиновых кислот согласно изобретению полученные в результате реплицированные нуклеиновая кислота, РНК, экспрессированный белок или полипептид входят в объем защиты настоящего изобретения в качестве продукта хозяйской клетки или организма. Продукт получают любым подходящим способом, известным в данной области.
Как только ген, соответствующий селективному полинуклеотиду, оказывается идентифицированным, его экспрессию можно регулировать в клетке, по отношению к которой этот ген является нативным. Например, экзогенный по отношению к клетке ген можно регулировать с помощью экзогенной регуляторной последовательности, включенной в геном клетки в такой локализации, которая позволит по меньшей мере увеличить экспрессию гена в этой клетке. Регуляторная последовательность может быть сконструирована таким образом, чтобы ее можно было интегрировать в геном путем гомологической рекомбинации, как описано в патентах США №№ 5641670 и 5733761, описание которых включено в настоящее описание в виде ссылки, или же может быть сконструирована таким образом, чтобы ее можно было интегрировать в геном путем негомологической рекомбинации, как описано в международной заявке WO 99/15650, описание которых включено в настоящее описание в виде ссылки. Настоящим изобретением охватывается также продукция белков согласно изобретению как таковых, т.е. чтобы, не обращаясь к кодирующей нуклеиновой кислоте как к таковой, производить вместо этого интеграцию регуляторной последовательности в геном клетки, которая уже содержит ген, кодирующий требуемый белок, как описано выше во включенных в настоящее описание патентных документах.
Предусматриваются также гомологи нуклеиновых кислот согласно изобретению. Гомологи идентифицируются любым из многочисленных способов. Фрагмент предусмотренной кДНК может быть использован в качестве гибридизационного зонда против библиотеки кДНК из представляющего интерес организма-мишени, при этом используются условия низкой жесткости. В качестве зонда можно использовать большой фрагмент или же один или более коротких вырожденных праймеров. Нуклеиновые кислоты, имеющие сходную последовательность, выявляются путем гибридизации в условиях низкой жесткости, например, при 50°C и 6×SSC (0,9 M хлорид натрия/0,09 M цитрат натрия), и остаются связанными при отмывании при 55°C в 1×SSC (0,15 M хлорид натрия/,015 M цитрат натрия). Идентичность последовательности можно определить путем гибридизации в жестких условиях, например при 50°C или выше и 0,1×SSC (15 мМ хлорид натрия/1,5 мМ цитрат натрия). Нуклеиновые кислоты, имеющие область существенной идентичности с предусмотренной последовательностью, например аллельные варианты, генетически измененные версии гена и т.д., связываются с предусмотренной последовательностью в жестких условиях гибридизации. С помощью зондов, в частности, меченых зондов ДНК-последовательностей, можно выделить гомологи или родственные гены.
Представляют интерес также промоторные элементы геномных последовательностей согласно изобретению, где в качестве промоторных элементов может быть использована последовательность 5'-фланкирующей области, включая сайты связывания энхансера, которые, например, обеспечивают регуляцию экспрессии в клетках/тканях, где экспрессируются белковые гены согласно изобретению.
Также предусмотрены малые фрагменты ДНК нуклеиновых кислот согласно изобретению, причем указанные фрагменты используются в качестве праймеров для ПЦР, зондов для гибридизационного скрининга и т.д. Более крупные фрагменты ДНК, например, насчитывающие более 100 н., как описано в предыдущем разделе, используются для продуцирования кодируемого полипептида. Для применения в реакциях геометрической амплификации, таких как геометрическая ПЦР, используется пара праймеров. Точный состав последовательностей праймера в данном изобретении не является критическим, но, как известно в данной области, для большинства применений праймеры должны гибридизоваться с последовательностью согласно изобретению в жестких условиях. Предпочтительно выбирать пару праймеров, которые будут вырабатывать продукт амплификации по меньшей мере размером приблизительно в 50 н., предпочтительно по меньшей мере приблизительно в 100 н. Алгоритмы для отбора последовательностей праймера, как правило, известны и доступны из коммерческих комплектов программного обеспечения. Амплификационные праймеры гибридизуются с комплементарными цепями ДНК и будут для каждой из них служить затравкой.
ДНК может быть использована также для идентификации экспрессии гена в биологическом образце. Способ, которым тестируют клетки на наличие конкретных нуклеотидных последовательностей, таких как геномная ДНК или РНК, достаточно хорошо представлен в литературе. Вкратце, ДНК или мРНК выделяли из клеточных образцов. мРНК может быть амплифицирована с помощью ОТ-ПЦР, с использованием обратной тарнскриптазы для образования комплементарной цепи ДНК, а затем - с помощью полимеразной цепной реакции амплификации, используя праймеры, специфичные в отношении последовательностей ДНК согласно изобретению. Альтернативно, образец мРНК выделяется с помощью гель-электрофореза, переносится на подходящий носитель, например нитроцеллюлозу, нейлон и т.д., а затем тестируется с помощью фрагмента ДНК согласно изобретению в качестве пробы. Могут найти свое применение также и другие технологии, такие как метод лигирования олигонуклеотида, гибридизация in situ и гибридизация с ДНК-зондами на твердом чипе. Выявление мРНК, гибридизующейся с последовательностью согласно изобретению, указывает на экспрессию гена белка Anthozoa в пробе.
Нуклеиновые кислоты согласно изобретению, включая фланкирующие промоторные участки и кодирующие области, могут быть подвергнуты мутированию различными путями, известными в данной области, для получения направленных изменений в интенсивности промотора, последовательности кодируемого белка, свойствах кодируемого белка, включая флуоресцентные свойства кодируемого белка, и т.д. ДНК-последовательность белкового продукта при такой мутации обычно по существу сходна с предусматриваемой здесь последовательностью, например, она может отличаться по меньшей мере одним нуклеотидом или соответственно аминокислотой, и может отличаться по меньшей мере двумя, но не более чем приблизительно десятью нуклеотидами или соответственно аминокислотами. Изменения в последовательности могут быть за счет замен, вставок, делеций или их комбинаций. Кроме того, делеции могут включать в себя более крупные изменения, такие как делеции домена или экзона, например, составляющие в длину порядка 10, 20, 50, 75, 100, 150 или более аминокислотных остатков. Технологии мутагенеза клонированных генов in vitro хорошо известны. Примеры методик сайт-направленного мутагенеза можно найти в публикациях Gustin et al. (1993), Biotechniques 14:22; Barany (1985), Gene 37:111-23; Colicelli et al. (1985), Mol. Gen. Genet. 199:537-9; и Prentki et al. (1984), Gene 29:303-13. Методы сайт-специфического мутагенеза можно найти в публикациях Sambrook et al., Molecular Cloning: A Laboratory Manual, CSH Press 1989, pp. 15.3-15. 108; Weiner et al. (1993), Gene 126:35-41; Sayers et al. (1992), Biotechniques 13:592-6; Jones and Winistorfer (1992), Biotechniques 12:528-30; Barton et al. (1990), NucleicAcids Res 18:7349-55; Marotti and Tomich (1989), Gene Anal.Tech. 6:67-70; и Zhu (1989), Anal Biochem 177:120-4. Такие мутированные производные нуклеиновых кислот могут быть использованы для изучения структурно-функциональных отношений конкретного хромо/флуоресцирующего белка или же для изменения свойств белка, которые влияют на его функцию или регуляцию.
Представляют интерес также гуманизированные варианты нуклеиновой кислоты согласно изобретению. Здесь термин "гуманизированный" относится к изменениям, произведенным в последовательности нуклеиновой кислоты, чтобы оптимизировать кодоны для экспрессии белка в клетках человека (Yang et al., Nucleic Acids Research 24 (1996), 4592-4593). См. также патент США № 5795737, в котором описано гуманизирование белков, описание которого включено в настоящее описание в виде ссылки.
БЕЛКОВЫЕ/ПОЛИПЕПТИДНЫЕ КОМПОЗИЦИИ
Здесь предусмотрены также композиции быстросозревающих хромо- и/или флуоресцирующих белков и их мутантов, а также родственных им полипептидов. Поскольку протеины согласно изобретению представляют собой хромопротеины, они являются окрашенными, цветными белками, которые могут быть флуоресцирующими, слабо флуоресцирующими или не флуоресцирующими. Здесь термин хромопротеин и флуоресцирующий белок не включает в себя люциферазы, такие как люцифераза морского пера Renilla, и относится к любому белку, который пигментирован или окрашен и/или флуоресцирует при облучении светом, например, белым светом или светом со специальной длиной волны (или узким пучком длин волн, таким как длина волны возбуждения). Используемый здесь термин полипептидная композиция относится как к полноразмерному белку, так и к его частям или фрагментам. Этим термином также охватываются вариации природно возникающего белка, причем такие вариации являются гомологичными или существенно сходными с природно возникающим белком, как более подробно описано ниже. Полипептиды согласно изобретению находятся в среде, отличающейся от их природного окружения.
Во многих воплощениях спектр возбуждения белков согласно изобретению обычно колеблется приблизительно от 300 до 700, обычно приблизительно от 350 до 650, а еще чаще - приблизительно от 400 до 600 нм, тогда как спектр излучения белков согласно изобретению как правило, колеблется приблизительно от 400 до 800, обычно приблизительно от 425 до 775, а еще чаще - приблизительно от 450 до 750 нм. Белки согласно изобретению обычно имеют максимальный коэффициент экстинкции, изменяющийся приблизительно от 10000 до 55000, обычно приблизительно от 15000 до 55000. Белки согласно изобретению обычно насчитывают в длину приблизительно от 150 до 300, обычно приблизительно от 200 до 300 аминокислотных остатков, и обычно имеют молекулярную массу приблизительно от 15 до 35 кДа, обычно приблизительно от 17,5 до 32,5 кДа.
В определенных воплощениях белки согласно изобретению являются светящимися, где под светящимися подразумевается то, что хромопротеины и их флуоресцирующие мутанты могут быть обнаружены общепринятыми способами (например, путем визуального скрининга, методом спектрофотометрии, спектрофлуорометрии, флуоресцентной микроскопии, методом FACS-анализа и т.д.) Флуоресцентное свечение отдельных флуоресцирующих белков определяется по их квантовому выходу, помноженному на максимальный коэффициент экстинкции. Свечение хромопротеина может быть выражено через его максимальный коэффициент экстинкции.
В определенных воплощениях белки согласно изобретению очень быстро вслед за их экспрессией в хозяйских клетках образуют свою пространственную упаковку (фолдинг). Под быстрым фолдингом подразумевается, что белки образуют свою третичную (пространственную) структуру, которая в очень короткий промежуток времени порождает их хромо- и/или флуоресцентные свойства. В этих воплощениях время пространственной упаковки белка соответствует периоду, обычно не превышающему примерно 3 дней, обычно не превышающему примерно 2 дней, а чаще не превышающему приблизительно 1 дня.
Специфические белки, вызывающие особый интерес, включают в себя быстросозревающие варианты DsRed, которые созревают по меньшей мере приблизительно в 5 раз быстрее, иногда по меньшей мере приблизительно в 10 раз быстрее, например, по меньшей мере приблизительно в 15 раз быстрее или скорее, чем соответствующий белок DsRed дикого типа. Примеры белков этого специфического воплощения включают в себя такие белки, которые описаны ниже в экспериментальной части описания, например, DsRed.T1, DsRed.T3 и DsRedT4.
Предусмотрены также гомологи белков (или их фрагментов), которые отличаются своей последовательностью от представленных выше аминокислотных последовательностей согласно изобретению. Под гомологами подразумевается белок, имеющий аминокислотную последовательность, по меньшей мере приблизительно на 10%, обычно по меньшей мере приблизительно на 20%, чаще по меньшей мере приблизительно на 30%, а во многих воплощениях по меньшей мере приблизительно на 35%, обычно по меньшей мере приблизительно на 40%, а чаще по меньшей мере приблизительно на 60% идентичную белку согласно изобретению, как определено с помощью алгоритма (clustal algorithm) MegAlign, DNAstar (1998), как описано в публикации D. G. Higgins and P. M. Sharp, "Fast and Sensitive multiple Sequence Alignments on a Microcomputer," (1989) CABIOS, 5:151-153. (Используемыми параметрами являются ktuple 1, gap penalty 3, window, 5 and diagonals saved 5). Во многих воплощениях представляющие интерес гомологи имеют значительно более высокую степень идентичности последовательностей, например 65%, 70%, 75%, 80%, 85%, 90% или выше.
Предусмотрены также белки, которые по существу идентичны специально описанным здесь белкам, где по существу идентичный означает, что белок имеет аминокислотную последовательность, идентичную аминокислотной последовательности белка, с которым проводится сравнение, по меньшей мере приблизительно на 65%, а чаще по меньшей мере приблизительно на 70%, причем в некоторых случаях идентичность может быть намного выше, например 75%, 80%, 85%, 90%, 95% или выше.
Во многих воплощениях гомологи согласно изобретению имеют структурные свойства, обнаруживаемые у предусмотренных выше специфических последовательностей, где такие структурные свойства включают в себя β-складчатую структуру, пространственно упакованную наподобие бочки.
Предусмотрены также белки, которые являются мутантами особо описанных здесь белков. Мутанты могут сохранять биологические свойства белков (например, природно возникшие) дикого типа или же могут иметь биологические свойства, которые отличаются от свойств белков дикого типа. Термин "биологическое свойство" белков согласно изобретению включает в себя, не ограничиваясь этим, спектральные свойства, такие как максимум поглощения, максимум излучения, максимальный коэффициент экстинкции, свечение (например, по сравнению с белком дикого типа или с другим белком, на который ссылаются, таким как зеленый флуоресцирующий белок из A. victoria) и тому подобное; стабильность in vivo и/или in vitro (например, время полу-жизни) и т.д. Мутанты включают в себя одиночные аминокислотные замены, делеции одной или более аминокислот, N-концевой процессинг (усечение), C-концевой процессинг, вставки и т.д.
Мутанты могут быть получены с помощью стандартных технологий молекулярной биологии, например, с помощью неспецифического мутагенеза и направленного мутагенеза. Здесь описано несколько мутантов. Используя руководство, представленное в Примерах, а также с помощью стандартных технологий специалисты в данной области могут с легкостью получить широкий спектр дополнительных мутантов и проверить, изменились ли их биологические свойства.
Например, интенсивность флуоресценции может быть измерена на спектрофотометре при различных длинах волн излучения.
Те белки согласно изобретению, которые являются природно возникшими белками, но находятся в неестественной для них среде, например отделены от их природного окружения. В определенных воплощениях белки согласно изобретению присутствуют в составе композиции, которая обогащена белком согласно изобретению, по сравнению с его природно возникшим окружением. Например, предусмотрен очищенный белок, где под очищенным белком подразумевается то, что белок присутствует в композиции, которая по существу свободна от представляющих интерес нехромо/флуоропротеиновых белков, где под по существу свободной подразумевается, что менее 90%, обычно менее 60%, а чаще менее 50% композиции состоит из представляющих интерес нехромопротеинов или их мутантов. Белки согласно изобретению могут быть представлены также в виде изолята, под которым подразумевается, что этот белок по существу свободен от других белков и других природно возникающих биологических молекул, таких как олигосахариды, полинуклеотиды и их фрагменты, и тому подобное, где термин "по существу свободен" в данном случае означает, что менее 70%, обычно менее 60%, а чаще менее 50% композиции, содержащей изолированный белок, составляет какая-нибудь другая природно возникающая биологическая молекула. В определенных воплощениях белки присутствуют по существу в чистом виде, где под "по существу в чистом виде" подразумевается по меньшей мере 95%, обычно по меньшей мере 97%, а чаще по меньшей мере 99% степень чистоты.
Вдобавок к специально описанным здесь белкам полипептиды, которые отличаются от этих белков, например, описанные выше мутантные белки, также предусмотрены данным изобретением. Обычно такие полипептиды включают в себя аминокислотную последовательность, кодируемую открытой рамкой считывания (ORF) гена, кодирующего белок дикого типа согласно изобретению, включая полноразмерный белок и его фрагменты, в частности биологически активные фрагменты и/или фрагменты, соответствующие функциональным доменам, и тому подобное; и включая слияния полипептидов согласно изобретению с другими белками или их частями. Представляющие интерес фрагменты обычно составляют в длину по меньшей мере приблизительно 10 аминокислотных остатков, обычно по меньшей мере приблизительно 50 аминокислотных остатков в длину, а могут и насчитывать в длину 300 или более аминокислотных остатков, но обычно количество аминокислотных остатков в них не превышает в длину 1000 aa, причем фрагмент будет иметь длину аминокислотной последовательности, идентичную белку согласно изобретению по меньшей мере приблизительно в 10 aa, обычно по меньшей мере приблизительно в 15 aa, а во многих воплощениях будет иметь в длину по меньшей мере приблизительно 50 aa. В некоторых воплощениях полипептиды согласно изобретению составляют в длину приблизительно 25 aa, приблизительно 50 aa, приблизительно 75 aa, приблизительно 100 aa, приблизительно 125 aa, приблизительно 150 aa, приблизительно 200 aa, приблизительно 210 aa, приблизительно 220 aa, приблизительно 230 aa или приблизительно 240 aa, вплоть до длины полного белка. В некоторых воплощениях белковый фрагмент сохраняет все или по существу все биологические свойства белка дикого типа.
Белки и полипептиды согласно изобретению могут быть получены из природно возникающих источников, или же они могут быть продуцированы искусственным путем. Например, белки дикого типа могут быть получены из биологических источников, которые экспрессируют белки, например, из небиолюминесцентных книдарий, например, из видов Anthozoa, таких как специфические виды, перечисленные выше. Белки согласно изобретению могут быть получены также синтетическим путем, например путем экспрессии рекомбинантного гена или нуклеиновой кислоты, кодирующей последовательность, кодирующую представляющий интерес белок в соответствующем хозяине, как описано выше. Могут быть использованы любые подходящие методы очистки белка, причем подходящие методологии очистки белка описаны в Руководстве по Очистке Белков (Deuthser ed.) (Academic Press, 1990). Например, из исходного источника может быть получен лизат, который затем будет очищен с помощью HPLC, хроматографии исключенного размера, гель-электрофореза, аффинной хроматографии и т.п.
КОМПОЗИЦИИ АНТИТЕЛ
Предусмотрены также антитела, которые специфически связываются с флуоресцирующими белками согласно изобретению. Соответствующие антитела получены путем иммунизации животного-хозяина пептидами, содержащими полностью или частично белок согласно изобретению. Соответствующие животные-хозяева включают в себя мышь, крысу, овцу, козу, хомячка, кролика и т.д. Источником белкового иммуногена обычно служат виды книдарии (Cnidaria), в частности, небиолюминесцентные виды книдарии, такие как виды Anthozoa или виды Anthozoa non-Petalucea. Животное-хозяин обычно должно принадлежать к другому виду, нежели тот, из которого получен иммуноген, например это могут быть мыши, и т.д.
Иммуноген может включать в себя полноразмерный белок или его фрагменты или производные. Предпочтительные иммуногены содержат белок полностью или частично, причем эти остатки содержат посттрансляционные модификации, в нативном белке-мишени. Иммуногены получают разными путями, известными в данной области, например, путем экспрессии клонированных генов с помощью обычных рекомбинантных технологий, выделения из исходных видов Anthozoa и т.д.
Для получения поликлональных антител первой стадией является иммунизация животного-хозяина белком-мишенью, где белок-мишень предпочтительно должен быть по существу в чистом виде, содержащем менее чем приблизительно 1% примесей. Иммуноген может содержать полный белок-мишень или его фрагменты или производные. Для повышения иммунного ответа животного-хозяина белок-мишень можно комбинировать с адъювантом, причем подходящие адъюванты включают в себя квасцы, декстран, сульфат, крупные полимерные анионы, водно-масляные эмульсии, например адъювант Фрейнда, полный адъювант Фрейнда, и тому подобное. Белок-мишень может быть конъюгирован также с искусственными белками-носителями или искусственными антигенами. Для получения моноклональных антител можно иммунизировать различных хозяев. Такие хозяева включают в себя кроликов, морских свинок, грызунов, например мышей, крыс, овец, коз и т.д. Белки-мишени вводят в организм хозяина обычно внутрикожно сначала в первичной дозировке, а затем в виде одной или более, обычно по меньшей мере двух дополнительных бустерных дозировок. После иммунизации собирают кровь животного-хозяина, затем сыворотку отделяют от клеток крови. Иммуноглобулин, присутствующий в результирующей антисыворотке, может быть в дальнейшем фракционирован с помощью известных методов, таких как фракционирование солью аммония, DEAE-хроматография и тому подобное.
Моноклональные антитела получают общепринятыми методами. Обычно в качестве источника плазматических клеток берут селезенку и/или лимфатические узлы иммунизированного животного-хозяина. Плазматические клетки иммортализуют путем слияния с клетками миеломы для получения гибридомных клеток. Культуральный супернатант от индивидуальных гибридом подвергают скринингу с помощью стандартных технологий для идентификации тех из них, которые продуцируют антитела требуемой специфичности. Подходящие животные для продуцирования моноклональных антител против человеческих белков включают в себя мышь, крысу, хомячка и т.д.
Для производства антител против мышиного белка в качестве животных обычно выбирают хомяка, морскую свинку, кролика и т.д. Антитело из супернатантов гибридомных клеток или асцитной жидкости может быть очищено с помощью общепринятых технологий, например аффинной хроматографии, с использованием белка, связанного с нерастворимым субстратом, протеин A сефарозы и т.д.
Вместо обычной мультимерной цепи антитело может быть получено в виде одинарной цепи. Одноцепочечные антитела описаны в публикациях Jost et al. (1994) J. B. C. 269:26267-73, и других. ДНК-последовательности, кодирующие вариабельную область тяжелой цепи и вариабельную область легкой цепи, лигируют со спейсером, кодирующим по меньшей мере приблизительно 4 аминокислоты из числа малых нейтральных аминокислот, включая глицин и/или серин. Белок, кодируемый в результате такого слияния, позволяет осуществиться сборке функциональной вариабельной области, которая сохраняет специфичность и аффинность исходного антитела.
В определенных воплощениях представляют интерес также гуманизированные антитела. Способы гуманизирования антител хорошо известны специалистам. Гуманизированное антитело может являться продуктом, полученным у животного, содержащего гены константной области трансгенного иммуноглобулина человека (см., например, международные патентные заявки WO 90/10077 и WO90/04036). Альтернативно представляющее интерес антитело может быть получено генно-инженерным путем с использованием технологий рекомбинантной ДНК с целью замены CH1, CH2, CH3, шарнирных доменов и/или домена рамки считывания соответствующей человеческой последовательностью (см. WO 92/02190).
Применение кДНК иммуноглобулина для конструкции химерных иммуноглобулиновых генов известно в данной области (Liu et al. (1987) P. N. A. S. 84:3439 и (1987) J. Immunol. 139:3521). мРНК выделяют из гибридомы или другой клетки, продуцирующей антитело, и используют для продуцирования кДНК. Представляющая интерес кДНК может быть амплифицирована с помощью полимеразной цепной реакции с использованием специфических праймеров (патенты США №№ 4683195 и 4683202). Альтернативно создается библиотека и осуществляется скрининг с целью выделения представляющей интерес последовательности. Последовательность ДНК, кодирующая вариабельную область антитела, затем сливают с последовательностью константной области человека. Последовательности генов константных областей человека можно найти у Kabat et al. (1991) Sequences of Proteins of Immunological Interest, N. I. H. публикация № 91-3242. Гены C-области человека легко можно получить из известных клонов. Выбор изотипа должен быть подсказан требуемыми эффекторными функциями, такими как фиксация комплемента или активность антитело-зависимой клеточной цитотоксичности. Предпочтительными изотипами являются IgG1, IgG3 и IgG4. Может быть использована любая из константных областей легкой цепи человека, каппа или лямбда. Затем химерное гуманизированное антитело экспрессируют общепринятыми методами.
Фрагменты антитела, такие как Fv, F(ab')2 и Fab, могут быть получены путем расщепления интактного белка, например, под действием протеиназы или путем химического расщепления. Альтернативно, конструируется укороченный ген. Например, чтобы получить усеченную молекулу, химерный ген, кодирующий часть F(ab')2-фрагмента, должен включать в себя последовательности ДНК, кодирующие домен CH1 и шарнирную часть цепи H, а затем трансляционный стоп-кодон.
Консенсусные последовательности J-районов H- и L-цепей могут быть использованы для конструирования олигонуклеотидов для их применения в качестве праймеров для введения эффективных сайтов рестрикции в J-район для последующего связывания сегментов V-района с человеческими сегментами C-района. кДНК C-района может быть модифицирована сайт-направленным мутагенезом, чтобы разместить рестрикционный сайт в аналогичном положении в человеческой последовательности.
Экспрессирующие векторы включают в себя плазмиды, ретровирусы, YACs, эписомы, полученные из EBV, и тому подобное. Подходящим вектором является такой вектор, который кодирует функционально полноценную CH- или CL-последовательность иммуноглобулина человека, с соответствующими рестрикционными сайтами, сконструированными таким образом, чтобы любая VH- или VL-последовательность могла быть легко встроена и экспрессирована: в таких векторах сплайсинг обычно имеет место между донорным сайтом сплайсинга в представляющем интерес J-районе и акцепторным сайтом сплайсинга, предшествующим человеческому C-району, а также в таких областях сплайсинга, которые имеют место в человеческих CH-экзонах. Полиаденилирование и терминация транскрипции в нативных хромосомах имеют место в сайтах ниже кодирующих областей. Полученное в результате химерное антитело может быть присоединено к любому сильному промотору, включая ретровирусные LTR (длинные повторы РНК), например, раннему промотору SV-40, (Okayama et al. (1983) Mol. Cell. Bio. 3:280), LTR вируса саркомы Рауса (Gorman et al. (1982) P. N. A. S. 79:6777), и LTR вируса мышиного лейкоза Молони (Grosschedl et al. (1985) Cell 41:885); нативные Ig-промоторы, и т.д.
ТРАНСГЕННЫЕ ОРГАНИЗМЫ
Нуклеиновые кислоты согласно изобретению могут быть использованы для выработки трансгенных растений и животных нечеловеческой природы или сайт-специфических генных модификаций в клеточных линиях. Трансгенные клетки согласно изобретению включают в себя одну или более нуклеиновых кислот согласно изобретению, присутствующих в качестве трансгена, причем это определение включает в себя родительские клетки, трансформированные с целью включения в них трансгена, и их потомство. Во многих воплощениях трансгенными клетками являются клетки, которые в норме не образуют скопления или содержат нуклеиновую кислоту согласно изобретению. В таких воплощениях, когда трансгенные клетки от природы содержат нуклеиновые кислоты согласно изобретению, нуклеиновая кислота будет присутствовать в клетке в ином положении, нежели то положение, которое соответствует ее природной локализации, т.e. интегрирована в геномный материал клетки с необычной для нее локализацией. Трансгенные животные могут быть получены путем гомологической рекомбинации, при которой изменен эндогенный локус. Альтернативно конструкция нуклеиновой кислоты случайным образом интегрирована в геном. Векторы для стабильной интеграции включают в себя плазмиды, ретровирусы и другие вирусы животных, YACs и тому подобное.
Трансгенные организмы согласно изобретению включают в себя клетки и многоклеточные организмы, например растения и животные, которые являются эндогенными «нокаутами», у которых экспрессия эндогенного гена по меньшей мере редуцирована, если не исключена полностью. Представляющие интерес трансгенные организмы включают в себя клетки и многоклеточные организмы, например, растения и животные, в которых белок или его варианты экспрессируются в клетках или тканях, в которых они в норме не экспрессированы, и/или экспрессируются на таком уровне, который в норме в этих тканях не встречается.
ДНК-конструкции для гомологичной рекомбинации будут содержать по меньшей мере часть гена согласно изобретению, причем ген содержит нужную генетическую модификацию (модификации) и включает в себя области, гомологичные локусу-мишени. Для случайной интеграции ДНК-конструкции не обязательно должны для опосредования рекомбинации включать в себя гомологичные участки. Для удобства включаются маркеры положительной и отрицательной селекции. Способы генерации клеток, имеющих нацеленные генные модификации, гомологической рекомбинации известны в данной области. По поводу различных технологий клеток млекопитающих см. Keown et al. (1990), Meth. Enzymol. 185:527-537.
Для эмбриональных стволовых клеток (ES-клетки) может быть использована клеточная линия ES, или же эмбриональные клетки могут быть получены в свежем виде у хозяина, например, у мыши, крысы, морской свинки и т.д. Такие клетки выращивают на соответствующем фидерном слое фибробластной подложки или выращивают в присутствии фактора, ингибирующего лейкемию (LIF). Когда клетки ES или эмбриональные клетки трансформированы, их можно использовать для продуцирования трансгенных животных. После трансформации клетки наносятся на фидерный слой в соответствующей среде. Клетки, содержащие конструкцию, могут быть выявлены с использованием селективной среды. По прошествии времени, необходимого для роста колоний, их собирают и анализируют на предмет образования гомологичной рекомбинации или интеграции конструкции. Те колонии, которые являются позитивными, могут быть затем использованы для эмбриоманипуляций и инъекции бластоциста. Бластоцисты получают из 4-6-недельных суперовулированных самок. ES-клетки обрабатывают трипсином, и модифицированные клетки инъецируют в бластоцель бластоциста. После инъекции бластоцисты возвращают в каждый из рогов матки псевдобеременных самок. Затем самки донашивают плод, и полученное в результате потомство подвергают скринингу на предмет наличия конструкции. Обеспечивая различный фенотип бластоциста и генетически модифицированных клеток, можно с легкостью обнаружить химерное потомство.
Химерных животных подвергают скринингу на предмет наличия модифицированного гена, и имеющие модификации самцы и самки спариваются с целью получения гомозиготного потомства. В случае если генетические изменения на той или иной стадии развития приводят к летальному исходу, ткани или органы могут быть сохранены в качестве аллогенных или конгенных протезов или трансплантатов, или же могут быть использованы в культуре in vitro. Трансгенное животное может быть любым млекопитающим, не являющимся человеком, таким как лабораторные животные, домашние животные и т.д. Трансгенные животные могут быть использованы для функциональных исследований, для скрининга лекарственных средств и т.д. Репрезентативные примеры использования трансгенных животных включают в себя те, которые описаны ниже.
Подобным же способом могут быть получены трансгенные растения. Способы получения трансгенных растительных клеток и растений описаны в патентах США №№ 5767367; 5750870; 5739409; 5689049; 5689045; 5674731; 5656466; 5633155; 5629470; 5595896; 5576198; 5538879; 5484956; описания всех этих патентов включены в настоящее описание в виде ссылки. Обзор способов продуцирования трансгенных растений приведен также в публикации Plant Biochemistry and Molecular Biology (eds Lea & Leegood, John Wiley & Sons) (1993) pp.275-295. Вкратце, в зависимости от природы видов растений, собирают подходящие клетки или ткани растения. В определенных случаях выделяют протопласты как таковые, причем такие протопласты могут быть выделены из разных тканей различных растений, например листьев, гипокотиля, корня и т.д. Для выделения протопласта собранные клетки инкубируют в присутствии целлюлаз для разрушения клеточной стенки, причем конкретные условия инкубирования варьируют в зависимости от типа растения и/или ткани, из которой получены клетки. Затем полученные протопласты отделяют от полученного клеточного дебриса путем просеивания и центрифугирования. Вместо протопластов для получения трансгенного хозяина могут быть использованы эмбриогенные эксплантаты, содержащие соматические клетки. После сбора клеток или тканей представляющая интерес экзогенная ДНК вводится в клетки растений, причем для такого введения может быть использован целый ряд различных доступных технологий. В случае выделенных протопластов создается благоприятная возможность использования методик ДНК-опосредованного переноса гена, включающих в себя инкубацию протопластов в присутствии оголенной ДНК, например, плазмид, содержащих представляющую интерес экзогенную кодирующую последовательность, в присутствии поливалентных катионов, например PEG или PLO; и электропорацию протопластов в присутствии оголенной ДНК, содержащей представляющую интерес экзогенную последовательность. Затем протопласты, которые успешно поглотили экзогенную ДНК, отбирают, выращивают в каллюсе, и в конце концов в трансгенном растении в контакте с соответствующими количествами стимулирующих факторов, взятых в определенных соотношениях, например ауксинов и цитокининов. В случае эмбриогенных эксплантатов подходящим способом введения экзогенной ДНК в соматические клетки-мишени является способ с применением метода ускорения частиц, или метода "генной пушки". Затем полученные эксплантаты выращивают в химерных растениях, скрещивают и получают трансгенное потомство. Помимо описанных выше подходов с использованием оголенной ДНК существует и другой удобный способ продуцирования трансгенных растений, связанный с трансформацией, опосредованной Agrobacterium. С помощью опосредованной Agrobacterium трансформации получают коинтегральные или бинарные векторы, содержащие экзогенную ДНК, которые затем вводятся в соответствующий штамм Agrobacterium, например, A. tumefaciens. Затем полученные в результате бактерии совместно инкубируют с полученными протопластами или тканевыми эксплантатами, например листьями, лиственными дисками, и получают каллус. Затем каллус выращивают в селективных условиях, отбирают и подвергают воздействию ростовой среды для индукции корневой системы и роста побегов, чтобы в конце концов получить трансгенное растение.
ПРИМЕНИМОСТЬ
Хромопротеины согласно изобретению и их флуоресцирующие мутанты находят целый ряд самых разных практических применений, причем эти применения обязательно отличаются, в зависимости от того, является ли белок хромопротеином или флуоресцирующим белком. Соответствующие применения каждого из этих типов белков будут описаны ниже, причем описанные ниже варианты применений носят скорее репрезентативный характер и никоим образом не предназначены для ограничения области применения белков согласно изобретению примерами применений, описанными ниже.
Хромопротеины
Хромопротеины согласно изобретению имеют самые разные применения. Одним из интересных применений белков согласно изобретению в качестве окрашивающих агентов, которые способны придать окраску или пигментирование конкретной композиции согласно изобретению. Особенно интересными в определенных воплощениях являются нетоксичные хромопротеины. Хромопротеины согласно изобретению могут быть включены во множество различных композиций согласно изобретению, где типичные композиции согласно изобретению включают в себя пищевые композиции, фармацевтические композиции, косметические композиции, живые организмы, например животные и растения, и тому подобное. При использовании в качестве окрашивающего агента или пигмента в композицию согласно изобретению включают достаточное количество хромопротеина, чтобы придать ей нужную степень окрашивания или пигментирования. Хромопротеин может быть включен в композицию согласно изобретению в соответствии с любой из известных методик, причем любая конкретно используемая методика будет обязательно зависеть, во всяком случае отчасти, от природы композиции согласно изобретению, которая подлежит окрашиванию.
Методики, которые могут быть использованы, включают в себя, не ограничиваясь перечисленным, смешивание, диффузию, растирание, распыление, впрыскивание, вколачивание и тому подобное.
Хромопротеины могут также найти применение в качестве меток, детектируемых в анализируемом веществе, например, при анализе представляющих интерес биологических веществ. Например, хромопротеины могут быть включены в продукты присоединения к анализируемым специфическим антителам или их связывающим фрагментам, а следовательно, могут быть использованы в иммуноанализах представляющих интерес анализируемых веществ в комплексном образце, как описано в патенте США № 4302536, описание которого включено в настоящее описание в виде ссылки. Вместо антител или их связывающих фрагментов хромопротеины или их хромогенные фрагменты согласно изобретению могут быть конъюгированы с лигандами, которые могут специфически связываться с представляющим интерес исследуемым веществом или иными фрагментами, факторами роста, гормонами и тому подобное, что с легкостью определяется любым специалистом в данной области.
В других воплощениях хромопротеины согласно изобретению могут быть использованы в качестве селектируемых маркеров в применениях в области рекомбинантных ДНК, например, при продуцировании трансгенных клеток и организмов, как описано выше. Можно создать как таковую пропись методики создания конкретного трансгенного продукта, в которой экспрессия хромопротеинов согласно изобретению будет использована в качестве селектируемого маркера, причем либо методики для случая, когда создание продукта прошло успешно, либо методики для случая, когда создание продукта прошло неуспешно. Таким образом, появление окраски хромопротеина согласно изобретению в фенотипе трансгенного организма, продуцируемого конкретным способом, может быть использовано для подтверждения того, что полученный этим способом конкретный организм включил в себя представляющий интерес трансген, чаще всего интегрируемый таким образом, что обеспечивает экспрессию трансгена в этом организме. При использовании селектируемого маркера нуклеиновая кислота, кодирующая хромопротеин согласно изобретению, может быть использована в способе получения трансгенного организма, причем этот способ более подробно описан выше. Конкретные представляющие интерес трансгенные организмы, где в качестве селектируемых маркеров могут быть использованы белки согласно изобретению, включают в себя трансгенные растения, животных, бактерии, грибы и тому подобное.
В других воплощениях хромопротеины (и флуоресцирующие белки) согласно изобретению находят свое применение в солнцезащитных фильтрах в качестве селективных фильтров, и т.д., по аналогии со способами применения белков, описанными в международной заявке WO 00/46233.
Флуоресцирующие белки
Флуоресцирующие белки согласно изобретению (так же как и другие описанные выше компоненты согласно изобретению) находят множество разнообразных применений, причем такие применения включают в себя, не ограничиваясь перечисленным, следующие применения. Первым интересным применением является применение белков согласно изобретению в приложении к переносу энергии флуоресцентного резонанса (FRET). В этих приложениях белки согласно изобретению служат в качестве донора и/или акцепторов в комбинации со вторым флуоресцирующим белком или красителем, например флуоресцирующим белком, как описано у Matz et al., Nature Biotechnology (October 1999) 17:969-973, зеленым флуоресцирующим белком из эквореи вида Aequoria victoria или ее флуоресцирующим мутантом, например, как описано в патентах США №№ 6066476; 6020192; 5985577; 5976796; 5968750; 5968738; 5958713; 5919445; 5874304, описание которых включено в настоящее описание в виде ссылок, другие флуоресцирующие красители, например кумарин и его производные, например 7-амино-4-метилкумарин, аминокумарин, красители bodipy, такие как Bodipy FL, cascade blue, флуоресцеин и его производные, например флуоресцеинизотиоцианат, Oregon green, родаминовые красители, например texas red, тетраметилродамин, эозины и эритрозины, цианиновые красители, например Cy3 и Cy5, макроциклические хелаты лантанидных ионов, например, quantum dye, и т.д., хемилюминесцентные красители, например люциферазы, включая и те, которые описаны в патентах США №№ 5843746; 5700673; 5674713; 5618722; 5418155; 5330906; 5229285; 5221623; 5182202, описание которых включено в настоящее описание в виде ссылок. Специфические примеры того, где могут быть использованы FRET-анализы с использованием флуоресцирующих белков согласно изобретению, включают в себя, не ограничиваясь перечисленным, детектирование белок-белковых взаимодействий, например двугибридную систему млекопитающих (двугибридные системы млекопитающих), димеризацию фактора транскрипции, мультимеризацию мембранных белков, образование мультибелковых комплексов и т.д., в качестве биосенсора целого ряда различных явлений, где пептид или белок ковалентно связывается с флуоресцентной комбинацией FRET, включающей в себя флуоресцирующие белки согласно изобретению и связывающий пептид или белок, который представляет собой, например, специфический протеиназный субстрат, например, для опосредованного каспазой расщепления, линкер, который подвергается конформационному изменению под действием воспринимаемого сигнала, который увеличивает или уменьшает FRET, например, регуляторный домен PKA (сенсор цАМФ), фосфорилирования, например, когда в линкере имеется сайт фосфорилирования или когда линкер имеет специфичность связывания с фосфорилированным/дефосфорилированным доменом другого белка, или когда линкер имеет Ca2+-связывающий домен. Типичные применения переноса энергии флуоресцентного резонанса, или FRET, в которых могут быть использованы белки согласно изобретению, включают в себя, не ограничиваясь перечисленным, те из них, которые описаны в патентах США №№ 6008373; 5998146; 5981200; 5945526; 5945283; 5911952; 5869255; 5866336; 5863727; 5728528; 5707804; 5688648; 5439797 описания которых включены в настоящее описание в виде ссылок.
Флуоресцирующие белки согласно изобретению находят также применение в качестве биосенсоров в прокариотических и эукариотических клетках, например, в качестве индикатора связывания иона Ca2+; в качестве pH-индикатора, в качестве индикатора фосфорилирования, в качестве индикатора других ионов, например, магния, натрия, калия, хлора и галогенов. Например, для детектирования иона Ca белки, содержащие мотив EF-hand, известны своей способностью при связывании Ca2+ транслоцироваться из цитозоля на мембраны. Эти белки содержат миристоильную группу, которая скрыта внутри молекулы гидрофобными взаимодействиями с другими участками белка. Связывание Ca2+ индуцирует конформационное изменение, в результате которого миристоильная группа обнажается и может включиться в липидный бислой (так называемое "Ca2+-переключение миристоила"). Слияние такого EF-hand-содержащего белка с флуоресцирующими белками (FP) может превратить его в индикатор внутриклеточного Ca2+ при условии мониторинга переноса из цитозоля на плазматическую мембрану с помощью конфокальной микроскопии. Белки, содержащие мотив EF-hand, подходяшие для применения в такой системе, включают в себя, не ограничиваясь перечисленным, рековерин (1-3), кальцинейрин B, тропонин C, визинин, нейрокальцин, кальмодулин, парвальбумин и прочие. Для pH может быть использована система на основе гисактофилинов. Гисактофилины представляют собой миристоилированные богатые гистидином белки, которые, как известно, содержатся у Dictyostelium. Их связывание с актином и кислыми липидами является четко pH-зависимым процессом в рамках изменений цитоплазматических значений pH. В живых клетках связывание с мембраной, как полагают, приводит к нарушению взаимодействия гисактофилинов с актиновыми филаментами. При pH ≤6,5 они локализованы на плазматической мембране и ядре. Наоборот, при pH 7,5 они четко распределены по всему цитоплазматическому пространству. Такое изменение распределения является обратимым и приписывается гистидиновым кластерам, экспонированным в петлях на поверхности молекулы. Обратимость внутриклеточного распределения в области изменений цитоплазматических значений pH согласуется со значениями pK 6,5 гистидиновых остатков. Клеточное распределение не зависит от миристоилирования белка. При слиянии флуоресцирующих белков (FP) с гисактофилином внутриклеточное распределение слитого белка можно проследить с помощью лазерного сканирования, конфокальной микроскопии или стандартной флуоресцентной микроскопии. Количественный анализ флуоресценции может быть предпринят с помощью чресклеточных линейных сканеров (конфокальная микроскопия лазерного сканирования) или данных других электронных анализов (например, с помощью программного обеспечения metamorph (Universal Imaging Corp) и усреднения данных, полученных на уровне клеточной популяции. Реальное pH-зависимое перераспределение слитого комплекса гисактофилин-FP из цитозоля на плазматическую мембрану происходит в течение 1-2 мин и достигает стабильного уровня через 5-10 мин. Обратная реакция происходит примерно в течение такого же промежутка времени. Как таковой слитый белок гисактофилин-флуоресцирующий белок, который действует аналогичным образом, может быть использован для мониторинга в реальном времени изменений pH в цитозоле в живых клетках млекопитающих. Такие методы применяются в анализах при скрининге большого количества пропускаемого материала, например, при измерении изменений pH как вследствие активации рецептора фактора роста (например, эпителиального или фактора роста, полученного из тромбоцитов), стимуляции хемотаксиса/движения клеток, при детектировании изменений внутриклеточных показателей pH как вторичного мессенджера, в мониторинге внутриклеточного pH в экспериментах, связанных с манипулированием pH, и так далее. Для детектирования активности PKC в репортерной системе используется тот факт, что молекула, называемая MARCKS (миристоилированный аланин-обогащенный субстрат киназы C), представляет собой субстрат PKC. Она заякорена на плазматической мембране в результате миристоилирования и растягивания положительно заряженных аминокислот (ED-домен), которые связываются с отрицательно заряженной плазматической мембраной посредством электростатических взаимодействий. При активации PKC ED-домен становится фосфорилированным под действием PKC, становясь, таким образом, отрицательно заряженным, и, как следствие электростатического отталкивания, MARCKS транслоцируется с плазматической мембраны в цитоплазму (так называемое "электростатическое переключение миристоила"). Слияние N-конца MARCKS, распространяющегося от миристоилированного мотива до ED-домена MARCKS, с флуоресцирующими белками согласно изобретению создает указанную выше детекторную систему для выявления активности PKC. При фосфорилировании под действием PKC слитый белок транслоцируется с плазматической мембраны в цитозоль. Указанная транслокация отслеживается с помощью стандартной флуоресцентной микроскопии или конфокальной микроскопии, например, с использованием технологии Cellomics или других систем скрининга высокого разрешения (например, Universal Imaging Corp./Becton Dickinson). Указанная выше репортерная система применяется в скрининге высокого разрешения, например, скрининге ингибиторов PKC, и, как индикатор активности PKC, во многих вариантах применения для скрининга потенциальных реагентов, создающих помехи этому пути передачи сигнала. Методы использования флуоресцирующих белков в качестве биосенсоров включают в себя также методы, описанные в патентах США №№ 972638; 5824485 и 5650135 (так же как и в ссылках, цитируемых выше), описание которых включено в настоящее описание в виде ссылок.
Флуоресцирующие белки согласно изобретению находят также применение в областях, включающих в себя автоматизированный скрининг элементов матрицы, экспрессирующих флуоресцирующие репортерные группы, с помощью микроскопического изображения и электронного анализа. Скрининг может быть использован для изучения лекарственных средств и в области функциональных геномов, например, когда белки согласно изобретению используются в качестве маркеров цельных клеток для детектирования изменений в мультиклеточной реорганизации и миграции, например, образования мультиклеточных трубочек (формирование кровеносных сосудов) эндотелиальными клетками, миграции клеток через систему Fluoroblok Insert System (Becton Dickinson Co.), ранозаживления, отрастания нейрона и т.д.; когда белки используются как маркеры, слитые с пептидами (например, нацеливающие последовательности), и белки, которые дают возможность детектирования изменения внутриклеточной локализации как индикатора клеточной активности, например, передачи сигнала, такого как транслокация киназы и фактора транскрипции под действием стимула, такого как протеинкиназа C, протеинкиназа A, фактор транскрипции NFkB, и NFAT; белки клеточного цикла, такие как циклин A, циклин B1 и циклин E; отщепление протеиназой с последующим передвижением отщепленного субстрата, фосфолипиды, с маркерами внутриклеточных структур, такими как эндоплазматический ретикулум, аппарат Гольджи, митохондрии, пероксисомы, ядро, ядрышко, плазматическая мембрана, гистоны, эндосомы, лизосомы, микротрубочки, актин) как средства для скрининга высокого разрешения: co-локализация других флуоресцирующих слитых белков с этими маркерами локализации в качестве индикаторов движений внутриклеточных флуоресцирующих слитых белков/пептидов или только в качестве маркеров; и тому подобное. Примеры применений, включающие в себя автоматизированный скрининг элементов матрицы, в котором находят применение флуоресцирующие белки согласно изобретению, включают в себя те, которые описаны в патенте США № 5989835; а также в международных заявках WO/0017624; WO 00/26408; WO 00/17643 и WO 00/03246, описание которых включено в настоящее описание в виде ссылок.
Флуоресцирующие белки согласно изобретению также находят применение в анализах при скрининге большого количества пропускаемого материала. Флуоресцирующие белки согласно изобретению являются стабильными белками со временем полужизни, составляющим более 24 ч. Предусмотрены также дестабилизированные варианты флуоресцирующих белков согласно изобретению с укороченными временами полужизни, которые могут быть использованы в качестве репортеров транскрипции при изучении лекарственных средств. Например, белок согласно изобретению может быть слит с предполагаемой протеолитической сигнальной последовательностью, полученной из белка с укороченным временем полужизни, например, последовательностью PEST из мышиного гена орнитиндекарбоксилазы, мышиным боксом деструкции циклина B1 и убиквитином, и т.д. Описание дестабилизированных белков и векторов, которые могут быть использованы для продуцирования перечисленного, см., например, в патенте США № 6130313, описание которого включено в настоящее описание в виде ссылки. Промоторы в путях передачи сигнала могут быть детектированы с помощью дестабилизированных версий флуоресцирующих белков согласно изобретению для скрининга лекарственных средств, например AP1, NFAT, NFkB, Smad, STAT, p53, E2F, Rb, myc, CRE, ER, GR и TRE и тому подобное.
Белки согласно изобретению могут быть использованы в качестве детекторов вторичных мессенджеров, например, путем слияния белков согласно изобретению со специфическими доменами, например, гамма-PKC Ca-связывающим доменом, гамма-PKC DAG-связывающим доменом, SH2-доменом и SH3-доменом, и т.д.
Секретируемые формы белков согласно изобретению могут быть получены, например, путем слияния секретируемых лидируемых последовательностей с белками согласно изобретению для конструирования секретируемых форм белков согласно изобретению, которые, в свою очередь, могут быть использованы для целого ряда различных применений.
Белки согласно изобретению находят также применение в методах сортинга клеток, активированных флуоресценцией. При таких применениях флуоресцирующий белок согласно изобретению используется в качестве метки для маркировки популяции клеток, и затем полученная в результате меченая популяция клеток сортируется с помощью известного в данной области устройства для сортинга клеток, активированных флуоресценцией. FACS-методы описаны в патентах США №№ 5968738 и 5804387, описания которых включены в настоящее описание в виде ссылок.
Белки согласно изобретению находят также применение в качестве маркеров у животных in vivo (например, трансгенных животных). Например, экспрессию белков согласно изобретению можно направлять с помощью специфических тканевых промоторов, причем такие методы находят применение при разработке генной терапии, например, помимо прочих применений, при тестировании эффективности трансгенной экспрессии. Типичное применение флуоресцирующих белков в трансгенных животных, которое иллюстрирует этот класс применений белков согласно изобретению, можно найти в международной заявке WO 00/02997, описание которой включено в настоящее описание в виде ссылки.
Дополнительные применения белков согласно изобретению включают в себя следующие: в качестве маркеров последующей инъекции в клетки или животных и в калибровке для количественных измерений (флуоресценции и белка); в качестве маркеров или репортеров в биосенсорных кислородных устройствах для мониторинга жизнеспособности клеток; в качестве маркеров или меток для животных, домашних животных-любимцев, игрушек, пищи и т.д.
Флуоресцирующие белки согласно изобретению находят также применение при изучении протеолитического расщепления. Например, флуоресцентные анализы инактивации при расщеплении можно развивать с помощью белков согласно изобретению, где белки согласно изобретению сконструированы таким образом, чтобы они включали в себя специфическую последовательность, расщепляемую протеиназой, и при этом чтобы флуоресцирующие свойства белков не нарушались. При расщеплении флуоресцирующего белка под действием активированной протеиназы флуоресценция будет резко уменьшаться в результате разрушения функционального хромофора. Альтернативно, активированная в результате расщепления флуоресценция может развиваться с помощью белков согласно изобретению, где белки согласно изобретению сконструированы таким образом, чтобы они включали в себя дополнительную спейсерную последовательность, в непосредственной близости/или внутри хромофора. В этом варианте флуоресцирующая активность будет существенно снижена, поскольку части функционального хромофора будут разделены спейсером. Спейсер будет состоять из двух идентичных сайтов специфического расщепления протеиназой. При расщеплении под действием активированной протеиназы спейсер окажется вырезанным, и две остаточные "субъединицы" флуоресцирующего белка будут готовы вновь соединиться с образованием функционального флуоресцирующего белка. Оба из описанных выше типов применения могут быть развиты при изучении целого ряда различных типов протеиназ, например каспаз, и т.д.
Белки согласно изобретению могут быть использованы в анализах по определению фосфолипидной композиции в биологических мембранах. Например, слитые белки белков согласно изобретению (или любой другой вид ковалентной или нековалентной модификации белков согласно изобретению), которые дают возможность связывания со специфическими фосфолипидами для локализации/визуализации характера распределения фосфолипидов в биологических мембранах, также допуская со-локализацию мембранных белков в специфических фосфолипидных скоплениях, могут быть получены с помощью белков согласно изобретению. Например, PH-домен GRP1 имеет высокое сродство к фосфатидилинозиттрифосфату (PIP3), но не к PIP2. Как таковой, слитый белок между PH-доменом GRP1 и белками согласно изобретению может быть сконструирован таким образом, чтобы специфически метить области биологических мембран, обогащенные PIP3.
Другим применением белков согласно изобретению является использование их в качестве флуоресцирующего таймера, в котором переключение одного флуоресцентного цвета на другой (например, с зеленого на красный), сопутствующее старению флуоресцирующего белка, используется для определения активации/инактивации экспрессии гена, например, развивающейся экспрессии гена, зависящей от клеточного цикла экспрессии гена, экспрессии гена, специфической в отношении циркадных ритмов, и тому подобное.
Антитела согласно изобретению, описанные выше, также находят целый ряд применений, включая дифференцирование белков согласно изобретению от остальных флуоресцирующих белков.
НАБОРЫ
Согласно изобретению предусмотрены также наборы для использования при воспроизводстве одного или более из описанных выше применений, причем наборы согласно изобретению обычно включают в себя элементы для создания белков согласно изобретению, например, конструкцию, содержащую вектор, который включает в себя кодирующую область белка согласно изобретению. Компоненты набора согласно изобретению обычно представлены в подходящей для хранения среде, например, забуференном растворе, обычно в соответствующем контейнере. В наборах согласно изобретению могут быть также предусмотрены и антитела против представленного белка. В определенных воплощениях набор содержит множество различных векторов, каждый из которых кодирует белок согласно изобретению, причем векторы сконструированы для экспрессии в различных средах и/или при различных условиях, например, для конститутивной экспрессии, когда вектор включает в себя сильный промотор для экспрессии в клетках млекопитающих, или же вектор без промотора, со множественным клонирующим сайтом для обычной инсерции промотора и расширенной экспрессии, и т.д.
Помимо указанных выше компонентов наборы согласно изобретению будут дополнительно содержать инструкции для осуществления способов согласно изобретению. Эти инструкции могут сопутствовать наборам согласно изобретению в различных формах, одна или более из которых могут быть включены конкретно в тот или иной набор. Одна из форм, в которой могут быть представлены указанные инструкции, представляет собой информацию, отпечатанную на соответствующей среде или субстрате, например куске или кусках бумаги, на которых напечатана эта информация, сверху на упаковке набора, внутри набора на упаковках вложенных в него компонентов, и т.д. Другим способом представления является информация, которая может быть расположена на вебсайте, по адресу которого можно через Интернет выйти на отдельный сайт, на котором эта информация представлена. В наборах может быть использован любой удобный способ предоставления необходимой информации.
Следующие примеры представлены в целях иллюстрации настоящего изобретения, но ни в коем случае не для его ограничения.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Введение
Красный флуоресцирующий белок DsRed имеет спектральные свойства, которые являются идеальными для постановки двухцветных экспериментов с зеленым флуоресцирующим белком (GFP). Однако белок DsRed дикого типа имеет целый ряд недостатков, включая медленное созревание хромофора и плохую растворимость. Для того, чтобы избежать медленное созревание, авторами изобретения был использован неспецифический и направленный мутагенез для создания вариантов DsRed, которые созревают в 10-15 раз быстрее, чем белок дикого типа. Замена аспарагина на глутамин в положении 42 значительно ускоряет созревание DsRed, однако при этом повышает также уровень излучения света в зеленой части спектра. Дополнительные аминокислотные замены тормозят указанную эмиссию в зеленой части спектра, дополнительно ускоряя созревание. Для повышения растворимости DsRed результирующий заряд вблизи N-конца белка был уменьшен. Полученные в результате варианты DsRed давали яркое флуоресцентное свечение даже в быстрорастущих организмах, таких как дрожжи.
II. Методика эксперимента
А. Мутагенез и скрининг.
Для мутагенеза мутантный ген DsRed или ген дикого типа, находящийся в векторе pDsRed1-N1 (Clontech, Palo Alto, CA), вырезали с помощью Nhel и Hpal и использовали в качестве матрицы для ПЦР с помощью полимеразы, склонной к ошибкам (Cadwell, R. C. & Joyce, G. F. In PCR Primer. A laboratory manual. (eds. Dieffenbach, C. W. & Dveksler, G. S.) 583-589 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; 1995)). Амплифицированный продукт подвергали расщеплению с помощью BamHl и BsaBl, гель-фильтрации и лигировали между сайтами BamHI и Ec113611 экспрессирующего вектора pQE31 (Qiagen, Valencia, CA), который кодирует N-концевую гексагистидиновую метку. Библиотеку мутированных генов DsRed трансформировали в штамм DH10B E.coli. За 1-3 цикла мутагенеза осуществили скрининг 50000-100000 колоний на предмет наличия флуоресцентного свечения с использованием описанной в тексте установки проектора слайдов. За четвертый цикл было собрано 4000 ярко флуоресцирующих колоний, которые были размещены в лунках 96-луночных планшетов, где их максимально выращивали, затем подвергали лизису с помощью реагента B-PER II (Pierce, Rockford, IL), а потом центрифугировали в течение 5 мин при 2500×g. Супернатанты переносили в другую партию 96-луночных планшетов, и сигналы флуоресценции, исходящие из осадков и супернатантов, визуально сравнивали, используя метод анализа с помощью проектора слайдов. Клоны, в которых было повышено отношение флуоресценции растворимой фракции к фракции осадка, изучали далее. За пятый цикл с трансформационных планшетов было получено 10 пулов по 10000 мутантных клонов в каждом, они были подвержены клеточному сортингу на проточном цитометре Becton Dickinson FACStarPlus. Сигналы флуоресценции измеряли одновременно в зеленой (FL-1) и красной (FL-2) областях и собирали клетки, которые флуоресцировали строго в красной области спектра и при этом имели ослабленную флуоресценцию в зеленой части спектра.
B. Очистка и спектральный анализ вариантов DsRed.
Для очистки белков, меченных гексагистидином, ген флуоресцирующего белка в векторе pQE31 трансформировали в клетках E. coli, несущих репрессорную плазмиду pREP4 (Qiagen). 250 мл культуры выращивали до показателя оптической плотности OD 600, соответствующего 0,5, а затем индуцировали с помощью 1 мМ изопропил-β-D-тиогалактазида (IPTG) в течение 6-8 ч при 37°C. Клетки лизировали, используя 10 мл B-PER II, и центрифугировали в течение 20 мин при 27000×g. Детергент удаляли из супернатанта путем добавления NaCl до 300 мМ, затем центрифугировали в течение 10 мин при 2500×g. После добавления 1 мл Ni2+-NTA-агарозных бусинок (Qiagen) содержимое пробирки перемешивали в течение 1 часа на установке, позволяющей переворачивать пробирку вверх дном. После этого бусинки трижды промывали 10 мл 300 мМ NaCl, 20 мМ имидазол-HCl, pH 7,4, 0,5% Тритон X-100, а затем трижды промывали таким же буфером, но без добавок Тритона X-100. Флуоресцирующий белок элюировали с использованием 2,5 мл 300 мМ имидазол-HCl, pH 7,4, и подвергали диализу в 50 мМ Na+-HEPES, pH 7,5, 100 мМ NaCl, 1 мМ EDTA.
Откорректированные спектры возбуждения и излучения очищенных вариантов DsRed, разбавленных до A558 <0,04 в Na+-HEPES, pH 7,5, 100 мМ NaCl, 1 мМ EDTA, были получены с помощью спектрофлуориметра Horiba FluoroMax-3. Сканирующие щели соответствовали 1 нМ. Излучение измеряли при 600 нм в спектре возбуждения, а максимум возбуждения наблюдался при 470 нм в спектре излучения. Для определения коэффициентов экстинкции определяли концентрации флуоресцирующего белка с использованием метода BCA (Pierce), а поглощение белков в точках, соответствующих максимуму их излучения, измеряли с помощью спектрофотометра Spectronic Unicam GENESYS 10 UV. Квантовые выходы определяли в соответствии с известными публикациями (Baird, et al., Proc. Natl. Acad. Sci. USA 97, 11984-11989 (2000); Lakowicz, J. R. Principles of fluorescence spectroscopy, Edn. 2. (Kluwer Academic/Plenum Publishers, New York, NY; 1999)) с использованием этанольного родамина 101 в качестве свидетеля; при этих измерениях длина волны возбуждения составляла 535 нм, а флуоресцентное излучение суммарно наблюдалось от 550 до 800 нм.
C. Измерение кинетик созревания.
Гены, кодирующие варианты DsRed, клонировали в экспрессирующий вектор pQE81 (Qiagen) и трансформировали в E.coli. Бактериальные культуры, растущие в условиях аэрации при 37°C, индуцировали с помощью 1 мМ IPTG в течение 30 мин для получения импульса экспрессии для каждого варианта DsRed. Затем инициировали чейз (отслеживание метки) путем ингибирования белкового синтеза под действием смеси 170 мкг/мл хлорамфеникола, 30 мкг/мл канамицина и 50 мкг/мл тетрациклина. В предопределенных временных точках отбирали аликвоты культур, доводили в них концентрацию глицерина до 15%, и замораживали при -80°C. Позже эти аликвоты подвергали быстрому оттаиванию и с использованием проточной цитометрии на установке Becton Dickinson FACScan определяли среднюю интенсивность красной флуоресценции (канал FL-2) на клетку. Часть каждой из аликвот осаждали с помощью трихлоруксусной кислоты, а затем подвергали SDS-электрофорезу на полиакриламидном геле и иммуноблоттингу в присутствии анти-гексагистидинового моноклонального антитела (Qiagen) для измерения общего количества полипептидов DsRed в культурах. Флуоресцентная микроскопия дрожжей. Использовали плазмиду CEN, полученную из pRS315 (Sikorski, Genetics 122, 19-27 (1989)) и несущую слитый ген pCox4-DsRed1 под контролем промотора ADH1. Производные этой плазмиды были созданы путем замены кодирующей последовательности DsRed1 кодирующей последовательностью DsRed.T3 или DsRed.T4. Эти плазмиды были внесены в штамм S. cerevisiae BGY101, который содержит хромосомный SEC7-EGFPx3 ген 20. Клетки полученных дрожжевых штаммов выращивали на минимальной среде с глюкозой и фиксировали, а затем получали проецируемые изображения флуоресценции, как описано в публикации Rossanese et al., J. Cell Biol. 145, 69-81 (1999).
III. Результаты и обсуждение
Описано семейство флуоресцирующих белков. Наиболее полезным из этих вновь открытых белков является белок DsRed, который получен из коралла Discosoma. DsRed флуоресцирует в оранжево-красной области с максимумом интенсивности излучения при 583 нм. В результате биофизических исследований и исследований с помощью рентгеновской кристаллографии выявлено, что DsRed образует стабильный тетрамер и что каждый мономер структурно весьма сходен с GFP. Смещенная в красную сторону спектра флуоресценция белка DsRed сходна с результатами на GFP, полученными из хромофора с более обширно конъюгированной π-системой. Флуоресценция DsRed имеет оптимум при длине волны возбуждения 558 нм, но может быть также возбуждена и под действием стандартного лазерного излучения при 488 нм, что дает возможность использовать DsRed в конфокальных лазерных микроскопах и в проточных цитометрах. DsRed является фотостабильным и имеет высокий квантовый выход. Указанные характеристики делают DsRed идеальным кандидатом, с помощью которого могут быть получены изображения флуоресценции, в частности, в разноцветных экспериментах с участием в них зеленого флуоресцирующего белка (GFP) и его вариантов. В настоящее время кодон-оптимизированная версия DsRed доступна под названием DsRed1.
Несмотря на все эти преимущества белок DsRed дикого типа имеет ряд недостатков с точки зрения возможностей его применения в качестве флуоресцирующего репортера. Когда DsRed слит с другим белком, тетрамеризация домена DsRed может вызвать изменение функции и локализации этого белка. Тетрамер DsRed способен также к самоассоциации с образованием агрегатов более высокой степени организации. Возможно, наиболее серьезной проблемой, связанной с белком DsRed, является медленное созревание хромофора, с периодом полужизни >24 ч при комнатной температуре. Новосинтезированные DsRed развивают притушенную зеленую флуоресценцию при формировании такого же самого хромофора, что и в белке GFP. Затем в результате реакции вторичного окисления вырабатывается красный хромофор. Такое медленное созревание используется в варианте DsRed, называемом "флуоресцентным таймером", в котором исходная видовая флуоресценция повышена. Однако для большинства применений медленное созревание DsRed является нежелательным признаком. В изображениях GFP с двойной меткой первоначальная зеленая флуоресценция белка DsRed продуцирует проступание в канал GFP. Чаще медленное созревание красной флуоресценции ограничивает интенсивность сигнала DsRed, в частности, в быстро растущих организмах, таких как дрожжи. Вариант, обозначаемый как DsRed2, созревает быстрее, чем DsRed1, но DsRed2 все еще требует многих часов для развития полноценной флуоресценции. Здесь использованы приемы неспецифического и направленного мутагенеза для создания улучшенных вариантов DsRed. Эти новые варианты быстро созревают, а кроме того, они являются более растворимыми, чем DsRed дикого типа.
Для идентификации быстросозревающих вариантов DsRed модифицировали ранее существующий способ визуализации флуоресценции GFP в колониях микроорганизмов. Меченный гексагистидином DsRed в больших количествах продуцировали в Escherichia coli. Флуоресценцию бактериальных колоний возбуждали, помещая их в полосу света 520±20 нм, получаемую при размещении фильтра над линзами слайдового проектора, и через очки, покрытые фильтром Kodak Wratten № 22, который пропускает длины волн >550 нм, детектировали излучение. Такая технология проста и эффективна.
Библиотеку мутантных экпрессирующих плазмид получали с помощью метода ПЦР с помощью полимеразы, склонной к ошибкам, для амплификации матрицы DsRed1. Эта библиотека была трансформирована в E. coli, и было обследовано свыше 100000 трансформированных колоний. Колониям, продуцирующим белок DsRed1 дикого типа, требовалось два дня для развития значительного уровня флуоресценции, однако в трех мутантных колониях развивалась интенсивная флуоресценция уже после одного дня роста. Путем секвенирования выяснили, что эти три мутантных плазмиды отличались друг от друга, но что все они содержали замену кодона N42H. Таким образом, авторы изобретения создали вариант, который имеет только одну замену N42H.
Вариант N42H был очищен параллельно с DsRed1, и оба белка были подвергнуты анализу методом спектрофлуориметрии. Как наблюдали ранее, спектры очищенных DsRed1 изменялись по прошествии нескольких дней после созревания белка (данные не приведены). И наоборот, спектры очищенного варианта N42H оставались стабильными во времени (данные не приведены) вследствие быстрого созревания. К сожалению, вдобавок к ускоренному созреванию, замена N42H способствовала изменению спектральных свойств зрелого белка (Фиг.1A). Зрелый DsRed1, по-видимому, представлял собой равновесную смесь молекул красной флуоресценции и некоторого числа молекул зеленой флуоресценции, которая спектрально подобна GFP. GFP-подобные виды излучают синий свет с максимумом при длине волны приблизительно 480 нм, а излучают зеленый свет с максимумом при длине волны приблизительно 500 нм; но DsRed является тетрамером, поэтому возбуждение зеленых молекул часто приводит к явлению переноса энергии флуоресцентного резонанса (FRET) на близлежащие красные молекулы с образованием красного излучения. Этот эффект FRET наряду с относительно низким содержанием в зрелом DsRed1 зеленых молекул приводит к образованию очень слабого пика зеленого излучения по сравнению с красным излучением (Фиг.1A). В варианте N42H максимумы возбуждения синего света и излучения зеленого света оказались исключительно повышенными (Фиг.1A), что указывает на смещение равновесия, такое, при котором содержание зрелых молекул, несущих зеленый хромофор, повышено.
Поскольку замена N42H существенно увеличивает размер боковой цепи, была произведена попытка получить также более консервативную замену N42Q. Эта мутация требовала замены двух оснований, и вероятнее всего, должна была бы отсутствовать в исходной коллекции мутантов. Вариант N42Q сохранял свойство быстрого созревания варианта N42H, однако имел значительно менее выраженные максимумы возбуждения синего света и излучения зеленого света (Фиг.1A). Вариант N42Q был, таким образом, выбран в качестве отправной точки для дальнейшего исследования.
Дополнительный мутагенез (см. ниже) привел к образованию вариантов DsRed, которые отличались даже более быстрым созреванием и более низким максимумом излучения зеленого света, чем исходный вариант N42Q. Через шесть циклов мутагенеза были отобраны три оптимизированных варианта, которые были обозначены как DsRed.T1, DsRed.T3 и DsRed.T4 (Таблица 1). Спектральные свойства DsRed.T4 (Фиг.1B) фактически идентичны таковым белка DsRed.T1 (данные не приведены) и очень сходны с таковыми у белка DsRed1 дикого типа (Фиг.1A). По сравнению с DsRed.T1 и DsRed.T4, белок DsRed.T3 несколько ярче (см. ниже), однако имеет значительно более высокий максимум возбуждения синего света и предельно высокий пик излучения зеленого света (Фиг.1 B).
Оптимизированные варианты DsRed были изучены in vivo и in vitro. Судя по флуоресценции колонии, размеру колонии и стабильности плазмиды, эти варианты оказались менее токсичными в отношении E.coli, чем DsRed1, и в них развивалась флуоресценция более эффективно при повышении температуры до 37°C и выше (данные не приведены). Подобно белку DsRed дикого типа, оптимизированные варианты, скорее всего, представляют собой тетрамер: в них имел место FRET между зелеными и красными молекулами (Фиг.1B), и при SDS-PAGE в неденатурирующих условиях они мигрировали наподобие тетрамеров (см. ниже). После очистки DsRed1 авторы изобретения измерили его коэффициент экстинкции, который составил 52000 M-1 см-1, и квантовый выход, который приблизительно составил 0,7 (Таблица 1).
Свойства зрелых вариантов DsRedа
(ч)с
1
2
T1
T3
T4
DsRed2: R2A, K5E, K9T, V105A, 1161T, S197A.
DsRed.T1: P(-4)L, R2A, K5E, N6D, T21S, H41T, N42Q, V44A, C117S, T217A.
DsRed.T3: P(-4)L, R2A, K5E, N6D, T21S, H41T, N42Q, V44A, A145P.
DsRed.T4: P(-4)L, R2A, K5E, N6D, T21S, H41T, N42Q, V44A, A145P, T217A.
b Яркость определяется коэффициентом экстинкции и квантовым выходом.
Относительную яркость вычисляют, принимая яркость DsRed1 за 1,00.
c Полувремена созревания устанавливали графически, используя методику эксперимента, результаты которого приведены на Фигуре 2. Приведенные значения представляют собой средние значения по результатам двух отдельных экспериментов; значения, полученные для каждого варианта DsRed в этих двух экспериментах, отличались друг от друга не более чем на 15%.
В прежнем исследовании DsRed дикого типа сообщалось о сходном квантовом выходе, но о более высоком коэффициенте экстинкции порядка 75000 М-1 см-1, причем причина такого расхождения непонятна. DsRed2 обнаруживает некоторое уменьшение как коэффициента экстинкции, так и квантового выхода, что выражается в значении относительной яркости 0,68 по сравнению с DsRed1 (Таблица 1). Яркость DsRed.T3 почти такая же, как и яркость DsRed1. Однако DsRed.T1 и DsRed.T4 представляют собой димеры с относительными яркостями 0,36-0,38 по отношению к DsRed1. Чтобы рассчитать кинетики созревания вариантов DsRed, был предпринят пульс-чейз-анализ in vivo на культурах E. coli, растущих при 37°C (Фиг.2). Через 30 мин после индукции импульсов добавляли ингибиторы белкового синтеза, и культуральные образцы отбирали при различных временах слежения за меткой. Среднюю клеточную флуоресценцию для каждого образца измеряли с помощью проточной цитометрии, используя для возбуждения лазерное излучение с длиной волны 488 нм.
На фигуре 2A показаны исходные данные, в то время как на фигуре 2B показаны нормализованные данные, т.е. отнесенные к значению максимальной флуоресценции, принятой за 100%. При этих условиях DsRed1 имеет полупериод созревания приблизительно 11 ч, хотя точные измерения в случае DsRed1 и затруднены, потому что значения флуоресценции не выходят на плато (Фиг.2), а также потому что белок DsRed1 подвергается деградации в течение периода слежения за меткой (данные не приведены). Иногда DsRed2 созревает немного быстрее с полупериодом созревания, приблизительно составляющим 6,5 ч. Скорости развития флуоресценции DsRed1 и DsRed2 увеличиваются после четко выраженного лаг-периода, что указывает на то, что в этом процессе участвует множество медленных стадий. Созревание DsRed.T3 происходит с коротким лаг-периодом, и период его полужизни приблизительно составляет 1,3 ч.
DsRed.T4 и DsRed.T1 созревают с неопределяемым лаг-периодом и с периодами полужизни порядка 0,7 ч приблизительно в 15 раз быстрее, чем DsRed1 (Фиг.2, данные не показаны). При такой методике пульс-чейза у различных вариантов DsRed воспроизводимо получали в области плато разные значения средней клеточной флуоресценции (Фиг.2A). Высокий сигнал у DsRed.T3 может быть объяснен относительно сильным возбуждением этого белка при 488 нм (см. Фиг.1 B). Все три варианта - DsRed 1, DsRed2 и DsRed.T4 - имеют сходные спектры флуоресценции, только плато флуоресценции белка DsRed.T4-экспрессирующих клеток в 4 раза выше, чем у DsRed1-экспрессирующих клеток, и в 10 раз выше, чем у DsRed2-экспрессирующих клеток. Этот результат является неожиданным, потому что очищенный dsRed.T4 менее яркий, чем DsRed1 или DsRed2 (Таблица 1). Авторы предполагают, что незрелый DsRed1 является нестабильным в E. coli и что нестабильность этого белка усугубляется белком DsRed2, так что большая часть новосинтезированных молекул DsRed1 и DsRed2 теряется в процессе агрегации и/или дезагрегации. Такое объяснение подтверждается предшествующим исследованием, в котором сообщалось, что большинство новосинтезированных молекул DsRed1 в клетках E. coli или Drosophila подвергаются деградации. Интересно, что в клетках млекопитающих DsRed2 испускает более яркий сигнал флуоресцентного излучения, чем DsRed1, что дает основания предполагать, что эффективность экспрессии того или иного варианта DsRed может зависеть от типа клеток.
Преимущества ускоренного созревания должны быть очевидны, в частности, в тех случаях, когда варианты DsRed продуцируются в быстрорастущих организмах. Для проверки такого предположения авторы изобретения нацелили различные варианты DsRed на митохондрии дрожжей. Родительские дрожжевые штаммы также содержали EGFP-меченный маркер для цистерн Гольджи. В клетках из растущих культур с нацеленным на митохондрии DsRed1 флуоресценция резко тускнела и становилась отчетливо заметной только в субпопуляции этих клеток, как только культуры достигали стационарной фазы роста (данные не приведены). И наоборот, нацеленный на митохондрии DsRed.T4 постоянно испускал сильный флуоресцирующий сигнал в клетках из растущих культур (Фиг.3). Как показано на картинке слияния, авторы изобретения не наблюдали сколько-нибудь заметного проступания флуоресценции DsRed.T4 в зеленую часть спектра, а флуоресценции EGFP - в красную часть. Подобные результаты были получены с нацеленным на митохондрии DsRed.T3 (данные не показаны). Однако в случае других слитых конструкций было обнаружено, что когда большое количество DsRed.T3 концентрировалось в малом объеме внутри клетки, наблюдалось некоторое проступание в зеленую область (не показано). Следовательно, DsRed.T4 относится к такому разряду белков, которые позволяют получить строгое разделение сигналов красной и зеленой флуоресценции.
Эти результаты подтверждают, что неспецифический мутагенез с последующим скринингом представляет собой превосходный метод для создания улучшенных флуоресцирующих белков. Ключевым открытием в данном изобретении явилось открытие того, что замены Asn42, такие как N42Q, исключительно ускоряют формирование хромофора.
Побочным эффектом замен Asn42 является четко выраженное увеличение поглощения возбуждающего синего света и излучение (испускание) зеленого света (Фиг.1A). Зрелый DsRed дикого типа, скорее всего, представляет собой равновесную смесь красных видов и зеленых видов, а замены Asn42, очевидно, смещают равновесие в сторону увеличения выхода зеленых видов. Производя серию дополнительных замен в исходном 42Q, авторам изобретения удалось подавить почти полностью поглощение синего света и испускание зеленого, что обусловливалось N42Q при предохранении от быстрого созревания (Фиг.1 и Таблица 1).
Другое усовершенствование свойств DsRed дикого типа было достигнуто путем уменьшения результирующего заряда вблизи N-конца. У полученных в результате вариантов DsRed была выявлена сниженная агрегация in vitro (см. ниже) и in vivo. DsRed дикого типа является необычно для него основным, с предсказанным pI порядка 8,0, и возможно, неспецифически ассоциированным с отрицательно заряженными клеточными компонентами. Кроме того, основные «пэтчи» на поверхности тетрамера DsRed могут взаимодействовать с кислотными «пэтчами» на втором тетрамере, вызывая агрегацию более высокого порядка. Такое взаимодействие DsRed с другими макромолекулами, очевидно, уменьшается при элиминации кластера положительных зарядов вблизи N-конца.
Конечным результатом настоящей работы является пара оптимизированных вариантов, обозначенных как DsRed.T3 и DsRed.T4. DsRed.T3 быстро созревает, и очищенный белок имеет почти такую же яркость, что и зрелый DsRed дикого типа (Таблица 1), что делает этот вариант очень подходящим для одноцветного изображения красной люминесценции. У DsRed.T3 наблюдается более высокий пик поглощения (возбуждения) в синей области и слегка более высокий максимум излучения в зеленой области, чем таковые у DsRed дикого типа (Фиг.1B), что в двухцветных экспериментах приводит к некоторому загрязнению сигнала GFP. Однако это загрязнение обычно носит минорный характер. Повышенное поглощение белком DsRed.T3 синего света может на самом деле быть полезным, если, например, флуоресценция вызвана лазерным излучением при 488 нм (Фиг.2). Спектры флуоресценции белка DsRed.T4 очень сходны с таковыми у DsRed дикого типа (Фиг.1 B), образуя на выходе нежелательное «загрязнение» сигнала GFP (Фиг.3). Хотя яркость свечения DsRed.T4 составляет приблизительно только половину таковой у DsRed.T3, in vivo этот эффект частично скомпенсирован, поскольку DsRed.T4 созревает приблизительно в два раза быстрее, чем DsRed.T3 (Таблица 1). Таким образом, DsRed.T4, возможно, является наилучшим вариантом для большинства применений. DsRed.T1 в основном идентичен DsRed.T4 (Таблица 1) за исключением того, что DsRed.T1 лишен цистеиновых остатков, а следовательно, его пространственная упаковка может более эффективно формироваться в окислительной среде секреторного пути.
DsRed.T4 является подходящей матрицей для дальнейшего мутагенеза для продуцирования дополнительных вариантов.
Выработка новых вариантов DsRed вероятнее всего связана как с неспецифическим, так и с направленным мутагенезом. Для изучения направленного мутагенеза важно иметь в виду, что в пяти из замен, имеющих место в DsRed.T4 (R2A, H41T, N42Q, A145P и T217A), данный остаток заменяется остатком, который в общем является более консервативным в семействе гомологов DsRed. Таким образом, на основании сравнения последовательностей DsRed и родственных ему последовательностей можно прогнозировать мутации, которые, вероятно, приведут к образованию полезных вариантов.
IV. Быстросозревающие варианты красного флуоресцирующего белка Discosoma (DsRed)
В этом разделе описана стратегия многоступенчатого мутагенеза, которая была использована для получения оптимизированных вариантов DsRed. Результаты сведены в дополнительную Таблицу 2.
Один из исходных N42H-содержащих мутантов продуцировал более яркую флуоресценцию в колонии, чем два других. Такое увеличение яркости происходило за счет второй мутации: H41L. В нескольких гомологах DsRed остаток 41 является треонином, и авторы изобретения обнаружили, что замена H41T приводит к несколько более яркому свечению колоний, чем H41L. В контексте N42Q, H41T вызывает незначительное изменение свойств очищенного белка (не показано), однако приводит к увеличению скорости созревания белка. Таким образом, после цикла 1 мутагенеза авторы изобретения включили две замены - H41T и N42Q (Дополнительная Таблица 1). Вариант цикла 1 дает диффузную полосу с высоким молекулярным весом при анализе методом SDS-PAGE в неденатурирующих условиях (Дополнительная Фиг.4A), что дает основания полагать, что это все еще тетрамерная структура.
Были предприняты дополнительные циклы мутагенеза для дальнейшего ускорения созревания. Используя вариант цикла 1 в качестве матрицы, авторы изобретения получили три мутанта, которые после одного дня роста продуцировали более яркие колонии E. coli. Все три мутанта содержали замену V44A. Помимо ускоренного образования хромофора у V44A по сравнению с вариантом цикла 1 были снижены интенсивность возбуждения синим светом и зеленое излучение (не показано). Один из трех мутантов V44A также содержал замену T21S, которая дополнительно уменьшает интенсивность возбуждения синим светом и зеленое излучение (не показано). Таким образом, вариант цикла 2 содержал четыре замены - T21S, H41T, N42Q и V44A (Дополнительная Таблица 2). В цикле 3 мутагенеза продуцировали несколько мутантов с дальнейшим увеличением флуоресцирования колоний. Неожиданно родственная мутация не вызвала каких-либо изменений белка DsRed как такового, однако вместо этого произошла замена пролинового кодона на лейциновый в положении 4 в линкере между гексагистидиновой меткой и инициирующим метионином. Этот результат указывает на то, что последовательности, добавленные к N-концу DsRed, могут повлиять на пространственную упаковку белка и/или созревание хромофора. Замена P(-4)L была включена в вариант, который был получен на выходе цикла 3 (Таблица 2).
При очистке флуоресцирующих белков авторы изобретения заметили, что DsRed и его варианты не поддавались эффективной экстракции из клеток E. coli в условиях лизиса, при которых экстрагировалось большинство из EGFP. Это наблюдение согласуется с сообщениями о том, что DsRed образует агрегаты внутри клеток. Цикл 4 мутагенеза был сконструирован таким образом, чтобы уменьшить такую агрегацию. Авторы изобретения придумали такой метод анализа, при котором мутантные бактериальные клоны выращивают в 96-луночных планшетах, лизируют в детергентном буфере и центрифугируют для отделения экстрагированных белков из бактериальных осадков. Представляющие интерес мутанты обнаруживают повышенное отношение красной флуоресценции в растворимой фракции к таковой в нерастворимой фракции. Более чем из 25 таких идентифицированных мутантных белков почти все характеризовались уменьшением результирующего заряда вблизи N-конца. После тестирования целого ряда мутантных комбинаций авторы изобретения включили три замены - R2A, K5E и N6D - для получения варианта на выходе цикла 4 (Таблица 2).
Для сравнения растворимостей различных вариантов DsRed авторы экспрессировали каждый белок в E. coli, лизировали клетки в детергентном буфере и подсчитывали процентную долю белковых молекул, которые были экстрагированы (Фиг.4B). Фактически 100% молекул EGFP экстрагируется в этих условиях. Только ˜25% молекул DsRed1 являются растворимыми. DsRed2 оказывается значительно более растворимым (-55%), чем DsRed1. Вариант цикла 3 также оказывается более растворимым (˜52%), чем DsRed1, но вариант цикла 4 оказался даже еще более высокорастворимым (˜73%). При анализе в неденатурирующем SDS-PAGE вариант цикла 3 давал диффузную полосу, которая может свидетельствовать об образовании олигомеров более высокого порядка, тогда как вариант цикла 4 давал тонкую полосу в положении, предсказываемом для тетрамера (Фиг.4A). Эти результаты дают основание предполагать, что уменьшение результирующего заряда вблизи N-конца DsRed препятствует агрегации тетрамеров.
Релевантные мутации в DsRed
мута-
генеза
H41L, H41T
H41T
T21S
T21S
K5E, K5Q, K5M
N6D
K5E
N6D
A145P, A145S
A145Pd
b Замена R2A вызывает также элиминацию дополнительного валинового кодона, который присутствует в DsRed1 после начального кодона.
с T217A присутствует в DsRed.T1 и DsRed.T4, но не в DsRed.T3.
d DsRed.T1 содержит C117S, но не содержат A145P, тогда как DsRed.T3 и DsRed.T4 содержат A145P, но не содержат C117S.
Вариант цикла 4 все еще дает проступающую флуоресценцию при использовании наборов фильтров GFP (не показано). В связи с этим авторы предприняли цикл 5 мутагенеза, чтобы еще более уменьшить зеленое излучение. Проточная цитометрия была использована для селекции клеток E. coli, в которых наблюдалась флуоресценция с повышенным отношением красного излучения к зеленому. Все семь из полученных в результате мутантов содержали замену T217A. Помимо снижения возбуждения в области синего света и излучения в зеленой области в мутантах T217A наблюдали некоторый возврат к слабому красному смещению спектра, наблюдаемому в N42Q (см. Фиг.4A в основном тексте). Мутация T217A была включена в вариант, получаемый на выходе из цикла 5 (Таблица 2).
Наконец, авторы изобретения решили воспользоваться преимуществом, выведенным из случайных наблюдений, полученных в не связанных с данным вопросом экспериментах по мутагенезу. Замена C117S приводила к еще большему снижению интенсивности возбуждения в синей области и излучения в зеленой области. В результате оптимизированный вариант, обозначаемый DsRed.T1, содержит следующие замены: P(-4)L, R2A, K5E, N6D, T21S, H41T, N42Q, V44A, C117S и T217A. Замена A145P подобна замене C117S в отношении влияния на флуоресцентные спектры, но у некоторых исходных мутантов DsRed флуоресценция колонии клеток слегка уменьшается под действием мутации C117S и слегка увеличивается под действием мутации A145P (не показано). Поэтому авторы создали второй оптимизированный вариант, называемый DsRed.T4, который идентичен DsRed.T1, за исключением того, что вместо замены C117S в нем была произведена замена A145P. При последующем анализе выявили, что преимущества, приобретенные в результате T217A, сопровождаются умеренным снижением яркости, поэтому авторы изобретения создали третий оптимизированный вариант, называемый DsRed.T3, который идентичен DsRed.T4, за исключением того, что в DsRed.T3 отсутствует замена T217A.
Возбуждение в синей области и излучение в зеленой области снижаются в результате замен T21S, V44A, C117S, A145P и T217A. Val44 и Thr217 обращены внутрь белка DsRed и закрыты для остатка 42, что указывает на то, что замены V44A и T217A облегчают стерические напряжения, вызываемые N42Q. И наоборот, Thr21, Cys117 и Ala145 обращены к наружной части мономера DsRed, поэтому замены T21S, C117S и A145P являются такими заменами, которые в целом изменяют пространственную упаковку белка. T21S и A145P могут оказывать влияние на структуру DsRed в результате модификаций, возникающих на границе тетрамеров.
Из приведенных выше результатов и обсуждения становится очевидно, что в настоящем изобретении представлен новый очень важный класс флуоресцирующих белков, которые способны к быстрому созреванию. Как таковое, настоящее изобретение представляет собой значительный вклад в данную область.
Все цитируемые в настоящем описании публикации и патентные заявки включены в настоящее описание в виде ссылки, как если бы каждая индивидуальная публикация или патентная заявка специально и индивидуально была бы отмечена как входящая в виде ссылки. Цитирование любой публикации прежде всего связано с раскрытием того, что эта публикация вышла в свет еще до даты подачи и не должно быть истолковано как допущение, что настоящее изобретение не вправе использовать такую публикацию в связи с тем, что оно является более ранним изобретением.
Несмотря на то что представленное выше изобретение описано в некоторых подробностях с целью его иллюстрации, а примеры приведены для того, чтобы облегчить его понимание, тем не менее специалистам в данной области в свете описанного изобретения совершенно очевидно, что определенные изменения и модификации, не выходящие за рамки объема прилагаемой формулы изобретения, являются вполне допустимыми.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЛУОРЕСЦИРУЮЩИЕ БЕЛКИ ИЗ ВЕСЛОНОГИХ РАКООБРАЗНЫХ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2003 |
|
RU2345137C2 |
| ФЛУОРЕСЦИРУЮЩИЕ БЕЛКИ И ХРОМОПРОТЕИНЫ ИЗ ВИДОВ HYDROZOA, НЕ ОТНОСЯЩИХСЯ К AEQUOREA, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2003 |
|
RU2338785C2 |
| НОВЫЕ ХРОМОФОРЫ/ФЛУОРОФОРЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2002 |
|
RU2303600C2 |
| НОВЫЕ ФЛУОРЕСЦЕНТНЫЕ БЕЛКИ ИЗ ENTACMAEA QUADRICOLOR И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2007 |
|
RU2395581C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ | 2015 |
|
RU2699715C2 |
| ПОЛИНУКЛЕОТИД, ОБЛАДАЮЩИЙ ПРОМОТОРНОЙ АКТИВНОСТЬЮ, И ПРИМЕНЕНИЕ ПОЛИНУКЛЕОТИДА ДЛЯ ПОЛУЧЕНИЯ АМИНОКИСЛОТЫ | 2021 |
|
RU2815942C1 |
| ГОМОЛОГИ (ATR) ПОЛИПЕПТИДА rad3, ПОЛИНУКЛЕОТИДЫ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ И МАТЕРИАЛЫ | 1996 |
|
RU2252256C2 |
| ПОЛИПЕПТИД С АСПАРТАТКИНАЗНОЙ АКТИВНОСТЬЮ И ЕГО ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ АМИНОКИСЛОТЫ | 2021 |
|
RU2821918C1 |
| ПОЛИПЕПТИДЫ С ПРОТИВОМИКРОБНОЙ АКТИВНОСТЬЮ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2005 |
|
RU2393224C2 |
| ПОЛИНУКЛЕОТИД НА ОСНОВЕ ГЕНА MDH, ОБЛАДАЮЩИЙ ПРОМОТОРНОЙ АКТИВНОСТЬЮ, И ЕГО ПРИМЕНЕНИЕ | 2022 |
|
RU2825450C2 |
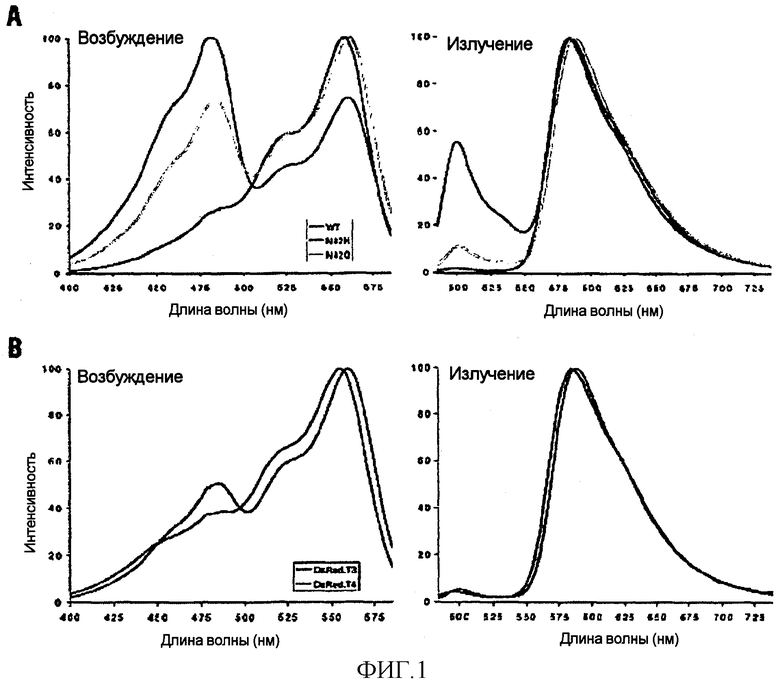
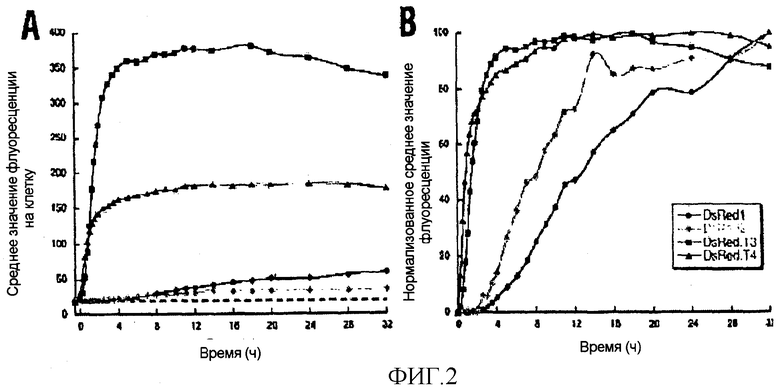

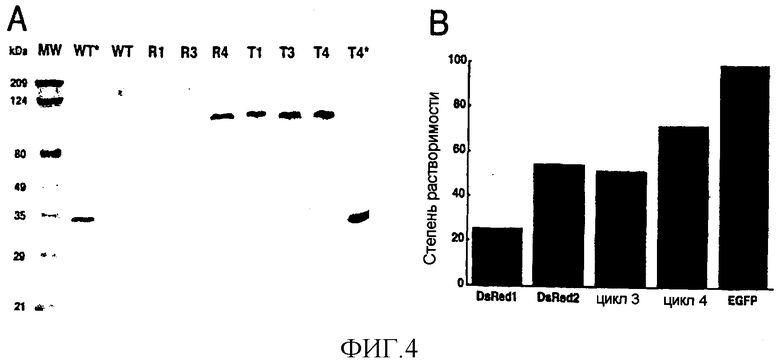
Изобретение относится к области биотехнологии, конкретно к нуклеиновым кислотам, кодирующим быстросозревающие флуоресцирующие белки, и может быть использовано в медицине и для диагностики. Получают полинуклеотид, кодирующий хромо- или флуоресцирующий мутант DsRed дикого типа (SEQ ID NO:2), где хромо- или флуоресцирующий мутант содержит замену в аминокислотном положении 42 SEQ ID NO:2, и необязательно одну или несколько замен в положениях аминокислот, выбранных из группы, состоящей из -4, 2, 5, 6, 21, 41, 44, 117, 217, 145. Полинуклеотидом в составе вектора трансформируют клетки-хозяева, которые экспрессируют хромо- или флуоресцирующий мутант DsRed. Изобретение позволяет получить хромо- или флуоресцирующий полипептид DsRed, который созревает быстрее, чем DsRed дикого типа. 7 н. и 19 з.п. ф-лы, 4 ил., 2 табл.
| US 5958713, 28.09.1999 | |||
| US 5976796, 02.11.1999 | |||
| US 6066476, 23.05.2000 | |||
| Способ получения люциферазы светляков | 1986 |
|
SU1339128A1 |

