ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПОДАННЫЙ В ВИДЕ ТЕКСТОВОГО ФАЙЛА ЧЕРЕЗ EFS WEB
[0001] Перечень последовательностей, записанный в файле 10335WO01_ST25.txt, имеет размер 88 килобайт, был создан 27 июня 2018 года и включен в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0002] Настоящее изобретение в данном документе в целом относится к рекомбинантным вирусным векторам с модифицированным тропизмом и композициям, содержащим их, применимым для нацеленного введения генетического материала в клетки и/или ткани.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0003] Доставка генов в конкретные клетки-мишени стала одной из важнейших технологий в современной медицине для диагностики и генной терапии различных хронических и генетических заболеваний. До настоящего времени прогресс в клиническом применении генной терапии ограничивался отсутствием идеальных сред-носителей для доставки генов. Для достижения терапевтического успеха среды-носители для доставки генов должны быть способны трансдуцировать клетки-мишени, избегая при этом трансдукции нецелевых клеток. В частности, когда нативный тропизм вируса не удовлетворяет текущим терапевтическим потребностям, существует потребность в рекомбинантных вирусных векторах, в которых естественный тропизм устранен или уменьшен и успешно сконструирован необходимый тропизм. (Buchholz et al.)
[0004] В последние годы наибольший прогресс в разработке векторов был достигнут с использованием вирусов без оболочки (например, вирусов, содержащих капсид, образованный вирусными капсидными белками без оболочки (например, липидного бислоя)), таких как аденоассоциированные вирусы (AAV) и аденовирусы (Ad), а также вирусов с оболочкой (например, вирусов, у которых капсид окружен липидным бислоем), таких как ретровирусы, лентивирусы и вирус простого герпеса. Векторы на основе AAV были предметом многих исследований, так как AAV представляют собой вирусы без оболочки, которые являются лишь слабо иммуногенными, но способны трансдуцировать широкий спектр видов и тканей in vivo без признаков токсичности.
[0005] AAV представляют собой небольшие однонитевые ДНК-вирусы без оболочки. Геном AAV составляет 4,7 т.о. и характеризуется двумя инвертированными концевыми повторами (ITR) и двумя открытыми рамками считывания, которые кодируют белки Rep и белки Сар, соответственно. Два ITR являются единственными цис-элементами, необходимыми для репликации, упаковки и интеграции AAV. Рамка считывания Rep кодирует четыре белка с молекулярной массой 78 кДа, 68 кДа, 52 кДа и 40 кДа. Эти белки функционируют, главным образом, при регуляции репликации AAV и интеграции AAV в хромосомы клетки-хозяина. Рамка считывания Сар кодирует три структурных (капсидных) вирусных белка (VP), имеющих молекулярную массу 83-85 кДа (VP1), 72-73 кДа (VP2) и 61-62 кДа (VP3). Более 80% всех белков в вирионе AAV составляет VP3; в зрелых вирионах VP1, VP2 и VP3 обнаруживаются при относительном содержании приблизительно 1:1:10. In vitro три белка спонтанно собираются в вирионоподобные структуры, например, вирусные капсиды. Таким образом, оказывается, что образование вирусного капсида в инфицированных клетках происходит независимо от синтеза вирусной ДНК (рассмотрено в Kotin et al. (1994) Hum. Gene Ther. 5:793).
[0006] Среди всех известных серотипов AAV, AAV2 является, пожалуй, наиболее хорошо охарактеризованным серотипом, поскольку его инфекционный клон был создан первым. (Samulski et al. (1982) Proc. Natl. Acad. Sci. USA 79:2077-2081). Впоследствии также были определены полные последовательности для AAV3A, AAV3B, AAV4 и AAV6. (Rutledge et al. (1998) J. Virol. 72:309-319; Chiorini et al. (1997) J. Virol. 71:6823-6833; S. Muramatsu et al. (1996) Virol. 221:208-217). Как правило, все AAV обладают более чем 80%-ой идентичностью нуклеотидной последовательности.
[0007] AAV является многообещающим вектором для генной терапии человека, поскольку, в отличие от других вирусных векторов, AAV, как было установлено, не связаны с каким-либо известным заболеванием человека и, как правило, не считаются патогенными. (Muzyczka et al. (1992) Current Topics in Microbiology and Immunology 158:97-129). Более того, AAV безопасно трансдупирует постмитотические ткани с относительно низкой иммуногенностью и способен интегрироваться в хромосомы хозяина сайт-специфическим образом и в клетки культивируемой ткани в хромосому 19, если белки Rep обеспечиваются с помощью другого вектора. (Kotin et al. (1990) Proc. Natl. Acad. Sci. USA 87:2211-2215; Samulski et al. (1991) EMBO J. 10(12):3941-3950; Balague et al. (1997) J. Virol. 71:3299-3306; Surosky et al. (1997) J. Virol. 71:7951-7959). Было показано, что интегрированные геномы AAV обеспечивают долгосрочную экспрессию генов в ряде тканей, включая мышцы, печень и головной мозг (Fisher (1997) Nature Med. 3(3):306-312; Snyder et al. (1997) Nature Genetics 16:270-276; Xiao et al. (1997) Experimental Neurology 144:113-124; Xiao et al. (1996) J. Virol. 70(11):8098-8108).
[0008] Ряд вирусов, включая AAV, инфицирует клетки посредством взаимодействия вирус/лиганд:клетка/рецептор, что в конечном итоге приводит к эндоцитозу вируса инфицированной клеткой. Это взаимодействие лиганд:рецептор является предметом большей части исследований вирусных векторов, например, им можно манипулировать, чтобы перенаправить естественный тропизм вируса с клетки, естественно пермиссивной к инфекции вирусом дикого типа, в клетку-мишень, например, через рецептор, который экспрессируется клеткой-мишенью.
[0009] Теоретически, перенацеливание вектора на любой белок или маркер клеточной поверхности должно привести к инфицированию клетки-мишени, так как большинство рецепторов или маркеров клеточной поверхности участвуют в путях эндоцитоза, либо конститутивных (например, для рециркуляции), либо индуцируемых лигандом (например, опосредованных рецептором). Эти рецепторы группируются в покрытых клатрином углублениях, попадают в клетку через покрытые клатрином везикулы, проходят через эндосому с повышенной кислотностью, в которой сортируются рецепторы, а затем либо возвращаются на поверхность клетки, остаются на хранение внутриклеточно, или разлагаются в лизосомах. Таким образом, платформы для перенацеливания вирусных векторов часто стремятся устранить естественный тропизм вирусного вектора и перенаправить вирусный вектор на рецептор или маркер, экспрессируемый исключительно или преимущественно клеткой-мишенью. Многие из достижений в направленной генной терапии с применением вирусных векторов можно кратко описать как нерекомбинаторную (отличную от генетической) или рекомбинаторную (генетическую) модификацию вирусного вектора, которая приводит к псевдотипированию, расширению тропизма и/или перенацеливанию естественного тропизма вирусного вектора, (рассмотрено в Nicklin and Baker (2002) Curr. Gene Ther. 2:273-93; Verheiji and Rottier (2012) Advances Virol 2012:1-15).
[0010] В подходах, отличных от генетических, обычно используют адаптер, который распознает как поверхностный белок вируса дикого типа (немодифицированный), так и клетку-мишень. Растворимые псевдорецепторы (для вируса дикого типа), полимеры, такие как полиэтиленгликоль, и антитела или их части, применяли в качестве вирус-связывающего домена адаптеров, тогда как для связывающего клетку домена адаптеров, описанных выше, применяли природные лиганды, представляющие собой пептиды или витамины, а также антитела и их части. При таком подходе перенацеливание вирусного вектора на клетку-мишень может быть выполнена после связывания комплекса вектор: адаптер с белком, экспрессируемым на поверхности клетки-мишени, например белком клеточной поверхности.
[0011] Такой подход применяли для AAV (Bartlett et al. (1999) Nat. Biotechnol. 74:2777-2785), аденовирусов (Hemminki et al. (2001) Cancer Res. 61: 6377-81; van Beusechem et al. (2003) Gene Therapy 10:1982-1991; Einfeld, et al. (2001) J. Virol. 75:11284-91; Glasgow et al. (2009) PLOS One 4:e8355), герпесвирусов (Nakano et al. (2005) Mol. Ther. 11:617-24) и парамиксовирусов (Bian et al. (2005) Cancer Gene Ther. 12:295-303; Bian et al. (2005) Int. J. Oncol. 29:1359-69), коронавирусов (Haijema et al. (2003) J. Virol. 77:4528-43] 8; Wurdinger et al. (2005) Gene Therapy 12:1394-1404).
[0012] Более популярным подходом являлась рекомбинаторная генетическая модификация вирусных капсидных белков и, таким образом, поверхности вирусного капсида. При непрямых рекомбинаторных подходах вирусный капсид модифицируется с помощью гетерологичного «каркаса», который затем присоединяется к адаптеру. Адаптер связывается с каркасом и клеткой мишенью. (Arnold et al. (2006) Mol. Ther. 5:125-132; Ponnazhagen et al. (2002) J. Virol. 76:12900-907; см. также WO 97/05266) Каркасы, такие как (1) Fc-связывающие молекулы (например, Fc-рецепторы, белок А и т.д.), которые связываются с Fc антител-адаптеров, (2) (стрепт)авидин, который связывается с биотинилированными адаптерами, (3) биотин, который связывается с адаптерами, слитыми с (стрепт)авидином, и (4) пары связывания белок: белок, которые образуют изометрические пептидные связи, такие как SpyCatcher, который связывает адаптер SpyTagged, были включены в Ad (Pereboeva et al. (2007) Gene Therapy 14:627-637; Park et al. (2008) Biochemical and Biophysical Research Communications 366:769-774; Henning et al. (2002) Human Gene Therapy 13:1427-1439; Banerjee et al. (2011) Bioorganic and Medicinal Chemistry Letters 21:4985-4988), AAV (Gigout et al. (2005) Molecular Therapy 11:856-865; Stachler et al. (2008) Molecular Therapy 16:1467-1473) и тогавирусы (Quetglas et al. (2010) Virus Research 153:179-196; Ohno et al. (1997) Nature Biotechnology 15:763-767; Klimstra et al. (2005) Virology 338:9-21).
[0013] При прямом рекомбинаторном подходе к нацеливанию нацеливающий лиганд непосредственно вставляется в вирусный капсид или соединяется с ним, то есть белковые вирусные капсиды модифицируются для экспрессии гетерологичного нацеливающего лиганда. Затем лиганд перенаправляет, например, связывает рецептор или маркер, преимущественно или исключительно экспрессируемый на клетке-мишени. (Stachler et al. (2006) Gene Ther. 13:926-931; White et al. (2004) Circulation 109:513-519.). Прямые рекомбинаторные подходы применяли для AAV (Park et al., (2007) Frontiers in Bioscience 13:2653-59; Girod et al. (1999) Nature Medicine 5:1052-56; Grifman et al. (2001) Molecular Therapy 3:964-75; Shi et al. (2001) Human Gene Therapy 12:1697-1711; Shi and Bartlett (2003) Molecular Therapy 7:515-525), ретровируса (Dalba et al. Current Gene Therapy 5:655-667; Tai and Kasahara (2008) Frontiers in Bioscience 13:3083-3095; Russell and Cosset (1999) Journal of Gene Medicine 1:300-311; Erlwein et al. (2002) Virology 302:333-341; Chadwick et al. (1999) Journal of Molecular Biology 285:485-494; Pizzato et al. (2001) Gene Therapy 8:1088-1096), поксвируса (Guse et al. (2011) Expert Opinion on Biological Therapy 11:595-608; Galmiche et al. (1997) Journal of General Virology 78:3019-3027; Paul et al. (2007) Viral Immunology 20:664-671), парамиксовируса (Nakamura and Russell (2004) Expert Opinion on Biological Therapy 4:1685-1692; Hammond et al. (2001) Journal of Virology 75:2087-2096; Galanis (2010) Clinical Pharmacology and Therapeutics 88:620-625; Blechacz and Russell (2008) Current Gene Therapy 8:162-175; Russell and Peng (2009) Current Topics in Microbiology and Immunology 330:213-241) и герпесвируса (Shah and Breakefield (2006) Current Gene Therapy 6:361-370; Campadelli-Fiume et al. (2011) Reviews in Medical Virology 21:213-226).
[0014] Каждый из трех подходов имеет преимущества и недостатки. Основным преимуществом прямого рекомбинаторного подхода является то, что специфичность вирусного вектора присуща вирусному геному и может сохраняться при репликации. Однако для прямого и непрямого рекомбинаторных подходов способность генетически модифицировать вирус требует сохранения структуры капсида, а нацеливающий лиганд или каркас должен располагаться в положении, которое будет допускать и надлежащим образом отображать нацеливающий лиганд или каркас, ограничивая таким образом репертуар соответствующих лигандов или каркасов, которые можно использовать. Как таковые, способы рекомбинаторного перенацеливания ограничены существующими в природе молекулами, полезными в качестве нацеливающих лигандов, что приводит к включению других связывающих лигандов, таких как антитела или их части. Как нерекомбинаторные, так и рекомбинаторные адаптерные платформы имеют преимущество в гибкости используемого адаптера. Однако с этими двухкомпонентными системами трудно достичь оптимальной эффективности трансдукции.
[0015] В данном документе предусмотрена стратегия перенацеливания вируса, которая решает проблемы, присущие предыдущим стратегиям рекомбинаторного перенацеливания, путем вставки гетерологичного эпитопа в вирусный капсид. Текущий рекомбинантный вирусный капсид проявляет сниженный естественный тропизм вплоть до устраненного, который восстанавливается и перенаправляется после комбинации с полиспецифической, необязательно биспецифической, связывающей молекулой, содержащей паратоп антитела, например, Fv, который специфически связывает гетер о логичный эпитоп, и перенацеливающий лиганд, который специфически связывает клетку-мишень, особенно в определенных соотношениях вирусного вектора:полиспепифической связывающей молекулы.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0016] В данном документе раскрыты рекомбинантные вирусные капсидные белки, вирусные капсиды, содержащие рекомбинантные вирусные капсидные белки, вирусные векторы, содержащие нуклеотид, представляющий интерес, заключенный в рекомбинантный вирусный капсид; где указанные капсидные белки, капсиды и вирусные векторы генетически модифицированы для включения (отображения) гетерологичного эпитопа, где гетерологичный эпитоп (его часть или в комбинации с вирусным капсидным белком) образует связывающую пару с паратопом антитела, и где рекомбинантный вирусный капсидный белок/капсид/вектор может дополнительно содержать мутацию, вставку или делецию в аминокислотном положении, которое участвует в связывании рецептора, например, (естественном) тропизме вирусного капсидного белка/капсида/вектора, так что рекомбинантный вирусный капсидный белок или вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, характеризуется пониженным вплоть до устраненного (естественным) тропизмом (например, обладает в отсутствие полиспецифической, необязательно биспецифической, связывающей молекулы меньшей эффективностью трансдукции, чем эффективность трансдукции эталонного вирусного капсидного белка/капсида/вектора, в котором отсутствует гетерологичный эпитоп, или неопределяемой эффективностью трансдукции в отсутствие полиспецифической, необязательно биспецифической, связывающей молекулы). Такой пониженный вплоть до устраненного (естественный) тропизм рекомбинантного вирусного капсидного белка/капсида/вектора можно усилить или восстановить в присутствии подходящего полиспецифического, необязательно биспецифического, связывающего фрагмента. Соответственно, также описаны композиции, содержащие (1) рекомбинантные вирусные векторы, имеющие капсид, содержащий рекомбинантный капсидный белок, описанный в данном документе, и (2) полиспецифическую, необязательно биспецифическую, связывающую молекулу, содержащую паратоп антитела и нацеливающий лиганд, включая композиции, содержащие определенные соотношения вирусного вектора: полиспецифической связывающей молекулы; и их пути применения для направления и/или введения генетического материала в клетку-мишень также описаны в данном документе. Также описаны способы перенацеливания рекомбинантного вирусного вектора, например, для нацеленной доставки нуклеотида, представляющего интерес, в клетку-мишень, включающие приведение в контакт рекомбинантного вирусного вектора с полиспецифической, необязательно биспецифической, связывающей молекулой, и способы получения рекомбинантного вирусного вектора также описаны.
[0017] В данном документе описаны рекомбинантные вирусные капсидные белки, содержащие эпитоп, который является гетерологичным по отношению к капсидному белку, где эпитоп или его часть специфически связывает паратоп антитела, и где рекомбинантный вирусный капсидный белок или вирусный капсид, содержащий рекомбинантный вирусный капсид, имеет пониженный вплоть до устраненного естественный тропизм, например, в отсутствие полиспецифического, необязательно биспецифического, связывающего фрагмента.
[0018] В некоторых вариантах осуществления гетерологичный эпитоп вставляется (отображается) рекомбинантным вирусным капсидным белком, так что вставка и/или отображение гетерологичного эпитопа понижает или устраняет (естественный) тропизм вирусного капсида по сравнению с эталонным вирусным капсидом, в котором отсутствует гетерологичный эпитоп, например, вирусный капсид содержит мутацию, предусматривающую вставку эпитопа в аминокислотном положении и/или замену аминокислоты эпитопом в аминокислотном положении, где мутация понижает или устраняет (естественный) тропизм капсидного белка, например, в отсутствие полиспецифического, необязательно биспецифического, связывающего фрагмента. В некоторых вариантах осуществления гетерологичный эпитоп вставлен (отображен) вирусным капсидным белком таким образом, что вставка и/или отображение частично понижает (естественный) тропизм рекомбинантного вирусного капсида по сравнению с эталонным вирусным капсидом, в котором отсутствует гетерологичный эпитоп, например, в отсутствие полиспецифического, необязательно биспецифического, связывающего фрагмента, и вирусный капсид дополнительно содержит дополнительную мутацию (например, замену, делецию, вставку, отличную от вставки гетерологичного эпитопа), которая дополнительно понижает и/или устраняет (естественный) тропизм рекомбинантного вирусного капсида или рекомбинантных вирусных векторов, содержащих их, например, в отсутствие полиспецифического, необязательно биспецифического, связывающего фрагмента, по сравнению с эталонным вирусным капсидом, в котором отсутствует эта мутация.
[0019] Как правило, рекомбинантные вирусные капсидные белки, описанные в данном документе, можно получить с помощью гена капсида, например, они кодируются геном капсида, модифицированным для экспрессии эпитопа, и/или представляют собой генетически модифицированный вирус без оболочки, который обычно инфицирует клетки человека, или серотипы вирусов без оболочки, которые обычно инфицируют клетки человека, например аденовирус, аденоассоциированный вирус и т.д. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в данном документе, получен с помощью гена капсида AAV, например, кодируется геном капсида, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок аденоассоциированного вируса (AAV) серотипа AAV, который инфицирует приматов, где необязательно AAV выбран из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, AAV6, AAV8 или AAV9, например, представляет собой генетически модифицированный капсидный белок AAV2, генетически модифицированный капсидный белок AAV6, генетически модифицированный капсидный белок AAV8 или генетически модифицированный капсидный белок AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, кодируется геном капсида AAV2, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV2, для которого аминокислотная последовательность белка VP1 AAV2 дикого типа представлена, соответственно, как SEQ ID NO: 1. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV6, например, кодируется геном капсида AAV6, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV6, для которого аминокислотная последовательность белка VP1 AAV6 дикого типа представлена под SEQ ID NO: 3. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV8, например, кодируется геном капсида AAV8, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV8, для которого аминокислотная последовательность белка VP1 AAV дикого типа представлена, соответственно, как SEQ ID NO: 21. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, кодируется геном капсида AAV9, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 или VP3 AAV9, для которого аминокислотная последовательность белка VP1 AAV9 дикого типа представлена, соответственно, как SEQ ID NO: 5. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, кодируется геном капсида AAV2, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV6, например, кодируется геном капсида AAV6, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV6. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV8, например, кодируется геном капсида AAV8, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV8. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, кодируется геном капсида AAV9, модифицированным для экспрессии эпитопа, и/или представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV9.
[0020] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью, например, кодируется химерным геном капсида AAV, модифицированным для экспрессии эпитопа, где химерный ген капсидного белка AAV содержит множество последовательностей нуклеиновых кислот, где каждая из множества последовательностей нуклеиновых кислот кодирует часть капсидного белка другого серотипа AAV, и где множество последовательностей нуклеиновых кислот вместе кодируют химерный капсидный белок AAV. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью химерного гена капсида AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью химерного гена капсида AAV6. В некоторых вариантах осуществления вирусный капсидный белок получен с помощью химерного гена капсида AAV8. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью химерного гена капсида AAV9.
[0021] Как правило, рекомбинантный вирусный капсидный белок, как описано в данном документе, содержит гетерологичный эпитоп, вставленный в рекомбинантный капсидный белок и/или отображенный на нем, так что сам гетерологичный эпитоп понижает и/или устраняет естественный тропизм рекомбинантного капсидного белка или капсида, содержащего его, по сравнению с эталонным капсидом, в котором отсутствует гетерологичный эпитоп, или капсидом, включающим эталонный капсид, соответственно. В некоторых вариантах осуществления гетерологичный эпитоп вставлен (отображен) в область капсидного белка, участвующую в естественном тропизме эталонного капсидного белка дикого типа, например, в область капсидного белка, участвующую в нацеливании на клетки. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в knob-домен волоконного белка Ad и/или отображен с его помощью. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в петлю HI волоконного белка Ad и/или отображен с ее помощью. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в сайт связывания гепарина капсидного белка AAV и/или отображен на нем. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в сайт связывания гепарина капсидного белка AAV2 и/или отображен на нем. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в сайт связывания гепарина капсидного белка AAV6 и/или отображен на нем. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в сайт связывания гепарина капсидного белка AAV8 и/или отображен на нем. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в сайт связывания гепарина капсидного белка AAV9 и/или отображен на нем. В некоторых вариантах осуществления (i) вирусный капсидный белок получен с помощью гена капсида AAV2 и эпитоп вставлен после аминокислоты в положении I453 или I587 капсидного белка VP1 AAV2 и/или аминокислот в соответствующих положениях капсидных белков VP2 и/или VP3 AAV2, и/или заменяет их; (ii) вирусный капсидный белок получен с помощью гена капсида AAV6 и эпитоп вставлен после аминокислоты в положении I585 капсидного белка VP1 AAV6 и/или аминокислот в соответствующих положениях капсидного белка VP2 и/или VP3 AAV6 и/или заменяет их; (iii) вирусный капсид получен с помощью гена капсида AAV8 и эпитоп вставлен после аминокислоты в положении I590 капсидного белка VP1 AAV8 и/или аминокислот в соответствующих положениях капсидных белков VP2 и/или VP3 AAV8 и/или заменяет их, или (iv) вирусный капсидный белок получен с помощью гена капсида AAV9 и эпитоп вставлен после аминокислоты в положении I453 или I589 капсидного белка VP1 AAV9 и/или аминокислот в соответствующих положениях капсидных белков VP2 и/или VP3 AAV9 и/или заменяет их. В некоторых вариантах осуществления гетерологичный эпитоп вставлен сразу после (например, слит с С-концом) аминокислоты, выбранной из группы, состоящей из G453 капсидного белка VP1 AAV2 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9), N587 капсидного белка VP1 AAV2 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9), Q585 капсидного белка VP1 AAV6 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV7, AAV8 и AAV9), N590 капсидного белка VP1 AAV8 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV9), G453 капсидного белка VP1 AAV9 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8) или А589 капсидного белка VP1 AAV9 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8). В некоторых вариантах осуществления гетерологичный эпитоп вставлен сразу после (например, слит с С-концом) G453 капсидного белка VP1 AAV2 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9). В некоторых вариантах осуществления гетерологичный эпитоп вставлен сразу после (например, слит с С-концом) N587 капсидного белка VP1 AAV2 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9). В некоторых вариантах осуществления гетерологичный эпитоп вставлен сразу после (например, слит с С-концом) Q585 капсидного белка VP1 AAV6 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV7, AAV8 и AAV9). В некоторых вариантах осуществления гетерологичный эпитоп вставлен сразу после (например, слит с С-концом) N590 капсидного белка VP1 AAV8 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV9). В некоторых вариантах осуществления гетерологичный эпитоп вставлен сразу после (например, слит с С-концом) G453 капсидного белка VP1 AAV9 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8). В некоторых вариантах осуществления гетерологичный эпитоп вставлен сразу после (например, слит с С-концом) А589 капсидного белка VP1 AAV9 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8). В некоторых вариантах осуществления гетерологичный эпитоп вставлен и/или отображен между аминокислотами N587 и R588 капсидного белка VP1 AAV2 (или соответствующими положениями VP2 и/или VP3 капсидов, кодируемых тем же геном капсида). В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, представленную под SEQ ID NO: 2. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, представленную под SEQ ID NO: 4. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 25. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 26. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 27.
[0022] В некоторых вариантах осуществления рекомбинантный капсидный белок, как описано в данном документе, содержит вторую и отличающуюся мутацию в дополнение к гетер о логичному эпитопу. Например, в некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, как описано в данном документе, может представлять собой генетически модифицированный капсидный белок AAV2, содержать гетерологичный эпитоп и может дополнительно содержать мутацию, например, мутацию R585A и/или R588A. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, представляет собой генетически модифицированный капсидный белок VP1 AAV2, содержит гетерологичный эпитоп, вставленный сразу после (например, слитый с С-концом) G453 белка VP1 AAV2 и дополнительно содержит мутацию, выбранную из группы, состоящей из R585A и/или R5889A. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, представляет собой генетически модифицированный капсидный белок VP1 AAV2, содержит гетерологичный эпитоп, вставленный сразу после (например, слитый с С-концом) N587 белка VP1 AAV2 и дополнительно содержит мутацию, выбранную из группы, состоящей из R585A и/или R588A. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, представляет собой генетически модифицированный капсидный белок VP1 AAV9, содержит гетерологичный эпитоп, вставленный сразу после (например, слитый с С-концом) G453 белка VP1 AAV9 и дополнительно содержит мутацию W503A. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, представляет собой генетически модифицированный капсидный белок VP1 AAV9, содержит гетерологичный эпитоп, вставленный сразу после (например, слитый с С-концом) А589 капсидного белка VP1 AAV9 и дополнительно содержит мутацию W503A.
[0023] Как правило, рекомбинантный вирусный капсидный белок и/или вирусный вектор, содержащий рекомбинантный вирусный капсид, содержит гетерологичный эпитоп, длина которого составляет по меньшей мере одну аминокислоту. В некоторых вариантах осуществления длина гетерологичного эпитопа может составлять от приблизительно 5 аминокислот до приблизительно 35 аминокислот, и он образует связывающую пару с паратопом антитела, например, вариабельным доменом иммуноглобулина. В некоторых вариантах осуществления длина гетерологичного эпитопа составляет по меньшей мере 10 аминокислот. В некоторых вариантах осуществления гетерологичный эпитоп содержит аффинную метку. В некоторых вариантах осуществления гетерологичный эпитоп и/или аффинная метка не образуют связывающую пару с константным доменом иммуноглобулина. В некоторых вариантах осуществления гетерологичный эпитоп и/или аффинная метка не образуют связывающую пару с ионом металла, например, Ni2+, Со2+, Cu2+, Zn2+, Fe3+ и т.д. В некоторых вариантах осуществления гетерологичный эпитоп не является полипептидом, выбранным из группы, состоящей из стрептавидина, Strep II, НА, L14, 4C-RGD, LH и белка А. В некоторых вариантах осуществления аффинная метка выбрана из группы, состоящей из FLAG (SEQ ID NO: 7), НА (SEQ ID NO: 8) и c-myc (EQKLISEEDL; SEQ ID NO: 6). В некоторых вариантах осуществления гетерологичный эпитоп содержит c-myc (EQKLISEEDL; SEQ ID NO: 6).
[0024] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок представляет собой генетически модифицированный капсидный белок VP1 AAV2 и содержит гетерологичный эпитоп, содержащий последовательность EQKLISEEDL (SEQ ID NO: 6), вставленную сразу после (например, слитую с С-концом) G453 капсидного белка VP1 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок (i) получен с помощью гена капсида AAV2, например, представляет собой генетически модифицированный капсидный белок VP1 AAV2, (ii) содержит гетерологичный эпитоп, который содержит последовательность EQKLISEEDL (SEQ ID NO: 6), и вставлен сразу после (например, слит с С-концом) G453 капсидного белка VP1 AAV2 и (ii) дополнительно содержит мутацию, выбранную из группы, состоящей из R585A и/или R5889A. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок представляет собой генетически модифицированный капсидный белок VP1 AAV2 и содержит гетерологичный эпитоп, содержащий последовательность EQKLISEEDL (SEQ ID NO: 6), вставленную сразу после (например, слитую с С-концом) N587 капсидного белка VP1 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок (i) получен с помощью гена капсида AAV2, например, представляет собой генетически модифицированный капсидный белок VP1 AAV2, (ii) содержит гетерологичный эпитоп, который содержит последовательность EQKLISEEDL (SEQ ID NO: 6), и вставлен сразу после (например, слит с С-концом) N587 капсидного белка VP1 AAV2 и (iii) дополнительно содержит мутацию, выбранную из группы, состоящей из R585A и/или R588A. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок (i) получен с помощью гена капсида AAV6, например, представляет собой генетически модифицированный капсидный белок VP1 AAV6, (ii) содержит гетерологичный эпитоп, который содержит последовательность EQKLISEEDL (SEQ ID NO: 6) и вставлен сразу после (например, слит с С-концом) Q585 капсидного белка VP1 AAV6. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок представляет собой генетически модифицированный капсидный белок VP1 AAV8 и содержит гетерологичный эпитоп, содержащий последовательность EQKLISEEDL (SEQ ID NO: 6), вставленную сразу после (например, слитую с С-концом) N590 капсидного белка VP1 AAV8. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок представляет собой генетически модифицированный капсидный белок VP1 AAV9 и содержит гетерологичный эпитоп, содержащий последовательность EQKLISEEDL (SEQ ID NO: 6), вставленную сразу после (например, слитую с С-концом) G453 капсидного белка VP1 AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок (i) получен с помощью гена капсида AAV9, например, представляет собой генетически модифицированный капсидный белок VP1 AAV9, (ii) содержит гетерологичный эпитоп, который содержит последовательность EQKLISEEDL (SEQ ID NO: 6) и вставлен сразу после (например, слит с С-концом) G453 капсидного белка VP1 AAV9, и (iii) дополнительно содержит мутацию W503A. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок представляет собой генетически модифицированный капсидный белок VP1 AAV9 и содержит гетерологичный эпитоп, содержащий последовательность EQKLISEEDL (SEQ ID NO: 6), вставленную сразу после (например, слитую с С-концом) А589 капсидного белка VP1 AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок (i) получен с помощью гена капсида AAV9, например, представляет собой генетически модифицированный капсидный белок VP1 AAV9, (ii) содержит гетерологичный эпитоп, который содержит последовательность EQKLISEEDL (SEQ ID NO: 6) и вставлен сразу после (например, слит с С-концом) А589 капсидного белка VP1 AAV9, и (iii) дополнительно содержит мутацию W503A.
[0025] В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV. В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I587 капсидного белка VP1 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между N587 и R588 капсидного белка VP1 AAV2, например, содержит аминокислотную последовательность, представленную под SEQ ID NO: 2.
[0026] В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV6. В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I585 капсидного белка VP1 AAV6. В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между Q585 и S586 капсидного белка VP1 AAV6, например, содержит аминокислотную последовательность, представленную под SEQ ID NO: 4.
[0027] В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами VP1 капсида AAV8. В некоторых вариантах осуществления рекомбинантный вирусный капсид содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I590 капсидного белка VP1 AAV8. В некоторых вариантах осуществления рекомбинантный вирусный капсид содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между N590 и Т591 капсидного белка VP1 AAV8, например, содержит аминокислотную последовательность, представленную под SEQ ID NO: 25.
[0028] В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсид содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I453 капсидного белка VP1 AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсид содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между G453 и S454 капсидного белка VP1 AAV9, например, содержит аминокислотную последовательность, представленную под SEQ ID NO: 26. В некоторых вариантах осуществления рекомбинантный вирусный капсид содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I589 капсидного белка VP1 AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсид содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между А589 и Q590 капсидного белка VP1 AAV9, например, содержит аминокислотную последовательность, представленную под SEQ ID NO: 27.
[0029] В некоторых вариантах осуществления гетерологичный эпитоп содержит аффинную метку и один или более линкеров. В некоторых вариантах осуществления гетерологичный эпитоп содержит аффинную метку, фланкированную линкером, например, гетерологичный эпитоп содержит от N-конца к С-концу первый линкер, аффинную метку и второй линкер. В некоторых вариантах осуществления первый и второй линкеры, каждый независимо, представляют собой полипептид длиной по меньшей мере 1 аминокислоту. В некоторых вариантах осуществления гетерологичный эпитоп, как описано в данном документе, например, аффинная метка сама по себе или в комбинации с одним или более линкерами, имеет длину от приблизительно 5 аминокислот до приблизительно 35 аминокислот. В некоторых вариантах осуществления первый и второй линкеры имеют одинаковую длину и/или содержат идентичные аминокислотные последовательности.
[0030] Как правило, в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, имеет пониженный вплоть до устраненного естественный тропизм, например, обладает пониженной способностью или неспособен нацелиться и связать эталонную клетку, естественно пермиссивную к трансдукции, по сравнению со способностью эталонного вирусного капсида, например капсида, содержащего эталонный вирусный капсидный белок, например, вирусный капсидный белок, который был бы идентичен рекомбинантному вирусному капсидному белку, но без гетерологичного эпитопа. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 10%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 20%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 30%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 40%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 50%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 60%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 70%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 75%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 80%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 85%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 90%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 95%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, проявляет по меньшей мере 99%-ное снижение эффективности трансдукции по сравнению с эталонным вирусным капсидом. В некоторых вариантах осуществления и в отсутствие подходящей полиспецифической, необязательно биспецифической, связывающей молекулы трансдукция контрольной клетки рекомбинантным вирусным капсидом, содержащим рекомбинантный вирусный капсидный белок, как описано в данном документе, устраняется, например, не обнаруживается.
[0031] В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, является мозаичным капсидом, например, содержит в определенном соотношении рекомбинантный вирусный капсидный белок, содержащий гетерологичный эпитоп, и эталонный капсидный белок, который не содержит гетерологичный эпитоп. В некоторых вариантах осуществления эталонный капсидный белок представляет собой эталонный капсидный белок дикого типа в том смысле, что он содержит аминокислотную последовательность капсидного белка дикого типа, имеющего тот же серотип, что и рекомбинантный вирусный капсидный белок. В некоторых вариантах осуществления эталонный капсидный белок представляет собой контрольный эталонный капсидный белок в том смысле, что он содержит аминокислотную последовательность рекомбинантного вирусного капсидного белка, за исключением того, что в контрольном эталонном капсидном белке отсутствует гетерологичный эпитоп. В некоторых вариантах осуществления эталонный капсидный белок представляет собой мутированный эталонный белок дикого типа в том смысле, что он содержит аминокислотную последовательность, по сути идентичную последовательности капсидного белка дикого типа, имеющего такой же серотип, что и рекомбинантный вирусный капсидный белок, за исключением мутации, (например, вставка аминокислотной последовательности, химеризация и т.д.), которая снижает тропизм капсидного белка дикого типа. В некоторых вариантах осуществления композиция, описанная в данном документе, содержит, или способ, описанный в данном документе, объединяет рекомбинантный вирусный капсидный белок и эталонный капсидный белок в соотношении, которое находится в диапазоне от 1:1 до 1:15. В некоторых вариантах осуществления соотношение составляет 1:2. В некоторых вариантах осуществления соотношение составляет 1:3. В некоторых вариантах осуществления соотношение составляет 1:4. В некоторых вариантах осуществления соотношение составляет 1:5. В некоторых вариантах осуществления соотношение составляет 1:6. В некоторых вариантах осуществления соотношение составляет 1:7. В некоторых вариантах осуществления соотношение составляет 1:8. В некоторых вариантах осуществления соотношение составляет 1:9. В некоторых вариантах осуществления соотношение составляет 1:10. В некоторых вариантах осуществления соотношение составляет 1:11. В некоторых вариантах осуществления соотношение составляет 1:12. В некоторых вариантах осуществления соотношение составляет 1:13. В некоторых вариантах осуществления соотношение составляет 1:14. В некоторых вариантах осуществления соотношение составляет 1:15.
[0032] Также в данном документе раскрыты нуклеиновые кислоты, которые кодируют рекомбинантный вирусный капсидный белок, описанный в данном документе, композиции, содержащие такие нуклеиновые кислоты (например, которые можно использовать в способах получения рекомбинантного вирусного капсида), и/или рекомбинантные вирусные капсидные белки (например, композиции, состоящие по существу из рекомбинантного вирусного капсидного белка, композиции, содержащие только вирусные векторы, заключенные в капсид, содержащий вирусный капсидный белок, описанный в данном документе, композиции, содержащие такие вирусные векторы и полиспецифическую, необязательно биспецифическую, связывающую молекулу (например, в определенных соотношениях вирусного вектора с полиспецифической, необязательно биспецифической, связывающей молекулой (молекула: молекула)), композиции, содержащие такие вирусные векторы, изменяющие мишень фрагменты и фармацевтически приемлемый носитель и т.д.). В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6) и нуклеотидную последовательность, которая кодирует по меньшей мере 5 смежных аминокислот капсидного белка аденовируса или аденоассоциированного вируса.
[0033] В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV2. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I587 капсидного белка VP1 AAV2. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между N587 и R588 капсидного белка VP1 AAV2, например, в некоторых вариантах осуществления нуклеиновая кислота, как описано, кодирует аминокислотную последовательность, предусматривающую аминокислотную последовательность, представленную под SEQ ID NO: 2.
[0034] В некоторых вариантах осуществления нуклеиновая кислота, как
описано в данном документе, содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV6. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I585 капсидного белка VP1 AAV6. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между Q585 и S586 капсидного белка VP1 AAV6, например, в некоторых вариантах осуществления нуклеиновая кислота, как описано, кодирует аминокислотную последовательность, предусматривающую аминокислотную последовательность, представленную под SEQ ID NO: 4.
[0035] В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV8. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I590 капсидного белка VP1 AAV8. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между N590 и Т591 капсидного белка VP1 AAV8, например, в некоторых вариантах осуществления нуклеиновая кислота, как описано, кодирует аминокислотную последовательность, предусматривающую аминокислотную последовательность, представленную под SEQ ID NO: 25.
[0036] В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV9. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I453 капсидного белка VP1 AAV9. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между G453 и S454 капсидного белка VP1 AAV9, например, в некоторых вариантах осуществления нуклеиновая кислота, как описано, кодирует аминокислотную последовательность, предусматривающую аминокислотную последовательность, представленную под SEQ ID NO: 26. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную в I589 капсидного белка VP1 AAV9. В некоторых вариантах осуществления нуклеиновая кислота, как описано в данном документе, содержит нуклеотидную последовательность, которая кодирует EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между А589 и Q590 капсидного белка VP1 AAV9, например, в некоторых вариантах осуществления нуклеиновая кислота, как описано, кодирует аминокислотную последовательность, предусматривающую аминокислотную последовательность, представленную под SEQ ID NO: 27.
[0037] Как правило, рекомбинантные вирусные векторы, как описано в данном документе, содержат вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, как описано в данном документе, где вирусный капсид инкапсулирует представляющий интерес нуклеотид. В некоторых вариантах осуществления представляющий интерес нуклеотид находится под контролем промотора, выбранного из группы, состоящей из вирусного промотора, бактериального промотора, промотора млекопитающего, промотора птицы, промотора рыбы, промотора насекомого и любой их комбинации. В некоторых вариантах осуществления представляющий интерес нуклеотид находится под контролем промотора, отличного от промотора человека. В некоторых вариантах осуществления промотор представляет собой промотор цитомегаловируса (CMV). В некоторых вариантах осуществления промотор представляет собой промотор EF1α.
[0038] Как правило, представляющий интерес нуклеотид может представлять собой один или более генов, которые могут кодировать выявляемый маркер, например, репортер, или терапевтический полипептид. В некоторых вариантах осуществления представляющим интерес нуклеотидом является репортерный ген. В некоторых вариантах осуществления представляющим интерес нуклеотидом является репортерный ген, который кодирует выявляемый маркер, выбранный из группы, состоящей из зеленого флуоресцентного белка, люциферазы, β-галактозидазы и т.д. В некоторых вариантах осуществления выявляемый маркер представляет собой зеленый флуоресцентний белок. В других вариантах осуществления представляющий интерес нуклеотид выбран из группы, состоящей из суицидного гена, нуклеотида, кодирующего антитело или его фрагмент, нуклеотида, кодирующего систему CRISPR/Cas или ее часть (части), нуклеотида, кодирующего антисмысловую РНК, нуклеотида, кодирующего siRNA, секретируемый фермент и т.д. В одном варианте осуществления представляющий интерес нуклеотид кодирует полидоменное терапевтическое средство, например, белок, который содержит по меньшей мере два домена, обеспечивающих две разные функции.
[0039] Композиции, описанные в данном документе, обычно содержат вирусный вектор, который содержит рекомбинантный вирусный капсидный белок, как описано в данном документе, например, содержит капсид, содержащий рекомбинантный вирусный капсидный белок, где капсид инкапсулирует представляющий интерес нуклеотид. В некоторых вариантах осуществления композиция, описанная в данном документе, содержит (1) вирусный вектор, имеющий капсид, содержащий рекомбинантный вирусный капсидный белок, генетически модифицированный для включения гетерологичного эпитопа, (2) полиспецифическую, необязательно биспецифическую, связывающую молекулу, содержащую (i) паратоп антитела, который специфически связывает эпитоп, и (ii) перенацеливающий лиганд, который специфически связывает рецептор, и, необязательно, (3) фармацевтически приемлемый носитель.
[0040] Паратоп антитела, как описано в данном документе, обычно содержит по меньшей мере определяющую комплементарность область (CDR), которая специфически распознает гетерологичный эпитоп, например, область CDR3 вариабельного домена тяжелой и/или легкой цепи. В некоторых вариантах осуществления полиспецифическая, необязательно биспецифическая, связывающая молекула содержит антитело (или его часть), которое содержит паратоп антитела, который специфически связывает гетерологичный эпитоп. Например, полиспецифическая, необязательно биспецифическая, связывающая молекула может содержать однодоменную вариабельную область тяжелой цепи или однодоменную вариабельную область легкой цепи, где однодоменная вариабельная область тяжелой цепи или однодоменная вариабельная область легкой цепи содержит паратоп антитела, который специфически связывает гетерологичный эпитоп. В некоторых вариантах осуществления полиспецифическая, необязательно биспецифическая, связывающая молекула может содержать Fv-область, например, полиспецифическая, необязательно биспецифическая, связывающая молекула может содержать scFv, содержащую паратоп антитела, который специфически связывает гетерологичный эпитоп. В некоторых вариантах осуществления полиспецифическая, необязательно биспецифическая, связывающая молекула содержит антитело (или его часть), содержащее паратоп антитела, который специфически связывает гетерологичный эпитоп, где антитело (или его часть) дополнительно содержит один или более константных доменов антитела (например, константный домен тяжелой цепи (например, СН1, шарнир, СН2, СН3, СН4 и т.д.) и/или константный домен легкой цепи (например, CL), где один или более константных доменов антитела не связываются с гетерологичным эпитопом.
[0041] Полиспецифическая, необязательно биспецифическая, связывающая молекула, как описано в данном документе, дополнительно содержит перенацеливающий лиганд в дополнение к паратопу (например, антителу или его части, содержащей паратоп), который специфически связывает гетерологичный эпитоп, вставленный в рекомбинантный вирусный капсидный белок/отображен с его помощью. В некоторых вариантах осуществления перенацеливающий лиганд специфически связывает рецептор на поверхности гранулы (например, для выделения и/или очищения рекомбинантного вирусного капсидного белка, как описано в данном документе). В некоторых вариантах осуществления перенацеливающий лиганд специфически связывает белок клеточной поверхности, например, рецептор, маркер клеточной поверхности и т.д., экспрессируемый на поверхности эукариотической клетки млекопитающего (например, человека), например, клетки-мишени. В некоторых вариантах осуществления перенацеливающий лиганд связывает (человеческую) клетку печени, (человеческую) клетку головного мозга, (человеческую) Т-клетку, (человеческую) клетку почки, (человеческую) клетку кишечника, (человеческую) клетку поджелудочной железы, (человеческую) раковую клетку и/или (человеческую) клетку, инфицированную гетерологичным патогеном. В некоторых вариантах осуществления перенацеливающий лиганд связывает специфический маркер (человеческой) клетки печени, специфический маркер (человеческой) клетки головного мозга, специфический маркер (человеческой) Т-клетки, специфический маркер (человеческой) клетки почки, специфический маркер (человеческой) клетки кишечника, специфический маркер (человеческой) клетки поджелудочной железы, специфический маркер (человеческой) опухолевой клетки и/или патогенный эпитоп.
[0042] В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый клеткой печени (человека), например, асиалогликопротеиновый рецептор, например, hASGR1. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый (человеческой) нервной клеткой, например, GABA, трансферрин и т.д. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый Т-клеткой (человека), например, CD3, например, CD3ε. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый (человеческой) гемопоэтической стволовой клеткой, например, CD34. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый клеткой почки (человека). В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый мышечной клеткой (человека), например, интегрин. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый (человеческой) раковой клеткой, например, опухолеассоциированный антиген, например, адипофилин, AIM-2, ALDH1A1, альфа-актинин-4, альфа-фетопротеин («AFP»), ARTC1, B-RAF, BAGE-1, BCLX (L), слитый белок BCR-ABL b3a2, бета-катенин, BING-4, СА-125, CALCA, онкоэмбриональный антиген («СЕА»), CASP-5, CASP-8, CD274, CD45, Cdc27, CDK12, CDK4, CDKN2A, СЕА, CLPP, СОА-1, CPSF, CSNK1A1, CTAG1, CTAG2, циклин D1, циклин-А1, слитый белок dek-can, DKK1, EFTUD2, фактор элонгации 2, ENAH (hMena), Ер-САМ, ЕрСАМ, EphA3, эпителиальный опухолевый антиген («ЕТА»), слитый белок ETV6-AML1, EZH2, Е6, Е7, FGF5, FLT3-ITD, FN1, G250/MN/CAIX, GAGE-1,2,8, GAGE-3,4,5,6,7, GAS7, глипикан-3, GnTV, gp100/Pme117, GPNMB, HAUS3, гепсин, HER-2/neu, HERV-K-MEL, HLA-A11, HLA-A2, HLA-DOB, hsp70-2, IDO1, IGF2B3, IL13Ralpha2, карбоксилэстеразу кишечника, K-ras, калликреин 4, KIF20A, KK-LC-1, KKLC1, KM-HN-1, KMHN1, также известный как CCDC110, LAGE-1, слитый белок LDLR-фукозилтрансферазу АS, ленгзин, M-CSF, MAGE-A1, MAGE-A10, MAGE-A12, MAGE-A2, MAGE-А3, MAGE-A4, MAGE-A6, MAGE-A9, MAGE-C1, MAGE-C2, маликфермент, маммаглобин-А, MART2, MATN, MC1R, MCSP, mdm-2, ME1, Melan-A/MART-1, Meloe, Midkine, MMP-2, MMP-7, MUC1, MUC5AC, муцин, MUM-1, MUM-2, MUM-3, миозин, миозин класса I, N-raw, NA88-A, нео-РАР, NFYC, NY-BR-1, NY-ESO-1/LAGE-2, OA1, OGT, OS-9, полипептид P, p53, PAP, PAX5, PBF, слитый белок pml-RARalpha, полиморфный эпителиальный муцин («РЕМ»), PPP1R3B, PRAME, PRDX5, PSA, PSMA, PTPRK, RAB38/NY-MEL-1, RAGE-1, RBAF600, RGS5, RhoC, RNF43, RU2AS, SAGE, сецернин 1, SIRT2, SNRPD1, SOX10, Sp17, SPA17, SSX-2, SSX-4, STEAP1, сурвивин, слитый белок SYT-SSX1 или -SSX2, TAG-1, TAG-2, теломеразу, TGF-betaRII, TPBG, TRAG-3, триозофосфатизомеразу, TRP-1/gp75, TRP-2, TRP2-INT2, тирозиназу, тирозиназу («TYR»), VEGF, WT1, XAGE-1b/GAGED2a, Kras, NY-ESO1, MAGE-A3, HPV E2, HPV E6, HPV E7, антиген WT-1 (при лимфоме и других солидных опухолях), рецепторы ErbB, Melan A [MART1], gp 100, тирозиназу, TRP-1/gp 75 и TRP-2 (при меланоме); MAGE-1 и MAGE-3 (при карциноме мочевого пузыря, головы и шеи и немелкоклеточной карциноме); белки EG и Е7 HPV (при раке шейки матки); муцин [MUC-1] (при раке молочной железы, поджелудочной железы, толстой кишки и предстательной железы); специфический антиген простаты [PSA] (при раке простаты); онкоэмбриональный антиген [СЕА] (при раке толстой кишки, молочной железы и желудочно-кишечного тракта) и такие общие опухолеспецифические антигены, как MAGE-2, MAGE-4, MAGE-6, MAGE-10, MAGE-12, BAGE-1, CAGE-1,2,8, CAGE-3-7, LAGE-1, NY-ESO-1/LAGE-2, NA-88, GnTV, TRP2-INT2 и т.д. В некоторых вариантах осуществления перенацеливающий лиганд связывает Е6 и/или Е7. В некоторых вариантах осуществления перенацеливающий лиганд связывает Her2. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор глюкагона человека (hGCGR). В некоторых вариантах осуществления перенацеливающий лиганд связывает эктонуклеозидтрифосфатдифосфогидролазу 3 (hENTPD3) человека.
[0043] В некоторых вариантах осуществления паратоп (например, антитело или его часть) и перенацеливающий лиганд непосредственно слиты друг с другом. В некоторых вариантах осуществления паратоп (например, антитело или его часть), который специфически связывает гетерологичный эпитоп и перенацеливающий лиганд, ковалентно присоединены друг к другу.
[0044] В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен содержит паратоп, который специфически связывает гетерологичный эпитоп, вставленный в рекомбинантный вирусный капсидный белок/отображенный с его помощью, и второй антигенсвязывающий домен, который специфически связывает белок клеточной поверхности, экспрессируемый клеткой-мишенью. В некоторых вариантах осуществления биспецифическая связывающая молекула представляет собой биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен содержит паратоп, который специфически связывает гетерологичный эпитоп, вставлен в рекомбинантный вирусный капсидный белок/отображенный с его помощью, и второй антигенсвязывающий домен специфически связывает рецептор, экспрессируемый клеткой-мишенью, где первый антигенсвязывающий домен функционально связан с первой областью тяжелой цепи, содержащей первый СН3-домен, где второй антигенсвязывающий домен функционально связан со второй областью тяжелой цепи, содержащей второй СН3-домен, где первый и второй CH3-домены Ig отличаются друг от друга по меньшей мере одной аминокислотой, и где по меньшей мере одно аминокислотное различие снижает связывание биспецифического антитела с белком А по сравнению с биспецифическим антителом, в котором отсутствует это аминокислотное различие. В одном варианте осуществления первый CH3-домен Ig связывает белок А, а второй CH3-домен Ig содержит мутацию, которая уменьшает или устраняет связывание белка А, такую как модификация H95R (нумерация экзонов согласно IMGT; H435R нумерация согласно EU). Второй CH3-домен может дополнительно содержать модификацию Y96F (по IMGT; Y436F по EU). Дополнительные модификации, которые можно обнаружить в пределах второго CH3-домена, включают: D16E, L18M, N44S, K52N, V57M и V82I (по IMGT; D356E, L358M, N384S, K392N, V397M и V422I по EU) в случае антител IgG1; N44S, K52N и V82I (IMGT; N384S, K392N и V422I по EU) в случае антител IgG2; и Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (по IMGT; Q355R, N384S, K392N, V397M, R409K, E419Q и V422I по EU) в случае антител IgG4.
[0045] В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например, биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен связывает аффинную метку, отображаемую рекомбинантным вирусным капсидным белком, как описано в данном документе, и где второй антигенсвязывающий домен связывает рецептор, экспрессируемый на поверхности клетки-мишени. В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например, биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6), отображаемую рекомбинантным вирусным капсидным белком, как описано в данном документе, и где второй антигенсвязывающий домен связывает рецептор, экспрессируемый на поверхности клетки-мишени. В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например, биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6), отображаемую рекомбинантным вирусным капсидным белком, как описано в данном документе, и где второй антигенсвязывающий домен связывает hASGR1. В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например, биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6), отображаемую рекомбинантным вирусным капсидным белком, как описано в данном документе, и где второй антигенсвязывающий домен связывает белок CD3, например, CD3ε. В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например, биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6), отображаемую рекомбинантным вирусным капсидным белком, как описано в данном документе, и где второй антигенсвязывающий домен связывает интегрин. В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например, биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6), отображаемую рекомбинантным вирусным капсидным белком, как описано в данном документе, и где второй антигенсвязывающий домен связывает интегрин, например, hGCGR. В некоторых вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическую связывающую молекулу, например, биспецифическое антитело, содержащее первый и второй антигенсвязывающие домены, где первый антигенсвязывающий домен связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6), отображаемую рекомбинантным вирусным капсидным белком, как описано в данном документе, и где второй антигенсвязывающий домен связывает ENTPD3.
[0046] Также в данном документе описаны способы получения и применения рекомбинантных вирусных капсидных белков, вирусных векторов, содержащих их, композиций и т.д. В некоторых вариантах осуществления способы перенаправления вируса, например, аденовируса, аденоассоциированного вируса и т.д.; доставки диагностического/терапевтического груза в клетку-мишень и т.д. включают объединение рекомбинантного вирусного вектора, содержащего рекомбинантный вирусный капсидный белок, как описано в данном документе, например, вирусный вектор, содержащий капсид, содержащий рекомбинантный вирусный капсид, отображающий гетерологичный эпитоп, с биспецифической связывающей молекулой, где биспецифическая связывающая молекула содержит (i) паратоп антитела, который специфически связывает эпитоп, и (ii) перенацеливающий лиганд, который специфически связывает рецептор. Такие способы могут включать в качестве первой стадии получение рекомбинантного вирусного вектора, например, культивирование упаковывающей клетки в условиях, достаточных для продуцирования вирусных векторов, где упаковывающая клетка содержит плазмиду, кодирующую капсидный белок, содержащий эпитоп. При доставке диагностического/терапевтического груза в клетку-мишень описанные в данном документе способы могут включать приведение клетки-мишени в контакт с комбинацией вирусного вектора, который содержит капсид, включая рекомбинантный вирусный капсид, отображающий гетерологичный эпитоп, и полиспецифическую связывающую молекулу, где полиспецифическая связывающая молекула содержит i) паратоп антитела, который специфически связывает эпитоп, и (ii) перенацеливающий лиганд, который специфически связывает рецептор, экспрессируемый клеткой-мишенью. В некоторых вариантах осуществления клетка-мишень находится в условиях in vitro. В других вариантах осуществления клетка-мишень находится в условиях in vivo у субъекта, например, человека.
[0047] В некоторых вариантах осуществления в композиции, описанной в данном документе, или в способе, описанном в данном документе, объединяют рекомбинантный вирусный вектор, содержащий представляющий интерес нуклеотид, заключенный в капсид, содержащий рекомбинантный капсидный белок, как описано в данном документе, и полиспецифическую связывающую молекулу в соотношении молекула:молекула, которое восстанавливает эффективность трансдукции вирусного вектора, сходную с эффективностью эталонного вирусного вектора. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула: молекула) находится в диапазоне от 1:0,5 до 1:100. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) находится в диапазоне от 1:4 до 1:20. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к биспецифической связывающей молекуле (молекула:молекула) находится в диапазоне от 1:8 до 1:15. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:4. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:8. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:15. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:20.
[0048] В данном документе также описаны способы инактивации вирусного капсида и/или получения вирусных векторов, при этом способы обычно включают (а) вставку нуклеиновой кислоты, кодирующей гетерологичный белок, в последовательность нуклеиновой кислоты, кодирующую вирусный капсидный белок, с образованием нуклеотидной последовательности, кодирующей генетически модифицированный капсидный белок, включая гетерологичный белок, и/или (b) культивирование упаковывающей клетки в условиях, достаточных для продуцирования вирусных векторов, где упаковывающая клетка содержит нуклеотидную последовательность. В некоторых вариантах осуществления упаковывающая клетка дополнительно содержит плазмиду-помощника и/или плазмиду для переноса, содержащую представляющий интерес нуклеотид. В некоторых вариантах осуществления способы дополнительно включают выделение самокомплементарных векторов на основе аденоассоциированного вируса из культурального супернатанта. В некоторых вариантах осуществления способы дополнительно включают лизис упаковывающей клетки и выделение однонитевых векторов на основе аденоассоциированного вируса из клеточного лизата. В некоторых вариантах осуществления способы дополнительно включают (а) удаление клеточного дебриса, (b) обработку супернатанта, содержащего вирусные векторы, ДНКазой I и MgCl2, (с) концентрирование вирусных векторов, (d) очистку вирусных векторов и (е) любую комбинацию (a)-(d). В данном документе также предусмотрен вирусный вектор, полученный в соответствии со способом, описанным в данном документе, и упаковывающая клетка, полезная при получении вирусного вектора, как описано в данном документе, например, упаковывающие клетки, содержащие плазмиду, кодирующую описанный рекомбинантный капсидный белок.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0049] Файл патента или приложения содержит по меньшей мере один графический материал, выполненный в цвете. Копии данного патента или публикации заявки на патент с цветным(-и) графическим(-и) материалом(-и) будут предоставлены Ведомством после запроса и уплаты необходимой пошлины.
[0050] На фигуре 1 представлены изображения иммунофлуоресцентной микроскопии (верхняя панель) или гистограммы, полученные при сортировке флуоресцентно-активированных клеток (FACS) (нижняя панель), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками HepG2, инкубированными с (А) вирусными векторами scAAV2-CMV-eGFP дикого типа отдельно; (В) вирусными векторами scAAV2-N587Myc-CMV-hrGFP отдельно; вирусными векторами scAAV2-N587Myc в смеси с биспецифическими антителами к Myc-ASGR1 при следующих соотношениях: (С) 1:0,5, (D) 1:1, (Е) 1:2, (F) 1:4, (G) 1:8, (Н) 1:15, (I) 1:20, (J) 1:50 или (K) 1:100; (L) или вирусными векторами scAAV-N587Myc в смеси с моноспецифическим антителом к Мус в соотношении 1:8.
[0051] На фигуре 2А представлены точечные графики, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 29T3-hASGR1, инкубированными с вирусными векторами на основе scAAV2 дикого типа отдельно (i), вирусными векторами scAAV2-N587Myc-CMV-eGFP отдельно (ii), вирусными векторами scAAV2-N587Myc-CMV-eGFP в смеси с биспецифическими антителами к Myc-ASGR1 при следующих соотношениях: 1:0,5 (iii), 1:1 (iv), 1:2 (v), 1:4 (vi), 1:8 (vii), 1:15 (viii), 1:20 (ix) или 1:100 (x), или вирусными векторами scAAV2-N587Myc-CMV-eGFP в смеси с нерелевантным биспецифическим антителом к Myc-GCGR в соотношении 1:8 (xii). Также показана экспрессия GFP клетками 29Т3, инкубированными с вирусными векторами scAAV-N587Myc в смеси с биспецифическими антителами к Myc-ASGR1 в соотношении 1:8 (xi). Для каждого эксперимента использовали 2×105 клеток и 5×109 вирусных векторов.
[0052] На фигуре 2В представлены гистограммы, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 29T3-hASGR1, инкубированными с немодифицированными вирусными векторами AAV9-CAGG-GFP отдельно (i) вирусными векторами AAV9-A589Myc-CAGG-eGFP отдельно (ii), вирусными векторами AAV9-A589Myc-CAGG-eGFP в смеси с биспецифическими антителами к Myc-ASGR1 при следующих соотношениях: 1:1 (iii), 1:2 (iv), 1:4 (v), 1:8 (vi), 1:20 (vii), 1:50 (viii) или 1:100 (ix). Для каждого эксперимента использовали 2×105 клеток и 1×1010 вирусных векторов (титрованных с помощью qPCR).
[0053] На фигуре 3 представлены точечные графики, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 29T3-hASGR1, предварительно инкубированными с бивалентным антителом к ASGR1 в концентрации 0 нМ (С), 50 нМ (D), 10 нМ (Е), 2 нМ (F), 0,4 нМ (G), 0,08 нМ (Н), 0,016 нМ (I) или 0,0032 нМ (J), и затем инфицированными вирусными векторами scAAV2-N587Myc-CMV-eGFP в смеси с биспецифическими антителами к Myc-ASGR1 в соотношении 1:8 (L). Клетки 293T3-hASGR1, инкубированные с вирусными векторами на основе scAAV дикого типа отдельно (А) или с вирусными векторами scAAV2-N587Myc-CMV-eGFP отдельно (В), выступали в качестве контроля. Для каждого эксперимента использовали 2×105 клеток и 5×109 вирусных векторов (титрованных с помощью qPCR).
[0054] На фигуре 4 представлены изображения иммунофлуоресцентной микроскопии для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 293T-hASGR1, инкубированными с вирусными векторами на основе scAAV дикого типа отдельно (А), вирусными векторами scAAV2-N587Myc-CMV-eGFP отдельно (В), или последовательно инкубированными с 1×109 (С), 2×109 (D), 4×109 (Е), 8×109 (F), 2×1010 (G), 1×1011 (Н), 1×1012 (I) биспецифических антител к Myc-ASGR1 и затем с 1×109 вирусных векторов scAAV2-N587Myc-CMV-eGFP. Также показаны изображения иммунофлуоресцентной микроскопии клеток 293Т, последовательно инкубированных с 1×1011 молекул антитела к myc-ASGR1 и затем с 1×109 вирусных векторов scAAV2-N587Myc-CMV-eGFP (J) и клеток 293T-hASGR1, последовательно инкубированных с 1×1011 молекул нерелевантного биспецифического антитела к Мус-GCGR и затем с 1×109 вирусных векторов scAAV2-N587Myc-CMV-eGFP (K).
[0055] На фигуре 5 представлены точечные графики, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 29T3-hASGR1, инкубированными с вирусными векторами на основе ssAAV дикого типа отдельно (А), вирусными векторами ssAAV2-N587Myc-CMV-hrGFP отдельно (В), вирусными векторами ssAAV2-N587Myc-CMV-hrGFP в смеси с биспецифическими антителами к Myc-ASGR1 при следующих соотношениях: 1:1 (С), 1:2 (D), 1:4 (Е), 1:8 (F), 1:20 (G), 1:100 (Н), 1:1000 (I), или с вирусными векторами ssAAV-N587Myc в смеси с нерелевантным биспецифическим антителом к Myc-GCGR в соотношении 1:8 (K). Также показана экспрессия GFP клетками 29Т3, инкубированными с вирусными векторами scAAV-N587Myc в смеси с биспецифическими антителами к MycASGR1 в соотношении 1:8 (J). Для каждого эксперимента использовали 2×105 клеток и 5×109 вирусных векторов.
[0056] На фигуре 6 представлены точечные графики, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 29T3-hGCGR, инкубированными с вирусными векторами на основе scAAV дикого типа отдельно (А), вирусными векторами scAAV2-N587Myc-CMV-eGFP отдельно (В), вирусными векторами scAAV2-N587Myc-CMV-eGFP в смеси с биспецифическими антителами к Myc-GCGR при следующих соотношениях: 1:0,5 (С), 1:1 (D), 1:2 (Е), 1:4 (F), 1:8 (G), 1:15 (Н), 1:20 (I), 1:50 (J) или 1:100 (K), или с вирусными векторами scAAV-2N587Myc-CMV-eGFP в смеси с нерелевантным моноспецифическим антителом к Мус (Regeneron Pharmaceuticals, Тэрритаун, Нью-Йорк) в соотношении 1:8 (L). Для каждого эксперимента использовали 2×105 клеток и 5×109 вирусных векторов.
[0057] На фигуре 7 представлены точечные графики, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками Jurkat отдельно (А) или клетками Jurkat, инкубированными с вирусными векторами scAAV6-EF1-eGFP на основе дикого типа отдельно (В), вирусными векторами AAV6-Q585Myc-EF1a-eGFP отдельно (С), вирусными векторами AAV6-Q585Myc-EF1a-eGFP в смеси с биспецифическими антителами к Myc-CD3 при следующих соотношениях: 1:1 (D), 1:5 (Е), 1:10 (F), 1:100 или (G), 1:1000 (Н). Для каждого эксперимента использовали 2×105 клеток и 1×109 вирусных векторов.
[0058] На фигуре 8А представлены изображения иммунофлуоресцентной микроскопии печени. На фигуре 8В представлены изображения иммунофлуоресцентной микроскопии селезенки. На фигуре 8С представлены изображения иммунофлуоресцентной микроскопии почки. Все образцы взяты у мышей C57BL/6, трансгенно модифицированных для экспрессии ASGR1 человека клетками печени (i-iv), или мышей C57BL/6 дикого типа (v-viii) через десять дней после внутривенной инъекции 1×1011 scAAV2-CMV-eGFP дикого типа (i, v), физиологического раствора (ii, vi), 1×1011 вирусных векторов scAAV2-N587myc-CMV-eGFP отдельно (iii, vii) или вирусных векторов scAAV2-N587myc-CMV-eGFP с биспецифическим антителом к myc-ASGR1 (iv, viii).
[0059] На фигуре 9 представлены изображения иммунофлуоресцентной микроскопии образцов печени, взятых у мышей C57BL/6, трансгенно модифицированных для экспрессии ASGR1 человека на клетках печени (D-F, J-L, Р-R), или мышей C57BL/6 дикого типа (А-С, G-I, М-О) через четыре недели после внутривенной инъекции 2,18×1011 ssAAV2-CAGG-eGFP дикого типа (В, С, Е, F), физиологического раствора (A, D), 2,18×1011 вирусных векторов ssAAV2-N587myc-CAGG-eGFP отдельно (G-I, J-L) или вирусных векторов ssAAV2-N587myc-CAGG-eGFP с биспецифическим антителом к myc-ASGR1 (М-О, P-R). Каждое изображение представляет одну мышь.
[0060] На фигуре 10 представлены изображения иммунофлуоресцентной микроскопии образцов печени, взятых у мышей C57BL/6, трансгенно модифицированных для экспрессии ASGR1 человека клетками печени, через десять дней после внутривенной инъекции (A) AAV9 дикого типа, (В) 250 нМ NaCl (С) вирусных частиц AAV9-A589myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-hCD3 или (D) вирусных частиц AAV9-A589myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1.
[0061] На фигуре 11 представлены иллюстративные, без соблюдения масштаба и не ограничивающие примеры форматов полиспецифических связывающих молекул, полезных в некоторых вариантах осуществления настоящего изобретения.
[0062] На фигуре 12 представлены точечные графики, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 29T3-hASGR1, инкубированными с (А)_ векторами на основе вируса AAV8 дикого типа отдельно, (С) вирусными векторами AA8-N590-myc отдельно или вирусными векторами pAAV RC8 N590myc в смеси с биспецифическими молекулами антитела к hASGR1-IgG4-Fc/антитела к myc при следующих соотношениях (D) 1:1, (Е) 1:2, (F) 1:4, (G) 1:8, (Н) 1:12, (I) 1:15, (J) 1:50 или (K) 1:100. Также показана экспрессия GFP ложно-трансфицированными клетками 29T3-hASGR1 (В). Для каждого эксперимента использовали 2×105 клеток и 1×109 вирусных векторов.
[0063] На фигуре 13 представлены изображения иммунофлуоресцентной микроскопии образцов печени, взятых у мышей C57BL/6, трансгенно модифицированных для экспрессии ASGR1 человека клетками печени, через десять дней после внутривенной инъекции (А)-(С) AAV8 дикого типа, (D)-(F) вирусных векторов AA8-N590-myc и контрольной биспецифической связывающей молекулы или (G)-(I) вирусных частиц AAV8 N590myc-CAGG-eGFP в комбинации с биспецифическими связывающими молекулами антитела к hASGR1-IgG4-Fc/антитела к myc.
[0064] На фигуре 14 представлены точечные графики, полученные при сортировке флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками 29T3-h ENTPD3, инкубированными с (А) вирусными векторами на основе AAV2 дикого типа отдельно, (В) вирусными векторами AAV2-N587Myc-CAGG-eGFP отдельно или вирусными векторами AAV2-N587Myc-CAGG-eGFP в смеси с биспецифическими молекулами антитела к hENTPD3-IgG4-Fc/антитела к myc при следующих соотношениях (С) 1:1, (D) 1:2, (Е) 1:4, (F) 1:8, (G) 1:20, (Н) 1:50, (I) 1:100 или (K) 1:200. Для каждого эксперимента использовали 2×105 клеток и 1×109 вирусных векторов.
[0065] На фигуре 15А представлены изображения иммунофлуоресцентной микроскопии образцов печени, на фигуре 15В представлены изображения иммунофлуоресцентной микроскопии образцов кишечника и на фигуре 15С представлены изображения иммунофлуоресцентной микроскопии образцов поджелудочной железы. Все образцы взяты у мышей C57BL/6 дикого типа через десять дней после внутривенной инъекции PBS (15A(i), 15B(i) и 15C(i)), 5×1011 AAV9 дикого типа (15A(ii), 15B(ii) и 15C(ii)), 5×1011 вирусных векторов AAV2-N587myc-CAGG-eGFP с 1×103 нерелевантных биспецифических связывающих белков IgG4-Fc/антител к myc (15A(iii), 15В(iii), и 15С(iii)) или 5×1011 вирусных векторов AAV2-N587myc-CAGG-eGFP с 1×1013 биспецифических связывающих белков hENTPD3-IgG4-Fc/антител к myc (15A(iv), 15B(iv) и 15C(iv)).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0066] Общей проблемой адапторных подходов с применением немодифицированных вирусных капсидов или вирусных капсидов с модифицированным каркасом, была субоптимальная эффективность трансдукции модифицированных капсидов. (Grifman et al. (2001) Mol. Ther. 3:964-75). Например, Curiel et al. описывают получение и характеристику рекомбинантного аденовирусного вектора, содержащего волокна с последовательностью RGD-4C, генетически включенной в петлю HI карбоксиконцевого knob-домена, и демонстрируют полезность петли HI в knob-домене волоконного белка как оптимального сайта для включения коротких пептидных лигандов. См., например, патент США №7297542; см., также, Beany and Curiel (2012) Adv. Cancer. Res. 115:39-67. Аналогично, вставка пептидов-лигандов в капсидные белки AAV приводила к образованию капсидов, которые были способны отображать лиганд на поверхности капсида и опосредовать трансдукцию путем взаимодействия лиганда с его рецептором, таким образом перенаправляя вирусный тропизм с помощью генетических модификаций капсида (Girod et al. (1999) Nat. Med. 5(9): 1052-6, 1438 (опечатки включены) (1999); Grifman et al. (2001) Mol. Ther. 3(6):964-75; Nicklin et al. (2001) Mol. Ther. 4(3): 174-81; Shi et al. (2001) Hum Gene Ther. 17(3):353-61 (2006); Wu et al. (2000) J. Virol. 74(18):8635-47). В частности, было продемонстрировано, что вставка связывающего интегрин мотива Arg-Gly-Asp (RGD) в сайт вставки 1-587 капсидного белка VP1 AAV позволяет векторам на основе вируса AAV трансдуцировать клетки с помощью интегринов αvβ1 (Girod et al. (1999) выше). В отличие от этого, хотя вставка пептида из 14 аминокислот L14 после аминокислоты R447 (1-447) приводила к образованию капсидов, все еще распознаваемых конформационно-чувствительным антителом А20, такие рекомбинантные вирусные векторы не могли трансдуцировать клетки, экспрессирующие рецептор L-14 (Girod et al., 1999; ср. Wu et al. (2000) (сообщается о вставке пептида гемагглютинина (НА) в положении 1-447 и успешной трансдукции клеток, экспрессирующих пептид НА). Вставка эпитопа Мус между Т448 и N449 распознавалась антителом к Мус и, следовательно, присутствовала на поверхности капсида, но приводила к образованию инактивированных вирусных векторов. (Grifman et al., 2001). В отличие от этого, об успешном перенацеливании снова сообщалось в отношении вставки мотива NGR после N587, но не для вставки с-myc после N587. (Grifman et al., 2001). В патенте США №9624274 описано I-453 капсидного белка AAV в качестве подходящего сайта вставки для гетерологичного эпитопа. Хотя эти исследования демонстрируют успешную вставку и отображение гетерологичного пептида, например, эпитопа, капсидными белками AAV, ни в одном из этих исследований не предполагается, что полиспецифическая связывающая молекула, например, биспецифическая связывающая молекула, такая как биспецифическое антитело, которое специфически связывает гетерологичный пептид и белок клеточной поверхности, может изменять мишень модифицированных вирусных векторов на клетки, экспрессирующие белок клеточной поверхности, и восстанавливать их эффективность трансдукции.
[0067] В данном документе раскрыты рекомбинантные вирусные капсидные белки, которые модифицированы гетерологичным эпитопом, который можно использовать в связи с полиспецифической связывающей молекулой, содержащей паратоп, например Fv-домен, который специфически связывает эпитоп, и лиганд, который связывает рецептор, экспрессируемый на поверхности клетки-мишени. Как показано в данном документе, приведение в контакт вирусных векторов, имеющих капсиды, образованные с помощью капсидных белков, описанных в данном документе, с полиспецифической связывающей молекулой при определенных соотношениях, восстанавливает эффективность трансдукции вирусного капсида до уровней, сопоставимых с вирусом дикого типа, см., например, пример 2. Как правило, капсидные белки, модифицированные гетерологичными эпитопами, как описано в данном документе, можно получить из вируса без оболочки, такого как, без ограничения, аденовирус (Ad) и аденоассоциированный вирус (AAV).
[0068] Хотя настоящее изобретение было конкретно показано и описано со ссылкой на ряд вариантов осуществления, специалистам в данной области техники должно быть понятно, что изменения в форме и деталях могут быть внесены в различные варианты осуществления, раскрытые в данном документе, без отступления от сущности и объема настоящего изобретения и что различные варианты осуществления, раскрытые в данном документе, не предназначены для ограничения объема формулы изобретения.
[0069] Хотя при практическом осуществлении или тестировании настоящего изобретения можно применять любые способы и материалы, аналогичные описанным в данном документе или эквивалентные им, в данном документе описаны лишь некоторые предпочтительные способы и материалы. Все публикации, упомянутые в данном документе, включены в данный документ посредством ссылки во всей своей полноте. Если не определено иное, то все технические и научные термины, используемые в данном документе, имеют такое же значение, как это обычно понимает специалист в данной области техники, к которой принадлежит настоящее изобретение.
Определения
[0070] Если не определено иное, то все технические и научные термины, используемые в данном документе, имеют такое же значение, как это обычно понимает специалист в данной области техники, к которой принадлежит настоящее изобретение.
[0071] Формы единственного числа включают ссылки на формы множественного числа, если контекст явно не указывает на иное. Таким образом, например, ссылка на «способ» включает один или более способов и/или стадий, описанных в данном документе, и/или которые станут очевидными специалистам в данной области техники при прочтении настоящего раскрытия.
[0072] Термин «антитело» предусматривает молекулы иммуноглобулинов, содержащие четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь содержит вариабельный домен тяжелой цепи (VH) и константную область тяжелой цепи (CH). Константная область тяжелой цепи содержит по меньшей мере три домена, CH1, CH2, CH3 и необязательно CH4. Каждая легкая цепь содержит вариабельный домен легкой цепи (CH) и константную область легкой цепи (CL). Вариабельные домены тяжелой цепи и легкой цепи могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), чередующиеся с более консервативными областями, называемыми остовными областями (FR). Каждый вариабельный домен тяжелой и легкой цепи содержит три CDR и четыре FR, расположенные от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи могут быть сокращены как HCDR1, HCDR2 и HCDR3; CDR легкой цепи могут быть сокращены как LCDR1, LCDR2 и LCDR3). Типичные антитела с тетрамерной структурой содержат два идентичных антигенсвязывающих домена, каждый из которых образован при ассоциации VH- и VL-доменов, и каждый из которых вместе с соответствующими CH- и СLдоменами образует Fv-область антитела. Однодоменные антитела содержат один антигенсвязывающий домен, например, VH или VL. Термин «антитело» охватывает моноклональные антитела, полиспецифические (например, биспецифические) антитела, человеческие антитела, гуманизированные антитела, химерные антитела, одноцепочечные Fv (scFv), одноцепочечные антитела, Fab-фрагменты, F(ab’)-фрагменты, связанные дисульфидом Fv (sdFv), интратела, минитела, диатела и антиидиотипические (anti-Id) антитела (включая, например, антитела anti-Id к антигенспецифичным TCR) и эпитоп-связывающие фрагменты любого из вышеперечисленного. Термины «антитело» и «антитела» также относятся к ковалентным диателам, таким как раскрытые в публикации заявки на патент США №2007/0004909 и Ig-DARTS, таким как раскрытые в публикации заявки на патент США №2009/0060910. Антитела включают молекулы иммуноглобулина и иммунологически активные фрагменты молекул иммуноглобулина, т.е. молекулы, которые содержат антигенсвязывающий участок. Молекулы иммуноглобулина могут относиться к любому типу (например, IgG, IgE, IgM, IgD, IgA и IgY), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу.
[0073] Антигенсвязывающий домен антитела, например часть антитела, которая распознает и связывается с эпитопом антигена, также называется «паратопом». Это небольшой участок (от 5 до 10 аминокислот) Fc-области антитела, часть антигенсвязывающего фрагмента (Fab-область), и может содержать части тяжелой и/или легкой цепей антитела. Паратоп специфически связывает эпитоп, когда паратоп связывает эпитоп с высокой аффинностью. Термин «высокоаффинное» антитело относится к антителу, которое характеризуется KD по отношению к своему целевому эпитопу, составляющему приблизительно 10-9 М или меньше (например, приблизительно 1×10-9 М, 1×10-10 М, 1×10-11 М или приблизительно 1×10-12 М). В одном варианте осуществления KD измеряют с помощью поверхностного плазмонного резонанса, например, BIACORE™; в другом варианте осуществления, KD измеряют с помощью ELISA.
[0074] Фраза «определяющая комплементарность область» или термин «CDR» включают аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулина организма, которая в норме (т.е. у животного дикого типа) находится между двух остовных областей в вариабельной области легкой или тяжелой цепи молекулы иммуноглобулина (например, антитела или Т-клеточного рецептора). CDR может кодироваться, например, последовательностью зародышевого типа либо перегруппированной или неперегруппированной последовательностью, и, например, непримированной или зрелой В-клеткой или Т-клеткой. CDR может быть подвергшейся соматической мутации (например, отличаться от последовательности, кодируемой в зародышевой линии животного), гуманизированной и/или модифицированной с помощью аминокислотных замен, добавлений или делеций. В некоторых случаях (например, в случае CDR3) CDR могут кодироваться двумя или более последовательностями (например, последовательностями зародышевой линии), которые не являются смежными (например, в случае неперегруппированной последовательности нуклеиновой кислоты), но они являются смежными в последовательности нуклеиновой кислоты в В-клетке, например, как результат сплайсинга или соединения последовательностей (например, V-D-J-рекомбинация с образованием CDR3 тяжелой цепи).
[0075] «Эпитоп» представляет собой часть макромолекулы, которая распознается иммунной системой, в частности антителами, В-клетками или цитотоксическими Т-клетками. Хотя обычно полагают, что эпитопы получены из чужих белков, последовательности, полученные от хозяина, которые могут быть распознаны, также классифицируются как эпитопы. Эпитопы имеют по меньшей мере 4 аминокислоты, предпочтительно от 4 до 30 аминокислот, более предпочтительно от 5 до 20 аминокислот, в частности от 5 до 15 аминокислот в длину. Эпитопы могут быть линейными или трехмерными, обычно образованными аминокислотами, которые удалены друг от друга в первичной структуре белка, но становятся близкородственными во вторичной и/или третичной структуре. Эпитопы, которые специфически распознаются В-клетками, называются В-клеточными эпитопами.
[0076] Фраза «инвертированный концевой повтор» или «ITR» включает симметричные последовательности нуклеиновых кислот в геноме аденоассоциированных вирусов, необходимые для эффективной репликации. Последовательности ITR расположены на каждом конце ДНК-генома AAV. ITR выступают источниками репликации для синтеза вирусной ДНК и являются важными цис-компонентами для получения интегрирующих векторов на основе AAV.
[0077] Фраза «легкая цепь» включает последовательность легкой цепи иммуноглобулина из любого организма, и, если не указано иное, включает κ и λ легкие цепи человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены легкой цепи обычно включают три CDR легкой цепи и четыре остовных (FR) области, если не указано иное. Как правило, полноразмерная легкая цепь включает от аминоконца к карбоксильному концу, вариабельный домен, который включает FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и константную область легкой цепи. Вариабельный домен легкой цепи кодируется последовательностью гена вариабельной области легкой цепи, которая обычно содержит сегменты VL и JL, полученные из репертуара сегментов V и J, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов V и J легкой цепи для различных организмов можно найти в базе данных IMGT, www.imgt.org. Легкие цепи включают, например, цепи, которые селективно не связывают ни первый, ни второй эпитоп, селективно связанный эпитоп-связывающим белком, в котором они встречаются. Легкие цепи также включают цепи, которые связывают и распознают, или помогают тяжелой цепи или другой легкой цепи связываться и распознавать один или более эпитопов, селективно связанных эпитоп-связывающим белком, в котором они встречаются. Обычные или универсальные легкие цепи включают цепи, полученные с помощью гена Vκ1-39Jκ человека или гена Vκ3-20Jκ человека, и включают их соматически мутированные (например, с созревшей аффинностью) версии. Иллюстративные VL-сегменты человека включают сегмент гена Vκ1-39 человека, сегмент гена Vκ3-20 человека, сегмент гена Vλ1-40 человека, сегмент гена Vλ1-44 человека, сегмент гена Yλ2-8 человека, сегмент гена Vλ2-14 человека, сегмент гена Vλ3-21 человека, и включают их соматически мутированные (например, с созревшей аффинностью) версии. Можно получить легкие цепи, которые включают вариабельный домен из одного организма (например, человека или грызуна, например, крысы или мыши; или птицы, например, курицы) и константную область из того же или другого организма (например, человека или грызуна, например, крысы или мыши; или птицы, например, курицы).
[0078] Термин «приблизительно» или «примерно» включает нахождение в пределах статистически значимого диапазона значения. Такой диапазон может быть в пределах порядка величины, предпочтительно в пределах 50%, более предпочтительно в пределах 20%, еще более предпочтительно в пределах 10% и еще более предпочтительно в пределах 5% от данного значения или диапазона. Допустимое отклонение, охватываемое терминами «приблизительно» или «примерно», зависит от конкретной исследуемой системы и может быть легко оценено специалистом в данной области техники.
[0079] Термин «аффинная метка» включает полипептидную последовательность, которая является представителем пары специфического связывания, например, которая специфически связывается с другой полипептидной последовательностью, например, паратопом антитела, с высокой аффинностью. Иллюстративные и неограничивающие аффинные метки включают гексагистидиновую метку, метку FLAG, метку Strep II, метку стрептавидин-связывающего пептида (SBP), кальмодулинсвязывающий пептид (СВР), глутатион-S-трансферазу (GST), мальтозосвязывающий белок (MBP), S-метку, метку НА и метку с-Мус. (Рассмотрено в Zhao et al. (2013) J. Analytical Meth. Chem. 1-8; включенной в данный документ посредством ссылки).
[0080] Термин «капсидный белок» включает белок, который является частью капсида вируса. Для аденоассоциированных вирусов капсидные белки обычно обозначаются как VP1, VP2 и/или VP3, и каждый кодируется единственным геном cap. Для AAV три капсидных белка AAV продуцируются перекрывающимся образом из открытой рамки считывания (ORF) для cap посредством альтернативного сплайсинга mRNA и/или альтернативного использования стартового кодона трансляции, хотя все три белка используют общий стоп-кодон. Warrington et al. (2004) J. Virol. 78:6595, включенная в данный документ посредством ссылки во всей своей полноте. VP1 AAV2 обычно транслируется со стартового кодона ATG (аминокислота M1) на mRNA длиной 2,4 т.о., в то время как VP2 и VP3 AAV2 появляются из mRNA меньшего размера 2,3 т.о. с использованием более слабого стартового кодона ACG для получения VP2 (аминокислота Т138) и со сквозной трансляцией до следующего доступного кодона ATG (аминокислота М203) для получения наиболее распространенного капсидного белка, VP3. Warrington, выше; Rutledge et al. (1998) J. Virol. 72:309-19, включенная в данный документ посредством ссылки во всей своей полноте. Аминокислотные последовательности капсидных белков аденоассоциированных вирусов хорошо известны в данной области и в целом являются консервативными, особенно в отношении депендопарвовирусов. См., Rutledge et al., выше. Например, Rutledge et al. (1998), выше, представляют на фигуре 4В выравнивания аминокислотных последовательностей для капсидных белков VP1, VP2 и VP3 AAV2, AAV3, AAV4 и AAV6, где сайты начала для каждого из капсидных белков VP1, VP2 и VP3 указаны стрелками, а вариабельные домены в рамке. Соответственно, хотя положения аминокислот, предусмотренные в данном документе, могут быть предоставлены в отношении капсидного белка VP1 AAV, и положения аминокислот, предусмотренные в данном документе, которые не указаны дополнительно, относятся к последовательности основного белка оболочки VP1 AAV2, представленной под SEQ ID NO: 1, квалифицированный специалист мог бы соответственно и легко определить положение той же аминокислоты в капсидном белке VP2 и/или VP3 AAV и соответствующее положение аминокислот в различных серотипах. Кроме того, специалист в данной области сможет поменять домены между капсидными белками различных серотипов AAV для образования «химерного капсидного белка».
[0081] Описан обмен доменами между двумя конструкциями капсидного белка AAV для получения «химерного капсидного белка AAV», см., например, Shen et al. (2007) Mol. Therapy 15(11):1955-1962, включенную в данный документ посредством ссылки во всей своей полноте. «Химерный капсидный белок AAV» включает капсидный белок AAV, который содержит аминокислотные последовательности, например домены, из двух или более различных серотипов AAV, и который способен образовывать и/или образует AAV-подобный вирусный капсид/вирусную частицу. Химерный капсидный белок AAV кодируется химерным геном капсида AAV, например, нуклеотидом, содержащим множество, например, по меньшей мере две последовательности нуклеиновой кислоты, каждая из которых идентична части гена капсида, кодирующего капсидный белок различных серотипов AAV, и это множество вместе кодирует функциональный химерный капсидный белок AAV. Ссылка на химерный капсидный белок в отношении конкретного серотипа AAV указывает на то, что капсидный белок содержит один или более доменов из капсидного белка этого серотипа и один или более доменов из капсидного белка другого серотипа. Например, химерный капсидный белок AAV2 включает капсидный белок, содержащий один или более доменов капсидного белка VP1, VP2 и/или VP3 AAV2 и один или более доменов капсидного белка VP1, VP2 и/или VP3 из другого AAV.
[0082] «Мозаичный капсид» содержит по меньшей мере два набора белков VP1, VP2 и/или VP3, где каждый из наборов кодируется другим геном cap.
[0083] В некоторых вариантах осуществления описанный в данном документе мозаичный капсид содержит рекомбинантные белки VP1, VP2, и/или VP3, кодируемые геном cap, генетически модифицированным вставкой последовательности нуклеиновой кислоты, кодирующей гетерологичный эпитоп, и дополнительно содержит белки VP1, VP2 и/или VP3, кодируемые эталонным геном cap, например, эталонным геном cap дикого типа, кодирующим белки VP1, VP2 и/или VP3 дикого типа того же серотипа AAV, что и рекомбинантные белки VP1, VP2 и/или VP3, контрольным эталонным геном cap, кодирующим белки VP1, VP2 и/или VP3, идентичные рекомбинантным белкам VP1, VP2 и VP3, но в отсутствие гетерологичного эпитопа, мутантным эталонным геном cap дикого типа, кодирующим белки VP1, VP2 и/или VP3 по сути дикого типа того же серотипа AAV, что и рекомбинантные белки VP1, VP2 и/или VP3, но с мутацией (например, вставкой, заменой, делецией), при этом мутация предпочтительно понижает тропизм белков VP1, VP2 и VP3 дикого типа. В некоторых вариантах осуществления эталонный капсидный белок представляет собой химерный эталонный белок, содержащий по меньшей мере один домен белков VP1, VP2 и/или VP3 того же серотипа AAV, что и рекомбинантные белки VP1, VP2 и/или VP3. В некоторых вариантах осуществления эталонный ген cap кодирует химерный белок VP1, VP2 и/или VP3.
[0084] Фраза «тяжелая цепь» или «тяжелая цепь иммуноглобулина» включает последовательность тяжелой цепи иммуноглобулина, включая последовательность константной области тяжелой цепи иммуноглобулина, из любого организма. Вариабельные домены тяжелой цепи включают три CDR тяжелой цепи и четыре FR-области, если не указано иное. Фрагменты тяжелых цепей включают CDR, CDR и FR и их комбинации. Типичная тяжелая цепь после вариабельного домена (от N-конца к С-концу) содержит СН1-домен, шарнир, СН2-домен и СН3-домен. Функциональный фрагмент тяжелой цепи включает фрагмент, который способен специфически распознавать эпитоп (например, распознавать эпитоп с KD в микромолярном, наномолярном или пикомолярном диапазоне), который способен к экспрессии и секреции из клетки и который содержит по меньшей мере одну CDR. Вариабельные домены тяжелой цепи кодируются нуклеотидной последовательностью вариабельной области, которая обычно содержит сегменты VH, DH и JH, полученные из репертуара сегментов VH, DH и JH, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов V, D и J тяжелой цепи для различных организмов можно найти в базе данных IMGT, которая доступна через Интернет во всемирной паутине (www) по адресу «imgt.org».
[0085] Термин «антитело только с тяжелой цепью», «антигенсвязывающий белок только с тяжелой цепью», «однодоменный антигенсвязывающий белок», «однодоменный связывающий белок» или т.п. относится к мономерной или гомодимерной молекуле иммуноглобулина, содержащей иммуноглобулиноподобную цепь, содержащую вариабельный домен, функционально связан с константной областью тяжелой цепи, которая не может связываться с легкой цепью, поскольку в константной области тяжелой цепи обычно отсутствует функциональный СН1-домен. Соответственно, термин «антитело только с тяжелой цепью», «антигенсвязывающий белок только с тяжелой цепью», «однодоменный антигенсвязывающий белок», «однодоменный связывающий белок» или т.п. охватывает как (i) мономерный однодоменный антигенсвязывающий белок, содержащий одну из иммуноглобулиноподобных цепей, содержащую вариабельный домен, функционально связан с константной областью тяжелой цепи, в которой отсутствует функциональный СН1-домен, так и (ii) гомодимерный одно доменный антигенсвязывающий белок, содержащий две иммуноглобулиноподобные цепи, каждая из которых содержит вариабельный домен, функционально связан с константной областью тяжелой цепи, в которой отсутствует функциональный СН1-домен. В различных аспектах гомодимерный однодоменный антигенсвязывающий белок содержит две идентичные иммуноглобулиноподобные цепи, каждая из которых содержит идентичный вариабельный домен, функционально связан с идентичной константной областью тяжелой цепи, в которой отсутствует функциональный СН1-домен. Кроме того, каждая иммуноглобулиноподобная цепь однодоменного антигенсвязывающего белка содержит вариабельный домен, который может быть получен из сегментов гена вариабельной области тяжелой цепи (например, VH, DH, JH), сегментов гена легкой цепи (например, VL, JL) или их комбинации, связанный с последовательностью гена константной области тяжелой цепи (СН), содержащей делецию или инактивирующую мутацию в последовательности, кодирующей СН1 (и, необязательно, шарнирной области), гена константной области тяжелой цепи, например, IgG, IgA, IgE, IgD или их комбинации. Однодоменный антигенсвязывающий белок, содержащий вариабельный домен, полученный из сегментов гена тяжелой цепи, может называться «VH-однодоменное антитело» или «VH-однодоменный антигенсвязывающий белок», см., например, патент США №8754287; публикации заявок на патент США №№20140289876; 20150197553; 20150197554; 20150197555; 20150196015; 20150197556 и 20150197557, каждая из которых включена посредством ссылки во всей своей полноте. Однодоменный антигенсвязывающий белок, содержащий вариабельный домен, полученный из сегментов гена легкой цепи, может называться «VL-однодоменный антигенсвязывающий белок», см., например, публикацию заявки на патент США №20150289489, включенную посредством ссылки во всей своей полноте.
[0086] Фраза «легкая цепь» включает последовательность легкой цепи иммуноглобулина из любого организма, и, если не указано иное, включает каппа (κ) и лямбда (λ) легкие цепи человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены легкой цепи обычно включают три CDR легкой цепи и четыре остовных (FR) области, если не указано иное. Как правило, полноразмерная легкая цепь включает от аминоконца к карбоксильному концу вариабельный домен, который включает FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и аминокислотную последовательность константной области легкой цепи. Вариабельные домены легкой цепи кодируются нуклеотидной последовательностью вариабельной области легкой цепи, которая обычно включает сегменты гена VL легкой цепи и JL легкой цепи, полученные из репертуара сегментов гена V и J легкой цепи, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов гена V и J легкой цепи для различных организмов можно найти в базе данных IMGT, которая доступна через Интернет во всемирной паутине (www) по адресу «imgt.org». Легкие цепи включают, например, цепи, которые селективно не связывают ни первый, ни второй эпитоп, селективно связанный эпитоп-связывающим белком, в котором они встречаются. Легкие цепи также включают цепи, которые связывают и распознают, или помогают тяжелой цепи связываться и распознавать один или более эпитопов, селективно связанных эпитоп-связывающим белком, в котором они встречаются. Легкие цепи также включают цепи, которые связывают и распознают, или помогают тяжелой цепи связываться и распознавать один или более эпитопов, селективно связанных эпитоп-связывающим белком, в котором они встречаются. Обычные или универсальные легкие цепи включают цепи, полученные с помощью гена Vκ1-39Jκ5 человека или гена Vκ3-20Jκ1 человека, и включают их соматически мутированные (например, с созревшей аффинностью) версии.
[0087] Используемая в данном документе фраза «функционально связан» включает физическое сопоставление (например, в трехмерном пространстве) компонентов или элементов, которые прямо или косвенно взаимодействуют друг с другом или иным образом координируют друг друга для участия в биологическом событии, и при таком сопоставлении достигается или разрешается такое взаимодействие и/или координация. В качестве только одного примера, говорят, что контролирующая последовательность (например, последовательность, контролирующая экспрессию) в нуклеиновой кислоте «функционально связана» с кодирующей последовательностью, когда она расположена относительно кодирующей последовательности так, что ее присутствие или отсутствие влияет на экспрессию и/или активность кодирующей последовательности. Во многих вариантах осуществления «функциональная связь» включает ковалентную связь соответствующих компонентов или элементов друг с другом. Специалистам в данной области техники должно быть понятно, однако, что в некоторых вариантах осуществления ковалентная связь не требуется для достижения эффективной функциональной связи. Например, в некоторых вариантах осуществления контролирующие последовательности нуклеиновых кислот, функционально связаны с кодирующими последовательностями, которые они контролируют, являются смежными с представляющим интерес нуклеотидом. Альтернативно или дополнительно, в некоторых вариантах осуществления одна или несколько таких контролирующих последовательностей действуют в транс-положении или на расстоянии для контроля представляющей интерес кодирующей последовательности. В некоторых вариантах осуществления используемый в данном документе термин «последовательность, контролирующая экспрессию» относится к полинуклеотидным последовательностям, которые необходимы и/или достаточны для осуществления экспрессии и процессинга кодирующих последовательностей, с которыми они лигированы. В некоторых вариантах осуществления последовательности, контролирующие экспрессию, могут представлять собой или содержать соответствующие последовательности инициации, терминации транскрипции, промотора и/или энхансера; сигналы эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, которые стабилизируют цитоплазматическую mRNA; последовательности, которые повышают эффективность трансляции (например, консенсусная последовательность Козак); последовательности, которые повышают стабильность белка; и/или, в некоторых вариантах осуществления, последовательности, которые усиливают секрецию белка. В некоторых вариантах осуществления одна или несколько контролирующих последовательностей предпочтительно или исключительно активны в конкретной клетке-хозяине или организме или их типе. В качестве примера, в прокариотах контролирующие последовательности обычно включают промотор, сайт связывания рибосомы и последовательность терминации транскрипции; у эукариот во многих вариантах осуществления контролирующие последовательности обычно включают промоторы, энхансеры и/или последовательности терминации транскрипции. Специалистам в данной области техники должно быть понятно из контекста, что во многих вариантах осуществления термин «контролирующие последовательности» относится к компонентам, присутствие которых является существенным для экспрессии и процессинга, и в некоторых вариантах осуществления включает компоненты, присутствие которых выгодно для экспрессии (включая например, лидерные последовательности, нацеливающие последовательности и/или последовательности партнеров по слиянию).
[0088] Термин «рекомбинантный капсидный белок» включает капсидный белок, который имеет по меньшей мере одну мутацию по сравнению с соответствующим капсидным белком вируса дикого типа, который может быть эталонным и/или контрольным вирусом для сравнительного исследования. Рекомбинантный капсидный белок включает капсидный белок, содержащий гетерологичный эпитоп, который может быть вставлен в капсидный белок и/или отображен с его помощью. «Гетерологичный» в данном контексте означает гетерологичный по сравнению с вирусом, из которого получают капсидный белок. Вставленные аминокислоты могут быть просто вставлены между двумя указанными аминокислотами капсидного белка. Вставка аминокислот может также сопровождаться делецией указанных аминокислот капсидного белка в месте вставки, например, 1 или более аминокислот капсидного белка замещают 5 или более гетерологичными аминокислотами).
[0089] Термины «полиспецифическая связывающая молекула» и «биспецифическая связывающая молекула» и т.п в целом и соответственно относятся к связывающей молекуле, содержащей по меньшей мере два и только два неидентичных связывающих компонента, причем каждый связывающий компонент специфически связывает другой эпитоп - либо на двух разных молекулах (например, разные эпитопы на двух разных иммуногенах), или на одной и той же молекуле (например, разные эпитопы на одном и том же иммуногене). Как правило, один из связывающих компонентов биспецифической связывающей молекулы в данном документе специфически связывает гетерологичный эпитоп, отображаемый вирусным капсидным белком, а второй связывающий компонент является специфическим для белка, например, маркера клеточной поверхности, экспрессируемого преимущественно и/или предпочтительно клеткой-мишенью, например, маркера Т-клеток (например, CD3, CD28 и т.д.). Биспепифические связывающие молекулы можно получить, например, путем объединения связывающих компонентов, которые распознают разные эпитопы одного и того же иммуногена. Например, последовательности нуклеиновой кислоты, кодирующие связывающие компоненты (например, вариабельные последовательности легкой или тяжелой цепи), которые распознают разные эпитопы, можно слить с последовательностями нуклеиновой кислоты, кодирующими одинаковые или разные константные области тяжелой цепи, одинаковые или разные константные области легкой цепи, или, соответственно, константную область тяжелой цепи и константную область легкой цепи, и такие последовательности могут экспрессироваться в клетке в виде полиспецифического антигенсвязывающего белка в формате, который аналогичен структуре Fab, структуре scFab, структуре диатела, структуре scFv, структуре scFv-Fc, структуре scFv-zipper, тетрамерной структуре, подобной типичному антителу, которое включает когнатную универсальную легкую цепь, тетрамерной структуре, содержащей типичное бивалентное антитело, которое включает когнатную универсальную легкую цепь и/или дополнительный связывающий компонент (например, scFv, scFv-zipper, scFab и т.д.), присоединенный к одной или обеим тяжелым цепям (например, на N- и/или С-конце) и/или к одной или обеим легким цепям (например, на N- и/или С-конце). Различные форматы полиспецифических, особенно биспецифических, связывающих молекул хорошо известны, см. например, Brinkmann and Konterman (2017) mAbs 9:182-212, включенную в данный документ посредством ссылки во всей своей полноте.
[0090] Иллюстративная полиспецифическая связывающая молекула имеет две тяжелые цепи, каждая из которых содержит CDR тяжелой цепи, за которыми следуют (от N-конца к С-концу) СН1-домен, шарнир, СН2-домен и СН3-домен, и легкую цепь иммуноглобулина, которая либо не придает эпитоп-связывающей специфичности, но может связываться с каждой тяжелой цепью (например, общая легкая цепь), или которая может связываться с каждой тяжелой цепью и может связывать один или более эпитопов, связанных эпитоп-связывающими областями тяжелой цепи, или которая может связываться с каждой тяжелой цепью и обеспечивать связывание одной или обеих тяжелых цепей с одним или обоими эпитопами. В некоторых вариантах осуществления полиспецифическая связывающая молекула содержит: (1) вариабельный домен тяжелой цепи иммуноглобулина, функционально связан с первой константной областью тяжелой цепи, содержащей первую аминокислотную последовательность СН3 человеческого IgG, выбранного из IgG1, IgG2, IgG4 и их комбинации; и (2) вариабельный домен тяжелой цепи иммуноглобулина, где второй вариабельный домен тяжелой цепи иммуноглобулина функционально связан со второй константной областью тяжелой цепи, содержащей вторую аминокислотную последовательность СН3 человеческого IgG, выбранного из IgG1, IgG2, IgG4 и их комбинации, где первый или второй вариабельные домены тяжелой цепи (с когнатной легкой цепью или без нее) связывают гетерологичный эпитоп, как описано в данном документе, а другой вариабельный домен тяжелой цепи (с когнатной легкой цепью или без нее) связывает рецептор на клетке-мишени, и где первая константная область тяжелой цепи связывается со второй константной областью цепи таким образом, который обеспечивает более легкое выделение полиспецифического связывающего белка, например, где первая и вторая константные области тяжелой цепи образуют формат выступ-во-впадину (KIH), или где вторая аминокислотная последовательность СН3 содержит модификацию, которая уменьшает или устраняет связывание второй аминокислотной последовательности СН3 с белком А (см., например, патент США №8586713, который включен в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления полиспецифическая связывающая молекула содержит: (1) вариабельный домен тяжелой цепи иммуноглобулина, функционально связан с первой константной областью тяжелой цепи, содержащей первую аминокислотную последовательность СН3 человеческого IgG, выбранного из IgG1, IgG2, IgG4 и их комбинации; и (2) вариабельный домен тяжелой цепи иммуноглобулина, где второй вариабельный домен тяжелой цепи иммуноглобулина функционально связан со второй константной областью тяжелой цепи, содержащей вторую аминокислотную последовательность СН3 человеческого IgG, выбранного из IgG1, IgG2, IgG4 и их комбинации, где первый и второй вариабельные домены тяжелой цепи (с когнатной легкой цепью или без нее) связывают одинаковые или разные антигены, при этом первая или вторая константные области тяжелой цепи модифицированы, чтобы дополнительно содержать дополнительный связывающий домен (например, scFv или Fv, который связывается с гетерологичным эпитопом, как описано в данном документе, например, где дополнительный связывающий домен присоединен к С-концу или N-концу одной или обеих тяжелых цепей), и где первая константная область тяжелой цепи связана со второй константной областью цепи таким образом, который обеспечивает более легкое выделение полиспецифического связывающего белка, например, где первая и вторая константные области тяжелой цепи образуют формат выступ-во-впадину (KIH), или где вторая аминокислотная последовательность СН3 содержит модификацию, которая уменьшает или устраняет связывание второй аминокислотной последовательности СН3 с белком А. В некоторых вариантах осуществления, в которых вторая аминокислотная последовательность СН3 имеет уменьшенное или устраненное связывание с белком А, вторая аминокислотная последовательность СН3 содержит модификацию H95R (нумерация экзонов согласно IMGT; H435R нумерация согласно EU). В одном варианте осуществления вторая аминокислотная последовательность СН3 дополнительно содержит модификацию Y96F (нумерация экзонов согласно IMGT; H436F по EU). В другом варианте осуществления вторая аминокислотная последовательность СН3 содержит как модификацию H95R (нумерация экзонов согласно IMGT; H435R нумерация согласно EU), так и модификацию Y96F (нумерация экзонов согласно IMGT; H436F согласно EU). В некоторых вариантах осуществления вторая аминокислотная последовательность СН3 получена из модифицированного IgG1 человека и дополнительно содержит мутацию, выбранную из группы, состоящей из D16E, L18M, N44S, K52N, V57M и V82I (IMGT; D356E, L38M, N384S, K392N, V397M и V422I по EU). В некоторых вариантах осуществления вторая аминокислотная последовательность СН3 получена из модифицированного IgG2 человека и дополнительно содержит мутацию, выбранную из группы, состоящей из N44S, K52N и V82I (IMGT: N384S, K392N и V422I по EU). В некоторых вариантах осуществления вторая аминокислотная последовательность СН3 получена из модифицированного IgG4 человека и дополнительно содержит мутацию, выбранную из группы, состоящей из Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (IMGT: Q355R, N384S, K392N, V397M, R409K, E419Q и V422I по EU). В некоторых вариантах осуществления аминокислотная последовательность константной области тяжелой цепи представляет собой аминокислотную последовательность константной области, отличной от человеческой, и эта аминокислотная последовательность константной области тяжелой цепи содержит одну или более модификаций любого типа, описанных выше.
[0091] В различных вариантах осуществления Fc-домены модифицированы для изменения связывания с Fc-рецептором, что, в свою очередь, влияет на эффекторную функцию. В некоторых вариантах осуществления сконструированная константная область тяжелой цепи (СН), которая включает Fc-домен, является химерной. Таким образом, химерная СН-область объединяет СН-домены, полученные из более чем одного изотипа иммуноглобулина. Например, химерная СН-область содержит часть или весь СН2-домен, полученный из молекулы IgG1 человека, IgG2 человека или IgG4 человека, в комбинации с частью или всем СН3-доменом, полученным из молекулы IgG1 человека, IgG2 человека или IgG4 человека. В некоторых вариантах осуществления химерная СН-область содержит химерную шарнирную область. Например, химерный шарнир может содержать «верхнюю шарнирную» аминокислотную последовательность (аминокислотные остатки с положения 216 по 227 согласно нумерации EU; аминокислотные остатки с положения 226 по 240 согласно нумерации Kabat), полученную из шарнирной области IgG1 человека, IgG2 человека или IgG4 человека, в комбинации с «нижней шарнирной» последовательностью (аминокислотные остатки с положения 228 по 236 согласно нумерации EU; аминокислотные остатки с положения 241 по 249 согласно нумерации Kabat), полученной из шарнирной области IgG1 человека, IgG2 человека или IgG4 человека. В некоторых вариантах осуществления химерная шарнирная область содержит аминокислотные остатки, полученные из верхней шарнирной последоательности IgG1 человека или IgG4 человека, и аминокислотные остатки, полученные из нижней шарнирной последовательности IgG2 человека.
[0092] В некоторых вариантах осуществления Fc-домен можно сконструировать для активации всех, некоторых или ни одной из эффекторных функций нормального Fc, не влияя на необходимые фармакокинетические свойства Fc-содержащего белка (например, антитела). Примеры белков, содержащих химерные СН-области и обладающих измененными эффекторными функциями, см. в WO 2014022540, которая включена в данный документ во всей своей полноте.
[0093] Термин «клетки-мишени» включает любые клетки, в которых экспрессия представляющего интерес нуклеотида является необходимой. Предпочтительно клетки-мишени демонстрируют белок, например рецептор, на своей поверхности, который позволяет нацелиться на клетку с помощью перенацеливающего лиганда, как описано ниже. Предпочтительно целевой белок, например рецептор, является специфическим для клетки-мишени, например, представляет собой «клеточно-специфический маркер», «клеточно-специфический антиген» или т.п. Термин «клеточно-специфический маркер», «клеточно-специфический антиген», «органоспецифический маркер», «тканеспецифический маркер» или т.п.относится к и включает те белки, экспрессия которых обогащена в клетке, ткани и/или органе, для которых он является специфическим маркером. «Обогащенный» в контексте экспрессии белка относится к и включает экспрессию или избыточную экспрессию белка, специфического для клетки/ткани/органа, преимущественно, предпочтительно или исключительно клеткой/тканью/органом, для которых белок является специфическим маркером, хотя такой маркер может также экспрессироваться другими клетками/тканями/или органами на минимальных уровнях. Атлас белков человека может использоваться для определения того, является ли белок специфическим для клетки/ткани/органа маркером, а также предоставляет хранилище специфичных для клетки/ткани/органа белков. См., www.proteinatlas.org; см., также Uhlen et al. (2010) Nat. Biotech. 28:1248-50, включенную в данный документ посредством ссылки во всей своей полноте.
[0094] Термин «трансдукция» или «инфицирование» или т.п. относится к введению нуклеиновой кислоты в клетку-мишень с помощью вирусного вектора. Термин эффективность в отношении трансдукции или т.п., например, «эффективность трансдукции», относится к доле (например, процентному содержанию) клеток, экспрессирующих представляющий интерес нуклеотид после инкубации с установленным количеством вирусных векторов, содержащих представляющий интерес нуклеотид. Хорошо известные способы определения эффективности трансдукции включают сортировку флуоресцентно-активированных клеток для клеток, трансдуцированных флуоресцентным репортерным геном, ПЦР для экспрессии представляющего интерес нуклеотида и т.д.
[0095] Используемый в данном документе термин «дикий тип» включает объект, обладающий структурой и/или активностью, обнаруживаемой в природе в «нормальном» (в противоположность мутантному, болезненному, измененному и т.д.) состоянии или окружении. Специалистам в данной области понятно, что вирусные векторы дикого типа, например, капсидные белки дикого типа, можно использовать в качестве эталонного вирусного вектора в сравнительных исследованиях. Как правило, эталонный вирусный капсидный белок/капсид/вектор идентичен тестируемому вирусному капсидному белку/капсиду/вектору за исключением изменения, для которого необходимо проверить эффект. Например, чтобы определить влияние, например, на эффективность трансдукции, вставки гетерологичного эпитопа в тестовый вирусный вектор, эффективность трансдукции тестового вирусного вектора (в отсутствие или в присутствии соответствующей полиспецифической связывающей молекулы) можно сравнить с эффективность трансдукции эталонного вирусного вектора (в отсутствие или в присутствии соответствующей полиспецифической связывающей молекулы, если необходимо), который идентичен тестовому вирусному вектору в каждом случае (например, дополнительные мутации, представляющий интерес нуклеотид, количество вирусных векторов и клеток-мишеней и т.д.), за исключением наличия гетерологичного эпитопа.
Рекомбинантные вирусные капсидные белки и вирусные векторы и нуклеиновые кислоты
[0096] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, как описано в данном документе, представляет собой капсидный белок Ad, например, капсидный белок серотипа Ad, выбранного из группы, состоящей из Ad1, Ad2, Ad3, Ad4, Ad5, Ad6 и Ad7. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида Ad2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида Ad5. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок Ad, как описано в данном документе, содержит гетерологичный эпитоп в домене волоконного белка, например, на карбокси-конце волоконного белка, в knob-домене волоконного белка и/или в петле HI knob-домена волоконного белка.
[0097] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в данном документе, получен с помощью гена капсида аденоассоциированного вируса (AAV), например, представляет собой генетически модифицированный капсидный белок серотипа AAV, выбранного из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, гена капсида AAV6, гена капсида AAV8 или гена капсида AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, представляет собой генетически модифицированный капсидный белок VP1 AAV2, аминокислотная последовательность для дикого типа которого представлена, соответственно, под SEQ ID NO: 1. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV8, например, представляет собой генетически модифицированный капсидный белок VP1 AAV8, аминокислотная последовательность для дикого типа которого представлена под SEQ ID NO: 21. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, представляет собой генетически модифицированный капсидный белок VP1 AAV9, аминокислотная последовательность для дикого типа которого представлена под SEQ ID NO: 5. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV6, например, представляет собой генетически модифицированный капсидный белок VP1 AAV6. В некоторых вариантах осуществления гетерологичный эпитоп вставляют в I-453 капсидного белка AAV9.
[0098] Как правило, рекомбинантный вирусный капсидный белок, как описано в данном документе, содержит гетерологичный эпитоп, вставленный в капсидный белок и/или отображенный с его помощью, так что гетерологичный эпитоп понижает и/или устраняет естественный тропизм капсидного белка или капсида, содержащего его. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в область капсидного белка, участвующую в естественном тропизме эталонного капсидного белка дикого типа, например, в область капсидного белка, связанную с рецептором клетки. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в knob-домен волоконного белка Ad и/или отображен с его помощью. В некоторых вариантах осуществления гетерологичный эпитоп вставлен в петлю HI волоконного белка Ad и/или отображен с ее помощью. В некоторых вариантах осуществления гетерологичный эпитоп вставлен после аминокислотного положения, выбранного из группы, состоящей из G453 капсидного белка VP1 AAV2, N587 капсидного белка VP1 AAV2, Q585 капсидного белка VP1 AAV6, G453 капсидного белка VP1 AAV9 и А589 капсидного белка VP1 AAV9. В некоторых вариантах осуществления гетерологичный эпитоп вставлен и/или отображен между аминокислотными положениями N587 и R588 VP1 капсида AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, представленную под SEQ ID NO: 2. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, представленную под SEQ ID NO: 4. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 25. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 26. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 27. Дополнительные подходящие сайты вставки, идентифицированные с использованием AAV2, хорошо известны из уровня техники (Wu et al. (2000) J. Virol. 74:8635-8647) и включают I-1, I-34, I-138, I-139, I-161, I-261, I-266, I-381, I-447, I-448, I-459, I-471, I-520, I-534, I-570, I-573, I-584, I-587, I-588, I-591, I-657, I-664, I-713 и I-716. Рекомбинантный вирусный капсидный белок, как описано в данном документе, может представлять собой капсидный белок AAV2, содержащий гетерологичный эпитоп, вставленный в положении, выбранном из группы, состоящей из I-1, I-34, I-138, I-139, I-161, I-261, I-266, I-381, I-447, I-448, I-459, I-471, I-520, I-534, I-570, I-573, I-584, I-587, I-588, I-591, I-657, I-664, I-713, I-716 и их комбинации. Дополнительные подходящие сайты вставки, идентифицированные с использованием дополнительных серотипов AAV, хорошо известны и включают I-587 (AAV1), I-589 (AAV1), I-585 (AAV3), I-585 (AAV4) и I-585 (AAV5). В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, как описано в данном документе, может представлять собой капсидный белок AAV2, содержащий гетерологичный эпитоп, вставленный в положении, выбранном из группы, состоящей из I-587 (AAV1), I-589 (AAV1), I-585 (AAV3), I-585 (AAV4), I-585 (AAV5) и их комбинации.
[0099] Используемая в данном документе номенклатура I-### относится к сайту вставки с ###, обозначающим номер аминокислоты относительно белка VP1 капсидного белка AAV, однако такая вставка может быть расположена непосредственно с N- или С-конца, предпочтительно с С-конца от одной аминокислоты в последовательности из 5 аминокислот с N- или С-конца от данной аминокислоты, предпочтительно 3, более предпочтительно 2, особенно 1 аминокислота с N- или С-конца от данной аминокислоты. Кроме того, положения, упоминаемые в данном документе, относятся к белку VP1, кодируемому геном капсида AAV, и соответствующие положения (и их мутации) можно легко идентифицировать для капсидных белков VP2 и VP3, кодируемых геном капсида, путем выполнения выравнивания последовательности белков VP1, VP2 и VP3, кодируемых эталонным геном капсида AAV.
[00100] Таким образом, вставка в соответствующее положение кодирующей нуклеиновой кислоты одного из этих сайтов гена cap приводит к вставке в VP1, VP2 и/или VP3, поскольку капсидные белки кодируются перекрывающимися рамками считывания с одного и того же гена со поочередными стартовыми кодонами. Следовательно, для AAV2, например, согласно этой номенклатуре вставки между аминокислотами 1 и 138 вставляются только в VP1, вставки между 138 и 203 вставляются в VP1 и VP2, а вставки между 203 и С-концом вставляются в VP1, VP2 и VP3, что, конечно, также относится и к сайту вставки I-587. Следовательно, настоящее изобретение охватывает структурные гены AAV с соответствующими вставками в белки VP1, VP2 и/или VP3.
[00101] Кроме того, из-за высокой консервативности по меньшей мере крупных участков и большого числа близкородственных представителей семейства соответствующие сайты встраивания для AAV, отличного от перечисленного AAV, можно идентифицировать путем выполнения аминокислотного выравнивания или сравнения структур капсида. См., например, Rutledge et al. (1998) J. Virol. 72:309-19 и патент США №9624274 для иллюстративных выравниваний капсидных белков различных AAV, каждый из этих источников включен в данный документ посредством ссылки во всей своей полноте.
[00102] В некоторых раскрытых в данном документе композициях, состоящих из рекомбинантного вирусного капсида (например, в отсутствие полиспецифической связывающей молекулы), рекомбинантный вирусный капсидный белок представляет собой капсидный белок VP1 AAV2 с гетерологичным эпитопом, вставленным в сайт 1587, где гетер о логичный эпитоп не содержит мотив Arg-Gly-Asp (RGD), мотив NGR или с-myc. В некоторых раскрытых в данном документе композициях, состоящих из рекомбинантного вирусного капсида (например, в отсутствие полиспецифической связывающей молекулы), рекомбинантный вирусный капсидный белок представляет собой капсидный белок VP1 с гетерологичным эпитопом, вставленным между Т448 и N449, где гетерологичный эпитоп не содержит с-myc. В некоторых раскрытых в данном документе композициях, состоящих из рекомбинантного вирусного капсида (например, в отсутствие полиспецифической связывающей молекулы), рекомбинантный вирусный капсидный белок представляет собой капсидный белок VP1 с гетерологичный эпитопом, вставленным в сайт I-447, где гетерологичный эпитоп не содержит L14 или НА.
[00103] В некоторых композициях, содержащих рекомбинантный вирусный капсид (например, дополнительно содержащих полиспецифическую связывающую молекулу), рекомбинантный вирусный капсидный белок представляет собой капсидный белок VP1 с гетерологичным эпитопом, вставленным в сайт I587, где гетерологичный эпитоп содержит мотив Arg-Gly-Asp (RGD), мотив NGR или с-myc. В некоторых раскрытых в данном документе композициях, содержащих рекомбинантный вирусный капсид (например, дополнительно содержащих полиспецифическую связывающую молекулу), вирусный капсид представляет собой капсид VP1, гетерологичный эпитоп содержит с-myc и гетерологичный эпитоп вставлен между Т448 и N449 или между N587 и R588. В некоторых раскрытых в данном документе композициях, содержащих рекомбинантный вирусный капсид (например, дополнительно содержащих полиспецифическую связывающую молекулу), рекомбинантный вирусный капсидный белок представляет собой капсидный белок VP1 с гетерологичным эпитопом, вставленным в сайт I-447, где гетерологичный эпитоп содержит L14 или НА. В некоторых раскрытых в данном документе композициях, содержащих рекомбинантный вирусный капсид (например, дополнительно содержащих полиспецифическую связывающую молекулу), рекомбинантный вирусный капсидный белок представляет собой капсидный белок VP1 с гетерологичным эпитопом, вставленным между Т448 и N449, где гетерологичный эпитоп содержит с-myc. В патенте США №9624274 описано I-453 капсидного белка AAV в качестве подходящего сайта вставки для гетерологичного эпитопа.
[00104] В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа устраняет естественный тропизм вирусного вектора, например, трансдукция клетки, естественным образом пермиссивной к инфекции эталонными вирусными векторами дикого типа, и/или клетки-мишени не обнаруживается в отсутствие подходящей полиспецифической связывающей молекулы. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора, например, по сравнению с трансдукцией клетки, естественным образом пермиссивной к инфекции эталонными вирусными векторами дикого типа. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 5%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 5%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 10%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 20%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 30%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 40%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 50%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 60%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 70%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 80%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 90%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 95%. В некоторых вариантах осуществления вставка (отображение) гетерологичного эпитопа понижает естественный тропизм вирусного вектора по меньшей мере на 90%. В этих вариантах осуществления, где вставка (отображение) гетерологичного эпитопа не устраняет естественный тропизм рекомбинантных вирусных капсидов, естественный тропизм таких рекомбинантных вирусных капсидов можно устранить с помощью второй и отличающейся мутации. Например, в одном варианте осуществления рекомбинантный вирусный капсидный белок, как описано в данном документе, может быть получен с помощью гена капсида AAV9, содержать гетерологичный эпитоп и может дополнительно содержать мутацию, например, мутацию W503A.
[00105] Это удаление целенаправленного воздействия вируса на его естественную клетку-хозяина является важным, особенно если предполагается системное введение по сравнению с локальным или локально-региональным введением вирусных векторов, поскольку поглощение вирусных векторов естественными клетками-хозяевами ограничивает эффективную дозу вирусных векторов. Сообщается, что в случае AAV2 и AAV6 HSPG является основным рецептором для захвата вируса в большом количестве клеток, особенно в клетках печени. Для AAV2 HSPG-связывающая активность зависит от группы из 5 основных аминокислот, R484, R487, R585, R588 и K532 (Kern et al., (2003) J Virol. 77(20): 11072-81). Недавно сообщалось, что аминокислотная замена лизина на глутамат K531E приводит к подавлению способности AAV6 связывать гепарин или HSPG ((Wu et al., 2006) J. of Virology 80(22):11393-11397). Соответственно, предпочтительными точечными мутациями являются те, которые снижают активность трансдукции вирусного вектора для данной клетки-мишени, опосредованной природным рецептором, по меньшей мере на 50%, предпочтительно по меньшей мере на 80%, особенно по меньшей мере на 95% в случае HSPG как основного рецептора связывания вирусных векторов с HSPG.
[00106] Следовательно, дополнительные мутации, предпочтительные для HSPG-связывающих вирусных векторов, представляют собой такие мутации, которые истощают или заменяют основную аминокислоту, такую как R, K или Н, предпочтительно R или K, которая участвует в связывании HSPG соответствующим вирусом, на аминокислоту, отличную от основной, такую как A, D, G, Q, S и Т, предпочтительно А, или аминокислоту, которая присутствует в соответствующем положении другого, но высоко консервативного серотипа AAV, в котором отсутствует такая основная аминокислота в этом положении. Следовательно, предпочтительными аминокислотными заменами являются R484A, R487A, R487G, K532A, K532D, R585A, R585S, R585Q, R585A или R588T, особенно R585A и/или R588A для AAV2, и K531A или K531E для AAV6. Одним особенно предпочтительным вариантом осуществления настоящего изобретения являются такие мутанты капсидного белка AAV2, которые дополнительно содержат две точечные мутации R585A и R588A, поскольку этих двух точечных мутаций достаточно для устранения активности связывания HSPG в значительной степени. Эти точечные мутации обеспечивают эффективное удаление целенаправленного воздействия с HSPG-экспрессирующих клеток, что для целенаправленного воздействия повышает специфичность соответствующего мутантного вируса в отношении его новой клетки-мишени.
[00107] Одним из вариантов осуществления настоящего изобретения является мультимерная структура, предусматривающая рекомбинантный вирусный капсидный белок по настоящему изобретению. Мультимерная структура предусматривает по меньшей мере 5, предпочтительно по меньшей мере 10, более предпочтительно по меньшей мере 30, наиболее предпочтительно по меньшей мере 60 рекомбинантных вирусных капсидных белков, содержащих гетерологичный эпитоп, как описано в данном документе. Они могут образовывать обычные вирусные капсиды (пустые вирусные частицы) или вирусные векторы (капсиды, инкапсулирующие представляющий интерес нуклеотид). Образование вирусных векторов, способных упаковывать вирусный геном, является весьма предпочтительным признаком для применения рекомбинантных вирусных капсидов, описанных в данном документе в качестве вирусных векторов.
[00108] Одним из вариантов осуществления настоящего изобретения является нуклеиновая кислота, кодирующая капсидный белок, как описано выше. Нуклеиновая кислота предпочтительно представляет собой вектор, содержащий заявленную последовательность нуклеиновой кислоты. Нуклеиновые кислоты, особенно векторы, необходимы для рекомбинантной экспрессии капсидных белков по настоящему изобретению.
[00109] Еще одним вариантом осуществления настоящего изобретения является применение по меньшей мере одного рекомбинантного вирусного капсидного белка и/или кодирующей его нуклеиновой кислоты, предпочтительно по меньшей мере одной мультимерной структуры (например, вирусного вектора), для изготовления и применения в качестве вектора для переноса гена.
Гетерологичные эпитопы
[00110] Как правило, рекомбинантный вирусный капсидный белок и/или вирусный вектор, содержащий рекомбинантный вирусный капсид, содержит гетерологичный эпитоп, который способен перенацеливать вирусный вектор, например, с помощью полиспецифической связывающей молекулы. В некоторых вариантах осуществления гетерологичный эпитоп представляет собой В-клеточный эпитоп, например, его длина составляет от приблизительно 1 аминокислоты до приблизительно 35 аминокислот, и образует связывающую пару с паратопом антитела, например, вариабельным доменом иммуноглобулина. В некоторых вариантах осуществления гетерологичный эпитоп содержит аффинную метку.
[00111] Большое количество аффинных меток известно из уровня техники. (См, например: Nilsson et al. (1997) "Affinity fusion strategies for detection, purification, and immobilization of recombinant proteins" Protein Expression and Purification 11:1-16, Terpe et al. (2003) "Overview of tag protein fusions: From molecular and biochemical fundamentals to commercial systems" Applied Microbiology and Biotechnology 60:523-533, и ссылки в них). Аффинные метки включают без ограничения полигистидиновую метку (например, метку His-6, His-8 или His-10), которая связывает иммобилизованные двухвалентные катионы (например, Ni2+), биотиновый фрагмент (например, на биотинилированной in vivo полипептидной последовательности), который связывает иммобилизованный авидин, последовательность GST (глутатион-8-трансфераза), которая связывает иммобилизованный глутатион, S-метку, которая связывает иммобилизованный белок S, антиген, который связывает иммобилизованное антитело или его домен или фрагмент (включая, например, метки Т7, myc, FLAG и В, которые связывают соответствующие антитела), метку FLASH (высокоаффинную метку, которая связывается со специфическими фрагментами на основе мышьяка), рецептор или рецепторный домен, который связывает иммобилизованный лиганд (или наоборот), белок А или его производное (например, Z), который связывает иммобилизованный IgG, мальтозосвязывающий белок (MBP), который связывает иммобилизованную амилозу, альбумин-связывающий белок, который связывает иммобилизованный альбумин, хитин-связывающий домен, который связывает иммобилизованный хитин, кальмодулин-связывающий пептид, который связывает иммобилизованный кальмодулин, и целлюлозо-связывающий домен, который связывает иммобилизованную целлюлозу. Другой иллюстративной меткой является SNAP-метка, коммерчески доступная от Covalys (www.covalys.com). В некоторых вариантах осуществления гетер о логичный эпитоп, раскрытый в данном документе, содержит аффинную метку, распознаваемую только паратопом антитела. В некоторых вариантах осуществления гетерологичный эпитоп, раскрытый в данном документе, содержит аффинную метку, распознаваемую паратопом антитела и другими парами специфического связывания.
[00112] В некоторых вариантах осуществления гетерологичный эпитоп и/или аффинная метка не образуют связывающую пару с константным доменом иммуноглобулина. В некоторых вариантах осуществления гетерологичный эпитоп и/или аффинная метка не образуют связывающую пару с ионом металла, например, Ni2+, Со2+, Cu2+, Zn2+, Fe3+ и т.д. В некоторых вариантах осуществления гетерологичный эпитоп не является полипептидом, выбранным из группы, состоящей из стрептавидина, Strep II, НА, L14, 4C-RGD, LH и белка А.
[00113] В некоторых вариантах осуществления аффинная метка выбрана из группы, состоящей из FLAG (SEQ ID NO: 7), НА (SEQ ID NO: 8) и c-myc (EQKLISEEDL; SEQ ID NO: 6). В некоторых вариантах осуществления гетерологичный эпитоп представляет собой с-myc.
[00114] В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV. В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит аминокислотную последовательность EQKLISEEDL (представленную под SEQ ID NO: 6), фланкированную и/или функционально связанную с по меньшей мере 5 смежными аминокислотами капсидного белка VP1 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсид, как описано в данном документе, содержит EQKLISEEDL (представленную под SEQ ID NO: 6), вставленную между N587 и R588 капсидного белка VP1 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, как описано в данном документе, содержит аминокислотную последовательность, представленную под SEQ ID NO: 2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, как описано в данном документе, содержит аминокислотную последовательность, представленную под SEQ ID NO: 4. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 25. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 26. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 27.
[00115] В некоторых вариантах осуществления гетерологичный эпитоп содержит аффинную метку и один или более линкеров. В некоторых вариантах осуществления гетерологичный эпитоп содержит аффинную метку, фланкированную линкером, например, гетерологичный эпитоп содержит от N-конца к С-концу первый линкер, аффинную метку и второй линкер. В некоторых вариантах осуществления длина каждого из первого и второго линкеров независимо составляет по меньшей мере одну аминокислоту. В некоторых вариантах осуществления первый и второй линкеры идентичны.
[00116] Как правило, гетерологичный эпитоп, как описано в данном документе, например, аффинная метка сама по себе или в комбинации с одним или более линкерами, имеет длину от приблизительно 5 аминокислот до приблизительно 35 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет по меньшей мере 5 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 6 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 7 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 8 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 9 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 10 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 11 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 12 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 13 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 14 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 15 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 16 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 17 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 18 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 19 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 20 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 21 аминокислоту. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 22 аминокислоты. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 23 аминокислоты. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 24 аминокислоты. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 25 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 26 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 27 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 28 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 29 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 30 аминокислот. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 31 аминокислоту. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 32 аминокислоты. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 33 аминокислоты. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 34 аминокислоты. В некоторых вариантах осуществления длина гетерологичного эпитопа (самого по себе или в комбинации с одним или более линкерами) составляет 35 аминокислот.
Перенацеливающие фрагменты
[00117] Вирусный вектор, описанный в данном документе, характеризуется пониженной вплоть до устраненной способностью к трансдукции в отсутствие полиспецифической связывающей молекулы, в частности, полиспецифической связывающей молекулы, содержащей (i) паратой антитела, который специфически связывает эпитоп, и (ii) перенацеливающий лиганд, специфически связывающий рецептор, который может быть конъюгирован с поверхностью гранулы (например, для очистки) или экспрессирован клеткой-мишенью. Соответственно, полиспецифическая связывающая молекула, содержащая (i) паратоп антитела, который специфически связывает эпитоп, и (ii) перенацеливающий лиганд, который специфически связывает рецептор, перенацеливает вирусный вектор. Такое «перенацеливание» или «перенаправление» может включать сценарий, в котором вирусный вектор дикого типа нацелен на несколько клеток в ткани и/или несколько органов в организме, это широкое нацеливание на ткань или органы понижают вплоть до устранения путем вставки гетерологичного эпитопа, и этого перенацеливания на более специфические клетки в ткани или более специфический орган в организме достигают с помощью полиспецифической связывающей молекулы. Такое перенацеливание или перенаправление может также включать сценарий, в котором вирусный вектор дикого типа нацелен на ткань, это нацеливание на ткань понижают вплоть до устранения путем вставки гетерологичного эпитопа, и этого перенацеливания на полностью другую ткань достигают с помощью полиспецифической связывающей молекулы. Паратоп антитела, как описано в данном документе, обычно содержит по меньшей мере определяющую комплементарность область (CDR), которая специфически распознает гетерологичный эпитоп, например, область CDR3 вариабельного домена тяжелой и/или легкой цепи. В некоторых вариантах осуществления полиспецифическая связывающая молекула содержит антитело (или его часть), которое содержит паратоп антитела, который специфически связывает гетерологичный эпитоп. Например, полиспецифическая связывающая молекула может содержать однодоменную вариабельную область тяжелой цепи или однодоменную вариабельную область легкой цепи, где однодоменная вариабельная область тяжелой цепи или однодоменная вариабельная область легкой цепи содержит паратоп антитела, который специфически связывает гетерологичный эпитоп. В некоторых вариантах осуществления полиспецифическая связывающая молекула может содержать Fv-область, например, полиспецифическая связывающая молекула может содержать scFv, содержащую паратоп антитела, который специфически связывает гетерологичный эпитоп. В некоторых вариантах осуществления полиспецифическая связывающая молекула, описанная в данном документе, содержит паратоп антитела, который специфически связывает с-myc.
[00118] В некоторых вариантах осуществления полиспецифическая связывающая молекула, описанная в данном документе, содержит паратоп антитела, который специфически связывает с-myc, причем этот паратоп содержит scFv, вариабельные домены тяжелой и легкой цепи scFv и/или аминокислотную последовательность набора из HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3, кодируемую последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 28, например, scFv, содержащий аминокислотную последовательность, представленную под SEQ ID NO: 37. В некоторых вариантах осуществления полиспецифическая связывающая молекула, описанная в данном документе, содержит Fv или scFv, кодируемый последовательностью нуклеиновой кислоты, представленной под SEQ ID NO: 28.
[00119] Соответственно, настоящее изобретение включает в качестве антител антигенсвязывающие фрагменты антител и полиспецифические связывающие белки, которые специфически связывают с-myc, где антитела, фрагменты антител и полиспецифические связывающие белки содержат паратоп, который содержит вариабельную область тяжелой цепи (HCVR), содержащую SEQ ID NO: 29, и вариабельную область легкой цепи (LCVR), содержащую SEQ ID NO: 30. Настоящее изобретение также включает антитела, антигенсвязывающие фрагменты антител и/или полиспецифические связывающие белки, которые содержат паратоп, который специфически связывает с-Мус, где паратоп содержит определяющую комплементарность область 1 тяжелой цепи (HCDR1), содержащую SEQ ID NO: 31, HCDR2, содержащую SEQ ID NO: 32, HCDR3, содержащую SEQ ID NO: 33, определяющую комплементарность область 1 легкой цепи (LCDR1), содержащую SEQ ID NO: 34, LCDR2, содержащую SEQ ID NO: 35, и LCDR3, содержащую SEQ ID NO: 36.
[00120] Настоящее изобретение предусматривает антитела, их антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит HCVR, содержащую аминокислотную последовательность, представленную под SEQ ID NO: 29, или по сути аналогичную последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности с ней.
[00121] Настоящее изобретение предусматривает антитела, антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит LCVR, содержащую аминокислотную последовательность, представленную под SEQ ID NO: 30, или по сути аналогичную последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности с ней.
[00122] Настоящее изобретение предусматривает антитела, их антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит пару аминокислотных последовательностей HCVR и LCVR (HCVR/LCVR), содержащую аминокислотную последовательность HCVR, представленную под SEQ ID NO: 29, в паре с аминокислотной последовательностью LCVR, представленной под SEQ ID NO: 30. В некоторых вариантах осуществления пара аминокислотных последовательностей HCVR/LCVR выбрана из группы, состоящей из SEQ ID NO: 29/30.
[00123] Настоящее изобретение предусматривает антитела, антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит CDR1 тяжелой цепи (HCDR1), содержащую аминокислотную последовательность, представленную под SEQ ID NO: 31, или по сути аналогичную ей последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности.
[00124] Настоящее изобретение предусматривает антитела, антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит CDR2 тяжелой цепи (HCDR2), содержащую аминокислотную последовательность, представленную под SEQ ID NO: 32, или по сути аналогичную ей последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности.
[00125] Настоящее изобретение предусматривает антитела, антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит CDR3 тяжелой цепи (HCDR3), содержащую аминокислотную последовательность, представленную под SEQ ID NO: 33, или по сути аналогичную ей последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности.
[00126] Настоящее изобретение предусматривает антитела, антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит CDR1 легкой цепи (LCDR1), содержащую аминокислотную последовательность, представленную под SEQ ID NO: 34, или по сути аналогичную ей последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности.
[00127] Настоящее изобретение предусматривает антитела, антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит CDR2 легкой цепи (LCDR2), содержащую аминокислотную последовательность, представленную под SEQ ID NO: 35, или по сути аналогичную ей последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности.
[00128] Настоящее изобретение предусматривает антитела, антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит CDR3 легкой цепи (LCDR3), содержащую аминокислотную последовательность, представленную под SEQ ID NO: 36, или по сути аналогичную ей последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности.
[00129] Настоящее изобретение предусматривает антитела, их антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит пару аминокислотных последовательностей HCDR3 и LCDR3 (HCDR3/LCDR3), содержащую аминокислотную последовательность HCDR3, представленную под SEQ ID NO: 33, в паре с аминокислотной последовательностью LCDR3, представленной под SEQ ID NO: 36. В некоторых вариантах осуществления пара аминокислотных последовательностей HCDR3/LCDR3 представлена под SEQ ID NO: 33/36.
[00130] Настоящее изобретение предусматривает антитела, их антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит набор из шести CDR (т.е., HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3), кодируемых нуклеотидной последовательностью, представленной под SEQ ID NO: 28. В определенных вариантах осуществления набор аминокислотных последовательностей HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3 представлен под SEQ ID NO: 31-32-33-34-35-36.
[00131] В родственном варианте осуществления настоящее изобретение предусматривает антитела, их антигенсвязывающие фрагменты и/или полиспецифические связывающие белки, содержащие паратоп, который содержит набор из шести CDR (т.е., HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3), содержащийся в паре аминокислотных последовательностей HCVR/LCVR, представленных под SEQ ID NO: 29/30. Способы и методики идентификации CDR в аминокислотных последовательностях HCVR и LCVR хорошо известны из уровня техники и могут быть использованы для идентификации CDR в указанных аминокислотных последовательностях HCVR и/или LCVR, раскрытых в данном документе. Иллюстративные подходы, которые можно использовать для определения границ CDR, включают, например, определение согласно Kabat, определение согласно Chothia и определение согласно AbM. В общих чертах, определение согласно Kabat основано на вариабельности последовательности, определение согласно Chothia основано на расположении структурных областей петли, а определение согласно AbM является компромиссным решением между подходами согласно Kabat и Chothia. См., например, Kabat, "Sequences of Proteins of Immunological Interest," National Institutes of Health, Бетесда, Мэриленд. (1991); Al-Lazikani et al., J. Mol. Biol. 273:927-948 (1997); и Martin et al., Proc. Natl. Acad. Sci. USA 56:9268-9272 (1989). Доступны также базы данных общего пользования для идентификации последовательностей CDR в антителе.
[00132] Настоящее изобретение также предусматривает молекулы нуклеиновой кислоты, кодирующие антитела к туе или их части.
[00133] Полиспецифическая связывающая молекула, как описано в данном документе, дополнительно содержит перенацеливающий лиганд в дополнение к паратопу (например, антителу или его части), который специфически связывает гетерологичный эпитоп, вставленный в рекомбинантный вирусный капсидный белок/отображенный с его помощью. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый на поверхности клетки, например, белок клеточной поверхности на (человеческой) эукариотической клетке, например, клетке-мишени. Существует большое количество подходящих белков клеточной поверхности, например, рецепторов клеточной поверхности, на которые можно направить перенацеливающий лиганд, и для которых изменяющие мишень лиганды, например, антитела или их части, уже доступны. Такие структуры включают без ограничения: антигены главного комплекса гистосовместимости класса I и класса II; рецепторы для различных цитокинов (например, рецепторы для IL-1, IL-4, IL-6, IL-13, IL-22, IL-25, IL-33 и т.д.), специфичные для типа клеток гормоны роста, нейротрофический фактор головного мозга (BDNF), цилиарный нейротрофический фактор (CTNF), колониестимулирующие факторы роста, эндотелиальные факторы роста, эпидермальные факторы роста, факторы роста фибробластов, глиальный нейротрофический фактор, глиальные факторы роста, gro-beta/mip 2, факторы роста гепатоцитов, инсулиноподобный фактор роста, интерфероны (α-IFN, β-IFN, γIFN, консенсусный IFN), интерлейкины (IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14), фактор роста кератиноцитов, факторы, ингибирующие лейкемию, фактор активации хемотаксиса макрофагов/моноцитов, фактор роста нервов, активирующий нейтрофилы белок 2, тромбоцитарный фактор роста, фактор стволовых клеток, трансформирующий фактор роста, факторы некроза опухолей, фактор роста эндотелия сосудов, липопротеины, включая дополнительные или другие трансмембранные рецепторы типа 1, такие как PRLR, рецепторы, связанные с G-белком, такие как GCGR, ионные каналы, такие как Nav1.7, ASIC1 или ASIC2; молекулы клеточной адгезии; транспортные молекулы для метаболитов, таких как аминокислоты; антигенные рецепторы В- и Т-лимфоцитов (например, В-клеточные рецепторы и ассоциированные белки (например, CD19, CD20 и т.д.) и Т-клеточные рецепторы и ассоциированные белки (например, CD3, CD4, CD8 и т.д.); тетраспаниновый белок (например, CD63). Описанный в данном документе рекомбинантный вирусный капсид обеспечивает специфическое инфицирование типа клеток путем использования полиспецифической связывающей молекулы, содержащей перенацеливающий лиганд, который связывает дифференцировочные антигены клеточной поверхности в качестве мишеней для комплекса вирусного вектора.
[00134] В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) клетками печени (человека), т.е. специфический для печени маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) клетками головного мозга (человека), т.е. специфический для клеток головного мозга маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) гемопоэтическими клетками (человека), т.е. специфический для гемопоэтических клеток маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) Т-клетками (человека), т.е. специфический для Т-клеток маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) В-клетками (человека), т.е. специфический для В-клеток маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) дендритными клетками (человека), т.е. специфический для дендритных клеток маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) макрофагами (человека), т.е. специфический для макрофагов маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) NK-клетками (человека), т.е. специфический для NK-клеток маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) клетками почки (человека), т.е. специфический для почки маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый главным образом (например, исключительно) клетками поджелудочной железы (человека), т.е. специфический для поджелудочной железы маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый главным образом (например, исключительно) клетками кишечника (человека), т.е. специфический для кишечника маркер. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) раковыми клетками (человека), т.е. опухолеассоциированный антиген. В некоторых вариантах осуществления перенацеливающий лиганд связывает белок, экспрессируемый главным образом (например, исключительно) клетками (человека), инфицированными гетерологичным патогеном. Белки, которые (1) специфически экспрессируются в клетке/ткани/органе, или экспрессия которых обогащена в них, и (2) распознаются антигенсвязывающим белком, применимым в качестве перенацеливающего лиганда, как описано в данном документе, хорошо известны и могут также быть обнаружены по адресу www.proteinatlas.org; см. также Uhlen et al. (2010) Nat. Biotech. 28:1248-50, включенную в данный документ посредством ссылки во всей своей полноте. В приведенной ниже таблице 1 представлены иллюстративные и неограничивающие маркеры, специфические для органа, для которых доступны антигенсвязывающие белки, которые могут быть полезны в качестве перенацеливающих лигандов, и клетки/ткань/орган, экспрессирующие такие маркеры.

[00135] В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый клеткой печени (человека), например, асиалогликопротеиновый рецептор, например, hASGR1. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый клеткой (человека) головного мозга. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый Т-клеткой (человека), например, CD3, например, CD3ε. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый клеткой почки (человека). В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый мышечной клеткой (человека), например, интегрин. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор, экспрессируемый раковой клеткой (человека), например, опухолеассоциированный антиген, например, Е6 и Е7. В некоторых вариантах осуществления перенацеливающий лиганд связывает рецептор глюкагона человека (hGCGR). В некоторых вариантах осуществления перенацеливающий лиганд связывает ENTPD3 человека.
[00136] В некоторых вариантах осуществления перенацеливающий лиганд связывает опухолеассоциированный антиген, экспрессируемый клеткой опухоли. Неограничивающие примеры специфических опухолеассоциированных антигенов включают, например, адипофилин, AIM-2, ALDH1A1, альфа-актинин-4, альфа-фетопротеин («AFP»), ARTC1, B-RAF, BAGE-1, BCLX (L), слитый белок BCR-ABL b3a2, бета-катенин, BING-4, СА-125, CALCA, онкоэмбриональный антиген («СЕА»), CASP-5, CASP-8, CD274, CD45, Cdc27, CDK12, CDK4, CDKN2A, СЕА, CLPP, СОА-1, CPSF, CSNK1A1, CTAG1, CTAG2, циклин D1, циклин-А1, слитый белок dek-can, DKK1, EFTUD2, фактор элонгации 2, ENAH (hMena), Ер-САМ, ЕрСАМ, EphA3, эпителиальный опухолевый антиген («ЕТА»), слитый белок ETV6-AML1, EZH2, Е6, Е7, FGF5, FLT3-ITD, FN1, G250/MN/CAIX, GAGE-1,2,8, GAGE-3,4,5,6,7, GAS7, глипикан-3, GnTV, gp100/Pme117, GPNMB, HAUS3, гепсин, HER-2/neu, HERV-K-MEL, HLA-A11, HLA-A2, HLA-DOB, hsp70-2, IDO1, IGF2B3, IL13Ralpha2, карбоксилэстеразу кишечника, K-ras, калликреин 4, KIF20A, KK-LC-1, KKLC1, KM-HN-1, KMHN1, также известный как CCDC110, LAGE-1, слитый белок LDLR-фукозилтрансферазу АS, ленгзин, M-CSF, MAGE-A1, MAGE-A10, MAGE-A12, MAGE-A2, MAGE-А3, MAGE-A4, MAGE-A6, MAGE-A9, MAGE-C1, MAGE-C2, малик-фермент, маммаглобин-А, MART2, MATN, MC1R, MCSP, mdm-2, МЕ1, Melan-A/MART-1, Meloe, Midkine, MMP-2, MMP-7, MUC1, MUC5AC, муцин, MUM-1, MUM-2, MUM-3, миозин, миозин класса I, N-raw, NA88-A, нео-РАР, NFYC, NY-BR-1, NY-ESO-1/LAGE-2, OA1, OGT, OS-9, полипептид P, p53, PAP, PAX5, PBF, слитый белок pml-RARalpha, полиморфный эпителиальный муцин («РЕМ»), PPP1R3B, PRAME, PRDX5, PSA, PSMA, PTPRK, RAB38/NY-MEL-1, RAGE-1, RBAF600, RGS5, RhoC, RNF43, RU2AS, SAGE, сецернин 1, SIRT2, SNRPD1, SOX10, Sp17, SPA17, SSX-2, SSX-4, STEAP1, сурвивин, слитый белок SYT-SSX1 или -SSX2, TAG-1, TAG-2, теломеразу, TGF-betaRII, TPBG, TRAG-3, триозофосфатизомеразу, TRP-1/gp75, TRP-2, TRP2-INT2, тирозиназу, тирозиназу («TYR»), VEGF, WT1, XAGE-1b/GAGED2a, Kras, NY-ESO1, MAGE-A3, HPV E2, HPV E6, HPV E7, антиген WT-1 (при лимфоме и других солидных опухолях), рецепторы ErbB, Melan A [MART1], gp 100, тирозиназу, TRP-1/gp 75 и TRP-2 (при меланоме); MAGE-1 и MAGE-3 (при карциноме мочевого пузыря, головы и шеи и немелкоклеточной карциноме); белки EG и Е7 HPV (при раке шейки матки); муцин [MUC-1] (при раке молочной железы, поджелудочной железы, толстой кишки и предстательной железы); специфический антиген простаты [PSA] (при раке простаты); онкоэмбриональный антиген [СЕА] (при раке толстой кишки, молочной железы и желудочно-кишечного тракта) и такие общие опухолеспецифические антигены, как MAGE-2, MAGE-4, MAGE-6, MAGE-10, MAGE-12, BAGE-1, CAGE-1,2,8, CAGE-3 TO 7, LAGE-1, NY-ESO-1/LAGE-2, NA-88, GnTV, TRP2-INT2. В некоторых вариантах осуществления опухолеассоциированный антиген представляет собой ErbB2/Her2. В некоторых вариантах осуществления опухолеассоциированный антиген представляет собой Е6 и/или Е7.
[00137] В некоторых вариантах осуществления перенацеливающий лиганд связывается с маркерами на основе CD, ассоциированными с иммунным ответом, например, CD3, CD4, CD8, CD19, CD20 и т.д. В некоторых вариантах осуществления маркер на основе CD представляет собой CD3.
[00138] В определенных иллюстративных вариантах осуществления полиспецифическая связывающая молекула представляет собой биспецифическое антитело. Каждый антигенсвязывающий домен биспецифического антитела содержит вариабельный домен тяжелой цепи (HCVR) и вариабельный домен легкой цепи (LCVR). В контексте биспецифической антигенсвязывающей молекулы, содержащей первый и второй антигенсвязывающий домен (например, биспецифического антитела), CDR первого антигенсвязывающего домена могут обозначаться с префиксом «А1» и CDR второго антигенсвязывающего домена могут обозначаться с префиксом «А2». Таким образом, CDR первого антигенсвязывающего домена могут упоминаться в данном документе как A1-HCDR1, A1-HCDR2 и A1-HCDR3; и CDR второго антигенсвязывающего домена могут упоминаться в данном документе как A2-HCDR1, A2-HCDR2 и A2-HCDR3.
[00139] Первый антигенсвязывающий домен и второй антигенсвязывающий домен могут быть прямо или косвенно соединены друг с другом с образованием биспецифической антигенсвязывающей молекулы по настоящему изобретению. Альтернативно, первый антигенсвязывающий домен и второй антигенсвязывающий домен, могут быть, каждый, соединены с отдельным доменом мультимеризации. Ассоциация одного домена мультимеризации с другим доменом мультимеризации облегчает ассоциацию между двумя антигенсвязывающими доменами, тем самым образуя биспецифическую антигенсвязывающую молекулу. Используемый в данном документе термин «домен мультимеризации» подразумевает любую макромолекулу, белок, полипептид, пептид или аминокислоту, которая обладает способностью связываться со вторым доменом мультимеризации той же или подобной структуры или состава. Например, домен мультимеризации может представлять собой полипептид, содержащий СН3-домен иммуноглобулина. Неограничивающим примером компонента мультимеризации является Fc-часть иммуноглобулина (содержащая СН2-СН3-домен), например, Fc-домен IgG, выбранного из изотипов IgG1, IgG2, IgG3 и IgG4, а также любого аллотипа в каждой изотипической группе.
[00140] Биспецифические антигенсвязывающие молекулы по настоящему изобретению обычно содержат два домена мультимеризации, например, два Fc-домена, каждый из которых по отдельности является частью отдельной тяжелой цепи антитела. Первый и второй домены мультимеризации могут быть одного и того же изотипа IgG, такого как, например, IgG1/IgG1, IgG2/IgG2, IgG4/IgG4. Альтернативно, первый и второй домены мультимеризации могут иметь разные изотипы IgG, такие как, например, IgG1/IgG2, IgG1/IgG4, IgG2/IgG4 и т.д.
[00141] В определенных вариантах осуществления домен мультимеризации представляет собой Fc-фрагмент или аминокислотную последовательность длиной от 1 до приблизительно 200 аминокислот, содержащую по меньшей мере один остаток цистеина. В других вариантах осуществления домен мультимеризации представляет собой остаток цистеина или короткий цистеинсодержащий пептид. Другие домены мультимеризации включают пептиды или полипептиды, содержащие или состоящие из лейциновой застежки, мотива спираль-петля или мотива суперспирали.
[00142] Любой формат или технологию биспецифического антитела можно использовать для получения биспецифических антигенсвязывающих молекул по настоящему изобретению. Например, антитело или его фрагмент, имеющий первую антигенсвязывающую специфичность, можно функционально связать (например, путем химического связывания, генетического слияния, нековалентной ассоциации или иным образом) с одним или более другими молекулярными объектами, таким как другое антитело или фрагмент антитела, имеющий вторую антигенсвязывающую специфичность, с получением биспецифической антигенсвязывающей молекулы. Конкретные иллюстративные биспецифические форматы, которые можно использовать в контексте настоящего изобретения, включают без ограничения, например, биспецифические форматы на основе scFv или диател, гибриды IgG-scFv, Ig с двойным вариабельным доменом (DVD)-Ig, квадрому, выступы-во-впадины, обычную легкую цепь (например, обычную легкую цепь с выступами-во-впадины и т.п.), CrossMab, CrossFab, (SEED)body, лейциновую застежку, DuoBody, IgG1/IgG2, Fab (DAF)-IgG двойного действия и биспецифические форматы Mab2 (см., например, Klein et al. 2012, mAbs 4:6, 1-11, и источники, упоминаемые в данном документе, для изучения вышеизложенных форматов; см. также Brinkmann and Konterman (2017) mAbs 9: 182-212; каждая из которых включена посредством ссылки во всей своей полноте).
[00143] Настоящее изобретение также включает биспецифические антигенсвязывающие молекулы, содержащие первый СН3-домен и второй СН3-домен Ig3, где первый и второй СН3-домены Ig3 отличаются друг от друга по меньшей мере одной аминокислотой, и где по меньшей мере одно различие в аминокислотах уменьшает связывание биспецифического антитела с белком А по сравнению с биспецифическим антителом, в котором отсутствует аминокислотное различие. В одном варианте осуществления первый СН3-домен Ig связывает белок А, а второй СН3-домен Ig содержит мутацию, которая уменьшает или устраняет связывание белка А, такую как модификация H95R (нумерация экзонов согласно IMGT; H435R нумерация согласно EU). Второй СН3 может дополнительно содержать модификацию Y96F (по IMGT; Y436F по EU). Дополнительные модификации, которые можно обнаружить в пределах второго СН3, включают: D16E, L18M, N44S, K52N, V57M и V82I (по IMGT; D356E, L358M, N384S, K392N, V397M и V422I по EU) в случае антител на основе IgG1; N44S, K52N и V82I (IMGT; N384S, K392N и V422I по EU) в случае антител на основе IgG2; и Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (по IMGT; Q355R, N384S, K392N, V397M, R409K, E419Q и V422I по EU) в случае антител на основе IgG4; см., например, WO 2010/151792.
[00144] В некоторых вариантах осуществления Fc-домен может быть химерным, объединяя последовательности Fc, полученные из более чем одного изотипа иммуноглобулина. Например, химерный Fc-домен может содержать часть или всю последовательность СН2, полученную из области СН2 IgG1 человека, IgG2 человека или IgG4 человека, и часть или всю последовательность СН3, полученную из IgG1 человека, IgG2 человека или IgG4 человека. Химерный Fc-домен также может содержать химерную шарнирную область. Например, химерный шарнир может содержать «верхнюю шарнирную» последовательность, полученную из шарнирной области IgG1 человека, IgG2 человека или IgG4 человека, в комбинации с «нижней шарнирной» последовательностью, полученной из шарнирной области IgG1 человека, IgG2 человека или IgG4 человека. Конкретный пример химерного Fc-домена, который может быть включен в любую из антигенсвязывающих молекул, представленных в данном документе, содержит от N- к С-концу: [CH1 IgG4] - [верхнюю шарнирную последовательность IgG4] - [нижнюю шарнирную последовательность IgG2] - [СН2 IgG4] - [CH3 IgG4]. Другой пример химерного Fc-домена, который может быть включен в любую из антигенсвязывающих молекул, представленных в данном документе, содержит от N- к С-концу: [CH1 IgG1] - [верхнюю шарнирную последовательность IgG1] - [нижнюю шарнирную последовательность IgG2] - [СН2 IgG4] - [СН3 IgG1]. Эти и другие примеры химерных Fc-доменов, которые могут быть включены в любую из антигенсвязывающих молекул по настоящему изобретению, описаны в заявке согласно РСТ № WO 2014/022540, включенной посредством ссылки во всей своей полноте. Химерные Fc-домены, содержащие эти общие структурные композиции и их варианты, могут иметь измененное связывание с Fc-рецептором, что, в свою очередь, влияет на эффекторную функцию Fc.
Способы применения и получения
[00145] Дополнительным вариантом осуществления рекомбинантных вирусных капсидных белков, описанных в данном документе, является их применение для доставки представляющего интерес нуклеотида, например репортерного гена или терапевтического гена, в клетку-мишень. Как правило, представляющим интерес нуклеотидом может быть плазмида для переноса, которая обычно может содержать 5'- и 3'-последовательности инвертированного концевого повтора (ITR), фланкирующие репортерный(-е) ген(ы) или терапевтический(-е) ген(ы), который(-е) может(-ут) находиться под контролем вирусного или невирусного промотора, когда он включен в вектор на основе AAV. В одном варианте осуществления представляющий интерес нуклеотид представляет собой плазмиду для переноса, содержащую от 5' к 3': 5'-ITR, промотор, ген (например, репортерный и/или терапевтический ген) и 3'-ITR.
[00146] Неограничивающие примеры полезных промоторов включают, например, цитомегаловирусный (CMV) промотор, промотор вируса некроза селезенки (SFFV), промотор фактора элонгации 1 альфа (EF1a) (EF1a-промотор размером 1,2 т.о. или EF1a-промотор размером 0,2 т.о.), химерный EF1a/IF4-промотор и промотор фосфоглицераткиназы (PGK). Внутренний энхансер может также присутствовать в вирусной конструкции для увеличения экспрессии представляющего интерес гена. Например, можно использовать энхансер CMV (Karasuyama et al. 1989. J. Exp. Med. 169:13, которая включена в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления энхансер CMV можно использовать в комбинации с промотором 13-актина курицы.
[00147] Разнообразные репортерные гены (или детектируемые фрагменты) можно заключать в мультимерную структуру, содержащую рекомбинантные вирусные капсидные белки, описанные в данном документе. Иллюстративные репортерные гены включают, например, β-галактозидазу (кодируемую геном lacZ), зеленый флуоресцентный белок (GFP), усиленный зеленый флуоресцентный белок (eGFP), MmGFP, синий флуоресцентный белок (BFP), усиленный синий флуоресцентный белок (eBFP), mPlum, mCherry, tdTomato, mStrawberry, J-Red, DsRed, mOrange, mKO, mCitrine, Venus, YPet, желтый флуоресцентный белок (YFP), усиленный желтый флуоресцентный белок (eYFP), Emerald, CyPet, голубой флуоресцентный белок (CFP), церулин, T-Sapphire, люциферазу, щелочную фосфатазу или их комбинацию. Способы, описанные в данном документе, демонстрируют построение целенаправленно воздействующих векторов, в которых используется репортерный ген, который кодирует зеленый флуоресцентный белок, однако специалисты в данной области техники поймут при прочтении данного раскрытия, что описанных в данном документе животных, отличных от человека, можно получить в отсутствие репортерного гена или с любым репортерным геном, известным из уровня техники.
[00148] Разнообразные терапевтические гены также можно заключать в мультимерную структуру, содержащую рекомбинантные вирусные капсидные белки, описанные в данном документе, например, как часть вектора для переноса. Неограничивающие примеры терапевтического гена включают те, которые кодируют токсин (например, суицидный ген), терапевтическое антитело или его фрагмент, систему CRISPR/Cas или ее часть (части), антисмысловую РНК, siRNA, shRNA и т.д.
[00149] Дополнительным вариантом осуществления настоящего изобретения является способ получения рекомбинантного капсидного белка, причем способ включает стадии:
a) экспрессии нуклеиновой кислоты, кодирующей рекомбинантный капсидный белок, в подходящих условиях, и
b) выделения экспрессированного капсидного белка со стадии а).
[00150] Дополнительные варианты осуществления настоящего изобретения представляют собой способ изменения тропизма вируса, включающий стадии: (а) вставки нуклеиновой кислоты, кодирующей гетерологичный эпитоп, в последовательность нуклеиновой кислоты, кодирующую вирусный капсидный белок, с образованием нуклеотидной последовательности, кодирующей генетически модифицированный капсидный белок, содержащий гетерологичный эпитоп, и/или (b) культивирования упаковывающей клетки в условиях, достаточных для продуцирования вирусных векторов, где упаковывающая клетка содержит нуклеотидную последовательность. Дополнительным вариантом осуществления настоящего изобретения является способ отображения гетерологичного эпитопа на поверхности капсидного белка, причем способ включает стадии: а) экспрессии нуклеиновой кислоты по данному изобретению в подходящих условиях и b) выделения экспрессированного капсидного белка со стадии а).
[00151] В некоторых вариантах осуществления упаковывающая клетка дополнительно содержит плазмиду-помощника и/или плазмиду для переноса, содержащую представляющий интерес нуклеотид. В некоторых вариантах осуществления способы дополнительно включают выделение самокомплементарных векторов на основе аденоассоциированного вируса из культурального супернатанта. В некоторых вариантах осуществления способы дополнительно включают лизис упаковывающей клетки и выделение однонитевых векторов на основе аденоассоциированного вируса из клеточного лизата. В некоторых вариантах осуществления способы дополнительно включают (а) удаление клеточного дебриса, (b) обработку супернатанта, содержащего вирусные векторы, ДНКазой I и MgCl2, (с) концентрирование вирусных векторов, (d) очистку вирусных векторов и (е) любую комбинацию (a)-(d).
[00152] Упаковывающие клетки, пригодные для продуцирования вирусных векторов, описанных в данном документе, включают, например, клетки животных, пермиссивные для вируса, или клетки, модифицированные таким образом, чтобы они были пермиссивными для вируса; или конструкцию упаковывающей клетки, например, с применением трансформирующего средства, такого как фосфат кальция. Неограничивающие примеры упаковывающих клеточных линий, полезных для получения вирусных векторов, описанных в данном документе, включают, например, клетки эмбриональной почки человека 293 (HEK-293) (например, Американская коллекция типовых культур [АТСС] № CRL-1573), клетки HEK-293, которые содержат большой Т-антиген SV40 (HEK-293Т или 293Т), клетки HEK293T/17, линию клеток саркомы человека НТ-1080 (CCL-121), линию лимфобластоподобных клеток Raji (CCL-86), линию эпителиоподобных клеток глиобластомы-астроцитомы U87-MG (НТВ-14), линию клеток Т-лимфомы HuT78 (TIB-161), клетки NIH/3T3, клетки яичника китайского хомячка (СНО) (например, АТСС № CRL9618, CCL61, CRL9096), клетки HeLa (например, АТСС № CCL-2), клетки Vero, клетки NIH 3Т3 (например, АТСС № CRL-1658), клетки Huh-7, клетки BHK (например, АТСС № CCL10), клетки РС12 (АТСС № CRL1721), клетки COS, клетки COS-7 (АТСС № CRL1651), клетки RATI, L-клетки мыши (АТСС № CCLI.3), клетки HLHepG2, клетки САР, клетки САР-Т и т.п.
[00153] Клетки L929, клеточная система упаковки вируса FLY, описанная в Cosset et al. (1995) J. Virol 69, 7430-7436, клетки NSO (мышиная миелома), человеческие амниоцитарные клетки (например, САР, САР-Т), клетки дрожжей (включая без ограничения S. cerevisiae, Pichia pastoris), растительные клетки (включая без ограничения Tobacco NT1, BY-2), клетки насекомых (включая без ограничения SF9, S2, SF21, Tni (например, High 5)) или бактериальные клетки (включая без ограничения Е. coli).
[00154] Для дополнительных упаковывающих клеток и систем, методик упаковки и векторов для упаковки генома в виде нуклеиновой кислоты в псевдотипированный вирусный вектор см., например, Polo et al., Proc Natl Acad Sci USA, (1999) 96:4598-4603. Способы упаковки включают использование упаковывающих клеток, которые постоянно экспрессируют вирусные компоненты, или при временной трансфекции клеток плазмидами.
[00155] Дополнительные варианты осуществления включают способы перенаправления вируса и/или доставки репортерного или терапевтического гена в клетку-мишень, где способ включает способ трансдукции клеток in vitro или in vivo, причем способ включает следующие стадии: приведение клетки-мишени в контакт с комбинацией вирусного вектора, который содержит капсид, включая рекомбинантный вирусный капсид, отображающий гетерологичный эпитоп, и полиспецифическую связывающую молекулу, где полиспецифическая связывающая молекула содержит i) паратоп антитела, который специфически связывает эпитоп, и (ii) перенацеливающий лиганд, который специфически связывает рецептор, экспрессируемый клеткой-мишенью. В некоторых вариантах осуществления в композиции, описанной в данном документе, или в способе, описанном в данном документе, объединяют рекомбинантный вирусный вектор и полиспецифическую связывающую молекулу в соотношении молекула:молекула, которое восстанавливает эффективность трансдукции вирусного вектора, сходную с эффективностью контрольного вирусного вектора дикого типа. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) находится в диапазоне от 1:0,5 до 1:100. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) находится в диапазоне от 1:4 до 1:20. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) находится в диапазоне от 1:8 до 1:15. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:4. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:8. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:15. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет 1:20. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет менее чем 1:100. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула: молекула) составляет менее чем 1:50. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет менее чем 1:20. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет менее чем 1:15. В некоторых вариантах осуществления соотношение рекомбинантного вирусного вектора к полиспецифической связывающей молекуле (молекула:молекула) составляет менее чем 1:10.
[00156] В некоторых вариантах осуществления клетка-мишень находится в условиях in vitro. В других вариантах осуществления клетка-мишень находится в условиях in vivo у субъекта, например, человека.
Клетки-мишени
[00157] Для доставки представляющего интерес нуклеотида с использованием рекомбинантного вирусного вектора, как раскрыто в данном документе, можно применять большое разнообразие клеток. Клетки-мишени обычно выбирают на основе представляющего интерес нуклеотида и необходимого эффекта.
[00158] В некоторых вариантах осуществления представляющий интерес нуклеотид может быть доставлен, чтобы позволить клетке-мишени продуцировать белок, который восполняет дефицит в организме, такой как ферментативный дефицит или иммунодефицит, такой как Х-сцепленный тяжелый комбинированный иммунодефицит. Таким образом, в некоторых вариантах осуществления клетки, которые обычно продуцируют белок у животного, являются мишенями. В других вариантах осуществления клетки в области, в которой белок был бы наиболее полезным, являются мишенями.
[00159] В других вариантах осуществления представляющий интерес нуклеотид, такой как ген, кодирующий siRNA, может ингибировать экспрессию конкретного гена в клетке-мишени. Представляющий интерес нуклеотид может, например, ингибировать экспрессию гена, вовлеченного в жизненный цикл патогена. Таким образом, клетки, восприимчивые к инфекции патогеном или инфицированные патогеном, могут быть мишенью. В других вариантах осуществления представляющий интерес нуклеотид может ингибировать экспрессию гена, который отвечает за продуцирование токсина в клетке-мишени.
[00160] В других вариантах осуществления представляющий интерес нуклеотид может кодировать токсичный белок, который приводит к гибели клеток, в которых он экспрессируется. В этом случае опухолевые клетки или другие нежелательные клетки могут быть мишенью.
[00161] В других вариантах осуществления можно использовать представляющий интерес нуклеотид, который кодирует белок, подлежащий сбору, такой как терапевтический белок, и клетки, которые способны продуцировать и секретировать белок, являются мишенью.
[00162] Как только идентифицирована определенная популяция клеток-мишеней, в которой необходима экспрессия представляющего интерес нуклеотида, выбирают рецептор-мишень, который специфически экспрессируется в этой популяции клеток-мишеней. Рецептор-мишень может экспрессироваться исключительно в этой популяции клеток или в большей степени в этой популяции клеток, чем в других популяциях клеток. Чем более специфичной является экспрессия, тем более специфично можно направить доставку в клетки-мишени. В зависимости от контекста желаемая величина специфичности маркера (и, следовательно, доставки гена) может варьироваться. Например, для введения токсичного гена высокая специфичность является наиболее предпочтительной, чтобы избежать гибели нецелевых клеток. Для экспрессии белка для сбора или экспрессии секретируемого продукта, когда необходимо глобальное воздействие, может потребоваться меньшая специфичность маркера.
[00163] Как обсуждалось выше, рецептором-мишенью может быть любой рецептор, для которого можно идентифицировать или создать перенацеливающий лиганд. Предпочтительно рецептор-мишень представляет собой пептид или полипептид, такой как рецептор. Однако в других вариантах осуществления рецептором-мишенью может быть углевод или другая молекула, которая может распознаваться партнером по связыванию. Если партнер по связыванию, например, лиганд, для рецептора-мишени уже известен, его можно использовать в качестве аффинной молекулы. Однако, если связывающая молекула неизвестна, антитела к рецептору-мишени можно получить с использованием стандартных процедур. Затем эти антитела можно использовать в качестве перенацеливающего лиганда как часть полиспецифической связывающей молекулы.
[00164] Таким образом, клетки-мишени можно выбрать на основе множества факторов, включая, например, (1) конкретное применение (например, терапия, экспрессия белка, подлежащего сбору, и придание устойчивости к заболеванию) и (2) экспрессию маркера с необходимой величиной специфичности.
[00165] Клетки-мишени никоим образом не ограничены и включают как клетки и линии клеток зародышевой линии, так и соматические клетки и линии клеток. Клетки-мишени могут быть стволовыми клетками любого происхождения. Когда клетки-мишени представляют собой клетки зародышевой линии, клетки-мишени предпочтительно выбирают из группы, состоящей из одноклеточных эмбрионов и эмбриональных стволовых клеток (ES).
Фармацевтические композиции, лекарственные формы и введение
[00166] В дополнительном варианте осуществления предусмотрен лекарственный препарат, содержащий по меньшей мере один рекомбинантный вирусный капсидный белок и соответствующую полиспецифическую связывающую молекулу согласно данному изобретению и/или нуклеиновую кислоту согласно данному изобретению, предпочтительно по меньшей мере одну мультимерную структуру согласно данному изобретению. Предпочтительно, чтобы такой лекарственный препарат был применимым в качестве вектора для переноса генов.
[00167] В данном документе также раскрыты фармацевтические композиции, содержащие вирусные векторы, описанные в данном документе, и фармацевтически приемлемый носитель и/или вспомогательное вещество. Кроме того, в данном документе раскрыты фармацевтические лекарственные формы, содержащие вирусный вектор, описанный в данном документе.
[00168] Как обсуждается в данном документе, вирусные векторы, описанные в данном документе, можно использовать для различных терапевтических применений (in vivo и ex vivo) и в качестве инструментов для исследования.
[00169] Фармацевтические композиции на основе вирусных векторов, раскрытых в данном документе, могут быть составлены любым общепринятым способом с использованием одного или более физиологически приемлемых носителей и/или вспомогательных веществ. Вирусный вектор можно составить для введения, например, путем инъекции, ингаляции или инстилляции (через рот или нос), либо путем перорального, буккального, парентерального или ректального введения, или путем введения непосредственно в опухоль.
[00170] Фармацевтические композиции можно составить для различных способов введения, включая системное, местное или локализованное введение. Методы и составы можно найти, например, в Remrnington's Pharmaceutical Sciences, Meade Publishing Co., Истон, Пенсильвания. Для системного введения предпочтительна инъекция, в том числе внутримышечная, внутривенная, внутрибрюшинная и подкожная. Для инъекции фармацевтические композиции можно составить в жидких растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хенкса или раствор Рингера. Кроме того, фармацевтические композиции можно составить в твердой форме и повторно растворить или суспендировать непосредственно перед применением. Лиофилизированные формы фармацевтической композиции также являются подходящими.
[00171] Для перорального введения фармацевтические композиции могут быть в форме, например, таблеток или капсул, приготовленных обычными способами с фармацевтически приемлемыми вспомогательными веществами, такими как связующие средства (например, предварительно желатинизированный кукурузный крахмал, поливинилпирролидон или гидроксипропилметилцеллюлоза); наполнители (например, лактоза, микрокристаллическая целлюлоза или гидрофосфат кальция); скользящие вещества (например, стеарат магния, тальк или диоксид кремния); разрыхлители (например, картофельный крахмал или крахмалгликолят натрия) или смачивающие средства (например, лаурилсульфат натрия). Таблетки могут также быть покрыты оболочкой с помощью способов, хорошо известных из уровня техники. Жидкие препараты для перорального введения могут быть в форме, например, растворов, сиропов или суспензий, или они могут быть представлены в виде сухого продукта для разведения водой или другой подходящей средой-носителем перед применением. Такие жидкие препараты можно получить обычными способами с фармацевтически приемлемыми добавками, такими как суспендирующие средства (например, сироп сорбита, производные целлюлозы или гидрогенизированные пищевые жиры); эмульгаторы (например, лецитин или аравийская камедь); неводные среды-носители (например, миндальное масло, масляные эфиры, этиловый спирт или фракционированные растительные масла) и консерванты (например, метил- или пропил-п-гидроксибензоаты или сорбиновая кислота). Препараты могут также содержать буферные соли, ароматизаторы, красители и подсластители, в случае необходимости.
[00172] Фармацевтические композиции можно составить для парентерального введения путем инъекции, например, путем болюсной инъекции или непрерывной инфузии. Составы для инъекций могут быть представлены в стандартной лекарственной форме, например, в ампулах или в многодозовых контейнерах необязательно с добавлением консерванта. Фармацевтические композиции могут быть дополнительно составлены в виде суспензий, растворов или эмульсий в масляных или водных средах-носителях и могут содержать другие средства, включая суспендирующие, стабилизирующие и/или диспергирующие средства.
[00173] Кроме того, фармацевтические композиции также можно составить в виде депо-препарата. Эти составы длительного действия можно вводить путем имплантации (например, подкожно или внутримышечно) или путем внутримышечной инъекции. Так, например, соединения могут быть составлены с подходящими полимерными или гидрофобными материалами (например, в виде эмульсии в приемлемом масле) или ионообменными смолами или в виде труднорастворимых производных, например, в виде труднорастворимой соли. Другие подходящие системы доставки включают микросферы, которые предлагают возможность локальной неинвазивной доставки лекарственных средств в течение продолжительного периода времени. Эта технология может включать микросферы, имеющие прекапиллярный размер, которые можно вводить через коронарный катетер в любую выбранную часть органа, не вызывая воспаления или ишемии. Вводимое терапевтическое средство медленно высвобождается из микросфер и поглощается окружающими клетками, присутствующими в выбранной ткани.
[00174] Системное введение также можно осуществлять трансмукозальным или трансдермальным способами. Для трансмукозального или трансдермального введения в составе используют проникающие вещества, подходящие для проникновения через барьер. Такие проникающие вещества обычно известны из уровня техники и включают, например, для трансмукозального введения соли желчных кислот и производные фузидиевой кислоты. Кроме того, можно использовать детергенты для облегчения проникновения. Трансмукозальное введение можно осуществлять с использованием назальных спреев или суппозиториев. Для местного введения вирусный вектор, описанный в данном документе, можно составить в виде мазей, бальзамов, гелей или кремов, общеизвестных из уровня техники. Раствор для промывания также можно использовать локально для лечения повреждения или воспаления с целью ускорения заживления.
[00175] Фармацевтические формы, подходящие для инъекционного применения, могут включать стерильные водные растворы или дисперсии; составы, включающие кунжутное масло, арахисовое масло или водный раствор пропиленгликоля; и стерильные порошки для получения стерильных инъекционных растворов или дисперсий для немедленного приема. Во всех случаях форма должна быть стерильной и должна быть жидкой. Она должна быть стабильной в условиях производства и при определенных параметрах хранения (например, охлаждение и заморозка) и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибы.
[00176] Если составы, раскрытые в данном документе, используют в качестве терапевтического средства для усиления иммунного ответа у субъекта, терапевтическое средство можно составить в композиции в нейтральной или солевой форме. Фармацевтически приемлемые соли включают соли присоединения кислоты (образованные с помощью свободных аминогрупп белка) и соли, которые образованы с неорганическими кислотами, такими как, например, хлористоводородная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная, миндальная и т.п. Соли, образованные со свободными карбоксильными группами, также можно получить из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и т.п.
[00177] Носителем также может быть растворитель или дисперсионная среда, содержащая, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п), их подходящие смеси и растительные масла. Подходящую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания требуемого размера вирусного вектора в случае дисперсии и путем применения поверхностно-активных веществ. Предотвращение действия микроорганизмов можно осуществить с помощью различных антибактериальных и противогрибковых средств, известных из уровня техники. Во многих случаях предпочтительно включать изотонические средства, например сахара или хлорид натрия. Длительной абсорбции инъекционных композиций можно достичь путем применения в композициях средств, замедляющих абсорбцию, например, моностеарата алюминия и желатина.
[00178] Стерильные инъекционные растворы можно получить путем включения активных соединений или конструкций в необходимом количестве в подходящем растворителе с различными другими ингредиентами, перечисленными выше, при необходимости, с последующей стерилизацией путем фильтрации.
[00179] После составления растворы можно вводить способом, соответствующим дозированному составу, и в таком количестве, которое является терапевтически эффективным. Составы можно легко вводить в различных лекарственных формах, таких как тип инъекционных растворов, описанных выше, но также можно использовать капсулы или микрочастицы и микросферы с замедленным высвобождением и т.п.
[00180] Например, для парентерального введения в водном растворе этот раствор должен быть подходящим образом забуферен, если необходимо, и жидкий разбавитель сначала должен быть сделан изотоническим с помощью достаточного количества физиологического раствора или глюкозы. Эти конкретные водные растворы особенно подходят для внутривенного, внутриопухолевого, внутримышечного, подкожного и внутрибрюшинного введения. В этом контексте стерильные водные среды, которые можно использовать, будут известны специалистам в данной области техники в свете настоящего раскрытия. Например, одну дозу можно растворить в 1 мл изотонического раствора NaCl и либо добавить к 1000 мл жидкости для гиподермоклизиса, либо ввести в предлагаемом месте инфузии.
[00181] Лицо, ответственное за введение, в любом случае определит подходящую дозу для индивидуального субъекта. Например, субъекту могут вводить вирусные векторы, описанные в данном документе, ежедневно или еженедельно в течение некоторого периода времени, или ежемесячно, два раза в год или ежегодно, в зависимости от потребности или воздействия патогенного организма или состояния у субъекта (например, рак).
[00182] В дополнение к соединениям, составленным для парентерального введения, такого как внутривенная, внутриопухолевая, внутрикожная или внутримышечная инъекция, другие фармацевтически приемлемые формы включают, например, таблетки или другие твердые вещества для перорального введения; липосомные составы; капсулы с замедленным высвобождением; биоразлагаемые и любые другие формы, используемые в настоящее время.
[00183] Можно также применять интраназальные или ингаляционные растворы или спреи, аэрозоли или ингаляторы. Назальные растворы могут быть водными растворами, предназначенными для введения в носовые ходы в виде капель или спреев. Назальные растворы можно получать так, что они во многом похожи на носовой секрет. Таким образом, водные назальные растворы обычно являются изотоническими и слегка забуференными для поддержания рН от 5,5 до 7,5. Кроме того, в состав можно включать противомикробные консерванты, подобные тем, которые применяют в офтальмологических препаратах, и соответствующие стабилизаторы лекарственных средств, если это необходимо. Известны различные коммерческие назальные препараты, которые могут включать, например, антибиотики и антигистаминные препараты, и применяются для профилактики астмы.
[00184] Составы для перорального применения могут включать вспомогательные вещества, такие как, например, маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлоза, карбонат магния и т.п. фармацевтической степени чистоты. Этим композициям придают форму растворов, суспензий, таблеток, пилюль, капсул, составов с замедленным высвобождением или порошков. В некоторых определенных вариантах осуществления фармацевтические композиции для перорального применения будут включать инертный разбавитель или усваиваемый съедобный носитель, или они могут быть заключены в желатиновую капсулу с твердой или мягкой оболочкой, или они могут быть спрессованы в таблетки, или они могут быть включены непосредственно в пищевой продукт, предусмотренный диетой. Для перорального терапевтического введения активные соединения можно включать со вспомогательными веществами и применять в форме принимаемых внутрь таблеток, буккальных таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, облаток и тому подобного.
[00185] Таблетки, пастилки, пилюли, капсулы и тому подобное могут также содержать следующее: связующее вещество, такое как трагакантовая камедь, аравийская камедь, кукурузный крахмал или желатин; вспомогательные вещества, такие как дикальция фосфат; разрыхлитель, такой как кукурузный крахмал, картофельный крахмал, альгиновая кислота и тому подобное; скользящее вещество, такое как стеарат магния; и может быть добавлен подсластитель, такой как сахароза, лактоза или сахарин, или вкусоароматическое средство, такое как мята перечная, масло грушанки или вишневый ароматизатор. Когда стандартная лекарственная форма представляет собой капсулу, она может содержать, помимо материалов вышеуказанного типа, жидкий носитель. Различные другие материалы могут присутствовать в качестве покрытий или иным образом модифицировать физическую форму лекарственной формы. Например, таблетки, пилюли или капсулы могут быть покрыты шеллаком, сахаром или и тем и другим. Сироп или эликсир может содержать активные соединения сахарозы как подсластитель, метил- и пропилпарабены в качестве консервантов, красителей и ароматизаторов, таких как вишневый или апельсиновый ароматизатор.
[00186] Другие варианты осуществления, раскрытые в данном документе, могут касаться наборов для применения в соответствии со способами и композициями. Наборы могут также включать подходящий контейнер, например флаконы, пробирки, мини- или микроцентрифужные пробирки, пробирку для испытания, колбу, сосуд, шприц или другой контейнер. Если предусмотрены дополнительные компонент или средство, то набор может содержать один или более дополнительных контейнеров, в которые можно поместить эти средство или компонент. Наборы в данном документе также обычно включают средства для герметичного содержания вирусных векторов и любых других контейнеров с реагентами для коммерческой продажи. Такие контейнеры могут включать пластмассовые контейнеры, полученные литьем под давлением или выдувным формованием, в которые помещают необходимые флаконы. Необязательно, для описанных композиций могут потребоваться одно или более дополнительных активных средств, таких как, например, противовоспалительные средства, противовирусные средства, противогрибковые или антибактериальные средства или противоопухолевые средства.
[00187] Диапазоны доз и частота введения могут варьироваться в зависимости от природы вирусных векторов и состояния здоровья, а также параметров конкретного пациента и используемого пути введения. В некоторых вариантах осуществления композиции с вирусными векторами можно вводить субъекту в дозе в диапазоне от приблизительно 1×105 бляшкообразующих единиц (БОЕ) до приблизительно 1×1015 БОЕ в зависимости от способа введения, пути введения, характера заболевания и состояния субъекта. В некоторых случаях композиции с вирусными векторами можно вводить в дозе в диапазоне от приблизительно 1×108 БОЕ до приблизительно 1×1015 БОЕ, или от приблизительно 1×1010 БОЕ до приблизительно 1×1015 БОЕ, или от приблизительно 1×108 БОЕ до приблизительно 1×1012 БОЕ. Более точная доза также может зависеть от субъекта, которому ее вводят. Например, более низкая доза может потребоваться, если субъект несовершеннолетний, и более высокая доза может потребоваться, если субъект является взрослым субъектом-человеком. В определенных вариантах осуществления более точная доза может зависеть от веса субъекта. В определенных вариантах осуществления, например, несовершеннолетний субъект-человек может получать от приблизительно 1×108 БОЕ до приблизительно 1×1010 БОЕ, в то время как взрослый субъект-человек может получать дозу от приблизительно 1×1010 БОЕ до приблизительно 1×1012 БОЕ.
[00188] Композиции, раскрытые в данном документе, можно вводить любым способом, известным в данной области. Например, композиции могут предусматривать введение субъекту внутривенно, внутриопухолево, внутрикожно, внутриартериально, внутрибрюшинно, внутрь повреждения, внутричерепно, внутрисуставно, внутрипростатически, внутриплеврально, внутритрахеально, интраназально, интравитреально, интравагинально, интраректально, местно, внутриопухолево, внутримышечно, интратекально, подкожно, субконъюнктивально, интравезикулярно, мукозально, интраперикардиально, внутрипуповинно, интраокулярно, перорально, локально, с помощью ингаляции, с помощью инъекции, с помощью инфузии, с помощью непрерывной инфузии, с помощью локальной перфузии, через катетер, с помощью промывания, в креме или в липидной композиции.
[00189] Любой способ, известный специалисту в данной области, можно применять для крупномасштабного производства вирусных векторов, упаковывающих клеток и векторных конструкций, описанных в данном документе. Например, основной и рабочий посевные материалы можно получать в условиях согласно GMP в отвечающих требованиям первичных CEF или другими способами. Упаковывающие клетки можно высевать в колбы с большой площадью поверхности, выращивать практически до конфлюентности и очищать вирусные векторы. Клетки можно собирать и вирусные векторы, высвобожденные в культуральную среду, выделять и очищать или внутриклеточные вирусные векторы, высвобожденные путем механического разрушения (клеточный дебрис можно удалять путем глубинной фильтрации с большими порами и расщепления ДНК клетки-хозяина эндонуклеазой). Вирусные векторы вирусов можно впоследствии очищать и концентрировать путем фильтрации в тангенциальном потоке с последующей диафильтрацией. Полученную концентрированную массу можно составлять путем разбавления буфером, содержащим стабилизаторы, наполнять флаконы и лиофилизировать. Композиции и составы можно хранить для последующего применения. Для применения лиофилизированные вирусные векторы можно восстанавливать путем добавления разбавителя.
[00190] Определенные дополнительные средства, применяемые в комбинированной терапии, можно составлять и вводить любым способом, известным из уровня техники.
[00191] Композиции, раскрытые в данном документе, могут также включать адъюванты, такие как соли алюминия и другие минеральные адъюванты, сурфактанты, бактериальные компоненты, среды-носители и цитокины. Адъюванты также могут обладать антагонистическими иммуномодулирующими свойствами. Например, адъюванты могут стимулировать Th1- или Th2-зависимый иммунитет. Композиции и способы, раскрытые в данном документе, также могут предусматривать адъювантную терапию.
ПРИМЕРЫ
[00192] Следующие примеры предоставлены лишь в иллюстративных целях и не предназначены для ограничения объема настоящего изобретения.
Пример 1. Получение векторов на основе аденоассоциированного вируса с гетерологичным эпитопом
[00193] Капсидные белки AAV модифицировали так, чтобы они содержали один из нескольких гетерологичных эпитопов: FLAG, c-myc, гексагистидин и т.д., с помощью ПЦР с получением плазмиды, кодирующей рекомбинантный капсидный белок. Кратко, последовательность, кодирующую FLAG, c-myc или гексагистидин, вставляли в рамку после ко дона, кодирующего N587 капсидного белка AAV2, Q585 капсидного белка VP1 AAV6, N590 капсидного белка VP1 AAV8, А589 капсидного белка VP1 AAV9 или G453 капсидного белка VP1 AAV9.
[00194] Выработку аденоассоциированных вирусов получали с использованием способа тройной трансфекции клеток HEK293 (см., например, Erik Arden and Joseph M. Metzger, J Biol Methods. 2016; 3(2)). Клетки высевали за один день до опосредованной PEFpro (Polyplus transfection, Нью-Йорк, штат Нью-Йорк) трансфекции соответствующими векторами:
плазмида-помощник, pHelper (Agilent, номер по каталогу 240074);
плазмида, кодирующая ген rep/cap AAV дикого типа или модифицированный (pAAV RC2 (Cell biolabs, номер по каталогу VPK-422), например pAAV RC2/6/9 (Cell Biolabs, номер по каталогу VPK-426), pAAV RC8, pAAV RC2-N587myc, pAAV RC2/6-Q585myc, pAAVRC8-N590myc, pAAV RC9-A589myc и т.д.; и
плазмида, кодирующая представляющую интерес нуклеотидную последовательность и последовательности ITR AAV, например pscAAV-CMV-eGFP, pAAV-CMVGFP (Agilent, номер по каталогу 240074), pAAV-EF1a-eGFP или pAAV-CAGG-eGFP и т.д.
[00195] Через семьдесят два часа после трансфекции среду собирали и клетки подвергали лизису в буфере [50 мМ Tris-HCl, 150 мМ NaCl и 0,5% дезоксихолата натрия (Sigma, номер по каталогу D6750-100G)]. Затем бензоназу (Sigma, Сент-Луис, штат Миссури) добавляли как к среде, так и к клеточному лизату до конечной концентрации 0,5 ед./мкл перед инкубацией при 37°С в течение 60 минут. Клеточный лизат центрифугировали при 4000 об./мин. в течение 30 минут. Клеточный лизат и среду объединяли и осаждали с помощью PEG 8000 (Teknova, номер по каталогу Р4340) в конечной концентрации 8%. Осадок ресуспендировали в 400 мМ NaCl и центрифугировали при 10000 g в течение 10 минут. Вирусы в надосадочной жидкости осаждали ультрацентрифугированием при 149000 g в течение 3 часов и титровали с помощью qPCR.
[00196] При qPCR для титрования геномов AAV образцы AAV обрабатывали ДНКазой I (Thermofisher Scientific, номер по каталогу EN0525) при 37°С в течение одного часа и подвергали лизису с использованием набора DNA extract All Reagents (Thermofisher Scientific, номер по каталогу 4403319). Заключенные в белковую оболочку вирусные геномы определяли количественно, используя систему для ПЦР в реальном времени QuantStudio 3 (Thermofisher Scientific) с применением праймеров для ITR AAV2. Последовательности праймеров для ITR AAV2 представляют собой 5'-GGAACCCCTAGTGATGGAGTT-3' (прямой праймер для ITR; SEQ ID NO: 9) и 5'-CGGCCTCAGTGAGCGA-3' (обратный праймер для ITR; SEQ ID NO: 10) (Aurnhammer et al., 2012), вырезают соответственно последовательность левого внутреннего инвертированного повтора (ITR) из AAV и последовательность правого внутреннего инвертированного повтора (ITR) из AAV. Последовательность зонда для ITR AAV2 представляет собой 5'-6-FAM-CACTCCCTCTCTGCGCGCTCG-TAMRA-3' (SEQ ID NO: 11) (Aurnhammer С., Haase M., Muether N., et al., 2012, Hum. Gene Ther. Methods 23, 18-28). После этапа активации при 95°С в течение 10 минут выполняли двухэтапный цикл ПЦР при 95°С в течение 15 секунд и 60°С в течение 30 секунд на протяжении 40 циклов. Универсальный мастер-микс для ПЦР TaqMan (Thermofisher Scientific, номер по каталогу 4304437) использовали для qPCR. ДНК-плазмиду (Agilent, номер по каталогу 240074) использовали в качестве стандарта для определения абсолютных титров.
[00197] Получали векторы на основе аденоассоциированного вируса, содержащие капсид с эпитопом с-myc. В этом примере эпитоп c-myc (EQKLISEEDL; SEQ ID NO: 6) вставляли между аминокислотами N587 и R588 капсидного белка VP1 AAV2 или между аминокислотами А589 и Q590 капсидного белка VP1 AAV9, т.е. нуклеотидную последовательность, кодирующую эпитоп c-myc (GAA САА AAA СТС АТС ТСА GAA GAG GAT CTG; SEQ ID NO: 12), вставляли в плазмиду pAAV RC2 (Cell Biolabs, Inc., Сан-Диего, штат Калифорния) или плазмиду pAAV RC2/9 и каждую из соответствующих модифицированных плазмид pAAV RC2-N587Myc, pAAV RC9-A589Myc, использовали для кодирования модифицированного капсидного белка для вирусных векторов на основе AAV с пониженным или устраненным тропизмом.
[00198] Для создания pAAV RC2-N587Myc получали первый продукт полимеразной цепной реакции (ПЦР), содержащий (от 5' к 3') сайт рестрикции BsiWl, нуклеотидную последовательность между положениями 3050 и 3773 pAAV RC2 и нуклеотидную последовательность эпитопа c-myc с «липкими» концами, и второй продукт ПЦР, содержащий (от 5' к 3') нуклеотидную последовательность эпитопа с-myc с «липкими» концами, нуклеотидную последовательность между положениями 3774 и 4370 pAAV RC2 и сайт рестрикции Pme1, с использованием праймеров, приведенных в таблице 2. Плазмиду pAAV RC2-N587Myc (т.е. плазмиду pAAV RC2, модифицированную для кодирования эпитопа с-myc между аминокислотами N587 и R588 капсидного белка VP1) создавали путем расщепления pAAV RC2 с помощью BsiW1 (New England Biolabs, R0553L) и Pmel (New England Biolabs, R0560L) и вставки двух продуктов ПЦР с помощью независимого от лигирования клонирования, описанного в (2012) Methods Mol. Biol. 52:51-9.
[00199] Для создания pAAV RC2/6-Q585Myc заказали фрагмент ДНК gblock, содержащий положения 3700 и 3940 pAAV RC2/6 с последовательностью эпитопа с-myc, вставленной между 3757 и 3758, от Integrated DNA Technologies (Коралвилль, штат Айова). Плазмиду pAAV RC2/6-Q585Myc создавали путем вставки фрагмента gblock в pAAV RC2/6, расщепленную с помощью MscI (New England Biolabs, номер по каталогу R0534L) и Aflll (New England Biolabs, номер по каталогу R0520L) с помощью независимого от лигирования клонирования, описанного в (2012) Methods Mol. Biol. 52:51-9.
[00200] Для создания pAAV RC9-A589Myc получали первый продукт полимеразной цепной реакции (ПЦР), содержащий (от 5' к 3') сайт рестрикции BsiW1, нуклеотидную последовательность между положениями 3052 и 3779 pAAV RC9 и нуклеотидную последовательность эпитопа с-myc с «липкими» концами, и второй продукт ПЦР, содержащий (от 5' к 3') нуклеотидную последовательность эпитопа с-myc с «липкими» концами, нуклеотидную последовательность между положениями 3779 и 4404 pAAV RC9 и сайт рестрикции Pme1, с использованием праймеров, приведенных в таблице 2.
[00201] Плазмиду pAAV RC9-A589Myc (т.е. плазмиду pAAV RC2/9, модифицированную для кодирования эпитопа с-myc между аминокислотами А589 и Q590 капсидного белка VP1) создавали путем расщепления pAAV RC9 с помощью BsiW1 (New England Biolabs, R0553L) и Pme1 (New England Biolabs, R0560L) и вставки двух продуктов ПЦР с помощью независимого от лигирования клонирования, описанного в (2012) Methods Mol. Biol. 52:51-9.
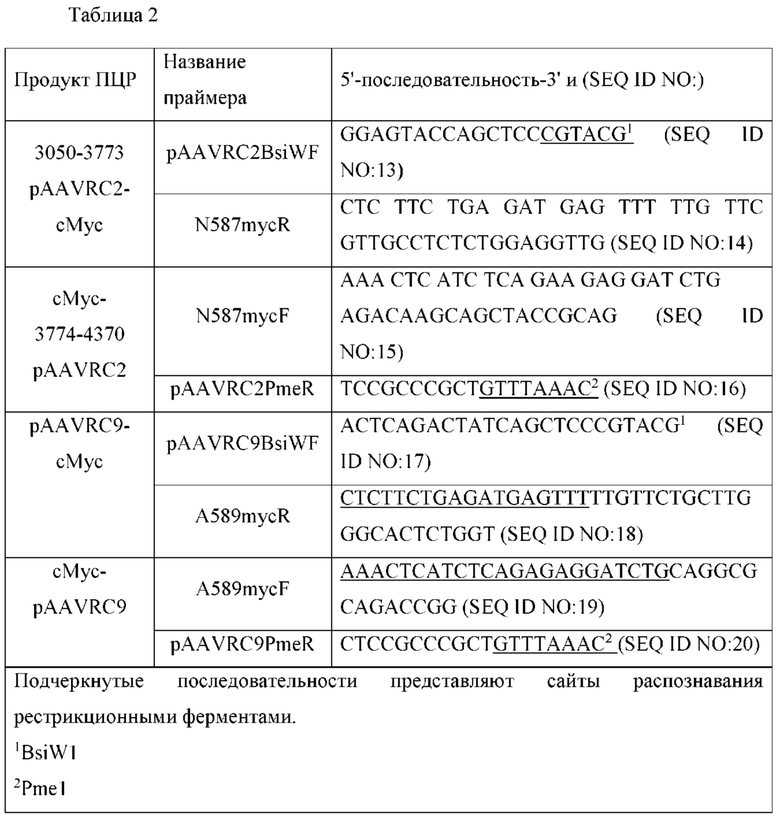
[00202] pscAAV-CMV-eGFP получали путем введения фрагмента GFP в вектор pscAAV MCS (Cell Biolabs, номер по каталогу VPK-430) с использованием сайтов рестрикции для BamHI и NotI. Плазмиду pAAV-EF1a-eGFP и pAAV-CAGG-eGFP синтезировали de novo в Thermofisher Scientific (Уолтем, штат Массачусетс).
Пример 2. Опосредованное антителами перенацеливание вирусных векторов на основе AAV in vitro
[00203] HepG2 представляет собой линию клеток рака печени человека, которая экспрессирует специфичный для печени маркерный асиалогликопротеиновый рецептор 1 (ASGR1). Чтобы проверить, могут ли вирусные векторы scAAV2-N587Myc-CMV-hrGFP инфицировать клетки HepG2, например, с помощью биспецифического антитела, которое распознает как c-myc, так и ASGR1 (антитело к myc-ASGR1), смеси, содержащие различные соотношения количества вирусных геномов (5е9 вирусных геномов) и количества молекул антитела (1:0,5, 1:1, 1:2, 1:4, 1:8, 1:15, 1:20, 1:50 или 1:100) scAAV2-N587Myc-CMV-eGFP, и биспецифическое антитело к myc-ASGR1 инкубировали при комнатной температуре в течение получаса, а затем добавляли к клеткам HepG2. Клетки HepG2 также инкубировали только с вирусными векторами scAAV2-CMV-eGFP дикого типа, только с вирусными векторами scAAV2-N587Myc-CMV-eGFP или вирусными векторами scAAV2-N587Myc-CMV-eGFP с моноспецифическим антителом к myc (Regeneron Pharmaceuticals, Inc., Тэрритаун, штат Нью-Йорк) в соотношении 1:8 в качестве контролей. Через три дня после инфицирования экспрессию GFP инфицированными клетками HepG2 подтверждали с помощью сортировки флуоресцентно-активированных клеток (FACS) (фигура 1). Экспрессию GFP также обнаруживали при сопоставимом процентном содержании клеток HepG2 при инкубации с scAAV2-CMV-eGFP дикого типа и при инкубации с вирусными векторами scAAV2-N587Myc-CMV-eGFP и биспецифическим антителом к myc-ASGR1 в соотношении 1:8 (44,6% и 44,1% соответственно, фигуры 1А и 1G, верхняя и нижняя панели). Более низкую экспрессию GFP обнаруживали с помощью FACS в клетках HepG2, инкубированных со смесью вирусных векторов scAAV2-N587Myc-CMV-eGFP и более высокого или более низкого количества биспецифического антитела к myc-ASGR1 (Regeneron Pharmaceuticals, Inc., Тэрритаун, штат Нью-Йорк) (фигура 1С, нижняя панель). Кроме того, GFP не обнаруживали в клетках HepG2, инфицированных scAAV-N587Myc-CMV-eGFP в отсутствие антитела к myc-ASGR1, или в клетках HepG2, инкубированных с scAAV2-N587Myc-CMV-eGFP и моноспецифическим антителом к myc.
[00204] Аналогично, клетки 293T-hASGR1, которые являются клетками 29Т3, генетически модифицированными для экспрессии человеческого (h) ASGR1 на поверхности клетки, инкубировали со смесями, содержащими различные соотношения вирусных геномов и количества молекул антитела (1:0,5, 1:1, 1:2, 1:4, 1:8, 1:15, 1:20 или 1:100) scAAV2-N587Myc-CMV-eGFP и биспецифического антитела к myc-ASGR1. Клетки 293Т дикого типа (которые не экспрессируют hASGR1), инкубированные со смесью scAAV2-N587Myc-CMV-eGFP и биспецифического антитела к myc-ASGR1 в соотношении 1:8, и клетки 293T-hASGR1, инкубированные с (а) только вирусными векторами scAAV дикого типа, (b) только вирусными векторами scAAV2-N587Myc-CMV-eGFP или (с) вирусными векторами scAAV2-N587Myc-CMV-eGFP с нерелевантным биспецифическим антителом к myc-GCGR, которое распознает с-myc и рецептор глюкагона (GCGR) (Regeneron Pharmaceuticals, Inc., Тэрритаун, штат Нью-Йорк), не экспрессируемый клетками 293Т, в соотношении 1:8, служили в качестве контролей. Через три дня после инфицирования клетки 293Т-hASGR1 или 293Т окрашивали с помощью человеческого антителом к hASGR1 (Regeneron Pharmaceuticals, Inc., Тэрритаун, штат Нью-Йорк), а затем конъюгированным с АРС антителом козы к антителу человека (Jackson ImmunoResearch laboratories Inc, номер по каталогу 109-136-098, Вест Грув, штат Пенсильвания) и экспрессию GFP анализировали с помощью FACS. Экспрессию hASGR1 обнаруживали на поверхности клеток 293T-hASGR1 (фигуры 2Ai-2Ax и 2Axii) но не клеток 293Т (фигура 2Axi). GFP-положительные клетки 293T-hASGR1 обнаруживали после инкубации со смесями scAAV2-N587Myc-CMV-eGFP и биспецифического антитела к myc-ASGR1 в соотношениях 1:0,5, 1:1, 1:2, 1:4, 1:8, 1:15, 1:20 и 1:50 на уровнях (56,9%-68,3%), сопоставимых с клетками 293T-hASGR1, инкубированными с scAAV дикого типа (56,7%) (фигуры 2Ai и 2Aiii-2Aix). Соотношение 1:100 для scAAV2-N587Myc-CMV-eGFP и биспецифического антитела к myc-ASGR1 снижало инфекционность (фигура 2Ах). Инкубация с scAAV2-N587Myc-CMV-eGFP только или scAAV-N587Myc с нерелевантным биспецифическим антителом к myc-GCGR не приводила к экспрессии GFP клетками 293T-hASGR1 (фигуры 2Aii и 2Axii).
[00205] В аналогичном эксперименте клетки 293T-hASGR1 инкубировали с клетками, инкубированными только с немодифицированными вирусными векторами AAV9-CAGG-GFP, только с вирусными векторами AAV9-A589Myc-CAGG-eGFP или с вирусными векторами AAV9-A589Myc-CAGG-eGFP в смеси с биспецифическими антителами к Myc-ASGR1 в разных соотношениях. Через три дня после инфицирования экспрессию GFP анализировали с помощью FACS. GFP-положительные клетки 293T-hASGR1 обнаруживали после инкубации со смесями AAV9-A589Myc-CAGG-eGFP и биспецифического антитела к myc-ASGR1 в соотношениях 1:1, 1:2, 1:4, 1:8, 1:20, 1:50 и 1:100 на уровнях (41,1%-91%), сопоставимых с клетками 293T-hASGR1, инкубированными с AAV9-CAGG-eGFP дикого типа (9,72%) (фигура 2B(i) и Фигуры 2B(iii)-(ix). Инкубация только с AAV9-A589Myc-CAGG-eGFP не приводила к экспрессии GFP клетками 293T-hASGR1 (фигура 2В(ii)).
[00206] Тестировали способность бивалентного антитела к hASGR1 (Regeneron Pharmaceuticals, Inc., Тэрритаун, штат Нью-Йорк) блокировать инфицирование клеток 293T-hASGR1. Клетки 293T-hASGR1 инкубировали с бивалентным антителом к hASGR1 в различных концентрациях в течение 1 часа при комнатной температуре, а затем инкубировали со смесью, содержащей в соотношении 1:8 scAAV2-N587Myc-CMV-eGFP и биспецифическое антитело к myc-ASGR1. Через три дня после инфицирования клетки фиксировали и анализировали с помощью FACS, как описано выше. На фигуре 3 представлены данные, показывающие, что поступление scAAV2-N587Myc-CMV-eGFP может блокироваться дозозависимым образом бивалентным антителом к hASGR1.
[00207] Чтобы определить, может ли последовательное введение биспецифического антитела и модифицированного AAV привести к опосредованному антителом изменению мишени для модифицированного AAV, различные количества молекул биспецифического антитела к myc-ASGR1 (в диапазоне от 1×109 до 1×1012 молекул) добавляли к 2×105 клеток 293T-hASGR1 на протяжении часа, после чего добавляли 1×109 вирусных векторов scAAV-N587Myc. Контроли включали (1) клетки 293T-hASGR1, инкубированные только с scAAV дикого типа, т.е. в отсутствие какого-либо антитела, (2) клетки 293T-hASGR1, последовательно инкубированные с 1×1011 молекул нерелевантного биспецифического антитела к myc-GCGR и 1×109 вирусных векторов scAAV2-N587Myc-CMV-eGFP, и (3) клетки 293Т, которые не экспрессируют hASGR1, последовательно инкубированные с 1×1011 молекул биспецифического антитела к myc-ASGR1 и 1×109 вирусных векторов scAAV-N587Myc. Через два дня после инфицирования экспрессию GFP инфицированными клеткам 293T-hASGR1 визуализировали с помощью микроскопии (фигура 4). Экспрессию GFP клетками 293T-hASGR1 наблюдали после инкубации только с scAAV дикого типа и после последовательной инкубации с молекулами антитела к myc-ASGR1 при всех концентрациях и scAAV2-N587Myc-CMV-eGFP (фигуры 4А, 4C-4I). GFP не обнаруживали в клетках 29T3-hASGR1 после инкубации только с scAAV2-N587Myc-CMV-eGFP (фигура 4 В), в клетках 293Т, которые не экспрессируют ASGR1, после последовательной инкубации с антителом к тус-hASGR1 и вирусными векторами scAAV2-N587Myc-CMV-eGFP (фигура 4J) или в клетках 29Т3-hASGR1 после последовательной инкубации с нерелевантным биспецифическим антителом к myc-GCGR и вирусными векторами scAAV2-N587Myc-CMV-eGFP (фигура 4K).
[00208] Как описано в данном документе, тропизм самокомплементарного AAV (scAAV) можно (1) инактивировать, например, путем модификации капсидного белка, например путем вставки эпитопа с-myc, и, необязательно, (2) перенаправить с использованием биспецифических антител, например с помощью биспецифического антитела, которое распознает эпитоп с-myc и лиганд, экспрессируемый клеткой-мишенью. Чтобы определить, можно ли тропизм одноцепочечного AAV (ssAAV) аналогичным образом (1) понизить или устранить и, необязательно, (2) перенаправить, клетки 293T-hASGR1 инкубировали с вирусными векторами ssAAV2-N587Myc-CMV-hrGFP, полученными, как описано в примере 1, в отсутствие или в присутствии биспецифического антитела к myc-hASGR1 при различных соотношениях вирусный вектор:антитело (1:0, 1:1, 1:2, 1:4, 1:8, 1:20, 1:100 или 1:1000). Контроли включали клетки 293T-hASGR1, инкубированные с ssAAV дикого типа, клетки, 293T-hASGR1, инкубированные с вирусными векторами ssAAV2-N587Myc-CMV-hrGFP и нерелевантным биспецифическим антителом к myc-GCGR при соотношении вирусный вектор:антитело 1:8, и клетки 293Т (которые не экспрессируют hASGR1), инкубированные с вирусными векторами ssAAV2-N587Myc-CMV-hrGFP и биспецифическим антителом к myc-hASGR1 при соотношении вирусный вектор:антитело 1:8. Через три дня после инфицирования клетки фиксировали и выявляли экспрессию GFP с помощью FACS (фигура 5). Экспрессию GFP обнаруживали в клетках 293T-hASGR1, инкубированных с ssAAV дикого типа и со смесями вирусных векторов ssAAV2-N587Myc-CMV-hrGFP и биспецифических антител к myc-ASGR1 при всех соотношениях (фигуры 5А, 5C-5I). Экспрессию GFP не обнаруживали на клетках 29Т3, инкубированных с вирусными векторами ssAAV2-N587Myc-CMV-hrGFP и биспепифическими антителами к myc-ASGR1 (фигура 5J), или на клетках 293T-hASGR1, инкубированных только с вирусными векторами ssAAV2-N587Myc-CMV-hrGFP или с вирусными векторами ssAAV-N587Myc и нерелевантным биспецифическим антителом к myc-GCGR (фигуры 5В, 5K).
[00209] Человеческий (h) рецептор глюкагона (GCGR) обычно не экспрессируется клетками 293Т. Однако 293T-hGCGR является стабильной линией клеток 293Т, генетически модифицированной для экспрессии hGCGR на клеточной поверхности. Чтобы проверить, может ли перенацеливание векторов scAAV2-N587Myc-CMV-eGFP опосредоваться биспецифическим антителом к myc-GCGR через рецептор hGCGR, клетки 293T-hGCGR инкубировали с различными смесями, содержащими разные соотношения scAAV2-N587Myc-CMV-eGFP:биспецифическое антитело к myc-GCGR. Вирусные векторы scAAV2-N587Myc-CMV-eGFP смешивали с биспецифическим антителом к myc-GCGR при соотношениях 1:0,5, 1:1, 1:2, 1:4, 1:8, 1:15, 1:20, 1:50 или 1:100 при комнатной температуре в течение получаса до добавления клеток 293T-hGCGR. Через три дня после инфицирования экспрессию GFP инфицированными клетками 293T-hCGCR подтверждали с помощью сортировки флуоресцентно-активированных клеток (FACS) (фигура 6). Значительное процентное содержание GFP-положительных клеток обнаруживали после инкубации с scAAV дикого типа (фигура 6А) или с scAAV2-N587Myc-CMV-eGFP/биспецифическое антитело к myc-GCGR при соотношении 1:0,5, 1:1, 1:2, 1:4, 1:8, 1:15, 1:20, 1:50 и 1:100 (фигуры 6С-6K). GFP не обнаруживали в клетках 293T-hGCGR, инкубированных только с вирусными векторами scAAV2-N587myc-CMV-eGFP или с вирусными векторами scAAV2-N587Myc-CMV-eGFP с моноспецифическим антителом к туе (фигуры 6В и 6L соответственно).
[00210] Jurkat представляет собой линию клеток острого Т-клеточного лейкоза человека, которая экспрессирует человеческий (h) CD3. Чтобы проверить, может ли перенацеливание вирусных векторов AAV6-Q585Myc-EF1a-eGFP опосредоваться биспецифическим антителом к myc-CD3 через рецептор hCD3, клетки Jurkat инкубировали с различными смесями, содержащими разные соотношения AAV6-Q585Myc-ЕР1а-eGFP:биспецифическое антитело к myc-CD3. Вирусные векторы AAV6-Q585Myc-EF1a-eGFP смешивали с биспецифическим антителом к myc-CD3 при соотношениях 1:1, 1:5, 1:10, 1:100, 1:1000 при комнатной температуре в течение получаса до добавления клеток Jurkat. Через три дня после инфицирования экспрессию GFP инфицированными клетками Jurkat подтверждали с помощью сортировки флуоресцентно-активированных клеток (FACS) (фигура 7). Значительное процентное содержание GFP-положительных клеток обнаруживали после инкубации с AAV6-EF1a-eGFP дикого типа (фигура 7В) или с AAV6-Q585Myc-EF1a-eGFP/биспецифическое антитело к myc-CD3 при соотношении 1:1, 1:5, 1:10 и 1:100 (фигуры 7D-7G). GFP не обнаруживали на клетках Jurkat, инкубированных с вирусными векторами AAV6-Q585Myc-EF1a-eGFP (фигуры 7С).
[00211] В этом примере описана инактивация природного тропизма нескольких серотипов самокомплементарных (sc) или одноцепочечных (ss) AAV, продемонстрированная с помощью неспособности scAAV2 или ssAAV2, генетически модифицированных с помощью эпитопа с-myc, вставленного между аминокислотами N587 и R588 капсидного белка VP1 (scAAV-N587myc или ssAAV-N587myc), AAV6 с эпитопом с-myc, вставленным между Q585 и S586, и AAV9 с инфицировать клетки, обычно инфицируемые AAV дикого типа (фигуры 1А-1В, 2А-2В, 3А-3В, 4А-4В, 5А-5В, 6А-6В и 7). В этом примере дополнительно продемонстрирована способность биспецифического антитела, которое распознает эпитоп с-myc и второй лиганд, экспрессируемый клеткой-мишенью (например, hASGR1, hGCGR или hCD3), изменять мишень тропизм модифицированного AAV и опосредовать инфицирование клетки-мишени с помощью псевдотипированного AAV специфичным для лиганда образом (фигуры 1-7). Кроме того, в данном примере показано, что одновременное введение генетически модифицированного sc- или ss-AAV и биспецифического антитела не является необходимым для инфицирования, т.е. последовательного введения достаточно для перенацеливания генетически модифицированного sc- или ss-AAV in vitro (фигура 4).
Пример 3. Опосредованное антителами перенаправление модифицированных вирусных векторов in vivo
Перенацеливание для вирусных векторов в отношении печени с помощью различных форматов полиспецифических связывающих молекул
[00212] Чтобы определить, могут ли биспецифическое антитело к myc-ASGR1 изменять мишень вирусных векторов scAAV2-N587myc-CMV-eGFP в отношении экспрессирующих hASGR1 клеток печени in vivo, мышам, генетически модифицированным так, что их клетки печени экспрессируют hASGR1 на фоне C57BL/6, и контрольным мышам C57BL/6 дикого типа вводили 1×1011 (титрованных в помощью qPCR) только scAAV2-CMV-eGFP дикого типа или вирусных векторов scAAV2-N587myc-CMV-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 в соотношении 1:8 вирусного генома и молекул антитела внутрисосудисто. Контроль включал мышей с инъекцией физиологического раствора [250 мМ NaCl] или только вирусного вектора scAAV2-N587myc-CMV-eGFP. Через десять дней после инъекции мышей умерщвляли и транскардиально перфузировали с использованием 4% PFA. Такие органы, как печень, почки и сердце, собирали и дегидратировали в 15% сахарозе, а затем в 30% сахарозе. Затем органы подвергали изготовлению срезов с замораживанием на предметных стеклах и окрашивали с помощью антитела цыпленка к EGFP (Jackson ImmunoResearch Labs, Inc., Вест Грув, штат Пенсильвания) и конъюгированным с Alexa-488 вторичным антителом к антителу цыпленка (Jackson ImmunoResearch Labs, Inc., Вест Грув, штат Пенсильвания). (Фигуры 8А-8С). GFP-положительные клетки обнаруживали в печени трансгенных животных, модифицированных для экспрессии ASGR1 в печени и получивших инъекцию scAAV2-CMV-eGFP дикого типа или scAAV2-N587myc-CMV-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигуры 8A(i) и 8A(iv)), и в печени мышей C57BL/6 дикого типа, получивших инъекцию scAAV2-CMV-eGFP дикого типа (фигура 8A(v)). GFP не обнаруживали ни в образцах селезенки или почек (фигуры 8В и 8С), ни в образцах печени, селезенки или почек от любого животного с инъекцией физиологического раствора или только вирусных векторов scAAV2-N587myc-CMV-eGFP (фигуры 8A(ii, iii, vi, vii)), ни в образцах печени, взятых у животных C57BL/6 дикого типа с инъекцией scAAV2-N587myc-CMV-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигура 8A(viii)). Таким образом, комбинация вирусных векторов scAAV2-N587myc-CMV-eGFP и биспецифического антитела к myc-ASGR1 инфицировала только те (печеночные) клетки, которые экспрессируют hASGR1, убедительно свидетельствуя о том, что вирусный вектор scAAV2-CMV-eGFP инактивируется модификацией капсидного белка, например, естественный тропизм вирусного вектора на основе scAAV можно понизить вплоть до устранения, например с помощью эпитопа с-myc, и такие вирусные векторы можно специфически реактивировать, например для них специфически изменяли мишень, например, в отношении клеток печени in vivo, например с помощью биспецифических антител к myc-ASGR1.
[00213] Аналогичным образом, чтобы определить, могут ли биспецифические антитела к myc-ASGR1 изменять мишень для вирусных векторов ssAAV2-N587myc-CAGG-eGFP в отношении экспрессирующих hASGR1 клеток печени in vivo, мышам, генетически модифицированным так, что их клетки печени экспрессируют hASGR1 на фоне C57BL/6, и контрольным мышам C57BL/6 дикого типа вводили 2,18×1011 (титрованных в помощью qPCR) только ssAAV2-CAGG-eGFP дикого типа или вирусных векторов ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 в соотношении 1:4 вирусного генома и молекул антитела внутрисосудисто. Контроли включали мышей с инъекцией PBS или только вирусного вектора ssAAV2-N587myc-CAGG-eGFP. Через четыре недели после инъекции мышей умерщвляли и транскардиально перфузировали с использованием 4% PFA. Такие органы, как печень, почки и сердце, собирали и дегидратировали в 15% сахарозе, а затем в 30% сахарозе. Затем органы подвергали изготовлению срезов с замораживанием на предметных стеклах и окрашивали с помощью антитела цыпленка к EGFP (Jackson ImmunoResearch Labs, Inc., Вест Грув, штат Пенсильвания) и конъюгированным с Alexa-488 вторичным антителом к антителу цыпленка (Jackson ImmunoResearch Labs, Inc., Вест Грув, штат Пенсильвания). GFP-положительные клетки обнаруживали в печени трансгенных животных, модифицированных для экспрессии ASGR1 в печени и получивших инъекцию ssAAV2-CAGG-eGFP дикого типа или ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигуры 9E-9F, 9P-9R), и в печени мышей C57BL/6 дикого типа, получивших инъекцию ssAAV2-CAGG-eGFP дикого типа (фигура 9В-9С). Удивительно, но эффективность инфицирования ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 намного выше, чем у WT ssAAV2-CAGG-GFP (фигуры 9E-9F, 9P-9R). GFP не обнаруживали или практически не обнаруживали ни в образцах печени от любого животного с инъекцией физиологического раствора или только вирусных векторов ssAAV2-N587myc-CAGG-eGFP (фигуры 9А, 9D, 9G-9L)), ни в образцах печени, взятых у животных C57BL/6 дикого типа с инъекцией ssAAV2-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигура 9М-9O). Таким образом, комбинация вирусных векторов ssAAV2-N587myc-CAGG-eGFP и биспецифического антитела к myc-ASGR1 инфицировала только те (печеночные) клетки, которые экспрессируют hASGR1, убедительно свидетельствуя о том, что вирусный вектор ssAAV2-N587myc-CAGG-eGFP инактивируется модификацией капсидного белка, например, естественный тропизм вирусного вектора на основе scAAV можно понизить вплоть до устранения, например с помощью эпитопа c-myc, и такие вирусные векторы можно специфически реактивировать, например для них специфически изменять мишень, например, в отношении клеток печени in vivo, например с помощью биспецифических антител к myc-ASGR1.
[00214] Дальнейшие эксперименты с вирусным вектором на основе псевдотипированного AAV9 проводили на мышах, генетически модифицированных таким образом, что их клетки печени экспрессируют hASGR1 на фоне C57BL/6. Этим мышам вводили инъекцию 1×1011 (титрованных с помощью qPCR) вирусных частиц AAV9-CAGG-eGFP дикого типа или AAV9-A589myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 в соотношении 1:100 вирусного генома и молекул антитела внутрисосудисто. Контроли включали мышей с инъекцией PBS или вирусных частиц AAV9-N587myc-CAGG-eGFP в комбинации с нерелевантным биспецифическим антителом к myc-hCD3. Через четыре недели после инъекции мышей умерщвляли. Печень фиксировали 10% формалином и отправляли в HistoWiz. Inc (Нью-Йорк, штат Нью-Йорк) для окрашивания в отношении GFP. GFP-положительные клетки обнаруживали в печенях животных, которым вводили вирусные частицы AAV9-CAGG-eGFP дикого типа или AAV9-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-ASGR1 (фигуры 10А и 10D). GFP не обнаруживали или практически не обнаруживали в образцах печени животных, которым вводили физиологический раствор или вирусные частицы AAV9-N587myc-CAGG-eGFP в комбинации с биспецифическим антителом к myc-hCD3 соответственно (фигуры 10В и 10С). Таким образом, аналогично AAV2, естественный тропизм AAV9 можно понизить вплоть до устранения путем модификации капсидного белка, например путем вставки эпитопа c-myc, и для такого модифицированного вирусного вектора, например AAV9-A589myc, можно изменять мишень в отношении специфических типов клеток с использованием соответствующего биспецифического связывающего белка к эпитопу и клеточно-специфичному маркеру, например биспецифического антитела к myc-ASGR1 в отношении клеток печени.
Fc-фирмат «выступ-во-впадину» на основе IgG4 к hASGR1 с добавлением scFc к myc
[00215] Чтобы определить, могут ли различные форматы полиспецифических связывающих молекул опосредовать инфицирование AAV, scFv к myc сливали с С-концом одной цепи антитела на основе IgG4 к hASGR1 формата «выступ-во-впадину» (см. фигуру 11А). В частности, gblock, кодирующий scFv к myc (SEQ ID NO: 28), фланкированный гомологичными группами к 1459-1478 и 1479-1498 pRG7078, кодирующей тяжелую цепь «выступа-невидимки» IgG4 к hASGR1, был предоставлен Integrated DNA technologies (Скоки, штат Иллинойс). Кодирующие нуклеотидные последовательности (NA) и аминокислотные (АА) последовательности HCVR, LCVR, HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 scFv (SEQ ID NO: 37), кодируемого SEQ ID NO: 28, представлены в таблице 3.
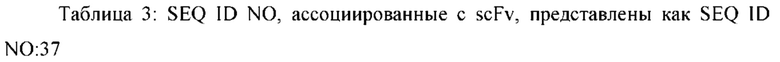
[00216] Gblock клонировали в pRG7078, кодирующую тяжелую цепь «выступа-невидимки» IgG4 к hASGR1, с получением pRG7078, кодирующей слитый полипептид «выступа-невидимки» IgG4 к hASGR1 и scFv к myc «биспецифическое антитело к hASGR1-IgG4-Fc/к myc». Плазмиду pRG7078, модифицированную с помощью с-myc, немодифицированную pRG7078 с «впадиной-невидимкой» IgG4 к hASGR1 и плазмиду, содержащую легкую цепь антитела, трансфицировали в клетки 293Т, культивируемые с OPTI-MEM (номер по каталогу 31985070, Thermo Fisher). Через четыре дня после трансфекции среду собирали и очищали биспецифическую связывающую молекулу к hASGR1-IgG4-Fc/к myc с использованием колонки с белком А (номер по каталогу 89948, Thermo Fisher).
[00217] Для создания вирусного вектора на основе псевдотипированного AAV8 pAAV RC8 N590myc, gblock с эпитопом c-myc (SEQ ID NO: 22), вставленным между N590 и Т591 капсидного белка VP1 AAV8, заказывали в Integrated DNA technologies (Скоки, штат Иллинойс). Плазмиду pAAV RC8 (SEQ ID NO: 23) расщепляли ферментами Mlu1 и Sbf1 и gblock клонировали в вектор с использованием SLIC для получения pAAV RC8 N590myc (SEQ ID NO: 24).
[00218] Клетки 293T-hASGR1, которые являются клетками 293Т, генетически модифицированными для экспрессии человеческого (h) ASGR1 на поверхности клетки, инкубировали либо со смесями, содержащими различные соотношения AAV8-N590Myc-CAGG-eGFP с молекулами биспецифической связывающей молекулы к hASGR1-IgG4-Fc/к myc (1:0, 1:1, 1:2, 1:4, 1:8, 1:12, 1:15, 1:50 или 1:100), либо с вирусными геномами AAV8-CAGG-eGFP. Через три дня после инфицирования экспрессию GFP анализировали с помощью FACS. GFP-положительные клетки 293T-hASGR1 обнаруживали после инкубации со смесями AAV8-N590Myc-CAGG-eGFP и биспецифических молекул к hASGR1-IgG4-Fc/к myc при соотношениях 1:1, 1:2, 1:4, 1:8, 1:12, 1:15, 1:50 или 1:100 (2,12%-22,2%; фигуры 12D-12K) сопоставимо с клетками 293T-hASGR1, инкубированными с AAV8-CAGG eGFP дикого типа (46,1%) (фигура 12А). Ни мнимо инфицированные клетки (Фигура 12В), ни инкубация только с AAV8-N590Myc-CAGG-eGFP не приводила к экспрессии GFP клетками 293Т hASGR1 (фигура 12С).
[00219] Чтобы определить, могут ли биспецифические связывающие белки к hASGR1-IgG4-Fc/к myc изменять мишень для вирусных частиц AAV8 N590myc-CAGG-eGFP в отношении экспрессирующих hASGR1 клеток печени in vivo, мышам, генетически модифицированным так, что их клетки печени экспрессируют hASGR1 на фоне C57BL/6, вводили внутрисосудисто 1×1011 (титрованных с помощью qPCR) вирусных частиц AAV8-CAGG-eGFP дикого типа или AAV8 N590myc-CAGG-eGFP в комбинации с биспецифическими связывающими молекулами к hASGR1-IgG4-Fc/к myc в соотношении 1:12 вирусного генома и молекул связывающего белка. Контрольным животным вводили вирусные частицы AAV8 N590myc-CAGG-eGFP в комбинации с нерелевантной биспецифической связывающей молекулой к hCD3 х к myc, имеющей сходный формат с биспецифической связывающей молекулой к hASGR1-IgG4-Fc/к myc. Через три недели после инъекции мышей умерщвляли и транскардиально перфузировали с использованием 4% PFA. Такие органы, как печень, почки и сердце, собирали и дегидратировали в 15% сахарозе, а затем в 30% сахарозе. Затем органы подвергали изготовлению срезов с замораживанием на предметных стеклах и окрашивали с помощью антитела цыпленка к EGFP (Jackson ImmunoResearch Labs, Inc., Вест Грув, штат Пенсильвания) и конъюгированным с Alexa-488 вторичным антителом к антителу цыпленка (Jackson ImmunoResearch Labs, Inc., Вест Грув, штат Пенсильвания). GFP-положительные клетки обнаруживали в печенях животных, которым вводили вирусные частицы AAV8-CAGG-eGFP дикого типа (фигуры 13А-13С) или AAV8 N590myc-CAGG-eGFP в комбинации с биспецифическими связывающими молекулами к hASGR1-IgG4-Fc/к myc (фигуры 13G-13I). GFP не обнаруживали или практически не обнаруживали в образцах печени животных, которым вводили вирусные частицы AAV8 N590myc-CAGG-eGFP в комбинации с нерелевантным связывающим белком к hCD3/myc (фигуры 13D-13F). Таким образом, для AAV8 с меткой эпитопа, например AAV8-N590myc, можно изменять мишень в отношении конкретного типа клеток с использованием биспецифической связывающей молекулы, которая специфически связывает эпитоп и клеточно-специфичный маркер, экспрессируемый целевым типом клеток.
Перенацеливание вирусных векторов в отношении кишечника и/или поджелудочной железы
[00220] Клетки 293Т, генетически модифицированные для экспрессии эктонуклеозидтрифосфат-дифосфогидролазы 3 человека (hENTPD3) на поверхности клетки («293T-ENTPD3»), инкубировали со смесями, содержащими различные соотношения вирусных геномов AAV2-N587Myc-CAGG-eGFP или AAV2-CAGG-GFP и биспецифической связывающей молекулы, образованной путем слияния scFv к myc с С-концом одной цепи антитела на основе IgG4 к hENTPD3 формата «выступ-во-впадину» (к hENTPD3-IgG4-Fc/к myc), в различных соотношениях (1:0, 1:1, 1:2, 1:4, 1:8, 1:20, 1:50, 1:100 или 1:200). Через три дня после инфицирования экспрессию GFP анализировали с помощью FACS. GFP-положительные клетки 293T-ENTPD3 обнаруживали после инкубации со смесями AAV2-N587Myc-CAGG-eGFP и молекул к hENTPD3-IgG4-Fc/к myc на уровнях (1,48%-16%; фигуры 14C-14J), сопоставимых с клетками 293T-ENTPD3, инкубированными с AAV2-CAGG-eGFP дикого типа (66%) (фигура 14А). Инкубация только с AAV2-N587Myc-CAGG-eGFP приводила к очень низкому уровню экспрессии GFP клетками 293T-ENTPD3 (фигура 14В).
[00221] Было определено, что ENTPD3 экспрессируется в слизистой оболочке кишечника (данные не показаны). Чтобы определить, может ли связывающий белок к hENTPD3-IgG4-Fc/к myc изменять мишень для вирусных частиц AAV2-N587myc-CAGG-eGFP в отношении экспрессирующих ENTPD3 клеток кишечника in vivo, мышам C57BL/6 вводили внутрисосудисто 5×1011 (титрованных в помощью qPCR) вирусных частиц AAV2-N587myc-CAGG-eGFP в комбинации со связывающими белками к hENTPD3-IgG4-Fc/к myc в соотношении 1:20 вирусного генома и молекул связывающего белка. Контрольным мышам вводили PBS, AAV9 дикого типа или вирусные частицы AAV2-N587myc-CAGG-eGFP в комбинации с нерелевантной молекулой связывающего белка. Через три недели после инъекции мышей умерщвляли. Печень, кишечник и поджелудочную железу фиксировали 10% формалином и отправляли в HistoWiz. Inc (Нью-Йорк, штат Нью-Йорк) для окрашивания в отношении GFP. GFP-положительные клетки обнаруживали у мышей с инъекцией AAV9 дикого типа (фигура 15В(ii)), но не в печени мышей с инъекцией PBS или вирусных частиц AAV2-N587myc-CAGG-eGFP в комбинации с нерелевантными связывающими молекулами или связывающими белками к hENTPD3-IgG4-Fc/к myc (фигура 15A(iii)-15A(iv), соответственно), указывая на то, что вирусные векторы AAV2-N587myc не инфицируют ENTPD3-отрицательные клетки печени даже при совместном введении со связывающими белками hENTPD3-IgG4-Fc/к myc. GFP обнаруживали в кишечнике только у мышей с инъекцией AAV9 дикого типа или вирусных частиц AAV2-N587myc-CAGG-eGFP в комбинации со связывающими белками hENTPD3-IgG4-Fc/к myc (фигуры 15B(i)-15B(iv)). GFP обнаруживали в островках поджелудочной железы мышей с инъекцией вирусных частиц AAV2-N587myc-CAGG-eGFP в комбинации со связывающими белками к hENTPD3-IgG4-Fc/к myc. (Фигура 15C(iv)). WT AAV9 инфицировал как островковые клетки, так и не островковые клетки поджелудочной железы (фигура 15C(ii)). GFP не обнаруживали в образцах поджелудочной железы мышей с инъекцией физиологического раствора или вирусных частиц AAV2-N587myc-CAGG-eGFP в комбинации с нерелевантными связывающими белками. (Фигуры 7C(i) и 7С(iii)). Таким образом, естественный тропизм AAV можно понизить вплоть до устранения путем введения гетерологичного эпитопа и перенацеливания в отношении тех же или других клеток с использованием полиспецифического связывающего белка, который связывает метку эпитопа и маркер, экспрессируемый клетками или тканями, в отношении которых изменяли мишень.
[00222] В данном примере показано, что вектор на основе нескольких различных серотипов аденоассоциированного вируса можно генетически модифицировать гетерологичным эпитопом (например, c-myc), как описано в данном документе, для инактивации инфекционности, и можно использовать биспецифическое антитело (независимо от биспецифических форматов), которое одновременно распознает гетерологичный эпитоп (например, c-myc) и маркер, экспрессируемый клеткой-мишенью, для перенацеливания вирусного вектора для доставки представляющего интерес нуклеотида.
Пример 4. Применение генетически модифицированных вирусных векторов AAV-N587Myc для доставки терапевтического груза в конкретные клетки
[00223] В данном примере показана способность вирусных векторов AAV-N587Myc специфически доставлять терапевтический груз, такой как один или более суицидных генов, биологическое терапевтическое средство (например, антитело), систему редактирования генов CRISPR/Cas, shRNA и т.д., в целевой тип клеток. В частности, в данном примере описана доставка суицидного гена, кодирующей последовательности антитела или системы редактирования гена CRISPR/Cas к клеткам, экспрессирующим целевой лиганд.
[00224] Доставка суицидного гена к клеткам, экспрессирующим целевой лиганд
[00225] Чтобы проверить способность вирусных векторов scAAV2-N587Myc доставлять суицидный ген в конкретную клетку, использовали модель ксенотрансплантата рака молочной железы HER2+ у бестимусных мышей, как описано в Wang et al. ((2010) Cancer Gene Therapy 17:559-570).
[00226] Вирусные векторы. Вирусные векторы scAAV2-N587Myc или ssAAV2-N587Myc, несущие репортерный ген EGFP (scAAV2-N587Myc-EGFP или ssAAV2-N587MycEGFP), получали, как описано в примере 1. Вирусные векторы scAAV2-N587Myc или ssAAV2-N587Myc, несущие суицидный ген (SG), получали аналогичным образом. Вкратце, клетки 293Т17 трансфицировали с помощью (1) pAd Helper и (2) pAAV RC2 (кодирующей капсид дикого типа) или pAAV RC2-N587Myc (кодирующей капсид, модифицированный эпитопом с-myc), как описано выше, и с помощью (3) вектора pAAV, несущего суицидный ген, например, ген цитозиндезаминазы, ген тимидинкиназы вируса простого герпеса, под контролем промотора, например CMV. Вирусные векторы ssAAV-N587MycSG или scAAV-N587SG выделяли и титровали, как описано в примере 1.
[00227] Линии клеток. Линии клеток рака молочной железы ВТ474, рака молочной железы SK-BR-3 и рака легкого Calu-3 представляют собой HER2-положительные линии опухолевых клеток человека (Bunn P.A. et al., (2001) Clin Cancer Res. 7:3239-3250; Pegram M, et al. (1999) Oncogene 18:2241-2251; Spiridon CI, et al., (2002) Clin Cancer Res. 8:1720-1730). Рабдомиосаркома A-673 и рак шейки матки HeLa представляют собой HER2-отрицательные линии опухолевых клеток человека, a BEAS-2b представляет собой иммортализованную HER2-отрицательную линию клеток эпителия бронхов (Jia LT et al. (2003) Cancer Res. 63:3257-3262; Kern JA, et al., (1993) Am J Respir Cell Mol Biol 9:448-454; Martinez-Ramirez A, et al., (2003) Cancer Genet Cytogenet. 2003; 141:138-142). Все эти линии клеток получены из Американской коллекции типовых культур (АТСС, Манассас, штат Вирджиния) и хранятся в среде, рекомендованной АТСС.
[00228] Мыши. Самок бестимусных мышей в возрасте от 6 до 8 недель получали и содержали в специальных свободных от патогенных микроорганизмов условиях. В день 0 мышам одновременно (1) вводили подкожно в правый бок 107 опухолевых клеток ВТ474, SK-BR-3, Calu-3, А-673 или HeLa и (2) внутривенно вводили биспецифическое антитело к myc-HER2 и вирусные векторы ss- или scAAV-N587Myc, несущие репортерный ген (например, EGFP) или суицидный ген. Контролями служили животные без обработки (животные с инъекцией только опухолевых клеток), животные с инъекцией вирусных векторов на основе ss- или scAAV дикого типа, несущих репортерный ген или суицидный ген, животные с инъекцией только биспецифического антитела к myc-HER2 или животные с инъекцией вирусных векторов ss- или scAAV-N587Myc, несущих только репортерный ген или суицидный ген. Всех животных обрабатывали соответствующим пролекарством через 1 день после инъекции и обработки. Размер каждой опухоли измеряют с помощью штангенциркуля 2 раза в неделю и объем опухоли рассчитывают как длина×ширина2×0,52. При болезненности, изъязвлении опухоли, диаметре опухоли 15 мм или объеме опухоли 1000 мм3 мышей умерщвляют и дату умерщвления записывают как дату смерти. Печень, селезенку, почки и опухоли животных с инъекцией вирусных векторов вирусов на основе ss- или scAAV дикого типа или ss- или scAAV-N587Myc, несущих репортерный ген, фиксируют и визуализируют экспрессию репортерного гена.
[00229] Хотя нацеленная доставка генов, индуцирующих самоубийство, была описана (Zarogoulidis P., et al. (2013) J. Genet. Syndr. Gene Ther. 4:16849), в данном примере описана доставка суицидного гена в клетку, экспрессирующую целевой лиганд HER2, с использованием описанных в данном документе вирусных векторов, содержащих гетерологичный эпитоп, например c-myc, и биспецифического антитела, которое специфически связывает целевой лиганд и гетерологичный эпитоп. В дополнительных экспериментах суицидный ген доставляют в другой тип клеток, экспрессирующих один или более других целевых лигандов, с использованием вирусного вектора, содержащего гетерологичный эпитоп, как описано в данном документе, и биспецифическое антитело, которое специфически связывает гетерологичный эпитоп (например, c-myc) и рецептор-мишень. Иллюстративные и неограничивающие примеры рецепторов, подходящих для нацеливания, включают те рецепторы, которые опосредуют эндоцитоз вирусного вектора, например, онкоэмбриональный антиген (СЕА) (Qiu Y, et al. (2012) Cancer Lett. 316:31-38) и рецептор фактора роста эндотелия сосудов (VEGFR) (Leng A, et al. (2013) Tumour Biol. 32:1103-1111; Liu T, et al. (2011) Exp Mol Pathol. 91:745-752). Дополнительные рецепторы, которые могут быть мишенью, включают: рецептор эпидермального фактора роста (EGFR) (Heimberger АВ, et al. (2009) Expert Opin Biol Ther. 9:1087-1098), кластер дифференцировки 44s (CD44s) (Heider KH, et al. (2004) Cancer Immunol Immunother. 53:567-579), кластер дифференцировки 133 (CD133, также известный как АС133) (Zhang SS, et al. (2012) BMC Med.; 10:85), фолатный рецептор (FR) (Duarte S, et al., (2011) J Control Release 149(3):264-72), рецептор трансферрина (TfR) или кластер дифференцировки 71 (CD71) (Habashy НО, et al., Breast Cancer Res Treat. 119(2):283-93), муцины (Torres MP, et al., (2012) Curr Pharm Des. 2012; 18(17):2472-81), стадиеспецифический эмбриональный антиген 4 (SSEA-4) (Malecki M., et al., (2012) J Stem Cell Res Ther. 2(5)), антиген опухолевой резистентности 1-60 (TRA-1-60) (Malecki M., et al., (2013) J Stem Cell Res Ther. 3:134).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Регенерон Фармасьютикалс, Инк.
Куратсос, Христос
Мерфи, Эндрю Дж
Ванг, Ченг
Сабин, Лия
<120> Рекомбинантные вирусные векторы с модифицированным тропизмом и пути
их применения для нацеленного введения генетического материала в клетки человека
<130> 009108.335WO1/10335WO01
<150> 62/525,704
<151> 2017-06-27
<160> 40
<170>PatentIn версия 3.5
<210> 1
<211> 735
<212> БЕЛОК
<213> аденоассоциированный вирус 2
<400> 1
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 2
<211> 745
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223>AAV2 VP1 N587Myc
<400> 2
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Glu Gln Lys Leu Ile
580 585 590
Ser Glu Glu Asp Leu Arg Gln Ala Ala Thr Ala Asp Val Asn Thr Gln
595 600 605
Gly Val Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu Gln
610 615 620
Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly His Phe His Pro
625 630 635 640
Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro Pro Gln Ile
645 650 655
Leu Ile Lys Asn Thr Pro Val Pro Ala Asn Pro Ser Thr Thr Phe Ser
660 665 670
Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val
675 680 685
Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg Trp
690 695 700
Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Asn Lys Ser Val Asn Val
705 710 715 720
Asp Phe Thr Val Asp Thr Asn Gly Val Tyr Ser Glu Pro Arg Pro Ile
725 730 735
Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 3
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 6
<400> 3
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Val Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 4
<211> 746
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AAV6 VP1 Q585Myc
<400> 4
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Glu Gln Lys Leu Ile Ser Glu
580 585 590
Glu Asp Leu Ser Ser Ser Thr Asp Pro Ala Thr Gly Asp Val His Val
595 600 605
Met Gly Ala Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu
610 615 620
Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly His Phe His
625 630 635 640
Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro Pro Gln
645 650 655
Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn Pro Pro Ala Glu Phe
660 665 670
Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln
675 680 685
Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg
690 695 700
Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn Tyr Ala Lys Ser Ala Asn
705 710 715 720
Val Asp Phe Thr Val Asp Asn Asn Gly Leu Tyr Thr Glu Pro Arg Pro
725 730 735
Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
740 745
<210> 5
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 9
<400> 5
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Gln Ala Gln
580 585 590
Thr Gly Trp Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Met
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asp Pro Pro Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Tyr Lys Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 6
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> c-myc
<400> 6
Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
1 5 10
<210> 7
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FLAG
<400> 7
Asp Tyr Lys Asp Asp Asp Asp Lys
1 5
<210> 8
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HA
<400> 8
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala
1 5
<210> 9
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер ITR
<400> 9
ggaaccccta gtgatggagt t 21
<210> 10
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер ITR
<400> 10
cggcctcagt gagcga 16
<210> 11
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд AAV2 ITR
<400> 11
cactccctct ctgcgcgctc g 21
<210> 12
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> c-myc
<400> 12
gaacaaaaac tcatctcaga agaggatctg 30
<210> 13
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAVRCBsiWF
<400> 13
ggagtaccag ctcccgtacg 20
<210> 14
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N587mycR
<400> 14
ctcttctgag atgagttttt gttcgttgcc tctctggagg ttg 43
<210> 15
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N587mycF
<400> 15
aaactcatct cagaagagga tctgagacaa gcagctaccg cag 43
<210> 16
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAVRCPmeR
<400> 16
tccgcccgct gtttaaac 18
<210> 17
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAVRC9BsiWF
<400> 17
actcagacta tcagctcccg tacg 24
<210> 18
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> A589mycR
<400> 18
ctcttctgag atgagttttt gttctgcttg ggcactctgg t 41
<210> 19
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> A589mycF
<400> 19
aaactcatct cagagaggat ctgcaggcgc agaccgg 37
<210> 20
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAVRC9PmeR
<400> 20
ctccgcccgc tgtttaaac 19
<210> 21
<211> 738
<212> БЕЛОК
<213> аденоассоциированный вирус 8
<400> 21
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Gln Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro
180 185 190
Pro Ala Ala Pro Ser Gly Val Gly Pro Asn Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val
225 230 235 240
Ile Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His
245 250 255
Leu Tyr Lys Gln Ile Ser Asn Gly Thr Ser Gly Gly Ala Thr Asn Asp
260 265 270
Asn Thr Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn
275 280 285
Arg Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn
290 295 300
Asn Asn Trp Gly Phe Arg Pro Lys Arg Leu Ser Phe Lys Leu Phe Asn
305 310 315 320
Ile Gln Val Lys Glu Val Thr Gln Asn Glu Gly Thr Lys Thr Ile Ala
325 330 335
Asn Asn Leu Thr Ser Thr Ile Gln Val Phe Thr Asp Ser Glu Tyr Gln
340 345 350
Leu Pro Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe
355 360 365
Pro Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn
370 375 380
Asn Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr
385 390 395 400
Phe Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Thr Tyr
405 410 415
Thr Phe Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser
420 425 430
Leu Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu
435 440 445
Ser Arg Thr Gln Thr Thr Gly Gly Thr Ala Asn Thr Gln Thr Leu Gly
450 455 460
Phe Ser Gln Gly Gly Pro Asn Thr Met Ala Asn Gln Ala Lys Asn Trp
465 470 475 480
Leu Pro Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Thr Thr Thr Gly
485 490 495
Gln Asn Asn Asn Ser Asn Phe Ala Trp Thr Ala Gly Thr Lys Tyr His
500 505 510
Leu Asn Gly Arg Asn Ser Leu Ala Asn Pro Gly Ile Ala Met Ala Thr
515 520 525
His Lys Asp Asp Glu Glu Arg Phe Phe Pro Ser Asn Gly Ile Leu Ile
530 535 540
Phe Gly Lys Gln Asn Ala Ala Arg Asp Asn Ala Asp Tyr Ser Asp Val
545 550 555 560
Met Leu Thr Ser Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr
565 570 575
Glu Glu Tyr Gly Ile Val Ala Asp Asn Leu Gln Gln Gln Asn Thr Ala
580 585 590
Pro Gln Ile Gly Thr Val Asn Ser Gln Gly Ala Leu Pro Gly Met Val
595 600 605
Trp Gln Asn Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile
610 615 620
Pro His Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe
625 630 635 640
Gly Leu Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val
645 650 655
Pro Ala Asp Pro Pro Thr Thr Phe Asn Gln Ser Lys Leu Asn Ser Phe
660 665 670
Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu
675 680 685
Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr
690 695 700
Ser Asn Tyr Tyr Lys Ser Thr Ser Val Asp Phe Ala Val Asn Thr Glu
705 710 715 720
Gly Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg
725 730 735
Asn Leu
<210> 22
<211> 453
<212> ДНК
<213> Искусственная последовательность
<220>
<223> AAV8 N590myc gblock
<400> 22
gaccctgtta ccgccaacaa cgcgtctcaa cgacaaccgg gcaaaacaac aatagcaact 60
ttgcctggac tgctgggacc aaataccatc tgaatggaag aaattcattg gctaatcctg 120
gcatcgctat ggcaacacac aaagacgacg aggagcgttt ttttcccagt aacgggatcc 180
tgatttttgg caaacaaaat gctgccagag acaatgcgga ttacagcgat gtcatgctca 240
ccagcgagga agaaatcaaa accactaacc ctgtggctac agaggaatac ggtatcgtgg 300
cagataactt gcagcagcaa aacgaacaaa aactcatctc agaagaggat ctgacggctc 360
ctcaaattgg aactgtcaac agccaggggg ccttacccgg tatggtctgg cagaaccggg 420
acgtgtacct gcagggtccc atctgggcca aga 453
<210> 23
<211> 7336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAV RC2/8
<400> 23
gcgcgccgat atcgttaacg ccccgcgccg gccgctctag aactagtgga tcccccggaa 60
gatcagaagt tcctattccg aagttcctat tctctagaaa gtataggaac ttctgatctg 120
cgcagccgcc atgccggggt tttacgagat tgtgattaag gtccccagcg accttgacga 180
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 240
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 300
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 360
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 420
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 480
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 540
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 600
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 660
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 720
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 780
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 840
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 900
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 960
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 1020
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 1080
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1140
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1200
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1260
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1320
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1380
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1440
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1500
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1560
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1620
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1680
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1740
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1800
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1860
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1920
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1980
catctttgaa caataaatga tttaaatcag gtatggctgc cgatggttat cttccagatt 2040
ggctcgagga caacctctct gagggcattc gcgagtggtg ggcgctgaaa cctggagccc 2100
cgaagcccaa agccaaccag caaaagcagg acgacggccg gggtctggtg cttcctggct 2160
acaagtacct cggacccttc aacggactcg acaaggggga gcccgtcaac gcggcggacg 2220
cagcggccct cgagcacgac aaggcctacg accagcagct gcaggcgggt gacaatccgt 2280
acctgcggta taaccacgcc gacgccgagt ttcaggagcg tctgcaagaa gatacgtctt 2340
ttgggggcaa cctcgggcga gcagtcttcc aggccaagaa gcgggttctc gaacctctcg 2400
gtctggttga ggaaggcgct aagacggctc ctggaaagaa gagaccggta gagccatcac 2460
cccagcgttc tccagactcc tctacgggca tcggcaagaa aggccaacag cccgccagaa 2520
aaagactcaa ttttggtcag actggcgact cagagtcagt tccagaccct caacctctcg 2580
gagaacctcc agcagcgccc tctggtgtgg gacctaatac aatggctgca ggcggtggcg 2640
caccaatggc agacaataac gaaggcgccg acggagtggg tagttcctcg ggaaattggc 2700
attgcgattc cacatggctg ggcgacagag tcatcaccac cagcacccga acctgggccc 2760
tgcccaccta caacaaccac ctctacaagc aaatctccaa cgggacatcg ggaggagcca 2820
ccaacgacaa cacctacttc ggctacagca ccccctgggg gtattttgac tttaacagat 2880
tccactgcca cttttcacca cgtgactggc agcgactcat caacaacaac tggggattcc 2940
ggcccaagag actcagcttc aagctcttca acatccaggt caaggaggtc acgcagaatg 3000
aaggcaccaa gaccatcgcc aataacctca ccagcaccat ccaggtgttt acggactcgg 3060
agtaccagct gccgtacgtt ctcggctctg cccaccaggg ctgcctgcct ccgttcccgg 3120
cggacgtgtt catgattccc cagtacggct acctaacact caacaacggt agtcaggccg 3180
tgggacgctc ctccttctac tgcctggaat actttccttc gcagatgctg agaaccggca 3240
acaacttcca gtttacttac accttcgagg acgtgccttt ccacagcagc tacgcccaca 3300
gccagagctt ggaccggctg atgaatcctc tgattgacca gtacctgtac tacttgtctc 3360
ggactcaaac aacaggaggc acggcaaata cgcagactct gggcttcagc caaggtgggc 3420
ctaatacaat ggccaatcag gcaaagaact ggctgccagg accctgttac cgccaacaac 3480
gcgtctcaac gacaaccggg caaaacaaca atagcaactt tgcctggact gctgggacca 3540
aataccatct gaatggaaga aattcattgg ctaatcctgg catcgctatg gcaacacaca 3600
aagacgacga ggagcgtttt tttcccagta acgggatcct gatttttggc aaacaaaatg 3660
ctgccagaga caatgcggat tacagcgatg tcatgctcac cagcgaggaa gaaatcaaaa 3720
ccactaaccc tgtggctaca gaggaatacg gtatcgtggc agataacttg cagcagcaaa 3780
acacggctcc tcaaattgga actgtcaaca gccagggggc cttacccggt atggtctggc 3840
agaaccggga cgtgtacctg cagggtccca tctgggccaa gattcctcac acggacggca 3900
acttccaccc gtctccgctg atgggcggct ttggcctgaa acatcctccg cctcagatcc 3960
tgatcaagaa cacgcctgta cctgcggatc ctccgaccac cttcaaccag tcaaagctga 4020
actctttcat cacgcaatac agcaccggac aggtcagcgt ggaaattgaa tgggagctgc 4080
agaaggaaaa cagcaagcgc tggaaccccg agatccagta cacctccaac tactacaaat 4140
ctacaagtgt ggactttgct gttaatacag aaggcgtgta ctctgaaccc cgccccattg 4200
gcacccgtta cctcacccgt aatctgtaat tgcctgttaa tcaataaacc gtttaattcg 4260
tttcagttga actttggtct ctgcgtattt ctttcttatc tagtttccat ggctacgtag 4320
ataagtagca tggcgggtta atcattaact acagcccggg cgtttaaaca gcgggcggag 4380
gggtggagtc gtgacgtgaa ttacgtcata gggttaggga ggtcctgtat tagaggtcac 4440
gtgagtgttt tgcgacattt tgcgacacca tgtggtctcg ctgggggggg gggcccgagt 4500
gagcacgcag ggtctccatt ttgaagcggg aggtttgaac gagcgctggc gcgctcactg 4560
gccgtcgttt tacaacgtcg tgactgggaa aaccctggcg ttacccaact taatcgcctt 4620
gcagcacatc cccctttcgc cagctggcgt aatagcgaag aggcccgcac cgatcgccct 4680
tcccaacagt tgcgcagcct gaatggcgaa tggaaattgt aagcgttaat attttgttaa 4740
aattcgcgtt aaatttttgt taaatcagct cattttttta accaataggc cgaaatcggc 4800
aaaatccctt ataaatcaaa agaatagacc gagatagggt tgagtgttgt tccagtttgg 4860
aacaagagtc cactattaag aacgtggact ccaacgtcaa agggcgaaaa accgtctatc 4920
agggcgatgg cccactacgt gaaccatcac cctaatcaag ttttttgggg tcgaggtgcc 4980
gtaaagcact aaatcggaac cctaaaggga gcccccgatt tagagcttga cggggaaagc 5040
cggcgaacgt ggcgagaaag gaagggaaga aagcgaaagg agcgggcgct agggcgctgg 5100
caagtgtagc ggtcacgctg cgcgtaacca ccacacccgc cgcgcttaat gcgccgctac 5160
agggcgcgtc aggtggcact tttcggggaa atgtgcgcgg aacccctatt tgtttatttt 5220
tctaaataca ttcaaatatg tatccgctca tgagacaata accctgataa atgcttcaat 5280
aatattgaaa aaggaagagt atgagtattc aacatttccg tgtcgccctt attccctttt 5340
ttgcggcatt ttgccttcct gtttttgctc acccagaaac gctggtgaaa gtaaaagatg 5400
ctgaagatca gttgggtgca cgagtgggtt acatcgaact ggatctcaac agcggtaaga 5460
tccttgagag ttttcgcccc gaagaacgtt ttccaatgat gagcactttt aaagttctgc 5520
tatgtggcgc ggtattatcc cgtattgacg ccgggcaaga gcaactcggt cgccgcatac 5580
actattctca gaatgacttg gttgagtact caccagtcac agaaaagcat cttacggatg 5640
gcatgacagt aagagaatta tgcagtgctg ccataaccat gagtgataac actgcggcca 5700
acttacttct gacaacgatc ggaggaccga aggagctaac cgcttttttg cacaacatgg 5760
gggatcatgt aactcgcctt gatcgttggg aaccggagct gaatgaagcc ataccaaacg 5820
acgagcgtga caccacgatg cctgtagcaa tggcaacaac gttgcgcaaa ctattaactg 5880
gcgaactact tactctagct tcccggcaac aattaataga ctggatggag gcggataaag 5940
ttgcaggacc acttctgcgc tcggcccttc cggctggctg gtttattgct gataaatctg 6000
gagccggtga gcgtgggtct cgcggtatca ttgcagcact ggggccagat ggtaagccct 6060
cccgtatcgt agttatctac acgacgggga gtcaggcaac tatggatgaa cgaaatagac 6120
agatcgctga gataggtgcc tcactgatta agcattggta actgtcagac caagtttact 6180
catatatact ttagattgat ttaaaacttc atttttaatt taaaaggatc taggtgaaga 6240
tcctttttga taatctcatg accaaaatcc cttaacgtga gttttcgttc cactgagcgt 6300
cagaccccgt agaaaagatc aaaggatctt cttgagatcc tttttttctg cgcgtaatct 6360
gctgcttgca aacaaaaaaa ccaccgctac cagcggtggt ttgtttgccg gatcaagagc 6420
taccaactct ttttccgaag gtaactggct tcagcagagc gcagatacca aatactgttc 6480
ttctagtgta gccgtagtta ggccaccact tcaagaactc tgtagcaccg cctacatacc 6540
tcgctctgct aatcctgtta ccagtggctg ctgccagtgg cgataagtcg tgtcttaccg 6600
ggttggactc aagacgatag ttaccggata aggcgcagcg gtcgggctga acggggggtt 6660
cgtgcacaca gcccagcttg gagcgaacga cctacaccga actgagatac ctacagcgtg 6720
agctatgaga aagcgccacg cttcccgaag ggagaaaggc ggacaggtat ccggtaagcg 6780
gcagggtcgg aacaggagag cgcacgaggg agcttccagg gggaaacgcc tggtatcttt 6840
atagtcctgt cgggtttcgc cacctctgac ttgagcgtcg atttttgtga tgctcgtcag 6900
gggggcggag cctatggaaa aacgccagca acgcggcctt tttacggttc ctggcctttt 6960
gctggccttt tgctcacatg ttctttcctg cgttatcccc tgattctgtg gataaccgta 7020
ttaccgcctt tgagtgagct gataccgctc gccgcagccg aacgaccgag cgcagcgagt 7080
cagtgagcga ggaagcggaa gagcgcccaa tacgcaaacc gcctctcccc gcgcgttggc 7140
cgattcatta atgcagctgg cacgacaggt ttcccgactg gaaagcgggc agtgagcgca 7200
acgcaattaa tgtgagttag ctcactcatt aggcacccca ggctttacac tttatgcttc 7260
cggctcgtat gttgtgtgga attgtgagcg gataacaatt tcacacagga aacagctatg 7320
accatgatta cgccaa 7336
<210> 24
<211> 7366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Плазмида paav RC2/8 myc
<400> 24
gcgcgccgat atcgttaacg ccccgcgccg gccgctctag aactagtgga tcccccggaa 60
gatcagaagt tcctattccg aagttcctat tctctagaaa gtataggaac ttctgatctg 120
cgcagccgcc atgccggggt tttacgagat tgtgattaag gtccccagcg accttgacga 180
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 240
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 300
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 360
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 420
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 480
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 540
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 600
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 660
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 720
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 780
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 840
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 900
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 960
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 1020
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 1080
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1140
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1200
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1260
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1320
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1380
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1440
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1500
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1560
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1620
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1680
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1740
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1800
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1860
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1920
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1980
catctttgaa caataaatga tttaaatcag gtatggctgc cgatggttat cttccagatt 2040
ggctcgagga caacctctct gagggcattc gcgagtggtg ggcgctgaaa cctggagccc 2100
cgaagcccaa agccaaccag caaaagcagg acgacggccg gggtctggtg cttcctggct 2160
acaagtacct cggacccttc aacggactcg acaaggggga gcccgtcaac gcggcggacg 2220
cagcggccct cgagcacgac aaggcctacg accagcagct gcaggcgggt gacaatccgt 2280
acctgcggta taaccacgcc gacgccgagt ttcaggagcg tctgcaagaa gatacgtctt 2340
ttgggggcaa cctcgggcga gcagtcttcc aggccaagaa gcgggttctc gaacctctcg 2400
gtctggttga ggaaggcgct aagacggctc ctggaaagaa gagaccggta gagccatcac 2460
cccagcgttc tccagactcc tctacgggca tcggcaagaa aggccaacag cccgccagaa 2520
aaagactcaa ttttggtcag actggcgact cagagtcagt tccagaccct caacctctcg 2580
gagaacctcc agcagcgccc tctggtgtgg gacctaatac aatggctgca ggcggtggcg 2640
caccaatggc agacaataac gaaggcgccg acggagtggg tagttcctcg ggaaattggc 2700
attgcgattc cacatggctg ggcgacagag tcatcaccac cagcacccga acctgggccc 2760
tgcccaccta caacaaccac ctctacaagc aaatctccaa cgggacatcg ggaggagcca 2820
ccaacgacaa cacctacttc ggctacagca ccccctgggg gtattttgac tttaacagat 2880
tccactgcca cttttcacca cgtgactggc agcgactcat caacaacaac tggggattcc 2940
ggcccaagag actcagcttc aagctcttca acatccaggt caaggaggtc acgcagaatg 3000
aaggcaccaa gaccatcgcc aataacctca ccagcaccat ccaggtgttt acggactcgg 3060
agtaccagct gccgtacgtt ctcggctctg cccaccaggg ctgcctgcct ccgttcccgg 3120
cggacgtgtt catgattccc cagtacggct acctaacact caacaacggt agtcaggccg 3180
tgggacgctc ctccttctac tgcctggaat actttccttc gcagatgctg agaaccggca 3240
acaacttcca gtttacttac accttcgagg acgtgccttt ccacagcagc tacgcccaca 3300
gccagagctt ggaccggctg atgaatcctc tgattgacca gtacctgtac tacttgtctc 3360
ggactcaaac aacaggaggc acggcaaata cgcagactct gggcttcagc caaggtgggc 3420
ctaatacaat ggccaatcag gcaaagaact ggctgccagg accctgttac cgccaacaac 3480
gcgtctcaac gacaaccggg caaaacaaca atagcaactt tgcctggact gctgggacca 3540
aataccatct gaatggaaga aattcattgg ctaatcctgg catcgctatg gcaacacaca 3600
aagacgacga ggagcgtttt tttcccagta acgggatcct gatttttggc aaacaaaatg 3660
ctgccagaga caatgcggat tacagcgatg tcatgctcac cagcgaggaa gaaatcaaaa 3720
ccactaaccc tgtggctaca gaggaatacg gtatcgtggc agataacttg cagcagcaaa 3780
acgaacaaaa actcatctca gaagaggatc tgacggctcc tcaaattgga actgtcaaca 3840
gccagggggc cttacccggt atggtctggc agaaccggga cgtgtacctg cagggtccca 3900
tctgggccaa gattcctcac acggacggca acttccaccc gtctccgctg atgggcggct 3960
ttggcctgaa acatcctccg cctcagatcc tgatcaagaa cacgcctgta cctgcggatc 4020
ctccgaccac cttcaaccag tcaaagctga actctttcat cacgcaatac agcaccggac 4080
aggtcagcgt ggaaattgaa tgggagctgc agaaggaaaa cagcaagcgc tggaaccccg 4140
agatccagta cacctccaac tactacaaat ctacaagtgt ggactttgct gttaatacag 4200
aaggcgtgta ctctgaaccc cgccccattg gcacccgtta cctcacccgt aatctgtaat 4260
tgcctgttaa tcaataaacc gtttaattcg tttcagttga actttggtct ctgcgtattt 4320
ctttcttatc tagtttccat ggctacgtag ataagtagca tggcgggtta atcattaact 4380
acagcccggg cgtttaaaca gcgggcggag gggtggagtc gtgacgtgaa ttacgtcata 4440
gggttaggga ggtcctgtat tagaggtcac gtgagtgttt tgcgacattt tgcgacacca 4500
tgtggtctcg ctgggggggg gggcccgagt gagcacgcag ggtctccatt ttgaagcggg 4560
aggtttgaac gagcgctggc gcgctcactg gccgtcgttt tacaacgtcg tgactgggaa 4620
aaccctggcg ttacccaact taatcgcctt gcagcacatc cccctttcgc cagctggcgt 4680
aatagcgaag aggcccgcac cgatcgccct tcccaacagt tgcgcagcct gaatggcgaa 4740
tggaaattgt aagcgttaat attttgttaa aattcgcgtt aaatttttgt taaatcagct 4800
cattttttta accaataggc cgaaatcggc aaaatccctt ataaatcaaa agaatagacc 4860
gagatagggt tgagtgttgt tccagtttgg aacaagagtc cactattaag aacgtggact 4920
ccaacgtcaa agggcgaaaa accgtctatc agggcgatgg cccactacgt gaaccatcac 4980
cctaatcaag ttttttgggg tcgaggtgcc gtaaagcact aaatcggaac cctaaaggga 5040
gcccccgatt tagagcttga cggggaaagc cggcgaacgt ggcgagaaag gaagggaaga 5100
aagcgaaagg agcgggcgct agggcgctgg caagtgtagc ggtcacgctg cgcgtaacca 5160
ccacacccgc cgcgcttaat gcgccgctac agggcgcgtc aggtggcact tttcggggaa 5220
atgtgcgcgg aacccctatt tgtttatttt tctaaataca ttcaaatatg tatccgctca 5280
tgagacaata accctgataa atgcttcaat aatattgaaa aaggaagagt atgagtattc 5340
aacatttccg tgtcgccctt attccctttt ttgcggcatt ttgccttcct gtttttgctc 5400
acccagaaac gctggtgaaa gtaaaagatg ctgaagatca gttgggtgca cgagtgggtt 5460
acatcgaact ggatctcaac agcggtaaga tccttgagag ttttcgcccc gaagaacgtt 5520
ttccaatgat gagcactttt aaagttctgc tatgtggcgc ggtattatcc cgtattgacg 5580
ccgggcaaga gcaactcggt cgccgcatac actattctca gaatgacttg gttgagtact 5640
caccagtcac agaaaagcat cttacggatg gcatgacagt aagagaatta tgcagtgctg 5700
ccataaccat gagtgataac actgcggcca acttacttct gacaacgatc ggaggaccga 5760
aggagctaac cgcttttttg cacaacatgg gggatcatgt aactcgcctt gatcgttggg 5820
aaccggagct gaatgaagcc ataccaaacg acgagcgtga caccacgatg cctgtagcaa 5880
tggcaacaac gttgcgcaaa ctattaactg gcgaactact tactctagct tcccggcaac 5940
aattaataga ctggatggag gcggataaag ttgcaggacc acttctgcgc tcggcccttc 6000
cggctggctg gtttattgct gataaatctg gagccggtga gcgtgggtct cgcggtatca 6060
ttgcagcact ggggccagat ggtaagccct cccgtatcgt agttatctac acgacgggga 6120
gtcaggcaac tatggatgaa cgaaatagac agatcgctga gataggtgcc tcactgatta 6180
agcattggta actgtcagac caagtttact catatatact ttagattgat ttaaaacttc 6240
atttttaatt taaaaggatc taggtgaaga tcctttttga taatctcatg accaaaatcc 6300
cttaacgtga gttttcgttc cactgagcgt cagaccccgt agaaaagatc aaaggatctt 6360
cttgagatcc tttttttctg cgcgtaatct gctgcttgca aacaaaaaaa ccaccgctac 6420
cagcggtggt ttgtttgccg gatcaagagc taccaactct ttttccgaag gtaactggct 6480
tcagcagagc gcagatacca aatactgttc ttctagtgta gccgtagtta ggccaccact 6540
tcaagaactc tgtagcaccg cctacatacc tcgctctgct aatcctgtta ccagtggctg 6600
ctgccagtgg cgataagtcg tgtcttaccg ggttggactc aagacgatag ttaccggata 6660
aggcgcagcg gtcgggctga acggggggtt cgtgcacaca gcccagcttg gagcgaacga 6720
cctacaccga actgagatac ctacagcgtg agctatgaga aagcgccacg cttcccgaag 6780
ggagaaaggc ggacaggtat ccggtaagcg gcagggtcgg aacaggagag cgcacgaggg 6840
agcttccagg gggaaacgcc tggtatcttt atagtcctgt cgggtttcgc cacctctgac 6900
ttgagcgtcg atttttgtga tgctcgtcag gggggcggag cctatggaaa aacgccagca 6960
acgcggcctt tttacggttc ctggcctttt gctggccttt tgctcacatg ttctttcctg 7020
cgttatcccc tgattctgtg gataaccgta ttaccgcctt tgagtgagct gataccgctc 7080
gccgcagccg aacgaccgag cgcagcgagt cagtgagcga ggaagcggaa gagcgcccaa 7140
tacgcaaacc gcctctcccc gcgcgttggc cgattcatta atgcagctgg cacgacaggt 7200
ttcccgactg gaaagcgggc agtgagcgca acgcaattaa tgtgagttag ctcactcatt 7260
aggcacccca ggctttacac tttatgcttc cggctcgtat gttgtgtgga attgtgagcg 7320
gataacaatt tcacacagga aacagctatg accatgatta cgccaa 7366
<210> 25
<211> 748
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pAAV8N590myc
<400> 25
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Gln Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro
180 185 190
Pro Ala Ala Pro Ser Gly Val Gly Pro Asn Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val
225 230 235 240
Ile Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His
245 250 255
Leu Tyr Lys Gln Ile Ser Asn Gly Thr Ser Gly Gly Ala Thr Asn Asp
260 265 270
Asn Thr Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn
275 280 285
Arg Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn
290 295 300
Asn Asn Trp Gly Phe Arg Pro Lys Arg Leu Ser Phe Lys Leu Phe Asn
305 310 315 320
Ile Gln Val Lys Glu Val Thr Gln Asn Glu Gly Thr Lys Thr Ile Ala
325 330 335
Asn Asn Leu Thr Ser Thr Ile Gln Val Phe Thr Asp Ser Glu Tyr Gln
340 345 350
Leu Pro Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe
355 360 365
Pro Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn
370 375 380
Asn Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr
385 390 395 400
Phe Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Thr Tyr
405 410 415
Thr Phe Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser
420 425 430
Leu Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu
435 440 445
Ser Arg Thr Gln Thr Thr Gly Gly Thr Ala Asn Thr Gln Thr Leu Gly
450 455 460
Phe Ser Gln Gly Gly Pro Asn Thr Met Ala Asn Gln Ala Lys Asn Trp
465 470 475 480
Leu Pro Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Thr Thr Thr Gly
485 490 495
Gln Asn Asn Asn Ser Asn Phe Ala Trp Thr Ala Gly Thr Lys Tyr His
500 505 510
Leu Asn Gly Arg Asn Ser Leu Ala Asn Pro Gly Ile Ala Met Ala Thr
515 520 525
His Lys Asp Asp Glu Glu Arg Phe Phe Pro Ser Asn Gly Ile Leu Ile
530 535 540
Phe Gly Lys Gln Asn Ala Ala Arg Asp Asn Ala Asp Tyr Ser Asp Val
545 550 555 560
Met Leu Thr Ser Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr
565 570 575
Glu Glu Tyr Gly Ile Val Ala Asp Asn Leu Gln Gln Gln Asn Glu Gln
580 585 590
Lys Leu Ile Ser Glu Glu Asp Leu Thr Ala Pro Gln Ile Gly Thr Val
595 600 605
Asn Ser Gln Gly Ala Leu Pro Gly Met Val Trp Gln Asn Arg Asp Val
610 615 620
Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly Asn
625 630 635 640
Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro
645 650 655
Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asp Pro Pro Thr
660 665 670
Thr Phe Asn Gln Ser Lys Leu Asn Ser Phe Ile Thr Gln Tyr Ser Thr
675 680 685
Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser
690 695 700
Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Tyr Lys Ser
705 710 715 720
Thr Ser Val Asp Phe Ala Val Asn Thr Glu Gly Val Tyr Ser Glu Pro
725 730 735
Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 26
<211> 746
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pAAV9A589myc
<400> 26
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Glu Gln Lys
580 585 590
Leu Ile Ser Glu Glu Asp Leu Gln Ala Gln Thr Gly Trp Val Gln Asn
595 600 605
Gln Gly Ile Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu
610 615 620
Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly Asn Phe His
625 630 635 640
Pro Ser Pro Leu Met Gly Gly Phe Gly Met Lys His Pro Pro Pro Gln
645 650 655
Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asp Pro Pro Thr Ala Phe
660 665 670
Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln
675 680 685
Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg
690 695 700
Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Tyr Lys Ser Asn Asn
705 710 715 720
Val Glu Phe Ala Val Asn Thr Glu Gly Val Tyr Ser Glu Pro Arg Pro
725 730 735
Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 27
<211> 746
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pAAV9G453myc
<400> 27
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Ser
450 455 460
Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser Val Ala Gly Pro Ser Asn
465 470 475 480
Met Ala Val Gln Gly Arg Asn Tyr Ile Pro Gly Pro Ser Tyr Arg Gln
485 490 495
Gln Arg Val Ser Thr Thr Val Thr Gln Asn Asn Asn Ser Glu Phe Ala
500 505 510
Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn Gly Arg Asn Ser Leu Met
515 520 525
Asn Pro Gly Pro Ala Met Ala Ser His Lys Glu Gly Glu Asp Arg Phe
530 535 540
Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly Lys Gln Gly Thr Gly Arg
545 550 555 560
Asp Asn Val Asp Ala Asp Lys Val Met Ile Thr Asn Glu Glu Glu Ile
565 570 575
Lys Thr Thr Asn Pro Val Ala Thr Glu Ser Tyr Gly Gln Val Ala Thr
580 585 590
Asn His Gln Ser Ala Gln Ala Gln Ala Gln Thr Gly Trp Val Gln Asn
595 600 605
Gln Gly Ile Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu
610 615 620
Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly Asn Phe His
625 630 635 640
Pro Ser Pro Leu Met Gly Gly Phe Gly Met Lys His Pro Pro Pro Gln
645 650 655
Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asp Pro Pro Thr Ala Phe
660 665 670
Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln
675 680 685
Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg
690 695 700
Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Tyr Lys Ser Asn Asn
705 710 715 720
Val Glu Phe Ala Val Asn Thr Glu Gly Val Tyr Ser Glu Pro Arg Pro
725 730 735
Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 28
<211> 814
<212> ДНК
<213> Искусственная последовательность
<220>
<223> cmyc-scfv-gblock
<400> 28
tctccctgtc tctgggtaaa ggcggaggtg gatccggagg gggcggatct gacatccaga 60
tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc atcacttgcc 120
gggcaagtca gagcattagc agctatttaa attggtatca acagaaacca gggaaagccc 180
ctaagctcct gatctatgct gtttccattt tgcaaagtgg ggtcccatca aggttcagtg 240
gcagtggatc tgggacagat ttcactctca ccatcaactc tctgcaacct gaagattttg 300
caacttactc ctgtcaacag acttacagta cccctccgat caccttcggc caagggacac 360
gactggagat taaaggaggg ggcggctcag gaggaggggg tagtggggga ggcggctccg 420
aggtgcagct ggtggagtct gggggaggct tggtacagcc tggagggtcc ctgagactct 480
cctgtgtcac ctctggattt cgttttagta gttatgaaat gaactgggtc cgccaggctc 540
cagggaaggg gctggaatgg atttcataca ttaacagaag tggttatacc atatactacg 600
cagcctctct gaagggccga ttcaccatct ccagagacaa cgccaagaac tcactgtatc 660
ttcaaatgaa cagcctgaga gacgaggaca cggctatcta ttactgtgcg agagggagta 720
tcgcccctcg gggggagatt cgacacatct ttgactactg gggccaggga accctggtca 780
ccgtctcctc atgatgagcg gccgctaatc agcc 814
<210> 29
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc HCVR AA
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Thr Ser Gly Phe Arg Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ser Tyr Ile Asn Arg Ser Gly Tyr Thr Ile Tyr Tyr Ala Ala Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ile Ala Pro Arg Gly Glu Ile Arg His Ile Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val
115 120
<210> 30
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc LCVR AA
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 31
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc HCDR1
<400> 31
Gly Phe Arg Phe Ser Ser Tyr Glu
1 5
<210> 32
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc HCDR2 AA
<400> 32
Ile Asn Arg Ser Gly Tyr Thr Ile
1 5
<210> 33
<211> 2
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc HCDR3
<400> 33
Ala Arg
1
<210> 34
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc LCDR1 AA
<400> 34
Gln Ser Ile Ser Ser Tyr
1 5
<210> 35
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc LCDR2
<400> 35
Ala Ala Ser
1
<210> 36
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> myc LCDR3 AA
<400> 36
Gln Gln Ser Tyr Ser Thr Pro
1 5
<210> 37
<211> 247
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> anti-myc scFv
<400> 37
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Val Ser Ile Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Ser Cys Gln Gln Thr Tyr Ser Thr Pro Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Gly Gly Gly Gly
100 105 110
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val
115 120 125
Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
130 135 140
Cys Val Thr Ser Gly Phe Arg Phe Ser Ser Tyr Glu Met Asn Trp Val
145 150 155 160
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Ser Tyr Ile Asn Arg
165 170 175
Ser Gly Tyr Thr Ile Tyr Tyr Ala Ala Ser Leu Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Asp Glu Asp Thr Ala Ile Tyr Tyr Cys Ala Arg Gly Ser Ile
210 215 220
Ala Pro Arg Gly Glu Ile Arg His Ile Phe Asp Tyr Trp Gly Gln Gly
225 230 235 240
Thr Leu Val Thr Val Ser Ser
245
<210> 38
<211> 774
<212> ДНК
<213> Искусственная последовательность
<220>
<223> scFv к myc NA
<400> 38
ggcggaggtg gatccggagg gggcggatct gacatccaga tgacccagtc tccatcctcc 60
ctgtctgcat ctgtaggaga cagagtcacc atcacttgcc gggcaagtca gagcattagc 120
agctatttaa attggtatca acagaaacca gggaaagccc ctaagctcct gatctatgct 180
gtttccattt tgcaaagtgg ggtcccatca aggttcagtg gcagtggatc tgggacagat 240
ttcactctca ccatcaactc tctgcaacct gaagattttg caacttactc ctgtcaacag 300
acttacagta cccctccgat caccttcggc caagggacac gactggagat taaaggaggg 360
ggcggctcag gaggaggggg tagtggggga ggcggctccg aggtgcagct ggtggagtct 420
gggggaggct tggtacagcc tggagggtcc ctgagactct cctgtgtcac ctctggattt 480
cgttttagta gttatgaaat gaactgggtc cgccaggctc cagggaaggg gctggaatgg 540
atttcataca ttaacagaag tggttatacc atatactacg cagcctctct gaagggccga 600
ttcaccatct ccagagacaa cgccaagaac tcactgtatc ttcaaatgaa cagcctgaga 660
gacgaggaca cggctatcta ttactgtgcg agagggagta tcgcccctcg gggggagatt 720
cgacacatct ttgactactg gggccaggga accctggtca ccgtctcctc atga 774
<210> 39
<211> 372
<212> ДНК
<213> Искусственная последовательность
<220>
<223> myc HCVR NA
<400> 39
gaggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggagggtc cctgagactc 60
tcctgtgtca cctctggatt tcgttttagt agttatgaaa tgaactgggt ccgccaggct 120
ccagggaagg ggctggaatg gatttcatac attaacagaa gtggttatac catatactac 180
gcagcctctc tgaagggccg attcaccatc tccagagaca acgccaagaa ctcactgtat 240
cttcaaatga acagcctgag agacgaggac acggctatct attactgtgc gagagggagt 300
atcgcccctc ggggggagat tcgacacatc tttgactact ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 40
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> myc LCVR NA
<400> 40
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattagc agctatttaa attggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccgtca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag agttacagta cccctccgat caccttcggc 300
caagggacac gactggagat taaa 324
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ЧАСТИЦЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА | 2018 |
|
RU2811426C2 |
| НОВЫЕ НАЦЕЛИВАЮЩИЕ НА ПЕЧЕНЬ АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ | 2019 |
|
RU2793735C2 |
| Выделенный модифицированный белок VPI капсида аденоассоциированного вируса 9 серотипа (AAV9), капсид и вектор на его основе | 2021 |
|
RU2825667C2 |
| ВАРИАНТ AAV, КОМПОЗИЦИИ И СПОСОБЫ, В КОТОРЫХ ОН ИСПОЛЬЗУЕТСЯ, А ТАКЖЕ СПОСОБЫ ЕГО ПРИМЕНЕНИЯ ДЛЯ ПЕРЕНОСА ГЕНОВ В КЛЕТКИ, ОРГАНЫ И ТКАНИ | 2014 |
|
RU2837753C2 |
| Выделенный модифицированный белок VP1 капсида аденоассоциированного вируса 5 серотипа (AAV5), капсид и вектор на его основе | 2019 |
|
RU2751592C2 |
| СПОСОБЫ ВЫЯВЛЕНИЯ AAV | 2017 |
|
RU2771622C2 |
| МОДИФИЦИРОВАННЫЙ ФАКТОР IX, А ТАКЖЕ КОМПОЗИЦИИ, СПОСОБЫ И ВАРИАНТЫ ПРИМЕНЕНИЯ ПЕРЕНОСА ГЕНОВ В КЛЕТКИ, ОРГАНЫ И ТКАНИ | 2016 |
|
RU2811445C2 |
| ВАРИАНТЫ, НА ОСНОВЕ АДЕНОАССОЦИИРОВАННОГО ВИРУСА, СОСТАВЫ И СПОСОБЫ ДЛЯ ПУЛЬМОНАЛЬНОЙ ДОСТАВКИ | 2021 |
|
RU2829874C1 |
| РЕКОМБИНАНТНЫЕ ВАКЦИНЫ ОТ FMDV И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2745373C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ПОВЫШЕНИЯ ИЛИ УСИЛЕНИЯ ТРАНСДУКЦИИ ВЕКТОРОВ ДЛЯ ГЕННОЙ ТЕРАПИИ И ДЛЯ УДАЛЕНИЯ ИЛИ СНИЖЕНИЯ ИММУНОГЛОБУЛИНОВ | 2021 |
|
RU2841237C1 |


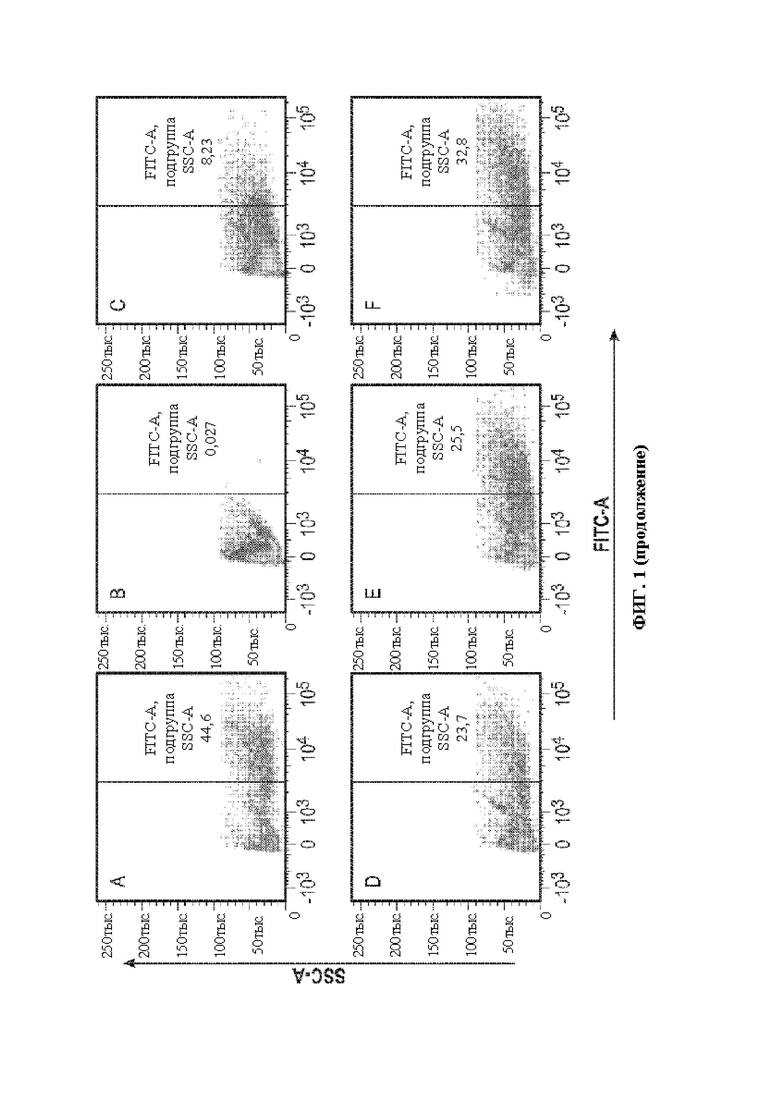
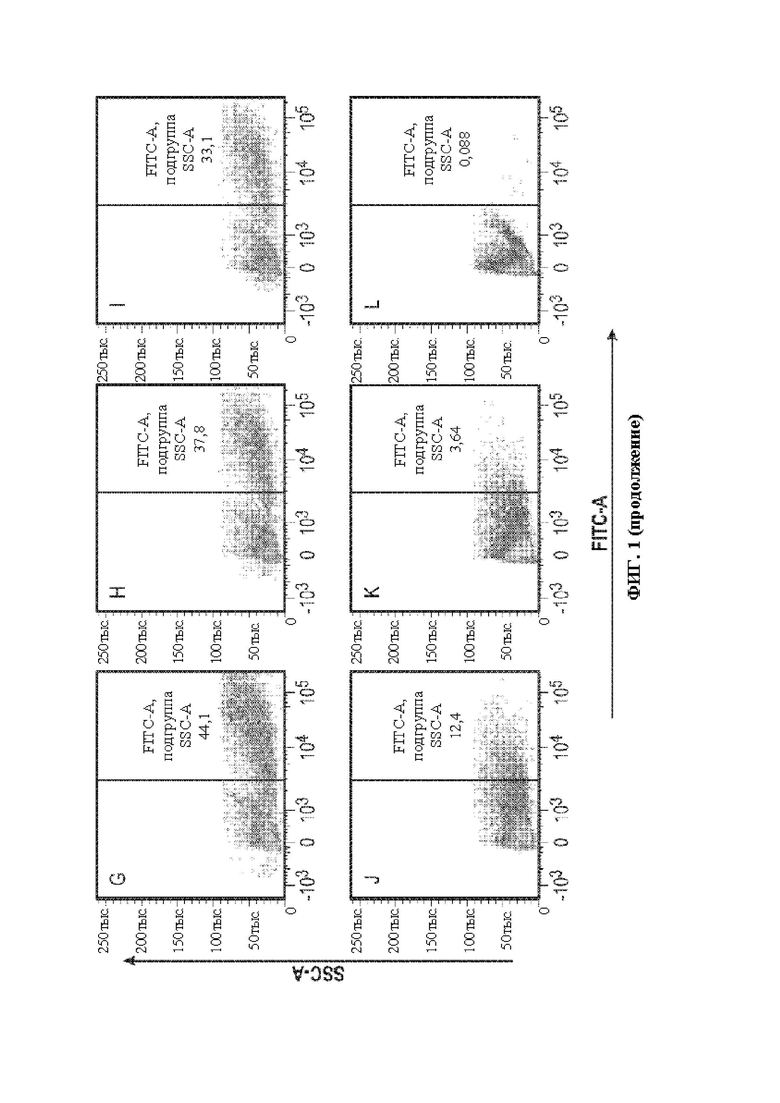
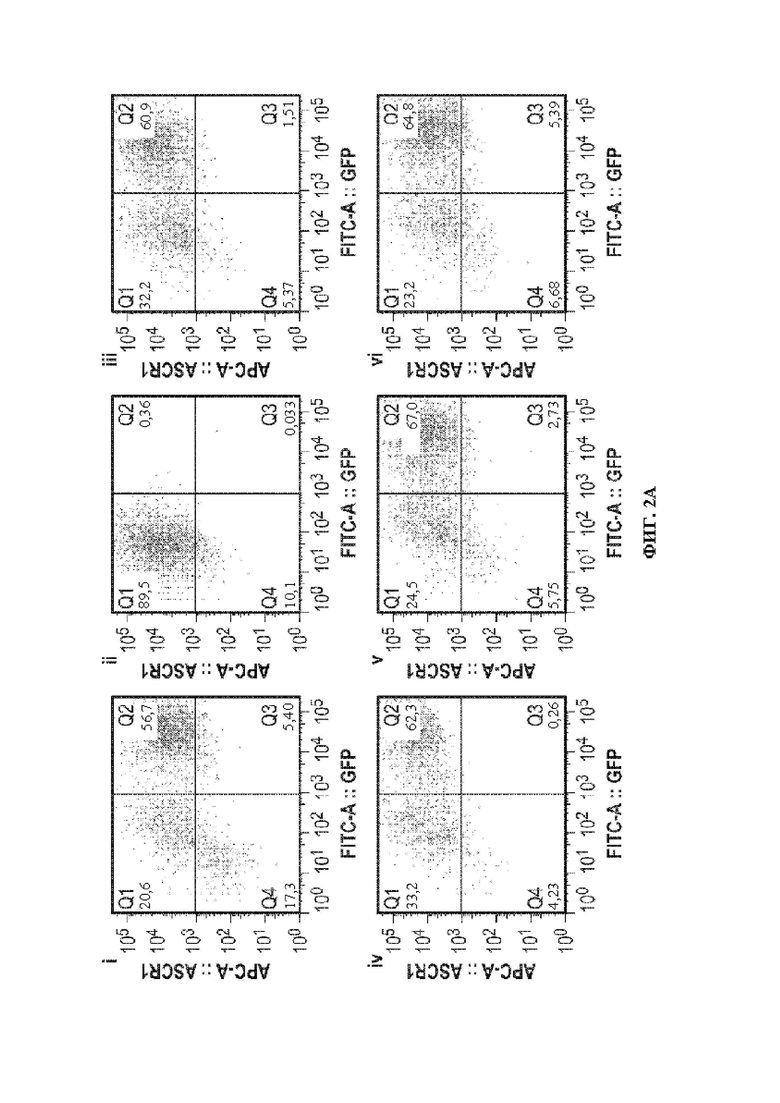
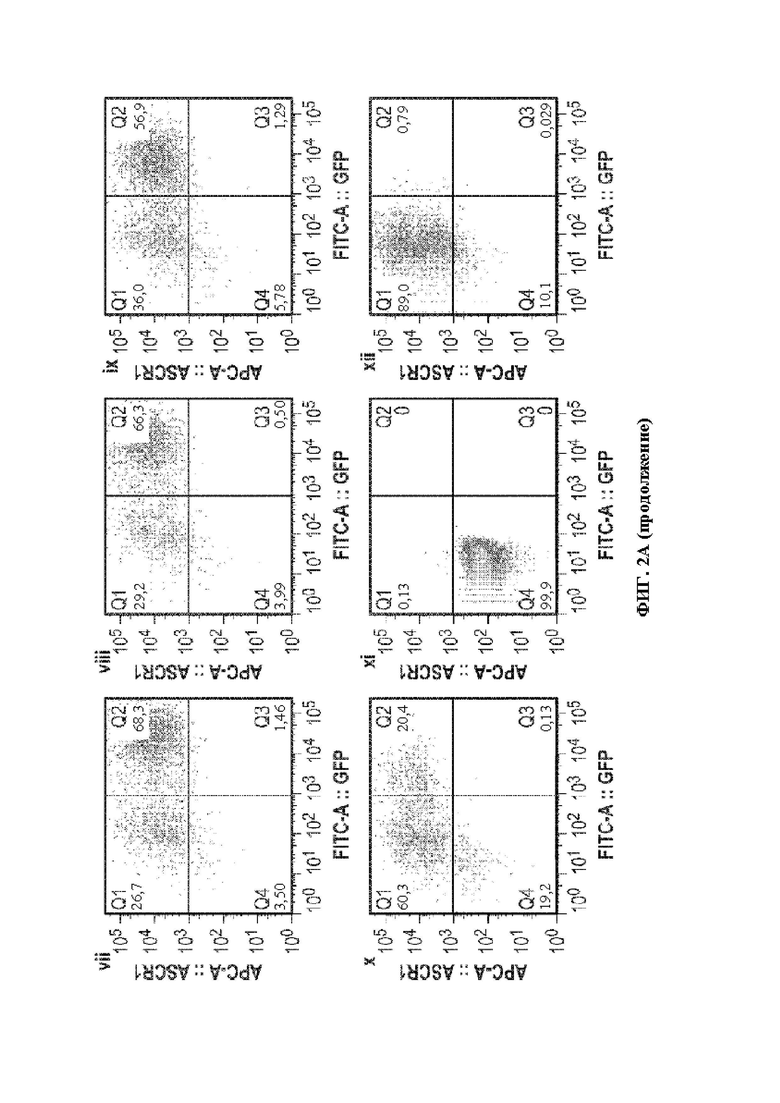
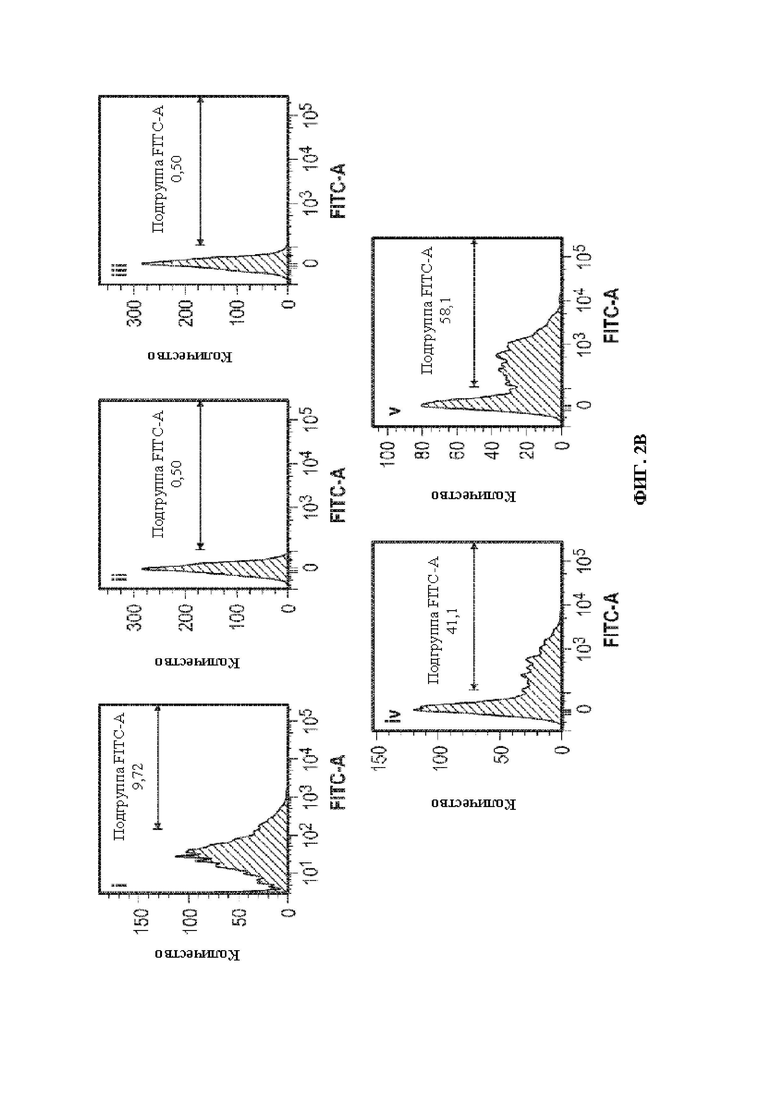
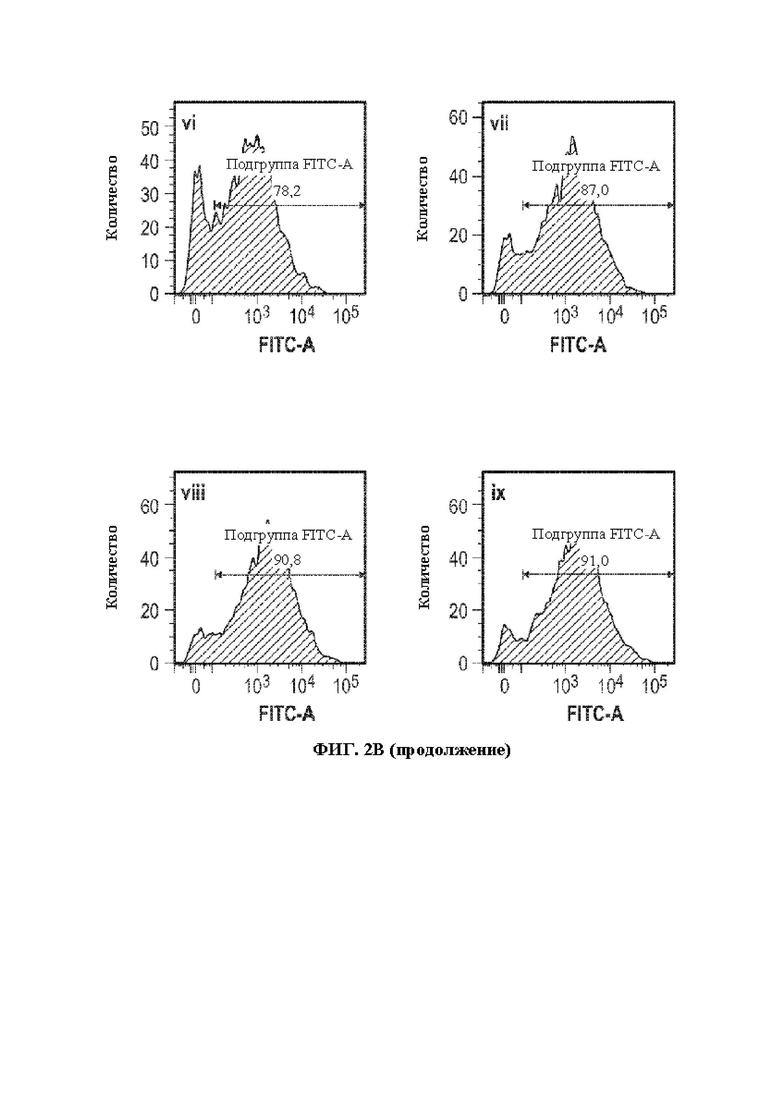
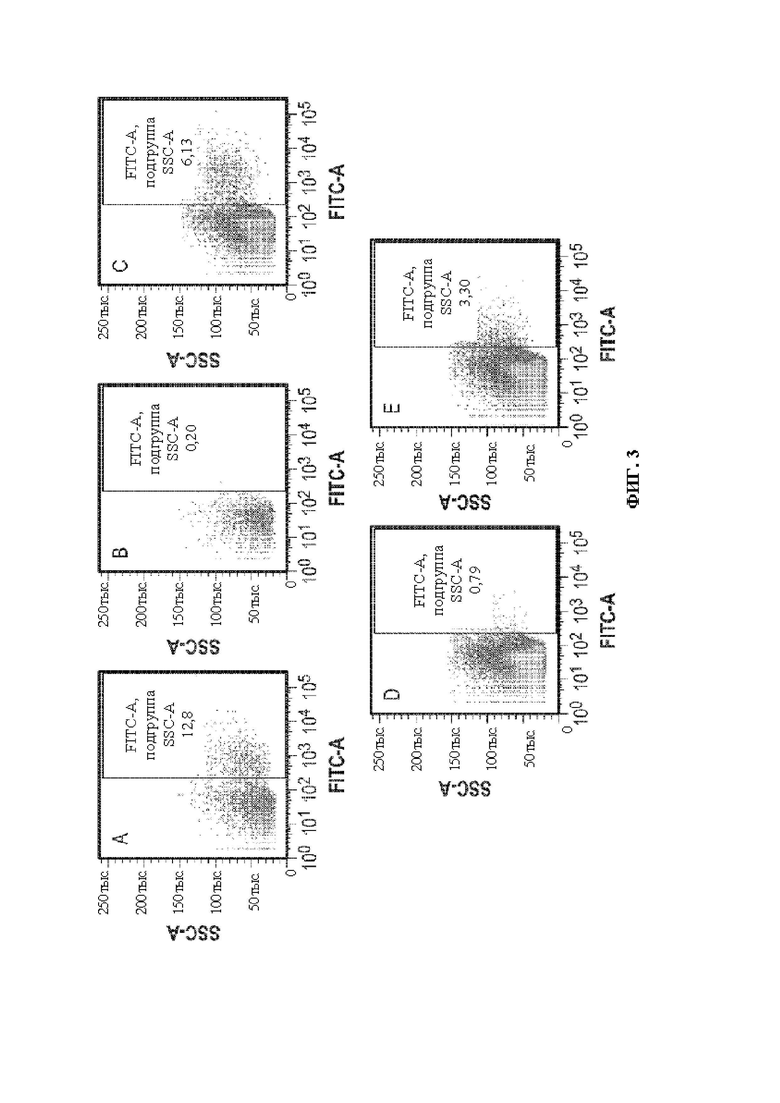
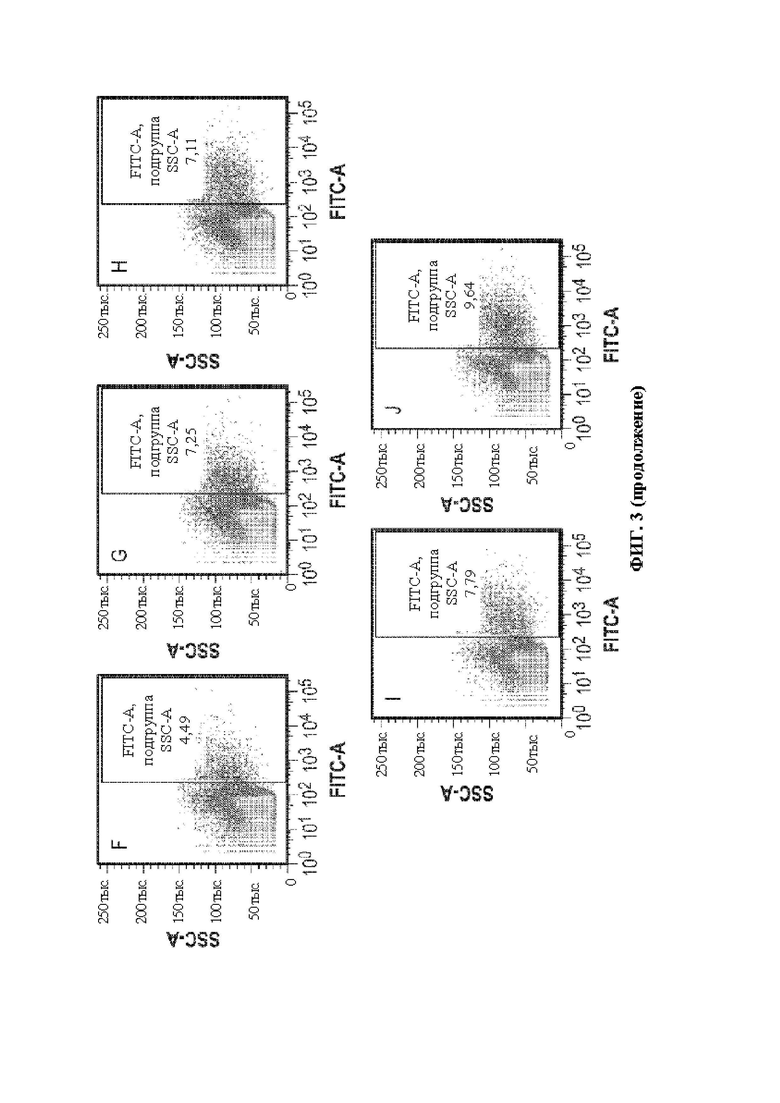

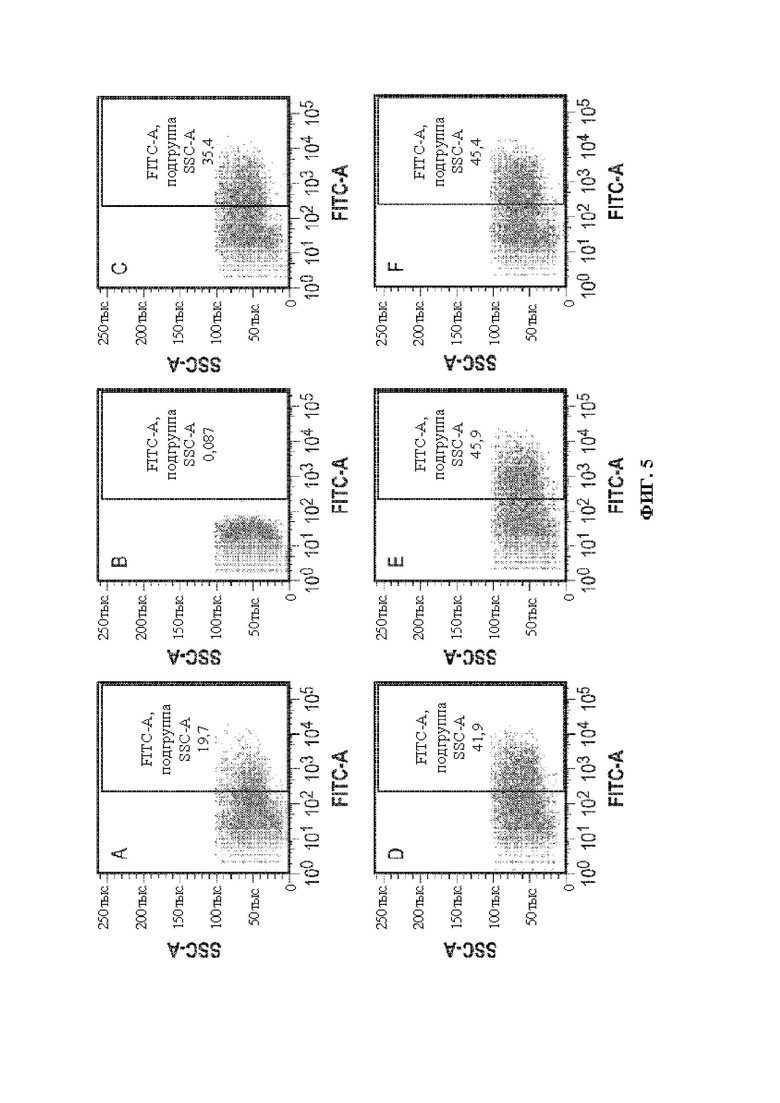
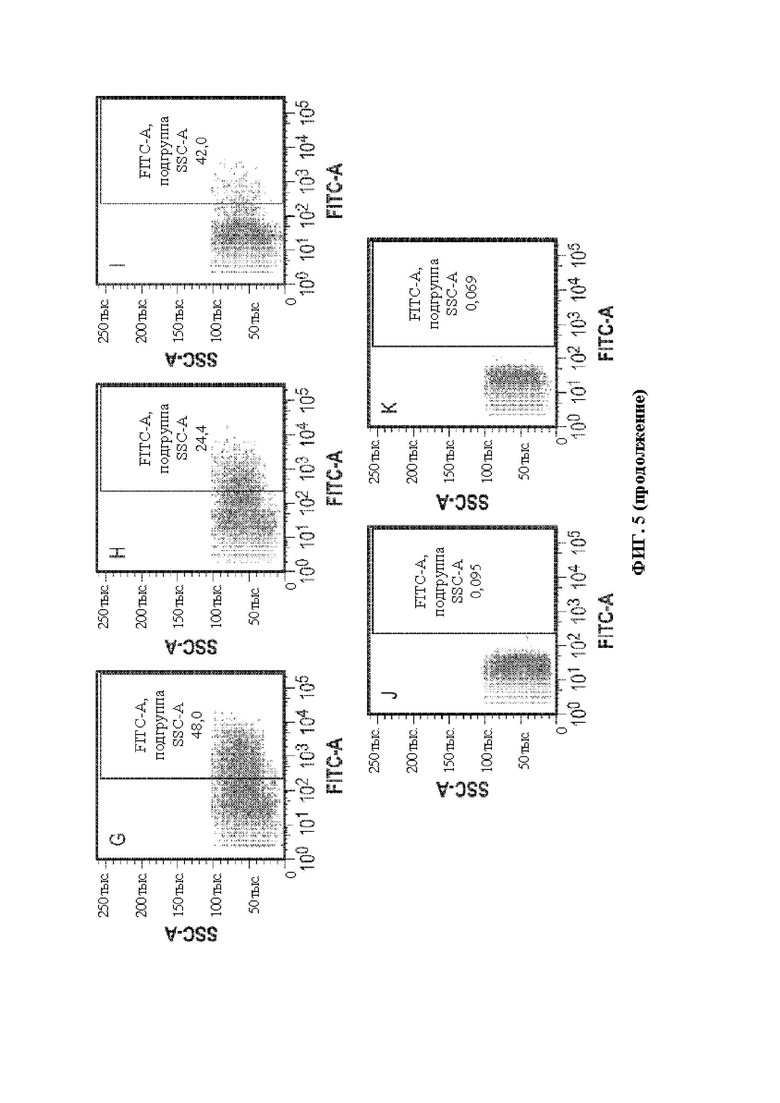
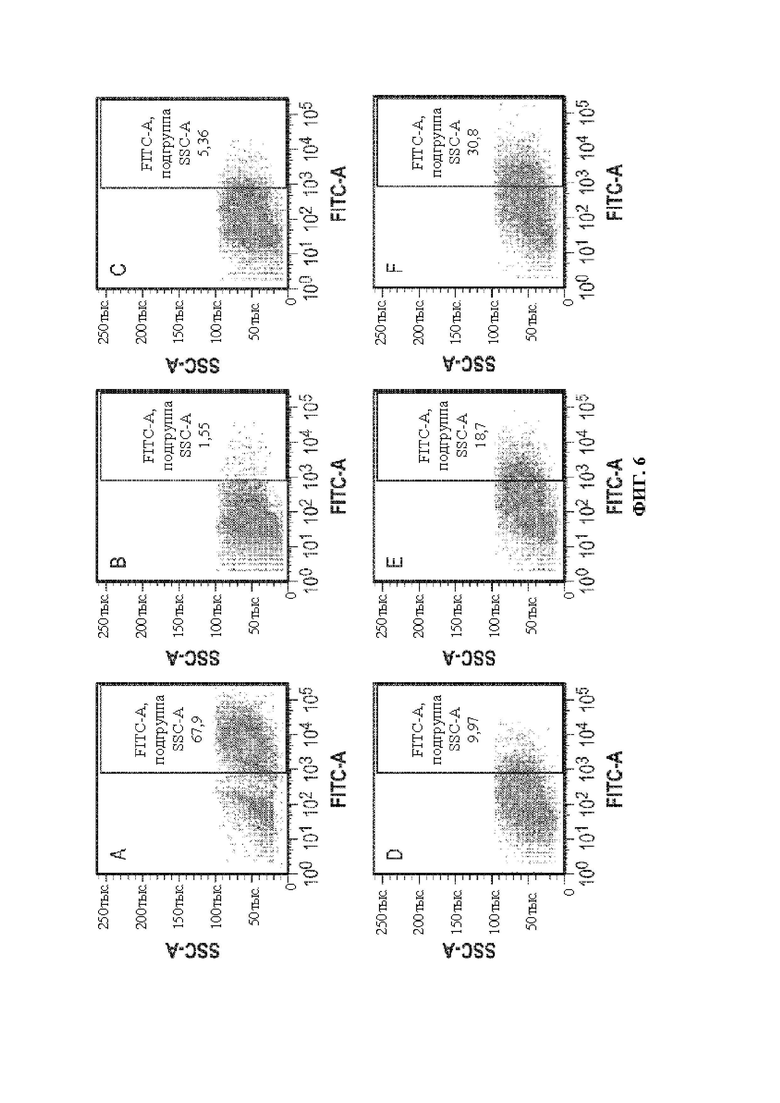
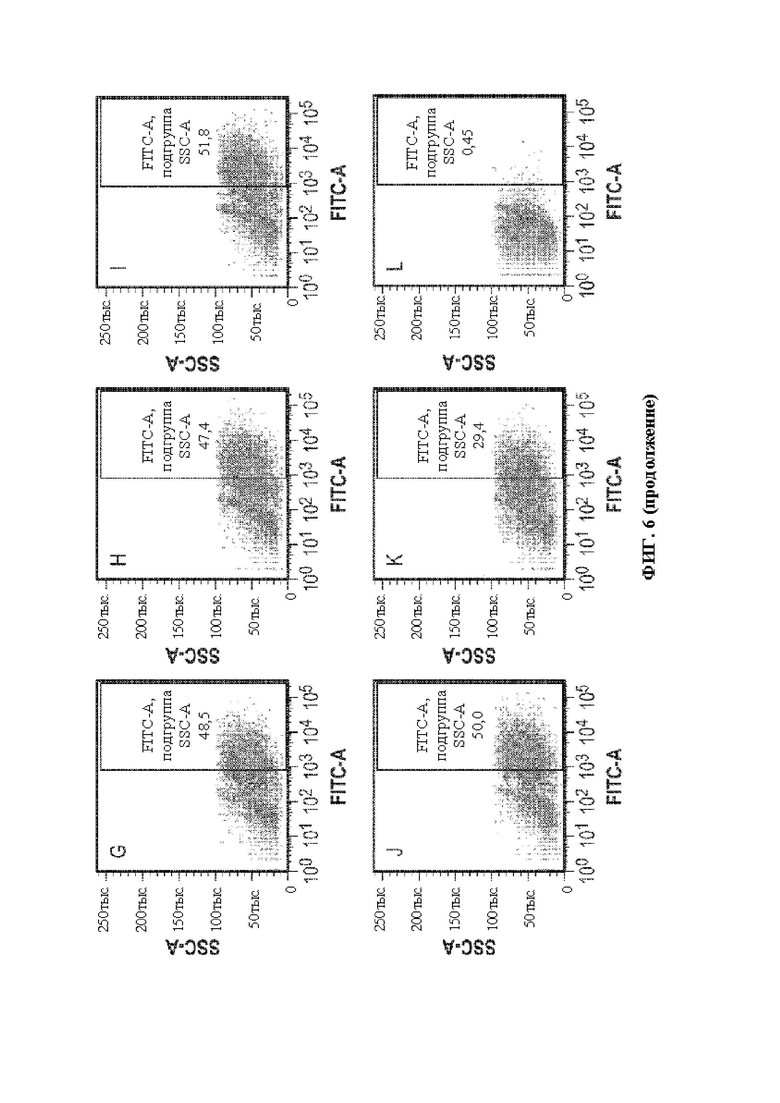
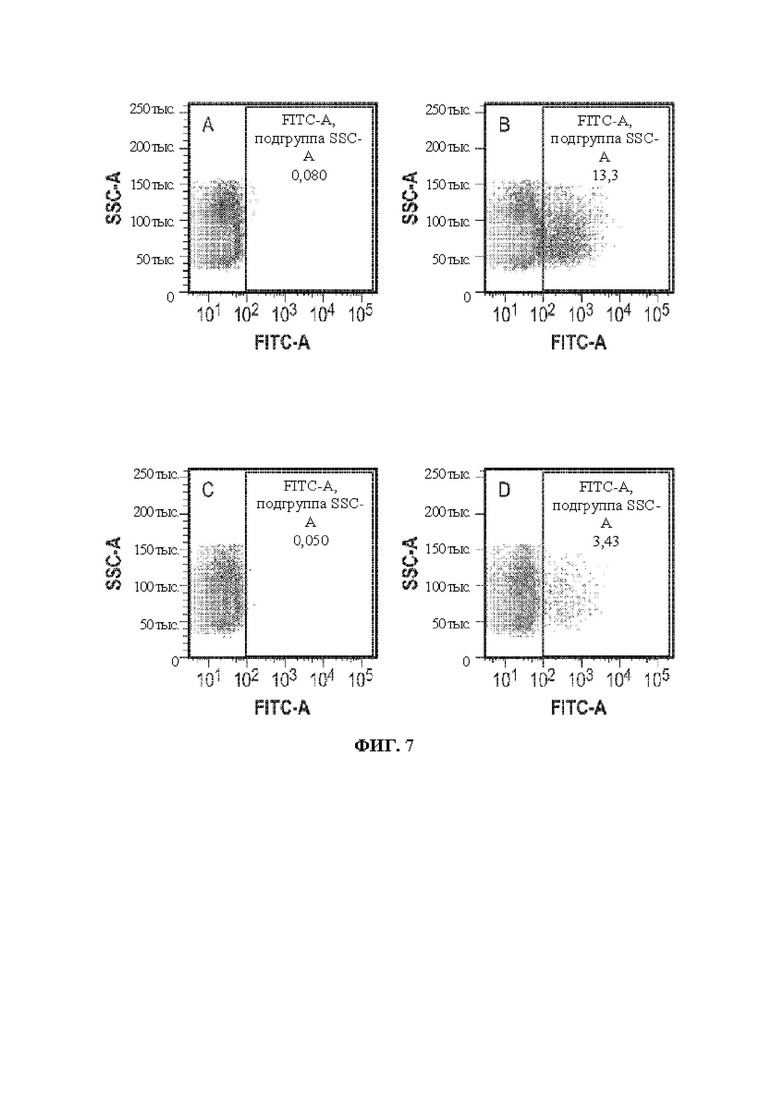
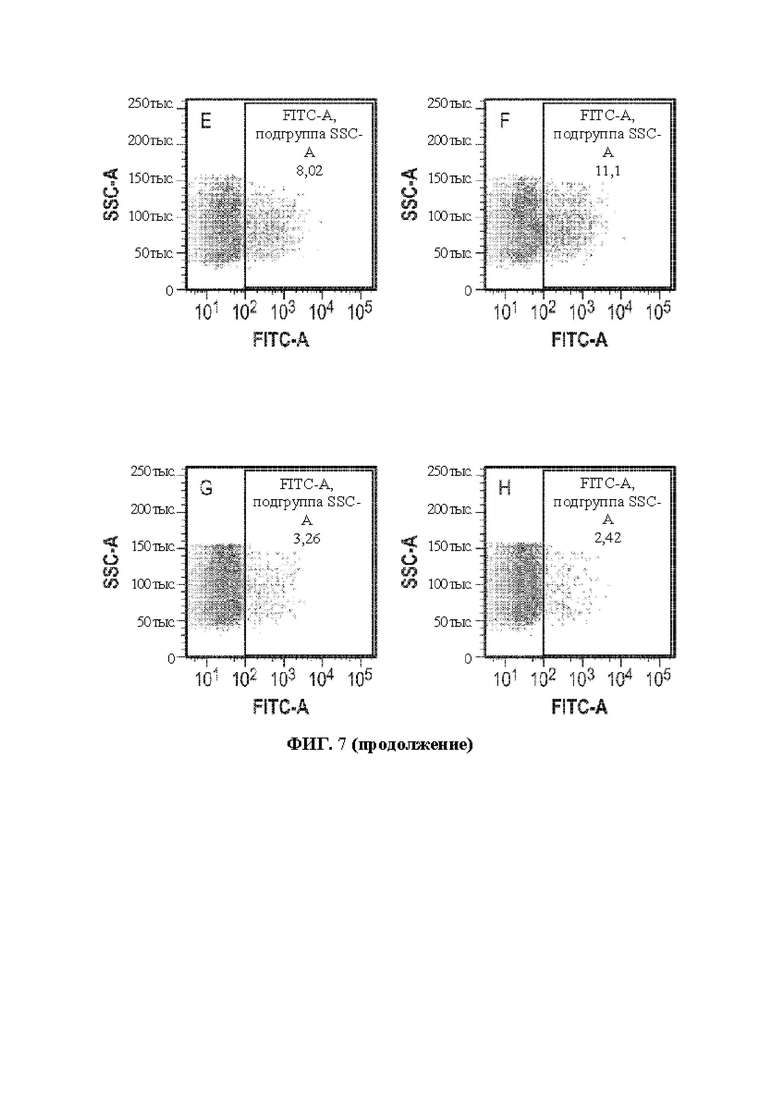





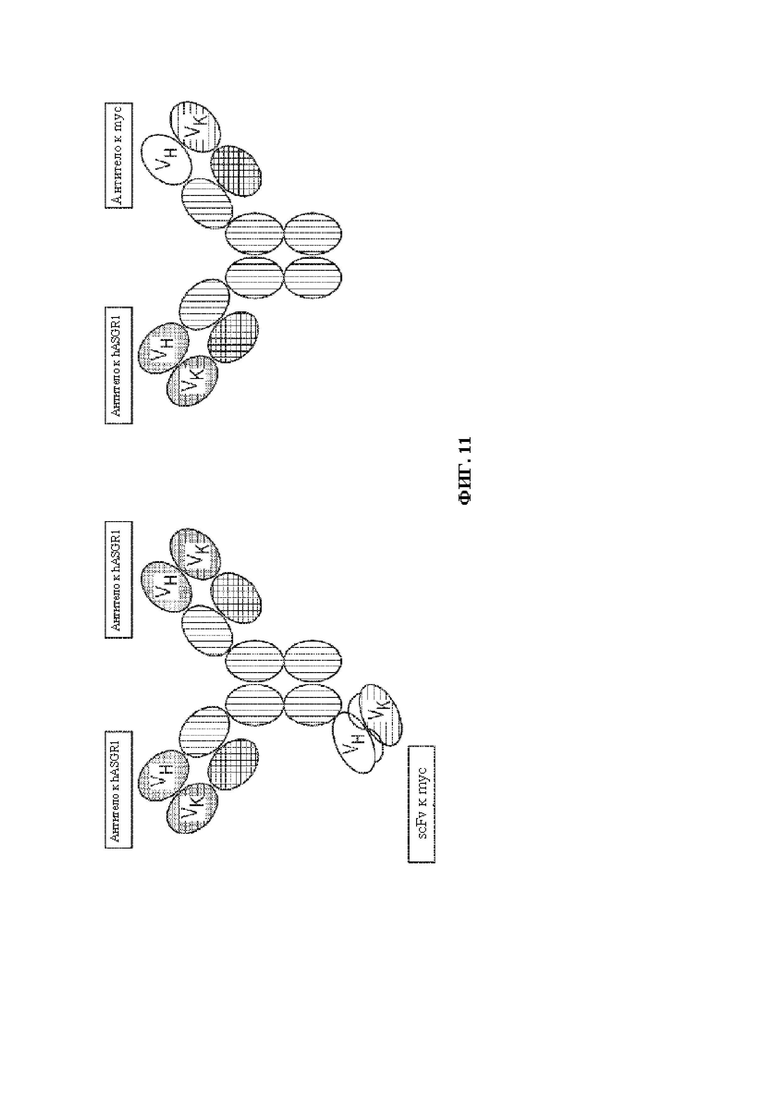
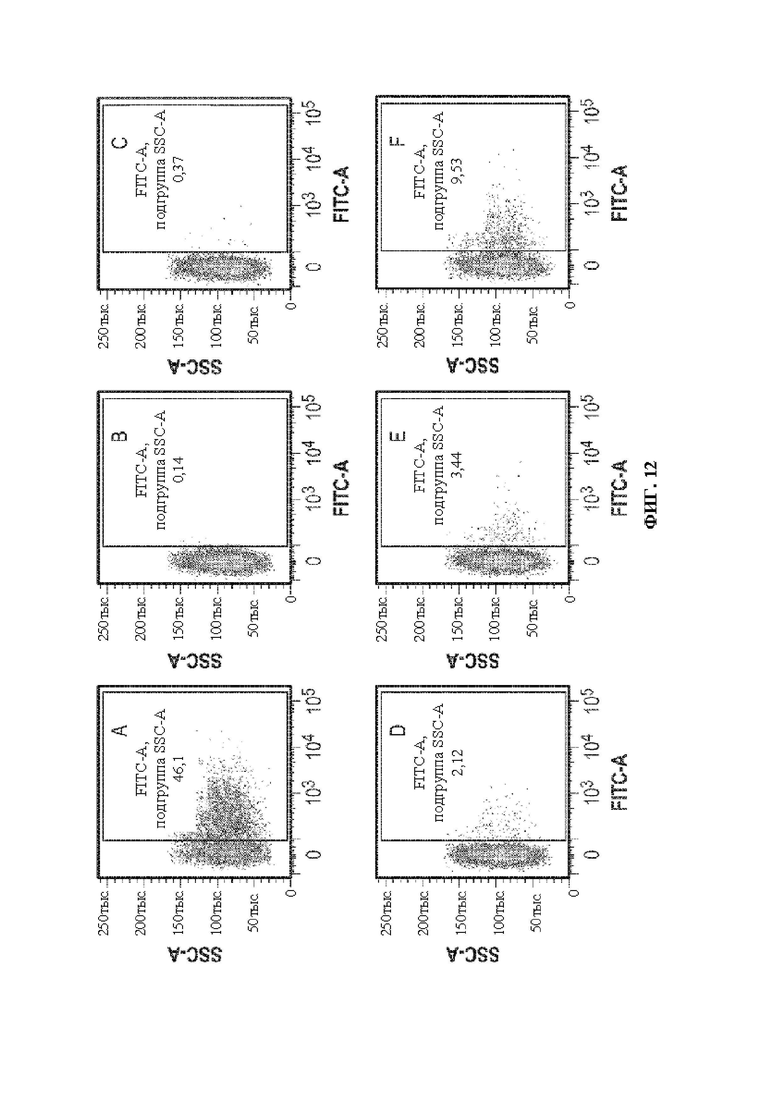
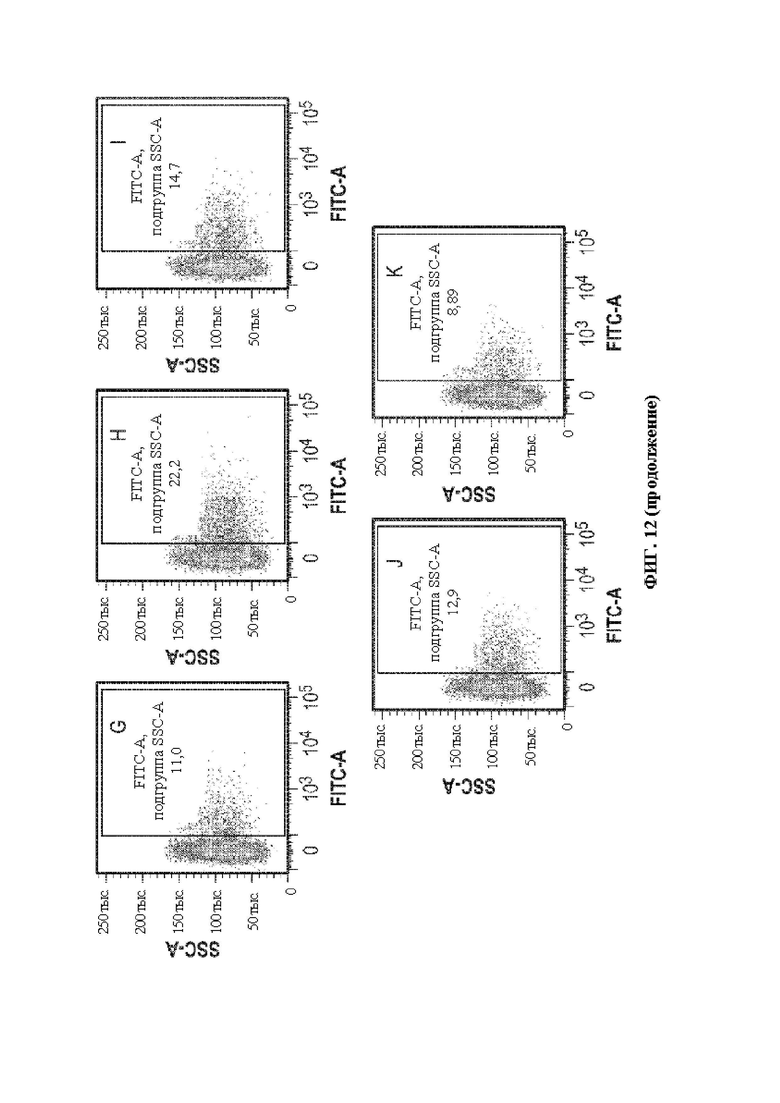


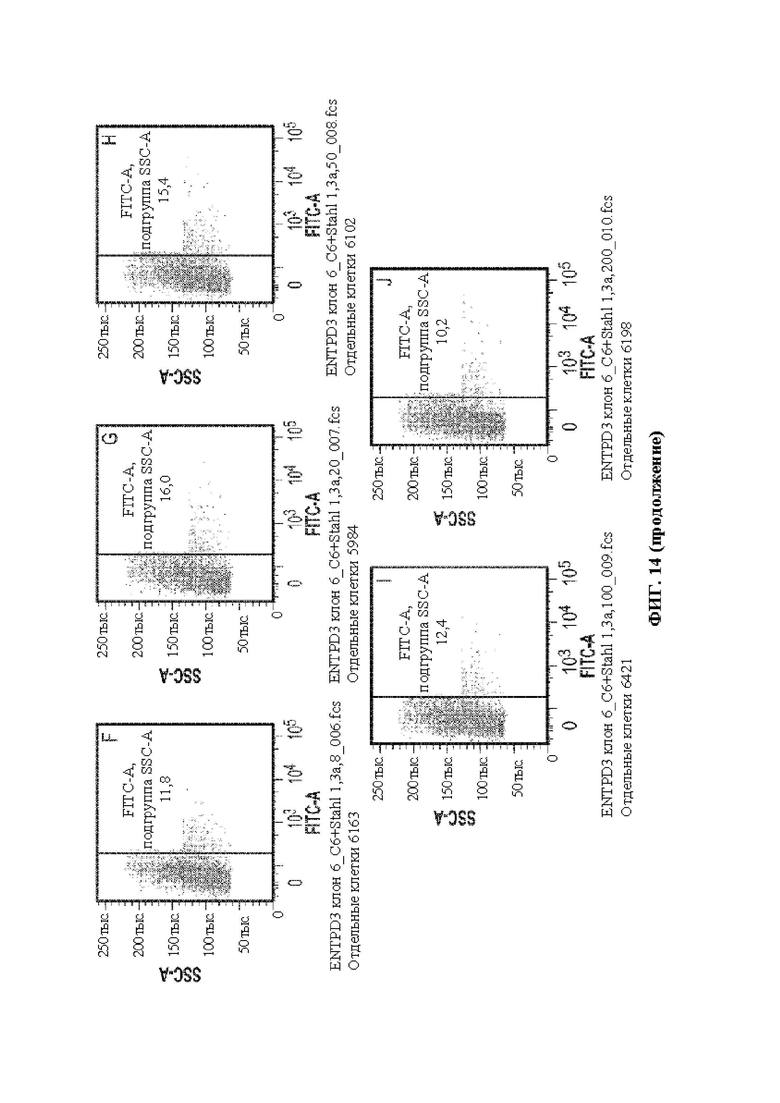



Изобретение относится к области биотехнологии, а именно к композиции, содержащей рекомбинантный аденоассоциированный-вирусный (AAV) вектор, содержащий капсид AAV, который инкапсулирует представляющий интерес нуклеотид, и полиспецифическую связывающую молекулу, содержащую паратоп антитела, который специфически связывает гетерологичный эпитоп, и перенацеливающий лиганд, который специфически связывает белок или маркер клеточной поверхности, а также к способу доставки представляющего интерес нуклеотида в клетку-мишень, экспрессирующую белок клеточной поверхности с ее использованием. Изобретение эффективно для нацеленного введения генетического материала в клетку. 2 н. и 41 з.п. ф-лы, 15 ил., 3 табл., 4 пр.
1. Композиция для нацеленного введения генетического материала в клетку, содержащая
(a) рекомбинантный аденоассоциированный-вирусный (AAV) вектор, содержащий капсид AAV, который инкапсулирует представляющий интерес нуклеотид, где капсид AAV содержит рекомбинантный капсидный белок AAV, модифицированный для включения гетерологичного эпитопа, где рекомбинантный капсидный белок AAV получен с помощью капсидного гена AAV, модифицированного для экспрессии гетерологичного эпитопа, и
(b) полиспецифическую связывающую молекулу, содержащую паратоп антитела, который специфически связывает гетерологичный эпитоп, и перенацеливающий лиганд, который специфически связывает белок или маркер клеточной поверхности, где соотношение рекомбинантного AAV вектора и полиспецифической связывающей молекулы (молекула:молекула) составляет от 1:0,5 до 1:100, и
где капсид AAV проявляет, по сравнению с капсидом AAV дикого типа того же серотипа:
сниженный или устраненный естественный тропизм в отсутствие полиспецифической связывающей молекулы, и
восстановленный и/или перенаправленный тропизм в комбинации с полиспецифической связывающей молекулой.
2. Композиция по п. 1, в которой рекомбинантный капсидный белок AAV содержит дополнительную мутацию в дополнение к гетерологичному эпитопу.
3. Композиция по п. 1 или 2, где гетерологичный эпитоп вставлен так, что вставка частично снижает тропизм капсида AAV по сравнению с капсидом AAV дикого типа того же серотипа, а рекомбинантный капсидный белок AAV включает дополнительную мутацию, необязательно, где дополнительная мутация включает замену, делецию или вставку, отличную от вставки гетерологичного эпитопа, в дополнение к вставке гетерологичного эпитопа, при этом мутация дополнительно снижает и/или устраняет тропизм рекомбинантного капсидного белка AAV по сравнению с капсидом AAV дикого типа того же серотипа.
4. Композиция по п. 2 или 3, где дополнительная мутация выбрана из группы, состоящей из R484A, R487A, R487G, K532A, K532D, R585A, R585S, R585Q, R585A или R588T.
5. Композиция по любому из предыдущих пунктов, где рекомбинантный AAV вектор и полиспецифическая связывающая молекула присутствуют в соотношении 1:4.
6. Композиция по любому из предыдущих пунктов, где паратоп антитела содержит Fv-домен.
7. Композиция по п. 6, где Fv-домен непосредственно слит с константным доменом тяжелой цепи.
8. Композиция по любому из предыдущих пунктов, где перенацеливающий лиганд содержит антитело или его антигенсвязывающую часть.
9. Композиция по любому из предыдущих пунктов, где полиспецифическая связывающая молекула содержит биспецифическое антитело, и
где каждый из паратопа антитела и перенацеливающего лиганда содержит отдельный Fv-домен, слитый с первым и вторым константным доменом тяжелой цепи.
10. Композиция по любому из пп. 1-8, где полиспецифическая связывающая молекула содержит биспецифическое антитело, и
где перенацеливающий лиганд содержит антитело с тетрамерной структурой, содержащее две идентичные тяжелые цепи иммуноглобулина и две идентичные легкие цепи, и
где паратоп антитела, который связывает гетерологичный эпитоп, присоединен к С-концу или N-концу одной или обеих тяжелых цепей.
11. Композиция по п. 10, где паратоп представляет собой scFv, где необязательно scFv содержит аминокислотную последовательность, представленную под SEQ ID NO: 37.
12. Композиция по любому из предыдущих пунктов, где
(i) гетерологичный эпитоп содержит аффинную метку и необязательно линкер и/или
(ii) рекомбинантный капсидный белок AAV содержит аминокислотную последовательность EQKLISEEDL (SEQ IDNO: 6) или ее часть, фланкированную и/или функционально связанную по меньшей мере с 5 смежными аминокислотами капсидного белка AAV,
и где паратоп антитела специфически связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6) или ее часть.
13. Композиция по любому из предыдущих пунктов, где гетерологичный эпитоп содержит аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6) или ее часть, и где паратоп антитела специфически связывает аминокислотную последовательность EQKLISEEDL (SEQ ID NO: 6) или ее часть.
14. Композиция по любому из предыдущих пунктов, где представляющий интерес нуклеотид:
(i) находится под контролем промотора, выбранного из группы, состоящей из вирусного промотора, бактериального промотора, промотора млекопитающего, птичьего промотора, промотора рыбы или промотора насекомого,
(ii) фланкирован последовательностями ITR AAV;
(iii) представляет собой репортерный ген, или
выбран из группы, состоящей из суицидного гена, нуклеотида, кодирующего антитело, или его связывающей части, нуклеотида, кодирующего систему CRISPR/Cas или ее часть(и), нуклеотида, кодирующего антисмысловую РНК, или нуклеотида, кодирующего siRNA; или
(iv) любая комбинация из (i) - (iii).
15. Композиция по любому из предыдущих пунктов, где представляющий интерес нуклеотид находится под контролем нечеловеческого промотора.
16. Композиция по п. 14 или 15, где репортерный ген кодирует зеленый флуоресцентный белок.
17. Композиция по любому из предыдущих пунктов, где AAV выбран из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9.
18. Композиция по любому из предыдущих пунктов, где гетерологичный эпитоп вставлен после и/или заменяет положение, выбранное из группы, состоящей из I-453 AAV2, I-587 AAV2, I-585 AAV6, I-590 AAV8, I-453 AAV9, I-589 AAV9 и любых соответствующих аминокислот серотипа AAV, инфицирующего приматов.
19. Композиция по любому из предыдущих пунктов, где капсид AAV представляет собой мозаичный капсид, который содержит эталонный капсидный белок AAV,
где эталонный капсидный белок AAV имеет тот же серотип, что и рекомбинантный капсидный белок AAV, и не включает гетерологичный эпитоп, и
где капсид AAV включает эталонный капсидный белок AAV и рекомбинантный капсидный белок AAV в соотношении от 1:1 до 15:1.
20. Композиция по любому из предыдущих пунктов, где рекомбинантный капсидный белок AAV получен с помощью химерного капсидного гена AAV, модифицированного для экспрессии гетерологичного эпитопа,
где химерный капсидный ген AAV включает множество последовательностей нуклеиновых кислот,
где каждое из множеств последовательностей нуклеиновых кислот кодирует часть капсидного белка другого серотипа AAV, и
где множество последовательностей нуклеиновых кислот вместе кодирует химерный капсидный белок AAV.
21. Композиция по любому из предыдущих пунктов, где белок или маркер клеточной поверхности содержит асиалогликопротеин 1 (ASGR1).
22. Композиция по любому из пп. 1-20, где белок или маркер клеточной поверхности содержит CD3.
23. Композиция по любому из пп. 1-20, где белок или маркер клеточной поверхности содержит ENTPD3.
24. Композиция по любому из пп. 1-20, где белок или маркер клеточной поверхности содержит CD71.
25. Композиция по любому из предыдущих пунктов, где белок или маркер клеточной поверхности представляет собой белок человека.
26. Композиция по любому из предыдущих пунктов, дополнительно содержащая фармацевтически приемлемый носитель.
27. Способ доставки представляющего интерес нуклеотида в клетку-мишень, экспрессирующую белок клеточной поверхности, включающий приведение в контакт клетки-мишени с композицией по любому из предыдущих пунктов, где клетка-мишень экспрессирует на своей поверхности белок или маркер клеточной поверхности.
28. Способ по п. 27, где клетка-мишень находится в условиях in vitro.
29. Способ по п. 27, где клетка-мишень находится в условиях in vivo в субъекте.
30. Способ по п. 29, где субъектом является человек.
31. Способ по любому из пп. 27-30, где клетка-мишень является клеткой человека.
32. Способ по любому из пп. 27-31, где клетка-мишень выбрана из группы, состоящей из клетки печени, клетки головного мозга, Т-клетки, клетки почки, клетки кишечника, клетки поджелудочной железы, раковой клетки, мышечной клетки, нервной клетки и клетки, инфицированной гетерологичным патогеном.
33. Способ по любому из пп. 27-32, где клетка-мишень является клеткой печени человека.
34. Способ по любому из пп. 27-33, где белок или маркер клеточной поверхности представляет собой асиалогликопротеиновый рецептор 1 человека (hASGR1).
35. Способ по любому из пп. 27-34, где клетка-мишень является Т-клеткой человека.
36. Способ по любому из пп. 27-32 и 35, где белок или маркер клеточной поверхности представляет собой CD3.
37. Способ по любому из пп. 27-32, где белок или маркер клеточной поверхности представляет собой рецептор глюкагона человека (hGCGR).
38. Способ по любому из пп. 27-32, где клетка-мишень является клеткой кишечника.
39. Способ по любому из пп. 27-32, где клетка-мишень является клеткой поджелудочной железы.
40. Способ по любому из пп. 27-32 и 38, 39, где белок или маркер клеточной поверхности представляет собой ENTPD3.
41. Способ по любому из пп. 27-32, где клетка-мишень представляет собой мышечную клетку.
42. Способ по любому из пп. 27-32, где клетка-мишень представляет собой нервную клетку.
43. Способ по любому из пп. 27-32 и 42, где белок или маркер клеточной поверхности представляет собой CD71.
| THOMAS J | |||
| WICKHAM et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| CHRISTIE L | |||
| BELL et al | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| ПЕРЕДВИЖНОЕ УСТРОЙСТВО ДЛЯ ОБСЛУЖИВАНИЯ ЛЕСНЫХ БИРЖ, СКЛАДОВ И Т. П. | 1927 |
|
SU7326A1 |
| MIRTA GRIFMAN et al | |||
| Incorporation of | |||
