ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка заявляет приоритет предварительной заявки США № 62/375314, поданной 15 августа 2016 года, которая включена в данный документ посредством ссылки во всей своей полноте.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Содержание нижеследующего представленного текстового файла ASCII включено в данный документ посредством ссылки во всей своей полноте: машиночитаемая форма (CRF) перечня последовательностей (название файла: 159792014140SEQLIST.txt, дата составления: 14 августа 2017 года, размер: 51 кбайт).
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Настоящее изобретение относится к способам серотипирования и/или определения гетерогенности вирусной частицы (например, частицы аденоассоциированого вируса (AAV)) с помощью определения массы, например, с помощью применения жидкостной хроматографии/масс-спектрометрии (LC/MS) или жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS). В некоторых аспектах настоящее изобретение относится к способам улучшения стабильности частиц AAV.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0004] Полная характеристика вирусных капсидных белков вирусных векторов (например, векторов на основе AAV), в том числе их последовательности и посттрансляционных модификаций, желательна в исследованиях и разработках, связанной с генной терапией, поскольку вирусные капсидные белки (VP) являются основными для инфекционности вирусов.
[0005] Продукты на основе вирусных векторов, такие как продукты на основе рекомбинантного аденоассоциированного вируса (rAAV) обычно идентифицируют с помощью молекулярных средств, нацеливающихся на трансген нуклеиновой кислоты. Эти способы могут включать методики полимеразной цепной реакции (ПЦР), нацеливающиеся на специфические в отношении трансгена последовательности, и полиморфмизма длины рестрикционных фрагментов (RFLP). По мере развития технологий на основе rAAV многие учреждения начинают изучать многочисленные серотипы капсида AAV, кодирующие их терапевтический трансген, в целях повышения нацеленного тропизма к ткани.
[0006] С помощью традиционных способов молекулярной идентификации определяют продукты, содержащие уникальные трансгены, но невозможно разграничить таковые, которые имеют различающиеся серотипы капсида AAV. В настоящее время большинство тестов для идентификации серотипа AAV основаны на характерах исчерченности SDS-PAGE, ELISA на основе антител или вестерн-блоттинге. Однако характеры исчерченности и антитела не являются достаточно специфическими для того, чтобы дифференцировать различные серотипы AAV. В качестве способа идентификации серотипа капсида была описана гель-LC/MS/MS. Однако этот способ включает несколько этапов, в том числе SDS-PAGE, расщепление в геле и LC/MS/MS, и, таким образом, требуется несколько дней для анализа, при этом обеспечивается ограниченный охват последовательностей. Способы идентификации векторов, таких как векторы на основе rAAV, представляют интерес в отношении векторов для генной терапии (см., например, публикацию PG США № US20110275529). Таким образом, было бы полезным улучшить способы характеристики вирусных частиц.
[0007] Все литературные источники, цитируемые в данном документе, в том числе патентные заявки и публикации, включены с помощью ссылки во всей своей полноте.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0008] С использованием rAAV в качестве примера в данном документе описано применение LC/MS в качестве аналитического средства для специфической идентификации различных серотипов капсидов вирусов (например, серотипов капсидов rAAV). В качестве части характеристики вируса можно применять LC/MS для дополнения способов молекулярной идентификации. Эта аналитическая комбинация может удовлетворять нормативным требованиям с помощью разграничения как особенностей терапевтического трансгена продукта, так и особенностей серотипа капсида. Этот способ можно применять, например, в качестве теста особенностей серотипа AAV или для контроля гетерогенности вирусных капсидных белков при разработке генной терапии на основе рекомбинантных AAV. Его также можно применять для подтверждения последовательностей VP в исследованиях по разработке капсидов. Помимо этого, эту методику можно применять для изучения влияния посттрансляционных модификаций, таких как N-концевое ацетилирование вирусных капсидных белков, в отношении эффективности трансфекции и миграции внутриклеточных белков.
[0009] Способы, описанные в данном документе, также можно применять для разработки частиц AAV в целях более высокой стабильности и/или повышенной эффективности трансдукции; например, с помощью изменения аминокислотного остатка в положении 2 VP1 и/или VP3 капсида AAV таким образом, что аминокислота в положении 2 является ацетилированной в большей степени по сравнению с капсидом AAV дикого типа. В некоторых вариантах осуществления способы можно применять для разработки частиц AAV со сниженной эффективностью трансдукции; например, с помощью изменения аминокислотного остатка в положении 2 VP1 и/или VP3 капсида AAV таким образом, что аминокислота в положении 2 является деацетилированной в большей или меньшей степени по сравнению с капсидом AAV дикого типа.
[0010] В некоторых аспектах в настоящем изобретении предусмотрен способ определения серотипа вирусной частицы, предусматривающий a) денатурирование вирусной частицы, b) подвергание денатурированной вирусной частицы жидкостной хроматографии/масс-спектрометрии (LC/MS) и c) определение масс одного или нескольких капсидных белков вирусной частицы; при этом конкретная комбинация масс одного или нескольких капсидных белков характеризует серотип вируса. В некоторых вариантах осуществления рассчитанные массы одного или нескольких капсидных белков сравнивают с теоретическими массами одного или нескольких капсидных белков одного или нескольких серотипов вируса.
[0011] В некоторых аспектах в настоящем изобретении предусмотрен способ определения гетерогенности вирусной частицы, предусматривающий a) денатурирование вирусной частицы, b) подвергание денатурированной вирусной частицы жидкостной хроматографии/масс-спектрометрии/масс-спектрометрии (LC/MS/MS), c) определение масс одного или нескольких капсидных белков вирусной частицы и d) сравнение масс из стадии c) с теоретическими массами одного или нескольких капсидных белков серотипа вируса; при этом отклонение одной или нескольких масс одного или нескольких капсидных белков характеризует гетерогенность капсидов вируса. В некоторых вариантах осуществления гетерогенность предусматривает одно или несколько из смешанных серотипов, вариантных капсидов, аминокислотных замен капсидов, усеченных капсидов или модифицированных капсидов.
[0012] В некоторых вариантах осуществления вышеуказанных аспектов жидкостная хроматография представляет собой обращенно-фазовую жидкостную хроматографию, эксклюзионную хроматографию, жидкостную хроматографию гидрофильных взаимодействий или катионообменную хроматографию. В некоторых вариантах осуществления вирусная частица содержит вирусный вектор, кодирующий гетерологичный трансген.
[0013] В некоторых аспектах в настоящем изобретении предусмотрен способ определения серотипа вирусной частицы, предусматривающий a) денатурирование вирусной частицы, b) подвергание денатурированной вирусной частицы восстановлению и/или алкилированию, c) подвергание денатурированной вирусной частицы расщеплению с образованием фрагментов одного или нескольких капсидных белков вирусной частицы, d) подвергание фрагментов одного или нескольких капсидных белков жидкостной хроматографии/масс-спектрометрии/масс-спектрометрии (LC/MS/MS), и e) определение масс фрагментов одного или нескольких капсидных белков вирусной частицы; при этом конкретная комбинация масс фрагментов одного или нескольких капсидных белков характеризует серотип вируса. В некоторых вариантах осуществления рассчитанные массы фрагментов одного или нескольких капсидных белков сравнивают с теоретическими массами фрагментов одного или нескольких капсидных белков одного или нескольких серотипов вируса.
[0014] В некоторых аспектах в настоящем изобретении предусмотрен способ определения гетерогенности серотипа вирусной частицы, предусматривающий a) денатурирование вирусной частицы, b) подвергание денатурированной вирусной частицы восстановлению и/или алкилированию, c) подвергание денатурированной вирусной частицы расщеплению с образованием фрагментов одного или нескольких капсидных белков вирусной частицы, d) подвергание фрагментов одного или нескольких капсидных белков жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS), e) определение масс фрагментов одного или нескольких капсидных белков вирусной частицы и f) сравнение масс из стадии e) с теоретическими массами фрагментов одного или нескольких капсидных белков серотипа вируса; при этом отклонение одной или нескольких масс одного или нескольких капсидных белков характеризует гетерогенность капсида вируса. В некоторых вариантах осуществления гетерогенность предусматривает одно или несколько из смешанных серотипов, вариантных капсидов, аминокислотных замен капсидов, усеченных капсидов или модифицированных капсидов. В некоторых вариантах осуществления жидкостная хроматография представляет собой обращенно-фазовую жидкостную хроматографию, эксклюзионную хроматографию, жидкостную хроматографию гидрофильных взаимодействий или катионообменную хроматографию.
[0015] Как показано в данном документе, способы можно осуществлять в отсутствие стадии разделения в геле (например, электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE)).
[0016] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления вирусная частица содержит вирусный вектор, кодирующий гетерологичный трансген. В некоторых вариантах осуществления вирусная частица принадлежит к семейству вирусов, выбранному из группы, состоящей из Adenoviridae, Parvoviridae, Retroviridae, Baculoviridae, и Herpesviridae. В некоторых вариантах осуществления вирусная частица принадлежит к роду вирусов, выбранному из группы, состоящей из Atadenovirus, Aviadenovirus, Ichtadenovirus, Mastadenovirus, Siadenovirus, Ambidensovirus, Brevidensovirus, Hepandensovirus, Iteradensovirus, Penstyldensovirus, Amdoparvovirus, Aveparvovirus, Bocaparvovirus, Copiparvovirus, Dependoparvovirus, Erythroparvovirus, Protoparvovirus, Tetraparvovirus, Alpharetrovirus, Betaretrovirus, Deltaretrovirus, Epsilonretrovirus, Gammaretrovirus, Lentivirus, Spumavirus, Alphabaculovirus, Betabaculovirus, Deltabaculovirus, Gammabaculovirus, Iltovirus, Mardivirus, Simplexvirus, Varicellovirus, Cytomegalovirus, Muromegalovirus, Proboscivirus, Roseolovirus, Lymphocryptovirus, Macavirus, Percavirus, и Rhadinovirus.
[0017] В некоторых аспектах в настоящем изобретении предусмотрен способ определения серотипа частицы аденоассоциированного вируса (AAV), предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV жидкостной хроматографии/масс-спектрометрии (LC/MS) и c) определение масс VP1, VP2 и VP3 частицы AAV; при этом конкретная комбинация масс VP1, VP2 и VP3 характеризует серотип AAV. В некоторых вариантах осуществления рассчитанные массы VP1, VP2 и VP3 сравнивают с теоретическими массами VP1, VP2 и VP3 одного или нескольких из серотипов AAV.
[0018] В некоторых аспектах в настоящем изобретении предусмотрен способ определения гетерогенности частицы AAV, предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV жидкостной хроматографии/масс-спектрометрии/масс-спектрометрии (LC/MS/MS), c) определение масс VP1, VP2 и VP3 частицы AAV и d) сравнение масс из стадии c) с теоретическими массами VP1, VP2 и VP3 частицы AAV; при этом отклонение одной или нескольких масс VP1, VP2 и VP3 характеризует гетерогенность капсида AAV. В некоторых вариантах осуществления гетерогенность предусматривает одно или несколько из смешанных серотипов, вариантных капсидов, аминокислотных замен капсидов, усеченных капсидов или модифицированных капсидов.
[0019] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления частицу AAV денатурируют уксусной кислотой, гуанидингидрохлоридом и/или органическим растворителем. В некоторых вариантах осуществления жидкостная хроматография представляет собой обращенно-фазовую жидкостную хроматографию, эксклюзионную хроматографию, жидкостную хроматографию гидрофильных взаимодействий или катионообменную хроматографию. В некоторых вариантах осуществления жидкостная хроматография представляет собой обращенно-фазовую жидкостную хроматографию. В некоторых вариантах осуществления обращенно-фазовая хроматография представляет собой хроматографию с обращенной фазой C4 или C8. В некоторых вариантах осуществления в хроматографии применяют подвижную фазу A, содержащую муравьиную кислоту в воде. В некоторых вариантах осуществления подвижная фаза A содержат приблизительно 0,1% муравьиной кислоты. В некоторых вариантах осуществления в хроматографии применяют подвижную фазу В, содержащую муравьиную кислоту в ацетонитриле. В некоторых вариантах осуществления подвижная фаза В содержат приблизительно 0,1% муравьиной кислоты. В некоторых вариантах осуществления долю подвижной фазы B в хроматографии увеличивают со временем. В некоторых вариантах осуществления долю подвижной фазы B в хроматографии увеличивают поэтапно. В некоторых вариантах осуществления подвижную фазу B увеличивают от приблизительно 10% до приблизительно 20%, от приблизительно 20% до приблизительно 30% и от приблизительно 30% до приблизительно 38%. В некоторых вариантах осуществления подвижную фазу B увеличивают от приблизительно 10% до приблизительно 20% в течение приблизительно 6 минут, от приблизительно 20% до приблизительно 30% в течение приблизительно 10 минут и от приблизительно 30% до приблизительно 38% в течение приблизительно 40 минут. В некоторых вариантах осуществления жидкостная хроматография представляет собой ультраэффективную хроматографию (UPLC).
[0020] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления масс-спектрометрия предусматривает капиллярное напряжение, составляющее приблизительно 3,5 кВ. В некоторых вариантах осуществления масс-спектрометрия предусматривает напряжение пробоотборного конуса, составляющее приблизительно 45 В. В некоторых вариантах осуществления масс-спектрометрия предусматривает калибровку, выполняемую с помощью дополнительного средства. В некоторых вариантах осуществления йодид натрия применяют в качестве калибранта.
[0021] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления N-конец VP1 и/или VP3 является ацетилированным. В некоторых вариантах осуществления частица AAV представляет собой частицу рекомбинантного AAV (rAAV). В некоторых вариантах осуществления частица AAV содержит капсид AAV1, капсид AAV2, капсид AAV3, капсид AAV4, капсид AAV5, капсид AAV6, капсид AAV7, капсид AAV8, капсид AAVrh8, капсид AAV9, капсид AAV10, капсид AAVrh10, капсид AAV11, капсид AAV12, капсид AAV LK03, капсид AAV2R471A, капсид AAV2/2-7m8, капсид AAV DJ, капсид AAV DJ8, капсид AAV2 N587A, капсид AAV2 E548A, капсид AAV2 N708A, капсид AAV V708K, капсид AAV козы, химерный капсид AAV1/AAV2, капсид AAV крупного рогатого скота или капсид AAV мыши rAAV2/HBoV1 (химерный AAV/вирус 1, относящийся к бокавирусам человека). В некоторых вариантах осуществления капсид AAV содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином. В некоторых вариантах осуществления массы VP1, VP2 и VP3 сравнивают с теоретическими массами одного или нескольких из капсида AAV1, капсида AAV2, капсида AAV3, капсида AAV4, капсида AAV5, капсида AAV6, капсида AAV7, капсида AAV8, капсида AAVrh8, капсида AAV9, капсида AAV10, капсида AAVrh10, капсида AAV11, капсида AAV12, капсида AAV LK03, капсида AAV2R471A, капсида AAV2/2-7m8, капсида AAV DJ, капсида AAV DJ8, капсида AAV2 N587A, капсида AAV2 E548A, капсида AAV2 N708A, капсида AAV V708K, капсида AAV козы, химерного капсида AAV1/AAV2, капсида AAV крупного рогатого скота или капсида AAV мыши rAAV2/HBoV1 (химерный AAV/вирус 1, относящийся к бокавирусам человека), капсида AAV2HBKO, капсида AAVPHP.B или капсида AAVPHP.eB.
[0022] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления вирусная частица содержит ITR AAV1, ITR AAV2, ITR AAV3, ITR AAV4, ITR AAV5, ITR AAV6, ITR AAV7, ITR AAV8, ITR AAVrh8, ITR AAV9, ITR AAV10, ITR AAVrh10, ITR AAV11 или ITR AAV12. В некоторых вариантах осуществления частица AAV содержит вектор на основе AAV, кодирующий гетерологичный трансген.
[0023] В некоторых аспектах в настоящем изобретении предусмотрен способ определения серотипа частицы аденоассоциированного вируса (AAV), предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV восстановлению и/или алкилированию, c) подвергание денатурированной частицы AAV расщеплению с образованием фрагментов VP1, VP2 и/или VP3 частицы AAV, d) подвергание фрагментов VP1, VP2 и/или VP3 жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS) и e) определение масс фрагментов VP1, VP2 и/или VP3 частицы AAV; при этом конкретная комбинация масс фрагментов VP1, VP2 и/или VP3 характеризует серотип AAV. В некоторых вариантах осуществления рассчитанные массы фрагментов VP1, VP2 и VP3 сравнивают с теоретическими массами фрагментов VP1, VP2 и/или VP3 одного или нескольких серотипов AAV.
[0024] В некоторых аспектах в настоящем изобретении предусмотрен способ определения гетерогенности серотипа частицы AAV, предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV восстановлению и/или алкилированию, c) подвергание денатурированной частицы AAV расщеплению с образованием фрагментов VP1, VP2 и/или VP3 частицы AAV, d) подвергание фрагментов VP1, VP2 и/или VP3 жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS), и e) определение масс фрагментов VP1, VP2 и VP3 частицы AAV; и f) сравнение масс из стадии e) с теоретическими массами фрагментов VP1, VP2 и VP3 серотипа AAV; при этом отклонение одной или нескольких масс VP1, VP2 или VP3 характеризует гетерогенность капсида AAV. В некоторых вариантах осуществления гетерогенность предусматривает одно или несколько из смешанных серотипов, вариантных капсидов, аминокислотных замен капсидов, усеченных капсидов или модифицированных капсидов. В некоторых вариантах осуществления восстановление осуществляют с помощью подвергания частицы AAV воздействию дитиотреитола, бета-меркаптоэтанола или трис(2-карбоксиэтил)фосфина (TCEP). В некоторых вариантах осуществления алкилирование осуществляют с помощью подвергания частицы AAV воздействию йодуксусной кислоты, йодацетамида или 4-винилпиридина. В некоторых вариантах осуществления расщепление представляет собой ферментативное расщепление или химическое расщепление. В некоторых вариантах осуществления ферментативное расщепление представляет собой расщепление с помощью эндопептидазы. В некоторых вариантах осуществления ферментативное расщепление представляет собой расщепление с помощью трипсина, расщепление с помощью LysC, расщепление с помощью Asp-N или расщепление с помощью Glu-C. В некоторых вариантах осуществления химическое расщепление представляет собой расщепление с помощью бромциана или кислотное расщепление. В некоторых вариантах осуществления частицу AAV денатурируют уксусной кислотой, гуанидингидрохлоридом и/или органическим растворителем.
[0025] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления жидкостная хроматография представляет собой обращенно-фазовую жидкостную хроматографию, эксклюзионную хроматографию, жидкостную хроматографию гидрофильных взаимодействий или катионообменную хроматографию. В некоторых вариантах осуществления жидкостная хроматография представляет собой обращенно-фазовую жидкостную хроматографию. В некоторых вариантах осуществления обращенно-фазовая хроматография представляет собой хроматографию с обращенной фазой C18. В некоторых вариантах осуществления в хроматографии применяют подвижную фазу A, содержащую муравьиную кислоту в воде. В некоторых вариантах осуществления подвижная фаза A содержат приблизительно 0,1% муравьиной кислоты. В некоторых вариантах осуществления в хроматографии применяют подвижную фазу В, содержащую муравьиную кислоту в ацетонитриле. В некоторых вариантах осуществления подвижная фаза В содержат приблизительно 0,1% муравьиной кислоты. В некоторых вариантах осуществления долю подвижной фазы B в хроматографии увеличивают со временем. В некоторых вариантах осуществления подвижную фазу B увеличивают от приблизительно 2% до приблизительно 60%. В некоторых вариантах осуществления подвижную фазу B увеличивают от приблизительно 2% до приблизительно 60% в течение приблизительно 121 минуты. В некоторых вариантах осуществления жидкостная хроматография представляет собой высокоэффективную жидкостную хроматографию (HPLC).
[0026] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления масс-спектрометрия предусматривает капиллярное напряжение, составляющее приблизительно 3,5 кВ. В некоторых вариантах осуществления масс-спектрометрия предусматривает напряжение пробоотборного конуса, составляющее приблизительно 45 В. В некоторых вариантах осуществления масс-спектрометрия предусматривает калибровку, выполняемую с помощью дополнительного средства. В некоторых вариантах осуществления йодид натрия применяют в качестве калибранта.
[0027] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления N-конец VP1 и/или VP3 является ацетилированным. В некоторых вариантах осуществления частица AAV представляет собой частицу рекомбинантного AAV (rAAV). В некоторых вариантах осуществления частица AAV содержит капсид AAV1, капсид AAV2, капсид AAV3, капсид AAV4, капсид AAV5, капсид AAV6, капсид AAV7, капсид AAV8, капсид AAVrh8, капсид AAV9, капсид AAV10, капсид AAVrh10, капсид AAV11, капсид AAV12, капсид AAV LK03, капсид AAV2R471A, капсид AAV2/2-7m8, капсид AAV DJ, капсид AAV DJ8, капсид AAV2 N587A, капсид AAV2 E548A, капсид AAV2 N708A, капсид AAV V708K, капсид AAV козы, химерный капсид AAV1/AAV2, капсид AAV крупного рогатого скота или капсид AAV мыши rAAV2/HBoV1 (химерный AAV/вирус 1, относящийся к бокавирусам человека). В некоторых вариантах осуществления капсид AAV содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином. В некоторых вариантах осуществления массы VP1, VP2 и VP3 сравнивают с теоретическими массами одного или нескольких из капсида AAV1, капсида AAV2, капсида AAV3, капсида AAV4, капсида AAV5, капсида AAV6, капсида AAV7, капсида AAV8, капсида AAVrh8, капсида AAV9, капсида AAV10, капсида AAVrh10, капсида AAV11, капсида AAV12, капсида AAV LK03, капсида AAV2R471A, капсида AAV2/2-7m8, капсида AAV DJ, капсида AAV DJ8, капсида AAV2 N587A, капсида AAV2 E548A, капсида AAV2 N708A, капсида AAV V708K, капсида AAV козы, химерного капсида AAV1/AAV2, капсида AAV крупного рогатого скота или капсида AAV мыши rAAV2/HBoV1 (химерный AAV/вирус 1, относящийся к бокавирусам человека).
[0028] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления вирусная частица содержит ITR AAV1, ITR AAV2, ITR AAV3, ITR AAV4, ITR AAV5, ITR AAV6, ITR AAV7, ITR AAV8, ITR AAVrh8, ITR AAV9, ITR AAV10, ITR AAVrh10, ITR AAV11 или ITR AAV12. В некоторых вариантах осуществления частица AAV содержит вектор на основе AAV, кодирующий гетерологичный трансген.
[0029] В некоторых вариантах осуществления в настоящем изобретении предусмотрена частица рекомбинантного AAV (rAAV), содержащая аминокислотную замену по аминокислотному остатку 2 VP1 и/или VP3; при этом аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления замена приводит к более высокой частоте N-концевого ацетилирования или более низкой частоте N-концевого ацетилирования. В некоторых вариантах осуществления частица rAAV содержит аминокислотную замену по аминокислотному остатку 2 VP1; при этом аминокислотная замена по аминокислотному остатку 2 VP1 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 частицы исходного AAV. В некоторых вариантах осуществления частица rAAV содержит аминокислотную замену по аминокислотному остатку 2 VP3; при этом аминокислотная замена по аминокислотному остатку 2 VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP3 частицы исходного AAV. В некоторых вариантах осуществления аминокислотный остаток 2 заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr. В некоторых вариантах осуществления аминокислотный остаток 2 заменен Ser, Asp или Glu.
[0030] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления частица AAV содержит капсид AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV DJ8, капсид серотипов AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерного AAV1/AAV2, AAV крупного рогатого скота, AAV мыши, rAAV2/HBoV1, AAV2HBKO, AAVPHP.B или AAVPHP.eB. В некоторых вариантах осуществления капсид AAV дополнительно содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином. В некоторых вариантах осуществления частица rAAV содержит вектор на основе rAAV. В некоторых вариантах осуществления вектор на основе rAAV содержит один или несколько ITR AAV. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV1, ITR AAV2, ITR AAV3, ITR AAV4, ITR AAV5, ITR AAV6, ITR AAV7, ITR AAV8, ITR AAVrh8, ITR AAV9, ITR AAV10, ITR AAVrh10, ITR AAV11 или ITR AAV12. В некоторых вариантах осуществления капсид AAV и ITR AAV получены из одного и того же серотипа. В некоторых вариантах осуществления капсид AAV и ITR AAV получены из разных серотипов. В некоторых вариантах осуществления частица AAV содержит вектор на основе AAV, кодирующий гетерологичный трансген, фланкированный одним или несколькими ITR AAV.
[0031] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления вектор на основе rAAV является самокомплементарным вектором. В некоторых вариантах осуществления вектор на основе rAAV содержит первую последовательность нуклеиновой кислоты, кодирующую трансген, и вторую последовательность нуклеиновой кислоты, кодирующую комплементарную трансгену последовательность, при этом первая последовательность нуклеиновой кислоты может образовывать внутринитевые пары оснований со второй последовательностью нуклеиновой кислоты по большей части или по всей ее длине. В некоторых вариантах осуществления первая последовательность нуклеиновой кислоты и вторая последовательность нуклеиновой кислоты соединены мутантным ITR AAV, где мутантный ITR AAV характеризуется делецией D-области и характеризуется мутацией последовательности концевого разрешения.
[0032] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления частицу rAAV получают с помощью трансфекции клетки-хозяина нуклеиновой кислотой, кодирующей вектор на основе rAAV, и нуклеиновой кислотой, кодирующей функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью трансфекции клетки-хозяина нуклеиновой кислотой, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью инфицирования клетки-хозяина вирусом-помощником AAV, который обеспечивает функциональные элементы помощника AAV. В некоторых вариантах осуществления вирус-помощник AAV представляет собой аденовирус, вирус простого герпеса или бакуловирус. В некоторых вариантах осуществления частицу rAAV получают с помощью клетки-продуцента AAV, содержащей нуклеиновую кислоту, кодирующую вектор на основе rAAV, и нуклеиновую кислоту, кодирующую функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления клетка-продуцент AAV содержит нуклеиновую кислоту, кодирующую функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью инфицирования клеток-продуцентов AAV вирусом-помощником AAV, который обеспечивает функциональные элементы помощника AAV. В некоторых вариантах осуществления вирус-помощник AAV представляет собой аденовирус, вирус простого герпеса или бакуловирус. В некоторых вариантах осуществления функциональные элементы cap AAV обеспечивают аминокислотную замену по аминокислотному остатку 2 VP1 и/или VP3; при этом аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 и/или VP3 частицы исходного AAV.
[0033] В некоторых аспектах в настоящем изобретении предусмотрена фармацевтическая композиция, содержащая частицу rAAV, описанную в данном документе. В некоторых аспектах в настоящем изобретении предусмотрен набор, содержащий частицу rAAV или фармацевтическую композицию, описанные в данном документе. В некоторых аспектах в настоящем изобретении предусмотрено изделие, содержащее частицу rAAV или фармацевтическую композицию, описанные в данном документе.
[0034] В некоторых аспектах в настоящем изобретении предусмотрен капсидный белок AAV, содержащий аминокислотную замену по аминокислотному остатку 2 капсидного белка исходного AAV; при этом аминокислотная замена по аминокислотному остатку 2 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 капсидного белка исходного AAV. В некоторых вариантах осуществления замена приводит к более высокой частоте N-концевого ацетилирования или более низкой частоте N-концевого ацетилирования. В некоторых вариантах осуществления капсидный белок AAV представляет собой VP1 или VP3. В некоторых вариантах осуществления аминокислотный остаток 2 капсидного белка AAV заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr. В некоторых вариантах осуществления аминокислотный остаток 2 капсидного белка AAV заменен Ser, Asp или Glu. В некоторых вариантах осуществления аминокислотная замена приводит к меньшей степени дезамидирования капсида AAV.
[0035] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления частица AAV содержит капсид AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV DJ8, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота, AAV мыши, rAAV2/HBoV1, капсид серотипов AAV2HBKO, AAVPHP.B или AAVPHP.eB. В некоторых вариантах осуществления капсид AAV дополнительно содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином.
[0036] В некоторых аспектах в настоящем изобретении предусмотрен способ повышения стабильности частицы rAAV, предусматривающий замену аминокислотного остатка 2 VP1 и/или VP3 исходного VP1 и/или VP3; при этом замена аминокислотного остатка 2 обеспечивает изменение N-концевого ацетилирования VP1 и/или VP3 по сравнению с аминокислотным остатком 2 исходного VP1 и/или VP3. В некоторых аспектах в настоящем изобретении предусмотрен способ улучшения сборки частиц rAAV в клетке, предусматривающий замену аминокислотного остатка 2 VP1 и/или VP3 исходного VP1 и/или VP3; при этом замена аминокислоты в положении 2 обеспечивает изменение N-концевого ацетилирования VP1 и/или VP3 по сравнению с аминокислотным остатком 2 исходного VP1 и/или VP3. В некоторых аспектах в настоящем изобретении предусмотрен способ улучшения трансдукции частиц rAAV в клетке, предусматривающий замену аминокислотного остатка 2 VP1 и/или VP3 исходного VP1 и/или VP3; при этом замена аминокислотного остатка 2 обеспечивает изменение N-концевого ацетилирования VP1 и/или VP3 по сравнению с аминокислотным остатком 2 исходного VP1 и/или VP3. В некоторых вариантах осуществления замена аминокислоты приводит к более высокой частоте N-концевого ацетилирования или более низкой частоте N-концевого ацетилирования. В некоторых вариантах осуществления аминокислотная замена является заменой по аминокислотному остатку 2 VP1. В некоторых вариантах осуществления аминокислотная замена является заменой по аминокислотному остатку 2 VP3. В некоторых вариантах осуществления аминокислотный остаток 2 заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr. В некоторых вариантах осуществления аминокислотный остаток 2 заменен Ser, Asp или Glu. В некоторых аспектах в настоящем изобретении предусмотрен способ уменьшения трансдукции частиц rAAV в клетке, предусматривающий замену аминокислотного остатка 2 VP1 и/или VP3; при этом замененная аминокислота в положении 2 обеспечивает изменение N-концевого ацетилирования VP1 и/или VP3 по сравнению с аминокислотным остатком 2 исходного VP1 и/или VP3.
[0037] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления частица AAV содержит капсид AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV DJ8, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота, AAV мыши, rAAV2/HBoV1, капсид серотипов AAV2HBKO, AAVPHP.B или AAVPHP.eB. В некоторых вариантах осуществления капсид AAV дополнительно содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином. В некоторых вариантах осуществления частица rAAV содержит вектор на основе rAAV. В некоторых вариантах осуществления вектор на основе rAAV содержит один или несколько ITR AAV. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV1, ITR AAV2, ITR AAV3, ITR AAV4, ITR AAV5, ITR AAV6, ITR AAV7, ITR AAV8, ITR AAVrh8, ITR AAV9, ITR AAV10, ITR AAVrh10, ITR AAV11 или ITR AAV12. В некоторых вариантах осуществления капсид AAV и ITR AAV получены из одного и того же серотипа. В некоторых вариантах осуществления капсид AAV и ITR AAV получены из разных серотипов. В некоторых вариантах осуществления частица AAV содержит вектор на основе AAV, кодирующий гетерологичный трансген, фланкированный одним или несколькими ITR AAV.
[0038] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления вектор на основе rAAV является самокомплементарным вектором. В некоторых вариантах осуществления вектор на основе rAAV содержит первую последовательность нуклеиновой кислоты, кодирующую трансген, и вторую последовательность нуклеиновой кислоты, кодирующую комплементарную трансгену последовательность, при этом первая последовательность нуклеиновой кислоты может образовывать внутринитевые пары оснований со второй последовательностью нуклеиновой кислоты по большей части или по всей ее длине. В некоторых вариантах осуществления первая последовательность нуклеиновой кислоты и вторая последовательность нуклеиновой кислоты соединены мутантным ITR AAV, где мутантный ITR AAV характеризуется делецией D-области и характеризуется мутацией последовательности концевого разрешения.
[0039] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления частицу rAAV получают с помощью трансфекции клетки-хозяина нуклеиновой кислотой, кодирующей вектор на основе rAAV, и нуклеиновой кислотой, кодирующей функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью трансфекции клетки-хозяина нуклеиновой кислотой, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью инфицирования клетки-хозяина вирусом-помощником AAV, который обеспечивает функциональные элементы помощника AAV. В некоторых вариантах осуществления вирус-помощник AAV представляет собой аденовирус, вирус простого герпеса или бакуловирус. В некоторых вариантах осуществления частицу rAAV получают с помощью клетки-продуцента AAV, содержащей нуклеиновую кислоту, кодирующую вектор на основе rAAV, и нуклеиновую кислоту, кодирующую функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления клетка-продуцент AAV содержит нуклеиновую кислоту, кодирующую функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью инфицирования клеток-продуцентов AAV вирусом-помощником AAV, который обеспечивает функциональные элементы помощника AAV. В некоторых вариантах осуществления вирус-помощник AAV представляет собой аденовирус, вирус простого герпеса или бакуловирус. В некоторых вариантах осуществления функциональные элементы cap AAV обеспечивают аминокислотную замену по аминокислотному остатку 2 VP1 и/или VP3; при этом аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 и/или VP3 частицы исходного AAV.
[0040] В некоторых аспектах в настоящем изобретении предусмотрена частица рекомбинантного AAV (rAAV), содержащая одну или несколько аминокислотных замен аминокислотного остатка A35, N57, G58, N382, G383, N511, G512, N715, или G716 VP1 или VP3 исходной частицы, где нумерация остатков приведена на основе VP1 AAV2; при этом одна или несколько аминокислотных замен обеспечивают изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления одна или несколько аминокислотных замен находятся по аминокислотному остатку A35, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 и обеспечивают изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления одна или несколько аминокислотных замен представляют собой замену на Asp по N57 VP1, N382 VP3, N511 VP3 или N715 VP3 и приводят к более высокой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления одна или несколько аминокислотных замен представляют собой замену в N57K или N57Q и приводят к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления одна или несколько аминокислотных замен представляют собой замену на Asp по A35 VP1 и приводят к более высокой частоте дезамидирования по сравнению с дезамидированием VP1 частицы исходного AAV. В некоторых вариантах осуществления одна или несколько аминокислотных замен находятся в G58 VP1, G383 VP3, G512 VP3 или G716 VP3 и приводят к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления G58 VP1 заменен на Asp. В некоторых вариантах осуществления частица rAAV представляет собой частицу AAV1 или частицу AAV2.
[0041] В некоторых аспектах в настоящем изобретении предусмотрены фармацевтические композиции, которые содержат частицы AAV, содержащие одну или несколько аминокислотных замен аминокислотного остатка A35, N57, G58, N382, G383, N511, G512, N715, или G716 VP1 или VP3, при этом нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых аспектах в настоящем изобретении предусмотрены наборы, содержащие частицы AAV или композиции, содержащие частицы AAV, при этом частицы AAV содержат одну или несколько аминокислотных замен аминокислотного остатка A35, N57, G58, N382, G383, N511, G512, N715, или G716 VP1 или VP3, при этом нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых аспектах в настоящем изобретении предусмотрены изделия, содержащие частицы AAV или композиции, содержащие частицы AAV, при этом частицы AAV содержат одну или несколько аминокислотных замен аминокислотного остатка A35, N57, G58, N382, G383, N511, G512, N715, или G716 VP1 или VP3, при этом нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых аспектах в настоящем изобретении предусмотрен капсидный белок AAV, содержащий аминокислотную замену капсидного белка исходного AAV; где аминокислотная замена обеспечивает изменение дезамидирования капсида по сравнению с капсидным белком исходного AAV.
[0042] В некоторых аспектах в настоящем изобретении предусмотрен способ повышения стабильности частицы rAAV, предусматривающий замену одного или нескольких аминокислотных остатков, при этом один или несколько аминокислотных остатков представляют собой остаток A35, N57, G58, N382, G383, N511, G512, N715, или G716, при этом нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых аспектах в настоящем изобретении предусмотрен способ улучшения сборки частиц rAAV в клетке, предусматривающий замену одного или нескольких аминокислотных остатков, при этом один или несколько аминокислотных остатков представляют собой остаток A35, N57, G58, N382, G383, N511, G512, N715, или G716, при этом нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых аспектах в настоящем изобретении предусмотрен способ улучшения трансдукции частиц rAAV в клетке, предусматривающий замену одного или нескольких аминокислотных остатков, при этом один или несколько аминокислотных остатков представляют собой остаток A35, N57, G58, N382, G383, N511, G512, N715, или G716, при этом нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления одна или несколько аминокислотных замен находятся в А35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3; где замена аминокислотного остатка обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления исходный остаток Ala в положении 35 VP1 заменен Asn. В некоторых вариантах осуществления исходный остаток Gly в положении 58 VP1 заменен Asp. В некоторых вариантах осуществления частица rAAV представляет собой частицу AAV1 или частицу AAV2.
[0043] В некоторых вариантах осуществления в настоящем изобретении предусмотрен способ улучшения стабильности, сборки и/или эффективности трансдукции частицы rAAV, предусматривающий замену одного или нескольких аминокислотных остатков, при этом один или несколько аминокислотных остатков представляют собой A35, N57, G58, N382, G383, N511, G512, N715 или G716, при этом нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV, описанной выше, при этом частица AAV содержит капсид AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV DJ8, капсид серотипов AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерного AAV1/AAV2, AAV крупного рогатого скота, AAV мыши или rAAV2/HBoV1. В некоторых вариантах осуществления капсид AAV дополнительно содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином. В некоторых вариантах осуществления частица rAAV содержит вектор на основе rAAV. В некоторых вариантах осуществления вектор на основе rAAV содержит один или несколько ITR AAV. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV1, ITR AAV2, ITR AAV3, ITR AAV4, ITR AAV5, ITR AAV6, ITR AAV7, ITR AAV8, ITR AAVrh8, ITR AAV9, ITR AAV10, ITR AAVrh10, ITR AAV11 или ITR AAV12. В некоторых вариантах осуществления капсид AAV и ITR AAV получены из одного и того же серотипа. В некоторых вариантах осуществления капсид AAV и ITR AAV получены из разных серотипов. В некоторых вариантах осуществления частица AAV содержит вектор на основе AAV, кодирующий гетерологичный трансген, фланкированный одним или несколькими ITR AAV.
[0044] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления вектор на основе rAAV является самокомплементарным вектором. В некоторых вариантах осуществления вектор на основе rAAV содержит первую последовательность нуклеиновой кислоты, кодирующую трансген, и вторую последовательность нуклеиновой кислоты, кодирующую комплементарную трансгену последовательность, при этом первая последовательность нуклеиновой кислоты может образовывать внутринитевые пары оснований со второй последовательностью нуклеиновой кислоты по большей части или по всей ее длине. В некоторых вариантах осуществления первая последовательность нуклеиновой кислоты и вторая последовательность нуклеиновой кислоты соединены мутантным ITR AAV, где мутантный ITR AAV характеризуется делецией D-области и характеризуется мутацией последовательности концевого разрешения.
[0045] В некоторых вариантах осуществления вышеуказанных аспектов и вариантов осуществления частицу rAAV получают с помощью трансфекции клетки-хозяина нуклеиновой кислотой, кодирующей вектор на основе rAAV, и нуклеиновой кислотой, кодирующей функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью трансфекции клетки-хозяина нуклеиновой кислотой, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью инфицирования клетки-хозяина вирусом-помощником AAV, который обеспечивает функциональные элементы помощника AAV. В некоторых вариантах осуществления вирус-помощник AAV представляет собой аденовирус, вирус простого герпеса или бакуловирус. В некоторых вариантах осуществления частицу rAAV получают с помощью клетки-продуцента AAV, содержащей нуклеиновую кислоту, кодирующую вектор на основе rAAV, и нуклеиновую кислоту, кодирующую функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV. В некоторых вариантах осуществления клетка-продуцент AAV содержит нуклеиновую кислоту, кодирующую функциональные элементы помощника AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV получают с помощью инфицирования клеток-продуцентов AAV вирусом-помощником AAV, который обеспечивает функциональные элементы помощника AAV. В некоторых вариантах осуществления вирус-помощник AAV представляет собой аденовирус, вирус простого герпеса или бакуловирус. В некоторых вариантах осуществления функциональные элементы cap AAV обеспечивают аминокислотную замену VP1 и/или VP3, при этом аминокислотная замена обеспечивала модулирование дезамидирования капсида по сравнению с частицей исходного AAV.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0046] На фиг. 1A-D представлены общие ионные хроматограммы VP AAV2, полученные с помощью LC/MS. Фиг. 1A: колонка BEH C4 длиной 10 см с градиентом 1,7%/мин.; фиг. 1B: колонка BEH C4 длиной 10 см с градиентом 0,5%/мин.; фиг. 1C: колонка BEH C4 длиной 15 см с градиентом 0,5%/мин.; фиг. 1D: колонка BEH C8 длиной 15 см с градиентом 0,5%/мин.
[0047] На фиг. 2A и В представлены спектры масс после деконволюции на основе пика 1 фиг. 1D (фиг. 2A) и пика 2 фиг. 1D (фиг. 2B).
[0048] На фиг. 3 представлен охват последовательности VP1 AAV2 (SEQ ID NO:3): зеленый - триптические пептиды, синий - Lys-C-пептиды, розовый - Asp-N-пептиды.
[0049] На фиг. 4A-4C представлены спектры MS/MS N-концевых пептидов VP AAV2. Фиг. 4A: N-концевой триптический пептид VP1 A(Ac)ADGYLPDWLEDTLSEGIR (SEQ ID NO: 4), фиг. 4B: N-концевой Asp-N-пептид VP2 APGKKRPVEHSPVEP (SEQ ID NO: 15). Фиг. 4C: полученный из N-концевого Asp-N-пептида VP-3 A(Ac)TGSGAPM (SEQ ID NO: 5).
[0050] На фиг. 5 представлено выравнивание последовательностей 13 серотипов AAV черными буквами на белом фоне - несходные; синими буквами на синем фоне - консервативные; черными буквами на зеленом фоне - блок сходных; красными буквами на желтом фоне - идентичные; зелеными буквами на белом фоне - в низкой степени сходные. AAVRh10 (SEQ ID NO: 17); AAV10 (SEQ ID NO: 18); AAV8 (SEQ ID NO: 19); AAV7 (SEQ ID NO: 20); AAV1 (SEQ ID NO: 21); AAV6 (SEQ ID NO: 22); AAV2 (SEQ ID NO: 23); AAV3 (SEQ ID NO: 24); AAV11 (SEQ ID NO: 25); AAV12 (SEQ ID NO: 26); AAV4 (SEQ ID NO: 27); AAV5 (SEQ ID NO: 28); AAV9 (SEQ ID NO: 29); консенсус (SEQ ID NO: 30).
[0051] На фиг. 6A и 6B показаны результаты анализа LC/MS/MS со сравнением процентной доли дезамидирования частиц AAV1 и AAV2, полученных с применением способов TTx и PCL. Пептид T9 YLGPFNGLDK (SEQ ID NO: 9) применяли для контроля потенциального участка дезамидирования N57 как в AAV1, так и в AAV2.
[0052] На фиг. 7A и 7B показаны результаты анализа LC/MS/MS со сравнением процентной доли дезамидирования частиц AAV1 и AAV2, полученных с применением способов TTx и PCL. Пептиды T49 YNLNGR (SEQ ID NO: 11) и YHLNGR (SEQ ID NO: 12) применяли для контроля потенциального участка дезамидирования N511 в AAV1 и AAV2 соответственно.
[0053] На фиг. 8A и 8B показаны результаты анализа LC/MS/MS со сравнением процентной доли дезамидирования частиц AAV1 и AAV2, полученных с применением способов TTx и PCL. Пептиды T67 SANVDFTVDNNGLYTEPR (SEQ ID NO: 13) и SVNVDFTVDTNGVYSEPR (SEQ ID NO: 14) применяли для контроля потенциального участка дезамидирования N715 в AAV1 и AAV2 соответственно.
[0054] На фиг. 9 показаны результаты анализа в белковом геле SYPRO в отношении получения и соотношения VP1:VP2:VP3 деацетилированных мутантных вариантов AAV5.
[0055] На фиг. 10 показан анализ трансдукции in vitro в целях исследования эффективности трансдукции деацетилированных вариантов AAV5.
[0056] На фиг. 11 показана эффективность вхождения в клетку указанных деацетилированных вариантов AAV5 или исходного немодифицированного AAV5, измеренная с помощью числа копий векторного генома/мкг белка. Применяли три клеточные линии: 293, HeLa и HuH7.
[0057] На фиг. 12 показана экспрессия eGFP (измеренная с помощью ELISA) клетками, трансдуцированными указанными деацетилированными вариантами AAV5, по сравнению с трансдукцией исходным немодифицированным AAV5. Применяли три клеточные линии: 293, HeLa и HuH7.
[0058] На фиг. 13 представлено выравнивание последовательностей 13 серотипов AAV, при этом выделен консервативный участок дезамидирования N57G58 и остаток A35 в AAV2. AAVRh10 (SEQ ID NO: 31); AAV10 (SEQ ID NO: 31); AAV8 (SEQ ID NO: 32); AAV7 (SEQ ID NO: 33); AAV1 (SEQ ID NO: 31); AAV6 (SEQ ID NO: 31); AAV2 (SEQ ID NO: 34); AAV3 (SEQ ID NO: 35); AAV11 (SEQ ID NO: 31); AAV12 (SEQ ID NO: 36); AAV4 (SEQ ID NO: 37); AAV5 (SEQ ID NO: 38); AAV9 (SEQ ID NO: 39); консенсус (SEQ ID NO: 40).
[0059] На фиг. 14 показаны капсидные белки VP1, VP2 и VP3 из частиц AAV1 или AAV2, полученных с помощью способа PCL или TTx, в белковом геле. *выделен усеченный белок VP1 (tVP1).
[0060] На фиг. 15 показаны результаты анализа LC/MS дезамидирования указанных мутантов AAV2 по сравнению с контрольными капсидами AAV2.
[0061] На фиг. 16 показаны результаты анализа в белковом геле SYPRO в отношении получения и соотношения VP1:VP2:VP3 мутантных по дезамидированию вариантов AAV2.
[0062] На фиг. 17 показан анализ трансдукции in vitro в целях исследования эффективности трансдукции дезамидированных вариантов AAV2.
[0063] На фиг. 18 показана эффективность вхождения в клетку указанных дезамидированных вариантов AAV2 или исходного немодифицированного AAV2, измеренная с помощью числа копий векторного генома/мкг белка. Применяли три клеточные линии: 293, HeLa и HuH7.
[0064] На фиг. 19 показана экспрессия eGFP (измеренная с помощью ELISA) клетками, трансдуцированными указанными дезамидированными вариантами AAV2, по сравнению с трансдукцией исходным немодифицированным AAV2. Применяли три клеточные линии: 293, HeLa и HuH7.
ПОДРОБНОЕ ОПИСАНИЕ
[0065] В некоторых аспектах в настоящем изобретении предусмотрен способ определения серотипа частицы (частиц) аденоассоциированного вируса (AAV), предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV жидкостной хроматографии/масс-спектрометрии (LC/MS) и c) определение масс VP1, VP2 и VP3 частицы AAV; при этом конкретная комбинация масс VP1, VP2 и VP3 характеризует серотип AAV.
[0066] В других аспектах в настоящем изобретении предусмотрен способ определения гетерогенности частицы AAV, предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV жидкостной хроматографии/масс-спектрометрии (LC/MS), а также c) определение масс VP1, VP2 и VP3 частицы AAV и сравнение масс из стадии c) с теоретическими массами VP1, VP2 и VP3 частицы AAV; при этом отклонение одной или нескольких масс VP1, VP2 или VP3 характеризует гетерогенность капсида AAV.
[0067] В других аспектах в настоящем изобретении предусмотрен способ определения серотипа частицы аденоассоциированного вируса (AAV), предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV восстановлению и/или алкилированию, c) подвергание денатурированной частицы AAV расщеплению с образованием фрагментов VP1, VP2 и/или VP3 частицы AAV, d) подвергание фрагментов VP1, VP2 и/или VP3 жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS) и e) определение масс фрагментов VP1, VP2 и/или VP3 частицы AAV; при этом конкретная комбинация масс фрагментов VP1, VP2 и/или VP3 характеризует серотип AAV.
[0068] В других аспектах в настоящем изобретении предусмотрен способ определения гетерогенности серотипа частицы AAV, предусматривающий a) денатурирование частицы AAV, b) подвергание денатурированной частицы AAV восстановлению и/или алкилированию, c) подвергание денатурированной частицы AAV расщеплению с образованием фрагментов VP1, VP2 и/или VP3 частицы AAV, d) подвергание фрагментов VP1, VP2 и/или VP3 жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS), e) определение масс фрагментов VP1, VP2 и VP3 частицы AAV и f) сравнение масс из стадии e) с теоретическими массами фрагментов VP1, VP2 и VP3 серотипа AAV; при этом отклонение одной или нескольких масс VP1, VP2 или VP3 характеризует гетерогенность капсида AAV.
[0069] В некоторых аспектах в настоящем изобретении предусмотрена частица рекомбинантного AAV (rAAV), содержащая аминокислотную замену по аминокислотному остатку 2 VP1 и/или VP3; при этом аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 и/или VP3 частицы исходного AAV.
[0070] В некоторых аспектах в настоящем изобретении предусмотрен способ улучшения сборки частиц rAAV в клетке, предусматривающий замену аминокислотного остатка 2 VP1 и/или VP3; при этом замененная аминокислота в положении 2 является N-ацетилированной с более высокой частотой, чем аминокислотный остаток 2 исходного VP1 и/или VP3. В некоторых аспектах в настоящем изобретении предусмотрен способ улучшения трансдукции частиц rAAV в клетке, предусматривающий замену аминокислотного остатка 2 VP1 и/или VP3; при этом замененная аминокислота в положении 2 является N-ацетилированной с более высокой частотой, чем аминокислотный остаток 2 исходного VP1 и/или VP3.
I. Общие методики
[0071] Методики и процедуры, описанные или упоминаемые в данном документе, обычно широко распространены и часто используются специалистами в данной области техники с использованием традиционной методологии, как например широко используемые методики, описанные в Molecular Cloning: A Laboratory Manual (Sambrook et al., 4th ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 2012); Current Protocols in Molecular Biology (F.M. Ausubel, et al. eds., 2003); серии Methods in Enzymology (Academic Press, Inc.); PCR 2: A Practical Approach (M.J. MacPherson, B.D. Hames and G.R. Taylor eds., 1995); Antibodies, A Laboratory Manual (Harlow and Lane, eds., 1988); Culture of Animal Cells: A Manual of Basic Technique and Specialized Applications (R.I. Freshney, 6th ed., J. Wiley and Sons, 2010); Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., Academic Press, 1998); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, Plenum Press, 1998); Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., J. Wiley and Sons, 1993-8); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds., 1996); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Ausubel et al., eds., J. Wiley and Sons, 2002); Immunobiology (C.A. Janeway et al., 2004); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane, Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995) и Cancer: Principles and Practice of Oncology (V.T. DeVita et al., eds., J.B. Lippincott Company, 2011).
II. Определения
[0072] Выражение "вектор", используемое в данном документе, относится к рекомбинантной плазмиде или вирусу, которые содержат нуклеиновую кислоту, которую необходимо доставить в клетку-хозяина либо in vitro, либо in vivo.
[0073] Термины "полинуклеотид" или "нуклеиновая кислота", используемые в данном документе, относятся к полимерной форме нуклеотидов любой длины, либо рибонуклеотидов, либо дезоксирибонуклеотидов. Таким образом, данный термин включает без ограничения одно-, двух- или многонитевые ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК или полимер, содержащий пуриновые и пиримидиновые основания или другие природные, химически или биохимически модифицированные, отличные от природных, или дериватизированные нуклеотидные основания. Остов нуклеиновой кислоты может содержать сахара и фосфатные группы (которые обычно могут обнаруживаться в РНК или ДНК) или модифицированные либо замещенные сахарные или фосфатные группы. В качестве альтернативы остов нуклеиновой кислоты может включать полимер из синтетических субъединиц, таких как фосфорамидаты и, таким образом, может представлять собой олигодезоксинуклеозидный фосфорамидат (P-NH2) или смешанный олигомер фосфорамидата-сложного фосфодиэфира. Кроме того, двухнитевая нуклеиновая кислота может быть получена из однонитевого полинуклеотидного продукта химического синтеза либо путем синтеза комплементарной нити и гибридизации нитей в соответствующих условиях, либо путем синтеза комплементарной нити de novo с использованием ДНК-полимеразы с соответствующим праймером.
[0074] Термины "полипептид" и "белок" используются взаимозаменяемо для обозначения полимера из аминокислотных остатков и не ограничены минимальной длиной. Такие полимеры из аминокислотных остатков могут содержать природные или отличные от природных аминокислотные остатки, и включают без ограничения пептиды, олигопептиды, димеры, тримеры и мультимеры из аминокислотных остатков. Данным определением охватываются как полноразмерные белки, так и их фрагменты. Термины включают также посттрансляционные модификации полипептида, например, гликозилирование, сиалирование, ацетилирование, фосфорилирование и т.п. Кроме того, применительно к целям настоящего изобретения выражение "полипептид" относится к белку, нативная последовательность которого содержит модификации, такие как делеции, добавления и замены (обычно консервативные по своей природе), при условии, что белок сохраняет требуемую активность. Данные модификации могут быть преднамеренными, как например полученными с помощью сайт-направленного мутагенеза, или могут быть случайными, как например возникшими вследствие мутаций у хозяев, которые продуцируют белки, или ошибок, обусловленных ПЦР-амплификацией.
[0075] Выражение "рекомбинантный вирусный вектор" относится к рекомбинантному полинуклеотидному вектору, содержащему одну или несколько гетерологичных последовательностей (т. е. последовательность нуклеиновой кислоты, не происходящая от вируса). В случае рекомбинантных векторов на основе AAV рекомбинантную нуклеиновую кислоту фланкируют с помощью по меньшей мере одного, например, двух, последовательностей инвертированных концевых повторов (ITR).
[0076] Выражение "рекомбинантный вектор на основе AAV (вектор на основе rAAV)" относится к полинуклеотидному вектору, содержащему одну или несколько гетерологичных последовательностей (т.е. последовательность нуклеиновой кислоты, не происходящую из AAV), которые фланкированы по меньшей мере одной, например, двумя последовательностями инвертированных концевых повторов (ITR). Такие векторы на основе rAAV могут реплицироваться и упаковываться в инфекционные вирусные частицы, когда они находятся в клетке-хозяине, которая была инфицирована подходящим вирусом-помощником (или которая экспрессирует подходящие хелперные функциональные элементы) и которая экспрессирует продукты генов rep и cap AAV (т.е. белки Rep и Cap AAV). Если вектор на основе rAAV встроен в более крупный полинуклеотид (например, в хромосому или в другой вектор, такой как плазмида, применяемая для клонирования или трансфекции), то вектор на основе rAAV можно называть "провектором", который может быть "спасен" с помощью репликации и заключения в капсид в присутствии упаковывающих функциональных элементов и подходящих функциональных элементов помощника AAV. Вектор на основе rAAV может находиться в любой из множества форм, в том числе без ограничения в форме плазмид, линейных искусственных хромосом, образующих комплексы с липидами, инкапсулированными в липосомах и, в вариантах осуществления, инкапсулированными в вирусной частице, в частности, частице AAV. Вектор на основе rAAV может быть упакован в капсид вируса AAV с образованием "частицы рекомбинантного аденоассоциированного вируса (частица rAAV)".
[0077] "Вирус rAAV" или "вирусная частица rAAV" относится к вирусной частице, состоящей по меньшей мере из одного капсидного белка AAV и инкапсидированного генома вектора на основе rAAV.
[0078] "Частица исходного AAV" и "капсидный белок исходного AAV", используемые в данном документе в контексте сравнения N-ацетилирования и/или дезамидирования, относится к частице или капсидному белку AAV, в которые вводят аминокислотные модификации для модуляции N-ацетилирования и/или дезамидирования (например, частице/капсидному белку AAV, которые являются аналогичными частице/капсиду AAV по настоящему изобретению или сходными с ними, но не содержат мутаций, которые обеспечивают модуляцию/изменение N-ацетилирования и/или дезамидирования, как описано в данном документе). В некоторых вариантах осуществления частица исходного AAV представляет собой частицу рекомбинантного AAV, содержащую геном рекомбинантного AAV. В некоторых вариантах осуществления капсидная частица исходного AAV или капсидный белок исходного AAV содержит аминокислотные замены, которые влияют в отношении других аспектов частицы AAV. Например, частица исходного AAV может содержать аминокислотные замены, которые оказывают влияние в отношении связывания AAV со своим рецептором, например, оказывают влияние в отношении связывания AAV2 с гепаринсульфатпротеогликаном (например, частицей AAV2 HBKO). Частицу AAV2 HBKO можно подвергнуть мутированию с введением аминокислотных замен, которые обеспечивают модуляцию N-ацетилирования и/или дезамидирования. Такую мутированную частицу AAV затем можно сравнить с частицей исходного AAV2 HBKO в аспектах настоящего изобретения, описанных в данном документе. Капсидный белок исходного AAV может включать исходный капсидный белок VP1, исходный капсидный белок VP2 или исходный капсидный белок VP3.
[0079] Используемый в данном документе термин "модулировать" или "изменять" в отношении к исходной молекуле означает изменять свойство исходной молекулы. Например, частица AAV с измененным N-ацетилированием может характеризоваться повышенной или сниженной степенью N-ацетилирования по сравнению с частицей исходного AAV, а частица AAV с измененным дезамидированием может характеризоваться повышенной или сниженной степенью дезамидирования по сравнению с частицей исходного AAV.
[0080] "Гетерологичный" означает полученный из объекта, генотипически отличающегося от остальной части объекта, с которым его сравнивают или в который его вводят или встраивают. Например, нуклеиновая кислота, введенная с помощью методик генетической инженерии в клетки другого типа, является гетерологичной нуклеиновой кислотой (и при экспрессии может кодировать гетерологичный полипептид). Аналогично, клеточная последовательность (например, ген или его часть), встроенная в вирусный вектор, является гетерологичной нуклеотидной последовательностью по отношению к вектору.
[0081] Термин "трансген" относится к нуклеиновой кислоте, вводимой в клетку и способной к транскрипции в РНК и необязательно к трансляции и/или экспрессии в соответствующих условиях. В некоторых аспектах он придает требуемое свойство клетке, в которую он был введен, или иным образом приводит к требуемому терапевтическому или диагностическому эффекту. В другом аспекте он может транскрибироваться в молекулу, которая опосредует РНК-интерференцию, такую как siRNA.
[0082] Термины "геномные частицы (gp)", "геномные эквиваленты" или "копии генома", используемые в отношении вирусного титра, относятся к числу вирионов, содержащих ДНК-геном рекомбинантного AAV, вне зависимости от инфекционности или функциональности. Число геномных частиц в конкретном препарате на основе векторов можно измерять с помощью процедур, таких как описанные в примерах в данном документе или, например, в Clark et al. (1999) Hum. Gene Ther., 10:1031-1039; Veldwijk et al. (2002) Mol. Ther., 6:272-278.
[0083] Термины "инфекционная единица (iu)", "инфекционная частица" или "единица репликации", используемые в отношении вирусного титра, относятся к числу инфекционных и репликационно компетентных частиц, представляющих собой вектор на основе рекомбинантного AAV, как измерено с помощью анализа инфекционных центров, также известного как анализ центров репликации, описанного например в McLaughlin et al. (1988) J. Virol., 62:1963-1973.
[0084] Термин "трансдуцирующая единица (tu)", используемый в отношении вирусного титра, относится к числу инфекционных частиц, представляющих собой вектор на основе рекомбинантного AAV, которые приводят к получению функционального трансгенного продукта, измеряемому в функциональных анализах, таких как описанные в примерах в данном документе или, например, в Xiao et al. (1997) Exp. Neurobiol., 144:113-124; или в Fisher et al. (1996) J. Virol., 70:520-532 (анализ LFU).
[0085] Последовательность "инвертированных концевых повторов" или "ITR" является термином, широко распространенным из уровня техники, и относится к относительно коротким последовательностям, встречающимся на концах вирусных геномов, которые имеют противоположную ориентацию.
[0086] Последовательность "инвертированного концевого повтора (ITR) AAV", термин, хорошо известный из уровня техники, обозначает последовательность из примерно 145 нуклеотидов, которая присутствует на обоих концах нативного однонитевого генома AAV. Крайние 125 нуклеотидов ITR могут присутствовать в одной из двух альтернативных ориентаций, что обуславливает гетерогенность между различными геномами AAV и между двумя концами одного генома AAV. Крайние 125 нуклеотидов также содержат несколько более коротких областей самокомплементарности (обозначаемых как A-, A'-, B-, B'-, C-, C'- и D-области), которые обеспечивают возможность образования внутринитевого спаривания оснований в пределах данной части ITR.
[0087] "Последовательность концевого разрешения" или "trs" представляет собой последовательность в D-области ITR AAV, которая отщепляется белками rep AAV в ходе репликации вирусной ДНК. Мутантная последовательность концевого разрешения устойчива к отщеплению белками rep AAV. "Функциональными элементами помощника AAV" называют функциональные элементы, которые обеспечивают возможность репликации и упаковки AAV в клетке-хозяине. Функциональные элементы помощника AAV могут быть представлены любой из множества форм, в том числе без ограничения вирусом-помощником или генами вируса-помощника, которые способствуют репликации и упаковке AAV. Из уровня техники известны другие функциональные элементы помощника AAV, такие как генотоксичные средства.
[0088] "Функциональными элементами помощника AAV" называют функциональные элементы, которые обеспечивают возможность репликации и упаковки AAV в клетке-хозяине. Функциональные элементы помощника AAV могут быть представлены любой из множества форм, в том числе без ограничения вирусом-помощником или генами вируса-помощника, которые способствуют репликации и упаковке AAV. Из уровня техники известны другие функциональные элементы помощника AAV, такие как генотоксичные средства.
[0089] Выражение "вирус-помощник" для AAV относится к вирусу, который обеспечивает возможность репликации и упаковки AAV (который является дефектным парвовирусом) в клетке-хозяине. Было идентифицировано множество таких вирусов-помощников, в том числе аденовирусы, герпесвирусы, поксвирусы, такие как вирус осповакцины и бакуловирус. Аденовирусы охватывают множество различных подгрупп, однако наиболее широко применяется аденовирус 5 типа подгруппы C (Ad5). Многочисленные аденовирусы, поражающие человека, млекопитающих, отличных от человека, и птиц, известны и доступны из депозитариев, таких как ATCC. Вирусы семейства герпесвирусов, которые также доступны из депозитариев, таких как ATCC, включают, например, вирусы простого герпеса (HSV), вирусы Эпштейна-Барр (EBV), цитомегаловирусы (CMV) и вирусы псевдобешенства (PRV). Бакуловирусы, доступные из депозитариев, включают вирус ядерного полиэдроза Autographa californica.
[0090] Выражение "процентная (%) идентичность последовательностей" относительно эталонной полипептидной последовательности или последовательности нуклеиновой кислоты определяется как процентная доля аминокислотных остатков или нуклеотидов в последовательности-кандидате, которые идентичны аминокислотным остаткам или нуклеотидам в эталонной полипептидной последовательности или последовательности нуклеиновой кислоты после выравнивания последовательностей и, при необходимости, введения гэпов для достижения максимальной процентной идентичности последовательностей, и не учитывая любые консервативные замены как часть идентичности последовательностей. Выравнивание для целей определения процентной идентичности аминокислотных последовательностей или последовательностей нуклеиновых кислот может быть достигнуто различными способами, которые находятся в пределах компетенции специалиста в данной области техники, например с помощью общедоступных компьютерных программ, например описанных в Current Protocols in Molecular Biology (Ausubel et al., eds., 1987), прилож. 30, раздел 7.7.18, таблица 7.7.1, и в том числе программного обеспечения BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Потенциальной программой выравнивания является ALIGN Plus (Scientific and Educational Software, Пенсильвания). Специалисты в данной области техники способны определять соответствующие параметры для оценки выравнивания, в том числе любые алгоритмы, необходимые для достижения максимального выравнивания по полной длине сравниваемых последовательностей. Для целей данного документа % идентичность аминокислотной последовательности, свойственная данной аминокислотной последовательности А, в отношении данной аминокислотной последовательности В, с ней или в сравнении с ней (что в качестве альтернативы можно перефразировать как данная аминокислотная последовательность A, которая характеризуется или обладает определенной % идентичностью аминокислотной последовательности в отношении данной аминокислотной последовательности В, с ней или в сравнении с ней) рассчитывается следующим образом: 100 умножить на частное от X/Y, где X представляет собой число аминокислотных остатков, учитываемых в качестве идентичных совпадений программой для выравнивания последовательностей в таком программном выравнивании A и B, и где Y представляет собой общее число аминокислотных остатков в B. Следует принимать во внимание, что если длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, то % идентичность аминокислотной последовательности A в отношении B не будет равна % идентичности аминокислотной последовательности B в отношении A. Для целей данного документа % идентичность последовательности нуклеиновой кислоты, свойственная данной последовательности нуклеиновой кислоты C, в отношении данной последовательности нуклеиновой кислоты D, с ней или в сравнении с ней (что в качестве альтернативы можно перефразировать как данная последовательность нуклеиновой кислоты C, которая характеризуется или обладает определенной % идентичностью последовательности нуклеиновой кислоты в отношении данной последовательности нуклеиновой кислоты D, с ней или в сравнении с ней) рассчитывается следующим образом: 100 умножить на частное от W/Z, где W представляет собой число нуклеотидов, учитываемых в качестве идентичных совпадений программой для выравнивания последовательностей в таком программном выравнивании C и D, и где Z представляет собой общее число нуклеотидов в D. Следует принимать во внимание, что если длина последовательности нуклеиновой кислоты C не равна длине последовательности нуклеиновой кислоты D, то % идентичность последовательности нуклеиновой кислоты C в отношении D не будет равна % идентичности последовательности нуклеиновой кислоты D в отношении C.
[0091] Выражение "выделенная" молекула (например, нуклеиновая кислота или белок) или клетка означает, что она была идентифицирована и отделена и/или извлечена из компонента своего природного окружения.
[0092] "Масс-спектрометрия" относится к методике аналитической химии, связанной с идентификацией количества и/или типа соединения (например, полипептида) путем измерения соотношения массы к заряду и избытка ионов газообразной фазы. Термин "масс-спектрометрия" может использоваться в данном документе взаимозаменяемо.
[0093] "Гетерогенность" при использовании в отношении капсида AAV относится к капсиду AAV, характеризующемуся одним или несколькими капсидными полипептидами, которые, как наблюдается, отклоняются в отношении эталонной массы полипептида VP1, VP2 и/или VP3 или его фрагмента. Эталонная масса может включать в себя без ограничения теоретическую, прогнозируемую или ожидаемую массу полипептида VP1, VP2 и/или VP3, например, известного серотипа AAV. Например, считается, что капсид AAV проявляет гетерогенность, если он характеризуется одним или несколькими из следующих свойств (без ограничения): смешанным серотипом, вариантным капсидом, аминокислотной заменой капсида, усеченным капсидом или модифицированным капсидом.
[0094] Ссылка на "приблизительно" в отношении значения или параметра в данном документе включает (и описывает) варианты осуществления, которые направлены на данное значение или параметр per se. Например, описание, относящееся к "приблизительно X", включает описание "X".
[0095] Формы единственного числа, используемые в данном документе, включают ссылки на множественное число, если не указано иное.
[0096] Понятно, что аспекты и варианты осуществления настоящего изобретения, описанные в данном документе, включают "содержащие", "состоящие из" и/или "состоящие по сути из" аспекты и варианты осуществления.
III. Способы
[0097] Определенные аспекты настоящего изобретения относятся к способам определения серотипа вирусной частицы. Другие аспекты настоящего изобретения относятся к способам определения гетерогенности вирусной частицы. Как описано ниже, точные массы VP1, VP2 и VP3 каждого серотипа AAV являются уникальными и могут применяться для идентификации или дифференциации серотипов капсидов AAV. Эти способы отчасти основаны на том открытии, описанном в данном документе, что прямую LC/MS различных типов AAV после денатурации можно применять для контроля последовательности белка и посттрансляционных модификаций с точным измерением масс на уровне интактного белка. Кроме того, случаи ацетилирования N-концов VP1 и VP3 также можно идентифицировать и/или контролировать в различных серотипах AAV. На основании этих результатов AAV и руководства, приведенного в данном документе, предполагается, что такие способы можно легко применять для получения профилей ряда вирусов, например, семейств, подсемейств и родов вирусов по настоящему изобретению. Эти способы по настоящему изобретению могут найти применение, например, в получении профилей VP для контроля процессов экспрессии, пострансляционных модификаций и усечений VP и для обеспечения однородности продуктов во время получения VLP, для подтверждения сайт-направленного мутагенеза или структурной характеристики применений, связанных с инженерией капсидных белков, и/или для контроля или выявления гетерогенности вирусной частицы или препарата.
[0098] В некоторых вариантах осуществления способы включают денатурирование вирусной частицы. В некоторых вариантах осуществления вирусную частицу, такую как частицу AAV, можно денатурировать с помощью детергента, тепла, высокой концентрации соли или буферизации при низком или высоком значении pH. В определенных вариантах осуществления частицу AAV можно денатурировать с применением уксусной кислоты или гуанидингидрохлорида. Специалист в данной области техники поймет, что в данной области техники доступен ряд способов, пригодных для способствования и/или контроля денатурации белка, и сможет подходящим образом выбрать способ денатурации, совместимый с жидкостной хроматографией/масс-спектрометрией. Например, если применяют тепловую денатурацию, то необходимо соблюдать осторожность с целью избежания осаждения белка и закупорки колонки с обращенной фазой. Аналогично денатурацию с помощью высокой концентрации соли можно сочетать с этапом обессоливания перед LC/MS или LC/MS/MS. В других вариантах осуществления применяют денатурацию при высоком значении pH, денатурацию при низком значении pH или денатурацию с помощью органических растворителей.
[0099] В некоторых вариантах осуществления способы предусматривают подвергание денатурированной вирусной частицы по настоящему изобретению жидкостной хроматографии/масс-спектрометрии (LC/MS). Как известно в данной области техники, при LC/MS применяют жидкостную хроматографию для физического разделения ионов и масс-спектрометрию для получения масс-спектральных данных на основании ионов. Такие масс-спектральные данные можно применять, например, для определения молекулярного веса или структуры, идентификации частиц по массе, количеству, чистоте и т.п. Эти данные могут представлять свойства выявляемых ионов, такие как сила сигнала (например, интенсивность) в течение времени (например, времени удержания) или относительная интенсивность относительно соотношения массы к заряду.
[0100] В некоторых вариантах осуществления жидкостная хроматография (например, применяемая в LC/MS, описанной в данном документе) представляет собой ультраэффективную жидкостную хроматографию (UPLC; термины "ультраэффективная жидкостная хроматография" или UHPLC могут быть использованы в данном документе взаимозаменяемо). UPLC известна в данной области техники в качестве методики LC, которая основана на применении колонки с уменьшенным размером частиц (например, менее 2 мкм) и повышенной скоростью потока для повышения хроматографического разрешения, эффективности, пиковой емкости и чувствительности (см., например, Plumb, R. et al. (2004) Rapid Commun. Mass Spectrom. 18:2331-2337). В некоторых вариантах осуществления UPLC относится к применению колонки с размером частиц менее 2 мкм в жидкостной хроматографии. В некоторых вариантах осуществления UPLC относится к применению высокой линейной скорости растворителя (например, наблюдаемой при функционировании при 6000 psi или выше) в жидкостной хроматографии. Иллюстративные устройства для UPLC являются коммерчески доступными (например, ACQUITY UPLC® от Waters; Милфорд, Массачусетс).
[0101] В некоторых вариантах осуществления масс-спектрометрия (например, применяемая при LC/MS, описанной в данном документе) может относится к масс-спектрометрии с ионизацией электрораспылением (ESI-MS). ESI-MS известна в данной области техники в качестве методики, при которой применяется электрическая энергия для анализа ионов, полученных из раствора с помощью масс-спектрометрии (см., например, Yamashita, M. and Fenn, J.B. (1984) J. Phys. Chem. 88:4451-4459). Ионные частицы (или нейтральные частицы, которые ионизированы в растворе или в газообразной фазе) переносят из раствора в газообразную фазу с помощью распыления в аэрозоле заряженных капель с последующим испарением растворителя, что снижает размер заряженных частиц и выброс ионов образца из заряженных капель по мере того, как раствор проходит через небольшой капилляр с напряжением по отношению к заземлению (например, стенке окружающей камеры). В некоторых вариантах осуществления капиллярное напряжение составляет от приблизительно 2 кВ до приблизительно 10 кВ или от приблизительно 2,5 кВ до приблизительно 6,0 кВ. В определенных вариантах осуществления при жидкостной хроматографии (например, применяемой при LC/MS, описанной в данном документе) используется капиллярное напряжение, составляющее приблизительно 3,5 кВ. В некоторых вариантах осуществления капиллярное напряжение находится в диапазоне от приблизительно 1 кВ до приблизительно 10 кВ. В других вариантах осуществления масс-спектрометрия (например, применяемая при LC/MS, описанной в данном документе) может относиться к лазерной десорбции-ионизации в присутствии матрицы (MALDI).
[0102] В некоторых вариантах осуществления при масс-спектрометрии (например, применяемой при LC/MS, описанной в данном документе) используют пробоотборный конус и/или конус скиммера, через который ионы могут проходить перед вхождением в анализатор. В некоторых вариантах осуществления, например, при применении напряжения по отношению к капилляру, описанному выше, пробоотборный конус удерживается при более низком напряжении, чем капиллярное напряжение. В определенных вариантах осуществления при жидкостной хроматографии (например, применяемой при LC/MS, описанной в данном документе) используется напряжение пробоотборного конуса, составляющее приблизительно 45 В. В некоторых вариантах осуществления напряжение пробоотборного конуса находится в диапазоне от приблизительно 0 В до приблизительно 200 В.
[0103] В некоторых вариантах осуществления при масс-спектрометрии (например, при LC/MS, применяемой в данном документе) применяют калибровку, выполняемую с помощью дополнительного средства. Калибровка при применении в отношении к масс-спектрометрии может включать введение одного или нескольких соединений, имеющих известную массу (например, стандарт) в целях калибровки устройства по отношению к выявлению масс (например, измерениям m/z). В некоторых вариантах осуществления калибровка, выполняемая с помощью дополнительного средства, может относиться к применению компьютерной программы для установления корреляции пика и/или положения известного стандарта (например, калибранта) с конкретным соотношением массы к заряду (m/z). После калибровки пользователь затем может осуществлять масс-спектрометрию образца, имеющего одно или несколько известных соединений или соединений, присутствующих при неизвестной концентрации, в пределах определенной степени точности или ошибки и/или необходимого уровня воспроизводимости, например, по сравнению с предыдущим или известным экспериментальным условием. Различные калибранты известны в данной области техники, в том числе без ограничения йодид натрия, йодид цезия, активированный натрием, полиэтиленгликоль и перфтортрибутиламин. В определенных вариантах осуществления йодид натрия используют в качестве калибранта. В некоторых вариантах осуществления калибранты представляют собой Glu-1-фибринопептид B и пептид лейцин-энкефалин для фиксации массы во время проведения LC/MS.
[0104] В некоторых вариантах осуществления способы включают подвергание денатурированной вирусной частицы по настоящему изобретению или подвергание расщепленных фрагментов денатурированной вирусной частицы по настоящему изобретению жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS). Как известно в данной области техники, в LC/MS/MS (термин "жидкостная хроматография-тандемная масс-спектрометрия" можно использовать взаимозаменяемо в данном документе) применяют жидкостную хроматографию для физического разделения ионов и масс-спектрометрию для получения масс-спектральных данных на основании ионов, при этом в масс-спектрометрии применяют несколько стадий разделения масс (например, m/z), обычно разделенных этапом фрагментации. Например, представляющие интерес ионы в диапазоне m/z можно разделить в первом цикле MS, фрагментировать и затем дополнительно разделить на основе отдельного m/z во втором цикле MS. Фрагментация ионов может включать без ограничения методику, такую как диссоциация, индуцированная столкновениями (CID), диссоциация, индуцированная столкновениями при повышенной энергии (HCD), электронно-захватная диссоциация (ECD) или диссоциация с переносом электронов (ETD).
[0105] В некоторых вариантах осуществления способы предусматривают подвергание денатурированной вирусной частицы по настоящему изобретению восстановлению и/или алкилированию. Средства для восстановления вирусной частицы включают без ограничения обработку дитиотреитолом, β-меркаптоэтанолом или трис(2-карбоксиэтил)фосфином (TCEP). Средства для алкилирования вирусной частицы включают без ограничения обработку частицы AAV йодуксусной кислотой, йодацетамидом или 4-винилпиридином.
[0106] В некоторых вариантах осуществления способы включают подвергание денатурированной вирусной частицы по настоящему изобретению расщеплению, например, с образованием фрагментов VP1, VP2 и/или VP3 частицы AAV. Например, денатурированную частицу AAV можно подвергать расщеплению с образованием пептидных фрагментов, которые можно анализировать, например, с помощью LC для разделения и MS/MS для анализа (см. ниже более подробное описание). В некоторых вариантах осуществления расщепление представляет собой ферментативное расщепление. В некоторых вариантах осуществления при расщеплении применяют химическое расщепление, такое как обработка CNBr инструментальной фрагментации (например, сверху вниз). В некоторых вариантах осуществления при расщеплении применяют химическое расщепление, такое как кислотное расщепление.
[0107] В некоторых вариантах осуществления ферментативное расщепление представляет собой расщепление с помощью эндопептидазы. Эндопептидаза может включать любую пептидазу, которая катализирует протеолиз пептидных связей неконцевых аминокислот полипептида. Известные пептидазы могут включать без ограничения трипсин, химотрипсин, AspN, Glu-C, LysC, пепсин, термолизин, глутамилэндопептидазу, эластазу и неприлизин. В определенных вариантах осуществления ферментативное расщепление представляет собой расщепление с помощью трипсина или расщепление с помощью LysC.
[0108] В некоторых вариантах осуществления жидкостная хроматография (например, применяемая в LC/MS или LC/MS/MS, описанных в данном документе) представляет собой обращенно-фазовую жидкостную хроматографию (термины "обратно-фазная жидкостная хроматография" или RPLC можно использовать в данном документе взаимозаменяемо с обращенно-фазовой жидкостной хроматографией). Как известно в данной области техники, обращенно-фазовая жидкостная хроматография может относиться к хроматографическому разделению с применением гидрофобной неподвижной фазы (например, подложки или субстрата, таких как колонка) с адсорбцией гидрофобных молекул в полярной подвижной фазе. В помощью снижения полярности подвижной фазы (например, путем добавления органического растворителя) можно достичь градиентного разделения молекул в зависимости от гидрофобности, поскольку более гидрофобные молекулы будут оставаться в колонке в более высоких концентрациях органического растворителя вследствие более сильных гидрофобных взаимодействий с колонкой. В некоторых вариантах осуществления разделение происходит с помощью капиллярного электрофореза (CE), эксклюзионной хроматографии (SEC), ионообменной хроматографии (IEC), такой как катионообменная хроматография, хроматографии с гидрофобным взаимодействием (HIC), жидкостной хроматографии гидрофильных взаимодействий (HILIC), без ограничения LC/MS он-лайн, такого как офф-лайн разделение до MS; например, наконечники, колонки; планшеты или картриджи.
[0109] Как правило, неподвижная фаза, подходящая для обращенно-фазовой жидкостной хроматографии (например, гидрофобный фрагмент) может быть соединена с подложкой, в том числе без ограничения колонкой или смолой, упакованной частицами или гранулами (например, частицами пористого кремнезема или полистирола). Ряд гидрофобных неподвижных фаз известны в данной области техники, в том числе без ограничения гидрофобные алкильные цепи, октил- или октадецилсилильные фрагменты, цианофрагменты и аминофрагменты. В некоторых вариантах осуществления неподвижная фаза может включать гидрофобную алкильную цепь определенной длины, такую как C4, C8 или C18. В определенных вариантах осуществления обращенно-фазовая хроматография представляет собой хроматографию с обращенной фазой C4 или C8 (например, обращенно-фазовую хроматографию с применением неподвижной фазы C4 или C8). Специалист в данной области техники может подходящим образом выбрать неподвижную фазы на основании молекулы, представляющей интерес (например, денатурированной частицы AAV или ее фрагмента).
[0110] Ряд подвижных фаз, подходящих для обращенно-фазовой жидкостной хроматографии, известны в данной области техники. Как описано выше, подвижная фаза обращенно-фазовой жидкостной хроматографии может включать смесь органических (например, гидрофобных) и водных (например, полярных) растворителей. Повышение доли органического растворителя повышает его силу элюирования гидрофобных соединений из неподвижной фазы. Удержание и/или селективность соединений можно изменять, например, с помощью изменения типа или воздействия неподвижной фазы, добавления полярных реагентов, таких как реагенты для кэпирования концевых групп, изменения температуры и/или изменения характеристик подвижной фазы, таких как доля органического растворителя, pH, буферы и тип используемого органического растворителя. В некоторых вариантах осуществления полярный компонент подвижной фазы может включать без ограничения воду или водный буфер. В некоторых вариантах осуществления полярный компонент подвижной фазы может включать без ограничения ацетонитрил, метанол, этанол, изопропиловый спирт, тетрагидрофуран (THF) и уксусную кислоту.
[0111] В некоторых вариантах осуществления можно применять две или более подвижных фаз (например, подвижная фаза А, подвижная фаза B и т. д.) в градиенте или пропорции, представляющих интерес. В определенных вариантах осуществления в хроматографии применяют подвижную фазу A, содержащую муравьиную кислоту в воде. В определенных вариантах осуществления подвижная фаза A содержит приблизительно 0,1% муравьиной кислоты. В определенных вариантах осуществления подвижная фаза A содержит от приблизительно 0,1% до приблизительно 5% муравьиной кислоты. В определенных вариантах осуществления в хроматографии применяют подвижную фазу B, содержащую муравьиную кислоту в ацетонитриле. В определенных вариантах осуществления подвижная фаза В содержит приблизительно 0,1% муравьиной кислоты.
[0112] В некоторых вариантах осуществления долю подвижной фазы B в хроматографии увеличивают со временем. Например, долю подвижной фазы B в хроматографии можно увеличивать поэтапно. В определенных вариантах осуществления подвижную фазу B увеличивают от приблизительно 10% до приблизительно 20%, от приблизительно 20% до приблизительно 30% и от приблизительно 30% до приблизительно 38%. В других вариантах осуществления подвижную фазу B увеличивают от приблизительно 2% до приблизительно 60%. В других вариантах осуществления подвижную фазу B увеличивают от приблизительно 2% до приблизительно 100% в течение периода от приблизительно 1 мин. до приблизительно 200 мин. В некоторых вариантах осуществления остаток подвижной фазы представляет собой вторую подвижную фазу по настоящему изобретению, например, подвижную фазу A. В определенных вариантах осуществления подвижную фазу B увеличивают от приблизительно 10% до приблизительно 20% в течение приблизительно 6 минут, от приблизительно 20% до приблизительно 30% в течение приблизительно 10 минут и от приблизительно 30% до приблизительно 38% в течение приблизительно 40 минут. В других вариантах осуществления подвижную фазу B увеличивают от приблизительно 2% до приблизительно 60% в течение приблизительно 121 минуты. Специалист в данной области техники может подходящим образом регулировать представляющую интерес подвижную фазу и время градиента на основании необходимого хроматографического разделения и/или аналита, представляющего интерес.
[0113] В некоторых вариантах осуществления жидкостная хроматография представляет собой высокоэффективную жидкостную хроматографию (HPLC). HPLC известна в данной области техники в качестве формы жидкостной хроматографии, в которой жидкий растворитель, содержащий образец, сжимают по мере того, как он проходит через колонку, содержащую твердую фазу. В то время как при традиционной LC или LC низкого давления можно применять силу тяжести для прохождения подвижной фазы через твердую фазу, при HPLC используют насосы для применения давления по отношению к подвижной фазе и обычно используют твердую фазу с более мелкими частицами для повышения разрешения. В некоторых вариантах осуществления при HPLC применяют давление от приблизительно 50 бар до приблизительно 350 бар. В некоторых вариантах осуществления обращенно-фазовую HPLC можно применять для концентрирования и/или обессоливания белков (например, капсидных белков AAV) для MS-анализа.
[0114] В некоторых вариантах осуществления один или несколько параметров, в том числе без ограничения напряжение источника, температуру капилляра, напряжение ESI (при применении ESI-MS), энергию CID и число событий MS/MS, можно регулировать, например, при LC/MS/MS, описанной в данном документе, на основании результатов, описанных в данном документе. В некоторых вариантах осуществления при масс-спектрометрии (например, применяемой при LC/MS/MS, описанной в данном документе) применяют напряжение источника (например, капиллярного напряжения), составляющее приблизительно 2,5 кВ. В некоторых вариантах осуществления при масс-спектрометрии (например, применяемой при LC/MS/MS, описанной в данном документе) применяют капиллярную температуру, составляющую приблизительно 275°C. В некоторых вариантах осуществления температура капилляра находится в диапазоне от приблизительно 20°C до приблизительно 400°C.
[0115] Ряд масс-анализаторов, подходящих для LC/MS и/или LC/MS/MS, известен в данной области техники, в том числе без ограничения времяпролетные (TOF) анализаторы, квадрупольные масс-фильтры, квадрупольные TOF (QTOF) и ионные ловушки (например, масс-спектрометр на основе преобразования Фурье или Orbitrap). В случае Orbitrap бочковидный внешний электрод при потенциале земли и веретенообразный центральный электрод применяют для захвата ионов в траекториях, вращающихся эллиптически вокруг центрального электрода с колебаниями вдоль центральной оси, ограниченными равновесием центробежных и электростатических сил. При применении таких устройств осуществляют операцию преобразования Фурье для преобразования временного сигнала (например, частоты) на основании выявления экранирующего тока в измерение массы высокого разрешения (например, нано-LC/MS/MS). Дополнительные описания и подробности можно найти, например, в Scheltema, R.A. et al. (2014) Mol. Cell Proteomics 13:3698-3708; Perry, R.H. et al. (2008) Mass. Spectrom. Rev. 27:661-699; и Scigelova, M. et al. (2011) Mol. Cell Proteomics 10:M111.009431.
[0116] Как описано выше, в некоторых вариантах осуществления MS включает нано-LC/MS/MS, например, применение масс-анализатора Orbitrap. В некоторых вариантах осуществления источник ионов может включать уложенный кольцом ионопровод или S-линзу. Как известно в области техники, S-линзу можно применять для фокусирования пучка ионов с помощью радиочастоты (RF) с повышением тем самым передачи ионов в устройство. Это может повышать чувствительность (например, в случае ионов низкой интенсивности) и/или повышать скорость сканирования. В определенных вариантах осуществления уровень RF S-линзы при масс-спектрометрии составляет приблизительно 55%. В определенных вариантах осуществления уровень RF S-линзы при масс-спектрометрии составляет от приблизительно 20% до приблизительно 100%.
[0117] В некоторых вариантах осуществления можно определять массы капсидных белков вирусов, например, на основании данных LC/MS и/или LC/MS/MS. В некоторых вариантах осуществления можно определять массы VP1, VP2 и VP3 частицы AAV или фрагментов VP1, VP2 и VP3 частицы AAV, например, на основании данных LC/MS и/или LC/MS/MS. Различные способы определения массы и/или идентичности белков на основании данных MS известны в области техники. Например, можно применять фингерпринтинг масс пептидов для определения белковой последовательности на основании данных MS или можно идентифицировать белки на основании данных MS/MS, относящихся к одному или нескольким составляющим их пептидам. При использовании тандемной MS сканирование ионов-продуктов можно применять для анализа данных m/z, относящихся к одному или нескольким пептидам белка, представляющего интерес. Можно применять компьютерные программы, известные в данной области техники, например, для приведения идентифицированных пиков в соответствие с эталонными или известными пиками, для группировки пиков в изотопомерные зоны и т.д. Значения масс пептидов можно сравнить с базой данных известных пептидных последовательностей. Например, можно применять Mascot для приведения наблюдаемых пептидов в соответствие с теоретическими пептидами из базы данных, например, в результате применения определенного паттерна расщепления к базе данных белков in silico. Другие подходящие компьютерные программы включают без ограничения Proteome Discoverer, ProteinProspector, X!Tandem, Pepfinder, Bonics или MassLynx™ (Waters). Другие компьютерные программы, подходящие для различных этапов анализа данных MS можно найти, например, по адресу www.ms-utils.org/wiki/pmwiki.php/Main/SoftwareList.
[0118] В некоторых вариантах осуществления определенную или рассчитанную массу по настоящему изобретению (например, определенную или рассчитанную массу VP1, VP2 и/или VP3 частицы AAV) можно сравнить с эталоном, например, с теоретической массой VP1, VP2 и/или VP3 одного или нескольких серотипов AAV. Эталон по настоящему изобретению может включать теоретическую массу VP1, VP2 и/или VP3 одного или нескольких серотипов AAV, описанных в данном документе. Например, в некоторых вариантах осуществления массы VP1, VP2 и/или VP3 сравнивают с теоретическими массами одного или нескольких из капсида AAV1, капсида AAV2, капсида AAV3, капсида AAV4, капсида AAV5, капсида AAV6, капсида AAV7, капсида AAV8, капсида AAVrh8, капсида AAV9, капсида AAV10, капсида AAVrh10, капсида AAV11, капсида AAV12, капсида AAV LK03 (см. патент США № 9169299), капсида AAV2R471A, капсида AAV2/2-7m8, капсида AAV DJ (см. патент США № 7588772), капсида AAV DJ8, капсида AAV2 N587A, капсида AAV2 E548A, капсида AAV2 N708A, капсида AAV V708K, капсида AAV козы, химерного капсида AAV1/AAV2, капсида AAV крупного рогатого скота или капсида AAV мыши rAAV2/HBoV1 (химерный AAV/вирус 1, относящийся к бокавирусам человека), капсида AAV2HBKO, капсида AAVPHP.B или капсида AAVPHP.eB. В некоторых вариантах осуществления определенную или рассчитанную массу по настоящему изобретению (например, определенную или рассчитанную массу VP1, VP2 и/или VP3 частицы AAV) можно сравнить с теоретической массой VP1, VP2 и/или VP3 соответствующего серотипа AAV.
[0119] В некоторых вариантах осуществления способы по настоящему изобретению могут включать определение гетерогенности частицы AAV. В некоторых вариантах осуществления отклонение одной или нескольких масс VP1, VP2 и/или VP3 (например, от эталонной массы, такой как теоретическая, прогнозируемая или ожидаемая масса) характеризует гетерогенность капсида AAV. В некоторых вариантах осуществления гетерогенность может включать без ограничения одно или несколько из следующего: смешанных серотипов, вариантных капсидов, аминокислотных замен капсидов, усеченных капсидов или модифицированных капсидов.
[0120] В некоторых вариантах осуществления можно комбинировать применение LC/MS и LC/MS/MS, описанных в данном документе. В некоторых вариантах осуществления способ определения серотипа частицы AAV может предусматривать подвергание денатурированной частицы AAV LC/MS (например, описанной в данном документе) и определение масс VP1, VP2 и VP3 частицы AAV; а также подвергание фрагментов VP1, VP2 и/или VP3 LC/MS/MS и определение масс фрагментов VP1, VP2 и VP3 частицы AAV (конкретная комбинация масс фрагментов VP1, VP2 и VP3 характеризует серотип AAV). В некоторых вариантах осуществления способ определения гетерогенности частицы AAV может предусматривать подвергание денатурированной частицы AAV LC/MS (например, описанной в данном документе), определение масс VP1, VP2 и VP3 частицы AAV и сравнение этих масс с теоретическими массами VP1, VP2 и VP3 серотипа AAV; а также подвергание фрагментов VP1, VP2 и/или VP3 LC/MS/MS, определение масс фрагментов VP1, VP2 и VP3 частицы AAV и сравнение этих масс с теоретическими массами VP1, VP2 и VP3 серотипа AAV (отклонение одной или нескольких масс VP1, VP2 или VP3 характеризует гетерогенность капсида AAV).
[0121] В некоторых вариантах осуществления частица AAV по настоящему изобретению может быть ацетилированной. Например, в некоторых вариантах осуществления N-конец VP1 и/или VP3 является ацетилированным. Как описано более подробно ниже, аминокислоту во 2-м положении по отношению к инициирующему метионину (iMet X) капсидного белка AAV можно мутировать в целях определения ее эффекта в отношении N-концевого (Nt-) ацетилирования, а также функциональных последствий влияния Nt-ацетилирования на миграцию, трансдукцию и/или посттрансляционную модификацию частицы AAV (например, гликозилирование, убиквитинирование и т.д.). В некоторых вариантах осуществления N-конец капсидного белка AAV (например, VP1 или VP3) может относиться к первой аминокислоте после инициирующего метионина, который в некоторых случаях может быть удален, например, с помощью Met-аминопептидазы.
[0122] В некоторых вариантах осуществления частица AAV по настоящему изобретению (например, частица рекомбинантного AAV или rAAV) содержит аминокислотную замену по аминокислотному остатку 2 VP1 и/или VP3. В некоторых вариантах осуществления аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 приводит к образованию VP1 и/или VP3 с другой частотой или долей N-концевого ацетилирования по сравнению с эталоном (например, частицей исходного AAV до аминокислотной замены или частицы AAV с другим аминокислотным остатком 2 VP1 и/или VP3). В некоторых вариантах осуществления аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 и/или VP3 частицы исходного AAV. Например, в определенных вариантах осуществления аминокислотная замена в положении аминокислотного остатка 2 VP1 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 частицы исходного AAV. В определенных вариантах осуществления аминокислотная замена в положении аминокислотного остатка 2 VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP3 частицы исходного AAV. В некоторых вариантах осуществления аминокислотная замена (например, аминокислотная замена по аминокислотному остатку 2 VP1 или VP3), которая "изменяет" N-концевое ацетилирование, приводит к более высокой частоте N-концевого ацетилирования или более низкой частоте N-концевого ацетилирования, например, по сравнению с VP1 или VP3 без замены, такими как исходный VP1 или VP3. VP1 и/или VP3 могут принадлежать к любому из иллюстративных серотипов AAV, описанных в данном документе, включая их варианты или гибриды (например, несущие мутацию по тирозину или мутации, влияющие на связывание с гепарином). Иллюстративные анализы N-концевого ацетилирования включают без ограничения масс-спектрометрию, введение изотопных меток (например, с помощью меченной изотопами ацетильной группы или ее предшественника), вестерн-блоттинг с помощью специфичного к ацетилированию антитела и т. д. В некоторых вариантах осуществления аминокислотный остаток 2 капсидного белка AAV (например, VP1 или VP3) заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr. В некоторых вариантах осуществления аминокислотная замена приводит к меньшей степени дезамидирования капсида AAV.
[0123] В некоторых вариантах осуществления частица AAV по настоящему изобретению может быть дезамидированной. Например, в некоторых вариантах осуществления N57 VP1 и/или N382, N511 и/или N715 VP3 являются дезамидированными. Как описано более подробно ниже, аминокислота, выбранная из A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 капсидного белка AAV (например, VP1 или VP3), может быть мутирована с целью определения ее эффекта в отношении дезамидирования, а также функциональных последствий влияния дезамидирования в отношении миграции, трансдукции и/или посттрансляционной модификации частицы AAV (например, гликозилирование, убиквитинирование и т..).
[0124] В некоторых вариантах осуществления частица AAV по настоящему изобретению (например, частица рекомбинантного AAV или rAAV) содержит аминокислотную замену в одном или нескольких аминокислотных остатках, выбранных из A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и G716 VP3. В некоторых вариантах осуществления аминокислотная замена A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и/или G716 VP3 приводит к образованию VP1/или VP3 с другой частотой или долей дезамидирования по сравнению с эталоном (например, частицей исходного AAV до введения аминокислотной замены или частицей AAV с другим соответствующим аминокислотным остатком 2). В некоторых вариантах осуществления аминокислотная замена (например, аминокислотная замена в A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3), которая обеспечивает "изменение" дезамидирования, приводит к более высокой частоте дезамидирования или более низкой частоте дезамидирования, например, по сравнению с VP1 или VP3 без замены, такими как исходные VP1 или VP3. VP1 и/или VP3 могут принадлежать к любому из иллюстративных серотипов AAV, описанных в данном документе, включая их варианты или гибриды (например, несущие мутацию по тирозину или мутации, влияющие на связывание с гепарином). Иллюстративные анализы дезамидирования включают без ограничения масс-спектрометрию, HPLC (см., например, набор для выявления изоаспартата ISOQUANT® от Promega) и т.д. В некоторых вариантах осуществления N57 VP1, N382 VP3, N511 VP3 и/или N715 VP3 заменены Asp, и аминокислотная замена приводит к более высокой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В других вариантах осуществления аминокислотная замена представляет собой N57K или N57Q. и аминокислотная замена приводит к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В еще других вариантах осуществления G58 VP1, G383 VP3, G512 VP3 и/или G716 VP3 заменены аминокислотой, которая не является Gly (например, Ala, Arg, Asn, Asp, Cys, Glu, Gln, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr, или Val), и аминокислотная замена приводит к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV. В еще других вариантах осуществления A35 VP1 заменен Asn и приводит к более высокой частоте дезамидирования по сравнению с VP1 исходной частицы.
[0125] Как используется в данном документе "N-ацетилирование" относится к процессу, в котором ацетильная группа ковалентно добавляется к аминогруппе N-концевой аминокислоты белка. Обычно N-концевые ацетилтрансферазы (NAT) переносят ацетильную группу от ацетилкофермента А (Ac-CoA) на α-аминогруппу первого аминокислотного остатка белка.
[0126] Как используется в данном документе, "дезамидирование" относится к химической реакции, в которой амидная функциональная группа в боковой цепи аспарагина или глутамина удаляется или превращается в другую функциональную группу. Например, аспарагин может превратиться в аспарагиновую кислоту или изоаспарагиновую кислоту. В других примерах глутамин превращается в глутаминовую кислоту или пироглутаминовую кислоту (5-оксопролин).
[0127] В некоторых вариантах осуществления частица AAV является N-ацетилированной до более высокой степени по сравнению с капсидным белком исходного AAV. В некоторых вариантах осуществления частица AAV содержит больше N-ацетильных групп на любое значение из более приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, или 100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV содержит больше N-ацетильных групп на любое значение в диапазонах 5%-10%, 10%-15%, 15%-20%, 20%-25%, 25%-30%, 30%-35%, 35%-40%, 40%-55%, 45%-50%, 50%-55%, 55%-60%, 60%-65%, 65%-70%, 70%-75%, 75%-80%, 80%-85%, 85%-90%, 90%-95%, 95%-100%, 5-25%, 25-50%, 50-75%, 75%-100%, 5-50% или 50%-100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV содержит больше N-ацетильных групп в любом из более приблизительно 2-кратного, 3-кратного, 4-кратного, 5-кратного, 10-кратного, 25-кратного, 50-кратного, 100-кратного, 500-кратного или 1000-кратного количества по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV содержит больше N-ацетильных групп в количестве в любом приблизительном диапазоне из от 2-кратного до 3-кратного, от 3-кратного до 4-кратного, от 4-кратного до 5-кратного, от 5-кратного до 10-кратного, от 10-кратного до 25-кратного, от 25-кратного до 50-кратного, от 50-кратного до 100-кратного, от 100-кратного до 500-кратного, от 500-кратного до 1000-кратного, от 2-кратного до 10-кратного, от 10-кратного до 100-кратного или от 100-кратного до 1000-кратного количества по сравнению с частицей исходного AAV.
[0128] В некоторых вариантах осуществления частица AAV является N-ацетилированной до более низкой степени по сравнению с капсидным белком исходного AAV. В некоторых вариантах осуществления частица AAV содержит меньше N-ацетильных групп на любое значение из более приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, или 100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV содержит меньше N-ацетильных групп на любое значение в диапазонах 5%-10%, 10%-15%, 15%-20%, 20%-25%, 25%-30%, 30%-35%, 35%-40%, 40%-55%, 45%-50%, 50%-55%, 55%-60%, 60%-65%, 65%-70%, 70%-75%, 75%-80%, 80%-85%, 85%-90%, 90%-95%, 95%-100%, 5-25%, 25-50%, 50-75%, 75%-100%, 5-50% или 50%-100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV содержит меньше N-ацетильных групп в любом из более приблизительно 2-кратного, 3-кратного, 4-кратного, 5-кратного, 10-кратного, 25-кратного, 50-кратного, 100-кратного, 500-кратного или 1000-кратного количества по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV содержит меньше N-ацетильных групп в количестве в любом приблизительном диапазоне из от 2-кратного до 3-кратного, от 3-кратного до 4-кратного, от 4-кратного до 5-кратного, от 5-кратного до 10-кратного, от 10-кратного до 25-кратного, от 25-кратного до 50-кратного, от 50-кратного до 100-кратного, от 100-кратного до 500-кратного, от 500-кратного до 1000-кратного, от 2-кратного до 10-кратного, от 10-кратного до 100-кратного или от 100-кратного до 1000-кратного количества по сравнению с частицей исходного AAV.
[0129] В некоторых вариантах осуществления частица AAV является дезамидированной до более высокой степени по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV является дезамидированной в большей степени на любое значение из более приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, или 100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV является дезамидированной в большей степени на любое значение в приблизительном диапазоне из 5%-10%, 10%-15%, 15%-20%, 20%-25%, 25%-30%, 30%-35%, 35%-40%, 40%-55%, 45%-50%, 50%-55%, 55%-60%, 60%-65%, 65%-70%, 70%-75%, 75%-80%, 80%-85%, 85%-90%, 90%-95%, 95%-100%, 5-25%, 25-50%, 50-75%, 75%-100%, 5-50% или 50%-100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV является дезамидированной в большей степени в любой из более приблизительно 2-кратной, 3-кратной, 4-кратной, 5-кратной, 10-кратной, 25-кратной, 50-кратной, 100-кратной, 500-кратной или 1000-кратной степени по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV является дезамидированной в большей степени в любом из приблизительных диапазонов от 2-кратной до 3-кратной, от 3-кратной до 4-кратной, от 4-кратной до 5-кратной, от 5-кратной до 10-кратной, от 10-кратного до 25-кратной, от 25-кратной до 50-кратной, от 50-кратной до 100-кратной, от 100-кратной до 500-кратной, от 500-кратной до 1000-кратной, от 2-кратной до 10-кратной, от 10-кратной до 100-кратной или от 100-кратной до 1000-кратной степени по сравнению с частицей исходного AAV.
[0130] В некоторых вариантах осуществления капсидный белок AAV является дезамидированным до более низкой степени по сравнению с капсидным белком исходного AAV. В некоторых вариантах осуществления частица AAV является в меньшей степени дезамидированной на любое значение из более приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, или 100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV является дезамидированной в меньшей степени на любое значение в приблизительном диапазоне из 5%-10%, 10%-15%, 15%-20%, 20%-25%, 25%-30%, 30%-35%, 35%-40%, 40%-55%, 45%-50%, 50%-55%, 55%-60%, 60%-65%, 65%-70%, 70%-75%, 75%-80%, 80%-85%, 85%-90%, 90%-95%, 95%-100%, 5-25%, 25-50%, 50-75%, 75%-100%, 5-50% или 50%-100% по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV является дезамидированной в меньшей степени в любой из более приблизительно 2-кратной, 3-кратной, 4-кратной, 5-кратной, 10-кратной, 25-кратной, 50-кратной, 100-кратной, 500-кратной или 1000-кратной степени по сравнению с частицей исходного AAV. В некоторых вариантах осуществления частица AAV является дезамидированной в меньшей степени в любом из приблизительных диапазонов от 2-кратной до 3-кратной, от 3-кратной до 4-кратной, от 4-кратной до 5-кратной, от 5-кратной до 10-кратной, от 10-кратного до 25-кратной, от 25-кратной до 50-кратной, от 50-кратной до 100-кратной, от 100-кратной до 500-кратной, от 500-кратной до 1000-кратной, от 2-кратной до 10-кратной, от 10-кратной до 100-кратной или от 100-кратной до 1000-кратной степени по сравнению с частицей исходного AAV.
[0131] В настоящем изобретении предусмотрена любая комбинация из N-ацетилирования и дезамидирования. Например, капсидный белок AAV может быть N-ацетилированным до более высокой степени, чем капсидный белок исходного AAV, и дезамидированным до более высокой степени, чем капсидный белок исходного AAV, капсидный белок AAV может быть N-ацетилированным до более высокой степени, чем капсидный белок исходного AAV, и дезамидированным до той же самой степени, что и капсидный белок исходного AAV, капсидный белок AAV может быть N-ацетилированным до более высокой степени, чем капсидный белок исходного AAV, и дезамидированным до более низкой степени, чем капсидный белок исходного AAV, капсидный белок AAV может быть N-ацетилированным до той же самой степени, что и капсидный белок исходного AAV, и дезамидированным до более высокой степени, чем капсидный белок исходного AAV, капсидный белок AAV может быть N-ацетилированным до той же самой степени, что и капсидный белок исходного AAV, и дезамидированным до той же самой степени, что и капсидный белок исходного AAV, капсидный белок AAV может быть N-ацетилированным до той же самой степени, что и капсидный белок исходного AAV, и дезамидированным до более низкой степени, чем капсидный белок исходного AAV, капсидный белок AAV может быть N-ацетилированным до более низкой степени, чем капсидный белок исходного AAV, и дезамидированным до более высокой степени, чем капсидный белок исходного AAV, капсидный белок AAV может быть N-ацетилированным до более низкой степени, чем капсидный белок исходного AAV, и дезамидированным до той же самой степени, что и капсидный белок исходного AAV, или капсидный белок AAV может быть N-ацетилированным до более низкой степени, чем капсидный белок исходного AAV, и дезамидированным до более низкой степени, чем капсидный белок исходного AAV.
IV. Векторы
[0132] В определенных аспектах настоящее изобретение относится к вирусным частицам, подходящим для применения в любом из способов, описанных в данном документе, которые могут содержать векторы на основе AAV (например, векторы на основе rAAV) или векторы, полученные из другого вируса. В некоторых вариантах осуществления вирусная частица содержит вектор, кодирующий гетерологичную нуклеиновую кислоту, например, гетерологичный трансген. В некоторых вариантах осуществления частица AAV содержит геном вектора на основе AAV, кодирующий гетерологичную нуклеиновую кислоту, например, гетерологичный трансген.
[0133] В настоящем изобретении предусмотрено применение рекомбинантного вирусного генома для введения одной или нескольких последовательностей нуклеиновой кислоты, кодирующих терапевтический полипептид и/или нуклеиновую кислоту, для упаковки в вирусную частицу rAAV. Рекомбинантный вирусный геном может содержать любой элемент для обеспечения экспрессии терапевтического полипептида и/или нуклеиновой кислоты, например, промотор, ITR по настоящему изобретению, элемент связывания рибосомы, терминатор, энхансер, селективный маркер, интрон, сигнал поли(А) и/или точку начала репликации.
[0134] В некоторых вариантах осуществления гетерологичная нуклеиновая кислота кодирует терапевтический полипептид. Терапевтический полипептид может, например, возмещать полипептид и/или ферментативную активность, которые отсутствуют или присутствуют в клетке или организме на сниженном уровне. Как альтернатива, терапевтический полипептид может возмещать полипептид и/или ферментативную активность, которые косвенно противодействуют дисбалансу в клетке или организме. Например, терапевтический полипептид, применяемый против нарушения, связанного с накоплением метаболита, что вызвано дефицитом метаболического фермента или активности, может возмещать недостающий метаболический фермент или активность, или он может обеспечивать другой метаболический фермент или активность, приводящие к снижению уровня метаболита. Терапевтический полипептид также можно применять для снижения активности полипептида (например, полипептида, который сверхэкспрессируется, активируется вследствие мутации приобретения функции, или полипептида, регуляция активности которого нарушена иным образом) за счет действия, например, в качестве полипептида с доминантно-отрицательным эффектом.
[0135] Нуклеиновые кислоты по настоящему изобретению могут кодировать полипептиды, которые представляют собой внутриклеточные белки, заякоренные в клеточной мембране, сохраняющиеся в клетке или секретируемые клеткой, трансдуцированной векторами по настоящему изобретению. В случае полипептидов, секретируемых клеткой, вмещающей вектор, полипептид может быть растворимым (т. е. не прикрепленным к клетке). Например, растворимые полипептиды лишены трансмембранной области и секретируются из клетки. Методики идентификации и удаления последовательностей нуклеиновой кислоты, которые кодируют трансмембранные домены, известны из уровня техники.
[0136] Нуклеиновые кислоты по настоящему изобретению (например, геном вектора на основе AAV) могут содержать в качестве трансгена нуклеиновую кислоту, кодирующую белок или функциональную РНК, которые обеспечивают модулирование или лечение нарушения, ассоциированного с ЦНС. Далее приведен неограничивающий перечень генов, связанных с нарушениями, ассоциированными с ЦНС: ген белка, ингибирующего апоптоз нейронов (NAIP), ген фактора роста нервов (NGF), ген глиального фактора роста (GDNF), ген мозгового фактора роста (BDNF), ген цилиарного нейротрофического фактора (CNTF), ген тирозингидроксилазы (TM), ген GTP-циклогидролазы (GTPCH), ген аспартоацилазы (ASPA), ген супероксиддисмутазы (SOD1), ген антиоксиданта, ген антиангиогенного полипептида, ген противовоспалительного полипептида и ген декарбоксилазы аминокислот (AADC). Например, трансген, применимый в лечении болезни Паркинсона, кодирует TH, которая представляет собой фермент, ограничивающий скорость синтеза дофамина. Трансген, кодирующий GTPCH, которая обеспечивает образование кофактора TH, тетрагидробиоптерина, также можно применять в лечении болезни Паркинсона. Трансген, кодирующий GDNF, или BDNF, или AADC, которая способствует превращению L-Dopa в DA, также можно применять для лечения болезни Паркинсона. Трансген, применимый для лечения ALS, может кодировать GDNF, BDNF или CNTF. Трансген, применимый для лечения ALS, также может кодировать функциональную РНК, например, shRNA, miRNA, которая ингибирует экспрессию SOD1. Трансген, применимый для лечения ишемии, может кодировать NAIP или NGF. Трансген, кодирующий бета-глюкуронидазу (GUS), может быть применимым для лечения определенных лизосомных болезней накопления (например, мукополисахаридоза VII типа (MPS VII)). Трансген, кодирующий продукт гена, активирующий пролекарство, например, тимидинкиназу HSV, превращающую ганцикловир в токсичный нуклеотид, который нарушает синтез ДНК и приводит к клеточной гибели, может быть применимым для лечения определенных форм рака, например при введении в комбинации с пролекарством. Трансген, кодирующий эндогенный опиоид, такой как β-эндорфин, может быть применимым для лечения боли. Примеры антиоксидантов включают без ограничения SOD1; SOD2; каталазу; сиртуины 1, 3, 4 или 5; NRF2; PGC1a; GCL (каталитическую субъединицу); GCL (модифицирующую субъединицу); адипонектин; глутатионпероксидазу 1 и нейроглобин. Примеры антиангиогенных полипептидов включают без ограничения ангиостатин, эндостатин, PEDF, растворимый рецептор VEGF и растворимый рецептор PDGF. Примеры противовоспалительных полипептидов включат без ограничения IL-10, растворимый IL17R, растворимый TNF-R, TNF-R-Ig, ингибитор IL-1 и ингибитор IL18. Другие примеры трансгенов, которые можно применять в векторах на основе rAAV по настоящему изобретению, будут очевидны специалисту в данной области (см., например, Costantini LC, et al., Gene Therapy (2000) 7, 93-109).
[0137] В некоторых вариантах осуществления гетерологичная нуклеиновая кислота кодирует терапевтическую нуклеиновую кислоту. В некоторых вариантах осуществления терапевтическая нуклеиновая кислота может включать без ограничения siRNA, shRNA, средство для RNAi, miRNA, антисмысловую РНК, рибозим или ДНК-фермент. Таким образом, терапевтическая нуклеиновая кислота может кодировать РНК, которая при транскрипции с нуклеиновых кислот вектора может обеспечивать лечение нарушения путем препятствования трансляции или транскрипции аномального или избыточного белка, ассоциированного с нарушением по настоящему изобретению. Например, нуклеиновые кислоты по настоящему изобретению могут кодировать РНК, которая обеспечивает лечение нарушения путем высокоспецифичного устранения или снижения уровня mRNA, кодирующей аномальные и/или избыточные белки. Последовательности терапевтических РНК включают средства для RNAi, малые ингибирующие РНК (siRNA), микроРНК (miRNA) и/или рибозимы (такие как рибозимы типа "головки молотка" и рибозимы, содержащие шпильки), которые могут обеспечивать лечение нарушений путем высокоспецифичного устранения или снижения уровня mRNA, кодирующей аномальные и/или избыточные белки.
[0138] В некоторых вариантах осуществления терапевтический полипептид или терапевтическую нуклеиновую кислоту применяют для лечения нарушения со стороны ЦНС. Не желая ограничиваться какой-либо теорией, полагают, что терапевтический полипептид или терапевтическую нуклеиновую кислоту можно применять для снижения или устранения экспрессии и/или активности полипептида, мутация приобретения функции которого была ассоциирована с нарушением, или для усиления экспрессии и/или активности полипептида с восполнением дефицита, который был ассоциирован с нарушением (например, мутацией гена, экспрессия которого проявляет сходную или связанную активность). Неограничивающие примеры нарушений по настоящему изобретению, лечение которых можно осуществлять с помощью терапевтического полипептида или терапевтической нуклеиновой кислоты по настоящему изобретению (иллюстративные гены, на которые можно нацеливаться или которые можно возмещать, приведены в скобках для каждого нарушения), включают инсульт (например, каспаза-3, Beclin1, Ask1, PAR1, HIF1α, PUMA и/или любой из генов, описанных в Fukuda, A.M. and Badaut, J. (2013) Genes (Basel) 4:435-456), болезнь Хантингтона (мутантный HTT), эпилепсию (например, SCN1A, NMDAR, ADK и/или любой из генов, описанных в Boison, D. (2010) Epilepsia 51:1659-1668), болезнь Паркинсона (ген альфа-синуклеина), болезнь Лу Герига (также известна как боковой амиотрофический склероз; SOD1), болезнь Альцгеймера (гены тау-белка, белка-предшественника амилоида), кортикобазальную дегенерацию или CBD (ген тау-белка), кортикобазальную ганглиозную дегенерацию или CBGD (ген тау-белка), лобно-височную деменцию или FTD (ген тау-белка), прогрессирующий надъядерный паралич или PSP (ген тау-белка), множественную системную атрофию или MSA (ген альфа-синуклеина), рак головного мозга (например, мутантный или сверхэкспрессируемый онкоген, вовлеченный в рак головного мозга) и лизосомные болезни накопления (LSD). Нарушения по настоящему изобретению могут включать нарушения, охватывающие большие зоны коры головного мозга, например, несколько функциональных зон коры головного мозга, несколько долей коры головного мозга и/или всю кору головного мозга. Другие неограничивающие примеры нарушений по настоящему изобретению, лечение которых можно осуществлять с помощью терапевтического полипептида или терапевтической нуклеиновой кислоты по настоящему изобретению, включают травматическое повреждение головного мозга, нарушения, предусматривающие ферментативную дисфункцию, нарушения психики (в том числе посттравматический стрессовый синдром), нейродегенеративные заболевания и когнитивные нарушения (в том числе формы деменции, аутизм и депрессию). Нарушения, предусматривающие ферментативную дисфункцию, включают без ограничения формы лейкодистрофии (в том числе болезнь Канавана) и любые лизосомные болезни накопления, описанные ниже.
[0139] В некоторых вариантах осуществления терапевтический полипептид или терапевтическую нуклеиновую кислоту применяют для лечения лизосомной болезни накопления. Как общеизвестно из уровня техники, лизосомные болезни накопления представляют собой редкие наследственные нарушения метаболизма, характеризующиеся дефектами функций лизосом. Такие нарушения часто вызваны дефицитом фермента, необходимого для правильного метаболизма мукополисахаридов, гликопротеинов и/или липидов, что приводит к патологическому накоплению хранящихся в лизосомах клеточных материалов. Неограничивающие примеры лизосомных болезней накопления по настоящему изобретению, лечение которых можно осуществлять с помощью терапевтического полипептида или терапевтической нуклеиновой кислоты по настоящему изобретению (иллюстративные гены, на которые можно нацеливаться или которые можно возмещать, приведены в скобках для каждого нарушения), включают болезнь Гоше 2 типа или 3 типа (ген кислой бета-глюкозидазы, GBA), GM1-ганглиозидоз (ген бета-галактозидазы-1, GLB1), болезнь Хантера (ген идуронат-2-сульфатазы, IDS), болезнь Краббе (ген галактозилцерамидазы, GALC), заболевание, представляющее собой α-маннозидоз (ген маннозидазы, такой как альфа-D-маннозидаза, MAN2B1), заболевание, представляющее собой β-маннозидоз (ген бета-маннозидазы, MANBA), заболевание, представляющее собой метахроматическую лейкодистрофию (ген псевдоарилсульфатазы A, ARSA), заболевание, представляющее собой муколипидоз II/III типа (ген N-ацетилглюкозамин-1-фосфотрансферазы, GNPTAB), болезнь Ниманна-Пика типа A (ген кислой сфингомиелиназы, ASM), болезнь Ниманна-Пика типа C (ген белка, связанного с болезнью Ниманна-Пика типа C, NPC1), болезнь Помпе (ген кислой альфа-1,4-глюкозидазы, GAA), болезнь Сандхоффа (ген бета-субъединицы гексозаминидазы, HEXB), болезнь Санфилиппо типа A (ген N-сульфоглюкозаминсульфогидролазы, MPS3A), болезнь Санфилиппо типа B (ген N-альфа-ацетилглюкозаминидазы, NAGLU), болезнь Санфилиппо типа C (ген гепарин-ацетил-CoA-альфа-глюкозаминид-N-ацетилтрансферазы, MPS3C), болезнь Санфилиппо типа D (ген N-ацетилглюкозамин-6-сульфатазы, GNS), болезнь Шиндлера (ген альфа-N-ацетилгалактозаминидазы, NAGA), болезнь Слая (ген бета-глюкуронидазы, GUSB), болезнь Тея-Сакса (ген альфа-субъединицы гексозаминидазы, HEXA) и болезнь Вольмана (ген лизосомной кислой липазы, LIPA).
[0140] Дополнительные лизосомные болезни накопления, а также дефектные ферменты, ассоциированные с каждым заболеванием, указаны в таблице 1 ниже. В некоторых вариантах осуществления заболевание, указанное в таблице ниже, лечат с помощью терапевтического полипептида или терапевтической нуклеиновой кислоты по настоящему изобретению, которые восполняют или иным образом компенсируют соответствующий дефект фермента.
Таблица 1. Лизосомные нарушения накопления и дефектные ферменты, ассоциированные с ними
[0141] В некоторых вариантах осуществления гетерологичная нуклеиновая кислота функционально связана с промотором. Иллюстративные промоторы включают без ограничения немедленно-ранний промотор цитомегаловируса (CMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40) и промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, LSP-промотор, химерные печень-специфические промоторы (LSP), промотор E2F, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (промотор CAG; Niwa et al., Gene, 1991, 108(2):193-9) и промотор фактора элонгации 1-альфа (EF1-альфа) (Kim et al., Gene, 1990, 91(2):217-23 и Guo et al., Gene Ther., 1996, 3(9):802-10). В некоторых вариантах осуществления промотор предусматривает промотор β-глюкуронидазы человека или промотор из энхансера цитомегаловируса, соединенного с промотором β-актина курицы (CBA). Промотор может представлять собой конститутивный, индуцируемый или репрессируемый промотор. В некоторых вариантах осуществления в настоящем изобретении предусмотрен рекомбинантный вектор, содержащий нуклеиновую кислоту, кодирующую гетерологичный трансген по настоящему изобретению, функционально связанный с промотором CBA. Иллюстративные промоторы и их описания можно найти, например, в заявке на патент США № 20140335054, опубликованной до выдачи патента.
[0142] Примеры конститутивных промоторов включают без ограничения ретровирусный LTR-промотор вируса саркомы Рауса (RSV) (необязательно с энхансером RSV), промотор цитомегаловируса (CMV) (необязательно с энхансером CMV) [см., например, Boshart et al., Cell, 41:521-530 (1985)], промотор SV40, промотор гена дигидрофолатредуктазы, промотор гена 13-актина, промотор гена фосфоглицеринкиназы (PGK) и промотор EF1a [Invitrogen].
[0143] Индуцируемые промоторы обеспечивают возможность регуляции экспрессии генов, и они могут регулироваться экзогенно вводимыми соединениями, факторами окружающей среды, такими как температура, или наличием конкретного физиологического состояния, например, острой фазы, определенного состояния дифференцировки клетки, или функционируют только в размножающихся клетках. Индуцируемые промоторы и индуцируемые системы доступны из ряда коммерческих источников, включающих без ограничения Invitrogen, Clontech и Ariad. Многие другие системы были описаны и могут быть без труда выбраны специалистом в данной области техники. Примеры индуцируемых промоторов, регулируемых экзогенно предоставляемыми соединениями, включают индуцируемый цинком промотор гена металлотионина овцы (MT), индуцируемый дексаметазоном (Dex) промотор вируса опухоли молочной железы у мышей (MMTV), промоторную систему полимеразы T7 (WO 98/10088); индуцируемый экдизоном промотор генов насекомых (No et al., Proc. Natl. Acad. Sci. USA, 93:3346-3351 (1996)), систему с тетрациклин-зависимой репрессией (Gossen et al., Proc. Natl. Acad. Sci. USA, 89:5547-5551 (1992)), систему с тетрациклин-зависимой индукцией (Gossen et al., Science, 268:1766-1769 (1995), см. также Harvey et al., Curr. Opin. Chem. Biol., 2:512-518 (1998)), систему с RU486-зависимой индукцией (Wang et al., Nat. Biotech., 15:239-243 (1997) и Wang et al., Gene Ther., 4:432-441 (1997)) и систему с рапамицин-зависимой индукцией (Magari et al., J. Clin. Invest., 100:2865-2872 (1997)). Несколько других типов индуцируемых промоторов, которые могут быть применимыми в данном контексте, представляют собой промоторы, регулируемые конкретным физиологическим состоянием, например, температурой, острой фазой, определенным состоянием дифференцировки клетки, или функционирующие только в размножающихся клетках.
[0144] В другом варианте осуществления для трансгена будет применяться нативный промотор или его фрагмент. Нативный промотор можно применять в том случае, если требуется, чтобы экспрессия трансгена имитировала экспрессию нативного гена. Нативный промотор можно применять в тех случаях, когда экспрессия трансгена должна регулироваться во времени, или в зависимости от стадии развития, или тканеспецифичным образом, или в ответ на конкретные транскрипционные стимулы. В дополнительном варианте осуществления для имитации экспрессии нативного гена также можно применять другие нативные элементы, осуществляющие контроль экспрессии, такие как энхансерные элементы, сайты полиаденилирования или консенсусные последовательности Козак.
[0145] В некоторых вариантах осуществления регуляторные последовательности придают способности к тканеспецифичной экспрессии генов. В некоторых случаях тканеспецифичные регуляторные последовательности связывают тканеспецифичные факторы транскрипции, которые индуцируют транскрипцию тканеспецифичным образом. Такие тканеспецифичные регуляторные последовательности (например, промоторы, энхансеры и т. д.) хорошо известны из уровня техники.
[0146] В некоторых вариантах осуществления вектор содержит интрон. Например, в некоторых вариантах осуществления интрон представляет собой химерный интрон, полученный из гена бета-актина цыпленка и бета-глобина кролика. В некоторых вариантах осуществления интрон представляет собой интрон мелкого вируса мышей (MVM).
[0147] В некоторых вариантах осуществления вектор содержит последовательность полиаденилирования (поли(A)). Многочисленные примеры последовательностей полиаденилирования известны из уровня техники, такие как последовательность поли(А) бычьего гормона роста (BGH) (см., например, номер доступа EF592533), последовательность полиаденилирования SV40 и последовательность полиаденилирования pA TK HSV.
V. Вирусные частицы и способы получения вирусных частиц
[0148] Определенные аспекты настоящего изобретения относятся к рекомбинантным вирусным частицам (например, частицам rAAV).
[0149] На основании руководства, представленного в данном документе, методики по настоящему изобретению могут быть подходящим образом адаптированы специалистом в данной области техники для применения с множеством различных вирусов.
[0150] В некоторых вариантах осуществления вирус относится к семейству Adenoviridae, которое включает безоболочечные вирусы, обычно известные как аденовирусы. В некоторых вариантах осуществления вирус относится к genus Atadenovirus, Aviadenovirus, Ichtadenovirus, Mastadenovirus, или Siadenovirus.
[0151] В некоторых вариантах осуществления вирус относится к семейству Parvoviridae, которое включает безоболочечные вирусы, такие как AAV и бокапарвовирус. В некоторых вариантах осуществления вирус относится к подсемейству Densovirinae. В некоторых вариантах осуществления вирус относится к роду Ambidensovirus, Brevidensovirus, Hepandensovirus, Iteradensovirus, или Penstyldensovirus. В некоторых вариантах осуществления вирус относится к подсемейству Parvovirinae. В некоторых вариантах осуществления вирус относится к роду Amdoparvovirus, Aveparvovirus, Bocaparvovirus, Copiparvovirus, Dependoparvovirus, Erythroparvovirus, Protoparvovirus, или Tetraparvovirus.
[0152] В некоторых вариантах осуществления вирус относится к семейству Retroviridae, которое включает оболочечные вирусы, в том числе лентивирус. В некоторых вариантах осуществления вирус относится к подсемейству Orthoretrovirinae. В некоторых вариантах осуществления вирус относится к роду Alpharetrovirus, Betaretrovirus, Deltaretrovirus, Epsilonretrovirus, Gammaretrovirus, или Lentivirus. В некоторых вариантах осуществления вирус относится к подсемейству Spumaretrovirinae. В некоторых вариантах осуществления вирус относится к роду Spumavirus.
[0153] В некоторых вариантах осуществления вирус относится к семейству Baculoviridae, которое включает оболочечные вирусы, в том числе альфа-бакуловирус. В некоторых вариантах осуществления вирус относится к роду Alphabaculovirus, Betabaculovirus, Deltabaculovirus,или Gammabaculovirus.
[0154] В некоторых вариантах осуществления вирус относится к семейству Herpesviridae, которое включает оболочечные вирусы, такие как вирусы простого герпеса HSV-1 и HSV-2. В некоторых вариантах осуществления вирус относится к подсемейству Alphaherpesvirinae. В некоторых вариантах осуществления вирус относится к роду Iltovirus, Mardivirus, Simplexvirus, или Varicellovirus. В некоторых вариантах осуществления вирус относится к подсемейству Betaherpesvirinae. В некоторых вариантах осуществления вирус относится к роду Cytomegalovirus, Muromegalovirus, Proboscivirus, или Roseolovirus. В некоторых вариантах осуществления вирус относится к подсемейству Gammaherpesvirinae. В некоторых вариантах осуществления вирус относится к роду Lymphocryptovirus, Macavirus, Percavirus, или Rhadinovirus.
[0155] В некоторых вариантах осуществления вирус представляет собой AAV вирус. В случае частицы AAV нуклеиновая кислота заключена в капсид частицы AAV. Частица AAV также содержит капсидные белки. В некоторых вариантах осуществления нуклеиновая кислота содержит гетерологичную нуклеиновую кислоту и/или одно или несколько из следующего: компоненты, функционально связанные в направлении транскрипции, последовательности, осуществляющие контроль, в том числе последовательности инициации и терминации, за счет чего обеспечивается образование кассеты экспрессии.
[0156] В некоторых вариантах осуществления вирусная частица содержит последовательность ITR AAV. Например, кассета экспрессии может быть фланкирована на 5'- и 3'-конце по меньшей мере одной функциональной последовательностью ITR AAV. Под "функциональными последовательностями ITR AAV" подразумевается, что последовательности ITR функционируют в качестве предполагаемых для спасения, репликации и упаковки вириона AAV. См. Davidson et al., PNAS, 2000, 97(7)3428-32; Passini et al., J. Virol., 2003, 77(12):7034-40; и Pechan et al., Gene Ther., 2009, 16:10-16, все из которых включены в данный документ посредством ссылки во всей своей полноте. Для осуществления на практике некоторых аспектов настоящего изобретения рекомбинантные векторы содержат по меньшей мере все последовательности AAV, необходимые для заключения в капсид, и физические структуры для инфицирования с помощью rAAV. ITR AAV для применения в векторах по настоящему изобретению не должны иметь нуклеотидную последовательность дикого типа (например, описанную в Kotin, Hum. Gene Ther., 1994, 5:793-801), и они могут быть изменены посредством вставки, делеции или замены нуклеотидов, или ITR AAV могут быть получены из любого из нескольких серотипов AAV. На сегодняшний день известно более 40 серотипов AAV, и новые серотипы и варианты существующих серотипов продолжают идентифицировать. См. Gao et al., PNAS, 2002, 99(18): 11854-6; Gao et al., PNAS, 2003, 100(10):6081-6; и Bossis et al., J. Virol., 2003, 77(12):6799-810. Применение любого серотипа AAV рассматривается как входящее в объем настоящего изобретения. В некоторых вариантах осуществления вектор на основе rAAV представляет собой вектор, полученный из серотипа AAV, в том числе без ограничения AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV DJ, AAV DJ8, AAV козы, AAV крупного рогатого скота или ITR AAV мыши или т. п. В некоторых вариантах осуществления нуклеиновая кислота в AAV (например, векторе на основе rAAV) содержит ITR AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV DJ, AAV DJ8, AAV козы, AAV крупного рогатого скота или ITR AAV мыши или т. п. В некоторых вариантах осуществления частица AAV содержит вектор на основе AAV, кодирующий гетерологичный трансген, фланкированный одним или несколькими ITR AAV.
[0157] В некоторых вариантах осуществления частица rAAV содержит инкапсулированный белок, выбранный из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6 (например, капсида AAV6 дикого типа или вариантного капсида AAV6, как, например, ShH10, описанный в заявке на патент США 2012/0164106, опубликованной до выдачи патента), AAV7, AAV8, AAVrh8, AAVrh8R, AAV9 (например, капсида AAV9 дикого типа или модифицированного капсида AAV9, описанных в публикации заявки на патент США 2013/0323226, опубликованной до выдачи патента), AAV10, AAVrh10, AAV11, AAV12, мутантного по тирозину капсида, мутантного по связыванию с гепарином капсида, капсида AAV2R471A, капсида AAVAAV2/2-7m8, капсида AAV LK03, капсида AAV DJ (например, капсида AAV-DJ/8, капсида AAV-DJ/9 или любого другого из капсидов, описанных в заявке на патент США 2012/0066783, опубликованной до выдачи патента), капсида AAV2 N587A, капсида AAV2 E548A, капсида AAV2 N708A, капсида AAV V708K, капсида AAV козы, химерного капсида AAV1/AAV2, капсида AAV крупного рогатого скота, капсида AAV мыши, капсида rAAV2/HBoV1, капсида AAV2HBKO, капсида AAVPHP.B или капсида AAVPHP.eB или капсида AAV, описанного в патенте США № 8283151 или в Международной публикации № WO/2003/042397. В дополнительных вариантах осуществления частица rAAV содержит капсидные белки серотипа AAV из клад A-F.
[0158] Определенные аспекты настоящего изобретения относятся к капсидному белку AAV (например, rAAV), содержащему аминокислотную замену по аминокислотному остатку 2. В некоторых вариантах осуществления аминокислотная замена по аминокислотному остатку 2 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 исходного капсидного белка AAV. Как описано в данном документе, аминокислоту во 2-м положении по отношению к инициирующему метионину (iMet X) капсидного белка AAV можно исследовать в отношении эффектов в отношении N-концевого ацетилирования миграции, трансдукции и/или другой (других) посттрансляционной(посттрансляционных) модификации(модификаций) (например, гликозилирования, убиквитирования и т.д.). Любой анализ, описанный в данном документе, для исследования ацетилирования или его последствия в отношении функций, связанных с частицами AAV, можно использовать для оценки N-концевого ацетилирования. В некоторых вариантах осуществления аминокислотный остаток 2 капсидного белка AAV (например, VP1 или VP3) заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr. В некоторых вариантах осуществления аминокислотная замена приводит к меньшей степени дезамидирования капсида AAV.
[0159] Другие аспекты настоящего раскрытия относятся к капсидному белку AAV (например, rAAV), содержащему аминокислотную замену, которая обеспечивает изменение дезамидирования. В некоторых вариантах осуществления аминокислотная замена (например, аминокислотная замена в A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3), которая обеспечивает "изменение" дезамидирования, приводит к более высокой частоте дезамидирования или более низкой частоте дезамидирования, например, по сравнению с VP1 или VP3 без замены, такими как исходные VP1 или VP3. Как описано в данном документе, потенциальный сайт дезамидирования капсидного белка AAV (например, VP1 или VP3) можно исследовать в отношении влияния на дезамидирование, миграцию, трансдукцию и/или другую (другие) посттрансляционную (посттрансляционные) модификацию (модификации) (например, гликозилирование, убиквитирование и т.д.). Любой анализ, описанный в данном документе, для исследования дезамидирования или его последствия в отношении функций, связанных с частицами AAV, можно использовать для оценки дезамидирования.
[0160] Несколько потенциальных сайтов дезамидирования описаны в данном документе. В некоторых вариантах осуществления аминокислотная замена, которая обеспечивает изменение дезамидирования, выбрана из A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3. Например, в некоторых вариантах осуществления N57 VP1, N382 VP3, N511 VP3 и/или N715 VP3 заменены Asp, и аминокислотная замена приводит к более высокой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 исходной частицы AAV. В других вариантах осуществления аминокислотная замена представляет собой N57K или N57Q, и аминокислотная замена приводит к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 исходной частицы AAV. В еще одних вариантах осуществления G58 VP1, G383 VP3, G512 VP3 и/или G716 VP3 заменены аминокислотой, которая не является Gly (например, Ala, Arg, Asn, Asp, Cys, Glu, Gln, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val), и аминокислотная замена приводит к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 исходной частицы AAV.
[0161] В некоторых вариантах осуществления капсидный белок AAV представляет собой VP1, VP2 или VP3. Частица AAV может предусматривать любой из иллюстративных серотипов капсида AAV, описанных в данном документе, таких как AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV DJ8, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота, AAV мыши или rAAV2/HBoV1. Капсидный белок AAV может дополнительно содержать любую из мутаций капсидного белка, описанных в данном документе, таких как мутации по тирозину и/или мутации, влияющие на связывание с гепарином.
[0162] Другие аспекты настоящего изобретения относятся к способам улучшения стабильности частицы rAAV. В некоторых вариантах осуществления способы предусматривают осуществление замены аминокислотного остатка 2 VP1 и/или VP3, например, как описано в данном документе. Например, в некоторых вариантах осуществления аминокислотный остаток 2 VP1 является замененным. В других вариантах осуществления аминокислотный остаток 2 VP3 является замененным. В некоторых вариантах осуществления замененная аминокислота в положении 2 подвергается N-ацетилированию с более высокой частотой, чем аминокислотный остаток 2 исходного VP1 и/или VP3, например, как описано в данном документе. В некоторых вариантах осуществления замена аминокислотного остатка 2 VP1 и/или VP3 обеспечивает улучшение стабильности частицы rAAV на по меньшей мере приблизительно 5%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 100%. В некоторых вариантах осуществления стабильность частицы rAAV с замененной аминокислотой в положении 2 можно сравнить с капсидом AAV дикого типа или исходным капсидом AAV, например, одного и того же серотипа. Например, в некоторых вариантах осуществления замена аминокислотного остатка 2 VP1 и/или VP3 обеспечивает улучшение стабильности частицы rAAV на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20%, например, по сравнению со стабильностью частицы rAAV, содержащей капсид дикого типа. Стабильность частицы AAV можно измерять с помощью различных анализов, известных из уровня техники, в том числе без ограничения дифференциальной сканирующей флуориметрии (DSF), дифференциальной сканирующей калориметрии (DSC), других анализов термической денатурации, восприимчивости к протеолизу, визуализации или структурного анализа с наблюдением денатурации (например, с помощью электронного микроскопа), эффективности трансдукции или другого функционального анализа в отношении композиций на основе частиц AAV, хранящихся в течение определенного периода времени при определенной температуре (например, комнатной температуре или 4°C в случае термоустойчивости) или обрабатываемых при определенном значении pH (например, стабильность в зависимости от значений pH) и т.п.
[0163] Другие аспекты настоящего изобретения относятся к способам улучшения сборки частицы rAAV. В некоторых вариантах осуществления способы предусматривают осуществление замены аминокислотного остатка 2 VP1 и/или VP3, например, как описано в данном документе. Например, в некоторых вариантах осуществления аминокислотный остаток 2 VP1 является замененным. В других вариантах осуществления аминокислотный остаток 2 VP3 является замененным. В некоторых вариантах осуществления замененная аминокислота в положении 2 подвергается N-ацетилированию с более высокой частотой, чем аминокислотный остаток 2 исходного VP1 и/или VP3, например, как описано в данном документе. В некоторых вариантах осуществления замена аминокислотного остатка 2 VP1 и/или VP3 обеспечивает улучшение сборки частицы rAAV на по меньшей мере приблизительно 5%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 100%. В некоторых вариантах осуществления сборку частицы rAAV с замененной аминокислотой в положении 2 можно сравнить с капсидом AAV дикого типа или исходным капсидом AAV, например, одного и того же серотипа. Например, в некоторых вариантах осуществления замена аминокислотного остатка 2 VP1 и/или VP3 обеспечивает улучшение сборки частицы rAAV на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20%, например, по сравнению со сборкой частицы rAAV, содержащей капсид дикого типа. Сборку частицы AAV можно измерять с помощью различных анализов, известных из уровня техники, в том числе без ограничения измерения количества и/или скорости образования частиц, количественной оценки образования капсидов (например, после очистки с помощью любого из способов, описанных в данном документе), анализа образования полных капсидов с векторами по сравнению с пустыми капсидами, измерения эффективности трансдукции, визуализации или структурного анализа в целях наблюдения образования частиц (например, с помощью электронного микроскопа), образования капсидных белков AAV (например, анализируемых с помощью вестерн-блоттинга) и т.п.
[0164] Другие аспекты настоящего изобретения относятся к способам улучшения трасндукции частицы rAAV. В некоторых вариантах осуществления способы предусматривают осуществление замены аминокислотного остатка 2 VP1 и/или VP3, например, как описано в данном документе. Например, в некоторых вариантах осуществления аминокислотный остаток 2 VP1 является замененным. В других вариантах осуществления аминокислотный остаток 2 VP3 является замененным. В некоторых вариантах осуществления замененная аминокислота в положении 2 подвергается N-ацетилированию с более высокой частотой, чем аминокислотный остаток 2 исходного VP1 и/или VP3, например, как описано в данном документе. В некоторых вариантах осуществления замена аминокислотного остатка 2 VP1 и/или VP3 обеспечивает улучшение трансдукции частицы rAAV на по меньшей мере приблизительно 5%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 100%. В некоторых вариантах осуществления трансдукцию частицы rAAV с замененной аминокислотой в положении 2 можно сравнить с капсидом AAV дикого типа или исходным капсидом AAV, например, одного и того же серотипа. Например, в некоторых вариантах осуществления замена аминокислотного остатка 2 VP1 и/или VP3 обеспечивает улучшение трансдукции частицы rAAV на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20%, например, по сравнению с трансдукцией частицы rAAV, содержащей капсид дикого типа. Трансдукцию частицы AAV можно измерять с помощью различных анализов, известных из уровня техники, в том числе без ограничения анализов эффективности трансдукции, описанных в данном документе. В некоторых вариантах осуществления в настоящем изобретении предусмотрены способы уменьшения трансдукции частицы rAAV; например, путем замены аминокислотного остатка 2 VP1 и/или VP3.
[0165] Другие аспекты настоящего изобретения относятся к способам повышения стабильности частицы rAAV. В некоторых вариантах осуществления способы предусматривают осуществление замены аминокислоты VP1 и/или VP3, которая обеспечивает изменение дезамидирования, например, как описано в данном документе. Например, в некоторых вариантах осуществления A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 являются замененными. В некоторых вариантах осуществления замененная аминокислота подвергается дезамидированию с более высокой частотой, чем аминокислотный остаток исходного VP1 и/или VP3, например, как описано в данном документе. В некоторых вариантах осуществления замена A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и/или G716 VP3 обеспечивает улучшение стабильности частицы rAAV на по меньшей мере приблизительно 5%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 100%. В некоторых вариантах осуществления стабильность частицы rAAV с замененными A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 можно сравнить с капсидом AAV дикого типа или исходным капсидом AAV, например, одного и того же серотипа. Например, в некоторых вариантах осуществления замена A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и/или G716 VP3 обеспечивает улучшение стабильности частицы rAAV на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20%, например, по сравнению со стабильностью частицы rAAV, содержащей капсид дикого типа. Стабильность частицы AAV можно измерять с помощью различных анализов, известных из уровня техники, в том числе без ограничения дифференциальной сканирующей флуориметрии (DSF), дифференциальной сканирующей калориметрии (DSC), других анализов термической денатурации, восприимчивости к протеолизу, визуализации или структурного анализа с наблюдением денатурации (например, с помощью электронного микроскопа), эффективности трансдукции или другого функционального анализа в отношении композиций на основе частиц AAV, хранящихся в течение определенного периода времени при определенной температуре (например, комнатной температуре или 4°C в случае термоустойчивости) или обрабатываемых при определенном значении pH (например, стабильность в зависимости от значений pH) и т.п.
[0166] Другие аспекты настоящего изобретения относятся к способам улучшения сборки частицы rAAV. В некоторых вариантах осуществления способы предусматривают осуществление замены аминокислоты VP1 и/или VP3, которая обеспечивает изменение дезамидирования, например, как описано в данном документе. Например, в некоторых вариантах осуществления A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 являются замененными. В некоторых вариантах осуществления замененная аминокислота подвергается дезамидированию с более высокой частотой, чем аминокислотный остаток исходного VP1 и/или VP3, например, как описано в данном документе. В некоторых вариантах осуществления замена A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и/или G716 VP3 обеспечивает улучшение сборки частицы rAAV на по меньшей мере приблизительно 5%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 100%. В некоторых вариантах осуществления стабильность частицы rAAV с замененными A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 можно сравнить с капсидом AAV дикого типа или исходным капсидом AAV, например, одного и того же серотипа. Например, в некоторых вариантах осуществления замена A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и/или G716 VP3 обеспечивает улучшение сборки частицы rAAV на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20%, например, по сравнению со сборкой частицы rAAV, содержащей капсид дикого типа. Сборку частицы AAV можно измерять с помощью различных анализов, известных из уровня техники, в том числе без ограничения измерения количества и/или скорости образования частиц, количественной оценки образования капсидов (например, после очистки с помощью любого из способов, описанных в данном документе), анализа образования полных капсидов с векторами по сравнению с пустыми капсидами, измерения эффективности трансдукции, визуализации или структурного анализа в целях наблюдения образования частиц (например, с помощью электронного микроскопа), образования капсидных белков AAV (например, анализируемых с помощью вестерн-блоттинга) и т.п.
[0167] Другие аспекты настоящего изобретения относятся к способам улучшения трасндукции частицы rAAV. В некоторых вариантах осуществления способы предусматривают осуществление замены аминокислоты VP1 и/или VP3, которая обеспечивает изменение дезамидирования, например, как описано в данном документе. Например, в некоторых вариантах осуществления A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 являются замененными. В некоторых вариантах осуществления замененная аминокислота подвергается дезамидированию с более высокой частотой, чем аминокислотный остаток исходного VP1 и/или VP3, например, как описано в данном документе. В некоторых вариантах осуществления замена A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и/или G716 VP3 обеспечивает улучшение трансдукции частицы rAAV на по меньшей мере приблизительно 5%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 100%. В некоторых вариантах осуществления стабильность частицы rAAV с замененными A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 можно сравнить с капсидом AAV дикого типа или исходным капсидом AAV, например, одного и того же серотипа. Например, в некоторых вариантах осуществления замена A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 и/или G716 VP3 обеспечивает улучшение трансдукции частицы rAAV на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20%, например, по сравнению с трансдукцией частицы rAAV, содержащей капсид дикого типа. Трансдукцию частицы AAV можно измерять с помощью различных анализов, известных из уровня техники, в том числе без ограничения анализов эффективности трансдукции, описанных в данном документе.
[0168] В некоторых аспектах в настоящем изобретении предусмотрены вирусные частицы, содержащие рекомбинантный самокомплементарный геном (например, самокомплементарный или самокомплементирующий вектор на основе rAAV). Вирусные частицы AAV с самокомплементарными геномами вектора и способы применения самокомплементарных геномов AAV описаны в патентах США №№ 6596535; 7125717; 7 465 583; 7785888; 7790154; 7846729; 8093054 и 8361457; и Wang Z., et al., (2003) Gene Ther 10:2105-2111, каждый из которых включен в данный документ посредством ссылки во всей своей полноте. rAAV, содержащий самокомплементарный геном, будет быстро образовывать двухцепочечную молекулу ДНК благодаря своим частично комплементарным последовательностям (например, комплементарным кодирующим и некодирующим цепям гетерологичной нуклеиновой кислоты). В некоторых вариантах осуществления вектор содержит первую последовательность нуклеиновой кислоты, кодирующую гетерологичную нуклеиновую кислоту, и вторую последовательность нуклеиновой кислоты, кодирующую комплементарную последовательность нуклеиновой кислоты, где первая последовательность нуклеиновой кислоты может образовывать внутрицепочечные пары оснований со второй последовательностью нуклеиновой кислоты вдоль большей части или всей ее длины.
[0169] В некоторых вариантах осуществления первая гетерологичная последовательность нуклеиновой кислоты и вторая гетерологичная последовательность нуклеиновой кислоты соединены с помощью мутантного ITR (например, правого ITR). В некоторых вариантах осуществления ITR содержит полинуклеотидную последовательность 5'-CACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCC GGGCGACCAAAGGTCGCCCACGCCCGGGCTTTGCCCGGGCG - 3' (SEQ ID NO: 8). Мутантный ITR характеризуется делецией D-области, содержащей последовательность концевого разрешения. В результате, при репликации вирусного генома AAV белки rep не будут расщеплять вирусный геном в месте мутантного ITR, и в связи с этим в вирусный капсид будет упакован рекомбинантный вирусный геном, содержащий следующее в порядке от 5'- к 3'-концу: ITR AAV, первую гетерологичную полинуклеотидную последовательность, в том числе регуляторные последовательности, мутантный ITR AAV, второй гетерологичный полинуклеотид в обратной ориентации по отношению к первому гетерологичному полинуклеотиду и третий ITR AAV.
[0170] Для оптимизации трансдукции конкретных клеток-мишеней или для нацеливания на конкретные типы клеток в конкретной ткани-мишени (например, пораженной заболеванием ткани) применяют различные серотипы AAV. Частица rAAV может содержать белки вируса и нуклеиновые кислоты вируса одного и того же серотипа или смешанного серотипа. Например, частица rAAV может содержать один или несколько ITR и капсид, полученные из одного и того же серотипа AAV, или частица rAAV может содержать один или несколько ITR, полученных из серотипа AAV, отличного от серотипа капсида частицы rAAV.
[0171] В некоторых вариантах осуществления капсид AAV содержит мутацию, например, капсид содержит мутантный капсидный белок. В некоторых вариантах осуществления мутация представляет собой мутацию по тирозину или мутацию, влияющую на связывание с гепарином. В некоторых вариантах осуществления мутантный капсидный белок сохраняет способность формировать капсид AAV. В некоторых вариантах осуществления частица rAAV содержит капсид AAV2 или AAV5, мутантный по тирозину (см., например, Zhong L. et al., (2008) Proc Natl Acad Sci U S A 105(22):7827-7832), например, с мутацией в Y444 или Y730 (нумерация в соответствии с AAV2). В дополнительных вариантах осуществления частица rAAV содержит капсидные белки серотипа AAV из клад A-F (Gao, et al., J. Virol. 2004, 78(12):6381).
[0172] В некоторых вариантах осуществления капсидный белок содержит одну или несколько аминокислотных замен в одном или нескольких положениях, которые участвуют во взаимодействии с гепаринсульфатпротеогликаном, или в одном или нескольких положениях, соответствующих аминокислотам 484, 487, 527, 532, 585 или 588, при этом нумерация приведена согласно нумерации VP1 AAV2. Из уровня техники известно, что гепарансульфатпротеогликан (HSPG) выступает в качестве клеточного рецептора частиц AAV2 (Summerford, C. and Samulski, R.J. (1998) J. Virol. 72(2):1438-45). Связывание между частицей AAV2 и HSPG на клеточной мембране необходимо для прикрепления частицы к клетке. Другие белки клеточной поверхности, такие как рецептор фактора роста фибробластов и интегрин αvβ5, также могут облегчать инфицирование клетки. После связывания частица AAV2 может проникать в клетку посредством механизмов, включающих рецептор-опосредованный эндоцитоз с помощью клатрин-окаймленных ямок. Частица AAV2 может высвобождаться из эндоцитозного пузырька при эндосомальном подкислении. Это способствует миграции частицы AAV2 к перинуклеарной области, а затем к клеточному ядру. Также известно, что частицы AAV3 связываются с гепарином (Rabinowitz, J.E., et al. (2002) J. Virol. 76(2):791-801).
[0173] Известно, что связывание между капсидными белками AAV2 и HSPG происходит за счет электростатических взаимодействий между основными остатками капсидных белков AAV2 и отрицательно зараженными остатками гликозаминогликанов (Opie, SR et al., (2003) J. Virol. 77:6995-7006; Kern, A et al., (2003) J. Virol. 77:11072-11081). Конкретные капсидные остатки, вовлеченные в эти взаимодействия, включают R484, R487, K527, K532, R585 и R588. Было показано, что мутации в этих остатках обеспечивают уменьшение связывания AAV2 с клетками Hela и самим гепарином (Opie, SR et al., (2003) J. Virol. 77:6995-7006; Kern, A et al., (2003) J. Virol. 77:11072-11081; WO 2004/027019 A2, патент США № 7629322). Кроме того, без ограничения какой-либо теорией, считается, что посредством аминокислотной (аминокислотных) замены (замен) в одном или нескольких остатках, соответствующих аминокислотам 484, 487, 527, 532, 585 или 588, нумерация которых приведена согласно нумерации VP1 AAV2, можно модулировать трансдукционные свойства типов капсидов AAV, которые не связываются с HSPG, или можно модулировать трансдукционные свойства типов капсидов AAV независимо от их способности к связыванию с HSPG. В некоторых вариантах осуществления одна или несколько аминокислотных замен предусматривает замену в положении R484, R487, K527, K532, R585 и/или R588 VP1, VP2 и/или VP3, нумерация которых приведена согласно нумерации VP1 AAV2.
[0174] В некоторых вариантах осуществления одна или несколько аминокислотных замен обеспечивают уменьшение связывания частицы rAAV с гепаринсульфатпротеогликаном на приблизительно по меньшей мере 10%, приблизительно по меньшей мере 25%, приблизительно по меньшей мере 50%, приблизительно по меньшей мере 75% или приблизительно по меньшей мере 100%. В некоторых вариантах осуществления одна или несколько аминокислотных замен обеспечивают уменьшение связывания частицы rAAV с гепаринсульфатпротеогликаном на приблизительно по меньшей мере 10%, приблизительно по меньшей мере 15%, приблизительно по меньшей мере 20%, приблизительно по меньшей мере 25%, приблизительно по меньшей мере 30%, приблизительно по меньшей мере 35%, приблизительно по меньшей мере 40%, приблизительно по меньшей мере 45%, приблизительно по меньшей мере 50%, приблизительно по меньшей мере 55%, приблизительно по меньшей мере 60%, приблизительно по меньшей мере 65%, приблизительно по меньшей мере 70%, приблизительно по меньшей мере 75%, приблизительно по меньшей мере 80%, приблизительно по меньшей мере 85%, приблизительно по меньшей мере 90%, приблизительно по меньшей мере 95% или приблизительно по меньшей мере 100% (по сравнению со связыванием частицы rAAV, содержащей капсид дикого типа). В некоторых вариантах осуществления одна или несколько аминокислотных замен обеспечивают уменьшение связывания частицы rAAV с гепаринсульфатпротеогликаном на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20% (по сравнению со связыванием частицы rAAV, содержащей капсид дикого типа). В некоторых вариантах осуществления одна или несколько аминокислотных замен приводят к отсутствию выявляемого связывания частицы rAAV с гепаринсульфатпротеогликаном по сравнению со связыванием частицы rAAV дикого типа. Способы измерения связывания частиц AAV с HSPG известны из уровня техники; например, связывание с хроматографической средой на основе гепаринсульфата или связывание с клеткой, которая, как известно, экспрессирует HSPG на своей поверхности. Например, см. Opie, SR et al., (2003) J. Virol. 77:6995-7006, и Kern, A et al., (2003) J. Virol. 77:11072-11081. В некоторых вариантах осуществления одна или несколько аминокислотных замен обеспечивают улучшение эффективности трансдукции частицы rAAV в клетку (например, клетку глаза или ЦНС) на приблизительно по меньшей мере 10%, приблизительно по меньшей мере 15%, приблизительно по меньшей мере 20%, приблизительно по меньшей мере 25%, приблизительно по меньшей мере 30%, приблизительно по меньшей мере 35%, приблизительно по меньшей мере 40%, приблизительно по меньшей мере 45%, приблизительно по меньшей мере 50%, приблизительно по меньшей мере 55%, приблизительно по меньшей мере 60%, приблизительно по меньшей мере 65%, приблизительно по меньшей мере 70%, приблизительно по меньшей мере 75%, приблизительно по меньшей мере 80%, приблизительно по меньшей мере 85%, приблизительно по меньшей мере 90%, приблизительно по меньшей мере 95% или приблизительно по меньшей мере 100% (по сравнению с эффективностью трансдукции частицы rAAV, содержащей капсид дикого типа). В некоторых вариантах осуществления одна или несколько аминокислотных замен обеспечивают улучшение эффективности трансдукции частицы rAAV в клетку (например, клетку глаза или ЦНС) на любую величину от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20% (по сравнению с эффективностью трансдукции частицы rAAV, содержащей капсид дикого типа). Способы измерения эффективности трансдукции частиц AAV в клетку (например, клетку в культуре или части ткани) известны из уровня техники. Например, популяция клеток (например, в культуре или части ткани) может быть инфицирована скоплением частиц rAAV, содержащих вектор, экспрессия в клетках которого обеспечивает образование репортера, который можно проанализировать (например, флуоресценция GFP, образование sFLT и т.д.).
Капсидные белки AAV
[0175] В некоторых аспектах в настоящем изобретении предусмотрен капсидный белок AAV, содержащий аминокислотную замену по аминокислотному остатку 2; при этом аминокислотная замена по аминокислотному остатку 2 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 исходного капсидного белка AAV. В некоторых вариантах осуществления капсидный белок AAV представляет собой VP1 или VP3. В некоторых вариантах осуществления аминокислотный остаток 2 капсидного белка AAV (например, VP1 или VP3) заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr. В некоторых вариантах осуществления аминокислотная замена приводит к меньшей степени дезамидирования капсидного белка AAV. Неограничивающие примеры капсидных белков AAV по настоящему изобретению включают VP1 и/или VP3 любого из следующих серотипов AAV: капсид серотипов AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV DJ8, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерного AAV1/AAV2, AAV крупного рогатого скота, AAV мыши или rAAV2/HBoV1. В некоторых вариантах осуществления капсид AAV дополнительно содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином.
Получение частиц AAV
[0176] Из уровня техники известны многочисленные способы получения векторов на основе rAAV, в том числе трансфекция, получение стабильных линий клеток и применение систем продуцирования на основе инфекционных гибридных вирусов, которые включают гибриды аденовирус-AAV, гибриды вирус герпеса-AAV (Conway, JE et al., (1997) J. Virology 71(11):8780-8789) и гибриды бакуловирус-AAV (Urabe, M. et al., (2002) Human Gene Therapy 13(16):1935-1943; Kotin, R. (2011) Hum Mol Genet. 20(R1): R2-R6). Для получения вирусных частиц rAAV всем культурам для продуцирования rAAV необходимы: 1) подходящие клетки-хозяева, 2) подходящий функциональный элемент вируса-помощника, 3) гены rep и cap AAV и продукты генов; 4) нуклеиновая кислота (такая как терапевтическая нуклеиновая кислота), фланкированная по меньшей мере одной последовательностью ITR AAV (например, генома AAV, кодирующего GNPTAB); и 5) подходящая среда и компоненты среды для поддержания продуцирования rAAV. В некоторых вариантах осуществления подходящая клетка-хозяин представляет собой клетку-хозяина, полученную от примата. В некоторых вариантах осуществления подходящая клетка-хозяин представляет собой линии клеток, полученные от человека, такие как клетки HeLa, A549, 293 или Perc.6. В некоторых вариантах осуществления подходящий функциональный элемент вируса-помощника обеспечивается аденовирусом дикого типа или мутантным аденовирусом (таким как термочувствительный аденовирус), вирусом герпеса (HSV), бакуловирусом или плазмидной конструкцией, обеспечивающей функциональные элементы помощника. В некоторых вариантах осуществления продукты генов rep и cap AAV могут происходить из любого серотипа AAV. В целом, но не обязательно, продукт гена rep AAV относится к тому же серотипу, что и ITR из генома вектора на основе rAAV, при условии, что продукты гена rep могут обеспечивать возможность репликации и упаковки генома rAAV. Для получения векторов на основе rAAV можно применять подходящие среды, известные из уровня техники. Эти среды включают без ограничения среды, производимые Hyclone Laboratories и JRH, в том числе модифицированную среду Игла (MEM), среду Игла, модифицированную по Дульбекко (DMEM), изготавливаемые по заказу составы, такие как описанные в патенте США №6566118, и среду Sf-900 II SFM, описанную в патенте США №6723551, каждый из которых включен в данный документ посредством ссылки во всей своей полноте, в особенности в отношении изготавливаемых по заказу составов сред для применения в получении рекомбинантных векторов на основе AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV представлены аденовирусом или HSV. В некоторых вариантах осуществления функциональные элементы помощника AAV представлены бакуловирусом, и клетка-хозяин представляет собой клетку насекомого (например, клетки Spodoptera frugiperda (Sf9)). В некоторых вариантах осуществления функциональные элементы cap AAV обеспечивают аминокислотную замену по аминокислотному остатку 2 VP1 и/или VP3; при этом аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 и/или VP3 частицы исходного AAV. В некоторых вариантах осуществления аминокислотный остаток 2 капсидного белка AAV (например, VP1 или VP3) заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr. В некоторых вариантах осуществления аминокислотная замена приводит к меньшей степени дезамидирования капсида AAV.
[0177] Один способ получения частиц rAAV представляет собой способ тройной трансфекции. Вкратце, плазмиду, содержащую ген rep и ген капсида, вместе с аденовирусной плазмидой-помощником можно ввести путем трансфекции (например, с применением способа с использованием фосфата кальция) в линию клеток (например, клетки HEK-293), и вирус можно собирать и необязательно очищать. Таким образом, в некоторых вариантах осуществления частица rAAV была получена с помощью введения путем тройной трансфекции нуклеиновой кислоты, кодирующей вектор на основе rAAV, нуклеиновой кислоты, кодирующей rep и cap AAV, и нуклеиновой кислоты, кодирующей функциональные элементы вируса-помощника AAV, в клетку-хозяина, при этом введение путем трансфекции нуклеиновых кислот в клетки-хозяева приводит к получению клетки-хозяина, способной продуцировать частицы rAAV.
[0178] В некоторых вариантах осуществления частицы rAAV можно получить с помощью способа с использованием линии клеток-продуцентов (см. Martin et al., (2013) Human Gene Therapy Methods 24:253-269, заявку на патент США № US 2004/0224411, опубликованную до выдачи патента; и Liu, X.L. et al. (1999) Gene Ther. 6:293-299). Вкратце, линию клеток (например, линию клеток HeLa, 293, A549 или Perc.6) можно стабильно трансфицировать плазмидой, содержащей ген rep, ген капсида и геном вектора, содержащий последовательность нуклеиновой кислоты с гетерологичным промотором (например, GNPTAB). Линии клеток можно подвергать скринингу для отбора ведущего клона для продуцирования rAAV, который можно затем размножать в производственном реакторе и инфицировать вирусом-помощником (например, аденовирусом или HSV) для инициации продуцирования rAAV. Затем вирус можно собрать, аденовирус можно инактивировать (например, под действием тепла) и/или удалить, а частицы rAAV можно очистить. В связи с этим, в некоторых вариантах осуществления частица rAAV была получена с помощью линии клеток-продуцентов, содержащих одну или несколько из нуклеиновой кислоты, кодирующей вектор на основе rAAV, нуклеиновой кислоты, кодирующей rep и cap AAV, и нуклеиновой кислоты, кодирующей функциональные элементы вируса-помощника AAV. Как описано в данном документе, способ с использованием линии клеток-продуцентов может быть преимущественным для получения частиц rAAV с увеличенным в размере геном по сравнению со способом тройной трансфекции.
[0179] В некоторых вариантах осуществления нуклеиновая кислота, кодирующая гены rep и cap AAV, и/или геном rAAV стабильно поддерживаются в линии клеток-продуцентов. В некоторых вариантах осуществления нуклеиновую кислоту, кодирующую гены rep и cap AAV, и/или геном rAAV вводят в линию клеток с помощью одной или нескольких плазмид с получением линии клеток-продуцентов. В некоторых вариантах осуществления rep AAV, cap AAV и геном rAAV вводят в клетку с помощью одной и той же плазмиды. В других вариантах осуществления rep AAV, cap AAV и геном rAAV вводят в клетку с помощью разных плазмид. В некоторых вариантах осуществления в линии клеток, стабильно трансформированной плазмидой, плазмида сохраняется в течение нескольких пассажей линии клеток (например, 5, 10, 20, 30, 40, 50 или более 50 пассажей клетки). Например, плазмида(плазмиды) может(могут) реплицироваться при делении клетки, или плазмида(плазмиды) может(могут) интегрироваться в геном клетки. Было идентифицировано множество последовательностей, которые обеспечивают возможность автономной репликации плазмиды в клетке (например, клетке человека) (см., например, Krysan, P.J. et al. (1989) Mol. Cell Biol. 9:1026-1033). В некоторых вариантах осуществления плазмида(плазмиды) может(могут) содержать селектируемый маркер (например, маркер устойчивости к антибиотикам), который обеспечивает возможность отбора клеток, содержащих плазмиду. Селектируемые маркеры, обычно используемые в клетках млекопитающих, включают без ограничения бластицидин, G418, гигромицин B, зеоцин, пуромицин и их производные. Способы введения нуклеиновых кислот в клетку известны из уровня техники и включают без ограничения вирусную трансдукцию, катионную трансфекцию (например, с использованием катионного полимера, такого как DEAE-декстран, или катионного липида, такого как липофектамин), трансфекцию с использованием фосфата кальция, микроинъекцию, бомбардировку частицами, электропорацию и трансфекцию с использованием наночастиц (для более подробной информации см., например, Kim, T.K. and Eberwine, J.H. (2010) Anal. Bioanal. Chem. 397:3173-3178).
[0180] В некоторых вариантах осуществления нуклеиновая кислота, кодирующая гены rep и cap AAV, и/или геном rAAV стабильно интегрируются в геном линии клеток-продуцентов. В некоторых вариантах осуществления нуклеиновую кислоту, кодирующую гены rep и cap AAV, и/или геном rAAV вводят в линию клеток с помощью одной или нескольких плазмид с получением линии клеток-продуцентов. В некоторых вариантах осуществления rep AAV, cap AAV и геном rAAV вводят в клетку с помощью одной и той же плазмиды. В других вариантах осуществления rep AAV, cap AAV и геном rAAV вводят в клетку с помощью разных плазмид. В некоторых вариантах осуществления плазмида (плазмиды) может (могут) содержать селектируемый маркер (например, маркер устойчивости к антибиотикам), который обеспечивает возможность отбора клеток, содержащих плазмиду. Способы стабильной интеграции нуклеиновых кислот во множество линий клеток-хозяев известны из уровня техники. Например, повторный отбор (например, посредством применения селектируемого маркера) можно использовать для отбора клеток, содержащих интегрированную нуклеиновую кислоту, содержащую селектируемый маркер (и гены cap и rep AAV и/или геном rAAV). В других вариантах осуществления нуклеиновые кислоты могут интегрироваться в линию клеток сайт-специфичным образом с получением линии клеток-продуцентов. Несколько систем сайт-специфичной рекомбинации известны из уровня техники, такие как FLP/FRT (см., например, O'Gorman, S. et al. (1991) Science 251:1351-1355), Cre/loxP (см., например, Sauer, B. and Henderson, N. (1988) Proc. Natl. Acad. Sci. 85:5166-5170) и phi C31-att (см., например, Groth, A.C. et al. (2000) Proc. Natl. Acad. Sci. 97:5995-6000).
[0181] В некоторых вариантах осуществления линия клеток-продуцентов получена из линии клеток приматов (например, линии клеток отличных от человека приматов, такой как линия клеток Vero или FRhL-2). В некоторых вариантах осуществления линия клеток получена из линии клеток человека. В некоторых вариантах осуществления линия клеток-продуцентов получена из клеток HeLa, 293, A549 или PERC.6® (Crucell). Например, перед введением и/или стабильным поддержанием/интеграцией нуклеиновой кислоты, кодирующей гены rep и cap AAV, и/или увеличенного в размере генома rAAV в линию клеток для получения линии клеток-продуцентов линия клеток представляет собой линию клеток HeLa, 293, A549 или PERC.6® (Crucell) или их производное.
[0182] В некоторых вариантах осуществления линия клеток-продуцентов адаптирована для роста в суспензии. Как известно из уровня техники, субстратзависимые клетки, как правило, не способны расти в суспензии без субстрата, такого как гранулы-микроносители. Адаптация линии клеток к росту в суспензии может предусматривать, например, выращивание линии клеток в культурах с постоянным перемешиванием с помощью перемешивающей лопасти, применение культуральной среды, не содержащей ионов кальция и магния с целью предотвращения агрегации (и необязательно противовспенивающего средства), применение сосуда для культивирования, покрытого силиконизирующим соединением, и отбор клеток в культуре (а не в больших скоплениях или на стенках сосуда) при каждом пассаже. Дополнительное описание см., например, в документе ATCC с часто задаваемыми вопросами (доступном во всемирной сети Интернет по адресу atcc.org/Global/FAQs/9/1/Adapting%20a%20monolayer%20cell%20line%
20to%20suspension-40.aspx) и ссылках, упоминаемых в данном документе.
[0183] Подходящие культуральные среды для продуцирования AAV по настоящему изобретению могут быть дополнены сывороткой крови или рекомбинантными белками, полученными из сыворотки крови, на уровне 0,5% - 20% (об./об. или вес/об.). В качестве альтернативы, как известно из уровня техники, векторы на основе AAV можно получать в условиях, предусматривающих отсутствие сыворотки крови, которые также могут означать применение среды, не содержащей продуктов животного происхождения. Специалист в данной области техники может понять, что коммерческие или изготавливаемые по заказу среды, разработанные для обеспечения продуцирования векторов на основе AAV, также могут быть дополнены одним или несколькими компонентами для культивирования клеток, известными из уровня техники, в том числе без ограничения глюкозой, витаминами, аминокислотами и/или факторами роста, в целях повышения титра AAV в культурах для продуцирования.
[0158] Культуры для продуцирования AAV можно выращивать при различных условиях (в широком диапазоне температур, в течение различных промежутков времени и т.п.), подходящих для конкретной используемой клетки-хозяина. Как известно из уровня техники, культуры для продуцирования AAV включают зависимые от прикрепления культуры, которые можно культивировать в подходящих сосудах для зависимых от прикрепления культур, таких как, например, роллерные флаконы, фильтры с полыми волокнами, микроносители и биореакторы с уплотненным слоем или псевдоожиженным слоем. Культуры для продуцирования векторов на основе AAV могут также включать клетки-хозяева, адаптированные к суспензии, такие как клетки HeLa, 293 и SF-9, которые можно культивировать с помощью множества способов, в том числе, например, с помощью флаконов с перемешиванием, биореакторов с механическим перемешиванием и одноразовых систем, таких как система в виде мешка Wave.
[0159] Частицы, представляющие собой вектор на основе AAV по настоящему изобретению, можно собирать из культур для продуцирования AAV посредством лизиса клеток-хозяев из культуры для продуцирования или посредством сбора отработанной среды из культуры для продуцирования, при условии, что клетки культивируют при условиях, которые, как известно из уровня техники, вызывают высвобождение частиц AAV из интактных клеток в среду, как описано более подробно в патенте США № 6566118. Подходящие способы лизиса клеток также известны из уровня техники и включают, например, несколько циклов замораживания/размораживания, обработку ультразвуком, микрофлюидизацию и обработку химическими веществами, такими как детергенты и/или протеазы.
[0160] В дополнительных вариантах осуществления частицы AAV являются очищенными. Термин "очищенный", используемый в данном документе, подразумевает препарат из частиц AAV, в котором отсутствуют по меньшей мере некоторые другие компоненты, которые могут также присутствовать в среде, где частицы AAV находятся в естественном состоянии или из которой они изначально получены. Таким образом, например, выделенные частицы AAV можно получать с помощью методики очистки для обогащения ими исходной смеси, такой как лизат культуры или надосадочная жидкость культуры для продуцирования. Обогащение можно измерять с помощью множества способов, таких как, например, по доле устойчивых к ДНКазе частиц (DRP) или копий генома (gc), присутствующих в растворе, или по инфекционности, либо его можно измерять по отношению ко второму потенциально мешающему веществу, присутствующему в исходной смеси, такому как контаминанты, в том числе контаминанты из культуры для продуцирования или контаминанты, образуемые в ходе продуцирования, в том числе вирус-помощник, компоненты среды и т. п.
[0161] В некоторых вариантах осуществления сбор культуры для продуцирования AAV очищают от примесей с удалением дебриса клеток-хозяев. В некоторых вариантах осуществления сбор культуры для продуцирования очищают от примесей путем фильтрации через серию пористых фильтров, включающих, например, модульный фильтр Millipore Millistak+ HC марки DOHC, модульный фильтр Millipore Millistak+ HC марки A1HC и фильтр с размером пор 0,2 мкм Opticap XL1O с гидрофильной мембраной фильтра Millipore Express SHC. Очистку от примесей также можно осуществлять с помощью ряда других стандартных методик, известных из уровня техники, таких как центрифугирование или фильтрация через любой фильтр из ацетата целлюлозы с размером пор 0,2 мкм или больше, известный из уровня техники.
[0162] В некоторых вариантах осуществления сбор культуры для продуцирования AAV дополнительно обрабатывают с помощью Benzonase® для расщепления любой высокомолекулярной ДНК, присутствующей в культуре для продуцирования. В некоторых вариантах осуществления расщепление с помощью Benzonase® осуществляют в стандартных условиях, известных из уровня техники, в том числе, например, при конечной концентрации 1-2,5 единиц Benzonase®/мл, при температуре, варьирующей в диапазоне от температуры окружающей среды до 37°C, в течение периода времени от 30 минут до нескольких часов.
[0163] Частицы AAV можно выделять или очищать с использованием одной или нескольких следующих стадий очистки: равновесного центрифугирования; проточной анионообменной фильтрации; тангенциальной поточной фильтрации (TFF) для концентрирования частиц AAV; захвата AAV посредством хроматографии на апатите; термоинактивации вируса-помощника; захвата AAV посредством хроматографии гидрофобных взаимодействий; замены буфера посредством эксклюзионной хроматографии (SEC); нанофильтрации и захвата AAV посредством анионообменной хроматографии, катионообменной хроматографии или аффинной хроматографии. Эти стадии можно использовать в отдельности, в различных комбинациях или в разном порядке. В некоторых вариантах осуществления способ включает все стадии в порядке, описанном ниже. Способы очистки частиц AAV встречаются, например, в Xiao et al., (1998) Journal of Virology 72:2224-2232; патентах США №№ 6989264 и 8137948 и WO 2010/148143.
Фармацевтические композиции
[0164] В некоторых вариантах осуществления частица AAV по настоящему изобретению (например, частица rAAV) находится в фармацевтической композиции. Фармацевтические композиции могут подходить для любого способа введения, описанного в данном документе или известного из уровня техники. В некоторых вариантах осуществления фармацевтическая композиция содержит частицы rAAV, модифицированные для улучшения стабильности и/или улучшения эффективности трансдукции частиц rAAV; например, для применения при осуществлении замены аминокислотного остатка в положении 2 VP1 и/или VP3 для улучшения ацетилирования капсидных белков rAAV. В некоторых вариантах осуществления фармацевтическая композиция содержит частицы rAAV, модифицированные для модулирования стабильности и/или эффективности трансдукции частиц rAAV (например, повышения стабильности и/или эффективности трансдукции или снижения стабильности и/или эффективности трансдукции), например, для применения при осуществлении замены аминокислотных остатков, которые обеспечивают модулирование дезамидирования (например, повышение степени дезамидирования или снижение степени дезамидирования).
[0165] В некоторых вариантах осуществления частица rAAV находится в фармацевтической композиции, содержащей фармацевтически приемлемое вспомогательное вещество. Как хорошо известно из уровня техники, фармацевтически приемлемые вспомогательные вещества представляют собой относительно инертные вещества, облегчающие введение фармакологически эффективного вещества, и могут предоставляться в виде жидких растворов или суспензий, в виде эмульсий или в виде твердых форм, подходящих для растворения или суспендирования в жидкости перед применением. Например, вспомогательное вещество может придавать форму или консистенцию или выступать в качестве разбавителя. Подходящие вспомогательные вещества включают без ограничения стабилизирующие средства, смачивающие и эмульгирующие средства, соли для изменения осмолярности, инкапсулирующие средства, буферные вещества для поддержания pH и буферы. Такие наполнители включают любое фармацевтическое средство, подходящее для непосредственной доставки в глаз, которое можно вводить без излишней токсичности. Фармацевтически приемлемые вспомогательные вещества включают без ограничения сорбит, любое из различных соединений TWEEN и жидкости, такие как вода, солевой раствор, глицерин и этанол. В них могут быть включены фармацевтически приемлемые соли, например, соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты, сульфаты и т.п.; и соли органических кислот, такие как ацетаты, пропионаты, малонаты, бензоаты и т.п. Всестороннее обсуждение фармацевтически приемлемых вспомогательных веществ доступно в REMINGTON'S PHARMACEUTICAL SCIENCES (Mack Pub. Co., N.J. 1991). В некоторых вариантах осуществления фармацевтическая композиция, содержащая частицу rAAV, описанную в данном документе, и фармацевтически приемлемый носитель, подходит для введения человеку. Такие носители хорошо известны из уровня техники (см., например, Remington's Pharmaceutical Sciences, 15th Edition, pp. 1035-1038 и 1570-1580).
[0166] Такими фармацевтически приемлемыми носителями могут быть стерильные жидкости, такие как вода и масло, в том числе масла нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло и т.п. В качестве жидких носителей, в частности для инъекционных растворов, также могут использоваться солевые растворы и водные растворы декстрозы, полиэтиленгликоля (PEG) и глицерина. Фармацевтическая композиция может дополнительно содержать дополнительные ингредиенты, например, консерванты, буферы, вещества, регулирующие тоничность, антиоксиданты и стабилизаторы, неионные смачивающие или осветляющие средства, загустители и т. п. Фармацевтические композиции, описанные в данном документе, могут быть упакованы в единичные дозированные формы или в многодозовые формы. Как правило, композиции составляют в виде стерильного или практически изотонического раствора.
Наборы и изделия
[0167] В настоящем изобретении также предусмотрены наборы или изделия, содержащие любые из частиц rAAV и/или фармацевтических композиций по настоящему изобретению. Наборы или изделия могут содержать любые из частиц rAAV или композиций на основе частиц rAAV по настоящему изобретению. В некоторых вариантах осуществления наборы применяют для улучшения стабильности и/или улучшения эффективности трансдукции частиц rAAV; например, для применения при осуществлении замены аминокислотного остатка в положении 2 VP1 и/или VP3 для улучшения ацетилирования капсидных белков rAAV. В некоторых вариантах осуществления наборы применяют для модулирования стабильности и/или эффективности трансдукции частиц rAAV (например, повышения стабильности и/или эффективности трансдукции или снижения стабильности и/или эффективности трансдукции), например, для применения при осуществлении замены аминокислотных остатков, которые обеспечивают модулирование дезамидирования (например, повышение степени дезамидирования или снижение степени дезамидирования).
[0168] В некоторых вариантах осуществления наборы или изделия дополнительно содержат инструкции по введению композиции на основе частиц rAAV. Наборы или изделия, описанные в данном документе, могут дополнительно содержать другие материалы, необходимые с коммерческой и потребительской точки зрения, в том числе другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши с инструкциями по выполнению любых способов, описанных в данном документе. Подходящие упаковочные материалы также могут быть включены и могут представлять собой любые упаковочные материалы, известные из уровня техники, включающие, например, колбы (такие как герметично закрытые колбы), сосуды, ампулы, бутылки, банки, гибкую упаковку (например, герметизированные пакеты Майлара или пластиковые пакеты) и т.п. Такие изделия можно дополнительно стерилизовать и/или герметизировать.
[0169] В некоторых вариантах осуществления наборы или изделия дополнительно содержат один или несколько буферов и/или фармацевтически приемлемых вспомогательных веществ, описанных в данном документе (например, описываемых в REMINGTON'S PHARMACEUTICAL SCIENCES (Mack Pub. Co., N.J. 1991). В некоторых вариантах осуществления наборы или изделия содержат один или несколько фармацевтически приемлемых вспомогательных веществ, носителей, растворов и/или дополнительных ингредиентов, описанных в данном документе. Наборы или изделия, описанные в данном документе, могут быть упакованы в единичные дозированные формы или в многодозовые формы. Содержимое наборов или изделий, как правило, составлено в стерильном виде, и оно может быть лиофилизированным или обеспечиваться в виде практически изотонического раствора.
ПРИМЕРЫ
[0170] Настоящее изобретение будет более понятным со ссылкой на следующие примеры. Тем не менее, их не следует истолковывать как ограничивающие объем настоящего изобретения. Понятно, что описанные в данном документе примеры и варианты осуществления служат только в качестве иллюстрации, и что различные модификации или изменения с их учетом будут понятны специалистам в данной области техники, и они должны находиться в пределах сути и содержания настоящей заявки, а также объема прилагаемой формулы изобретения.
Пример 1. Прямая LC/MS и LC/MS/MS для полной характеристики вирусного капсидного белка рекомбинантного AAV
[0171] Рекомбинантные аденоассоциированные вирусы (rAAV) стали распространенными векторами для генной терапии благодаря своей непатогенной природе, способности инфицировать как делящиеся, так и неделящиеся клетки и длительному периоду экспрессии генов. В настоящее время виды генной терапии на основе AAV используются в клинических исследованиях для многочисленных целевых заболеваний, таких как мышечная дистрофия, гемофилия, болезнь Паркинсона, врожденный амавроз Лебера и макулярная дегенерация.
[0172] AAV представляет собой небольшой и не имеющий оболочки парвовирус с геномом на основе одноцепочечной ДНК, инкапсулированным в икосаэдрическую оболочку. Каждый капсид включает шестьдесят копий трех вирусных капсидных белков VP1 (87 кДа), VP2 (73 кДа) и VP3 (62 кДА) в примерном соотношении 1:1:10. Все три вирусных капсидных белка экспрессируются из одной и той же открытой рамки считывания посредством альтернативного сплайсинга и атипичного старт-кодона и, таким образом, имеют перекрывающиеся последовательности. VP1 имеет ~137 дополнительных N-концевых аминокислотных остатков по сравнению с VP3, в то время как VP2 имеет ~65 дополнительных N-концевых аминокислотных остатков по сравнению с VP3. По меньшей мере 13 серотипов AAV и ~150 генных последовательностей выделили из тканей человека и приматов, отличных от человека; серотипы AAV отличаются аминокислотной последовательностью вирусных капсидных белков и их соответствующих клеточных рецепторов и корецепторов для нацеливания.
[0173] Капсид AAV, помимо защиты генома изнутри, играет важную роль в опосредовании связывания с рецепторами, ускользании вируса от эндосомы и транспорте вирусной ДНК в ядро в цикле инфицирования вируса, тем самым, непосредственно влияя на инфекционность вируса. Было показано, что N-конец VP1 содержит домен фосфолипазы PLA2 (аминокислоты 52-97), который является основным при эндосомальном ускользании вируса [1-3]. N-концы VP1 и VP2 также содержат три основных аминокислотных кластера в качестве сигналов внутриядерной локализации. Эти последовательности являются высококонсервативными среди различных серотипов AAV. Было показано, что мутации этих аминокислот снижают или полностью устраняют инфекционность [4]. Помимо этого, каждый серотип AAV характеризуется соответствующими специфичными по отношению к последовательности рецепторами и корецепторами. Например, протеогликан гепарин-сульфат идентифицировали в качестве основного рецептора AAV2 и нескольких других корецепторов, в том числе интегрина αVβ5, идентифицировали рецептор 1 фактора роста фибробластов и рецептор фактора роста гепатоцитов [5-8]. В результате мутационного анализа капсидных белков AAV2 идентифицировали группу основных аминокислот (аргинин 484, 487, 585 и лизин 532) в качестве гепарин-связывающего мотива, который способствует связыванию гепарина и клеток HeLa [9]. Домен NGR AAV2 идентифицировали в качестве домена, связывающегося с интегрином α5β1, который является необходимым для вхождения вируса в клетку [10]. Таким образом, последовательности вирусных капсидных белков являются важными при клеточном нацеливании и миграции в цикле вирусной инфекции. Поскольку различные условия получения могут вызывать различные уровни экспрессии вирусных капсидных белков, посттрансляционных модификаций и усечений, вирусные капсидные белки необходимо характеризовать и контролировать в целях обеспечения однородности продукта в программах разработки генной терапии.
[0174] Традиционно для характеристики вирусных капсидных белков AAV использовали SDS-PAGE, обеспечивающий примерную информацию о молекулярном весе, такую как 87 кДа, 73 кДа и 62 кДа. Исходя из секвенирования по Эдману, информация о последовательностях не была получена, возможно, в связи с блокированными N-концами вирусных капсидных белков, кроме VP2. Несмотря на то, что были определены рентгеновские структуры многочисленных AAV, лишь последовательность участка VP3 наблюдали в кристаллических структурах. Пятнадцать N-концевых аминокислотных остатков VP3 по-прежнему отсутствовали в рентгеновской структуре, по-видимому, в связи с ее внутренним нарушением [11-13]. По-видимому, отсутствие информации о N-концевых участках VP1 и VP2 в атомной структуре могло быть связано с низкой стехиометрией VP1 и VP2 в капсиде. Кроме этого, N-концы VP1 и VP2 погружены внутрь капсида и недоступны для антител в нативном состоянии, как описано в некоторых литературных источниках [3, 14, 15]. Обычно при характеристике VP использовали способ Gel-LC/MS (SDS-PAGE, триптическое переваривание в геле и LC/MS/MS) [16-18]. Однако с использованием этого подхода не были определены N-концы VP1, VP2 и VP3, поскольку с помощью этого способа не удалось получить 100% охват последовательностей VP в связи с ограниченным извлечением пептида из геля.
[0175] Прямой анализ с помощью MALDI-TOF MS был описан для нескольких вирусных капсидных белков, в том числе вируса табачной мозаики U2 после диссоциации с органической кислотой [19]. После амидно-водородного обмена и масс-спектрометрии использовали прямое пептидное картирование для изучения pH-индуцированных структурных изменений капсида вируса мозаики костра (BMV) [20]. Поскольку AAV представляют собой не имеющие оболочки вирусы, содержащие только капсидные белки и геном, капсиды AAV можно непосредственно анализировать с помощью RP-LC/MS белков и LC/MS/MS пептидного картирования для достижения 100% охвата последовательностей после диссоциации капсида без разделения с помощью SDS-PAGE. Фрагменты ДНК можно элюировать в "мертвом объеме" и тем самым избегать интерференции при выявлении белков/пептидов с помощью LC/MS. Для изучения этих способов прямую LC/MS различных типов AAV после денатурации использовали для контроля последовательности белка и посттрансляционных модификаций капсидных белков AAV. Как описано в данном документе, N-концы VP1, VP2 и VP3 AAV определяли с помощью масс-спектрометрии. Ацетилирования N-концов VP1 и VP3 идентифицировали в другом серотипе AAV. Также разрабатывали прямое пептидное картирование AAV с помощью LC/MS/MS для обеспечения охвата последовательностей VP1, VP2 и VP3 и подтверждения N-концевого ацетилирования VP1 и VP3.
Способы
Материалы и реагенты
[0176] Дитиотреитол (DTT), 4-винилпиридин, ультрачистую муравьиную кислоту, уксусную кислоту, гуанидин-HCl, Трис-HCl и Трис-основание приобретали у Sigma Chemicals (Сент-Луис, Миссури). Фильтры Amicon ultra-4 приобретали у Millipore (Биллерика, Массачусетс). Свиной трипсин со степенью чистоты, пригодной для секвенирования, приобретали у Promega (Милуоки, Висконсин). Эндопротеиназу Lys-C и Asp-N приобретали у Roche (Германия). Кассеты Slide-A-Lyzer с 10000 MWCO приобретали у Pierce (Рокфорд, Иллинойс).
Получение и очистка векторов
[0177] Векторы AAV получали с помощью способа тройной транзиентной трансфекции, описанного ранее (Xiao, 1998 #123). Вкратце, клетки HEK293 трансфицировали с использованием полиэтиленимина, PEI и трех плазмид в соотношении 1:1:1 (вектор на основе ITR, AAV rep/cap и плазмида-помощник Ad). Векторная плазмида содержит векторный геном CBA-EGFP и последовательности ITR из AAV2. Экспрессия EGFP находилась под управлением гибридного промотора (CBA) на основе промотора бета-актина курицы и энхансера CMV, как описано (Miyazaki, 1989 #124) (Niwa, 1991 #125). Хелперные элементы rep/cap AAV содержали последовательности rep из AAV2 и специфичные в отношении серотипа капсидные последовательности с обозначением rep2/cap2, rep2/cap5, rep2/cap7 и т. д. Используемым хелперным элементом pAd был pHelper (Stratagene/Agilent Technologies, Санта-Клара, Калифорния). Очистку AAV выполняли, как описано Qu et al. (2007, J. Virol. Methods 140:183-192).
Анализ интактных белков с помощью LC/MS
[0178] Вирионы AAV концентрировали с помощью фильтра Amicon ultra-4 (10 кДа MWCO) и денатурировали 10% уксусной кислотой с последующим прямым анализом в устройстве Acquity UPLC -Xevo® QTOF MS (Waters, Милфорд, Массачусетс). Разделения осуществляли с помощью колонки С4 или С8 UPLC BEH (1,7 мкм, 2,1 мм внутр. диам.) при расходе 0,25 мл/мин. Подвижная фаза A представляла собой 0,1% муравьиную кислоту в воде, в то время как подвижная фаза B представляла собой 0,1% муравьиную кислоту в ацетонитриле. Конечный градиент был следующим: от 10% B до 20% B в течение 6 минут, от 20% B до 30% B в течение 10 минут, затем от 30% до 38% B в течение 40 минут. В случае MS капиллярное напряжение и напряжение пробоотборного конуса устанавливали на 3,5 кВ и 45 В соответственно. Масс-спектры получали в режиме чувствительности с диапазоном m/z, составляющим 500-4000. Калибровку, выполняемую с помощью дополнительного средства, с применением йодида натрия в качестве калибрующего вещества осуществляли в случае калибровки по массе. MaxEnt1 в компьютерной программе Masslynx использовали для деконволюции белков.
Ферментативные переваривания VP AAV2
[0179] Концентрированные вирионы AAV2 денатурировали 6 M гуанидин-HCl, 0,1 M Трис при значении pH 8,5. Белки восстанавливали с помощью 30 мM DTT при 55°C в течение 1 часа в темноте и алкилировали 0,07% 4-винилпиридином при комнатной температуре в течение 2 часов. Реакции гасили посредством добавления 1 M DTT. Образцы диализировали с помощью кассет Slide-A-Lyzer (10000 MWCO) против 25 мM Трис-буфера при значении pH 8,5 в течение ~18 часов. После диализа образцы разделяли на три аликвоты. Каждую аликвоту переваривали с помощью трипсина при соотношении фермент:белок (вес./вес.) 1:25 или Lys-C - при 1:50 или Asp-N - при 1:100 в течение 18 часов при 37°C соответственно.
Пептидное картирование с помощью LC/MS/MS
[0180] Нано-LC/MS/MS осуществляли с помощью системы NanoAcquity HPLC (Waters, Милфорд, Массачусетс) в сочетании с масс-спектрометром Orbitrap Velos (Thermo-Fisher Scientific, Волтгем, Массачусетс) с использованием упакованной в заводских условиях колонки для нано-LC (75 мкм × 10 мм) с Magic C18 с упаковочным материалом (5 мкм, Bruker, Биллерика, Массачусетс) при расходе 300 нл/мин. Подвижные фазы A и B представляли собой 0,1% муравьиную кислоту в воде и ацетонитрил соответственно. Градиент составлял от 2% B до 60% B в течение 121 мин.
[0181] Параметры источника в случае Velos были следующими: напряжение источника: 2,5 кВ, температура капилляра 275°C; уровень RF S-линзы: 55%. Данные получали с помощью информационно-зависимого способа первой десятки с точностью MS при разрешении 60000 и 10 MS/MS в ионной ловушке. Для поиска в базах данных использовали Mascot против последовательностей капсидных вирусных белков AAV2. Устойчивость MS 10 ppm и устойчивость ms/ms 0,8 Да использовали для поиска в базе данных.
Пептидное картирование с помощью UPLC/MS/MS
[0182] Переваренные белки также анализировали с помощью UPLC/MS/MS в Acquity UPLC-Xevo qTOF MS. Колонку C18 BEH300 (2,1×150 мм) использовали в подвижных фазах с 0,1% муравьиной кислотой в градиенте вода/ацетонитрил с расходом 0,25 мл/мин. Масс-спектры получали в положительном режиме MSe в диапазоне масс, составляющим 200-2000.
Результаты
Способ денатурации AAV
[0183] AAV можно денатурировать с помощью ряда способов с использованием детергента, посредством нагревания, с помощью высокого содержания соли или буфера с низкими или высокими значениями pH. Тепловая денатурация может приводить к преципитации белка и в результате этого колонки с обращенной фазой легко закупориваются и чрезмерно сжимаются. Денатурация с помощью высокой концентрации соли требует дополнительного этапа обессоливания перед анализом с помощью LC/MS. Денатурацию с помощью 10% уксусной кислоты использовали для анализа интактных белков с помощью LC/MS, поскольку она обеспечивала чистый масс-спектр. В случае пептидного картирования 0,1% RapiGest или 6 M гуанидин HCl можно использовать в качестве денатурирующего реагента.
Разработка способа анализа интактных белков
[0184] Исходный анализ интактных белков AAV2 осуществляли с помощью колонки C4 UPLC BEH при быстром градиенте. При этом условии наблюдали только один пик в общей ионной хроматограмме с массой, соответствующей VP3 (фиг. 1A). Без привязки к какой-либо теории, считается, что отсутствие VP1 и VP2, возможно, связано с низкой стехиометрией VP1 и VP2 или подавлением сигналов VP1 и VP2 с помощью VP3, если все VP элюируют совместно. Предпринимали попытку повышения ввода пробы или длины колонки с использованием пологого градиента и с помощью альтернативных колонок в целях выявления VP1 и VP2. Более высокая нагрузка (1,7 мкг) с использованием пологого градиента при 0,5% B/мин. приводила к плечевому пику слева (фиг. 1B). Увеличение длины колонки от 10 см до 15 см не приводило к усилению разделения плечевого пика (фиг. 1C). Однако плечевой пик дополнительно разделяли от основного пика с помощью колонки C8 BEH, при этом наблюдали повышенные интенсивности сигналов (фиг. 1D).
[0185] В результате массы VP1 и VP2 получали в этом плечевом пике при интенсивностях сигналов, показанных на фиг. 2A. Массы VP1 и VP3 соответствуют аминокислотам 2-735 (ацетилирование) и аминокислотам 204-735 (ацетилирование) соответственно (фиг. 2A и 2B). В случае VP2 (аминокислоты 139-735) ацетилирования не наблюдали. Кроме того, наблюдали небольшой пик с массой, меньшей VP3, при этом масса соответствовала аминокислотной последовательности 212-735 с одним ацетилированием (фиг. 2B). Эти данные согласуются с последовательностями ДНК, поскольку VP3 содержит два кодона инициации ATG в AAV2: ATGGCTACAGGCAGTGGCGCACCAATGGCAGAC (SEQ ID NO:1), приводя к образованию двух возможных N-концов (подчеркнуто): MATGSGAPMAD (SEQ ID NO:2). N-концевые метиониновые остатки не присутствовали как в VP1, так и в VP3, как измеряли с помощью анализа интактных белков. Ацетилирование VP1 и VP3 не является артефактом, вызванным способом (денатурация AAV 10% уксусной кислотой), поскольку ацетилирование VP1 и VP3 также наблюдали при получении AAV с помощью альтернативного способа денатурации без использования уксусной кислоты. Данные об интактных белках также подтвердили, что гликозилирование не присутствовало в вирусных капсидных белках, даже несмотря на то, что присутствовали несколько N-связанных консенсусных последовательностей [16].
Пептидное картирование с помощью LC/MS/MS
[0186] Для дополнительного подтверждения N-концов и ацетилирования, наблюдаемого в анализе интактных белков, пептидное картирование осуществили с помощью нескольких ферментов и анализировали с помощью нескольких устройств. Оценивали различные способы получения образцов, в том числе способы денатурации и этапы обессоливания. Конечный способ переваривания, предусматривающий денатурацию с помощью 6 M гуанидин-HCl, восстановление и алкилирование 4-винилпиридином, а также диализ с помощью Slide-A-lyzer с последующим ферментативным перевариванием приводили к чистому пептидному картированию с низкими искусственными модификациями в течение процесса переваривания. Исследовали всего 5 мкг исходного материала, что приводило к полному охвату последовательностей с помощью нано-LC/MS/MS и UPLC/MS/MS.
[0187] Поиск Mascot триптических гидролизатов в результате нано-LC/MS/MS в отдельности приводил к охвату 78% последовательностей с граничным ионным значением 13, показанным на фиг. 3. Два больших отсутствующих триптических пептида, T27 и T38 (заключенных в рамку) на основании нано-LC/MS/MS обнаружили в LC/MS в Xevo TOF MS с использованием колонки C18 BEH (фиг. 3). Кроме того, большинство из пептидных последовательностей T27 и T38 дополнительно подтверждали с помощью нано-LC/MS/MS гидролизатов Asp-N, показанных курсивом на фиг. 3. Полные N-концевые и C-концевые пептиды охватывали гидролизатами Lys-C, подчеркнутыми на фиг. 3. Таким образом, 100% охват последовательности VP1 достигали посредством нескольких ферментативных перевариваний и с помощью двух способов LC/MS/MS.
[0188] С помощью LC/MS/MS подтверждали N- и C-концы VP1, VP2 и VP3 и N-концевое ацетилирование VP1 и VP3, наблюдаемое при анализе интактных белков. На фиг. 4A-4C показан спектр MS/MS N-концевого триптического пептида VP1 A(Ac)ADGYLPDWLEDTLSEGIR (SEQ ID NO:4) (фиг. 4A), полученного из N-концевого Asp-N пептида VP2 (APGKKRPVEHSPVEP) (SEQ ID NO:15) (фиг. 4B) и N-концевого Asp-N пептида VP3 A(Ac)TGSGAPM (SEQ ID NO:5) (фиг. 4C). С помощью MS/MS подтвердили положение ацетилирования в N-концевых аланиновых остатках как в пептидах VP1, так и в пептидах VP3. Наличие немодифицированных y18- и y17-ионов, а также все детектируемые b-ионы со сдвигом массы 42 Да на фиг. 4A указывает на то, что модификация 42 Да расположена на N-конце VP1. Аналогично наличие немодифицированых y3-y8-ионов на фиг. 4C подтвердило положение ацетилирования в N-концевом аланиновом остатке.
Сравнение N-концов VP AAV
[0189] Помимо AAV2, AAV1, AAV5, AAV7, AAV9 и AAV Rh10 также анализировали с помощью анализа интактных белков. Теоретические и прогнозируемые массы VP AAV показаны в табл. 2.
Таблица 2. Теоретическая масса и экспериментальная масса VP AAV
тельность
тельность
Ms (Да)
тальная Ms (Да)
[0190] N-концы, а также их посттрансляционные модификации, являются высококонсервативными среди анализируемых серотипов AAV, даже несмотря на то, что AAV5 описан в качестве наиболее разнообразной последовательности серотипа AAV, как показано при выравниваниях последовательностей на фиг. 5. В 11 из 13 серотипов AAV N-концы VP1 имеют идентичную последовательность из 13 аминокислот (MAADGYLPDWLED) (SEQ ID NO:6), в то время как во всех 13 серотипах AAV идентифицировали TAP… N-концевые последовательности в VP2 (фиг. 5). LC/MS AAV2 указывала, что T отсутствует в VP2 на уровне белка. N-концы VP3 являются наиболее разнообразными среди всех трех вирусных капсидных белков, при этом 8 из 13 серотипов AAV имеют MA… N-концевую последовательность. Аналогично AAV2, AAV1 и AAV Rh10 также имеют два кодона инициации ATG, при этом первый в качестве преобладающего N-конца на основе анализа интактных белков с помощью LC/MS. Интересно, что несмотря на то, что AAV7 имеет два потенциальных кодона инициации (GTGGCTGCAGGCGGTGGCGCACCAATGGCAGACAATAAC…) (SEQ ID NO:7), второй кодон инициации (ATG) является предпочтительным, исходя из анализа интактных белков: VP3` с 213(ac)-737 был преобладающим пиком, в то время как VP3 с 203(ac)-737 был минорным пиком.
Выводы
Применения анализа интактных белков с помощью LC/MS и пептидного картирования с помощью LC/MS/MS VP AAV при исследовании и разработке генной терапии
[0191] Эти результаты показывают, что прямая LC/MS различных типов AAV после денатурации оказалась простым и эффективным способом контроля белковой последовательности и посттрансляционных модификаций с точным измерением масс на уровне интактного белка. N-концы VP1, VP2 и VP3 AAV определяли с помощью масс-спектрометрии. Ацетилирования N-концов VP1 и VP3 идентифицировали в других серотипах AAV. Также разрабатывали прямое пептидное картирование AAV с помощью LC/MS/MS для обеспечения 100% охвата последовательностей VP1, VP2 и VP3 и подтверждения N-концевого ацетилирования VP. Теоретические массы прогнозируемых последовательностей 13 серотипов AAV на основе выравнивания последовательностей и анализа интактных белков нескольких серотипов AAV показаны в табл. 3.
Таблица 3. Прогнозируемые последовательности и массы
руемая последовательность VP1
руемая последовательность VP2
руемая последовательность VP3
[0192] Точные массы VP1, VP2 и VP3 каждого серотипа являются уникальными и, таким образом, анализ интактных белков может быть использован в качестве теста на идентичность для дифференциации серотипов капсидов AAV. В табл. 4-6 показаны различия масс VP среди 13 распространенных серотипов AAV. Обычным шрифтом показаны дельта-массы более 10, в то время как дельта-массы менее 10 показаны жирным шрифтом.
Таблица 4. Различия масс VP1 среди 13 изотипов AAV
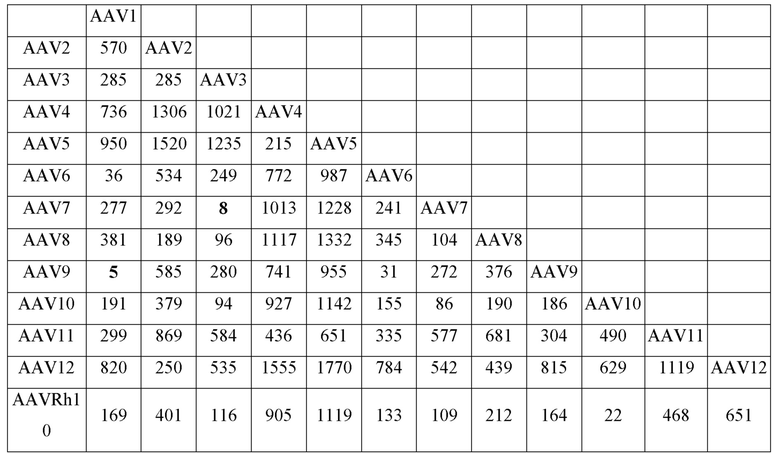
Таблица 5. Различия масс VP2 среди 13 изотипов AAV
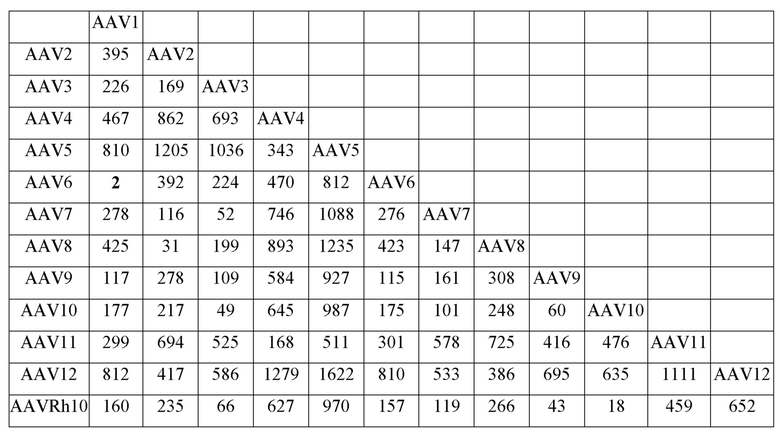
Таблица 6. Различия масс VP3 среди 13 изотипов AAV
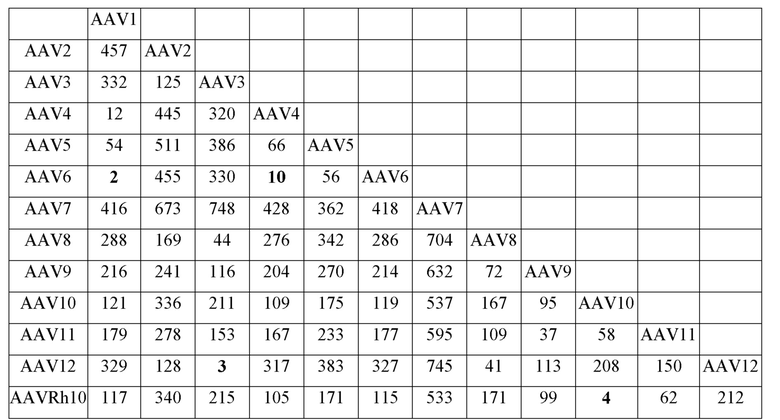
[0193] Различий масс в пределах 10 Да всех трех VP между двумя изотипами не наблюдали. Даже несмотря на то, что VP2 и VP3 имеют разницу лишь в 2 Да между AAV1 и AAV6, разница масс VP1 между AAV1 и AAV6 составляет 36, что является достаточно значительным для различения с помощью измерения точных масс. Таким образом, измерение интактных белков VP1, VP2 и VP3 является высокоспецифичным в качестве теста на идентичность.
[0194] Эти результаты показывают, что анализ интактных белков и LC/MS/MS могут быть использованы для профилирования VP для контроля экспрессий, посттрансляционных модификаций и усечений VP и для обеспечения однородности продукта во время получения VLP. Эти два анализа также могут быть использованы для подтверждения сайт-направленного мутагенеза или структурной характеристики в случае применений инженерии капсидных белков.
Пример 2. Роль N-концевого ацетилирования капсидных белков AAV
[0195] Химические модификации клеточных белков являются распространенным средством контроля их функций (Arnesen, T. (2006) Virology 353(2): 283-293). N-концевое ацетилирование (Nt-ацетилирование), которое включает в себя перенос ацетильной группы от ацетил-кофермента A к α-аминогруппе первого аминокислотного остатка белка (Brown, J.L. and Roberts, W.K. (1976) J Biol Chem 251: 1009-1014; Arnesen, T. et al. (2009) Proc Natl Acad Sci USA 106: 8157-8162), является наиболее распространенной из модификаций белка. В отличие от большинства других модификаций белка, Nt-ацетилирование является необратимым; оно происходит главным образом во время синтеза белка, катализируемого N-концевыми ацетилтрансферазами (NAT), ассоциированными с рибосомами (Gautschi, M. et al. (2003) Mol Cell Biol 23: 7403-7414; Pestana, A. and Pitot, H.C. (1975) Biochemistry 14: 1404-1412; Polevoda, B. et al. (2003) J Biol Chem 278: 30686-97). У эукариот существует несколько различных NAT -NatA-NatF- каждая из них состоит из одной или нескольких субъединиц и каждая из них ацетилирует конкретную подгруппу N-конца в зависимости от аминокислотной последовательности первых нескольких аминокислот (Jornvall, H. (1975) J Theor Biol 55: 1-12; Persson, B. et al. (1985) Eur J Biochem 152: 523-527).
[0196] Экспериментальные данные указывают на то, что белки с ацетилированным N-концом являются наиболее стабильными in vivo, чем неацетилированные белки; т.е., Nt-ацетилирование защищает белки от распада (Hershko, A. et al. (1984) Proc Natl Acad Sci U S A 81: 7021-7025). Одним объяснением этого может быть открытие в 2004 году того, что другая модификация N-конца, убиквитинирование, которая включает непосредственное прикрепление малого белка убиквитина к N-концевому аминокислотному остатку, способствует последующему разрушению белка (Ben Saadon, R. et al. (2004) J Biol Chem 279: 41414-41421). В отличие от этого, сигналы Nt-ацетилирования также могут быть частью механизма контроля качества в целях разрушения несвернутых или неправильно свернутых белков для регуляции стехиометрий белка in vivo (Hwang, C.S. et al. (2010) Science 327: 973-977).
[0197] Систематический анализ прогнозируемого N-концевого процессинга цитозольных белков по сравнению с таковыми, которые предназначены для сортировки в секреторный путь, показал, что цитозольные белки были значительно смещены в сторону процессинга, однако существует равноценное и противоположное смещение против такой модификации в случае секреторных белков (Forte, G.M.A. et al. (2011) PLoS Biology, 4 May 2011, Volume 9). Мутации секреторных сигнальных последовательностей, которые способствуют их ацетилированию, приводят к нарушению сортировки в цитозоле с помощью способа, зависимого от механизма N-концевого процессинга. В связи с этим N-концевое ацетилирование представляет этап раннего определения клеточной сортировки образующихся полипептидов, которые представляют повышенный уровень жесткости в целях обеспечения того, что белки, которые предназначены остаться в цитозоле, фактически находятся в цитозоле. Эукариотическая клетка содержит несколько различных компартментов, называемых органеллами, необходимых для осуществления конкретных функций. Белки в этих компартментах синтезируются в цитоплазме и таким образом требуют сложных механизмов сортировки для обеспечения своей доставки в соответствующую органеллу. Белки модифицируются путем ацетилирования своего аминоконца на очень ранней стадии своего синтеза. Существует выраженное отличие между вероятностью такой модификации в цитоплазматических белках и таковых, которые предназначены для одной из основных органелл, эндоплазматической сети (ER): в то время как цитоплазматические белки обычно являются ацетилированными, таковые, привязанные к ER, являются главным образом немодифицированными. Более того, при разработке конкретных белков для ER для индуцирования их ацетилирования, их нацеливание на ER было ингибировано (Forte, G.M.A. et al. (2011) PLoS Biology, 4 May 2011, Volume 9).
[0198] Было показано, что сократительные белки актин и тропомиозин требуют NatB-опосредованного Nt-ацетилирования для соответствующей функции, в частности, включающей связывание актина и тропомиозина и регуляцию актомиозина (Coulton, A.T. et al. (2010) J Cell Sci 123: 3235-3243; Polevoda, B. et al. (2003) J Biol Chem 278: 30686-97). Таким образом, Nt-ацетилирование капсидных белков AAV может иметь значение при трансдукционном потенциале векторов на основе rAAV. Если векторы AAV не могут получить доступ в ядро, впоследствии они не могут трансдуцировать клетки. Роль актиновых филаментов и FKBP52 (FK506-связывающего белка p52) в транслокации капсидов AAV из эндосомы в ядро является четко определенной (Zhao, W. et al. (2006) Virology 353(2): 283-293). Важно, что Nt-ацетилирование является необходимым для функционирования актиновых филаментов в результате модулирования белок-белковых взаимодействий (Coulton, A.T. et al. (2010) J Cell Sci 123: 3235-3243; Polevoda, B. et al. (2003) J Biol Chem 278: 30686-97).
[0199] Несмотря на то, что N-концевое ацетилирование белков является широко известным явлением, биологическая значимость Nt-ацетилирования капсидных белков AAV не является в достаточной мере понимаемой. Оба прогнозируемых N-конца VP1 и VP3 на основании секвенирования ДНК представляют собой метионин и следующий за ним аланин. Было описано, что удаление N-концевого метионина Met-аминопептидазами часто приводит к Nt-ацетилированию образующихся N-концевых остатков аланина, валина, серина, треонина и цистеина, и что ацетилирование N-конца выступает в качестве потенциального сигнала разрушения [21]. Убиквитинирование вирусных капсидных белков предполагалось в качестве потенциального сигнала процессинга капсида во время разборки вириона [22]. Связь между N-ацетилированием VP1 и VP3 и разрушением и сбрасыванием оболочки вирусного капсида перед вхождением в ядро исследуют дополнительно.
[0200] Для понимания функциональных последствий N-концевого ацетилирования в отношении капсидных белков AAV, для генерации мутантов AAV используется сайт-направленный мутагенез N-концевых кодонов инициации VP3.
Способы
[0201] Капсидные белки AAV получают с различными аминокислотами в положении 2 по отношению к инициаторному метионину (iMet X) для определения того, является ли Nt-ацетилирование ингибированным или ослабленным, а затем определяются функциональные последствия. Определяется способность капсидных белков мигрировать внутриклеточно и/или приобретать посттрансляционные модификации, такие как гликозилирование, а также затем определяется, влияет ли эта способность на инфекционность собранной частицы AAV. Помимо этого, определяется влияние ацетилирования на убиквитинирование/разрушение и нацеливание на лизосому, ER, аппарат Гольджи или внутреннюю ядерную мембрану.
[0202] Например, для анализа миграции или нацеливания частицы AAV с капсидными белками, имеющими мутантное положение 2 (например, iMet X), метят флуоресцентным путем и используют для инфицирования клеток (например, клеток HeLa). Такие частицы AAV анализируют в отношении одного или нескольких из: времени захвата вирусных частиц, колокализации частиц AAV с конкретными маркерами компартментов (например, аппаратом Гольджи, ER или лизосомальными белками или другими маркерами), накопления в ядре (например, при анализе путем колокализации с ядерным маркером или красителем) и/или чувствительности миграции к конкретным ингибиторам раннего эндосомального ускользания (такого как бафиломицин A или хлорид аммония), по сравнению с мечеными флуоресцентным путем частицами AAV дикого типа, используемыми для инфицирования той же самой линии клеток (см., например, Bartlett, J.S. et al. (2000) J. Virol. 74:2777-2785, касательно описания таких анализов).
[0203] Для анализа инфекционности частицы AAV с капсидными белками, имеющими мутированное положение 2 (например, iMet X), используют для инифицирования клеток (например, клеток HeLa), а их эффективность трансдукции сравнивают с частицами AAV дикого типа (например, имеющими тот же самый серотип AAV и инфицирующими тот же самый тип клеток).
[0204] Для анализа гликозилирования частицы AAV с капсидными белками, имеющими мутантное положение 2 (например, iMet X), используют для инфицирования клеток (например, клеток HeLa). Частицы AAV из инфицированных клеток подвергают одному или нескольким анализам, в том числе без ограничения химическому выявлению гликозилирования (например, применению коммерчески доступного набора для выявления дигоксигенингликана (DIG) и/или набора для выявления флуоресцентного гликопротеина в отношении денатурированных и электрофоретически разделенных капсидных белков) и масс-спектрометрии (например, FT-ICR MS), для сравнения с частицами AAV дикого типа, используемыми для инфицирования той же самой линии клеток (см., например, Murray, S. et al. (2006) J. Virol. 80:6171-6176, касательно описания таких анализов).
[0205] Для анализа убиквитинирования, частицы AAV с капсидными белками, имеющие мутантное положение 2, используют для инфицирования клеток (например, клеток HeLa). Частицы AAV иммунопреципитируют из инфицированных клеток с помощью антитела к капсиду, затем подвергают вестерн-блоттингу с помощью антитела к убиквитину и сравнивают с частицами AAV дикого типа, используемыми для инфицирования клеток таким же самым способом. Мутантные частицы AAV также могут быть использованы для анализов убиквитинирования in vitro для сравнения с частицами AAV дикого типа (см., например, Yan, Z. et al. (2002) J. Virol. 76:2043-2053).
Пример 3. Роль дезамидирования капсидных частиц AAV2
[0206] Анализ последовательностей капсидного белка AAV2 показал потенциальные сайты дезамидирования, подчеркнутые в следующей аминокислотной последовательности: MAADGYLPDWLEDTLSEGIRQWWKLKPGPPPPKPAERHKDDSRGLVLPGYKYLGPFNG LDKGEPVNEADAAALEHDKAYDRQLDSGDNPYLKYNHADAEFQERLKEDTSFGGNLG RAVFQAKKRVLEPLGLVEEPVKTAPGKKRPVEHSPVEPDSSSGTGKAGQQPARKRLNFG QTGDADSVPDPQPLGQPPAAPSGLGTNTMATGSGAPMADNNEGADGVGNSSGNWHCD STWMGDRVITTSTRTWALPTYNNHLYKQISSQSGASNDNHYFGYSTPWGYFDFNRFHC HFSPRDWQRLINNNWGFRPKRLNFKLFNIQVKEVTQNDGTTTIANNLTSTVQVFTDSEY QLPYVLGSAHQGCLPPFPADVFMVPQYGYLTLNNGSQAVGRSSFYCLEYFPSQMLRTG NNFTFSYTFEDVPFHSSYAHSQSLDRLMNPLIDQYLYYLSRTNTPSGTTTQSRLQFSQAG ASDIRDQSRNWLPGPCYRQQRVSKTSADNNNSEYSWTGATKYHLNGRDSLVNPGPAM ASHKDDEEKFFPQSGVLIFGKQGSEKTNVDIEKVMITDEEEIRTTNPVATEQYGSVSTNL QRGNRQAATADVNTQGVLPGMVWQDRDVYLQGPIWAKIPHTDGHFHPSPLMGGFGLK HPPPQILIKNTPVPANPSTTFSAAKFASFITQYSTGQVSVEIEWELQKENSKRWNPEIQYTS NYNKSVNVDFTVDTNGVYSEPRPIGTRYLTRNL (SEQ ID NO: 3).
[0207] В частности, потенциальный сайт дезамидирования обнаружен в N57/G58 в домене фосфолипазы A2 (Ca++ связывающий сайт), приведенный жирным курсивом в вышеуказанной последовательности. Следующие эксперименты были направлены на исследование того, может ли дезамидирования в N57 приводить к сниженной активности и/или усечению AAV2, а также могут ли оказывать различные способы получения AAV различные эффекты на дезамидирование. Например, с помощью способа с использованием линии клеток-продуцентов (см. Martin et al., (2013) Human Gene Therapy Methods 24:253-269; публикацию заявки на патент США № US 2004/0224411, опубликованной до выдачи патента; и Liu, X.L. et al. (1999) Gene Ther. 6:293-299) можно индуцировать более высокий уровень дезамидирования в N57 по сравнению с тройным способом трансфекции. В соответствии с кристаллической структурой AAV2, N57 не показана; однако N382 и N511 находятся под частичным воздействием, а N715 находится под полным воздействием.
Способы
Ферментативные переваривания VP AAV1 и AAV2
[0208] 10 мкг каждого материала AAV1-EGFP или AAV2-EGFP (образованного с помощью тройной трансфекции, а также способа с использованием линии клеток-продуцентов) концентрировали с помощью фильтров Amicon (10 кДа MWCO), денатурировали 6 M гуанидин-HCl, 50 мM Трис при значении pH 8,5. Белки восстанавливали с помощью 5 мM DTT при 60°C в течение 30 минут в темноте, алкилировали 15 мM йодацетамидом при комнатной температуре в течение 30 минут и затем буфер заменяли на 25 мM Трис, pH 7,1, для переваривания с использованием микроколонок Bio-Spin® 6 Трис. После замены буфера образцы разделяли на две аликвоты. Каждую аликвоту переваривали трипсином при соотношении фермент:белок (вес./вес.) 1:25 или Asp-N - при 1:50 в течение 2 часов при 37°C соответственно.
Пептидное картирование с помощью UPLC/MS/MS
[0209] Переваренные белки также анализировали с помощью UPLC/MS/MS в Acquity UPLC-Xevo qTOF MS. Колонку C18 BEH300 (2,1×150 мм) использовали для разделения в подвижных фазах с 0,1% муравьиной кислотой в градиенте вода/ацетонитрил с расходом 0,25 мл/мин. Масс-спектры получали в положительном режиме разрешения MSe в диапазоне масс, составляющим 50-2000.
Определение уровней дезамидирования в VP AAV
[0210] Экстрагированные ионные хроматограммы (XIC) пептидов, содержащие сайты NG (T9, T49 и T67 в VP AA1 и AAV2) и их соответствующие дезамидированные молекулы, использовали для расчета уровней дезамидирования.
[0211] Для сравнения векторов на основе AAV, полученных с помощью способов тройной трансфекции (TTx) и с использованием линии клеток-продуцентов (PCL), AAV1 или AAV2, меченые EGFP, получали с помощью способа TTx или PCL. Было обнаружено, что усеченный VP1 (tVP1) присутствует в AAV2-EGFP, полученных с помощью PCL, но не в AAV2-EGFP, полученном с помощью TTx. Было обнаружено, что AAV1-EGFP не имеет tVP1, вне зависимости от способа получения. Также было обнаружено, что эффективность AAV2 in vitro, полученного с помощью способа PCL, снижена по сравнению с AAV2, полученного с помощью TTx. Также было обнаружено, что мутантные по N57K и N57Q частицы AAV2, имеют сниженную активность и нарушение связывания с Са++.
[0212] В следующей таблице представлены триптические пептиды, которые анализировали для исследования каждого потенциального сайта дезамидирования, а также соответствующего остатка.
Таблица 7. Триптические пептиды, содержащие сайты NG
[0213] Как показано в табл. 7, пептид T9 YLGPFNGLDK (SEQ ID NO: 9) использовали для контроля N57, пептид T38 EVTQNDGTTTIANNLTSTVQVFTDSEYQLPYVLGSAHQGCLPPFPADVFMVPQYGYLTL NNGSQAVGRSSFYCLEYFPSQMLR (SEQ ID NO: 10) использовали для контроля N382, пептиды T49 YNLNGR (SEQ ID NO: 11) и YHLNGR (SEQ ID NO: 12) использовали для контроля N511 в AAV1 или AAV2 (соответственно) и пептиды T67 SANVDFTVDNNGLYTEPR (SEQ ID NO: 13) и SVNVDFTVDTNGVYSEPR (SEQ ID NO: 14) использовали для контроля N715 в AAV1 или AAV2 (соответственно).
[0214] Анализ с помощью LC/MS/MS использовали для сравнения процента дезамидирования в частицах AAV1 и AAV2, полученных с помощью способов TTx и PCL. Результаты в отношении пептида T9 показаны на фиг. 6A и 6B. Результаты в отношении пептида T49 показаны на фиг. 7A и 7B. Результаты в отношении пептида T67 показаны на фиг. 8A и 8B. Эти результаты обобщены в табл. 8. Пептид T38 не был выявлен из-за его размера.
Таблица 8. Обобщение результатов LC/MS/MS
[0215] В частности, AAV2, получаемый с помощью PCL, характеризовался почти 3-кратным повышением дезамидирования по сравнению с AAV2, получаемым с помощью TTx. Эти результаты указывают на то, что дезамидирование снижает активность AAV, также как снижается активность AAV2 in vitro, полученного с помощью PCL.
Выводы
[0216] В совокупности примеры 1-3 показывают способы анализа интактных белков вирусных частиц (например, капсидных белков AAV) с помощью LC/MS. Молекулярный вес измеряли точно, и эти методики также могут быть использованы для определения N-концов и/или модификаций вирусных капсидных белков. Более того, эти способы можно адаптировать в качестве анализов идентичности серотипов капсидов, пригодных в генной терапии, например, в качестве аналитической платформы. Эти результаты дополнительно определяют связь между структурой (например, усечениями, дезамидированием и т. д.) и активностью капсидных белков, свидетельствуя о том, что точечные мутации в основных сайтах могут быть использованы для разработки более эффективных векторов.
Пример 4. Определение роли N-концевого ацетилирования капсидных белков AAV
[0217] Как обсуждалось выше, N-концы капсидных белков AAV являются высококонсервативными в пределах серотипов (фиг. 5). Методики, описанные в примере 1, позволяют изучать экспрессию и посттрансляционные модификации VP. Далее изучали роль и биологическое значение N-концевого ацетилирования капсидных белков AAV.
Результаты
[0218] Для определения потенциальной роли деацетилирования капсидных белков AAV исследовали варианты деацетилирования AAV5. Частицу AAV5, экспрессирующую eGFP с использованием промотора CBA (AAV5-CBA-Egfp), сравнивали с вариантами AAV5 с аминокислотой, прилегающей к инициирующему метионину (iMET), мутировавшему в случае VP1 и VP3 (deAC-AAV5-CBA-eGFPs). Три аминокислоты, которые согласно прогнозам, имеют низкую вероятность ацетилирования NatA, NatC или NatD, выбирали для образования вариантов: Gly, Leu и Pro, как показано в табл. 9 ниже.
Таблица 9. Частота N-концевого ацетилирования
Обычно встречается в VP1 и VP3
Обычно встречается в VP1 и VP3 в случае AAV5
[0219] Образовывали следующие деацетилированные мутанты AAV5 (deAC):
S2GVP1 - Ser заменен на Gly в положении 2 в AAV5VP1
S2LVP1 - Ser заменен на Leu в положении 2 в AAV5VP1
S2PVP1 - Ser заменен на Pro в положении 2 в AAV5VP1
S2GVP3 - Ser заменен на Gly в положении 2 в AAV5VP3
S2LVP3 - Ser заменен на Leu в положении 2 в AAV5VP3
S2PVP3 - Ser заменен на Pro в положении 2 в AAV5VP3
S2PVP1/VP3 - Ser заменен на Pro в положении 2 как в VP1, так и в VP3 AAV5
S2GVP1/VP3 - Ser заменен на Gly в положении 2 как в VP1, так и в VP3 AAV5
S2LVP1/VP3 - Ser заменен на Leu в положении 2 как в VP1, так и в VP3 AAV5
[0220] Эти варианты образовывали с помощью способа TTX, описанного выше. Все варианты AAV5 характеризовались высокой продуктивностью, при этом выход составлял более 1013 суммарных VG. Все варианты AAV5 также характеризовались прогнозируемым соотношением белков VP1:VP2:VP3 с помощью анализа белкового геля SYPRO (фиг. 9). После этого использовали LC/MS для подтверждения того, что все варианты AAV5 характеризовались сниженным ацетилированием, как показано в табл. 10.
Таблица 10. Анализ LC/MS в случае ацетилирования варианта AAV5
н.о. = не определено.
[0221] Эти анализы с помощью LC/MS подтверждали, что варианты AAV5 были деацетилированными. Все варианты S2LVP1, S2LVP3 и S2LVP1/VP3 характеризовались повышенной массой (повышенной от 173 до 182) в белках VP1 и VP3, указывая на то, что изменение второй N-концевой аминокислоты на лейцин в VP1 или VP3 изменяет белок, приводя к повышению массы.
[0222] После этого варианты AAV5 анализировали в анализе трансдукции in vitro с помощью eGFP в качестве репортерного гена (фиг. 10). Анализ разрабатывали для оценки трансдукции с помощью деацетилированных мутантных вариантов AAV5 при множественности заражения 106 (MOI), сравнивая каждый вариант с исходной, немодифицированной частицей AAV5. Применяли три клеточные линии: клетки 293, HuH7 и HeLa. После инфицирования клетки анализировали для определения числа копий генома вектора (vg/мкг клеточного белка) и экспрессии eGFP (с помощью ELISA). Число копий генома вектора (vg/мкг белка) представляет эффективность, при которой вариант AAV5 входит в клетку, а eGFP представляет эффективность внутриклеточной миграции капсида, поскольку для экспрессии трансгенов требуется, чтобы ДНК капсида/вектора эффективно мигрировала в ядро (фиг. 10). Геномы векторов количественно оценивали с помощью анализа TaqMan.
[0223] На фиг. 11 показано, что на основании анализа геномов векторов деацетилированные мутантные векторы AAV5 инфицировали все три линии исследуемых клеток при аналогичных, но сниженных уровнях по сравнению с исходными немодифицированными частицами AAV5. На фиг. 12 показано, что все деацетилированные мутантные векторы AAV5 приводили к сниженной экспрессии eGFP во всех трех линиях клеток, по сравнению с трансдукцией исходным немодифицированным AAV5.
Выводы
[0224] Согласно прогнозам, ацетилирование в мутантных вариантах, в которых N-концевой Ser заменен на Pro/Leu/Gly, при исследовании с помощью LC/MS не наблюдалось. Варианты deAC AAV5 характеризовались устойчивым продуцированием векторов, а варианты deAC AAV5 инфицировали клетки при уровнях, сопоставимых с исходным AAV5. Однако функциональные уровни белков в клетках, инфицированных вариантами deAC, были значительно снижены по сравнению с исходным AAV5. Эти данные указывают на то, что тропизм является минимально нарушенным в результате отсутствия N-концевого деацетилирования в VP1/VP3, однако последующий процессинг (например, миграция и/или разрушение) нарушен значительно. Поскольку исследуемые варианты характеризовались сниженной активностью in vitro, специалисту в данной области будет понятно, что можно использовать варианты, характеризуемые сниженным или устраненным ацетилированием, среди прочего, если сниженные уровни трансдукции являются желательными.
Пример 5. Оценка дезамидирования капсидных белков AAV
[0225] Примеры 1 и 3 показывают методики, которые позволяют изучать посттрансляционные модификации капсидных белков AAV и исследовать роль дезамидирования капсида AAV2. В следующем примере исследовали, снижает ли дезамидирование активность и/или индуцирует ли усечение капсидных белков, и могут ли различные способы производства влиять на уровни дезамидирования.
Способы
Частицы AAV образовывали и статус дезамидирования анализировали, как описано в примере 3.
Результаты
[0226] Как описано в примере 3, потенциальный сайт дезамидирования обнаружен в N57/G58 в домене фосфолипазы A2 (Ca++ связывающий сайт) в VP1 капсида AAV2. Мотив N57/G58 является консервативным в пределах серотипов AAV (фиг. 13). В примере 3 показано, что AAV2, получаемый с помощью PCL, характеризовался почти 3-кратным повышением дезамидирования по сравнению с AAV2, получаемым с помощью TTx (см. фиг. 6A и 6B и табл. 8).
[0227] При исследовании получения VP1, VP2 и VP3 с помощью белковых гелей усеченный белок VP1 (tVP1) выявляли только в капсидных белках AAV2, получаемых с помощью способа PCL (фиг. 14).
[0228] После этого образовывали ряд мутантов по дезамидированию AAV2. Эти мутанты нацеливались на остаток Gly в канонической последовательности NG. Также образовывали мутации, нацеливающиеся на остаток A35 (см. фиг. 13), N-концевую аминокислоту в случае tVP1, как показано в табл. 11. Мутанты pAF277 и pAF279, несущие несколько мутаций, не упаковывались.
Таблица 11. Мутанты по дезамидированию
K = положительный заряд (основная);
D = отрицательный заряд (кислая);
Q = полярная.
[0229] Дезамидирование вариантов затем анализировали с помощью LC/MS, как описано в примере 3 выше. Варианты AAV2A35N и AAV2G58D характеризовались измененным дезамидированием по сравнению с исходным AAV2 (фиг. 15). В частности, мутант AAV2A35N характеризовался повышенным дезамидированием (17,8%) по сравнению с исходным AAV2 (5,7%). Вариант AAV2G58D характеризовался сниженным дезамидированием (1,1%) по сравнению с исходным AAV2. Анализ в белковом геле SYPRO показал, что мутанты по дезамидированию AAV2 характеризовались надлежащим соотношением VP1:VP2:VP2 (фиг. 16).
[0230] После этого варианты дезамидирования AAV2 анализировали в анализе трансдукции in vitro с помощью eGFP в качестве репортерного гена (фиг. 17). Анализ разрабатывали для оценки трансдукции мутантными вариантами по дезамидированию AAV2 при множественности заражения 106 (MOI), сравнивая каждый вариант с исходной, немодифицированной частицей AAV2. Применяли три клеточные линии: клетки 293, HuH7 и HeLa. После инфицирования клетки анализировали для определения числа копий генома вектора (vg/мкг клеточного белка) и экспрессии eGFP (с помощью ELISA). Число копий генома вектора (vg/мкг белка) представляет эффективность, при которой вариант AAV2 входит в клетку, а eGFP представляет эффективность внутриклеточной миграции капсида, поскольку для экспрессии трансгенов требуется, чтобы ДНК капсида/вектора эффективно мигрировала в ядро (фиг. 17). Геномы векторов количественно оценивали с помощью анализа TaqMan.
[0231] Анализ векторных геномов указывал на то, что мутантные варианты по дезамидированию AAV2 инфицировали все тестируемые линии клеток при уровнях, сопоставимых с исходными векторами AAV2 (фиг. 18). Важно, что было обнаружено, что вариант AAV2A35N является более активным, чем исходный вектор AAV2 для трансдукции во все линии клеток (фиг. 19). Было обнаружено, что вариант AAV2G58D является более активным, чем исходный вектор AAV2 в клетках HuH7 (фиг. 19).
Выводы
[0232] В заключение следует отметить, что мутантные векторы по дезамидированию AAV2 инфицируют клетки при уровнях, сопоставимых с исходными частицами AAV2 (например, сопоставимых с vg/мкг клеточного белка). Однако на основании анализа уровней eGFP в трансдуцированных клетках вариант AAV2A35N характеризовался более высокой активностью, чем исходный AAV2 во всех исследуемых линиях клеток, и вариант AAV2G58D характеризовался более высокой активностью, чем исходный AAV2 в клетках HuH7 (линия клеток печеночного происхождения). Эти результаты указывают на то, что мутация A35N может быть эффективной в повышении активности вектора в случае трансдукции в различные типы клеток, и что мутация G58D также может быть эффективной в повышении активности в определенные типы клеток, например, клетки печени.
Литературные источники
1. Girod, A., et al., The VP1 capsid protein of adeno-associated virus type 2 is carrying a phospholipase A2 domain required for virus infectivity. J Gen Virol, 2002. 83(Pt 5): p. 973-8.
2. Stahnke, S., et al., Intrinsic phospholipase A2 activity of adeno-associated virus is involved in endosomal escape of incoming particles. Virology, 2011. 409(1): p. 77-83.
3. Bleker, S., F. Sonntag, and J.A. Kleinschmidt, Mutational analysis of narrow pores at the fivefold symmetry axes of adeno-associated virus type 2 capsids reveals a dual role in genome packaging and activation of phospholipase A2 activity. J Virol, 2005. 79(4): p. 2528-40.
4. Popa-Wagner, R., et al., Impact of VP1-specific protein sequence motifs on adeno-associated virus type 2 intracellular trafficking and nuclear entry. J Virol, 2012. 86(17): p. 9163-74.
5. Kashiwakura, Y., et al., Hepatocyte growth factor receptor is a coreceptor for adeno-associated virus type 2 infection. J Virol, 2005. 79(1): p. 609-14.
6. Qing, K., et al., Human fibroblast growth factor receptor 1 is a co-receptor for infection by adeno-associated virus 2. Nat Med, 1999. 5(1): p. 71-7.
7. Sanlioglu, S., et al., Endocytosis and nuclear trafficking of adeno-associated virus type 2 are controlled by rac1 and phosphatidylinositol-3 kinase activation. J Virol, 2000. 74(19): p. 9184-96.
8. Summerford, C., J.S. Bartlett, and R.J. Samulski, AlphaVbeta5 integrin: a co-receptor for adeno-associated virus type 2 infection. Nat Med, 1999. 5(1): p. 78-82.
9. Kern, A., et al., Identification of a Heparin-Binding Motif on Adeno-Associated Virus Type 2 Capsids. J Virol, 2003. 77(20): p. 11072-11081.
10. Asokan, A., et al., Adeno-associated virus type 2 contains an integrin alpha5beta1 binding domain essential for viral cell entry. J Virol, 2006. 80(18): p. 8961-9.
11. Xie, Q., et al., The atomic structure of adeno-associated virus (AAV-2), a vector for human gene therapy. Proc Natl Acad Sci U S A, 2002. 99(16): p. 10405-10.
12. DiMattia, M.A., et al., Structural insight into the unique properties of adeno-associated virus serotype 9. J Virol, 2012. 86(12): p. 6947-58.
13. Nam, H.J., et al., Structure of adeno-associated virus serotype 8, a gene therapy vector. J Virol, 2007. 81(22): p. 12260-71.
14. Kronenberg, S., et al., A conformational change in the adeno-associated virus type 2 capsid leads to the exposure of hidden VP1 N termini. J Virol, 2005. 79(9): p. 5296-303.
15. Sonntag, F., et al., Adeno-associated virus type 2 capsids with externalized VP1/VP2 trafficking domains are generated prior to passage through the cytoplasm and are maintained until uncoating occurs in the nucleus. J Virol, 2006. 80(22): p. 11040-54.
16. Murray, S., et al., Characterization of the capsid protein glycosylation of adeno-associated virus type 2 by high-resolution mass spectrometry. J Virol, 2006. 80(12): p. 6171-6.
17. Salganik, M., et al., Evidence for pH-dependent protease activity in the adeno-associated virus capsid. J Virol, 2012. 86(21): p. 11877-85.
18. Van Vliet, K., et al., Adeno-associated virus capsid serotype identification: Analytical methods development and application. J Virol Methods, 2009. 159(2): p. 167-77.
19. Thomas, J.J., et al., Viral characterization by direct analysis of capsid proteins. Anal Chem, 1998. 70(18): p. 3863-7.
20. Wang, L., L.C. Lane, and D.L. Smith, Detecting structural changes in viral capsids by hydrogen exchange and mass spectrometry. Protein Sci, 2001. 10(6): p. 1234-43.
21. Hwang, C.S., A. Shemorry, and A. Varshavsky, N-terminal acetylation of cellular proteins creates specific degradation signals. Science, 2010. 327(5968): p. 973-7.
22. Yan, Z., et al., Ubiquitination of both Adeno-Associated Virus Type 2 and 5 Capsid Proteins Affects the Transduction Efficiency of Recombinant Vectors. J Virol, 2002. 76(5): p. 2043-2053.
ПОСЛЕДОВАТЕЛЬНОСТИ
Все полипептидные последовательности представлены в направлении от N-конца к C-концу, если не указано иное.
Все последовательности нуклеиновых кислот представлены в направлении от 5'- к 3'-концу, если не указано иное.
Нуклеотидные последовательности потенциальных кодонов инициации VP3 AAV2 (кодоны ATG подчеркнуты)
ATGGCTACAGGCAGTGGCGCACCAATGGCAGAC (SEQ ID NO: 1)
Полипептидная последовательность, соответствующая потенциальным кодонам инициации VP3 AAV2 (метионины подчеркнуты)
MATGSGAPMAD (SEQ ID NO: 2)
Полипептидная последовательность VP1 AAV2
MAADGYLPDWLEDTLSEGIRQWWKLKPGPPPPKPAERHKDDSRGLVLPGYKYL GPFNGLDKGEPVNEADAAALEHDKAYDRQLDSGDNPYLKYNHADAEFQERLKEDTSF GGNLGRAVFQAKKRVLEPLGLVEEPVKTAPGKKRPVEHSPVEPDSSSGTGKAGQQPAR KRLNFGQTGDADSVPDPQPLGQPPAAPSGLGTNTMATGSGAPMADNNEGADGVGNSS GNWHCDSTWMGDRVITTSTRTWALPTYNNHLYKQISSQSGASNDNHYFGYSTPWGYF DFNRFHCHFSPRDWQRLINNNWGFRPKRLNFKLFNIQVKEVTQNDGTTTIANNLTSTVQ VFTDSEYQLPYVLGSAHQGCLPPFPADVFMVPQYGYLTLNNGSQAVGRSSFYCLEYFPS QMLRTGNNFTFSYTFEDVPFHSSYAHSQSLDRLMNPLIDQYLYYLSRTNTPSGTTTQSRL QFSQAGASDIRDQSRNWLPGPCYRQQRVSKTSADNNNSEYSWTGATKYHLNGRDSLV NPGPAMASHKDDEEKFFPQSGVLIFGKQGSEKTNVDIEKVMITDEEEIRTTNPVATEQYG SVSTNLQRGNRQAATADVNTQGVLPGMVWQDRDVYLQGPIWAKIPHTDGHFHPSPLM GGFGLKHPPPQILIKNTPVPANPSTTFSAAKFASFITQYSTGQVSVEIEWELQKENSKRWN PEIQYTSNYNKSVNVDFTVDTNGVYSEPRPIGTRYLTRNL (SEQ ID NO: 3)
N-концевой триптический пептид VP1 (N-концевой аланин является ацетилированным)
AADGYLPDWLEDTLSEGIR (SEQ ID NO: 4)
N-концевой Asp-N-пептид VP3 (N-концевой аланин является ацетилированным)
ATGSGAPM (SEQ ID NO: 5)
Обычная N-концевая последовательность VP1
MAADGYLPDWLED (SEQ ID NO: 6)
Нуклеотидные последовательности потенциальных кодонов инициации VP3 AAV7 (старт-кодоны подчеркнуты)
GTGGCTGCAGGCGGTGGCGCACCAATGGCAGACAATAAC (SEQ ID NO:7)
Нуклеотидная последовательность мутированного ITR
CACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTC GCCCACGCCCGGGCTTTGCCCGGGCG (SEQ ID NO: 8)
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> JIN, Xiaoying
O'RIORDAN, Catherine
LIU, Ling
Zhang, Kate
<120> СПОСОБЫ ВЫЯВЛЕНИЯ AAV
<130> 159792014140
<140> Пока еще не назначен
<141> Одновременно с этим
<150> 62/375,314
<151> 2016-08-15
<160> 40
<170> FastSEQ для Windows версии 4.0
<210> 1
<211> 33
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 1
Ala Thr Gly Gly Cys Thr Ala Cys Ala Gly Gly Cys Ala Gly Thr Gly
1 5 10 15
Gly Cys Gly Cys Ala Cys Cys Ala Ala Thr Gly Gly Cys Ala Gly Ala
20 25 30
Cys
<210> 2
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 2
Met Ala Thr Gly Ser Gly Ala Pro Met Ala Asp
1 5 10
<210> 3
<211> 735
<212> БЕЛОК
<213> Аденоассоциированный вирус 2
<400> 3
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 4
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> АЦЕТИЛИРОВАНИЕ
<222> 1
<400> 4
Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser Glu
1 5 10 15
Gly Ile Arg
<210> 5
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> АЦЕТИЛИРОВАНИЕ
<222> 1
<400> 5
Ala Thr Gly Ser Gly Ala Pro Met
1 5
<210> 6
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 6
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp
1 5 10
<210> 7
<211> 39
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 7
Gly Thr Gly Gly Cys Thr Gly Cys Ala Gly Gly Cys Gly Gly Thr Gly
1 5 10 15
Gly Cys Gly Cys Ala Cys Cys Ala Ala Thr Gly Gly Cys Ala Gly Ala
20 25 30
Cys Ala Ala Thr Ala Ala Cys
35
<210> 8
<211> 78
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 8
Cys Ala Cys Thr Cys Cys Cys Thr Cys Thr Cys Thr Gly Cys Gly Cys
1 5 10 15
Gly Cys Thr Cys Gly Cys Thr Cys Gly Cys Thr Cys Ala Cys Thr Gly
20 25 30
Ala Gly Gly Cys Cys Gly Gly Gly Cys Gly Ala Cys Cys Ala Ala Ala
35 40 45
Gly Gly Thr Cys Gly Cys Cys Cys Ala Cys Gly Cys Cys Cys Gly Gly
50 55 60
Gly Cys Thr Thr Thr Gly Cys Cys Cys Gly Gly Gly Cys Gly
65 70 75
<210> 9
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 9
Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys
1 5 10
<210> 10
<211> 83
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 10
Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu Thr
1 5 10 15
Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr Val
20 25 30
Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp Val
35 40 45
Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser Gln
50 55 60
Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser Gln
65 70 75 80
Met Leu Arg
<210> 11
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 11
Tyr Asn Leu Asn Gly Arg
1 5
<210> 12
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 12
Tyr His Leu Asn Gly Arg
1 5
<210> 13
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 13
Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu Tyr Thr Glu
1 5 10 15
Pro Arg
<210> 14
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 14
Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr Ser Glu
1 5 10 15
Pro Arg
<210> 15
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 15
Ala Pro Gly Lys Lys Arg Pro Val Glu His Ser Pro Val Glu Pro
1 5 10 15
<210> 16
<211> 735
<212> БЕЛОК
<213> Аденоассоциированный вирус 2
<400> 16
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 17
<211> 239
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 17
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro
180 185 190
Pro Ala Gly Pro Ser Gly Leu Gly Ser Gly Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg
225 230 235
<210> 18
<211> 239
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 18
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Glu Ser Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro
180 185 190
Pro Ala Gly Pro Ser Gly Leu Gly Ser Gly Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg
225 230 235
<210> 19
<211> 239
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 19
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Gln Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro
180 185 190
Pro Ala Ala Pro Ser Gly Val Gly Pro Asn Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg
225 230 235
<210> 20
<211> 239
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 20
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asn Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Ala Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro
180 185 190
Pro Ala Ala Pro Ser Ser Val Gly Ser Gly Thr Val Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn
210 215 220
Ala Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg
225 230 235
<210> 21
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 21
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg
225 230 235
<210> 22
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 22
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg
225 230 235
<210> 23
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 23
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg
225 230 235
<210> 24
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 24
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Val Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Arg Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Ile Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Gly
130 135 140
Ala Val Asp Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Val Gly
145 150 155 160
Lys Ser Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Thr Ser Leu Gly Ser Asn Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg
225 230 235
<210> 25
<211> 232
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 25
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Leu Glu Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly Lys
145 150 155 160
Lys Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Glu Glu Asp Thr
165 170 175
Gly Ala Gly Asp Gly Pro Pro Glu Gly Ser Asp Thr Ser Ala Met Ser
180 185 190
Ser Asp Ile Glu Met Arg Ala Ala Pro Gly Gly Asn Ala Val Asp Ala
195 200 205
Gly Gln Gly Ser Asp Gly Val Gly Asn Ala Ser Gly Asp Trp His Cys
210 215 220
Asp Ser Thr Trp Ser Glu Gly Lys
225 230
<210> 26
<211> 241
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 26
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Lys Gln Leu Glu Gln Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Gln Arg Leu Ala Thr Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Ile Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Leu Glu Lys Thr Pro Asn Arg Pro Thr Asn Pro Asp Ser Gly Lys
145 150 155 160
Ala Pro Ala Lys Lys Lys Gln Lys Asp Gly Glu Pro Ala Asp Ser Ala
165 170 175
Arg Arg Thr Leu Asp Phe Glu Asp Ser Gly Ala Gly Asp Gly Pro Pro
180 185 190
Glu Gly Ser Ser Ser Gly Glu Met Ser His Asp Ala Glu Met Arg Ala
195 200 205
Ala Pro Gly Gly Asn Ala Val Glu Ala Gly Gln Gly Ala Asp Gly Val
210 215 220
Gly Asn Ala Ser Gly Asp Trp His Cys Asp Ser Thr Trp Ser Glu Gly
225 230 235 240
Arg
<210> 27
<211> 232
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 27
Met Thr Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser Glu
1 5 10 15
Gly Val Arg Glu Trp Trp Ala Leu Gln Pro Gly Ala Pro Lys Pro Lys
20 25 30
Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro Gly
35 40 45
Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro Val
50 55 60
Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp Gln
65 70 75 80
Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala Asp
85 90 95
Ala Glu Phe Gln Gln Arg Leu Gln Gly Asp Thr Ser Phe Gly Gly Asn
100 105 110
Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro Leu
115 120 125
Gly Leu Val Glu Gln Ala Gly Glu Thr Ala Pro Gly Lys Lys Arg Pro
130 135 140
Leu Ile Glu Ser Pro Gln Gln Pro Asp Ser Ser Thr Gly Ile Gly Lys
145 150 155 160
Lys Gly Lys Gln Pro Ala Lys Lys Lys Leu Val Phe Glu Asp Glu Thr
165 170 175
Gly Ala Gly Asp Gly Pro Pro Glu Gly Ser Thr Ser Gly Ala Met Ser
180 185 190
Asp Asp Ser Glu Met Arg Ala Ala Ala Gly Gly Ala Ala Val Glu Gly
195 200 205
Gly Gln Gly Ala Asp Gly Val Gly Asn Ala Ser Gly Asp Trp His Cys
210 215 220
Asp Ser Thr Trp Ser Glu Gly His
225 230
<210> 28
<211> 228
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 28
Met Ser Phe Val Asp His Pro Pro Asp Trp Leu Glu Glu Val Gly Glu
1 5 10 15
Gly Leu Arg Glu Phe Leu Gly Leu Glu Ala Gly Pro Pro Lys Pro Lys
20 25 30
Pro Asn Gln Gln His Gln Asp Gln Ala Arg Gly Leu Val Leu Pro Gly
35 40 45
Tyr Asn Tyr Leu Gly Pro Gly Asn Gly Leu Asp Arg Gly Glu Pro Val
50 55 60
Asn Arg Ala Asp Glu Val Ala Arg Glu His Asp Ile Ser Tyr Asn Glu
65 70 75 80
Gln Leu Glu Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala Asp
85 90 95
Ala Glu Phe Gln Glu Lys Leu Ala Asp Asp Thr Ser Phe Gly Gly Asn
100 105 110
Leu Gly Lys Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro Phe
115 120 125
Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Thr Gly Lys Arg Ile
130 135 140
Asp Asp His Phe Pro Lys Arg Lys Lys Ala Arg Thr Glu Glu Asp Ser
145 150 155 160
Lys Pro Ser Thr Ser Ser Asp Ala Glu Ala Gly Pro Ser Gly Ser Gln
165 170 175
Gln Leu Gln Ile Pro Ala Gln Pro Ala Ser Ser Leu Gly Ala Asp Thr
180 185 190
Met Ser Ala Gly Gly Gly Gly Pro Leu Gly Asp Asn Asn Gln Gly Ala
195 200 205
Asp Gly Val Gly Asn Ala Ser Gly Asp Trp His Cys Asp Ser Thr Trp
210 215 220
Met Gly Asp Arg
225
<210> 29
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 29
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg
225 230 235
<210> 30
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> ВАРИАНТ
<222> 24, 148, 151, 152, 154, 155, 168, 169, 170, 171, 172, 208,
209
<223> Xaa = любая аминокислота
<400> 30
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Xaa Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Xaa Ser Pro Xaa Xaa Gln Xaa Xaa Pro Asp Ser Ser Ser
145 150 155 160
Gly Ile Gly Lys Lys Gly Gln Xaa Xaa Xaa Xaa Xaa Gln Pro Ala Lys
165 170 175
Lys Arg Leu Asn Phe Gly Gln Thr Gly Asp Ser Glu Ser Val Pro Asp
180 185 190
Pro Gln Pro Leu Gly Glu Pro Pro Ala Ala Pro Ser Gly Leu Gly Xaa
195 200 205
Xaa Thr Met Ala Ala Gly Gly Gly Ala Pro Met Ala Asp Asn Asn Glu
210 215 220
Gly Ala Asp Gly Val Gly Asn Ala Ser Gly Asn Trp His Cys Asp Ser
225 230 235 240
Thr Trp Leu Gly Asp Arg
245
<210> 31
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 31
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys
50 55 60
<210> 32
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 32
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys
50 55 60
<210> 33
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 33
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asn Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys
50 55 60
<210> 34
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 34
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys
50 55 60
<210> 35
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 35
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Val Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Arg Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys
50 55 60
<210> 36
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 36
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Lys Ala Asn Gln Gln His Gln Asp Asn Gly Arg Gly Leu Val Leu
35 40 45
Pro Gly Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys
50 55 60
<210> 37
<211> 60
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 37
Met Thr Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser Glu
1 5 10 15
Gly Val Arg Glu Trp Trp Ala Leu Gln Pro Gly Ala Pro Lys Pro Lys
20 25 30
Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro Gly
35 40 45
Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys
50 55 60
<210> 38
<211> 60
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 38
Met Ser Phe Val Asp His Pro Pro Asp Trp Leu Glu Glu Val Gly Glu
1 5 10 15
Gly Leu Arg Glu Phe Leu Gly Leu Glu Ala Gly Pro Pro Lys Pro Lys
20 25 30
Pro Asn Gln Gln His Gln Asp Gln Ala Arg Gly Leu Val Leu Pro Gly
35 40 45
Tyr Asn Tyr Leu Gly Pro Gly Asn Gly Leu Asp Arg
50 55 60
<210> 39
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 39
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys
50 55 60
<210> 40
<211> 61
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> ВАРИАНТ
<222> 24
<223> Xaa = любая аминокислота
<400> 40
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Xaa Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys
50 55 60
<---
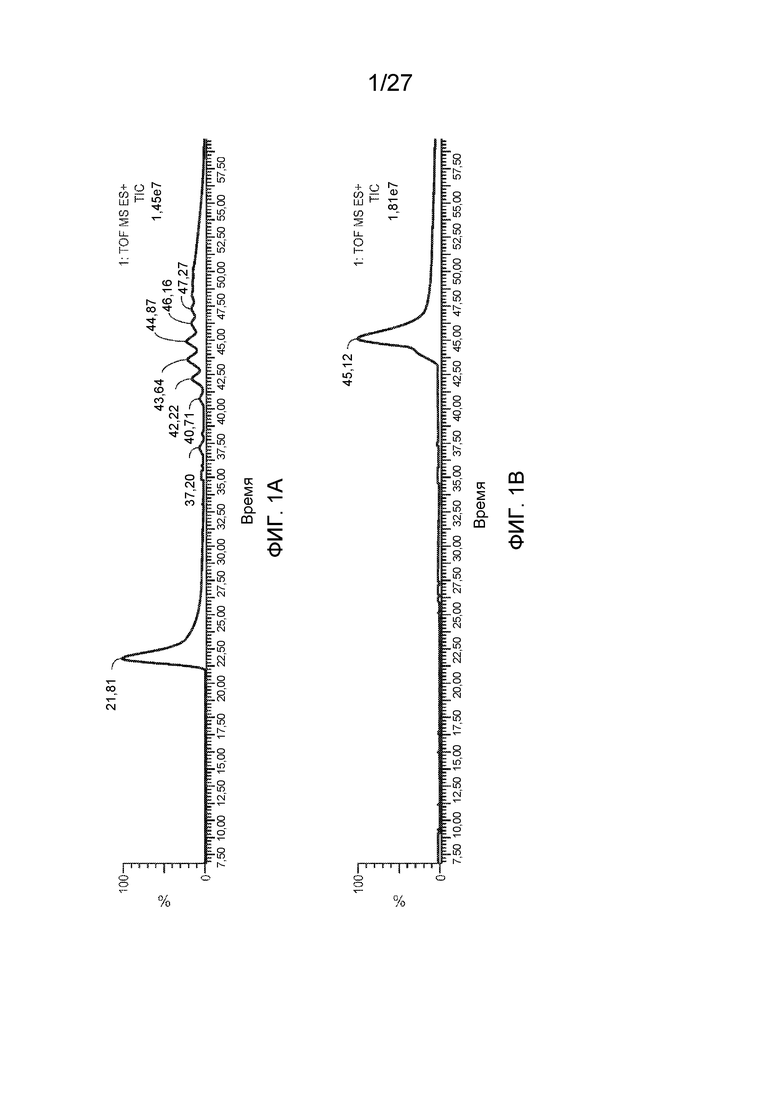
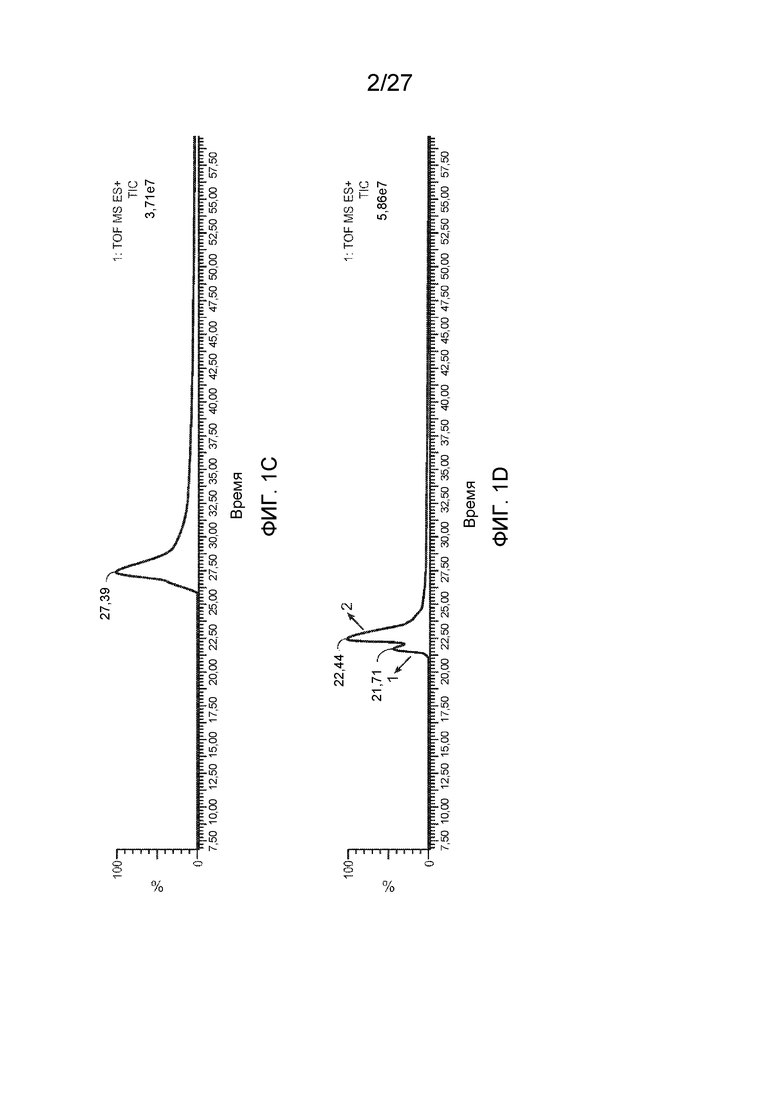


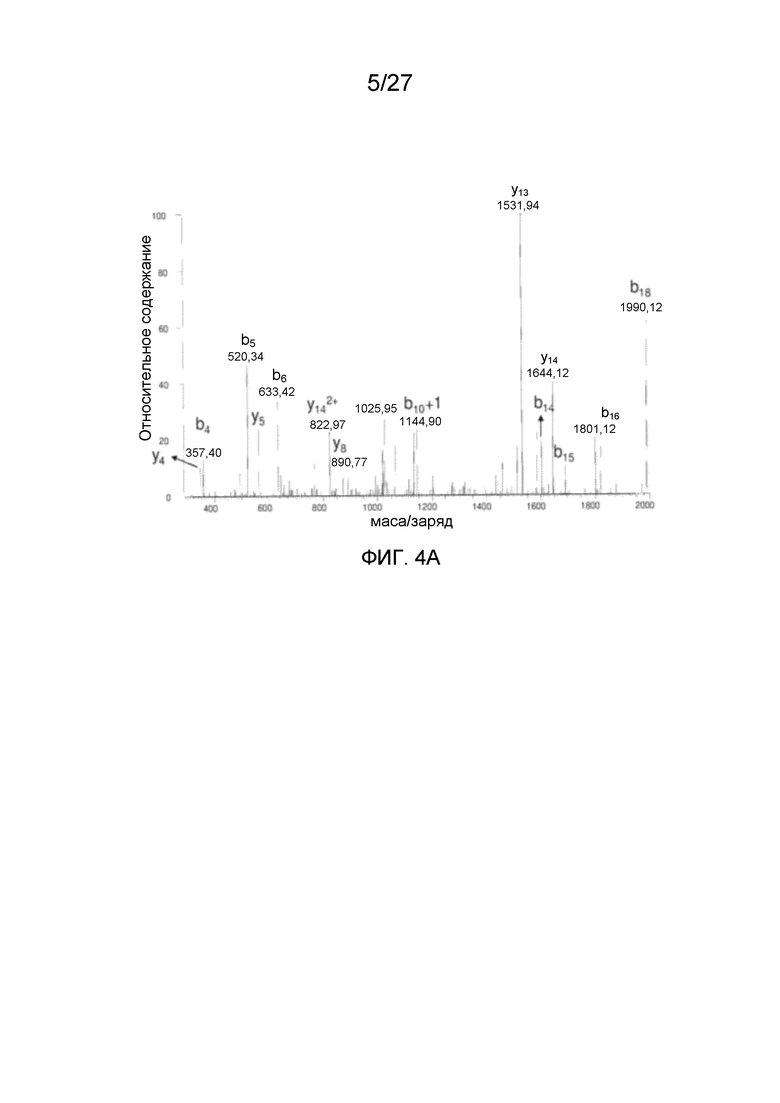
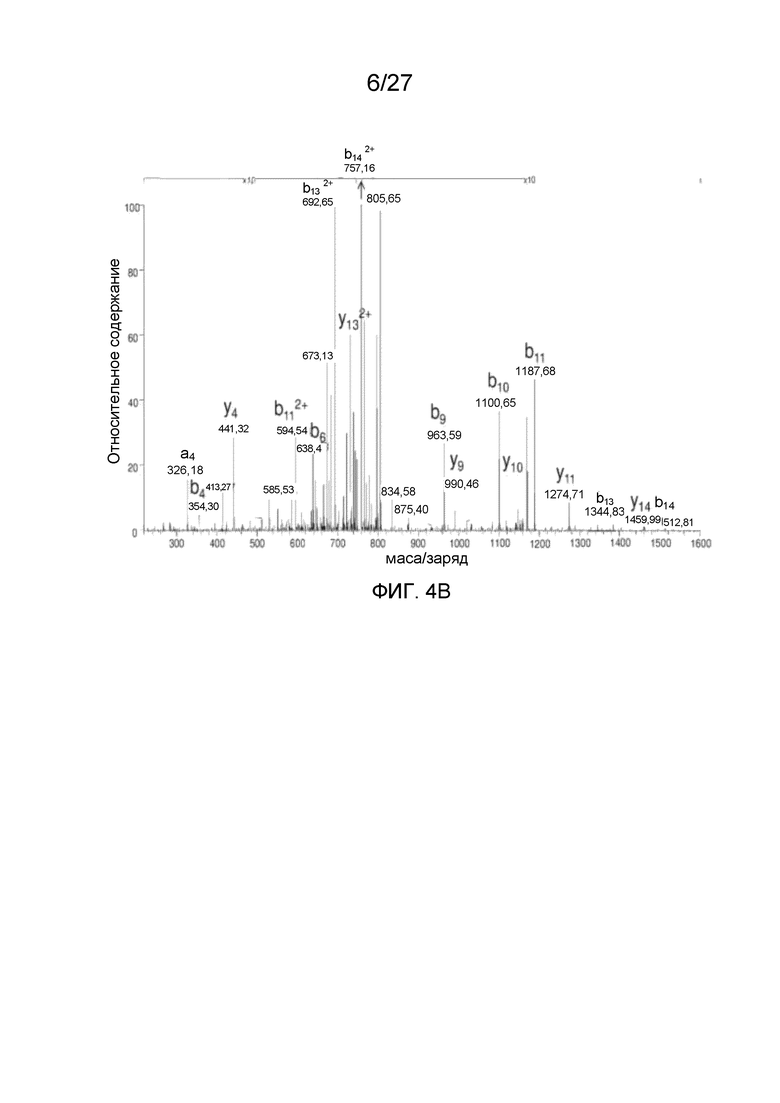
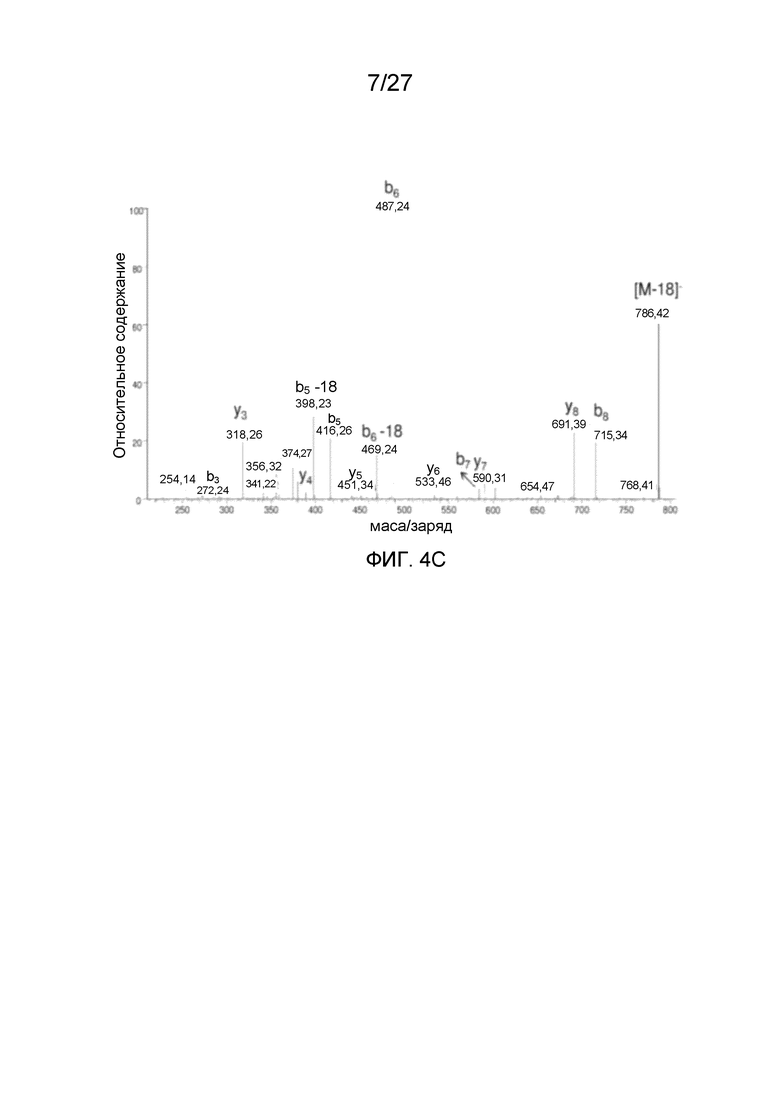
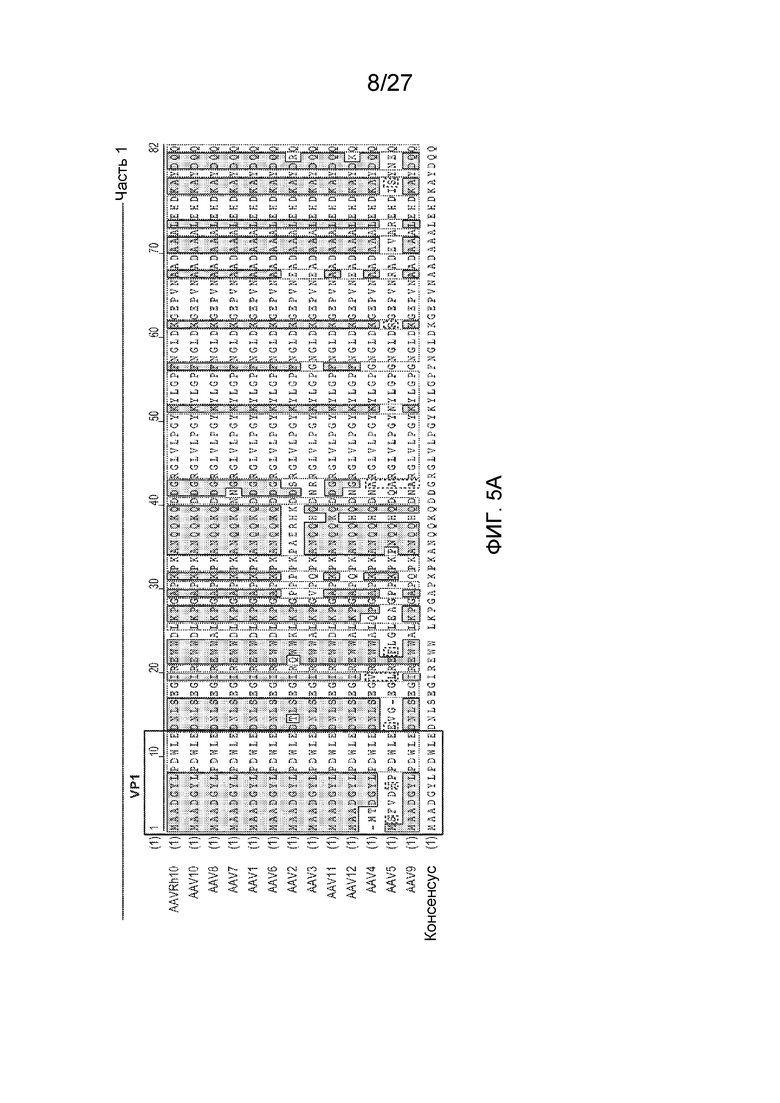

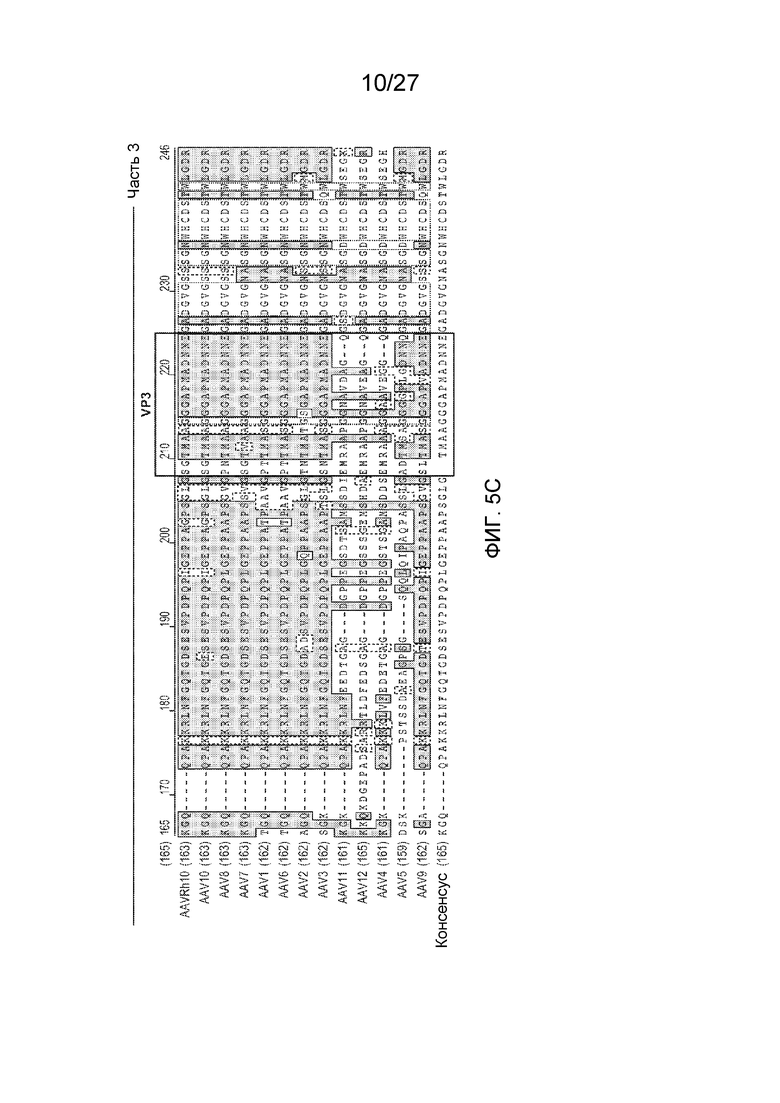
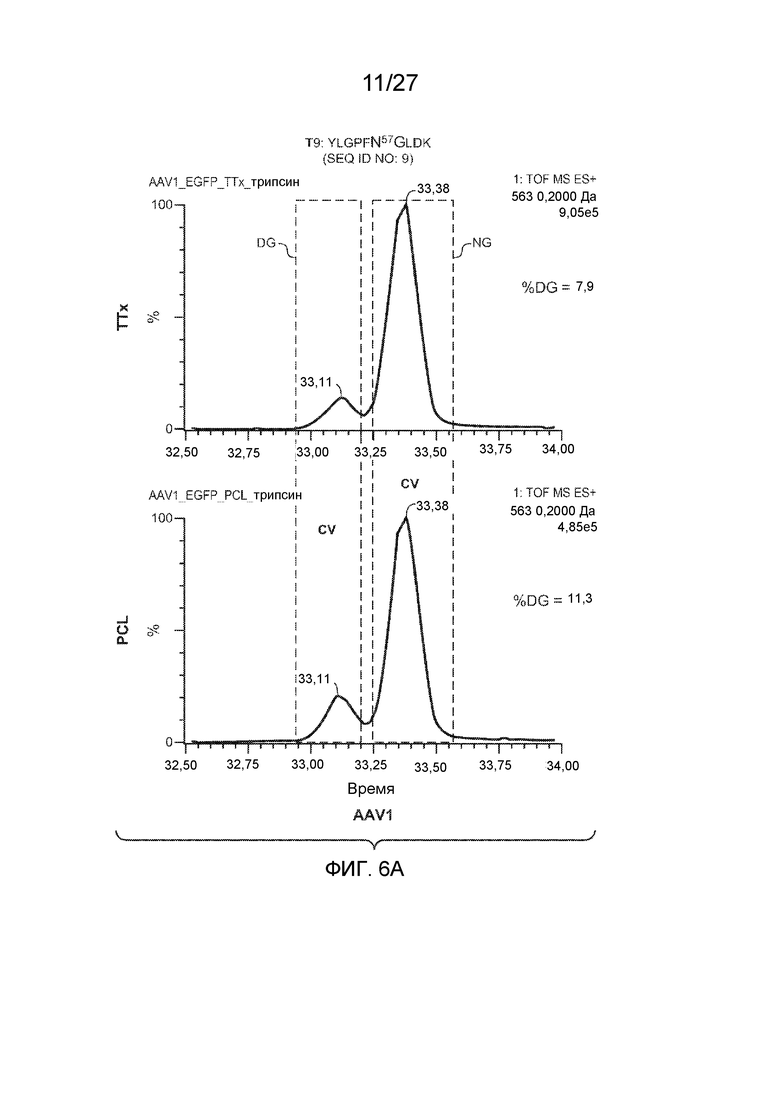
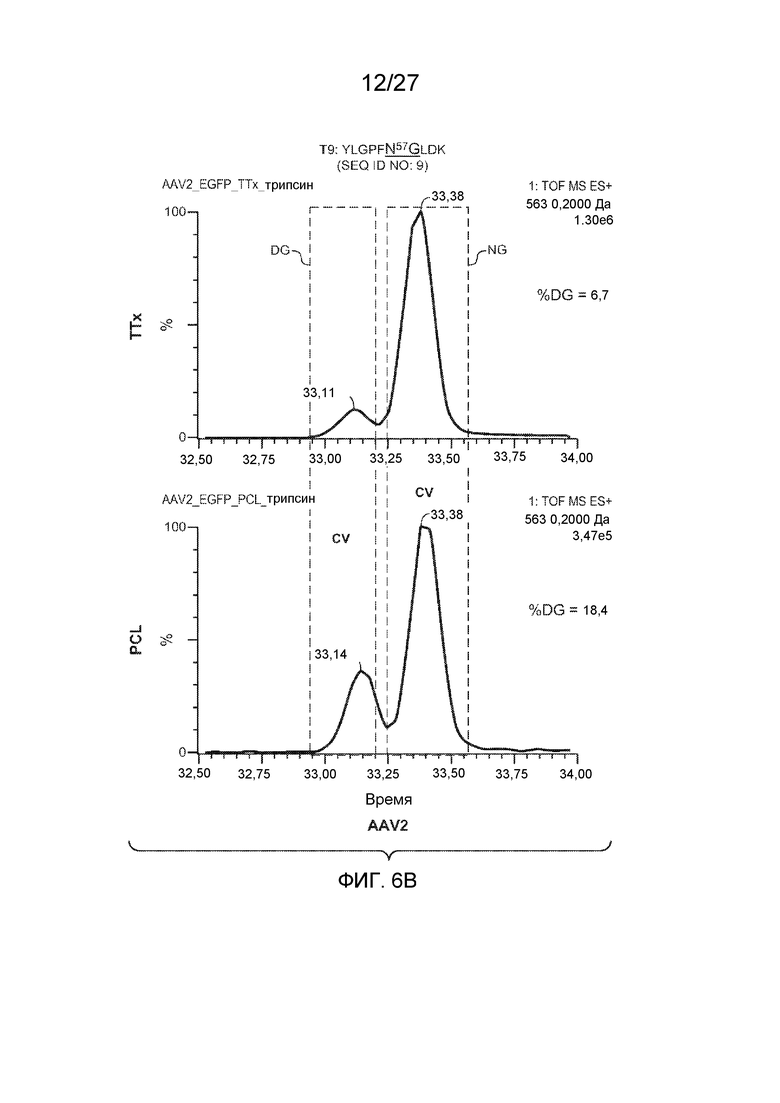
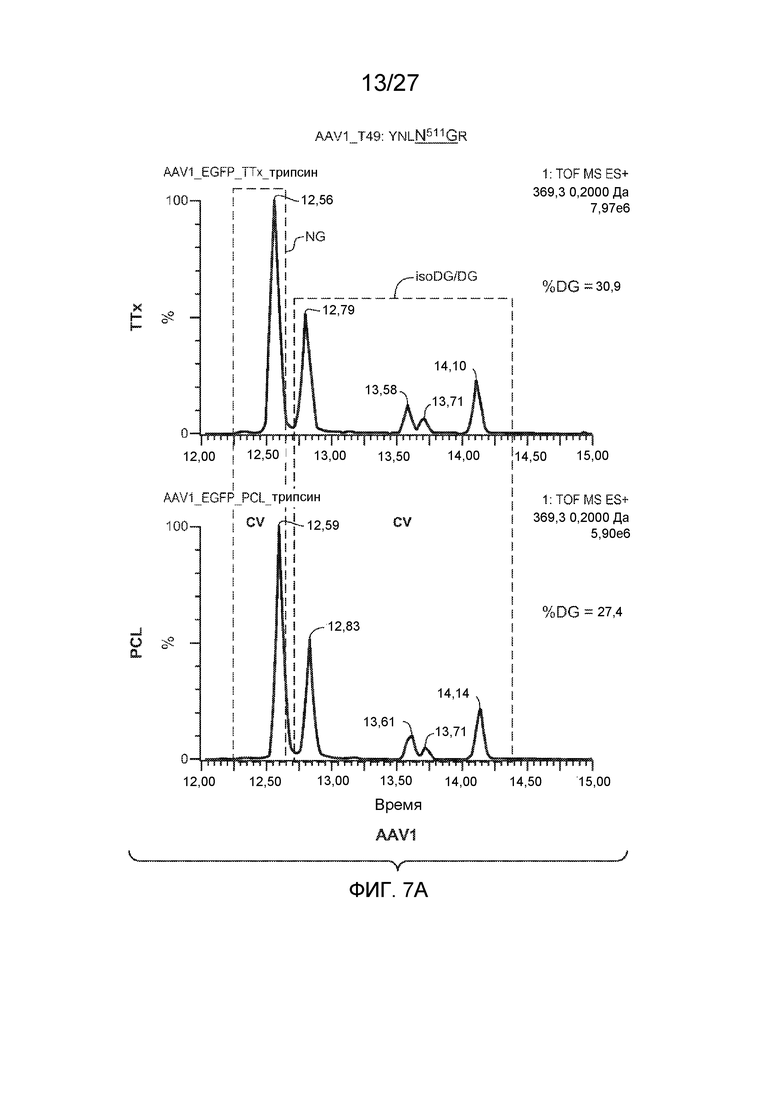
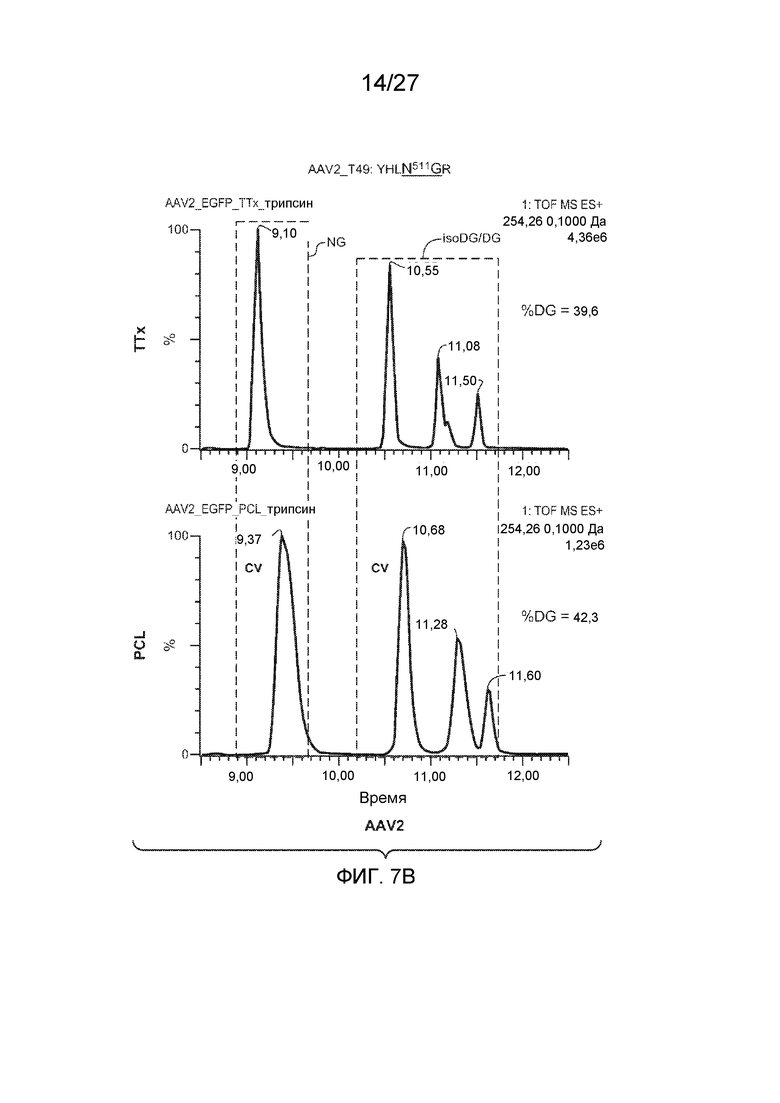
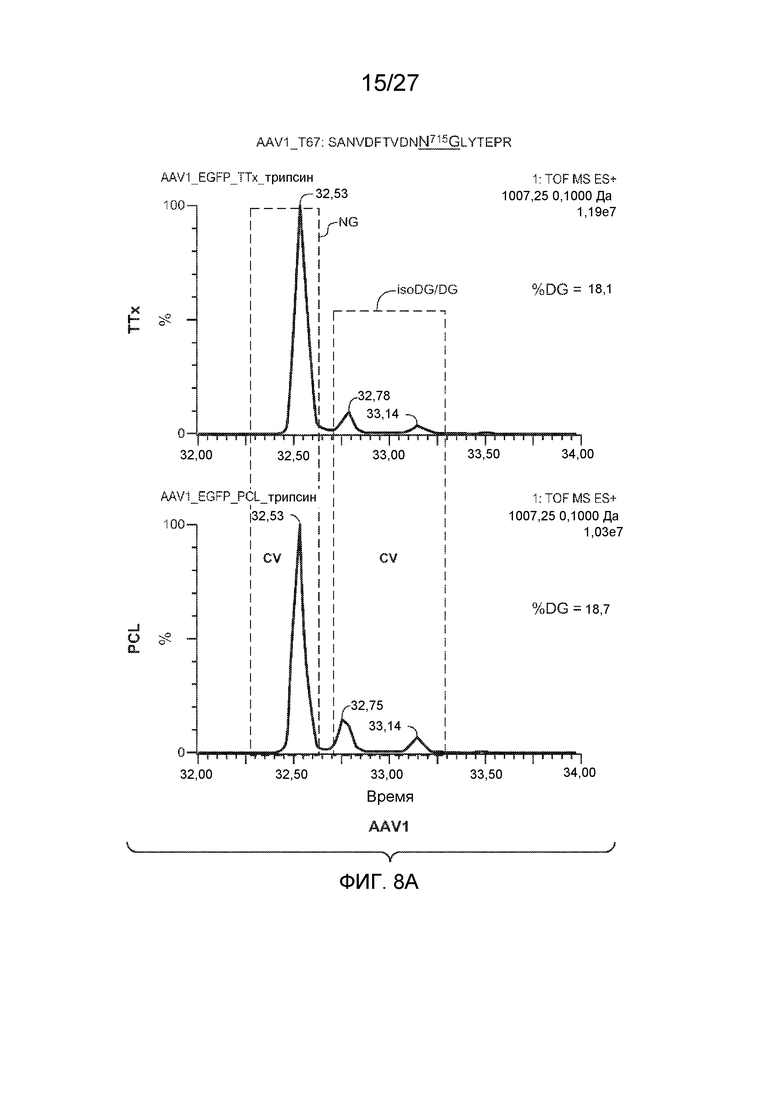
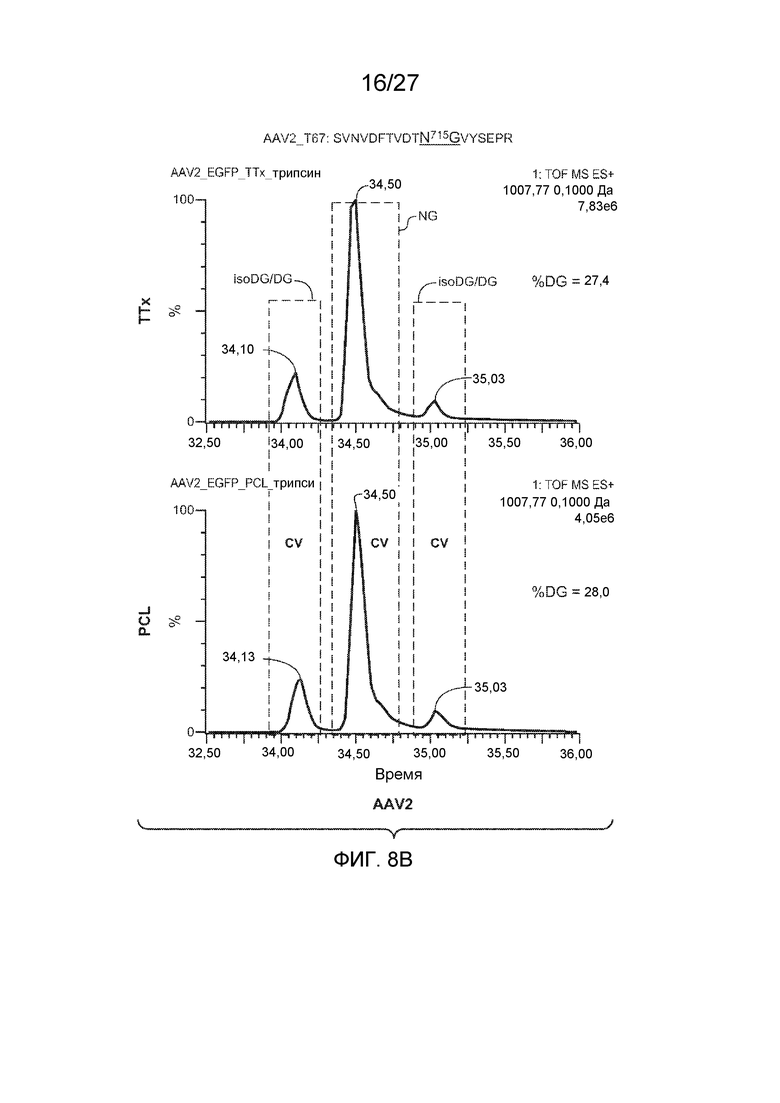

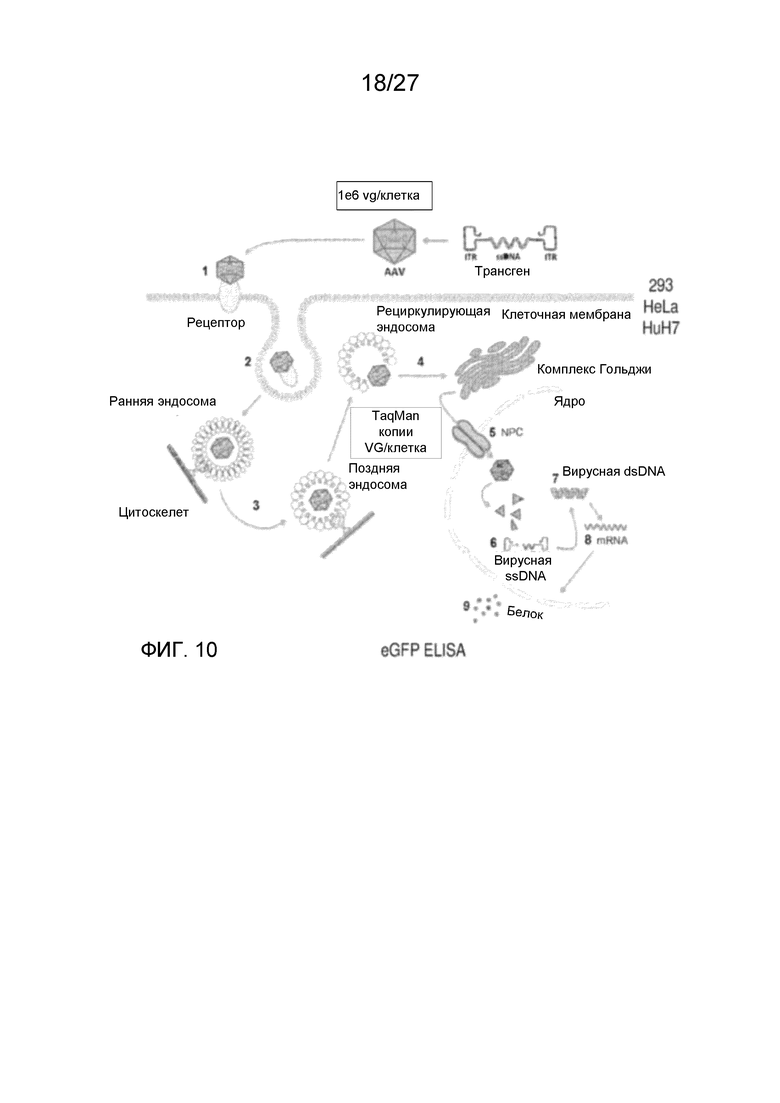
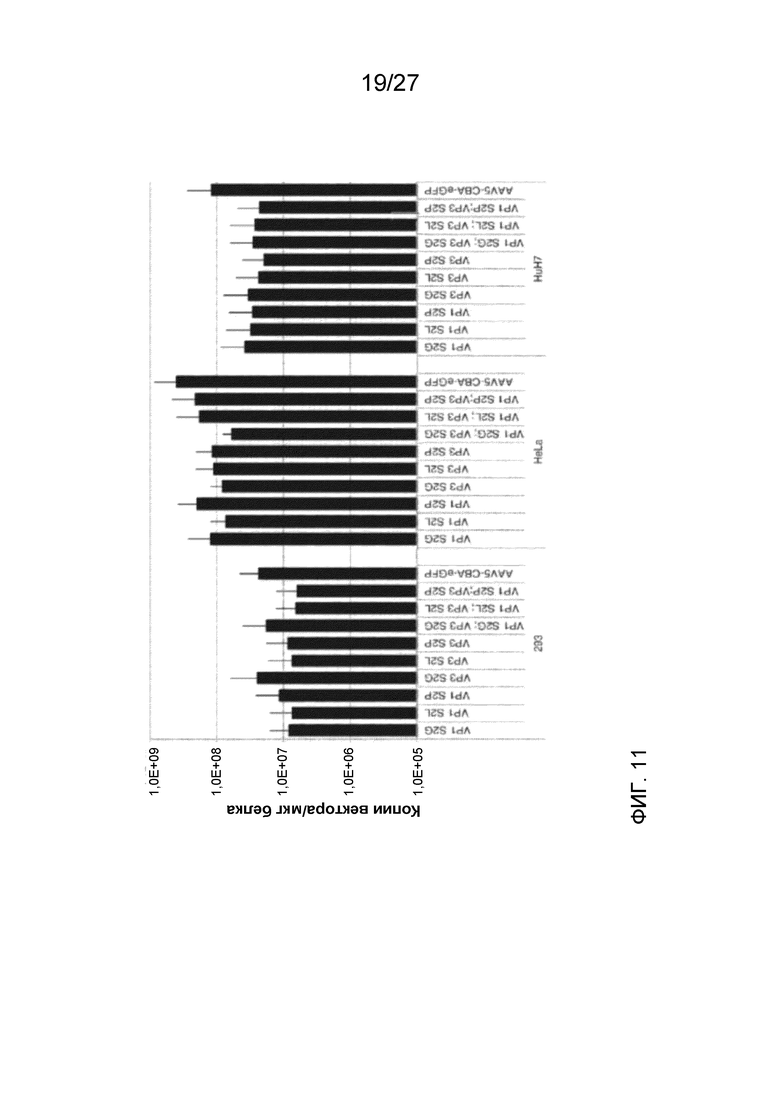
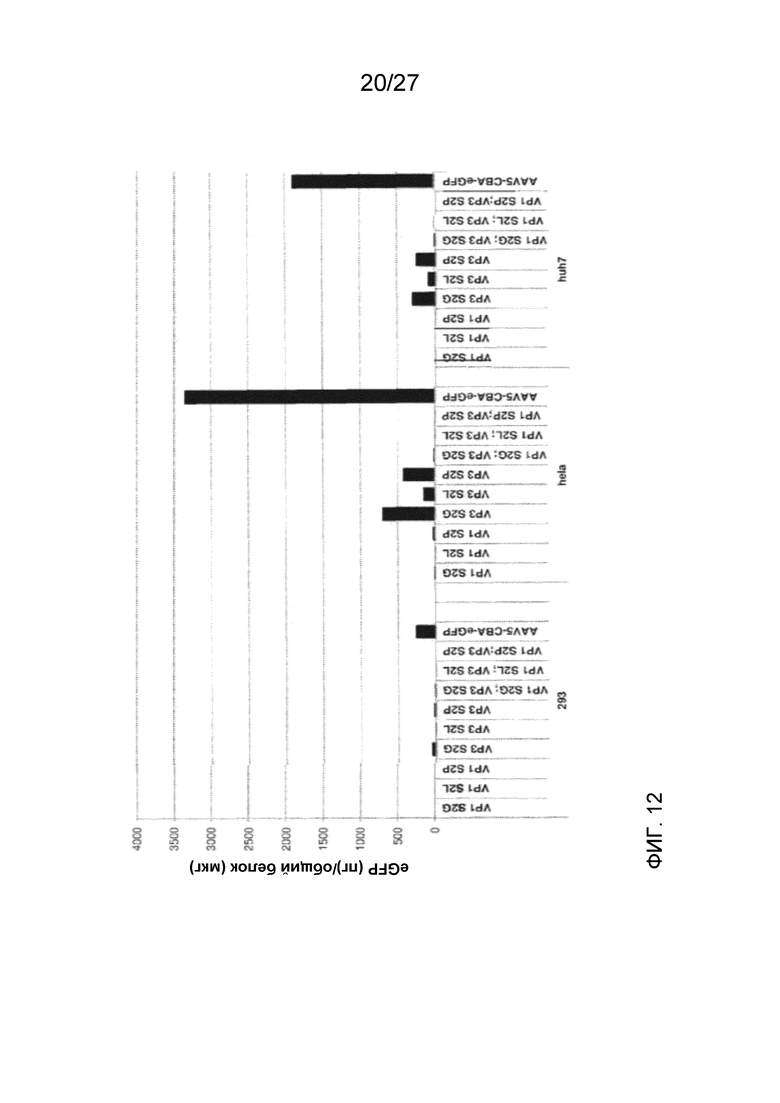


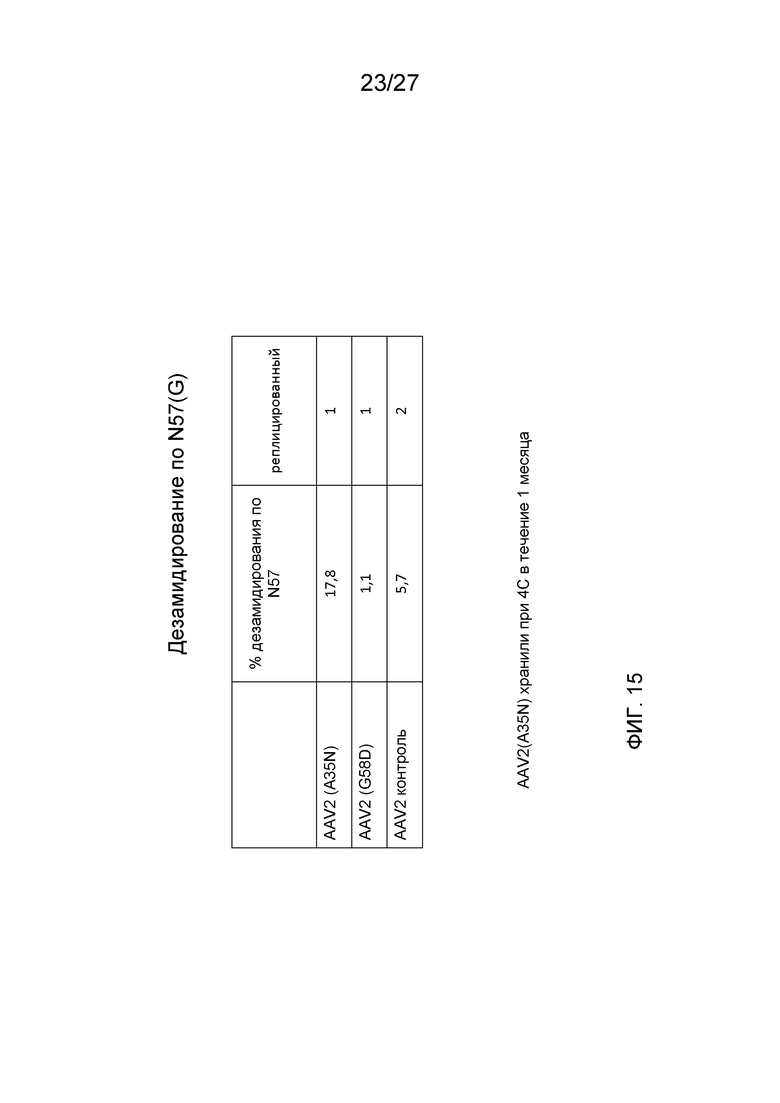

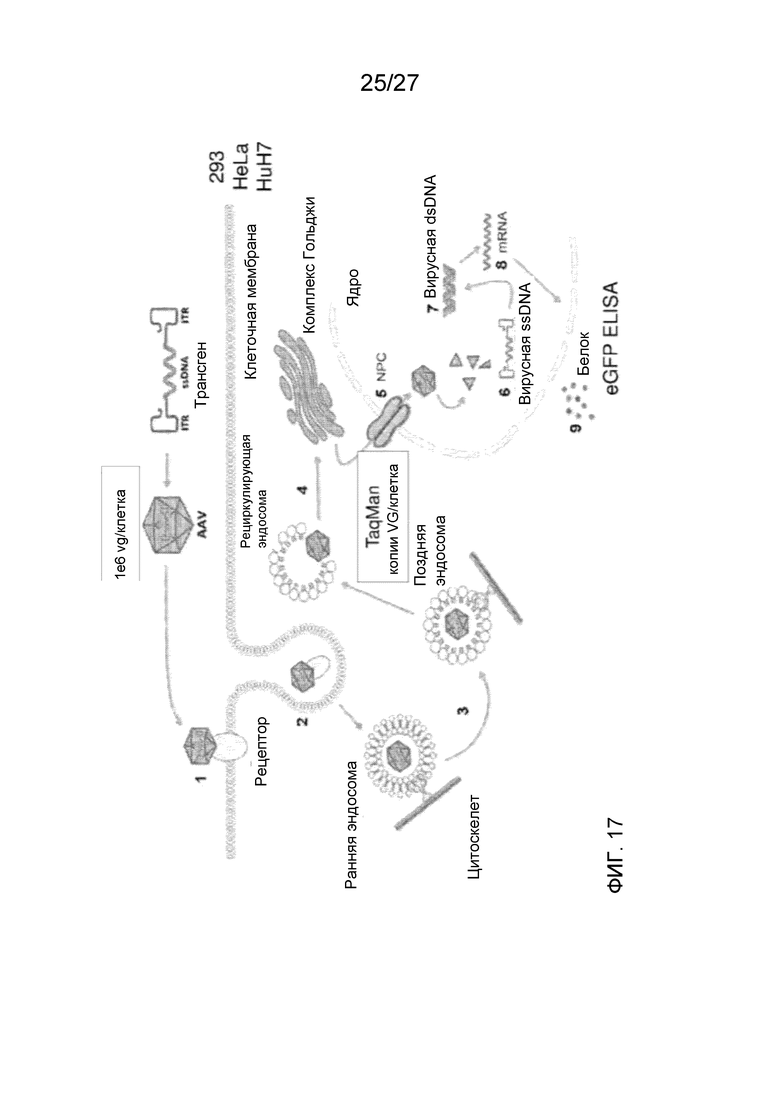
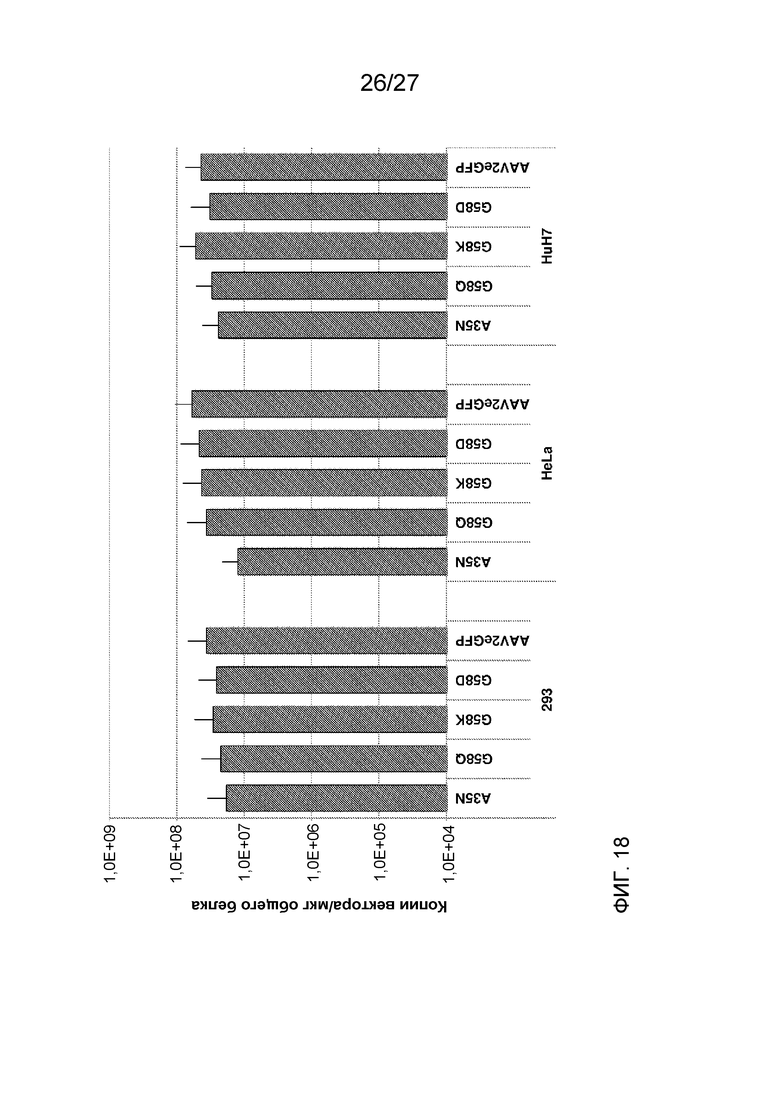
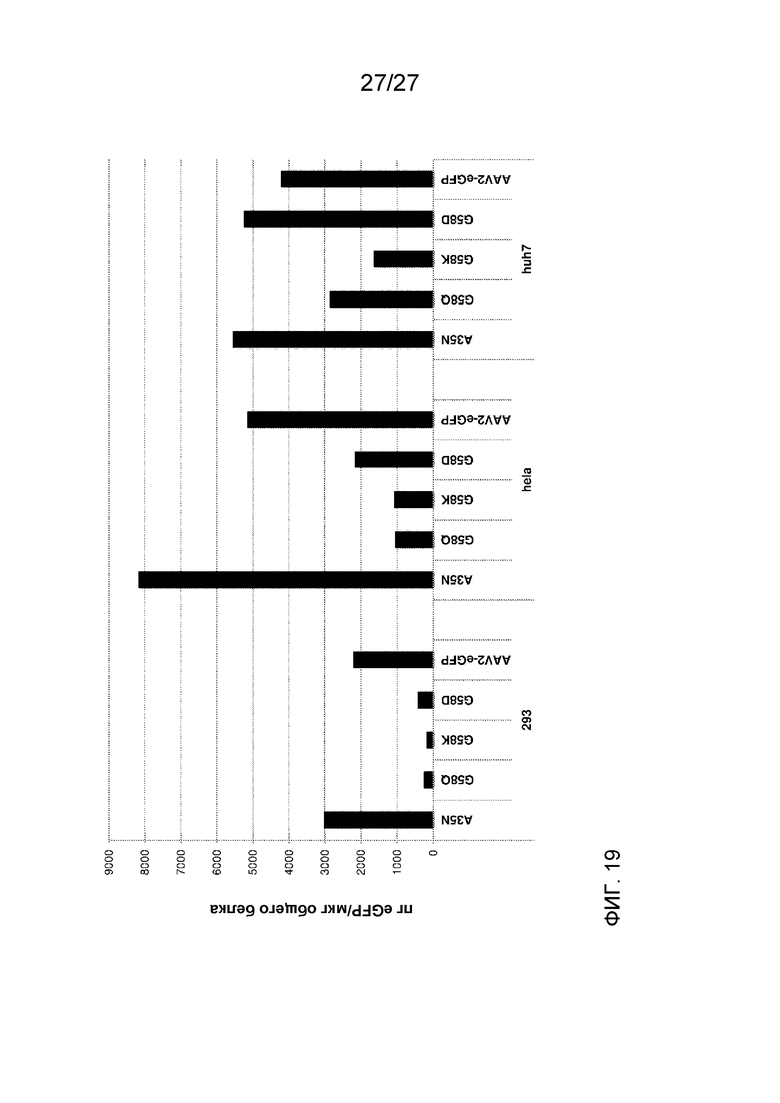
Изобретение относится к области биотехнологии и вирусологии. Описаны способы серотипирования и/или определения гетерогенности вирусной частицы с помощью определения массы, в частности, с помощью применения жидкостной хроматографии/масс-спектрометрии (LC/MS) или жидкостной хроматографии/масс-спектрометрии-масс-спектрометрии (LC/MS/MS). Описаны также способы улучшения стабильности частиц rAAV, улучшения сборки частиц rAAV в клетке или улучшения трансдукции частиц rAAV в клетке с помощью повышения степени ацетилирования и/или дезамидирования капсидных белков. Группа изобретений также относится к капсидному белку AAV для получения частицы AAV и частице рекомбинантного AAV, фармацевтической композиции, набору и изделию для доставки нуклеиновой кислоты в клетку-хозяин. Группа изобретений позволяет улучшить способы характеристики вирусных частиц. 15 н. и 49 з.п. ф-лы, 30 ил., 11 табл., 5 пр.
1. Способ определения серотипа вирусной частицы, включающий:
a) денатурацию вирусной частицы;
b) непосредственное введение денатурированной вирусной частицы в анализ интактного белка путем жидкостной хроматографии/масс-спектрометрии (LC/MS); и
c) определение масс одного или более капсидных белков вирусной частицы;
где конкретная комбинация масс одного или более капсидных белков характеризует серотип вируса, и где способ осуществляют в отсутствие стадии разделения на геле.
2. Способ по п.1, в котором рассчитанные массы одного или более капсидных белков сравнивают с теоретическими массами одного или более капсидных белков одного или более серотипов вируса.
3. Способ определения серотипа вирусной частицы, включающий:
a) денатурацию вирусной частицы;
b) восстановление и/или алкилирование денатурированной вирусной частицы;
c) расщепление денатурированной вирусной частицы с образованием фрагментов одного или более капсидных белков вирусной частицы;
d) жидкостную хроматографию/масс-спектрометрию/масс-спектрометрию (LC/MS/MS) фрагментов одного или более капсидных белков; и
e) определение масс фрагментов одного или более капсидных белков вирусной частицы;
где конкретная комбинация масс фрагментов одного или более капсидных белков характеризует серотип вируса, и где способ осуществляют в отсутствие стадии разделения на геле.
4. Способ по п.3, в котором рассчитанные массы фрагментов одного или более капсидных белков сравнивают с теоретическими массами фрагментов одного или более капсидных белков одного или более серотипов вируса.
5. Способ определения гетерогенности вирусной частицы, включающий:
a) денатурацию вирусной частицы;
b) непосредственное введение денатурированной вирусной частицы в анализ интактного белка путем жидкостной хроматографии/масс-спектрометрии/масс-спектрометрии (LC/MS/MS);
c) определение масс одного или более капсидных белков вирусной частицы и
d) сравнение масс со стадии c) с теоретическими массами одного или более капсидных белков серотипа вируса;
где отклонение одной или более масс одного или более капсидных белков характеризует гетерогенность капсида вируса, и где способ осуществляют в отсутствие стадии разделения на геле.
6. Способ определения гетерогенности серотипа вирусной частицы, включающий:
a) денатурацию вирусной частицы;
b) восстановление и/или алкилирование денатурированной вирусной частицы;
c) расщепление денатурированной вирусной частицы с образованием фрагментов одного или более капсидных белков вирусной частицы;
d) жидкостную хроматографию/масс-спектрометрию/масс-спектрометрию (LC/MS/MS) фрагментов одного или более капсидных белков;
e) определение масс фрагментов одного или более капсидных белков вирусной частицы и
f) сравнение масс со стадии e) с теоретическими массами фрагментов одного или более капсидных белков серотипа вируса;
где отклонение одной или более масс одного или более капсидных белков характеризует гетерогенность капсида вируса, и где способ осуществляют в отсутствие стадии разделения на геле.
7. Способ по п.5 или 6, в котором гетерогенность включает одно или более из смешанных серотипов, вариантных капсидов, аминокислотных замен капсидов, усеченных капсидов или модифицированных капсидов.
8. Способ по любому из пп. 1-4, где
(i) масс-спектрометрия включает:
(1) капиллярное напряжение, составляющее 3,5 кВ, и/или
(2) напряжение пробоотборного конуса, составляющее 45 B, и/или
(3) калибровку, выполняемую с помощью дополнительного средства; и/или
(ii) жидкостная хроматография представляет собой ультраэффективную жидкостную хроматографию (UPLC) и/или обращенно-фазовую жидкостную хроматографию.
9. Способ по п.8, в котором масс-спектрометрия включает калибровку, выполняемую с помощью дополнительного средства, где в качестве калибранта используют йодид натрия.
10. Способ по п.8 или 9, в котором жидкостная хроматография представляет собой хроматографию с обращенной фазой C4 или C8.
11. Способ по п.10, в котором:
(1) в хроматографии используют подвижную фазу A, содержащую муравьиную кислоту в воде; и/или
(2) в хроматографии используют подвижную фазу B, содержащую муравьиную кислоту в ацетонитриле.
12. Способ по п.11, в котором в хроматографии используют подвижную фазу A, содержащую 0,1% муравьиной кислоты в воде, и/или подвижную фазу B, содержащую 0,1% муравьиной кислоты в ацетонитриле.
13. Способ по п.12, в котором долю подвижной фазы B в хроматографии увеличивают со временем, увеличивают поэтапно, увеличивают от 10% до 20%, увеличивают от 20% до 30%, увеличивают от 30% до 38%, увеличивают от 10% до 20% в течение 6 минут, увеличивают от 20% до 30% в течение 10 минут или увеличивают от 30% до 38% в течение 40 минут.
14. Способ по любому из пп. 5-7, где
(i) масс-спектрометрия включает:
(1) напряжение источника, составляющее 2,5 кВ, и/или
(2) температуру капилляра, составляющую 275°C, и/или
(3) уровень RF S-линзы, составляющий 55%; и/или
(ii) жидкостная хроматография представляет собой высокоэффективную жидкостную хроматографию (HPLC) и/или обращенно-фазовую жидкостную хроматографию.
15. Способ по п.14, в котором жидкостная хроматография представляет собой жидкостную хроматографию с обращенной фазой, причем жидкостная хроматография с обращенной фазой представляет собой хроматографию с обращенной фазой C18.
16. Способ по п.15, в котором:
(1) в хроматографии используют подвижную фазу A, содержащую муравьиную кислоту в воде; и/или
(2) в хроматографии используют подвижную фазу B, содержащую муравьиную кислоту в ацетонитриле.
17. Способ по п.16, в котором в хроматографии используют подвижную фазу A, содержащую 0,1% муравьиной кислоты в воде, и/или подвижную фазу B, содержащую 0,1% муравьиной кислоты в ацетонитриле.
18. Способ по любому из пп. 14-17, в котором долю подвижной фазы B в хроматографии увеличивают со временем, увеличивают от 2% до 60% или увеличивают от 2% до 60% в течение 121 минуты.
19. Способ по п.3 или 6, где вирусная частица представляет собой частицу AAV, и где
(i) восстановление осуществляют воздействием на частицу AAV дитиотреитола, бета-меркаптоэтанола или трис(2-карбоксиэтил)фосфина (TCEP); и/или
(ii) алкилирование осуществляют воздействием на частицу AAV йодуксусной кислоты, йодацетамида или 4-винилпиридина; и/или
(iii) расщепление представляет собой ферментативное расщепление или химическое расщепление.
20. Способ по п.19, в котором ферментативное расщепление представляет собой расщепление с помощью эндопептидазы.
21. Способ по п.20, в котором расщепление с помощью эндопептидазы представляет собой расщепление с помощью трипсина, расщепление с помощью LysC, расщепление с помощью Asp-N или расщепление с помощью Glu-C.
22. Способ по п.19, в котором химическое расщепление представляет собой расщепление с помощью бромциана или кислотное расщепление.
23. Способ по любому из пп. 1-22, в котором вирусная частица включает вирусный вектор, кодирующий гетерологичный трансген.
24. Способ по любому из пп. 1-23, в котором
(i) вирусную частицу денатурируют уксусной кислотой, гуанидин гидрохлоридом и/или органическим растворителем; и/или
(ii) жидкостная хроматография представляет собой жидкостную хроматографию с обращенной фазой, эксклюзионную хроматографию, жидкостную хроматографию гидрофильных взаимодействий или катионообменную хроматографию.
25. Способ по любому из пп. 1-24, в котором вирусная частица представляет собой частицу аденоассоциированного вируса (AAV), и один или более капсидных белков представляют собой VP1, VP2 и VP3.
26. Способ по п.25, в котором (A) частица AAV представляет собой частицу рекомбинантного AAV (rAAV) и/или (B) N-конец VP1 и/или VP3 является ацетилированным.
27. Способ определения серотипа частицы AAV, включающий способ по п.1 в комбинации со способом по п.3.
28. Способ определения гетерогенности частицы AAV, включающий способ по п.5 в комбинации со способом по п.6.
29. Способ по любому из пп. 1-28, где вирусная частица представляет собой частицу AAV, и частица AAV содержит:
(i) капсид AAV1, капсид AAV2, капсид AAV3, капсид AAV4, капсид AAV5, капсид AAV6, капсид AAV7, капсид AAV8, капсид AAVrh8, капсид AAV9, капсид AAV10, капсид AAVrh10, капсид AAV11, капсид AAV12, капсид AAV LK03, капсид AAV2R471A, капсид AAV2/2-7m8, капсид AAV DJ, капсид AAV DJ8, капсид AAV2 N587A, капсид AAV2 E548A, капсид AAV2 N708A, капсид AAV V708K, капсид AAV козы, химерный капсид AAV1/AAV2, капсид AAV крупного рогатого скота, капсид AAV мыши rAAV2/HBoV1, капсид AAV2HBKO, капсид AAVPHP.B или капсид AAVPHP.eB, и/или капсид AAV содержит мутацию по тирозину, мутацию, влияющую на связывание с гепарином, или мутацию HBKO; и/или
(ii) ITR AAV1, ITR AAV2, ITR AAV3, ITR AAV4, ITR AAV5, ITR AAV6, ITR AAV7, ITR AAV8, ITR AAVrh8, ITR AAV9, ITR AAV10, ITR AAVrh10, ITR AAV11 или ITR AAV12.
30. Способ по любому из пп. 1-29, где массы VP1, VP2 и VP3 сравнивают с теоретическими массами одного или нескольких из капсида AAV1, капсида AAV2, капсида AAV3, капсида AAV4, капсида AAV5, капсида AAV6, капсида AAV7, капсида AAV8, капсида AAVrh8, капсида AAV9, капсида AAV10, капсида AAVrh10, капсида AAV11, капсида AAV12, капсида AAV LK03, капсида AAV2R471A, капсида AAV2/2-7m8, капсида AAV DJ, капсида AAV DJ8, капсида AAV2 N587A, капсида AAV2 E548A, капсида AAV2 N708A, капсида AAV V708K, капсида AAV козы, химерного капсида AAV1/AAV2, капсида AAV крупного рогатого скота, капсида AAV мыши rAAV2/HBoV1, капсида AAV2HBKO, капсида AAVPHP.B или капсида AAVPHP.eB.
31. Способ по любому из пп. 1-18, где вирусная частица принадлежит к семейству вирусов, выбранному из группы, состоящей из Adenoviridae, Parvoviridae, Retroviridae, Baculoviridae и Herpesviridae.
32. Способ по п.31, в котором вирусная частица принадлежит к роду вирусов, выбранному из группы, состоящей из Atadenovirus, Aviadenovirus, Ichtadenovirus, Mastadenovirus, Siadenovirus, Ambidensovirus, Brevidensovirus, Hepandensovirus, Iteradensovirus, Penstyldensovirus, Amdoparvovirus, Aveparvovirus, Bocaparvovirus, Copiparvovirus, Dependoparvovirus, Erythroparvovirus, Protoparvovirus, Tetraparvovirus, Alpharetrovirus, Betaretrovirus, Deltaretrovirus, Epsilonretrovirus, Gammaretrovirus, Lentivirus, Spumavirus, Alphabaculovirus, Betabaculovirus, Deltabaculovirus, Gammabaculovirus, Iltovirus, Mardivirus, Simplexvirus, Varicellovirus, Cytomegalovirus, Muromegalovirus, Proboscivirus, Roseolovirus, Lymphocryptovirus, Macavirus, Percavirus и Rhadinovirus.
33. Способ повышения стабильности частицы rAAV, улучшения сборки частиц rAAV в клетке или улучшения трансдукции частиц rAAV в клетке, включающий замену по аминокислотному остатку 2 VP1 и/или VP3 исходного VP1 и/или VP3; где замена аминокислоты в положении 2 обеспечивает изменение N-концевого ацетилирования VP1 и/или VP3 по сравнению с аминокислотным остатком 2 исходного VP1 и/или VP3.
34. Способ по п.33, в котором замена по аминокислотному остатку 2 приводит к более высокой частоте N-концевого ацетилирования или более низкой частоте N-концевого ацетилирования.
35. Способ по п.33 или 34, в котором аминокислотный остаток 2 заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr.
36. Способ по п.35, в котором аминокислотный остаток 2 заменен Ser, Asp или Glu.
37. Способ повышения стабильности частицы rAAV, улучшения сборки частиц rAAV в клетке или улучшения трансдукции частиц rAAV в клетке, включающий замену по одному или более аминокислотным остаткам VP1 и/или VP3 исходного VP1 и/или VP3, где один или более аминокислотных остатков представляют собой A35, N57, G58, N382, G383, N511, G512, N715 или G716, причем нумерация остатков приведена на основе VP1 AAV2; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с аминокислотным остатком 2 исходного VP1 и/или VP3.
38. Способ по п.37, в котором одна или более аминокислотных замен осуществляются по A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3; где аминокислотная замена обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV.
39. Способ по п.37 или 38, в котором исходный остаток Ala в положении 35 VP1 заменен Asn, и/или исходный остаток Gly в положении 58 VP1 заменен Asp.
40. Способ по любому из пп. 33-39, где частицу rAAV получают с помощью
a) трансфекции клетки-хозяина нуклеиновой кислотой, кодирующей вектор на основе rAAV, и нуклеиновой кислотой, кодирующей функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV; или
b) клетки-продуцента AAV, содержащей нуклеиновую кислоту, кодирующую вектор на основе rAAV, и нуклеиновую кислоту, кодирующую функциональные элементы rep и cap AAV, и получения нуклеиновой кислоты, кодирующей функциональные элементы помощника AAV.
41. Способ по п.40, в котором функциональные элементы cap AAV обеспечивают аминокислотную замену VP1 и/или VP3, где аминокислотная замена VP1 и/или VP3 обеспечивает изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV.
42. Способ по любому из пп. 33-41, где частица AAV содержит:
(i) капсид AAV1, капсид AAV2, капсид AAV3, капсид AAV4, капсид AAV5, капсид AAV6, капсид AAV7, капсид AAV8, капсид AAVrh8, капсид AAV9, капсид AAV10, капсид AAVrh10, капсид AAV11, капсид AAV12, капсид AAV LK03, капсид AAV2R471A, капсид AAV2/2-7m8, капсид AAV DJ, капсид AAV DJ8, капсид AAV2 N587A, капсид AAV2 E548A, капсид AAV2 N708A, капсид AAV V708K, капсид AAV козы, химерный капсид AAV1/AAV2, капсид AAV крупного рогатого скота, капсид AAV мыши rAAV2/HBoV1 (химерный AAV/вирус 1, относящийся к бокавирусам человека), капсид AAV2HBKO, капсид AAVPHP.B или капсид AAVPHP.eB; и/или
(ii) вектор на основе rAAV.
43. Способ по п.42, в котором капсид AAV содержит мутацию по тирозину, мутацию, влияющую на связывание с гепарином, или мутацию HBKO.
44. Способ по п.42 или 43, в котором вектор на основе rAAV представляет собой самокомплементарный вектор.
45. Частица рекомбинантного AAV (rAAV) для доставки нуклеиновой кислоты в клетку-хозяин, содержащая аминокислотную замену по аминокислотному остатку 2 VP1 и/или VP3 или исходного полипептида; где аминокислотная замена по аминокислотному остатку 2 VP1 и/или VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 и/или VP3 частицы исходного AAV.
46. Частица rAAV по п.45, в которой
a) замена по аминокислотному остатку 2 приводит к более высокой частоте N-концевого ацетилирования или более низкой частоте N-концевого ацетилирования;
b) частица rAAV содержит аминокислотную замену по аминокислотному остатку 2 VP1, и при этом аминокислотная замена по аминокислотному остатку 2 VP1 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP1 частицы исходного AAV;
c) частица rAAV содержит аминокислотную замену по аминокислотному остатку 2 VP3; и при этом аминокислотная замена по аминокислотному остатку 2 VP3 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 VP3 частицы исходного AAV; и/или
d) аминокислотный остаток 2 заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr.
47. Частица rAAV по п.46, в которой аминокислотный остаток 2 заменен Ser, Asp или Glu.
48. Частица rAAV по любому из пп. 45-47, где частица rAAV содержит:
(i) капсид AAV1, капсид AAV2, капсид AAV3, капсид AAV4, капсид AAV5, капсид AAV6, капсид AAV7, капсид AAV8, капсид AAVrh8, капсид AAV9, капсид AAV10, капсид AAVrh10, капсид AAV11, капсид AAV12, капсид AAV LK03, капсид AAV2R471A, капсид AAV2/2-7m8, капсид AAV DJ, капсид AAV DJ8, капсид AAV2 N587A, капсид AAV2 E548A, капсид AAV2 N708A, капсид AAV V708K, капсид AAV козы, химерный капсид AAV1/AAV2, капсид AAV крупного рогатого скота, капсид AAV мыши rAAV2/HBoV1 (химерный AAV/вирус 1, относящийся к бокавирусам человека), капсид AAV2HBKO, капсид AAVPHP.B или капсид AAVPHP.eB, и/или капсид AAV содержит мутацию по тирозину, мутацию, влияющую на связывание с гепарином, или мутацию HBKO; и/или
(ii) вектор на основе rAAV.
49. Частица rAAV по п.48, где частица rAAV содержит геном rAAV, причем геном rAAV содержит:
(1) один или более ITR AAV; и/или
(2) ITR AAV1, ITR AAV2, ITR AAV3, ITR AAV4, ITR AAV5, ITR AAV6, ITR AAV7, ITR AAV8, ITR AAVrh8, ITR AAV9, ITR AAV10, ITR AAVrh10, ITR AAV11 или ITR AAV12; и/или
(3) кодируемый гетерологичный трансген, фланкированный одним или более ITR AAV.
50. Частица rAAV по п.48 или 49, где вектор на основе rAAV представляет собой самокомплементарный вектор.
51. Частица rAAV по п.48, где вектор на основе rAAV содержит первую последовательность нуклеиновой кислоты, кодирующую трансген, и вторую последовательность нуклеиновой кислоты, кодирующую комплементарную трансгену последовательность, где первая последовательность нуклеиновой кислоты образует внутринитевые пары оснований со второй последовательностью нуклеиновой кислоты по большей части или по всей ее длине.
52. Частица rAAV по п.51, где первая последовательность нуклеиновой кислоты и вторая последовательность нуклеиновой кислоты соединены мутантным ITR AAV, и при этом мутантный ITR AAV характеризуется делецией D-области и характеризуется мутацией последовательности концевого разрешения.
53. Частица рекомбинантного AAV (rAAV) для доставки нуклеиновой кислоты в клетку-хозяин, содержащая одну или более аминокислотных замен по аминокислотному остатку A35, N57, G58, N382, G383, N511, G512, N715 или G716 VP1 или VP3 частицы исходного AAV, где нумерация остатков приведена на основе VP1 AAV2, и одна или несколько аминокислотных замен обеспечивают изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV.
54. Частица rAAV по п.53, где частица rAAV представляет собой частицу AAV1 или частицу AAV2.
55. Частица rAAV по п.53 или 54, в которой
a) одна или более аминокислотных замен осуществляются по аминокислотному остатку A35 VP1, N57 VP1, G58 VP1, N382 VP3, G383 VP3, N511 VP3, G512 VP3, N715 VP3 или G716 VP3 и обеспечивают изменение дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV; или
b) одна или более аминокислотных замен включают замену Asp по N57 VP1, N382 VP3, N511 VP3 или N715 VP3 и приводят к более высокой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV; или
c) одна или более аминокислотных замен включают замену N57K или N57Q и приводят к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV; или
d) одна или более аминокислотных замен включают замену Asn по A35 VP1 и приводят к более высокой частоте дезамидирования по сравнению с дезамидированием VP1 частицы исходного AAV; или
e) одна или более аминокислотных замен осуществляются по G58 VP1, G383 VP3, G512 VP3 или G716 VP3 и приводят к более низкой частоте дезамидирования по сравнению с дезамидированием VP1 и/или VP3 частицы исходного AAV.
56. Частица rAAV по п.55, где G58 VP1 заменен на Asp.
57. Фармацевтическая композиция для доставки нуклеиновой кислоты в клетку-хозяин, содержащая частицу rAAV по любому из пп. 45-56.
58. Набор для доставки нуклеиновой кислоты в клетку-хозяин, содержащий частицу rAAV по любому из пп. 45-56 или фармацевтическую композицию по п.57.
59. Изделие для доставки нуклеиновой кислоты в клетку-хозяин, содержащее частицу rAAV по любому из пп. 45-56 или фармацевтическую композицию по п.57.
60. Капсидный белок AAV для получения частицы AAV, содержащий аминокислотную замену по аминокислотному остатку 2 капсидного белка исходного AAV; при этом аминокислотная замена по аминокислотному остатку 2 обеспечивает изменение N-концевого ацетилирования по сравнению с N-концевым ацетилированием по аминокислотному остатку 2 капсидного белка исходного AAV.
61. Капсидный белок AAV по п.60, где:
a) замена приводит к более высокой частоте N-концевого ацетилирования или более низкой частоте N-концевого ацетилирования; и/или
b) капсидный белок AAV представляет собой VP1 или VP3; и/или
c) аминокислотный остаток 2 заменен Cys, Ser, Thr, Val, Gly, Asn, Asp, Glu, Ile, Leu, Phe, Gln, Lys, Met, Pro или Tyr; и/или
d) аминокислотная замена приводит к меньшей степени дезамидирования капсида AAV.
62. Капсидный белок AAV по п.61, где аминокислотный остаток 2 заменен Ser, Asp или Glu.
63. Капсидный белок AAV по любому из пп. 60-62, где капсидный белок относится к серотипу AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV LK03, AAV2R471A, AAV2/2-7m8, AAV DJ, капсиду AAV DJ8, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерного AAV1/AAV2, AAV крупного рогатого скота, AAV мыши, rAAV2/HBoV1, AAV2HBKO, AAVPHP.B или AAVPHP.eB; и/или капсидный белок AAV дополнительно содержит мутацию по тирозину или мутацию, влияющую на связывание с гепарином.
64. Капсидный белок AAV для получения частицы AAV, содержащий аминокислотную замену капсидного белка исходного AAV; где аминокислотная замена обеспечивает изменение дезамидирования капсида по сравнению с капсидным белком исходного AAV.
| VLIET K.V | |||
| et al., "Adeno-associated virus capsid serotype identification: Analytical methods development and application" //JOURNAL OF VIROLOGICAL METHODS, 159, 2009, pp.167-177 | |||
| STEVEN J | |||
| BARK et al., "High-Temperature Protein Mass Mapping Using a Thermophilic Protease" // JOURNAL OF THE AMERICAN CHEMICAL SOCIETY, vol | |||
| Устройство для разметки подлежащих сортированию и резанию лесных материалов | 1922 |
|
SU123A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |

