ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ПРИЛОЖЕНИЯ
[0001] По этой заявке испрашивается приоритет временной заявки на патент США № 63/016246, поданной 27 апреля 2020 г., и временной заявки на патент США № 63/088432, поданной 6 октября 2020 г., полное раскрытие которых включено в настоящее описание посредством ссылки.
Подача списка последовательностей через EFS-WEB
[0002] Машиночитаемый текстовый файл под названием «090400-5013-WO-Sequence-Listing», созданный 26 апреля 2021 г. или приблизительно того, с размером файла приблизительно 186 КБ, содержит список последовательностей для настоящей заявки, который в полном объеме включен в настоящее описание посредством ссылки.
Уровень техники
[0003] Векторы для доставки генов на основе аденоассоциированных вирусов (AAV) продемонстрировали многообещающие результаты как на доклинических моделях заболеваний, так и в недавних клинических испытаниях на людях для лечения нескольких целевых заболеваний. Векторы на основе AAV по существу безопасны, поскольку AAV дикого типа является непатогенным и не имеет этиологической ассоциации с какими-либо известными заболеваниями. Кроме того, AAV обеспечивает возможность высокоэффективной доставки генов и устойчивой экспрессии трансгенов во многих тканях, включая печень, мышцы, легкие, сетчатку и головной мозг.
[0004] AAV представляет собой вирус, содержащий одноцепочечную ДНК, который содержит две открытые рамки считывания, rep и cap. Первый ген кодирует четыре белка, необходимых для репликации генома (Rep78, Rep68, Rep52 и Rep40), и второй экспрессирует три структурных белка (VP1-3), которые собираются с формированием вирусного капсида. Как следует из названия, активная репликация AAV зависит от присутствия вируса-помощника, такого как аденовирус или вирус герпеса. В отсутствии помощника он переходит в латентное состояние, в котором его геном сохраняется эписомально или интегрируется в хромосому хозяина. Были идентифицированы многочисленные гомологичные серотипы AAV приматов и многочисленные серотипы приматов, отличных от человека. AAV2 лучше всего охарактеризован в качестве носителя для доставки генов.
[0005] По состоянию на 2010 г. имеется 75 текущих клинических испытаний, в которых AAV использовался в качестве носителя для доставки генов. Однако высокое распространение антикапсидных нейтрализующих антител в результате широкой экспозиции многочисленных вариантов и серотипов AAV в человеческой популяции снижает эффективность генной терапии с использованием AAV. Такой ранее развившийся иммунитет, а также последующее развитие иммунитета за счет введения вектора могут препятствовать более широкому внедрению генной терапии с использованием AAV. Например, к настоящему времени AAV был наиболее успешным в клинических исследованиях, включающих доставку в иммуннопривилегированные области.
[0006] Недавний анализ показал, что распространение IgG анти-AAV антител у людей было самым высоким для AAV2 (72%) и AAV1 (67%), но антитела к AAV9 (47%), AAV6 (46%), AAV5 (40%), и AAV8 (38%) также присутствовали у значительной части исследованной популяции. Результаты нескольких исследований показали, что гуморальный иммунитет к капсиду AAV во время генной терапии можно предотвратить, снизив количество доставляемых частиц rAAV. К сожалению, введение малых доз вектора приводит к низкой трансдукции и, следовательно, к низкой экспрессии терапевтического гена.
[0007] В данной области техники существует потребность в разработке новых вариантов AAV, устойчивых к нейтрализации анти-AAV антителами.
Сущность изобретения
[0008] В некоторых вариантах осуществления настоящее изобретение относится к вектору на основе рекомбинантного аденоассоциированного вируса (rAAV), содержащему (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую в направлении от 5' к 3': (a) концевой повтор AAV2, (b) промотор, (c) нуклеотидную последовательность, кодирующую белок регулятора трансмембранной проводимости при муковисцидозе человека (CFTR) или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека, (d) последовательность полиаденилирования и (e) концевой повтор AAV2.
[0009] В родственных вариантах осуществления нуклеотидная последовательность, кодирующая CFTR человека или его биологически активный фрагмент, кодирует нативный белок CFTR человека и имеет следующую последовательность или последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей:
ATGCAGAGGTCGCCTCTGGAAAAGGCCAGCGTTGTCTCCAAACTTTTTTTCAGCTGGACCAGACCAATTTTGAGGAAAGGATACAGACAGCGCCTGGAATTGTCAGACATATACCAAATCCCTTCTGTTGATTCTGCTGACAATCTATCTGAAAAATTGGAAAGAGAATGGGATAGAGAGCTGGCTTCAAAGAAAAATCCTAAACTCATTAATGCCCTTCGGCGATGTTTTTTCTGGAGATTTATGTTCTATGGAATCTTTTTATATTTAGGGGAAGTCACCAAAGCAGTACAGCCTCTCTTACTGGGAAGAATCATAGCTTCCTATGACCCGGATAACAAGGAGGAACGCTCTATCGCGATTTATCTAGGCATAGGCTTATGCCTTCTCTTTATTGTGAGGACACTGCTCCTACACCCAGCCATTTTTGGCCTTCATCACATTGGAATGCAGATGAGAATAGCTATGTTTAGTTTGATTTATAAGAAGACTTTAAAGCTGTCAAGCCGTGTTCTAGATAAAATAAGTATTGGACAACTTGTTAGTCTCCTTTCCAACAACCTGAACAAATTTGATGAAGGACTTGCATTGGCACATTTCGTGTGGATCGCTCCTTTGCAAGTGGCACTCCTCATGGGGCTAATCTGGGAGTTGTTACAGGCGTCTGCCTTCTGTGGACTTGGTTTCCTGATAGTCCTTGCCCTTTTTCAGGCTGGGCTAGGGAGAATGATGATGAAGTACAGAGATCAGAGAGCTGGGAAGATCAGTGAAAGACTTGTGATTACCTCAGAAATGATTGAAAATATCCAATCTGTTAAGGCATACTGCTGGGAAGAAGCAATGGAAAAAATGATTGAAAACTTAAGACAAACAGAACTGAAACTGACTCGGAAGGCAGCCTATGTGAGATACTTCAATAGCTCAGCCTTCTTCTTCTCAGGGTTCTTTGTGGTGTTTTTATCTGTGCTTCCCTATGCACTAATCAAAGGAATCATCCTCCGGAAAATATTCACCACCATCTCATTCTGCATTGTTCTGCGCATGGCGGTCACTCGGCAATTTCCCTGGGCTGTACAAACATGGTATGACTCTCTTGGAGCAATAAACAAAATACAGGATTTCTTACAAAAGCAAGAATATAAGACATTGGAATATAACTTAACGACTACAGAAGTAGTGATGGAGAATGTAACAGCCTTCTGGGAGGAGGGATTTGGGGAATTATTTGAGAAAGCAAAACAAAACAATAACAATAGAAAAACTTCTAATGGTGATGACAGCCTCTTCTTCAGTAATTTCTCACTTCTTGGTACTCCTGTCCTGAAAGATATTAATTTCAAGATAGAAAGAGGACAGTTGTTGGCGGTTGCTGGATCCACTGGAGCAGGCAAGACTTCACTTCTAATGGTGATTATGGGAGAACTGGAGCCTTCAGAGGGTAAAATTAAGCACAGTGGAAGAATTTCATTCTGTTCTCAGTTTTCCTGGATTATGCCTGGCACCATTAAAGAAAATATCATCTTTGGTGTTTCCTATGATGAATATAGATACAGAAGCGTCATCAAAGCATGCCAACTAGAAGAGGACATCTCCAAGTTTGCAGAGAAAGACAATATAGTTCTTGGAGAAGGTGGAATCACACTGAGTGGAGGTCAACGAGCAAGAATTTCTTTAGCAAGAGCAGTATACAAAGATGCTGATTTGTATTTATTAGACTCTCCTTTTGGATACCTAGATGTTTTAACAGAAAAAGAAATATTTGAAAGCTGTGTCTGTAAACTGATGGCTAACAAAACTAGGATTTTGGTCACTTCTAAAATGGAACATTTAAAGAAAGCTGACAAAATATTAATTTTGCATGAAGGTAGCAGCTATTTTTATGGGACATTTTCAGAACTCCAAAATCTACAGCCAGACTTTAGCTCAAAACTCATGGGATGTGATTCTTTCGACCAATTTAGTGCAGAAAGAAGAAATTCAATCCTAACTGAGACCTTACACCGTTTCTCATTAGAAGGAGATGCTCCTGTCTCCTGGACAGAAACAAAAAAACAATCTTTTAAACAGACTGGAGAGTTTGGGGAAAAAAGGAAGAATTCTATTCTCAATCCAATCAACTCTATACGAAAATTTTCCATTGTGCAAAAGACTCCCTTACAAATGAATGGCATCGAAGAGGATTCTGATGAGCCTTTAGAGAGAAGGCTGTCCTTAGTACCAGATTCTGAGCAGGGAGAGGCGATACTGCCTCGCATCAGCGTGATCAGCACTGGCCCCACGCTTCAGGCACGAAGGAGGCAGTCTGTCCTGAACCTGATGACACACTCAGTTAACCAAGGTCAGAACATTCACCGAAAGACAACAGCATCCACACGAAAAGTGTCACTGGCCCCTCAGGCAAACTTGACTGAACTGGATATATATTCAAGAAGGTTATCTCAAGAAACTGGCTTGGAAATAAGTGAAGAAATTAACGAAGAAGACTTAAAGGAGTGCTTTTTTGATGATATGGAGAGCATACCAGCAGTGACTACATGGAACACATACCTTCGATATATTACTGTCCACAAGAGCTTAATTTTTGTGCTAATTTGGTGCTTAGTAATTTTTCTGGCAGAGGTGGCTGCTTCTTTGGTTGTGCTGTGGCTCCTTGGAAACACTCCTCTTCAAGACAAAGGGAATAGTACTCATAGTAGAAATAACAGCTATGCAGTGATTATCACCAGCACCAGTTCGTATTATGTGTTTTACATTTACGTGGGAGTAGCCGACACTTTGCTTGCTATGGGATTCTTCAGAGGTCTACCACTGGTGCATACTCTAATCACAGTGTCGAAAATTTTACACCACAAAATGTTACATTCTGTTCTTCAAGCACCTATGTCAACCCTCAACACGTTGAAAGCAGGTGGGATTCTTAATAGATTCTCCAAAGATATAGCAATTTTGGATGACCTTCTGCCTCTTACCATATTTGACTTCATCCAGTTGTTATTAATTGTGATTGGAGCTATAGCAGTTGTCGCAGTTTTACAACCCTACATCTTTGTTGCAACAGTGCCAGTGATAGTGGCTTTTATTATGTTGAGAGCATATTTCCTCCAAACCTCACAGCAACTCAAACAACTGGAATCTGAAGGCAGGAGTCCAATTTTCACTCATCTTGTTACAAGCTTAAAAGGACTATGGACACTTCGTGCCTTCGGACGGCAGCCTTACTTTGAAACTCTGTTCCACAAAGCTCTGAATTTACATACTGCCAACTGGTTCTTGTACCTGTCAACACTGCGCTGGTTCCAAATGAGAATAGAAATGATTTTTGTCATCTTCTTCATTGCTGTTACCTTCATTTCCATTTTAACAACAGGAGAAGGAGAAGGAAGAGTTGGTATTATCCTGACTTTAGCCATGAATATCATGAGTACATTGCAGTGGGCTGTAAACTCCAGCATAGATGTGGATAGCTTGATGCGATCTGTGAGCCGAGTCTTTAAGTTCATTGACATGCCAACAGAAGGTAAACCTACCAAGTCAACCAAACCATACAAGAATGGCCAACTCTCGAAAGTTATGATTATTGAGAATTCACACGTGAAGAAAGATGACATCTGGCCCTCAGGGGGCCAAATGACTGTCAAAGATCTCACAGCAAAATACACAGAAGGTGGAAATGCCATATTAGAGAACATTTCCTTCTCAATAAGTCCTGGCCAGAGGGTGGGCCTCTTGGGAAGAACTGGATCAGGGAAGAGTACTTTGTTATCAGCTTTTTTGAGACTACTGAACACTGAAGGAGAAATCCAGATCGATGGTGTGTCTTGGGATTCAATAACTTTGCAACAGTGGAGGAAAGCCTTTGGAGTGATACCACAGAAAGTATTTATTTTTTCTGGAACATTTAGAAAAAACTTGGATCCCTATGAACAGTGGAGTGATCAAGAAATATGGAAAGTTGCAGATGAGGTTGGGCTCAGATCTGTGATAGAACAGTTTCCTGGGAAGCTTGACTTTGTCCTTGTGGATGGGGGCTGTGTCCTAAGCCATGGCCACAAGCAGTTGATGTGCTTGGCTAGATCTGTTCTCAGTAAGGCGAAGATCTTGCTGCTTGATGAACCCAGTGCTCATTTGGATCCAGTAACATACCAAATAATTAGAAGAACTCTAAAACAAGCATTTGCTGATTGCACAGTAATTCTCTGTGAACACAGGATAGAAGCAATGCTGGAATGCCAACAATTTTTGGTCATAGAAGAGAACAAAGTGCGGCAGTACGATTCCATCCAGAAACTGCTGAACGAGAGGAGCCTCTTCCGGCAAGCCATCAGCCCCTCCGACAGGGTGAAGCTCTTTCCCCACCGGAACTCAAGCAAGTGCAAGTCTAAGCCCCAGATTGCTGCTCTGAAAGAGGAGACAGAAGAAGAGGTGCAAGATACAAGGCTTTAG (SEQ ID NO:42).
[0010] В предпочтительных вариантах осуществления нуклеотидная последовательность, кодирующая человеческий CFTR или биологически активный усеченный белок CFTR, содержит следующую нуклеотидную последовательность или последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей:
ATGCAGCGCAGCCCACTGGAGAAGGCAAGCGTGGTGTCCAAGCTGTTCTTTTCCTGGACCAGGCCTATCCTGAGGAAGGGATACAGGCAGCGGCTGGAGCTGAGCGACATCTATCAGATCCCTTCTGTGGACAGCGCCGATAATCTGTCCGAGAAGCTGGAGAGAGAGTGGGATAGGGAGCTGGCCTCTAAGAAGAACCCAAAGCTGATCAATGCCCTGCGGAGATGCTTCTTTTGGCGGTTCATGTTCTACGGCATCTTCCTGTATCTGGGCGAGGTGACCAAGGCCGTGCAGCCACTGCTGCTGGGCAGAATCATCGCCTCTTACGACCCCGATAACAAGGAGGAGAGGAGCATCGCCATCTATCTGGGCATCGGCCTGTGCCTGCTGTTTATCGTGAGGACACTGCTGCTGCACCCAGCCATCTTCGGCCTGCACCACATCGGCATGCAGATGAGAATCGCCATGTTCAGCCTGATCTACAAGAAGACCCTGAAGCTGAGCTCCAGGGTGCTGGACAAGATCTCCATCGGCCAGCTGGTGTCCCTGCTGTCTAACAATCTGAACAAGTTTGATGAGGGACTGGCCCTGGCACACTTCGTGTGGATCGCACCACTGCAGGTGGCCCTGCTGATGGGCCTGATCTGGGAGCTGCTGCAGGCAAGCGCCTTTTGCGGACTGGGCTTCCTGATCGTGCTGGCCCTGTTCCAGGCAGGACTGGGACGCATGATGATGAAGTACAGAGACCAGAGGGCCGGCAAGATCTCTGAGCGGCTGGTCATCACCAGCGAGATGATCGAGAACATCCAGTCCGTGAAGGCCTATTGTTGGGAGGAGGCCATGGAGAAGATGATCGAGAATCTGCGCCAGACAGAGCTGAAGCTGACCAGAAAGGCCGCCTACGTGAGGTACTTCAACTCTAGCGCCTTCTTTTTCTCTGGCTTTTTCGTGGTGTTCCTGAGCGTGCTGCCATACGCCCTGATCAAGGGCATCATCCTGCGGAAGATCTTTACCACAATCTCCTTCTGCATCGTGCTGAGAATGGCCGTGACAAGGCAGTTTCCCTGGGCCGTGCAGACCTGGTATGACTCTCTGGGCGCCATCAATAAGATCCAGGATTTCCTGCAGAAGCAGGAGTACAAGACACTGGAGTATAACCTGACCACAACCGAGGTGGTCATGGAGAATGTGACCGCCTTCTGGGAGGAGGGCTTTGGCGAGCTGTTCGAGAAGGCCAAGCAGAACAATAACAATCGCAAGACATCTAACGGCGACGATAGCCTGTTTTTCAGCAATTTTTCCCTGCTGGGCACCCCCGTGCTGAAGGACATCAACTTCAAGATCGAGAGGGGACAGCTGCTGGCAGTGGCAGGCTCCACAGGCGCCGGCAAGACCTCTCTGCTGATGATGATCATGGGCGAGCTGGAGCCAAGCGAGGGCAAGATCAAGCACTCCGGCCGGATCTCTTTTTGCAGCCAGTTCTCCTGGATCATGCCCGGCACCATCAAGGAGAATATCATCTTTGGCGTGTCCTACGATGAGTACAGATATAGGTCTGTGATCAAGGCCTGTCAGCTGGAGGAGGACATCAGCAAGTTCGCCGAGAAGGATAACATCGTGCTGGGCGAGGGCGGCATCACACTGAGCGGAGGACAGAGGGCAAGGATCTCCCTGGCCAGAGCCGTGTACAAGGACGCCGATCTGTATCTGCTGGACAGCCCCTTTGGCTATCTGGATGTGCTGACCGAGAAGGAGATCTTCGAGTCCTGCGTGTGCAAGCTGATGGCCAATAAGACAAGGATCCTGGTGACCTCTAAGATGGAGCACCTGAAGAAGGCCGACAAGATCCTGATCCTGCACGAGGGCTCCTCTTACTTTTATGGCACATTCAGCGAGCTGCAGAATCTGCAGCCTGACTTCAGCTCCAAGCTGATGGGCTGTGACTCCTTTGATCAGTTCTCTGCCGAGAGGCGCAACTCCATCCTGACAGAGACCCTGCACAGATTCTCTCTGGAGGGCGACGCACCCGTGAGCTGGACAGAGACCAAGAAGCAGTCCTTTAAGCAGACCGGCGAGTTCGGCGAGAAGAGGAAGAATTCTATCCTGAACCCTATCAATAGCACACTGCAGGCCCGGAGAAGGCAGTCTGTGCTGAACCTGATGACCCACAGCGTGAACCAGGGCCAGAATATCCACAGAAAGACAACCGCCAGCACAAGGAAGGTGTCCCTGGCACCTCAGGCAAACCTGACCGAGCTGGACATCTACTCCCGCCGGCTGTCTCAGGAGACCGGACTGGAGATCTCTGAGGAGATCAATGAGGAGGATCTGAAGGAGTGCTTTTTCGACGATATGGAGAGCATCCCAGCCGTGACAACCTGGAACACATACCTGCGCTATATCACCGTGCACAAGTCCCTGATCTTTGTGCTGATCTGGTGTCTGGTCATCTTCCTGGCAGAGGTGGCAGCATCTCTGGTGGTGCTGTGGCTGCTGGGCAACACACCCCTGCAGGACAAGGGCAATTCTACCCACAGCCGCAACAATTCCTACGCCGTGATCATCACATCTACCTCTAGCTACTACGTGTTCTACATCTATGTGGGCGTGGCCGATACACTGCTGGCCATGGGCTTTTTCCGGGGCCTGCCCCTGGTGCACACACTGATCACCGTGAGCAAGATCCTGCACCACAAGATGCTGCACAGCGTGCTGCAGGCCCCTATGTCCACACTGAACACCCTGAAGGCCGGCGGCATCCTGAATCGGTTTTCCAAGGACATCGCCATCCTGGACGATCTGCTGCCTCTGACCATCTTTGATTTCATCCAGCTGCTGCTGATCGTGATCGGAGCAATCGCAGTGGTGGCCGTGCTGCAGCCTTACATCTTCGTGGCCACAGTGCCAGTGATCGTGGCCTTTATCATGCTGCGCGCCTATTTCCTGCAGACCAGCCAGCAGCTGAAGCAGCTGGAGAGCGAGGGCCGGTCCCCTATCTTTACACACCTGGTGACCTCCCTGAAGGGACTGTGGACACTGAGGGCCTTCGGCCGGCAGCCATACTTTGAGACCCTGTTCCACAAGGCCCTGAACCTGCACACAGCCAATTGGTTTCTGTATCTGAGCACCCTGCGCTGGTTTCAGATGCGGATCGAGATGATCTTCGTGATCTTTTTCATCGCCGTGACCTTCATCTCCATCCTGACAACCGGAGAGGGAGAGGGAAGAGTGGGAATCATCCTGACACTGGCCATGAACATCATGTCTACCCTGCAGTGGGCCGTGAATTCCTCTATCGACGTGGATAGCCTGATGAGATCTGTGAGCAGGGTGTTTAAGTTCATCGACATGCCCACAGAGGGCAAGCCTACAAAGAGCACCAAGCCATACAAGAACGGCCAGCTGTCCAAAGTGATGATCATCGAGAATTCTCACGTGAAGAAGGACGATATCTGGCCATCCGG (SEQ ID NO:43)
[0011] SEQ ID NO:43 представляет собой нуклеотидную последовательность, которая была кодон-оптимизирована для экспрессии у человека и кодирует биологически активный усеченный белок CFTR человека, в котором отсутствуют аминокислоты 708-759. В некоторых вариантах осуществления изобретение относится к выделенной нуклеиновой кислоте, содержащей нуклеотидную последовательность SEQ ID NO:43 или последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей, где необязательно нуклеотидная последовательность функционально связана с регуляторной последовательностью контроля экспрессии. Настоящее изобретение также относится к плазмидам и векторам, содержащим последовательность нуклеиновой кислоты SEQ ID NO:43 или последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей, и клеткам-хозяевам, содержащим такие плазмиды и векторы. Также настоящее изобретение относится к применению нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:43 или последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей, где необязательно нуклеотидная последовательность функционально связана с регуляторной последовательностью контроля экспрессии, для лечения муковисцидоза или заболевания легких, ассоциированного с ним, как описано в настоящем изобретении, или к применению в получении лекарственного средства для лечения муковисцидоза или заболевания легких, ассоциированного с ним.
[0012] В некоторых аспектах промотор представляет собой конститутивный промотор, необязательно усеченный непосредственный/ранний энхансер/промотор цитомегаловируса (CMVie) и функционально связанный с нуклеотидной последовательностью, кодирующей человеческий CFTR или его биологически активный фрагмент.
[0013] В еще одних аспектах промотор представляет собой тканеспецифический промотор, где предпочтительно промотор направляет предпочтительную экспрессию нуклеиновой кислоты в клетке легкого и функционально связан с нуклеотидной последовательностью, кодирующей человеческий CFTR или его биологически активный фрагмент.
[0014] В предпочтительных вариантах осуществления промотор представляет собой усеченный промотор CMVie и функционально связан с нуклеотидной последовательностью, кодирующей человеческий CFTR или его биологически активный фрагмент. В особенно предпочтительном варианте осуществления промотор CMVie представляет собой CMV173, имеющий следующую последовательность или последовательность, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей:
ACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAATAACCCCGCCCCGTTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCGTTTAGTGAACCGT (SEQ ID NO:44)
[0015] В особенно предпочтительном варианте осуществления вектор rAAV содержит (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую в направлении от 5' к 3': (a) концевой повтор AAV2 (b) промотор (c) нуклеотидную последовательность, кодирующую белок регулятора трансмембранной проводимости при муковисцидозе человека (CFTR) или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека, (d) последовательность полиаденилирования и (e) концевой повтор AAV2, где нуклеиновая кислота содержит в направлении от 5' к 3' следующую последовательность или последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей:
TTGGCCACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCTTTGCCCGGGCGGCCTCAGTGAGCGAGCGAGCGCGCAGAGAGGGAGTGGCCAACTCCATCACTAGGGGTTCCTGCGGCCGCACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAATAACCCCGCCCCGTTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCGTTTAGTGAACCGTCAGAATTCTCGAGTGATCGAAAGAGCCTGCTAAAGCAAAAAAGAAGTCACCATGCAGCGCAGCCCACTGGAGAAGGCAAGCGTGGTGTCCAAGCTGTTCTTTTCCTGGACCAGGCCTATCCTGAGGAAGGGATACAGGCAGCGGCTGGAGCTGAGCGACATCTATCAGATCCCTTCTGTGGACAGCGCCGATAATCTGTCCGAGAAGCTGGAGAGAGAGTGGGATAGGGAGCTGGCCTCTAAGAAGAACCCAAAGCTGATCAATGCCCTGCGGAGATGCTTCTTTTGGCGGTTCATGTTCTACGGCATCTTCCTGTATCTGGGCGAGGTGACCAAGGCCGTGCAGCCACTGCTGCTGGGCAGAATCATCGCCTCTTACGACCCCGATAACAAGGAGGAGAGGAGCATCGCCATCTATCTGGGCATCGGCCTGTGCCTGCTGTTTATCGTGAGGACACTGCTGCTGCACCCAGCCATCTTCGGCCTGCACCACATCGGCATGCAGATGAGAATCGCCATGTTCAGCCTGATCTACAAGAAGACCCTGAAGCTGAGCTCCAGGGTGCTGGACAAGATCTCCATCGGCCAGCTGGTGTCCCTGCTGTCTAACAATCTGAACAAGTTTGATGAGGGACTGGCCCTGGCACACTTCGTGTGGATCGCACCACTGCAGGTGGCCCTGCTGATGGGCCTGATCTGGGAGCTGCTGCAGGCAAGCGCCTTTTGCGGACTGGGCTTCCTGATCGTGCTGGCCCTGTTCCAGGCAGGACTGGGACGCATGATGATGAAGTACAGAGACCAGAGGGCCGGCAAGATCTCTGAGCGGCTGGTCATCACCAGCGAGATGATCGAGAACATCCAGTCCGTGAAGGCCTATTGTTGGGAGGAGGCCATGGAGAAGATGATCGAGAATCTGCGCCAGACAGAGCTGAAGCTGACCAGAAAGGCCGCCTACGTGAGGTACTTCAACTCTAGCGCCTTCTTTTTCTCTGGCTTTTTCGTGGTGTTCCTGAGCGTGCTGCCATACGCCCTGATCAAGGGCATCATCCTGCGGAAGATCTTTACCACAATCTCCTTCTGCATCGTGCTGAGAATGGCCGTGACAAGGCAGTTTCCCTGGGCCGTGCAGACCTGGTATGACTCTCTGGGCGCCATCAATAAGATCCAGGATTTCCTGCAGAAGCAGGAGTACAAGACACTGGAGTATAACCTGACCACAACCGAGGTGGTCATGGAGAATGTGACCGCCTTCTGGGAGGAGGGCTTTGGCGAGCTGTTCGAGAAGGCCAAGCAGAACAATAACAATCGCAAGACATCTAACGGCGACGATAGCCTGTTTTTCAGCAATTTTTCCCTGCTGGGCACCCCCGTGCTGAAGGACATCAACTTCAAGATCGAGAGGGGACAGCTGCTGGCAGTGGCAGGCTCCACAGGCGCCGGCAAGACCTCTCTGCTGATGATGATCATGGGCGAGCTGGAGCCAAGCGAGGGCAAGATCAAGCACTCCGGCCGGATCTCTTTTTGCAGCCAGTTCTCCTGGATCATGCCCGGCACCATCAAGGAGAATATCATCTTTGGCGTGTCCTACGATGAGTACAGATATAGGTCTGTGATCAAGGCCTGTCAGCTGGAGGAGGACATCAGCAAGTTCGCCGAGAAGGATAACATCGTGCTGGGCGAGGGCGGCATCACACTGAGCGGAGGACAGAGGGCAAGGATCTCCCTGGCCAGAGCCGTGTACAAGGACGCCGATCTGTATCTGCTGGACAGCCCCTTTGGCTATCTGGATGTGCTGACCGAGAAGGAGATCTTCGAGTCCTGCGTGTGCAAGCTGATGGCCAATAAGACAAGGATCCTGGTGACCTCTAAGATGGAGCACCTGAAGAAGGCCGACAAGATCCTGATCCTGCACGAGGGCTCCTCTTACTTTTATGGCACATTCAGCGAGCTGCAGAATCTGCAGCCTGACTTCAGCTCCAAGCTGATGGGCTGTGACTCCTTTGATCAGTTCTCTGCCGAGAGGCGCAACTCCATCCTGACAGAGACCCTGCACAGATTCTCTCTGGAGGGCGACGCACCCGTGAGCTGGACAGAGACCAAGAAGCAGTCCTTTAAGCAGACCGGCGAGTTCGGCGAGAAGAGGAAGAATTCTATCCTGAACCCTATCAATAGCACACTGCAGGCCCGGAGAAGGCAGTCTGTGCTGAACCTGATGACCCACAGCGTGAACCAGGGCCAGAATATCCACAGAAAGACAACCGCCAGCACAAGGAAGGTGTCCCTGGCACCTCAGGCAAACCTGACCGAGCTGGACATCTACTCCCGCCGGCTGTCTCAGGAGACCGGACTGGAGATCTCTGAGGAGATCAATGAGGAGGATCTGAAGGAGTGCTTTTTCGACGATATGGAGAGCATCCCAGCCGTGACAACCTGGAACACATACCTGCGCTATATCACCGTGCACAAGTCCCTGATCTTTGTGCTGATCTGGTGTCTGGTCATCTTCCTGGCAGAGGTGGCAGCATCTCTGGTGGTGCTGTGGCTGCTGGGCAACACACCCCTGCAGGACAAGGGCAATTCTACCCACAGCCGCAACAATTCCTACGCCGTGATCATCACATCTACCTCTAGCTACTACGTGTTCTACATCTATGTGGGCGTGGCCGATACACTGCTGGCCATGGGCTTTTTCCGGGGCCTGCCCCTGGTGCACACACTGATCACCGTGAGCAAGATCCTGCACCACAAGATGCTGCACAGCGTGCTGCAGGCCCCTATGTCCACACTGAACACCCTGAAGGCCGGCGGCATCCTGAATCGGTTTTCCAAGGACATCGCCATCCTGGACGATCTGCTGCCTCTGACCATCTTTGATTTCATCCAGCTGCTGCTGATCGTGATCGGAGCAATCGCAGTGGTGGCCGTGCTGCAGCCTTACATCTTCGTGGCCACAGTGCCAGTGATCGTGGCCTTTATCATGCTGCGCGCCTATTTCCTGCAGACCAGCCAGCAGCTGAAGCAGCTGGAGAGCGAGGGCCGGTCCCCTATCTTTACACACCTGGTGACCTCCCTGAAGGGACTGTGGACACTGAGGGCCTTCGGCCGGCAGCCATACTTTGAGACCCTGTTCCACAAGGCCCTGAACCTGCACACAGCCAATTGGTTTCTGTATCTGAGCACCCTGCGCTGGTTTCAGATGCGGATCGAGATGATCTTCGTGATCTTTTTCATCGCCGTGACCTTCATCTCCATCCTGACAACCGGAGAGGGAGAGGGAAGAGTGGGAATCATCCTGACACTGGCCATGAACATCATGTCTACCCTGCAGTGGGCCGTGAATTCCTCTATCGACGTGGATAGCCTGATGAGATCTGTGAGCAGGGTGTTTAAGTTCATCGACATGCCCACAGAGGGCAAGCCTACAAAGAGCACCAAGCCATACAAGAACGGCCAGCTGTCCAAAGTGATGATCATCGAGAATTCTCACGTGAAGAAGGACGATATCTGGCCATCCGGAGGACAGATGACCGTGAAGGATCTGACAGCCAAGTATACCGAGGGCGGCAACGCCATCCTGGAGAATATCTCCTTTTCTATCAGCCCTGGACAGAGGGTGGGACTGCTGGGACGGACAGGCTCCGGCAAGTCTACCCTGCTGAGCGCCTTCCTGAGGCTGCTGAATACAGAGGGCGAGATCCAGATCGACGGCGTGAGCTGGGATTCCATCACCCTGCAGCAGTGGAGAAAGGCCTTTGGCGTGATCCCTCAGAAGGTGTTTATCTTCTCCGGCACCTTCAGGAAGAACCTGGACCCATACGAGCAGTGGTCTGATCAGGAGATCTGGAAGGTGGCCGACGAAGTGGGCCTGAGATCTGTGATCGAGCAGTTTCCAGGCAAGCTGGACTTCGTGCTGGTGGATGGAGGATGCGTGCTGAGCCACGGACACAAGCAGCTGATGTGCCTGGCCAGGTCTGTGCTGAGCAAGGCCAAGATCCTGCTGCTGGACGAGCCAAGCGCCCACCTGGATCCCGTGACATACCAGATCATCAGAAGGACCCTGAAGCAGGCCTTTGCCGATTGCACCGTGATCCTGTGCGAGCACCGCATCGAGGCCATGCTGGAGTGCCAGCAGTTCCTGGTCATCGAGGAGAACAAGGTGCGGCAGTATGACAGCATCCAGAAGCTGCTGAATGAGCGGAGCCTGTTTCGGCAGGCCATCTCCCCCTCTGATCGCGTGAAGCTGTTCCCTCACCGGAACAGCTCCAAGTGTAAGTCCAAGCCCCAGATCGCCGCCCTGAAGGAGGAGACAGAGGAGGAGGTGCAGGACACCAGACTGTGAAATAAAACATCTTTATTTTCATTACATCTGTGTGTTGGTTTTTTGTGTGAACAACGGCCGGCCGGAGGAACCCCTAGTGATGGAGTTGGCCACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGGTCGCCCGGCCTCAGTGAGCGAGCGAGCGCGCAGAGAGGGAGTGGCCAA (SEQ ID NO: 45).
[0016] Нуклеиновая кислота, имеющая нуклеотидную последовательность SEQ ID NO:45, содержит в направлении от 5' к 3': (a) концевой повтор AAV2, (b) промотор CMV173 с SEQ ID NO:44, (c) кодон-оптимизированную нуклеотидную последовательность, кодирующую биологически активный усеченный белок CFTR человека, в котором отсутствуют аминокислоты 708-759 SEQ ID NO: 43, (d) последовательность полиаденилирования и (e) концевой повтор AAV2.
[0017] Также настоящее изобретение относится к способам лечения муковисцидоза у субъекта, нуждающегося в этом, включающим введение субъекту терапевтически эффективного количества инфекционного rAAV, содержащего (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую белок регулятора трансмембранной проводимости при муковисцидозе человека (CFTR) или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека, функционально связанную с промотором. В предпочтительных вариантах осуществления нуклеотидная последовательность, кодирующая CFTR, имеет последовательность SEQ ID NO: 43, и/или промотор имеет последовательность SEQ ID NO: 44, и/или нуклеиновая кислота содержит последовательность SEQ ID NO: 45. В некоторых аспектах субъекту вводят количество rAAV, эффективное для ослабления одного или более признаков муковисцидоза, не ограничивающие примеры которых включают воспаление верхних и нижних дыхательных путей, аберрантную передачу сигналов цитокинов в эпителии и повышенные уровни IgE.
[0018] В еще одних аспектах настоящее изобретение относится к способам лечения заболевания легких, ассоциированного с муковисцидозом, включая, не ограничиваясь этим, заболевание верхних дыхательных путей, заболевание нижних дыхательных путей, заболевание носоглотки, синусит и/или заболевание слюнных желез, ассоциированное с муковисцидозом, включающим введение субъекту терапевтически эффективного количества инфекционного rAAV, содержащего (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую белок регулятора трансмембранной проводимости при муковисцидозе человека (CFTR) или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека, функционально связанную с промотором. В предпочтительных вариантах осуществления нуклеотидная последовательность, кодирующая CFTR, имеет последовательность SEQ ID NO: 43, и/или промотор имеет последовательность SEQ ID NO: 44, и/или нуклеиновая кислота содержит последовательность SEQ ID NO: 45.
[0019] Векторы rAAV для генной терапии по настоящему изобретению, содержащие капсид, содержащий капсидный белок с SEQ ID NO:12 и последовательности нуклеиновой кислоты, кодирующие CFTR или его биологически активный фрагмент (например, содержащие нуклеиновую кислоту, содержащую нуклеотидную последовательность SEQ ID NO: 43, необязательно связанную с промотором с SEQ ID NO: 44 и/или содержащую нуклеотидную последовательность SEQ ID NO: 45), можно вводить пациенту различными путями для достижения и поддержания терапевтически эффективного уровня CFTR или его фрагмента, для лечения муковисцидоза или заболевания легких, ассоциированного с ним.
[0020] В некоторых аспектах инфекционный rAAV вводят субъекту с муковисцидозом в одной или более дозах, каждая из которых включает от приблизительно 1×1013 до приблизительно 1×1015 векторных геномов (вг), от приблизительно 1×1013 до приблизительно 1×1014 вг, от приблизительно 1×1014 до приблизительно 1×1015 вг или от приблизительно 1×1015 до приблизительно 5×1015 вг. В некоторых предпочтительных аспектах каждая доза содержит приблизительно 1×1014 вг или приблизительно 1×1015 вг rAAV.
[0021] В некоторых аспектах лечение включает введение субъекту не более одной дозы и является эффективным для достижения длительной и сохраняемой терапевтической концентрации CFTR или его биологически активного фрагмента. В родственных аспектах лечение включает введение не более чем одной дозы путем ингаляции от приблизительно 1×1013 до приблизительно 1×1015 бляшкообразующих единиц (БОЕ), вирусных частиц (вч) или вирусных геномов (вг) rAAV, содержащих капсидный белок SEQ ID NO:12 и нуклеиновую кислоту SEQ ID NO:45 человеку с муковисцидозом. В других аспектах дозированное лечение может представлять собой схему многократного введения.
[0022] Способы, относящиеся к введению векторов AAV людям, ранее были описаны в публикации Kay et al. (2000, Nat. Genet., 24:257-261), полное содержание которой включено в настоящее описание посредством ссылки. В некоторых предпочтительных вариантах осуществления инфекционный rAAV вводят субъекту пульмональным, эндобронхиальным, интраназальным, интратрахеальным и/или внутрибронхиальным введением. В некоторых предпочтительных вариантах осуществления инфекционный rAAV вводят с помощью распылителя.
[0023] В родственных аспектах настоящее изобретение относится к инфекционному rAAV, содержащему (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую трансмембранный регулятор проводимости при муковисцидозе человека (CFTR)) белок или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека, функционально связанную с промотором, для применения в лечении муковисцидоза или для применения в получении лекарственного средства для лечения муковисцидоза. В некоторых предпочтительных вариантах осуществления нуклеотидная последовательность, кодирующая белок CFTR человека, содержит или состоит из последовательности SEQ ID NO: 43 или последовательности, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей, функционально связанную с промотором, содержащим последовательность SEQ ID NO:44 или последовательность, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную ей. В особенно предпочтительных вариантах осуществления rAAV содержит нуклеиновую кислоту, содержащую или состоящую из нуклеотидной последовательности SEQ ID NO:45.
[0024] В еще одних вариантах осуществления настоящее изобретение относится к фармацевтической композиции, подходящей для ингаляции, содержащей (i) rAAV, инфекционный rAAV, содержащий (a) капсид, содержащий капсидный белок SEQ ID NO:12, и (b) нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую один или более генных продуктов в (ii) буфере, содержащем от приблизительно 10 мМ до приблизительно 50 мМ цитрата, от приблизительно 70 мМ до приблизительно 150 мМ NaCl и необязательно поверхностно-активное вещество, предпочтительно неионогенное поверхностно-активное вещество, такое как Плюроник F-68, более предпочтительно приблизительно 0,005% Плюроника F68, и имеющем рН от 5 до 7, предпочтительно имеющем рН приблизительно 6,0. В некоторых предпочтительных аспектах фармацевтическая композиция содержит от приблизительно 20 мМ до приблизительно 50 мМ цитрата, от приблизительно 85 мМ до приблизительно 125 мМ NaCl и приблизительно 0,005% Плюроника F68 и имеет рН приблизительно 6,0. В некоторых особенно предпочтительных аспектах фармацевтическая композиция содержит приблизительно 20 мМ цитрата, приблизительно 125 мМ NaCl и приблизительно 0,005% Плюроника F68 и имеет рН приблизительно 6,0. В предпочтительных вариантах осуществления фармацевтическая композиция содержит rAAV, содержащий (а) капсид, содержащий капсидный белок SEQ ID NO:12, и (b) нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую белок регулятора трансмембранной проводимости при муковисцидозе человека (CFTR) или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека, функционально связанную с промотором. В родственных вариантах осуществления rAAV содержит нуклеиновую кислоту, содержащую или состоящую из нуклеотидной последовательности SEQ ID NO:45.
[0025] В некоторых аспектах фармацевтическая композиция содержит от 1011 до 1014 векторных геномов (вг) на мл. В некоторых предпочтительных вариантах осуществления фармацевтическая композиция содержит от приблизительно 1×1013 до приблизительно 9×1013 вг/мл, предпочтительно от приблизительно 2×1013 до 6×1013 гв/мл. В других предпочтительных вариантах осуществления фармацевтический препарат содержит приблизительно 1×1013 вг/мл, приблизительно 2×1013 вг/мл, приблизительно 3×1013 вг/мл, приблизительно 4×1013 вг/мл, приблизительно 5×1013 вг/мл, приблизительно 6×1013 вг/мл, приблизительно 7×1013 вг/мл, приблизительно 8× 1013 вг/мл или приблизительно 9×1013 вг/мл. В особенно предпочтительном воплощении фармацевтическая композиция содержит от приблизительно 2×1013 вг/мл до приблизительно 5×1013 вг/мл.
[0026] В некоторых вариантах осуществления фармацевтическая композиция составлена в виде жидкости/суспензии, подходящей для аэрозольной доставки. В родственных вариантах осуществления фармацевтическая композиция составлена в виде аэрозоля и/или представляет собой дозированную форму для ингаляций.
[0027] Также настоящее изобретение относится к способам доставки гетерологичной нуклеиновой кислоты в клетку легкого, включающим приведение клетки легкого в контакт с вирионом rAAV, содержащим (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую один или более генных продуктов. В некоторых вариантах осуществления гетерологичная нуклеиновая кислота кодирует белок и/или малую интерферирующую РНК. В некоторых предпочтительных вариантах клетка легкого представляет собой любую клетку легкого или трахеи. В других предпочтительных вариантах осуществления клетка легкого представляет собой эпителиальную клетку дыхательных путей, включая, не ограничиваясь этим, клетку альвеолярного эпителия, бронхиальную (первичную, вторичную или третичную) эпителиальную клетку или трахеальную эпителиальную клетку. В некоторых предпочтительных аспектах клетка легкого представляет собой реснитчатую эпителиальную клетку дыхательных путей. В некоторых предпочтительных аспектах клетка легкого представляет собой клетку альвеолярного эпителия легких типа 1 (AECI) или типа 2 (AECII). В других вариантах осуществления клетка легкого представляет собой гладкомышечную или эндотелиальную клетку. В еще одних вариантах осуществления клетка легкого представляет собой базальную клетку, бокаловидную клетку или ооцит. В особенно предпочтительных вариантах осуществления rAAV содержит нуклеиновую кислоту, содержащую или состоящую из нуклеотидной последовательности SEQ ID NO:45.
[0028] Также настоящее изобретение относится к способам доставки гетерологичной нуклеиновой кислоты в легкие субъекта (например, человека), включающим введение субъекту вириона rAAV, содержащего (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую один или более генных продуктов. В некоторых вариантах осуществления гетерологичная нуклеиновая кислота кодирует белок и/или малую интерферирующую РНК. В родственных вариантах осуществления способы доставки гетерологичной нуклеиновой кислоты в верхние дыхательные пути, носоглотку, пазухи, ротовую/щечную область и/или слюнные железы субъекта (например, человека), включают введение субъекту вириона rAAV, содержащего (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую один или более генных продуктов. В родственных аспектах rAAV или содержащую его фармацевтическую композицию вводят субъекту пульмональным, эндобронхиальным, интраназальным, интратрахеальным и/или внутрибронхиальным введением. В особенно предпочтительных вариантах осуществления rAAV содержит нуклеиновую кислоту, содержащую или состоящую из нуклеотидной последовательности SEQ ID NO:45.
[0029] Также настоящее изобретение относится к способам лечения заболевания легких, включающим введение субъекту, нуждающемуся в этом, терапевтически эффективного количества рекомбинантного AAV (rAAV), содержащего (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую один или более генных продуктов, где один или более генных продуктов функционально связаны с промотором. В некоторых аспектах гетерологичная нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую несколько генных продуктов, и в этом случае экспрессия нескольких генных продуктов (например, 2) может быть опосредована несколькими (например, 2) независимыми промоторами или может быть опосредована одним промотором, где несколько трансгенов разделены внутренним сайтом посадки рибосомы (IRES) или последовательностью пептида 2А. В предпочтительных вариантах осуществления гетерологичная нуклеиновая кислота кодирует терапевтический белок и/или терапевтическую малую интерферирующую РНК. В родственных аспектах генный продукт(ы), доставляемый rAAV, снижает уровень продукта мешающего гена и/или вводит или дополняет уровень продукта поддерживающего гена. В особенно предпочтительных вариантах осуществления rAAV содержит нуклеиновую кислоту, содержащую или состоящую из нуклеотидной последовательности SEQ ID NO:45. В других предпочтительных вариантах осуществления rAAV содержит нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую альфа-1-антитрипсин.
[0030] В некоторых аспектах заболевание легких выбрано из легочной артериальной гипертензии, легочной гипертензии, рака легкого (первичного, вторичного и метастатического), дефицита сурфактанта, вирусной и/или бактериальной инфекции, муковисцидоза, острого бронхита, пневмонии (включая вирусную, бактериальную и грибковую пневмонию), инфекции дыхательных путей (включая фарингит, круп, аспергиллез, кокцидиомикоз, хантавирусный пульмональный синдром и гистоплазмоз), химического и гиперчувствительного пневмонита, туберкулеза и других микобактериальных инфекций (включая, помимо прочего, Mycobacterium avium), саркоидоза, инфекции, вызванной респираторно-синцитиальным вирусом, отека легкого, острого респираторного дистресс-синдрома (ARDS), пневмокониоза (включая черную болезнь легких, асбестоз и силикоз), интерстициального заболевания легких (включая саркоидоз и аутоиммунное заболевание), легочной эмболии, плеврального выпота, плеврита, мезотелиомы, пневмоторакса, острого бронхита, бронхиолита (включая облитерирующий бронхиолит), синдрома внезапной детской смерти, апноэ во сне, бронхоэктазов, бронхолегочной дисплазии, криптогенной организующейся пневмонии, повреждения легких, связанного с использованием электронных сигарет или вейпинга (EVALI), ближневосточного респираторного синдрома (MERS), первичной цилиарной дискинезии, тяжелого острого респираторного синдрома (SARS), дефицита альфа-1-антитрипсина, астмы, интерстициального заболевания легких и COVID-19 (короновирусная инфекция 2019). В других аспектах заболевание легких представляет собой хроническую обструктивную болезнь легких (COPD) или идиопатический легочный фиброз (IPF). В связанных аспектах обеспечивается способ лечения COVID-19, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества рекомбинантного AAV (rAAV), содержащего (i) капсид, содержащий капсидный белок SEQ ID NO:12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую один или более генных продуктов, функционально связанных с одним или более промоторами, или фармацевтическую композицию, содержащую rAAV, где генный продукт(ы) обеспечивает нокдаун, модифицирует и/или сверхэкспрессирует вирусный продукт гена или гена клетки-хозяина для снижения или элиминации вирусной патогенности или репликации либо в легких, либо в носоглотке, и/или экспрессирует нейтрализующее антитело против эпитопа на вирусе.
[0031] В некоторых аспектах гены, на которые может быть направлено нацеливание для лечения IPF, включают, не ограничиваясь этим, SFTPA1 (сурфактант А1) и кавеолин-1. Гены, на которые может быть направлено нацеливание для лечение COPD, включают, помимо прочего, альфа-1-антитрипсин, альфа-1-антихимотрипсин, альфа-1-макроглобулин, матриксную металлопротеиназу 1 (ММР1), матриксную металлопротеиназу 12 (ММР12), микросомальные эпоксидгидролиаза, CYP1A1, глутатион S-трансфераза, гемоксигеназу-1, TGF-бета-1, TNF-альфа, комплекс IL-1, IL-8, IL-13, человеческий лейкоцитарный антиген (HLA-B7 и Bw16), витамин D-связывающий белок и бета-2-адренорецептор.
[0032] В родственных аспектах rAAV или фармацевтическую композицию вводят пульмональным, эндобронхиальным, интраназальным, интратрахеальным и/или внутрибронхиальным введением для лечения заболевания легких у субъекта, нуждающегося в этом. В некоторых предпочтительных вариантах осуществления инфекционный rAAV вводят с помощью распылителя.
[0033] В еще одних аспектах субъекту для лечения легочного заболевания вводят по меньшей мере одну дозу из от приблизительно 1012 до 1014 векторных геномов (вг)/кг rAAV. В родственных аспектах субъекту вводят от приблизительно 1×1011 до приблизительно 1×1014 вг/кг, от приблизительно 1×1012 до приблизительно 9×1013 вг/кг, от приблизительно 1×1012 вг/кг до приблизительно 9×1012 вг/кг, предпочтительно от приблизительно 2×1012 вг/кг до приблизительно 3×1012 вг/кг, более предпочтительно приблизительно 2,6×1012 вг/кг, приблизительно 2,7×1012 вг/кг, приблизительно 2,8×1012 вг/кг, приблизительно 2,9×1012 вг/кг, приблизительно 3,0×1012 вг/кг или приблизительно 3,1×1012 вг/кг. В предпочтительных вариантах осуществления субъекту вводят одну или более доз в одной или более дозировках, каждая доза включает от приблизительно 1×1013 до приблизительно 1×1015 векторных геномов (вг), от приблизительно 1×1013 до приблизительно 1×1014 вг, от приблизительно 1×1014 и приблизительно 1×1015 вг, или приблизительно 1×1015 и приблизительно 5×1015 вг rAAV. В некоторых предпочтительных аспектах каждая доза содержит приблизительно 1×1014 вг или приблизительно 1×1015 вг rAAV.
[0034] Настоящее изобретение дополнительно относится к инфекционным вирионам рекомбинантного аденоассоциированного вируса (rAAV), которые содержат вариантный капсидный белок и гетерологичную нуклеиновую кислоту. Изобретение также относится к капсидным белкам вариантного аденоассоциированного вируса (AAV) (и/или нуклеиновой кислоте, кодирующей капсидные белки вариантного AAV), которые придают инфекционному вириону rAAV повышенную устойчивость к нейтрализующим анти-AAV антителам человека. Настоящее изобретение дополнительно относится к клеткам-хозяевам, содержащим инфекционный вирион rAAV и/или нуклеиновую кислоту, кодирующую вариантный капсидный белок AAV по настоящему изобретению. Настоящее раскрытие дополнительно обеспечивает библиотеки вышеуказанных вирионов, капсидных белков, нуклеиновых кислот и/или клеток-хозяев; где вариантный капсидный белок AAV по меньшей мере одного члена библиотеки содержит аминокислотную последовательность, имеющую по меньшей мере одну аминокислотную замену относительно аминокислотной последовательности, показанной в одной из SEQ ID NO: 10-13 и 26-33.
[0035] Настоящее изобретение также относится к способам доставки гетерологичной нуклеиновой кислоты в клетку-мишень, где клетку-мишень приводят в контакт с заявленным инфекционным вирионом rAAV. Настоящее изобретение дополнительно относится к способам доставки генного продукта субъекту, где способы, как правило, включают введение эффективного количества заявленного вириона rAAV субъекту, нуждающемуся в этом. Также настоящее изобретение относится к композициям и наборам для применения в заявленных способах.
[0036] Признаки настоящего изобретения включают инфекционный вирион рекомбинантного аденоассоциированного вируса (rAAV), содержащий (а) вариантный капсидный белок аденоассоциированного вируса (AAV), содержащий аминокислотную последовательность, имеющую по меньшей мере приблизительно 90% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в одной из SEQ ID NO: 11-13 и 26-33; и (b) гетерологичную нуклеиновую кислоту. В некоторых случаях вариантный капсидный белок AAV содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в одной из SEQ ID NO: 11-13 и 26-33. В некоторых случаях вариантный капсидный белок AAV содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 11-13 и 26-33.
[0037] Признаки настоящего изобретения включают инфекционный вирион рекомбинантного аденоассоциированного вируса (rAAV), содержащий (а) вариантный капсидный белок аденоассоциированного вируса (AAV), который содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 10, и содержит аминокислотные замены N312K, N449D, D472N, N551S, 1698V и L735Q относительно SEQ ID NO: 2; и (b) гетерологичную нуклеиновую кислоту. В некоторых случаях вариантный капсидный белок AAV содержит аминокислотную последовательность, показанную в SEQ ID NO: 10. В некоторых случаях rAAV проявляет повышенную устойчивость к нейтрализующим анти-AAV антителам человека по сравнению с устойчивостью, проявляемой AAV2 (AAV дикого типа серотипа 2). В некоторых случаях rAAV проявляет, по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 30 раз и т. д.) большую устойчивость к нейтрализующим анти-AAV антителам человека, чем устойчивость, проявляемая AAV2. В некоторых случаях rAAV проявляет повышенную трансдукцию клеток млекопитающих в присутствии нейтрализующих анти-AAV антител человека по сравнению с трансдукцией клеток млекопитающих, проявляемой серотипом 2 AAV дикого типа (AAV2). В некоторых случаях клетки млекопитающих представляют собой клетки печени, клетки поджелудочной железы, клетки скелетной мышцы, клетки сердечной мышцы, фибробласты, ретинальные клетки, синовиальные клетки суставов, клетки легкого, Т-клетки, нейроны, глиальные клетки, стволовые клетки (например, гемопоэтические стволовые клетки, гемопоэтические клетки-предшественники, нейральные стволовые клетки, нейральные клетки-предшественники, стволовые клетки нервного гребня, эмбриональные стволовые клетки, индуцированные плюрипотентные стволовые клетки (iPS), мезенхимальные стволовые клетки, мезодермальные стволовые клетки, стволовые клетки печени, стволовые клетки поджелудочной железы, клетки-предшественники поджелудочной железы, мышечные стволовые клетки, ретинальные стволовые клетки и т.п.), эндотелиальные клетки или опухолевые клетки. В некоторых случаях гетерологичная нуклеиновая кислота содержит РНКи-агент. В некоторых случаях гетерологичная нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую полипептид.
[0038] Признаки настоящего изобретения включают выделенную нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая кодирует вариантный капсидный белок аденоассоциированного вируса (AAV), содержащий аминокислотную последовательность, имеющую по меньшей мере приблизительно 90% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в одной из SEQ ID NO: 11-13 и 26-33. В некоторых случаях кодируемый вариантный капсидный белок AAV содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в одной из SEQ ID NO: 11-13 и 26-33. В некоторых случаях кодируемый вариантный капсидный белок AAV содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 11-13 и 26-33.
[0039] Признаки настоящего изобретения включают выделенную нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая кодирует вариантный капсидный белок аденоассоциированного вируса (AAV), который содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 10, и включает аминокислотные замены N312K, N449D, D472N, N551S, I698V и L735Q относительно SEQ ID NO: 2.
[0040] В некоторых случаях кодируемый вариантный капсидный белок AAV (кодируемый выделенной нуклеиновой кислотой) придает инфекционному вириону рекомбинантного аденоассоциированного вируса (rAAV) повышенную устойчивость к нейтрализующим анти-AAV антителам человека по сравнению с устойчивостью, проявляемой AAV2 (дикий тип AAV серотипа 2). В некоторых случаях повышенная устойчивость по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 30 раз и т. д.) выше, чем устойчивость, проявляемая AAV2. В некоторых случаях кодируемый вариантный капсидный белок AAV (кодируемый выделенной нуклеиновой кислотой) придает инфекционному вириону рекомбинантного аденоассоциированного вируса (rAAV) повышенную трансдукцию клеток млекопитающих в присутствии нейтрализующих анти-AAV антител человека по сравнению с трансдукцией, проявляемой ААВ2.
[0041] Признаки настоящего изобретения включают выделенную клетку-хозяин, содержащую заявленную нуклеиновую кислоту, как описано выше. В некоторых случаях клетка-хозяин стабильно трансфектируется нуклеиновой кислотой. В некоторых случаях клетка-хозяин дополнительно содержит нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую белок rep AAV. В некоторых случаях клетка-хозяин дополнительно содержит рекомбинантный вектор AAV.
[0042] Признаки настоящего изобретения включают способ доставки гетерологичной нуклеиновой кислоты в клетку-мишень, включающий приведение в контакт клетки-мишени с заявленным вирионом (описанным выше). В некоторых случаях клетка-мишень представляет собой клетку печени, клетку поджелудочной железы, клетку скелетной мышцы, клетку сердечной мышцы, фибробласт, ретинальную клетку, синовиальную клетку суставов, клетку легкого, Т-клетку, нейрон, глиальную клетку, стволовую клетку (например, гемопоэтическую стволовую клетку, гемопоэтическую клетку-предшественник, нейральную стволовую клетку, нейральную клетку-предшественник, стволовую клетку нервного гребня, эмбриональную стволовую клетку, индуцированную плюрипотентную стволовую клетку (iPS), мезенхимальную стволовую клетку, мезодермальную стволовую клетку, стволовую клетку печени, стволовую клетку поджелудочной железы, клетку-предшественник поджелудочной железы, мышечную стволовую клетку, ретинальную стволовую клетку и т.п.), эндотелиальную клетку или опухолевую клетку. В некоторых случаях клетка-мишень находится in vitro. В некоторых случаях клетка-мишень находится in vivo.
[0043] Признаки настоящего изобретения включают способ доставки генного продукта субъекту, нуждающемуся в этом, где способ включает введение субъекту эффективного количества заявленного инфекционного вириона рекомбинантного аденоассоциированного вируса (rAAV) (описанного выше). В некоторых случаях гетерологичная нуклеиновая кислота вириона rAAV содержит РНК-интерферирующий агент. В некоторых случаях гетерологичная нуклеиновая кислота вириона rAAV содержит нуклеотидную последовательность, кодирующую полипептид. В некоторых случаях стадия введения включает опосредованную доставку инфекционного вириона rAAV. В некоторых случаях стадия введения включает прямую доставку инфекционного вириона rAAV.
[0044] Признаки настоящего изобретения включают вариантный капсидный белок аденоассоциированного вируса (AAV), содержащий аминокислотную последовательность, имеющую по меньшей мере приблизительно 90% идентичность с аминокислотной последовательностью, показанной в одной из SEQ ID NO: 11-13 и 26-33. В некоторых случаях капсидный белок AAV содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в одной из SEQ ID NO: 11-13 и 26-33. В некоторых случаях капсидный белок AAV содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 11-13 и 26-33.
[0045] Признаки настоящего изобретения включают вариантный капсидный белок аденоассоциированного вируса (AAV), который содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 10, и содержит аминокислотные замены N312K, N449D, D472N, N551S, 1698V и L735Q относительно SEQ ID NO: 2. В некоторых случаях вариантный капсидный белок AAV содержит аминокислотную последовательность, показанную в SEQ ID NO: 10. В некоторых случаях вариантный капсидный белок AAV придает инфекционному вириону рекомбинантного аденоассоциированного вируса (rAAV) повышенную устойчивость к нейтрализующим анти-AAV антителам человека по сравнению с устойчивостью, проявляемой AAV2. В некоторых случаях повышенная устойчивость по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 30 раз и т. д.) выше, чем устойчивость, проявляемая AAV2. В некоторых случаях вариантный капсидный белок AAV придает инфекционному вириону рекомбинантного аденоассоциированного вируса (rAAV) повышенную трансдукцию клеток млекопитающих в присутствии нейтрализующих анти-AAV антител человека по сравнению с трансдукцией, проявляемой AAV2.
[0046] Признаки настоящего изобретения включают библиотеку, содержащую по меньшей мере одно из: (i) двух или более инфекционных вирионов rAAV, каждый из которых содержит вариантный капсидный белок аденоассоциированного вируса (AAV) и гетерологичную нуклеиновую кислоту; (ii) двух или более выделенных нуклеиновых кислот, каждая из которых содержит нуклеотидную последовательность, кодирующую вариантный капсидный белок AAV; (iii) двух или более клеток-хозяев, каждая из которых содержит нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую вариантный капсидный белок AAV; и (iv) двух или более вариантных капсидных белков AAV; где вариантный капсидный белок AAV по меньшей мере одного члена библиотеки содержит аминокислотную последовательность, имеющую по меньшей мере одну аминокислотную замену относительно аминокислотной последовательности, показанной в одной из SEQ ID NO: 10-13 и 26-33.
[0047] Признаки настоящего изобретения включают способ получения и идентификации модифицированного инфекционного вириона rAAV, который проявляет измененное свойство инфицирования по сравнению с исходным (родительским) вирионом, содержащим исходный капсидный белок, где способ включает: (a) получение вариантных капсидных белков аденоассоциированного вируса (AAV) из исходного капсидного белка, где исходный капсидный белок содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33, и где каждый вариантный капсидный белок AAV содержит по меньшей мере одну аминокислотную замену относительно исходного капсидного белка; (b) получение вариантных вирионов AAV, каждый из которых содержит вариантный капсидный белок AAV, полученный на стадии (a); и (c) анализ вариантных вирионов AAV, полученных на стадии (b), на измененное свойство инфицирования для идентификации модифицированного инфекционного вириона rAAV. В некоторых случаях создание библиотеки вариантных капсидных белков AAV включает метод мутагенеза, выбранный из группы, состоящей из: мутагенеза с использованием полимеразной цепной реакции, олигонуклеотид-направленного мутагенеза, мутагенеза с насыщением, мутагенеза с заменой петель, мутагенеза с шаффлингом фрагментов и их комбинации. В некоторых случаях измененным свойством инфицирования является повышенная устойчивость к нейтрализующим анти-AAV антителам человека по сравнению с устойчивостью, проявляемой исходным вирионом. В некоторых случаях измененным свойством инфицирования является повышенная трансдукция клеток млекопитающих в присутствии нейтрализующих анти-AAV антител человека по сравнению с трансдукцией, проявляемой исходным вирионом. В некоторых случаях модифицированный инфекционный вирион rAAV содержит модифицированный капсидный белок AAV, содержащий аминокислотную последовательность, имеющую по меньшей мере приблизительно 90% идентичность аминокислотной последовательности с исходным капсидным белком.
[0048] Признаки настоящего изобретения включают способ получения вариантного капсидного белка AAV из исходного капсидного белка, где способ включает: подвергание нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую исходный капсидный белок, типу мутагенеза, выбранному из группы, состоящей из: мутагенеза с использованием полимеразной цепной реакции, олигонуклеотид-направленного мутагенеза, мутагенеза с насыщением, мутагенеза с заменой петель, мутагенеза с шаффлингом фрагментов и их комбинации; где исходный капсидный белок содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33.
Краткое описание фигур
[0049] На фиг. 1A-B показана направленная эволюция AAV для повышения уклонения от антител.
[0050] На фиг. 2A-2B показаны профили нейтрализации вариантов, уклоняющихся от антител, с использованием человеческого IVIG.
[0051] На фиг. 3A-3C приведены профили нейтрализации вариантов, уклоняющихся от антител, с использованием человеческой сыворотки, полученной от субъектов, которые были исключены из клинических испытаний с гемофилией B за счет наличия высоких титров нейтрализующих анти-AAV антител.
[0052] На фиг. 4A-4B приведены аминокислотные последовательности клонов с заменой петель/шаффингом и мутагенезом с насыщением.
[0053] На фиг. 5 показан тропизм вариантов AAV in vitro.
[0054] На фиг. 6A-6B показана локализация и нейтрализация новых вариантов AAV in vivo.
[0055] На фиг. 7A-7D показано количество вариантов с уклонением от антител.
[0056] На фиг. 8A-8I приведена последовательность капсидного белка Shuffle 100-1 (SEQ ID NO: 11), выровненная с последовательностями капсидного белка AAV1-9 дикого типа (SEQ ID NO: 1-9).
[0057] На фиг. 9A-9I приведена последовательность капсидного белка Shuffle 100-3 (SEQ ID NO: 12), выровненная с последовательностями капсидного белка AAV1-9 дикого типа (SEQ ID NO: 1-9).
[0058] На фиг. 10A-I приведена последовательность капсидного белка Shuffle 100-7 (SEQ ID NO: 13), выровненная с последовательностями капсидного белка AAV1-9 дикого типа (SEQ ID NO: 1-9).
[0059] На фиг. 11 показаны титры нейтрализующих антител библиотечных клонов и исходных серотипов в сыворотке иммунизированных мышей.
[0060] На фиг. 12 показан процесс направленной эволюции, использованный для идентификации капсидного варианта «А101» (содержащего капсидный белок с SEQ ID NO:12) с повышенной доставкой гена в легкие в присутствии нейтрализующих антител человека.
[0061] Фиг. 13A-13B: на фиг. 13A показано предполагаемое генетическое разнообразие библиотек капсидов, используемых для процесса направленной эволюции. Общее разнообразие библиотек составляет более 1 млр. генетических вариантов. На фиг. 13В показана продуктивность библиотек капсидов. Все капсидные библиотеки были генерированы на уровне, достаточном для получения материала для исследования в программе эволюции терапевтического вектора in vivo. Введенные вирусные геномы (вг) представляют собой целевую дозу без учета потерь, связанных с устройством для доставки и путем введения.
[0062] Фиг. 14A-14B: на фиг. 14A показана внешняя ПЦР-амплификация вирусных геномов из выделенных клеток AT II после а) введения AeroProbe® или b) введения с использованием распылителя в первом раунде селекции. Полосы в синих прямоугольниках представляют собой успешную амплификацию вирусных геномов. Температурный градиент представляет собой температуры отжига, используемые во время ПЦР, соответствующие каждой дорожке в геле. На фиг. 14В показана внутренняя ПЦР-амплификация вирусных геномов из выделенных клеток AT II после а) введения AeroProbe® или b) введения с использованием распылителя в первом раунде селекции. Полосы в синих прямоугольниках представляют собой успешную амплификацию вирусных геномов.
[0063] Фиг. 15A-15B: на фиг. 15A показана частота химерного мотива в анализе секвенирования для исследования. Анализ секвенирования основан на общей частоте в секвенированной популяции для обоих устройств для доставки AeroProbe и Распылитель. На фиг. 15B показана частота варианта A101 в химерном мотиве для исследования. Анализ секвенирования на основе общей частоты в секвенированной популяции для устройств для доставки AeroProbe и Распылитель.
[0064] На фиг. 16 приведена схема отбора образцов легких (примеры 3 и 7). Схематическое представление отбора образцов трахеи и легких. Кружки в правом легком представляют смежные образцы, полученные для выделения ДНК и белка. Образцы, ориентированные вдоль длинной и короткой оси для разреза ткани, представлены квадратами.
[0065] На фиг. 17 представлен вариантный капсид (содержащий капсидный белок с SEQ ID NO:12). Трансдукция с образцами сыворотки NHP при разведении сыворотки 1:10. Образцы сыворотки от NHP, отобранных для включения в исследование, анализировали на наличие нейтрализующих анти-AAV антител. Трансдукция в присутствии разведения сыворотки 1:10 (по сравнению с трансдукцией в отсутствии сыворотки) имеет место у всех NHP. NHP, отобранные для включения в исследование, обозначены желтыми полосами. Столбцы ошибок = стандартное отклонение, n=3 (внутренние повторы).
[0066] На фиг. 18 представлено биораспределения геномов, опосредованное вариантным капсидом. Количественная оценка вирусных геномов в легких и дополнительных системных органах с использованием кПЦР с использованием праймеров и зонда против трансгена EGFP. Вирусные геномы детектировались во всех 48 образцах (n=16 образцов на NHP; n=3 NHP). Все тестированные образцы скелетных мышц (трехглавая мышца плеча, латеральная широкая мышца бедра), диафрагмы, почек, селезенки, головного и спинного мозга были ниже предела количественного определения. Среднее ± стандартная ошибка; n=3 NHP (n=16 участков биопсии на легкое на каждого NHP, n=10 участков биопсии на печень на каждого NHP, n=15 участков биопсии на сердце на каждого NHP, n=9 участков биопсии на скелетную мышцу на каждого NHP, n=2 образца на почку на каждого NHP, n=1 образец на селезенку на NHP, n=8 участков биопсии на головной мозг на каждого NHP, n=3 участка биопсии на спинной мозг на каждого NHP).
[0067] На фиг. 19 показана экспрессия белка, опосредованная вариантным капсидом, в легких. Количественная оценка экспрессии белка EGFP в легких с использованием ELISA против белка EGFP. Экспрессию EGFP наблюдали во всех 48 образцах легких (n=16 образцов на каждого NHP; n=3 NHP). Экспрессию EGFP наблюдали в 10 образцах печени, положительных на вирусные геномы (n=10 образцов на каждого NHP; n=3 NHP). Среднее ± стандартная ошибка.
[0068] На фиг. 20 показана локализация белка, опосредованная вариантным капсидом, в легких. Репрезентативные изображения экспрессии EGFP в трахее (a-b), бронхах (c, e, g) и альвеолах (d, f, h) у NHP V002969. Секции, обозначенные белыми прямоугольниками в трахее (b), альвеолах (d) и бронхах (e), представлены в виде увеличенных изображений на i, j и k соответственно. Примерное расположение изображений обозначено пурпурными прямоугольниками на схеме. Экспрессия EGFP определяется антителом против GFP (красный цвет) на всех изображениях. Ядра доокрашивали DAPI (синий цвет).
[0069] На фиг. 21А-21D приведена характеристика клеток альвеолярного эпителия типа II примата, отличного от человека. Клетки NHP AECII были более чем на 90% положительными на окрашивание LysoTracker, что было показано с помощью флуоресцентной микроскопии (фиг. 17А) и количественно определено проточной цитометрией (фиг. 21В). Сурфактантный белок С, зрелый маркер клеток AECII, был наглядно виден на сутки 1 и 5 после посева (фиг. 21С). Клетки AECII снижали скорость пролиферации в культуре во времени, о чем свидетельствует включение EdU (фиг. 21D). EdU=5-этинил-2'-дезоксиуридин, столбцы ошибок = стандартное отклонение, n=3 внутренних повтора.
[0070] На фиг. 22А-22В приведена характеристика клеточного вектора для альвеолярного эпителия типа II «нечеловеческих» приматов. Капсид rAAV с капсидом, содержащим капсидный белок с SEQ ID NO:12 (4D-A101), показал более высокую скорость трансдукции, чем капсид AAV5, которые оба несут CAG-eGFP в культурах ALI клеток AECII NHP. Количественная оценка eGFP-положительных клеток проточной цитометрией (фиг. 22А). Репрезентативные изображения ICC положительных клеток eGFP (фиг. 22B). Время после заражения 3 дня, всего 5 дней в культуре. Столбцы ошибок = стандартное отклонение, n=3 внутренних повтора. t-критерий Стьюдента, p<0,05 в сравнении с AAV5.
[0071] На фиг. 23А-23D приведена характеристика клеток человека альвеолярного эпителия типа II. Клетки AECII человека были приблизительно на 80% положительными на окрашивание LysoTracker до суток 11 в культуре, когда их количество снизилось до 50%, как показано с использованием флуоресцентной микроскопии (фиг. 23А) и количественно определено проточной цитометрией (фиг. 23В). Сурфактантный белок С, зрелый маркер клеток AECII, был наглядно виден на сутки 5 и сутки 11 после посева (фиг. 23С). Клетки AECII снижали скорость пролиферации во времени в культуре, о чем свидетельствовало включение EdU (фиг. 23D). EdU=5-этинил-2'-дезоксиуридин, столбцы ошибок = стандартное отклонение, n=3 внутренних повтора.
[0072] На фиг. 24 приведена характеристика векторов для клеток альвеолярного эпителия человека типа II. Капсид, содержащий капсидный белок с SEQ ID NO:12 (4D-A101), показал более высокую скорость трансдукции, чем капсид AAV5, которые оба несут CAG-eGFP в культурах ALI клеток человека AECII. Репрезентативные изображения ICC eGFP-положительных клеток. Время после инифицирования 6 и 10 суток, всего 7 и 11 суток в культуре.
[0073] На фиг. 25 приведены профили нейтрализации in vitro AAV1, AAV2, AAV5, AAV8, AAV9 и rAAV дикого типа, содержащих капсид, содержащий капсидный белок с SEQ ID NO:12 (4D-A101). rAAV, содержащий капсид, содержащий капсидный белок с SEQ ID NO:12, продемонстрировал превосходную способность улоняться от нейтрализующих анти-AAV антител в человеческом IVIG по сравнению с AAV дикого типа. Векторы AAV.CAG.Luciferase инкубировали с разведениями IVIG до инфицирования клеток 2V6.11 при MOI 1000. Векторы, способные уклоняться от антител, трансдуцировали клетки, и активность люциферазы измеряли через 48 ч после инфицирования. IVIG=иммуноглобин для внутривенного введения, столбцы ошибок=стандартное отклонение, n=3, внутренние повторы. * p <0,05 для 4D-A101 по сравнению с AAV1, AAV2, AAV8 и AAV9, † p <0,05 для 4D-A101 по сравнению с AAV5.
[0074] На фиг. 26 представлен график зависимости суммарного заряда от pH для капсидных белков A101 VP1 и VP3.
[0075] На фиг. 27 представлен график рН-растворимости A101-GFP после 1-суточного хранения при комнатной температуре.
[0076] На фиг.28А-С показано, что трансдукция приводит к надежной экспрессии белка и мембранной локализации в клетках HEK2v6.11. HEK2v6.11 трансдуцировали с 4D-710 и исследовали вестерн-блоттингом (фиг. 28А) с анти-CFTR антителом (фиг. 28А). Репрезентативные изображения (фиг. 28B) показывают клетки, анализированные с помощью иммуноцитохимии, анти-CFTR (красный), F-актин (зеленый), DAPI, ядерный (синий). Шкала столбцов составляет 100 мкМ (фиг. 28B) и 25 мкМ (фиг. 28C).
[0077] На фиг. 29A-B показана трансдукция клеток 16HBE14o-G542X с 4D-710. Цифровую капельную ПЦР (ddPCR) с обратной транскрипцией-ddPCR (RT-ddPCR) проводили на РНК, выделенной из культур HBE после трансдукции 4D-710 при повышении MOI (фиг. 29A). Уровни экзогенного транскрипта CFTRΔR определяли и количественно оценивали в виде количества копий/мкл выше установленного порога и наносили на линейную шкалу. BLQ, ниже предела количественного определения. NT, нетрансдуцированный. Иммуноцитохимия культур HBE после трансдукции при множественности инфекции 35000 и 50000 (фиг. 29B). Синий цвет представляет DAPI, и красный цвет представляет белок CFTR. Масштаб 100 мкм.
[0078] На фиг. 30 показана трансдукция культур здоровых легких ALI ex vivo с 4D-710. ddPCR проводили на кДНК, полученной из РНК, экстрагированной из культур после трансдукции 4D-710. Приготовили два набора праймеров/зондов для специфической дифференциации трансгена CFTRΔR человека с оптимизированными кодонами от эндогенного гена CFTR человека. Количественный анализ анализировал количество капель выше установленного порога, содержащих транскрипт исследуемого набора праймеров/зондов. BLQ, ниже предела количественного определения. NT, нетрансдуцированный.
[0079] На фиг. 31 приведена трансдукция с 4D-A101 образцов сыворотки NHP при разведении сыворотки 1:10. Образцы сыворотки от NHP, подходящих для включения в исследование, анализировали на наличие нейтрализующих анти-AAV антител к капсиду 4D-710 (4D-A101, содержащий капсидный белок с SEQ ID NO:12). Трансдукция в присутствии разведения сыворотки 1:10 (по сравнению с трансдукцией в отсутствии сыворотки) имела место у всех NHP. Столбцы ошибок=стандартное отклонение, n=3 (внутренние повторы).
[0080] На фиг. 32А-32С приведена количественная оценка вирусных геномов с использованием кПЦР с использованием праймеров и зонда против трансгена CFTRΔR. На фиг. 32А вирусные геномы стабильно детектировались в образцах легких, распределенные по всему правому легкому. На фиг. 32B отдельные образцы легких животных обозначены примерной областью и долей легкого: альвеолы (зеленый цвет), первичные/вторичные бронхи (синий цвет), третичные/нижние бронхи (красный цвет), верхняя доля (круг), средняя доля (квадрат), нижняя доля (треугольник), добавочная доля (ромб). На фиг. 32C показано, что вирусные геномы, количественно определенные у животных, получавших дозу 3×1013 вг, во всех тестированных образцах сердца, печени, головного мозга, скелетных мышц (трехглавая мышца плеча, латеральная широкая мышца бедра, диафрагма), спинного мозга, поджелудочной железы, почек и яичка находились на уровне ниже предела количественного определения. У всех трех животных детектировались вирусные геномы в трахеобронхиальном (TB) лимфатическом узле, и у одного животного вирусные геномы детектировались в селезенке. Среднее значение ± стандартное отклонение.
[0081] На фиг. 33A-33B показана экспрессия трансгенного транскрипта 4D-710 в легких. Количественная оценка транскрипта CFTRΔR с использованием ОТ-кПЦР с использованием праймеров и зонда против трансгена 4D-710. На фиг. 33A показано, что транскрипты детектировались в образцах правого легкого, распределенные по долям, у животных, получавших 3×1013 вг, и у всех животных, получавших носитель, уровни транскриптов были на уровне ниже BLQ. На фиг. 33B показаны отдельные образцы легких животных, которым ввели дозу 3×1013 вг, обозначены примерной областью и долей легкого: альвеолы (зеленый цвет), первичные/вторичные бронхи (синий цвет), третичные/нижние бронхи (красный цвет), верхняя доля (круг), средняя доля (квадрат), нижняя доля (треугольник), добавочная доля (ромб). Среднее значение ± стандартное отклонение.
[0082] На фиг. 34A-B показана 4D-710 экспрессия белка в легких. Экспрессия белка CFTR в легких показана с использованием иммуногистохимического окрашивания. На фиг. 34A показана экспрессия CFTR в срезах эпителия трахеи, бронхиального эпителия и альвеол в каждой группе обработки, репрезентативные изображения. На фиг. 34B показана экспрессия белка CFTR в эпителии трахеи, эпителии бронхов и срезах альвеол 3×1013 вг обработанных животных (показаны отдельные животные), репрезентативные изображения.
[0083] На фиг. 35 приведен график зависимости растворимости A101-Luc от рН.
Подробное описание изобретения
[0084] Определения
[0085] Аденоассоциированный вирус представляет собой непатогенный парвовирус, состоящий из генома одноцепочечной ДНК длиной 4,7 т.п.н. в икосаэдрическом капсиде без оболочки. «AAV» является аббревиатурой для аденоассоциированного вируса и может использоваться для обозначения самого вируса или его производных. Геном содержит три открытые рамки считывания (ORF), окруженные инвертированными концевыми повторами (ITR), которые функционируют в качестве ориджина репликации вируса и сигнал упаковки. ORF rep кодирует четыре неструктурных белка, которые играют роль в репликации вируса, регуляции транскрипции, сайт-специфической интеграции и сборке вириона. ORF cap кодирует три структурных белка (VP1-3), которые собираются с образованием 60-мерного вирусного капсида. Наконец, ORF, присутствующая в качестве альтернативной рамки считывания в гене cap, продуцирует белок, активирующий сборку (AAP), вирусный белок, который локализует капсидные белки AAV в ядрышке и участвует в процессе сборки капсида.
[0086] Существует несколько встречающихся в природе серотипов и более 100 вариантов AAV, каждый из которых отличается аминокислотной последовательностью, в частности, в гипервариабельных областях капсидных белков, и, таким образом, своими свойствами доставки генов. Не было выявлено связи между каким-либо AAV и каким-либо заболеванием человека, что делает рекомбинантный AAV привлекательным для клинических применений.
[0087] В рамках настоящего изобретения, термин «AAV» охватывает все подтипы и как встречающиеся в природе, так и рекомбинантные формы, за исключением случаев, когда требуется иное. Термин «AAV» включает AAV типа 1 (AAV-1 или AAV1), AAV типа 2 (AAV-2 или AAV2), AAV типа 3 (AAV-3 или AAV3), AAV типа 4 (AAV-4 или AAV4), AAV типа 5 (AAV-5 или AAV5), AAV типа 6 (AAV-6 или AAV6), AAV типа 7 (AAV-7 или AAV7), AAV типа 8 (AAV-8 или AAV8), AAV типа 9 (AAV-9 или AAV9), птичий AAV, бычий AAV, собачий AAV, лошадиный AAV, AAV приматов, AAV животных, отличных от приматов, и овечий AAV. «AAV приматов» относится к AAV, которые инфицируют приматов, «AAV животных, отличных от приматов» относится к AAV, которые инфицируют млекопитающих, не являющихся приматами, «бычий AAV» относится к AAV, которые инфицируют млекопитающих, относящихся к подсемейству бычьих, и т. д.
[0088] В рамках настоящего изобретения, термин «4D-A101» или «A101» относится к капсиду AAV, содержащему капсидный белок с последовательностью SEQ ID NO:12.
[0089] В рамках настоящего изобретения, термин «4D-710» относится к рекомбинантному AAV, содержащему (i) капсид, содержащий капсидный белок с SEQ ID NO:12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность SEQ ID NO:45.
[0090] Геномные последовательности различных серотипов AAV, а также последовательности нативных концевых повторов (TR), белков Rep и капсидных субъединиц известны в данной области. Такие последовательности можно найти в литературе или в общедоступных базах данных, таких как GenBank. См., например, идентификационные номера в GenBank NC-002077.1 (AAV-1), AF063497.1 (AAV-1), NC-001401.2 (AAV-2), AF043303.1 (AAV-2), J01901.1 (AAV-2), U48704.1 (AAV-3), NC-001729.1 (AAV-3), NC-001829.1 (AAV-4), U89790.1 (AAV-4), NC-006152.1 (AAV-5), AF085716.1 (ААВ-5), AF028704.1 (ААВ-6), NC-006260.1 (ААВ-7), AF513851.1 (ААВ-7), AF513852.1 (ААВ-8) NC-006261.1 (ААВ-8), и AY530579.1 (ААВ-9); описание которых включено в настоящее описание посредством ссылки для описания последовательностей нуклеиновых кислот и аминокислот AAV. См. также, например, Srivistava et al. (1983) J. Virology 45:555; Chiorini et al. (1998) J. Virology 71:6823; Chiorini et al. (1999) J. Virology 73:1309; Bantel-Schaal et al. (1999) J. Virology 73:939; Xiao et al. (1999) J. Virology 73:3994; Muramatsu et al. (1996) Virology 221:208; Shade et al., (1986) J. Virol. 58:921; Gao et al. (2002) Proc. Nat. Acad. Sci. USA 99:11854; Moris et al. (2004) Virology 33:375-383; публикации международных заявок WO 00/28061, WO 99/61601, WO 98/11244; и патент США № 6156303.
[0091] Последовательности природных cap (капсидных) белков, связанных с серотипами AAV, известны в данной области и включают: AAV1 (SEQ ID NO: 1), AAV2 (SEQ ID NO: 2), AAV3 (SEQ ID NO: 3)), AAV4 (SEQ ID NO: 4), AAV5 (SEQ ID NO: 5), AAV6 (SEQ ID NO: 6), AAV7 (SEQ ID NO: 7), AAV8 (SEQ ID NO: 8) и AAV9 (SEQ ID NO: 9). Термин «вариантный капсидный белок AAV» представляет собой капсидный белок AAV, содержащий аминокислотную последовательность, которая включает, по меньшей мере одну замену (включая делецию, инсерцию и т. д.) относительно одной из существующих в природе последовательностей капсидного белка AAV, показанных в SEQ ID. №: 1-9.
[0092] «Вирион AAV» или «вирусная частица AAV» относится к вирусной частице, состоящей по меньшей мере из одного капсидного белка AAV и инкапсулированного полинуклеотида AAV.
[0093] «Рекомбинантный» применительно к полинуклеотиду означает, что полинуклеотид является продуктом различных комбинаций стадий клонирования, рестрикции или лигирования и других процедур, в результате которых получается конструкция, отличная от полинуклеотида, встречающегося в природе. Рекомбинантный вирус представляет собой вирусную частицу, содержащую рекомбинантный полинуклеотид. Термины соответственно включают репликаты исходной полинуклеотидной конструкции и потомство исходной конструкции вируса.
[0094] Если вирион AAV содержит гетерологичный полинуклеотид (т.е. полинуклеотид, отличный от генома AAV дикого типа, например, трансген, который необходимо доставить в клетку-мишень, РНКи-агент или агент CRISPR, который нужно доставить в клетку-мишень, и т. д.), то обычно его называют «рекомбинантным вирионом AAV (rAAV)» или «вирусной частицей rAAV». Как правило, гетерологичный полинуклеотид фланкирован по меньшей мере одной, и обычно двумя последовательностями инвертированных концевых повторов (ITR) AAV.
[0095] Термин «вектор rAAV» охватывает вирионы rAAV (т.е. вирусные частицы rAAV) (например, инфекционный вирион rAAV), которые по определению включают полинуклеотид rAAV; и также включает полинуклеотиды, кодирующие rAAV (например, одноцепочечный полинуклеотид, кодирующий rAAV (ss-rAAV); двухцепочечный полинуклеотид, кодирующий rAAV (ds-rAAV), например, плазмиды, кодирующие rAAV, и т.п.).
[0096] Термин «упаковка» относится к серии внутриклеточных событий, которые приводят к сборке и инкапсулированию частицы AAV.
[0097] Гены «rep» и «cap» AAV относятся к полинуклеотидным последовательностям, кодирующим белки репликации и инкапсидирования аденоассоциированного вируса. AAV rep и cap называются здесь «упаковочными генами» AAV.
[0098] «Вирус-помощник» для AAV относится к вирусу, который позволяет AAV (например, AAV дикого типа) реплицироваться и упаковываться с помощью клетки млекопитающего. Ряд таких вирусов-помощников для AAV известны в данной области техники, включая аденовирусы, герпевирусы и проксивирусы, такие как вирус коровьей оспы. Аденовирусы охватывают много различных подгрупп, хотя обычно используется аденовирус 5 типа, подгруппы С. Известны многочисленные аденовирусы человека, млекопитающих, отличных от человека, и птичьего происхождения и доступные из хранилищ, таких как АТСС. Вирусы семейства герпеса включают, например, вирусы герпеса простого (HSV) и вирусы Эпштейн-Барра (EBV), а также цитомегаловирусы (CMV) и вирусы псевдобешенства (PRV); которые также доступны из хранилищ АТСС.
[0099] «Функция(и) вируса-помощника» относится к функции(ям), закодированной в геноме вируса-помощника, которая обеспечивает репликацию и упаковку AAV (в сочетании с другими необходимыми условиями для репликации и упаковывания, описанными в настоящем изобретении). Как здесь описано, «функция вируса-помощника» может обеспечиваться несколькими путями, в том числе путем обеспечения вируса-помощника или обеспечения, например, полинуклеотидных последовательностей, кодирующих необходимую(ие) функцию(и) клетки-продуцента в транс-положении. Например, плазмидный или другой экспрессионный вектор, содержащий нуклеотидные последовательности, кодирующие один или более аденовирусных белков, трансфектируют в клетку-продуцент вместе с вектором rAAV.
[00100] Термин «инфекционный» вирус или вирусная частица представляет собой вирус, который содержит правильно собранный вирусный капсид и способен доставить полинуклеотидный компонент в клетку, для которой вирусный вид является тропным. Термин не обязательно подразумевает любую репликационную способность вируса. Методы подсчета инфекционных вирусных частиц описаны в другом разделе в настоящем описании и в документах в данной области техники. Вирусная инфекционность может быть выражена как отношение инфекционных вирусных частиц ко всем вирусным частицам. Способы определения отношения инфекционных вирусных частиц ко всем вирусным частицам известны в данной области техники. См., например, Grainger et al. (2005) Mol. Ther. 11:S337. См. также примеры.
[00101] В рамках настоящего изобретения, термин «тропизм» относится к преимущественному нацеливанию вируса (например, AAV) на определенные виды-хозяев или определенные типы клеток внутри видов-хозяев. Например, вирус, способный инфицировать клетки сердца, легких, печени и мышц, обладает более широким (т.е. повышенным) тропизмом по сравнению с вирусом, способным поражать только клетки легких и мышц. Тропизм также может включать зависимость вируса от определенных типов молекул клеточной поверхности хозяина. Например, некоторые вирусы могут инфицировать только клетки с поверхностными гликозаминогликанами, тогда как другие вирусы могут инфицировать только клетки с сиаловой кислотой (такие зависимости можно тестировать, используя различные клеточные линии, дефицитные по определенным классам молекул, в качестве потенциальных клеток-хозяев для вирусной инфекции). В некоторых случаях тропизм вируса описывает относительные предпочтения вируса. Например, первый вирус может быть способен заражать все типы клеток, но на гораздо более высоком уровне заражает клетки с поверхностными гликозаминогликанами. Можно считать, что второй вирус обладает таким же (или идентичным) тропизмом, что и первый вирус, если второй вирус также предпочитает те же характеристики (например, второй вирус также на более высоком уровне инфицирует клетки с поверхностными гликозаминогликанами), даже если абсолютная эффективность трансдукции неодинакова. Например, второй вирус может быть более эффективным, чем первый вирус, при инфицировании каждого данного тестируемого типа клеток, но если относительные предпочтения сходны (или идентичны), можно считать, что второй вирус имеет аналогичный (или идентичный) тропизм, как первый вирус. В некоторых вариантах осуществления тропизм вириона, содержащего рассматриваемый вариантный капсидный белок AAV по настоящему изобретению, не изменяется по сравнению с встречающимся в природе вирионом. В некоторых вариантах осуществления тропизм вириона, содержащего вариантный капсидный белок AAV по настоящему изобретению, увеличивается (т.е. расширяется) по сравнению с встречающимся в природе вирионом. В некоторых вариантах осуществления тропизм вириона, содержащего указанный вариантный капсидный белок AAV, снижен по сравнению с встречающимся в природе вирионом.
[00102] «Репликационно-компетентный» вирус (например, репликационно-компетентный AAV) относится к фенотипически дикому вирусу, который является инфекционным, и также способен к репликации в инфицированной клетке (т.е. в присутствии вируса-помощника или функции вируса-помощника). В случае AAV способность к репликации обычно требует присутствия функциональных генов упаковки AAV. Как правило, векторы rAAV, описанные в настоящем изобретении, являются репликационно-некомпетентными в клетках млекопитающих (особенно в клетках человека) за счет отсутствия одного или более генов упаковки AAV. Как правило, в таких векторах rAAV отсутствуют какие-либо последовательности генов упаковки AAV для сведения к минимуму возможности того, что репликационно-компетентные AAV будут генерироваться путем рекомбинации между генами упаковки AAV и входящим вектором rAAV. Во многих вариантах осуществления препараты векторов rAAV, описанные в настоящем изобретении, содержат мало или вообще не содержат репликационно-компетентных AAV (rcAAV, также называемых RCA) (например, ниже приблизительно 1 rcAAV на 102 частиц rAAV, ниже приблизительно 1 rcAAV на 104 rAAV частиц, ниже приблизительно 1 rcAAV на 108 частиц rAAV, ниже приблизительно 1 rcAAV на 1012 частиц rAAV, или не содержат rcAAV).
[00103] Термин «полинуклеотид» относится к полимерной форме нуклеотидов любой длины, включая дезоксирибонуклеотиды или рибонуклеотиды или их аналоги. Полинуклеотиды могут содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и нуклеотидные аналоги, и могут прерываться ненуклеотидными компонентами. Если имеются, то модификации нуклеотидной структуре могут придаваться до или после сборки полимера. В рамках настоящего изобретения, термин полинуклеотид относится взаимозаменяемо к дву- и одноцепочечным молекулам. Если не определено и не требуется иное, то любой вариант осуществления изобретения, описанного здесь, которое является полинуклеотидом, охватывает как двуцепочечную форму, так и каждую из двух комплементарных одноцепочечных форм, известных или прогнозированных для составления двуцепочечной формы.
[00104] Полинуклеотид или полипептид имеет определенную процентную «идентичность последовательности» с другим полинуклеотидом или полипептидом, означает что, при выравнивании этот процент оснований или аминокислот является таким же при сравнении этих двух последовательностей. Сходство последовательностей можно определить с использованием ряда различных способов. Для определения идентичности последовательности можно выравнить с использованием методов и компьютерных программ, включая BLAST, доступный из всемирной компьютерной сети с ncbi.nlm.nih.gov/BLAST/. Другим алгоритмом выравнивания является FASTA, доступный из пакета Genetics Computing Group (GCG) из Madison, Wisconsin, USA, находящегося в полной собственности филиала Oxford Molecular Group, Inc. Другие методы выравнивания описаны в монографии Methods in Enzymology, vol. 266: Computer Methods for Macromolecular Sequence Analysis (1996), ed. Doolittle, Academic Press, Inc., a division of Harcourt Brace & Co., San Diego, California, USA. Особый интерес представляют программы выравнивания, которые допускают гэпы в последовательности. Алгоритм Смита-Уотермана является одним из типов алгоритма, который допускают гэпы при выравнивании последовательностей. Смотри Meth. Mol. Biol. 70:173-187 (1997). Также, программу GAP, в которой используется метод выравнивания Нидлмана-Вунша, можно применить для выравнивания последовательностей. См. J. Mol. Biol., 48:443-453 (1970).
[00105] «Ген» относится к полинуклеотиду, который выполняет какую-либо функцию в клетке. Например, ген может содержать открытую рамку считывания, способную кодировать определенный белок после его транскрипции и трансляции. С другой стороны, ген может кодировать нетранслируемый продукт функциональной РНК (например, аптамер, интерферирующую РНК, рибосомальную РНК (рРНК), транспортную РНК (тРНК) и т. д.).
[00106] «Продукт экспрессии гена» или «генный продукт» представляет собой молекулу, полученную в результате экспрессии конкретного гена, как определено выше. Продукты генной экспрессии включают, например, полипептид, аптамер, интерферирующую РНК, информационную РНК (мРНК), рРНК, тРНК, некодирующую РНК (нкРНК) и т.п.
[00107] «РНК-интерферирующий агент» или «РНКи-агент» включает любой агент (или полинуклеотид, кодирующий такой агент), который можно использовать для изменения экспрессии гена (как определено выше). Примеры РНКи-агентов, известных специалистам в данной области, включают, не ограничиваясь этим, (i) миРНК агенты; (ii) антисмысловую РНК; (iii) агенты CRISPR; (iv) агенты нуклеазы цинковых пальцев и (v) агенты на основе нуклеаза эффектор-подобного активатора транскрипции (TALEN).
[00108] (i) агент миРНК («малая интерферирующая» или «короткая интерферирующая РНК» (или миРНК)) представляет собой РНК-дуплекс нуклеотидов, нацеленный на интересующий ген («ген-мишень»). «РНК-дуплекс» относится к структуре, образованной путем комплементарного спаривания двух участков молекулы РНК с образованием области двухцепочечной РНК (дцРНК). миРНК «нацелена» на ген, когда нуклеотидная последовательность дуплексной области миРНК комплементарна нуклеотидной последовательности гена-мишени. В некоторых вариантах осуществления длина дуплекса миРНК составляет менее 30 нуклеотидов. В некоторых вариантах осуществления дуплекс может иметь длину 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11 или 10 нуклеотидов. В некоторых вариантах осуществления длина дуплекса составляет 19-25 нуклеотидов. Дуплексная область РНК в миРНЕ может быть частью шпилькообразной структуры. Агенты миРНК, которые содержат шпильку, также могут называться «агентами shRNA (короткая шпилька РНК)». В дополнение к дуплексной части структура шпильки может содержать петлевую область, расположенную между двумя последовательностями, образующими дуплекс. Петля может быть разной длины. В некоторых вариантах осуществления петля имеет длину 5, 6, 7, 8, 9, 10, 11, 12 или 13 нуклеотидов. Структура шпильки также может содержать 3'- или 5'-выступы. В некоторых вариантах осуществления выступ представляет собой 3'- или 5'-выступ длиной 0, 1, 2, 3, 4 или 5 нуклеотидов. Как правило, уровень продукта экспрессии (например, мРНК, полипептида и т. д.) гена-мишени снижается под действием агента миРНК (например, миРНК, кшРНК и т. д.), который содержит специфические двухцепочечные нуклеотидные последовательности, комплементарные друг другу, по меньшей мере к сегменту длиной 19-25 нуклеотидов (например, последовательности из 20-21 нуклеотида) транскрипта гена-мишени, включая 5'-нетранслируемую (UT) область, ORF или 3'-UT область. В некоторых вариантах осуществления малые интерферирующие РНК имеют длину приблизительно 19-25 нуклеотидов. См., например, заявки РСТ WO0/44895, WO99/32619, WO01/75164, WO01/92513, WO01/29058, WO01/89304, WO02/16620 и WO02/29858; и публикацию патента США № 20040023390 для описания технологии миРНК. миРНК и/или shRNA могут кодироваться последовательностью нуклеиновой кислоты, и последовательность нуклеиновой кислоты может также включать промотор. Последовательность нуклеиновой кислоты может также включать сигнал полиаденилирования. В некоторых вариантах осуществления сигнал полиаденилирования представляет собой синтетический минимальный сигнал полиаденилирования.
[00109] (ii) антисмысловая РНК представляет собой РНК, комплементарную продукту экспрессии гена. Например, антисмысловая РНК, нацеленная на конкретную мРНК, представляет собой агент на основе РНК (или может представлять собой модифицированную РНК), комплементарную мРНК, где гибридизация антисмысловой РНК с мРНК изменяет экспрессию мРНК (например, путем изменения стабильности РНК, изменения трансляции РНК и т. д.). В понятие «антисмысловая РНК» также включаются нуклеиновые кислоты, кодирующие антисмысловую РНК.
[00110] (iii) агенты CRISPR. Системы CRISPR (короткие палиндромные повторы, регулярно расположенные кластерами)/CRISPR-ассоциированные (Cas) обеспечивают бактериям и археям адаптивный иммунитет против вирусов и плазмид за счет использования РНК CRISPR (crРНК) для сайленсинга чужеродных нуклеиновых кислот. Белок Cas 9 (или его функциональный эквивалент и/или вариант, т. е. Cas9-подобный белок) в природе обладает ДНК-эндонуклеазной активностью, которая зависит от ассоциации белка с двумя встречающимися в природе или синтетическими молекулами РНК, называемыми crРНК и tracrRNA (также называемыми направляющими РНК). В некоторых случаях две молекулы ковалентно связываются с образованием одной молекулы (также называемой одиночной направляющей РНК («sgRNA»). Таким образом, Cas9 или Cas9-подобный белок связывается с ДНК-нацеливающей РНК (этот термин охватывает как конфигурацию направляющей РНК с двумя молекулами, так и конфигурацию направляющей РНК с одной молекулой), которая активирует Cas9 или Cas9-подобный белок и направляет белок к последовательности-мишени нуклеиновой кислоты. Если Cas9 или Cas9-подобный белок сохраняет свою естественную ферментативную функцию, то он будет расщеплять ДНК-мишень, создавая двухцепочечный разрыв, что может привести к изменению генома (т. е. редактированию: делеции, инсерции (при наличии донорного полинуклеотида), замене и т. д.), тем самым изменяя экспрессию гена. Некоторые варианты Cas9 (которые охватываются термином Cas9-подобные) были изменены таким образом, что они приобретают сниженную активность в расщеплении ДНК (в некоторых случаях они расщепляют одну цепь вместо обеих цепей ДНК-мишени, в то время как в других случаях их активность в расщеплении ДНК резко снижается или отсутствует). Cas9-подобные белки со сниженной активностью расщепления ДНК (даже без активности расщепления ДНК) все еще могут направляться к ДНК-мишени и могут блокировать активность РНК-полимеразы. Таким образом, ферментативно неактивные Cas9-подобные белки могут направляться в определенное место в ДНК-мишени с помощью ДНК-нацеливающей РНК, чтобы блокировать транскрипцию ДНК-мишени. Подробную информацию об агентах CRISPR можно найти, например, в (a) Jinek et. al., Science, 2012 Aug. 17; 337(6096):816-21: «A programmed dual-RNA-guided DNA endonuclease in adaptive bacterial immunity»; (b) Qi et al., Cell, 2013 Feb. 28; 152(5):1173-83: «Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression» и (c) заявке на патент США сер. № 13/842859 и заявке РСТ PCT/US13/32589; которые все в полном объеме включены в настоящее описание посредством ссылки. Таким образом, используемый здесь термин «агент CRISPR» охватывает любой агент (или нуклеиновую кислоту, кодирующую такой агент), содержащий встречающиеся в природе и/или синтетические последовательности, которые можно использовать в системе на основе Cas9 (например, Cas9 или Cas9-подобный белок; любой компонент ДНК-нацеливающей РНК, например, crРНК-подобная РНК, tracrRNA-подобная РНК, одиночная направляющая РНК и т. д.; донорный полинуклеотид и т. п.).
[00111] (iv) Агенты на основе нуклеазы цинкового пальца (ZFN). Нуклеазы цинкового пальца (ZFN) представляют собой искусственные ДНК-эндонуклеазы, генерируемые путем слияния ДНК-связывающего домена с цинковыми пальцами с доменом расщепления ДНК. ZFN можно сконструировать для нацеливания на желаемые последовательности ДНК, и это позволяет нуклеазам цинковых пальцев расщеплять уникальные последовательности-мишени. При введении в клетку ZFN можно использовать для редактирования ДНК-мишени в клетке (например, генома клетки) путем индукции двухцепочечных разрывов. Для получения дополнительной информации об использовании ZFN см., например: Asuri et al., Mol Ther. 2012 February; 20(2):329-38; Bibikova et al. Science. 2003 May 2; 300(5620):764; Wood et al. Science. 2011 Jul. 15; 333(6040):307; Ochiai et al. Genes Cells. 2010 August; 15(8):875-85; Takasu et. al., Insect Biochem Mol Biol. 2010 October; 40(10):759-65; Ekker et al, Zebrafish 2008 Summer; 5(2):121-3; Young et al, Proc Natl Acad Sci USA. 2011 Apr. 26; 108(17):7052-7; Goldberg et al, Cell. 2010 Mar. 5; 140(5):678-91; Geurts et al, Science. 2009 Jul. 24; 325(5939):433; Flisikowska et al, PLoS One. 2011; 6(6):e21045. doi: 10.1371/journal.pone.0021045. Epub 2011 Jun. 13; Hauschild et al, Proc Natl Acad Sci USA. 2011 Jul. 19; 108(29):12013-7; и Yu et al, Cell Res. 2011 November; 21(11):1638-40; которые все включены здесь посредством ссылки для их идей, связанных с ZFN. Термин «средство ZFN» включает нуклеазу цинкового пальца и/или полинуклеотид, содержащий нуклеотидную последовательность, кодирующую нуклеазу цинкового пальца.
[00112] (v) Агенты на основе нуклеаза эффектор-подобного активатора транскрипции (TALEN). Эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), представляют собой искусственные ДНК-эндонуклеазы, генерируемые путем слияния эффекторного ДНК-связывающего домена TAL (подобного активатору транскрипции) с доменом расщепления ДНК. TALEN можно быстро сконструировать для связывания практически любой желаемой последовательности ДНК, и при введении в клетку TALEN можно использовать для редактирования ДНК-мишени в клетке (например, клеточного генома) путем индукции двухцепочечных разрывов. Для получения дополнительной информации об использовании TALEN см., например: Hockemeyer et al. Nat Biotechnol. 2011 Jul. 7; 29(8):731-4; Wood et al. Science. 2011 Jul. 15; 333(6040):307; Tesson et al. Nat Biotechnol. 2011 Aug. 5; 29(8):695-6; and Huang et. al., Nat Biotechnol. 2011 Aug. 5; 29(8):699-700; которые все включены здесь посредством ссылки для их идей, связанных с TALEN. Термин «агент TALEN» включает TALEN и/или полинуклеотид, содержащий нуклеотидную последовательность, кодирующую TALEN.
[00113] Термин «регуляторный элемент» или «регуляторная последовательность» представляет собой нуклеотидную последовательность, участвующую во взаимодействии молекул, которое способствует функциональной регуляции полинуклеотида, включая репликацию, дупликацию, транскрипцию, сплайсинг, трансляцию или деградацию полинуклеотида. Регуляция может оказывать влияние на частоту, скорость или специфичность процесса и может носить повышающий или ингибирующий характер. Регуляторные элементы, известные в данной области, включают, например, регуляторные последовательности транскрипции, такие как промоторы и энхансеры. Промотор представляет собой участок ДНК, способный в определенных условиях связывать РНК-полимеразу и инициировать транскрипцию кодирующего участка, обычно расположенного ниже (в 3'-направлении) от промотора.
[00114] «Оперативно связанные» или «функционально связанные» относятся к сопоставлению генетических элементов, где элементы находятся во взаимоотношениях, позволяющих им действовать ожидаемым образом. Например, промотор функционально связан с кодирующей областью, если промотор помогает инициировать транскрипцию кодирующей последовательности. Между промотором и кодирующей областью могут находиться промежуточные остатки при условии, что сохраняется эта функциональная взаимосвязь.
[00115] «Экспрессионный вектор» представляет собой вектор, содержащий область, которая кодирует представляющий интерес полипептид, и используется для осуществления экспрессии белка в предполагаемой клетке-мишени. Экспрессионный вектор также содержит регуляторные элементы, оперативно связанные с кодирующей областью для облегчения экспрессии белка в мишени. Комбинация регуляторных элементов и гена или генов, с которыми они функционально связаны для экспрессии, иногда называют «экспрессионной кассетой», большое количество которых известно и доступно в данной области или может быть легко сконструировано из компонентов, которые доступны в данной области техники.
[00116] «Гетерологичный» означает полученный из молекулы, генотипически отличной от остальной части молекулы, с которой его сравнивают. Например, полинуклеотид, введенный методами генной инженерии в плазмиду или вектор, полученный из другого вида, является гетерологичным полинуклеотидом. Промотор, удаленный из его нативной кодирующей последовательности и оперативно связанный с кодирующей последовательностью, с которой он в природе не связан, является гетерологичным промотором. Таким образом, например, rAAV, который включает гетерологичную нуклеиновую кислоту, кодирующую продукт гетерологичного гена, представляет собой rAAV, который включает нуклеиновую кислоту, обычно не входящую в состав встречающегося в природе AAV дикого типа, и кодируемый продукт гетерологичного гена представляет собой продукт гена, который обычно не кодируется встречающимся в природе AAV дикого типа.
[00117] «Пептид 2А» относится к «саморасщепляющимся» пептидам приблизительно из 20 аминокислот, которые продуцируют эквимолярные уровни нескольких генов из одной и той же мРНК и могут использоваться вместо элементов IRES в мультицистронных векторах. Неограничивающие примеры включают последовательности пептидов T2A, P2A, E2A и F2A.
[00118] Термины «генетическое изменение» и «генетическая модификация» (и их грамматические варианты) используются в настоящем изобретении взаимозаменяемо для обозначения процесса, при котором генетический элемент (например, полинуклеотид) вводят в клетку иначе, чем путем митоза или мейоза. Элемент может быть гетерологичным для клетки или может быть дополнительной копией или улучшенной версией элемента, уже находящегося в клетке. Генетическое изменение может быть осуществлено, например, путем трансфекции клетки рекомбинантной плазмидой или другим полинуклеотидом любым способом, известным в данной области, таким как электропорация, осаждение фосфатом кальция или контактирование с комплексом полинуклеотид-липосома. Генетическое изменение также можно осуществить, например, путем трансдукции или инфицирования ДНК- или РНК-вирусом или вирусным вектором. Как правило, генетический элемент вводится в хромосому или мини-хромосому в клетке; но любое изменение, которое изменяет фенотип и/или генотип клетки и ее потомства, включается в этот термин.
[00119] Клетка была «генетически модифицирована», или «трансформирована», или «трансфектирована» экзогенной ДНК (например, с помощью рекомбинантного вируса), когда такая ДНК была введена в клетку. Наличие экзогенной ДНК приводит к постоянному или транзиентному генетическому изменению. Трансформирующая ДНК может быть или не быть интегрирована (ковалентно связана) в геном клетки. «Клон» представляет собой популяцию клеток, полученных из одной клетки или общего предка путем митоза. «Клеточная линия» представляет собой клон первичной клетки, способный к стабильному росту in vitro в течение многих поколений.
[00120] Клетка считается «стабильно» измененной, трансдуцированной, генетически модифицированной или трансформированной с помощью генетической последовательности, если последовательность доступна для выполнения своей функции во время длительного культивирования клетки in vitro и/или в течение длительного периода времени в естественных условиях. Как правило, такая клетка является «наследственно» измененной (генетически модифицированной) в том смысле, что в нее вводится генетическое изменение, которое также наследуется потомством измененной клетки.
[00121] Термины «полипептид», «пептид» и «белок» используются здесь взаимозаменяемо для обозначения полимеров из аминокислот любой длины. Термины также охватывают аминокислотный полимер, который был модифицирован; например, посредством образования дисульфидной связи, гликозилирования, липидирования, фосфорилирования или конъюгирования с метящим компонентом. Полипептиды, такие как антиангиогенные полипептиды, нейропротекторные полипептиды и т.п., при обсуждении в контексте доставки генного продукта субъекту-млекопитающему и их композиции, относятся к соответствующему интактному полипептиду или любому его фрагменту или сконструированному производному, который сохраняет желаемую биохимическую функцию интактного белка. Аналогично, ссылки на нуклеиновые кислоты, кодирующие антиангиогенные полипептиды, нуклеиновые кислоты, кодирующие нейропротекторные полипептиды, и другие подобные нуклеиновые кислоты для применения при доставке генного продукта субъекту-млекопитающему (которые могут называться «трансгенами», которые должны быть доставлены субъекту-млекопитающему клетка-реципиент), включают полинуклеотиды, кодирующие интактный полипептид или любой фрагмент или сконструирвоанное производное, обладающее желаемой биохимической функцией.
[00122] «Выделенная» плазмида, нуклеиновая кислота, вектор, вирус, вирион, клетка-хозяин, белок или другое вещество относится к препарату вещества, по меньшей мере без некоторых других компонентов, которые также могут присутствовать там, откуда вещество или подобное вещество происходит или изначально получено. Так, например, выделенное вещество может быть получено с использованием метода очистки для обогащения его из исходной смеси. Обогащение можно измерить в абсолютном выражении, например, по массе на объем раствора, или его можно измерить по отношению ко второму, потенциально интерферирующему веществу, присутствующему в исходной смеси. Увеличение обогащения вариантов осуществления настоящего раскрытия становится все более выделенными. Выделенная плазмида, нуклеиновая кислота, вектор, вирус, клетка-хозяин или другое вещество в некоторых вариантах осуществления очищены, например, от приблизительно 80% до приблизительно 90% чистоты, по меньшей мере до приблизительно 90% чистоты, по меньшей мере до приблизительно 95% чистоты, по меньшей мере до приблизительно 98% чистоты или по меньшей мере приблизительно до 99% или выше.
[00123] В рамках настоящего изобретения, термины «лечение», «терапия» и т.п. относятся к получению желаемого фармакологического и/или физиологического эффекта. Эффект может быть профилактическим с точки зрения полного или частичного предотвращения заболевания или его симптома и/или может быть терапевтическим с точки зрения частичного или полного излечения заболевания и/или неблагоприятного эффекта, ассоциированного с заболеванием. «Лечение», как здесь используется, охватывает любое лечение заболевания у млекопитающего, в частности у человека, и включает: (а) предотвращение развития заболевания (и/или симптомов, вызванных заболеванием) у субъекта, который может быть предрасположен к заболеванию или находится в группе риска по заболеванию, но еще не были диагностированы как имеющие его; (b) ингибирование заболевания (и/или симптомов, вызванных заболеванием), т.е. остановку его развития; и (c) облегчение заболевания (и/или симптомов, вызванных заболеванием), т.е. обеспечение регрессии заболевания (и/или симптомов, вызванных заболеванием).
[00124] Термины «индивидуум», «хозяин», «субъект» и «пациент» используются здесь взаимозаменяемо и относятся к млекопитающим, включая, не ограничиваясь ими, людей; приматов, отличных от человека, в том числе обезьян; спортивных животных млекопитающих (например, лошадей); сельскохозяйственных животных млекопитающих (например, овец, коз и т.д.); домашних животных млекопитающих (собак, кошек и т.д.); и грызунов (например, мышей, крыс и т.д.).
[00125] В некоторых вариантах осуществления субъектом является человек, который ранее подвергался естественному воздействию AAV и, как следствие, содержит анти-AAV антитела (т.е. нейтрализующие антитела к AAV). В некоторых вариантах осуществления субъектом является человек, которому ранее вводили AAV-вектор (и в результате он может содержать антитела против AAV), и которому требуется повторное введение вектора для лечения другого патологического состояния или для дальнейшего лечения того же состояния. Основываясь на положительных результатах клинических испытаний, связанных с доставкой гена AAV, например, в печень, мышцы и сетчатку - все ткани, пораженные нейтрализующими антителами против этого носителя, - существует множество таких терапевтических применений/мишеней для лечения заболеваний.
[00126] В рамках настоящего изобретения, термин «эффективное количество» представляет собой количество, достаточное для достижения полезных или желаемых клинических результатов. Эффективное количество можно вводить в одно или несколько введений. Для целей настоящего изобретения эффективное количество соединения (например, инфекционного вириона rAAV) представляет собой количество, достаточное для облегчения, ослабления, стабилизации, реверсии, предотвращения, замедления или задержки прогрессирования (и/или симптомов, ассоциированных с) конкретного болезненного состояния (например, рака). Соответственно, эффективное количество инфекционного вириона rAAV представляет собой количество инфекционного вириона rAAV, которое способно уклоняться от нейтрализующей активности индивидуальных антител против AAV, таким образом, эффективно доставляя гетерологичную нуклеиновую кислоту в клетку-мишень (или клетки-мишени) субъекта.
[00127] Перед дальнейшим описанием настоящего изобретения следует понимать, что это изобретение не ограничивается конкретными описанными вариантами осуществления, поскольку они, конечно, могут варьировать. Также следует понимать, что используемая здесь терминология предназначена только для целей описания конкретных вариантов осуществления и не предназначена для ограничения, поскольку объем настоящего изобретения будет ограничен только прилагаемой формулой изобретения.
[00128] Если указан диапазон значений, то подразумевается, что каждое промежуточное значение, с точностью до десятой доли единицы нижнего предела, если контекст явно не указывает иное, между верхним и нижним пределом этого диапазона и любым другим указанным или промежуточное значение в указанном диапазоне охватывается изобретением. Верхний и нижний пределы этих меньших диапазонов могут независимо включаться в меньшие диапазоны, а также охватываются изобретением с учетом любого специально исключенного предела в указанном диапазоне. Если указанный диапазон включает один или оба предела, диапазоны, исключающие один или оба из этих включенных пределов, также включены в изобретение.
[00129] Если не указано иное, то все технические и научные термины, используемые в настоящем изобретении, имеют то же значение, которое обычно понимается специалистами в области техники, к которой относится данное изобретение. Хотя любые способы и материалы, подобные или эквивалентные описанным здесь, также могут быть использованы при практическом применении или тестировании настоящего изобретения, теперь описаны предпочтительные способы и материалы. Все упомянутые здесь публикации включены сюда в качестве ссылки для раскрытия и описания способов и/или материалов, в связи с которыми цитируются публикации.
[00130] Следует отметить, что используемые здесь и в прилагаемой формуле изобретения формы единственного числа включают ссылки на множественное число, если контекст явно не указывает иное. Так, например, ссылка на «инфекционный вирион рекомбинантного аденоассоциированного вируса (rAAV)» включает множество таких вирионов, и ссылка на «инфекционный вирион рекомбинантного аденоассоциированного вируса (rAAV)» включает ссылку на один или более таких вирионов и их эквиваленты, известные специалистам в данной области, и так далее. Далее следует отметить, что формула изобретения может быть составлена таким образом, чтобы исключить любой необязательный элемент. Как таковое, это заявление предназначено служить предшествующей основой для использования такой исключительной терминологии, как «исключительно», «только» и т.п., в связи с перечислением элементов формулы изобретения или использованием «негативного» ограничения.
[00131] Понятно, что некоторые признаки изобретения, которые для ясности описаны в контексте отдельных вариантов осуществления, также могут быть представлены в комбинации в одном варианте осуществления. И наоборот, различные признаки изобретения, которые для краткости описаны в контексте одного варианта осуществления, также могут быть предоставлены по отдельности или в любой подходящей комбинации. Все комбинации вариантов осуществления, относящихся к изобретению, охватываются настоящим изобретением и раскрываются здесь точно так же, как если бы каждая комбинация была раскрыта отдельно и явно. Кроме того, все подкомбинации различных вариантов осуществления и их элементы также конкретно охватываются настоящим изобретением и раскрыты здесь точно так же, как если бы каждая такая подкомбинация была отдельно и явно раскрыта здесь.
[00132] Публикации, цитированные здесь, представлены исключительно для их раскрытия до даты подачи настоящей заявки. Ничто в настоящем изобретении не должно быть истолковано как допущение того, что настоящее изобретение не имеет права предшествуть такой публикации в силу предшествующего изобретения. Кроме того, указанные даты публикации могут отличаться от фактических дат публикации, которые могут потребовать независимого подтверждения.
[00133] Настоящее изобретение относится к инфекционным вирионам рекомбинантного аденоассоциированного вируса (rAAV), которые содержат вариантный капсидный белок и гетерологичную нуклеиновую кислоту. Изобретение также относится к вариантным капсидным белкам аденоассоциированного вируса (AAV) (и/или нуклеиновой кислоте, кодирующей вариантные капсидные белки AAV), которые придают инфекционному вириону rAAV повышенную устойчивость к нейтрализующим анти-AAV антителам человека. Настоящее раскрытие дополнительно относится к клеткам-хозяевам, содержащим инфекционный вирион rAAV и/или нуклеиновую кислоту, кодирующую заявленный вариантный капсидный белок AAV. Настоящее раскрытие дополнительно относится к библиотекам указанных выше вирионов, капсидных белков, нуклеиновых кислот и/или клеток-хозяев; где вариантный капсидный белок AAV по меньшей мере одного члена библиотеки содержит аминокислотную последовательность, имеющую по меньшей мере одну аминокислотную замену относительно аминокислотной последовательности, показанной в одной из SEQ ID NO: 10-13 и 26-33.
[00134] Настоящее изобретение также относится к способам доставки гетерологичной нуклеиновой кислоты в клетку-мишень, где клетку-мишень подвергают контактиррованию с инфекционным вирионом rAAV субъекта. Также настоящее изобретение относится к способам доставки генного продукта субъекту, где способы, как правило, включают введение эффективного количества вириона rAAV субъекта индивидууму, нуждающемуся в этом. Также настоящее изобретение относится к композициям и наборам для применения в заявленных способах. Во многих вариантах осуществления заявленный инфекционный вирион rAAV, заявленную нуклеиновую кислоту, заявленный вариантный капсидный белок AAV, заявленную клетку-хозяина и т.д. выделяют.
[00135] Вариантные капсидные полипептиды AAV
[00136] Вариантный капсидный полипептид AAV по настоящему изобретению (или вариантный капсидный белок AAV, кодируемый нуклеиновой кислотой по настоящему изобретению) придает вириону инфекционного rAAV, содержащему вариантный капсидный полипептид AAV, повышенную устойчивость к нейтрализующим антителам человека AAV по сравнению с устойчивостью, проявляемой AAV дикого типа (например, AAV2 (AAV дикого типа серотипа 2)) или AAV, содержащий капсидный белок дикого типа. В некоторых вариантах осуществления повышенная устойчивость по меньшей мере приблизительно 1,5 раза (например, по меньшей мере приблизительно в 1,5 раза, по меньшей мере приблизительно в 2 раза, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 4 раза, по меньшей мере приблизительно в 5 раз), по меньшей мере приблизительно в 7,5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 12 раз, по меньшей мере приблизительно в 15 раз, по меньшей мере приблизительно в 17 раз, по меньшей мере приблизительно в 20 раз, по меньшей мере приблизительно в 25 раз, по меньшей мере приблизительно в 30 раз, по меньшей мере приблизительно в 40 раз, по меньшей мере приблизительно в 50 раз, по меньшей мере приблизительно в 75 раз, по меньшей мере приблизительно в 100 раз, по меньшей мере приблизительно в 150 раз, по меньшей мере приблизительно в 200 раз, по меньшей мере приблизительно в 250 раз, по меньшей мере приблизительно в 300 раз и т. д.) выше, чем устойчивость, проявляемая AAV дикого типа (например, AAV2 (AAV дикого типа серотипа 2)) или AAV, содержащего капсидный белок дикого типа.
[00137] Можно сказать, что вариантный капсидный белок AAV по настоящему изобретению (или вариантный капсидный белок AAV, кодируемый нуклеиновой кислотой по настоящему изобретению) придает вириону инфекционного rAAV повышенную трансдукцию клеток млекопитающих в присутствии человеческих нейтрализующих антител к AAV по сравнению с трансдукцией, проявляемой AAV дикого типа (например, AAV2 (AAV дикого типа серотипа 2)) или AAV, содержащим капсидный белок дикого типа. В некоторых вариантах осуществления повышенная трансдукция по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 1,5 раза, по меньшей мере приблизительно в 2 раза, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 4 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 7,5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 12 раз, по меньшей мере приблизительно в 15 раз, по меньшей мере приблизительно в 17 раз, по меньшей мере приблизительно в 20 раз, по меньшей мере приблизительно в 25 раз, по меньшей мере приблизительно в 30 раз, по меньшей мере приблизительно в 40 раз, по меньшей мере приблизительно в 50 раз, по меньшей мере приблизительно в 75 раз, по меньшей мере приблизительно в 100 раз, по меньшей мере приблизительно в 150 раз, по меньшей мере приблизительно в 200 раз, по меньшей мере приблизительно в 250 раз, по меньшей мере приблизительно в 300 раз и т. д.) выше, чем трансдукция, проявляемая AAV дикого типа (например, AAV2 (AAV дикого типа серотипа 2)) или AAV, содержащим капсидный белок дикого типа.
[00138] В некоторых вариантах осуществления вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый нуклеиновой кислотой по настоящему изобретению) проявляет пониженное связывание с нейтрализующим антителом, которое связывает капсидный белок AAV дикого типа. Например, вариантный капсидный белок AAV может проявлять по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 1,5 раза, по меньшей мере приблизительно в 2 раза, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 4 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 7,5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 12 раз, по меньшей мере приблизительно в 15 раз, по меньшей мере приблизительно в 17 раз, по меньшей мере приблизительно в 20 раз, по меньшей мере приблизительно в 25 раз, по меньшей мере приблизительно в 30 раз, по меньшей мере приблизительно в 40 раз, по меньшей мере приблизительно в 50 раза, по меньшей мере приблизительно в 75 раз, по меньшей мере приблизительно в 100 раз, по меньшей мере приблизительно в 150 раз, по меньшей мере приблизительно в 200 раз, по меньшей мере приблизительно в 250 раз, по меньшей мере приблизительно в 300 раз и т. д.) более низкое связывание (например, пониженную аффинность) с нейтрализующим антителом, которое связывает капсидный белок AAV дикого типа, по сравнению с аффинностью связывания антитела к капсидному белку AAV дикого типа.
[00139] В некоторых вариантах осуществления нейтрализующее анти-AAV антитело связывается с вариантным капсидным белком AAV по настоящему изобретению (или вариантным капсидным белком AAV, кодируемым нуклеиновой кислотой по настоящему изобретению) с аффинностью ниже чем приблизительно 10-7 М, ниже чем приблизительно 5×10-6 М, ниже приблизительно 10-6 М, ниже приблизительно 5×10-5 М, ниже приблизительно 10-5 М, ниже приблизительно 10-4 М или ниже.
[00140] Термин «вариантный капсидный белок» не охватывает капсидные белки AAV дикого типа. «Вариантный капсидный белок AAV» не содержит аминокислотную последовательность, присутствующую в природном капсидном белке AAV. Например, вариантный капсидный белок не содержит аминокислотную последовательность, которая на 100% идентична любой из последовательностей, показанных в SEQ ID NO:1-9. Другими словами, указанный вариантный капсидный белок не содержит аминокислотную последовательность, показанную ни в одной из SEQ ID NO: 1-9. Вариантный капсидный белок может отличаться по аминокислотной последовательности от «исходного» или «родительского» капсидного белка AAV, где родительский капсидный белок AAV может представлять собой капсидный белок AAV дикого типа или капсидный белок AAV недикого типа.
[00141] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 90% (например, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотами 203-736 аминокислотной последовательности, показанной в одной из SEQ ID NO: 10-13 и 26-33.
[00142] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый указанной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 90% (например, по меньшей мере приблизительно 92%, по меньшей мере приблизительно 95%%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в одной из SEQ ID NO: 10-13 и 26-33.
[00143] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый указанной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотами 203-736 аминокислотной последовательности, показанной в SEQ ID NO:10, и включает аминокислотные замены N312K, N449D, D472N, N551S, I698V и L735Q относительно капсидного белка AAV AAV2 (например, SEQ ID NO: 2) или соответствующие положения в другом исходном серотипе AAV.
[00144] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:10, и включает аминокислотные замены N312K, N449D, D472N, N551S, I698V и L735Q относительно капсидного белка AAV AAV2 (например, SEQ ID NO: 2) или соответствующие положения в другом родительском серотипе AAV.
[00145] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотами 203-736 аминокислотной последовательности, показанной в SEQ ID NO:31, и включает аминокислотные замены N312K, N449D, N551S и I698V относительно капсидного белка AAV AAV2 (например, SEQ ID NO:2) или соответствующие положения в другом исходном серотипе AAV.
[00146] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:31, и включает аминокислотные замены N312K, N449D, N551S и I698V относительно капсидного белка AAV AAV2 (например, SEQ ID NO:2) или соответствующие положения в другом родительском серотипе AAV.
[00147] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотами 203-736 аминокислотной последовательности, показанной в SEQ ID NO:32, и включает аминокислотные замены D180N, N312K, Q385R, N449D, N551S, I698V и S721T относительно капсидного белка AAV AAV2 (например, SEQ ID NO:2) или соответствующие положения в другом родительском серотипе AAV.
[00148] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:32, и включает аминокислотные замены D180N, N312K, Q385R, N449D, N551S, I698V и S721T относительно капсидного белка AAV AAV2 (например, SEQ ID NO:2) или соответствующие положения в другом родительском серотипе AAV.
[00149] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере примерна 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотами 203-736 аминокислотной последовательности, показанной в SEQ ID NO:33, и включает аминокислотные замены N312K, N449D, T450A, N551S и I698V относительно капсидного белка AAV AAV2 (например, SEQ ID NO:2) или соответствующие положения в другом родительском серотипе AAV.
[00150] В некоторых вариантах осуществления заявленный вариантный капсидный белок AAV (или вариантный капсидный белок AAV, кодируемый заявленной нуклеиновой кислотой) содержит аминокислотную последовательность, имеющую по меньшей мере приблизительно 95% (например, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, по меньшей мере приблизительно 99,5% или 100%) идентичность аминокислотной последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:33, и включает аминокислотные замены N312K, N449D, T450A, N551S и I698V относительно капсидного белка AAV AAV2 (например, SEQ ID NO:2) или соответствующие положения в другом исходном серотипе AAV.
[00151] Иллюстративные вариантные капсидные белки AAV включают, не ограничиваясь этим (см. фиг. 8-10 для выбранных примеров выравнивания последовательностей):
MAADGYLPDWLEDTLSEGIRQWWKLKPGPPPPKPAERHKDDSRGLVLPGYKYLGPFNGLDKGEPVNEADAAALEHDKAYDQQLKAGDNPYLKYNHADAEFQQRLQGDTSFGGNLGRAVFQAKKRVLEPLGLVEQAGETAPGKKRPLIESPQQPDSSTGIGKKGKQPAKKRLNFGQTGDSESVPDPQPLGEPPATPAAVGPTTMASGGGAPMADNNEGADGVGNASGNWHCDSTWLGDRVITTSTRTWALPTYNNHLYKQISSASTGASNDNHYFGYSTPWGYFDFNRFHCHFSPRDWQRLINNNWGFRPKRLNFKLFNIQVKEVTTNDGVTTIANNLTSTVQVFSDSDYQLPYVLGSAHEGCLPPFPADVFMVPQYGYLTLNNGSQAVGRSSFYCLEYFPSQMLRTGNNFTFSYTFEDVPFHSSYAHSQSLDRLMNPLIDQYLYYLNRTQNQSGSAQNKDLLFSRGSPTGMSVQPKNWLPGPCYRQQRVSKTKTDNNNSNFTWTGASKYNLNGRESIINPGTAMASHKDDKDKFFPMSGVMIFGKESAGASNTALDNVMITDEEEIKATNPVATERFGTVAVNLQSSSTDPATGDVHAMGALPGMVWQDRDVYLQGPIWAKIPHTDGHFHPSPLMGGFGLKNPPPQILIKNTPVPANPPAEFSATKFASFITQYSTGQVSVEIEWELQKENSKRWNPEVQYTSNYAKSANVDFTVDNNGLYTEPRPIGTRYLTRPL;
[00154] Shuffle 100-3 (нуклеотидная последовательность) (SEQ ID NO: 24):
atggctgctgatggttatcttccagattggctcgaggacactctctctgaaggaataagacagtggtggaagctcaaacctggcccaccaccaccaaagcccgcagagcggcataaggacgacagcaggggtcttgtgcttcctgggtacaagtacctcggacccttcaacggactcgacaagggagagccggtcaacgaggcagacgcagcggccctcgagcacgacaaggcctacgaccagcagctcaaggccggtgacaacccctacctcaagtacaaccacgccgacgcggagttccagcagcggcttcagggcgacacatcgtttgggggcaacctcggcagagcagtcttccaggccaaaaagagggttcttgaacctcttggtctggttgagcaagcgggtgagacggctcctggaaagaagagaccgttgattgaatccccccagcagcccgactcctccacgggtatcggcaaaaaaggcaagcagccggctaaaaagagactcaattttggtcagactggcgactcagagtcagtccccgacccacaacctctcggagaacctccagcaacccccgctgctgtgggacctactacaatggcttcaggtggtggcgcaccaatggcagacaataacgaaggcgccgacggagtgggtaatgcctcaggaaattggcattgcgattccacatggctgggcgacagagtcatcaccaccagcacccgcacctgggccttgcccacctacaataaccacctctacaagcaaatctccagtgcttcaacgggggccagcaacgacaaccactacttcggctacagcaccccctgggggtattttgacttcaacagattccactgccacttttcaccacgtgactggcagcgactcatcaacaacaattggggattccggcccaagagactcaacttcaaactcttcaacatccaagtcaaggaggtcacgacgaatgatggcgtcacaaccatcgctaataaccttaccagcacggttcaagtcttctcggactcagactatcagctcccgtacgtgctcgggtcggctcacgagggctgcctcccgccgttcccagcagacgtcttcatggtgccacagtatggatacctcaccctgaacaacgggagtcaggcagtaggacgctcttcattttactgcctggagtactttccttctcagatgctgcgtaccggaaacaactttaccttcagctacacttttgaggacgttcctttccacagcagctacgctcacagccagagtctggaccgtctcatgaatcctctcatcgaccagtacctgtattacctgaacagaactcagaatcagtccggaagtgcccaaaacaaggacttgctgtttagccgggggtctccaactggcatgtctgttcagcccaaaaactggctacctggaccctgttatcggcagcagcgcgtttctaaaacaaaaacagacaacaacaacagcaactttacctggactggtgcttcaaaatataaccttaatgggcgtgaatctataatcaaccctggcactgctatggcctcacacaaagacgacaaagacaagttctttcccatgagcggtgtcatgatttttggaaaggagagcgccggagcttcaaacactgcattggacaatgtcatgatcacagacgaagaggaaatcaaagccactaaccccgtggccactgaaagatttgggactgtggcagtcaatctccagagcagcagcacagaccctgcgaccggagatgtgcatgccatgggagccttacctggaatggtgtggcaagacagagacgtatacctgcagggtcctatttgggccaaaattcctcacacggatggacactttcacccgtctcctctcatgggcggctttggactcaagaacccgcctcctcagatcctcatcaaaaacacgcctgttcctgcgaatcctccggcggagttttcagctacaaagtttgcttcattcatcacccagtattccacaggacaagtgagcgtggagattgaatgggagctgcagaaagaaaacagcaaacgctggaatcccgaagtgcagtatacatctaactatgcaaaatctgccaacgttgatttcactgtggacaacaatggactttatactgagcctcgccccattggcacccgttacctcacccgtcccctgtaa.
[00155] Shuffle 100-7 (аминокислотная последовательность) (SEQ ID NO: 13); Shuffle 100-7 (нуклеотидная последовательность) (SEQ ID NO: 25); Shuffle 10-2 (аминокислотная последовательность) (SEQ ID NO: 26); Shuffle 10-2 (нуклеотидная последовательность) (SEQ ID NO: 34); Shuffle 10-6 (аминокислотная последовательность) (SEQ ID NO: 27); Shuffle 10-6 (нуклеотидная последовательность) (SEQ ID NO: 35); Shuffle 10-8 (аминокислотная последовательность) (SEQ ID NO: 28); Shuffle 10-8 (нуклеотидная последовательность) (SEQ ID NO: 36); Shuffle 100-2 (аминокислотная последовательность) (SEQ ID NO: 29); Shuffle 100-2 (нуклеотидная последовательность) (SEQ ID NO: 37); SM 10-1 (аминокислотная последовательность) (SEQ ID NO: 30); SM 10-1 (нуклеотидная последовательность) (SEQ ID NO: 38); SM 10-8 (аминокислотная последовательность) (SEQ ID NO: 31); SM 10-8 (нуклеотидная последовательность) (SEQ ID NO: 39); SM 100-3 (аминокислотная последовательность) (SEQ ID NO: 32); SM 100-3 (нуклеотидная последовательность) (SEQ ID NO: 40); SM 100-10 (аминокислотная последовательность) (SEQ ID NO: 33); и SM 100-10 (нуклеотидная последовательность) (SEQ ID NO: 41).
[00156] Нуклеиновые кислоты и клетки-хозяева
[00157] Настоящее изобретение относится к нуклеиновым кислотам, содержащим нуклеотидные последовательности, кодирующие вариантный капсидный белок AAV (как описано выше), а также к клеткам-хозяевам, содержащим нуклеиновую кислоту по настоящему изобретению. Нуклеиновые кислоты и клетки-хозяева пригодны для получения вирионов rAAV (как описано ниже).
[00158] Настоящее изобретение относится к клеткам-хозяевам, например, выделенным клеткам-хозяевам, содержащим нуклеиновую кислоту по настоящему изобретению. Клетка-хозяин по настоящему изобретению может относиться к «генетически модифицированной клетке-хозяину» и обычно представляет собой выделенную клетку, например, клетку в культуре in vitro. Клетка-хозяин по настоящему изобретению является пригодной для получения вириона rAAV по настоящему изобретению, как описано ниже. Когда указанная клетка-хозяин используется для получения указанного вириона rAAV, то ее называют «упаковывающей клеткой». В некоторых вариантах осуществления указанная клетка-хозяин является стабильно генетически модифицированной (т.е. стабильно трансфектированной) нуклеиновой кислотой по настоящему изобретению. В других вариантах осуществления указанная клетка-хозяин является транзиентно генетически модифицированной (т.е. транзиентно трансфектированной) нуклеиновой кислотой по настоящему изобретению.
[00159] Нуклеиновую кислоту по настоящему изобретению стабильно или транзиентно вводят в клетку-хозяин с использованием хорошо известных способов, включая, не ограничиваясь этим, электропорацию, преципитацию фосфатом кальция, липосома-опосредованную трансфекцию и т.п. Для стабильной трансформации нуклеиновая кислота по настоящему изобретению обычно дополнительно включает селектируемый маркер, например, любой из нескольких хорошо известных селектируемых маркеров, таких как устойчивость к неомицину и т.п.
[00160] Клетку-хозяина по настоящему изобретению получают введения заявленной нуклеиновой кислоты в любую из множества клеток, например, клетки млекопитающих, включая, например, мышиные клетки и клетки приматов (например, клетки человека). Подходящие клетки млекопитающих включают, не ограничиваясь этим, первичные клетки и клеточные линии, где подходящие клеточные линии включают, помимо прочего, клетки 293, клетки COS, клетки HeLa, клетки Vero, мышиные фибробласты 3T3, фибробласты C3H10T1/2, клетки CHO и т.п.
[00161] В некоторых вариантах осуществления клетка-хозяин по настоящему изобретению включает, в дополнение к нуклеиновой кислоте, содержащей нуклеотидную последовательность, кодирующую мутантный капсидный белок, нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую один или более белков rep AAV. В других вариантах осуществления рассматриваемая клетка-хозяин дополнительно содержит вектор rAAV, как описано ниже. Как более подробно описано ниже, вирион rAAV создается с использованием клетки-хозяина по настоящему изобретению.
[00162] Инфекционные вирионы rAAV
[00163] Инфекционный вирион rAAV субъекта содержит вариантный капсидный белок AAV и гетерологичную нуклеиновую кислоту (более подробно описанную ниже) и проявляет повышенную устойчивость к нейтрализующим анти-AAV антитела человека по сравнению с устойчивостью, проявляемой AAV дикого типа (например, AAV2 (AAV дикого типа серотипа 2)) или AAV, содержащий капсидный белок дикого типа. Под «повышенной устойчивостью» подразумевается, что инфекционный вирион rAAV субъекта проявляет повышенную инфекционность в присутствии человеческих анти-AAV антител. Как описано выше, инфекционность вируса можно выразить как отношение инфекционных вирусных частиц к общему количеству вирусных частиц. Таким образом, повышенная инфекционность означает повышенное отношение инфекционных вирусных частиц к общему количеству вирусных частиц. Для определения устойчивости AAV к человеческим анти-AAV антителам, инфекционность AAV измеряют в присутствии различных концентраций человеческих анти-AAV антител, чтобы получить концентрацию антител (например, концентрацию в сыворотке, концентрацию IVIG и т. д.) (мг/мл), необходимых для снижения эффективности доставки гена (т.е. инфекционности) до 50% от эффективности в отсутствии анти-AAV антител человека. Считается, что вирус, для которого требуется более высокая концентрация антител для снижения эффективности доставки гена до 50% по сравнению с отсутствием анти-AAV антител человека, обладает повышенной устойчивостью к нейтрализации антителами. Таким образом, двукратное увеличение устойчивости означает двукратное увеличение концентрации антител, необходимой для снижения эффективности доставки гена до 50% по сравнению с отсутствием анти-AAV антител человека. В некоторых вариантах осуществления инфекционный вирион rAAV субъекта проявляет более высокую устойчивость к нейтрализующим анти-AAV антителам человека, по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 1,5 раза, по меньшей мере приблизительно в 2 раза, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 4 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 7,5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 12 раз, по меньшей мере приблизительно в й5 раз, по меньшей мере приблизительно в 17 раз, по меньшей мере приблизительно в 20 раз, по меньшей мере приблизительно в 25 раз, по меньшей мере приблизительно в 30 раз, по меньшей мере приблизительно в 40 раз, по меньшей мере приблизительно в 50 раз, по меньшей мере приблизительно в 75 раз, по меньшей мере приблизительно в 100 раз, по меньшей мере приблизительно в 150 раз, по меньшей мере приблизительно в 200 раз, по меньшей мере приблизительно в 250 раз, по меньшей мере приблизительно в 300 раз и т. д.), чем устойчивость, проявляемая AAV дикого типа (например, AAV2 (AAV дикого типа серотипа 2)) или AAV содержащий капсидный белок дикого типа.
[00164] Можно сказать, что инфекционный вирион rAAV по настоящему изобретению проявляет повышенную трансдукцию клеток млекопитающих в присутствии антител, нейтрализующих AAV человека. В некоторых вариантах осуществления инфекционный вирион rAAV по настоящему изобретению проявляет более высокую трансдукцию клеток млекопитающих, по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 1,5 раза, по меньшей мере приблизительно в 2 раза, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 4 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 7,5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 12 раз, по меньшей мере приблизительно в 15 раз, по меньшей мере приблизительно в 17 раз, по меньшей мере приблизительно в 20 раз, по меньшей мере приблизительно в 25 раз, по меньшей мере приблизительно в 30 раз, по меньшей мере приблизительно в 40 раз, по меньшей мере приблизительно в 50 раз, по меньшей мере приблизительно в 75 раз, по меньшей мере приблизительно в 100 раз, по меньшей мере приблизительно в 150 раз, по меньшей мере приблизительно в 200 раз, по меньшей мере приблизительно в 250 раз, по меньшей мере приблизительно в 300 раз и т. д.) в присутствии антител, нейтрализующих AAV человека, чем трансдукция, проявляемая AAV дикого типа (например, AAV2 (серотип AAV дикого типа) 2)) или AAV, содержащий капсидный белок дикого типа.
[00165] В некоторых вариантах осуществления инфекционный вирион rAAV субъекта демонстрирует пониженное связывание с нейтрализующим антителом, которое связывает капсидный белок AAV дикого типа. Например, инфекционный вирион rAAV субъекта может иметь пониженное связывание (например, пониженную аффинность) по меньшей мере приблизительно в 1,5 раза (например, по меньшей мере приблизительно в 1,5 раза, по меньшей мере приблизительно в 2 раза, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 4 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 7,5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 12 раз, по меньшей мере приблизительно в 15 раз, по меньшей мере приблизительно в 17 раз, по меньшей мере приблизительно в 20 раз, по меньшей мере приблизительно в 25 раз, по меньшей мере приблизительно в 30 раз, по меньшей мере приблизительно в 40 раз, по меньшей мере приблизительно в 50 раз, по меньшей мере приблизительно в 75 раз, по меньшей мере приблизительно в 100 раз, по меньшей мере приблизительно в 150 раз, по меньшей мере приблизительно в 200 раз кратно, по меньшей мере приблизительно в 250 раз, по меньшей мере приблизительно в 300 раз и т. д.) с нейтрализующим антителом, которое связывает капсидный белок AAV дикого типа, по сравнению с аффинностью связывания антитела с капсидным белком AAV дикого типа.
[00166] В некоторых вариантах осуществления нейтрализующее анти-AAV антитело связывается с инфекционным вирионом rAAV субъекта с аффинностью ниже приблизительно 10-7 М, ниже приблизительно 5×10-6 М, ниже приблизительно 10-6 М, ниже приблизительно 5×10-5 М, ниже приблизительно 10-5 М, ниже приблизительно 10-4 М или ниже.
[00167] В некоторых вариантах осуществления инфекционный вирион rAAV по настоящему изобретению демонстрирует увеличенное время нахождения in vivo по сравнению с AAV дикого типа. Например, инфекционный вирион rAAV по настоящему изобретению демонстрирует время нахождения, которое составляет, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 100%, по меньшей мере приблизительно в 3 раза, по меньшей мере приблизительно в 5 раз, по меньшей мере приблизительно в 10 раз, по меньшей мере приблизительно в 25 раз, по меньшей мере приблизительно в 50 раз, по меньшей мере приблизительно в 100 раз или более дольше, чем время нахождения AAV дикого типа.
[00168] Насколько заявленный инфекционный вирион rAAV субъекта проявляет пониженное связывание с нейтрализующим антителом и/или повышенную устойчивость к нейтрализующему антителу, можно определить с помощью любого подходящего анализа, известного специалисту в данной области.
[00169] В некоторых вариантах осуществления инфекционный вирион rAAV по настоящему изобретению содержит белки Rep78, Rep68, Rep52 и Rep40 дикого типа. В других вариантах осуществления инфекционный вирион rAAV по настоящему изобретению содержит, в дополнение к одному или более вариантным капсидных белкам, одну или более мутаций в одном или более белках Rep78, Rep68, Rep52 и Rep40.
[00170] Гетерологичные нуклеиновые кислоты
[00171] Подходящей гетерологичной молекулой ДНК (также называемой в настоящем изобретении «гетерологичной нуклеиновой кислотой») для применения в заявленном векторе rAAV (например, заявленном инфекционном вирионе rAAV) может быть любая гетерологичная нуклеиновая кислота. В некоторых вариантах осуществления гетерологичная нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую полипептид (например, белок, который придает клетке-мишени определенные желаемые характеристики, например, флуоресцентный белок, позволяющий анализировать клетки, фермент, который обеспечивает активность, отсутствующую или измененную в клетке-мишени и т. д.). В некоторых вариантах осуществления гетерологичная нуклеиновая кислота содержит агент РНК-интерференции, (как определено выше).
[00172] Заявленная гетерологичная нуклеиновая кислота, как правило, будет иметь размер ниже приблизительно 5 т.п.н. и будет включать, например, ген (нуклеотидную последовательность), который кодирует белок, дефектный или отсутствующий у реципиента или в клетке-мишени; ген, который кодирует белок, обладающий желаемым биологическим или терапевтическим эффектом (например, антибактериальной, противовирусной или противоопухолевой/противораковой активностью); нуклеотидную последовательность, которая кодирует РНК, которая ингибирует или снижает продукцию вредного или иным образом нежелательного белка (например, нуклеотидная последовательность, которая кодирует агент РНК-интерференции, как определено выше); и/или нуклеотидную последовательность, которая кодирует антигенный белок.
[00173] Подходящие гетерологичные нуклеиновые кислоты включают, не ограничиваясь этим, такие, которые кодируют белки, используемые для лечения эндокринных, метаболических, гематологических, сердечно-сосудистых, неврологических заболеваний, болезней костно-мышечной системы, урологических, легочных и иммунных заболеваний, включая такие заболевания, как воспалительные заболевания, аутоиммунные хронические и инфекционные заболевания, такие как синдром приобретенного иммунодефицита (СПИД), рак, гиперхолестеринемия, лизосомные болезни накопления, такие как дефицит активатора/GM2-ганглиозидоз, альфа-маннозидоз, аспаргалглюкозаминурия, болезнь накопления эфиров холестерина, хронический дефицит гексозаминидазы А, цистиноз, болезнь Данона, болезнь Фабри, болезнь Фарбера, фукозидоз, галактосиалидоз, болезнь Гоше, GM1-ганглиозидоз, I-клеточная болезнь/муколипидоз II, инфантильная болезнь накопления свободных сиаловых кислот/ISSD, ювенильная форма дефицит гексозаминидазы А, болезнь Краббе, дефицит лизосомальной кислой липазы, метахроматическая лейкодистрофия, мукополисахаридозы (включая псевдо-полидистрофию Херлер/муколипидоз IIIA, МПС I синдром Гурлера, МПС I синдром Шейе, МПС I синдром Гурлера-Шейе, МПС I II синдром Хантера, синдром Санфилиппо типа A/МПС III A, синдром Санфилиппо типа B/МПС III B, синдром Санфилиппо типа C/МПС III C, синдром Санфилиппо типа D/МПС III D, синдром Моркио типа A/МПС IVA, синдром Моркио типа B/МПС IVB, МПС IX/дефицит гиалуронидазы, МПС VI синдром Марко-Лами, МПС VII синдром Слая, муколипидоз I/сиалидоз, муколипидоз типа IIIC и муколипидоз типа IV), множественный дефицит сульфатазы, болезнь Ниманна-Пика, нейрональные цероидные липофусцинозы, болезнь Помпе/болезнь накопления гликогена типа II, пикнодизостоз, болезнь Сандхоффа с началом во взрослом возрасте/GM2-ганглиозидоз, болезнь Сандхоффа с началом в детском возрасте/GM2-ганглиозидоз, болезнь Сандхоффа с началом в юношеском возрасте/GM2-ганглиозидоз, болезнь Шиндлера, болезнь Салла/болезнь накопления сиаловой кислоты, болезнь Тея-Сакса/GM2-ганглиозидоз и болезнь Вольмана, нарушения инсулина, такие как диабет, нарушения роста, различные заболевания крови, включая различные анемии, талассемию и гемофилию; генетические дефекты, такие как муковисцидоз, болезнь Гоше, синдром Герлера, дефицит аденозиндезаминазы (ADA), эмфизема и т.п.
[00174] Подходящие гетерологичные нуклеиновые кислоты включают, не ограничиваясь этим, такие, которые кодируют любой из множества белков, включая, не ограничиваясь этим: интерферон (например, IFN-γ, IFN-α, IFN-β, IFN-ω; IFN-τ); инсулин (например, новолин, хумулин, хумалог, лантус, ультраленте и т.д.); эритропоэтин («EPO»; например, Procrit®, Eprex® или Epogen® (эпоэтин-α); Aranesp® (дарбэпоэтин-α); NeoRecormon®, Epogin® (эпоэтин-α) и т.п.); антитело (например, моноклональное антитело) (например, Rituxan® (ритуксимаб); Remicade® (инфликсимаб); Herceptin® (трастузумаб); Humira™ (адалимумаб); Xolair® (омализумаб); Bexxar® (тозитумомаб); Raptiva™ (эфализумаб), Erbitux™ (цетуксимаб), Avastin® (бевацизумаб) и т.п.), включая антигенсвязывающий фрагмент моноклонального антитела (например, Lucentis® (ранибизумаб)); фактор крови (например, Activase® (алтеплаза) тканевой активатор плазминогена; NovoSeven® (рекомбинантный человеческий фактор VIIa); фактор VIIa; фактор VIII (например, Kogenate®); фактор IX; β-глобин; гемоглобин и т.п.); колониестимулирующий фактор (например, Neupogen® (филграстим; G-CSF); Neulasta (пегфилграстим); гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарно-моноцитарный колониестимулирующий фактор, макрофагальный колониестимулирующий фактор, мегакариоцитарный колониестимулирующий фактор и тому подобное; гормон роста (например, соматотропин, например, Genotropin®, Nutropin®, Norditropin®, Saizen®, Serostim®, Humatrope® и т.д.; гормон роста человека и тому подобное); интерлейкин (например, IL-1; IL-2, включая, например, Proleukin®; IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9 и т.д.); фактор роста (например, Regranex® (беклапермин; PDGF); Fiblast® (трафермин; bFGF); Stemgen® (анцестим; фактор стволовых клеток); фактор роста кератиноцитов; кислый фактор роста фибробластов, фактор стволовых клеток, основный фактор роста фибробластов, фактор роста гепатоцитов и тому подобное); растворимый рецептор (например, растворимый рецептор, связывающий TNF-α, такой как Enbrel® (этанерцепт); растворимый рецептор VEGF; растворимый рецептор интерлейкина; растворимый рецептор γ/σ Т-клеток и тому подобное); фермент (например, α-глюкозидаза; Cerazyme® (имиглюкараза; β-глюкоцереброзидаза, Ceredase® (алглюцераза); активатор фермента (например, тканевой активатор плазминогена); хемокин (например, IP-10; Mig; Groα/ IL-8, RANTES; MIP-1α; MIP-1β; MCP-1; PF-4 и т.п.); ангиогенный агент (например, фактор роста эндотелия сосудов (VEGF); антиангиогенный агент (например, растворимый рецептор VEGF); белковая вакцина; нейроактивный пептид, такой как брадикинин, холецистокинин, гастин, секретин, окситоцин, гонадотропин-рилизинг-гормон, бета-эндорфин, энкефалин, субстанция Р, соматостатин, пролактин, галанин, гормон роста-рилизинг-гормон, бомбезин, динорфин, нейротензин, мотилин, тиротропин, нейропептид Y, лютеинизирующий гормон, кальцитонин, инсулин, глюкагон, вазопрессин, ангиотензин II, тиреотропин-рилизинг-гормон, вазоактивный интестинальный пептид, дельта-сон индуцирующий пептид и тому подобное; другие белки, такие как тромболитический агент, предсердный натрийуретический пептид, костный морфогенный белок, тромбопоэтин, релаксин, глиальный фибриллярный кислый белок, фолликулостимулирующий гормон, человеческий альфа-1-антитрипсин, фактор, ингибирующий лейкемии, трансформирующий фактор роста, инсулиноподобный фактор роста, лютеинизирующий гормон, фактор активации макрофагов, фактор некроза опухоли, фактор хемотаксиса нейтрофилов, фактор роста нервов, тканевой ингибитор металлопротеиназ; вазоактивный интестинальный пептид, ангиогенин, ангиотропин, фибрин; гирудин; фактор ингибирования лейкемии; антагонист рецептора IL-1 (например, Kineret® (анакинра)); ионный канал, например регулятор трансмембранной проводимости при муковисцидозе (CFTR); дистрофин; атрофин, супрессор опухоли; лизосомальный фермент аналог кислой α-глюкозидазы (GAA) и тому подобное. Подходящие нуклеиновые кислоты также включают такие, которые кодируют функциональный фрагмент любого из вышеуказанных белков; и нуклеиновые кислоты, которые кодируют функциональные варианты любого из вышеуказанных белков.
[00175] Подходящие гетерологичные нуклеиновые кислоты также включают такие, которые кодируют антигенные белки. Заявленный вектор rAAV, содержащий гетерологичную нуклеиновую кислоту, кодирующую антигенный белок, подходит для стимуляции иммунного ответа на антигенный белок у млекопитающего-хозяина. Антигенный белок происходит из аутоантигена, аллергена, опухоль/рак-ассоциированного антигена, патогенного вируса, патогенной бактерии, патогенного простейшего, патогенного гельминта или любого другого патогенного организма, инфицирующего хозяина-млекопитающего. В рамках настоящего изобретения, термин «нуклеиновая кислота, кодирующая антигенный белок, полученный из» включает нуклеиновые кислоты, кодирующие антигенные белки дикого типа, например, нуклеиновую кислоту, выделенную из патогенного вируса, которая кодирует вирусный белок; синтетические нуклеиновые кислоты, полученные в лаборатории, которые кодируют антигенные белки, аминокислотная последовательность которых идентична природному антигенному белку; синтетические нуклеиновые кислоты, полученные в лаборатории, которые кодируют антигенные белки, отличающиеся по аминокислотной последовательности (например, от одной аминокислоты до приблизительно 15 аминокислот) от природного антигенного белка, но, тем не менее, индуцирующие иммунный ответ на соответствующие встречающийся в природе антигенный белок; синтетические нуклеиновые кислоты, полученные в лаборатории, которые кодируют фрагменты антигенных белков (например, фрагменты приблизительно из от 5 аминокислот до приблизительно 50 аминокислот, такие фрагменты содержат один или более антигенных эпитопов), такие фрагменты индуцируют иммунный ответ на соответствующие встречающийся в природе антигенный белок; и т.п.
[00176] Аналогично, антигенный белок, который «происходит из» аутоантигена, аллергена, опухоль/рак-ассоциированного антигена, патогенного вируса, патогенной бактерии, патогенного простейшего, патогенного гельминта или любого другого патогенного организма, инфицирующего млекопитающего-хозяина, включает белки, которые идентичны по аминокислотной последовательности природному антигенному белку, и белки, которые отличаются по аминокислотной последовательности (например, от одной аминокислоты до приблизительно 15 аминокислот) от природного антигенного белка, но которые, тем не менее, индуцируют иммунный ответ на соответствующий встречающийся в природе антигенный белок; и фрагменты антигенных белков (например, фрагменты от приблизительно 5 до приблизительно 100 аминокислот, например, от приблизительно 5 до приблизительно 50 аминокислот, где фрагменты содержат один или более антигенных эпитопов), такие фрагменты индуцируют иммунный ответ на соответствующий природный антигенный белок.
[000177] В некоторых вариантах осуществления иммунный ответ на антигенный белок, кодируемый заявленным вектором rAAV, будет индуцировать защитный иммунный ответ на патогенный организм, который экспонирует антигенный белок или антигенный эпитоп (или белок или эпитоп, который является перекрестно-реактивным) с кодируемым rAAV антигенным белком или антигенными эпитопами) у млекопитающего-хозяина. В некоторых вариантах осуществления у млекопитающего-хозяина индуцируется ответ цитотоксических Т-лимфоцитов (CTL) на антигенный белок, кодируемый rAAV. В еще одних вариантах осуществления у млекопитающего-хозяина индуцируется гуморальный ответ на кодируемый rAAV антигенный белок, и в результате продуцируются антитела, специфичные к антигенному белку. Во многих вариантах осуществления у млекопитающего-хозяина будет индуцироваться TH1 иммунный ответ на антигенный белок, кодируемый rAAV. Подходящие антигенные белки включают опухоль/рак-ассоциированные антигены, вирусные антигены, бактериальные антигены и протозойные антигены; и их антигенные фрагменты. В некоторых вариантах осуществления антигенный белок происходит от внутриклеточного патогена. В других вариантах осуществления антигенный белок представляет собой аутоантиген. В еще одних вариантах осуществления антигенный белок представляет собой аллерген.
[00178] Опухоль/рак-специфические антигены включают, не ограничиваясь этим, любой из различных MAGE (меланома-ассоциированный антиген E), включая MAGE 1 (например, с идентификационным номером GenBank M77481), MAGE 2 (например, с идентификационным номером GenBank U03735), MAGE 3, MAGE 4 и т. д.; любую из различных тирозиназ; мутантный ras; мутантный p53 (например, с идентификационным номером GenBank X54156 и AA494311); и антиген меланомы p97 (например, с идентификационным номером GenBank M12154). Другие опухоль/рак-специфические антигены, включают пептид Ras и пептид p53, ассоциированные с раком на поздних стадиях, антигены HPV 16/18 и E6/E7, ассоциированные с раком шейки матки, антиген MUCI1-KLH, ассоциированный с карциномой молочной железы (например, с идентификационным номером GenBank J03651), CEA (карциноэмбриональный антиген), ассоциированный с колоректальным раком (например, с идентификационным номером GenBank X98311), gp100 (например, с идентификационныым номером GenBank S73003) или антигены MART1, ассоциированные с меланомой, и антиген PSA, ассоциированный с раком предстательной железы (например, с идентификационным номером GenBank X14810). Последовательность гена р53 известна (см., например, Harris et al. (1986) Mol. Cell. Biol., 6:4650-4656) и депонирована в GenBank с идентификационным номером M14694. Таким образом, белки, нуклеиновые кислоты и/или вирионы по настоящему изобретению можно использовать в качестве иммунотерапевтических средств для лечения рака, включая, не ограничиваясь этим, рак шейки матки, рак молочной железы, колоректальный рак, рак предстательной железы, легких и меланому.
[00179] Вирусные антигены происходят от известных возбудителей, ответственных за заболевания, включая, не ограничиваясь этим, корь, эпидемический паротит, краснуху, полиомиелит, гепатит А, В (например, с идентификационным номером GenBank E02707) и C (например, с идентификационным номером GenBank E06890), а также другие вирусы гепатита, вирус гриппа, аденовирусы (например, типы 4 и 7), вирус бешенства (например, с идентификационным номером GenBank M34678), желтой лихорадки, японского энцефалита (например, с идентификационным номером GenBank E07883), лихорадки денге (например, с идентификационным номером GenBank M24444), хантавирус и вирус иммунодефицита человека (например, с идентификационным номером GenBank U18552).
[00180] Подходящие бактериальные и паразитарные антигены включают антигены, происходящие от известных возбудителей, ответственных за заболевания, включая, помимо прочего, дифтерию, коклюш (например, с идентификационным номером GenBank M35274), столбняк (например, с идентификационным номером GenBank M64353), туберкулез, бактериальные и грибковые пневмонии (например, Haemophilus influenzae, Pneumocystis carinii и т. д.), холеру, брюшной тиф, чуму, шигеллез, сальмонеллез (например, с идентификационным номером GenBank L03833), болезнь легионеров, болезнь Лайма (например, с идентификационным номером GenBank U59487), малярию (например, с идентификационным номером GenBank X53832), анкилостомоз, онхоцеркоз (например, с идентификационным номером GenBank M27807), шистосомоз (например, с идентификационным номером GenBank L08198), трипаносомоз, лейшманиоз, лямблиоз (например, с идентификационным номером GenBank M33641), амебиаз, филяриатоз (например, с идентификационным номером GenBank J03266), боррелиоз и трихинеллез.
[00181] Подходящие гетерологичные нуклеиновые кислоты, которые кодируют продукты гетерологичных генов, включают нетранслируемые РНК, такие как РНКи агент (как более подробно описано выше) (например, антисмысловая РНК; миРНК; кшРНК; двухцепочечная РНК (дцРНК)); CRISPR-агент, например, Cas9 или Cas9-подобный белок, crРНК-подобная РНК, tracrRNA-подобная РНК, одиночная направляющая РНК и/или донорный полинуклеотид и т.п.), рибозим и т.д. РНК-и агенты можно использовать для ингибирования экспрессии генов. Некоторые агенты РНК-интерференции предоставляют инструмент, который впоследствии можно использовать для ингибирования экспрессии генов (например, агент CRISPR, такой как cas9 или cas9-подобный белок).
[00182] Целевые гены включают любой ген, кодирующий продукт целевого гена (РНК или белок), который является вредным (например, патологическим), например, продукт целевого гена, который имеет нарушенную функцию (например, за счет мутации в кодированной белковой последовательности, за счет мутации в некодирующих последовательностях, которые контролируют устойчивый уровень продукта гена и т. д.). Продукты целевого гена включают, не ограничиваясь этим, хантигтин; вирус гепатита С; вирус иммунодефицита человека; белок-предшественник амилоида; тау; белок, который содержит полиглутаминовые повторы; вирус герпеса (например, вирус варицелла-зостер); любой патологический вирус; и тому подобное.
[00183] Таким образом, rAAV по настоящему изобретению, который включает гетерологичную нуклеиновую кислоту, кодирующую РНК-и агент, пригоден для лечения различных расстройств и патологических состояний, включая, не ограничиваясь этим, нейродегенеративные заболевания, например, болезни экспансии тринуклеотидных повторов, такое как заболевание, связанное с полиглутаминовыми повторами, например болезнь Хантингтона, спиноцеребеллярную атаксию, спинальную и бульбарную мышечную атрофию (SBMA), дентато-рубро-паллидо-льюисову атрофию (DRPLA) и т.д.; приобретенную патологию (например, заболевание или синдром, проявляющийся аномальным физиологическим, биохимическим, клеточным, структурным или молекулярно-биологическим состоянием), такую как вирусная инфекция, например, гепатит, который возникает или может возникнуть в результате инфекции HCV, приобретенный синдром иммунодефицита, возникающий в результате ВИЧ-инфекции; рак; и тому подобное.
[00184] Во многих вариантах осуществления гетерологичная нуклеиновая кислота, кодирующая РНКи агент, функционально связана с промотором. Подходящие промоторы известны специалистам в данной области и включают промотор любого гена, кодирующего белок, например, эндогенно регулируемого гена или конститутивно экспрессируемого гена. Например, промоторы генов, регулируемых клеточными физиологическими событиями, например, тепловым шоком, уровнями кислорода и/или уровнями окиси углерода, например, при гипоксии, могут быть функционально связаны с нуклеиновой кислотой, кодирующей миРНК.
[00185] Выбранная гетерологичная нуклеотидная последовательность, такая как EPO-кодирующая или представляющая интерес нуклеиновая кислота, функционально связана с регуляторными элементами, которые регулируют транскрипцию или экспрессию нуклеотидной последовательности in vivo. Такие регуляторные элементы могут содержать регуляторные последовательности, обычно связанные с выбранным геном (например, эндогенные клеточные регуляторные элементы). Альтернативно можно использовать гетерологичные регуляторные последовательности. Пригодные гетерологичные регуляторные последовательности обычно включают последовательности, полученные из последовательностей, кодирующих гены млекопитающих или вирусы. Примеры включают, не ограничиваясь этим, ранний промотор SV40, промотор длинного концевого повтора (LTR) вируса опухоли молочной железы мышей; главный поздний промотор аденовируса (Ad MLP); промотор вируса простого герпеса (HSV), эндогенный клеточный промотор, гетерологичный для представляющего интерес гена, промотор цитомегаловируса (CMV), такой как область непосредственно раннего промотора CMV (CMVIE), промотор вируса саркомы Рауса (RSV), синтетический промоторы, гибридные промоторы и т.п. Кроме того, здесь также найдут применение последовательности, полученные из невирусных генов, таких как ген металлотионеина мыши. Такие промоторные последовательности коммерчески доступны, например, от Stratagene (San Diego, Калифорния).
[00186] В некоторых вариантах осуществления клеточно-специфический или тканеспецифический промотор будет функционально связан с гетерологичной нуклеиновой кислотой, кодирующей продукт гетерологичного гена, так что продукт гена продуцируется селективно или предпочтительно в конкретном(ых) типе(ах) клеток или ткань(и). В некоторых вариантах осуществления индуцируемый промотор будет функционально связан с гетерологичной нуклеиновой кислотой.
[00187] Например, мышечноспецифические и индуцибельные промоторы, энхансеры и т.п. можно использовать для доставки генного продукта в мышечную клетку. Такие регуляторные элементы включают, не ограничиваясь этим, элементы, происходящие из семейств генов актина и миозина, например, из семейства генов myoD; фактор связывания миоцит-специфического энхансера MEF-2; регуляторные элементы, полученные из гена скелетного актина человека и гена кардиального актина; элементы последовательности мышечной креатинкиназы и элемент энхансера мышиной креатинкиназы (mCK); регуляторные элемент, происходящие из гена быстрого скелетного тропонина С, гена медленного кардиального тропонина С и гена медленного тропонина I; ядерные факторы, индуцируемые гипоксией; индуцируемые стероидами элементы и промоторы, такие как глюкокортикоидный ответный элемент (GRE); слитый консенсусный элемент для индукции RU486; и элементы, которые обеспечивают регулируемую тетрациклином экспрессию генов.
[00188] Экспрессионный вектор AAV, который содержит представляющую интерес молекулу ДНК (гетерологичную ДНК), связанную ITR AAV, можно сконструировать путем прямой инсерцией выбранной(ых) последовательности(ей) в геном AAV, который имеет основные открытые рамки считывания AAV («ОРС»), вырезанные из них. Другие части генома AAV также могут быть делецированы при условии, что остается достаточная часть ITR для обеспечения функций репликации и упаковки. Такие конструкции могут быть разработаны с использованием методов, хорошо известных в данной области техники. См., например, патент США № 5173414 и 5139941; международные публикации WO 92/01070 (опубликована 23 января 1992 г.) и WO 93/03769 (опубликована 4 марта 1993 г.); Lebkowski et al. (1988) Molec. Cell. Biol., 8:3988-3996; Vincent et al. (1990) Vaccines, 90 (Cold Spring Harbor Laboratory Press); Carter, B. J. (1992) Current Opinion in Biotechnology, 3:533-539; Muzyczka N. (1992) Current Topics in Microbiol. and Immunol. 158:97-129; Kotin, R. M. (1994) Human Gene Therapy, 5:793-801; Shelling and Smith (1994) Gene Therapy, 1:165-169; и Zhou et al. (1994) J. Exp. Med., 179:1867-1875.
[00189] Алтернативно ITR AAV могут быть вырезаны из вирусного генома или из вектора AAV, содержащего такие же и слитые 5' и 3' выбранной конструкции нуклеиновой кислоты, которая присутствует в другом векторе, с использованием любого подходящего метода, известного специалистам а данной области. Например, в одном подходящем подходе используются стандартные методики лигирования, такие как описанные Sambrook et al., см. выше. Например, лигирование можно проводить в 20 мМ Tris-Cl pH 7,5, 10 мМ MgCl2, 10 мМ DTT, 33 мкг/мл BSA, 10-50 мМ NaCl и либо 40 мкМ АТФ, 0,01-0,02 единицы (Вейсса). ДНК-лигаза Т4 при температуре от 0°С до 16°С (для лигирования по «липким концам») или 1 мМ АТФ, 0,3-0,6 единиц (Вейсса) ДНК-лигаза Т4 при 14°С (для лигирования по «тупым концам»). Межмолекулярное лигирование «липких концов» обычно проводят при общей концентрации ДНК 30-100 мкг/мл (общая концевая концентрация 5-100 нМ). Векторы AAV, которые содержат ITR, описаны, например, в патентах США No. № 5 139 941. В частности, в нем описано несколько векторов AAV, которые доступны в Американской коллекции типовых культур («ATCC») под регистрационными номерами 53222, 53223, 53224, 53225 и 53226.
[00190] Кроме того, химерные гены могут быть получены синтетическим путем, чтобы включать последовательности ITR AAV, расположенные на 5' и 3' одной или нескольких выбранных последовательностях нуклеиновой кислоты. Можно использовать предпочтительные кодоны для экспрессии последовательности химерного гена в мышечных клетках млекопитающих. Полная химерная последовательность собирается из перекрывающихся олигонуклеотидов, полученных стандартными методами. См., например, Edge, Nature (1981) 292:756; Nambair et al. Science (1984) 223:1299; Jay et al. J. Biol. Chem. (1984) 259:6311.
[00191] Поучение инфекционных вирионов rAAV по настоящему изобретению
[00192] В качестве введения, обычно используют клетку-хозяин или «клетку-продуцент» для репликации и упаковки вектора rAAV. Такая клетка-продуцент (обычно клетка-хозяин млекопитающего) обычно содержит или модифицирована для включения нескольких различных типов компонентов для продукции rAAV. Первый компонент представляет собой геном вектора рекомбинантного аденоассоциированного вируса (rAAV) (или «провектор rAAV»), который может реплицироваться и упаковываться в векторные частицы упаковочной клеткой-хозяином. Провектор rAAV обычно содержит гетерологичный полинуклеотид (или «трансген»), с помощью которого желательно генетически изменить другую клетку в контексте генной терапии (поскольку может быть эффективно использована упаковка такого трансгена в частицы вектора rAAV) для доставки трансгена в различные клетки млекопитающих). Трансген обычно фланкирован двумя инвертированными концевыми повторами (ITR) AAV, которые содержат последовательности, распознаваемые во время вырезания, репликации и упаковки вектора AAV, а также во время интеграции вектора в геном клетки-хозяина.
[00193] Второй компонент представляет собой вирус-помощник, который может обеспечивать вспомогательные функции для репликации AAV. Хотя обычно используется аденовирус, другие вирусы-помощники также могут быть использованы, как известно в данной области. Альтернативно, необходимые функции вируса-помощника можно выделить генетически из вируса-помощника, и кодирующие гены можно использовать для обеспечения функций вируса-помощника в транс-положении. Векторные элементы AAV и вирус-помощник (или функции вируса-помощника) могут быть введены в клетку-хозяин либо одновременно, либо последовательно в любом порядке.
[00194] Конечными компонентами для продукции AAV, которые должны обеспечиваться в клетке-продуценте, являются «гены упаковки AAV», такие как гены rep и cap AAV, которые обеспечивают белки репликации и инкапсидирования, соответственно. Могут быть предоставлены несколько различных вариантов генов упаковки AAV (включая кассеты rep-cap и отдельные кассеты rep и/или cap, в которых гены rep и/или cap могут находиться под контролем нативных промоторов или функционально связаны с гетерологичными промоторами. Такие гены упаковки AAV могут быть транзиентно или стабильно введены в упаковочную клетку-хозяина, как известно в данной области и более подробно описано ниже.
[00195] 1. Вектор rAAV
[00196] Вирион субъекта rAAV по настоящему изобретению, включающий представляющую интерес гетерологичную ДНК (где «представляющая интерес гетерологичная ДНК» также относится в настоящем изобретении к «гетерологичной нуклеиновой кислоте»), может быть получен с использованием стандартной методики, известной специалистам в данной области. Способы обычно включают следующие стадии: (1) введение вектора rAAV по настоящему изобретению в клетку-хозяин; (2) введение вспомогательной конструкции AAV в клетку-хозяин, где вспомогательная конструкция включает кодирующие области AAV, способные экспрессироваться в клетке-хозяине, чтобы дополнить вспомогательные функции AAV, отсутствующие в векторе AAV; (3) введение одного или более вирусов-помощников и/или векторов со вспомогательными функциями в клетку-хозяин, где вирус-помощник и/или векторы со вспомогательной функцией обеспечивают дополнительные функции, способные обеспечивать эффективную продукцию рекомбинантного вириона AAV («rAAV») в клетке-хозяине; и (4) культивирование клетки-хозяина для получения вирионов rAAV. Экспрессионный вектор AAV, вспомогательная конструкция AAV и вирус-помощник или вектор(ы) со вспомогательной функцией могут быть введены в клетку-хозяин либо одновременно, либо последовательно, с использованием стандартных методов трансфекции.
[00197] Экспрессионные векторы AAV конструируют с использованием известных методов для обеспечения, по меньшей мере оперативно связанных компонентов в направлении транскрипции, регуляторные элементы, включая область инициации транскрипции, представляющую интерес ДНК и область терминации транскрипции. Регуляторные элементы контроля транскрипции выбраны таким образом, чтобы они функционировали в мышечной клетке млекопитающего. Полученная конструкция, которая содержит функционально связанные компоненты, связана (5' и 3') с функциональными последовательностями ITR AAV.
[00198] Нуклеотидные последовательности областей ITR AAV известны. См., например, Kotin, R. M. (1994) Human Gene Therapy 5:793-801; Berns, K. I. «Parvoviridae and their Replication» in Fundamental Virology, 2nd Edition, (B. N. Fields and D. M. Knipe, eds.) для последовательности AAV-2. ITR AAV, используемые в векторах по изобретению, не обязательно должны иметь нуклеотидную последовательность дикого типа, и они могут быть изменены, например, путем инсерции, делеции или замены нуклеотидов. Кроме того, ITR AAV могут быть получены из любого из нескольких серотипов AAV, включая, помимо прочего, AAV-1, AAV-2, AAV-3, AAV-4, AAV-5, AAV-7 и т. д. Кроме того, 5' и 3' ITR, которые фланкируют выбранную нуклеотидную последовательность в экспрессионном векторе AAV, необязательно должны быть идентичными или происходить из одного и того же серотипа или изолята AAV, при условии, что они функционируют, как предполагалось, позволяя вырезать или спасти представляющую интерес последовательность из гена клетки-хозяина или вектора, и обеспечивая возможность интеграции молекулы ДНК в геном клетки-реципиента, когда в клетке присутствуют продукты гена AAV Rep. ITR позволяют векторной последовательности реплицироваться в присутствии соответствующей смеси белков Rep. ITR также позволяют включать векторную последовательность в капсид для генерации частицы AAV.
[00199] Для получения вирионов rAAV экспрессионный вектор AAV вводят в подходящую клетку-хозяин с использованием известных методов, таких как трансфекция. Ряд методов трансфекции общеизвестен в данной области. См., например, Graham et al. (1973) Virology, 52:456, Sambrook et al. (1989) Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratories, New York, Davis et al. (1986) Basic Methods in Molecular Biology, Elsevier, and Chu et al. (1981) Gene 13:197. Особенно подходящие методы трансфекции включают преципитацию фосфатом кальция (Graham et al. (1973) Virol. 52:456-467), прямую микроинъекцию в культивируемые клетки (Capecchi, M.R. (1980) Cell 22:479-488), электропорацию. (Shigekawa et al. (1988) BioTechniques 6:742-751), липосома-опосредованный перенос генов (Mannino et al. (1988) BioTechniques, 6:682-690), липид-опосредованную трансдукцию (Felgner et al. (1987) Proc. Natl.Acad.Sci.USA, 84:7413-7417) и доставку нуклеиновой кислоты с использованием высокоскоростных микрочастиц (Klein et al. (1987) Nature 327:70-73).
[00200] Для целей настоящего изобретения подходящие клетки-хозяева для продукции вирионов rAAV включают микроорганизмы, дрожжевые клетки, клетки насекомых и клетки млекопитающих, которые могут использоваться или использовались в качестве реципиентов гетерологичной молекулы ДНК. Термин включает потомство исходной клетки, которая была трансфектирована. Таким образом, «клетка-хозяин» для продукции вирионов rAAV обычно относится к клетке, которая была трансфектирована экзогенной последовательностью ДНК. Клетки из стабильной клеточной линии человека 293 (легкодоступные, например, из Американской коллекции типовых культур с идентификационным номеров в ATCC CRL1573) используются во многих вариантах осуществления. В частности, линия клеток человека 293 представляет собой линию клеток эмбриональной почки человека, которая была трансформирована фрагментами ДНК аденовируса типа 5 (Graham et al. (1977) J. Gen. Virol., 36:59) и экспрессирует аденовирусные гены E1a и E1b (Aiello et al. (1979) Virology, 94:460). Клеточная линия 293 легко подвергается трансфекции и обеспечивает особенно удобную платформу для получения вирионов rAAV.
[00201] 2. Вспомогательные функции AAV
[00202] Клетки-хозяева, содержащие вышеописанные экспрессионные векторы AAV, должны быть наделены способностью обеспечивать вспомогательные функции AAV для обеспечения репликации и инкапсидирования нуклеотидных последовательностей, фланкированных ITR AAV, для продукции вирионов rAAV. Вспомогательные функции AAV, как правило, представляют собой происходящие от AAV кодирующие последовательности, которые могут экспрессироваться с получением генных продуктов AAV, которые, в свою очередь, функционируют в транс-положении для продуктивной репликации AAV. Вспомогательные функции AAV используются здесь для дополнения необходимых функций AAV, которые отсутствуют в экспрессионных векторах AAV. Таким образом, вспомогательные функции AAV включают одну или обе основные ORF AAV, а именно кодирующие области rep и cap, или их функциональные гомологи. рамках настиящего изобретения, функции cap включают один или более мутантных капсидных белков, где по меньшей мере один капсидный белок содержит по меньшей мере одну мутацию, как описано выше.
[00203] Под термином «кодирующая область rep AAV» подразумевается известная в данной области область генома AAV, которая кодирует белки репликации Rep 78, Rep 68, Rep 52 и Rep 40. Было показано, что эти продукты экспрессии Rep обладают многими функциями, включая распознавание, связывание и никинг ориджина репликации ДНК AAV, активность ДНК-геликазы и модуляцию транскрипции с AAV (или других гетерологичных) промоторов. Продукты экспрессии Rep в совокупности необходимы для репликации генома AAV. Для описания кодирующей области rep AAV см., например, Muzyczka, N. (1992) Current Topics in Microbiol. and Immunol. 158:97-129; и Kotin, R. M. (1994) Human Gene Therapy 5:793-801. Подходящие гомологи области, кодирующей rep AAV, включают ген rep вируса герпеса человека 6 (HHV-6), который также известен как опосредующий репликацию ДНК AAV-2 (Thomson et al. (1994) Virology, 204:304-311).
[00204] Белки cap AAV включают VP1, VP2 и VP3, где по меньшей мере один из VP1, VP2 и VP3 содержит, по меньшей мере одну мутацию, как описано выше.
[00205] Вспомогательные функции AAV вводят в клетку-хозяин путем трансфекции клетки-хозяина вспомогательной конструкцией AAV либо до, либо одновременно с трансфекцией экспрессионного вектора AAV. Таким образом, вспомогательные конструкции AAV используются для обеспечения по меньшей мере транзиентной экспрессии генов rep и/или cap AAV для дополнения отсутствующих функций AAV, которые необходимы для продуктивного инфицирования AAV. Вспомогательные конструкции AAV не имеют ITR AAV и не могут ни реплицироваться, ни упаковываться. Эти конструкции могут находиться в форме плазмиды, фага, транспозона, космиды, вируса или вириона. Описан ряд вспомогательных конструкций AAV, таких как обычно используемые плазмиды pAAV/Ad и pIM29+45, которые кодируют продукты экспрессии как Rep, так и Cap. См., например, Samulski et al. (1989) J. Virol., 63:3822-3828; и McCarty et al. (1991) J. Virol., 65:2936-2945. Описан ряд других векторов, которые кодируют продукты экспрессии Rep и/или Cap. См., например, патент США № 5139941.
[00206] Как экспрессионные векторы AAV, так и вспомогательные конструкции AAV могут быть сконструированы таким образом, чтобы они содержали один или более необязательных селектируемых маркеров. Подходящие маркеры включают гены, которые придают устойчивость или чувствительность к антибиотикам, придают окраску или изменяют антигенные свойства тех клеток, которые были трансфектированы нуклеиновокислотной конструкцией, содержащей селектируемый маркер, при культивировании клеток в соответствующей селективной среде. Несколько селектируемых маркерных генов, которые можно использовать при практическом применении способов по настоящему изобретению, включают ген устойчивости к гигромицину В (кодирующий аминогликозидфосфотрансферазу (АРН)), который обеспечивает селекцию клеток млекопитающих путем придания устойчивости к гигромицину; ген неомицинфосфотрансферазы (кодирующий неомицинфосфотрансферазу), который обеспечивает селекцию клеток млекопитающих путем придания устойчивости к G418; и тому подобное. Другие подходящие маркеры известны специалистам в данной области.
[00207] 3. Дополнительные функции AAV
[00208] Клетка-хозяин (или упаковочная клетка) также должна быть наделена способностью обеспечивать функции, не связанные с AAV, или «дополнительные функции», чтобы продуцировать вирионы rAAV. Дополнительные функции представляют собой вирусные и/или клеточные функции, происходящие не от AAV, но от которых зависит репликация AAV. Таким образом, дополнительные функции включают по меньшей мере такие белки и РНК, отличные от AAV, которые необходимы для репликации AAV, в том числе такие, которые участвуют в активации транскрипции гена AAV, стадия-специфическом сплайсинге мРНК AAV, репликации ДНК AAV, синтезе продуктов экспрессии Cap и сборке капсида AAV. Дополнительные функции, основанные на вирусах, могут быть получены от любого из известных вирусов-помощников.
[00209] В частности, дополнительные функции могут быть введены и затем экспрессированы в клетках-хозяевах с использованием способов, известных специалистам в данной области. Обычно дополнительные функции обеспечиваются инфицированием клеток-хозяев неродственным вирусом-помощником. Известен ряд подходящих вирусов-помощников, включая аденовирусы; вирусы герпеса, такие как вирус простого герпеса типов 1 и 2; и вирусы коровьей оспы. Здесь также найдут применение невирусные дополнительные функции, такие как функции, обеспечиваемые синхронизацией клеток с использованием любого из различных известных агентов. См., например, Buller et al. (1981) J. Virol., 40:241-247; McPherson et al. (1985) Virology, 147:217-222; Schlehofer et al. (1986) Virology, 152:110-117.
[00210] Альтернативно дополнительные функции могут быть обеспечены с использованием вектора с дополнительными функциями. Векторы с дополнительной функций включают нуклеотидные последовательности, которые обеспечивают одну или более дополнительных функций. Вектор с дополнительной функцией можно ввести в подходящую клетку-хозяин для обеспечения эффективной продукции вириона AAV в клетке-хозяине. Векторы с дополнительными функциями могут находиться в форме плазмиды, фага, транспозона, космиды или другого вируса. Векторы с дополнительной функцией также могут находиться в форме одного или более линеаризованных фрагментов ДНК или РНК, которые при связывании с соответствующими регуляторными элементами и ферментами могут транскрибироваться или экспрессироваться в клетке-хозяине для обеспечения дополнительных функций.
[00211] Последовательности нуклеиновых кислот, обеспечивающие дополнительные функции, могут быть получены из природных источников, таких как геном аденовирусной частицы, или сконструированы с использованием рекомбинантных или синтетических способов, известных в данной области. В связи с этим дополнительные функции, происходящие от аденовирусов, были широко изучены, и ряд генов аденовирусов, участвующих в дополнительных функциях, был идентифицирован и частично охарактеризован. См., например, Carter, B. J. (1990) “Adeno-Associated Virus Helper Functions,” in CRC Handbook of Parvoviruses, vol. I (P. Tijssen, ed.), и Muzyczka, N. (1992) Curr. Topics. Microbiol. and Immun. 158:97-129. В частности, полагается, что участки ранних аденовирусных генов E1a, E2a, E4, РНК VAI и, возможно, E1b участвуют в процессе обеспечения дополнительных функций. Janic et al. (1981) Proc. Natl. Acad. Sci. USA, 78:1925-1929. Описаны дополнительные функции вируса герпеса. См., например, Young et al. (1979) Prog. Med. Virol., 25:113. Также были описаны дополнительные функции, происходящие от вируса коровьей оспы. См., например, Carter, B.J. (1990), выше, Schlehofer et al. (1986) Virology, 152:110-117.
[00212] В результате инфицирования клетки-хозяина вирусом-помощником или трансфекции клетки-хозяина вектором с дополнительной функцией экспрессируются дополнительные функции, которые трансактивируют вспомогательную конструкцию AAV с образованием белков Rep и/или Cap AAV. Продукты экспрессии Rep вырезают рекомбинантную ДНК (включая представляющую интерес ДНК, например, гетерологичную нуклеиновую кислоту) из экспрессионного вектора AAV. Белки Rep также служат для копирования генома AAV. Экспрессированные белки Cap собираются в капсиды, и рекомбинантный геном AAV упаковывается в капсиды. Таким образом, происходит продуктивная репликация AAV, и ДНК упаковывается в вирионы rAAV.
[00213] После репликации рекомбинантного AAV вирионы rAAV можно выделить из клетки-хозяина с использованием различных традиционных способов очистки, таких как очистка в градиенте CsCl, аффинная хроматография и ионообменная хроматография. Кроме того, если для обеспечения дополнительных функций используется инфицирование, то остаточный вирус-помощник можно инактивировать с использованием известных способов. Например, аденовирус можно инактивировать нагреванием до температуры приблизительно 60°С в течение, например, 20 мин или более. Такая обработка эффективно инактивирует только вирус-помощник, поскольку AAV является очень термостабильным, а аденовирус-помощник является термолабильным.
[00214] Полученные вирионы rAAV затем готовы к использованию для доставки ДНК, например, в приложениях генной терапии или для доставки генного продукта хозяину-млекопитающему.
[00215] Доставка гетерологичной нуклеиновой кислоты
[00216] Настоящее изобретение также относится к способам доставки гетерологичной нуклеиновой кислоты в клетку-мишень и/или субъекту, нуждающемуся в этом. В некоторых вариантах осуществления субъект, нуждающийся в этом, представляет собой человека, который ранее подвергался естественному воздействию AAV и, как следствие, содержит анти-AAV антитела (т.е. нейтрализующие антитела к AAV). Основываясь на положительных результатах клинических испытаний, включающих доставку гена AAV, например, в печень, мышцы и сетчатку - все ткани, содержащие нейтрализующие антитела против этого носителя, - существует множество таких терапевтических применений/мишеней для лечения заболеваний.
[00217] Способ по настоящему изобретению обычно включает: (i) введение эффективного количества заявленного вириона rAAV субъекту и/или (ii) приведение клетки-мишени в контакт с заявленным вирионом. Как правило, вирионы rAAV вводят субъекту с использованием методов трансдукции либо in vivo («прямая»), либо in vitro («опосредованно»). При трансдукции in vitro («опосредованно») желаемая клетка-реципиент (т.е. «клетка-мишень») может быть выделена из организма субъекта, трансдуцирована вирионами rAAV и повторно введена субъекту. В качестве альтернативы можно использовать сингенные или ксеногенные клетки, если только эти клетки не индуцируют неадекватного иммунного ответа у субъекта.
[00218] Описаны подходящие способы доставки и введения трансдуцированных клеток-мишеней субъекту. Например, клетки можно трансдуцировать in vitro путем объедирнения рекомбинантных вирионов AAV с клетками, например, в соответствующих средах, и скрининга тех клеток, которые содержат представляющую интерес ДНК, с использованием обычных методов, таких как саузерн-блоттинг и/или ПЦР, или с использованием селектируемых маркеров. Затем трансдуцированные клетки можно формулировать в виде фармацевтических композиций, более подробно описанных ниже, и композиция может быть введена субъекту различными методами, такими как внутримышечная, внутривенная, подкожная и внутрибрюшинная инъекция.
[00219] Для доставки in vivo (т. е. «прямой») вирионы rAAV формулируют в виде фармацевтических композиций и, как правило, вводят парентеральным путем введения (например, путем внутримышечного, подкожного, внутриопухолевого, чрескожного, интратекального, внутривенного и т.д.).
[00220] Фармацевтические композиции должны содержать достаточное количество генетического материала для получения терапевтически эффективного количества представляющего интерес продукта экспрессии гена, т.е. количества, достаточного для подавления или облегчения симптомов указанного болезненного состояния, или количества, достаточного для получения желаемого полодительного эффекта. Фармацевтические композиции также будут содержать фармацевтически приемлемый эксципиент. Такие эксципиенты включают любой фармацевтический агент, который сам по себе не индуцирует выработку антител, вредных для субъекта, получающего композицию, и который можно вводить без проявления чрезмерной токсичности. Фармацевтически приемлемые эксципиенты включают, не ограничиваясь этим, жидкости, такие как вода, физиологический раствор, глицерин и этанол. Сюда могут быть включены фармацевтически приемлемые соли, например, соли неорганических кислот, такие как гидрохлориды, гидробромиды, фосфаты, сульфаты и т.п.; и соли органических кислот, такие как ацетаты, пропионаты, малонаты, бензоаты и т.п. Кроме того, в таких носителях могут присутствовать вспомогательные вещества, такие как смачивающие или эмульгирующие вещества, вещества, регулирующие рН, и т.п. В данной области техники известно большое разнообразие фармацевтически приемлемых эксципиентов, и нет необходимости подробно здесь их обсуждать. Фармацевтически приемлемые эксципиенты подробно описаны в различных публикациях, включая, например, A. Gennaro (2000) «Remington: The Science and Practice of Pharmacy,» 20th edition, Lippincott, Williams, & Wilkins Pharmaceutical Dosage Forms and Drug Delivery Systems (1999) H. C. Ansel et al., eds., 7th ed., Lippincott, Williams, & Wilkins и Handbook of Pharmaceutical Excipients (2000) A. H. Kibbe et al., eds., 3rd ed. Amer. Pharmaceutical Assoc.
[00221] Соответствующие дозы будут зависеть от млекопитающего, который подвергается лечению (например, человека или примата, отличного от человека, или другого млекопитающего), возраста и общего состояния здоровья субъекта, подлежащего лечению, тяжести патологического состояния, подлежащего лечению, конкретного рассматриваемого терапевтического белка, способ его введения, среди прочих других факторов. Подходящее эффективное количество может легко определить специалист в данной области.
[00222] Таким образом, «терапевтически эффективное количество» будет находиться в относительно широком диапазоне, который может быть определен в клинических испытаниях. Например, для инъекции in vivo, т.е. инъекции непосредственно в скелетную или сердечную мышцу, терапевтически эффективная доза будет составлять порядка от приблизительно 106 до приблизительно 1015 вирионов rAAV, например, от приблизительно 108 до 1012 вирионов rAAV. Для трансдукции in vitro эффективное количество вирионов rAAV, которые должны быть доставлены в клетки, будет находиться в диапазоне от приблизительно 108 до приблизительно 1013 вирионов rAAV. Другие эффективные дозировки могут легко установить специалисты в данной области посредством стандартных испытаний, устанавливающих кривые доза-эффект.
[00223] Лечение введением доз может представлять собой схему с однократным введением дозы или схему с многократным введением дозы. Более того, субъекту можно вводить столько доз, сколько это необходимо. Специалист в данной области может легко определить подходящее количество доз.
[00224] Представляющие интерес клетки (т.е. «клетки-мишени») обычно представляют собой клетки млекопитающих, где термин относится к любому животному, классифицируемому как млекопитающее, включая человека, домашних и сельскохозяйственных животных, а также зоопарковых животных, лабораторных животных, спортивных или домашних животных, таких как собаки, лошади, кошки, коровы, мыши, крысы, кролики и т.д. В некоторых вариантах осуществления клетка-мишень представляет собой клетку человека.
[00225] Представляющие интерес клетки-мишени включают любые клетки, восприимчивые к инфицированию вирионом rAAV по настоящему изобретению. В некоторых случаях, например, когда способ представляет собой способ доставки гетерологичной нуклеиновой кислоты в клетку-мишень, где клетка-мишень может представлять собой клетку, отобранную у субъекта (например, «первичную» клетку), или клетка-мишень может быть тканевой культуральной клеткой (например, из хорошо известной клеточной линии).
[00226] Примеры клеток-мишеней включают, не ограничиваясь этим, клетки печени, клетки поджелудочной железы (например, клетки островков Лангерганса: альфа-клетки, бета-клетки, дельта-клетки, гамма-клетки и/или эпсилон-клетки), скелеьные мышечные клетки, клетки сердечной мышцы, фибробласты, ретинальные клетки, синовиальные клетки суставов, клетки легких, Т-клетки, нейроны, глиальные клетки, стволовые клетки, гемопоэтические клетки-предшественники, нейральные клетки-предшественники, эндотелиальные клетки и опухолевые клетки. Типичные стволовые клетки-мишени включают, не ограничиваются этим, гемопоэтические стволовые клетки, стволовые клетки нервного гребня, эмбриональные стволовые клетки, индуцированные плюрипотентные стволовые клетки (iPS), мезенхимальные стволовые клетки, мезодермальные стволовые клетки, стволовые клетки печени, стволовые клетки поджелудочной железы, мышечные стволовые клетки и ретинальные стволовые клетки.
[00227] Термин «стволовая клетка» используется в настоящем изобретении для обозначения клетки млекопитающего, которая обладает способностью как к самообновлению, так и к генерации дифференцированного потомства (см., например, Morrison et al. (1997) Cell, 88:287- 298). Как правило, стволовые клетки также обладают одним или более из следующих свойств: способностью к асинхронной или симметричной репликации, когда две дочерние клетки после деления могут иметь разные фенотипы; высокой способностью к самообновлению; способностью к существованию в митотически покоящейся форме; и к клональной регенерации всей ткани, в которой они находятся, например, способность гемопоэтических стволовых клеток восстанавливать все гемопоэтические линии. Как понятно специалистам в данной области, «клетки-предшественники» отличаются от стволовых клеток тем, что обычно они не обладают высокой способностью к самообновлению и часто могут генерировать более ограниченное подмножество клонов в ткани, из которой они происходят, например, только из лимфоидных или эритроидных клонов в гематопоэтическом окружении. В рамках настоящего изобретения, термин «стволовая клетка» охватывает как «стволовые клетки», так и «клетки-предшественники», которые имеют значения, определенные выше.
[00228] Стволовые клетки могут характеризоваться как наличием маркеров, ассоциированных со специфическими эпитопами, идентифицируемыми с помощью антител, так и отсутствием определенных маркеров, что определяется по отсутствию связывания специфическими антителами. Стволовые клетки также можно идентифицировать с помощью функциональных анализов как in vitro, так и in vivo, в частности, анализов, касающихся способности стволовых клеток давать многочисленное дифференцированное потомство.
[00229] Подходящие представляющие интерес стволовые клетки включают, не ограничиваясь этим: гемопоэтические стволовые клетки и полученные из них клетки-предшественники (патент США № 5061620); стволовые клетки нервного гребня (см. Morrison et al. (1999) Cell, 96:737-749); нейральные стволовые клетки и нейральные клетки-предшественники; эмбриональные стволовые клетки; мезенхимальные стволовые клетки; мезодермальные стволовые клетки; стволовые клетки печени, мышечные стволовые клетки, ретинальные стволовые клетки, индуцированные плюрипотентные стволовые клетки (iPS-клетки) и т. д. Другие представляющие интерес гемопоэтические клетки-предшественники включают клетки, характерные для лимфоидных линий, например незрелые популяции Т-клеток и В-клеток.
[00230] Можно использовать очищенные популяции стволовых клеток или клеток-предшественников. Например, гемопоэтические стволовые клетки человека могут подвергнуться положительной селекции с использованием антител, специфичных к CD34, thy-1; или могут подвергнуться отрицательной селекции с использованием специфичных для линии дифференцировки маркеров, которые могут включать гликофорин А, CD3, CD24, CD16, CD14, CD38, CD45RA, CD36, CD2, CD19, CD56, CD66a и CD66b; Т-клеточные маркеры, опухоль/рак-специфичные маркеры и т. д. Маркеры, пригодные для выделения мезодермальных стволовых клеток, включают FcγRII, FcγRIII, Thy-1, CD44, VLA-4a, LFA-113, HSA, ICAM-1, CD45, Aa4.1, Sca-1 и т. д. Стволовые клетки нервного гребня можно подвергнуть положительной селекции с использованием антител, специфичных к рецептору фактора роста нервов с низким сродством (LNGFR), и подвергнуть отрицательной селекции по маркерам сульфатида, глиального фибриллярного кислого белка (GFAP)), миелинового белка Ро, периферина и нейрофиламента. Мезенхимальные стволовые клетки человека можно выделить положительной селекцией с использованием маркеров SH2, SH3 и SH4.
[00231] Используемые клетки-мишени могут быть свежевыделенными, замороженными или подвергнутыми предварительному культивированию. Они могут представлять собой клетки эмбрионов, новорожденных и взрослых. Гемопоэтические клетки могут быть получены из фетальной печени, костного мозга, крови, в частности, из мобилизованного G-CSF или GM-CSF периферической крови, или из любого другого обычного источника. Способ, которым стволовые клетки отделяют от других клеток гемапоэтической или другой линии, не имеет решающего значения для настоящего раскрытия. Как описано выше, по существу однородная популяция стволовых клеток или клеток-предшественников может быть получена путем селективного выделения клеток, свободных от маркеров, связанных с дифференцированными клетками, при этом демонстрирующих эпитопные характеристики, связанные со стволовыми клетками.
[00232] Нуклеиновые кислоты, которые могут быть доставлены субъекту, включают любую из вышеуказанных гетерологичных нуклеиновых кислот. Белки, которые можно доставить с использованием заявленного способа, также включают функциональный фрагмент любого из вышеуказанных белков; и функциональные варианты любого из вышеуказанных белков.
[00233] В некоторых вариантах осуществления терапевтически эффективное количество белка продуцируется у хозяина-млекопитающего. Насколько имеет место продукция терапевтически эффективного количества определенного белка у хозяина-млекопитающего с использованием заявленного способа, легко определить с помощью анализов, соответствующих определенному белку. Например, если белок представляет собой EPO, то измеряют гематокрит.
[00234] Если rAAV кодирует антигенный белок, то подходящие антигенные белки, которые могут быть доставлены субъекту с использованием способа по настоящему изобретению, включают, не ограничиваясь этим, опухоль/рак-ассоциированные антигены, аутоантигены («собственные» антигены), вирусные антигены, бактериальные антигены, протозойные антигены и аллергены; и их антигенные фрагменты. В некоторых вариантах осуществления у хозяина-млекопитающего индуцируется ответ цитотоксических Т-лимфоцитов (CTL) на антигенный белок, кодируемый rAAV. В других вариантах осуществления у хозяина-млекопитающего индуцируется гуморальный ответ на кодируемый rAAV антигенный белок, так что продуцируются антитела, специфичные к антигенному белку. Во многих вариантах осуществления у хозяина-млекопитающего будет индуцироваться TH1 иммунный ответ на антигенный белок, кодируемый rAAV. Насколько развился иммунный ответ на антигенный белок, легко определить с помощью хорошо известных методов. Например, твердофазный иммуноферментный анализ можно использовать для определения того, образовалось ли антитело к антигенному белку. Способы детектирования антигенспецифических CTL хорошо известны в данной области. Например, детектируемо меченую клетку-мишень, экспрессирующую антигенный белок на своей поверхности, используют для анализа на наличие антигенспецифических CTL в образце крови.
[00235] Насколько терапевтически эффективное количество гетерологичной нуклеиновой кислоты (например, нуклеиновой кислоты, кодирующей полипептид, агент для РНК-интерференции и т. д.) доставлено хозяину-млекопитающему с использованием заявленного способа, легко определить с использованием любого подходящего анализа. Например, если продукт гена представляет собой РНКи агент, который ингибирует ВИЧ, можно определить вирусную нагрузку.
[00236] Способы создания и идентификации модифицированных вирионов rAAV
[00237] Настоящее изобретение относится к способу создания и идентификации модифицированного инфекционного вириона рекомбинантного аденоассоциированного вируса (rAAV), который содержит вариантный капсидный белок, содержащий аминокислотную последовательность по меньшей мере с одной аминокислотной заменой (включая делеции, инсерции и т. д.) в сравнении с исходным капсидным белком AAV. Исходный капсидный белок AAV содержит аминокислотную последовательность, представленную в одной из SEQ ID NO: 10-13 и 26-33.
[00238] Способ обычно включает создание библиотеки мутантных вирионов rAAV; и селекцию библиотеки на модифицированные вирионы rAAV с измененными свойствами по сравнению с исходным вирионом rAAV. Исходный вирион rAAV содержит вариантный капсидный белок AAV, который содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33. Настоящее раскрытие дополнительно относится к библиотеке и композициям, содержащим библиотеки.
[00239] В некоторых вариантах осуществления указанную стадию селекции повторяют два, три, четыре или более раз для обогащения рассматриваемой библиотеки AAV измененными свойствами вириона. В некоторых вариантах осуществления после выбора библиотеки AAV выделяют и секвенируют отдельные клоны.
[00240] Создание библиотеки мутантных AAV
[00241] Создают библиотеку мутантных AAV, которая содержит одну или более мутаций относительно исходного гена cap AAV. Исходный гена cap представляет собой cap, содержащий нуклеотидную последовательность, которая кодирует вариантный капсидный белок AAV, содержащий аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33. Мутации в гене cap rAAV генерируют любым известным способом. Подходящие способы мутагенеза исходного гена cap AAV включают, не ограничиваются этим, метод, основанный на полимеразной цепной реакции (ПЦР), олтгонуклеотид-направленный мутагенез, мутагенез с насыщением, мутагенез с заменой петель, мутагенез с шаффлингом фрагментов (ДНК-шаффлинг и т.п.) и тому подобное. Способы получения мутаций хорошо описаны в данной области. См., например, Zhao et al. Nat. Biotechnol., 1998 March; 16(3):234-5; Koerber et. al.; Mol. Ther., 2008 October; 16(10):1703-9; Koerber et. al.; Mol. Ther., 2009 December; 17(12):2088-95; патент США № 6579678; патент США № 6573098; и патент США № 6582914; которые все включены в данный документ посредством ссылки в отношении идей изобретения, относящихся к мутагенезу.
[00242] В некоторых вариантах осуществления библиотека мутантных AAV, содержащая мутации в гене cap, будет создана с использованием процесса постадийного удлинения. Процесс постадийного удлинения включает амплификацию гена cap с использованием метода, основанного на ПЦР. Матрица гена cap праймируется с использованием специфических праймеров для ПЦР с последующими повторными циклами денатурации и очень коротким отжигом/катализируемым полимеразой удлинением. В каждом цикле растущие фрагменты отжигаются с разными матрицами на основе комплементарности последовательностей и удлиняются далее. Циклы денатурации, отжига и удлинения повторяются до образования полноразмерных последовательностей. Полученные полноразмерные последовательности включают по меньшей мере одну мутацию в гене cap в сравнении с геном cap AAV дикого типа.
[00243] Продукты ПЦР, содержащие последовательности cap AAV, которые включают одну или более мутаций, встраивают в плазмиду, содержащую геном AAV дикого типа. Результатом является библиотека мутантов cap AAV. Таким образом, настоящее изобретение относится к библиотеке мутантных генов cap AAV, содержащей от приблизительно 10 до приблизительно 1010 членов и включающей мутации в гене cap AAV. Данный член библиотеки имеет от приблизительно одной до приблизительно 50 мутаций в гене cap AAV. Библиотека по настоящему изобретению включает от приблизительно 10 до приблизительно 109 различных членов, каждый из которых имеет различную(ые) мутацию(и) в гене cap AAV.
[00244] Как только библиотека мутантов cap создана, то продуцируются вирусные частицы, которые затем можно селектировать на основе измененных свойств капсида. Библиотеку плазмидной ДНК трансфектируют в подходящую клетку-хозяина (например, клетки 293) с последующим введением в клетку вируса-помощника. Собирают вирусные частицы, продуцированные трансфектированными клетками-хозяевами (частицы библиотеки rAAV).
[00245] Селекция библиотеки
[00246] После создания библиотеки ее селектируют по определенному свойству вириона (т.е. измененному свойству инфицирования). Вирусные частицы генерируются, как обсуждалось выше (таким образом создается библиотека модифицированных вирионов rAAV) и подвергаются одной или более стадиям селекции для идентификации модифицированного вириона rAAV с измененным свойством инфицирования (по сравнению с инфекционным вирионом rAAV, содержащим вариантный капсидный белок), который содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33). Свойства инфицирования, по которым проходила селекция, могут включать, не ограничиваясь этим: 1) измененное связывание (например, пониженное связывание) с нейтрализующими анти-AAV антителами; 2) повышенное уклонение от нейтрализующих анти-ААВ антител; 3) повышенная инфекционность для клетки, устойчивой к заражению ААВ; и 4) изменение связывания гепарина.
[00247] 1. Селекция на пониженное связывание с нейтрализующими анти-AAV антителами
[00248] В некоторых вариантах осуществления библиотеку AAV по настоящему изобретению селектируют на измененное (например, сниженное) связывание с нейтрализующими антителами, которые связываются и нейтрализуют вирионы AAV дикого типа, по сравнению со связыванием таких антител с вирионами AAV дикого типа и нейтрализацией вирионов AAV дикого типа (или относительно инфекционного вириона rAAV, содержащего вариантный капсидный белок, который содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33). Частицы библиотеки AAV (вирион библиотеки AAV) подвергают контактированию с нейтрализующими антителами и тестируют на способность частиц библиотеки AAV инфицировать пермиссивную клетку-хозяина. Обычно частицы библиотеки AAV подвергают контактированию с различными концентрациями нейтрализующих антител. Чем выше концентрация нейтрализующих антител, необходимая для снижения инфекционности частиц библиотеки AAV, тем более устойчивы частицы AAV к нейтрализации. Любой удобный анализ, известный специалисту в данной области, можно использовать для непосредственного измерения связывания (например, измерения аффинности связывания) вириона библиотеки AAV с нейтрализующими анти-AAV антителами.
[00249] 2. Селекция на повышенное уклонение от нейтрализующих анти-AAV антител
[00250] В некоторых вариантах осуществления библиотеку AAV по настоящему изобретению селектируют на повышенную способность уклоняться от нейтрализующих антител (т.е. повышенную устойчивость к нейтрализующим анти-AAV антителам человека) по сравнению с инфекционным вирионом rAAV, содержащим вариантный капсидный белок, который содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33. Частицы библиотеки AAV подвергают контактированию с клетками-мишенями в присутствии нейтрализующих антител к AAV (обычно нейтрализующих анти-AAV человека). По прошествии подходящего периода времени, необходимого для инфицирования клеток частицами библиотеки AAV, добавляют вирус-помощник и собирают частицы библиотеки AAV, которые успешно инфицировали клетку (клетки). В некоторых вариантах осуществления инфекционность измеряют (например, как описано выше) для тех вирионов, которые проявляют успешное инфицирование. В некоторых вариантах осуществления цикл инфицирования, добавления вируса-помощника и сбора частиц AAV повторяют один, два, три или более раз. Селекцию может иметь место с различными количествами (концентрациями) нейтрализующих антител к AAV для отбора различных степеней уклонения (например, в каждом повторном раунде может использоваться более высокая концентрация антител по сравнению с предыдущим раундом).
[00251] 3. Селекция на повышенную инфекционность для непермиссивных клеток
[00252] В некоторых вариантах осуществления библиотеку AAV по настоящему изобретению селектируют на повышенную инфекционность для непермиссивных клеток (по сравнению с инфекционным вирионом rAAV, содержащим вариантный капсидный белок, который содержит аминокислотную последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33). Частицы библиотеки AAV подвергают контактированию с непермиссивной клеткой (например, популяцией непермиссивных клеток). По прошествии подходящего периода времени, необходимого для инфицирования клеток частицами библиотеки AAV, добавляют вирус-помощник и собирают частицы библиотеки AAV, которые успешно инфицировали непермиссивную(ые) клетку(и). В некоторых вариантах осуществления цикл инфицирования, добавления вируса-помощника и сбора частиц AAV повторяют один, два, три или более раз.
[0253] 4. Селекция на измененное связывание с гепарином
[00254] В некоторых вариантах осуществления библиотеку селектируют на измененное связывание с гепарином, включая повышенное связывание с гепарином и пониженное связывание с гепарином по сравнению со связыванием гепарина вириона AAV дикого типа (или по сравнению с инфекционным вирионом rAAV, содержащим вариантный капсидный белок, который содержит аминокислотная последовательность, показанную в одной из SEQ ID NO: 10-13 и 26-33). Частицы библиотеки AAV подвергают контактированию с аффинной матрицей с гепарином. Например, частицы библиотеки AAV загружают на колонку для аффинной хроматографии с гепарином в условиях, которые обеспечивают связывание частиц библиотеки AAV с гепарином. Типичные условия включают уравновешивание колонки 0,15 М NaCl и 50 мМ Трис при рН 7,5. После предоставления частице библиотеки AAV возможности связываться с аффинной матрицей с гепарином комплекс частица библиотеки AAV/гепарин на аффинной матрице промывают объемами буфера, содержащего постепенно возрастающие концентрации NaCl, и при каждой концентрации NaCl собирают элюированные частицы библиотеки AAV. Например, после связывания комплекса частицы библиотеки AAV/гепарин на аффинной матрице промывают объемом 50 мМ Трис-буфера, pH 7,5, содержащим 200 мМ NaCl, и собирают элюированные частицы библиотеки AAV. Стадию элюирования повторяют с 50 мМ Трис-буфером, рН 7,5, содержащим приблизительно 250 мМ NaCl, приблизительно 300 мМ NaCl, приблизительно 350 мМ, приблизительно 400 мМ NaCl, приблизительно 450 мМ NaCl, приблизительно 500 мМ NaCl, приблизительно 550 мМ NaCl, приблизительно 600 мМ NaCl, приблизительно 650 мМ NaCl, приблизительно 700 мМ NaCl или приблизительно 750 мМ NaCl.
[00255] Частицы библиотеки AAV, которые элюируются при концентрациях NaCl ниже приблизительно 450 мМ NaCl, проявляют пониженные свойства связывания с гепарином по сравнению с AAV дикого типа. Частицы библиотеки AAV, которые элюируются при концентрациях NaCl выше приблизительно 550 мМ NaCl, проявляют повышенные свойства связывания с гепарином по сравнению с AAV дикого типа.
[00256] В некоторых вариантах осуществления элюированные частицы библиотеки AAV амплифицируют путем коинфицирования пермиссивных клеток вирусом-помощником и рефракционируют на аффинной матрице с гепарином. Эту стадию можно повторить несколько раз, чтобы обогатить частицы библиотеки AAV с измененными свойствами в отношении связывания с гепарином.
[00257] В способах по настоящему одна или более стадий селекции могут следовать за созданием частиц библиотеки AAV. Например, в некоторых вариантах осуществления способ включает селекцию на повышенное связывание с гепарином с последующей селекцией на пониженное связывание с нейтрализующими антителами. В других вариантах осуществления способ включает селекцию на пониженное связывание с нейтрализующими антителами с последующй селекцией на повышенное связывание с гепарином. В еще одних вариантах осуществления способ включает селекцию на пониженное связывание с гепарином с последующей селекцией на пониженное связывание с нейтрализующими антителами. В еще одних вариантах осуществления способ включает селекцию на пониженное связывание с нейтрализующими антителами с последующей селекцией на пониженноое связывание с гепарином. В еще одних вариантах осуществления способ включает селекцию на пониженное связывание с нейтрализующими антителами с последующей селекцией на повышенную инфекционность для стволовой клетки. В еще одних вариантах осуществления способ включает селекцию на пониженное связывание с нейтрализующими антителами с последующей селекцией на повышенное уклонению от нейтрализующих антител. В еще одних вариантах осуществления способ включает селекцию на повышенное уклонение от нейтрализующих антител с последующей селекцией на пониженное связывание с нейтрализующими антителами.
[00258] Таким образом, настоящее изобретение относится к библиотеке аденоассоциированных вирусов (AAV), включающей множество нуклеиновых кислот, каждая из которых включает нуклеотидную последовательность, кодирующую вариантный капсидный белок AAV. Кодируемый вариантный капсидный белок AAV включает по меньшей мере одну аминокислотную замену относительно последовательности, показанной в одной из SEQ ID NO: 10-13 и 26-33. Настоящее изобретение относится к библиотеке частиц мутантного аденоассоциированного вируса (AAV), включающей множество частиц AAV, каждая из которых включает капсидный белок AAV, который содержит по меньшей мере одну аминокислотную замену относительно последовательности, показанной в одной из SEQ ID NO: 10-13 и 26-33. Нуклеиновые кислоты, кодирующие мутантные капсидные белки AAV, описаны выше, как и свойства кодированных мутантных капсидных белков AAV.
[00259] Настоящее изобретение также относится к библиотеке, включающей по меньшей мере одно из следующего: (i) два или более инфекционных вирионов rAAV, каждый из которых содержит вариантный капсидный белок аденоассоциированного вируса (AAV) и гетерологичную нуклеиновую кислоту; (ii) две или более выделенных нуклеиновых кислот, каждая из которых содержит нуклеотидную последовательность, кодирующую вариантный капсидный белок AAV; (iii) две или более клеток-хозяев, каждая из которых содержит нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую вариантный капсидный белок AAV; и (iv) два или более вариантных капсидных белков AAV; где вариантный капсидный белок AAV по меньшей мере одного члена библиотеки содержит аминокислотную последовательность, имеющую по меньшей мере одну аминокислотную замену относительно аминокислотной последовательности, показанной в одной из SEQ ID NO: 10-13 и 26-33.
[00260] Композиции и наборы
[00261] Также настоящее изобретение относится к композициям и наборам для применения в способах по настоящему изобретению. Композиции и наборы по настоящему изобретению включают по меньшей мере одно из следующего: заявленный инфекционный вирион rAAV, заявленный вектор rAAV, заявленную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую заявленный вариантный капсидный белок AAV, выделенную клетку-хозяин, содержащую заявленную нуклеиновую кислоту (т.е. заявленную генетически модифицированную клетку-хозяин, содержащую нуклеиновую кислоту, которая содержит нуклеотидную последовательность, кодирующую заявленный вариантный капсидный белок AAV); заявленную библиотеку (например, любую из вышеописанных библиотек); и заявленный вариантный капсидный белок AAV. Композиция или набор могут включать любую удобную комбинацию вышеуказанного. Композиция или набор также могут включать вирус-помощник и/или нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая кодирует вирус-помощник. Набор также может включать реагенты для получения нуклеиновых кислот (т. е. «мутантных» нуклеиновых кислот), кодирующих модифицированные вариантные капсидные белки AAV.
[00262] В дополнение к вышеуказанным компонентам наборы по настоящему изобретению могут дополнительно включать (в некоторых вариантах осуществления) инструкции по применению способов по настоящему изобретению. Эти инструкции могут находиться в заявленных наборах в различных формах, одна или несколько из которых могут находиться в наборе. Одна из форм, в которой могут находиться эти инструкции, представляет собой печатную информацию на подходящем носителе или подложке, например, листе или листах бумаги, на которых напечатана информация, в упаковке набора, вкладыше в упаковку и т.п. Еще одной формой этих инструкций является машиночитаемый носитель, например дискета, компакт-диск (CD), флэш-накопитель и т.п., на котором записана информация. Еще одна форма этих инструкций, которая может присутствовать, представляет собой адрес веб-сайта, который может использоваться через Интернет для доступа к информации на удаленном сайте.
[00263] Теперь, когда изобретение полностью описано, специалисту в данной области будет очевидно, что различные изменения и модификации могут быть сделаны без отклонения от сущности или объема изобретения.
Примеры
[00264] Пример 1
[00265] К настоящему времени векторы для генной терапии на основе аденоассоциированного вируса (AAV) продемонстрировали многообещающие результаты в нескольких клинических испытаниях. Однако циркулирующие анти-AAV антитела, продуцированные в результате экспозиции в детстве или предшествующего введения вектора AAV, препятствуют проведению генной терапии AAV для многих потенциальных пациентов. Авторы изобретения выделили новые варианты AAV, которые способны к повышенному уклонению от анти-AAV антител, как in vitro, так и in vivo. Жесткое давление, возникающее в результате селекции с использованием пулов сывороток человека с низкой и высокой эффективностью и IVIG человека, привело к появлению вариантов AAV, способных уклоняться от нейтрализации антителами из индивидуальных сывороток человека, IVIG человека и мышиных сывороток, представляющих собой наиболее широко уклоняющиеся варианты на сегодняшний день.
[00266] Материалы и методы
[00267] Клеточные линии
[00268] Клеточные линии культивировали при 37°C и 5% CO2 и, если не указано иное, то их получали из Американской коллекции типовых культур (Manassas, VA). Клетки HEK293T, HeLa и HT1080 культивировали в среде Игла, модифицированной по способу Дульбекко, с добавлением 10% фетальной бычьей сыворотки (Gibco, Carlsbad, Calif.) и 1% пенициллина/стрептомицина (Invitrogen, Carlsbad, Calif.). Клетки CHO K1 и CHO pgsA культивировали в среде F-12K (ATCC) с добавлением 10% фетальной бычьей сыворотки (Gibco) и 1% пенициллина/стрептомицина (Invitrogen). Клетки Pro5 и Lec1 культивировали в среде MEM-альфа (Gibco) с добавлением 10% фетальной бычьей сыворотки (Gibco) и 1% пенициллина/стрептомицина (Invitrogen).
[00269] Пулы сывороток человека для селекции
[00270] Восемнадцать индивидуальных образцов человеческой сыворотки получали из Innovative Research, Inc. (Southfield, Mich.), и для каждого образца определяли титр нейтрализующих антител к AAV2 дикого типа (таблица 2). Поскольку индивидуальные образцы, вероятно, обладают вариациями как по аффинности, так и по эпитопной специфичности антител, то создали три эффективных пула сывороток (α=A+F+G, β=B+H+M и γ=I+J+N) смешиванием равных объемов индивидуальных образцов сывороток. Селекция в присутствии этих вариаций антител должна приводить к общему повышению устойчивости ко многим ранее существовавшим человеческим антителам. Более поздние селекции проводили в присутствии Гамимуна Н, 10% человеческого IVIG (Bayer, Elkhart Ind.) для селекции на устойчивость к еще более широкому диапазону антител.
[00271] Таблица 2: титры нейтрализующих антител в индивидуальных образцах сыворотки человека
Титры нейтрализующих антител (NAb) для каждого образца представлены в виде обратной величины объема фракции сыворотки, необходимой для снижения инфекционности до значения 37%, измеренного в отсутствии сыворотки. Затем были созданы три пула сывороток (α=A+F+G, β=B+H+M и γ=I+J+N) смешиванием эквиобъемных количеств трех индивидуальных образцов сыворотки.
[00272]
[00273] Создание библиотеки и получение вируса
[00274] Для создания библиотеки мутагенеза с насыщением была создана библиотека cap AAV2 с использованием ПЦР пониженной точности с последующим процессом постадийного удлинения, описанным Zhao et al. с использованием 5′-GCGGAAGCTTCGATCAACTACGC-3′ (SEQ ID NO: 14) и 5′-GGGGCGGCCGCAATTACAGATTACGAGTCAGGTATCTGGTG-3′ (SEQ ID NO: 15) в качестве прямого и обратного праймеров соответственно. Селекции с использованием объединенных индивидуальных сывороток человека выявили вариант, содержащий четыре точечные мутации (описаны в разделе «Результаты»), которые послужили основой для создания библиотеки мутагенеза с насыщением. Ген cap для этого варианта был подвергнут дальнейшему мутагенезу путем замены аминокислот в определенных сайтах. Праймер 5'-cattNNKgaccagtctaggaactgg-3' (SEQ ID NO: 16) и соответствующий комплементарный обратный праймер использовали для мутагенеза аминокислотного сайта R471. Праймер 5' gccacaaggacgatgaagaaNNKttttttcctcagagcggggttctcatctttgggaagcaaggctcaNNKaaaacaagt gtggacattg-3' (SEQ ID NO: 17) и соответствующий комплементарный обратный праймер использовали для мутагенизации аминокислотных сайтов K532 и E548. Праймер 5'-ccaacctccagagaggcNNKagacaagcagctacc-3' (SEQ ID NO: 18) и соответствующий комплементарный обратный праймер использовали для мутагенеза аминокислотного сайта N587. Праймер 5'-ccaactacaacaagtctNNKaatgtggactttactgtggacNNKaatggcgtgtatt-3' (SEQ ID NO: 19) и соответствующий комплементарный обратный праймер использовали для мутагенизации аминокислотных сайтов V708 и T716. Библиотеку, состоящую из AAV2, содержащего рандомизированные области петли cap, и библиотеку, содержащую перетасованную ДНК из генов cap AAV1, AAV2, AAV4, AAV5, AAV6, AAV8, AAV9 дикого типа, упаковывали и объединяли для начальных стадий селекции (Koerber et. al.; Mol. Ther., 2008 October, 16(10):1703-9 и Koerber et al., Mol. Ther, 2009, December, 17(12):2088-95, которые обе в полном объеме включены в настоящее описание посредством ссылки).
[00275] Для второго и третьего раундов эволюции создавали библиотеки случайного мутагенеза посредством подвергания генов cap из библиотеки замены петель/щаффлинга и библиотеки мутагенеза с насыщением с использованием ПЦР пониженной точности с использованием 5'-CATGGGAAAGGTGCCAGACG-3' (SEQ ID NO: 20) и 5'-ACCATCGGCAGCCATACCTG-3' (SEQ ID NO: 21) в качестве прямого и обратного праймеров соответственно, как описано выше. Репликационно-компетентные библиотеки AAV и рекомбинантные векторы AAV, экспрессирующие GFP под контролем промотора CMV, упаковывали с использованием клеток HEK293T (ATCC) с использованием метода трансфекции фосфатом кальция, и вирусы очищали центрифугированием в градиенте йодиксанола. Рекомбинантные векторы AAV, экспрессирующие GFP или люциферазу под контролем промотора CMV, для применения in vivo дополнительно очищали фильтрацией на устройстве Amicon. Геномные титры устойчивости к ДНКазе определяли с помощью количественной ПЦР (Excoffon et al., Proc. Natl. Acad. Sci. USA, 2009 Mar. 10; 106(10):3865-70; и Maheshri et al., Nat. Biotechnol., 2006 February; 24(2):198-204; которые обе в полном объеме включены здесь посредством ссылки).
[00276] Селекция и эволюция библиотеки
[00277] Один раунд селекции определяется как инфицирование клеток HEK293T с использованием исходной библиотеки AAV (инкубация в течение 30 мин при комнатной температуре для объединенных индивидуальных сывороток человека или в течение 1 ч при 37°C с инактивированным нагреванием IVIG перед инфицированием), с последующим спасением аденовируса и сбором успешных вариантов. Каждый раунд эволюции состоит из мутагенеза гена cap для создания исходной библиотеки и трех раундов селекции. С каждой библиотекой было проведено три раунда эволюции с проведением клонального анализа между каждым раундом эволюции. Исходные библиотеки для каждого раунда эволюции были созданы, как описано выше. После третьего раунда селекции гены cap AAV выделяли из пула успешных вариантов AAV и амплифицировали с использованием ПЦР. Гены сap встраивали в рекомбинантную упаковочную плазмиду AAV pXX2 с использованием NotI и HindIII. Затем гены cap секвенировали в лаборатории секвенирования ДНК Калифорнийского университета в Беркли и анализировали с использованием программного обеспечения Geneious (Biomatters, Auckland, Новая Зеландия). Трехмерные модели капсида AAV2 (идентификационный номер в банке данных белков 1LP3) визуализировали в системе Pymol (DeLano Scientific, San Carlos, Калифорния).
[00278] Анализ трансдукции in vitro вариантов, ускользающих от антител
[00279] HEK293T высевали при плотности 3×104 клеток/лунку за 24 ч до инфицирования. Варианты инкубировали при 37°С в течение 1 ч с инактивированным нагреванием IVIG, индивидуальными человеческими сыворотками или индивидульными мышиными сыворотками до инфицирования, и затем клетки инфицировали rAAV-GFP при геномной множественности инфекции 2000. Процент GFP-позитивных клеток оценивали через 48 ч после инфицирования с использованием системы визуализации ImageXpress Micro Cellular Imaging и Analysis System (Molecular Devices, Sunnyvale, Калифорния) и программного обеспечения MetaXpress Image Analysis, версия 3.1.0, Multi Wavelength Cell Scoring Application Module (Molecular Devices).
[00280] Анализ трансдукции in vitro
[00281] Для определения относительной эффективности трансдукции селектированные мутанты сравнивали с родительскими серотипами AAV дикого типа, клетки HEK293T, CHO K1, CHO pgsA (у всех отсутствуют поверхностные гликозаминогликаны), CHO Pro5 (родительская линия для нескольких мутантов по гликозилированию, включая клетки Lec1), CHO Lec1 (дефектные по гликозилированию), HeLa и HT1080 (клеточная линия фибросаркомы человека) высевали при плотности 2,5×104 клеток на лунку за 24 ч до инфицирования. Клетки инфицировали rAAV1-GFP, rAAV2-GFP, rAAV6-GFP, Shuffle 100.1-GFP, Shuffle 100.3-GFP, SM 10.2-GFP или Shuffle 100.7-GFP в диапазоне значений MOI 100-1000. Процент GFP-позитивных клеток оценивали через 48 ч после инфицирования с использованием проточного цитометра Beckman-Coulter Cytomics FC500 (Beckman-Coulter, Brea, Калифорния).
[00282] Анализ in vivo вариантов, ускользающих от антител
[00283] Для анализа экспрессии генов in vivo восьминедельных мышей самок Balb/c праймировали IVIG из расчета 4 мг на мышь или забуференным фосфатом физиологическим раствором (контрольных мышей) введением в хвостовую вену за 24 ч до введения рекомбинантного Shuffle 100-3 (см. SEQ ID NO: 12), SM 10-2 (см. SEQ ID NO: 10) или векторов AAV2. Мышей инфицировали 1011 вирусных геномов рекомбинантных векторов AAV, кодирующих люциферазу под контролем промотора CMV, введением в хвостовую вену. Для биолюминесцентной визуализации мышей анестезировали смесью 2% изофлурана и кислорода. Субстрат D-люциферин (GOLD Biotechnology, St. Louis, Mo.) вводили внутрибрюшинно в дозе 500 мкг/г массы тела. Изображения получали с использованием устройства для визуализации VivoVision IVIS Lumina (Xenogen, Alameda, Калифорния). Для каждой мыши делали вентральные изображения через 7-10 мин после инъекции субстрата раз в неделю в течение четырех недель. Через пять недель после инфицирования отбирали кровь сердечной пункцией и готовили сыворотку, и затем мышей перфузировали 0,9% физиологическим раствором. Отбирали образцы сердца, печени, легких, почек, селезенки, головного мозга, спинного мозга и мышц задних конечностей и замораживали. Образцы замороженной ткани гомогенизировали и ресуспендировали в лизирующем буфере для репортерного теста (Promega, Mannheim, Германия) для анализа люциферазы in vitro. Лизат, содержащий люциферазу, осветляли центрифугированием в течение 10 мин при 10000 g. Для анализа образцов 20 мкл лизата добавляли к 100 мкл буфера для анализа люциферазы, перемешивали, инкубировали в течение 5 мин и помещали в люминометр. Сигнал интегрировали в течение 30 с с 2-с интервалом и выражали в относительных световых единицах (RLU), определенных люминометром TD 20/20 (Turner Designs, Sunnyvale, CA). Люциферазный сигнал нормализовали к общему содержанию белка, определенному с помощью анализа с бицинхониновой кислотой (Pierce).
[00284] Результаты
[00285] Полученные результаты демонстрируют, что AAV может эволюционировать, чтобы существенно преодолеть нейтрализацию анти-AAV антителами, как in vitro, так и in vivo. Были выделены новые варианты AAV, для которых требовались титры нейтрализующих антител в 2-35 раз выше (с использованием IVIG человека), чем для AAV дикого типа in vitro. Их способность подвергаться нейтрализации антителами также проявлялась в усилении трансдукции in vivo в присутствии нейтрализующих антител. Выделение таких новых клонов, устойчивых к анти-AAV антителам, позволяет более широко применять методы лечения, основанные на использовании AAV в качестве вектора для доставки нуклеиновой кислоты (включая субъектов с высокими титрами антител, которые в настоящее время не подходят для генной терапии AAV).
[00286] Создание и селекция библиотеки AAV посредством направленной эволюции
[00287] На фиг. 1а приведена схема подхода направленной эволюции, используемой для выделения новых вариантов AAV, способных «ускользнуть» от нейтрализации человеческими антителами. Библиотеки вирусов создавали с использованием методов мутагенеза ДНК, описанных в следующих параграфах (фиг. 1а, стадии 1 и 2). Во время начальной селекции пулы вирусных библиотек, полученные из мутаций с использованием ПЦР пониженной точности, генов cap AAV2, инкубировали с различными разведениями α-пула сывороток человека с низкой активностью в течение 30 мин при комнатной температуре перед инфицированием клеток HEK293T (стадия 3). После трех раундов селекции против α-пула человеческих сывороток с низкой активностью (фиг. 1а, стадии 4 и 5) было получено несколько вариантов с повышенной устойчивостью к этому пулу нейтрализующих сывороток (фиг. 1а, стадия 6, фиг. 7а). Вариант 1.45 содержал две точечные мутации (N312K, N449D), которые приводили к более чем в 10 раз высокой устойчивости к нейтрализации α-пулом по сравнению с AAV2 дикого типа.
[00288] Ген cap из варианта 1.45 подвергали дополнительному случайному мутагенезу, и полученную библиотеку селектировали в трех дополнительных циклах селекции против β- и γ-пулов параллельно. Поскольку наблюдали лишь незначительное усиление в уклонении от антител (данные не показаны), то восстановленные гены cap объединяли и подвергали дополнительной диверсификации с помощью шаффлинга ДНК и EP-ПЦР. Еще три раунда селекции с возрастающим количеством сывороток как из β-пула, так и из γ-пула приводили к значительному увеличению количества восстановленного вируса из вирусной библиотеки по сравнению с AAV2 дикого типа (фиг. 7b, c). Секвенирование успешных генов cap из обоих пулов выявило несколько мутантов на низкой частоте и один доминантный мутант, вариант γ4.3, который содержал четыре точечные мутации (N312K, N449D, N551S и 1698V), присутствующие в обеих библиотеках. В присутствии IVIG человека вариант 1.45 показал умеренную повышенную в 1,2 раза устойчивость к нейтрализации, тогда как вариант γ4.3 продемонстрировал повышенную в 3,1 раза устойчивость к нейтрализации (фиг. 7d). Это наблюдение подтверждает гипотезу о том, что пулы индивидуальных сывороток человека можно использовать для выделения вариантов AAV, способных к усиленному уклонению от антител, присутствующих в общей популяции человека.
[00289] Средний успех варианта γ4.3 в устойчивости к нейтрализации анти-AAV антителами побудил к разработке библиотеки на основе гена cap γ4.3. Аминокислотные сайты R471, K532, E548, N587, V708, T716, которые ранее были определены как иммуногенные сайты в капсиде AAV2, подвергали мутагенезу с насыщением в попытке найти аминокислотные мутации, которые могут привести к повышению устойчивости γ4.3 к антителам. Эта библиотека «насыщающего мутагенеза» вместе с «перетасованной» библиотекой, состоящей из случайных химер cap 7 родительских серотипов AAV, и библиотекой «перестановки петель», состоящей из cap AAV2 с замещенными петлевыми областями, подвергали трем дополнительным раундам селекции в котором пулы вирусных библиотек инкубировали с различными разведениями человеческого IVIG в течение одного часа при 37°С до инфицирования клеток НЕК293Т. После инфицирования библиотеками AAV и амплификации инфекционных вариантов AAV посредством аденовирусной суперинфекции количество вирусных геномов или титр вируса в каждой библиотеке определяли количественно и сравнивали с титрами AAV2 дикого типа в качестве метода определения успеха селекции (фиг. 1b). Для каждого раунда селекции с использованием библиотек мутагенеза с насыщением и замены петель/шаффлинга вирусные пулы из условий разведения 1:10 и 1:100 IVIG, которые давали более высокие титры вируса, чем AAV2 дикого типа, использовали в качестве отправной точки для последующего раунда селекции. После трех раундов селекции успешные вирусные гены cap выделяли и тестировали по отдельности, чтобы определить вирус с наиболее эффективной доставкой генов. Кроме того, гены cap, выделенные из третьего раунда селекции, подвергали дополнительным раундам мутагенеза с ПЦР пониженной точности, и этот процесс повторяли для итеративного повышения пригодности вируса.
[00290] На фиг. 1 представлена направленная эволюция AAV для обеспечения повышенного уклонения от антител. (а) Схема направленной эволюции. 1). Вирусную библиотеку создают генетической диверсификацией гена cap с использованием нескольких взаимодополняющих подходов. 2) Вирусы упаковывают в клетки HEK293T с использованием плазмидной трансфекции, затем собирают и очищают. 3) Вирусную библиотеку инкубируют с человеческим IVIG в нескольких концентрациях и вводят в клетки HEK293T in vitro. 4) Успешные вирусы амплифицируют и выделяют с помощью аденовирусной суперинфекции. 5) Успешные клоны обогащают за счет повторных селекций при более низких значениях MOI. 6) Выделенная вирусная ДНК выявляет успешные гены cap. 7) Успешные гены cap снова мутируют, чтобы служить новой отправной точкой для селекции. (b) Отбор мутантов, уклоняющихся от антител, из библиотек замены петель/шаффлинга и мутагенеза с насыщением. Клетки HEK293T инфицировали вирусными библиотеками в течение 24 ч. Вирусные частицы, которые продуктивно инфицировали клетки, амплифицировали с помощью аденовирусной инфекции, и спасенный AAV определяли количественно с помощью кПЦР (количественная полимеразная цепная реакция). Разведение IVIG 1:10 соответствует концентрации IVIG 10 мг/мл. Столбцы ошибок указывают на стандартное отклонение (n=3).
[00291] На фиг. 7 показано создание вариантов, ускользающих от антител человека, на основе AAV2. (а) Четыре вирусных клона, селектированных после трех раундов селекции против α-пула низкой жесткости, демонстрируют повышенную устойчивость к 1 мкл α-сыворотки при множественности инфекции 1. Два дополнительных раунда диверсификации (т.е. мутагенеза и ДНК-шаффлинга) и селекции (3 раунда с увеличением числа сывороток) приводило к значительному повышению выделения вируса в присутствии больших количеств сильнодействующих (b) β- и (c) γ-пулов. (d) Кроме того, два вирусных клона (1,45 и γ4,3) показали повышенную в 1,23 и 3,10 раза резистентность к высоко разнообразному пулу ранее продуцированных антител, присутствующих с объединенным иммуноглобулином человека для внутривенного ввведения (IVIg) от ~100000 субъектов по сравнению с дикими типа ААВ2.
[00292] Повышенное уклонение от антител новых эволюционировавших вариантов AAV in vitro
[00293] Из двенадцати клонов, отобранных и упакованных для индивидуального анализа из библиотек мутагенеза с насыщением и замены петель/шаффлинга после девяти раундов скрининга против человеческого IVIG, для всех двенадцати требовались более высокие титры нейтрализующих антител, чем для AAV1 и AAV2 дикого типа (на фиг. 2а и таблица 1). Вариант Shuffle 100-3 (см. SEQ ID NO: 12), для нейтрализации которого требовалась в 35 раз более высокая концентрация IVIG in vitro, чем для AAV2 дикого типа, по-прежнему был способен трансдуцировать приблизительно 10% клеток в присутствии 1 мг/мл IVIG (фиг. 2b). Кроме того, для варианта SM 10-2 из библиотеки мутагенеза с насыщением AAV2 требовалась для нейтрализации в 7,5 раз более высокая концентрация WIG in vitro, чем для AAV2 дикого типа. Кроме того, варианты Shuffle 100-3 и SM 10-2 (см. SEQ ID NO: 10) показали усиленную трансдукцию в присутствии образцов сыворотки от отдельных пациентов, исключенных из клинического исследования c гемофилией B (фиг. 3) (Nathwani et al., N. Engl. J. Med., 2011, Dec. 22, 365(25):2357-65).
[00294] На фиг. 2 показаны профили нейтрализации вариантов, уклоняющихся от антител. Гены cap мутантов, уклоняющихся от антител, выделенных после трех раундов эволюции, использовали для упаковки рекомбинантного AAV, кодирующего GFP, и инкубировали с человеческим IVIG перед инфицированием клеток HEK293T. Фракцию оставшихся инфекционных частиц определяли с использованием высококонцентрированной флуоресцентной визуализации и нормализовали к инфекционному титру в отсутствии IVIG. Показаны два клона из каждой библиотеки с устойчивостью к IVIG. Данные для других анализированных клонов представлены в таблице 1. (а) Кривые нейтрализации. Столбцы ошибок указывают на стандартное отклонение (n=3). (b) Репрезентативные флуоресцентные изображения из нескольких разведений IVIG показывают, что мутанты способны к трансдукции HEK293T в присутствии высоких концентраций нейтрализующих антител.
[00295] На фиг. 3 показаны профили нейтрализации вариантов, ускользающих от антител. Сыворотки человека получали от субъектов, которые были исключены из клинических испытаний с гемофилией В за счет наличия высоких титров нейтрализующих анти-AAV антител. Рекомбинантный AAV, кодирующий GFP, инкубировали с индивидуальными образцами сыворотки человека перед инфицированием клеток HEK293T. Фракцию оставшихся инфекционных частиц определяли с использованием флуоресцентной микроскопии и нормализовали к инфекционному титру в отсутствии сыворотки человека. Столбцы ошибок указывают на стандартное отклонение (n=3).
[00296] Анализ последовательности двенадцати клонов показал, что два варианта с самой высокой устойчивостью к нейтрализующим антителам, Shuffle 100-3 (см. SEQ ID NO: 12) и Shuffle 100-1 (см. SEQ ID NO: 11), почти идентичны по перетасованным капсидам, содержащим фрагменты AAV1-4, AAV6 и AAV9 (фиг. 4). Различия в аминокислотах 469 (от остатка AAV6 до остатка AAV7) и 598 (от остатка AAV6 до остатка AAV1) между двумя вариантами приводят к почти 3-кратному повышению титра нейтрализующих антител для Shuffle 100-3 (см. SEQ ID NO: 12) (таблица 1). Вариант Shuffle 100-7 (см. SEQ ID NO: 13), который имел четвертое место по устойчивости к нейтрализующим антителам (таблица 1), также представляет собой перетасованный капсид, содержащий фрагменты AAV1, AAV6 и AAV8 (фиг. 4), что хорошо согласуется с опубликованными данными, показывающими, что AAV1 и AAV8 дикого типа способны уклоняться от анти-AAV2 антител. Интересно, что вариант SM 10-2 (SEE SEQ ID NO: 10) сохранил точечные мутации, приобретенные вариантом γ4.3, и также сохранил остатки дикого типа в сайтах мутагенеза с насыщением. Вариант SM 10-2 (СМ. SEQ ID NO: 10) приобрел дополнительные точечные мутации в поверхностном остатке D472N и внутреннем остатке L735Q. На фиг. 4 приведены аминокислотные последовательности клонов с мутагенезом с заменой петель/шаффлингом и насыщением. (а) Схемы капсидного белка показаны для двух клонов из каждой библиотеки с самыми высокими нейтрализующими концентрациями IVIG. Каждая область затенина в соответствии с родительским серотипом, из которого она получена. Черные стрелки обозначают (слева направо) исходные кодоны капсидных белков VP1, VP2 и VP3. Серые стрелки обозначают (слева направо) участки поверхностной петли I, II, III, IV и V на основе капсида AAV2. (b) Молекулярные модели полного капсида AAV2, основанные на разрешенной структуре, показаны для двух клонов из каждой библиотеки с самыми высокими нейтрализующими концентрациями IVIG. Каждая область затемнена в соответствии с родительским серотипом, из которого она получена. Для варианта Shuffle 100-3 (см. SEQ ID NO: 12): черные стрелки указывают на отличия от варианта Shuffle 100-1 (см. SEQ ID NO: 11). Для варианта SM 10-2 (см. SEQ ID NO: 10): мутации N449D, D472N, N551S и 1698V представляют собой поверхностные мутации (черные).
[00297] Таблица 1: титры нейтрализующих IVIG антител для библиотек клонов и исходных серотипов
[00298] IVIG человека использовали для нейтрализации рекомбинантных векторов AAV-GFP с капсидами из AAV1, AAV2, AAV8 дикого типа и вариантов, извлеченных из библиотек мутагенеза с заменой петель/шаффлингом и насыщением. Показана концентрация IVIG (мг/мл), необходимая для снижения эффективности доставки гена до 50% при отсутствии IVIG, в сравнении с концентрацией, необходимой для снижения доставки AAV2. Для всех анализированных вариантов требовались более высокие концентраций IVIG, чем для AAV1 и AAV2 дикого типа. Титр нейтрализующих антител определяли, подгоняя кривые на фиг. 2 к экспоненциальной кривой. SEQ ID NO указаны как «аминокислота, нуклеотид».
[00299]
концентрация мг/мл
[00300] Варианты Shuffle 100-3 (см. SEQ ID NO: 12), Shuffle 100-1 (см. SEQ ID NO: 11) и Shuffle 100-7 (см. SEQ ID NO: 13) имеют профили трансдукции, которые сходны с профилями трансдукции родительских серотипов AAV1 и AAV6 (фиг. 5). Кроме того, мутации в SM 10-2 (см. SEQ ID NO: 10) не предотвращают зависимость от гепарина (как это наблюдается у родительского серотипа AAV2), что приводит к профилю, сходному с AAV2 (фиг. 5).
[00301] На фиг. 5 показан тропизм новых вариантов aav in vitro. Рекомбинантные векторы AAV, экспрессирующие зеленый флуоресцентный белок, использовали для трансдукции ряда клеточных линий: CHO, pgsA (все с отсутствием поверхностных гликозаминогликанов), Pro5, Lec1 (с отсутствием сиаловой кислоты), HEK293T, HeLa и HT1080 (линия клеток фибросаркомы человека) для получения профиля трансдукционных свойств новых вариантов AAV. Столбцы ошибок указывают на стандартное отклонение (n=3).
[00302] Повышенное уклонение от антител новых эволюционировавших вариантов AAV in vivo
[00303] Для определения паттерна локализации вариантов Shuffle 100-3 и Shuffle 100-7, определяли активность фермента люциферазы в различных тканях наивных мышей, которым вводили AAV2, Shuffle 100-3 или Shuffle 100-7 (фиг. 6А). Вариант Shuffle 100-7 показал тропизм in vivo, аналогичный AAV2, за исключением того, что трансдукция клеток в сердце была в 7 раз выше, трансдукция в легких в 5 раз выше, и трансдукция в печени была в 4,5 раза ниже. Вариант Shuffle 100-3 проявил более чем в 4 раза более высокую трансдукцию клеток в головном мозге, более чем в 3 раза более высокую трансдукцию в легких и в 27 раз более высокую трансдукцию в мышцах, чем AAV2. Анализ сыворотки от этих мышей показал, что для варианта Shuffle 100-3 требуются равные или более высокие концентрации в сыворотке in vitro для нейтрализации, чем AAV1 и AAV8 для сыворотки от мышей, которым вводили векторы для доставки генов AAV1, AAV2, AAV8 или Shuffle 100-3 (фиг. 11). Для нейтрализации Shuffle 100-7 требовались равные или более высокие концентрации в сыворотке in vitro, чем AAV1 для сыворотки мышей, которым вводили векторы для доставки генов AAV1, AAV2, AAV8, Shuffle 100-3 или SM 10-2 (фиг. 11). Кроме того, оба варианта в меньшей степени подвергались нейтрализации сывороткой мышей, которым вводили векторы для доставки гена AAV2, чем все тестированные серотипы AAV дикого типа. Интересно отметить, что вариант Shuffle 100-3 также в меньшей степени подвергался нейтрализации сывороткой мышей, иммунизированных против него, чем любой из других испытанных серотипов или вариантов (фиг. 11). Полученные данные иллюстрируют возможность того, что эти варианты можно использовать в комбинации с серотипами AAV дикого типа или другим вариантом в применениях, для которых требуется многократное введение вектора.
[00304] На фиг. 11 показаны титры нейтрализующих антител библиотечных клонов и исходных серотипов в сыворотке иммунизированных мышей. Сыворотки мышей, которым вводили библиотечные клоны или AAV дикого типа, использовали для нейтрализации рекомбинантных векторов AAV-GFP с капсидами из AAV1, AAV2, AAV8 дикого типа и вариантов, выделенных из библиотек мутагенеза с заменой петель/шаффлингом и насыщением. Показано разведение сыворотки, необходимое для снижения эффективности доставки генов до 50% в отсутствии сыворотки.
[00305] Для определения способности вариантов Shuffle 100-7 и Shuffle 100-3 уклоняться от нейтрализации антителами in vivo, мышей пассивно иммунизировали человеческим IVIG перед введением AAV. Вариант Shuffle 100-7 имел достоверно более высокую трансдукцию клеток сердца, печени и мышц, чем AAV2, что измеряли по активности фермента люциферазы (фиг. 6b). Вариант Shuffle 100-3 имел достоверно более высокую трансдукцию клеток в сердце и мышцах по сравнению с AAV2 (фиг. 6b).
[00306] На фиг. 6 показана локализация и нейтрализация новых вариантов AAV in vivo. (а) Рекомбинантные векторы AAV, кодирующие люциферазу, вводили в хвостовую вену мышам самкам BALB/c. Через 5 недель определяли уровни активности люциферазы и нормализовали их к общему белку для каждого анализированного образца. (b) Рекомбинантные векторы AAV, экспрессирующие люциферазу, вводили в хвостовую вену мышам самкам BALB/c через 24 ч после инъекции в хвостовую вену 4 мг человеческого IVIG. Через 5 недель уровни экспрессии люциферазы нормализовали к общему белку для каждого анализированного образца. Столбцы ошибок указывают на стандартное отклонение (n=3), *=p<0,05. RLU, относительная световая единица.
[00307] Вариант γ4.3, выделенный из библиотеки, пролученной ПЦР пониженной точности, на основе AAV2, выбранной из пула индивидуальных сывороток человека, содержал четыре точечные мутации (N312K, N449D, N551S и I698V). Интересно отметить, что два из этих положений (N449 и N551) ранее были идентифицированы в качестве иммуногенных остатков с использованием других пулов сывороток человека, свидетельствуя о том, что антигенные эпитопы, включающие эти сайты, являются мишенями для многих различных нейтрализующих антител. Таким образом, эти сайты представляют собой интересные и ценные мишени для мутаций. Сочетание направленной эволюции и рационального дизайна в получении библиотеки мутагенеза с насыщением привело к выделению варианта SM 10-2, который был способен к более высокой устойчивости к антителам, чем как AAV1, так и AAV2 in vitro. Вариант SM 10-2 включает две дополнительные точечные мутации (D472N и L735Q) по сравнению с обнаруженными в варианте γ4.3. Ранее было показано, что мутация D472N повышает уровень синтеза капсида в клетках HEK293. Аналогично замена положительно заряженной боковой цепи лизина в положении аминокислоты 735 на незаряженную боковую цепь глутамина может способствовать стабилизации капсида, поскольку она также присутствует в варианте Shuffle 100-7, несмотря на то, что расположена внутри собранного капсида (фиг. 4).
[00308] Создание химерных капсидов AAV позволяет генерировать вирусные варианты, которые могут объединять желаемые свойства нескольких серотипов AAV. Несмотря то, что также было показано, что AAV8 и AAV9 существенно более устойчивы к нейтрализации под действием IVIG, чем AAV2, аминокислоты, специфичные для этих капсидов, присутствовали только в небольших участках на поверхности полученных шаффлингом вариантов, выделенных во время селекции (фиг. 4). Вариант, демонстрирующий более эффективное уклонение от нейтрализации антителами in vitro, Shuffle 100-3, проявлял in vitro тропизм, сходный с родительскими серотипами AAV1 и AAV6, но с более высокой степенью инфекционности, чем любой из серотипов дикого типа. Различия в аминокислотах 469 и 598 между вариантами Shuffle 100-1 и Shuffle 100-3 приводят к почти 3-кратному увеличению титра нейтрализующих антител для Shuffle 100-3. Результаты исследования Lochrie et al. свидетельствуют о том, что иммуногенные остатки, распознаваемые сывороткой человека и IVIG, различны, что позволяет предположить, что у разных людей могут продуцироваться различные нейтрализующие антитела к разным наборам эпитопов на капсиде AAV, и полностью избежать нейтрализации является непростой задачей (Lochrie et al., J. Virol., 2006 January; 80(2):821-34). Работа авторов изобретения показывает, что использование нескольких раундов направленной эволюции с использованием нескольких различных пулов сыворотки, содержащих различное количество и различающихся по эффективности антител против AAV, приведет к выделению новых вариантов AAV, которые способны к повышенной клеточной трансдукции, как in vitro, так и in vivo, в присутствии нескольких пулов анти-AAV антител.
[00309] Адаптивные иммунные ответы на компоненты вектора AAV у животных и человека часто препятствуют повторному введению векторов AAV одного и того же серотипа, что затрудняет применение с доставкой генов, требующее многократного введения вектора. Анализ нейтрализации in vitro с использованием сыворотки мышей, использованных в исследованиях биораспределения, показывают, что варианты подвергаются нейтрализации этими сыворотками в меньшей степени, чем AAV дикого типа (фиг. 11), что может быть пригодно для стратегий генной терапии, в которых необходимо повторное введение вектора. Например, Shuffle 100-3 не нейтрализуется сывороткой мышей, которым инъецировали AAV2, и AAV2 не нейтрализуется сывороткой мышей, которым ввели Shuffle 100-3, что позволяет предположить, что этот вариант можно использовать в комбинации с серотипами AAV дикого типа или в применениях, в которых требуется введение нескольких векторов. В заключение, авторы изобретения использовали направленную эволюцию для выделения новых вариантов AAV, которые способны к пониженной нейтрализации анти-AAV антителами, полученными от отдельных пациентов, объединенной сыворотки человека и сыворотки мыши, как in vitro, так и in vivo.
Пример 2
Идентификация варианта капсида, подходящего для применения в генной терапии, направленной в легкие приматов
[00310] Введение
[00311] Стратегию направленной эволюции использовали для идентификации вариантов капсида AAV с повышенной эффективностью доставки генов в клетки альвеолярного эпителия легкого примата, отличного от человека (NHP), типа II (AT II) после интратрахеального аэрозольного введения в присутствии нейтрализующих антител человека (NAbs)). Вкратце, гены cap аденоассоциированного вируса (AAV) дикого типа были диверсифицированы с использованием нескольких подходов для создания больших генетических библиотек, которые были упакованы для генерации библиотек вирусных частиц, и затем применяли селективное давление для выделения новых вариантов, которые могут преодолеть барьеры доставки генов, включая, помимо прочего, антикапсидный иммунный ответ, ограниченную трансдукцию клеток в определенных тканях и невозможность адресной доставки к определенным типам клеток.
[00312] Методы
[00313] Клеточные линии и получение библиотек
[00314] Клетки HEK293T получали из Американской коллекции типовых культур (Manassas, VA). Клетки культивировали при 37°C и 5% CO2 в среде Игла, модифицированной по способу Дульбекко, с добавлением 10% фетальной бычьей сыворотки (Gibco, Carlsbad, Калифорния) и 1% пенициллина/стрептомицина (Invitrogen, Carlsbad, Калифорния). Вирусные библиотеки получали в клетках HEK293T с использованием тройной трансфекции, и вирусы очищали центрифугированием в градиенте йодиксанола и фильтрацией через устройство Amicon. Геномные титры устойчивости к ДНКазе определяли с помощью количественной ПЦР (кПЦР).
[00315] Внутритрахеальная инъекция и забор тканей
[00316] Для каждого устройства для доставки, использованного в селекции, вводили дозу одному яванскому макаку самцу (macaca fascicularis) в возрасте 4-6 лет и с массой тела 5,5-6,0 кг. Животных анестезировали кетамином в дозе 10 мг/кг и дексмедетомидином в дозе 15 мкг/кг внутримышечно (в/м). Пять мл библиотеки предварительно смешивали с 1,75 мг/мл человеческого иммуноглобулина для внутривенного введения (IVIG) и вводили, как описано ниже. Каждое животное интубировали эндотрахеальной трубкой диаметром 5 мм, при этом конец трубки располагали на уровне ключицы (приблизительно на 5 см выше киля), и его положение подтверждали рентгеноскопией.
[00317] Распылительное устройство соединяли с дистальным концом эндотрахеальной трубки, и респиратор для птиц использовали для обеспечения вдохов со скоростью 15±1 вдохов/мин при давлении 20 см H2O. Катетер AeroProbe® (Trudell Medical International) был снабжен отрезком звездообразной трубки диаметром 0,144 дюйма для облегчения правильного размещения внутри эндотрахеальной трубки. Кончик катетера располагали непосредственно над концом эндотрахеальной трубки. Катетер AeroProbe® подсоединяли к системе управления катетером AeroProbe, и дыхательный аппарат (Harvard Appartatus) использовали для обеспечения дыхания со скоростью 20 вдохов/мин при давлении 18-20 см H2O. После завершения введения дозы каждое животное экстубировали и внутримышечно вводили 0,15 мг/кг атипамезола для отмены седации. Животные находились под визуальным контролем до тех пор, пока они полностью не восстанавливались от анестезии, прежде чем их возвращали в клетки, в которых их содержались.
[00318] Эвтаназию проводил квалифицированный ветеринарный персонал с использованием 100 мг/кг пентобарбитала натрия, введенного внутривенно на сутки 15±1. Легкие, включая трахею, извлекали и рассекали, как подробно описано ниже. ДНК выделяли из клеток АТ II и хранили при -20°С до амплификации вирусного генома.
[00319] Выделение клеток альвеолярного эпителия типа II (AT II)
[00320] Клетки AT II выделяли из легких приматов, отличных от человека, как описано Fang et al., Measurement of Protein Permeability and Fluid Transport of Human Alveolar Epithelial Type II Cells Under Pathological Conditions. Humana Press, New York, NY, 2018, pp. 121-128. Вкратце, легкие промывали 500 мл PBS, содержащего 5 мМ ЭДТА и 5 мМ ЭГТА, с помощью шприца, введенного в трахею. Затем легкие заполняли 250 мл раствора эластазы с концентрацией 1,2 мг/мл, и инкубировали в течение 1 ч при 37°С. Легкие гомогенизировали для высвобождения клеток, выстилающих легкие, и удаляли дыхательные пути. Клетки AT II выделяли после серии стадий включения и исключения, включая градиент Percoll и магнитные частицы CD14/CD45 Dynabeads. Клетки AT II помещали на вставки, покрытые коллагеном IV, на 24 ч перед выделением ДНК. Проводили анализ с использованием красителя Lysotracker и сурфактантного белка C с использованием проточной цитометрии и иммуноцитохимии, чтобы гарантировать чистоту клеточного изолята.
[00321] Эволюция терапевтического вектора
[00322] Используемый процесс эволюции вектора показан на фиг. 12. Вкратце, создавали библиотеку вирусных капсидов, содержащую запатентованные комбинации методов мутации ДНК и генов cap (а). Затем вирусы упаковывали (b) таким образом, чтобы каждая частица состояла из мутантного капсида, окружающего ген cap, кодирующий этот капсид, и очищали. Капсидную библиотеку помещали под селективное давление in vivo. Представляющий интерес тип ткани или клеток собирали для выделения вариантов AAV, которые успешно локализовались в мишени. Успешные вирусы извлекали с использованием ПЦР-амплификации. Успешные клоны обогащали путем повторной селекции (стадия I - (c)). Затем селектированные гены cap подвергали запатентованной повторной диверсификации и обогащали посредством дальнейших стадий селекции для итерационного повышения приспособленности вируса (стадия 2 - (d)). Варианты, идентифицированные как наиболее успешные на стадиях 1 и 2 селекции вектора, оценивали для выявления вариантов капсида с желаемыми свойствами (e).
[00323] Мотивы были признаны «попадаемыми», когда они соответствовали следующим критериям: 1) мотив представляет приблизительно 5% от секвенированной популяции в двух или более последовательных раундах селекции; или 2) мотив представляет, по меньшей мере 10% от секвенированной популяции в одном или нескольких раундах селекции.
[00324] Результаты
[00325] Пилотные исследования параметров устройства для доставки и выделение клеток AT II
[00326] Два устройства для доставки, катетер AeroProbe® (Trudell Medical International) и распылитель собственного производства CRO, использовали для обеспечения последующей совместимости с несколькими клинически применимыми устройствами. Оба устройства для доставки оценивали в пилотных исследованиях с доставкой красителя синего Эванса для гарантии того, что параметры вентиляции приводят к адекватному распределению во все доли легких и альвеолярные мешочки. Оба устройства доставки продемонстрировали хорошее распределение во всех долях, включая альвеолярный компартмент, с более интенсивным окрашиванием, наблюдаемым в зависимых долях.
[00327] Протокол выделения клеток AT II оптимизировали с использованием в целом 6 легких от NHP. Оптимизация протокола привела к высокому выходу и чистоте клеток AT II, выделенных из обоих легких NHP, которые использовали во время эволюции терапевтического вектора.
[00328] Эволюция терапевтического вектора
[00329] До начала раунда 1 программы эволюции терапевтического вектора было синтезировано, получено и охарактеризовано 37 векторных библиотек. Как показано на фиг. 13А, разнообразие плазмидных библиотек оценивается как включающее от приблизительно 1×106 до >1×108 отдельных уникальных вариантов на библиотеку. Она представляет собой высококачественную и очень разнообразную исходную библиотеку вариантов AAV. Затем завершали получение каждой отдельной библиотеки для получения достаточного материала для первого раунда селекции. Как показано на фиг. 13В, все библиотеки получали на уровне, достаточном для генерации материала для проведения селекции в процессе эволюции терапевтического вектора in vivo.
[00330] Все 37 библиотек объединяли и успешно вводили NHP однократным аэрозольным введением с использованием либо AeroProbe®, либо распылителя. Перед введением библиотеки инкубировали с человеческим иммуноглобулином для внутривенного введения в концентрации 1,75 мг/мл. Это представляет собой высокую, но физиологически релевантную концентрацию человеческих NAb на слизистых легких. Доза библиотеки 1,7×1012 вг на NHP представляет собой дозу, которая приблизительно в 10 раз ниже, чем реальная максимально возможная доза, исходя из получениинных соображений. Следовательно, это представляет собой жесткое селективное давление, чтобы сделать возможным открытие вектора, способного трансдуцировать клетки AT II в альвеолярном пространстве в присутствии NAb. Легкие NHP извлекали через две недели после введения. Клетки AT II выделяли из легких, и ДНК выделяли из клеток AT II.
[00331] Успешная амплификация геномов капсида AAV
[00332] Амплификация генов капсида из ткани представляет собой успешную локализацию векторов библиотеки в представляющем интерес типе клеток. Капсиды, амплифицированные с использованием каждого устройства для доставки (фиг. 14A-B), клонировали в упаковочную плазмиду библиотеки AAV, для анализа последовательности и инициации последующего цикла селекции (при необходимости).
[00333] Анализ последовательности
[00334] Секвенирование проводили на отдельных клонах в библиотеке для определения частоты вариантов в популяции. Было выполнено секвенирование как минимум 90 клонов для каждого устройства для доставки. Варианты оценивали на наличие мотивов в данных секвенирования. Варианты были сгруппированы в мотивы на основе наличия объединяющей вариации (например, специфической точечной мутации или специфической последовательности вставки пептида в постоянном сайте внутри капсида), которая имела место во многих последовательностях. Мотив подвергался дальнейшей оценке только в том случае, если он представлял, по меньшей мере 5% от секвенированной популяции в двух или более последовательных раундах селекции или по меньшей мере 10% от секвенированной популяции в одном или более раундах селекции. Мотив, отвечающий последнему критерию, представлен на фиг. 15А-15В. Селекцию считали завершенной после первого раунда, поскольку при использовании обоих устройств для доставки отмечали высокую конвергенцию с вариантом A101 (содержащим капсидный белок SEQ ID NO:12).
[00335] На основе вышеприведенных критериев ранжирования вариант капсида A101, содержащий капсидный белок с SEQ ID NO:12, был идентифицирован как обеспечивающий повышенную эффективность доставки генов в легкие приматов после интратрахеального аэрозольного введения в присутствии нейтрализующих антител человека (NAb). A101 представляет собой химеру, состоящую в основном из AAV1, но также включающую аминокислоты из AAV2, AAV4, AAV6 и AAV9.
Пример 3
[00336] Несмотря на то, что первоначальные попытки разработки генной терапии на основе AAV для легких продемонстрировали клиническую безопасность, использование вектора AAV2 в конечном итоге не привело к эффективной трансдукции клеток легких, чтобы продемонстрировать клиническую пользу при муковисцидозе. Позднее дополнительные серотипы AAV, включая AAV1 и AAV5, показали улучшенную, но все еще не оптимальную трансдукцию клеток легких приматов после аэрозольного введения. Приведенные ниже экспериментальные данные подтверждают удивительную пригодность rAAV, содержащего капсид, содержащий капсидный белок с SEQ ID NO:12, в качестве носителя для эффективной доставки трансгенов, таких как CFTR человека, в легкое примата в присутствии нейтрализующих антител человека.
[00337] Доставка гена рекомбинантного AAV (rAAV), содержащего (i) капсид, содержащий капсидный белок с SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую репортерный трансген (GFP или EGFP), функционально связанную с промотором CAG, была охарактеризована (включая гистопатологический анализ) после введения аэрозоля трем приматам, отличным от человека (NHP). Доставка rAAV распылителем привела к надежной доставке вирусных геномов во все области легкого, включая периферические (бронхиоальвеолярные области), с минимальным системным биораспределением, и rAAV опосредовал экспрессию белка во всех областях легкого, включая альвеолы.
[00338] Материалы и методы
[00339] Анализ нейтрализующих антител
[00340] Клетки HEK2v6.11 (полученные из Университета Джона Хопкинса) высевали в черные непрозрачные 96-луночные планшеты при плотности клеток 30000 клеток/лунку в среде Игла, модифицированной по способу Дульбекко (DMEM; Corning) с 1% инактивированной нагреванием фетальной бычьей сыворотки (FBS; GE Healthcare Life Sciences) и 1% пенициллина/стрептомицина (Invitrogen). Клеткам давали прикрепиться к планшету в течение 24 ч до начала эксперимента.
[00341] Каждый образец сыворотки NHP анализировали в разведениях 1:10, 1:25, 1:50. Каждый планшет содержал положительные и отрицательные контроли для трансдукции. Образцы сыворотки NHP инкубировали с rAAV, содержащим (i) капсид, содержащий капсидный белок с SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую промотор CAG, функционально связанную с геном люцерифазы, при MOI 1000 при 37°C в течение 1 ч. После инкубации в течение 1 ч каждое разведение образца сыворотки NHP плюс rAAV добавляли в отдельные лунки черных непрозрачных 96-луночных планшетов, содержащих клетки 2V6.11. Люциферазу определяли с помощью набора для анализа люциферазы ONE-Glo EX (Promega) через 48 ч после трансдукции. При добавлении ONE-Glo EX клетки лизировали, и к клеткам добавляли субстрат люциферазы в одну стадию. Люминесценцию считывали с помощью микропланшетного ридера Cytation 3 (BioTek).
[00342] Коэффициенты вариации (CV) и стандартные отклонения рассчитывали для всех разведений образцов сыворотки NHP и каждой точки на стандартной кривой. Образцы сыворотки NHP нормализовали к положительному контролю трансдукции. Для каждого NHP определяли титр нейтрализующих антител. Титр нейтрализующих антител для каждого образца сыворотки NHP определяли как самое низкое разведение сыворотки, при котором наблюдалась трансдукция ≥50%. NHP, для которых отмечали трансдукцию ≥50% при разведении сыворотки 1:10, рассматривались как подходящие для включения в исследование.
[00343] Тест-система и иммуносупрессия
[00344] В исследование включали три яванского макака самца. Животные были в возрасте от 4 лет 10 месяцев до 9 лет 8 месяцев и имели массу тела от 4,73 кг до 7,09 кг. Животным проводили иммуносупрессию метилпреднизолоном (20 мг/кг, внутримышечно) один раз в неделю, начиная с суток -7.
[00345] Приготовление и введение тестируемого образца
[00346] Партии тестируемых образцов rAAV, содержащих (i) капсид, содержащий капсидный белок с SEQ ID NO:12, и (ii) нуклеиновую кислоту, содержащую промотор CAG, функционально связанную с EGFP, оттаивали на льду, объединяли вместе и разводили в буфере для формуляции для доставки конечной дозы 2,80×1012 вг/кг в 5 мл каждому NHP. Разведения испытуемого образца для каждого животного приведены в таблице 3. Животным проводили седацию кетамином и дексмедетомидином. Животных интубировали таким образом, чтобы конец интубационной трубки располагался приблизительно на 5 см выше киля. Животных помещали в кресло в сидячем положении для введения. Для каждого животного 5 мл разведенного тестируемого образца загружали в резервуар для распылителя AeroEclipseII (Trudell Medical). Во время введения давление доводили до 15-20 см H2O. Тестируемый образец вводили со скоростью 12-24 вдоха/мин до тех пор, пока в течение 10 импульсов не образовывался видимый туман (животным вводили дозу непрерывно в течение <40 мин). После завершения введения животных экстубировали, и седацию снимали.
[00347]
Тестирование: подготовка дозы образца
(вг)
(мл)
(мл)
4DER000043.01
4DER000043.01
4DER000043.01
[00348] Прижизненные наблюдения, некропсия и сбор тканей
[00349] Наблюдения за животными, находящимися в клетках, проводили два раза в день персоналом CRO с суток -7 до времени проведения некропсии. Массу тела определяли еженедельно, и образцы крови собирали на определенные временные точки для гематологического и биохимического анализа. Эвтаназию проводил на сутки 57±1 квалифицированный ветеринарный персонал с помощью внутривенной инъекции пентобарбитала натрия (100 мг/кг) с последующей двусторонней торакотомией и транскардиальной перфузией гепаринизированным забуференным фосфатом физиологическим раствором. После перфузии отбирали образцы легких (включая трахею), головного мозга, спинного мозга (шейный, грудной и поясничный отделы), сердца (желудочковый и предсердный отделы), печени, селезенки, скелетных мышц (трехглавая мышца плеча, латеральная широкая мышца бедра), диафрагмы и почки. Образцы тканей отбирали и быстро замораживали для последующего выделения ДНК и белка. Дополнительные образцы отбирали и фиксировали в 4% параформальдегиде (легкие и нервные ткани) или 10% нейтральном забуференном формалине (периферические ткани) для последующей заливки в парафин и приготовления срезов для иммунофлуоресценции.
[00350] Из трахеи и легких отбирали большие образцы, чтобы получить несколько образцов для каждого анализа. Легкие извлекали и пережимали как можно выше над трахеей. Правое легкое пережимали дважды приблизительно на 1 мм от главного бронха. Правое легкое извлекали разрезом между зажимами. Шестнадцать образцов для выделения ДНК и белка были отобраны из областей правого легкого, охватывающих первичные/вторичные бронхи, третичные бронхи и альвеолы, как показано на фиг. 16. Трахею и левое легкое наполняли 4% параформальдегидом и фиксировали в 10-кратном объеме 4% параформальдегида. Затем готовили срезы трахеи и левого легкого с охватом образцов трахеи, первичных/вторичных бронхов, третичных бронхов и альвеол, как показано на фиг. 16.
[00351] Биораспределение вирусного генома
[00352] Биораспределение вирусного генома определяли в Маттаванском центре Charles River Laboratories с использованием подходящего анализа вирусных геномов AAV, содержащих трансгенную последовательность EGFP. Общую ДНК экстрагировали из образцов тканей с использованием станции QIAsymphony (Qiagen) и соответствующего мини-набора DSP DNA. Реакции количественной ПЦР проводили в 96-луночных планшетах, где для каждого планшета была стандартная кривая, набор образцов для контроля качества и образцы для исследования. Дублирующие образцы для контроля качества готовили при высоком, среднем и низком числе копий/реакцию на фоне 1000 нг матричной ДНК NHP на реакцию. По возможности образцы тканевой ДНК тестировали из расчета 1000 нг на реакцию. Если было невозможно загрузить количество, указанное выше для конкретного образца (за счет слишком низкой концентрации ДНК или ограничения объема образца), то анализировали меньшее количество ДНК образца.
[00353] Все реакции с образцами проводили в трех повторах, и в третью реакцию вносили 200 копий ДНК pAAV-CAG-EGFP-SV40 для оценки потенциального ингибирования кПЦР. Если наблюдалось ингибирование кПЦР, что было показано измеренным значением в третьей лунке с внесением менее 110 копий ДНК-мишени, то образец ДНК повторно анализировали в меньшем количестве.
[00354] Биораспределение экспрессии белка
[00355] Общий белок экстрагировали из образцов тканей с использованием гомогенизатора тканей softMACS (Miltenyi Biotec) и соответствующих реагентов. EGFP количественно определяли с использованием набора GFP ELISA (Abcam), и общий белок определяли количественно с использованием набора для анализа белка Pierce BCA (ThermoFisher). Как для GFP, так и для общего белка реакции проводили в трех повторах, и для каждого набора была стандартная кривая.
[00356] Иммунофлуоресцентная визуализация
[00357] Ткани обрабатывали для получения парафиновых блоков с использованием Sakura VIP 5, используя стандартную программу для тканей собак, NHP и свиней, разработанную лабораторией Seventh Wave. Готовили срезы толщиной 10 мкм в лаборатории Seventh Wave, где они хранились при 4°С. Иммуногистохимический анализ проводили на 6 срезах легких (включая альвеолярную и бронхиальную области) и 2 срезах трахеи от каждого опытного животного (n=3) и дополнительного контрольного животного. Парафиновые срезы дегидратировали с использованием стандартного протокола окрашивания парафиновых срезов антителами. Вкратце, срезы депарафинизировали ксилолом и регидратировали с уменьшающимися концентрациями этанола в воде (100%, 90%, 70%, 50% и 30%) с последующей промывкой PBS. Извлечение антигена выполняли до окрашивания антителом с использованием комбинации теплового индуцированного извлечения (HEIR) и давления. Срезы инкубировали в течение 10 мин в кипящем цитратно-натриевом буфере, затем инкубировали в течение 3 мин под давлением. Срезы охлаждали до комнатной температуры перед окрашиванием антителами. После извлечения антигена срезы окрашивали первичным куриным поликлональным анти-GFP антителом (Abcam #13970) в разведении 1:1000, вторичным козьим антителом против куриного IgY (Abcam #175779) в разведении 1:1000 и DAPI. Флуоресцентную визуализацию проводили с использованием микроскопа Zeiss AxioObserver. Сигнал анти-GFP получали в крайнем красном канале (647 нм) через 1000 мс. Сигнал DAPI детектировали в синем канале (355 нм) при 100 мс. Все изображения обрабатывали с помощью программного обеспечения Zeiss ZenPro с использованием тех же параметров и значений интенсивности пикселей в канале анти-GFP.
[00358] Гистопатологический анализ
[00359] Обработку, заливку, приготовление срезов ткани и окрашивание гематоксилин-эозином проводили в лаборатории Seventh Wave. Ткани обрабатывали для получения парафиновых блоков с использованием Sakura VIP 5, используя стандартную программу для тканей собак, NHP и свиней. Готовили срезы толщиной 4 мкм и окрашивали гематоксилин-эозином (H&E) с использованием автоматического аппарата для окрашивания Leica XL. Гистопатологический анализ проводили на 16 срезах легкого, 4 срезах трахеи и 1 срезе киля от каждого опытного животного (n=3) и дополнительного контрольного животного, при этом гистопатолог работал «вслепую» и не знал об условии обработки. Срезы оценивали по характеру и степени показателей с использованием стандартной оценочной шкалы.
[00360] Компьютеризированные системы
[00361] Для скрининга сывороточных нейтрализующих антител данные получали и анализировали с использованием микропланшетного ридера Cytation 3 (Biotek), программного обеспечения Gen5plus версии 3.03.14 и Microsoft Excel версии 15.32.
[00362] Для количественного определения вирусных геномов в образцах тканей данные получали и анализировали с использованием системы ПЦР в режиме реального времени QuantStudio 7 Flex, программного обеспечения для ПЦР в реальном времени QuantStudio v1.4, Microsoft Excel и GraphPad Prism версии 8.1.2.
[00363] Для количественного определения экспрессии EGFP в образцах тканей данные получали и анализировали с использованием микропланшетного ридера Cytation 3 (Biotek), программного обеспечения Gen5plus версии 3.03.14, Microsoft Excel версии 15.32 и GraphPad Prism версии 8.1.2.
[00364] Для репрезентативной иммунофлуоресцентной визуализации изображения получали с использованием микроскопа Zeiss Axio Observer z1 и программного обеспечения ZenPro. Изображения процессировали с помощью программного обеспечения ZenPro. Изображения переносили в Microsoft PowerPoint версии 15.32 для презентации.
[00365] Весь анализ данных и компиляцию проводили на Macbook Pro под управлением OSX (10.12.6).
[00366] Результаты и обсуждение
[00367] Скрининг нейтрализующих анти-AAV антител идентифицирует NHP для включения в исследование
[00368] Анализ нейтрализующих антител использовали для оценки уровней нейтрализующих антител против капсида AAV, содержащего капсидный белок с SEQ ID NO:12, в сыворотке приматов, отличных от человека (NHP). Для каждого образца сыворотки NHP определяли титр нейтрализующих антител. Животное считали серонегативным и соответствующим критериям включения в исследование, если наблюдали трансдукцию на уровне ≥50% при разведении сыворотки 1:10.
[00369] В целом оценивали 20 образцов сыворотки NHP в четырех 96-луночных планшетах. Критерии приемлемости анализа были установлены для 1) коэффициента вариации (CV) для стандартной кривой, 2) CV для неизвестных образцов сыворотки и 3) процентного отклонения от фактического входного белка для стандартной кривой. Приемлемые значения CV для стандартной кривой были установлены на уровне < 25%, но фактические значения CV не превышали 5%. Приемлемые CV для неизвестных образцов сыворотки в пределах количественного определения были установлены на уровне < 30%, но фактические значения CV не превышали 19%. Приемлемое процентное отклонение от входного белка для стандартной кривой было установлено на уровне < 25%, но фактическое процентное отклонение не превышало 19%. Все планшеты соответствовали всем критериям приемлемости анализа, и данные, полученные с этими планшетами, использовали для оценки. В целом, 11 (55%) анализированных образцов сыворотки NHP были серонегативными для капсида, содержащего капсидный белок с SEQ ID NO:12 (фиг. 17). Для включения в исследование были отобраны NHP с тремя самыми высокими процентами трансдукции при разведении сыворотки 1:10. Все выбранные NHP продемонстрировали трансдукцию выше трансдукции, наблюдаемой в отсутствие сыворотки NHP. Это наблюдение было отмечено в предыдущих исследованиях и, вероятно, является результатом благоприятного взаимодействия с неизвестным белком сыворотки. Номера отобранных NHP и процент трансдукции при разведении сыворотки 1:10 представлены в таблице 4.
[00370]
NHP, включенные в исследование
[00371] Доставка с распылителем вариантного капсида, содержащего капсидный белок с SEQ ID NO:12, хорошо переносится NHP
[00372] Схема исследования представлена в таблице 5. Все животные восстановились до нормы после введения тестируемого образца и оставались живыми до запланированных дат проведения некропсии. Никакой значимой клинической патологии не было зарегистрировано ни у одного животного на любую временную точку во время прижизненной части исследования. У некоторых животных на некоторые временные точки отмечали незначительные изменения, выходящие за пределы референтных диапазонов для гематологических и биохимических показателей, но эти изменения интерпретировались как нормальные физиологические колебания. Никакой серьезной патологии не было зарегистрировано во время макроскопического исследования при вскрытии.
[00373]
Обобщенный дизайн исследования
(AeroEclipse II)
вг/кг
сердце
печень
скелетные мышцы (трехглавая мышца плеча, четырехглавая мышца, диафрагма)
почка
селезенка
ЦНС (головной мозг, спинной мозг)
ELISA (легкое & кПЦР+ ткани)
IHC (легкое & ELISA+ ткани)
[00374] Вариантный капсид, содержащий капсидный белок с SEQ ID NO:12, опосредует надежную доставку гена во все области легкого
[00375] Вирусные геномы количественно определяли с использованием кПЦР для всех образцов, полученных при некропсии, для определения геномного биораспределения rAAV, содержащего вариантный капсид (содержащего капсидный белок SEQ ID NO:12) и нуклеиновую кислоту, содержащую трансген EGFP, функционально связанную с промотором CAG.
[00376] Большое количество вирусных геномов, варьирующее от приблизительно 104 до 105 вг/мкг, наблюдали во всех 48 образцах легких (n=16 образцов на NHP; n=3 NHP), которые представляли собой образцы из альвеолярных мешочков, третичных бронхов и первичных/вторичных бронхов (фиг. 18). Для всех трех животных не было отмечено значимых различий в количестве вирусных геномов в разных долях легких или разных областях легких. Небольшое количество образцов сердца и печени, 3 (из 15) образца сердца от NHP V003424 и 10 (из 10) образцов печени от NHP V002969, содержали детектированные вирусные геномы. Однако количество присутствующих вирусных геномов в расчете на мкг ДНК было в 1000-10000 раз ниже, чем количество вирусных геномов, находящихся в легких. Содержание геномов во всех тестированных образцах скелетных мышц (трехглавая мышца плеча, латеральная широкая мышца бедра), диафрагмы, почек, селезенки, головного и спинного мозга были ниже предела количественного определения (BLQ). Таким образом, доставка rAAV распылителем приводит к надежной доставке вирусных геномов во все области легкого с минимальной системной экспозицией.
[00377] Вариантный капсид (содержащий капсидный белок с SEQ ID NO:12) опосредует экспрессию белка во всех областях легкого и трансдуцирует многочисленные типы клеток
[00378] Экспрессию белка EGFP количественно определяли для всех образцов, для которых результаты кПЦР продемонстрировали присутствие вирусных геномов выше нижнего предела количественного определения анализа. Данный набор образцов включал все образцы легких от всех трех NHP, 3 (из 15) образца сердца от NHP V003424 и 10 (из 10) образцов печени от NHP V002969. Экспрессию EGFP отмечали во всех 48 образцах легких (n=16 образцов на NHP; n=3 NHP), которые представляли собой образцы из альвеолярных мешочков, третичных бронхов и первичных/вторичных бронхов (фиг. 19). Экспрессия была самой высокой для животного V002969 во всех областях и во всех долях легкого. Для всех трех животных не было отмечено существенных различий в уровне экспрессии белка EGFP в разных долях легких. В целом образцы из альвеолярных областей содержали среднее или выше среднего количество EGFP, но эта тенденция не была статистически значимой. В образцах печени кПЦР+ от NHP V002969 детектировалась экспрессия белка EGFP, но уровни экспрессии были на несколько порядков ниже, чем уровни экспрессии в легких.
[00379] Полученные результаты согласуются с данными по биораспределению генома, демонстрирующими, что локализация генома в печени была на несколько порядков ниже, чем локализация генома в легких (фиг. 18). В образцах сердца кПЦР+ от NHP V003424 не было обнаружено детектируемой экспрессии белка EGFP. Полученные результаты свидетельствуют о том, что доставка rAAV с помощью распылителя, содержащего капсид с капсидным белком с SEQ ID NO:12 и нуклеиновой кислотой, кодирующей трансген, опосредует экспрессию белка во всех областях легкого.
[00380] Иммунофлуоресцентную визуализацию выполняли для дальнейшего определения степени трансдукции клеток в различных областях легкого. Были сканированы срезы, представляющие 6 секций легкого (включая альвеолярную и бронхиальную области) и 2 среза трахеи от каждого опытного животного (n=3) и дополнительного контрольного животного, и для каждой области были получены репрезентативные изображения. В целом, экспрессия EGFP была самой высокой у животного V002969 во всех областях и во всех долях легкого, что соответствует наблюдаемой относительной экспрессии у животных, определенной с использованием ELISA. В трахее и бронхах экспрессия EGFP наблюдалась преимущественно в клетках реснитчатого эпителия (фиг. 20). В альвеолах наблюдали высокую экспрессию EGFP (фиг. 20), но клетки ATI и ATII невозможно определить без использования специфических клеточных маркеров. Эти результаты демонстрируют, что доставка с распылителем rAAV, содержащего капсид, содержащий капсидный белок SEQ ID NO:12 и нуклеиновую кислоту, кодирующую трансген, опосредует экспрессию белка во всех областях легкого.
[00381] Введение rAAV, содержащего капсид, содержащий капсидный белок с SEQ ID NO:12, является безопасным и не приводит к воспалению легочной ткани
[00382] Наблюдения за животными, находящимися в клетках, в которых их содержали, проводили два раза в сутки на протяжении всей прижизненной части исследования, начиная за неделю до введения дозы. Отсутствовали какие-либо существенные клинические признаки патологии у всех животных на протяжении всей прижизненной части исследования. Гематологический и биохимический анализы образцов крови проводили раз в две недели на протяжении всей прижизненной части исследования и за одну неделю до введения дозы. Хотя некоторые индивидуальные гематологические и/или биохимические показатели выходили за пределы референтного диапазона, эти значения в значительной степени интерпретировались как физиологические колебания.
[00383] После некропсии проводили гистопатологический анализ легочной ткани и сравнивали с контрольным животным (без введения). Очаговое кровоизлияние, черный пигмент и минимальные инфильтраты мононуклеарных клеток в альвеолярных пространствах наблюдали у всех животных, и они являются обычными случайными находками у обезьян. Кроме того, подслизистые лимфоидные инфильтраты в трахее и воспаление на слизистой поверхности киля также, вероятно, носят случайный характер и не связаны с введением тестируемого образца. Ни одно из наблюдений не было признано неблагоприятным. Никакие результаты у обработанных животных не отличались или не были более серьезными, чем те, которые наблюдались у контрольного животного.
[00384] Выводы
rAAV, содержащий капсид с вариантным капсидным белком SEQ ID NO:12 и нуклеиновую кислоту, кодирующую репортерный трансген (EGFP), характеризовали при аэрозольной доставке репортерного гена яванским макакам. Сыворотки предварительно подвергали скринингу для выявления животных, которые были серонегативными в отношении ранее продуцированных нейтрализующих антител к капсидам тестируемого образца. Животные (n=3) получали максимально возможную дозу rAAV, доставляемую с использованием устройства AeroEclipseII, клинически значимого действующего распылителя.
[00385] Большое количество вирусных геномов (по данным кПЦР) и в результате высокую экспрессию EGFP (по данным ELISA и иммуноокрашивания) наблюдали во всех образцах легких от всех 3 NHP, включенных в исследовании, которые представляли собой образцы из альвеолярных мешочков, третичных бронхов и первичных /вторичных бронхов. Небольшое количество образцов сердца и печени имело низкий, но поддающийся детектированию уровень вирусного генома, и во всех образцах всех других тканей вирусный геном не детектировался. Полученные данные демонстрируют, что доставка rAAV через распылитель приводит к надежной доставке вирусных геномов во все области легкого с минимальным системным биораспределением, и rAAV опосредует экспрессию белка во всех областях легкого, включая альвеолы. Доставка rAAV и экспрессия EGFP были стабильными среди животных с равномерным распределением на нескольких уровнях бронхов и альвеол и даже распределением по верхним, средним и нижним отделам. Данные об экспрессии белка согласуются с данными по биораспределению генома, демонстрирующими, что локализация генома в печени была на несколько порядков ниже, чем локализация генома в легких.
[00386] Дополнительные эксперименты проводили для определения эффективности и специфичности для клеток AECII в легких приматов. Если желательна дополнительная специфичность помимо того, что является присущим для rAAV, содержащему капсид, содержащий капсидный белок с SEQ ID NO: 12, то для направления экспрессии представляющего интерес трансгена применяют клеточно-специфический промотор. Эксперименты, описанные здесь, показывают, что rAAV, содержащий капсидный белок с SEQ ID NO:12, можно использовать для безопасной и эффективной доставки трансгена CFTR в легкие субъектов с муковисцидозом и для доставки терапевтических генов для лечения других легочных заболеваний.
Пример 4
[00387] Резюме
[00388] Была создана клеточная культуральная модель клеток альвеолярного эпителия легкого типа II (AECII) из свежевыделенных легких приматов, отличных от человека, и легких человека-донора, отклоненных для трансплантации. Данную модель использовали для характеристики rAAV, содержащего капсид, содержащий вариантный капсидный белок SEQ ID NO:12, идентифицированный как мишень для клеток AECII. Эффективность трансдукции rAAV и природного серотипа AAV5 оценивали в культуре клеток AECII на границе раздела воздух-жидкость (ALI). rAAV, содержащий капсид, содержащий вариантный капсидный белок SEQ ID NO:12, после апикальной трансдукции с множественностью инфекции (MOI) 35000 показал усиленную трансдукцию клеток AECII по сравнению с AAV5. rAAV также проявил высокую устойчивость к человеческим анти-AAV антителам.
[00389] Для подтверждения того, что капсид rAAV нацелен на клетки AECII и что инфекционность перносится на клетки AECII человека, клетки AECII выделяли из легких NHP и легких человека-донора, отклоненных для трансплантации, и культивировали на границе раздела воздух-жидкость для имитации среды легких. Эффективность трансдукции rAAV определяли по экспрессии репортерного зеленого флуоресцентного белка с усиленными флуоресцентными свойствами (eGFP), управляемой промотором CAG, на определенные временные точки после инфицирования и по сравнению с AAV5.CAG-eGFP. Полученные данные демонстрируют, что варианты rAAV, содержащие капсид, содержащий вариантный капсидный белок SEQ ID NO:12, являются более инфекционными, чем встречающиеся в природе серотипы (т.е. являются более эффективными трансдукторами, чем капсид AAV5), потенциально обеспечивая усовершенствование лечение генетических заболеваний.
[00390] Одна общая проблема в доклинических и клинических исследованиях генной терапии с AAV заключается в том, что ранее продуцированные нейтрализующие антитела могут ингибировать успешную трансдукцию. Для оценки способности rAAV, содержащего капсид, содержащий вариантный капсидный белок SEQ ID NO:12, уклоняться от нейтрализующих антител в человеческой популяции по сравнению с AAV дикого типа, rAAV и AAV1, AAV2, AAV5, AAV8 и AAV9 дикого типа анализировали против человеческого IVIG в люциферазном анализе in vitro. Данные, приведенные здесь, свидетельствуют о том, что rAAV, содержащий капсид, содержащий вариантный капсидный белок с SEQ ID NO:12, обладает устойчивостью к нейтрализующим антителам при воздействии человеческого IVIG (в 4 раза больше, чем AAV5, и в 32 раза больше, чем AAV2), критический компонент для обработки с аэрозольной доставкой и селективного давления, используемых в процессе эволюции терапевтического вектора.
[00391] Материалы и методы
[00392] Выделение клеток альвеолярного эпителия II типа легких
[00393] Легкие приматов, отличных от человека (NHP, Cynomolgus macaque, CRO), или человеческие донорские легкие, отклоненные для трансплантации (Donor Network West, номер донора: AGES430), использовали для выделения клеток альвеолярного эпителия II типа (AECII), как описано Fang et al.1,2. Для выделения клеток NHP использовали все легкое полностью; для донорских легких человека вырезали правую среднюю долю и использовали для выделения клеток. Бронхи промывали PBS, не содержащим кальций или магний (ThermoFisher), содержащим ЭДТА (Sigma) и ЭГТА (Sigma), с последующим заполнением эластазой (Worthington Chemicals). Ткань инкубировали в течение 1 ч при 37°С. Легкое гомогенизировали, трахею и бронхи удаляли. Клеточный гомогенат пропускали через слои марли для удаления крупных оставшихся кусочков ткани. Клетки последовательно пропускали через сита 100 мкм и 20 мкм. Клетки загружали в двухступенчатый градиент плотности Percoll (ThermoFisher) (70% и 30%) и центрифугировали при 1800 об/мин в течение 20 мин. Промежуточный слой удаляли, центрифугировали и дважды промывали PBS, не содержащим кальций или магний. Моноциты и макрофаги удаляли с помощью магнитных частиц CD14 и CD45 Dynabeads (ThermoFisher). Оставшиеся клетки инкубировали в течение ночи при 37°С в планшетах, покрытых IgG, для удаления Т-клеток. На следующий день неприкрепленные клетки собирали с планшетов, центрифугировали и помещали в гипотонический раствор для лизиса эритроцитов (ACK Lysis Buffer 1:10, ThermoFisher). Полученные клетки высевали на вставки, покрытые человеческим коллагеном IV (Sigma, человеческий плацентарный коллаген IV, 18-24 ч при 25°C) при плотности 1,5×106 клеток/см2 и инкубировали при 37°C, 5% CO2. Клетки культивировали в базальной среде для эпителиальных клеток дыхательных путей (ATCC) с коммерческими добавками, фетальной бычьей сывороткой (10%, HyClone, ThermoFisher) и инсулином, трансферрином и селеном (1:200, ThermoFisher). Через сутки после посева вкладыши дважды промывали PBS для удаления неадгезированных клеток. Верхняя часть вставки оставалась сухой, чтобы обеспечить формирование границы раздела воздух-жидкость (ALI).
[00394] Окрашивание красителем LysoTracker
[00395] Клетки, достигшие ALI, анализировали окрашиванием LysoTracker в культуре. LysoTracker (ThermoFisher) представляет собой индикаторный краситель, который поглощается сильнокислотными компонентами живых клеток, такими как лизосомы и пластинчатые тела в клетках AECII. Он обычно используется для маркировки клеток AECII.
[00396] LysoTracker на адгезированных клетках и микроскопия
[00397] Клетки на вкладышах, предназначенные для окрашивания, дважды промывали PBS. Концентрат LysoTracker разводили средой (1:1000). Во вставку для окрашивания живых клеток добавляли 100 мкл разбавленного LysoTracker. Клетки инкубировали в течение 5 мин при 37°С. После инкубации вставку трижды промывали PBS и визуализировали на флуоресцентном микроскопе Zeiss Axio Observer D.1. Неокрашенную лунку использовали в качестве контроля и для установки экспозиции.
[00398] LysoTracker в клеточной суспензии и проточная цитометрия
[00399] Клетки инкубировали с 0,05% трипсин-ЭДТА (ThermoFisher) в течение 10 мин при 37°C. Трипсин инактивировали соевым ингибитором трипсина (ThermoFisher), клетки собирали со вставок и центрифугировали при 300×g в течение 4 мин. Концентрат LysoTracker разводили в среде (1:1000). К каждому клеточному осадку добавляли 100 мкл разбавленного LysoTracker и перемешивали на вортексе на половинной скорости. Образец неокрашенных клеток использовали в качестве контроля и для гейтирования в проточной цитометрии. Клетки инкубировали в течение 5 мин при 37°С. После инкубации клетки центрифугировали и дважды промывали PBS. Клетки ресуспендировали в PBS и анализировали на проточном цитометре BD Accuri C6 Plus. LysoTracker-положительная популяция была идентифицирована как популяция со сдвигом вправо по сравнению с неокрашенным контролем.
[00400] Включение EdU
[00401] Пролиферацию клеток определяли с использованием набора Click-iT EdU Alexa Fluor (ThermoFisher) в соответствии с инструкциями производителя. Вкратце, клетки обрабатывали EdU в течение 2 ч, дважды промывали PBS и фиксировали 4% параформальдегидом (15 мин при 4°C). Готовили реакционную смесь Click-iT, включающую азид Alexa Fluor, и инкубировали с клетками в течение 30 мин при 25°C. После проведения реакции клетки дважды промывали PBS и докрашивали DAPI в течение 10 мин при 25°C. Визуализацию клеток проводили на флуоресцентном микроскопе Zeiss Axio Observer D.1.
[00402] Трансдукция вектором
[00403] Клетки NHP AECII трансдуцировали через 2 суток после посева. Клетки AECII человека трансдуцировали через сутки после посева. В день проведения трансдукции три вставки инкубировали с 0,05% трипсин-ЭДТА (ThermoFisher) в течение 10 мин при 37°С. Трипсин инактивировали с использованием соевого ингибитора трипсина (ThermoFisher). Клетки собирали со вставок и подсчитывали на гемоцитометре. Среднее количество клеток определяли в расчете на вкладыш и использовали для расчета общего количества вирусных геномов, необходимых для каждой вставки. Множественность инфекции (MOI) 35000 использовали для всех экспериментов в общем объеме 100 мкл на вставку. Клетки обрабатывали апикально в течение 48 ч rAAV, содержащим капсид с капсидным белком с SEQ ID NO:12 и геном GFP, функционально связанным с промотором CAG, или нативным AAV серотипа 5, содержащим ген GFP, функционально связанный с промотором CAG. Через 2 суток после инфицирования вирус удаляли со вставок, чтобы восстановить границу раздела воздух-жидкость. Через 3 суток после инфицирования клетки NHP собирали для анализа, всего пять дней в культуре. Через 6 и 10 суток после инфицирования человеческие клетки собирали для анализа, в общей сложности семь и одиннадцать суток в культуре.
[00404] Иммуноцитохимия (ICC)
[00405] Клетки дважды промывали PBS и фиксировали 4% параформальдегидом (15 мин при 4°C). Клетки блокировали 5% козьей сывороткой и 2% бычьим сывороточным альбумином (BSA) в 0,2% Тритон X-100 в PBS в течение 30 мин. Клетки инкубировали с антителом к сурфактантному белку С или контрольным IgG (MilliporeSigma, 1:100) в течение 2 ч при 25°С. Первичное антитело трижды отмывали 0,2% Тритоном в PBS с последующей инкубацией с вторичным антителом (козье антикроличье, конъюгированное с Alexa Fluor 555, 1:500) в течение 30 мин при 25°C. Клетки докрашивали DAPI в течение 10 мин при 25°C и трижды промывали PBS. Клетки визуализировали на флуоресцентном микроскопе Zeiss Axio Observer D.1 с контрольным IgG для установки экспозиции.
[00406] Проточная цитометрия
[00407] Клетки после трансдукции снимали с использованием трипсина-ЭДТА 0,05% (ThermoFisher) в течение 10 мин при 37°C. Трипсин инактивировали соевым ингибитором трипсина (ThermoFisher), клетки собирали со вставок и центрифугировали при 300×g в течение 4 мин. Клетки ресуспендировали в PBS и анализировали на проточном цитометре BD Accuri C6 Plus. Трансдуцированные (положительные на eGFP) клетки идентифицировали как популяцию со сдвигом вправо по сравнению с нетрансдуцированным контролем.
[00408] Анализ устойчивости к нейтрализующим антителам
[00409] Клетки HEK 2V6.11 (полученные из Университета Джона Хопкинса) высевали на 96-луночные планшеты с плотностью 3×104 клеток на лунку. Через 24 ч после посева rAAV, содержащий (i) капсид, содержащий капсидный белок с SEQ ID NO:12 («A101»), и (ii) трансген люциферазы, функционально связанный с промотором CAG, и AAV дикого типа (серотипы AAV1, AAV2 , AAV5, AAV8 и AAV9, несущие репортерный трансген люциферазы), инкубировали при 37°C в течение 1 ч с пятью разведениями 1:50, 1:100, 1:200, 1:400, 1:800, 1:1600 иммуноглобулина человека для внутривенного введения (IVIG) до инфицирования. Затем клетки инфицировали rAAV и AAV дикого типа при MOI 1000. Каждый планшет содержал положительные и отрицательные контроли для трансдукции. Положительным контролем был либо rAAV, либо AAV дикого типа в отсутствии IVIG. Отрицательным контролем для трансдукции служила среда без сыворотки или AAV.CAG-люциферазы. Активность люциферазы измеряли через 48 ч после инфицирования с использованием планшетного ридера Cytation 3 (Biotek).
[00410] Компьютеризированные системы
[00411] Программное обеспечение FloJo, LLC использовали для анализа данных проточной цитометрии.
[00412] Microsoft Excel использовали для расчета средних значений и стандартных отклонений от выходных данных FloJo и построения гистограмм.
[00413] Для скрининга нейтрализующих антител сыворотки данные получали и анализировали с использованием микропланшетного ридера Cytation 3 (Biotek), программного обеспечения Gen5plus версии 3.03.14 и Microsoft Excel.
[00414] Результаты и обсуждение
[00415] Характеристика культуры AECII ALI приматов, отличных от человека
[00416] Клетки альвеолярного эпителия легкого типа II (AECII) выделяли из легкого примата, отличного от человека (NHP), и культивировали на границе раздела воздух-жидкость (ALI). Клетки культивировали на вставках, покрытых коллагеном, и анализировали на наличие специфических маркеров AECII, краситель LysoTracker и поверхностно-активного белка-C (SPC). Клетки на сутки 1 и 5 содержали более 90% положительных на LysoTracker клеток, что было показано ICC и количественно проточной цитометрией (фиг. 21A и 21B). Культивированные клетки AECII также анализировали на наличие SPC, зрелого маркера AECII, на сутки 1 и 5 культивирования. Клетки экспрессировали SPC, что было определено ICC на временные точки (фиг. 21C). Чтобы дополнительно охарактеризовать систему культивирования AECII, исследовали во времени клеточный митоз. Митоз анализировали по включению EdU с последующей реакцией Click-iT с азид-содержащим флуоресцентным красителем.
[00417] Митоз анализировали с суток 2 по сутки 5 флуоресцентной микроскопией и окрашиванием ядер (фиг. 21D). Митоз был самым высоким через 2 суток после посева; по мере поддержания клеток в культуре количество клеток с митозом снижалось, о чем свидетельствует отсутствие включения EdU в ядра.
[00418] Характеристика rAAV, содержащего капсид с вариантным капсидным белком с SEQ ID NO:12, в системе культивирования AECII ALI приматов, отличных от человека
[00419] Через 2 суток после посева клеток AECII, выделенных от NHP, их трансдуцировали rAAV, содержащим капсид с вариантным капсидным белком с SEQ ID NO:12 и трансген GFP под контролем промотора CAG, или AAV, содержащим капсид AAV5 дикого типа и трансген GFP под контролем промотора CAG при множественности инфекции 35000. Через 3 суток после инфицирования, в целом 5 суток в культуре, клетки анализировали на экспрессию eGFP с использованием ICC и проточной цитометрии. rAAV, содержащий капсид, содержащий капсидный белок с SEQ ID NO:12, давал достоверно более высокий процент eGFP-положительных клеток (~30%) по сравнению с AAV5 (~8%), что указывает на более высокую эффективность трансдукции (фиг. 22A и 22B).
[00420] Характеристика культуры AECII ALI человека
[00421] Правую среднюю долю легкого человека, полученную из Donor Network West (номер донора: AGES430), вырезали из оставшихся легких и выделяли клетки AECII. Клетки культивировали на вставках, покрытых коллагеном, и получали ALI. Для определения чистоты исследовали два маркера для AECII, LysoTracker и экспрессию белка SPC. LysoTracker анализировали во времени от свежевыделенного изолята (сутки 0) до суток 11 в культуре. LysoTracker-положительные клетки составляли приблизительно 80% до суток 7. Популяция LysoTracker-положительных клеток снижалась приблизительно до 50% на сутки 11, поскольку AECII трансдифференцируется в AECI (фиг. 23A и 23B). Клетки также демонстрировали высокую экспрессию SPC на анализированные временные точки (фиг. 23C).
[00422] Митоз анализировали во времени, как в системе NHP AECII. Аналогичный результат наблюдали, когда наибольшее количество клеток с митозом появлялось в течение первых 3 суток в культуре и снижалось во времени (фиг. 23D).
[00423] Характеристика 4D-A101 в системе культивирования AECII ALI человека
[00424] Через 1 сутки после посева клетки AECII человека инфицировали rAAV, содержащим капсид, содержащий капсидный белок с SEQ ID NO:12 или AAV5, содержащий трансген eGFP, с множественностью инфекции 35000. Через 6 и 10 суток после инфицирования клетки визуализировали на предмет экспрессии eGFP в качестве показателя эффективности трансдукции. rAAV, содержащий капсид, содержащий капсидный белок SEQ ID NO:12, как на сутки 6, так и на сутки 10 после инфицирования показал более высокий уровень eGFP-позитивных клеток по сравнению с AAV5 (фиг. 24). Клетки сохраняли окрашивание на LysoTracker, указывающее на фенотип клеток AECII.
[00425] Анализ устойчивости к нейтрализующим антителам
[00426] Относительные световые единицы (RLU) каждого вектора люциферазы AAV в каждом разведении IVIG нормализовали относительно положительного контроля трансдукции, только вектора люциферазы AAV. После нормализации каждый вектор при каждом разведении IVIG преобразовывали и выражали в виде процента трансдукции. Высокий процент трансдукции коррелирует с низкой нейтрализацией, и низкий процент трансдукции коррелирует с высокой нейтрализацией. Затем для каждого вектора люциферазы AAV определяли титр нейтрализующего антитела, определяемый как наименьшее разведение, при котором он достигает более чем 50% трансдукции. В целом, 4D-A101 продемонстрировал повышение устойчивости к нейтрализующим антителам по сравнению с каждым тестированным AAV дикого типа (фиг. 25, иллюстрирующая процентную трансдукцию клеток HEK2v6.11 в присутствии человеческого IVIG). 4D-A101 продемонстрировал более чем 32-кратное повышение уклонения от антител по сравнению с AAV1, AAV2, AAV8 и AAV9 дикого типа и 4-кратное повышение уклонения от антител по сравнению с AAV5 дикого типа (таблица 6).
[00427] Таблица 6. Титры нейтрализующих антител для AAV1, AAV2, AAV5, AAV8, AAV9 и 4D-A101 дикого типа. Показано разведение IVIG, необходимое для снижения активности люциферазы до 50% по сравнению с отсутствием IVIG.
[00428] Выводы
[00429] Клетки AECII выделяли и культивировали в ALI, и они сохраняли высокую чистоту, определяемую окрашиванием Lysotracker и экспрессией SPC. rAAV, содержащий капсид, содержащий капсидный белок с SEQ ID NO:12, продемонстрировал более высокую эффективность трансдукции по сравнению с AAV5 как в культуральных системах NHP, так и AECII ALI человека.
[00430] Была показана способность rAAV, содержащего капсид, содержащий капсидный белок SEQ ID NO:12, уклоняться от нейтрализующих антител, находящихся в пуле IVIG человека, что повышает его терапевтический потенциал для пациентов. Основываясь на этих результатах, вполне вероятно, что гораздо меньшая часть целевой популяции пациентов будет исключаться из лечения по причине наличия ранее продуцированных нейтрализующих антител.
Пример 5
[00431] Исследования по скринингу формуляции были проведены для выбора и характеристики оптимальных буферов для формуляции в отношении растворимости, доставки и сохранности рекомбинантных частиц AAV, содержащих капсид, содержащий капсидный белок VP1 SEQ ID NO:12, капсидный белок VP2, содержащий аминокислоты 138- 736 SEQ ID NO:12 и капсид VP3, содержащий аминокислоты 203-736 SEQ ID NO:12 и нуклеиновой кислоты, кодирующей трансген (например, репортерный ген, такой как люцифераза или зеленый флуоресцентный белок, или терапевтический трансген, такой как CFTR).
[00432] Все исследования проводили с капсидами (содержащими VP1 SEQ ID NO:12), очищенными аффинной (AVB) и анионообменной (CIMQA) хроматографией. Первоначальные исследования проводились с очищенным материалом, предварительно формулированным в DPBS (фосфатно-солевой буфер Дульбекко) с 0,005% Плюроника. Эти партии либо разбавляли, либо подвергали замене на соответствующие буферные системы. Более поздние исследования были проведены с партиями, буферы которых были заменены непосредственно на соответствующие буферные системы сразу после очистки CIMQA.
[00433] Стабильность в циклах замораживания-оттаивания
[00434] Небольшие аликвоты образцов (≤ 100 мкл), содержащие ~2-3×1013 вирусных геномов на мл, переносили в полипропиленовые пробирки емкостью 1,5 мл и подвергали 5 или 10 циклам быстрого замораживания (т.е. резкому охлаждению при -80°C) с последующим оттаиванием при комнатной температуре до исчезновения кристаллов льда. В отдельном исследовании объем ≥ 5 мл, содержащий приблизительно ~6×1011 вирусных геномов/мл, переносили в полипропиленовую пробирку Falcon емкостью 15 мл и помещали при температуре -80°C приблизительно на 1 ч (до замораживания), и затем оттаивали в течение приблизительно 1 ч при комнатной температуре до тех пор, пока при осторожном перемешивании не обнаруживали кристаллов льда. Данный процесс повторяли еще 2 раза (3X FT). Образец хранили в течение ночи при комнатной температуре после третьего цикла замораживания-оттаивания.
[00435] Ускоренная оценка стабильности при 5°C, комнатной температуре, 37°C и 40°C/75% относительной влажности
[00436] Различные образцы с концентрацией от приблизительно 1×1012 до 6×1013 вирусных геномов на мл разделяли на аликвотные порции (≤ 100 мкл), которые переносили в полипропиленовые пробирки емкостью 1,5 мл или полипропиленовые криопробирки емкостью 2,0 мл и хранили при 5°C, комнатной температуре, 37°C и 40°C/75% относительной влажности в течение заданного времени в каждом протоколе исследования. Пробирки с образцами помещали в вентилируемые коробки для хранения, и обычно использовали Парафильм, чтобы свести к минимуму влияние испарения во время хранения образцов.
[00437] Стабильность при перемешивании
[00438] Небольшие аликвотные порции образцов (≤ 100 мкл), содержащие ~2-3×1013 вирусных геномов на мл, переносили в полипропиленовые пробирки емкостью 1,5 мл и помещали в коробку для образцов, которая была присоединена к вихревому смесителю, установленному на 1500 об/мин. Образцы перемешивали при комнатной температуре в течение 2-4 суток.
[00439] Моделирование состояния заряда
[00440] Калькулятор состояния заряда на основе Excel (онлайн-загрузка; Gale Rhodes, University of Southern Maine) модифицировали для автоматического извлечения и количественного определения ионизируемых и лабильных остатков. Суммарный заряд мономера капсидного белка (например, VP1 и VP3) определяли путем добавления частичных отрицательных и частичных положительных зарядов для каждого ионизируемого вещества в каждом интервале значений pH (≤ 1 единица pH):
[00441] Вклад отрицательного заряда = количество остатков * -1 * (10^-(pKa-pH)/((10^-(pKa-pH))+1)
[00442] Вклад положительного заряда = количество остатков * (10^(pKa-pH)/((10^(pKa-pH))+1)
[00443] Указанная изоэлектрическая точка (pI) представляет собой значение pH, при котором суммарный заряд был математически определен как самый близкий к 0.
[00444] Коэффициент экстинкции при 280 нм (теоретический)
[00445] ProtParam (web.expasy.org/protparam/) использовали для определения молярной массы белков VP1, VP2 и VP3 на основе известных первичных последовательностей. Молярный коэффициент экстинкции при 280 нм рассчитывали по следующему уравнению: (M-1 см-1)=(#Trp)(5500) + (#Tyr)(1490) + (#цистин)(125). Примечание: все VP1, VP2 и VP3 содержат 5 остатков цистеина, но, согласно литературным данным, цистеины AAV не образуют дисульфидных связей (SS=цистин). Abs280нм0,1% (или единицы поглощения для раствора с концентрацией 1 мг/мл) затем рассчитывали делением молярного коэффициента экстинкции на молярную массу.
[00446] Поглощение при 280 нм (А280) и мутность по данным УФ-спектрофотометрии
[00446] Приблизительно 30 мкл образца (или буфера) переносили в 3 мм кварцевую кювету и снимали УФ-спектры (250-400 нм; интервал 1 нм) на спектрофотометре NanoDrop. Альтернативно несколько испытуемых образцов (или буферов) переносили в 96-луночный титрационный микропланшет UV Star и снимали УФ-спектры (250-400 нм; интервал 1 нм) на планшетном ридере Cytation. Длину оптического пути лунок с образцами титрационного микропланшета определяли определяли эмпирически измерением оптической плотности при 977 и 900 нм и использовали следующее уравнение:

[00448] Спектры буфера и образца нормализовали к ячейке с длиной оптического пути 1 см делением оптической плотности (O.D.), измеренной при каждой длине волны, на длину оптического пути кюветы или лунки для образца. Нормализованные спектры усредняли, при этом анализировали дубликаты. Спектры образцов были скорректированы на фон вычитанием значений нормализованной оптической плотности для соответствующего контрольного холостого буфера. Вклад светорассеяния при 280 нм (LS280) рассчитывали с использованием логарифмической экстраполяции, полученной для непоглощающей УФ-области (~300-400 нм) и вычитали из O.D.280 -LS280=A280. Светорассеяние с поправкой при A280 умножали на коэффициент разбавления образца (при необходимости) и делили на теоретический коэффициент экстинкции VP3 (Abs280нм0,1%=1,71), с получением расчетной концентрации капсидного белка в мг/мл белка. Однако для данного метода свойственна завышенная оценка капсидного белка за счет поглощения ДНК. Альтернативно значения при A280 представлены в этом отчете без дополнительного преобразования, чтобы избежать завышенной оценки капсидного белка за счет поглощения ДНК. Значения O.D.350 первоначально использовали для полуколичественного мониторинга изменений светорассеяния или мутности, возникающих в результате самоассоциации. Однако в этом отчете также представлены значения 350/A280 для нормализации рассеяния к количеству капсида в растворимой фракции.
[00449] рН
[00450] Для измерения рН образца или буфера соответственно использовали рН-зонды малого или большого объема. pH-зонды были откалиброваны с использованием предварительно упакованных стандартов pH Mettler Toledo с pH 4,01, 7,01, 10,01.
[00451] Осмоляльность при депрессии точки замерзания
[00452] 15 мкл референсного раствора (например, 290 мОсм/кг) или образца переносили в морозильную камеру осмометра по депрессии точки замерзания Advanced Instruments. Образец переохлаждали до замораживания и контролировали температуру до тех пор, пока не наблюдалось плато. Температуру плато использовали для расчета осмоляльности в соответствии со следующим уравнением: 1 мОсм растворенного вещества/кг воды=1,858 миллиградусов (м°C) ↓ в точке замерзания.
[00453] Гидродинамический размер и полидисперсность с использованием динамического рассеяния света
[00454] Приблизительно 30 мкл образца загружали в чистую 3-мм кювету (ZN2112) и выполняли несколько сканирований (n=2 или 3) на Malvern Zetasizer Ultra (диспергатор=вода, тип образца = белок). Данные сканирования усредняли для построения графиков интенсивности (%) и объема (%). Первый использовался во время раннего скрининга формуляции как низкоуровневый, виды HMW легче наблюдать по интенсивности (LS ~ диаметр6 ~ Mw2). Данные сканирования также усреднялись для получения значений полидисперсности с помощью Cumulants Fits, среднего размера по интенсивности (10-100 днм), среднего размера по объему (10-100 днм), % площади по интенсивности (10-100 днм) и % площади по объему (10-100 днм), где это уместно.
[00455] Определение размера субвидимых частиц с помощью фоновой мембранной визуализации Horizon (BMI)
[00456] Фоновую мембранную визуализацию (BMI) выполняли с помощью системы для анализа частиц Horizon. Пустой 96-луночный планшет загружали в систему Horizon и получали фоновое изображение. Затем планшет переносили в вакуумный сборник. В каждую лунку загружали приблизительно 20-30 мкл образца. Промокательную бумагу и адаптер для промокательной бумаги прикрепляли к вакуумному сборнику, и планшет с образцами перемещали в верхнюю часть стопки. Включали вакуум для удаления любой оставшейся жидкости на дне лунок. Затем планшет с образцами переносили в прибор Horizon. Получали значения количества частиц и изображения (2-10 мкм, 10-25 мкм, > 25 мкм в целом).
[00457] Титр с использованием капельной цифровой полимеразной цепной реакции (ddPCR)
[00458] Замороженные аликвоты объемом 5-10 мкл использовали для определения концентрации генома AAV с помощью ddPCR в соответствии с SOP-AD-018. Образцы rAAV, содержащие капсидный белок с SEQ ID NO:12, разводили в фосфатно-солевом буфере Дульбекко с кальцием и магнием, содержащем 0,02% неионогенного поверхностно-активного вещества Плюроник™ F-68 (DPBS+0,02% F68), смешанного с ферментом ДНКазой I, для расщепления любой неинкапсулированной ДНК, и дополнительно разбавляли DPBS+0,02% F68, чтобы привести тестируемый образец в динамический диапазон анализа. Затем образец, обработанный ДНКазой, смешивали с праймерами ddPCR Supermix и SV40 (или CFTR)/зондами, меченными FAM. Затем 20 мкл реакционной смеси разделяли на капли с помощью генератора капель Bio-Rad QX200 Auto DG, подвергали ПЦР, затем считывали на ридере для капель Bio-Rad QX200, который измеряет флуоресцентный сигнал каждой капли отдельно. Данные анализировали с использованием программного обеспечения Bio-Rad QuantaSoft, в котором используется статистический анализ Пуассона положительных и отрицательных капель для обеспечения абсолютного количественного определения последовательностей-мишеней. Контроли без матрицы использовали для установки отрицательного базового уровня для образцов. Также на отдельных образцах проводили ddPCR без обработки ферментом ДНКазой I.
[00459] Химическая чистота с использованием электрофореза в полиакриламидном геле (PAGE)
[00460] Замороженные аликвоты объемом 5-10 мкл подвергали анализу химической чистоты в соответствии с SOP-AD-028 Rev. 00 (Определение чистоты капсида с использованием SDS-PAGE с окрашиванием криптоном™) или SOP-AD-002 (Определение чистоты капсида с окрашиванием серебром LDS-PAGE). Вкратце, приблизительно 1,00×1010 (SOP-AD-028) или 1,00×109 (SOP-AD-002) вирусных геномов смешивали с 4× буфером для образцов LDS, 10× восстанавливающим агентом для образцов, водой и нагревали при 95°C в течение 10 мин. Гель NuPAGE™ 4-12% Bis-Tris помещали в коробку для геля, содержащую верхнюю камеру (1× SDS буфер+антиоксидант) и нижнюю камеру (1× SDS буфер). На гель наносили денатурированные нагреванием (и восстановленные) реакционные смеси. Крышку коробки с гелем прикрепляли, и электроды подключали к внешнему источнику питания. Затем подавали мощность в соответствии с инструкцией производителя. Поток тока останавливали, когда фронт красителя образца составлял > ¾ длины полного геля. Гели промывали и окрашивали в соответствии с инструкциями производителя. Визуализацию геля проводили на приборе ChemiDoc MP Imager.
[00461] Функциональная активность по экспрессии зеленого флуоресцентного белка (GFP)
[00462] 15 мкл замороженных аликвот анализировали на экспрессию зеленого флуоресцентного белка (GFP). Клетки HEK2v6.11 трансдуцировали при 10000 MOI и собирали через 72 ч после трансдукции. Экспрессию GFP визуализировали флуоресцентной микроскопией и количественно определяли проточной цитометрией.
[00463] Значительную потерю (~50%) rAAV, содержащего капсид с VP1 SEQ ID NO:12, наблюдали во время концентрирования до ≥ 1×1013 вирусных геномов/мл (вг/мл) и замены буфера очищенных AAV в буфер для формуляции на основе фосфатно-солевого буфера Дульбекко (фосфатно-солевой буфер Дульбекко с кальцием и магнием, содержащий 0,05% неионогенного поверхностно-активного вещества Плюроник™ F-68 (DPBS+0,05% F68)). В экспериментах, описанных здесь, определили первопричину потерь, и описан улучшенный буфер для формуляции.
[00464] Установленное значение pI для VP1 SEQ ID NO:12, VP2 и VP3 составляет 6,7, 7,4 и 6,8 соответственно. VP1 имеет большее количество заряженных остатков по сравнению с VP2 и VP3, что, вероятно, объясняет теоретические различия в суммарном заряде, наблюдаемые при pH ниже 5 и выше pH 9 (фиг. 26). Однако суммарный заряд между значениями pH 5 и pH 9, по-видимому, в значительной степени одинаков для VP1 и VP3, в то время как для VP2 прогнозированы некоторые тонкие различия. Мономеры VP1, VP2 и VP3 содержат остатки аспарагиновой кислоты (D), аспарагина (N), метионина (M) и свободного цистеина (C). Следовательно, предполагается, что мономеры VP1 и VP3 чувствительны к шаффлингу аспарагиновой кислоты при низком pH, дезамидированию при нейтральном/основном pH, окислению и шаффлингу дисульфидов при высоком pH.
[00465] pH DBPS составляет приблизительно 7,0. Данное значение очень близко к значению pI или теоретическому минимуму растворимости для белков VP1 и VP3. Следовательно, предполагалось, что рН может играть роль в физической нестабильности, наблюдаемой во время концентрирования и замены буфера при фильтрации с тангенциальным потоком.
[00466] рН против растворимости
[00467] Исходный материал rAAV, содержащий капсид с VP1 SEQ ID NO:12 и нуклеиновую кислоту, кодирующую GFP (~2×1013 вг/мл в DPBS+0,005% Плюроника F68), оттаивали и разбавляли в 10 раз, приблизительно до 2×1012 вг/мл в различных буферах (pH 4-8) как с низкой ионной силой (~14 мМ NaCl), так и с физиологической ионной силой (~150 мМ NaCl). Составы с низкой ионной силой показали зависимое от рН увеличение высокомолекулярной массы (HMW) при увеличении рН с 4 до 8. Эта зависимость от рН достоверно снижалась в присутствии 150 мМ NaCl.
[00467] Затем образцы выдерживали при комнатной температуре, и затем анализировали с помощью УФ-спектрофотометрии на следующий день (T=1 день при комнатной температуре). УФ-поглощение при 280 нм (A280) представлено в зависимости от pH на фиг. 27. Образцы с низкой ионной силой показали очень резкое снижение сигнала A280 (т. е., что свидетельствует о сниженной концентрации rAAV) выше pH 5, что согласуется с физической нестабильностью, наблюдаемой с помощью DLS. Все образцы, содержащие 150 мМ NaCl, показали одинаковые значения A280. Полученные данные показывают, что более низкое значение рН и добавление соли приводят к повышению растворимости. Детали по формуляции, значениям pH образца и поглощения при A280 (с поправкой на светорассеяние и длину пути) можно найти ниже:
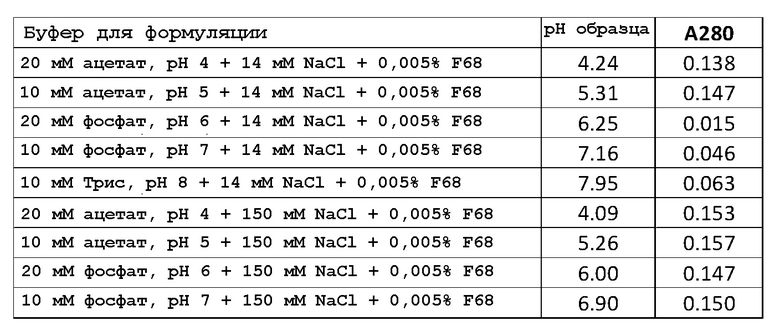
[00469] Эти результаты позволяют предположить, что как ионная сила, так и pH влияют на поведение rAAV в растворенном состоянии. Поэтому было проведено последующее исследование для дальнейшей оценки пределов растворимости rAAV при значениях pH от 5 до 8 в присутствии хлорида натрия (ионная сила ~0,15M) или цитрата тринатрия (высокая ионная сила). Исходный материал rAAV, содержащий капсид с VP1 SEQ ID NO:12 и нуклеиновую кислоту, кодирующую люциферазу (1,76×1013 гв/мл в DPBS+0,005% Плюроника F68), размораживали, заменяли буфер и концентрировали в различных буферах (таблица 7) до целевого уровня приблизительно 3×1013 вг/мл. Выходы процесса определяли делением извлеченных вирусных геномов на вирусные геномы в исходном материале. Снижение извлечения наблюдали для составов DPBS+0,005% F68 и 10 мМ Трис, pH 8+150 мМ NaCl+0,005% F68. Такие более низкие выходы, по-видимому, также соответствовали более медленной скорости буферного обмена и концентрирования.
Таблица 7
Обощенные результаты исследования pH против растворимости A101-Luc (после фильтрации через фильтр 0,2 мкм)
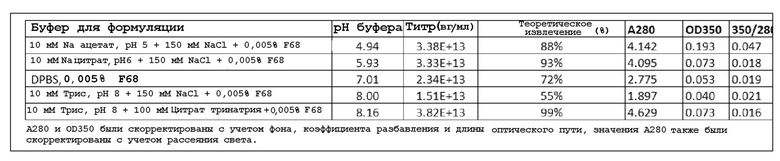
[00470] Образцы, содержащие хлорид натрия, продемонстрировали аналогичную зависимость от рН, т.е. ↓ растворимости при ↑ рН, по титру и поглощению при A280. Состав с высокой ионной силой (10 мМ трис, рН 8+100 мМ цитрата натрия+0,005% F68) показал самую высокую растворимость среди испытанных составов, что дополнительно демонстрирует влияние ионной силы. Для образцов, профильрованных через фильтр с размером пор 0,2 мкм, наблюдали зависимость от pH, т. е. ↑ O.D.350 при более низком pH. Однако по значениям 350/A280 был построен график, чтобы представить физическую нестабильность относительно количества rAAV в растворимой фракции. Значение 350/A280 показало нестабильность при рН 5. Однако при pH 6 наблюдали перегиб (10 мМ цитрата, pH 6+150 мМ NaCl+0,005% F68), который, по-видимому, представляет собой баланс между высокой растворимостью и физической стабильностью. Состав с высокой ионной силой показал мутность, сравнимую с составом с рН 6. Как показано на фиг. 35 (соответствует данным в таблице 7), более низкий рН показывает повышенную растворимость в присутствии 0,15 М NaCl (растворимость выше при рН 5 и 6 по сравнению с рН 7 и 8, когда концентрация соли составляет ~150 мМ NaCl) и ясно демонстрирует, что для «восстановления» растворимости при более высоком pH требуется повышенная ионная сила. Полученные результаты в конечном итоге привели к использованию состава с цитратом с pH 6.
[00471] DLS (динамическое рассеяние света) показало различные уровни высокомолекулярных частиц в концентрированных образцах, за исключением состава с высокой ионной силой (10 мМ Трис, pH 8+100 мМ цитрата тринатрия+0,005% F68). Тем не менее, количество высокомолекулярных частиц в значительной степени уменьшалось после фильтрации через фильтр 0,2 мкм, за исключением устойчивых субмикронных частиц, наблюдаемых в составе с 10 мМ ацетата натрия, рН 5+150 мМ NaCl+0,005% F68. Это согласуется с увеличением показателя 350/A280, наблюдаемым для того же образца.
[00472] Результаты данного исследования позволяют предположить, что можно формулировать rAAV на уровне ≥ 3×1013 вг/мл, когда pH ≤ 6 или когда ионная сила увеличивается выше 0,15M. Однако при низком pH (например, при pH 5) может иметь место риск появления растворимых высокомолекулярных частиц, а для составов с более высоким pH может потребоваться концентрация модификатора ионной силы (например, цитрата натрия), который превышает количество, присутствующее в других одобренных продуктов для ингаляционного применения в соответствии с базой данных неактивных ингредиентов FDA. Поэтому следующее исследование было направлено на сужение диапазона рН состава (6-8), а также на оценку воздействия цитрата натрия в диапазоне концентраций от 20 до 100 мМ.
[00473] Исходный материал rAAV, содержащий капсид с VP1 SEQ ID NO:12 и нуклеиновую кислоту, кодирующую GFP (8,75×1012 гв/мл в DPBS+0,005% Плюроника F68), размораживали, заменяли буфер и концентрировали в различных буферах (таблица 8; все составы также включают 0,005% F68) до целевого уровня приблизительно 3×1013 вг/мл. Значения рН и осмоляльности буфера (без rAAV) можно найти в таблице 9. Формулированные образцы фильтровали через фильтр 0,2 мкм и аликвоты распределяли в полипропиленовые пробирки. 1 аликвотную порцию каждого состава использовали для измерения на T=0, и затем подвергали 4-суточному перемешиванию при комнатной температуре при 1500 об/мин. Отдельные аликвоты использовали для оценки влияния циклов замораживания-оттаивания (5/10× циклов FT), хранения при комнатной температуре (T=13 суток) и 28-суточного хранения (15 суток при 2-8°C и затем 13 суток при комнатной температуре). Исходный материал rAAV, 8,46×1012 вг/мл в DPBS+0,005% F68, оценивали в выбранных условиях.
[00474]
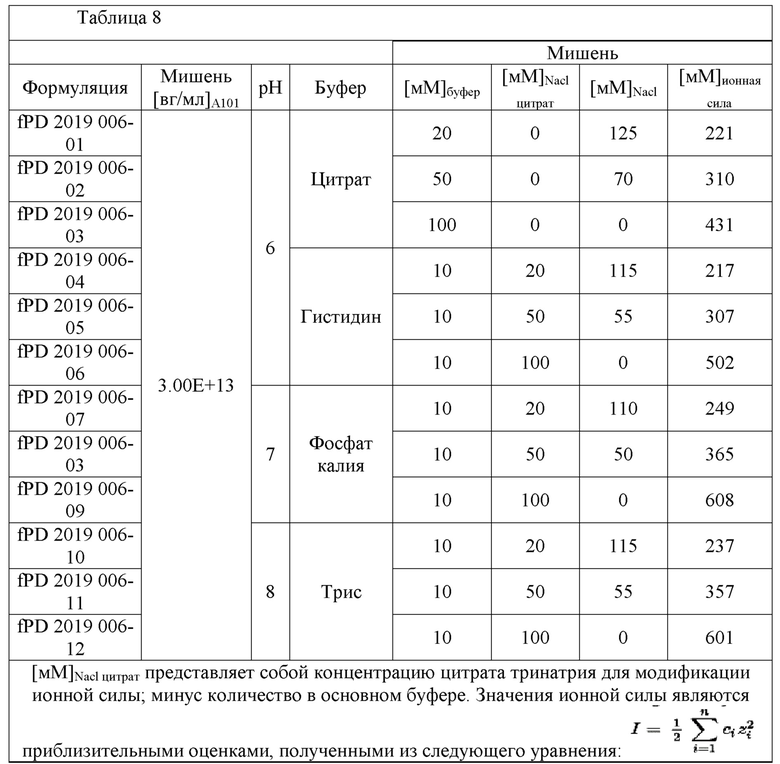
[00475]
Таблица 9
pH и осмоляльность буфера (мОсм/кг)
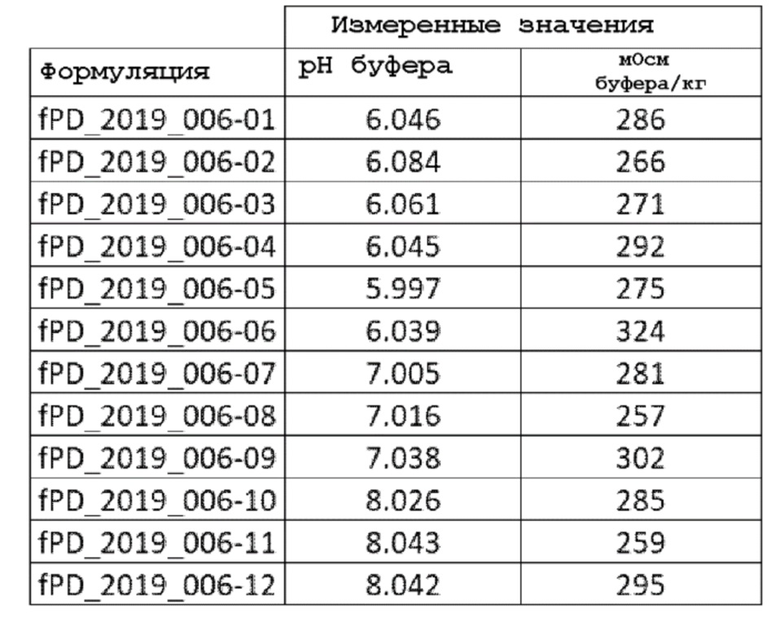
[00476] Анализ титра с помощью ddPCR проводили в небольшом объеме за счет большого количества составов и условий, подлежащих скринингу. Однако на Т=0 титры образцов варьировались от приблизительно 2,3 до 2,9×1013 вг/мл (таблица 10). Предположили, что такая низкая изменчивость связана с небольшим масштабом процесса, так как большинство условий соответствовали или превышали степень извлечения > 90%. Исключением являются fPD_2019_006_003, fPD_2019_006_005 и fPD_2019_006_007, которые показали более низкое извлечение. Титр образца также измеряли после 28-суточного хранения (15 суток при 2-8°С, затем 13 суток при комнатной температуре). Все составы сохраняли титр ≥ 100% относительно T=0.
[00477]
Таблица 10
Титр на T=0 и извлечения (%)
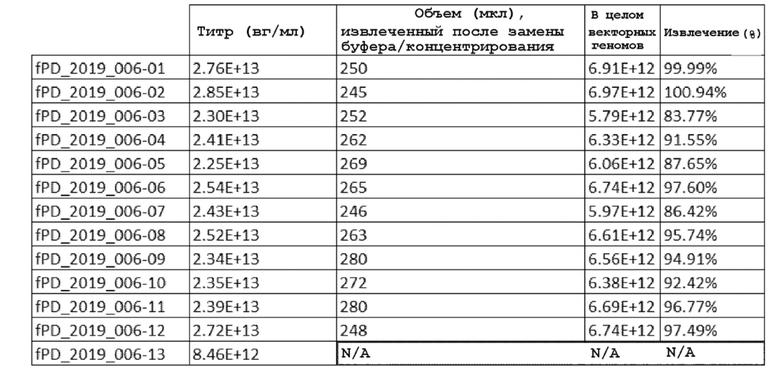
N/A - отсутствует
[00478] Для любого из составов в тестированных условиях наблюдали незначительное изменение гидродинамического размера в объемных % или его отсутствие. Следовательно, были созданы совмещения для гидродинамического размера в процентах интенсивности, которые обычно более чувствительны к присутствию низкомолекулярных частиц. fPD_2019_006-05 и fPD_2019_006-10 показали увеличение ширины пика после 4-суточного перемешивания при 1500 об/мин. Это также можно было наблюдать на графике полидисперсности образца. Полидисперсность на T=0 исходного материала (fPD_2019_006-13) была выше, чем у любого из 12 скринированных составов, что еще больше указывает на улучшение по сравнению с исходным составом DPBS.
[00479] Таблица 9 также включает значения A280, а также % оставшегося сигнала при A280 по отношению к T=0. Большинство составов сохраняли ≥ 95% соответствующих сигналов при A280, за исключением fPD_2019_006-11 и fPD_2019_006-13 (контроль DPBS) после 4-суточного перемешивания при 1500 об/мин, и fPD_2019_006-03 и fPD_2019_006-08, которые показали небольшое снижение после 28-суточного хранения.
[00480] Для подвергшихся перемешиванию образцов наблюдалось значительно большее количество частиц по сравнению с другими условиями испытаний, означая, что продолжительность перемешивания могла быть чрезмерно агрессивной. Тем не менее, составы, по-видимому, различались по своей реакции на различные применяемые стрессоры. Например, fPD_2019_006-05 содержал повышенное количество частиц > 10 мкм после перемешивания, в то время как fPD_2019_006-09 и fPD_2019_006-10 показали повышенное количество частиц после 10-кратного замораживания-оттаивания.
[00481] Результаты по титру и физической стабильности, полученные в исследовании с fPD_2019_006, оценивали с помощью системы полуколичественного взвешивания, и было решено, что fPD_2019_006-01 (20 мМ цитрата, pH 6+125 мМ NaCl+0,005% F68), fPD_2019_006-02 (50 мМ цитрата, pH 6+70 мМ NaCl+0,005% F68), fPD_2019_006-08 (10 мМ фосфата, pH 5+ 50 мМ NaCl+50 мМ тринатрийцитрата+0,005% F68) и fPD_2019_006-12 (10 мМ Трис, pH 8+100 мМ цитрата тринатрия+0,005% F68), которые все показали благоприятные характеристики, нужно исследовать дополнительно. После принятия этого решения оставшиеся аликвоты 28-суточных образцов анализировали с помощью электрофореза с окрашиванием криптоном. VP1, VP2 и VP3 имели место во всех тестированных образцах. Полосы с низкой молекулярной массой (LMW) также наблюдались в различной степени; наиболее заметным из них является fPD_2019_006-01. К сожалению, значения на T=0 не были доступны для сравнения, поэтому было неясно, являются ли эти полосы LMW продуктами разложения или технологическими примесями. Поэтому химическую стабильность оценивали в последующих исследованиях.
[00482] rAAV, содержащий капсид с VP1 SEQ ID NO:12 и нуклеиновую кислоту, кодирующую пул CFTR, CIMQA (партия № dPD_2020_001, 7,92×1011 вг/мл), подвергали буферному обмену и концентрировали непосредственно в 4 буферах, указанных в fPD_2019_006 до целевого уровня, составляющего приблизительно 3×1013 вг/мл. rAAV, содержащий капсид с VP1 SEQ ID NO:12, и нуклеиновую кислоту, кодирующую GFP (партия № 4DER000057 г/мл), в DPBS+0,005% F68, подвергли буферному обмену и концентрировали в 20 мМ цитрата, pH 6+125 мМ NaCl+0,005% F68 до целевого уровня приблизительно 6E13 вг/мл. Затем композиции rAAV-CFTR и rAAV-GFP разделяли на аликвоты и помещали в полипропиленовые пробирки и подвергали 10 циклам замораживания-оттаивания, 40-ч перемешиванию при 1500 об/мин или 40-ч хранению при 40°C.
[00483]
Таблица 11
Дизайн исследования скрининга формуляции
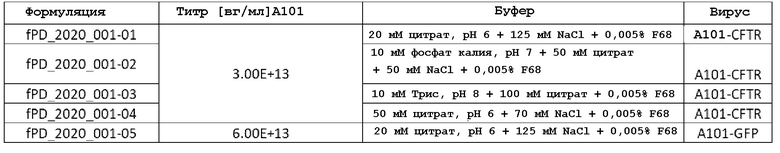
[00484] Титры rAAV-CFTR, T=0, варьировались от 1,7 до 1,9×1013 вг/мл. Они были на ≥ 30% ниже, чем целевое значение 3× 1013 вг/мл, что было связано с малым рабочим объемом, а не с ограничением растворимости. Составы с pH 7 и pH 8 (fPD_2020_001-02 и fPD_2020_001-03) продемонстрировали приблизительно потери на уровне 4% при воздействии стресса при перемешивании, в то время как для составов с pH 6 потерь не наблюдалось (fPD_2020_001-01 и fPD_2020_001-04). Эта тенденция к стабильности значительно усилилась при 40°C, поскольку составы с pH 7 и pH 8 показали потери ≥ 90%, тогда как потери при pH 6 составили <30%. Все 4 препарата rAAV-CFTR сохраняли титр ≥ 100% после 10 циклов замораживания-оттаивания. rAAV-GFP (fPD_2020_001-05, 5,3E13 вг/мл на T=0) оказался высокостабильным при pH 6 без потерь при перемешивании и <3% потерь при хранении при 40°C в течение 10 циклов замораживания-оттаивания. Предполагается, что разница в относительной стабильности между rAAV-CFTR и rAAV-GFP, формулированными в одном и том же буфере, связана с размером трансгена. Однако в будущем потребуются дополнительные исследования.
[00485] fPD_2020_001-02 (pH 7) и fPD_2020_001-03 (pH 8) показали более высокую потерю сигнала при A280 по сравнению с образцами с pH 6 во время хранения при 40°C. Это согласуется с тенденцией, наблюдаемой для титра, но в меньшей степени. Возможно, это связано с увеличением количества пустых капсидов (и свободной ДНК) при хранении при 40°С. Пустые капсиды и ДНК по-прежнему будут поглощать УФ-свет, но последние будут восприимчивы к обработке ДНКазой. Результаты A280 для состава rAAV-GFP (fPD_2020_001-05) также согласуются с анализом титра; от минимального до отсутствия изменений.
[00486]
Таблица 12
Поглощение при 280 нм (A280)
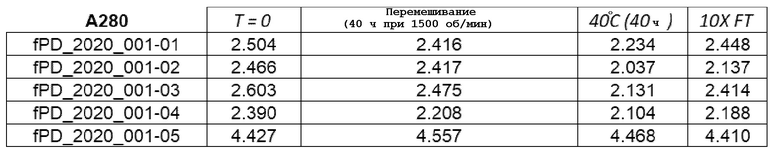
[00487] Графики гидродинамического размера по интенсивности показали увеличение ширины пика для всех образцов rAAV-CFTR при 40°C. В fPD_2020_001-01 на низком уровне были обнаружены высокомолекулярные частицы. Однако значительные различия в титре и УФ-излучении могут свидетельствовать об отсутствии этого пика в менее стабильных препаратах за счет преципитации, поверхностной адсорбции или изменения соотношения пустой/полный капсид. Различия в размерах были менее очевидны по объему, но полидисперсность также оказалась очень чувствительной к незначительным различиям. В соответствии с другими методами тестирования, для rAAV-GFP практически отсутствовали изменения.
[00488] Хранение при 40°С привело к тому, что оказалось высоким число частиц с размером 2-10 мкм в составе A101-GFP (fPD_2020_001-05). Предполагалось, что это явление зависит от концентрации (т. е. титр rAAV-GFP был приблизительно в 3 раза выше, чем у образцов rAAV-CFTR). Интересно отметить, что этот образец показал сравнительно небольшое количество частиц > 10 мкм, что, возможно, свидетельствует о том, что формуляция предотвращает образование более крупных агрегатов. rAAV-CFTR, приготовленный в том же буфере (fPD_2020_001-01), проявил чувствительность к перемешиванию и нагреванию, но, по-видимому, отсутствовало образование частиц > 25 мкм. Другие составы rAAV-CFTR показали небольшую тенденцию к частицам > 25 мкм.
[00489] Никаких существенных различий не наблюдали на Т=0, 40 ч при 1500 об/мин или после 10X FT. Однако составы с pH 7 (fPD_2020_001-02) и pH 8 (fPD_2020_001-03) показали повышенное содержание низкомолекулярных частиц и оказались перегруженными. Напротив, fPD_2020_001-01 показал присутствие некоторых высокомолекулярных полос после хранения при 40°C. Однако было неясно, был ли это артефакт, связанный с подготовкой образца или процедурой окрашивания (например, окрашивание серебром считается неколичественным анализом). К счастью, на тех же образцах выполняли анализ вестерн-блоттингом. Результаты вестерн-блоттинга четко показали, что образцы с pH 7 и pH 8 при 40°C были перегружены (загружены на основе титра). Это еще раз подтверждает идею о том, что непропорциональная потеря титра по сравнению с поглощением при A280, вероятно, связана с увеличением количества пустых капсидов. Для образца 40°C fPD_2020_001-01 с помощью вестерн-блоттинга не наблюдалось высокомолекулярных полос.
[00490] Большая часть усилий по скринингу состава была сосредоточена на улучшении физической стабильности rAAV и, в меньшей степени, на мониторинге химической стабильности. Однако важная часть все еще не была рассмотрена, а именно функциональная активность. Поэтому функциональную активность rAAV-GFP оценивали в трех составах, тестированных в исследовании fPD_2020_001. rAAV-GFP (1,58 a 1013 вг/мл) в DPBS+0,005% F68 подвергали буферному обмену и концентрировали в 20 мМ цитрата, pH 6+125 мМ NaCl+0,005% F68 (fPD_2020_002-01), 10 мМ фосфата калия, pH 7+50 мМ цитрата+50 мМ NaCl+0,005% F68 (fPD_2020_002-02) или 50 мМ цитрата, pH 6+70 мМ NaCl+0,005% F68 (fPD_2020_002-03) до целевого уровня приблизительно 3×1013 вг/мл. Затем составы rAAV-GFP аликвотировали в полипропиленовые пробирки и хранили при комнатной температуре в течение 13 суток. Титр измеряли на Т=0 (таблица 12), и значения применяли как к Т=0, так и к Т=13-суточным образцам (т. е. одинаковый объем нагрузки для функционального тестирования).
[00491]
Таблица 13
fPD_2020_002: исследование функциональной активности A101-GFP

[00492] Гидродинамический размер оценивали на Т=0. Для трех тестированных составов не наблюдали никаких различий. При хранении в течение 13 суток при комнатной температуре потери сигнала при A280 (таблица 14) не отмечали.
[00493]
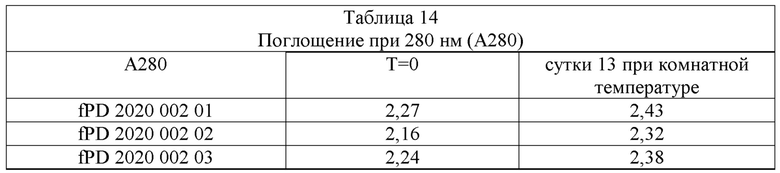
[00494] Результаты флуоресцентной микроскопии и проточной цитометрии показали, что все образцы rAAV-GFP демонстрировали экспрессию зеленого флуоресцентного белка, в то время как для растворителя флуоресценция не наблюдалась. В % GFP-положительных клеток после 13-суточного хранения при комнатной температуре различий не наблюдалось, что означает, что все три препарата сохранили функциональную активность rAAV.
[00495] На основе результатов исследований fPD_2020_001 и fPD_2020_002 было принято решение провести окончательную оценку двух составов с цитратом, рН 6. Очищенный rAAV-CFTR (4D130109), содержащий 20 мМ цитрата, pH 6+125 мМ NaCl+0,005% F68 (fPD_2020_003-01, номер партии dPD_2020_007) и 50 мМ цитрата, pH 6+85 мМ NaCl+0,005% F68 (fPD_3-3-2020_007) 02, номер партии dPD_2020_007) размораживали и распределяли на аликвоты (~100 мкл) в полипропиленовые пробирки. Две аликвоты каждой композиции (n=2) использовали для измерений на T=0. Остальные флаконы помещали при 40°С/75% относительной влажности и извлекали через 4 ч (n=2), 20 ч (n=2) или 44 ч (n=2). В составе с 20 мМ цитрата, pH 6 (fPD_2020_003-01) использовали тот же буфер, который показал хорошую стабильность в предыдущих исследованиях. Состав с 50 мМ цитрата был подобен составу с 50 мМ цитрата, pH 6, определенному в предыдущих исследованиях, с небольшим изменением; концентрацию NaCl повышали с 70 мМ до 85 мМ для увеличения тоничности раствора. Осмоляльность 20 мМ и 50 мМ цитратных буферов, рН 6, составляла 289 и 295 мОсм/кг соответственно.
[00496]

[00497] Анализ титра с использованием ddPCR (n=2) проводили с и без предварительной обработки ДНКазой. Между двумя методами наблюдалась измеримая разница, предполагающая либо наличие свободной капсидной ДНК, либо чувствительных к ДНКазе капсидов (например, нарушенных или химически поврежденных). Однако потеря титра при 40°С/75% относительной влажности была сопоставима для двух составов.
[00498] Повышенная потеря сигнала при A280 наблюдалась для fPD_2020_003-02 (состав с 50 мМ цитрата, pH 6), что, возможно, свидетельствует о высаливании (или осаждении) некоторой части растворимой фракции. Интересно, что мутность увеличилась в fPD_2020_003-01, что еще больше указывает на присутствие «нарушенных» видов, которые сохранялись в растворе при более низкой концентрации цитрата.
[00499] Анализ результатов DLS коррелировал с данными по мутности. Образец fPD_2020_003-01 показал обнаруживаемые уровни высокомолекулярных частиц по интенсивности через 20 и 44 ч при 40°C/75% относительной влажности, в то время как в fPD_2020_003-02 эти виды отсутствовали. Этот пик объясняет различия, наблюдаемые в % площади пика и полидисперсности.
[00500] Образцы также анализировали на химическую чистоту с использованием PAGE. Наблюдения были следующими. Продукты расщепления обнаруживаются на Т=0 для обоих составов. Предположительно, это технологические примеси, унесенные в процессе последующей очистки. Продукты расщепления низкого уровня начинают формироваться в течение длительного времени при 40°С/75% относительной влажности. Химическая чистота обоих составов оказалась сопоставимой.
[00501] На основе этих результатов было определено, что повышенный уровень цитрата может приводить к небольшому повышению физической стабильности rAAV-CFTR при 40°C/75% относительной влажности. Однако было неясно, был ли это истинный защитный эффект или одновременная потеря в A280 предполагала, что физически нестабильные виды подверглись высаливанию. Кроме того, 50 мМ цитрат не показал снижения потери титра по сравнению с 20 мМ цитратной композицией, и обе композиции, по-видимому, имели одинаковое увеличение низкомолекулярных частиц, что было установлено PAGE. Таким образом, было установлено, что состав с 50 мМ цитрата не обеспечивает достаточного улучшения, чтобы гарантировать более высокую концентрацию цитрата, которая может быть менее переносимой в ингаляционном лекарственном продукте.
[00502] Таким образом, состав с 20 мМ цитрата, pH 6+125 мМ NaCl+0,005% F68 был признан предпочтительным составом для пилотного исследования токсичности/установления дозы на NHP. Предварительные требования к пробным образцам для испытаний на токсичность можно найти в таблице 16. Было проведено исследование (fPD_2020_006) для оценки стабильности низких доз (~6×1011 вг/мл) при замораживании-оттаивании.
[00503]
Таблица 16
Требования к опытному тестируемому образцу для пилотного исследования токсичности на NHP
[00504] rAAV-CFTR (4D130109, партия № 4DER000060.02, ~3×1013 вг/мл) в 20 мМ цитрате, pH 6+125 мМ NaCl+0,005% F68, разбавляли в том же буфере для формуляции до целевого значения приблизительно 6×1011 вг/мл (низкая доза при общем объеме 5,5 мл). Аликвоту объемом 100 мкл отбирали для титра T=0 и DLS. Оставшийся материал помещали в морозильную камеру при -80°С на 1 ч. Пробирку извлекали и оставляли оттаивать при комнатной температуре на 1 ч. Размороженную пробирку осторожно переворачивали для перемешивания и отбирали аликвоту 100 мкл для DLS и титра (1X замораживание-оттаивание). Данный процесс повторяли еще два раза. После третьего цикла замораживания-оттаивания материал оставляли на ночь при комнатной температуре.
[00505] Титр показал очень небольшое изменение в течение 3 циклов замораживания-оттаивания (3× FT), в то время как небольшое снижение титра (~10%) наблюдалось, когда образец 3X FT хранили при комнатной температуре в течение ночи. Однако это значение все еще находилось в пределах вариации анализа ddPCR и, следовательно, рассматривалось в качестве наихудшего сценария.
[00506] Для образцов методом замораживания-оттаивания с использованием DLS не наблюдалось существенной разницы в физической стабильности. Меньшие аликвоты (~50 мкл) образцов с T=0, 2X и 3X FT также измеряли после еще одного часа хранения при комнатной температуре (2 ч после полного оттаивания). Никаких изменений в этих образцах не наблюдали.
Пример 6
In Vitro и Ex Vivo характеристика 4D-710 в клетках человека
[00507] Оценивали способность rAAV, содержащего (i) капсид SEQ ID NO:12 и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность SEQ ID NO:45 (4D-710), трансдуцировать клетки человека и доставлять кодон-оптимизированный трансген CFTR человека с SEQ ID NO: 43 (cohCFTRΔR, кодирующий функциональный ген CFTR с делецированными аминокислотами 708-759). Клетки человека представляли собой культуры HEK2v6.11, 16HBE14o-G542X (доступные через Фонд муковисцидоза) и ex vivo культуры ALI без CF. Оценивали экспрессию белка, локализацию белка в мембране и экспрессию мРНК кассеты терапевтического трансгена.
[00508] Материалы и методы - SDS-PAGE и вестерн-блоттинг. Клетки лизировали для анализа вестерн-блоттингом в PBS, содержащем 1% об./об. NP-40, в течение 30 мин при 4°С перед удалением нерастворимого материала. Для вестерн-блоттинга равные объемы лизата, содержащие краситель 1Х и восстановитель (Thermo Bolt), нагревали при 50°C в течение 10 мин, затем загружали на 4%-12% бис-трис-акриламидный гель (Thermo Bolt) и анализировали при 250 В в течение 45 мин. Гель полусухим переносили на 0,2 мкМ нитроцеллюлозу (BIO-RAD) при настройке переноса «High MW» BIO-RAD Trans-Blot Turbo (10 мин, 1,3A). Затем блот быстро промывали сверхчистой водой, и затем блокировали раствором 1X iBind Flex (Thermo) в течение 10 мин. Затем блот инкубировали в 1X растворе iBindFlex, содержащем антитело к CFTR (Ab660, CFF/UNC 1:500) или антитело к тубулину (E-7, DSHB 1:1000), в течение 2 ч при комнатной температуре при осторожном встряхивании. Затем блоты промывали PBS (Corning), содержащим 0,01% Твина-20 (Sigma), в течение 5 мин при встряхивании; эту стадию промывания повторяли еще два раза. Затем блоты инкубировали во вторичном антителе против мышиного HRP (R&D systems, 1:5000) в растворе 1X iBind Flex в течение 1 ч при комнатной температуре. Затем повторяли стадии промывания PBS-0,01% Твина-20. Затем промытые блоты покрывали DuraWest HRP Detection Substrate (Thermo) на 5 мин, и затем блоты визуализировали с помощью системы визуализации BIO-RAD ChemiDoc.
[00509] Материалы и методы - Иммуноцитохимия. Для иммуноцитохимии клетки 2v6.11 культивировали на стеклянных предметных стеклах с лунками, покрытыми поли-L-лизином, 16HBE14o- культивировали на вкладышах Transwell, покрытых как бычьим коллагеном I, так и фибронектином плазмы человека. Через 2 суток (2v6.11) или 4 суток (16HBE14o-) после трансдукции клетки фиксировали в 4% PFA в течение 20 мин при комнатной температуре в темноте. Затем клетки пермеабилизировали 0,2% Triton-X в PBS (без кальция и магния) и инкубировали при 4°C в течение ночи с антителом CFTR MM13-4 (EMD Millipore, 1:100) в блокирующем буфере. Блокирующий буфер состоял из 0,2% Тритона-X, 5% козьей сыворотки и 2% бычьего сывороточного альбумина в PBS. После вымывания антител с помощью PBS-Тритона в блокирующий буфер на один час при комнатной температуре в темноте добавляли конъюгат вторичного козьего антимышиного антитела с Alexa Fluor-647 (Invitrogen, 1:500). Затем вымывали вторичное антитело. Ядра окрашивали 4',6-диамидино-2-фенилиндолом (DAPI) в PBS, затем покрывали покровным стеклом Prolong Gold (Invitrogen) и затем визуализировали. Клетки визуализировали с помощью флуоресцентного микроскопа Zeiss Axio Observer.D1. Обработку изображений проводили с использованием программного обеспечения Zeiss Zen 2 (Carl Zeiss Microscopy LLC, White Plains, NY).
[00510] Материалы и методы - экстракция РНК и капельная цифровая ПЦР. РНК выделяли из клеток с использованием набора RNeasy Micro Qiagen (Thermo Fisher Scientific), и кДНК получали с использованием набора для синтеза кДНК Maxima H minus (Thermo Fisher). Образцы кДНК готовили приготовлением серийных разведений в 1× буфере ТЕ (10 мМ Трис, 0,1 мМ ЭДТА, рН 8,0). Затем образцы помещали в мастер-микс, содержащий супермикс ddPCR и либо набор праймеров/зондов, нацеленных на CFTRΔR (SEQ ID NO:43), либо на эндогенный ген CFTR (Thermo Fisher). Затем образцы разделяли на нанолитровые капли с помощью автоматического генератора капель (BIO-RAD), подвергали ПЦР с анализом результатов «по конечной точке» в сенсорном термоциклере C1000 (BIO-RAD) и считывали на ридере для считывания капель QX200 (BIO-RAD), который измеряет каждую каплю индивидуально по флуоресцентному сигналу. Затем данные анализировали с использованием программного обеспечения BIO-RAD QuantaSoft, в котором используется статистический анализ Пуассона положительных (капли, содержащие флуоресцентный сигнал выше порогового значения) и отрицательных (капли с флуоресцентным сигналом ниже порогового значения) капель для обеспечения абсолютного количественного определения целевой последовательности (последовательностей). Дальнейший анализ проводили с использованием Microsoft Excel.
[00511] Результаты
[00512] По прнципу «проверки концепции» первоначальную экспериментальную экспрессию трансгена и локализацию белка в мембране после трансдукции с 4D-710 оценивали путем трансдукции клеточной линии HEK2v6.11 - относительно простой системы (по сравнению с более сложной моделью человеческой клетки на границе воздух-жидкость). Клетки HEK2v6.11 получали из линии почек эмбриона человека, HEK-293T, с экспрессией белка ORF6 человеческого аденовируса E4, индуцированной понастероном А. Многочисленные серотипы AAV способны к высокой эффективности трансдукции в данной клеточной линии. Вкратце, HEK2v6.11 культивировали в среде DMEM (Gibco) с 10% FBS и 1% пенициллина/стрептомицина. После слияния клетки обрабатывали при различных значениях MOI в течение 48 ч. Понастерон добавляли до концентрации 1 мкг/мл для увеличения экспрессии из вектора AAV. Клетки анализировали вестерн-блоттингом или иммуноцитохимией и трансдуцировали с 4D-710 в течение 24 ч и анализировали вестерн-блоттингом или иммуноцитохимией (ICC) через 48 ч после трансдукции. Дозозависимая трансдукция 4D-710 в клетках HEK2v6.11 была продемонстрирована вестерн-блоттингом при увеличении множественности инфекции (MOI) (фиг. 28A). Можно увидеть полосу B, которая соответствует ER-кор-гликозилированной форме белка (150 кДа), и полосу C, отражающую полностью гликозилированную зрелую форму CFTR (170-180 кДа), что предполагает правильную трансляцию белка и клеточный процессинг. Репрезентативные иммунофлуоресцентные изображения дополнительно демонстрируют дозозависимую трансдукцию (фиг. 28В), цитоплазматическую (фиг. 28В) и мембранную (фиг. 28С) экспрессию белка CFTR. Нетрансдуцированные контроли показывают, что эндогенный белок CFTR не детектируется, что указывает на то, что обнаруженный белок является трансдуцированным трансгеном CFTRΔR. Таким образом, не только образуются незрелые и зрелые формы CFTRΔR, но и экспрессия белка CFTRΔR на мембране наблюдалась с использованием ICC (с использованием анти-CFTR антитела и анти- F-актин антитела для окрашивания клеточной мембраны).
[00513] Оценивали трансдукцию 4D-710 линии клеток легких человека. Клетки 16HBE14o-G542X представляют собой иммортализованную клеточную линию бронхиального эпителия (HBE) человека (первоначально иммортализованную с помощью плазмиды SV40, дефектной в отношении ориджина репликации), содержащую нулевую мутацию CFTR (G542X, созданную с помощью редактирования генов на основе CRISPR) и не экспрессирующую CFTR. Клетки 16HBE14o-G542X культивировали в MEM (Gibco 11095-072) с добавлением 10% FBS (Gibco 26140-079) и 1% пенициллина/стрептомицина (Gibco 15140-122). Клетки высевали на вкладыши Transwell по 9000 клеток на вкладыш. Среду добавляли как в верхнюю часть (100 мкл), так и нижнюю часть (500 мкл). На сутки 3 после посева клетки обрабатывали при различных значениях MOI апикально в течение 72 ч. Клетки анализировали иммуноцитохимией или капельной цифровой полимеразной цепной реакцией с обратной транскрипцией (RT-ddPCR) с использованием праймеров и зондов, специфичных для кодон-оптимизированной мРНК CFTRΔR (трансген 4D-710). Для детектирования экзогенного белка CFTR трансдукцию проводили параллельно с обработкой доксорубицином (1 мкМ) в течение 24 ч в культуральной среде. Для оценки уровней транскриптов кодон-оптимизированного трансгена CFTRΔR после трансдукции 4D-710 в клетки бронхиального эпителия человека культуры 16HBE14o-G542X трансдуцировали 4D-710 при повышении значений MOI. Для дальнейшего подтверждения трансдукции HBEs с 4D-710 проводили ОТ-ddPCR для определения количества копий мРНК CFTRΔR. На фиг. 29A показана кривая зависимости от дозы для количества копий мРНК CFTRΔR для увеличения множественности заражения. В присутствии доксорубицина трансдуцированные клетки идентифицировали с использованием иммуноцитохимии (фиг. 29B), где большее количество клеток трансдуцировалось при MOI 50000 по сравнению с MOI 35000. Таким образом, была продемонстрирована зависимая от дозы трансдукция клеток бронхиального эпителия человека и стабильная экспрессия трансгена CFTR в этих клетках человека, имеющих отношение к заболеванию.
[00514] Клеточные культуры на поверхности раздела воздух-жидкость (ALI) получали из здоровых легких (без CF) и, как таковые, имеют эндогенную экспрессию CFTR. Эндогенный CFTR отличали от экспрессии усеченного трансгена 4D-710 CFTR с использованием ОТ-ddPCR с использованием праймеров и зондов, специфичных для эндогенного CFTR или специфичных для CFTRΔR мРНК FTRΔR. Стандартное культивирование ALI клеток легких ex vivo без муковисцидоза представляет собой наилучшую модель in vitro легочной системы in vivo (эти культуры представляют собой комплексы с негомогенными типами клеток (такими как легкие in vivo)). Вкратце, ex vivo ALI культуры легких человека без CF высевали на покрытые коллагеном типа IV вкладыши Corning Transwell, культивировали базально в среде PneumaCult ALI (StemCell Technologies 05040) с включением гена Pen/Strep (Gibco 15140-122), гентамицина и амфотерицин В по протоколу производителя. Через 30 суток после посева, когда ALI культуры созревали, культуры обрабатывали при различных значениях MOI апикально в присутствии 0,625 мкМ идарубицина в течение 24 ч в культуральной среде. Через 7 суток после трансдукции клетки анализировали с помощью ОТ-ddPCR с использованием праймеров и зондов, специфичных для эндогенного CFTR или специфичных для мРНК CFTRΔR. Для оценки уровней транскриптов кодон-оптимизирвоанного трансгена CFTRΔR после трансдукции в ex vivo здоровое легкое ALI культуры ALI трансдуцировали с помощью 4D-710 при MOI 50000 и 100000 в присутствии идарубицина. РНК выделяли через 7 суток после трансдукции и синтезировали кДНК. ОТ-ddPCR проводили на подготовленных образцах, и уровни транскриптов в расчете на каплю определяли как значение копий/мкл. Количественный анализ давал число капель выше установленного порога, содержащих транскрипт исследуемого набора праймеров/зондов. Было созданы два набора праймеров/зондов для специфической дифференциации кодон-оптимизированного трансгена CFTRΔR человека от эндогенного гена CFTR человека. Экспрессия нетрансдуцированных ALI культур была ниже предела уровней количественного определения транскрипта CFTRΔR, как и ожидалось (фиг. 30). После трансдукции 4D-710 клетки показали дозозависимое увеличение транскрипта CFTRΔR (фиг. 30). При 50000 MOI уровень транскрипта CFTRΔR составлял ~80% от уровня эндогенного CFTR (фиг. 30). При 100000 MOI транскрипт CFTRΔR достигал уровня эндогенного CFTR, и в некоторых случаях превышал его (фиг. 30). Эти данные показывают, что 4D-710 может трансдуцировать ex vivo легочные ALI культуры с дозозависимой экспрессией трансгена CFTRΔR и при высоком значении MOI наблюдается увеличение мРНК CFTRΔR по сравнению с эндогенным CFTR.
[00515] Выводы. 4D-710 представляет собой рекомбинантный продукт генной заместительной терапии AAV, предназначенный для лечения муковисцидоза, вызванного мутациями CFTR, респираторного заболевания легких, предпочтительно путем доставки однократной дозы в легкие в виде аэрозоля. 4D-710 содержит капсидный белок с SEQ ID NO:12, идентифицированный направленной эволюцией как неожиданно пригодный для доставки терапевтических генных продуктов в легкие, и гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность SEQ ID NO:45 (нуклеотидная последовательность SEQ ID NO:45 содержит кодон-оптимизированную полезную нагрузку для генной терапии муковисцидоза человека (SEQ ID NO:43), функционально связанную с промотором CMV173 (SEQ ID NO:44)). Полученные данные in vitro и ex vivo для клеток человека показали, что 4D-710 восстанавливает транскрипт CFTR человека и экспрессию трансгена в клетках бронхиального эпителия человека, содержащих мутацию класса I муковисцидоза, и в ALI культурах здоровых легких. Кроме того, белок CFTR, экспрессированный после трансдукции 4D-710, был экспрессирован, посттрансляционно гликозилирован и локализован в цитоплазме и клеточной мембране в клетках HEK2v6.11. Эти данные демонстрируют, что 4D-710 способен трансдуцировать линию клеток легких человека и доставлять детектируемую мРНК CFTRΔR, что приводит к дозозависимой продукции белка CFTR и локализации экспрессированного белка CFTR на мембране. В сочетании с данными примера 3 и примера 7, демонстрирующими безопасную, надежную и широко распространенную трансдукцию и экспрессию трансгена в легких приматов после доставки в виде аэрозоля с минимальным системным воздействием, эти данные представляют собой существенный прогресс по сравнению с существующими серотипами AAV для разработки генной терапии для муковисцидоза и других заболеваний легких.
Пример 7
Диапазон доз и пилотное исследование безопасности аэрозольной доставки 4D-710 в легкие яванских макак
[00516] Было инициировано исследование in vivo для проверки безопасности и трансдукционной активности терапевтического продукта 4D-710 в экспериментальном исследовании на яванских макаках для оценки способности сконструированных вирусных векторов трансдуцировать клетки и экспрессировать трансген CFTRΔR в дыхательных путях, легких и других тканях после однократного аэрозольного введения в диапазоне доз. Как обсуждалось в примере 6, 4D-710 представляет собой rAAV, содержащий капсидный белок с SEQ ID NO:12 и гетерологичную нуклеиновую кислоту SEQ ID NO:45. Компонент гетерологичной нуклеиновой кислоты 4D-710 содержит нуклеотидную последовательность SEQ ID NO:43 - кодон-оптимизированный трансген CFTRΔR человека, функционально связанный с промотором CMV173 SEQ ID NO:44. Экспрессию и функцию предполагаемой кассеты терапевтического трансгена характеризовали in vivo у приматов, яванских макак. Сыворотку предварительно подвергали скринингу для выявления животных, которые были серонегативными в отношении ранее продуцированных нейтрализующих антител к капсидам тестируемого препарата. Животные получали носитель (только буфер для формуляции), 3×1012 вг 4D-710 или 3×1013 вг 4D-710, доставленные эндоскопически непосредственно ниже гортани с использованием устройства AeroEclipseII (Trudell Medical) для обеспечения оптимальной доставки в дистальный отдел легкого. Через 8 недель для анализа отбирали выбранные ткани (легкие, сердце, скелетные мышцы, печень, почки, поджелудочная железа, селезенка, головной мозг, спинной мозг, трахеобронхиальные лимфатические узлы и яички). Вирусные геномы были количественно определены с помощью количественной ПЦР. Образцы тканей, которые содержали детектируемые вирусные геномы, оценивали на транскрипт CFTRΔR с использованием ОТ-кПЦР, и делали срезы образцов легких и визуализировали на предмет экспрессии белка CFTR с помощью IHC.
[00517] Данные, приведенные ниже, демонстрируют, что доставка 4D-710 через распылитель приводила к надежной доставке вирусных геномов во все области легкого, включая периферические (бронхиоальвеолярные) области, с минимальным системным биораспределением, и 4D-710 опосредовал транскрипт CFTRΔR и экспрессию белка во всех областях легких. Отсутствовали сообщения о нежелательных явлениях, связанных с безопасностью, у животных, которым вводили носитель или испытуемый препарат 4D-710 (3×1012 и 3×1013 вг).
[00518] Материалы и методы
[00519] Анализ нейтрализующих антител
[00520] Клетки HEK2v6.11 (полученные из Университета Джона Хопкинса) высевали в черные непрозрачные 96-луночные планшеты при плотности клеток 30000 клеток/лунку в среде Игла, модифицированной по способу Дульбекко (DMEM; Corning) с 1% инактивированной нагреванием фетальной бычьей сыворотки (FBS; GE Healthcare Life Sciences) и 1% пенициллина/стрептомицина (Invitrogen). Клеткам давали прикрепиться к планшету в течение 24 ч до начала эксперимента.
[00521] Каждый образец сыворотки «нечеловеческого» примата (NHP) анализировали в разведениях 1:10, 1:25, 1:50. Каждый планшет содержал положительные и отрицательные контроли для трансдукции. Образцы сыворотки NHP инкубировали с вирусом при 37°C в течение 1 ч. Каждую лунку инфицировали 4D-A101.CAG-люциферазой (rAAV, содержащей капсидный белок SEQ ID NO:12 и гетерологичную нуклеиновую кислоту, кодирующую люциферазу, функционально связанную с промотором CAG) при множественности инфекции на уровне 1000. После инкубации в течение 1 ч каждый образец сыворотки NHP плюс разведение 4D-A101.CAG-люциферазы добавляли в отдельные лунки черных непрозрачных 96-луночных планшетов, содержащих клетки HEK2v6.11. Люциферазу определяли с помощью набора для анализа люциферазы ONE-Glo EX (Promega) через 48 ч после трансдукции. При добавлении ONE-Glo EX клетки лизировали, и к клеткам добавляли субстрат люциферазы в одну стадию. Люминесценцию считывали с помощью микропланшетного ридера Cytation 3 (BioTek).
[00522] Коэффициенты вариации (CV) и стандартные отклонения рассчитывали для всех разведений образцов сыворотки NHP и каждой точки стандартной кривой. Образцы сыворотки NHP нормализовали к положительному контролю трансдукции. Каждому NHP был присвоен титр нейтрализующих антител. Титр нейтрализующих антител для каждого образца сыворотки NHP определяли как самое низкое разведение сыворотки, при котором наблюдали трансдукцию ~ 50%. NHP, для которых наблюдалась трансдукция ~ 50% при разведении сыворотки 1:10, рассматривались для включения в исследование.
[00523] Тест-система и иммуносупрессия
[00524] В исследование было включено семь яванских макак самцов. Животные имели возраст от 4,7 до 4,9 лет и массу тела от 3,8 до 6,0 кг. Животные получали иммуносупрессию метилпреднизолоном (10 мг/кг, внутримышечно) один раз в неделю, начиная с суток -7, и до одной недели до эвтаназии и отбора образцов тканей (сутки 49). Дополнительные детали эксперимента представлены в отчете об исследовании контрактной исследовательской организации (CRO) (отчет о доклиническом исследовании находится на рассмотрении).
[00525] Приготовление и введение тестируемого образца
[00526] Партии испытуемого образца (ТА) 4D-710 и носителя разводили в буфере для формуляции для доставки конечных доз 3×1012 вг и 3×1013 вг 5 мл. ТА оттаивали при комнатной температуре в течение >1 ч и пробирки осторожно переворачивали 5 раз перед добавлением 5 мл в устройство для доставки (AeroEclipseII).
[00527] Животных анестезировали телозолом (в/м, 7-8 мг/кг) и эндотрахеальную трубку с манжетой (размер 4,5) вводили в трахею непосредственно под гортанью и закрепляли, чтобы избежать выскальзывания во время переноса и размещения животного в кресле. После этого животных помещали в кресло в вертикальном положении, чтобы соединить эндотрахеальную трубку с системой генерации аэрозоля. Животных укрывали одеялами и с помощью устройства Bair Hugger для внешнего нагревания, и частоту сердечных сокращений и насыщение кислородом контролировали во время экспозиции и выхода из анестезии.
[00528] В системе генерирования и доставки аэрозоля для генерирования аэрозоля использовался аэрозольный распылитель AeroEclipse II (BAN, Trudell Medical International, Канада). В общей сложности пять (5) мл контрольного или испытуемого препарата вводили анестезированным животным через эндотрахеальную трубку. Распылитель работал в режиме непрерывной генерации аэрозоля посредством переключения ручки на крышке распылителя. Кроме того, вентиляционные отверстия внутри крышки распылителя были заглушены.
[00529] Наблюдения за животными
[00530] Дважды в день наблюдения у клетки, в которой содержались животные, проводили в соответствии с СОП «Процедуры ухода и содержания домашних приматов, отличных от человека». В дни обработки и проведения процедуры животные находились под постоянным наблюдением исследовательского персонала. Основные наблюдения включали, помимо прочего, признаки тошноты, вялость, респираторный дистресс и цианоз, обесцвечивание слизистых оболочек, рвоту и нерегулярные выделения из физиологических отверстий или наличие крови в фекалиях/моче.
[00531] Отбор крови
[00532] Животных не кормили в течение ночи со свободным доступом к питьевой воде перед любым забором крови и кормили сразу после проведения процедуры. Образцы крови для клинической патологии и анализа титра антител и иммуногенности собирали путем венепункции бедренной вены. Образцы (1 мл в ЭДТА) для гематологии и полного подсчета клеток отправляли во внешнюю референс-лабораторию (IDEXX) в день отбора для анализа в течение 24-48 ч после отбора. Образцы для клинической химии (1 мл в SST) были проанализированы в лаборатории патологии CRO в день отбора.
[00533] Бронхоальвеолярные лаважи (БАЛ)
[00534] Животных выдерживали натощак в течение ночи и анестезировали кетамином гидрохлоридом с последующей ингаляцией изофлураном в соответствии с СОП «Анестезия приматов, отличных от человека». Эндотрахеальную трубку с манжетой надлежащего размера вводили непосредственно проксимальнее киля, чтобы можно было ввести волоконно-оптический бронхоскоп, применяемый в педиатрии, для выполнения БАЛ в соответствии с СОП ACL-1536 «Процедуры проведения лаважа легких и/или бронхоскопии у приматов, отличных от человека». Образцы с обеих сторон легкого отбирали в начале исследования (сутки -7) и через 4 недели после обработки. Три аликвоты по 10 мл PBS Дульбекко вводили в соответствующую сторону легкого через бронхоскоп и последовательно аспирировали. Лаважные изоляты из первого промывания хранили отдельно, и 2 и 3 промывание объединяли во время сбора и хранили на влажном льду до обработки в течение не более 2 ч после отбора. Подсчитывали объединенные клетки из всех промываний и готовили предметные стекла для проведения дифференциального подсчета клеток с использованием морфологических критериев в CRO.
[00535] Эвтаназия и некропсия
[00536] Для эвтаназии животных анестезировали кетамином (от 11,0 до 12,4 мг/кг, в/м) с последующей внутривенной инъекцией раствора для эвтаназии в соответствии с CRO СОП «Эвтаназия крупных животных». После подтверждения наступления смерти обученный персонал проводил вскрытие в соответствии с CRO СОП «Процедура вскрытия негрызунов», и ткани были собраны, взвешены, сохранены и исследованы. Ткани обрабатывали соответствующим образом на ДНК и РНК или готовили для гистопатологического анализа.
[00537] Животных перфузировали гепаринизированным физиологическим раствором перед сбором любой ткани. Были отобраны образцы легких и трахеи, скелетных мышц (диафрагмы, трехглавой мышцы плеча и латеральной широкой мышцы бедра), сердца, печени, селезенки, поджелудочной железы, яичка, почек, головного мозга, трахеобронхиальных лимфатических узлов и спинного мозга. Каждую ткань разделяли на разные области, и из каждой области было отобрано несколько образцов. Проводили макроскопическое исследование основных периферических органов и образцов тканей, отобранных из любых выявленных патологических зон. Образцы тканей собирали и быстро замораживали для последующего анализа ДНК или хранили в RNALater для последующего выделения РНК. Отбирали дополнительные образцы и фиксировали в 10% нейтральном забуференном формалине для последующей заливки в парафин и приготовления срезов для иммуногистохимии.
[00538] Из трахеи и легких отбирали больште образцы, чтобы получить несколько образцов для каждого анализа. Легкие отбирали и зажимали как можно выше над трахеей. Правое легкое было дважды пережато с интервалом приблизительно 1 мм на главных бронхах. Правое легкое удаляли разрезом между зажимами. Шестнадцать образцов для выделения ДНК и РНК были собраны из областей правого легкого, охватывающих первичные/вторичные бронхи, третичные бронхи и альвеолы, как показано на фиг. 16. Трахею и левое легкое заполняли фиксатором и фиксировали в 10% нейтральном растворе забуференного формалина. Затем готовили срезы трахеи и левого легкого, чтобы охватить образцы трахеи, первичных/вторичных бронхов, третичных бронхов и альвеол, как описано на фиг. 16.
[00539] Биораспределение вирусного генома и экспрессия трансгена
[00540] Метод количественной полимеразной цепной реакции в режиме реального времени (кПЦР) и метод количественной полимеразной цепной реакции в режиме реального времени с обратной транскрипцией (ОТ-кПЦР) были разработаны для количественного анализа в тканях NHP. Количественную оценку вирусных геномов в легких NHP из каждой группы проводили с помощью количественной ПЦР и ОТ-кПЦР с использованием праймеров и зондов против (и специфичных для) кодон-оптимизированного трансгена (SEQ ID NO:43). Титры вирусов, измеренные в ткани, выражали в виде число копий на мкг ДНК (BLQ=50), и трансгенный транскрипт экспрессируется как число копий на реакцию с 250 нг РНК (BLQ=25).
[00541] Иммуногистохимию проводили с образцами ткани трахеи и левого легкого, включая образцы трахеи, первичных/вторичных бронхов, третичных бронхов и альвеол, как показано на фиг. 16, от всех животных. Анализ CFTR IHC выполняли с использованием автоматического красителя Roche Discovery ULTRA и реагентов для обнаружения, протокола извлечения антигена CC1S и набора для амплификации тирамида, а также мышиного моноклонального антитела против CFTR (Abcam ab270238 M3A7).
[00542] Результаты
[00543] Скрининг нейтрализующих анти-AAV антител идентифицирует NHP для включения в исследование
[00544] Анализ нейтрализующих антител использовали для оценки уровней нейтрализующих антител против капсида 4D-A101 (содержащего капсидный белок SEQ ID NO:12) в сыворотке NHP. Для каждого образца сыворотки NHP определяли титр нейтрализующих антител. Животное считали серонегативным, и оно соответствовало критериям включения в исследование, если наблюдалась трансдукция на уровне ≥50% при разведении сыворотки 1:10.
[00545] В целом было анализировано 50 образцов сыворотки NHP. Критерии приемлемости анализа были установлены для 1) коэффициента вариации (CV) стандартной кривой, 2) CV неизвестных образцов сыворотки и 3) процентного отклонения от фактического входного белка для стандартной кривой. Приемлемые значения коэффициентов вариации для стандартной кривой были определены на уровне <25%. Приемлемые CV для неизвестных образцов сыворотки в пределах количественного определения были определены на уровне <30%. Приемлемое процентное отклонение от входного белка для стандартной кривой было определено на уровне <25%. Все планшеты соответствовали всем критериям приемлемости анализа, и данные, полученные на этих планшетах, использовались для оценки. В целом, 19 (38%) анализированных образцов сыворотки NHP были серонегативными в отношении 4D-A101 (фиг. 31). Для включения в исследование были отобраны NHP с самым высоким процентом трансдукции при разведении сыворотки 1:10 и прошедшие скрининг состояния здоровья у поставщика. Выбранные идентификаторы NHP и процент трансдукции при разведении 1:10 приведены в таблице 17:
NHP, включенные в исследование
анализа
[00546] Доставка 4D-710 с использованием распылителя хорошо переносится у NHP
[00547] Дизайн исследования представлен в таблице 18:
Обобщенный дизайн исследования
Трахеобронхивальный лимфатический узел
Сердце
Печень
Скелетная мышца (трехглавая мышца плеча, четырехглавая мышца, диафрагма)
Почка
Селезенка
Поджелудочная железа
Головной мозг
Спинной мозг
ОТ-кПЦР (легкое & кПЦР+ ткань)
IHC (легкое)
[00548] Ни одно животное не пало или не было преждевременно подвергнуто эвтаназии во время проведения данного исследования. Ни у одного животного не было каких-либо признаков дистресса или проблем со здоровьем, связанных с воздействием ТА в течение всего периода исследования. Во время обработки и сбора образцов не наблюдали потери аппетита и изменений в пищевом поведении. Во время этого исследования отсутстовали какие-либо изменения массы тела, связанные с лечением.
[00549] Образцы для гематологии и полного подсчета клеток были отправлены во внешнюю референс-лабораторию (IDEXX) в день отбора, и анализ был проведен не более чем через 48 ч после отбора. Образцы для конечных точек биохимического анализа были анализированы лабораторией патологии CRO в день отбора. Некоторые из биохимических показателей изменялись во времени, но не наблюдалось никакой разницы между группой с носителем по сравнению с обоими уровнями доз группы, получавшей 4D-710, на все временные точки. Аналогично через 8 недель после ингаляционной обработки не наблюдали серьезных изменений в гематологических показателях и лейкоцитарной формуле по сравнению с исходными уровнями.
[00550] Дифференциальное соотношение клеток и количество клеток в лаважной жидкости определяли для правой и левой стороны лаважа, и все данные представлены в виде средних значений для обеих сторон. Общее количество клеток и дифференциальное соотношение клеток в BALF не различалось между группами обработки, измеренными на сутки 28, по сравнению с исходными уровнями, и за счет вариабельности и небольшого размера выборки невозможно сделать дополнительные выводы.
[00551] Массу органов и массу органов по отношению к массе тела определяли в день эвтаназии для всех групп обработки. Массы органов и массы, нормализованные к массе тела, не различались ни в одной из групп обработки.
[00552] Во время макроскопического исследования при вскрытии не сообщалось о серьезных патологиях. Микроскопическое исследование показало, что в исследованных тканях не было никаких наблюдений, связанных с обработкой.
[00553] 4D-710 опосредует надежную доставку генов во все области легкого
[00554] Вирусные геномы количественно определяли многоуровневым подходом для образцов, полученных во время некропсии, для определения геномного биораспределения 4D-710. Анализировали все образцы легких, полученные от животных, получавших носитель, 3×1012 вг и 3×1013 вг. Головной мозг, спинной мозг (шейный, грудной и поясничный отделы), сердце, печень, селезенка, поджелудочная железа, почки, скелетные мышцы (трехглавая мышца плеча, латеральная широкая мышца бедра, диафрагма), трахеобронхиальные лимфатические узлы и яички, от всех животные после введения из 3×1013 вг. Ткани, которые имели уровни вирусных геномов выше BLQ, затем анализировали при более низкой дозе 3×1012 вг (данные ожидаются). Данные представлены в виде вирусных геномов на мкг ДНК.
[00555] Результаты анализа кПЦР указывают на равномерную доставку вируса в различные области легких, которые представляют собой образцы из альвеолярных мешочков (дистальные), третичных бронхов (медиальные) и первичных/вторичных бронхов (проксимальные), за исключением образцов с маркировкой R6. Данный образец было трудно отобрать за счет близости к килю и, следовательно, места для разделения и связывания правой и левой сторон легкого (см. фиг. 16). Среднее количество копий для остальных положений образцов для групп с дозой 3×1012 вг и 3×1013 вг составляло 104 и 105 соответственно (фиг. 32A). По меньшей мере 10-кратная разница между группами с низкой и высокой дозой хорошо соответствует 10-кратной разнице в дозе обработки. Для всех животных в одной и той же группе обработки не было отмечено существенных различий в количестве вирусных геномов в разных долях легких или разных областях легких (фиг. 32B).
[00556] Системное воздействие помимо легких оценивали у животных, которым вводили 3×1013 вг 4D-710 (фиг. 32C). Животные имели детектируемые вирусные геномы ~104 вг/мкг, присутствующие в трахеобронхиальных лимфатических узлах (фиг. 32C). Патологические показатели у этих животных для этой ткани являются нормальными. У одного NHP были детектированы вирусные геномы в селезенке (фиг. 32C), при этом патология считалась нормальной. Во всех других тестированных образцах головного и спинного мозга, сердца, печени, поджелудочной железы, почек, скелетных мышц и яичек уровень вирусных геномов был ниже нижнего предела количественного определения (фиг. 32C). О патологиях, связанных с тестируемым образцом, не сообщалось. Таким образом, доставка 4D-710 с распылителем приводит к безопасной и надежной доставке вирусных геномов во все области легкого с минимальным внелегочным системным воздействием.
[00557] 4D-710 опосредует экспрессию трансгена во всех областях легкого
[00558] Транскрипт CFTRΔR количественно определяли многоуровневым подходом для образцов, полученных во время некропсии, для определения экспрессии трансгена 4D-710. Были анализированы все образцы легких, полученные от животных, получавших носитель в дозе 3×1012 вг и 3×1013 вг. Анализировали образцы головного мозга, спинного мозга, сердца, печени, селезенки, поджелудочной железы, почек, скелетных мышц, трахеобронхиальных лимфатических узлов и яичек, отобранные у животных, получавших дозу 3×1013 вг (данные ожидаются). Ткани, которые имели уровни транскриптов выше BLQ, затем анализировали при более низкой дозе 3×1012 вг (данные ожидаются). Данные представлены в виде копий на реакцию 250 нг РНК и представлены на графике в виде копий на мкг РНК.
[00559] Результаты анализа ОТ-кПЦР указывают на успешную трансдукцию и экспрессию транскрипта в легких. У животных, которым вводили дозу 3×1013 вг, 44 из 48 образцов легких превышали BLQ с ~103 копиями на мкг РНК без образцов носителя выше BLQ (фиг. 33A). Три из четырех образцов BLQ были отобраны из одного и того же места отбора проб R6, где сбор был технически сложным, и у одного NHP был дополнительный образец BLQ. Трансдукцию отмечали в различных областях легких (фиг. 33B), которые представляли образцы из альвеолярных мешочков (дистальные), третичных бронхов (медиальные) и первичных/вторичных бронхов (проксимальные), за исключением образцов с маркировкой R6 (см. обоснование выше). Напротив, в образцах легких от животных с более низкой дозой 3×1012 вг 41 из 48 образцов представляли собой BLQ. Эти данные свидетельствуют о том, что доза 3×1013 вг 4D-710 способна количественно преобразовывать легочную ткань NHP и экспрессировать терапевтический транскрипт CFTRΔR.
[00560] Экспрессия белка CFTR, детектированная с использованием иммуногистохимии, показывает повышенную экспрессию CFTR в обработанном 4D-710 легком у NHP по сравнению с носителем в эпителии трахеи, бронхиальном эпителии и срезах альвеол (фиг. 34A). Высокое повышение экспрессии CFTR наблюдали у всех животных, получавших 3×1013 вг (фиг. 34B), по сравнению с носителем (фиг. 34A). Эти результаты демонстрируют, что доставка 4D-710 через распылитель опосредует экспрессию белка во всех областях легкого.
[00561] Выводы
[00562] Терапевтический продукт 4D-710 характеризовали при аэрозольной доставке яванским макакам. Сыворотку предварительно подвергали скринингу для выявления животных, которые были серонегативными в отношении ранее продуцированных нейтрализующих антител к капсиду тестируемого образца 4D-A101. Животные получали либо носитель (буфер для формуляции), либо дозу 4D-710 3×1012 вг, либо дозу 4D-710 3×1013 вг, доставляемую с помощью устройства AeroEclipseII.
[00563] В исследовании наблюдали большое количество вирусных геномов и в результате экспрессию транскрипта CFTRΔR и экспрессию белка CFTR в образцах легких у NHP, которые представляли собой образцы из альвеолярных мешочков, третичных бронхов и первичных/вторичных бронхов. В одном образце селезенки находилось небольшое количество вирусных геномов, но они поддавались детектированию, а во всех образцах из всех других тканей детектируемые вирусные геномы отсутствовали. Полученные данные демонстрируют, что доставка 4D-710 с помощью распылителя приводит к безопасной и надежной доставке вирусных геномов во все области легких приматов (трахею, альвеолы и бронхиальный эпителий) с минимальным системным биораспределением. Эти данные также демонстрируют, что 4D-710 опосредует стабильную экспрессию мРНК CFTRΔR и белкового продукта во всех областях легких, включая альвеолы, что подтверждает пригодность rAAV с капсидами, содержащими капсидный белок SEQ ID NO:12, для доставки в легкие вектора для лечения различных заболеваний легких и, в частности, поддерживают применение 4D-710 (содержащего капсидный белок с SEQ ID NO:12 и гетерологичную нуклеиновую кислоту SEQ ID NO:45) для доставки биологически активного терапевтического трансгена CFTR для лечения муковисцидоза у пациентов-людей.
[00564] Несмотря на то что настоящее изобретение было описано со ссылкой на его конкретные варианты осуществления, специалистам в данной области должно быть понятно, что могут быть внесены различные изменения и эквиваленты могут быть заменены без отклонения от истинной сущности и объема изобретения. Кроме того, можно сделать множество модификаций, чтобы адаптировать конкретную ситуацию, материал, состав вещества, процесс, стадию или стадии способа к цели, сущности и объему настоящего изобретения. Предполагается, что все такие модификации входят в объем прилагаемой формулы изобретения.
--->
Список последовательностей
<110> 4D Molecular Therapeutics, Inc.
<120> ВАРИАНТЫ, СОСТАВЫ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННОГО ВИРУСА И СПОСОБЫ ДЛЯ
ПУЛЬМОНАЛЬНОЙ ДОСТАВКИ
<130> 090400-5013 WO
<150> US 63/016,246
<151> 2020-04-27
<150> US 63/088,432
<151> 2020-10-06
<160> 45
<170> PatentIn version 3.5
<210> 1
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 1
<400> 1
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Glu Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Phe Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Ala Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys Asn Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 2
<211> 735
<212> БЕЛОК
<213> аденоассоциированный вирус 2
<400> 2
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 3
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 3
<400> 3
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Val Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Arg Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Ile Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Gly
130 135 140
Ala Val Asp Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Val Gly
145 150 155 160
Lys Ser Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Thr Ser Leu Gly Ser Asn Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Lys Leu Ser Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Arg Gly Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg Thr
435 440 445
Gln Gly Thr Thr Ser Gly Thr Thr Asn Gln Ser Arg Leu Leu Phe Ser
450 455 460
Gln Ala Gly Pro Gln Ser Met Ser Leu Gln Ala Arg Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Leu Ser Lys Thr Ala Asn Asp Asn
485 490 495
Asn Asn Ser Asn Phe Pro Trp Thr Ala Ala Ser Lys Tyr His Leu Asn
500 505 510
Gly Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Glu Lys Phe Phe Pro Met His Gly Asn Leu Ile Phe Gly
530 535 540
Lys Glu Gly Thr Thr Ala Ser Asn Ala Glu Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln
565 570 575
Tyr Gly Thr Val Ala Asn Asn Leu Gln Ser Ser Asn Thr Ala Pro Thr
580 585 590
Thr Gly Thr Val Asn His Gln Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Met Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Thr Thr Phe Ser Pro Ala Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 4
<211> 734
<212> БЕЛОК
<213> аденоассоциированный вирус 4
<400> 4
Met Thr Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser Glu
1 5 10 15
Gly Val Arg Glu Trp Trp Ala Leu Gln Pro Gly Ala Pro Lys Pro Lys
20 25 30
Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro Gly
35 40 45
Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro Val
50 55 60
Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp Gln
65 70 75 80
Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala Asp
85 90 95
Ala Glu Phe Gln Gln Arg Leu Gln Gly Asp Thr Ser Phe Gly Gly Asn
100 105 110
Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro Leu
115 120 125
Gly Leu Val Glu Gln Ala Gly Glu Thr Ala Pro Gly Lys Lys Arg Pro
130 135 140
Leu Ile Glu Ser Pro Gln Gln Pro Asp Ser Ser Thr Gly Ile Gly Lys
145 150 155 160
Lys Gly Lys Gln Pro Ala Lys Lys Lys Leu Val Phe Glu Asp Glu Thr
165 170 175
Gly Ala Gly Asp Gly Pro Pro Glu Gly Ser Thr Ser Gly Ala Met Ser
180 185 190
Asp Asp Ser Glu Met Arg Ala Ala Ala Gly Gly Ala Ala Val Glu Gly
195 200 205
Gly Gln Gly Ala Asp Gly Val Gly Asn Ala Ser Gly Asp Trp His Cys
210 215 220
Asp Ser Thr Trp Ser Glu Gly His Val Thr Thr Thr Ser Thr Arg Thr
225 230 235 240
Trp Val Leu Pro Thr Tyr Asn Asn His Leu Tyr Lys Arg Leu Gly Glu
245 250 255
Ser Leu Gln Ser Asn Thr Tyr Asn Gly Phe Ser Thr Pro Trp Gly Tyr
260 265 270
Phe Asp Phe Asn Arg Phe His Cys His Phe Ser Pro Arg Asp Trp Gln
275 280 285
Arg Leu Ile Asn Asn Asn Trp Gly Met Arg Pro Lys Ala Met Arg Val
290 295 300
Lys Ile Phe Asn Ile Gln Val Lys Glu Val Thr Thr Ser Asn Gly Glu
305 310 315 320
Thr Thr Val Ala Asn Asn Leu Thr Ser Thr Val Gln Ile Phe Ala Asp
325 330 335
Ser Ser Tyr Glu Leu Pro Tyr Val Met Asp Ala Gly Gln Glu Gly Ser
340 345 350
Leu Pro Pro Phe Pro Asn Asp Val Phe Met Val Pro Gln Tyr Gly Tyr
355 360 365
Cys Gly Leu Val Thr Gly Asn Thr Ser Gln Gln Gln Thr Asp Arg Asn
370 375 380
Ala Phe Tyr Cys Leu Glu Tyr Phe Pro Ser Gln Met Leu Arg Thr Gly
385 390 395 400
Asn Asn Phe Glu Ile Thr Tyr Ser Phe Glu Lys Val Pro Phe His Ser
405 410 415
Met Tyr Ala His Ser Gln Ser Leu Asp Arg Leu Met Asn Pro Leu Ile
420 425 430
Asp Gln Tyr Leu Trp Gly Leu Gln Ser Thr Thr Thr Gly Thr Thr Leu
435 440 445
Asn Ala Gly Thr Ala Thr Thr Asn Phe Thr Lys Leu Arg Pro Thr Asn
450 455 460
Phe Ser Asn Phe Lys Lys Asn Trp Leu Pro Gly Pro Ser Ile Lys Gln
465 470 475 480
Gln Gly Phe Ser Lys Thr Ala Asn Gln Asn Tyr Lys Ile Pro Ala Thr
485 490 495
Gly Ser Asp Ser Leu Ile Lys Tyr Glu Thr His Ser Thr Leu Asp Gly
500 505 510
Arg Trp Ser Ala Leu Thr Pro Gly Pro Pro Met Ala Thr Ala Gly Pro
515 520 525
Ala Asp Ser Lys Phe Ser Asn Ser Gln Leu Ile Phe Ala Gly Pro Lys
530 535 540
Gln Asn Gly Asn Thr Ala Thr Val Pro Gly Thr Leu Ile Phe Thr Ser
545 550 555 560
Glu Glu Glu Leu Ala Ala Thr Asn Ala Thr Asp Thr Asp Met Trp Gly
565 570 575
Asn Leu Pro Gly Gly Asp Gln Ser Asn Ser Asn Leu Pro Thr Val Asp
580 585 590
Arg Leu Thr Ala Leu Gly Ala Val Pro Gly Met Val Trp Gln Asn Arg
595 600 605
Asp Ile Tyr Tyr Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp
610 615 620
Gly His Phe His Pro Ser Pro Leu Ile Gly Gly Phe Gly Leu Lys His
625 630 635 640
Pro Pro Pro Gln Ile Phe Ile Lys Asn Thr Pro Val Pro Ala Asn Pro
645 650 655
Ala Thr Thr Phe Ser Ser Thr Pro Val Asn Ser Phe Ile Thr Gln Tyr
660 665 670
Ser Thr Gly Gln Val Ser Val Gln Ile Asp Trp Glu Ile Gln Lys Glu
675 680 685
Arg Ser Lys Arg Trp Asn Pro Glu Val Gln Phe Thr Ser Asn Tyr Gly
690 695 700
Gln Gln Asn Ser Leu Leu Trp Ala Pro Asp Ala Ala Gly Lys Tyr Thr
705 710 715 720
Glu Pro Arg Ala Ile Gly Thr Arg Tyr Leu Thr His His Leu
725 730
<210> 5
<211> 724
<212> БЕЛОК
<213> аденоассоциированный вирус 5
<400> 5
Met Ser Phe Val Asp His Pro Pro Asp Trp Leu Glu Glu Val Gly Glu
1 5 10 15
Gly Leu Arg Glu Phe Leu Gly Leu Glu Ala Gly Pro Pro Lys Pro Lys
20 25 30
Pro Asn Gln Gln His Gln Asp Gln Ala Arg Gly Leu Val Leu Pro Gly
35 40 45
Tyr Asn Tyr Leu Gly Pro Gly Asn Gly Leu Asp Arg Gly Glu Pro Val
50 55 60
Asn Arg Ala Asp Glu Val Ala Arg Glu His Asp Ile Ser Tyr Asn Glu
65 70 75 80
Gln Leu Glu Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala Asp
85 90 95
Ala Glu Phe Gln Glu Lys Leu Ala Asp Asp Thr Ser Phe Gly Gly Asn
100 105 110
Leu Gly Lys Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro Phe
115 120 125
Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Thr Gly Lys Arg Ile
130 135 140
Asp Asp His Phe Pro Lys Arg Lys Lys Ala Arg Thr Glu Glu Asp Ser
145 150 155 160
Lys Pro Ser Thr Ser Ser Asp Ala Glu Ala Gly Pro Ser Gly Ser Gln
165 170 175
Gln Leu Gln Ile Pro Ala Gln Pro Ala Ser Ser Leu Gly Ala Asp Thr
180 185 190
Met Ser Ala Gly Gly Gly Gly Pro Leu Gly Asp Asn Asn Gln Gly Ala
195 200 205
Asp Gly Val Gly Asn Ala Ser Gly Asp Trp His Cys Asp Ser Thr Trp
210 215 220
Met Gly Asp Arg Val Val Thr Lys Ser Thr Arg Thr Trp Val Leu Pro
225 230 235 240
Ser Tyr Asn Asn His Gln Tyr Arg Glu Ile Lys Ser Gly Ser Val Asp
245 250 255
Gly Ser Asn Ala Asn Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr
260 265 270
Phe Asp Phe Asn Arg Phe His Ser His Trp Ser Pro Arg Asp Trp Gln
275 280 285
Arg Leu Ile Asn Asn Tyr Trp Gly Phe Arg Pro Arg Ser Leu Arg Val
290 295 300
Lys Ile Phe Asn Ile Gln Val Lys Glu Val Thr Val Gln Asp Ser Thr
305 310 315 320
Thr Thr Ile Ala Asn Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp
325 330 335
Asp Asp Tyr Gln Leu Pro Tyr Val Val Gly Asn Gly Thr Glu Gly Cys
340 345 350
Leu Pro Ala Phe Pro Pro Gln Val Phe Thr Leu Pro Gln Tyr Gly Tyr
355 360 365
Ala Thr Leu Asn Arg Asp Asn Thr Glu Asn Pro Thr Glu Arg Ser Ser
370 375 380
Phe Phe Cys Leu Glu Tyr Phe Pro Ser Lys Met Leu Arg Thr Gly Asn
385 390 395 400
Asn Phe Glu Phe Thr Tyr Asn Phe Glu Glu Val Pro Phe His Ser Ser
405 410 415
Phe Ala Pro Ser Gln Asn Leu Phe Lys Leu Ala Asn Pro Leu Val Asp
420 425 430
Gln Tyr Leu Tyr Arg Phe Val Ser Thr Asn Asn Thr Gly Gly Val Gln
435 440 445
Phe Asn Lys Asn Leu Ala Gly Arg Tyr Ala Asn Thr Tyr Lys Asn Trp
450 455 460
Phe Pro Gly Pro Met Gly Arg Thr Gln Gly Trp Asn Leu Gly Ser Gly
465 470 475 480
Val Asn Arg Ala Ser Val Ser Ala Phe Ala Thr Thr Asn Arg Met Glu
485 490 495
Leu Glu Gly Ala Ser Tyr Gln Val Pro Pro Gln Pro Asn Gly Met Thr
500 505 510
Asn Asn Leu Gln Gly Ser Asn Thr Tyr Ala Leu Glu Asn Thr Met Ile
515 520 525
Phe Asn Ser Gln Pro Ala Asn Pro Gly Thr Thr Ala Thr Tyr Leu Glu
530 535 540
Gly Asn Met Leu Ile Thr Ser Glu Ser Glu Thr Gln Pro Val Asn Arg
545 550 555 560
Val Ala Tyr Asn Val Gly Gly Gln Met Ala Thr Asn Asn Gln Ser Ser
565 570 575
Thr Thr Ala Pro Ala Thr Gly Thr Tyr Asn Leu Gln Glu Ile Val Pro
580 585 590
Gly Ser Val Trp Met Glu Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp
595 600 605
Ala Lys Ile Pro Glu Thr Gly Ala His Phe His Pro Ser Pro Ala Met
610 615 620
Gly Gly Phe Gly Leu Lys His Pro Pro Pro Met Met Leu Ile Lys Asn
625 630 635 640
Thr Pro Val Pro Gly Asn Ile Thr Ser Phe Ser Asp Val Pro Val Ser
645 650 655
Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val Thr Val Glu Met Glu
660 665 670
Trp Glu Leu Lys Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln
675 680 685
Tyr Thr Asn Asn Tyr Asn Asp Pro Gln Phe Val Asp Phe Ala Pro Asp
690 695 700
Ser Thr Gly Glu Tyr Arg Thr Thr Arg Pro Ile Gly Thr Arg Tyr Leu
705 710 715 720
Thr Arg Pro Leu
<210> 6
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 6
<400> 6
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Val Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 7
<211> 735
<212> БЕЛОК
<213> аденоассоциированный вирус 7
<400> 7
Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser Glu
1 5 10 15
Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro Lys
20 25 30
Ala Asn Gln Gln Lys Gln Asp Asn Gly Arg Gly Leu Val Leu Pro Gly
35 40 45
Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro Val
50 55 60
Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp Gln
65 70 75 80
Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala Asp
85 90 95
Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly Asn
100 105 110
Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro Leu
115 120 125
Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Ala Lys Lys Arg Pro
130 135 140
Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile Gly
145 150 155 160
Lys Lys Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Ser Val Gly Ser Gly Thr Val Ala Ala Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Glu Thr Ala Gly Ser Thr Asn Asp Asn Thr
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Lys Leu Arg Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Ile Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ser Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Glu Phe Ser Tyr Ser Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ala Arg
435 440 445
Thr Gln Ser Asn Pro Gly Gly Thr Ala Gly Asn Arg Glu Leu Gln Phe
450 455 460
Tyr Gln Gly Gly Pro Ser Thr Met Ala Glu Gln Ala Lys Asn Trp Leu
465 470 475 480
Pro Gly Pro Cys Phe Arg Gln Gln Arg Val Ser Lys Thr Leu Asp Gln
485 490 495
Asn Asn Asn Ser Asn Phe Ala Trp Thr Gly Ala Thr Lys Tyr His Leu
500 505 510
Asn Gly Arg Asn Ser Leu Val Asn Pro Gly Val Ala Met Ala Thr His
515 520 525
Lys Asp Asp Glu Asp Arg Phe Phe Pro Ser Ser Gly Val Leu Ile Phe
530 535 540
Gly Lys Thr Gly Ala Thr Asn Lys Thr Thr Leu Glu Asn Val Leu Met
545 550 555 560
Thr Asn Glu Glu Glu Ile Arg Pro Thr Asn Pro Val Ala Thr Glu Glu
565 570 575
Tyr Gly Ile Val Ser Ser Asn Leu Gln Ala Ala Asn Thr Ala Ala Gln
580 585 590
Thr Gln Val Val Asn Asn Gln Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asn Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Glu Val Phe Thr Pro Ala Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Phe Glu Lys Gln Thr Gly Val Asp Phe Ala Val Asp Ser Gln Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn
725 730 735
<210> 8
<211> 738
<212> БЕЛОК
<213> аденоассоциированный вирус 8
<400> 8
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Gln Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Pro Ser Pro Gln Arg Ser Pro Asp Ser Ser Thr Gly Ile
145 150 155 160
Gly Lys Lys Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln
165 170 175
Thr Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro
180 185 190
Pro Ala Ala Pro Ser Gly Val Gly Pro Asn Thr Met Ala Ala Gly Gly
195 200 205
Gly Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser
210 215 220
Ser Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val
225 230 235 240
Ile Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His
245 250 255
Leu Tyr Lys Gln Ile Ser Asn Gly Thr Ser Gly Gly Ala Thr Asn Asp
260 265 270
Asn Thr Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn
275 280 285
Arg Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn
290 295 300
Asn Asn Trp Gly Phe Arg Pro Lys Arg Leu Ser Phe Lys Leu Phe Asn
305 310 315 320
Ile Gln Val Lys Glu Val Thr Gln Asn Glu Gly Thr Lys Thr Ile Ala
325 330 335
Asn Asn Leu Thr Ser Thr Ile Gln Val Phe Thr Asp Ser Glu Tyr Gln
340 345 350
Leu Pro Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe
355 360 365
Pro Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn
370 375 380
Asn Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr
385 390 395 400
Phe Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Thr Tyr
405 410 415
Thr Phe Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser
420 425 430
Leu Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu
435 440 445
Ser Arg Thr Gln Thr Thr Gly Gly Thr Ala Asn Thr Gln Thr Leu Gly
450 455 460
Phe Ser Gln Gly Gly Pro Asn Thr Met Ala Asn Gln Ala Lys Asn Trp
465 470 475 480
Leu Pro Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Thr Thr Thr Gly
485 490 495
Gln Asn Asn Asn Ser Asn Phe Ala Trp Thr Ala Gly Thr Lys Tyr His
500 505 510
Leu Asn Gly Arg Asn Ser Leu Ala Asn Pro Gly Ile Ala Met Ala Thr
515 520 525
His Lys Asp Asp Glu Glu Arg Phe Phe Pro Ser Asn Gly Ile Leu Ile
530 535 540
Phe Gly Lys Gln Asn Ala Ala Arg Asp Asn Ala Asp Tyr Ser Asp Val
545 550 555 560
Met Leu Thr Ser Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr
565 570 575
Glu Glu Tyr Gly Ile Val Ala Asp Asn Leu Gln Gln Gln Asn Thr Ala
580 585 590
Pro Gln Ile Gly Thr Val Asn Ser Gln Gly Ala Leu Pro Gly Met Val
595 600 605
Trp Gln Asn Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile
610 615 620
Pro His Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe
625 630 635 640
Gly Leu Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val
645 650 655
Pro Ala Asp Pro Pro Thr Thr Phe Asn Gln Ser Lys Leu Asn Ser Phe
660 665 670
Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu
675 680 685
Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr
690 695 700
Ser Asn Tyr Tyr Lys Ser Thr Ser Val Asp Phe Ala Val Asn Thr Glu
705 710 715 720
Gly Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg
725 730 735
Asn Leu
<210> 9
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 9
<400> 9
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Gln Ala Gln
580 585 590
Thr Gly Trp Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Met
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asp Pro Pro Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Tyr Lys Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 10
<211> 735
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 10
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Lys Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asp Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asn Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Ser Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Gln
725 730 735
<210> 11
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 11
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Gln Arg Leu Gln Gly Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Gln Ala Gly Glu Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Leu Ile Glu Ser Pro Gln Gln Pro Asp Ser Ser Thr Gly Ile Gly
145 150 155 160
Lys Lys Gly Lys Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Asp Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Val Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys Asn Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 12
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 12
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Gln Arg Leu Gln Gly Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Gln Ala Gly Glu Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Leu Ile Glu Ser Pro Gln Gln Pro Asp Ser Ser Thr Gly Ile Gly
145 150 155 160
Lys Lys Gly Lys Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Asp Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Thr Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Ala Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys Asn Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 13
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 13
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Ser Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Glu Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Phe Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Ala Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys Asn Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Ile Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Gln
725 730 735
<210> 14
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Artificial nucleic acid sequence
<400> 14
gcggaagctt cgatcaacta cgc 23
<210> 15
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 15
ggggcggccg caattacaga ttacgagtca ggtatctggt g 41
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<220>
<221> misc_feature
<222> (5)..(6)
<223> n представляет собой a, c, g или t
<400> 16
cattnnkgac cagtctagga actgg 25
<210> 17
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<220>
<221> misc_feature
<222> (21)..(22)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (69)..(70)
<223> n представляет собой a, c, g или t
<400> 17
gccacaagga cgatgaagaa nnkttttttc ctcagagcgg ggttctcatc tttgggaagc 60
aaggctcann kaaaacaagt gtggacattg 90
<210> 18
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<220>
<221> misc_feature
<222> (18)..(19)
<223> n представляет собой a, c, g или t
<400> 18
ccaacctcca gagaggcnnk agacaagcag ctacc 35
<210> 19
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<220>
<221> misc_feature
<222> (18)..(19)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (42)..(43)
<223> n представляет собой a, c, g или t
<400> 19
ccaactacaa caagtctnnk aatgtggact ttactgtgga cnnkaatggc gtgtatt 57
<210> 20
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 20
catgggaaag gtgccagacg 20
<210> 21
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 21
accatcggca gccatacctg 20
<210> 22
<211> 2208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 22
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctatgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggaacgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaagttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggtaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacagacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cggaaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca agtgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc agagaggcaa cagacaagca gctaccgcag atgtcaacac acaaggcgtt 1800
cttccaggca tggtctggca ggacagagat gtgtaccttc aggggcccat ctgggcaaag 1860
attccacaca cggacggaca ttttcacccc tctcccctca tgggtggatt cggacttaaa 1920
caccctcctc cacagattct catcaagaac accccggtac ctgcgaatcc ttcgaccacc 1980
ttcagtgcgg caaagtttgc ttccttcatc acacagtact ccacgggaca ggtcagcgtg 2040
gagatcgagt gggagctgca gaaggaaaac agcaaacgct ggaatcccga agttcagtac 2100
acttccaact acaacaagtc tgttaatgtg gactttactg tggacactaa tggcgtgtat 2160
tcagagcctc gccccattgg caccagatac ctgactcgta atcagtaa 2208
<210> 23
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 23
atggctgctg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgca gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggtga caacccctac ctcaagtaca accacgccga cgcggagttc 300
cagcagcggc ttcagggcga cacatcgttt gggggcaacc tcggcagagc agtcttccag 360
gccaaaaaga gggttcttga acctcttggt ctggttgagc aagcgggtga gacggctcct 420
ggaaagaaga gaccgttgat tgaatccccc cagcagcccg actcctccac gggtatcggc 480
aaaaaaggca agcagccggc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctccagcaa cccccgctgc tgtgggacct 600
actacaatgg cttcaggtgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgcacctg ggccttgccc acctacaata accacctcta caagcaaatc 780
tccagtgctt caacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg acttcaacag attccactgc cacttttcac cacgtgactg gcagcgactc 900
atcaacaaca attggggatt ccggcccaag agactcaact tcaaactctt caacatccaa 960
gtcaaggagg tcacgacgaa tgatggcgtc acaaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc agactatcag ctcccgtacg tgctcgggtc ggctcacgag 1080
ggctgcctcc cgccgttccc agcagacgtc ttcatggtgc cacagtatgg atacctcacc 1140
ctgaacaacg ggagtcaggc agtaggacgc tcttcatttt actgcctgga gtactttcct 1200
tctcagatgc tgcgtaccgg aaacaacttt accttcagct acacttttga ggacgttcct 1260
ttccacagca gctacgctca cagccagagt ctggaccgtc tcatgaatcc tctcatcgac 1320
cagtacctgt attacctgaa cagaactcag aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgggggtc tccagctggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt atcggcagca gcgcgtttct aaaacaaaaa cagacaacaa caacagcaac 1500
tttacctgga ctggtgcttc aaaatataac cttaatgggc gtgaatctat aatcaaccct 1560
ggcactgcta tggcctcaca caaagacgac aaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaggagag cgccggagct tcaaacactg cattggacaa tgtcatgatc 1680
acagacgaag aggaaatcaa agccactaac cccgtggcca ccgaaagatt tgggactgtg 1740
gcagtcaatc tccagagcag cagcacagac cctgcgaccg gagatgtgca tgttatggga 1800
gccttacctg gaatggtgtg gcaagacaga gacgtatacc tgcagggtcc catttgggcc 1860
aaaattcctc acacagatgg acactttcac ccgtctcctc ttatgggcgg ctttggactc 1920
aagaacccgc ctcctcagat cctcatcaaa aacacgcctg ttcctgcgaa tcctccggcg 1980
gagttttcag ctacaaagtt tgcttcattc atcacccaat actccacagg acaagtgagt 2040
gtggaaattg aatgggagct gcagaaagaa aacagcaagc gctggaatcc cgaagtgcag 2100
tacacatcca attatgcaaa atctgccaac gttgatttta ctgtggacaa caatggactt 2160
tatactgagc ctcgccccat tggcacccgt tacctcaccc gtcccctgta a 2211
<210> 24
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 24
atggctgctg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgca gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggtga caacccctac ctcaagtaca accacgccga cgcggagttc 300
cagcagcggc ttcagggcga cacatcgttt gggggcaacc tcggcagagc agtcttccag 360
gccaaaaaga gggttcttga acctcttggt ctggttgagc aagcgggtga gacggctcct 420
ggaaagaaga gaccgttgat tgaatccccc cagcagcccg actcctccac gggtatcggc 480
aaaaaaggca agcagccggc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctccagcaa cccccgctgc tgtgggacct 600
actacaatgg cttcaggtgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgcacctg ggccttgccc acctacaata accacctcta caagcaaatc 780
tccagtgctt caacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg acttcaacag attccactgc cacttttcac cacgtgactg gcagcgactc 900
atcaacaaca attggggatt ccggcccaag agactcaact tcaaactctt caacatccaa 960
gtcaaggagg tcacgacgaa tgatggcgtc acaaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc agactatcag ctcccgtacg tgctcgggtc ggctcacgag 1080
ggctgcctcc cgccgttccc agcagacgtc ttcatggtgc cacagtatgg atacctcacc 1140
ctgaacaacg ggagtcaggc agtaggacgc tcttcatttt actgcctgga gtactttcct 1200
tctcagatgc tgcgtaccgg aaacaacttt accttcagct acacttttga ggacgttcct 1260
ttccacagca gctacgctca cagccagagt ctggaccgtc tcatgaatcc tctcatcgac 1320
cagtacctgt attacctgaa cagaactcag aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgggggtc tccaactggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt atcggcagca gcgcgtttct aaaacaaaaa cagacaacaa caacagcaac 1500
tttacctgga ctggtgcttc aaaatataac cttaatgggc gtgaatctat aatcaaccct 1560
ggcactgcta tggcctcaca caaagacgac aaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaggagag cgccggagct tcaaacactg cattggacaa tgtcatgatc 1680
acagacgaag aggaaatcaa agccactaac cccgtggcca ctgaaagatt tgggactgtg 1740
gcagtcaatc tccagagcag cagcacagac cctgcgaccg gagatgtgca tgccatggga 1800
gccttacctg gaatggtgtg gcaagacaga gacgtatacc tgcagggtcc tatttgggcc 1860
aaaattcctc acacggatgg acactttcac ccgtctcctc tcatgggcgg ctttggactc 1920
aagaacccgc ctcctcagat cctcatcaaa aacacgcctg ttcctgcgaa tcctccggcg 1980
gagttttcag ctacaaagtt tgcttcattc atcacccagt attccacagg acaagtgagc 2040
gtggagattg aatgggagct gcagaaagaa aacagcaaac gctggaatcc cgaagtgcag 2100
tatacatcta actatgcaaa atctgccaac gttgatttca ctgtggacaa caatggactt 2160
tatactgagc ctcgccccat tggcacccgt tacctcaccc gtcccctgta a 2211
<210> 25
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 25
atggctgccg atggttatct tccagattgg ctcgaggaca acctctctga gggcattcgc 60
gagtggtggg cgctgaaacc tggagccccg aagcccaaag ccaaccagca aaagcaggac 120
gacggccggg gtctggtgct tcctggctac aagtacctcg gacccttcaa cggactcgac 180
aagggggagc ccgtcaacgc ggcggatgca gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aagcgggtga caatccgtac ctgcggtata accacgccga cgccgagttt 300
caggagcgtc tgcaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaagaagc gggttctcga acctctcggt ctggttgagg aaggcgctaa gacggctcct 420
ggaaagaaac gtccggtaga gcaatcgcca caagagccag actcctcctc gggcatcggc 480
aagacaggcc agcagcccgc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctccagcaa cccccgctgc tgtgggacct 600
actacaatgg cttcaggcgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgaacatg ggccttgccc acctataaca accacctcta caagcaaatc 780
tccagtgctt cgacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg actttaacag attccactgc cacttttcac cacgtgactg gcagcgactc 900
atcaacaaca actggggatt ccggcccaag agactcagct tcaagctctt caacatccag 960
gtcaaggagg tcacgacgaa tgatggcgtc acaaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc ggagtaccag cttccgtacg tcctcggctc tgcgcaccag 1080
ggctgcctcc ctccgttccc ggcggacgtg ttcatgattc cgcaatacgg ctacctgacg 1140
ctcaacaatg gcagccaagc cgtgggacgt tcatcctttt actgcctgga atatttccct 1200
tctcagatgc tgagaacggg caacaacttt accttcagct acacctttga ggaagtgcct 1260
ttccacagca gctacgcgca cagccagagc ctggaccggc tgatgaatcc tctcatcgat 1320
caatacctgt attacctgaa cagaactcaa aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgtgggtc tccagctggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt atcggcagca gcgcgtttct aaaacaaaaa cagacaacaa caacagcaat 1500
tttacctgga ctggtgcttc aaaatataac ctcaatgggc gtgaatccat catcaaccct 1560
ggcactgcta tggcctcaca taaagacgac gaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaagagag cgccggagct tcaaacactg cattggacaa tgtcatgatt 1680
acagacgaag aggaaattaa agccactaac cctgtggcca ccgaaagatt tgggaccgtg 1740
gcagtcaatt tccagagcag cagcacagac cctgcgaccg gagatgtgca tgctatggga 1800
gcattacctg gcatggtgtg gcaagataga gacgtgtacc tgcagggtcc catttgggcc 1860
aaaattcctc acacagatgg acactttcac ccgtctcctc ttatgggcgg ctttggactc 1920
aagaacccgc ctcctcagat cctcatcaaa aacacgcctg ttcctgcgaa tcctccggcg 1980
gagttttcag ctacaaagtt tgcttcattc atcacccaat actccacagg acaagtgagc 2040
gtggagattg aatgggagct gcagaaagaa aacagcaaac gctggaatcc cgaagtgcag 2100
tatacatcta actatgcaaa atctgccaac attgatttca ctgtggacaa caatggactt 2160
tatactgagc ctcgccccat tggcacccgt tacctcaccc gtccccagta a 2211
<210> 26
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 26
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Glu Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Cys Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Val Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys Asn Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 27
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 27
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Val Asn Gln Gln Lys Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Thr Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Ala Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 28
<211> 723
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 28
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Val Asn Gln Gln Lys Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Gln Val
305 310 315 320
Lys Glu Thr Thr Asp Val Thr Thr Ile Ala Asn Asn Leu Thr Ser Thr
325 330 335
Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro Tyr Val Leu Gly
340 345 350
Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp Val Phe Met
355 360 365
Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser Gln Ala Val
370 375 380
Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser Gln Met Leu
385 390 395 400
Arg Thr Gly Asn Asn Phe Thr Ser Tyr Thr Phe Glu Asp Val Pro Phe
405 410 415
His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg Leu Met Asn Pro
420 425 430
Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg Thr Gln Asn Gln Ser
435 440 445
Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser Arg Gly Ser Pro Thr
450 455 460
Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro Gly Pro Cys Tyr Arg
465 470 475 480
Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn Asn Asn Ser Asn Phe
485 490 495
Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn Gly Arg Glu Ser Ile
500 505 510
Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys Asp Asp Glu Asp Lys
515 520 525
Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly Lys Glu Ser Ala Gly
530 535 540
Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile Thr Asp Glu Glu Ala
545 550 555 560
Thr Asn Pro Val Ala Thr Glu Arg Phe Gly Thr Val Ala Val Asn Leu
565 570 575
Gln Ser Ser Pro Ala Thr Asp Val His Ala Met Gly Ala Leu Pro Gly
580 585 590
Met Val Trp Gln Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala
595 600 605
Lys Ile Pro His Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly
610 615 620
Gly Phe Gly Leu Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr
625 630 635 640
Pro Val Pro Ala Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala
645 650 655
Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu
660 665 670
Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln
675 680 685
Tyr Thr Ser Asn Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp
690 695 700
Asn Asn Gly Leu Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu
705 710 715 720
Thr Arg Pro
<210> 29
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 29
Met Ala Ser Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Arg Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Val Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 30
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 30
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Ser Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Glu Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Phe Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Ala Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys Asn Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 31
<211> 735
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 31
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Lys Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asp Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Ser Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 32
<211> 735
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 32
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asn Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Lys Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Arg Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asp Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Ser Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 33
<211> 735
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая аминокислотная последовательность
<400> 33
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Lys Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asp Ala Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Ser Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 34
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 34
atggctgccg atggttatct tccagattgg ctcgaggaca acctctctga gggcattcgc 60
gagtggtggg acttgaaacc tggagccccg aaacccaaag ccaaccagca aaagcaggac 120
gacggccggg gtctggtgct tcctggctac aagtacctcg gacccttcaa cggactcgac 180
aagggggagc ccgtcaacgc ggcggatgca gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aagcgggtga caatccgtac cttcggtata accacgccga cgccgagttt 300
caggagcgtc tgcaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga gggttctcga acctctcggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaac gtccggtaga gcagtcgcca caagagccag actcctcctc gggcattggc 480
aagacaggcc agcagcccgc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgcacctg ggccttgccc acctacaata accacctcta caagcaaatc 780
tccagtgctt caacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg acttcaacag attccactgc cacttttcac cacgtgactg gcaaagactc 900
atcaacaaca attggggatt ccggcccaag agactcaact tcaagctctt caacatccaa 960
gtcaaggagg tcacgacgaa tgatggcgtc acgaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc ggagtaccag ttgccgtacg tcctcggctc tgcgcaccag 1080
ggctgcctcc ctccgttccc ggcggacgtg ttcatgattc cgcagtacgg ctacctaacg 1140
ctcaacaatg gcagccaggc agtgggacgg tcatcctttt actgcctgga atatttccca 1200
tcgcagatgc tgagaacggg caacaacttt accttcagct acacctttga ggaagtgcct 1260
ttccacagca gctacgcgca cagccagagc ctggaccggc tgatgaatcc tctcatcgac 1320
cagtacctgt attacctgaa cagaactcaa aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgtgggtc tccagctggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt accggcagca gtgcgtttct aaaacaaaaa cagacaacaa caacagcaac 1500
tttacctgga ctggtgcttc aaaatataac cttaatgggc gtgaatctat aatcaaccct 1560
ggcactgcta tggcctcaca caaagacgac aaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaggagag cgccggagct tcaaacactg cattggacaa tgtcatgatc 1680
acagacgaag aggaaatcaa agccactaac cccgtggcca ccgaaagatt tgggactgtg 1740
gcagtcaatc tccagagcag cagcacagac cctgcgaccg gagatgtgca tgttatggga 1800
gccttacctg gaatggtgtg gcaagacaga gacgtatacc tgcagggtcc tatttgggcc 1860
aaaattcctc acacagatgg acactttcac ccgtctcctc ttatgggcgg ctttggactc 1920
aagaacccgc ctcctcagat cctcatcaaa aacacgcctg ttcctgcgaa tcctccggcg 1980
gagttttcag ctacaaagtt tgcttcattc atcacccaat actccacagg acaagtgagc 2040
gtggagattg aatgggagct gcagaaagaa aacagcaagc gctggaatcc cgaagtgcag 2100
tacacatcca attatgcaaa atctgccaac gttgatttca ctgtggacaa caatggactt 2160
tatactgagc ctcgccccat tggcacccgt tacctcaccc gtcccctgta a 2211
<210> 35
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 35
atggctgccg atggttatct tccagattgg ctcgaggaca acctctctga gggcattcgc 60
gaatggtggg acttgaaacc tggagccccg aaacccaaag tcaaccagca aaagcaggac 120
aacgctcggg gtcttgtgct tccgggttac aaatacctcg gacccttcaa cggactcgac 180
aagggggagc ccgtcaacgc ggcggacgca gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aagcgggtga caatccgtac cttcggtata accacgccga cgccgagttt 300
caggagcgtc tgcaagaaga tacgtctttt gggggcaacc ttggacgagc agtcttccag 360
gccaagaaga gggttctcga accttttggt ctggttgagg aaggtgctaa gacggctcct 420
ggaaagaaac gtccggtaga gcagtcgcca caagagccag actcctcctc gggcattggc 480
aagacaggcc agcagcccgc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctccagcaa cccccgctgc tgtgggacct 600
actacaatgg cttcaggcgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgcacctg ggccttgccc acctacaata accacctcta caagcaaatc 780
tccagtgctt caacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg acttcaacag attccactgc cacttttcac cacgtgactg gcaaagactc 900
atcaacaaca attggggatt ccggcccaag agactcaact tcaagctctt caacatccaa 960
gtcaaggagg tcacgacgaa tgatggcgtc acgaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc ggagtaccag ttgccgtacg tcctcggctc tgcgcaccag 1080
ggctgcctcc ctccgttccc ggcggacgtg ttcatgattc cgcaatacgg ctacctgacg 1140
ctcaacaatg gcagccaggc agtgggacgg tcatcctttt actgcctgga atatttccca 1200
tcgcagatgc tgagaacggg caataacttt accttcagct acacttttga ggacgttcct 1260
ttccacagca gctacgctca cagccagagc ctggaccggc tgatgaatcc tctcatcgac 1320
cagtacctgt attacctgaa cagaactcag aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgtgggtc tccaactggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt atcggcagca gcgcgtttct aaaacaaaaa cagacaacaa caacagcaac 1500
tttacctgga ctggtgcttc aaaatataac cttaatgggc gtgaatctat aatcaaccct 1560
ggcactgcta tggcctcaca caaagacgac gaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaggagag cgccggagct tcaaacactg cattggacaa tgtcatgatc 1680
acagacgaag aggaaatcaa agccactaac cccgtggcca ctgaaagatt tgggactgtg 1740
gcagtcaatc tccagagcag cagcacagac cctgcgaccg gagatgtgca tgccatggga 1800
gccttacctg gaatggtgtg gcaagacaga gacgtatacc tgcagggtcc tatttgggcc 1860
aaaattcctc acacggatgg acactttcac ccgtctcctc tcatgggcgg ctttggactt 1920
aagcacccgc ctcctcagat cctcatcaaa aacacgcctg ttcctgcgaa tcctccggca 1980
gagttttcgg ctacaaagtt tgcttcattc atcacccagt attccacagg acaagtgagc 2040
gtggagattg aatgggagct gcagaaagaa aacagcaaac gctggaatcc cgaagtgcag 2100
tatacatcta actatgcaaa atctgccaac gttgatttca ctgtggacaa caatggactt 2160
tatactgagc ctcgccccat tggcacccgt tacctcaccc gtcccctgta a 2211
<210> 36
<211> 2208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<220>
<221> misc_feature
<222> (955)..(955)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (971)..(972)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (981)..(981)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (986)..(986)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (1235)..(1235)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (1694)..(1696)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (1698)..(1698)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (1763)..(1765)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (1778)..(1778)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (2204)..(2204)
<223> n представляет собой a, c, g или t
<400> 36
atggctgccg atggttatct tccagattgg ctcgaggaca acctctctga gggcattcgc 60
gaatggtggg acttgaaacc tggagccccg aaacccaaag tcaaccagca aaagcaggac 120
aacgctcggg gtcttgtgct tccgggttac aaatacctcg gacccttcaa cggactcgac 180
aagggggagc ccgtcaacgc ggcggacgca gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aagcgggtga caatccgtac cttcggtata accacgccga cgccgagttt 300
caggagcgtc tgcaagaaga tacgtctttt gggggcaacc ttggacgagc agtcttccag 360
gccaagaaga gggttctcga accttttggt ctggttgagg aaggtgctaa gacggctcct 420
ggaaagaaac gtccggtaga gcagtcgcca caagagccag actcctcctc gggcattggc 480
aagacaggcc agcagcccgc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctccagcaa cccccgctgc tgtgggacct 600
actacaatgg cttcaggcgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgaacatg ggccttgccc acctataaca accacctcta caagcaaatc 780
tccagtgctt caacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg atttcaacag attccactgc cacttttcac cacgtgactg gcagcgactc 900
atcaataaca attggggatt ccggcccaag agactcaact tcaaactctt caacntccaa 960
gtcaaggagg nnacgacgaa ngatgncgtc acaaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc ggagtaccag cttccgtacg tcctcggctc tgcgcaccag 1080
ggctgcctcc ctccgttccc ggcggacgtg ttcatgattc cgcaatacgg ctacctgacg 1140
ctcaacaatg gcagccaggc agtgggacgg tcatcctttt actgcctgga atatttccca 1200
tcgcagatgc tgagaacggg caataacttt acctncagct acacttttga ggacgttcct 1260
ttccacagca gctacgctca cagccagagc ctggaccggc tgatgaatcc tctcatcgac 1320
cagtacctgt attacctgaa cagaactcag aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgtgggtc tccaactggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt atcggcagca gcgcgtttct aaaacaaaaa cagacaacaa caacagcaac 1500
tttacctgga ctggtgcttc aaaatataac cttaatgggc gtgaatctat aatcaaccct 1560
ggcactgcta tggcctcaca caaagacgac gaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaggagag cgccggagct tcaaacactg cattggacaa tgtcatgatc 1680
acagacgaag agannncnaa gccactaacc ccgtggccac tgaaagattt gggactgtgg 1740
cagtcaatct ccaagcagca cannnaccct gcgaccgnag atgtgcatgc catgggagcc 1800
ttacctggaa tggtgtggca agacagagac gtatacctgc agggtcctat ttgggccaaa 1860
attcctcaca cggatggaca ctttcacccg tctcctctca tgggcggctt tggacttaag 1920
cacccgcctc ctcagatcct catcaaaaac acgcctgttc ctgcgaatcc tccggcagag 1980
ttttcggcta caaagtttgc ttcattcatc acccagtatt ccacaggaca agtgagcgtg 2040
gagattgaat gggagctgca gaaagaaaac agcaaacgct ggaatcccga agtgcagtat 2100
acatctaact atgcaaaatc tgccaacgtt gatttcactg tggacaacaa tggactttat 2160
actgagcctc gccccattgg cacccgttac ctcacccgtc cccngtaa 2208
<210> 37
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 37
atggcttccg atggttatct tccagattgg ctcgaggaca acctctctga gggcatccgc 60
gagtggtggg acttgaaacc tggagccccg aaacccaaag ccaaccagca aaagcaggac 120
gacggccggg gtctggtgct tcctggctac aagtacctcg gacccttcaa cggactcgac 180
aagggggagc ccgtcaacgc ggcggatgca gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca gagcgggtga caatccgtac ctgcggtata accacgccga cgccgagttt 300
caggagcgtc tgcaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaagaaga gggttctcga accttttggt ctggttgagg aaggtgctaa gacggctcct 420
ggaaagaaac gtccggtaga gcagtcgcca caagagccag actcctcctc gggcattggc 480
aagacaggcc agcagcccgc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctccagcaa cccccgctgc tgtgggacct 600
actacaatgg cttcaggcgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgaacatg ggccttgccc acctataaca accacctcta caagcaaatc 780
tccagtgctt caacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg atttcaacag attccactgc catttctcac cacgtgactg gcagcgactc 900
atcaacaaca attggggatt ccggcccaag agactcaact tcaaactctt caacatccaa 960
gtcaaggagg tcacgacgaa tgatggcgtc acaaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc ggagtaccag cttccgtacg tcctcggctc tgcgcaccag 1080
ggctgcctcc ctccgttccc ggcggacgtg ttcatgattc cgcagtacgg ctacctaacg 1140
ctcaacaatg gcagccaggc agtgggacgg tcatcctttt actgcctgga atatttccca 1200
tcgcagatgc tgagaacggg caataacttt accttcagct acaccttcga ggacgtgcct 1260
ttccacagca gctacgcgca cagccagagc ctggaccggc tgatgaatcc tctcatcgac 1320
cagtacctgt attacctgaa cagaactcag aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgggggtc tccagctggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt accggcagca gcgcgtttct aaaacaaaaa cagacaacaa caacagcaac 1500
tttacctgga ctggtgcttc aaaatataac cttaatgggc gtgaatctat aatcaaccct 1560
ggcactgcta tggcctcaca caaagacgac aaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaggagag cgccggagct tcaaacactg cattggacaa tgtcatgatc 1680
acagacgaag aggaaatcaa agccactaac cccgtggcca ccgaaagatt tgggactgtg 1740
gcagtcaatc tccagagcag cagcacagac cctgcgaccg gagatgtgca tgttatggga 1800
gccttacctg gaatggtgtg gcaagacaga gacgtatacc tgcagggtcc catttgggcc 1860
aaaattcctc acacagatgg acactttcac ccgtctcctc ttatgggcgg ctttggactt 1920
aagcacccgc ctcctcagat cctcatcaaa aacacgcctg ttcctgcgaa tcctccggca 1980
gagttttcgg ctacaaagtt tgcttcattc atcacccagt attctactgg ccaagtcagc 2040
gtggagattg aatgggagct gcagaaagaa aacagcaaac gctggaatcc cgaagtgcag 2100
tatacatcta actatgcaaa atctgccaac gttgatttca ctgtggacaa caatggactt 2160
tatactgagc ctcgtcccat tggcacccgt tacctcaccc gtcccctgta a 2211
<210> 38
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 38
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aagcgggtga caatccgtac ctgcggtata accacgccga cgccgagttt 300
caggagcgtc tgcaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaagaagc gggttctcga acctctcggt ctggttgagg aaggcgctaa gacggctcct 420
ggaaagaaac gtccggtaga gcagtcgcca caagagccag actcctcctc gggcatcggc 480
aagacaggcc agcagcccgc taaaaagaga ctcaattttg gtcagactgg cgactcagag 540
tcagtccccg acccacaacc tctcggagaa cctccagcaa cccccgctgc tgtgggacct 600
actacaatgg cttcaggcgg tggcgcacca atggcagaca ataacgaagg cgccgacgga 660
gtgggtaatg cctcaggaaa ttggcattgc gattccacat ggctgggcga cagagtcatc 720
accaccagca cccgaacatg ggccttgccc acctataaca accacctcta caagcaaatc 780
tccagtgctt cgacgggggc cagcaacgac aaccactact tcggctacag caccccctgg 840
gggtattttg actttaacag attccactgc cacttttcac cacgtgactg gcagcgactc 900
atcaacaata actggggatt ccggcccaag agactcagct tcaagctctt caacatccag 960
gtcaaggagg tcacgacgaa tgatggcgtc acaaccatcg ctaataacct taccagcacg 1020
gttcaagtct tctcggactc ggagtaccag cttccgtacg tcctcggctc tgcgcaccag 1080
ggctgcctcc ctccgttccc ggcggacgtg ttcatgattc cgcaatacgg ctacctgacg 1140
ctcaacaatg gcagccaagc cgtgggacgt tcatcctttt actgcctgga atatttccct 1200
tctcagatgc tgagaacggg caacaacttt accttcagct acacctttga ggaagtgcct 1260
ttccacagca gctacgcgca cagccagagc ctggaccggc tgatgaatcc tctcatcgat 1320
caatacctgt attacctgaa cagaactcaa aatcagtccg gaagtgccca aaacaaggac 1380
ttgctgttta gccgtgggtc tccagctggc atgtctgttc agcccaaaaa ctggctacct 1440
ggaccctgtt atcggcagca gcgcgtttct aaaacaaaaa cagacaacaa caacagcaat 1500
tttacctgga ctggtgcttc aaaatataac ctcaatgggc gtgaatccat catcaaccct 1560
ggcactgcta tggcctcaca caaagacgac gaagacaagt tctttcccat gagcggtgtc 1620
atgatttttg gaaaagagag cgccggagct tcaaacactg cattggacaa tgtcatgatt 1680
acggacgaag aggaaattaa agccactaac cctgtggcca ccgaaagatt tgggaccgtg 1740
gcagtcaatt tccagagcag cagcacagac cctgcgaccg gagatgtgca tgctatggga 1800
gcattacctg gcatggtgtg gcaagataga gacgtgtacc tgcagggtcc catttgggcc 1860
aaaattcctc acacagatgg acactttcac ccgtctcctc ttatgggcgg ctttggactc 1920
aagaacccgc ctcctcagat cctcatcaaa aacacgcctg ttcctgcgaa tcctccggcg 1980
gagttttcag ctacaaagtt tgcttcattc atcactcaat actccacagg acaagtgagc 2040
gtggaaattg aatgggagct gcagaaagaa aacagcaaac gctggaatcc cgaagtgcag 2100
tatacatcta actatgcaaa atctgccaac gttgatttca ctgtggacaa caatggactt 2160
tatactgagc ctcgccccat tggcacccgt tacctcaccc gtcccctgta a 2211
<210> 39
<211> 2208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 39
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctatgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg atccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaagttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtatgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggtaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacagacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca agtgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc agagaggcaa cagacaagca gctaccgcag atgtcaacac acaaggcgtt 1800
cttccaggca tggtctggca ggacagagat gtgtaccttc aggggcccat ctgggcaaag 1860
attccacaca cggacggaca ttttcacccc tctcccctca tgggtggatt cggacttaaa 1920
caccctcctc cacagattct catcaagaac accccggtac ctgcgaatcc ttcgaccacc 1980
ttcagtgcgg caaagtttgc ttccttcatc acacagtact ccacgggaca ggtcagcgtg 2040
gagatcgagt gggagctgca gaaggaaaac agcaaacgct ggaatcccga agttcagtac 2100
acttccaact acaacaagtc tgttaatgtg gactttactg tggacactaa tggcgtgtat 2160
tcagagcctc gccccattgg caccagatac ctgactcgta atctgtaa 2208
<210> 40
<211> 2208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 40
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctatgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcaaac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaagttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcgggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggtaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacagacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca agtgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac gaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc agagaggcaa cagacaagca gctaccgcag atgtcaacac acaaggcgtt 1800
cttccaggca tggtctggca ggacagagat gtgtaccttc aggggcccat ctgggcaaag 1860
attccacaca cggacggaca ttttcacccc tctcccctca tgggtggatt cggacttaaa 1920
caccctcctc cacagattct catcaagaac accccggtac ctgcgaatcc ttcgaccacc 1980
ttcagtgcgg caaagtttgc ttccttcatc acacagtact ccacgggaca ggtcagcgtg 2040
gagatcgagt gggagctgca gaaggaaaac agcaaacgct ggaatcccga agttcagtac 2100
acttccaact acaacaagtc tgttaatgtg gactttactg tggacactaa tggcgtgtat 2160
acagagcctc gccccattgg caccagatac ctgactcgta atctgtaa 2208
<210> 41
<211> 2208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 41
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctatgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggtc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaagttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggtaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacagacgct ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca agtgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc agagaggcaa cagacaagca gctaccgcag atgtcaacac acaaggcgtt 1800
cttccaggca tggtctggca ggacagagat gtgtaccttc aggggcccat ctgggcaaag 1860
attccacaca cggacggaca ttttcacccc tctcccctca tgggtggatt cggacttaaa 1920
caccctcctc cacagattct catcaagaac accccggtac ctgcgaatcc ttcgaccacc 1980
ttcagtgcgg caaagtttgc ttccttcatc acacagtact ccacgggaca ggtcagcgtg 2040
gagatcgagt gggagctgca gaaggaaaac agcaaacgct ggaatcccga agttcagtac 2100
acttccaact acaacaagtc tgttaatgtg gactttactg tggacactaa tggcgtgtat 2160
tcagagcctc gccccattgg caccagatac ctgactcgta atctgtaa 2208
<210> 42
<211> 4443
<212> ДНК
<213> Homo sapiens
<400> 42
atgcagaggt cgcctctgga aaaggccagc gttgtctcca aacttttttt cagctggacc 60
agaccaattt tgaggaaagg atacagacag cgcctggaat tgtcagacat ataccaaatc 120
ccttctgttg attctgctga caatctatct gaaaaattgg aaagagaatg ggatagagag 180
ctggcttcaa agaaaaatcc taaactcatt aatgcccttc ggcgatgttt tttctggaga 240
tttatgttct atggaatctt tttatattta ggggaagtca ccaaagcagt acagcctctc 300
ttactgggaa gaatcatagc ttcctatgac ccggataaca aggaggaacg ctctatcgcg 360
atttatctag gcataggctt atgccttctc tttattgtga ggacactgct cctacaccca 420
gccatttttg gccttcatca cattggaatg cagatgagaa tagctatgtt tagtttgatt 480
tataagaaga ctttaaagct gtcaagccgt gttctagata aaataagtat tggacaactt 540
gttagtctcc tttccaacaa cctgaacaaa tttgatgaag gacttgcatt ggcacatttc 600
gtgtggatcg ctcctttgca agtggcactc ctcatggggc taatctggga gttgttacag 660
gcgtctgcct tctgtggact tggtttcctg atagtccttg ccctttttca ggctgggcta 720
gggagaatga tgatgaagta cagagatcag agagctggga agatcagtga aagacttgtg 780
attacctcag aaatgattga aaatatccaa tctgttaagg catactgctg ggaagaagca 840
atggaaaaaa tgattgaaaa cttaagacaa acagaactga aactgactcg gaaggcagcc 900
tatgtgagat acttcaatag ctcagccttc ttcttctcag ggttctttgt ggtgttttta 960
tctgtgcttc cctatgcact aatcaaagga atcatcctcc ggaaaatatt caccaccatc 1020
tcattctgca ttgttctgcg catggcggtc actcggcaat ttccctgggc tgtacaaaca 1080
tggtatgact ctcttggagc aataaacaaa atacaggatt tcttacaaaa gcaagaatat 1140
aagacattgg aatataactt aacgactaca gaagtagtga tggagaatgt aacagccttc 1200
tgggaggagg gatttgggga attatttgag aaagcaaaac aaaacaataa caatagaaaa 1260
acttctaatg gtgatgacag cctcttcttc agtaatttct cacttcttgg tactcctgtc 1320
ctgaaagata ttaatttcaa gatagaaaga ggacagttgt tggcggttgc tggatccact 1380
ggagcaggca agacttcact tctaatggtg attatgggag aactggagcc ttcagagggt 1440
aaaattaagc acagtggaag aatttcattc tgttctcagt tttcctggat tatgcctggc 1500
accattaaag aaaatatcat ctttggtgtt tcctatgatg aatatagata cagaagcgtc 1560
atcaaagcat gccaactaga agaggacatc tccaagtttg cagagaaaga caatatagtt 1620
cttggagaag gtggaatcac actgagtgga ggtcaacgag caagaatttc tttagcaaga 1680
gcagtataca aagatgctga tttgtattta ttagactctc cttttggata cctagatgtt 1740
ttaacagaaa aagaaatatt tgaaagctgt gtctgtaaac tgatggctaa caaaactagg 1800
attttggtca cttctaaaat ggaacattta aagaaagctg acaaaatatt aattttgcat 1860
gaaggtagca gctattttta tgggacattt tcagaactcc aaaatctaca gccagacttt 1920
agctcaaaac tcatgggatg tgattctttc gaccaattta gtgcagaaag aagaaattca 1980
atcctaactg agaccttaca ccgtttctca ttagaaggag atgctcctgt ctcctggaca 2040
gaaacaaaaa aacaatcttt taaacagact ggagagtttg gggaaaaaag gaagaattct 2100
attctcaatc caatcaactc tatacgaaaa ttttccattg tgcaaaagac tcccttacaa 2160
atgaatggca tcgaagagga ttctgatgag cctttagaga gaaggctgtc cttagtacca 2220
gattctgagc agggagaggc gatactgcct cgcatcagcg tgatcagcac tggccccacg 2280
cttcaggcac gaaggaggca gtctgtcctg aacctgatga cacactcagt taaccaaggt 2340
cagaacattc accgaaagac aacagcatcc acacgaaaag tgtcactggc ccctcaggca 2400
aacttgactg aactggatat atattcaaga aggttatctc aagaaactgg cttggaaata 2460
agtgaagaaa ttaacgaaga agacttaaag gagtgctttt ttgatgatat ggagagcata 2520
ccagcagtga ctacatggaa cacatacctt cgatatatta ctgtccacaa gagcttaatt 2580
tttgtgctaa tttggtgctt agtaattttt ctggcagagg tggctgcttc tttggttgtg 2640
ctgtggctcc ttggaaacac tcctcttcaa gacaaaggga atagtactca tagtagaaat 2700
aacagctatg cagtgattat caccagcacc agttcgtatt atgtgtttta catttacgtg 2760
ggagtagccg acactttgct tgctatggga ttcttcagag gtctaccact ggtgcatact 2820
ctaatcacag tgtcgaaaat tttacaccac aaaatgttac attctgttct tcaagcacct 2880
atgtcaaccc tcaacacgtt gaaagcaggt gggattctta atagattctc caaagatata 2940
gcaattttgg atgaccttct gcctcttacc atatttgact tcatccagtt gttattaatt 3000
gtgattggag ctatagcagt tgtcgcagtt ttacaaccct acatctttgt tgcaacagtg 3060
ccagtgatag tggcttttat tatgttgaga gcatatttcc tccaaacctc acagcaactc 3120
aaacaactgg aatctgaagg caggagtcca attttcactc atcttgttac aagcttaaaa 3180
ggactatgga cacttcgtgc cttcggacgg cagccttact ttgaaactct gttccacaaa 3240
gctctgaatt tacatactgc caactggttc ttgtacctgt caacactgcg ctggttccaa 3300
atgagaatag aaatgatttt tgtcatcttc ttcattgctg ttaccttcat ttccatttta 3360
acaacaggag aaggagaagg aagagttggt attatcctga ctttagccat gaatatcatg 3420
agtacattgc agtgggctgt aaactccagc atagatgtgg atagcttgat gcgatctgtg 3480
agccgagtct ttaagttcat tgacatgcca acagaaggta aacctaccaa gtcaaccaaa 3540
ccatacaaga atggccaact ctcgaaagtt atgattattg agaattcaca cgtgaagaaa 3600
gatgacatct ggccctcagg gggccaaatg actgtcaaag atctcacagc aaaatacaca 3660
gaaggtggaa atgccatatt agagaacatt tccttctcaa taagtcctgg ccagagggtg 3720
ggcctcttgg gaagaactgg atcagggaag agtactttgt tatcagcttt tttgagacta 3780
ctgaacactg aaggagaaat ccagatcgat ggtgtgtctt gggattcaat aactttgcaa 3840
cagtggagga aagcctttgg agtgatacca cagaaagtat ttattttttc tggaacattt 3900
agaaaaaact tggatcccta tgaacagtgg agtgatcaag aaatatggaa agttgcagat 3960
gaggttgggc tcagatctgt gatagaacag tttcctggga agcttgactt tgtccttgtg 4020
gatgggggct gtgtcctaag ccatggccac aagcagttga tgtgcttggc tagatctgtt 4080
ctcagtaagg cgaagatctt gctgcttgat gaacccagtg ctcatttgga tccagtaaca 4140
taccaaataa ttagaagaac tctaaaacaa gcatttgctg attgcacagt aattctctgt 4200
gaacacagga tagaagcaat gctggaatgc caacaatttt tggtcataga agagaacaaa 4260
gtgcggcagt acgattccat ccagaaactg ctgaacgaga ggagcctctt ccggcaagcc 4320
atcagcccct ccgacagggt gaagctcttt ccccaccgga actcaagcaa gtgcaagtct 4380
aagccccaga ttgctgctct gaaagaggag acagaagaag aggtgcaaga tacaaggctt 4440
tag 4443
<210> 43
<211> 3464
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 43
atgcagcgca gcccactgga gaaggcaagc gtggtgtcca agctgttctt ttcctggacc 60
aggcctatcc tgaggaaggg atacaggcag cggctggagc tgagcgacat ctatcagatc 120
ccttctgtgg acagcgccga taatctgtcc gagaagctgg agagagagtg ggatagggag 180
ctggcctcta agaagaaccc aaagctgatc aatgccctgc ggagatgctt cttttggcgg 240
ttcatgttct acggcatctt cctgtatctg ggcgaggtga ccaaggccgt gcagccactg 300
ctgctgggca gaatcatcgc ctcttacgac cccgataaca aggaggagag gagcatcgcc 360
atctatctgg gcatcggcct gtgcctgctg tttatcgtga ggacactgct gctgcaccca 420
gccatcttcg gcctgcacca catcggcatg cagatgagaa tcgccatgtt cagcctgatc 480
tacaagaaga ccctgaagct gagctccagg gtgctggaca agatctccat cggccagctg 540
gtgtccctgc tgtctaacaa tctgaacaag tttgatgagg gactggccct ggcacacttc 600
gtgtggatcg caccactgca ggtggccctg ctgatgggcc tgatctggga gctgctgcag 660
gcaagcgcct tttgcggact gggcttcctg atcgtgctgg ccctgttcca ggcaggactg 720
ggacgcatga tgatgaagta cagagaccag agggccggca agatctctga gcggctggtc 780
atcaccagcg agatgatcga gaacatccag tccgtgaagg cctattgttg ggaggaggcc 840
atggagaaga tgatcgagaa tctgcgccag acagagctga agctgaccag aaaggccgcc 900
tacgtgaggt acttcaactc tagcgccttc tttttctctg gctttttcgt ggtgttcctg 960
agcgtgctgc catacgccct gatcaagggc atcatcctgc ggaagatctt taccacaatc 1020
tccttctgca tcgtgctgag aatggccgtg acaaggcagt ttccctgggc cgtgcagacc 1080
tggtatgact ctctgggcgc catcaataag atccaggatt tcctgcagaa gcaggagtac 1140
aagacactgg agtataacct gaccacaacc gaggtggtca tggagaatgt gaccgccttc 1200
tgggaggagg gctttggcga gctgttcgag aaggccaagc agaacaataa caatcgcaag 1260
acatctaacg gcgacgatag cctgtttttc agcaattttt ccctgctggg cacccccgtg 1320
ctgaaggaca tcaacttcaa gatcgagagg ggacagctgc tggcagtggc aggctccaca 1380
ggcgccggca agacctctct gctgatgatg atcatgggcg agctggagcc aagcgagggc 1440
aagatcaagc actccggccg gatctctttt tgcagccagt tctcctggat catgcccggc 1500
accatcaagg agaatatcat ctttggcgtg tcctacgatg agtacagata taggtctgtg 1560
atcaaggcct gtcagctgga ggaggacatc agcaagttcg ccgagaagga taacatcgtg 1620
ctgggcgagg gcggcatcac actgagcgga ggacagaggg caaggatctc cctggccaga 1680
gccgtgtaca aggacgccga tctgtatctg ctggacagcc cctttggcta tctggatgtg 1740
ctgaccgaga aggagatctt cgagtcctgc gtgtgcaagc tgatggccaa taagacaagg 1800
atcctggtga cctctaagat ggagcacctg aagaaggccg acaagatcct gatcctgcac 1860
gagggctcct cttactttta tggcacattc agcgagctgc agaatctgca gcctgacttc 1920
agctccaagc tgatgggctg tgactccttt gatcagttct ctgccgagag gcgcaactcc 1980
atcctgacag agaccctgca cagattctct ctggagggcg acgcacccgt gagctggaca 2040
gagaccaaga agcagtcctt taagcagacc ggcgagttcg gcgagaagag gaagaattct 2100
atcctgaacc ctatcaatag cacactgcag gcccggagaa ggcagtctgt gctgaacctg 2160
atgacccaca gcgtgaacca gggccagaat atccacagaa agacaaccgc cagcacaagg 2220
aaggtgtccc tggcacctca ggcaaacctg accgagctgg acatctactc ccgccggctg 2280
tctcaggaga ccggactgga gatctctgag gagatcaatg aggaggatct gaaggagtgc 2340
tttttcgacg atatggagag catcccagcc gtgacaacct ggaacacata cctgcgctat 2400
atcaccgtgc acaagtccct gatctttgtg ctgatctggt gtctggtcat cttcctggca 2460
gaggtggcag catctctggt ggtgctgtgg ctgctgggca acacacccct gcaggacaag 2520
ggcaattcta cccacagccg caacaattcc tacgccgtga tcatcacatc tacctctagc 2580
tactacgtgt tctacatcta tgtgggcgtg gccgatacac tgctggccat gggctttttc 2640
cggggcctgc ccctggtgca cacactgatc accgtgagca agatcctgca ccacaagatg 2700
ctgcacagcg tgctgcaggc ccctatgtcc acactgaaca ccctgaaggc cggcggcatc 2760
ctgaatcggt tttccaagga catcgccatc ctggacgatc tgctgcctct gaccatcttt 2820
gatttcatcc agctgctgct gatcgtgatc ggagcaatcg cagtggtggc cgtgctgcag 2880
ccttacatct tcgtggccac agtgccagtg atcgtggcct ttatcatgct gcgcgcctat 2940
ttcctgcaga ccagccagca gctgaagcag ctggagagcg agggccggtc ccctatcttt 3000
acacacctgg tgacctccct gaagggactg tggacactga gggccttcgg ccggcagcca 3060
tactttgaga ccctgttcca caaggccctg aacctgcaca cagccaattg gtttctgtat 3120
ctgagcaccc tgcgctggtt tcagatgcgg atcgagatga tcttcgtgat ctttttcatc 3180
gccgtgacct tcatctccat cctgacaacc ggagagggag agggaagagt gggaatcatc 3240
ctgacactgg ccatgaacat catgtctacc ctgcagtggg ccgtgaattc ctctatcgac 3300
gtggatagcc tgatgagatc tgtgagcagg gtgtttaagt tcatcgacat gcccacagag 3360
ggcaagccta caaagagcac caagccatac aagaacggcc agctgtccaa agtgatgatc 3420
atcgagaatt ctcacgtgaa gaaggacgat atctggccat ccgg 3464
<210> 44
<211> 173
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 44
actcacgggg atttccaagt ctccacccca ttgacgtcaa tgggagtttg ttttggcacc 60
aaaatcaacg ggactttcca aaatgtcgta ataaccccgc cccgttgacg caaatgggcg 120
gtaggcgtgt acggtgggag gtctatataa gcagagctcg tttagtgaac cgt 173
<210> 45
<211> 4874
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновокислотная последовательность
<400> 45
ttggccactc cctctctgcg cgctcgctcg ctcactgagg ccgggcgacc aaaggtcgcc 60
cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc gagcgcgcag agagggagtg 120
gccaactcca tcactagggg ttcctgcggc cgcactcacg gggatttcca agtctccacc 180
ccattgacgt caatgggagt ttgttttggc accaaaatca acgggacttt ccaaaatgtc 240
gtaataaccc cgccccgttg acgcaaatgg gcggtaggcg tgtacggtgg gaggtctata 300
taagcagagc tcgtttagtg aaccgtcaga attctcgagt gatcgaaaga gcctgctaaa 360
gcaaaaaaga agtcaccatg cagcgcagcc cactggagaa ggcaagcgtg gtgtccaagc 420
tgttcttttc ctggaccagg cctatcctga ggaagggata caggcagcgg ctggagctga 480
gcgacatcta tcagatccct tctgtggaca gcgccgataa tctgtccgag aagctggaga 540
gagagtggga tagggagctg gcctctaaga agaacccaaa gctgatcaat gccctgcgga 600
gatgcttctt ttggcggttc atgttctacg gcatcttcct gtatctgggc gaggtgacca 660
aggccgtgca gccactgctg ctgggcagaa tcatcgcctc ttacgacccc gataacaagg 720
aggagaggag catcgccatc tatctgggca tcggcctgtg cctgctgttt atcgtgagga 780
cactgctgct gcacccagcc atcttcggcc tgcaccacat cggcatgcag atgagaatcg 840
ccatgttcag cctgatctac aagaagaccc tgaagctgag ctccagggtg ctggacaaga 900
tctccatcgg ccagctggtg tccctgctgt ctaacaatct gaacaagttt gatgagggac 960
tggccctggc acacttcgtg tggatcgcac cactgcaggt ggccctgctg atgggcctga 1020
tctgggagct gctgcaggca agcgcctttt gcggactggg cttcctgatc gtgctggccc 1080
tgttccaggc aggactggga cgcatgatga tgaagtacag agaccagagg gccggcaaga 1140
tctctgagcg gctggtcatc accagcgaga tgatcgagaa catccagtcc gtgaaggcct 1200
attgttggga ggaggccatg gagaagatga tcgagaatct gcgccagaca gagctgaagc 1260
tgaccagaaa ggccgcctac gtgaggtact tcaactctag cgccttcttt ttctctggct 1320
ttttcgtggt gttcctgagc gtgctgccat acgccctgat caagggcatc atcctgcgga 1380
agatctttac cacaatctcc ttctgcatcg tgctgagaat ggccgtgaca aggcagtttc 1440
cctgggccgt gcagacctgg tatgactctc tgggcgccat caataagatc caggatttcc 1500
tgcagaagca ggagtacaag acactggagt ataacctgac cacaaccgag gtggtcatgg 1560
agaatgtgac cgccttctgg gaggagggct ttggcgagct gttcgagaag gccaagcaga 1620
acaataacaa tcgcaagaca tctaacggcg acgatagcct gtttttcagc aatttttccc 1680
tgctgggcac ccccgtgctg aaggacatca acttcaagat cgagagggga cagctgctgg 1740
cagtggcagg ctccacaggc gccggcaaga cctctctgct gatgatgatc atgggcgagc 1800
tggagccaag cgagggcaag atcaagcact ccggccggat ctctttttgc agccagttct 1860
cctggatcat gcccggcacc atcaaggaga atatcatctt tggcgtgtcc tacgatgagt 1920
acagatatag gtctgtgatc aaggcctgtc agctggagga ggacatcagc aagttcgccg 1980
agaaggataa catcgtgctg ggcgagggcg gcatcacact gagcggagga cagagggcaa 2040
ggatctccct ggccagagcc gtgtacaagg acgccgatct gtatctgctg gacagcccct 2100
ttggctatct ggatgtgctg accgagaagg agatcttcga gtcctgcgtg tgcaagctga 2160
tggccaataa gacaaggatc ctggtgacct ctaagatgga gcacctgaag aaggccgaca 2220
agatcctgat cctgcacgag ggctcctctt acttttatgg cacattcagc gagctgcaga 2280
atctgcagcc tgacttcagc tccaagctga tgggctgtga ctcctttgat cagttctctg 2340
ccgagaggcg caactccatc ctgacagaga ccctgcacag attctctctg gagggcgacg 2400
cacccgtgag ctggacagag accaagaagc agtcctttaa gcagaccggc gagttcggcg 2460
agaagaggaa gaattctatc ctgaacccta tcaatagcac actgcaggcc cggagaaggc 2520
agtctgtgct gaacctgatg acccacagcg tgaaccaggg ccagaatatc cacagaaaga 2580
caaccgccag cacaaggaag gtgtccctgg cacctcaggc aaacctgacc gagctggaca 2640
tctactcccg ccggctgtct caggagaccg gactggagat ctctgaggag atcaatgagg 2700
aggatctgaa ggagtgcttt ttcgacgata tggagagcat cccagccgtg acaacctgga 2760
acacatacct gcgctatatc accgtgcaca agtccctgat ctttgtgctg atctggtgtc 2820
tggtcatctt cctggcagag gtggcagcat ctctggtggt gctgtggctg ctgggcaaca 2880
cacccctgca ggacaagggc aattctaccc acagccgcaa caattcctac gccgtgatca 2940
tcacatctac ctctagctac tacgtgttct acatctatgt gggcgtggcc gatacactgc 3000
tggccatggg ctttttccgg ggcctgcccc tggtgcacac actgatcacc gtgagcaaga 3060
tcctgcacca caagatgctg cacagcgtgc tgcaggcccc tatgtccaca ctgaacaccc 3120
tgaaggccgg cggcatcctg aatcggtttt ccaaggacat cgccatcctg gacgatctgc 3180
tgcctctgac catctttgat ttcatccagc tgctgctgat cgtgatcgga gcaatcgcag 3240
tggtggccgt gctgcagcct tacatcttcg tggccacagt gccagtgatc gtggccttta 3300
tcatgctgcg cgcctatttc ctgcagacca gccagcagct gaagcagctg gagagcgagg 3360
gccggtcccc tatctttaca cacctggtga cctccctgaa gggactgtgg acactgaggg 3420
ccttcggccg gcagccatac tttgagaccc tgttccacaa ggccctgaac ctgcacacag 3480
ccaattggtt tctgtatctg agcaccctgc gctggtttca gatgcggatc gagatgatct 3540
tcgtgatctt tttcatcgcc gtgaccttca tctccatcct gacaaccgga gagggagagg 3600
gaagagtggg aatcatcctg acactggcca tgaacatcat gtctaccctg cagtgggccg 3660
tgaattcctc tatcgacgtg gatagcctga tgagatctgt gagcagggtg tttaagttca 3720
tcgacatgcc cacagagggc aagcctacaa agagcaccaa gccatacaag aacggccagc 3780
tgtccaaagt gatgatcatc gagaattctc acgtgaagaa ggacgatatc tggccatccg 3840
gaggacagat gaccgtgaag gatctgacag ccaagtatac cgagggcggc aacgccatcc 3900
tggagaatat ctccttttct atcagccctg gacagagggt gggactgctg ggacggacag 3960
gctccggcaa gtctaccctg ctgagcgcct tcctgaggct gctgaataca gagggcgaga 4020
tccagatcga cggcgtgagc tgggattcca tcaccctgca gcagtggaga aaggcctttg 4080
gcgtgatccc tcagaaggtg tttatcttct ccggcacctt caggaagaac ctggacccat 4140
acgagcagtg gtctgatcag gagatctgga aggtggccga cgaagtgggc ctgagatctg 4200
tgatcgagca gtttccaggc aagctggact tcgtgctggt ggatggagga tgcgtgctga 4260
gccacggaca caagcagctg atgtgcctgg ccaggtctgt gctgagcaag gccaagatcc 4320
tgctgctgga cgagccaagc gcccacctgg atcccgtgac ataccagatc atcagaagga 4380
ccctgaagca ggcctttgcc gattgcaccg tgatcctgtg cgagcaccgc atcgaggcca 4440
tgctggagtg ccagcagttc ctggtcatcg aggagaacaa ggtgcggcag tatgacagca 4500
tccagaagct gctgaatgag cggagcctgt ttcggcaggc catctccccc tctgatcgcg 4560
tgaagctgtt ccctcaccgg aacagctcca agtgtaagtc caagccccag atcgccgccc 4620
tgaaggagga gacagaggag gaggtgcagg acaccagact gtgaaataaa acatctttat 4680
tttcattaca tctgtgtgtt ggttttttgt gtgaacaacg gccggccgga ggaaccccta 4740
gtgatggagt tggccactcc ctctctgcgc gctcgctcgc tcactgaggc cgcccgggca 4800
aagcccgggc gtcgggcgac ctttggtcgc ccggcctcag tgagcgagcg agcgcgcaga 4860
gagggagtgg ccaa 4874
<---
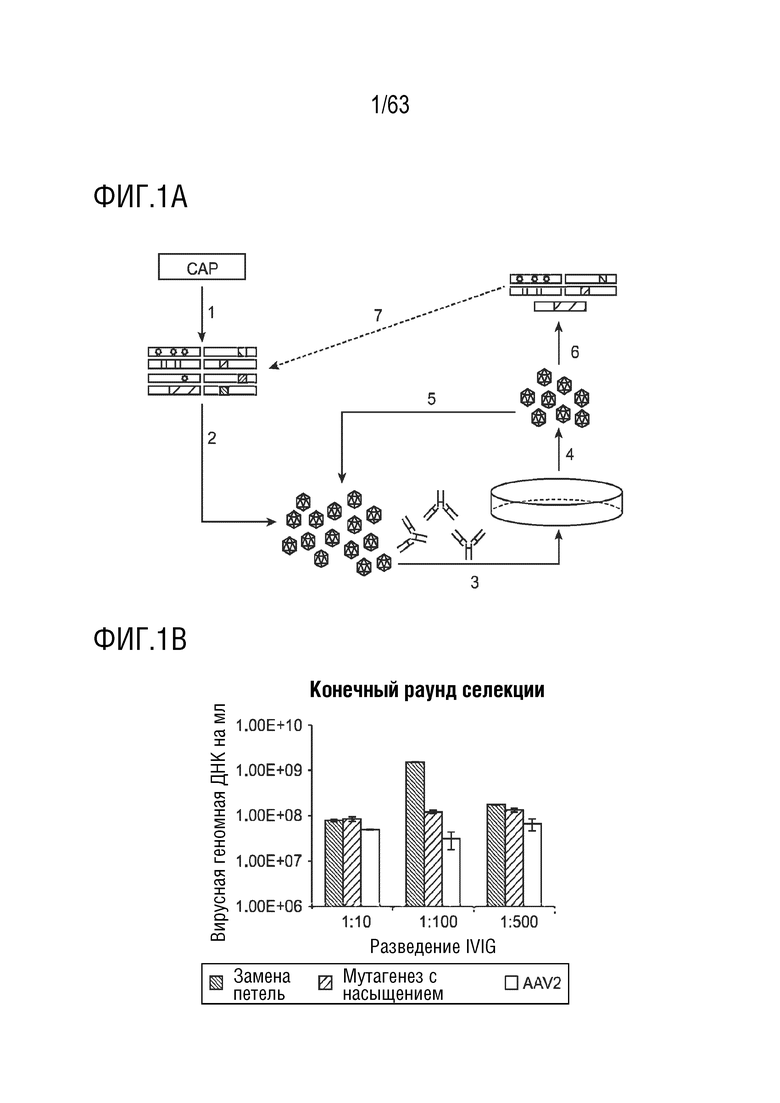
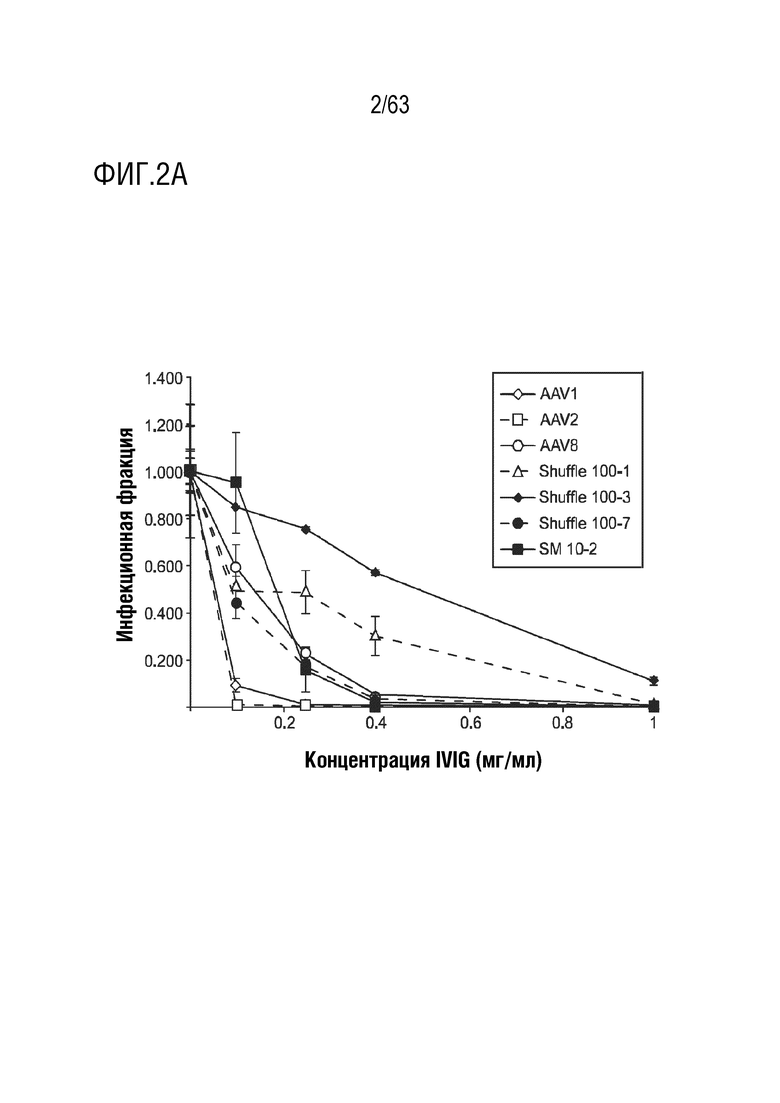

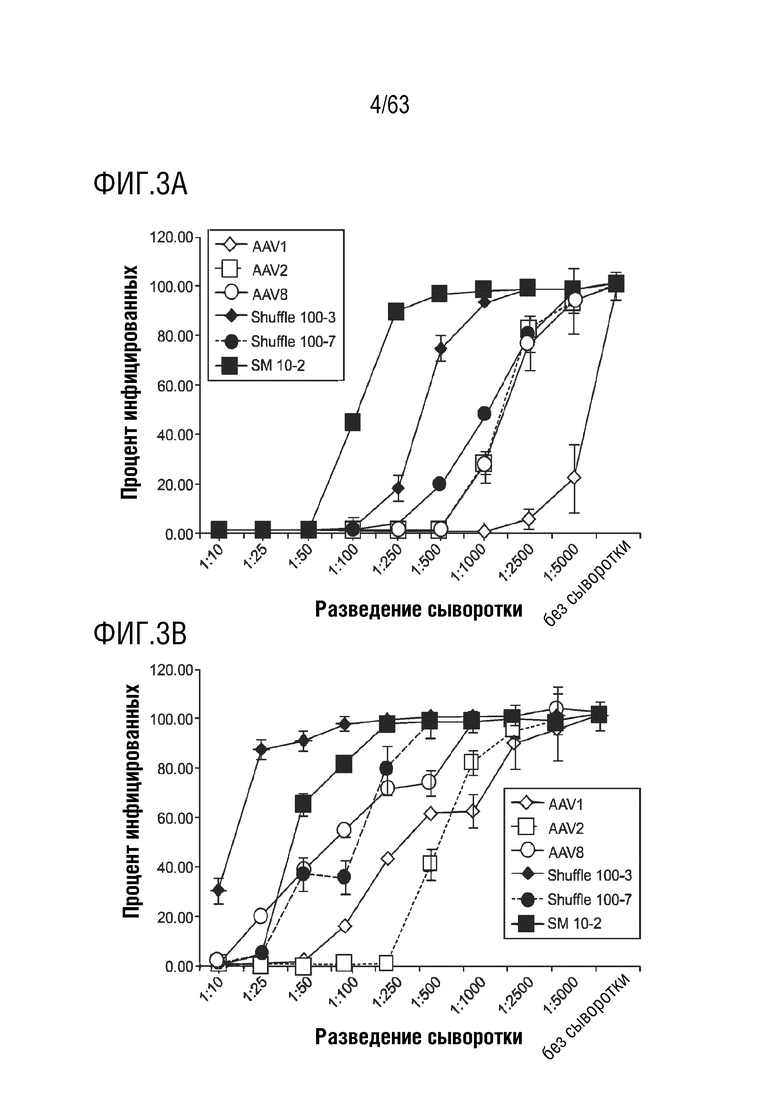
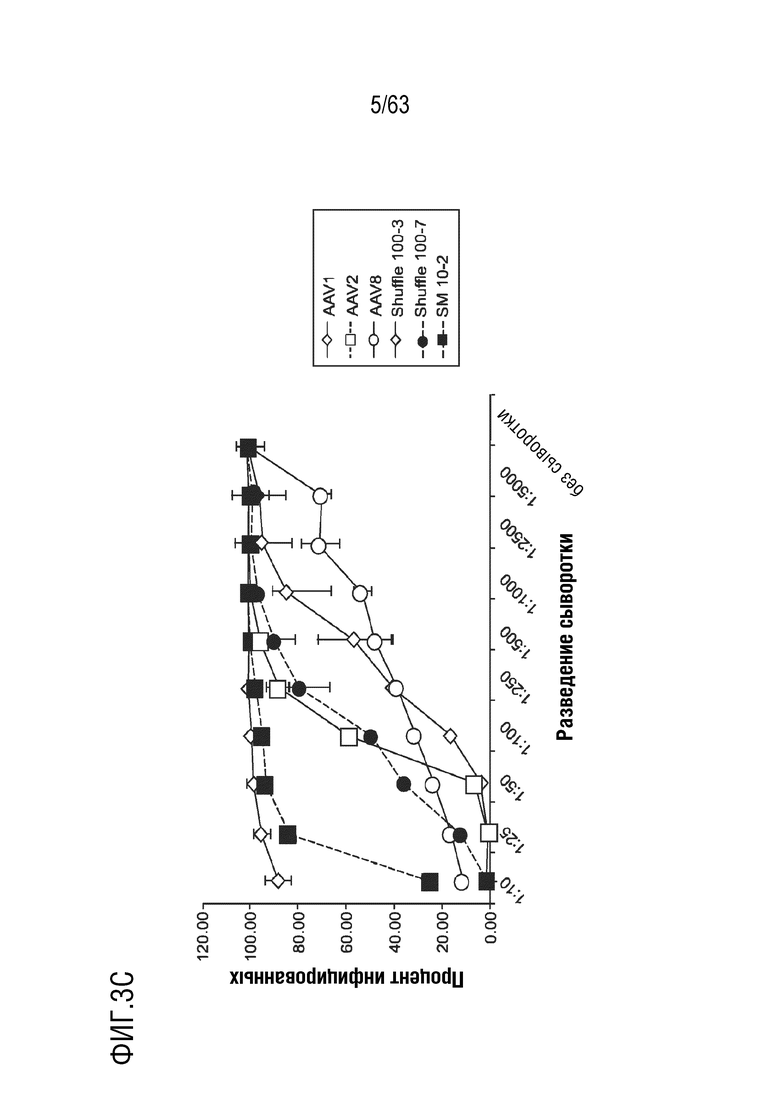

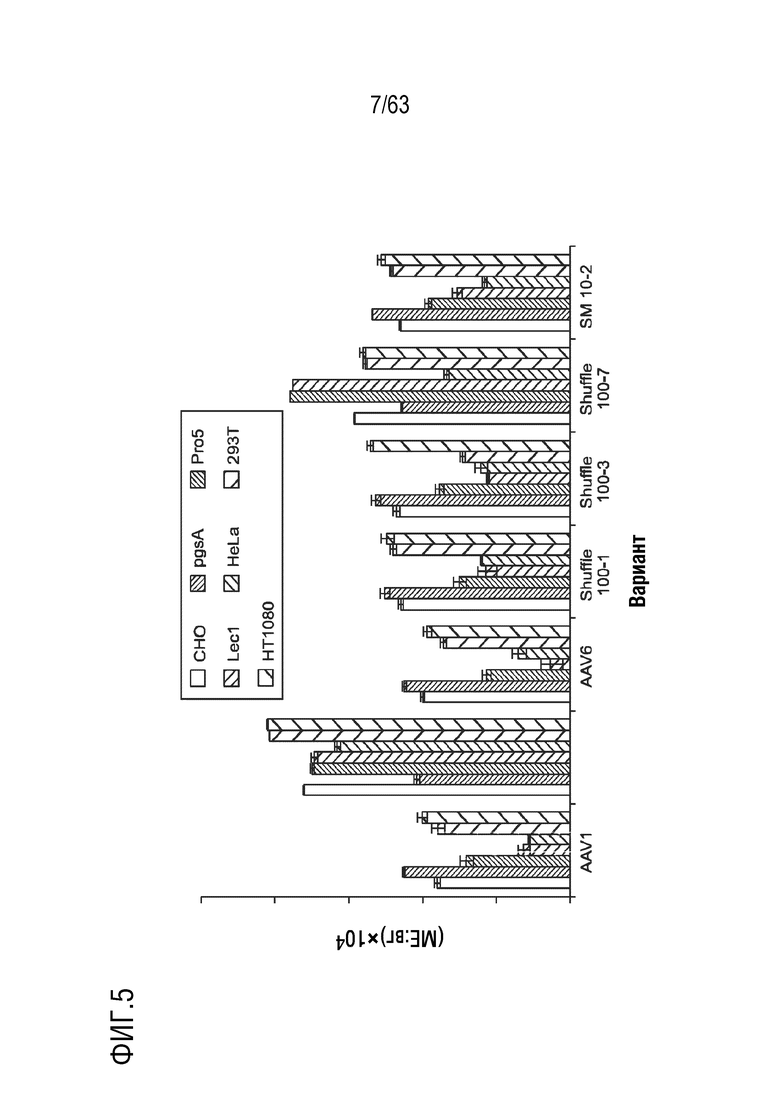
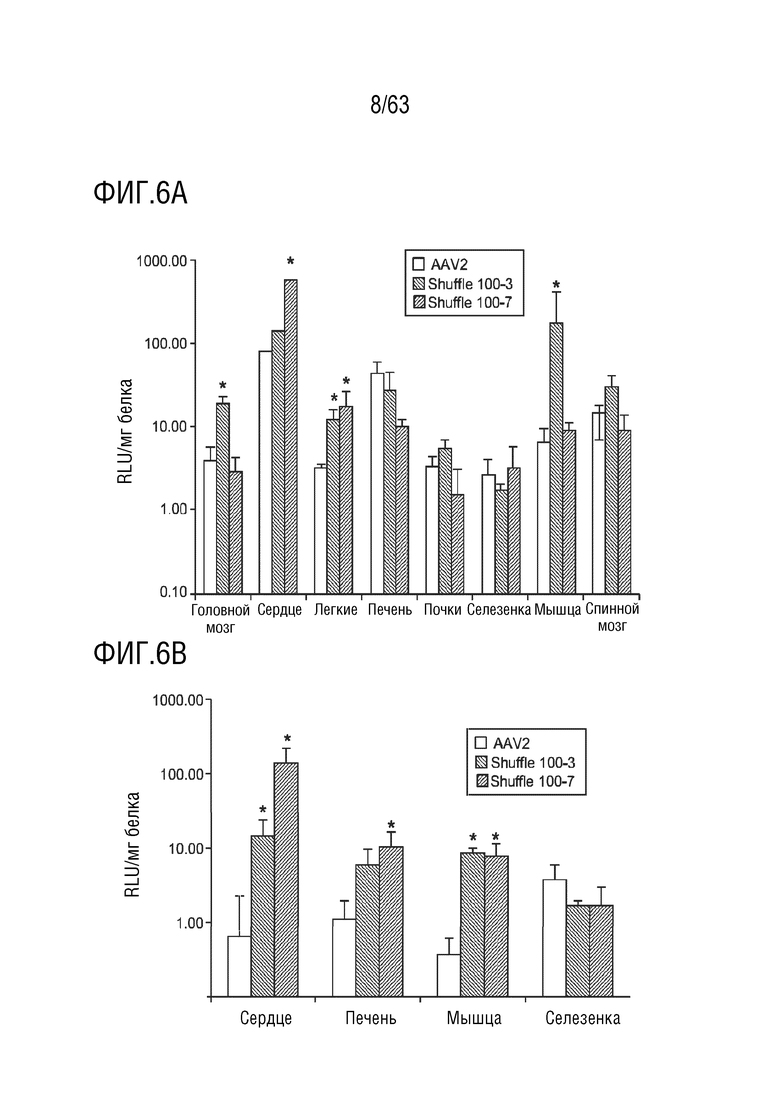
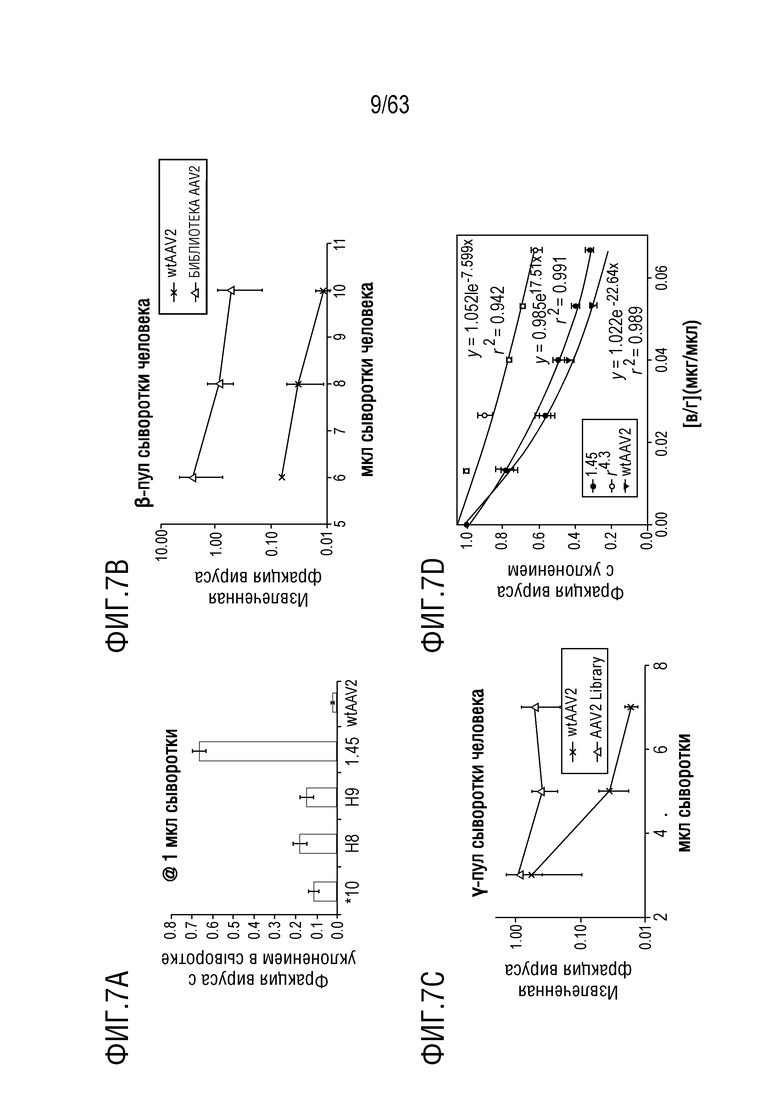









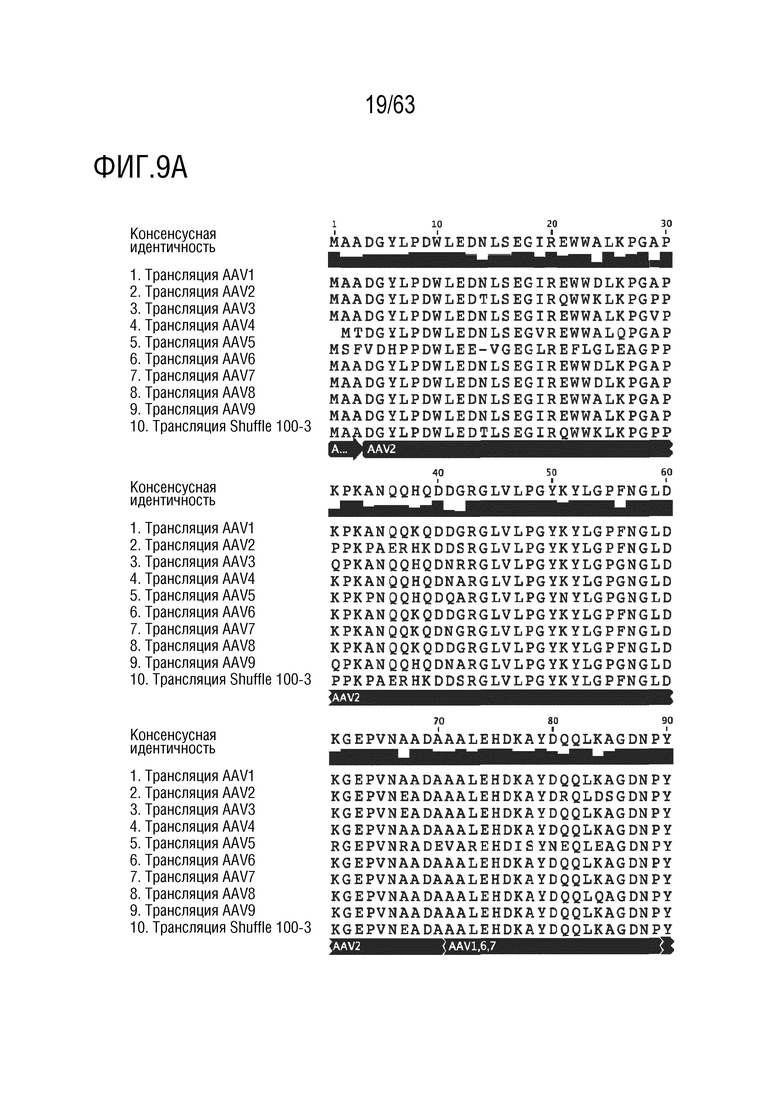













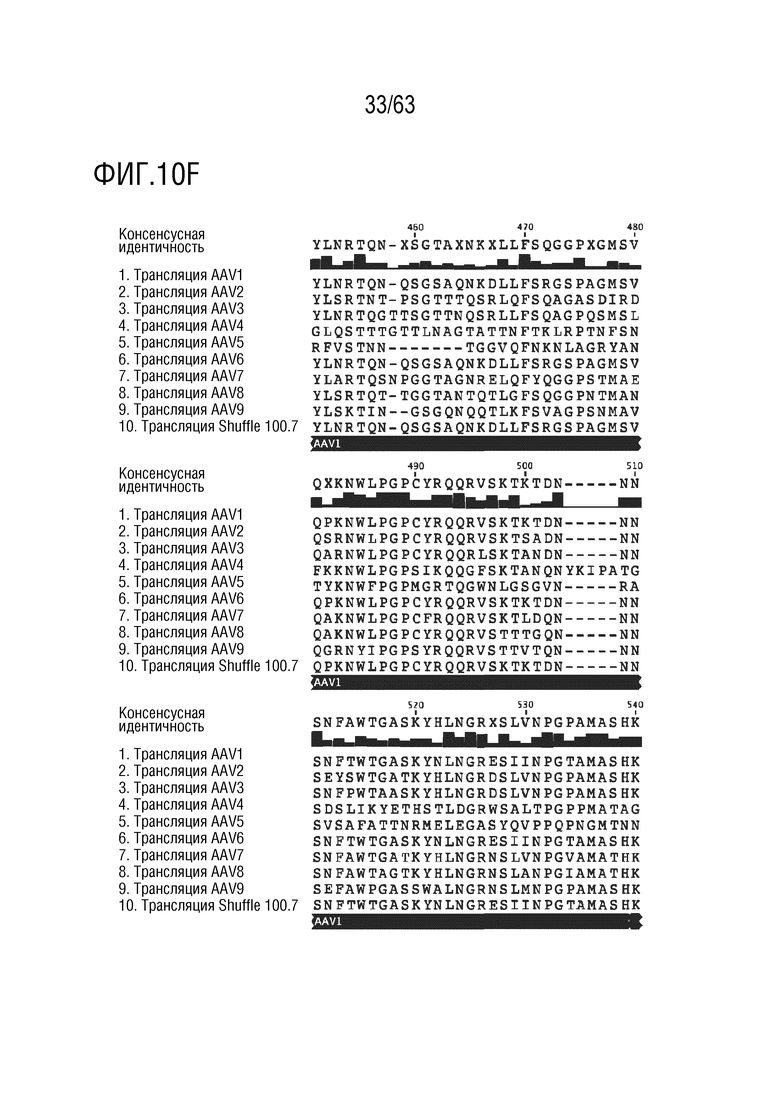



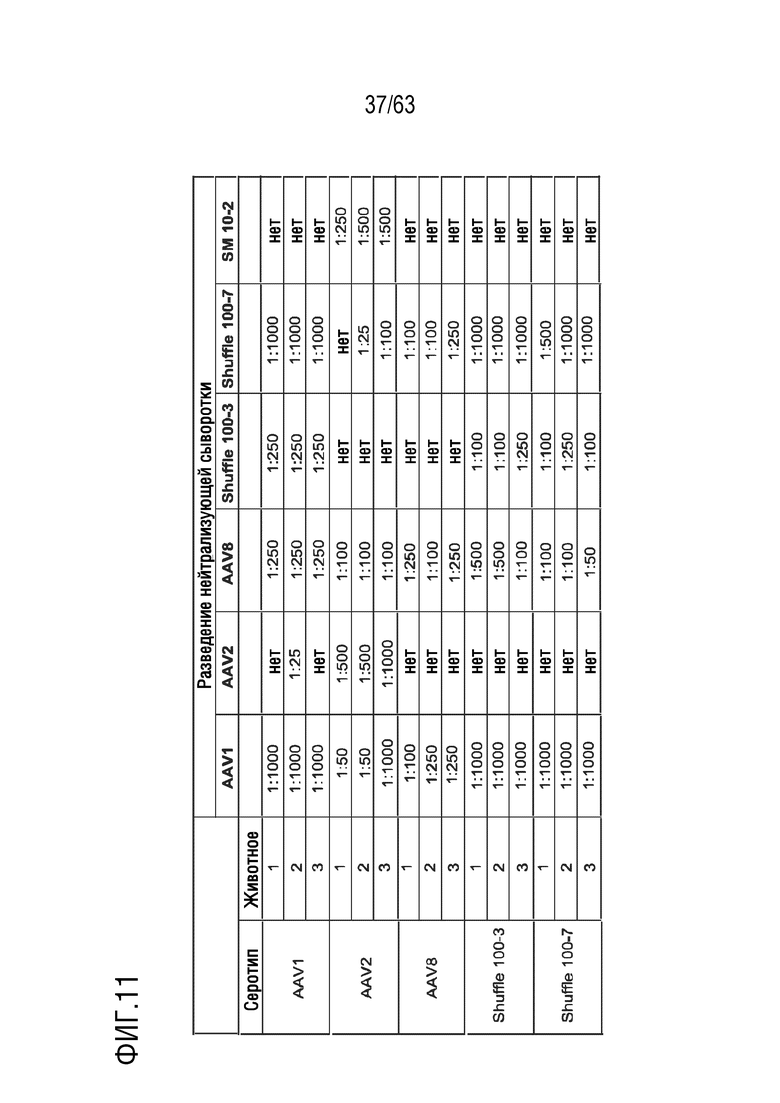
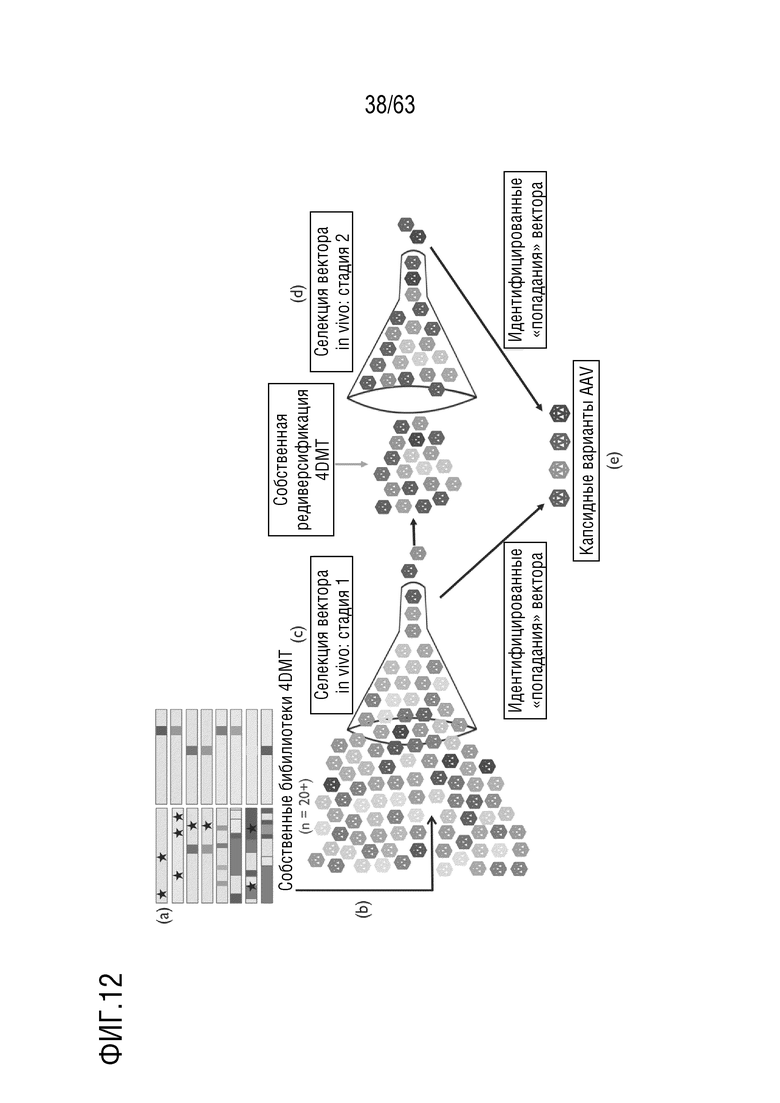
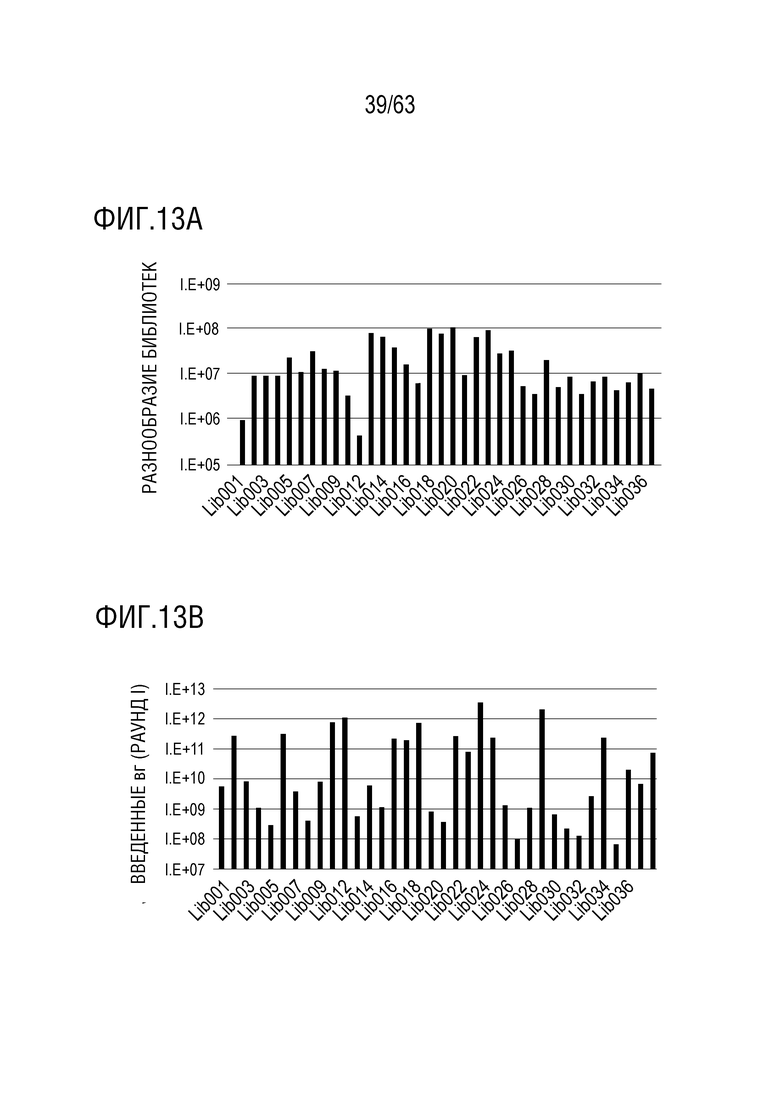
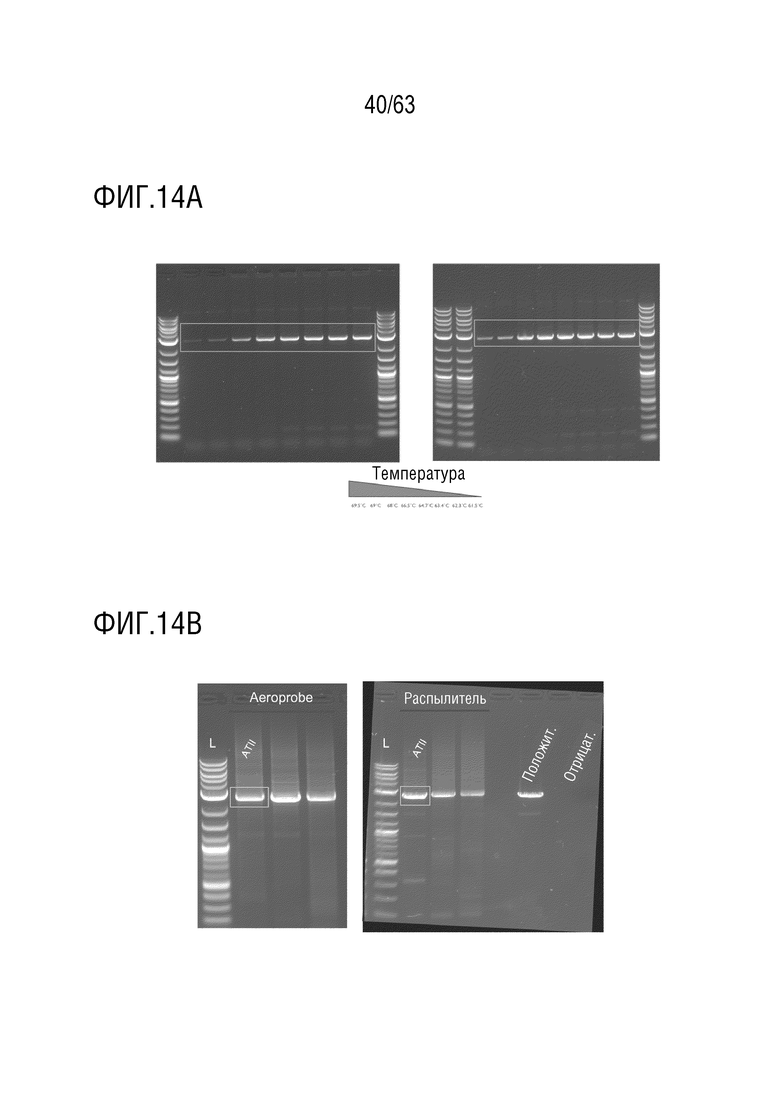

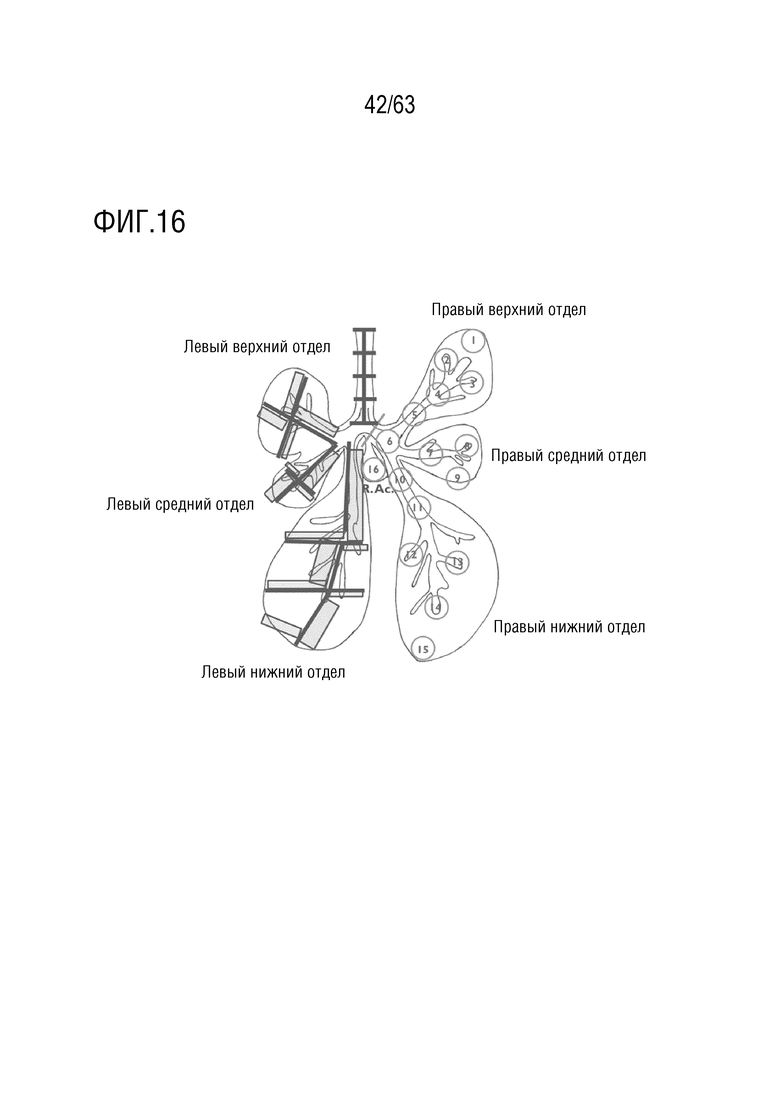
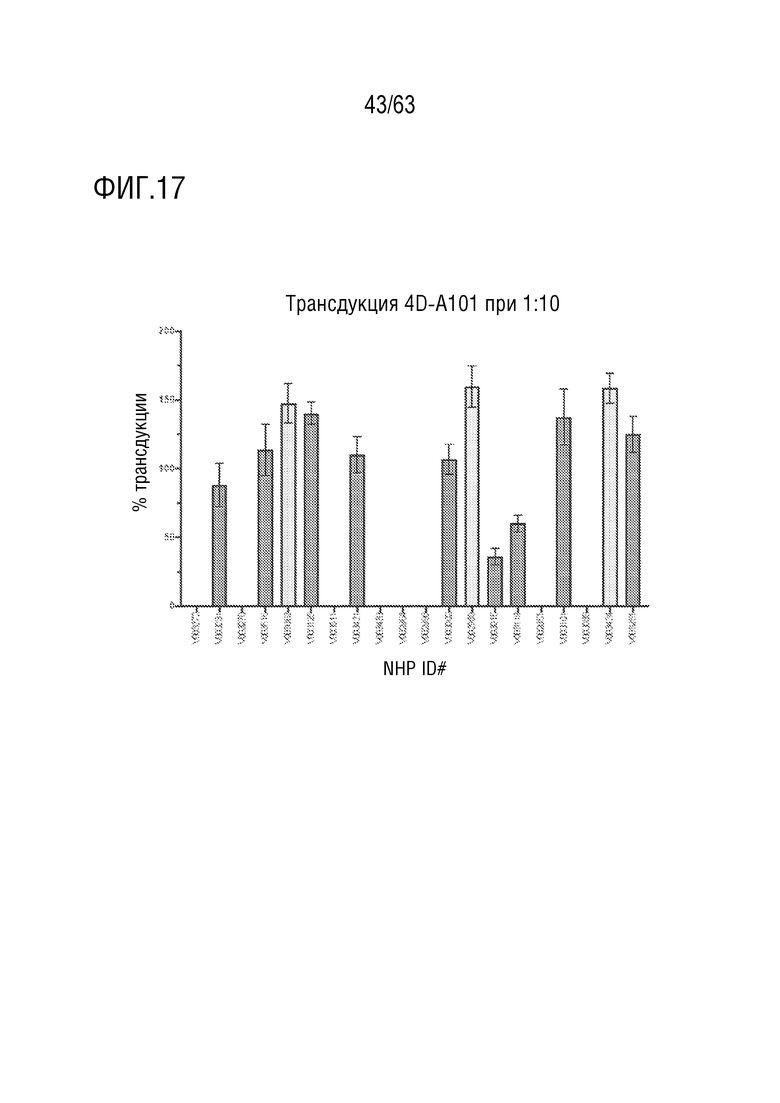
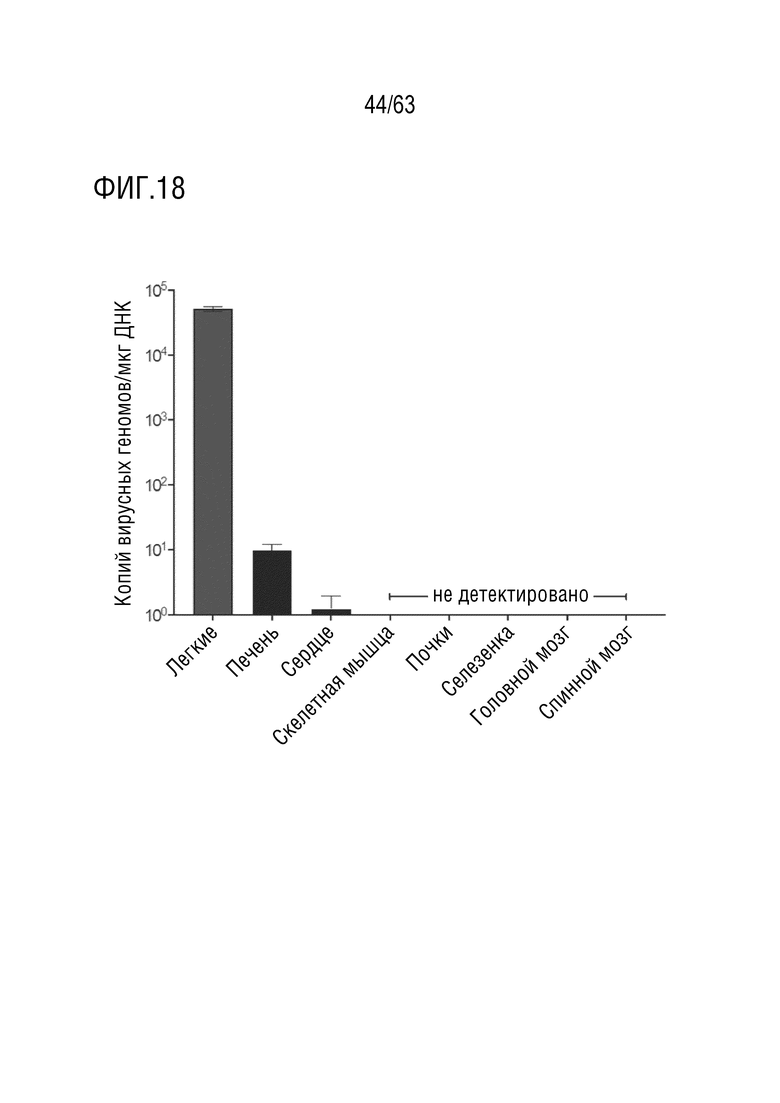
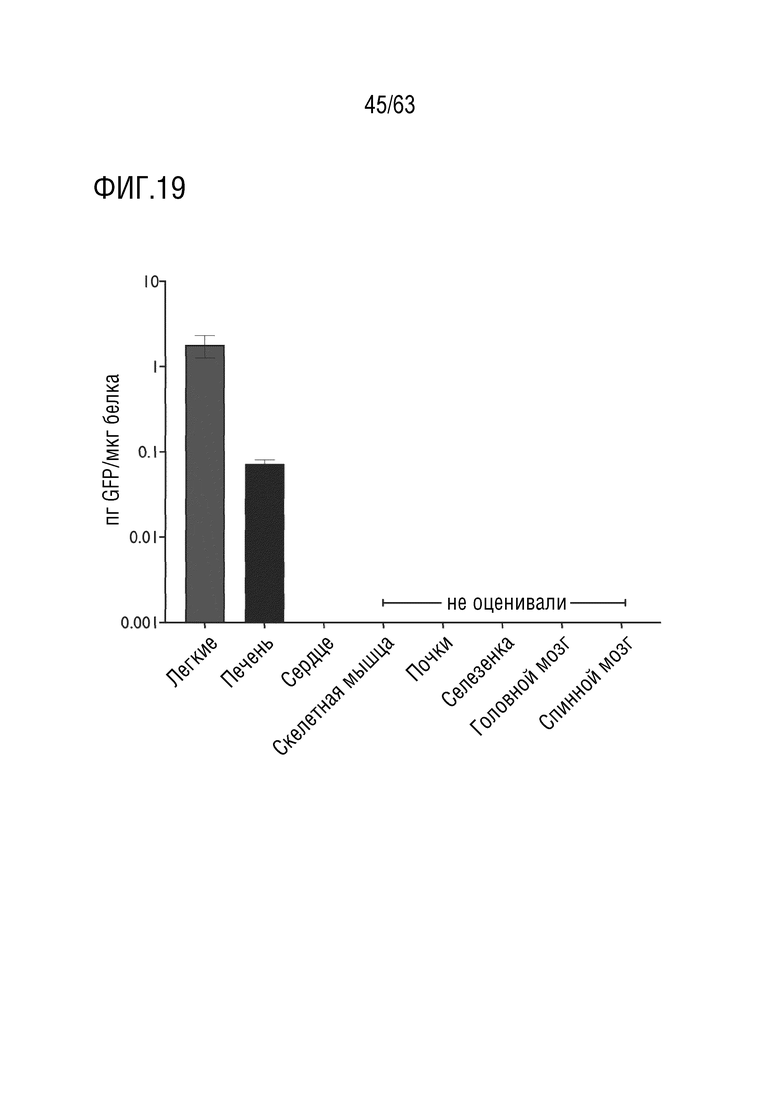
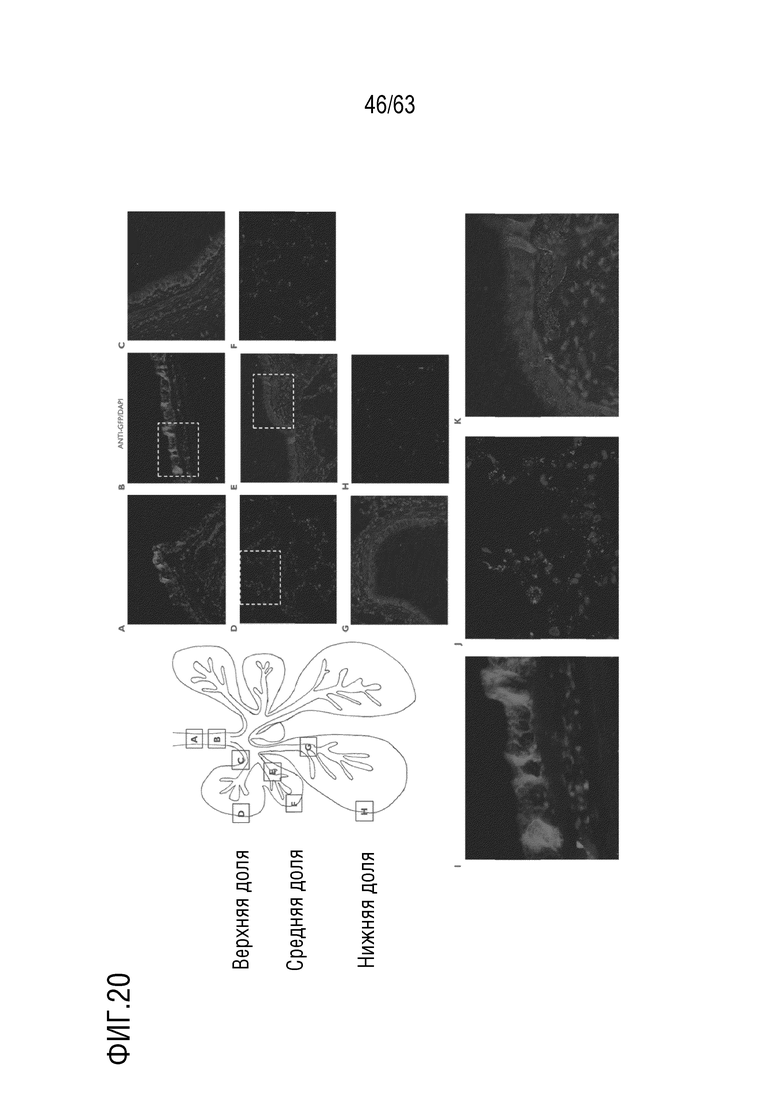
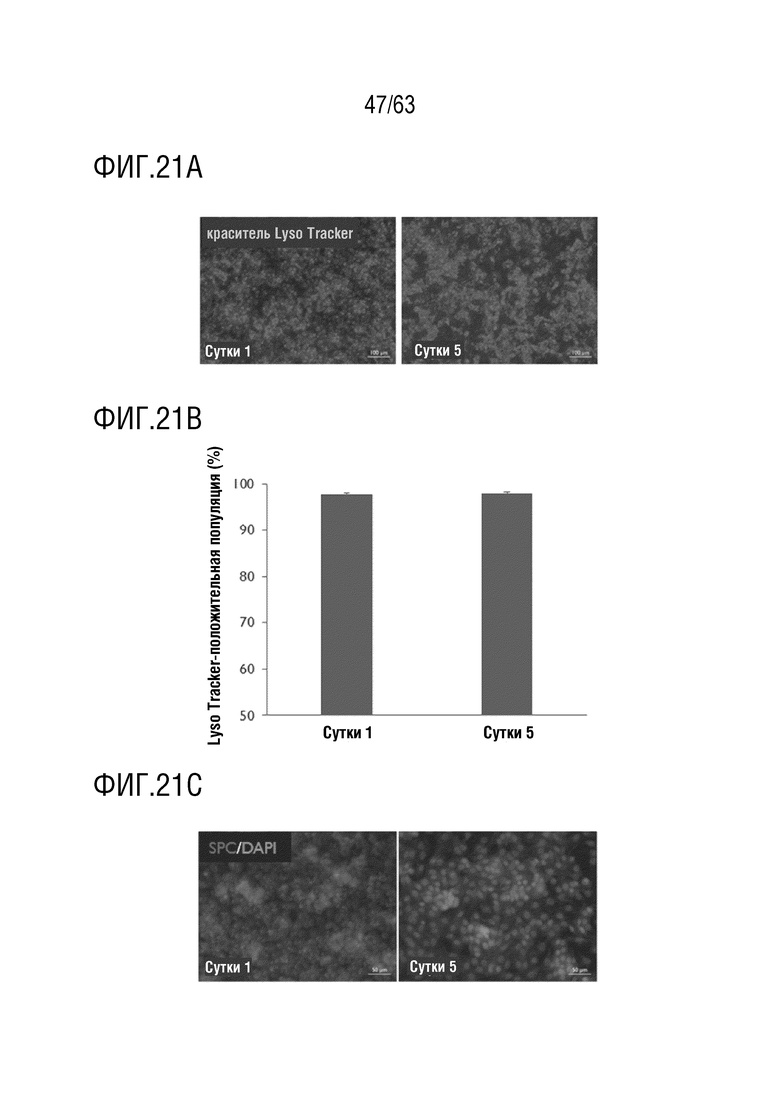

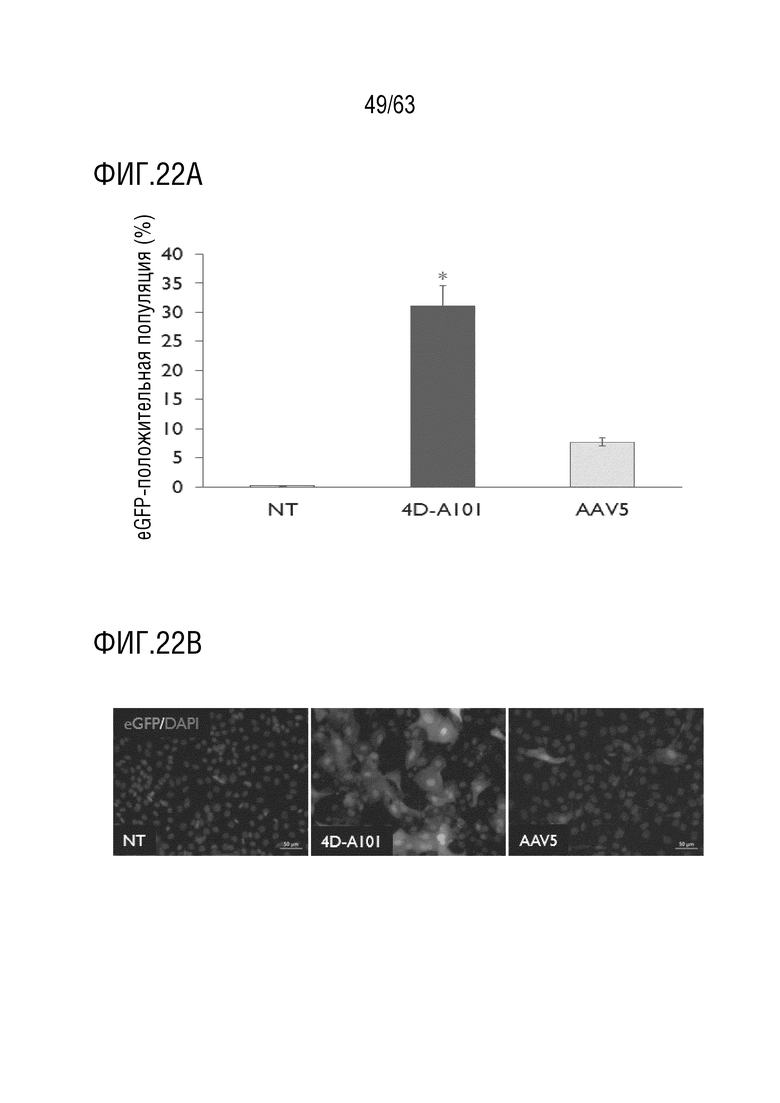
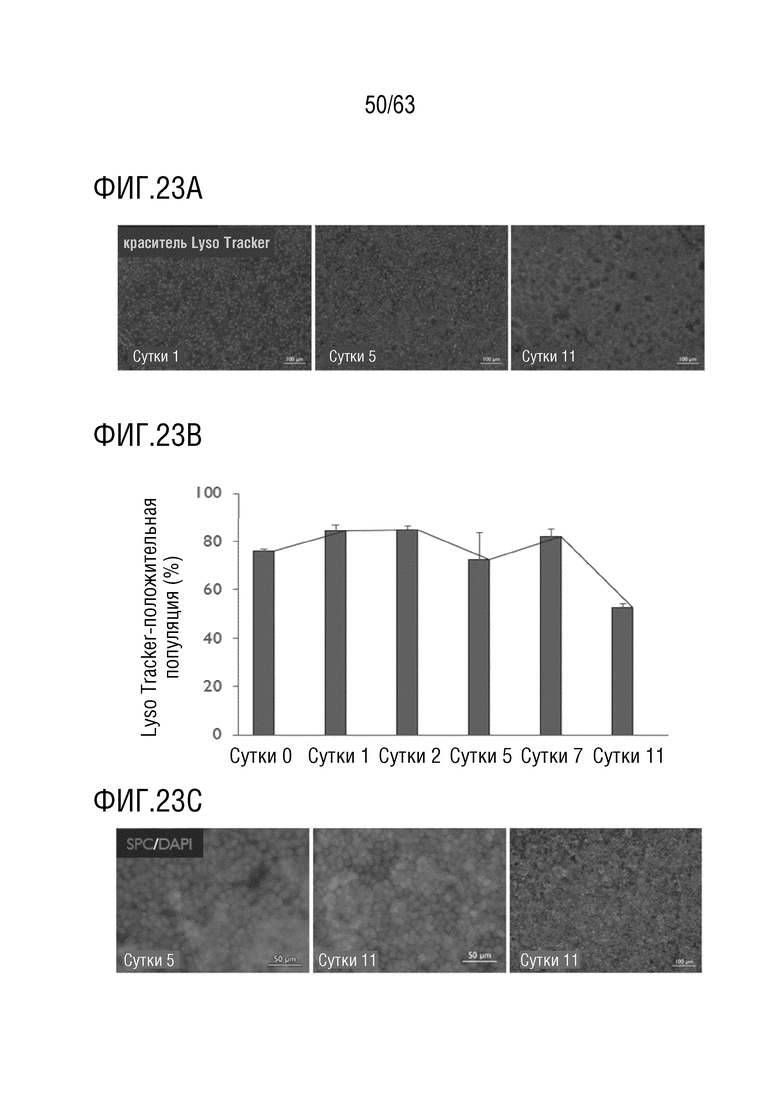

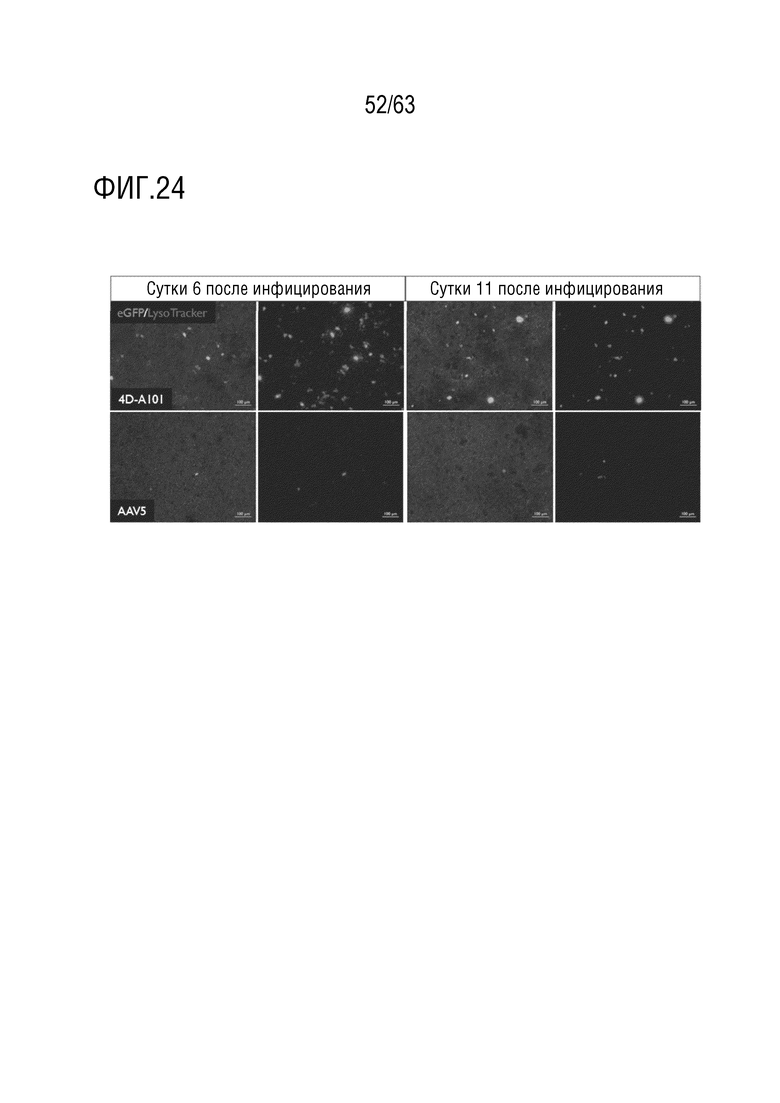
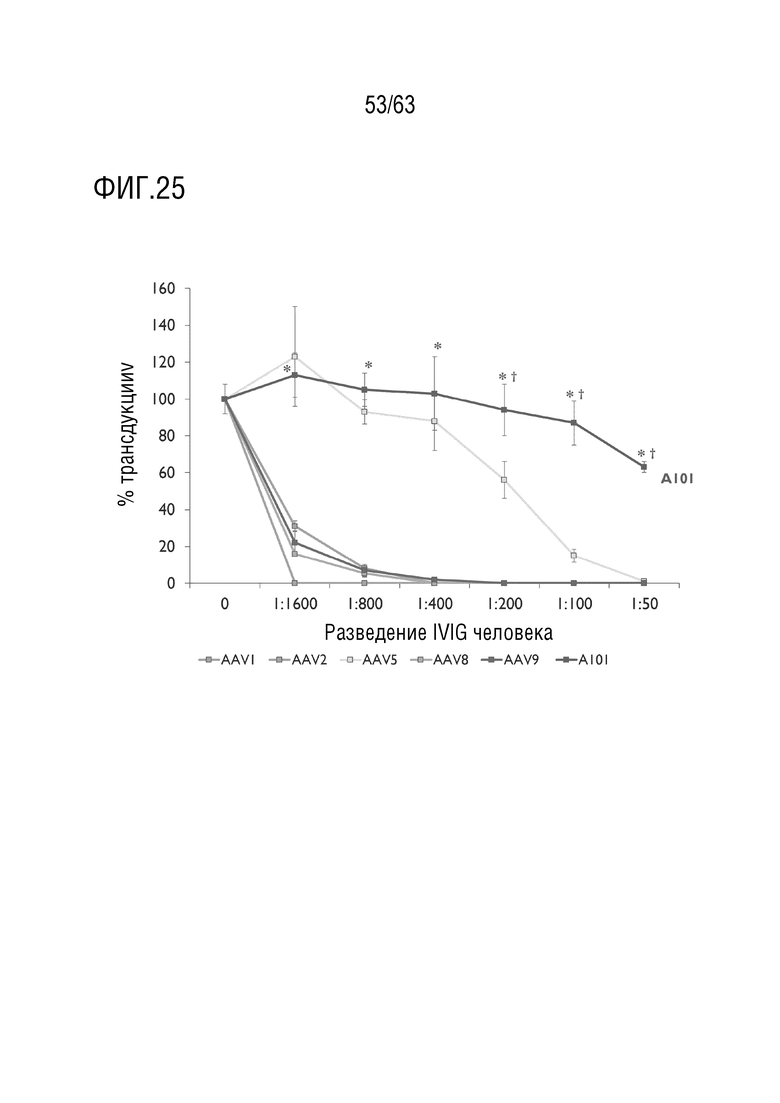
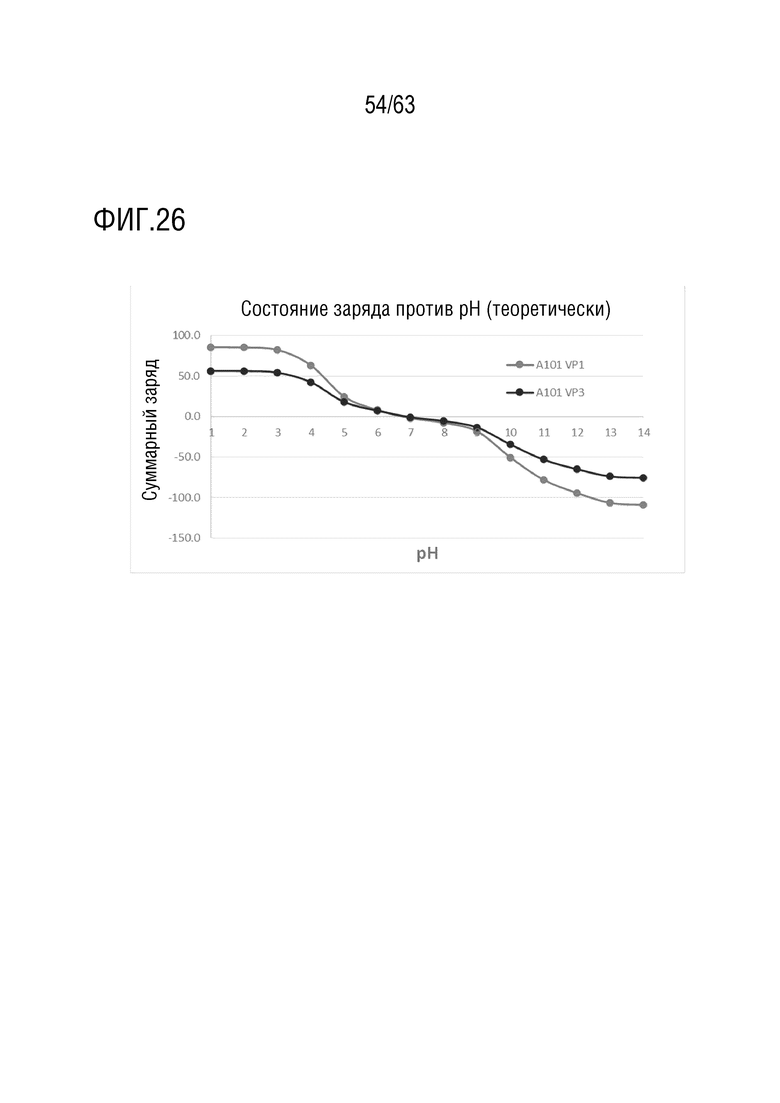
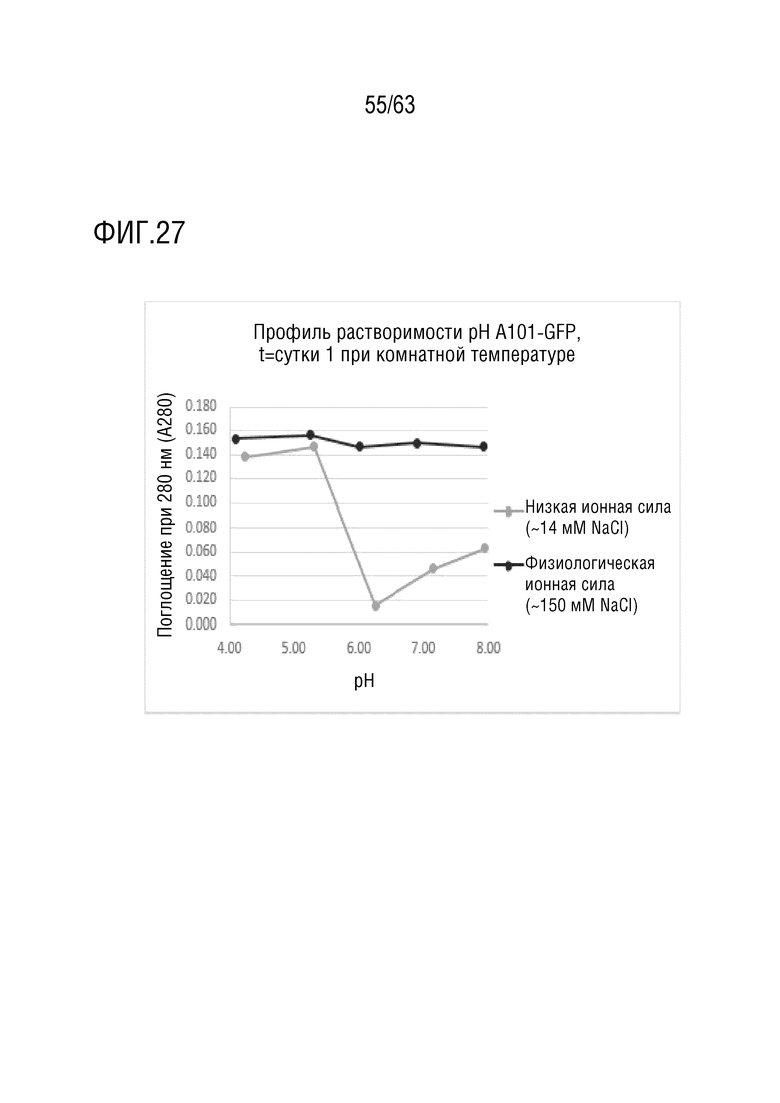
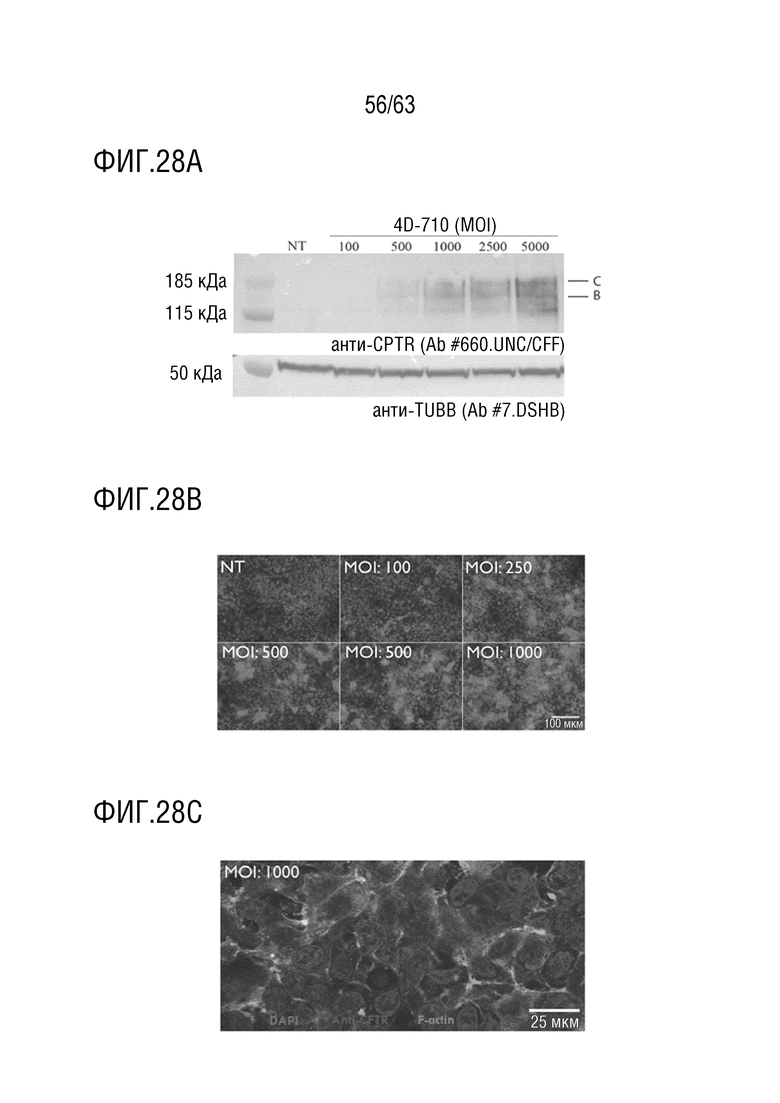
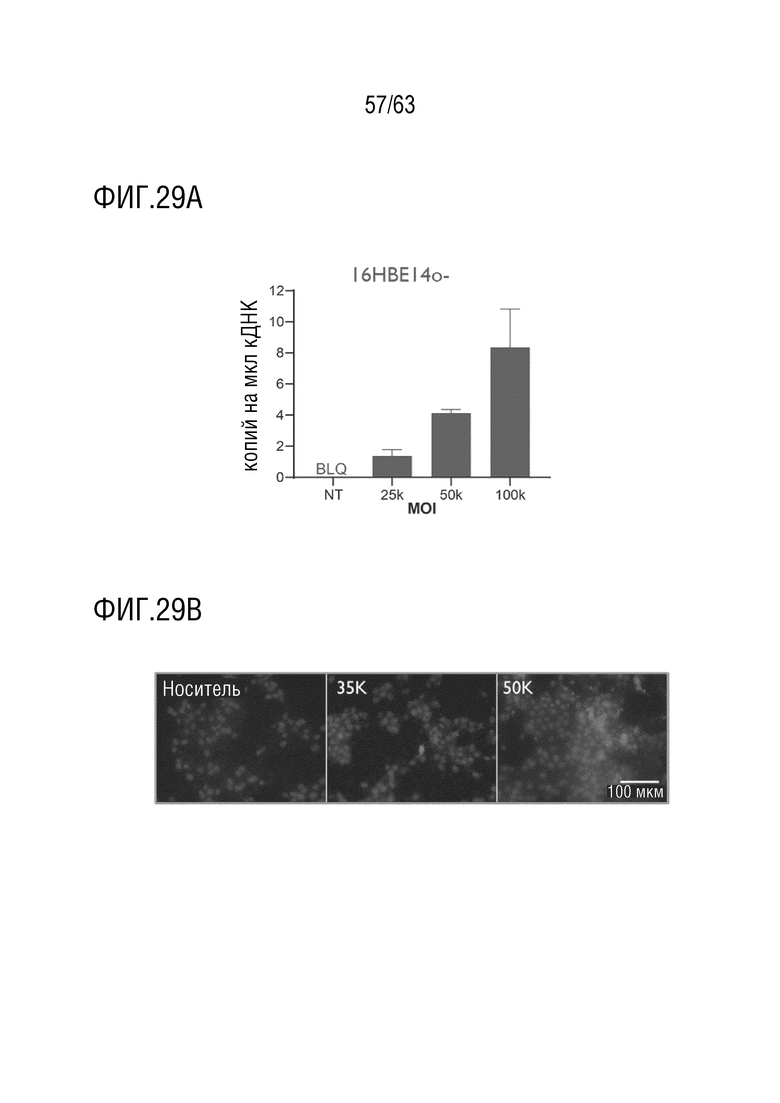
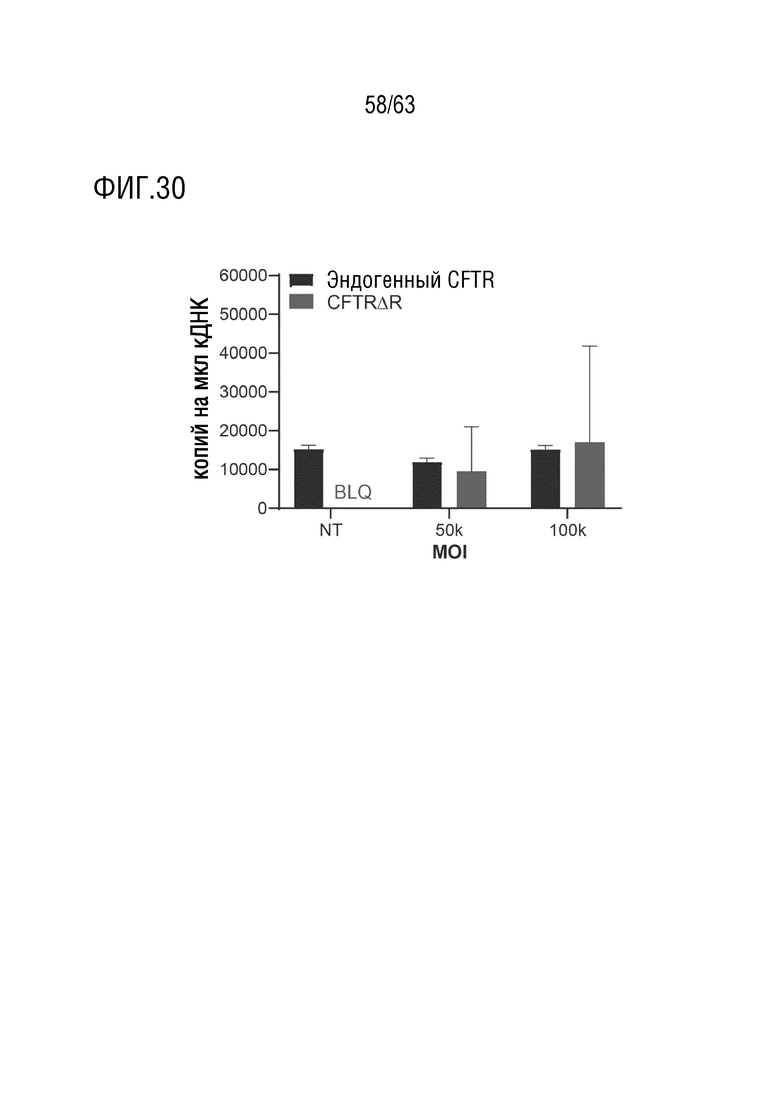
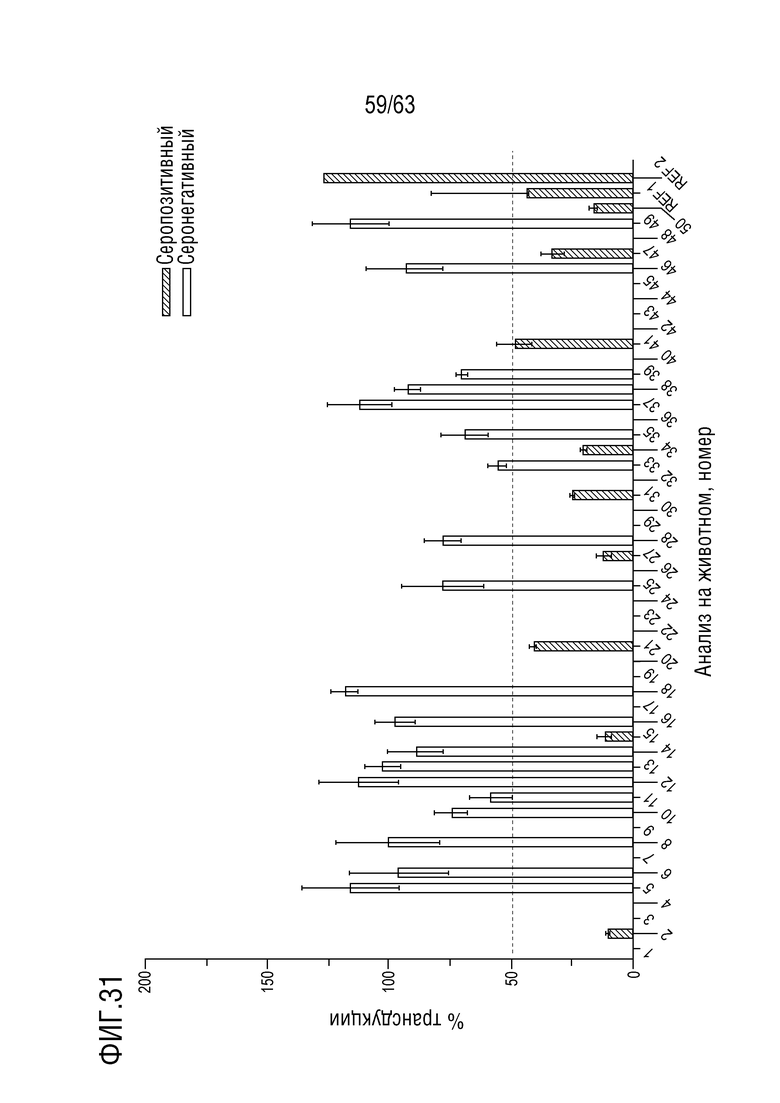
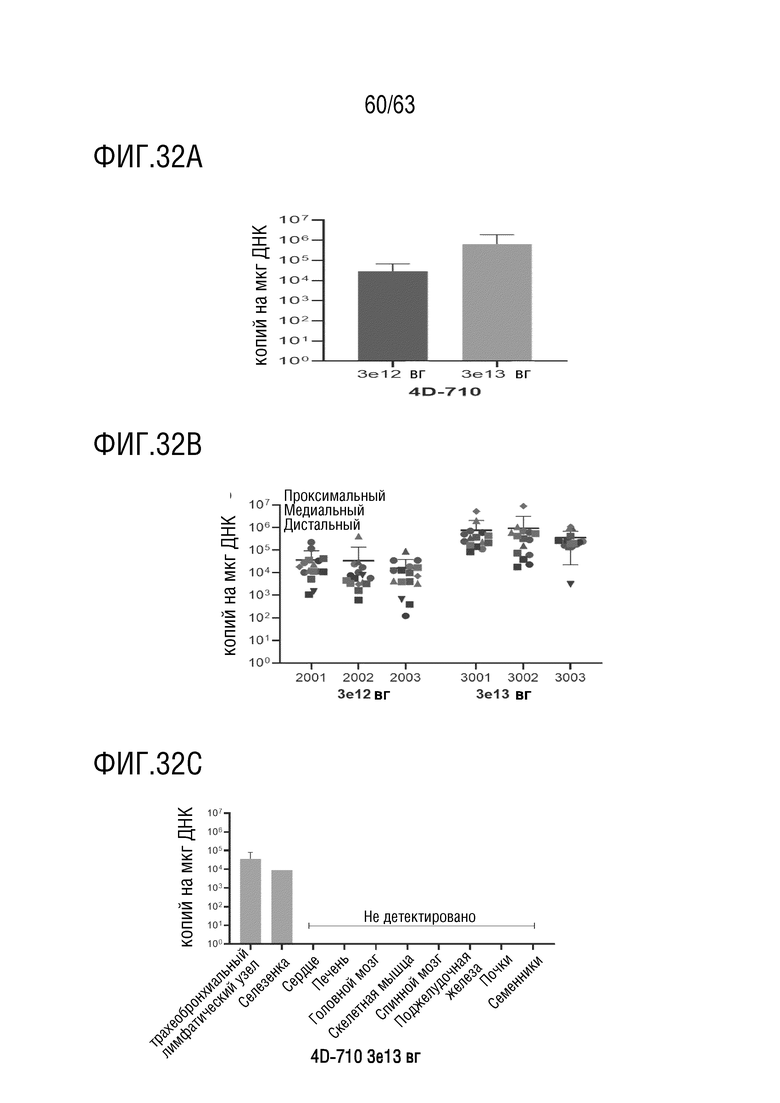
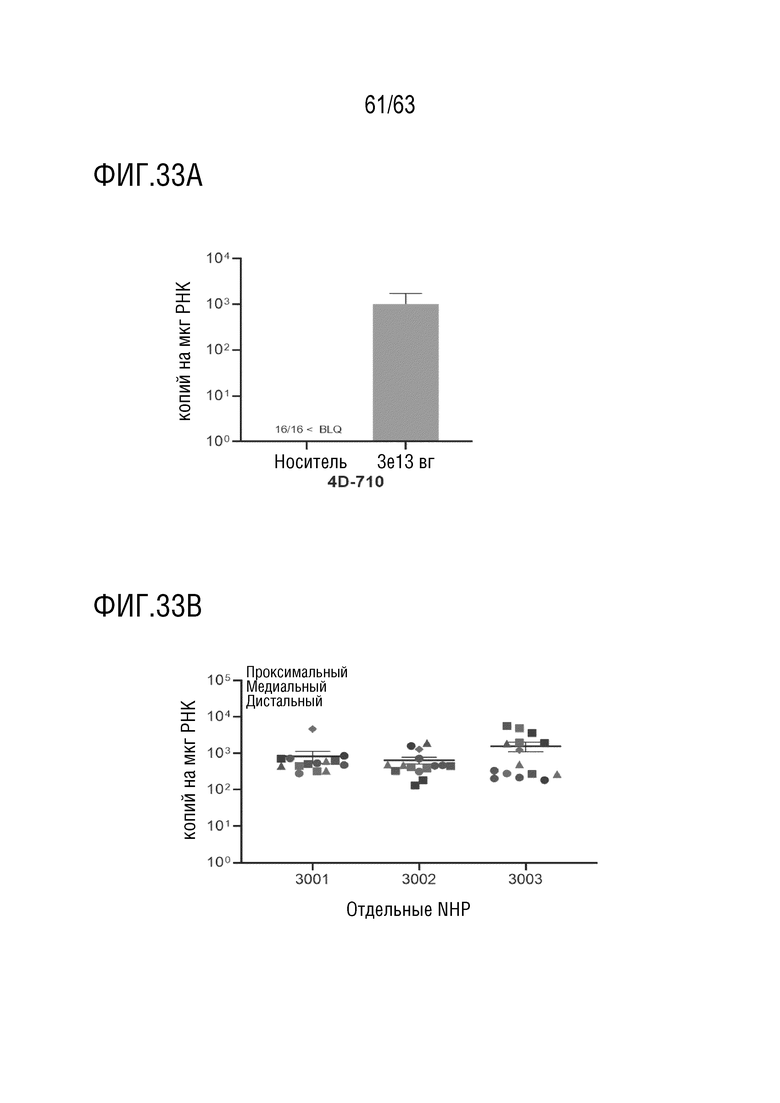
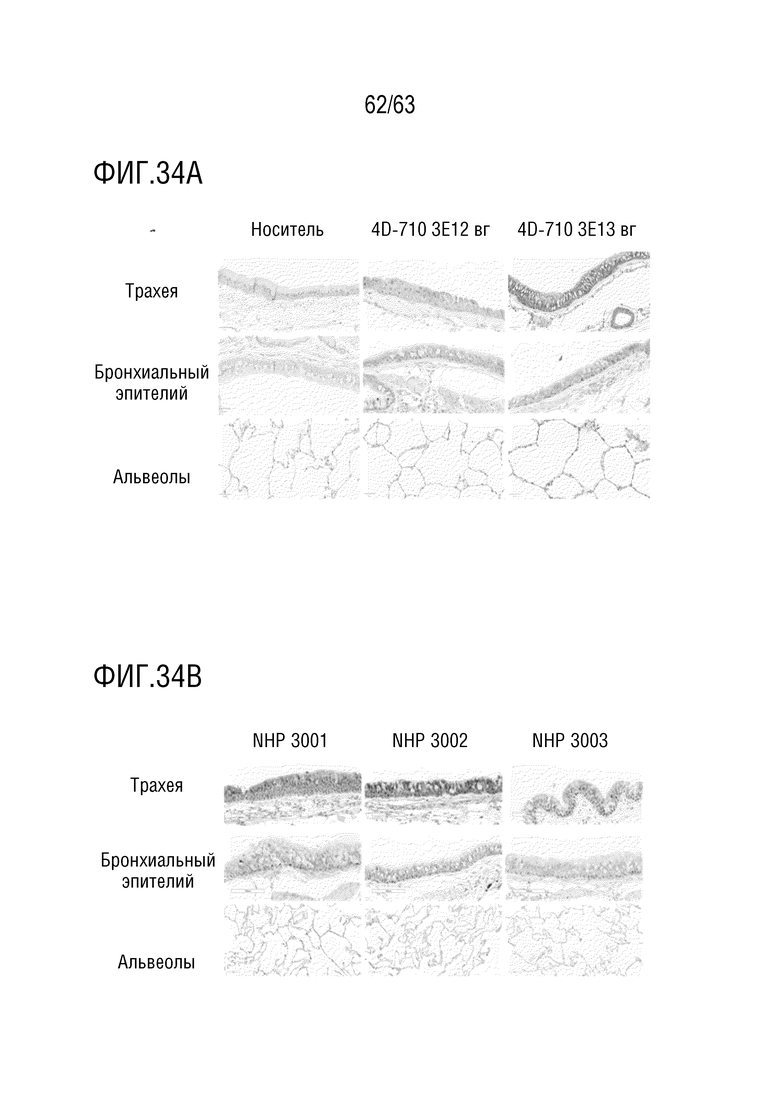
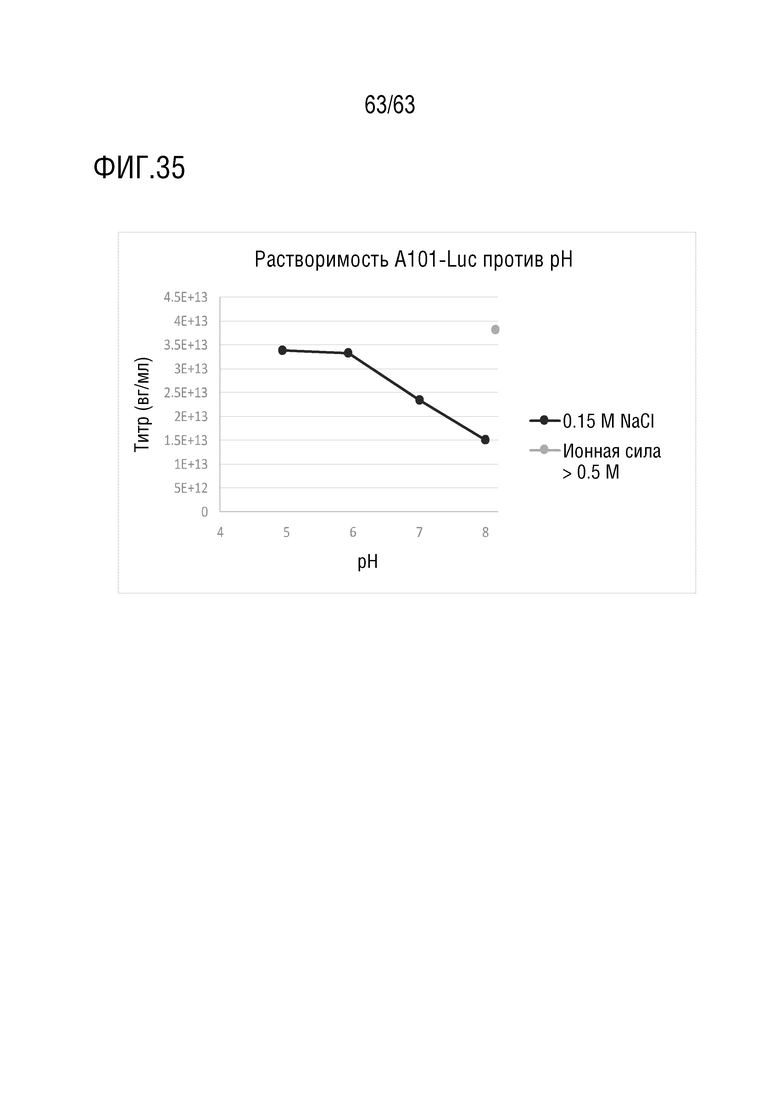
Изобретение относится к биохимии. Описан способ доставки гетерологичной нуклеиновой кислоты в клетку легкого у субъекта, включающий введение субъекту композиции, содержащей рекомбинантный вектор на основе аденоассоциированного вируса (rAAV). Причем указанный вектор rAAV содержит (i) капсид, содержащий капсидный белок, содержащий аминокислотную последовательность, представленную как SEQ ID NO: 12, или аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 12 и содержащую Thr по аминокислоте 469 и Ala по аминокислоте 598, исходя из нумерации аминокислот, представленной в SEQ ID NO: 12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт. Введение включает пульмональное, эндобронхиальное, интраназальное, интратрахеальное и/или внутрибронхиальное введение. Описана соответствующая композиция, содержащая указанный вектор. Кроме того, описан способ лечения муковисцидоза у субъекта, включающий введение субъекту указанной композиции. Применение указанной композиции в получении лекарственного средства для лечения муковисцидоза, где лекарственное средство предназначено для пульмонального, эндобронхиального, интраназального, внутритрахеального или внутрибронхиального введения. Изобретение расширяет арсенал средств для лечения легочных заболеваний. 5 н. и 25 з.п. ф-лы, 35 ил., 18 табл., 7 пр.
1. Способ доставки гетерологичной нуклеиновой кислоты в клетку легкого у субъекта, включающий введение субъекту композиции, содержащей рекомбинантный вектор на основе аденоассоциированного вируса (rAAV), причем указанный вектор rAAV содержит (i) капсид, содержащий капсидный белок, содержащий аминокислотную последовательность, представленную как SEQ ID NO: 12, или аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 12 и содержащую Thr по аминокислоте 469 и Ala по аминокислоте 598, исходя из нумерации аминокислот, представленной в SEQ ID NO: 12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт, причем стадия введения включает пульмональное, эндобронхиальное, интраназальное, интратрахеальное и/или внутрибронхиальное введение.
2. Способ по п. 1, в котором клетка легкого выбрана из эпителиальной клетки дыхательных путей, гладкомышечной клетки и эндотелиальной клетки.
3. Способ по п. 2, в котором эпителиальная клетка дыхательных путей представляет собой базальную клетку, бокаловидную клетку или реснитчатую клетку.
4. Способ по п. 3, в котором эпителиальная клетка дыхательных путей представляет собой альвеолярную эпителиальную клетку легкого 1 типа (AECI), альвеолярную клетку легкого 2 типа (AECII), эпителиальную клетку бронхов или эпителиальную клетку трахеи.
5. Способ по любому из пп. 1-4, где композиция составлена в виде аэрозоля.
6. Способ по п. 5, включающий введение композиции распылителем.
7. Способ по любому из пп. 1-6, где композиция содержит от 1011 до 1014 векторных геномов (вг) rAAV на 1 мл.
8. Способ по любому из пп. 1-7, где нуклеотидная последовательность, кодирующая генный продукт, функционально связана с промотором.
9. Способ по п. 8, где промотор представляет собой конститутивный промотор.
10. Способ по п. 8, где промотор представляет собой тканеспецифичный промотор.
11. Способ по любому из пп. 1-10, где гетерологичная нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую генный продукт, выбранный из белка регулятора трансмембранной проводимости при муковисцидозе человека (CFTR) или его биологически активного фрагмента, SFTPA1 (сурфактанта А1), кавеолина-1, альфа-1-антитрипсина, альфа-1-антихимотрипсина, альфа-1-макроглобулина, матриксной металлопротеиназы-1 (MMP1), матриксной металлопротеиназы-12 (MMP12), микросомальной эпоксидгидролиазы, CYP1A1, глутатион-S-трансферазы, гемоксигеназы-1, TGF-бета-1, TNF-альфа, комплекса IL-1, IL-8, IL-13, человеческого лейкоцитарного антигена (HLA-B7 и Bw16), белка, связывающего витамин D, и бета-2-адренергического рецептора.
12. Способ по п. 11, где генный продукт представляет собой белок регулятора трансмембранной проводимости при муковисцидозе человека (CFTR) или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека.
13. Способ по п. 12, в котором генный продукт представляет собой биологически усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека.
14. Способ по п. 13, где гетерологичная нуклеиновая кислота содержит нуклеотидную последовательность, представленную в SEQ ID NO: 43, или последовательность, идентичную ей по меньшей мере на 80%.
15. Фармацевтическая композиция для лечения муковисцидоза, содержащая вектор rAAV, причем указанный вектор rAAV содержит (i) капсид, содержащий капсидный белок с SEQ ID NO: 12 или аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 12, и содержащий Thr по аминокислоте 469 и Ala по аминокислоте 598, исходя из нумерации аминокислот, указанной в SEQ ID NO: 12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую CFTR, или биологически активный усеченный белок CFTR, в котором отсутствуют аминокислоты 708-759 последовательности белка CFTR человека, причем указанная нуклеотидная последовательность функционально связана с последовательностью контроля экспрессии.
16. Фармацевтическая композиция по п. 15, где нуклеотидная последовательность, кодирующая белок CFTR, по меньшей мере на 80% идентична нуклеотидной последовательности, представленной как SEQ ID NO: 43.
17. Фармацевтическая композиция по п. 16, где нуклеотидная последовательность, кодирующая CFTR, содержит нуклеотидную последовательность, представленную как SEQ ID NO: 43.
18. Фармацевтическая композиция по любому из пп. 15-17, где последовательность контроля экспрессии содержит универсальный промотор, предпочтительно, где универсальный промотор представляет собой CMV173.
19. Фармацевтическая композиция по любому из пп. 15-18, где композиция составлена для ингаляции или пригодна для ингаляции.
20. Фармацевтическая композиция по любому из пп. 15-19, содержащая от приблизительно 20 до приблизительно 50 мМ цитрата, от приблизительно 85 до приблизительно 125 мМ NaCl и приблизительно 0,005% плюроника F68 и имеющая pH приблизительно 6,0.
21. Фармацевтическая композиция по п. 20, содержащая приблизительно 20 мМ цитрата, приблизительно 125 мМ NaCl и приблизительно 0,005% плюроника F68 и имеющая pH приблизительно 6,0.
22. Способ лечения муковисцидоза у субъекта, включающий введение субъекту фармацевтической композиции по любому из пп. 15-21, где стадия введения включает пульмональное, эндобронхиальное, интраназальное, внутритрахеальное и/или внутрибронхиальное введение.
23. Применение фармацевтической композиции по любому из пп. 15-21 в получении лекарственного средства для лечения муковисцидоза, где лекарственное средство предназначено для пульмонального, эндобронхиального, интраназального, внутритрахеального или внутрибронхиального введения.
24. Композиция, содержащая рекомбинантный вектор на основе аденоассоциированного вируса (rAAV) для лечения заболевания легких путем доставки гетерологичной нуклеиновой кислоты в клетку легкого у субъекта, где указанный вектор rAAV содержит (i) капсид, содержащий капсидный белок, содержащий аминокислотную последовательность, представленную как SEQ ID NO: 12, или аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 12 и содержащую Thr по аминокислоте 469 и Ala по аминокислоте 598, исходя из нумерации аминокислот, представленной в SEQ ID NO: 12, и (ii) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт, где композицию вводят пульмональным, эндобронхиальным, интраназальным, внутритрахеальным или внутрибронхиальным введением.
25. Композиция по п. 24, где композицию используют в лечении муковисцидоза, легочного заболевания или связанного с ним заболевания легких.
26. Композиция по п. 25, где композицию используют в лечении муковисцидоза.
27. Способ по п. 1, где капсидный белок содержит аминокислотную последовательность, по меньшей мере на 96% идентичную SEQ ID NO: 12.
28. Способ по п. 1, где капсидный белок содержит аминокислотную последовательность, по меньшей мере на 97% идентичную SEQ ID NO: 12.
29. Фармацевтическая композиция по п. 15, где капсидный белок содержит аминокислотную последовательность, по меньшей мере на 98% идентичную SEQ ID NO: 12.
30. Фармацевтическая композиция по п. 15, где капсидный белок содержит аминокислотную последовательность, по меньшей мере на 99% идентичную SEQ ID NO: 12.
| US 20160017295 A1, 21.01.2016 | |||
| US 20080003204 A1, 03.01.2008 | |||
| RU 2017131875 A, 14.03.2019. |

