ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ,
ПОДАННЫЙ ЧЕРЕЗ EFS-WEB В ВИДЕ ТЕКСТОВОГО ФАЙЛА
[0001] Перечень последовательностей, записанный в файле 10359WO01_ST25.txt, имеет размер 183 килобайта, создан 27 июня 2018 года и включен в настоящий документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0002] Настоящее изобретение в данном документе в целом относится к рекомбинантным вирусным частицам с модифицированным тропизмом и композициям, содержащим их, которые применимы для нацеленного введения генетического материала в клетки.
ПРЕДПОСЫЛКИ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
[0003] Доставка генов в конкретные клетки-мишени стала одной из важнейших технологий в современной медицине для потенциального лечения множества хронических и генетических заболеваний. До настоящего времени прогресс в клиническом применении генной терапии ограничивался отсутствием идеальных носителей для доставки генов. С целью достижения терапевтического успеха носители для доставки генов должны характеризоваться способностью трансдуцировать клетки-мишени с избежанием при этом трансдукции нецелевых клеток. В частности, если нативный тропизм вируса не достигает желаемых в терапевтическом отношении тканей-мишеней или клеточных типов-мишеней, существует потребность в рекомбинантных вирусных частицах, где природный тропизм устранен или уменьшен и успешно сконструирован желаемый тропизм (Buchholz et al.).
[0004] В последние годы наибольший прогресс в разработке векторов был достигнут при применении вирусов без оболочки (например, вирусов, содержащих капсид, образованный вирусными капсидными белками без оболочки (например, липидный бислой)), таких как аденоассоциированные вирусы (AAV) и аденовирусы (Ad), а также вирусов с оболочкой (например, вирусов, у которых капсид окружен липидным бислоем), таких как ретровирусы, лентивирусы и вирус простого герпеса. Векторы на основе AAV являлись предметом многих исследований, поскольку AAV лишь в незначительной степени иммуногенны и способны к трансдукции широкого спектра видов и тканей in vivo без признаков токсичности.
[0005] AAV представляют собой небольшие однонитевые ДНК-вирусы без оболочки. Геном AAV составляет 4,7 т.п.о. и характеризуется двумя инвертированными концевыми повторами (ITR) и двумя открытыми рамками считывания, которые кодируют белки Rep и белки Сар соответственно. Два ITR являются единственными цис-элементами, необходимыми для репликации и капсидизации AAV. Рамка считывания Rep кодирует четыре белка с молекулярной массой 78 кДа, 68 кДа, 52 кДа и 40 кДа. Эти белки функционируют главным образом при регуляции транскрипции и репликации генома AAV. Рамка считывания Сар кодирует три структурных (капсидных) вирусных белка (VP), имеющих молекулярные массы 83-85 кДа (VP1), 72-73 кДа (VP2) и 61-62 кДа (VP3). Более 80% всех белков в вирионе AAV составляет VP3; в зрелых вирионах VP1, VP2 и VP3 обнаруживаются при относительном содержании примерно 1:1:10. In vitro три белка самопроизвольно собираются в вирионоподобные структуры, например, вирусные капсиды. Таким образом, оказывается, что образование вирусного капсида в инфицированных клетках происходит независимо от синтеза вирусной ДНК (рассмотрено в Kotin et al. (1994) Hum. Gene Ther. 5:793).
[0006] Среди всех известных серотипов AAV AAV2 является,
пожалуй, наиболее хорошо охарактеризованным серотипом, поскольку его инфекционный клон был получен первым (Samulski et al. (1982) Proc. Natl. Acad. Sci. USA, 79:2077-2081). Впоследствии также были определены полные последовательности для нескольких серотипов AAV. (см., например, Rutledge et al. (1998) J. Virol., 72:309-319; Gao et al. (2005) Curr. Gen. Ther. 5(3)285-97; Chiorini et al. (1997) J. Virol., 71:6823-6833; S. Muramatsu et al., (1996) Virol., 221:208-217). Как правило, все AAV обладают более 80% общей идентифицированной нуклеотидной последовательности.
[0007] Вектор на основе AAV является многообещающим вектором для генной терапии человека, поскольку, в отличие от других вирусных векторов, AAV, как было установлено, не связаны с каким-либо известным заболеванием человека и, как правило, не считаются патогенными (Muzyczka, et al. (1992) Current Topics in Microbiology and Immunology, 158:97-129). Более того, AAV безопасно трансдуцирует постмитотические ткани с относительно низкой иммуногенностью, и хотя вирус может иногда интегрироваться в хромосомы хозяина, он очень редко делает это в локусе safe-harbor в хромосоме 19 человека, и только когда белки Rep обеспечены в транс-форме. Геномы AAV быстро циркулируют и конкатемеризируются в инфицированных клетках и существуют в стабильном эписомальном состоянии в инфицированных клетках, обеспечивая долгосрочную стабильную экспрессию своих полезных нагрузок.
[0008] Ряд вирусов, в том числе AAV, инфицируют клетки посредством взаимодействия вирус/лиганд:клетка/рецептор, что в конечном итоге приводит к эндоцитозу вируса инфицированной клеткой. Такое взаимодействие лиганд:рецептор является предметом большей части исследований вирусных векторов, которым можно манипулировать с целью перенаправления природного тропизма вируса от естественным образом пермиссивного состояния клетки к состоянию инфицирования ненативной клетки-мишени вирусом дикого типа, например, посредством рецептора, экспрессируемого клеткой-мишенью.
[0009] Теоретически, перенацеливание вектора на любой белок или маркер клеточной поверхности должно привести к инфекции, поскольку большинство рецепторов клеточной поверхности вовлечены в пути эндоцитоза, либо конститутивно (например, для рециркуляции), либо посредством индукции лигандом (например, опосредованно с помощью рецептора). Эти рецепторы группируются в ямках, окаймленных клатрином, попадают в клетку посредством клатрин-окаймленных везикул, проходят через подкисленную эндосому, в которой происходит сортировка рецепторов, а затем либо рециркулируют к поверхности клетки при последующем внутриклеточном хранении или разлагаются в лизосомах. Таким образом, платформы для перенацеливания вирусных векторов часто направлены на устранение природного тропизма вирусного вектора и перенаправление вирусного вектора на рецептор или маркер, экспрессируемый исключительно или преимущественно клеткой-мишенью. Многие из преимуществ таргетной генной терапии с применением вирусных векторов можно кратко описать как нерекомбинаторную (негенетическую) или рекомбинаторную (генетическую) модификацию вирусного вектора, которая приводит к псевдотипированию, расширению тропизма и/или перенацеливанию природного тропизма вирусного вектора. (Рассмотрено в Nicklin and Baker (2002) Curr. Gene Ther. 2:273-93; Verheiji and Rottier (2012) Advances Virol 2012:1-15).
[0010] Наиболее популярный подход представляет собой рекомбинаторную генетическую модификацию вирусных капсидных белков и, таким образом, поверхности вирусного капсида. С другой стороны, в непрямых рекомбинаторных подходах вирусный капсид модифицируется с помощью гетерологичного «каркаса», который затем соединяется с адаптером. Адаптер связывается и с каркасом, и с клеткой-мишенью. В прямом рекомбинаторном подходе нацеливания нацеливающий лиганд вставляется непосредственно в вирусный капсид или соединяется с ним, т.е. белковые вирусные капсиды модифицируются с обеспечением экспрессии гетерологичного лиганда. Затем лиганд перенаправляет, например, связывает рецептор или маркер, преимущественно или исключительно экспрессируемый на клетке-мишени.
[0011] Каждый из подходов имеет свои преимущества и недостатки. Возможность генетической модификации вируса требует сохранения структуры капсида, и нацеливающий лиганд или каркас должен быть размещен в положении внутри капсидного белка, который будет переносить и соответствующим образом презентировать нацеливающий лиганд или каркас. Например, нацеливающий лиганд или каркас, введенный в вирусный белок, должен будет соответствовать ограничениям по размеру, чтобы не нарушать структуру модифицированного капсида, что обеспечивает вероятность того, что направляющий лиганд или каркас может быть представлен капсидом неправильно и/или возникнут ограничения в отношении спектра существующих в природе молекул, доступных для применения в качестве нацеливающих лигандов или каркасов. Кроме того, применение нацеливающих лигандов, вставленных непосредственно в вирусный капсид, не является модульным и должно подвергаться переконструированию для каждой мишени. Хотя платформа на основе каркаса имеет преимущества в гибкости модульной природе используемого адаптера, каркас на вирусной частице и адаптер взаимодействуют посредством ионных связей и остаются двумя отдельными объектами, и внутренняя нестабильность их взаимодействий может ограничивать их применимость in vivo. Оптимальные значения эффективности трансдукции могут быть трудно достижимыми посредством таких двухкомпонентных систем.
[0012] Очевидно, остается потребность в вирусных векторных системах, которые сохраняют целостность модифицированной вирусной структуры, в то же время оставаясь адаптируемыми для нацеленного переноса представляющих интерес нуклеиновых кислот в различные клетки-мишени.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0013] В данном документе описана стратегия перенацеливания вирусов, которая обеспечивает решение проблем, присущих предыдущим стратегиям перенацеливания, путем применения первого партнера и второго родственного партнера специфической связывающейся пары, причем первый партнер и второй родственный партнер специфически взаимодействуют с образованием химической, предпочтительно ковалентной, связи. Первый партнер, в случае отображения на капсидном белке, действует как каркас для какого-либо нацеливающего лиганда, слитого со вторым родственным партнером, но после связывания первого партнера и второго родственного партнера образуется изопептидная связь, и рекомбинантная вирусная частица действует как однокомпонентный нацеливающий вектор.
[0014] В данном документе предусмотрена рекомбинантная вирусная частица (например, рекомбинантный вирусный капсидный белок, рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, и/или рекомбинантный вирусный вектор, содержащий рекомбинантный вирусный капсид, который инкапсулирует представляющий интерес нуклеотид), которая генетически модифицирована с обеспечением отображения гетерологичной аминокислотной последовательности, содержащей первого партнера специфической связывающейся пары, например, пептидную метку, где аминокислотная последовательность имеет длину менее 50 аминокислот, и где рекомбинантный вирусный белок капсида/частицы проявляет природный тропизм, находящийся в пределах от уменьшенного до устраненного. Тропизм рекомбинантного вирусного капсидного белка/капсида/вектора может быть восстановлен и/или перенаправлен после образования изопептидной связи с родственным вторым партнером специфической связывающейся пары, причем этот родственный второй партнер слит с нацеливающим лигандом, который специфически связывается с клеткой-мишенью. Такое связывание приводит к рекомбинантному вирусному капсидному белку/капсиду/вектору, отображающему нацеливающий лиганд. Значения эффективности и специфичности трансдукции неожиданно повышаются, в случае когда нацеливающий лиганд отображен посредством рекомбинантного вирусного капсида/вектора с помощью линкера и/или в ограниченных количествах на поверхности вирусного капсида. Такие вирусные частицы, композиции, содержащие их, и способы их получения и применения предусмотрены в данном документе.
[0015] Соответственно, в данном документе описан рекомбинантный вирусный капсидный белок, содержащий пептидную метку, функционально связанную (например, ковалентно связанную) с капсидным белком, где вирусный капсидный белок получен с помощью гена капсида вируса, который инфицирует эукариотические клетки, и где пептидная метка является первым партнером специфической связывающейся пары, который образует изопептидную связь со вторым родственным партнером специфической связывающейся пары. В некоторых вариантах осуществления рекомбинантный капсидный белок (который может быть получен с помощью гена капсида вируса, который инфицирует эукариотические клетки, например, представляет собой генетически модифицированный капсидный белок вируса, который инфицирует эуркариотические клетки), описанный в настоящем документе, содержит первого партнера специфической связывающейся пары (т.е. пептидную метку), функционально связанный с капсидным белком, и дополнительно содержит второго родственного партнера специфической связывающейся пары, где первый и второй партнеры специфической связывающейся пары связаны ковалентной (изопептидной) связью, например, капсидный белок содержит первого партнера, функционально связанный с капсидным белком, и дополнительно содержит второго родственного партнера специфической связывающейся пары, ковалентно связанного с первым партнером. В некоторых вариантах осуществления рекомбинантный капсидный белок (который может быть получен с помощью гена капсида вируса, который инфицирует эукариотические клетки, например, представляет собой генетически модифицированный капсидный белок вируса, который инфицирует эуркариотические клетки), описанный в настоящем документе, содержит первого партнера специфической связывающейся пары (т.е. пептидную метку), функционально связанного с капсидным белком, и дополнительно содержит второго родственного партнера специфической связывающейся пары, слитого с нацеливающим лигандом, где первый и второй партнеры специфической связывающейся пары связаны посредством ковалентной (изопептидной) связи, например, капсидный белок содержит первого партнера специфической связывающейся пары, функционально связанного с капсидным белком, и дополнительно содержит второго родственного партнера специфической связывающейся пары, ковалентно связанного с первым партнером, и где второй родственный партнер специфической связывающейся пары функционально связан с нацеливающим лигандом, который специфически связывается с маркером клеточной поверхности (например, олигосахаридом клеточной поверхности, рецептором клеточной поверхности и/или маркером клеточной поверхности и т.д.) на клетке-мишени. Также в данном документе описаны вирусные капсиды, содержащие рекомбинантные вирусные капсидные белки, и вирусные векторы, содержащие нуклеотид, представляющий интерес, заключенные в описанные в данном документе вирусные капсиды. Также описаны композиции, содержащие рекомбинантные вирусные частицы, описанные в данном документе (например, рекомбинантные вирусные капсидные белки, рекомбинантные вирусные капсиды и/или рекомбинантные вирусные векторы), способы их применения для нацеленной доставки нуклеотида, представляющего интерес, и способы их получения.
[0016] В некоторых вариантах осуществления пептидная метка (первый партнер специфической связывающейся пары) функционально связана с капсидным белком (транслируется в рамке считывания с ним, химически присоединена к нему и/или воспроизводится с помощью него) посредством первого или второго линкера, например, аминокислотного спейсера, длина которого составляет по меньшей мере одну аминокислоту. В некоторых вариантах осуществления пептидная метка (первый партнер) фланкирована первым и/или вторым линкером, например, первым и/или вторым аминокислотным спейсером, длина каждого из которых составляет по меньшей мере одну аминокислоту.
[0017] В некоторых вариантах осуществления первый и/или второй линкеры не являются идентичными. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну или две аминокислоты. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две или три аминокислоты. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три или четыре аминокислоты. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре или пять аминокислот. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре или пять аминокислот. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре, пять или шесть аминокислот. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре, пять, шесть или семь аминокислот. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре, пять, шесть, семь или восемь аминокислот. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре, пять, шесть, семь, восемь или девять аминокислот. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре, пять, шесть, семь, восемь, девять или десять аминокислот. В некоторых вариантах осуществления длина каждого из первого и/или второго линкера независимо составляет одну, две, три, четыре, пять, шесть, семь, восемь, девять, десять или больше аминокислот.
[0018] В некоторых вариантах осуществления первый и второй линкеры являются идентичными по последовательности и/или по длине, и длина каждого составляет одну аминокислоту. В некоторых вариантах осуществления первый и второй линкеры являются идентичными по длине, и длина каждого составляет одну аминокислоту. В некоторых вариантах осуществления первый и второй линкеры являются идентичными по длине, и длина каждого составляет две аминокислоты. В некоторых вариантах осуществления первый и второй линкеры являются идентичными по длине, и длина каждого составляет три аминокислоты. В некоторых вариантах осуществления первый и второй линкеры идентичны по длине, и длина каждого составляет четыре аминокислоты, например, линкер представляет собой GLSG (SEQ ID NO: 40). В некоторых вариантах осуществления первый и второй линкеры являются идентичными по длине, и длина каждого составляет пять аминокислот. В некоторых вариантах осуществления первый и второй линкеры идентичны по длине, и длина каждого составляет шесть аминокислот, например, каждый из первого и второго линкеров содержит последовательность GLSGSG (SEQ ID NO: 41). В некоторых вариантах осуществления первый и второй линкеры являются идентичными по длине, и длина каждого составляет семь аминокислот. В некоторых вариантах осуществления первый и второй линкеры идентичны по длине, и длина каждого составляет восемь аминокислот, например, каждый из первого и второго линкеров содержит последовательность GLSGLSGS (SEQ ID NO: 42). В некоторых вариантах осуществления первый и второй линкеры являются идентичными по длине, и длина каждого составляет девять аминокислот. В некоторых вариантах осуществления первый и второй линкеры идентичны по длине, и длина каждого составляет восемь аминокислот, например, каждый из первого и второго линкеров содержит последовательность GLSGLSGLSG (SEQ ID NO: 43) или GLSGGSGLSG (SEQ ID NO: 55). В некоторых вариантах осуществления первый и второй линкеры являются идентичными по длине, и длина каждого составляет более чем десять аминокислот.
[0019] Как правило, длина пептидной метки и необязательно одного или более линкеров, описанных в данном документе, например, пептидной метки самой по себе или в комбинации с одним или более линкерами, составляет от приблизительно 5 аминокислот до приблизительно 50 аминокислот. В некоторых вариантах осуществления длина пептидной метки сама по себе или в комбинации с одним или более линкерами составляет по меньшей мере 5 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 6 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 7 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 8 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 9 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 10 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 11 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 12 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 13 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 14 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 15 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 16 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 17 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 18 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 19 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 20 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 21 аминокислоту. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 22 аминокислоты. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 23 аминокислоты. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 24 аминокислоты. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 25 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 26 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 27 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 28 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 29 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 30 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 31 аминокислоту. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 32 аминокислоты. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 33 аминокислоты. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 34 аминокислоты. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 35 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 36 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 37 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 38 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 39 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет 40 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет более 40 аминокислот. В некоторых вариантах осуществления длина пептидной метки самой по себе или в комбинации с одним или более линкерами составляет не более 50 аминокислот.
[0020] Как правило, рекомбинантные вирусные капсидные белки, описанные в данном документе, могут быть получены с помощью гена капсида вируса без оболочки, например, кодируются геном cap, модифицированным с обеспечением экспрессии генетически модифицированного капсидного белка вируса без оболочки, где вирус без оболочки инфицирует клетки человека, или серотипов вирусов без оболочки, которые, как правило, инфицируют клетки человека, например, аденовируса, аденоассоциированного вируса и т.д. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в данном документе, получен с помощью гена капсида AAV, который кодирует капсидные белки VP1, VP2 и/или VP3 AAV (или части капсидных белков VP1, VP2 и/или VP3) например, кодируется геном cap, модифицированным с обеспечением кодирования генетически модифицированного капсидного белка VP1, VP2 и/или VP3 аденоассоциированного вируса (AAV), например, генетически модифицированного капсидного белка серотипа AAV, который инфицирует человека, выбранного из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2 или AAV9, который соответственно кодирует капсидный белок VP1, VP2 и/или VP3 AAV2 или капсидный белок VP1, VP2 и/или VP3 AAV9, например, кодируется геном cap AAV2 или AAV9, модифицированным с обеспечением кодирования соответственно генетически модифицированного капсидного белка VP1, VP2 и/или VP3 AAV2 или генетически модифицированного капсидного белка VP1, VP2 и/или VP3 AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV6, например, кодируется геном cap AAV6, модифицированным с обеспечением кодирования генетически модифицированного капсидного белка VP1, VP2 и/или VP3 AAV6, аминокислотная последовательность дикого типа капсидного белка VP1 AAV6 которого изложена соответственно под SEQ ID NO: 51. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, кодируется геном cap AAV2, модифицированным с обеспечением кодирования генетически модифицированного капсидного белка VP1, VP2 и/или VP3 AAV2, аминокислотная последовательность дикого типа капсидного белка VP1 AAV2 которого изложена соответственно под SEQ ID NO: 9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, кодируется геном cap AAV9, модифицированным с обеспечением кодирования генетически модифицированного капсидного белка VP1, VP2 и/или VP3 AAV9, аминокислотная последовательность дикого типа капсидного белка VP1 AAV9 которого изложена соответственно под SEQ ID NO: 31.
[0021] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью (кодируется) химерным геном капсида AAV, где химерный ген капсида содержит множество последовательностей нуклеиновой кислоты, где каждая из множества последовательностей нуклеиновой кислоты кодирует часть капсидного белка другого серотипа AAV, и где множество последовательностей нуклеиновых кислот вместе кодирует химерный капсидный белок AAV. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью химерного гена капсида AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью химерного гена капсида AAV6. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью химерного гена капсида AAV9.
[0022] Как правило, рекомбинантный вирусный капсидный белок, описанный в настоящем документе, модифицируют таким образом, чтобы он содержал пептидную метку (первого партнера связывающейся пары белок : белок), функционально связанную, необязательно посредством линкера, с рекомбинантным капсидным белком (например, вставленную в него и/или отображаемую с помощью него), таким образом, что пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер сами по себе обеспечивает уменьшение и/или устранение природного тропизма рекомбинантного капсидного белка или капсида, содержащего его, по сравнению с эталонным капсидным белком, в котором отсутствуют пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер, или капсидом, содержащим эталонный капсидный белок, соответственно. В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) функционально связана, необязательно с помощью линкера, с областью капсидного белка (например, вставлена в нее и/или отображена с помощью нее), участвующей в природном тропизме эталонного капсидного белка дикого типа, например, областью капсидного белка, участвующей в нацеливании на клетку. В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер функционально связаны, необязательно посредством линкера, с доменом «выступа» белка фибера Ad (например, вставлены в него и/или отображены с помощью него). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) функционально связана, необязательно посредством линкера, с HI-петлей белка фибера Ad (например, вставлена в нее и/или отображена с помощью нее). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) функционально связана, необязательно посредством линкера, с открытой вариабельной петлей в капсидном белке AAV (например, вставлена в нее и/или отображена с помощью нее). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) функционально связана, необязательно посредством линкера, с открытой вариабельной петлей в капсидном белке AAV2 (например, вставлена в нее и/или отображена с помощью нее). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) функционально связана, необязательно посредством линкера, с открытой вариабельной петлей в капсидном белке AAV9 (например, вставлена в нее и/или отображена с помощью нее).
[0023] В некоторых вариантах осуществления (i) вирусный капсидный белок получен с помощью гена капсида AAV2, который кодирует капсидный белок VP1, VP2 и/или VP3 AAV2, и пептидная метка функционально связана, необязательно с помощью линкера, с аминокислотой (например, вставленав нее и/или отображена с помощью нее) в положении 1453 или 1587 капсидного белка VP1 AAV2 (или в соответствующих положениях капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2, и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9); (ii) вирусный капсидный белок получен с помощью гена капсида AAV6, и пептидная метка (первый партнер связывающейся пары белок : белок) функционально связана, необязательно с помощью линкера, с аминокислотой (например, вставленав нее и/или отображена с помощью нее) в положении 1585 капсидного белка VP1 AAV6 (или соответствующих положениях капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV7, AAV8 и AAV9); или (iii) вирусный капсидный белок получен с помощью гена капсида AAV9, который кодирует капсидный белок VP1, VP2 и/или VP3 AAV9, и пептидная метка функционально связана, необязательно с помощью линкера, с аминокислотой (например, вставленав нее и/или отображена с помощью нее) в положении 1453 или 1589 капсидного белка VP1 AAV9 (или соответствующих положениях капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8).
[0024] В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) функционально связана, необязательно с помощью линкера, с аминокислотой в положении, выбранном из группы, состоящей из 453 капсидного белка VP1 AAV2, 587 капсидного белка VP1 AAV2, 585 капсидного белка VP1 AAV6, 453 капсидного белка VP1 AAV9 и 589 капсидного белка VP1 AAV9 (или соответствующих положений капсидных белков VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислотных положений капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9), например, слита с С-концом аминокислоты в положении, выбранном из группы, состоящей из 453 капсидного белка VP1 AAV2, 587 капсидного белка VP1 AAV2, 585 капсидного белка VP1 AAV6, 453 капсидного белка VP1 AAV9 и 589 VP1 капсидного белка AAV9 (или соответствующих положений капсидных белков VP2 и/ ли VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер вставлены непосредственно после (например, слиты с С-концом) аминокислоты в положении 453 капсидного белка VP1 AAV2 (или соответствующих положениях в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер вставлены непосредственно после (например, слиты с С-концом) аминокислоты в положении 587 капсидного белка VP1 AAV2 (или соответствующих положениях в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер вставлены непосредственно после (например, слиты с С-концом) аминокислоты в положении 585 капсидного белка VP1 AAV6 (или соответствующих положениях в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV7, AAV8 и AAV9). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер вставлены непосредственно после (например, слиты с С-концом) аминокислоты в положении 453 капсидного белка VP1 AAV9 (или соответствующих положениях в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер вставлены непосредственно после (например, слиты с С-концом) аминокислоты в положении 589 капсидного белка VP1 AAV9 (или соответствующих положениях в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида, или соответствующих аминокислот капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8). В некоторых вариантах осуществления пептидная метка (первый партнер связывающейся пары белок : белок) и необязательный линкер вставлены и/или отображены между положениями 587 и 588 капсидного белка VP1 AAV2 (или соответствующими положениями в капсидных белках VP2 и/или VP3, кодируемыми тем же геном капсида, или соответствующими аминокислотами капсидных белков VP1, VP2 и/или VP3 из другого AAV, который инфицирует человека, например, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8).
[0025] В некоторых вариантах осуществления рекомбинантный капсидный белок, описанный в данном документе, содержит (вторую и отличную) мутацию, которая может являться дополнением к пептидной метке (первому партнеру связывающейся пары белок : белок) и необязательному линкеру. В некоторых вариантах осуществления (вторая и отличная мутация) предусматривает вставку гетерологичного пептида в капсидный белок, замену одной или более аминокислот капсидного белка одной или несколькими гетерологичными аминокислотами, делецию одной или более аминокислот из капсидного белка или их комбинацию. Например, в некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в настоящем документе, может быть получен с помощью гена капсида AAV2 (например, является генетически модифицированным капсидным белком VP1, VP2 и/или VP3 AAV2), содержит пептидную метку (первого партнера связывающейся пары белок : белок) и необязательный линкер и может дополнительно содержать мутацию, например, мутацию R585A и/или R588A, в капсидном белке VP1 AAV2 (или соответствующую мутацию в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида AAV2). В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV2, содержит пептидную метку (первого партнера связывающейся пары белок : белок) и необязательный линкер, вставленные непосредственно после (например, слиты с С-концом) аминокислоты в положении 453 белка VP1 AAV2 (или аминокислот в соответствующих положениях капсидных белков VP2 и/или VP3 AAV2, кодируемых геном капсида AAV2) и дополнительно содержит мутацию, выбранную из группы, состоящей из R585A и/или R588A (или соответствующих мутаций в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида AAV2). В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2, например, представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV2, содержит пептидную метку (первого партнера связывающейся пары белок : белок) и необязательный линкер, вставленные непосредственно после (например, слиты с С-концом) аминокислоты в положении 587 капсидного белка VP1 AAV2 (или аминокислот в соответствующих положениях капсидных белков VP2 и/или VP3 AAV2, кодируемых тем же геном капсида AAV2), и дополнительно содержит мутацию, выбранную из группы, состоящей из R585A, R588A, и/или соответствующих мутаций в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида AAV2.
[0026] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV9, содержит пептидную метку (первого партнера связывающейся пары белок : белок) и необязательный линкер, вставленные непосредственно после (например, слиты с С-концом) аминокислоты в положении 453 белка VP1 AAV9 (или аминокислоты в соответствующих положениях капсидных белков VP2 или VP3 AAV9, кодируемых тем же геном капсида AAV9), и дополнительно содержит мутацию W503A (или соответствующую мутацию в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида AAV2). В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV9, например, представляет собой генетически модифицированный капсидный белок VP1, VP2 и/или VP3 AAV9, содержит пептидную метку (первого партнера связывающейся пары белок : белок) и необязательный линкер, вставленные непосредственно после (например, слиты с С-концом) аминокислоты в положении 589 белка VP1 AAV9 (или аминокислот в соответствующих положениях капсидных белков VP2 и/или VP3 AAV9, кодируемых тем же геном капсида AAV9) и дополнительно содержит мутацию W503A (или соответствующую мутацию в капсидных белках VP2 и/или VP3, кодируемых тем же геном капсида AAV2).
[0027] В некоторых вариантах осуществления связывающаяся пара белок : белок может быть выбрана из группы, состоящей из SpyTag:SpyCatcher, SpyTag002:SpyCatcher002, SpyTag:KTag, Isopeptag:пилин-C и SnoopTag:SnoopCatcher. В некоторых вариантах осуществления, где пептидная метка (первый партнер) представляет собой SpyTag (или его биологически активную часть), белок (второй родственный партнер) представляет собой SpyCatcher (или его биологически активную часть). В некоторых вариантах осуществления, где пептидная метка (первый партнер) представляет собой SpyTag (или его биологически активную часть), белок (второй родственный партнер) представляет собой KTag (или его биологически активную часть). В некоторых вариантах осуществления, где пептидная метка (первый партнер) представляет собой KTag (или его биологически активную часть), белок (второй родственный партнер) представляет собой SpyTag (или его биологически активную часть). В некоторых вариантах осуществления, где пептидная метка (первый партнер) представляет собой SnoopTag (или его биологически активную часть), белок (второй родственный партнер) представляет собой SnoopCatcher (или его биологически активную часть). В некоторых вариантах осуществления, где пептидная метка (первый партнер) представляет собой Isopeptag (или его биологически активную часть), белок (второй родственный партнер) представляет собой пилин-С (или его биологически активную часть). В некоторых вариантах осуществления, где пептидная метка (первый партнер) представляет собой SpyTag002 (или его биологически активную часть), белок (второй родственный партнер) представляет собой SpyCatcher002 (или его биологически активную часть).
[0028] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок содержит SpyTag. В некоторых вариантах осуществления рекомбинантный вирусный капсид, рекомбинантный вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид или вирусный вектор, содержат аминокислотную последовательность, изложенную под любым из SEQ ID NO, перечисленных в таблице 1, в качестве аминокислотной последовательности рекомбинантного вирусного капсидного белка. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 13. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 15. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 17. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 19. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 21. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 23. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 25. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 27. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 29. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантныи вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 35. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 37. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 39.
[0029] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в данном документе, содержит первого партнера специфической связывающейся пары (например, пептидную метку), ковалентно связанный со вторым родственным белковым партнером специфической связывающейся пары. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в данном документе, содержит пептидную метку (первого партнера), ковалентно связанную с адаптерным полипептидом, содержащим родственный белок (второго партнера), функционально связанный с нацеливающим лигандом. В некоторых вариантах осуществления нацеливающий лиганд функционально связан с белком (вторым партнером), например, слит с белком, необязательно с помощью линкера. Обычно нацеливающий лиганд может представлять собой связывающий фрагмент, например, природный лиганд, антитело, полиспецифическую связывающую молекулу и т.д. В некоторых вариантах осуществления нацеливающий лиганд представляет собой антитело или его часть. В некоторых вариантах осуществления нацеливающий лиганд представляет собой антитело, содержащее вариабельный домен, который связывает белок клеточной поверхности на клетке-мишени, и константный домен тяжелой цепи. В некоторых вариантах осуществления нацеливающий лиганд представляет собой антитело, содержащее вариабельный домен, который связывает белок клеточной поверхности на клетке-мишени, и константный домен тяжелой цепи IgG. В некоторых вариантах осуществления нацеливающий лиганд представляет собой антитело, содержащее вариабельный домен, который связывает белок клеточной поверхности на клетке-мишени, и константный домен тяжелой цепи IgG, где константный домен тяжелой цепи IgG функционально связан, например, посредством линкера, с белком (например, вторым партнером связывающейся пары белок : белок), который образует изопептидную ковалентную связь с пептидной меткой. В некоторых вариантах осуществления рекомбинантный капсидный белок, описанный в данном документе, содержит SpyTag, функционально связанный с вирусным капсидным белком, и ковалентно связанный с SpyTag адаптерный полипептид, содержащий SpyCatcher, связанный с нацеливающим лигандом, содержащим вариабельный домен антитела и домен тяжелой цепи IgG, где SpyCatcher и домен тяжелой цепи IgG связаны посредством аминокислотного линкера, например, GSGESG (SEQ ID NO: 48). В некоторых вариантах осуществления адаптерный полипептид содержит последовательность, изложенную под SEQ ID NO: 46, которая содержит часть тяжелой цепи человеческого IgG4, при этом указанная часть IgG4 имеет последовательность, изложенную под SEQ ID NO: 49, связанную посредством линкера (SEQ ID NO: 48) с SpyCatcher (SEQ ID NO: 3).
[0030] В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат SpyTag, ковалентно связанный с SpyCatcher, слитым с нацеливающим лигандом. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под любым из SEQ ID NO, перечисленных в таблице 1, в качестве кодирующей рекомбинантный вирусный капсидный белок, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат рекомбинантный вирусный капсид, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 13, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 15, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 17, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 19, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 21, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 23, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 25, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 27, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 29, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 35, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 37, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 39, и полипептидный адаптер, содержащий аминокислотную последовательность, изложенную под SEQ ID NO: 46.
[0031] Как правило, нацеливающий лиганд специфически связывается с молекулой клеточной поверхности, например олигосахарид, рецептор, маркер клеточной поверхности и т.д., экспрессируемую на поверхности эукариотической клетки млекопитающего (например, человека), например, клетки-мишени. В некоторых вариантах осуществления нацеливающий лиганд связывается с клеткой печени (человека), клеткой мозга (человека), Т-клеткой (человека), клеткой почки (человека), клеткой кишечника (человека), клеткой легкого (человека), раковой клеткой (человека) или клеткой (человека), инфицированной гетерологичным патогеном.
[0032] В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым клеткой печени (человека), например, асиалогликопротеиновым рецептором, например, hASGR1. В некоторых вариантах осуществления нацеливающий лиганд связывается с молекулой, экспрессируемой нейронной клеткой (человека), например, GABA, трансферрином и т.д. В некоторых вариантах осуществления нацеливающий лиганд связывается с молекулой, экспрессируемую Т-клеткой (человека), например, CD3, например, CD3ε. В некоторых вариантах осуществления нацеливающий лиганд связывается с CD63. В некоторых вариантах осуществления нацеливающий лиганд связывается с молекулой, экспрессируемой гемопоэтической стволовой клеткой (человека), например, CD34. В некоторых вариантах осуществления нацеливающий лиганд связывается с молекулой, экспрессируемой клеткой почки (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с молекулой, экспрессируемой мышечной клеткой (человека), например, интегрином. В некоторых вариантах осуществления нацеливающий лиганд связывается с молекулой, экспрессируемой раковой клеткой (человека), например, опухолеассоциированным антигеном, например, адипофилином, AIM-2, ALDH1A1, альфа-актинином-4, альфа-фетопротеином («AFP»), ARTC1, B-RAF, BAGE-1, BCLX (L), слитым белком BCR-ABL b3a2, бета-катенином, BING-4, СА-125, CALCA, карциноэмбриональным антигеном («СЕА»), CASP-5, CASP-8, CD274, CD45, Cdc27, CDK12, CDK4, CDKN2A, СЕА, CLPP, СОА-1, CPSF, CSNK1A1, CTAG1, CTAG2, циклином D1, Cyclin-A1, слитым белком dek-can, DKK1, EFTUD2, фактором элонгации 2, ENAH (hMena), Ер-САМ, ЕрСАМ, ЕрпА3, эпителиальным опухолевым антигеном («ЕТА»), слитым белком ETV6-AML1, EZH2, Е6, Е7, FGF5, FLT3-ITD, FN1, G250/MN/CAIX, GAGE-1,2,8, GAGE-3,4,5,6,7, GAS7, глипиканом-3, GnTV, gp100/Pme117, GPNMB, HAUS3, гепсином, HER-2/neu, HERV-K-MEL, HLA-A11, HLA-A2, HLA-DOB, hsp70-2, ID01, IGF2B3, IL13Ralpha2, кишечной карбоксилэстеразой, K-ras, Kallikrein 4, KIF20A, KK-LC-1, KKLC1, KM-HN-1, KMHN1 также известным как CCDC110, LAGE-1, слитым белком LDLR-фукозилтрансфераза-AS, ленгсином, M-CSF, MAGE-A1, MAGE-A10, MAGE-A12, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A6, MAGE-A9, MAGE-C1, MAGE-C2, маликферментом, маммаглобином-А, MART2, MATN, MC1R, MCSP, mdm-2, ME1, Melan-A/MART-1, Meloe, Midkine, MMP-2, MMP-7, MUC1, MUC5AC, муцином,MUM-1, MUM-2, MUM-3, миозином, миозином I класса, N-raw, NA88-A, нео-РАР, NFYC, NY-BR-1, NY-ESO-1/LAGE-2, ОА1, OGT, OS -9, полипептидом Р, р53, РАР, РАХ5, PBF, слитым белком pml-RAR-альфа, полиморфным эпителиальным муцином («РЕМ»), PPP1R3B, PRAME, PRDX5, PSA, PSMA, PTPRK, RAB38/NY-MEL-1, RAGE-1, RBAF600, RGS5, RhoC, RNF43, RU2AS, SAGE, сецернином 1, SIRT2, SNRPD1, SOX10, Sp17, SPA17, SSX-2, SSX-4, STEAP1, сурвивином, слитым белком SYT-SSX1 или -SSX2, TAG-1, TAG-2, теломеразой, TGF-бетаRII, TPBG, TRAG-3, триозофосфатизомеразой, TRP-1/gp75, TRP-2, TRP2-INT2, тирозиназой, тирозиназой («TYR»), VEGF, WT1, XAGE-1b/GAGED2a, Kras, NY-ESO1, MAGE-A3, HPV E2, HPV E6, HPV E7, антигеном WT-1 (при лимфомах и других солидных опухолях), рецепторами ErbB, Melan A [MART1], gp 100, тирозиназой, TRP-1/gp 75 и TRP-2 (при меланоме); MAGE-1 и MAGE-3 (при раке мочевого пузыря, головы и шеи, а также немелкоклеточной карциноме); белками EG и Е7 HPV (при раке шейки матки); муцином [MUC-1] (при раке молочной железы, поджелудочной железы, толстой кишки и простаты); простатоспецифическим антигеном [PSA] (при раке предстательной железы); карциноэмбриональным антигеном [СЕА] (при раке толстой кишки, молочной железы и желудочно-кишечного тракта) и такими общими опухолеспецифическими антигенами, как MAGE-2, MAGE-4, MAGE-6, MAGE-10, MAGE-12, BAGE-1, CAGE-1,2,8, CAGE-3-7, LAGE-1, NY-ESO-1/LAGE-2, NA-88, GnTV, TRP2-INT2 и др. В некоторых вариантах осуществления нацеливающий лиганд связывается с Е6 и/или Е7. В некоторых вариантах осуществления нацеливающий лиганд связывается с Her2. В некоторых вариантах осуществления нацеливающий лиганд связывается с CD63. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором глюкагона человека (hGCGR). В некоторых вариантах осуществления перенацеливающий лиганд связывается с эктонуклеозидтрифосфатдифосфогидролазой 3 человека (hENTPD3).
[0033] Как правило, вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, не способен инфицировать клетку-мишень в отсутствие нацеливающего лиганда, например, второго партнера, функционально связанного с нацеливающим лигандом. Как правило, в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в настоящем документе, характеризуется природным тропизмом, находящимся в пределах от уменьшенного до устраненного, например, характеризуется уменьшенной способностью или отсутствием способности нацеливаться и связываться с эталонной клеткой, естественным образом пермиссивной для трансдукции, по сравнению со способностью эталонного вирусного капсида, например, капсида, содержащего эталонный вирусный капсидный белок, например, контрольный вирусный капсидный белок дикого типа или вирусный капсидный белок, который был бы идентичен рекомбинантному вирусному капсидному белку, кроме отсутствия нацеливающего лиганда и необязательно одного или обоих партнеров связывающейся пары белок : белок. В некоторых вариантах осуществления эффективность трансдукции рекомбинантного вирусного капсидного белка, содержащего SpyTag, уменьшена или устранена по сравнению с контрольным вирусным капсидным белком дикого типа.
[0034] В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 10% по сравнению с соответствующим контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 20% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 30% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 40% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 50% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 60% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 70% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 75% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 80% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 85% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 90% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 95% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, демонстрирует снижение эффективности трансдукции на по меньшей мере 99% по сравнению с контрольным вирусным капсидом дикого типа, например, представленным в таблице 1. В некоторых вариантах осуществления и в отсутствие подходящего нацеливающего лиганда трансдукция контрольной клетки вирусным капсидом, содержащим рекомбинантный вирусный капсидный белок, описанный в настоящем документе, устранена, например, не обнаруживается, например, с помощью способов измерения экспрессии представляющего интерес нуклеотида, например, с помощью репортерных анализов и т.д.
[0035] Наоборот, вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, содержащий пептидную метку, ковалентно связанную с соответствующим адаптерным полипептидом, например, родственным белком, функционально связанным с нацеливающим лигандом, способен инфицировать клетку-мишень, например, обладает частично или полностью восстановленной способностью нацеливаться и связываться с эталонной клеткой, естественным образом пермиссивной для трансдукции, по сравнению со способностью эталонного вирусного капсида, например, капсида, содержащего эталонный вирусный капсидный белок, например, контрольный вирусный капсидный белок дикого типа. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 10% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 20% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 30% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 40% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 50% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 60% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 70% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 75% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 80% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 85% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 90% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 95% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая составляет по меньшей мере 99% от эффективности трансдукции соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, идентичную таковой соответствующего контрольного вирусного капсида дикого типа, например, приведенного в таблице 1.
[0036] Аналогично, вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, содержащий пептидную метку, ковалентно связанную с соответствующим адаптерным полипептидом, например, родственным белком, функционально связанным с нацеливающим лигандом, способен инфицировать клетку-мишень, например, обладает повышенной способностью нацеливаться и связываться с эталонной клеткой, естественным образом пермиссивной для трансдукции, по сравнению с таковой эталонного вирусного капсида, который идентичен рекомбинантному вирусному капсидному белку, кроме отсутствия в нем одного или обоих партнеров связывающейся пары белок : белок, например, содержит эталонный капсидный белок. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 10% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 20% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 30% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 40% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с соответствующим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 50% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 60% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 70% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 75% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 80% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 85% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 90% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 95% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1. В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, например, приведенный в таблице 1, ковалентно связанный с подходящим адаптерным полипептидом, демонстрирует эффективность трансдукции, которая на 99% превышает эффективность трансдукции соответствующего контрольного эталонного вирусного капсида, например, приведенного в таблице 1.
[0037] В некоторых вариантах осуществления вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, представляет собой мозаичный капсид, например, содержит по меньшей мере два набора белков VP1, VP2 и/или VP3, каждый набор из которых кодируется различным геном cap, например, содержит рекомбинантный вирусный капсидный белок, содержащий пептидную метку, и эталонный капсидный белок, который не содержит пептидной метки, в определенном соотношении. В некоторых вариантах осуществления эталонный капсидный белок представляет собой эталонный капсидный белок дикого типа в том смысле, что он содержит аминокислотную последовательность капсидного белка дикого типа, имеющего тот же серотип, что и рекомбинантный вирусный капсидный белок. В некоторых вариантах осуществления эталонный капсидный белок представляет собой контрольный эталонный капсидный белок в том смысле, что он содержит аминокислотную последовательность рекомбинантного вирусного капсидного белка, кроме того, что контрольный эталонный капсидный белок не содержит пептидной метки. В некоторых вариантах осуществления эталонный капсидный белок представляет собой мутированный эталонный белок дикого типа в том смысле, что он содержит аминокислотную последовательность, по существу идентичную последовательности капсидного белка дикого типа, имеющего такой же серотип, что и рекомбинантный вирусный капсидный белок, но содержит мутацию (например, делецию аминокислотной последовательности, вставку аминокислотной последовательности, химеризацию и т.д.), которая обеспечивает уменьшение тропизма капсидного белка дикого типа. В некоторых вариантах осуществления композиция, описанная в настоящем документе, содержит, или в способе, описанном в настоящем документе, объединяют рекомбинантный вирусный капсидный белок и эталонный капсидный белок в соотношении, которое находится в диапазоне от 1:1 до 1:15. В некоторых вариантах осуществления соотношение составляет 1:2. В некоторых вариантах осуществления соотношение составляет 1:3. В некоторых вариантах осуществления соотношение составляет 1:4. В некоторых вариантах осуществления соотношение составляет 1:5. В некоторых вариантах осуществления соотношение составляет 1:6. В некоторых вариантах осуществления соотношение составляет 1:7. В некоторых вариантах осуществления соотношение составляет 1:8. В некоторых вариантах осуществления соотношение составляет 1:9. В некоторых вариантах осуществления соотношение составляет 1:10. В некоторых вариантах осуществления соотношение составляет 1:11. В некоторых вариантах осуществления соотношение составляет 1:12. В некоторых вариантах осуществления соотношение составляет 1:13. В некоторых вариантах осуществления соотношение составляет 1:14. В некоторых вариантах осуществления соотношение составляет 1:15.
[0038] В некоторых вариантах осуществления композиция, описанная в настоящем документе, содержит, или в способе, описанном в настоящем документе, объединяют рекомбинантный вирусный капсидный белок, приведенный в таблице 1, и его соответствующий эталонный капсидный белок (или комбинацию его эталонных капсидных белков), также приведенный в таблице 1, в соотношении (рекомбинантный капсидный белок: эталонный(-ые) капсидный(-ые) белок(белки)), которое находится в диапазоне от 1:1 до 1:15. В некоторых вариантах осуществления соотношение составляет 1:2. В некоторых вариантах осуществления соотношение составляет 1:3. В некоторых вариантах осуществления соотношение составляет 1:4. В некоторых вариантах осуществления соотношение составляет 1:5. В некоторых вариантах осуществления соотношение составляет 1:6. В некоторых вариантах осуществления соотношение составляет 1:7. В некоторых вариантах осуществления соотношение составляет 1:8. В некоторых вариантах осуществления соотношение составляет 1:9. В некоторых вариантах осуществления соотношение составляет 1:10. В некоторых вариантах осуществления соотношение составляет 1:11. В некоторых вариантах осуществления соотношение составляет 1:12. В некоторых вариантах осуществления соотношение составляет 1:13. В некоторых вариантах осуществления соотношение составляет 1:14. В некоторых вариантах осуществления соотношение составляет 1:15.
[0039] В таблице 1 приведены идентификационные номера последовательностей (SEQ ID NO), под которыми изложены аминокислотные последовательности (1) иллюстративных и неограничивающих рекомбинантных вирусных капсидных белков, содержащих описанную в данном документе пептидную метку, (2) иллюстративных и неограничивающих примеров соответствующих контрольных (С) вирусных капсидных белков дикого типа, которые необязательно можно применять в качестве эталонных для определения уменьшения или устранения эффективности трансдукции рекомбинантного капсидного белка, содержащего ковалентную белковую метку в отсутствие нацеливающего вектора, и (3) иллюстративных и неограничивающих примеров соответствующих эталонных вирусных капсидных белков для получения мозаичных капсидов и/или применения их в качестве эталонных для определения восстановления эффективности трансдукции рекомбинантного капсидного белка, содержащего связывающуюся пару белок : белок и нацеливающий лиганд.

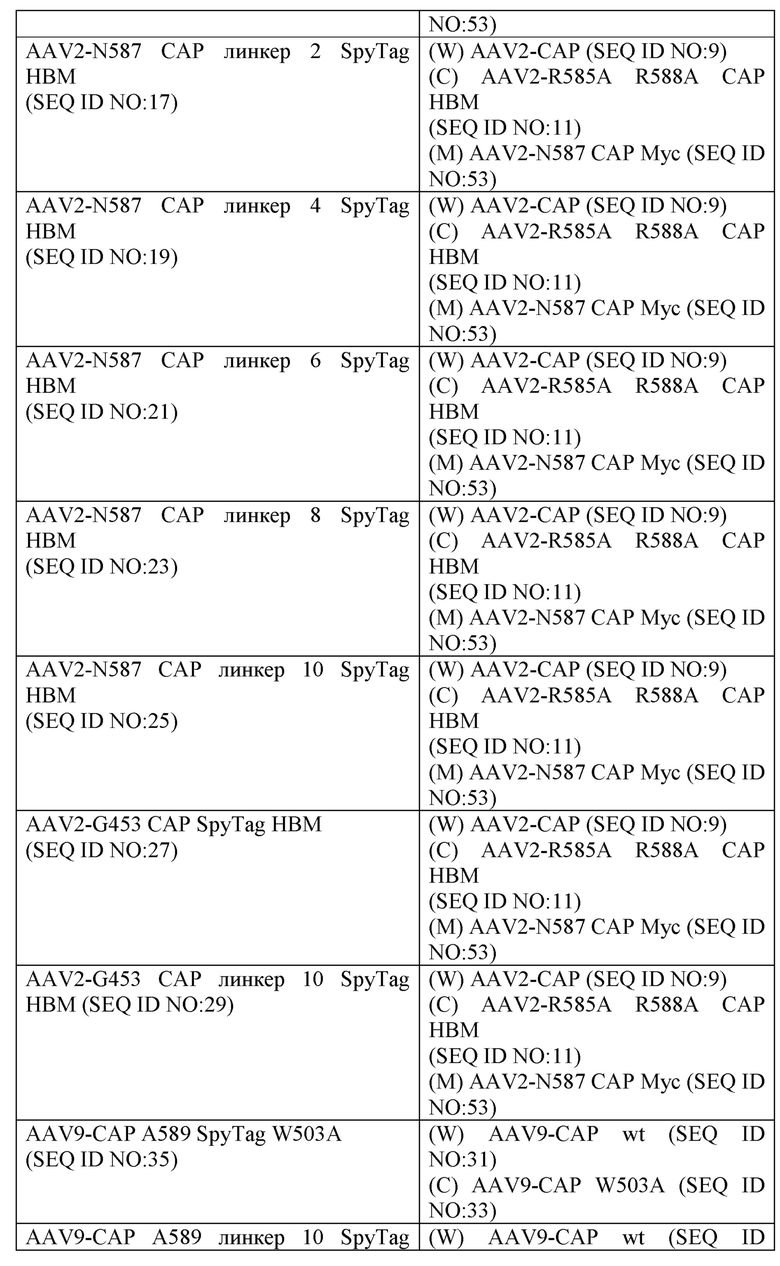

[0040] Как правило, рекомбинантные вирусные векторы, описанные в настоящем документе, содержат вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, в том числе мозаичные вирусные капсиды, где вирусный капсид инкапсулирует представляющий интерес нуклеотид. В некоторых вариантах осуществления представляющий интерес нуклеотид находится под контролем промотора, выбранного из группы, состоящей из вирусного промотора, бактериального промотора, промотора млекопитающего, промотора птицы, промотора рыбы, промотора насекомого и любой их комбинации. В некоторых вариантах осуществления представляющий интерес нуклеотид находится под контролем промотора, отличного от человеческого. В некоторых вариантах осуществления промотор представляет собой промотор цитомегаловируса (CMV). В некоторых вариантах осуществления промотор представляет собой промотор EF1α. В некоторых вариантах осуществления промотор представляет собой промотор CAGG. В некоторых вариантах осуществления промотор представляет собой промотор убиквитина С (UbC).
[0041] Как правило, представляющий интерес нуклеотид может представлять собой один или более генов, которые могут кодировать детектируемый маркер, например, репортер, или терапевтический полипептид. В некоторых вариантах осуществления представляющий интерес нуклеотид представляет собой репортерный ген. В некоторых вариантах осуществления представляющий интерес нуклеотид представляет собой репортерный ген, который кодирует детектируемый маркер, выбранный из группы, состоящей из зеленого флуоресцентного белка, люциферазы, β-галактозидазы и т.д. В некоторых вариантах осуществления детектируемый маркер представляет собой зеленый флуоресцентный белок. В других вариантах осуществления представляющий интерес нуклеотид выбран из группы, состоящей из суицидального гена, нуклеотида, кодирующего антитело или его фрагмент, нуклеотида, кодирующего систему CRISPR/Cas или ее часть(части), нуклеотида, кодирующего антисмысловую РНК, нуклеотида, кодирующего siRNA, секретируемый фермент, ген, кодирующий терапевтический белок, и т.д. В одном варианте осуществления представляющий интерес нуклеотид кодирует многодоменное терапевтическое средство, например, белок, который содержит по меньшей мере два домена, обеспечивающих две разные функции.
[0042] Композиции, описанные в данном документе, как правило, содержат вирусный вектор, который содержит рекомбинантный вирусный капсидный белок, описанный в данном документе, например, содержит капсид, содержащий рекомбинантный вирусный капсидный белок (в том числе мозаичный капсид), где капсид инкапсулирует представляющий интерес нуклеотид. В некоторых вариантах осуществления композиция, описанная в настоящем документе, содержит (1) вирусный вектор, имеющий капсид, содержащий рекомбинантный вирусный капсидный белок, описанный в данном документе, и (2)) фармацевтически приемлемый носитель.
[0043] Также в данном документе описаны способы получения и применение рекомбинантных вирусных капсидных белков, вирусных векторов, содержащих их, композиций и т.д. В некоторых вариантах осуществления способы перенаправления вируса, например, аденовируса, аденоассоциированного вируса и т.д., доставка диагностической/терапевтической нагрузки в клетку-мишень и т.д. предусматривают приведение в контакт клетки-мишени (которая может пребывать in vitro или in vivo, например, у человека) с рекомбинантным вирусным вектором, содержащим рекомбинантный вирусный капсидный белок, описанный в настоящем документе, где вирусный капсид или вирусный вектор содержит нацеливающий лиганд, который специфически связывает белок, экспрессируемый на поверхности клетки-мишени. Такие способы могут включать в качестве первой стадии получение рекомбинантного вирусного вектора, например, культивирование упаковывающей клетки в условиях, достаточных для продуцирования вирусных векторов, где упаковывающая клетка содержит плазмиду, кодирующую капсидный белок, содержащий пептидную метку (первого партнера), в отсутствие или в присутствии плазмиды, кодирующей эталонный капсидный белок, инкубирование рекомбинантного капсидного белка со вторым родственным партнером, функционально связанным с нацеливающим лигандом, и т.д. В некоторых вариантах осуществления клетка-мишень представляет собой клетку печени (человека), и (мозаичный) рекомбинантный вирусный вектор содержит нацеливающий лиганд, который специфически связывается с асиалогликопротеиновым рецептором, например, (h)ASGR1. В некоторых вариантах осуществления клетка-мишень представляет собой нейронную клетку (человека), и (мозаичный) рекомбинантный вирусный вектор содержит нацеливающий лиганд, который специфически связывается с GABA, рецептором трансферрина и т.д. В некоторых вариантах осуществления клетка-мишень представляет собой Т-клетку (человека), и (мозаичный) рекомбинантный вирусный вектор содержит нацеливающий лиганд, который специфически связывается с CD3, например, CD3ε. В некоторых вариантах осуществления клетка-мишень представляет собой гемопоэтическую стволовую клетку (человека), и (мозаичный) рекомбинантный вирусный вектор содержит нацеливающий лиганд, который специфически связывается с CD34. В некоторых вариантах осуществления клетка-мишень представляет собой клетку почки (человека). В некоторых вариантах осуществления клетка-мишень представляет собой мышечную клетку (человека), и (мозаичный) рекомбинантный вирусный вектор содержит нацеливающий лиганд, который специфически связывается с интегрином. В некоторых вариантах осуществления клетка-мишень представляет собой раковую клетку (человека), и (мозаичный) рекомбинантный вирусный вектор содержит нацеливающий лиганд, который специфически связывается с опухолеассоциированным антигеном, например, Е6 и Е7, Her2 и т.д. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором глюкагона человека (hGCGR).
[0044] В данном документе также описаны способы инактивации вирусного капсида и/или получения вирусных векторов, при этом способы, как правило, предусматривают (а) осуществление вставки нуклеиновой кислоты, кодирующей гетерологичный белок, в последовательность нуклеиновой кислоты, кодирующую вирусный капсидный белок, с образованием нуклеотидной последовательности, кодирующей генетически модифицированный капсидный белок, содержащий пептидную метку (в отсутствие или в присутствии плазмиды, кодирующей эталонный капсидный белок), и/или (b) культивирование упаковывающей клетки в условиях, достаточных для продуцирования вирусных векторов, где упаковывающая клетка содержит нуклеотидную последовательность. В некоторых вариантах осуществления упаковывающая клетка дополнительно содержит хелперную плазмиду и/или плазмиду для переноса, содержащую представляющий интерес нуклеотид. В некоторых вариантах осуществления способы дополнительно предусматривают выделение самокомплементарных векторов на основе аденоассоциированного вируса из культурального супернатанта. В некоторых вариантах осуществления способы дополнительно предусматривают лизирование упаковывающей клетки и выделение однонитевых векторов на основе аденоассоциированного вируса из клеточного лизата. В некоторых вариантах осуществления способы дополнительно предусматривают (а) удаление клеточного дебриса, (b) обработку супернатанта, содержащего вирусные векторы, нуклеазами, например, ДНКазой I, в присутствии MgCl2, (с) концентрирование вирусных векторов, (d) очистку вирусных векторов и (е) любую комбинацию (a)-(d). В данном документе также предусмотрены вирусный вектор, полученный в соответствии со способом, описанным в данном документе, и упаковывающая клетка, применимая для получения вирусного вектора, описанного в настоящем документе, например, упаковывающие клетки, содержащие плазмиду, кодирующую описанный рекомбинантный капсидный белок.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0045] Файл патента или заявки содержит по меньшей мере один графический материал, выполненный в цвете. Копии данного патента или публикации заявки на патент с цветным(-и) графическим(-и) материалом(-и) будут предоставлены Ведомству после запроса и уплаты необходимой пошлины.
[0046] На фигуре 1 представлены диаграммы рассеяния, полученные в ходе сортировки флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) HER2-положительными (+) 293 hErbB2 или HER2-отрицательными (-) исходными клетками 293, либо «неинфицированными», либо клетками, инфицированными частицами «N587 AAV2-SpyTag», или клетками, инфицированными частицами «N587 AAV2-SpyTag + С6.5-SpyC». Капсиды обоих вирусов содержат следующие мутации: R585A, delR588 и вставку пептида SpyTag (SEQ ID NO: 1) непосредственно после остатка N587 (SEQ ID NO: 13). Частицы C6.5-SpyC были конъюгированы с scFv к HER2 (С6.5), слитым с SpyCatcher (SEQ ID NO: 3) посредством SpyTag. Вирусы экспрессируют GFP как маркер трансдукции.
[0047] На фигуре 2 представлены диаграммы рассеяния, полученные в ходе сортировки флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) HER2-положительными (+) 293 hErbB2 или HER2-отрицательными (-) исходными клетками 293, либо «неинфицированными», либо клетками, инфицированными частицами «N587 AAV2-SpyTag», или клетками, инфицированными частицами «N587 AAV2-SpyTag + SpyC-антитело-к-HER2». Капсиды обоих вирусов содержат следующие мутации: R585A, delR588 и вставку пептида SpyTag (SEQ ID NO: 1) непосредственно после остатка N587 (SEQ ID NO: 13). Частицы SpyC-антитело-к-HER2 были конъюгированы с антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3) посредством SpyTag. Вирусы экспрессируют GFP как маркер трансдукции.
[0048] На фигуре 3 представлены диаграммы рассеяния, полученные в ходе сортировки флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) HER2-положительными (+) 293 hErbB2 или HER2-отрицательными (-) исходными клетками 293, инфицированными частицами «AAV2 дикого типа», или клетками, инфицированными частицами «G453 AAV2-SpyTag + SpyC-антитело-к-HER2». Капсиды «AAV2 дикого типа» не имеют мутаций или модификаций (SEQ ID NO: 9), тогда как капсид «G453 AAV2-SpyTag» представляет собой мозаичную вирусную частицу, состоящую, при соотношении 1:7, капсидных белков «SpyTag», где SpyTag вставлен непосредственно после остатка G453, фланкированного линкером из 10 аминокислот (SEQ ID NO: 29), и капсидов «AAV2 НВМ» без SpyTag, но содержащих мутации R585A и R588A (SEQ ID NO: 11). Мозаичные частицы G453 AAV2-SpyTag были конъюгированы с антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3) посредством SpyTag. Вирусы экспрессируют GFP как маркер трансдукции.
[0049] На фигуре 4А представлен вестерн-блоттинг с применением антитела В1, которое распознает линейный эпитоп, общий для капсидных белков VP1, VP2 и VP3 AAV2, с анализом реакции между scFv к HER2, слитым с SpyCatcher, «SpyC-scFv-к-Her2», и панелью вирусных частиц AAV2, состоящей из капсидов со следующими мутациями: R585A, delR588 и вставкой пептида SpyTag непосредственно после остатка N587, фланкированного аминокислотными линкерами различной длины (линкер 1, линкер 2, линкер 4, линкер 6, линкер 8 и линкер 10) (SEQ ID NO: 13, 15, 17, 19, 21, 23, 25). На фигуре 4В представлено процентное содержание клеток HER2+ (серый, ось у), которые экспрессируют GFP, по сравнению с процентным содержанием клеток HER2-, которые экспрессируют GFP (черный, ось у), через 5 дней после инфицирования вирусными частицами AAV2, состоящими из капсидов со следующими мутациями: R585A, delR588 и N587-SpyTag, фланкированной аминокислотными линкерами указанной длины (линкер 1, линкер 2, линкер 4, линкер 6, линкер 8 и линкер 10) (SEQ ID NO: 13, 15, 17, 19, 21, 23 и 25 соответственно) (ось х). Частицы N587 AAV2 SpyTag были конъюгированы с scFv к HER2, слитым с SpyCatcher (SEQ ID NO: 3).
[0050] На фигуре 5А представлен вестерн-блоттинг с применением антитела В1, которое распознает линейный эпитоп, общий для капсидных белков VP1, VP2 и VP3 AAV2, с анализом реакции между антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher, «SpyC-антитело-к-Her2», и панелью вирусных частиц AAV2, состоящей из капсидов со следующими мутациями: R585A, delR588 и вставкой пептида SpyTag непосредственно после остатка N587, фланкированного аминокислотными линкерами различной длины (линкер 1, линкер 2, линкер 4, линкер 6, линкер 8 и линкер 10) (SEQ ID NO: 13, 15, 17, 19, 21, 23 и 25 соответственно). На фигуре 5 В представлено процентное содержание клеток HER2+ (серый, ось у), которые экспрессируют GFP, по сравнению с процентным содержанием клеток HER2-, которые экспрессируют GFP (черный, ось у), через 5 дней после инфицирования частицами AAV2 (wt) дикого типа или вирусными частицами AAV2, состоящими из капсидов, со следующими мутациями: R585A, delR588 и N587-SpyTag, фланкированного аминокислотными линкерами указанной длины (без линкера, линкер 1, линкер 2, линкер 4, линкер 6, линкер 8 и линкер 10) (SEQ ID NO: 13, 15, 17, 19, 21, 23 и 25 соответственно) (ось х). Частицы N587 AAV2 SpyTag были конъюгированы с антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3).
[0051] На фигуре 6А представлен вестерн-блоттинг с применением антитела В1, которое распознает линейный эпитоп, общий для капсидных белков VP1, VP2 и VP3 AAV2, с анализом реакции между scFv-к-HER2 С6.5, слитым с SpyCatcher (SEQ ID NO: 3), «SpyC-scFv-к-HER2», и панелью мозаичных вирусных частиц AAV2, состоящих из смесей капсидных белков «SpyTag», содержащих мутации R585A, delR588 и N587-линкер 10-SpyTag (SEQ ID NO: 25), а также капсидных белков «НВМ», содержащих мутации R585A и R588A, но без SpyTag (SEQ ID NO: 11). Капсидные белки «SpyTag» и «НВМ» смешивали в различных соотношениях (1:0, 1:1 и 1:3). На фигуре 6 В представлено процентное содержание клеток 293 HER2+ hErbB2 (серые столбцы) и исходных клеток 293 HER2- (черные столбцы), которые экспрессируют GFP (ось у), через 5 дней после инфицирования мозаичными вирусными частицами AAV2 (ось х), состоящими из смесей капсидных белков «линкер 10», содержащих мутации R585A, delR588 и №87-линкер 10-SpyTag (SEQ ID NO: 25), и капсидных белков «НВМ», содержащих мутации R585A и R588A, но без SpyTag (SEQ ID NO: 11). Капсидные белки «линкер 10» и «НВМ» смешивали в различных соотношениях (1:0, 3:1, 1:1 и 1:3) и конъюгировали с scFv к HER2, слитым с SpyCatcher (SEQ ID NO: 3).
[0052] На фигуре 7А представлен вестерн-блоттинг с применением антитела В1, которое распознает линейный эпитоп, общий для капсидных белков VP1, VP2 и VP3 AAV2, с анализом реакции между антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3), «SpyC-антитело к HER2», и панелью мозаичных вирусных частиц AAV2, состоящих из смесей капсидных белков «SpyTag», содержащих мутации R585A, delR588 и N587-линкер 10-SpyTag (SEQ ID NO: 25), а также капсидных белков «НВМ», содержащих мутации R585A и R588A, но без SpyTag (SEQ ID NO: 11). Капсидные белки «SpyTag» и «НВМ» смешивали в различных соотношениях (1:0, 3:1, 1:1 и 1:3). На фигуре 7 В представлено процентное содержание клеток 293 HER2+ hErbB2 (серые столбцы) и исходных клеток 293 HER2-(черные столбцы), которые экспрессируют GFP (ось у), через 5 дней после инфицирования мозаичными вирусными частицами AAV2 (ось х), состоящими из смесей капсидных белков «линкер 10», содержащих мутации R585A, delR588 и N587-линкер 10-SpyTag (SEQ ID NO: 25), и капсидных белков «НВМ», содержащих мутации R585A и R588A, но без SpyTag (SEQ ID NO: 11). Капсидные белки «линкер 10» и «НВМ» смешивали в различных соотношениях (1:0, 3:1, 1:1 и 1:3) и конъюгировали с антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3).
[0053] На фигуре 8А представлен вестерн-блоттинг с применением антитела В1, которое распознает линейный эпитоп, общий для капсидных белков VP1, VP2 и VP3 AAV2, с анализом реакции между антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3), «SpyC-антитело-к-HER2», и панелью мозаичных вирусных частиц AAV2, состоящих из смесей капсидных белков «SpyTag» и капсидных белков «НВМ», содержащих мутации R585A и R588A, но без SpyTag (SEQ ID NO: 11). Капсидные белки «SpyTag» включают «G453 SpyTag», который содержит мутации R585A, R588A и вставку пептида SpyTag непосредственно после остатка G453 (SEQ ID NO: 27), и «G453 линкер 10 SpyTag», который содержит мутации R585A, R588A и вставку пептида SpyTag непосредственно после остатка G453 и фланкированного с одной из сторон 10 линкерными аминокислотами (SEQ ID NO: 29). Указанные капсиды «G453 SpyTag» и «G453 линкер 10 SpyTag» смешивали с капсидами «НВМ» в различных соотношениях (1:0, 1:3 и 1:7). На фигуре 8 В представлено процентное содержание клеток 293 HER2+ hErbB2 (серые столбцы) и исходных клеток 293 HER2- (черные столбцы), которые экспрессируют GFP (ось у), через 5 дней после инфицирования вирусными частицами дикого типа «wt» или мозаичными вирусными частицами AAV2 (ось х), состоящими из смесей капсидных белков «G453 SpyTag», содержащих мутации R585A, R588A и вставку пептида SpyTag непосредственно после остатка G453 (SEQ ID NO: 27), или капсидных белков «G453 линкер 10 SpyTag», содержащих мутации R585A, R588A и вставку пептида SpyTag непосредственно после остатка G453 и фланкированного с любой из сторон 10 линкерными аминокислотами (SEQ ID NO: 29), и капсидных белков «НВМ», содержащих мутации R585A и R588A, но без SpyTag (SEQ ID NO: 11). Капсиды «G453 SpyTag» или «G453 линкер 10 SpyTag» и «НВМ» смешивали в различных соотношениях (1:0 «чистый», 1:3 и 1:7) и конъюгировали с антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3).
[0054] На фигуре 9 представлены диаграммы рассеяния, полученные в ходе сортировки флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками, положительными (+) в отношении экспрессии ASGR1 после инфекции частицами «AAV2 wt», частицами «AAV2 SpyTag без антитела» или частицами «AAV2 SpyTag антитело-к-ASGR1». Капсиды «AAV2 wt» представляют собой капсиды дикого типа без мутаций или модификаций (SEQ ID NO: 9), тогда как капсид «AAV2 SpyTag» содержит следующие мутации: R585A, delR588 и вставку пептида SpyTag непосредственно после остатка N587 (SEQ ID NO: 13). Частицы «AAV2 SpyTag антитело-к-ASGR1» были конъюгированы со слитым с SpyCatcher антителом, которое специфически связывает ASGR1. Также представлены диаграммы рассеяния, полученные в ходе сортировки флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками, положительными (+) в отношении экспрессии CD63, после инфекции частицами «AAV2 wt», частицами «AAV2 SpyTag без антитела» или частицами «AAV2 SpyTag антитело-к-CD63». Капсиды «AAV2 wt» представляют собой капсиды дикого типа без мутаций или модификаций (SEQ ID NO: 9), тогда как капсид «AAV2 SpyTag» содержит следующие мутации: R585A, delR588 и вставку пептида SpyTag непосредственно после остатка N587 (SEQ ID NO: 13). Частицы «AAV2 SpyTag антитело-к-CD63» были конъюгированы со слитым с SpyCatcher антителом, которое специфически связывает CD63. Вирусы экспрессируют GFP как маркер трансдукции. Также представлены диаграммы рассеяния, полученные в ходе сортировки флуоресцентно-активированных клеток (FACS), для оценки экспрессии зеленого флуоресцентного белка (GFP) клетками, положительными (+) в отношении экспрессии PTPRN, после инфекции частицами «AAV9 wt», частицами «AAV2 SpyTag нерелевантное-антитело» или частицами «AAV2 SpyTag антитело-к-PTPRN». Капсиды «AAV9 wt» представляют собой капсиды дикого типа без мутаций или модификаций (SEQ ID NO: 31), в то время как капсиды «AAV2 SpyTag» представляют собой мозаичные вирусные частицы, состоящие из соотношения капсидных белков 1:7 «SpyTag», где SpyTag вставлен непосредственно после остатка G453, фланкированного линкером из 10 аминокислот (SEQ ID NO: 29), и капсидов без SpyTag, но содержащих аминокислотную последовательность Мус-метки, вставленную непосредственно после остатка N587 (SEQ ID NO: 53), что уменьшает природное связывание с рецептором. Частицы «AAV2 SpyTag нерелевантное-антитело» были конъюгированы со слитым с SpyCatcher антителом, которое не связывает PTPRN. Частицы «AAV2 SpyTag антитело-к-PTPRN» были конъюгированы со слитым с SpyCatcher антителом, которое специфически связывает PTPRN. Вирусы экспрессируют GFP как маркер трансдукции. Также показаны результаты анализа люциферазы, в котором оценивали экспрессию люциферазы светлячка клетками, положительными (+) в отношении hENTPD3 после инфекции частицами «AAV2 wt», частицами «AAV2 + нерелевантное mAb», частицами «AAV2 + антитело к ENTPD3» и «AAV2 + антитело к hCD20». Капсиды «AAV2 wt» представляют собой капсиды дикого типа без мутаций или модификаций (SEQ ID NO: 9), тогда как капсид «AAV2 + антитело» содержит следующие мутации: R585A, delR588 и вставку пептида SpyTag непосредственно после остатка N587 (SEQ ID NO: 13). Частицы «AAV2 + нерелевантное антитело» были конъюгированы со слитым с SpyCatcher антителом, которое не связывает hENTPD3. Частицы «AAV2 + антитело к hCD20» были конъюгированы с антителом, слитым с SpyCatcher, которое специфически связывает hCD20, который не экспрессируется на клетках hENTPD3+ и служит как дополнительный отрицательный контроль. Частицы «AAV2 + антитело к ENTPD3» были конъюгированы со слитым с SpyCatcher антителом, которое специфически связывает hENTPD3. Вирусы экспрессируют люциферазу светлячка как маркер трансдукции. Также показаны результаты анализа люциферазы, в котором оценивали экспрессию люциферазы светлячка клетками, положительными (+) в отношении hCD20 после инфекции частицами «AAV2 wt», частицами «AAV2 + нерелевантное mAb», частицами «AAV2 + антитело к ENTPD3» и «AAV2 + антитело к hCD20». Капсиды «AAV2 wt» представляют собой капсиды дикого типа без мутаций или модификаций (SEQ ID NO: 9), тогда как капсид «AAV2 + антитело» содержит следующие мутации: R585A, delR588 и вставку пептида SpyTag непосредственно после остатка N587 (SEQ ID NO: 13). Частицы «AAV2 + нерелевантное антитело» были конъюгированы со слитым с SpyCatcher антителом, которое не связывает hENTPD3. Частицы «AAV2 + антитело к ENTPD3» были конъюгированы с антителом, слитым с SpyCatcher, которое специфически связывает hENTPD3, который не экспрессируется на клетках hCD20+ и служит как дополнительный отрицательный контроль. Частицы «AAV2 + антитело к hCD20» были конъюгированы со слитым с SpyCatcher антителом, которое специфически связывает hCD20. Вирусы экспрессируют люциферазу светлячка как маркер трансдукции.
[0055] На фигуре 10А представлен вестерн-блоттинг с применением антитела В1, которое распознает линейный эпитоп, общий для капсидных белков VP1, VP2 и VP3 AAV9, с анализом реакции между антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (SEQ ID NO: 3), «SpyC-герцептин», и панелью вирусных частиц AAV9. Эти вирусные частицы AAV9 состоят из капсидов, содержащих мутацию W503A, которая обеспечивает уменьшение связывания рецептора, и вставку пептида SpyTag непосредственно после остатка А589 или G453 без линкера или фланкированного линкером из 10 аминокислот, или мозаичных вирусных частиц AAV9, состоящих из соотношения 1:7 капсидных белков «SpyTag», содержащих мутации W503A и или А589-линкер 10-SpyTag, или С453-линкер 10-SpyTag, и капсидных белков «W503A», содержащих мутацию W503A, но без SpyTag. На фигуре 10 В представлено процентное содержание клеток 293 HER2 + hErbB2 (серые столбцы) и исходных клеток 293 HER2- (черные столбцы) (ось у), которые экспрессируют GFP, через пять дней после инфицирования вирусными частицами AAV9, конъюгированными с антителом к HER2 (HERCEPTIN®), слитым с SpyCatcher (ось х). Эти вирусные частицы AAV9 состоят из капсидов, содержащих мутацию W503A, которая обеспечивает уменьшение связывания рецептора, и вставку пептида SpyTag непосредственно после остатка А589 (или G453) без линкера или фланкированного линкером из 10 аминокислот, или мозаичных вирусных частиц AAV9, состоящих из соотношения 1:7 капсидных белков «SpyTag», содержащих мутации W503A и или А589-линкер 10-SpyTag, или С453-линкер 10-SpyTag, и капсидных белков «W503A», содержащих мутацию W503A, но без SpyTag. Вирусы экспрессируют GFP как маркер трансдукции.
[0056] На фигуре 11 представлены изображения иммунофлуоресцентной микроскопии образцов печени, взятых у мышей C57BL/6, трансгенно модифицированных с обеспечением экспрессии человеческого ASGR1 на клетках печени. Образцы собирали через десять дней после внутривенной инъекции фосфатно-солевого буферного раствора (PBS) или 2,5×1011 вирусных геномов (vg)/животное SpyTag-меченых частиц AAV2, несущих представляющий интерес нуклеотид CAGG eGFP, и модифицированных посредством (1) SpyCatcher-антитело-к-CD3 человека (AAV SpyT-антитело-к-hCD3 CAGG eGFP) или (2) SpyCatcher-антитело-к-ASGR1 человека (AAV SpyT-антитело-к-hASGR1 CAGG eGFP). Мышей умерщвляли и транскардиально перфузировали 4% PFA. Органы печени, почек и сердца собирали и дегидратировали в 15% сахарозе, а затем в 30% сахарозе. Затем органы подвергали изготовлению срезов с замораживанием на предметных стеклах и окрашивали куриным антителом к EGFP (Jackson ImmunoResearch Labs, Inc. Уэст-Гроув, Пенсильвания) и конъюгированным с Alexa-488 вторичным куриным антителом (Jackson ImmunoResearch Labs, Inc. Уэст-Гроув, Пенсильвания). Каждое изображение представляет одну мышь. «SpyTag-меченый капсид AAV2» содержит следующие мутации: R585A, delR588 и вставку пептида SpyTag непосредственно после остатка N587 (SEQ ID NO: 13). Вирусы экспрессируют eGFP как маркер трансдукции.
[0057] На фигуре 12 представлены изображения люминесценции от отдельных мышей, которые не экспрессируют ASGR1 человека на клетках печени (контроль), и генетически модифицированных мышей, которые экспрессируют ASGR1 человека на клетках печени (гуманизированные по ASGR1 мыши) через 14 дней после внутривенной инъекции фосфатно-солевого буфера (PBS) или 3,0×1011 вирусных геномов (vg)/животное частиц AAV2 дикого типа (wt) или SpyTag-меченых частиц AAV2, несущих представляющий интерес нуклеотид люциферазы светлячка и модифицированных посредством (1) SpyCatcher-антитело-к-CD63 человека или (2) SpyCatcher-антитело-к-ASGR1 человека. Такие вирусные частицы AAV2 представляют собой мозаичные вирусные частицы, состоящие из соотношения капсидных белков 1:7«SpyTag», где SpyTag вставлен непосредственно после остатка G453, фланкированного линкером из 10 аминокислот (SEQ ID NO: 29), и капсидов без SpyTag, но содержащих аминокислотную последовательность Мус-метки, вставленную непосредственно после остатка N587 (SEQ ID NO: 53), что уменьшает природное связывание с рецептором. Вирусы экспрессируют люциферазу светлячка как маркер трансдукции. Мышей анестезировали, используя изофлуран, инъецировали субстратом люциферина и визуализировали через 10 минут с применением системы визуализации IVIS Spectrum In Vivo (PerkinElmer).
[0058] На фигуре 13 представлены изображения иммуногистохимии образцов печени и поджелудочной железы, взятых у мышей C57BL/6 через 4 недели после внутривенной инъекции физиологического раствора с фосфатным буфером (PBS) или 1,0×1012 вирусных геномов (vg)/животное частиц AAV9 дикого типа (wt) или 1,0×1013 вирусных геномов (vg)/животное SpyTag-меченых частиц AAV2, несущих представляющий интерес нуклеотид CMG eGFP и модифицированных посредством (1) SpyCatcher-антитело-к-ASGR1 человека (AAV2 SpyTag + нерелевантное mAb) или (2) SpyCatcher-антитело-к-ENTPD3 человека (AAV2 SpyTag + антитело к ENTPD3). Мышей умерщвляли, а печень и поджелудочную железу собирали и фиксировали в 10% нейтральном забуференном формалине. Затем органы фиксировали и подвергали изготовлению срезов с замораживанием на предметных стеклах и окрашивали антителами к GFP. Каждое изображение представляет одну мышь. Такие вирусные частицы AAV2 представляют собой мозаичные вирусные частицы, состоящие из соотношения капсидных белков 1:7 «SpyTag», где SpyTag вставлен непосредственно после остатка G453, фланкированного линкером из 10 аминокислот (SEQ ID NO: 29), и капсидов без SpyTag, но содержащих аминокислотную последовательность Мус-метки, вставленную непосредственно после остатка N587 (SEQ ID NO: 53), что уменьшает природное связывание с рецептором. Вирусы экспрессируют eGFP как маркер трансдукции. Сообщается, что ENTPD3 экспрессируется в островковых клетках поджелудочной железы и в языке, среди других типов клеток, кроме печени. SpyTag-меченые частицы AAV2 характеризовались отменой нацеливания на печень; экспрессия eGFP не наблюдалась в органах печени мышей, которым инъецировали частицы «AAV2 SpyTag + нерелевантное mAb» или «AAV2 SpyTag + антитело к ENTPD3». Положительное окрашивание в островковых клетках поджелудочной железы было обнаружено в образце поджелудочной железы одной из мышей, которой инъецировали частицы «AAV2 SpyTag + антитело к ENTPD3».
[0059] На фигуре 14 представлены изображения иммуногистохимии образцов печени и языка, взятых у мышей C57BL/6 через 14 дней после внутривенной инъекции физиологического раствора с фосфатным буфером (PBS) или 2,0×1012 вирусных геномов (vg)/животное частиц AAV9 дикого типа (wt) или SpyTag-меченых частиц AAV2, несущих представляющую интерес нуклеиновую кислоту CMV eGFP и модифицированных посредством (1) SpyCatcher-aHTKreno-к-ASGR1 человека (AAV2 SpyTag + нерелевантное mAb) или (2) SpyCatcher-антитело-к-ENTPD3 человека, которое связывается с ENTPD3 мыши (AAV2 SpyTag + антитело к ENTPD3). Мышей умерщвляли, а образцы печени и языка собирали и фиксировали в 10% нейтральном забуференном формалине. Затем органы фиксировали и подвергали изготовлению срезов с замораживанием на предметных стеклах и окрашивали антителами к GFP. Каждое изображение представляет одну мышь; трех мышей из групп «AAV2 SpyTag + нерелевантное mAb» и «AAV2 SpyTag + антитело к ENTPD3» подвергали инъекции и анализу, и все они показали сходные характеристики экспрессии GFP. Такие вирусные частицы AAV2 представляют собой мозаичные вирусные частицы, состоящие из соотношения капсидных белков 1:7«SpyTag», где SpyTag вставлен непосредственно после остатка G453, фланкированного линкером из 10 аминокислот (SEQ ID NO: 29), и капсидов без SpyTag, но содержащих аминокислотную последовательность Мус-метки, вставленную непосредственно после остатка N587 (SEQ ID NO: 52), что уменьшает природное связывание с рецептором. Вирусы экспрессируют eGFP как маркер трансдукции. Сообщается, что ENTPD3 экспрессируется в языке мыши, но не в печени (данные получены из общедоступных баз данных GenePaint.org http://www.informatics.jax.org/assay/MGI:5423021 и проекта Riken FANTOM5, набор данных для взрослых мышей). Экспрессия eGFP не наблюдалась в органах печени мышей, которым инъецировали частицы «AAV2 SpyTag + нерелевантное mAb» или «AAV2 SpyTag + антитело к ENTPD3», но была обнаружена в языке всех трех мышей, которым инъецировали частицы «AAV2 SpyTag + антитело к ENTPD3».
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0060] В WO 201611291 описано применение специфической связывающейся пары (SpyCatcher: SpyTag) для получения вирусоподобных частиц (VLP) из модифицированного бактериофага АР205, отображающего иммуногенные антигены с высокой плотностью на капсиде АР205 для целей вакцинации. Теоретически, для целей перенацеливания вирусного вектора такая высокая степень модификации вирусного капсида может быть желательной для обеспечения того, чтобы природный тропизм вирусного вектора был существенно уменьшен или устранен. Однако такое отображение нацеливающих лигандов с высокой плотностью на вирусной поверхности может мешать показателям эффективности трансдукции. См. примеры 4 и 5. С целью достижения оптимальных показателей эффективности трансдукции было обнаружено, что оптимальные показатели эффективности трансдукции имеют место, когда степень модификации вирусной поверхности с помощью партнера снижена. Таким образом, в настоящем документе предусмотрены генетически модифицированные вирусные частицы, композиции, содержащие их, и способы их получения и применение.
[0061] Если не определено иное, то все технические и научные термины, используемые в данном документе, имеют такое же значение, как это обычно понимает специалист в данной области техники, к которой принадлежит настоящее изобретение.
[0062] Формы единственного числа включают определяемые объекты во множественном числе, если из контекста явно не следует иное. Таким образом, например, ссылка на «способ» включает один или более способов и/или стадий, описанных в данном документе, и/или которые станут очевидными специалистам в данной области техники после прочтения настоящего раскрытия.
[0063] Термин «антитело» включает молекулы иммуноглобулинов, содержащие четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь содержит вариабельный домен тяжелой цепи (VH) и константную область тяжелой цепи (CH). Константная область тяжелой цепи содержит по меньшей мере три домена, CH1, CH2, CH3, и необязательно CH4. Каждая легкая цепь содержит вариабельный домен легкой цепи (VH) и константную область легкой цепи (CL). Вариабельные домены легких и тяжелых цепей могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), чередующиеся с более консервативными областями, называемыми каркасными областями (FR). Каждый вариабельный домен тяжелой и легкой цепей содержит три CDR и четыре FR, расположенных от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи могут сокращенно называться HCDR1, HCDR2 и HCDR3; CDR легкой цепи могут сокращенно называться LCDR1, LCDR2 и LCDR3. Типичные тетрамерные структуры антитела содержат два идентичных антигенсвязывающих домена, каждый из которых образован ассоциацией доменов VH и VL, и каждый из которых вместе с соответствующими доменами CH и CL образует Fv-область антитела. Однодоменные антитела содержат один антигенсвязывающий домен, например, VH или VL. Антигенсвязывающий домен антитела, например, часть антитела, которая распознает и связывается с первым партнером специфической связывающейся пары антигена, также называется «паратопом». Он представляет собой небольшую область (от 5 до 10 аминокислот) Fc-области антитела, часть антигенсвязывающего фрагмента (Fab-область) и может содержать части тяжелой и/или легкой цепей антитела. Паратоп специфически связывает первого партнера специфической связывающейся пары, в случае когда паратоп связывает первого партнера специфической связывающейся пары с высокой аффинностью. Термин «высокоаффинное» антитело относится к антителу, которое имеет KD по отношению к своему первому партнеру-мишени специфической связывающейся пары, составляющую приблизительно 10-9 М или меньше (например, приблизительно 1×10-9 М, 1×10-10 М, 1×10-11 М или приблизительно 1×10-12 М). В одном варианте осуществления KD измеряют посредством поверхностного плазмонного резонанса, например, BIACORE™; в другом варианте осуществления KD измеряют при помощи ELISA.
[0064] Фраза «определяющая комплементарность область» или термин «CDR» включают аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулина организма, которая в норме (т.е. у животного дикого типа) находится между двух каркасных областей в вариабельной области легкой или тяжелой цепи молекулы иммуноглобулина (например, антитела или Т-клеточного рецептора). CDR может кодироваться, например, последовательностью зародышевого типа, либо перегруппированной или неперегруппированной последовательностью, и, например, непримированной или зрелой В-клеткой или Т-клеткой. CDR может быть подвергшейся соматической мутации (например, отличаться от последовательности, кодируемой зародышевым типом животного), гуманизированной и/или модифицированной с помощью аминокислотных замен, добавлений или делеций. В некоторых случаях (например, в случае CDR3) CDR могут кодироваться двумя или более последовательностями (например, последовательностями зародышевого типа), которые не являются смежными (например, в случае неперегруппированной последовательности нуклеиновой кислоты), но они являются смежными в последовательности нуклеиновой кислоты в В-клетке, например, как результат сплайсинга или соединения последовательностей (например, V-D-J-рекомбинапия с образованием CDR3 тяжелой цепи).
[0065] Фраза «инвертированный концевой повтор» или «ITR» включает симметричные последовательности нуклеиновой кислоты в геноме аденоассоциированных вирусов, необходимые для эффективной репликации. Последовательности ITR расположены на каждом конце ДНК-генома AAV. ITR служат в качестве точек начала репликации для синтеза вирусной ДНК и являются важными цис-компонентами для получения векторов на основе AAV.
[0066] Фраза «легкая цепь» подразумевает последовательность легкой цепи иммуноглобулина из любого организма и, если не указано иное, предусматривает легкие κ и λ цепи человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены легкой цепи обычно содержат три CDR легкой цепи и четыре каркасные (FR) области, если не указано иное. В целом, полноразмерная легкая цепь включает, от аминоконца к карбоксильному концу, вариабельный домен, который включает FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и константный домен легкой цепи. Вариабельный домен легкой цепи кодируется последовательностью гена вариабельной области тяжелой цепи, который, как правило, содержит генные сегменты VL и JL, полученные из репертуара сегментов V и J, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатуру для сегментов V и J легкой цепи у различных организмов можно просмотреть в базе данных IMGT, www.imgt.org. Легкие цепи охватывают, например, те, которые не обеспечивают селективное связывание либо первого, либо второго первого партнера в специфической связывающейся паре, селективно связанной с первым партнером в белке, связывающем специфическую связывающуюся пару, в котором они обнаруживаются. Легкие цепи также охватывают те, которые обеспечивают связывание или распознавание или способствуют тяжелой цепи или другой легкой цепи при связывании или распознавании одного или более первых партнеров в специфических связывающихся парах, селективно связанных с первым партнером в белке, связывающем специфическую связывающуюся пару, в котором они обнаруживаются. Традиционные или универсальные легкие цепи включают цепи, полученные из гена VκI-39Jκ человека или гена Vκ3-20Jκ человека, и включают их варианты, подвергшиеся соматической мутации (например, созреванию аффинности). Иллюстративные сегменты VL включают сегмент гена Vκ1-39 человека, сегмент гена Vκ3-20 человека, сегмент гена Vλ1-40 человека, сегмент гена Vλ1-44 человека, сегмент гена Vλ2-8 человека, сегмент гена Vλ2-14 человека, а также сегмент гена Yλ3-21 человека и включают их варианты, подвергшиеся соматической мутации (например, созреванию аффинности). Можно получать легкие цепи, которые содержат вариабельный домен из одного организма (например, человека или грызуна, например, крысы или мыши; или птицы, например, курицы) и константную область из того же или другого организма (например, человека или грызуна, например, крысы или мыши; или птицы, например, курицы).
[0067] Термин «приблизительно» или «примерно» включает расположение в пределах статистически значимого диапазона значения. Такой диапазон может быть в пределах порядка величины, предпочтительно в пределах 50%, более предпочтительно в пределах 20%, еще более предпочтительно в пределах 10% и еще более предпочтительно в пределах 5% от данного значения или диапазона. Допустимое отклонение, охватываемое терминами «приблизительно» или «примерно», зависит от конкретной исследуемой системы и может быть легко оценено специалистом в данной области техники.
[0068] Термин «капсидный белок» включает белок, который является частью капсида вируса. Для аденоассоциированных вирусов капсидные белки обычно обозначаются как VP1, VP2 и/или VP3 и кодируются одним геном cap. Для AAV три капсидных белка AAV продуцируются перекрывающимся образом из открытой рамки считывания cap (ORF) посредством альтернативного сплайсинга mRNA и/или альтернативного применения стартового кодона трансляции, при этом все три белка используют общий стоп-кодон. Warrington et al. (2004) J. Virol. 78:6595, включенный в данный документ посредством ссылки во всей своей полноте. VP1 AAV2, как правило, транслируется со стартового кодона ATG (аминокислота M1) на mRNA размером 2,4 т.п.о., в то время как VP2 и VP3 AAV2 образуются с помощью mRNA меньшего размера в 2,3 т.п.о. с применением более слабого стартового кодона ACG для продуцирования VP2 (аминокислота Т138) и сквозной трансляции к следующему доступному ко дону ATG (аминокислота М203) для продуцирования наиболее многочисленного капсидного белка, VP3. Warrington, выше; Rutledge et al. (1998) J. Virol. 72:309-19, включенный в данный документ посредством ссылки во всей своей полноте. Аминокислотные последовательности капсидных белков аденоассоциированных вирусов хорошо известны в данной области и в целом консервативны, особенно в пределах депендопарвовирусов. См. Rutledge et al., выше. Например, в Rutledge et al. (1998), выше, на фигуре 4В приведены выравнивания аминокислотных последовательностей для капсидных белков VP1, VP2 и VP3 для AAV2, AAV3, AAV4 и AAV6, где стартовые сайты для каждого из капсидных белков VP1, VP2 и VP3 указаны стрелками, а вариабельные домены заключены в рамку. Соответственно, несмотря на то, что положения аминокислот, представленные в данном документе, могут быть предоставлены в отношении капсидного белка VP1 AAV, и положения аминокислот, представленные в данном документе, которые не указаны дополнительно, относятся к последовательности AAV2 основного белка оболочки VP1, изложенной под SEQ ID NO: 1, квалифицированный специалист сможет соответственно и легко определить положение той же аминокислоты в капсидном белке VP2 и/или VP3 AAV и соответствующее положение аминокислот среди различных серотипов. Соответственно, несмотря на то, что положения аминокислот, предусмотренные в данном документе, могут быть предоставлены в отношении капсидного белка VP1 AAV, и положения аминокислот, представленные в данном документе, которые не указаны дополнительно, относятся к последовательности AAV2 основного белка оболочки VP1, изложенной под SEQ ID NO: 9, квалифицированный специалист сможет соответственно и легко определить положение той же аминокислоты в капсидном белке VP2 и/или VP3 AAV и соответствующее положение аминокислоты среди различных серотипов AAV. Кроме того, квалифицированный специалист сможет произвести обмен доменов между капсидными белками различных серотипов AAV с образованием «химерного капсидного белка».
[0069] Был описан обмен доменами между двумя конструкциями капсидного белка AAV с получением «химерного капсидного белка AAV», см., например, Shen et al. (2007) Mol. Therapy 15(11): 1955-1962, включенном в данный документ посредством ссылки во всей своей полноте. «Химерный капсидный белок AAV» включает капсидный белок AAV, который содержит аминокислотные последовательности, например, домены, из двух или более различных серотипов AAV и который способен образовывать и/или образует AAV-подобные вирусный капсид/вирусную частицу. Химерный капсидный белок AAV кодируется химерным геном капсида AAV, например, нуклеотидом, содержащим множество, например, по меньшей мере две последовательности нуклеиновой кислоты, каждая из которых идентична части гена капсида, кодирующего капсидный белок различных серотипов AAV, и это множество совместно кодирует функциональный химерный капсидный белок AAV. Ссылка на химерный капсидный белок в отношении конкретного серотипа AAV указывает на то, что капсидный белок содержит один или более доменов из капсидного белка данного серотипа и один или более доменов из капсидного белка другого серотипа. Например, химерный капсидный белок AAV2 включает капсидный белок, содержащий один или более доменов капсидного белка VP1, VP2 и/или VP3 AAV2 и один или более доменов капсидного белка VP1, VP2 и/или VP3 другого AAV.
[0070] «Мозаичный капсид» содержит по меньшей мере два набора белков VP1, VP2 и/или VP3, каждый из которых кодируется другим геном cap.
[0071] В некоторых вариантах осуществления описанный в данном документе мозаичный капсид содержит рекомбинантные белки VP1, VP2 и/или VP3, кодируемые геном cap, генетически модифицированным посредством вставки последовательности нуклеиновой кислоты, кодирующей гетерологичный эпитоп, и дополнительно содержит белки VP1, VP2 и/или VP3, кодируемые эталонным геном cap, например, эталонным геном cap дикого типа, кодирующим белки VP1, VP2 и/или VP3 дикого типа того же серотипа AAV, что и рекомбинантные белки VP1, VP2 и/или VP3, контрольным эталонным геном cap, кодирующим белки VP1, VP2 и VP3, идентичные рекомбинантным белкам VP1, VP2 и/или VP3, но с отсутствием гетерологичного эпитопа, мутированным эталонным геном cap дикого типа, кодирующим по сути белки VP1, VP2 и/или VP3 дикого типа того же серотипа AAV, что и рекомбинантные белки VP1, VP2 и/или VP3, но с мутацией (например, вставкой, заменой, делецией), которая предпочтительно обеспечивает уменьшение тропизма белков VP 1, VP2 и VP3 дикого типа. В некоторых вариантах осуществления эталонный ген cap кодирует химерный белок VP1, VP2 и/или VP3.
[0072] Фраза «тяжелая цепь» или «тяжелая цепь иммуноглобулина» подразумевает последовательность тяжелой цепи иммуноглобулина, в том числе последовательность константной области тяжелой цепи иммуноглобулина, из любого организма. Вариабельные домены тяжелой цепи содержат три CDR тяжелой цепи и четыре FR-области, если не указано иное. Фрагменты тяжелых цепей включают CDR, CDR и FR, а также их комбинации. Типичная тяжелая цепь после вариабельного домена (от N-конца к С-концу) содержит CH1-домен, шарнир, CH2-домен и CH3-домен. Функциональный фрагмент тяжелой цепи включает фрагмент, который способен специфически распознавать первого партнера специфической связывающейся пары {например, распознавать первого партнера специфической связывающейся пары с KD в микромолярном, наномолярном или пикомолярном диапазонах), который способен экспрессироваться и секретироваться клеткой и который содержит по меньшей мере одну CDR. Вариабельные домены тяжелой цепи кодируются нуклеотидной последовательностью вариабельной области, которая, как правило, содержит сегменты VH, DH и JH, полученные из репертуара сегментов VH, DH и JH, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатура для сегментов тяжелых цепей V, D и J для различных организмов можно найти в базе данных IMGT, которая доступна через Интернет во всемирной сети (www) по адресу «imgt.org».
[0073] Термин «антитело, содержащее только тяжелую цепь», «антигенсвязывающий белок, содержащий только тяжелую цепь», «однодоменный антигенсвязывающий белок», «однодоменный связывающий белок» или тому подобное относится к мономерной или гомодимерной молекуле иммуноглобулина, содержащей иммуноглобулиноподобную цепь, содержащую вариабельный домен, функционально связанный с константной областью тяжелой цепи, который не может ассоциироваться с легкой цепью, поскольку в константной области тяжелой цепи, как правило, отсутствует функциональный домен CH1. Соответственно, термин «антитело, содержащее только тяжелую цепь», «антигенсвязывающий белок, содержащий только тяжелую цепь», «однодоменный антигенсвязывающий белок», «однодоменный связывающий белок» или тому подобное охватывает и (i) мономерный однодоменный антигенсвязывающий белок, содержащий одну из иммуноглобулиноподобной цепей, содержащих вариабельный домен, функционально связанный с константной областью тяжелой цепи, в которой отсутствует функциональный домен CH1, и (ii) гомодимерный одно доменный антигенсвязывающий белок, содержащий две иммуноглобулиноподобные цепи, каждая из которых содержит вариабельный домен, функционально связанный с константной областью тяжелой цепи, в которой отсутствует функциональный домен CH1. В различных аспектах гомодимерный одно доменный антигенсвязывающий белок содержит две идентичные иммуноглобулиноподобные цепи, каждая из которых содержит идентичный вариабельный домен, функционально связанный с идентичной константной областью тяжелой цепи, в которой отсутствует функциональный домен CH 1. Кроме того, каждая иммуноглобулиноподобная цепь одно доменного антигенсвязывающего белка содержит вариабельный домен, который может быть получен из сегментов гена вариабельной области тяжелой цепи (например, VH, DH, JH), сегментов гена легкой цепи (например, VL, JL) или их комбинации, связанный с последовательностью гена константной области (CH ) тяжелой цепи, содержащей делецию или инактивирующую мутацию в кодирующей последовательности CH1 (и необязательно шарнирную область) гена константной области тяжелой цепи, например, IgG, IgA, IgE, IgD или их комбинации. Однодоменный антигенсвязывающий белок, содержащий вариабельный домен, полученный из сегментов генов тяжелой цепи, может называться «VH-однодоменное антитело» или «VH-однодоменный антигенсвязывающий белок», см., например, патент США №8754287; патентные публикации США №№20140289876; 20150197553; 20150197554; 20150197555; 20150196015; 20150197556 и 20150197557, каждая из которых полностью включена посредством ссылки во всей своей полноте. Однодоменный антигенсвязывающий белок, содержащий вариабельный домен, полученный из сегментов генов легкой цепи, может называться «VL-однодоменный антигенсвязывающий белок», см., например, публикацию патента США №20150289489, включенную посредством ссылки во всей своей полноте.
[0074] Фраза «легкая цепь» подразумевает последовательность легкой цепи иммуноглобулина из любого организма и, если не указано иное, предусматривает легкие каппа (κ) и лямбда (λ) цепи человека и VpreB, а также суррогатные легкие цепи. Вариабельные домены легкой цепи обычно содержат три CDR легкой цепи и четыре каркасные (FR) области, если не указано иное. В целом, полноразмерная легкая цепь включает, от аминоконца к карбоксильному концу, вариабельный домен, который включает FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и аминокислотную последовательность константной области легкой цепи. Вариабельные домены легкой цепи кодируется нуклеотидной последовательностью вариабельной области легкой цепи, которая, как правило, содержит сегменты генов VL и JL легких цепей, полученные из репертуара сегментов генов VL и JL легких цепей, присутствующих в зародышевой линии. Последовательности, местоположения и номенклатура для сегментов генов легких цепей V и J для различных организмов можно найти в базе данных IMGT, которая доступна через Интернет во всемирной сети (www) по адресу «imgt.org». Легкие цепи охватывают, например, те, которые не обеспечивают селективное связывание либо первого, либо второго первого партнера в специфической связывающейся паре, селективно связанной с первым партнером в белке, связывающем специфическую связывающуюся пару, в котором они обнаруживаются. Легкие цепи также охватывают те, которые обеспечивают связывание или распознавание или способствуют тяжелой цепи при связывании или распознавании одного или более первых партнеров в специфических связывающихся парах, селективно связанных с первым партнером в белке, связывающем специфическую связывающуюся пару, в котором они обнаруживаются. Легкие цепи также охватывают те, которые обеспечивают связывание или распознавание или способствуют тяжелой цепи при связывании или распознавании одного или более первых партнеров в специфических связывающихся парах, селективно связанных с первым партнером в белке, связывающем специфическую связывающуюся пару, в котором они обнаруживаются. Традиционные или универсальные легкие цепи включают цепи, полученные из гена Vκ1-39Jκ5 человека или гена Vκ3-20Jκ1 человека, и включают их варианты, подвергшиеся соматической мутации {например, созреванию аффинности).
[0075] Фраза «функционально связанный», используемая в данном документе, включает в себя физическое положение (например, в трехмерном пространстве) компонентов или элементов, которые прямо или косвенно взаимодействуют друг с другом или иным образом координируют друг друга для участия в биологическом событии, при этом такое положение достигает или разрешает такое взаимодействие и/или координацию. В качестве лишь одного примера, говорят, что контрольная последовательность (например, контрольная последовательность экспрессии) в нуклеиновой кислоте «функционально связана» с кодирующей последовательностью, когда она расположена относительно кодирующей последовательности таким образом, что ее присутствие или отсутствие влияет на экспрессию и/или активность кодирующей последовательности. Во многих вариантах осуществления «функциональная связь» включает ковалентную связь соответствующих компонентов или элементов друг с другом. Специалисты в данной области легко поймут, что в некоторых вариантах осуществления ковалентная связь не требуется для достижения эффективной функциональной связи. Например, в некоторых вариантах осуществления контролирующие последовательности нуклеиновой кислоты, которые функционально связаны с кодирующими последовательностями, которые они контролируют, являются смежными с представляющим интерес нуклеотидом. Альтернативно или дополнительно, в некоторых вариантах осуществления одна или более таких контролирующих последовательностей действуют в транс-положении или на расстоянии для осуществления контроля представляющей интерес кодирующей последовательности. В некоторых вариантах осуществления термин «последовательность, контролирующая экспрессию», используемый в данном документе, относится к полинуклеотидным последовательностям, необходимым и/или достаточным для осуществления экспрессии и процессинга кодирующих последовательностей, с которыми они лигированы. В некоторых вариантах осуществления «последовательности, контролирующие экспрессию» могут представлять собой или включают соответствующие последовательности для инициации и терминации транскрипции, промоторные и/или энхансерные последовательности; сигналы для эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, стабилизирующие цитоплазматическую mRNA; последовательности, повышающие эффективность трансляции (например, консенсусную последовательность Козак); последовательности, повышающие стабильность белка; и в некоторых вариантах осуществления последовательности, усиливающие секрецию белка. В некоторых вариантах осуществления одна или более контролирующих последовательностей предпочтительно или исключительно активны в конкретной клетке-хозяине или организме или их типе. В качестве лишь примера, у прокариот контролирующие последовательности обычно включают промотор, участок связывания рибосом и последовательность терминации транскрипции; у эукариот во многих вариантах осуществления контролирующие последовательности обычно включают промоторы, энхансеры и/или последовательности терминации транскрипции. Специалистам в данной области техники должно быть понятно из контекста, что во многих вариантах осуществления термин «контролирующие последовательности» относится к компонентам, присутствие которых является существенным для экспрессии и процессинга, и в некоторых вариантах осуществления включает компоненты, присутствие которых является преимущественным для экспрессии (в том числе, например, лидерные последовательности, нацеливающие последовательности и/или последовательности партнеров по слиянию).
[0076] Термин «рекомбинантный капсидный белок» включает капсидный белок, который характеризуется по меньшей мере одной мутацией по сравнению с соответствующим капсидным белком вируса дикого типа, который может быть эталонным и/или контрольным вирусом для сравнительного исследования. Рекомбинантный капсидный белок предусматривает капсидный белок, который содержит гетерологичную аминокислотную последовательность, которая может быть встроена и/или отображена с помощью капсидного белка. «Гетерологичный» в данном контексте означает гетерологичный по сравнению с вирусом, из которого получен капсидный белок. Вставленные аминокислоты могут быть просто вставлены между двумя указанными аминокислотами капсидного белка. Вставка аминокислот также может сопровождаться делецией указанных аминокислот капсидного белка в участке вставки, например, 1 или более аминокислот капсидного белка заменены на 5 или больше гетерологичных аминокислот).
[0077] «Перенацеливание» или «перенаправление» может предусматривать сценарий, при котором вектор дикого типа нацеливается на несколько клеток в ткани и/или несколько органов в организме, причем общее нацеливание на ткань или органы находится в пределах от уменьшенного до устраненного, путем введения гетерологичного эпитопа, и при этом перенацеливание на более конкретную клетку в ткани или конкретном органе в организме достигнуто с помощью нацеливающего лиганда, который связывается с маркером, экспрессируемым конкретной клеткой. Такое перенацеливание или перенаправление может также предусматривать сценарий, при котором вектор дикого типа нацеливается на ткань, причем нацеливание на ткань находится в пределах от уменьшенного до устраненного, путем введения гетерологичного эпитопа, и при этом перенацеливание на совершенно другую ткань достигнуто с помощью нацеливающего лиганда.
[0078] «Специфическая связывающаяся пара», «связывающаяся пара белок: белок» и тому подобное включает два белка (например, первого партнера (например, первый полипептид) и второго родственного партнера (например, второй полипептид)), которые взаимодействуют с образованием ковалентной изопептидной связи в условиях, которые обеспечивают или облегчают образование изопептидной связи, где термин «родственный» относится к компонентам, которые функционируют совместно, т.е. взаимодействуют друг с другом с образованием изопептидной связи. Таким образом, два белка, которые эффективно реагируют друг с другом с образованием изопептидной связи в условиях, которые обеспечивают или облегчают образование изопептидной связи, также могут называться «комплементарной» парой пептидных линкеров. Специфические связывающиеся пары, способные взаимодействовать с образованием ковалентной изопептидной связи, рассмотрены в Veggiani et al. (2014) Trends Biotechnol. 32:506, и включают связывающиеся пары пептид: пептид, такие как SpyTag: SpyCatcher, SpyTag002:SpyCatcher002; SpyTag:KTag; isopeptag:пилин С, SnoopTag:SnoopCatcher и т.д. Как правило, пептидная метка относится к партнеру связывающейся пары белок: белок, длина которой обычно составляет менее 30 аминокислот, и которая образует ковалентную изопептидную связь со вторым родственным белком, где второй родственный белок, как правило, больше, но также может иметь длину менее 30 аминокислот, например, в системе SpyTag:KTag.
[0079] Термин «изопептидная связь» относится к амидной связи между карбоксильной или карбоксамидной группой и аминогруппой, по меньшей мере одна из которых не происходит из основной цепи белка или, альтернативно, не является частью остова белка. Изопептидная связь может образовываться в пределах одного белка или может иметь место между двумя пептидами или пептидом и белком. Таким образом, изопептидная связь может образовываться внутримолекулярно в пределах одного белка или межмолекулярно, т.е. между двумя молекулами пептид/белок, например, между двумя пептидными линкерами. Как правило, изопептидная связь может находиться между остатком лизина и остатком аспарагина, аспарагиновой кислоты, глутамина или глутаминовой кислоты или концевой карбоксильной группой белка или пептидной цепи или может находиться между альфа-аминоконцом белка или пептидной цепи и аспарагином, аспарагиновой кислотой, глутамином или глутаминовой кислотой. Каждый остаток пары, участвующей в изопептидной связи, упоминается в данном документе как реакционноспособный остаток. В предпочтительных вариантах осуществления настоящего изобретения изопептидная связь может образовываться между остатком лизина и остатком аспарагина или между остатком лизина и остатком аспарагиновой кислоты. В частности, изопептидные связи могут находиться между аминной группой боковой цепи лизина и карбоксамидной группой аспарагина или карбоксильной группой аспартата.
[0080] Система SpyTag: SpyCatcher описана в патенте США №9547003 и Zakeri et al. (2012) PNAS 109:E690-E697, каждый из которых полностью включен в настоящее описание посредством ссылки, и получена из домена CnaB2 фибронектинсвязывающего белка FbaB Streptococcus pyogenes. Путем расщепления домена Zakeri et al. получили пептид «SpyTag», имеющий последовательность AHIVMVDAYKPTK (SEQ ID NO: 1), который образует амидную связь с его родственным белком «SpyCatcher», полипептидом из 112 аминокислот, имеющим аминокислотную последовательность, изложенную под SEQ ID NO: 3. (Zakeri (2012), выше). Дополнительной специфической связывающейся парой, полученной из домена CnaB2, является система SpyTag:KTag, которая образует изопептидную связь в присутствии SpyLigase. (Fierer (2014) PNAS 111:Е1176-1181) SpyLigase была сконструирована путем вырезания β-нити из SpyCatcher, которая содержит реакционноспособный лизин, с получением в результате KTag, пептидной метку из 10 остатков, имеющей аминокислотную последовательность ATHIKFSKRD (SEQ ID NO: 2). Система SpyTag002:SpyCatcher002 описана в Keeble et al (2017) Angew Chem Int Ed Engl 56:1652I-25, включенном в данный документ посредством ссылки во всей своей полноте. SpyTag002 имеет аминокислотную последовательность VPTIVMVDAYKRYK, изложенную под SEQ ID NO: 54, и связывается с SpyCatcher002 (SEQ ID NO: 55).
[0081] Система SnoopTag:SnoopCatcher описана в Veggiani (2016) PNAS 113:1202-07. Ig-подобный домен D4 RrgA, отвечающий за адгезию у Streptococcus pneumoniae, расщепляли с образованием SnoopTag (остатки 734-745; SEQ ID NO: 5) и SnoopCatcher (остатки 749-860). Инкубация SnoopTag и SnoopCatcher приводит к спонтанному образованию изопептидной связи, которая является специфической в отношении комплементарных белков. Veggiani (2016)), выше.
[0082] Специфическая связывающаяся пара isopeptag:пилин-C была получена из основного белка пилина Spy0128 из Streptococcus pyogenes. (Zakeir and Howarth (2010) J. Am. Chem. Soc. 132:4526-27). Isopeptag имеет аминокислотную последовательность TDKDMTITFTNKKDAE, изложенную под SEQ ID NO: 7, и связывается с пилином-С (остатки 18-299 из Spy0128). Инкубация SnoopTag и SnoopCatcher приводит к спонтанному образованию изопептидной связи, которая является специфической в отношении комплементарных белков. Zakeir and Howarth (2010), выше.
[0083] Термин «пептидная метка» включает полипептиды, которые (1) гетерологичны белку, который помечен с помощью пептидной метки, (2) являются партнером специфической связывающейся пары белок: белок, способной образовывать изопептидную связь, и (3) их длина составляет не более 50 аминокислот.
[0084] Термин «клетки-мишени» включает любые клетки, в которых желательна экспрессия представляющего интерес нуклеотида. Предпочтительно клетки-мишени демонстрируют рецептор на своей поверхности, который обеспечивает нацеливание на клетку с помощью нацеливающего лиганда, как описано ниже.
[0085] Термин «трансдукция» или «инфекция» или тому подобное относится к введению нуклеиновой кислоты в ядро клетки-мишени с помощью вирусного вектора. Термин эффективность в отношении трансдукции или т.п., например, «эффективность трансдукции», относится к фракции (например, процентному содержанию) клеток, экспрессирующих представляющий интерес нуклеотид после инкубации с установленным количеством вирусных векторов, содержащих представляющий интерес нуклеотид. Хорошо известные способы определения эффективности трансдукции включают в себя флуоресцентную сортировку клеток, трансдуцированных флуоресцентным репортерным геном, ПЦР в отношении экспрессии представляющего интерес нуклеотида и т.д.
[0086] Термин «дикий тип», используемый в данном документе, включает объект, обладающий структурой и/или активностью, обнаруживаемой в природе в «нормальном» (в противоположность мутантному, болезненному, измененному и т.д.) состоянии или окружении. Специалистам в данной области будет понятно, что вирусные векторы дикого типа, например, капсидные белки дикого типа, могут быть использованы как эталонный вирусный вектор в сравнительных исследованиях. Как правило, эталонный вирусный капсидный белок/капсид/вектор идентичен исследуемому вирусному капсидному белку/капсиду/вектору, за исключением изменения, в отношении влияния которого проводят исследование. Например, для определения влияния, например, на эффективность трансдукции, вставки первого партнера специфической связывающейся пары в исследуемый вирусный вектор, показатели эффективности трансдукции исследуемого вирусного вектора (в отсутствие или в присутствии соответствующего нацеливающего лиганда) можно сравнить с эффективностью трансдукции эталонного вирусного вектора (в отсутствие или в присутствии соответствующего нацеливающего лиганда, если необходимо), который идентичен исследуемому вирусному вектору по всем параметрам (например, дополнительные мутации, представляющий интерес нуклеотид, количество вирусных векторов и клетки-мишени и т.д.), за исключением присутствия первого партнера специфической связывающейся пары.
[0087] Стратегия перенацеливания, описанная в данном документе, обеспечивает преимущества как каркасного, так и прямого рекомбинаторного подходов, описанных выше, а также устраняет многие недостатки, присущие обоим. В стратегии применяется специфическая связывающаяся пара, в которой первый партнер и второй родственный партнер специфически связываются друг с другом и при связывании образуют ковалентную связь, которая перманентно соединяет вирусную частицу с любым нацеливающим лигандом, который слит с родственным партнером. В случае такой генетически модифицированной вирусной частицы тропизм сохраняется до тех пор, пока вирусный капсид остается интактным, например, одним из преимуществ системы, представленной в настоящем документе, по сравнению с другими каркасными подходами является постоянство, с которым «адаптер», например, нацеливающий лиганд, связан с рекомбинантной вирусной частицей, аналогично прямому рекомбинаторному подходу. Однако, в отличие от прямого рекомбинаторного подхода, в системе, описанной в настоящем документе, сохраняется гибкость подходов каркасного адаптера в том, что рекомбинантная вирусная частица может оставаться постоянной при изменчивости, обнаруживаемой в адаптере, например, родственный партнер может быть слит с разными нацеливающими лигандами, и затем различные слитые белки связаны с вирусной частицей в соответствии с клеткой-мишенью.
[0088] Рекомбинантные вирусные капсидные белки, а также вирусные векторы и нуклеиновые кислоты
[0089] В данном документе предусмотрена рекомбинантная вирусная частица (например, вирусный капсидный белок, рекомбинантный вирусный капсид и/или рекомбинантный вирусный вектор, содержащий рекомбинантный вирусный капсидный белок), которая генетически модифицирована с обеспечением отображения гетерологичной аминокислотной последовательности, содержащей первого партнера специфической связывающейся пары, например, где длина аминокислотной последовательности составляет менее 50 аминокислот, и где рекомбинантный вирусный белок капсида/частицы проявляет природный тропизм, находящийся в пределах от уменьшенного до устраненного. В некоторых вариантах осуществления вирусная частица дополнительно содержит второго родственного партнера специфической связывающейся пары, где первый и второй партнеры ковалентно связаны, и где второй партнер слит с нацеливающим лигандом.
[0090] В некоторых вариантах осуществления гетерологичная аминокислотная последовательность содержит первого партнера специфической связывающейся пары и один или более линкеров. В некоторых вариантах осуществления гетерологичная аминокислотная последовательность содержит первого партнера специфической связывающейся пары, фланкированный линкером, например, гетерологичная аминокислотная последовательность содержит, в направлении от N-конца к С-концу, первый линкер, первого партнера специфической связывающейся пары и второй линкер. В некоторых вариантах осуществления длина каждого из первого и второго линкеров независимо составляет по меньшей мере одну аминокислоту. В некоторых вариантах осуществления первый и второй линкеры являются идентичными.
[0091] Как правило, длина гетерологичной аминокислотной последовательности, описанной в настоящем документе, например, содержащей первого партнера специфической связывающейся пары сам по себе или в комбинации с одним или более линкерами, составляет от приблизительно 5 до приблизительно 50 аминокислот.В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательность составляет по меньшей мере 5 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательностм составляет 6 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 7 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 8 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 9 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 10 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 11 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 12 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 13 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 14 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 15 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 16 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 17 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 18 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 19 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 20 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 21 аминокислоту. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 22 аминокислоты. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 23 аминокислоты. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 24 аминокислоты. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 25 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 26 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 27 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотн ой последовательности составляет 28 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 29 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 30 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 31 аминокислоту. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 32 аминокислоты. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 33 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 34 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 35 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 36 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 37 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 38 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 39 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 40 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 41 аминокислоту. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 42 аминокислоты. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 43 аминокислоты. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 44 аминокислоты. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 45 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 46 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 47 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 48 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 49 аминокислот. В некоторых вариантах осуществления длина гетерологичной аминокислотной последовательности составляет 50 аминокислот.
[0092] В некоторых вариантах осуществления специфическая связывающаяся пара представляет собой связывающуюся пару SpyTag:SpyCatcher, где первым партнером является SpyTag, а вторым родственным партнером является SpyCatcher. В некоторых вариантах осуществления специфическая связывающаяся пара представляет собой SpyTag:KTag, где первым партнером является SpyTag, а вторым родственным партнером является KTag. В некоторых вариантах осуществления специфическая связывающаяся пара представляет собой SpyTag:KTag, где первым партнером является KTag, а вторым родственным партнером является SpyTag. В некоторых вариантах осуществления специфическая связывающаяся пара представляет собой isopeptag:пилин-C, где первым партнером является isopeptag, а вторым родственным партнером является пилин-С или его часть. В некоторых вариантах осуществления специфическая связывающаяся пара представляет собой SnoopTag:SnoopCatcher, и первым партнером является SnoopTag, а вторым родственным партнером является SnoopCatcher.
[0093] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в настоящем документе, представляет собой капсидный белок Ad, например, капсидный белок серотипа Ad, выбранный из группы, состоящей из Ad1, Ad2, Ad3, Ad4, Ad5, Ad6 и Ad7. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида Ad2. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида Ad5. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок Ad, описанный в настоящем документе, предусматривает первого партнера специфической связывающейся пары в домене белка фибера, например, на карбоксиконце белка фибера, в «выступе» фибера и/или HI-петле «выступа» фибера.
[0094] В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в настоящем документе, получен с помощью гена капсидного белка аденоассоциированного вируса (AAV), например, гена капсида серотипа AAV, выбранного из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок получен с помощью гена капсида AAV2 или гена капсида AAV9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок представляет собой генетически модифицированный капсидный белок VP1 AAV2, аминокислотная последовательность дикого типа которого изложена под SEQ ID NO: 9. В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок представляет собой генетически модифицированный капсидный белок VP1 AAV9, аминокислотная последовательность дикого типа которого изложена под SEQ ID NO: 31.
[0095] Как правило, рекомбинантный вирусный капсидный белок, описанный в настоящем документе, содержит первого партнера специфической связывающейся пары, вставленный в капсидный белок и/или отображенный с помощью него таким образом, что первый партнер специфической связывающейся пары обеспечивает уменьшение и/или устранение природного тропизм капсидного белка или капсида, содержащего его. В некоторых вариантах осуществления первый партнер специфической связывающейся пары вставлен в область капсидного белка, участвующую в природном тропизме эталонного капсидного белка дикого типа, например, в область капсидного белка, связанную с клеточным рецептором. В некоторых вариантах осуществления первый партнер специфической связывающейся пары вставлен в домен «выступа» белка фибера Ad и/или отображен с помощью него. В некоторых вариантах осуществления первый партнер специфической связывающейся пары вставлен в HI-петлю белка фибера Ad и/или отображен с помощью нее. В некоторых вариантах осуществления первый партнер специфической связывающейся пары вставлен после аминокислотного положения, выбранного из группы, состоящей из G453 капсидного белка VP1 AAV2, N587 капсидного белка VP1 AAV2, G453 капсидного белка VP1 AAV9 и А589 капсидного белка VP1 AAV9. В некоторых вариантах осуществления первый партнер специфической связывающейся пары вставлен и/или отображен между аминокислотами N587 и R588 капсида VP1 AAV2. В некоторых вариантах осуществления рекомбинантный вирусный капсид, вирусный вектор, содержащий рекомбинантный вирусный капсид, и/или композиции, содержащие рекомбинантный вирусный капсид, содержат аминокислотную последовательность, изложенную под SEQ ID NO: 13, 15, 17, 19, 21, 23, 25, 27, 29, 35, 37 или 39. Дополнительные подходящие участки вставки, идентифицированные с помощью AAV2, хорошо известны в данной области (Wu et al. (2000) J. Virol. 74:8635-8647) и включают I-1, I-34, I-138, I-139, I-161, I-261, I-266, I-381, I-447, I-448, I-459, I-471, I-520, I-534, I-570, I-573, I-584, I-587, I-588, I-591, I-657, I-664, I-713 и I-716. Рекомбинантный вирусный капсидный белок, описанный в данном документе, может представлять собой капсидный белок AAV2, содержащий первого партнера специфической связывающейся пары, вставленный в положение, выбранное из группы, состоящей из I-1, I-34, I-138, I-139, I-161, I-261, I-266, I-381, I-447, I-448, I-459, I-471, I-520, I-534, I-570, I-573, I-584, I-587, I-588, I-591, I-657, I-664, I-713, I-716 и их комбинации. Дополнительные подходящие участки вставки, идентифицированные с помощью дополнительных серотипов AAV, хорошо известны и включают I-587 (AAV1), I-589 (AAV1), I-585 (AAV3), I-585 (AAV4) и I-585 (AAV5). В некоторых вариантах осуществления рекомбинантный вирусный капсидный белок, описанный в данном документе, может представлять собой капсидный белок AAV2, содержащий первого партнера специфической связывающейся пары, вставленного в положение, выбранное из группы, состоящей из I-587 (AAV1), I-589 (AAV1), I-585 (AAV3), I-585 (AAV4), I-585 (AAV5) и их комбинации.
[0096] Используемая номенклатура I-### в данном документе относится к участку вставки с ### обозначением номера аминокислоты относительно белка VP1 капсидного белка AAV, однако такая вставка может быть расположена непосредственно на N- или С-конце, предпочтительно на С-конце одной аминокислоты в последовательности из 5 аминокислот, расположенных в направлении N- или С-конца от данной аминокислоты, предпочтительно 3, более предпочтительно 2, особенно 1 аминокислоты (аминокислот) в направлении к N- или С-концу от данной аминокислоты. Кроме того, положения, на которые ссылаются в данном документе, находятся относительно белка VP1, кодируемого геном капсида AAV, и соответствующие положения (и их мутации) могут быть легко идентифицированы для капсидных белков VP2 и VP3, кодируемых геном капсида, посредством выполнения выравнивания последовательностей белков VP1, VP2 и VP3, кодируемых эталонным геном капсида AAV.
[0097] Соответственно, вставка в соответствующее положение кодирующей нуклеиновой кислоты одного из этих участков гена cap приводит к вставке в VP1, VP2 и/или VP3, поскольку капсидные белки кодируются с помощью перекрывающихся рамок считывания того же гена с поочередными стартовыми кодонами. Следовательно, для AAV2, например, согласно этой номенклатуре вставки между аминокислотами 1 и 138 осуществлены только в VP1, вставки между 138 и 203 осуществлены в VP1 и VP2, а вставки между 203 и С-концом осуществлены в VP1, VP2 и VP3, что, конечно, также относится и к участку вставки I-587. Следовательно, настоящее изобретение охватывает структурные гены AAV с соответствующими вставками в белки VP1, VP2 и/или VP3.
[0098] Кроме того, вследствие высокой степени консервативности по меньшей мере крупных участков и большого партнера близкородственных партнеров семейства соответствующие участки вставки для AAV, отличные от перечисленных для AAV, можно идентифицировать путем выполнения выравнивания аминокислот или сравнения капсидных структур. См., например, Rutledge et al. (1998) J. Virol. 72:309-19 и патент США №9624274 для иллюстративных выравниваний различных капсидных белков AAV, каждый из которых включен в настоящий документ посредством ссылки во всей своей полноте.
[0099] В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение или устранение природного тропизма вирусного вектора, например, трансдукция клетки, естественным образом пермиссивной для инфекции с помощью эталонных вирусных векторов дикого типа и/или клетки-мишени не обнаруживается в отсутствие ковалентной связи со вторым родственным партнером связывающейся пары, который слит с соответствующим нацеливающим лигандом. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора, например, по сравнению с трансдукцией клетки, естественным образом пермиссивной для инфекции эталонными вирусными векторами дикого типа. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 5%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 5%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 10%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 20%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 30%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 40%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 50%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 60%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 70%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 80%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 90%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 95%. В некоторых вариантах осуществления вставка (отображение) первого партнера специфической связывающейся пары обеспечивает уменьшение природного тропизма вирусного вектора на по меньшей мере 90%. В вариантах осуществления, где вставка (отображение) первого партнера специфической связывающейся пары не обеспечивает полное устранение природного тропизма рекомбинантных вирусных капсидов, природный тропизм таких рекомбинантных вирусных капсидов может быть дополнительно уменьшен с помощью второй и другой мутации. Например, в одном варианте осуществления рекомбинантный вирусный капсидный белок, описанный в данном документе, может быть получен из серотипа AAV9, и может содержать первого партнера специфической связывающейся пары, и может дополнительно содержать мутацию, например, мутацию W503A.
[00100] Такая отмена нацеливания вируса в отношении его природной клетки-хозяина важна, особенно если предполагается системное, а не локальное или локально-региональное введение вирусных векторов, поскольку поглощение вирусных векторов природными клетками-хозяевами ограничивает эффективную дозу вирусных векторов. Сообщается, что в случае AAV2 и AAV6 HSPG является основным рецептором для поглощения вируса в большом количестве клеток, особенно клеток печени. Для AAV2 HSPG-связывающая активность зависит от группы из 5 основных аминокислот, R484, R487, R585, R588 и К532 (Kern et al., (2003) J Virol. 77(20): 11072-81). Соответственно, предпочтительными точечными мутациями являются те, которые обеспечивают снижение трансдуцирующей активности вирусного вектора в отношении данной клетки-мишени, опосредованной природным рецептором, на по меньшей мере 50%, предпочтительно на по меньшей мере 80%, особенно предпочтительно на по меньшей мере 95%, в случае HSPG как первичного рецептора для связывания вирусных векторов с клетками-мишенями.
[00101] Следовательно, дополнительные мутации, предпочтительные для HSPG-связывающих вирусных векторов, представляют собой такие мутации, при которых удалена или заменена основная аминокислота, такая как R, K или Н, предпочтительно R или K, которая участвует в связывании HSPG соответствующего вируса, на неосновную аминокислоту, такую как A, D, G, Q, S и Т, предпочтительно А или аминокислоту, которая присутствует в соответствующем положении другого, но высококонсервативного серотипа AAV, в котором отсутствует такая основная аминокислота в данном положении. Следовательно, предпочтительными аминокислотными заменами являются R484A, R487A, R487G, К532А, K532D, R585A, R585S, R585Q, R585A или R588T, особенно R585A и/или R588A в случае AAV2 и К531А или К531Е в случае AAV6. Одним особенно предпочтительным вариантом осуществления настоящего изобретения являются такие мутанты капсидного белка AAV2, которые дополнительно содержат две точечные мутации R585A и R588A, поскольку этих двух точечных мутаций достаточно для удаления активности связывания HSPG в значительной степени. Эти точечные мутации обеспечивают эффективную отмену нацеливания в отношении HSPG-экспрессирующих клеток, что, в целях нацеливания, повышает специфичность соответствующего мутантного вируса к его новой клетке-мишени.
Нацеливающие лиганды
[00102] Описанная в данном документе вирусная частица может дополнительно содержать второго партнера специфической связывающейся пары, который специфически образует ковалентную связь с первым партнером специфической связывающейся пары, который вставлен в рекомбинантный вирусный капсидный белок/отображен с помощью него, где второй партнер слит с нацеливающим лигандом. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым на поверхности клетки, например, белком клеточной поверхности на эукариотической клетке (человека), например, клетке-мишени. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) клетками печени (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) клетками мозга (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) лимфоцитами (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) Т-клетками (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) В-клетками (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) дендритными клетками (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) макрофагами (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) NK-клетками (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) клетками почки (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) раковой клеткой (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) клеткой (человека), инфицированной гетерологичным патогеном.
[00103] Существует большое количество белков клеточной поверхности, например, рецепторов клеточной поверхности, на которые подходящим образом можно нацеливаться с помощью нацеливающего лиганда, и для которых нацеливающий лиганд, например, антитела или их части, уже являются доступными. Такие структуры включают без ограничения антигены главного комплекса гистосовместимости класса I и класса II; рецепторы для различных цитокинов (например, рецепторы для IL-1, IL-4, IL-6, IL-13, IL-22, IL-25, IL-33 и т.д.), специфические для клеточного типа гормоны роста, нейротрофический фактор мозга (BDNF), цилиарный нейротрофический фактор (CTNF), колониестимулирующие факторы роста, эндотелиальные факторы роста, эпидермальные факторы роста, факторы роста фибробластов, нейротрофический фактор глиальных клеток, глиальные факторы роста, gro-beta/mip 2, факторы роста гепатоцитов, инсулиноподобный фактор роста, интерфероны (α-IFN, β-IFN, γIFN, консенсусный IFN), интерлейкины (IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14), фактор роста кератиноцитов, факторы, подавляющие лейкоз, фактор активации хемотаксиса макрофагов/моноцитов, фактор роста нервной ткани, активирующий нейтрофилы белок 2, фактор роста тромбоцитов, фактор роста стволовых клеток, трансформирующий фактор роста, факторы некроза опухолей, фактор роста эндотелия сосудов, липопротеины, в том числе дополнительные или другие трансмембранные рецепторы типа 1, такие как PRLR, рецепторы, связанные с G-белком, такие как GCGR, ионные каналы, такие как Nav1.7, ASIC1 или ASIC2; молекулы клеточной адгезии; транспортные молекулы для метаболитов, таких как аминокислоты; антигенные рецепторы В- и Т-лимфоцитов (например, рецепторы В-клеток и ассоциированных белков (например, CD 19, CD20 и т.д.) и рецепторы Т-клеток и ассоциированных белков (например, CD3, CD4, CD8 и т.д.); белок тетраспанин (например, CD63). Рекомбинантный вирусный капсид, описанный в настоящем документе, обеспечивает специфическую инфекцию клеточного типа путем применения нацеливающего лиганда, который связывается с дифференцирующими антигенами клеточной поверхности в качестве мишеней для комплекса вирусного вектора.
[00104] В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) клетками печени человека, т.е. специфическим для печени маркером. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) клетками мозга (человека), специфическим маркером клеток головного мозга. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) гемопоэтическими клетками (человека), т.е. специфическим маркером гемопоэтических клеток. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) Т-клетками (человека), т.е. специфическим для Т-клеток маркером. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) В-клетками (человека), т.е. специфическим для В-клеток маркером. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) дендритными клетками (человека), т.е. специфическим маркером дендритных клеток. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) макрофагами (человека), т.е. специфическим для макрофагов маркером. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) NK-клетками (человека), т.е. специфическим маркером NK-клеток. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) клетками почки (человека), т.е. специфическим для клеток почки маркером. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) клетками поджелудочной железы (человека), т.е. специфическим для клеток поджелудочной железы маркером. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым преимущественно (например, исключительно) клетками кишечника (человека), т.е. специфическим для клеток кишечника маркером. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) раковой клеткой (человека), т.е. опухолеассоциированным антигеном. В некоторых вариантах осуществления нацеливающий лиганд связывается с белком, экспрессируемым преимущественно (например, исключительно) клеткой (человека), инфицированной гетерологичным патогеном. Белки, которые (1) специфически экспрессируются, или в случае которых экспрессия в клетке/ткани/органе является обогащенной, и (2) распознаются антигенсвязывающим белком, применимым как нацеливающий лиганд, описанный в данном документе, хорошо известны и также могут быть найдены на www.proteinatlas.org; см. также Uhlen et al. (2010) Nat. Biotech. 28:1248-50, включенный в данный документ посредством ссылки во всей своей полноте. В приведенной ниже таблице 2 представлены иллюстративные и неограничивающие маркеры, специфические для органа, для которых доступны антигенсвязывающие белки, которые могут быть применимы как нацеливающие лиганды, и клетки/ткани/органы, экспрессирующие такие маркеры.
[00105] В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым клеткой печени (человека), например, асиалогликопротеиновым рецептором, например, hASGR1. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым клеткой мозга (человека). В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым Т-клеткой (человека), например, CD3, например, CD3ε. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым клеткой почки (человека), например. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым мышечной клеткой (человека), например, интегрином. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором, экспрессируемым раковой клеткой (человека), например, опухолеассоцированным антигеном, например, Е6 и Е7. В некоторых вариантах осуществления нацеливающий лиганд связывается с рецептором глюкагона человека (hGCGR).
[00106] В некоторых вариантах осуществления нацеливающий лиганд связывается с опухолеассоциированным антигеном, экспрессируемым опухолевой клеткой. Неограничивающие примеры специфических опухолеассоциированных антигенов включают, например, AFP, ALK, белки BAGE, β-катенин, brc-abl, BRCA1, BORIS, СА9, карбоангидразу IX, каспазу-8, CCR5, CD19, CD20, CD30, CD40, CDK4, СЕА, CTLA4, циклин-В1, CYP1B1, EGFR, EGFRvIII, ErbB2/Her2, ErbB3, ErbB4, ETV6-AML, ЕрСАМ, EphA2, Fra-1, FOLR1, белки GAGE (например, GAGE-1, -2), GD2, GD3, GloboH, глипикан-3, GM3, gp100, Her2, HLA/B-raf, HLA/k-ras, HLA/MAGE-A3, hTERT, LMP2, белки MAGE (например, MAGE-1, -2, -3, -4, -6 и -12), MART-1, мезотелин, ML-IAP, Muc1, Muc2, Muc3, Muc4, Muc5, Muc16 (CA-125), MUM1, NA17, NY-BR1, NY-BR62, NY-BR85, NY-ESO1, OX40, p15, p53, PAP, PAX3, PAX5, PCTA-1, PLAC1, PRLR, PRAME, PSMA (FOLH1), белки RAGE, Ras, RGS5, Rho, SART-1, SART-3, Steap-1, Steap-2, сурвивин, TAG-72, TGF-β, TMPRSS2, Tn, TRP-1, TRP-2, тирозиназу и уроплакин-3.
[00107] В некоторых вариантах осуществления нацеливающий лиганд связывается с маркерами CD, связанными с иммунным ответом, например, CD3, CD4, CD8, CD19, CD20 и т.д.
[00108] Одним из вариантов осуществления настоящего изобретения является мультимерная структура, содержащая рекомбинантный вирусный капсидный белок по настоящему изобретению. Мультимерная структура содержит по меньшей мере 5, предпочтительно по меньшей мере 10, более предпочтительно по меньшей мере 30, наиболее предпочтительно по меньшей мере 60 рекомбинантных вирусных капсидных белков, содержащих первого партнера специфической связывающейся пары, описанной в настоящем документе. Они могут образовывать обычные вирусные капсиды (пустые вирусные частицы) или вирусные векторы (капсиды, инкапсулирующие представляющий интерес нуклеотид). Образование вирусных векторов, способных упаковывать вирусный геном, является весьма предпочтительным признаком при применении рекомбинантных вирусных капсидов, описанных в данном документе в качестве вирусных векторов.
[00109] Одним вариантом осуществления настоящего изобретения является нуклеиновая кислота, кодирующая капсидный белок, описанный выше. Нуклеиновая кислота предпочтительно представляет собой вектор, содержащий заявленную нуклеиновую. Нуклеиновые кислоты, в частности, векторы, необходимы для рекомбинантной экспрессии капсидных белков по настоящему изобретению.
[00110] Еще одним вариантом осуществления настоящего изобретения является применение по меньшей мере одного рекомбинантного вирусного капсидного белка и/или нуклеиновой кислоты, кодирующей его, предпочтительно по меньшей мере одной мультимерной структуры (например, вирусного вектора), для изготовления и применения в качестве вектора для переноса генов.
Способы применения и получения
[00111] Дополнительный вариант осуществления описанных в данном документе рекомбинантных вирусных капсидных белков предусматривает их применение для доставки представляющего интерес нуклеотида, например, репортерного гена или терапевтического гена, в клетку-мишень. Как правило, представляющим интерес нуклеотидом может быть плазмида для переноса, которая обычно может содержать последовательности 5' и 3' инвертированного концевого повтора (ITR), фланкирующие репортерный(-ые) ген(ы) или терапевтический(-ие) ген(ы) (который может находиться под контролем вирусного или отличного от вирусного промотора при включении в вектор AAV). В одном варианте осуществления представляющим интерес нуклеотидом является плазмида для переноса, содержащая в направлении от 5' до 3': 5'-ITR, промотор, ген (например, репортерный и/или терапевтический ген) и 3'-ITR.
[00112] Неограничивающие примеры применимых промоторов включают, например, промотор цитомегаловируса (CMV), промотор вируса некроза селезенки (SFFV), промотор фактора элонгации 1 альфа (EF1a) (промотор EFla 1,2 т.п.о. или промотор EFla 0,2 т.п.о.), химерный промотор EF 1 a/IF4 и фосфоглицераткиназный (PGK) промотор. В вирусной конструкции также может присутствовать внутренний энхансер для увеличения экспрессии представляющего интерес гена. Например, можно применять энхансер CMV (Karasuyama et al. 1989. J. Exp. Med. 169:13, который включен в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления энхансер CMV можно применять в комбинации с промотором гена β-актина курицы.
[00113] Разнообразные репортерные гены (или выявляемые фрагменты) могут быть заключены в мультимерную структуру, содержащую рекомбинантные вирусные капсидные белки, описанные в данном документе. Иллюстративные репортерные гены включают, например, β-галактозидазу (кодируемую геном lacZ), зеленый флуоресцентный белок (GFP), усиленный зеленый флуоресцентный белок (eGFP), MmGFP, синий флуоресцентный белок (BFP), усиленный синий флуоресцентный белок (eBFP), mPlum, mCherry, tdTomato, mStrawberry, J-Red, DsRed, mOrange, mKO, mCitrine, Venus, YPet, желтый флуоресцентный белок (YFP), усиленный желтый флуоресцентный белок (eYFP), Emerald, CyPet, голубой флуоресцентный белок (CFP), Cerulean, T-Sapphire, люциферазу, щелочную фосфатазу или их комбинацию. Способы, описанные в данном документе, демонстрируют конструкцию нацеливающих векторов, в которой задействовано применение репортерного гена, который кодирует зеленый флуресцентный белок, однако специалистам в данной области при ознакомлении с настоящим изобретением будет понятно, что животных, отличных от человека, описанных в данном документе, можно создавать при отсутствии репортерного гена или с любым репортерным геном, известным из уровня техники.
[00114] Разнообразные терапевтические гены также могут быть заключены, могут быть заключены в мультимерную структуру, содержащую рекомбинантные вирусные капсидные белки, описанные в данном документе, например, как часть вектора для переноса. Неограничивающие примеры терапевтического гена включают гены, которые кодируют токсин (например, суицидальный ген), терапевтическое антитело или его фрагмент, систему CRISPR/Cas или ее часть(части), антисмысловую РНК, siRNA, shRNA и т.д.
[00115] Еще одним вариантом осуществления настоящего изобретения является способ получения рекомбинантного капсидного белка, при этом способ предусматривает следующие стадии:
a) осуществление экспрессии нуклеиновой кислоты, кодирующей рекомбинантный капсидный белок в подходящих условиях и
b) выделение экспрессированного капсидного белка со стадии а).
[00116] В некоторых вариантах осуществления вирусная частица, описанная в данном документе, содержит мозаичный капсид, например, капсид, содержащий капсидные белки, генетически модифицированные, как описано в настоящем документе (в отсутствие или в присутствии ковалентной связи с нацеливающим лигандом) в определенном соотношении с эталонными капсидными белками. Способ получения такой мозаичной вирусной частицы предусматривает:
a) осуществление экспрессии нуклеиновой кислоты, кодирующей рекомбинантный капсидный белок, и нуклеотида, кодирующего эталонный капсидный белок, в соотношении (вес/вес) 1:1 и 10:1 в подходящих условиях и
b) выделение экспрессированного капсидного белка со стадии а).
[00117] Говоря в общем, будет считаться, что мозаичный капсид, образованный в соответствии с этим способом, характеризуется соотношением модифицированный капсидный белок:эталонный капсидный белок, аналогичным соотношению (вес:вес) нуклеиновых кислот, кодирующих их, применяемых для получения мозаичного капсида. Соответственно, в некоторых вариантах осуществления композиция, описанная в настоящем документе, содержит, или в способе, описанном в настоящем документе, объединяют рекомбинантный вирусный капсидный белок и эталонный капсидный белок (или комбинацию эталонных капсидных белков) в соотношении, которое находится в диапазоне от 1:1 до 1:15. В некоторых вариантах осуществления соотношение составляет 1:2. В некоторых вариантах осуществления соотношение составляет 1:3. В некоторых вариантах осуществления соотношение составляет 1:4. В некоторых вариантах осуществления соотношение составляет 1:5. В некоторых вариантах осуществления соотношение составляет 1:6. В некоторых вариантах осуществления соотношение составляет 1:7. В некоторых вариантах осуществления соотношение составляет 1:8. В некоторых вариантах осуществления соотношение составляет 1:9. В некоторых вариантах осуществления соотношение составляет 1:10. В некоторых вариантах осуществления соотношение составляет 1:11. В некоторых вариантах осуществления соотношение составляет 1:12. В некоторых вариантах осуществления соотношение составляет 1:13. В некоторых вариантах осуществления соотношение составляет 1:14. В некоторых вариантах осуществления соотношение составляет 1:15.
[00118] Дополнительные варианты осуществления настоящего изобретения представляют собой способ изменения тропизма вируса, предусматривающий стадии (а) осуществления вставки нуклеиновой кислоты, кодирующей гетерологичную аминокислотную последовательность, в последовательность нуклеиновой кислоты, кодирующую вирусный капсидный белок, с образованием нуклеотидной последовательности, кодирующей генетически модифицированный капсидный белок, содержащий гетерологичную аминокислотную последовательность, и/или (b) культивирования упаковывающей клетки в условиях, достаточных для продуцирования вирусных векторов, где упаковывающая клетка содержит нуклеиновую кислоту. Еще один вариант осуществления настоящего изобретения представляет собой способ отображения нацеливающего лиганда на поверхности капсидного белка, при этом способ предусматривает стадии (а) осуществления экспрессии нуклеиновой кислоты, кодирующей рекомбинантный вирусный капсидный белок, описанный в настоящем документе (и необязательно с нуклеотидом, кодирующим эталонный капсидный белок), в подходящих условиях, где нуклеиновая кислота кодирует капсидный белок, содержащий первого партнера специфической связывающейся пары, (b) выделения экспрессированного капсидного белка, содержащего первого партнера специфической связывающейся пары, со стадии (а) или капсида, содержащий его, и (с) инкубирования капсидного белка или капсида со вторым родственным партнером специфической связывающейся пары в условиях, подходящих для обеспечения возможности образования изопептидной связи между первым и вторым партнером, где второй родственный партнер специфической связывающейся пары слит с нацеливающим лигандом.
[00119] В некоторых вариантах осуществления упаковывающая клетка дополнительно содержит хелперную плазмиду и/или плазмиду для переноса, содержащую представляющий интерес нуклеотид. В некоторых вариантах осуществления способы дополнительно предусматривают выделение самокомплементарных векторов на основе аденоассоциированного вируса из культурального супернатанта. В некоторых вариантах осуществления способы дополнительно предусматривают лизирование упаковывающей клетки и выделение однонитевых векторов на основе аденоассоциированного вируса из клеточного лизата. В некоторых вариантах осуществления способы дополнительно предусматривают (а) удаление клеточного дебриса, (b) обработку супернатанта, содержащего вирусные векторы, нуклеазами, например ДНКазой I и MgCl2, (с) концентрирование вирусных векторов, (d) очистку вирусных векторов и (е) любую комбинацию (a)-(d).
[00120] Упаковывающие клетки, пригодные для продуцирования вирусных векторов, описанных в настоящем документе, включают, например, клетки животных, пермиссивные для вируса, или клетки, модифицированные с обеспечением пермиссивности для вируса; или конструкцию упаковывающей клетки, например, с применением трансформирующего средства, такого как фосфат кальция. Неограничивающие примеры линий упаковывающих клеток, применимых для продуцирования вирусных векторов, описанных в данном документе, включают, например, эмбриональные клетки почки 293 человека (HEK-293) (например, под № Американской коллекции типовых культур [АТСС] CRL-1573), клетки HEK-293, которые содержат большой Т-антиген SV40 (HEK-293Т или 293Т), клетки HEK293T/17, клеточную линию саркомы человека НТ-1080 (CCL-121), лимфобластоподобную клеточную линию Raji (CCL-86), эпителиоподобную клеточную линию глиобластомы-астроцитомы U87-MG (НТВ-14), клеточную линию Т-лимфомы HuT78 (TIB-161), клетки NIH/3T3, клетки яичника китайского хомячка (СНО) (например, под №№ АТСС CRL9618, CCL61, CRL9096), клетки HeLa (например, №АТСС CCL-2), клетки Vero, клетки NIH 3Т3 (например, №АТСС CRL-1658), клетки Huh-7, клетки BHK (например, № АТСС CCL10), клетки PC12 (№ АТСС CRL1721), клетки COS, клетки COS-7 (№ АТСС CRL1651), клетки RATI, мышиные L-клетки (№ АТСС CCLI.3), клетки HLHepG2, клетки САР, клетки САР-Т и т.п.
[00121] Клетки L929, вирусная система упаковывающих клеток FLY, описанная в Cosset et al. (1995) J. Virol 69, 7430-7436, клетки NS0 (мышиной миеломы), человеческие амниоцитарные клетки (например, САР, САР-Т), дрожжевые клетки (в том числе без ограничения S. cerevisiae, Pichia pastoris), растительные клетки (в том числе без ограничения Tobacco NT1, BY-2), клетки насекомых (в том числе без ограничения SF9, S2, SF21, Tni (например, High 5)) или бактериальные клетки (в том числе без ограничения Е. coli).
[00122] Для дополнительных упаковывающих клеток и систем, методик упаковки и векторов для упаковки генома нуклеиновой кислоты в псевдотипированный вирусный вектор см., например, Polo et al., Proc Natl Acad Sci USA, (1999) 96:4598-4603. Способы упаковки предусматривают применение упаковывающих клеток, которые постоянно экспрессируют вирусные компоненты, или путем временной трансфекции клеток плазмидами.
[00123] Дополнительные варианты осуществления предусматривают способы перенаправления вируса и/или доставки репортерного или терапевтического гена в клетку-мишень, при этом способ предусматривает способ трансдукции клеток in vitro или in vivo, при этом способ предусматривает стадии приведения в контакт клетки-мишени с вирусным вектором, содержащим капсид, описанный в данном документе, где капсид содержит нацеливающий лиганд, который специфически связывается с рецептором, экспрессируемым клеткой-мишенью. В некоторых вариантах осуществления клетка-мишень находится in vitro. В других вариантах осуществления клетка-мишень находится in vivo в организме субъекта, например, человека.
Клетки-мишени
[00124] Для доставки представляющего интерес нуклеотида с применением рекомбинантного вирусного вектора, описанного в настоящем документе, можно нацеливаться на широкий ряд клеток. Клетки-мишени обычно будут выбирать на основе представляющего интерес нуклеотида и желаемого эффекта.
[00125] В некоторых вариантах осуществления представляющий интерес нуклеотид можно доставлять с целью обеспечения клетке-мишени возможности продуцировать белок, который восполняет дефицит в организме, такой как ферментативный дефицит или иммунный дефицит, такой как тяжелый комбинированный иммунодефицит, связанный с Х-хромосомой. Таким образом, в некоторых вариантах осуществления клетки животных, которые обычно продуцируют белок, являются мишенью. В других вариантах осуществления мишенью являются клетки в участке, в котором белок был бы наиболее полезным.
[00126] В других вариантах осуществления представляющий интерес нуклеотид, такой как ген, кодирующий siRNA, может подавлять экспрессию конкретного гена в клетке-мишени. Представляющий интерес нуклеотид может, например, подавлять экспрессию гена, вовлеченного в жизненный цикл патогена. Таким образом, клетки, восприимчивые к инфекции патогеном или инфицированные патогеном, могут являться мишенью. В других вариантах осуществления представляющий интерес нуклеотид может подавлять экспрессию гена, который отвечает за продуцирование токсина в клетке-мишени.
[00127] В других вариантах осуществления представляющий интерес нуклеотид может кодировать токсичный белок, который обеспечивает гибель клеток, в которых он экспрессируется. В этом случае опухолевые клетки или другие нежелательные клетки могут являться мишенью.
[00128] В еще других вариантах осуществления представляющий интерес нуклеотид кодирует терапевтический белок.
[00129] Как только определенная популяция клеток-мишеней, в которой желательна экспрессия представляющего интерес нуклеотида, идентифицирована, выбирают рецептор-мишень, который специфически экспрессируется в этой популяции клеток-мишеней. Рецептор-мишень может экспрессироваться исключительно в этой популяции клеток или в большей степени в этой популяции клеток, чем в других популяциях клеток. Чем более специфична экспрессия, тем более специфичная доставка может быть направлена к клеткам-мишеням. В зависимости от контекста желаемый уровень специфичности маркера (и, следовательно, доставки гена) может варьироваться. Например, при введении токсичного гена высокая специфичность наиболее предпочтительна во избежание гибели нецелевых клеток. Для экспрессии белка для сбора или экспрессии секретируемого продукта, когда требуется глобальное воздействие, может потребоваться меньшая специфичность маркера.
[00130] Как обсуждалось выше, рецептор-мишень может представлять собой любой рецептор, для которого может быть идентифицирован или создан нацеливающий лиганд. Предпочтительно рецептор-мишень представляет собой пептид или полипептид, такой как рецептор. Однако в других вариантах осуществления рецептор-мишень может представлять собой углевод или другую молекулу, которую может распознавать партнер по связыванию. Если партнер по связыванию, например, лиганд, для рецептора-мишени уже известен, его можно использовать как аффинную молекулу. Однако, если связывающая молекула неизвестна, могут быть получены антитела к рецептору-мишени с применением стандартных процедур. Затем антитела можно применять в качестве нацеливающего лиганда.
[00131] Таким образом, клетки-мишени могут быть выбраны на основе множества факторов, в том числе, например, (1) конкретного применения (например, терапия, экспрессия белка для сбора и придание устойчивости к заболеваниям) и (2) экспрессии маркера с желаемым уровнем специфичности.
[00132] Клетки-мишени никоим образом не ограничены и включают как клетки и клеточные линии зародышевой линии, так и соматические клетки и клеточные линии. Клетки-мишени могут быть стволовыми клетками любого происхождения. Когда клетки-мишени представляют собой клетки зародышевой линии, клетки-мишени предпочтительно выбраны из группы, состоящей из одноклеточных эмбрионов и эмбриональных стволовых клеток (ES).
Фармацевтические композиции, лекарственные формы и введение
[00133] Дополнительный вариант осуществления предусматривает лекарственный препарат, содержащий по меньшей мере один рекомбинантный вирусный капсидный белок и соответствующий нацеливающий лиганд по настоящему изобретению и/или нуклеиновую кислоту по настоящему изобретению. Предпочтительно такой лекарственный препарат применим в качестве частицы для переноса генов.
[00134] Также в данном документе раскрыты фармацевтические композиции, содержащие описанные в данном документе вирусные частицы и фармацевтически приемлемый носитель и/или вспомогательное вещество. Кроме того, в данном документе раскрыты фармацевтические лекарственные формы, содержащие вирусную частицу, описанную в данном документе.
[00135] Как обсуждено в настоящем документе, вирусные частицы, описанные в данном документе, можно использовать для различных терапевтических путей применения (in vivo и ex vivo) и как инструменты для исследования.
[00136] Фармацевтические композиции на основе вирусных частиц, раскрытых в настоящем документе, могут быть составлены любым общепринятым способом с применением одного или более физиологически приемлемых носителей и/или вспомогательных веществ. Вирусные частицы могут быть составлены для введения, например, путем инъекции, ингаляции или инсуляции (либо через рот, либо через нос), либо путем перорального, трансбуккального, парентерального или ректального введения, либо путем введения непосредственно в опухоль.
[00137] Фармацевтические композиции могут быть составлены для различных способов введения, в том числе системного, местного или локального введения. Методики и составы можно найти, например, в Remrnington's Pharmaceutical Sciences, Meade Publishing Co., Easton, Pa. Для системного введения предпочтительна инъекция, в том числе внутримышечная, внутривенная, внутрибрюшинная и подкожная. Для целей инъекции фармацевтические композиции могут быть составлены в жидких растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хенка или раствор Рингера. Кроме того, фармацевтические композиции могут быть составлены в твердой форме и повторно растворены или суспендированы непосредственно перед применением. Лиофилизированные формы фармацевтической композиции также являются подходящими.
[00138] Для перорального введения фармацевтические композиции могут находиться в форме, например, таблеток или капсул, полученных традиционными способами, с фармацевтически приемлемыми вспомогательными веществами, такими как связующие средства (например, предварительно желатинизированный маисовый крахмал, поливинилпирролидон или гидроксипропилметилцеллюлоза); наполнители (например, лактоза, микрокристаллическая целлюлоза или гидрофосфат кальция); смазывающие вещества (например, стеарат магния, тальк или диоксид кремния); разрыхлители (например, картофельный крахмал или гликолят крахмала натрия) или смачивающие средства (например, лаурилсульфат натрия). Таблетки также могут быть покрыты оболочкой с помощью способов, хорошо известных в данной области. Жидкие препараты для перорального введения могут находится в форме, например, растворов, сиропов или суспензий, или они могут быть представлены в виде сухого продукта для разведения водой или другой подходящей средой-носителем перед применением. Такие жидкие препараты можно получать с помощью общепринятых способов с фармацевтически приемлемыми добавками, такими как суспендирующие средства (например, сорбитный сироп, производные целлюлозы или гидрогенизированные пищевые жиры); эмульгирующие средства (например, лецитин или аравийская камедь); неводные среды-носители (например, эфирное масло, масляные сложные эфиры, этиловый спирт или фракционированные растительные масла) и консерванты (например, метил- или пропил-п-гидроксибензоаты или сорбиновая кислота). Препараты могут также содержать при необходимости буферные соли, ароматизирующие, красящие и подслащивающие средства.
[00139] Фармацевтические композиции могут быть составлены для парентерального введения путем инъекции, например, путем болюсной инъекции или непрерывной инфузии. Составы для инъекций могут быть представлены в виде стандартной лекарственной формы, например, в ампулах или в многодозовых контейнерах с необязательно добавленным консервантом. Фармацевтические композиции могут быть дополнительно составлены в виде суспензий, растворов или эмульсий в масляных или водных средах-носителях и могут содержать другие средства, в том числе суспендирующие, стабилизирующие и/или диспергирующие средства.
[00140] Кроме того, фармацевтические композиции также могут быть составлены в виде депо-препарата. Эти составы длительного действия можно вводить путем имплантации (например, подкожно или внутримышечно) или посредством внутримышечной инъекции. Так, например, соединения могут быть составлены с подходящими полимерными или гидрофобными материалами (например, в виде эмульсии в приемлемом масле) или ионообменными смолами или в виде труднорастворимых производных, например, в виде труднорастворимой соли. Другие подходящие системы доставки включают микросферы, которые обеспечивают возможность локальной неинвазивной доставки лекарственных средств в течение продолжительного периода времени. Эта технология может включать микросферы, имеющие прекапиллярный размер, которые можно вводить через коронарный катетер в любую выбранную часть органа, не вызывая воспаления или ишемии. Введенное терапевтическое средство медленно высвобождается из микросфер и поглощается окружающими клетками, присутствующими в выбранной ткани.
[00141] Системное введение также можно осуществлять чрезслизистым или чрескожным способами. В случае чрезслизистого или чрескожного введения в составе используют пенетранты, соответствующие барьеру, через который необходимо проникнуть. Такие пенетранты в целом известны из уровня техники и включают, например, в случае чрезслизистого введения, соли желчных кислот и производные фузидовой кислоты. Кроме того, для облегчения проникновения можно использовать детергенты. Чрезслизистое введение можно осуществлять посредством применения назальных спреев или суппозиториев. В случае местного применения вирусные частицы, описанные в данном документе, можно составлять в виде мазей, бальзамов, гелей или кремов, как в целом известно из уровня техники. Так же локально для лечения травмы или воспаления с целью ускорения заживления можно применять раствор для промывания.
[00142] Фармацевтические формы, подходящие для инъекционного применения, включают стерильные водные растворы или дисперсии, составы, включающие кунжутное масло, арахисовое масло или водный раствор пропиленгликоля; а также стерильные порошки для приготовления стерильных инъекционных растворов или дисперсий для немедленного применения. Во всех случаях форма должна быть стерильной и должна быть текучей. Она должна быть стабильной в условиях производства и при определенных параметрах хранения (например, охлаждение и заморозка) и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибы.
[00143] Если составы, раскрытые в данном документе, применяют как терапевтическое средство для усиления иммунного ответа у субъекта, терапевтическое средство может быть составлено в композицию в нейтральной или солевой форме. Фармацевтически приемлемые соли включают соли присоединения кислот (образованные с помощью свободных аминогрупп белка) и которые образуются с помощью неорганических кислот, таких как, например, хлористоводородная или фосфорная кислоты, или таких органических кислот, как уксусная, щавелевая, винная, миндальная и т.п. Соли, образованные с помощью свободных карбоксильных групп, также могут быть получены с помощью неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и т.п.
[00144] Носитель также может представлять собой растворитель или дисперсионную среду, содержащие, например, воду, этанол, многоатомный спирт (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.), их подходящие смеси и растительные масла. Надлежащую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Предотвращение действия микроорганизмов можно осуществлять с помощью различных антибактериальных и противогрибковых средств, известных в области техники. Во многих случаях будет предпочтительно включать изотонические средства, например, сахара или хлорид натрия. Пролонгированного всасывания инъекционных композиций можно добиться путем применения в композициях средств, задерживающих всасывание, например, моностеарата алюминия и желатина.
[00145] Стерильные инъекционные растворы можно получать путем включения активных соединений или конструкций в требуемом количестве в соответствующий растворитель вместе с различными другими ингредиентами, перечисленными выше, в случае необходимости, с последующей стерилизацией путем фильтрации.
[00146] После составления растворы можно вводить способом, совместимым с дозированным составом, и в таком количестве, которое является терапевтически эффективным. Составы без труда вводят в разнообразных лекарственных формах, как, например, типа инъекционных растворов, описанных выше, однако также можно применять капсулы или микрочастицы или микросферы с медленным высвобождением и т.п.
[00147] Для парентерального введения в виде водного раствора, например, раствор следует надлежащим образом забуферить при необходимости, а жидкий разбавитель вначале сделать изотоническим с помощью достаточного количества солевого раствора или глюкозы. Данные конкретные водные растворы особенно подходят для внутривенного, внутриопухолевого, внутримышечного, подкожного и внутрибрюшинного введения. В этом контексте стерильные водные среды, которые можно применять, будут известны специалистам в данной области техники в свете настоящего изобретения. Например, одну дозу можно растворить в 1 мл изотонического раствора NaCl и либо добавить к 1000 мл жидкости для гиподермоклизиса, либо инъецировать в предполагаемое место инфузии.
[00148] Лицо, ответственное за введение, в любом случае будет определять соответствующую дозу для отдельного субъекта. Например, субъекту можно вводить описанные в данном документе вирусные частицы один раз в день или один раз в неделю в течение определенного периода времени или один раз в месяц, два раза в год или один раз в год, в зависимости от необходимости или воздействия патогенного организма или состояния у субъекта (например, рака).
[00149] В дополнение к соединениям, составленным для парентерального введения, такого как внутривенная, внутриопухолевая, внутрикожная или внутримышечная инъекция, другие фармацевтически приемлемые формы включают, например, таблетки или другие твердые вещества для перорального введения; липосомные составы; капсулы с замедленным высвобождением; биоразлагаемую и любую другую форму, применяемую в настоящее время.
[00150] Можно также применять интраназальные или ингаляционные растворы или спреи, аэрозоли или ингаляторы. Назальные растворы могут быть водными растворами, предназначенными для введения в носовые ходы в виде капель или спреев. Назальные растворы могут быть получены таким образом, что они во многом сходны с носовыми выделениями. Таким образом, водные назальные растворы обычно являются изотоническими и слегка забуференными с поддержанием рН от 5,5 до 7,5. Кроме того, в состав могут быть включены противомикробные консерванты, подобные тем, которые применяют в офтальмологических препаратах, и соответствующие стабилизаторы лекарственного средства, если это необходимо. Известны различные коммерческие назальные препараты, которые могут включать, например, антибиотики и антигистаминные препараты, и применяются для профилактики астмы.
[00151] Составы для перорального применения могут включать вспомогательные вещества, такие как, например, маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлоза, карбонат магния и т.п. фармацевтической степени чистоты. Этим композициям придают форму растворов, суспензий, таблеток, пилюль, капсул, составов с замедленным высвобождением или порошков. В некоторых определенных вариантах осуществления фармацевтические композиции для перорального применения будут включать инертный разбавитель или усваиваемый съедобный носитель, или они могут быть заключены в желатиновую капсулу с твердой или мягкой оболочкой, или они могут быть спрессованы в таблетки, или они могут быть включены непосредственно в пищевой продукт, предусмотренный диетой. Для перорального терапевтического введения активные соединения можно соединять со вспомогательными веществами и применять в форме принимаемых внутрь таблеток, буккальных таблеток, троше, капсул, эликсиров, суспензий, сиропов, облаток и тому подобного.
[00152] Таблетки, троше, пилюли, капсулы и тому подобное также могут содержать следующее: связующее вещество, такое как трагакантовая камедь, аравийская камедь, кукурузный крахмал или желатин; вспомогательные вещества, такие как дикальцийфосфат; разрыхлитель, такой как кукурузный крахмал, картофельный крахмал, альгиновая кислота и тому подобное; смазывающее вещество, такое как стеарат магния; и может быть добавлен подсластитель, такой как сахароза, лактоза или сахарин, или ароматизатор, такой как мята перечная, масло грушанки или вишневый ароматизатор. Когда стандартная лекарственная форма представляет собой капсулу, она может содержать, помимо веществ вышеуказанного типа, жидкий носитель. Различные другие вещества могут присутствовать в качестве покрытий или иным образом модифицировать физическую форму лекарственной формы. Например, таблетки, пилюли или капсулы могут быть покрыты шеллаком, сахаром или и тем и другим. Сироп или эликсир может содержать активные соединения сахарозы как подсластитель, метил- и пропилпарабены как консерванты, красители и ароматизаторы, такие как вишневый или апельсиновый ароматизатор.
[00153] Другие варианты осуществления, раскрытые в данном документе, могут касаться наборов для применения в соответствии со способами и композициями. Наборы могут также включать подходящую емкость, например, флаконы, пробирки, мини- или микроцентрифужные пробирки, пробирку для испытания, колбу, флакон, шприц или другую емкость. Если предоставляется дополнительный компонент или средство, набор может содержать одну или более дополнительных емкостей, в которые может быть помещено это средство или компонент. Наборы в данном документе, как правило, также включают средства для содержания вирусных частиц и любые другие емкости с реагентами, плотно упакованные для коммерческой продажи. Такие емкости могут включать пластмассовые контейнеры, полученные путем литья под давлением или выдувного формования, в которые помещаются желаемые флаконы. Необязательно для описанных композиций может потребоваться одно или более дополнительных активных средств, таких как, например, противовоспалительные средства, противовирусные средства, противогрибковые или антибактериальные средства или противоопухолевые средства.
[00154] Композиции, раскрытые в данном документе, можно вводить любым способом, известным в данной области. Например, композиции могут предусматривать введение субъекту внутривенно, внутриопухолево, внутрикожно, внутриартериально, внутрибрюшинно, внутриочагово, внутричерепно, внутриартикулярно, внутрипростатически, внутриплеврально, внутритрахеально, интраназально, интравитреально, интравагинально, интраректально, местным нанесением, внутриопухолево, интрамускулатурно, интратекально, подкожно, субконъюнктивально, интравезикулярно, мукозально, интраперикардиально, внутрибрюшинно, интраокулярно, перорально, локально, путем ингаляции, путем инъекции, путем инфузии, непрерывной инфузии, посредством локальной перфузии, посредством катетера, лаважа, с помощью крема или в составе липидной композиции.
[00155] Для крупномасштабного производства вирусных частиц, упаковывающих клеток и конструкций частиц, описанных в данном документе, можно применять любой способ, известный специалисту в данной области. Например, основные и рабочие материалы для посева могут быть подготовлены в условиях GMP в квалифицированных первичных CEF или с помощью других способов. Упаковывающие клетки можно помещать в колбы с большой площадью поверхности, выращивать практически до достижения конфлюентности и выполнять очистку вирусных частиц. Клетки можно собирать и выделять и очищать вирусные частицы в культуральную среду, или высвобождать внутриклеточные вирусные частицы путем механического разрушения (клеточный дебрис можно удалять с помощью глубинной фильтрации с крупными порами и расщепления ДНК клетки-хозяина эндонуклеазой). Вирусные частицы могут быть впоследствии очищены и сконцентрированы путем фильтрации в тангенциальном потоке с последующей диафильтрацией. Полученную концентрированную массу можно составлять путем разбавления буфером, содержащим стабилизаторы, заполнить им флаконы и лиофилизировать. Композиции и составы могут храниться для последующего применения. Для применения лиофилизированные вирусные частицы могут быть восстановлены путем добавления разбавителя.
[00156] Определенные дополнительные средства, применяемые в комбинированных средствах терапии, могут быть составлены и введены любым способом, известным в данной области.
[00157] Композиции, раскрытые в данном документе, также могут включать адъюванты, такие как соли алюминия и другие минеральные адъюванты, тензоактивные средства, производные бактерий, носители и цитокины. Адъюванты также могут обладать антагонистическими иммуномодулирующими свойствами. Например, адъюванты могут стимулировать Th1- или Th2-иммунитет. Композиции и способы, раскрытые в данном документе, также могут предусматривать адъювантную терапию.
ПРИМЕРЫ
[00158] Следующие примеры приведены только в иллюстративных целях и не предназначены для ограничения объема настоящего изобретения.
Материалы и способы
[00159] Клеточные линии и антитела
[00160] Все клеточные линии 293 содержали в DMEM с добавлением 10% FBS, 1% пен./стреп. и 1% L-глютамина. Клеточные линии 293 hErbB2 и 293hASGR1/2 получали посредством лентивирусной трансдукции исходной клеточной линии 293 вектором, экспрессирующим соответствующую cDNA. Все клеточные линии получали из профильного центра Regeneron ТС. Антитело В1 распознает линейный эпитоп, общий для VP1, VP2 и VP3 AAV.
[00161] Конструкции капсидного белка AAV
[00162] GeneBlock, кодирующие нужные вставки SpyTag, фланкирующие линкерные аминокислоты и дополнительные мутации, приобретали у IDT и клонировали в расщепленные BsiWI и XcmI pAAV2-CAP wt или pAAV9-CAP wt с помощью Gibson Assembly в соответствии с протоколом производителя (NEB).
[00163] Слияние SpyCatcher с антителами
[00164] GeneBlock, кодирующие SpyCatcher, приобретали у IDT и использовали Gibson Assembly для клонирования кодирующей последовательности в рамке в плазмиды экспрессии для scFv или тяжелых цепей антитела на С-конце каждой конструкции, отделенной гибким аминокислотным линкером GSGESG (SEQ ID NO: 48).
[00165] Получение вирусных векторов на основе AAV
[00166] Вирус получали путем трансфекции упаковывающих клеток 293Т с применением PEI Pro следующими плазмидами: pAd Helper, AAV2-ITR-содержащей геномной плазмидой, кодирующей репортерный белок, и плазмидой pAAV-CAP, кодирующей гены Rep и Cap AAV, либо в присутствии, либо в отсутствие дополнительных плазмид, кодирующих либо scFv, либо тяжелые и легкие цепи антитела. Все конструкции scFv и тяжелой цепи антитела слиты с SpyCatcher на своем С-конце, как описано выше. Трансфекцию проводили в OptiMEM, а среду заменяли на DMEM, дополненную 10% FBS, 1% пен./стреп. и 1% L-глют., через 8 часов.
[00167] Трансфицированные упаковывающие клетки инкубировали в течение 3 дней при 37°С, затем вирус собирали из клеточных лизатов с применением стандартного протокола замораживания-оттаивания. Вкратце, упаковывающие клетки снимали с помощью соскоба и гранулировали. Супернатант удаляли и клетки ресуспендировали в растворе 50 мМ Tris-HCl; 150 мМ NaCl и 2 мМ MgCl2 [рН 8,0]. Внутриклеточные вирусные частицы высвобождали путем индуцирования лизиса клеток посредством трех последовательных циклов замораживания-оттаивания, состоящих из возвратно-поступательных перемещений суспензии клеток между баней с сухим льдом/этанолом и водяной баней при 37°С при энергичном встряхивании. Вязкость снижали путем обработки лизата с помощью EMD Millipore Benzonase (50 Ед/мл клеточного лизата) в течение 60 мин. при 37°С при периодическом перемешивании. Затем дебрис осаждали центрифугированием, а полученный супернатант фильтровали через PVDF-фильтр Millex-GV 0,22 мкм непосредственно в верхнюю камеру центробежного фильтра Amicon Ultra-15 с фильтрующим картриджем с мембраной Ultracel-100 (100 кДа MWCO). Фильтрующий блок центрифугировали с интервалами 5-10 минут, пока в верхней камере не был достигнут желаемый объем, затем концентрированный неочищенный вирус пипетировали в пробирку с незначительным связыванием с белками и хранили при 4С. Титр (вирусные геномы на миллилитр, vg/мл) определяли с помощью qPCR с применением стандартной кривой вируса известной концентрации.
[00168] Клеточная инфекция/трансдукция и анализ посредством проточной цитометрии
[00169] Для инфекции клеток вирусные частицы добавляли непосредственно в среду клеток в культуре, а смесь инкубировали в течение ночи при 37°С. Среду в каждой лунке заменяли через 24 часа и клетки инкубировали в течение 5 дней. На 5-й день после инфекции клетки трипсинизировали, ресуспендировали в PBS с 2% FBS, а процентное содержание клеток GFP+ собирали на проточном питометре BD FACSCanto и анализировали с применением программного обеспечения FlowJo.
[00170] Вестерн-блоттинг анализ
Реакцию между SpyTag-мечеными белками VP1, VP2 и VP3 AAV и SpyCatcher-мечеными антителами или scFv контролировали с помощью вестерн-блоттинг анализа. Буфер для образцов Novex® Tris-глицин SDS с восстанавливающим средством добавляли к равным объемам неочищенных вирусных препаратов, и образцы нагревали до 85°С в течение 5 минут, затем охлаждали до комнатной температуры и загружали в предварительно подготовленный 4-12% Tris-глициновый гель (Invitrogen). Белки разделяли путем восстановления при SDS-PAGE и подвергали блоттингу на PVDF с помощью влажного переноса. Мембраны блокировали блокирующим буфером Li-Cor Odyssey TBS и зондировали с помощью моноклонального антитела В1 мыши (ARP American Research Products, Inc.) при разведении 1:100 в TBST в течение ночи при 4°С. Блоты промывали в TBST, зондировали вторичным антителом против мыши, конъюгированным с инфракрасным красителем, и визуализировали на Li-Cor Odyssey.
Пример 1. Конъюгирование scFv с пептидом, вставленным в капсид AAV2 по остатку N587, направляет антигенспецифическое нацеливание
[00171] Каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293 Т следующими плазмидами и при следующих количествах:
В ПРИСУТСТВИИ или В ОТСУТСТВИЕ
[00172] Клетки, инкубированные с вирусными частицами, описанными выше, оценивали с помощью проточной цитометрии для выявления инфекции.
[00173] AAV2, несущий мутации, влияющие на связывание с гепарином (НВМ), R585A и R588A, а также пептид SpyTag в положении N587 капсида, получали в присутствии или в отсутствие C6.5-SpyCatcher, scFv, который связывает HER2 и слит с SpyCatcher на его С-конце. AAV2, конъюгированный с HER2-нацеливающимся scFv, специфически инфицирует клетки HER2+ и демонстрирует очень незначительную фоновую инфекцию клеток HER2-. Фигура 1.
Пример 2. Конъюгирование антитела с пептидом, вставленным в капсид AAV2 по остатку N587, направляет антигенспецифическое нацеливание
[00174] Каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами и при следующих количествах:
ИЛИ
В ПРИСУТСТВИИ ИЛИ В ОТСУТСТВИЕ
[00175] AAV2 дикого типа и AAV2, несущий мутации, влияющие на связывание с гепарином (НВМ), R585A и R588A, а также пептид SpyTag в положении капсида N587, получали в присутствии или в отсутствие тяжелых и легких цепей антитела, кодирующих SpyCatcher-герцептин, антитело, которое связывает HER2 и которое слито с SpyCatcher на С-конце тяжелой цепи. Клетки, инфицированные вирусными частицами, описанными выше, оценивали с помощью проточной цитометрии для контроля трансдукции. AAV2, конъюгированный с HER2-нацеливающим антителом, специфически инфицирует клетки HER2+ и демонстрирует очень незначительную фоновую инфекцию клеток HER2-. Фигура 2.
Пример 3. Конъюгирование антитела с пептидом, вставленным в капсид AAV2 по остатку G453, направляет антигенспецифическое нацеливание.
[00176] Каждый из остатков N587 и G453 лежит на открытых участках капсида AAV2, которые образуют шипы белка, которые выступают на поверхности вириона. Остатки лежат на двух разных шипах, в связи с этим исследовали, будет ли SpyTag, вставленный после остатка G453, функционировать так же, как SpyTag, вставленный после остатка N587. Каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами и при следующих количествах:
ИЛИ
В ПРИСУТСТВИИ или В ОТСУТСТВИЕ
[00177] AAV2 дикого типа и AAV2, несущий мутации, влияющие на связывание с гепарином (HBM), R585A и R588A, а также пептид SpyTag в положении капсида G453, получали в присутствии или в отсутствие тяжелых и легких цепей антитела, кодирующих SpyCatcher-герцептин, антитело, которое связывает HER2 и которое слито с SpyCatcher на С-конце тяжелой цепи. Вирус получали как мозаичный путем смешивания капсидов, экспрессирующих SpyTag, с капсидами НВМ. Клетки, инфицированные вирусными частицами, описанными выше, оценивали с помощью проточной цитометрии для контроля трансдукции. AAV2, конъюгированный с HER2- нацеливающим антителом, специфически инфицирует клетки HER2+ и демонстрирует очень незначительную фоновую инфекцию клеток HER2-. Фигура 3
Пример 4. Увеличение степени модификации вирионов AAV с помощью scFvs снижает их инфекционность
[00178] В попытках оптимизировать эффективность реакции SpyTag-SpyCatcher, доступность SpyTag на поверхности вируса улучшали путем фланкирования пептидной метки гибкими линкерными аминокислотами с каждой стороны. Получали панель инсерционных мутантов SpyTag N587, фланкированных с помощью линкеров увеличенной длины, и получали вирус с применением этих конструкций Rep-Cap AAV2 в присутствии или в отсутствие C6.5-SpyCatcher, scFv, который связывает HER2 и который слит с SpyCatcher на его С-конце. Реакцию между SpyTag-мечеными белками AAV2, VP1, VP2 и VP3, и SpyCatcher-меткой С6.5 контролировали с помощью вестерн-блоттинга; SpyTag-меченые капсидные белки, которые реагировали с меченым SpyCatcher scFv, демонстрируют увеличение размера с помощью SDS-PAGE. Клетки, инфицированные вирусными частицами, описанными выше, оценивали с помощью проточной цитометрии для измерения трансдукции.
[00179] Каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами и при следующих количествах:
[00180] Конструкции pAAV2-N587 CAP линкер SpyTag включают:
pAAV2-N587 CAP линкер 1 SpyTag HBM
pAAV2-N587 CAP линкер 2 SpyTag HBM
pAAV2-N587 CAP линкер 4 SpyTag HBM
pAAV2-N587 CAP линкер 6 SpyTag HBM
pAAV2-N587 CAP линкер 8 SpyTag HBM
pAAV2-N587 CAP линкер 10 SpyTag HBM
[00181] В случае, когда SpyTag не был фланкирован линкерными аминокислотами, комплексы VP-SpyTag-SpyCatcher-scFv не обнаруживались с помощью вестерн-блоттинга, и вирусы достигали низких уровней специфической трансдукции клеток HER2+. Фигура 4. Когда длина линкера увеличилась (1-6 аминокислот), комплексы VP-SpyTag-SpyCatcher-scFv начинали обнаруживаться с помощью вестерн-блоттинга, и эффективность трансдукции вирусов увеличивалась. Фигура 4. Однако, когда SpyTag был фланкирован двумя наиболее длинными линкерами (8-10 аминокислот), почти все белки VP реагировали с SpyCatcher-Vh при применении вестерн-блоттинга, но такие полностью отделанные вирусы больше не трансдуцировали клетки эффективно. Фигура 4. Таким образом, оказалось, что чрезмерная модификация частиц AAV с помощью scFv отрицательно сказывается их способности трансдуцировать клетки-мишени, и для перенацеливания вируса на клетки-мишени требуется лишь небольшое количество конъюгированных scFv
Пример 5. Увеличение степени модификации вирионов AAV с помощью антител снижает их инфекционность
[00182] С применением панели инсерционных мутантов SpyTag N587, фланкированных с помощью линкеров увеличенной длины, получали вирус с применением этих конструкций Rep-Cap AAV2 в присутствии или в отсутствие тяжелых и легких цепей антитела, кодирующих SpyCatcher-герцептин, антитела, которое связывает HER2 и которое слито с SpyCatcher на С-конце тяжелой цепи. Реакцию между SpyTag-мечеными белками AAV, VP1, VP2 и VP3, и SpyCatcher-меченой тяжелой цепью герцептина (Vh) контролировали с помощью вестерн-блоттинга; SpyTag-меченые капсидные белки, которые реагировали с мечеными SpyCatcher антителами, будут демонстрировать сдвиг размера с помощью SDS-PAGE.
[00183] Каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами и при следующих количествах:
[00184] Конструкции pAAV2-N587 CAP линкер SpyTag включают:
pAAV2-N587 CAP SpyTag HBM
pAAV2-N587 CAP линкер 1 SpyTag HBM
pAAV2-N587 CAP линкер 2 SpyTag HBM
pAAV2-N587 CAP линкер 4 SpyTag HBM
pAAV2-N587 CAP линкер 6 SpyTag HBM
pAAV2-N587 CAP линкер 8 SpyTag HBM
pAAV2-N587 CAP линкер 10 SpyTag HBM
[00185] Когда SpyTag не был фланкирован линкерными аминокислотами или фланкирован очень короткими аминокислотами, комплексы VP-SpyTag-SpyCatcher-Vh не обнаруживались при помощи вестерн-блоттинга, но вирусы специфически инфицировали клетки HER2+ с эффективностью, близкой к уровням дикого типа. Фигура 5. Наоборот, когда SpyTag был фланкирован двумя самыми длинными линкерами (6 или более аминокислот), почти все белки VP реагировали со SpyCatcher-Vh с помощью вестерн-блоттинга, но такие полностью отделанные вирусы больше не являлись инфекционными. Фигура 5. Увеличение степени модификации частиц AAV с помощью антител отрицательно сказывается на их способности трансдуцировать клетки-мишени, и для перенацеливания вируса на клетки-мишени требуется лишь небольшое количество конъюгированных антител.
Пример 6. Мозаичность модулирует эффективность трансдукции вирусных частиц
[00186] Поскольку длинные гибкие линкеры обеспечивают эффективную реакцию SpyTag-меченых капсидов AAV с scFv, слитыми с SpyCatcher, но чрезмерная модификация частиц AAV с помощью scFv отрицательно сказывается на их способности трансдуцировать клетки-мишени, количество SpyTag на каждом вирионе уменьшали, а доступность SpyTag и эффективную реакционноспособность сохраняли путем получения мозаичных частиц AAV, которые представляют собой смесь при различных соотношениях высокореакционноспособных SpyTag-меченых конструкций с конструкциями, отличными от SpyTag-меченых капсидных конструкций, все из которых несут мутацию R585A R588A, влияющую на связывание с гепарином (НВМ). Каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами и при следующих количествах:
ПРИ РАЗЛИЧНЫХ СООТНОШЕНИЯХ:
pAAV2-N587 САР линкер 10 SpyTag HBM Xug
pAAV2-R585A R588A CAP Xug
или
ПРИ РАЗЛИЧНЫХ СООТНОШЕНИЯХ:
pAAV2-N587 САР линкер 10 SpyTag HBM Xug
pAAV2-R585A R588A CAP Xug
или
ПРИ РАЗЛИЧНЫХ СООТНОШЕНИЯХ:
pAAV2-G453 САР линкер X SpyTag НВМ Xug
pAAV2-R585A R588A CAP Xug
[00187] Соотношение плазмид pAAV2-N587 САР линкер 10 SpyTag HBM и плазмид pAAV2-R585A R588A CAP составляло 1:0 (4 мкг:0 мкг), что представляет чистые вирионы pAAV2-N587 САР линкер 10 SpyTag НВМ, 3:1 (3 мкг:1 мкг), 1:1 (2 мкг:2 мкг) или 1:3 (1 мкг:3 мкг) в смеси для трансфекции. Соотношение плазмид pAAV2-G453 САР линкер 10 SpyTag НВМ и плазмид pAAV2-R585A R588A САР составляло 1:0 (4 мкг:0 мкг), что представляет чистые вирионы pAAV2-G453 САР линкер 10 SpyTag НВМ, 1:3 (1 мкг:3 мкг) или 1:7 (0,5 мкг:3,5 мкг) в смеси для трансфекции. Реакцию между SpyTag-мечеными белками AAV, VP1, VP2 и VP3, и SpyCatcher-меченым scFv к HER или SpyCatcher-меченой тяжелой цепью герцептина (Vh) контролировали с помощью вестерн-блоттинга; SpyTag-меченые капсидные белки, которые реагировали с SpyCatcher-мечеными scFv или антителами, будут демонстрировать сдвиг размера с помощью SDS-PAGE. Реакция панели N587 SpyTag линкер с SpyCatcher-scFv к HER2 показана на фигуре 6. Реакция панели N587 SpyTag линкер с SpyCatcher-антитело к HER2 показана на фигуре 7. Клетки, инфицированные мозаичными вирусными частицами, описанными выше, оценивали с помощью проточной цитометрии для измерения трансдукции, фигуры 6-7. По мере того, как количество высокореакционносособных капсидов SpyTag, фланкированных линкером 10, уменьшалось, а количество капсидов, не меченых SpyTag, увеличивалось, с помощью вестерн-блоттинга наблюдали уменьшение количества комплексов VP-SpyTag-SpyCatcher-scFv в сочетании с увеличением эффективности трансдукции вируса, фигуры 6-7. Была продемонстрирована обратная зависимость между количеством антител, которыми отделан вирион, и эффективностью трансдукции с помощью перенацеленного вируса.
[00188] Реакция G453 SpyTag НВМ и G453 линкер 10 SpyTag НВМ с SpyCatcher-антитело к HER2 показана на фигуре 8. Обе вставки SpyTag по G453 либо с помощью SpyTag отдельно, либо с помощью SpyTag, фланкированного линкером 10, очень эффективно реагировали с SpyCatcher-меченым белком с герцептином, что измеряли с помощью вестерн-блоттинга, что позволяет предположить, что участок вставки G453, естественно, более доступен, чем N587, который нелегко реагирует в отсутствие фланкирования линкерными аминокислотами. В случае линкерной панели N587 наблюдалось, что если вирусы были в высокой степени модифицированы антителом герцептином с SpyCatcher, вирусы больше не являлись инфекционными. Следовательно, оказалось, что высокие уровни модификации частиц AAV с помощью антител отрицательно сказываются на их способности трансдуцировать клетки-мишени, и можно сделать вывод, что SpyTag, вставленный после G453, естественным образом более доступен, чем SpyTag, вставленный по N587.
Пример 7. Систему SpyTag-SpyCatcher можно применять с дополнительными парами антитело-мишень для достижения специфического перенацеливания in vitro
[00189] Исследовали эффективность подхода SpyTag-SpyCatcher для перенацеливания AAV на мишени, отличные от HER2. SpyCatcher-меченые антитела, нацеливающиеся на дополнительные белки клеточной поверхности, клонировали и исследовали способность этих антител перенацеливать SpyTag-меченый AAV к типам клеток, экспрессирующим эти дополнительные мишени. Для экспериментов по нацеливанию на ASGR1 и CD63 каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами при следующих количествах:
В ПРИСУТСТВИИ ИЛИ В ОТСУТСТВИЕ
[00190] Для экспериментов по нацеливанию на PTPRN каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами при следующих количествах:
В ПРИСУТСТВИИ ИЛИ В ОТСУТСТВИЕ
[00191] Для экспериментов по нацеливанию на ENTPD3 и CD20 каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами при следующих количествах:
В ПРИСУТСТВИИ ИЛИ В ОТСУТСТВИЕ
[00192] Исследовали плазмиды SpyCatcher-Vh и Vk, которые кодируют тяжелые и легкие цепи антител, которые распознают человеческие белки ASGR1, CD63, PTPRN, ENTPD3 и CD20. Для получения мозаичных частиц AAV с небольшим количеством открытых SpyTag плазмиды SpyTag и плазмиды без SpyTag присутствовали в соотношении 1:7 в смеси для трансфекции, которое ранее было определено как идеальное соотношение капсидов с SpyTag и капсидов без SpyTag, для перенацеливания AAV с помощью антител. Клетки, экспрессирующие ASGR1, CD63 или PTPRN, инфицировали мозаичными частицами AAV2, описанными выше, и измеряли трансдукцию с помощью проточной цитометрии. AAV2, конъюгированная с антителами, специфичными к ASGR1, CD63 и PTPRN, была способна специфически инфицировать родственные клетки-мишени, экспрессирующие ASGR1, CD63 и PTPRN соответственно, и демонстрировала очень низкий фоновый уровень инфицирования в отсутствие антитела. Клетки, экспрессирующие ENTPD3 или CD20, инфицировали частицами AAV2, описанными выше, и измеряли трансдукцию с помощью люциферазного анализа с применением стандартных протоколов. AAV2, конъюгированная с антителами, специфичными к ENTPD3 и CD20, была способна специфически инфицировать родственные клетки-мишени, экспрессирующие ENTPD3 и CD20 соответственно, и демонстрировала очень низкий фоновый уровень инфицирования в отсутствие антитела. Фигура 9
Пример 8. Система SpyTag-SpyCatcher может быть адаптирована для перенацеливания AAV9
[00193] Исследовали адаптивность системы SpyTag-SpyCatcher к другим серотипам AAV. AAV9 является широко применяемым серотипом, который обеспечивает высокий титр вируса и очень эффективен при трансдукции мышиных тканей. Остатки, важные для связывания рецептора, отличаются у AAV2 и AAV9, поскольку AAV2 связывается с гепаринсульфатпротеогликанами, a AAV9 связывается с галактозой. Остатки, которые, как известно, важны для связывания с рецептором, были определены из доступной литературы (Bell, С.L., Gurda, В.L., Van Vliet, K., Agbandje-McKenna, М., & Wilson, J. М. (2012). Identification of the galactose binding domain of the adeno-associated virus serotype 9 capsid. Journal of Virology, 86(13), 7326-7333. http://doi.org/10.1128/JVI.00448-12) и включают N470, D271, N272, Y446 и W503. Мутация W503A была выбрана в качестве мутации, влияющей на связывание с рецептором, для применения при создании мутантных конструкций, поскольку такая одиночная аминокислотная мутация обеспечивала сильное уменьшение связывания рецептора. Области капсида AAV9, которые ортологичны по двум проекциям (переменные петли), в которых лежат N587 AAV2 и G453, также были идентифицированы; соответствующие остатки в AAV9 представляют собой А589 и G453. SpyTag вставляли в эти два участка в пределах капсида AAV9, с фланкирующими линкерными аминокислотами и без них и в комбинации с мутацией W503A, влияющей на связывание с рецептором.
[00194] Каждый вирус получали, как описано выше, путем трансфекции в одном 15 см планшете упаковывающих клеток 293Т следующими плазмидами и при следующих количествах:
[00195] Для AAV9 wt, AAV9-CAP A589SpyT_W503A, AAV9-A589 CAP линкер 10 SpyT_W503A и AAV9-G453 CAP линкер 10 SpyT_W503A использовали по 4 мкг каждой плазмиды для каждой трансфекции.
[00196] Для мозаичных вирусов использовали 3,5 мкг pAAV9-W503A САР и 0,5 мкг либо pAAV9-A589 САР линкер 10 SpyT_W503A, либо pAAV9-G453 САР линкер 10 SpyT_W503A для каждой трансфекции с достижением соотношения 1:7 плазмид Rep-Cap с SpyTag и без SpyTag.
[00197] Клеточную трансдукцию, анализ проточной цитометрии и вестерн-блоттинг проводили, как описано выше.
[00198] Вставки SpyTag А589 и G453 AAV9 RC обеспечивали поддержание реакции с SpyCatcher-герцептин и опосредование специфической трансдукции клеток HER2+. Фигура 10. Подобно AAV2, SpyTag, вставленный по А589 AAV9 RC без фланкирующих линкеров, не являлся хорошо доступным и плохо реагировал с SpyCatcher. Фигура 10. Наоборот, добавление аминокислотных линкеров с любой из сторон SpyTag обеспечило более устойчивую реакционную способность с SpyCatcher, и мозаичные частицы с несколькими высокореакционноспособными SpyTag были очень эффективными при трансдукции клеток-мишеней. Фигура 10.
Пример 9. Перенацеливание комплексов SpyTag-меченая частица AAV-SpyCatcher-Vh in vivo
[00199] Чтобы определить, возможно ли перенацелить VP-SpyTag-SpyCatcher-Vh на клетки печени, экспрессирующие hASGR1 in vivo, мышам, генетически модифицированным таким образом, что их клетки печени экспрессируют hASGRl с фоновым C57BL/6, и контрольным мышам дикого типа внутривенно вводили AAV дикого типа отдельно или вирусные частицы VP-SpyTag-SpyCatcher-Vh (в виде чистых или мозаичных частиц, в присутствии или в отсутствие аминокислотного линкера), несущие репортерный ген, например, зеленый флуоресцентный белок или люциферазу светлячка. Контроли включают мышей, которым инъецировали физиологический раствор с фосфатным буфером (PBS). Чтобы определить, можно ли отменить нацеливание на печень VP-SpyTag-SpyCatcher-Vh и перенацелить на другие органы, мышам дикого типа внутривенно вводили AAV дикого типа отдельно или вирусные частицы VP-SpyTag-SpyCatcher-Vh (в виде мозаичных частиц), несущие репортерный ген, например, зеленый флуоресцентный белок. Для демонстрации отмены нацеливания на печень и перенацеливания на другой орган выбрали белок ENTPD3, поскольку он не экспрессируется в печени, но экспрессируется в других органах, таких как островковые клетки поджелудочной железы (Syed et al. 2013, Am J Physiol Endocrinol Metab 305: E1319-E1326, 2013) и язык, в соответствии с общедоступными базами данных (данные получены из GenePaint.org http://www.informatics.jax.org/assay/MGI:5423021 и проекта Riken FANTOM5, набор данных для взрослых мышей).
[00200] SpyCatcher-меченые антитела, нацеливающиеся на CD3 человека, CD63 человека, ASGR1 человека (ни один из которых не экспрессируется у мышей дикого типа) или ENTPD3 человека (который также распознает белок мыши), клонировали и исследовали способность этих антител перенацеливать SpyTag-меченый AAV2, несущий либо eGFP, либо люциферазу светлячка, in vivo. Каждый вирус получали, как описано выше, путем трансфекции в 15 см планшетах упаковывающих клеток 293Т следующими плазмидами и при следующих количествах:
Антитело к CD3 человека/антитело к ASGR1 человека, GFP
В ПРИСУТСТВИИ ИЛИ В ОТСУТСТВИЕ
Антитело к CD63 человека/антитело к ASGR1 человека, люцифераза
В ПРИСУТСТВИИ или В ОТСУТСТВИЕ
Антитело к ENTPD3 человека/антитело к ASGR1 человека, GFP
В ПРИСУТСТВИИ или В ОТСУТСТВИЕ
[00201] Плазмиды SpyCatcher-Vh и Vk, которые кодируют тяжелые и легкие цепи антител, которые распознают человеческие белки ASGR1, CD3 или CD63, исследовали на мышах, генетически модифицированных таким образом, что их клетки печени экспрессируют hASGR1 с фоновым C57BL/6. Плазмиды SpyCatcher-Vh и Vk, которые кодируют антитело, которое распознает человеческий белок ASGR1 (как ненацеливающийся контроль) или мышиный и человеческий белок ENTPD3, исследовали на мышах дикого типа. Через десять дней после инфекции мышей умерщвляли и исследовали экспрессию репортерного гена; печени, селезенки и почки фиксировали и окрашивали куриным антителом к EGFP (Jackson ImmunoResearch Labs, Inc. Уэст-Гроув, Пенсильвания) и конъюгированным с Alexa-488 куриным вторичным антителом (Jackson ImmunoResearch Labs, Inc. Уэст-Гроув, Пенсильвания). Для исследования люминисценции у животных через 14 дней после инфекции живых животных анестезировали с помощью изофлурана, инъецировали субстрат люциферина и визуализировали через 10 минут с применением системы визуализации IVIS Spectrum In Vivo (PerkinElmer). На фигурах 11 и 12 показано, что инфекция комплексами AAV2 -SpyTag-SpyCatcher-Vh обнаруживалась только в печени мышей, экспрессирующих hASGR1, которым инъецировали перенацеленный на hASGR1 AAV, и не обнаруживалась в печени мышей дикого типа, которые не экспрессируют hASGR1. В других органах положительный результат EGF не обнаруживался (данные не показаны).
[00202] На фигуре 13 показано иммуногистохимическое окрашивание в отношении экспрессии eGFP в печени и органах поджелудочной железы мышей дикого типа после инъекции AAV, конъюгированных с антителами, нацеливающимися на ENTPD3 или, в качестве ненацеливающегося контроля, на hASGR1, поскольку hASGR1 не экспрессируется у мышей дикого типа. Через четыре недели после инфекции органы собирали у инфицированных животных и фиксировали в 10% нейтральном забуференном формалине в течение 48 часов, затем окрашивали в отношении eGFP с помощью иммуногистохимического окрашивания. На фигуре 13 показано, что в отношении комплексов AAV2-SpyTag-SpyCatcher-Vh отменено нацеливание на печень мышей дикого типа; у всех мышей, которым инъецировали AAV, конъюгированные с антителами, нацеливающимися на ENTPD3 и hASGR1, выявлено сходное отсутствие экспрессии eGFP в печени. Мышь, которой инъецировали AAV, конъюгированные с антителами, нацеливающимися на ENTPD3, имела клетки, экспрессирующие eGFP в островках поджелудочной железы, где, как полагают, экспрессируется ENTPD3.
[00203] На фигуре 14 показано иммуногистохимическое окрашивание в отношении экспрессии eGFP в печени и языке мышей дикого типа после инъекции AAV, конъюгированных с антителами, нацеливающимися на ENTPD3 или hASGR1 в качестве ненецеливающегося контроля, поскольку hASGR1 не экспрессируется у мышей дикого типа. Через 14 дней после инфицирования органы собирали у инфицированных животных и фиксировали в 10% нейтрально забуференном формалине в течение 48 часов, затем окрашивали в отношении eGFP посредством иммуногистохимического окрашивания.
На фигуре 14 показано, что в отношении комплексов AAV2-SpyTag-SpyCatcher-Vh отменено нацеливание на печень мышей дикого типа; у всех мышей, которым инъецировали AAV, конъюгированные с антителами, нацеливающимися на ENTPD3 и hASGR1, выявлено сходное отсутствие экспрессии eGFP в печени. У всех трех мышей, которым инъецировали AAV, конъюгированные с ненацеливающимся нерелевантным антителом (антителом к ASGR1), не наблюдалось окрашивания в языке, в то время как у всех трех мышей, которым инъецировали AAV, конъюгированные с антителом к ENTPD3, обнаруживались клетки, экспрессирующие eGFP, в языке, где, как полагают, экспрессируется ENTPD3.
Пример 10. Доставка суицидального гена к клеткам, экспрессирующим целевой лиганд
[00204] Также описана способность комплексов VP-SpyTag-SpyCatcher-Vh специфически доставлять терапевтическую нагрузку, такую как один или более суицидальных генов, биологическое терапевтическое средство (например, антитело), систему редактирования генов CRISPR/Cas, shRNA и т.д., к целевому типу клеток.
[00205] С целью исследовать способность комплексов VP-SpyTag-SpyCatcher-Vh доставлять суицидальный ген в конкретную клетку, применяли ксенотрансплантатную модель рака молочной железы HER2+ у голых мышей, как описано в Wang et al. ((2010) Cancer Gene Therapy 17:559-570).
[00206] Комплексы VP-SpyTag-SpyCatcher-Vh, несущие суицидальный ген (SG), получают аналогично тому, как описано в разделе «Материалы и способы».
[00207] Клеточные линии. Клеточные линии рака молочной железы ВТ474, рака молочной железы SK-BR-3 и рака легких Calu-З представляют собой HER2-положительные линии опухолевых клеток человека (Bunn P.A. et al., (2001) Clin Cancer Res. 7:3239-3250; Pegram M, et al. (1999) Oncogene 18:2241-2251; Spiridon CI, et al., (2002) Clin Cancer Res. 8:1720-1730). Клетки рабдомиосаркомы A-673 и рака шейки матки HeLa представляют собой HER2-отрицательные линии опухолевых клеток человека, a BEAS-2b представляет собой иммортализованную HER2-отрицательную клеточную линию эпителия бронхов (Jia LT et al. (2003) Cancer Res. 63:3257-3262; Kern JA, et al., (1993) Am J Respir Cell Mol Biol 9:448-454; Martinez-Ramirez A, et al., (2003) Cancer Genet Cytogenet. 2003; 141:138-142). Все эти клеточные линии получены из Американской коллекции типовых культур (АТСС, Манассас, Виргиния) и хранятся в среде, рекомендованной АТСС.
[00208] Мыши. Самок голых мышей возрастом от 6 до 8 недель получают и содержат в специфических условиях без патогенов. В день 0 мышам одновременно (1) вводят подкожно в правый бок 107 опухолевых клеток ВТ474, SK-BR-3, Calu-3, А-673 или HeLa и (2) внутривенно вводят комплексы VP-SpyTag-SpyCatcher-Vh, несущие репортерный (например, EGFP) или суицидальный ген. В качестве контроля служат необработанные животные (животные, которым вводят только опухолевые клетки), животные, которым вводят частицы AAV дикого типа, несущие репортерный или суицидальный ген, животные, которым вводят только вирусные частицы с SpyTag и т.д. Всех животных подвергают обработке соответствующим пролекарством через 1 день после инъекции и обработки. Размер каждой опухоли измеряют с помощью штангенциркуля 2 раза в неделю, а объем опухоли рассчитывают как длина×ширина2×0,52. При заболеваемости, изъязвлении опухоли, диаметре опухоли 15 мм или объеме опухоли 1000 мм3 мышей умерщвляют, а дату умерщвления записывают как дату смерти. Органы печени, селезенки, почки и опухоли животных, которым инъецировали вирусные частицы, несущие репортерный ген, фиксируют и визуализируют экспрессию репортерного гена.
[00209] Хотя нацеленная доставка генов, индуцирующих суицид, была описана (Zarogoulidis P., et al. (2013) J. Genet. Syndr. Gene Ther. 4:16849), в этом примере описана доставка суицидального гена в клетку, экспрессирующую нацеливающий лиганд HER2, с применением описанных в данном документе вирусных частиц. В дополнительных экспериментах обеспечивают доставку суицидального гена в другой тип клеток, экспрессирующих один или более других лигандов-мишеней, с применением описанной в данном документе вирусной частицы. Иллюстративные и неограничивающие примеры рецепторов, подходящих для нацеливания, включают те рецепторы, которые опосредуют эндоцитоз вирусной частицы, например, карцино-эмбриональный антиген (СЕА) (Qiu Y, et al. (2012) Cancer Lett. 316:31-38) и рецептор фактора роста эндотелия сосудов (VEGFR) (Leng A, et al. (2013) Tumour Biol. 32:1103-1111; Liu T, et al. (2011) Exp Mol Pathol. 91:745-752). Дополнительные рецепторы, на которые можно нацеливаться, включают рецептор эпидермального фактора роста (EGFR)(Heimberger АВ, et al. (2009) Expert Opin Biol Ther. 9:1087-1098), дифференцировочный кластер 44s (CD44s) (Heider KH, et al. (2004) Cancer Immunol Immunother. 53:567-579), дифференцировочный кластер 133 (CD133 aka AC133) (Zhang SS, et al. (2012) BMC Med.; 10:85), фолатный рецептор (FR) (Duarte S, et al., (2011) J Control Release 149(3):264-72), рецептор трансферрина (TfR) или дифференцировочный кластер 71 (CD71) (Habashy НО, et al., Breast Cancer Res Treat. 119(2):283-93), муцины (Torres MP, et al., (2012) Curr Pharm Des. 2012; 18(17):2472-81), стадиеспецифический эмбриональный антиген 4 (SSEA-4) (Malecki M., et al., (2012) J Stem Cell Res Ther. 2(5)), антиген опухолевой резистентности 1-60 (TRA-1-60) (Malecki M., et al., (2013) J Stem Cell Res Ther. 3:134).
[00210] Хотя настоящее изобретение было конкретно представлено и описано со ссылкой на некоторые его варианты осуществления, специалистам в данной области будет понятно, что изменения в форме и деталях могут быть внесены в различные варианты осуществления, раскрытые в данном документе, без отступления от сущности и объема настоящего изобретения и что различные варианты осуществления, раскрытые в данном документе, не предназначены для ограничения объема формулы изобретения.
[00211] Хотя при практическом осуществлении или исследовании настоящего изобретения можно применять любые способы и материалы, аналогичные описанным в данном документе или эквивалентные им, в данном документе описаны лишь некоторые предпочтительные способы и материалы. Все публикации, упомянутые в данном документе, включены в данный документ посредством ссылки во всей своей полноте. Если не определено иное, то все технические и научные термины, используемые в данном документе, имеют такое же значение, как это обычно понимает специалист в данной области техники, к которой принадлежит настоящее изобретение.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Редженерон Фармасьютикалз, Инк.
Сабин, Леа
Шенгерр, Кристофер
Экономидес, Арис Н.
Киратсоус, Кристос
Мерфи, Эндрю Дж.
<120> РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ЧАСТИЦЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА
<130> 009108.359WO1/10359WO01
<150> 62525708
<151> 27-06-2017
<160> 55
<170>PatentIn, версия 3.5
<210> 1
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> SpyTag
<400> 1
Ala His Ile Val Met Val Asp Ala Tyr Lys Pro Thr Lys
1 5 10
<210> 2
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> KTag
<400> 2
Ala Thr His Ile Lys Phe Ser Lys Arg Asp
1 5 10
<210> 3
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Spycatcher
<400> 3
Val Asp Thr Leu Ser Gly Leu Ser Ser Glu Gln Gly Gln Ser Gly Asp
1 5 10 15
Met Thr Ile Glu Glu Asp Ser Ala Thr His Ile Lys Phe Ser Lys Arg
20 25 30
Asp Glu Asp Gly Lys Glu Leu Ala Gly Ala Thr Met Glu Leu Arg Asp
35 40 45
Ser Ser Gly Lys Thr Ile Ser Thr Trp Ile Ser Asp Gly Gln Val Lys
50 55 60
Asp Phe Tyr Leu Tyr Pro Gly Lys Tyr Thr Phe Val Glu Thr Ala Ala
65 70 75 80
Pro Asp Gly Tyr Glu Val Ala Thr Ala Ile Thr Phe Thr Val Asn Glu
85 90 95
Gln Gly Gln Val Thr Val Asn Gly Lys Ala Thr Lys Gly Asp Ala His
100 105 110
Ile
<210> 4
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223>SpyLigase
<400> 4
Met Ser Tyr Tyr His His His His His His Asp Tyr Asp Gly Gln Ser
1 5 10 15
Gly Asp Gly Lys Glu Leu Ala Gly Ala Thr Met Glu Leu Arg Asp Ser
20 25 30
Ser Gly Lys Thr Ile Ser Thr Trp Ile Ser Asp Gly Gln Val Lys Asp
35 40 45
Phe Tyr Leu Tyr Pro Gly Lys Tyr Thr Phe Val Glu Thr Ala Ala Pro
50 55 60
Asp Gly Tyr Glu Val Ala Thr Ala Ile Thr Phe Thr Val Asn Glu Gln
65 70 75 80
Gly Gln Val Thr Val Asn Gly Lys Ala Thr Lys Gly Gly Ser Gly Gly
85 90 95
Ser Gly Gly Ser Gly Glu Asp Ser Ala Thr His Ile
100 105
<210> 5
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> SnoopTag
<400> 5
Lys Leu Gly Asp Ile Glu Phe Ile Lys Val Asn Lys
1 5 10
<210> 6
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> SnoopCatcher
<400> 6
Met Gly Ser Ser His His His His His His Ser Ser Gly Leu Val Pro
1 5 10 15
Arg Gly Ser His Met Lys Pro Leu Arg Gly Ala Val Phe Ser Leu Gln
20 25 30
Lys Gln His Pro Asp Tyr Pro Asp Ile Tyr Gly Ala Ile Asp Gln Asn
35 40 45
Gly Thr Tyr Gln Asn Val Arg Thr Gly Glu Asp Gly Lys Leu Thr Phe
50 55 60
Lys Asn Leu Ser Asp Gly Lys Tyr Arg Leu Phe Glu Asn Ser Glu Pro
65 70 75 80
Ala Gly Tyr Lys Pro Val Gln Asn Lys Pro Ile Val Ala Phe Gln Ile
85 90 95
Val Asn Gly Glu Val Arg Asp Val Thr Ser Ile Val Pro Gln Asp Ile
100 105 110
Pro Ala Thr Tyr Glu Phe Thr Asn Gly Lys His Tyr Ile Thr Asn Glu
115 120 125
Pro Ile Pro Pro Lys
130
<210> 7
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Isopeptag
<400> 7
Thr Asp Lys Asp Met Thr Ile Thr Phe Thr Asn Lys Lys Asp Ala Glu
1 5 10 15
<210> 8
<211> 2208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-CAPwt
<400> 8
atggctgccgatggttatcttccagattggctcgaggacactctctctgaaggaataaga 60
cagtggtggaagctcaaacctggcccaccaccaccaaagcccgcagagcggcataaggac 120
gacagcaggggtcttgtgcttcctgggtacaagtacctcggacccttcaacggactcgac 180
aagggagagccggtcaacgaggcagacgccgcggccctcgagcacgacaaagcctacgac 240
cggcagctcgacagcggagacaacccgtacctcaagtacaaccacgccgacgcggagttt 300
caggagcgccttaaagaagatacgtcttttgggggcaacctcggacgagcagtcttccag 360
gcgaaaaagagggttcttgaacctctgggcctggttgaggaacctgttaagacggctccg 420
ggaaaaaagaggccggtagagcactctcctgtggagccagactcctcctcgggaaccgga 480
aaggcgggccagcagcctgcaagaaaaagattgaattttggtcagactggagacgcagac 540
tcagtacctgacccccagcctctcggacagccaccagcagccccctctggtctgggaact 600
aatacgatggctacaggcagtggcgcaccaatggcagacaataacgagggcgccgacgga 660
gtgggtaattcctcgggaaattggcattgcgattccacatggatgggcgacagagtcatc 720
accaccagcacccgaacctgggccctgcccacctacaacaaccacctctacaaacaaatt 780
tccagccaatcaggagcctcgaacgacaatcactactttggctacagcaccccttggggg 840
tattttgacttcaacagattccactgccacttttcaccacgtgactggcaaagactcatc 900
aacaacaactggggattccgacccaagagactcaacttcaagctctttaacattcaagtc 960
aaagaggtcacgcagaatgacggtacgacgacgattgccaataaccttaccagcacggtt 1020
caggtgtttactgactcggagtaccagctcccgtacgtcctcggctcggcgcatcaagga 1080
tgcctcccgccgttcccagcagacgtcttcatggtgccacagtatggatacctcaccctg 1140
aacaacgggagtcaggcagtaggacgctcttcattttactgcctggagtactttccttct 1200
cagatgctgcgtaccggaaacaactttaccttcagctacacttttgaggacgttcctttc 1260
cacagcagctacgctcacagccagagtctggaccgtctcatgaatcctctcatcgaccag 1320
tacctgtattacttgagcagaacaaacactccaagtggaaccaccacgcagtcaaggctt 1380
cagttttctcaggccggagcgagtgacattcgggaccagtctaggaactggcttcctgga 1440
ccctgttaccgccagcagcgagtatcaaagacatctgcggataacaacaacagtgaatac 1500
tcgtggactggagctaccaagtaccacctcaatggcagagactctctggtgaatccgggc 1560
ccggccatggcaagccacaaggacgatgaagaaaagttttttcctcagagcggggttctc 1620
atctttgggaagcaaggctcagagaaaacaaatgtggacattgaaaaggtcatgattaca 1680
gacgaagaggaaatcaggacaaccaatcccgtggctacggagcagtatggttctgtatct 1740
accaacctccagagaggcaacagacaagcagctaccgcagatgtcaacacacaaggcgtt 1800
cttccaggcatggtctggcaggacagagatgtgtaccttcaggggcccatctgggcaaag 1860
attccacacacggacggacattttcacccctctcccctcatgggtggattcggacttaaa 1920
caccctcctccacagattctcatcaagaacaccccggtacctgcgaatccttcgaccacc 1980
ttcagtgcggcaaagtttgcttccttcatcacacagtactccacgggacaggtcagcgtg 2040
gagatcgagtgggagctgcagaaggaaaacagcaaacgctggaatcccgaaattcagtac 2100
acttccaactacaacaagtctgttaatgtggactttactgtggacactaatggcgtgtat 2160
tcagagcctcgccccattggcaccagatacctgactcgtaatctgtaa 2208
<210> 9
<211> 735
<212> БЕЛОК
<213> аденоассоциированный вирус 2
<400> 9
MetAlaAlaAspGlyTyrLeuProAspTrpLeuGluAspThrLeuSer
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 10
<211> 2208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-R585A R588A CAP (HBM)
<400> 10
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cgcgcaagca gctaccgcag atgtcaacac acaaggcgtt 1800
cttccaggca tggtctggca ggacagagat gtgtaccttc aggggcccat ctgggcaaag 1860
attccacaca cggacggaca ttttcacccc tctcccctca tgggtggatt cggacttaaa 1920
caccctcctc cacagattct catcaagaac accccggtac ctgcgaatcc ttcgaccacc 1980
ttcagtgcgg caaagtttgc ttccttcatc acacagtact ccacgggaca ggtcagcgtg 2040
gagatcgagt gggagctgca gaaggaaaac agcaaacgct ggaatcccga aattcagtac 2100
acttccaact acaacaagtc tgttaatgtg gactttactg tggacactaa tggcgtgtat 2160
tcagagcctc gccccattgg caccagatac ctgactcgta atctgtaa 2208
<210> 11
<211> 735
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-R585A R588A CAP (HBM)
<400> 11
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Ala Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 12
<211> 2244
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-N587 CAP SpyTag HBM
<400> 12
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cgcccacatc gtgatggtgg acgcctacaa gccgacgaag 1800
caagcagcta ccgcagatgt caacacacaa ggcgttcttc caggcatggt ctggcaggac 1860
agagatgtgt accttcaggg gcccatctgg gcaaagattc cacacacgga cggacatttt 1920
cacccctctc ccctcatggg tggattcgga cttaaacacc ctcctccaca gattctcatc 1980
aagaacaccc cggtacctgc gaatccttcg accaccttca gtgcggcaaa gtttgcttcc 2040
ttcatcacac agtactccac gggacaggtc agcgtggaga tcgagtggga gctgcagaag 2100
gaaaacagca aacgctggaa tcccgaaatt cagtacactt ccaactacaa caagtctgtt 2160
aatgtggact ttactgtgga cactaatggc gtgtattcag agcctcgccc cattggcacc 2220
agatacctga ctcgtaatct gtaa 2244
<210> 13
<211> 747
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-N587 CAP SpyTag HBM
<400> 13
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Ala His Ile Val Met
580 585 590
Val Asp Ala Tyr Lys Pro Thr Lys Gln Ala Ala Thr Ala Asp Val Asn
595 600 605
Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr
610 615 620
Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly His Phe
625 630 635 640
His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro Pro
645 650 655
Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn Pro Ser Thr Thr
660 665 670
Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln Tyr Ser Thr Gly
675 680 685
Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys
690 695 700
Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Asn Lys Ser Val
705 710 715 720
Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr Ser Glu Pro Arg
725 730 735
Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 14
<211> 2253
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-N587 CAP Линкер 1 SpyTag HBM
<400> 14
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cgcgagcgcc cacatcgtga tggtggacgc ctacaagccg 1800
acgaaggcgc aagcagctac cgcagatgtc aacacacaag gcgttcttcc aggcatggtc 1860
tggcaggaca gagatgtgta ccttcagggg cccatctggg caaagattcc acacacggac 1920
ggacattttc acccctctcc cctcatgggt ggattcggac ttaaacaccc tcctccacag 1980
attctcatca agaacacccc ggtacctgcg aatccttcga ccaccttcag tgcggcaaag 2040
tttgcttcct tcatcacaca gtactccacg ggacaggtca gcgtggagat cgagtgggag 2100
ctgcagaagg aaaacagcaa acgctggaat cccgaaattc agtacacttc caactacaac 2160
aagtctgtta atgtggactt tactgtggac actaatggcg tgtattcaga gcctcgcccc 2220
attggcacca gatacctgac tcgtaatctg taa 2253
<210> 15
<211> 750
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-N587 CAP Линкер 1 SpyTag HBM
<400> 15
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Ala Ser Ala His Ile
580 585 590
Val Met Val Asp Ala Tyr Lys Pro Thr Lys Ala Gln Ala Ala Thr Ala
595 600 605
Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp Arg
610 615 620
Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp
625 630 635 640
Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His
645 650 655
Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn Pro
660 665 670
Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln Tyr
675 680 685
Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu
690 695 700
Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Asn
705 710 715 720
Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr Ser
725 730 735
Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745 750
<210> 16
<211> 2256
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-N587 CAP Линкер 2 SpyTag HBM
<400> 16
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cggcagcgcc cacatcgtga tggtggacgc ctacaagccg 1800
acgaagggca gccaagcagc taccgcagat gtcaacacac aaggcgttct tccaggcatg 1860
gtctggcagg acagagatgt gtaccttcag gggcccatct gggcaaagat tccacacacg 1920
gacggacatt ttcacccctc tcccctcatg ggtggattcg gacttaaaca ccctcctcca 1980
cagattctca tcaagaacac cccggtacct gcgaatcctt cgaccacctt cagtgcggca 2040
aagtttgctt ccttcatcac acagtactcc acgggacagg tcagcgtgga gatcgagtgg 2100
gagctgcaga aggaaaacag caaacgctgg aatcccgaaa ttcagtacac ttccaactac 2160
aacaagtctg ttaatgtgga ctttactgtg gacactaatg gcgtgtattc agagcctcgc 2220
cccattggca ccagatacct gactcgtaat ctgtaa 2256
<210> 17
<211> 751
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-N587 CAP Линкер 2 SpyTag HBM
<400> 17
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Gly Ser Ala His Ile
580 585 590
Val Met Val Asp Ala Tyr Lys Pro Thr Lys Gly Ser Gln Ala Ala Thr
595 600 605
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
610 615 620
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
625 630 635 640
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
645 650 655
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
660 665 670
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
675 680 685
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
690 695 700
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
705 710 715 720
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
725 730 735
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745 750
<210> 18
<211> 2268
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-N587 CAP Линкер 4 SpyTag HBM
<400> 18
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cggcctgagc ggcgcccaca tcgtgatggt ggacgcctac 1800
aagccgacga agggcctgag cggccaagca gctaccgcag atgtcaacac acaaggcgtt 1860
cttccaggca tggtctggca ggacagagat gtgtaccttc aggggcccat ctgggcaaag 1920
attccacaca cggacggaca ttttcacccc tctcccctca tgggtggatt cggacttaaa 1980
caccctcctc cacagattct catcaagaac accccggtac ctgcgaatcc ttcgaccacc 2040
ttcagtgcgg caaagtttgc ttccttcatc acacagtact ccacgggaca ggtcagcgtg 2100
gagatcgagt gggagctgca gaaggaaaac agcaaacgct ggaatcccga aattcagtac 2160
acttccaact acaacaagtc tgttaatgtg gactttactg tggacactaa tggcgtgtat 2220
tcagagcctc gccccattgg caccagatac ctgactcgta atctgtaa 2268
<210> 19
<211> 755
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-N587 CAP Линкер 4 SpyTag HBM
<400> 19
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Gly Leu Ser Gly Ala
580 585 590
His Ile Val Met Val Asp Ala Tyr Lys Pro Thr Lys Gly Leu Ser Gly
595 600 605
Gln Ala Ala Thr Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met
610 615 620
Val Trp Gln Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys
625 630 635 640
Ile Pro His Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly
645 650 655
Phe Gly Leu Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro
660 665 670
Val Pro Ala Asn Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser
675 680 685
Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp
690 695 700
Glu Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr
705 710 715 720
Thr Ser Asn Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr
725 730 735
Asn Gly Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr
740 745 750
Arg Asn Leu
755
<210> 20
<211> 2280
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-N587 CAP Линкер 6 SpyTag HBM
<400> 20
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cggcctgagc ggcagcggcg cccacatcgt gatggtggac 1800
gcctacaagc cgacgaaggg cctgagcggc agcggccaag cagctaccgc agatgtcaac 1860
acacaaggcg ttcttccagg catggtctgg caggacagag atgtgtacct tcaggggccc 1920
atctgggcaa agattccaca cacggacgga cattttcacc cctctcccct catgggtgga 1980
ttcggactta aacaccctcc tccacagatt ctcatcaaga acaccccggt acctgcgaat 2040
ccttcgacca ccttcagtgc ggcaaagttt gcttccttca tcacacagta ctccacggga 2100
caggtcagcg tggagatcga gtgggagctg cagaaggaaa acagcaaacg ctggaatccc 2160
gaaattcagt acacttccaa ctacaacaag tctgttaatg tggactttac tgtggacact 2220
aatggcgtgt attcagagcc tcgccccatt ggcaccagat acctgactcg taatctgtaa 2280
<210> 21
<211> 759
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-N587 CAP Линкер 6 SpyTag HBM
<400> 21
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Gly Leu Ser Gly Ser
580 585 590
Gly Ala His Ile Val Met Val Asp Ala Tyr Lys Pro Thr Lys Gly Leu
595 600 605
Ser Gly Ser Gly Gln Ala Ala Thr Ala Asp Val Asn Thr Gln Gly Val
610 615 620
Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu Gln Gly Pro
625 630 635 640
Ile Trp Ala Lys Ile Pro His Thr Asp Gly His Phe His Pro Ser Pro
645 650 655
Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro Pro Gln Ile Leu Ile
660 665 670
Lys Asn Thr Pro Val Pro Ala Asn Pro Ser Thr Thr Phe Ser Ala Ala
675 680 685
Lys Phe Ala Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val
690 695 700
Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro
705 710 715 720
Glu Ile Gln Tyr Thr Ser Asn Tyr Asn Lys Ser Val Asn Val Asp Phe
725 730 735
Thr Val Asp Thr Asn Gly Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr
740 745 750
Arg Tyr Leu Thr Arg Asn Leu
755
<210> 22
<211> 2292
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-N587 CAP Линкер 8 SpyTag HBM
<400> 22
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cggcctgagc ggcctgagcg gcagcgccca catcgtgatg 1800
gtggacgcct acaagccgac gaagggcctg agcggcctga gcggcagcca agcagctacc 1860
gcagatgtca acacacaagg cgttcttcca ggcatggtct ggcaggacag agatgtgtac 1920
cttcaggggc ccatctgggc aaagattcca cacacggacg gacattttca cccctctccc 1980
ctcatgggtg gattcggact taaacaccct cctccacaga ttctcatcaa gaacaccccg 2040
gtacctgcga atccttcgac caccttcagt gcggcaaagt ttgcttcctt catcacacag 2100
tactccacgg gacaggtcag cgtggagatc gagtgggagc tgcagaagga aaacagcaaa 2160
cgctggaatc ccgaaattca gtacacttcc aactacaaca agtctgttaa tgtggacttt 2220
actgtggaca ctaatggcgt gtattcagag cctcgcccca ttggcaccag atacctgact 2280
cgtaatctgt aa 2292
<210> 23
<211> 763
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-N587 CAP Линкер 8 SpyTag HBM
<400> 23
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Gly Leu Ser Gly Leu
580 585 590
Ser Gly Ser Ala His Ile Val Met Val Asp Ala Tyr Lys Pro Thr Lys
595 600 605
Gly Leu Ser Gly Leu Ser Gly Ser Gln Ala Ala Thr Ala Asp Val Asn
610 615 620
Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr
625 630 635 640
Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly His Phe
645 650 655
His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro Pro
660 665 670
Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn Pro Ser Thr Thr
675 680 685
Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln Tyr Ser Thr Gly
690 695 700
Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys
705 710 715 720
Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Asn Lys Ser Val
725 730 735
Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr Ser Glu Pro Arg
740 745 750
Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
755 760
<210> 24
<211> 2304
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-N587 CAP Линкер 10 SpyTag HBM
<400> 24
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc aggcgggcaa cggcctgagc ggcctgagcg gcctgagcgg cgcccacatc 1800
gtgatggtgg acgcctacaa gccgacgaag ggcctgagcg gcctgagcgg cctgagcggc 1860
caagcagcta ccgcagatgt caacacacaa ggcgttcttc caggcatggt ctggcaggac 1920
agagatgtgt accttcaggg gcccatctgg gcaaagattc cacacacgga cggacatttt 1980
cacccctctc ccctcatggg tggattcgga cttaaacacc ctcctccaca gattctcatc 2040
aagaacaccc cggtacctgc gaatccttcg accaccttca gtgcggcaaa gtttgcttcc 2100
ttcatcacac agtactccac gggacaggtc agcgtggaga tcgagtggga gctgcagaag 2160
gaaaacagca aacgctggaa tcccgaaatt cagtacactt ccaactacaa caagtctgtt 2220
aatgtggact ttactgtgga cactaatggc gtgtattcag agcctcgccc cattggcacc 2280
agatacctga ctcgtaatct gtaa 2304
<210> 25
<211> 767
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-N587 CAP Линкер 10 SpyTag HBM
<400> 25
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Gly Leu Ser Gly Leu
580 585 590
Ser Gly Leu Ser Gly Ala His Ile Val Met Val Asp Ala Tyr Lys Pro
595 600 605
Thr Lys Gly Leu Ser Gly Leu Ser Gly Leu Ser Gly Gln Ala Ala Thr
610 615 620
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
625 630 635 640
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
645 650 655
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
660 665 670
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
675 680 685
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
690 695 700
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
705 710 715 720
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
725 730 735
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
740 745 750
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
755 760 765
<210> 26
<211> 2247
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-G453 CAP SpyTag HBM
<400> 26
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggag cccacatcgt gatggtggac 1380
gcctacaagc cgacgaagac caccacgcag tcaaggcttc agttttctca ggccggagcg 1440
agtgacattc gggaccagtc taggaactgg cttcctggac cctgttacgc ccagcaggcc 1500
gtatcaaaga catctgcgga taacaacaac agtgaatact cgtggactgg agctaccaag 1560
taccacctca atggcagaga ctctctggtg aatccgggcc cggccatggc aagccacaag 1620
gacgatgaag aagccttttt tcctcagagc ggggttctca tctttgggaa gcaaggctca 1680
gagaaaacaa atgtggacat tgaaaaggtc atgattacag acgaagagga aatcaggaca 1740
accaatcccg tggctacgga gcagtatggt tctgtatcta ccaacctcca ggccggcaac 1800
gcccaagcag ctaccgcaga tgtcaacaca caaggcgttc ttccaggcat ggtctggcag 1860
gacagagatg tgtaccttca ggggcccatc tgggcaaaga ttccacacac ggacggacat 1920
tttcacccct ctcccctcat gggtggattc ggacttaaac accctcctcc acagattctc 1980
atcaagaaca ccccggtacc tgcgaatcct tcgaccacct tcagtgcggc aaagtttgct 2040
tccttcatca cacagtactc cacgggacag gtcagcgtgg agatcgagtg ggagctgcag 2100
aaggaaaaca gcaaacgctg gaatcccgaa attcagtaca cttccaacta caacaagtct 2160
gttaatgtgg actttactgt ggacactaat ggcgtgtatt cagagcctcg ccccattggc 2220
accagatacc tgactcgtaa tctgtaa 2247
<210> 27
<211> 748
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-G453 CAP SpyTag HBM
<400> 27
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Ala His Ile Val Met Val Asp Ala Tyr Lys Pro
450 455 460
Thr Lys Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln Ala Gly Ala
465 470 475 480
Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly Pro Cys Tyr
485 490 495
Ala Gln Gln Ala Val Ser Lys Thr Ser Ala Asp Asn Asn Asn Ser Glu
500 505 510
Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly Arg Asp Ser
515 520 525
Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp Asp Glu Glu
530 535 540
Ala Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys Gln Gly Ser
545 550 555 560
Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr Asp Glu Glu
565 570 575
Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr Gly Ser Val
580 585 590
Ser Thr Asn Leu Gln Ala Gly Asn Ala Gln Ala Ala Thr Ala Asp Val
595 600 605
Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val
610 615 620
Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly His
625 630 635 640
Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro
645 650 655
Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn Pro Ser Thr
660 665 670
Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln Tyr Ser Thr
675 680 685
Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser
690 695 700
Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Asn Lys Ser
705 710 715 720
Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr Ser Glu Pro
725 730 735
Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 28
<211> 2307
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV2-G453 CAP Линкер10 SpyTag HBM
<400> 28
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggag gcctgagcgg cctgagcggc 1380
ctgagcggcg cccacatcgt gatggtggac gcctacaagc cgacgaaggg cctgagcggc 1440
ctgagcggcc tgagcggcac caccacgcag tcaaggcttc agttttctca ggccggagcg 1500
agtgacattc gggaccagtc taggaactgg cttcctggac cctgttacgc ccagcaggcc 1560
gtatcaaaga catctgcgga taacaacaac agtgaatact cgtggactgg agctaccaag 1620
taccacctca atggcagaga ctctctggtg aatccgggcc cggccatggc aagccacaag 1680
gacgatgaag aagccttttt tcctcagagc ggggttctca tctttgggaa gcaaggctca 1740
gagaaaacaa atgtggacat tgaaaaggtc atgattacag acgaagagga aatcaggaca 1800
accaatcccg tggctacgga gcagtatggt tctgtatcta ccaacctcca ggccggcaac 1860
gcccaagcag ctaccgcaga tgtcaacaca caaggcgttc ttccaggcat ggtctggcag 1920
gacagagatg tgtaccttca ggggcccatc tgggcaaaga ttccacacac ggacggacat 1980
tttcacccct ctcccctcat gggtggattc ggacttaaac accctcctcc acagattctc 2040
atcaagaaca ccccggtacc tgcgaatcct tcgaccacct tcagtgcggc aaagtttgct 2100
tccttcatca cacagtactc cacgggacag gtcagcgtgg agatcgagtg ggagctgcag 2160
aaggaaaaca gcaaacgctg gaatcccgaa attcagtaca cttccaacta caacaagtct 2220
gttaatgtgg actttactgt ggacactaat ggcgtgtatt cagagcctcg ccccattggc 2280
accagatacc tgactcgtaa tctgtaa 2307
<210> 29
<211> 768
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV2-G453 CAP Линкер 10 SpyTag HBM
<400> 29
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Gly Leu Ser Gly Leu Ser Gly Leu Ser Gly Ala
450 455 460
His Ile Val Met Val Asp Ala Tyr Lys Pro Thr Lys Gly Leu Ser Gly
465 470 475 480
Leu Ser Gly Leu Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser
485 490 495
Gln Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro
500 505 510
Gly Pro Cys Tyr Ala Gln Gln Ala Val Ser Lys Thr Ser Ala Asp Asn
515 520 525
Asn Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn
530 535 540
Gly Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys
545 550 555 560
Asp Asp Glu Glu Ala Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly
565 570 575
Lys Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile
580 585 590
Thr Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln
595 600 605
Tyr Gly Ser Val Ser Thr Asn Leu Gln Ala Gly Asn Ala Gln Ala Ala
610 615 620
Thr Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln
625 630 635 640
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
645 650 655
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
660 665 670
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
675 680 685
Asn Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr
690 695 700
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
705 710 715 720
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
725 730 735
Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val
740 745 750
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
755 760 765
<210> 30
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV9-CAP wt
<400> 30
atggctgccg atggttatct tccagattgg ctcgaggaca accttagtga aggaattcgc 60
gagtggtggg ctttgaaacc tggagcccct caacccaagg caaatcaaca acatcaagac 120
aacgctcgag gtcttgtgct tccgggttac aaataccttg gacccggcaa cggactcgac 180
aagggggagc cggtcaacgc agcagacgcg gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggaga caacccgtac ctcaagtaca accacgccga cgccgagttc 300
caggagcggc tcaaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga ggcttcttga acctcttggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaga ggcctgtaga gcagtctcct caggaaccgg actcctccgc gggtattggc 480
aaatcgggtg cacagcccgc taaaaagaga ctcaatttcg gtcagactgg cgacacagag 540
tcagtcccag accctcaacc aatcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca gtggcagaca ataacgaagg tgccgatgga 660
gtgggtagtt cctcgggaaa ttggcattgc gattcccaat ggctggggga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca atcacctcta caagcaaatc 780
tccaacagca catctggagg atcttcaaat gacaacgcct acttcggcta cagcaccccc 840
tgggggtatt ttgacttcaa cagattccac tgccacttct caccacgtga ctggcagcga 900
ctcatcaaca acaactgggg attccggcct aagcgactca acttcaagct cttcaacatt 960
caggtcaaag aggttacgga caacaatgga gtcaagacca tcgccaataa ccttaccagc 1020
acggtccagg tcttcacgga ctcagactat cagctcccgt acgtgctcgg gtcggctcac 1080
gagggctgcc tcccgccgtt cccagcggac gttttcatga ttcctcagta cgggtatctg 1140
acgcttaatg atggaagcca ggccgtgggt cgttcgtcct tttactgcct ggaatatttc 1200
ccgtcgcaaa tgctaagaac gggtaacaac ttccagttca gctacgagtt tgagaacgta 1260
cctttccata gcagctacgc tcacagccaa agcctggacc gactaatgaa tccactcatc 1320
gaccaatact tgtactatct ctcaaagact attaacggtt ctggacagaa tcaacaaacg 1380
ctaaaattca gtgtggccgg acccagcaac atggctgtcc agggaagaaa ctacatacct 1440
ggacccagct accgacaaca acgtgtctca accactgtga ctcaaaacaa caacagcgaa 1500
tttgcttggc ctggagcttc ttcttgggct ctcaatggac gtaatagctt gatgaatcct 1560
ggacctgcta tggccagcca caaagaagga gaggaccgtt tctttccttt gtctggatct 1620
ttaatttttg gcaaacaagg aactggaaga gacaacgtgg atgcggacaa agtcatgata 1680
accaacgaag aagaaattaa aactactaac ccggtagcaa cggagtccta tggacaagtg 1740
gccacaaacc accagagtgc ccaagcacag gcgcagaccg gctgggttca aaaccaagga 1800
atacttccgg gtatggtttg gcaggacaga gatgtgtacc tgcaaggacc catttgggcc 1860
aaaattcctc acacggacgg caactttcac ccttctccgc tgatgggagg gtttggaatg 1920
aagcacccgc ctcctcagat cctcatcaaa aacacacctg tacctgcgga tcctccaacg 1980
gccttcaaca aggacaagct gaactctttc atcacccagt attctactgg ccaagtcagc 2040
gtggagatcg agtgggagct gcagaaggaa aacagcaagc gctggaaccc ggagatccag 2100
tacacttcca actattacaa gtctaataat gttgaatttg ctgttaatac tgaaggtgta 2160
tatagtgaac cccgccccat tggcaccaga tacctgactc gtaatctgta a 2211
<210> 31
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 9
<400> 31
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Gln Ala Gln
580 585 590
Thr Gly Trp Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Met
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asp Pro Pro Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Tyr Lys Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 32
<211> 2211
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV9-W503A CAP
<400> 32
atggctgccg atggttatct tccagattgg ctcgaggaca accttagtga aggaattcgc 60
gagtggtggg ctttgaaacc tggagcccct caacccaagg caaatcaaca acatcaagac 120
aacgctcgag gtcttgtgct tccgggttac aaataccttg gacccggcaa cggactcgac 180
aagggggagc cggtcaacgc agcagacgcg gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggaga caacccgtac ctcaagtaca accacgccga cgccgagttc 300
caggagcggc tcaaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga ggcttcttga acctcttggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaga ggcctgtaga gcagtctcct caggaaccgg actcctccgc gggtattggc 480
aaatcgggtg cacagcccgc taaaaagaga ctcaatttcg gtcagactgg cgacacagag 540
tcagtcccag accctcaacc aatcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca gtggcagaca ataacgaagg tgccgatgga 660
gtgggtagtt cctcgggaaa ttggcattgc gattcccaat ggctggggga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca atcacctcta caagcaaatc 780
tccaacagca catctggagg atcttcaaat gacaacgcct acttcggcta cagcaccccc 840
tgggggtatt ttgacttcaa cagattccac tgccacttct caccacgtga ctggcagcga 900
ctcatcaaca acaactgggg attccggcct aagcgactca acttcaagct cttcaacatt 960
caggtcaaag aggttacgga caacaatgga gtcaagacca tcgccaataa ccttaccagc 1020
acggtccagg tcttcacgga ctcagactat cagctcccgt acgtgctcgg gtcggctcac 1080
gagggctgcc tcccgccgtt cccagcggac gttttcatga ttcctcagta cgggtatctg 1140
acgcttaatg atggaagcca ggccgtgggt cgttcgtcct tttactgcct ggaatatttc 1200
ccgtcgcaaa tgctaagaac gggtaacaac ttccagttca gctacgagtt tgagaacgta 1260
cctttccata gcagctacgc tcacagccaa agcctggacc gactaatgaa tccactcatc 1320
gaccaatact tgtactatct ctcaaagact attaacggtt ctggacagaa tcaacaaacg 1380
ctaaaattca gtgtggccgg acccagcaac atggctgtcc agggaagaaa ctacatacct 1440
ggacccagct accgacaaca acgtgtctca accactgtga ctcaaaacaa caacagcgaa 1500
tttgctgctc ctggagcttc ttcttgggct ctcaatggac gtaatagctt gatgaatcct 1560
ggacctgcta tggccagcca caaagaagga gaggaccgtt tctttccttt gtctggatct 1620
ttaatttttg gcaaacaagg aactggaaga gacaacgtgg atgcggacaa agtcatgata 1680
accaacgaag aagaaattaa aactactaac ccggtagcaa cggagtccta tggacaagtg 1740
gccacaaacc accagagtgc ccaagcacag gcgcagaccg gctgggttca aaaccaagga 1800
atacttccgg gtatggtttg gcaggacaga gatgtgtacc tgcaaggacc catttgggcc 1860
aaaattcctc acacggacgg caactttcac ccttctccgc tgatgggagg gtttggaatg 1920
aagcacccgc ctcctcagat cctcatcaaa aacacacctg tacctgcgga tcctccaacg 1980
gccttcaaca aggacaagct gaactctttc atcacccagt attctactgg ccaagtcagc 2040
gtggagatcg agtgggagct gcagaaggaa aacagcaagc gctggaaccc ggagatccag 2100
tacacttcca actattacaa gtctaataat gttgaatttg ctgttaatac tgaaggtgta 2160
tatagtgaac cccgccccat tggcaccaga tacctgactc gtaatctgta a 2211
<210> 33
<211> 736
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV9-W503A CAP
<400> 33
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Ala Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Gln Ala Gln
580 585 590
Thr Gly Trp Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Met
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asp Pro Pro Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Tyr Lys Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 34
<211> 2250
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV9-A589 CAP SpyTag W503A
<400> 34
atggctgccg atggttatct tccagattgg ctcgaggaca accttagtga aggaattcgc 60
gagtggtggg ctttgaaacc tggagcccct caacccaagg caaatcaaca acatcaagac 120
aacgctcgag gtcttgtgct tccgggttac aaataccttg gacccggcaa cggactcgac 180
aagggggagc cggtcaacgc agcagacgcg gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggaga caacccgtac ctcaagtaca accacgccga cgccgagttc 300
caggagcggc tcaaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga ggcttcttga acctcttggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaga ggcctgtaga gcagtctcct caggaaccgg actcctccgc gggtattggc 480
aaatcgggtg cacagcccgc taaaaagaga ctcaatttcg gtcagactgg cgacacagag 540
tcagtcccag accctcaacc aatcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca gtggcagaca ataacgaagg tgccgatgga 660
gtgggtagtt cctcgggaaa ttggcattgc gattcccaat ggctggggga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca atcacctcta caagcaaatc 780
tccaacagca catctggagg atcttcaaat gacaacgcct acttcggcta cagcaccccc 840
tgggggtatt ttgacttcaa cagattccac tgccacttct caccacgtga ctggcagcga 900
ctcatcaaca acaactgggg attccggcct aagcgactca acttcaagct cttcaacatt 960
caggtcaaag aggttacgga caacaatgga gtcaagacca tcgccaataa ccttaccagc 1020
acggtccagg tcttcacgga ctcagactat cagctcccgt acgtgctcgg gtcggctcac 1080
gagggctgcc tcccgccgtt cccagcggac gttttcatga ttcctcagta cgggtatctg 1140
acgcttaatg atggaagcca ggccgtgggt cgttcgtcct tttactgcct ggaatatttc 1200
ccgtcgcaaa tgctaagaac gggtaacaac ttccagttca gctacgagtt tgagaacgta 1260
cctttccata gcagctacgc tcacagccaa agcctggacc gactaatgaa tccactcatc 1320
gaccaatact tgtactatct ctcaaagact attaacggtt ctggacagaa tcaacaaacg 1380
ctaaaattca gtgtggccgg acccagcaac atggctgtcc agggaagaaa ctacatacct 1440
ggacccagct accgacaaca acgtgtctca accactgtga ctcaaaacaa caacagcgaa 1500
tttgctgctc ctggagcttc ttcttgggct ctcaatggac gtaatagctt gatgaatcct 1560
ggacctgcta tggccagcca caaagaagga gaggaccgtt tctttccttt gtctggatct 1620
ttaatttttg gcaaacaagg aactggaaga gacaacgtgg atgcggacaa agtcatgata 1680
accaacgaag aagaaattaa aactactaac ccggtagcaa cggagtccta tggacaagtg 1740
gccacaaacc accagagtgc ccaagcagcc cacatcgtga tggtggacgc ctacaagccg 1800
acgaagcagg cgcagaccgg ctgggttcaa aaccaaggaa tacttccggg tatggtttgg 1860
caggacagag atgtgtacct gcaaggaccc atttgggcca aaattcctca cacggacggc 1920
aactttcacc cttctccgct gatgggaggg tttggaatga agcacccgcc tcctcagatc 1980
ctcatcaaaa acacacctgt acctgcggat cctccaacgg ccttcaacaa ggacaagctg 2040
aactctttca tcacccagta ttctactggc caagtcagcg tggagatcga gtgggagctg 2100
cagaaggaaa acagcaagcg ctggaacccg gagatccagt acacttccaa ctattacaag 2160
tctaataatg ttgaatttgc tgttaatact gaaggtgtat atagtgaacc ccgccccatt 2220
ggcaccagat acctgactcg taatctgtaa 2250
<210> 35
<211> 749
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV9-A589 CAP SpyTag W503A
<400> 35
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Ala Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Ala His Ile
580 585 590
Val Met Val Asp Ala Tyr Lys Pro Thr Lys Gln Ala Gln Thr Gly Trp
595 600 605
Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp Gln Asp Arg Asp
610 615 620
Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly
625 630 635 640
Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Met Lys His Pro
645 650 655
Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asp Pro Pro
660 665 670
Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr Gln Tyr Ser
675 680 685
Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn
690 695 700
Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Tyr Lys
705 710 715 720
Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly Val Tyr Ser Glu
725 730 735
Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 36
<211> 2310
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV9-A589 CAP Линкер 10 SpyTag W503A
<400> 36
atggctgccg atggttatct tccagattgg ctcgaggaca accttagtga aggaattcgc 60
gagtggtggg ctttgaaacc tggagcccct caacccaagg caaatcaaca acatcaagac 120
aacgctcgag gtcttgtgct tccgggttac aaataccttg gacccggcaa cggactcgac 180
aagggggagc cggtcaacgc agcagacgcg gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggaga caacccgtac ctcaagtaca accacgccga cgccgagttc 300
caggagcggc tcaaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga ggcttcttga acctcttggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaga ggcctgtaga gcagtctcct caggaaccgg actcctccgc gggtattggc 480
aaatcgggtg cacagcccgc taaaaagaga ctcaatttcg gtcagactgg cgacacagag 540
tcagtcccag accctcaacc aatcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca gtggcagaca ataacgaagg tgccgatgga 660
gtgggtagtt cctcgggaaa ttggcattgc gattcccaat ggctggggga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca atcacctcta caagcaaatc 780
tccaacagca catctggagg atcttcaaat gacaacgcct acttcggcta cagcaccccc 840
tgggggtatt ttgacttcaa cagattccac tgccacttct caccacgtga ctggcagcga 900
ctcatcaaca acaactgggg attccggcct aagcgactca acttcaagct cttcaacatt 960
caggtcaaag aggttacgga caacaatgga gtcaagacca tcgccaataa ccttaccagc 1020
acggtccagg tcttcacgga ctcagactat cagctcccgt acgtgctcgg gtcggctcac 1080
gagggctgcc tcccgccgtt cccagcggac gttttcatga ttcctcagta cgggtatctg 1140
acgcttaatg atggaagcca ggccgtgggt cgttcgtcct tttactgcct ggaatatttc 1200
ccgtcgcaaa tgctaagaac gggtaacaac ttccagttca gctacgagtt tgagaacgta 1260
cctttccata gcagctacgc tcacagccaa agcctggacc gactaatgaa tccactcatc 1320
gaccaatact tgtactatct ctcaaagact attaacggtt ctggacagaa tcaacaaacg 1380
ctaaaattca gtgtggccgg acccagcaac atggctgtcc agggaagaaa ctacatacct 1440
ggacccagct accgacaaca acgtgtctca accactgtga ctcaaaacaa caacagcgaa 1500
tttgctgctc ctggagcttc ttcttgggct ctcaatggac gtaatagctt gatgaatcct 1560
ggacctgcta tggccagcca caaagaagga gaggaccgtt tctttccttt gtctggatct 1620
ttaatttttg gcaaacaagg aactggaaga gacaacgtgg atgcggacaa agtcatgata 1680
accaacgaag aagaaattaa aactactaac ccggtagcaa cggagtccta tggacaagtg 1740
gccacaaacc accagagtgc ccaagcaggc ctgagcagcg gcagcggcct gagcggcgcc 1800
cacatcgtga tggtggacgc ctacaagccg acgaagggcc tgagcggcgg cagcggcctg 1860
agcggccagg cgcagaccgg ctgggttcaa aaccaaggaa tacttccggg tatggtttgg 1920
caggacagag atgtgtacct gcaaggaccc atttgggcca aaattcctca cacggacggc 1980
aactttcacc cttctccgct gatgggaggg tttggaatga agcacccgcc tcctcagatc 2040
ctcatcaaaa acacacctgt acctgcggat cctccaacgg ccttcaacaa ggacaagctg 2100
aactctttca tcacccagta ttctactggc caagtcagcg tggagatcga gtgggagctg 2160
cagaaggaaa acagcaagcg ctggaacccg gagatccagt acacttccaa ctattacaag 2220
tctaataatg ttgaatttgc tgttaatact gaaggtgtat atagtgaacc ccgccccatt 2280
ggcaccagat acctgactcg taatctgtaa 2310
<210> 37
<211> 769
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV9-A589 CAP Линкер 10 SpyTag W503A
<400> 37
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Ala Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Gly Leu Ser
580 585 590
Ser Gly Ser Gly Leu Ser Gly Ala His Ile Val Met Val Asp Ala Tyr
595 600 605
Lys Pro Thr Lys Gly Leu Ser Gly Gly Ser Gly Leu Ser Gly Gln Ala
610 615 620
Gln Thr Gly Trp Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp
625 630 635 640
Gln Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro
645 650 655
His Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly
660 665 670
Met Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro
675 680 685
Ala Asp Pro Pro Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile
690 695 700
Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu
705 710 715 720
Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser
725 730 735
Asn Tyr Tyr Lys Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly
740 745 750
Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn
755 760 765
Leu
<210> 38
<211> 2310
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК pAAV9-G453 CAP Линкер 10 SpyTag W503A
<400> 38
atggctgccg atggttatct tccagattgg ctcgaggaca accttagtga aggaattcgc 60
gagtggtggg ctttgaaacc tggagcccct caacccaagg caaatcaaca acatcaagac 120
aacgctcgag gtcttgtgct tccgggttac aaataccttg gacccggcaa cggactcgac 180
aagggggagc cggtcaacgc agcagacgcg gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggaga caacccgtac ctcaagtaca accacgccga cgccgagttc 300
caggagcggc tcaaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga ggcttcttga acctcttggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaga ggcctgtaga gcagtctcct caggaaccgg actcctccgc gggtattggc 480
aaatcgggtg cacagcccgc taaaaagaga ctcaatttcg gtcagactgg cgacacagag 540
tcagtcccag accctcaacc aatcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca gtggcagaca ataacgaagg tgccgatgga 660
gtgggtagtt cctcgggaaa ttggcattgc gattcccaat ggctggggga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca atcacctcta caagcaaatc 780
tccaacagca catctggagg atcttcaaat gacaacgcct acttcggcta cagcaccccc 840
tgggggtatt ttgacttcaa cagattccac tgccacttct caccacgtga ctggcagcga 900
ctcatcaaca acaactgggg attccggcct aagcgactca acttcaagct cttcaacatt 960
caggtcaaag aggttacgga caacaatgga gtcaagacca tcgccaataa ccttaccagc 1020
acggtccagg tcttcacgga ctcagactat cagctcccgt acgtgctcgg gtcggctcac 1080
gagggctgcc tcccgccgtt cccagcggac gttttcatga ttcctcagta cgggtatctg 1140
acgcttaatg atggaagcca ggccgtgggt cgttcgtcct tttactgcct ggaatatttc 1200
ccgtcgcaaa tgctaagaac gggtaacaac ttccagttca gctacgagtt tgagaacgta 1260
cctttccata gcagctacgc tcacagccaa agcctggacc gactaatgaa tccactcatc 1320
gaccaatact tgtactatct ctcaaagact attaacggtg gcctgagcag cggcagcggc 1380
ctgagcggcg cccacatcgt gatggtggac gcctacaagc cgacgaaggg cctgagcggc 1440
ggcagcggcc tgagcggctc tggacagaat caacaaacgc taaaattcag tgtggccgga 1500
cccagcaaca tggctgtcca gggaagaaac tacatacctg gacccagcta ccgacaacaa 1560
cgtgtctcaa ccactgtgac tcaaaacaac aacagcgaat ttgctgctcc tggagcttct 1620
tcttgggctc tcaatggacg taatagcttg atgaatcctg gacctgctat ggccagccac 1680
aaagaaggag aggaccgttt ctttcctttg tctggatctt taatttttgg caaacaagga 1740
actggaagag acaacgtgga tgcggacaaa gtcatgataa ccaacgaaga agaaattaaa 1800
actactaacc cggtagcaac ggagtcctat ggacaagtgg ccacaaacca ccagagtgcc 1860
caagcacagg cgcagaccgg ctgggttcaa aaccaaggaa tacttccggg tatggtttgg 1920
caggacagag atgtgtacct gcaaggaccc atttgggcca aaattcctca cacggacggc 1980
aactttcacc cttctccgct gatgggaggg tttggaatga agcacccgcc tcctcagatc 2040
ctcatcaaaa acacacctgt acctgcggat cctccaacgg ccttcaacaa ggacaagctg 2100
aactctttca tcacccagta ttctactggc caagtcagcg tggagatcga gtgggagctg 2160
cagaaggaaa acagcaagcg ctggaacccg gagatccagt acacttccaa ctattacaag 2220
tctaataatg ttgaatttgc tgttaatact gaaggtgtat atagtgaacc ccgccccatt 2280
ggcaccagat acctgactcg taatctgtaa 2310
<210> 39
<211> 769
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA pAAV9-RC G453 Линкер 10 SpyTag W503A
<400> 39
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Gly Leu Ser Ser Gly Ser Gly Leu Ser Gly Ala
450 455 460
His Ile Val Met Val Asp Ala Tyr Lys Pro Thr Lys Gly Leu Ser Gly
465 470 475 480
Gly Ser Gly Leu Ser Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe
485 490 495
Ser Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile
500 505 510
Pro Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln
515 520 525
Asn Asn Asn Ser Glu Phe Ala Ala Pro Gly Ala Ser Ser Trp Ala Leu
530 535 540
Asn Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His
545 550 555 560
Lys Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe
565 570 575
Gly Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met
580 585 590
Ile Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu
595 600 605
Ser Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Gln Ala
610 615 620
Gln Thr Gly Trp Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp
625 630 635 640
Gln Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro
645 650 655
His Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly
660 665 670
Met Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro
675 680 685
Ala Asp Pro Pro Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile
690 695 700
Thr Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu
705 710 715 720
Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser
725 730 735
Asn Tyr Tyr Lys Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly
740 745 750
Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn
755 760 765
Leu
<210> 40
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер 4
<400> 40
Gly Leu Ser Gly
1
<210> 41
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер 6
<400> 41
Gly Leu Ser Gly Ser Gly
1 5
<210> 42
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер 8
<400> 42
Gly Leu Ser Gly Leu Ser Gly Ser
1 5
<210> 43
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер 10
<400> 43
Gly Leu Ser Gly Leu Ser Gly Leu Ser Gly
1 5 10
<210> 44
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер 10 SEQ ID NO:44
<400> 44
Gly Leu Ser Gly Gly Ser Gly Leu Ser Gly
1 5 10
<210> 45
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь IgG4 человека - линкер - SpyCatcher
<400> 45
gcctccacca agggcccatc ggtcttcccc ctggcgccct gctccaggag cacctccgag 60
agcacagccg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacgaagacc 240
tacacctgca acgtagatca caagcccagc aacaccaagg tggacaagag agttgagtcc 300
aaatatggtc ccccatgccc accctgccca gcacctgagt tcctgggggg accatcagtc 360
ttcctgttcc ccccaaaacc caaggacact ctcatgatct cccggacccc tgaggtcacg 420
tgcgtggtgg tggacgtgag ccaggaagac cccgaggtcc agttcaactg gtacgtggat 480
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagttcaa cagcacgtac 540
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaacggcaa ggagtacaag 600
tgcaaggtct ccaacaaagg cctcccgtcc tccatcgaga aaaccatctc caaagccaaa 660
gggcagcccc gagagccaca ggtgtacacc ctgcccccat cccaggagga gatgaccaag 720
aaccaggtca gcctgacctg cctggtcaaa ggcttctacc ccagcgacat cgccgtggag 780
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 840
gacggctcct tcttcctcta cagcaggctc accgtggaca agagcaggtg gcaggagggg 900
aatgtcttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagtcc 960
ctctccctgt ctctgggtaa aggaagcggc gaaagcggcg tggataccct gtccggactg 1020
agcagtgagc aaggccagtc cggagatatg acaattgaag aagatagcgc cacccatatt 1080
aaattctcca aaagagatga ggacggcaaa gagctggctg gagcaacaat ggagctgaga 1140
gattcctctg gaaagactat tagtacatgg atctctgatg gccaagtgaa agatttctat 1200
ctgtatccag gaaagtacac atttgtcgaa accgctgcac cagacggata tgaggtggct 1260
acagctatta cctttacagt gaatgagcaa ggacaggtga ctgttaatgg caaagctact 1320
aaaggagacg ctcatattta a 1341
<210> 46
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> SEQ ID NO:46
<400> 46
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys Gly Ser Gly Glu Ser Gly Val Asp Thr
325 330 335
Leu Ser Gly Leu Ser Ser Glu Gln Gly Gln Ser Gly Asp Met Thr Ile
340 345 350
Glu Glu Asp Ser Ala Thr His Ile Lys Phe Ser Lys Arg Asp Glu Asp
355 360 365
Gly Lys Glu Leu Ala Gly Ala Thr Met Glu Leu Arg Asp Ser Ser Gly
370 375 380
Lys Thr Ile Ser Thr Trp Ile Ser Asp Gly Gln Val Lys Asp Phe Tyr
385 390 395 400
Leu Tyr Pro Gly Lys Tyr Thr Phe Val Glu Thr Ala Ala Pro Asp Gly
405 410 415
Tyr Glu Val Ala Thr Ala Ile Thr Phe Thr Val Asn Glu Gln Gly Gln
420 425 430
Val Thr Val Asn Gly Lys Ala Thr Lys Gly Asp Ala His Ile
435 440 445
<210> 47
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК IgG-Линкер SpyCatcher
<400> 47
ggaagcggcg aaagcggc 18
<210> 48
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA IgG-Линкер SpyCatcher
<400> 48
Gly Ser Gly Glu Ser Gly
1 5
<210> 49
<211> 1020
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IgG
<400> 49
gcctccacca agggcccatc ggtcttcccc ctggcgccct gctccaggag cacctccgag 60
agcacagccg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacgaagacc 240
tacacctgca acgtagatca caagcccagc aacaccaagg tggacaagag agttgagtcc 300
aaatatggtc ccccatgccc accctgccca gcacctgagt tcctgggggg accatcagtc 360
ttcctgttcc ccccaaaacc caaggacact ctcatgatct cccggacccc tgaggtcacg 420
tgcgtggtgg tggacgtgag ccaggaagac cccgaggtcc agttcaactg gtacgtggat 480
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagttcaa cagcacgtac 540
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaacggcaa ggagtacaag 600
tgcaaggtct ccaacaaagg cctcccgtcc tccatcgaga aaaccatctc caaagccaaa 660
gggcagcccc gagagccaca ggtgtacacc ctgcccccat cccaggagga gatgaccaag 720
aaccaggtca gcctgacctg cctggtcaaa ggcttctacc ccagcgacat cgccgtggag 780
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 840
gacggctcct tcttcctcta cagcaggctc accgtggaca agagcaggtg gcaggagggg 900
aatgtcttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagtcc 960
ctctccctgt ctctgggtaa aggaagcggc gaaagcggcg tggataccct gtccggactg 1020
<210> 50
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AA IgG
<400> 50
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 51
<211> 736
<212> БЕЛОК
<213> аденоассоциированный вирус 6
<400> 51
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Asp Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Phe Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Ile Gly
145 150 155 160
Lys Thr Gly Gln Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Thr Pro Ala Ala Val Gly Pro Thr Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ala
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Ala Ser Thr Gly Ala Ser Asn Asp Asn His
260 265 270
Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe
275 280 285
His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn
290 295 300
Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln
305 310 315 320
Val Lys Glu Val Thr Thr Asn Asp Gly Val Thr Thr Ile Ala Asn Asn
325 330 335
Leu Thr Ser Thr Val Gln Val Phe Ser Asp Ser Glu Tyr Gln Leu Pro
340 345 350
Tyr Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala
355 360 365
Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly
370 375 380
Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro
385 390 395 400
Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe
405 410 415
Glu Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp
420 425 430
Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg
435 440 445
Thr Gln Asn Gln Ser Gly Ser Ala Gln Asn Lys Asp Leu Leu Phe Ser
450 455 460
Arg Gly Ser Pro Ala Gly Met Ser Val Gln Pro Lys Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Lys Thr Asp Asn
485 490 495
Asn Asn Ser Asn Phe Thr Trp Thr Gly Ala Ser Lys Tyr Asn Leu Asn
500 505 510
Gly Arg Glu Ser Ile Ile Asn Pro Gly Thr Ala Met Ala Ser His Lys
515 520 525
Asp Asp Lys Asp Lys Phe Phe Pro Met Ser Gly Val Met Ile Phe Gly
530 535 540
Lys Glu Ser Ala Gly Ala Ser Asn Thr Ala Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Lys Ala Thr Asn Pro Val Ala Thr Glu Arg
565 570 575
Phe Gly Thr Val Ala Val Asn Leu Gln Ser Ser Ser Thr Asp Pro Ala
580 585 590
Thr Gly Asp Val His Val Met Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Ala Glu Phe Ser Ala Thr Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Val Gln Tyr Thr Ser Asn
690 695 700
Tyr Ala Lys Ser Ala Asn Val Asp Phe Thr Val Asp Asn Asn Gly Leu
705 710 715 720
Tyr Thr Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<210> 52
<211> 2238
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодирует pAAV2N587Myc
<400> 52
atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga aggaataaga 60
cagtggtgga agctcaaacc tggcccacca ccaccaaagc ccgcagagcg gcataaggac 120
gacagcaggg gtcttgtgct tcctgggtac aagtacctcg gacccttcaa cggactcgac 180
aagggagagc cggtcaacga ggcagacgcc gcggccctcg agcacgacaa agcctacgac 240
cggcagctcg acagcggaga caacccgtac ctcaagtaca accacgccga cgcggagttt 300
caggagcgcc ttaaagaaga tacgtctttt gggggcaacc tcggacgagc agtcttccag 360
gcgaaaaaga gggttcttga acctctgggc ctggttgagg aacctgttaa gacggctccg 420
ggaaaaaaga ggccggtaga gcactctcct gtggagccag actcctcctc gggaaccgga 480
aaggcgggcc agcagcctgc aagaaaaaga ttgaattttg gtcagactgg agacgcagac 540
tcagtacctg acccccagcc tctcggacag ccaccagcag ccccctctgg tctgggaact 600
aatacgatgg ctacaggcag tggcgcacca atggcagaca ataacgaggg cgccgacgga 660
gtgggtaatt cctcgggaaa ttggcattgc gattccacat ggatgggcga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca accacctcta caaacaaatt 780
tccagccaat caggagcctc gaacgacaat cactactttg gctacagcac cccttggggg 840
tattttgact tcaacagatt ccactgccac ttttcaccac gtgactggca aagactcatc 900
aacaacaact ggggattccg acccaagaga ctcaacttca agctctttaa cattcaagtc 960
aaagaggtca cgcagaatga cggtacgacg acgattgcca ataaccttac cagcacggtt 1020
caggtgttta ctgactcgga gtaccagctc ccgtacgtcc tcggctcggc gcatcaagga 1080
tgcctcccgc cgttcccagc agacgtcttc atggtgccac agtatggata cctcaccctg 1140
aacaacggga gtcaggcagt aggacgctct tcattttact gcctggagta ctttccttct 1200
cagatgctgc gtaccggaaa caactttacc ttcagctaca cttttgagga cgttcctttc 1260
cacagcagct acgctcacag ccagagtctg gaccgtctca tgaatcctct catcgaccag 1320
tacctgtatt acttgagcag aacaaacact ccaagtggaa ccaccacgca gtcaaggctt 1380
cagttttctc aggccggagc gagtgacatt cgggaccagt ctaggaactg gcttcctgga 1440
ccctgttacc gccagcagcg agtatcaaag acatctgcgg ataacaacaa cagtgaatac 1500
tcgtggactg gagctaccaa gtaccacctc aatggcagag actctctggt gaatccgggc 1560
ccggccatgg caagccacaa ggacgatgaa gaaaagtttt ttcctcagag cggggttctc 1620
atctttggga agcaaggctc agagaaaaca aatgtggaca ttgaaaaggt catgattaca 1680
gacgaagagg aaatcaggac aaccaatccc gtggctacgg agcagtatgg ttctgtatct 1740
accaacctcc agagaggcaa cgaacaaaaa ctcatctcag aagaggatct gagacaagca 1800
gctaccgcag atgtcaacac acaaggcgtt cttccaggca tggtctggca ggacagagat 1860
gtgtaccttc aggggcccat ctgggcaaag attccacaca cggacggaca ttttcacccc 1920
tctcccctca tgggtggatt cggacttaaa caccctcctc cacagattct catcaagaac 1980
accccggtac ctgcgaatcc ttcgaccacc ttcagtgcgg caaagtttgc ttccttcatc 2040
acacagtact ccacgggaca ggtcagcgtg gagatcgagt gggagctgca gaaggaaaac 2100
agcaaacgct ggaatcccga aattcagtac acttccaact acaacaagtc tgttaatgtg 2160
gactttactg tggacactaa tggcgtgtat tcagagcctc gccccattgg caccagatac 2220
ctgactcgta atctgtaa 2238
<210> 53
<211> 745
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность pAAV2N587 Myc
<400> 53
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Glu Pro Asp Ser Ser Ser Gly Thr Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Gln Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Thr Asn Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Glu Gln Lys Leu Ile
580 585 590
Ser Glu Glu Asp Leu Arg Gln Ala Ala Thr Ala Asp Val Asn Thr Gln
595 600 605
Gly Val Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu Gln
610 615 620
Gly Pro Ile Trp Ala Lys Ile Pro His Thr Asp Gly His Phe His Pro
625 630 635 640
Ser Pro Leu Met Gly Gly Phe Gly Leu Lys His Pro Pro Pro Gln Ile
645 650 655
Leu Ile Lys Asn Thr Pro Val Pro Ala Asn Pro Ser Thr Thr Phe Ser
660 665 670
Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val
675 680 685
Ser Val Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg Trp
690 695 700
Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr Asn Lys Ser Val Asn Val
705 710 715 720
Asp Phe Thr Val Asp Thr Asn Gly Val Tyr Ser Glu Pro Arg Pro Ile
725 730 735
Gly Thr Arg Tyr Leu Thr Arg Asn Leu
740 745
<210> 54
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> SpyTag002
<400> 54
Val Pro Thr Ile Val Met Val Asp Ala Tyr Lys Arg Tyr Lys
1 5 10
<210> 55
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> SpyCatcher002
<400> 55
Val Thr Thr Leu Ser Gly Leu Ser Gly Glu Gln Gly Pro Ser Gly Asp
1 5 10 15
Met Thr Thr Glu Glu Asp Ser Ala Thr His Ile Lys Phe Ser Lys Arg
20 25 30
Asp Glu Asp Gly Arg Glu Leu Ala Gly Ala Thr Met Glu Leu Arg Asp
35 40 45
Ser Ser Gly Lys Thr Ile Ser Thr Trp Ile Ser Asp Gly His Val Lys
50 55 60
Asp Phe Tyr Leu Tyr Pro Gly Lys Tyr Thr Phe Val Glu Thr Ala Ala
65 70 75 80
Pro Asp Gly Tyr Glu Val Ala Thr Ala Ile Thr Phe Thr Val Asn Glu
85 90 95
Gln Gly Gln Val Thr Val Asn Gly Glu Ala Thr Lys Gly Asp Ala His
100 105 110
Thr
<---
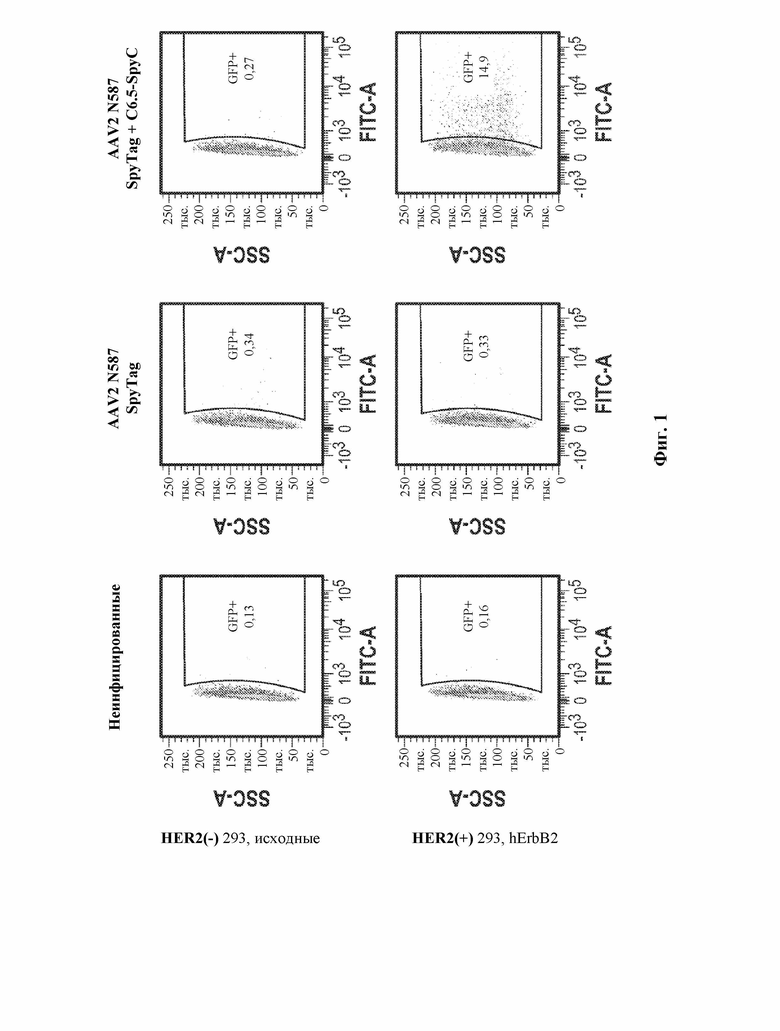
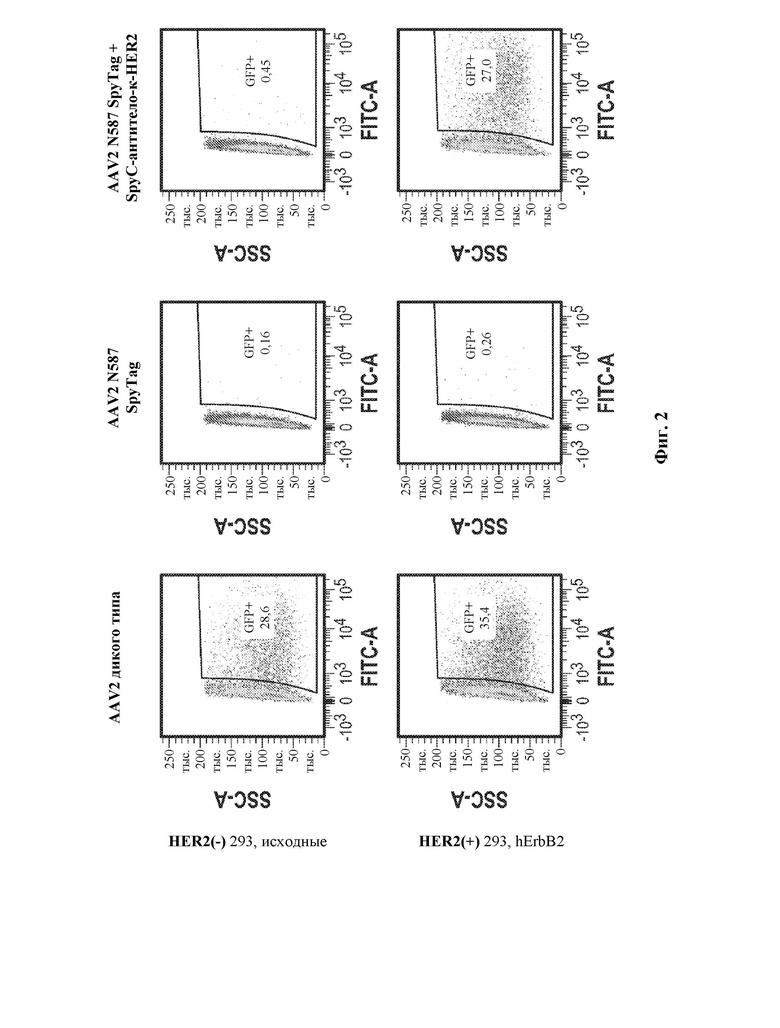
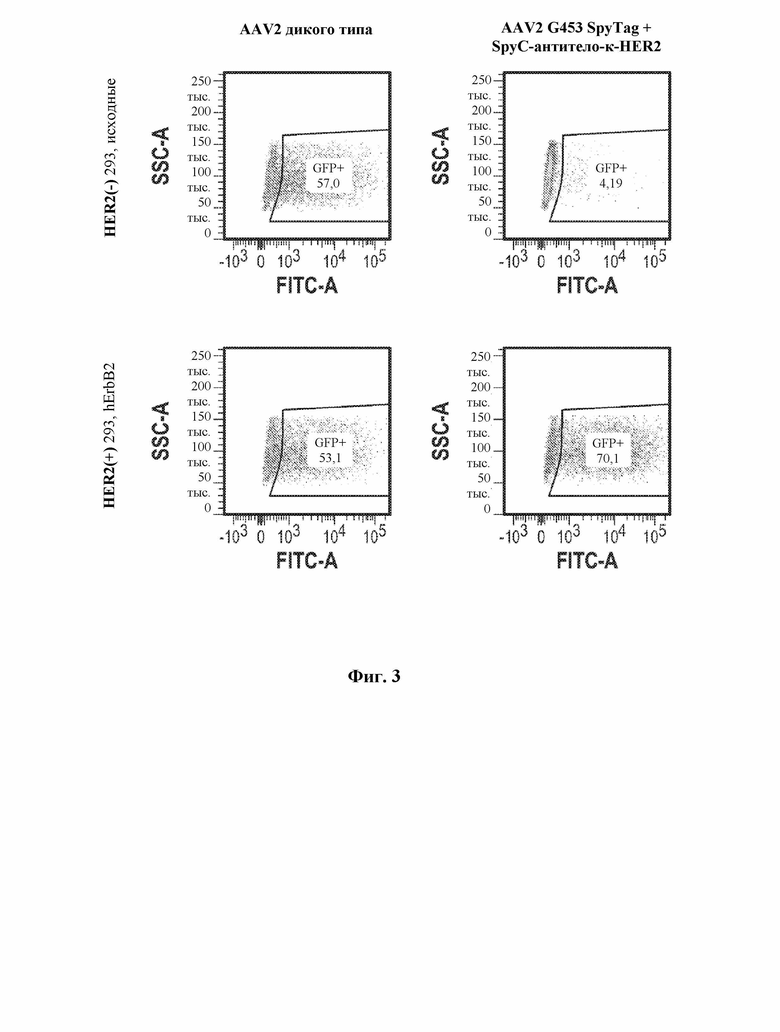
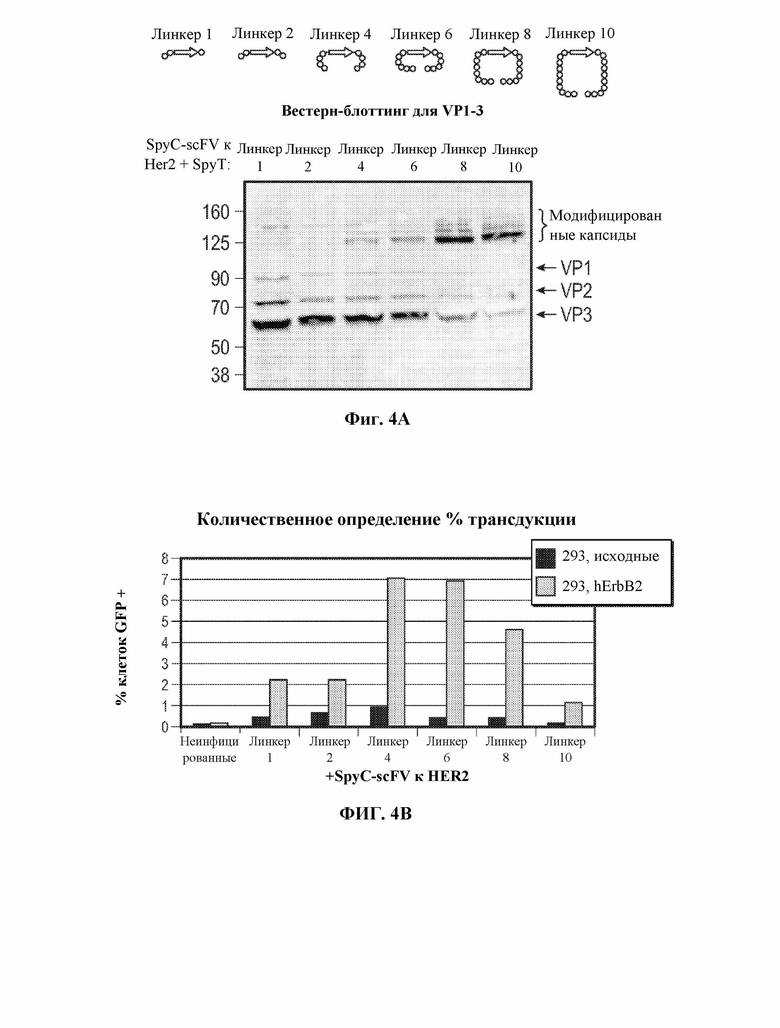
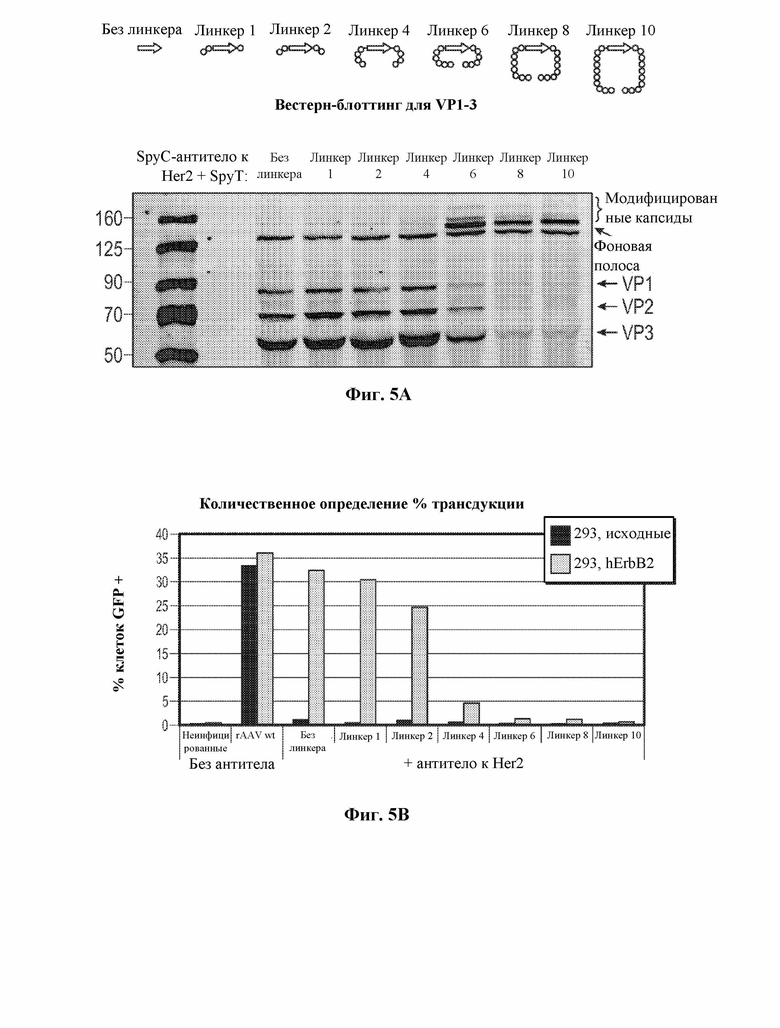
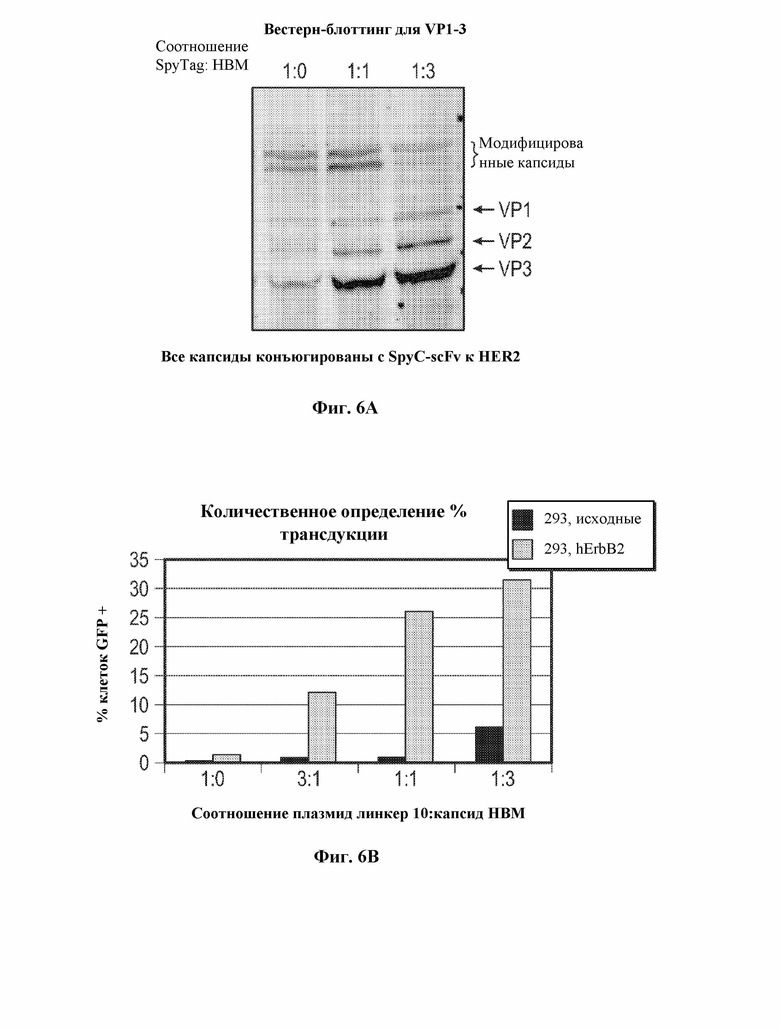
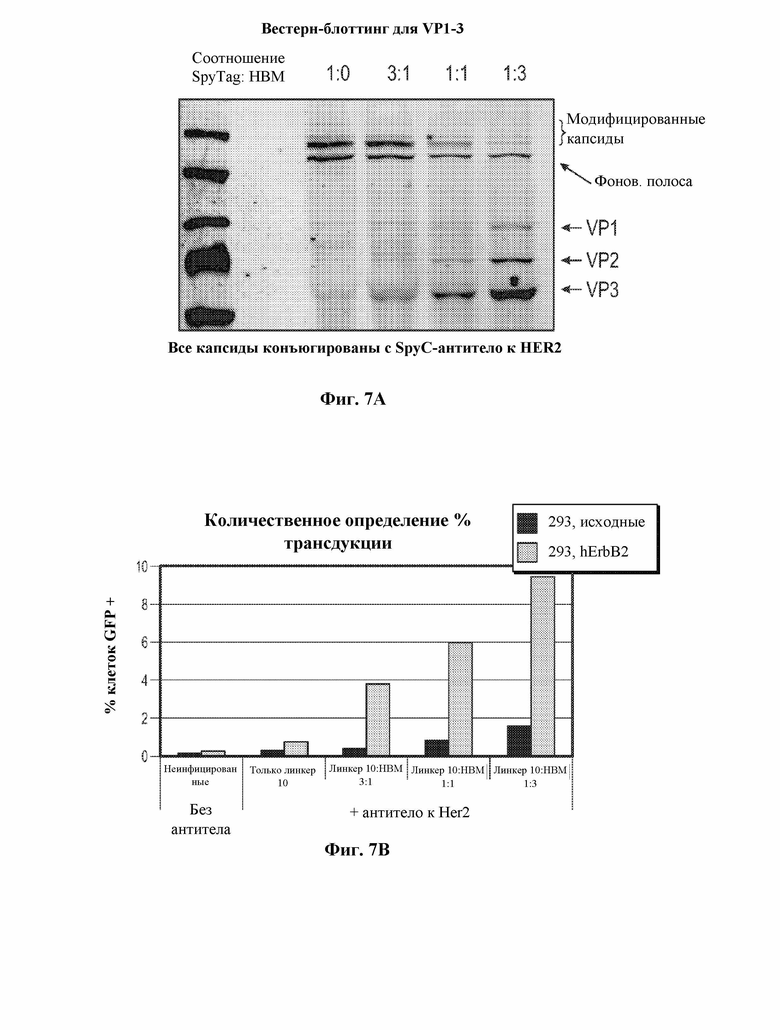
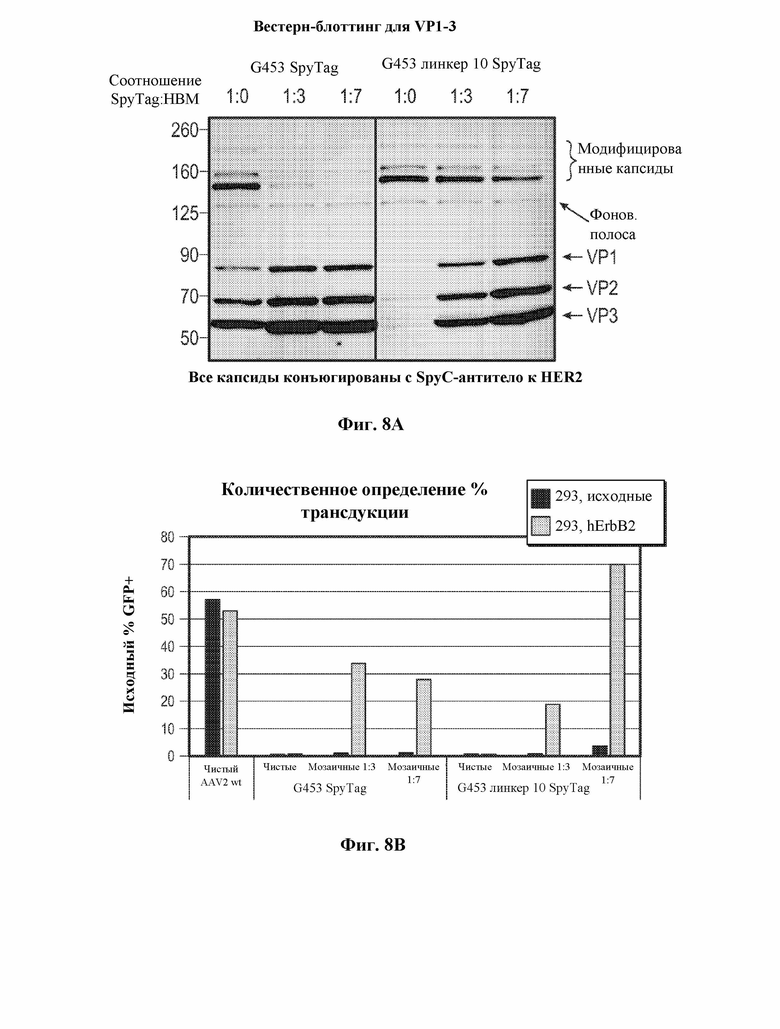
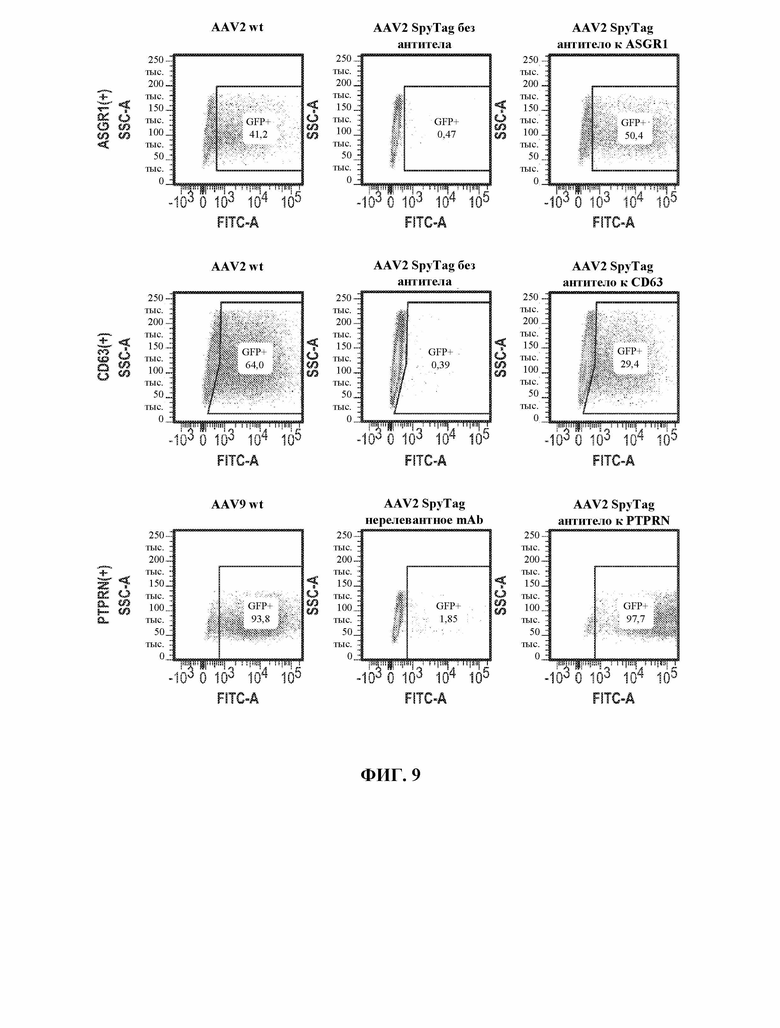
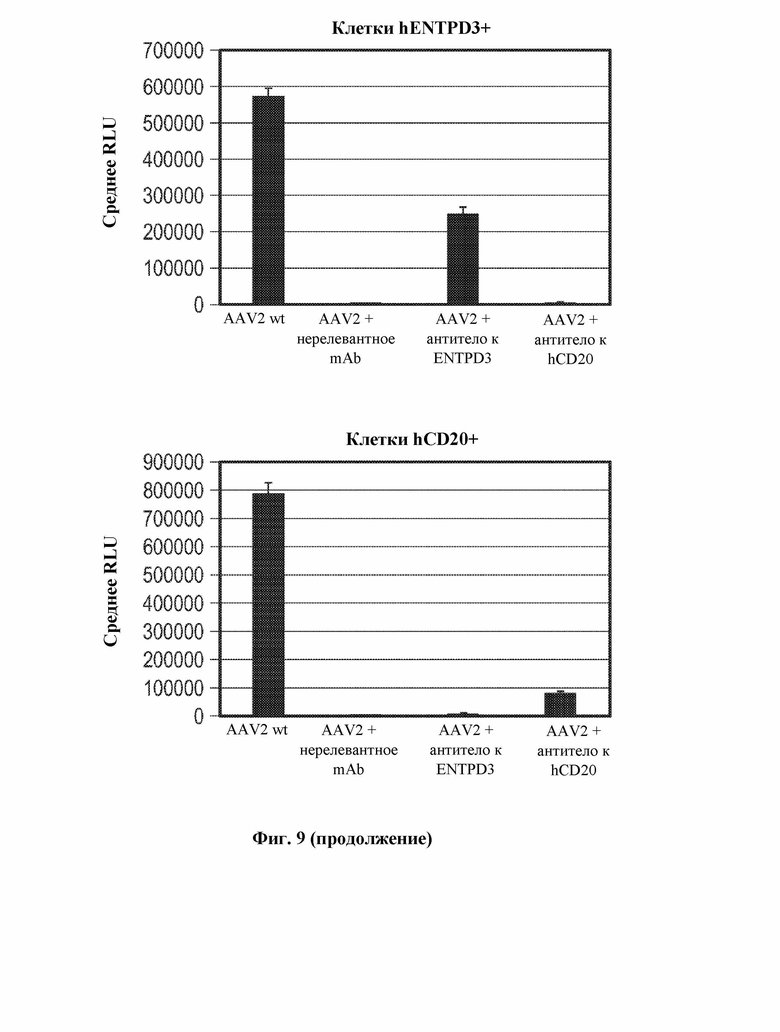

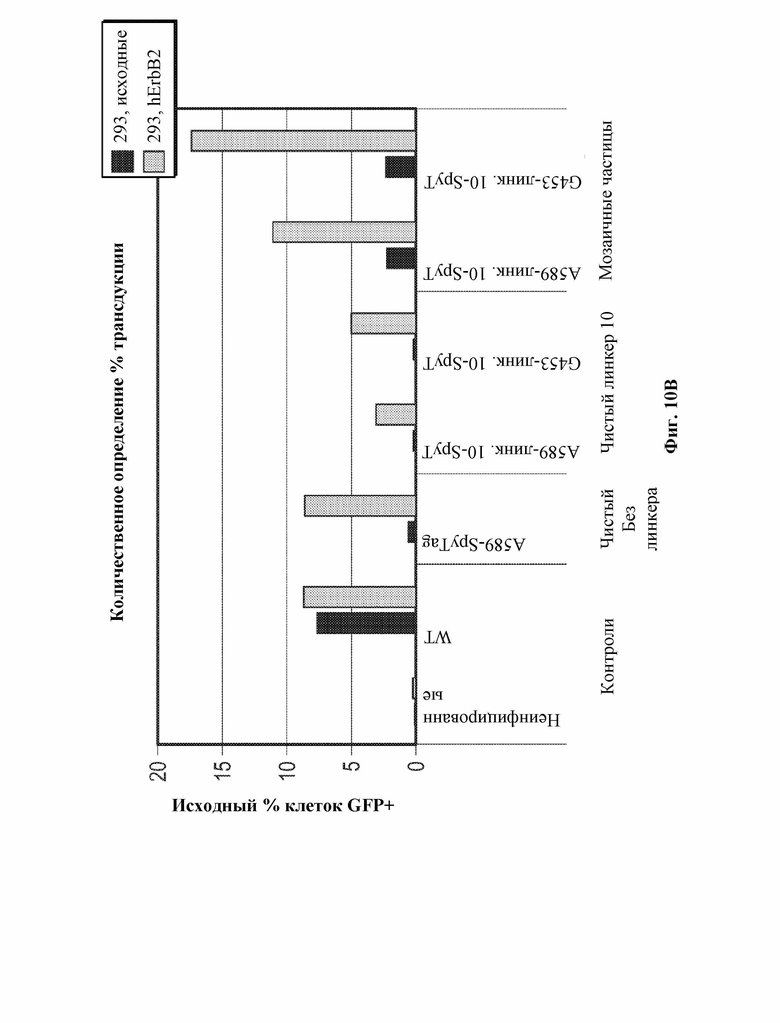

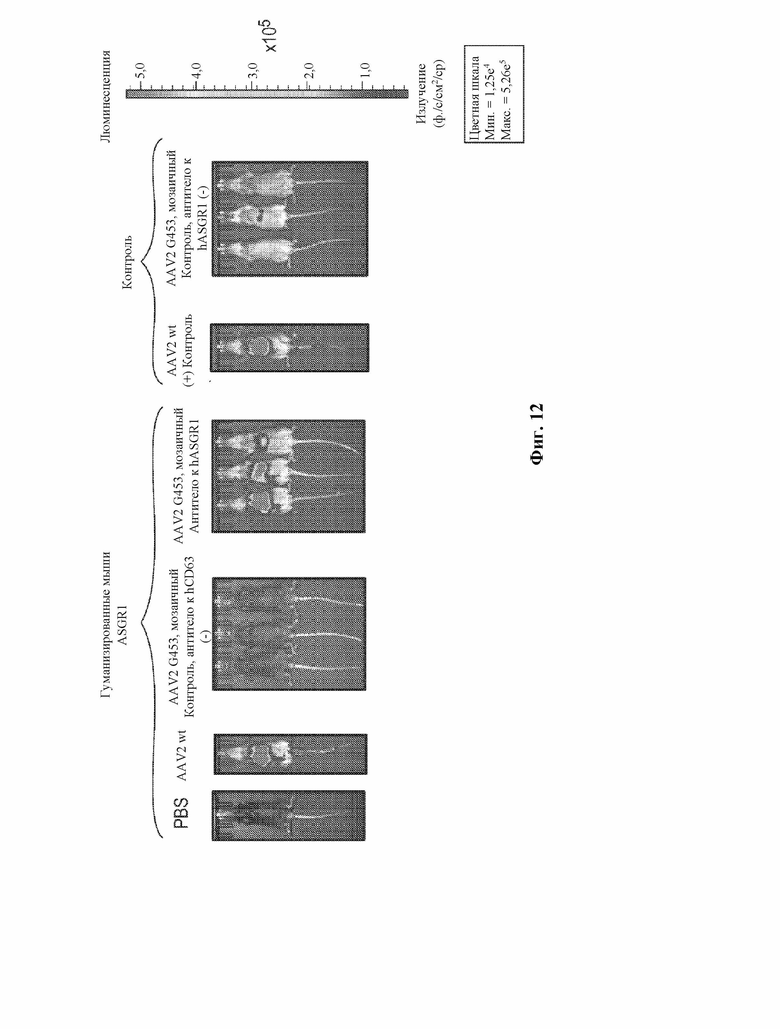


Изобретение относится к области биотехнологии. Описана группа изобретений, включающая рекомбинантный вирусный капсидный белок для отображения гетерологичной аминокислотной последовательности, рекомбинантный вирусный капсид для отображения гетерологичной аминокислотной последовательности, содержащий вышеуказанный рекомбинантный вирусный капсидный белок, рекомбинантную вирусную частицу для отображения гетерологичной аминокислотной последовательности, композицию для отображения гетерологичной аминокислотной последовательности, способ доставки представляющего интерес нуклеотида в клетку-мишень. Изобретение расширяет арсенал рекомбинантных вирусных капсидных белков для отображения гетерологичной аминокислотной последовательности. 5 н. и 58 з.п. ф-лы, 14 ил., 2 табл., 10 пр.
1. Рекомбинантный вирусный капсидный белок для отображения гетерологичной аминокислотной последовательности, где рекомбинантный вирусный капсидный белок содержит первого партнера связывающейся пары белок:белок, вставленного в рекомбинантный вирусный капсидный белок,
где рекомбинантный вирусный капсидный белок получен с помощью гена cap аденоассоциированного вируса (AAV), который кодирует капсидный белок VP1, VP2 и/или VP3 AAV,
где первый партнер связывающейся пары белок:белок содержит пептидную метку, длина которой составляет менее 50 аминокислот, и способен образовывать изопептидную связь со вторым родственным партнером связывающейся пары белок:белок.
2. Рекомбинантный вирусный капсидный белок по п. 1, дополнительно содержащий второй родственный партнер связывающейся пары белок:белок, где первый партнер связывающейся пары белок:белок связан со вторым родственным партнером связывающейся пары белок:белок ковалентной связью.
3. Рекомбинантный вирусный капсидный белок по п. 2, где ковалентная связь представляет собой изопептидную связь.
4. Рекомбинантный вирусный капсидный белок по любому из пп. 2, 3, где второй родственный партнер функционально связан с нацеливающим лигандом направленным на клетку-мишень, где необязательно нацеливающий лиганд представляет собой связывающий фрагмент.
5. Рекомбинантный вирусный капсидный белок по любому из пп. 1-4, где первый партнер фланкирован первым и/или вторым линкером, который(ые) соединяет(ют) первого партнера с рекомбинантным вирусным капсидным белком, и где длина каждого из первого и/или второго линкера независимо составляет по меньшей мере одну аминокислоту.
6. Рекомбинантный вирусный капсидный белок по п. 5, где первый и второй линкеры не являются идентичными.
7. Рекомбинантный вирусный капсидный белок по п. 5, где первый и второй линкеры являются идентичными, и их длина составляет 10 аминокислот.
8. Рекомбинантный вирусный капсидный белок по любому из пп. 1-7, где
(a) первый партнер связывающейся пары белок:белок обеспечивает уменьшение или устранение природного тропизма рекомбинантного вирусного капсидного белка; и/или
(b) рекомбинантный вирусный капсидный белок дополнительно содержит мутацию в дополнение к первому партнеру связывающейся пары белок:белок, которая обеспечивает уменьшение или устранение природного тропизма рекомбинантного вирусного капсидного белка,
необязательно, где:
мутация находится в аминокислотном положении, участвующем в связывании рекомбинантного вирусного капсидного белка с его природным рецептором, и/или
мутация предусматривает вставку гетерологичного пептида в рекомбинантный вирусный капсидный белок, замену аминокислоты рекомбинантного вирусного капсидного белка на гетерологичный пептид, делецию аминокислоты рекомбинантного вирусного капсидного белка или их комбинацию.
9. Рекомбинантный вирусный капсидный белок по любому из пп. 4-8, где эффективность трансдукции рекомбинантного вирусного капсидного белка в отношении клетки-мишени уменьшается или устраняется в отсутствие нацеливающего лиганда, необязательно, где эффективность трансдукции в отношении клетки-мишени в отсутствие нацеливающего лиганда:
(i) снижена на 10%,
(ii) снижена на 20%,
(iii) снижена на 30%,
(iv) снижена на 40%,
(v) снижена на 50%,
(vi) снижена на 60%,
(vii) снижена на 70%,
(viii) снижена на 80%, или
(ix) снижена на 90%.
10. Рекомбинантный вирусный капсидный белок по любому из пп. 1-9, где AAV выбран из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и AAV9.
11. Рекомбинантный вирусный капсидный белок по п. 10, где AAV представляет собой AAV2.
12. Рекомбинантный вирусный капсидный белок по п. 10, где AAV представляет собой AAV9.
13. Рекомбинантный вирусный капсидный белок по любому из пп. 1-12, где связывающаяся пара белок:белок представляет собой
(i) SpyTag:SpyCatcher,
(ii) SpyTag:KTag,
(iii) Isopeptag:пилин-C,
(iv) SnoopTag:SnoopCatcher или
(v) SpyTag002:SpyCatcher002.
14. Рекомбинантный вирусный капсидный белок по любому из пп. 1-13, где совокупная длина первого партнера и любого линкера составляет не более приблизительно 50 аминокислот.
15. Рекомбинантный вирусный капсидный белок по любому из пп. 1-14, где первый партнер представляет собой SpyTag, содержащий последовательность, изложенную под SEQ ID NO: 1.
16. Рекомбинантный вирусный капсидный белок по любому из пп. 2-15, где второй родственный партнер представляет собой SpyCatcher, содержащий последовательность, изложенную под SEQ ID NO: 3.
17. Рекомбинантный вирусный капсидный белок по любому из пп. 2-15, где второй родственный партнер представляет собой KTag, содержащий последовательность, изложенную под SEQ ID NO: 2.
18. Рекомбинантный вирусный капсидный белок по любому из пп. 2-15, где первый партнер представляет собой KTag, содержащий последовательность, изложенную под SEQ ID NO: 2, а второй родственный партнер представляет собой Spy Tag, содержащий последовательность, изложенную под SEQ ID NO: 1.
19. Рекомбинантный вирусный капсидный белок по любому из пп. 2-14, где
первый партнер представляет собой SnoopTag, содержащий последовательность, изложенную под SEQ ID NO: 5, а второй родственный партнер представляет собой SnoopCatcher.
20. Рекомбинантный вирусный капсидный белок по любому из пп. 2-14, где
первый партнер представляет собой isopeptag, содержащий последовательность, изложенную под SEQ ID NO: 7, а второй родственный партнер представляет собой пилин-С.
21. Рекомбинантный вирусный капсидный белок по любому из пп. 2-14, где первый партнер представляет собой SpyTag002, а второй родственный партнер представляет собой SpyCatcher002.
22. Рекомбинантный вирусный капсидный белок по любому из пп. 4-21, где нацеливающий лиганд представляет собой антитело или его часть.
23. Рекомбинантный вирусный капсидный белок по п. 22, где антитело или его часть слиты с SpyCatcher, содержащим аминокислотную последовательность, изложенную под SEQ ID NO: 3.
24. Рекомбинантный вирусный капсидный белок по п. 23, где антитело или его часть слиты с линкером на С-конце, и линкер слит с SpyCatcher на С-конце линкера.
25. Рекомбинантный вирусный капсидный белок по п. 24, где линкер содержит аминокислотную последовательность, изложенную под SEQ ID NO: 48.
26. Рекомбинантный вирусный капсидный белок по любому из пп. 4-25, где нацеливающий лиганд содержит аминокислотную последовательность, изложенную под SEQ ID NO: 50.
27. Рекомбинантный вирусный капсидный белок по любому из пп. 4-26, где нацеливающий лиганд специфически связывается с молекулой клеточной поверхности, где необязательно молекула клеточной поверхности представляет собой
(i) асиалогликопротеин 1 (ASGR1),
(ii) эктонуклеозидтрифосфатдифосфогидролаза 3 (ENTPD3),
(iii) тирозиновая протеинфосфатаза N рецепторного типа (PTPRN),
(iv) дифференцировочный кластер 20 (CD20),
(v) дифференцировочный кластер 63 (CD63),
(vi) рецептор эпидермального фактора роста человека типа 2 (Her2), или
(vii) дифференцировочный кластер 71 (CD71).
28. Рекомбинантный вирусный капсидный белок по любому из пп. 4-26, где:
(i) клетка-мишень представляет собой клетку печени человека;
(ii) клетка-мишень представляет собой нейронную клетку человека;
(iii) клетка-мишень представляет собой Т-клетку человека;
(iv) клетка-мишень представляет собой гемопоэтическую клетку человека;
(vi) клетка-мишень представляет собой клетку почки человека;
(vii) клетка-мишень представляет собой раковую клетку человека; или
(viii) клетка-мишень представляет собой мышечную клетку человека.
29. Рекомбинантный вирусный капсидный белок по любому из пп. 1-26, где cap ген кодирует генетически модифицированный капсидный белок AAV VP1, содержащий аминокислотную последовательность, изложенную в SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 35, SEQ ID NO: NO: 37 или SEQ ID NO: 39.
30. Рекомбинантный вирусный капсидный белок по любому из пп. 2-29, где второй родственный партнер, слитый с нацеливающим лигандом, содержит аминокислотную последовательность, изложенную в SEQ ID NO: 46.
31. Рекомбинантный вирусный капсид для отображения гетерологичной аминокислотной последовательности, содержащий первого партнера связывающейся пары белок:белок, где рекомбинантный вирусный капсид содержит рекомбинантный вирусный капсидный белок по любому из пп. 1-30.
32. Рекомбинантный вирусный капсид по п. 31, дополнительно содержащий эталонный вирусный капсидный белок, в котором отсутствует какой-либо партнер связывающейся пары белок:белок.
33. Рекомбинантный вирусный капсид по п. 32, где каждый из рекомбинантного вирусного капсидного белка и эталонного вирусного капсидного белка содержит мутацию остатка, участвующего в связывании вирусного капсида с его природным рецептором.
34. Рекомбинантный вирусный капсид по любому из пп. 32, 33, содержащий рекомбинантный вирусный капсидный белок и эталонный вирусный капсидный белок в соотношении от 1:1 до 1:15.
35. Рекомбинантная вирусная частица для отображения гетерологичной аминокислотной последовательности, содержащая первого партнера связывающейся пары белок:белок, где рекомбинантная вирусная частица содержит представляющий интерес нуклеотид, заключенный в рекомбинантный вирусный капсид по любому из пп. 31-34.
36. Рекомбинантная вирусная частица по п. 35, где представляющий интерес нуклеотид находится под контролем промотора, выбранного из группы, состоящей из вирусного промотора, бактериального промотора, промотора млекопитающего, промотора птицы, промотора рыбы, промотора насекомого и любой их комбинации.
37. Рекомбинантная вирусная частица по п. 35, где представляющий интерес нуклеотид находится под контролем человеческого промотора.
38. Рекомбинантная вирусная частица по п. 35, где представляющий интерес нуклеотид находится под контролем промотора, отличного от человеческого.
39. Рекомбинантная вирусная частица по любому из пп. 35-38, где представляющий интерес нуклеотид представляет собой репортерный ген.
40. Рекомбинантная вирусная частица по п. 39, где репортерный ген кодирует зеленый флуоресцентный белок.
41. Рекомбинантная вирусная частица по п. 39, где репортерный ген кодирует зеленый флуоресцентный белок, β-галактозидазу, усиленный зеленый флуоресцентный белок (eGFP), MmGFP, синий флуоресцентный белок (BFP), усиленный синий флуоресцентный белок (eBFP), mPlum, mCherry, tdTomato, mStrawberry, J-Red, DsRed, mOrange, mKO, mCitrine, Venus, YPet, желтый флуоресцентный белок (YFP), усиленный желтый флуоресцентный белок (eYFP), Emerald, CyPet, голубой флуоресцентный белок (CFP), Cerulean, T-Sapphire, люциферазу, щелочную фосфатазу или их комбинацию.
42. Рекомбинантная вирусная частица по любому из пп. 35-38, где представляющий интерес нуклеотид выбран из группы, состоящей из гена, кодирующего терапевтический белок, суицидального гена, нуклеотида, кодирующего антитело или его фрагмент, нуклеотида, кодирующего систему CRISPR/Cas или ее часть(части), нуклеотида, кодирующего антисмысловую РНК, и нуклеотида, кодирующего shRNA.
43. Композиция для отображения гетерологичной аминокислотной последовательности, содержащая первого партнера связывающейся пары белок:белок, где композиция содержит (а) рекомбинантный вирусный капсид по любому из пп. 31-34 или рекомбинантную вирусную частицу по любому из пп. 35-42 и (b) фармацевтически приемлемый носитель.
44. Способ доставки представляющего интерес нуклеотида в клетку-мишень, предусматривающий приведение клетки-мишени в контакт с:
(a) рекомбинантной вирусной частицей, содержащей:
(i) рекомбинантный вирусный капсид, содержащий рекомбинантный вирусный капсидный белок, содержащий первого партнера связывающейся пары белок:белок, вставленного в рекомбинантный вирусный капсидный белок,
где рекомбинантный вирусный капсидный белок получен с помощью гена cap аденоассоциированного вируса (AAV), который кодирует капсидный белок VP1, VP2 и/или VP3 AAV, и
где первый партнер связывающейся пары белок:белок содержит пептидную метку, длина которой составляет менее 50 аминокислот, и способен образовывать изопептидную связь со вторым родственным партнером связывающейся пары белок:белок; и
(ii) представляющий интерес нуклеотид, заключенный в рекомбинантный вирусный капсид; или
(b) фармацевтической композицией, содержащей рекомбинантную вирусную частицу из (а),
где рекомбинантный вирусный капсид содержит нацеливающий лиганд, который специфически связывается с белком, экспрессируемым на поверхности клетки-мишени.
45. Способ по п. 44, где клетка-мишень находится in vitro.
46. Способ по п. 44, где клетка-мишень находится in vivo у субъекта.
47. Способ по п. 46, где субъектом является человек.
48. Способ по любому из пп. 44-47, где клетка-мишень представляет собой клетку-мишень человека.
49. Способ по п. 48, где клетка-мишень представляет собой клетку печени человека, и где нацеливающий лиганд связывается с асиалогликопротеиновым рецептором человека (ASGR1).
50. Способ по п. 48, где клетка-мишень представляет собой нейронную клетку человека, и где нацеливающий лиганд связывается с GABA.
51. Способ по п. 48, где клетка-мишень представляет собой Т-клетку человека, и где нацеливающий лиганд связывается с CD3, необязательно CD3ε.
52. Способ по п. 48, где нацеливающий лиганд связывается с PTPRN.
53. Способ по п. 48, где клетка-мишень представляет собой гемопоэтическую клетку человека, и где нацеливающий лиганд связывается с CD34.
54. Способ по п. 48, где клетка-мишень представляет собой клетку почки человека.
55. Способ по п. 48, где клетка-мишень представляет собой раковую клетку человека, и где нацеливающий лиганд связывается с опухолеассоциированным антигеном.
56. Способ по п. 55, где опухолевый антиген представляет собой Е6, Е7 или Her2.
57. Способ по п. 48, где нацеливающий лиганд связывается с CD20.
58. Способ по п. 48, где нацеливающий лиганд связывается с рецептором глюкагона человека.
59. Способ по любому из пп. 44-48, где нацеливающий лиганд специфически связывается с CD63.
60. Способ по любому из пп. 44-48, где нацеливающий лиганд специфически связывается с эктонуклеозидтрифосфатдифосфогидролазой 3 человека (hENTPD3).
61. Способ по п. 48, где клетка-мишень представляет собой нейронную клетку.
62. Способ по п. 48, где клетка-мишень представляет собой мышечную клетку.
63. Способ по п. 48, где нацеливающий лиганд специфически связывается с CD71.
| WO 2001021216 A1, 29.03.2001 | |||
| WO 2013037961 A1, 21.03.2013 | |||
| WO 2010138263 A2, 02.12.2010 | |||
| RU 2017116353 A, 22.11.2018 | |||
| J.S | |||
| Bartlett, J | |||
| Kleinschmidt, R.C | |||
| Boucher, R.J | |||
| Samulski | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Nature Biotechnology, 1999, 17(2), 181-186 | |||

