[0001] Содержание любых и всех заявок, для которых иностранный или внутренний приоритет заявлен в информационном листке заявки, поданном вместе с настоящей заявкой, включено посредством ссылки в соответствии с 37 CFR §1.57.
Перечень последовательностей
[0002] Настоящая заявка подается вместе с перечнем последовательностей в электронном виде. Указанный перечень последовательностей предоставлен в виде файла под названием «KDIAK044.txt», созданного 1 марта 2019 года, размер которого составляет 395430 байтов. Полная информация, содержащаяся в перечне последовательностей в электронном виде, включена в настоящее описание посредством ссылки.
ПОЛОЖЕНИЕ О ДЕПОНИРОВАНИИ БИОЛОГИЧЕСКИХ ОБРАЗЦОВ
[0003] В некоторых вариантах реализации предложены анти-IL-6 антитела, их слитые конструкции и конъюгаты, которые депонированы в Американской коллекции типовых культур (АТСС) в соответствии с Будапештским договором под номерами ____ (дата) _____.
[0004] Эти депонирования осуществлены в соответствии с положениями Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры и инструкциями к нему (Будапештский договор). Это обеспечивает сохранение депонированного образца в течение 30 лет с даты депонирования. Депонированный образец будет доступен в ATCC в соответствии с условиями Будапештского договора и по соглашению между заявителем и ATCC, что обеспечивает постоянную и неограниченную общедоступность депонированного образца после выдачи соответствующего патента США или после публикации любой заявки на патент США или иностранной заявки на патент в зависимости от того, что произойдет раньше, и обеспечивает доступность депонированного образца для лиц, определенных уполномоченным США по патентам и товарным знакам как имеющих на это право согласно 35 U.S.C. (Свода законов США) § 122 и соответствующим правилам уполномоченного (в том числе 37 C.F.R. (Свод федеральных нормативных актов США) § 1.14). Доступность депонированного биологического материала не должна рассматриваться как разрешение осуществление изобретения в нарушение прав, предоставленных уполномоченным органом любого правительства в соответствии с его патентным законодательством.
ОБЛАСТЬ ТЕХНИКИ
[0005] Настоящее изобретение, в целом, относится к слитым конструкциям, которые связываются с IL-6 и/или VEGF.
УРОВЕНЬ ТЕХНИКИ
[0006] Фактор роста эндотелия сосудов-А (VEGF-A) представляет собой сигнальный белок, который опосредует проангиогенные функции, такие как выживание, пролиферация, миграция клеток эндотелия и межклеточная проницаемость. Было показано, что его активность способствует прогрессированию заболеваний сетчатки, таких как хориоидальная неоваскуляризация (ХНВ) при возрастной дегенерации желтого пятна (ВДЖП) и диабетическом отеке желтого пятна (ДОЖП). Доказано, что ингибирование передачи сигнала VEGF-A является эффективным способом остановки прогрессирования неоваскулярных заболеваний сетчатки (Ferrara et al., Retina, 2006). Для ингибирования функции VEGF были разработаны различные терапевтические молекулы. Было показано, что среди указанных молекул такие моноклональные анти-VEGF антитела, как ранибизумаб и бевацизумаб, представляют собой безопасное и эффективное лечение патологического ангиогенеза. В последнее время полученные рекомбинантным способом слитые белки VEGFR (рецептора фактора роста сосудистого эндотелия - ФРЭС), такие как афлиберцепт (Эйлеа) и конберцепт (Китай), которые действуют как «ловушки» для VEGF, оказываются более эффективными и демонстрируют более длительное действие, чем их конкуренты-антитела.
[0007] Воспаление вовлечено в патогенез заболеваний сетчатки, и такие виды противовоспалительной терапии, как стероиды, эффективны в лечении увеита и диабетического отека желтого пятна (ДОЖП). Тщательные исследования воспаления и инфекции в глазу показали, что во внутриглазной жидкости пациентов с рефрактерным/хроническим увеитом уровень провоспалительного цитокина, интерлейкина-6 (IL-6), значительно повышен, и ингибирование IL-6 в моделях у животных препятствует возникновению увеита. Высокие уровни IL-6 во внутриглазной жидкости также обнаруживают у пациентов с ДОЖП и окклюзией вены сетчатки. Более того, клетки хронического воспаления наблюдали на поверхности мембраны Бруха в глазах с неоваскулярной ВДЖП, и пациенты с ВДЖП, по сообщениям, демонстрировали повышенные уровни IL-6 в сыворотке. Интересно, что согласно наблюдениям IL-6 также стимулировал нарушение ангиогенеза. Было показано, что в дополнение к аутоиммунным нарушениям, таким как ревматоидный артрит, лечение, направленное против IL-6, позволяет эффективно лечить увеит и отек желтого пятна при увеите.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0008] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело, которое специфично связывается с IL-6, конъюгированное с полимером.
[0009] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело. Указанное антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи, составляющую тяжелую цепь, в таблице 1; и аминокислотную последовательность вариабельной области легкой цепи, составляющую легкую цепь, в таблице 2.
[0010] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело. Указанное антитело содержит: вариабельную область тяжелой цепи (VH), содержащую определяющий комплементарность область (участок) (CDR) 1 (CDR1) VH, CDR2 VH и CDR3 VH, имеющую аминокислотную последовательность, выбранную из группы, состоящей из CDR в таблице 1; и вариабельную область легкой цепи (VL), содержащую CDR1 VL, CDR2 VL и CDR3 VL, имеющую аминокислотную последовательность, выбранную из группы, состоящей из CDR в таблице 2.
[0011] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело, которое связывается с IL-6. Указанное антитело содержит по меньшей мере одну из следующих мутаций согласно нумерации EU (ЕС): L234A, L235A и G237A.
[0012] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело, которое связывается с IL-6. Указанное антитело содержит CDRH1, которая представляет собой CDRH1 в таблице 1; CDRH2, которая представляет собой CDRH2 в Таблице 1; CDRH3, которая представляет собой CDRH3 в Таблице 1; CDRL1, которая представляет собой CDRL1 в таблице 2; CDRL2, которая представляет собой CDRL2 в таблице 2; CDRL3, которая представляет собой CDRL3 в таблице 2; по меньшей мере одну из следующих мутаций (нумерация EU): L234A, L235A и G237A; и по меньшей мере одну из следующих мутаций (нумерация EU): Q347C или L443C.
[0013] В соответствии с некоторыми аспектами предложен слитый белок, содержащий выделенное антагонистическое антитело; и фрагмент VEGF-связывающего белка.
[0014] В соответствии с некоторыми аспектами предложен конъюгат, содержащий любое из выделенных антагонистических антител согласно настоящему изобретению и полимер. Указанный полимер ковалентно присоединен к антителу.
[0015] В соответствии с некоторыми аспектами предложен конъюгат. Указанный конъюгат содержит слитый белок, предложенный согласно настоящему изобретению, и полимер, при этом указанный полимер ковалентно присоединен к слитому белку.
[0016] В соответствии с некоторыми аспектами предложена выделенная линия клеток, продуцирующая выделенное антагонистическое антитело или слитый белок.
[0017] В соответствии с некоторыми аспектами предложена выделенная нуклеиновая кислота, кодирующая выделенное антагонистическое антитело или слитый белок.
[0018] В соответствии с некоторыми аспектами предложен рекомбинантный вектор экспрессии. Указанный вектор содержит нуклеиновую кислоту, кодирующую любую из этих конструкций, предложенных согласно настоящему изобретению.
[0019] В соответствии с некоторыми аспектами предложена клетка-хозяин, содержащая вектор экспрессии, предложенный согласно настоящему изобретению.
[0020] В соответствии с некоторыми аспектами предложен способ получения антитела-антагониста IL-6 или его слитого белка. Указанный способ включает: культивирование линии клеток, которая рекомбинантно продуцирует выделенное антагонистическое антитело или слитый белок в условиях, в которых продуцируется антитело; и извлечение антитела.
[0021] В соответствии с некоторыми аспектами предложена фармацевтическая композиция. Она содержит выделенное антагонистическое антитело или слитый белок и/или конъюгат, предложенный согласно настоящему изобретению, и фармацевтически приемлемый носитель.
[0022] В соответствии с некоторыми аспектами предложен способ лечения или профилактики заболевания у нуждающегося в этом пациента. Указанный способ включает введение пациенту выделенного антитела-антагониста или слитого белка и/или конъюгата, предложенного согласно настоящему изобретению.
[0023] В соответствии с некоторыми аспектами предложен способ лечения или профилактики заболевания у нуждающегося в этом пациента. Указанный способ включает идентификацию пациента с гиперактивностью IL-6 и/или VEGF; и введение указанному пациенту выделенного антитела-антагониста или слитого белка и/или конъюгата, предложенного согласно настоящему изобретению.
[0024] В соответствии с некоторыми аспектами предложен слитый белок, содержащий IL-6 VH, IL-6 VL, IL-6 Fc и ловушку VEGF, при этом указанная ловушка VEGF слита с IL-6 одним из следующих способов: 1) с N-концом тяжелой цепи, содержащей IL-6 VH; или 2) между шарнирной областью и после CH1-домена тяжелой цепи, содержащей IL-6 VH.
[0025] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело к IL-6, содержащее вариабельную область тяжелой цепи, которая содержит тяжелую цепь, которая имеет последовательность по меньшей мере одной из SEQ ID NO: 7-13, 19-27, 89, 90, 256-262; и вариабельную область легкой цепи, которая содержит легкую цепь, которая имеет последовательность по меньшей мере одной из SEQ ID NO: 91-93, 28-30.
[0026] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело к IL-6, содержащее: вариабельную область тяжелой цепи (VH), содержащую 3 определяющие комплементарность области: CDR1 VH, CDR2 VH и CDR3 VH, имеющие аминокислотную последовательность из CDR, перечисленных в SEQ ID NO: 256; и вариабельную область легкой цепи (VL), содержащую CDR1 VL, CDR2 VL и CDR3 VL, имеющие аминокислотную последовательность, выбранную из группы CDR, перечисленных в SEQ ID NO: 91-93.
[0027] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело, которое связывается с IL-6. Указанное антитело содержит по меньшей мере одну из следующих мутаций согласно нумерации EU: L234A, L235A и G237A.
[0028] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, которое связывается с IL-6, и оно содержит: CDRH1, представляющую собой CDRH1 в SEQ ID NO: 172; CDRH2, представляющую собой CDRH2 в SEQ ID NO: 173; CDRH3, представляющую собой CDRH3 в SEQ ID NO: 174; CDRL1, представляющую собой CDRL1 в SEQ ID NO: 199; CDRL2, представляющую собой CDRL2 в SEQ ID NO: 200; CDRL3, представляющую собой CDRL3 в SEQ ID NO: 201; по меньшей мере одну из следующих мутаций (нумерация EU): L234A, L235A и G237A; и по меньшей мере одну из следующих мутаций (нумерация EU): Q347C или L443C.
[0029] В соответствии с некоторыми аспектами предложен слитый белок, содержащий: выделенное антагонистическое антитело из любых, предложенных согласно настоящему изобретению (включая их фрагменты), и ловушку VEGF.
[0030] В соответствии с некоторыми аспектами предложен конъюгат, содержащий: любое из выделенных антагонистических антител (включая фрагменты), предложенных согласно настоящему изобретению; и полимер, при этом указанный полимер ковалентно присоединен к антителу.
[0031] В соответствии с некоторыми аспектами предложен конъюгат, содержащий слитый белок, предложенный согласно настоящему изобретению; и полимер, при этом указанный полимер ковалентно присоединен к слитому белку.
[0032] В соответствии с некоторыми аспектами предложен двойной ингибитор VEGFR-анти-IL-6. Указанный двойной ингибитор VEGFR-анти-IL-6 содержит слитую конструкцию ловушка-антитело из анти-IL6 антитела и анти-VEGF ловушки (VEGFR1/2), при этом двойной ингибитор содержит по меньшей мере одну точечную мутацию в последовательности VEGFR для уменьшения расщепления белка VEGFR.
[0033] В соответствии с некоторыми аспектами предложена белковая конструкция, содержащая: по меньшей мере 3 CDR тяжелой цепи; по меньшей мере 3 CDR легкой цепи; последовательность ловушки VEGF; и линкерную последовательность, при этом каждая из указанных последовательностей выбрана из соответствующей последовательности на фиг.18-25 или из соответствующих последовательностей из таблиц в настоящем описании (включая, например, 1, 2, 6, 7, 8 и 9).
[0034] В соответствии с некоторыми аспектами предложен слитый белок, содержащий IL-6 VH; IL-6 VL; IL-6 Fc; и ловушку VEGF, при этом указанный слитый белок изменяет пролиферацию эндотелиальных клеток пупочной вены человека (ЭКПВЧ). В некоторых вариантах реализации изменение пролиферации ЭКПВЧ ингибирует пролиферацию, опосредуемую VEGF/IL6.
[0035] В соответствии с некоторыми аспектами предложен слитый белок, содержащий последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 263 и по меньшей мере на 80% идентична SEQ ID NO: 117, при этом указанный слитый белок дополнительно конъюгирован с полимером.
[0036] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело, которое связывается с IL-6. Указанное антитело содержит по меньшей мере одну из следующих мутаций согласно нумерации EU: L234A, L235A и G237A.
[0037] В соответствии с некоторыми аспектами предложено выделенное антагонистическое антитело, которое связывается с IL-6, при этом указанное антитело содержит: CDRH1, представляющую собой CDRH1 в SEQ ID NO: 172; CDRH2, представляющую собой CDRH2 в SEQ ID NO: 173; CDRH3, представляющую собой CDRH3 в SEQ ID NO: 174; CDRL1, представляющую собой CDRL1 в SEQ ID NO: 199; CDRL2, представляющую собой CDRL2 в SEQ ID NO: 200; CDRL3, представляющую собой CDRL3 в SEQ ID NO: 201; по меньшей мере одну из следующих мутаций (нумерация EU): L234A, L235A и G237A; и по меньшей мере одну из следующих мутаций (нумерация EU): Q347C или L443C.
[0038] В соответствии с некоторыми аспектами двойной ингибитор VEGFR-анти-IL-6 содержит последовательности анти-IL-6 вариабельной области тяжелой цепи, выбранные из SEQ ID NO: 7-13, 89, 90 и/или 256-262.
[0039] В соответствии с некоторыми аспектами двойной ингибитор VEGFR-анти-IL-6 содержит последовательности ловушки VEGF, выбранные из по меньшей мере одной из SEQ ID NO: 145, 15, 16 или 17.
[0040] В соответствии с некоторыми аспектами линкерная последовательность представляет собой SEQ ID NO: 18.
[0041] В соответствии с некоторыми аспектами предложена последовательность тяжелой цепи для анти-IL-6 молекулы, выбранная из по меньшей мере одной из SEQ ID NO: 19-27 или содержащая по меньшей мере последовательность в одной из SEQ ID NO: 89, 90, 256-262.
[0042] В соответствии с некоторыми аспектами предложена последовательность легкой цепи для анти-IL-6 молекулы, содержащая по меньшей мере 1, 2 или 3 CDR легкой цепи из по меньшей мере одной из SEQ ID NO: 76-84.
[0043] В соответствии с некоторыми аспектами предложена последовательность тяжелой цепи для анти-IL-6 молекулы, содержащая по меньшей мере 1, 2 или 3 CDR тяжелой цепи из по меньшей мере одной из SEQ ID NO: 49-75.
[0044] В соответствии с некоторыми аспектами предложен двойной ингибитор VEGFR-анти-IL-6, содержащий последовательность VEGFR-Fc из по меньшей мере одной из SEQ ID NO: 85-88.
[0045] В соответствии с некоторыми аспектами предложен двойной ингибитор VEGFR-анти-IL-6, содержащий одну или более последовательностей в любых одной или более SEQ ID NO: 7-13, 145, 15-17, 18-84.
[0046] В некоторых вариантах реализации предложен двойной ингибитор VEGFR-анти-IL-6, содержащий IL-6 VH; IL-6 VL; IL-6 Fc; ловушку VEGF; и линкер. В некоторых вариантах реализации IL-6 VH содержит последовательность из последовательности IL6 VH в любой из SEQ ID NO: 19-27, 31-39, 89, 90 или 256-262. В некоторых вариантах реализации IL-6 VL содержит последовательность из последовательности IL6 VL в любой из SEQ ID NO: 28-30 или 91-93. В некоторых вариантах реализации Fc содержит последовательность из последовательности Fc в любой из SEQ ID NO: 40-48. В некоторых вариантах реализации ловушка VEGF содержит последовательность из последовательности ловушки VEGF в любой из SEQ ID NO: 145, 15, 16 или 17.
[0047] В соответствии с некоторыми аспектами предложен слитый белок, содержащий IL-6 VH; IL-6 VL; IL-6 Fc; и ловушку VEGF, при этом указанный слитый белок изменяет пролиферацию ЭКПВЧ. В некоторых вариантах реализации изменение пролиферации ЭКПВЧ ингибирует пролиферацию, опосредуемую VEGF/IL6.
[0048] В соответствии с некоторыми аспектами предложен слитый белок, содержащий последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 263 и по меньшей мере на 80% идентична SEQ ID NO: 117. Указанный слитый белок дополнительно конъюгирован с полимером. В некоторых вариантах реализации слитый белок по меньшей мере на 95% идентичен SEQ ID NO: 263 и по меньшей мере на 95% идентичен SEQ ID NO: 117. В некоторых вариантах реализации белок содержит по меньшей мере а) SEQ ID NO: 172, 173, 174, 199, 200 и 201 или b) замены 1, 2 или 3 аминокислот в SEQ ID NO: 172, 173, 174, 199, 200 и/или 201, при этом указанная замена представляет собой консервативную замену.
[0049] В соответствии с некоторыми аспектами антитело из любых предложенных согласно настоящему изобретению содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
[0050] В соответствии с некоторыми аспектами слитый белок из любых предложенных согласно настоящему изобретению содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
[0051] В соответствии с некоторыми аспектами конъюгат из любых предложенных согласно настоящему изобретению содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
[0052] В соответствии с некоторыми аспектами двойной ингибитор VEGFR-анти-IL-6 из любых предложенных согласно настоящему изобретению содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
[0053] В соответствии с некоторыми аспектами белок из любых предложенных согласно настоящему изобретению содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0054] На фиг.1 представлена последовательность IL-6.
[0055] На фиг.2A показано Соединение L.
[0056] На фиг.2B показано Соединение K.
[0057] На фиг.2C представлен синтез OG1802 из R3707.
[0058] На фиг.2D показан OG1786.
[0059] На фиг.2E представлен синтез OG1546 из OG1550.
[0060] На фиг.2F представлен синтез OG1784 из OG1546 и OG1563.
[0061] На фиг.2G представлен синтез OG1405 из OG1784.
[0062] На фиг.2H представлен синтез OG1785 из OG1405.
[0063] На фиг.2I представлен синтез OG1786 из OG1785.
[0064] На фиг.2J показан OG1801.
[0065] На фиг.2K показан OG1802.
[0066] На фиг.2L показано Соединение Е.
[0067] На фиг.3 представлена схема выбора и оптимизации антитела.
[0068] На фиг.4 представлены данные ELISA, демонстрирующие, что анти-IL-6 моноклональное антитело (mAb, МАТ) связывается с IL-6, но не с комплексом IL-6/IL-6R, и что МАТ к IL-6 ингибирует образование комплекса IL-6/IL-6R.
[0069] На фиг.5 представлены некоторые варианты реализации вариабельных областей тяжелой и легкой цепи антитела (Ab, АТ) к IL-6. Варианты реализации CDR показаны в заключенных в рамку областях. Эти последовательности могут быть также использованы в слитой конструкции АТ к IL-6-ловушка VEGF.
[0070] На фиг.6 представлены некоторые варианты реализации слитого белка IL-6-ловушка VEGF. Домены ловушки VEGF расположены либо на N-конце, непосредственно предшествующем вариабельному домену (слева), либо расположены между Fab-областью и шарнирной областью антитела (справа).
[0071] На фиг.7 представлены сенсограммы и таблица, которые демонстрируют, что молекулы двойного ингибитора VEGFR-антиIL-6 и антиIL-6-VEGFR связываются со схожей аффинностью с VEGF-A, что и антитело анти-VEGF, OG1950, и Эйлеа. Таким образом, положение ловушки VEGF не изменяет ее аффинность к мишени.
[0072] На фиг.8 представлены сенсограммы независимого или комбинированного связывания VEGF-A и IL-6 с двойными ингибиторами. Сенсограммы IL-6 для смешанных мишеней сравнивали с теоретической кривой (сумма отдельных сенсограмм IL-6 и VEGF-A). Результаты демонстрируют наложение теоретической и экспериментальной кривых, что качественно указывает на то, что обе мишени могут связываться с молекулой двойного ингибитора и не влияют на связывание друг друга.
[0073] На фиг.9 представлены результаты при ОП450 нм из различных ELISA-анализов IL-6. В мостиковом ELISA (вверху) оба двойных ингибитора связывали мостиком btVEGF с IL-6, что указывает на то, что обе конфигурации могут связываться с обеими мишенями. EC50 конструкции ловушка VEGF-антиIL-6=0,079 нМ и антиIL-6-ловушка VEGF=0,026 нМ. Эйлеа и анти-IL-6 служили в качестве отрицательных контролей. На фиг.9 также представлены результаты ELISA для комплекса IL-6/IL-6R (посередине). Двойные ингибиторы и антиIL-6 ингибировали образование комплекса IL-6/IL-6R в равной степени. Значения IC50: антиIL-6=0,36 нМ, конструкция ловушка VEGF-антиIL-6=0,47 нМ и конструкция антиIL-6-ловушка VEGF=0,32 нМ. Эйлеа служило в качестве отрицательного контроля. На фиг.9 также представлены результаты конкурентного ELISA для VEGF/VEGFR (внизу). Двойные ингибиторы, Эйлеа и OG1950 ингибировали связывание VEGF с VEGFR в различной степени. Эйлеа и конструкция антиIL-6-ловушка VEGF сопоставимы (4,24 нМ относительно 4,53 нМ), тогда как конструкция ловушка VEGF-антиIL-6 была в ~2 раза эффективнее (1,74 нМ), и OG1950 демонстрировало максимальное ингибирование, превосходящее другие ингибиторы (1,55 нМ).
[0074] На фиг.10 представлены некоторые варианты реализации способа получения конъюгата антитела (который можно также применять для конъюгата АТ-ловушка или ловушка-АТ). Хотя изображено в виде антитела, для специалиста в этой области техники в контексте настоящего изобретения очевидно, что изображение АТ на фиг.10 можно заменить на слитую конструкцию ловушки (показанную на фиг.6). Для простоты обобщенное антитело, изображенное на этой фигуре, представляет собой как варианты в виде антитела, так и варианты в виде слитой структуры в контексте слитой конструкции ловушки (если, конечно, оно уже не изображено в виде слитой конструкции).
[0075] На фиг.11А-11 В представлены полосы электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-ПААГ) для стандарта SeeBlue®Plus2, анти-IL-6, конструкции анти-IL-6-VEGFR и VEGFR-анти-IL-6. На фиг.11С изображена конъюгированная конструкция VEGFR-анти-IL6, которая представляет собой слитую конструкцию анти-VEGF (VEGFR1/2) и анти-IL-6, конъюгированную с полимером на основе фосфорилхолина.
[0076] На фиг.12 представлены результаты переноса на PVDF-мембрану, N-концевого секвенирования по Эдману, которые демонстрируют, что расщепленные продукты содержат одинаковую N-концевую последовательность (LTHRQT), что показывает, что сайт расщепления расположен в области ловушки VEGF.
[0077] На фиг.13А-В представлены полосы SDS-ПААГ для стандарта SeeBlue®Plus2 и 19 конструкций ловушки VEGF.
[0078] На фиг.13С-13Е представлены перечни последовательностей для: VEGF_ловушка_вариант_1, VEGF_ловушка_вариант_2 и VEGF_ловушка_вариант_3.
[0079] На фиг.13F-13G представлены полосы SDS-ПААГ для стандарта SeeBlue®Plus2 и 4 вариантов ловушки VEGF, включая VEGFR_вариант_3, содержащий двойные точечные мутации T94I и H95I.
[0080] На фиг.14A-14F представлены результаты анализа Biacore и измерения аффинности к VEGF-A.
[0081] На фиг.15 представлен репортерный анализ VEGF-стимулируемого VEGFR в клеточной системе.
[0082] На фиг.16A представлен анализ ингибирования VEGF/IL6-опосредуемого образования трубочек эндотелиальными клетками пупочной вены человека («ЭКПВЧ»).
[0083] На фиг.16B-16C представлена статистика анализов образования трубочек с различными параметрами.
[0084] На фиг.17 представлены результаты анализа пролиферации ЭКПВЧ.
[0085] На фиг.18 показаны варианты последовательностей анти-IL-6 вариабельной области тяжелой цепи. CDR подчеркнуты.
[0086] На фиг.19 показаны различные варианты последовательностей ловушки VEGF. Участок, варьирующийся среди указанных последовательностей, выделен жирным шрифтом и подчеркнут.
[0087] На фиг.20 показаны некоторые варианты линкерной (GS) последовательности. Линкер может присутствовать в виде линкера Gly-Gly-Gly-Gly-Ser (GS) с двойным повтором.
[0088] На фиг.21 показаны некоторые варианты последовательности тяжелой цепи для анти-IL-6 молекул. CDR подчеркнуты. Фиг.21 включает как фиг.21A, так и фиг.21B, на которых представлены различные варианты CDR в указанных последовательностях.
[0089] На фиг.22 показаны некоторые варианты последовательностей легкой цепи для анти-IL-6 молекул. CDR подчеркнуты. Фиг.22 включает как фиг.22A, так и фиг.22B, на которых представлены различные варианты CDR в указанных последовательностях.
[0090] На фиг.23 показаны некоторые варианты последовательностей тяжелой цепи для анти-IL-6 молекул. CDR подчеркнуты. Фиг.23 включает как фиг.23A, так и фиг.23B, на которых представлены различные варианты CDR в указанных последовательностях.
[0091] На фиг.24A-24B показаны некоторые варианты комбинаций CDR с фиг.21-23.
[0092] На фиг.25 показаны некоторые варианты последовательности VEGFR-Fc. Участок, варьирующийся среди указанных последовательностей, выделен жирным шрифтом и подчеркнут.
[0093] На фиг.26A-26С представлены данные аффинности связывания.
[0094] На фиг.27 представлены последовательности некоторых вариантов VEGFR-антиIL6-последовательностей. CDR (определенные согласно Kabat) подчеркнуты. Участки, окрашенные в серый цвет, обозначают конструкции VEGFR. Жирным шрифтом выделен линкерный участок. Мутации L234A, L235A, G237A и L443C (нумерация EU) подчеркнуты двойной линией. Каждый из этих участков может быть заменен на другие соответствующие участки, представленные в настоящем описании (например, альтернативные линкеры или CDR и т.д.).
[0095] На фиг.28 представлен SDS-ПААГ продуктов реакции восстановления конструкции VEGFR-антиIL6 (Cys - деблокирование (decapping)). Дорожки следующие: 1. VEGFR-антиIL6; 2. VEGFR-антиIL6 - полностью восстановленная (TCEP); 3. стандарт Novex sharp pre-stained protein standard (предварительно окрашенный белковый стандарт Novex); 4. VEGFR-антиIL6+30x TCEP, исходная точка; 5. VEGFR-антиIL6+30x TCEP, через 30 минут; 6. VEGFR-антиIL6+30x TCEP, через 60 минут; 7. VEGFR-антиIL6, обработанная TCEP, буфер заменен; 8. VEGFR-антиIL6+15x dHAA, исходная точка; 9. VEGFR-антиIL6+15x dHAA, через 30 минут; 10. VEGFR-антиIL6+15x dHAA, через 60 минут; 11. VEGFR-антиIL6, деблокированная; 12. VEGFR-антиIL6, деблокированная, полностью восстановленная (TCEP); Гель: Бис-Трис NuPAGE 4-12% Количество белка: 4 мкг/дорожка; 30x TCEP=30-кратный молярный избыток TCEP, 15x dHAA=15-кратный молярный избыток dHAA.
[0096] На фиг.29 представлен SDS-ПААГ для катионообменной хроматографии (CEX) конъюгата VEGFR-антиIL6-OG1802. Невосстанавливающий гель: Бис-Трис NuPAGE 4-12%. Буфер A: 20 мМ ацетат натрия, pH 5,5. Буфер В: 20 мМ ацетат натрия, pH 5,5, 500 мM NaCl. Дорожки следующие: 1. VEGFR-антиIL6; 2. VEGFR-антиIL6-OG1802 (загрузка); 3. Стандарт Novex sharp pre-stained protein standard (предварительно окрашенный белковый стандарт Novex); 4. Проточная фракция; 5. Чейз; 6. 30% буфер B - аликвота 1; 7. 30% буфер B - аликвота 2; 8. 30% буфер B - аликвота 3; 9. 30% буфер B - аликвота; 10. 40% буфер B - аликвота 1; 11. 40% буфер B - аликвота 2; 12. 40% буфер B - аликвота 3; 13. 40% буфер B - аликвота 4; 14. 60% буфер B - аликвота 1; 15. 60% буфер B - аликвота 2; 16. 60% буфер B - аликвота 3; 17. 100% буфер B - стрип. Дорожки 6-13 демонстрируют конъюгированный белковый материал, конъюгат не может проникать в гель из-за большого размера. Дорожки 14-17 демонстрируют смесь белков, возможно содержащую агрегированный, конъюгированный и неконъюгированный белковый материал.
[0097] На фиг.30 представлен SDS-ПААГ конъюгата белок-полимер относительно неконъюгированного белкового материала. Гель-анализ демонстрирует соотношение 57% интенсивности полосы тяжелой цепи и 96% интенсивности полосы легкой цепи при сравнении биоконъюгата (дорожка 3) с эталонным стандартом VEGFR-антиIL6 (дорожка 2), что указывает на присутствие одного полимера OG1802 на молекулу VEGFR-антиIL6. Восстанавливающий гель представляет собой: Бис-Трис NuPAGE 4-12%. Восстанавливающий гель представляет собой: Бис-Трис NuPAGE 4-12%.
[0098] На фиг.31 представлена эксклюзионная хроматография с многоугловым светорассеянием (SEC-MALS) для конструкции VEGFR-антиIL6-OG1802. Молекулярную массу конъюгата VEGFR-антиIL6 определяли с помощью интегрированной эксклюзионной хроматографии (Shodex-SB806M-HQ) и светорассеяния (MALS). Верхний график. Хроматограмма показывает наличие единственного пика элюирования. Отсутствие дополнительных пиков и плеча позволяет предположить отсутствие агрегатов и распавшегося вещества после конъюгации и последующих этапов разделения путем CEX. Нижний график. Анализ конъюгата белка выбранного пика демонстрировал экспериментально измеренную среднюю молекулярную массу (Mw, ММ), равную 983 кДа для биоконъюгата VEGFR-антиIL6-OG1802. Это значение является результатом конъюгации одной молекулы VEGFR-антиIL6 (ММ ~ 189 кДа) и одного полимера OG1802 (ММ ~ 794 кДа).
[0099] На фиг.32 представлены результаты CEX-хроматографии конструкции VEGFR-антиIL6. Гель: представляет собой 8-16% Трис-Глицин Novex (восстанавливающие условия), M=стандарт SeeBlue®Plus2. Дорожки D12-F11 соответствуют образцам, поделенным на аликвоты при различных концентрациях буфера B, как показано на хроматограмме. Показаны интактные (I) и расщепленные (C) тяжелые цепи. В колонке Porus XS буфер A представляет собой 20 мМ фосфат натрия, pH 6, и буфер B представляет собой 20 мМ фосфат натрия, pH 6, 1 М NaCl.
[0100] На фиг.33 представлены результаты хроматографии гидрофобного взаимодействия (HIC) для конструкции VEGFR-антиIL6. Гель: представляет собой 8-16% Трис-Глицин Novex/восстанавливающие условия, M=стандарт SeeBlue®Plus2. L=конструкция VEGFR-антиIL6, интактная и расщепленная смесь (загрузка). Остальные дорожки соответствуют образцам, поделенным на аликвоты при различных концентрациях буфера B, как показано на хроматограмме. Показаны интактные (I) и расщепленные (C) тяжелые цепи. В колонке Hi Trap Butly HP буфер A представляет собой: 20 мM фосфат натрия pH 6, 1 M сульфат аммония. Буфер B представляет собой: 20 мМ фосфат натрия, pH 6.
[0101] На фиг.34 представлены результаты конкурентного ELISA для VEGF/VEGFR.
[0102] На фиг.35 представлены результаты ELISA для комплекса IL6/IL6R.
[0103] На фиг.36 представлены результаты репортерного анализа VEGF-стимулируемого VEGFR в клеточной системе.
[0104] На фиг.37 представлено стимулируемое липополисахаридом образование трубочек в ЭКПВЧ.
[0105] На фиг.38 представлено стимулируемое липополисахаридом образование трубочек в ЭКПВЧ.
[0106] На фиг.39 представлено стимулируемое липополисахаридом образование трубочек в ЭКПВЧ.
[0107] На фиг.40А представлено ингибирование VEGF/IL6-опосредуемой пролиферации ЭКПВЧ.
[0108] На фиг.40В представлено ингибирование пролиферации ЭКПВЧ с увеличением количества клеток на лунку.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0109] Для непосредственного уменьшения сопутствующего воспаления и нарушенного ангиогенеза, которые управляют патогенезом неоваскулярных патологий сетчатки, согласно настоящему изобретению предложены сконструированные молекулы, которые одновременно блокируют функции провоспалительного цитокина IL-6 и проангиогенного сигнального белка VEGF. Эти молекулы состоят из (1) моноклонального анти-IL-6 антитела, слитого с (2) двумя VEGF-связывающими доменами рецепторов VEGF (VEGFR). Анти-IL-6 фрагмент, специфично связывает IL-6 и ингибирует его взаимодействие с рецептором IL-6 (IL-6R). Фрагмент, представляющий собой ловушку VEGF, содержит слияние двух VEGF-связывающих доменов (домена 2 VEGFR1, домена 3 VEGFR2), которые действуют как ловушка VEGF и предотвращают связывание VEGF с рецепторами VEGF. В дополнение к этому, в некоторых вариантах реализации каждая из этих молекул двойного ингибитора «снабжена» неспаренным цистеином на С-конце, который может быть конъюгирован с биополимером на основе фосфорилхолина, увеличивающим период полувыведения.
[0110] В различных вариантах реализации, представленных в настоящем описании, применяют, если не указано иное, традиционные методы молекулярной биологии (включая рекомбинантные методы), микробиологии, биологии клетки, биохимии и иммунологии, которые находятся в рамках компетенции специалиста в этой области техники. Такие методы полностью описаны в литературе, например, в Molecular Cloning: A Laboratory Manual, second edition (Sambrook et al., 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995).
[0111] Следует понимать, что следующие термины, если не указано иное, имеют следующие значения: термин «выделенная молекула» относится к молекуле (при этом указанная молекула представляет собой, например, полипептид, полинуклеотид или антитело), которая в силу своего происхождения или источника получения (1) не связана с естественным образом связанными с ней компонентами, которые сопровождают ее в нативном состоянии, (2) по существу свободна от других молекул из того же источника, например, вида, клетки, в которой она экспрессируется, библиотеки и т.д., (3) экспрессируется клеткой другого вида или (4) не встречается в природе. Таким образом, молекула, химически синтезированная или экспрессированная в клеточной системе, отличной от системы, из которой она происходит в природе, будет «отделена (выделена)» от естественным образом связанных с ней компонентов. Молекула также может быть по существу освобождена от естественным образом связанных с ней компонентов путем выделения с применением методов очистки, хорошо известных в этой области техники. Анализ чистоты или гомогенности молекул можно осуществлять рядом способов, хорошо известных в этой области техники. Например, чистоту образца полипептида можно анализировать с применением электрофореза в полиакриламидном геле и окрашивания геля для визуализации указанного полипептида с применением методов, хорошо известных в этой области техники. Для некоторых целей может быть обеспечено более высокое разрешение путем применения ВЭЖХ или других способов очистки, хорошо известных в этой области техники.
[0112] В настоящем описании, если не указано иное, термин «IL-6» (ИЛ-6) или «IL6» (ИЛ6) относится к IL-6 человека. В некоторых вариантах реализации предусмотрены другие формы IL-6 и они будут обозначены посредством конкретной ссылки на другие организмы, например, собаку, кошку, лошадь и быка. Один из типичных IL-6 человека имеет номер доступа UniProt P05231.
[0113] Анти-IL-6 антитела или другие биопрепараты, описанные в настоящем документе, как правило, представлены в выделенном виде. Это означает, что антитело является, как правило, по меньшей мере на 50 масс.% чистым и не содержит нежелательных белков и других загрязняющих примесей, возникающих в результате его получения или очистки, но не исключает возможности того, что указанное антитело комбинировано с фармацевтически приемлемым вспомогательным веществом, предназначенным для облегчения его применения. Иногда антитела являются по меньшей мере на 60, 70, 80, 90, 95 или 99 масс.% чистыми и не содержат нежелательных белков и загрязняющих примесей в результате получения или очистки. Зачастую антитело (или конъюгат антитела) является преобладающим видом макромолекулы, который остается после его очистки.
[0114] «Антитело» представляет собой молекулу иммуноглобулина, способную специфично связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и т.д., посредством по меньшей мере одного сайта распознавания антигена, расположенного в вариабельной области молекулы иммуноглобулина. В настоящем описании указанный термин включает не только интактные поликлональные или моноклональные антитела, а также, если не указано иное, любую их антигенсвязывающую часть, которая конкурирует с интактным антителом за специфическое связывание, слитые белки, содержащие антигенсвязывающую часть, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена. Антигенсвязывающие части включают, например, Fab, Fab’, F(ab’)2, Fd, Fv, доменные антитела (dAb (дАТ), например, антитела акулы и верблюдовых), фрагменты, содержащие определяющие комплементарность области (CDR), антитела, представляющие собой одноцепочечные вариабельные фрагменты (scFv), макситела, минитела, интратела, диатела, триатела, тетратела, v-NAR и бис-scFv, и полипептиды, которые содержат по меньшей мере часть иммуноглобулина, достаточную для придания полипептиду свойства специфического связывания антигена. Антитело включает антитело любого класса, например, IgG, IgA или IgM (или их подкласса), и антитело не обязательно должно относиться к какому-либо конкретному классу. В зависимости от аминокислотной последовательности константной области тяжелых цепей антитела иммуноглобулины могут быть отнесены к разным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно поделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные области тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, обозначаются альфа, дельта, эпсилон, гамма и мю соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
[0115] Термин «вариабельная область» антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела либо отдельно, либо в комбинации. Как известно в этой области техники, каждая вариабельная область тяжелых и легких цепей состоит из четырех каркасных областей (FR), соединенных тремя определяющими комплементарность областями (CDR), также известными как гипервариабельные области, и вносит вклад в образование антигенсвязывающего центра антител. Если необходимы варианты рассматриваемой вариабельной области, в частности, содержащие замену аминокислотных остатков вне CDR-области (т.е. в каркасной области), соответствующую замену аминокислоты, предпочтительно консервативную замену аминокислоты, можно идентифицировать путем сравнения рассматриваемой вариабельной области с вариабельными областями других антител, которые содержат последовательности CDR1 и CDR2, в том же каноническом классе, что и рассматриваемая вариабельная область (Chothia and Lesk, J Mol Biol 196(4): 901-917, 1987).
[0116] В некоторых вариантах реализации точное определение CDR и идентификацию остатков, составляющих центр связывания антитела, осуществляют путем определения структуры антитела и/или определения структуры комплекса антитело-лиганд. В некоторых вариантах реализации это можно осуществлять любым из различных способов, известных специалисту в этой области техники, таким как рентгеновская кристаллография. В некоторых вариантах реализации для идентификации или аппроксимации CDR-областей можно применять различные методы анализа. В некоторых вариантах реализации для идентификации или аппроксимации CDR-областей можно применять различные методы анализа. Примеры таких методов включают, но не ограничиваются ими, определение согласно Kabat, определение согласно Chotia, подход IMGT (Lefranc et al., 2003) Dev Comp Immunol. 27:55-77), компьютерные программы, такие как Paratome (Kunik et al., 2012, Nucl Acids Res. W521-4), определение AbM (АтМ) и конформационное определение.
[0117] Определение согласно Kabat является стандартом для нумерации остатков в антителе и его, как правило, используют для идентификации CDR-областей. См., например, Johnson & Wu, 2000, Nucleic Acids Res., 28: 214-8. Определение согласно Chotia подобно определению согласно Kabat, но определение согласно Chotia учитывает положения некоторых структурных петлевых областей. См., например, Chothia et al., 1986, J. Mol. Biol., 196: 901-17; Chothia et al., 1989, Nature, 342: 877-83. В определении AbM используют комплексный пакет компьютерных программ, разработанных Oxford Molecular Group, которые моделируют структуру антитела. См., например, Martin et al., 1989, Proc Natl Acad Sci (USA), 86:9268-9272; “AbM™, A Computer Program for Modeling Variable Regions of Antibodies,” Oxford, UK; Oxford Molecular, Ltd. Определение AbM позволяет моделировать третичную структуру антитела на основе первичной последовательности с использованием комбинации баз знаний и методов ab initio, таких как описаны Samudrala et al., 1999, “Ab Initio Protein Structure Prediction Using a Combined Hierarchical Approach,” в PROTEINS, Structure, Function and Genetics Suppl., 3:194-198. Контактное определение основано на анализе доступных сложных кристаллических структур. См., например, MacCallum et al., 1996, J. Mol. Biol., 5:732-45. Согласно другому подходу, называемому в настоящем описании «конформационным определением» CDR, положения CDR могут быть определены как остатки, которые вносят энтальпийный вклад в связывание антигена. См., например, Makabe et al., 2008, Journal of Biological Chemistry, 283:1156-1166. Другие определения границ CDR могут не следовать строго одному из вышеуказанных подходов, но тем не менее они будут перекрываться по меньшей мере с частью CDR согласно Kabat, хотя они могут быть укорочены или удлинены ввиду прогнозирования или экспериментальных данных о том, что конкретные остатки или группы остатков не оказывают значительного влияния на связывание антигена. В настоящем описании термин «CDR» может относиться к CDR-областям, определенным посредством любого подхода, известного в этой области техники, в том числе комбинаций подходов. В способах, применяемых согласно настоящему изобретению, можно использовать CDR, определенные в соответствии с любым из этих подходов. Для любого конкретного варианта реализации, включающего более одной CDR, CDR могут быть определены в соответствии с любым из: определений согласно Kabat, Chotia, расширенного определения, определения IMGT, Paratome, AbM и/или конформационного определения, или комбинацией любых из вышеуказанных определений.
[0118] Как известно в этой области техники, термин «константная область» антитела относится к константной области легкой цепи антитела или константной области тяжелой цепи антитела либо отдельно, либо в комбинации.
[0119] В настоящем описании термин «моноклональное антитело» относится к антителу, полученному из популяции по существу однородных антител, т.е. отдельные антитела, составляющие эту популяцию, являются идентичными за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, будучи направленными против одной антигенной детерминанты. Более того, в отличие от препаратов поликлональных антител, которые, как правило, содержат разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Определение «моноклональное» указывает на характер антитела как антитела, полученного по существу из однородной популяции антител, и его не следует рассматривать как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела для применения согласно настоящему изобретению могут быть получены способом на основе гибридом, впервые описанным Kohler and Milstein, 1975, Nature 256:495, или могут быть получены способами на основе рекомбинантных ДНК, такими как описаны в патенте США №4816567. Моноклональные антитела могут быть также выделены из фаговых библиотек, полученных с применением методик, описанных, например, в McCafferty et al., 1990, Nature 348:552-554. В настоящем описании термин «гуманизированное» антитело относится к формам антител нечеловеческого происхождения (например, мыши), которые представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), которые содержат минимальную последовательность, полученную из иммуноглобулина нечеловеческого происхождения. Предпочтительно, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из CDR реципиентного антитела заменены остатками из CDR вида, не относящегося к человеку (донорское антитело), такого как мышь, крыса или кролик, обладающими желаемой специфичностью, аффинностью и способностью. Гуманизированное антитело может содержать остатки, не встречающиеся ни в реципиентном антителе, ни в вводимых CDR или каркасных последовательностях, но они включены для дополнительного улучшения и оптимизации эффективности антитела.
[0120] «Антитело человека» представляет собой антитело, содержащее аминокислотную последовательность, которая соответствует последовательности антитела, продуцируемого человеком, и/или которое получено с применением любого из способов получения антител человека, описанных в настоящем документе. Это определение антитела человека, в частности, исключает гуманизированное антитело, содержащее антигенсвязывающие остатки нечеловеческого происхождения.
[0121] Термин «химерное антитело» относится к антителам, в которых последовательности вариабельной области получены из одного вида, а последовательности константной области получены из другого вида, таким как антитело, в котором последовательности вариабельной области получены из антитела мыши, а последовательности константной области получены из антитела человека. Термин «эпитоп» относится к той части молекулы, которая может быть распознана и связана антителом в одной или более антигенсвязывающих областях указанного антитела. Эпитопы часто состоят из поверхностной группы молекул, таких как аминокислоты или боковые цепи сахаров, и обладают конкретными характеристиками трехмерной структуры, а также конкретными характеристиками заряда. В некоторых вариантах реализации эпитоп может представлять собой белковый эпитоп.Белковые эпитопы могут быть линейными или конформационными. В линейном эпитопе все точки взаимодействия между белком и взаимодействующей молекулой (такой как антитело) расположены линейно вдоль первичной аминокислотной последовательности белка. «Нелинейный эпитоп» или «конформационный эпитоп» содержит несмежные полипептиды (или аминокислоты) в пределах антигенного белка, с которыми связывается антитело, специфичное в отношении этого эпитопа. В настоящем описании термин «антигенный эпитоп» определен как часть антигена, с которой антитело может специфично связываться, что определяют любым способом, хорошо известным в этой области техники, например, путем традиционных иммуноанализов. После определения желаемого эпитопа на антигене можно получить антитела к этому эпитопу, например, с применением методик, описанных в настоящем документе. В качестве альтернативы, в ходе процесса обнаружения получение и определение характеристик антител могут дать информацию о желаемых эпитопах. Затем на основании этой информации можно проводить конкурентный скрининг антител в отношении связывания с одинаковым эпитопом. Подход для достижения этого заключается в проведении исследований конкуренции и перекрестной конкуренции для обнаружения антител, которые конкурируют или перекрестно конкурируют друг с другом за связывание с IL-6, например, антитела конкурируют за связывание с указанным антигеном.
[0122] В настоящем описании термин «конкурировать» применительно к антителу означает, что первое антитело или его антигенсвязывающая часть связывается с эпитопом способом, достаточно схожим со связыванием второго антитела или его антигенсвязывающей части, так, что результат связывания первого антитела с его распознанным эпитопом детектируемо снижается в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствии второго антитела. В качестве альтернативы, может происходить, но не обязательно, также детектируемое уменьшение связывания второго антитела с эпитопом в присутствии первого антитела. То есть первое антитело может ингибировать связывание второго антитела с эпитопом без ингибирования вторым антителом связывания первого антитела с соответствующим эпитопом. Однако, когда каждое антитело детектируемо ингибирует связывание другого антитела с его распознанным эпитопом или лигандом, независимо от того, в той же, большей или меньшей степени, говорят, что указанные антитела «перекрестно конкурируют» друг с другом за связывание с соответствующим эпитопом (эпитопами). Настоящее изобретение включает как конкурирующие, так и перекрестно конкурирующие антитела. Независимо от механизма, по которому происходит такая конкуренция или перекрестная конкуренция (например, стерическое препятствие, конформационное изменение или связывание с общим эпитопом или его частью), специалисту в этой области техники понятно на основании идей, представленных в настоящем описании, что такие конкурирующие и/или перекрестно конкурирующие антитела включены и могут подходить для способов, описанных в настоящем документе.
[0123] В настоящем описании антитело «взаимодействует с» IL-6, когда равновесная константа диссоциации равна или меньше 20 нМ, предпочтительно меньше примерно 6 нМ, более предпочтительно меньше примерно 1 нМ, наиболее предпочтительно меньше примерно 0,75 нМ. В некоторых вариантах реализации аффинность антитела составляет от 400 до 800 пМ, например, 450-700 или 500-600 пM.
[0124] антагонистическое антитело к IL-6 включает антитела, которые блокируют, противодействуют, подавляют или снижают (в любой степени, в том числе в значительной степени) биологическую активность IL-6, такую как связывание с IL-6R, связывание комплекса IL-6/IL-6R с gp130, фосфорилирование и активация Stat3, пролиферация клеток и стимуляция IL-6-опосредуемых воспалительных или проангиогенных путей. Для цели настоящего изобретения очевидно, что термин «антагонистическое антитело к IL-6» включает все ранее определенные термины, названия и функциональные состояния, и характеристики, благодаря которым сам IL-6, биологическая активность IL-6 или последствия указанной биологической активности по существу исчезают, уменьшаются или нейтрализуются в любой значимой степени. В некоторых вариантах реализации антагонистическое антитело к IL-6 связывает IL-6. В настоящем описании приведены примеры антител-антагонистов IL-6.
[0125] Антитело, которое «предпочтительно связывается» или «специфично связывается» (взаимозаменяемы в настоящем описании) с эпитопом, является термином, хорошо известным в этой области техники, и способы определения такого специфического или предпочтительного связывания также хорошо известны в этой области техники. Говорят, что молекула демонстрирует «специфическое связывание» или «предпочтительное связывание», если она реагирует или связывается чаще и/или быстрее, и/или с большей продолжительностью, и/или с большей аффинностью с конкретной клеткой или веществом, чем с альтернативными клетками или веществами. Антитело «специфично связывается» или «предпочтительно связывается» с мишенью, если оно связывается с большей аффинностью и/или авидностью, и/или легче, и/или с большей продолжительностью, чем оно связывается с другими веществами. Например, антитело, которое специфично или предпочтительно связывается с эпитопом IL-6, представляет собой антитело, которое связывает этот эпитоп с большей аффинностью и/или авидностью, и/или легче, и/или с большей продолжительностью, чем оно связывается с другими эпитопами IL-6 или эпитопами не IL-6. Из этого определения также понятно, что, например, антитело (или фрагмент, или эпитоп), которое специфично или предпочтительно связывается с первой мишенью, может специфично или предпочтительно связываться со второй мишенью или может не связываться специфично или предпочтительно со второй мишенью. Соответственно, «специфическое связывание» или «предпочтительное связывание» необязательно требует (хотя оно может включать) исключительного связывания. Как правило, но не обязательно, указание на связывание означает предпочтительное связывание.
[0126] В настоящем описании термин «по существу чистый» относится к веществу, которое является по меньшей мере на 50% чистым (т.е. не содержащим загрязняющих примесей), более предпочтительно по меньшей мере на 90% чистым, более предпочтительно по меньшей мере на 95% чистым, более предпочтительно по меньшей мере на 98% чистым и наиболее предпочтительно по меньшей мере на 99% чистым.
[0127] Термин «клетка-хозяин» включает отдельную клетку или культуру клеток, которая может являться или являлась реципиентом для вектора (векторов) для включения полинуклеотидных встраиваний. Клетки-хозяева включают потомство одной клетки-хозяина и это потомство не обязательно может быть полностью идентичным (по морфологии или по комплементарной последовательности геномной ДНК) исходной родительской клетке вследствие естественной, случайной или намеренной мутации. Клетка-хозяин включает клетки, трансфицированные in vivo полинуклеотидом (полинуклеотидами) согласно настоящему изобретению.
[0128] Как известно в этой области техники, термин «Fc-область» используют для определения C-концевой области тяжелой цепи иммуноглобулина. «Fc-область» может представлять собой Fc-область с нативной последовательностью или вариантную Fc-область. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьироваться, обычно определяют, что Fc-область тяжелой цепи IgG человека простирается от аминокислотного остатка в положении Cys226 или от Pro230 до карбоксильного конца. Нумерация остатков в Fc-области представляет собой нумерацию согласно индексу EU, как у Kabat. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md., 1991. Fc-область иммуноглобулина, как правило, содержит два константных домена, CH2 и CH3. Как известно в этой области техники, Fc-область может присутствовать в форме димера или мономера.
[0129] В этой области техники термины «Fc-рецептор» и «FcR» описывают рецептор, который связывается с Fc-областью антитела. Предпочтительный FcR представляет собой FcR человека с нативной последовательностью. Более того, предпочтительный FcR представляет собой FcR, который связывает антитело IgG (гамма-рецептор), и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и формы этих рецепторов, полученные в результате альтернативного сплайсинга. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIВ («ингибирующий рецептор»), которые содержат схожие аминокислотные последовательности, которые отличаются главным образом своими цитоплазматическими доменами. FcR рассмотрены в Ravetch and Kinet, 1991, Ann. Rev. Immunol., 9:457-92; Capel et al., 1994, Immunomethods, 4:25-34; и de Haas et al., 1995, J. Lab. Clin. Med., 126:330-41. Термин «FcR» также включает неонатальный рецептор, FcRn, ответственный за передачу материнских IgG плоду (Guyer et al., 1976, J. Immunol., 117:587; and Kim et al., 1994, J. Immunol., 24:249).
[0130] «Функциональная Fc-область» обладает по меньшей мере одной эффекторной функцией Fc-области с нативной последовательностью. Типичные «эффекторные функции» включают связывание C1q; комплементзависимую цитотоксичность; связывание Fc-рецептора; антителозависимую клеточноопосредованную цитотоксичность; фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, рецептора B-клетки) и т.д. Для таких эффекторных функций, как правило, необходимо комбинирование Fc-области со связывающим доменом (например, вариабельным доменом антитела), и их можно оценить с использованием различных анализов, известных в этой области техники для оценки таких эффекторных функций антитела.
[0131] «Fc-область с нативной последовательностью» содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. «Вариантная Fc-область» содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности Fc-области с нативной последовательностью по меньшей мере одной модификацией аминокислоты, но тем не менее сохраняет по меньшей мере одну эффекторную функцию Fc-области с нативной последовательностью. Предпочтительно, вариантная Fc-область содержит по меньшей мере одну замену аминокислоты по сравнению с Fc-областью с нативной последовательностью или с Fc-областью исходного полипептида, например, от примерно одной до примерно десяти замен аминокислот и предпочтительно от примерно одной до примерно пяти замен аминокислот в Fc-области с нативной последовательностью или в Fc-области исходного полипептида. Вариантная Fc-область согласно настоящему изобретению предпочтительно обладает по меньшей мере примерно 80% идентичностью последовательности по отношению к Fc-области с нативной последовательностью и/или Fc-области исходного полипептида, и наиболее предпочтительно по меньшей мере примерно 90% идентичностью последовательности по отношению к ней, более предпочтительно по меньшей мере примерно 95%, по меньшей мере примерно 96%, по меньшей мере примерно 97%, по меньшей мере примерно 98%, по меньшей мере примерно 99% идентичностью последовательности по отношению к ней.
[0132] В настоящем описании «лечение» представляет собой подход для получения полезных или желаемых клинических результатов.
[0133] В настоящем описании «нарушения, связанные с IL-6 и/или VEGF» включают, например, глазные нарушения и системные нарушения. Глазные нарушения включают такие глазные нарушения, как офтальмологическое воспалительное заболевание - склерит, непролиферативная диабетическая ретинопатия, пролиферативная диабетическая ретинопатия, диабетический отек желтого пятна, предотвращение диабетического отека желтого пятна, предотвращение пролиферативной диабетической ретинопатии, влажная форма возрастной дегенерации желтого пятна, предотвращение влажной формы возрастной дегенерации желтого пятна, сухая форма возрастной дегенерации желтого пятна, венозная, артериальная окклюзия или другая окклюзия кровеносных сосудов глаза или сетчатки с отеком сетчатки или без него, передний и задний увеит, отек желтого пятна при увеите и внутриглазные опухоли. Нарушения, связанные с IL-6, также включают нарушения, при которых наблюдают повышенный уровень активности IL-6 из-за взаимодействия IL-6 с IL-6R или растворимым IL-6R (sIL-6R). В некоторых вариантах реализации любой один или более слитых белков, предложенных согласно настоящему изобретению, и/или любой один или более конъюгатов, предложенных согласно настоящему изобретению, можно применять для лечения или предотвращения любого одного или более нарушений, связанных с IL-6 и/или VEGF. В некоторых вариантах реализации указанные нарушения включают системные заболевания, поражающие глаз, такие как болезнь Грейвса или нейромиелит зрительного нерва, или системные заболевания, не поражающие глаз, такие как рассеянный склероз, ревматоидный артрит.В некоторых вариантах реализации нарушения включают синдром высвобождения цитокинов после CAR-T или подобных иммуноонкологических лекарственных средств.
[0134] В дополнение к этому, анти-IL-6 молекулы подавляют индукцию экспрессии IL-6, наблюдаемую после лечения анти-PD-1/PD-L1 молекулами (Tsukamoto et al, Cancer Res; 2018 78(17); 5011-22). Также было показано, что блокада передачи сигнала VEGF может повышать эффективность анти-PD-L1 лечения (Allen et al, Sci Transl Med 2017 April 12: 9(385)). Таким образом, эти двойные ингибиторы можно применять в комбинации с модуляторами PD-1/PDL-1 и/или другими ингибиторами иммунных контрольных точек для синергетического лечения рака. Другие нарушения могут включать отек мозга при глиобластоме, при котором анти-IL6 терапия может демонстрировать полезные эффекты в дополнение к лечению анти-VEGF. Другие нарушения включают нарушения, связанные с солидными опухолями.
[0135] В настоящем описании термин «улучшение» означает ослабление или облегчение одного или более симптомов по сравнению с отсутствием введения антитела к IL-6, слитой конструкции антитело к IL-6-ловушка VEGF, конъюгата антитела к IL-6 и/или конъюгата слитой конструкции антитело к IL-6-ловушка VEGF. Термин «улучшение» также включает сокращение или уменьшение продолжительности симптома.
[0136] В настоящем описании термин «ловушка VEGF» или аналогичный термин означает VEGF-связывающие домены (домен 2 VEGFR1, домен 3 VEGFR2). Этот фрагмент позволяет белку действовать как ловушка VEGF и предотвращать связывание VEGF с рецепторами VEGF, экспрессируемыми клетками. Пример этой последовательности можно найти в таблице 10. В некоторых вариантах реализации ловушка VEGF содержит только домен 2 VEGFR1, домен 3 VEGFR2. В этой области техники известны различные варианты белков-ловушек и их можно найти, например, в публикации США №20150376271, полное содержание которой в отношении различных вариантов ловушек VEGF (которые представляют собой белки VEGFR или их фрагменты) и их слитых конструкций включено в настоящее описание посредством ссылки. В некоторых вариантах реализации термин «ловушка VEGF» или аналогичный термин относится к полноразмерной внеклеточной области или любой ее части, или комбинации частей из различных рецепторов VEGF, которые могут противодействовать передаче сигнала между по меньшей мере одним VEGF и VEGFR.
[0137] В настоящем описании термины «слитая конструкция антитело к IL-6-ловушка VEGF», «антитело к IL-6-ловушка VEGF», « АТ к IL-6-ловушка VEGF», «антиIL-6-ловушка VEGF», «VEGFR-антиIL6», «VEGFR-антиIL-6», «слитая конструкция ловушка VEGF-анти-IL6 антитело(TAF)», «ловушка VEGF-IL6», «VEGFR IL-6», «IL6-VEGFR» или аналогичный термин, или термины наоборот (например, «ловушка VEGF- АТ к IL-6, «слитая конструкция ловушка VEGF-антитело к IL-6» и т.д.) означают слияние антитела IL-6 и ловушки VEGF. Варианты представлены на фиг.6. При использовании в общем порядок двух терминов можно менять. При использовании в конкретном случае порядок двух терминов означает относительное положение компонентов в конструкции. Термин «ТА-ловушка», «IL-6 АТ-ловушка VEGF», « АТ к IL-6-ловушка VEGF» или « АТ к IL-6-ловушка», или «антиIL-6 -ловушка VEGF», «ловушка-АТ», «антиIL-6-VEGFR», «антиIL6-VEGFR» или другой аналогичный термин или термины наоборот (например, «ловушка VEGF- АТ к IL-6», «слитая конструкция ловушка VEGF-антитело к IL-6» и т.д.) означает расположение АТ, слитого с соответствующими доменами VEGF-связывающего белка таким образом, чтобы получить ловушку VEGF. Как отмечено выше, этот участок VEGF-связывающего белка представляет собой участок, который предотвращает связывание VEGF с рецепторами VEGF. Как описано в настоящем документе, расположение (порядок) участков, представляющих собой ловушку и антитело, можно варьировать. Таким образом, если явным образом не указано иное или в контексте не указано иное, термины, употребляемые в настоящем описании в отношении слитых конструкций АТ-ловушка (или Il-6/ловушка VEGF и т.д.), означают все описанные варианты расположения антитела и ловушки. Таким образом, если не указано иное, термин АТ-ловушка (или Il-6/ловушка VEGF и т.д.) означает левый вариант на фиг.6 и правый вариант на фиг.6, и оба варианта на фиг.6. Таким образом, это общее выражение описывает для удобства все три варианта. Если обозначена конкретная ориентация, она может быть обозначена, например, путем указания, что «расположение» может быть одним из: ловушка-АТ, ловушка АТ к IL-6, ловушка VEGF АТ к IL-6, ловушка VEGF АТ к IL6. Подобным образом очевидно, что в контексте некоторых примеров в настоящем описании указана конкретная ориентация или расположение молекул, обозначенное в контексте примеров. Оба расположения (альтернативное и комбинированное) явным образом предусмотрены для всего описания слитых белков, предложенных согласно настоящему изобретению. Кроме того, благодаря расположению в определенном порядке очевидно, что термин « АТ к IL-6» при использовании в контексте слитого белка, включает как вариант, когда антитело является смежным, фиг.6, левая сторона, так и вариант, когда ловушка расположена «в пределах» АТ (фиг.6, правая сторона). Снова, термин «АТ» (Ab) или «антитело» при использовании в контексте слитого белка (или другой подобный термин) включает все три варианта (левая сторона фиг.6, правая сторона фиг.6 и оба варианта), если не указано иное. В некоторых вариантах реализации ловушка VEGF слита с IL-6 одним из следующих способов: с N-концом тяжелой цепи, содержащей VH кIL-6; или между шарнирной областью и после CH1-домена тяжелой цепи, содержащей VH к IL-6. Нет различия между обозначениями «АТ» (Ab), «антитело», «анти» или другим подобным термином при использовании в названии для обозначения антитела или его фрагмента. Нет различия между обозначениями «Il-6» или «IL6», или «IL-6». В настоящем описании при ссылке на слитую конструкцию с IL-6 термины «VEGF», «VEGFR», «ловушка VEGF», «ловушка VEGFR» являются взаимозаменяемыми. Указанные термины могут иметь разные значения при употреблении отдельно от расположения в слитой конструкции с IL-6, которые будут зависеть от контекста рассматриваемого термина.
[0138] В настоящем описании термин «биополимер» означает, что полимер связан с белком, представляющим интерес.Указанный термин может быть также определен как «конъюгированная» форма белка. Это можно осуществлять для всех белков, описанных в настоящем документе. Таким образом, для всех таких АТ к IL-6 и конструкций антитело к IL-6-ловушка VEGF, предложенных согласно настоящему изобретению, предусмотрены биополимеры АТ к IL-6 и биополимеры конструкции антитело к IL-6-ловушка VEGF. В дополнение к этому также предложены биополимеры ловушки VEGF.
[0139] В настоящем описании термин «антагонистическое антитело» означает антитело, которое блокирует одну или более функций, или активность молекулы, с которое связывается указанное антитело.
[0140] В настоящем описании термин «эффективная доза» или «эффективное количество» лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для обеспечения любого одного или более полезных или желаемых результатов. В соответствии с более конкретными аспектами эффективное количество предотвращает, облегчает или устраняет симптомы заболевания и/или увеличивает продолжительность жизни вылечиваемого субъекта. В случае профилактического применения полезные или желаемые результаты включают устранение или снижение риска, снижение тяжести или отсрочивание начала заболевания, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические проявления, присутствующие во время развития заболевания. В случае терапевтического применения полезные или желаемые результаты включают клинические результаты, такие как уменьшение одного или более симптомов заболевания, такого как, например, ВДЖП, включая, например, без исключения, сухую форму ВДЖП и влажную форму ВДЖП, снижение дозы других лекарственных средств, необходимых для лечения указанного заболевания, усиление действия другого лекарственного средства и/или замедление прогрессирования ВДЖП у пациентов. Эффективную дозу можно вводить в виде одного или более введений. Для целей настоящего изобретения эффективная доза лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для осуществления профилактического или терапевтического лечения либо напрямую, либо опосредовано. Как понятно в клиническом контексте, эффективная доза лекарственного средства, соединения или фармацевтической композиции может быть или не быть достигнута в сочетании с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, «эффективную дозу» можно рассматривать в контексте введения одного или более терапевтических агентов и можно рассматривать введение одного агента в эффективном количестве, если в сочетании с одним или более другими агентами может быть достигнут или достигается желаемый результат.
[0141] Анти-IL-6 антитела вводят в эффективном режиме, означающем дозу, путь введения и частоту введения, который отсрочивает возникновение, уменьшает тяжесть, препятствует дальнейшему ухудшению и/или устраняет по меньшей мере один признак или симптом нарушения. Если пациент уже страдает нарушением, режим может называться терапевтически эффективным режимом. Если пациент подвержен повышенному риску возникновения нарушения относительно общей популяции, но еще не испытывает симптомов, режим может называться профилактически эффективным режимом. В некоторых случаях терапевтическую или профилактическую эффективность можно наблюдать у отдельного пациента относительно исторического контроля или прошлого опыта у этого же пациента. В других случаях терапевтическая или профилактическая эффективность может быть продемонстрирована в доклиническом или клиническом исследовании в популяции пациентов, получавших лечение, относительно контрольной популяции пациентов, не получавших лечение.
[0142] Конструкции анти-IL-6-ловушка VEGF вводят в эффективном режиме, означающем дозу, путь введения и частоту введения, который отсрочивает возникновение, уменьшает тяжесть, препятствует дальнейшему ухудшению и/или устраняет по меньшей мере один признак или симптом нарушения. Если пациент уже страдает нарушением, режим может называться терапевтически эффективным режимом. Если пациент подвержен повышенному риску возникновения нарушения относительно общей популяции, но еще не испытывает симптомов, режим может называться профилактически эффективным режимом. В некоторых случаях терапевтическую или профилактическую эффективность можно наблюдать у отдельного пациента относительно исторического контроля или прошлого опыта у этого же пациента. В других случаях терапевтическая или профилактическая эффективность может быть продемонстрирована в доклиническом или клиническом исследовании в популяции пациентов, получавших лечение, относительно контрольной популяции пациентов, не получавших лечение.
[0143] «Биологический период полувыведения» вещества представляет собой фармакокинетический параметр, который обозначает время, необходимое для выведения из организма половины вещества после введения вещества в указанный организм.
[0144] Термин «предотвращение» или «предотвращать» относится к (а) предотвращению возникновения нарушения, (b) отсрочиванию возникновения нарушения или возникновения симптомов нарушения, или (с) замедлению прогрессирования существующего состояния. Если не указано иное, термин «предотвращение» не предусматривает полное недопущение возникновения явления.
[0145] «Индивидуум» или «субъект» представляет собой млекопитающее или птицу, более предпочтительно человека. Млекопитающие также включают, но не ограничиваются ими, сельскохозяйственных животных (например, коров, свиней, лошадей, цыплят и т.д.), спортивных животных, домашних животных, приматов, лошадей, собак, кошек, мышей и крыс.
[0146] В настоящем описании термин «вектор» означает конструкцию, способную доставлять и предпочтительно экспрессировать один или более генов, или последовательностей, представляющих интерес, в клетке-хозяине. Примеры векторов включают, но не ограничиваются ими, вирусные векторы, векторы экспрессии на основе депротеинизированной ДНК или РНК, плазмидные, космидные или фаговые векторы, ДНК- или РНК-векторы экспрессии, ассоциированные с катионными конденсирующими агентами, ДНК- или РНК-векторы экспрессии, инкапсулированные в липосомах, и некоторые эукариотические клетки, такие как клетки-продуценты.
[0147] В настоящем описании термин «последовательность контроля экспрессии» означает последовательность нуклеиновой кислоты, которая направляет транскрипцию нуклеиновой кислоты. Последовательность контроля экспрессии может представлять собой промотор, такой как конститутивный или индуцируемый промотор, или энхансер. Последовательность контроля экспрессии функционально связана с транскрибируемой последовательностью нуклеиновой кислоты.
[0148] В настоящем описании термин «фармацевтически приемлемый носитель» или «фармацевтически приемлемое вспомогательное вещество» включает любое вещество, которое при комбинировании с активным ингредиентом позволяет указанному ингредиенту сохранять биологическую активность и которое не реагирует с иммунной системой субъекта. Примеры включают, но не ограничиваются ими, любой из стандартных фармацевтических носителей, таких как фосфатный буферный раствор, вода, эмульсии, такие как эмульсия масло-в-воде, различных типов смачивающих агентов, детергентов, таких как полисорбат 20, для предотвращения агрегации и сахаров, таких как сахароза, в качестве криопротектора. Предпочтительные разбавители для аэрозольного или парентерального введения представляют собой фосфатный буферный раствор (ФБР) или изотонический (0,9%) раствор. Композиции, содержащие такие носители, изготавливают хорошо известными традиционными способами (см., например, Remington's Pharmaceutical Sciences, 18th edition, A. Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000).
[0149] В настоящем описании термин «kon» относится к константе скорости ассоциации антитела (или биоконъюгата) с антигеном. В частности, константы скорости (kon и koff) и равновесные константы диссоциации измеряют с использованием полноразмерных антител и/или Fab-фрагментов антител (т.е. одновалентных) и IL-6.
[0150] В настоящем описании термин «koff» относится к константе скорости диссоциации антитела (или биоконъюгата) из комплекса антитело/антиген.
[0151] В настоящем описании термин «KD» относится к равновесной константе диссоциации взаимодействия антитело-антиген (или биоконъюгат-антиген).
[0152] Термин «примерно» применительно к значению или параметру в настоящем описании включает (и описывает) варианты реализации, которые относятся к этому значению или параметру per se. Например, описание, относящееся к «примерно X», включает описание «X». Числовые диапазоны включают числа, определяющие этот диапазон.
[0153] Термин «пациент» включает человека и других субъектов (включая млекопитающих), которые получают либо профилактическое, либо терапевтическое лечение.
[0154] Для целей классификации замен аминокислот как консервативные или неконсервативные, аминокислоты группируют следующим образом: группа I (гидрофобные боковые цепи): met, ala, val, leu, ile; группа II (нейтральные гидрофильные боковые цепи): cys, ser, thr; группа III (кислотные боковые цепи): asp, glu; группа IV (основные боковые цепи): asn, gln, his, lys, arg; группа V (остатки, влияющие на ориентацию цепи): gly, pro; и группа VI (ароматические боковые цепи): trp, tyr, phe. Консервативные замены включают замены между аминокислотами в одном и том же классе. Неконсервативные замены представляют собой замену представителя одного из этих классов на представитель другого.
[0155] Процент идентичности последовательностей определяют для последовательностей антител, максимально выровненных согласно системе нумерации Kabat для вариабельной области или нумерации EU для константной области. После выравнивания, если область рассматриваемого антитела (например, всю зрелую вариабельную область тяжелой или легкой цепи) сравнивают с той же областью эталонного антитела, процент идентичности последовательностей между указанными областями рассматриваемого и эталонного антитела представляет собой количество положений, занимаемых одной и той же аминокислотой в области как рассматриваемого, так и эталонного антитела, деленное на общее количество выровненных положений двух областей без учета гэпов, умноженное на 100 для перевода в проценты. Идентичность последовательностей в случае других последовательностей может быть определена путем выравнивания последовательностей с применением таких алгоритмов, как BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, версия 7.0, Genetics Computer Group, 575 Science Dr., Мэдисон, Висконсин, с использованием параметров гэпа по умолчанию, или визуально, и наилучшего выравнивания (т.е. обеспечивающего наибольший процент сходства последовательностей в окне сравнения). Процент идентичности последовательностей рассчитывают путем сравнения двух оптимально выровненных последовательностей в окне сравнения, определения количества положений, в которых в обеих последовательностях встречаются идентичные остатки, с получением количества совпавших положений, деления количества совпавших положений на общее количество положений в окне сравнения (т.е. размер окна) и умножения результата на 100 с получением процента идентичности последовательностей.
[0156] Термин «антителозависимая клеточная цитотоксичность» или АЗКЦ представляет собой механизм индукции гибели клеток, который зависит от взаимодействия клеток-мишеней, покрытых антителом (т.е. клеток со связанным антителом), с иммунными клетками, обладающими литической активностью (также называемыми эффекторными клетками). Такие эффекторные клетки включают естественные клетки-киллеры, моноциты/макрофаги и нейтрофилы. К запуску АЗКЦ приводят взаимодействия между Fc-областью антитела, связанного с клеткой, и рецепторами Fcγ, в частности FcγRI и FcγRIII, на иммунных эффекторных клетках, таких как нейтрофилы, макрофаги и естественные клетки-киллеры. Клетка-мишень удаляется путем фагоцитоза или лизиса в зависимости от типа опосредующей эффекторной клетки. Гибель клетки-мишени, покрытой антителом, происходит в результате активности эффекторной клетки.
[0157] Гуманизированное антитело представляет собой генетически сконструированное антитело, в котором CDR из «донорского» антитела нечеловеческого происхождения «привиты» в последовательности «акцепторного» антитела человека (см., например, Queen, US 5530101 и 5585089; Winter, US 5225539, Carter, US 6407213, Adair, US 5859205 6881557, Foote, US 6881557). Последовательности акцепторного антитела могут представлять собой, например, последовательность зрелого антитела человека, композит таких последовательностей, консенсусную последовательность антитела человека или последовательность зародышевой линии. Таким образом, гуманизированное антитело представляет собой антитело, содержащее некоторые или все CDR полностью или по существу из донорского антитела и каркасные последовательности вариабельной области и константные области, если присутствуют, полностью или по существу из последовательностей антитела человека. Подобным образом, гуманизированная тяжелая цепь содержит по меньшей мере одну, две и обычно все три CDR полностью или по существу из тяжелой цепи донорского антитела, и каркасную последовательность вариабельной области тяжелой цепи и константную область тяжелой цепи, если присутствует, по существу из каркасных последовательностей вариабельной области тяжелой цепи и последовательностей константной области человека. Подобным образом, гуманизированная легкая цепь содержит по меньшей мере одну, две и обычно все три CDR полностью или по существу из легкой цепи донорского антитела, и каркасную последовательность вариабельной области легкой цепи и константную область легкой цепи, если присутствует, по существу из каркасных последовательностей вариабельной области легкой цепи и последовательностей константной области человека. За исключением нанотел и dAb гуманизированное антитело содержит гуманизированную тяжелую цепь и гуманизированную легкую цепь. CDR в гуманизированном антителе получена по существу из соответствующей CDR в антителе нечеловеческого происхождения, когда по меньшей мере 85%, 90%, 95% или 100% соответствующих остатков (определенных согласно Kabat) идентичны между соответствующими CDR. Каркасные последовательности вариабельной области цепи антитела или константная область цепи антитела получены по существу из каркасной последовательности вариабельной области человека или константной области человека соответственно, когда по меньшей мере 85%, 90%, 95% или 100% соответствующих остатков, определенных согласно Kabat, являются идентичными.
[0158] Хотя гуманизированные антитела часто содержат все шесть CDR (предпочтительно определенные согласно Kabat) из антитела мыши, они могут быть также получены с меньшим количеством, чем все CDR (например, по меньшей мере 3, 4 или 5 CDR из антитела мыши) (например, Pascalis et al, J. Immunol. 169:3076, 2002; Vajdos et al., Journal of Molecular Biology, 320: 415-428, 2002; Iwahashi et al., Mol. Immunol. 36: 1079-1091, 1999; Tamura et al, Journal of Immunology, 164: 1432-1441, 2000).
[0159] Химерное антитело представляет собой антитело, в котором зрелые вариабельные области легкой и тяжелой цепей антитела нечеловеческого происхождения (например, мыши) комбинированы с константными областями легкой и тяжелой цепи человека. Такие антитела по существу или полностью сохраняют специфичность связывания, характерную для антитела мыши, и примерно на две трети представляют собой последовательность человека.
[0160] Венированное антитело представляет собой тип гуманизированного антитела, в котором сохранены некоторые и, как правило, все CDR, и некоторые остатки каркасного участка вариабельной области антитела нечеловеческого происхождения, но заменены другие остатки каркасного участка вариабельной области, которые могут вносить вклад в связывание с В- или Т-клеточными эпитопами, например, экспонированные остатки (Padlan, Mol. Immunol. 28:489, 1991) заменены на остатки из соответствующих положений в последовательности антитела человека. Результатом является антитело, в котором CDR получены полностью или по существу из антитела нечеловеческого происхождения, а степень гуманизированности каркасных участков вариабельных областей антитела нечеловеческого происхождения повышена посредством указанных замен. Антитело человека может быть выделено от человека или иным образом получено в результате экспрессии генов иммуноглобулина человека (например, у трансгенной мыши, in vitro или посредством фагового дисплея). Способы получения антител человека включают способ на основе триом согласно Oestberg et al., Hybridoma 2:361-367 (1983); Oestberg, патент США №4634664; и Engleman et al., патент США 4634666, использование трансгенных мышей, содержащих гены иммуноглобулина человека (см. например, Lonberg et al., W093/12227 (1993); US 5877397, US 5874299, US 5814318, US 5789650, US 5770429, US 5661016, US 5633425, US 5625126, US 5569825, US 5545806, Nature 148, 1547-1553 (1994), Nature Biotechnology 14, 826 (1996), Kucherlapati, WO 91/10741 (1991) и способы фагового дисплея (см., например, Dower et al., WO 91/17271 и McCafferty et al., WO 92/01047, US 5877218, US 5871907, US 5858657, US 5837242, US 5733743 и US 5565332.
[0161] «Полимер» представляет собой молекулу, состоящую из множества повторяющихся субъединиц. Указанные субъединицы, также иногда называемые «мономерами», могут быть одинаковыми или разными. Существуют как природные, так и полученные путем синтеза полимеры. Примерами природных полимеров являются ДНК, белок и сложные углеводы. Примерами полученных путем синтеза полимеров являются полистирол и полиакриламид. Полимер, состоящий из повторяющихся звеньев одного мономера, называется гомополимером. Полимер, состоящий из двух или более мономеров, называется сополимером или иногда гетерополимером. Сополимер, в котором определенные типы мономеров сгруппированы вместе, иногда называют блок-сополимером. Полимеры могут быть линейными или разветвленными. Когда полимер разветвлен, полимерные цепи, имеющие общее начало, иногда называют ответвлением (ответвлениями) полимера.
[0162] «Инициатор» представляет собой соединение, способное служить субстратом, по которому может происходить одна или более полимеризаций с использованием мономеров или сомономеров, описанных в настоящем документе. Полимеризация может представлять собой традиционную свободнорадикальную полимеризацию или предпочтительно контролируемую/«живущую» радикальную полимеризацию, такую как радикальная полимеризация с переносом атома (ATRP), полимеризация с обратимым присоединением-фрагментацией-терминацией (RAFT) или нитроксид-опосредованная полимеризация (NMP). Полимеризация может представлять собой «псевдо» контролируемую полимеризацию, такую как вырожденная передача. Инициаторы, подходящие для ATRP, содержат одну или более лабильных связей, которые могут гомолитически расщепляться с образованием фрагмента инициатора, I, представляющего собой радикал, способный инициировать радикальную полимеризацию, и акцептора радикалов, I', реагирующего с радикалом растущей полимерной цепи с обратимой терминацией полимеризации. Указанный акцептор радикалов I’, как правило, представляет собой галоген, но может также представлять собой органический фрагмент, такой как нитрил. В некоторых вариантах реализации настоящего изобретения инициатор содержит одну или более 2-бромизобутиратных групп в качестве центров для полимеризации путем ATRP.
[0163] Термин «химический линкер» относится к химическому фрагменту, который связывает две группы, такие как фрагмент, увеличивающий период полувыведения, и белок. Линкер может быть расщепляемым или нерасщепляемым. Расщепляемые линкеры могут представлять собой в том числе гидролизуемые, ферментативно расщепляемые, pH-чувствительные, фотолабильные или дисульфидные линкеры. Другие линкеры включают гомобифункциональные и гетеробифункциональные линкеры. «Связывающая группа» представляет собой функциональную группу, способную образовывать ковалентную связь, состоящую из одной или более связей, с биоактивным агентом. Неограничивающие примеры включают те, которые представлены в таблице 1 в WO2013059137 (содержание которого включено посредством ссылки).
[0164] Термин «реакционноспособная группа» относится к группе, которая способна реагировать с другой химической группой с образованием ковалентной связи, т.е. ковалентно реакционноспособна в подходящих условиях реакции, и, как правило, представляет собой точку присоединения другого вещества. Реакционноспособная группа представляет собой группу, такую как малеимид или сложный сукцинимидиловый эфир, способную химически реагировать с функциональной группой в другом фрагменте с образованием ковалентной связи. Реакционноспособные группы, как правило, включают нуклеофилы, электрофилы и фотоактивируемые группы.
[0165] В настоящем описании термин «фосфорилхолин», также обозначаемый «PC», относится к следующему:
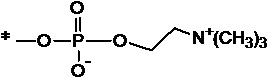
[0166] где * показывает точку присоединения. Фосфорилхолин представляет собой цвиттерионную группу и включает соли (такие как внутренние соли) и их протонированные и депротонированные формы.
[0167] В настоящем описании «полимер на основе фосфорилхолина» представляет собой полимер, содержащий фосфорилхолин. Термин «цвиттерион-содержащий полимер» относится к полимеру, который содержит цвиттерион.
[0168] Термин «полимер, содержащий поли(акрилоилоксиэтилфосфорилхолин)» относится к полимеру, содержащему в качестве мономера 2-(акрилоилокси)этил-2-(триметиламмоний)этилфосфат.
[0169] Термин «полимер, содержащий поли(метакрилоилоксиэтилфосфорилхолин)» относится к полимеру, содержащему в качестве мономера 2-(метакрилоилокси)этил-2-(триметиламмоний)этилфосфат.
[0170] В настоящем описании «молекулярная масса» в контексте полимера может быть выражена в виде либо среднечисловой молекулярной массы, либо среднемассовой молекулярной массы, либо пиковой молекулярной массы. Если не указано иное, все упоминания молекулярной массы в настоящем описании относятся к пиковой молекулярной массе. Эти определения молекулярной массы, среднечисловой (Mn), среднемассовой (Mw) и пиковой (Mp), могут быть выполнены с использованием эксклюзионной хроматографии или других методов жидкостной хроматографии. Можно также использовать другие способы измерения значений молекулярной массы, такие как применение анализа концевых групп или измерение коллигативных свойств (например, понижение температуры замерзания, повышение температуры кипения или осмотическое давление) для определения среднечисловой молекулярной массы или применение методов светорассеяния, ультрацентрифугирования или вискозиметрии для определения среднемассовой молекулярной массы. В предпочтительном варианте реализации настоящего изобретения молекулярную массу измеряют путем SEC-MALS (эксклюзионная хроматография - многоугловое светорассеяние). Полимерные реагенты согласно настоящему изобретению, как правило, являются полидисперсными (т.е. среднечисловая молекулярная масса и среднемассовая молекулярная масса полимеров не равны). Коэффициент полидисперсности (PDI) представляет меру дисперсности полимеров в смеси. PDI определяется формулой Mw/Mn. В этом отношении гомогенный белок будет иметь PDI, равный 1,0 (Mn совпадает с Mw). PDI для полимеров, как правило, будет выше 1,0. Полимеры согласно настоящему изобретению предпочтительно имеют относительно низкие значения полидисперсности (PDI), составляющие, например, менее примерно 1,5, по оценке, например, путем SEC-MALS. В других вариантах реализации значения полидисперсности (PDI) более предпочтительно находятся в диапазоне от примерно 1,4 до примерно 1,2, составляют более предпочтительно менее примерно 1,15 и более предпочтительно менее примерно 1,10, более предпочтительно менее примерно 1,05 и наиболее предпочтительно менее примерно 1,03.
[0171] В настоящем описании термины «защищенный», «защищенная форма», «защищающая группа» и «защитная группа» относятся к присутствию группы (т.е. защитной группы), которая предотвращает или блокирует реакцию конкретной химически активной функциональной группы в молекуле в определенных условиях реакции. Защитные группы варьируются в зависимости от типа защищаемой химически активной группы, а также используемых условий реакции и присутствия дополнительных реакционноспособных или защитных групп в молекуле, при наличии таковых. Подходящие защитные группы включают группы, такие как описаны в трактате Greene et al., “Protective Groups In Organic Synthesis,” 3rd Edition, John Wiley and Sons, Inc., New York, 1999.
[0172] В настоящем описании термин «алкил» относится к линейному или разветвленному насыщенному алифатическому радикалу, содержащему указанное число атомов углерода. Например, C1-C6 алкил включает, но не ограничивается ими, метил, этил, пропил, изопропил, бутил, изобутил, втор-бутил, трет-бутил, пентил, изопентил, гексил и т.д. Другие алкильные группы включают, но не ограничиваются ими, гептил, октил, нонил, децил и т.д. Алкил может содержать любое число атомов углерода, например, 1-2, 1-3, 1-4, 1-5, 1-6, 1-7, 1-8, 1-9, 1-10, 2-3, 2-4, 2-5, 2-6, 3-4, 3-5, 3-6, 4-5, 4-6 и 5-6 атомов углерода.
[0173] Термин «низший», упоминаемый выше и ниже в настоящем описании в сочетании с органическими радикалами или соединениями соответственно, определяет соединение или радикал, который может быть разветвленным или неразветвленным и содержать до 7 включительно, предпочтительно до 4 включительно и (как неразветвленный) один или два атома углерода.
[0174] В настоящем описании термин «алкилен» относится к алкильной группе, определенной выше, связывающей по меньшей мере две другие группы, т.е. двухвалентному углеводородному радикалу. Указанные две группы, связанные с алкиленом, могут быть связаны с одним и тем же атомом или разными атомами алкилена. Например, линейный алкилен может представлять собой двухвалентный радикал -(CH2)n,, где n равен 1, 2, 3, 4, 5 или 6. Алкиленовые группы включают, но не ограничиваются ими, метилен, этилен, пропилен, изопропилен, бутилен, изобутилен, втор-бутилен, пентилен и гексилен.
[0175] Заместители для алкильных, алкенильных, алкиленовых, гетероалкильных, гетероалкиленовых, гетероалкенильных, алкинильных, циклоалкильных, гетероциклоалкильных, циклоалкенильных и гетероциклоалкенильных радикалов могут представлять собой одну или более из различных групп, выбранных из, но не ограниченных ими: -OR’,=O,=NR’,=N-OR’, -NR’R”, -SR’, -галогена, -SiR’R”R”’, -OC(O)R’, -C(O)R’, -CO2R’, -CONR’R”, -OC(O)NR’R”, -NR”C(O)R’, -NR’-C(O)NR”R”’, -NR”C(O)2R’,-NR-C(NR’R”R’”)=NR””, -NR-C(NR’R”)=NR’”, -S(O)R’, -S(O)2R’, -S(O)2NR’R”, -NRSO2R’, -CN и -NO2, в количестве в диапазоне от 1 до (2m’+1), где m’ представляет собой общее число атомов углерода в таком радикале. Каждый из R’, R”, R”’ и R”” независимо относится к водороду, содержащему заместители или не содержащему заместителей гетероалкилу, содержащему заместители или не содержащему заместителей арилу, например, арилу, содержащему в качестве заместителей 1-3 атома галогена, содержащему заместители или не содержащему заместителей алкилу, алкокси- или тиоалкоксигруппе, или арилалкильным группам. В случае, когда R’ и R” присоединены к одному атому азота, они могут образовывать вместе с указанным атомом азота 5-, 6- или 7-членное кольцо. Например, -NR’R” включает, но не ограничивается ими, 1-пирролидинил и 4-морфолинил.
[0176] В настоящем описании термин «алкокси» относится к алкильной группе, присоединенной к атому кислорода, и образует радикал -O-R, где R представляет собой алкил. Алкоксигруппы включают, например, метокси, этокси, пропокси, изопропокси, бутокси, 2-бутокси, изобутокси, втор-бутокси, трет-бутокси, пентокси, гексокси и т.д. Алкоксигруппы могут дополнительно содержать различные заместители, описанные в настоящем документе. Например, алкоксигруппы могут содержать в качестве заместителей атомы галогена с образованием «галогеналкокси» группы.
[0177] В настоящем описании термин «карбоксиалкил» означает алкильную группу (определенную в настоящем описании), содержащую в качестве заместителей карбоксигруппу. Термин «карбоксициклоалкил» означает циклоалкильную группу (определенную в настоящем описании), содержащую в качестве заместителей карбоксигруппу. Термин «алкоксиалкил» означает алкильную группу (определенную в настоящем описании), содержащую в качестве заместителей алкоксигруппу. В настоящем описании термин «карбокси» относится к карбоновым кислотам и их сложным эфирам.
[0178] В настоящем описании термин «галогеналкил» относится к алкилу, определенному выше, в котором некоторые или все атомы водорода замещены атомами галогена. Галоген (галогено) предпочтительно представляет собой хлор или фтор, но может также представлять собой бром или йод. Например, галогеналкил включает трифторметил, фторметил, 1,2,3,4,5-пентафторфенил и т.д. Термин «перфтор» определяет соединение или радикал, в котором все доступные атомы водорода заменены на фтор. Например, перфторфенил относится к 1,2,3,4,5-пентафторфенилу, перфторметил относится к 1,1,1-трифторметилу, и перфторметокси относится к 1,1,1-трифторметокси. Галогеналкил может также называться алкилом, содержащим в качестве заместителей галоген, таким как алкил, содержащий в качестве заместителей фтор.
[0179] В настоящем описании «цитокин» в контексте настоящего изобретения является членом группы белковых сигнальных молекул, которые могут участвовать в межклеточной коммуникации в иммунных и воспалительных ответах. Цитокины представляют собой, как правило, малые водорастворимые гликопротеиды, масса которых составляет примерно 8-35 кДа.
[0180] В настоящем описании термин «циклоалкил» относится к насыщенной моно- или полициклической алифатической кольцевой системе, которая содержит от примерно 3 до 12, от 3 до 10, от 3 до 7 или от 3 до 6 атомов углерода. В случае, когда циклоалкильная группа состоит из двух или более колец, указанные кольца могут быть соединены друг с другом с образованием конденсированного кольца или спирокольцевой структуры. В случае, когда циклоалкильная группа состоит из трех или более колец, указанные кольца могут быть также соединены друг с другом с образованием мостиковой кольцевой структуры. Моноциклические кольца включают, например, циклопропил, циклобутил, циклопентил, циклогексил и циклооктил. Бициклические и полициклические кольца включают, например, бицикло[1.1.1]пентан, бицикло[2.1.1]гептан, норборнан, декагидронафталин и адамантан. Например, C3-8 циклоалкил включает циклопропил, циклобутил, циклопентил, циклогексил, циклооктил и норборнан.
[0181] В настоящем описании термин «эндоциклический» относится к атому или группе атомов, составляющих часть циклической кольцевой структуры.
[0182] В настоящем описании термин «экзоциклический» относится к атому или группе атомов, которые присоединены, но не определяют циклическую кольцевую структуру.
[0183] В настоящем описании термин «циклический алкиловый эфир» относится к 4- или 5-членной циклической алкильной группе, содержащей 3 или 4 эндоциклических атома углерода и 1 эндоциклический атом кислорода или серы (например, оксетан, тиетан, тетрагидрофуран, тетрагидротиофен); или 6-7-членной циклической алкильной группе, содержащей 1 или 2 эндоциклических атома кислорода или серы (например, тетрагидропиран, 1,3-диоксан, 1,4-диоксан, тетрагидротиопиран, 1,3-дитиан, 1,4-дитиан, 1,4-оксатиан).
[0184] В настоящем описании термин «алкенил» относится либо к линейному, либо к разветвленному углеводороду из 2-6 атомов углерода, содержащему по меньшей мере одну двойную связь. Примеры алкенильных групп включают, но не ограничиваются ими, винил, пропенил, изопропенил, 1-бутенил, 2-бутенил, изобутенил, бутадиенил, 1-пентенил, 2-пентенил, изопентенил, 1,3-пентадиенил, 1,4-пентадиенил, 1-гексенил, 2-гексенил, 3-гексенил, 1,3-гексадиенил, 1,4-гексадиенил, 1,5-гексадиенил, 2,4-гексадиенил или 1,3,5-гексатриенил. Алкенильные группы могут также содержать от 2 до 3, от 2 до 4, от 2 до 5, от 3 до 4, от 3 до 5, от 3 до 6, от 4 до 5, от 4 до 6 и от 5 до 6 атомов углерода.
[0185] В настоящем описании термин «алкенилен» относится к алкенильной группе, определенной выше, связывающей по меньшей мере две другие группы, т.е. двухвалентному углеводородному радикалу. Указанные две группы, связанные с алкениленом, могут быть связаны с одним и тем же атомом или разными атомами алкенилена. Алкениленовые группы включают, но не ограничиваются ими, этенилен, пропенилен, изопропенилен, бутенилен, изобутенилен, втор-бутенилен, пентенилен и гексенилен.
[0186] В настоящем описании термин «алкинил» относится либо к линейному, либо к разветвленному углеводороду из 2-6 атомов углерода, содержащему по меньшей мере одну тройную связь. Примеры алкинильных групп включают, но не ограничиваются ими, ацетиленил, пропинил, 1-бутинил, 2-бутинил, изобутинил, втор-бутинил, бутадиинил, 1-пентинил, 2-пентинил, изопентинил, 1,3-пентадиинил, 1,4-пентадиинил, 1-гексинил, 2-гексинил, 3-гексинил, 1,3-гексадиинил, 1,4-гексадиинил, 1,5-гексадиинил, 2,4-гексадиинил или 1,3,5-гексатриинил. Алкинильные группы могут также содержать от 2 до 3, от 2 до 4, от 2 до 5, от 3 до 4, от 3 до 5, от 3 до 6, от 4 до 5, от 4 до 6 и от 5 до 6 атомов углерода.
[0187] В настоящем описании термин «алкинилен» относится к алкинильной группе, определенной выше, связывающей по меньшей мере две другие группы, т.е. двухвалентному углеводородному радикалу. Указанные две группы, связанные с алкиниленом, могут быть связаны с одним и тем же атомом или разными атомами алкинилена. Алкиниленовые группы включают, но не ограничиваются ими, этинилен, пропинилен, бутинилен, втор-бутинилен, пентинилен и гексинилен.
[0188] В настоящем описании термин «циклоалкилен» относится к циклоалкильной группе, определенной выше, связывающей по меньшей мере две другие группы, т.е. двухвалентному углеводородному радикалу. Указанные две группы, связанные с циклоалкиленом, могут быть связаны с одним и тем же атомом или разными атомами циклоалкилена. Циклоалкиленовые группы включают, но не ограничиваются ими, циклопропилен, циклобутилен, циклопентилен, циклогексилен и циклооктилен.
[0189] В настоящем описании термин «гетероциклоалкил» относится к кольцевой системе, содержащей от 3 атомов кольца до примерно 20 атомов кольца и от 1 до примерно 5 гетероатомов, таких как N, О и S. Также подходящими могут быть дополнительные гетероатомы, включая, но не ограничиваясь ими, B, Al, Si и P. Гетероатомы могут быть также окисленными, например, но не ограничиваясь ими, -S(O)- и -S(O)2-. Например, гетероцикл включает, но не ограничивается ими, тетрагидрофуранил, тетрагидротиофенил, морфолино, пирролидинил, пирролинил, имидазолидинил, имидазолинил, пиразолидинил, пиразолинил, пиперазинил, пиперидинил, индолинил, хинуклидинил и 1,4-диокса-8-аза-спиро[4.5]дец-8-ил.
[0190] В настоящем описании термин «гетероциклоалкилен» относится к гетероциклоалкильной группе, определенной выше, связывающей по меньшей мере две другие группы. Указанные две группы, связанные с гетероциклоалкиленом, могут быть связаны с одним и тем же атомом или разными атомами гетероциклоалкилена.
[0191] В настоящем описании термин «арил» относится к моноциклической или полициклической (например, конденсированной бициклической, трициклической или с бóльшим количеством циклов) ароматической кольцевой системе, содержащей от 6 до 16 атомов углерода. Например, арил может представлять собой фенил, бензил или нафтил, предпочтительно представляет собой фенил. Арильные группы могут быть моно-, ди- или тризамещенными и содержать в качестве заместителей один, два или три радикала, выбранные из алкила, алкокси, арила, гидрокси, галогена, циано, амино, аминоалкила, трифторметила, алкилендиокси и окси-C2-C3-алкилена; все из которых возможно также содержат заместители, например, определенные выше; или 1- или 2-нафтила; или 1- или 2-фенантренила. Алкилендиокси представляет собой двухвалентный заместитель, присоединенный к двум смежным атомам углерода фенила, например, метилендиокси или этилендиокси. Окси-C2-C3-алкилен также представляет собой двухвалентный заместитель, присоединенным к двум смежным атомам углерода фенила, например, оксиэтилен или оксипропилен. Примером окси-C2-C3-алкиленфенила является 2,3-дигидробензофуран-5-ил.
[0192] Предпочтительный арил представляет собой нафтил, фенил или фенил, моно- или дизамещенный алкокси, фенилом, галогеном, алкилом или трифторметилом, особенно фенил или фенил, моно- или дизамещенный алкокси, галогеном или трифторметилом, и, в частности, фенил.
[0193] Примерами содержащих заместители фенильных групп в качестве R являются, например, 4-хлорфен-1-ил, 3,4-дихлорфен-1-ил, 4-метоксифен-1-ил, 4-метилфен-1-ил, 4-аминометилфен-1-ил, 4-метоксиэтиламинометилфен-1-ил, 4-гидроксиэтиламинометилфен-1-ил, 4-гидроксиэтил-(метил)-аминометилфен-1-ил, 3-аминометилфен-1-ил, 4-N-ацетиламинометилфен-1-ил, 4-аминофен-1-ил, 3-аминофен-1-ил, 2-аминофен-1-ил, 4-фенил-фен-1-ил, 4-(имидазол-1-ил)-фенил, 4-(имидазол-1-илметил)-фен-1-ил, 4-(морфолин-1-ил)-фен-1-ил, 4-(морфолин-1-илметил)-фен-1-ил, 4-(2-метоксиэтиламинометил)-фен-1-ил и 4-(пирролидин-1-илметил)-фен-1-ил, 4-(тиофенил)-фен-1-ил, 4-(3-тиофенил)-фен-1-ил, 4-(4-метилпиперазин-1-ил)-фен-1-ил и 4-(пиперидинил)-фенил, и 4-(пиридинил)-фенил, возможно содержащие заместители в гетероциклическом кольце.
[0194] В настоящем описании термин «арилен» относится к арильной группе, определенной выше, связывающей по меньшей мере две другие группы. Указанные две группы, связанные с ариленом, связаны с разными атомами арилена. Ариленовые группы включают, но не ограничиваются ими, фенилен.
[0195] В настоящем описании термин «ариленокси» относится к ариленовой группе, определенной выше, при этом одна из групп, связанных с ариленом, связана через атом кислорода. Ариленоксигруппы включают, но не ограничиваются ими, фениленокси.
[0196] Подобным образом, заместители для арильных и гетероарильных групп варьируются и выбраны из: -галогена, -OR’, -OC(O)R’, -NR’R”, -SR’, -R’, -CN, -NO2, -CO2R’, -CONR’R”, -C(O)R’, -OC(O)NR’R”, -NR”C(O)R’, -NR”C(O)2R’, -NR’-C(O)NR”R”’, -NH-C(NH2)=NH, -NR’C(NH2)=NH, -NH-C(NH2)=NR’, -S(O)R’, -S(O)2R’, -S(O)2NR’R”, -N3, -CH(Ph)2, перфтор(C1-C4)алкокси и перфтор(C1-C4)алкила в количестве в диапазоне от нуля до общего числа свободных валентностей в ароматической кольцевой системе; и где R’, R” и R”’ независимо выбраны из водорода, (C1-C8)алкила и гетероалкила, не содержащего заместителей арила и гетероарила, (не содержащий заместителей арил)-(C1-C4)алкила и (не содержащий заместителей арил)окси-(C1-C4)алкила.
[0197] Два из указанных заместителей у смежных атомов арильного или гетероарильного кольца могут быть заменены на заместитель формулы -T-C(O)-(CH2)q-U-, где T и U независимо представляют собой -NH-, -O-, -CH2- или одинарную связь, и q представляет собой целое число от 0 до 2. В качестве альтернативы, два из заместителей у смежных атомов арильного или гетероарильного кольца могут быть заменены на заместитель формулы -A-(CH2)r-B-, где А и В независимо представляют собой -CH2-, -O-, -NH-, -S-, -S(O)-, -S(O)2-, -S(O)2NR’- или одинарную связь, и r представляет собой целое число от 1 до 3. Одна из одинарных связей нового кольца, образованного таким образом, может быть заменена на двойную связь. В качестве альтернативы, два из заместителей у смежных атомов арильного или гетероарильного кольца могут быть заменены на заместитель формулы -(CH2)s-X-(CH2)t-, где s и t независимо представляют собой целые числа от 0 до 3, и X представляет собой -O-, -NR’-, -S-, -S(O)-, -S(O)2- или -S(O)2NR’-. Заместитель R’ в -NR’- и -S(O)2NR’- выбран из водорода или не содержащего заместителей (C1-C6)алкила.
[0198] В настоящем описании термин «гетероарил» относится к моноциклической или конденсированной бициклической или трициклической ароматической кольцевой системе, содержащей от 5 до 16 атомов кольца, где каждый из 1-4 атомов кольца представляет собой гетероатом N, O или S. Например, гетероарил включает пиридил, индолил, индазолил, хиноксалинил, хинолинил, изохинолинил, бензотиенил, бензофуранил, фуранил, пирролил, тиазолил, бензотиазолил, оксазолил, изоксазолил, триазолил, тетразолил, пиразолил, имидазолил, тиенил или любые другие радикалы, содержащие в качестве заместителей (особенно моно- и дизамещенные), например, алкил, нитро или галоген. Пиридил представляет собой 2-, 3- или 4-пиридил, предпочтительно 2- или 3-пиридил. Тиенил представляет собой 2- или 3-тиенил. Хинолинил предпочтительно представляет собой 2-, 3- или 4-хинолинил. Изохинолинил предпочтительно представляет собой 1-, 3- или 4-изохинолинил. Бензопиранил, бензотиопиранил предпочтительно представляют собой 3-бензопиранил или 3-бензотиопиранил соответственно. Тиазолил предпочтительно представляет собой 2- или 4-тиазолил и наиболее предпочтительно 4-тиазолил. Триазолил предпочтительно представляет собой 1-, 2- или 5-(1,2,4-триазолил). Тетразолил предпочтительно представляет собой 5-тетразолил.
[0199] Предпочтительно, гетероарил представляет собой пиридил, индолил, хинолинил, пирролил, тиазолил, изоксазолил, триазолил, тетразолил, пиразолил, имидазолил, тиенил, фуранил, бензотиазолил, бензофуранил, изохинолинил, бензотиенил, оксазолил, индазолил или любой из содержащих заместители радикалов, особенно моно- или дизамещенных.
[0200] Термин «гетероалкил» относится к алкильной группе, содержащей от 1 до 3 гетероатомов, таких как N, O и S. Также подходящими могут быть дополнительные гетероатомы, включая, но не ограничиваясь ими, B, Al, Si и P. Гетероатомы могут быть также окисленными, например, но не ограничиваясь ими, -S(O)- и -S(O)2-. Например, гетероалкил может включать эфиры, тиоэфиры, алкиламины и алкилтиолы.
[0201] Термин «гетероалкилен» относится к гетероалкильной группе, определенной выше, связывающей по меньшей мере две другие группы. Указанные две группы, связанные с гетероалкиленом, могут быть связаны с одним и тем же атомом или разными атомами гетероалкилена.
[0202] В настоящем описании термин «электрофил» относится к иону или атому, или группе атомов, которые могут быть ионными, содержащих электрофильный центр, т.е. электроноакцепторный центр, способный реагировать с нуклеофилом. Электрофил (или электрофильный реагент) представляет собой реагент, который образует связь со своим партнером по реакции (нуклеофилом) путем акцептирования обоих связывающих электронов от этого партнера по реакции.
[0203] В настоящем описании термин «нуклеофил» относится к иону или атому, или группе атомов, которые могут быть ионными, содержащих нуклеофильный центр, т.е. центр, «ищущий» электрофильный центр или способный реагировать с электрофилом. Нуклеофил (или нуклеофильный реагент) представляет собой реагент, который образует связь со своим партнером по реакции (электрофилом) путем донирования обоих связывающих электронов. Термин «нуклеофильная группа» относится к нуклеофилу после того, как он прореагировал с реакционноспособной группой. Неограничивающие примеры включают амино, гидроксил, алкокси, галогеналкокси и т.п.
[0204] В настоящем описании термин «малеимидо» относится к пиррол-2,5-дион-1-ильной группе, имеющей структуру:
[0205] которая при реакции с сульфгидрилом (например, тиоалкилом) образует -S-малеимидогруппу, имеющую структуру
[0206] где «•» показывает точку присоединения малеимидогруппы, и « » показывает точку присоединения атома серы тиола к оставшейся части исходной сульфгидрил-содержащей группы.
» показывает точку присоединения атома серы тиола к оставшейся части исходной сульфгидрил-содержащей группы.
[0207] Для цели настоящего изобретения «встречающиеся в природе аминокислоты», обнаруживаемые в белках и полипептидах, представляют собой L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин, L-глутамин, L-глутаминовую кислоту, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и/или L-валин. «Не встречающиеся в природе аминокислоты», обнаруживаемые в белках, представляют собой любую аминокислоту кроме тех, которые перечислены в качестве встречающихся в природе аминокислот.Не встречающиеся в природе аминокислоты включают, без ограничения, D-изомеры встречающихся в природе аминокислот и смеси D- и L-изомеров встречающихся в природе аминокислот.Другие аминокислоты, такие как 4-гидроксипролин, десмозин, изодесмозин, 5-гидроксилизин, эпсилон-N-метиллизин, 3-метилгистидин, хотя и обнаруживаются во встречающихся в природе белках, считаются не встречающимися в природе аминокислотами, обнаруживаемыми в белках, для цели настоящего изобретения, поскольку их, как правило, вводят способами, отличными от рибосомальной трансляции мРНК.
[0208] В настоящем описании термин «линейный» применительно к геометрии, строению или общей структуре полимера относится к полимеру, содержащему одно ответвление полимера.
[0209] В настоящем описании термин «разветвленный» применительно к геометрии, строению или общей структуре полимера относится к полимеру, содержащему 2 или более ответвлений полимера, тянущихся от коровой структуры, содержащейся в инициаторе. Инициатор можно использовать в реакции радикальной полимеризации с переносом атома (ATRP). Разветвленный полимер может содержать 2 полимерные цепи (ответвления полимера), 3 ответвления полимера, 4 ответвления полимера, 5 ответвлений полимера, 6 ответвлений полимера, 7 ответвлений полимера, 8 ответвлений полимера, 9 ответвлений полимера или более. Каждое ответвление полимера тянется от центра инициации синтеза полимера. Каждый центр инициации синтеза полимера может являться центром роста полимерной цепи путем присоединения мономеров. Например, но не в качестве ограничения, при использовании ATRP центр инициации синтеза полимера на инициаторе представляет собой, как правило, органический галогенид, подвергающийся обратимому окислительно-восстановительному процессу, катализируемому соединением переходного металла, таким как галогенид меди. Предпочтительно, галогенид представляет собой бром.
[0210] В настоящем описании термин «фармацевтически приемлемое вспомогательное вещество» относится к вспомогательному веществу, которое может быть включено в композиции согласно настоящему изобретению и которое не оказывает значительного нежелательного токсикологического действия на пациента и одобрено или может быть одобрено FDA для терапевтического применения, в частности, у людей. Неограничивающие примеры фармацевтически приемлемых вспомогательных веществ включают воду, NaCl, физиологические растворы, раствор Рингера с лактатом, стандартный раствор сахарозы, стандартный раствор глюкозы и т.п.
[0211] В настоящем описании «OG1786» представляет собой инициатор с 9 ответвлениями, используемый для синтеза полимера, со структурой, показанной на фиг.2D, на которой представлена солевая форма OG1786 с трифторуксусной кислотой. OG1786 можно применять в соответствии с настоящим изобретением в виде других солей или в виде свободного основания.
[0212] В настоящем описании «OG1801» представляет собой полимер массой приблизительно (+/- 15%) 750 кДа (либо Mn, либо Mp), полученный с использованием OG1786 в качестве инициатора для синтеза путем ATRP с использованием мономера HEMA-PC. Структура OG1801 представлена на фиг.2J.
[0213] В настоящем описании «OG1802» представляет собой OG1801 с присоединенной функциональной малеимидной группой и он имеет структуру, представленную на фиг.2K, где каждый из n1, n2, n3, n4, n5, n6, n7, n8 и n9 представляет собой целое число (положительное) (от 0 до примерно 3000), так что общая молекулярная масса указанного полимера составляет (Mw) 750000±15% Дальтон. При употреблении термина «OG1802» для модификации термина белка (такого как ловушка VEGF или анти-IL6 антитело) он означает, что указанный белок представляет собой конъюгированный белок.
[0214] Если не определено иное, все технические и научные термины, используемые в настоящем описании, имеют те же значения, которые обычно понятны специалисту в области техники, к которой относится настоящее изобретение. В случае противоречия настоящее описание, включая определения, будет иметь преимущественную силу. На протяжении всего настоящего описания и формулы изобретения термин «содержать (включать)» или такие вариации, как «содержит (включает)» или «содержащий (включающий)», означают включение указанного целого числа или группы целых чисел, но не исключение какого-либо другого целого числа или группы целых чисел. Если по контексту не требуется иное, термины в единственном числе включают множественное число, а термины во множественном числе включают единственное число. Любой пример (примеры), следующий за термином «например», не является исчерпывающим или ограничивающим.
[0215] Термин, называющий объект в единственном числе, относится к одному или более из таких объектов; например, термин «соединение» относится к одному или более соединениям или по меньшей мере одному соединению. Таким образом, термины в единственном числе, «один или более» и «по меньшей мере один» могут являться взаимозаменяемыми в настоящем описании.
[0216] В настоящем описании термин «примерно» означает вариацию, которую можно наблюдать при измерениях, производимых различными инструментами, в разных образцах и при получении образцов.
[0217] В настоящем описании положения CDR следуют в порядке их появления в вариабельном домене при описании. Например, положения в тяжелой цепи могут быть описаны как S35H или G66D. В настоящем описании положения Fc следуют нумерации EU при описании или указании, что они приведены согласно нумерации EU. Например, мутации антител могут быть описаны как L234A или L235A, что соответствует нумерации EU. Положения могут быть также определены в соответствии с указанными положениями в конкретной SEQ ID или последовательности, представленной в настоящем описании.
[0218] Многоугловое светорассеяние (MALS) представляет собой метод анализа макромолекул, в котором лазерный свет падает на молекулу, осциллирующее электрическое поле света индуцирует в ней осциллирующий диполь. Этот осциллирующий диполь будет обратно излучать свет и его можно измерить с использованием детектора MALS, такого как miniDawn TREOS Wyatt. Интенсивность излучаемого света зависит от величины диполя, индуцированного в макромолекуле, которая в свою очередь пропорциональна поляризуемости макромолекулы, чем больше индуцированный диполь, тем, следовательно, больше интенсивность рассеянного света. Соответственно, для анализа рассеяния из раствора таких макромолекул необходимо знать их поляризуемость относительно окружающей среды (например, растворителя). Она может быть определена в результате измерения изменения, Δn, коэффициента преломления раствора n при изменении молекулярной концентрации, Δc, путем измерения значения dn/dc (=Δn/Δc) с использованием дифференциального рефрактометра Optilab T-rEX Wyatt. Два параметра молярной массы, которые используют при определении MALS, представляют собой среднечисловую молекулярную массу (Mn) и среднемассовую молекулярную массу (Mw), при этом коэффициент полидисперсности (PDI) равен Mw, деленной на Mn. SEC также позволяет определить еще одну среднюю молекулярную массу - пиковую молекулярную массу Mp, которую определяют как молекулярную массу наивысшего пика в SEC.
[0219] PDI используют в качестве меры широты распределения молекулярных масс для полимера и биоконъюгата, полученного в результате конъюгации отдельного белка с полидисперсным биополимером (например, OG1802). Для образца белка полидисперсность близка к 1,0 поскольку он представляет собой продукт трансляции, при которой ожидают, что каждая молекула белка в растворе имеет почти одинаковую длину и молярную массу. Напротив, из-за полидисперсной природы биополимера, когда во время процесса полимеризации синтезируются полимерные цепи различной длины, очень важное значение имеет определение PDI образца как одного из показателей его качества для узкого распределения молекулярной массы.
[0220] Эксклюзионная хроматография (SEC) представляет собой метод хроматографии, в котором молекулы в растворе разделяют по их размеру. Как правило, для перемещения образца через колонку, заполненную смолами с разным размером пор, используют водный раствор. Ожидают, что смола будет инертной по отношению к аналиту при прохождении через колонку, и аналиты будут отделяться друг от друга на основе их уникального размера и характеристик размера пор выбранной колонки.
[0221] Сочетание SEC с MALS или SEC/MALS обеспечивает точное распределение молярной массы и размера (среднеквадратичный радиус) в отличие от опоры на набор калибровочных стандартов SEC. Этот тип устройства имеет много преимуществ по сравнению с традиционными способами калибровки колонки. Поскольку светорассеяние и концентрацию измеряют для каждой элюируемой фракции, молярная масса и размер могут быть определены независимо от положения элюирования. Это особенно важно для веществ с макромолекулами неглобулярной формы, таких как биополимеры (OG1802) или биоконъюгаты; такие вещества, как правило, не элюируются способом, который можно описать набором стандартов для калибровки колонок.
[0222] В некоторых вариантах реализации анализ SEC/MALS включает систему ВЭЖХ Waters с модулем подачи растворителя Alliance 2695 и фотодиодным матричным детектором Waters 2996, оснащенным колонкой SEC-ВЭЖХ Shodex (7,8x300 мм). Ее подключают онлайн к miniDawn TREOS Wyatt и дифференциальному рефрактометру Optilab T-rEX Wyatt. Программное обеспечение Empower от Waters можно использовать для контроля системы ВЭЖХ Waters, а программное обеспечение ASTRA V 6.1.7.16 от Wyatt можно использовать для получения данных MALS от miniDawn TREOS Wyatt, данных dn/dc от детектора T-rEX и данных о степени извлечения по массе с использованием сигнала поглощения A280 от фотодиодного матричного детектора Waters 2996. SEC можно проводить со скоростью 1 мл/мин в 1xФБР pH 7,4, после ввода образца сигналы MALS и RI можно анализировать посредством программного обеспечения ASTRA для определения абсолютной молярной массы (Mp, Mw, Mn) и коэффициента полидисперсности (PDI). В дополнение к этому расчет также включает входные значения dn/dc для полимера и белка, такие как 0,142 и 0,183 соответственно. Для биоконъюгатов значение dn/dc рассчитывают на основе определенной молекулярной массы полимера и белка с использованием формулы ниже, и оно составляет примерно 0,148.
dn/dc конъюгата=0,142 x [MWполимера /(MWполимера+MWбелка)]+0,183 x [MWбелка/(MWполимера+MWбелка)]
[0223] Когда MWполимера для OG1802, измеренная путем SEC-MALS, составляет примерно 800 кДа, и MWбелка для анти-IL-6, измеренная путем SEC-MALS, составляет примерно 145 кДа, ожидаемая общая молекулярная масса биоконъюгата, измеренная путем SEC-MALS, составляет примерно 1000 кДа. MWбелка для конструкции антиIL-6-ловушка VEGF или ловушка VEGF-антиIL-6 составляет примерно 192 кДа, и ожидаемая общая молекулярная масса биоконъюгата составляет 1000-1100 кДа.
[0224] В настоящем документе описаны типичные способы и вещества, хотя при практическом применении или тестировании настоящего изобретения можно также использовать способы и вещества, аналогичные или эквивалентные описанным в настоящем документе. Указанные вещества, способы и примеры приведены только в качестве иллюстрации и не являются ограничивающими.
I. АНТИТЕЛА-АНТАГОНИСТЫ IL-6, КОНСТРУКЦИИ AB IL-6-ЛОВУШКА VEGF И/ИЛИ ИХ КОНЪЮГАТЫ
[0225] Согласно настоящему изобретению предложены анти-IL-6 антитела, которые блокируют, подавляют или снижают (в том числе значительно снижают) биологическую активность IL-6, включая нисходящие (downstream) явления, опосредуемые IL-6. В некоторых вариантах реализации антагонистическое антитело к IL-6 будет содержать одну или более последовательностей CDR, представленных в настоящем описании.
[0226] В некоторых вариантах реализации выделенное антагонистическое антитело специфично связывается с IL-6.
[0227] В некоторых вариантах реализации указанное антитело предпочтительно реагирует с IL-6 таким образом, что ингибирует функцию передачи сигнала IL-6. В некоторых вариантах реализации антагонистическое антитело к IL-6 специфично связывает IL-6 примата.
[0228] Антитела, подходящие для настоящего изобретения, могут включать моноклональные антитела, поликлональные антитела, фрагменты антител (например, Fab, Fab’, F(ab’)2, Fv, Fc и т.д.), химерные антитела, биспецифические антитела, гетероконъюгатные антитела, одноцепочечные фрагменты (ScFv), их мутанты, слитые белки, содержащие часть, представляющую собой антитело (например, доменное антитело), гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена необходимой специфичности, включая гликозилированные варианты антител, варианты аминокислотной последовательности антител и ковалентно модифицированные антитела. Антитела могут представлять собой антитела мыши, крысы, человека или быть любого другого происхождения (включая химерные или гуманизированные антитела). В некоторых вариантах реализации антагонистическое антитело к IL-6 представляет собой моноклональное антитело. В некоторых вариантах реализации антитело представляет собой антитело человека или гуманизированное антитело.
[0229] В некоторых вариантах реализации антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи, представленную в таблицах 1, 2, 6, 7, 8 и/или 9, или на фиг.5. В некоторых вариантах реализации выделенное антагонистическое антитело содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область 1 (CDR1) VH, CDR2 VH и CDR3 VH, имеющую аминокислотную последовательность из представленных в таблицах 1, 2, 6, 7, 8 и/или 9, и вариабельную область легкой цепи (VL), содержащую CDR1 VL, CDR2 VL и CDR3 VL, имеющую аминокислотную последовательность из представленных в таблице.
[0230] В некоторых вариантах реализации предложено выделенное антагонистическое антитело к IL-6. Указанное антитело содержит константный домен тяжелой цепи, содержащий одну или более мутаций для снижения эффекторной функции. В некоторых вариантах реализации указанная одна или более мутаций снижают эффекторные функции антитела, связанные с системой комплемента, например, снижают активацию системы комплемента. В некоторых вариантах реализации снижение эффекторной функции составляет по меньшей мере примерно 50%.
[0231] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, которое специфично связывается с IL-6, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), при этом указанное антитело содержит мутации L234A, L235A и G237A (согласно нумерации EU). В некоторых вариантах реализации выделенное антагонистическое антитело содержит мутации L234A, L235A и G237A. В некоторых вариантах реализации выделенное антагонистическое антитело, содержащее мутации, демонстрирует минимизированное связывание с гамма-рецепторами FC или C1q. В некоторых вариантах реализации выделенное антагонистическое антитело, содержащее мутации L234A, L235A и G237A, демонстрирует минимизированное связывание с гамма-рецепторами FС или C1q. В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, в котором мутация (мутации) расположена в одном или более из следующих положений аминокислот (нумерация EU): E233, L234, L235, G236, G237, A327, A330 и P331. В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, в котором мутация (мутации) выбрана из группы, состоящей из E233P, L234V, L234A, L235A, G237A, A327G, A330S и P331S.
[0232] В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело. Константный домен тяжелой цепи дополнительно содержит остаток цистеина, введенный методом рекомбинантных ДНК. В некоторых вариантах реализации указанный остаток цистеина выбран из группы, состоящей из Q347C и L443C (нумерация EU). В некоторых вариантах реализации остаток цистеина представляет собой L443C (нумерация EU).
[0233] В некоторых вариантах реализации антитело содержит все три следующие мутации (нумерация EU): L234A, L235A и G237A, и антитело содержит L443C (нумерация EU). В некоторых вариантах реализации антитело представляет собой IgG1 человека, и константный домен тяжелой цепи антитела содержит одну или более мутаций, снижающих иммуноопосредованную эффекторную функцию.
[0234] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, связывающее эпитоп на IL-6 человека, который является таким же или перекрывается с эпитопом, распознаваемым антителом, содержащим аминокислотные последовательности в любых одной или более таблицах: 1 и 2 и/или 6-9. В некоторых вариантах реализации предложено антитело к IL-6, содержащее cys и связанное через этот цистеин с полимером (как показано в Формуле 17 в настоящем описании).
[0235] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, которое связывается с IL-6. В некоторых вариантах реализации указанное выделенное антагонистическое антитело, которое связывается с IL-6, содержит тяжелую цепь, содержащую аминокислотную последовательность, представленную в таблицах 1, 2, 6, 7, 8 и/или 9, с С-концевым лизином или без него, и легкую цепь, содержащую аминокислотную последовательность, представленную в таблицах 1, 2, 6, 7, 8 и/или 9.
[0236] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, которое связывается с IL-6. Указанное антитело содержит VH, содержащую аминокислотную последовательность, представленную в таблицах 1, 2, 6, 7, 8 и/или 9, или последовательность, которая по меньшей мере на 90% ей идентична, содержащую замены аминокислот в остатках, не входящих в CDR. В некоторых вариантах реализации антитело содержит одну или более из: HCDR1: на фиг.5, HCDR2: на фиг.5, HCDR3: на фиг.5; LCDR1: на фиг.5, LCDR2: на фиг.5, LCDR3: на фиг.5, например, 1, 2, 3, 4, 5 или все 6 CDR.
[0237] В некоторых вариантах реализации предложено антитело, которое связывается с IL-6, при этом указанное антитело содержит CDRH1, представляющую собой CDRH1 в таблицах 1, 2, 6, 7, 8 и/или 9, CDRH2, представляющую собой CDRH2 в таблицах 1, 2, 6, 7, 8 и/или 9, CDRH3, представляющую собой CDRH3 в таблицах 1, 2, 6, 7, 8 и/или 9, CDRL1, представляющую собой CDRL1 в таблицах 1, 2, 6, 7, 8 и/или 9, CDRL2, представляющую собой CDRL2 в таблицах 1, 2, 6, 7, 8 и/или 9, CDRL3, представляющую собой CDRL3 в таблицах 1, 2, 6, 7, 8 и/или 9; по меньшей мере одну из следующих мутаций: L234A, L235A и G237A согласно нумерации EU и по меньшей мере одну из следующих мутаций: Q347C или L443C согласно нумерации EU.
[0238] В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело. Вариабельная область тяжелой цепи указанного антитела содержит три определяющие комплементарность области (CDR), содержащие аминокислотные последовательности, представленные в таблице 1. В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом вариабельная область легкой цепи указанного антитела содержит три определяющие комплементарность области (CDR), содержащие аминокислотные последовательности, представленные в таблице 2. В некоторых вариантах реализации указанное антитело представляет собой антитело, которое содержит одну или более последовательностей, определенных на фиг.5, например, одну или более CDR (включая 2, 3, 4, 5 или 6 CDR, заключенных в рамку) и/или целые вариабельные области тяжелой и легкой цепи.
[0239] В некоторых вариантах реализации выделенное антагонистическое анти IL-6 антитело содержит вариабельную область тяжелой цепи (VH), которая содержит три CDR, содержащие аминокислотные последовательности, представленные таблице 1, и вариабельную область легкой цепи (VL) антитела, которая содержит три CDR, содержащие аминокислотные последовательности, представленные в таблице 2.
[0240] В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело. VH содержит аминокислотные последовательности, представленные таблице 1, и вариабельная область легкой цепи антитела содержит три CDR, содержащие аминокислотные последовательности, представленные в таблице 2.
[0241] В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом указанное антитело содержит VL, содержащую аминокислотную последовательность, представленную в таблице 2, или ее вариант, содержащий одну замену аминокислоты в аминокислотах, не входящих в CDR. В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом указанное антитело содержит VH, содержащую аминокислотную последовательность, представленную в таблице 1, или ее вариант, содержащий несколько замен аминокислот в аминокислотах, не входящих в CDR.
[0242] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, при этом указанное антитело содержит тяжелую цепь, содержащую аминокислотную последовательность, представленную в таблице 1, с С-концевым лизином, и легкую цепь, содержащую аминокислотную последовательность, представленную в таблице 2. В некоторых вариантах реализации предложено выделенное антагонистическое антитело, при этом указанное антитело содержит тяжелую цепь, содержащую аминокислотную последовательность, представленную в таблице 1, без С-концевого лизина, и легкую цепь, содержащую аминокислотную последовательность, представленную в таблице 2.
[0243] Антитела-антагонисты IL-6 могут быть получены любым способом, известным в этой области техники. Общие способы получения антител человека и мыши известны в этой области техники и/или описаны в настоящем документе.
[0244] Антитела-антагонисты IL-6 могут быть идентифицированы или охарактеризованы с применением способов, известных в этой области техники, посредством которых детектируют и/или измеряют снижение, устранение или нейтрализацию биологической активности IL-6. В некоторых вариантах реализации антагонистическое антитело к IL-6 идентифицируют путем инкубации антитела-кандидата совместно с IL-6 и мониторинга связывания с IL-6R или связывания IL-6/IL-6R с gp130, и/или сопутствующего снижения или нейтрализации биологической активности IL-6. Анализ связывания можно проводить, например, с использованием очищенного полипептида (полипептидов) IL-6 или клеток, естественным образом экспрессирующих (например, различных штаммов) или трансфицированных для экспрессии полипептида (полипептидов) IL-6. В одном из вариантов реализации анализ связывания представляет собой анализ конкурентного связывания, в котором оценивают способность антитела-кандидата конкурировать с известным антителом-антагонистом IL-6 за связывание с IL-6. Указанный анализ можно проводить в различных форматах, включая формат ELISA.
[0245] После первоначальной идентификации активность антитела-антагониста IL-6, являющегося кандидатом, может быть дополнительно подтверждена и уточнена путем биоанализов, известных для тестирования биологической активности-мишени. В некоторых вариантах реализации для получения дополнительных характеристик антитела-антагониста IL-6, являющегося кандидатом, используют клеточный анализ in vitro.
[0246] Характеристики антител-антагонистов IL-6 могут быть получены с применением способов, хорошо известных в этой области техники. Например, один из способов заключается в идентификации эпитопа, с которым оно связывается, или в «картировании эпитопов». Существует много способов картирования и определения расположения эпитопов на белках, известных в этой области техники, включая определение кристаллической структуры комплекса антитело-антиген, конкурентные анализы, анализы экспрессии фрагментов гена и анализы на основе синтетических пептидов, описанные, например, в главе 11 в Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999. В дополнительном примере картирование эпитопов можно использовать для определения последовательности, с которой связывается антагонистическое антитело к IL-6. Картирование эпитопов для антитела-антагониста IL-6 коммерчески доступно из различных источников, например, Pepscan Systems (Edelhertweg 15, 8219 PH Лелистад, Нидерланды). Эпитоп может представлять собой линейный эпитоп, т.е. содержащийся в одном участке аминокислот, или конформационный эпитоп, образованный в результате трехмерного взаимодействия аминокислот, которые могут необязательно содержаться в одном участке. Пептиды различной длины (например, длиной по меньшей мере 4-6 аминокислот) можно выделять или синтезировать (например, рекомбинантно) и использовать для анализов связывания с антителом-антагонистом IL-6. В другом примере эпитоп, с которым связывается антагонистическое антитело к IL-6, может быть определен в систематическом скрининге путем использования перекрывающихся пептидов, полученных из последовательности IL-6, и определения связывания антителом-антагонистом IL-6. В соответствии с анализами экспрессии фрагментов гена открытую рамку считывания, кодирующую IL-6, фрагментируют либо произвольно, либо посредством конкретных генетических конструкций, и определяют реакционную способность экспрессированных фрагментов IL-6 в отношении тестируемого антитела. Фрагменты гена могут быть получены, например, путем ПЦР, а затем транскрибированы и транслированы в белок in vitro в присутствии радиоактивных аминокислот. Затем определяют связывание антитела с радиоактивно мечеными фрагментами IL-6 путем иммунопреципитации и гель-электрофореза. Некоторые эпитопы можно также идентифицировать путем использования больших библиотек случайных пептидных последовательностей, представленных на поверхности фаговых частиц (фаговые библиотеки) или дрожжей (дрожжевой дисплей). В качестве альтернативы, определенную библиотеку перекрывающихся пептидных фрагментов можно тестировать на предмет связывания с тестируемым антителом в простых анализах связывания. В дополнительном примере можно осуществлять мутагенез антигена, эксперименты по обмену доменов и аланин-сканирующий мутагенез для идентификации остатков, требуемых, достаточных и/или необходимых для связывания эпитопа. Например, эксперименты по аланин-сканирующему мутагенезу можно проводить с использованием мутантного IL-6, в котором различные остатки в полипептиде IL-6 заменены на аланин. Путем оценки связывания антитела с мутантным IL-6 можно оценить важное значение конкретных остатков IL-6 для связывания с антителом.
[0247] В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом указанное антитело связывает IL-6 человека с аффинностью от примерно 0,01 пМ до примерно 10 нМ. В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом указанное антитело связывает IL-6 человека с аффинностью от примерно 0,1 пМ до примерно 2 нМ. В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом указанное антитело связывает IL-6 человека с аффинностью, равной примерно 0,01, 0,05, 0,1, 0,5, 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 пМ.
[0248] Аффинность связывания (KD) антитела-антагониста IL-6 в отношении IL-6 может составлять от примерно 0,001 до примерно 200 нМ. В некоторых вариантах реализации аффинность связывания имеет любое значение из: примерно 200 нМ, примерно 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ, примерно 60 пМ, примерно 50 пМ, примерно 20 пМ, примерно 15 пМ, примерно 10 пМ, примерно 5 пМ, примерно 2 пМ или примерно 1 пМ. В некоторых вариантах реализации аффинность связывания меньше любого значения из: примерно 250 нМ, примерно 200 нМ, примерно 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ, примерно 50 пМ, примерно 20 пМ, примерно 10 пМ, примерно 5 пМ, примерно 2 пМ, примерно 1 пМ, примерно 0,5 пМ, примерно 0,1 пМ, примерно 0,05 пМ, примерно 0,01 пМ, примерно 0,005 пМ или примерно 0,001 пМ.
[0249] В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом указанное антитело связывает IL-6 человека с koff, составляющей по меньшей мере 5,0E-03 при 37 градусах. В некоторых вариантах реализации koff составляет 5E-04. В некоторых вариантах реализации предложено выделенное антагонистическое анти IL-6 антитело, при этом указанное антитело связывает IL-6 человека с koff, которая больше 5,0E-04 при 37 градусах.
[0250] В некоторых вариантах реализации аффинность связывания может быть определена в терминах одной или более из: константы ассоциации (ka), константы диссоциации (kd) и концентрации аналита, обеспечивающей полумаксимальную связывающую способность (KD). В некоторых вариантах реализации ka может находиться в диапазоне от примерно 0,50E+05 до примерно 5,00E+08. В некоторых вариантах реализации kd может находиться в диапазоне от примерно 0,50E-06 до примерно 5,00E-03. В некоторых вариантах реализации KD может находиться в диапазоне от примерно 0,50E-12 до примерно 0,50E-07.
[0251] В некоторых вариантах реализации предложена фармацевтическая композиция, содержащая любое из антител, описанных в настоящем документе. В некоторых вариантах реализации предложена фармацевтическая композиция, содержащая любой из конъюгатов, описанных в настоящем документе. В некоторых вариантах реализации указанная фармацевтическая композиция содержит один или более фармацевтически приемлемых носителей. В некоторых вариантах реализации фармацевтическая композиция представляет собой жидкость. В некоторых вариантах реализации фармацевтическая композиция имеет уровень эндотоксинов ниже примерно 0,2 ЕЭ/мл. В некоторых вариантах реализации фармацевтическая композиция представляет собой жидкость и имеет уровень эндотоксинов ниже примерно 0,2 ЕЭ/мл. В некоторых вариантах реализации фармацевтическая композиция представляет собой жидкость и имеет уровень эндотоксинов ниже примерно 2,0, 1, 0,5, 0,2 ЕЭ/мл. В некоторых вариантах реализации, например, при интравитреальной инъекции предел эндотоксинов составляет 0,01-0,02 ЕЭ/инъекция/глаз.
[0252] В некоторых вариантах реализации предложено любое из нижеследующего или композиции (включая фармацевтические композиции), содержащие антитело, содержащее частичную последовательность легкой цепи и частичную последовательность тяжелой цепи, приведенную в таблицах 1 и 2, или их варианты. В таблицах 1 и 2 подчеркнутые последовательности представляют собой некоторые варианты последовательностей CDR, представленных в настоящем описании.
Таблица 1. Последовательности анти-IL-6 вариабельной области тяжелой цепи. CDR подчеркнуты.
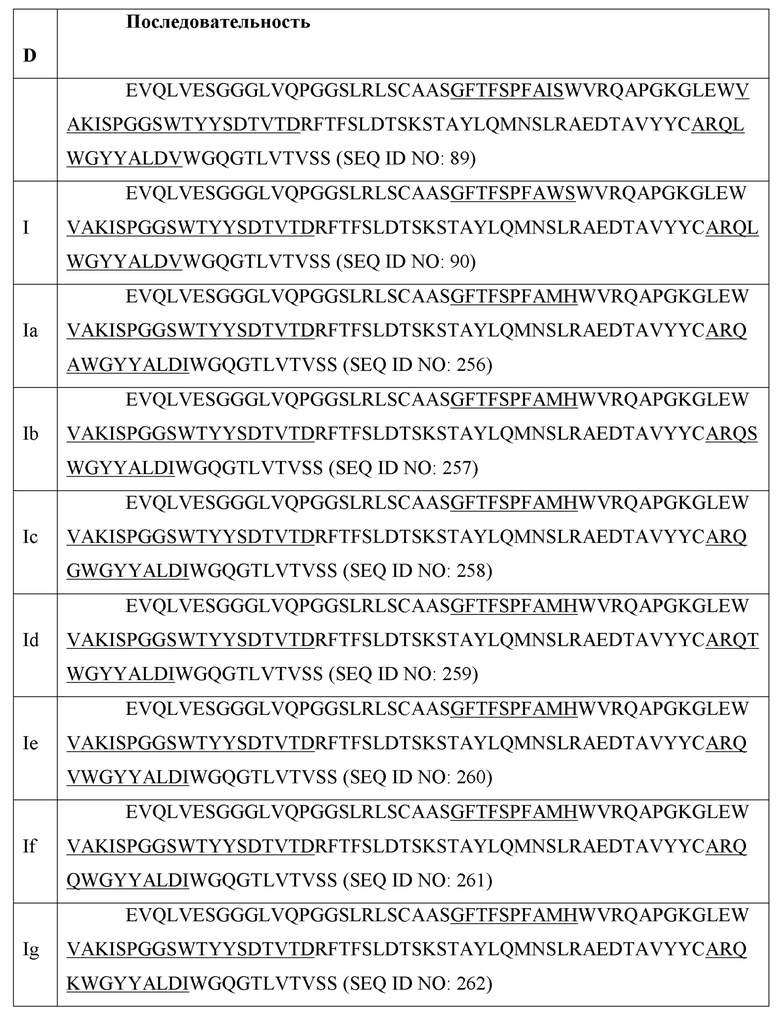
Таблица 2. Последовательности анти-IL-6 вариабельной области легкой цепи. CDR подчеркнуты.
[0253] В некоторых вариантах реализации антитело не содержит одной или более (или никаких) следующих CDR, таблицы 3, 4 и/или 5.
Таблица 3
Таблица 4
Таблица 5
[0254] В некоторых вариантах реализации композиция, описанная в настоящем документе, содержит антитело, содержащее частичную или полную последовательность легкой цепи и частичную или полную последовательность тяжелой цепи из любых предложенных в таблицах 1, 2, 6, 7, 8 и/или 9, или их варианты. В некоторых вариантах реализации антитело (или его связывающий фрагмент) может содержать любую одну или более CDR, представленных в таблицах 1, 2, 6, 7, 8 и/или 9. В некоторых вариантах реализации антитело (или его связывающий фрагмент) может содержать любые три или более CDR, представленных в таблицах 1, 2, 6, 7, 8 и/или 9. В некоторых вариантах реализации антитело (или его связывающий фрагмент) может содержать любые или все шесть CDR, представленных в таблицах 1, 2, 6, 7, 8 и/или 9. В некоторых вариантах реализации тяжелая и/или легкая цепь может представлять собой любую одну или более из других конструкций антител, предложенных согласно настоящему изобретению, включая, например, те, которые представлены на фиг.5, 18 и/или 21-24 и в таблицах 1, 3, 4, 5, 6, 7, 8 и/или 9.
[0255] В некоторых вариантах реализации композиция, описанная в настоящем документе, содержит антитело, содержащее частичную или полную последовательность CDR легкой цепи и частичную или полную последовательность CDR тяжелой цепи из любых вариантов, предложенных в таблицах 1, 2, 6, 7, 8 и/или 9.
[0256] В некоторых вариантах реализации также предложены CDR-части антител-антагонистов IL-6. Определение CDR-областей хорошо известно специалисту в этой области техники. Понятно, что в некоторых вариантах реализации CDR могут представлять собой комбинацию CDR согласно IMGT и Paratome (также называемые «комбинированными CDR» или «расширенными CDR»). Определение CDR хорошо известно специалисту в этой области техники. В некоторых вариантах реализации CDR представляют собой CDR согласно IMGT. В некоторых вариантах реализации CDR представляют собой CDR согласно Paratome. В других вариантах реализации CDR представляют собой расширенные CDR, AbM, конформационные CDR, CDR согласно Kabat или Chotia. В вариантах реализации с более чем одной CDR эти CDR могут представлять собой любую из: CDR согласно IMGT, Paratome, расширенной CDR, CDR согласно Kabat, Chotia, AbM, конформационной CDR или их комбинаций. В некоторых вариантах реализации могут быть также использованы другие определения CDR. В некоторых вариантах реализации используют только остатки, которые являются общими для 2, 3, 4, 5, 6 или 7 определений, приведенных выше (что обуславливает более короткую последовательность). В некоторых вариантах реализации может быть использован любой остаток в любом из 2, 3, 4, 5, 6 или 7 определений, приведенных выше (что обуславливает более длинную последовательность).
[0257] В некоторых вариантах реализации антагонистическое антитело к IL-6 содержит три CDR любой из вариабельных областей тяжелой цепи, представленных в таблицах 1, 2, 6, 7, 8 и/или 9. В некоторых вариантах реализации антитело содержит три CDR любой из вариабельных областей легкой цепи, представленных в таблицах 1, 2, 6, 7, 8 и/или 9. В некоторых вариантах реализации антитело содержит три CDR любой из вариабельных областей тяжелой цепи, представленных в таблице 1, и три CDR любой из вариабельных областей легкой цепи, представленных в таблице 2. В некоторых вариантах реализации CDR представляют собой одну или более указанных в таблицах 6 и/или 7, или 8 и/или 9 ниже:
Таблица 6. ПОСЛЕДОВАТЕЛЬНОСТИ CDR АНТИ-IL-6 ТЯЖЕЛОЙ ЦЕПИ
Таблица 7. ПОСЛЕДОВАТЕЛЬНОСТИ CDR АНТИ-IL-6 ТЯЖЕЛОЙ ЦЕПИ. Kabat
Таблица 8. ПОСЛЕДОВАТЕЛЬНОСТИ CDR АНТИ-IL-6 ЛЕГКОЙ ЦЕПИ
Таблица 9. ПОСЛЕДОВАТЕЛЬНОСТИ CDR АНТИ-IL-6 ЛЕГКОЙ ЦЕПИ
[0258] В некоторых вариантах реализации антитело, применяемое для связывания с IL-6, может представлять собой антитело, содержащее одну или более последовательностей в таблицах 1, 2, 6, 7, 8 и/или 9. В некоторых вариантах реализации антитело, применяемое для связывания с IL-6, может представлять собой антитело, содержащее три или более последовательностей в таблицах 1, 2, 6, 7, 8 и/или 9. В некоторых вариантах реализации антитело, применяемое для связывания с IL-6, может представлять собой антитело, содержащее шесть последовательностей в любой из таблиц 1, 2, 6 и/или 7, 8 и/или 9. В некоторых вариантах реализации антитело, которое связывается с IL-6, может представлять собой антитело, конкурирующее за связывание с антителом, содержащим 6 из CDR, указанных в любой из таблиц 1, 2, 6 и/или 7, 8 и/или 9.
[0259] В некоторых вариантах реализации антитело может быть связано или подвергнуто слиянию с последовательностью ловушки VEGF. В некоторых вариантах реализации эта последовательность ловушки может быть такой, как показано в таблице 10. В некоторых вариантах реализации указанная последовательность по меньшей мере на 80% идентична последовательности, представленной в таблице 10, например, по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% идентична последовательности, представленной в таблице 10. В некоторых вариантах реализации настоящего изобретения может быть использована любая из молекул-ловушек VEGF из публикации США №20150376271. В некоторых вариантах реализации последовательность ловушки VEGF слита с IL-6 одним из следующих способов: с N-концом тяжелой цепи, содержащей IL-6 VH (фиг.6 слева), или между шарнирной областью и после CH1-домена тяжелой цепи, содержащей IL-6 VH (фиг.6 справа). Если не указано иное, в вариантах реализации, представленных в настоящем описании, в которых рассмотрена слитая конструкция АТ-ловушка, предусмотрены оба варианта в качестве альтернативы и вместе. В некоторых вариантах реализации термин «ловушка» относится к полноразмерной внеклеточной области или любой ее части, или комбинации частей из различных рецепторов VEGF, которые могут противодействовать передаче сигнала между по меньшей мере одним VEGF и VEGFR. Предпочтительно, внеклеточный сегмент ловушки содержит по меньшей мере один домен из одного из VEGFR-1, -2 или -3, и более предпочтительно по меньшей мере два смежных домена, таких как D2 и D3. Внеклеточный домен возможно содержит по меньшей мере один домен из по меньшей мере двух различных VEGFR. Предпочтительный внеклеточный домен содержит или по существу состоит из D2 из VEGFR-1 и D3 из VEGFR-2.
Таблица 10. Слитая последовательность домена 2 VEGFR1 и домена 3 VEGFR2
SDTGRPFVEMYSEIPEIIHMTEGRELVIPCRVTSPNITVTLKKFPLDTLIPDGKRIIWDSRKGFIISNATYKEIGLLTCEATVNGHLYKTNYLTHRQTNTIIDVVLSPSHGIELSVGEKLVLNCTARTELNVGIDFNWEYPSSKHQHKKLVNRDLKTQSGSEMKKFLSTLTIDGVTRSDQGLYTCAASSGLMTKKNSTFVRVHEK (SEQ ID NO: 114)
[0260] В некоторых вариантах реализации конструкция АТ к IL-6-ловушка VEGF может содержать любую из последовательностей, представленных в таблице 11. В некоторых вариантах реализации указанная конструкция может быть по меньшей мере идентична последовательностям в таблице 11, например, на 80, 85, 90, 95, 96, 97, 98, 99% или больше. В некоторых вариантах реализации слитый белок может соответствовать этим процентам за исключением того, что домен антитела к IL-6 не содержит одной или более CDR в таблицах 3, 4 и/или 5. В некоторых вариантах реализации слитый белок представляет собой слитый белок, который содержит одну или более последовательностей, определенных на фиг.5, например, одну или более CDR (включая 2, 3, 4, 5 или 6 CDR, заключенных в рамку) и/или целые вариабельные области тяжелой и легкой цепи совместно с последовательностью ловушки VEGF (например, таблица 10). В некоторых вариантах реализации указанные последовательности могут быть непосредственно подвергнуты слиянию друг с другом. В некоторых вариантах реализации может быть использована одна или более гибких связывающих последовательностей или участков. Связывающая последовательность может быть расположена между последовательностью АТ и последовательностью ловушки VEGF. Длина этих последовательностей может составлять от 5 до 30 аминокислот. В некоторых вариантах реализации связывающая последовательность может содержать G и S в соотношении примерно 4:1. В некоторых вариантах реализации линкер содержит следующую последовательность: GGGGSGGGGS (SEQ ID NO: 115). В некоторых вариантах реализации может быть использован любой гибкий линкер. В некоторых вариантах реализации Fc-часть АТ к Il-6 представляет собой IgG1.
Таблица 11. Последовательности тяжелой и легкой цепи для молекул двойного ингибитора. CDR в тяжелой и легкой цепях подчеркнуты, последовательность ловушки VEGF выделена черным жирным шрифтом, линкер Gly-Ser выделен курсивом
[0261] Для экспрессии анти-IL-6 антител и/или конструкций IL-6-ловушка VEGF, предложенных согласно настоящему изобретению, сначала могут быть получены фрагменты ДНК, кодирующие описанные VH- и VL-области. В последовательности ДНК могут быть также введены различные модификации, например, мутации, делеции и/или присоединения с применением стандартных способов, известных специалисту в этой области техники. Например, мутагенез можно осуществлять с применением стандартных способов, таких как ПЦР-опосредованный мутагенез, в котором мутированные нуклеотиды включают в праймеры для ПЦР, в результате чего продукт ПЦР содержит желаемые мутации, или сайт-направленный мутагенез.
[0262] Настоящее изобретение включает модификации вариабельных областей, представленных в настоящем описании. Например, настоящее изобретение включает антитела, содержащие функционально эквивалентные вариабельные области и CDR, которые не влияют в значительной степени на их свойства, а также варианты, обладающие повышенной или пониженной активностью и/или аффинностью. Например, аминокислотная последовательность может быть мутирована с получением антитела, обладающего желаемой аффинностью связывания в отношении IL-6. В этой области техники модификация полипептидов является рутинной практикой и нет необходимости в ее подробном описании в настоящем документе. Примеры модифицированных полипептидов включают полипептиды, содержащие консервативные замены аминокислотных остатков, одну или более делеций, или присоединений аминокислот, которые не оказывают значительного отрицательного влияния и не изменяют функциональную активность или которые приводят к созреванию (повышению) аффинности полипептида в отношении своего лиганда, или использование химических аналогов.
[0263] Встраивания аминокислотных последовательностей включают амино- и/или карбоксиконцевые слияния, длина которых находится в диапазоне от одного остатка до полипептидов, содержащих сто или более остатков, а также встраивания одного или нескольких аминокислотных остатков внутри последовательности. Примеры концевого встраивания включают антитело с N-концевым метионильным остатком или антитело, подвергнутое слиянию с эпитопной меткой. Другие варианты встраивания для молекулы антитела включают слияние с N- или С-концом антитела фермента или полипептида, увеличивающего период полувыведения антитела из кровообращения.
[0264] У вариантов, содержащих замену, по меньшей мере один аминокислотный остаток в молекуле антитела удален и на его место встроен другой остаток. Сайты, представляющие наибольший интерес для мутагенеза с заменой, включают гипервариабельные области, но также предусмотрены изменения в каркасной области. Консервативные замены показаны в таблице 12 под заголовком «консервативные замены». Если такие замены приводят к изменению биологической активности, то могут быть внесены более значительные изменения, называемые «типичными заменами» в таблице 12 или дополнительно описанные ниже применительно к классам аминокислот, и проведен скрининг продуктов.
Таблица 12. Замены аминокислот
[0265] Значительные модификации биологических свойств антитела осуществляют путем выбора замен, которые значительно отличаются своим влиянием на сохранение (а) структуры полипептидного остова в области замены, например, в виде β-складчатого слоя или спиральной конформации, (b) заряда или гидрофобности молекулы в сайте-мишени или (с) объема боковой цепи. Встречающиеся в природе остатки делятся на группы на основе общих свойств боковой цепи:
[0266] (1) Неполярные: норлейцин, Met, Ala, Val, Leu, Ile;
[0267] (2) Полярные незаряженные: Cys, Ser, Thr, Asn, Gln;
[0268] (3) Кислотные (отрицательно заряженные): Asp, Glu;
[0269] (4) Основные (положительно заряженные): Lys, Arg;
[0270] (5) Остатки, влияющие на ориентацию цепи: Gly, Pro; и
[0271] (6) Ароматические: Trp, Tyr, Phe, His.
[0272] Неконсервативные замены осуществляют путем замены представителя одного из этих классов на представитель из другого класса.
[0273] Один из типов замены, например, который может быть произведен, заключается в замене одного или более остатков цистеина в антителе, которые могут быть химически активными, на другой остаток, такой как, без ограничения, аланин или серин. Например, может быть осуществлена замена неканонического цистеина. Замена может быть осуществлена в CDR или каркасной области вариабельного домена или в константной области антитела. В некоторых вариантах реализации цистеин является каноническим. Любой остаток цистеина, не вовлеченный в сохранение подходящей конформации антитела, также может быть заменен, как правило, на серин для повышения окислительной стабильности молекулы и предотвращения нарушения перекрестного сшивания. В антитело может быть, напротив, введена цистеиновая связь (связи) для повышения его стабильности, в частности, когда указанное антитело представляет собой фрагмент антитела, такой как Fv-фрагмент.
[0274] Антитела могут быть также модифицированы, например, в вариабельных доменах тяжелых и/или легких цепей, например, для изменения свойства связывания антитела. Изменения в вариабельной области могут изменять аффинность и/или специфичность связывания. В некоторых вариантах реализации в пределах CDR-домена осуществляют не более одной-пяти консервативных замен аминокислот.В других вариантах реализации в пределах CDR-домена осуществляют не более одной-трех консервативных замен аминокислот.Например, мутация может быть произведена в одной или более CDR-областях для увеличения или уменьшения KD антитела для IL-6, для увеличения или уменьшения koff или для изменения специфичности связывания антитела. Методики сайт-направленного мутагенеза хорошо известны в этой области техники. См., например, Sambrook et al. and Ausubel et al., выше.
[0275] В соответствии с одним из аспектов IgG-домен антитела-антагониста IL-6 может представлять собой IgG1, IgG2, IgG3 или IgG4. В соответствии с другим аспектом IgG-домен может представлять собой составной домен, в котором константные области образованы из более чем одного из вышеуказанных изотипов (например, CH1-область из IgG2 или IgG4, шарнирная область, CH2- и CH3-области из IgG1). В этой области техники известно, что при выборе изотипа изотипы IgGl и IgG3 человека обладают комплемент-опосредованной цитотоксичностью, тогда как изотипы IgG2 и IgG4 человека обладают слабой комплемент-опосредованной цитотоксичностью или не обладают ею. В некоторых вариантах реализации изотип антитела-антагониста IL-6 представляет собой IgG1.
[0276] Константная область легкой цепи может относиться либо к лямбда, либо к каппа человека. В некоторых вариантах реализации антагонистическое антитело к IL-6 содержит константную область легкой каппа-цепи человека.
[0277] Константные области человека демонстрируют аллотипическую вариацию и изоаллотипическую вариацию у разных индивидуумов, т.е. константные области могут различаться у разных индивидуумов по одному или более полиморфным положениям. Изоаллотипы отличаются от аллотипов тем, что белки сыворотки, распознающие изоаллотип, связываются с неполиморфной областью одного или более других изотипов. Ссылка на константную область человека включает константную область с любым природным аллотипом или любой перестановкой остатков, занимающих полиморфные положения в природных аллотипах, или содержащую до 3, 5 или 10 замен для снижения или повышения эффекторной функции, как описано ниже.
[0278] Одна или несколько аминокислот на амино- или карбоксиконце легких и/или тяжелых цепей, такая как С-концевой лизин тяжелой цепи, может отсутствовать или быть дериватизирована в части или во всех молекулах.
[0279] Замены могут быть осуществлены в константных областях для снижения или повышения эффекторной функции, такой как комплемент-опосредованная цитотоксичность (КОЦ), антителозависимая клеточноопосредованная цитотоксичность (АЗКЦ) (см., например, Winter et al., патент США №5624821; Tso et al., патент США №5834597; и Lazar et al., Proc. Natl. Acad. Sci. USA 103:4005, 2006), или для увеличения периода полувыведения у людей (см., например, Hinton et al., J. Biol. Chem. 279:6213, 2004).
[0280] В некоторых вариантах реализации антитела-антагонисты IL-6, предложенные согласно настоящему изобретению, содержат одну или более замен, снижающих комплемент-опосредованную цитотоксичность. Снижение комплемент-опосредованной цитотоксичности может быть достигнуто с уменьшением связывания Fc-рецептора или без него в зависимости от природы мутации (мутаций). Антитела со сниженной комплемент-опосредованной цитотоксичностью, но с небольшим уменьшением или отсутствием уменьшения связывания Fc-рецептора, обеспечивают желаемый эффект Fc-опосредованного фагоцитоза iC3b без активации комплемента, которая может вносить вклад в побочные эффекты. Типичные мутации в константных областях человека, которые, как известно, снижают комплемент-опосредованную цитотоксичность, включают мутации в положениях 241, 264, 265, 270, 296, 297, 322, 329 и 331 согласно нумерации EU. Существуют данные о том, что мутации в положениях 318, 320 и 322 в антителах мыши уменьшают активацию комплемента. Предпочтительным остатком для того, чтобы занять эти положения в мутированной константной области является аланин. Некоторые типичные мутации антитела человека, которые использовали, включают F241A, V264A, D265A, V296A, N297A, K322A и P331S в IgG3 человека и D270A или E, N297Q, K322A, P329A и P331S в IgGl человека (нумерация EU).
[0281] Здесь, как и в других местах настоящего описания, для нумерации аминокислот в константной области антитела используют схему нумерации EU. Когда в настоящем описании приведена ссылка на остаток в вариабельной области (если не указано иное), нумерация остатков соответствует вариабельному домену (или, если указано, SEQ ID NO). Замена в любом или во всех положениях 234, 235, 236 и/или 237 снижает аффинность к рецепторам Fcγ, в частности, рецептору FcγRI, а также уменьшает связывание и активацию комплемента (см., например, US 6624821 WO/2009/052439). Замена на аланин в положениях 234, 235 и 237 снижает эффекторные функции, в частности, в контексте IgGl человека. Возможно, положения 234, 236 и/или 237 в IgG2 человека заменяют на аланин, а положение 235 - на глутамин (см., например, US 5624821) для уменьшения связывания с Fc-рецептором. Типичные замены для увеличения периода полувыведения включают Gln в положении 250 и/или Leu в положении 428. В соответствии с одним из аспектов настоящего изобретения, когда представленное анти-IL6 антитело имеет изотип IgG1 человека, предпочтительно, чтобы указанное антитело содержало по меньшей мере одну мутацию в константной области. Предпочтительно, указанная мутация уменьшает фиксацию или активацию комплемента константной областью. В соответствии с наиболее предпочтительными аспектами настоящего изобретения антитело содержит одну или более мутаций в положениях E233, L234, L235, G236, G237, A327, A330 и P331 согласно нумерации EU. Более предпочтительно, указанные мутации представляют собой одну или более из следующих: E233P, L234V, L234A, L235A, G237A, A327G, A330S и P331S согласно нумерации EU. В самых предпочтительных вариантах реализации IgG1 человека содержит следующие мутации: L234A, L235A и G237A согласно нумерации EU.
Конъюгаты
[0282] Период полувыведения антител-антагонистов IL-6 и/или конструкций АТ к IL-6-ловушка VEGF может быть увеличен путем присоединения «фрагментов, увеличивающих период полувыведения» или «групп, увеличивающих период полувыведения»; указанные термины являются взаимозаменяемыми в настоящем описании и относятся к одной или более химическим группам, присоединенным к одной или более функциональным группам боковой цепи аминокислот, таким как -SH, -OH, -COOH, -CONH2, -NH2, или одной или более структурам N- и/или O-гликанов, и которые могут увеличивать период полувыведения белков/пептидов из крови in vivo при конъюгировании с этими белками/пептидами. Примеры фрагментов, увеличивающих период полувыведения, включают полимеры, описанные в настоящем документе, в частности, полимеры из цвиттерионных мономеров, например, HEMA-фосфорилхолин, ПЭГ, биосовместимые жирные кислоты и их производные, гидроксиалкилкрахмал (HAS), например, гидроксиэтилкрахмал (HES), полиэтиленгликоль (ПЭГ), поли(Glyx-Sery) (HAP), гиалуроновую кислоту (HA), гепарозановые полимеры (HEP), флексимеры, декстран, полисиаловые кислоты (PSA), Fc-домены, трансферрин, 25 альбумин, эластиноподобные пептиды (ELP), полимеры XTEN, полимеры пролин-аланин-серин (ПАС), полиамидные (ПА) полимеры, связывающие альбумин пептиды, пептиды CTP, FcRn-связывающие пептиды и любую их комбинацию.
[0283] В некоторых вариантах реализации антитело конъюгировано с полимером, содержащим фосфорилхолин. В некоторых вариантах реализации антитело конъюгировано с полимером, содержащим поли(акрилоилоксиэтилфосфорилхолин), таким как полимер акриловой кислоты, содержащий по меньшей мере один мономер акрилоилоксиэтилфосфорилхолина, такой как 2-метакрилоилоксиэтилфосфорилхолин (т.е. 2-метакрилоил-2'-триметиламмоний-этилфосфат).
[0284] В некоторых вариантах реализации антитело и/или слитая конструкция антитело-ловушка VEGF конъюгирована с водорастворимым полимером, который относится к полимеру, растворимому в воде. Раствор водорастворимого полимера может пропускать по меньшей мере примерно 75%, более предпочтительно по меньшей мере примерно 95% света, пропускаемого этим же раствором после фильтрации. По массе водорастворимый полимер или его сегмент может быть по меньшей мере примерно на 35%, по меньшей мере примерно на 50%, примерно на 70%, примерно на 85%, примерно на 95% или 100% (по массе сухого полимера) растворимым в воде.
[0285] В одном из вариантов реализации фрагмент, увеличивающий период полувыведения, может быть конъюгирован с антителами-антагонистами IL-6 и/или конструкцией АТ к IL-6-ловушка VEGF через свободные аминогруппы белка с использованием сложных эфиров N-гидроксисукцинимида (NHS). Реагенты, направленные на конъюгацию с аминогруппами, могут случайным образом реагировать с ϵ-аминогруппой остатков лизина, α-аминогруппой N-концевых аминокислот и δ-аминогруппой остатков гистидина.
[0286] Однако антитела-антагонисты IL-6 согласно настоящему изобретению содержат много аминогрупп, доступных для конъюгации с полимером. Таким образом, конъюгация полимеров со свободными аминогруппами может отрицательно влиять на способность антитела связываться с эпитопом.
[0287] В другом варианте реализации фрагмент, увеличивающий период полувыведения, подвергают реакции сочетания с одной или более свободными SH-группам с использованием любой подходящей тиол-активной химии, включая, без ограничения, химию малеимида или сочетание полимерных гидразидов или полимерных аминов с углеводными фрагментами антител-антагонистов IL-6 и/или конструкций АТ к IL-6-ловушка VEGF после предварительного окисления. Применение сочетания с использованием малеимида представляет собой наиболее предпочтительный вариант реализации настоящего изобретения. Сочетание предпочтительно происходит по остаткам цистеина, присутствующим естественным образом или введенным посредством генной инженерии.
[0288] В некоторых вариантах реализации полимеры ковалентно присоединяют к остаткам цистеина, введенным в антитела-антагонисты IL-6 и/или конструкции АТ к IL-6-ловушка VEGF путем сайт-направленного мутагенеза. В некоторых вариантах реализации можно использовать остатки цистеина в Fc-части антитела-антагониста IL-6 и/или конструкции АТ к IL-6-ловушка VEGF. В некоторых вариантах реализации сайты для введения остатков цистеина в Fc-область представлены в WO 2013/093809, US 7521541, WO 2008/020827, US 8008453, US 8455622 и US2012/0213705, содержание которых включено в настоящее описание посредством ссылки для всех целей. В некоторых вариантах реализации мутации цистеина представляют собой Q347C и L443C, относящиеся к тяжелой цепи IgG человека согласно EU-индексу Kabat. В некоторых вариантах реализации цистеин, присоединенный путем направленного мутагенеза для последующего присоединения полимера, представляет собой L443C. В некоторых вариантах реализации стехиометрия антитела-антагониста IL-6 и полимера составляет 1:1; другими словами, конъюгат состоит по существу из молекул, каждая из которых содержит одну молекулу антитела-антагониста IL-6 и/или конструкции АТ к IL-6-ловушка VEGF, конъюгированную с одной молекулой полимера. В некоторых вариантах реализации сочетание может происходить по одному или более остаткам лизина.
[0289] В некоторых вариантах реализации конъюгат содержит выделенное антагонистическое антитело, которое специфично связывается с IL-6, или конструкцию АТ к IL-6-ловушка VEGF, конъюгированную с полимером. В некоторых вариантах реализации полимер содержит цвиттерионный мономер. В некоторых вариантах реализации указанный цвиттерионный мономер без ограничений представляет собой HEMA-фосфорилхолин, ПЭГ, биосовместимые жирные кислоты и их производные, гидроксиалкилкрахмал (HAS), например, гидроксиэтилкрахмал (HES), полиэтиленгликоль (ПЭГ), поли(Glyx-Sery) (HAP), гиалуроновую кислоту (HA), гепарозановые полимеры (HEP), флексимеры, декстран, полисиаловые кислоты (PSA), Fc-домены, трансферрин, 25 альбумин, эластиноподобные пептиды (ELP), полимеры XTEN, полимеры ПАС, ПА-полимеры, связывающие альбумин пептиды, пептиды CTP или FcRn-связывающие пептиды. В некоторых вариантах реализации полимер, содержащий цвиттерионный мономер, представляет собой фрагмент, увеличивающий период полувыведения
[0290] В некоторых вариантах реализации антагонистическое антитело к IL-6 и/или конструкция АТ к IL-6-ловушка VEGF может содержать присоединенный фрагмент, увеличивающий период полувыведения. В некоторых вариантах реализации фрагмент, увеличивающий период полувыведения, представляет собой цвиттерионный полимер, но в качестве альтернативы можно использовать ПЭГ или другие агенты, увеличивающие период полувыведения, рассмотренные ниже. В некоторых вариантах реализации цвиттерионный полимер образован мономерами, содержащими фосфорилхолиновую группу. В некоторых вариантах реализации мономер представляет собой 2-(акрилоилоксиэтил)-2’-(триметиламмонийметил)фосфат. В некоторых вариантах реализации мономер представляет собой 2-(метакрилоилоксиэтил)-2’-(триметиламмонийметил)фосфат (HEMA-PC).
[0291] В некоторых вариантах реализации полимер, конъюгированный с антителом-антагонистом IL-6 и/или конструкцией АТ к IL-6-ловушка VEGF, содержит по меньшей мере 2 или 3, или более ответвлений. Некоторые полимеры содержат 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 ответвлений. В некоторых вариантах реализации полимер содержит 3, 6 или 9 ответвлений. В некоторых вариантах реализации полимер содержит 9 ответвлений. В некоторых вариантах реализации пиковая молекулярная масса полимера составляет от 300000 до 1750000 Да. В некоторых вариантах реализации полимер имеет пиковую молекулярную массу от 500000 до 1000000 Да. В некоторых вариантах реализации полимер имеет пиковую молекулярную массу от 600000 до 800000 Да.
[0292] В некоторых вариантах реализации предложен конъюгат антагонистического анти-IL-6 антитела и/или конструкции АТ к IL-6-ловушка VEGF и полимера. В некоторых вариантах реализации полимер имеет пиковую молекулярную массу от 300000 до 1750000 Дальтон, измеренную путем эксклюзионной хроматографии с многоугловым светорассеянием (далее в настоящем описании «SEC-MALS»). В некоторых вариантах реализации полимер имеет пиковую молекулярную массу от 500000 до 1000000 Дальтон, измеренную путем SEC-MALS. В некоторых вариантах реализации полимер имеет пиковую молекулярную массу от 600000 до 800000 Дальтон, измеренную путем SEC-MALS.
[0293] В некоторых вариантах реализации фрагмент, увеличивающий период полувыведения, может быть конъюгирован со встречающимся в природе остатком цистеина антитела-антагониста IL-6 и/или конструкции АТ к IL-6-ловушка VEGF, предложенной согласно настоящему изобретению. В некоторых вариантах реализации фрагмент, увеличивающий период полувыведения, конъюгирован с цистеином, присоединенным путем сайт-направленного мутагенеза. В некоторых вариантах реализации цистеин присоединяют путем применения метода рекомбинантных ДНК для присоединения пептида SGGGC или CAA к С-концу либо легкой, либо тяжелой цепи. В некоторых вариантах реализации пептид присоединяют к тяжелой цепи. В некоторых вариантах реализации остаток цистеина, введенный методом рекомбинантных ДНК, выбран из группы, состоящей из (нумерация EU) Q347C и L443C.
[0294] В соответствии с другим аспектом настоящего изобретения представлена фармацевтическая композиция, содержащая антагонистическое антитело к IL-6 и/или конструкцию АТ к IL-6-ловушка VEGF, и фармацевтически приемлемое вспомогательное вещество.
[0295] Антитела-антагонисты IL-6 и/или конструкции АТ к IL-6-ловушка VEGF (или другие конструкции, предложенные согласно настоящему изобретению) могут быть получены путем рекомбинантной экспрессии, включающей (i) получение рекомбинантной ДНК посредством генной инженерии, (ii) введение рекомбинантной ДНК в прокариотические или эукариотические клетки, например и без ограничения, путем трансфекции, электропорации или микроинъекции, (iii) культивирование трансформированных клеток, (iv) экспрессию анти-IL-6 антител, например, конститутивную или при индукции и (v) выделение анти-IL-6 антитела, например, из культуральной среды или путем сбора трансформированных клеток с (vi) получением очищенного анти-IL-6 антитела.
[0296] В некоторых вариантах реализации предложен конъюгат, содержащий анти-IL6 антитело и/или конструкцию АТ к IL-6-ловушка VEGF и полимер (а также другие конструкции, предложенные согласно настоящему изобретению, такие как биоконъюгат). Антитело и/или конструкция АТ-ловушка представляет собой любое из антител и/или ловушек, описанных в настоящем документе. В некоторых вариантах реализации антитело представляет собой IgG. В некоторых вариантах реализации антитело представляет собой IgA, IgE, IgD или IgM. В некоторых вариантах реализации полимер ковалентно связан с сульфгидрильной группой. В некоторых вариантах реализации полимер ковалентно связан с сульфгидрильной группой остатка цистеина. В некоторых вариантах реализации полимер ковалентно связан с сульфгидрильной группой остатка цистеина в тяжелой цепи. В некоторых вариантах реализации полимер ковалентно связан с сульфгидрильной группой остатка цистеина в тяжелой цепи IgG. В некоторых вариантах реализации антитело содержит остаток цистеина в положении 347 или 443 (нумерация EU). В некоторых вариантах реализации полимер ковалентно связан с сульфгидрильной группой остатка цистеина в положении 347 или 443 (нумерация EU).
[0297] Антитела-антагонисты IL-6 и/или конструкция АТ к IL-6-ловушка VEGF могут быть получены путем экспрессии в подходящей прокариотической или эукариотической системе-хозяине, характеризующейся продуцированием фармакологически приемлемой молекулы анти-IL-6 антитела. Примерами эукариотических клеток являются клетки млекопитающих, такие как CHO, COS, HEK 293, BHK, SK-Hip и HepG2. Другие подходящие системы экспрессии представляют собой прокариотические клетки (например, E.coli с системой экспрессии pET/BL21), клетки дрожжей (системы Saccharomyces cerevisiae и/или Pichia pastoris) и клетки насекомых. В некоторых вариантах реализации предложена выделенная линия клеток, продуцирующая любое из антител и/или конструкций АТ-ловушка, описанных в настоящем документе. В некоторых вариантах реализации указанная выделенная линия клеток выбрана, без ограничений, из одной или более из: CHO, k1SV, XCeed, CHOK1SV, GS-KO.
[0298] В некоторых вариантах реализации предложена выделенная нуклеиновая кислота, кодирующая любое из антител и/или конструкций АТ-ловушка, описанных в настоящем документе. В некоторых вариантах реализации предложен рекомбинантный вектор экспрессии, содержащий выделенную нуклеиновую кислоту. В некоторых вариантах реализации клетка-хозяин содержит вектор экспрессии.
[0299] Для получения антител-антагонистов IL-6 и/или конструкций АТ к IL-6-ловушка VEGF можно использовать широкий спектр векторов и они могут быть выбраны из векторов эукариотической и прокариотической экспрессии. Примеры векторов для прокариотической экспрессии включают плазмиды, такие как, без ограничения, preset, pet и pad, при этом промоторы, используемые в векторах прокариотической экспрессии, включают один или более из, без ограничения, lac, trc, trp, recA или araBAD. Примеры векторов для эукариотической экспрессии включают без ограничения: (i) для экспрессии в дрожжах: векторы, такие как и без ограничения, pAO, pPIC, pYES или pMET, с использованием промоторов, таких как и без ограничения, AOX1, GAP, GAL1 или AUG1; (ii) для экспрессии в клетках насекомых: векторы, такие как и без ограничения, pMT, pAc5, pIB, pMIB или pBAC, с использованием промоторов, таких как и без ограничения, PH, p10, MT, Ac5, OpIE2, gp64 или polh, и (iii) для экспрессии в клетках млекопитающих: векторы, такие как и без ограничения, pSVL, pCMV, pRc/RSV, pcDNA3 или pBPV, и векторы, полученные в соответствии с одним из аспектов из вирусных систем, таких как и без ограничения, вирус коровьей оспы, аденоассоциированные вирусы, вирусы герпеса или ретровирусы, с использованием промоторов, таких как и без ограничения, CMV, SV40, EF-1, UbC, RSV, ADV, BPV и бета-актин.
[0300] В некоторых вариантах реализации предложен способ получения антитела-антагониста IL-6 и/или конструкции АТ к IL-6-ловушка VEGF. В некоторых вариантах реализации указанный способ включает культивирование линии клеток, рекомбинантно продуцирующей любое из антител и/или конструкций АТ-ловушка, описанных в настоящем документе, в условиях, в которых антитело продуцируется и его извлекают.В некоторых вариантах реализации предложен способ получения антитела-антагониста IL-6 и/или конструкции АТ к IL-6-ловушка VEGF. В некоторых вариантах реализации указанный способ включает культивирование линии клеток, содержащей нуклеиновую кислоту, кодирующую антитело и/или конструкцию АТ-ловушка, содержащую тяжелую цепь, содержащую аминокислотную последовательность, представленную в таблице 1, и легкую цепь, содержащую аминокислотную последовательность, представленную в таблице 2, в условиях, в которых антитело и/или конструкция АТ к IL-6-ловушка VEGF продуцируется и ее извлекают.В некоторых вариантах реализации тяжелая и легкая цепи антитела и/или конструкции АТ к IL-6-ловушка VEGF кодируются отдельными векторами. В некоторых вариантах реализации тяжелая и легкая цепи антитела и/или конструкции АТ к IL-6-ловушка VEGF кодируются одним вектором.
[0301] В соответствии с другим аспектом настоящего изобретения предложены способы синтеза конъюгатов цвиттерионный полимер-антагонистическое антитело к IL-6 и/или АТ к IL-6-ловушка VEGF, при этом указанный конъюгат содержит один или более функциональных агентов и одно или более ответвлений полимера, при этом каждое из ответвлений полимера содержит одно или более мономерных звеньев, при этом по меньшей мере одно из указанных звеньев содержит цвиттерион. Например, такой способ может включать этапы:
a. получения инициатора, содержащего один или более центров полимеризации мономера и первый линкер, содержащий аминогруппу, при этом указанный инициатор представляет собой соль трифторуксусной кислоты;
b. получения одного или более мономеров, подходящих для полимеризации, при этом по меньшей мере один из указанных мономеров является цвиттерионным;
c. проведения реакции мономеров с инициатором с образованием одного или более ответвлений полимера, каждое из которых соответствует центрам полимеризации мономера, с получением конъюгата инициатор-полимер, содержащего первый линкер с аминогруппой;
d. получения второго линкера, содержащего по меньшей мере вторую и третью реакционноспособные группы;
e. сочетания одной из указанных второй и третьей реакционноспособных групп второго линкера с аминогруппой первого линкера конъюгата инициатор-полимер с получением конъюгата линкер-инициатор-полимер, содержащего одну или более реакционноспособные групп, не использованных на этапе сочетания; и
f. сочетания одного или более функциональных агентов с одной или более непрореагировавшими реакционноспособными группами конъюгата линкер-инициатор-полимер с получением конъюгата полимер-функциональный агент.
[0302] В некоторых вариантах реализации предложен конъюгат, содержащий выделенное антагонистическое антитело и/или конструкцию АТ к IL-6-ловушка VEGF, специфично связывающуюся с IL-6, и полимер, содержащий фосфорилхолин. В некоторых вариантах реализации указанный полимер ковалентно связан с антителом и/или конструкцией АТ-ловушка. В некоторых вариантах реализации полимер нековалентно связан с антителом и/или конструкцией АТ-ловушка.
[0303] В некоторых вариантах реализации предложен конъюгат, содержащий анти-IL6 антитело и/или конструкцию АТ к IL-6-ловушка VEGF и полимер, содержащий фосфорилхолин. В некоторых вариантах реализации указанный полимер ковалентно связан с антителом за пределами вариабельной области антитела. В некоторых вариантах реализации предложен конъюгат, содержащий анти-IL6 антитело и/или конструкцию АТ к IL-6-ловушка VEGF и полимер, содержащий фосфорилхолин. Указанный полимер ковалентно связан с антителом в остатке цистеина за пределами вариабельной области антитела. В некоторых вариантах реализации предложен конъюгат, содержащий анти-IL6 антитело и/или конструкцию АТ к IL-6-ловушка VEGF и полимер, содержащий фосфорилхолин, при этом указанный полимер ковалентно связан с антителом в остатке цистеина за пределами вариабельной области антитела, при этом указанный цистеин был присоединен методом рекомбинантных ДНК. В некоторых вариантах конъюгата полимер содержит мономеры, представляющие собой 2(метакрилоилокси)этил (2-(триметиламмоний)этил)фосфат (MPC).
[0304] В соответствии с другим аспектом настоящего изобретения предложен способ, в котором после синтеза полимера присоединяют группу для конъюгации (например, малеимид). Его иногда называют «стратегией пристегивания (snap-on strategy)» или «универсальной стратегией получения полимера». См., например, заявку на патент США №14/916180 (опубликованную в виде публикации заявки на патент США №20160199501), полное содержание которой включено в настоящее описание посредством ссылки. В некоторых вариантах реализации один инициаторный фрагмент можно использовать для крупномасштабного синтеза полимера. Таким образом, могут быть разработаны условия для увеличения масштаба оптимального синтеза полимера. Такие полимеры можно затем адаптировать к различным типам функциональных агентов путем «пристегивания» различных типов линкеров. Например, если необходимо конъюгировать более крупный функциональный агент с полимером согласно настоящему изобретению, такой как антитело или даже Fab-фрагмент, к указанному полимеру может быть «пристегнута» более длинная линкерная последовательность. Для более мелких функциональных агентов, напротив, могут быть необходимы относительно более короткие линкерные последовательности.
[0305] В некоторых вариантах способов инициатор содержит примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 центров инициации синтеза полимера. В некоторых вариантах реализации инициатор содержит примерно 3, примерно 6 или примерно 9 центров инициации синтеза полимера.
[0306] В соответствии с другим аспектом настоящего изобретения второй линкер содержит вторую, третью, четвертую, пятую и шестую реакционноспособные группы. Более предпочтительно, второй линкер содержит только вторую и третью реакционноспособные группы.
[0307] В соответствии с одним из аспектов настоящего изобретения каждое ответвление полимера содержит от примерно 20 до примерно 2000 мономерных звеньев. Предпочтительно, каждое ответвление содержит от примерно 100 до 500 мономерных звеньев или от примерно 500 до 1000 мономерных звеньев, или от примерно 1000 до 1500 мономерных звеньев, или от примерно 1500 до 2000 мономерных звеньев.
[0308] В соответствии с одним из аспектов настоящего изобретения пиковая молекулярная масса конъюгата полимер-функциональный агент составляет от примерно 100000 до 1500000 Да. Предпочтительно, пиковая молекулярная масса конъюгата полимер-функциональный агент составляет от примерно 200000 до примерно 300000 Да, от примерно 400000 до примерно 600000 Да или от примерно 650000 до примерно 850000 Да.
[0309] В соответствии с другим аспектом настоящего изобретения первый линкер предпочтительно представляет собой алкил, содержащий заместители алкил, алкилен, алкокси, карбоксиалкил, галогеналкил, циклоалкил, циклический алкиловый эфир, алкенил, алкенилен, алкинил, алкинилен, циклоалкилен, гетероциклоалкил, гетероциклоалкилен, арил, арилен, ариленокси, гетероарил, амино, амидо или любую их комбинацию. Более предпочтительно, первый линкер имеет формулу:
Формула (1)
где m равен от 1 до 10. В некоторых вариантах реализации первый линкер имеет приведенную выше формулу (Формула (1)), и m равен 4.
[0310] В некоторых вариантах реализации инициатор предпочтительно содержит структуру, выбранную из группы, состоящей из:
Формула (2)
Формула (3) и
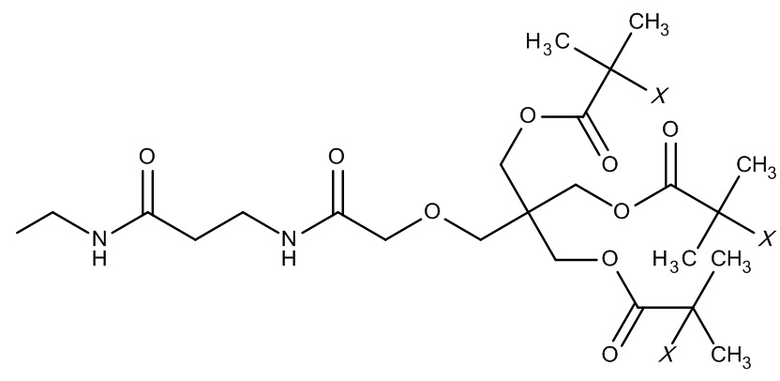
Формула (4)
где X выбран из группы, состоящей из NCS, F, Cl, Br и I. Более предпочтительно, X в Формуле (2), Формуле (3) и/или Формуле (4) представляет собой Br.
[0311] В некоторых вариантах реализации мономер выбран из группы, состоящей из:
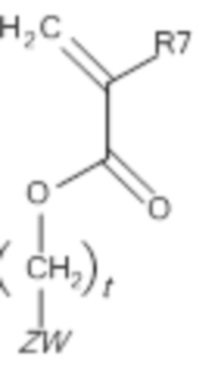
Формула (5)
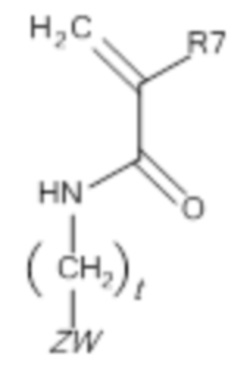
Формула (6)
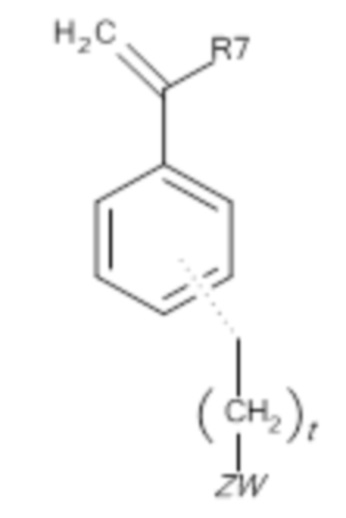
Формула (7)
Формула (8) и
Формула (9)
где R7 представляет собой H или C1-6 алкил, и t равен от 1 до 6.
[0312] Более предпочтительно, мономер выбран из группы, состоящей из 2-(метакрилоилоксиэтил)-2’-(триметиламмонийметил)фосфата (HEMA-PC) и 2-(акрилоилоксиэтил)-2’-(триметиламмонийметил)фосфата.
[0313] Наиболее предпочтительно, мономер представляет собой 2-(метакрилоилоксиэтил)-2’-(триметиламмонийметил)фосфат.
[0314] Второй линкерный фрагмент предпочтительно содержит активированный сложный эфир, имеющий структуру
Формула (10)
где R8 выбран из группы, состоящей из:
 и
и  ,
,
и R9 представляет собой
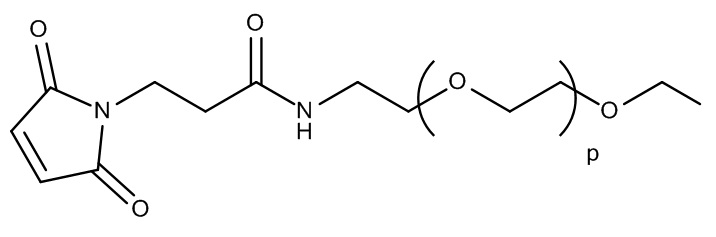
Формула (11)
где p равен от 1 до 12.
[0315] В более предпочтительных вариантах реализации настоящего изобретения полимер содержит 9 ответвлений, m равен 2-4, R9 представляет собой
Формула (11)
где p равен от 4 до 15. Более предпочтительно, m равен 4, и p равен 12.
[0316] В некоторых вариантах реализации радикально полимеризуемый мономер представляет собой
Формула (12)
где R1 представляет собой H или C1-6 алкил, R2, R3, R4 являются одинаковыми или разными и представляют собой H или C1-4 алкил, и X и Y являются одинаковыми или разными и представляют собой целые числа 1-6. В некоторых вариантах реализации каждый из R1, R2, R3 и R4 представляет собой метил, и каждый из X и Y равен 2 в Формуле (12).
[0317] В некоторых вариантах реализации радикально полимеризуемый мономер представляет собой
Формула (13)
где R1 представляет собой H или C1-6 алкил, R2 и R3 являются одинаковыми или разными и представляют собой H или C1-4 алкил, R4 представляет собой PO4-, SO3- или CO2-, и X и Y являются одинаковыми или разными и представляют собой целые числа 1-6. В некоторых вариантах реализации R1, R2 и R3 представляют собой метил, R4 представляет собой PO4-, и каждый из X и Y равен 2 в Формуле (13).
[0318] В некоторых вариантах реализации мономер представляет собой
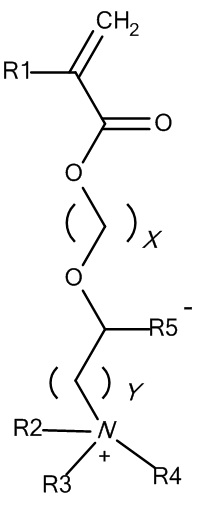
Формула (14)
где R1 представляет собой H или C1-6 алкил, R2, R3 и R4 являются одинаковыми или разными и представляют собой H или C1-4 алкил, R5 представляет собой PO4-, SO3- или CO2-, и X и Y являются одинаковыми или разными и представляют собой целые числа 1-6. В некоторых вариантах реализации R1, R2, R3 и R4 представляют собой метил, R5 представляет собой PO4-, и X и Y равны 2 в Формуле (14).
[0319] Когда полимер конъюгируют через цистеин (или другой указанный остаток), полимер может быть связан с указанным остатком напрямую или опосредованно (например, с помощью промежуточного инициатора и/или спейсера и т.п.).
[0320] В некоторых вариантах реализации полимер, содержащий фосфорилхолин, содержит мономеры, представляющие собой 2-(метакрилоилоксиэтил)-2'-(триметиламмоний)этилфосфат (MPC), как представлено ниже:
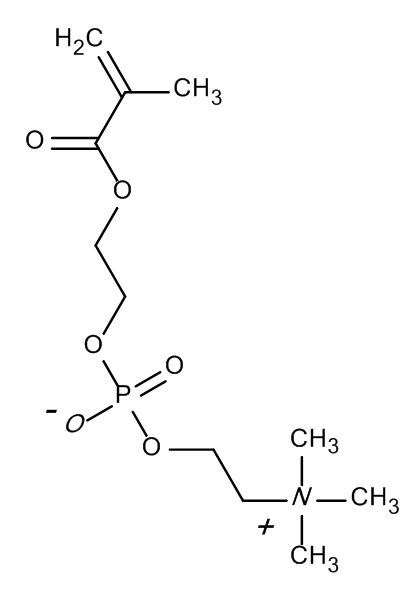
Формула (15),
так, что указанный полимер содержит следующие повторяющиеся звенья:
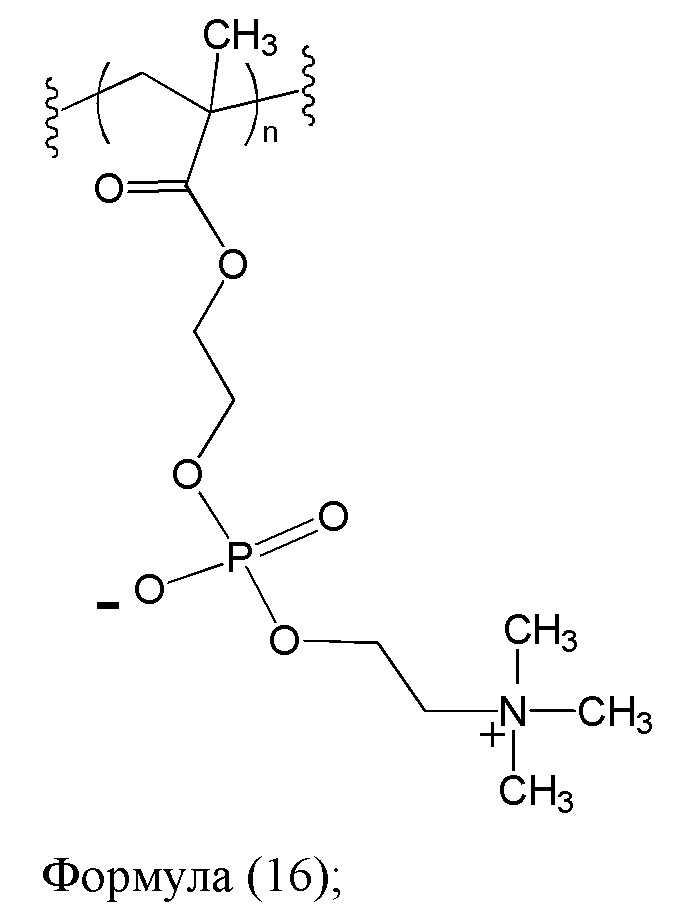
где n представляет собой целое число от 1 до 3000, а волнистые линии показывают точки присоединения между мономерными звеньями в полимере.
[0321] В некоторых вариантах реализации полимер содержит три или более ответвлений, или синтезирован с использованием инициатора, содержащего 3 или более центров инициации синтеза полимера. В некоторых вариантах реализации полимер содержит 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 ответвлений или синтезирован с использованием инициатора, содержащего 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 центров инициации синтеза полимера. Более предпочтительно, полимер содержит 3, 6 или 9 ответвлений или синтезирован с использованием инициатора, содержащего 3, 6 или 9 центров инициации синтеза полимера. В некоторых вариантах реализации полимер содержит 9 ответвлений или синтезирован с использованием инициатора, содержащего 9 центров инициации синтеза полимера.
[0322] В некоторых вариантах реализации присоединяемый полимер имеет молекулярную массу от примерно 300000 до примерно 1750000 Да (SEC-MALs). В некоторых вариантах реализации полимер имеет молекулярную массу от примерно 500000 до примерно 1000000 Да. В некоторых вариантах реализации полимер имеет молекулярную массу от примерно 600000 до примерно 900000 Да. В некоторых вариантах реализации полимер имеет молекулярную массу от примерно 750000 до примерно 850000 Да. В некоторых вариантах реализации полимер имеет молекулярную массу от примерно 800000 до примерно 850000 Да. В некоторых вариантах реализации полимер имеет молекулярную массу от примерно 750000 до примерно 800000 Да.
[0323] В некоторых вариантах реализации любое из антител и/или конструкций АТ-ловушка, описанных в настоящем документе, может быть дополнительно конъюгировано с полимером с образованием биоконъюгата. Молекулярная масса указанного биоконъюгата (общая, SEC-MALs) может составлять от примерно 350000 до 2000000 Дальтон, например, от примерно 450000 до 1900000 Дальтон, от примерно 550000 до 1800000 Дальтон, от примерно 650000 до 1700000 Дальтон, от примерно 750000 до 1600000 Дальтон, от примерно 850000 до 1500000 Дальтон, от примерно 900000 до 1400000 Дальтон, от примерно 950000 до 1300000 Дальтон, от примерно 900000 до 1000000 Дальтон, от примерно 1000000 до 1300000 Дальтон, от примерно 850000 до 1300000 Дальтон, от примерно 850000 до 1000000 Дальтон и от примерно 1000000 до 1200000 Дальтон. В некоторых вариантах реализации биоконъюгат имеет молекулярную массу от примерно 350000 до 1900000 Дальтон.
[0324] В некоторых вариантах реализации антитело и/или конструкцию АТ-ловушка очищают.В некоторых вариантах реализации полимер в аспекте конъюгата антитела и/или конструкции АТ-ловушка является полидисперсным, т.е. PDI полимера не равен 1,0. В некоторых вариантах реализации PDI меньше 1,5. В некоторых вариантах реализации PDI меньше 1,4. В некоторых вариантах реализации PDI меньше 1,3. В некоторых вариантах реализации PDI меньше 1,2. В некоторых вариантах реализации PDI меньше 1,1. В некоторых вариантах реализации PDI конъюгата равен или меньше 1,5.
[0325] В некоторых вариантах реализации конъюгат антитела и/или конструкции АТ-ловушка содержит анти-IL-6 иммуноглобулин G (IgG), связанный с полимером, при этом указанный полимер содержит мономеры MPC, при этом последовательность анти-IL-6 тяжелой цепи представлена в таблице 1, а последовательность анти-IL-6 легкой цепи представлена в таблице 2, и при этом антитело и/или конструкция АТ-ловушка связана с полимером только в C442. В некоторых вариантах реализации полимер содержит 9 ответвлений и имеет молекулярную массу от примерно 600000 до примерно 1000000 Да.
[0326] В некоторых вариантах реализации конъюгат антитела и/или конструкции АТ-ловушка содержит анти-IL-6 иммуноглобулин G (IgG), связанный с полимером, при этом указанный полимер содержит мономеры MPC, при этом последовательность анти-IL-6 тяжелой цепи содержится в таблице 1, а последовательность анти-IL-6 легкой цепи содержится в таблице 2, и при этом антитело связано с полимером только в C443 (нумерация EU). В некоторых вариантах реализации полимер содержит 9 ответвлений и имеет молекулярную массу от примерно 600000 до примерно 1000000 Да. В некоторых вариантах реализации конъюгат содержит полимер, содержащий 9 ответвлений, и указанный полимер имеет молекулярную массу от примерно 600000 до примерно 900000 Да.
[0327] В некоторых вариантах реализации конъюгат антитела имеет следующую структуру:
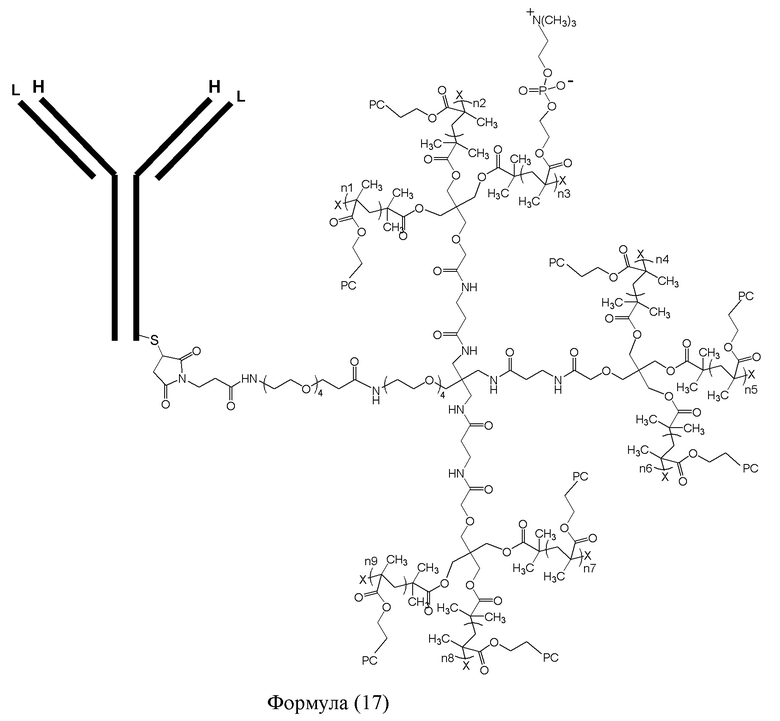
где: каждая анти-IL-6 тяжелая цепь антитела обозначена буквой H и каждая легкая цепь анти-IL-6 антитела обозначена буквой L (которые в некоторых вариантах реализации могут быть также связаны со структурой АТ-ловушка, представленной на фиг.6);
полимер связан с анти-IL-6 антителом (и/или конструкцией АТ-ловушка) через сульфгидрильную группу C443 (нумерация EU); указанная связь изображена на одной из тяжелых цепей; PC представляет собой
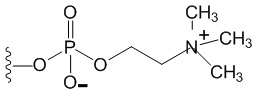 ,
,
где волнистая линия показывает точку присоединения к остальной части полимера; где X=a) OR, где R=H, метил, этил, пропил, изопропил, b) H или c) любой галогенид, включая Br; и n1, n2, n3, n4, n5, n6, n7, n8 и n9 являются одинаковыми или разными так, что сумма n1, n2, n3, n4, n5, n6, n6, n7, n8 и n9 равна 2500 плюс или минус 10%. В некоторых вариантах реализации n1, n2, n3, n4, n5, n6, n7, n8 и n9 являются одинаковыми или разными и представляют собой целые числа от 0 до 3000. В некоторых вариантах реализации n1, n2, n3, n4, n5, n6, n7, n8 и n9 являются одинаковыми или разными и представляют собой целые числа от 0 до 500. В некоторых вариантах реализации X=OR, где R представляет собой сахар, аминоалкил, монозамещенные, полизамещенные или не содержащие заместителей варианты следующих остатков: насыщенный C1 -C24 алкил, ненасыщенный C2 -C24 алкенил или C2 -C24 алкинил, ацил, ацилокси, алкилоксикарбонилокси, арилоксикарбонилокси, циклоалкил, циклоалкенил, алкокси, циклоалкокси, арил, гетероарил, арилалкоксикарбонил, алкоксикарбонилацил, амино, аминокарбонил, аминокарбоилокси, нитро, азидо, фенил, гидрокси, алкилтио, арилтио, оксисульфонил, карбокси, циано и галогенированный алкил, включая полигалогенированный алкил, --CO--O--R7, карбонил --CCO--R7, --CO--NR8R9, --(CH2)n--COOR7, --CO--(CH) n--COOR7, --(CH2) n--NR8R9, сложный эфир, алкоксикарбонил, арилоксикарбонил, где n представляет собой целое число от 1 до 6, где каждый из R7, R8 и R9 отдельно выбран из группы, состоящей из атома водорода, атома галогена, монозамещенных, полизамещенных или не содержащих заместителей вариантов следующих остатков: насыщенный C1 -C24 алкил, ненасыщенный C2 -C24 алкенил или C2 -C24 алкинил, ацил, ацилокси, алкилоксикарбонилокси, арилоксикарбонилокси, циклоалкил, циклоалкенил, алкокси, циклоалкокси, арил, гетероарил, арилалкоксикарбонил, алкоксикарбонилацил, амино, аминокарбонил, аминокарбоилокси, нитро, азидо, фенил, гидрокси, алкилтио, арилтио, оксисульфонил, карбокси, циано и галогенированный алкил, включая полигалогенированный алкил, 5-членное кольцо и 6-членное кольцо. В некоторых вариантах реализации Формула 17 может представлять собой часть слитого белка, который будет дополнительно содержать часть или всю последовательность из таблицы 10 для получения таким образом слитого белка АТ к IL-6-ловушка VEGF. В некоторых вариантах реализации слитую конструкцию на фиг.27 можно использовать в формуле 17, при этом указанная слитая конструкция заменяет изображенное антитело.
[0328] В некоторых вариантах реализации конъюгат антитела имеет следующую структуру:
где:
каждая тяжелая цепь антитела обозначена буквой H, и каждая легкая цепь анти-IL-6 антитела обозначена буквой L (которые в некоторых вариантах реализации могут быть также связаны с расположением АТ-ловушка, как показано на фиг.6);
полимер связан с антителом через сульфгидрильную группу C443 (нумерация EU); указанная связь изображена на одной из тяжелых цепей;
PC представляет собой
, где волнистая линия показывает точку присоединения к остальной части полимера, где X=a) OR, где R=H, метил, этил, пропил, изопропил, b) H или c) любой галогенид, включая Br; и
n1, n2, n3, n4, n5, n6, n7, n8 и n9 являются одинаковыми или разными так, что сумма n1, n2, n3, n4, n5, n6, n6, n7, n8 и n9 равна 2500 плюс или минус 15%. В некоторых вариантах реализации сумма n1, n2, n3, n4, n5, n6, n6, n7, n8 и n9 составляет от примерно 1500 до примерно 3500 плюс или минус примерно 10% - примерно 20%. В некоторых вариантах реализации слитую конструкцию на фиг.27 можно использовать в формуле 17, при этом указанная слитая конструкция заменяет изображенное антитело.
[0329] В некоторых вариантах реализации конъюгат антитела и/или конструкции АТ-ловушка находится в жидком составе. В некоторых вариантах реализации конъюгат антитела и/или конструкции АТ-ловушка комбинируют с фармацевтически приемлемым носителем.
[0330] В некоторых вариантах реализации изотип тяжелой цепи анти-IL-6 антитела (и/или конструкции АТ к IL-6-ловушка VEGF) представляет собой IgG1 и содержит CH1-домен, шарнирный домен, CH2- и CH3-домены. В некоторых вариантах реализации изотип легкой цепи представляет собой каппа-изотип.
[0331] В некоторых вариантах реализации IgG1-домен анти-IL-6 антитела (и/или конструкции АТ к IL-6-ловушка VEGF) содержит одну или более мутаций для модуляции эффекторной функции, такой как АЗКЦ, антителозависимый клеточный фагоцитоз (АЗКФ) и КОЦ. В некоторых вариантах реализации указанные мутации IgG1 снижают эффекторную функцию. В некоторых вариантах реализации аминокислоты, используемые для мутаций для эффекторной функции, включают (нумерация EU) E233X, L234X, L235X, G236X, G237X, G236X, D270X, K322X, A327X, P329X, A330X, A330X, P331X, и P331X, где X представляет собой любую природную или неприродную аминокислоту. В некоторых вариантах реализации мутации включают собой одну или более из следующих: E233P, L234V, L234A, L235A, G237A, A327G, A330S и P331S (нумерация EU). В некоторых вариантах реализации тяжелая цепь анти-IL-6 содержит следующие мутации (нумерация EU): L234A, L235A и G237A. В некоторых вариантах реализации количество мутаций для эффекторной функции относительно природной последовательности IgG1 человека составляет не более 10. В некоторых вариантах реализации количество мутаций для эффекторной функции относительно природной последовательности IgG1 человека составляет не более 5, 4, 3, 2 или 1. В некоторых вариантах реализации антитело (и/или АТ к IL-6-ловушка VEGF) демонстрирует уменьшение связывания Fc-гамма и/или связывания C1q комплемента, благодаря чему снижается способность указанного антитела обуславливать эффекторную функцию. Это может быть особенно предпочтительным при офтальмологических симптомах/нарушениях.
[0332] В некоторых вариантах реализации анти-IL6 антитело (и/или АТ к IL-6-ловушка VEGF) содержит одну или более следующих мутаций аминокислот: L234A, L235A, G237A и L443C (нумерация EU, или 451A, 452A, 454A и 660C в SEQ ID NO: 170, как показано двойным подчеркиванием на фиг.27).
[0333] В некоторых вариантах реализации анти-IL6 антитело (и/или АТ к IL-6-ловушка VEGF) представляет собой иммуноглобулин G (IgG1) человека или является его частью.
[0334] В некоторых вариантах реализации антитело к IL-6 (и/или АТ к IL-6-ловушка VEGF) содержит константный домен тяжелой цепи, содержащий одну или более мутаций, снижающих иммуноопосредованную эффекторную функцию.
[0335] В некоторых вариантах реализации анти-IL-6 тяжелая цепь содержит остаток цистеина, присоединенный в качестве мутации методом рекомбинантных ДНК, который можно использовать для конъюгации фрагмента, увеличивающего период полувыведения. В некоторых вариантах реализации указанная мутация представляет собой Q347C (нумерация EU) и/или L443C (нумерация EU). В некоторых вариантах реализации мутация представляет собой L443C (нумерация EU). В некоторых вариантах реализации стехиометрия антитела и полимера составляет 1:1; другими словами, конъюгат содержит одну молекулу антитела, конъюгированную с одной молекулой полимера.
[0336] Период полувыведения анти-IL-6 антител может быть увеличен путем присоединения «фрагментов, увеличивающих период полувыведения» или «групп, увеличивающих период полувыведения». Фрагменты, увеличивающие период полувыведения, включают пептиды и белки, которые могут быть экспрессированы в одной рамке с рассматриваемым биологическим лекарственным средством (или химически конъюгированы в зависимости от ситуации), и различные полимеры, которые могут быть присоединены или конъюгированы с одной или более функциональными группами боковых цепей аминокислот или концевыми функциональными группами, такими как -SH, -OH, -COOH, -CONH2, -NH2, или одной или более структурами N- и/или O-гликанов. Фрагменты, увеличивающие период полувыведения, как правило, действуют с увеличением периода полувыведения биологических лекарственных средств из крови in vivo.
[0337] Примеры пептидных/белковых фрагментов, увеличивающих период полувыведения, включают слияние с Fc (Capon DJ, Chamow SM, Mordenti J, et al. Designing CD4 immunoadhesions for AIDS therapy. Nature. 1989. 337:525-31), слияние с сывороточным альбумином человека (HAS) (Yeh P, Landais D, Lemaitre M, et al. Design of yeast-secreted albumin derivatives for human therapy: biological and antiviral properties of a serum albumin-CD4 genetic conjugate. Proc Natl Acad Sci USA. 1992. 89:1904-08), слияние с карбоксиконцевым пептидом (CTP) (Fares FA, Suganuma N. Nishimori K., et al. Design of a long-acting follitropin agonist by fusing the C-terminal sequence of the chorionic gonadotropin beta subunit to the follitropin beta subunit. Proc Natl Acad Sci USA. 1992. 89:4304-08), слияние с неточной повторяющейся пептидной последовательностью методом генной инженерии (XTEN) (Schellenberger V, Wang CW, Geething NC, et al. A recombinant polypeptide extends the in vivo half-life of peptides and proteins in a tunable manner. Nat Biotechnol. 2009. 27:1186-90), эластиноподобный пептид (ELP) (слияние с ELP) (MCpherson DT, Morrow C, Minehan DS, et al. Production and purification of a recombinant elastomeric polypeptide, G(VPGVG19-VPGV, from Escheriachia coli. Biotechnol Prog. 1992. 8:347-52), слияние с трансферрином человека (Prior CP, Lai C-H, Sadehghi H et al. Modified transferrin fusion proteins. патент WO2004/020405. 2004), пролин-аланин-серин (ПАСилирование) (Skerra A, Theobald I, Schlapsky M. Biological active proteins having increased in vivo and/or vitro stability. Патент WO2008/155134 A1. 2008), гомоаминокислотный полимер (ГАПилирование) (Schlapschy M, Theobald I, Mack H, et al. Fusion of a recombinant antibody fragment with a homo-amino acid polymer: effects on biophysical properties and prolonged plasma half-life. Protein Eng Des Sel. 2007. 20:273-84) и слияние с желатиноподобным белком (GLK) (Huang Y-S, Wen X-F, Zaro JL, et al. Engineering a pharmacologically superior form of granulocyte-colony-stimulating-factor by fusion with gelatin-like protein polymer. Eur J. Pharm Biopharm. 2010. 72:435-41).
[0338] Примеры представляющих собой полимер фрагментов, увеличивающих период полувыведения, включают полиэтиленгликоль (ПЭГ), разветвленный ПЭГ, PolyPEG® (Warwick Effect Polymers; Ковентри, Великобритания), полисиаловую кислоту (PSA), крахмал, гидроксилэтилкрахмал (HES), гидроксиалкилкрахмал (HAS), углевод, полисахариды, пуллулан, хитозан, гиалуроновую кислоту, хондроитинсульфат, дерматансульфат, декстран, карбоксиметилдекстран, полиалкиленоксид (PAO), полиалкиленгликоль (PAG), полипропиленгликоль (PPG), полиоксазолин, полиакрилоилморфолин, поливиниловый спирт (PVA), поликарбоксилат, поливинилпирролидон, полифосфазен, полиоксазолин, сополимер этилена и ангидрида малеиновой кислоты, сополимер стирола и ангидрида малеиновой кислоты, поли(1-гидроксиметэтиленгидроксиметилформаль) (PHF), цвиттерионный полимер, полимер, содержащий фосфорилхолин, и полимер, содержащий MPC, поли(Glyx-Sery), гиалуроновую кислоту (HA), гепарозановые полимеры (HEP), флексимеры, декстран и полисиаловые кислоты (PSA).
[0339] В одном из вариантов реализации фрагмент, увеличивающий период полувыведения, может быть конъюгирован с антителом через свободные аминогруппы белка с использованием сложных эфиров N-гидроксисукцинимида (NHS). Реагенты, направленные на конъюгацию с аминогруппами, могут случайным образом реагировать с ε-аминогруппой остатков лизина, α-аминогруппой N-концевых аминокислот и δ-аминогруппой остатков гистидина.
[0340] Однако анти-IL-6 антитела (и/или конструкции АТ к IL-6-ловушка VEGF), описанные в настоящем документе, могут содержать много аминогрупп, доступных для конъюгации с полимером. Таким образом, конъюгация полимеров со свободными аминогруппами может отрицательно влиять на способность антител-белков связываться с IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF).
[0341] В некоторых вариантах реализации фрагмент, увеличивающий период полувыведения, подвергают реакции сочетания с одной или более свободными SH-группам с использованием любой подходящей тиол-активной химии, включая, без ограничения, химию малеимида или сочетание полимерных гидразидов или полимерных аминов с углеводными фрагментами антитела после предварительного окисления. В некоторых вариантах реализации используют сочетание с использованием малеимида. В некоторых вариантах реализации сочетание происходит по остаткам цистеина, присутствующим естественным образом или введенным посредством генной инженерии.
[0342] В некоторых вариантах реализации предложены конъюгаты антитела и высокомолекулярных полимеров, служащих в качестве агентов, увеличивающих период полувыведения. В некоторых вариантах реализации конъюгат содержит антитело (и/или конструкцию АТ к IL-6-ловушка VEGF), подвергнутое реакции сочетания с цвиттерионным полимером, при этом указанный полимер образован из одного или более мономерных звеньев, и при этом по меньшей мере одно мономерное звено содержит цвиттерионную группу. В некоторых вариантах реализации указанная цвиттерионная группа представляет собой фосфорилхолин.
[0343] В некоторых вариантах реализации одно из мономерных звеньев представляет собой HEMA-PC. В некоторых вариантах реализации полимер синтезируют из одного мономера, который представляет собой HEMA-PC.
[0344] В некоторых вариантах реализации некоторые конъюгаты антитела (и/или конструкции АТ к IL-6-ловушка VEGF) содержат 2, 3 или более ответвлений полимера, в которых мономер представляет собой HEMA-PC. В некоторых вариантах реализации конъюгаты содержат 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 ответвлений полимера, в которых мономер представляет собой HEMA-PC. В некоторых вариантах реализации конъюгаты содержат 3, 6 или 9 ответвлений. В некоторых вариантах реализации конъюгат содержит 9 ответвлений.
[0345] В некоторых вариантах реализации конъюгаты полимер-антитело (и/или полимер- АТ к IL-6-ловушка VEGF) содержат полимерную часть с молекулярной массой от 100000 до 1500000 Да. В некоторых вариантах реализации конъюгат содержит полимерную часть с молекулярной массой от 500000 до 1000000 Да. В некоторых вариантах реализации конъюгат содержит полимерную часть с молекулярной массой от 600000 до 800000 Да. В некоторых вариантах реализации конъюгат содержит полимерную часть с молекулярной массой от 600000 до 850000 Да и содержит 9 ответвлений. Когда молекулярная масса приведена для антитела, конъюгированного с полимером, указанная молекулярная масса складывается из молекулярной массы белка, включая любые связанные с ним углеводные фрагменты, и молекулярной массы полимера.
[0346] В некоторых вариантах реализации предложено анти-IL6 антитело (и/или конструкция АТ к IL-6-ловушка VEGF), которое содержит полимер HEMA-PC, имеющий молекулярную массу, измеренную как Mw, составляющую от примерно 100 кДа до 1650 кДа. В некоторых вариантах реализации молекулярная масса полимера, измеренная как Mw, составляет от примерно 500 кДа до 1000 кДа. В некоторых вариантах реализации молекулярная масса полимера, измеренная как Mw, составляет от примерно 600 кДа до примерно 900 кДа. В некоторых вариантах реализации молекулярная масса полимера, измеренная как Mw, составляет 750 или 800 кДа плюс или минус 15%.
[0347] В некоторых вариантах реализации полимер получают из инициатора, подходящего для ATRP, содержащего один или более центров инициации синтеза полимера. В некоторых вариантах реализации центр инициации синтеза полимера содержит 2-бромизобутиратный центр. В некоторых вариантах реализации инициатор содержит 3 или более центров инициации синтеза полимера. В некоторых вариантах способов инициатор содержит 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 центров инициации синтеза полимера. В некоторых вариантах реализации инициатор содержит 3, 6 или 9 центров инициации синтеза полимера. В некоторых вариантах реализации инициатор содержит 9 центров инициации синтеза полимера. В некоторых вариантах реализации инициатор представляет собой OG1786.
[0348] Анти-IL-6 антитела (и/или конструкции АТ к IL-6-ловушка VEGF) могут быть получены путем рекомбинантной экспрессии, включающей (i) получение рекомбинантной ДНК посредством генной инженерии, (ii) введение рекомбинантной ДНК в прокариотические или эукариотические клетки, например и без ограничения, путем трансфекции, электропорации или микроинъекции, (iii) культивирование трансформированных клеток, (iv) экспрессию антитела, например, конститутивную или при индукции и (v) выделение антитела, например, из культуральной среды или путем сбора трансформированных клеток с (vi) получением очищенного антитела.
[0349] Анти-IL-6 антитела (и/или конструкции АТ к IL-6-ловушка VEGF) могут быть получены путем экспрессии в подходящей прокариотической или эукариотической системе-хозяине, характеризующейся продуцированием фармакологически приемлемой молекулы антитела. Примерами эукариотических клеток являются клетки млекопитающих, такие как CHO, COS, HEK 293, BHK, SK-Hip и HepG2. Другие подходящие системы экспрессии представляют собой прокариотические клетки (например, E.coli с системой экспрессии pET/BL21), клетки дрожжей (системы Saccharomyces cerevisiae и/или Pichia pastoris) и клетки насекомых.
[0350] Для получения антител (и/или конструкции АТ к IL-6-ловушка VEGF), описанных в настоящем документе, можно использовать широкий спектр векторов, и они выбраны из векторов эукариотической и прокариотической экспрессии. Примеры векторов для прокариотической экспрессии включают плазмиды, такие как, без ограничения, preset, pet и pad, при этом промоторы, используемые в векторах прокариотической экспрессии, включают один или более из, без ограничения, lac, trc, trp, recA или araBAD. Примеры векторов для эукариотической экспрессии включают: (i) для экспрессии в дрожжах: векторы, такие как и без ограничения, pAO, pPIC, pYES или pMET, с использованием промоторов, таких как и без ограничения, AOX1, GAP, GAL1 или AUG1; (ii) для экспрессии в клетках насекомых: векторы, такие как и без ограничения, pMT, pAc5, pIB, pMIB или pBAC, с использованием промоторов, таких как и без ограничения, PH, p10, MT, Ac5, OpIE2, gp64 или polh, и (iii) для экспрессии в клетках млекопитающих: векторы, такие как и без ограничения, pSVL, pCMV, pRc/RSV, pcDNA3 или pBPV, и векторы, полученные в соответствии с одним из аспектов из вирусных систем, таких как и без ограничения, вирус коровьей оспы, аденоассоциированные вирусы, вирусы герпеса или ретровирусы, с использованием промоторов, таких как и без ограничения, CMV, SV40, EF-1, UbC, RSV, ADV, BPV и бета-актин.
[0351] В некоторых вариантах реализации на белок приходится по меньшей мере один полимер. В некоторых вариантах реализации соотношение белка и полимера составляет от 1:4 до 1:6. В некоторых вариантах реализации соотношение белка (Ab, АТ) и полимера представляет собой молярное соотношение 1:5. В некоторых вариантах реализации количество белка составляет 5, 4, 3, 2 или примерно 2,4 мг/мл.
[0352] В некоторых вариантах реализации очистка после комбинирования полимера и антитела также включает использование катионообменной колонки.
[0353] В некоторых вариантах реализации присутствие полимера, присоединенного к антителам-антагонистам IL-6 и/или конструкциям АТ к IL-6-ловушка VEGF, может обеспечивать более высокую эффективность благодаря присутствию самого полимера. В некоторых вариантах реализации присутствие полимера при использовании в комбинации с гепарин-связывающим доменом (таким как в VEGF) позволяет получить конъюгированную молекулу с более высокой эффективностью. В некоторых вариантах реализации в соответствии с данными ЭКПВЧ, приведенными в примерах ниже, указанная молекула обладает превосходной эффективностью при ангиогенезе. В некоторых вариантах реализации это различие заключается в природе молекулы конъюгата, а не просто в степени (например, сравнение конъюгата с неконъюгированными молекулами). Более того, как показано в анализах пролиферации ЭКПВЧ в примерах ниже, в некоторых вариантах реализации присутствие полимера обеспечивает синергетическое ингибирование.
[0354] Кроме того, в некоторых вариантах реализации по мере увеличения плотности клеток (например, с 4000 до 6000) эффективность конъюгата может также повышаться.
[0355] В некоторых вариантах реализации количество (по массе белка) применяемого конъюгата составляет более 50 мг/мл. Оно может составлять, например, 55, 60, 65, 70 мг/мл или более (по массе белка до конъюгации).
Способ конъюгации белков с полимерами
[0356] В некоторых вариантах реализации представлен способ получения конъюгата терапевтический белок-фрагмент, увеличивающий период полувыведения, включающий этап конъюгации терапевтического белка, содержащего остаток цистеина, присоединенный методом рекомбинантных ДНК, с фрагментом, увеличивающим период полувыведения, содержащим сульфгидрил-специфичную реакционноспособную группу, выбранную из группы, состоящей из малеимида, винилсульфонов, ортопиридил-дисульфидов и йодацетамидов, с получением конъюгата терапевтический белок-фрагмент, увеличивающий период полувыведения.
[0357] В некоторых вариантах реализации предложен способ получения конъюгата антитела (и/или конструкции АТ к IL-6-ловушка VEGF). Как показано на фиг.10, указанный способ включает восстановление белка с использованием 30х молярного избытка восстановителя TCEP (фиг.10). После восстановления антитело окисляют с получением деблокированного антитела, в котором образованы дисульфидные связи между и внутри легкой и тяжелой цепей, естественным образом встречающиеся в антителе, но искусственно сконструированный цистеин в положении L443C тяжелой цепи (нумерация EU) еще предстоит деблокировать (фиг.10). Затем антитело конъюгируют путем добавления вспомогательного вещества и добавления 2-10х молярного избытка малеимидного биополимера (фиг.10). Указанный биополимер связывается с антителом ковалентной тиолэфирной связью (фиг.10). После конъюгации конъюгат антитела очищают с удалением как неконъюгированного антитела, так и полимера (фиг.10). В вариантах реализации, в которых белок представляет собой конструкцию АТ-ловушка, можно использовать это же положение на антителе и применять этот же подход.
[0358] Белок и способ, описанный выше, можно также варьировать. Таким образом, в некоторых вариантах реализации предложен способ получения конъюгированного белка (который не обязательно должен представлять собой антитело или анти-IL6 антитело). Указанный способ включает восстановление одного или более остатков цистеина в белке с образованием деблокированного белка в растворе. После восстановления указанного одного или более остатков цистеина деблокированный белок повторно окисляют с восстановлением по меньшей мере одной дисульфидной связи в восстановленном белке и с сохранением при этом искусственно сконструированного остатка цистеина в белке в форме свободного тиола с образованием повторно окисленного деблокированного белка в растворе. Затем к раствору добавляют по меньшей мере одно вспомогательное вещество. Указанное вспомогательное вещество уменьшает осаждение белка, индуцированное полимером. После добавления вспомогательного вещества к раствору добавляют полимер, который конъюгируется с повторно окисленным деблокированным белком в искусственно сконструированном остатке цистеина с образованием конъюгированного белка.
[0359] В некоторых вариантах реализации молярный избыток восстановителя может быть изменен на любое значение, которое выполняет свою функцию. В некоторых вариантах реализации можно использовать 10, 20, 30, 40, 50, 60, 70, 80, 90х молярный избыток восстановителя (который не обязательно должен представлять собой TCEP во всех вариантах реализации). В некоторых вариантах реализации восстановитель может представлять собой тринатрий 3,3',3''-фосфинтриилтрис(бензол-1-сульфонат) (TPPTS). В некоторых вариантах реализации можно использовать любое антитело (терапевтическое или иное). В некоторых вариантах реализации можно использовать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15х молярный избыток малеимидного биополимера. В некоторых вариантах реализации присутствует избыток деблокированного белка относительно полимера. В некоторых вариантах реализации количество восстановленного белка меньше количества полимера. В некоторых вариантах реализации количество восстановленного белка составляет 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10%, 5%, 4%, 3%, 2%, 1% от количества полимера. В некоторых вариантах реализации полимера используют в 10-15 раз больше, чем белка. В некоторых вариантах реализации количество восстановленного антитела больше количества полимера. В некоторых вариантах реализации количество полимера больше количества восстановленного антитела (и/или конструкции АТ к IL-6-ловушка VEGF).
[0360] В некоторых вариантах реализации этап очистки является необязательным.
[0361] В некоторых вариантах реализации способ получения конъюгата антитела (и/или конструкции АТ-ловушка) включает конъюгацию анти-IL-6 антитела (и/или конструкции АТ к IL-6-ловушка VEGF) с полимером, содержащим фосфорилхолин. В некоторых вариантах реализации способ включает этапы конъюгации анти-IL-6 антитела с полимером, содержащим фосфорилхолин. Анти-IL6 антитело содержит аминокислотный остаток, присоединенный методом рекомбинантных ДНК. В некоторых вариантах реализации указанный присоединенный аминокислотный остаток представляет собой остаток цистеина. В некоторых вариантах реализации указанный остаток цистеина присоединяют за пределами вариабельной области антитела. Остаток цистеина может быть присоединен либо к тяжелой цепи, либо к легкой цепи антитела.
[0362] В некоторых вариантах реализации полимер содержит или состоит из полимера, содержащего фосфорилхолин. В некоторых вариантах реализации полимер, содержащий фосфорилхолин, содержит сульфгидрил-специфичную реакционноспособную группу, выбранную из группы, состоящей из малеимида, винилсульфона, ортопиридил-дисульфида и йодацетамида. В некоторых вариантах реализации указанная сульфгидрил-специфичная реакционноспособная группа в полимере, содержащем фосфорилхолин, реагирует с остатком цистеина в анти-IL-6 антителе с образованием конъюгата антитела (и/или конструкции АТ к IL-6-ловушка VEGF).
[0363] В некоторых вариантах реализации конъюгируемый белок может представлять собой антитело, слитый с антителом белок (например, конструкцию АТ к IL-6-ловушка VEGF) или его связывающий фрагмент.В некоторых вариантах реализации белок не является антителом, а представляет собой фермент, лиганд, рецептор или другой белок, или их мутанты или варианты. В некоторых вариантах реализации нативный белок содержит по меньшей мере одну дисульфидную связь и по меньшей мере один ненативный цистеин.
[0364] В некоторых вариантах реализации вспомогательное вещество может представлять собой кислоту или основание. В некоторых вариантах реализации вспомогательное вещество представляет собой детергент, сахар или заряженную аминокислоту. В некоторых вариантах реализации вспомогательное вещество помогает удерживать белок в растворе во время конъюгации с полимером. В некоторых вариантах реализации вспомогательное вещество добавляют к раствору, содержащему белок, до добавления полимера к раствору, содержащему белок.
[0365] В некоторых вариантах реализации реакцию проводят в водных условиях, pH от примерно pH 5 до примерно pH 9. В некоторых вариантах реализации реакцию проводят от 6,0 до 8,5, от 6,5 до 8,0 или от 7,0 до 7,5.
[0366] В некоторых вариантах реализации полимер конъюгируют с белком при 2-37 градусах Цельсия. В некоторых вариантах реализации конъюгацию осуществляют при 0-40 градусах Цельсия, 5-35 градусах Цельсия, 10-30 градусах Цельсия и 15-25 градусах Цельсия.
[0367] В некоторых вариантах реализации конъюгированные белки, описанные в настоящем документе, могут быть приведены в контакт с ионообменной средой или средой для хроматографии гидрофобного взаимодействия, или аффинной хроматографии для очистки (для отделения конъюгированного от неконъюгированного). В некоторых вариантах реализации указанная ионообменная среда, среда для хроматографии гидрофобного взаимодействия и/или аффинной хроматографии позволяет отделить конъюгированный белок от неконъюгированного свободного полимера и от неконъюгированного повторно окисленного деблокированного белка.
[0368] В некоторых вариантах реализации способы, описанные в настоящем документе и представленные в общих чертах на фиг.10, включают вспомогательное вещество, которое может способствовать и/или поддерживать систему растворимости. В некоторых вариантах реализации способ обеспечивает поддержание раствором растворимости двух взаимодействующих компонентов. Растворимость может включать растворимость белка и полимера, а затем также конечного конъюгата. В некоторых вариантах реализации без применения подхода с использованием вспомогательного вещества проблема может заключаться в том, что хотя белок растворим, при добавлении биополимера растворимость в растворе (например, белка) падает, и он выпадает из раствора в осадок. Конечно, когда белок выпадает в осадок, его нельзя эффективно конъюгировать с биополимером. Таким образом, можно использовать вспомогательное вещество для поддержания растворимости белка в присутствии биополимера, чтобы эти два вещества могли соединиться с образованием конъюгата белка (или, как показано на фиг.10, конъюгата антитела). Это также позволяет поддерживать растворимость конъюгата.
[0369] В некоторых вариантах реализации полимеры, описанные в настоящем документе, могут содержать один или более из следующего: цвиттерион, фосфорилхолин или линкер ПЭГ, соединяющий мостиком центр точки разветвления полимера с малеимидной функциональной группой. В некоторых вариантах реализации любой из полимеров, предложенных согласно настоящему изобретению, может быть присоединен к белку способами, предложенными согласно настоящему изобретению.
[0370] В некоторых вариантах реализации любой из белков, предложенных согласно настоящему изобретению, может быть конъюгирован с любым из полимеров, предложенных согласно настоящему изобретению, одним или более способами, предложенными согласно настоящему изобретению.
[0371] В некоторых вариантах реализации указанный способ (способы), предложенный согласно настоящему изобретению, позволяет осуществлять более крупномасштабный процесс получения и очистки конъюгатов белка и/или антитела. В некоторых вариантах реализации используемый объем составляет по меньшей мере 1 литр, например, 1, 10, 100, 1000, 5000, 10000 литров или более. В некоторых вариантах реализации количество полученного и/или очищенного конъюгата антитела может составлять 0,1, 1, 10, 100, 1000 или более граммов.
[0372] В некоторых вариантах реализации терапевтический белок может представлять собой любое из анти-IL-6 антител (и/или конструкций АТ к IL-6-ловушка VEGF), описанных в настоящем документе, содержащее остаток цистеина, присоединенный методом рекомбинантных ДНК. В некоторых вариантах реализации тяжелая цепь анти-IL-6 антитела содержит следующие CDR: CDRH1: в таблице 1, CDRH2: в таблице 1 и CDRH3: в таблице 1. Тяжелая цепь может также содержать аргинин (R) в положении 84 (согласно последовательной нумерации). В некоторых вариантах реализации анти-IL-6 легкая цепь содержит следующие CDR: CDRL1: в таблице 2, CDRL2: в таблице 2 и CDRL3: в таблице 2.
[0373] В некоторых вариантах реализации анти-IL6 антитело (и/или АТ к IL-6-ловушка VEGF) содержит IgG1. В некоторых вариантах реализации тяжелая цепь содержит одну или более мутаций для модуляции эффекторной функции. В некоторых вариантах реализации указанные мутации относятся к одному или более следующим положениям аминокислот (нумерация EU): E233, L234, L235, G236, G237, A327, A330 и P331. В некоторых вариантах реализации мутации выбраны из группы, состоящей из: E233P, L234V, L234A, L235A, G237A, A327G, A330S и P331S (нумерация EU). В некоторых вариантах реализации мутации представляют собой (нумерация EU) L234A, L235A и G237A.
[0374] В некоторых вариантах реализации остаток цистеина, присоединенный к терапевтическому белку методом рекомбинантных ДНК, не должен быть вовлечен в образование дисульфидных связей Cys-Cys. В этом отношении терапевтические белки могут быть димерными. Так, например, интактное анти-IL6 антитело (и/или АТ к IL-6-ловушка VEGF) содержит две легкие цепи и две тяжелые цепи. Если остаток Cys вводят в тяжелую цепь, например, интактное антитело будет содержать два таких введенных остатка цистеина в идентичных положениях, и существует вероятность того, что эти остатки цистеина будут образовывать внутрицепочечные дисульфидные связи. Если введенные остатки цистеина образуют дисульфидные связи Cys-Cys или склонны к этому, этот введенный остаток Cys не будет подходить для конъюгации. В этой области техники известно как избежать положений в тяжелой и легкой цепях, которые приводят к образованию внутрицепочечных дисульфидных связей. См., например, заявку на патент США №2015/0158952.
[0375] В некоторых вариантах реализации остаток цистеина, введенный методом рекомбинантных ДНК, выбран из группы, состоящей из (нумерация EU) Q347C и L443C. В некоторых вариантах реализации остаток цистеина представляет собой L443C (нумерация EU). В некоторых вариантах реализации тяжелая цепь антитела содержит аминокислотную последовательность, представленную в таблице 1, и легкая цепь содержит аминокислотную последовательность из таблицы 2.
[0376] В некоторых вариантах реализации сульфгидрил-специфичная реакционноспособная группа представляет собой малеимид.
[0377] В некоторых вариантах реализации фрагмент, увеличивающий период полувыведения, выбран из группы, состоящей из полиэтиленгликоля (ПЭГ), разветвленного ПЭГ, PolyPEG ® (Warwick Effect Polymers; Ковентри, Великобритания), полисиаловой кислоты (PSA), крахмала, гидроксилэтилкрахмала (HES), гидроксиалкилкрахмала (HAS), углевода, полисахаридов, пуллулана, хитозана, гиалуроновой кислоты, хондроитинсульфата, дерматансульфата, декстрана, карбоксиметилдекстрана, полиалкиленоксида (PAO), полиалкиленгликоля (PAG), полипропиленгликоля (PPG), полиоксазолина, полиакрилоилморфолина, поливинилового спирта (PVA), поликарбоксилата, поливинилпирролидона, полифосфазена, полиоксазолина, сополимера этилена и ангидрида малеиновой кислоты, сополимера стирола и ангидрида малеиновой кислоты, поли(1-гидроксиметэтиленгидроксиметилформаля) (PHF), цвиттерионного полимера, полимера, содержащего фосфорилхолин, и полимера, содержащего 2-метакрилоилокси-2’-этилтриметиламмонийфосфат (MPC).
[0378] В некоторых вариантах реализации фрагмент, увеличивающий период полувыведения, представляет собой цвиттерионный полимер. В некоторых вариантах реализации цвиттерион представляет собой фосфорилхолин, т.е. полимер, содержащий фосфорилхолин. В некоторых вариантах реализации указанный полимер состоит из звеньев MPC.
[0379] В некоторых вариантах реализации полимер MPC содержит три или более ответвлений. В некоторых вариантах реализации полимер MPC содержит 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 ответвлений. В некоторых вариантах реализации полимер MPC содержит 3, 6 или 9 ответвлений. В некоторых вариантах реализации полимер MPC содержит 9 ответвлений. В некоторых вариантах реализации полимер синтезирован с использованием инициатора, содержащего 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более центров инициации синтеза полимера.
[0380] В некоторых вариантах реализации полимер MPC имеет молекулярную массу от примерно 300000 до 1750000 Да. В некоторых вариантах реализации полимер MPC имеет молекулярную массу от примерно 500000 до 1000000 Да или от примерно 600000 до 900000 Да.
[0381] В некоторых вариантах реализации способ получения конъюгата терапевтический белок-фрагмент, увеличивающий период полувыведения, включает дополнительный этап приведения терапевтического белка в контакт с тиоловым восстановителем в условиях, позволяющих получить восстановленную сульфгидрильную группу цистеина. Как рассмотрено выше, предпочтительно, чтобы остатки цистеина, присоединенные методом рекомбинантных ДНК, были неспаренными, т.е. не были вовлечены во внутрицепочечные дисульфидные связи Cys-Cys или по существу не были вовлечены в образование таких связей. Однако известно, что остатки Cys, которые не вовлечены в образование таких дисульфидных связей Cys-Cys и свободны для конъюгации, реагируют со свободным цистеином в культуральной среде с образованием дисульфидных аддуктов. См., например, WO 2009/052249. Дериватизированный таким образом цистеин будет недоступен для конъюгации. Для высвобождения вновь присоединенного цистеина из дисульфидного аддукта белок после очистки обрабатывают восстановителем, например, дитиотреитолом. Однако такая обработка восстановителем приведет к восстановлению всех остатков цистеина в терапевтическом белке, включая нативные остатки цистеина, многие из которых вовлечены в меж- и внутрицепочечные дисульфидные связи Cys-Cys. Нативные дисульфиды Cys-Cys, как правило, имеют критическое значение для стабильности и активности белка, и они должны быть заново сформированы. В некоторых вариантах реализации заново формируют все нативные (например, меж- и внутрицепочечные) дисульфиды Cys-Cys.
[0382] Для того, чтобы заново сформировать нативные меж- и внутрицепочечные дисульфидные остатки, после восстановления с удалением дисульфидных аддуктов цистеина терапевтический белок подвергают воздействию окислительных условий и/или окислителей в течение установленного периода времени, например, в течение ночи. В некоторых вариантах реализации для формирования заново нативных дисульфидных связей можно использовать воздействие воздухом окружающей среды в течение ночи. В некоторых вариантах реализации для возвращения нативных дисульфидных связей используют окислитель. В некоторых вариантах реализации указанный окислитель выбран из группы, состоящей из водного CuSO4 и дегидроаскорбиновой кислоты (DHAA). В некоторых вариантах реализации окислитель представляет собой DHAA. В некоторых вариантах реализации используемая DHAA находится в диапазоне 5-30 эквивалентов. В некоторых вариантах реализации указанный диапазон составляет 10-30 эквивалентов. В некоторых вариантах реализации диапазон составляет 15 эквивалентов.
[0383] В некоторых вариантах реализации тиоловый восстановитель выбран из группы, состоящей из: 3,3',3''-фосфантриилтрипропановой кислоты (TCEP), дитиотреитола (DTT), дитиоэритритола (DTE), боргидрида натрия (NaBH4), цианоборгидрида натрия (NaCNBH3), β-меркаптоэтанола (BME), гидрохлорида цистеина, тринатрий 3,3',3''-фосфинтриилтрис(бензол-1-сульфоната) (TPPTS) и цистеина. В некоторых вариантах реализации тиоловый восстановитель представляет собой TCEP.
[0384] В некоторых вариантах реализации концентрация тиолового восстановителя представляет собой 1-100-кратный молярный избыток относительно концентрации терапевтического белка. В некоторых вариантах реализации концентрация тиолового восстановителя представляет собой 20-50-кратный молярный избыток относительно концентрации терапевтического белка. В некоторых вариантах реализации тиоловый восстановитель удаляют после инкубации совместно с терапевтическим белком до окисления терапевтического белка.
[0385] В некоторых вариантах реализации способ конъюгации терапевтического белка с фрагментом, увеличивающим период полувыведения, включает дополнительный этап очистки конъюгата терапевтического белка после конъюгации. В некоторых вариантах реализации конъюгат терапевтического белка очищают с использованием метода, выбранного из группы, состоящей из ионообменной хроматографии, хроматографии гидрофобного взаимодействия, эксклюзионной хроматографии и аффинной хроматографии или их комбинаций.
[0386] В некоторых вариантах реализации конъюгат терапевтического белка сохраняет по меньшей мере 20% биологической активности относительно неконъюгированного терапевтического белка. В некоторых вариантах реализации конъюгат терапевтического белка сохраняет по меньшей мере 50% биологической активности относительно неконъюгированного терапевтического белка. В некоторых вариантах реализации конъюгат терапевтического белка сохраняет по меньшей мере 90% биологической активности относительно нативного терапевтического белка.
[0387] В некоторых вариантах реализации конъюгат терапевтического белка демонстрирует увеличенный период полувыведения относительно неконъюгированного терапевтического белка. В некоторых вариантах реализации конъюгат терапевтического белка демонстрирует по меньшей мере 1,5-кратное увеличение периода полувыведения относительно неконъюгированного терапевтического белка. В некоторых вариантах реализации конъюгат терапевтического белка демонстрирует по меньшей мере 1,5-, 2-, 2,5-, 3-, 3,5-, 4-, 4,5- или 5-кратное увеличение периода полувыведения относительно неконъюгированного терапевтического белка.
[0388] В некоторых вариантах реализации цвиттерионный полимер в способе конъюгации терапевтического белка с фрагментом, увеличивающим период полувыведения, представляет собой радикально полимеризуемый мономер, содержащий цвиттерионную группу, и способ включает дополнительный этап полимеризации указанного свободнорадикально-полимеризуемого цвиттерионного мономера в среде для полимеризации с получением полимера, при этом указанная среда содержит: радикально полимеризуемый цвиттерионный мономер; катализатор на основе переходного металла Mt(q-1)+, где Mt представляет собой переходный металл, q представляет собой более высокую степень окисления металла, и q-1 представляет собой более низкую степень окисления металла, при этом металлический катализатор представлен в виде соли формы Mt(q-1)+X’(q-1), где X' представляет собой противоион или группу, или катализатор на основе переходного металла получают in situ путем использования соли неактивного металла в более высокой степени окисления Mtq+X’q совместно с восстановителем, который способен восстановить переходный металл из окисленного неактивного состояния в восстановленное активное состояние; лиганд; и инициатор.
[0389] Для выполнения функции катализатора на основе переходного металла для ATRP переходный металл должен иметь по меньшей мере две легко доступные степени окисления, отделенные одним электроном, более высокую степень окисления и более низкую степень окисления. В ATRP обратимая окислительно-восстановительная реакция приводит к циклическому переходу катализатора на основе переходного металла между более высокой степенью окисления и более низкой степенью окисления, в то время как полимерные цепи проходят цикл между концами растущей цепи и концами спящей цепи. См., например, патент США №7893173.
[0390] В некоторых вариантах реализации радикально полимеризуемый цвиттерионный мономер выбран из группы, состоящей из:
Формула (18)
Формула (19)

Формула (20) и
Формула (21)
где R1 представляет собой H или C1-6 алкил, ZW представляет собой цвиттерион, и n представляет собой целое число 1-6.
[0391] В некоторых вариантах реализации радикально полимеризуемый мономер представляет собой
Формула (12)
где R1 представляет собой H или C1-6 алкил, R2, R3, R4 являются одинаковыми или разными и представляют собой H или C1-4 алкил, и X и Y являются одинаковыми или разными и представляют собой целые числа 1-6. В некоторых вариантах реализации каждый из R1, R2, R3 и R4 представляет собой метил, и каждый из X и Y равен 2 в Формуле (12).
[0392] В некоторых вариантах реализации радикально полимеризуемый мономер представляет собой
Формула (13)
где R1 представляет собой H или C1-6 алкил, R2 и R3 являются одинаковыми или разными и представляют собой H или C1-4 алкил, R4 представляет собой PO4-, SO3- или CO2-, и X и Y являются одинаковыми или разными и представляют собой целые числа 1-6. В некоторых вариантах реализации R1, R2 и R3 представляют собой метил, R4 представляет собой PO4-, и каждый из X и Y равен 2 в Формуле (13).
[0393] В некоторых вариантах реализации мономер представляет собой
Формула (14)
где R1 представляет собой H или C1-6 алкил, R2, R3 и R4 являются одинаковыми или разными и представляют собой H или C1-4 алкил, R5 представляет собой PO4-, SO3- или CO2-, и X и Y являются одинаковыми или разными и представляют собой целые числа 1-6. В некоторых вариантах реализации R1, R2, R3 и R4 представляют собой метил, R5 представляет собой PO4-, и X и Y равны 2 в Формуле (14).
[0394] В некоторых вариантах реализации переходный металл Mt выбран из группы, состоящей из Cu, Fe, Ru, Cr, Mo, W, Mn, Rh, Re, Co, V, Zn, Au и Ag. В некоторых вариантах реализации металлический катализатор представлен в виде соли формы Mt(q-1)+X’(q-1). Mt(q-1)+ выбран из группы, состоящей из Cu1+, Fe2+, Ru2+, Cr2+, Mo2+, W2+, Mn3+, Rh3+, Re2+, Co+, V2+, Zn+, Au+и Ag+, и X’ выбран из группы, состоящий из галогена, C1-6 алкокси, (SO4)1/2, (PO4)1/3, (R7PO4)1/2, (R72PO4), трифлата, гексалуорофосфата, метансульфоната, арилсульфоната, CN и R7CO2, где R7 представляет собой H или линейную, или разветвленную C1-6 алкильную группу, которая может содержать в качестве заместителей галоген от 1 до 5 раз. В некоторых вариантах реализации Mt(q-1)+ представляет собой Cu1+, и X’ представляет собой Br.
[0395] В некоторых вариантах реализации Mt(q-1)+ получают in situ. В некоторых вариантах реализации Mtq+Xq представляет собой CuBr2. В некоторых вариантах реализации восстановитель представляет собой неорганическое соединение. В некоторых вариантах реализации восстановитель выбран из группы, состоящей из соединения серы с низкой степенью окисления, гидросульфита натрия, неорганической соли, содержащей ион металла, металла, гидразин-гидрата и производных таких соединений. В некоторых вариантах реализации восстановитель представляет собой металл. В некоторых вариантах реализации восстановитель представляет собой Cu0.
[0396] В некоторых вариантах реализации восстановитель представляет собой органическое соединение. В некоторых вариантах реализации указанное органическое соединение выбрано из группы, состоящей из алкилтиолов, меркаптоэтанола или карбонильных соединений, которые могут быть легко енолизированы, аскорбиновой кислоты, ацетилацетоната, камфорсульфоновой кислоты, гидроксиацетона, восстанавливающих сахаров, моносахаридов, глюкозы, альдегидов и производных таких органических соединений.
[0397] В некоторых вариантах реализации лиганд выбран из группы, состоящей из 2,2'-бипиридина, 4,4'-ди-5-нонил-2,2'-бипиридина, 4,4-динонил-2,2'-дипиридила, 4,4',4''-трис(5-нонил)-2,2':6',2''-терпиридина, N,N,N',N',N''-пентаметилдиэтилентриамина, 1,1,4,7,10,10-гексаметилтриэтилентетрамина, трис(2-диметиламиноэтил)амина, N,N-бис(2-пиридилметил)октадециламина, N,N,N',N'-тетра[(2-пиридал)метил]этилендиамина, трис[(2-пиридил)метил]амина, трис(2-аминоэтил)амина, трис(2-бис(3-бутокси-3-оксопропил)аминоэтил)амина, трис(2-бис(3-(2-этилгексокси)-3-оксопропил)аминоэтил)амина и трис(2-бис(3-додекокси-3-оксопропил)аминоэтил)амина. В некоторых вариантах реализации лиганд представляет собой 2,2’-бипиридин.
[0398] В некоторых вариантах реализации инициатор имеет структуру:
Формула (22)
где R1 представляет собой нуклеофильную реакционноспособную группу, R2 содержит линкер, и R3 содержит фрагмент инициатора синтеза полимера, имеющий структуру
Формула (23)
где R4 и R5 являются одинаковыми или разными и выбраны из группы, состоящей из алкила, содержащего заместители алкила, алкилена, алкокси, карбоксиалкила, галогеналкила, циклоалкила, циклического алкилового эфира, алкенила, алкенилена, алкинила, алкинилена, циклоалкилена, гетероциклоалкила, гетероциклоалкилена, арила, арилена, ариленокси, гетероарила, амино, амидо или любой их комбинации; Z представляет собой галоген или CN; и s представляет собой целое число от 1 до 20.
[0399] В некоторых вариантах реализации Z в Формуле (23) представляет собой Br, и каждый из R4 и R5 представляет собой метил. В некоторых вариантах реализации R1 в Формуле (22) выбран из группы, состоящей из NH2-, OH- и SH-.
[0400] В некоторых вариантах реализации R2 в Формуле (22) представляет собой алкил, содержащий заместители алкил, алкилен, алкокси, карбоксиалкил, галогеналкил, циклоалкил, циклический алкиловый эфир, алкенил, алкенилен, алкинил, алкинилен, циклоалкилен, гетероциклоалкил, гетероциклоалкилен, арил, арилен, ариленокси, гетероарил, амино, амидо или любую их комбинацию. В некоторых вариантах реализации R2 в Формуле (22) представляет собой
Формула (24)
где X и Y являются одинаковыми или разными и представляют собой целые числа 1-20. В некоторых вариантах реализации каждый из X и Y равен 4.
[0401] В некоторых вариантах реализации R3 в Формуле (22) представляет собой
Формула (25)
где R6, R7 и R8 являются одинаковыми или разными и выбраны из группы, состоящей из
Формула (26),
Формула (27) и

Формула (28)
где Z представляет собой NCS, F, Cl, Br или I. В некоторых вариантах реализации Z в Формуле (26), Формуле (27) и/или Формуле (28) представляет собой Br, и каждый из R6, R7 и R8 в Формуле (25) представляет собой
Формула (29)
[0402] В некоторых вариантах реализации инициатор имеет структуру:
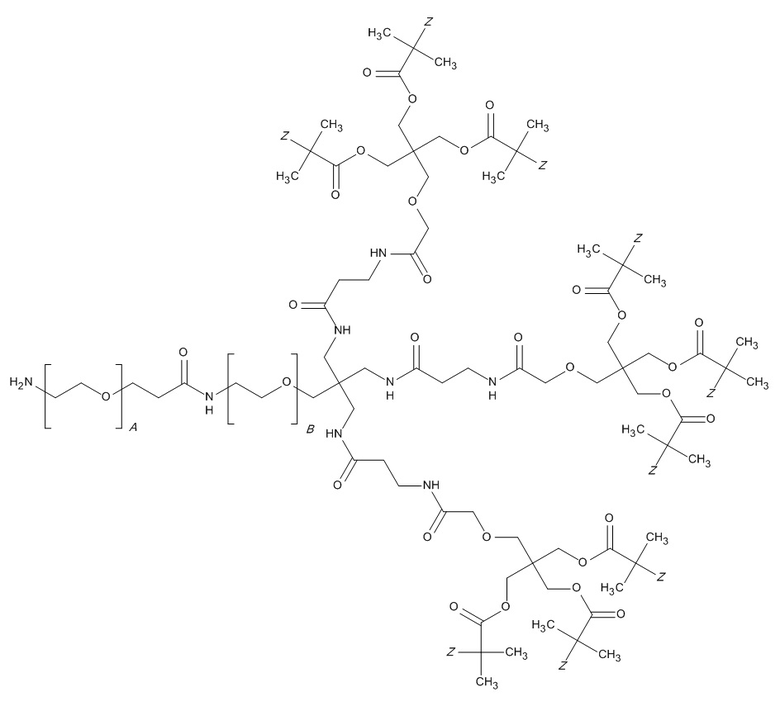
Формула (30)
где A и B являются одинаковыми или разными и представляют собой целые числа от 2 до 12, и Z представляет собой любой галогенид, например, Br. В некоторых вариантах реализации каждый из A и B равен 4 в Формуле (30).
[0403] В некоторых вариантах реализации способ дополнительно включает этап проведения реакции полимера с малеимидным реагентом с получением полимера, содержащего концевой малеимид. В некоторых вариантах реализации указанное малеимидное соединение представляет собой
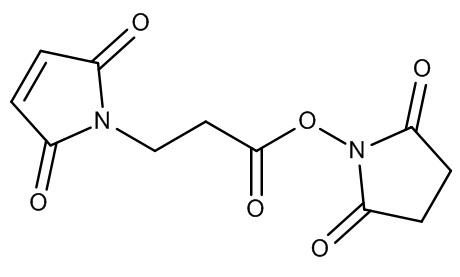
Формула (31).
[0404] Модификацию или мутацию можно также осуществлять в каркасной области или константной области для увеличения периода полувыведения антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF). См., например, публикацию PCT №WO 00/09560. Можно также осуществлять мутацию в каркасной области или константной области для изменения иммуногенности антитела, получения сайта для ковалентного или нековалентного связывания с другой молекулой или для изменения таких свойств, как фиксация комплемента, связывание FcR и антителозависимая клеточноопосредованная цитотоксичность. В некоторых вариантах реализации в пределах каркасной области или константной области осуществляют не более одной-пяти консервативных замен аминокислот.В других вариантах реализации в пределах каркасной области или константной области осуществляют не более одной-трех консервативных замен аминокислот.Согласно настоящему изобретению одно антитело может содержать мутации в любых одной или более CDR или каркасных областях вариабельного домена, или в константной области.
[0405] Модификации также включают гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, гликозилирование различными сахарами, ацетилирование и фосфорилирование. Антитела подвергают гликозилированию в консервативных положениях в константных областях (Jefferis and Lund, 1997, Chem. Immunol. 65:111-128; Wright and Morrison, 1997, TibTECH 15:26-32). Олигосахаридные боковые цепи иммуноглобулинов влияют на функцию белка (Boyd et al., 1996, Mol. Immunol. 32:1311-1318; Wittwe and Howard, 1990, Biochem. 29:4175-4180) и внутримолекулярное взаимодействие между частями гликопротеида, которое может влиять на конформацию и представляемую трехмерную поверхность гликопротеида (Jefferis and Lund выше; Wyss and Wagner, 1996, Current Opin. Biotech. 7:409-416). Олигосахариды могут также служить для направления конкретного гликопротеида на определенные молекулы на основе структур для специфичного распознавания. Существуют также данные о том, что гликозилирование антител влияет на антителозависимую клеточную цитотоксичность (АЗКЦ). Существуют данные о том, что антитела, продуцируемые клетками СНО с тетрациклин-регулируемой экспрессией β(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII), гликозилтрансферазы, катализирующей образование разветвляющегося GlcNAc, обладают повышенной АЗКЦ-активностью (Umana et al., 1999, Nature Biotech. 17:176-180).
[0406] Гликозилирование антител является, как правило, либо N-связанным, либо O-связанным. N-связанное относится к присоединению углеводного фрагмента к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-X-серин, аспарагин-X-треонин и аспарагин-X-цистеин, где X представляет собой любую аминокислоту за исключением пролина, являются распознаваемыми последовательностями для ферментативного присоединения углеводного фрагмента к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает возможный центр гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров: N-ацетилгалактозамина, галактозы или ксилозы к гидроксиаминокислоте, чаще всего серину или треонину, хотя может быть также использован 5-гидроксипролин или 5-гидроксилизин.
[0407] Добавление центров гликозилирования в антитело (и/или конструкцию АТ-ловушка) удобным образом осуществляют путем изменения аминокислотной последовательности таким образом чтобы она содержала одну или более вышеописанных трипептидных последовательностей (для центров N-связанного гликозилирования). Указанное изменение можно также осуществлять путем присоединения или замены на один или более остатков серина или треонина в последовательности исходного антитела (для центров О-связанного гликозилирования).
[0408] Схему гликозилирования антител (и/или конструкций АТ-ловушка) можно также изменять без изменения основной нуклеотидной последовательности. Гликозилирование в значительной степени зависит от клетки-хозяина, используемой для экспрессии антитела. Поскольку тип клеток, используемый для экспрессии рекомбинантных гликопротеидов, например, антител в качестве потенциальных терапевтических средств, редко представляет собой нативную клетку, можно ожидать вариации в схеме гликозилирования антител (см., например, Hse et al., 1997, J. Biol. Chem. 272:9062-9070).
[0409] В дополнение к выбору клеток-хозяев факторы, влияющие на гликозилирование во время рекомбинантного получения антител (и/или конструкций АТ-ловушка), включают режим роста, состав среды, плотность культуры, оксигенацию, рН, схемы очистки и т.п.Были предложены различные способы изменения схемы гликозилирования, осуществляемого в конкретном организме-хозяине, включая введение или сверхэкспрессию некоторых ферментов, вовлеченных в получение олигосахаридов (патенты США №№5047335; 5510261 и 5278299). Гликозилирование или некоторые типы гликозилирования могут быть ферментативно удалены из гликопротеида, например, с использованием эндогликозидазы Н (Endo Н), N-гликозидазы F, эндогликозидазы F1, эндогликозидазы F2, эндогликозидазы F3. Более того, рекомбинантная клетка-хозяин может быть генетически сконструирована таким образом чтобы она демонстрировала нарушение процессинга некоторых типов полисахаридов. Эти и подобные методики хорошо известны в этой области техники.
[0410] Некоторые варианты реализации, представленные в настоящем описании, можно применять в качестве терапевтического средства в случае воспалительных заболеваний сетчатки и/или везикулярных заболеваний сетчатки с воспалительным компонентом. В дополнение к ангиогенезу в патогенез этих заболеваний сетчатки вовлечено воспаление. Виды противовоспалительной терапии, такие как стероиды, эффективны в лечении как увеита (спектр заболеваний, определяющей характеристикой которых является внутриглазное воспаление), так и диабетического отека желтого пятна (ДОЖП). Подобным образом генетически унаследованные полиморфизмы IL-6 связывали с более высокой частотой возникновения пролиферативной диабетической ретинопатии (ПДР) у пациентов с диабетом 2 типа. Более того, существуют данные о том, что прогрессирование заболевания в случае ВДЖП, диабетической ретинопатии (ДР) и окклюзии вены сетчатки (ОВС) связано с повышенными уровнями IL-6 в сыворотке и/или глазах. Кроме того, клетки хронического воспаления наблюдали на поверхности мембраны Бруха в глазах при неоваскулярной ВДЖП. IL-6 вовлечен в резистентность к анти-VEGF лечению у пациентов с ДОЖП. Полагают, это отчасти является косвенным результатом IL-6-опосредованной повышающей регуляции экспрессии VEGF [ссылка], а также более непосредственных VEGF-независимых ангиогенных функций, опосредованных передачей сигнала IL-6, которые осуществляются в присутствии ингибиторов VEGF. В некоторых вариантах реализации любое одно или более этих состояний можно лечить и/или предотвращать одной или более композициями, предложенными согласно настоящему изобретению.
[0411] Фиг.11С иллюстрирует некоторые варианты конструкции биоконъюгата, представляющего собой слитую конструкцию ловушка-антитело (TAF) для ловушки VEGF (VEGFR1/2) и анти-IL-6 антитела, конъюгированную с биополимером на основе фосфорилхолина. В некоторых вариантах реализации биоконъюгат анти-VEGF/анти-IL-6 может иметь молекулярную массу 1,0 МДа с клинической дозой 6,0 мг в некоторых вариантах реализации. Эквивалентная молярная доза анти-VEGF части, в биоконъюгате может быть в 6 раз больше, чем в случае клинической дозы ранибизумаба, в то время как период полувыведения из глаза может быть в 3-5 раз больше, чем у ранибизумаба. CH представляет собой константную область тяжелой цепи, CL представляет собой константную область легкой цепи, Fab представляет собой антигенсвязывающий фрагмент, Fc представляет собой кристаллизующийся фрагмент, VEGFR представляет рецептор фактора роста эндотелия сосудов, VH представляет собой вариабельную область тяжелой цепи, VL представляет собой вариабельную область легкой цепи, и * представляют собой CDR-области. Эквивалентные значения приведены в виде кратных изменений относительно ранибизумаба.
[0412] Может быть использовано различное расположение последовательностей в конструкции анти-IL6 антитело/ловушка VEGF (VEGFR1/2) или указанные последовательности могут быть представлены в различном расположении. Фиг.21 иллюстрирует некоторые варианты возможных последовательностей вариабельной области анти-IL-6 тяжелой цепи. CDR подчеркнуты.
[0413] На основе результатов в случае вариантов ловушки VEGF и улучшенных анти-IL-6 паратопов конструировали и получали 216 молекул в двух разных конфигурациях, которые содержали комбинации последовательностей, представленных на фиг.19, 20, 21, 22 и 23. В случае первой конфигурации (VEGFR-анти-IL-6) ловушку VEGF, представленную последовательностями 1A-1D на фиг.19, располагали в начале белка с последующим двойным повтором линкера Gly-Gly-Gly-Gly-Ser (GS), представленного последовательностью 2A на фиг.20, который соединяет ловушку с N-концом тяжелой цепи анти-IL-6, представленной последовательностями 3A-3I на фиг.21. Эти конструкции могут быть спарены с легкими цепями, перечисленными в виде последовательностей 4A-4C на фиг.22. В некоторых вариантах реализации ловушку VEGF (на фиг.19) можно комбинировать с любой из легких цепей и/или анти-IL-6 тяжелых цепей, приведенных на любой из фигур или таблиц, представленных в настоящем описании.
[0414] В некоторых вариантах реализации любой из вариантов ловушки VEGF, представленных в настоящем описании, может быть спарен с любой из: последовательностей тяжелой и легкой цепи антител к IL-6, представленных в настоящем описании, и может быть дополнительно модифицирован любым из полимеров, представленных в настоящем описании. В некоторых вариантах реализации любой из вариантов ловушки VEGF, представленных в настоящем описании, может быть спарен с любой из: последовательностей вариабельной области тяжелой и легкой цепи антител к IL-6, представленных в настоящем описании, и может быть дополнительно модифицирован любым из полимеров, представленных в настоящем описании. В некоторых вариантах реализации любой из вариантов ловушки VEGF, представленных в настоящем описании, может быть спарен с любой из: 3 CDR тяжелой и легкой цепи (HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3) антител к IL-6, представленных в настоящем описании, и может быть дополнительно модифицирован любым из полимеров, представленных в настоящем описании. В некоторых вариантах реализации слитая конструкция IL-6-ловушка VEGF содержит компоненты на фиг.27.
[0415] В некоторых вариантах реализации последовательность конструкции анти-IL-6-ловушка VEGF (или двойного ингибитора, слитого белка или его конъюгата) содержит SEQ ID NO: 169 в качестве легкой цепи, SEQ ID NO: 170 в качестве тяжелой цепи, подвергнутой слиянию с VEGFR-ловушкой через линкер. В некоторых вариантах реализации последовательность конструкции анти-IL-6-ловушка VEGF (или двойного ингибитора, или слитого белка, или его конъюгата) по меньшей мере на 80% идентична молекуле, содержащей SEQ ID NO: 169 и 170, включая, например, по меньшей мере 85, 90, 95, 96, 97, 98, 99% или бóльшую идентичность комбинации SEQ ID NO: 170 и 169. В некоторых вариантах реализации последовательность конструкции анти-IL-6-ловушка VEGF (или ее конъюгата) по меньшей мере на 80% идентична молекуле, содержащей SEQ ID NO: 169, и по меньшей мере на 80% идентична молекуле, содержащей SEQ ID NO: 170, включая, например, по меньшей мере 85, 90, 95, 96, 97, 98, 99% или бóльшую идентичность SEQ ID NO: 170, и по меньшей мере 85, 90, 95, 96, 97, 98, 99% или бóльшую идентичность 169. В некоторых вариантах реализации CDR остаются такими, как изображены в любой из CDR, представленных в настоящем описании для анти-IL-6 антитела (включая указанные области в SEQ ID NO: 170 и 169), в то время как остальные последовательности (тяжелая и легкая цепь, вариабельные области тяжелой и легкой цепи и/или полные слитые последовательности (такие как SEQ ID NO 169 и 170)) можно варьировать таким образом, чтобы полная последовательность была по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% или больше идентична исходной последовательности. В некоторых вариантах реализации CDR можно варьировать путем 1, 2 или 3 консервативных изменений, в то время как остальные последовательности (тяжелая и легкая цепь, вариабельные области тяжелой и легкой цепи и/или полные слитые последовательности (такие как SEQ ID NO 169 и 170)) можно варьировать таким образом, чтобы полная последовательность была по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% или больше идентична исходной последовательности. В некоторых вариантах реализации последовательность ловушки (показанная серым цветом на фиг.27 или на фиг.19) может сохранять по меньшей мере 80, 85, 90, 95, 96, 97, 98, 99 процентов идентичности или больше, и CDR можно варьировать путем 1, 2 или 3 консервативных изменений, в то время как остальные последовательности (тяжелая и легкая цепь, вариабельные области тяжелой и легкой цепи и/или полные слитые последовательности (такие как SEQ ID NO 169 и 170)) можно варьировать таким образом, чтобы полная последовательность была по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99% или больше идентична исходной последовательности.
[0416] В некоторых вариантах реализации слитый белок (или двойной ингибитор) или его конъюгат содержит 1, 2, 3, 4, 5 или все 6 CDR, представленных в настоящем описании. В некоторых вариантах реализации слитый белок или его конъюгат содержит по меньшей мере SEQ ID NO: 49, 50 и 51 (или вариант, содержащий 1, 2 или 3 консервативные замены). В некоторых вариантах реализации слитый белок (или двойной ингибитор) или его конъюгат содержит по меньшей мере SEQ ID NO: 172, 173 и 174 (или вариант, содержащий 1, 2 или 3 консервативные замены). В некоторых вариантах реализации слитый белок или его конъюгат содержит по меньшей мере SEQ ID NO: 49, 50 и 51 (или вариант, содержащий 1, 2 или 3 консервативные замены) и по меньшей мере SEQ ID NO: 172, 173 и 174 (или вариант, содержащий 1, 2 или 3 консервативные замены). В некоторых вариантах реализации эти 6 CDR (или их варианты) содержатся в каркасной области антитела человека. В некоторых вариантах реализации CDR расположены в каркасной области любого из антител, представленных в настоящем описании, или их вариантов, которые по меньшей мере на 80, 85, 90, 95, 96, 97, 98, 99 процентов или больше идентичны вариабельным областям тяжелой и/или легкой цепи, представленным в настоящем описании, включая, без ограничения, те, которые приведены в таблицах 1 для тяжелой цепи и 2 для легкой цепи. В некоторых вариантах реализации слитый белок (или двойной ингибитор) или его конъюгат содержит по меньшей мере SEQ ID NO: 49, 50 и 51 (или вариант, содержащий 1, 2 или 3 консервативные замены) и по меньшей мере SEQ ID NO: 172, 173 и 174 (или вариант, содержащий 1, 2 или 3 консервативные замены), и эти CDR (включая варианты) заменены на CDR в одной или более последовательностях в таблице 11 или в варианте в таблице 11 (где без учета CDR в таблице 11, остальная последовательность по меньшей мере на 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99,5 процентов или больше идентична участку, содержащему тяжелую цепь, и участку легкой цепи в таблице 11). Таким образом, в некоторых вариантах реализации эти 6 CDR (и их варианты) могут быть заменены не только на CDR последовательностей, указанных в таблицах 1 и 2, но также на их варианты. В некоторых вариантах реализации CDR, представленные в таблицах 3, 4, 5, исключены как возможные области в вариантах возможных CDR. В некоторых вариантах реализации антитело (включая его фрагменты) представляет собой антитело, которое по меньшей мере на 80% идентично антителу, содержащему вариабельные области тяжелой и легкой цепи, представленные на фиг.27, при сохранении конкретных CDR на фиг.27 (таким образом в этом варианте реализации отсутствует вариация в CDR-участке).
[0417] В некоторых вариантах реализации предложено выделенное антагонистическое антитело к IL-6. Оно может содержать вариабельную область тяжелой цепи, которая содержит тяжелую цепь, которая имеет последовательность по меньшей мере одной из SEQ ID NO: 7-13, 19-27, 89, 90, 256-262; и вариабельную область легкой цепи, которая содержит легкую цепь, которая имеет последовательность по меньшей мере одной из SEQ ID NO: 91-93, 28-30.
[0418] В некоторых вариантах реализации предложено выделенное антагонистическое антитело к IL-6, содержащее вариабельную область тяжелой цепи (VH), содержащую 3 определяющие комплементарность области: CDR1 VH, CDR2 VH и CDR3 VH, имеющие аминокислотную последовательность из CDR, перечисленных в SEQ ID NO: 256; и вариабельную область легкой цепи (VL), содержащую CDR1 VL, CDR2 VL и CDR3 VL, имеющие аминокислотную последовательность, выбранную из группы CDR, перечисленных в SEQ ID NO: 91-93.
[0419] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, которое связывается с IL-6. Указанное антитело содержит по меньшей мере одну из следующих мутаций согласно нумерации EU: L234A, L235A и G237A.
[0420] В некоторых вариантах реализации предложено выделенное антагонистическое антитело, которое связывается с IL-6, при этом указанное антитело содержит: CDRH1, представляющую собой CDRH1 в SEQ ID NO: 172; CDRH2, представляющую собой CDRH2 в SEQ ID NO: 173; CDRH3, представляющую собой CDRH3 в SEQ ID NO: 174; CDRL1, представляющую собой CDRL1 в SEQ ID NO: 199; CDRL2, представляющую собой CDRL2 в SEQ ID NO: 200; CDRL3, представляющую собой CDRL3 в SEQ ID NO: 201; по меньшей мере одну из следующих мутаций (нумерация EU): L234A, L235A и G237A; и по меньшей мере одну из следующих мутаций (нумерация EU): Q347C или L443C.
[0421] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит последовательности анти-IL-6 вариабельной области тяжелой цепи, выбранные из SEQ ID NO: 7-13, 89, 90 и/или 256-262.
[0422] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит последовательности ловушки VEGF, выбранные из по меньшей мере одной из SEQ ID NO: 145, 15, 16 или 17.
[0423] В некоторых вариантах реализации линкерная последовательность представляет собой SEQ ID NO: 18.
[0424] В некоторых вариантах реализации последовательность тяжелой цепи для анти-IL-6 молекул выбрана из по меньшей мере одной из SEQ ID NO: 19-27 или содержит по меньшей мере последовательность в одной из SEQ ID NO: 89, 90, 256-262.
[0425] В некоторых вариантах реализации последовательность легкой цепи для анти-IL-6 молекулы содержит по меньшей мере 1, 2 или 3 CDR легкой цепи из по меньшей мере одной из SEQ ID NO: 76-84.
[0426] В некоторых вариантах реализации последовательность тяжелой цепи для анти-IL-6 молекулы содержит по меньшей мере 1, 2 или 3 CDR тяжелой цепи из по меньшей мере одной из SEQ ID NO: 49-75.
[0427] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит последовательность VEGFR-Fc из по меньшей мере одной из SEQ ID NO: 85-88.
[0428] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит одну или более последовательностей в любых одной или более из SEQ ID NO: 7-13, 145, 15-17, 18-84.
[0429] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит IL-6 VH; IL-6 VL; IL-6 Fc; ловушку VEGF; и линкер. В некоторых вариантах реализации IL-6 VH содержит последовательность из последовательности IL6 VH в любой из SEQ ID NO: 19-27, 31-39, 89, 90 или 256-262. В некоторых вариантах реализации IL-6 VL содержит последовательность из последовательности IL6 VL в любой из SEQ ID NO: 28-30 или 91-93. В некоторых вариантах реализации Fc содержит последовательность из последовательности Fc в любой из SEQ ID NO: 40-48. В некоторых вариантах реализации ловушка VEGF содержит последовательность из последовательности ловушки VEGF в любой из SEQ ID NO: 145, 15, 16 или 17.
[0430] В некоторых вариантах реализации предложен слитый белок, содержащий IL-6 VH; IL-6 VL; IL-6 Fc; и ловушку VEGF, при этом указанный слитый белок изменяет пролиферацию ЭКПВЧ. В некоторых вариантах реализации изменение пролиферации ЭКПВЧ ингибирует пролиферацию, опосредуемую VEGF/IL6.
[0431] В некоторых вариантах реализации предложен слитый белок, содержащий последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 263 и по меньшей мере на 80% идентична SEQ ID NO: 117. Указанный слитый белок дополнительно конъюгирован с полимером. В некоторых вариантах реализации слитый белок по меньшей мере на 95% идентичен SEQ ID NO: 263 и по меньшей мере на 95% идентичен SEQ ID NO: 117. В некоторых вариантах реализации белок содержит по меньшей мере а) SEQ ID NO: 172, 173, 174, 199, 200 и 201 или b) замены 1, 2 или 3 аминокислот в SEQ ID NO: 172, 173, 174, 199, 200 и/или 201, при этом указанная замена представляет собой консервативную замену.
[0432] В некоторых вариантах реализации любая из конструкций, представленных в настоящем описании, может содержать одну или более мутаций в положениях 94 и/или 95 последовательности VEGFR. В некоторых вариантах реализации указанная мутация (мутации) может представлять собой T94I и H95I. В некоторых вариантах реализации она уменьшает любое расщепление белка VEGFR (как подробно описано в примерах ниже).
[0433] В некоторых вариантах реализации (в которых ориентация представляет собой анти-IL-6-VEGFR) вариабельный и константный домены тяжелой цепи, представленные тяжелой цепью - Fab, последовательностями 5A-5I на фиг.23, соединяют с ловушкой VEGF, представленной последовательностями 1A-1D на фиг.19, через линкер GS, представленный последовательностью 2A на фиг.20, а затем Fc-домен, представленный тяжелой цепью - Fc, последовательностями 5A-5I на фиг.23, подвергают слиянию с C-концевой областью ловушки VEGF. Таким образом, ловушка VEGF «зажата» между Fab- и Fc-областями антитела. В некоторых вариантах реализации эти конструкции могут быть спарены с легкими цепями, перечисленными в виде последовательностей 4A-4C на фиг.22.
[0434] На фиг.19 показаны некоторые варианты последовательностей ловушки VEGF. Вариации между указанными последовательностями подчеркнуты и выделены жирным шрифтом. В некоторых вариантах реализации любая из слитых конструкций, представленных в настоящем описании, может содержать любые из выделенных жирным шрифтом вариаций, представленных на этой фигуре или на любой из фигур в настоящем описании.
[0435] На фиг.20 показаны некоторые варианты линкерной (GS) последовательности Gly-Gly-Gly-Gly-Ser с двойным повтором. В некоторых вариантах реализации можно использовать более одной единицы последовательности (например, можно использовать две для двойного повтора, три или более). В некоторых вариантах реализации могут быть использованы другие линкерные последовательности. В некоторых вариантах реализации могут быть использованы альтернативные линкерные последовательности.
[0436] На фиг.21 представлены последовательности тяжелой цепи для некоторых вариантов анти-IL-6 молекул. CDR подчеркнуты. В некоторых вариантах реализации любая из последовательностей, представленных в настоящем описании, может быть использована вместо других последовательностей тяжелой цепи, которые являются частью слитых конструкций в настоящем описании.
[0437] На фиг.22 показаны некоторые варианты последовательностей легкой цепи для анти-IL-6 молекул. CDR подчеркнуты. В некоторых вариантах реализации любая из последовательностей, представленных в настоящем описании, может быть использована вместо других последовательностей легкой цепи, которые являются частью слитых конструкций в настоящем описании.
[0438] На фиг.23 показаны некоторые варианты последовательностей тяжелой цепи (разделенной на Fab и Fc) для анти-IL-6 молекул. CDR подчеркнуты. В некоторых вариантах реализации любая из последовательностей, представленных в настоящем описании, может быть использована вместо других последовательностей тяжелой цепи, которые являются частью слитых конструкций в настоящем описании.
[0439] На фиг.24A-24B представлены комбинации CDR с фиг.21-23. На фиг.24A представлены последовательности CDR тяжелой цепи, определенные на фиг.21 и 23. На фиг.24B представлены последовательности CDR легкой цепи, определенные на фиг.22. В некоторых вариантах реализации вместо использования в конструкции целой вариабельной области тяжелой и/или легкой цепи можно использовать только CDR-области с одной или более фигур, представленных в настоящем описании.
[0440] В некоторых вариантах реализации слитая конструкция VEGFR-Fc представляет собой часть искусственно сконструированной каркасной области IgG1. В дополнение к конструкциям VEGFR-анти-IL-6 и анти-IL-6-VEGFR может быть получена молекула анти-VEGF (VEGFR-Fc), содержащая ловушку VEGF с устойчивыми к расщеплению вариантами, описанными выше, и искусственно сконструированный Fc-домен IgG1, представленный последовательностями 6A-6D на фиг.25. Указанный Fc-домен для этих молекул может содержать замены L234A, L235A и G237A, которые минимизируют эффекторную функцию, и L443C, которая обеспечивает сайт-специфичную конъюгацию с биополимером на основе фосфорилхолина, увеличивающим период полувыведения (положения остатков соответствуют нумерации EU). На фиг.25 показаны некоторые варианты последовательности VEGFR-Fc. Вариация между указанными последовательностями подчеркнута и выделена жирным шрифтом. Следует отметить, что эти вариации и/или комбинации могут обеспечивать биотерапевтическое средство, которое еще больше превосходит и/или является альтернативой другим соединениям, применяемым в настоящее время в терапии.
[0441] Как показано на фиг.26A-26C, VEGFR-Fc и Эйлеа связываются с VEGF со схожей аффинностью. Фиг.26A иллюстрирует анализы Biacore для VEGFR-Fc. Фиг.26B иллюстрирует анализы Biacore для Эйлеа. На фиг.26C представлены результаты экспериментов (Ka, Kd, KD и Rmax) для VEGFR-Fc и Эйлеа.
[0442] В некоторых вариантах реализации любая одна или более последовательностей для указанной аминокислотной последовательности на любой одной или более фиг.18-23 и 25 могут быть заменены на соответствующую структуру любого из других вариантов реализации, представленных в настоящем описании, или заменены любой из других последовательностей, представленных в настоящем описании. Например, любую из последовательностей на фиг.13C-13E, 19 или 25 можно использовать с любой из тяжелых цепей IL-6 на фиг.18, 21 или 23, с любым из линкеров, представленных в настоящем описании (например, фиг.20), с любой из легких цепей (фиг.22). В некоторых вариантах реализации конструкция представляет собой конфигурацию VEGFR-анти-IL-6 и содержит комбинацию одной из последовательностей 1A-1D (фиг.19), связанной с линкером (например, фиг.20, последовательность 2A), связанным с последовательностью тяжелой цепи IL-6 (например, фиг.21, последовательность 3A или 3B), связанной с последовательностью легкой цепи (например, фиг.22, последовательность 4A). В некоторых вариантах реализации любую из последовательностей 1A-1D (фиг.19) можно комбинировать с линкером (фиг.20) и с последовательностью анти-IL-6 тяжелой цепи (фиг.21, последовательности 3A-3I) и с последовательностью анти-IL-6 легкой цепи (фиг.22, последовательность 4A-4C). В некоторых вариантах реализации любая из других соответствующих последовательностей для любой конкретной единицы конструкции, представленной в настоящем описании, может быть заменена на любую из этих единиц или быть вместо нее.
[0443] В некоторых вариантах реализации двойной ингибитор обеспечивает один или более синергетических результатов. В некоторых вариантах реализации двойной ингибитор синергетически превосходит каждый его участок, вводимый в виде монотерапии (например, комбинация терапии IL-6 и терапии VEGF). В некоторых вариантах реализации комбинированная конструкция VEGFR-IL-6 обеспечивает синергетический результат, продемонстрированный результатами анализа пролиферации (например, в то время как двойной ингибитор оказывает очевидное влияние на пролиферацию ЭКПВЧ, важный компонент ангиогенеза, ни Эйлеа, ни анти-IL-6 не обеспечивают никакого изменения). Таким образом, в некоторых вариантах реализации предложен способ контроля пролиферации ЭКПВЧ, в котором применяют конструкцию VEGFR-анти-IL-6, предложенную согласно настоящему изобретению.
[0444] В других вариантах реализации слитая конструкция просто обеспечивает дополнительный полезный эффект за счет достижения как результатов ловушки VEGF, так и результатов для анти-IL-6.
[0445] В некоторых вариантах реализации любой один или более компонентов в любых одной или более таблицах, представленных в настоящем описании, могут быть комбинированы с получением таким образом конструкции VEGFR-анти-IL-6 или с получением таким образом конструкции анти-IL-6-VEGFR. В некоторых вариантах реализации указанная конструкция будет также содержать Fc-область. В некоторых вариантах реализации указанная Fc-область может представлять собой Fc-область человека.
[0446] В некоторых вариантах реализации одни только CDR (с любым конкретным каркасным участком или без него) можно использовать в качестве участка, представляющего собой антитело. Таким образом, любую одну или более (например, 3 или 6) CDR в каждой цепи или паре цепей (тяжелой и легкой) можно использовать из любой из таблиц или фигур, представленных в настоящем описании, в комбинации с любым из других компонентов (VEGFR и/или линкеры). CDR и/или 1, 2 или 3 CDR тяжелой цепи из любой из конструкций IL-6, представленных в настоящем описании, можно комбинировать с любой одной или более последовательностями VEGFR, представленными в настоящем описании. В некоторых вариантах реализации любую вариабельную область тяжелой и/или легкой цепи из любой из конструкций IL-6, представленных в настоящем описании, можно комбинировать с любой одной или более последовательностями VEGFR, представленными в настоящем описании. В некоторых вариантах реализации в конечной конструкции также может быть представлен любой из линкеров.
[0447] В некоторых вариантах реализации также может быть получена любая последовательность нуклеиновой кислоты, кодирующая любую одну или более аминокислотных последовательностей, представленных в настоящем описании (например, выделенная, в векторе или в клетке и т.д.).
[0448] В некоторых вариантах реализации любой один или более полимеров для конъюгации с образованием биоконъюгата можно комбинировать с любой из конструкций VEGFR-IL6, представленных в настоящем описании.
[0449] В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1A-1D (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3A или 3B на фиг.21), связанной с легкой цепью 4А на фиг.22.
[0450] В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1A (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3A на фиг.21), связанной с легкой цепью 4А на фиг.22. В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1B (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3A на фиг.21), связанной с легкой цепью 4А на фиг.22. В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1C (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3A на фиг.21), связанной с легкой цепью 4А на фиг.22. В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1D (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3A на фиг.21), связанной с легкой цепью 4А на фиг.22.
[0451] В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1A (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3B на фиг.21), связанной с легкой цепью 4А на фиг.22. В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1B (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3B на фиг.21), связанной с легкой цепью 4А на фиг.22. В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1C (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3B на фиг.21), связанной с легкой цепью 4А на фиг.22. В некоторых вариантах реализации комбинация VEGFR-анти-IL-6 представляет собой последовательность 1D (фиг.19) для VEGFR, связанную с линкером (2A на фиг.20), связанным с тяжелой цепью (3B на фиг.21), связанной с легкой цепью 4А на фиг.22.
[0452] В некоторых вариантах реализации любую из последовательностей антитела к IL-6 можно использовать с любой из последовательностей VEGFR, представленных в настоящем описании. В некоторых вариантах реализации любые последовательности 1, 2, 3, 4, 5 или 6 CDR (1, 2 или 3 легкой цепи и 1, 2 или 3 тяжелой цепи) анти-IL-6 можно использовать с VEGFR в любом из расположений.
[0453] В некоторых вариантах реализации можно использовать любую легкую и/или тяжелую цепь анти-IL-6 антитела, содержащую любую одну или более точечных мутаций, представленных в любых одной или более таблицах 30-33. В некоторых вариантах реализации в слитой конструкции используют анти-IL-6 конструкцию, содержащую точечную мутацию в любом одном или более положениях, определенных в любых одной или более таблицах 30-33. В некоторых вариантах реализации измененные положения представляют собой положения, подчеркнутые и выделенные жирным шрифтом на соответствующих фигурах как показывающие измененные остатки. В некоторых вариантах реализации конструкция представляет собой конструкцию, содержащую изменения в тяжелой цепи, включающие одно или более из: S35H, G66D, L100A или L100S, и изменения в легкой цепи, включающие одно или более из: M32L, M50D, N52S или M88Q. При описании положения CDR следуют в порядке их появления в вариабельном домене (например, S35H, G66D…). При описании положения Fc соответствуют нумерации EU (например, L234A, L235A…). В некоторых вариантах реализации конструкция (например, антитело к IL-6 и/или IL-6-ловушка VEGF, и/или биоконъюгат (с одним или более биополимерами, представленными в настоящем описании) конструкции IL-6-ловушка VEGF) содержит в тяжелой и легкой цепи 1, 2, 3, 4, 5, 6, 7 или 8 из следующего: S35H, G66D, L100A и/или L100S в тяжелой цепи и M32L, M50D, N52S и/или M88Q в легкой цепи. В некоторых вариантах реализации эти точечные мутации можно использовать в любом из антител к Il-6 или конструкций, содержащих антитела к Il-6, представленные в настоящем описании.
[0454] Как очевидно для специалиста в этой области техники, настоящее изобретение предусматривает множество конструкций. В целом, эти конструкции можно подразделить на: антитела к IL-6 и/или конструкции анти-IL-6-ловушка VEGF (слияние антитела и ловушки VEGF), и/или биоконъюгат конструкции IL-6-ловушка VEGF (слияние с одним или более биополимерами, представленными в настоящем описании), и/или конструкции ловушки VEGF, и/или биополимеры ловушки VEGF. Таким образом, любой из отдельных компонентов, представленных в настоящем описании (и их вариантов), можно использовать отдельно или в комбинации друг с другом. В некоторых вариантах реализации конструкции любой из этих групп (антитела к IL-6 и/или конструкции анти-IL-6-ловушка VEGF (слияние антитела и ловушки VEGF), и/или биоконъюгат конструкции IL-6-ловушка VEGF (слияние с одним или более биополимерами, представленными в настоящем описании), и/или конструкции ловушки VEGF, и/или биополимеры ловушки VEGF) можно использовать для любого из способов или других композиций, представленных в настоящем описании. Например, любой из этих вариантов можно использовать для любого из способов лечения, описанных в отношении терапии конструкцией IL-6-ловушка VEGF и/или терапии IL-6, и/или биоконъюгатами конструкции IL-6-ловушка VEGF. Как очевидно, характеристики и свойства каждой конструкции могут быть различными для каждого лечения в соответствии со свойствами компонентов, подробно описанных в настоящем документе. Таким образом, приведенное в настоящем документе описание в отношении, например, линий клеток, нуклеиновых кислот, видов терапии и других вариантов реализации предусмотрено не только именно для конструкции анти-IL-6-ловушка VEGF и/или ее биоконъюгатов, но также для антител (и, например, их фрагментов), биоконъюгатов антител и конструкций ловушки VEGF отдельно от конструкций анти-IL-ловушка VEGF и/или биоконъюгатов. Например, в некоторых вариантах реализации любую из точечных мутаций (или их комбинаций) для IL-6 можно использовать в антителе (или его связывающем фрагменте) для лечения, которое основано только на антителе, а не на антителе и ловушке VEGF. Таким образом, точечные мутации для ловушки VEGF и антител к IL-6 не ограничиваются контекстом слитых структур (хотя они предложены), поскольку контекст также описывает такие конструкции (и их применение, и способ получения) отдельно от слитых структур. Следует отметить, что вместо повтора всех таких вариантов реализации отдельно, любое такое описание для одной из структур (антитела к IL-6 и/или конструкции анти-IL-6-ловушка VEGF (слияние антитела и ловушки VEGF), и/или биоконъюгат конструкции IL-6-ловушка VEGF (слияние с одним или более биополимерами, представленными в настоящем описании), и/или конструкции ловушки VEGF, и/или биополимеры ловушки VEGF) применимо и определяет варианты для всех остальных таких структур (антитела к IL-6 и/или конструкции анти-IL-6-ловушка VEGF (слияние антитела и ловушки VEGF), и/или биоконъюгат конструкции IL-6-ловушка VEGF (слияние с одним или более биополимерами, представленными в настоящем описании), и/или конструкции ловушки VEGF, и/или биополимеры ловушки VEGF).
[0455] В некоторых вариантах реализации любые из конструкций ловушки VEGF предусмотрены для применения в качестве композиций, компонентов или видов терапии, включая, например, те, что на фиг.13C-13E, 19 и 25 из вариантов реализации, представленных в настоящем описании.
[0456] В некоторых вариантах реализации любые из конструкций анти-IL-6 антител предусмотрены для применения в качестве композиций, компонентов или видов терапии, включая, например, те, что в таблицах 1 и 3, 4, 5, и на фиг.5, 18, 21, 22, 23 и 24 из вариантов реализации, представленных в настоящем описании.
[0457] В некоторых вариантах реализации слитый белок содержит мутацию в положении 94 в последовательности ловушки VEGF, в положении 95 в последовательности ловушки VEGF или в T94I и H95I в последовательности ловушки VEGF.
[0458] В некоторых вариантах реализации предложен двойной ингибитор VEGFR-анти-IL-6. Указанный двойной ингибитор VEGFR-анти-IL-6 содержит слитую конструкцию ловушка-антитело из анти-IL6 антитела и ловушки VEGF (VEGFR1/2), при этом двойной ингибитор содержит по меньшей мере одну точечную мутацию в последовательности VEGFR для уменьшения расщепления последовательности VEGFR. В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 имеет молекулярную массу 1,0 МДа.
[0459] В некоторых вариантах реализации двойной ингибитор VEGR-анти-IL-6 обеспечивает терапию воспалительных заболеваний сетчатки.
[0460] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит константную область тяжелой цепи, константную область легкой цепи, антигенсвязывающий фрагмент, кристаллизующийся фрагмент (Fc), рецептор фактора роста эндотелия сосудов (VEGFR), вариабельную область тяжелой цепи, вариабельную область легкой цепи и CDR-области.
[0461] В некоторых вариантах реализации последовательности анти-IL-6 могут быть выбраны из вариантов VI, VII, VIII, IX, X, XI или XII на фиг.18.
[0462] В некоторых вариантах реализации последовательность ловушки VEGF может быть выбрана из по меньшей мере одного из вариантов 1A, 1B, 1C или 1D на фиг.19.
[0463] В некоторых вариантах реализации для двойного ингибитора VEGR-анти-IL-6 последовательность тяжелой цепи для анти-IL-6 молекул может быть выбрана из по меньшей мере одного из вариантов 3A, 3B, 3C, 3D, 3E, 3F, 3G, 3H или 3I на фиг.21.
[0464] В некоторых вариантах реализации для двойного ингибитора VEGFR-анти-IL-6 последовательность легкой цепи для анти-IL-6 молекул содержит по меньшей мере 1, 2 или 3 CDR легкой цепи из по меньшей мере одного из вариантов 4A, 4B или 4C на фиг.24 В.
[0465] В некоторых вариантах реализации для двойного ингибитора VEGFR-анти-IL-6 последовательность тяжелой цепи для анти-IL-6 молекулы содержит по меньшей мере 1, 2 или 3 CDR тяжелой цепи из по меньшей мере одного из вариантов 5A, 5B, 5C, 5D, 5E, 5F, 5G, 5H или 5I на фиг.24А.
[0466] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит последовательность VEGFR-Fc из по меньшей мере одного из вариантов 6A, 6B, 6C или 6D на фиг.25.
[0467] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит одну или более последовательностей на фиг.18-25.
[0468] В некоторых вариантах реализации двойной ингибитор VEGFR-анти-IL-6 содержит IL-6 VH, IL-6 VL, IL-6 Fc, ловушку VEGF и линкер. В некоторых вариантах реализации IL-6 VH содержит последовательность из последовательности IL6 VH на фиг.21 или 23. В некоторых вариантах реализации IL-6 VL содержит последовательность из последовательности IL6 VL на фиг.22. В некоторых вариантах реализации Fc содержит последовательность из последовательности Fc на фиг.23. В некоторых вариантах реализации ловушка VEGF содержит последовательность из последовательности ловушки VEGF.
[0469] В некоторых вариантах реализации предложена белковая конструкция, содержащая: по меньшей мере 3 CDR тяжелой цепи; по меньшей мере 3 CDR легкой цепи; последовательность ловушки VEGF; и линкерную последовательность, при этом каждая из указанных последовательностей выбрана из последовательности на фиг.18-25.
[0470] В некоторых вариантах реализации предложен слитый белок, содержащий: IL-6 VH, IL-6 VL, IL-6 Fc, ловушку VEGF, и при этом указанный слитый белок изменяет пролиферацию ЭКПВЧ. В некоторых вариантах реализации каждая последовательность выбрана из последовательности на фиг.18-25.
Полинуклеотиды, векторы и клетки-хозяева
[0471] Согласно настоящему изобретению также предложены полинуклеотиды, кодирующие любое из антител (и/или конструкций АТ к IL-6-ловушка VEGF), включая фрагменты антител и модифицированные антитела, описанные в настоящем документе. В соответствии с другим аспектом настоящего изобретения предложен способ получения любого из полинуклеотидов, описанных в настоящем документе. Полинуклеотиды можно получать и экспрессировать способами, известными в этой области техники. Соответственно, согласно настоящему изобретению предложены полинуклеотиды или композиции, в том числе фармацевтические композиции, содержащие полинуклеотиды, кодирующие любое из антител-антагонистов IL-6 (и/или конструкций АТ к IL-6-ловушка VEGF), представленных в настоящем описании.
[0472] Настоящее изобретение также включает полинуклеотиды, комплементарные любым таким последовательностям. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут представлять собой молекулы ДНК (геномной, кДНК или синтетической) или РНК. Молекулы РНК включают молекулы гетерогенной ядерной РНК (гяРНК), которые содержат интроны и соответствуют молекуле ДНК один к одному, и молекулы мРНК, которые не содержат интронов. В полинуклеотиде согласно настоящему изобретению могут, но не обязательно, присутствовать дополнительные кодирующие или некодирующие последовательности, и полинуклеотид может быть, но не обязательно, связан с другими молекулами и/или подложкой.
[0473] Полинуклеотиды могут содержать нативную последовательность (т.е. эндогенную последовательность, кодирующую антитело или его фрагмент) или могут содержать вариант такой последовательности. Варианты полинуклеотидов содержат одну или более замен, присоединений, делеций и/или встраиваний таким образом, что иммунореактивность кодируемого полипептида не снижается относительно нативной иммунореактивной молекулы. Влияние на иммунореактивность кодируемого полипептида можно, в целом, оценить, как описано в настоящем документе. Варианты предпочтительно демонстрируют по меньшей мере примерно 70% идентичность, более предпочтительно по меньшей мере примерно 80% идентичность, более предпочтительно по меньшей мере примерно 90% идентичность и наиболее предпочтительно по меньшей мере примерно 95% идентичность последовательности полинуклеотида, кодирующей нативное антитело или его фрагмент.
[0474] Говорят, что две полинуклеотидные или полипептидные последовательности «идентичны», если последовательность нуклеотидов или аминокислот в этих двух последовательностях одинакова при выравнивании для максимального соответствия, как описано ниже. Сравнение двух последовательностей, как правило, осуществляют путем сравнения указанных последовательности в окне сравнения для идентификации и сравнения локальных областей сходства последовательностей. В настоящем описании термин «окно сравнения» относится к сегменту по меньшей мере из примерно 20 следующих друг за другом положений, как правило, от 30 до примерно 75 или от 40 до примерно 50, в котором последовательность можно сравнить с эталонной последовательностью из такого же числа следующих друг за другом положений после оптимального выравнивания этих двух последовательностей.
[0475] Оптимальное выравнивание последовательностей для сравнения может быть выполнено с использованием программы MegAlign® в пакете программного обеспечения для биоинформатики Lasergene® (DNASTAR®, Inc., Мэдисон, Висконсин) с использованием параметров по умолчанию. Эта программа включает несколько схем выравнивания, описанных в следующих источниках: Dayhoff, M.O., 1978, A model of evolutionary change in proteins - Matrices for detecting distant relationships. In Dayhoff, M.O. (ed.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Washington DC Vol.5, Suppl. 3, pp.345-358; Hein J., 1990, Unified Approach to Alignment and Phylogenes pp.626-645 Methods in Enzymology vol. 183, Academic Press, Inc., San Diego, CA; Higgins, D.G. and Sharp, P.M., 1989, CABIOS 5:151-153; Myers, E.W. and Muller W., 1988, CABIOS 4:11-17; Robinson, E.D., 1971, Comb. Theor. 11:105; Santou, N., Nes, M., 1987, Mol. Biol. Evol. 4:406-425; Sneath, P.H.A. and Sokal, R.R., 1973, Numerical Taxonomy the Principles and Practice of Numerical Taxonomy, Freeman Press, San Francisco, CA; Wilbur, W.J. and Lipman, D.J., 1983, Proc. Natl. Acad. Sci. USA 80:726-730.
[0476] Предпочтительно, «процент идентичности последовательностей» определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения по меньшей мере из 20 положений, при этом часть полинуклеотидной или полипептидной последовательности в указанном окне сравнения может содержать присоединения или делеции (т.е. гэпы), составляющие 20 процентов или менее, как правило, от 5 до 15 процентов или от 10 до 12 процентов по сравнению с эталонными последовательностями (которые не содержат присоединений или делеций), для оптимального выравнивания двух последовательностей. Процент рассчитывают путем определения количества положений, в которых в обеих последовательностях встречаются идентичные основания нуклеиновых кислот или аминокислотные остатки, с получением количества совпавших положений, деления количества совпавших положений на общее количество положений в эталонной последовательности (т.е. размер окна) и умножения результатов на 100 с получением процента идентичности последовательностей.
[0477] Варианты могут быть также или в качестве альтернативы по существу гомологичны нативному гену или его части, или набору. Такие варианты полинуклеотидов способны гибридизоваться в умеренно жестких условиях со встречающейся в природе последовательностью ДНК, кодирующей нативное антитело (или комплементарной последовательностью).
[0478] Подходящие «умеренно жесткие условия» включают предварительную промывку в растворе 5 X SSC, 0,5% SDS, 1,0 мM ЭДТА (pH 8,0); гибридизацию при 50°C-65°C, 5 X SSC в течение ночи; с последующей промывкой два раза при 65°C в течение 20 минут каждым из 2X, 0,5X и 0,2X SSC, содержащим 0,1% SDS.
[0479] В настоящем описании «очень жесткие условия» или «условия высокой жесткости» представляют собой условия, в которых: (1) используют низкую ионную силу и высокую температуру для промывки, например, 0,015 М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50°С; (2) во время гибридизации используют денатурирующий агент, такой как формамид, например, 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1% Фиколлом/0,1% поливинилпирролидоном/50 мМ натрий-фосфатным буфером при рН 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°С; или (3) используют 50% формамид, 5 x SSC (0,75 М NaCl, 0,075 М цитрат натрия), 50 мМ фосфат натрия (рН 6,8), 0,1% пирофосфат натрия, 5 х раствор Денхардта, обработанную ультразвуком ДНК из молок лососевых (50 мкг/мл), 0,1% SDS и 10% сульфат декстрана при 42°С, с промывками при 42°С в 0,2 х SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°С с последующей промывкой высокой жесткости, состоящей из 0,1 х SSC, содержащего ЭДТА, при 55°С. Специалисту известно как корректировать температуру, ионную силу и т.д. при необходимости, для согласования с такими факторами, как длина зонда и т.п.
[0480] Для специалиста в этой области техники очевидно, что из-за вырожденности генетического кода существует много нуклеотидных последовательностей, кодирующих полипептид, описанный в настоящем документе. Некоторые из этих полинуклеотидов имеют минимальную гомологию с нуклеотидной последовательностью какого-либо нативного гена. Однако полинуклеотиды, которые варьируются вследствие различий в использовании кодонов, в частности, предусмотрены настоящим изобретением. Более того, в объем настоящего изобретения включены аллели генов, содержащих полинуклеотидные последовательности, представленные в настоящем описании. Аллели представляют собой эндогенные гены, измененные в результате одной или более мутаций, таких как делеции, присоединения и/или замены нуклеотидов. Полученные мРНК и белок могут, но не обязательно, иметь измененную структуру или функцию. Аллели могут быть идентифицированы с применением стандартных методов (таких как гибридизация, амплификация и/или сравнение последовательностей базы данных).
[0481] Полинуклеотиды согласно настоящему изобретению могут быть получены с применением химического синтеза, рекомбинантных методов или ПЦР. Способы химического синтеза полинуклеотидов хорошо известны в этой области техники и не требуют подробного описания в настоящем документе. Специалист в этой области техники может использовать последовательности, представленные в настоящем описании, и коммерческий ДНК-синтезатор для получения желаемой последовательности ДНК.
[0482] Для получения полинуклеотидов с применением рекомбинантных методов полинуклеотид, содержащий желаемую последовательность, может быть встроен в подходящий вектор, а вектор в свою очередь может быть введен в подходящую клетку-хозяина для репликации и амплификации, как описано ниже в настоящем документе. Полинуклеотиды могут быть введены в клетки-хозяева любыми способами, известными в этой области техники. Клетки трансформируют путем введения экзогенного полинуклеотида посредством прямого поглощения, эндоцитоза, трансфекции, F-спаривания или электропорации. После введения экзогенный полинуклеотид может содержаться в клетке в виде неинтегрированного вектора (такого как плазмида) или может быть интегрирован в геном клетки-хозяина. Полинуклеотид, амплифицированный таким образом, может быть выделен из клетки-хозяина способами, хорошо известными в этой области техники. См., например, Sambrook et al., 1989.
[0483] В качестве альтернативы, ПЦР позволяет репродуцировать последовательности ДНК. Технология ПЦР хорошо известна в этой области техники и описана в патентах США №№4683195, 4800159, 4754065 и 4683202, а также в PCR: The Polymerase Chain Reaction, Mullis et al. eds., Birkauswer Press, Boston, 1994.
[0484] РНК может быть получена путем использования выделенной ДНК в подходящем векторе и ее введения в подходящую клетку-хозяина. Когда указанная клетка реплицируется и ДНК транскрибируется в РНК, РНК может быть затем выделена с применением способов, хорошо известных специалисту в этой области техники, как указано, например, в Sambrook et al., 1989 выше.
[0485] Подходящие клонирующие векторы могут быть сконструированы в соответствии со стандартными способами или могут быть выбраны из большого числа клонирующих векторов, доступных в этой области техники. Хотя выбранный клонирующий вектор может варьироваться в зависимости от используемой клетки-хозяина, подходящие клонирующие векторы, в целом, обладают способностью к саморепликации, могут иметь одну мишень для конкретной рестрикционной эндонуклеазы и/или могут нести гены маркера, который можно использовать при отборе клонов, содержащих вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например, pUC18, pUC19, Bluescript (например, pBS SK+) и его производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговые ДНК и челночные векторы, такие как pSA3 и pAT28. Эти и многие другие клонирующие векторы доступны от коммерческих поставщиков, таких как BioRad, Strategene и Invitrogen.
[0486] Также предложены векторы экспрессии. Векторы экспрессии, как правило, представляют собой реплицируемые полинуклеотидные конструкции, которые содержат полинуклеотид согласно настоящему изобретению. Подразумевается, что вектор экспрессии должен реплицироваться в клетках-хозяевах либо в виде эписом, либо в виде составной часть хромосомной ДНК. Подходящие векторы экспрессии включают, но не ограничиваются ими, плазмиды, вирусные векторы, включая аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды и вектор (векторы) экспрессии, описанный в публикации PCT №WO 87/04462. Компоненты вектора могут, в целом, включать, но не ограничиваются ими, один или более следующих компонентов: сигнальную последовательность; точку начала репликации; один или более маркерных генов; подходящие элементы, контролирующие транскрипцию (такие как промоторы, энхансеры и терминатор). Для экспрессии (т.е. трансляции) также, как правило, необходим один или более элементов, контролирующих трансляцию, таких как сайты связывания рибосом, сайты инициации трансляции и стоп-кодоны.
[0487] Векторы, содержащие полинуклеотиды, представляющие интерес, могут быть введены в клетку-хозяина любым из ряда подходящих способов, включая электропорацию, трансфекцию с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию; и инфицирование (например, когда вектор представляет собой инфекционный агент, такой как вирус коровьей оспы). Выбор вводимых векторов или полинуклеотидов часто зависит от особенностей клетки-хозяина.
[0488] Согласно настоящему изобретению также предложены клетки-хозяева, содержащие любой из полинуклеотидов, описанных в настоящем документе. Для выделения генов, кодирующих представляющее интерес антитело, полипептид или белок, можно использовать любые клетки-хозяева, способные сверхэкспрессировать гетерологичные ДНК. Неограничивающие примеры клеток-хозяев млекопитающих включают, но не ограничиваются ими, клетки COS, HeLa и CHO. См. также публикацию PCT №WO 87/04462. Подходящие клетки-хозяева, не относящиеся к млекопитающим, включают прокариоты (такие как E.coli или B. subtillis) и дрожжи (такие как S. cerevisae, S. pombe; или K. lactis). Предпочтительно, клетки-хозяева экспрессируют кДНК на уровне, который примерно в 5 раз выше, более предпочтительно в 10 раз выше, более предпочтительно в 20 раз выше уровня для соответствующего эндогенного антитела или белка, представляющего интерес, если присутствует, в клетках-хозяевах. Скрининг клеток-хозяев в отношении специфического связывания с IL-6 осуществляют путем иммуноанализа или FACS. Может быть идентифицирована клетка, сверхэкспрессирующая антитело или белок, представляющий интерес.
[0489] Вектор экспрессии можно использовать для направления экспрессии антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF). Специалисту в этой области техники известно введение векторов экспрессии для достижения экспрессии экзогенного белка in vivo. См., например, патенты США №№6436908; 6413942; и 6376471. Введение векторов экспрессии включает местное или системное введение, включая инъекцию, пероральное введение, генную пушку или введение через катетер, и топическое введение. В другом варианте реализации вектор экспрессии вводят непосредственно в симпатический ствол или нервный узел, или в коронарную артерию, предсердие, желудочек или перикард.
[0490] Можно также использовать направленную доставку терапевтических композиций, содержащих вектор экспрессии или субгеномные полинуклеотиды. Способы опосредованной рецепторами доставки ДНК описаны, например, в Findeis et al., Trends Biotechnol., 1993, 11:202; Chiou et al., Gene Therapeutics: Methods And Applications Of Direct Gene Transfer, J.A. Wolff, ed., 1994; Wu et al., J. Biol. Chem., 1988, 263:621; Wu et al., J. Biol. Chem., 1994, 269:542; Zenke et al., Proc. Natl. Acad. Sci. USA, 1990, 87:3655; Wu et al., J. Biol. Chem., 1991, 266:338. Терапевтические композиции, содержащие полинуклеотид, вводят в диапазоне от примерно 100 нг до примерно 200 мг ДНК в случае местного введения в протоколе генной терапии. В протоколе генной терапии можно также использовать диапазоны концентрации от примерно 500 нг до примерно 50 мг, от примерно 1 мкг до примерно 2 мг, от примерно 5 мкг до примерно 500 мкг и от примерно 20 мкг до примерно 100 мкг ДНК. Терапевтические полинуклеотиды и полипептиды могут быть доставлены с использованием средств доставки гена. Указанное средство доставки гена может быть вирусного или невирусного происхождения (в целом, см. Jolly, Cancer Gene Therapy, 1994, 1:51; Kimura, Human Gene Therapy, 1994, 5:845; Connelly, Human Gene Therapy, 1995, 1:185; и Kaplitt, Nature Genetics, 1994, 6:148). Экспрессию таких кодирующих последовательностей можно индуцировать с использованием эндогенных промоторов млекопитающего или гетерологичных промоторов. Экспрессия кодирующей последовательности может быть либо конститутивной, либо регулируемой.
[0491] Векторы на основе вирусов для доставки желаемого полинуклеотида и экспрессии в желаемой клетке хорошо известны в этой области техники. Типичные средства доставки на основе вирусов включают, но не ограничиваются ими, рекомбинантные ретровирусы (см., например, публикации PCT №№WO 90/07936; WO 94/03622; WO 93/25698; WO 93/25234; WO 93/11230; WO 93/10218; WO 91/02805; патенты США №№5219740 и 4777127; патент Великобритании №2200651; и Европейский патент №0345242), векторы на основе альфавирусов (например, векторы на основе вируса Синдбис, вирус леса Семлики (ATCC VR-67; ATCC VR-1247), вирус Росс-Ривер (ATCC VR-373; ATCC VR-1246) и вирус венесуэльского энцефалита лошадей (ATCC VR-923; ATCC VR-1250; ATCC VR 1249; ATCC VR-532)) и векторы на основе аденоассоциированного вируса (AAV) (см., например, публикации PCT №№WO 94/12649, WO 93/03769; WO 93/19191; WO 94/28938; WO 95/11984 и WO 95/00655). Можно также использовать введение ДНК, связанной с инактивированным аденовирусом, как описано в Curiel, Hum. Gene Ther., 1992, 3:147.
[0492] Можно также использовать невирусные средства и способы доставки, включая, но не ограничиваясь ими, поликатионную конденсированную ДНК, связанную или несвязанную только с инактивированным аденовирусом (см., например, Curiel, Hum. Gene Ther., 1992, 3:147); ДНК, связанную с лигандом (см., например, Wu, J. Biol. Chem., 1989, 264:16985); средства доставки на основе эукариотических клеток (см., например, патент США №5814482; публикации PCT №№WO 95/07994; WO 96/17072; WO 95/30763; и WO 97/42338) и нейтрализацию заряда нуклеиновой кислоты или слияние с клеточными мембранами. Можно также использовать депротеинизированную ДНК. Типичные способы введения депротеинизированной ДНК описаны в публикации PCT №WO 90/11092 и патенте США №5580859. Липосомы, которые могут выступать в качестве средств доставки гена, описаны в патенте США №5422120; публикациях PCT №№WO 95/13796; WO 94/23697; WO 91/14445; и EP 0524968. Дополнительные подходы описаны в Philip, Mol. Cell Biol., 1994, 14:2411 и в Woffendin, Proc. Natl. Acad. Sci., 1994, 91:1581.
Композиции
[0493] Согласно настоящему изобретению также предложены фармацевтические композиции, содержащие эффективное количество конъюгата антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF), описанного в настоящем документе. Примеры таких композиций, а также то, как их изготавливать, также описано в настоящем документе. В некоторых вариантах реализации композиция содержит одно или более антител-антагонистов IL-6 (и/или конструкций АТ к IL-6-ловушка VEGF) и/или конъюгатов антитела (и/или конструкции АТ-ловушка). В других вариантах реализации указанное антагонистическое антитело к IL-6 (и/или конструкция АТ к IL-6-ловушка VEGF) распознает IL-6 человека. В других вариантах реализации антагонистическое антитело к IL-6 представляет собой антитело человека. В других вариантах реализации антагонистическое антитело к IL-6 представляет собой гуманизированное антитело. В некоторых вариантах реализации антагонистическое антитело к IL-6 содержит константную область, которая не запускает неблагоприятный или нежелательный иммунный ответ, такой как антителоопосредованный лизис или АЗКЦ. В других вариантах реализации антагонист IL-6 (и/или конструкция АТ к IL-6-ловушка VEGF) содержит одну или более CDR антитела (например, одну, две, три, четыре, пять или в некоторых вариантах реализации все шесть CDR).
[0494] Понятно, что указанные композиции могут содержать более одного антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) (например, смесь антител к IL-6, которые распознают разные эпитопы IL-6). Другие типичные композиции содержат более одного антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF), распознающего один и тот же эпитоп (эпитопы), или различные виды антител-антагонистов IL-6 (и/или конструкций АТ к IL-6-ловушка VEGF), которые связываются с разными эпитопами IL-6. В некоторых вариантах реализации композиции содержат смесь антител-антагонистов IL-6 (и/или конструкций АТ к IL-6-ловушка VEGF), распознающих различные варианты IL-6. В дополнение к этому в некоторых вариантах реализации композиция может также или в качестве альтернативы содержать более одной ловушки VEGF. Эти альтернативные части, представляющие собой ловушку VEGF, могут включать различные последовательности для части, представляющей собой ловушку VEGF, если указанные части по меньшей мере демонстрируют одинаковую или схожую функциональность, указанную в настоящем описании.
[0495] Композиция, применяемая согласно настоящему изобретению, может дополнительно содержать фармацевтически приемлемые носители, вспомогательные вещества или стабилизаторы (Remington: The Science and practice of Pharmacy 20th Ed., 2000, Lippincott Williams and Wilkins, Ed. K. E. Hoover) в форме лиофилизированных составов или водных растворов. Приемлемые носители, вспомогательные вещества или стабилизаторы нетоксичны для реципиентов в указанных дозах и концентрациях, и могут включать буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензетония хлорид; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстраны; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннитол, трегалоза или сорбитол; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как Полисорбат, ПЛЮРОНИК™ или полиэтиленгликоль (ПЭГ). Фармацевтически приемлемые вспомогательные вещества описаны ниже.
[0496] Фармацевтические композиции для парентерального введения включают водный и неводный стерильный раствор для инъекций, который может содержать антиоксиданты, буферы, бактериостатические агенты и растворенные вещества, которые делают состав по существу изотоничным с кровью предполагаемого реципиента; и водные и неводные стерильные суспензии, которые могут содержать суспендирующие агенты и загустители. Вспомогательные вещества, которые можно использовать для растворов для инъекций, включают воду, спирты, полиолы, глицерин и растительные масла, например. Композиции могут быть представлены в однодозовых или многодозовых емкостях, например, герметичных ампулах и флаконах, и могут храниться в высушенном методом сублимации (лиофилизированном) состоянии, требующем только добавления стерильного жидкого носителя, например, воды для инъекций, непосредственно перед применением. Растворы и суспензии для инъекций для немедленного применения могут быть приготовлены из стерильных порошков, гранул и таблеток. Фармацевтические композиции могут быть по существу изотоническими, что подразумевает осмоляльность, равную примерно 250-350 мОсм/кг воды.
[0497] Фармацевтические композиции могут содержать консерванты, солюбилизирующие агенты, стабилизаторы, смачивающие агенты, эмульгаторы, подсластители, красители, отдушки, соли (вещества согласно настоящему изобретению могут быть сами представлены в форме фармацевтически приемлемой соли), буферы, покрывающие агенты или антиоксиданты. Они могут также содержать терапевтически активные агенты в дополнение к веществу согласно настоящему изобретению. Фармацевтические композиции согласно настоящему изобретению можно применять в комбинации с одним или более фармацевтически приемлемыми вспомогательными веществами. Такие вспомогательные вещества могут включать, но не ограничиваются ими, физиологический раствор, буферный физиологический раствор (такой как фосфатный буферный раствор), декстрозу, липосомы, воду, глицерин, этанол и их комбинации.
[0498] Антитела-антагонисты IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF), конъюгаты антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) и фармацевтические композиции согласно настоящему изобретению можно применять отдельно или совместно с другими соединениями, такими как терапевтические соединения или молекулы, например, противовоспалительные лекарственные средства, анальгезирующие средства или антибиотики. Такое введение совместно с другими соединениями может быть одновременным, раздельным или последовательным. Компоненты могут быть представлены в форме набора, который может содержать инструкции при необходимости.
Способы предотвращения или лечения офтальмологических нарушений
[0499] В некоторых вариантах реализации антитела (и/или конструкции АТ к IL-6-ловушка VEGF) и конъюгаты антитела (и/или конструкции АТ к IL-6-ловушка VEGF) подходят для различного применения, включая, но не ограничиваясь ими, способы терапевтического лечения.
[0500] В некоторых вариантах реализации предложен способ лечения заболевания глаз, такого как, например, ВДЖП. В некоторых вариантах реализации способ лечения ВДЖП у субъекта включает введение нуждающемуся в этом субъекту эффективного количества композиции (например, фармацевтической композиции), содержащей любое из анти-IL-6 или конструкций антител АТ к IL6-ловушка VEGF, описанных в настоящем документе. В настоящем описании ВДЖП включает сухую форму ВДЖП и влажную форму ВДЖП. В некоторых вариантах реализации предложен способ лечения ВДЖП у субъекта, включающий введение нуждающемуся в этом субъекту эффективного количества композиции, содержащей антитела-антагонисты IL-6 или конструкции АТ к IL6-ловушка VEGF, или их конъюгаты, описанные в настоящем документе. В некоторых вариантах реализации антитело к IL-6 или конструкцию АТ к IL6-ловушка VEGF и/или их конъюгаты можно применять для лечения и/или предотвращения, и/или снижения риска возникновения влажной формы дегенерации желтого пятна или неоваскулярной дегенерации желтого пятна, влажной формы возрастной дегенерации желтого пятна или неоваскулярной возрастной дегенерации желтого пятна, сухой формы возрастной дегенерации желтого пятна, венозной, артериальной окклюзии или другой окклюзии кровеносных сосудов глаза и/или сетчатки с отеком сетчатки или без него, переднего и заднего увеита, отека желтого пятна при увеите, диабетической ретинопатии, пролиферативной диабетической ретинопатии, диабетического отека желтого пятна, непролиферативного диабетического отека желтого пятна и внутриглазных опухолей. В некоторых вариантах реализации антитело или конструкцию АТ к IL6-ловушка VEGF можно применять для: (i) пациентов, резистентных к терапии VEGF, (ii) пациентов, у которых происходит потеря эффективности анти-VEGF агентов из-за прогрессирования дополнительных базовых патологий, таких как фиброз, заживление раны и атрофия, и/или (iii) при недостаточном лечении из-за необходимости в частых интравитреальных инъекциях.
[0501] В некоторых вариантах реализации предложен способ лечения заболевания глаз, такого как системное заболевание. В некоторых вариантах реализации системные заболевания, поражающие глаз, такие как болезнь Грейвса или нейромиелит зрительного нерва, или системные заболевания, не поражающие глаз, такие как рассеянный склероз, ревматоидный артрит, можно лечить способами и композициями, предложенными согласно настоящему изобретению.
[0502] В некоторых вариантах реализации композиции, предложенные согласно настоящему изобретению, можно применять в комбинации с иммуномодуляторами, такими как PD-1/PDL-1, при показаниях в случае онкологического заболевания.
[0503] В некоторых вариантах реализации способы, описанные в настоящем документе, дополнительно включают этап лечения субъекта одной или более дополнительными формами терапии. В некоторых вариантах реализации указанная дополнительная форма терапии представляет собой дополнительную терапию ВДЖП, включая, но не ограничиваясь ими, Визудин®, лазерную фотокоагуляцию или интравитреальную инъекцию, например, Луцентиса®, Макуджена®, Эйлеа®, Озурдекса®, Илювьена®, Триесенса® (Triesence®) или Тривариса®.
[0504] Применительно ко всем способам, описанным в настоящем документе, ссылка на антитела-антагонисты IL-6 (и/или конструкцию АТ к IL6-ловушка VEGF) также включает композиции, содержащие один или более дополнительных агентов. Эти композиции могут дополнительно содержать подходящие вспомогательные вещества, такие как фармацевтически приемлемые вспомогательные вещества, включая буферы, которые хорошо известны в этой области техники. Настоящее изобретение можно применять отдельно или в комбинации с другими способами лечения.
[0505] В некоторых вариантах реализации предложен способ лечения или профилактики заболевания у нуждающегося в этом пациента. В некоторых вариантах реализации указанный способ включает введение пациенту любого из выделенных антител-антагонистов (и/или конструкций АТ к IL-6-ловушка VEGF), описанных в настоящем документе. В некоторых вариантах реализации способ включает введение пациенту любого из конъюгатов, описанных в настоящем документе. В некоторых вариантах реализации способ включает введение пациенту любой из композиций, описанных в настоящем документе. В некоторых вариантах реализации способ включает идентификацию пациента с гиперактивностью IL-6, и введение указанному пациенту любого из выделенных антител-антагонистов, описанных в настоящем документе. В некоторых вариантах реализации способ включает идентификацию пациента с гиперактивностью IL-6, и введение указанному пациенту любого из конъюгатов, описанных в настоящем документе. В некоторых вариантах реализации способ включает идентификацию пациента с гиперактивностью IL-6, и введение указанному пациенту любой из композиций, описанных в настоящем документе. В некоторых вариантах реализации у пациента наблюдают повышенную активность VEGF вместо повышенной активности IL-6 или в дополнение к ней.
[0506] В некоторых вариантах реализации нарушение выбрано из группы, состоящей из влажной формы дегенерации желтого пятна или неоваскулярной дегенерации желтого пятна, влажной формы возрастной дегенерации желтого пятна или неоваскулярной возрастной дегенерации желтого пятна, сухой формы возрастной дегенерации желтого пятна, венозной, артериальной окклюзии или другой окклюзии кровеносных сосудов глаза и/или сетчатки с отеком сетчатки или без него, переднего и заднего увеита, отека желтого пятна при увеите, диабетической ретинопатии, пролиферативной диабетической ретинопатии, диабетического отека желтого пятна, непролиферативного диабетического отека желтого пятна и внутриглазных опухолей.
[0507] В некоторых вариантах реализации выделенное антитело (и/или конструкцию АТ к IL-6-ловушка VEGF), конъюгат (и/или конъюгат конструкции АТ к IL-6-ловушка VEGF), композицию или их комбинацию вводят не чаще одного раза в месяц. В некоторых вариантах реализации выделенное антитело, конъюгат, композицию или их комбинацию вводят не чаще одного раза в два месяца. В некоторых вариантах реализации выделенное антитело (и/или конструкцию АТ к IL-6-ловушка VEGF), конъюгат (и/или конъюгат конструкции АТ к IL-6-ловушка VEGF), композицию или их комбинацию вводят не чаще одного раза в три месяца. В некоторых вариантах реализации выделенную конструкцию (например, АТ или АТ-ловушка с полимером или без него) вводят с частотой от одного раза в год до одного раза в два месяца. В некоторых вариантах реализации лечение может представлять собой однократное применение антитела. В некоторых вариантах реализации лечение может представлять собой столько применений антитела, сколько необходимо или требуется для достижения результата.
[0508] антагонистическое антитело к IL-6 (и/или конструкцию АТ к IL6-ловушка VEGF) или его конъюгат можно вводить субъекту любым подходящим путем. Для специалиста в этой области техники очевидно, что примеры, описанные в настоящем документе, не ограничивают, а иллюстрируют доступные способы. Соответственно, в некоторых вариантах реализации антагонистическое антитело к IL-6 (и/или конструкцию АТ к IL6-ловушка VEGF) вводят субъекту в соответствии с известными способами, такими как внутривенное введение, например, в виде болюсного введения или путем непрерывной инфузии в течение некоторого периода времени, внутримышечный, интраперитонеальный, интрацереброспинальный, трансдермальный, подкожный, внутрисуставной, сублингвальный, интрасиновиальный путь, путем инсуффляции, интратекальный, пероральный, ингаляционный или топический путь. Введение может быть системным, например, внутривенное введение, или местным. Для введения подходят коммерчески доступные небулайзеры для жидких составов, включая струйные небулайзеры и ультразвуковые небулайзеры. Жидкие составы можно непосредственно распылять, а лиофилизированный порошок можно распылять после восстановления. В качестве альтернативы, антагонистическое антитело к IL-6 (и/или конструкцию АТ к IL6-ловушка VEGF) можно распылять в виде аэрозоля с использованием фторуглеродного состава и ингалятора отмеренных доз или вводить путем ингаляции в виде лиофилизированного и молотого порошка.
[0509] В некоторых вариантах реализации антитела-антагонисты IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF), конъюгаты антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) и фармацевтические композиции, описанные в настоящем документе, применяют для профилактики или лечения заболевания или состояния глаз. Применяемые таким образом конъюгаты, как правило, изготавливают в форме для такого введения и вводят путем глазной, внутриглазной и/или интравитреальной инъекции, и/или околосклеральной инъекции, и/или субтеноновой инъекции, и/или супрахориоидальной инъекции, и/или топического введения в форме глазных капель и/или мази. Такие антитела-антагонисты IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF), конъюгаты антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) и композиции можно доставлять различными способами, например, интравитреально в виде устройства и/или депо, обеспечивающего медленное высвобождение соединения в стекловидное тело, включая те, которые описаны в таких источниках, как Intraocular Drug Delivery, Jaffe, Jaffe, Ashton, and Pearson, editors, Taylor & Francis (March 2006). В одном из примеров устройство может иметь форму системы минимальной и/или матричной, и/или пассивной диффузии, и/или инкапсулированных клеток, высвобождающих соединение в течение длительного периода времени (Intraocular Drug Delivery, Jaffe, Jaffe, Ashton, and Pearson, editors, Taylor & Francis (March 2006). В терапии или в качестве профилактического средства активный агент можно вводить индивидууму в виде композиции для инъекций, например, в виде стерильной водной дисперсии, предпочтительно изотонической или по существу изотонической.
[0510] Составы для глазного, внутриглазного или интравитреального введения могут быть получены способами и с использованием ингредиентов, известных в этой области техники. Для эффективного лечения необходимо подходящее проникновение в глаз. В отличие от заболеваний передней части глаза, когда лекарственные средства можно доставлять топически, для заболеваний сетчатки необходим более специфичный для этой области подход. Глазные капли и мази редко проникают в заднюю часть глаза, а гематоофтальмический барьер препятствует проникновению лекарственных средств, вводимых системно, в ткань глаза. В некоторых вариантах реализации предпочтительный способ доставки лекарственного средства для лечения заболевания сетчатки, такого как ВДЖП и ХНВ, представляет собой прямую интравитреальную инъекцию. В некоторых вариантах реализации интравитреальные инъекции повторяют с интервалами, которые зависят от состояния пациента и свойств, и периода полувыведения доставляемого лекарственного средства.
[0511] Ожидают, что для введения млекопитающим и, в частности, людям доза активного агента составляет от 0,01 мг/кг массы тела до, как правило, примерно 1 мг/кг в случае системного введения. Для заболеваний глаз, при которых необходимо местное введение (например, интравитреальное, супрахориоидальное, периокулярное и т.д.), доза составляет, как правило, от 0,1 мг/глаз/доза до 10 мг/глаз/доза или более. В некоторых вариантах реализации доза составляет 100 мкл/доза/глаз. В некоторых вариантах реализации для введения указанной дозы можно использовать иглу. Игла может представлять собой, например, иглу ½ дюйма 30 калибра или иглу ½ дюйма калибра 27G или 29G. Врач может определить фактическую дозу, наиболее подходящую для индивидуума, которая зависит от факторов, включающих возраст, массу тела, пол и ответ индивидуума, вылечиваемое заболевание или нарушение и возраст, и состояние индивидуума, которого лечат.Вышеприведенные дозы являются типичными для среднего случая. Конечно, могут быть случаи, когда необходимы более высокие или более низкие дозы.
[0512] Эту дозу можно повторять с любой необходимой периодичностью (например, еженедельно, раз в две недели, ежемесячно, ежеквартально). Если развиваются побочные эффекты, величину и/или частоту введения дозы можно уменьшать в соответствии с обычной клинической практикой. В одном из вариантов реализации фармацевтическую композицию можно вводить один раз в один-тридцать дней.
[0513] Антитела-антагонисты IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) согласно настоящему изобретению можно применять в соответствии с настоящим изобретением путем экспрессии таких полипептидов у пациента in vivo, т.е. генной терапии. Существуют два основных подхода к введению нуклеиновой кислоты (возможно содержащейся в векторе) в клетки пациента: in vivo и ex vivo. Для доставки in vivo нуклеиновую кислоту вводят путем инъекции непосредственно пациенту, как правило, в участках, где необходим терапевтический белок, т.е. где необходима биологическая активность терапевтического белка. Для лечения ex vivo извлекают клетки пациента, в эти выделенные клетки вводят нуклеиновую кислоту и модифицированные клетки вводят пациенту либо напрямую, либо, например, инкапсулированными в пористые мембраны, которые имплантируют пациенту (см., например, патенты США №№4892538 и 5283187). Существуют различные способы введения нуклеиновых кислот в жизнеспособные клетки. Эти способы варьируются в зависимости от того, переносят ли нуклеиновую кислоту в культивированные клетки in vitro или переносят в клетки предполагаемого хозяина in vivo. Способы, подходящие для переноса нуклеиновой кислоты в клетки млекопитающего in vitro, включают использование липосом, электропорацию, микроинъекцию, трансдукцию, слияние клеток, использование DEAE-декстрана, способ осаждения фосфатом кальция и т.д. Трансдукция включает связывание дефектной по репликации рекомбинантной вирусной (предпочтительно ретровирусной) частицы с клеточным рецептором с последующим введением нуклеиновых кислот, содержащихся в указанной частице, в клетку. Вектор, обычно используемый для доставки гена ex vivo, представляет собой ретровирус.
[0514] В настоящее время предпочтительные способы переноса нуклеиновой кислоты in vivo включают трансфекцию вирусными или невирусными векторами (например, аденовирус, лентивирус, вирус простого герпеса I или аденоассоциированный вирус (AAV)) и системами на основе липидов (липиды, подходящие для липид-опосредованного переноса гена, представляют собой, например, DOTMA, DOPE и DC-Chol; см., например, Tonkinison et al., Cancer Investigation, 14(1): 54-65 (1996)). Наиболее предпочтительные векторы для применения в генной терапии представляют собой вирусы, наиболее предпочтительно аденовирусы, AAV, лентивирусы или ретровирусы. Вирусный вектор, такой как ретровирусный вектор, содержит по меньшей мере один промотор/энхансер транскрипции или локус-определяющий элемент (элементы), или другие элементы, которые контролируют экспрессию гена другими способами, такими как альтернативный сплайсинг, ядерный экспорт РНК или посттрансляционная модификация мессенджера. В дополнение к этому, вирусный вектор, такой как ретровирусный вектор, содержит молекулу нуклеиновой кислоты, которая когда транскрибирована в присутствии гена, кодирующего терапевтический белок, функционально связана с ним и выступает в качестве последовательности инициации трансляции. Такие векторные конструкции также содержат сигнал упаковки, длинные концевые повторы (LTR) или их части и участки связывания праймера положительной и отрицательной цепи, подходящие для используемого вируса (если они уже не присутствуют в вирусном векторе). В дополнение к этому, такой вектор, как правило, содержит сигнальную последовательность для секреции прополипептида из клетки-хозяина, в которую он помещен. Предпочтительно, указанная сигнальная последовательность для этой цели представляет собой сигнальную последовательность млекопитающего, наиболее предпочтительно нативную сигнальную последовательность для терапевтического белка. Необязательно, векторная конструкция может также содержать сигнальную последовательность, которая направляет полиаденилирование, а также один или более сайтов рестрикции и последовательность терминации транскрипции. Например, такие векторы, как правило, содержат 5' LTR, сайт связывания тРНК, сигнал упаковки, точку начала синтеза второй цепи ДНК и 3' LTR или его часть. Можно использовать другие векторы, которые являются невирусными, такие как катионные липиды, полилизин и дендримеры.
[0515] В некоторых ситуациях желательно обеспечить источник нуклеиновой кислоты агентом, который направленно воздействует на клетки-мишени, таким как антитело, специфичное в отношении мембранного белка клеточной поверхности или клетки-мишени, лиганд для рецептора на клетке-мишени и т.д. При использовании липосом белки, которые связываются с мембранным белком клеточной поверхности, ассоциированным с эндоцитозом, можно использовать для направленного воздействия и/или для облегчения поглощения, например, капсидные белки или их фрагменты, тропные к конкретному типу клеток, антитела к белкам, которые подвергаются интернализации при прохождении цикла, и белки, которые направленно воздействуют на внутриклеточную локализацию и увеличивают внутриклеточный период полувыведения. Методика эндоцитоза, опосредованного рецепторами, описана, например, в Wu et al., J. Biol. Chem.,262: 4429-4432 (1987); и Wagner et al., Proc. Natl. Acad. Sci. USA, 87: 3410-3414(1990). Обзор известных в настоящее время протоколов маркирования генов и генной терапии см. в Anderson et al., Science, 256: 808-813 (1992). Также см. WO 93/25673 и указанные там источники.
[0516] Подходящую генную терапию и способы получения ретровирусных частиц и структурных белков можно найти, например, в патенте США №5681746.
[0517] В соответствии с некоторыми аспектами представлен способ лечения или профилактики заболевания глаз у млекопитающего, в котором вводят молекулу нуклеиновой кислоты, кодирующую антагонистическое антитело к IL-6 (и/или конструкцию АТ к IL-6-ловушка VEGF).
[0518] В некоторых вариантах реализации эти молекулы двойного ингибитора IL6/VEGF можно применять для лечения глазных нарушений, таких как офтальмологическое воспалительное заболевание - склерит, непролиферативная диабетическая ретинопатия, пролиферативная диабетическая ретинопатия (ПДР), диабетический отек желтого пятна, предотвращение диабетического отека желтого пятна, предотвращение пролиферативной диабетической ретинопатии, влажная форма возрастной дегенерации желтого пятна, предотвращение влажной формы возрастной дегенерации желтого пятна, увеит и отек желтого пятна при увеите.
[0519] Лекарственные средства VEGFR-антиIL6 можно также применять для лечения системных заболеваний, поражающих глаз, таких как болезнь Грейвса или нейромиелит зрительного нерва, или системных заболеваний, не поражающих глаз, таких как рассеянный склероз, ревматоидный артрит.
[0520] Эти молекулы можно также применять для лечения синдрома высвобождения цитокинов после CAR-T или подобных иммуноонкологических лекарственных средств. В дополнение к этому, анти-IL-6 молекулы подавляют индукцию экспрессии IL-6, наблюдаемую после лечения анти-PD-1/PD-L1 молекулами (Tsukamoto et al, 2018). В дополнение к этому, блокада передачи сигнала VEGF может также повышать эффективность лечения анти-PD-L1 средствами (Allen et al., 2017). Таким образом, эти двойные ингибиторы можно применять в комбинации с ингибиторами иммунных контрольных точек (такими как модуляторы PD-1/PDL-1) для синергетического лечения рака. Кроме того, ингибирование VEGF оказывает дополнительный полезный эффект. Еще одним видом применения указанных молекул является применение при отеке мозга при глиобластоме, когда анти-IL6 терапия (терапия анти-IL6 средствами) может демонстрировать дополнительные к лечению анти-VEGF полезные эффекты. Биополимер может также обеспечивать увеличение распределения в тканях, что благоприятно при проникновении в солидные опухоли.
[0521] В некоторых вариантах реализации конъюгат (или различные белки) подходит для предотвращения любого одного или более нарушений, представленных в настоящем описании.
Составы
[0522] Терапевтические составы антител-антагонистов IL-6 (и/или конструкций АТ к IL-6-ловушка VEGF) и конъюгатов антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF), применяемые согласно настоящему изобретению, получают путем смешивания антитела, демонстрирующего желаемую степень чистоты, с возможными фармацевтически приемлемыми носителями, вспомогательными веществами или стабилизаторами (Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000) для хранения в форме лиофилизированных составов или водных растворов. Приемлемые носители, вспомогательные вещества или стабилизаторы нетоксичны для реципиентов в используемых дозах и концентрациях, и могут включать буферы, такие как фосфат, цитрат и другие органические кислоты; соли, такие как хлорид натрия; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензетония хлорид; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннитол, трегалоза или сорбитол; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как ТВИН™, ПЛЮРОНИК™ или полиэтиленгликоль (ПЭГ).
[0523] Липосомы, содержащие антагонистическое антитело к IL-6 (и/или конструкцию АТ к IL-6-ловушка VEGF) и/или конъюгат антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF), получают способами, известными в этой области техники, такими как описаны в Epstein, et al., Proc. Natl. Acad. Sci. USA 82:3688 (1985); Hwang, et al., Proc. Natl Acad. Sci. USA 77:4030 (1980); и патентах США №№4485045 и 4544545. Липосомы с увеличенным временем циркуляции описаны в патенте США №5013556. Наиболее подходящие липосомы могут быть получены методом обращенно-фазового выпаривания с использованием липидной композиции, содержащей фосфатидилхолин, холестерин и ПЭГ-дериватизированный фосфатидилэтаноламин (ПЭГ-ФЭ). Липосомы подвергают экструзии через фильтры с определенным размером пор с получением липосом с желаемым диаметром.
[0524] Активные ингредиенты могут быть также заключены в микрокапсулы, полученные, например, методами коацервации или путем полимеризации на границе фаз, например, в микрокапсулы из гидроксиметилцеллюлозы или желатина и микрокапсулы из поли-(метилметакрилата) соответственно, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методики описаны в Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing (2000).
[0525] Могут быть получены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащие антитело, при этом указанные матрицы имеют определенную форму изделий, например, пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и 7 этил-L-глутамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (микросферы для инъекций, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида), изобутират ацетата сахарозы и поли-D-(-)-3-гидроксимасляную кислоту.
[0526] Составы, применяемые для введения in vivo, должны быть стерильными. Стерилизацию легко осуществлять путем, например, фильтрации через стерильные фильтрационные мембраны. Терапевтические композиции антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) и/или конъюгата антитела (и/или конструкции АТ к IL-6-ловушка VEGF), как правило, помещают в емкость со стерильным портом доступа, например, пакет или флакон с раствором для внутривенного введения с пробкой, прокалываемой гиподермальной иглой для инъекций. В некоторых вариантах реализации композиции антитела и/или конъюгата антитела помещают в шприц с получением предварительно заполненного шприца.
[0527] В некоторых вариантах реализации композиция представляет собой не содержащую пирогенов композицию, которая по существу не содержит эндотоксинов и/или родственных пирогенных веществ. Эндотоксины включают токсины, которые заключены внутри микроорганизма и высвобождаются только тогда, когда эти микроорганизмы разрушаются или погибают.Пирогенные вещества также включают индуцирующие лихорадку термостабильные вещества из наружной мембраны бактерий и других микроорганизмов. Оба эти вещества могут вызывать лихорадку, гипотензию и шок при введении людям. Из-за возможных вредных воздействий даже низкие количества эндотоксинов должны быть удалены из растворов лекарственных средств, вводимых внутривенно. Для системной инъекции, такой как внутривенная или интраперитонеальная, Управление США по контролю за качеством пищевых продуктов и лекарственных средств («FDA») установило верхний предел в 5 единиц эндотоксина (ЕЭ) на дозу на килограмм массы тела в один одночасовой период для внутривенного введения лекарственных средств (The United States Pharmacopeial Convention, Pharmacopeial Forum 26 (1):223 (2000)). Когда терапевтические белки вводят в количестве нескольких сотен или тысяч миллиграммов на килограмм массы тела, как может быть в случае представляющих интерес белков (например, антител), должны быть удалены даже следовые количества вредного и опасного эндотоксина. В некоторых вариантах реализации уровни эндотоксинов и пирогенов в композиции составляют менее 10 ЕЭ/мг или менее 5 ЕЭ/мг, или менее 1 ЕЭ/мг, или менее 0,1 ЕЭ/мг, или менее 0,01 ЕЭ/мг или менее 0,001 ЕЭ/мг.В некоторых вариантах реализации в композициях или способах, представленных в настоящем описании, допускается 0,1 ЕЭ/глаз/инъекция. В некоторых вариантах реализации в композициях или способах, представленных в настоящем описании, допускается 0,05 ЕЭ/глаз/инъекция. В некоторых вариантах реализации в композициях или способах, представленных в настоящем описании, допускается 0,02 ЕЭ/глаз/инъекция. В некоторых вариантах реализации в композициях или способах, представленных в настоящем описании, допускается 0,01 ЕЭ/глаз/инъекция.
[0528] Композиции согласно настоящему изобретению могут быть представлены в виде единичных лекарственных форм, таких как таблетки, пилюли, капсулы, порошки, гранулы, растворы или суспензии, или суппозитории, для перорального, парентерального или ректального введения, или введения путем ингаляции или инсуффляции.
[0529] Для получения твердых композиций, таких как таблетки, основной активный ингредиент смешивают с фармацевтическим носителем, например, с традиционными ингредиентами для получения таблеток, такими как кукурузный крахмал, лактоза, сахароза, сорбитол, тальк, стеариновая кислота, стеарат магния, дикальцийфосфат или камеди, и с другими фармацевтическими разбавителями, например, водой, с образованием твердой композиции предварительного состава, содержащей гомогенную смесь соединения согласно настоящему изобретению или его нетоксичной фармацевтически приемлемой соли. Когда эти композиции предварительного состава называют гомогенными, это означает, что активный ингредиент равномерно диспергирован по всей композиции так, что указанная композиция может быть легко разделена на одинаково эффективные единичные лекарственные формы, такие как таблетки, пилюли и капсулы. Эту твердую композицию предварительного состава затем разделяют на единичные лекарственные формы описанного выше типа, содержащие от примерно 0,1 до примерно 500 мг активного ингредиента согласно настоящему изобретению. Таблетки или пилюли из новой композиции могут иметь покрытие или могут быть приготовлены иным образом для получения лекарственной формы, обеспечивающей преимущество пролонгированного действия. Например, таблетка или пилюля может содержать компонент внутренней дозы и внешней дозы, при этом последний находится в форме оболочки над первым. Указанные два компонента могут быть разделены энтеросолюбильным слоем, служащим для препятствования распаду в желудке и обеспечивающим прохождение внутреннего компонента в интактном состоянии в двенадцатиперстную кишку или его отсроченное высвобождение. Для таких энтеросолюбильных слоев или покрытий можно использовать различные вещества; такие вещества включают ряд полимерных кислот и смесей полимерных кислот с такими веществами, как шеллак, цетиловый спирт и ацетат целлюлозы.
[0530] Подходящие поверхностно-активные агенты включают, в частности, неионные агенты, такие как Полисорбат или полиоксиэтиленсорбитаны (например, Твин™ 20, 40, 60, 80 или 85) и другие сорбитаны (например, Спан™ 20, 40, 60, 80 или 85). Композиции с поверхностно-активным агентом обычно содержат от 0,01% до 5% поверхностно-активного агента и могут содержать от 0,01 до 0,02% или от 0,1 до 2,5% (полисорбат 20 или 80). Очевидно, что могут быть добавлены другие ингредиенты, например, маннитол, или другие фармацевтически приемлемые носители при необходимости.
[0531] Подходящие эмульсии могут быть получены с использованием коммерчески доступных жировых эмульсий, таких как Интралипид™, Липозин™ (Liposyn™), Инфонутрол™, Липофундин™ и Липифизан™. Активный ингредиент может быть либо растворен в предварительно смешанной композиции эмульсии, либо в качестве альтернативы он может быть растворен в масле (например, соевом масле, сафлоровом масле, хлопковом масле, кунжутном масле, кукурузном масле или миндальном масле) и эмульсии, образованной при смешивании с фосфолипидом (например, с фосфолипидами яйца, соевыми фосфолипидами или соевым лецитином) и водой. Очевидно, что могут быть добавлены другие ингредиенты, например, глицерин или глюкоза, для регулирования тоничности эмульсии. Подходящие эмульсии, как правило, содержат до 20% масла, например, от 5 до 20%. Жировая эмульсия может содержать мелкие капли жира от 0,1 до 1,0 мкм, в частности, от 0,1 до 0,5 мкм, и иметь рН в диапазоне от 5,5 до 8,0.
[0532] Композиции эмульсий могут представлять собой композиции, полученные путем смешивания антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) с Интралипидом™ или его компонентами (соевое масло, фосфолипиды яйца, глицерин и вода).
[0533] Композиции для ингаляции или инсуффляции включают растворы и суспензии в фармацевтически приемлемых водных или органических растворителях или их смесях, и порошки. Жидкие или твердые композиции могут содержать подходящие фармацевтически приемлемые вспомогательные вещества, указанные выше. В некоторых вариантах реализации композиции вводят пероральным или назальным дыхательным путем для оказания местного или системного действия. Композиции в предпочтительно стерильных фармацевтически приемлемых растворителях можно распылять путем использования газов. Распыляемые растворы можно вдыхать непосредственно из распыляющего устройства или указанное распыляющее устройство может быть подсоединено к маске для лица, палатке или дыхательному аппарату с перемежающимся положительным давлением. Композиции в форме раствора, суспензии или порошка можно вводить, предпочтительно перорально или назально, из устройств, доставляющих состав подходящим способом.
Наборы
[0534] Согласно настоящему изобретению также предложены наборы, содержащие любое из антител, описанных в настоящем документе, или все антитела. Наборы согласно настоящему изобретению содержат одну или более емкостей, содержащих антагонистическое антитело к IL-6 (и/или конструкцию АТ к IL-6-ловушка VEGF) или конъюгат, описанный в настоящем документе, и инструкции по применению в соответствии с любым из способов согласно настоящему изобретению, описанных в настоящем документе. Как правило, эти инструкции содержат описание введения антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) или конъюгата для терапевтического лечения, описанного выше. В некоторых вариантах реализации предложены наборы для получения дозы для однократного введения. В некоторых вариантах реализации набор может содержать как первую емкость, содержащую высушенный белок, так и вторую емкость, содержащую водный состав. В некоторых вариантах реализации включены наборы, содержащие одно- и многокамерные предварительно заполненные шприцы (например, шприцы с жидкостью и шприцы с лиофилизатом).
[0535] В некоторых вариантах реализации антитело представляет собой антитело человека. В некоторых вариантах реализации антитело представляет собой гуманизированное антитело. В некоторых вариантах реализации антитело представляет собой моноклональное антитело. Инструкции, касающиеся применения антитела-антагониста IL-6 (и/или конструкции АТ к IL-6-ловушка VEGF) или конъюгата, как правило, содержат информацию о дозе, режиме дозирования и пути введения для предполагаемого лечения. Емкости могут представлять собой однократные дозы, нефасованные упаковки (например, упаковки, содержащие несколько доз) или субъединичные дозы. Инструкции, предоставленные в наборах согласно настоящему изобретению, как правило, представляют собой письменные инструкции на этикетке или листке-вкладыше в упаковке (например, лист бумаги, включенный в набор), а также приемлемыми являются машиночитаемые инструкции (например, инструкции, содержащиеся на диске магнитного или оптического запоминающего устройства).
[0536] Наборы согласно настоящему изобретению находятся в подходящей упаковке. Подходящая упаковка включает, но не ограничивается ими, флаконы, бутылки, банки, гибкую упаковку (например, герметичные майларовые или пластиковые пакеты) и т.п.Также предусмотрена упаковка для применения в комбинации с конкретным устройством, таким как предварительно заполненный шприц, ингалятор, устройство для введения в нос (например, распылитель) или инфузионное устройство, такое как мининасос.Набор может иметь стерильный порт доступа (например, емкость может представлять собой пакет или флакон с раствором для внутривенного введения с пробкой, прокалываемой гиподермальной иглой для инъекций). Емкость может также иметь стерильный порт доступа (например, емкость может представлять собой пакет или флакон с раствором для внутривенного введения с пробкой, прокалываемой гиподермальной иглой для инъекций). По меньшей мере один активный агент в композиции представляет собой антагонистическое антитело к IL-6 или его конъюгат.Емкость может дополнительно содержать второй фармацевтически активный агент.
[0537] В наборах могут быть предоставлены дополнительные компоненты, такие как буферы и пояснительная информация. В некоторых вариантах реализации наборы могут содержать дополнительный шприц и иглу, используемые для заполнения дозирующего шприца. Обычно набор содержит емкость и этикетку или листок (листки)-вкладыш в упаковке на емкости или соединенный с емкостью.
[0538] Следующие примеры предложены только в целях иллюстрации и никоим образом не ограничивают объем настоящего изобретения. Действительно, различные модификации настоящего изобретения в дополнение к тем, которые представлены и описаны в настоящем документе, будут очевидны для специалиста в этой области техники исходя из описания, приведенного выше, и входят в объем прилагаемой формулы изобретения.
ПРИМЕРЫ
[0539] Несмотря на терапевтический успех биологических лекарственных средств, направленных против фактора роста эндотелия сосудов (VEGF, ФРЭС), для лечения неоваскулярных заболеваний сетчатки по-прежнему существует неудовлетворенная потребность из-за наличия: (i) пациентов, резистентных к терапии VEGF, (ii) пациентов, у которых происходит потеря эффективности анти-VEGF агентов из-за прогрессирования дополнительных базовых патологий, таких как фиброз, заживление раны и атрофия, и/или (iii) недостаточного лечения из-за необходимости в частых интравитреальных инъекциях. Ниже приведены результаты, относящиеся к новым биспецифическим биологическим агентам, разработанным для ингибирования как VEGF, так и IL-6.
[0540] Способы. Разрабатывали двойные ингибиторы и получали характеристики их аффинности связывания в отношении соответствующих мишеней путем поверхностного плазмонного резонанса (SPR). Функциональные аспекты молекул тестировали: (1) путем ELISA-анализов, направленных на конкурентное блокирование каждой мишени для соответствующего рецептора (2) могут быть выполнены анализы VEGF- и/или IL-6-стимулируемой пролиферации первичных клеток эндотелия микрососудов сетчатки (HRMVEC), стимулированные и (3) могут быть выполнены анализы трехмерной сокультуры первичных клеток для оценки ингибирования разрастания кровеносных сосудов.
[0541] Конструкции АТ к IL-6-ловушка VEGF связывались с пM аффинностью с VEGF и IL-6. Кинетика связывания каждой мишени не зависела от присутствия другой, что указывает на то, что обе мишени могут одновременно связываться без препятствий. Связывание этих молекул с каждой из их мишеней ингибировало взаимодействия с соответствующими распознаваемыми рецепторами.
[0542] При комбинировании в рамках конъюгата с биополимером эти молекулы двойного ингибитора обеспечивают дополнительный терапевтический агент для следующего поколения лечения и предотвращения заболеваний сетчатки.
Пример 1 - получение и скрининг антител
Способы/результаты:
Anti-IL-6
[0543] В примерах в настоящем документе кратко описано конструирование молекулы двойного анти-IL-6 и анти-VEGF ингибитора, содержащей антитело к IL-6 в искусственно сконструированной каркасной области IgG1 человека и Fc-домен IgG1, подвергнутый слиянию с анти-VEGF ловушкой. Указанный Fc-домен для этих молекул содержит замены 234A, L235A и G237A, которые минимизируют эффекторную функцию, и L443C, которая обеспечивает сайт-специфичную конъюгацию с биополимером на основе фосфорилхолина, увеличивающим период полувыведения (положения остатков соответствуют нумерации EU).
[0544] Анти-IL-6 паратоп в искусственно сконструированной каркасной области IgG1 человека подвергали аффинному созреванию с использованием комбинации сканирующего мутагенеза библиотеки в E.coli с последующим сайт-направленным мутагенезом в системе млекопитающего. Конструирование плазмидной системы для двойной экспрессии, содержащей Fab тяжелой и легкой цепи с последовательностями анти-IL-6 CDR, осуществляли в E.coli. На фиг.3 представлена схема этого процесса, который приводил к получению вариабельных областей тяжелой и легкой цепи антитела на фиг.5.
[0545] Конструировали библиотеки Fab, содержащие случайные единичные точечные мутации для всего 71 положения CDR за исключением замен для Cys, Met и Asn из-за химической неустойчивости, связанной с этими аминокислотами. Для скрининга библиотек Fab использовали мелкомасштабную экспрессию в формате 96-луночных планшетов. Колонии E.coli (клетки TG1, Zymo Research), содержащие случайные единичные точечные мутации, собирали в 96-луночные планшеты (1,5 мл) и выращивали в течение ночи при 30°C в LB Amp+2% глюкоза. На следующий день формировали отдельный 96-луночный планшет путем удаления 100 мкл культуры и добавления 100 мкл 30% глицерина, затем хранили при -80°C. Оставшуюся культуру центрифугировали в течение 20 минут, а затем ресуспендировали в 1,5 мл LB Amp, содержащей 1 мМ IPTG, для индукции экспрессии белка при 30°C. Через 5 часов клетки центрифугировали, ресуспендировали в 500 мкл буфера HBS-EP+и лизировали путем использования одного цикла замораживания/оттаивания. Лизаты центрифугировали в течение 30 минут и подвергали вакуумной фильтрации (200 мкл) в круглодонные 96-луночные планшеты для анализа Biacore.
[0546] Аффинность анти-IL-6 Fab определяли с использованием Biacore T200 (GE Healthcare) при 25°C. Чипы CM5 активировали EDC и NHS, рекомбинантный IL-6 человека (R&D Systems) разводили в 10 мМ ацетате натрия, pH 5, до концентрации 0,1 мг/мл и вводили на активированный чип. Получали три диапазона антигенной плотности (100-150 RU, 300-400 RU, 400-600 RU) путем использования различного времени потока для каждого канала чипа, затем блокированного этаноламином. Надосадочные жидкости фильтрованных лизатов культуры из 96 лунок вводили со скоростью 50 мкл/мин в течение 120 секунд. Подгонку кривых диссоциации (300 секунд) к одиночной экспоненциальной кривой спада использовали для определения скоростей диссоциации (koff). Поверхности чипов регенерировали 10 мМ раствором глицина pH 1,7, вводимым со скоростью 50 мкл/мин в течение 60 секунд после каждого цикла связывания и диссоциации образца. Клоны, демонстрировавшие схожие или улучшенные значения koff по сравнению с диким типом, подвергали секвенированию ДНК для идентификации мутации.
[0547] Данные скрининга показали, что из общего количества 71 положения остатков CDR, 15 остатков имели критическое значение для связывания IL-6 и не могли быть заменены никакой другой аминокислотой. Замены в допустимых положениях приводили к значениям koff, схожим со значениями для исходной молекулы (таблица 13). Большую часть из этих единичных замен затем вводили в гены полных тяжелых и легких цепей, клонированные в векторах экспрессии млекопитающих, которые котрансфицировали в 3 мл культуры клеток Expi293 для продуцирования белка. В пятый день эти культуры центрифугировали и надосадочные жидкости, содержащие секретированные антитела, собирали и разводили в подвижном буфере HBS-EP+(соотношение 1:20). Точечные мутанты представлены в таблицах 13-18.
Таблица 13
CDR1 тяжелой цепи
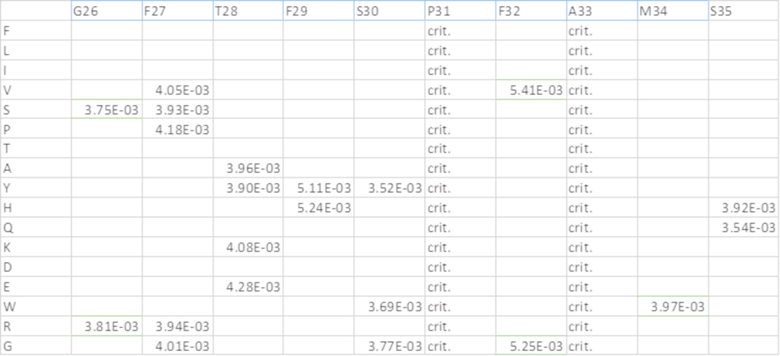
[0548] В таблице представлены значения koff (s-1) для каждой допустимой замены в пределах определенной CDR. В общем, мутанты демонстрировали koff, схожую со значением для исходной молекулы (4,94E-03±7,67E-04). Остатки, имеющие критическое значение для связывания IL-6 и не допускающие замен, обозначены как «крит.».
Таблица 14
CDR2 тяжелой цепи
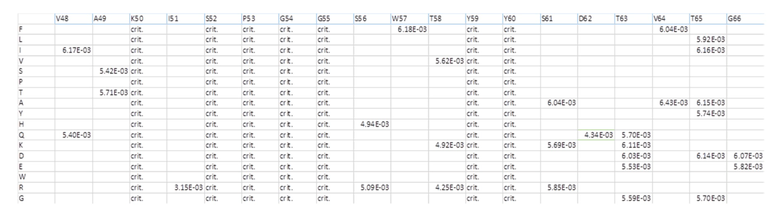
[0549] В таблице представлены значения koff (s-1) для каждой допустимой замены в пределах определенной CDR. В общем, мутанты демонстрировали koff, схожую со значением для исходной молекулы (4,94E-03±7,67E-04). Остатки, имеющие критическое значение для связывания IL-6 и не допускающие замен, обозначены как «крит.».
Таблица 15
CDR3 тяжелой цепи
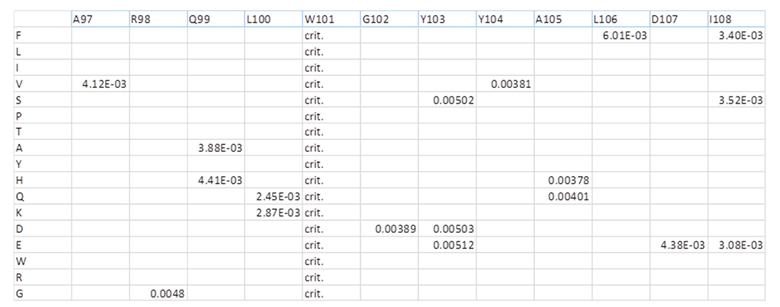
[0550] В таблице представлены значения koff (s-1) для каждой допустимой замены в пределах определенной CDR. В общем, мутанты демонстрировали koff, схожую со значением для исходной молекулы (4,94E-03±7,67E-04). Остатки, имеющие критическое значение для связывания IL-6 и не допускающие замен, обозначены как «крит.».
Таблица 16
CDR1 тяжелой цепи

[0551] В таблице представлены значения koff (s-1) для каждой допустимой замены в пределах определенной CDR. В общем, мутанты демонстрировали koff, схожую со значением для исходной молекулы (4,94E-03±7,67E-04). Остатки, имеющие критическое значение для связывания IL-6 и не допускающие замен, обозначены как «крит.».
Таблица 17
CDR2 тяжелой цепи
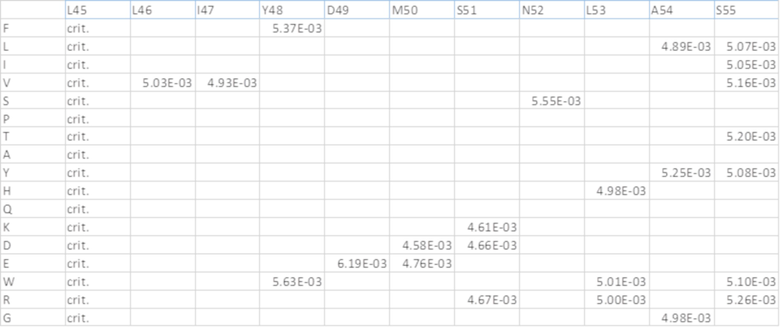
[0552] В таблице представлены значения koff (s-1) для каждой допустимой замены в пределах определенной CDR. В общем, мутанты демонстрировали koff, схожую со значением для исходной молекулы (4,94E-03±7,67E-04). Остатки, имеющие критическое значение для связывания IL-6 и не допускающие замен, обозначены как «крит.».
Таблица 18
CDR3 тяжелой цепи
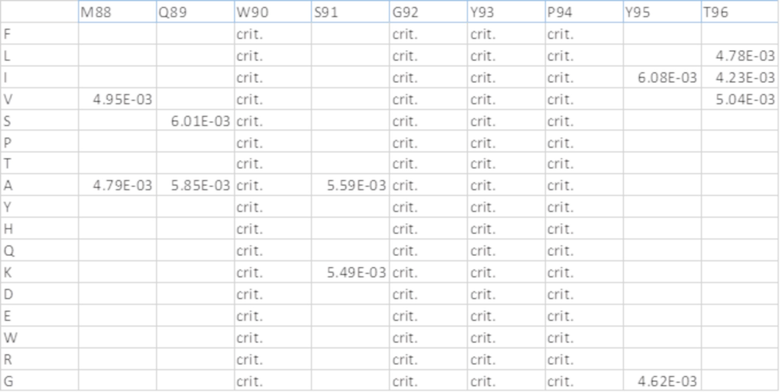
[0553] В таблице представлены значения koff (s-1) для каждой допустимой замены в пределах определенной CDR. В общем, мутанты демонстрировали koff, схожую со значением для исходной молекулы (4,94E-03±7,67E-04). Остатки, имеющие критическое значение для связывания IL-6 и не допускающие замен, обозначены как «крит.».
[0554] Кинетику связывания этих молекул с одной мутацией в отношении IL-6 измеряли при 25°C с использованием Biacore T200. Антитела захватывали на чип с белком А (GE) путем введения надосадочной жидкости клеток в разведении 1:20 со скоростью 10 мкл/мин в течение 60 секунд. Через захваченные антитела пропускали пять концентраций (0,56, 1,67, 5, 15 и 45 нМ) IL-6 в течение 60 секунд со скоростью потока 30 мкл/мин и подвергали диссоциации в течение 180 секунд в формате анализа кинетики Biacore с одним циклом. Сенсорный чип регенерировали путем 60-секундного введения 10 мМ глицина, pH 1,7, со скоростью потока 50 мкл/мин. Аналогичную систему анализа использовали для мониторинга взаимодействий между очищенными образцами антитела и IL-6 при 37°C, где применимо. Для этой системы 1 мкг/мл очищенного антитела вводили на чип с белком A со скоростью 10 мкл/мин в течение 25-28 секунд. Анализ данных для надосадочной жидкости и очищенных образцов проводили с использованием программного обеспечения BIAevaluation (GE). Из всех сенсограмм вычитали два эталонных сигнала и осуществляли подгонку с использованием модели связывания Ленгмюра 1:1.
[0555] Из всех тестируемых единичных точечных мутаций (таблица 19) мутация G66D в CDR2 тяжелой цепи (последовательная нумерация вариабельного домена антитела) приводила к заметному увеличению аффинности (~2-кратное уменьшение KD по сравнению с диким типом: 6,95x10-11 M относительно 1,58x10-10 M соответственно). G66D сохраняли, а остальные остатки CDR тяжелой цепи и легкой цепи, которые могут быть химически неустойчивыми при производстве (Cys, Met, Asn), подвергали мутагенезу. Исходя из этого положения получали два варианта тяжелой цепи, содержащие тройные мутации (таблица 21): I) M34I/G66D/I108V, II) M34W/G66D/I108V, и три варианта легкой цепи, содержащие тройные и четверные мутации (таблица 22): III) M32L/M50D/N52S/M88Q, IV) M32L/M50A/N52S/M88Q, V) M32L/M50D/M88Q (положения остатков соответствуют последовательной нумерации вариабельной области тяжелой цепи антитела). Авторы настоящего изобретения оценивали связывание IL-6 со всеми возможными комбинациями среди этих вариантов с использованием такой же системы анализа Biacore, которая описана для единичных точечных мутаций. Результаты, полученные авторами настоящего изобретения, продемонстрировали, что эти мутации противодействовали эффекту единичной точечной мутации G66D и сохраняли при этом аффинность к IL-6, более близкую к исходной последовательности CDR (таблица 20). На основе этих результатов эти модифицированные CDR включали в создание анти-IL-6 и конструкции АТ к IL-6-ловушка VEGF.
Таблица 19
[0556] Кинетика Biacore для единичных точечных мутаций анти-IL-6. Данные получали для образцов до (надосадочная жидкость (надос.)) при 25°C и после очистки при 37°C. Мутации, выбранные для конечных комбинаций CDR (таблица 19), выделены курсивом. Положения CDR, которые не допускали мутаций при бактериальном скрининге, обозначены как критические.
Таблица 20
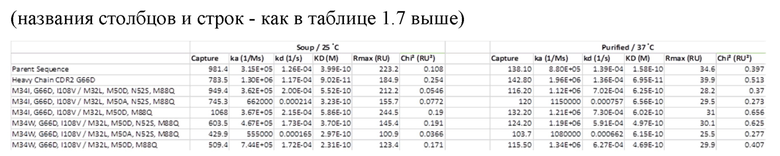
[0557] Данные кинетики Biacore для вариантов анти-IL-6 CDR. Кинетические данные получали для надосадочных жидкостей культур клеток (надос.) при 25°C и очищенных образцов при 37°C. Мутации CDR тяжелой и легкой цепи показаны с левой и правой стороны соответственно (тяжелая/легкая). В таблице приведены скорости захвата, kon (ka), koff (kd) и константы диссоциации.
Таблица 21. Последовательности анти-IL-6 вариабельной области тяжелой цепи. Некоторые варианты CDR подчеркнуты
Таблица 22. Последовательности анти-IL-6 вариабельной области легкой цепи. Некоторые варианты CDR подчеркнуты
Пример 2
Разработка и конструирование молекулы ab анти-IL-6-ловушка VEGF
[0558] Фрагмент, связывающий VEGF, в конструкции АТ к IL-6-ловушка VEGF состоял из слияния домена 2 VEGFR1 и домена 3 VEGFR2 (таблица 23). Это слияние действует как ловушка VEGF и предотвращает связывание VEGF с клеточными рецепторами VEGF.
Таблица 23. Последовательность ловушки VEGF
[0559] Тестировали 12 конструкций, содержащих ловушку VEGF в двух разных конфигурациях (таблица 24). В случае первой конфигурации ловушку VEGF располагали в начале белка с последующей двойной последовательностью линкера Gly-Gly-Gly-Gly-Ser (GS), соединяющего ловушку VEGF с N-концом анти-IL-6 тяжелой цепи (ловушка VEGF-анти-IL-6; таблица 24, молекулы A-F). В случае второй конфигурации вариабельный и константный домены тяжелой цепи (Fab-область) соединяли с ловушкой VEGF через линкер GS, а затем Fc-домен подвергали слиянию с C-концом ловушки VEGF. Таким образом, в этой конфигурации ловушка VEGF оказывалась «зажата» между Fab- и Fc-областями антитела (антиIL-6-ловушка VEGF; таблица 24, молекулы G-L).
Таблица 24. Последовательности тяжелой и легкой цепи для молекул двойного ингибитора. Некоторые варианты CDR подчеркнуты в тяжелой и легкой цепях, последовательность ловушки VEGF выделена жирным шрифтом, линкер Gly-Ser выделен курсивом (и жирным шрифтом)
[0560] Кинетические данные Biacore (система, описанная в предыдущем анализе) показали, что хотя значения KD среди всех вариантов сопоставимы, существует различие в значениях захвата для образцов надосадочной жидкости (таблица 25). Этот параметр чувствителен к количеству антитела, присутствующего в растворе, что позволило предположить варьирование уровня экспрессии белка среди этих молекул вследствие различных композиций CDR. С другой стороны, два тестируемых положения ловушки VEGF не демонстрировали заметного влияния на кинетические данные. Эксперименты по мониторингу уровней экспрессии (данные не представлены) показали, что конфигурация IL-6-ловушка VEGF обеспечивала примерно на 20% более высокий уровень экспрессии по сравнению с конфигурацией ловушка VEGF-антиIL-6. Предварительные данные Biacore также указывают на то, что конфигурация антиIL-6-ловушка VEGF прочнее связывается с гомологом VEGF, фактором роста плаценты (PLGF) (данные не представлены).
Таблица 25
[0561] Данные кинетики Biacore для вариантов A-L двойного ингибитора, связывающихся с IL-6. Кинетические данные получали для образцов надосадочной жидкости культур клеток (надос.) при 25°C и очищенных образцов при 37°C. Приведены скорости захвата, kon (ka), koff (kd) и константы диссоциации.
Пример 3
Связывание двойного ингибитора с VEGF по данным анализа Biacore
[0562] Для оценки кинетики связывания различных конфигураций двойного ингибитора с VEGF проводили серию анализов Biacore с измерением аффинности среди этих молекул. Кинетику связывания измеряли при 37°C в буфере HBS-EP+, содержащем 1 мг/мл БСА. Анти-VEGF агенты (конструкция ловушка VEGF-антиIL-6 (5 мкг/мл), конструкция антиIL-6-ловушка VEGF (5 мкг/мл), OG1950 (1 мкг/мл) и Эйлеа (0,5 мкг/мл)) захватывали на чип с белком A (GE) со скоростью 10 мкл/мин в течение 25 секунд, пять концентраций VEGF-A165 (0,19, 0,56, 1,67, 5, 15 нМ) пропускали через захваченные антитела в течение 120 секунд со скоростью 30 мкл/мин и подвергали диссоциации в течение 30 минут.Поверхность сенсорного чипа регенерировали путем 60-секундного введения 10 мМ глицина, pH 1,7, со скоростью потока 50 мкл/мин. Из всех сенсограмм вычитали два эталонных сигнала и осуществляли подгонку с использованием модели связывания Ленгмюра 1:1.
[0563] Данные показывают, что компонент в молекуле двойного ингибитора, представляющий собой ловушку VEGF, прочно связывается с VEGF-A независимо от конфигурации. Анти-IL-6 компонент молекулы, по-видимому, не влиял на связывание VEGF-A с ловушкой VEGF, поскольку молекулы двойного ингибитора демонстрируют аффинность, схожую с аффинностью к VEGF для другой молекулы на основе ловушки VEGF (Эйлеа). (Фиг.7).
Связывание IL-6 и VEGF-A с молекулами двойного ингибитора
[0564] Для непосредственного мониторинга двойного связывания IL-6 и VEGF-A проводили анализ Biacore. Молекулы двойного ингибитора (2 мкг/мл) захватывали на чип с белком A (GE) со скоростью 10 мкл/мин в течение 25 секунд, пять концентраций IL-6 и/или VEGF-A165 (0,19, 0,56, 1,67, 5, 15 нМ) пропускали по отдельности или в смеси через захваченные антитела в течение 120 секунд со скоростью 30 мкл/мин и подвергали диссоциации в течение 30 минут. Поверхность сенсорного чипа регенерировали путем 60-секундного введения 10 мМ глицина, pH 1,7, со скоростью потока 50 мкл/мин. Из всех данных вычитали данные двух контролей и сумму отдельных сенсограмм IL-6 и VEGF-A использовали для построения теоретической кривой, которую сравнивали с сенсограммами смешанных образцов.
[0565] Результаты показали, что теоретические и экспериментальные кривые практически совпадают при наложении, что в высокой степени подтверждает модель, в которой обе мишени связываются с молекулами двойного ингибитора и не влияют на их соответствующие взаимодействия (Фиг.8).
ELISA-анализ связывания анти-IL-6 с комплексом IL-6/IL-6-R
[0566] Комплексы IL-6/IL-6R могут связываться с мембраносвязанным рецептором gp130, и этот тримерный комплекс стимулирует передачу сигнала IL-6, такую как пролиферация и повышающая регуляция VEGF. Ингибирование либо связывания IL-6 с IL-6R, либо связывания IL-6/IL-6R с gp130 блокирует IL-6-опосредуемые нисходящие пути передачи сигнала. Для определения, блокируют ли анти-IL-6 антитела любой из этих процессов связывания, сначала осуществляли ELISA для оценки, могут ли они связываться с IL-6, когда он находится в комплексе с рецептором IL-6.
[0567] Nunc MaxiSorp ELISA покрывали 100 мкл 1 мкг/мл рекомбинантного IL-6R человека (R&D Systems) в течение ночи. 10 нМ IL-6 (R&D Systems) смешивали с серией 3-кратных разведений анти-IL-6 (конечные концентрации от 30 нМ до 10 пМ). После блокирования и промывки смесь добавляли в планшеты, покрытые IL-6R. Этот анализ также выполняли сначала путем инкубации планшета, покрытого IL-6R, совместно с 10 нМ IL-6, промывки, а затем добавления анти-IL-6 антитела. Для обоих способов связанные комплексы антитело/IL-6 детектировали с помощью HRP-антитела к IgG человека и проявляли с помощью TMB (KPL) в течение 10 минут. На планшет-ридере SpectraMax Plus (Molecular Devices) измеряли поглощение при 450 нМ и наносили на график в зависимости от концентрации антитела с использованием программного обеспечения GraphPad Prism и приводили к кривой нелинейной регрессии.
[0568] Nunc MaxiSorp ELISA покрывали 100 мкл 1 мкг/мл рекомбинантного IL-6 человека (R&D systems) в течение ночи. Осуществляли серию 3-кратных разведений анти-IL-6 (R&D Systems; конечные концентрации от 30 нМ до 10 пМ) и растворы добавляли в блокированные планшеты, покрытые IL-6. Комплексы антитело/IL-6 детектировали с помощью HRP-антитела к IgG человека и проявляли с помощью TMB (KPL) в течение 10 минут.На планшет-ридере SpectraMax Plus (Molecular Devices) измеряли поглощение при 450 нМ и наносили на график в зависимости от концентрации антитела с использованием программного обеспечения GraphPad Prism и приводили к кривой нелинейной регрессии.
[0569] В тестируемых условиях анти-IL6 антитело, описанное в настоящем документе, специфично связывается с IL-6, но не когда оно находится в комплексе с рецептором IL-6, что позволяет предположить, что антитело и IL-6R имеют общий эпитоп на IL-6. Результаты представлены на фиг.4.
Конкурентный ELISA-анализ IL-6/IL-6R
[0570] Затем проводили ELISA для определения, блокирует ли анти-IL6 антитело и двойные ингибиторы IL-6/VEGF связывание IL-6 с IL-6R.
[0571] Nunc MaxiSorp ELISA покрывали 100 мкл 1 мкг/мл антитела к комплексу IL-6/IL-6R (R&D systems; IL-6/IL-6R DuoSet) в течение ночи. Блокировали планшет 2% бычьим сывороточным альбумином (БСА) в растворе ФБР-Твин (ФБР-T) в течение 2 часов. Во время блокирования 6 нМ IL-6 (R&D Systems), 1 нМ IL-6R (R&D Systems) и серию 3-кратных разведений для получения 8 точек разведения двойных ингибиторов или одной группы контролей (начиная с 30 нМ) инкубировали при комнатной температуре. Затем эту смесь добавляли в планшет для ELISA после этапа промывки. Через 1 час планшет промывали и инкубировали совместно с биотинилированным антителом к комплексу IL-6/IL-6R (R&D Duoset). Комплексы IL-6/IL-6R детектировали с помощью стрептавидин-HRP (разведение 1:2000; R&D Systems) и проявляли с использованием реагента TMB (KPL) в течение 10 минут. На планшет-ридере SpectraMax Plus (Molecular Devices) измеряли поглощение при 450 нМ и наносили на график в зависимости от концентрации антитела с использованием программного обеспечения GraphPad Prism и приводили к кривой нелинейной регрессии.
[0572] В этих условиях анти-IL6 антитело и двойные ингибиторы IL-6/VEGF эффективно конкурируют с IL-6R за связывание с IL-6 и, следовательно, ингибируют взаимодействие IL-6/IL-6R. Значения IC50 для молекул двойного ингибитора сопоставимы (анти-IL-6=0,36 нМ, конструкция ловушка VEGF-анти-IL-6=0,47 нМ и конструкция анти-IL-6-ловушка VEGF=0,32 нМ), что указывает на то, что конфигурация молекулы не влияет на связывание или ингибирование IL-6. Эйлеа служило в качестве отрицательного контроля. (Фиг.9).
Конкурентный ELISA-анализ VEGF/VEGFR
[0573] Для оценки функции ловушки VEGF способность двойных ингибиторов IL-6/VEGF блокировать связывание VEGF с рецептором (VEGFR) тестировали путем конкурентного ELISA.
[0574] Nunc MaxiSorp ELISA покрывали 100 мкл 1 мкг/мл рекомбинантного VEGFR-Fc человека (R&D systems) в течение ночи. Блокировали планшет 2% бычьим сывороточным альбумином (БСА) в растворе ФБР-Твин (ФБР-T) в течение 2 часов. Во время блокирования 100 пМ биотинилированный VEGF (btVEGF; R&D Systems) инкубировали совместно с возрастающими концентрациями конструкции анти-IL-6-ловушка VEGF, конструкции ловушка VEGF-анти-IL-6 или двухвалентного анти-VEGF антитела, OG1950. Связанный bt-VEGF детектировали с помощью стрептавидин-HRP (разведение 1:2000; R&D Systems) и проявляли с использованием реагента TMB (KPL) в течение 10 минут.На планшет-ридере SpectraMax Plus (Molecular Devices) измеряли поглощение при 450 нМ и наносили на график в зависимости от концентрации антитела с использованием программного обеспечения GraphPad Prism и приводили к кривой нелинейной регрессии.
[0575] В этих условиях двойные ингибиторы IL-6/VEGF связываются с VEGF и ингибируют его связывание с рецептором. Когда ловушка VEGF расположена на N-конце анти-IL-6 антитела, ингибирование VEGF примерно в 2,5 раза сильнее, чем в случае молекулы, в которой ловушка VEGF расположена между Fab- и Fc-доменами анти-IL-6 антитела (IC50 1,74 нМ относительно 4,53 нМ соответственно), вероятно, из-за изменения валентности с 2 центров связывания на 1 центр связывания. Результаты представлены на фиг.9.
Пример 4
Мостиковый ELISA-анализ IL-6-VEGF
[0576] Для подтверждения того, что двойные ингибиторы могут одновременно связываться как с IL-6, так и с VEGF, их подвергали мостиковому ELISA, в котором их сначала подвергали взаимодействию с IL-6, связанным с планшетом, а затем инкубировали совместно с биотинилированным VEGF (btVEGF). Детектирование btVEGF посредством стрептавидин-HRP показывает, что молекулы, связанные с IL-6, также способны связывать btVEGF.
[0577] Nunc MaxiSorp ELISA покрывали 100 мкл 1 мкг/мл рекомбинантного IL-6 человека (Tonbo Sciences) в течение ночи. Блокировали планшет 2% бычьим сывороточным альбумином (БСА) в растворе ФБР-Твин (ФБР-T) в течение 2 часов. Во время блокирования 100 пМ btVEGF инкубировали совместно с возрастающими концентрациями конструкции анти-IL-6-ловушка VEGF, конструкции ловушка VEGF-анти-IL-6 или двухвалентного антитела антиVEGF, OG1950, в качестве отрицательного контроля. Связанный btVEGF детектировали с помощью стрептавидин-HRP (разведение 1:2000; R&D systems) и проявляли с использованием реагента TMB (KPL) в течение 10 минут.На планшет-ридере SpectraMax Plus (Molecular Devices) измеряли поглощение при 450 нМ и наносили на график в зависимости от концентрации антитела с использованием программного обеспечения GraphPad Prism и приводили к кривой нелинейной регрессии.
[0578] Этот анализ подтвердил, что двойные ингибиторы IL-6/VEGF одновременно связывают IL-6 и btVEGF. Ориентация анти-IL-6 и ловушки VEGF демонстрирует слегка вариабельное связывание, при этом ориентация анти-IL-6-ловушка VEGF является более подходящей в этих условиях (EC50 конструкции ловушка VEGF-антиIL-6=0,079 нМ и конструкции антиIL-6-ловушка VEGF=0,026 нМ). Эйлеа и анти-IL-6 служили в качестве отрицательных контролей. Результаты представлены на фиг.9.
Пример 5
Путь 1. Синтез OG1802
[0579] Первый путь синтеза OG1802 является следующим. Сначала синтезировали инициатор - соль ТФУ/амин (Соединение L), имеющий структуру, представленную на фиг.2А, следующим образом.
[0580] Сначала синтезировали Соединение K, имеющее структуру, представленную на фиг.2B, следующим образом. В 200 мл круглодонную колбу в атмосфере азота помещали Соединение J (OG1563) (1,9 г, 2,67 ммоль, 3,3 экв.)
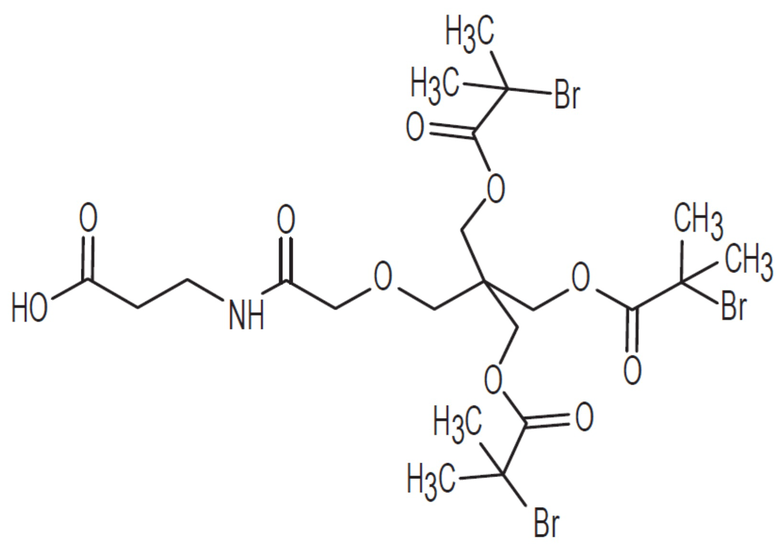
СОЕДИНЕНИЕ J
и Соединение E (0,525 г, 0,81 ммоль, 1,0 экв.) (см. фиг.2L), а затем диметилформамид (10 мл), затем диизопропилэтиламин (2,5 мл, 14,6 ммоль, 18 экв.). Колбу охлаждали до 0°C с использованием ледяной бани. К этому добавляли раствор пропилфосфонового ангидрида (50 масс.% в этилацетате, 2,5 мл, 4,04 ммоль, 5 экв.) в течение ~6 минут.
[0581] Реакционную смесь нагревали до комнатной температуры и перемешивали в течение 15 минут. Реакцию гасили путем добавления воды (20 мл), насыщенного водного бикарбоната натрия (20 мл) и этилацетата (100 мл). Отделяли органическую фазу и водную фазу экстрагировали этилацетатом (75 мл). Объединенные органические фазы промывали насыщенным водным бикарбонатом натрия (30 мл), 0,5 М водной лимонной кислотой (40 мл), водой (25 мл) и насыщенным водным хлоридом натрия (40 мл), затем сушили (сульфат натрия), фильтровали и концентрировали в вакууме. Остаток, который использовали без дополнительной очистки, позволял получить 2,0 г (0,80 ммоль, 99%) Соединения K.
1H ЯМР (400 МГц ДМСО-d6): (=1,36 (s, 9H, OCCH3), 1,90 (s, 54H, CC(CH3)2Br), 2,31 (t, J=7,2 Гц, 6H, CCH2CH2NH), 2,98 (d, J=5,6 Гц, 6H, CCH2NH), 3,04 (q, J=6,0 Гц, 2H, OCH2CH2NH), 3,18 (s, 2H, OCH2C), 3,3-3,37 (m, 8H, CH2), 3,47-3,55 (m, 12H, CH2), 3,58 (s, 6H, OCH2C), 3,87 (s, 6H, O=CCH2O), 4,27 (s, 18H, CCH2OC=O), 6,74 (br t, 1H, CH2NHC=O), 7,69 (t, J=6,8 Гц, 3H, CH2NHC=O), 7,84 (t, J=6,0 Гц, 3H, CH2NHC=O).
ЖХ-МС (ES, m/z): [(M+2H-boc)/2]+рассчит. для (C84H136Br9N7O33+2H-Boc)/2=1196,6; Обнаружено 1196,6.
[0582] Затем синтезировали Соединение L (фиг.2A) следующим образом: в 100 мл круглодонную колбу в атмосфере азота добавляли Соединение K (2,0 г, 0,8 ммоль), дихлорметан (10 мл), а затем трифторуксусную кислоту (5 мл). Реакционную смесь перемешивали при комнатной температуре в течение 30 минут. Реакционную смесь концентрировали в вакууме. Реакционную смесь разбавляли с использованием дихлорметана (10 мл) и концентрировали в вакууме. Остаток растворяли с использованием ацетонитрила (10 мл), фильтровали через шприцевой фильтр (Acrodisc CR25, PN 4225T) и загружали в колонку для препаративной ВЭЖХ, и элюировали 60% ацетонитрилом в воде (с 0,1% трифторуксусной кислотой) - 98% ацетонитрилом (с 0,1% трифторуксусной кислотой). Пробирки, содержащие продукт, объединяли в пул, концентрировали в вакууме, замораживали и помещали в лиофилизатор. В результате получали 990 мг (0,4 ммоль, 50% за 2 этапа) Соединения L в виде белого порошка.
1H ЯМР (400 Мгц ДМСО-d6): (=1,90 (s, 54H, CC(CH3)2Br), 2,31 (t, J=7,2 Гц, 6H, CCH2CH2NH), 2,97-3,0 (m, 8H, CCH2NH и OCH2CH2NH), 3,17 (s, 2H, OCH2C), 3,3 (q, 6H, CH2CH2NHC=O), 3,4-3,59 (m, 20H, CH2), 3,87 (s, 6H, O=CCH2O), 4,27 (s, 18H, CCH2OC=O), 7,69-7,84 (m, 9H, как CH2NHC=O, так и NH3+).
ЖХ-МС (ES, m/z): [(M+2H)/2]+рассчит.для (C84H136Br9N7O33+2H)/2=1196,6; обнаружено 1197,4.
[0583] Затем Соединение L (фиг.2A) использовали в качестве инициатора для синтеза полимера MPC. Инициатор, как правило, получают в виде исходного раствора в ДМФА примерно 100 мг/мл. Инициатор и лиганд (2,2’-бипиридил) вводили в сосуд Шленка. Полученный раствор охлаждали до -78°C с использованием смеси сухой лед/ацетон и дегазировали в вакууме в течение 10 минут. Сосуд снова заполняли в атмосфере аргона, и катализатор (CuBr, если не указано иное), хранимый в атмосфере аргона, вводили в сосуд Шленка (молярное соотношение атома брома в инициаторе/катализаторе (CuBr)/лиганде поддерживали на уровне 1/1/2). Раствор сразу становился темно-коричневым. Сосуд Шленка герметично закрывали и сразу же продували путем использования короткого цикла вакуум/аргон. Получали раствор HEMA-PC путем смешивания определенного количества мономера, полученного в перчаточном боксе в атмосфере азота, с дегазированным этанолом крепости 200 пруф. Раствор мономера по каплям добавляли в сосуд Шленка (через канюлю) (и гомогенизировали путем легкого перемешивания). Температуру поддерживали на уровне -78°C. Реакционную смесь подвергали воздействию полного вакуума в течение по меньшей мере 10-15 минут до прекращения выделения пузырьков из раствора. Затем сосуд снова заполняли аргоном и нагревали до комнатной температуры. Раствор перемешивали и по мере протекания полимеризации раствор становился вязким. Через 3-8 часов или после того как просто оставляли на ночь, реакцию гасили путем прямого воздействия воздуха для окисления Cu (I) до Cu (II), смесь приобретала сине-зеленый цвет и ее пропускали через колонку с диоксидом кремния с удалением медного катализатора. Собранный раствор концентрировали путем ротационного выпаривания и полученную смесь либо осаждали тетрагидрофураном, либо подвергали диализу против воды с последующей сушкой методом сублимации с получением сыпучего белого порошка. В таблице 26 ниже представлены данные для полимера, в котором соединение L использовано в качестве инициатора.
Таблица 26
(кДа)
[0584] Затем «пристегивали» сложный эфир малеимида Mal-PEG4-PFP (как показано на фиг.2C) к указанному выше полимеру массой 750 кДа с получением OG1802. В 20 мл флакон помещали полимер R3707 (полимер массой 750 кДа, полученный с использованием соединения L в качестве инициатора, 515 мг) и растворяли с использованием этанола (4,0 мл) после перемешивания в течение 40 минут.Добавляли 1% раствор 4-метилморфолина в ацетонитриле (22 мкл). В отдельном флаконе растворяли Mal-PEG4-PFP (1,97 мг) в ацетонитриле (1,0 мл), и этот раствор добавляли к раствору полимера в течение ~2 минут при комнатной температуре, и полученный раствор перемешивали в течение ночи. Реакционную смесь разбавляли 0,1% водной трифторуксусной кислотой (2 мл) (pH ~5), затем водой (~12 мл), фильтровали через шприцевой фильтр (Acrodisc Supor, PN 4612) и поровну помещали в 3 центрифужные пробирки с диализной мембраной Amicon (30000 номинальное отсечение по молекулярной массе (mwco)). Указанные пробирки разбавляли и смешивали с водой (~5 мл каждая), помещали в центрифугу (3200 об/мин) на 25 минут. Фильтрат извлекали для анализа, а ретентат разбавляли и смешивали с водой (~10 мл/пробирка). Процедуру центрифугирования повторяли еще 5 раз, после чего ретентат извлекали и помещали во флакон. Пробирки с мембраной Amicon промывали водой (2 × ~2 мл каждую пробирку) и объединяли с ретентатом. Раствор ретентата фильтровали через шприцевой фильтр (Acrodisc Supor, PN 4612), замораживали и помещали в лиофилизатор. В результате получали 485 мг белого порошка.
Пример 6. Синтез инициатора OG1786
[0585] OG1786 представляет собой инициатор с девятью ответвлениями для синтеза полимера, используемый в качестве предшественника в синтезе OG1802. Каждое ответвление заканчивается 2-бромизобутиратом, который способен инициировать полимеризацию у условиях ATRP. OG1786 представляет собой соль трифторуксусной кислоты (ТФУ), показанную на фиг.2D. OG1786 получали следующим образом. Сначала OG1550 подвергали реакции с ТФУ (трифторуксусной кислотой) с получением OG1546, как показано на фиг.2E.
[0586] В 1 л круглодонную колбу, оснащенную магнитной мешалкой и капельной воронкой, добавляли OG1550 (14,8 г), метил-трет-бутиловый эфир (MTBE) (350 мл) и воду (30 мл). Смесь перемешивали с растворением OG1550, затем охлаждали на ледяной бане. К этой смеси по каплям добавляли раствор трифторуксусной кислоты (4,9 мл) в воде (90 мл) в течение 90 минут. После завершения добавления смесь перемешивали в течение еще 15 минут, затем снимали с ледяной бани и оставляли нагреваться до комнатной температуры. Смесь перемешивали (после снятия с ледяной бани) в течение еще 4-5 часов, пока тонкослойная хроматография (ТСХ) не показала, что остается ~5% исходного вещества, и pH водной смеси не составил от 3 до 4 (индикаторная бумага для определения pH).
[0587] Смесь разделяли. Фазу MTBE промывали водой (30 мл). Водные фазы объединяли, затем экстрагировали МТВЕ (150 мл). Вторую фазу MTBE промывали водой (30 мл). Объединенные водные фазы промывали третьей порцией МТВЕ (100 мл). Третью фазу MBTE промывали водой (25 мл). Водные фазы снова объединяли (~250 мл, pH ~4 по индикаторной бумаге для определения pH).
[0588] Продукт собирали путем лиофилизации. Получали 11,5 г белого твердого вещества. Указанное вещество чрезвычайно гигроскопично, поэтому манипуляции с ним лучше всего проводить в атмосфере азота. Продукт подтверждали путем ЖХ-МС.
[0589] Затем полученный OG1546 подвергали реакции с OG1563 с получением OG1784 (как показано на фиг.2F).
[0590] В 250 мл колбу в атмосфере азота, оснащенную мешалкой, добавляли OG1546 (гигроскопичный, 9,0 г), а затем N,N-диметилформамид (110 мл). Смесь перемешивали при комнатной температуре до растворения всего OG1546 (примерно 15 минут), затем добавляли OG1563 (29,9 г) и смесь перемешивали в течение еще 3 минут также до растворения OG1563. Полученный раствор охлаждали на ледяной бане и добавляли N,N-диизопропилэтиламин (37,6 мл) в течение 3 минут, а затем по каплям пропилфосфоновый ангидрид (T3P), 50% в этилацетате (34,5 мл) в течение 5 минут (добавление T3P является экзотермическим). После завершения добавления T3P колбу снимали с охлаждающей бани и оставляли нагреваться до комнатной температуры. Затем брали образцы для анализа ЖХ-МС с 5-минутными интервалами. Реакционная смесь демонстрировала очень светлый желто-коричневый цвет.
[0591] Через 20 минут реакционную смесь снова охлаждали на ледяной бане и добавляли 5 мл воды. Затем смесь снимали с охлаждающей бани и добавляли дополнительную 50 мл порцию воды, а затем 50 мл 0,5 М лимонной кислоты, затем изопропилацетат (300 мл). Смесь разделяли. Водную фазу (~300 мл) экстрагировали дополнительным количеством изопропилацетата (150 мл). Водная фаза представляла собой AQ1 для тестирования путем ВЭЖХ. Объединенные органические фазы промывали водной лимонной кислотой (115 мл, 65 мМ, которая представляла собой смесь 15 мл 0,5 М лимонной кислоты плюс 100 мл воды), и водная фаза представляла собой AQ2 (pH~3). Органическую фазу промывали водой/насыщенным хлоридом натрия (100 мл/25 мл), и водная фаза представляла собой AQ3 (pH~3). Органическую фазу, наконец, промывали насыщенным хлоридом натрия (100 мл), и водная фаза представляла собой AQ4. Ни одна из водных (AQ) фракций не содержала никакого значимого продукта (данные не представлены). Подтверждали продукт в органической фазе путем ЖХ-МС.Продукт сушили над сульфатом натрия (80 г), фильтровали и промывали изопропилацетатом (75 мл), и концентрировали на ротационном испарителе с получением желтовато-коричневого масла (33,2 г). Неочищенный продукт хранили в течение ночи в атмосфере азота.
[0592] На следующий день неочищенный продукт оставляли дойти до комнатной температуры, затем растворяли в ацетонитриле/воде (46 мл/12 мл) и фильтровали с использованием фильтровального диска ВЭЖХ (Cole-Parmer PTFE 0,2 мкм, номер продукта 02915-20). Фильтрат делили на три равные части и очищали в трех сериях.
[0593] Фильтрат загружали в колонку C18 RediSep Rf Gold (275 г, SN 69-2203-339, серия # 24126-611Y), уравновешенную смесью 50% ацетонитрил/вода. Вещество элюировали со скоростью 100 мл/мин с использованием следующего градиента (растворитель A: вода, растворитель B: ацетонитрил). Все соответствующие фракции проверяли путем ВЭЖХ. Фракции, которые, как было установлено, являлись достаточно чистыми, объединяли в пул (из всех трех серий) и концентрировали (температуру бани поддерживали на уровне примерно 20°C) на ротационном испарителе, затем разделяли между дихлорметаном (100 мл) и водой (5 мл)/насыщенным хлоридом натрия (25 мл). Водную фазу еще два раза экстрагировали дихлорметаном (2 × 30 мл). Объединенные органические фазы сушили над сульфатом натрия (35 г), фильтровали, промывали дихлорметаном (ДХМ) (30 мл) и концентрировали. Продукт и чистоту подтверждали методами ЖХ-МС. Фактический выход и чистота партий R5172 и R5228 представлены в Таблице 27.
Таблица 27
[0594] Затем получали OG1405 из OG1784, как показано на фиг.2G. В 500 мл круглодонную колбу, оснащенную магнитной мешалкой, добавляли OG1784 (20,9 г), затем дихлорметан (50 мл), затем трифторуксусную кислоту (20 мл). Смесь перемешивали при комнатной температуре, и анализ ВЭЖХ демонстрировал полное снятие защитных групп через 23 минуты. Смесь концентрировали на ротационном испарителе, снова растворяли в дихлорметане (25 мл) и снова концентрировали, затем снова растворяли в ацетонитриле (25 мл) и снова концентрировали. Продукт подтверждали путем ЖХ-МС. Указанное выше вещество (OG1405, 34,5 г, предполагаемый количественный выход 21,0 г) использовали в виде неочищенного масла на следующем этапе. В очистке не было необходимости.
[0595] Затем OG1405 подвергали реакции с OG1402 с получением OG1785, как показано на фиг.2H. В 500 мл колбу в атмосфере азота, оснащенную мешалкой, помещали OG1402 (5,5 г), затем ацетонитрил (70 мл), затем N,N-диизопропилэтиламин (26,3 мл) и раствор T3P (см. выше) (7,9 мл). Раствор перемешивали при комнатной температуре в течение 30 минут, затем охлаждали на бане с ледяной водой и добавляли раствор OG1405 (неочищенное масло с этапа выше, 34,5 г) в ацетонитриле (70 мл). Смесь нагревали до комнатной температуры. Через 20 минут реакционную смесь охлаждали на бане с ледяной водой и гасили водой (5 мл). Затем смесь концентрировали в вакууме с использованием ротационного испарителя до половины объема. Брали образцы для ЖХ-МС.
[0596] Добавляли еще воду (50 мл), затем 0,5 М лимонную кислоту (75 мл) и изопропилацетат (175 мл). Через 5 минут смесь разделяли. Водную фазу экстрагировали дополнительным количеством изопропилацетата (50 мл). Объединенные органические фазы промывали водной лимонной кислотой (0,13 М, 30 мл, состояла из 10 мл 0,5 М лимонной кислоты и 20 мл воды). Затем органические фазы промывали смесью насыщенного хлорида натрия (25 мл) и воды (25 мл), затем, наконец, промывали насыщенным хлоридом натрия (25 мл). Затем их сушили над сульфатом натрия (124 г), фильтровали и промывали изопропилацетатом (30 мл), и концентрировали на ротационном испарителе с получением желтовато-коричневого масла (27,3 г). Брали образцы для анализа ЖХ-МС.
[0597] Указанное масло растворяли в смеси ацетонитрил/вода (3:1, 15 мл/5 мл), фильтровали через фильтровальный диск ВЭЖХ (мембрана PTFE Cole-Parmer 0,2 мкм, номер продукта 02915-20) и делили на три равные порции, каждую из которых отдельно очищали следующим образом.
[0598] Порции загружали в колонку C18 Redi-Sep Gold (275 г, SN - 69-2203-339, серия 241234-611W), уравновешенную 50% растворителем B (ацетонитрил)/50% растворителем A (вода). Затем вещество очищали путем обращенно-фазовой ВЭЖХ с использованием градиента растворитель А: вода/растворитель В: ацетонитрил. Соответствующие фракции объединяли в пул и разделяли между дихлорметаном (150 мл) и водой (5 мл)/насыщенным хлоридом натрия (25 мл). Водную фазу два раза экстрагировали дихлорметаном (2 х 50 мл). Объединенные органические фазы сушили над сульфатом натрия (60 г), фильтровали и промывали дихлорметаном (40 мл), и концентрировали. Структуру и чистоту подтверждали различными методами анализа, включая ЖХ-МС: OG1785 выделяли в виде пенообразного твердого вещества (R5329, 19,0 г, выход 83%, чистота 95,1% (a/a 210 нм), хранили в атмосфере азота при 4°C.
[0599] Затем удаляли трет-бутилоксикарбонильную защитную группу в OG1785 с использованием трифторуксусной кислоты (ТФУ) с получением OG1786, как показано на фиг.2I.
Пример 7. Синтез полимера OG1801
[0600] Полимер OG1801 сначала получали из инициатора OG1786. OG1801 содержит функциональную аминогруппу, которая более стабильна (чем малеимид) во время синтеза полимера. Для синтеза полимера OG1801 использовали модифицированный вариант ATRP, в котором соединение меди (Cu(I)) получали in situ путем добавления металлической меди к Cu (II). Исходные вещества и реагенты, необходимые в этой реакции, рассчитывали на основе загружаемой партии мономера (HEMA-PC) OG47, а также целевой молекулярной массы (ММ).
[0601] В перчаточном боксе взвешивали 50 г мономера OG47 и добавляли 200 мл дегазированного EtOH с растворением указанного мономера при комнатной температуре; брали образцы для тестирования концентрации мономера. Взвешивали Cu (II), Bpy, Cu(0) и помещали в 500 мл колбу; продували аргоном при добавлении раствора мономера в колбу; колбу закрывали пробкой и подвергали воздействию вакуумом в течение 25 минут до исчезновения пузырьков. Реакционная смесь постепенно меняла цвет от светло-зеленого до темно-зеленого, затем до светло-коричневого; в перчаточном боксе взвешивали ~200 мг инициатора OG1786 и растворяли в ~2000 мкл ДМФА при комнатной температуре с получением исходного раствора 100 мг/мл; брали образцы для тестирования концентрации и чистоты инициатора; добавляли раствор инициатора в колбу в атмосфере аргона. Со временем реакционный раствор становился темно-коричневым и начинал густеть; систему закрывали и оставляли реакцию протекать в течение 2 дней.
[0602] Затем OG1801 готовили к добавлению малеимида и удаляли катализатор (медь) следующим образом: для удаления катализатора использовали заполненную колонку с нормальной фазой с диоксидом кремния RediSep® Rf. Размер колонки выбирали исходя из количества меди в реакционной смеси. Например, для 50 г партии OG1801 использовали колонку 330 г (кат.№69-2203-330, размер колонки 330 г, объем колонки (ОК)=443 мл). Для соединения использовали тефлоновую трубку, поскольку растворитель для элюирования представлял собой EtOH.
[0603] После удаления меди все фракции партиями переносили в круглодонную колбу и досуха выпаривали EtOH посредством ротационного испарителя при 45-50°C при пониженном давлении. На этом этапе контролировали объем EtOH, собранный в результате конденсации, чтобы убедиться, что удаление EtOH составляло>90%. Полимер растворяли в 250 мл воды для инъекций и фильтровали с использованием 0,2 мкм фильтра. В результате получали прозрачный-светло-желтый раствор полимера в концентрации ~150 мг/мл. Указанный раствор можно хранить при 2-8°C до 3 месяцев перед применением.
Пример 8. Синтез полимера OG1802
[0604] Исходные вещества и реагенты, необходимые в этой реакции, рассчитывали на основе загружаемой партии OG1801. Линкер представляет собой сложный NHS-эфир 3-малеимидопропионовой кислоты. К 50 г раствора полимера (~150 мг/мл) добавляли 30 мл 0,5 М фосфата натрия (в воде для инъекций, pH 8). Оставляли перемешиваться в течение 1 минуты; pH, измеренный индикаторной бумагой для определения pH, составлял 8,0. Взвешивали 204,8 мг линкера и растворяли в 4,1 мл ДМФА с получением исходного раствора 50 мг/мл. К раствору полимера по каплям добавляли раствор линкера со скоростью 815 мкл в минуту при энергичном перемешивании. Добавление 4095 мкл раствора линкера занимало 5 минут. Реакцию проводили в течение 30 минут при комнатной температуре. Гасили реакцию 20 мл 5% уксусной кислоты с достижением конечного pH, равного 5. Раствор фильтровали с использованием 1 л вакуумного фильтра (0,2 мкм).
[0605] Затем очищали OG1802 (представленный на фиг.2K) следующим образом: использовали кассеты фильтрации в перекрестном потоке Millipore для очистки полимера в водной системе. Начинали с концентрирования раствора полимера до 250 мл (~ 200 мг/мл). Добавляли свежую воду для инъекций из резервуара и регулировали скорость потока свежей подаваемой воды для инъекций до уровня пермеата (~2 мл/мин). УФ/ДФ устанавливали на 2-8°C на ночь. Как правило, использовали 2,5 л воды для инъекций (10х объемное отношение к раствору полимера). Брали образец ретентата для тестирования чистоты. Целевая чистота составляла>98%. Раствор полимера фильтровали посредством 1 л фильтр-флакона 0,2 мкМ. Раствор полимера можно хранить при 2-8°C до 3 месяцев перед конъюгацией.
Пример 9. Альтернативные полимеры, содержащие фосфорилхолин
[0606] Полимер HEA-PC синтезировали, как описано ниже. HEA-PC (2-(акрилоилокси)этил-2-(триметиламмоний)этилфосфат), который представляет собой акрилат, в отличие от метакрилата HEMA-PC, описанного выше, имеет следующую структуру:
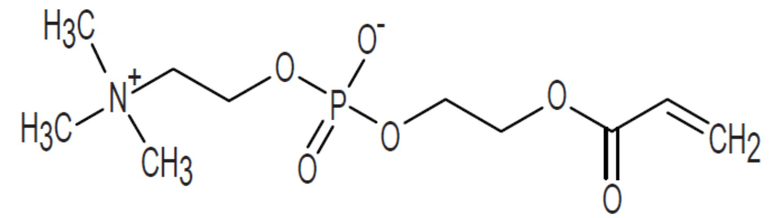
HEA-PC
[0607] HEA-PC полимеризовали с инициатором, представленным в примере 5 как соединение L.
Таблица 28
(Me6TREN)
[0608] Готовили исходный раствор инициатора 200 мг/мл путем растворения 2,2 мг инициатора в 11 мкл сухого ДМФА и раствор лиганда 200 мг/мл путем растворения 4,6 мг Me6TREN в 23 мкл сухого ДМФА. В пробирку отмеряли 8,25 мкл исходного раствора инициатора и 13,6 мкл лиганда. Дегазировали при -78°C в течение 5 минут, затем снова заполняли аргоном и добавляли 1,2 мг CuBr. Дегазировали и снова заполняли аргоном. Добавляли исходный раствор HEA-PC в метаноле (взвешивали 0,461 г HEA-PC и растворяли его в 0,5 мл метанола) к раствору внутри реактора при -78°C. Флакон промывали 200 мкл метанола и добавляли в реактор при -78°C, а затем 0,5 мл дистиллированной воды, затем еще 200 мкл воды. Тщательно дегазировали до отсутствия выделения пузырьков и исчезновения всей неоднородности (растворения или исчезновения твердых частиц). Снова заполняли 4 фунт/кв.дюйм аргона и оставляли реакцию протекать при комнатной температуре в течение часа. Реакционная смесь была уже вязкой. Реакцию оставляли протекать в течение примерно одного часа. Добавляли раствор бипириндина в метаноле (5 мг в 0,5 мкл). Добавляли еще 2-3 мл метанола и катализатор оставляли окисляться в течение ночи при 4°C. Превращение, определенное путем 1H ЯМР, составляло 94%.
[0609] На следующий день полимер подвергали диализу и анализу SEC/MALS с использованием колонки SB806M_HQ Shodex (7,8x300 мм) в 1x ФБР, pH 7,4 со скоростью 1 мл/мин, с получением PDI, составляющего 1,157, Mn, равной 723,5 кДа, Mp, равной 820,4 кДа, и Mw, равной 837,2 кДа (до диализа PDI составлял 1,12, Mn=695 кДа, Mp=778 кДа). Затем к полимеру присоединяли малеимидную функциональную группу, чтобы его можно было конъюгировать с белком.
[0610] Затем малеимидный сложный эфир Mal-PEG4-PFP (см. пример 20 выше) «пристегивали» к полимеру HEA-PC, как показано в примере 20. Полученный полимер HEA-PC, содержащий малеимидную функциональную группу, можно затем конъюгировать с сульфгидрильными группами, как рассмотрено в настоящем описании для полимеров HEMA-PC.
[0611] Полимер акриламида, содержащий PC, также получали с использованием мономера 2-(акриламил)этил-2-(триметиламмоний)этилфосфата (Am-PC), имеющего следующую структуру:
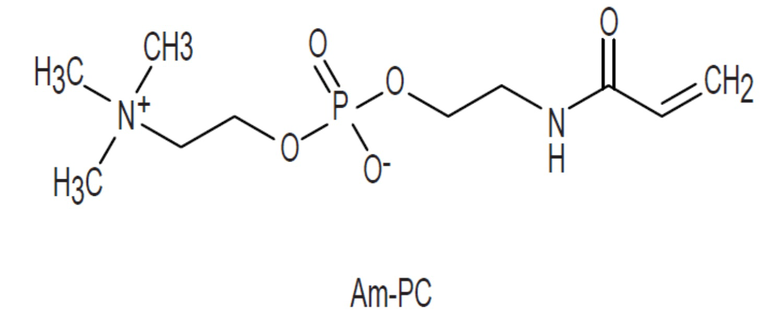
[0612] Am-PC использовали для полимеризации с использованием инициатора с 3 ответвлениями (соль ТФУ), имеющего структуру:
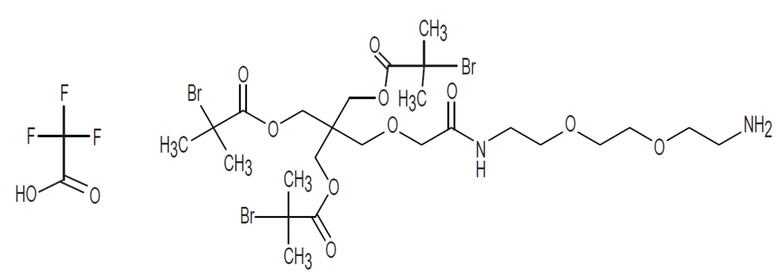
[0613] Синтез полимера Am-PC осуществляли следующим образом:
Таблица 29
(Me6TREN)
[0614] Готовили исходный раствор лиганда с концентрацией 200 мг/мл путем растворения 9 мг Me6TREN в 45 мкл сухого ДМФА. Добавляли 19,7 мкл исходного раствора в реакционный сосуд. Готовили исходный раствор инициатора с концентрацией 200 мг/мл путем растворения 6,5 мг вещества в 32,5 мкл сухого ДМФА. Добавляли 11 мкл исходного раствора инициатора к лиганду, полученному выше. Дегазировали в течение 5 минут. Добавляли 1 мг CuBr. Готовили исходный раствор CuBr2 с концентрацией 200 мг/мл путем растворения 4 мг CuBr2 в 20 мкл сухого ДМФА. Добавляли 0,5 г мономера (AmPC) к 1 мл метанола (медленное растворение/вязкий раствор), а затем 1 мкл исходного раствора CuBr2. К реакционной смеси, полученной выше, по каплям добавляли раствор мономера. Промывали 1 мл воды. Реакционную смесь тщательно дегазировали (замораживание-оттаивание). Реакцию оставляли протекать в течение 24 часов.
[0615] Затем полимер Am-PC можно подвергать диализу. Молекулярную массу полимера, указанного выше, определяли путем SEC/MALS: Mn составляла 215 кДа, Mp: 250 кДа, PDI составлял 1,17. Превращение определяли путем 1H ЯМР и оно составляло 94%. Малеимидная функциональная группа может быть присоединена к полимеру Am-PC, как рассмотрено выше для HEMA-PC и HEA-PC. Полимер Am-PC, содержащий малеимидную функциональную группу, может быть конъюгирован с белком, как описано выше.
Пример 10. Обратный анализ Эллмана для расчета свободного малеимида в соединении
[0616] После присоединения малеимидной функциональной группы к полимеру OG1801 с образованием OG1802 (см. выше) использовали анализ Эллмана для определения количества функционального малеимида, выраженного в виде функции в процентах (т.е. конъюгируемого), в образце. Тиол превращал реагент Эллмана (DTNB) в TNB-, затем в TNB2- в воде при нейтральном и щелочном pH, что обеспечивало желтый цвет (измерение при 412 нм). Строили стандартную кривую с цистеином. Поскольку малеимид реагирует с тиолом, этот анализ позволял фактически измерить оставшийся тиол (цистеин). Ингибирование рассчитывали в виде соотношений молярности (исходный тиол - тиол, оставшийся после присоединения малеимида к полимеру)/(исходный тиол) и выражали в процентах, при этом чем выше процент, тем выше функционализация малеимидом.
[0617] Реагенты, используемые в анализе: стандартную кривую получали с использованием цистеина от 62,5 мкМ до 2 мкМ. Исходные растворы полимера получали путем растворения порошка в 1xФБР pH 7,4 (реакционный буфер) и тщательного перемешивания. Смешивали растворы полимера и цистеина с одинаковой молярностью и оставляли для протекания реакции при 27°C в течение 30 минут. Добавляли 150 мкМ раствора DTNB к стандартным растворам цистеина и реакционной смеси полимер/цистеин, и цвет проявлялся при 27°C в течение 5 минут. ОП при 412 нм считывали на планшет-ридере Spectramax и процент ингибирования рассчитывали с помощью программного обеспечения Softmax Pro и стандартной кривой для цистеина.
Пример 11. Очистка и деблокирование молекул антиIL-6-ловушка VEGF
[0618] Тяжелые и легкие цепи клонировали в плазмиды экспрессии и трансфицировали в клетки СНО. Клетки выращивали в подходящих средах и собирали. Антитело (или, в качестве альтернативы, конструкцию АТ-ловушка) очищали с использованием захвата и элюирования на колонке для аффинной хроматографии на основе белка А. Остаток цистеина антитела (или, в качестве альтернативы, конструкции АТ-ловушка) в положении 443 (L443C (нумерация EU)), как правило, «блокируется» или окисляется химическими веществами в средах для культивирования клеток и недоступен для конъюгации. Поэтому очищенное антитело (или, в качестве альтернативы, конструкцию АТ-ловушка) подвергали процессу деблокирования (т.е. восстановления) для снятия блокировки и обеспечения конъюгации свободного (т.е. не вовлеченного в дисульфидные связи Cys-Cys) остатка цистеина с малеимидной функциональной группой полимера. Деблокирование осуществляли путем смешивания очищенного белка, представляющего собой антитело (или, в качестве альтернативы, конструкцию АТ-ловушка), с 30х молярным избытком восстановителя, такого как TCEP (3,3',3''-фосфантриилтрипропановая кислота), тринатрий 3,3',3''-фосфинтриилтрис(бензол-1-сульфонат) (TPPTS) и т.п., в течение 1 часа при температуре окружающей среды. Мониторинг реакции восстановления осуществляли путем SDS-ПААГ. После восстановления осуществляли замену буфера белка, представляющего собой антитело (или, в качестве альтернативы, конструкцию АТ-ловушка), с использованием кассеты XL Ultrafiltration Cassette Pellicon на буфер 20 мМ Трис pH 7,5, 100 мМ NaCl, 0,5 мМ TCEP для снятия блокировки. Затем реагент TCEP удаляли в той же системе для замены буфера с 20 мМ Трис pH 7,5, 100 мМ NaCl. Восстановленное антитело (или, в качестве альтернативы, конструкцию АТ-ловушка) подвергали повторному окислению с использованием 15х молярного избытка окислителя DHAA (дегидроксиаскорбиновой кислоты) в течение 1 часа при температуре окружающей среды, которое снова контролировали путем анализа SDS-ПААГ. Затем реагент DHAA удаляли в той же системе для замены буфера с 20 мМ Трис pH 7,5, 100 мМ NaCl.
Пример 12A. Конъюгация антитела (или, в качестве альтернативы, конструкции АТ-ловушка) с полимером MPC
[0619] Деблокированное антитело (или, в качестве альтернативы, конструкцию АТ-ловушка) конъюгировали с полимером OG1802 с получением биоконъюгата. Использовали избыток OG1802 (3-20-кратный молярный избыток). Осуществляли мониторинг конъюгации путем катионообменной ВЭЖХ-хроматографии и доводили почти до завершения. Конъюгат антитела (или, в качестве альтернативы, конструкции АТ-ловушка) очищали путем катионообменной хроматографии и заменяли буфер на буфер для получения состава путем ультрафильтрации/диафильтрации (УФ/ДФ). Конъюгат антитела (или, в качестве альтернативы, конструкции АТ-ловушка) очищали путем хроматографии, как описано выше.
Пример 12B. Реакция конъюгации конструкции VEGFR-антиIL6 с OG1802
[0620] Способ конъюгации конструкции VEGFR-антиIL6 с OG1802 включал два этапа: первоначальную реакцию восстановления с удалением защитных групп цистеина S (деблокирование) с последующей конъюгацией с биополимером OG1802 посредством химического взаимодействия малеимид-цистеин. Деблокирование осуществляли сначала путем восстановления (30-60 минут) конструкции VEGFR-антиIL6 30х молярным избытком 3,3',3''-фосфантриилтрипропановой кислоты (TCEP) с последующей заменой буфера на 20 мМ Трис-HCl pH 7,5, 100 мМ NaCl со снятием блокировки цистеина и удалением TCEP (фиг.28). В качестве альтернативы, для этапа деблокирования также можно использовать другие восстановители, такие как тринатриевая соль 3',3''-фосфантриилтрис(бензолсульфоновой кислоты) (TPPTS).
[0621] Затем восстановленную конструкцию VEGFR-антиIL6 окисляли (30-60 минут) 15х молярным избытком дегидроаскорбиновой кислоты (dHAA) с последующей заменой буфера на 20 мМ Трис pH 7,5, 100 мМ NaCl с удалением окислителя (фиг.28). Суммарный выход для процесса деблокирования составлял 89% (восстановление TCEP - 94%, окисление dHAA - 95%).
[0622] Процесс конъюгации осуществляли путем смешивания конструкции VEGFR-антиIL6 в конечной концентрации 2,4 мг/мл (или 12,5 мкМ) с 5х молярным избытком биополимера OG1802 при pH 8,0. Процесс конъюгации протекал при 2-8°C в течение периода от 2 до 3 дней. Конъюгированную конструкцию VEGFR-анти-IL6 очищали от неконъюгированного вещества путем катионообменной хроматографии (CEX) со смолой Poros XS с использованием ступенчатого градиента возрастающих концентраций соли (0-500 мМ NaCl) в буфере, содержащем 20 мМ ацетат натрия, pH 5,5 (фиг.29). Фракции, содержащие биоконъюгат VEGFR-антиIL6-OG1802, объединяли в пул и заменяли буфер на фосфатный буферный раствор (ФБР), pH 7,4. Эффективность конъюгации составляла приблизительно 60% (определено путем SDS-ПААГ в невосстанавливающих условиях и анализа ImageJ). Суммарный выход от исходного белкового вещества до биоконъюгата составлял приблизительно 27%.
[0623] Анализ SDS-ПААГ (фиг.30) и SEC-MALS (фиг.31) демонстрирует одну молекулу VEGFR-антиIL6 (~189 кДа), конъюгированную с одной молекулой полимера OG1802 (~794 кДа), что позволило получить биоконъюгат приблизительно ~983 кДа. Проводили эксперимент SEC-MALS путем введения 10 мкг (масса белка) со скоростью потока 1 мл/мин в колонку для эксклюзионной хроматографии (Shodex-SB806M-HQ), предварительно уравновешенную буфером ФБР, наряду с детекторами рассеяния света (miniDAWN TREOS) и показателя преломления (Optilab T-rEX) (Wyatt Technology). Анализ данных и конъюгата белка проводили в ASTRA 7 (Wyatt Technology).
Пример 13. Определение и устранение расщепления ловушки VEGF
[0624] Как показано на фиг.11A и 11B, SDS-ПААГ очищенных белком A восстановленных продуктов VEGFR-анти-IL-6 и анти-IL-6-VEGFR, полученных из клеток ExpiCHO-S, демонстрировал неожиданные полосы, размер которых зависел от ориентаций указанной конструкции. Эти полосы указывали на то, что часть молекул двойного ингибитора подвергалась расщеплению во время получения. В этом примере конструкция VEGFR-антиIL6 соответствует молекуле тяжелой цепи, содержащей последовательности 1A-2A-3H, спаренные с легкой цепью, содержащей последовательность 4A. Конструкция антиIL-6-VEGFR соответствует тяжелой цепи, содержащей последовательности 5H(Fab)-2A-1A-5H(Fc), спаренные с легкой цепью, содержащей последовательность 4A.
[0625] Как показано на фиг.12, эти полосы белка переносили на PVDF-мембрану и первые шесть остатков полос белка секвенировали путем N-концевого секвенирования по Эдману (устройство Procise 494HT). Результаты показали, что расщепленные продукты содержали общую N-концевую последовательность (93LTHRQT), которая расположена в домене 2 VEGFR1 в области ловушки VEGF.
[0626] На фиг.13A и 13B представлены результаты мутаций в области, фланкирующей сайт расщепления (K89TNY/LTHR96), которые препятствовали расщеплению белка при сохранении общей стабильности белка и свойств связывания с VEGF. Эти мутации выбирали на основе сохранения последовательностей, для этой цели использовали средство поиска основного локального выравнивания (BLAST) для поиска сходства последовательностей и выравнивания между последовательностью домена 2 VEGFR1 и гомологами белка в пределах класса млекопитающих. Выбирали 500 лучших результатов на основе их Е-величины BLAST. Из этого начального кластера, содержащего 500 последовательностей (кластер_1), выбирали последовательность с наилучшими показателями для каждого отдельного вида, сокращая его до набора из 106 типичных последовательностей (кластер_2). На основе вариаций последовательности в области, фланкирующей сайт расщепления, наблюдаемых в кластере_2, получали 16 мутантов конструкции анти-IL-6-VEGFR с единичными точечными мутациями и оценивали их по расщеплению белка. Эти мутации в начале вводили только в конструкцию анти-IL-6-VEGFR, поскольку она демонстрирует лучшее разделение по размеру между расщепленными и интактными продуктами из-за различий в их молекулярной массе (~35 кДа).
[0627] На фиг.13A и 13B представлены результаты SDS-ПААГ мутантов конструкции анти-IL-6-VEGFR, демонстрирующие, что модификации в сайте расщепления способствовали уменьшению количества расщепленного продукта (показано стрелками). Мутации T94I и H95I (нумерация остатков согласно положению в последовательности ловушки VEGF) заметно уменьшали расщепление ловушки VEGF. В частности, на фиг.13A и 13B показано, что большая часть мутаций уменьшала расщепление белка, а мутации T94I (VEGFR_вариант_1), представленная на фиг.13C, и H95I (VEGFR_вариант_2), представленная на фиг.13D, оказывали более заметный эффект. Единичную точечную мутацию (T94I, H95I) и двойную точечную мутацию (T94I/H95I, VEGFR_вариант_3), представленные на фиг.13E, затем вводили в конструкцию VEGFR-анти-IL-6. На фиг.13C указано положение T94I (подчеркнуто). На фиг.13D указано положение H95I (подчеркнуто). На фиг.13E указано положение T94I и H95I (подчеркнуто). Как показано на фиг.13F и 13G, SDS-ПААГ демонстрировал схожее уменьшение расщепления белка, и этот эффект дополнительно усиливался, когда вводили двойную мутацию.
Пример 14. Связывание вариантов двойного ингибитора VEGFR с VEGF по данным анализа Biacore
[0628] На фиг.14A-14F представлены результаты серии анализов Biacore, используемых для определения аффинности между агентами анти-VEGF и VEGF. Кинетику связывания между молекулами измеряли с использованием Biacore T200 (GE Healthcare) при 37°C в буфере HBS-EP+, содержащем 1 мг/мл БСА. Агенты анти-VEGF (конструкция VEGFR-анти-IL-6 (5 мкг/мл), конструкция VEGFR_вариант_1-анти-IL-6 (5 мкг/мл), конструкция VEGFR_вариант_2-анти-IL-6 (5 мкг/мл), конструкция VEGFR_вариант_3-анти-IL-6 (5 мкг/мл) и Эйлеа (3 мкг/мл)) захватывали на чип с белком A (GE) со скоростью 10 мкл/мин в течение 25 секунд, пять концентраций VEGF-A165 (0,19, 0,56, 1,67, 5, 15 нМ) пропускали через захваченные антитела в течение 120 секунд со скоростью 30 мкл/мин и подвергали диссоциации в течение 60 минут. Поверхность сенсорного чипа регенерировали путем 60-секундного введения 10 мМ глицина, pH 1,7, со скоростью потока 50 мкл/мин. Из всех сенсограмм вычитали два эталонных сигнала и осуществляли подгонку с использованием модели связывания Ленгмюра 1:1. На фиг.14A-14F показано, что введение единичных точечных мутаций T94I (конструкция VEGFR_вариант_1-анти-IL-6), H95I (конструкция VEGFR_вариант_2-анти-IL-6) и двойной мутации T94I/H95I (конструкция VEGFR_вариант_3-анти-IL-6) минимально влияет на связывание с VEGF. Как видно на фиг.14F, сенсограммы демонстрируют, что молекулы двойного ингибитора VEGFR-анти-IL-6 и варианты прочно связываются с VEGF-A, как и Эйлеа. В этом примере молекулы VEGFR-антиIL6 соответствуют вариантам VEGFR (последовательности 1A-1D), слитым с фрагментом, представляющим собой антиIL-6, через линкер GS (последовательности 2A-3H), спаренный с легкой цепью, содержащей последовательность 4A.
Пример 15. Репортерный анализ VEGF-стимулируемого VEGFR в клеточной системе
[0629] Результаты репортерного анализа VEGF-стимулируемого VEGFR в клеточной системе представлены на фиг.15. Двойной ингибитор VEGFR-антиIL6, как указано выше, блокировал связывание VEGF с VEGFR и стимуляцию нисходящей передачи сигнала в той же степени, что и Эйлеа (IC50 0,18 нМ относительно 0,11 нМ), тогда как анти-IL6 антитело не оказывало никакого влияния.
[0630] Для этих экспериментов клетки HEK293 KDR/VEGF Promega, готовые к применению после оттаивания (CS181405; Promega), подвергали оттаиванию и добавляли к 4,6 мл среды для анализа (10% FBS в DMEM). 25 мкл клеток переносили в каждую лунку 96-луночного планшета. VEGF (конечная концентрация 500 пМ) смешивали с серией разведений ингибиторов 1:3 для получения 10 точек разведения, начиная с 100 нМ (конечная концентрация), и инкубировали в течение 30 минут перед добавлением 50 мкл к высеянным клеткам. Через 6 часов к клеткам добавляли 75 мкл приготовленного реагента Bio-Glo (Promega) и инкубировали в течение 3-10 минут при комнатной температуре. Люминесценцию измеряли на планшет-ридере iD3 (Molecular Devices). Относительные единицы люминесценции приводили к самой низкой концентрации ингибитора для каждой кривой и наносили на график в виде среднего значения для повторностей со стандартной ошибкой в зависимости от логарифма концентрации антитела с использованием программного обеспечения GraphPad Prism и подгоняли к кривой нелинейной регрессии. В этом примере конструкция VEGFR-антиIL6 соответствует молекуле тяжелой цепи, содержащей последовательности 1A-2A-3H, спаренные с легкой цепью, содержащей последовательность 4A.
Пример 16. Анализ образования трубочек ЭКПВЧ
[0631] На фиг.16A-16C представлены результаты анализа образования трубочек ЭКПВЧ.
[0632] ЭКПВЧ-клетки 2 или 3 пересева культивировали в базальных средах для клеток сосудов с добавлением 1x набора Bovine Brain Extract endothelial cell growth kit (АТСС; питательные среды) в 5% CO2 при 37°C. По достижении клетками 70% конфлюэнтности, клетки отделяли от планшета с использованием трипсина (0,05%; ATCC) и 0,75x10^6 клеток высевали в колбу Т25 в питательных средах. На следующий день среды заменяли на среды для анализа (2% FBS в базальной среде для клеток сосудов). Через 6 часов в среды непосредственно добавляли 5 мкг/мл Кальцеина AM и инкубировали в течение 30 минут при 37°C. Одновременно в каждую лунку стеклянного 8-луночного предметного стекла добавляли 200 мкл предварительно размороженного Матригеля (с пониженным содержанием факторов роста; Corning) и инкубировали при 37°C в течение 30 минут.Во время инкубации 25 нМ каждого ингибитора (конечные концентрации) смешивали с 3 нМ VEGF (R&D), 12,5 нМ IL6 (Tonbo) и 25 нМ IL6R (Tonbo) (все конечные концентрации). После 30-минутной инкубации совместно с Кальцеином AM клетки промывали 3х ФБР, а затем обрабатывали трипсином. Собранные клетки центрифугировали в течение 3 минут при 1500×g и ресуспендировали в среде для анализа c плотностью 500000 клеток/мл. Затем 100 мкл клеток смешивали со 100 мкл приготовленных растворов ингибитор/комплекс, а затем переносили на предметные стекла, покрытые Матригелем, и инкубировали в течение ночи. На следующий день трубочки визуализировали с использованием 10x линзы объектива и GFP-фильтра на флуоресцентном микроскопе EVOS (Life Technologies). Для каждого условия получали по меньшей мере 2 типовых изображения, а затем анализировали с использованием Angiogenesis Analyzer Plugin для ImageJ (NIH), который оценивает и количественно определяет 20 параметров образования сети трубочек.
[0633] На фиг.16A-16C представлены данные в виде среднего значения со стандартной ошибкой из 4 независимых экспериментов. Статистическую значимость определяли для всех повторностей 4 независимых экспериментов с использованием непарного двустороннего t-критерия Стьюдента. Как показано на фиг.16C, конструкция VEGFR-антиIL6 значительно ингибировала параметры ангиогенеза: 17/20 параметров по сравнению с контролем. Эйлеа, напротив, ингибировало 4/20 параметра, а анти-IL-6 ингибировало 7/20 параметров. Конструкция VEGFR-антиIL6 была значительно эффективнее, чем Эйлеа и анти-IL-6, по 12/20 параметрам, как показано звездочками на фиг.16B. Кроме того, статистика с t-критерием представлена на фиг.16С. В этом примере конструкция VEGFR-анти-IL6 соответствует молекуле тяжелой цепи, содержащей последовательности 1A-2A-3H, спаренные с легкой цепью, содержащей последовательность 4A.
[0634] В некоторых вариантах реализации конструкция VEGFR-анти-IL6 обеспечивает синергетические механизмы для повышения эффективности и длительности в конкретных подверженных риску популяциях. Подверженные риску популяции включают тех, кто подвержен риску или у кого диагностирована диабетическая ретинопатия, или одно или более воспалительных заболеваний сетчатки.
Пример 17. Анализ пролиферации ЭКПВЧ
[0635] На фиг.17 представлены результаты анализа пролиферации ЭКПВЧ. ЭКПВЧ-клетки (2-5 пересевы) культивировали в базальных средах для клеток сосудов с добавлением 1x набора Bovine Brain Extract endothelial cell growth kit (АТСС; питательные среды) в 5% CO2 при 37 градусах. По достижении клетками 70% конфлюэнтности, клетки отделяли от планшета с использованием трипсина (0,05%; ATCC) и 3500 клеток/лунка высевали в центральные 60 лунок 96-луночного планшета, покрытого желатином, в 100 мкл сред для анализа (базальные среды для клеток сосудов с добавлением l-глутамина, аскорбиновой кислоты, гепарансульфата, гидрокортизона и 1% FBS) (Corning). После 24 часов нахождения в минимальной среде клетки стимулировали VEGF (1,5 нМ), IL-6 (5 нМ) и IL6R (10 нМ), смешанными с серией разведений ингибиторов 1:3 для получения 10 точек разведения, начиная с 100 нМ. Эти концентрации VEGF и IL-6 индуцировали пролиферацию на субмаксимальных уровнях при добавлении независимо друг от друга, а при комбинировании демонстрировали кривую синергетического роста. Через 48 часов жизнеспособность клеток измеряли с использованием реагента CellTiterBlue (Promega) в соответствии с инструкциями изготовителя на планшет-ридере iD3 (Molecular Devices). Для измерений поглощения определяли соотношение A570/A605 и данные приводили к контрольным (без ингибитора) клеткам для каждой повторности. Измерения флуоресценции (555ex/605em) приводили к контрольным клеткам для каждой повторности.
[0636] Как показано на фиг.17, приведенные значения наносили на график в виде среднего значения для повторностей со стандартной ошибкой в зависимости от концентрации антитела с использованием программного обеспечения GraphPad Prism и приводили к кривой нелинейной регрессии.
[0637] Как видно на фиг.17, двойной ингибитор VEGFR-антиIL6 блокировал синергетическую стимуляцию IL-6 и VEGF пролиферации ЭКПВЧ, тогда как ни Эйлеа, ни анти-Il-6 не влияли на рост ЭКПВЧ в этих условиях.
Пример 18. Получение улучшенных анти-IL-6 паратопов
[0638] В настоящем примере описано получение улучшенных анти-IL-6 паратопов. Результаты кинетики Biacore представлены в таблицах 30, 31, 32 и 33.
[0639] Для получения лучшего анти-IL-6 паратопа в искусственно сконструированной каркасной области IgG1 человека, мутант тяжелой цепи G66D (CDR2) подвергали созреванию аффинности. В начале конструировали библиотеки полных тяжелых цепей G66D, содержащие случайные единичные точечные мутации в положениях S35, I51, T63 и T65 за исключением замен на Cys, Met и Asn из-за химической неустойчивости, связанной с этими аминокислотами. Эти четыре положения выбирали исходя из допустимости для мутаций, наблюдаемых при скрининге бактерий и млекопитающих. Векторы экспрессии млекопитающих, содержащие мутации тяжелой цепи, котрансфицировали с легкой цепью дикого типа и вариантом M32L/M50D/N52S/M88Q в 3 мл культуры клеток Expi293 и инкубировали в течение пяти дней. Эти культуры центрифугировали и надосадочные жидкости, содержащие секретированные антитела, собирали и разводили в подвижном буфере HBS-EP+(соотношение 1:20).
[0640] Различные конструкции и получаемые кинетические свойства представлены ниже в таблицах 30-33.
Таблица 30
Кинетика Biacore для двойных мутантов анти-IL-6 тяжелой цепи
(названия столбцов и строк - как в таблице 1.7 выше)
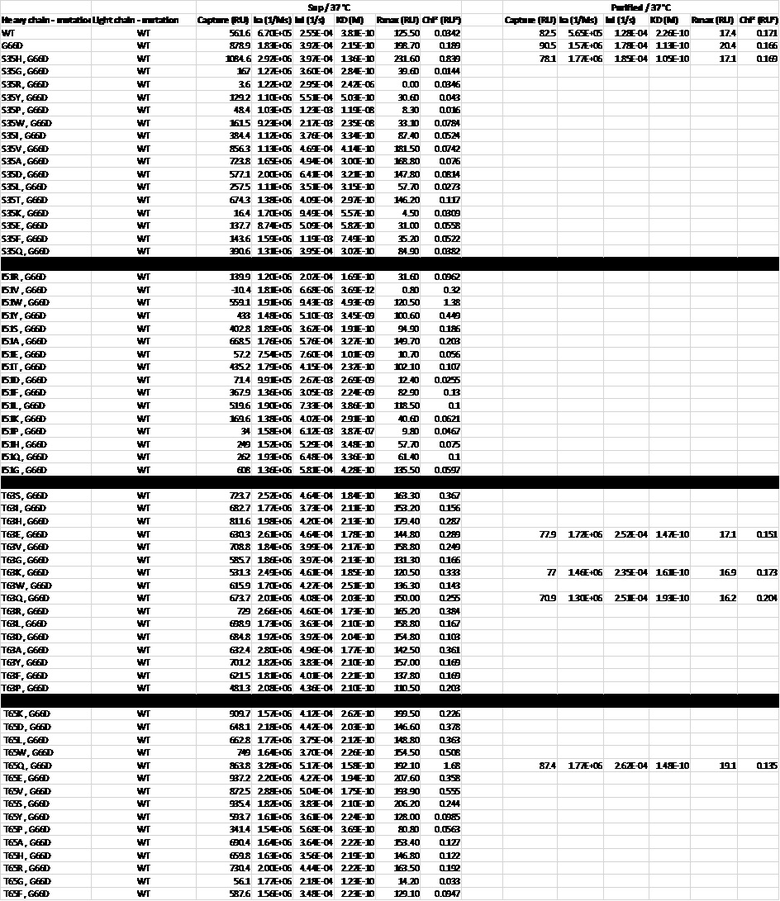
Таблица 31
Кинетика Biacore для двойных мутантов анти-IL-6 тяжелой цепи с подвергнутой мутагенезу легкой цепью
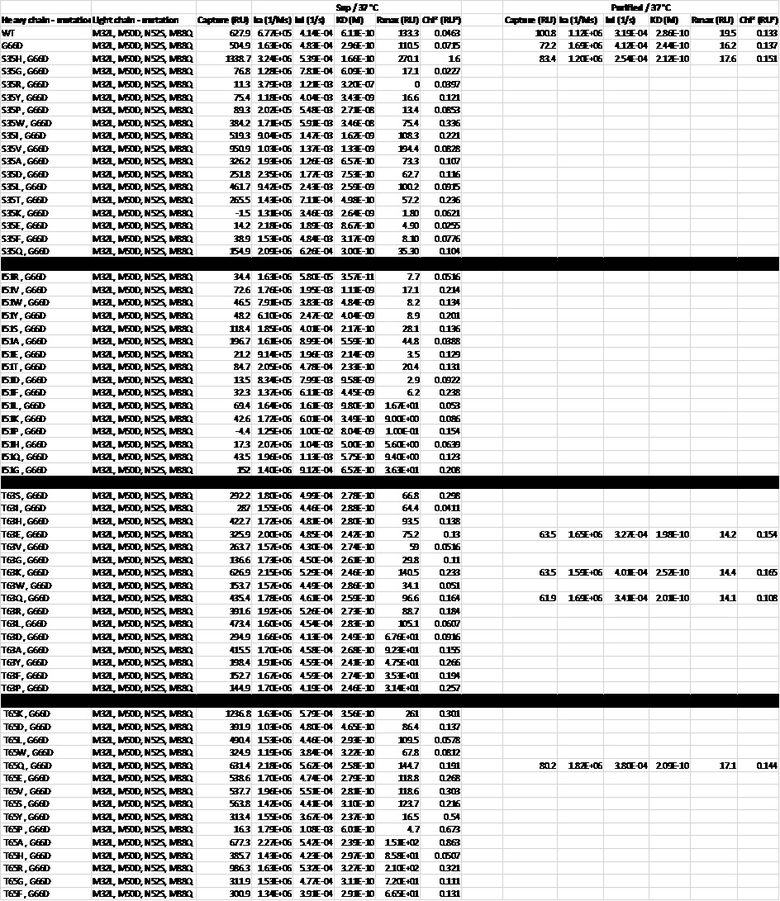
Таблица 32
Кинетика Biacore для тройных мутантов анти-IL-6 тяжелой цепи
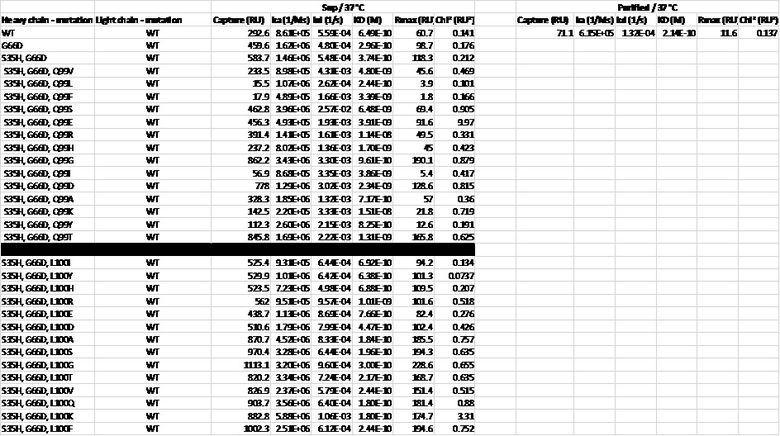
Таблица 33
Кинетика Biacore для тройных мутантов анти-IL-6 тяжелой цепи, спаренных с подвергнутой мутагенезу легкой цепью
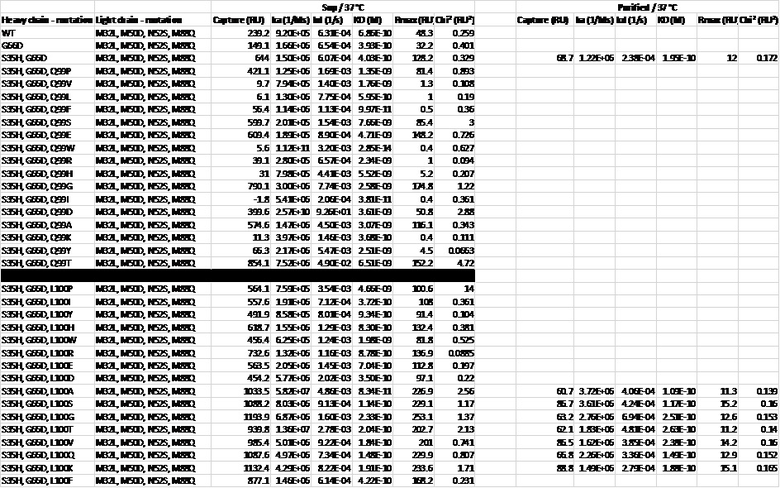
[0641] Кинетику связывания этих мутантных молекул в отношении IL-6 измеряли при 37°C с использованием Biacore T200. Антитела захватывали на чип с белком А (GE) путем введения надосадочной жидкости клеток в разведениях 1:20 со скоростью 10 мкл/мин в течение 60 секунд. Через захваченные антитела пропускали пять концентраций (0,56, 1,67, 5, 15 и 45 нМ) IL-6 в течение 60 секунд со скоростью потока 30 мкл/мин и подвергали диссоциации в течение 180 секунд в формате анализа кинетики Biacore с одним циклом. Сенсорный чип регенерировали путем 60-секундного введения 10 мМ глицина, pH 1,7, со скоростью потока 50 мкл/мин. Аналогичную систему анализа использовали для мониторинга взаимодействий между очищенными образцами антитела и IL-6 при 37°C, где применимо. Для этой системы 1 мкг/мл очищенного антитела вводили на чип с белком A со скоростью 10 мкл/мин в течение 25 секунд. Через захваченные антитела пропускали пять концентраций (0,56, 1,67, 5, 15 и 45 нМ) IL-6 в течение 60 секунд со скоростью потока 30 мкл/мин и подвергали диссоциации в течение 1800 секунд в формате анализа кинетики Biacore с одним циклом. Сенсорный чип регенерировали путем 60-секундного введения 10 мМ глицина, pH 1,7, со скоростью потока 50 мкл/мин. Анализ данных для надосадочной жидкости и очищенных образцов проводили с использованием программного обеспечения BIAevaluation (GE). Из всех сенсограмм вычитали два эталонных сигнала и осуществляли подгонку с использованием модели связывания Ленгмюра 1:1.
[0642] Как показано в таблицах 30 и 31, тяжелая цепь двойного мутанта S35H G66D демонстрировала усиление экспрессии белка, на что указывает приблизительно двукратное повышение уровня захвата относительно тяжелой цепи дикого типа. Этот мутант также демонстрировал небольшое увеличение аффинности в зависимости от спаренной легкой цепи (дикий тип: в ~2 раза, M32L/M50D/N52S/M88Q: в ~1,3 раза). В таблице 30 показана G66D-оптимизация кинетики Biacore легкой цепи дикого типа для двойных мутантов анти-IL-6 тяжелой цепи. Данные получали для образцов до (надос.) и после очистки, в обоих случаях при 37°C. В таблице 31 показана G66D-оптимизация кинетики Biacore мутанта легкой цепи для двойных мутантов анти-IL-6 тяжелой цепи, спаренных с подвергнутой мутагенезу легкой цепью. Данные получали для образцов до (надос.) и после очистки, в обоих случаях при 37°C.
[0643] Таким образом, этот двойной мутант выбирали в качестве исходной точки для другого этапа, на котором допустимые для замен положения Q99 и L100 в CDR3 подвергали случайному мутагенезу, экспрессировали и оценивали на предмет связывания IL-6, как описано для этапа скрининга G66D. Как показано в таблицах 32 и 33, замены L100 на Ala, Ser, Gly, Thr, Gln, Lys демонстрировали лучшую экспрессию по сравнению с диким типом, на что указывают уровни захвата. Соответственно, этих мутантов выбирали для очистки и оценки связывания IL-6. Измерения аффинности в случае очищенного вещества также демонстрировали лучшее связывание IL-6, как показано в таблицах 32 и 33. На основе этих результатов эти дополнительные модифицированные CDR, показанные на фиг.18, включали в создание молекул двойного анти-IL-6 и анти-VEGF ингибитора.
[0644] В таблице 32 представлена кинетика Biacore для модификации тяжелой цепи G66D/S35H, спаренной с легкой цепью дикого типа, для тройных мутантов анти-IL-6 тяжелой цепи. Данные получали для образцов до (надос.) и после очистки, в обоих случаях при 37°C. В таблице 33 представлена кинетика Biacore для модификации тяжелой цепи G66D/S35H, спаренной с мутантной легкой цепью, для тройных мутантов анти-IL-6 тяжелой цепи, спаренных с подвергнутой мутагенезу легкой цепью. Данные получали для образцов до (надос.) и после очистки, в обоих случаях при 37°C.
Пример 19. Варианты двойных анти-IL-6-анти-VEGF ингибиторов
[0645] На основе результатов в случае вариантов ловушки VEGF и улучшенных анти-IL-6 паратопов конструировали 216 молекул в двух разных конфигурациях, которые содержали комбинации последовательностей, как показано на фиг.19, 20, 21, 22 и 23. В случае первой конфигурации (VEGFR-анти-IL-6) ловушку VEGF, представленную последовательностями 1A-1D на фиг.19, располагали в начале белка с последующим двойным повтором линкера Gly-Gly-Gly-Gly-Ser (GS), представленного последовательностью 2A на фиг.20, который соединяет ловушку с N-концом анти-IL-6 тяжелой цепи, представленной последовательностями 3A-3I на фиг.21. Эти конструкции могут быть спарены с легкими цепями, перечисленными в виде последовательностей 4A-4C на фиг.22. В случае второй конфигурации (анти-IL-6-VEGFR) вариабельный и константный домены тяжелой цепи, представленные тяжелой цепью - Fab, последовательностями 5A-5I на фиг.23, соединяли с ловушкой VEGF, представленной последовательностями 1A-1D на фиг.19, через линкер GS, представленный последовательностью 2A на фиг.20, а затем Fc-домен, представленный тяжелой цепью - Fc, последовательностями 5A-5I на фиг.23, подвергали слиянию с C-концом ловушки VEGF. Таким образом, ловушка VEGF оказывалась «зажата» между Fab- и Fc-областями антитела.
[0646] На фиг.19 показаны последовательности ловушки VEGF. Вариация последовательности подчеркнута и выделена жирным шрифтом. На фиг.20 показана линкерная (GS) последовательность Gly-Gly-Gly-Gly-Ser с двойным повтором. На фиг.21 показаны последовательности тяжелой цепи для анти-IL-6 молекул. CDR подчеркнуты. На фиг.22 показаны последовательности легкой цепи для анти-IL-6 молекул. CDR подчеркнуты. На фиг.23 показаны последовательности тяжелой цепи (разделенной на Fab и Fc) для анти-IL-6 молекул. CDR подчеркнуты.
[0647] В некоторых вариантах реализации любой из вариантов ловушки VEGF, в котором она «зажата» между Fab- и Fc-областями антитела, может быть спарен с легкими цепями, перечисленными в виде последовательностей 4A-4C на фиг.22. Таким образом, в некоторых вариантах реализации конструкции, представленные на фиг.19 (последовательности VEGFR), можно использовать для любого из расположений ловушки VEGF, представленных в настоящем описании, или самостоятельно.
Пример 20. Кинетика связывания между слитой конструкцией VEGFR-ловушка-Fc и VEGF
[0648] На фиг.26A-26C представлены результаты анализов Biacore, используемых для определения аффинности между анти-VEGF и VEGF агентами. Кинетику связывания измеряли с использованием Biacore T200 при 37°C в буфере HBS-EP+, содержащем 1 мг/мл БСА. VEGFR-Fc (1 мкг/мл) и Эйлеа (1 мкг/мл) захватывали на чип с белком A (GE) со скоростью 10 мкл/мин в течение 25 секунд, пять концентраций VEGF-A165 (0,19, 0,56, 1,67, 5, 15 нМ) пропускали через захваченные анти-VEGF молекулы в течение 120 секунд со скоростью 30 мкл/мин и подвергали диссоциации в течение 30 минут. Поверхность сенсорного чипа регенерировали путем 60-секундного введения 10 мМ глицина, pH 1,7, со скоростью потока 50 мкл/мин. Из всех сенсограмм вычитали два эталонных сигнала и осуществляли подгонку с использованием модели связывания Ленгмюра 1:1. Как показано на фиг.26A-26C, VEGFR-Fc и Эйлеа связываются с VEGF со схожей аффинностью. В этом примере VEGFR-Fc содержит последовательность 6A.
Пример 21. Удаление продукта расщепления
[0649] Как рассмотрено в примере 13, конструкция VEGFR-антиIL6 демонстрирует незначительную степень расщепления в домене 2 VEGFR1 в области ловушки VEGF. Авторы настоящего изобретения разработали два метода ортогональной хроматографии для отделения интактного белка от расщепленного вещества после этапа очистки с белком А: i) катионообменную хроматографию (CEX) и ii) хроматографию гидрофобного взаимодействия (HIC).
[0650] Для метода CEX pH раствора элюата из колонки с белком А VEGFR-антиIL6 доводили до 6, затем вводили в колонку Porous XS, предварительно уравновешенную 20 мМ фосфатом натрия, pH 6. Интактный белок элюировался при меньшей ионной силе, чем расщепленное вещество, и почти полностью отделялся в пределах линейного градиента возрастающей ионной силы (0-1 М хлорид натрия) (фиг.32). Фракции, обогащенные интактным белком, объединяли в пул и общий выход после извлечения составлял приблизительно 57%.
[0651] Для метода HIC смесь интактного белка VEGFR-антиIL6, обогащенную расщепленным веществом, вводили в колонку Butyl HP, предварительно уравновешенную 20 мМ фосфатом натрия, pH 6, 500 мМ-1 М сульфатом аммония. Интактный и расщепленный белки разделяли в пределах линейного градиента понижающейся ионной силы (1-0 М сульфат аммония), при этом интактный белок элюировался при меньшей ионной силе, чем расщепленное вещество, и мог быть частично или полностью отделен в зависимости от объединенных фракций (фиг.33).
Пример 22. Аффинность конструкции VEGFR-антиIL6 к IL6 и VEGF-A, измеренная путем KinExA
[0652] Систему анализа кинетического исключения (KinExA®) 3200 (Sapidyne Instruments Inc., Бойсе, Айдахо) использовали для измерения аффинности и кинетики равновесного связывания между конструкцией VEGFR-антиIL6 или VEGFR-антиIL6-OG1802 (белковый компонент которой представлен на фиг.27) и VEGF или IL6 в растворе. Гранулы азлактона, покрытые VEGF или IL6, использовали для захвата фракции свободной конструкции VEGFR-антиIL6 или VEGFR-антиIL6-OG1802 из уравновешенного образца VEGFR-антиIL6 или VEGFR-антиIL6-OG1802 и VEGF или IL6. Захваченные молекулы двойного ингибитора детектировали с помощью флуоресцентно меченого поликлонального антитела к иммуноглобулину человека (антитело козы к IgG человека Alexa 647).
[0653] Аффинность между молекулами двойного ингибитора и VEGF определяли по 2-3 кривым титрования в возрастающих концентрациях двойного ингибитора и соответствующих серийных разведениях VEGF для титрования (указаны в скобках), перечисленных ниже: I) конструкция VEGFR-антиIL6: 1 нМ (20 нМ, 2-кратное разведение, 12 разведений), инкубация 3,5 часа; 50 пМ (500 пМ, 2-кратное разведение, 14 разведений), инкубация 2,5 часа; 10 пМ (1 нМ, 2-кратное разведение, 13 разведений), инкубация 23 часа; II) конструкция VEGFR-антиIL6-OG1802: 2 нМ (30 нМ, 2-кратное разведение, 12 разведений), инкубация 45 минут; 100 пМ (15 нМ, 2-кратное разведение, 14 разведений), инкубация 6 часов; 7,5 пМ (1 нМ, 2-кратное разведение, 14 разведений), инкубация 48 часов. Аналогичные измерения проводили с IL6 в следующих условиях: III) конструкция VEGFR-антиIL6: 1 нМ (20 нМ, 2-кратное разведение, 13 разведений), инкубация 7 часов; 10 пМ (1 нМ, 2-кратное разведение, 13 разведений), инкубация 42 часа; 100 пМ (1 нМ, 2-кратное разведение, 13 разведений), инкубация 2,5 часа; IV) конструкция VEGFR-антиIL6-OG1802: 1 нМ (22,5 нМ, 2-кратное разведение, 13 разведений), инкубация 5,5 часов; 100 пМ (22,5 нМ, 2-кратное разведение, 14 разведений), инкубация 12 часов.
[0654]Скорости ассоциации (kon, M-1 с-1) между молекулами двойного ингибитора и VEGF или IL6 непосредственно определяли в результате экспериментов по измерению кинетики (1-2 кривые для каждой анализируемой пары), тогда как скорости диссоциации (koff, с-1) рассчитывали на основе следующего уравнения: koff=Kd x kon. Измерения проводили на четырех препаратах, содержащих i) 100 пМ VEGFR-антиIL6 и 86,4 пМ VEGF, ii) 217 пМ VEGFR-антиIL6-OG1802 и 200 пМ VEGF, iii) 100 пМ VEGFR-антиIL6 и 84,3 пМ IL6, iv) 1000 пМ VEGFR-антиIL6 и 900 пМ IL6.
[0655] Все измерения осуществляли при 37°C с использованием 1х подвижного буфера ФБР (pH 7,4) и образцов, полученных в 1xФБР (pH 7,4) с 1 мг/мл БСА. Анализ данных проводили с использованием программного обеспечения KinExA Pro 4.3.11.
[0656] В тестируемых условиях конструкция VEGFR-антиIL6-OG1802 связывалась с VEGF и IL6 сопоставимо с конструкцией VEGFR-антиIL6 со значениями Kd, составляющими 2,10 пМ для конструкции VEGFR-антиIL6 и VEGF, 1,02 пМ для конструкции VEGFR-антиIL6-OG1802 и VEGF, 21,10 пМ для конструкции VEGFR-антиIL6 и IL6, 12,10 пМ для конструкции VEGFR-антиIL6-OG1802 и IL6 (см. таблицы 34 и 35).
Таблица 35. IL-6
[0657] Измеренная аффинность конструкций VEGFR-антиIL6 и VEGFR-антиIL6-OG1802 была в пределах ошибки как для IL6, так и для VEGF; таким образом, эти результаты демонстрируют, что биополимер не влиял на связывание двойных ингибиторов с их мишенями.
Пример 23. Конкурентный ELISA-анализ VEGF/VEGFR
[0658] Этот анализ выполняли, как описано в примере 3. В этом анализе все из: конструкции VEGFR-антиIL6, VEGFR-антиIL6-OG1802 (как показано на фиг.27) и Эйлеа в различной степени ингибировали связывание VEGF с VEGFR (фиг.34). Конструкция VEGFR-антиIL6-OG1802 демонстрировала большее ингибирование (IC50=0,99 нМ) по сравнению с конструкцией VEGFR-анти-IL6 (IC50=1,98 нМ) и Эйлеа (IC50=1,38 нМ). Кроме того, конструкция VEGFR-антиIL6 демонстрировала максимальное ингибирование, превосходящее ингибирование в случае Эйлеа, тогда как конструкция VEGFR-антиIL6-OG1802 демонстрировала еще более выраженное максимальное ингибирование (VEGFR-антиIL6-OG1802=79,5%, VEGFR-анти-IL6=67,0% относительно Эйлеа=49,6%). Эти данные показывают, что присоединение биополимера OG1802 к конструкции VEGFR-антиIL6 обуславливает дополнительные ингибирующие способности в отношении связывания VEGF с VEGFR. Соответственно, указанный биополимер усиливает функцию молекулы.
Пример 24. Конкурентный ELISA-анализ IL-6/IL-6R
[0659] Этот анализ выполняли, как описано в примере 3.
[0660] В этих условиях конструкция VEGFR-антиIL6-OG1802 эффективно блокировала взаимодействие IL-6/IL-6R сопоставимо с конструкцией VEGFR-антиIL6 со значениями IC50, составляющими 0,26 нМ для VEGFR-антиIL6 и 0,23 нМ для VEGFR-антиIL6-OG1802 (фиг.35). Эти результаты показывают, что биополимер не изменял ингибирующее действие двойного ингибитора на связывание IL6 с его рецептором. Эйлеа служило в качестве отрицательного контроля.
Пример 25. Репортерный анализ VEGF-стимулируемого VEGFR в клеточной системе
[0661] Этот анализ выполняли, как описано в примере 15.
[0662] В тестируемых условиях конструкция VEGFR-антиIL6-OG1802 ингибировала VEGF-стимулированные VEGFR-репортерные клетки сопоставимо с конструкцией VEGFR-антиIL6 и Эйлеа со средними значениями IC50, составляющими 0,35±0,10 нМ, 0,51±0,07 нМ и 0,35±0,03 нМ соответственно (фиг.36). Может существовать тенденция к тому, что конструкция VEGR-антиIL6-OG1802 (белковый компонент представлен на фиг.27) и Эйлеа умеренно более эффективны, чем конструкция VEGFR-антиIL6, но это не является значимым обнаружением и приходится на погрешность анализа. На основе этих данных можно сделать вывод, что биоконъюгат ингибирует передачу сигнала VEGFR2 аналогично своему белку-предшественнику.
Пример 26. Стимулируемое липополисахаридом образование трубочек в ЭКПВЧ
[0663] ЭКПВЧ-клетки 2 пересева культивировали в базальных средах для клеток сосудов с добавлением 1x набора Bovine Brain Extract endothelial cell growth kit (АТСС; питательные среды) в 5% CO2 при 37 градусах. По достижении клетками 70% конфлюэнтности, клетки отделяли от планшета с использованием трипсина (0,05%; ATCC) и 0,75x106 клеток высевали в колбу Т25 в питательных средах.
[0664] На следующий день среды заменяли на среды для анализа (2% FBS в базальной среде для клеток сосудов). Через 6 часов в среды непосредственно добавляли 5 мкг/мл Кальцеина AM и инкубировали в течение 30 минут при 37 градусах.
[0665] Одновременно в каждую лунку стеклянного 8-луночного предметного стекла добавляли 200 мкл предварительно размороженного Матригеля (с пониженным содержанием факторов роста; Corning) и инкубировали при 37 градусах в течение 30 минут.
[0666] Во время инкубации ингибиторы (конечная концентрация 37,5 нМ) смешивали с липополисахаридом (LPS; Sigma; конечная концентрация 1 мкг/мл).
[0667] После 30-минутной инкубации совместно с Кальцеином AM клетки промывали 3х ФБР, а затем обрабатывали трипсином. Собранные клетки центрифугировали в течение 3 минут при 1500×g и ресуспендировали в среде для анализа c плотностью 500000 клеток/мл. Затем 150 мкл клеток смешивали со 100 мкл приготовленных растворов ингибитор/комплекс, а затем переносили на предметные стекла, покрытые Матригелем, и инкубировали в течение ночи.
[0668] На следующий день трубочки визуализировали с использованием 10x линзы объектива и GFP-фильтра на флуоресцентном микроскопе EVOS (Life Technologies). Для каждого условия получали по меньшей мере 2 типовых изображения, а затем анализировали с использованием Angiogenesis Analyzer Plugin для ImageJ (NIH), который оценивает и количественно определяет 20 параметров образования сети трубочек.
[0669] Статистическую значимость определяли для всех повторностей независимых экспериментов с использованием непарного двустороннего t-критерия Стьюдента.
[0670] В этих условиях конструкция VEGFR-антиIL6 демонстрировала статистически значимое ингибирование 15/20 измеренных параметров ангиогенеза по сравнению с контролем, тогда как Эйлеа значительно ингибировало 5/20, и анти-IL6 ингибировало 3/20 по сравнению с клетками, обработанными контролем. В целом, конструкция VEGFR-антиIL6 значительно ингибировала все параметры, кроме одного, на которые Эйлеа или анти-IL6 были склонны влиять или значительно влияли. Более того, было несколько параметров, в отношении которых конструкция VEGFR-антиIL6 демонстрировала значительно лучшее ингибирование, чем Эйлеа (7/20) и анти-IL6 (10/20), 3 из которых конструкция VEGFR-антиIL6 значительно лучше ингибировала, чем Эйлеа и анти-IL6. (Фиг.37 и фиг.38 и 11)
[0671] Конструкция VEGFR-антиIL6-OG1802 эффективно ингибировала образование трубочек ЭКПВЧ и, что интересно, конструкция VEGFR-антиIL6-OG1802 демонстрировала тенденцию к усилению ингибирования 9/20 параметров по сравнению с конструкцией VEGFR-антиIL6. (Фиг.39). Это означает, что конструкция VEGFR-антиIL6-OG1802 имеет дополнительное преимущество по сравнению с ее неконъюгированным аналогом.
Пример 27. Анализ пролиферации ЭКПВЧ
[0672] Этот анализ выполняли, как описано в примере 17.
[0673] В тестируемых условиях конструкция VEGFR-антиIL6-OG1802 (белковый компонент показан на фиг.27) ингибировала VEGF/IL6-стимулируемую пролиферацию ЭКПВЧ сопоставимо с конструкцией VEGFR-антиIL6, тогда как Эйлеа не оказывало влияния (фиг.40A).
[0674] Интересно, что конструкция VEGFR-антиIL6-OG1802 демонстрировала повышенную эффективность по сравнению с конструкцией VEGFR-антиIL6, когда в начале эксперимента клетки высевали в лунки с разной плотностью. Как показано на фиг.40B, при плотности 2000 клеток на лунку конструкции VEGFR-антиIL6 и VEGFR-антиIL6-OG1802 демонстрировали практически одинаковое ингибирование пролиферации ЭКПВЧ. При плотности 4000 клеток на лунку конструкция VEGFR-антиIL6-OG1802 демонстрировала чуть большее максимальное ингибирование, чем конструкция VEGFR-антиIL6 (40% относительно 30% соответственно), и при плотности 6000 клеток на лунку конструкция VEGFR-антиIL6-OG1802 ингибировала пролиферацию на 25% относительно контроля, тогда как конструкция VEGFR-антиIL6 не оказывала влияния.
[0675] Как показано в примерах выше, не ограничиваясь рамками какой-либо теории, оказывается, что биополимер помогает доставлять белок, с которым он связан, на поверхность планшета и клеток. Это позволяет лекарственному средству прежде всего ингибировать пулы VEGF и IL6, которые с наибольшей вероятностью активируют передачу сигнала в клетке, и избежать насыщения VEGF и IL6, находящимися на более высоком уровне в жидкой фазе, и с меньшей вероятностью влияют на передачу сигнала в клетке. По мере увеличения количества клеток этот эффект будет более выраженным, поскольку для воздействия на большее количество клеток необходимо большее количество лекарственного средства. Соответственно, в некоторых вариантах реализации указанный конъюгат обеспечивает превосходный продукт, поскольку он может способствовать распределению молекулы.
[0676] Полное содержание всех заявок на патент, веб-сайтов, других публикаций, номеров доступа и т.п., указанных выше или ниже, включено в настоящее описание посредством ссылки для всех целей так же, как если бы содержание каждого индивидуального элемента из этого перечня было специально включено отдельно. Если различные варианты последовательности ассоциированы с номером доступа в разное время, подразумевается вариант, ассоциированный с номером доступа на действительную дату подачи настоящей заявки. Действительная дата подачи означает более раннюю из действительной даты подачи или даты подачи приоритетной заявки, относящейся к номеру доступа, если применимо. Подобным образом, если различные версии публикации, веб-сайта и т.п. опубликованы в разное время, подразумевается версия, опубликованная последней на действительную дату подачи заявки, если не указано иное. Любой признак, этап, элемент, вариант реализации или аспект настоящего изобретения может быть использован в сочетании с любым другим, если специально не указано иное. Хотя настоящее изобретение подробно описано для иллюстрации и примера для его ясного и полного понимания, очевидно, что могут быть произведены некоторые изменения и модификации в пределах объема прилагаемой формулы изобретения.
--->
АНТИТЕЛА К IL-6 И ИХ СЛИТЫЕ КОНСТРУКЦИИ И КОНЪЮГАТЫ
<110>КОДИАК САЙЕНСЕС ИНК.
ДЖЕЙКОБСОН, Рэйчел Д.
КОРРЕА, Фернандо
ЛЯН, Хун
ПЕРЛРОТ, Д. Виктор
<120>АНТИТЕЛА К IL-6 И ИХ СЛИТЫЕ КОНСТРУКЦИИ И КОНЪЮГАТЫ
<130>KDIAK.044WO
<150>62/637575
<151>2018-03-02
<150>62/727950
<151>2018-09-06
<160>269
<170>PatentIn, верисия 3.5
<210>1
<211>183
<212>PRT/БЕЛОК
<213>Homo sapiens
<400>1
Val Pro Pro Gly Glu Asp Ser Lys Asp Val Ala Ala Pro His Arg Gln
1 5 10 15
Pro Leu Thr Ser Ser Glu Arg Ile Asp Lys Gln Ile Arg Tyr Ile Leu
20 25 30
Asp Gly Ile Ser Ala Leu Arg Lys Glu Thr Cys Asn Lys Ser Asn Met
35 40 45
Cys Glu Ser Ser Lys Glu Ala Leu Ala Glu Asn Asn Leu Asn Leu Pro
50 55 60
Lys Met Ala Glu Lys Asp Gly Cys Phe Gln Ser Gly Phe Asn Glu Glu
65 70 75 80
Thr Cys Leu Val Lys Ile Ile Thr Gly Leu Leu Glu Phe Glu Val Tyr
85 90 95
Leu Glu Tyr Leu Gln Asn Arg Phe Glu Ser Ser Glu Glu Gln Ala Arg
100 105 110
Ala Val Gln Met Ser Thr Lys Val Leu Ile Gln Phe Leu Gln Lys Lys
115 120 125
Ala Lys Asn Leu Asp Ala Ile Thr Thr Pro Asp Pro Thr Thr Asn Ala
130 135 140
Ser Leu Leu Thr Lys Leu Gln Ala Gln Asn Gln Trp Leu Gln Asp Met
145 150 155 160
Thr Thr His Leu Ile Leu Arg Ser Phe Lys Glu Phe Leu Gln Ser Ser
165 170 175
Leu Arg Ala Leu Arg Gln Met
180
<210>2
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>2
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>3
<211>106
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>3
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210>4
<211>205
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>4
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Ile His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200 205
<210>5
<211>204
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>5
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr Ile Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr Glu
115 120 125
Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys His
130 135 140
Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly Ser
145 150 155 160
Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr Arg
165 170 175
Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met Thr
180 185 190
Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200
<210>6
<211>205
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>6
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Ile Ile Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200 205
<210>7
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>7
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>8
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>8
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ser Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>9
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Gly Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>10
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Thr Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>11
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>11
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Val Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>12
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>12
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Gln Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>13
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>13
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Lys Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>14
<211>205
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>14
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200 205
<210>15
<211>205
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>15
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Ile His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200 205
<210>16
<211>204
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>16
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr Ile Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr Arg
165 170 175
Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met Thr
180 185 190
Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200
<210>17
<211>205
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>17
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Ile Ile Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200 205
<210>18
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>linker
<400>18
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210>19
<211>453
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>19
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Trp Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Leu Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Cys Ser Pro Gly Lys
450
<210>20
<211>450
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>20
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ser Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Leu Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Val Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Ile His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro
435 440 445
Gly Lys
450
<210>21
<211>444
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Thr Phe Ser Pro Phe Ala
20 25 30
Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
35 40 45
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
50 55 60
Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gln Gly Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Arg Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
435 440
<210>22
<211>448
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Thr Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Glu Ala Ala Gly Ala Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Leu Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
435 440 445
<210>23
<211>448
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>23
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Val Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
435 440 445
<210>24
<211>449
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>24
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Gln Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly
435 440 445
Lys
<210>25
<211>448
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>25
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Lys Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
435 440 445
<210>26
<211>449
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>26
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly
435 440 445
Lys
<210>27
<211>447
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
35 40 45
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
50 55 60
Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
435 440 445
<210>28
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>28
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>29
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>29
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>30
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>30
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>31
<211>223
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>31
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Ile Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>32
<211>222
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>32
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ser Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>33
<211>221
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>33
Glu Val Gln Leu Val Glu Ser Gly Gly Leu Val Gln Pro Gly Gly Ser
1 5 10 15
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe Ala
20 25 30
Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
35 40 45
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
50 55 60
Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gln Gly Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>34
<211>222
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>34
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Thr Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>35
<211>222
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>35
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Ile Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Val Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>36
<211>222
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>36
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Ile Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Gln Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>37
<211>222
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>37
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Lys Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>38
<211>222
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>38
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>39
<211>222
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>39
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Pro Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210>40
<211>225
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>40
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Asp Val Ser His Glu
35 40 45
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
50 55 60
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
65 70 75 80
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
85 90 95
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
100 105 110
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
115 120 125
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
130 135 140
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
145 150 155 160
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
165 170 175
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
180 185 190
Lys Ser Arg Trp Gln Gln Gly Asn Phe Ser Cys Ser Val Met His Glu
195 200 205
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly
210 215 220
Lys
225
<210>41
<211>225
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>41
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
20 25 30
Ser Arg Thr Pro Glu Val Thr Cys Val Val Asp Val Ser His Glu Asp
35 40 45
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
50 55 60
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
65 70 75 80
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
85 90 95
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
100 105 110
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
115 120 125
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
130 135 140
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
145 150 155 160
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
165 170 175
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
180 185 190
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
195 200 205
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly
210 215 220
Lys
225
<210>42
<211>226
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>antobyd
<400>42
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ile Glu
100 105 110
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
115 120 125
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
130 135 140
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
145 150 155 160
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
165 170 175
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
180 185 190
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
195 200 205
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro
210 215 220
Gly Lys
225
<210>43
<211>227
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>43
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser
210 215 220
Pro Gly Lys
225
<210>44
<211>227
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>44
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser
210 215 220
Pro Gly Lys
225
<210>45
<211>227
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>45
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser
210 215 220
Pro Gly Lys
225
<210>46
<211>227
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>46
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser
210 215 220
Pro Gly Lys
225
<210>47
<211>227
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>47
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser
210 215 220
Pro Gly Lys
225
<210>48
<211>225
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>48
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Val Glu Val His
50 55 60
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
65 70 75 80
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
85 90 95
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
100 105 110
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
115 120 125
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
130 135 140
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
145 150 155 160
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
165 170 175
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
180 185 190
Lys Ser Arg Trp Gln Gln Gly Asn Phe Ser Cys Ser Val Met His Glu
195 200 205
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly
210 215 220
Lys
225
<210>49
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>49
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>50
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>50
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>51
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>51
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>52
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>52
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>53
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>53
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>54
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>54
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>55
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>55
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>56
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>56
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>57
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>57
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>58
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>58
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>59
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>59
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>60
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>60
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>61
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>61
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>62
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>62
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>63
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>63
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>64
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>64
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>65
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>65
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>66
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>66
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>67
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>67
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>68
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>68
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>69
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>69
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>70
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>70
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser
1 5 10
<210>71
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>71
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>72
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>72
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>73
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>73
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser
1 5 10
<210>74
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>74
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>75
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>75
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>76
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>76
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>77
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>77
Leu Leu Ile Tyr Asp Asp Ser Ser Leu Ala Ser
1 5 10
<210>78
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>78
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>79
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>79
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>80
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>80
Leu Leu Ile Tyr Asp Asp Ser Ser Leu Ala Ser
1 5 10
<210>81
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>81
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>82
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>82
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>83
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>83
Leu Leu Ile Tyr Asp Asp Ser Ser Leu Ala Ser
1 5 10
<210>84
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>84
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>85
<211>431
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>85
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Thr Leu Lys Lys Phe Pro Leu Asp
35 40 45
Thr Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly
50 55 60
Phe Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys
65 70 75 80
Glu Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His
85 90 95
Arg Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly
100 105 110
Ile Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg
115 120 125
Thr Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser
130 135 140
Lys His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser
145 150 155 160
Gly Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val
165 170 175
Thr Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu
180 185 190
Met Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Asp Lys
195 200 205
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
210 215 220
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
225 230 235 240
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
245 250 255
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
260 265 270
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
275 280 285
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
290 295 300
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
305 310 315 320
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
325 330 335
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
340 345 350
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
355 360 365
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
370 375 380
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
385 390 395 400
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Val Met His Glu Ala Leu
405 410 415
His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
420 425 430
<210>86
<211>432
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>86
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Ile His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Asp Lys Thr
195 200 205
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser
210 215 220
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
225 230 235 240
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
245 250 255
Glu Val Lys Phe Asn Trp Tyr Tyr Asp Gly Val Glu Val His Asn Ala
260 265 270
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
275 280 285
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
290 295 300
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
305 310 315 320
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
325 330 335
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
340 345 350
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
355 360 365
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
370 375 380
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
385 390 395 400
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
405 410 415
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
420 425 430
<210>87
<211>432
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>87
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Leu Thr Ile
85 90 95
Arg Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly
100 105 110
Ile Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg
115 120 125
Thr Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser
130 135 140
Lys His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser
145 150 155 160
Gly Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val
165 170 175
Thr Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu
180 185 190
Met Thr Lys Lys Ser Thr Phe Val Arg Val His Glu Lys Asp Lys Thr
195 200 205
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser
210 215 220
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
225 230 235 240
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
245 250 255
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
260 265 270
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
275 280 285
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
290 295 300
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
305 310 315 320
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
325 330 335
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
340 345 350
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
355 360 365
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
370 375 380
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
385 390 395 400
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
405 410 415
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
420 425 430
<210>88
<211>430
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>88
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Leu Asp Thr Leu
35 40 45
Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe Ile
50 55 60
Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu Ala
65 70 75 80
Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Ile Ile Arg Gln
85 90 95
Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile Glu
100 105 110
Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr Glu
115 120 125
Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys His
130 135 140
Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly Ser
145 150 155 160
Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr Arg
165 170 175
Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Thr Lys
180 185 190
Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Asp Lys Thr His Thr
195 200 205
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
210 215 220
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
225 230 235 240
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
245 250 255
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
260 265 270
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
275 280 285
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
290 295 300
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
305 310 315 320
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
325 330 335
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
340 345 350
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
355 360 365
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
370 375 380
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
385 390 395 400
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
405 410 415
Asn His Tyr Thr Gln Lys Ser Leu Ser Cys Ser Pro Gly Lys
420 425 430
<210>89
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>89
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>90
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>90
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>91
<211>106
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>91
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210>92
<211>106
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>92
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210>93
<211>106
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>93
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210>94
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>94
Gly Phe Thr Phe Ser Pro Phe Ala Met Ser
1 5 10
<210>95
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>95
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Gly
<210>96
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>96
Ser Ala Ser Ile Ser Val Ser Tyr Met Tyr
1 5 10
<210>97
<211>7
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>97
Asp Met Ser Asn Leu Ala Ser
1 5
<210>98
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>98
Met Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>99
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>artifical
<400>99
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser
1 5 10
<210>100
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>100
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>101
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>101
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>102
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>102
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser
1 5 10
<210>103
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>103
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>104
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>104
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>105
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>105
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>106
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>106
Leu Leu Ile Tyr Asp Asp Ser Ser Leu Ala Ser
1 5 10
<210>107
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>107
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>108
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>108
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>109
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>109
Leu Leu Ile Tyr Asp Ala Ser Ser Leu Ala Ser
1 5 10
<210>110
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>110
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>111
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>111
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>112
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>112
Leu Leu Ile Tyr Asp Asp Ser Asn Leu Ala Ser
1 5 10
<210>113
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>113
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>114
<211>205
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>114
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200 205
<210>115
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>115
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210>116
<211>666
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>fuison
<400>116
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys Gly Lys
660 665
<210>117
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>117
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>118
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>118
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>119
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>119
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>120
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>120
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>121
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>121
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>122
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>122
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>123
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>123
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>124
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>124
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>125
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>125
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>126
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>126
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>127
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>127
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>128
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>128
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>129
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>129
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>130
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>130
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>131
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>131
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>132
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>132
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>133
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>133
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>134
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>134
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>135
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>135
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>136
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>136
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>137
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>137
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>138
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>138
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>139
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>139
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>140
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>140
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>141
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>141
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>142
<211>106
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>142
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210>143
<211>106
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>143
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210>144
<211>106
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>144
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210>145
<211>205
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>145
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys
195 200 205
<210>146
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>146
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>147
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>147
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>148
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>148
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>149
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>149
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>150
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>150
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>151
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>151
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>152
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>152
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>153
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>153
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>154
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>154
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>155
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>155
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>156
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>156
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>157
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>157
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>158
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>158
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>159
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>159
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>160
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>160
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>161
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>161
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>162
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>162
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>163
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>163
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>164
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>164
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>165
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>165
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>166
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>166
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Ala Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>167
<211>667
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>167
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Trp Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys Pro Gly Lys
660 665
<210>168
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>легкая цепь
<400>168
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>169
<211>213
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>169
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ile Ser Val Ser Tyr Leu
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Asp Ser Ser Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210>170
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>170
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Met His Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Ala Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Ile Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>171
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>171
Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>172
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>172
Pro Phe Ala Met His
1 5
<210>173
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>173
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>174
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>174
Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>175
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>175
Pro Phe Ala Met His
1 5
<210>176
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>176
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>177
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>177
Gln Ser Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>178
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>178
Pro Phe Ala Met His
1 5
<210>179
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>179
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>180
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>180
Gln Gly Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>181
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>181
Pro Phe Ala Met His
1 5
<210>182
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>182
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>183
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>183
Gln Thr Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>184
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>184
Pro Phe Ala Met His
1 5
<210>185
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>185
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>186
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>186
Gln Val Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>187
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>187
Pro Phe Ala Met His
1 5
<210>188
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>188
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>189
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>189
Gln Gln Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>190
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>190
Pro Phe Ala Met His
1 5
<210>191
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>191
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>192
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>192
Gln Lys Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>193
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>193
Pro Phe Ala Ile Ser
1 5
<210>194
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>194
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>195
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>195
Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>196
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>196
Pro Phe Ala Trp Ser
1 5
<210>197
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>197
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Asp
<210>198
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>198
Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>199
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>199
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>200
<211>7
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>200
Asp Asp Ser Ser Leu Ala Ser
1 5
<210>201
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>201
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>202
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>202
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>203
<211>7
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>203
Asp Ala Ser Ser Leu Ala Ser
1 5
<210>204
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>204
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>205
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>205
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>206
<211>7
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>206
Asp Asp Ser Asn Leu Ala Ser
1 5
<210>207
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>207
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>208
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>208
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>209
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>209
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>210
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>210
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>211
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>211
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>212
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>212
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>213
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>213
Ala Arg Gln Ser Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>214
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>214
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>215
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>215
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>216
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>216
Ala Arg Gln Gly Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>217
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>217
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>218
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>218
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>219
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>219
Ala Arg Gln Thr Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>220
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>220
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>221
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>221
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>222
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>222
Ala Arg Gln Val Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>223
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>223
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>224
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>224
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>225
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>225
Ala Arg Gln Gln Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>226
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>226
Gly Phe Thr Phe Ser Pro Phe Ala Met His
1 5 10
<210>227
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>227
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>228
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>228
Ala Arg Gln Lys Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>229
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>229
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser
1 5 10
<210>230
<211>19
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>230
Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr
1 5 10 15
Val Thr Asp
<210>231
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>231
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>232
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>232
Gly Phe Thr Phe Ser Pro Phe Ala Trp Ser
1 5 10
<210>233
<211>20
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>233
Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp
1 5 10 15
Thr Val Thr Asp
20
<210>234
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>234
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Val
1 5 10
<210>235
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>235
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>236
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>236
Leu Leu Ile Tyr Asp Asp Ser Ser Leu Ala Ser
1 5 10
<210>237
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>237
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>238
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>238
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>239
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>239
Leu Leu Ile Tyr Asp Ala Ser Ser Leu Ala Ser
1 5 10
<210>240
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>240
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>241
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>241
Ser Ala Ser Ile Ser Val Ser Tyr Leu Tyr
1 5 10
<210>242
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>242
Leu Leu Ile Tyr Asp Asp Ser Asn Leu Ala Ser
1 5 10
<210>243
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>243
Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>244
<211>5
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>244
Pro Phe Ala Met Ser
1 5
<210>245
<211>17
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>245
Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val Thr
1 5 10 15
Gly
<210>246
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>246
Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>247
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>247
Ser Ala Ser Ile Ser Val Ser Tyr Met Tyr
1 5 10
<210>248
<211>7
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>248
Asp Met Ser Asn Leu Ala Ser
1 5
<210>249
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>249
Met Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>250
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>250
Gly Phe Thr Phe Ser Pro Phe Ala Met Ser
1 5 10
<210>251
<211>20
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>251
Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp
1 5 10 15
Thr Val Thr Gly
20
<210>252
<211>12
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>252
Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Ile
1 5 10
<210>253
<211>10
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>253
Ser Ala Ser Ile Ser Val Ser Tyr Met Tyr
1 5 10
<210>254
<211>11
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>254
Leu Leu Ile Tyr Asp Met Ser Asn Leu Ala Ser
1 5 10
<210>255
<211>9
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>255
Met Gln Trp Ser Gly Tyr Pro Tyr Thr
1 5
<210>256
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>256
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>257
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>257
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ser Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>258
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>258
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Gly Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>259
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>259
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Thr Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>260
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>260
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Val Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>261
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>261
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Gln Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>262
<211>119
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>антитело
<400>262
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Lys Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210>263
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>263
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Met His Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Ala Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Ile Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>264
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>264
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Met His Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Ala Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Ile Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>265
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>ловушка
<400>265
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Met His Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Ala Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Ile Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>266
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>266
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>267
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>267
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>268
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>268
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Lys Ile Ser Pro Gly Gly Ser Trp Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Thr Asp Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Ala Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Gly Gly Gly Gly Ser Ser Asp Thr Gly Arg Pro Phe Val
225 230 235 240
Glu Met Tyr Ser Glu Ile Pro Glu Ile Ile His Met Thr Glu Gly Arg
245 250 255
Glu Leu Val Ile Pro Cys Arg Val Thr Ser Pro Asn Ile Thr Val Thr
260 265 270
Leu Lys Lys Phe Pro Leu Asp Thr Leu Ile Pro Asp Gly Lys Arg Ile
275 280 285
Ile Trp Asp Ser Arg Lys Gly Phe Ile Ile Ser Asn Ala Thr Tyr Lys
290 295 300
Glu Ile Gly Leu Leu Thr Cys Glu Ala Thr Val Asn Gly His Leu Tyr
305 310 315 320
Lys Thr Asn Tyr Leu Thr His Arg Gln Thr Asn Thr Ile Ile Asp Val
325 330 335
Val Leu Ser Pro Ser His Gly Ile Glu Leu Ser Val Gly Glu Lys Leu
340 345 350
Val Leu Asn Cys Thr Ala Arg Thr Glu Leu Asn Val Gly Ile Asp Phe
355 360 365
Asn Trp Glu Tyr Pro Ser Ser Lys His Gln His Lys Lys Leu Val Asn
370 375 380
Arg Asp Leu Lys Thr Gln Ser Gly Ser Glu Met Lys Lys Phe Leu Ser
385 390 395 400
Thr Leu Thr Ile Asp Gly Val Thr Arg Ser Asp Gln Gly Leu Tyr Thr
405 410 415
Cys Ala Ala Ser Ser Gly Leu Met Thr Lys Lys Asn Ser Thr Phe Val
420 425 430
Arg Val His Glu Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<210>269
<211>664
<212>PRT/БЕЛОК
<213>Искусственная последовательность
<220>
<223>слитая
<400>269
Ser Asp Thr Gly Arg Pro Phe Val Glu Met Tyr Ser Glu Ile Pro Glu
1 5 10 15
Ile Ile His Met Thr Glu Gly Arg Glu Leu Val Ile Pro Cys Arg Val
20 25 30
Thr Ser Pro Asn Ile Thr Val Thr Leu Lys Lys Phe Pro Leu Asp Thr
35 40 45
Leu Ile Pro Asp Gly Lys Arg Ile Ile Trp Asp Ser Arg Lys Gly Phe
50 55 60
Ile Ile Ser Asn Ala Thr Tyr Lys Glu Ile Gly Leu Leu Thr Cys Glu
65 70 75 80
Ala Thr Val Asn Gly His Leu Tyr Lys Thr Asn Tyr Leu Thr His Arg
85 90 95
Gln Thr Asn Thr Ile Ile Asp Val Val Leu Ser Pro Ser His Gly Ile
100 105 110
Glu Leu Ser Val Gly Glu Lys Leu Val Leu Asn Cys Thr Ala Arg Thr
115 120 125
Glu Leu Asn Val Gly Ile Asp Phe Asn Trp Glu Tyr Pro Ser Ser Lys
130 135 140
His Gln His Lys Lys Leu Val Asn Arg Asp Leu Lys Thr Gln Ser Gly
145 150 155 160
Ser Glu Met Lys Lys Phe Leu Ser Thr Leu Thr Ile Asp Gly Val Thr
165 170 175
Arg Ser Asp Gln Gly Leu Tyr Thr Cys Ala Ala Ser Ser Gly Leu Met
180 185 190
Thr Lys Lys Asn Ser Thr Phe Val Arg Val His Glu Lys Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly
210 215 220
Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser
225 230 235 240
Gly Phe Thr Phe Ser Pro Phe Ala Ile Ser Trp Val Arg Gln Ala Pro
245 250 255
Gly Lys Gly Leu Glu Trp Val Ala Lys Ile Ser Pro Gly Gly Ser Trp
260 265 270
Thr Tyr Tyr Ser Asp Thr Val Thr Asp Arg Phe Thr Phe Ser Leu Asp
275 280 285
Thr Ser Lys Ser Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
290 295 300
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala
305 310 315 320
Leu Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
325 330 335
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
340 345 350
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
355 360 365
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
370 375 380
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
385 390 395 400
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
405 410 415
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
420 425 430
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
435 440 445
Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
450 455 460
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
465 470 475 480
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
485 490 495
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
500 505 510
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
515 520 525
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
530 535 540
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
545 550 555 560
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
565 570 575
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
580 585 590
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
595 600 605
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
610 615 620
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
625 630 635 640
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
645 650 655
Ser Leu Ser Cys Ser Pro Gly Lys
660
<---

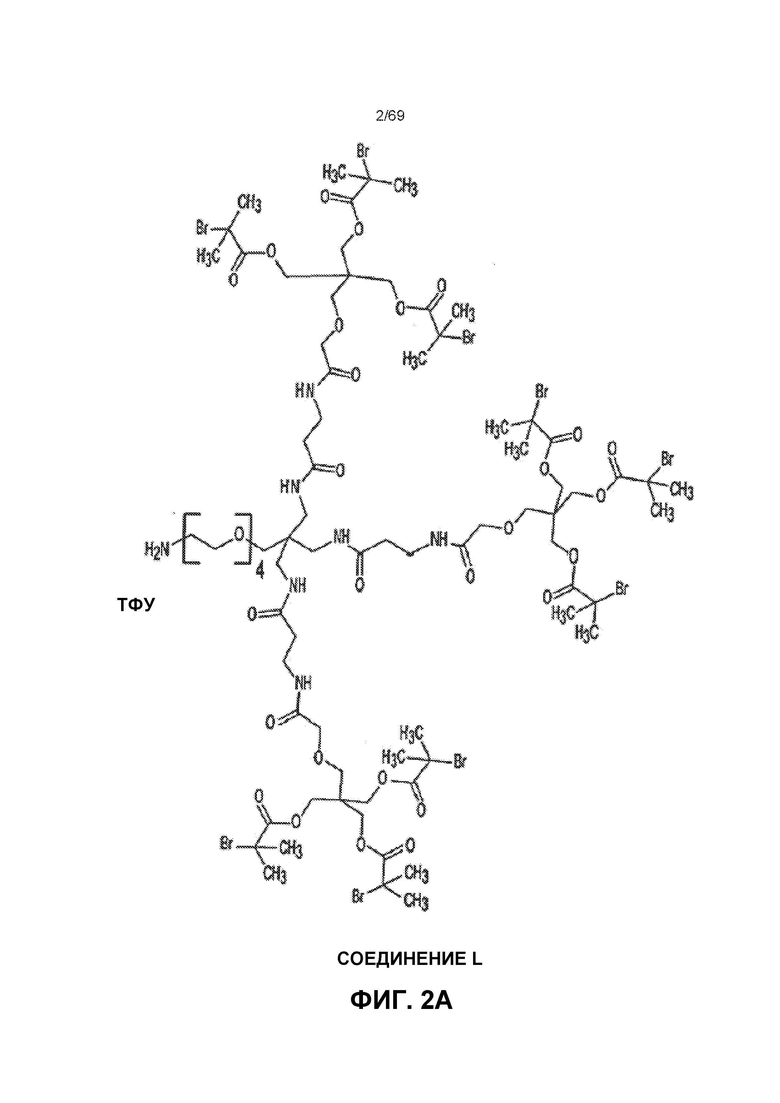
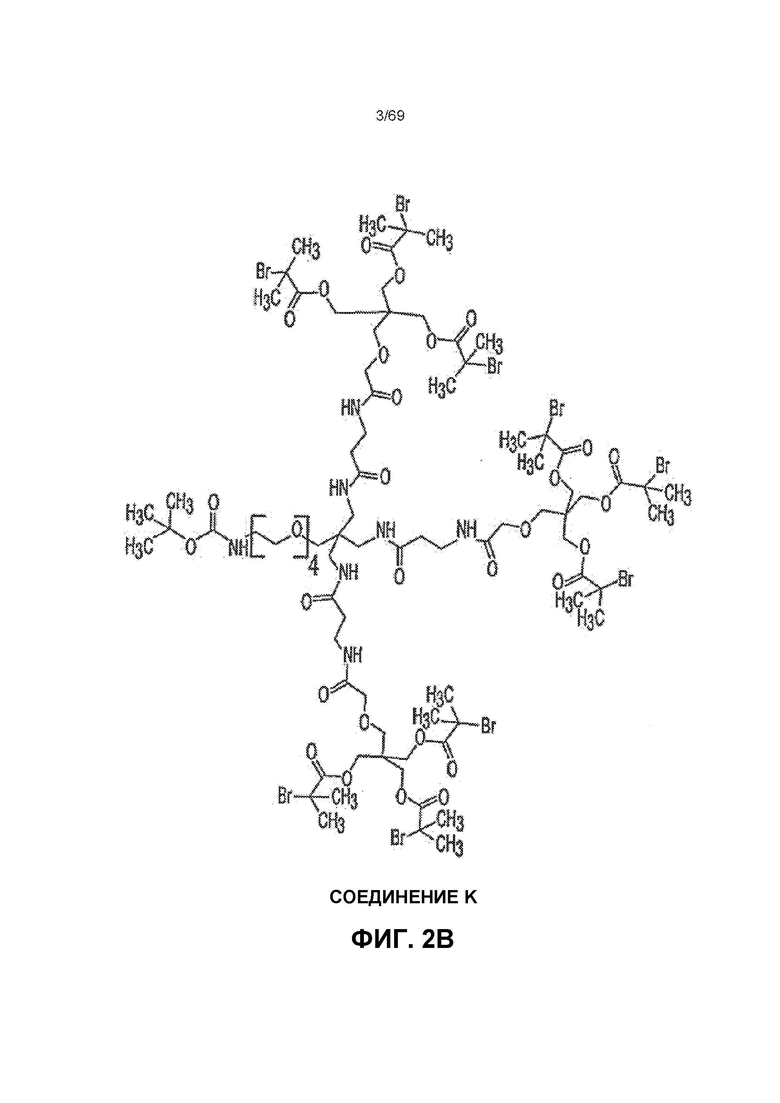
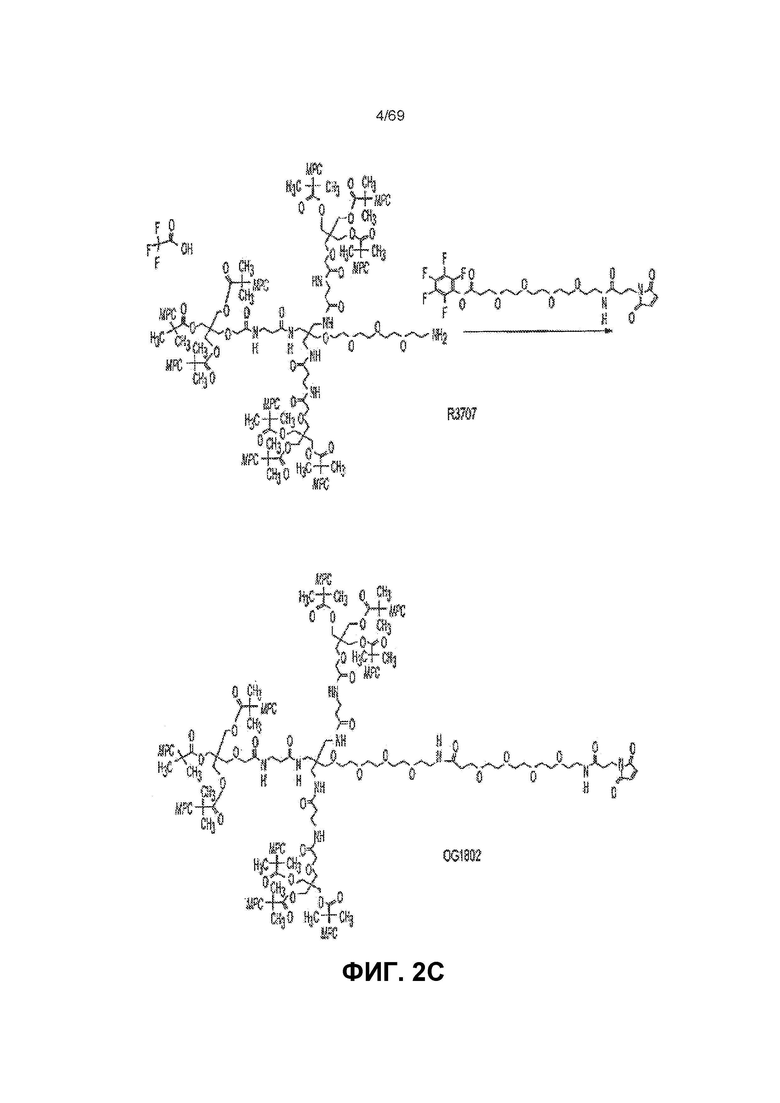
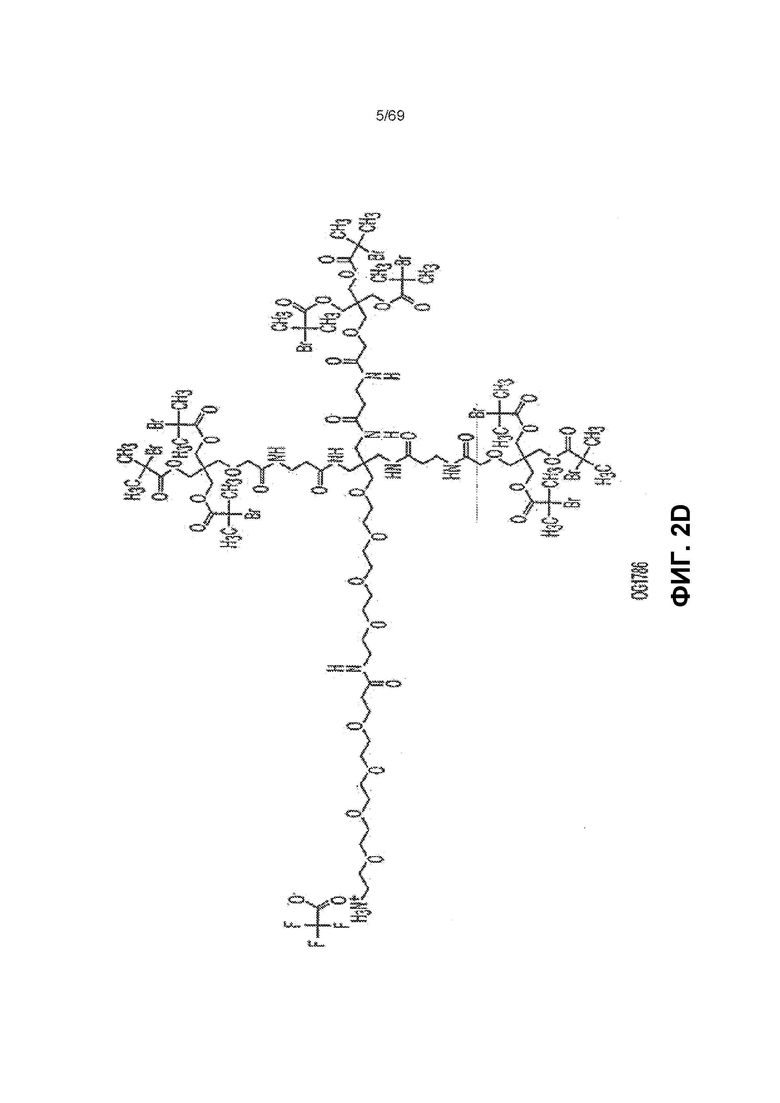
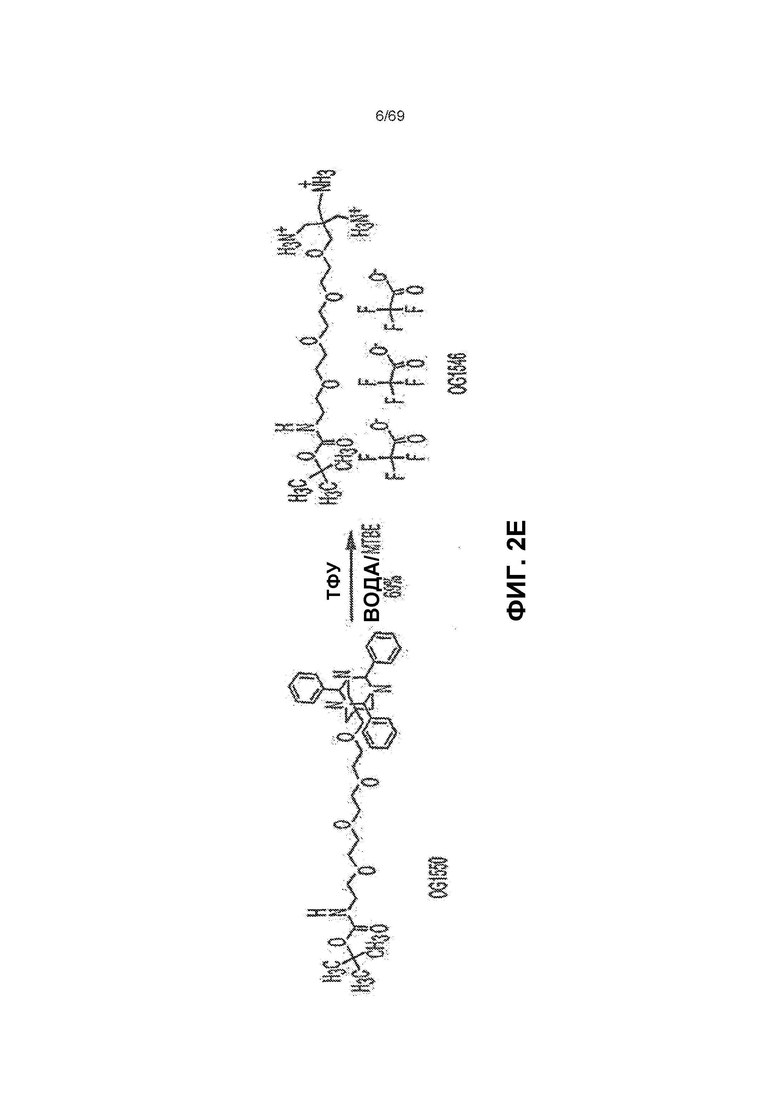
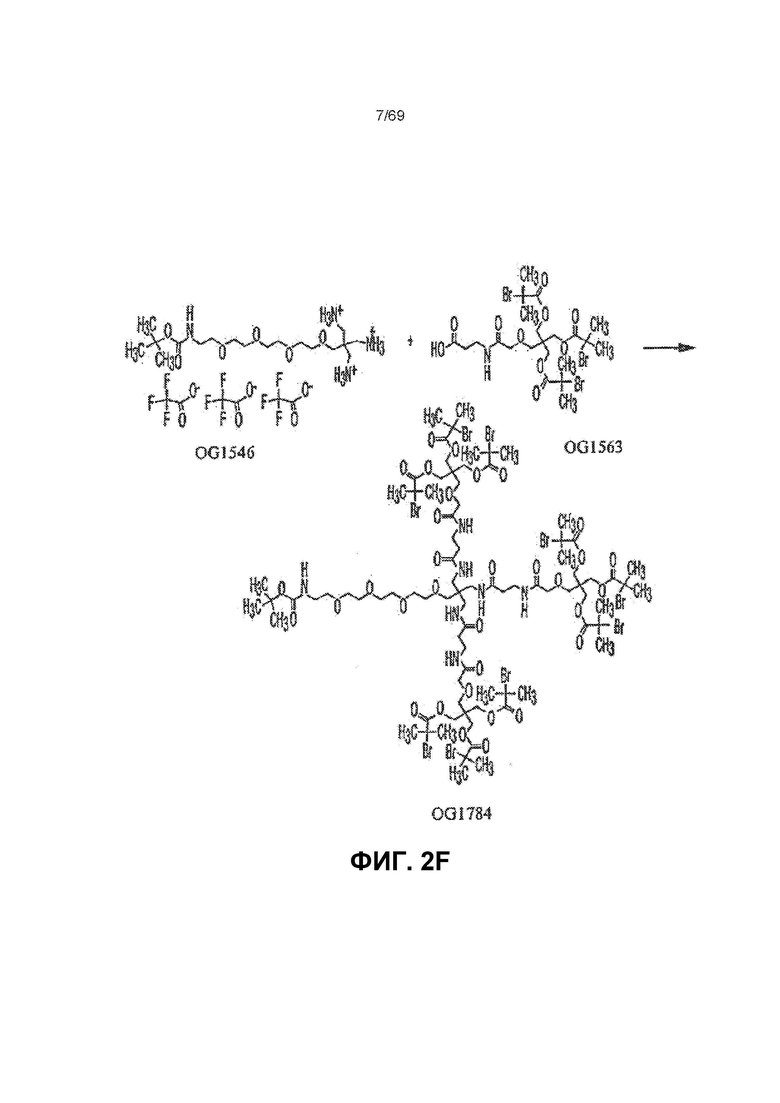
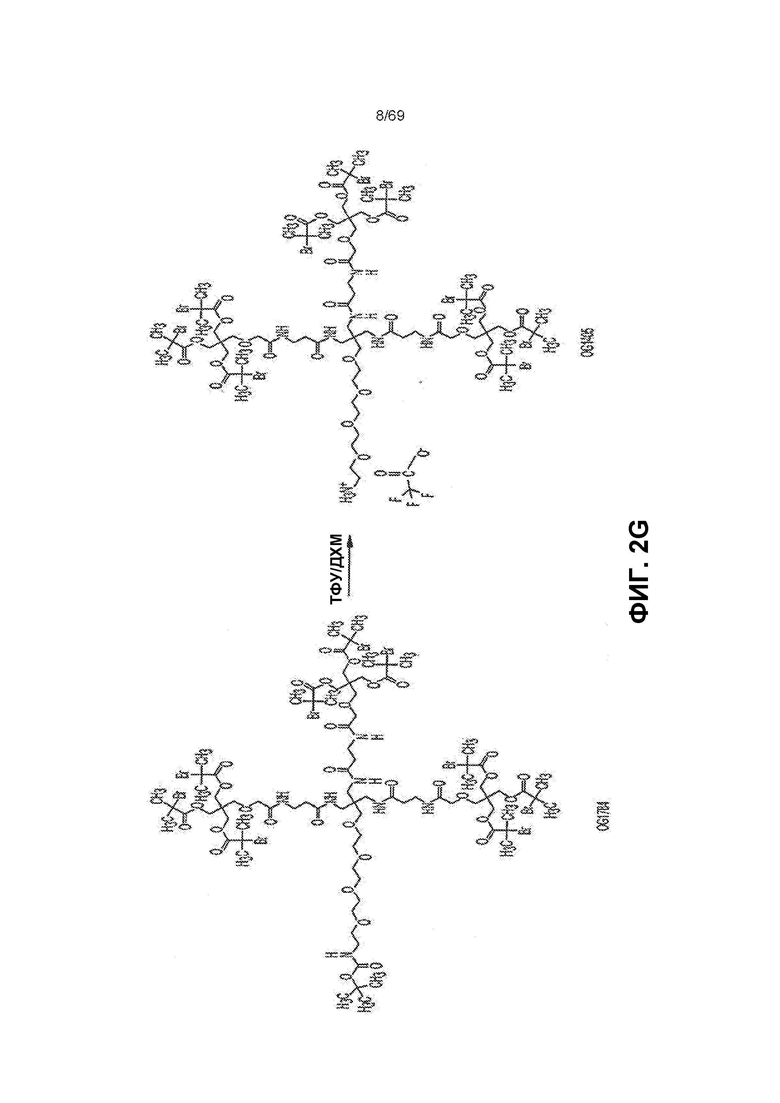
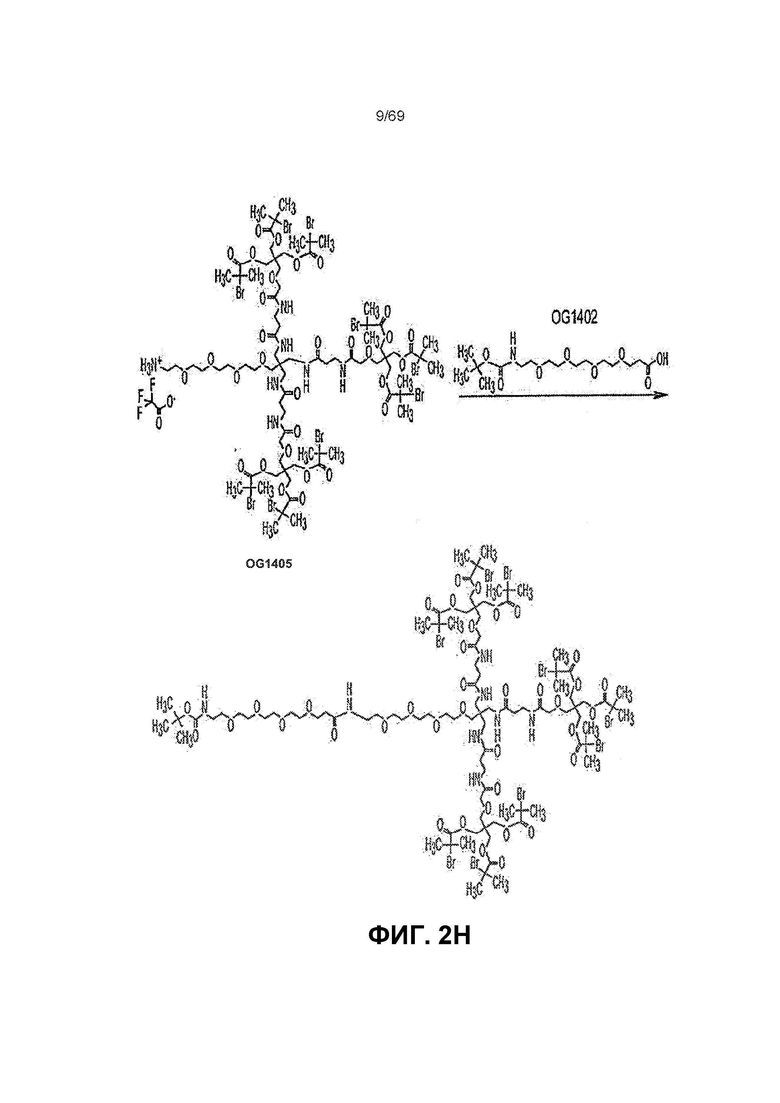
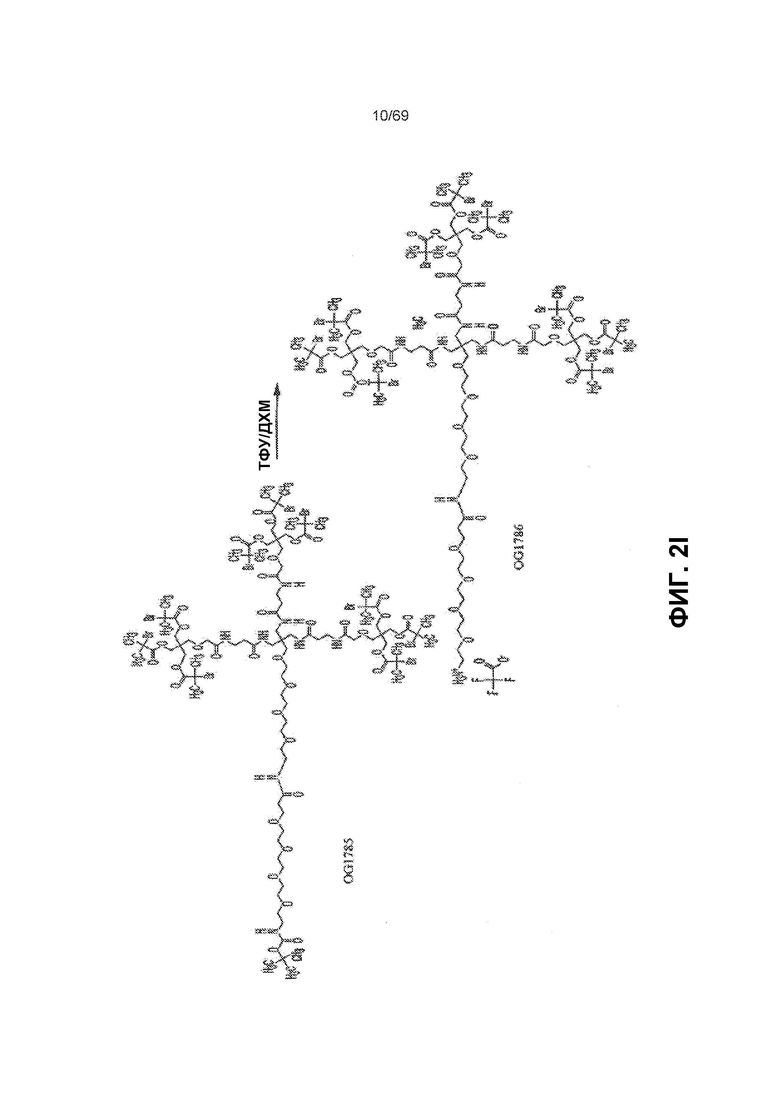
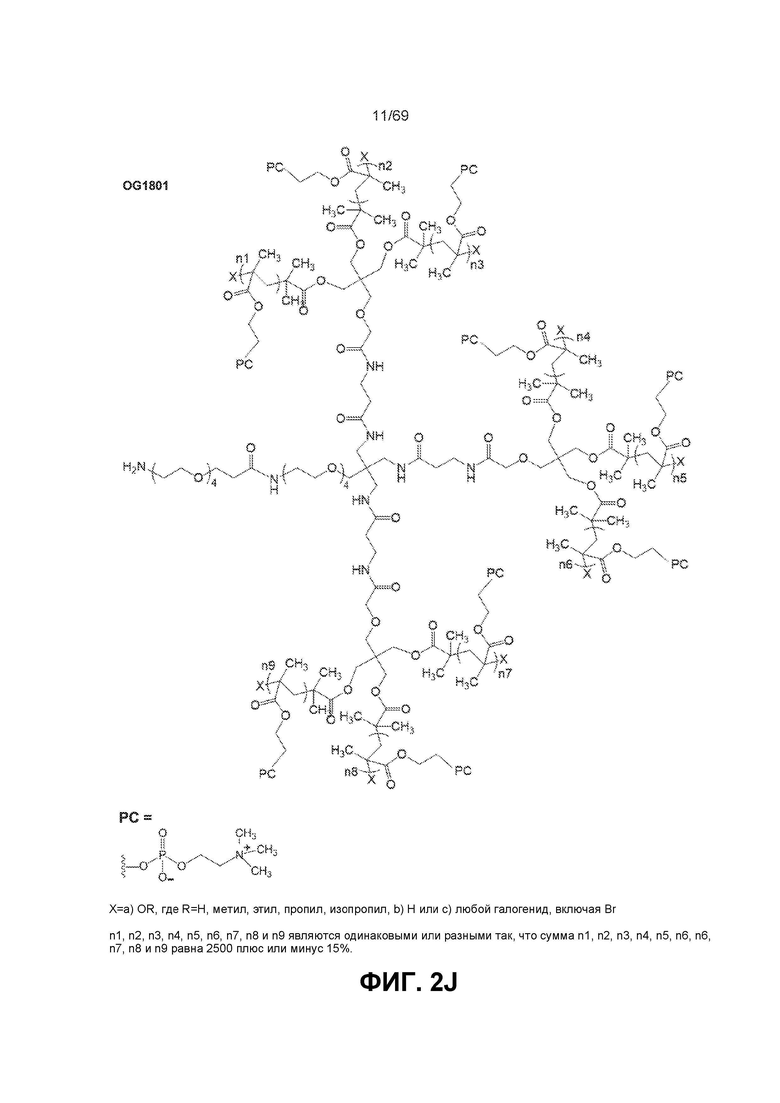
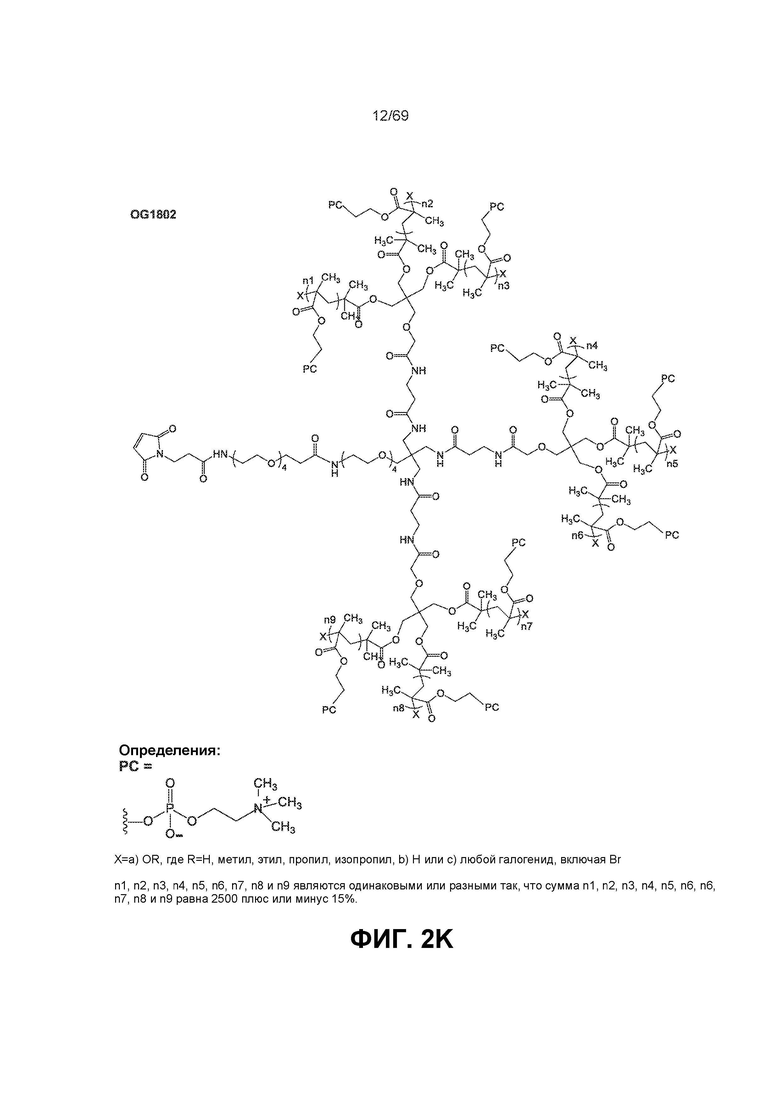
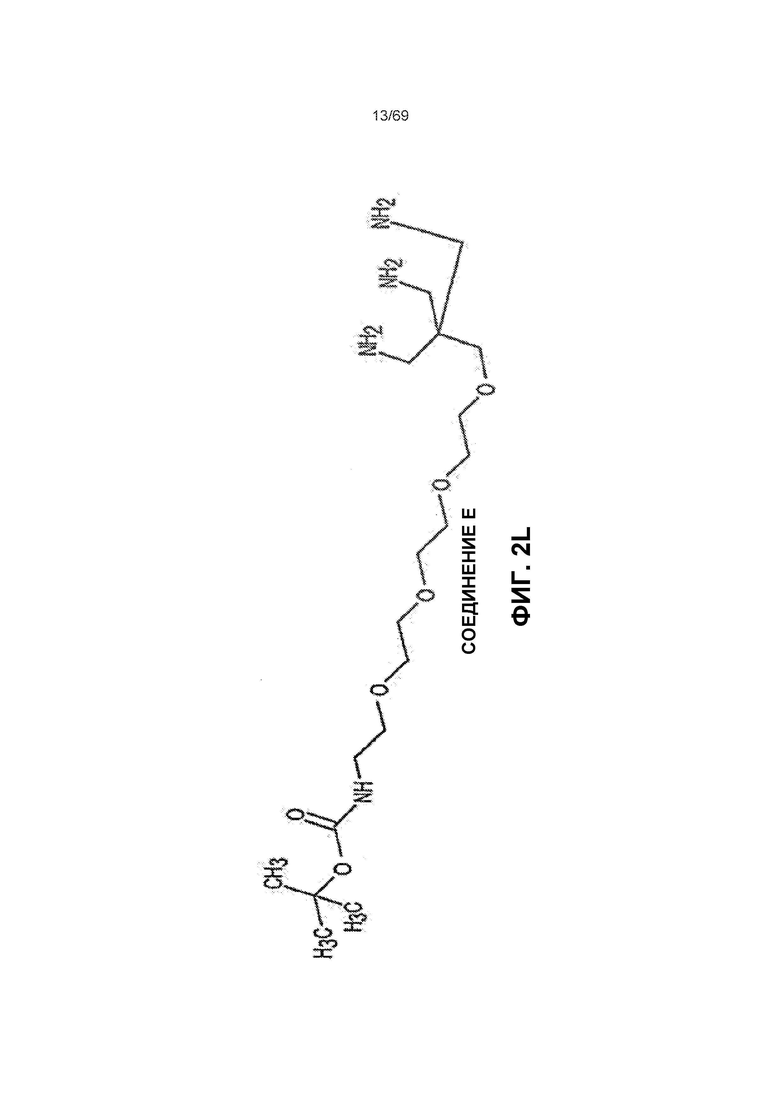

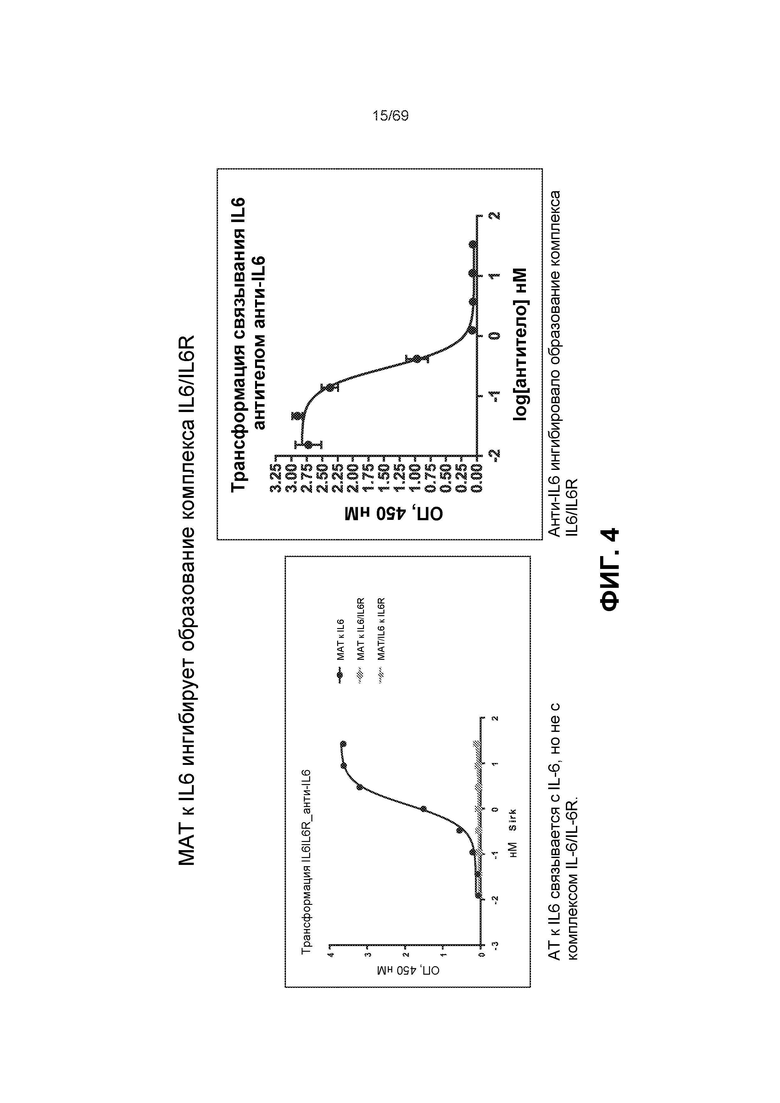
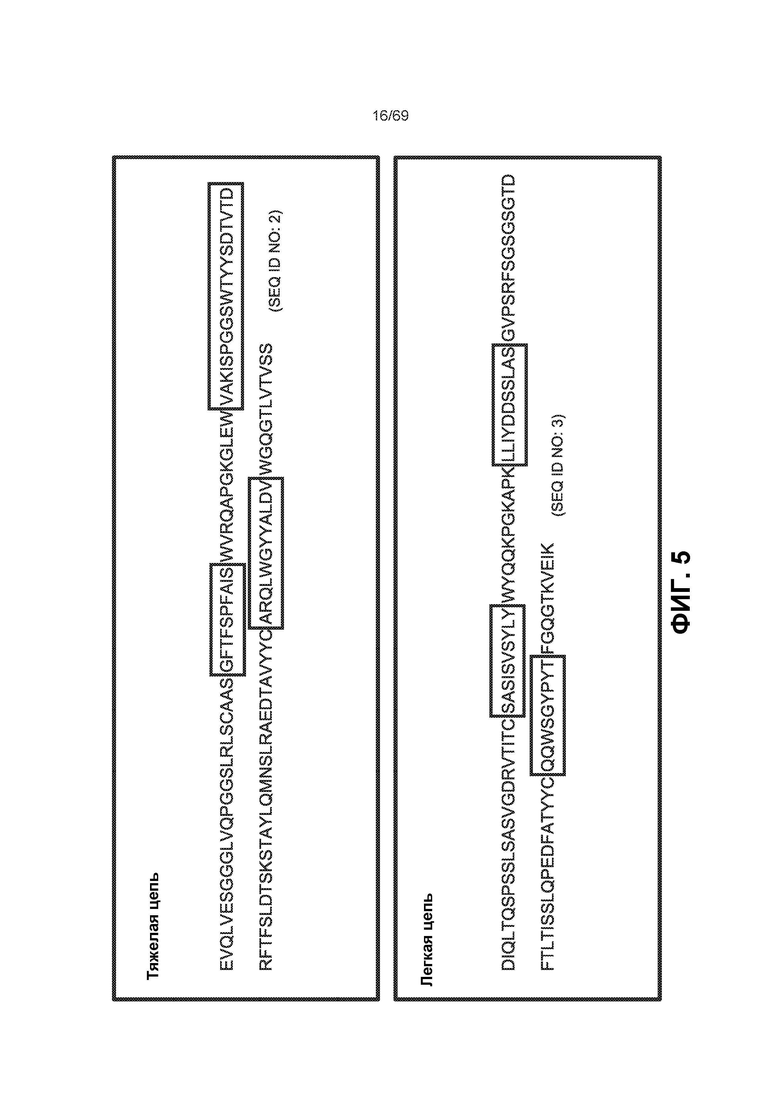
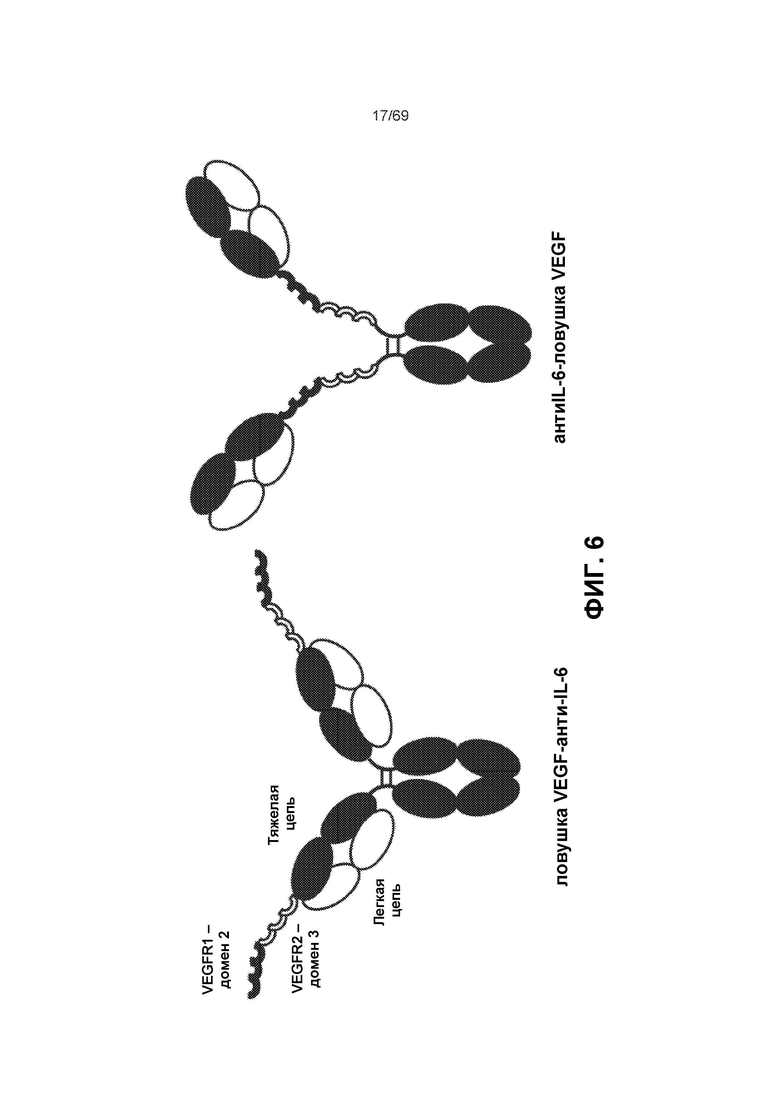
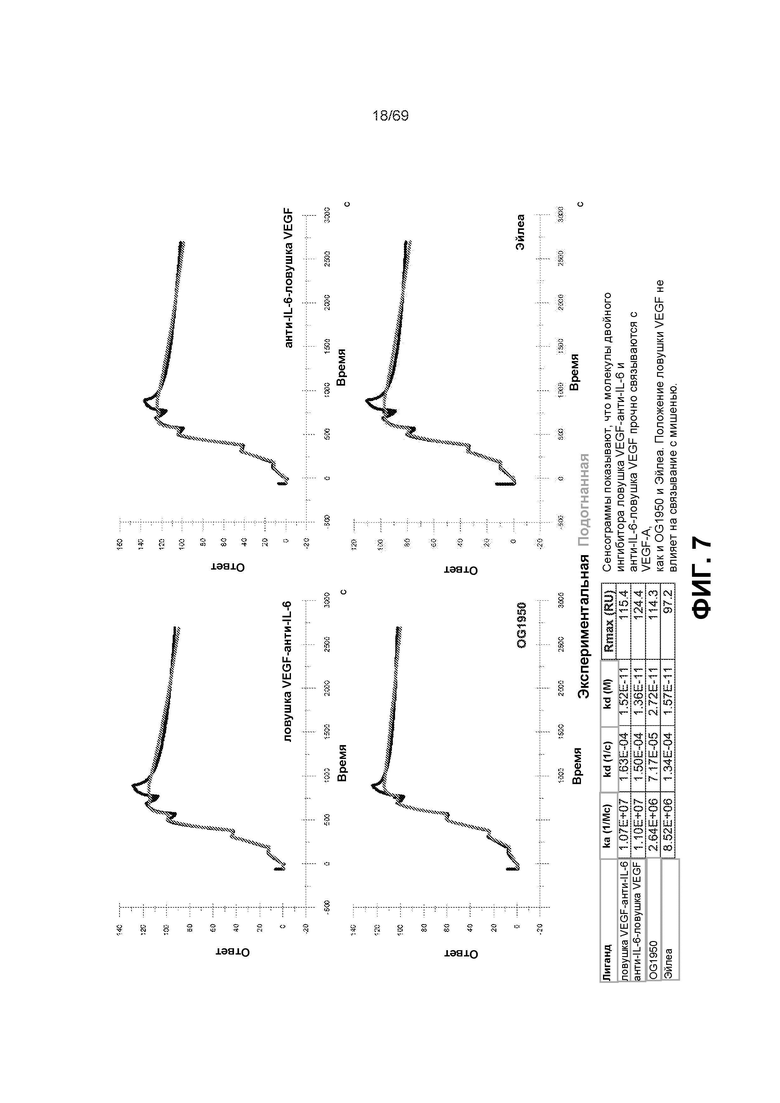
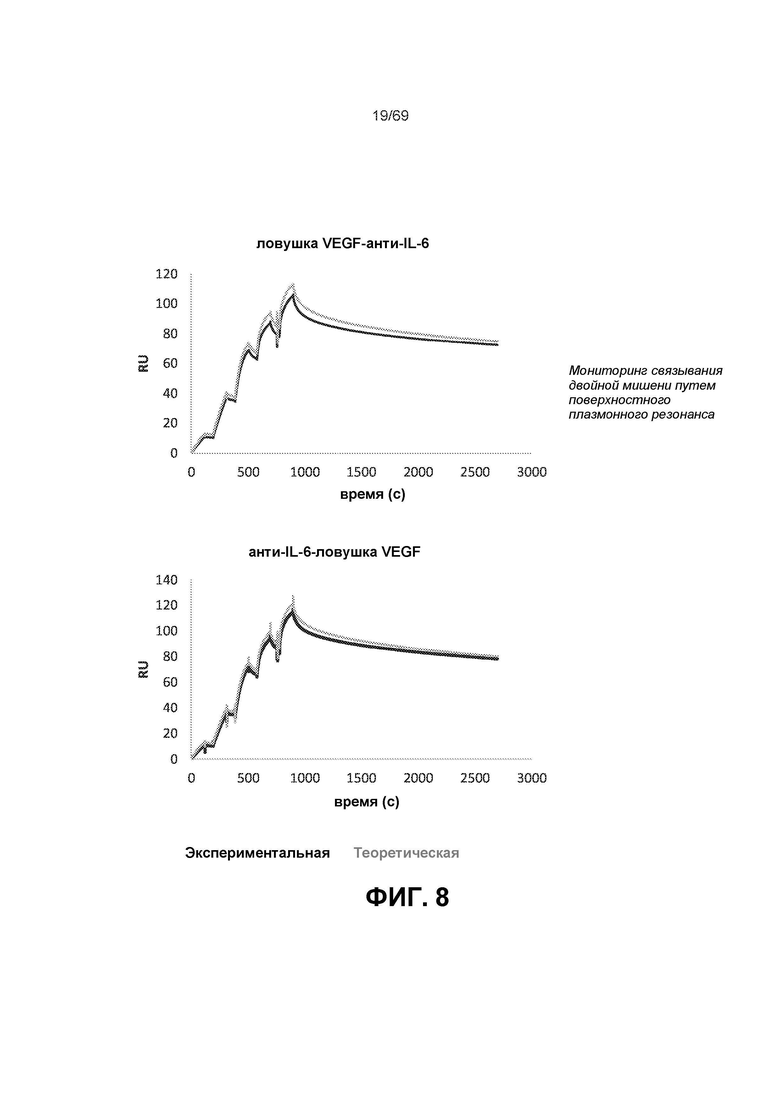
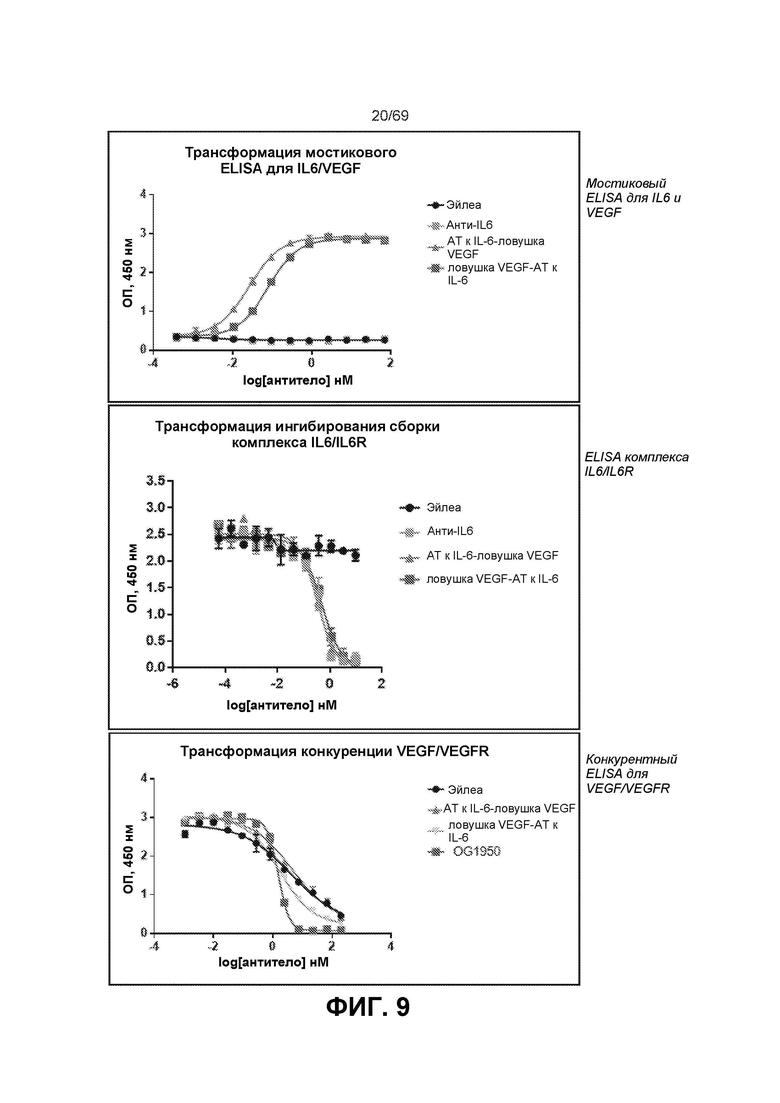
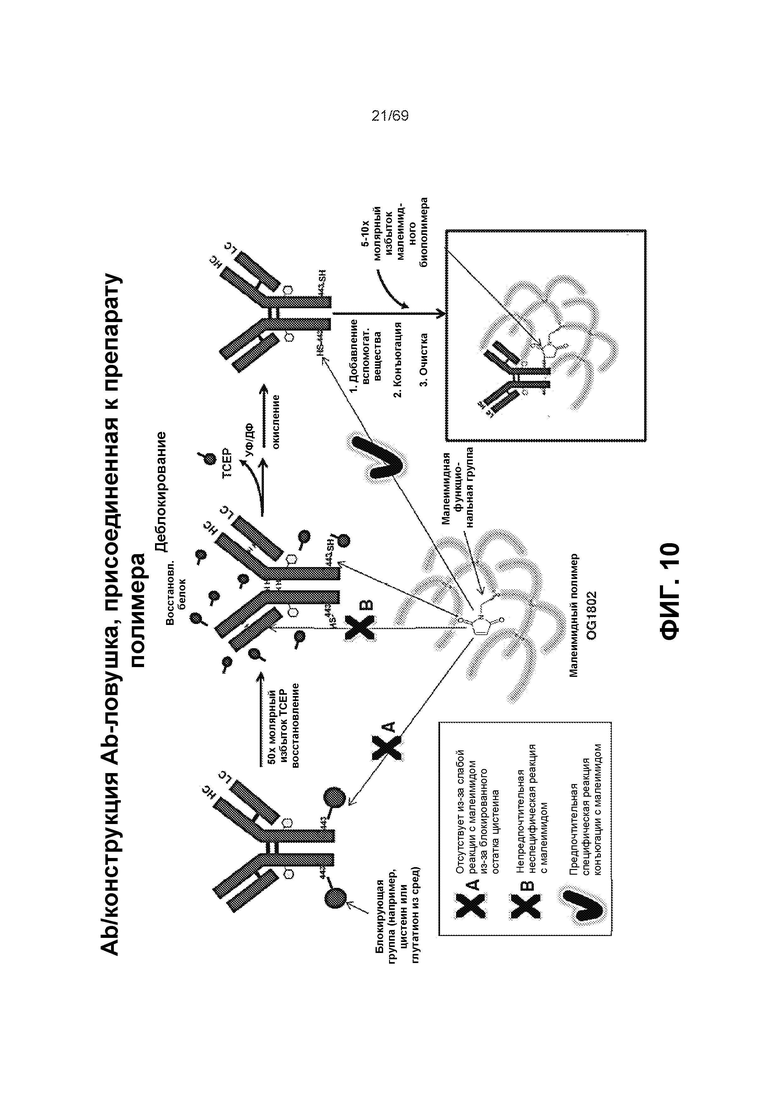
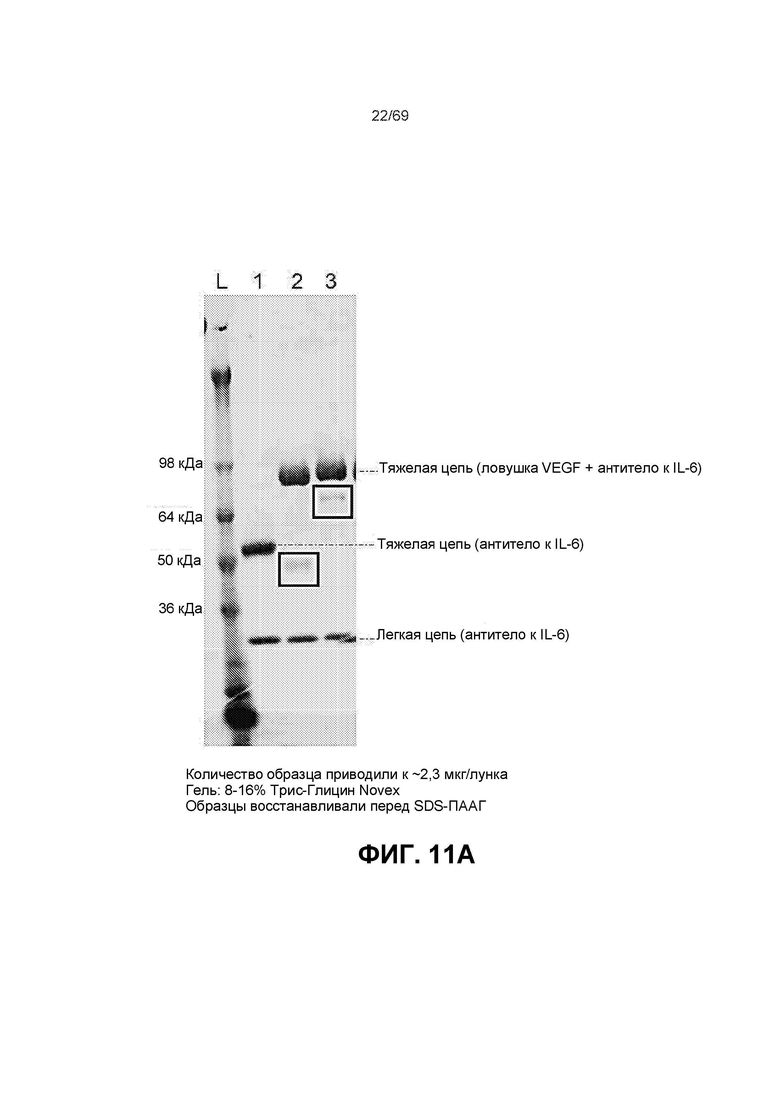

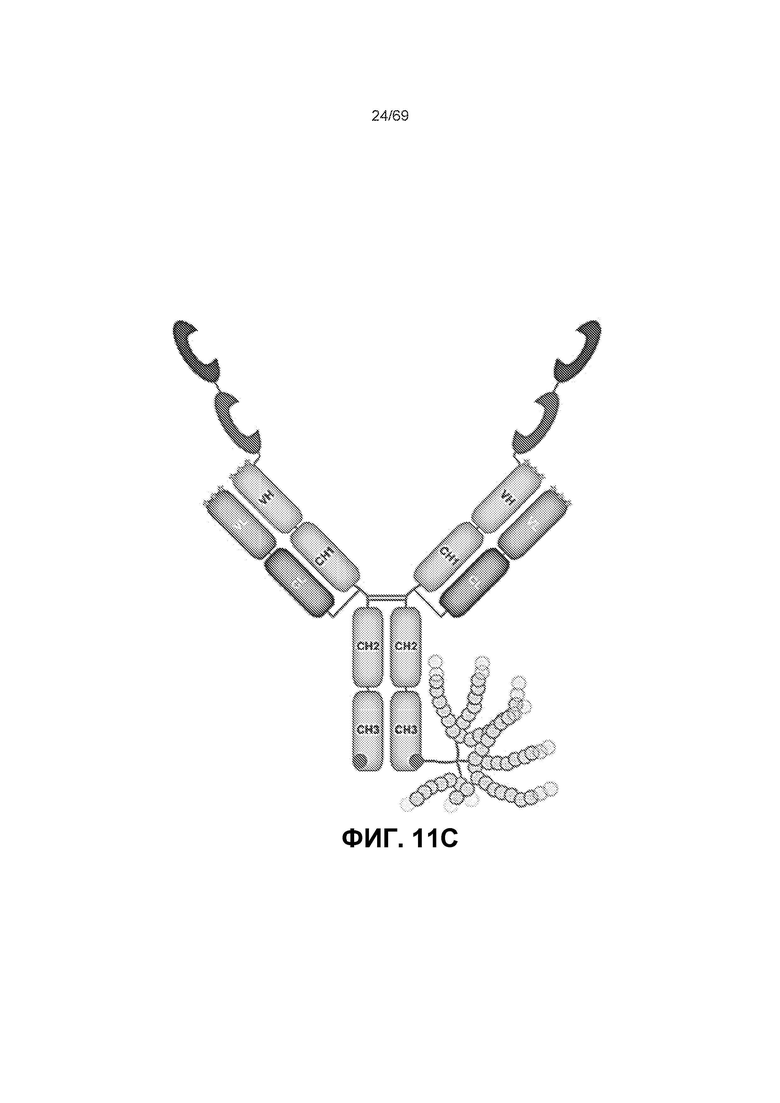
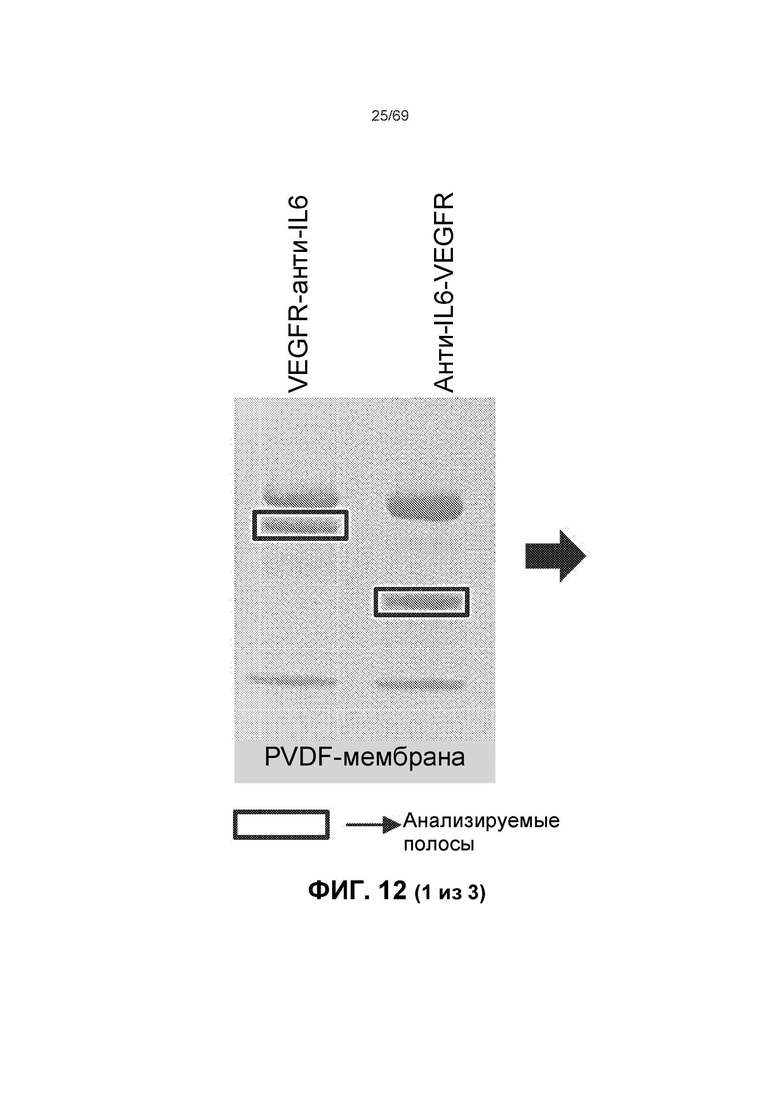
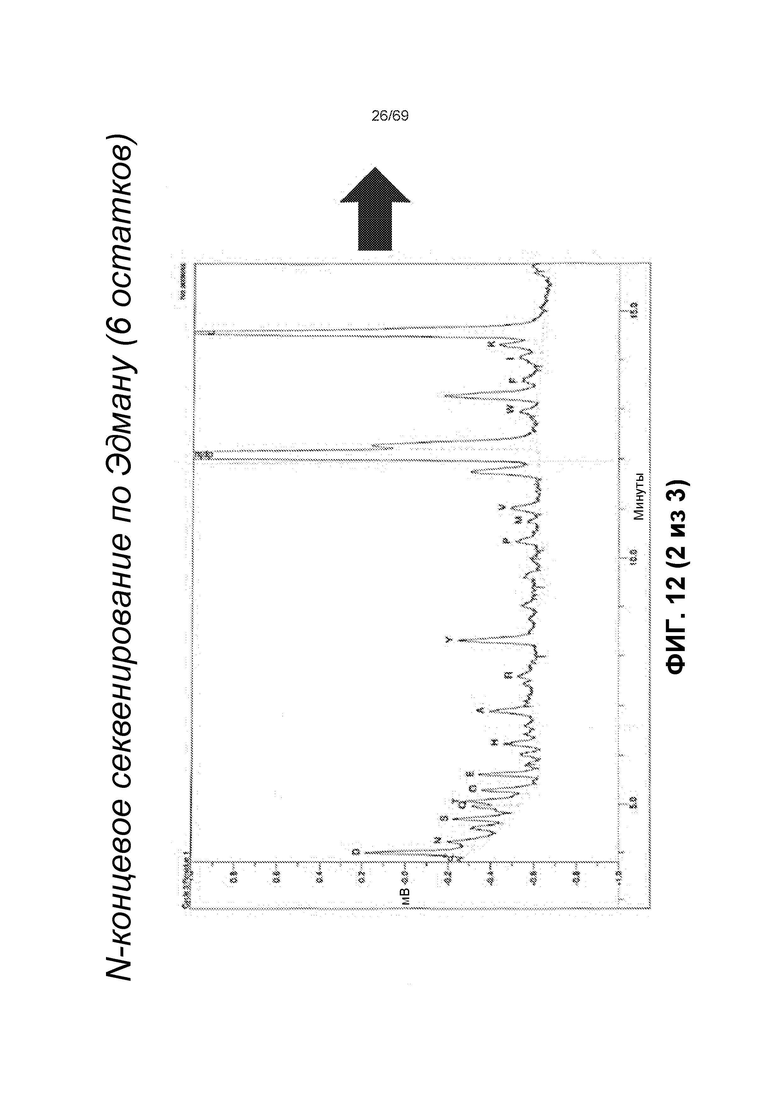





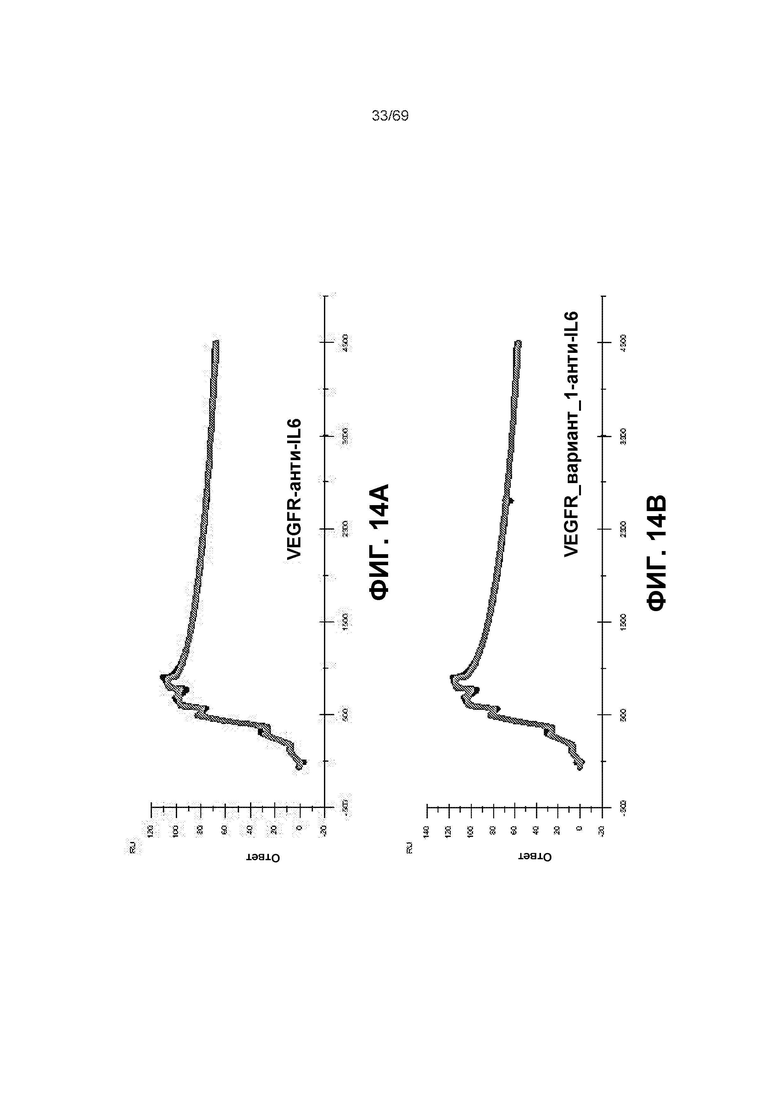

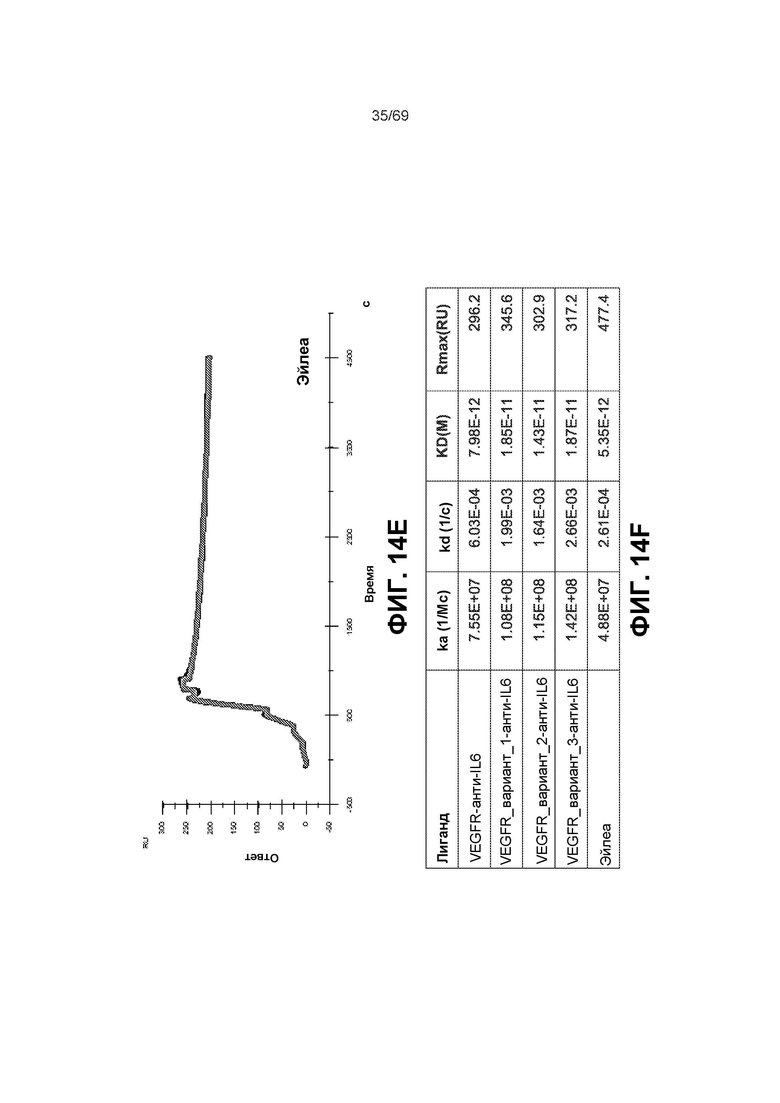
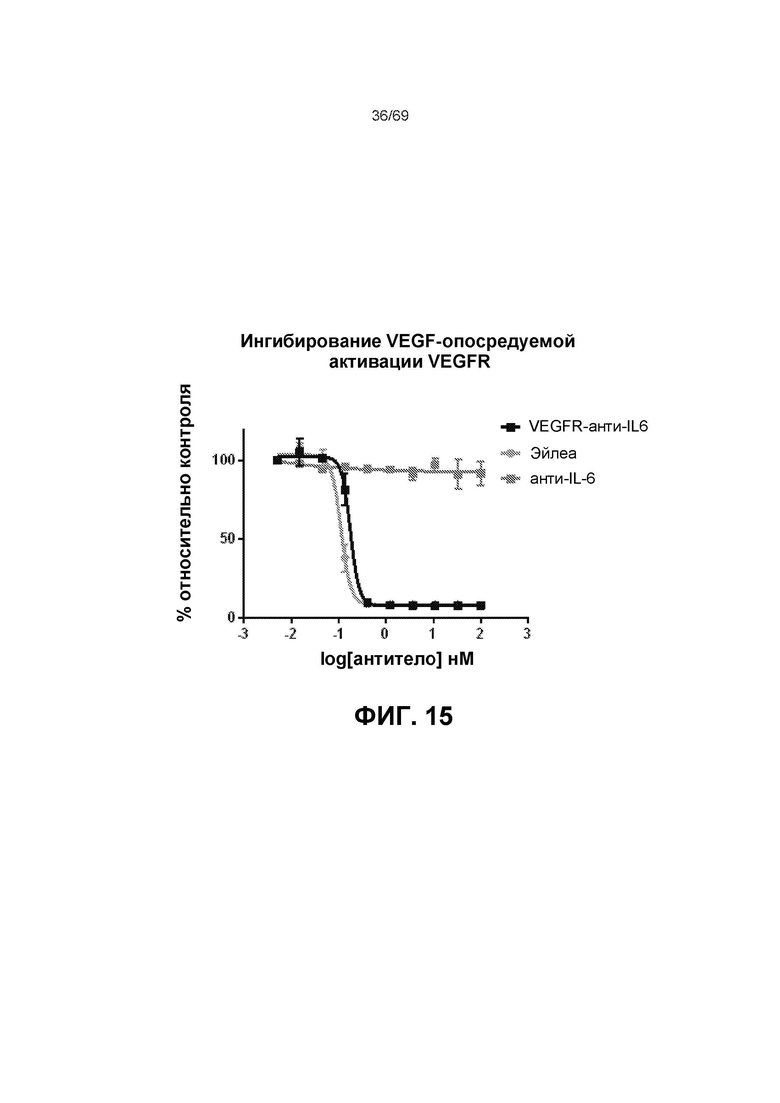
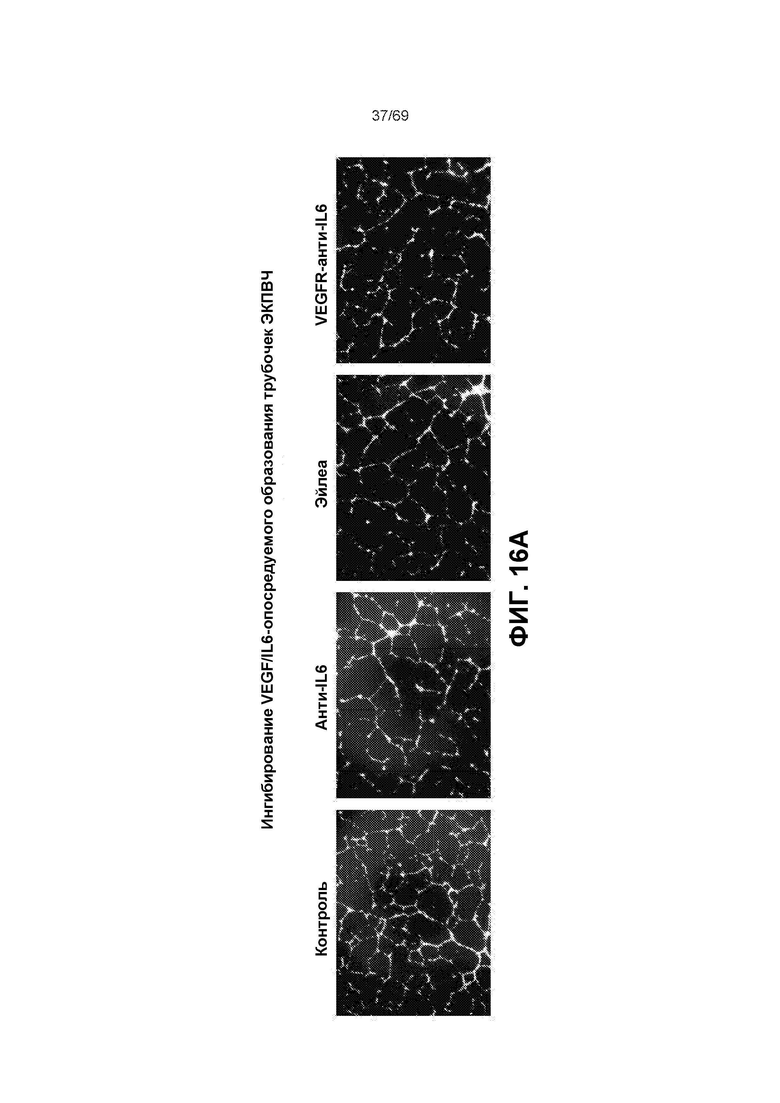
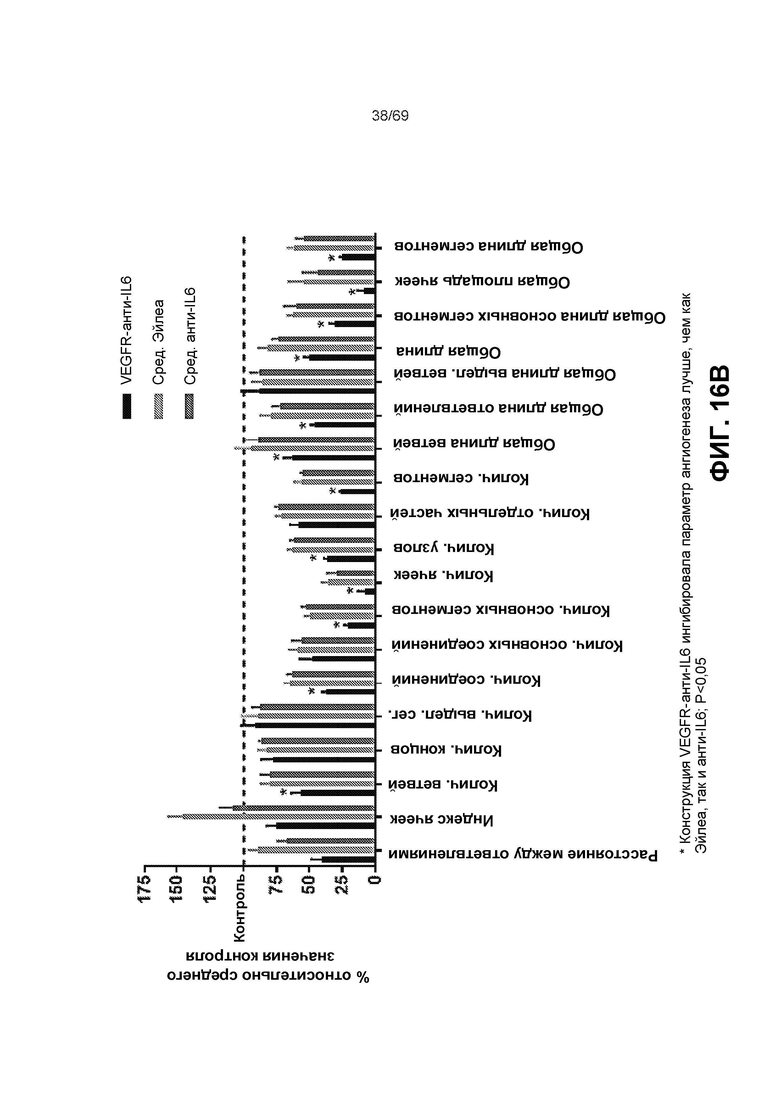
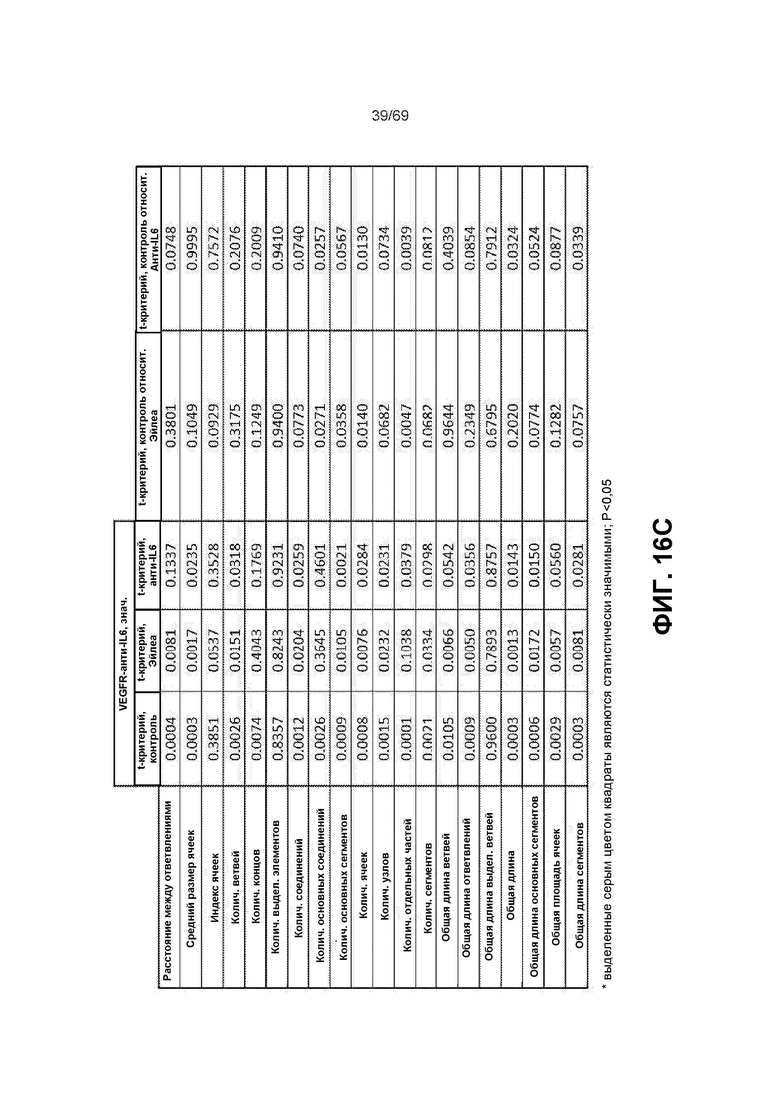
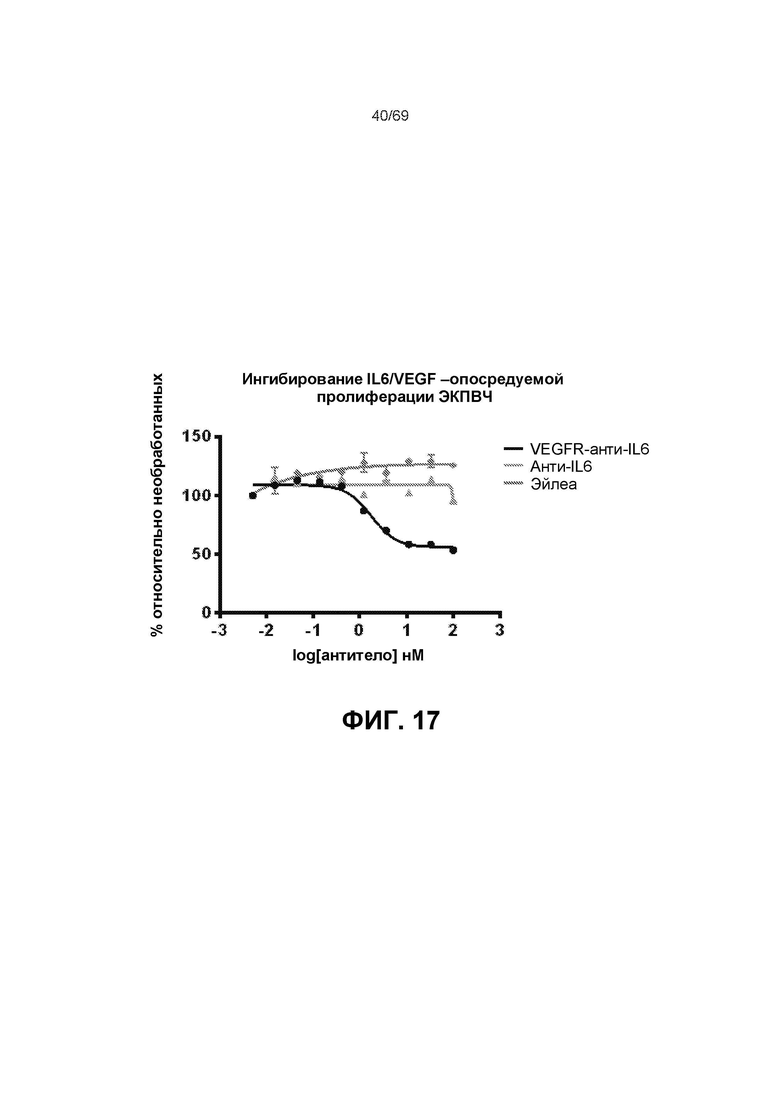
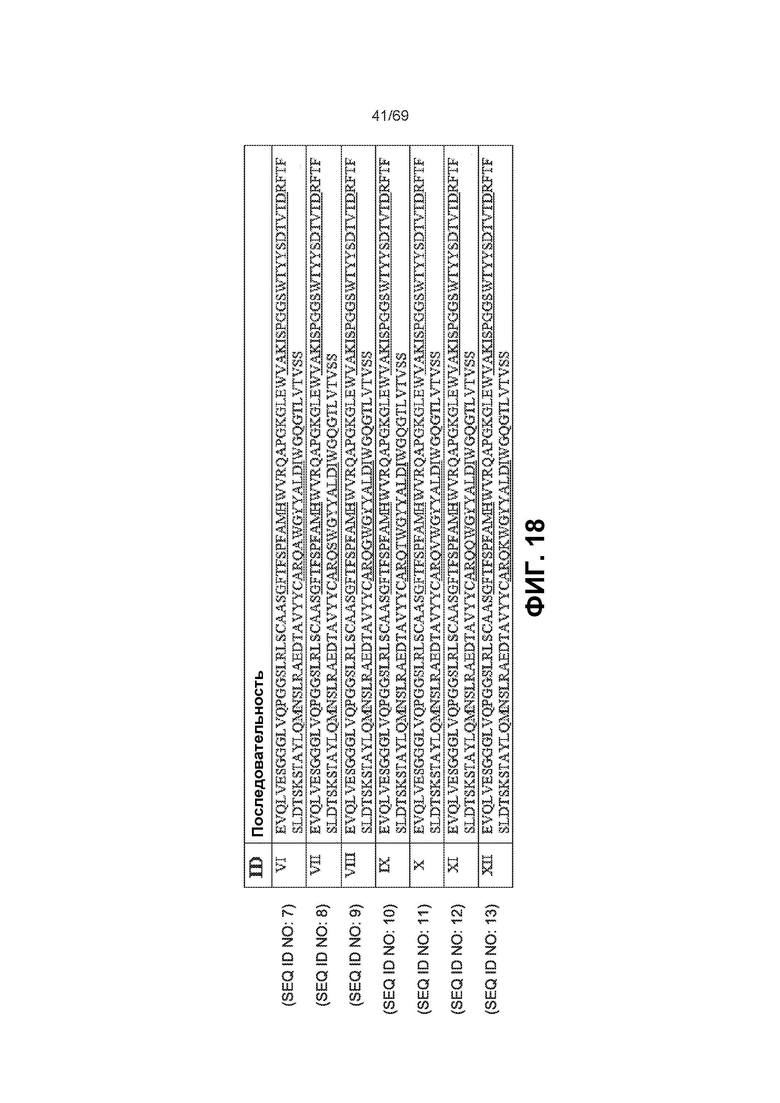

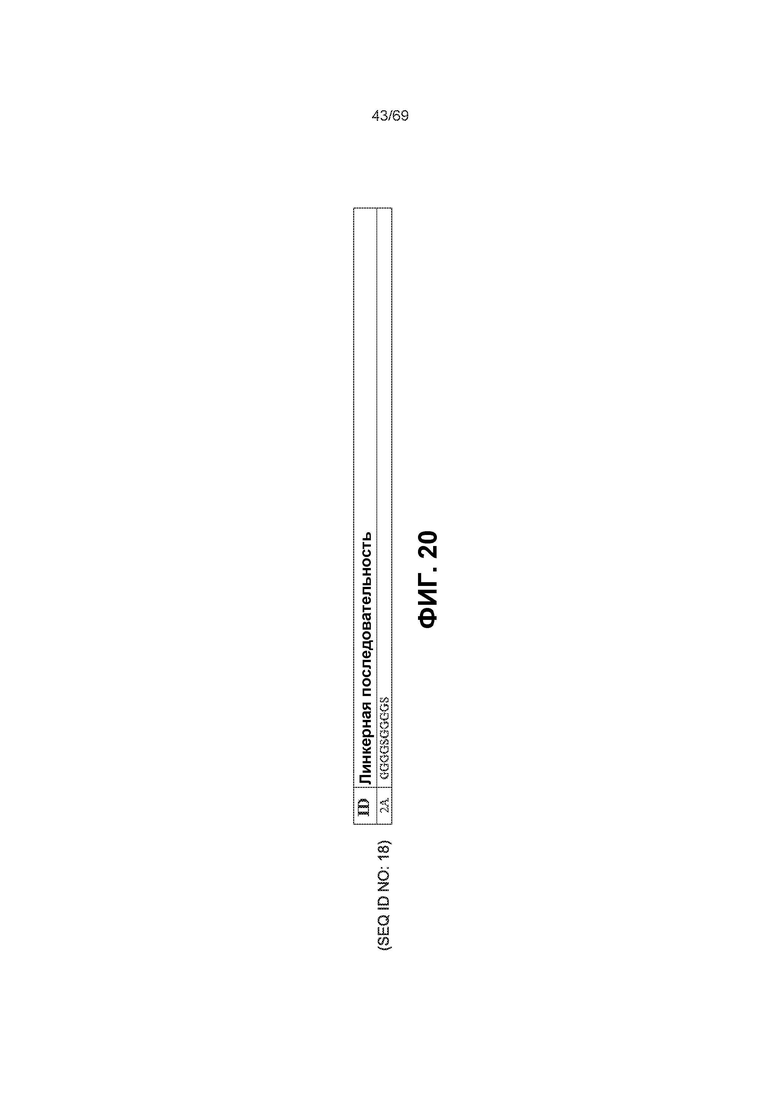
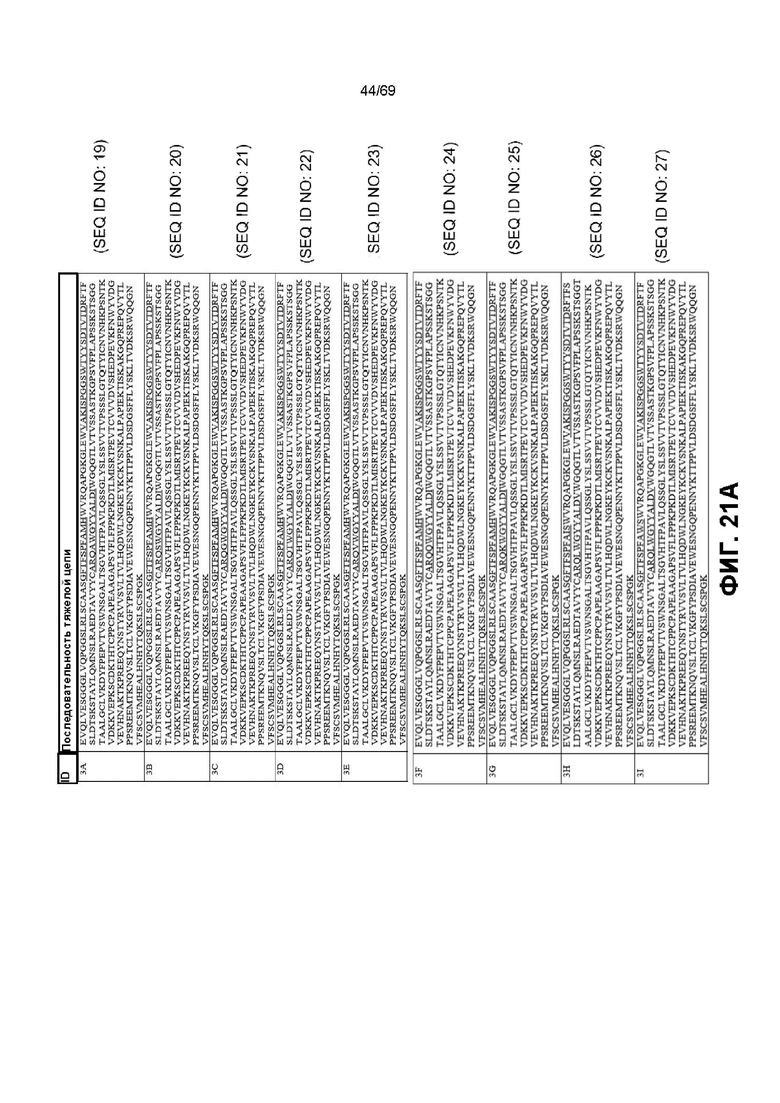




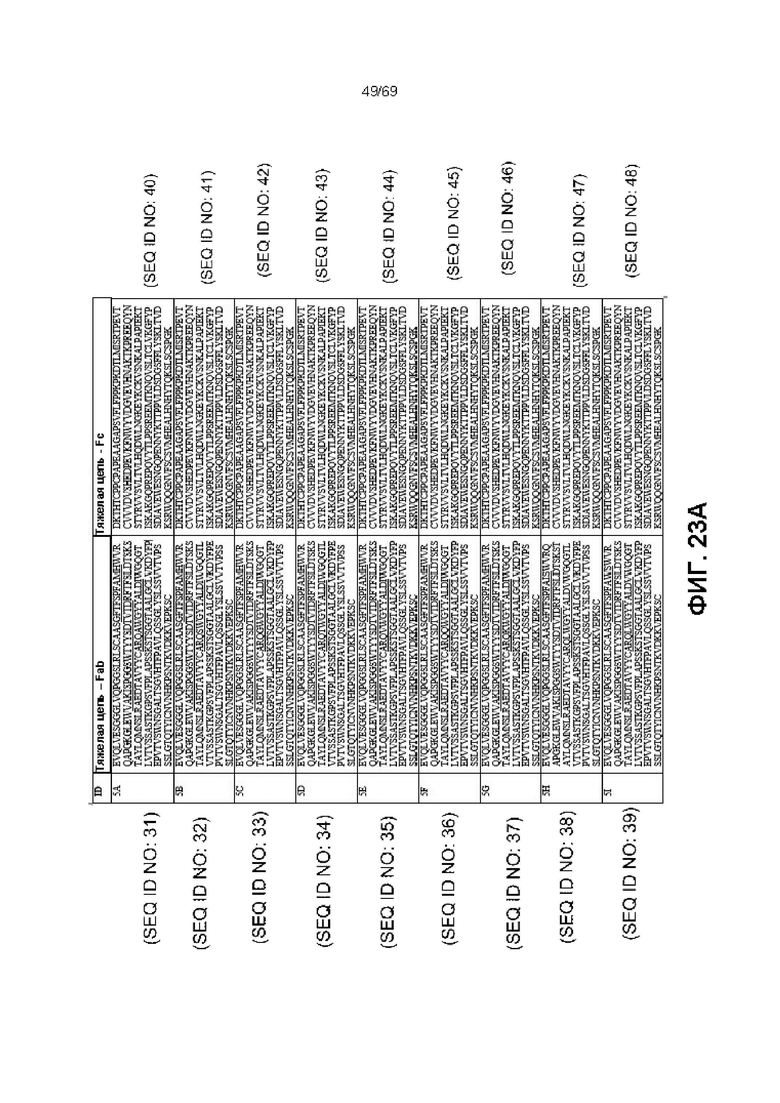


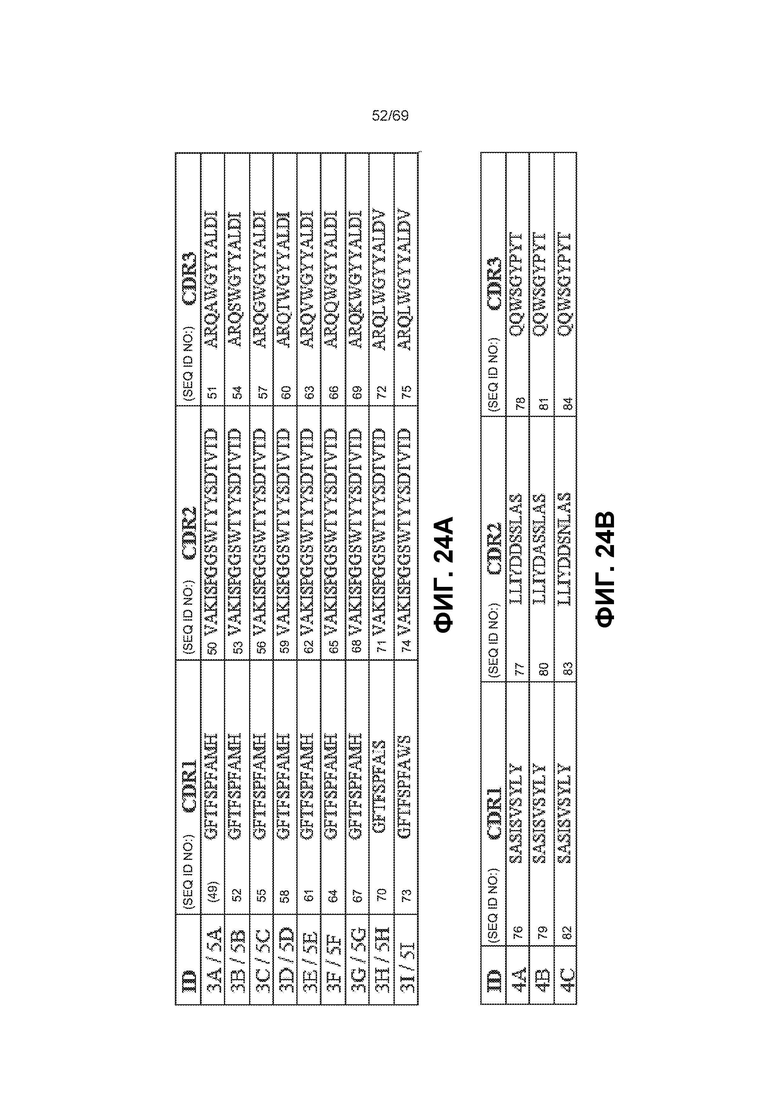

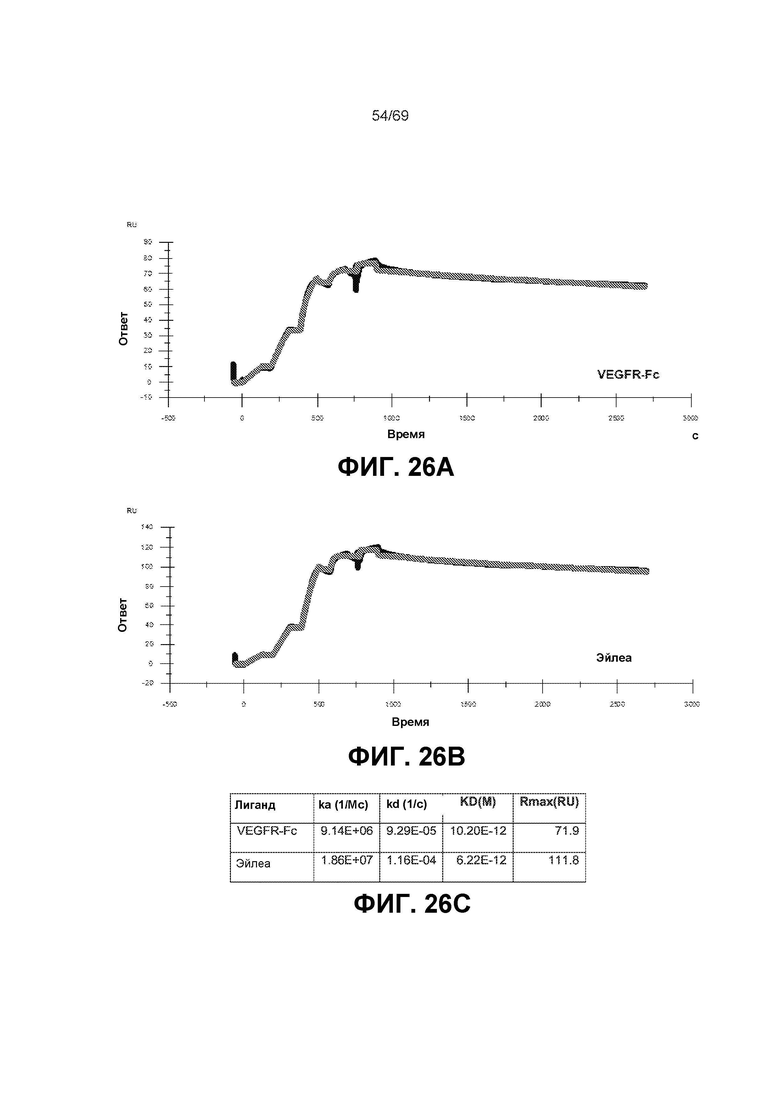
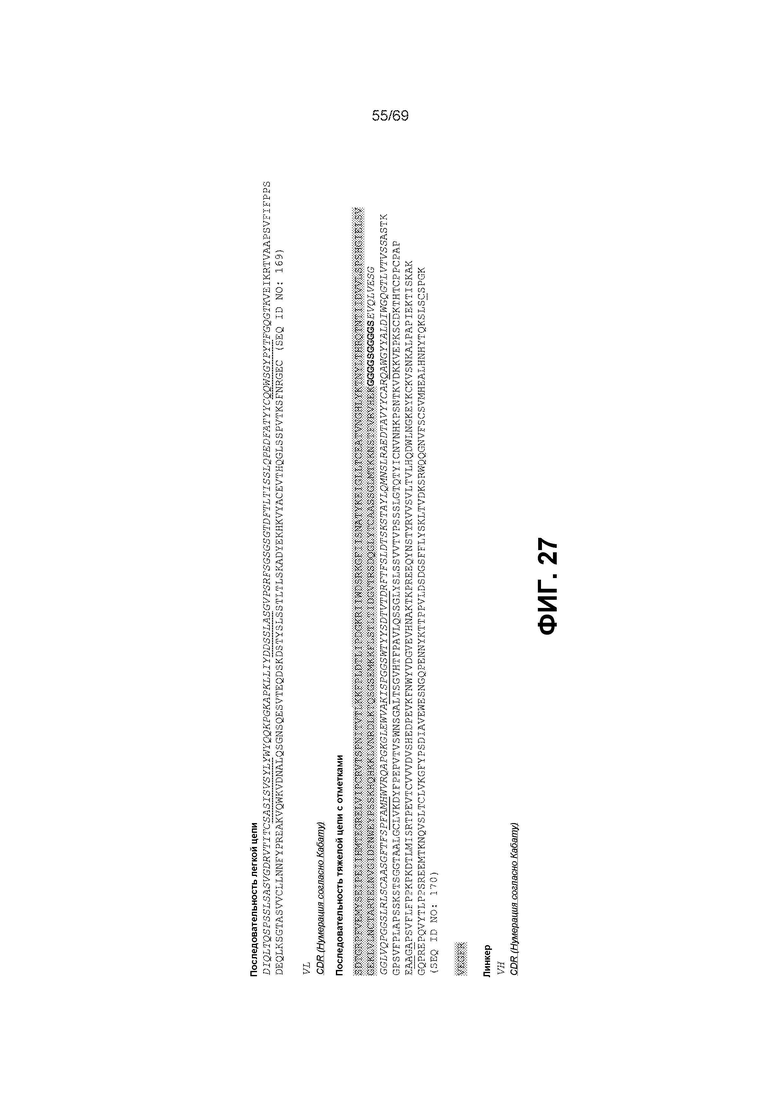
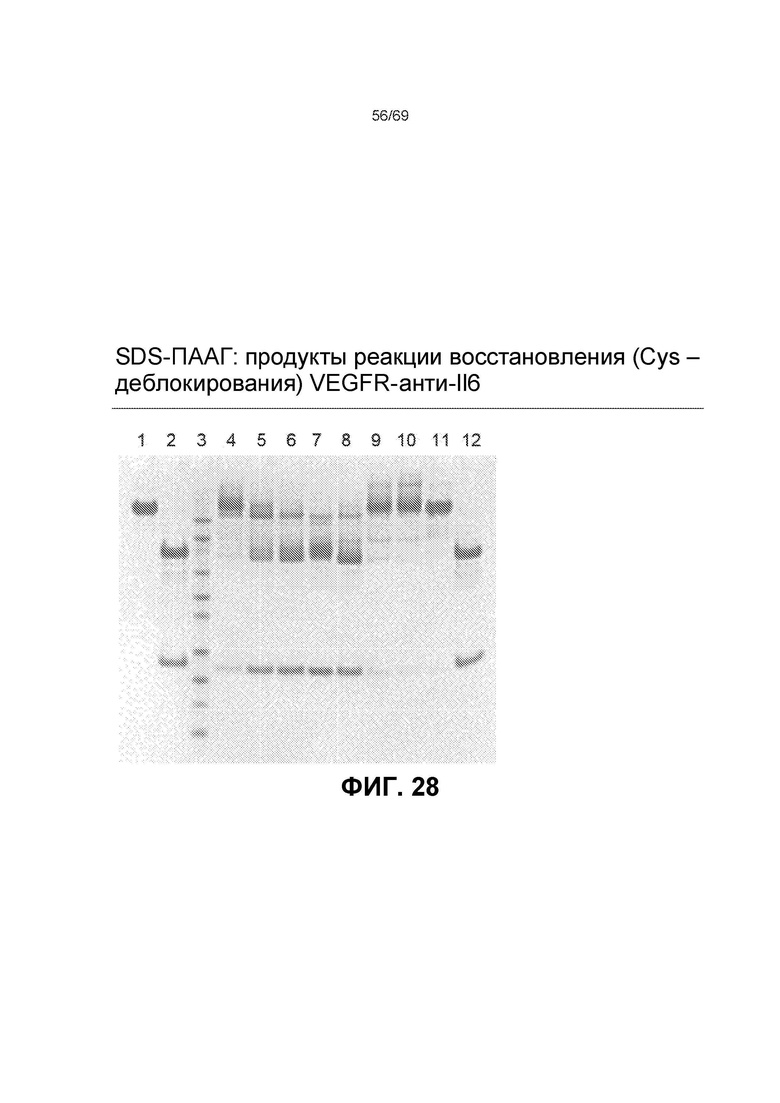


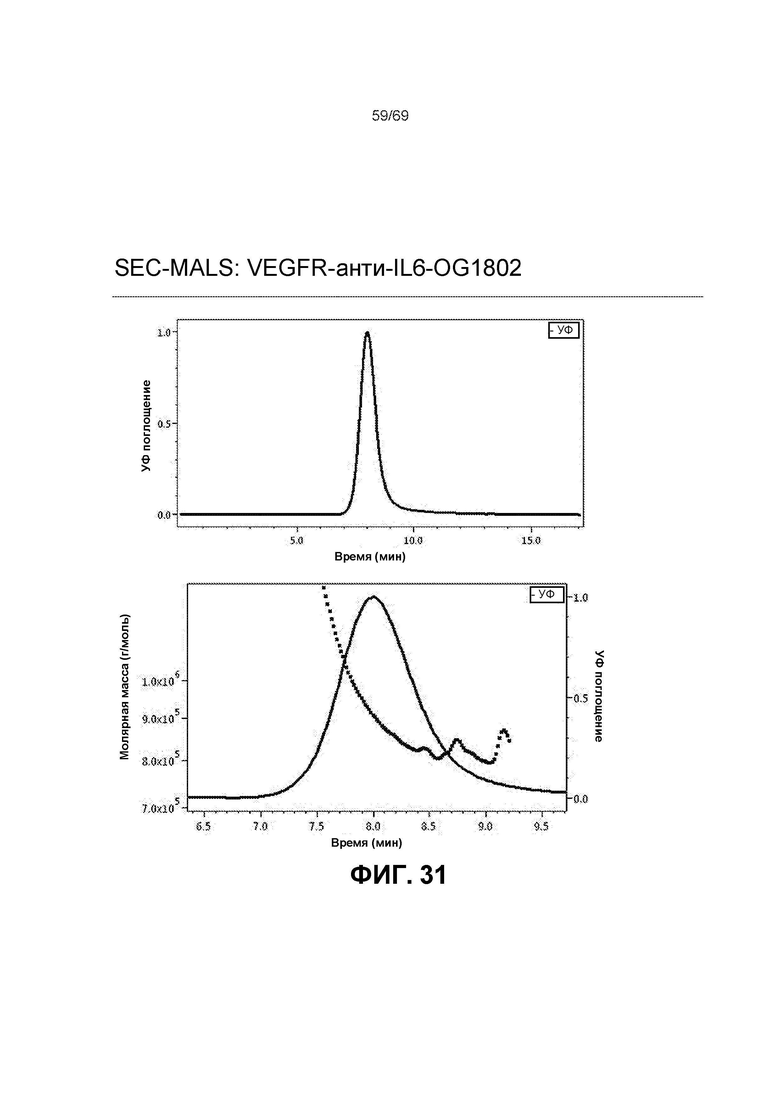
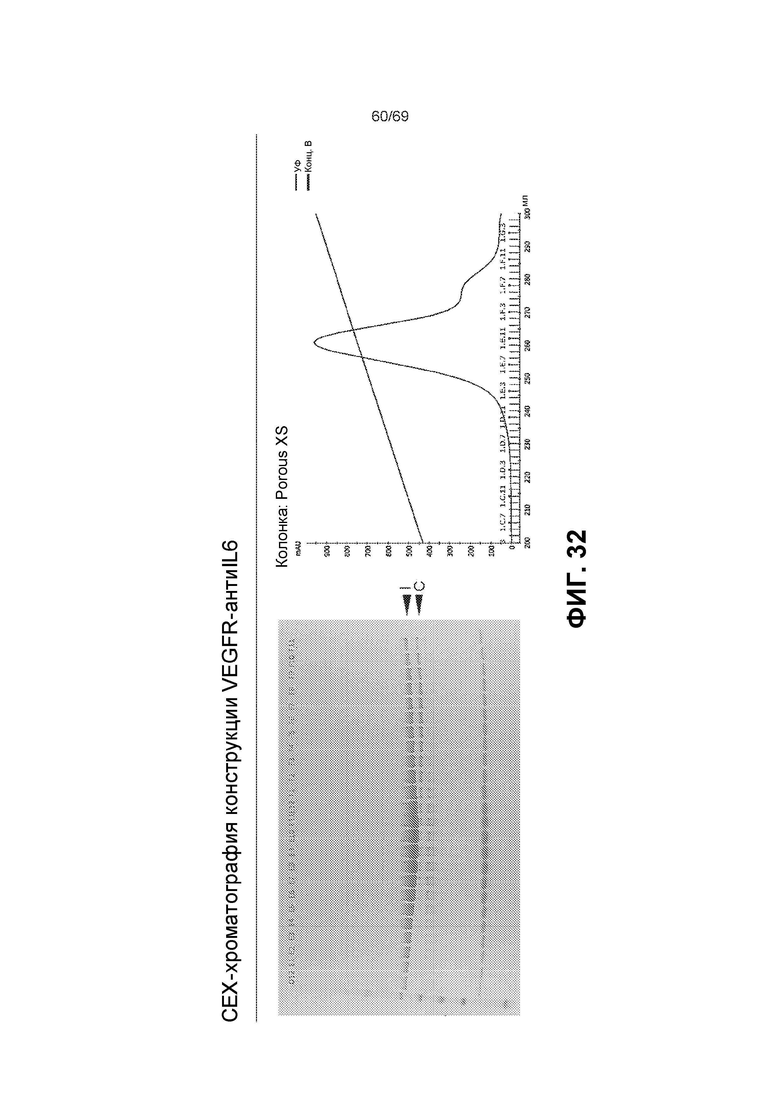
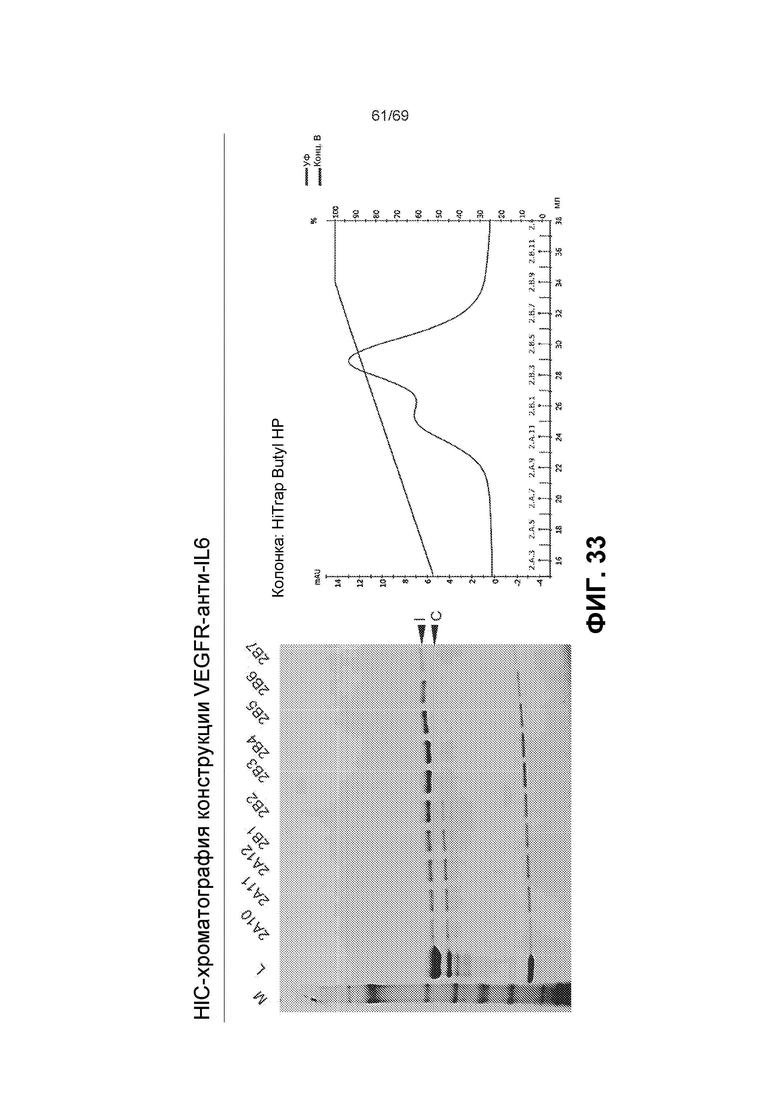
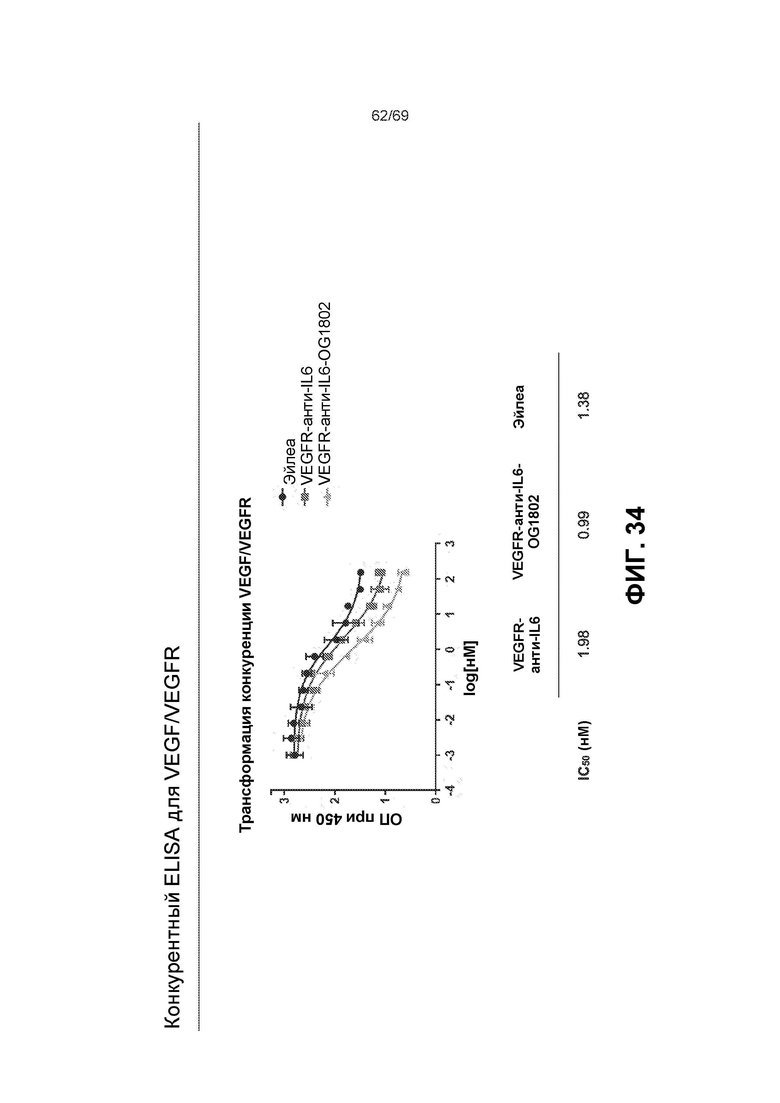
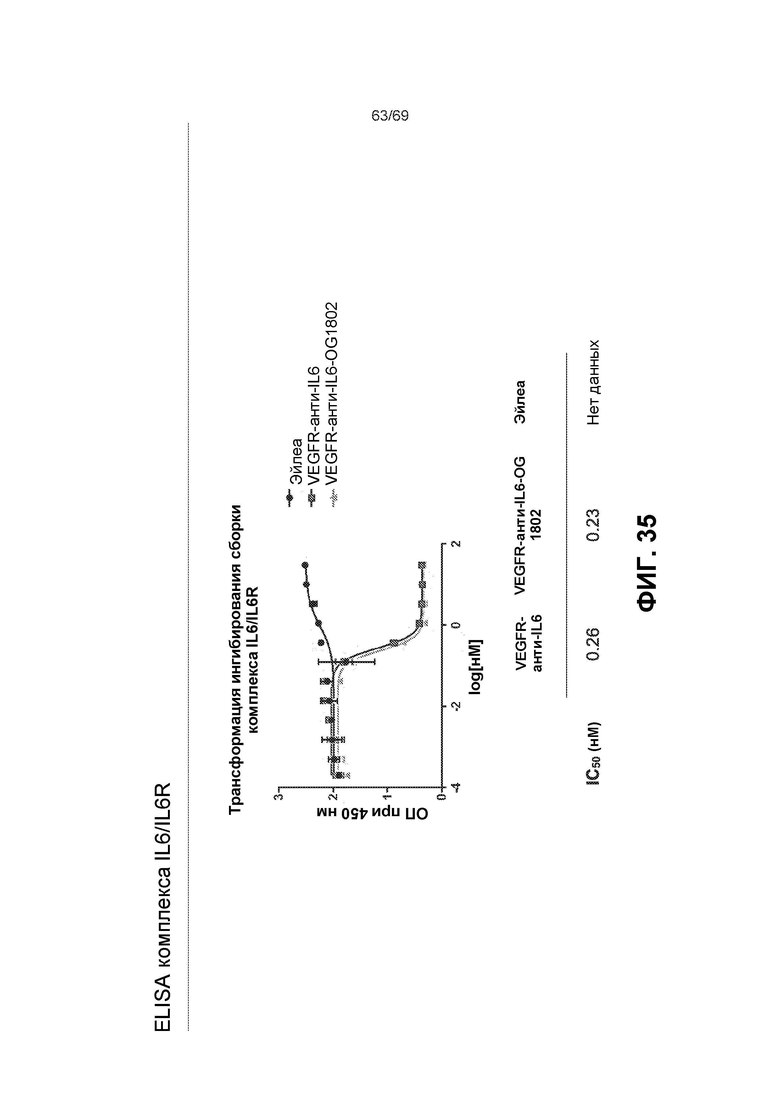
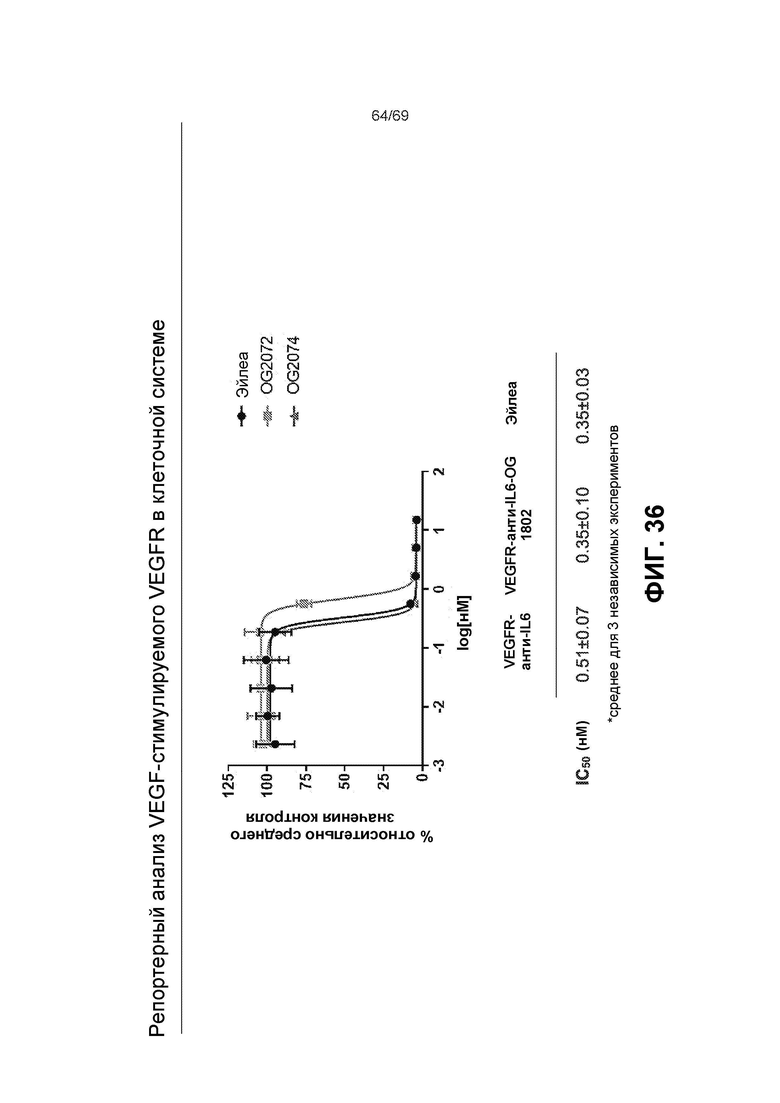
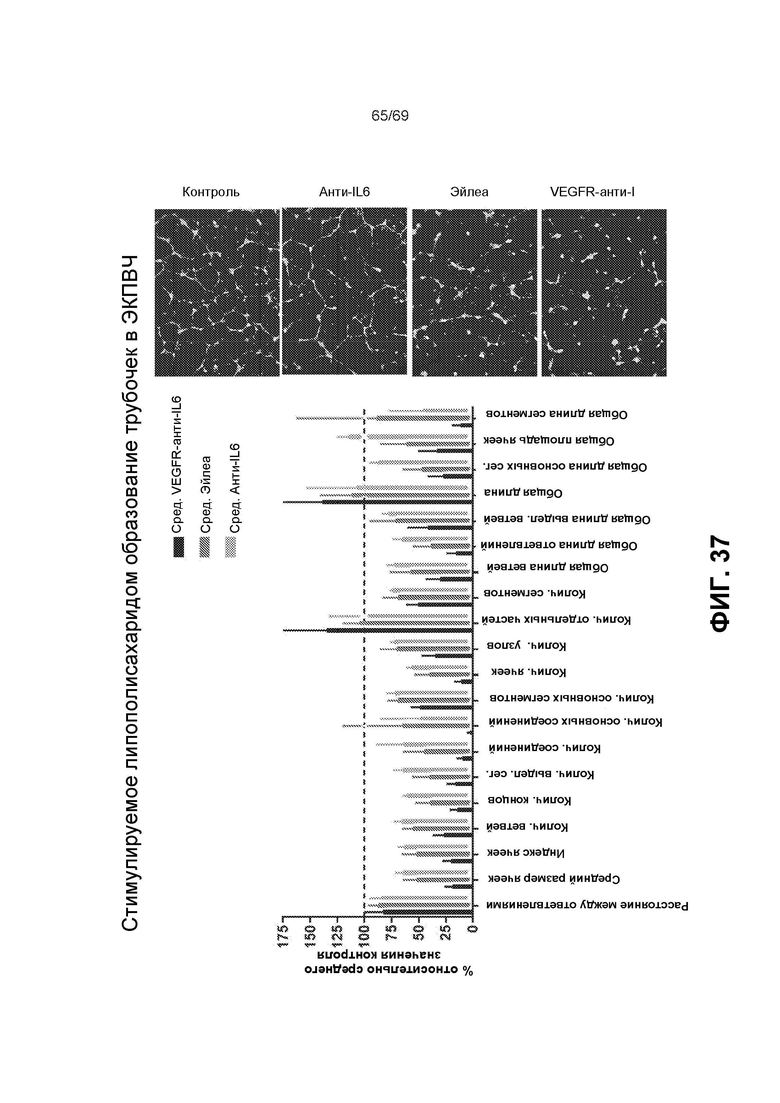
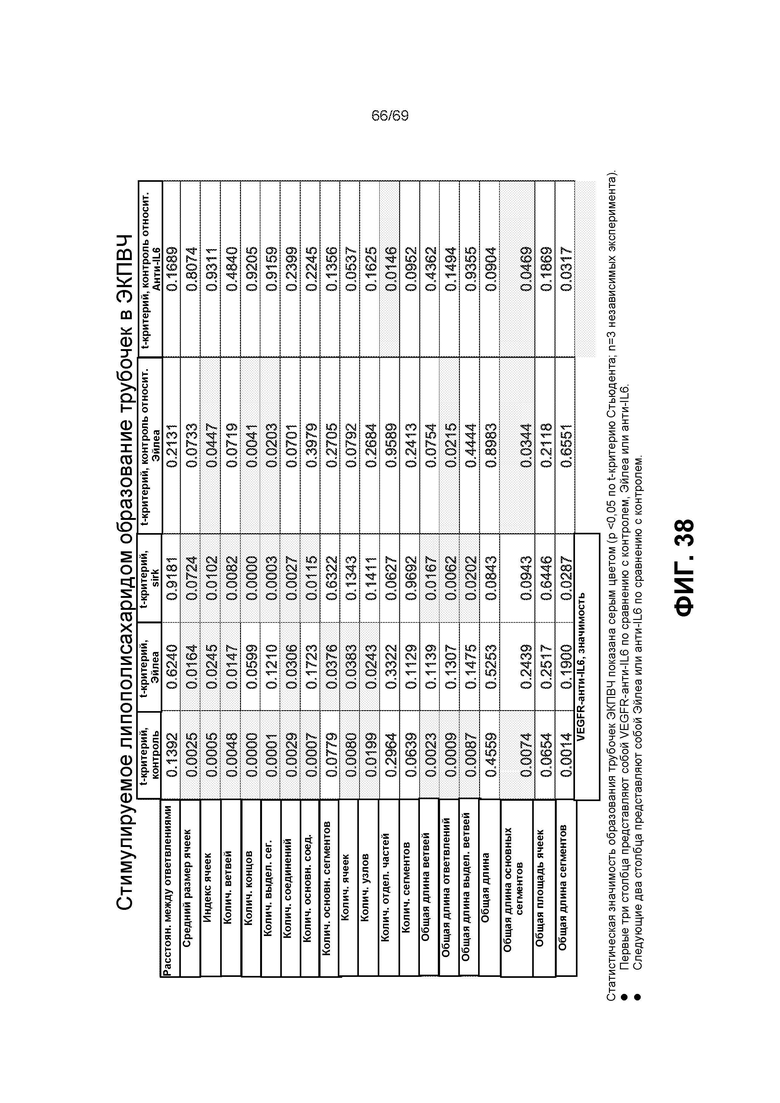
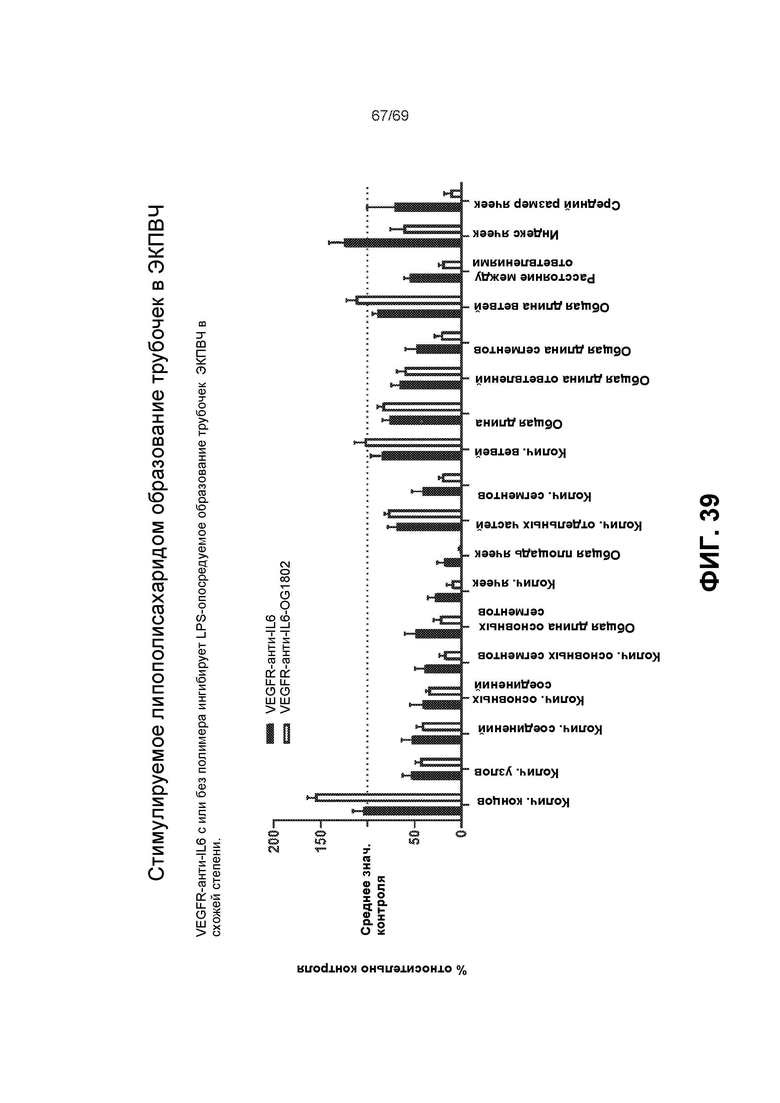
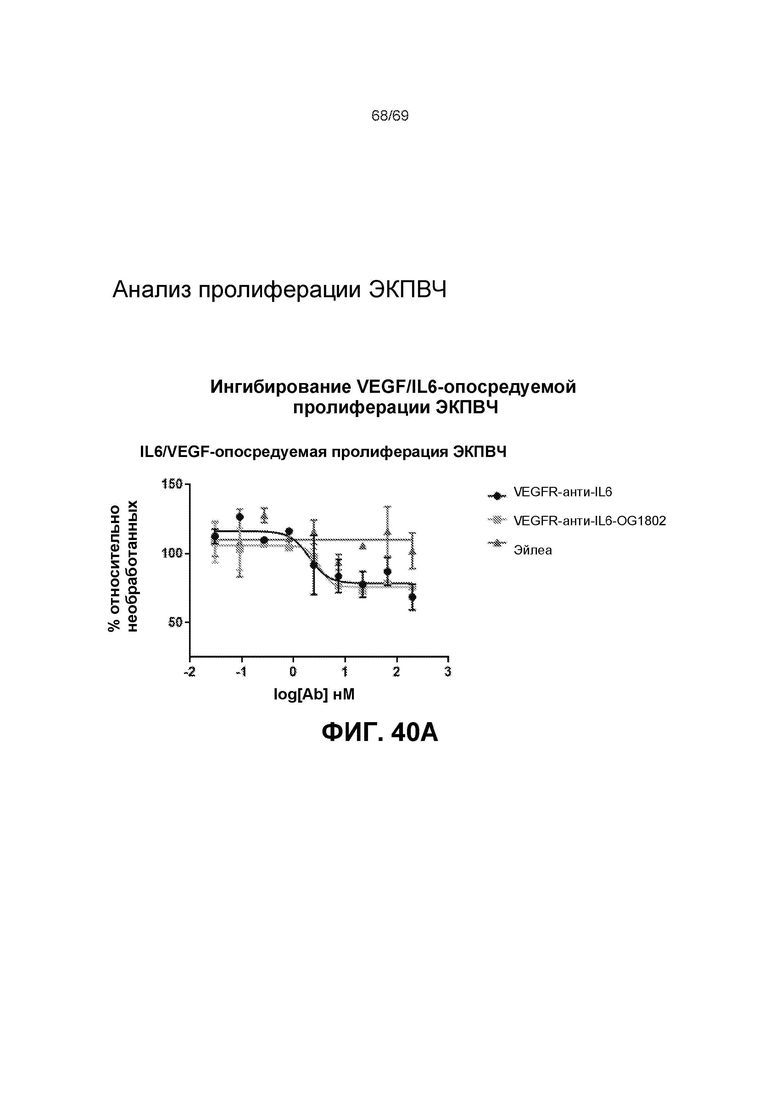
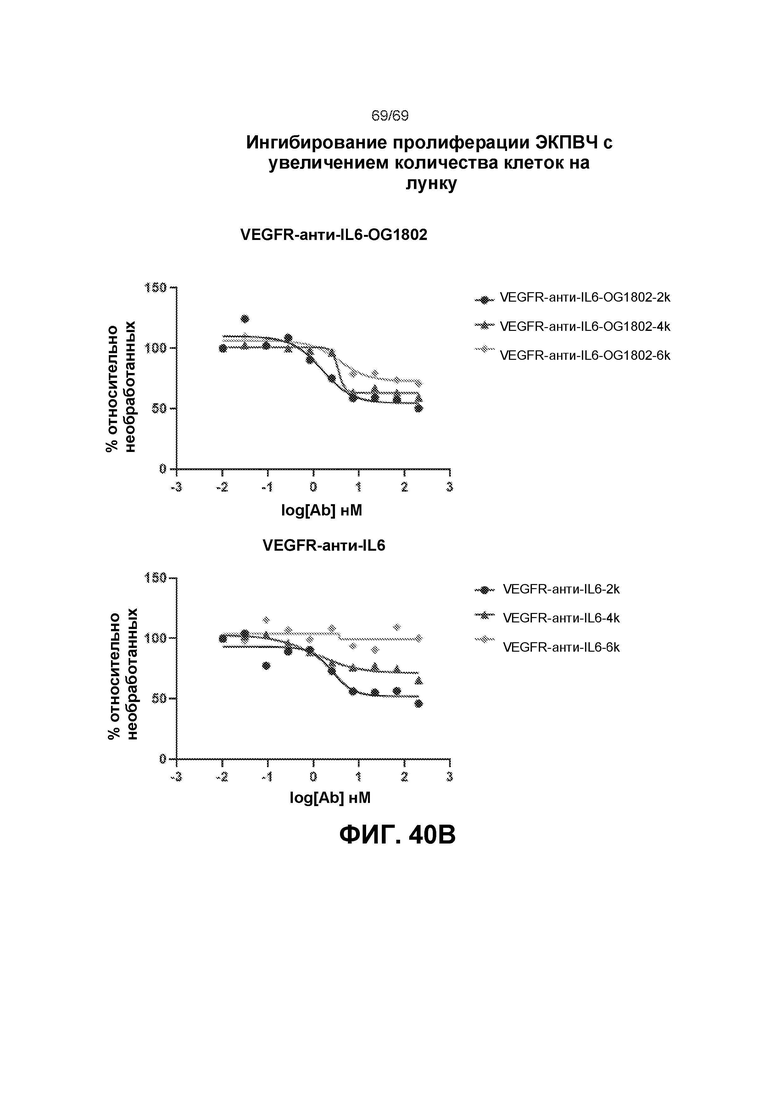
Изобретение относится к области биотехнологии. Предложены слитые белки для связывания VEGF и IL-6, каждый из которых содержит антагонистическое антитело, связывающееся с IL-6, и ловушку VEGF. Ловушка VEGF содержит домен 2 VEGFR1 и домен 3 VEGFR2. Также предложены конъюгаты указанных слитых белков. Кроме того, изобретение относится к кодирующим слитые белки нуклеиновым кислотам, векторам экспрессии, клеткам для продуцирования слитых белков, способам получения и применения указанных слитых белков. Изобретение может найти дальнейшее применение в терапии. 16 н. и 57 з.п. ф-лы, 77 ил., 35 табл., 27 пр.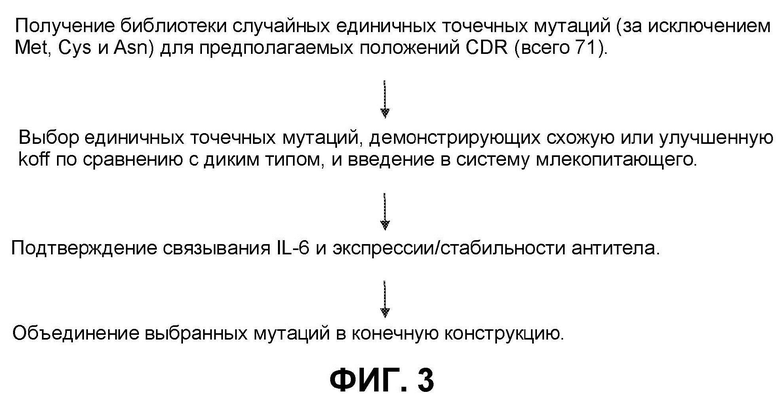
1. Слитый белок для связывания VEGF и IL-6, содержащий:
выделенное антагонистическое антитело, которое специфично связывается с IL-6, где указанное антитело содержит тяжелую цепь и легкую цепь; и
ловушку VEGF (фактора роста эндотелия сосудов), где указанная ловушка VEGF содержит домен 2 VEGFR1 и домен 3 VEGFR2,
причем указанная ловушка VEGF расположена либо
на N-конце указанной тяжелой цепи, содержащей IL-6 VH, с линкером, связывающим указанную ловушку VEGF и указанную тяжелую цепь; либо
между шарнирной областью и CH1-доменом тяжелой цепи, содержащей IL-6 VH, с линкером, связывающим указанный CH1-домен и указанную ловушку VEGF.
2. Слитый белок по п. 1, характеризующийся тем, что ловушка VEGF расположена на N-конце тяжелой цепи, содержащей IL-6 VH.
3. Слитый белок по п. 1, характеризующийся тем, что ловушка VEGF расположена между шарнирной областью и после CH1-домена тяжелой цепи, содержащей IL-6 VH.
4. Слитый белок по п. 1, характеризующийся тем, что ловушка VEGF состоит из последовательности SEQ ID NO: 114.
5. Слитый белок по любому из пп. 1-4, характеризующийся тем, что указанное антитело связывает IL-6 человека с аффинностью от примерно 10 пМ до примерно 1 нМ.
6. Слитый белок по любому из пп. 1-5, содержащий константный домен тяжелой цепи, содержащий ненативный остаток цистеина.
7. Слитый белок по п. 6, характеризующийся тем, что указанный ненативный остаток цистеина расположен в положении, выбранном из группы, состоящей из 347 и 443, в нумерации EU.
8. Слитый белок по п. 7, характеризующийся тем, что указанный ненативный остаток цистеина расположен в положении 443, в нумерации EU.
9. Слитый белок по любому из пп. 1-8, характеризующийся тем, что
(i) указанная тяжелая цепь содержит аминокислотную последовательность любой из SEQ ID NO: 19-27, или содержит вариабельную область тяжелой цепи, (VH), содержащую аминокислотную последовательность любой из SEQ ID NO: 7-13, 89, 90, 256-262; и легкую цепь, которая содержит аминокислотную последовательность любой из SEQ ID NO: 28-30, или содержит вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность любой из SEQ ID NO: 91-93; или
(ii) указанная тяжелая цепь содержит вариабельную область тяжелой цепи (VH), содержащую 3 определяющие комплементарность области (CDR): VH CDR1, VH CDR2 и VH CDR3, имеющие аминокислотные последовательности из CDR1, CDR2 и CDR3, соответственно, в SEQ ID NO: 256; и легкая цепь содержит вариабельную область легкой цепи (VL), содержащую VL CDR1, VL CDR2 и VL CDR3, имеющие аминокислотные последовательности из CDR1, CDR2 и CDR3, соответственно, в одной из SEQ ID NO: 91-93; или
(iii) указанное антитело содержит:
CDRH1, содержащую аминокислотную последовательность в SEQ ID NO: 172;
CDRH2, содержащую аминокислотную последовательность в SEQ ID NO: 173;
CDRH3, содержащую аминокислотную последовательность в SEQ ID NO: 174;
CDRL1, содержащую аминокислотную последовательность в SEQ ID NO: 199;
CDRL2, содержащую аминокислотную последовательность в SEQ ID NO: 200;
CDRL3, содержащую аминокислотную последовательность в SEQ ID NO: 201; и
Fc-домен, содержащий:
по меньшей мере одну из следующих мутаций (нумерация EU): L234A, L235A и G237A; и/или
по меньшей мере одну из следующих мутаций (нумерация EU): Q347C или L443C.
10. Конъюгат для связывания VEGF и IL-6, содержащий:
слитый белок по любому из пп. 1-9; и
полимер, содержащий фосфорилхолин, при этом указанный полимер ковалентно присоединен к тяжелой цепи выделенного антагонистического антитела.
11. Конъюгат по п. 10, характеризующийся тем, что полимер содержит цвиттерионный мономер, при этом указанный цвиттерионный мономер выбран из группы, состоящей из HEMA-фосфорилхолина, ПЭГ, биосовместимых жирных кислот и их производных, гидроксиалкилкрахмала (HAS), гидроксиэтилкрахмала (HES), полиэтиленгликоля (ПЭГ), поли(Glyx-Sery) (HAP), гиалуроновой кислоты (HA), гепарозановых полимеров (HEP), флексимеров, декстрана, полисиаловых кислот (PSA), Fc-доменов, трансферрина, 25 альбумина, эластиноподобных пептидов (ELP), полимеров XTEN, полимеров ПАС, ПА-полимеров, связывающих альбумин пептидов, пептидов CTP и FcRn-связывающих пептидов.
12. Конъюгат по любому из пп. 10-11, характеризующийся тем, что полимер содержит мономеры 2(метакрилоилокси)этил-(2-(триметиламмоний)этил)фосфата (MPC).
13. Конъюгат по любому из пп. 10-12, характеризующийся тем, что полимер имеет пиковую молекулярную массу от 300000 до 1750000 Дальтон, измеренную путем эксклюзионной хроматографии с многоугловым светорассеянием (далее «SEC-MALS»).
14. Конъюгат по любому из пп. 10-13, характеризующийся тем, что полимер имеет пиковую молекулярную массу от 500000 до 1000000 Дальтон, измеренную путем SEC-MALS.
15. Конъюгат по п. 14, характеризующийся тем, что полимер имеет пиковую молекулярную массу от 600000 до 800000 Дальтон, измеренную путем SEC-MALS.
16. Конъюгат по любому из пп. 10-15, характеризующийся тем, что полимер содержит 2 или более ответвлений.
17. Конъюгат по любому из пп. 10-16, характеризующийся тем, что полимер содержит 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 ответвлений.
18. Конъюгат по п. 17, характеризующийся тем, что полимер содержит 9 ответвлений.
19. Конъюгат по любому из пп. 10-18, характеризующийся тем, что антитело представляет собой IgG.
20. Конъюгат по п. 19, характеризующийся тем, что полимер ковалентно связан с сульфгидрильной группой остатка цистеина в тяжелой цепи IgG.
21. Конъюгат по любому из пп. 10-20, характеризующийся тем, что полимер ковалентно связан с сульфгидрильной группой остатка цистеина в положении 347 или 443, в нумерации EU.
22. Конъюгат по любому из пп. 10-21, характеризующийся тем, что антитело дополнительно конъюгировано с полимером с образованием биоконъюгата и при этом указанный биоконъюгат имеет молекулярную массу от примерно 350000 до 1900000 Дальтон.
23. Конъюгат по любому из пп. 10-22, характеризующийся тем, что коэффициент полидисперсности (PDI) равен или меньше 1,5.
24. Конъюгат по любому из пп. 10-23, характеризующийся тем, что
полимер содержит 9 ответвлений; и
полимер имеет молекулярную массу от примерно 600000 до примерно 900000 Да.
25. Конъюгат по любому из пп. 10-24, содержащий следующую структуру:
где:
каждая тяжелая цепь анти-IL-6 антитела обозначена буквой H и каждая легкая цепь анти-IL-6 антитела обозначена буквой L;
полимер связан с антителом через сульфгидрильную группу C443, в нумерации EU; указанная связь изображена на одной из тяжелых цепей;
PC представляет собой 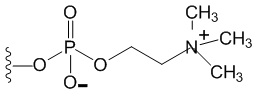 , где волнистая линия показывает точку присоединения к остальной части полимера, где X=a) OR, где R=H, метил, этил, пропил, изопропил, b) H или c) любой галогенид, включая Br; и
, где волнистая линия показывает точку присоединения к остальной части полимера, где X=a) OR, где R=H, метил, этил, пропил, изопропил, b) H или c) любой галогенид, включая Br; и
n1, n2, n3, n4, n5, n6, n7, n8 и n9 являются одинаковыми или разными так, что сумма n1, n2, n3, n4, n5, n6, n7, n8 и n9 равна 2500 плюс или минус 15%.
26. Выделенная линия клеток, продуцирующая слитый белок, где клетки указанной линии клеток содержат выделенную нуклеиновую кислоту, кодирующую слитый белок по любому из пп. 1-9.
27. Выделенная линия клеток по п. 26, характеризующаяся тем, что указанная линия клеток выбрана из группы, состоящей из CHO, k1SV, XCeed, CHOK1SV и GS-KO.
28. Выделенная нуклеиновая кислота, кодирующая слитый белок по любому из пп. 1-9.
29. Рекомбинантный вектор экспрессии, содержащий нуклеиновую кислоту по п. 28.
30. Клетка-хозяин для продукции слитого белка, содержащая вектор экспрессии по п. 29.
31. Способ получения слитого белка, при этом указанный способ включает: культивирование линии клеток, которая рекомбинантно продуцирует слитый белок по любому из пп. 1-9, в условиях, в которых продуцируется слитый белок; и извлечение слитого белка.
32. Фармацевтическая композиция для лечения глазного нарушения или для ингибирования передачи сигнала VEGF и IL-6 у пациента, нуждающегося в этом, содержащая слитый белок по любому из пп. 1-9 и/или конъюгат по любому из пп. 10-25, и фармацевтически приемлемый носитель.
33. Фармацевтическая композиция по п. 32, характеризующаяся тем, что указанная композиция представляет собой жидкость и имеет уровень эндотоксинов ниже примерно 0,2 ЕЭ/мл.
34. Фармацевтическая композиция по п. 33, характеризующаяся тем, что указанная композиция представляет собой жидкость и имеет уровень эндотоксинов ниже примерно 2,0, 1, 0,5 или 0,2 ЕЭ/мл.
35. Способ лечения или профилактики заболевания у нуждающегося в этом пациента, включающий введение пациенту слитого белка по любому из пп. 1-9 и/или конъюгата по любому из пп. 10-25, где указанное заболевание представляет собой глазное нарушение, выбранное из влажной формы дегенерации желтого пятна или неоваскулярной дегенерации желтого пятна, влажной формы возрастной дегенерации желтого пятна или неоваскулярной возрастной дегенерации желтого пятна, сухой формы возрастной дегенерации желтого пятна, венозной, артериальной окклюзии или другой окклюзии кровеносных сосудов глаза и/или сетчатки с отеком сетчатки или без него, переднего и заднего увеита, отека желтого пятна при увеите, диабетической ретинопатии, непролиферативной диабетической ретинопатии, диабетического отека желтого пятна, непролиферативного диабетического отека желтого пятна, офтальмологического воспалительного заболевания - склерита и внутриглазных опухолей.
36. Способ ингибирования передачи сигнала VEGF и IL-6 у пациента, нуждающегося в этом, включающий:
идентификацию пациента с гиперактивностью IL-6 и/или VEGF; и
введение указанному пациенту слитого белка по любому из пп. 1-9 и/или конъюгата по любому из пп. 10-25.
37. Способ по п. 36, характеризующийся тем, что пациент нуждается в лечении глазного нарушения посредством ингибирования передачи сигнала VEGF и IL-6.
38. Способ по п. 37, характеризующийся тем, что указанное глазное нарушение представляет собой по меньшей мере одно из: влажной формы дегенерации желтого пятна или неоваскулярной дегенерации желтого пятна, влажной формы возрастной дегенерации желтого пятна или неоваскулярной возрастной дегенерации желтого пятна, сухой формы возрастной дегенерации желтого пятна, венозной, артериальной окклюзии или другой окклюзии кровеносных сосудов глаза и/или сетчатки с отеком сетчатки или без него, переднего и заднего увеита, отека желтого пятна при увеите, диабетической ретинопатии, непролиферативной диабетической ретинопатии, диабетического отека желтого пятна, непролиферативного диабетического отека желтого пятна, офтальмологического воспалительного заболевания - склерита и внутриглазных опухолей.
39. Способ по любому из пп. 35-38, характеризующийся тем, что слитый белок и/или конъюгат вводят не чаще одного раза в месяц.
40. Способ по любому из пп. 35-38, характеризующийся тем, что слитый белок и/или конъюгат вводят не чаще одного раза в два месяца.
41. Способ по любому из пп. 35-38, характеризующийся тем, что слитый белок и/или конъюгат вводят не чаще одного раза в три месяца.
42. Способ по любому из пп. 35-38, характеризующийся тем, что слитый белок и/или конъюгат вводят с частотой от одного раза в год до одного раза в два месяца.
43. Слитый белок для связывания VEGF и IL-6, содержащий:
IL-6 VH;
IL-6 VL;
Fc-область;
ловушку VEGF, где указанная ловушка VEGF содержит домен 2 VEGFR1 и домен 3 VEGFR2,
при этом указанная ловушка VEGF слита с IL-6 одним из следующих способов:
c N-концом тяжелой цепи, содержащей IL-6 VH, линкером, связывающим указанную ловушку VEGF и указанную тяжелую цепь; или
между шарнирной областью и после CH1-домена тяжелой цепи, содержащей IL-6 VH, с линкером, связывающим указанный CH1-домен и указанную ловушку VEGF.
44. Слитый белок по любому из пп. 1-9 или конъюгат по любому из пп. 10-25, содержащий мутацию в положении 94 в последовательности ловушки VEGF.
45. Слитый белок по любому из пп. 1-9 или конъюгат по любому из пп. 10-25, содержащий мутацию в положении 95 в последовательности ловушки VEGF.
46. Слитый белок по пп. 1-9 или конъюгат по любому из пп. 10-25, содержащий мутации T94I и/или H95I в последовательности ловушки VEGF.
47. Двойной ингибитор передачи сигнала VEGFR и IL-6, где указанный двойной ингибитор содержит слитую конструкцию ловушка-антитело из анти-IL6 антитела и ловушки VEGF, причем указанное анти-IL 6 антитело содержит IL-6 VH и IL-6 VL, причем указанная ловушка VEGF содержит домен 2 VEGFR1 и домен 3 VEGFR2, причем указанный двойной ингибитор содержит по меньшей мере одну точечную мутацию в последовательности VEGFR для уменьшения расщепления белка VEGFR,
причем указанная ловушка VEGF расположена либо
на N-конце тяжелой цепи, содержащей IL-6 VH, с линкером, связывающим указанную ловушку VEGF и указанную тяжелую цепь; либо
между шарнирной областью и CH1-доменом тяжелой цепи, содержащей IL-6 VH, с линкером, связывающим указанный CH1-домен и указанную ловушку VEGF.
48. Двойной ингибитор по п. 47, характеризующийся тем, что молекулярная масса указанного двойного ингибитора составляет 1,0 МДа.
49. Двойной ингибитор по п. 47, характеризующийся тем, что указанный двойной ингибитор обеспечивает терапию воспалительных заболеваний сетчатки.
50. Двойной ингибитор по п. 47, содержащий последовательность вариабельной области тяжелой цепи, выбранную из опций SEQ ID NO: 7-13, 89, 90 и/или 256-262.
51. Двойной ингибитор по любому из пп. 47-50, характеризующийся тем, что ловушка VEGF содержит последовательность, выбранную из по меньшей мере одной из SEQ ID NO: 15, 16 или 17.
52. Двойной ингибитор по любому из пп. 47-51, характеризующийся тем, что линкерная последовательность представляет собой SEQ ID NO: 18.
53. Двойной ингибитор по любому из пп. 47-52, характеризующийся тем, что последовательность тяжелой цепи выбрана из по меньшей мере одной из SEQ ID NO: 19-27 или содержит по меньшей мере последовательность в одной из SEQ ID NO: 89, 90, 256-262.
54. Двойной ингибитор по любому из пп. 47-53, содержащий последовательность легкой цепи, содержащую по меньшей мере 3 CDR легкой цепи из по меньшей мере одной из SEQ ID NO: 76-84.
55. Двойной ингибитор по любому из пп. 47-54, характеризующийся тем, что последовательность тяжелой цепи для анти-IL-6 молекулы содержит по меньшей мере 3 CDR тяжелой цепи из по меньшей мере одной из SEQ ID NO: 49-75.
56. Двойной ингибитор по любому из пп. 47-55, содержащий последовательность VEGFR-Fc из по меньшей мере одной из SEQ ID NO: 85-88.
57. Двойной ингибитор по любому из пп. 47-56, содержащий одну или более последовательностей в любых одной или более из SEQ ID NO: 7-13, 145, 15-17, 18-84.
58. Двойной ингибитор по п. 47, характеризующийся тем, что IL-6 VH содержит последовательность из последовательности IL6 VH в любой из SEQ ID NO: 19-27, 31-39, 89, 90 или 256-262.
59. Двойной ингибитор по п. 47 или 58, характеризующийся тем, что IL-6 VL содержит последовательность из последовательности IL6 VL в любой из SEQ ID NО: 28-30 или 91-93.
60. Двойной ингибитор по любому из пп. 47-59, содержащий Fc-область, содержащую последовательность из последовательности Fc в любой из SEQ ID NO: 40-48.
61. Белковая конструкция для связывания VEGF и IL-6, содержащая:
тяжелую цепь, содержащую по меньшей мере 3 CDR тяжелой цепи, выбранных из соответствующих CDR тяжелой цепи в любой из SEQ ID NO:7-13, 19-27, 31-39, 49-75;
легкую цепь, содержащую по меньшей мере 3 CDR легкой цепи, выбранных из соответствующих CDR легкой цепи в любой из SEQ ID NO:28-30, 76-84;
последовательность ловушки VEGF, выбранную из соответствующей последовательности ловушки VEGF любой из SEQ ID NO: 14-17, 85-88; и
линкерную последовательность SEQ ID NO:18,
причем указанная ловушка VEGF расположена либо
на N-конце указанной тяжелой цепи, с линкером, связывающим указанную ловушку VEGF и указанную тяжелую цепь; либо
между шарнирной областью и CH1-доменом указанной тяжелой цепи, с линкером, связывающим указанный CH1-домен и указанную ловушку VEGF.
62. Слитый белок для связывания VEGF и IL-6, содержащий:
IL-6 VH;
IL-6 VL;
Fc-область; и
ловушку VEGF, причем указанная ловушка VEGF, содержит домен 2 VEGFR1 и домен 3 VEGFR2,
причем указанная ловушка VEGF расположена либо
на N-конце указанной тяжелой цепи, содержащей IL-6 VH, с линкером, связывающим указанную ловушку VEGF и указанную тяжелую цепь; либо
между шарнирной областью и CH1-доменом тяжелой цепи, содержащей IL-6 VH, с линкером, связывающим указанный CH1-домен и указанную ловушку VEGF,
при этом указанный слитый белок ингибирует пролиферацию эндотелиальных клеток пупочной вены человека (ЭКПВЧ).
63. Слитый белок по п. 62, характеризующийся тем, что изменение пролиферации ЭКПВЧ ингибирует пролиферацию, опосредуемую VEGF/IL6.
64. Конъюгат для связывания VEGF и IL-6, содержащий слитый белок, конъюгированный с полимером, содержащим фосфорилхолин, причем указанный слитый белок содержит (i) последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 263, причем указанная последовательность содержит последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 114, и последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 256, и (ii) последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 117, причем указанная последовательность содержит последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 91.
65. Конъюгат по п. 64, характеризующийся тем, что указанный слитый белок по меньшей мере на 95% идентичен SEQ ID NO: 263 и по меньшей мере на 95% идентичен SEQ ID NO: 117.
66. Конъюгат по п. 64 или 65, характеризующийся тем, что указанный белок содержит по меньшей мере а) SEQ ID NO: 172, 173, 174, 199, 200 и 201 или b) SEQ ID NO: 172, 173, 174, 199, 200 и/или 201, где содержится до 3 замен аминокислот, при этом указанная замена представляет собой консервативную замену.
67. Слитый белок по любому из пп. 1-9 или 43-46, характеризующийся тем, что указанный слитый белок содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
68. Конъюгат по любому из пп. 10-25, характеризующийся тем, что указанный конъюгат содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
69. Двойной ингибитор по любому из пп. 47-60, характеризующийся тем, что указанный двойной ингибитор содержит аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
70. Белковая конструкция по п. 61, характеризующаяся тем, что указанная конструкция содержит а) аминокислотную последовательность SEQ ID NO: 169 и 170 или b) аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 169 и по меньшей мере на 80% идентична SEQ ID NO: 170.
71. Конъюгат для связывания VEGF и IL-6, содержащий:
белковую конструкцию, содержащую две тяжелых цепи и две легких цепи, причем каждая из тяжелых цепей содержит аминокислотную последовательность SEQ ID NO: 170 с С-концевым лизином или без него, и каждая из легких цепей содержит аминокислотную последовательность SEQ ID NO: 169; и
полимер,
причем конъюгат содержит следующую структуру:
где:
каждая тяжелая цепь белковой конструкции обозначена буквой H и каждая легкая цепь белковой конструкции обозначена буквой L;
полимер связан с белковой конструкцией через сульфгидрильную группу C443 (нумерация EU), указанная связь изображена на одной из тяжелых цепей;
PC представляет собой , где волнистая линия показывает точку присоединения к остальной части полимера; где X=a) OR, где R=H, метил, этил, пропил, изопропил, b) H, или c) любой галогенид, включая Br; и
при этом n1, n2, n3, n4, n5, n6, n7, n8 и n9 каждый представляет собой число n следующих повторяющихся звеньев в формуле (17):
где волнистые линии показывают точки присоединения между мономерными звеньями в полимере, и
при этом n1, n2, n3, n4, n5, n6, n7, n8 и n9 каждый представляет собой целое число и являются одинаковыми или разными так, что сумма n1, n2, n3, n4, n5, n6, n7, n8 и n9 составляет 2500 плюс или минус 15%.
72. Конъюгат по п. 70, где каждая тяжелая цепь не содержит C-концевой лизин.
73. Слитый белок по п. 67, конъюгат по п. 68, двойной ингибитор по п. 69 или белковая конструкция по п. 70, где указанная аминокислотная последовательность, которая по меньшей мере на 80% идентична SEQ ID NO: 170, не содержит C-концевой лизин.

