ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0001] Настоящее изобретение относится к области биофармацевтических технологий, в частности к связывающей молекуле для IL-5 и ее способу получения и применению.
УРОВЕНЬ ТЕХНИКИ
[0002] Интерлейкин-5 человека, также называемый IL-5, наряду с IL-13 и гранулоцитарно-макрофагальным колониестимулирующим фактором (GM-CSF) является цитокином, участвующим в гемопоэзе и воспалении. Все эти три цитокина могут стимулировать образование, функционирование и выживание эозинофилов, и поэтому они способны влиять на воспалительные заболевания. Например, эозинофилы главным образом функционируют в качестве эффекторов при астме, атопическом дерматите и аллергическом рините.
[0003] Внимание большинства исследователей было сосредоточено на IL-5 в качестве цитокина, специфично воздействующего на эозинофилы, при этом обнаружено повышение уровней мРНК и белка IL-5 в легочных тканях и жидкости бронхоальвеолярного лаважа (BAL) у пациентов с симптоматической астмой. Исследователи также наблюдали, что существует корреляция между уровнями IL-5 и провокацией аллергенами и активностью заболевания. Однако, очевидно, что в дополнение к тому, что IL-5, GM-CSF и IL-3 функционируют для обеспечения образования и активации эозинофилов при астме, было продемонстрировано, что GM-CSF и IL-3 синтезируются в случае наличия аллергического воспаления. Экспрессия таких цитокинов может способствовать увеличению общего количества инфильтрирующих эозинофилов и степени активации эозинофилов. Также возможно, что эти цитокины функционируют для обеспечения эозинофильной инфильтрации на различных стадиях. Согласно недавним данным кинетического анализа от пациентов, получивших стимуляцию антигеном, было показано, что уровень IL-5 повышается от дня 2 до дня 7, а уровень GM-CSF достигает своего пика в день 2 и продолжает расти в день 16.
[0004] IL-5, GM-CSF и IL-3, связываясь с рецепторами клеточной поверхности, стимулируют эозинофилы, а также другие нормальные и раковые клетки. Эти рецепторы клеточной поверхности содержат лиганд-специфическую α-цепь и цепь (βc), общую для трех рецепторов. Связывание с α-цепью каждого рецептора является начальной стадией активации рецептора. Однако, одного связывания с α-цепью недостаточно для активации. Затем лиганд привлекает βc, после чего следует стадия с двумя основными функциональными последствиями. Во-первых, это позволяет связыванию IL-5, GM-CSF и IL-3 становиться практически необратимым; и кроме того, это приводит к полной активации рецептора. В качестве основного компонента передачи сигнала в этих рецепторах βc приводит к активации JAK-2, STAT-5 и других сигнальных молекул и в конечном счете приводит к избыточной клеточной активности, в нормальных условиях ассоциированной с IL-5, стимуляцией GM-CSF и IL-3, такой как адгезия эозинофилов, что приводит к дегрануляции и цитотоксичности, а также к продлению жизнеспособности клеток.
[0005] Исследователями было опробовано три основных способа блокирования цитокинов, активирующих эозинофилы, или оказания антагонистического действия на них. Один из них предусматривает применение антител к вовлеченным цитокинам. Например, антитело к IL-5 применяется в животной модели аллерген-индуцированной астмы. Данный способ продемонстрировал относительно длительный эффект предупреждения сильного ответа вследствие притока эозинофилов в дыхательные пути и бронхиолы. Однако по-прежнему существует недостаток высокоаффинных IL-5-специфичных антител или лекарственных средств, нацеленных на IL-5. С учетом вышеизложенного представлено настоящее изобретение.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0006] Целью настоящего изобретения является обеспечение IL-5-связывающей молекулы, а также ее способа получения и применения.
[0007] Настоящее изобретение реализуется следующим образом.
[0008] В соответствии с первым аспектом в варианте осуществления настоящего изобретения предложена IL-5-связывающая молекула, способная специфично связываться с IL-5 и содержащая по меньшей мере один из одиночных вариабельных доменов иммуноглобулина, содержащих определяющие комплементарность области CDR1, CDR2 и CDR3.
[0009] Аминокислотная последовательность CDR1 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 43-49; аминокислотная последовательность CDR2 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 50-56; и аминокислотная последовательность CDR3 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 57-62.
[0010] В соответствии со вторым аспектом в варианте осуществления настоящего изобретения предложена выделенная нуклеиновая кислота, кодирующая IL-5-связывающую молекулу согласно предыдущему варианту осуществления.
[0011] В соответствии с третьим аспектом в варианте осуществления настоящего изобретения предложен рекомбинантный вектор, содержащий выделенную нуклеиновую кислоту согласно предыдущему варианту осуществления.
[0012] В соответствии с четвертым аспектом в варианте осуществления настоящего изобретения предложена клетка-хозяин, содержащая рекомбинантный вектор согласно предыдущему варианту осуществления.
[0013] В соответствии с пятым аспектом в варианте осуществления настоящего изобретения предложен способ получения IL-5-связывающей молекулы, включающий культивирование клетки-хозяина согласно предыдущему варианту осуществления с получением IL-5-связывающей молекулы.
[0014] В соответствии с шестым аспектом в варианте осуществления настоящего изобретения предложен конъюгат для связывания белка IL-5, содержащий конъюгирующий компонент и IL-5-связывающую молекулу согласно предыдущим вариантам осуществления, где
[0015] конъюгирующий компонент конъюгирован с IL-5-связывающей молекулой; и конъюгирующий компонент включает в себя маркер и/или соединение для выявления.
[0016] В соответствии с седьмым аспектом в варианте осуществления настоящего изобретения предложен набор для выявления IL-5, содержащий IL-5-связывающие молекулы согласно предыдущему варианту осуществления.
[0017] В соответствии с восьмым аспектом в варианте осуществления настоящего изобретения предложено применение IL-5-связывающей молекулы согласно вышеописанным вариантам осуществления при получении лекарственных средств, нацеленных на IL-5, для лечения заболевания.
[0018] Настоящее изобретение имеет следующие благоприятные эффекты.
[0019] В вариантах осуществления настоящего изобретения предложена IL-5-связывающая молекула, а также ее способ получения и применение. Данная связывающая молекула способна специфично связываться с IL-5 и содержит по меньшей мере один из одиночных вариабельных доменов иммуноглобулина, содержащих определяющие комплементарность области CDR1, CDR2 и CDR3. Аминокислотная последовательность CDR1 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 43-49; аминокислотная последовательность CDR2 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 50-56; и аминокислотная последовательность CDR3 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 57-62. Данная связывающая молекула способна специфично связываться с IL-5 и эффективно блокировать клеточную пролиферацию, индуцируемую IL-5, и ее можно применять при предупреждении, диагностике и/или лечении заболеваний, связанных с IL-5.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0020] Для более четкого описания технических решений вариантов осуществления настоящего изобретения ниже приведено краткое описание сопутствующих графических материалов, необходимых для описания вариантов осуществления. Следует понимать, что на следующих сопутствующих графических материалах показаны только некоторые варианты осуществления настоящего изобретения, которые не следует считать ограничивающими объем настоящего изобретения. Средний специалист в данной области все равно может вывести другие сопутствующие графические материалы на основе этих сопутствующих графических материалов без творческих усилий.
[0021] На фиг. 1 показаны результаты гель-электрофореза рекомбинантного белка IL-5 человека из примера 1;
[0022] на фиг. 2 показано обогащение рекомбинантным белком IL-5 после скрининга библиотеки наноантител из примера 1; P/N = число моноклональных бактерий, выросших после инфицирования бактерий TG1 фагами, элюированными из положительных лунок в ходе биопэннинга/число моноклональных бактерий, выросших после инфицирования бактерий TG1 фагами, элюированными из отрицательных лунок, где данный параметр постепенно увеличивается в ходе обогащения; I/E = общее количество бактериофагов, добавленных в положительные лунки в каждом цикле биопэннинга/общее количество фагов, элюированных из положительных лунок в ходе биопэннинга, где данный параметр постепенно приближается к 1 в ходе обогащения;
[0023] на фиг. 3 показаны соответствующие результаты выравнивания гуманизированных вариантов антител клонов 1B3 и 2B3 из примера 2;
[0024] на фиг. 4 показаны результаты анализа связывания 12 клонов антител, экспрессируемых в E. coli, полученных в примере 1, с IL-5 согласно проверочному примеру 1;
[0025] на фиг. 5 изображен график, на котором показана зависимость «доза-эффект» для связывания Tab1 и Tab2 с IL-5 из проверочного примера 2;
[0026] на фиг. 6 изображен график, на котором показана зависимость «доза-эффект» для индуцируемой IL-5 пролиферации клеток TF-1, нейтрализуемой контрольным антителом 1 (Tab1) и контрольным антителом 2 (Tab2) в проверочном примере 2;
[0027] на фиг. 7 изображен график, на котором показана зависимость «доза-эффект» для индуцируемой IL-5 пролиферации TF-1, нейтрализуемой IL-5-специфичным слитым с Fc однодоменным антителом, полученным в примере 3, из проверочного примера 3; и
[0028] на фиг. 8 изображен график, на котором показана зависимость «доза-эффект» для индуцируемой IL-5 пролиферации TF-1, нейтрализуемой различными гуманизированными слитыми с Fc однодоменными антителами, полученными в примере 3, из проверочного примера 3.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[0029] Для более ясного понимания целей, технических решений и преимуществ вариантов осуществления настоящего изобретения далее приводится четкое и полное описание технических решений в вариантах осуществления настоящего изобретения. Если в примерах не приведены конкретные условия, то следуют общепринятым условиям или условиям, рекомендуемым производителем. Все используемые реагенты и приборы, для которых не указаны производители, являются широко распространенными продуктами, коммерчески доступными на рынке.
[0030] Определения терминов
[0031] В данном документе «однодоменное антитело» (sdAb), также известное как наноантитело, означает антитело, в естественных условиях не имеющее легких цепей и содержащее только один вариабельный домен тяжелой цепи (VHH).
[0032] В данном документе «гуманизированное антитело» относится к антителу, полученному путем слияния вариабельного домена тяжелой цепи целевого антитела (такого как антитело животного) с константным доменом антитела человека, либо антителу, полученному путем пересадки определяющей комплементарность области (последовательностей CDR 1-3) целевого антитела в вариабельную область антитела человека, либо антителу, полученному путем осуществления аминокислотной мутации в целевом антителе в соответствии с характеристиками каркасной области (FR1-4) антитела человека. Гуманизированное антитело может быть получено путем синтеза или сайт-направленного мутагенеза.
[0033] В данном документе «диатело» означает небольшой двухвалентный и биспецифический фрагмент антитела, способный одновременно распознавать два антигена. Hollinger и др. с помощью коротких пептидных молекул соединяли гены вариабельного домена легкой цепи антитела к антигену A (VLA) и вариабельного домена тяжелой цепи антитела к антигену B (VHB); а также соединяли VHA и VLB и вставляли две группы химерных генов в бицистронную экспрессионную плазмиду для конструирования плазмиды для экспрессии диатела. После экспрессии VLA-VHB сшивали с VHA-VLB с образованием биспецифического антитела. В настоящем изобретении одним из антигенов, которые способно распознавать двухвалентное антитело, является белок IL-5, а другой антиген, который способно распознавать двухвалентное антитело, может быть выбран из любого из существующих антигенов.
[0034] В данном документе «поливалентное антитело», также называемое полиантителом, относится к антителу с модифицированной структурой (сходной с диателом), которое способно одновременно распознавать различные антигены (аналогично диателу). В настоящем изобретении одним из антигенов, которые способно распознавать поливалентное антитело, является белок IL-5.
[0035] «CDR», упоминаемая в настоящем описании, означает определяющую комплементарность область антитела. Антитело обычно содержит две вариабельные области - вариабельный домен тяжелой цепи и вариабельный домен легкой цепи. Вариабельный домен тяжелой цепи или вариабельный домен легкой цепи обычно содержит три CDR.
[0036] Варианты осуществления
[0037] В варианте осуществления настоящего изобретения предложена IL-5-связывающая молекула. IL-5-связывающая молекула способна специфично связываться с IL-5 и содержит по меньшей мере один из одиночных вариабельных доменов иммуноглобулина, содержащих определяющие комплементарность области CDR1, CDR2 и CDR3;
[0038] где аминокислотная последовательность CDR1 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 43-49; аминокислотная последовательность CDR2 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 50-56; и аминокислотная последовательность CDR3 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 57-62.
[0039] Предпочтительно аминокислотные последовательности определяющих комплементарность областей CDR1, CDR2 и CDR3 приведены в любом из (1)-(13):
[0040] (1) SEQ ID NO: 43, 56 и 58;
[0041] (2) SEQ ID NO: 49, 51 и 60;
[0042] (3) SEQ ID NO: 47, 56 и 57;
[0043] (4) SEQ ID NO: 45, 54 и 57;
[0044] (5) SEQ ID NO: 46, 50 и 61;
[0045] (6) SEQ ID NO: 47, 56 и 62;
[0046] (7) SEQ ID NO: 48, 56 и 57;
[0047] (8) SEQ ID NO: 45, 53 и 57;
[0048] (9) SEQ ID NO: 44, 55 и 57;
[0049] (10) SEQ ID NO: 43, 56 и 59;
[0050] (11) SEQ ID NO: 43, 52 и 59;
[0051] (12) SEQ ID NO: 47, 56 и 59; а также
[0052] (13) SEQ ID NO: 47, 52 и 59.
[0053] В необязательном варианте осуществления одиночные вариабельные домены иммуноглобулина дополнительно содержат каркасную область, содержащую FR1, FR2, FR3 и FR4. Структура однодоменного антитела является следующей: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
[0054] Аминокислотная последовательность FR1 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 63-68; аминокислотная последовательность FR2 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 69-72; аминокислотная последовательность FR3 выбрана из любой из последовательностей, приведенных под SEQ ID NO: 73-86; а аминокислотная последовательность FR4 приведена под SEQ ID NO: 87.
[0055] Предпочтительно каркасные области FR1, FR2 и FR3 одиночных вариабельных доменов иммуноглобулина имеют последовательности, приведенные в любом из (14)-(28):
[0056] (14) SEQ ID NO: 65, 72 и 85;
[0057] (15) SEQ ID NO: 65, 72 и 82;
[0058] (16) SEQ ID NO: 64, 71 и 86;
[0059] (17) SEQ ID NO: 65, 72 и 73;
[0060] (18) SEQ ID NO: 65, 72 и 84;
[0061] (19) SEQ ID NO: 66, 69 и 83;
[0062] (20) SEQ ID NO: 68, 72 и 76;
[0063] (21) SEQ ID NO: 66, 69 и 79;
[0064] (22) SEQ ID NO: 63, 72 и 85;
[0065] (23) SEQ ID NO: 67, 70 и 80;
[0066] (24) SEQ ID NO: 65, 72 и 78;
[0067] (25) SEQ ID NO: 65, 72 и 77;
[0068] (26) SEQ ID NO: 67, 70 и 81;
[0069] (27) SEQ ID NO: 65, 72 и 74; а также
[0070] (28) SEQ ID NO: 65, 72 и 75.
[0071] Предпочтительно одиночный вариабельный домен иммуноглобулина представляет собой VHH.
[0072] Предпочтительно аминокислотная последовательность VHH выбрана из любой из последовательностей, приведенных под SEQ ID NO: 1-12.
[0073] Предпочтительно VHH представляет собой гуманизированный VHH.
[0074] Предпочтительно последовательность гуманизированного VHH выбрана из любой из последовательностей, приведенных под SEQ ID NO: 13-21.
[0075] Предпочтительно связывающая молекула дополнительно содержит Fc-область иммуноглобулина, соединенную с VHH.
[0076] Предпочтительно Fc-область иммуноглобулина представляет собой Fc-область иммуноглобулина человека.
[0077] Предпочтительно Fc-область иммуноглобулина человека представляет собой Fc-область IgG4 человека.
[0078] В варианте осуществления настоящего изобретения предложена выделенная нуклеиновая кислота, кодирующая IL-5-связывающую молекулу согласно любому из предыдущих вариантов осуществления.
[0079] Предпочтительно последовательность нуклеиновой кислоты выбрана из любой из последовательностей, приведенных под SEQ ID NO: 22-42.
[0080] В варианте осуществления настоящего изобретения предложен рекомбинантный вектор, содержащий выделенную нуклеиновую кислоту согласно любому из предыдущих вариантов осуществления. В необязательном варианте осуществления рекомбинантный вектор может представлять собой плазмиду, бактериофаг или вирусный вектор.
[0081] В варианте осуществления настоящего изобретения предложена клетка-хозяин, содержащая рекомбинантный вектор согласно предыдущему варианту осуществления. В альтернативном варианте осуществления клетка-хозяин может являться прокариотической клеткой или эукариотической клеткой.
[0082] В варианте осуществления настоящего изобретения предложен способ получения IL-5-связывающей молекулы, включающий культивирование клетки-хозяина согласно предыдущему варианту осуществления с получением IL-5-связывающей молекулы.
[0083] Следует отметить, что связывающая молекула, описанная в любом из приведенных выше вариантов осуществления, может быть получена путем искусственного синтеза или может быть получена путем вначале синтеза кодирующего ее гена, а затем осуществления биологической экспрессии.
[0084] В варианте осуществления настоящего изобретения предложен конъюгат для связывания белка IL-5, содержащий конъюгирующий компонент и IL-5-связывающую молекулу согласно любому из предыдущих вариантов осуществления. Конъюгирующий компонент конъюгирован с IL-5-связывающей молекулой; и конъюгирующий компонент включает в себя маркер и/или соединение для выявления.
[0085] Предпочтительно маркер для выявления представляет собой радиоактивный элемент.
[0086] В варианте осуществления настоящего изобретения предложен набор для выявления IL-5, содержащий IL-5-связывающую молекулу согласно любому из предыдущих вариантов осуществления.
[0087] В дополнение, в варианте осуществления настоящего изобретения предложено применение IL-5-связывающей молекулы при получении лекарственных средств, нацеленных на IL-5, для лечения заболевания.
[0088] Предпочтительно заболевание выбрано из любого из астмы, аллергического дерматита, экземы, артрита, герпеса, хронической первичной крапивницы, склеродермии, гипертрофических рубцов, хронического обструктивного заболевания легких, атопического дерматита, идиопатического легочного фиброза, болезни Кавасаки, серповидноклеточной анемии, болезни Грейвса, синдрома Шегрена, аутоиммунного лимфопролиферативного синдрома, аутоиммунной гемолитической анемии, пищевода Барретта, аутоиммунного увеита, туберкулеза и заболевания почек.
[0089] Ниже проводится дополнительное подробное описание признаков и характеристик настоящего изобретения со ссылкой на примеры.
[0090] Пример 1
[0091] Получение однодоменного антитела к белку IL-5
[0092] A. Конструирование вектора для экспрессии рекомбинантного белка IL-5 человека
[0093] Последовательность, кодирующую IL-5, получали путем информационного поиска в NCBI, и ее номером доступа является NM_000879.2. Кодирование кодирующей последовательностью обеспечивало получение аминокислотной последовательности с номером доступа NP_000870.1. В аминокислотной последовательности, соответствующей NP_000870.1, на веб-сайтах ™HMM и SMART соответственно проводился анализ в отношении трансмембранных областей и внеклеточных концов. Результаты анализа продемонстрировали, что белок IL-5 является секреторным белком без трансмембранной области и имеет 134 аминокислоты в своей полной длине с сигнальным пептидом в положениях аминокислот с номерами 1-19. Посредством синтеза генов нуклеотидную последовательность, кодирующую аминокислоты с номерами 20-134 белка IL-5, клонировали в вектор pcDNA3.4. Сконструированный вектор подвергали секвенированию по Сэнгеру и выравнивали с исходной последовательностью. При успешном подтверждении рекомбинантные плазмиды экстрагировали партиями, чтобы удалить из них эндотоксины, и суспендированные клетки 293F трансфицировали для экспрессии и очистки целевого белка. Результаты анализа очищенного рекомбинантного белка IL-5 человека методом SDS-PAGE показаны на фиг. 1 (где маркер - градиент молекулярной массы стандартного белка; hrIL-5-cHis - рекомбинантный белок IL-5 человека с гистидиновой меткой на карбокси-конце).
[0094] Как видно из фиг. 1, чистота экспрессированного и очищенного рекомбинантного белка IL-5 человека составляет приблизительно 90%. Полученный в данном примере белок IL-5 применяется для иммунизации верблюдов и скрининга антител.
[0095] B. Конструирование библиотеки однодоменных антител к белку IL-5
[0096] 600 мкг очищенного рекомбинантного белка IL-5 человека, полученного на стадии A, смешивали с полным адъювантом Фрейнда в равном объеме для иммунизации алашаньского двугорбого верблюда из Внутренней Монголии один раз в неделю в общей сложности семь раз. За исключением первой иммунизации, остальные шесть иммунизаций проводили с использованием смеси 300 мкг рекомбинантного белка IL-5 и неполного адъюванта Фрейнда в равном объеме для продуцирования у верблюда антитела к IL-5.
[0097] После иммунизации у верблюда отбирали 100 мл периферической крови, и из лимфоцитов экстрагировали РНК. Экстрагированную общую РНК использовали для синтеза кДНК, и вариабельную область тяжелой цепи антитела (VHH) амплифицировали посредством вложенной ПЦР с кДНК в качестве матрицы. Затем вектор pMECS и амплифицированный VHH-фрагмент расщепляли соответственно с применением рестрикционной эндонуклеазы, и расщепленные фрагменты и вектор соединяли; соединенные фрагменты вводили путем трансформации в компетентные клетки TG1 для конструирования фаг-дисплейной библиотеки белка IL-5, и объем библиотеки (рекомбинантных клеток TG1) определяли как составляющий приблизительно 1×109.
[0098] C. Скрининг однодоменного антитела к белку IL-5
[0099] 200 мкл рекомбинантных клеток TG1, полученных на стадии B, инокулировали в 2× среду TY для инкубирования. В ходе инкубирования добавляют 40 мкл фага-помощника VCSM13 для инфицирования клеток TG1, и их культивируют в течение ночи для амплификации фага. На следующий день фаг осаждали с помощью PEG/NaCl, и амплифицированный фаг собирали путем центрифугирования с получением библиотеки амплифицированных фагов.
[0100] 500 мкг белка IL-5, разбавленного в NaHCO3 (100 мМ, pH 8,3), наносили на поверхность планшета для ELISA при 4°C на ночь. Также были представлены лунки отрицательного контроля. В день 2 добавляют 200 мкл 3% обезжиренного молока для блокирования при комнатной температуре в течение 2 ч. После блокирования добавляют 100 мкл библиотеки амплифицированных фагов (приблизительно 2×1011 фаговых частиц) для воздействия при комнатной температуре в течение 1 ч. После 1 часа воздействия планшет промывают пять раз с помощью PBS и 0,05% Tween-20, чтобы смыть несвязавшиеся фаги. Фаг, специфично связывающийся с белком IL-5, подвергали диссоциации с помощью трипсина в конечной концентрации 2,5 мг/мл для инфицирования клеток TG1 E. coli в фазе логарифмического роста. Инфицированные клетки TG1 E. coli культивировали при 37°C в течение 1 ч. для продуцирования и сбора фагов для следующего скрининга. Тот же самый процесс скрининга повторяли в течение еще одного цикла для обеспечения постепенного обогащения. Для кратности обогащения, составляющей более 10x, эффект обогащения показан на фиг. 2 и в таблице 1.
[0101] Из результатов можно видеть, что для обогащения в цикле 3 P/N составляет 2500, а I/E составляет 4.
[0102] D. Скрининг конкретных положительных клонов против IL-5 методом ELISA бактериофагов.
[0103] Если кратность обогащения составляет более 10, из подвергнутых скринингу положительных клонов отбирают 400 отдельных колоний и инокулируют в среду TB, содержащую 100 мкг/мл ампициллина, в 96-луночном планшете с глубокими лунками. Также был представлен холостой контроль. Содержимое планшета культивировали при 37°C до логарифмической фазы, и затем добавляли IPTG в конечной концентрации 1 мМ для культивирования в течение ночи при 28°C.
[0104] Неочищенный экстракт антитела получали путем осмотического взрыва. Рекомбинантный белок IL-5 человека разбавляли в NaHCO3 (100 мМ pH 8,3), и 100 мкг рекомбинантного белка IL-5 наносили на поверхность планшета для ELISA при 4°C на ночь. 100 мкл полученного неочищенного экстракта антитела переносили на планшет для ELISA, в который был добавлен антиген (рекомбинантный белок IL-5), и инкубировали при комнатной температуре в течение 1 ч. Несвязавшиеся антитела смывали с помощью PBST. Добавляли 100 мкл антител мыши к HA-метке (Thermo Fisher) в разведении 1:2000 и инкубировали при комнатной температуре в течение 1 ч. Несвязавшиеся антитела смывали с помощью PBST. Добавляли 100 мкл антитела к иммуноглобулину кролика, конъюгированного с HRP (антитела козы к иммуноглобулину кролика, меченного пероксидазой хрена, Thermo Fisher), в разведении 1:20000, и инкубировали при комнатной температуре в течение 1 ч. Несвязавшиеся антитела смывали с помощью PBST. Добавляли раствор для проявления цвета пероксидазы хрена для осуществления реакции при 37°C в течение 15 мин., добавляли останавливающий раствор, и измеряли поглощение при длине волны 450 нм в микропланшет-ридере. Если оптическая плотность (OD) в лунках с образцом в более чем 5 раз превышает таковую в контрольных лунках, то лунки с образцом определяют как лунки с положительными клонами. Бактерии из лунок с положительными клонами переносили в среду LB, содержащую 100 мкг/мкл ампициллина, на шейкере для экстракции и секвенирования плазмид. Последовательности генов каждого клона анализировали согласно программному обеспечению для выравнивания последовательностей Vector NTI. Клоны с одинаковыми последовательностями CDR1, CDR2 и CDR3 считаются одним и тем же клоном, а клоны с разными последовательностями считаются разными клонами, и в конечном счете получают однодоменное антитело, специфичное к белку IL-5. Однодоменное антитело имеет аминокислотную последовательность FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, которая образует целый VHH. Полученная рекомбинантная плазмида для экспрессии однодоменного антитела (положительная плазмида, целевая последовательность) может экспрессироваться в прокариотической системе с получением в конечном счете белкового однодоменного антитела.
[0105] E. Очистка и экспрессия однодоменного антитела, специфичного к белку IL-5, в клетках-хозяевах E. coli.
[0106] Положительные плазмиды (pMECS-VHH) различных клонов, полученные посредством анализа методом секвенирования на стадии D, подвергали электропорации в HB2151 E. coli, распределяли по культуральной чашке с LB+ампициллин+глюкоза (т.е. содержащей ампициллин и глюкозу) и инкубировали при 37°C в течение ночи. Отдельную колонию отбирали и инокулировали в 5 мл среды LB, содержащей ампициллин, и инкубировали в шейкере при 37°C в течение ночи. 1 мл суточной культуры инокулировали в 330 мл среды TB и инкубировали в шейкере при 37°C. Поглощение измеряли при длине волны 600 нм с помощью спектрометра и регистрировали как OD600. Если измеряемое значение OD600 находилось в диапазоне 0,6-0,9, то добавляли 1M IPTG для инкубирования в шейкере при 28°C в течение ночи. E. coli собирали путем центрифугирования. Неочищенный экстракт антитела получали путем осмотического взрыва. Антитело очищали посредством аффинной хроматографии на колонке с Ni. Очищенные однодоменные антитела показаны в таблице 2.
[0107] Примечание: в таблице 2 числа, соответствующие CDR 1-3 и FR 1-4, являются их SEQ ID NO.
[0108] Пример 2
[0109] Гуманизация однодоменного антитела к IL-5
[0110] Гуманизацию осуществляли путем гуманизации аминокислот на поверхности белка и пересадки универсальной каркасной области для гуманизации однодоменных антител.
[0111] Гуманизацию аминокислот на поверхности белка проводили в следующих стадиях. Гомологичное моделирование проводили для клонов антител 1B3 и 2B3 с использованием программного обеспечения для моделирования Discovery Studio с антителом NbBcII10 (номер в PDB: 3DWT), имеющим гомологичную последовательность, в качестве эталона, и относительную доступность аминокислот для растворителя рассчитывали согласно трехмерной структуре белка.
[0112] Пересадку универсальной каркасной области для гуманизации VHH проводили в следующих конкретных стадиях. Вначале разрабатывали и получали универсальную каркасную область h-NbBcII10FGLA (№ доступа в PDB: 3EAK) для гуманизации VHH в соответствии с гомологией последовательностей и на основе наноантитела NbBcII10 (№ доступа в PDB: 3DWT) путем гуманизации аминокислот на поверхности белка со ссылкой на гуманизированное антитело DP-47 и модификации некоторых аминокислот каркасной области 2 VHH как FGLA. В качестве каркасной области непосредственно используется h-NbBcII10FGLA, в котором CDR заменяются на CDR клонов антител 1B3 и 2B3 соответственно с достижением гуманизации антитела.
[0113] Клоны антител 1B3 и 2B3 были гуманизированы с получением пяти вариантов гуманизированных клонов антител. В таблице 3 и таблице 4 показаны номера последовательностей и изменения аминокислот для этих гуманизированных вариантов. Аминокислотные остатки были пронумерованы в соответствии с системой нумерации по Kabat. Знаком « » в таблице 3 и таблице 4 указано, что в данном положении в клоне наноантитела имеется мутация (замена). В частности, например, «S11L» означает, что в положении 11 S (серин) заменен на L (лейцин); и «S66T» означает, что в положении 66 S (серин) заменен на T (треонин). На фиг. 3 показаны результаты выравнивания гуманизированных последовательностей.
» в таблице 3 и таблице 4 указано, что в данном положении в клоне наноантитела имеется мутация (замена). В частности, например, «S11L» означает, что в положении 11 S (серин) заменен на L (лейцин); и «S66T» означает, что в положении 66 S (серин) заменен на T (треонин). На фиг. 3 показаны результаты выравнивания гуманизированных последовательностей.
[0114] Пример 3
[0115] F. Конструирование вектора экспрессии в эукариотических клетках для слитого белка на основе Fc и однодоменного антитела к белку IL-5
[0116] Нуклеотидную последовательность однодоменного антитела, подвергнутого скринингу на стадии D в примере 1, получали путем секвенирования по Сэнгеру. Вышеуказанную нуклеотидную последовательность, подвергнутую оптимизации кодонов посредством синтеза последовательности, вставляли в модифицированный вектор RJK-V4-hFC.
[0117] RJK-V4-hFC представляет собой универсальный целевой вектор для экспрессии наноантитела, полученный путем слияния Fc-фрагмента последовательности, кодирующей тяжелую цепь (номер доступа в NCBI: AB776838.1) IgG человека, с коммерческим вектором pCDNA3.4 (информация о векторе доступна по адресу https://assets.thermofisher.com/TFS-Assets/LSG/manuals/pcdna3_4_topo_ta_cloning_kit_man.pdf) от Invitrogen. Данный вектор содержит шарнирные области, CH2 и CH3 тяжелой цепи IgG. Ниже показана специфическая модификация.
[0118] В pcDNA3.4 были выбраны сайты разрезания рестрикционными ферментами XbaI и AgeI. Сайт множественного клонирования (MCS) и 6×His-метку вводили соответственно на 5’- и 3’-концы последовательности, кодирующей Fc-фрагмент, посредством ПЦР с перекрывающимися праймерами. Вышеописанный фрагмент амплифицировали посредством ПЦР с использованием пары праймеров с сайтами разрезания ферментами XbaI и AgeI соответственно. pcDNA3.4 и амплифицированный фрагмент с сайтами разрезания ферментами XbaI и AgeI из праймеров расщепляли соответственно с применением рестрикционных эндонуклеаз XbaI и AgeI. Расщепленный вектор и вставленный фрагмент соединяли с помощью лигазы T4, после чего продукт, полученный в результате такого соединения, вводили путем трансформации в E. coli, амплифицировали и подтверждали путем секвенирования с получением рекомбинантного вектора экспрессии в эукариотических клетках.
[0119] Сконструированный рекомбинантный вектор экспрессии в эукариотических клетках вводили путем трансформации в DH5α E. coli и инкубировали для экстракции плазмид в максимасштабе и удаления эндотоксинов. Плазмиду, экстрагированную в максимасштабе, секвенировали для идентификации. Подтвержденный рекомбинантный вектор использовали для последующей трансфекции и экспрессии в эукариотических клетках.
[0120] G. Экспрессия слитого белка на основе Fc и однодоменного антитела, специфичного к белку IL-5, в суспендированных клетках ExpiCHO-S.
[0121] За 3 дня до трансфекции пассировали и размножали клетки ExpiCHO-S™ в концентрации 2,5×105/мл. Клетки в рассчитанном требуемом объеме переносили во встряхиваемую колбу на 500 мл, содержащую 120 мл (конечный объем) среды для экспрессии ExpiCHO™. Клетки инкубировали до концентрации, составляющей приблизительно от 4×106 до 6×106 живых клеток/мл. За день до трансфекции клетки ExpiCHO-S™ разбавляли до 3,5×106 живых клеток/мл и культивировали в течение ночи. В день трансфекции определяли плотность клеток и процентную долю живых клеток. Перед трансфекцией плотность клеток должна достигать приблизительно от 7×106 до 10×106 живых клеток/мл. Клетки разбавляли до 6×106 живых клеток/мл в свежей среде для экспрессии ExpiCHO™, предварительно нагретой до 37°C. Клетки в рассчитанном требуемом объеме переносили во встряхиваемую колбу на 500 мл, содержащую 100 мл (конечный объем) свежей предварительно нагретой среды для экспрессии ExpiCHO™. Реагент ExpiFectamine™CHO равномерно перемешивали путем осторожного переворачивания вверх дном и разбавляли в 3,7 мл среды OptiPRO™, встряхиваемой или перемешиваемой равномерно. Плазмидную ДНК (полученную на стадии F) разбавляли в 4 мл охлажденной среды OptiPRO™, встряхивали и перемешивали равномерно. Смесь ExpiFectamine CHO/плазмидная ДНК инкубировали при комнатной температуре в течение 3 мин., а затем осторожно добавляли к полученной суспензии клеток, в ходе чего встряхиваемую колбу осторожно встряхивали. Клетки культивировали со встряхиванием на увлаженном воздухе с 8% CO2 при 37°C. В день 1 после трансфекции (спустя 18-22 часа) добавляли 600 мкл усилителя ExpiFectamine™CHO и 24 мл подпитки ExpiCHO. Через приблизительно 8 дней после трансфекции (когда жизнеспособность клеток составляла менее 70%) собирали надосадочную жидкость.
[0122] H. Экспрессия слитого белка на основе Fc и однодоменного антитела к белку IL-5 в суспендированных клетках 293F.
[0123] За 3 дня до трансфекции пассировали и размножали клетки 293F в концентрации 2,5×105/мл. Клетки в рассчитанном требуемом объеме переносили во встряхиваемую колбу на 500 мл, содержащую 120 мл (конечный объем) свежей предварительно нагретой среды OPM-293 CD05. Клетки инкубировали до концентрации, составляющей приблизительно от 2×106 до 3×106 живых клеток/мл. В день трансфекции определяли плотность клеток и процентную долю живых клеток. Перед трансфекцией плотность клеток должна достигать приблизительно от 2×106 до 3×106 живых клеток/мл. Клетки разбавляли до концентрации 1×106 живых клеток/мл в предварительно нагретой среде OPM-293 CD05. Клетки в рассчитанном требуемом объеме переносили во встряхиваемую колбу на 500 мл, содержащую 100 мл (конечный объем) свежей предварительно нагретой среды. Реагент PEI (1 мг/мл) разбавляли в 4 мл среды Opti-MEM и встряхивали или перемешивали равномерно путем пипетирования. Плазмидную ДНК (полученную на стадии F) разбавляли в 4 мл среды Opti-MEM, встряхивали и перемешивали равномерно, фильтровали с помощью головки фильтра с размером пор 0,22 мкм и инкубировали при комнатной температуре в течение 5 мин. Разбавленный реагент PEI добавляли к разбавленной ДНК и переворачивали вверх дном для равномерного перемешивания. Смесь PEI/плазмидная ДНК инкубировали при комнатной температуре в течение 15-20 мин., а затем осторожно добавляли к полученной суспензии клеток, в ходе чего встряхиваемую колбу осторожно встряхивали. Клетки культивировали со встряхиванием при 37°C в 5% CO2 при 120 об./мин. Через 24 ч. и 72 ч. после трансфекции добавляли 5 мл подпитки OPM-CHO PFF05. Через приблизительно 7 дней после трансфекции (когда жизнеспособность клеток составляла менее 70%) собирали надосадочную жидкость.
[0124] F. Очистка слитого белка на основе Fc и однодоменного антитела к белку IL-5
[0125] Полученную на стадии G или H надосадочную жидкость с экспрессируемым слитым белком на основе Fc фильтровали с помощью одноразовой головки фильтра с размером пор 0,45 мкм для удаления нерастворимых примесей. Фильтрат очищали посредством аффинной хроматографии с использованием устройства для очистки белков с агарозным наполнителем, конъюгированным с белком A, по принципу связывания слитого белка на основе Fc человека с белком A. Фильтрату позволяли проходить через предварительно упакованную колонку с белком A при скорости потока 1 мл/мин. На данной стадии целевой белок в фильтрате связывается с наполнителем. Примесные белки, связывающиеся с колонкой, смывали с помощью низкосолевых и высокосолевых буферных растворов. Целевой белок, связывающийся с колонкой, элюировали буферным раствором с низким pH. Элюированный раствор быстро добавляли в раствор Tris-HCl с pH 9,0 для нейтрализации.
[0126] Нейтрализованный раствор белка подвергали диализу и затем подвергали анализу методом SDS-PAGE. Белок, имеющий чистоту 95% или больше и концентрацию 0,5 мг/мл или больше, был криоконсервирован и готов к применению.
[0127] Проверочный пример 1
[0128] Определение зависимости «доза-эффект» для связывания однодоменного антитела, специфичного к белку IL-5, представленного в примере 1
[0129] 50 мкл 1 мкг/мл белка IL-5 наносили на поверхность планшета для ELISA при 4°C на ночь. Планшет промывали. 200 мкл 5% молока добавляли для блокирования при 37°C в течение 1 ч. Однодоменное антитело (VHH), специфичное к белку IL-5, полученное в примере 1, разбавляли до 2 мкг/мл, а затем разбавляли с 5-кратным градиентом в общей сложности в 8 градиентах концентрации. Планшет промывали. 50 мкл антитела добавляли в дублирующие лунки и затем инкубировали при 37°C в течение 1 ч. Планшет промывали. Добавляли 50 мкл вторичного антитела мыши к HA-метке, конъюгированного с HRP, и затем инкубировали при 37°C в течение 30 мин. Планшет промывали (несколько раз). 50 мкл ™B, который был возвращен к комнатной температуре, добавляли для осуществления реакции при комнатной температуре в темноте в течение 15 мин. Добавляли 50 мкл останавливающего раствора (1 н. HCl). Результаты измеряли с помощью микропланшет-ридера и затем регистрировали. Кривую откладывали на графике. Концентрация для 50% от максимального эффекта (EC50), которая является концентрацией, вызывающей 50% от максимального эффекта, была рассчитана и показана на фиг. 4 и в таблице 5.
[0130] Проверочный пример 2
[0131] Экспрессия и очистка антитела Tool (Tab), нацеливающегося на белок IL-5 человека
[0132] Были получены Tab1 (контрольное антитело 1) и Tab2 (контрольное антитело 2). Tab1 представляет собой меполизумаб, имеющий последовательность, приведенную в патенте US7982005B2. Tab2 представляет собой реслизумаб, имеющий последовательность, приведенную в патенте US6056957.
[0133] Искомые последовательности Tab1 и Tab2 были переданы в General Biol (Anhui) Co., Ltd. для оптимизации кодонов в системе экспрессии в клетках млекопитающих и клонированы в вектор pcDNA3.1 соответственно.
[0134] Бактерии, положительные по плазмидам и отсеянные при скрининге в отношении устойчивости, амплифицировали, и плазмиды экстрагировали с применением набора для экстракции плазмид в среднем масштабе (Macherey Nagel, № по кат. 740412.50). На каждые 100 мл клеток добавляли 100 мкг плазмид (50 мкг тяжелых цепей и 50 мкг легких цепей), которые подвергали транзиентной трансфекции и экспрессировали в клетках 293F (среда для экспрессии: FreeStyle 293, Thermo, № по кат. 12338026+F-68, Thermo, № по кат. 24040032) с использованием PEI. Через 6-24 ч. после трансфекции добавляли 5% объем 10% пептона (Sigma, № по кат. P0521-100G) и культивировали в 8% CO2 при 130 об./мин. в течение приблизительно 7-8 дней. Когда жизнеспособность клеток падала до 50%, надосадочную жидкость с экспрессируемым белком собирали и очищали с помощью гравитационной колонки с белком A (GE, № по кат. 17-5438-02). После диализа против PBS определяли концентрацию с помощью NanoDrop, идентифицировали чистоту с помощью SEC и подтверждали связывание с помощью непрямого ELISA. Антитела Tab, полученные посредством данного способа, имеют концентрацию не менее 2 мг/мл и чистоту более 94%. EC50 для связывания с белком IL-5 (Novoprotein, № по кат. CS 33) показана на фиг. 5 и в таблице 6.
[0135] Эксперимент по пролиферации клеток TF1, индуцируемой рекомбинантным белком IL-5 человека и нейтрализуемой антителом Tool (Tab)
[0136] Эксперимент по пролиферации клеток TF1, индуцируемой рекомбинантным белком IL-5 человека. Клетки TF-1, пассированные в течение 3-4 пассажей после реанимации, инокулировали в 96-луночный планшет при 10000 клеток/лунка. Белок IL-5 человека составляли в виде раствора с максимальной концентрацией 500 нг/мл и разбавляли с 5-кратным градиентом. Разбавленный с градиентом раствор белка IL-5 добавляли в лунки для культивирования клеток в объеме, равном объему среды для культивирования клеток. После инкубирования в течение 72 ч. выявляли жизнеспособность клеток с помощью набора для люминесцентного анализа жизнеспособности клеток. EC80 для пролиферации клеток TF-1, индуцируемой IL-5, была рассчитана согласно результатам выявления. EC80 (концентрация для 80% от максимального эффекта) является концентрацией, вызывающей 80% от максимального эффекта.
[0137] Эксперимент по пролиферации клеток TF1, индуцируемой IL-5 человека и нейтрализуемой Tab. Клетки TF-1, пассированные в течение 3-4 пассажей после реанимации, инокулировали в 96-луночный планшет при 10000 клеток/лунка. Tab1 и Tab2 составляли в виде 10 мкг/мл раствора и разбавляли с 5-кратным градиентом. Разбавленное с градиентом Tab смешивали с IL-5 при концентрации EC80, полученной в эксперименте по пролиферации, в соотношении 1:1 с получением смешанного раствора. Смешанный раствор добавляли в лунки для культивирования клеток в объеме, равном объему среды для культивирования клеток. После инкубирования в течение 72 ч. выявляли жизнеспособность клеток с помощью набора для люминесцентного анализа жизнеспособности клеток. EC50 Tab1 и Tab2 для нейтрализации пролиферации клеток TF-1, индуцируемой IL-5, была рассчитана согласно результатам выявления и показана на фиг. 6 и в таблице 7.
[0138] Выявление нейтрализации пролиферации клеток TF-1, индуцируемой IL-5, под действием слитого белка на основе Fc и однодоменного антитела к белку IL-5
[0139] Клетки TF-1, пассированные в течение 3-4 пассажей после реанимации, инокулировали в 96-луночный планшет при 10000 клеток/лунка. Tab1, Tab2 и слитый белок на основе Fc и однодоменного антитела, представленные в примере 3-I, составляли в виде 10 мкг/мл раствора и разбавляли с 5-кратным градиентом. Разбавленное с градиентом Tab и однодоменное антитело смешивали соответственно с белком IL-5 при концентрации EC80, полученной в эксперименте по пролиферации, в соотношении 1:1 с получением смешанного раствора. Смешанный раствор добавляли в лунки для культивирования клеток в объеме, равном объему среды для культивирования клеток. После инкубирования в течение 72 ч. выявляли жизнеспособность клеток с помощью набора для люминесцентного анализа жизнеспособности клеток. EC50 различных однодоменных антител для нейтрализации пролиферации клеток TF-1, индуцируемой IL-5, была рассчитана согласно результатам выявления и показана на фиг. 7-8 и в таблицах 8-9.
[0140] Выявление кинетики аффинности IL-5-связывающей молекулы
Составление буферного раствора SD. Надлежащее количество бычьего сывороточного альбумина и Tween-20 растворяли в 1×PBS (pH 7,4), так чтобы массовые (или объемные) фракции бычьего сывороточного альбумина и Tween-20 составляли 0,1% и 0,02% соответственно. IL-5-связывающую молекулу составляли в буферном растворе SD до концентрации 10 мкг/мл. Составление рабочего раствора антигена: антиген составляли в буферном растворе SD до 200 нМ, а затем разбавляли с 2-кратным градиентом в общей сложности в 5 градиентах концентрации. В дополнение также был представлен холостой раствор в виде буферного раствора SD. Надлежащее количество 0,1 M исходного раствора глицина растворяли в 10 раз в деионизированной воде и равномерно перемешивали с получением регенерирующего раствора. Запускали Octet 96 и программное обеспечение для сбора данных на компьютере, поддерживающем работу с ними. Донную и боковые поверхности зонда для сбора данных очищали с помощью протирочной ткани для оптических стекол с надлежащим количеством 75% этанола. Прибор предварительно нагревали в течение 15 мин. или больше. Предварительное увлажнение датчика: датчик погружали в буферный раствор SD на 10 мин. или дольше перед анализом. Затем машинную процедуру настраивали в соответствии со следующим порядком: исходный уровень → антитело → исходный уровень → связывание антигена → диссоциация антигена → регенерация датчика для проведения анализа.
[0141] Специально тестируемые параметры кинетики аффинности IL-5-связывающей молекулы показаны в таблице 10.
[0142] KD - константа аффинности, выраженная в молях (M).
[0143] Ka - константа скорости ассоциации, выраженная величиной, обратной молям на секунду (1/М⋅с).
[0144] Kd - константа скорости диссоциации, выраженная величиной, обратной времени.
[0145] R2 - степень соответствия, иными словами, степень соответствия между кривой, построенной по результатам измерений, и аппроксимированной кривой. R2 ближе к 1 указывает на то, что подобранное значение ближе к измеренному значению, и в такой системе R2 должна составлять по меньшей мере > 0,95.
[0146] X2 - характеристика статистического параметра значений, измеренных в системе, которая должна составлять < 3, и меньшее измеренное значение является более надежным.
[0147] Другие значения погрешностей являются значениями погрешностей соответствующих параметров, которые должны быть более чем на порядок величины (в 10 раз) меньше параметров, которым они соответствуют.
[0148] Результаты продемонстрировали, что по сравнению с антителом Tool клон 1B3 обладает незначительно меньшей кинетической константой аффинности, а остальные клоны обладают незначительно большей кинетической константой аффинности, но среди тестируемых образцов не наблюдается значимое различие кинетических констант аффинности и отсутствует статистически значимое различие.
[0149] В вышеприведенном описании представлены лишь предпочтительные примеры согласно настоящему изобретению, которые не предполагаются как ограничивающие настоящее изобретение. Специалист в данной области может производить различные изменения и вариации настоящего изобретения. Любые модификации, эквивалентные замены или улучшения, выполненные в рамках сущности и принципа настоящего изобретения, должны находиться в пределах объема правовой охраны настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> РЕГЕНЕКОР БИОТЕХ КО., ЛТД
<120> IL-5-СВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ЕЕ ПРИМЕНЕНИЕ
<130> PCT/CN2021/077650
<150> 2020108435011
<151> 2020-08-20
<160> 87
<170> PatentIn, версия 3.5
<210> 1
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 1
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Ala Lys Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 2
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 2
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Gln Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Ala Lys Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 3
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 3
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Ala Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Leu Asp Leu Met
20 25 30
Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala Gly
35 40 45
Ile Tyr Ser Ser Gly Thr Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg
50 55 60
Phe Thr Val Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr Leu Gln Met
65 70 75 80
Asn Ser Leu Lys Pro Glu Asp Ala Ala Met Tyr Tyr Cys Ala Ala Lys
85 90 95
Val Ser Trp Tyr Gly Arg Trp Tyr Gln Ser Glu Thr Tyr Glu Tyr Trp
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 4
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 4
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Phe Ser Phe Ser
20 25 30
Thr Tyr Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu
35 40 45
Gly Ser Leu Ala Thr Ile Tyr Asp Gly Ser Thr Ala Tyr Ala Gly Ser
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu
65 70 75 80
Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr
85 90 95
Cys Ala Ala Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 5
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 5
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Lys Pro Ala Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Ala Lys Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 6
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 6
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ser Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Gly Asn Ile Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 7
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 7
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Leu Thr Leu Asn Asp Val
20 25 30
Ser Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Glu Val
35 40 45
Ala Gly Ile Asp Ile Asp Gly Leu Ile Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Asn Ala Gln Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Ser Cys Ala
85 90 95
Ala Val Arg Leu Tyr Phe Ser Ser Ser Arg Asn Gln Tyr Lys Tyr Glu
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 8
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 8
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Phe Ser Phe Ser
20 25 30
Thr Tyr Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu
35 40 45
Gly Ser Leu Ala Thr Ile Tyr Asp Gly Ser Thr Ala Tyr Ala Gly Ser
50 55 60
Val Lys Gly Arg Phe Thr Val Ser Val Asp Asn Ala Lys Asn Ile Leu
65 70 75 80
Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr
85 90 95
Cys Ala Ser Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 9
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 9
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Leu Thr Leu Asn Asp Val
20 25 30
Ser Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Glu Val
35 40 45
Ala Gly Ile Asp Ile Asp Gly Leu Ile Ser Tyr Ala Asp Pro Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Asn Ala Gln Asn Thr Met Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Met Tyr Ser Cys Ala
85 90 95
Ala Val Arg Leu Tyr Phe Ser Ser Ser Arg Asn Gln Tyr Lys Tyr Glu
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 10
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 10
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Phe Ser Thr Ser
20 25 30
Thr Tyr Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu
35 40 45
Gly Ser Leu Ala Thr Ile Tyr Asp Gly Ser Thr Ala Tyr Ala Gly Ser
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu
65 70 75 80
Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr
85 90 95
Cys Ala Ala Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 11
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 11
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ser Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Gly Ala Ile Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 12
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> вариабельный домен тяжелой цепи
<400> 12
Gln Val Gln Leu Val Glu Ser Glu Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Thr Ser Ser Thr Ala
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Gly Ser Ile Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 13
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 13
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Gly Leu Glu Ala Val
35 40 45
Ala Ala Thr Ile Tyr Asp Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 14
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 14
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Ala Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 15
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 15
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Ala Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 16
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 16
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Ala Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 17
<211> 120
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 17
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Ala Ser Thr Ser
20 25 30
Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser
35 40 45
Leu Ala Thr Ile Tyr Asp Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp Gly Gln
100 105 110
Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 18
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 18
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Phe Ser Phe Ser
20 25 30
Thr Tyr Cys Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Gly Leu Glu
35 40 45
Ala Val Ala Ala Thr Ile Tyr Asp Gly Ser Thr Tyr Tyr Ala Gly Ser
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu
65 70 75 80
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 19
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 19
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Phe Ser Phe Ser
20 25 30
Thr Tyr Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu
35 40 45
Gly Ser Leu Ala Thr Ile Tyr Asp Ala Ser Thr Ala Tyr Ala Gly Ser
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ile Leu
65 70 75 80
Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 20
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 20
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Phe Ser Phe Ser
20 25 30
Thr Tyr Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu
35 40 45
Gly Ser Leu Ala Thr Ile Tyr Asp Ala Ser Thr Ala Tyr Ala Gly Ser
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ala Lys Asn Ile Leu
65 70 75 80
Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp Thr Ala Met Tyr Tyr
85 90 95
Cys Ala Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 21
<211> 122
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> гуманизированный VHH
<400> 21
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Asn Thr Phe Ser Phe Ser
20 25 30
Thr Tyr Cys Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu
35 40 45
Gly Ser Leu Ala Thr Ile Tyr Asp Ala Ser Thr Ala Tyr Ala Gly Ser
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ile Leu
65 70 75 80
Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp
100 105 110
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 22
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 22
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atggcagcac cagctatgcg 180
gatagcgtga aaggccgttt taccattagc gtggataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgaaacc ggaagatacc gcgatgtatt attgcgcggc ggcgaaatat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 23
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 23
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atggcagcac cagctatgcg 180
gatagcgtga aaggccagtt taccattagc gtggataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgaaacc ggaagatacc gcgatgtatt attgcgcggc ggcgaaatat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 24
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 24
caggtgcagc tggtggaaag cggcggcggc agcgcgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa caccctggat ctgatggcgt ggtttcgtca ggcgccgggc 120
aaagaacgtg aaggcgtggc gggcatttat agcagcggca ccacctatta tgcggatagc 180
gtgaaaggcc gttttaccgt gagccgtgat aacgcgaaaa acaccgtgta tctgcagatg 240
aacagcctga aaccggaaga tgcggcgatg tattattgcg cggcgaaagt gagctggtat 300
ggccgttggt atcagagcga aacctatgaa tattggggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 25
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 25
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa cacctttagc tttagcacct attgcatggg ctggtttcgt 120
cagagcccgg gcaaagaacg tgaaggcagc ctggcgacca tttatgatgg cagcaccgcg 180
tatgcgggca gcgtgaaagg ccgttttacc attagcgtgg ataacgcgaa aaacattctg 240
tatctgcaga tgaacaacct gaaaccggaa gataccgcga tgtattattg cgcggcggcg 300
aaatattgca tgttttggag cgatccgagc tattggggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 26
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 26
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atggcagcac cagctatgcg 180
gatagcgtga aaggccgttt taccattagc gtggataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgaaacc ggcggatacc gcgatgtatt attgcgcggc ggcgaaatat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 27
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 27
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccagcagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atggcaacat tagctatgcg 180
gatagcgtga aaggccgttt taccattagc gtggataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgaaacc ggaagatacc gcgatgtatt attgcgcggc ggcgaaatat 300
tgcatgtttt ggagcgatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 28
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 28
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgtgg cgagcggcct gaccctgaac gatgtgagca tggcgtggtt tcgtcaggcg 120
ccgggcaaag aacgtgaaga agtggcgggc attgatattg atggcctgat tagctatgcg 180
gatagcgtga aaggccgttt taccattagc aaagataacg cgcagaacac cctgtatctg 240
cagatgaaca gcctgaaacc ggaagatacc gcgatgtata gctgcgcggc ggtgcgtctg 300
tattttagca gcagccgtaa ccagtataaa tatgaaggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 29
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 29
caggtgcagc tggtggaaag cggcggcggc agcgtgcagc cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa cacctttagc tttagcacct attgcatggg ctggtttcgt 120
cagagcccgg gcaaagaacg tgaaggcagc ctggcgacca tttatgatgg cagcaccgcg 180
tatgcgggca gcgtgaaagg ccgttttacc gtgagcgtgg ataacgcgaa aaacattctg 240
tatctgcaga tgaacaacct gaaaccggaa gataccgcga tgtattattg cgcgagcgcg 300
aaatattgca tgttttggag cgatccgagc tattggggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 30
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 30
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgtgg cgagcggcct gaccctgaac gatgtgagca tggcgtggtt tcgtcaggcg 120
ccgggcaaag aacgtgaaga agtggcgggc attgatattg atggcctgat tagctatgcg 180
gatccggtga aaggccgttt taccattagc aaagataacg cgcagaacac catgtatctg 240
cagatgaaca gcctgaaacc ggaagatacc gcgatgtata gctgcgcggc ggtgcgtctg 300
tattttagca gcagccgtaa ccagtataaa tatgaaggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 31
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 31
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa cacctttagc accagcacct attgcatggg ctggtttcgt 120
cagagcccgg gcaaagaacg tgaaggcagc ctggcgacca tttatgatgg cagcaccgcg 180
tatgcgggca gcgtgaaagg ccgttttacc attagcgtgg ataacgcgaa aaacattctg 240
tatctgcaga tgaacaacct gaaaccggaa gataccgcga tgtattattg cgcggcggcg 300
aaatattgca tgttttggag cgatccgagc tattggggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 32
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 32
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccagcagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atggcgcgat tagctatgcg 180
gatagcgtga aaggccgttt taccattagc gtggataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgaaacc ggaagatacc gcgatgtatt attgcgcggc ggcgaaatat 300
tgcatgtttt ggagcgatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 33
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 33
caggtgcagc tggtggaaag cgaaggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg tgagcggcat taccagcagc accgcgtgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atggcagcat tagctatgcg 180
gatagcgtga aaggccgttt taccattagc gtggataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgaaacc ggaagatacc gcgatgtatt attgcgcggc ggcgaaatat 300
tgcatgtttt ggagcgatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 34
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 34
caggtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcaggcg 120
ccgggcaaag gcctggaagc ggtggcggcg accatttatg atggcagcac ctattatgcg 180
gatagcgtga aaggccgttt taccattagc cgtgataaca gcaaaaacac cctgtatctg 240
cagatgaaca gcctgcgtgc ggaagatacc gcggtgtatt attgcgcggc ggcgcgttat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca ccctggtgac cgtgagcagc 360
<210> 35
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 35
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atgcgagcac cagctatgcg 180
gatagcgtga aaggccgttt taccattagc cgtgataaca gcaaaaacat tctgtatctg 240
cagatgaaca acctgcgtgc ggaagatacc gcggtgtatt attgcgcggc ggcgcgttat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 36
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 36
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atgcgagcac cagctatgcg 180
gatagcgtga aaggccgttt taccattagc gtggataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgaaacc ggaagatacc gcgatgtatt attgcgcggc ggcgcgttat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 37
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 37
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atgcgagcac cagctatgcg 180
gatagcgtga aaggccgttt taccattagc cgtgataacg cgaaaaacat tctgtatctg 240
cagatgaaca acctgcgtgc ggaagatacc gcggtgtatt attgcgcggc ggcgcgttat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 38
<211> 360
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 38
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcat taccgcgagc accagctgca tgggctggtt tcgtcagagc 120
ccgggcaaag aacgtgaagg cagcctggcg accatttatg atggcagcac cagctatgcg 180
gatagcgtga aaggccgttt taccattagc cgtgataaca gcaaaaacat tctgtatctg 240
cagatgaaca acctgcgtgc ggaagatacc gcggtgtatt attgcgcggc ggcgcgttat 300
tgcatgtttt ggagccatcc gagctattgg ggccagggca cccaggtgac cgtgagcagc 360
<210> 39
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 39
caggtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa cacctttagc tttagcacct attgcatggg ctggtttcgt 120
caggcgccgg gcaaaggcct ggaagcggtg gcggcgacca tttatgatgg cagcacctat 180
tatgcgggca gcgtgaaagg ccgttttacc attagccgtg ataacagcaa aaacaccctg 240
tatctgcaga tgaacagcct gcgtgcggaa gataccgcgg tgtattattg cgcggcggcg 300
cgttattgca tgttttggag ccatccgagc tattggggcc agggcaccct ggtgaccgtg 360
agcagc 366
<210> 40
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 40
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa cacctttagc tttagcacct attgcatggg ctggtttcgt 120
cagagcccgg gcaaagaacg tgaaggcagc ctggcgacca tttatgatgc gagcaccgcg 180
tatgcgggca gcgtgaaagg ccgttttacc attagccgtg ataacagcaa aaacattctg 240
tatctgcaga tgaacaacct gcgtgcggaa gataccgcgg tgtattattg cgcggcggcg 300
cgttattgca tgttttggag ccatccgagc tattggggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 41
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 41
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa cacctttagc tttagcacct attgcatggg ctggtttcgt 120
cagagcccgg gcaaagaacg tgaaggcagc ctggcgacca tttatgatgc gagcaccgcg 180
tatgcgggca gcgtgaaagg ccgttttacc attagcgtgg ataacgcgaa aaacattctg 240
tatctgcaga tgaacaacct gaaaccggaa gataccgcga tgtattattg cgcggcggcg 300
cgttattgca tgttttggag ccatccgagc tattggggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 42
<211> 366
<212> ДНК
<213> искусственная последовательность
<220>
<223> нуклеиновая кислота, кодирующая IL-5-связывающую молекулу
<400> 42
caggtgcagc tggtggaaag cggcggcggc agcgtgcagg cgggcggcag cctgcgtctg 60
agctgcgcgg cgagcggcaa cacctttagc tttagcacct attgcatggg ctggtttcgt 120
cagagcccgg gcaaagaacg tgaaggcagc ctggcgacca tttatgatgc gagcaccgcg 180
tatgcgggca gcgtgaaagg ccgttttacc attagccgtg ataacgcgaa aaacattctg 240
tatctgcaga tgaacaacct gcgtgcggaa gataccgcgg tgtattattg cgcggcggcg 300
cgttattgca tgttttggag ccatccgagc tattggggcc agggcaccca ggtgaccgtg 360
agcagc 366
<210> 43
<211> 8
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR1
<400> 43
Gly Ile Thr Ala Ser Thr Ser Cys
1 5
<210> 44
<211> 8
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR1
<400> 44
Gly Ile Thr Ser Ser Thr Ala Cys
1 5
<210> 45
<211> 8
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR1
<400> 45
Gly Ile Thr Ser Ser Thr Ser Cys
1 5
<210> 46
<211> 8
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR1
<400> 46
Gly Leu Thr Leu Asn Asp Val Ser
1 5
<210> 47
<211> 10
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR1
<400> 47
Gly Asn Thr Phe Ser Phe Ser Thr Tyr Cys
1 5 10
<210> 48
<211> 10
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR1
<400> 48
Gly Asn Thr Phe Ser Thr Ser Thr Tyr Cys
1 5 10
<210> 49
<211> 6
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR1
<400> 49
Gly Asn Thr Leu Asp Leu
1 5
<210> 50
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR2
<400> 50
Ile Asp Ile Asp Gly Leu Ile
1 5
<210> 51
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR2
<400> 51
Ile Tyr Ser Ser Gly Thr Thr
1 5
<210> 52
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR2
<400> 52
Thr Ile Tyr Asp Ala Ser Thr
1 5
<210> 53
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR2
<400> 53
Thr Ile Tyr Asp Gly Ala Ile
1 5
<210> 54
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR2
<400> 54
Thr Ile Tyr Asp Gly Asn Ile
1 5
<210> 55
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR2
<400> 55
Thr Ile Tyr Asp Gly Ser Ile
1 5
<210> 56
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR2
<400> 56
Thr Ile Tyr Asp Gly Ser Thr
1 5
<210> 57
<211> 15
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR3
<400> 57
Ala Ala Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp
1 5 10 15
<210> 58
<211> 15
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR3
<400> 58
Ala Ala Ala Lys Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp
1 5 10 15
<210> 59
<211> 15
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR3
<400> 59
Ala Ala Ala Arg Tyr Cys Met Phe Trp Ser His Pro Ser Tyr Trp
1 5 10 15
<210> 60
<211> 19
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR3
<400> 60
Ala Ala Lys Val Ser Trp Tyr Gly Arg Trp Tyr Gln Ser Glu Thr Tyr
1 5 10 15
Glu Tyr Trp
<210> 61
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR3
<400> 61
Ala Ala Val Arg Leu Tyr Phe Ser Ser Ser Arg Asn Gln Tyr Lys Tyr
1 5 10 15
Glu
<210> 62
<211> 15
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> определяющая комплементарность область CDR3
<400> 62
Ala Ser Ala Lys Tyr Cys Met Phe Trp Ser Asp Pro Ser Tyr Trp
1 5 10 15
<210> 63
<211> 25
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR1
<400> 63
Gln Val Gln Leu Val Glu Ser Glu Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser
20 25
<210> 64
<211> 25
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR1
<400> 64
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Ala Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 65
<211> 25
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR1
<400> 65
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 66
<211> 25
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR1
<400> 66
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser
20 25
<210> 67
<211> 25
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR1
<400> 67
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 68
<211> 25
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR1
<400> 68
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 69
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR2
<400> 69
Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Glu Val Ala
1 5 10 15
Gly
<210> 70
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR2
<400> 70
Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Gly Leu Glu Ala Val Ala
1 5 10 15
Ala
<210> 71
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR2
<400> 71
Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Gly Val Ala
1 5 10 15
Gly
<210> 72
<211> 17
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR2
<400> 72
Met Gly Trp Phe Arg Gln Ser Pro Gly Lys Glu Arg Glu Gly Ser Leu
1 5 10 15
Ala
<210> 73
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 73
Ala Tyr Ala Gly Ser Val Lys Gly Arg Phe Thr Ile Ser Val Asp Asn
1 5 10 15
Ala Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp
20 25 30
Thr Ala Met Tyr Tyr Cys
35
<210> 74
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 74
Ala Tyr Ala Gly Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ser Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 75
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 75
Ala Tyr Ala Gly Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ala Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 76
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 76
Ala Tyr Ala Gly Ser Val Lys Gly Arg Phe Thr Val Ser Val Asp Asn
1 5 10 15
Ala Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp
20 25 30
Thr Ala Met Tyr Tyr Cys
35
<210> 77
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 77
Ser Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ala Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 78
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 78
Ser Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ser Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 79
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 79
Ser Tyr Ala Asp Pro Val Lys Gly Arg Phe Thr Ile Ser Lys Asp Asn
1 5 10 15
Ala Gln Asn Thr Met Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp
20 25 30
Thr Ala Met Tyr Ser Cys
35
<210> 80
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 80
Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 81
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 81
Tyr Tyr Ala Gly Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 82
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 82
Ser Tyr Ala Asp Ser Val Lys Gly Gln Phe Thr Ile Ser Val Asp Asn
1 5 10 15
Ala Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp
20 25 30
Thr Ala Met Tyr Tyr Cys
35
<210> 83
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 83
Ser Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Lys Asp Asn
1 5 10 15
Ala Gln Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp
20 25 30
Thr Ala Met Tyr Ser Cys
35
<210> 84
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 84
Ser Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Val Asp Asn
1 5 10 15
Ala Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Lys Pro Ala Asp
20 25 30
Thr Ala Met Tyr Tyr Cys
35
<210> 85
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 85
Ser Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Val Asp Asn
1 5 10 15
Ala Lys Asn Ile Leu Tyr Leu Gln Met Asn Asn Leu Lys Pro Glu Asp
20 25 30
Thr Ala Met Tyr Tyr Cys
35
<210> 86
<211> 38
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR3
<400> 86
Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Val Ser Arg Asp Asn
1 5 10 15
Ala Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp
20 25 30
Ala Ala Met Tyr Tyr Cys
35
<210> 87
<211> 10
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> каркасная область FR4
<400> 87
Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ IL-17A И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2816204C2 |
| ГУМАНИЗИРОВАННОЕ ОДНОДОМЕННОЕ АНТИТЕЛО К IL-4Rα И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2816867C1 |
| ГУМАНИЗИРОВАННОЕ ОДНОДОМЕННОЕ АНТИТЕЛО К IL-4Rα И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2836057C2 |
| KIR3DL3 - ИНГИБИРУЮЩИЙ РЕЦЕПТОР ИММУННОЙ СИСТЕМЫ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2827900C1 |
| АНТИТЕЛО К GIPR И ЕГО СЛИТЫЙ С GLP-1 БЕЛОК, А ТАКЖЕ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800370C2 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| ЛЕЧЕНИЕ И ИНГИБИРОВАНИЕ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ ЛЕГКИХ У ПАЦИЕНТОВ С АЛЛЕЛЯМИ РИСКА В ГЕНАХ, КОДИРУЮЩИХ IL33 И IL1RL1 | 2018 |
|
RU2776241C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ИНТЕРЛЕЙКИНА-33 (IL-33) | 2015 |
|
RU2829846C2 |
| АНТИ-CEACAM1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2754876C2 |
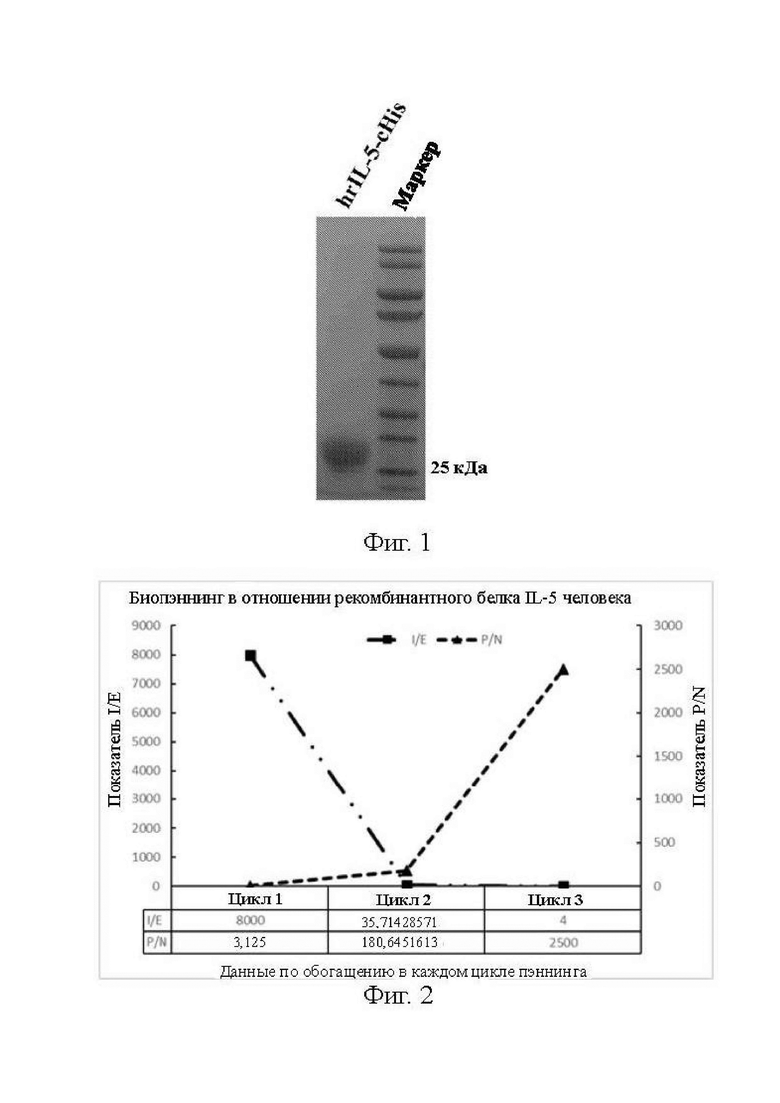
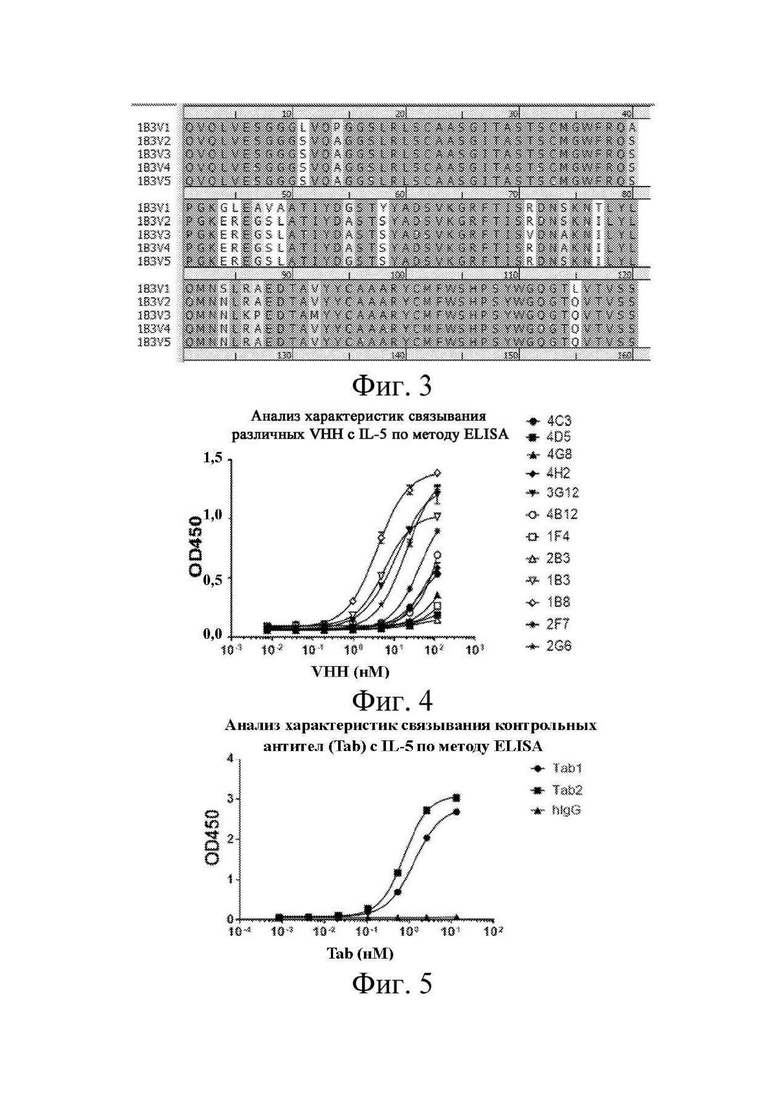
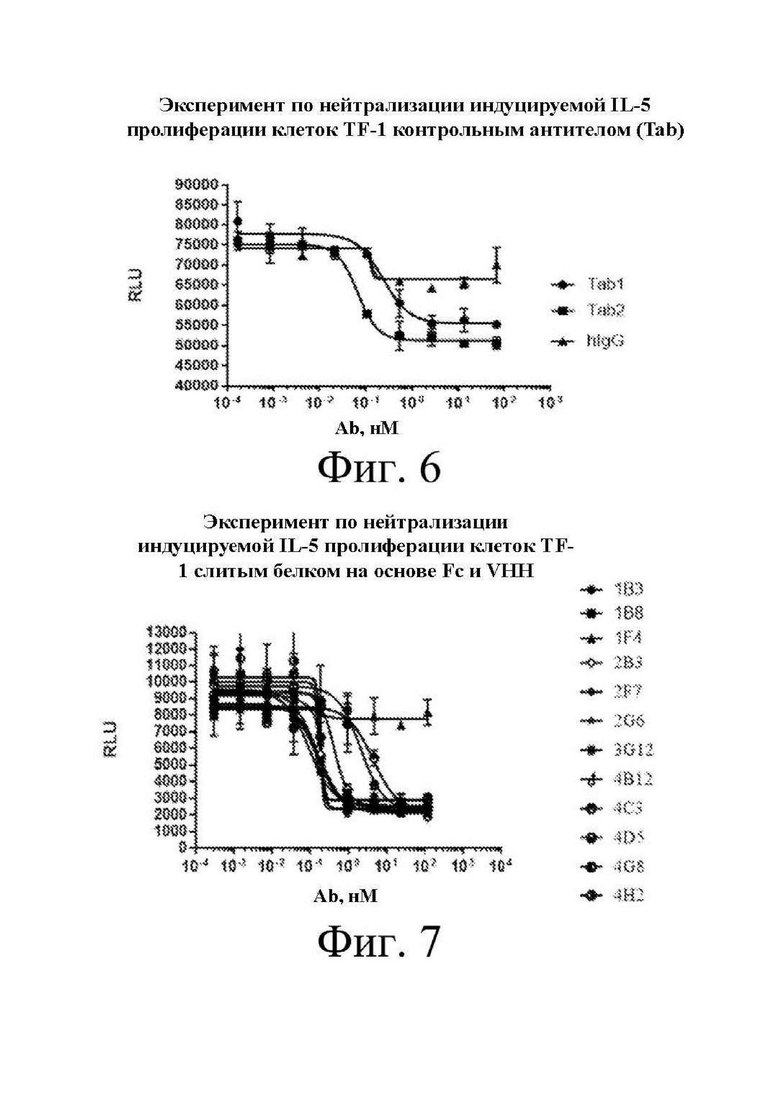
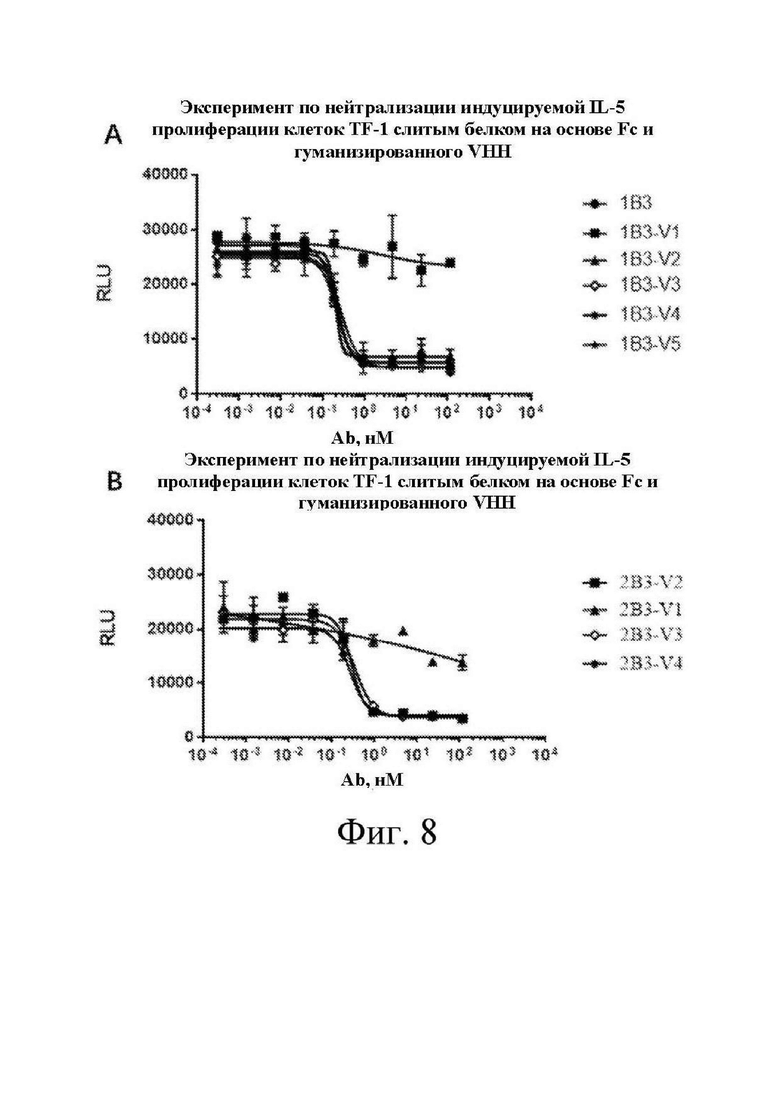
Изобретение относится к области биотехнологии, конкретно к рекомбинантным IL-5-связывающим молекулам, и может быть использовано в медицине при получении лекарственных средств, нацеленных на IL-5, для лечения лейкемии. Предложена IL-5-связывающая молекула, способная специфично связываться с интерлейкином-5 (IL-5) и содержащая одиночный вариабельный домен иммуноглобулина с определенным набором определяющих комплементарность областей CDR1, CDR2 и CDR3. Изобретение позволяет получить молекулу, которая способна специфично связываться с IL-5 и эффективно блокировать клеточную пролиферацию, индуцируемую IL-5. 6 н. и 8 з.п. ф-лы, 8 ил., 10 табл., 5 пр.
1. IL-5-связывающая молекула, способная специфично связываться с IL-5 и содержащая одиночный вариабельный домен иммуноглобулина, содержащий определяющие комплементарность области CDR1, CDR2 и CDR3, имеющие аминокислотные последовательности, приведенные в любом из (1)-(13):
(1) SEQ ID NO: 43, 56 и 58;
(2) SEQ ID NO: 49, 51 и 60;
(3) SEQ ID NO: 47, 56 и 57;
(4) SEQ ID NO: 45, 54 и 57;
(5) SEQ ID NO: 46, 50 и 61;
(6) SEQ ID NO: 47, 56 и 62;
(7) SEQ ID NO: 48, 56 и 57;
(8) SEQ ID NO: 45, 53 и 57;
(9) SEQ ID NO: 44, 55 и 57;
(10) SEQ ID NO: 43, 56 и 59;
(11) SEQ ID NO: 43, 52 и 59;
(12) SEQ ID NO: 47, 56 и 59; а также
(13) SEQ ID NO: 47, 52 и 59.
2. IL-5-связывающая молекула по п. 1, где одиночный вариабельный домен иммуноглобулина дополнительно содержит каркасную область, содержащую FR1, FR2, FR3 и FR4, имеющие аминокислотные последовательности, приведенные в любом из (14)-(28):
(14) SEQ ID NO: 65, 72 и 85;
(15) SEQ ID NO: 65, 72 и 82;
(16) SEQ ID NO: 64, 71 и 86;
(17) SEQ ID NO: 65, 72 и 73;
(18) SEQ ID NO: 65, 72 и 84;
(19) SEQ ID NO: 66, 69 и 83;
(20) SEQ ID NO: 68, 72 и 76;
(21) SEQ ID NO: 66, 69 и 79;
(22) SEQ ID NO: 63, 72 и 85;
(23) SEQ ID NO: 67, 70 и 80;
(24) SEQ ID NO: 65, 72 и 78;
(25) SEQ ID NO: 65, 72 и 77;
(26) SEQ ID NO: 67, 70 и 81;
(27) SEQ ID NO: 65, 72 и 74; а также
(28) SEQ ID NO: 65, 72 и 75.
3. IL-5-связывающая молекула по п. 2, где одиночный вариабельный домен иммуноглобулина представляет собой VHH.
4. IL-5-связывающая молекула по п. 3, где VHH имеет аминокислотную последовательность, выбранную из любой из последовательностей, приведенных под SEQ ID NO: 1-12.
5. IL-5-связывающая молекула по п. 3, где VHH представляет собой гуманизированный VHH.
6. IL-5-связывающая молекула по п. 5, где гуманизированный VHH имеет последовательность, выбранную из любой из последовательностей, приведенных под SEQ ID NO: 13-21.
7. IL-5-связывающая молекула по п. 3, дополнительно содержащая Fc-область иммуноглобулина, соединенную с VHH.
8. IL-5-связывающая молекула по п. 7, где Fc-область иммуноглобулина представляет собой Fc-область иммуноглобулина человека.
9. IL-5-связывающая молекула по п. 8, где Fc-область иммуноглобулина человека представляет собой Fc-область IgG4 человека.
10. Выделенная нуклеиновая кислота для кодирования IL-5-связывающей молекулы по любому из пп. 1-9;
при этом нуклеиновая кислота имеет последовательность, выбранную из любой из последовательностей, приведенных под SEQ ID NO: 22-42.
11. Рекомбинантный вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 10.
12. Клетка-хозяин для экспрессии IL-5-связывающей молекулы по любому из пп. 1-9, содержащая рекомбинантный вектор экспрессии по п. 11.
13. Способ получения IL-5-связывающей молекулы, включающий культивирование клетки-хозяина по п. 12 в среде для получения IL-5-связывающей молекулы и выделение IL-5-связывающей молекулы из среды.
14. Применение IL-5-связывающей молекулы по любому из пп. 1-5 при получении лекарственных средств, нацеленных на IL-5, для лечения лейкемии.
| WO 2019062831 A1, 04.04.2019 | |||
| WO 2018119016 A1, 28.06.2018 | |||
| EA 201890572 A1, 31.10.2018 | |||
| Прибор для установления наличия и развития стереоскопичности зрения | 1930 |
|
SU27160A1 |
| DAVIES J | |||
| et al., Affinity improvement of single antibody VH domains: residues in all three hypervariable regions affect antigen binding, Immunotechnology, 1996, v.2, p.169-179 | |||
| RIECHMANN L | |||
| et al., Reshaping human antibodies for | |||
