Область техники
[0001] Настоящее изобретение относится к способу продукции 3'-фосфоаденозин-5'-фосфосульфата (далее в настоящем описании обозначенного как PAPS) из недорогих сырьевых материалов, таких как глюкоза и аденин, с использованием бактерии из рода Corynebacterium, экспрессирующей ATP-сульфурилазу и APS-киназу (аденилилсульфат-киназу), и к способу продукции сульфатированного полисахарида с использованием бактерии из рода Corynebacterium и микроорганизма, принадлежащего к прокариотам, экспрессирующего различные ферменты сульфатирования.
Уровень техники
[0002] PAPS представляет собой кофермент, который широко распространен от микроорганизмов до высших организмов и функционирует в качестве донора сульфатной группы in vivo. У высших организмов, PAPS является необходимым для биосинтеза гликозаминогликанов, таких как хондроитинсульфат и гепарансульфат. Известно, что сульфатированные in vivo метаболиты имеют различные физиологические функции, и ожидают использование собственно PAPS в качестве средства для восстановления волос (PTL 1), препарата для наружного применения для кожи, имеющего увлажняющий эффект (PTL 2), и т.п.
[0003] В качестве способа продукции PAPS, известны способ использования ATP-сульфурилазы, происходящей из очищенных дрожжей, APS-киназы, происходящей из голубой плесени, и пирофосфатазы, происходящей из Escherichia coli, с использованием ATP в качестве сырьевого материала (NPL 1); способ с использованием термостабильной ATP-сульфурилазы, происходящей из термофильной бактерии, APS-киназы и пирофосфатазы, происходящих из дрожжей, с использованием ATP в качестве сырьевого материала, как в вышеуказанном способе (PTL 3); способ с использованием ATP-сульфурилазы, происходящей из термостабильной бактерии, APS-киназы и полифосфат-киназы, происходящих из Pseudomonas aeruginosa, с использованием аденозин-5'-монофосфата (далее в настоящем описании обозначенного как AMP) в качестве сырьевого материала (PTL 4); и т.п.
[0004] Тем не менее, известен способ промышленной продукции ATP с использованием мутантного штамма Corynebacterium ammoniagenes, в котором мембрану бактериальных клеток обрабатывают с использованием ксилола или поверхностно-активного вещества, для придания ей проницаемости, с использованием недорогих сырьевых материалов, таких как глюкоза и аденин (NPL 2).
[0005] PAPS представляет собой очень нестабильное соединение. Даже в способе с использованием неочищенного фермента, PAPS продуцируют посредством получения неочищенного фермента из APS-киназы и ATP-сульфурилазы, происходящих из термостабильной бактерии, которые рекомбинантным способом экспрессируют посредством Escherichia coli в качестве хозяина, с последующей обработкой нагреванием до реакции, таким образом, супрессируя активность контаминирующих ферментов, происходящих из Escherichia coli (PTL 4).
[0006] В PTL8 описано, что PAPS подвергается деградации в неочищенном растворе ферментов из Escherichia coli. Escherichia coli имеет некоторые ферменты, связанные с деградацией PAPS, и является возможным ингибирование деградации PAPS посредством делеции гена таких ферментов.
[0007] Гепарин, представляющий собой сульфатированный полисахарид, представляет собой ведущее антикоагулянтное средство, и его используют для лечения тромбоэмболии и синдрома диссеминированного внутрисосудистого свертывания, и для предотвращения свертывания во время искусственного диализа и при экстракорпоральном кровообращении, и т.п. В промышленности, большинство используемого гепарина выделено и очищено из слизистой оболочки кишечника свиней. Поскольку случай со смертельным исходом произошел в 2008 г. из-за контаминации гепарина свиного происхождения примесями, проводили исследования и разработку гепарина, не из животных источников, с контролем производства/с контролем качества.
[0008] В качестве конкретного примера, способ, в котором ферментно полученный и очищенный N-ацетилгепаросан (далее в настоящем описании обозначенный как гепаросан), представляющий собой капсульный полисахарид из некоторых грамотрицательных микроорганизмов, химически N-деацетилируют и N-сульфатируют, с последующим ферментными эпимеризацией и сульфатированием для получения гепарина, имеющего такую же структуру и антикоагулянтную активность, как происходящие из свиней (PTL 5-7, и NPL 3 и 4).
[0009] Хондроитинсульфат известен как другой сульфатированный полисахарид, который можно использовать, и его использовали в качестве лекарственного средства против боли в суставах и в качестве глазных капель для защиты поверхностного слоя роговицы. Использовали хондроитинсульфат, выделенный и очищенный из различных тканей животных, однако, недавно опубликован способ с использованием фермента для сульфатирования, с использованием капсульного полисахарида, происходящего из Escherichia coli, в качестве сырьевого материала, таким же способом, как гепарин (NPL 5 и 6).
[0010] В способе осуществления сульфатирования с использованием очищенного фермента, с использованием полисахарида, происходящего из капсулы микроорганизма в качестве сырьевого материала, PAPS служит в качестве донора сульфатной группы в реакции фермента сульфатирования и является необходимым в качестве кофермента. В вышеупомянутом способе продукции сульфатированного полисахарида с использованием капсульного полисахарида, происходящего из капсулы микроорганизма, в качестве сырьевого материала, 3'-фосфоаденозин-5'-фосфат (далее в настоящем описании обозначенный как PAP) образуется посредством переноса сульфатной группы PAPS на субстратный полисахарид, сульфатная группа п-нитрофенилсульфата ферментно переносится на PAP, таким образом, регенерируя PAPS, и реакция ферментного сульфатирования полисахарида продолжается (PTL 6 и NPL 3).
Список литературы
Патентные документы
[0011] [PTL 1] WO2012/057336
[PTL 2] JP-A-2012-201665
[PTL 3] JP-A-H5-137588
[PTL 4] Японский патент No. 4505011
[PTL 5] Японский патент No. 5830464
[PTL 6] Патент США No. 8771995
[PTL 7] WO 2018/048973
[PTL 8] WO 2020/013346
Непатентные документы
[0012] [NPL 1] Glycobiology vol. 21, no. 6, pp. 771-780, 2011
[NPL 2] Biosci. Biotechnol. Biochem. 65 (3), 644-650, 2001
[NPL 3] Carbohydrate Polymers 122 (2015) 399-407
[NPL 4] Advanced Drug Delivery Reviews 97 (2016) 237-249
[NPL 5] Metabolic Engineering 27 (2015) 92-100
[NPL 6] Biotechnology and Bioengineering 115 (2018) 1561-1570
Сущность изобретения
Техническая проблема
[0013] В каждом из общепринятых способов продукции PAPS, описанных в NPL 1, и PTL 1 и 2, используют относительно дорогие нуклеотиды, такие как ATP и AMP, в качестве сырьевых материалов, и использование фермента или неочищенного фермента, который получают посредством культивирования бактерий, с последующим отделением бактериальных клеток, разрушения бактериальных клеток посредством обработки ультразвуком или т.п., и центрифугирования. Таким образом, не является простым осуществление этих способов в промышленном масштабе.
[0014] С другой стороны, как описано выше, для продукции PAPS, известен способ с использованием очищенного фермента или неочищенного фермента, с использованием ATP или AMP в качестве сырьевого материала, однако, пример продукции PAPS с использованием собственно микроорганизма, не известен. Кроме того, как описано выше, PAPS представляет собой очень нестабильное соединение, и на предшествующем уровне техники, в случае, в котором используют неочищенный фермент, осуществляют сложные стадии, такие как супрессия активности контаминирующих ферментов посредством термической обработки.
[0015] С учетом вышеуказанного, является сложным прогнозировать, что PAPS можно продуцировать посредством способа, сходного со способом промышленной продукции ATP, просто посредством экспрессии ATP-сульфурилазы и APS-киназы в бактерии из рода Corynebacterium в качестве хозяина. Это из-за того, что, как в случае Escherichia coli, род Corynebacterium также имеет некоторые ферменты, связанные с деградацией PAPS.
[0016] Кроме того, как описано выше, в общепринятом способе ферментного сульфатирования полисахаридов используют множество ферментов сульфатирования, очищенных из лизата клеток, которые получают посредством культивирования микроорганизмов, экспрессирующих фермент сульфатирования, с последующим сбором и обработкой ультразвуком или т.п. Не являлось простым осуществление этих способов в промышленном масштабе. Кроме того, дорогостоящие PAPS и п-нитрофенилсульфат являлись необходимыми в качестве сырьевых материалов.
[0017] Соответственно, целью настоящего изобретения является предоставление способа простой продукции сульфатированных полисахаридов посредством реакции системы продукции/регенерации PAPS с использованием метаболической активности микроорганизма или его обработанного материала, с использованием микроорганизма, экспрессирующего фермент сульфатирования, или его обработанного материала или экстракта, при смешивании с недорогими сырьевыми материалами, такими как сульфат магния. Другой целью настоящего изобретения является предоставление практического способа продукции PAPS из недорогих сырьевых материалов.
Решение проблемы
[0018] Авторы настоящего изобретения завершили настоящее изобретение на основании следующих обнаружений (1) и (2).
(1) PAPS можно продуцировать более просто и недорого, по сравнению со способами в данной области, посредством культивирования штамма, в котором активности ATP-сульфурилазы и APS-киназы усилены посредством способов рекомбинантной ДНК, с использованием бактерии из рода Corynebacterium, имеющей способность к продукции ATP, в качестве хозяина, придавая ей проницаемость мембраны с использованием поверхностно-активных веществ или т.п., и добавления сырьевых материалов, таких как глюкоза и аденин.
(2) Сульфатированные полисахариды можно продуцировать с использованием бактериальных клеток и недорогих сырьевых материалов, таких как сульфат магния, без очистки фермента или добавления PAPS, посредством использования штамма с усиленной активностью ATP-сульфурилазы и APS-киназы, описанного в (1), в качестве клетки бактериального микроорганизма, ответственной за реакцию продукции/регенерации PAPS, придания проницаемости мембраны микроорганизму, принадлежащему к прокариотам, который экспрессирует эпимеразу и/или ферменты сульфатирования, и проведения смешанной реакции.
[0019] Таким образом, настоящее изобретение представляет собой следующее.
1. Способ продукции сульфатированного полисахарида, включающий следующие стадии (1-1) и (1-2):
(1-1) получение трансформанта (a) бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта (a); и
(1-2) проведение реакции для продукции PAPS посредством использования реакционного раствора, содержащего ATP или источник ATP, источник иона сульфата, и трансформант (a) или его обработанный материал.
2. Способ продукции сульфатированного полисахарида в соответствии с 1, дополнительно включающий следующие стадии (2-1) и (2-2):
(2-1) получение трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b); и
(2-2) проведение C5-эпимеризации посредством включения трансформанта (b) или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
3. Способ продукции сульфатированного полисахарида в соответствии с 1 или 2, включающий проведение сульфатирования посредством трансформанта, содержащего ген, кодирующий сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта.
4. Способ продукции сульфатированного полисахарида в соответствии с 3, дополнительно включающий следующие стадии (3-1) и (3-2):
(3-1) получение трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c); и
(3-2) проведение 2-O-сульфатирования посредством включения трансформанта (c), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
5. Способ продукции сульфатированного полисахарида в соответствии с 3 или 4, дополнительно включающий следующие стадии (3'-1) - (3'-3):
(3'-1) получение трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b);
(3'-2) получение трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c); и
(3'-3) проведение C5-эпимеризации и 2-O-сульфатирования посредством включения трансформанта (b) или его обработанного материала или экстракта, и трансформанта (c) или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
6. Способ продукции сульфатированного полисахарида в соответствии с любым из 3-5, дополнительно включающий следующие стадии (4-1) и (4-2):
(4-1) получение трансформанта (d) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 6-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (d); и
(4-2) проведение 6-O-сульфатирования посредством включения трансформанта (d), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
7. Способ продукции сульфатированного полисахарида в соответствии с любым из 3-6, дополнительно включающий следующие стадии (5-1) и (5-2):
(5-1) получение трансформанта (e) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 3-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (e); и
(5-2) проведение 3-O-сульфатирования посредством включения трансформанта (e), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
8. Способ продукции сульфатированного полисахарида, включающий получение сульфатированного полисахарида посредством включения в реакционный раствор, в присутствии ATP или источника ATP, источника иона сульфата и N-сульфогепаросана, трансформанта (a) бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта (a), и
по меньшей мере одного, выбранного из:
трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b),
трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c),
трансформанта (d) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 6-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (d), и
трансформанта (e) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 3-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (e).
9. Способ продукции сульфатированного полисахарида в соответствии с 7, дополнительно включающий получение сульфатированного полисахарида посредством включения в реакционный раствор, в присутствии ATP или источника ATP, источника иона сульфата и N-сульфогепаросана, трансформанта (a) или его обработанного материала, и трансформантов (b) - (e), или их обработанных материалов или экстрактов.
10. Способ продукции сульфатированного полисахарида в соответствии с любым из 1-9, предназначенный для продукции гепарина.
11. Способ продукции PAPS, включающий следующие стадии (i) и (ii):
(i) получение трансформанта бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта; и
(ii) проведение реакции для продукции PAPS посредством использования реакционного раствора, содержащего ATP или источник ATP, источник иона сульфата, и трансформанта, полученного на стадии (i), или его обработанного материала.
Обеспечивающие преимущество эффекты изобретения
[0020] В соответствии со способом продукции сульфатированного полисахарида по настоящему изобретению, сульфатированные полисахариды можно продуцировать просто и недорого посредством использования системы продукции/регенерации PAPS, использующей метаболическую активность микроорганизма, и микроорганизма, экспрессирующего фермент сульфатирования, или его обработанного материала или экстракта. Кроме того, в соответствии со способом продукции PAPS по настоящему изобретению, PAPS можно продуцировать просто и недорого посредством использования метаболической активности микроорганизма или его обработанного материала. Кроме того, посредством использования рода Corynebacterium в способе по настоящему изобретению, не является необходимой модификация генов комплексным способом для избегания деградации PAPS.
Краткое описание чертежей
[0021] [Фиг. 1] На ФИГ. 1 показана схематическая диаграмма кластера генов синтеза гепаросана на хромосоме Escherichia coli штамма K5.
[Фиг. 2] На ФИГ. 2 показана схематическая диаграмма ферментов, вовлеченных в продукцию гепаросана.
[Фиг. 3] На ФИГ. 3 показана схематическая диаграмма биосинтетического пути гепаросана.
[Фиг. 4] ФИГ. 4 представляет собой график, показывающий зависимые от времени изменения концентрации PAPS, содержащегося в супернатанте реакционного раствора, в тестировании реакции 2-O-сульфатирования N-сульфогепаросана.
[Фиг. 5] ФИГ. 5 представляет собой график, показывающий зависимые от времени изменения соотношения площади дельта-UA, 2S-GlcNS в анализе HPLC ненасыщенного дисахарида, в тестировании реакции 2-O-сульфатирования N-сульфогепаросана.
[Фиг. 6] ФИГ. 6 представляет собой график, показывающий зависимые от времени изменения концентрации PAPS, содержащегося в супернатанте реакционного раствора, в тестировании реакции 6-O-сульфатирования 2-O-сульфатированного N-сульфогепаросана.
[Фиг. 7] ФИГ. 7 представляет собой график, показывающий зависимые от времени изменения соотношения площади дельта-UA, 2S-GlcN, 6S в анализе HPLC ненасыщенного дисахарида, в тестировании реакции 6-O-сульфатирования 2-O-сульфатированного N-сульфогепаросана.
[Фиг. 8] ФИГ. 8 представляет собой график, показывающий зависимые от времени изменения концентрации PAPS, содержащегося в супернатанте реакционного раствора, в тестировании реакции сульфатирования в 6-O-положении и 3-O-положении 2-O-сульфатированного N-сульфогепаросана.
[Фиг. 9] ФИГ. 9 представляет собой график, показывающий зависимые от времени изменения концентрации PAPS, содержащегося в супернатанте реакционного раствора, в тестировании продукции PAPS.
Описание вариантов осуществления
[0022] Трансформант
Способ продукции сульфатированного полисахарида по настоящему изобретению отличается тем, что стадию 1) снабжения/регенерации PAPS проводят посредством реакции с использованием бактериальных клеток. В способе по настоящему изобретению, предпочтительно, по меньшей мере одну из стадий 2) C5-эпимеризации, 3) 2-O-сульфатирования, 4) 6-O-сульфатирования и 5) 3-O-сульфатирования также проводят посредством реакции с использованием бактериальных клеток. Далее в настоящем описании будут описаны трансформанты (a) - (e), используемые в каждой реакции.
[0023] Трансформант (a): снабжение/регенерация PAPS
В способе продукции сульфатированного полисахарида по настоящему изобретению, снабжение/регенерацию PAPS проводят с использованием трансформанта (a) микроорганизма, принадлежащего к роду Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта (a).
[0024] Примеры видов бактерий из рода Corynebacterium включают Corynebacterium ammoniagenes, Corynebacterium acetoacidophilum, Corynebacterium acetoglutamicum, Corynebacterium alkanolyticum, Corynebacterium callunae, Corynebacterium crenatum, Corynebacterium glutamicum, Corynebacterium lilium, Corynebacterium melassecola, Corynebacterium thermoaminogenes (Corynebacterium efficiens) и Corynebacterium herculis.
[0025] Примеры штаммов из рода Corynebacterium включают Corynebacterium ammoniagenes (Corynebacterium stationis) ATCC 6871 и ATCC 6872, Corynebacterium acetoacidophilum ATCC 13870, Corynebacterium acetoglutamicum ATCC 15806, Corynebacterium alkanolyticum ATCC 21511, Corynebacterium callunae ATCC 15991, Corynebacterium crenatum AS1.542, Corynebacterium glutamicum ATCC 13020, ATCC 13032, ATCC 13060, ATCC 13869 и FERM BP-734, Corynebacterium lilium ATCC 15990, Corynebacterium melassecola ATCC 17965, Corynebacterium efficiens (Corynebacterium thermoaminogenes) AJ12340 (FERM BP-1539), Corynebacterium herculis ATCC 13868, Brevibacterium divaricatum (Corynebacterium glutamicum) ATCC 14020, Brevibacterium flavum (Corynebacterium glutamicum) ATCC 13826, ATCC 14067 и AJ12418 (FERM BP-2205), и Brevibacterium lactofermentum (Corynebacterium glutamicum) ATCC 13869.
[0026] Бактерии, принадлежащие к роду Corynebacterium, также включают бактерии, которые были традиционно классифицированы в род Brevibacterium, но в настоящее время интегрированы в род Corynebacterium (Int Int. J. Syst. Bacteriol., 41, 255 (1991)). Кроме того, Corynebacterium stationis также включает бактерии, которые были традиционно классифицированы как Corynebacterium ammoniagenes, но в настоящее время переклассифицированы как Corynebacterium stationis по анализу нуклеотидной последовательности 16S рРНК или т.п. [Int. J Syst. Evol. Microbiol., 60, 874-879 (2010)].
[0027] Эти штаммы являются доступными, например, из Американской коллекции типовых культур (адрес: 12301 Parklawn Drive, Rockville, Maryland 20852, P.O. Box 1549, Manassas, VA 20108, United States of America). То есть, регистрационный номер присвоен каждому штамму, и штаммы могут быть получены с использованием этого регистрационного номера (ссылка https://www.atcc.org/). Регистрационный номер, соответствующий каждому штамму, описан в каталоге Американской коллекции типовых культур. Кроме того, эти штаммы являются доступными, например, из депозитария, в котором депонирован каждый штамм. Бактерия из рода Corynebacterium может представлять собой штамм дикого типа, его мутантный штамм или искусственный рекомбинант.
[0028] По настоящему изобретению, ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, вводят в бактерию из рода Corynebacterium подходящим для экспрессии образом.
[0029] ATP-сульфурилаза образует аденозин-5'-фосфосульфат (APS) из сульфата посредством следующей формулы реакции.
[0030] Химическая формула 1
ATP+SO42- → APS+PPi
(В вышеуказанной формуле, ATP представляет собой аденозин-5'-трифосфат.)
[0031] Одним аспектом ATP-сульфурилазы является MET3. Источник ATP-сульфурилазы не является конкретно ограниченным, и ее примеры включают ATP-сульфурилазы, происходящие из Saccharomyces cerevisiae, Candida albicans, Shizosaccharomyces pombe, Yarrowia lipolytica, Neurospora crassa, Penicillium chrysogenum, Kluyveromyces lactis, Fusarium fujikuroi, Aspergillus oryzae или Ashbya gossypii, и среди них, ATP-сульфурилаза, происходящая из Saccharomyces cerevisiae, является предпочтительной.
[0032] Примеры гена, кодирующего ATP-сульфурилазу, включают нуклеотидную последовательность, показанную в SEQ ID NO: 22. Его примеры дополнительно включают ДНК, кодирующую полипептид, имеющий активность ATP-сульфурилазы, и содержащую нуклеотидную последовательность, которая является, предпочтительно, на 80% или более, более предпочтительно, на 90% или более, даже более предпочтительно, на 95% или более, и особенно предпочтительно, на 98% или более идентичной нуклеотидной последовательности, показанной в SEQ ID NO: 22; и ДНК, кодирующую полипептид, имеющий активность ATP-сульфурилазы, и содержащую нуклеотидную последовательность, которая гибридизуется, в строгих условиях, с нуклеотидной последовательностью, показанной в SEQ ID NO: 22, или нуклеотидную последовательность, комплементарную вышеуказанной нуклеотидной последовательности. Термин «строгие условия» относится к условиям, в которых образуются так называемые специфические гибриды, и неспецифические гибриды не образуются. Их примеры включают условия, в которых ДНК, идентичные друг другу на более высоком уровне, например, ДНК, которые являются на 80% или более, предпочтительно, на 90% или более, более предпочтительно, на 95% или более, даже более предпочтительно, на 97% или более, и особенно предпочтительно, на 99% или более идентичными друг другу, гибридизуются друг с другом, и ДНК, идентичные друг другу на более низком уровне, не гибридизуются друг с другом; или условия, представляющие собой условия для отмывки при нормальной Саузерн-гибридизации, в которых отмывку проводят один раз, предпочтительно, от двух до трех раз, при концентрации соли и температуре, соответствующих 60 градусам C, 1 x SSC и 0,1% SDS, предпочтительно, 60 градусам C, 0,1 x SSC, и 0,1% SDS, и более предпочтительно, 68 градусам C, 0,1 x SSC и 0,1% SDS.
[0033] Активность ATP-сульфурилазы в трансформанте проверяют по увеличению значений активности ATP-сульфурилазы в жидком экстракте клеток трансформанта. Активность ATP-сульфурилазы можно проверять посредством способа, описанного в литературе [Medina DC et al., Temperature effects on the allosteric transition of ATP sulfurylase from Penicillium chrysogenum. Arch. Biochem. Biophys. 1; 393 (1): 51-60 (2001)].
[0034] Числовые значения, применительно к идентичности по настоящему изобретению, могут представлять собой числовые значения, рассчитанные с использованием программы поиска гомологии, известной специалисту в данной области, если не указано иное. Примеры числовых значений для нуклеотидных последовательностей включают числовые значения, рассчитанные с использованием параметров по умолчанию в BLAST [J. Mol. Biol., 215, 403 (1990)], и примеры числовых значений для аминокислотных последовательностей включают числовые значения, рассчитанные с использованием параметров по умолчанию в BLAST2 [Nucleic Acids Res., 25, 3389 (1997), Genome Res., 7, 649 (1997), http://www.ncbi.nlm.nih.gov/Education/BLASTinfo/information3.htmL].
[0035] APS-киназа образует PAPS из APS в соответствии со следующей формулой реакции.
[0036] Химическая формула 2
APS+ATP → PAPS+ADP
(В вышеуказанной формуле, ADP представляет собой аденозин-5'-дифосфат.)
[0037] Один аспект APS-киназы включает MET14. Источник APS-киназы не является конкретно ограниченным, и ее примеры включают APS киназы, происходящие из Saccharomyces cerevisiae, Candida albicans, Shizosaccharomyces pombe, Yarrowia lipolytica, Neurospora crassa, Penicillium chrysogenum, Kluyveromyces lactis, Fusarium fujikuroi, Aspergillus oryzae или Ashbya gossypii, и среди них, APS-киназа, происходящая из Saccharomyces cerevisiae, является предпочтительной.
[0038] Примеры гена, кодирующего APS-киназу, включают нуклеотидную последовательность, показанную в SEQ ID NO: 23. Его примеры дополнительно включают ДНК, кодирующую полипептид, имеющий активность APS-киназы, и содержащую нуклеотидную последовательность, которая является предпочтительно на 80% или более, более предпочтительно, на 90% или более, даже более предпочтительно, на 95% или более, и особенно предпочтительно, на 98% или более идентичной нуклеотидной последовательности, показанной в SEQ ID NO: 23; и ДНК, кодирующую полипептид, имеющий активность APS-киназы, и содержащую нуклеотидную последовательность, которая гибридизуется, в строгих условиях, с нуклеотидной последовательностью, показанной в SEQ ID NO: 23, или нуклеотидную последовательность, комплементарную вышеуказанной нуклеотидной последовательности.
[0039] Активность APS-киназы в трансформанте проверяют по увеличению значений активности APS-киназы в жидком экстракте клеток трансформанта. Активность APS-киназы можно проверять посредством способа, описанного в литературе [Renosto F. et al., Adenosine 5'-phosphosulfate kinase from Penicillium chrysogenum. Purification and kinetic characterization. J. Biol. Chem. 259 (4): 2113-2123 (1984)].
[0040] Введение гена, кодирующего ATP-сульфурилазу, и гена, кодирующего APS-киназу, в бактерию из рода Corynebacterium можно проводить, соответственно, посредством введения вышеописанных ДНК в хромосому хозяина, или посредством клонирования вышеописанных ДНК в соответствующий плазмидный вектор, который может амплифицироваться в хозяине, и введения вектора в хозяина.
[0041] Плазмидный вектор может представлять собой любой плазмидный вектор, при условии, что он имеет ген, контролирующий функцию автономной репликации в бактериях из рода Corynebacterium. Его конкретные примеры включают pAM330, происходящую из Brevibacterium lactofermentum 2256 [JP-A-S58-67699], [Miwa, K. et al., Cryptic plasmids in glutamic acid-producing bacteria. Agric. Biol. Chem. 48: 2901-2903 (1984)], и [Yamaguchi, R. et al., Determination of the complete nucleotide sequence of the Brevibacterium lactofermentum plasmid pAM330 and the analysis of its genetic information. Nucleic Acids Symp. Ser. 16: 265-267 (1985)]; pHM1519 [Miwa, K. et al., Cryptic plasmids in glutamic acid-producing bacteria. Agric. Biol. Chem. 48: 2901-2903 (1984)] и pCRY30 [Kurusu, Y. et al., Identification of plasmid partition function in coryneform bacteria. Appl. Environ. Microbiol. 57: 759-764 (1991)], происходящие из Corynebacterium glutamicum ATCC 3058; pCG4 [JP-A-S57-183799] и [Katsumata, R. et al., Protoplast transformation of glutamate-producing bacteria with plasmid DNA. J. Bacteriol., 159: 306-311 (1984)], pAG1, pAG3, pAG14 и pAG50 [JP-A-S62-166890], и pEK0, pEC5 и pEKEx1 [Eikmanns, B.J. et al., A family of Corynebacterium glutamicum/Escherichia coli shuttle vectors for cloning, controlled gene expression, and promoter probing. Gene, 102: 93-98 (1991)], происходящие из Corynebacterium glutamicum T250, и т.п.
[0042] Промотор может представлять собой промотор, происходящий из хозяина, или гетерологичный промотор. Его примеры включают промотор PgapA гена глицеральдегид-3-фосфат-дегидрогеназы A (gapA), промотор Pmdh гена малат-дегидрогеназы (mdh), промотор PldhA гена лактат-дегидрогеназы A (ldhA) и т.п., происходящие из Corynebacterium glutamicum R.
[0043] Примеры терминатора включают терминатор rrnB T1T2 оперона рРНК из Escherichia coli, терминатор trpA из Escherichia coli, терминатор trp из Brevibacterium lactofermentum и т.п.
[0044] В качестве одного аспекта настоящего изобретения, экспрессия гена фермента, связанного с деградацией PAPS, не является ослабленной. Примеры гена фермента, связанного с деградацией PAPS, включают ген cysQ и ген CP01850. В качестве одного аспекта трансформанта (a), экспрессия по меньшей мере одного гена, выбранного из гена cysQ и гена CP01850, не является ослабленной.
[0045] Трансформант (b): C5-эпимеризация
В способе продукции сульфатированного полисахарида в соответствии с настоящим изобретением, C5-эпимеризацию N-сульфогепаросана проводят с использованием трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b).
[0046] C5-эпимераза не является конкретно ограниченной, при условии, что она может катализировать изомеризацию остатков глюкуроновой кислоты (GlcUA) до остатков идуроновой кислоты (IdoA). C5-эпимераза может являться происходящей из любого из животных, растений, микроорганизмов и т.п. Например, C5-эпимеразу человека можно использовать в качестве C5-эпимеразы.
[0047] Примеры гена, кодирующего C5-эпимеразу, включают нуклеотидную последовательность, показанную в SEQ ID NO: 24. Его примеры дополнительно включают ДНК, кодирующую полипептид, имеющий активность C5-эпимеразы, и содержащую нуклеотидную последовательность, которая является, предпочтительно, на 80% или более, более предпочтительно, на 90% или более, даже более предпочтительно, на 95% или более, и особенно предпочтительно, на 98% или более идентичной нуклеотидной последовательности, показанной в SEQ ID NO: 24, и ДНК, кодирующую полипептид, имеющий активность C5-эпимеразы, и содержащую нуклеотидную последовательность, которая гибридизуется, в строгих условиях, с нуклеотидной последовательностью, показанной в SEQ ID NO: 24, или нуклеотидную последовательность, комплементарную вышеуказанной нуклеотидной последовательности.
[0048] Активность C5-эпимеразы в трансформанте проверяют по увеличению значений активности C5-эпимеразы в жидком экстракте клеток трансформанта. Активность C5-эпимеразы можно измерять посредством способа, описанного в литературе [Babu P. et al., A rapid, nonradioactive assay for measuring heparan sulfate C-5 epimerase activity using hydrogen/deuterium exchange-mass spectrometry. Methods. Mol. Biol. 1229: 209-219 (2015)].
[0049] Далее в настоящем описании будет описан трансформант, экспрессирующей сульфотрансферазу. В способе продукции сульфатированного полисахарида по настоящему изобретению, сульфатирование проводят с использованием трансформанта, содержащего ген, кодирующий сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта. По настоящему изобретению, сульфотрансфераза не является конкретно ограниченной, при условии, что она переносит группу сульфата на полисахарид для получения сульфатированного полисахарида. Примеры сульфотрансферазы включают 2-O-сульфотрансферазу (2-OST), 6-O-сульфотрансферазу (6-OST) и 3-O-сульфотрансферазу (3-OST).
[0050] Трансформант (c): 2-O-сульфатирование
В способе продукции сульфатированного полисахарида по настоящему изобретению, 2-O-сульфатирование проводят с использованием трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу (2-OST), который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c).
[0051] 2-OST не является конкретно ограниченной, при условии, что она может катализировать сульфатирование остатков IdoA в положении O-2. Кроме того, 2-OST может являться происходящей из любого из животных, растений, микроорганизмов и т.п. Например, 2-OST, происходящую из хомяка, можно использовать в качестве 2-OST.
[0052] Примеры гена, кодирующего 2-OST, включают нуклеотидную последовательность, показанную в SEQ ID NO: 19. Его примеры дополнительно включают ДНК, кодирующую полипептид, имеющий активность 2-OST, и содержащую нуклеотидную последовательность, которая является, предпочтительно, на 80% или более, более предпочтительно, на 90% или более, даже более предпочтительно, на 95% или более, и особенно предпочтительно, на 98% или более идентичной нуклеотидной последовательности, показанной в SEQ ID NO: 19; и ДНК, кодирующую полипептид, имеющий активность 2-OST, и содержащую нуклеотидную последовательность, которая гибридизуется, в строгих условиях, с нуклеотидной последовательностью, показанной в SEQ ID NO: 19, или нуклеотидную последовательность, комплементарную вышеуказанной нуклеотидной последовательности.
[0053] Активность 2-OST в трансформанте проверяют по увеличению значений активности 2-OST в жидком экстракте клеток трансформанта. Активность 2-OST можно проверять посредством способа, описанного в литературе [Zhang J. et al., High cell density cultivation of recombinant Escherichia coli strains expressing 2-O-sulfotransferase and C5-epimerase for the production of bioengineered heparin. Appl. Biochem. Biotechnol. 175(6): 2986-2995 (2015)].
[0054] Трансформант (d): 6-O-сульфатирование
В способе продукции сульфатированного полисахарида по настоящему изобретению, 6-O-сульфатирование проводят с использованием трансформанта (d) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 6-O-сульфотрансферазу (6-OST), который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (d).
[0055] 6-OST не является конкретно ограниченной при условии, что она может катализировать сульфатирование остатков N-сульфатированного глюкозамина (GlcNS) в положении O-6. 6-OST может являться происходящей из любого из животных, растений, микроорганизмов и т.п. Примеры 6-OST включают 6-OST-1, происходящую из хомяка, и 6-OST-3, происходящую из мыши.
[0056] Примеры гена, кодирующего 6-OST, включают нуклеотидную последовательность, показанную в SEQ ID NO: 25. Его примеры дополнительно включают ДНК, кодирующую полипептид, имеющий активность 6-OST, и содержащую нуклеотидную последовательность, которая является, предпочтительно, на 80% или более, более предпочтительно, на 90% или более, даже более предпочтительно, на 95% или более, и особенно предпочтительно, на 98% или более идентичной нуклеотидной последовательности, показанной в SEQ ID NO: 25; и ДНК, кодирующую полипептид, имеющий активность 6-OST, и содержащую нуклеотидную последовательность, которая гибридизуется, в строгих условиях, с нуклеотидной последовательностью, показанной в SEQ ID NO: 25, или нуклеотидную последовательность, комплементарную вышеуказанной нуклеотидной последовательности.
[0057] Активность 6-OST в трансформанте проверяют по увеличению значений активности 6-OST в жидком экстракте клеток трансформанта. Активность 6-OST можно проверять посредством способа, описанного в литературе [Zhang J. et al., High cell density cultivation of a recombinant Escherichia coli strain expressing a 6-O-sulfotransferase for the production of bioengineered heparin. J. Appl. Microbiol. 118(1): 92-98 (2015)].
[0058] Трансформант (e): 3-O-сульфатирование
В способе продукции сульфатированного полисахарида по настоящему изобретению, 3-O-сульфатирование проводят с использованием трансформанта (e) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 3-O-сульфотрансферазу (3-OST), который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (e).
[0059] 3-OST не является конкретно ограниченной, при условии, что она может катализировать сульфатирование остатков N-сульфатированного/6-O-сульфатированного глюкозамина в положении O-3. 3-OST может являться происходящей из любого из животных, растений, микроорганизмов и т.п. Например, 3-OST-1, происходящую из мыши, можно использовать в качестве 3-OST.
[0060] Примеры гена, кодирующего 3-OST, включают нуклеотидную последовательность, показанную в SEQ ID NO: 26. Его примеры дополнительно включают ДНК, кодирующую полипептид, имеющий активность 3-OST, и содержащую нуклеотидную последовательность, которая является, предпочтительно, на 80% или более, более предпочтительно, на 90% или более, даже более предпочтительно, на 95% или более, и особенно предпочтительно, на 98% или более идентичной нуклеотидной последовательности, показанной в SEQ ID NO: 26; и ДНК, кодирующую полипептид, имеющий активность 3-OST, и содержащую нуклеотидную последовательность, которая гибридизуется, в строгих условиях, с нуклеотидной последовательностью, показанной в SEQ ID NO: 26, или нуклеотидную последовательность, комплементарную вышеуказанной нуклеотидной последовательности.
[0061] Активность 3-OST в трансформанте проверяют по увеличению значений активности 3-OST в жидком экстракте клеток трансформанта. Активность 3-OST можно проверять посредством способа, описанного в литературе [Jin W. et al., Increased soluble heterologous expression of a rat brain 3-O-sulfotransferase 1 - A key enzyme for heparin biosynthesis. Protein Expr. Purif. 151:23-29 (2018)].
[0062] Микроорганизм, принадлежащий к прокариотам
Микроорганизм, принадлежащий к прокариотам, используемый в качестве хозяина трансформанта по настоящему изобретению, не является конкретно ограниченным, при условии, что он может экспрессировать предопределенный ген по настоящему изобретению. Его примеры включают бактерии, такие как микроорганизмы, принадлежащие к роду Escherichia, роду Serratia, роду Bacillus, роду Corynebacterium, роду Microbacterium и роду Pseudomonas, и предпочтительно, используют бактерию из рода Escherichia.
[0063] Бактерия, принадлежащая к роду Escherichia, не является конкретно ограниченной, и ее примеры включают бактерию, классифицированную в род Escherichia, по классификации, известной экспертам в микробиологии. Примеры бактерий, принадлежащих к роду Escherichia, включают бактерии, описанные в литературе [Backmann, BJ 1996. Derivations and Genotypes of some mutant derivatives of Escherichia coli K-12, p. 2460-2488. Таблица 1. In F.D. Neidhardt (ed.), Escherichia coli and Salmonella Cellular and Molecular Biology/Second Edition, American Society for Microbiology Press, Washington, DC].
[0064] Примеры бактерий, принадлежащих к роду Escherichia, включают Escherichia coli. Примеры Escherichia coli включают Escherichia coli штаммов K-12, таких как штамм W3110 (ATCC 27325) и штамм MG1655 (ATCC 47076); Escherichia coli штамма K5 (ATCC 23506); Escherichia coli штаммов B, таких как штамм BL21 (DE3); Escherichia coli штамма Nissle 1917 (DSM 6601); и их производные штаммы.
[0065] Эти штаммы являются доступными, например, из Американской коллекции типовых культур (адрес: 12301 Parklawn Drive, Rockville, Maryland 20852, P.O. Box 1549, Manassas, VA 20108, United States of America). То есть, регистрационный номер присвоен каждому штамму, и штаммы могут быть получены с использованием этого регистрационного номера (ссылка https://www.atcc.org/). Регистрационный номер, соответствующий каждому штамму, описан в каталоге Американской коллекции типовых культур. Кроме того, штамм BL21 (DE3) является доступным, например, из Life Technologies (номер продукта C6000-03).
[0066] Для цели введения генов подходящим для экспрессии образом, можно использовать вектор, который может автономно реплицироваться в хозяине, в качестве вектора, используемого для трансформации микроорганизма, принадлежащего к прокариотам. Вектор, предпочтительно, представляет собой мультикопийный вектор. Кроме того, вектор, предпочтительно, имеет маркер, такой как ген устойчивости к антибиотику или другой ген, описанный в литературе [Karl Friehs, Plasmid Copy Number and Plasmid Stability, Adv Biochem Engin/Biotechnol 86: 47-82 (2004)], для отбора трансформанта. Кроме того, вектор может иметь промотор или терминатор для экспрессии вставленного гена. Примеры вектора включают вектор, происходящий из бактериальной плазмиды, вектор, происходящий из дрожжевой плазмиды, вектор, происходящий из бактериофага, космиды, фагмиды и т.п.
[0067] Конкретные примеры векторов, способных к автономной репликации в бактериях из рода Escherichia coli, включают pUC19, pUC18, pHSG299, pHSG399, pHSG398, pBR322 и pSTV29 (все из Takara Bio Inc.), pACYC184 и pMW219 (Nippon Gene), pTrc99A (Pharmacia), вектор pPROK (Clontech), pKK233-2 (Clontech), вектор pET (Novagen), вектор pQE (Qiagen) и вектор с широким спектром хозяев RSF1010.
[0068] Промотор может представлять собой промотор, происходящий из хозяина, или гетерологичный промотор. Промотор может представлять собой собственный промотор гена, подлежащего введению, или промотор других генов.
[0069] Примеры терминаторов включают терминатор T7, терминатор T4, терминатор фага fd, терминатор tet и терминатор trpA.
[0070] В вышеупомянутых трансформантах (b) - (e), два или более из генов, введенных в микроорганизм, принадлежащий к прокариотам, могут быть введены в один трансформант. Более конкретно, например, ген, кодирующий C5-эпимеразу, и ген, кодирующий 2-O-сульфотрансферазу, могут быть введены в один микроорганизм, принадлежащий к прокариотам, для получения одного трансформанта.
[0071] Обработанный материал трансформанта
Обработанный материал трансформанта по настоящему изобретению относится к материалу, в котором клеточная плазматическая мембрана бактериальной клетки является проницаемой. По настоящему изобретению, когда клеточная плазматическая мембрана обозначена как являющаяся проницаемой для веществ, это означает, что различные малые (ионы и т.п.) и большие (белки и т.п.) молекулы проникают через клеточную мембрану посредством диффузии и таким образом, могут свободно входить в и покидать клеточную мембрану. Обработанный материал трансформанта по настоящему изобретению, предпочтительно, представляет собой покоящуюся бактериальную клетку, утратившую свою способность к росту из-за обработки для придания проницаемости мембраны.
[0072] Примеры обработанного материала трансформанта включают обработанный поверхностно-активным веществом материал бактериальной клетки, представляющей собой трансформант, обработанный растворителем материал бактериальной клетки, обработанный ферментом материал бактериальной клетки, обработанный материал, содержащий живые бактериальные клетки, которые сохраняют такую же функцию, как культура бактериальных клеток, в качестве источника фермента, такой как иммобилизованный продукт бактериальной клетки, обработанный ультразвуком материал бактериальных клеток и механически измельченный обработанный материал бактериальной клетки.
[0073] Примеры способов сделать клеточную плазматическую мембрану проницаемой для веществ включают химическую обработку и механическую обработку. В способе продукции по настоящему изобретению, временные рамки, в которых клеточную плазматическую мембрану трансформанта делают проницаемой для веществ, не являются конкретно ограниченными, при условии, что проявляются эффекты по настоящему изобретению. Клеточную плазматическую мембрану каждого трансформанта можно сделать проницаемой для веществ заранее, или это можно сделать, когда реакцию проводят посредством приведения трансформантов, используемых в реакции, в контакт друг с другом.
[0074] Примеры химической обработки включают способ с использованием поверхностно-активного вещества, способ с использованием органического растворителя и способ с использованием фермента. В качестве поверхностно-активного вещества, неионное поверхностно-активное вещество является предпочтительным, поскольку его действие на белки или т.п. является более мягким (по сравнению с ионными поверхностно-активными веществами). Примеры поверхностно-активных веществ включают дигитонин, сапонин, Triton X100, Triton X114, Tween 20, Tween 80, N, N-бис(3-D-глюконамидопропил)холамид [BIGCHAP], N, N-бис(3-D-глюконамидопропил)дезоксихоламид [дезокси-BIGCHAP], NIKKOLBL-9EX [полиоксиэтилен(9)лауриловый эфир], октаноил-N-метилглюкамид [MEGA-8], хлорид бензалкония и т.п.
[0075] Примеры органических растворителей включают бензол, толуол, ксилол, другие спирты и т.п. Примеры ферментов включают лизоцим, ахромопептидазу и т.п.
[0076] Условия, такие как концентрация, температура, время, и т.п., обработки с использованием вышеупомянутых веществ, отличаются в зависимости от типа клеток, и необходимо устанавливать соответствующие условия для проведения желательного анализа, однако, обычная концентрация при обработке составляет 10-1000 микрограмм/мл, и более обычно, 20-200 микрограмм/мл при температуре 2 градуса C - 37 градусов C и времени 1-30 минут.
[0077] Примеры механической обработки включают обработку ультразвуком и обработку механическим измельчением.
[0078] Экстракт трансформанта
Примеры экстрактов трансформанта по настоящему изобретению включают неочищенный экстракт фермента, полученный из бактериальной клетки, представляющей собой трансформант, очищенный фермент, полученный из бактериальной клетки, обработанной, как выше (химически или механически, например), концентрат культуры, полученной посредством культивирования вышеописанного трансформанта или его обработанного материала, и высушенный продукт культуры. Примеры экстракта трансформанта также включают бактериальные клетки, полученные посредством центрифугирования или фильтрации культуры, высушенные бактериальные клетки и лиофилизированные бактериальные клетки.
[0079] Способ трансформации и способ культивирования трансформанта
Можно использовать известные способы трансформации, без ограничения. Примеры таких известных способов включают способ с использованием хлорида кальция/хлорида рубидия, способ с использованием фосфата кальция, опосредованную DEAE-декстраном трансфекцию, способ с использованием электрических импульсов и т.п. Среди них, способ с использованием электрических импульсов является пригодным для коринебактерий, и способ с использованием электрических импульсов можно осуществлять посредством известного способа [Kurusu, Y. et al., Electroporation-transformation system for Coryneform bacteria by auxotrophic complementation. Agric Biol. Chem. 54: 443-447 (1990)].
[0080] Является предпочтительным выращивание трансформанта посредством культивирования с использованием среды, обычно используемой для культивирования микроорганизма, перед каждой реакцией. В качестве среды, можно в общем использовать природную среду или синтетическую среду, содержащую источник углерода, источник азота, неорганические соли и другие питательные вещества.
[0081] Источник углерода может представлять собой источник ATP. Примеры источников углерода включают углеводы или сахарные спирты, такие как глюкоза, фруктоза, сахароза, манноза, мальтоза, маннит, ксилоза, арабиноза, галактоза, крахмал, меласса, сорбит и глицерин; органические кислоты, такие как уксусная кислота, лимонная кислота, молочная кислота, фумаровая кислота, малеиновая кислота и глюконовая кислота; и спирты, такие как этанол и пропанол. В качестве источника углерода, один вид можно использовать отдельно, или два или более их видов можно смешивать. Как правило, является достаточным, чтобы концентрация этих источников углерода в среде составляла приблизительно 0,1-10 (масс./об.%).
[0082] Примеры источников азота включают неорганические или органические соединения аммония, такие как хлорид аммония, сульфат аммония, нитрат аммония и ацетат аммония, мочевина, водный аммиак, нитрат натрия, нитрат калия и т.п. Кроме того, можно использовать также жидкий кукурузный экстракт, мясной экстракт, пептон, NZ-амин, гидролизат белка, содержащие азот органические соединения, такие как аминокислоты, и т.п. В качестве источника азота, один вид можно использовать отдельно, или два или более их видов можно смешивать и использовать. Концентрация источников азота в среде меняется в зависимости от используемого соединения азота, но, как правило, составляет приблизительно 0,1-10 (масс./об.%).
[0083] Примеры неорганических солей включают одноосновный фосфат калия, двухосновный фосфат калия, нитрат железа, хлорид натрия, карбонат кальция и т.п., в дополнение к источникам ионов сульфата, таким как сульфат магния, сульфат марганца, сульфат цинка и сульфат кобальта. В качестве неорганических солей, один вид можно использовать отдельно, или два или более их видов можно смешивать и использовать. Концентрация неорганических солей в среде меняется в зависимости от используемой неорганической соли, но, как правило, составляет приблизительно 0,01-1 (масс./об.%).
[0084] Примеры питательных веществ включают мясной экстракт, пептон, полипептон, дрожжевой экстракт, сухие дрожжи, жидкий кукурузный экстракт, порошок обезжиренного молока, обезжиренный гидрохлорид гидролизата сои, экстракты клеток животных и растений или бактериальных микроорганизмов, и продукты их деградации, и т.п., и их концентрация, как правило, составляет приблизительно 0,1-10 (масс./об.%). Кроме того, витамины можно добавлять, по необходимости. Примеры витаминов включают биотин, тиамин (витамин B1), пиридоксин (витамин B6), пантотеновую кислоту, инозитол, никотиновую кислоту и т.п. pH среды, предпочтительно, составляет приблизительно 6-8.
[0085] Предпочтительные примеры культуральных сред для микроорганизмов включают среду A [Inui, M. et al., Metabolic analysis of Corynebacterium glutamicum during lactate and succinate productions under oxygen deprivation conditions. J. Mol. Microbiol. Biotechnol. 7:182-196 (2004)], среду BT [Omumasaba, C.A. et al., Corynebacterium glutamicum glyceraldehyde-3-phosphate dehydrogenase isoforms with opposite, ATP-dependent regulation. J. Mol. Microbiol. Biotechnol. 8:91-103 (2004)] и т.п. В качестве конкретных условий культивирования, например, температура культивирования составляет приблизительно 15 градусов C - 45 градусов C, и время культивирования составляет приблизительно 1-7 суток.
[0086] Способ продукции сульфатированного полисахарида
Один аспект способа продукции сульфатированного полисахарида, в соответствии с настоящим изобретением, представляет собой способ продукции сульфатированного полисахарида, включающий получение сульфатированного полисахарида посредством включения в реакционный раствор, в присутствии ATP или источника ATP, источника иона сульфата и N-сульфогепаросана, трансформанта (a) или его обработанного материала, и по меньшей мере одного, выбранного из трансформантов (b) - (e) или их обработанных материалов или экстрактов.
[0087] В качестве одного аспекта способа продукции сульфатированного полисахарида, в соответствии с настоящим изобретением, его примеры включают способ продукции сульфатированного полисахарида, включающий получение сульфатированного полисахарида посредством использования реакционного раствора, содержащего трансформант (a) или его обработанный материал, и трансформанты (b) - (e) или их обработанные материалы или экстракты, в присутствии ATP или источника ATP, источника иона сульфата и N-сульфогепаросана.
[0088] Кроме того, один аспект способа продукции сульфатированного полисахарида, в соответствии с настоящим изобретением, представляет собой способ продукции сульфатированного полисахарида, включающий получение сульфатированного полисахарида посредством включения по меньшей мере одного, выбранного из трансформантов (b) - (e) или их обработанных материалов или экстрактов, в реакционный раствор, в присутствии PAPS и N-сульфогепаросана.
[0089] В способе продукции по настоящему изобретению, каждый трансформант можно первоначально добавлять в реакционный раствор, можно последовательно добавлять в него, или можно первоначально и последовательно добавлять в него. Конкретные примеры аспектов первоначального добавления каждого трансформанта в реакционный раствор включают аспект, в котором сульфатированные полисахариды получают посредством первоначального добавления трансформанта (a) или его обработанного материала, и трансформантов (b) - (e), или их обработанных материалов или экстрактов в реакционный раствор, в присутствии ATP или источника ATP, источника иона сульфата и N-сульфогепаросана.
[0090] В способе продукции сульфатированного полисахарида по настоящему изобретению, с использованием трансформанта (a) или его обработанного материала и трансформантов (b) - (e), или их обработанных материалов или экстрактов, является возможным осуществлять 1) снабжение/регенерацию PAPS с использованием трансформанта (a) или его обработанного материала, 2) C5-эпимеризацию с использованием трансформанта (b), или его обработанного материала или экстракта, 3) 2-O-сульфатирование с использованием трансформанта (c), или его обработанного материала или экстракта, 4) 6-O-сульфатирование с использованием трансформанта (d), или его обработанного материала или экстракта, и 5) 3-O-сульфатирование с использованием трансформанта (e), или его обработанного материала или экстракта.
[0091] Далее в настоящем описании, каждая из стадий будет описана отдельно, но порядок 2) C5-эпимеризация, 3) 2-O-сульфатирование, 4) 6-O-сульфатирование и 5) 3-O-сульфатирование не является конкретно ограниченным, при условии, что желательные сульфатированные полисахариды могут быть получены.
[0092] Кроме того, далее в настоящем описании, каждая реакция из 1) снабжения/регенерации PAPS, 2) C5-эпимеризации, 3) 2-O-сульфатирования, 4) 6-O-сульфатирования и 5) 3-O-сульфатирования будет описана отдельно, но две или более из этих реакций можно проводить в одно и то же время. Например, эти реакции можно проводить в одно и то же время с использованием реакционного раствора, содержащего все из трансформанта (a) или его обработанного материала и трансформантов (b) - (e), или их обработанных материалов или экстрактов.
[0093] Примеры сульфатированного полисахарида, продуцированного посредством способа продукции сульфатированного полисахарида по настоящему изобретению, включают гепарин.
[0094] Снабжение/регенерация PAPS
Способ продукции сульфатированного полисахарида по настоящему изобретению отличается включением следующих стадий (1-1) и (1-2):
(1-1) получение трансформанта (a) бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта (a); и
(1-2) проведение реакции для продукции PAPS посредством использования реакционного раствора, содержащего ATP или источник ATP, источник иона сульфата и трансформант (a) или его обработанный материал.
[0095] ATP-сульфурилаза и APS-киназа, экспрессированные посредством трансформанта (a) или его обработанного материала, вступают в реакцию с ATP или источником ATP и источником иона сульфата в трансформанте (a) или его обработанном материале, и таким образом, продуцируется PAPS. PAPS функционирует в качестве донора сульфатной группы в продукции сульфатированных полисахаридов. Кроме того, PAPS, продуцированный посредством трансформанта (a) или его обработанного материала, используют в способе продукции сульфатированного полисахарида, и таким образом, получают PAP. PAPS можно продуцировать из этого PAP посредством трансформанта (a) или его обработанного материала. Соответственно, можно осуществлять снабжение/регенерацию PAPS посредством включения трансформанта (a) или его обработанного материала в реакционный раствор.
[0096] C5-эпимеризация
Способ продукции сульфатированного полисахарида по настоящему изобретению, предпочтительно, включает следующие стадии (2-1) и (2-2):
(2-1) получение трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b); и
(2-2) проведение C5-эпимеризации посредством включения трансформанта (b), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
[0097] На стадии (2-2), C5-эпимеризацию N-сульфогепаросана проводят посредством трансформанта (b), экспрессирующего C5-эпимеразу, или его обработанного материала или экстракта.
[0098] 2-O-сульфатирование
Способ продукции сульфатированного полисахарида по настоящему изобретению, предпочтительно, включает следующие стадии (3-1) и (3-2):
(3-1) получение трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c); и
(3-2) проведение 2-O-сульфатирования посредством включения трансформанта (c), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
[0099] На стадии (3-2), 2-O-сульфатирование N-сульфогепаросана проводят посредством трансформанта (c), экспрессирующего 2-O-сульфотрансферазу, или его обработанного материала или экстракта, с использованием PAPS, продуцированного посредством трансформанта (a) или его обработанного материала, в качестве донора сульфатной группы. Когда сульфатированный полисахарид, продуцированный на стадии (2-2), существует в реакционном растворе, проводят 2-O-сульфатирование сульфатированного полисахарида, продуцированного на этой стадии. Например, когда существует N-сульфогепаросан с C5-эпимеризацией, проводят 2-O-сульфатирование N-сульфогепаросана с C5-эпимеризацией.
[0100] Вышеупомянутые C5-эпимеризацию и 2-O-сульфатирование можно проводить в одно и то же время. Таким образом, один аспект способа продукции сульфатированного полисахарида по настоящему изобретению, предпочтительно, включает следующие стадии (3'-1) - (3'-3):
(3'-1) получение трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b); (3'-2) получение трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c); и (3'-3) проведение C5-эпимеризации и 2-O-сульфатирования посредством включения трансформанта (b), или его обработанного материала или экстракта, и трансформанта (c), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
[0101] 6-O-сульфатирование
Способ продукции сульфатированного полисахарида по настоящему изобретению, предпочтительно, включает следующие стадии (4-1) и (4-2):
(4-1) получение трансформанта (d) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 6-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (d); и
(4-2) проведение 6-O-сульфатирования посредством включения трансформанта (d), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
[0102] На стадии (4-2), 6-O-сульфатирование N-сульфогепаросана проводят посредством трансформанта (d), экспрессирующего 6-O-сульфотрансферазу, или его обработанного материала или экстракта, с использованием PAPS, продуцированного посредством трансформанта (a) или его обработанного материала, в качестве донора сульфатной группы.
[0103] Когда сульфатированный полисахарид, продуцированный на стадии (2-2), (3-2) или (3'-3), существует в реакционном растворе, проводят 6-O-сульфатирование сульфатированного полисахарида, продуцированного на этой стадии.
[0104] Например, когда существует N-сульфогепаросан с C5-эпимеризацией, проводят 6-O-сульфатирование N-сульфогепаросана с C5-эпимеризацией. Когда существует N-сульфогепаросан с 2-O-сульфатированием, проводят 6-O-сульфатирование N-сульфогепаросана с 2-O-сульфатированием.
[0105] Когда существует N-сульфогепаросан с C5-эпимеризацией и 2-O-сульфатированием, проводят 6-O-сульфатирование N-сульфогепаросана с C5-эпимеризацией и 2-O-сульфатированием.
[0106] 3-O-сульфатирование
Способ продукции сульфатированного полисахарида по настоящему изобретению, предпочтительно, включает следующие стадии (5-1) и (5-2):
(5-1) получение трансформанта (e) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 3-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (e); и
(5-2) проведение 3-O-сульфатирования посредством включения трансформанта (e), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
[0107] На стадии (5-2), 3-O-сульфатирование N-сульфогепаросана проводят посредством трансформанта (e), экспрессирующего 3-O-сульфотрансферазу, или его обработанного материала или экстракта, с использованием PAPS, продуцированного посредством трансформанта (a) или его обработанного материала, в качестве донора сульфатной группы.
[0108] Когда сульфатированный полисахарид, продуцированный на стадии (2-2), (3-2), (3'-3) или (4-2), существует в реакционном растворе, проводят 3-O-сульфатирование сульфатированного полисахарида, продуцированного на этой стадии.
[0109] Например, когда существует N-сульфогепаросан с C5-эпимеризацией, проводят 3-O-сульфатирование N-сульфогепаросана с C5-эпимеризацией. Когда существует N-сульфогепаросан с 2-O-сульфатированием, проводят 3-O-сульфатирование N-сульфогепаросана с 2-O-сульфатированием. Когда существует N-сульфогепаросан с 6-O-сульфатированием, проводят 3-O-сульфатирование N-сульфогепаросана с 6-O-сульфатированием.
[0110] Когда существует N-сульфогепаросан с C5-эпимеризацией и 2-O-сульфатированием, проводят 3-O-сульфатирование N-сульфогепаросана с C5-эпимеризацией и 2-O-сульфатированием. Когда существует N-сульфогепаросан с C5-эпимеризацией и 6-O-сульфатированием, проводят 3-O-сульфатирование N-сульфогепаросана с C5-эпимеризацией и 6-O-сульфатированием. Когда существует N-сульфогепаросан с 2-O-сульфатированием и 6-O-сульфатированием, проводят 3-O-сульфатирование N-сульфогепаросана с 2-O-сульфатированием и 6-O-сульфатированием.
[0111] Когда существует N-сульфогепаросан с C5-эпимеризацией, 2-O-сульфатированием и 6-O-сульфатированием, проводят 3-O-сульфатирование N-сульфогепаросана с C5-эпимеризацией, 2-O-сульфатированием и 6-O-сульфатированием.
[0112] Условия реакции
Условия реакции для вышеупомянутых 1) снабжения/регенерации PAPS, 2) C5-эпимеризации, 3) 2-O-сульфатирования, 4) 6-O-сульфатирования и 5) 3-O-сульфатирования будут описаны ниже.
[0113] pH реакционного раствора, предпочтительно, составляет приблизительно 6-8. Во время реакции, является предпочтительным проведение реакции с использованием водного раствора аммиака, водного раствора гидроксида натрия, гидроксида калия или т.п. со средством для контроля pH, для контроля pH реакционного раствора, близкого к нейтральному, в частности, приблизительно 7.
[0114] Температура реакции, то есть, температура выживаемости трансформанта, во время реакции, составляет, предпочтительно, 20 градусов C - 50 градусов C, и более предпочтительно, 25 градусов C - 47 градусов C. Время реакции составляет, предпочтительно, приблизительно 1-7 суток и более предпочтительно, приблизительно 1-3 суток. Культивирование может представлять собой любой из периодического типа, периодического типа с подпиткой и непрерывного типа. Среди них, периодический тип является предпочтительным.
[0115] Условия аэрации реакции можно осуществлять в восстанавливающих условиях или микроаэробных условиях. Восстанавливающие условия определяют посредством окислительно-восстановительного потенциала реакционного раствора. Окислительно-восстановительный потенциал реакционного раствора составляет, предпочтительно, приблизительно -200 мВ - -500 мВ и более предпочтительно, -250 мВ - -500 мВ.
[0116] Состояние восстановления реакционного раствора можно легко оценивать с использованием индикатора резазурина, и можно точно измерять с использованием окислительно-восстановительного потенциометра. В качестве способа получения реакционного раствора в восстанавливающих условиях, можно использовать известный способ, без ограничения.
[0117] Конкретно, водный раствор для реакционного раствора в восстанавливающих условиях можно получать посредством удаления растворенного газа посредством подвергания дистиллированной воды или т.п. термической обработке или обработке пониженным давлением. Кроме того, подходящее восстанавливающее средство (например, тиогликолевую кислоту, аскорбиновую кислоту, гидрохлорид цистина, меркаптоуксусную кислоту, тиолуксусную кислоту, глутатион, сульфид натрия или т.п.) можно добавлять для получения водного раствора для реакционного раствора в восстанавливающих условиях. Подходящая комбинация этих способов также является эффективным способом получения водного раствора для реакционного раствора в восстанавливающих условиях.
[0118] В случае, когда восстанавливающие условия поддерживают во время реакции, является желательным предотвращение смешивания с кислородом извне реакционной системы, настолько строго, насколько возможно, и конкретные примеры такого способа включают способ, в котором реакционная система отгорожена с использованием инертного газа, такого как газ азот или газ диоксид углерода.
[0119] В случае, когда микроаэробные условия поддерживают во время реакции, реакцию можно проводить в условиях, в которых скорость аэрации установлена на низкое значение 0,5 об./об./мин или т.п. или более низкое значение, и скорость перемешивания установлена на низкое значение 500 об./мин или т.п., или более низкое значение. В некоторых случаях, реакцию можно проводить в комбинации с состоянием, в котором аэрацию останавливают с подходящим расписанием после начала реакции, и степень анаэробного состояния увеличивается в условиях скорости перемешивания 100 об./мин или менее.
[0120] Сбор сульфатированного полисахарида
Сульфатированный полисахарид продуцируют в реакционный раствор посредством культивирования, как описано выше. Сульфатированный полисахарид можно собирать посредством выделения реакционного раствора, и кроме того, сульфатированный полисахарид можно отделять от реакционного раствора посредством известного способа. Примеры таких известных способов включают способ дистилляции, способ пропускания через мембрану, способ экстракции органическим растворителем и т.п.
[0121] Получение N-сульфогепаросана
N-сульфогепаросан, используемый в способе продукции сульфатированного полисахарида по настоящему изобретению, получают посредством деацетилирования, деполимеризации и N-сульфатирования гепаросана. Получение гепаросана и получение N-сульфогепаросана из гепаросана можно проводить посредством известного способа (например, WO2018/048973).
[0122] Примеры способа продукции гепаросана включают способ, включающий культивирование микроорганизма, принадлежащего к прокариотам, который имеет генетическую модификацию из следующей (a1) и имеет способность к продукции гепаросана, в среде для продукции гепаросана, и сбор гепаросана из среды:
(a1) генетической модификации, увеличивающей уровень экспрессии гена kpsS.
Микроорганизм, принадлежащий к прокариотам, может дополнительно иметь по меньшей мере одну из следующих генетических модификаций (a2) и (a3), в дополнение к (a1):
(a2) генетической модификации, увеличивающей уровень экспрессии по меньшей мере одного гена, выбранного из генов kfiA, kfiB, kfiC и kfiD; и
(a3) генетической модификации, вызывающей потерю функции гена yhbJ
[0123] Ген kpsS представляет собой ген, кодируемый областью I, среди группы генов из области I, области II и области III. kpsS вовлечен в инициацию синтеза гепаросана. В продукции гепаросана, kpsS, вместе с kpsC, играет роль в добавлении множества линкеров Kdo к фосфатидилглицерину на внутренней мембране.
[0124] В качестве гена kpsS, ген kpsS, происходящий из рода Escherichia, является предпочтительным. Его конкретные примеры включают ген kpsS из Escherichia coli штамма K5. Нуклеотидная последовательность гена kpsS из Escherichia coli штамма K5 и аминокислотная последовательность белка, кодируемого геном, могут быть получены из публичных баз данных. Ген kpsS из Escherichia coli штамма K5 зарегистрирован как номер доступа в GenBank CAA52659.1.
[0125] Примеры гена kpsS включают ДНК, имеющую нуклеотидную последовательность, показанную в SEQ ID NO: 27, или ДНК имеющую свойство увеличения способности к продукции гепаросана микроорганизма, когда уровень экспрессии увеличен в микроорганизме, принадлежащем к прокариотам, имеющем нуклеотидную последовательность, на 90% или более идентичную нуклеотидной последовательности, показанной в SEQ ID NO: 27, и имеющем способность к продукции гепаросана.
[0126] kfiA, kfiB, kfiC и kfiD представляют собой гены, кодированные областью II, среди группы генов из области I, области II и области III. Как показано на ФИГ. 2, kfiA, kfiB, kfiC и kfiD вовлечены в синтез гепаросана, и играют роль добавления сахарида и таким образом, синтеза гепаросана.
[0127] В качестве гена kfiA, kfiB, kfiC или kfiD, ген kfiA, kfiB, kfiC или kfiD, происходящий из рода Escherichia, является предпочтительным. Его конкретные примеры включают ген kfiA, kfiB kfiC или kfiD из Escherichia coli штамма K5. Нуклеотидная последовательность гена kfiA, kfiB, kfiC или kfiD из Escherichia coli штамма K5 и аминокислотная последовательность белка, кодированного геном, могут быть получены из публичных баз данных. kfiA зарегистрирован как номер доступа в GenBank CAA54711.1; kfiB зарегистрирован как номер доступа в GenBank CAE55824.1; kfiC зарегистрирован как номер доступа в GenBank CAA54709.1; и kfiD зарегистрирован как номер доступа в GenBank CAA54708.1.
[0128] Примеры гена kfiA включают ДНК, имеющую нуклеотидную последовательность, показанную в SEQ ID NO: 28, или ДНК, имеющую свойство увеличения способности к продукции гепаросана микроорганизма, когда уровень экспрессии увеличен в микроорганизме, принадлежащем к прокариотам, имеющем нуклеотидную последовательность, на 90% или более идентичную нуклеотидной последовательности, показанной в SEQ ID NO: 28, и имеющем способность к продукции гепаросана.
[0129] Примеры гена kfiB включают ДНК, имеющую нуклеотидную последовательность, показанную в SEQ ID NO: 29, или ДНК, имеющую свойство увеличения способности к продукции гепаросана микроорганизма, когда уровень экспрессии увеличен в микроорганизме, принадлежащем к прокариотам, имеющем нуклеотидную последовательность, на 90% или более идентичную нуклеотидной последовательности, показанной в SEQ ID NO: 29, и имеющем способность к продукции гепаросана.
[0130] Примеры гена kfiC включают ДНК, имеющую нуклеотидную последовательность, показанную в SEQ ID NO: 30, или ДНК, имеющую свойство увеличения способности к продукции гепаросана микроорганизма, когда уровень экспрессии увеличен в микроорганизме, принадлежащем к прокариотам, имеющем нуклеотидную последовательность, на 90% или более идентичную нуклеотидной последовательности, показанной в SEQ ID NO: 30, и имеющем способность к продукции гепаросана.
[0131] Примеры гена kfiD включают ДНК, имеющую нуклеотидную последовательность, показанную в SEQ ID NO: 31, или ДНК, имеющую свойство увеличения способности к продукции гепаросана микроорганизма, когда уровень экспрессии увеличен в микроорганизме, принадлежащем к прокариотам, имеющем нуклеотидную последовательность, на 90% или более идентичную нуклеотидной последовательности, показанной в SEQ ID NO: 31, и имеющем способность к продукции гепаросана.
[0132] На ФИГ. 3 показана схематическая диаграмма биосинтетического пути гепаросана. GlmS представляет собой первый фермент в пути снабжения UDP-N-ацетилглюкозамином, являющимся предшественником гепаросана, и представляет собой фермент, который катализирует реакцию из фруктоза-6-фосфата до глюкозамин-6-фосфата. YhbJ представляет собой фермент, отрицательно контролирующий GlmS.
[0133] В качестве гена yhbJ, ген yhbJ, происходящий из рода Escherichia, является предпочтительным. Его конкретные примеры включают ген yhbJ из Escherichia coli штамма K-12. Нуклеотидная последовательность гена yhbJ из Escherichia coli штамма K-12 и аминокислотная последовательность белка, кодированного геном, могут быть получены из публичных баз данных. Ген yhbJ из Escherichia coli штамма K-12 зарегистрирован как номер доступа в GenBank BAE77249.1.
[0134] Примеры гена yhbJ включают ДНК, имеющую нуклеотидную последовательность, показанную в SEQ ID NO: 32, или ДНК, имеющую свойство увеличения способности к продукции гепаросана микроорганизма, когда уровень экспрессии уменьшен в микроорганизме, принадлежащем к прокариотам, имеющем нуклеотидную последовательность, на 90% или более идентичную нуклеотидной последовательности, показанной в SEQ ID NO: 32, и имеющем способность к продукции гепаросана.
[0135] Каждый из генов в (a1) - (a3) выше может быть легко получен из публичных баз данных посредством, например, поиска BLAST или поиска FASTA с использованием нуклеотидной последовательности каждого гена, описанной выше. Кроме того, гомолог каждого гена может быть получен, например, посредством ПЦР с использованием хромосомы микроорганизма, такого как бактерия, в качестве матрицы, и с использованием олигонуклеотида, полученного на основе этих известных последовательностей генов, в качестве праймера.
[0136] Каждый из генов в (a1) - (a3) выше могут представлять собой варианты генов, при условии, что исходные функции (например, активность или свойство) белка, кодированного генами, сохраняется. Можно проверять, сохраняет или нет белок, кодированный вариантами генов, свою исходную функцию; конкретно, например, когда исходной функцией является улучшение способности к продукции гепаросана, посредством введения варианта гена в микроорганизм, принадлежащий к прокариотам, имеющий способность к продукции гепаросана.
[0137] Варианты каждого из генов в (a1) - (a3) выше могут быть получены в соответствии со способом сайт-направленного мутагенеза, посредством модификации кодирующей области гена таким образом, что аминокислотные остатки в специфических положениях кодированного белка заменены, делетированы, вставлены или добавлены. Кроме того, варианты каждого из генов в (a1) - (a3) выше могут быть также получены, например, посредством обработки для мутагенеза.
[0138] При условии, что их исходные функции сохраняются, каждый из генов в (a1) - (a3) выше могут представлять собой гены, кодирующие белок, имеющий аминокислотную последовательность, в которой одна или несколько аминокислот в одном или нескольких положениях заменены, делетированы, вставлены или добавлены. Например, в кодированном белке, его N-конец и/или C-конец может быть удлинен или укорочен. Фраза «одна или несколько» отличается, в зависимости от положения и типа аминокислотного остатка в трехмерной структуре белка. Конкретные примеры этого включают 1-50, 1-40, 1-30, и это составляет, предпочтительно, 1-20, более предпочтительно, 1-10, даже более предпочтительно, 1-5, и особенно предпочтительно, 1-3.
[0139] Замена, делеция, вставка или добавление одной или нескольких аминокислот, как описано выше, представляет собой консервативную мутацию, которая нормально сохраняет функцию белка. Репрезентативными консервативными мутациями являются консервативные замены. Консервативная замена представляет собой мутацию, в которой замена возникает между Phe, Trp и Tyr, в случае, когда участок замены представляет собой ароматическую аминокислоту, замена возникает между Leu, Ile и Val, в случае, когда участок замены представляет собой гидрофобную аминокислоту, замена возникает между Gln и Asn, в случае, когда участок замены представляет собой полярную аминокислоту, замена возникает между Lys, Arg и His, в случае, когда участок замены представляет собой основную аминокислоту, замена возникает между Asp и Glu, в случае, когда участок замены представляет собой кислую аминокислоту, и замена возникает между Ser и Thr, в случае, когда участок замены представляет собой аминокислоту, имеющую гидроксильные группы. Конкретные примеры замен, рассматриваемых как консервативная замена, включают замену Ala на Ser или Thr; замену Arg на Gln, His или Lys; замену Asn на Glu, Gln, Lys, His или Asp; замену Asp на Asn, Glu или Gln; замену Cys на Ser или Ala; замену Gln на Asn, Glu, Lys, His, Asp или Arg; замену Glu на Gly, Asn, Gln, Lys или Asp; замену Gly на Pro; замену His на Asn, Lys, Gln, Arg или Tyr; замену Ile на Leu, Met, Val или Phe; замену Leu на Ile, Met, Val или Phe; замену Lys на Asn, Glu, Gln, His, или Arg; замену Met на Ile, Leu, Val или Phe; замену Phe на Trp, Tyr, Met, Ile или Leu; замену Ser на Thr или Ala; замену Thr на Ser или Ala; замену Trp на Phe или Tyr; замену Tyr на His, Phe или Trp; и замену Val на Met, Ile или Leu. Кроме того, замены, делеции, вставки, добавления аминокислот, как описано выше, их инверсии и т.п. включают замены, делеции, вставки и добавления, инверсии, и т.п., которые вызваны мутациями (мутанты или варианты), возникающими естественным образом, такими как мутации, основанные на индивидуальных различиях или видовых различиях в организме, из которого происходит ген.
[0140] Кроме того, при условии, что их исходные функции сохраняются, каждый из генов в (a1) - (a3) выше могут представлять собой гены, кодирующие белок, имеющий 80% или более, предпочтительно, 90% или более, более предпочтительно, 95% или более, даже более предпочтительно, 97% или более, и особенно предпочтительно, 99% или более идентичность с полной аминокислотной последовательностью.
[0141] Кроме того, при условии, что их исходные функции сохраняются, каждый из генов (a1) - (a3) выше может представлять собой ДНК, которая гибридизуется, в строгих условиях, с зондом, который может быть получен из известных последовательностей гена, таких как последовательность, комплементарная полной или части нуклеотидной последовательности. Термин «строгие условия» относится к условиям, в которых образуются так называемые специфические гибриды, и неспецифические гибриды не образуются. Их примеры включают условия, в которых ДНК, идентичные друг другу на более высоком уровне, например, ДНК, которые являются на 80% или более, предпочтительно, на 90% или более, более предпочтительно, на 95% или более, даже более предпочтительно, на 97% или более, и особенно предпочтительно, на 99% или более идентичными друг другу, гибридизуются друг с другом, и ДНК, идентичные друг другу на более низком уровне, не гибридизуются друг с другом; или условия, представляющие собой условия для отмывки при нормальной Саузерн-гибридизации, в которых отмывку проводят один раз, предпочтительно, от двух до трех раз, при концентрации соли и температуре, соответствующих 60 градусам C, 1 x SSC и 0,1% SDS, предпочтительно, 60 градусам C, 0,1 x SSC и 0,1% SDS, и более предпочтительно, 68 градусам C, 0,1 x SSC и 0,1% SDS.
[0142] Зонд, используемый для вышеуказанной гибридизации, может являться частью последовательности, комплементарной каждому гену. Такой зонд может быть получен посредством ПЦР с использованием олигонуклеотида, полученного на основе известной последовательности гена, в качестве праймера, и с использованием фрагмента ДНК, содержащего каждый из генов в (a1) - (a3) выше, в качестве матрицы. Например, фрагмент ДНК, имеющий длину приблизительно 300 п.о., можно использовать в качестве зонда. В случае, когда фрагмент ДНК, имеющий длину приблизительно 300 п.о., используют в качестве зонда, примеры условий для отмывки при гибридизации включают условия 50 градусов C, 2 x SSC и 0,1% SDS.
[0143] Кроме того, поскольку вырожденность кодонов отличается, в зависимости от хозяев, каждый из генов в (a1) - (a3) выше, могут представлять собой гены, полученные посредством замены любого кодона на эквивалентный кодон, при условии, что их исходные функции сохраняются. Например, гены в таблицах 1-3 можно модифицировать таким образом, чтобы они имели оптимальные кодоны, в зависимости от частоты использования кодонов для используемого хозяина.
[0144] Примеры обработок для мутагенеза включают способ обработки in vitro молекулы ДНК, имеющей нуклеотидную последовательность каждого из генов в (a1) - (a3) выше, с использованием гидроксиламина или т.п.; способ обработки микроорганизмов, несущих каждый из генов в (a1) - (a3) выше, с использованием рентгеновского излучения, ультрафиолетового излучения или мутагена, такого как N-метил-N'-нитро-N-нитрозогуанидин (NTG), этилметансульфонат (EMS) и метилметансульфонат (MMS); и т.п.
[0145] Генетическая модификация для увеличения уровня экспрессии гена
Фраза «увеличение экспрессии гена» означает, что экспрессия гена увеличена, по сравнению с экспрессией для немодифицированного штамма. Примеры одного аспекта увеличения экспрессии гена включают аспект, в котором экспрессия гена, предпочтительно, увеличена в 1,5 раза или более, более предпочтительно, увеличена в 2 раза или более, и даже более предпочтительно, увеличена в 3 раза или более, по сравнению с экспрессией для немодифицированного штамма.
[0146] Кроме того, фраза «экспрессия гена увеличена» означает не только увеличение экспрессии целевого гена в штамме, в котором целевой ген исходно экспрессируется, но также означает, что целевой ген экспрессируется в штамме, в котором целевой ген исходно не экспрессируется. То есть, фраза «экспрессия гена увеличена» включает, например, случай, в котором целевой ген вводят в штамм, не имеющий целевого гена, и целевой ген экспрессируется в нем. Кроме того, фраза «экспрессия гена увеличена» также обозначена как фразы «экспрессия гена усилена» или «экспрессия гена повышена».
[0147] Увеличения экспрессии гена можно достигать, например, посредством увеличения количества копий гена. Увеличения количества копий гена можно достигать посредством введения гена в хромосому хозяина. Введение гена в хромосому можно проводить с использованием, например, гомологичной рекомбинации (Miller I, J.H. Experiments in Molecular Genetics, 1972, Cold Spring Harbor Laboratory). Можно вводить только одну копию гена, или можно вводить две или более его копий.
[0148]
Например, множество копий гена можно вводить в хромосому посредством проведения гомологичной рекомбинации с нацеливанием в то же время на последовательность, имеющую множество копий на хромосоме. Примеры последовательности, имеющей множество копий на хромосоме, включают повторяющуюся последовательность ДНК (повторяющуюся ДНК) и инвертированные повторы, присутствующие на обоих концах транспозона.
[0149] Альтернативно, гомологичную рекомбинацию можно проводить с нацеливанием в то же время на соответствующую последовательность на хромосоме, такую как ген, не являющийся необходимым для продукции целевого вещества. Гомологичную рекомбинацию можно проводить, например, посредством способа с использованием линейной ДНК, способа с использованием плазмиды, содержащей термочувствительную точку начала репликации, способа с использованием плазмиды, способной к конъюгативному переносу, способа с использованием суицидного вектора, не имеющего точку начала репликации, которая функционирует в хозяине, или способа трансдукции с использованием фага. Кроме того, ген можно также случайным образом вводить в хромосому с использованием транспозона или Mini-Mu (JP-A-H2-109985).
[0150] Введен ли целевой ген в хромосому, можно проверять посредством Саузерн-гибридизации с использованием зонда, имеющего последовательность, комплементарную всей или части гена, ПЦР с использованием праймера, сконструированного на основе последовательности гена, или т.п.
[0151] Кроме того, количество копий гена также можно увеличивать посредством введения вектора, содержащего ген, в хозяина. Например, является возможным увеличивать количество копий гена посредством конструирования экспрессирующего вектора для гена, посредством лигирования фрагмента ДНК, содержащего целевой ген, с вектором, который функционирует в хозяине, и трансформации хозяина с использованием экспрессирующего вектора. Фрагмент ДНК, содержащий целевой ген, можно получать, например, посредством ПЦР с использованием геномной ДНК микроорганизма, имеющего целевой ген, в качестве матрицы. Способ трансформации не является конкретно ограниченным, и можно использовать общеизвестный способ.
[0152] В качестве вектора, можно использовать вектор, который может автономно реплицироваться в клетке-хозяине. Вектор, предпочтительно, представляет собой мультикопийный вектор. Кроме того, вектор, предпочтительно, имеет маркер, такой как ген устойчивости к антибиотику или другой ген, описанный в литературе [Karl Friehs, Plasmid Copy Number and Plasmid Stability, Adv Biochem Engin/Biotechnol 86: 47-82 (2004)], для отбора трансформанта. Кроме того, вектор может иметь промотор или терминатор для экспрессии вставленного гена. Примеры вектора включают вектор, происходящий из бактериальной плазмиды, вектор, происходящий из дрожжевой плазмиды, вектор, происходящий из бактериофага, космиды, фагмиды и т.п.
[0153] Конкретные примеры векторов, способных к автономной репликации в бактериях из числа Enterobacteriaceae, таких как Escherichia coli, включают pUC19, pUC18, pHSG299, pHSG399, pHSG398, pBR322 и pSTV29 (все из Takara Bio Inc.), pACYC184 и pMW219 (Nippon Gene), pTrc99A (Pharmacia), вектор pPROK (Clontech), pKK233-2 (Clontech), вектор pET (Novagen), вектор pQE (Qiagen) и вектор с широким спектром хозяев RSF1010.
[0154] В случае введения гена, является достаточным, чтобы ген сохранялся в микроорганизме, принадлежащем к прокариотам, имеющем генетическую модификацию по настоящему изобретению. Конкретно, является достаточным, чтобы ген был введен таким образом, чтобы он экспрессировался под контролем промоторной последовательности, которая функционирует в бактерии по настоящему изобретению. Промотор может представлять собой промотор, происходящий из хозяина, или гетерологичный промотор. Промотор может представлять собой собственный промотор гена, подлежащего введению, или промотор других генов. В качестве промотора, например, можно использовать более сильный промотор, который будет описан позже.
[0155] Терминатор для терминации транскрипции может быть расположен ниже гена. Терминатор не является конкретно ограниченным, при условии, что он функционирует в бактерии по настоящему изобретению. Терминатор может представлять собой терминатор, происходящий из хозяина, или гетерологичный терминатор. Терминатор может представлять собой терминатор, специфический для гена, подлежащего введению, или может представлять собой терминатор из других генов. Конкретные примеры терминаторов включают терминатор T7, терминатор T4, терминатор фага fd, терминатор tet и терминатор trpA.
[0156] Векторы, промоторы и терминаторы, которые можно использовать в различных микроорганизмах, подробно описаны, например, в «Basic Lecture 8 on Microbiology, Gene Engineering, KYORITSU SHUPPAN, 1987», и их можно использовать.
[0157] Кроме того, в случае, когда вводят два или более генов, является достаточным, чтобы каждый ген сохранялся в бактерии по настоящему изобретению подходящим для экспрессии образом. Например, каждый ген из всех может сохраняться в отдельном экспрессирующем векторе, или все могут сохраняться на хромосоме. Кроме того, каждый ген может отдельно сохраняться на множестве экспрессирующих векторов, или может отдельно сохраняться на одном или множестве экспрессирующих векторов и на хромосоме. Кроме того, два или более генов могут формировать оперон и быть введены. Примеры «случаев, когда вводят два или более генов», включают случай введения генов, соответственно, кодирующих два или более ферментов, случай введения генов, соответственно, кодирующих две или более субъединиц, которые формируют один фермент, и их комбинации.
[0158] Ген, подлежащий введению, не является конкретно ограниченным, при условии, что он кодирует белок, который функционирует в хозяине. Ген, подлежащий введению, может представлять собой ген, происходящий из хозяина, или гетерологичный ген. Ген, подлежащий введению, может быть получен, например, посредством ПЦР с использованием праймеров, сконструированных на основе нуклеотидной последовательности гена, и с использованием геномной ДНК из организма, имеющего ген, или плазмиды или т.п., несущей ген, в качестве матрицы. Кроме того, ген, подлежащий введению, можно полностью синтезировать, например, на основании нуклеотидной последовательности гена [Gene, 60 (1), 115-127 (1987)].
[0159] Кроме того, увеличения экспрессии гена можно достигать посредством улучшения эффективности транскрипции гена. Улучшения эффективности транскрипции гена можно достигать, например, посредством замены промотора гена на хромосоме на более сильный промотор. «Более сильный промотор» относится к промотору, который усиливает транскрипцию гена, по сравнению с природным промотором дикого типа.
[0160] Примеры «более сильных промоторов» включают известные промоторы с высокой экспрессией, такие как промотор uspA, промотор T7, промотор trp, промотор lac, промотор thr, промотор tac, промотор trc, промотор tet, промотор araBAD, промотор rpoH, промотор PR и промотор PL.
[0161] Кроме того, в качестве более сильного промотора, общепринятый промотор высоко активного типа может быть получен с использованием различных репортерных генов. Например, активность промотора можно увеличивать посредством переноса областей -35 и -10 в промоторной области ближе к консенсусной последовательности (WO 2000/18935).
[0162] Примеры промоторов высоко активного типа включают различные tac-подобные промоторы (Katashkina JI et al. Патентная заявка Российской Федерации 2006134574) и промотор pnlp8 (Международная заявка WO 2010/027045). Способы оценки силы промотора и примеры сильных промоторов описаны в известных литературных источниках [Prokaryotic promoters in biotechnology. Biotechnol. Annu. Rev., 1, 105-128 (1995) и т.п.].
[0163] Кроме того, увеличения уровня экспрессии гена можно достигать посредством улучшения эффективности трансляции гена. Улучшения эффективности трансляции гена можно достигать, например, посредством замены последовательности Шайна-Дальгарно (SD) (также называемой участок связывания рибосомы (RBS)) гена на хромосоме на более сильную последовательность SD.
[0164] «Более сильная последовательность SD» относится к последовательности SD, для которой трансляция мРНК является улучшенной, по сравнением с природной последовательностью SD дикого типа. Примеры более сильной последовательности SD включают RBS гена 10 из фага T7 [Olins P.O. et al, Gene, 1988, 73, 227-235]. Кроме того, известно, что замена, вставка или делеция нескольких нуклеотидов в спейсерной области между RBS и инициирующим кодоном, в частности, в последовательности, непосредственно выше инициирующего кодона (5'-UTR), значительно влияет на стабильность и эффективность трансляции мРНК, и таким образом, эффективность трансляции гена можно улучшать посредством их модификации.
[0165] По настоящему изобретению, участки, влияющие на экспрессию гена, такие как промотор, последовательность SD и спейсерная область между RBS и инициирующим кодоном, также в совокупности обозначены как «области контроля экспрессии». Области контроля экспрессии можно определять с использованием программного обеспечения для поиска генов, такого как vector или GENETYX для поиска промоторов. Модификацию этих областей контроля экспрессии можно проводить, например, посредством способа с использованием термочувствительного вектора или способа направляемой Red интеграции (WO2005/010175).
[0166] Улучшения эффективности трансляции генов можно также достигать, например, посредством модификации кодонов. Конкретно, например, в случае, когда проводят гетерологичную экспрессию гена, и т.п., эффективность трансляции гена можно улучшать посредством замены редких кодонов, присутствующих в гене, на синонимические кодоны, используемые более часто.
[0167] Замену кодонов можно проводить, например, посредством способа сайт-направленного мутагенеза, в котором целевую мутацию вводят в целевой участок в ДНК. Примеры способов сайт-направленного мутагенеза включают способ с использованием ПЦР [Higuchi, R., 61, in PCR technology, Erlich, H.A. Eds., Stockton press (1989); Carter, P., Meth. In Enzymol., 154, 382 (1987)], и способ с использованием фага [Kramer, W. and Frits, H.J., Meth. In Enzymol., 154, 350 (1987); Kunkel, T.A. et al., Meth. In Enzymol., 154, 367 (1987)]. Альтернативно, фрагмент гена, в котором кодоны заменены, можно полностью синтезировать. Частота использования кодонов в различных организмах описана в «базе данных использования кодонов» [http://www.kazusa.or.jp/codon; Nakamura, Y. et al, Nucl. Acids Res., 28, 292 (2000)].
[0168] Кроме того, экспрессию гена можно также увеличивать посредством амплификации регулятора, который увеличивает экспрессию гена, или посредством делеции или ослабления регулятора, который уменьшает экспрессию гена. Такие способы увеличения экспрессии гена, как описано выше, можно использовать отдельно или можно использовать в любой комбинации.
[0169] Увеличение экспрессии гена можно проверять, например, посредством проверки увеличения уровня транскрипции гена или посредством проверки увеличения количества белка, экспрессированного с гена. Кроме того, увеличение экспрессии гена можно проверять, например, посредством проверки увеличения активности белка, экспрессированного с гена.
[0170] Проверку увеличения уровня транскрипции гена можно проводить посредством сравнения количества мРНК, транскрибированной с гена, с немодифицированным штаммом, таким как штамм дикого типа или родительский штамм. Примеры способов оценки количества мРНК включают Нозерн-гибридизацию, RT-ПЦР и т.п. [Sambrook, J., et al., Molecular Cloning A Laboratory Manual/Third Edition, Cold spring Harbor Laboratory Press, Cold spring Harbor (USA), 2001]. Увеличение количества мРНК относится, например, к случаю, в котором количество мРНК, предпочтительно, увеличено в 1,5 раза или более, более предпочтительно, увеличено в 2 раза или более, и даже более предпочтительно, увеличено в 3 раза или более, по сравнению с количеством в немодифицированном штамме.
[0171] Увеличение количества белка можно проверять, например, посредством Вестерн-блоттинга с использованием антитела. Увеличение количества белка относится, например, к случаю, в котором количество белка, предпочтительно, увеличено в 1,5 раза или более, более предпочтительно, увеличено в 2 раза или более, и даже более предпочтительно, увеличено в 3 раза или более, по сравнению с количеством в немодифицированном штамме.
[0172] Увеличение активности белка можно проверять, например, посредством измерения активности белка. Увеличение активности белка относится, например, к случаю, в котором активность белка, предпочтительно, увеличена в 1,5 раза или более, более предпочтительно, увеличена в 2 раза или более, и даже более предпочтительно, увеличена в 3 раза или более, по сравнению с активностью в немодифицированном штамме.
[0173] Вышеописанные способы увеличения экспрессии гена можно использовать для усиления экспрессии каждого из генов (a1) и (a2) выше.
[0174] Генетическая модификация, которая увеличивает экспрессию гена kpsS, предпочтительно, представляет собой по меньшей мере одну из модификации областей контроля экспрессии гена kpsS и генетической модификации, увеличивающей количество копий. Ген kpsFEDUCS присутствует в качестве группы генов продукции гепаросана, но как будет описано позднее в примерах, авторы настоящего изобретения обнаружили, что эффект особенного улучшения продукции гепаросана получен посредством увеличения экспрессии только гена kpsS среди них. Соответственно, в качестве генетической модификации, которая увеличивает экспрессию гена kpsS, генетическая модификация для увеличения количества копий гена kpsS является особенно предпочтительной.
[0175] Генетическая модификация, которая увеличивает экспрессию по меньшей мере одного гена, выбранного из генов kfiA, kfiB, kfiC и kfiD, предпочтительно, представляет собой по меньшей мере одну из модификации областей контроля экспрессии по меньшей мере одного гена, выбранного из генов kfiA, kfiB, kfiC и kfiD, и увеличения количества копий генов. Как показано на ФИГ. 1, гены kfiA, kfiB, kfiC и kfiD составляют, оперон. Генетическая модификация, которая усиливает весь оперон, составляемый генами kfiA, kfiB, kfiC и kfiD, является предпочтительной, и модификация областей контроля экспрессии генов kfiA, kfiB, kfiC и kfiD, является более предпочтительной.
[0176] Генетическая модификация, которая вызывает потерю функции гена
Примеры генетических модификаций, которые вызывают потерю функции гена yhbJ из (a3) выше, включают генетическую модификацию, при которой функция белка, кодированного частью, соответствующей yhbJ, уменьшена или полностью остановлена посредством модификации ДНК, кодирующей часть, соответствующую yhbJ, в геномной ДНК микроорганизма, принадлежащего к прокариотам, в качестве хозяина.
[0177] В способе по настоящему изобретению, форма модификации, подлежащей добавлению к ДНК, кодирующей часть, соответствующую yhbJ, не является конкретно ограниченной, при условии, что функция белка, кодированного частью, соответствующей yhbJ, уменьшена или полностью остановлена, и можно соответствующим образом использовать известный способ.
[0178] Примеры форм, которые уменьшают или полностью останавливают функцию белка, кодированного частью, соответствующей yhbJ, включают любую из следующих модификаций (I) - (III):
(I) Вся или часть ДНК, кодирующей часть, соответствующую yhbJ, удалена.
(II) Одна или несколько замен, делеций или добавлений внесены в ДНК, кодирующую часть, соответствующую yhbJ.
(III) ДНК, кодирующая часть, соответствующую yhbJ, заменена на последовательность ДНК, имеющую менее чем 80% идентичность с последовательностью ДНК до модификации.
[0179] Примеры потери функции гена yhbJ включают случай, в котором активность гена yhbJ составляет, предпочтительно, 20% или менее, более предпочтительно, 10% или менее, и даже более предпочтительно, 5% или менее, по сравнению с активностью немодифицированного штамма. Активность yhbJ можно проверять посредством проверки уровня экспрессии glmS посредством способа Нозерн-блоттинга, способа Вестерн-блоттинга или т.п. [Kalamorz F. et al, (2007) «Feedback control of glucosamine-6-phosphate synthase GlmS expression depends on the small RNA GlmZ and involves the novel protein YhbJ in Escherichia coli». Mol Microbiol. 65 (6):1518-33].
[0180] В способе продукции сульфатированного полисахарида по настоящему изобретению, можно использовать, для реакционного раствора, N-сульфогепаросан, полученный посредством химической модификации гепаросана посредством известного способа, где гепаросан получен посредством вышеописанного способа продукции гепаросана.
[0181] Способ продукции PAPS
Способ продукции PAPS по настоящему изобретению отличается тем, что реакцию продукции PAPS проводят посредством включения источника ATP, источника иона сульфата, вышеописанного трансформанта (a) или его обработанного материала в реакционный раствор. Таким образом, способ продукции PAPS по настоящему изобретению включает следующие стадии (i) и (ii):
(i) получение трансформанта бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, в котором клеточная плазматическая мембрана трансформанта является проницаемой для веществ, или обработанного материала трансформанта; и
(ii) проведение реакции для продукции PAPS посредством использования реакционного раствора, содержащего источник ATP, источник иона сульфата и трансформант, полученный на стадии (i), или его обработанный материал.
[0182] В соответствии со способом продукции PAPS по настоящему изобретению, PAPS можно продуцировать посредством реакции бактериальной клетки с использованием трансформанта бактерии из рода Corynebacterium, экспрессирующего ATP-сульфурилазу и APS-киназу, или его обработанного материала.
[0183]
ПРИМЕРЫ
Примеры показаны ниже, но настоящее изобретение не является ограниченным следующими примерами.
Пример 1
[0184] Конструирование плазмиды Corynebacterium ammoniagenes DE3
Плазмиду для вставки лямбда-DE3, содержащего ген РНК-полимеразы T7, между gltD-purT на хромосоме Corynebacterium ammoniagenes дикого типа штамма ATCC 6872 конструировали следующим образом. Конструировали два вида праймеров для амплификации стороны gltD [gltD-purT_1 (SEQ ID NO: 1) и gltD-purT_2BX (SEQ ID NO: 2)], и два вида праймеров для амплификации стороны purT [gltD-purT_3BX] (SEQ ID NO: 3) и gltD-purT_4 (SEQ ID NO: 4)].
[0185] В этот раз, для проведения второй ПЦР (ПЦР для слияния) для связывания фланкирующего фрагмента gltD и фланкирующего фрагмента purT, амплифицированных в первой ПЦР, последовательность, которая является комплементарной последовательности приблизительно 25 оснований на 3'-стороне 5' праймера (gltD-purT_4; SEQ ID NO: 4) для амплификации фланкирующего фрагмента purT, добавляли на 5'-стороне 3'-праймера (gltD-purT_2BX; SEQ ID NO: 2) для амплификации фланкирующего фрагмента gltD. Кроме того, последовательность для клонирования In-Fusion добавляли к 5'-стороне 5'-праймера (gltD-purT_1) для амплификации фланкирующего фрагмента gltD и 3'-праймера (gltD-purT_4) для амплификации фланкирующего фрагмента purT. Кроме того, последовательности узнавания BglII и XhoI добавляли к связывающей части на стороне gltD и на стороне purT для вставки лямбда-DE3.
[0186] Хромосомную ДНК Corynebacterium ammoniagenes штамма ATCC 6872 (далее в настоящем описании обозначенного как ATCC 6872), представляющего собой штамм дикого типа Corynebacterium ammoniagenes, получали в соответствии со способом из Saito et al. [Biochim. Biophys. Acta 72, 619 (1963)].
[0187] С использованием хромосомной ДНК в качестве матрицы, проводили первую ПЦР для амплификации фланкирующего фрагмента gltD и фланкирующего фрагмента purT, и таким образом, получали фрагмент ДНК приблизительно 0,8 т.п.о., фланкирующий gltD, и фрагмент ДНК приблизительно 0,8 т.п.о., фланкирующий purT. Затем проводили вторую ПЦР для связывания этих фланкирующего фрагмента gltD и фланкирующего фрагмента purT, и таким образом, получали фрагмент ДНК приблизительно 1,6 т.п.о. (gltD-BX-purT).
[0188] Плазмиду pESB30, в которой PstI-фрагмент ДНК 2,6 т.п.о. [Mol. Microbiol., 6, 1195 (1992)], содержащий ген левансахаразы sacB из Bacillus subtilis, содержится в участке расщепления PstI вектора pHSG299 Escherichia coli [Gene, 61, 63, (1987)], имеющего ген устойчивости к канамицину, разрезали с использованием BamHI. Затем фрагмент gltD-BX-purT, полученный выше, связывали с использованием набора для клонирования In-Fusion (Takara Bio Inc.). С использованием продукта реакции, трансформировали Escherichia coli DH5-альфа [полученную в TOYOBO CO., LTD.], в соответствии с общепринятым способом [Molecular cloning: a laboratory manual, 3rd ed., 2001, Cold Spring Harbor Laboratory Press].
[0189] Полученный штамм культивировали на среде LB с агаром, содержащей 20 микрограмм/мл канамицина [среде, которая содержит 10 г бактотриптона (изготовленного в Difco), 5 г дрожжевого экстракта (изготовленного в Difco), 10 г хлорида натрия и 16 г бактоагара (изготовленного в Difco) в 1 л воды, и pH которой доводили до pH 7,0], и отбирали трансформированный штамм. После отбора целевого клона посредством ПЦР колоний, трансформированный штамм инокулировали в среду LB, содержащую 20 микрограмм/мл канамицина (среду, которая имеет такой же состав, как среда LB с агаром, за исключением того, что не содержит агара), и культивировали в течение ночи, и таким образом, плазмиду получали из полученного культурального бульона с использованием набора QIAprep Spin Miniprep (Qiagen). Анализ нуклеотидной последовательности подтвердил, что плазмида имеет структуру, в которой фрагмент gltD-BX-purT приблизительно 1,6 т.п.о. вставлен в pESB30.
[0190] Затем, фрагмент лямбда-DE3 4,5 т.п.о. амплифицировали посредством ПЦР с использованием хромосомной ДНК, выделенной из Escherichia coli BL21 (DE3), в качестве матрицы и с использованием праймеров DE3-for_Xho (SEQ ID NO: 5) и DE3-rev_Bgl (SEQ ID NO: 6). Последовательность узнавания XhoI добавляли к DE3-for_Xho, и последовательность узнавания BglII добавляли к DE3-rev_Bgl. Этот фрагмент и плазмиду, имеющую структуру, в которой фрагмент gltD-BX-purT приблизительно 1,6 т.п.о. вставлен в pESB30, расщепляли с использованием XhoI и BglII, и затем связывали с использованием набора для лигирования ДНК (Takara Bio Inc.).
[0191] Escherichia coli DH5 альфа трансформировали таким же способом, как описано выше, полученный таким образом штамм культивировали на среде LB с агаром, содержащей 20 микрограмм/мл канамицина, и отбирали трансформированный штамм. Трансформированный штамм инокулировали в среду LB, содержащую 20 микрограмм/мл канамицина, и культивировали в течение ночи, и таким образом, плазмиду получали из полученного культурального бульона с использованием набора набора QIAprep Spin Miniprep (Qiagen). Анализ нуклеотидной последовательности подтвердил, что плазмида имеет структуру, в которой фрагмент лямбда-DE3 4,5 т.п.о. вставлен между приблизительно 1,6 т.п.о. gltD-purT в pESB30. Эта плазмида обозначена как pC-DE3.
Пример 2
[0192] Конструирование штамма Corynebacterium ammoniagenes с вставкой лямбда-DE3
PC-DE3 вводили в штамм ATCC 6872 посредством электропорации, в соответствии со способом Rest et al. [Appl. Microbiol. Biotech., 52, 541 (1999)], и отбирали устойчивые к канамицину штаммы. Когда структуру хромосомы, полученной из одного из устойчивых к канамицину штаммов, проверяли посредством Саузерн-гибридизации, [Molecular cloning: a laboratory manual, 3rd ed., 2001, Cold Spring Harbor Laboratory Press], подтвердили, что pC-DE3 была интегрирована в хромосому посредством гомологичной рекомбинации типа Кэмпбелла.
[0193]
Трансформированный штамм (с одиночной рекомбинацией) наносили на среду Suc с агаром [среду, которая содержит 100 г сахарозы, 7 г мясного экстракта, 10 г пептона, 3 г хлорида натрия, 5 г дрожжевого экстракта (изготовленного в Difco) и 15 г бактоагара (изготовленного в Difco) в 1 л воды, и pH которой доводили до pH 7,2], и культивировали при 30 градусах C в течение 1 суток, и отбирали растущие колонии. Штамм, в котором присутствовал ген sacB, не мог расти на этой среде, поскольку он превращает сахарозу в суицидный субстрат [J. Bacteriol., 174, 5462 (1991)]. С другой стороны, штамм, в котором ген sacB был делетирован посредством второй гомологичной рекомбинации между лямбда-DE3 типа вставки и дикого типа, присутствующего около хромосомы, мог расти на этой среде без образования суицидного субстрата. Во время этой гомологичной рекомбинации, либо структура дикого типа, либо штамм, в котором фрагмент лямбда-DE3 был вставлен между gltD-purT, подвергались удалению вместе с sacB. В то же время, в штамме, в котором структура дикого типа подвергалась удалению вместе с sacB, происходила замена гена на лямбда-DE3 типа вставки.
[0194] Штамм, в котором лямбда-DE3 вставлен между gltD-purT ATCC 6872, получали посредством ПЦР колоний с использованием второго рекомбинанта, полученного таким образом, с использованием праймеров DE3-for_Xho и DE3-rev_Bgl. Этот штамм обозначен как ATCC 6872 (DE3).
Пример 3
[0195] Конструирование плазмиды, имеющей фрагмент ДНК для экспрессии MET3-MET14
Плазмиду pCS299P [Appl. Microbiol. Biotech., 63, 592 (2004)] расщепляли с использованием BamHI. ПЦР проводили с использованием pCET_Fw2 (SEQ ID NO: 7) и pCET_Rv2 (SEQ ID NO: 8) и с использованием pET21b в качестве матрицы для получения фрагмента ДНК, содержащего lacI-PT7. Их очищали посредством набора для очистки продуктов ПЦР QIAquick (Qiagen) и связывали с использованием набора для клонирования In-Fusion (Takara Bio Inc.). С использованием продукта реакции, Escherichia coli DH5 альфа (полученную в TOYOBO CO., LTD.), трансформировали в соответствии с общепринятым способом и культивировали на среде LB с агаром, содержащей 20 микрограмм/мл канамицина, и отбирали трансформированный штамм. После отбора целевого клона посредством ПЦР колоний, трансформированный штамм инокулировали в среду LB, содержащую 20 микрограмм/мл канамицина, и культивировали в течение ночи, и таким образом, плазмиду получали из полученного культурального бульона с использованием набора для получения препаратов в минимасштабе QIAprep Spin Miniprep (Qiagen). Анализ нуклеотидной последовательности подтвердил, что плазмида представляла собой плазмиду, имеющую структуру, в которой фрагмент ДНК lacI-PT7 приблизительно 1,9 т.п.о., происходящий из pET21b, вставлен в pCS299P. Эта плазмида обозначена как pCET212.
[0196] Затем конструировали плазмиду, в которой MET3 и MET14, происходящие из Saccharomyces cerevisiae, были вставлены ниже промотора T7 pCET212. Конструировали два вида праймеров (MET3_1 (SEQ ID NO: 9) и MET3_2 (SEQ ID NO: 10)) для амплификации MET3 с хромосомной ДНК Saccharomyces cerevisiae штамма S288C (далее в настоящем описании обозначенного как S288C), и два вида праймеров (MET14_3 (SEQ ID NO: 11) и MET14_4 (SEQ ID NO: 12)) для амплификации MET14.
[0197] В этот раз, для проведения второй ПЦР (ПЦР для слияния), которая связывает фрагмент MET3 и фрагмент MET14, амплифицированные в первой ПЦР, последовательность, которая является комплементарной последовательности приблизительно 15 оснований на 5'-стороне 5' праймера (MET14_3; SEQ ID NO: 11) для амплификации MET14, добавляли к 5'-стороне 3'-праймера (MET3_2; SEQ ID NO: 10) для амплификации MET3, и последовательность, которая является комплементарной последовательности приблизительно 15 оснований на 5'-стороне MET3_3, добавляли к 5'-стороне MET14_3. Кроме того, последовательность для клонирования In-Fusion добавляли к 5'-стороне 5' праймера (MET3_1) для амплификации MET3 и 3'-праймера (MET14_4) для амплификации MET13. С использованием хромосомной ДНК штамма S288C в качестве матрицы, проводили первую ПЦР для амплификации MET3 и MET14, и таким образом, получали фрагмент ДНК MET3 приблизительно 1,5 т.п.о. и фрагмент ДНК приблизительно 0,6 т.п.о. из нижележащей области. Затем проводили вторую ПЦР для связывания этих фрагмента MET3 и фрагмента MET14, и таким образом, получали фрагмент ДНК (MET3-MET14) приблизительно 2,1 т.п.о. Этот фрагмент ДНК очищали с использованием набора для очистки продуктов ПЦР QIAquick (Qiagen) и затем лигировали с pCET212, расщепленной с использованием NdeI и XhoI, с использованием набора для клонирования In-Fusion (Takara Bio Inc.). С использованием продукта реакции, Escherichia coli DH5 альфа (полученную в TOYOBO CO., LTD.) трансформировали в соответствии с общепринятым способом. Полученный таким образом штамм культивировали на среде LB с агаром, содержащей 20 микрограмм/мл канамицина, и отбирали трансформированный штамм. После отбора целевого клона посредством ПЦР колоний, трансформированный штамм инокулировали в среду LB, содержащую 20 микрограмм/мл канамицина, и культивировали в течение ночи, и таким образом, плазмиду получали из полученного культурального бульона с использованием набора QIAprep Spin Miniprep (Qiagen). Анализ нуклеотидной последовательности подтвердил, что плазмида имеет структуру, в которой фрагмент MET3-MET14 приблизительно 2,1 т.п.о. вставлен в pCET212. Эта плазмида обозначена как pSC-3-13. Посредством трансформации pSC-3-13 в ATCC 6872 (DE3), получали продуцирующий PAPS штамм Corynebacterium ammoniagenes ATCC 6872 (DE3)/pSC-3-13.
Пример 4
[0198] Конструирование плазмиды, имеющей фрагмент ДНК для экспрессии белка шаперона
ПЦР проводили с использованием Gro_F (SEQ ID NO: 13) и Gro_R (SEQ ID NO: 14), и с использованием хромосомной ДНК Escherichia coli штамма BL21 (DE3) в качестве матрицы, и таким образом, получали фрагмент ДНК, содержащий GroES-GroEL. Затем, ПЦР проводили с использованием pKD46 [Datsenko, K.A., Warner, B.L., Proceedings of the National Academy of Science of the United States of America, Vol. 97. 6640-6645 (2000)] в качестве матрицы, и с использованием AraC_ParaB_F (SEQ ID NO: 15) и AraC_ParaB_R (SEQ ID NO: 16), и таким образом, получали фрагмент ДНК, содержащий AraC_ParaB. ПЦР проводили с использованием pCDF-SmOri-F (SEQ ID NO: 17) и pCDF-SmOri-R (SEQ ID NO: 18), и с использованием pCDF-Duet1 (Novagen) в качестве матрицы, и таким образом, получали фрагмент ДНК, содержащий ген устойчивости к стрептомицину и точку начала репликации ColdDF. Их очищали посредством набора для очистки продуктов ПЦР QIAquick (Qiagen) и связывали с использованием набора для клонирования In-Fusion (Takara Bio Inc.). С использованием продукта реакции, Escherichia coli DH5 альфа (полученную в TOYOBO CO., LTD.) трансформировали в соответствии с общепринятым способом и культивировали на среде LB с агаром, содержащей 50 микрограмм/мл стрептомицина, и отбирали трансформированный штамм. После отбора целевого клона посредством ПЦР колоний, трансформированный штамм инокулировали в среду LB, содержащую 50 микрограмм/мл стрептомицина, и культивировали в течение ночи, и таким образом, плазмиду получали из полученного культурального бульона с использованием набора QIAprep Spin Miniprep (Qiagen). Анализ нуклеотидной последовательности подтвердил, что плазмида имеет структуру, в которой фрагмент ДНК AraC-ParaB приблизительно 1,2 т.п.о., происходящий из pKD46, и фрагмент ДНК GroES-GroEL приблизительно 2,0 т.п.о., происходящий из BL21 (DE3), вставлены в pCDF-Duet1. Плазмида обозначена как pGro (Sm).
Пример 5
[0199] Конструирование Escherichia coli, экспрессирующей C5-эпимеразу, 2OST, 6OST-3 и 3OST-1
Область каталитического домена C5-эпимеразы человека (E53-N609) клонировали в вектор pMAL-C2X (New England Biolabs), в соответствии со способом, описанным в литературе [Biochemical and Biophysical Research Communications Volume 339, Issue 2, 13 January 2006, pages 597-602], для конструирования MBP-C5. MBP-C5 трансформировали в Origami-B (DE3) (Novagen) вместе с pGro7 (Takara Bio Inc.), и таким образом, конструировали штамм Escherichia coli Origami-B (DE3)/MBP-C5_pGro7, экспрессирующий C5-эпимеразу.
[0200] В SEQ ID NO: 19 показана нуклеотидная последовательность каталитического домена (R51-N356) изоформы 1 2-O-сульфотрансферазы, происходящая из китайского хомяка и подвергнутая оптимизации кодонного состава для экспрессии в Escherichia coli. ПЦР проводили с использованием искусственно синтезированной последовательности, описанной выше, в качестве матрицы, и праймеров из SEQ ID NO: 20 и 21. Этот фрагмент ПЦР клонировали в pET-His6-MBP-TEV-LIC (Addgene) посредством способа безлигазного клонирования [Methods Mol Biol. 2009; 498: 105-115] для конструирования H-MBP-2OST. H-MBP-2OST трансформировали в Origami-B (DE3) (Novagen) с pGro (Sm), и таким образом, конструировали штамм Escherichia coli Origami-B (DE3)/H-MBP-2OST_pGro (Sm), экспрессирующий 2OST.
[0201] Каталитический домен (P120-L424) изоформы 3 6-O-сульфотрансферазы, происходящей из мыши, клонировали в вектор pMAL-C2X (New England Biolabs) посредством способа, описанного в литературе [Chemistry & Biology Volume 14, Issue 9, 21 September 2007, pages 986-993], для конструирования MBP-6OST3. MBP-6OST3 трансформировали в Origami-B (DE3) (Novagen) вместе с pGro7 (Takara Bio Inc.), и таким образом, конструировали штамм Escherichia coli Origami-B (DE3)/MBP-6OST3_pGro7, экспрессирующий 6OST-3.
[0202] Каталитический домен (G48-H311) 3-O сульфотрансферазы, происходящей из мыши, клонировали в вектор pET28a (Novagen) посредством способа, описанного в литературе [J Biol Chem. 2004 Jun 11; 279 (24): 25789-97], для конструирования HIS-3OST1. HIS-3OST1 трансформировали в BL21-CodonPlus (DE3) -RIL (Agilent Technologies) вместе с pGro (Sm), и таким образом, конструировали штамм Escherichia coli RIL/HIS-3OST1_pGro(Sm), экспрессирующий 3OST1.
Пример 6
[0203] Получение N-сульфогепаросана и 2-O-сульфатированного N-сульфогепаросана Гепаросан получали посредством ферментации с использованием Escherichia coli штамма K5 или штамма Nissle, в соответствии со способом, описанным в Патентном документе [WO2018/048973 A1]. В соответствии с этим документом, полученный гепаросан подвергали химическому деацетилированию и деполимеризации, и затем химическому N-сульфатированию. Затем проводили фракционирование посредством преципитации этанолом для получения N-сульфогепаросана. Полученный N-сульфогепаросан подвергали эпимеризации положения C5 и сульфатированию положения 2-O в остатках уроновой кислоты посредством ферментной реакции, в соответствии с тем же документом, и затем подвергали фракционированию посредством преципитации этанолом, и таким образом, получали 2-O-сульфатированный N-сульфогепаросан.
Пример 7
[0204] Культивирование ATCC 6872 (DE3)/pSC-3-13
Штамм ATCC 6872 (DE3)/pSC-3-13, полученный в примере 3, инокулировали на среду BY-глюкоза с агаром, содержащую 50 микрограмм/мл канамицина [среду, содержащую 10 г глюкозы, 20 г обычной бульонной среды (изготовленной в Kyokuto Pharmaceutical Industrial Co., Ltd.), 5 г дрожжевого экстракта (изготовленного в Difco) и 20 г бактоагара (изготовленного в Difco) в 1 л воды], и культивировали при 30 градусах C в течение ночи.
[0205] Бактериальные клетки с двух чашек инокулировали в 350 мл среды для первичного посева [среды, которая содержит глюкозу, 50 г/л, полипептон высокой молекулярной массы (Nihon Pharmaceutical Co., Ltd.) 10 г/л, дрожжевой экстракт (Asahi), 10 г/л, KH2PO4, 1 г/л, K2HPO4, 1 г/л, (NH4)2SO4, 0,5 г/л, мочевину, 0,5 г/л, L-цистин, 0,03 г/л, MgSO4-7H2O, 1 г/л, CaCl2-2H2O, 0,1 г/л, ZnSO4-7H2O, 0,01 г/л, FeSO4-7H2O, 0,01 г/л, MnSO4-5H2O, 0,02 г/л, D-пантотенат кальция, 0,01 г/л, биотин, 40 микрограмм/л, гидрохлорид тиамина, 0,005 г/л, и никотиновую кислоту, 0,005 г/л, pH которой доводили до pH 7,2 с использованием гидроксида натрия, и к которой L-цистеин, 0,1 г/л, тиосульфат натрия, 1 г/л, и канамицин, 0,1 г/л, добавляли отдельно после стерилизации при 122 градусах C в течение 20 минут с использованием автоклава] в 2 л колбе Эрленмейера, и культивировали при 30 градусах C, при скорости перемешивания 220 об./мин в течение 24 часов.
[0206] В 1700 мл среды для вторичного посева [среды, которая содержит глюкозу, 100 г/л, фруктозу, 4 г/л, дрожжевой экстракт (изготовленный в Asahi), 10 г/л, KH2PO4, 1,25 г/л, K2HPO4, 1 г/л, моногидрат глутамата натрия, 2,1 г/л, L-цистин, 0,02 г/л, MgSO4-7H2O, 1,25 г/л, CaCl2-2H2O, 0,1 г/л, CuSO4-5H2O, 0,002 г/л, ZnSO4-7H2O, 0,01 г/л, FeSO4-7H2O, 0,02 г/л, MnSO4-5H2O, 0,02 г/л, D-пантотенат кальция, 0,015 г/л, биотин, 40 микрограмм/л, никотиновую кислоту, 0,005 г/л, и ADEKA NOL LG-109 (изготовленный в ADEKA), 1 мл/л, pH которой доводили до 7,2 с использованием гидроксида натрия, которую стерилизовали в автоклаве при 122 градусов C в течение 20 минут после отдельного добавления мочевины до 3,2 г/л, и к которой гидрохлорид тиамина 0,1 г/л, L-цистеин, 0,3 г/л, тиосульфат натрия 2,5 г/л, и канамицин 0,2 г/л добавляли отдельно] в 6 л культуральном резервуаре, инокулировали 300 мл вышеуказанного культурального раствора и культивировали в течение 24 часов в условиях культивирования 30 градусов C, скорость перемешивания 650 об./мин и скорость аэрации 2 л/мин, с доведением в то же время ее pH до 6,8 с использованием 18% водного аммиака.
[0207] В 1700 мл среды для основного культивирования [среды, которая содержит KH2PO4, 10 г/л, K2HPO4, 10 г/л, моногидрат глутамата натрия, 1 г/л, L-цистин, 0,02 г/л, CaCl2-2H2O, 0,1 г/л, CuSO4-5H2O, 0,005 г/л, ZnSO4-7H2O, 0,01 г/л, FeSO4-7H2O, 0,02 г/л, биотин, 150 микрограмм/л, никотиновую кислоту, 0,005 г/л, мочевину, 2 г/л, и ADEKA NOL LG-109 (изготовленный в ADEKA), 1 мл/л, и в которую глюкозу, 125 г/л, фруктозу, 25 г/л, MgSO4-7H2O, 10 г/л, MnSO4-5H2O, 0,02 г/л, D-пантотенат кальция, 0,015 г/л, гидрохлорид тиамина 0,005 г/л, L-цистеин, 0,15 г/л, тиосульфат натрия, 2,5 г/л и канамицин, 0,2 г/л, добавляли отдельно после стерилизации при 122 градусах C в течение 20 минут в автоклаве] в 6 л культуральном резервуаре, инокулировали 300 мл вышеуказанного культурального раствора и культивировали в течение 25 часов в условиях культивирования 30 градусов C и скорость аэрации 2 л/мин, с доведением в то же время скорости перемешивания до между 650 об./мин и 900 об./мин, так чтобы количество растворенного кислорода не падало ниже 1 м.д., и с доведением в то же время ее pH до 6,8 с использованием 18% водного аммиака.
[0208] В ходе этого периода, IPTG добавляли таким образом, чтобы конечная концентрация становилась 1 мМ через 6 часов от начала культивирования, и биотин, 100 микрограмм/л, никотиновую кислоту, 0,015 г/л, D-пантотенат кальция, 0,015 г/л, и гидрохлорид тиамина, 0,005 г/л дополнительно добавляли через 10 часов от начала культивирования. После завершения культивирования, культуральный раствор разделяли на бактериальные клетки и культуральный супернатант посредством центрифуги, и таким образом, осадок получали в форме влажных бактериальных клеток и замораживали при -80 градусов C.
Пример 8
[0209] Культивирование Origami-B (DE3)/MBP-C5_pGro7, Origami-B (DE3)/H-MBP-2OST_pGro (Sm), Origami-B (DE3)/MBP-6OST3_pGro7 и RIL/HIS-3OST1_pGro (Sm)
Штамм Origami-B (DE3)/MBP-C5_pGro7, полученный в примере 5, инокулировали в большую пробирку, содержащую 5 мл среды TB, содержащей 50 микрограмм/мл ампициллина, 20 микрограмм/мл хлорамфеникола, 15 микрограмм/мл тетрациклина и 15 микрограмм/мл канамицина, и культивировали при 30 градусах C в течение 16 часов. 1,2% культуральный раствор инокулировали в колбу Эрленмейера, содержащую 500 мл среды TB, содержащей 50 микрограмм/мл ампициллина, 20 микрограмм/мл хлорамфеникола, 15 микрограмм/мл тетрациклина и 15 микрограмм/мл канамицина, и культивировали с встряхиванием при 37 градусах C в течение 6 часов. Затем в него добавляли IPTG при конечной концентрации 1 мМ и арабинозу при конечной концентрации 4 мМ, и культивирование проводили при 28 градусах C в течение 20 часов. Затем культуральный раствор центрифугировали, и таким образом, осадок получали в форме влажных бактериальных клеток и замораживали при -80 градусов C.
[0210] Штамм Origami-B(DE3)/H-MBP-2OST_pGro(Sm), полученный в примере 5, инокулировали в большую пробирку, содержащую 5 мл среды TB, содержащей 50 микрограмм/мл ампициллина, 50 микрограмм/мл стрептомицина, 15 микрограмм/мл тетрациклина и 15 микрограмм/мл канамицина, и культивировали при 30 градусах C в течение 16 часов. 1,2% культуральный раствор инокулировали в колбу Эрленмейера, содержащую 500 мл среды TB, содержащей 50 микрограмм/мл ампициллина, 50 микрограмм/мл стрептомицина, 15 микрограмм/мл тетрациклина и 15 микрограмм/мл канамицина, и культивировали с встряхиванием при 30 градусах C в течение 12 часов. Затем в него добавляли IPTG при конечной концентрации 1 мМ и арабинозу при конечной концентрации 4 мМ, и культивирование проводили при 28 градусах C в течение 20 часов. Затем культуральный раствор центрифугировали, и таким образом, осадок получали в форме влажных бактериальных клеток и замораживали при -80 градусов C.
[0211] Штамм Origami-B(DE3)/MBP-6OST3_pGro7, полученный в примере 5, инокулировали в большую пробирку, содержащую 5 мл среды TB, содержащей 50 микрограмм/мл ампициллина, 20 микрограмм/мл хлорамфеникола, 15 микрограмм/мл тетрациклина и 15 микрограмм/мл канамицина, и культивировали при 30 градусах C в течение 16 часов. 1,2% культуральный раствор инокулировали в колбу Эрленмейера, содержащую 500 мл среды TB, содержащей 50 микрограмм/мл ампициллина, 20 микрограмм/мл хлорамфеникола, 15 микрограмм/мл тетрациклина и 15 микрограмм/мл канамицина, и культивировали с встряхиванием при 37 градусах C в течение 4 часов. Затем в него добавляли IPTG при конечной концентрации 1 мМ и арабинозу при конечной концентрации 4 мМ, и культивирование проводили при 28 градусах C в течение 20 часов. Затем культуральный раствор центрифугировали, и таким образом, осадок получали в форме влажных бактериальных клеток и замораживали при -80 градусов C.
[0212] Штамм RIL/HIS-3OST1_pGro(Sm), полученный в примере 5, инокулировали в большую пробирку, содержащую 5 мл среды TB, содержащей 50 микрограмм/мл стрептомицина, 15 микрограмм/мл канамицина, и культивировали при 30 градусах C в течение 16 часов. 1,2% культуральный раствор инокулировали в колбу Эрленмейера, содержащую 500 мл среды TB, содержащей 50 микрограмм/мл стрептомицина и 15 микрограмм/мл канамицина, и культивировали с встряхиванием при 37 градусах C в течение 4 часов. Затем в него добавляли IPTG при конечной концентрации 1 мМ и арабинозу при конечной концентрации 4 мМ, и культивирование проводили при 28 градусах C в течение 20 часов. Затем культуральный раствор центрифугировали, и таким образом, осадок получали в форме влажных бактериальных клеток и замораживали при -80 градусов C.
Пример 9
[0213] Тестирование реакции сульфатирования в положении 2-O N-сульфогепаросана с использованием ATCC 6872 (DE3)/pSC-3-13, Origami-B (DE3)/MBP-C5_pGro7 и Origami-B (DE3)/H-MBP-2OST_pGro (Sm)
Каждые из замороженных бактериальных клеток ATCC 6872 (DE3)/pSC-3-13, Origami-B (DE3)/MBP-C5_pGro7 и Origami-B (DE3)/H-MBP-2OST_pGro (Sm), полученных в примерах 7 и 8, суспендировали в дистиллированной воде таким образом, чтобы масса замороженных бактериальных клеток стала 333 г/л, для получения суспензии бактериальных клеток. Полученные таким образом 30 мл суспензии бактериальных клеток ATCC 6872 (DE3)/pSC-3-13, 6 мл суспензии бактериальных клеток Origami-B (DE3)/MBP-C5_pGro7 и 6 мл суспензии бактериальных клеток Origami-B (DE3)/H- MBP-2OST_pGro (Sm) добавляли к 18 мл реакционного раствора [водного раствора, содержащего глюкозу, 60 г/л, KH2PO4, 8,75 г/л, K2HPO4, 15 г/л, MgSO4-7H2O, 20 г/л, D-пантотенат кальция, 0,3 г/л, никотиновую кислоту, 0,2 г/л, аденин, 4,05 г/л, хлорид бензалкония, 1,25 г/л, ADEKA NOL LG-109 (изготовленный в ADEKA), 1 мл/л, и N-сульфогепаросан, 1 г/л (продуцированный в примере 6)] в 250 мл биореактор, и подвергали реакции в течение 22 часов в условиях культивирования 37 градусов C, скорость перемешивания 500 об./мин и скорость аэрации 0,75 мл/мин, с доведением в то же время его pH до 6,5 с использованием 2,8% водного раствора аммиака. Во время этого периода, глюкозу, 60 г/л, MgSO4-7H2O, 5,0 г/л, и аденин, 2,7 г/л, дополнительно добавляли 6 часов после начала реакции.
[0214] Отбор образцов проводили соответствующим образом во время реакции для получения реакционного раствора. Полученный реакционный раствор соответствующим образом разводили и затем центрифугировали, и PAPS детектировали и количественно оценивали посредством измерения оптической плотности при 254 нм с использованием УФ-детектора, с использованием устройства для HPLC, изготовленного в Shimadzu Corporation. Результаты показаны на ФИГ. 4. Кроме того, получение ненасыщенного дисахарида посредством ферментного расщепления и анализа посредством HPLC проводили в соответствии с Патентным документом [WO2018/048973 A1].
[0215] То есть, полученный реакционный раствор центрифугировали, и полученный супернатант нагревали и поддерживали при 80 градусах C в течение 10 минут для денатурации белка. Раствор после денатурации белка центрифугировали, и полученный супернатант подвергали обессоливанию посредством ультрафильтрации с использованием устройства для фильтрации, для молекулярной массы 3000 (изготовленного в Merck). Раствор после обессоливания вводили в реакционный раствор для гепариназы [состоящий из 50 мМ ацетата аммония и 2 мМ хлорида кальция], содержащий 0,5 ед./мл каждой из гепариназ I, II и III (изготовленных в Sigma, но можно использовать также другие гепариназы), и подвергали ферментному расщеплению при 35 градусах C в течение 2 часов. Гепариназы инактивировали посредством поддержания раствора, после ферментного расщепления, при 95 градусах C в течение 15 минут.
[0216] Анализ ненасыщенных дисахаридов проводили посредством подвергания раствора после инактивации гепариназы анализу, в режиме градиентной элюции, подвижной фазы A [водного раствора, который содержит 1,8 мМ дигидрофосфат натрия, и pH которого доводили до pH 3,0 с использованием фосфорной кислоты] и подвижной фазы B [водного раствора, который содержит 1,8 мМ дигидрофосфат натрия и 1 M перхлорат натрия, и pH которого доводили до pH 3,0 с использованием фосфорной кислоты], с использованием Shimadzu HPLC и колонки для обмена сильных анионов (хроматографической колонки spherisorb-SAX, 4,0×250 мм, 5 микрометров, изготовленной в Waters).
[0217] Ненасыщенный дисахарид детектировали посредством измерения оптической плотности при 232 нм с использованием УФ-детектора. Время удержания дельта-UA-GlcNS и дельта-UA, 2S-GlcNS проверяли посредством сравнения со стандартным реагентом ненасыщенного дисахарида (изготовленным в Iduron). 2-O-сульфатирование проверяли посредством получения соотношения площади для дельта-UA, 2S-GlcNS, относительно общей площади для детектированных дельта-UA-GlcNS и дельта-UA, 2S-GlcNS. Результаты показаны на ФИГ. 5.
Пример 10
[0218] Тестирование реакции сульфатирования в положении 6-O 2-O-сульфатированного N-сульфогепаросана с использованием ATCC 6872 (DE3)/pSC-3-13 и Origami-B (DE3)/MBP-6OST3_pGro7
Каждые из замороженных бактериальных клеток ATCC 6872(DE3)/pSC-3-13 и Origami-B(DE3)/MBP-6OST3_pGro7, полученных в примерах 7 и 8, суспендировали в дистиллированной воде таким образом, чтобы масса замороженных бактериальных клеток стала 333 г/л, для получения суспензии бактериальных клеток. Полученные таким образом 30 мл суспензии бактериальных клеток ATCC 6872 (DE3)/pSC-3-13 и 6 мл суспензии бактериальных клеток Origami-B(DE3)/MBP-6OST3_pGro7, так же как 6 мл дистиллированной воды, добавляли к 18 мл реакционного раствора [водного раствора, содержащего глюкозу, 60 г/л, KH2PO4 8,75 г/л, K2HPO4 15 г/л, MgSO4-7H2O, 20 г/л, D-пантотенат кальция, 0,3 г/л, никотиновую кислоту, 0,2 г/л, аденин, 4,05 г/л, хлорид бензалкония, 1,25 г/л, ADEKA NOL LG-109 (изготовленный в ADEKA), 1 мл/л, и 2-O-сульфатированный N-сульфогепаросан, 0,5 г/л (продуцированный в примере 6)], в 250 мл биореакторе, и подвергали реакции в течение 22 часов в условиях культивирования 32 градусов C, скорость перемешивания 500 об./мин, и скорость аэрации 0,75 мл/мин, с доведением в то же время его pH до 7,4 с использованием водного раствора 2Н гидроксида калия.
[0219] Во время этого периода, глюкозу, 60 г/л, MgSO4-7H2O, 5,0 г/л, и аденин, 2,7 г/л дополнительно добавляли 6 часов после начала реакции. Отбор образцов проводили соответствующим образом во время реакции для получения реакционного раствора. С использованием такого же способа, как в примере 9, проводили детекцию и количественную оценку PAPS. Результаты показаны на ФИГ. 6.
[0220] Кроме того, состав сульфатирования, содержащегося в сахарной цепи после реакции, анализировали посредством такого же способа, как в примере 9. Время удержания дельта-UA, 2S-GlcNS и дельта-UA, 2S-GlcN, 6S проверяли посредством сравнения со стандартным реагентом ненасыщенного дисахарида (изготовленном в Iduron). 6-O-сульфатирование проверяли посредством получения соотношения площади для дельта-UA, 2S-GlcN, 6S, относительно общей площади для детектированных дельта-UA, 2S-GlcNS и дельта-UA, 2S-GlcN, 6S. Результаты показаны на ФИГ. 7.
Пример 11
[0221] Тестирование реакции сульфатирования в положении 6-O и положении 3-O 2-O-сульфатированного N-сульфогепаросана с использованием ATCC 6872 (DE3)/pSC-3-13, Origami-B (DE3)/MBP-6OST3_pGro7 и RIL/HIS-3OST1_pGro (Sm)
Реакцию проводили в течение 22 часов в условиях, описанных в примере 10. Во время этого периода, 3 часа после начала реакции, замороженные бактериальные клетки IL/HIS-3OST1_pGro(Sm), полученные в примере 8, суспендировали в дистиллированной воде таким образом, чтобы масса замороженных бактериальных клеток стала 333 г/л, и добавляли 6 мл полученной суспензии бактериальных клеток. Отбор образцов проводили соответствующим образом во время реакции для получения реакционного раствора. С использованием такого же способа, как в примере 9, проводили детекцию и количественную оценку PAPS. Результаты показаны на ФИГ. 8.
[0222] После завершения реакции, реакционный раствор центрифугировали, полученный супернатант соответствующим образом разводили, и затем, активность anti-IIa измеряли с использованием набора для измерения ANTI-IIa BIOPHEN (торговое наименование) (изготовленного в Hyphen Biomed), для проверки 3-O-сульфатирования. Кроме того, активность anti-IIa раствора после реакции из примера 10 также измеряли в качестве отрицательного контроля, в который не добавляли RIL/HIS-3OST1_pGro (Sm). Калибровочную кривую получали с использованием калибратора UFH BIOPHEN (торговое наименование) (изготовленного в Hyphen Biomed). Результаты показаны в таблице 1.
[0223]
Пример 12
[0224] Замороженные бактериальные клетки ATCC 6872(DE3)/pSC-3-13, полученные в примере 7, суспендировали в дистиллированной воде таким образом, чтобы масса замороженных бактериальных клеток стала 333 г/л, для получения суспензии бактериальных клеток. 30 мл полученной таким образом суспензии бактериальных клеток добавляли к 30 мл реакционного раствора [водного раствора, содержащего глюкозу, 60 г/л, KH2PO4, 8,75 г/л, K2HPO4, 15 г/л, MgSO4-7H2O, 20 г/л, D-пантотенат кальция, 0,3 г/л, никотиновую кислоту, 0,2 г/л, аденин, 4,05 г/л, хлорид бензалкония, 0,625 г/л, ADEKA NOL LG-109 (изготовленный в ADEKA), 1 мл/л], в 250 мл биореакторе, и подвергали реакции в течение 22 часов в условиях культивирования 32 градусов C, скорость перемешивания 500 об./мин и скорость аэрации 0,75 мл/мин, с доведением в то же время его pH до 7,4 с использованием водного раствора 2Н гидроксида калия.
[0225] Во время этого периода, глюкозу, 60 г/л, KH2PO4, 2,19 г/л, K2HPO4, 3,75 г/л, MgSO4-7H2O, 5,0 г/л, и аденин, 2,7 г/л, дополнительно добавляли 6 часов после начала реакции. Отбор образцов проводили соответствующим образом во время реакции для получения реакционного раствора.
[0226] Полученный реакционный раствор соответствующим образом разводили и затем центрифугировали, и PAPS детектировали и количественно оценивали посредством измерения оптической плотности при 254 нм с использованием УФ-детектора, с использованием устройства для HPLC, изготовленного в Shimadzu Corporation. Результаты показаны на ФИГ. 9.
[0227] Как описано выше, бактерию из рода Corynebacterium, способную к продукции и регенерации PAPS, и микроорганизм, принадлежащий к прокариотам, экспрессирующий фермент сульфатирования, добавляли и подвергали реакции с сырьевыми материалами, такими как глюкоза, аденин и сульфат магния, и таким образом, сульфатированные полисахариды эффективно получали из полисахаридов.
[0228] В то время как изобретение описано подробно и со ссылкой на конкретные варианты его осуществления, специалисту в данной области очевидно, что можно осуществлять его различные изменения и модификации, без отклонения от его содержания и объема. Полное содержание всех ссылок, процитированных в настоящем описании, приведено в настоящем описании в качестве ссылки. Эта заявка основана на Международной заявке No. PCT/JP2020/015388, поданной 3 апреля 2020 г., полное содержание которой приведено в настоящем описании в качестве ссылки.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> RENSSELAER POLYTECHNIC INSTITUTE
OTSUKA PHARMACEUTICAL FACTORY, INC
KIRIN BIOMATERIALS CO., LTD.
<120> Способ продукции сульфатированного полисахарида и способ продукции PAPS
<130> W531376
<150> PCT/JP2020/015388
<151> 2020-04-03
<160> 32
<170> PatentIn версии 3.5
<210> 1
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 1
cggtacccgg ggatccgact ggccagtaca ggctg 35
<210> 2
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 2
agcgactcga gtttagatct tcaatgctca acccgacctg 40
<210> 3
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 3
attgaagatc taaactcgag tcgcttgacg atgttcaact 40
<210> 4
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 4
cgactctaga ggatcgtttg tcgcgggcga cgata 35
<210> 5
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 5
tttctcgaga actgcgcaac tcgtgaaagg 30
<210> 6
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 6
tttagatctg ttacgcgaac gcgaagtc 28
<210> 7
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 7
cggtacccgg ggatcgctta atgcgccgct acaggg 36
<210> 8
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 8
atgcctgcag gtcgaggagc tgactgggtt gaagg 35
<210> 9
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 9
gaaggagata tacatatgcc tgctcctcac ggtgg 35
<210> 10
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 10
gttgtgtctc ctctattaaa atacaaaaaa gccat 35
<210> 11
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 11
tagaggagac acaacatggc tactaatatt acttg 35
<210> 12
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 12
ggtggtggtg ctcgattaca aatgcttacg gatga 35
<210> 13
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 13
ttgggaattc gagctctaag gaggttataa aaaatgaata ttcgtccatt gcatgatcgc 60
g 61
<210> 14
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 14
ttacatcatg ccgcccatgc cacc 24
<210> 15
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 15
ttatgacaac ttgacggcta catcattcac 30
<210> 16
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 16
cgcgatcatg caatggacga atattcattt tttataacct ccttagagct cgaattccca 60
a 61
<210> 17
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 17
ggcggcatga tgtaattatt tgccgactac cttggtgatc tc 42
<210> 18
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 18
gtcaagttgt cataatctag agcggttcag tagaaaagat caaag 45
<210> 19
<211> 918
<212> ДНК
<213> Chinese hamster
<400> 19
cgcgaaatcg aacagcgtca cacgatggat ggcccgcgcc aagatgcaac cctggacgaa 60
gaagaagata tggttatcat ctacaaccgc gtcccgaaaa ccgcttcaac gtcgtttacc 120
aatattgcgt acgacctgtg cgccaaaaac aaatatcatg tgctgcacat caataccacg 180
aaaaacaatc cggttatgag cctgcaggat caagtccgtt ttgtgaaaaa catcacctct 240
tggaaagaaa tgaaaccggg cttctaccat ggtcacgtta gttatctgga ctttgccaaa 300
ttcggcgtga agaaaaaacc gatctacatc aacgttatcc gtgatccgat cgaacgcctg 360
gtctcttatt actattttct gcgtttcggc gatgactatc gcccgggtct gcgtcgccgt 420
aaacagggtg acaagaaaac ctttgatgaa tgcgtcgcag aaggcggttc agattgtgct 480
ccggaaaaac tgtggctgca gatcccgttt ttctgcggcc atagctctga atgttggaac 540
gtgggttcgc gctgggcaat ggaccaagct aaatacaacc tgatcaacga atattttctg 600
gtgggtgtta ccgaagaact ggaagatttc atcatgctgc tggaagccgc actgccgcgc 660
tttttccgtg gcgcgacgga actgtaccgt accggcaaaa aatctcatct gcgcaaaacc 720
acggagaaaa aactgccgac gaaacagacc attgcgaaac tgcagcaatc cgacatctgg 780
aaaatggaaa acgaattcta cgaattcgcc ctggaacagt ttcaattcat tcgtgcgcac 840
gccgttcgcg aaaaagatgg tgacctgtat atcctggcac aaaacttctt ctatgaaaaa 900
atctatccga aatcaaat 918
<210> 20
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 20
tacttccaat ccaatcgcga aatcgaacag cgtca 35
<210> 21
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 21
ttatccactt ccaatatttg atttcggata gattt 35
<210> 22
<211> 1536
<212> ДНК
<213> Saccharomyces cerevisiae
<400> 22
atgcctgctc ctcacggtgg tattctacaa gacttgattg ctagagatgc gttaaagaag 60
aatgaattgt tatctgaagc gcaatcttcg gacattttag tatggaactt gactcctaga 120
caactatgtg atattgaatt gattctaaat ggtgggtttt ctcctctgac tgggtttttg 180
aacgaaaacg attactcctc tgttgttaca gattcgagat tagcagacgg cacattgtgg 240
accatcccta ttacattaga tgttgatgaa gcatttgcta accaaattaa accagacaca 300
agaattgccc ttttccaaga tgatgaaatt cctattgcta tacttactgt ccaggatgtt 360
tacaagccaa acaaaactat cgaagccgaa aaagtcttca gaggtgaccc agaacatcca 420
gccattagct atttatttaa cgttgccggt gattattacg tcggcggttc tttagaagcg 480
attcaattac ctcaacatta tgactatcca ggtttgcgta agacacctgc ccaactaaga 540
cttgaattcc aatcaagaca atgggaccgt gtcgtagctt tccaaactcg taatccaatg 600
catagagccc acagggagtt gactgtgaga gccgccagag aagctaatgc taaggtgctg 660
atccatccag ttgttggact aaccaaacca ggtgatatag accatcacac tcgtgttcgt 720
gtctaccagg aaattattaa gcgttatcct aatggtattg ctttcttatc cctgttgcca 780
ttagcaatga gaatgagtgg tgatagagaa gccgtatggc atgctattat tagaaagaat 840
tatggtgcct cccacttcat tgttggtaga gaccatgcgg gcccaggtaa gaactccaag 900
ggtgttgatt tctacggtcc atacgatgct caagaattgg tcgaatccta caagcatgaa 960
ctggacattg aagttgttcc attcagaatg gtcacttatt tgccagacga agaccgttat 1020
gctccaattg atcaaattga caccacaaag acgagaacct tgaacatttc aggtacagag 1080
ttgagacgcc gtttaagagt tggtggtgag attcctgaat ggttctcata tcctgaagtg 1140
gttaaaatcc taagagaatc caacccacca agaccaaaac aaggtttttc aattgtttta 1200
ggtaattcat taaccgtttc tcgtgagcaa ttatccattg ctttgttgtc aacattcttg 1260
caattcggtg gtggcaggta ttacaagatc tttgaacaca ataataagac agagttacta 1320
tctttgattc aagatttcat tggttctggt agtggactaa ttattccaaa tcaatgggaa 1380
gatgacaagg actctgttgt tggcaagcaa aacgtttact tattagatac ctcaagctca 1440
gccgatattc agctagagtc agcggatgaa cctatttcac atattgtaca aaaagttgtc 1500
ctattcttgg aagacaatgg cttttttgta ttttaa 1536
<210> 23
<211> 609
<212> ДНК
<213> Saccharomyces cerevisiae
<400> 23
atggctacta atattacttg gcatccaaat cttacttacg acgaacgcaa ggcattgaga 60
aaacaggacg gttgtactat ttggttaaca ggtctaagtg cgtcaggtaa aagtacaatc 120
gcctgtgcgc tagaacagtt actgctccaa aaaaacttgt ctgcatatag attggatggt 180
gacaacattc gttttggatt gaacaaggat ttgggtttct cagaaaagga cagaaatgaa 240
aacattcgta gaattagcga agtttctaag ctatttgctg attcatgtgc tatttcaatc 300
acctcattta tctctccata cagagttgac agagatagag ctcgtgaact acataaggag 360
gctggtttga agttcattga aatatttgtt gatgttccat tagaagtcgc tgagcaaagg 420
gaccctaagg gtttatacaa gaaagctagg gagggtgtaa tcaaggagtt tacaggtatt 480
tctgccccat atgaagcgcc aaaagctcca gagctacatt tgagaaccga ccagaagacg 540
gttgaagaat gtgctaccat tatttatgag tacttaatca gtgaaaaaat catccgtaag 600
catttgtaa 609
<210> 24
<211> 1695
<212> ДНК
<213> Homo sapiens
<400> 24
gaaaaaagag cagcagcatc tgagagtaac aactatatga accacgtggc caaacaacag 60
tctgaggaag cattccctca ggaacagcag aaagcacccc ctgttgttgg gggcttcaat 120
agcaatgtgg gaagtaaggt gttagggctc aaatatgaag aaattgactg tctcataaat 180
gatgaacaca caattaaagg gagacgagag gggaacgaag tctttcttcc attcacttgg 240
gttgagaaat attttgatgt ttatggaaag gtggttcagt atgatggcta tgatcggttt 300
gaattctctc atagctattc caaagtctat gcacagagag ccccctatca ccccgatggt 360
gtgtttatgt cttttgaagg ctacaatgtg gaagtccgag acagagtcaa gtgcataagt 420
ggggttgaag gtgtgccatt atctacacaa tggggacctc aaggctattt ctatccaatc 480
cagattgcac agtatggatt aagtcattac agcaagaatc taactgagaa acctcctcac 540
atagaggtat atgaaacagc agaagacaga gacaaaaaca agcctaatga ctggactgtg 600
ccaaagggct gctttatggc gaatgtggct gataagtcta gattcaccaa tgtcaaacag 660
tttattgcac cagaaaccag tgaaggtgta tccttgcaac tgggaaacac aaaagatttt 720
attatttcat ttgacctcaa gttcttgaca aatggaagtg tgtccgtggt tctagagacc 780
acagaaaaga atcagctctt cactatacat tatgtctcaa atgctcagct aattgctttt 840
aaagaaagag atatatacta tggcattggg cccagaactt catggagcac agttaccagg 900
gacctggtca ctgacctcag gaaaggagtg ggtctttcaa acacaaaagc tgtcaagcca 960
accaaaataa tgcccaagaa ggtggttagg ttgattgcaa aaggtaaggg attcctcgac 1020
aacattacca tctctaccac agcccacatg gctgcatttt ttgctgctag tgattggcta 1080
gtaaggaacc aggatgagaa aggtggctgg ccaattatgg tgacccgtaa gttaggggaa 1140
gggttcaagt ctttagagcc aggatggtat tctgccatgg cccaagggca agccatttct 1200
acattagtca gggcctatct gttaacaaaa gaccatatat tcctcaattc agctttaagg 1260
gcaacagccc cttataagtt tctatctgag cagcatggag ttaaagctgt gtttatgaat 1320
aaacatgact ggtatgaaga atatccaacc acacctagct cttttgtttt aaatggcttt 1380
atgtattctt taattgggct gtatgactta aaagaaactg caggggaaaa actcggaaaa 1440
gaagcaaggt ccttgtatga gcgtggcatg gaatctctta aagccatgct gcccttgtat 1500
gacactggct caggaaccat ctatgacctc cgtcacttca tgcttggcat cgctcctaac 1560
ctggctcgct gggactatca taccacccac atcaatcagt tgcagctact cagtaccatt 1620
gatgagtccc caatcttcaa agaatttgtc aagaggtgga aaagctacct taaaggcagc 1680
agggcaaagc acaac 1695
<210> 25
<211> 1695
<212> ДНК
<213> Mus musculus
<400> 25
gaaaaaagag cagcagcatc tgagagtaac aactatatga accacgtggc caaacaacag 60
tctgaggaag cattccctca ggaacagcag aaagcacccc ctgttgttgg gggcttcaat 120
agcaatgtgg gaagtaaggt gttagggctc aaatatgaag aaattgactg tctcataaat 180
gatgaacaca caattaaagg gagacgagag gggaacgaag tctttcttcc attcacttgg 240
gttgagaaat attttgatgt ttatggaaag gtggttcagt atgatggcta tgatcggttt 300
gaattctctc atagctattc caaagtctat gcacagagag ccccctatca ccccgatggt 360
gtgtttatgt cttttgaagg ctacaatgtg gaagtccgag acagagtcaa gtgcataagt 420
ggggttgaag gtgtgccatt atctacacaa tggggacctc aaggctattt ctatccaatc 480
cagattgcac agtatggatt aagtcattac agcaagaatc taactgagaa acctcctcac 540
atagaggtat atgaaacagc agaagacaga gacaaaaaca agcctaatga ctggactgtg 600
ccaaagggct gctttatggc gaatgtggct gataagtcta gattcaccaa tgtcaaacag 660
tttattgcac cagaaaccag tgaaggtgta tccttgcaac tgggaaacac aaaagatttt 720
attatttcat ttgacctcaa gttcttgaca aatggaagtg tgtccgtggt tctagagacc 780
acagaaaaga atcagctctt cactatacat tatgtctcaa atgctcagct aattgctttt 840
aaagaaagag atatatacta tggcattggg cccagaactt catggagcac agttaccagg 900
gacctggtca ctgacctcag gaaaggagtg ggtctttcaa acacaaaagc tgtcaagcca 960
accaaaataa tgcccaagaa ggtggttagg ttgattgcaa aaggtaaggg attcctcgac 1020
aacattacca tctctaccac agcccacatg gctgcatttt ttgctgctag tgattggcta 1080
gtaaggaacc aggatgagaa aggtggctgg ccaattatgg tgacccgtaa gttaggggaa 1140
gggttcaagt ctttagagcc aggatggtat tctgccatgg cccaagggca agccatttct 1200
acattagtca gggcctatct gttaacaaaa gaccatatat tcctcaattc agctttaagg 1260
gcaacagccc cttataagtt tctatctgag cagcatggag ttaaagctgt gtttatgaat 1320
aaacatgact ggtatgaaga atatccaacc acacctagct cttttgtttt aaatggcttt 1380
atgtattctt taattgggct gtatgactta aaagaaactg caggggaaaa actcggaaaa 1440
gaagcaaggt ccttgtatga gcgtggcatg gaatctctta aagccatgct gcccttgtat 1500
gacactggct caggaaccat ctatgacctc cgtcacttca tgcttggcat cgctcctaac 1560
ctggctcgct gggactatca taccacccac atcaatcagt tgcagctact cagtaccatt 1620
gatgagtccc caatcttcaa agaatttgtc aagaggtgga aaagctacct taaaggcagc 1680
agggcaaagc acaac 1695
<210> 26
<211> 792
<212> ДНК
<213> Mus musculus
<400> 26
ggcacagcat ccaatggttc cacacagcag ctgccacaga ccatcatcat tggggtgcgc 60
aagggtggta cccgagccct gctagagatg ctcagcctgc atcctgatgt tgctgcagct 120
gaaaacgagg tccatttctt tgactgggag gagcattaca gccaaggcct gggctggtac 180
ctcacccaga tgcccttctc ctcccctcac cagctcaccg tggagaagac acccgcctat 240
ttcacttcgc ccaaagtgcc tgagagaatc cacagcatga accccaccat ccgcctgctg 300
cttatcctga gggacccatc agagcgcgtg ctgtccgact acacccaggt gttgtacaac 360
caccttcaga agcacaagcc ctatccaccc attgaggacc tcctaatgcg ggacggtcgg 420
ctgaacctgg actacaaggc tctcaaccgc agcctgtacc atgcacacat gctgaactgg 480
ctgcgttttt tcccgttggg ccacatccac attgtggatg gcgaccgcct catcagagac 540
cctttccctg agatccagaa ggtcgaaaga ttcctgaagc tttctccaca gatcaacgcc 600
tcgaacttct actttaacaa aaccaagggc ttctactgcc tgcgggacag tggcaaggac 660
cgctgcttac acgagtccaa aggccgggcg cacccccagg tggatcccaa actacttgat 720
aaactgcacg aatactttca tgagccaaat aagaaatttt tcaagctcgt gggcagaaca 780
ttcgactggc ac 792
<210> 27
<211> 1170
<212> ДНК
<213> Escherichia coli
<400> 27
atgcaaggta atgcactaac cgttttatta tccggtaaaa aatatctgct attgcagggg 60
ccaatgggac cctttttcag tgatgttgcc gagtggctag agtcattagg tcgtaacgct 120
gtgaatgttg tattcaacgg tggggatcgt ttttactgcc gccatcgaca atacctagct 180
tactaccaga caccgaaaga gtttcccgga tggttacggg atctccaccg gcaatatgac 240
tttgacacaa tcctctgctt tggcgactgc cgcccattgc ataaagaagc aaaacgctgg 300
gcaaagtcga aagggatccg cttcctggca tttgaagaag gatatttacg cccgcaattt 360
attaccgttg aagaaggcgg agtgaacgca tattcatcgc taccgcgcga tccggatttt 420
tatcgtaagt taccagatat gcctacgccg cacgttgaga acttaaaacc ttcaacgatg 480
aaacgtatag gccatgctat gtggtattac ctgatgggct ggcattaccg tcatgagttt 540
cctcgctacc gccaccacaa atcattttcc ccctggtatg aagcacgttg ctgggttcgt 600
gcatactggc gcaagcaact ttacaaggta acacagcgta aggtattacc gaggttaatg 660
aacgaactgg accagcgtta ttatcttgct gttttgcagg tgtataacga tagccagatt 720
cgtaaccaca gcagttataa cgatgtgcgt gactatatta atgaagtcat gtactcattt 780
tcgcgtaaag cgccgaaaga aagttatttg gtgatcaaac atcatccgat ggatcgtggt 840
cacagactct atcgaccatt aattaaacgg ttgagtaagg aatatggctt aggtgagcga 900
atcctttatg tgcacgatct cccgatgccg gaattattac gccatgcaaa agcggtggtg 960
acgattaaca gtacggcggg gatctctgcg ctgattcata acaaaccact caaagtgatg 1020
ggcaatgccc tgtacgacat caagggcttg acgtatcaag ggcatttgca ccagttctgg 1080
caggctgatt ttaaaccaga tatgaaactg tttaagaagt ttcgtgggta tttattggtg 1140
aagacgcagg ttaatgcggt ttattattaa 1170
<210> 28
<211> 717
<212> ДНК
<213> Escherichia coli
<400> 28
atgattgttg caaatatgtc atcataccca cctcgaaaaa aagagttggt gcattctata 60
caaagtttac atgctcaagt agataaaatt aatctttgcc tgaatgagtt tgaagaaatt 120
cctgaggaat tagatggttt ttcaaaatta aatccagtta ttccagataa agattataag 180
gatgtgggca aatttatatt tccttgcgct aaaaatgata tgatcgtact tacagatgat 240
gatattattt accctcccga ttatgtagaa aaaatgctca atttttataa ttcctttgca 300
atattcaatt gcattgttgg gattcatggc tgtatataca tagatgcatt tgatggagat 360
cagtctaaaa gaaaagtatt ttcatttact caagggctat tgcgaccgag agttgtaaat 420
caattaggta cagggactgt ttttcttaag gcagatcaat taccatcttt aaaatatatg 480
gatggttctc aacgattcgt cgatgttaga ttttctcgct atatgttaga gaatgaaatt 540
ggtatgatat gtgttcccag agaaaaaaac tggctaagag aggtctcatc aggttcaatg 600
gaaggacttt ggaacacatt tacaaaaaaa tggcctttag acatcataaa agaaacacaa 660
gcaatcgcag gatattcaaa acttaacctc gaattagtgt ataatgtgga agggtaa 717
<210> 29
<211> 1689
<212> ДНК
<213> Escherichia coli
<400> 29
atgaataaat tagtgctagt cggacatcct ggctcaaagt atcagatagt tgaacatttt 60
ttgaaagaaa ttggcatgaa ctcaccaaat tattctacaa gtaataaaat ttccccagaa 120
tatatcaccg cttcattatg tcaattttat caaacaccag aagttaatga tgtagtagat 180
gagagagaat tctcagctgt tcaagtctca accatgtggg atagcatggt tcttgaacta 240
atgatgaaca atctaaataa caaactttgg gggtgggcag atccatctat aatatttttt 300
cttgattttt ggaaaaatat agataaaagc ataaaattca tcatgatata tgatcaccct 360
aaatataatt taatgcgttc agtaaataat gcccctctct ctttaaatat aaataatagt 420
gtagataact ggattgcata taataaaaga ttgcttgatt tttttttgga gaataaagaa 480
cgatgtgtgt tgattaattt tgaggcgttt caaagcaata agaaaaatat tataaagcca 540
ttgagtaata ttataaaaat agataatcta atgtctgcgc attacaaaaa ttcaatattg 600
tttgatgtgg ttgagaataa tgattataca aaatcaaatg aaattgccct gcttgaaaaa 660
tatacaactt tattttcttt aagtgcaaat gagactgaaa ttacatttaa tgatacaaag 720
gttagtgagt acttagtatc tgaattaata aaagaaagaa ccgaggttct gaagctttat 780
aatgagttac aagcctatgc aaacctacct tatatagaaa catcgaaaga taacgtttcg 840
gctgaggctg cattatggga ggtagtcgaa gagagaaatt ctatcttcaa tattgtatct 900
catttggtgc aagagtcaaa aaagaaggat gcagatattg aattgactaa atctatattt 960
aagaaaagac aatttttatt attgaacagg attaatgagc taaaaaaaga aaaggaagag 1020
gtaattaaac tttcaaaaat aaatcacaac gatgttgtga gacaagaaaa atatccagat 1080
gatattgaaa aaaaaataaa tgacatacag aaatatgaag aagagataag cgaaaaagaa 1140
tcaaaactca ctcaggcaat atcagaaaaa gaacagattt taaaacaatt gcataaatat 1200
gaagaagaga taagcgaaaa agaatcaaaa ctcactcagg caatatcaga aaaagaacag 1260
attttaaaac aattgcatat agtgcaagag cagttggaac actattttat agaaaatcag 1320
gaaattaaaa agaaacttcc acctgtgcta tatggagcag ctgagcagat aaaacaagag 1380
ttaggttatc gacttggtta tattatagtc tcgtattcta aatccctcaa ggggattatt 1440
accatgccat ttgcacttat ccgtgagtgt gtttttgaaa aaaaacgtaa gaagagttat 1500
ggcgttgatg tgccactcta tttatatgct gatgctgata aggctgaaag agttaagaaa 1560
catttatctt atcaattagg gcaggctatt atctccagtg ctaattcgat atttggattc 1620
attacccttc catttaagtt aattgttgtt gtttataaat ataggagagc taaaatcaag 1680
ggctgttaa 1689
<210> 30
<211> 1563
<212> ДНК
<213> Escherichia coli
<400> 30
atgaacgcag aatatataaa tttagttgaa cgtaaaaaga aattagggac aaatattggt 60
gctcttgatt ttttattatc aattcataag gagaaagttg atcttcaaca taaaaactcg 120
cctttaaaag gtaacgataa ccttattcac aaaagaataa acgaatacga caatgtactt 180
gaactatcta agaatgtatc agctcagaat tctggcaatg agttttctta tttattggga 240
tatgcagatt ctcttagaaa agttggtatg ttggatactt atattaaaat tgtttgttat 300
ctaacaattc aatctcgtta ttttaaaaat ggcgaacgag ttaagctttt tgaacatata 360
agtaacgctc tacggtattc aaggagtgat tttctcatta atcttatttt tgaacgatat 420
atcgaatata taaaccatct aaaattgtcg cccaaacaaa aagattttta tttttgtacg 480
aagttttcaa aatttcatga ttatactaaa aatggatata aatatttagc atttgataat 540
caagccgatg cagggtatgg cctgacttta ttattaaatg caaacgatga tatgcaagat 600
agttataatc tactccctga gcaagaactt tttatttgta atgctgtaat agataatatg 660
aatatttata ggagtcaatt taacaaatgt ctacgaaaat acgatttatc agaaataact 720
gatatatacc caaataaaat tatattgcaa ggaattaagt tcgataagaa aaaaaatgtt 780
tatggaaaag atcttgttag tataataatg tcagtattca attcagaaga tactattgca 840
tactcattac attcattgtt gaatcaaaca tatgaaaata ttgaaattct cgtgtgcgat 900
gattgttcat cggacaaaag ccttgaaata attaagagca tagcttattc tgattcaaga 960
gtgaaagtat atagctcacg aaaaaaccaa ggcccttata atataagaaa tgagctaata 1020
aaaaaagcac acggtaattt catcaccttt caagatgcag atgatctttc tcatccggag 1080
agaatacaaa gacaagttga ggttcttcgc aataataagg ctgtaatctg tatggctaac 1140
tggatccgtg ttgcgtcaaa tggaaaaatt caattcttct atgatgataa agccacaaga 1200
atgtctgttg tatcgtcaat gataaaaaaa gatatttttg cgacagttgg tggctataga 1260
caatctttaa ttggtgcaga tacggagttt tatgaaacag taataatgcg ttatgggcga 1320
gaaagtattg taagattact gcagccattg atattggggt tatggggaga ctccggactt 1380
accaggaata aaggaacaga agctctacct gatggatata tatcacaatc tcgaagagaa 1440
tatagtgata tcgcggcaag acaacgagtg ttagggaaaa gtatcgtaag tgataaagat 1500
gtacgtggtt tattatctcg ctatggtttg tttaaagatg tatcaggaat aattgaacaa 1560
tag 1563
<210> 31
<211> 1179
<212> ДНК
<213> Escherichia coli
<400> 31
atgttcggaa cactaaaaat aactgtttca ggcgctggtt acgttgggct ttcaaatgga 60
attctaatgg ctcaaaatca tgaagtggtt gcatttgata cccatcaaaa aaaagttgac 120
ttacttaatg ataaactctc tcctatagag gataaggaaa ttgaaaatta tctttcaact 180
aaaatactta attttcgcgc aactactaac aaatatgaag cctataaaaa tgccaattac 240
gttattattg ctacaccaac gaattatgac ccaggttcaa attactttga tacatcaagc 300
gttgaagctg tcattcgtga cgtaacggaa atcaacccaa acgcaattat ggtggttaaa 360
tctacggtcc cagtaggttt cacaaaaaca attaaagaac atttaggtat taataatatt 420
atcttctctc cagaattttt acgagaagga agagccctat acgataatct ccatccatct 480
cgcattatta tcggtgaatg ttctgaacgg gcagaacgtt tggcagtgtt atttcaggaa 540
ggagcgatta aacaaaatat acccgtttta tttacagatt ctacggaagc ggaagcgatt 600
aagttatttt caaatactta tttggctatg cgagttgcat tttttaatga attggatagt 660
tacgcagaaa gttttggtct gaatacgcgt cagattattg acggtgtttg tttggatccg 720
cgcattggta attactacaa taatccttct tttggttatg gtggctactg tttgccaaaa 780
gataccaagc aattattagc caactatcag tctgttccga ataaacttat atctgcaatt 840
gttgatgcta accgtacacg taaggacttt atcactaatg ttattttgaa acatagacca 900
caagttgtgg gggtttatcg tttgattatg aaaagtggtt cagataattt tagagattct 960
tctattcttg gtattataaa gcgtatcaag aaaaaaggcg tgaaagtaat tatttatgag 1020
ccgcttattt ctggagatac attctttaac tcacctttgg aacgggagct ggcgatcttt 1080
aaagggaaag ctgatattat tatcactaac cgaatgtcag aggagttgaa cgatgtggtc 1140
gacaaagtct atagtcgcga tttgtttaaa tgtgactaa 1179
<210> 32
<211> 855
<212> ДНК
<213> Escherichia coli
<400> 32
atggtactga tgatcgtcag cggacgttca ggttcaggta aatctgtcgc cctgcgtgcg 60
ctggaagata tgggttttta ctgcgtggat aaccttcccg tagtgttgtt acccgatctg 120
gctcgaactc tggccgatcg agagatttct gccgccgtca gcattgatgt tcgtaatatg 180
ccggagtcac cagaaatatt cgaacaggcg atgagtaacc tgcctgacgc tttctcaccg 240
caactactgt tcctggatgc cgaccgtaat accttaattc gtcgttacag tgacacgcgc 300
cgactgcatc cgctttccag caaaaacctg tcgctggaaa gtgctatcga caaagaaagc 360
gatttgctgg agcctctgcg ttcgcgagcg gatctgattg tcgacacctc agaaatgtcc 420
gttcacgagc tggcagaaat gctgcgtacc cgtctgctgg gtaaacgtga acgtgaactg 480
accatggtct ttgagtcttt cggcttcaaa cacggtatcc ctatcgatgc agattacgtc 540
tttgacgtgc gcttcttgcc gaacccgcac tgggatccga aactgcgtcc aatgacaggt 600
cttgataaac ctgtcgccgc gttcctcgac cgccacacag aagtacacaa ttttatctac 660
cagacgcgaa gctatcttga gctatggtta cctatgctgg aaaccaacaa ccgtagctac 720
ctgacggtcg ccattggttg taccggcggg aagcaccgtt cggtgtatat tgcagagcaa 780
ctggcagact acttccgctc gcgcggtaaa aacgtccagt cacgccatcg gacgctggaa 840
aaacgtaaac catga 855
<---
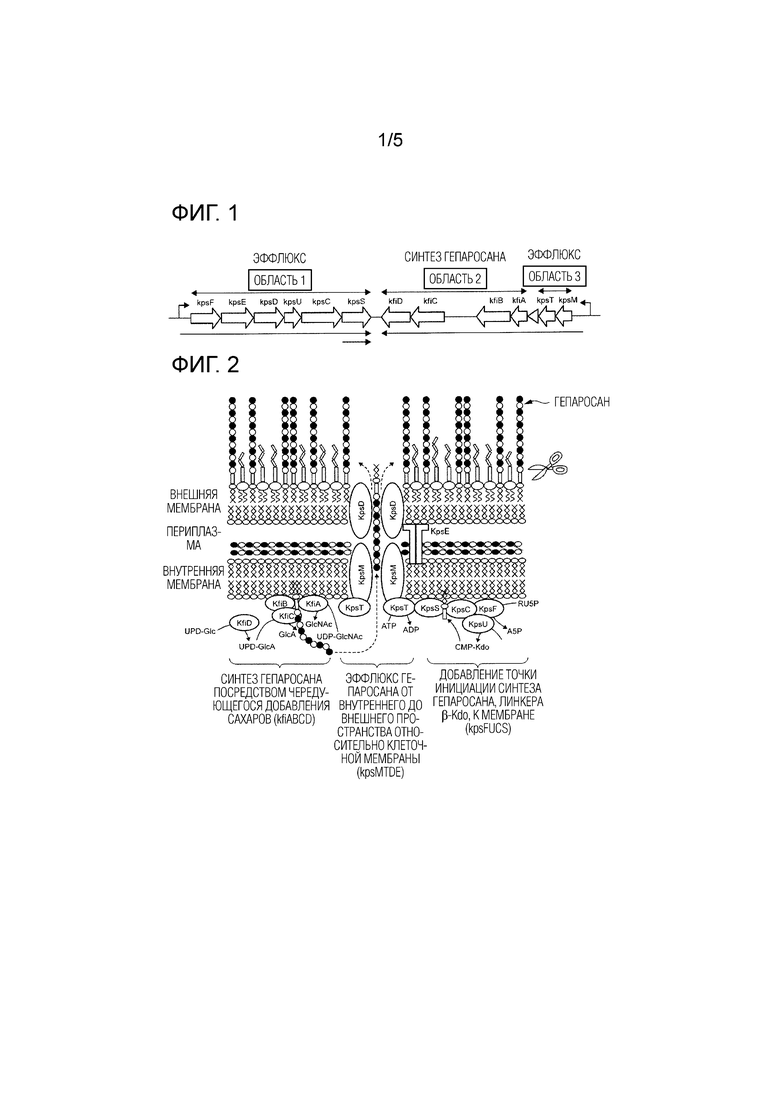
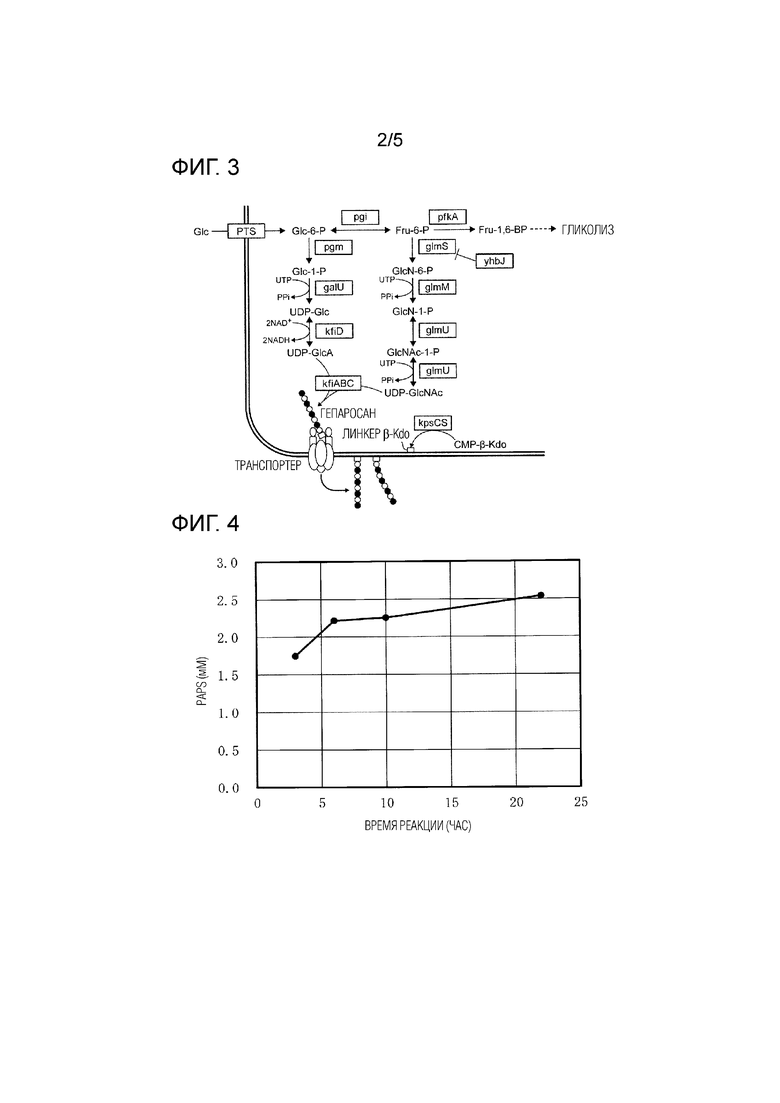
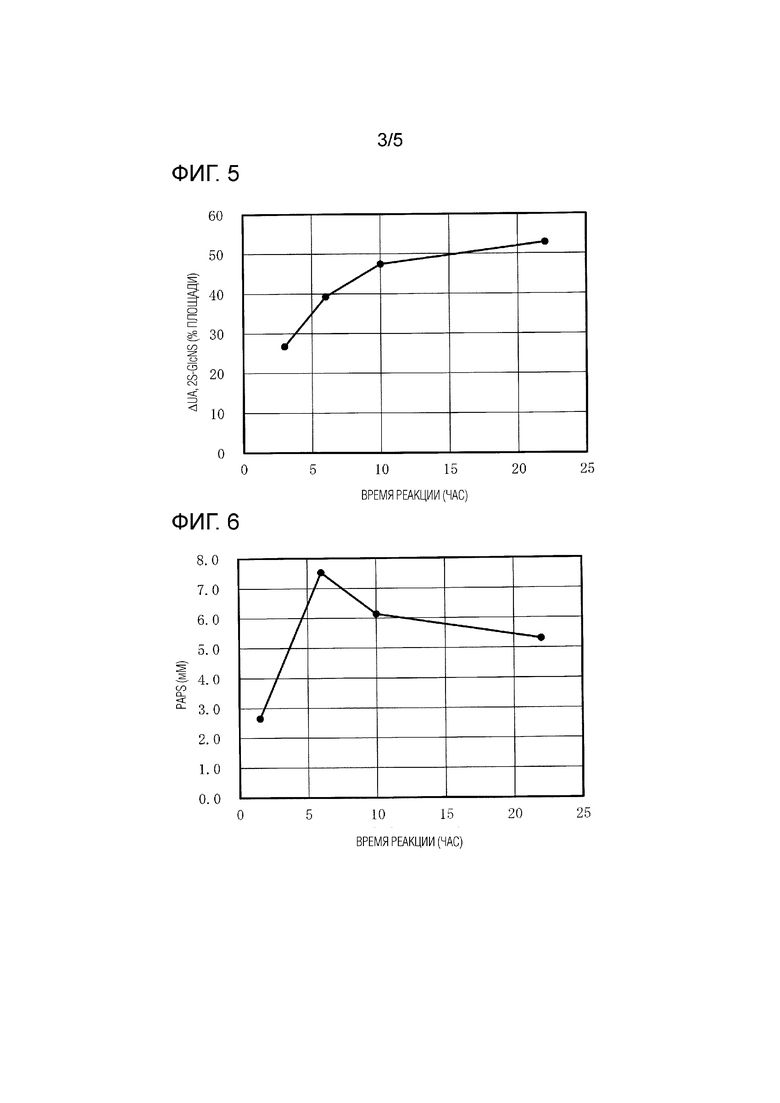
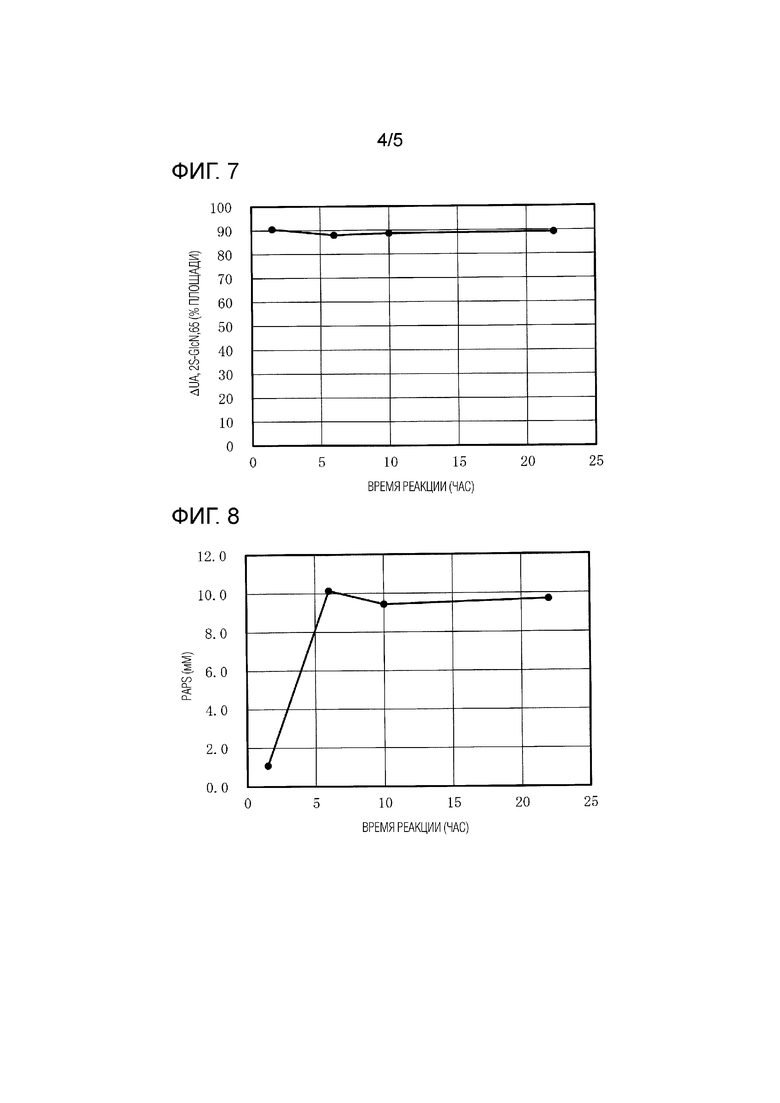
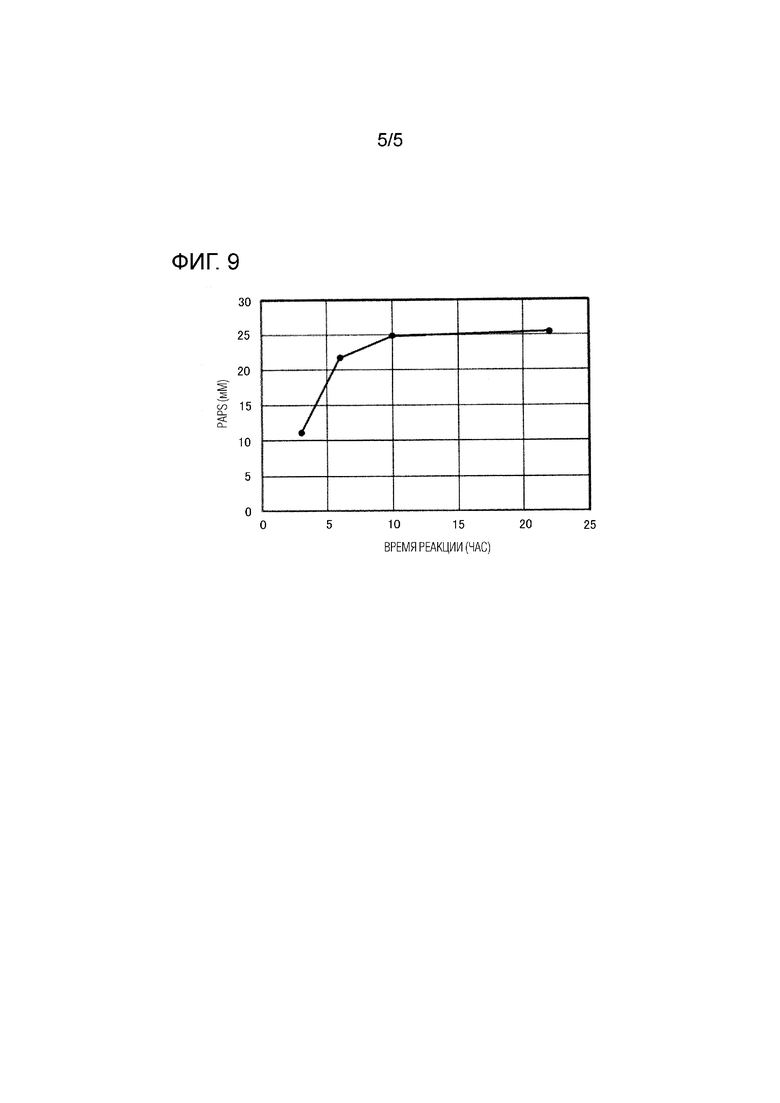
Группа изобретений относится к биотехнологии. Предложены способы продукции сульфатированного полисахарида и продукции 3'-фосфоаденозин-5'-фосфосульфата (PAPS). Способы включают стадию получения трансформанта из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу. Затем проводят реакцию для продукции PAPS в реакционном растворе, содержащем ATP или источник ATP, источник иона сульфата и трансформант или его обработанный материал. Сульфатирование проводят посредством трансформанта, содержащего ген, кодирующий сульфотрансферазу. Изобретения позволяют продуцировать сульфатированные полисахариды с использованием бактериальных клеток и недорогих сырьевых материалов, таких как сульфат магния, без очистки фермента или добавления PAPS. 3 н. и 9 з.п. ф-лы, 9 ил., 1 табл., 12 пр.
1. Способ продукции сульфатированного полисахарида, включающий следующие стадии:
(1-1) получение трансформанта (a) бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта (a); и
(1-2) проведение реакции для продукции 3'-фосфоаденозин-5'-фосфосульфата (PAPS) посредством использования реакционного раствора, содержащего ATP или источник ATP, источник иона сульфата и трансформант (a) или его обработанный материал;
дополнительно включающий проведение сульфатирования посредством трансформанта, содержащего ген, кодирующий сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта.
2. Способ по п.1, дополнительно включающий следующие стадии:
(2-1) получение трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b); и
(2-2) проведение C5-эпимеризации посредством включения трансформанта (b), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
3. Способ по п.1 или 2, дополнительно включающий следующие стадии:
(3-1) получение трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c); и
(3-2) проведение 2-O-сульфатирования посредством включения трансформанта (c), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
4. Способ по любому из пп.1-3, дополнительно включающий следующие стадии:
(3'-1) получение трансформанта (b) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (b);
(3'-2) получение трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c); и
(3'-3) проведение C5-эпимеризации и 2-O-сульфатирования посредством включения трансформанта (b), или его обработанного материала или экстракта, и трансформанта (c), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
5. Способ по любому из пп. 1-4, дополнительно включающий следующие стадии:
(4-1) получение трансформанта (d) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 6-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (d); и
(4-2) проведение 6-O-сульфатирования посредством включения трансформанта (d), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
6. Способ по любому из пп. 1-5, дополнительно включающий следующие стадии:
(5-1) получение трансформанта (e) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 3-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (e); и
(5-2) проведение 3-O-сульфатирования посредством включения трансформанта (e), или его обработанного материала или экстракта в реакционный раствор, в присутствии N-сульфогепаросана.
7. Способ продукции сульфатированного полисахарида, включающий получение сульфатированного полисахарида посредством включения в реакционный раствор, в присутствии ATP или источника ATP, источника иона сульфата и N-сульфогепаросана,
трансформанта (a) бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта (a), и
по меньшей мере одного, выбранного из:
трансформанта (c) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 2-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (c),
трансформанта (d) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 6-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (d), и
трансформанта (e) микроорганизма, принадлежащего к прокариотам, содержащего по меньшей мере ген, кодирующий 3-O-сульфотрансферазу, который введен в него подходящим для экспрессии образом, или обработанного материала или экстракта трансформанта (e).
8. Способ по п.7, отличающийся тем, что в реакционный раствор дополнительно включен трансформант (b) микроорганизма, принадлежащего к прокариотам, содержащий по меньшей мере ген, кодирующий C5-эпимеразу, который введен в него подходящим для экспрессии образом, или обработанный материал или экстракт трансформанта (b).
9. Способ по п.8, отличающийся тем, что в реакционный раствор последовательно добавляют в этом порядке трансформант (b) или обработанный материал или его экстракт, и трансформант (с) или обработанный материал или его экстракт.
10. Способ по любому из пп.7–9, включающий получение сульфатированного полисахарида посредством включения в реакционный раствор, в присутствии ATP или источника ATP, источника иона сульфата и N-сульфогепаросана, трансформанта (a) или его обработанного материала, и трансформантов (b) - (e) или их обработанных материалов или экстрактов.
11. Способ по любому из пп. 1-10, предназначенный для продукции гепарина.
12. Способ продукции PAPS, включающий следующие стадии (i) и (ii):
(i) получение трансформанта бактерии из рода Corynebacterium, содержащего по меньшей мере ген, кодирующий ATP-сульфурилазу, и ген, кодирующий APS-киназу, которые введены в него подходящим для экспрессии образом, или обработанного материала трансформанта; и
(ii) проведение реакции для продукции PAPS посредством использования реакционного раствора, содержащего ATP или источник ATP, источник иона сульфата и трансформант, полученный на стадии (i), или его обработанный материал.
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| US 8735106 B2, 27.05.2014 | |||
| ФЕРМЕНТАЦИЯ И ОЧИСТКА ГЕПАРОСАНА К5 | 2010 |
|
RU2564566C2 |
