Настоящая заявка испрашивает приоритет по предварительной заявке на патент США №61/275, 675, поданной 1 сентября 2009 года, которая включена в настоящую заявку посредством ссылки в полном объеме.
ЛИЦЕНЗИОННЫЕ ПРАВА ГОСУДАРСТВА
Настоящее изобретение было осуществлено при поддержке Федерального правительства США по грантам GM38060 и HL096972, присужденным Национальным институтом здоровья. Федеральное правительство США имеет определенные права на настоящее изобретение.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Гепарин и гепарансульфат представляют собой биологически важные молекулы, вовлеченные в процесс антикоагуляции крови, вирусные и бактериальные инфекции, ангиогепез, воспаление, рак и в процессы развития. Linhardt (2003) "Heparin: structure and activity," J Med Chem, 46: 2551-2554. Linhardt RJ, Toida T. (2004) "Role of glycosaminoglycans in cellular communication," Ace Chem Res, 37: 431-438. Гепарин находит широкий спектр применения, в том числе в хирургии, в процессе оксигенации в аппарате искусственного кровообращения и при почечном диализе, при лечении тромбоза глубоких вен и острого коронарного синдрома. Linhardt "Heparin: an important drug enters its seventh decade." Chem. Ind. 2, 45-50 (1991); Agnelli et al. "Enoxaparin plus compression stockings compared with compression stockings alone in the prevention of venous thromboembolism after elective neurosurgery" N Engl J Med 339 (2), 80-5 (1998). Гепарином также покрывают поверхности емкостей для крови и медицинского оборудования, в том числе пробирок и гемодиализного аппарата, для формирования антикоагулянтной поверхности. Гепарин в настоящее время получают из тканей животных в количестве примерно 100 тонн/год, но такой гепарин может быть загрязнен другими биологическими продуктами. Linhardt Chem. Ind. 2, 45-50 (1991). Гепарин, загрязненный сверхсульфатированным хондроитинсульфатом, стал причиной гибели примерно 100 американцев в 2008 году. Guerrini, et al. "Oversulfated chondroitin sulfate is a contaminant in heparin associated with adverse clinical events." Nature Biotechnology 26, 669-675 (2008).
Природный предшественник биосинтеза эукариотического гепарина, гепаросан, представляет собой полисахарид с повторяющимся дисахаридным блоком [→ 4) β-D-глюкуроновая кислота (GlcA) (1→4) N-ацетил-α-D-глюкозамин (GlcNAc) (1→]n, показанным на Фиг.1.
Гепаросан также синтезируются в виде полисахаридиой капсулы у бактерий, в том числе у Escherichia coli K5 и Pasteurella multicida. Lindahl U et al. (1998) "Regulated diversity of heparan sulfate" J Biol Chem 273(39):24979-24982. Синтез гепаросана K5 по имеющимся данным начинается с 2-кето-3-дезоксиоктулозоновой кислоты. Finke A et al. 5 (1991) "Biosynthesis of the Escherichia coli K5 polysaccharide, a representative of group II capsular polysaccharides: polymerization in vitro and characterization of the product" J Bacterial 173(13):4088-94. Затем гепаросан K5 удлиняется за счет альтернативного действия гликотрансфераз KfiA и KfiC, которые присоединяют GlcNAc и GlcA к невосстанавливающему концу растущей полисахаридной цепи. Hodson N et al. (2000) "Identification that KfiA, a protein essential for the biosynthesis of the Escherichia coli K5 capsular polysaccharide, is an alpha -UDP-GlcNAc glycosyltransferase. The formation of a membrane-associated K5 biosynthetic complex requires KfiA, KfiB, and KfiC." J Biol Chem 275(35):27311-5. После синтеза цепь гепаросана транспортируется на поверхность клетки посредством пути, в который вовлечены шесть белков: KpsC, KpsD, KpsE, KpsM, KpsS и KpsT. McNulty С et al. (2006) "The cell surface expression of group 2 capsular polysaccharides in Escherichia coli: the role of KpsD, RhsA and a multi-protein complex at the pole of the cell" Mol Microbiol 59(3):907-22. Считается, что цепь гепаросана K5 заякорена на поверхности клетки через замещение липидной группы на молекулу фосфатидной кислоты на редуцирующем конце полисахарида в наружной мембране Е. coli. Jann В, Jann K. (1990) "Structure and biosynthesis of the capsular antigens of Escherichia coli" Curr Top Microbiol Immunol 150:19-42. Фрагменты полисахарида гепаросана могут высвобождаться из клеток Е. coli K5 за счет действия гепаросаплиазы К5, фермента бактериофагального происхождения, который расщепляет гепаросановую цепь посредством механизма β-элиминирования. Manzoni М et al. (1996) "Production of K5 polysaccharides of different molecular weight by Escherichia coli" Journal of Bioactive and Compatible Polymers 11 (4):301 -311. Manzoni М, et al. (2000) "Influence of the culture conditions on extracellular lyase activity related to K5 polysaccharide." Biotechnology Letters 22(1):81-85. Активность лиазы K5 может оказывать влияние на количество гепаросана, выделяемого в культуральную среду, а также на структуру и характеристику молекулярного веса гепаросана, как находящегося в клетках, так и высвобождаемого в среду (Фиг.16). Гепаросан К5, но оценкам, имеет молекулярный вес (Mw) 20000 и состоит из двух основных субкомпонентов с молекулярным весом (Mw) 16000 и 1500. Соотношение двух субкомпонентов соответствует общему молекулярному весу и зависит от активности лиазы K5. Vann WF et al./ (1981) "The structure of the capsular Polysaccharide (K5 Antigen) of Urinary-Tract-Infective Escherichia-Coli 010-K5-H4 - a Polymer Similar to Desulfo-Heparin" European Journal of Biochemistry 116(2):359-364; Manzoni (2000) Biotechnology Letters 22(1):81-85.
Как показали лабораторные исследования, гепаросап со средним молекулярным весом (Mw)>10000, полученный из штамма Е. coli K5, может быть ферментативно превращен в полисахарид-антикоагулянт, схожий с гепарином. Lindahl et al. (2005) "Generation of "Neoheparin" from E coli K5 capsular polysaccharide"./ Med Chem 48(2):349-352; Zhang et al. (2008) "Solution structures of chemoenzymatically synthesized heparin and its precursors" Journal of the American Chemical Society 130(39): 12998-13007. Гепаросан также можно применять для различных целей (WO 2009/014559).
Настоящее изобретение описывает способ ферментации Е. coli K5 с высоким выходом гепаросана и эффективного выделения гепаросана высокой степени чистоты, применимый для промышленного производства гепаросана.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к усовершенствованному способу производства гепаросана с помощью ферментации Е. coli K5, выделения полисахарида K5, и очистки.
В одном из вариантов реализации указанный способ включает (а) культивирование E.coli K5 в среде заданного состава с глюкозой в качестве основного источника углерода (б) осуществление связывания гепаросана с твердофазным носителем с последующим элюированием, и (в) осаждение гепаросана из элюата. Способ пригоден для продукции по существу чистого гепаросана, степень чистоты которого составляет не менее 90%.
В соответствующих вариантах реализации указанный способ включает в себя две стадии культивирования, а именно периодическое культивирование и периодическое культивирование с подпиткой, причем (а) среда, применяемая на стадии периодического культивирования, включает (на 1 литр) примерно 20 г глюкозы, 10-300 мг тиамина, примерно 13,5 г KН2РО4, примерно 4,0 г (NH4)2HPO4, примерно 1,4 г MgSO4·7H2O, 1,7 г лимонной кислоты, и примерно 10,0 мл раствора микроэлементов (на литр), при этом раствор микроэлементов включает (на 1 л 5 М НСl) 10,0 г FeSO4·7H2O, 2,0 г CaCl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г CuSO4 ·5H2O, 0,1 г (NH4)6МO7O24·4Н2O и 0,02 г Na2B4O7·10H2O и где (б) раствор для подпитки, используемый на стадии культивирования с подпиткой, включает (на 1 литр): 250-1000 г глюкозы, примерно 20 г MgSO4·7H2O и 0,15-0,5 г тиамина, и может содержать примерно 47 г KН2РO4.
В соответствующих вариантах реализации поступление кислорода обеспечивают путем барботирования воздухом. Воздух может быть дополнен кислородом. Предпочтительно концентрацию растворенного кислорода поддерживают на уровне примерно 20%. В некоторых вариантах реализации применяют чистый кислород. В других соответствующих вариантах реализации условия культивирования включают температуру, поддерживаемую па уровне 37°С, и рН, поддерживаемый па уровне примерно 7. рН можно поддерживать путем добавления раствора аммиака или газообразного аммиака. В некоторых вариантах, раствор аммиака составляет примерно 25-35%, в частности, примерно 30%, например, 29%.
В конкретных вариантах реализации среду для подпитки, применяемую на стадии культивирования с подпиткой, подают со скоростью, определяемой по формуле:
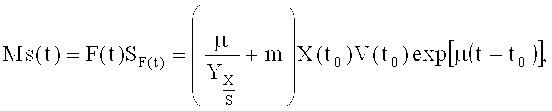
где MS представляет собой скорость потока источника углерода (г/ч), F - скорость подачи раствора для подпитки (л/ч); SF представляет собой концентрацию источника углерода в подпитке (г/л); Х - концентрация клеток (г/л с.м.к.), m - удельный коэффициент поддержания (г/г DCW/ч), V - объем культуры (л); t0 - время начала подпитки; t - время процесса; µ - удельная скорость роста (ч-1) и YX/s - выход клеток на количество источника углерода (г/г).
Указанный способ также включает осуществление связывания и элюирования гепаросана из культуральной среды. В одном варианте реализации гепаросан получают из бесклеточного супернатанта. В другом варианте реализации гепаросан получают из клеток путем их обработки детергентом, таким как ДСН (например, 1% ДСН), при перемешивании.
После удаления клеток связывание и элюирование может включать смешивание анионообменных смол с супернатантом культуры с последующим удалением супернатанта, промыванием смолы с 50 мМ раствором хлорида натрия в натрий-ацетатном буфере при рН 4, элюированием раствором 1 М хлорида натрия в натрий-ацетатном буфере при рН 4.
Кроме того, реакцию связывания и элюирования можно проводить с помощью хитозана. В одном из вариантов реализации супернатант культуры смешивают с раствором хитозана для осаждения. Затем отделяют осадок, промывают его, например, водой или каким-либо разбавленным буфером и элюируют с помощью сильного основания, такого как 1М NaOH. Связывание и элюирование с применением хитозана можно проводить в дополнение с анионным обменом. После осуществления связывания и элюирования гепаросан осаждают из элюата, например, с помощью этанола или метанола. В одном из вариантов реализации используют 3 объема этанола. Полученный осадок обычно промывают и высушивают.
Этот способ также включает дополнительную стадию депирогенизации, например, путем окисления, в том числе с помощью перекиси водорода.
Способ настоящего изобретения позволяет получить высокий выход гепаросана с высокой степенью чистоты из небольшого объема культуры за короткий период времени и с низким содержанием загрязняющих веществ. Таким образом, настоящее изобретение можно применять для промышленного производства гепаросана. Выход продукта гепаросана может быть получен при ферментации менее чем за 60 часов, менее чем за 48 часов и менее чем 40 часов без учета роста стартовой культуры. В других вариантах реализации первоначальный объем культуры составляет 3 л на этапе периодического культивирования до 7 литров на стадии подпитки, и подобные соотношения. В соответствующих вариантах реализации, таким образом, из культуры получают по меньшей мере 10 г/л, по меньшей мере 11 г/л, по меньшей мере 12 г/л, по меньшей мере 13 г/л, по меньшей мере 14 г/л, а по меньшей мере 15 г/л гепаросана. Степень чистоты по существу чистого гепаросана составляет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, или по меньшей мере 99% чисты. В соответствующих вариантах реализации гепаросан содержит менее 1% ДНК и менее 2% белка. Гепаросан можно применять для переработки в гепарин. В одном варианте реализации гепаросан имеет среднечисленный молекулярный вес по меньшей мере 10, 20, 30, 40, 50 или 60 кДа, например примерно 58 кДа, средневесовой молекулярный вес по меньшей мере 20, 30, 40, 50, 60, 70, 80 или 90 кДа, например примерно 84 кДа, и индекс полидисперсности (ИПД) менее 2,0, например, меньше 1,9, 1,8, 1,7, 1,6, 1,5, 1,4 или 1,3, в том числе примерно 1,4.
В других вариантах реализации настоящее изобретение включает применение гепаросана, полученного описанным выше способом, при приготовлении фармацевтических препаратов, например для нанесения покрытия на медицинские изделия. В схожих вариантах реализации гепаросан применяют для производства гепарина. Соответственно, изобретение представляет собой способ применения гепаросана или гепарина, полученного вышеизложенным способом, для изготовления лекарственных средств.
В других вариантах реализации изобретение относится к гепаросану, полученному вышеизложенным способом, в том числе гепаросану со степенью чистоты по меньшей 5 мере 90%. по меньшей мере 95% и более. В схожих вариантах реализации изобретение относится к гепарину, полученному из указанного гепаросана.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1. Структура гепаросана. А. Повторяющийся дисахаридный блок гепаросана В. Структура цепи гепаросана в результате действия K5 гепаросанлиазы.
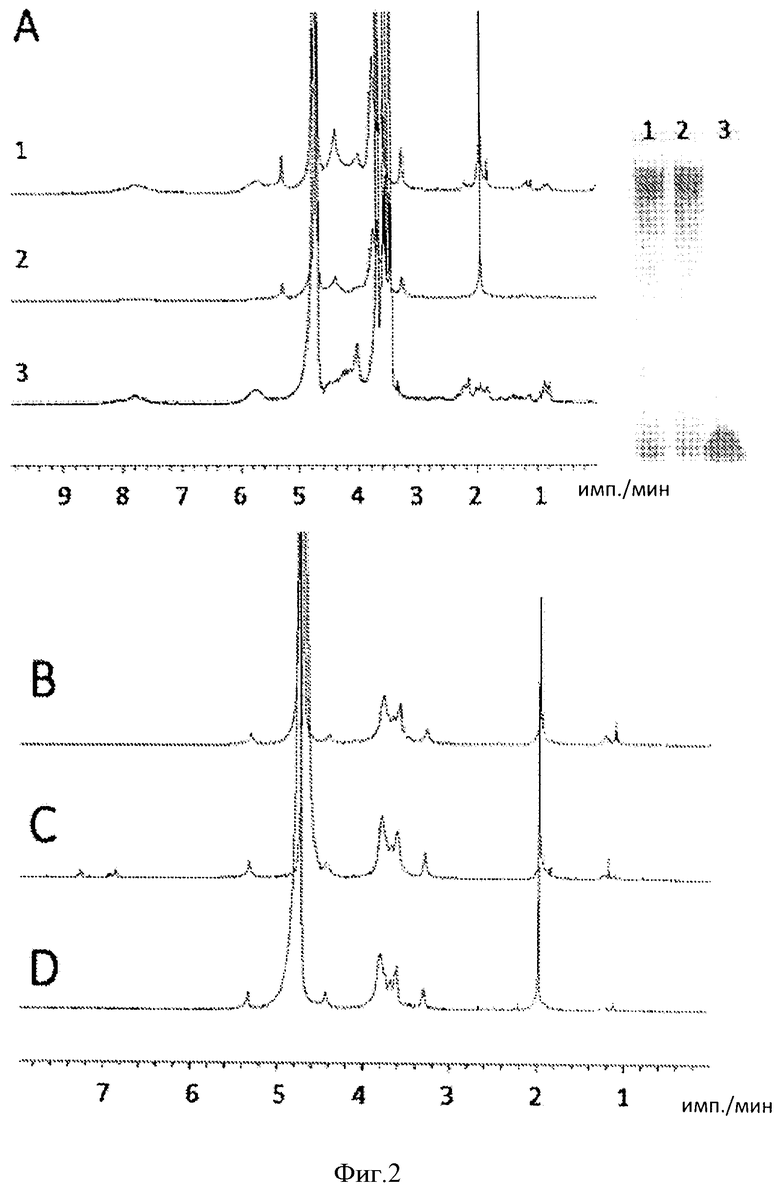
Фигура 2. Спектр 1Н-ЯМР (600 МГц) гепаросана, полученного из культуры, выращенной во встряхиваемой колбе. А. Стек-диаграмма, показывающая количество гепаросана, полученного из: 1. среды LB, 2. среды LB с МWсредн>3000, 3. среды LB с МWсредн<3000; на вставке представлен анализ в образцов 1, 2 и 3 с помощью электрофореза в ПААГ. В. Гепаросан, полученный из среды М9. С. Гепаросан, полученный из среды с заданным содержанием глицерина. D. Гепаросан, полученный из среды заданного состава с глюкозой.
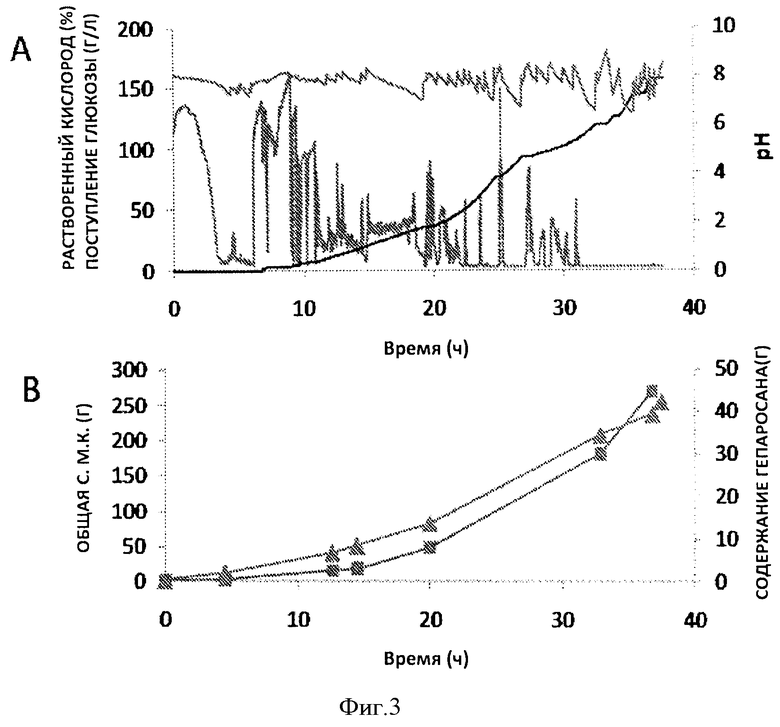
Фигура 3. Продукция гепаросана в ферментере объемом 7 л. На Панели А показана кривая подачи глюкозы (черный цвет), кривая изменения рН (синяя) и кривая содержания 20 растворенного кислорода (% DO) (красная) в зависимости от времени ферментации (часы). На Панели В показана кривая роста клеток (общ. с.м.к. г,▲) и выработки гепаросана (г,▪) в зависимости от времени ферментации (часы).
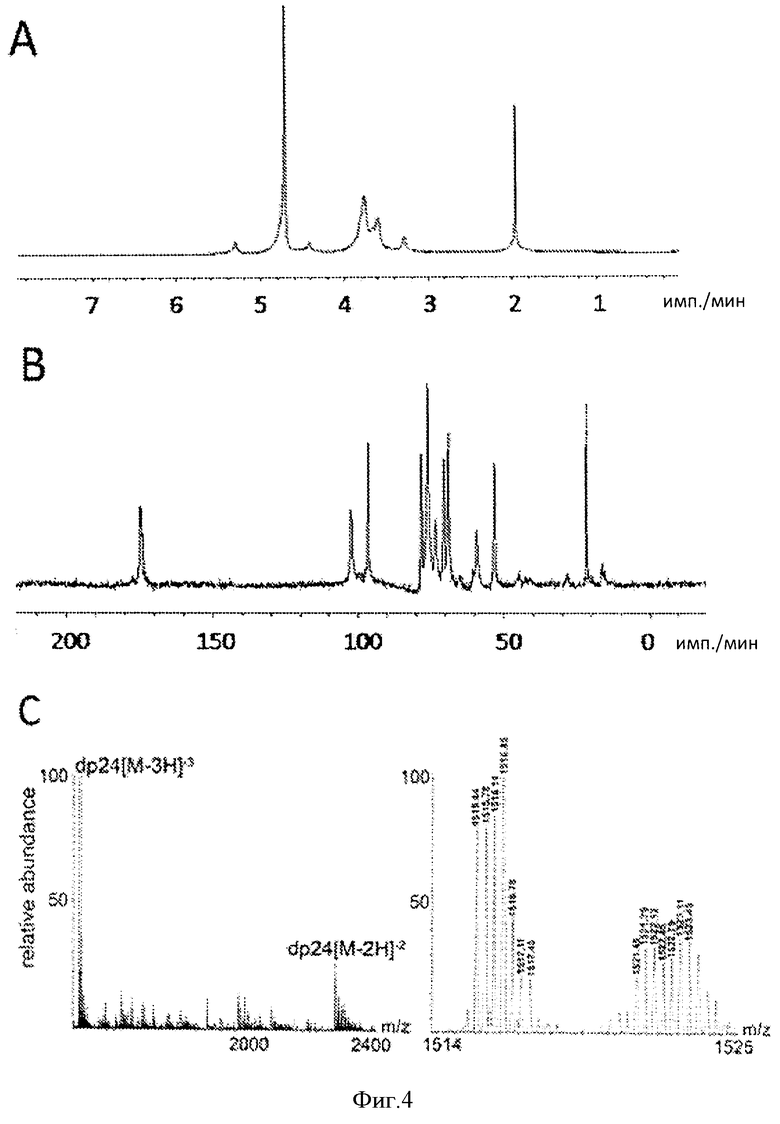
Фигура 4. Характеристика гепаросана, очищенного из супернатанта 7 л культуральной среды. А. 1Н-ЯМР (600 МГц) (600 МГц) гепаросана. В. 13С-ЯМР 25 гепаросана, полученного из среды М9, содержащей 13C-глюкозу и 15N-сульфат аммония. С. Анализ с помощью Фурье-МС цепи гепаросана со средней молекулярной массой 4551,81 (степень полимеризации=24), очищенного с помощью препаративного электрофореза в ПААГ (Ly et al., 2010).
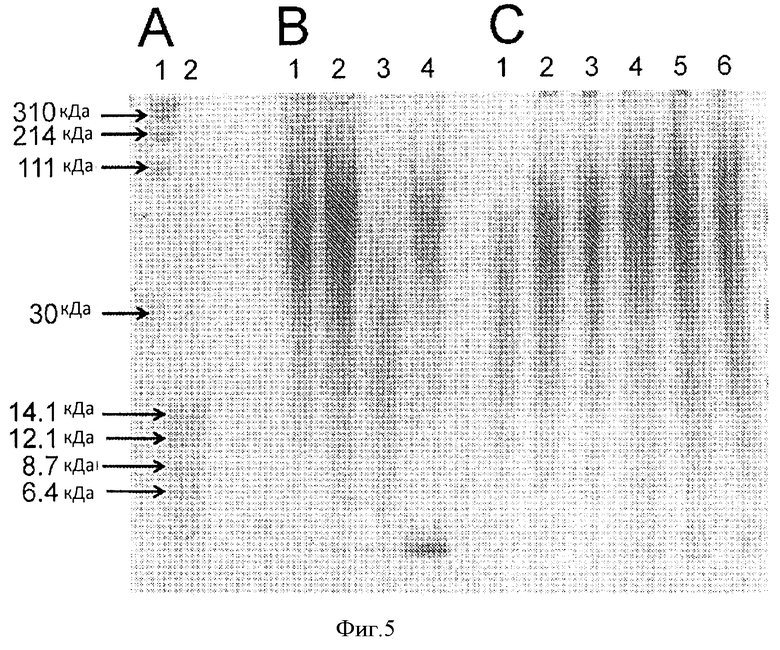
Фигура 5. Анализ молекулярной массы гепаросана с помощью электрофореза в 30 ПААГ, окрашенном красителем альциановый синий. А. Молекулярные стандарты, охватывающие диапазон градиентного геля. На дорожках представлены: 1. Молекулярные маркеры ГК (30 кДа-310 кДа), 2. Молекулярные маркеры гепаросана (6,4 кДа-14.1 кДа). В. Гепаросан, полученный при культивировании во встряхиваемой колбе в различных средах. На дорожках представлены: 1. Гепаросан из среды М9, 2. Гепаросан из синтетической среды с глицерином, 3. Гепаросан из синтетической среды с глюкозой, и 4. Гепаросан из среды LB. С. Пробы гепаросана, производимого в 7 л ферментере, взятые на разных сроках. На дорожках представлены: 1-6. Пробы гепаросана из ферментера взятые через 4,5 ч, 12,6 ч, 14,5 ч, 20 ч, 32,9 ч и 37,6 ч после начала культивирования.
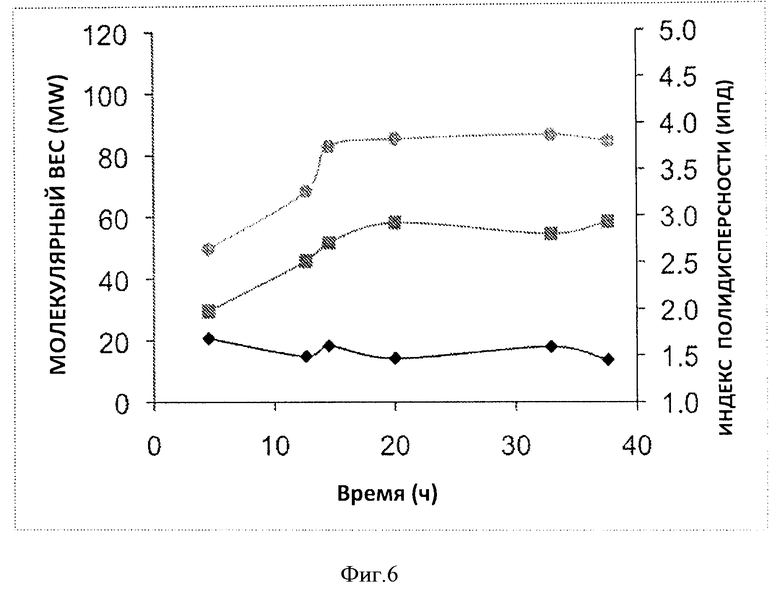
Фигура 6. Временная динамика характеристики молекулярного веса гепаросана, производимого в 7 л ферментере. Показаны изменения среднечисленного молекулярного веса MN(■), средневесового молекулярного веса MW (●) и индекса полидисперсности PDIС(▲).
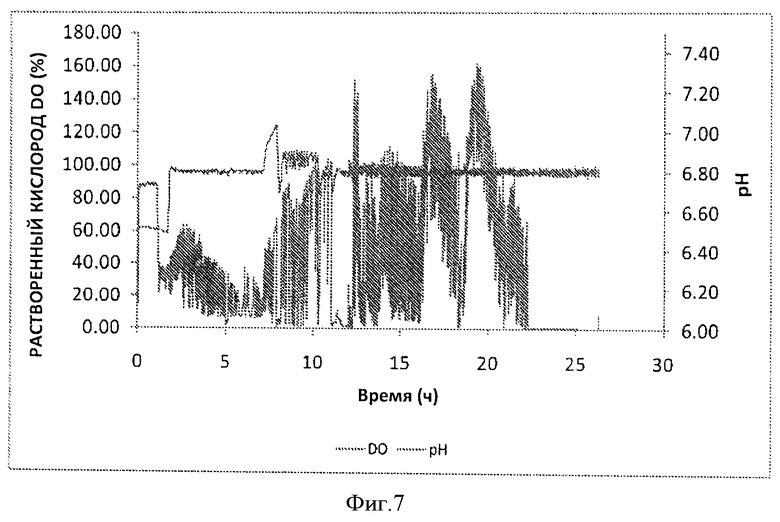
Фигура 7. Изменение содержания рН (■) и растворенного кислорода (% DO) (♦) в зависимости от времени ферментации (часы) в 20 л ферментере.
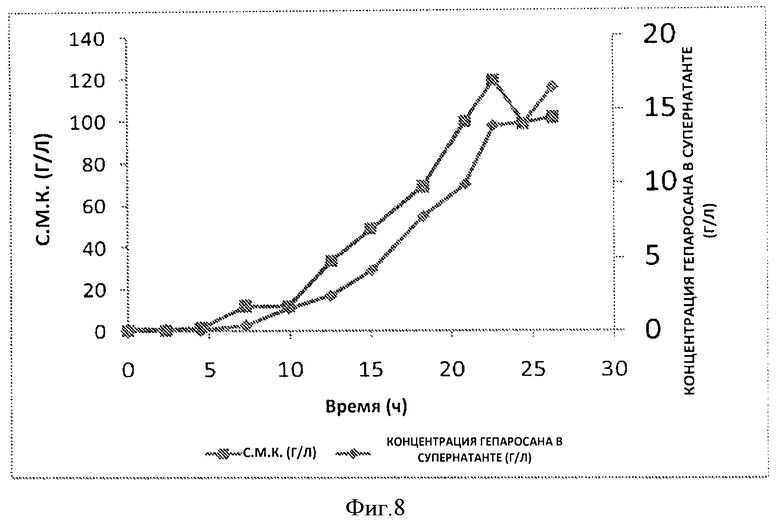
Фигура 8. Временная динамика роста клеток (с.м.к. г/л, (■)) и концентрации гепаросана в супернатанте культуральной среды (г/л, (♦)) в 20 л ферментере.
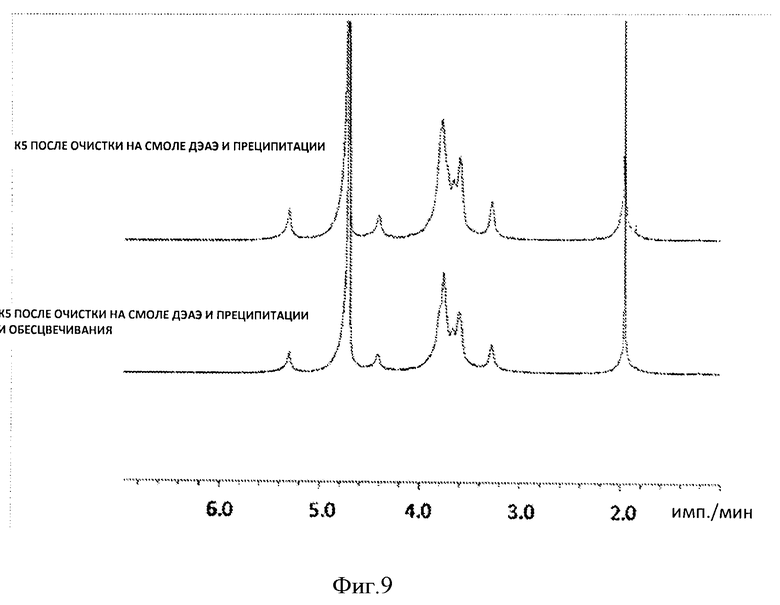
Фигура 9. 1Н-ЯМР-спектр образцов очищенного гепаросана.
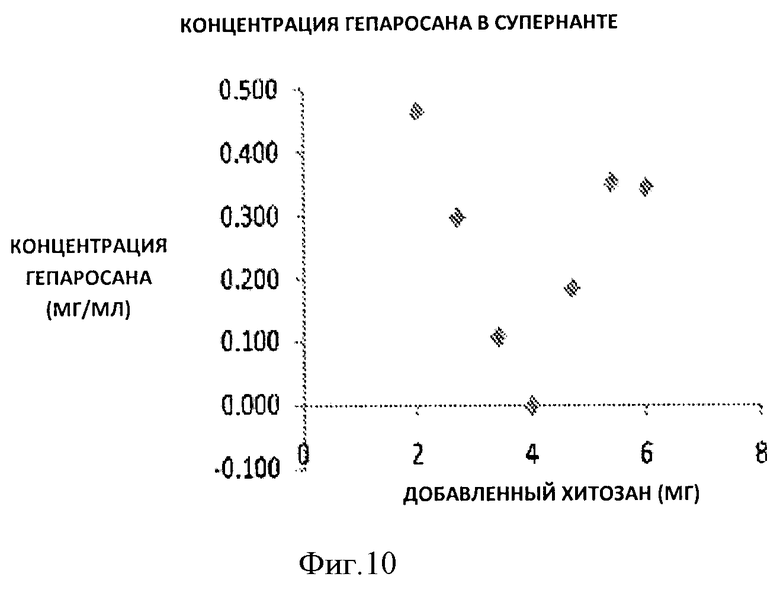
Фигура 10. Концентрация гепаросана в супернатанте после осаждения с 15 использованием различного количества хитозана из 10 мл раствора чистого гепаросана 1 мг/мл.
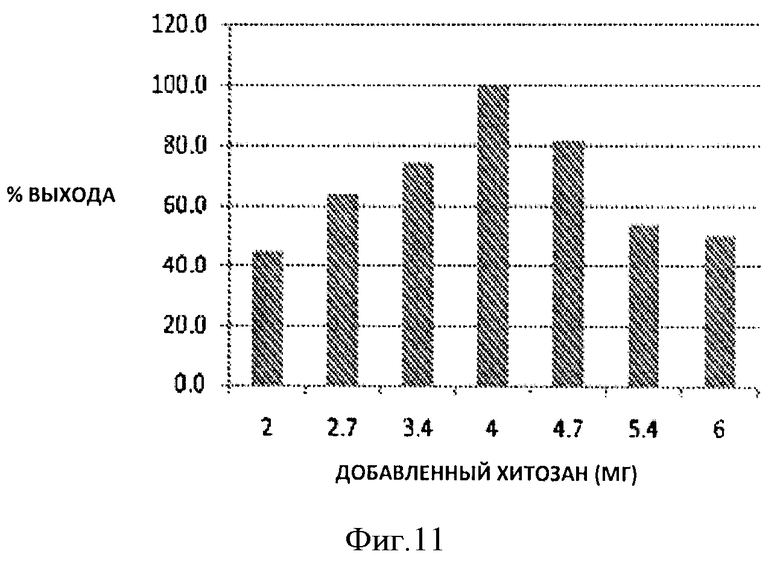
Фигура 11. Процент выхода с использованием различного количества хитозана из 10 мл раствора чистого гепаросана 1 мг/мл.
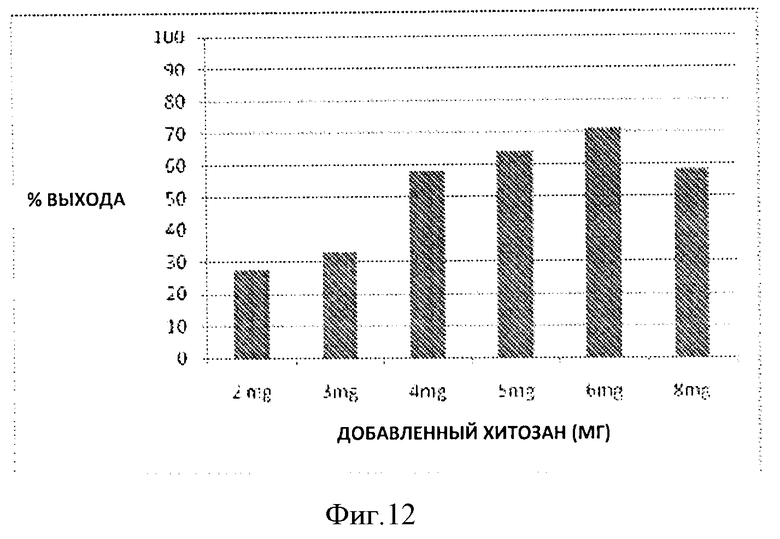
Фигура 12. Процент выхода с использованием различного количества хитозана из 20 10 мл проб разбавленной в 5 раз культуральной среды.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ОПРЕДЕЛЕНИЯ
Гепаросан представляет собой полисахарид с повторяющимся дисахаридным блоком [GlcAα-(1-4)GlcNAcR(1-4)]n. Поскольку экзополисахарид K5 образуется из гепаросана, который выделяют и очищают в соответствии с настоящим изобретением, «гепаросан» в определенном контексте может относиться к гепаросану, связанному с бактериями, присутствующему в культуральной среде, и к гепаросану на стадии выделения и очистки. Значение термина «гепаросан» в соответствующем контексте будет 30 понятно специалисту в данной области.
Escherichia coli K5 относится к вариантам Е. coli, которые продуцируют экзополисахарид К5. Подходящие для применения в настоящем изобретении штаммы Е. coli K5 могут быть получены из государственных коллекций, таких как АТСС (Американская коллекция типовых культур, США), например штамм Е. coli ATCC23506.
Штаммы Escherichia coli K5 могут быть выделены из клинических источников и/или могут быть генетически модифицированными.
При использовании в настоящей заявке термин «ферментация» относится к размножению бактерий и продукции экзополисахарида, в частности экзополисахарида K5. При использовании в настоящей заявке термин «изолированный/выделенный» означает, что гепаросан отделен от питательной среды и бактериальных клеток и присутствует в количестве, достаточном для его идентификации или применения.
При использовании в настоящей заявке термин «по существу чистый» означает, что гепаросан по существу не содержит других веществ до такой степени, что он подходит для практического применения по назначению. По существу чистый гепаросан является по меньшей мере на 90% чистым. Предпочтительно, вещество свободно от примесей более чем на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или даже более чем 99%. Степень чистоты можно оценить с помощью способов, известных в данной области.
При использовании в настоящей заявке объекта в единственном числе, подразумевается «один или более», если особым образом не оговаривается, что подразумевается «только один».
Термин «примерно/примерно» используется в соответствии с пониманием специалиста в данной области в том контексте, к которому относится указанный термин. Для численных значений предполагается, что термин «примерно/примерно» охватывает изменения указанного значения в пределах 10%.
ФЕРМЕНТАЦИЯ Е. coli K5 ДЛЯ ПРОИЗВОДСТВА ГЕПАРОСАНА
Для определения надлежащих условий ферментации необходимо учитывать многие факторы. Е. coli, в целом, может расти на самых разных средах и при различных значениях рН, температуры, О2 и других условиях. Однако в отличие от лабораторных штаммов, штамм Е. coli K5 не подвергался обширному генетическому исследованию, изучению его роста и функций.
Изменения условий роста могут оказывать влияние на (а) скорость роста бактерий, (б) максимальную плотность клеток (в) производство экзополисахаридных капсул K5, (г) степени модификации капсулы Е. coli K5, на состояние резидентного бактериофага в Е. coli K5, и другие факторы, (д) высвобождение экзополисахаридной капсулы K5 в среду, (е) соответствие диапазона размеров образующегося полисахарида гепаросана возможности его переработки в гепарин, и (г) количество и тип примесей. Например, условия, которые способствуют лизису клеток, могут увеличить выход экзополисахарида K5 в супернатант культуры, но также могут увеличить деградацию экзополисахаридной капсулы К5, а также количество и объем примесей в супернатанте.
Важным моментом является то, насколько легко какая-либо из примесей может быть удалена на последующих стадиях очистки и обработки, влияет ли эта примесь на последующую переработку гепаросана в гепарин, и представляет ли примесь опасность для людей и животных. Е. coli K5 является известным патогеном, и, следовательно, токсины и другие факторы, связанные с вирулентностью, должны быть удалены. Липополисахариды (ЛПС) являются широко распространенными загрязнителями экстрактов грамотрицательных бактерий, и они известны как мощные иммуностимулирующие агенты и токсины. Нуклеиновые кислоты также могут обладать иммуностимулирующим эффектом, как ЛПС, так и нуклеиновые кислоты имеют полисахаридное ядро, и поэтому могут выделяться совместно с гепаросаном. Другие примеси могут включать в себя полисахариды, отличные от гепаросана, или производные гепаросана.
Для производства фармацевтической и медицинской продукции источник и характеристики питательной среды могут иметь большое значение. Например, комплексная среда, полученная путем гидролиза животных или растительных белков, отлично подходит для роста и продукции К5, но для такой среды характерны различия между партиями и она может содержать аитигенные и другие загрязняющие примеси.
Для производства гепаросана в промышленных масштабах необходимо учитывать дополнительные факторы, в том числе объем культуры, время и температуру ферментации. Таким образом, являются предпочтительными меньшие объемы питательной среды, и условия, способствующие более быстрому росту. Также актуальным является выявление питательной среды, в которой применяются широкодоступные, стандартизированные и недорогие исходные вещества.
Кроме того, условия культивирования должны быть масштабируемыми. Так, выявленные оптимальные условия для культуры 5 мл должны также подтверждаться при культивировании в более крупных масштабах, например, 5 л, 20 л, 100 л и более.
Соответственно, процесс культивирования должен быть надежными и воспроизводимым, и, таким образом, не должен быть чувствительным к небольшим изменениям условий.
В связи с вышеизложенным, выявление условий, подходящих для промышленного производства гепаросана, не является тривиальной задачей. Авторы настоящего изобретения определили подходящие условия для роста и культивирования Е. coli K5 для облегчения производства, выделения и очистки гепаросана, которые могут быть адаптированы для промышленного производства.
Среда для культивирования согласно настоящему изобретению представляет собой среду с заданным составом с глюкозой в качестве источника углерода, варьирующую в зависимости от фазы роста. Состав среды для периодического культивирования является следующим на литр: примерно 20 г глюкозы, 10-300 мг тиамина, примерно 13,5 г KH2PO4, примерно 4,0 г (NH4)2HPO4, примерно 1,4 г MgSO4·7H2O, примерно 1,7 г лимонной кислоты и примерно 10,0 мл раствора микроэлементов. Раствор микроэлементов включал (на 1 л 5М НСl) 10,0 г FeSO4·7H2O, 2,0 г СаСl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г 10 CuSO4·5H2O, 0,1 г (N4)6МO7O24·4Н2O и 0,02 г Na2B4O7·10H2O. Раствор для подпитки, используемый на стадии культивирования с подпиткой, включал (на 1 л): 250-1000 г глюкозы, примерно 20 г MgSO4·7H2O и 0,15-0,5 г тиамина, и мог содержать примерно 47 г KН2РO4. В некоторых вариантах реализации раствор для подпитки содержал 700 г глюкозы, 20 г MgSO4·7H2O и 0,2 г тиамина. В другом варианте реализации раствор для подпитки содержал 700 г глюкозы, 20 г MgSO4·7H2O 47 г KН2РO4 и 0,4 г тиамина.
Культуру для инокуляции получали, выращивая штамм Е. coli K5 в течение ночи при 37°С в конической колбе. Затем культурой для инокуляции инокулировали ферментер. Процесс ферментации в ферментере включает две фазы: фазу периодического роста, и фазу, на которой осуществляют подпитку согласно комбинации стратегий экспоненциальной подпитки (exponential feeding) и подпитки с учетом содержания растворенного кислорода (DO-stat feeding). Фаза периодического роста начинается после посева культуры для инокуляции в ферментер. Температура поддерживается при 37°С, а уровень рН поддерживается между 6 и 8, путем добавления NH4OH при снижении рН.
Второй этап ферментации начинается после того, как концентрация растворенного кислорода резко увеличивается, а содержание глюкозы в среде истощается. Скорость подачи глюкозы рассчитывается в целом по формуле, но с возможными отклонениями:
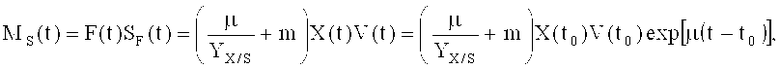
где MS представляет собой скорость потока источника углерода (г/ч), F - скорость подачи раствора для подпитки (л/ч); Sp представляет собой концентрацию источника углерода в подпитке (г/л); Х - концентрация клеток (г/л с.м.к.), m - удельный коэффициент поддержания (г/г DCW/ч), V - объем культуры (л); t0 - время начала подпитки; t - время процесса; µ - удельная скорость роста (ч-1) и YX/S - выход клеток на количество источника углерода (г/г).
В течение всего процесса подпитки р. находится между 0,1 и 0,4. В некоторых вариантах реализации в процессе подпитки возможны паузы, используемые как контрольные точки для проверки того, не превышен ли уровень глюкозы, и для проверки того, что уровень токсичных веществ, таких как ацетат, в культуральной среде является низким. В таких контрольных точках подпитку возобновляют после резкого повышения концентрации растворенного кислорода, что указывает па истощение глюкозы в питательной среде. При крупномасштабном культивировании такие наблюдения проводят непрерывно с помощью датчиков или путем автоматического отбора проб и анализа в процессе ферментации.
Как правило, ферментацию продолжают до тех пор, пока плотность клеток не перестанет увеличиваться. Ферментацию затем останавливают, и мешалку ферментера оставляют еще на 30-60 минут для того, чтобы с поверхности клеток и в среду могло бы высвободиться большее количество полисахаридных капсул К5.
ОЧИСТКА ГЕПАРОСАНА ИЗ КУЛЬТУРЫ ПОСЛЕ ФЕРМЕНТАЦИИ
В одном из примеров вариантов реализации настоящего изобретения очистка гепаросана может включать (а) этап получения супернатанта культуры или фильтрата, (б) осуществление связывания гепаросана с твердофазным носителем, таким как смола, и элюирование с него гепаросана; (с) осаждение с помощью спирта и (г) депирогенизацию. Могут быть добавлены дополнительные этапы связывания, преципитации и депирогенизации.
На этапе (а) культуру центрифугируют или фильтруют для отделения супернатанта от осадка клеток. Для инактивации бактериальных клеток к культуре могут быть добавлены формалин или другие агенты. Как правило, гепаросан выделяют из супернатанта, но также его можно выделять, промывая клетки, в том числе с помощью детергентов, таких как додецилсульфат натрия, и механического перемешивания с последующим центрифугированием с получением осадка и экстрагирования гепаросана из супернатанта.
На этапе (б) гепаросан связывают с твердофазным носителем, который преимущественно связывает гепаросан. Например, к супернатанту после стадии (а) добавляют смолы, например, анионообменную смолу. Смесь перемешивают, чтобы связать гепаросан в супернатанте со смолой. Затем смесь фильтруют для отделения смолы от раствора. Твердофазный носитель затем промывают, чтобы смыть не связавшиеся примеси, и элюируют.
В случае анионообменных смол промывку осуществляют с помощью раствора с низкой концентрацией соли для смыва не связавшихся примесей и элюируют с помощью 1-2 М раствора хлорида натрия. Для связывания гепаросана также можно применять хитозан. Поскольку хитозан можно добавлять в виде раствора, и таким образом, не в «твердой фазе», он преципитирует и таким образом «выводит» гепаросан из супернатанта. Кроме того, гепаросан можно пропускать через соответствующую колонку, содержащую твердофазный носитель, промывать и элюировать.
На этапе (в) элюат с гепаросаном, образовавшийся на этапе (б), преципитируют, например, путем добавления 1-5 частей по объему этанола или метанола. Этот этап не только позволяет сконцентрировать гепаросан, но и избирательно удалить примеси. Осадок отделяют центрифугированием и промывают 50-80% спиртом.
В стадии (г) преципитат растворяют в воде и подвергают депирогенизации для 15 инактивации оставшегося липополисахарида и других загрязнений. Обычно, для этого применяют окисляющий агент, такой как перекись водорода, но также известны другие пероксиды или обесцвечивающие агенты. Образующийся в результате гепаросан вновь преципитируют, как указано выше, и высушивают.
АНАЛИЗ
Полученный на предыдущем этапе гепаросан анализируют для определения его выхода, степени чистоты и пригодности для дальнейшей обработки. Можно применять несколько аналитических инструментальных подходов.
Дисахаридный состав можно проанализировать с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) с масс-спектрометрией (MS) с ионизацией электрораспылением (ESI, electrospray ionization), после частичного гидролиза гепаросана. Bhattacharyya S et al. (2010) '-Cell-bound IL-8 increases in bronchial epithelial cells after arylsulfatase В silencing due to sequestration with chondroitin-4-sulfate," Am J Respir Cell and Molec Biol 42, 51-61. Также можно применять 1Н-ЯМР и 13C-ЯМР для анализа негидролизованного гепаросана, кроме того, этот метод позволяет определить концентрацию и чистоты гепаросана. Wang Z, Zhang Z, McCallum SA, Linhardt RJ. 2009. Nuclear magnetic resonance quantification for monitoring heparosan K5 capsular polysaccharide production. Anal Biochem. 398(2):275-7. Гепаросан также можно проанализировать с помощью анализа с карбазолом, как описано в работе Bitter Т, Muir HM. (1962) “А Modified Uronic Acid Carbazole Reaction." Analytical Biochemistry 4(4):330.
Содержание ДИК можно определить по УФ-поглощению. Содержание белка можно определить с помощью хорошо известных наборов, таких как набор Micro BCA Protein Assay Kit согласно инструкциям производителя.
Профиль молекулярного веса очищенного гепаросана определяют путем сравнения с лестницей гепаросана или гиалуроновой кислоты с известным молекулярным весом, и на основании его рассчитывают средний молекулярный вес, количество и распределение гепаросана.
Содержание пирогена можно определить с помощью анализа с применением лизата амебоцитов Limulus (LAL).
Изобретение можно лучше понять со ссылкой на следующие примеры, которые приведены для иллюстрации некоторых вариантов реализации изобретения, которые не являются ограничивающими. Как понятно специалисту в данной области, возможны 15 изменения без отступления от сущности изобретения.
ПРИМЕРЫ
ПРИМЕР 1. Материалы
В качестве ферментера применяли автоклавируемый стеклянный биореактор объемом 7 л от Applikon (Schiedam, Netherlands). Для управления ферментеров и сбора данных использовали программу BioXpert VI.5. Среда Difco™ LB в сухом виде приобретена у компании BD (Franklin Lakes, NJ). Большинство химических веществ, используемых для приготовления синтетической среды приобретены у Sigma-Aldrich (St Louis, МО). Antifoam 204 приобретен у Sigma-Aldrich (St Louis, МО). Встряхиваемые колбы с перегородками приобретены у Coming (Corning, NY). Для очистки гепаросана использовали быстропроточную ДЭАЭ-сефарозу DEAE от GE Healthcare (Piscataway, NJ). Колонки Vivapure D Mini H приобретены у Sartorius Stedim Biotech (Aubagne, France). Набор Micro BCA Protein Assay Kit - от Thermo Scientific (Rockford, IL). Ферменты, используемые для гидролитического расщепления гепаросана для дисахаридного анализа, нарабатывали и очищали в нашей лаборатории, как описано Man et al.(2009) "Structural snapshots of heparin depolymerization by heparin lyase I"./ Biol Chem 284(49):34019-27. ВЭЖХ-МС проводили с помощью прибора Agilent 1100 (Santa Clara, California). Планшеты для ТСХ с силикагелем получали от EMD (Gibbstown, NJ). Вещества для подготовки лестницы молекулярных весов гепаросана и определения молекулярного веса гепаросана были подробно описаны в работе Ly М, et al. (2010) "Analysis of E.coli K5 capsular polysaccharide heparosan" Anal Bioanal Chem (DOI: 10.1007/s00216-010-3679-7). Набор маркера молекулярных весов гиалуропана Select_HA™ LoLadder приобретен у Hyalose (Oklahoma City, OK).
Рост E.coli K5 во встряхиваемой колбе объемом 2,8 л
Штамм E.coli K5 из Американской коллекции типовых культур (АТСС U 23506) хранили замороженными в 1 мл среды М9, LB, среды с глицерином или среды с глюкозой, содержащей с 25% глицерин. Во встряхиваемую колбу объемом 250 мл, содержащую 25 мл соответствующей среды, вносили 0,5 мл размороженной культуры E.coli. Культуру собирали в конце экспоненциальной стадии роста при плотности клеток 1,1 г сухой массы клеток (с.м.к.) на л для культуры в среде М9, 5,4 г с.м.к./л для синтетической среды с глицерином, 5,6 г с.м.к./л в случае синтетической среды с глюкозой, и 1,9 г с.м.к./л для среды LB. Культуру E.coli K5 (300 мл) затем выращивали во встряхиваемой колбе объемом 2,8 л, инокулированной 5% по объему клетками, взятыми в конце стадии экспоненциального роста. Культуры выращивали при скорости покачивания 220 оборотов в минуту и при 37°С до 1-4 ч после того, как рост достигал стационарной фазы, а затем культуру собирали для выделения гепаросана. Среда, используемая для ферментации в колбе 2,8 л, включала следующие среды. 1. Среду LB: Difco™ LB broth, Lennox, 20 г/л 2. среда М9: 2 г/л глюкозы, 0,12 г/л MgSO4, 0,011 г/л CaCl2, 0,337 г/л тиамина-HCl, 6 г/л Na2HPO4, KH2PO4 3 г/л, 0,5 г/л NaCl, 1 г/л NH4Cl. 3. Среда с заданным содержанием глицерина: 20 г/л глицерина, 20 мг/л тиамина, 13,5 г КН2РO4, 4,0 г (NH4)2HPO4, 1,4 г MgSO4·7H2O, 1,7 г лимонной кислоты и 10,0 мл раствора микроэлементов. Раствор микроэлементов включал (на 1 л 5М НСl) 10,0 г FeSO4·7H2O, 2,0 г CaCl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г CuSO4, 0,1 г (NН4)6МO7O24·4Н2O и 0,02 г Nа2 В4O7·10Н2O Wang FL, Lee SY (1998). См. "High cell density culture of metabolically engineered Escherichia coli for the production of poly(3-hydroxybutyrate) in a defined medium" Biotechnology and Bioengineering 58(2-3):325-328. 4. Среда заданного состава с глюкозой содержала: на литр 20 г глюкозы, 20 мг тиамина, 13,5 г KН2РO4, 4,0 г (NH4)2HPO4, 1,4 г MgSO4·7H2O, 1,7 г лимонной кислоты и 10,0 мл раствора микроэлементов. Раствор микроэлементов включал (на 1 л 5М НСl) 10,0 г FeSO4·7H2O, 2,0 г СаСl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г CuSO4·5H2O, 0,1 г (Na4)6МO7O24·4Н2О и 0,02 г Na2B4O7·10H2O (см. Wang FL, Lee SY (1998), выше).
Рост Е. coli K5 в ферментере объемом 7 л
Такая ферментация включает стадию периодического культивирования и стадию периодического культивирования с подпиткой. Среда для периодического культивирования включала: (на 1 литр) 20 г глюкозы, 20 мг тиамина, 13,5 г KН2РO4, 4,0 г (NH4)2HPO4, 1,4 г MgSO4·7H2O, 1,7 г лимонной кислоты, и 10,0 мл раствора микроэлементов. Раствор микроэлементов включал (на 1 л 5М НСl) 10,0 г FeSO4·7H2O, 2,0 г СаСl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г CuSO4H2O, 0,1 г (NH2)6МO7O24·4Н2O и 0,02 г Na2B4O7·10H2O. Раствор для подпитки, используемый на стадии культивирования с подпиткой, включал (на 1 л): 700 г глюкозы, 20 г MgSO4·7H2O и 0,2 г тиамина (см. Wang FL, Lee SY (1998), выше).
Стадия периодического культивирования начиналась с посева культуры для инокуляции (300 мл на 5,6 г/л с.м.к.), выращенной во встряхиваемой колбе и взятой в конце фазы экспоненциального роста. Температура поддерживали при ~ 37°С, а рН поддерживали примерно 7 (путем добавления 29% раствора аммиака). Для подачи кислорода барботировали воздух в ферментер, а скорость перемешивания составляла до 520 об/мин.
Второй этап ферментации начинали после того, как уровень глюкозы в питательной среде периодической культуры истощался, а содержание растворенного кислорода резко увеличивалось. После этого в культуру подавали раствор для подпитки экспоненциально согласно формуле 1:
(1)
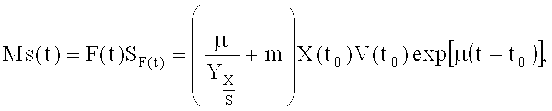
где Ms представляет собой скорость потока источника углерода (г/ч), F - скорость подачи раствора для подпитки (л/ч); SF представляет собой концентрацию источника углерода в подпитке (г/л); Х - концентрация клеток (г/л с.м.к.), m - удельный коэффициент поддержания (г/г DCW/ч), V - объем культуры (л); to - время начала подпитки; t - время процесса; µ - удельная скорость роста (ч-1) и Yx/s - выход клеток на количество источника углерода (г/г).
См. Lee SY. 1996. "High cell-density culture of Escherichia coli". Trends Biotechnol 14(3):98-105.
В данном исследовании использовали значение µ в пределах от 0,10-0,15 ч для обеспечения достаточного размножения клеток, избегая при этом накопления токсичных побочных продуктов за счет более высокой скорости роста. «Кислотный насос», прикрепленный к ферментеру, использовали для выполнения функции подачи глюкозы, а не для добавления реальной кислоты, а основной насос использовали для добавления 29% раствора аммиака для поддержания стабильного рН и обеспечения культуры источником азота. Третий насос использовали для добавления в культуру реагента Antifoam 204, когда вспенивание превышало заданный уровень, который отслеживался с помощью обратной связи через цифровой блок управления ферментера. Скорость подачи глюкозы регулировали путем программирования «кислотного насоса» на время вкл./выкл. в течение одной минуты. Уравнение Твкл=0,72* exp[0.0023*(t-420)] использовали между 7 ч и 24 ч по истечении времени ферментации, где Твкл - это время, в течение которого «кислотный насос» был включен на одну секунду во время периода, равного 1 минуте. Скорость подачи глюкозы снижали через 24 часа после того, как наблюдали недостаток кислорода. рН контролировали вручную, используя основной насос для добавления 29% раствора гидроксида аммония. Для более точного контроля рН применяли с помощью специализированной программы, написанной с помощью программного обеспечения BioXpert VI.5. Скорость перемешивания и/или расход воздуха увеличивали, когда уровень растворенного кислорода опускался ниже 20% насыщения воздуха. Чистый кислород смешивался с воздухом для обеспечения достаточного количества растворенного кислорода после 24 ч ферментации. Подачу глюкозу периодически прекращали, для того, чтобы удостовериться, что клетки потребили всю глюкозу, и что в среде не накопились токсичные побочные продукты (например, ацетат). Подпитку возобновляли после всплеска уровня растворенного кислорода, что указывало на истощение глюкозы и образование токсичных побочных продуктов. Johnston WA, et al. (2003) "Tracking the acetate threshold using DO-transient control during medium and high cell density cultivation of recombinant Escherichia coli in complex media" Biolechnol Bioeng 84(3):314-23. Образцы периодически собирали из ферментера. Взятые пробы центрифугировали при 12000 × g в течение 30 мин для отделения супернатанта от клеток E.coli.
Определение концентрации гепаросана в супсрнатанте культуры после ферментации
Концентрацию гепаросана в супернатанте культуры после ферментации оценивали с помощью анализа с использование карбазола и ЯМР-анализа. В анализе гепаросана с использованием карбазола на обессоливающую центрифужную колонку Millipore YM-3 (1200×g) наносили 0,5 мл супернатанта ферментационной культуры для удаления солей и низкомолекулярных соединений. Получали концентрат, содержащий неочищенный сырой гепаросан, и лиофилизировали его. Высушенный неочищенный гепаросан восстанавливали с помощью 0,5 мл дистиллированной воды и анализировали с помощью карбазола. Концентрацию гепаросана рассчитывали по стандартной кривой, полученной с использованием чистого гепаросана. В анализе методом ЯМР гепаросан (1 мл) аналогичным способом извлекали из супернатанта культуры, лиофилизировали, а затем растворяли в 400 мкл D2O, содержащей 71 мкг полиэтилентерефталата натрия (внутренний стандарт), а затем переносили в пробирку для ЯМР для анализа 1Н-ЯМР при частоте 600 МГц. Концентрацию гепаросана рассчитывали из соотношения интегрированной области N-ацетильной группы гепаросана и таковой для внутреннего стандарта.
Быстрое выделение образцов гепаросана в процессе ферментации для анализа
Гепаросан выделяли быстрым способом из супернатанта культуры после ферментации и частично очищали с помощью цептрифужных колонок Vivapure D Mini H. Супернатант (1 мл) отделяли от клеток путем центрифугирования (12000 мкг), смешивали с 1 мл буфера А (50 мМ хлорида натрия, 20 мМ ацетата натрия, рН 4). Подводили рН смеси до значения 4, и наносили смесь на предварительно уравновешенную колонку Vivapure D Mini. После этого колонку промывали буфером А и элюировали буфером В (1 М хлорида натрия, 20 мМ ацетата натрия, рН 4). Элюированные образцы затем осаждали с помощью 3 объемов этанола, оставляя из на ночь при -20°С (во взрывобезопасной морозильной камере) и полученный преципитат промывали с помощью 75% этанола, растворяли в воде и лиофилизировали. Поле этого чистоту полученных образцов анализировали с помощью 1Н-ЯМР.
Очистка гепаросана
Гепаросан выделяли по окончании ферментации либо во встряхиваемой колбе или в ферментере объемом 7 л путем центрифугирования культуры при 12000 × g в течение 30 мин. Доводили рН полученного супернатанта до значения 4, добавляли ледяную уксусную кислоту, а затем фильтровали через воронку Бюхнера Руrех с пористым диском (размер пор 40-60 мкм). Быстропроточной смолой ДЭАЭ-сефарозой заполняли колонку соответствующего размера (<20 мг гепаросана/мл набухшей смолы). Колонку сначала уравновешивали 50 мМ хлоридом натрия в 20 мМ буфере ацетата натрия при рН 4. После этого на колонку наносили супернатант ферментационной культуры, и промывали колонку 3 объемами 50 мМ хлорида натрия в 20 мМ натрий-ацетатном буфере при рН 4. После этого гепаросан элюировали с колонки с 1 М хлоридом натрия в 20 мМ натрий-ацетатном буфере при рН 4. Гепаросан, элюированный с колонки, осаждали путем добавления 3-х объемов этанола, и оставляли такой раствор в течение ночи при -20°С во взрывобезопасной морозильной камере. В результате центрифугирования при 12000 g в течение 30 мин получали осадок. Осадок промывали 75% этанолом, снова центрифугировали и полученный осадок либо лиофилизировали для хранения, либо использовали непосредственно на стадии обесцвечивания. Гепаросан также может быть очищен от осадка клеток путем добавления 0,02% ДСН с последующим интенсивным перемешиванием, центрифугированием и хроматографией с помощью ДЭАЭ.
На этапе обесцвечивания гепаросан сначала растворяли в 1 М хлорида натрия в концентрации ~ 15 г/л. Доводили рН раствора до 9,5 с помощью 1 М NaOH, и добавляли перекись водорода (30%) до конечной концентрации 1,5%. Смесь инкубировали в течение ночи при комнатной температуре, после чего гепаросан осаждали путем добавления 3-х объемов этанола в течение ночи при температуре -20° (во взрывобезопасной морозильной камере). Полученный осадок промывали 75% этанолом, растворяли в воде и высушивали.
Дисахаридный анализ гепаросана
Гепаросан (10 мг/мл) растворяли в 200 мМ натрий-фосфатном буфере при рН 7 и обрабатывали 1 мЕд каждой из гепарин-лиаз 1, 2 и 3 при 30°С в течение ночи. Образующийся дисахаридный продукт подвергали высокоэффективной жидкостной хроматографии (ВЭЖХ) с масс-спектрометрией (MS) с ионизацией электрораспылением (ESI, electrospray ionization) с помощью ионной ловушки Agilent Ion trap, и определяли дисахаридный состав. Анализ методом ЖХ-МС проводили с помощью системы LC/MSD trap MS; Agilent, Santa Clara, CA. В качестве растворов А и В для высокоэффективной жидкостной хроматографии применяли 15% и 65% ацетонитрил, соответственно, содержащие одну и ту же концентрацию 37,5 мм NH4НСО3 и 11,25 мМ трибутиламин.
Разделение проводили па колонке С-18 (Agilent) с помощью раствора А в течение 20 мин, затем линейного градиента от 20 до 45 мин 0-50% раствора В. Сток с колонки попадал в источник для масс-спектрометрии (MS) с ионизацией электрораспылением (ESI-MS), для непрерывной детекции с помощью MS. Интерфейс электрораспыления был установлен в режим отрицательной ионизации с потенциалом скиммера 400 В, капиллярным выходом 240,0 В, и источником температуры 325°С для получения максимального количества ионов в спектре при полном сканировании (150-1500 Да, 10 полных спектров/сек). Азот использовали в качестве высушивающего (5 л/мин) и распыляющего газа (20 фунт/кВ. дюйм). Bhattacharyya S, Am J Respir Cell and Molec Biol 42, 51-61.
Анализ гепаросана с помощью ЯМР
1Н-ЯМР и 13H-ЯМР проводили на спектрометре Bruker 600 МГц. Образцы гепаросана готовили в концентрации 2 мг/мл D2O (99,99+атом. %), лиофилизировали, удаляли обменные протоны, вновь растворяли в D2O и переносили в стандартные 5 мм 10 пробирки для ЯМР. Спектры регистрировали с помощью программы TOPSPIN 2,0. Все спектры получали при температуре 298 K.
Анализ содержания белка и ЛНК
Содержание ДНК в конечном продукте гепаросана определяли путем измерения УФ-поглощения 0,1 мг/мл раствора гепаросан при 260 им и 320 нм. Концентрацию ДНК рассчитывали как концентрацию (мкг/мл)=(значение А260 - значение А320) × коэффициент разведения × 50 мкг/мл. Содержание белка определяли с помощью набора для определения белка Micro BCA Protein Assay Kit согласно инструкциям производителя.
Анализ молекулярной массы гепаросана
Лестница стандартов гепаросана с известным молекулярным весом была изготовлена из обесцвеченного полисахарида гепаросана K5 путем непрерывного препаративного электрофореза в полиакриламидном геле с использованием аппарата Mini Prep Cell (Biorad, Hercules, CA) аппарат (Ly et al., 2010). Фракции гепаросана анализировали методом масс-спектрометрии с Фурье-преобразованием (FT-MS), а вновь вмешивали для получения лестницы стандартов молекулярного веса для определения молекулярного веса гепаросана (Ly et al., 2010). Молекулярные маркеры для гиалуроновой кислоты (ГК), которая также представляет собой линейный полисахарид с той же плотностью заряда, как и у N-ацетилгепаросана, применяли в качестве стандартов для верхнего диапазона образцов.
Маркеры молекулярного веса и для ГК, и для гепаросана использовали в качестве набора стандартов для анализа молекулярного веса гепаросана, полученного на различных культуральных средах и при различных сроках ферментации. Для анализа молекулярноговеса гепаросана использовали полиакриламидиый гель (4-15%) размером 0,75 мм×6,8 см×8,6 см. Образцы гепаросана (25 мкг) загружали в гель, затем подвергали электрофорезу (200В в течение 20 мин) и окрашивали альциановым синим в течение 1 ч, а затем обесцвечивают с 25% этанолом/10% уксусной кислотой (Ly et al., 2010). Гели 5 сканировали, цифровые изображения анализировали с помощью компьютерной программы UN-SCANIT. Строили графики зависимости плотности изображения от расстояния миграции в геле. На основании полученных данных определяли молекулярный вес образцов гепаросана с помощью стандартной кривой.
Результаты
Ферментация Е. coli K5 во встряхиваемой колбе колбы для получения гепаросана Культуру Е. coli K5 выращивали во встряхиваемой колбе до 1-4 ч после достижения стационарной фазы роста. Ферментацию продолжали после стационарной фазы для накопления максимального количества гепаросана в среде, в соответствии известными данными о том, что накопление гепаросана в среде отстает от роста клеток Manzoni M et al. (1993) Journal of Bioactive and Compatible Polymers 8(3):251-257. Выхода гепаросан из культур, выращенных во встряхиваемой колбе, варьировал от 70 до 500 мг/л. Степень чистоты гепаросана, выделенного из таких культур, во всех случаях составила>85%, как было показано с помощью ЯМР Wang Z et al (2009) Anal Biochem. 398(2):275-7.
Zhang ZQ et al/ (2008) "Solution structures of chemoenzymatically synthesized heparin and its precursors" Journal of the American Chemical Society 130(39):12998-13007. Гепаросан, выделенный из синтетических сред, в том числе среды М9, среды с глюкозой и среды с глицерином, характеризовался более высоким уровнем чистоты (>95%), но сравнению с гепаросаном, выделенным из среды LB, который демонстрировал дополнительные пики в ЯМР-анализе, что соответствовало ~85% чистоты (Фиг.2). Дальнейший анализ гепаросана, полученного в среде LB, с помощью электрофореза в полиакриламидном геле (ПААГ) показал наличие низкомолекулярной полосы, соответствующей компонентам среды, выделяемых совместно с гепаросаном. Такие примеси можно удалить с помощью спин-колонок с показателем отсекаемой молекулярной массы 3K (molecular weight cut-off, 30 MWCO) (Фиг.2А).
Дисахаридный состав гепаросана определяли с помощью ВЭЖХ-МС (данные не представлены) после полного ферментативного гидролиза с гепаринлиазой 1, 2 и 3, и было показано наличие одного дисахарида м/з 378,2. Такой дисахарид соответствует ΔUA(1→4)-α-D-GlcNAc, что согласуется со стандартным повторяющимся структурным элементом гепаросана, → 4)-β-D-GlcA (1 → 4)-α-D-GlcNAc (1 →. Для подтверждения такой структуры получали ^С- и ^М-меченный гепаросан при культивировании Е. coli K5 в среде М9, содержащей равномерно меченую 13C-глюкозу и 15N-аммония хлорид. Структура полученного равномерно меченого гепаросана 13С-, 15N- была подтверждена путем полного ферментативного гидролиза с гепаринлиазой 1, 2 и 3 с последующим анализом с помощью ВЭЖХ-МС, показавшим наличие одного дисахарида м/з 392,9. Разница в 15 а.е.м. между м/з 378,2 и 392,9 соответствует полному 13С и 15N изотопному обогащению гепаросана и подтверждает повторяющуюся структуру гепаросана.
Ферментация Е. coli K5 в ферментере объемом 7 л
Скорость подачи подпитки экспоненциально увеличивалась для поддержания экспоненциального роста Е. coli. На стадии подпитки было несколько пауз, чтобы исключить накопление глюкозы в среде, что в свою очередь может привести к накоплению ацетата и ингибированию роста Е. coli. Подпитку возобновляли только после увеличения содержания растворенного кислорода, подтверждающего истощение глюкозы (Фиг.3). На поздней стадии ферментации (после 22 ч), растворенный кислород становился ограниченным. Для увеличения до максимальной величины концентрации растворенного кислорода в культуре увеличивали скорость перемешивания и вводили чистый кислород в культуру. Несмотря такие усилия, уровень растворенного кислорода по-прежнему оставался довольно низким после 31 часов. Скорость добавления глюкозы уменьшали, когда кислород становился лимитирующим элементом. В ходе ферментации рН поддерживали между 6 и 8, добавляя NH4ОH. После 37,5 ч ферментации плотность клеток достигла 85 г с.м.к./л, а концентрация гепаросана в супернатанте культуры после ферментации составила 15 г/л. Общий показатель роста культуры составил 0,12 ч-1, общий уровень продукции составил 1,2 г/ч, а объемная скорость продукции составила 0,4 г/л Гепаросан, очищенный из культуры после ферментации характеризовался высокой чистотой, согласно 1Н-ЯМР-спектру (Фиг.4а). Кроме того, анализ содержания ДНК и анализ содержания белка с помощью бицинхониновой кислоты (ВСА) показал, что в конечном веществе гепаросана присутствует<1% ДНК и<2% белка.
Структурная характеристика гепаросана
1Н-ЯМР 13C-ЯМР, ВЭЖХ-МС и МС с Фурье-преобразованием (FT-MS) (Фиг.4) подтвердили, что общая структура гепаросана является следующей: → 4)-β-D-GlcA (1 → 4)-α-D- GlcNAc (1 →(Фиг.1А). Фракция гепаросана в FT-MS со степенью полимеризации (СП), равной 24, полученная с помощью препаративного электрофореза, показана на Фигуре 4С. Этот спектр также указывает на наличие остатков ненасыщенных уронатов ΔUA (Фиг.1 В) на невосстанавливающем конце некоторых из полисахаридных цепей. Наличие концевых остатков ΔUA согласуется с действием K5 гепаросанлиазой, также присутствующей в штамме Е. coli K5. Анализ с помощью FT-MS показал, что гепаросан, выделенный из супернатанта культуры после ферментации, содержит смесь гепаросана с концевыми остатками ΔUA и остатками GlcA. Такой результат согласуется с тем, что часть гепаросана в среде отделялась от поверхности клеток за счет действия поперечной силы, и некоторая часть гепаросана высвобождалась в среду в результате ферментативного гидролиза с участием гепаросанлиазы. Гепаросан, выделенный из супернатанта, обрабатывали экзолитическим ферментом β-глюкуронидазой, способной удалять GlcA, но не DUA, с невосстанавливающего конца гепаросановой цепочки. Выявление высвобожденного GlcA с помощью тонкослойной хроматографии (данные не представлены) подтверждает, что некоторая часть гепаросановых цепочек, высвобождаемых в супернатант, содержала на конце GlcA.
Характеристика молекулярного веса гепаросана
Гепаросан, выделенный из культуры Е. coli K5, выращенной во встряхиваемой колбе, анализировали с помощью электрофореза в ПААГ в сравнении с лестницей стандартов очищенного гепаросана (Фиг.5 и Таблица 1). Пробы гепаросана, выделенного из супернатанта культур, выращенных в среде М9, синтетической среде с глицерином, комплексной среде LB, показали небольшие различия в Мn или MW. Гепаросан, выделенный из супернатанта синтетической среды с глюкозой, характеризовался наиболее низкими значениями Мn и MW. Гепаросан, выделенный из осадка, характеризовался более высоким MW, no сравнению с гепаросаном, выделенным из сунернатанта культуры после ферментации (данные не представлены).
Затем из ферментера на разных сроках в ходе ферментации отбирали пробы, и гепаросан очищали с помощью спин-колонок Vivapure D Mini H. Чистота полученных образцов составила >85% для образцов, выделенных из супернатантов культур, что было показано с помощью 1Н-ЯМР. Анализ а ПААГ (Фиг.5Б) показал начальное увеличение молекулярного веса гепаросана, после чего по мере увеличения времени ферментации молекулярный вес не изменялся (Фиг.6). Индекс полидисперсности гепаросана (polydispcrsity index, PDI=Mw/Mn) мало изменялся в ходе ферментации.
Обсуждение
В настоящем исследовании был достигнут высокий выход гепаросана (15 г/л) из супернатанта культуры, выращенной в ферментере, при периодическом культивировании с подпиткой в среде заданного состава с глюкозой. Это выгодно отличается от выхода гепаросана, который составляет 10,2 г/л, полученного от этого же организма, выращенного в среде с заданным содержанием глицерина (публикация заявки на Патент США №2008/0032349) Объемная скорость продукции в настоящем исследовании также увеличена, по сравнению с указанным предыдущим сообщением. Кроме того, глюкоза как менее дорогой источник углерода, по сравнению с глицерином, делает такой процесс ферментации более экономичным. Способ ферментации при периодическом культивировании с подпиткой, разработанный для объема в 3 л, может служить прототипом ферментации в больших объемах при лабораторном производстве, и в конечном счете, позволяет осуществить производство гепаросана в промышленных масштабах. Исследования по увеличению масштаба производства в настоящее время проводятся в нашей лаборатории, позволяющие перейти от рабочего объема 3 л к 750 л, что будет служить основой для увеличения объема до ~100000 л, необходимого для крупномасштабного (100 метрических тонн) производства гепаросана, поставляемого в качестве исходного вещества для биоинженерного синтеза гепарина. Изучается возможность применения более высокой скорости подачи глюкозы при ферментации и более высокого содержания кислорода в ферментере для увеличения скорости продукции гепаросана.
Применение среды с заданным составом, содержащей глюкозу в качестве источника углерода, снижает как стоимость ферментации, так и уменьшает комплексность среды, что делает процесс очистки более простым. Гепаросан, выделенный из комплексной среды LB, характеризовался меньшей степенью чистоты из-за примесей компонентов комплексной среды, и для него необходимы дополнительные стадии очистки, чем для гепаросана, выделенный из среды заданного состава с глюкозой.
Гепаросан, выделенный из супернатанта культуры, накапливается в результате его высвобождения в среду в ходе ферментации, вероятно, благодаря совместному действию лиазы K5 и поперечной силы. Увеличение высвобождения гепаросана в среду увеличивает выход гепаросана в супернатанте, снижает стоимость последующих этапов очистки.
Наличие лиазы K5 способствует желаемому высвобождению гепаросановых капсул в среду, увеличивая выход гепаросана. В то же время, наличие гепаросанлиазы K5 приводит к появлению неприродного сахаридного остатка, ΔUA, на невосстапавливающем конце цепочки, что может требовать последующей обработки. Гепаросанлиаза K5 также увеличивает индекс полидисперсности гепаросана, что может усложнить последующую его переработку в гепарин. Характеристики молекулярного веса полученного биоинженерным способом гепарина неразрывно связаны с таковыми 5 характеристиками гепаросана. Данный способ позволяет получить гепаросан со свойствами, подходящими для его переработки в гепарин. Таким образом, авторы настоящего изобретения показали, что в присутствии лиазы все-таки возможно получение гепаросана с желаемыми характеристиками молекулярного веса при тщательном контроле условий ферментации и времени процесса ферментации.
В будущем контроль активности лиазы K5 может представлять собой подход для точного контроля продукции гепаросана.
Среднечисленный молекулярный вес (MN), средневесовой молекулярный вес (MW) и индекс полидисперсности (ИПД) гепаросана, выделенного из 15 различных культуральных сред во встряхиваемой колбе объемом 2,8 л.
ПРИМЕР 2. Протокол ферментации 20 л отчет (рН стат федеральным партии брожения)
Среда
Среда с заданным составом глюкозы (на литр), 20 г глюкозы, 20 мг тиамина, 13,5 г KН2РO4, 4,0 г (NH4)2HPO4, 1,4 г MgSO4·7H2O, 1,7 г лимонной кислоты и 10,0 мл раствора микроэлементов. Раствор микроэлементов включал (на 1 л 5М НСl) 10,0 г FeSO4·7H2O, 2,0 г CaCl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г CuSO4·5H2O, 0,1 г (NH4)6MO7O24·4H2O и 0,02 г Na2B4O7·10H2O. Раствор для подпитки, используемый на стадии культивирования с подпиткой, включал (на 1 л): 700 г глюкозы, 20 г MgSO4·7H2O, 47 г KН2РO4 и 0,4 г тиамина. Среду R, которая представляет собой глюкозосодержащую среду, применяемую в процессе ферментации, получали следующим образом:
Компонент А среды R: (NH4)2HPO4, KH2PO4, и лимонную кислоту добавляли примерно в 8 литров воды с магнитной мешалкой, добавляли раствор микроэлементов (100х кратный сток), подводили рН до 6,80, затем добавляли воду до 9 литров. Компонент А стерилизовали в ферментере при температуре 121°С в течение 45 минут.
Компонент В среды R: Готовили растворы MgSO4 и глюкозы в виде 10× стоков и стерилизовали их по отдельности в автоклаве при 110°С в течение 20 минут (также возможна стерилизация путем фильтрации).
Раствор для подпитки: Глюкоза, MgSO4·7H2O и 47 г KH2PO4 растворяли в 1 л воды при нагревании и перемешивания, стерилизовали в автоклаве при 115°С в течение минут. Тиамин растворяли в воде, стерилизовали путем фильтрации.
Протокол
День 1:
В четыре культуральные пробирки 5 мл, содержащие синтетическую среду с глюкозой, описанную выше (среда R) утром засевали штамм Е. coli K5 (штамм хранили в морозильной камере при -80°С), выращивали в инкубаторе при температуре 37°С, при скорости покачивания 220 об/мин в течение 10 часов. Указанные четыре культуры вечером переносили в 500 мл среды во встряхиваемые колбы с перегородками 2,8 л и выращивали при 37°С, покачивании 220 об/мин в течение 10 часов.
День 2: Датчик рН и датчик уровня растворенного кислорода (DO, dissolved oxygen) стерилизовали и калибровали при температуре культивации (37°С). Устанавливали три насоса: насос 1 - NH4OH для поддержания рН; насос 2 - для подачи раствора для подпитки, насос 3 - для пеногасителя.
Компонент В среды R добавляли в ферментер, смешивая с Компонентом А, и инокулировали его с помощью 500 мл культуры. рН поддерживали на уровне 6,8, путем закачки в NH4OH. Добавление раствора для подпитки осуществляли, когда наблюдали всплеск уровня рН и DO. Насос для подачи подпитки был запрограммирован таким образом, что он работал па 100% мощность (выброс ~ 6,7 г глюкозы) при рН>6,8. Скорость перемешивания и/или скорость воздушного потока увеличивали для поддержания DO выше 20%. Поток чистого кислорода добавляли, когда DO не удерживался па уровне выше 20% в результате изменения скорости перемешивания и воздушного потока.
День 3: Ферментацию прекращали, когда значение оптической плотности культуры OD600 начинало уменьшаться. Культуры собирали и центрифугировали. Гепаросан выделяли из супернатанта.
Синяя кривая на Фигуре 7 представляет собой изменение DO (растворенного кислорода в культуре). Из-за высокой плотности клеток, потребляемых кислород очень быстро, DO снижалась практически до 0 за несколько часов перед сбором культуры, тогда как подача чистого кислорода не прекращался. Ферментацию останавливали через несколько часов после того, когда OD600 (оптическая плотность культуры при длине волны 600 нм) начинала снижаться. Ферментацию останавливали отключением подачи 10 кислорода, остановкой перемешивания, а затем собирали культуру.
Взятие проб: Примерно 40 мл культуры отбирали для измерения оптической плотности при 600 нм, а затем центрифугировали при 12000 × g в течение 30 мин для отделения супернатанта от клеток Е. coli. Супернатант и осадок клеток замораживали отдельно.
Анализ проб после ферментации: Сухую массу клеток (с.м.к.) измеряли, взвешивая высушенный осадок клеток на весах. Концентрацию гепаросана в супернатанте измеряли методом количественной оценки с помощью ЯМР или с помощью карбазола, после чего проводили осаждение этанолом Song J-M et al. (2009) "A simple method for hyaluronic acid quantification in culture broth" Carbohydrate Polymers 78 (2009) 633-634. Wang Z et al. (2009) Anal Biochem. 398(2):275-7.
Результаты приведены на Фигурах 7, 8 и 9.
Гепаросан, полученный на этой стадии ферментации, имел среднечисленный молекулярный вес примерно 58000 Да, средневесовой молекулярный вес примерно 84000 25 Да, и индекс полидисперсности (ИПД) примерно 1,4. Степень чистоты составила 95% или выше.
ПРИМЕР 3. Очистка гепаросана с помощью хроматографии с использованием хитозана.
Хитозап представляет собой линейный полисахарид, состоящий из беспорядочно распределенный β-(1-4)-связанных D-глюкозамина и Н-ацетил-D-глюкозамина, и он является альтернативой анионообменной смоле для хроматографической очистки гепаросапа. Аминогруппы хитозана (~рКа 6,5) делают его положительно заряженным и растворимым в кислых условиях, и таким образом, подходящим для аффинной очистки отрицательно заряженного гепаросана. Очистку гепаросана с помощью хитозана можно легко контролировать с помощью концентрации хитозана и начальном значении рН. Кроме того, хитозан является недорогим, доступным в изобилии и биодеградируемым. В этом примере описан способ разделения с помощью растворимого хитозана для эффективного извлечения гепаросана непосредственно из культуральной среды. Такое разделение на основе нейтрализации заряда с применением хитозана потенциально можно применять для очистки других отрицательно заряженных макромолекул из сложных ферментационных смесей. Культуральную среду после ферментации Escherichia coli IC5 в 7 л, центрифугировали при 7000 об/мин в течение 30 мин для удаления клеток.
1. Проводили количественную оценку с помощью ЯМР содержания гепаросана в супернатанте.
2. Подводили рН супернатанта до 4.0 с помощью раствора НСl с последующим центрифугированием при 7000 об/мин в течение 1 часа для удаления все примесей, выпадающих в осадок.
3. После этого супернатант разбавляли в 5 раз с помощью рН 4,0 ДИ воды.
4. Готовили раствор хитозана 10 мг/мл путем растворения хитозана в 1% растворе уксусной кислоты. На каждые 10 мг гепаросана добавляли 2,5 мг хитозана к супернатанту, используя раствор хитозана 10 мг/мл.
5. Затем смесь перемешивали в течение 10 мин при комнатной температуре для обеспечения надлежащего смешивания полученного раствора. После этого инкубировали смесь при 4°С в течение ночи.
6. Полиэлектролитный комплекс хитозан-гепаросан формировал преципитат, 25 который оседал, который собирали с помощью центрифугирования при 8000 об/мин в течение 1 часа.
7. После этого промывали осадок ДИ водой с рH 4,0 и центрифугировали еще раз при 8000 об/мин в течение 1 часа.
8. Хитозан теряет катионные свойства при щелочных условиях, что приводит к разрушению электростатического комплекса, в результате чего в растворе образуется свободный гепаросан и нерастворимый хитозан. Промытый осадок помещали в 1М раствор NaOH и инкубировали в течение ночи для извлечения гепаросана в раствор. Нерастворимый хитозан удаляли либо путем фильтрации, либо путем центрифугирования при 10000 об/мин в течение 2 часов. Эта стадия может быть объединена вместе с катионообменной или анионообменной стадией для удаления других примесей, что приводит к дополнительной очистки полученного гепаросана.
Пример 3А.
С применением описанной выше процедуры гепаросан выделяли из ферментационной среды с помощью колонок с быстропроточной смолой ДЭАЭ-сефарозой с последующим осаждением с четырьмя объемами этанола. Полученный гепаросан подвергали диализу с помощью мембраны 3500 MWCO мембраны против ДИ воды. Диализированный раствор затем высушивали сублимацией с последующим сокращением объема с помощью роторного испарителя.
После этого готовили раствор гепаросана, таким образом очищенного, концентрацией 1 мг/мл, растворяя его в деионизированной воде (ДИ) при рН 4,0. Готовили шесть различных образцов с использованием 10 мл раствора 1 мг/мл и добавляли различное количество хитозана в диапазоне 2-6 мг с использованием 10 мг/мл раствора хитозана в 1% уксусной кислоте. Образцы инкубировали при 4°С в течение ночи. Осажденный полиэлектролитный комплекс собирали путем центрифугирования образцов при 8000 об/мин в течение 1 часа. После этого к осадкам добавляли 1М раствор NaOH, не промывая их ДИ водой при рН 4,0. Нерастворимый хитозан удаляли центрифугированием при 10000 об/мин в течение 2 часов. После этого полученные образцы и супернатант анализировали на наличие гепаросана с помощью анализа с использованием карбазола.
По данным анализа с использованием карбазола 100% извлечения гепаросана достигали при рН 4,0 добавлением 4 мг хитозана к 10 мл 1 мг/мл раствора гепаросана.
Пример 3 В.
В дополнительном эксперименте, супернатант, получены в результате центрифугирования после ферментации, кондиционировали добавлением соляной кислоты до рН 4,0. После этого центрифугировали при 8000 об/мин в течение 1 ч для удаления примесей, выпадающих в осадок. рН супернатанта затем вновь подводили до 4,0 с помощью 1 М раствора NaOH. Количественная оценка методом ЯМР показала, что в таких образцах с подведенным значением рН не наблюдали потери гепаросана.
Полученные образцы затем разбавляли в пять раз с использованием ДИ воды рН 4,0. После этого готовили шесть образцов, по 10 мл каждый, путем добавления хитозана в диапазоне 2-8 мг в образец с использованием 10 мг/мл раствора хитозана в 1% уксусной кислоте. Образцы инкубировали при 4°С в течение ночи. Осажденный полиэлектролитный комплекс собирали путем центрифугирования образцов при 8000 об/мин в течение 1 часа. Затем к полученным осадкам добавляли 1М раствор NaOH без промывки ДИ водой рН 4,0. Нерастворимый хитозан удаляли центрифугированием при 10000 об/мин в течение 2 часов. После этого полученные образцы и супернатант анализировали на наличие гепаросана с помощью анализа с использованием карбазола. По данным анализа с использованием карбазола показано ~70% очистка гепаросан из ферментационной среды.
Изобретение обеспечивает эффективный способ очистки гепаросана из ферментационной среды. Хитозан, применяемый в данном способе, представляет собой дешевое, доступное, биосовместимое и биоразлагаемое вещество. Эти свойства делают его идеальным для применения в качестве преципитанта для очистки гепаросана, который может быть ферментативно модифицирован для возможности внутривенной доставки гепарина. Настоящее изобретение также обеспечивает критическую стадию концентрирования, таким образом, уменьшая рабочие объемы в данном способе, по сравнению с обычно применяемыми стадиями осаждения этанолом. Рабочие объемы для осаждения этанолом, например, в некоторых случаях превышает объемы ферментации. Биосовместимая и биоразлагаемая природа хитозана делает его гораздо более предпочтительными, по сравнению с другими поликатионами, которые являются токсичными, такими как, цетилпиридиний хлорид (ЦПХ) и поли (диаллилдиметилхлорид аммония). Кроме того, способ настоящего изобретения потенциально применим для очистки других подобных полисахаридов, таких как гиалуроновая кислота и кислые белки, а также ДНК.
Настоящее изобретение также обеспечивает экономически эффективное выделение бактериального капсульного полисахарида, гепаросана K5, из ферментационной среды, с помощью осаждения, для потенциального применении в продукции гепарина биоинженерным способом. В способе применяются природные поликатионами, которые являются биосовместимыми и биоразлагаемыми, и таким образом обеспечивают эффективную и безопасную технологию очистки препаратов для внутривенной доставки, таких как гепарин. Высокий выход продукта, полученный с помощью такого способа, делают идеальным его применение па стадии концентрирования, тем самым уменьшая рабочий объем. Легкость извлечения гепаросана на этом этапе и его совместимость с дальнейшими этапами в химико-ферментативного биоинженерного синтеза гепарин делает его идеальным для применения на этапе очистки. Способ настоящего изобретения потенциально применим для удаления других анионных полисахаридов и белков, полученных после бактериальной ферментации, а также молекул с высоким отрицательным зарядом, таких как ДНК.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОМЫШЛЕННОГО КУЛЬТИВИРОВАНИЯ ШТАММОВ E.coli, ПОЛУЧЕННЫХ НА ОСНОВЕ ШТАММА BL21(DE3), НЕСУЩЕГО ГЕН T7 RNA ПОЛИМЕРАЗЫ ПОД КОНТРОЛЕМ lacUV5 ПРОМОТОРА, С ПОВЫШЕННЫМ СИНТЕЗОМ БИОМАССЫ И ВЫХОДОМ ЦЕЛЕВОГО БЕЛКА В ТЕЛЬЦАХ ВКЛЮЧЕНИЯ | 2011 |
|
RU2473683C1 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ АМИНОКИСЛОТ И ПРОИЗВОДНЫХ АМИНОКИСЛОТ ИЗ СЕМЕЙСТВА ФОСФОГЛИЦЕРАТОВ | 2003 |
|
RU2346038C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА | 2011 |
|
RU2631001C2 |
| Рекомбинантный штамм бактерии Escherichia coli - продуцент L-треонина | 2018 |
|
RU2697219C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА SAV-RGD | 2014 |
|
RU2577138C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА SAV-RGD, СПЕЦИФИЧЕСКИ УЗНАЮЩЕГО КЛЕТКИ МЕЛАНОМЫ | 2013 |
|
RU2563540C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЧУЖЕРОДНОГО ПРОТЕИНА В КЛЕТКАХ E.COLI, ПЛАЗМИДНЫЙ ВЕКТОР И ТРАНСФОРМИРОВАННЫЙ ШТАММ Е.COLI ДЛЯ ЭКСПРЕССИИ ЧУЖЕРОДНОГО ПРОТЕИНА | 1996 |
|
RU2201455C2 |
| СПОСОБ ПОЛУЧЕНИЯ БАКТЕРИОХЛОРОФИЛЛА А | 2014 |
|
RU2671158C2 |
| СПОСОБ ПРОДУКЦИИ СУЛЬФАТИРОВАННОГО ПОЛИСАХАРИДА И СПОСОБ ПРОДУКЦИИ PAPS | 2021 |
|
RU2811941C1 |
| ВЫСОКОЭФФЕКТИВНЫЙ СПОСОБ ПОЛУЧЕНИЯ БАКТЕРИАЛЬНОГО ПОЛИСАХАРИДА, ПОЛУЧЕНИЕ КОНЪЮГАТА НА ЕГО ОСНОВЕ И ИММУНОГЕННАЯ КОМПОЗИЦИЯ | 2013 |
|
RU2627156C2 |
Способ получения по существу чистого гепаросана из E.coli K5 предусматривает культивирование клеток E.coli K5 в среде, содержащей глюкозу в качестве основного источника углерода, осуществление связывания гепаросана с твердофазным носителем с последующим элюированием и осаждение гепаросана из элюата. При этом указанное культивирование включает фазу периодического культивирования и фазу периодического культивирования с подпиткой. Среда, применяемая на стадии периодического культивирования, содержит (на литр) примерно 20 г глюкозы, 10-300 мг тиамина, примерно 13,5 г KH2PO4, примерно 4,0 г (NH4)2HPO4, примерно 1,4 г MgSO4·7H2O, примерно 1,7 г лимонной кислоты, и примерно 10,0 мл раствора микроэлементов, раствор микроэлементов по существу состоит из (на 1 л 5М HCl) 10,0 г FeSO4·7H2O, 2,0 г CaCl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г CuSO4·5H2O, 0,1 г (NH4)6Mo7O24·4H2O и 0,02 г Na2B4O7·10H2O. Среда для подпитки, применяемая на стадии периодического культивирования с подпиткой, содержит (на литр): 250-1000 г глюкозы, 20 г MgSO4, 0,15-0,5 г тиамина и, возможно, 47 г KH2PO4. Снабжение кислородом осуществляют барботированием воздуха с дополнительной подачей или без дополнительной подачи кислорода. Способ позволяет получить по меньшей мере на 90% чистый гепаросан. 13 з.п. ф-лы, 12 ил., 1 табл., 3 пр.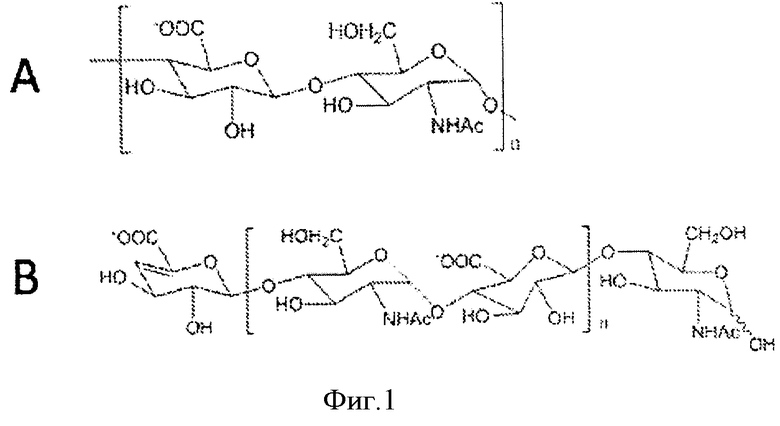
1. Способ получения по существу чистого гепаросана из E.coli K5, включающий:
(а) культивирование клеток E.coli K5 в заданной среде, содержащей глюкозу в качестве основного источника углерода, при этом указанное культивирование включает фазу периодического культивирования и фазу периодического культивирования с подпиткой, при этом
(I) среда, применяемая на стадии периодического культивирования, содержит (на литр) примерно 20 г глюкозы, 10-300 мг тиамина, примерно 13,5 г KH2PO4, примерно 4,0 г (NH4)2HPO4, примерно 1,4 г MgSO4·7H2O, примерно 1,7 г лимонной кислоты, и примерно 10,0 мл раствора микроэлементов, при этом раствор микроэлементов по существу состоит из (на 1 л 5М HCl) 10,0 г FeSO4·7H2O, 2,0 г CaCl2, 2,2 г ZnSO4·7H2O, 0,5 г MnSO4·4H2O, 1,0 г CuSO4·5H2O, 0,1 г (NH4)6Mo7O24·4H2O и 0,02 г Na2B4O7·10H2O и при этом;
(II) среда для подпитки, применяемая на стадии периодического культивирования с подпиткой, содержит (на литр): 250-1000 г глюкозы, 20 г MgSO4, 0,15-0,5 г тиамина, и возможно 47 г KH2PO4 и
(III) при этом снабжение кислородом осуществляют путем барботирования воздуха с дополнительной подачей или без дополнительной подачи кислорода;
(б) осуществление связывания гепаросана с твердофазным носителем с последующим элюированием; и
(в) осаждение гепаросана из элюата;
при этом указанный по существу чистый гепаросан является по меньшей мере на 90% чистым.
2. Способ по п.1, отличающийся тем, что содержание растворенного кислорода поддерживают на уровне примерно 20%.
3. Способ по п.1, отличающийся тем, что температуру поддерживают на уровне примерно 37°C, а pH поддерживают на уровне примерно 7.
4. Способ по п.3, отличающийся тем, что pH поддерживают путем добавления 29% раствора аммиака.
5. Способ по п.1, отличающийся тем, что среду, применяемую на стадии периодического культивирования с подпиткой, подают со скоростью, определяемой по формуле:

где MS представляет собой скорость потока источника углерода (г/ч), F - скорость подачи раствора для подпитки (л/ч); SF представляет собой концентрацию источника углерода в подпитке (г/л); X - концентрация клеток (г/л с. м.к.), m - удельный коэффициент поддержания (г/г DCW/ч), V - объем культуры (л); t0 - время начала подпитки; t - время процесса; µ - удельная скорость роста (ч-1) и YX/S - выход клеток на количество источника углерода (г/г).
6. Способ по п.1, отличающийся тем, что выход гепаросана составляет более 12 г на литр супернатанта культуры.
7. Способ по п.6, отличающийся тем, что ферментацию проводят в течение менее 48 часов, без учета роста стартовой культуры.
8. Способ по п.1, отличающийся тем, что стадия осуществления связывания и элюирования включает (I) удаление клеток, (II) смешивание анионной смолы с супернатантом культуры и удаление супернатанта, (III) промывку смолы с помощью 50 мМ хлорида натрия в натрий-ацетатном буфере при pH 4, (IV) элюирование 1 М хлорида натрия в натрий-ацетатном буфере при pH 4.
9. Способ по п.1, отличающийся тем, что стадия осуществления связывания и элюирования включает (I) удаление клеток, (II) смешивание раствора хитозана с супернатантом культуры, (III) осаждение хитозана и выделение осадка; (IV) промывку хитозана, (V) элюирование гепаросана с помощью примерно 1М раствора NaOH.
10. Способ по п.1, отличающийся тем, что осаждение гепаросана из элюата включает осаждение с помощью этанола.
11. Способ по п.1, дополнительно включающий депирогенизацию с помощью перекиси водорода.
12. Способ по п.1, отличающийся тем, что указанный гепаросан содержит менее 1% ДНК и менее 2% белка.
13. Способ по п.1, отличающийся тем, что указанный гепаросан имеет среднечисленный молекулярный вес примерно 58000 Да, средневесовой молекулярный вес 84000 Да и индекс полидисперсности (ИПД) примерно 1,4.
14. Способ по п.1, отличающийся тем, что указанный гепаросан является по меньшей мере на 95% чистым.
| US 20080207895 A1, 28.08.2008 | |||
| JP 2004018840 А, 22.01.2004 | |||
| WO 2005014656 A1, 17.02.2005 | |||
| МАЛЕР Г., КОРДЕС Ю | |||
| Основы биологической химии | |||
| М.: Издательство "МИР", 1970, с | |||
| Приспособление для уменьшения дымовой тяги паровоза | 1920 |
|
SU270A1 |
| LEGOUX R | |||
| ET AL | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ДВЕРНОЙ ЗАМОК С ДВУМЯ КЛЮЧАМИ | 1926 |
|
SU7260A1 |

