Область техники
Настоящее изобретение относится к мультиспецифическим антигенсвязывающим молекулам, обладающим способностью замещать функциональную активность кофактора коагулирующего фактора крови VIII (FVIII), и фармацевтическим композициям, содержащим указанную молекулу. Изобретение также относится к антигенсвязывающим молекулам, в которых регулируется ассоциация между легкой и тяжелой цепями, способам получения антигенсвязывающих молекул, в которых регулируется ассоциация между легкой и тяжелой цепями, и к способам регулирования ассоциации между легкой и тяжелой цепью в антигенсвязывающих молекулах.
Уровень техники
Гемофилия А представляет собой заболевание, характеризующееся ненормальными кровотечениями, которое характеризуется наследственным снижением функциональной активности или дефицитом функциональной активности коагулирующего фактора крови VIII (FVIII). Пациентам с гемофилией А обычно вводят препараты FVIII для прекращения кровотечений (применение по требованию). Недавно препараты FVIII стали также использовать профилактически для предотвращения эпизодов кровотечений (превентивное применение; непатентные источники информации 1 и 2). Период полувыведения FVIII в крови составляет примерно от 12 до 16 часов. Поэтому для постоянного предупреждения кровотечений препараты FVIII следует вводить пациентам три раза в неделю (непатентные документы 3 и 4). Для применения по требованию препараты FVIII также дополнительно вводят, когда существует необходимость в регулярном предотвращении повторных кровотечений в определенные промежутки времени. Следует также отметить, что препараты FVIII применяют внутривенно. Таким образом, существует острая необходимость в разработке фармацевтических агентов, которые обеспечивают меньшую нагрузку на пациентов, чем препараты FVIII.
Случается, что антитела к FVIII (ингибиторы) образуются в организме больных гемофилией. Такие ингибиторы отменяют действие препаратов FVIII. Для остановки кровотечений у пациентов, в организме которых образовались такие ингибиторы, используются «обходные» (шунтирующие) препараты. Механизмы их действия не зависит от функционирования FVIII, а именно, они связаны с возможностью катализировать активацию коагулирующего фактора крови X (FX) путем активации коагулирующего фактора крови IX (FIXa). Таким образом, в некоторых случаях «обходные» препараты неспособны в достаточной степени остановить кровотечение. Соответственно, существует значительная потребность в создании фармацевтических агентов, действие которых не зависит от присутствия ингибиторов, и которые функционируют как заместители FVIII.
Сообщалось, что в качестве средств решения указанной проблемы могут быть использованы биспецифические антитела, замещающие функцию FVIII (патентные документы 1, 2, 3 и 4). Биспецифические антитела к FIXa и FX способны замещать функцию FVIII путем позиционирования этих двух факторов в непосредственной близости друг от друга для проявления ими функциональной замещающей активности кофактора FVIII (непатентный документ 5). Сообщалось, что функциональная замещающая активность кофактора FVIII, свойственная антителам, может быть усилена путем оптимизации аффинности и их пространственной ориентации по отношению к FIXa и FX (непатентный документ 6). Более того, известно, что антительная функциональная замещающая активность кофактора FVIII определяется изотипом IgG, паттерном дисульфидной связи, аминокислотной последовательностью шарнирного участка и наличием или отсутствием сахарных цепей в Fc-области (непатентный документ 7). Было показано, что АСЕ910 (Эмицизумаб), имеющий высокую функциональную замещающую активность кофактора FVIII, который представляет собой одно из указанных антител, обладает гемостатическими эффектами на модельных обезьянах с гемофилией (непатентные документы 8 и 9). Также клинические исследования АСЕ910 (Эмицизумаб) на здоровых субъектах обнаружили наличие у него превосходной фармакокинетики (времени полужизни) и переносимости (непатентный документ 10), а в клинических исследованиях на пациентах, больных гемофилией А, в организме которых как образуются, так и не образуются ингибиторы, было показано, что введение АСЕ910 (Эмицизумаб) значимо редуцирует интенсивность кровотечения по сравнению с тем, что наблюдалось до введения этого препарата (непатентный документ 11).
Как указано выше, в клинических исследованиях АСЕ910 (Эмицизумаб) наблюдался эффект снижения интенсивности кровотечения. Однако, в in vitro исследованиях генерации тромбина с использованием дефицитной по FVIII плазмы улучшенное влияние АСЕ910 (Эмицизумаб) на максимальное образование количества тромбина (тромбиновый пик) было ниже по сравнению с количеством тромбина, образовавшегося в присутствии нормального уровня FVIII, которое составляет 100 Ед/дл (непатентный документ 8). Таким образом, было желательным повышать эффективность соответствующих лекарственных препаратов. Дополнительно, принимая во внимание соображения комфорта для больных гемофилией А, существовала потребность в биспецифических антителах, имеющих функциональную замещающую активность кофактора F VIII, за счет одновременного повышения специфической активности как таковой.
В целом, известны случаи, когда антитело фармацевтически действует как антиген и индуцирует образование анти-антител (ААТ) (непатентный документ 12). Поскольку постоянное применение АСЕ910 (Эмицизумаб) вызывает трудности у пациентов с гемофилией, в организме которых образуются ААТ [(анти-идиотипические антитела к АСЕ910 (Эмицизумаб)], то имеется необходимость в разработке биспецифических антител, имеющих функциональную замещающую активность кофактора FVIII, которые могут вводиться таким пациентам.
АСЕ910 (Эмицизумаб) представляет собой биспецифическое антитело, которое оптимизировано по многим параметрам за счет включения многочисленных аминокислотных замен в исходное антитело hBS1. Исходное антитело hBS1 было сконструировано путем гуманизации биспецифического антитела, полученного иммунизацией животных, которое распознает FIX и/или FIXa, а также FX. АСЕ910 (Эмицизумаб) имеет высокую функциональную замещающую активность кофактора FVIII (непатентные документы 4 и 6). Однако для увеличения лекарственной эффективности и улучшения специфической активности необходимо биспецифическое антитело, замещающее функцию FVIII, которое будет иметь максимальную активность (максимальную функциональную замещающую активность кофактора FVIII) по сравнению с активностью АСЕ910 (Эмицизумаб) и сможет проявлять функциональную замещающую активность кофактора FVIII при более низких концентрациях по сравнению с АСЕ910 (Эмицизумаб). Однако, на сегодняшнее время, не существует сообщений о получении биспецифических антител, имеющих значительно более высокую функциональную замещающую активность кофактора FVIII, чем АСЕ910 (Эмицизумаб), как с точки зрения используемой концентрации, так и с точки зрения максимальной активности (патентные документы 4 и 5).
Описаны различные способы получения биспецифических антител, относящихся к классу IgG, содержащих константные участки антитела человека (антитела IgG, содержащие константные участки антитела человека, которые имеют специфическую связывающую активность антигена А на одном плече и специфическую связывающую активность антигена В на другом). В принципе, биспецифические антитела IgG состоят из двух типов Н-цепей (называемых Н-цепью для антигена А и Н-цепью для антигена В) и двух типов L-цепей (называемых L-цепью для антигена А и L-цепью для антигена В). Когда такие биспецифические антитела IgG экспрессируются, возможно образование десяти типов комбинаций H2L2, поскольку синтезируются два типа Н-цепей и два типа L-цепей. Среди всех антител существует один тип комбинации, который имеет желательную связывающую активность (IgG, имеющие специфическую активность по отношению к антигену А на одном плече и специфическую активность по отношению к антигену В на другом плече). Соответственно, помимо получения нужной биспецифической активности антитела необходимо выделить представляющий интерес тип антител из смеси десяти типов антител, что имеет достаточно низкую эффективность и чрезвычайно трудно.
Способы, которые известны для решения данной проблемы, включают предпочтительную секрецию антител IgG, имеющих гетерологичную комбинацию Н-цепи для антигена А и Н-цепи для антигена В, за счет замещения аминокислотных остатков в СН3-участке Н-цепи IgG (патентные документы 6, 7, 8 и 9 и непатентные документы 13 и 14). Среди указанных способов описаны способы, которые используют физические препятствия в форме «выпуклость» и «впадина», а также взаимное отталкивание электрических зарядов.
Кроме того, сообщалось о способе эффективного получения нужной молекулы, который использует общую L-цепь, в которой L-цепь для антигена А и L-цепь для антигена В входят в состав одной и той же аминокислотной последовательности (патентные документы 10 и 11). Однако, поскольку использование общей L-цепи способно значительно снизить аффинность связывания с антигеном, данный способ не является оптимально необходимым. Соответственно, наряду с получением биспецифического антитела, связывающего два антигена с высокой аффинностью, предпочтительно, чтобы были ассоциированы только L-цепь и Н-цепь для антигена А и только L-цепь и Н-цепь для антигена В. Более того, указанный способ позволяет Н-цепям и L-цепям для каждого антигена ассоциировать независимо от вариабельных участков, которые содержат замещения аминокислот в СН1 и CL-доменах, являющихся константными участками, вместо таковых в вариабельных участках (патентные документы 7, 12 и 13). Однако, данный способ оставляет возможность для значительного улучшения с целью эффективного получения биспецифического антитела, представляющего интерес.
Список процитированных документов
Патентные документы
Патентный документ 1 WO 2005/035754
Патентный документ 2 WO 2005/035756
Патентный документ 3 WO 2006/109592
Патентный документ 4 WO 2012/067176
Патентный документ 5 WO 2017/110980
Патентный документ 6 WO 1996/027011
Патентный документ 7 WO 2006/106905
Патентный документ 8 WO 2009/089004
Патентный документ 9 WO 2010/129304
Патентный документ 10 WO 1998/050431
Патентный документ 11 WO 2006/109592
Патентный документ 12 WO 2007/147901
Патентный документ 13 WO 2013/065708
Непатентные документы
Непатентный документ 1 Blood 58,1-13 (1981)
Непатентный документ 2 Nature 312, 330-337 (1984)
Непатентный документ 3 Nature 312, 337-342 (1984)
Непатентный документ 4 Biochim. Biophys. Acta 871, 268-278 (1986)
Непатентный документ 5 Nat. Med. 2012 Oct; 18 (10):1570-4.
Непатентный документ 6 PLoS One. 2013; 8 (2): e57479.
Непатентный документ 7 MAbs. 2015;7 (1): 120-8.
Непатентный документ 8 J Thromb Haemost. 2014 Feb; 12 (2): 206-213.
Непатентный документ 9 Blood. 2014 Nov 13; 124 (20):3165-71.
Непатентный документ 10 Blood. 2016, Vol. 127, 13
Непатентный документ 11 New Eng J Med 2016, 374; 21, 2044-2053
Непатентный документ 12 Self/Nonself Volume 1, 2010- Issue 4
Непатентный документ 13 Protein Engineering. 1996, Vol. 9:617-621
Непатентный документ 14 Nature Biotechnology. 1998, Vol. 16:677-681
Краткое описание изобретения
Проблемы, решаемые настоящим изобретением
Настоящее изобретение было создано с учетом следующих аспектов. Задачей настоящего изобретения является разработка мультиспецифических антигенсвязывающих молекул, имеющих функциональную замещающую активность кофактора коагулирующего фактора крови VIII (FVIII), а также фармацевтических композиций, содержащих такие молекулы в качестве активного ингредиента.
Другой задачей настоящего изобретения является создание антител с регулируемой ассоциацией тяжелой и легкой цепей, способов получения указанных антител и способов регулирования ассоциации между тяжелой и легкой цепями в указанных антителах.
Способы решения указанных выше задач
Для создания антител с высокой специфической активностью и максимальной функциональной замещающей активностью кофактора FVIII авторы настоящего изобретения получили из библиотеки антител человека новые последовательности легких цепей, имеющие последовательности, отличные от таковых АСЕ910 (Эмицизумаб), которые имеют функциональную замещающую активность кофактора FVIII, и сконструировали биспецифические антитела, в которых аминокислотные замены были введены в различные сайты легких цепей и тяжелых цепей АСЕ910 (Эмицизумаб). Затем авторы обнаружили, что у данных антител активирующая-ингибирующая активность коагулирующего фактора крови IX (FIX) увеличивается с увеличением функциональной замещающей активности кофактора FVIII.
В результате объявленного тестирования было показано, что авторы настоящего изобретения преуспели в создании биспецифических антител, чья активирующая-ингибирующая активность FIX не увеличивается, а функциональная замещающая активность кофактора FVIII увеличивается.
Более того, авторы настоящего изобретения отобрали константный участок СН1 тяжелой цепи и константный участок CL легкой цепи в качестве участков тяжелой и легкой цепей, ассоциация между которыми подлежит регулированию, и провели объявленные исследования по регулированию ассоциации между СН1 и CL. В результате авторы с успехом обнаружили, что нежелательная ассоциация между СН1 и CL может быть супрессирована путем замещений специфических аминокислотных остатков, находящихся в промежуточном пространстве (интерфейсе) между СН1 и CL, на аминокислотные остатки, которые электростатистически отталкивают друг друга, и гетерогенные молекулы образуются более эффективно чем в случае использования описанных выше модификаций, которые подразумевают только введение выпуклости и впадины в СН3.
Настоящее изобретение было создано на основе таких находок и специфически обеспечивает следующие указанные ниже аспекты от [1] до [25]: [1] мультиспецифическая антигенсвязывающая молекула, обладающая способностью замещать функцию коагулирующего фактора крови VIII, содержащая первый антигенсвязывающий участок, который связывает коагулирующий фактор крови IX и/или активированный коагулирующий фактор крови IX, и второй антигенсвязывающий участок, который связывает коагулирующий фактор крови X,
где первый антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи (Q499) включает HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 1, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 2, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3, и вариабельный домен легкой цепи (QNK131) включает HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 162, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 163, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 164, и где второй антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи (J327) включает HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 4, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 5, и HVR-H3, содержащий аминокислотную последовательность S
EQ ID NO: 6, и вариабельный домен легкой цепи (JNL095) включает HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 165, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 166, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 167; причем один или более аминокислотных остатков замещены другими аминокислотными остатками, делетированы или встроены по крайней мере в одном из участков HVRs;
[2] мультиспецифическая антигенсвязывающая молекула [1], в которой
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 97, 98, 100, 100а, 100b и 100е в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене тяжелой цепи первого антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 53, 55, 92, 93 и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или встроен в вариабельном домене легкой цепи первого антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 51, 56, 57, 59, 61, 62, 65 и 102 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком в вариабельном домене тяжелой цепи второго антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 24, 26, 27, 29, 30, 31, 32, 50, 92, 94, 95, 95а и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене легкой цепи второго антигенсвязывающего участка;
[3] мультиспецифическая антигенсвязывающая молекула [1] или [2], в которой:
в вариабельном домене тяжелой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин или аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 26 представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене тяжелой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин или аминокислотный остаток в положении 102 представляет собой валин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин, где указанные положения соответствуют нумерации по Кабату;
[4] любая мультиспецифическая антигенсвязывающая молекула от [1] до [3], где первый антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 168, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 169, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 170 (QH01);
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 171, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 172, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 173 (QH02);
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 174, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 175, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 176 (QH03);
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 177, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 178, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 179 (QH04);
5) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 180, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 181, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 182 (QH06); или
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 183, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 184, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 185 (QH07); и
вариабельный домен легкой цепи включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 186, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 187, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 188 (QL21);
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 189, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 190, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 191 (QL22);
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 192, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 193, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 194 (QL23);
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 195, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 196, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 197 (QL24);
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 198, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 199, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 200 (QL25);
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 201, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 202, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 203 (QL26);
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 204, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 205, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 206 (QL28);
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 207, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 208, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 209 (QL29);
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 210, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 211, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 212 (QL30);
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 213, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 214, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 215 (QL31);
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 216, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 217, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 218 (QL32);
12) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 219, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 220, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 221 (QL33); и
где второй антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 222, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 223, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 224 (JH01);
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 225, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 226, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 227 (JH02);
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 228, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 229, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 230 (JH03);
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 231, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 232, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 233 (JH04);
5) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 234, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 235, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 236 (JH05);
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 237, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 238, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 239 (JH06);
7) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 240, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 241, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 242 (JH07);
8) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 243, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 244, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 245 (JH08);
9) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 246, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 247, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 248 (JH09);
10) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 249, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 250, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 251 (JH10);
11) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 252, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 253, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 254 (JH11); и
вариабельный домен легкой цепи включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 255, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 256, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 257 (JL01);
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 258, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 259, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 260 (JL02);
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 261, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 262, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 263 (JL03);
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 264, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 265, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 266 (JL04);
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 267, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 268, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 269 (JL05);
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 270, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 271, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 272 (JL06);
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 273, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 274, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 275 (JL07);
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 276, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 277, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 278 (JL08);
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 279, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 280, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 281 (JL09);
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 282, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 283, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 284 (JL10); или
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 285, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 286, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 287 (JL11);
[5] мультиспецифическая антигенсвязывающая молекула [1], в которой первый антигенсвязывающий участок содержит вариабельный домен тяжелой цепи SEQ ID NO: 45 (Q499) и вариабельный домен легкой цепи SEQ ID NO: 13 (QNK131), и второй антигенсвязывающий участок содержит вариабельный домен тяжелой цепи SEQ ID NO: 46 (J327) и вариабельный домен легкой цепи SEQ ID NO: 31 (JNL095),
причем один или более аминокислотных остатков замещены другими аминокислотами, делетированы или встроены по крайней мере в одном из вариабельных доменов тяжелой или легкой цепей;
[6] мультиспецифическая антигенсвязывающая молекула [5], в которой
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 39, 97, 98, 100, 100а, 100b и 100е в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене тяжелой цепи первого антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 38, 45, 53, 55, 60, 70, 76, 79, 80, 83, 85, 92, 93, 95 и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или встроен в вариабельном домене легкой цепи первого антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 28, 31, 39, 51, 56, 57, 59, 61, 62, 65, 67, 73, 82b и 102 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком в вариабельном домене тяжелой цепи второго антигенсвязывающего участка и
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 3, 8, 15, 24, 26, 27, 29, 30, 31, 32, 38, 48, 49, 50, 79, 92, 94, 95, 95а и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене легкой цепи второго антигенсвязывающего участка; [7] мультиспецифическая антигенсвязывающая молекула [5] или [6], в которой:
в вариабельном домене тяжелой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 39 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин или аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 26 представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой лизин, аминокислотный остаток в положении 45 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 60 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 70 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 76 представляет собой аспарагин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 80 представляет собой пролин или аланин, аминокислотный остаток в положении 83 представляет собой метионин или аланин, аминокислотный остаток в положении 85 представляет собой треонин, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене тяжелой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 28 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 39 представляет собой лизин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин, аминокислотный остаток в положении 67 представляет собой лейцин, аминокислотный остаток в положении 73 представляет собой изолейцин, аминокислотный остаток в положении 82b представляет собой глутаминовую кислоту или аминокислотный остаток в положении 102 представляет собой валин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 3 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 8 представляет собой пролин, аминокислотный остаток в положении 15 представляет собой лейцин, аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 3 1 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 48 представляет собой изолейцин, аминокислотный остаток в положении 49 представляет собой тирозин, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин, где указанные положения соответствуют нумерации по Кабату;
[8] любая мультиспецифическая антигенсвязывающая молекула от [1] до [7] в которой первый антигенсвязывающий участок содержит: вариабельный домен тяжелой цепи (QH), содержащий аминокислотную последовательность SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59 или SEQ ID NO: 60, и
вариабельный домен легкой цепи (QL), содержащий аминокислотную последовательность SEQ ID NO: 61 SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72; и
второй антигенсвязывающий участок содержит вариабельный домен тяжелой цепи (JH), содержащий аминокислотную последовательность SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82, или SEQ ID NO: 83, и
вариабельный домен легкой цепи (JL), содержащий аминокислотную последовательность SEQ ID NO: 84 SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID N091, SEQ ID NO: 92, SEQ ID NO: 93 или SEQ ID NO: 94;
[9] любая мультиспецифическая антигенсвязывающая молекула от [1] до [8] в которой первый антигенсвязывающий участок содержит константные домены, содержащие аминокислотные последовательности (1) и (2), указанные ниже, и второй антигенсвязывающий участок содержит константные домены, содержащие аминокислотные последовательности (1) и (2), указанные ниже, которые отличаются от константных участков первого антигенсвязывающего участка:
(1) SEQ ID NO: 119 относится к константному участку тяжелой цепи и SEQ ID NO: 100 относится к константному участку легкой цепи;
(2) SEQ ID NO: 118 относится к константному участку тяжелой цепи и SEQ ID NO: 102 относится к константному участку легкой цепи;
[10] любая мультиспецифическая антигенсвязывающая молекула от [1] до [9], которая представляет собой мультиспецифическое антитело или биспецифическое антитело;
[11] биспецифическое антитело, содержащее тяжелую цепь первого антитела и легкую цепь первого антитела, которые связываются с коагулирующим фактором крови IX и/или активированным коагулирующим фактором крови IX, и тяжелую цепь второго антитела и легкую цепь второго антитела, которые связываются с коагулирующим фактором крови X, причем биспецифическое антитело представляет собой любое из антител от (а) до (v), как указано ниже:
(а) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 120, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 126, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 138, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 149 (QH01/QL21/JH01/JL01);
(б) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 138, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 149 (QH02/QL22//JH01/JL01);
(в) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 122, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 128, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150 (QH03/QL23//JH02/JL02);
(г) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 122, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 129, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150 (QH03/QL24//JH02/JL02);
(д) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 140, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 151 (QH02/QL22//JH03/JL03);
(е) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 141, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 152 (QH02/QL22//JH04/JL04);
(ж) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150 (QH02/QL22//JH02/JL02);
(з) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 130, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150 (QH04/QL25//JH02/JL02);
(и) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 131, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150 (QH04/QL26//JH02/JL02);
(к) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 131, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153 (QH04/QL26//JH05/JL05);
(л) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 132, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153 (QH04/QL28//JH05/JL05);
(м) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 132, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 143, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 154 (QH04/QL28//JH06/JL06);
(н) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 133, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153 (QH04/QL29//JH05/JL05);
(о) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 133, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 143, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 154 (QH04/QL29//JH06/JL06);
(п) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 134, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 144, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 155 (QH06/QL30//JH07/JL07);
(р) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 135, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 145, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 156 (QH04/QL31//JH08/JL08);
(с) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 136, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 144, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 155 (QH06/QL32//JH07/JL07);
(т) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 136, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 146, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 157 (QH06/QL32//JH09/JL09);
(у) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 134, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 147, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 158 (QH06/QL30//JH10/JL10);
(ф) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 125, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 137, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 148, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 159 (QH07/QL33//JH11/JL11);
(х) биспецифическое антитело, которое связывается с эпитопами, идентичными эпитопам коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопу коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (ф);
(ц) биспецифическое антитело, которое конкурирует за связывание с эпитопами коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопом коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (ф);
[12] антигенсвязывающая молекула, в которой регулируется ассоциация между тяжелой цепью и легкой цепью, причем один, или два, или более наборов аминокислотных остатков, выбранных из группы, содержащей наборы от (а) до (в), как показано ниже, в тяжелой цепи и в легкой цепи указанной антигенсвязывающей молекулы представляют собой аминокислотные остатки, которые электростатически отталкиваются друг от друга:
(а) аминокислотный остаток в константном домене тяжелой цепи (СН1) в положении 175 согласно нумерации EU и аминокислотный остаток в константном домене легкой цепи (CL) в положении 180 согласно нумерации по Кабату;
(б) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 131 согласно нумерации по Кабату;
(в) аминокислотные остатки в СН1 в положениях 147 и 175 согласно нумерации EU и аминокислотные остатки в CL в положениях 131 и 180 согласно нумерации по Кабату;
[13] антигенсвязывающая молекула [12] в которой также два или более двух аминокислотных остатков, которые формируют интерфейс между вариабельным доменом тяжелой цепи и вариабельным доменом легкой цепи, представляют собой аминокислотные остатки, которые электростатически отталкиваются друг от друга;
[14] антигенсвязывающая молекула [13], в которой аминокислотные остатки, которые электростатически отталкиваются друг от друга, представляют собой один или два набора аминокислотных остатков, выбранных из группы, содержащей наборы аминокислотных остатков от (а) до (б), как показано ниже:
(а) аминокислотный остаток в вариабельном домене тяжелой цепи в положении 39 согласно нумерации по Кабату и аминокислотный остаток в вариабельном домене легкой цепи в положении 38 согласно нумерации по Кабату;
(б) аминокислотный остаток в вариабельном домене тяжелой цепи в положении 45 согласно нумерации по Кабату и аминокислотный остаток в вариабельном домене легкой цепи в положении 44 согласно нумерации по Кабату;
[15] любая антигенсвязывающая молекула от [12] до [14], в которой аминокислотные остатки, которые электростатически отталкиваются друг от друга, выбраны из аминокислотных остатков, включенных в наборы (X) или (Y), как показано ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D);
(Y) лизин (К), аргинин (R) и гистидин (Н);
[16] любая антигенсвязывающая молекула от [12] до [15], представляющая собой биспецифическое антитело;
[17] способ получения антигенсвязывающей молекулы, в которой регулируется ассоциация между тяжелой цепью и легкой цепью, включающий осуществление стадий от (1) до (3), приведенных ниже:
(1) модификация нуклеиновой кислоты (кислот), кодирующей константный домен тяжелой цепи (СН1) и константный домен легкой цепи (CL) таким образом, что один набор, или два, или более наборов аминокислотных остатков, выбранных из группы, содержащей наборы аминокислотных остатков от (а) до (в), как показано ниже, электростатически отталкиваются друг от друга:
(а) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 180 согласно нумерации по Кабату;
(б) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 131 согласно нумерации по Кабату;
(в) аминокислотные остатки в СН1 в положениях 147 и 175 согласно нумерации EU и аминокислотные остатки в CL в положениях 131 и 180 согласно нумерации по Кабату;
(2) введение модифицированной нуклеиновой кислоты (кислот) в клетку-хозяин и культивирование клетки-хозяина в условиях, обеспечивающих экспрессию указанной нуклеиновой кислоты (кислот);
(3) выделение антигенсвязывающей молекулы из культуры клеток-хозяев;
[18] способ регулирования ассоциации между тяжелой цепью и легкой цепью в антигенсвязывающей молекуле, включающий модификацию нуклеиновой кислоты таким образом, что один набор, или два, или более наборов аминокислотных остатков, выбранных из группы, содержащей наборы аминокислотных остатков от (а) до (с), как показано ниже, электростатически отталкиваются друг от друга:
(а) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 180 согласно нумерации по Кабату;
(б) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 131 согласно нумерации по Кабату;
(в) аминокислотные остатки в СН1 в положениях 147 и 175 согласно нумерации EU и аминокислотные остатки в CL в положениях 131 и 180 согласно нумерации по Кабату;
[19] изолированная нуклеиновая кислота, кодирующая любую мультиспецифическую антигенсвязывающую молекулу от [1] до [9], мультиспецифическое антитело [10], биспецифическое антитело [10], [11] или [16], любую антигенсвязывающую молекулу от [12] до [15];
[20] клетка-хозяин, содержащая нуклеиновую кислоту [19];
[21] способ получения мультиспецифической антигенсвязывающей молекулы, мультиспецифического антитела, биспецифического антитела или антигенсвязывающей молекулы, включающий культивирование клетки-хозяина [20] в условиях, обеспечивающих продуцирование мультиспецифической антигенсвязывающей молекулы, мультиспецифического антитела, биспецифического антитела или антигенсвязывающей молекулы;
[22] фармацевтическая композиция, содержащая любую мультиспецифическую антигенсвязывающую молекулу от [1] до [9], мультиспецифическое антитело [10], биспецифическое антитело [10], [11] или [16] или любую антигенсвязывающую молекулу от [12] до [15] и фармацевтически приемлемый носитель;
[23] фармацевтическая композиция [22], предназначенная для применения для профилактики и/или лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением;
[24] фармацевтическая композиция [23], где кровотечение, заболевание, в которое вовлечено кровотечение, или заболевание, вызываемое кровотечением, представляет собой заболевание, развитие и/или прогрессивное течение которого обусловлено уменьшением или дефицитом активности коагулирующего фактора крови VIII и/или активированного коагулирующего фактора крови VIII;
[25] фармацевтическая композиция [24], где заболевание, развитие и/или прогрессивное течение которого обусловлено уменьшением или дефицитом активности коагулирующего фактора крови VIII и/или активированного коагулирующего фактора крови VIII, представляет собой гемофилию А, заболевание, при котором непредвиденно образуется ингибитор коагулирующего фактора крови VIII и/или активированного коагулирующего фактора крови VIII, приобретенную гемофилию или болезнь Виллебранда;
Также настоящее изобретение относится к следующим объектам:
[26] любая мультиспецифическая антигенсвязывающаямолекула от [1] до [9], мультиспецифическое антитело [10], биспецифическое антитело [10], [11] или [16] или любая антигенсвязывающая молекула от [12] до [15], которые используются в качестве медикамента;
[27] любая мультиспецифическая антигенсвязывающая молекула от [1] до [9], мультиспецифическое антитело [10] или биспецифическое антитело [10] или [11], которые используются для профилактики и/или лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением;
[28] любая мультиспецифическая антигенсвязывающая молекула от [1] до [9], мультиспецифическое антитело [10] или биспецифическое антитело [10] или [11], которые используются для замещения функциональной активности кофактора FVIII;
[29] применение любой мультиспецифической антигенсвязывающей молекулы от [1] до [9], мультиспецифического антитела [10] или биспецифического антитела [10] или [11] в производстве медикамента для лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением;
[30] применение любой мультиспецифической антигенсвязывающей молекулы от [1] до [9], мультиспецифического антитела [10] или биспецифического антитела [10] или [11] в производстве медикамента для замещения функциональной активности кофактора FVIII;
[31] способ лечения индивидуума, страдающего от кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением, включающий применение по отношению к индивидууму эффективного количества любой мультиспецифической антигенсвязывающей молекулы от [1] до [9], мультиспецифического антитела [10] или биспецифического антитела [10] или [11] и
[32] способ замещения функциональной активности кофактора FVIII у индивидуума, включающий применение по отношению к индивидууму эффективного количества любой мультиспецифической антигенсвязывающей молекулы от [1] до [9], мультиспецифического антитела [10] или биспецифического антитела [10] или [11] для замещения функциональной активности кофактора FVIII.
Также настоящее изобретение относится к следующим объектам:
[33] антигенсвязывающая молекула, полученная способом [17], который включает на стадии (1) модификацию нуклеиновой кислоты таким образом, что аминокислотные остатки, которые электростатически отталкиваются друг от друга, выбираются из аминокислотных остатков, включенных в наборы (X) или (Y), как показано ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D);
(Y) лизин (К), аргинин (R) и гистидин (Н);
[34] антигенсвязывающая молекула, полученная способом [17] или [33], который также включает на стадии (1) модификацию нуклеиновой кислоты таким образом, что два или более наборов аминокислотных остатков, которые образуют интерфейс между вариабельным доменом тяжелой цепи и вариабельным доменом легкой цепи, представляют собой аминокислотные остатки, которые электростатически отталкиваются друг от друга;
[35] антигенсвязывающая молекула, полученная способом [34], в которой аминокислотные остатки, которые электростатически отталкиваются друг от друга, представляют собой один любой набор аминокислотных остатков, выбранных из группы, содержащей наборы аминокислотных остатков от (а) до (б), как показано ниже:
(а) аминокислотный остаток в вариабельном домене тяжелой цепи в положении 39 согласно нумерации по Кабату и аминокислотный остаток в вариабельном домене легкой цепи в положении 38 согласно нумерации по Кабату;
(б) аминокислотный остаток в вариабельном домене тяжелой цепи в положении 45 согласно нумерации по Кабату и аминокислотный остаток в вариабельном домене легкой цепи в положении 44 согласно нумерации по Кабату;
[36] антигенсвязывающая молекула, полученная способом [34] или [35], в которой аминокислотные остатки, которые электростатически отталкиваются друг от друга, выбраны из аминокислотных остатков, включенных в наборы (X) или (Y), как показано ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D);
(Y) лизин (К), аргинин (R) и гистидин (Н);
[37] способ [18], где аминокислотные остатки, которые электростатически отталкиваются друг от друга, выбраны из аминокислотных остатков, включенных в наборы (X) или (Y), как показано ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D);
(Y) лизин (К), аргинин (R) и гистидин (Н);
[38] способ [18] или [37], где также два или более аминокислотных остатков, которые образуют интерфейс между вариабельным доменом тяжелой цепи и вариабельным доменом легкой цепи, представляют собой аминокислотные остатки, которые электростатически отталкиваются друг от друга;
[39] способ [38], где аминокислотные остатки, которые электростатически отталкиваются друг от друга, представляют собой один любой набор аминокислотных остатков, выбранных из группы, содержащей наборы аминокислотных остатков от (а) до (b), как показано ниже:
(a) аминокислотный остаток в вариабельном домене тяжелой цепи в положении 39 согласно нумерации по Кабату и аминокислотный остаток в вариабельном домене легкой цепи в положении 38 согласно нумерации по Кабату;
(b) аминокислотный остаток в вариабельном домене тяжелой цепи в положении 45 согласно нумерации по Кабату и аминокислотный остаток в вариабельном домене легкой цепи в положении 44 согласно нумерации по Кабату; и
[40] способ [38] или [39], где аминокислотные остатки, которые электростатически отталкиваются друг от друга, выбраны из аминокислотных остатков, включенных в наборы (X) или (Y), как показано ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D);
(Y) лизин (К), аргинин (R) и гистидин (Н).
Настоящее изобретение также относится к следующим объектам:
[41] любая мультиспецифическая антигенсвязывающая молекула от [1] до [7], которая представляет собой любую из молекул от (а) до (ц), как указано ниже:
(а) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 56, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 61, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84 (QH01/QL21//JH01/JL01);
(б) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84 (QH02/QL22//JH01/JL01);
(в) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 63, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH03/QL23//JH02/JL02);
(г) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 64, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID N0:74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH03/QL24//JH02/JL02);
(д) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 75, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 86 (QH02/QL22//JH03/JL03);
(е) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 76, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 87 (QH02/QL22//JH04/JL04);
(ж) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH02/QL22//JH02/JL02);
(з) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 65, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH04/QL25//JH02/JL02);
(и) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID N0:74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH04/QL26//JH02/JL02);
(к) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88 (QH04/QL26//JH05/JL05);
(л) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88 (QH04/QL28//JH05/JL05);
(м) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89 (QH04/QL28//JH06/JL06);
(н) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88 (QH04/QL29//JH05/JL05);
(о) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89 (QH04/QL29//JH06/JL06);
(п) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90 (QH06/QL30//JH07/JL07);
(р) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 70, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 80, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 91 (QH04/QL31//JH08/JL08);
(с) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 71, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90 (QH06/QL32//JH07/JL07);
(т) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 71, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 92 (QH06/QL32//JH09/JL09);
(у) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 82, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 93 (QH06/QL30//JH10/JL10);
(ф) мультиспецифическая антигенсвязывающая молекула, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 105, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 72, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 83, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 94 (QH07/QL33//JH11/JL11);
(х) мультиспецифическая антигенсвязывающая молекула, которая связывается с эпитопами, идентичными эпитопам коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопу коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (Ф);
(ц) мультиспецифическая антигенсвязывающая молекула, которая конкурирует за связывание с эпитопами коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопом коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (ф); и
[42] биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела и вариабельный домен легкой цепи первого антитела, которые связываются с коагулирующим фактором крови IX и/или активированным коагулирующим фактором крови IX, и вариабельный домен тяжелой цепи второго антитела и вариабельный домен легкой цепи второго антитела, которые связываются с коагулирующим фактором крови X, причем биспецифическое антитело представляет собой любое из антител от (а) до (ц), как указано ниже:
(а) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 56, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 61, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84 (QH01/QL21//JH01/JL01);
(б) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84 (QH02/QL22//JH01/JL01);
(в) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 63, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH03/QL23//JH02/JL02);
(г) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 64, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH03/QL24//JH02/JL02);
(д) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 75, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 86 (QH02/QL22//JH03/JL03);
(е) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 76, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 87 (QH02/QL22//JH04/JL04);
(ж) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH02/QL22//JH02/JL02);
(з) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 65, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH04/QL25//JH02/JL02);
(и) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85 (QH04/QL26//JH02/JL02);
(к) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88 (QH04/QL26//JH05/JL05);
(л) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88 (QH04/QL28//JH05/JL05);
(м) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89 (QH04/QL28//JH06/JL06);
(н) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88 (QH04/QL29//JH05/JL05);
(о) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89 (QH04/QL29//JH06/JL06);
(п) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90 (QH06/QL30//JH07/JL07);
(р) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 70, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 80, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 91 (QH04/QL31//JH08/JL08);
(с) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 71, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90 (QH06/QL32//JH07/JL07);
(т) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO:60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO:71, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 81, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO:92 (QH06/QL32//JH09/JL09);
(у) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO:60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO:69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 82, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO:93 (QH06/QL30//JH10/JL10);
(ф) биспецифическое антитело, содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO:105, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO:72, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 83, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO:94 (QH07/QL33//JH11/JL11);
(х) биспецифическое антитело, которое связывается с эпитопами, идентичными эпитопам коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопу коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (t); (ц) биспецифическое антитело, которое конкурирует за связывание с эпитопами коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопом коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (t).
Краткое описание фигур
На Фиг. 1. показано взаимодействие антитела АСЕ910 (Эмицизумаб) с идиотипическим антителом к биспецифическому антителу, имеющему новые L-цепи.
На Фиг. 2 показан эффект, который оказывает активность, ингибирующая активацию FIX, на активацию FX.
На Фиг. 3 приведены графические диаграммы, иллюстрирующие взаимодействие активности, ингибирующей активацию FIX, с функционально замещающей активностью кофактора FVIII, что обусловлено новыми модификациями L-цепей и новыми модификациями Н-цепей.
На Фиг. 4-1 приведена концентрационная зависимость функционально замещающей активности кофактора FVIII, свойственной биспецифическому антителу, имеющему комбинации модификаций L-цепей и модификаций Н-цепей.
На Фиг. 4-2 приведена концентрационная зависимость функционально замещающей активности кофактора FVIII, свойственной биспецифическому антителу, имеющему комбинации модификаций L-цепей и модификаций Н-цепей.
На Фиг. 5 показано максимальные количества образования тромбина (длины пиков) под влиянием биспецифического антитела, имеющего комбинации модификаций L-цепей и модификаций Н-цепей, в анализе образования тромбина.
На Фиг. 6 показано ЕСМ-связывание для полученных биспецифических антител.
На Фиг. 7 показана РК антитела для полученных биспецифических антител.
На Фиг. 8 приведены типы взаимных пар доменов легких и тяжелых цепей, которые могут быть экспрессированы.
На Фиг. 9-1 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-2 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-3 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-4 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-5 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-6 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-7 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-8 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-9 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-10 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-11 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-12 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-13 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-14 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-15 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-16 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-17 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-18 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-19 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 9-20 приведены данные CIEX анализа для полученного биспецифического антитела.
На Фиг. 10 приведены результаты измерения функциональной замещающей активности кофактора FVIII для биспецифических антител, имеющих новые L-цепи.
Способ осуществления изобретения
I. Определения
"Акцепторный каркасный участок человека" для целей настоящего изобретения означает каркасный участок, содержащий аминокислотную последовательность вариабельного домена легкой цепи (VL) или аминокислотную последовательность вариабельного домена тяжелой цепи (VH), происходящие из каркасного участка иммуноглобулина человека или консенсусного каркасного участка человека, как определено ниже. Акцепторный каркасный участок человека «происходящий из» каркасного участка иммуноглобулина человека или консенсусного каркасного участка человека, может включать ту же самую аминокислотную последовательность или может содержать изменения в аминокислотной последовательности. В некоторых случаях число аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В определенных воплощениях изобретения последовательность VL акцепторного каркасного участка человека идентична последовательности VL каркасного участка иммуноглобулина человека или последовательности консенсусного каркасного участка человека.
"Аффинность" относится к величине тотального суммарного нековалентного взаимодействия между единичным антигенсвязывающим участком молекулы (например, антитела) и соответствующим партнером по связыванию (например, антигеном). Если не указано иное, как здесь используется, «связывающая аффинность» означает внутреннюю связывающую активность, отражающую взаимодействие между партнерами связывающейся пары 1:1 (например, между антителом и антигеном). Аффинность молекулы X по отношению к своему связывающему партнеру Y, в основном, определяется константой диссоциации. Аффинность может быть измерена общими методами, известными из уровня техники, включая описанные здесь. Специфические иллюстративные примеры осуществления измерения связывающей активности описаны ниже.
"Аффинно-зрелое" антитело относится к антителу с одним или более изменениями в одном или более гипервариабельном участке (HVRs) по сравнению с родительским антителом, которое не имеет таких изменений, причем эти изменения приводят к улучшению аффинности антитела по отношению к антигену.
Мультиспецифические антигенсвязывающие молекулы, описанные здесь, включают первый антигенсвязывающий участок и второй антигенсвязывающий участок, которые могут специфически связываться по крайней мере с двумя различными типами антигенов.
В одном варианте осуществления изобретения требуется, чтобы первый антигенсвязывающий участок и второй антигенсвязывающий участок имели способность к связыванию с коагулирующим фактором крови IX (FIX) и/или активированным коагулирующим фактором крови IX (FIXa) и коагулирующим фактором крови X (FX), соответственно. Примеры включают участки, необходимые для связывания с антигенами, такие как антитела, скаффолд молекулы (молекулы, подобные антителам), или пептиды, или фрагменты, содержащие такие участки. Скаффолд молекулы представляют собой молекулы, которые обнаруживают функцию связывания с молекулами-мишенями, и в их качестве могут использоваться какие-либо полипептиды, если только они являются конформационно стабильными полипептидами, способными связываться по крайней мере с одним антигеном-мишенью. Примеры таких полипептидов включают вариабельные домены антител, фибронектин (WO 2002/032925), домен белка A (WO 1995/002937), LDL-рецептор домена белка А (WO 2004/044011, WO 2005/040229), анкирин ((WO 2002/020565) и т.д., а также молекулы, описанные в документах Nygren et al. (Current Opinion in Structural Biology, 7:463-469 (1997) и Journal of Immunol Methods, 290:3-28 (2004)), Binz et al. (Nature Biotech 23:1257-1266 (2005) и Hosse et al. (Protein Science 15:14-27 (2006). Более того, как указано в Curr Opin Mol Ther. 2010 Aug; 12(4):487-95 и Drugs. 2008; 68(7):901-12, могут быть использованы пептидные молекулы, способные связываться с антигенами-мишенями.
Под полипептидами в описании настоящего изобретения, в основном, понимаются белки и пептиды, имеющие в длину примерно десять аминокислотных остатков или более. Полипептиды обычно происходят из живых организмов, но этим их происхождение не ограничивается, и, например, они могут состоять из искусственно созданных последовательностей. Полипептиды могут являться природными полипептидами, синтетическими полипептидами, рекомбинантными полипептидами и т.п. Дополнительно, фрагменты описанных выше полипептидов также входят в состав полипептидов по настоящему изобретению.
В одном воплощении осуществления изобретения примеры мультиспецифических антигенсвязывающих молекул включают антитела, которые могут специфически связываться по крайней мере с двумя различными антигенами или двумя различными эпитопами одного и того же антигена.
В еще одном воплощении осуществления изобретения мультиспецифические антитела по настоящему изобретению представляют собой биспецифические антитела (BsAbs) (их также называют антителами с двойной специфичностью).
В настоящем описании термины «функциональная замещающая активность кофактора FVIII», «функциональная замещающая активность FVIII» и «активность по замещению функции FVIII» используются как синонимы и относятся к активности связывания FIX, и/или FIXa, и FX для инициации активации FX (инициация образования активированного коагулирующего фактора крови X (FXa)).
В одном варианте осуществления изобретения мультиспецифические антигенсвязывающие молекулы, обладающие функциональной замещающей активностью кофактора FVIII, представляют собой мультиспецифические антигенсвязывающие молекулы, которые связывают FIX, и/или FIXa, и FX.
В некоторых вариантах осуществления изобретения мультиспецифические антигенсвязывающие молекулы, обладающие функциональной замещающей активностью кофактора FVIII, представляют собой мультиспецифические антитела, которые связывают FIX, и/или FIXa, и FX. В другом специфическом варианте осуществления изобретения мультиспецифические антитела, которые связывают FIX, и/или FIXa, и FX, представляют собой биспецифические антитела, которые связывают FIX, и/или FIXa, и FX.
Термин «биспецифические антитела, которые связывают FIX, и/или FIXa, и FX», относится к биспецифическим антителам, которые связывают FIX, и/или FIXa, и FX с достаточной аффинностью, такой, что антитела полезны для использования в качестве диагностических и/или терапевтических агентов в нацеливании на FIX, и/или FIXa, и FX. В одном варианте осуществления изобретения величина связывания биспецифического антитела, связывающего FIX, и/или FIXa, и FX, с неродственным белком, а именно с не FIX-белком, и/или не FIXa-белком, или не FX-белком менее 10% измеренной величины связывания данного антитела с FIX, и/или FIXa, и FX (например, как определяется радио-иммунологическим методом (РИА)). В отдельных вариантах осуществления изобретения антитело, связывающее FIX, и/или FIXa, и FX, имеет константу диссоциации (Kd) 100 мкм или менее, 10 мкм или менее, 1 мкм или менее, 100 нМ или менее, 10 нМ или менее, 1 нМ или менее, 0,1 нМ или менее, 0, 01 нМ или менее, или 0, 001 нМ или менее, или, например, 10-5 М или менее, например, от 10-5 М до 10-10 М, например, от 10-6 М до 10-10 М). В некоторых вариантах осуществления изобретения биспецифическое антитело, которое связывается с FIX, и/или FIXa, и FX, связывается с эпитопами FIX, и/или FIXa, и FX, которые являются консервативными по отношению к FIX, и/или FIXa, и FX различных видов, соответственно.
В настоящем изобретении в определенных вариантах осуществления изобретения термин «антитело» используется синонимически с термином «антигенсвязывающая молекула». В настоящем изобретении термины «антитело» и «антигенсвязывающая молекула» используются в наиболее широком смысле и относятся к моноклональным антителам, поликлональным антителам и вариантам антител [(таким, как химерные антитела, гуманизированные антитела, минитела (антитела с низкой молекулярной массой), фрагменты антител, к которым могут быть необязательно добавлены другие молекулы и мультиспецифические антитела] при условии, что они проявляют желаемую антигенсвязывающую активность или биологическую активность. Примерами антитела или антигенсвязывающей молекулы в настоящем изобретении могут служить также молекулы, в которых HAS связывающая скэффолд активность добавлена к Fab (антитело, в котором только Fab-фрагмент является нормальным). Более того, в настоящем изобретении понятие «антитело» также относится к полипептиду или гетеромерному мультимеру. Предпочтительно использование моноклональных антител, химерных антител, гуманизированных антител, антител человека, Fc-слитых антител и минител так же, как и фрагментов антител.
Как используется в настоящем описании, термин «антитело» относится к связывающему белку, содержащему антигенсвязывающий участок. Понятия «связывающий участок» и «антигенсвязывающий участок» характеризуют здесь область молекулы антитела, с которой реально связывается антиген. Термин «антигенсвязывающий участок» относится к вариабельному домену тяжелой цепи (VH) и вариабельному домену легкой цепи (VL) (паре VH/VL).
Термин «фрагмент антитела» относится к молекуле, которая не является интактным антителом, и включает участок интактного антитела, связывающий антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают без ограничений Fv, Fab, Fab', Fab' - SH, F(ab')2, диатела, линейные антитела, одноцепочечные антительные молекулы (например, scFv) и мультиспецифические антитела, созданные из фрагментов антител.
Термин «антитело, которое связывается с тем же самым эпитопом что референсное антитело», относится антителу, которое блокирует связывание референсного антигена со своим антигеном в конкурентном методе исследования на 50% или более, и, наоборот, референсное антитело блокирует связывание указанного антитела со своим антигеном в конкурентном методе исследования на 50% или более. Примеры конкурентного анализа приведкны в описании.
Термин «химерное антитело» относится к антителу, в котором участок тяжелой и/или легкой цепи происходит из определенного источника или организма, в то время как оставшийся участок антитела происходит из другого источника или организма.
Понятие «класс антитела» относится к типу константного домена или константного участка тяжелой цепи. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM и некоторые из них могут далее подразделяться на подклассы (изотипы), например, IgGl, IgG2, IgG3, IgG4, IgAl и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются альфа, дельта, эпсилон, гамма и мю, соответственно.
Термин «эффективное количество агента», например, фармацевтической композиции, относится количеству, которое эффективно в определенных дозах в необходимые временные интервалы, для обеспечения желаемого терапевтического или профилактического результата.
Термин «Fc-участок» используется здесь для обозначения С-концевой области тяжелой цепи иммуноглобулина, который содержит по меньшей мере часть константного участка. Этот термин относится к нативной последовательности Fc-участка и к вариантам Fc-участка. В одном варианте осуществления изобретения Fc-участок тяжелой цепи IgG имеет длину от Cys226 или от Pro230 до С-концевой области тяжелой цепи. Однако, С-концевой лизин (Lys447) или глицин-лизин (Gly446-Lys447) Fc-участка может присутствовать или отсутствовать. Если специально не указано иное, нумерация аминокислотных остатков Fc-участка или константного участка соответствует нумерации EU и также называется индексом EU, как описано Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
Термин «каркасный участок» или «FR» относится к аминокислотным остаткам вариабельного домена за пределами остатков гипервариабельного участка (HVR). FR вариабельного домена традиционно состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, HVR и FR последовательности обычно располагаются в VH (или VL) следующим образом: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Понятия «антитело полной длины», «интактное антитело» и «целое антитело» используются здесь взаимозаменяемо и относятся к антителу, имеющему структуру, по существу, аналогичную структуре нативного антитела или имеющему тяжелые цепи, содержащие Fc-участок, определенный в настоящем описании.
Термины «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и относятся к клеткам, в которые была введена экзогенная нуклеиновая кислота, а также к потомству указанных клеток. Клетки-хозяева могут называться «трансформантами» и «трансформированными клетками» и включают как первично трансформированные клетки, так и потомство указанных клеток, полученное из первично трансформированных клеток путем определенного количества пассажей. Потомство может содержать экзогенную нуклеиновую кислоту, не полностью идентичную нуклеиновой кислоте, содержащейся в родительских клетках, поскольку экзогенная нуклеиновая кислота потомства может включать мутации. Мутантное потомство, которое имеет ту же биологическую функцию или биологическую активность, как определено скринингом или отбором, что и первично трансформированные клетки, также включено в объем настоящего изобретения.
Термин «антитело человека», в частности, касается антитела, которое имеет аминокислотную последовательность, соответствующую таковой антитела, образующегося в организме человека, или продуцируемого клетками человека, или происходящего из источника, не имеющего отношения к человеку, но использующему репертуар последовательностей антитела человека или других последовательностей, кодирующих антитело человека. Это определение антитела человека специальным образом включает и гуманизированное антитело, содержащее антигенсвязывающие аминокислотные остатки нечеловеческого происхождения.
Термин «человеческий консенсусный каркасный участок» относится к каркасному участку, который представлен наиболее часто встречающимися аминокислотными остатками в отобранных последовательностях каркасного участка VH и VL иммуноглобулина человека. В целом, отбор последовательностей VH и VL иммуноглобулина человека происходит из подгруппы последовательностей вариабельного домена. Обычно, указанная подгруппа последовательностей представляет собой подгруппу, описанную Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3. В одном варианте осуществления изобретения подгруппа VL является подгруппой каппа I согласно Kabat et al., (см. выше). В другом варианте осуществления изобретения подгруппа для VH является подгруппой III согласно Kabat et al., (см. выше).
Термин «гуманизированное антитело» относится к химерному антителу, содержащему аминокислотные остатки из HVRs нечеловеческого происхождения и аминокислотные остатки FRs человеческого происхождения. В некоторых вариантах осуществления изобретения гуманизированное антитело может содержать по существу все из по крайней мере одного, обычно двух вариабельных доменов, в которых все или по существу все HVRs (например, CDRs) соответствуют таковым антитела нечеловеческого происхождения и все или по существу все FRs соответствуют таковым антитела человека. Гуманизированное антитело необязательно может содержать по крайней мере участок константного участка антитела, имеющего человеческое происхождение. Понятие «гуманизированная форма антитела», например, антитела нечеловеческого происхождения, относится к антителу, которое подверглось гуманизации.
Термин «гипервариабельный участок» или «HVR» используется здесь для обозначения каждого из участков вариабельного домена антитела, последовательности которых являются гипервариабельными (участки, определяющие комплементарность связывания с антигеном или CDRs) и/или которые формируют определенные петли (гипервариабельные петли) и/или содержат аминокислотные остатки, контактирующие с антигеном (антигенные контакты). В целом, антитела содержат шесть HVRs: три в VH (H1, Н2, Н3) и три в VL (L1, L2, L3). Примеры HVRs здесь включают:
(а) гипервариабельные петли, образованные аминокислотными остатками 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987));
(б) CDRs, образованные аминокислотными остатками 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991));
(в) антигенные контакты, образованные аминокислотными остатками 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum et al, J. Mol. Biol. 262:732-745 (1996)); и
(г) комбинации (a), (b) и/или (с), включая аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3.
Если не указано иное, остатки HVR и другие аминокислотные остатки вариабельного домена (например, остатки FR) пронумерованы здесь согласно Kabat et al., см. выше).
Термин «иммуноконъюгат» относится к антителу, конъюгированному с одной или более гетерологичными молекулами, в том числе, без ограничений указанным, с цитотоксическим агентом.
Понятия «индивидуум» или «субъект» относятся к млекопитающему. Млекопитающие включают, без ограничений указанным, домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, человека и приматов, не относящихся к человеку, скажем, обезьян), кроликов и грызунов (например, мышей и крыс). В некоторых случаях осуществления изобретения индивидуум или субъект представляет собой человека.
Термин «изолированное антитело» означает антитело, которое было выделено из своего природного окружения. В некоторых вариантах осуществления изобретения антитело может быть очищено со степенью более 95% или 99%, как определяется, например, методом электрофореза [в том числе, SDS-PAGE, изоэлектрофокусированием (IEF), капиллярным электрофорезом] или хроматографией (например, ионообменной или обратно-фазовой хроматографией). Обзор методов очистки антител приведен, например, Flatman et al., J. Chromatogr. 848:79-87 (2007).
Термин «изолированная нуклеиновая кислота» относится к молекуле нуклеиновой кислоты, которая была выделена из своего природного окружения. Изолированная нуклеиновая кислота включает молекулу нуклеиновой кислоты, которая содержалась в клетках, которые обычно содержат молекулу нуклеиновой кислоты, но молекула нуклеиновой кислоты присутствует в клетках экстрахромосомально или же ее локализация в хромосоме отличается от природной хромосомной локализации.
Термин «изолированная нуклеиновая кислота, кодирующая биспецифическое антитело, которое связывается с FIX, и/или FIXa и FX», относится к одной или более молекулам нуклеиновой кислоты, кодирующей тяжелые и легкие цепи антитела (или их фрагменты), включая молекулу (молекулы) нуклеиновой кислоты, содержащиеся в одном векторе или отдельных векторах, и молекулу (молекулы), имеющие однк или более локализаций в клетке-хозяине.
Термин «моноклональное антитело» здесь означает антитело, полученное из популяции по существу гомогенных антител, т.е. индивидуальные антитела, образующие популяцию, являются идентичными и/или связываются с одним и тем же эпитопом, за исключением возможных вариантов антител, например, антител, содержащих природные мутации или мутации, возникающие в процессе получения препаратов моноклональных антител, однако такие варианты обычно обнаруживаются в минимальных количествах. В противоположность препаратам поликлональных антител, которые обычно представляют собой смесь различных антител, направленных против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклональных антител направлено против одной и той же детерминанты антигена. Таким образом, определение «моноклональный» обозначает характеристику антитела, которое было получено из практически гомогенной популяции антител, и не подразумевает получение антитела каким-либо определенным методом. Например, моноклональные антитела, которые используются в соответствии с настоящим изобретением, могут быть получены с помощью различных технологий, включая, без ограничений указанным, гибридомный метод, методы рекомбинантных ДНК, фагово-дисплейные методами и методы, использующие трансгенных животных, содержащих все или части иммуноглобулиновых локусов. Такие методы и другие возможные способы получения моноклональных антител описаны в настоящей заявке.
Термин «нативные антитела» относится к природным молекулам иммуноглобулинов различной структуры. Например, нативные IgG антитела представляют собой гетеротетрамерные гликопротеины с мол. массой около 150 000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидными связями. В направлении от N- до С-концевого участка каждая тяжелая цепь содержит вариабельный участок (VH), также называемый вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которым следуют три константных домена (CH1, СН2 и СН3). Аналогичным образом, в направлении от N- до С-концевого участка каждая легкая цепь содержит вариабельный участок (VL), также называемый вариабельным легким доменом или вариабельным доменом легкой цепи, за которым следует константный домен (CL). Легкая цепь антитела может быть представлена одним из двух типов, называемых каппа (к) и лямбда (к), основанным на аминокислотной последовательности соответствующего константного домена.
Термин «вкладыш в упаковку» используется здесь по отношению к инструкциям, традиционно включаемым в коммерческие упаковки терапевтических продуктов, которые содержат информацию о сфере применения, форме выпуска, способах применения, дозах, комбинационной терапии, противопоказаниях и/или побочных эффектах используемых терапевтических продуктов.
Понятие «процент (%) идентичности аминокислотной последовательности с референсной полипептидной последовательностью» определяет процент аминокислотных остатков в тестируемой последовательности, которые идентичны аминокислотным остаткам в референсной полипептидной последовательности, что определяется после выравнивания последовательностей и введения промежутков, если необходимо, с достижением максимальной процентной идентичности последовательностей, без учета каких-либо консервативных замещений как составляющей идентичности последовательностей. Выравнивание для целей определения процента идентичности аминокислотных последовательностей может быть обеспечено различными методами, широко известными из уровня техники, например, использованием публично доступного компьютерного обеспечения, такого, как BLAST, BLAST-2, ALIGN, Megalign (DNASTAR) или GENETYX (зарегистрированный товарный знак) (Genetyx Co., Ltd.). Специалист в данной области знаний может определить подходящие параметры для выравнивания последовательностей, включая алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
Компьютерная программа для выравнивания последовательностей ALIGN-2 авторизована Genentech Inc., и код источника включен в документацию пользователя в Агентстве Авторских Прав США, Вашингтон, D.С., 20559, где программа зарегистрирована под номером TXU510087. Программа ALIGN-2 публично доступна в компании Genentech Inc., South San Francisco, California, или может быть получена посредством кода источника. Программа ALIGN-2 может быть доступна для использования в операционной системе UNIX, включая цифровую систему UNIX V4.0D. Все параметры сравнения последовательностей закреплены программой ALIGN-2 и не могут быть изменены. В случаях, когда ALIGN-2 используется для сравнения аминокислотных последовательностей, % идентичности заданной аминокислотной последовательности А по отношению, с или в сравнении с аминокислотной последовательностью В (или, альтернативно, % идентичности заданной аминокислотной последовательности В по отношению, с или в сравнении с аминокислотной последовательностью А) вычисляют следующим образом:
100-кратное увеличение фракции X/Y
где X представляет собой число аминокислотных остатков, подсчитанных в идентичных участках совпадения в программе выравнивания последовательностей ALIGN-2, когда выравниваются последовательности А и В, и Y представляет собой общее число аминокислотных остатков в последовательности В. Следует иметь в виду, что длина аминокислотной последовательности А не эквивалентна длине аминокислотной последовательности В, а % идентичности аминокислотной последовательности А по отношению к последовательности В не эквивалентен % идентичности аминокислотной последовательности В по отношению к последовательности А. Если специальным образом не указано иное, все значения % идентичности аминокислотных последовательностей, используемые здесь, получены, как описано в данном параграфе с помощью компьютерной программы ALIGN-2.
Термин «фармацевтическая композиция» относится к препарату, который находится в такой форме, которая обеспечивает эффективную биологическую активность действующего начала композиции, причем композиция не содержит дополнительных компонентов, которые являются неприемлемо токсичными для индивидуума, по отношению к которому применяется композиция.
Термин «фармацевтически приемлемый носитель» относится к ингредиенту фармацевтической композиции, который не является активным ингредиентом композиции, причем указанный носитель не токсичен для индивидуума. Фармацевтически приемлемый носитель включает, без ограничений указанным, буфер, эксципиент, стабилизатор или консервант.
Используемые здесь обозначения «FIX», «FIXa» и «FX» относятся к нативному FIX, нативному FIXa и нативному FX, соответственно, полученным из любого источника, относящегося к позвоночным животным, включая млекопитающих, например, приматов (в частности, человека) и грызунов (в частности, мышей и крыс), если не указано иное. Эти обозначения охватывают полноразмерные (полной длины) непроцессированные FIX, FIXa и FX, любые формы FIX, FIXa и FX, полученные процессингом полноразмерных факторов в клетке, а также природно существующие варианты FIX, FIXa и FX, например, варианты сплайсинга или аллельные варианты.
Используемое здесь понятие «лечение» («терапия») относится к клиническому вмешательству с целью попытки изменить естественное состояние индивидуума, подлежащего лечению, и может осуществляться как в профилактических целях, так и в процессе течения клинической патологии. Желательные эффекты лечения включают, без ограничений указанным, предотвращение первичного возникновения заболевания или рецидива заболевания, смягчение симптомов заболевания, уменьшение любых прямых или опосредованных патологических последствий заболевания, предотвращение метастаз, уменьшение скорости прогрессивного развития заболевания, улучшение или облегчение имеющегося заболевания и ремиссию заболевания или улучшение прогноза. В некоторых воплощениях осуществления изобретения заявленные антитела используются для замедления развития заболевания или снижения уровня его прогрессии.
Термин «вариабельный участок» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который вовлечен в связывание антитела с антигеном. Вариабельные домены тяжелой и легкой цепей (VH и VL, соответственно) нативного антитела обычно имеют сходную структуру, причем каждый домен содержит четыре консервативных каркасных участка (FRs) и три гипервариабельных участка (HVRs). (См., например, Kindt et al. Kuby Immunology, 6th., W.H. Freeman and Co., стр. 91 (2007).) Одного VH- или VL-домена может быть достаточно для обеспечения антигенсвязывающей специфичности. Более того, антитела, которые связывают определенный антиген, могут быть изолированы при использовании VH- или VL-домена из антитела, которое связывает антиген, путем скрининга библиотеки комплементарных VH- или VL-доменов, соответственно. См., например, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
Используемый здесь термин «вектор» относится к молекуле нуклеиновой кислоты, способной к увеличению числа копий молекулы другой нуклеиновой кислоты, которая связана с вектором. Этот термин включает вектор как самореплицирующуюся структуру нуклеиновой кислоты, так же, как и вектор, включенный в геном клетки-хозяина, в которую он был интродуцирован. Некоторые векторы способны к направлению экспрессии нуклеиновых кислот, с которыми они оперативно связаны. Такие векторы называются здесь «векторами экспрессии».
II. Композиции и способы
Согласно одному аспекту, настоящее изобретение частично основано на мультиспецифических антигенсвязывающих молекулах, имеющих функциональную замещающую активность кофактора фактора VIII. В определенных вариантах осуществления изобретения обеспечиваются мультиспецифические антигенсвязывающие молекулы, которые связываются с FIX, и/или FIXa и FX. Мультиспецифические антигенсвязывающие молекулы в соответствии с настоящим изобретением полезны, например, для лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением.
А. Примеры мультиспецифических антигенсвязывающих молекул, которые связываются с FIX, и/или FIXa и FX.
Согласно одному аспекту, настоящее изобретение обеспечивает изолированные мультиспецифические антигенсвязывающие молекулы, которые связываются с FIX, и/или FIXa и FX. В определенных вариантах осуществления изобретения мультиспецифические антигенсвязывающие молекулы представляют собой биспецифические антитела, которые связываются с FIX, и/или FIXa и FX, причем такие биспецифические антитела имеют функциональную замещающую активность кофактора фактора VIII.
Биспецифическое антитело, которое связывается с FIX, и/или FIXa и FX, АСЕ910 (Эмицизумаб), является антителом, описанным в патентном документе WO 2012/067176, и охарактеризовано ниже.
Антитело представляет собой биспецифическое антитело, в котором ассоциированы первая полипептидная цепь и третья полипептидная цепь, а также вторая полипептидная цепь и четвертая полипептидная цепь. Данное биспецифическое антитело содержит первый полипептид, который является Н-цепью, содержащей HVR-H1, HVR-H2 и HVR-H3 аминокислотные последовательности SEQ ID Nos:1, 2 и 3 (CDRs Н-цепи Q499), соответственно; второй полипептид, который является Н-цепью, содержащей HVR-H1, HVR-H2 и HVR-H3 аминокислотные последовательности SEQ ID Nos:4, 5 и 6 (CDRs Н-цепи J327), соответственно; и третий и четвертый полипептиды, которые обычно являются L-цепями, содержащими HVR-L1, HVR-L2 и HVR-L3 аминокислотные последовательности SEQ ID Nos:7,8 и 9 (CDRs L-цепи L404), соответственно.
Более точно, антитело представляет собой биспецифическое антитело, в котором ассоциированы первая полипептидная цепь и третья полипептидная цепь, а также вторая полипептидная цепь и четвертая полипептидная цепь. Данное биспецифическое антитело содержит первый полипептид, который является Н-цепью, содержащей вариабельный участок с аминокислотной последовательностью SEQ ID NO:45; второй полипептид, который является Н-цепью, содержащей вариабельный участок с аминокислотной последовательностью SEQ ID NO:46, и третий и четвертый полипептиды, которые обычно являются L-цепями, содержащими вариабельный домен с аминокислотной последовательностью SEQ ID NO:47.
Еще более точно, антитело представляет собой биспецифическое антитело, в котором ассоциированы первая полипептидная цепь и третья полипептидная цепь, а также вторая полипептидная цепь и четвертая полипептидная цепь. Данное биспецифическое антитело содержит первый полипептид, который является Н-цепью, состоящей из аминокислотной последовательности SEQ ID NO:10; второй полипептид, который является Н-цепью, состоящей из аминокислотной последовательности SEQ ID NO: 11, и третий и четвертый полипептиды, которые обычно являются L-цепями, состоящими из аминокислотной последовательности SEQ ID NO:12 (Q499-z121/J327-zll9/L404-k).
Согласно настоящему изобретению мультиспецифические антигенсвязывающие молекулы представляют собой мультиспецифические антигенсвязывающие молекулы, которые имеют функциональную замещающую активность FVIII, причем такие молекулы содержат первый антигенсвязывающий участок, который связывается с FIX, и/или FIXa и FX, и второй антигенсвязывающий участок, который связывается с FX, а первый антигенсвязывающий участок содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
где в указанном участке вариабельный домен тяжелой цепи (Q499) содержит HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:1, HVR-Н2, содержащий аминокислотную последовательность SEQ ID NO:2, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:3, и вариабельный домен легкой цепи (QNK131) содержит HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 162, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 163, HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 164, и второй антигенсвязывающий участок содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, где в указанном участке вариабельный домен тяжелой цепи (J327) содержит HVR-H1, содержащий аминокислотную последовательность SEQ ID NO:4, HVR-Н2, содержащий аминокислотную последовательность SEQ ID NO:5, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO:6, и вариабельный домен легкой цепи (JNL095) содержит HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 165, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 166, HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 167, причем один или более аминокислотных остатков замещены другими аминокислотными остатками, делетированы или встроены по крайней мере в одном из HVRs.
В некоторых вариантах осуществления изобретения заявленные мультиспецифические антигенсвязывающие молекулы представляют собой антигенсвязывающие молекулы, которые имеют функциональную замещающую активность FVIII.
Далее, согласно другим аспектам изобретения, заявленные мультиспецифические антигенсвязывающие молекулы обладают пониженной способностью взаимодействовать с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом по сравнению с АСЕ910 (Эмицизумаб). В настоящем описании выражение «обладают пониженной способностью взаимодействовать с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом по сравнению с АСЕ910 (Эмицизумаб)» означает, что величина связывания (%) между анти-идиотипическим антителом и меченным АСЕ910, измеряется с помощью метода, описанного в примере 1, и если концентрация антитела составляет 100 мкг/мл, то величина связывания предпочтительно уменьшается на 10% или более, более предпочтительно, на 20% или более, и еще более предпочтительно, на 30% или более.
Кроме того, в других вариантах осуществления изобретения мультиспецифические антигенсвязывающие молекулы представляют собой мультиспецифические антигенсвязывающие молекулы, которые имеют усиленную функциональную замещающую активность FVIII и более высокую биологическую активность при более низкой концентрации антитела по сравнению с АСЕ910 (Эмицизумаб).
В других вариантах осуществления изобретения мультиспецифические антигенсвязывающие молекулы, по существ, не имеют активирующей-ингибирующей активности FIX и имеют повышенную функциональную замещающую активность кофактора FVIII по сравнению с АСЕ910 (Эмицизумаб). В настоящем описании выражение «по существу не имеют активирующей-ингибирующей активности FIX» означает, что значения OD измеряются с помощью методов, описанных в примерах, и уменьшение значения OD по сравнению с контрольным значением OD составляет 0,025 или менее, предпочтительно, 0,02 или менее или, более предпочтительно, 0,01 или менее.
В одном варианте осуществления изобретения вариабельный домен тяжелой цепи первого антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы согласно настоящему изобретению представляет собой вариабельный домен тяжелой цепи первого антигенсвязывающего участка, причем по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 97, 98, 100, 100а, 100b и 100е, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене тяжелой цепи первого антигенсвязывающего участка.
В одном варианте осуществления изобретения вариабельный домен легкой цепи первого антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы согласно настоящему изобретению представляет собой вариабельный домен легкой цепи первого антигенсвязывающего участка, причем по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 53, 55, 92, 93, 95 и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене легкой цепи первого антигенсвязывающего участка.
В одном варианте осуществления изобретения вариабельный домен тяжелой цепи второго антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы согласно настоящему изобретению представляет собой вариабельный домен тяжелой цепи второго антигенсвязывающего участка, причем по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 51, 56, 57, 59, 61, 62, 65 и 102, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене тяжелой цепи второго антигенсвязывающего участка.
В одном варианте осуществления изобретения вариабельный домен легкой цепи второго антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы согласно настоящему изобретению представляет собой вариабельный домен легкой цепи второго антигенсвязывающего участка, причем по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 24, 26, 27, 29, 30, 31, 32, 50, 92, 94, 95, 95а и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене легкой цепи второго антигенсвязывающего участка.
В одном варианте осуществления изобретения заявленная мультиспецифическая антигенсвязывающая молекула представляет собой мультиспецифическую антигенсвязывающую молекулу, где:
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 97, 98, 100, 100а, 100b и 100е, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене тяжелой цепи первого антигенсвязывающего участка;
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 53, 55, 92, 93, 95 и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или встроен в вариабельном домене легкой цепи первого антигенсвязывающего участка;
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 51, 56, 57, 59, 61, 62, 65 и 102, согласно нумерации по Кабату, замещен другим аминокислотным остатком в вариабельном домене тяжелой цепи второго антигенсвязывающего участка; и
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 24, 26, 27, 29, 30, 31, 32, 50, 92, 94, 95, 95а и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене легкой цепи второго антигенсвязывающего участка.
В одном варианте осуществления изобретения, вариабельный домен тяжелой цепи первого антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен тяжелой цепи первого антигенсвязывающего участка, причем:
в вариабельном домене тяжелой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин или аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин, где указанные положения соответствуют нумерации по Кабату;
В одном варианте осуществления изобретения, вариабельный домен легкой цепи первого антигенсвязывающего участка, согласно изобретению, представляет собой вариабельный домен легкой цепи первого антигенсвязывающего участка, причем:
в вариабельном домене легкой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 26 представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин, где указанные положения соответствуют нумерации по Кабату.
В одном варианте осуществления изобретения, вариабельный домен тяжелой цепи второго антигенсвязывающего участка, согласно изобретению, представляет собой вариабельный домен тяжелой цепи второго антигенсвязывающего участка, причем:
в вариабельном домене тяжелой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин или аминокислотный остаток в положении 102 представляет собой валин, где указанные положения соответствуют нумерации по Кабату.
В одном варианте осуществления изобретения, вариабельный домен легкой цепи второго антигенсвязывающего участка, согласно изобретению, представляет собой вариабельный домен легкой цепи второго антигенсвязывающего участка, причем:
в вариабельном домене легкой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин, где указанные положения соответствуют нумерации по Кабату.
В одном варианте осуществления изобретения мультиспецифическая антигенсвязывающая молекула, согласно настоящему изобретению, представляет собой мультиспецифическую антигенсвязывающую молекулу, причем:
в вариабельном домене тяжелой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин или аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 26 представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин, где указанные положения соответствуют нумерации по Кабату; в вариабельном домене тяжелой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин или аминокислотный остаток в положении 102 представляет собой валин, где указанные положения соответствуют нумерации по Кабату; и
в вариабельном домене легкой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин, где указанные положения соответствуют нумерации по Кабату.
В одном варианте осуществления изобретения вариабельный домен тяжелой цепи первого-антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен тяжелой цепи, который включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 168, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 169, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 170 (QH01);
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 171, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 172, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 173 (QH02);
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 174, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 175, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 176 (QH03);
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 177, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 178, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 179 (QH04);
5) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 180, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 181, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 182 (QH06); или
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 183, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 184, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 185 (QH07).
В одном варианте осуществления изобретения вариабельный домен легкой цепи первого-антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен легкой цепи, который включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 186, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 187, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 188 (QL21);
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 189, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 190, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 191 (QL22);
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 192, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 193, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 194 (QL23);
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 195, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 196, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 197 (QL24);
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 198, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 199, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 200 (QL25);
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 201, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 202, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 203 (QL26);
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 204, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 205, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 206 (QL28);
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 207, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 208, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 209 (QL29);
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 210, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 211, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 212 (QL30);
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 213, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 214, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 215 (QL31);
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 216, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 217, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 218 (QL32);
12) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 219, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 220, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 221 (QL33).
В одном варианте осуществления изобретения вариабельный домен тяжелой цепи второго-антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен тяжелой цепи, который включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 222, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 223, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 224 (JH01);
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 225, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 226, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 227 (JH02);
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 228, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 229, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 230 (JH03);
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 231, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 232, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 233 (JH04);
5) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 234, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 235, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 236 (JH05);
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 237, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 238, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 239 (JH06);
7) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 240, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 241, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 242 (JH07);
8) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 243, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 244, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 245 (JH08);
9) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 246, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 247, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 248 (JH09);
10) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 249, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 250, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 251 (JH10);
11) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 252, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 253, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 254 (JH11).
В одном варианте осуществления изобретения вариабельный домен легкой цепи второго антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен легкой цепи, который включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 255, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 256, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 257 (JL01);
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 258, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 259, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 260 (JL02);
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 261, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 262, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 263 (JL03);
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 264, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 265, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 266 (JL04);
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 267, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 268, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 269 (JL05);
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 270, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 271, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 272 (JL06);
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 273, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 274, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 275 (JL07);
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 276, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 277, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 278 (JL08);
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 279, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 280, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 281 (JL09);
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 282, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 283, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 284 (JL10); или
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 285, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 286, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 287 (JL11).
В одном варианте осуществления изобретения мультиспецифическая антигенсвязывающая молекула, согласно настоящему изобретению, представляет собой мультиспецифическую антигенсвязывающую молекулу, в которой первый антигенсвязывающий участок содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, причем вариабельный домен тяжелой цепи включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 168, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 169, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 170 (QH01);
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 171, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 172, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 173 (QH02);
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 174, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 175, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 176 (QH03);
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 177, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 178, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 179 (QH04);
5) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 180, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 181, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 182 (QH06); или
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 183, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 184, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 185 (QH07) и
вариабельный домен легкой цепи включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 186, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 187, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 188 (QL21);
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 189, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 190, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 191 (QL22);
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 192, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 193, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 194 (QL23);
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 195, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 196, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 197 (QL24);
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 198, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 199, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 200 (QL25);
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 201, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 202, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 203 (QL26);
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 204, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 205, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 206 (QL28);
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 207, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 208, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 209 (QL29);
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 210, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 211, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 212 (QL30);
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 213, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 214, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 215 (QL31);
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 216, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 217, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 218 (QL32);
12) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 219, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 220, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 221 (QL33), причем
второй антигенсвязывающий участок мультиспецифической антигенсвязывающей молекулы содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
где вариабельный домен тяжелой цепи включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 222, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 223, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 224 (JH01);
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 225, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 226, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 227 (JH02);
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 228, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 229, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 230 (JH03);
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 231, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 232, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 233 (JH04);
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 234, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 235, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 236 (JH05);
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 237, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 238, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 239 (JH06);
7) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 240, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 241, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 242 (JH07);
8) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 243, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 244, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 245 (JH08);
9) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 246, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 247, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 248 (JH09);
10) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 249, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 250, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 251 (JH10);
11) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 252, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 253, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 254 (JH11) или
вариабельный домен легкой цепи включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 255, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 256, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 257 (JL01);
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 258, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 259, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 260 (JL02);
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 261, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 262, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 263 (JL03);
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 264, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 265, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 266 (JL04);
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 267, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 268, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 269 (JL05);
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 270, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 271, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 272 (JL06);
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 273, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 274, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 275 (JL07);
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 276, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 277, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 278 (JL08);
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 279, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 280, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 281 (JL09);
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 282, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 283, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 284 (JL10); или
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 285, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 286, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 287 (JL11).
В одном варианте осуществления настоящего изобретения мультиспецифическая антигенсвязывающая молекула представляет собой мультиспецифическую антигенсвязывающую молекулу, имеющая функцию замещения функциональной активности коагулирующего фактора крови VIII, причем молекула содержит первый антигенсвязывающий участок, который связывается с коагулирующим фактором крови IX и/или активированным коагулирующим фактором крови IX, и второй антигенсвязывающий участок, который связывается с коагулирующим фактором крови X, где первый антигенсвязывающий участок содержит вариабельный домен тяжелой цепи (Q499) с последовательностью SEQ ID NO: 45 и вариабельный домен легкой цепи (QNK131) с последовательностью SEQ ID NO: 13, а второй антигенсвязывающий участок содержит вариабельный домен тяжелой цепи (J327) с последовательностью SEQ ID NO: 46 и вариабельный домен легкой цепи (JNL095) с последовательностью SEQ ID NO: 31 и где один или более аминокислотных остатков замещены другими аминокислотными остатками или делетированы по крайней мере в одном вариабельном домене тяжелой цепи или в одном вариабельном домене легкой цепи.
В одном варианте осуществления изобретения в вариабельном домене тяжелой цепи первого антигенсвязывающего участка, согласно настоящему изобретению, по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 39, 97, 98, 100, 100а, 100b и 100е, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован.
В одном варианте осуществления изобретения в вариабельном домене легкой цепи первого антигенсвязывающего участка, согласно настоящему изобретению, по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 38, 45, 53, 55, 60, 70, 76, 79, 80, 83, 85, 92, 93, 95 и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или встроен в указанном выше вариабельном домене легкой цепи первого антигенсвязывающего участка.
В одном варианте осуществления изобретения в вариабельном домене тяжелой цепи второго антигенсвязывающего участка, согласно настоящему изобретению, по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 28, 31, 39, 51, 56, 57, 59, 61, 62, 65, 67, 73, 82b и 102, согласно нумерации по Кабату, замещен другим аминокислотным остатком.
В одном варианте осуществления изобретения в вариабельном домене легкой цепи второго антигенсвязывающего участка, согласно настоящему изобретению, по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 3, 8, 15, 24, 26, 27, 29, 30, 31, 32, 38, 48, 49, 50, 79, 92, 94, 95, 95а и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован.
В одном варианте осуществления изобретения в заявленной мультиспецифической антигенсвязывающей молекуле, согласно настоящему изобретению: по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 39, 97, 98, 100, 100а, 100b и 100е, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в указанном выше вариабельном домене тяжелой цепи первого антигенсвязывающего участка; по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 38, 45, 53, 55, 60, 70, 76, 79, 80, 83, 85, 92, 93, 95 и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или встроен в указанном выше вариабельном домене легкой цепи первого антигенсвязывающего участка; по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 28, 31, 39, 51, 56, 57, 59, 61, 62, 65, 67, 73, 82b и 102, согласно нумерации по Кабату, замещен другим аминокислотным остатком в указанном выше вариабельном домене тяжелой цепи второго антигенсвязывающего участка; и по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 3, 8, 15, 24, 26, 27, 29, 30, 31, 32, 38, 48, 49, 50, 79, 92, 94, 95, 95а и 96, согласно нумерации по Кабату, замещен другим аминокислотным остатком или делетирован в указанном выше вариабельном домене легкой цепи второго антигенсвязывающего участка.
В одном варианте осуществления изобретения, в вариабельном домене тяжелой цепи первого антигенсвязывающего участка, согласно настоящему изобретению, аминокислотный остаток в положении 31 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 39 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин и аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин.
В одном варианте осуществления изобретения, в вариабельном домене легкой цепи первого антигенсвязывающего участка, согласно изобретению, аминокислотный остаток в положении 26 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой лизин, аминокислотный остаток в положении 45 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 60 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 70 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 76 представляет собой аспарагин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 80 представляет собой пролин или аланин, аминокислотный остаток в положении 83 представляет собой метионин или аланин, аминокислотный остаток в положении 85 представляет собой треонин, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин.
В одном варианте осуществления изобретения в вариабельном домене тяжелой цепи второго антигенсвязывающего участка, согласно изобретению, аминокислотный остаток в положении 28 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 39 представляет собой лизин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин, аминокислотный остаток в положении 67 представляет собой лейцин, аминокислотный остаток в положении 73 представляет изолейцин, аминокислотный остаток в положении 82b представляет собой глутаминовую кислоту или аминокислотный остаток в положении 102 представляет собой валин.
В одном варианте осуществления изобретения, в вариабельном домене легкой цепи второго антигенсвязывающего участка, согласно изобретению, аминокислотный остаток в положении 3 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой глутаминовую кислоту, аминокислотный остаток в положении 8 представляет собой пролин, аминокислотный остаток в положении 15 представляет собой лейцин, аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аргинин, 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 48 представляет собой изолейцин, аминокислотный остаток в положении 49 представляет собой тирозин, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин.
В одном варианте осуществления изобретения в мультиспецифической антигенсвязывающей молекуле, согласно настоящему изобретению, аминокислотный остаток в положении 31 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 39 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин или аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин в указанном выше вариабельном домене тяжелой цепи первого антигенсвязывающего участка; аминокислотный остаток в положении 26 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой лизин, аминокислотный остаток в положении 45 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 60 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 70 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 76 представляет собой аспарагин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 80 представляет собой пролин или аланин, аминокислотный остаток в положении 83 представляет собой метионин или аланин, аминокислотный остаток в положении 85 представляет собой треонин, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин в указанном выше вариабельном домене легкой цепи первого антигенсвязывающего участка; аминокислотный остаток в положении 28 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 39 представляет собой лизин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин, аминокислотный остаток в положении 67 представляет собой лейцин, аминокислотный остаток в положении 73 представляет изолейцин, аминокислотный остаток в положении 82b представляет собой глутаминовую кислоту или аминокислотный остаток в положении 102 представляет собой валин в указанном выше вариабельном домене тяжелой цепи второго антигенсвязывающего участка; аминокислотный остаток в положении 3 (согласно нумерации по Кабату, что справедливо и для указанного ниже) представляет собой глутаминовую кислоту, аминокислотный остаток в положении 8 представляет собой пролин, аминокислотный остаток в положении 15 представляет собой лейцин, аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аргинин, 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 48 представляет собой изолейцин, аминокислотный остаток в положении 49 представляет собой тирозин, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин в указанном выше вариабельном домене легкой цепи второго антигенсвязывающего участка.
В одном варианте осуществления изобретения вариабельный домен тяжелой цепи первого антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59 или SEQ ID NO: 60.
В одном варианте осуществления изобретения вариабельный домен легкой цепи первого антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72.
В одном варианте осуществления изобретения вариабельный домен тяжелой цепи второго антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82 или SEQ ID NO: 83.
В одном варианте осуществления изобретения вариабельный домен легкой цепи второго антигенсвязывающего участка мультиспецифической антигенсвязывающей молекулы, согласно настоящему изобретению, представляет собой вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87 или SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93 или SEQ ID NO: 94.
В одном варианте осуществления изобретения мультиспецифическая антигенсвязывающая молекула, согласно настоящему изобретению, представляет собой мультиспецифическую антигенсвязывающую молекулу, в которой первый антигенсвязывающий участок содержит:
вариабельный домен тяжелой цепи (QH) с аминокислотной последовательностью SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59 или SEQ ID NO: 60; и
вариабельный домен легкой цепи (QL) с аминокислотной последовательностью SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72; и
второй антигенсвязывающий участок содержит:
вариабельный домен тяжелой цепи (JH), содержащий аминокислотную последовательность SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82 или SEQ ID NO: 83; и
вариабельный домен легкой цепи (JL), содержащий аминокислотную последовательность SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93 или SEQ ID NO: 94.
В одном варианте осуществления изобретения мультиспецифическая антигенсвязывающая молекула содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи первого антитела, который связывается с FIX и/или FIXa, и вариабельный домен тяжелой цепи и вариабельный домен легкой цепи второго антитела, который связывается с FX, причем указанная молекула является любой из молекул от (а) до (v), как указано ниже:
(а) мультиспецифическая антигенсвязывающая молекула (QH01/QL21//JH01/JL01), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 56, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 61, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84;
(б) мультиспецифическая антигенсвязывающая молекула (QH02/QL22//JH01/JL01), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84;
(в) мультиспецифическая антигенсвязывающая молекула (QH03/QL23//JH02/JL02), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 63, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(г) мультиспецифическая антигенсвязывающая молекула (QH03/QL24//JH02/JL02), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 64, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(д) мультиспецифическая антигенсвязывающая молекула (QH02/QL22//JH03/JL03), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 75, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 86;
(е) мультиспецифическая антигенсвязывающая молекула (QH02/QL22//JH04/JL04), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 76, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 87;
(ж) мультиспецифическая антигенсвязывающая молекула (QH02/QL22//JH02/JL02), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(з) мультиспецифическая антигенсвязывающая молекула (QH04/QL25//JH02/JL02), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 65, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(и) мультиспецифическая антигенсвязывающая молекула (QH04/QL26//JH02/JL02), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(к) мультиспецифическая антигенсвязывающая молекула (QH04/QL26//JH05/JL05), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88;
(л) мультиспецифическая антигенсвязывающая молекула (QH04/QL28//JH05/JL05), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88;
(м) мультиспецифическая антигенсвязывающая молекула (QH04/QL28//JH06/JL06), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89;
(н) мультиспецифическая антигенсвязывающая молекула (QH04/QL29//JH05/JL05), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88;
(о) мультиспецифическая антигенсвязывающая молекула (QH04/QL29//JH06/JL06), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89;
(п) мультиспецифическая антигенсвязывающая молекула (QH06/QL30//JH07/JL07), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90;
(р) мультиспецифическая антигенсвязывающая молекула (QH04/QL31//JH08/JL08), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 70, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 80, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 91;
(с) мультиспецифическая антигенсвязывающая молекула (QH06/QL32//JH07/JL07), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 71, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90;
(т) мультиспецифическая антигенсвязывающая молекула (QH06/QL32//JH09/JL09), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 71, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 81, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 92;
(у) мультиспецифическая антигенсвязывающая молекула (QH06/QL30//JH10/JL10, содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 82, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 93;
(ф) мультиспецифическая антигенсвязывающая молекула (QH07/QL33//JH11/JL11), содержащая вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 105, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 72, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 83, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 94;
(х) мультиспецифическая антигенсвязывающая молекула, которая связывается с эпитопами, идентичными эпитопам FIX и/или FIXa, и эпитопу FX, которые распознаются любым антителом из антител от (а) до (ф);
(ц) мультиспецифическая антигенсвязывающая молекула, которая конкурирует за связывание с эпитопами FIX и/или FIXa, и эпитопом FX, которые распознаются любым антителом из антител от (а) до (ф).
В одном варианте осуществления изобретения константный участок тяжелой цепи заявленной мультиспецифической антигенсвязывающей молекулы представляет собой константный участок тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 118 или SEQ ID NO: 119.
В одном варианте осуществления изобретения константный участок легкой цепи заявленной мультиспецифической антигенсвязывающей молекулы представляет собой константный участок легкой, содержащий аминокислотную последовательность SEQ ID NO: 100 или SEQ ID NO: 102.
В одном варианте осуществления изобретения заявленная мультиспецифическая антигенсвязывающая молекула представляет собой мультиспецифическую антигенсвязывающую молекулу, в которой первый антигенсвязывающий участок содержит константный участок (1) или (2), как указано ниже, и второй антигенсвязывающий участок содержит константный участок (1) или (2), как указано ниже, который не является константным участком, содержащемся в первом антигенсвязывающем участке:
(1) SEQ ID NO: 119 для константного участка тяжелой цепи и SEQ ID NO: 100 для константного участка легкой цепи;
(2) SEQ ID NO: 118 для константного участка тяжелой цепи и SEQ ID NO: 102 для константного участка легкой цепи.
В одном варианте осуществления изобретения заявленная мультиспецифическая антигенсвязывающая молекула представляет собой мультиспецифическое антитело. Еще в одном варианте осуществления изобретения мультиспецифическое антитело представляет собой биспецифическое антитело.
Согласно одному аспекту настоящее изобретение относится к биспецифическому антителу, содержащему вариабельный домен тяжелой цепи первого антитела и вариабельный домен легкой цепи первого антитела, которые связываются с FIX и/или FIXa, и вариабельный домен тяжелой цепи второго антитела и вариабельный домен легкой цепи второго антитела, которые связываются с FX, причем биспецифическое антитело представляет собой любое из антител от (а) до (v), как указано ниже:
(а) биспецифическое антитело (QH01/QL21//JH01/JL01), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 56, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 61, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84;
(б) биспецифическое антитело (QH02/QL22//JH01/JL01), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 73, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 84;
(в) биспецифическое антитело (QH03/QL23//JH02/JL02), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 63, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(г) биспецифическое антитело (QH03/QL24//JH02/JL02), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 58, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 64, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(д) биспецифическое антитело (QH02/QL22//JH03/JL03), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 75, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 86;
(е) биспецифическое антитело (QH02/QL22//JH04/JL04), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 76, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 87;
(ж) биспецифическое антитело (QH02/QL22//JH02/JL02), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 57, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 62, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(з) биспецифическое антитело (QH04/QL25//JH02/JL02), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 65, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(и) биспецифическое антитело (QH04/QL26//JH02/JL02), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 74, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 85;
(к) биспецифическое антитело (QH04/QL26//JH05/JL05), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 66, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88;
(л) биспецифическое антитело (QH04/QL28//JH05/JL05), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88;
(м) биспецифическое антитело (QH04/QL28//JH06/JL06), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 67, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89;
(н) биспецифическое антитело (QH04/QL29//JH05/JL05), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 77, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 88;
(o) биспецифическое антитело (QH04/QL29//JH06/JL06), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 68, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 78, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 89;
(п) биспецифическое антитело (QH06/QL30//JH07/JL07), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90;
(р) биспецифическое антитело (QH04/QL31//JH08/JL08), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 59, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 70, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 80, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 91;
(с) биспецифическое антитело (QH06/QL32//JH07/JL07), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 71, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 79, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 90;
(т) биспецифическое антитело (QH06/QL32//JH09/JL09), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 71, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 81, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 92;
(у) биспецифическое антитело (QH06/QL30//JH10/JL10), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 60, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 69, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 82, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 93;
(ф) биспецифическое антитело (QH07/QL33//JH11/JL11), содержащее вариабельный домен тяжелой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 105, вариабельный домен легкой цепи первого антитела, содержащий аминокислотную последовательность SEQ ID NO: 72, вариабельный домен тяжелой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 83, и вариабельный домен легкой цепи второго антитела, содержащий аминокислотную последовательность SEQ ID NO: 94;
(х) биспецифическое антитело, которое связывается с эпитопами, идентичными эпитопам коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопу коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (ф);
(ц) биспецифическое антитело, которое конкурирует за связывание с эпитопами коагулирующего фактора крови IX и/или активированного коагулирующего фактора крови IX, и эпитопом коагулирующего фактора крови X, которые распознаются любым антителом из антител от (а) до (ф).
Согласно одному аспекту настоящее изобретение относится к биспецифическому антителу, содержащему тяжелую цепь первого антитела и легкую цепь первого антитела, которые связываются с FIX и/или FIXa, и тяжелую цепь второго антитела и легкую цепь второго антитела, которые связываются с FX, причем биспецифическое антитело представляет собой любое из антител от (а) до (ц), как указано ниже:
(а) биспецифическое антитело (QH01/QL21//JH01/JL01), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 120, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 126, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 138, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 149;
(б) биспецифическое антитело (QH02/QL22//JH01/JL01), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 138, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 149;
(в) биспецифическое антитело (QH03/QL23//JH02/JL02), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 122, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 128, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(г) биспецифическое антитело (QH03/QL24//JH02/JL02), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 122, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 129, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(д) биспецифическое антитело (QH02/QL22//JH03/JL03), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 140, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 151;
(е) биспецифическое антитело (QH02/QL22//JH04/JL04), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 141, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 152;
(ж) биспецифическое антитело (QH02/QL22//JH02/JL02), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(з) биспецифическое антитело (QH04/QL25//JH02/JL02), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 130, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(и) биспецифическое антитело (QH04/QL26//JH02/JL02), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 131, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(к) биспецифическое антитело (QH04/QL26//JH05/JL05), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 131, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153;
(л) биспецифическое антитело (QH04/QL28//JH05/JL05), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 132, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153;
(м) биспецифическое антитело (QH04/QL28//JH06/JL06), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 132, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 143, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 154;
(н) биспецифическое антитело (QH04/QL29//JH05/JL05), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 133, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153;
(о) биспецифическое антитело (QH04/QL29//JH06/JL06), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 133, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 143, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 154;
(п) биспецифическое антитело (QH06/QL30//JH07/JL07), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 134, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 144, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 155;
(р) биспецифическое антитело (QH04/QL31//JH08/JL08), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 135, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 145, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 156;
(с) биспецифическое антитело (QH06/QL32//JH07/JL07), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 136, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 144, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 155;
(т) биспецифическое антитело (QH06/QL32//JH09/JL09), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 136, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 146, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 157;
(у) биспецифическое антитело (QH06/QL30//JH10/JL10), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 143, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 147, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 158;
(ф) биспецифическое антитело (QH07/QL33//JH11/JL11), содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 125, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 137, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 148, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 159;
(х) биспецифическое антитело, которое связывается с эпитопами, идентичными эпитопам FIX и/или FIXa, и эпитопу FX, которые распознаются любым антителом из антител от (а) до (t);
(ц) биспецифическое антитело, которое конкурирует за связывание с эпитопами FIX и/или FIXa, и эпитопом FX, которые распознаются любым антителом из антител от (а) до (t).
Настоящее изобретение также обеспечивает мультиспецифические антигенсвязывающие молекулы, которые имеют функциональную замещающую активность коагулирующего фактора крови VIII, причем такие молекулы содержат первый антигенсвязывающий участок, который связывается с коагулирующим фактором крови IX, и/или активированным коагулирующим фактором крови IX, и второй антигенсвязывающий участок, который связывается с коагулирующим фактором крови X,
причем первый антигенсвязывающий участок содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
где вариабельный домен тяжелой цепи (Q499) содержит HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 1, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 2, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3, и вариабельный домен легкой цепи (QNK131) содержит HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 162, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 163, HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 164, и
второй антигенсвязывающий участок содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
где вариабельный домен тяжелой цепи (J327) содержит HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 4, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 5, HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 6,
и вариабельный домен легкой цепи (JNL095) содержит HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 165, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 166, HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 167.
В некоторых вариантах осуществления изобретения описанные выше антитела от (а) до (v) обладают функциональной замещающей активностью FVIII.
Далее, согласно другим аспектам изобретения, описанные выше антитела от (а) до (ц) представляют собой антитела, которые обладают пониженной способностью взаимодействовать с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом по сравнению с АСЕ910 (Эмицизумаб). В настоящем описании выражение «обладают пониженной способностью взаимодействовать с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом по сравнению с АСЕ910 (Эмицизумаб)» означает, что величина связывания (%) между анти-идиотипическим антителом и меченным АСЕ910, измеряется с помощью раскрытых в описании методов, и если тестируемое вещество добавлено в концентрации 100 мкг/мл, то величина взаимодействия предпочтительно уменьшается на 10% или более, более предпочтительно, на 20% или более, и еще более предпочтительно, на 30% или более.
Анти-АСЕ910 (Эмицизумаб) идиотипические антитела могут быть получены, например, следующим способом. F(ab')2 - фрагменты) Q499/L404, которые распознают FIX(a), и F(ab')2- фрагменты J327/L404, которые распознают FX, полученные из АСЕ910 (Эмицизумаб), вводят кроликам и получают сыворотку, содержащую антитела кролика к F(ab')2- фрагментам Q499/L404, и сыворотку, содержащую антитела кролика к F(ab')2- фрагментам J327/L404. Фракции указанных сывороток в сульфате аммония наносят на колонку для удаления IgG-реакционноспособных антител человека. Далее, проводят аффинную очистку на колонке, связанной с Q499/L404 (J327/L404), и удаляют взаимодействующие с J327/L404 (Q499/L404) антитела при использовании колонки, связанной с J327/L404 (Q499/L404). Таким образом, получают поликлональные анти-идиотипические антитела, специфически связывающиеся с Q499/L404 (J327/L404).
Величину связывания (%) анти-идиотипических антител с меченным АСЕ910 (Эмицизумаб) можно измерить, например, в электрохимическом люминесцентном иммуноанализе, что позволяет оценить ингибирование связывания тестируемым веществом. Когда используются анти-идиотипические антитела, полученные, как указано выше, то добавляют тестируемое вещество (биспецифическое антитело) в концентрации 0, 1, 3, 10, 30 или 100 мкг/мл к смеси анти-идиотипических антител к Q499/L404, анти-идиотипических антител к J327/L404, меченного биотином АСЕ910 (Эмицизумаб) и меченного SULFO-TAG ACE910 (Эмицизумаб). Полученный перемешанный раствор инкубируют в течение ночи в холодильнике. Затем, перемешанный раствор вносят в лунки 96-луночного планшета для микротитрования, отмывают и определяют величину связывания электрохимическим люминесцентным методом.
В других вариантах осуществления изобретения описанные выше антитела от (а) до (ц), по существу, не имеют активирующей-ингибирующей активности FIX и имеют повышенную функциональную замещающую активность кофактора FVIII по сравнению с АСЕ910 (Эмицизумаб). В настоящем описании выражение «по существу не имеют активирующей-ингибирующей активности FIX» означает, что значения OD измеряются с помощью методов, раскрытых в настоящем описании, и уменьшение значения OD по сравнению с контрольным значением OD составляет 0,025 или менее, предпочтительно, 0,02 или менее или, более предпочтительно, 0,01 или менее.
Аминокислоты, входящие в состав описанных здесь аминокислотных последовательностей, могут быть подвергнутыми пост-трансляционным модификациям (например, модификация N-концевого глутамина в пироглутаминовую кислоту путем пироглутамилирования хорошо известна специалистам в данной области техники). Соответственно, такие последовательности, содержащие модифицированные аминокислоты, подвергнутые пост-трансляционным модификациям, также включены в объем настоящего изобретения.
Согласно другому аспекту, настоящее изобретение обеспечивает мультиспецифические антигенсвязывающие молекулы или биспецифические антитела, которые связываются с тем же самым эпитопом, что и антигенсвязывающие молекулы или биспецифические антитела, которые связываются с FIX и/или FIXa, и FX и описаны здесь. Например, в некоторых воплощениях изобретение обеспечивает мультиспецифические антигенсвязывающие молекулы или биспецифические антитела, которые связываются с тем же самым эпитопом, что и мультиспецифические антигенсвязывающие молекулы или биспецифические антитела, охарактеризованные выше в подразделах от (а) до (t).
Согласно еще одному варианту осуществления изобретения описанные выше биспецифические антитела, которые связываются с FIX и/или FIXa, и FX, представляют собой моноклональные антитела, включая химерные, гуманизированные и человеческие антитела. В одном случае биспецифические антитела, которые связываются с FIX и/или FIXa, и FX, представляют собой фрагменты антител, например, (scFv)2, диатела или F(ab')2-фрагменты. В другом случае антитела являются полноразмерными антителами, например, интактными IgG4-антителами или антителами другого класса или изотипа, как описано здесь.
Далее, описанные выше биспецифические антитела, которые связываются с FIX и/или FIXa, и FX, могут обладать любым из признаков, по отдельности или в комбинации, приведенных ниже в подразделах 1-7:
1. Аффинность антител
В некоторых вариантах осуществления изобретения заявленные антитела имеют константу диссоциации (Kd) 100 мкМ или менее, 10 мкМ или менее, 1 мкМ или менее, 100 нМ или менее, 10 нМ или менее, 1 нМ или менее, 0,1 нМ или менее, 0,01 нМ или менее, 0,001 нМ или менее (например, 10-5 М или менее, скажем, от 10-5 М до 10-10 М, например, от 10-6 М до 10-10 М).
Согласно одному варианту осуществления изобретения Kd измеряют радиоиммунологическим анализом связывания антигена (RIA). В частности, RIA осуществляют при использовании Fab-версии представляющего интерес антитела и антигена, с которым связывается это антитело. Например, величину аффинности связывания с антигеном Fabs-фрагментов измеряют уравновешиванием Fabs с минимальной концентрацией (125I)-меченного антигена в присутствии серийных титрований немеченого антигена и дальнейшим захватом связавшегося антигена в лунках планшета для микротитрования, покрытых анти-Fab антителами (см., например, Chen et al., J. Mot Biol. 293: 865-881 (1999)). Для обеспечения условий анализа MICROTITER (зарегистрированный товарный знак) микролуночные планшеты (Thermo Scientific) в течение ночи покрывают 5 мкг/мл захватывающими антиген анти-Fab антителами (Cappel Labs) в 50 мМ карбонате натрия (рН 9,6) и затем блокируют 2%-ным (вес на объем) бычьим сывороточным альбумином в PBS в течение двух-пяти часов при комнатной температуре (примерно, 23°С). В неабсорбирующем планшете (Nunc#269620) 100 пкМ или 26 пкМ (125I)-меченного антигена смешивают с серийными разведениями представляющих интерес Fab-фрагментов (например, что соответствует оценке аффинности анти-VEGF антитела, Fab-12, как описано Presta et al., Cancer Res. 57:4593-4599 (1997)). Представляющие интерес Fab-фрагменты затем инкубируют в течение ночи, однако инкубация может продолжаться и дольше (например, около 65 часов), чтобы убедиться в том, что достигнуто уравновешивание. Далее смеси переносят в планшеты с захватывающим антигеном для инкубирования при комнатной температуре (например, в течение часа). Затем раствор удаляют и планшеты отмывают восемь раз 0,1%-ным полисорбатом 20 (TWEEN-20 (зарегистрированный товарный знак)) в PBS. После этого планшеты высушивают добавляют 150 мкл на лунку сцинтиллятора MICROSCINT-20™, Packard, и планшеты просчитывают в гамма-счетчике TOPCOUNT™ (Packard) в течение десяти минут. Концентрацию каждого Fab, который обнаруживает меньшую или эквивалентную 20% от максимальной связывающую активность, выбирают для использования в конкурентном связывающем анализе.
Согласно другому варианту осуществления изобретения Kd измеряют с использованием BIACORE (зарегистрированный товарный знак) методом поверхностного плазмонного резонанса. Например, метод, использующий BIACORE (зарегистрированный товарный знак) 2000 или BIACORE (зарегистрированный товарный знак) 3000 (BIAcore, Inc., Piscataway, NJ), осуществляют при 25°С с помощью микрочипов СМ5 с иммобилизованным антигеном примерно при 10 отвечающих единицах (RU). В одном случае карбоксиметилированные декстрановые биосенсорные чипы (СМ5, BIACORE, Inc.) активируют М-этил-N'-(3-диметиламинопропил)-карбодиимид гидрохлоридом (NHS) в соответствии с инструкциями производителя. Антиген разводят 10 мМ ацетатом натрия, рН 4,8, до концентрации 5 мкг/мл (примерно 0,2 мкМ) до введения со скоростью потока 5 мкл/мин до достижения приблизительно 10 отвечающих единиц (RU) связанного белка. Вслед за инъекцией антигена инъецируют 1 М этаноламина для блокирования непрореагировавших групп. Для измерения кинетики двукратные последовательные разведения Fab (от 0,78 нМ до 500 нМ) инъецируют в PBS вместе с 0,05%-ным полисорбатным 20 (TWEEN-20™) сурфактантом (PBST) при 25°С при скорости потока примерно 25 мкл/мин. Скорости ассоциации (kon) и диссоциации (koff) вычисляют при использовании простой «один к одному» связывающей модели Langmuir ((BIACORE (зарегистрированный товарный знак) версия Evaluation Software 3.2)) путем одновременного изучения сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляют как отношение kon/koff. См., например, Chen et al., J. Mol. Biol. 293: 865-881 (1999). Если скорость ассоциации превышает 106M-1s-1, как определяется указанным выше методом поверхностного плазмонного резонанса, то скорость ассоциации может быть установлена при использовании технологии флуоресцентного гашения, которая измеряет увеличение или уменьшение флуоресцентной интенсивности эмиссии (возбуждение=295 нм; эмиссия=340 нм, 16 нм полоса пропуска) при 25°С 20 нМ антитела к антигену (Fab) в PBS, рН 7,2, в присутствии увеличивающихся концентраций антигена, что измеряют в спектрометре, таком, как спектрофотометр, оснащенный возможностью остановки потока (Aviv Instruments) или 8000-серийный спектрофотометр SLM-AMINCO™ (ThermoSpectronic) в кювете с перемешиванием.
2. Фрагменты антитела
В некоторых вариантах осуществления изобретения обеспечиваемые антитела представляют собой антигенные фрагменты. Фрагменты антител включают, без ограничений указанным (scFv)2, диатела, F(ab')2 и другие фрагменты, описанные ниже. Обзор определенных фрагментов антител приведен Hudson et al. Nat. Med. 9: 129-134 (2003). Обзор scFv-фрагментов приведен, например,  в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore., (Springer- Verlag, New York), стр. 269-315 (1994); см. также WO 93/16185 и патенты US 5571894 и 5587458. F(ab')2-фрагменты, содержащие аминокислотные остатки, взаимодействующие с рецептор-связывающим эпитопом реутилизации и имеющие увеличенное время полужизни in vivo, описаны в патенте US 5869046.
в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore., (Springer- Verlag, New York), стр. 269-315 (1994); см. также WO 93/16185 и патенты US 5571894 и 5587458. F(ab')2-фрагменты, содержащие аминокислотные остатки, взаимодействующие с рецептор-связывающим эпитопом реутилизации и имеющие увеличенное время полужизни in vivo, описаны в патенте US 5869046.
Диатела представляют собой антительные фрагменты с двумя антигенсвязывающими участками, которые могут быть бивалентными или биспецифическими. См., например, ЕР 404097; WO 1993/01161; Hudson et al. Nat. Med. 9:129-134 (2003); и Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993).
Фрагменты антител могут быть получены с помощью различных технологий, включая без ограничений указанным, протеолитическое расщепление интактного антитела и продукцию фрагментов рекомбинантными клетками-хозяевами (например, фагами или Е. coli), как здесь описано.
3. Химерные или гуманизированные антитела
В некоторых вариантах осуществления изобретения обеспечиваемые антитела представляют собой химерные антитела. Некоторые химерные антитела описаны, например, в патенте US 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984). Один пример относится к химерному антителу, содержащему вариабельный участок нечеловеческого антитела (например, вариабельный участок, происходящий из антитела мыши, крысы, хомяка, кролика или примата, не являющегося человеком, таким, как обезьяна) и константный участок антитела человека. Еще один пример касается химерного антитела, класс или подкласс которого может быть изменен по сравнению с родительским антителом («класс-переключаемое» антитело). Химерные антитела включают также антигенсвязывающие фрагменты указанных выше антител.
В некоторых вариантах осуществления изобретения химерное антитело является гуманизированным антителом. Обычно, антитело не человека гуманизируют для снижения иммуногенности по отношению к человеку, при этом не затрагивая специфичности и аффинности родительского антитела нечеловеческого происхождения. В целом, гуманизированные антитела содержат один или более вариабельных доменов, в которых HVRs, например, CDRs (или их участки) происходят из антитела не человека, a FRs (или их участки) происходят из последовательностей антитела человека. Гуманизированное антитело необязательно также может содержать по меньшей мере часть константного домена антитела человека. В определенных случаях некоторые FR аминокислотные остатки в гуманизированном антителе замещены соответствующими остатками антитела не человека (например, антитела, из которого получены остатки HVR), в частности, для восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения описаны, например, в обзоре Almagro and Fransson, Front. Biosci. 13: 1619-1633 (2008) и далее, например, в обзоре Riechmann et al., Nature 332: 323-329 (1988); Queen et al., Proc. Natl. Acad. Sci. USA 86: 10029-10033 (1989); патентах US 5821337, 7527791, 6982321 и 7087409; Kashmiri et al., Methods 36: 25-34 (2005) (описана «прививка» участков, определяющих специфичность (SDR)); Padlan, Mol. Immunol. 28: 489-498 (1991) (описана «перестройка» поверхностей); Dall'Acqua et al., Methods 36: 43-60 (2005) (описана перестановка FR); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83: 252-260 (2000) (описан подход направленной селекции для перестановки FR).
Каркасные участки антитела человека, которые могут быть использованы для гуманизации, включают, без ограничений указанным: каркасные участки, отобранные методом «наилучшей подгонки» (см., например, Sims et al., J. Immunol. 151: 2296 (1993)); каркасные участки, полученные из консенсусной последовательности вариабельных участков антител человека определенной подгруппы легкой или тяжелой цепи (см., например, Carter et al., Proc. Natl. Acad. Sci. USA, 89: 4285 (1992); и Presta et al., J. Immunol. 151: 2296 (1993)); каркасные участки зрелых антител человека (соматически мутированные) или каркасные участки антител человека зародышевой линии (см., например, Almagro and Fransson, Front. Biosci. 13: 1619-1633 (2008)); и каркасные участки, полученные из скринированных FR-библиотек (см., например, Baca et al., J. Biol. Chem. 272: 10678-10684 (1997) и Rosok et al., J. Biol. Chem. 271: 22611-22618 (1996).
В некоторых вариантах осуществления изобретения обеспечиваемые антитела являются антителами человека. Антитела человека могут быть получены различными способами, известными из уровня техники. Такие антитела, в целом, описаны в работах van DiJk and van Winkel, Curr. Opin. Pharmacol. 5: 368-74 (2001) и Lonberg, Curr. Opin. Pharmacol. 20: 450-459 (2008).
Антитела человека могут быть получены путем введения иммуногена трансгенным животным, которые были модифицированы для продуцирования интактных антител человека или интактных антител с вариабельными доменами антитела человека в ответ на введение антигена. Такие животные обычно содержат все или часть иммуноглобулиновых человеческих локусов, которые замещают эндогенные иммуноглобулиновые локусы, или которые присутствуют экстрахромосомально или выборочно интегрированы в хромосомы животного. У такого рода трансгенных мышей эндогенные иммуноглобулиновые локусы обычно инактивированы. Обзор методов получения антител человека при использовании трансгенных животных опубликован Lonberg, Nat. Biotech. 23: 1117-1125 (2005). См. также, например, патенты US 6075181 и 6150584, описывающие технологию XENOMOUSE™, патент US 5770429, описывающий технологию HuMAB (зарегистрированный товарный знак); патент US 7041870, описывающий технологию K-М MOUSE (зарегистрированный товарный знак), и заявку US 2007/0061900, описывающую технологию VELOCIMOUSE (зарегистрированный товарный знак). Вариабельные участки антител человека из интактных антител, полученные при использовании трансгенных животных, могут быть далее модифицированы, например, путем объединения с константными участками различных антител человека.
Антитела человека также могут быть получены методом гибридомной технологии. Описаны клеточные линии миеломы человека и гетеромиеломы «человек-мышь» для продуцирования моноклональных антител человека. (См., например, Kozbor, J. Immunol., 133: 3001 (1984); Brodeur et al., Технологии получения и применения моноклональных антител, стр. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol., 147: 86 (1991). Антитела человека, полученные посредством технологии В-клеточных гибридом, также описаны Li et al., Proc. Natl. Acad. Sci. USA, 103: 3557-3562 (2006). Другие методы включают, например, описанные в патенте US 7189826 (раскрывается получение моноклональных антител человека IgM с помощью гибридомных клеточных линий) и в работе Ni, Xiandai Mianyixue, 26(4): 265-268 (2006) (описываются гибридомы «человек-человек»). Гибридомная технология для получения антител человека (триомная технология) также описана в работах Vollmers and Brandlein, Histology and Histopathology, 20(3): 927-937 (2005) и Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3): 185-91 (2005).
Антитела человека также могут быть получены путем выделения Fv-клона последовательностей вариабельного домена, отобранных из фагово-дисплейных библиотек человеческого происхождения. Такие последовательности вариабельного домена могут быть затем скомбинированы с нужным константным доменом человеческого происхождения. Технологии отбора антител человека из антительных библиотек описаны ниже.
5. Антитела, происходящие из библиотек
Антитела, согласно изобретению, могут быть выделены путем скрининга комбинаторных антительных библиотек на антитела нужной активности или активностей. Например, из уровня техники известны различные методы получения фагово-дисплейных библиотек и скрининга таких библиотек на антитела, обладающие желательными характеристиками связывания. Такие методы, например, описаны в обзорах Hoogenboon et al., Methods in Molecular Biology, 178: 1-37 (O'Brien et al., Human Press, Totowa, NJ, 2001) и далее, например, в работах McCafferty et al., Nature 348: 552-554; Clackson et al., Nature, 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Marks and Bradbury, Methods in Molecular Biology 248: 161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34): 12467-12472 (2004); Lee et al., J. Immunol. Methods 284(1-2): 119-132 (2004).
В некоторых фагово-дисплейных методах репертуар VH- и VL-генов по отдельности клонируют с помощью полимеразной цепной реакции (ПЦР) и выборочно рекомбинируют в фаговых библиотеках, которые затем могут быть скринированы на антигенсвязывающие фаги, как описано Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). Обычно на поверхности фага экспонируются фрагменты антител, такие, как одноцепочечные Fv (scFv)-фрагменты или Fab-фрагменты. Библиотеки из иммунизированных источников обеспечивают получение высокоаффинных антител к иммуногену, что не требует конструирования гибридом. Альтернативно, наивный репертуар может быть клонирован (например, из организма человека) с обеспечением конкретного источника антител к широкому спектру как несобственных, так и собственных антигенов без иммунизации, как описано Griffiths et al., EMBO J. 12: 725-734 (1993). И, наконец, наивные библиотеки могут быть созданы синтетическим путем клонирования не подвергнутых реарранжировке сегментов V-генов из стволовых клеток и при использовании ПЦР-праймеров, содержащих выборочные последовательности для кодирования высоко-вариабельных участков CDRs и достижения реарранжировки in vitro, как описано Hoogenboom and Winter, J. Mol. Biol, 227: 381-388 (1992). Патентные документы, раскрывающие фаговые библиотеки антител человека, включают, например, патент US 5750373 и патентные заявки US 2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Под антителами или фрагментами антител, выделенных из антительных библиотек человека, подразумеваются описанные здесь антитела человека или их фрагменты.
6. Мультиспецифические антитела
В контексте настоящего изобретения термин «мультиспецифическое антитело» относится к антителу, которое может специфически связываться с различными типами эпитопов. Более точно, мультиспецифические антитела представляют собой антитела, обладающие специфичностью по крайней мере к двум различным типам эпитопов, антитела, распознающие различные антигены, а также антитела, распознающие различные эпитопы одного и того же антигена. (Например, когда антигены представляют собой гетерологичные рецепторы, мультиспецифические антитела связываются с различными доменами, образующими гетерологичные рецепторы; альтернативно, когда антигены представляют собой мономеры, мультиспецифические антитела связываются с множественными сайтами на мономерных антигенах). Обычно, такие молекулы связываются с двумя антигенами (биспецифические антитела), но они даже могут иметь специфичность по отношению к большему числу антигенов (например, к трем). В специфических случаях один их антигенов представляет собой FIX и/или FIXa, а другой представляет собой FX. Биспецифические антитела могут быть получены в формате полноразмерных антител или фрагментов антител.
Технологии получения мультиспецифических антител включают, без ограничений указанным, рекомбинантную совместную экспрессию двух иммуноглобулиновых пар тяжелой и легкой цепей, имеющих различные специфичности (см. Milstein and Cuello, Nature, 305: 537 (1983)), WO 93/08829 и Traunecker et al., EMBO J. 10: 3655 (1991)), технология «выступ в полости» (см., например, патент US 5731168). Мультиспецифические антитела также могут быть получены с помощью технологии создания направленных электростатических эффектов для получения антительных Fc-гетеродимерных молекул (WO 2009/089004 А1); перекрестным связыванием двух или более антител или их фрагментов (см., например, патент US 4676980 и Brennan et al., Science, 229: 81 (1985)); использованием лейциновых застежек для получения биспецифических антител (см., например, Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992)); использованием технологии диател для получения фрагментов биспецифических антител (см., например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)); использованием одноцепочечных Fv (scFv)-димеров (см., например, Gruber et al., J. Immunol., 152: 5368 (1994)); и получением триспецифических антител, как описано, например, Tutt et al., J. Immunol, 147: 60 (1991).
Изобретение также включает антитела с тремя или более функциональными антигенсвязывающими участками, в том числе «октопус-антитела» (см., например, заявку US 2006/0025576А1).
7. Варианты антител
В некоторых вариантах изобретение охватывает аминокислотные последовательности вариантов антител. Например, это может быть нужным для улучшения связывающей аффинности и/или других биологических свойств антител. Аминокислотные последовательности вариантов антител могут быть получены путем введения подходящих модификаций в нуклеотидные последовательности, кодирующие исходные антитела, или путем пептидного синтеза. Такие модификации включают, например, делеции, и/или вставки, и/или замещения аминокислотных остатков в последовательностях антитела. Любые комбинации делеций, вставок или замещений могут быть сделаны для создания конечной конструкции с обеспечением того, что такая конструкция обладает желательными свойствами, например, антигенсвязывающими.
а) Варианты с делециями, вставками и замещениями
Согласно некоторым вариантам осуществления изобретение обеспечивает варианты антител, содержащие одно или более аминокислотных замещений. Представляющие интерес сайты для замещающего мутагенеза включают HVRs и FRs. Консервативные замены показаны в Таблице 1 в графе «Предпочтительные замены». Более существенные замены показаны в Таблице 1 в графе «Примеры замен» и далее описаны ниже со ссылками на классы боковых цепей антител. Аминокислотные замены могут быть введены в представляющие интерес антитела и полученные продукты скринированы на желательную активность, например, сохраняемую/улучшенную антигенсвязывающую способность, повышенную иммуногенность или улучшенную ADCC или CDC.

Аминокислоты могут быть сгруппированы в соответствии с общими боковыми цепями:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Thr, Tyr, Phe.
Неконсервативные замены будут обеспечивать изменение принадлежности аминокислоты к какому-либо из указанных классов на принадлежность к другому классу.
Один тип варианта с аминокислотными заменами включает замены одного или более аминокислотных остатков гипервариабельного участка родительского антитела (например, гуманизированного или человеческого антитела). В целом, полученный вариант (полученный варианты), отобранные для дальнейшего изучения, должны иметь модификации (например, улучшения) определенных биологических свойств (например, улучшенную аффинность, сниженную иммуногенность) по отношению к родительскому антителу. Примером варианта с заменами служит аффинно зрелое антитело, которое может быть получено любым удобным способом, например, при использовании технологии основанной на фаговом дисплее созревании аффинности, такой, как раскрывается в настоящем описании. Коротко говоря, один или более HVR-остатков подвергают мутагенезу и варианты антител, экспонируемые на поверхности фага, скринируют на определенную биологическую активность (например, аффинность связывания).
Изменения (например, замены) могут быть сделаны в HVRs, например, для улучшения аффинности антител. Такие изменения могут быть сделаны в «горячих точках» HVRs, т.е. затрагивать остатки, кодируемые кодонами, которые подвергаются мутациям с высокой частотой в процессе соматического мутагенеза (см., например, Chowdhury, Methods. Mol. Biol. 207: 179-196 (2008)), и/или остатки, которые контактируют с антигеном, с получением вариантов VH или VL, которые далее будут тестированы на аффинность связывания. Описана технология аффинного созревания из вторичных библиотек (см., например, Hoogenboom et al., Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, (2001).) В некоторых воплощениях аффинного созревания в генах вариабельных участков, отобранных для созревания, генерируют разнообразие любым из широко известных способов (например, ПЦР с введением ошибок, перестановка цепей или направленный мутагенез олигонуклеотидов). Далее создают вторичную библиотеку. Затем библиотеку скринируют для идентификации антител с желательной активностью. Другой способ генерации разнообразия включает HVR-направленные походы, в которых некоторые HVR-остатки (например, 4-6 в одно время) рандомизируют. HVR-остатки, вовлеченные в связывание с антигеном, могут быть специфическим образом идентифицированы, например, при использовании аланин-сканирующего мутагенеза или моделирования. Мишенями, в частности, часто служат CDR-H3 и CDR-L3.
В некоторых вариантах осуществления изобретения замены, вставки или делеции аминокислотных остатков могут происходить в пределах одного или более HVRs так, чтобы такие изменения существенно не уменьшали способность антител связывать антиген. Например, консервативные замены (в частности, замены, указанные выше), которые не снижают значительным образом аффинность связывания, могут быть сделаны в HVRs. Такие замены, могут быть генерированы, например, вне остатков HVRs, которые контактируют с антигеном. В некоторых воплощениях изобретения в вариантных последовательностях VH и VL, описанных выше, каждый HVR также может быть не изменен, или может содержать не более одной, двух или трех аминокислотных замен.
Подходящий метод для идентификации остатков или участков антитела, которые могут являться мишенями мутагенеза, называется «аланин-сканирующим мутагенезом», как описано Cunningham and Wells (1989) Science, 244: 1081-1085. Согласно данному методу, остаток или группу остатков, являющихся мишенями (например, заряженные остатки, такие, как Arg, Asp, His, Lys и Glu), идентифицируют и замещают нейтральными или отрицательно заряженными аминокислотами (например, аланином или полиаланином) для определения того, нарушается ли взаимодействие антитела с антигеном. Далее, замены могут быть введены в локализации аминокислот, демонстрирующих функциональную чувствительность к первоначальным заменам. Альтернативно или дополнительно может быть проанализирована кристаллическая структура комплекса «антиген-антитело» для идентификации точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки могут рассматриваться в качестве мишеней или быть элиминированы как кандидаты для замен. Варианты могут быть скринированы для определения того, обладают ли они желательными свойствами.
Вставки в аминокислотную последовательность включают амино- и/или карбокси-концевые слияния различной длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутрь последовательности одной или более аминокислот. Примеры терминальных вставок включают антитела с N-концевым метионином. Другие варианты вставок в молекулу антитела включают их слияние с ферментом (например, ADEPT) или с полипептидом, который увеличивает время полужизни антитела в плазме крови, по N- или С-концевому участку антитела.
б) Гликозилированные варианты
В некоторых вариантах осуществления изобретения обеспечиваемые антитела изменены для увеличения или уменьшения степени гликозилирования. Дополнение или делетирование сайтов гликозилирования в молекуле антитела может быть достигнуто путем изменения аминокислотной последовательности таким образом, что один или более сайтов гликозилирования создается или удаляется.
Когда антитела содержат Fc-участок, могут быть изменены углеводы, которые прикреплены к этому участку. Нативные антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленные олигосахариды, которые традиционно присоединены с помощью N-связи к Asn297 СН2-домена Fc-участка. См., например, Wright et al. TIBTECH 15: 26-32 (1997). Олигосахариды могут включать различные углеводы, например, маннозу, N-ацетил-глюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, прикрепленную к GlcNAc в «стволе» разветвленной структуры олигосахарида. В некоторых случаях модификации олигосахарида в молекуле антитела, согласно изобретению, могут быть сделаны для получения вариантов антител с определенными улучшенными свойствами.
в) Варианты Fc-участка
В некоторых вариантах осуществления изобретения одна или более аминокислотных модификаций могут быть введены в Fc-участок обеспечиваемых антител с получением варианта Fc-участка. Вариант Fc-участка может содержать последовательность Fc-участка человеческого происхождения (например, участок IgG1, IgG2, IgG3 или IgG4 человека), включающий аминокислотные модификации (например, замены) в одном или более положений аминокислот.
В некоторых вариантах осуществления изобретение охватывает варианты антител, обладающие некоторыми, но не всеми эффекторными функциями, что делает их желательными кандидатами для применения в тех случаях, когда время полужизни антител in vivo является важным, в то время как определенные эффекторные функции (такие, как связывание комплемента или ADCC), являются нежелательными или неприемлемыми. In vitro и/или in vivo исследования цитотоксичности могут быть сделаны для определения уменьшения/потери CDC и/или ADCC активностей. Например, исследования связывания Fc-рецептора (FcR) могут осуществляться для подтверждения того, что антитело утратило способность связывания с Fc-гамма рецептором (и вследствие этого, вероятно, утратило ADCC-активность), но сохранило способность связываться с FcRn. Первичные клетки, опосредующие ADCC, а именно, NK-киллеры, экспрессируют только Fc-гамма III-рецептор, в то время как моноциты экспрессируют Fc-гамма I-рецептор, Fc-гамма II-рецептор и Fc-гамма III-рецептор. Экспрессия FcR на гематопоэтических клетках суммирована в Таблице 3 на стр. 464 работы Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-492 (1991). Неограничивающие примеры исследований in vitro для определения ADCC-активности представляющей интерес молекулы приведены в патенте US 5500362 (см., например, Hellstrom, I. et al. Proc. Natl. Acad. Sci. USA 83: 7059-7063 (1986) и Hellstrom, I. et al. Proc. Natl. Acad. Sci. USA 82: 1499-1502 (1985); патенте US 5821337 (см. Bruggemann, M. et al., J. Exp. Med. 166: 1351-1361 (1987)). Альтернативно, могут быть использованы нерадиоактивные методы (см., например, АСТ1™ - нерадиоактивное исследование цитотоксичности для проточной цитометрии (CellTechnology, Inc. Mountain View, CA; и Cyto Tox 96 (зарегистрированный товарный знак)-нерадиоактивное исследование цитотоксичности (Promega, Madison, WI). Полезные эффекторные клетки для такого рода исследований включают мононуклеарные клетки периферической крови (РВМС) и природные киллеры (NK). Альтернативно или дополнительно ADCC-активность представляющей интерес молекулы может быть определена in vivo, например, на животных моделях, как описано в работе Clynes et al. Proc. Natl. Acad. Sci. USA 95: 652-656 (1998). Исследования связывания также могут быть выполнены для подтверждения того, что антитело неспособно связывать C1q и поэтому утратило CDC-активность. См., например, ELISA для оценки связывания C1q- и С3с в WO 2006/029879 и WO 2005/100402. С целью выявления активации комплемента может быть использован анализ CDC (см., например, Gazzano-Santoro et al J. Immunol Methods 202: 163 (1996); Cragg, M.S. et al., Blood 101: 1045-1052 (2003); и Cragg, M.S. and M. J. Glennie, Blood 103: 2738-2743 (2004)). Связывание EcRn и исследование in vivo клиренса/времени полужизни молекулы также могут быть осуществлены методами, известными из уровня техники (см., например, Petkova, S.B. et al., Intl. Immunol. 18(12): 1759-1769 (2006).
Описаны некоторые варианты антител с увеличенной или пониженной способностью к связыванию с FcRs. (См., например, патент US 6737056; WO 2004/056312 и Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001).)
Антитела с увеличенным временем полужизни и усиленной способностью к связыванию с неонатальным Fc-рецептором (FcRn), которые ответственны за перенос материнских IgGs к плоду (Guyer et al., J. Immunol. 117: 587 (1976) и Kim et al., J. Immunol. 24: 249 (1994)), описаны в заявке US 2005/0014934 A1 (Hinton et al.). Указанные антитела содержат Fc-участок с одной или более аминокислотными заменами, которые усиливают связывание Fc-участка с FcRn. Такие Fc-варианты имеют одну или более замен в Fc-участке в следующих положениях: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, замену в положении 434 (патент US 7371826).
См. также Duncan & Winter, Nature 322: 738-40 (1998); патент US 5648260; патент US 5624821; и WO 94/29351, описывающие другие примеры Fc-вариантов.
г) Варианты антител на основе инжиниринга цистеина
В некоторых вариантах осуществления изобретения может быть желательным создать антитела на основе инжиниринга цистеинов, например, «тиоMABs», в которых один или более аминокислотных остатков заменен на остатки цистеина. В частных вариантах воплощения изобретения заменяемые остатки находятся в доступных участках антитела. Для замены этих остатков на остатки цистеина в доступные сайты антитела вводят реакционноспособные тиоловые группы, которые могут быть использованы для конъюгации антитела с другими веществами, такими, как молекулы лекарственных средств или молекулы линкеров, связанных с молекулами лекарственных средств, с целью получения конъюгатов, как описано ниже. В некоторых случаях один или более следующих остатков могут быть заменены на цистеин: V205 (нумерация по Кабату) в легкой цепи; А118 (EU нумерация) тяжелой цепи; и S400 (EU нумерация) Fc-участка тяжелой цепи. Антитела на основе инжиниринга цистеина могут быть созданы, как раскрыто, например, в патенте US 7521541.
д) Производные антител
В некоторых вариантах осуществления изобретения обеспечиваемые антитела могут быть далее модифицированы для включения в их состав дополнительных соединений небелковой природы, которые хорошо известны и постоянно доступны. Соединения, подходящие для дериватизации антител, включают без ограничений указанным, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксалан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (как гомополимеры, так и рандомизированные сополимеры), декстран или поли(п-
винилпирролидон)полиэтиленгликоль, гомополимеры полипропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерол) или их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества в приготовлении благодаря его стабильности в воде. Полимер может быть любой мол. массы, причем как разветвленным, так и неразветвленным. К антителу может быть прикреплено различное число полимеров и, если более, чем один полимер прикреплен к антителу, это могут быть одинаковые или разные молекулы. В целом, число и/или тип полимеров, используемых для дериватизации антитела, могут определяться, без ограничений указанным, свойствами или функциями антитела, которые подлежат улучшению, возможностью использования антитела в терапии при определенных условиях, и т.д.
В другом варианте осуществления изобретения обеспечиваются конъюгаты антитела и компонентов небелковой природы, которые могут быть селективно нагреты путем подвергания радиоактивному облучению. Частным случаем компонента небелковой природы служит углеродная нанотрубка (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Радиоактивное облучение может иметь волны различной длины и включает, без ограничений, волны, которые не повреждают обычные клетки, но нагревают небелковый компонент до температуры, при которой погибают клетки в проксимальной ориентации по отношению к антителу в конъюгате «антитело-небелковый компонент».
В. Рекомбинантные способы и средства
Антитела могут быть получены с помощью рекомбинантных способов и средств, например, как описано в патенте US 4816567. В одном варианте осуществления изобретения обеспечивается изолированная нуклеиновая кислота, кодирующая описанное здесь мультиспецифическое антитело, которое связывается с FIX и/или FIXa и FX. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В другом варианте осуществления изобретения обеспечивается один или более векторов (например, векторов экспрессии), содержащих указанную молекулу нуклеиновой кислоты. Изобретение также касается клетки-хозяина, содержащей указанную молекулу нуклеиновой кислоты. В частности, клетка-хозяин содержит (например, она трансформирована): (1) вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, кодирующую аминокислотную последовательность, содержащую VL антитела. Согласно одному аспекту изобретения, клетка-хозяин является эукариотической клеткой, например, клеткой яичника китайского хомячка (СНО) или лимфоидной клеткой (например, Y0, NS0, Sp2/0). Согласно другому аспекту изобретения, обеспечивается способ получения мультиспецифического антитела, которое связывается с FIX и/или FIXa и FX, причем данный способ включает культивирование описанной выше клетки-хозяина, содержащей нуклеиновую кислоту, в условиях, обеспечивающих экспрессию мультиспецифического антитела, которое связывается с FIX и/или FIXa и FX, и, необязательно, выделение антитела из клетки-хозяина (или культуральной среды клетки-хозяина).
Для рекомбинантного получения мультиспецифического антитела, которое связывается с FIX и/или FIXa и FX, нуклеиновую кислоту, кодирующую антитело, например, описанную выше, выделяют и встраивают в один или более векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такая нуклеиновая кислота может быть без затруднений выделена и секвенирована при использовании соответствующих процедур (например, путем использования олигонуклеотидных проб, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи антитела).
Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитело, включают описанные здесь прокариотические или эукариотические клетки. Например, антитела могут быть получены в бактериях, в частности, когда не требуются гликозилирование и Fc-эффекторные функции. Экспрессия фрагментов антител и полипептидов в бактериях раскрыта, например, в патентах US 5648237, 5789199 и 5840523. (См. также работу Charlton, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ, 2003), стр. 245-254, в которой описана экспрессия фрагментов антител в Е. coli). После экспрессии антитело может быть переведено в раствор из гомогената бактериальных клеток и далее очищено.
В дополнение к прокариотическим клеткам, эукариотические микроорганизмы, такие, как нитевидные грибы или дрожжи, являются подходящими для клонирования или экспрессии антитело-кодирующих векторов в качестве клеток-хозяев, в том числе, штаммы грибов и дрожжей, пути гликозилирования в которых могут быть «гуманизированы», что приводит к продуцированию антитела, имеющего профиль гликозилирования, частично или полностью совпадающий с таковым антитела человека. См. Gerngross, Nat. Biotech. 22: 1409-1414 (2004) и Li et al., Nat. Biotech. 24: 210-215 (2006).
Подходящими клетками-хозяевами для экспрессии гликозилированного антитела также являются клетки многоклеточных организмов (позвоночных и не позвоночных). Примерами клеток не позвоночных являются клетки растений и насекомых. Идентифицированы многочисленные бакуловирусные штаммы, которые могут быть использованы совместно с клетками насекомых, в особенности, для трансфекции клеток Spodoptera frugiperda.
В качестве клеток-хозяев могут быть использованы и клетки растений. См., например, патенты US 5959177, 6040498, 6420548 и 6417429 (описывающие технологию PLANTIBODIES™ для продуцирования антител в трансгенных растениях).
В качестве клеток-хозяев могут быть использованы и клетки позвоночных. Например, могут применяться линии клеток млекопитающих, адаптированные для роста в суспензии. Другие примеры полезных линий клеток-хозяев млекопитающих включают линию клеток почки обезьяны CV1, трансформированную SV40 (COS-7); линию клеток почки человека (293 или клетки 293, как описано Graham et al., J. Gen. Virol. 36: 59 (1977)); клетки почки детеныша хомячка (ВНК); клетки мышей сертоли (ТМ4 клетки, как описано, например, в работе Mather, Biol. Reprod. 23: 243-251 (1980)); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки почки крыс буффало (BRL 3А); клетки легкого человека (W 138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, как описано, например, Mather et al., Annals N. Y. Acad. Sci. 383: 44-68 (1982); клетки MRC 5 и клетки FS4. Другие полезные линии клеток-хозяев млекопитающих включают клетки яичника китайского хомячка (СНО), включая DHFR- СНО (Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 4216 (1980)); и миеломные клеточные линии, такие, как Y0, NS0 и Sp2/0. Обзор некоторых линий клеток-хозяев млекопитающих, подходящих для продуцирования антител см. у Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), стр. 255-268 (2003).
С. Методы исследований
Мультиспецифические антигенсвязывающие молекулы, которые связывают FIX и/или FIXa и FX, обеспечиваемые настоящим изобретением, могут быть идентифицированы, скринированы или охарактеризованы с целью определения их физико-химических свойств и/или биологических активностей с помощью различных способов, известных из уровня техники.
1. Исследования связывания и другие методы
Согласно одному аспекту осуществления изобретения, обеспечиваемое антитело тестируют на активность связывания с антигеном, например, известными методами, такими, как ELISA, Вестерн-блоттинг и т.д.
Согласно другому аспекту осуществления изобретения, для идентификации антитела, которое конкурирует с каким-либо из описанных здесь мультиспецифических антител, связывающих FIX и/или FIXa и FX, за связывание с FIX и/или FIXa и FX, используют методы конкурентного анализа. В некоторых случаях такое конкурирующее антитело связывается с тем же самым эпитопом (например, линейным или конформационным эпитопом), который связывается описанными здесь мультиспецифическими антителами, связывающими FIX и/или FIXa и FX. Подробные примеры методов картирования эпитопа, с которым связываются антитела, приведены в работе Morris (1996) «Протоколы эпитопного картирования» в Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ).
Один пример конкурентного анализа включает инкубацию иммобилизованных FIX, и/или FIXa, или FX в растворе, содержащем первое меченное антитело, которое связывается с FIX, и/или FIXa, или FX (например, любое из описанных здесь антител), и второго немеченого антитела, которое тестируется на способность конкурировать с первым антителом за связывание с FIX, и/или FIXa, или FX. Второе антитело может находиться в гибридомном супернатанте. В качестве контроля иммобилизованные FIX, и/или FIXa, или FX инкубируют в растворе, содержащем первое меченное антитело, но не второе немеченое антитело. После инкубации в условиях, пермиссивных для связывания первого антитела с FIX, и/или FIXa, или FX, удаляют избыток несвязавшегося антитела, и измеряют количество метки, связанной с иммобилизованными FIX, и/или FIXa, или FX. Если количество метки, связанной с иммобилизованными FIX, и/или FIXa, или FX, существенно снижено в тестируемом образце по сравнению с контрольным, то это свидетельствует о том, что второе антитело конкурирует с первым за связывание с FIX, и/или FIXa, или FX. См. Harlow and Lane (1988) Антитела: лабораторный учебник, гл. 14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NJ).
2. Исследования активности
Согласно одному аспекту изобретения, обеспечиваются исследования для идентификации биологической активности мультиспецифической антигенсвязывающей молекулы, которая связывается с FIX, и/или FIXa, или FX и обладает биологической активностью. Биологическая активность может включать, например, способность обеспечивать образование FXa. Мультиспецифические антигенсвязывающие молекулы, имеющие такую биологическую активность in vivo и/или in vitro, также входят в объем настоящего изобретения.
В некоторых вариантах осуществления изобретения обеспечиваемое антитело тестируют на указанную выше биологическую активность. Выражение «замещающая функциональная активность кофактора FVIII» означает, например, способность увеличивать поглощение в тесте на реакцию активации FX под действием FIXa при использовании метода количественной колориметрии и способность повышать количество образуемого тромбина, что вычисляется в исследовании синтеза тромбина в сыворотке больных гемофилией А.
При использовании методов количественной колориметрии специфически может быть определена замещающая функциональная активность кофактора FVIII в измерительной системе, содержащей мультиспецифическую антигенсвязывающую молекулу, согласно изобретению, и, например, синтетический субстрат S-2222 (синтетический субстрат FXa), FIXa, и фосфолипид. Например, измерение может быть осуществлено следующим способом. Все реакции проводят при комнатной температуре. 5 мкл раствора антитела разводят физиологическим раствором в Трис-буфере, содержащем 0,1% бычьего сывороточного альбумина (далее обозначается как TBSB), смешивают с 5 мкл фактора 1Ха бета человека концентрации 150 нг/мл (Enzyme Research Laboratories) и полученную смесь инкубируют в 384-луночном планшете в течение 30 мин при комнатной температуре. Энзиматическую реакцию в этой смеси инициируют добавлением 5 мкл фактора X человека в концентрации 24, 7 мкг/мл (Enzyme Research Laboratories) и через четыре минуты останавливают добавлением 5 мкл 0,5 М ЭДТА. Хромогенную реакцию инициируют добавлением 5 мкл раствора хромогенного субстрата. Через 30 мин хромогенной реакции измеряют изменение поглощения при 405 нм с помощью SpectroMax 340РС384 (Molecular Devices). Растворителем для фактора IXa бета человека и фактора X человека является TBSB, содержащий 4,0 мкМ раствора фосфолипида (SYSMEX СО.) и 1,5 мМ CaCl2. S-2222 (SEKISUI MEDICAL) растворяют в очищенной воде с получением раствора хромогенного субстрата до концентрации 1, 47 мг/мл, и в описанном исследовании используют этот раствор. Измерительная система показывает корреляцию между тяжестью заболевания и клиническими симптомами в случае гемофилии A (Rosen S, Andersson М, Blomback М et al. Клинические применения метода хромогенного субстрата для определения активности FVIII. Thromb Haemost 1985; 54:811-23).
При использовании теста на образование тромбина замещающая функциональная активность кофактора FVIII может быть специфически оценена с помощью измерительной системы, содержащей мультиспецифическую антигенсвязывающую молекулу, согласно настоящему изобретению, и, например, FVIII-дефицитную плазму, активированный коагулирующий фактор крови XI, фосфолипид, Флуо-буфер, Флуо-субстрат (FluCa-Kit; синтетический субстрат тромбина). Например, измерение может быть осуществлено следующим образом. 8 мкл биспецифического антитела, разведенного TBSB, добавляют к 72 мкл FVIII-дефицитной плазмы (George King) и смесь инкубируют в течение 30 мин или более при комнатной температуре. Далее добавляют 20 мкл триггерного раствора, содержащего 20 мкМ фосфолипида и 5 нг/мл фактора XIa (Enzyme Research Laboratories). Затем инициируют реакцию коагуляции добавлением 20 мкл смеси раствора Флуо-буфера и Флуо-субстрата FluCa-Kit (Thrombinospore). Количество образовавшегося тромбина может быть определено при использовании системы флуоресцентного измерения и анализа синтеза тромбина (Thrombinospore). Тест на образование тромбина, использующий плазму больных гемофилией А, показывает общую коагулирующую активность в случае гемофилии А и корреляцию с клиническими симптомами заболевания (Shima М, Matsumoto Т & Ogiwara К. Новые исследования мониторинга лечения гемофилии. Haemophilia 2008; 14:83-92).
Термин «активирующая-ингибирующая активность FIX» относится к увеличению поглощения в реакции активации FIX под действием FXIa при использовании количественного колориметрического метода. Специфически, применяется следующая процедура. 5 мкл раствора антитела, разбавленного TBSB, смешивают с 5 мкл фактора IX человека в концентрации 3 Ед/мл (Christmassin М, Japan Blood Products Organization) и полученную смесь инкубируют в 384-луночном планшете в течение 30 мин при комнатной температуре. Энзиматическую реакцию в смеси инициируют добавлением 5 мкл фактора Х1а человека концентрации 90 нг/мл (Enzyme Research Laboratories) и через 60 минут останавливают добавлением 5 мкл 0,5 М ЭДТА. Хромогенную реакцию инициируют добавлением 10 мкл раствора хромогенного субстрата. Через 60 мин хромогенной реакции измеряют изменение поглощения при 405 нм с помощью SpectroMax 340РС384 (Molecular Devices). Растворителем для фактора IX и фактора ХГа человека является TBSB, содержащий 6,0 мкМ раствора фосфолипида (SYSMEX СО.) и 1,5 мМ CaCl2. Спектрозим FIXa (Sekisui Diagnostics) растворяют в очищенной воде с получением раствора хромогенного субстрата до концентрации 6, 7 мг/мл, смешивают с этиленгликолем в соотношении 5:8 и в описанном исследовании используют этот раствор.
Согласно патентной заявке US 2014/0080153 (WO 2012/093704), связывание антитела с ВМ (внеклеточным матриксом) может быть определено следующим образом. ВМ со свободным феноловым красным (BD Matrigel # 6137013) разводят TBS до концентрации 2 мг/мл и далее 5 мкл полученного раствора по каплям добавляют в центр каждой лунки планшета для ECL измерения (L15XB-6, MSD высокого связывания), размещенного на льду. Затем запечатывают планшет пленкой и выдерживают в течение ночи при 4°С. После этого образец антитела разводят до концентрации 9 мкг/мл ACES-T (20 mM ACES, 150 тМ NaCl, рН 7,4 или 5,8, с добавлением 0,01% Tween 20). Второе антитело разводят до концентрации 2 мг/мл ECLDB (ACES с добавлением 0,1% БСА и 0,01% Tween 20). 25 мкл раствора антитела добавляют круглодонный планшет, в каждую лунку которого внесено по 50 мкл ECLDB. Блокирующий буфер ECL удаляют из ЕСМ-планшета, содержащего Блокирующий буфер ECL, и 50 мкл каждого раствора антитела, указанного выше, добавляют в планшет. Далее, смесь перемешивают в течение часа при комнатной температуре. После удаления образца 50 мкл 0,25%-ного глутеральдегида (приготовленного с помощью ACES-Т) добавляют в каждую лунку и оставляют на 10 мин при комнатной температуре. Каждую лунку отмывают трижды PBS-T (PBS с добавлением 0,05% Tween 20), 50 мкл второго антитела, разведенного до концентрации 1 мкг/мл ECLDB, добавляют в каждую лунку и смесь перемешивают в течение часа при комнатной температуре на свету. Затем 150 мкл READ-буфера (MSD) добавляют в каждую лунку и детектируют сигнал световой эмиссии по сульфо-метке при использовании MESO SECTOR S600 (Meso Scale Discovery). F. Фармацевтические композиции
Описанные здесь фармацевтические композиции, содержащие мультиспецифическую антигенсвязывающую молекулу, которая связывается с FIX и/или FIXa и FX, приготавливают путем смешивания мультиспецифической антигенсвязывающей молекулы, имеющей нужную степень очистки, с одним или более необязательными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences, 16 издание, Osol, A. Ed. (1980)) в лиофилизированной форме или в форме водного раствора. Фармацевтически приемлемые носители обычно нетоксичны для реципиентов в используемых дозах и концентрациях и включают, без ограничений указанным, буферы, такие, как фосфатный, цитратный и на основе других органических кислот; антиоксиданты, в том числе, аскорбиновая кислота и метионин; консерванты (такие, как хлорид октадецилдиметилбензил аммония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкил-парабены, такие, как метил- или пропил-парабен; катехол; резорцинол; циклогексанол; 3-пентанол; m-крезол); полипептиды с низкой мол. массой (менее 10 аминокислотных остатков); белки, такие, как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие, как поливинилпирролидон; аминокислоты, такие, как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстраны; желирующие агенты, такие, как EDTA; сахара, такие, как сахароза, маннитол, трегалоза или сорбитол; образующие соль противоионы, такие, как натрий; комплексы металлов (например, Zn-белковые комплексы) и/или неионные сурфактанты, такие, как полиэтиленгликоль (ПЭГ). Примеры используемых здесь фармацевтически приемлемых носителей также включают агенты лекарственной дисперсии, такие, как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, растворимый гликопротеин гиалуронидазы человека РН-20, такой, как rHuPH20 (HYLENEX) (зарегистрированный товарный знак), Baxter International, Inc)). Некоторые примеры sHASEGP и методы его использования, включая rHuPH20, описаны в патентных заявках US 2005/0260186 и 2006/0104968. В одном аспекте sHASEGP объединяют с одной или более дополнительными глюкозаминоглюканазами, например, хондроитиназами.
Примерами лиофилизированных антительных композиций описаны в патенте US 6267958. Водные антительные композиции, например, описаны в патенте US 6171586 и WO 2006/044908 (раскрытые в этом документе композиции содержат гистидин-ацетатный буфер).
Обеспечиваемые изобретением композиции могут также содержать более одного активного ингредиента, если это необходимо для лечения определенного заболевания, предпочтительно, ингредиенты с комплементарной активностью и не оказывающие побочных эффектов друг на друга. Например, это может быть желательным для обеспечения FVIII, FVII, FIX, TFPI ингибитора, антитромбиновой нацеливающей siRNA, более специфически, Advate, Adynovate, Feiba, NovoSeven, NovoEight, N8-GP, N9-GP, Concizumab, Elocta и Fitusiran. Такие активные ингредиенты обычно присутствуют в комбинации в количествах, которые эффективны для обеспечения заданных целей. FVIII, FVII и FIX могут включать их слияния с Fc и ПЭГ.
Активные ингредиенты могут быть заключены в микрокапсулы, например, путем технологии коацервации или межповерхностной полимеризации, в частности, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметакрилатные) микрокапсулы, соответственно, в коллоидных системы доставки лекарств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие технологии описаны в Remington's Pharmaceutical Sciences, 16 издание, Osol, А. Ed. (1980).
Могут быть получены препараты с отложенным высвобождением. Подходящие примеры препаратов с отложенным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащие мультиспецифическую антигенсвязывающую молекулу, причем такие матрицы могут быть в виде оформленных частиц, например, пленок или микрокапсул.
Препараты для использования in vivo обычно являются стерильными. Стерильность может быть обеспечена, например, фильтрованием через стерильные фильтрующие мембраны.
G. Терапевтические способы и композиции
Любая обеспечиваемая изобретением мультиспецифическая антигенсвязывающая молекула, которая связывается с FIX и/или FIXa и FX, может быть использована в терапевтических способах.
Согласно одному аспекту, настоящее изобретение относится к мультиспецифической антигенсвязывающей молекуле, которая связывается с FIX и/или FIXa и FX, для применения в качестве лекарственного средства. Согласно другим аспектам, настоящее изобретение относится к мультиспецифической антигенсвязывающей молекуле, которая связывается с FIX и/или FIXa и FX, для применения для лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением. Согласно еще одним аспектам, настоящее изобретение относится к мультиспецифической антигенсвязывающей молекуле, которая связывается с FIX и/или FIXa и FX, для применения в способе лечения. Изобретение также относится к мультиспецифической антигенсвязывающей молекуле, которая связывается с FIX и/или FIXa и FX, для применения в способе лечения индивидуума, имеющего кровотечение, заболевание, в которое вовлечено кровотечение, или заболевание, вызываемое кровотечением, причем указанный способ включает введение индивидууму эффективного количества мультиспецифической антигенсвязывающей молекулы, которая связывается с FIX и/или FIXa и FX. В одном варианте осуществления изобретения указанный способ также включает введение индивидууму эффективного количества по крайней мере одного дополнительного терапевтического агента, например, как описано ниже. Согласно другим вариантам осуществления, настоящее изобретение относится к мультиспецифической антигенсвязывающей молекуле, которая связывается с FIX и/или FIXa и FX, для применения в целях замещения функции кофактора FVIII. Согласно некоторым вариантам осуществления, настоящее изобретение относится к мультиспецифической антигенсвязывающей молекуле, которая связывается с FIX и/или FIXa и FX, для применения в способе замещения функции кофактора FVIII у индивидуума, причем указанный способ включает введение индивидууму эффективного количества мультиспецифической антигенсвязывающей молекулы, которая связывается с FIX и/или FIXa и FX, для замещения функции кофактора FVIII. Под «индивидуумом» согласно любому из приведенных выше аспектов или вариантов осуществления изобретения предпочтительно понимается человек. Согласно настоящему изобретению, «кровотечение, заболевание, в которое вовлечено кровотечение, или заболевание, вызываемое кровотечением», предпочтительно, представляет собой заболевание, развитие и/или прогрессивное течение которого обусловлено снижением или дефицитом активности FVIII и/или активированного коагулирующего фактора крови VIII (FVIIIa). Такие заболевания включают указанную выше гемофилию А, гемофилию В, гемофилию С, заболевание, при котором непредвиденно образуется ингибитор FVIII/FVIIIa, приобретенную гемофилию и болезнь Виллебранда, без ограничений указанными.
Согласно еще одному аспекту, изобретение относится к применению мультиспецифической антигенсвязывающей молекулы, которая связывается с FIX и/или FIXa и FX, в производстве или приготовлении лекарственного средства. В одном случае лекарственное средство предназначено для лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением. В другом случае лекарственное средство предназначено для применения в способе лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением, причем указанный способ включает введение индивидууму, имеющему кровотечение, заболевание, в которое вовлечено кровотечение, или заболевание, вызываемое кровотечением, эффективного количества лекарственного средства. В одном варианте осуществления изобретения указанный способ также включает введение индивидууму эффективного количества по крайней мере одного дополнительного терапевтического агента, например, как описано ниже. Согласно другому варианту осуществления изобретения, лекарственное средство предназначено замещения функции кофактора FVIII. Согласно другому варианту осуществления изобретения, лекарственное средство предназначено для применения в способе замещения функции кофактора FVIII у индивидуума, причем указанный способ включает введение индивидууму эффективного количества лекарственного средства для замещения функции кофактора FVIII. Под «индивидуумом» согласно любому из приведенных выше аспектов или вариантов осуществления изобретения предпочтительно понимается человек. Согласно настоящему изобретению, «кровотечение, заболевание, в которое вовлечено кровотечение, или заболевание, вызываемое кровотечением», предпочтительно, представляет собой заболевание, развитие и/или прогрессивное течение которого обусловлено снижением или дефицитом активности FVIII и/или FVIIIa. Такие заболевания включают указанную выше гемофилию А, гемофилию В, гемофилию С, заболевание, при котором непредвиденно образуется ингибитор FVIII/FVIIIa, приобретенную гемофилию и болезнь Виллебранда, без ограничений указанными.
Согласно еще одному аспекту, изобретение относится к способу лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением. В одном варианте осуществления изобретения указанный способ включает введение индивидууму, имеющему кровотечение, заболевание, в которое вовлечено кровотечение, или заболевание, вызываемое кровотечением, эффективного количества мультиспецифической антигенсвязывающей молекулы, которая связывается с FIX и/или FIXa и FX. В другом варианте осуществления изобретения указанный способ также включает введение индивидууму эффективного количества по крайней мере одного дополнительного терапевтического агента, например, как описано ниже. Под «индивидуумом» согласно любому из приведенных выше аспектов или вариантов осуществления изобретения предпочтительно понимается человек.
Согласно одному аспекту, изобретение относится к способу замещения функции кофактора FVIII у индивидуума. Согласно другому аспекту изобретения, способ включает введение индивидууму эффективного количества мультиспецифической антигенсвязывающей молекулы, которая связывается с FIX и/или FIXa и FX, для замещения функции кофактора FVIII. В одном варианте осуществления изобретения индивидуумом является человек.
Один вариант осуществления изобретения относится к фармацевтическим композициям, содержащим любую из обеспечиваемых изобретением мультиспецифических антигенсвязывающих молекул, которые связываются с FIX и/или FIXa и FX, например, для применения в любом из описанных выше терапевтических способов. Согласно одному аспекту изобретения, фармацевтическая композиция содержит любую из обеспечиваемых изобретением мультиспецифических антигенсвязывающих молекул, которые связываются с FIX и/или FIXa и FX, и фармацевтически приемлемый носитель. Согласно другому аспекту изобретения, фармацевтическая композиция содержит любую из обеспечиваемых изобретением мультиспецифических антигенсвязывающих молекул, которые связываются с FIX и/или FIXa и FX, и по крайней мере один дополнительный терапевтический агент, например, как описано ниже.
Обеспечиваемая изобретением мультиспецифическая антигенсвязывающая молекула может быть использована в терапии как сама по себе, так и в комбинации с другими агентами. Например, обеспечиваемое изобретением антитело может совместно использоваться по крайней мере с одним дополнительным терапевтическим агентом. В некоторых вариантах осуществления изобретения дополнительный терапевтический агент представляет собой, например, FVIII, FVII, FIX, TFPI ингибитор, антитромбиновую нацеливающую siRNA, более специфически, Advate, Adynovate, Feiba, NovoSeven, NovoEight, N8-GP, N9-GP, Concizumab, Elocta и Fitusiran. FVIII, FVII и FIX могут включать их слияния с Fc и ПЭГ.
Описанные выше способы комбинированной терапии включают комбинированное введение (когда два или более терапевтических агента включены в одну или отдельные композиции) и отдельное введение, что подразумевает введение обеспечиваемой изобретением мультиспецифической антигенсвязывающей молекулы до, одновременно и/или после введения дополнительного терапевтического агента или агентов. Согласно одному аспекту осуществления изобретения, введение мультиспецифической антигенсвязывающей молекулы, которая связывается с FIX и/или FIXa и FX, и введение дополнительного терапевтического агента осуществляют примерно в течение месяца, двух или трех месяцев, или примерно в течение одной, двух или трех недель, или примерно в течение одного, двух, трех, четырех, пяти или шести дней.
Введение обеспечиваемой изобретением мультиспецифической антигенсвязывающей молекулы и любого дополнительного терапевтического агента может осуществляться любым подходящим образом, включая парентеральное, внутрилегочное, интраназальное и, если желательно, локальное применение и введение в очаг поражения. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, интраперитонеальное или подкожное введение. Дозирование может осуществляться любым подходящим способом, например, путем инъекций, таких, как внутривенные или подкожные инъекции, частично, в зависимости от того, является ли введение кратковременным или постоянным. Различные схемы дозирования включают, без ограничений указанным, однократное или многократное введение в различные временные точки, болюсное введение и пульсовую инфузию, что включено в объем настоящего изобретения.
Обеспечиваемая изобретением мультиспецифическая антигенсвязывающая молекула может быть сформирована, дозирована и введена таким образом, чтобы соответствовать наилучшей медицинской практике. Факторы, которые влияют на этот выбор, включают конкретное заболевание, подлежащее лечению, вид млекопитающего, подлежащего лечению, клиническое состояние индивидуального пациента, причину заболевания, место доставки средства, способ применения, схему введения и другие факторы, известные практикующим специалистам в области медицины. Мультиспецифическая антигенсвязывающая молекула необязательно может быть объединена с одним или более агентами, традиционно используемыми для предотвращения или лечения определенного нарушения. Эффективное количество таких других агентов зависит от количества мультиспецифической антигенсвязывающей молекулы, входящей в состав фармацевтической композиции, типа нарушения или вида терапии, а также от других факторов, которые приведены выше. Такие агенты обычно применяются в соответствии со способами и в дозах, описанных в настоящей заявке, или в количестве примерно от 1 до 99% описанных здесь доз, или в любой дозе и любым способом, которые эмпирически/клинически определены как подходящие.
Для предотвращения или лечения заболевания подходящая доза обеспечиваемой изобретением мультиспецифической антигенсвязывающей молекулы (когда она используется самостоятельно или в комбинации с одним или более другими дополнительными терапевтическими агентами) будет зависеть от вида заболевания, подлежащего лечению, типа мультиспецифической антигенсвязывающей молекулы, тяжести и причины заболевания, используется ли мультиспецифическая антигенсвязывающая молекула в профилактических или терапевтических целях, предшествующего лечения, клинической истории пациента, отвечаемости на антитело и благоразумия лечащего врача. Мультиспецифическая антигенсвязывающая молекула вводится пациенту однократно или путем серийных применений. В зависимости от типа и тяжести заболевания первоначальная кандидатная доза для введения пациенту может составлять от около 1 мкг/кг до 15 мкг/кг (например, 0,1 мг/кг 10 мг/кг), например, как при одном или более однократных применениях, так и при постоянной инфузии. Типичная однократная ежедневная доза может варьировать от около 1 мкг/кг до 100 мкг/кг или более, в зависимости от факторов, описанных выше. Для повторяющихся введений в течение нескольких дней или более, в зависимости от состояния, лечение будет продолжаться до тех пор, пока не будет достигнута желательная супрессия симптомов заболевания. В качестве примера дозы мультиспецифической антигенсвязывающей молекулы может быть доза от около 0,05 мг/кг до около 10 мг/кг. Таким образом, одна или более доз, составляющих примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любая их комбинация) могут быть введены пациенту. Такие дозы могут применяться с перерывами, например, каждую неделю или каждые три недели (например, таким образом, что пациент получает от около двух до около двадцати, или, например, около шести доз мультиспецифической антигенсвязывающей молекулы). Может быть использована первоначальная доза с более высокой нагрузкой, за которой следуют одна или более доз с меньшей нагрузкой. Однако, и другой дозовый режим может оказаться подходящим. Оказалась ли терапия успешной или нет, можно легко определить путем мониторинга с помощью подходящих технологий и исследований.
Следует понимать, что любая композиция или любой способ, описанные выше, могут включать использование иммуноконъюгатов, входящих в объем настоящего изобретения, вместо или дополнительно к мультиспецифической антигенсвязывающей молекуле, которая связывается с FIX и/или FIXa и FX.
Н. Упаковочные средства
Согласно другому аспекту осуществления, изобретение обеспечивает упаковочные средства, содержащие материалы, используемые для лечения, профилактики и/или диагностики описанных выше заболеваний. Такое средство включает контейнер и наклейку на контейнер или упаковочный вкладыш, ассоциированный с контейнером. Подходящие контейнеры включают, например, флаконы, ампулы, шприцы, пакеты с растворами и т.д. Контейнеры могут быть выполнены из различных материалов, таких, как стекло или пластик. Контейнер может содержать как отдельную композицию, так и композицию, объединенную с другим составом, эффективным для лечения, профилактики и/или диагностики заболевания, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного введения или флакон, прокалываемую пробку иглой для подкожной инъекции). По крайней мере один активный ингредиент композиции представляет собой обеспечиваемую изобретением мультиспецифическую антигенсвязывающую молекулу. Наклейка или упаковочный вкладыш указывают на то, что композиция используется для лечения выбранного заболевания. Более того, упаковочное средство может содержать (а) первый контейнер с помещенной внутрь композицией, причем композиция содержит обеспечиваемую изобретением мультиспецифическую антигенсвязывающую молекулу, и (b) второй контейнер с помещенной внутрь композицией, причем композиция содержит цитотоксический агент или другой терапевтический агент. Такое упаковочное средство может содержать и упаковочный вкладыш, указывающий на то, что композиции используются для лечения определенного состояния. Альтернативно или дополнительно упаковочное средство может содержать второй (или третий) контейнер, содержащий фармацевтически-приемлемый буфер, такой, как бактериостатическая вода для инъекции (BWFI), фосфатно-буферный раствор, раствор Рингера и раствор декстрозы. Средство также может включать другие материалы, нужные с точки зрения коммерческого производителя или частного потребителя для применения в соответствующих целях, в том числе другие буферы, растворители, фильтры, иглы и шприцы.
Другой вариант осуществления изобретения относится к антигенсвязывающей молекуле, в которой регулируется ассоциация между тяжелой и легкой цепями, способу получения антигенсвязывающей молекулы, в которой регулируется ассоциация между тяжелой и легкой цепями, и способу регулирования ассоциации между тяжелой и легкой цепями в антигенсвязывающей молекуле.
Антигенсвязывающая молекула, обеспечиваемая изобретением, относится к антигенсвязывающей молекуле, в которой регулируется ассоциация между тяжелой и легкой цепями, причем тяжелая цепь и легкая цепь, образующие антигенсвязывающую молекулу, представляют собой комбинацию представляющих интерес тяжелой и легкой цепей, и в которой аминокислотные остатки в определенных положениях константного участка тяжелой цепи (СН1) и константного участка легкой цепи (CL) представляют собой взаимно электростатически-отталкивающиеся остатки (имеющие одинаковый заряд).
В настоящем изобретении при переводе аминокислотных остатков в определенных положениях СН1 и CL нежелательной комбинации тяжелой и легкой цепей в аминокислотные остатки, которые взаимно электростатически отталкиваются (т.е. имеют одинаковый заряд), образование нежелательной комбинации тяжелой и легкой цепей может быть предотвращено за счет указанного отталкивания зарядов и в результате образуется нужная комбинация тяжелой и легкой цепей.
В настоящем изобретении выражения «регулировать ассоциацию» и «ассоциация регулируется» относятся к регулированию для достижения желательной ассоциации и, более специфически, к такому регулированию, когда нежелательная ассоциация между тяжелой и легкой цепями не образуется.
В настоящем изобретении понятие «интерфейс», в целом, относится к поверхности ассоциации, которая является результатом ассоциации (взаимодействия), и аминокислотные остатки, которые формируют интерфейс, обычно представляют собой один или более аминокислотных остатков, включенных в полипептидные участки, которые принимают участие в ассоциации, и, более предпочтительно, представляют собой аминокислотные остатки, которые приближаются друг к другу в процессе ассоциации и вовлечены во взаимодействие друг с другом. Более точно, такое взаимодействие включает, например, расстояние, на которое тесно сближаются аминокислотные остатки во время ассоциации с формированием водородных связей между ними, электростатических взаимодействий или солевых мостиков.
В настоящем изобретении понятие «аминокислотные остатки, формирующие интерфейс», более специфически, относится к аминокислотным остаткам, включенным в полипептидные участки, которые образуют интерфейс. Например, полипептидные участки, образующие интерфейс, относятся к полипептидным участкам, ответственным за селективное связывание между молекулами, такими, как антигенсвязывающие молекулы (например, антитела), лиганды, рецепторы или субстраты. Более точно, в антигенсвязывающих молекулах такие участки включают, например, константные участки тяжелой цепи, вариабельные участки тяжелой цепи, константные участки легкой цепи и вариабельные участки легкой цепи.
Согласно настоящему изобретению, модификации аминокислотных остатков относятся к замене оригинальных аминокислотных остатков (аминокислотного остатка) на другие аминокислотные остатки (аминокислотный остаток), делетированию оригинальных аминокислотных остатков (аминокислотного остатка), добавлению новых аминокислотных остатков (аминокислотного остатка), но, предпочтительно, относятся к замене одного или более оригинальных аминокислотных остатков на другие аминокислотные остатки.
В предпочтительном варианте осуществления антигенсвязывающей молекулы согласно настоящему изобретению, такая молекула имеет аминокислотные остатки в заданных положениях СН1 и CL нежелательной комбинации тяжелой и легкой цепей до регулирования ассоциации, которые электростатически взаимно отталкиваются (имеют одинаковый заряд).
При модификации аминокислотных остатков в указанной выше антигенсвязывающей молекуле с целью получения остатков, которые электростатически взаимно отталкиваются (имеют одинаковый заряд), считается, что ассоциация этих аминокислотных остатков ингибируется за счет репульсивной силы электрических зарядов.
Таким образом, в указанной выше антигенсвязывающей молекуле модифицированные аминокислотные остатки представляют собой, предпочтительно, аминокислотные остатки, которые приближены друг к другу при ассоциации в полипептидных участках, формирующих интерфейс.
Аминокислотные остатки, которые способны к ассоциации друг с другом, могут быть определены, например, при анализе трехмерной структуры полипептида и исследовании аминокислотных последовательностей полипептидных участков, которые формируют интерфейс в процессе полипептидной ассоциации. Аминокислотные остатки в области интерфейса, которые взаимно приближены друг к другу, являются предпочтительными мишенями «модификации» в обеспечиваемой изобретением антигенсвязывающей молекуле.
Известно, что некоторые аминокислоты имеют электрический заряд. В целом, считается, что лизин (K), аргинин (R) и гистидин (Н) являются аминокислотами, имеющими положительный заряд (положительно заряженные аминокислоты). Аспарагиновая кислота (D), глутаминовая кислота (Е) представляют собой аминокислоты, имеющие отрицательный заряд (отрицательно заряженные аминокислоты). Далее, аланин (А), аспарагин (N), цистеин (С), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (М), фенилаланин (F), пролин (Р), серин (S), треонин (Т), триптофан (W), тирозин (Y), валин (V) и подобные им аминокислоты являются аминокислотами, которые не имеют заряда (неполярные аминокислоты).
Таким образом, аминокислоты, которые взаимно электростатически отталкиваются (имеют одинаковый заряд), согласно настоящему изобретению, относятся к следующим аминокислотам:
аминокислоты, одна из которых является положительно заряженной аминокислотой, и другие аминокислоты также являются положительно заряженными аминокислотами и
аминокислоты, одна из которых является отрицательно заряженной аминокислотой, и другие аминокислоты также являются отрицательно заряженными аминокислотами и
Аминокислоты могут быть модифицированы в соответствии с различными методами, известными из уровня техники. Примеры таких методов включают, без ограничений указанным, сайт-направленный мутагенез (Hashimoto-Gotoh, Т., Mizuno, Т., Ogasahara, Y. and Nakagawa, М. (1995), Олигодезоксирибонуклеотид-направленный двойной амбер-метод для сайт-направленного мутагенеза, Gene 152, 271-275; Zoller, М.J. and Smith, M. (1983), Олигонуклеотид-направленный мутагенез ДНК-фрагментов, клонированных в М13 векторах, Methods Enzymol. 100, 468-500; Kramer, W., Drutsa, V., Jansen, H.W., Kramer, В., Pflugfelder, M. and Fritz, H.J. (1984), Дуплексный ДНК-метод с пробелами для олигонуклеотид-направленного создания мутаций, Nucleic Acids Res. 12, 9441-9456; Kramer, W. and Fritz, H.J. (1987) Олигонуклеотид-направленное создание мутаций посредством дуплексного ДНК-метода с пробелами, Methods Enzymol. 154, 350-367; Kunkel, Т. А. (1985) Быстрый и эффективный сайт-специфический мутагенез без селекции фенотипа, Proc. Natl. Acad. Sci. USA 82, 488-492); ПЦР-мутагенез; кассетный мутагенез и т.д.
Примеры модификаций аминокислот включают модификацию незаряженной аминокислоты или неполярной аминокислоты в положительно заряженную аминокислоту, модификацию незаряженной аминокислоты или неполярной аминокислоты в отрицательно заряженную аминокислоту, модификацию положительно заряженной аминокислоты в отрицательно заряженную аминокислоту и модификацию отрицательно заряженной аминокислоты в положительно заряженную аминокислоту. Кроме того, модификация незаряженной аминокислоты или неполярной аминокислоты в другую незаряженную или неполярную аминокислоту, модификация положительно заряженной аминокислоты в другую положительно заряженную и модификация отрицательно заряженной аминокислоты в другую отрицательно заряженную аминокислоту также включены в обеспечиваемые настоящим изобретением аминокислотные модификации.
Модификации аминокислот, согласно настоящему изобретению, включают создание одной модификации в каждой из тяжелой и легкой цепей или создание множественных модификаций в каждой из тяжелой и легкой цепей. В дополнение, число модификаций, создаваемых в тяжелой и легкой цепи, может быть одинаковым или различным.
Модификации аминокислот, согласно настоящему изобретению, включают множественные модификации с созданием положительно заряженных аминокислот в тяжелой или легкой цепи и множественные модификации с созданием отрицательно заряженных аминокислот в одной цепи. Более того, множественные модификации с получением положительно заряженных аминокислот, так же, как и множественные модификации с получением отрицательно заряженных аминокислот могут быть осуществлены в одной и той же тяжелой или легкой цепи. При указанных модификациях модификации в незаряженные или неполярные аминокислоты, а также модификации незаряженных или неполярных аминокислот могут быть скомбинированы.
Согласно настоящему изобретению, например, аминокислотные остатки одной из цепей могут использоваться, как если бы они не были модифицированы, и в таких случаях как тяжелая, так и легкая цепи не нуждаются в модификации, и только одна из цепей может быть модифицирована.
Несмотря на то, что практически не существует ограничений в количестве аминокислотных остатков, подвергаемых модификации в антигенсвязывающей молекуле, согласно изобретению, например, когда модифицируется константный участок антитела, то чтобы не происходило снижения связывающей активности по отношению к антигену и усиления иммуногенности, предпочтительно модифицировать так мало аминокислотных остатков, как это возможно. Приведенное выше слово «несколько» относится, например, к количеству, составляющему около 1-30, предпочтительно, около 1-20, даже более предпочтительно к количеству около 1-15, и наиболее предпочтительно, к 1-5.
Константный участок легкой цепи антигенсвязывающей молекулы, согласно изобретению, предпочтительно, представляет собой константный участок легкой цепи человеческого происхождения. Примеры константного участка легкой цепи антитела человека включают константные участки IgK (каппа), IgL1, IgL2, IgL3, IgL6 и IgL7 (лямбда). Константный участок легкой цепи антигенсвязывающей молекулы, согласно изобретению, практически не ограничен определенным типом; используемые легкие цепи могут принадлежать к различным типам, например, каппа и лямбда. Некоторые аллотипные последовательности, полученные путем генетического полиморфизма, описаны в Списке последовательностей белков, представляющих иммунологический интерес, публикация NIH No. 91-3242, как константный участок IgK (каппа) антитела человека и константный участок IgL7 (лямбда) антитела человека, и любой из них может использоваться в настоящем изобретении.
Константные участки антитела, в частности, константные участки тяжелой цепи могут быть модифицированы, если необходимо улучшить функциональную активность или стабильность антигенсвязывающей молекулы. Примерами модификаций, улучшающих функциональную активность антигенсвязывающей молекулы, являются модификации, которые усиливают или ослабляют связывание между антигенсвязывающей молекулой и Fc-гамма рецептором, модификации, которые усиливают или ослабляют связывание между антигенсвязывающей молекулой и FcRn, модификации, которые усиливают или ослабляют цитотоксическую активность (например, ADCC и CDC активность) антигенсвязывающей активности и подобные указанным. Дополнительно, в объем изобретения включены модификации, которые улучшают гетерогенность антигенсвязывающей молекулы, и модификации, которые улучшают ее иммуногенность и/или фармакокинетику.
Более того, сообщалось о снижении гетерогенности С-концевой последовательности тяжелой цепи IgG-антитела путем амидирования С-концевой карбоксильной группы за счет делеции С-концевой аминокислоты, а именно, лизина, или путем делеции двух С-концевых аминокислотных остатков, глицина и лизина в (Anal. Biochem. 2007 Jan 1:360 (1):75-83).
Таким образом, в настоящем изобретении для уменьшения гетерогенности С-концевого участка тяжелой цепи предпочтительно использовать IgG, в котором С-концевой лизин или С-концевые глицин и лизин делетированы.
Поскольку антигенность химерных и гуманизированных антител в организме человека снижена, химерные и гуманизированные антитела, содержащие последовательности человеческого происхождения, как ожидается, являются полезными для применения по отношению к человеку в терапевтических и подобных им целях.
Предпочтительным примером антигенсвязывающей молекулы, согласно изобретению, является гетеродимерный мультимер, содержащий два или более типов СН1 и два или более типов CL. Такой гетеродимерный мультимер, предпочтительно, связывается с двумя или более типами эпитопов, и примером его является мультиспецифическое антитело.
Предпочтительным примером мультиспецифического антитела, согласно изобретению, является биспецифическое антитело. Таким образом, предпочтительным вариантом антигенсвязывающей молекулы, согласно изобретению, является биспецифическое антитело, состоящее из двух типов тяжелых цепей (первая тяжелая цепь и вторая тяжелая цепь) и двух типов легких цепей (первая легкая цепь и вторая легкая цепь).
При более точном описании биспецифических антител как предпочтительных вариантов обеспечиваемых изобретением антигенсвязывающих молекул следует отметить, что указанная выше «первая тяжелая цепь» относится к одной из двух тяжелых цепей (Н цепей), формирующих биспецифическое антитело, а «вторая тяжелая цепь» относится к другой тяжелой цепи, которая отличается от первой Н цепи. Соответственно, из двух Н цепей одна может быть произвольно обозначена как первая Н цепь, а другая может быть обозначена как вторая Н цепь. Сходным образом, «первая легкая цепь» относится к одной из двух легких цепей (L цепей), формирующих биспецифическое антитело, а «вторая легкая цепь» относится к другой легкой цепи, которая отличается от первой L цепи. Из двух L цепей одна может быть произвольно обозначена как первая L цепь, а другая может быть обозначена как вторая L цепь. Обычно, первая L цепь и первая Н цепь получены из одного и того же антитела, которое связывается с конкретным антигеном (или эпитопом), а вторая L цепь и вторая H цепь также получены из одного и того же антитела, которое связывается с конкретным антигеном (или эпитопом). В настоящем описании пара L цепь - H цепь, образованная первой H цепью и первой L цепью, называется первой парой; а пара L цепь - Н цепь, образованная второй Н цепью и второй L цепью, называется второй парой. Антиген (или эпитоп), используемый для получения антитела, из которого происходит вторая пара, предпочтительно, отличается от антигена, используемого для получения антитела, из которого происходит первая пара. Более точно, антигены, распознаваемые первой парой и второй парой, могут быть одинаковыми, но, предпочтительно, пары связываются с различными антигенами (или эпитопами). В этом случае Н цепи и L цепи первой пары и второй пары, предпочтительно, имеют аминокислотные последовательности, которые отличаются друг от друга. Когда первая пара и вторая пара связываются с различными эпитопами, первая пара и вторая пара могут распознавать полностью различные антигены, или же могут распознавать различные сайты (различные эпитопы) одного и того же антигена. Кроме того, одна из них может распознавать антиген, представляющий собой белок, пептид, ген или сахар, а другая пара может распознавать цитотоксическое вещество, такое, как радиоактивные соединения, химиотерапевтические агенты или происходящие из клеток токсины. Однако, когда желательно получить антитело, содержащее пары, образованные специфическими комбинациями Н цепей и L цепей, такие специфические Н цепи и L цепи могут быть произвольно определены как первая пара и вторая пара.
Ниже более подробно описано биспецифическое антитело IgG класса, имеющее два типа константных участков СН1 тяжелой цепи (СН1-А и СН1-В) и два типа константных участков легкой цепи (CL-A и CL-B); однако настоящее изобретение также обеспечивает и другие антитела.
Если желательно получить биспецифическое антитело, которое будет распознавать один эпитоп за счет первого СН1-А и первого CL-A, и связывать другой эпитоп за счет второго СН1-В и второго CL-B, теоретически, существует возможность, что 10 типов антительных молекул могут быть получены, когда каждый из четырех типов цепей экспрессируется для производства антитела.
В этом случае нужные молекулы антитела могут быть преимущественным образом произведены, если, например, регулируется ассоциация между цепями, так, что ассоциация СН1-А и CL-B и/или между СН1-В и CL-A ингибируется.
Примером является модифицирование аминокислотных остатков, формирующих интерфейс между СН1-А и CL-B, в положительно заряженные аминокислотные остатки и модифицирование аминокислотных остатков, формирующих интерфейс между СН1-В и CL-A, в отрицательно заряженные аминокислотные остатки. В результате таких модификаций нежелательная ассоциация между СН1-А и CL-B ингибируется, поскольку все аминокислотные остатки, формирующие интерфейс, являются положительно заряженными, и ассоциация между СН1-В и CL-A также ингибируется, поскольку все аминокислотные остатки, формирующие интерфейс, являются отрицательно заряженными. Таким образом, нежелательная ассоциация между СН1-А и CL-B и ассоциация между СН1-В и CL-A ингибируется, поскольку аминокислотные остатки, формирующие интерфейс, взаимно имеют одинаковый заряд. Таким образом, антитела, имеющие желательную ассоциацию между СН1-А и CL-A и желательную ассоциацию между СН1-В и CL-B, могут быть успешно получены. Более того, обеспечивается преднамеренная ассоциация между СН1-А и CL-A, поскольку аминокислотные остатки, формирующие интерфейс, имеют различные заряды по отношению друг к другу и преднамеренная ассоциация между СН1-В и CL-B также обеспечивается, поскольку аминокислотные остатки, формирующие интерфейс, имеют различные заряды по отношению друг к другу. Соответственно, могут быть успешно получены антитела, имеющие желательную ассоциацию цепей.
Другим примером является модифицирование аминокислотных остатков, формирующих интерфейс между СН1-А и CL-B, в положительно заряженные аминокислотные остатки, когда аминокислотные остатки, формирующие интерфейс между СН1-В и CL-A являются взаимно незаряженными или неполярными аминокислотами. В результате такой модификации нежелательная ассоциация между СН1-А и CL-B ингибируется, поскольку все аминокислотные остатки, формирующие интерфейс, являются положительно заряженными. С другой стороны, поскольку аминокислотные остатки, формирующие интерфейс, являются аминокислотными остатками, которые взаимно электростатически не отталкиваются, желательная ассоциация между СН1-А и CL-A и желательная ассоциация между СР1-В и CL-B будет происходить более легко в том случае, когда аминокислотные остатки электростатически отталкиваются. Соответственно, антитела, имеющие желательную ассоциацию между СН1-А и CL-A и желательную ассоциацию между СН1-В и CL-B, могут быть успешно получены. В то же время в данном примере в том случае, когда аминокислотные остатки, формирующие интерфейс между CL-A и СН1-В, не являются взаимно незаряженными или неполярными аминокислотами, они могут быть модифицированы таким образом, что становятся взаимно незаряженными или неполярными аминокислотами.
Более того, в другом примере, когда аминокислотные остатки, формирующие интерфейс между CL-B и СН1-В, являются незаряженными или неполярными аминокислотами в СН1-В, один из аминокислотных остатков, формирующих интерфейс между СН1-А и CL-A, является модифицированным в положительно заряженный аминокислотный остаток, в то время как другой аминокислотный остаток является модифицированным в отрицательно заряженный аминокислотный остаток, а аминокислотные остатки, формирующие интерфейс между CL-B и СН1-В в CL-B, являются модифицированными таким образом, что имеют тот же самый заряд, что модифицированные остатки в СН1-А. В результате такой модификации при том, что желательная ассоциация между СН1-А и CL-A промотирутся, поскольку аминокислотные остатки, формирующие интерфейс, представляют собой комбинацию положительно заряженных и отрицательно заряженных остатков, желательная ассоциация между СН1-В и CL-B не ингибируется, поскольку аминокислотные остатки, формирующие интерфейс, представляют собой аминокислотные остатки, которые взаимно электростатически не отталкиваются. В итоге может быть успешно получено антитело, имеющее преднамеренную ассоциацию между СН1-А и CL-A и преднамеренную ассоциацию между СН1-В в CL-B. Там не менее, в этом примере, в том случае, когда аминокислотные остатки, формирующие интерфейс между CL-B и СН1-В, не являются незаряженными или неполярными аминокислотами в СН1-В, они могут быть модифицированы таким образом, что становятся незаряженными или неполярными аминокислотами.
Дополнительно, использование регулирования ассоциации, согласно настоящему изобретению, делает возможным супрессировать ассоциацию между CH1s (СН1-А и СН1-В) или ассоциацию между CLs (CL-A и CL-B). Специалисты в данной области техники будут способны без затруднений установить типы аминокислотных остатков, которые тесно приближаются друг к другу в процессе ассоциации в интерфейсе СН1 и CL в нужном полипептиде, для которых регулирование ассоциации, согласно настоящему изобретению, является нужным.
Далее, специалисты в данной области техники также без затруднений будут способны выявить последовательности, которые могут быть использованы как СН1 или CL антитела в организме человека, обезьяны, мыши, кролика и других подобных организмах, путем применения доступных баз данных. Более точно, информация об аминокислотных последовательностях СН1 или CL может быть получена с помощью средств, описанных ниже в Примерах.
Например, с точки зрения биспецифических антител, описанных ниже в Примерах, специфические примеры аминокислотных остатков, которые тесно приближаются друг к другу (находятся в непосредственной близости или в контакте) в процессе ассоциации в интерфейсе СН1 и CL, включают следующие комбинации:
- глутамин (Q) в положении 175, согласно нумерации EU, в СН1 и контактирующий треонин (Т) или серин (S) в положении 180, согласно нумерации по Кабату, в CL;
- глутамин (Q) в положении 175, согласно нумерации EU, в СН1 и контактирующий треонин (Т) или серин (S) в положении 131, согласно нумерации по Кабату, в CL;
- глутамин (Q) в положении 175, согласно нумерации EU, в СН1 и контактирующий серин (S) или треонин (Т) в положении 131 и серин (S) или треонин (Т) в положении 180, согласно нумерации по Кабату, в CL; и
- лизин (К) в положении 147 и глутамин (Q) в положении 175, согласно нумерации EU, в СН1 и контактирующий серин (S) или треонин (Т) в положении 131 и серин (S) или треонин (Т) в положении 180, согласно нумерации по Кабату, в CL.
В настоящем изобретении номера положений, согласно нумерации EU, указаны в соответствии с EU-нумерацией (Последовательности белков, представляющих иммунологический интерес, NIH Публикация No. 91-3242). В настоящем изобретении выражения «аминокислотный остаток в положении X согласно нумерации EU» и «аминокислота в положении X согласно нумерации EU» (где X является произвольным номером) также могут быть прочитаны, как выражения «аминокислотный остаток, который соответствует положению X, согласно нумерации EU» и «аминокислота, которая соответствует положению X, согласно нумерации EU». Как указано в Примерах, изложенных ниже, желательные антигенсвязывающие молекулы могут быть преимущественно получены путем модификации этих аминокислотных остатков при использовании методов, раскрытых в настоящем изобретении.
В одном варианте осуществления настоящее изобретение обеспечивает антигенсвязывающую молекулу, в которой регулируется ассоциация между тяжелой цепью и легкой цепью, причем один, или два, или более наборов аминокислотных остатков, выбранных из группы, содержащей наборы от (а) до (в), как показано ниже, в тяжелой цепи и в легкой цепи указанной антигенсвязывающей молекулы представляют собой аминокислотные остатки, которые взаимно электростатически отталкиваются друг от друга:
(а) аминокислотный остаток в константном домене тяжелой цепи (СН1) в положении 17, согласно нумерации EU, и аминокислотный остаток в константном домене легкой цепи (CL) в положении 180, согласно нумерации по Кабату;
(б) аминокислотный остаток в СН1 в положении 175, согласно нумерации EU, и аминокислотный остаток в CL в положении 131, согласно нумерации по Кабату;
(в) аминокислотные остатки в СН1 в положениях 147 и 175, согласно нумерации EU, и аминокислотные остатки в CL в положениях 131 и 180, согласно нумерации по Кабату.
В указанной выше антигенсвязывающей молекуле «аминокислотные остатки, которые взаимно электростатически отталкиваются друг от друга» или «аминокислотные остатки, имеющие одинаковый заряд», предпочтительно, выбраны из аминокислотных остатков, включенных, например, в наборы (X) или (Y), как показано ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D);
(Y) лизин (К), аргинин (R) и гистидин (Н).
В указанной выше антигенсвязывающей молекуле специфические примеры наборов аминокислотных остатков, которые взаимно электростатически отталкиваются друг от друга, включают наборы аминокислотных остатков, указанные ниже:
(а) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, и аминокислотный остаток, содержащийся в CL в положении 180, согласно нумерации EU;
(б) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, и аминокислотный остаток, содержащийся в CL в положении 131, согласно нумерации по Кабату;
(в) аминокислотные остатки, содержащиеся в СН1 в положениях 147 и 175, согласно нумерации EU, и аминокислотные остатки, содержащиеся в CL в положениях 131 и 180, согласно нумерации по Кабату;
(г) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, и аминокислотный остаток, содержащийся в CL в положениях 131 и 180, согласно нумерации по Кабату.
Настоящее изобретение обеспечивает антигенсвязывающую молекулу, в которой один, или два, или более наборов аминокислотных остатков, выбранных из группы, содержащей наборы от (a1) до (с2), как показано ниже, в тяжелой цепи и в легкой цепи указанной антигенсвязывающей молекулы представляют собой следующие аминокислотные остатки, которые взаимно электростатически отталкиваются друг от друга:
(a1) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D) и аминокислотный остаток, содержащийся в CL в положении 180, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D);
(а2) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R) и аминокислотный остаток, содержащийся в CL в положении 180, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R);
(б1) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D) и аминокислотный остаток, содержащийся в CL в положении 131, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D);
(б2) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R) и аминокислотный остаток, содержащийся в CL в положении 131, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R);
(в1) аминокислотные остатки, содержащиеся в СН1 в положениях 147 и 175, согласно нумерации EU, каждый представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D) и аминокислотные остатки, содержащиеся в CL в положениях 131 и 180, согласно нумерации EU, каждый представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D);
(в2) аминокислотные остатки, содержащиеся в СН1 в положениях 147 и 175, согласно нумерации EU, каждый представляет собой лизин (К), гистидин (Н) или аргинин (R) и аминокислотные остатки, содержащиеся в CL в положениях 131 и 180, согласно нумерации EU, каждый представляет собой лизин (К), гистидин (Н) или аргинин (R).
В указанной выше антигенсвязывающей молекуле специфические примеры аминокислотных остатков, которые взаимно электростатически отталкиваются друг от друга, включают аминокислоты, указанные ниже:
(a1) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D) и аминокислотный остаток, содержащийся в CL в положении 180, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D);
(а2) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R) и аминокислотный остаток, содержащийся в CL в положении 180, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R);
(б1) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D) и аминокислотный остаток, содержащийся в CL в положении 131, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D);
(б2) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R) и аминокислотный остаток, содержащийся в CL в положении 131, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R);
(в1) аминокислотные остатки, содержащиеся в СН1 в положениях 147 и 175, согласно нумерации EU, каждый представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D) и аминокислотные остатки, содержащиеся в CL в положениях 131 и 180, согласно нумерации EU, каждый представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D);
(в2) аминокислотные остатки, содержащиеся в СН1 в положениях 147 и 175, согласно нумерации EU, каждый представляет собой лизин (К), гистидин (Н) или аргинин (R) и аминокислотные остатки, содержащиеся в CL в положениях 131 и 180, согласно нумерации EU, каждый представляет собой лизин (К), гистидин (Н) или аргинин (R);
(г1) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D) и аминокислотные остатки, содержащиеся в CL в положениях 131 и 180, согласно нумерации EU, каждый представляет собой глутаминовую кислоту (Е) или аспарагиновую кислоту (D);
(г2) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, представляет собой лизин (К), гистидин (Н) или аргинин (R) и аминокислотные остатки, содержащиеся в CL в положениях 131 и 180, согласно нумерации EU, каждый представляет собой лизин (К), гистидин (Н) или аргинин (R).
В дополнение к указанному выше, технология ингибирования нежелательной ассоциации CH1/CL путем введения отталкивания электрического заряда в интерфейс между СН1 и CL (WO 2013/065708) может быть также применена по отношению к обеспечиваемой изобретением антигенсвязывающей молекуле. Более точно, настоящее изобретение относится к антигенсвязывающей молекуле, имеющей СН1 и CL, причем один, или два, или более наборов аминокислотных остатков, которые взаимно электростатически отталкиваются друг от друга, выбраны из группы, включающей наборы аминокислотных остатков от (а) до (d), указанные ниже:
(а) аминокислотный остаток, содержащийся в константном участке тяжелой цепи (СН1) в положении 147, согласно нумерации EU, и аминокислотный остаток, содержащийся в константном участке легкой цепи (CL) в положении 180, согласно нумерации EU;
(б) аминокислотный остаток, содержащийся в СН1 в положении 147, согласно нумерации EU, и аминокислотный остаток, содержащийся в CL в положении 131, согласно нумерации EU;
(в) аминокислотный остаток, содержащийся в СН1 в положении 175, согласно нумерации EU, и аминокислотный остаток, содержащийся в CL в положении 160, согласно нумерации EU;
(г) аминокислотный остаток, содержащийся в СН1 в положении 213, согласно нумерации EU, и аминокислотный остаток, содержащийся в CL в положении 123, согласно нумерации EU.
Методика введения электростатического отталкивания в интерфейс второго константного участка тяжелой цепи (СН2) или третьего константного участка тяжелой цепи (СН3) для подавления нежелательной ассоциации между тяжелыми цепями, методика введения электростатического отталкивания в интерфейс вариабельного участка тяжелой цепи и вариабельного участка легкой цепи для подавления нежелательной ассоциации между тяжелой и легкой цепями или методика модифицирования аминокислотных остатков, формирующих гидрофобное ядро, находящееся в интерфейсе вариабельного участка тяжелой цепи и вариабельного участка легкой цепи, в полярные аминокислоты, имеющие электрический заряд, для подавления нежелательной ассоциации между тяжелой цепью и легкой цепью, также могут быть применены по отношению к антигенсвязывающиммолекулам, обеспечиваемым изобретением (см. WO 2006/106905).
В технологии, которая супрессирует нежелательную ассоциацию между тяжелыми цепями путем введения электростатического отталкивания в интерфейсе СН2 и СН3, примеры аминокислотных остатков, которые находятся в контакте в зоне интерфейса других константных участков тяжелой цепи, включают участки, соответствующие положению 356 (нумерация EU) и положению 439 (нумерация EU), положению 357 (нумерация EU) и положению 370 (нумерация EU) и положению 399 (нумерация EU) и положению 409 (нумерация EU) в СН3 участке. С нумерацией константных участков антитела можно ознакомиться в публикации Kabat et al. (Kabat, E.A., et al., 1991, Белковые последовательности, представляющие иммунологический интерес, NIH); для ознакомления с нумерацией константных участков тяжелой цепи следует обратиться к EU нумерации.
Более точно, например, в антигенсвязывающей молекуле, содержащей два типа СН3 участков тяжелой цепи, от одного до трех наборов аминокислотных остатков в первом СН3 участке тяжелой цепи, которые выбраны из наборов аминокислотных остатков от (1) до (3), могут быть получены как взаимно электростатически отталкивающиеся:
(1) аминокислотные остатки, содержащиеся в СН3 участке тяжелой цепи в положении 356 и в положении 439, согласно нумерации EU;
(2) аминокислотные остатки, содержащиеся в СН3 участке тяжелой цепи в положении 357 и в положении 370, согласно нумерации EU;
(3) аминокислотные остатки, содержащиеся в СН3 участке тяжелой цепи в положении 399 и в положении 400, согласно нумерации EU.
Более того, антитело может представлять собой антитело, имеющее набор аминокислотных остатков во втором СН3 участке тяжелой цепи, которые отличаются от указанных выше аминокислотных остатков в первом СН3 участке тяжелой цепи, причем набор аминокислотных остатков выбран из наборов аминокислотных остатков от (1) до (3), как указано выше, и причем от одного до трех наборов аминокислотных остатков, которые соответствуют наборам (1)-(3), как указано выше, которые взаимно электростатически отталкиваются в первом СН3 участке тяжелой цепи, электростатически не отталкиваются от соответствующих аминокислотных остатков в первом СН3 участке тяжелой цепи.
Аминокислотные остатки, приведенные в указанных выше наборах от (1) до (3), приближаются друг к другу в процессе ассоциации. Специалист в данной области знаний будет способен определить сайты, соответствующие аминокислотным остаткам, указанным в наборах от (1) до (3), для желательного СН3 участка тяжелой цепи или константного участка тяжелой цепи путем гомологичного моделирования и использования коммерчески доступного программного обеспечения, а также для подходящей модификации аминокислотных остатков в указанных сайтах.
Применительно к указанной выше антигенсвязывающей молекуле выражения «электростатическое отталкивание» и «имеющие одинаковый заряд» относятся, например, к любым двум или более аминокислотным остаткам, которые приведены выше в любой из групп от (X) до (Y). В предпочтительном варианте воплощения указанной выше
антигенсвязывающей молекулы первый СН3 участок тяжелой цепи и второй СН3 участок тяжелой цепи могут быть перекрестно-связаны друг с другом за счет дисульфидных связей.
В настоящем изобретении аминокислотный остаток, подвергающийся модификации, не ограничен аминокислотным остатком вариабельного участка антигенсвязывающей молекулы или константного участка антитела, указанным выше. Специалист в данной области знаний будет способен определить аминокислотные остатки, которые формируют интерфейс в варианте полипептида или гетеромерном мультимере, путем гомологичного моделирования и использования коммерчески доступного программного обеспечения для модификации аминокислотных остатков в указанных сайтах с целью регулирования ассоциации. Гомологичное моделирование представляет собой технологию для предсказания трехмерной структуры белка с помощью коммерчески доступных компьютерных программ. При установлении строения белка неизвестной трехмерной структуры, в первую очередь следует обнаружить белок, трехмерная структура которого является высоко гомологичной структуре искомого белка. Далее, при использовании трехмерной структуры в качестве матрицы следует сконструировать строение белка с неизвестной пространственной структурой и затем оптимизировать строение искомого белка с помощью методов молекулярной динамики и подобных им методов для предсказания его трехмерной структуры.
В технологии введения электростатического отталкивания в интерфейс вариабельного участка тяжелой цепи и вариабельного участка легкой цепи для подавления нежелательной ассоциации тяжелой цепи и легкой цепи примеры аминокислотных остатков, которые контактируют в интерфейсе вариабельного участка тяжелой цепи (VH) и вариабельного участка легкой цепи (VL), включают глутамин (Q) в положении 39 согласно нумерации по Кабату в VH (FR2-участок) и контактирующий глутамин (Q) в положении 38 согласно нумерации по Кабату в VL (FR2-участок). Более того, предпочтительным примером является лейцин (L) в положении 45 согласно нумерации по Кабату в VH (FR2-участок) и контактирующий пролин (Р) в положении 44 согласно нумерации по Кабату в VL (FR2-участок). Нумерация этих сайтов сделана в соответствии с опубликованной нумерацией по Кабату (Kabat, Е.А., et al., 1991, Белковые последовательности, представляющие иммунологический интерес, NIH).
Поскольку известно, что аминокислотные остатки являются высоко консервативными у человека и мыши (J. Mol. Recognit. 2003; 16:113-120), ассоциация вариабельных участков антигенсвязывающих молекул может быть регулирована для ассоциации VH-VL других антигенсвязывающих молекул помимо указанных в Примерах, путем модификации аминокислотных остатков, соответствующих указанным выше аминокислотным остаткам.
Специфический пример относится к антигенсвязывающей молекуле, в которой два или более аминокислотных остатка, формирующих интерфейс VH и VL, являются аминокислотными остатками, которые взаимно электростатически отталкиваются. Более точно, примеры включают антигенсвязывающую молекулу с одним или более наборами аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, приведенных ниже в (а) или (б):
(а) аминокислотный остаток, содержащийся в VH в положении 39 согласно нумерации по Кабату, и аминокислотный остаток, содержащийся в VL в положении 38 согласно нумерации по Кабату; или
(б) аминокислотный остаток, содержащийся в VH в положении 45 согласно нумерации по Кабату, и аминокислотный остаток, содержащийся в VL в положении 44 согласно нумерации по Кабату.
Каждый из аминокислотных остатков, указанный выше в (а) или (б), приближается к другому в процессе ассоциации. Специалист в данной области техники будет способен определить сайты, которые соответствуют аминокислотным остаткам, приведенным выше в (а) или (б), в желательных VH или VL путем гомологичного моделирования и использования коммерчески доступного программного обеспечения, для подходящей модификации аминокислотных остатков в указанных сайтах.
В указанной выше антигенсвязывающей молекуле «аминокислотные остатки, которые взаимно электростатически отталкиваются друг от друга» или «аминокислотные остатки, имеющие одинаковый заряд», предпочтительно, выбраны из аминокислотных остатков, включенных, например, в наборы (X) или (Y), как показано ниже:
(X) глутаминовая кислота (Е) и аспарагиновая кислота (D);
(Y) лизин (К), аргинин (R) и гистидин (Н).
В технологии модификации аминокислотных остатков, формирующих гидрофобное ядро, находящееся в интерфейсе VH или VL, в полярные аминокислотные остатки, имеющие электрический заряд, для подавления нежелательной ассоциации тяжелой цепи и легкой цепи предпочтительные примеры аминокислотных остатков, которые способны сформировать гидрофобное ядро в интерфейсе VH и VL, включают лейцин (L) в положении 45 согласно нумерации по Кабату в VH (FR2-участок) и контактирующий пролин (Р) в положении 44 согласно нумерации по Кабату в VL (FR2-участок). Нумерация этих сайтов сделана в соответствии с публикацией Кабата (Kabat, Е.А., et al., 1991, Белковые последовательности, представляющие иммунологический интерес, NIH), которая включена в настоящее описание в качестве ссылки.
В целом, термин «гидрофобное ядро» относится к структуре, образованной боковыми цепями гидрофобных аминокислот во внутренней области ассоциированных полипептидов. Примеры гидрофобных аминокислот включают аланин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан и валин. Более того, аминокислотные остатки, иные чем гидрофобные аминокислоты (например, тирозин) могут быть вовлечены в формирование гидрофобного ядра. Это гидрофобное ядро вместе с гидрофильной поверхностью, в которой гидрофильные боковые цепи аминокислот экспонированы во внешнюю часть, становится направляющей силой для обеспечения ассоциации водорастворимых полипептидов. Когда гидрофобные аминокислоты двух различных доменов присутствуют на поверхности молекулы и экспонированы к молекулам воды, энтропия, как и свободная энергия, возрастает. Соответственно, два домена будут ассоциировать друг с другом с уменьшением свободной энергии и становятся стабильными, а гидрофобные аминокислоты в интерфейсе будут проникать во внутреннюю часть молекулы с формированием гидрофобного ядра.
Считается, что, когда происходит ассоциация полипептидов, формирование гидрофобного ядра ингибируется за счет модификации гидрофобных аминокислот, образующих гидрофобное ядро, в полярные аминокислоты, имеющие электрический заряд, и, соответственно, ассоциация полипептидов подавляется.
Специалист в данной области техники будет способен установить присутствие или отсутствие гидрофобного ядра, образование сайта (участка) и т.п. для анализа аминокислотных последовательностей желательной антигенсвязывающей молекулы. Антигенсвязывающая молекула, обеспечиваемая настоящим изобретением, представляет собой антигенсвязывающую молекулу, характеризующуюся тем, что в этой молекуле аминокислотные остатки, способные формировать гидрофобное ядро в интерфейсе, являются модифицированными аминокислотными остатками, имеющими электрический заряд. Более точно, примеры включают антигенсвязывающую молекулу, в которой аминокислотные остатки, показанные ниже в (1) или (2), являются аминокислотными остатками, имеющими электрический заряд. Боковые цепи аминокислотных остатков, приведенных в (1) и (2), примыкают друг к другу и могут сформировать гидрофобное ядро: аминокислотный остаток, содержащийся в VH в положении 45 согласно нумерации по Кабату, и
аминокислотный остаток, содержащийся в VL в положении 44 согласно нумерации по Кабату.
Предпочтительные примеры аминокислотных остатков, имеющих электрический заряд, в указанной выше антигенсвязывающей молекуле включают глутаминовую кислоту(Е), аспарагиновую кислоту (D), лизин (К), аргинин (R) и гистидин (Н). Более предпочтительные примеры включают глутаминовую кислоту (Е) и лизин (К).
В целом, аминокислотные остатки человека и мыши, описанные выше в (1) и (2), представляют собой:
лейцин (L) и
пролин (Р).
Таким образом, в предпочтительном варианте осуществления изобретения указанные аминокислотные остатки подвергаются модификации (таким, как замены на аминокислотные остатки, имеющие электрический заряд). Более того, типы указанных выше аминокислотных остатков в (1) и (2) необязательно ограничены указанными выше аминокислотными остатками, а могут представлять собой и другие аминокислоты, эквивалентные указанным аминокислотным остаткам.
Другие известные технологии могут быть приведены по отношению к обеспечиваемой изобретением антигенсвязывающей молекуле. Например, для обеспечения ассоциации первого VH (VH1) и первого VL (VL1) и/или второго VH (VH2) и второго VL (VL2) аминокислоты, находящиеся в вариабельном участке одной из Н цепей, могут быть заменены на аминокислоты с более крупными боковыми цепями (выпуклыми), и аминокислоты, находящиеся в противоположном вариабельном участке другой Н цепи, могут быть заменены на аминокислоты с меньшими боковыми цепями (формирующими впадину), так что выпуклость может быть размещена во впадине и обеспечивается ассоциация VH1 и VL1 и/или VH2 и VL2 и, соответственно, ассоциация VH1 и VL2 и/или VH2 и VL1 может быть супрессирована (WO 1996/027011; Ridgway, J.B., et al., Белковая инженерия (1996) 9, 617-621; Merchant, A.M., et al., Nature Biotechnology (1998) 16, 677-681).
Например, в случае IgG1 человека для замены аминокислоты в СН3 участке Н цепи на аминокислоту с более крупной боковой цепью (формирование выпуклости) осуществляют модификации Y349C и T366W, а для замены аминокислоты в СН3 участке другой Н цепи на аминокислоту с меньшей боковой цепью (формирование впадины) осуществляют модификации D356C, T336S, L368A и Y407V.
Другие известные технологии также могут быть применены по отношению к обеспечиваемой изобретением антигенсвязывающей молекуле. Нацеленная на мишень антигенсвязывающая молекула может быть успешно получена комплементарной ассоциацией СН3 за счет использования инженерии обмена цепей в домене СН3, причем участок СН3 одной из Н цепей антигенсвязывающей молекулы замещается последовательностью, происходящей из IgA и соответствующей указанному участку, а в комплементарный участок СН3 другой Н цепи вводится последовательность, происходящая из IgA и соответствующая указанному комплементарному участку (Protein Engineering Desin & Selection, 23:195-202, 2010).
Существуют и другие известные технологии, которые также могут быть применены по отношению к обеспечиваемой изобретением антигенсвязывающей молекуле. При производстве биспецифических антител нацеленное на мишень биспецифическое антитело может быть получено, например, изменением изоэлектрической точки за счет внесения различных аминокислотных модификаций в каждый вариабельный участок каждой из двух типов Н цепей, принимая во внимание, что измененная изоэлектрическая точка позволит более эффективно очищать антитело ион-обменной хроматографией (WO 2007/114325).
Технология модификации аминокислотного остатка в положении 435 согласно нумерации EU, который имеет отношение к сайту связывания между IgG и белком А, в аминокислоту, имеющую другую величину связывания с белком А, такую, как аргинин, также может быть использована по отношению к обеспечиваемой изобретением антигенсвязывающей молекуле в комбинации с указанными выше подходами. При использовании указанной технологии взаимодействие между Н цепью и белком А может быть изменено и только гетеродимерные антигенсвязывающие молекулы могут быть эффективно очищены на колонке с белком А. Указанная технология также может быть использована независимо от комбинирования с описанными выше методиками. В соответствии с настоящим изобретением могут быть осуществлены различные модификации антигенсвязывающих молекул, такие, как одна из описанных ниже модификаций. Например, может быть получена антигенсвязывающая молекула, имеющая структуру, в которой для обеспечения ассоциации первого VH (VH1) и первого VL (VL1) и/или второго VH (VH2) и второго VL (VL2) VH1 связан с Fc-участком посредством первого СН1 и VL1 связан с первым CL, a VH2 связан с другим Fc-участком посредством второго CL и VL2 связан со вторым CHI (WO 09/80254).
Более того, например, две или более указанных выше технологий могут быть использованы совместно по отношению к обеспечиваемой изобретением антигенсвязывающей молекуле. Так, в соответствии с изобретением, антигенсвязывающая молекула может быть получена на основании антитела, которое было модифицировано с помощью указанных выше известных подходов.
Описанные ниже способы регулирования ассоциации цепей, согласно настоящему изобретению, позволяют, например, эффективно производить антигенсвязывающие молекулы, которые являются активными. Примеры активности включают связывающую активность, нейтрализующую активность, цитотоксическую активность, агонистическую активность, антагонистическую активность, энзиматическую активность и т.д. Агонистическая активность представляет собой активность, которая индуцирует определенный вид изменений в физиологической активности посредством связывания антигенсвязывающей молекулы с антигеном, таким, как рецептор, который вызывает передачу сигнала в клетки. Примерами физиологической активности являются ростовая активность, выживаемость, дифференцировка, транскрипционная активность, активность мембранного транспорта, связывающая активность, протеолитическая активность, фосфорилирующая/дефосфорилирующая активность, окислительно-восстановительная активность, трансферная активность, нуклеолитическая активность, дегидратационная активность, активность индукции клеточной смерти и апоптоз-индуцирующая активность, без ограничений указанным.
Антигенсвязывающие молекулы, которые связываются с желательными антигенами или с желательными рецепторами, могут быть успешно получены с помощью методов, описанных в настоящем изобретении.
В соответствии с настоящим изобретением типы антигенов практически не ограничены и может быть использован любой антиген. Примеры антигенов включают рецепторы или их фрагменты, раковые антигены, антигены МНС, дифференцировочные антигены и другие подобные указанным антигены, без ограничений указанными.
Примеры рецепторов, согласно настоящему изобретению, включают рецепторы, принадлежащие к семейству рецепторов гематопоэтического фактора, семейству цитокинового рецептора, семейству рецептора тирозинкиназы, семейству рецептора типа серин/треонинкиназа, семейству рецептора TNF, семейству рецептора связанного G-белка, семейству рецептора заякоренного GPI, семейству рецептора типа тирозинфосфатаза, семейству рецептора фактора адгезии, семейству гормонального рецептора и т.п. Обзоры, посвященные рецепторам, принадлежащим указанным семействам, и их характеристики, приведены в различных публикациях, например, Cooke В.А., King R.J.В., van der Molen H.J. ed. Новая усовершенствованная биохимия, Vol. 18В «Гормоны и их действие», часть II, стр. 1-46 (1998) Elsevier Science Publishers BV., New York, USA; Patthy L. (1990) Cell, 61:13-14; Ullrich A., et al. (1990) Cell, 61: 203-212; Massagul J. (1992) Cell, 69: 1067-1070; Miyajima A., et a/.(1992) Annu. Rev. Immunol., 10: 295-331; Taga T. and Kishimoto T. (1992) FASEB J., 7: 3387-3396; Fantl W.I. et al. (1993) Annu. Rev. Biochem., 62: 453-481; Smith., et al. (1994) Cell, 76: 959-962; Flower D.R. (1999) Biochim. Biophys. Acta, 1422: 207-234; Miyasaka M. ed. Cell Technology, Серия справочников «Справочник по факторам адгезии» (1994) Shujunsha, Tokyo, Japan; и т.д. Примеры специфических рецепторов, принадлежащим указанным выше семействам рецепторов, включают рецептор эритропоэтина (ЭПО) человека или мыши, рецептор гранулоцитарного колоние-стимулирующего фактора (G-CSF) человека или мыши, рецептор тромбопоэтина (ТПО) человека или мыши, рецептор инсулина человека или мыши, рецептор лиганда Flt-3 человека или мыши, рецептор происходящего из тромбоцитов ростового фактора (PDGF) человека или мыши, рецептор интерферона (ИФР) альфа или,TNF человека или мыши, рецептор лептина человека или мыши, рецептор гормона роста (ГР) человека или мыши, рецептор интерлейкина-10 (ИЛ-10) человека или мыши, рецептор инсулин-подобного ростового фактора (ИГФ) I человека или мыши, рецептор ингибирующего лейкоз (ЛИФ) фактора человека или мыши и рецептор цилиарного нейротрофического фактора (ЦНТФ) человека или мыши (hEPOR: Simon, S. et al. (1990) Blood 76, 31-35; mEPOR: D'Andrea, A.D. et al. (1989) Cell 57, 277-285; hG-CSFR: Fukunaga, R. et. al. (1990) Proc. Natl. Acad. Sci. USA. 87, 8702-8706; mG-CSFR: Fukunaga, R. et. al. (1990) Cell 61, 341-350; hTPOR: Vigon, I. et. al. (1992) 89, 5640-5644; mTPOR: Skoda, R.C. et. al. (1993) 12, 2645-2653; hlnsR: Ullrich, A. et. al. (1985) Nature 313, 756-761; hFlt-3: Small, D. et. al. (1994) Proc. Natl. Acad. Sci. USA. 91, 459-463; hPDGFR: Gronwald, RGK. et. al. (1998) Proc. Natl. Acad. Sci. USA. 85, 3435-3439; hIFN альфа/бета рецептор: Uze, G. et. al. (1990) Cell 60, 225-234; и Novick, D. et. al. (1994) Cell 77, 391-400).
Раковые антигены представляют собой антигены, которые экспрессируются после злокачественной трансформацией клетки, и также называются опухолево-специфическими антигенами. Кроме того, необычные сахарные цепи, которые появляются на поверхности клетки или на белковой молекуле, когда клетка становится злокачественной, также считаются раковыми антигенами и могут называться антигенами раковой сахарной цепи. Примеры раковых антигенов включают ЕрСАМ, который экспрессируется при множестве раковых опухолей, включая рак легких (Proc. Natl. Acad. Sci. USA (1989) 86 (1), 27-31) (полинуклеотидная последовательность антигена указана как RefSeq номер доступа NM 002354.2 и полипептидная последовательность антигена указана как RefSeq номер доступа NP_002345.2), СА19-9, СА15-3, сиалилSSЕА-1 (SLX) и др.
Антигены МНС в широком смысле классифицируются на антигены МНС класса I и антигены МНС класса II. Антигены МНС класса I включают HLA-A, -В, -С, -Е, -F, -G и -Н, а антигены МНС класса II включают HLA-DR, -DQ и -DP. Антигены дифференцировки включают CD1, CD2, CD3, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD49e, CD49f, CD51, CD54, CD55, CD56, CD57, CD58, CD61, CD62E, CD62L, CD62P, CD69, CD71, CD73, CD95, CD102, CD106, CD122, CD126 и CDw130.
Антигенсвязывающая молекула, согласно настоящему изобретению, может представлять собой биспецифическое антитело; в этом случае два антигена (или эпитопа), распознаваемые этим антителом, могут быть произвольно выбраны из указанных выше рецепторов или их фрагментов, раковых антигенов, антигенов МНС, антигенов дифференцировки и др. Например, два антигена могут быть выбраны из рецепторов или их фрагментов, раковых антигенов, антигенов МНС и антигенов дифференцировки. Кроме того, каждый один антиген может быть выбран из двух антигенов, произвольно выбранных, например, из рецепторов или их фрагментов, раковых антигенов, антигенов МНС и антигенов дифференцировки.
Настоящее изобретение также обеспечивает способ получения антигенсвязывающей молекулы, в которой регулируется ассоциация между тяжелой цепью и легкой цепью. Предпочтительный вариант указанного выше способа по настоящему изобретению относится к способу получения антигенсвязывающей молекулы, в которой регулируется ассоциация между тяжелой и легкой цепью, причем указанный способ включает:
(1) модификацию нуклеиновых кислот, кодирующих СН1 и CL, так, что один набор, или два, или более наборов аминокислотных остатков, выбранных из группы, включающей наборы аминокислотных остатков, показанные ниже в наборах от (а) до (в), принадлежат аминокислотным остаткам, которые электростатически отталкиваются друг от друга:
(а) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 180 согласно нумерации по Кабату;
(б) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 131 согласно нумерации по Кабату; и
(в) аминокислотные остатки в СН1 в положениях 147 и 175 согласно нумерации EU и аминокислотные остатки в CL в положениях 131 и 180 согласно нумерации по Кабату;
(2) введение модифицированных нуклеиновых кислот в клетку-хозяин и культивирование клетки-хозяина таким образом, что указанные нуклеиновые кислоты экспрессируются; и
(3) сбор антигенсвязывающей молекулы из культуральной среды клетки-хозяина.
Дополнительно, настоящее изобретение относится к способу, включающему на указанной выше стадии (а) модификацию нуклеиновых кислот так, что аминокислотные остатки, которые электростатически отталкиваются друг от друга, выбраны из аминокислотных остатков, входящих в состав каждой из указанных выше групп (X) и (Y).
Настоящее изобретение также относится к способу получения, содержащему указанную выше стадию (1), включающую модификацию нуклеиновых кислот таким образом, что два или более аминокислотных остатка, формирующих интерфейс между VH и VL, представляют собой аминокислотные остатки, которые электростатически отталкиваются друг от друга. Предпочтительно, аминокислотные остатки, которые электростатически отталкиваются друг от друга, принадлежат любому набору остатков, выбранному из группы, содержащей, например, наборы аминокислотных остатков (а) и (б), как показано ниже:
(а) аминокислотный остаток в VH в положении 39 согласно нумерации по Кабату и аминокислотный остаток в VL в положении 38 согласно нумерации по Кабату или (б) аминокислотный остаток в VH в положении 45 согласно нумерации по Кабату и аминокислотный остаток в VL в положении 44 согласно нумерации по Кабату.
Указанные выше аминокислотные остатки, которые электростатически отталкиваются друг от друга, предпочтительно, выбраны из аминокислотных остатков, входящих в любой из наборов (X) и (Y), указанных выше.
Кроме того, настоящее изобретение относится к способу регулирования ассоциации между тяжелой и легкой цепями антигенсвязывающей молекулы. Предпочтительный вариант способа регулирования ассоциации, согласно настоящему изобретению, относится к способу регулирования ассоциации между тяжелой и легкой цепями антигенсвязывающей молекулы, включающему модификацию нуклеиновых кислот таким образом, что один, или два, или более наборов аминокислотных остатков, выбранных из группы, содержащей наборы аминокислотных остатков от (а) до (с), как показано ниже, представляют собой аминокислотные остатки, которые электростатически отталкиваются друг от друга:
(а) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 180 согласно нумерации EU;
(б) аминокислотный остаток в СН1 в положении 175 согласно нумерации EU и аминокислотный остаток в CL в положении 131 согласно нумерации EU;
(в) аминокислотные остатки в СН1 в положениях 147 и 175 согласно нумерации EU и аминокислотные остатки в CL в положениях 131 и 180 согласно нумерации EU.
Настоящее изобретение также относится к способу регулирования ассоциации, содержащему указанную выше стадию (1), включающую модификацию нуклеиновых кислот таким образом, что аминокислотные остатки, которые электростатически отталкиваются друг от друга, принадлежат любому набору остатков, выбранному из указанной выше группы, содержащей наборы аминокислотных остатков (X) или (Y).
Настоящее изобретение также относится к способу регулирования ассоциации, содержащему указанную выше стадию (1), включающую модификацию нуклеиновых кислот таким образом, что два или более аминокислотных остатка, формирующих интерфейс между VH и VL, представляют собой аминокислотные остатки, которые электростатически отталкиваются друг от друга. Предпочтительно, аминокислотные остатки, которые электростатически отталкиваются друг от друга, принадлежат любому набору остатков, выбранному из группы, содержащей, например, наборы аминокислотных остатков (а) и (б), как показано ниже:
(а) аминокислотный остаток в VH в положении 39 согласно нумерации по Кабату и аминокислотный остаток в VL в положении 38 согласно нумерации по Кабату,
(б) аминокислотный остаток в VH в положении 45 согласно нумерации по Кабату и аминокислотный остаток в VL в положении 44 согласно нумерации по Кабату.
Согласно способу регулирования ассоциации в соответствии с настоящим изобретением, нужное биспецифическое антитело может быть успешно и преимущественно получено, как описано ранее. Конкретно, желательный гетеромерный мультимер в форме биспецифического антитела может быть эффективно сформирован из смеси мономеров.
Выражение «модификация нуклеиновых кислот», приведенное применительно к описанным выше способам, составляющим предмет настоящего изобретения, относится к такой модификации, что нуклеиновые кислоты обеспечивают интродуцирование аминокислотных остатков за счет указанной модификации, сделанной в соответствии с изобретением. Более точно, это выражение касается модификации нуклеиновых кислот, кодирующих оригинальные (пре-модифицированные) аминокислотные остатки, в нуклеиновые кислоты, кодирующие аминокислотные остатки, которые подлежат интродуцированию в результате модификации. Обычно, указанная выше процедура означает осуществление генных манипуляций или введение мутаций, что способно привести к вставке, делеции или замене, по крайней мере, одного нуклеотида в оригинальной нуклеиновой кислоте, в результате чего формируются кодоны, кодирующие представляющие интерес аминокислотные остатки. В частности, кодоны, кодирующие оригинальные аминокислотные остатки, замещаются кодонами, кодирующими аминокислотные остатки, которые должны быть интродуцированы в результате модификации. Такая модификация нуклеиновой кислоты может быть сделана без затруднений специалистом в данной области техники с использованием известных методов, таких, как сайт-специфический мутагенез и ПЦР-мутагенез.
Дополнительно, настоящее изобретение обеспечивает нуклеиновые кислоты, кодирующие антигенсвязывающую молекулу, согласно изобретению. Кроме того, векторы, содержащие такие нуклеиновые кислоты, также включены в объем настоящего изобретения.
Настоящее изобретение также относится к фармацевтическим композициям, содержащим антигенсвязывающую молекулу, согласно изобретению, и фармацевтически приемлемый носитель. В настоящем изобретении фармацевтические композиции обычно относятся к фармацевтическим агентам для лечения или предотвращения, или тестирования и диагностики заболеваний.
Фармацевтические композиции, согласно настоящему изобретению, могут быть получены с помощью способов, известных специалистам в данной области знаний. Более того, антигесвязывающая молекула, согласно настоящему изобретению, если требуется, может находиться в комбинации с другими фармацевтическими субстанциями. Например, они могут быть использованы парентерально в форме инъекции, или стерильного раствора, или суспензии в воде или в другой фармацевтически приемлемой жидкости. В частности, они могут быть представлены единичными дозами, которые удовлетворяют требованиям к приготовлению фармацевтических средств путем соответствующего комбинирования с фармацевтически приемлемыми носителями или средами, например, со стерильной водой, физиологическим раствором, растительным маслом, эмульгатором, суспензией, детергентом, стабилизатором, агентом, улучшающим вкус и запах, эксципиентом, переносчиком, консервантом, связующим и т.д. В таких препаратах количество активного ингредиента подбирают таким образом, что доза попадает в подходящий, предварительно определенный интервал.
Аминокислоты, содержащиеся в аминокислотных последовательностях, согласно настоящему изобретению, могут быть пост-трансляционно модифицированы (например, модификация N-концевого глутамина в пироглутаминовую кислоту хорошо известна специалистам в данной области знаний). Соответственно, такие пост-трансляционно модифицированные аминокислоты включены в аминокислотные последовательности, обеспечиваемые настоящим изобретением.
Все документы предшествующего уровня техники, процитированные в настоящем описании, включены в него в качестве ссылок.
Примеры
Ниже приведены примеры способов и композиций, обеспечиваемых изобретением. Следует понимать, что в объем изобретения включены и различны другие варианты объектов изобретения, практическое воплощение которых определяется описанием изобретения.
Несмотря на то, что изобретение описано детально иллюстративным путем и за счет приведения примеров в целях ясности его понимания, описание и примеры не должны пониматься как ограничивающие объем изобретения. Описания всех патентов и научной литературы, приведенные здесь, включены в изобретение исключительно в качестве ссылок.
Пример 1. Получение новых L-цепей, совместимых с каждой Н-цепью АСЕ910 (Эмицизумаб)
АСЕ910 (Эмицизумаб) представляет собой гуманизированное IgG4 антитело, состоящее из анти-FIX(a) и анти-FX, и проявляет замещающую функциональную активность кофактора FVIII. Это антитело состоит из двух различных тяжелых цепей (Q499 и J327), которые связываются с FIX(a) и FX, соответственно, и общей L-цепи (L404) (тяжелые цепи: SEQ ID NOs: 10 и 11; легкая цепь: SEQ ID NO: 12). Возможный способ редуцирования взаимодействия с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом и улучшение активности, замещающей функцию кофактора FVIII, заключался в получении из библиотеки антител человека новой L-цепи с последовательностью, полностью отличающейся от обычной последовательности L-цепи, соединенной с каждой из Н-цепей анти-FIX(a) антитела и анти-FX антитела (Q499 и J327). Таким образом, авторы настоящего изобретения получили новые L-цепи, приведенные на Фиг. 10 и в Таблице 7 в соответствии со ссылочным примером 1. В настоящих примерах FIX(a) относится к FIX и/или FIXa.
Эти антитела были протестированы на взаимодействие с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом. Анти-АСЕ910 (Эмицизумаб) идиотипическое антитело было получено следующим способом. Сначала, F(ab')2 Q499/L404 и F(ab')2 J327/L404 АСЕ910 (Эмицизумаб), которые распознают FIX(a) и FX, соответственно, были введены кроликам, и получили сыворотку кролика, содержащую анти-Q499/L404 F(ab')2, и сыворотку кролика, содержащую анти-J327/L404 F(ab')2. Фракции этих сывороток в сульфате аммония пропустили через колонку со связанным IgG человека для удаления антител, взаимодействующих с IgG человека. Далее, осуществляли аффинную очистку с помощью колонки, связанной с Q499/L404 (или J327/L404), и пропускали фракцию через колонку, связанную с J327/L404 (или Q499/L404), для удаления антител, взаимодействующих с J327/L404 (или Q499/L404) так, что в результате были получены поликлональные анти-идиотипические антитела, специфически связывающиеся с Q499/L404 (или J327/L404).
Полученное анти-идиотипическое антитело было использовано для измерения величины его связывания с меченным АСЕ910 (Эмицизумаб) путем электрохемилюминесцентного иммуноанализа, причем оценивали ингибирование связывания тестируемым веществом. Сначала тестируемое вещество (биспецифическое антитело) добавляли в концентрации 0, 1, 3, 10, 30 или 100 мкг/мл к смеси Q499/L404-идиотипического антитела, J327/L404-идиотипического антитела, меченного биотином АСЕ910 (Эмицизумаб) и SULFO-TAG-меченного АСЕ910 (Эмицизумаб). Полученную смесь инкубировали в течение ночи в холодильнике. Затем смесь внесли в лунки 96-луночного планшета. Далее планшет отмывали и величину связывания измеряли электрохемилюминесцентным методом.
В качестве примера полученное биспецифическое антитело Q499/QNK131//J327/JNL095 (вариабельные участки: SEQ ID NOs: 45/13//46/31; константные участки: SEQ ID NOs: 95/99//97/101) приведено в Таблице 2 и биспецифическое антитело Q499/QL20//J327/JL01 (вариабельные участки: SEQ ID NOs: 45/43//46/44; константные участки: SEQ ID NOs: 95/99//97/101), полученное согласно Примеру 3, приведено на Фиг. 1. АСЕ910 (Эмицизумаб) снижает величину связывания между меченным АСЕ910 (Эмицизумаб) и анти-идиотипическим антителом в зависимости от концентрации добавленного антитела, таким образом, проявляя ингибирующий эффект. С другой стороны, обнаружено, что антитела с новыми L-цепями показывают значительное снижение связывания с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом.
Таким образом, авторы изобретения с успехом обнаружили новые L-цепи, которые, даже будучи спаренными с одинаковыми Н-цепями АСЕ910 (Эмицизумаб) (SEQ ID NOs: 45 и 46), проявляют значительное снижение связывания с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом и обладают замещающей функциональной активностью кофактора FVIII. Соответственно, ожидается, что такие биспецифические антитела могут быть применены даже у тех пациентов с гемофилией А, у которых индуцируется образование анти-АСЕ910 (Эмицизумаб) идиотипического антитела.

Пример 2. Получение вариантов и вариантов Н-цепи биспецифических антител, имеющих новые L-цепи
Для того, чтобы улучшить функциональную замещающую активность кофактора FVIII биспецифических антител, имеющих новые L-цепи, полученные согласно ссылочному Примеру 1, QNK131 (SEQ ID NO: 13), были отобраны новая L-цепь анти-FIX(a) антитела, и JNL095 (SEQ ID NO:31), новая L-цепь анти-FX антитела, и их аминокислотные последовательности были подвергнуты всестороннему мутагенезу с помощью методов, известных специалистам в данной области знаний, например, ПЦР-мутагенезу. Мутантные последовательности далее обширно скринировали на наличие функциональной замещающей активности кофактора FVIII и таким образом были получены варианты с аминокислотными заменами, обладающие улучшенной функциональной замещающей активностью кофактора FVIII.
В то же время, при использовании новых L-цепей были получены варианты с аминокислотными замещениями, в которых все CDRs Q499 и J327 были подвергнуты всестороннему мутагенезу путем замены всех аминокислот, за исключением цистеина. Варианты далее широкомасштабно скринировали на наличие функциональной замещающей активности кофактора FVIII для поиска аминокислотных замен, улучшающих функциональную замещающую активность кофактора FVIII.
Как описано в WO 2012/067176, были успешно получены биспецифические антитела, чья функциональная замещающая активность кофактора FVIII улучшена, а ингибирующий эффект на способность FIXa активировать FX при использовании FVIIIa в качестве кофактора не увеличен. Способ оптимизации, согласно настоящему изобретению, также улучшает функциональную замещающую активность кофактора FVIII без усиления ингибирующего эффекта на способность FIXa активировать FX при использовании FVIIIa в качестве кофактора, однако новым является обнаружение способности воздействия FXIa на активацию FIX (Фиг. 3(1)). Функциональную замещающую активность кофактора FVIII, приведенную на Фиг. 3(1), Фиг. 3(2) и Фиг. 4, измеряли способом, описанным в ссылочном Примере 1, за исключением того, что концентрация фактора X человека была изменена и составила 22 мкг/мл, а концентрация фосфолипида составила 6,0 мкМ. Кроме того, активность ингибирования активации FIX определяется по уменьшению поглощения в реакции активации FIX под действием FXIa, как определяется колориметрическим методом. Более конкретно, измерение осуществляли следующим способом. Пять мкл раствора антитела, разведенного TBSB, смешивали с 5 мкл фактора IX человека с концентрацией 3 Ед/мл (Christmassin М, Japan Blood Products Organization) и инкубировали в 384-луночном планшете при комнатной температуре в течение 30 мин. Энзиматическую реакцию в указанной смеси инициировали добавлением 5 мкл фактора XIa человека с концентрацией 90 нг/мл (Enzyme Research Laboratories). Через 60 мин реакцию останавливали добавлением 5 мкл 0,5М ЭДТА. Цветную реакцию инициировали добавлением 10 мкл раствора окрашивающего субстрата. Через 60 мин цветной реакции измеряли изменение поглощения при 405 нм при использовании SpectroMax 340РС384 (Molecular Devices). Растворителем фактора IX человека и фактора XIa человека являлся TBSB, содержащий 6,0 мкл раствора фосфолипида (SYSMEX СО.) и 1,5 мМ CaCl2. Для этого исследования использовали раствор окрашивающего субстрата Spectrozyme FIX (Sekisui Diagnostics) после растворения субстрата в очищенной воде с достижением концентрации 6,7 мМ и далее смешивали с этиленгликолем в соотношении 5:8. В результате указанные выше образцы показывали увеличение значения OD в сравнении с раствором, не содержащим антитело, что свидетельствует о снижении образования FIX. Конечная концентрация антитела, обеспечивающая функциональную замещающую активность кофактора FVIII и ингибирующую активность активации FIX, приведенные на Фиг. 3, составляет 100 мкг/мл. Конечная концентрация антитела относится к концентрации в смешанном растворе антитела, фактора IX человека и фактора XIa человека. В реакции коагуляции in vivo FIXa образуется посредством активации FIX под действием FXIa. Соответственно, ингибирование этого активационного процесса приводит к уменьшению образования FIXa, что далее оказывает негативное влияние на величину возрастания опосредованной FIXa активации FX биспецифическим антителом, имеющим функциональную замещающую активность кофактора FVIII (Фиг. 2). Поэтому для получения биспецифического антитела с высокой функциональной замещающей активностью кофактора FVIII, является предпочтительным, чтобы его ингибирующая активность по отношению к активации FIX была как можно меньшей.
Соответственно, на основании результатов указанного выше широкомасштабного скрининга на наличие функциональной замещающей активности кофактора FVIII авторы настоящего изобретения обнаружили многочисленные аминокислотные модификации, которые значительно увеличивают функциональную замещающую активность кофактора FVIII по сравнению с увеличением ингибирующей активности по отношению к активации FIX. Более точно, этими модификациями являются S30R, S31R, T53R (нумерация по Кабату) и др. Такие многочисленные модификации были введены в QNK131 (SEQ ID NO: 13) с получением QL20 (QAL201) (SEQ ID NO:43). Далее, варианты с аминокислотными заменами были получены путем усовершенствованного введения множественных CDR-аминокислотных замен в вариант, имеющий QL20 в качестве L-цепи анти-FIX(a) антитела. Далее варианты подвергали широкомасштабному скринингу на наличие функциональной замещающей активности кофактора FVIII и ингибирующей активности по отношению к активации FIX. В результате были обнаружены комбинации модификаций, которые снижают ингибирующую активность по отношению к активации FIX, в то время как сохраняется функциональная замещающая активность кофактора FVIII. Конкретно, эти комбинации представляли собой комбинации основных и кислых остатков, такие как S30R/S31R/N32D, S30R/S31R/N32E и Q27R/R93D (все согласно нумерации по Кабату), в L-цепи анти-FIX(a) антитела.
Дополнительно, варианты с аминокислотными заменами были также получены путем усовершенствованного введения аминокислотных замен в FRs, сайты, не контактирующие с антигеном, и далее эти варианты широкомасштабно скринировали. В результате были обнаружены многочисленные модификации, которые усиливают функциональную замещающую активность кофактора FVIII, в то время как сохраняется ингибирующая активность по отношению к активации FIX. Конкретно, варианты с аминокислотными заменами, в которых Phe83 в L-цепи анти-FIX(a) антитела был замещен на остаток Met, остаток Ala или подобные им остатки, или остаток Arg45 был замещен на остаток Glu, обнаруживали увеличенную функциональную замещающую активность кофактора FVIII.
Для оптимизации комбинации указанных выше модификаций и также баланса антигенсвязывающей активности (ассоциация и диссоциация) в плече анти-FIX(a) и в плече анти-FX, были получены исчерпывающие пары анти-FIX(a) и анти-FX антител. Более точно, для анти-FIX(a) антитела Н-цепь была модифицирована путем введения Y100E, Y100eI, или делеции G100a, или комбинации указанных модификаций, и L-цепь была модифицирована путем введения А55Е, K92R, вставки Pro в положение 95, или L96G, или комбинации указанных модификаций. Для анти-FX антитела Н-цепь была модифицирована путем введения T28E/D31H или D31N/Q, 151S, G56T/R, S57V, Y59S, E61R, E62K, D65N/Q, V67L, K73I, S82bE, E102V или комбинации указанных модификаций, и L-цепь, JNL095 (SEQ ID NO:31) была модифицирована путем введения Е24Т, N26E, H27Q, G29S, D30S/Q/E, K31R, H32E/Q, R50Q, D92A, S94D, S95D/A, A95aY, V96T или комбинации указанных модификаций (все согласно нумерации по Кабату) с оптимизацией антигенсвязывающей аффинности.
В результате авторы настоящего изобретения успешно получили варианты, чья ингибирующая активность по отношению к активации FIX не была повышена, однако их функциональная замещающая активность кофактора FVIII была существенно улучшена (биспецифические антитела, состоящие из комбинации константного участка Н-цепи, имеющего любую из SEQ ID NOs: 95-98, и константного участка L-цепи, имеющего любую из SEQ ID NOs: 99-104, и комбинации вариабельных участков, показанных в Таблице 4) в сравнении с АСЕ910 (Эмицизумаб) и в сравнении с вариантами, которые, как показано в WO 2012/067176, имеют более высокую функциональную замещающую активность кофактора FVIII, чем АСЕ910 (Эмицизумаб) (биспецифические антитела, состоящие из константного участка Н-цепи, имеющего SEQ ID NO: 95 или 97, и константного участка L-цепи, имеющего SEQ ID NO: 99, и комбинации вариабельных участков, показанных в Таблице 3) (Фиг. 3(2)). Более того, как показано на Фиг. 4, было обнаружено, что эти варианты имеют значительно усиленный максимум активности и специфической активности в сравнении с АСЕ910 (Эмицизумаб). Функциональную замещающую активность кофактора FVIII, приведенную на Фиг. 4, измеряли способом, описанным в ссылочном Примере 1, за исключением того, что концентрация фактора X человека была изменена и составляла 22, 9 мкг/мл, а концентрация фосфолипида составляла 6,0 мкМ.
Далее, для подтверждения того, что функциональная замещающая активность кофактора FVIII обеспечивает образование тромбина в плазме, проводили тестирование генерации тромбина способом, известным специалистам в данной области техники. Более точно, 8 мкл биспецифического антитела, разведенного TBSB, добавляют к 72 мкл FVIII-дефицитной плазмы (George King) и смесь инкубируют в течение 30 мин или более при комнатной температуре. Далее добавляют 20 мкл триггерного реагента, содержащего 20 мкМ фосфолипида и фактор XIa человека в концентрации 5 нг/мл (Enzyme Research Laboratories). Затем инициируют реакцию коагуляции добавлением 20 мкл смеси растворов Флуо-буфера и Флуо-субстрата компании FluCa-Kit (Thrombinospore). Количество образовавшегося тромбина определяют при использовании системы флуоресцентного измерения и анализа синтеза тромбина (Thrombinospore). После этого добавляют антитело до обеспечения конечной концентрации 10 мкг/мл. Конечная концентрация антитела относится к его концентрации в смеси дефицитной по FVIII плазмы и раствора антитела. Когда высота пика во время добавления антитела используется в качестве индекса уровня генерации тромбина, полученные результаты показывают, что все специфические антитела из Таблицы 5 также увеличивают уровень образования тромбина по сравнению с АСЕ910 (Эмицизумаб, Q499/L404//J327/L404). Более того, при сравнении с добавленным FVIII в качестве положительного контроля (Kogenate-FS BIO-SET; Bayer) многие варианты (QH01/QL21//JH01/JL01, QH02/QL22//JH01/JL01, QH03/QL3//JH02/JL02, QH03/QL24//JH02/JL02, QH02/QL22//JH0/JL04, QH04/QL26//JH02/JL02, QH04/QL26//JH05/JL05, QH06/QL30//JH07/JL11) показали более высокий уровень генерации тромбина по сравнению с тем, когда добавляют FVIII в количестве 40 Ед/дл, что является уровнем здоровых индивидуумов (Фиг. 5).

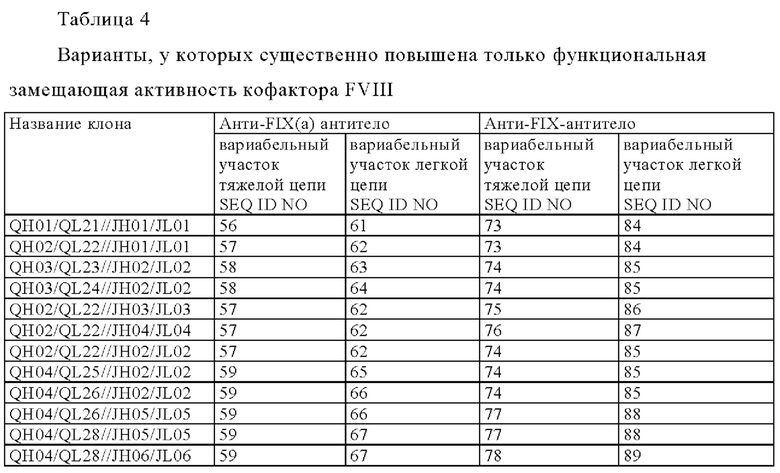
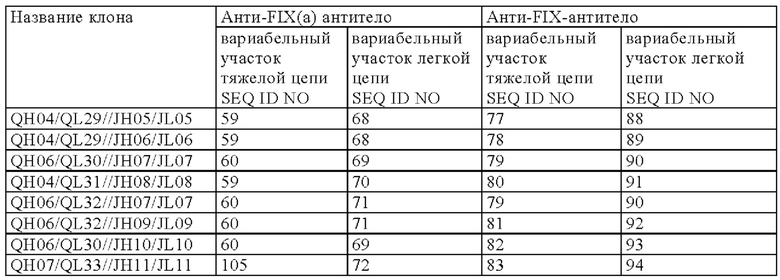
Пример 3. Фармакокинетика (ФК) полученных биспецифических антител
Для успешной медикаментозной терапии больных гемофилией А предпочтительно, чтобы вводимое антитело имело как можно более долгое время полужизни для снижения частоты применения. Основные способы улучшения ФК антитела включают способ увеличения времени его циркуляции в крови посредством FcRn и способ уменьшения клеточного захвата посредством неспецифического связывания (ADME и Трансляционная фармакокинетика/фармакодинамика терапевтически активных белков (2015) стр. 25-37).
Как показано в Примере 2, авторы настоящего изобретения успешно получили биспецифические антитела со значительно улучшенной функциональной замещающей активностью кофактора FVIII и в то же время не имеющие увеличенной ингибирующей активности по отношению к активации FIX. В способе получения таких антител авторы изобретения также предприняли попытку улучшения неспецифического связывания, что способно повлиять на ФК антител.
Более точно, система исследования связывания с внеклеточным матриксом (ЕСМ), которая известна как система определения неспецифического связывания in vitro, была использована для оценки антител (заявка US 2014/0080153). В результате ЕСМ-связывание варианта Q499/QL20//J327/JL01 (вариабельные участки: SEQ ID NOs:45/43/46/44), который обнаружил высокую функциональную замещающую активность кофактора FVIII в Примере 2, было очень высоким по сравнению с АСЕ910 (Эмицизумаб). Однако, для вариантов, в которых Н-цепь анти-FIX(a) антитела была модифицирована за счет G97D, Y100D, Y100E и подобных им замен, и L-цепь анти-FIX(a) антитела была модифицирована за счет N32D, N32E, А55Е и подобных им замен (состоящих из комбинации константного участка Н-цепи, имеющего любую из SEQ ID NOs:95-98, и константного участка L-цепи, имеющего любую из SEQ ID NOs: 99-104, и комбинации вариабельных участков, приведенных в Таблице 5), ЕСМ-связывание было значительно снижено (Фиг. 6).
Далее, варианты, полученные путем введения ACT-Fc модификаций (Mabs, (2017) Vol. 9, No. 5, 844-853), которые улучшают циркуляцию в крови за счет FcRn, в константные участки указанных выше вариантов, тестировали путем внутривенного введения мышам. После введения антитела собирали образцы крови и получали плазму. Затем измеряли концентрацию введенного антитела в плазме с помощью метода LC-MS/MS. Данные по концентрации антитела в зависимости от времени перемещения в плазме анализировали на приборе WinNonlin версия 7.0 (Certara) для вычисления клиренса антитела (CL). В результате при сравнении с вариантом без ACT-Fc модификаций, т.е. с вариантом QH01/QL21//JH01/JL01 (вариабельные участки: SEQ ID NOs: 56/61//73/84) или с вариантом QH02/QL22//JH02/JL02 (вариабельные участки: SEQ ID NOs: 57/62//74/85) клиренс антитела CL был существенно повышен у вариантов с ACT-Fc модификациями, а именно QH04/QL28//JH05/JL05 (вариабельные участки: SEQ ID NOs: 59/67//77/88), QH04/QL28//JH06/JL06 (вариабельные участки: SEQ ID NOs: 59/67//78/89), QH04/QL29//JH05/JL05 (вариабельные участки: SEQ ID NOs: 59/68//77/88) и QH04/QL29//JH06/JL06 (вариабельные участки: SEQ ID NOs: 59/68//78/9) (Фиг. 7).
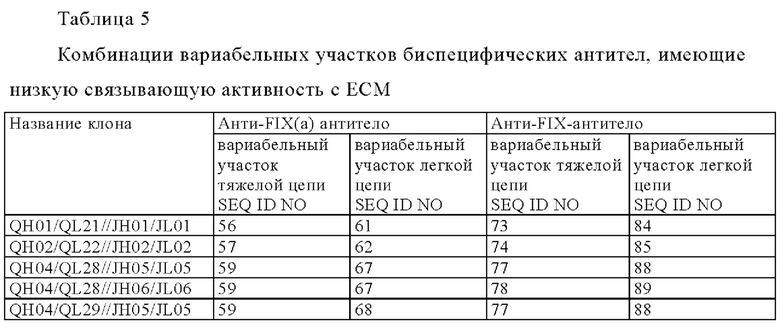
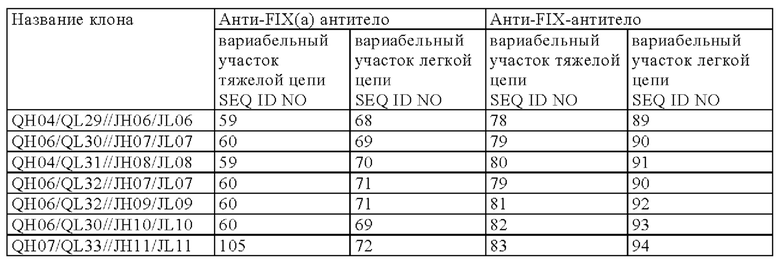
Пример 4. Получение биспецифических антител
Как описано выше, авторы настоящего изобретения ввели модификации в две различные тяжелые цепи (Н-цепи) и в две различные легкие цепи (L-цепи) биспецифических антител для получения антител с преимущественной эффективностью и свойствами.
Когда экспрессируется биспецифическое антитело, экспрессируются две различные тяжелые цепи (Н-цепи) и две различные легкие цепи (L-цепи). Соответственно, возможно получение десяти различных комбинаций. Поскольку только одна из этих комбинаций обладает нужной биспецифичностью, получение биспецифического антитела, представляющего интерес, требует выделения этого антитела из десяти различных антител, что крайне неэффективно и вызывает значительные трудности. В качестве средства, решающего данную проблему, может быть использован известный способ, в котором аминокислотные замены вводятся в СН3-домен Н-цепей IgG, так что преимущественно секретируется IgG с комбинацией из двух различных Н-цепей и, для эффективного получения представляющей интерес молекулы используются известные аминокислотные замены и их комбинации в вариабельных участках и в интерфейсе CH1-CL домена для обеспечения желательной ассоциации между Н-цепью и L-цепью (WO 2013/065708).
Полученные антитела и способы получения
В приведенном ниже описании тяжелая цепь и легкая цепь анти-FIX(a) антитела обозначены как Q и k, соответственно, а тяжелая цепь и легкая цепь анти-FX антитела обозначены как J и λ, соответственно.
Биспецифическое антитело
Q1014-G4T1k.LG.A5/AL869AE.F83M- kT0//J1494-G4T1h.LG.A5/YL681K27Q-lam1NL95 (вариабельные участки: SEQ ID NOs: 106/108//109/111, константные участки: SEQ ID NOs: 107/99//110/101) были использованы в качестве матричной последовательности. Комбинации модификаций, приведенные в Таблице 6, были введены в указанное антитело с получением вариантов.
Целью эксперимента было обеспечение регулирования ассоциации между Н-цепью и L-цепью. Однако, когда одна тяжелая цепь имеет константный участок, который облегчает гомологичную ассоциацию, экспрессируются все девять типов неправильно спаренных молекул, включающих не только те, которые имеют две различные тяжелые цепи, ассоциированные гетерологично, как показано на Фиг. 8(8)-(10), но и те, которые показаны на Фиг. 8(2)-(7). Это создает трудности для анализа и оценки представляющей интерес молекулы (Фиг. 8(1)). Соответственно, для того, чтобы вывести одну тяжелую цепь из гомологичной ассоциации, модификации типа «выпуклость» и «впадина» (KiH: Knobs into Hole) были введены в константные участки тяжелых цепей (Q и J), соответственно (SEQ ID NOs: 107 и 110), таким образом, что количество потенциально экспрессируемых неправильно спаренных молекул антител, уменьшается с 9 типов до 3 типов (Фиг. 8(8)-(10)), согласно проведенным исследованиям.
Во время трансфекции четыре цепи были экспрессированы в трех плазмидах при количественных соотношениях (Q:k:J:λ=1:3:1:1, 1:1:1:1 и 1:1:1:3) и очищены способами, известными специалистам в данной области техники. При экспрессии этих цепей в условиях, когда легкая цепь избыточна, недостаточное регулирование ассоциации между тяжелой и легкой цепями будет вызывать формирование неправильных пар. Наоборот, если представляющее интерес биспецифическое антитело образуется с высокой скоростью при всех трех условиях, можно предположить, что пара с введенной модификацией будет иметь более высокую способность к регулированию.
Дополнительно, поскольку референсные стандарты неправильного спаривания используются в CIEX (катион-обменная хроматография)-анализе, референсные стандарты с общей легкой цепью (Q/k//J/k и Q/λ/J/λ) были получены путем экспрессии трех цепей Q/k/J или Q/λ/J.
Анализ и способы анализа
Полученные антитела были проанализированы с помощью метода CIEX с помощью системы Alliance (Waters). Осуществляли метод двух-жидкостного градиента при использовании анализирующей колонки YMC-BioPro SP-F, 4,6 × 100 мм, СХ-1 рН градиентного буфера А, рН 5,6 (Thermo), в качестве подвижной фазы А и СХ-1 рН градиентного буфера В, рН 10,2 (Thermo), в качестве подвижной фазы В. Измерение осуществляли при длине волны 280 нм. Данные анализировали при использовании Empower3 (Waters) и вычисляли пропорцию каждого детектируемого пика. В случаях, когда наблюдался только единичный пик при экспрессии четырех цепей, этот пик считали основным, т.е. пиком, соответствующим биспецифическому антителу. Когда наблюдались множественные пики, пик, который не перекрывался с пиками референсных стандартов неправильного спаривания, имеющих общую легкую цепь, считали основным пиком.
Результаты и дискуссия
Пропорция пиков биспецифического антитела, которую вычисляли на основании данных анализа, приведена в Таблице 6, а каждая хроматограмма приведена на Фиг. 9.
Было обнаружено, что новые модифицированные пары No. 14 (пара из аминокислотного остатка в положении 175 (нумерация EU) в СН1 и аминокислотного остатка в положении 180 (нумерация по Кабату) в CL) и No. 15 (пара из аминокислотного остатка в положении 175 (нумерация EU) в СН1 и аминокислотного остатка в положении 131 (нумерация по Кабату) в CL), обладают способностью к регулированию по сравнению с парами No. 10 (пара из аминокислотного остатка в положении 147 (нумерация EU) в СН1 и аминокислотного остатка в положении 180 (нумерация по Кабату) в CL) и No. 11 (пара из аминокислотного остатка в положении 147 (нумерация EU) в СН1 и аминокислотного остатка в положении 131 (нумерация по Кабату) в CL), которые относятся к известным модификациям, описанным в WO 2013/065708.
Более того, было показано, что новая модифицированная пара No. 9 (пара из аминокислотных остатков в положении 147 (нумерация EU) и положении 175 (нумерация EU) в CH1 и аминокислотных остатков в положении 131 (нумерация по Кабату) и в положении 180 (нумерация по Кабату) в CL) обладает такой же высокой способностью к регулированию, что и известная комбинация No. 6 (пара из аминокислотных остатков в положении 147 (нумерация EU) и положении 175 (нумерация EU) в СН1 и аминокислотных остатков в положении 131 (нумерация по Кабату) и в положении 160 (нумерация по Кабату) в CL).
Далее, известно, что специфическая ассоциация между тяжелой и легкой цепями обеспечивается путем модификации пары из аминокислотного остатка в положении 39 (нумерация по Кабату) в VH и аминокислотного остатка в положении 38 (нумерация по Кабату) в VL (WO 2013/065708). Соответственно, эта модифицированная пара была введена в пару No. 9 с получением No. 18 и No. 19 и полученные пары были оценены. В результате было сделано наблюдение о том, что все модификации приводят к высокой способности к регулированию.
Дополнительно, когда, согласно настоящему изобретению, обнаруженная новая модифицированная пара No. 9 была применена к матрице, имеющей пару модификаций (Е356К-К439Е) для обеспечения ассоциации Н-цепи, отличной от KiH (Q1014Q39E-G4T1ALG409K.E356K/AL869Q38KAE.F83M- kT0//J1494Q39K-G4T1A5LG409K.K439E/YL681.K27Q.Q38E-lam1NL95 (вариабельные участки: SEQ ID NOs: 112/113//114/115, константные участки: SEQ ID NOs: 117/99//116/101)), также наблюдалась высокая способность к регулированию (No. 20:Q1014Q39E-G4T1th2A5LG409K.E356K/AL869Q38KAE.F83M- k0MTtl17E160Q//J1494Q39K-G4Tlth13LG409K.K439E/YL681.K27Q.Q38E-lam1p9 (вариабельные участки: SEQ ID NOs: 112/113//114/115, константные участки: SEQ ID NOs: 119/100//118/102).

«-» обозначает, что модификации не были введены. Номер каждого остатка обозначен в соответствии с нумерацией по Кабату для вариабельного участка Н-цепи, вариабельного участка L-цепи и константного участка L-цепи и в соответствии с нумерацией EU для константного участка Н-цепи.
Согласно настоящему изобретению, обнаружены новые L-цепи, которые уменьшают взаимодействие с анти-АСЕ910 (Эмицизумаб) идиотипическим антителом и проявляют функциональную замещающую активность кофактора FVIII, а также обнаружены аминокислотные замены и их комбинации для тяжелой и легкой цепей, которые усиливают функциональную замещающую активность кофактора FVIII биспецифических антител, имеющих эти легкие цепи, и при этом не увеличивается ингибирующая активность по отношению к активации FIX указанных антител. Эти аминокислотные замены и их комбинации полезны для создания биспецифических антител, имеющих превосходящую функциональную замещающую активность кофактора FVIII по отношению к АСЕ910 (Эмицизумаб).
Ссылочный Пример 1
Возможный способ улучшения активности АСЕ910 для замещения функции кофактора FVIII заключался в получении из библиотеки антител человека новой L-цепи с последовательностью, полностью отличающейся от обычной L-цепи, соединенной с каждой из Н-цепей анти-FIX(a) антитела и анти-FX антитела (Q499 и J327).
Обратившись к способам, известным специалисту в данной области техники, в особенности, к Сообщениям о биохимических и биофизических исследованиях (2000), 275, 2, 553-557, и подобным публикациям, авторы изобретения создали новую библиотеку путем замены L-цепи анти-АСЕ910 (Эмицизумаб) идиотипического антитела на L-цепь библиотеки человека и осуществили пэннинг против меченного биотином FIXa человека или меченного биотином FX человека для получения антител с новыми легкими цепями, обладающих функциональной замещающей активностью кофактора FVIII.
В результате QNK131 (SEQ ID NO: 13), QNK284 (SEQ ID NO: 14), QNK315 (SEQ ID NO: 15), QNL182 (SEQ ID NO: 16), QNL492 (SEQ ID NO: 17) и QNL576 (SEQ ID NO: 18) были обнаружены как вариабельные участки L-цепи для антитела против FIX(a) человека биспецифических антител, обладающих функциональной замещающей активностью кофактора FVIII, и JNK131 (SEQ ID NO: 19), JNK163 (SEQ ID NO: 20), JNK252 (SEQ ID NO: 21), JNK263 (SEQ ID NO: 22), JNK339 (SEQ ID NO: 23), (SEQ ID NO: 24), JNK351 (SEQ ID NO: 25), JNK360 (SEQ ID NO: 26), JNK378 (SEQ ID NO: 27), JNK382 (SEQ ID NO: 28), JNL036 (SEQ ID NO: 29), JNL036 (SEQ ID NO: 29), JNL072 (SEQ ID NO: 30), JNL095 (SEQ ID NO: 31), JNL176 (SEQ ID NO: 32), JNL208 (SEQ ID NO: 33), JNL224 (SEQ ID NO: 34), JNL260 (SEQ ID NO: 35), JNL056 (SEQ ID NO: 36), JNL059 (SEQ ID NO: 37), JNL226 (SEQ ID NO: 38), JNL250 (SEQ ID NO: 39), JNL263 (SEQ ID NO: 40) и JNL281 (SEQ ID NO: 41) были обнаружены как вариабельные участки L-цепи для антитела против FX человека.
Различные биспецифические антитела, содержащие указанные новые L-цепи, были экспрессированы и очищены способами, известными для специалиста в данной области знаний. Полученные антитела приведены в Таблице 7 (показаны обозначения клонов, SEQ ID NOs для вариабельных участков тяжелой цепи и SEQ ID NOs для вариабельных участков легкой цепи). Новый вариабельный участок L-цепи был использован только для одной из цепей. Для другой цепи был использован вариабельный участок L404, обычной L-цепи АСЕ910 (SEQ ID NO: 45). Для Н-цепи были использованы вариабельный участок Q499 (SEQ ID NO: 45) и вариабельный участок J327 (SEQ ID NO: 46). Для удобства обозначения новых L-цепей используются как обозначения клонов. Для анти-FIX(a) антитела обозначения QC1 (SEQ ID NO: 95) и CL1 (SEQ ID NO: 99) были использованы для константных участков Н-цепи и L-цепи, соответственно. Для анти-FX антитела обозначения JC1 (SEQ ID NO: 97) и CL3 (SEQ ID NO: 101) были использованы для константных участков Н-цепи и L-цепи, соответственно.
Каждое очищенное биспецифическое антитело было исследовано на наличие функциональной замещающей активностью кофактора FVIII способом, известным специалисту в данной области техники. Более точно, измерение проводили следующим способом. Все реакции осуществляли при комнатной температуре. Пять мкл раствора антитела, разведенного физиологическим раствором с Трис-буфером, содержащим 0,1% бычьего сывороточного альбумина (далее - TBSB), смешивали с фактором IXa бета человека (Enzyme Research Laboratories) в концентрации 150 нг/мл и инкубировали в 384-луночном планшете при комнатной температуре в течение 30 мин. Энзиматическую реакцию в смеси инициировали добавлением 5 мкл фактора X человека (Enzyme Research Laboratories) с концентрацией 24,7 мкг/мл. Через 4 минуты реакцию останавливали добавлением 5 мкл 0,5 М ЭДТА. Цветную реакцию инициировали добавлением 5 мкл раствора окрашивающего субстрата. Через 30 мин цветной реакции измеряли изменение поглощения при 405 нм при использовании SpectroMax 340РС384 (Molecular Devices). Растворителем фактора IXa бета человека и фактора X человека являлся TBSB, содержащий 4,0 мкл раствора фосфолипида (SYSMEX СО.) и 1,5 мМ CaCl2. Для этого исследования использовали раствор окрашивающего субстрата Spectrozyme FIX (Sekisui Diagnostics) после растворения субстрата в очищенной воде с достижением концентрации 1,47 мг/мл. Результаты измерения функциональной замещающей активности кофактора FVIII каждого очищенного биспецифического антитела приведены на Фиг. 10. Конечная концентрация антитела, используемого для измерения функциональной замещающей активности кофактора FVIII, показанная на Фиг. 10, составляла 66,7 мкг/мл для антител, образованных с помощью новой анти- FXI(a) легкой цепи, или 100 мкг/мл для антител, образованных с помощью новой анти- FX легкой цепи. Конечная концентрация антитела относится к концентрации в смеси раствора антитела, фактора IXa бета человека и фактора X человека. Было обнаружено, что все биспецифические антитела, образованные любой из новых L-цепей, обладают функциональной замещающей активностью кофактора FVIII.

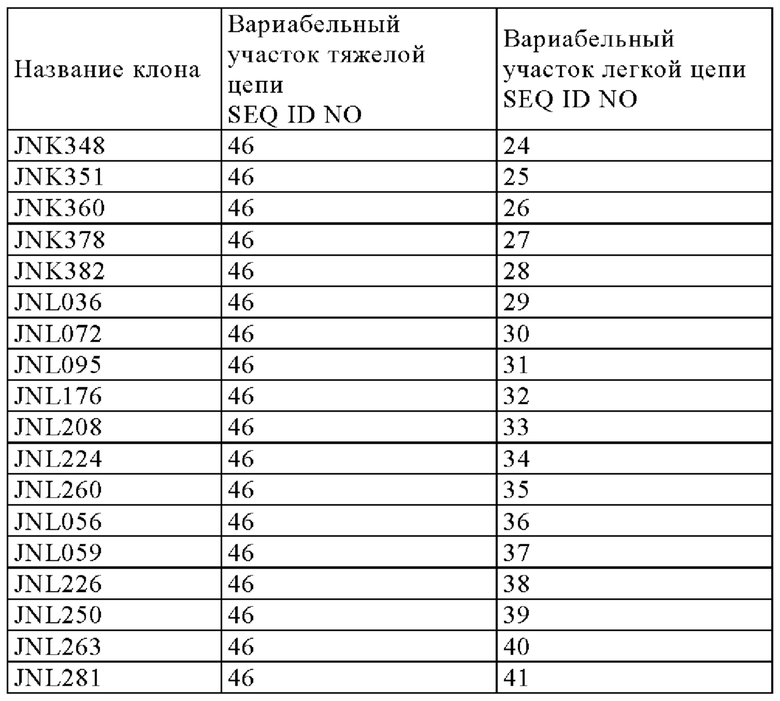
Ссылочный Пример 2
Были получены варианты с замещениями, в которых все CDRs Q499 и J327, Н-цепей АСЕ910, были избыточно мутированы путем замены всех аминокислот, за исключением цистеина. Был осуществлен широкомасштабный скрининг этих вариантов и новых L-цепей, полученных в Ссылочном Примере 1, на наличие функциональной замещающей активности кофактора FVIII для поиска аминокислотных замен, которые обеспечивают повышенную функциональную замещающую активность кофактора FVIII.
Например, аминокислотные замены были введены в QNK131, новую L-цепь для анти-FIX(а) антитела (вариабельный участок: SEQ ID NO: 13), с получением QAL187 (вариабельный участок: SEQ ID NO: 42) и QAL201 (вариабельный участок: SEQ ID NO: 43). Сходным образом, аминокислотные замены были введены в JNL095, новую L-цепь для анти-FX антитела (вариабельный участок: SEQ ID NO: 31), с получением JVL280 (вариабельный участок: SEQ ID NO: 44). Для анти-FIX(a) антитела QC1 (SEQ ID NO: 95) и CL1 (SEQ ID NO: 99) были использованы в качестве константных участков Н-цепи и L-цепи, соответственно. Для анти-FX антитела JC1 (SEQ ID NO: 97) и CL3 (SEQ ID NO: 101) были использованы в качестве константных участков Н-цепи и L-цепи, соответственно.
Результаты исследования показали, что некоторые аминокислотные замены, показанные в Таблице 8 ниже, способны улучшить функциональную замещающую активность кофактора FVIII, свойственную АСЕ910 (Эмицизумаб). При измерении функциональной замещающей активностью кофактора FVIII способ, описанный в Ссылочном Примере 1, был использован для вариантов с модифицированной Н-цепью для анти-FX антитела. В случае вариантов с модифицированной Н-цепью для анти-FX антитела измерения осуществляли при использовании способа, описанного в Ссылочном Примере 1, за исключением того, концентрация фактора IХа бета человека была изменена и составляла 450 нг/мл. В Таблице 8 показана специфическая активность каждого варианта с аминокислотной заменой относительно АСЕ910 (Эмицизумаб) (каждое значение показано в таблице). В Таблице 8 (-) означает, что уровень экспрессии антитела был низким.
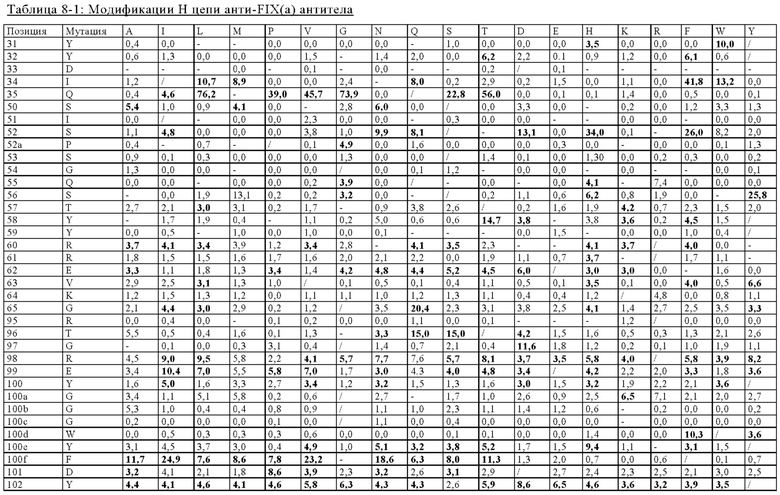
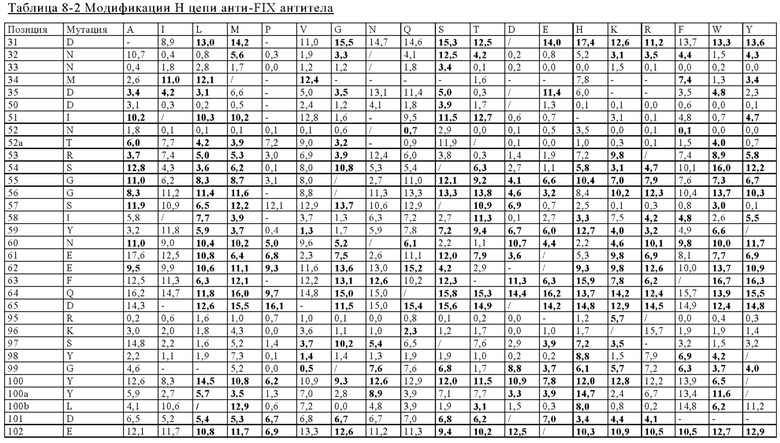
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЧУГАИ СЕЙЯКУ КАБУСИКИ КАЙСЯ
<120> МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ
ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО
ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ
УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО
ИНГРЕДИЕНТА
<130> C1-A1716P
<150> JP 2017-189647
<151> 2017-09-29
<160> 287
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 1
Tyr Tyr Asp Ile Gln
1 5
<210> 2
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 2
Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val Lys
1 5 10 15
Gly
<210> 3
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 3
Arg Thr Gly Arg Glu Tyr Gly Gly Gly Trp Tyr Phe Asp Tyr
1 5 10
<210> 4
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 4
Asp Asn Asn Met Asp
1 5
<210> 5
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 5
Asp Ile Asn Thr Arg Ser Gly Gly Ser Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 6
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 6
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu
1 5 10
<210> 7
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 7
Lys Ala Ser Arg Asn Ile Glu Arg Gln Leu Ala
1 5 10
<210> 8
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 8
Gln Ala Ser Arg Lys Glu Ser
1 5
<210> 9
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 9
Gln Gln Tyr Ser Asp Pro Pro Leu Thr
1 5
<210> 10
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 10
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Tyr Gly Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Lys Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 11
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 11
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Glu Ser Leu Ser Leu Ser Pro
435 440
<210> 12
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 12
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Arg Asn Ile Glu Arg Gln
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Arg Lys Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Asp Pro Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 13
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 13
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Lys Arg Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 14
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 14
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Ser Tyr Leu Gly
85 90 95
Ile Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 15
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 15
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Val Ser Cys Lys Ser Ser Gln Ser Leu Leu Arg Thr
20 25 30
Asp Gly Lys Ala Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Arg
85 90 95
Ile Gln Ala Leu Ser Phe Gly Gly Gly Thr Lys Val Asp Ile Lys
100 105 110
<210> 16
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 16
Ser Ser Gly Leu Thr Gln Pro Pro Ser Leu Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly His Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Ala Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Val
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Val Pro Asn Ser Gly Tyr
85 90 95
Val Phe Gly Thr Gly Thr Gln Val Thr Val Val
100 105
<210> 17
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 17
Gln Ser Val Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Ser Tyr
20 25 30
Asn Leu Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Ser
85 90 95
Ser Thr Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 18
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 18
Gln Ser Ala Leu Thr Gln Pro Pro Ser Leu Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly His Lys Val Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Ala Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Val
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Val Pro Asn Ser Gly Tyr
85 90 95
Val Phe Gly Thr Gly Thr Gln Val Thr Val Val
100 105
<210> 19
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 19
Asn Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Thr Val Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Ser Asp Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Arg Pro Pro Lys Val Leu Ile
35 40 45
Tyr Arg Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Ile Tyr Tyr Cys Gln Gln Ala Lys Asn Phe Pro Trp
85 90 95
Ala Phe Gly Gln Gly Thr Lys Val Glu Phe Lys
100 105
<210> 20
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 20
Asn Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Arg Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Phe Thr Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Ala Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr His Cys Gln Gln Ser Tyr Arg Ile Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 21
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 21
Asn Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Lys Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Gly Thr Ser
20 25 30
Leu Asn Trp Tyr Gln Gln Arg Pro Gly Thr Ala Pro Lys Leu Leu Ile
35 40 45
Phe Asp Thr Ser Asn Leu Glu Lys Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Lys Ser Glu Thr Tyr Phe Thr Phe Ala Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Asn Ile Pro Trp
85 90 95
Thr Phe Gly Arg Gly Thr Lys Val Glu Ile Thr
100 105
<210> 22
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 22
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Phe Thr Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Gly Gly Ser Gly Ala Glu Tyr Thr Leu Thr Ile Ser Asn Leu Gln Pro
65 70 75 80
Asp Asp Ser Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 23
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 23
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Gln Tyr Ile Gly Thr Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Asn Ser Val Ser Arg Leu Gln Thr Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Gly Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Tyr Ser Thr Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Arg Val Glu Ile Lys
100 105
<210> 24
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 24
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Arg Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Gln Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Val Ser Ser Arg Phe Ser Gly Val Pro
50 55 60
Glu Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Glu Ile
65 70 75 80
Ser Arg Val Glu Ala Asp Asp Val Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Leu His Leu Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Val Lys
100 105 110
<210> 25
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 25
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Ser Gly Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Thr Arg
20 25 30
Tyr Leu Ala Trp Tyr Tyr Gln Arg Pro Gly Gln Val Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Thr Ser Asn Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Gly Asp Ser Gly Val Tyr Tyr Cys Gln Gln Ser Arg Ser Ser Gln
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 26
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 26
Val Ile Trp Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Asn Ser Arg
20 25 30
Leu Val Trp Tyr Gln Gln Lys Pro Gly Thr Ala Pro Lys Val Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Phe Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Val Lys
100 105
<210> 27
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 27
Asn Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Thr Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Arg Ala Pro Arg Ser Leu Ile
35 40 45
Tyr Thr Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Thr Asn Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala His Ser Phe Pro Trp
85 90 95
Ser Phe Gly Pro Gly Thr Asn Val Glu Ile Lys
100 105
<210> 28
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 28
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Lys Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Asp Ser Ala Thr Asn Leu Arg Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Leu Ala Phe Tyr Tyr Cys Gln Gln Gly Lys Ser Phe Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 29
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 29
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Ser Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 30
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 30
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Pro Cys Gly Gly Asn Asn Ile Gly Asn Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Thr Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly His Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Ser Cys Gln Val Trp Asp Ser Gly Thr Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 31
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 31
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn His Ile Gly Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Ala Val
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 32
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 32
Ser Val Leu Thr Gln Pro Pro Ser Leu Ser Ala Ala Pro Gly Gln Arg
1 5 10 15
Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn His Leu
20 25 30
Val Ser Trp His Gln Gln Phe Pro Gly Thr Ala Pro Lys Ala Leu Ile
35 40 45
Tyr Asp Asn Asp Arg Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Lys Ser Gly Thr Ser Ala Thr Leu Asp Ile Thr Gly Leu Gln Thr
65 70 75 80
Gly Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Ala Ser Leu Arg
85 90 95
Ala Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 33
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 33
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Val Ser Cys Asn Gly Gly Ser Ser Asn Ile Gly Thr Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Ile
35 40 45
Val Ile Phe Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Gly Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Val Ile Ala Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Arg Ser
85 90 95
Leu Ser Gly Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 34
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 34
Ser Tyr Val Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Arg Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Arg Pro Gly Lys Ala Pro Lys Val
35 40 45
Met Ile Tyr Asp Val Ile Lys Arg Pro Ser Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ala Gly Ala
85 90 95
Ser Ser Phe Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 35
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 35
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Asn Ile Ser Cys Ser Gly Ser Arg Ser Asn Ile Ala Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln His Leu Pro Gly Thr Val Pro Lys Val Leu
35 40 45
Ile Ser Asp Asn Asp Gln Arg Ser Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Arg Met
85 90 95
Arg Gly Phe Val Phe Gly Ser Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 36
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 36
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Ser Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 37
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 37
Asn Phe Met Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Arg Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Arg
20 25 30
Ala Val Ser Trp Tyr Gln His Val Pro Gly Lys Pro Pro Arg Leu Ile
35 40 45
Val Tyr His Asp Asp Val Leu Ser Ser Gly Val Ser Gly Arg Phe Ser
50 55 60
Ala Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Phe Cys Ala Ala Trp Asp Ala Arg Leu
85 90 95
Asn Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 38
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 38
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Leu Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105
<210> 39
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 39
Gln Ser Val Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Thr Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Ser Ser Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Thr Ser
85 90 95
Gly Thr Tyr Val Phe Gly Thr Gly Thr Thr Val Thr Val Leu
100 105 110
<210> 40
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 40
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Arg
85 90 95
Arg Gly Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 41
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 41
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Ala Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Lys Tyr Val Ser Trp Tyr Gln Gln Tyr Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Ser
85 90 95
Asn Thr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105
<210> 42
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 42
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Lys Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 43
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 43
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Arg Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Lys Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 44
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 44
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn His Ile Gly Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 45
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 45
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Tyr Gly Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 46
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 46
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 47
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 47
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Arg Asn Ile Glu Arg Gln
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Arg Lys Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Asp Pro Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 48
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 48
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ser Gly Arg Glu Tyr Gly Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 49
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 49
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ser Gly His Asn Tyr Gly Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 50
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 50
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr His Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 51
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 51
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Gly Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 52
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 52
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Arg Asn Ile Glu Arg Gln
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Arg Lys Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Pro Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 53
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 53
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Lys Asn Ile Glu Arg Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Arg Ala Asp Arg Lys Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Pro Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 54
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Arg Asn Ile Glu Arg Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Arg Ala Asp Arg Lys Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Pro Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 55
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 55
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Arg Asn Ile Glu Arg Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Arg Lys Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Arg Tyr Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Pro Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 56
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 56
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Tyr Asp Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 57
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 57
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ala Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Ser Glu Asp Gly Ala Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 58
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 58
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Asp Gly Gly Trp His Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 59
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 59
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Glu Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 60
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 60
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 61
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 61
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Lys Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 62
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 62
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Gln Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 63
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 63
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Gln Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 64
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 64
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 65
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 65
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 66
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 66
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 67
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 67
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Arg Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Glu Thr Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 68
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 68
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 69
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 69
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 70
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 70
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 71
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 71
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 72
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 72
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Arg Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 73
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 73
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ser Asn Thr Arg Ser Gly Thr Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 74
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 74
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asn Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 75
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 75
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Arg Val Ile Ser Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 76
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 76
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Gln Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 77
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 77
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 78
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 78
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Thr His Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Arg Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Ile Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 79
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 79
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 80
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 80
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 81
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 81
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Lys Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Glu Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 82
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 82
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Thr His Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Arg Ser Ile Tyr Asn Arg Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Ile Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 83
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 83
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Glu Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 84
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 84
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn His Ile Gly Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 85
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 85
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn His Ile Gly Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Gln Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 86
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 86
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Thr Gly Asn His Ile Ser Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Gln Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Asp Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 87
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 87
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn Gln Ile Ser Gln Lys Gln Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 88
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 88
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 89
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 89
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn Gln Ile Gly Ser Arg Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Ala Ser Asp Ala Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 90
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 90
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 91
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 91
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Ile Tyr
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 92
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 92
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Ile Tyr
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 93
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 93
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn Gln Ile Gly Glu Lys Glu Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Ala Ser Asp Ala Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 94
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 94
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 95
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 95
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Arg Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Lys Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 96
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 96
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Lys Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 97
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 97
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Arg Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 98
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 98
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Glu Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Glu Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 99
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 99
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 100
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 100
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 101
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 101
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 102
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 102
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 103
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 103
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Glu Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 104
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 104
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Arg Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 105
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 105
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Asp Arg Glu Asp His Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 106
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 106
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 107
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 107
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 108
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 108
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 109
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 109
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 110
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Cys Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 111
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 111
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 112
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 112
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Glu Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 113
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 113
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 114
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 114
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 115
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 115
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val
100 105
<210> 116
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 116
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Glu Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 117
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 117
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Lys Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 118
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 118
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Glu Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Glu Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Glu Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 119
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 119
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Lys Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu
305 310 315 320
Leu Ser Leu Ser Pro
325
<210> 120
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 120
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Tyr Asp Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Lys Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His
420 425 430
Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 121
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 121
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ala Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Ser Glu Asp Gly Ala Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Lys Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His
420 425 430
Glu Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 122
<211> 447
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 122
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Asp Gly Gly Trp His Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Lys Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 123
<211> 447
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 123
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Glu Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Lys Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 124
<211> 447
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 124
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Lys Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 125
<211> 447
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 125
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Asp Arg Glu Asp His Gly Trp Ile Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Lys Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Lys Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ala His Tyr Thr Arg Lys Glu Leu Ser Leu Ser Pro
435 440 445
<210> 126
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 126
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Lys Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 127
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 127
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Gln Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 128
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 128
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Gln Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Ser Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 129
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 129
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 130
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 130
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 131
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 131
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 132
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 132
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Arg Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Glu Thr Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 133
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 133
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 134
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 134
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Met Ala Val Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 135
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 135
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Lys Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 136
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 136
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Arg Arg Glu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 137
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 137
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Thr Arg Ser Val Arg Arg Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Arg Glu Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Asp Pro Pro Gly
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Glu Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Glu Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 138
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 138
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ser Asn Thr Arg Ser Gly Thr Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 139
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 139
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asn Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 140
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 140
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Arg Val Ile Ser Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 141
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 141
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Gln Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 142
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 142
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 143
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 143
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Thr His Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Arg Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Ile Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 144
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 144
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 145
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 145
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 146
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 146
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Lys Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Glu Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 147
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 147
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Glu Phe Thr His Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Arg Ser Ile Tyr Asn Arg Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Ile Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 148
<211> 444
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 148
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gln Asn
20 25 30
Asn Met Asp Trp Val Arg Lys Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Leu Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Glu Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Glu Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His
420 425 430
Ala His Tyr Thr Arg Glu Glu Leu Ser Leu Ser Pro
435 440
<210> 149
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 149
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn His Ile Gly Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 150
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 150
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn His Ile Gly Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Gln Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 151
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 151
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Thr Gly Asn His Ile Ser Asp Lys His Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Gln Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Asp Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 152
<211> 212
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 152
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Arg Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn Gln Ile Ser Gln Lys Gln Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 153
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 153
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 154
<211> 212
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 154
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn Gln Ile Gly Ser Arg Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Ala Ser Asp Ala Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 155
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 155
Ser Tyr Val Leu Thr Gln Pro Val Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 156
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 156
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Ile Tyr
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 157
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 157
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Ile Tyr
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 158
<211> 212
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 158
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Asn Gln Ile Gly Glu Lys Glu Val
20 25 30
His Trp Tyr His Gln Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Ala Ser Asp Ala Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 159
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 159
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Glu Gly Glu Gln Ile Gly Ser Lys Glu Val
20 25 30
His Trp Tyr His Glu Arg Pro Gly Gln Ala Pro Ile Leu Val Met Phe
35 40 45
Arg Asp Ala Arg Arg Pro Ser Gly Ile Pro Glu Arg Leu Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Ala Glu Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Tyr Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Val Thr Val Val Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Lys Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Lys Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 160
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 160
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Tyr Gly Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 161
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 161
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Asn
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Asp Ile Asn Thr Arg Ser Gly Gly Ser Ile Tyr Asn Glu Glu Phe
50 55 60
Gln Asp Arg Val Ile Met Thr Val Asp Lys Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr His Cys
85 90 95
Ala Arg Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 162
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 162
Arg Ala Ser Gln Ser Val Ser Ser Asn Leu Ala
1 5 10
<210> 163
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 163
Gly Ala Ser Thr Arg Ala Thr
1 5
<210> 164
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 164
Gln Gln Tyr Lys Arg Pro Leu Thr
1 5
<210> 165
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 165
Glu Gly Asn His Ile Gly Asp Lys His Val His
1 5 10
<210> 166
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 166
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 167
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 167
Gln Val Trp Asp Ser Ser Ser Ala Val Val
1 5 10
<210> 168
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 168
Tyr Tyr Asp Ile Gln
1 5
<210> 169
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 169
Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val Lys
1 5 10 15
Gly
<210> 170
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 170
Arg Thr Gly Arg Glu Tyr Asp Gly Gly Trp Tyr Phe Asp Tyr
1 5 10
<210> 171
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 171
Tyr Tyr Asp Ala Gln
1 5
<210> 172
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 172
Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val Lys
1 5 10 15
Gly
<210> 173
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 173
Arg Thr Gly Ser Glu Asp Gly Ala Gly Trp Tyr Phe Asp Tyr
1 5 10
<210> 174
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 174
Tyr Tyr Asp Ile Gln
1 5
<210> 175
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 175
Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val Lys
1 5 10 15
Gly
<210> 176
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 176
Arg Thr Gly Arg Glu Asp Gly Gly Trp His Phe Asp Tyr
1 5 10
<210> 177
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 177
Tyr Tyr Asp Ile Gln
1 5
<210> 178
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 178
Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val Lys
1 5 10 15
Gly
<210> 179
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 179
Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr
1 5 10
<210> 180
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 180
Tyr Tyr Asp Ile Gln
1 5
<210> 181
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 181
Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val Lys
1 5 10 15
Gly
<210> 182
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 182
Arg Thr Gly Arg Glu Glu Gly Gly Trp Ile Phe Asp Tyr
1 5 10
<210> 183
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 183
His Tyr Asp Ile Gln
1 5
<210> 184
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 184
Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val Lys
1 5 10 15
Gly
<210> 185
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 185
Arg Thr Asp Arg Glu Asp His Gly Trp Ile Phe Asp Tyr
1 5 10
<210> 186
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 186
Arg Ala Ser Gln Ser Val Arg Arg Asp Leu Ala
1 5 10
<210> 187
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 187
Gly Ala Ser Arg Arg Ala Thr
1 5
<210> 188
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 188
Gln Gln Tyr Lys Ser Pro Leu Thr
1 5
<210> 189
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 189
Arg Ala Thr Gln Ser Val Arg Arg Asp Leu Ala
1 5 10
<210> 190
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 190
Gly Ala Ser Arg Arg Ala Thr
1 5
<210> 191
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 191
Gln Gln Tyr Arg Ser Pro Leu Thr
1 5
<210> 192
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 192
Arg Ala Thr Gln Ser Val Arg Arg Asp Leu Ala
1 5 10
<210> 193
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 193
Gly Ala Ser Arg Arg Ala Thr
1 5
<210> 194
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 194
Gln Gln Tyr Arg Ser Pro Leu Thr
1 5
<210> 195
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 195
Arg Ala Ser Arg Ser Val Arg Arg Glu Leu Ala
1 5 10
<210> 196
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 196
Gly Ala Ser Thr Arg Glu Thr
1 5
<210> 197
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 197
Gln Gln Tyr Arg Asp Pro Leu Thr
1 5
<210> 198
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 198
Arg Ala Ser Arg Ser Val Arg Arg Glu Leu Ala
1 5 10
<210> 199
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 199
Gly Ala Ser Thr Arg Glu Thr
1 5
<210> 200
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 200
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 201
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 201
Arg Ala Ser Arg Ser Val Arg Arg Glu Leu Ala
1 5 10
<210> 202
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 202
Gly Ala Ser Thr Arg Glu Thr
1 5
<210> 203
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 203
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 204
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 204
Arg Ala Thr Arg Ser Val Arg Arg Asp Leu Ala
1 5 10
<210> 205
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 205
Gly Ala Ser Arg Arg Glu Thr
1 5
<210> 206
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 206
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 207
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 207
Arg Ala Ser Arg Ser Val Arg Arg Glu Leu Ala
1 5 10
<210> 208
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 208
Gly Ala Ser Thr Arg Glu Thr
1 5
<210> 209
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 209
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 210
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 210
Arg Ala Ser Arg Ser Val Arg Arg Glu Leu Ala
1 5 10
<210> 211
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 211
Gly Ala Ser Thr Arg Glu Thr
1 5
<210> 212
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 212
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 213
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 213
Arg Ala Ser Arg Ser Val Arg Arg Glu Leu Ala
1 5 10
<210> 214
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 214
Gly Ala Ser Thr Arg Glu Thr
1 5
<210> 215
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 215
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 216
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 216
Arg Ala Ser Arg Ser Val Arg Arg Glu Leu Ala
1 5 10
<210> 217
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 217
Gly Ala Ser Thr Arg Glu Thr
1 5
<210> 218
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 218
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 219
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 219
Arg Ala Thr Arg Ser Val Arg Arg Asp Leu Ala
1 5 10
<210> 220
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 220
Gly Ala Ser Arg Arg Glu Thr
1 5
<210> 221
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 221
Gln Gln Tyr Arg Asp Pro Pro Gly Thr
1 5
<210> 222
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 222
Asp Asn Asn Met Asp
1 5
<210> 223
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 223
Asp Ser Asn Thr Arg Ser Gly Thr Ser Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 224
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 224
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu
1 5 10
<210> 225
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 225
Asn Asn Asn Met Asp
1 5
<210> 226
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 226
Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asn
<210> 227
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 227
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val
1 5 10
<210> 228
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 228
Asp Asn Asn Met Asp
1 5
<210> 229
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 229
Asp Ile Asn Thr Arg Ser Gly Arg Val Ile Ser Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 230
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 230
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu
1 5 10
<210> 231
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 231
Gln Asn Asn Met Asp
1 5
<210> 232
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 232
Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Gln
<210> 233
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 233
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val
1 5 10
<210> 234
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 234
Gln Asn Asn Met Asp
1 5
<210> 235
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 235
Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 236
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 236
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val
1 5 10
<210> 237
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 237
His Asn Asn Met Asp
1 5
<210> 238
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 238
Asp Ile Asn Thr Arg Ser Gly Arg Ser Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 239
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 239
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val
1 5 10
<210> 240
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 240
Gln Asn Asn Met Asp
1 5
<210> 241
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 241
Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 242
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 242
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val
1 5 10
<210> 243
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 243
Gln Asn Asn Met Asp
1 5
<210> 244
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 244
Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 245
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 245
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu
1 5 10
<210> 246
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 246
Gln Asn Asn Met Asp
1 5
<210> 247
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 247
Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Lys Phe Gln
1 5 10 15
Asp
<210> 248
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 248
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Val
1 5 10
<210> 249
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 249
His Asn Asn Met Asp
1 5
<210> 250
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 250
Asp Ile Asn Thr Arg Ser Gly Arg Ser Ile Tyr Asn Arg Glu Phe Gln
1 5 10 15
Asp
<210> 251
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 251
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu
1 5 10
<210> 252
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 252
Gln Asn Asn Met Asp
1 5
<210> 253
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 253
Asp Ile Asn Thr Arg Ser Gly Gly Val Ile Tyr Asn Glu Glu Phe Gln
1 5 10 15
Asp
<210> 254
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 254
Arg Lys Ser Tyr Gly Tyr Tyr Leu Asp Glu
1 5 10
<210> 255
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 255
Glu Gly Asn His Ile Gly Asp Lys His Val His
1 5 10
<210> 256
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 256
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 257
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 257
Gln Val Trp Asp Ser Ser Ser Tyr Thr Val
1 5 10
<210> 258
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 258
Glu Gly Asn His Ile Gly Asp Lys His Val His
1 5 10
<210> 259
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 259
Gln Asp Ala Arg Arg Pro Ser
1 5
<210> 260
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 260
Gln Val Trp Asp Ser Ser Ser Tyr Thr Val
1 5 10
<210> 261
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 261
Thr Gly Asn His Ile Ser Asp Lys His Val His
1 5 10
<210> 262
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 262
Gln Asp Ala Arg Arg Pro Ser
1 5
<210> 263
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 263
Gln Val Trp Asp Ser Ser Asp Tyr Thr Val
1 5 10
<210> 264
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 264
Glu Gly Asn Gln Ile Ser Gln Lys Gln Val His
1 5 10
<210> 265
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 265
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 266
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 266
Gln Val Trp Asp Ser Ser Ala Val Val
1 5
<210> 267
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 267
Glu Gly Glu Gln Ile Gly Ser Lys Glu Val His
1 5 10
<210> 268
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 268
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 269
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 269
Gln Val Trp Asp Ser Ser Ser Tyr Thr Val
1 5 10
<210> 270
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 270
Glu Gly Asn Gln Ile Gly Ser Arg Glu Val His
1 5 10
<210> 271
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 271
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 272
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 272
Gln Val Trp Ala Ser Asp Ala Val Val
1 5
<210> 273
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 273
Glu Gly Glu Gln Ile Gly Ser Lys Glu Val His
1 5 10
<210> 274
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 274
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 275
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 275
Gln Val Trp Asp Ser Ser Ser Tyr Thr Val
1 5 10
<210> 276
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 276
Glu Gly Glu Gln Ile Gly Ser Lys Glu Val His
1 5 10
<210> 277
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 277
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 278
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 278
Gln Val Trp Asp Ser Ser Ser Tyr Thr Val
1 5 10
<210> 279
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 279
Glu Gly Glu Gln Ile Gly Ser Lys Glu Val His
1 5 10
<210> 280
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 280
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 281
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 281
Gln Val Trp Asp Ser Ser Ser Tyr Thr Val
1 5 10
<210> 282
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 282
Glu Gly Asn Gln Ile Gly Glu Lys Glu Val His
1 5 10
<210> 283
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 283
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 284
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 284
Gln Val Trp Ala Ser Asp Ala Val Val
1 5
<210> 285
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 285
Glu Gly Glu Gln Ile Gly Ser Lys Glu Val His
1 5 10
<210> 286
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 286
Arg Asp Ala Arg Arg Pro Ser
1 5
<210> 287
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 287
Gln Val Trp Asp Ser Ser Ser Tyr Thr Val
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО, ОБЛАДАЮЩЕЕ ПОВЫШЕННОЙ АКТИВНОСТЬЮ, АЛЬТЕРНАТИВНОЙ ФУНКЦИИ КОФАКТОРА FVIII | 2017 |
|
RU2821642C2 |
| ЛИГАНДСВЯЗЫВАЮЩАЯ МОЛЕКУЛА С РЕГУЛИРУЕМОЙ ЛИГАНДСВЯЗЫВАЮЩЕЙ АКТИВНОСТЬЮ | 2017 |
|
RU2803067C2 |
| БИСПЕЦИФИЧНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2800779C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| АНТИТЕЛА К ВИРУСУ ДЕНГЕ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2758596C2 |
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732591C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2789884C2 |
| АНТИТЕЛА К ВИРУСУ ДЕНГЕ, ОБЛАДАЮЩИЕ ПЕРЕКРЕСТНОЙ РЕАКТИВНОСТЬЮ С ВИРУСОМ ЗИКА | 2019 |
|
RU2811697C2 |
| МЕЖВИДОВЫЕ АНТИТЕЛА К ЛАТЕНТНОМУ TGF-БЕТА 1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2837609C2 |
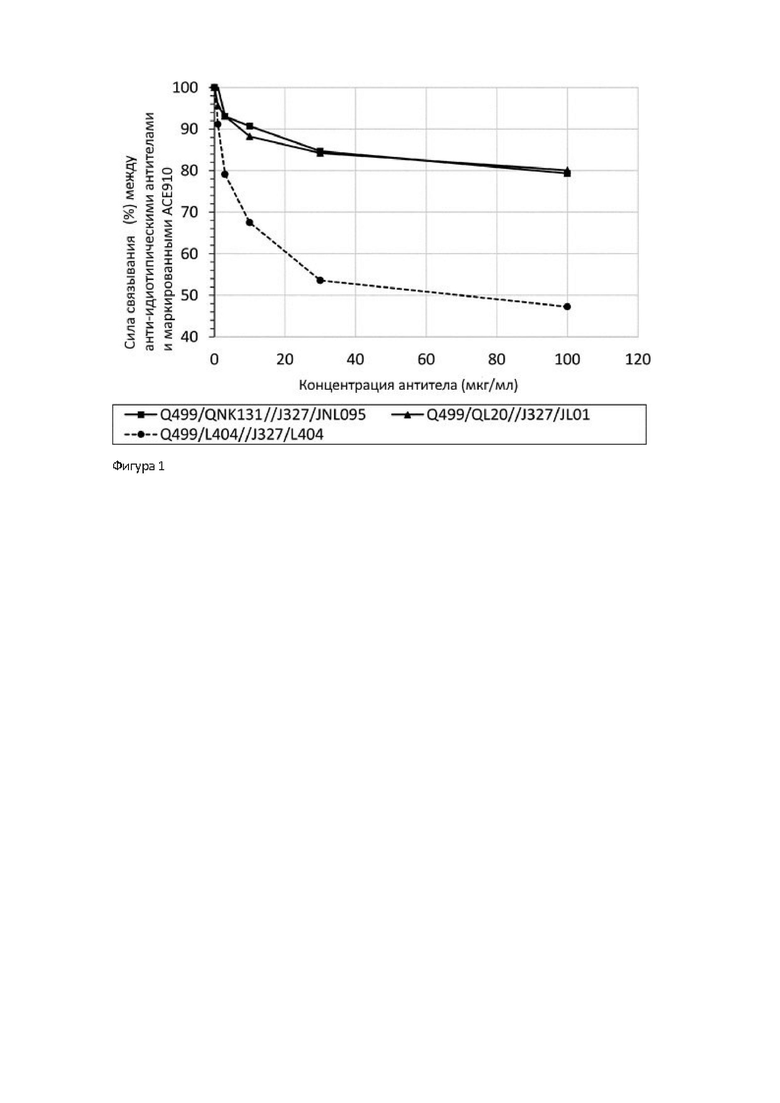
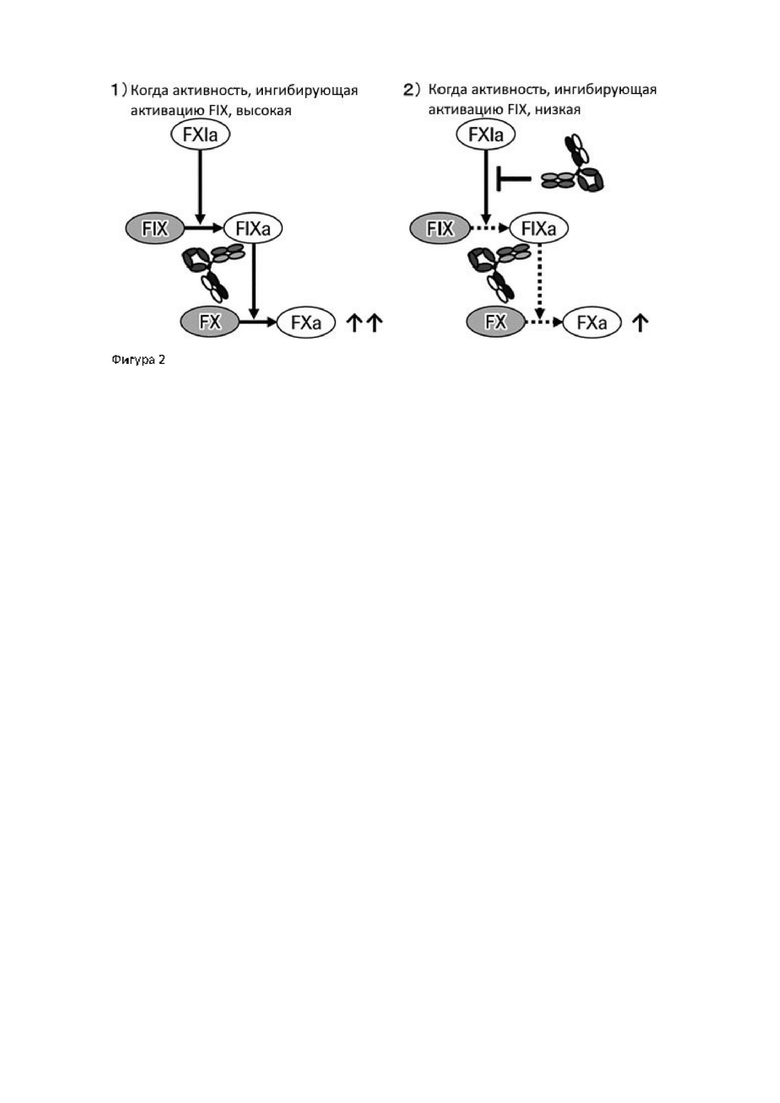

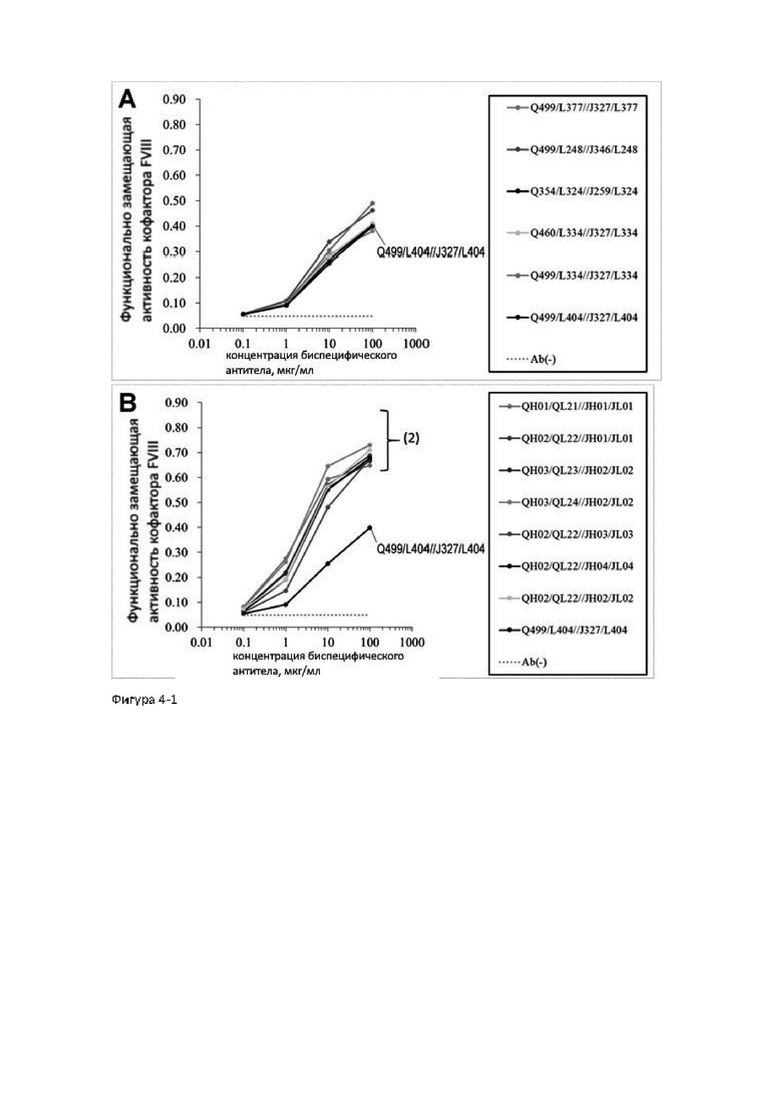
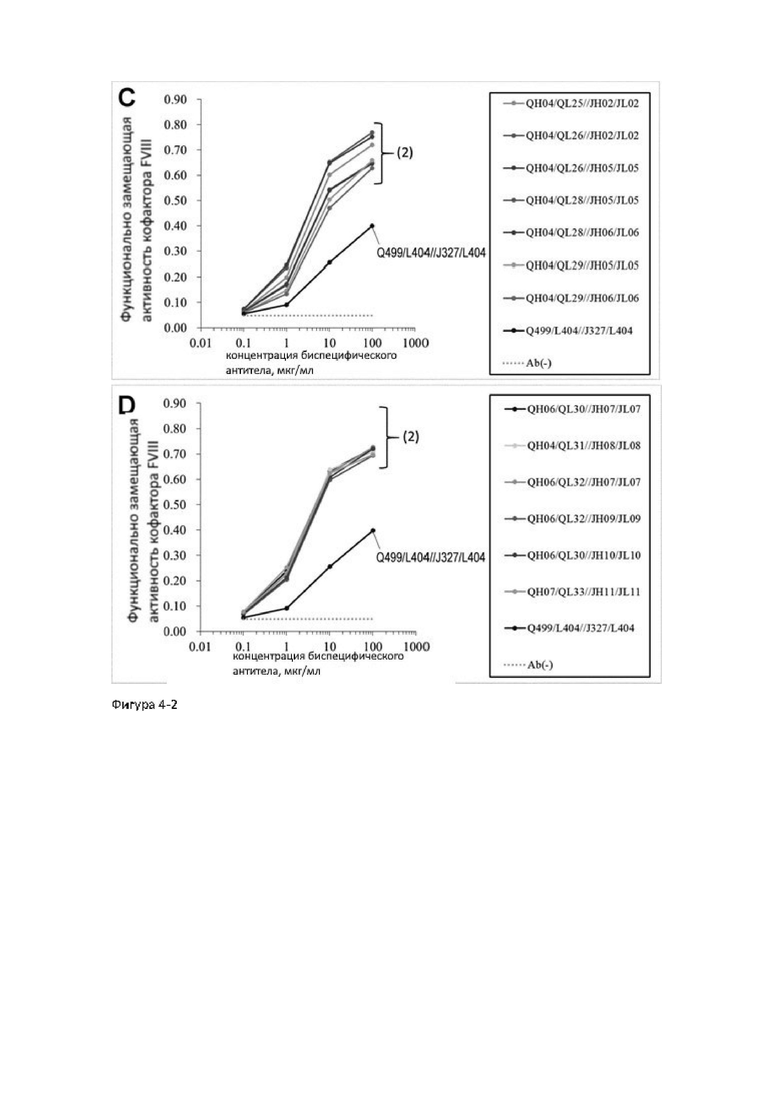

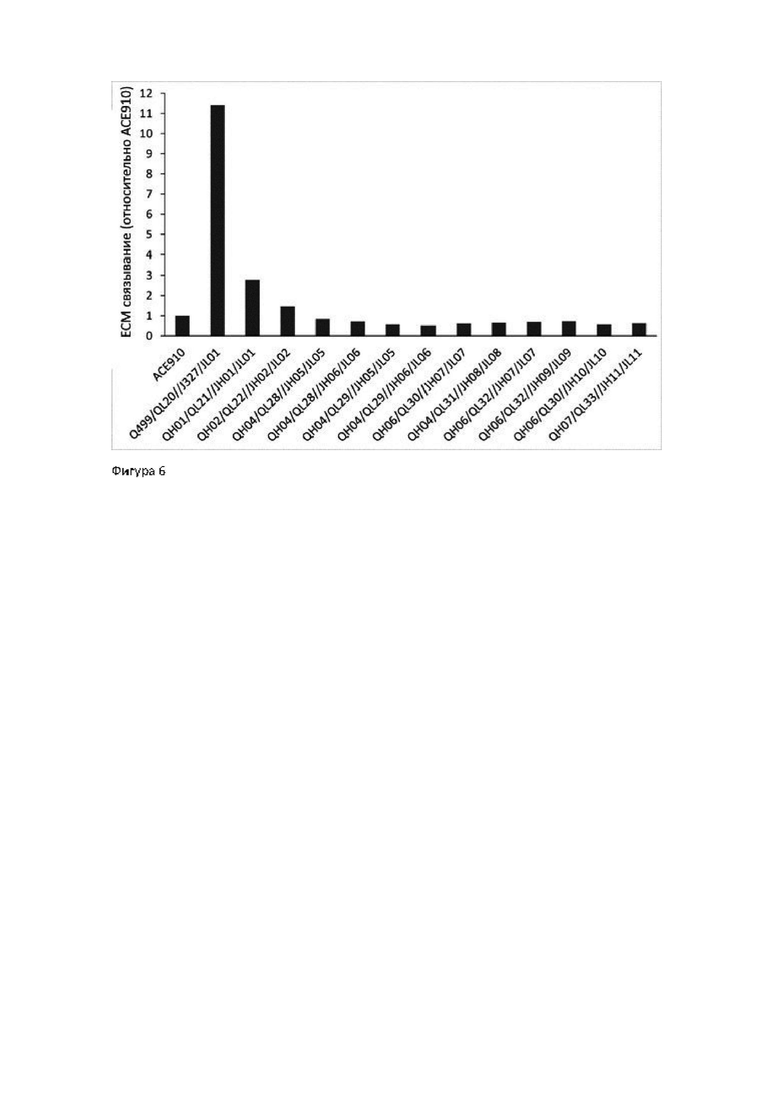
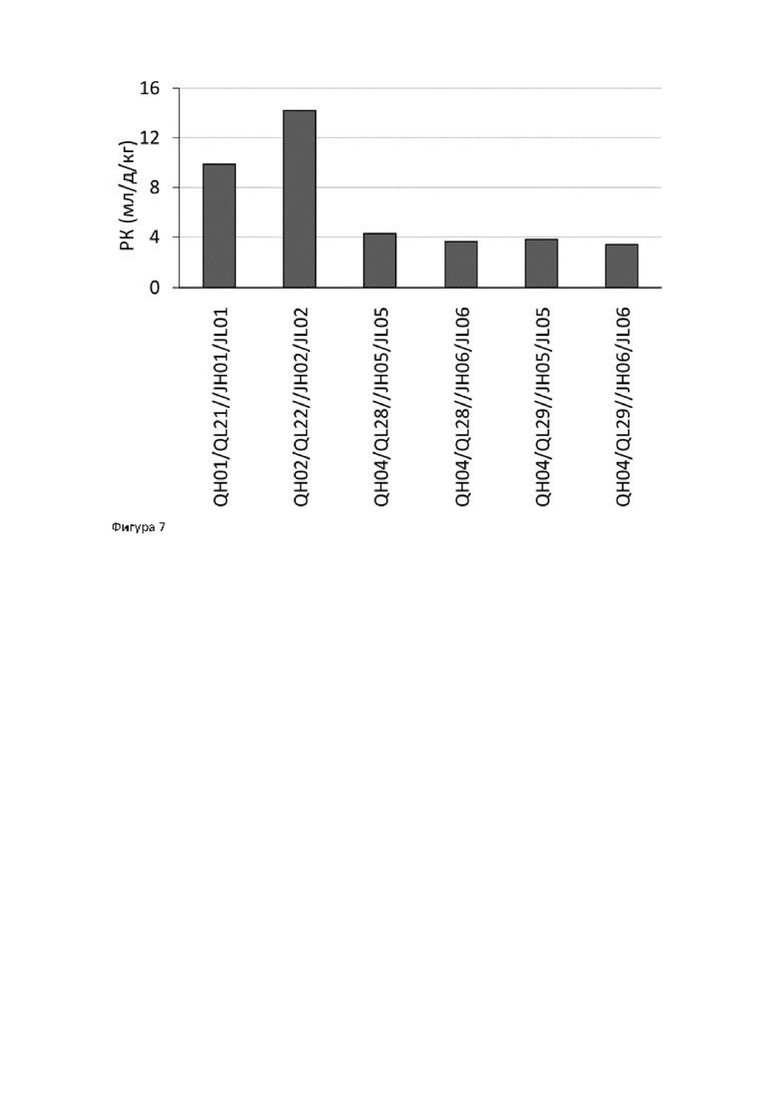
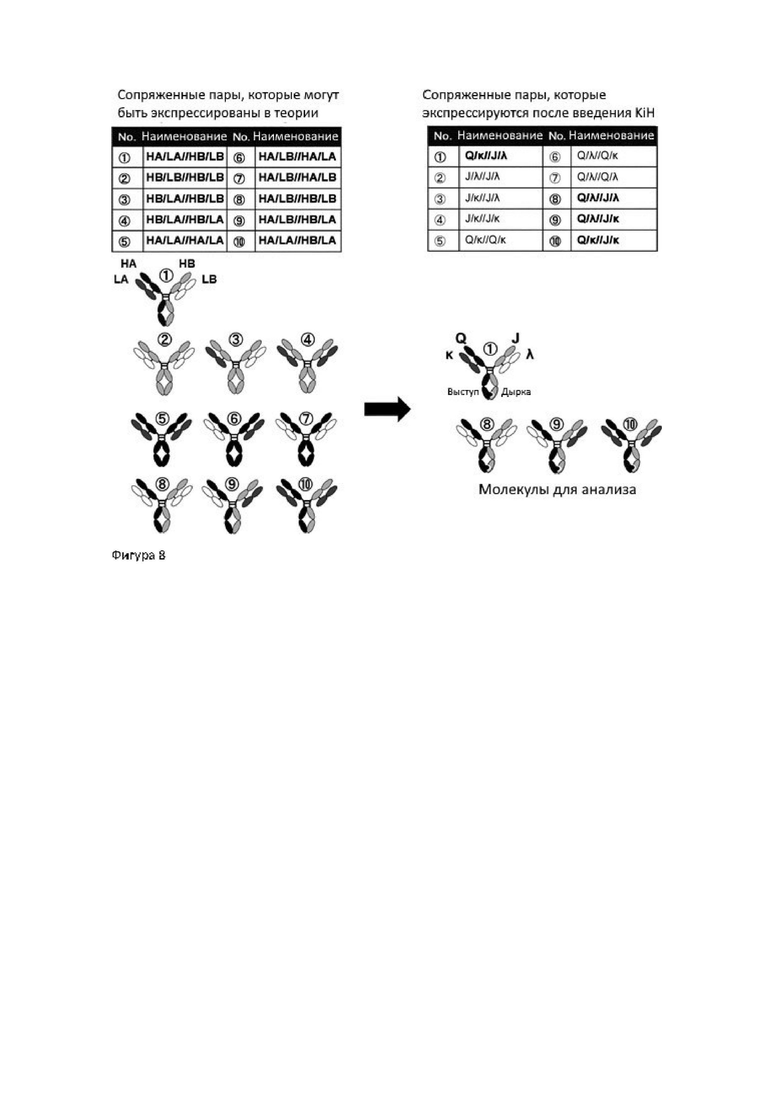
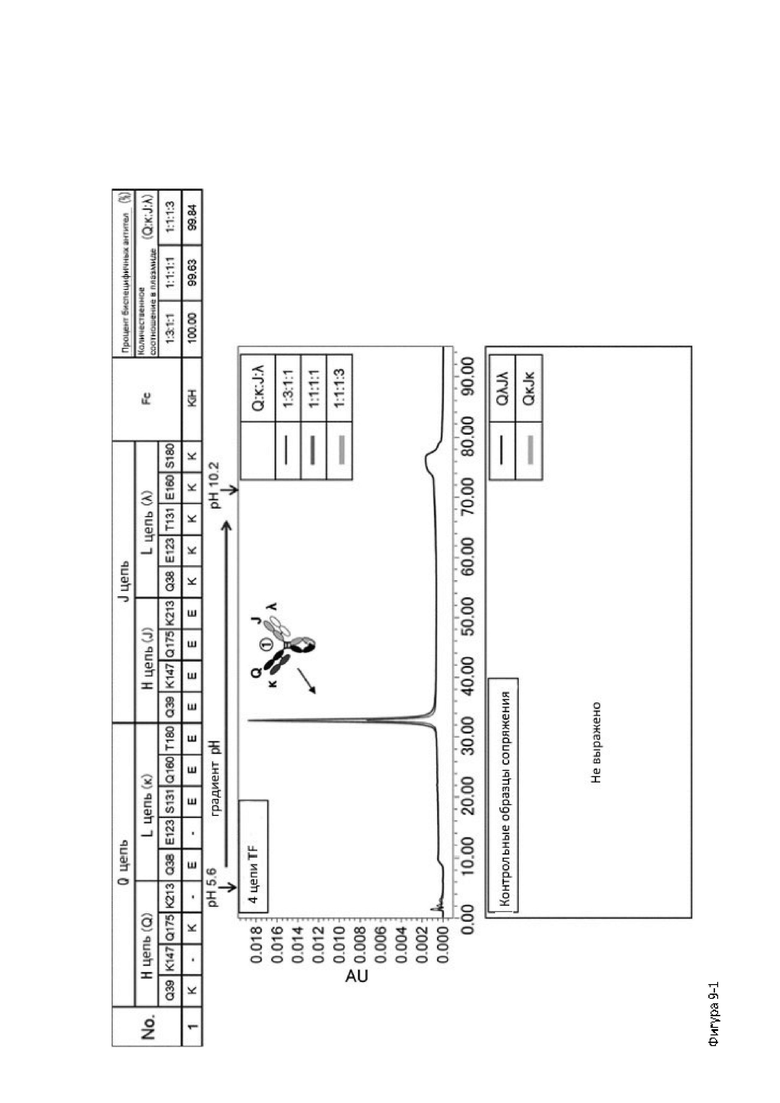
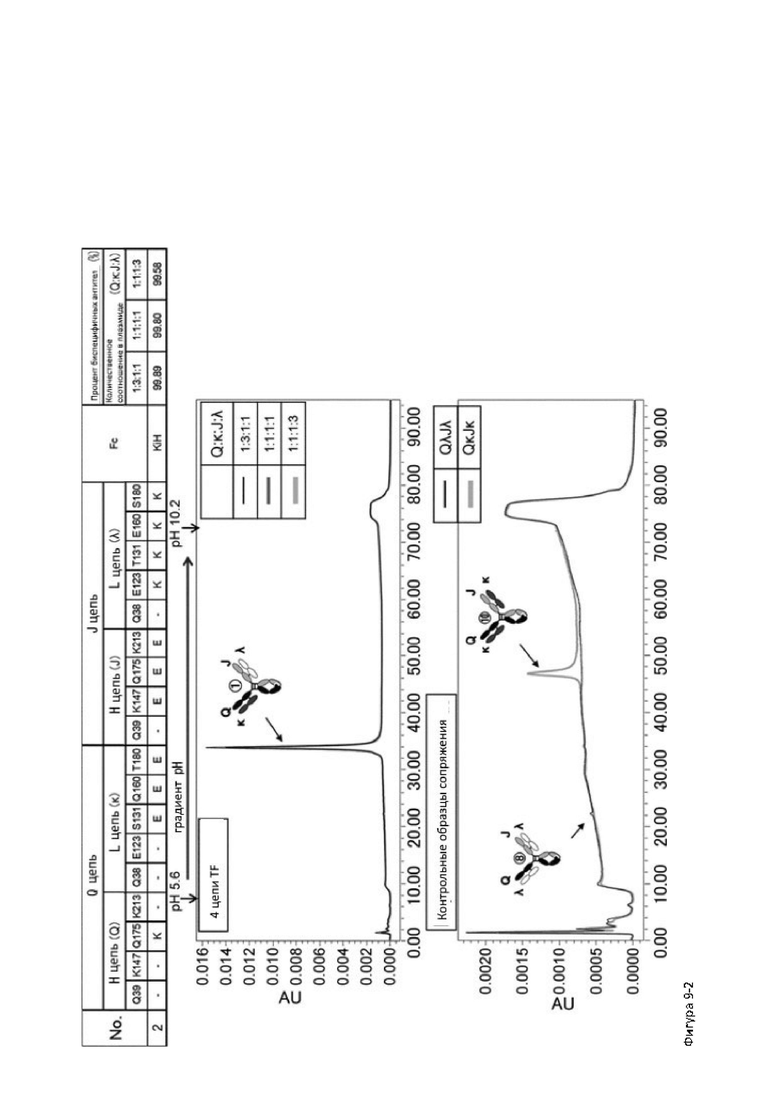
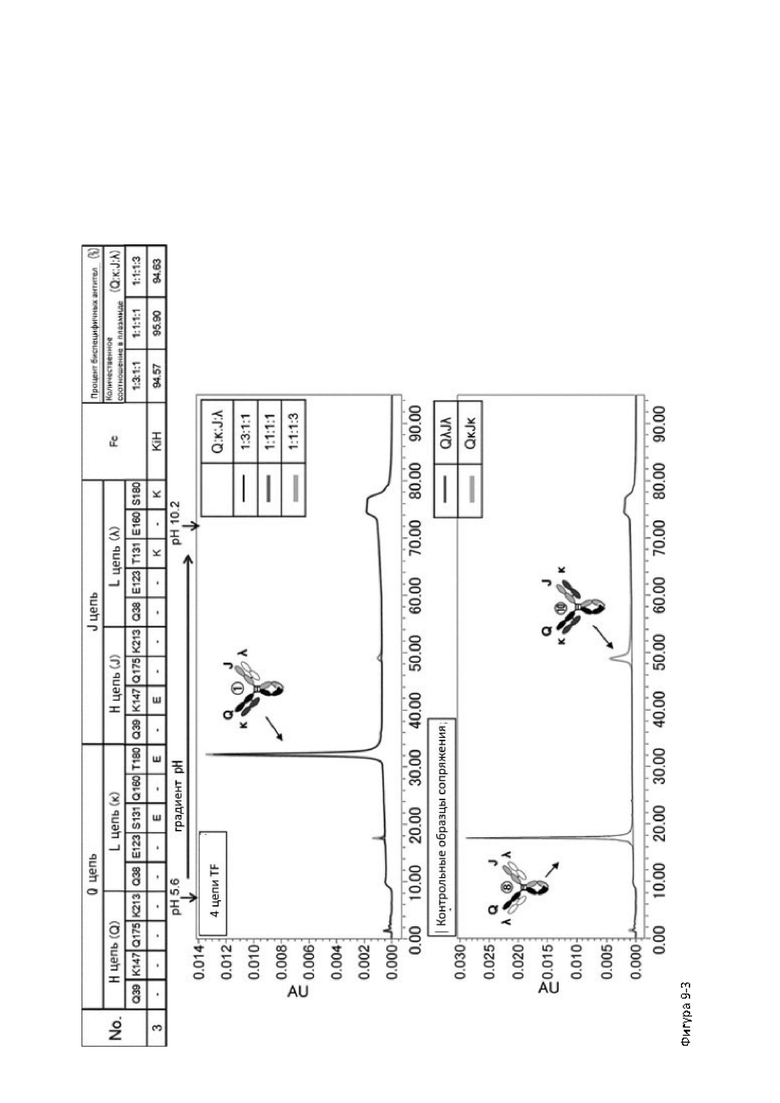
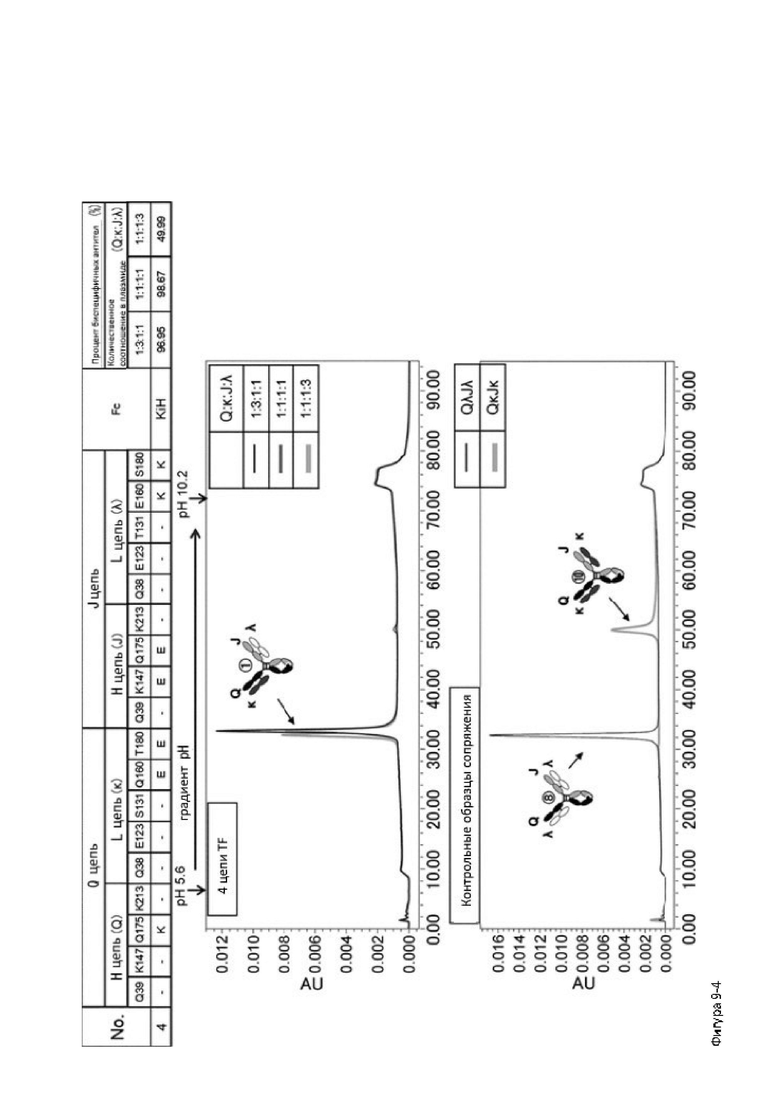
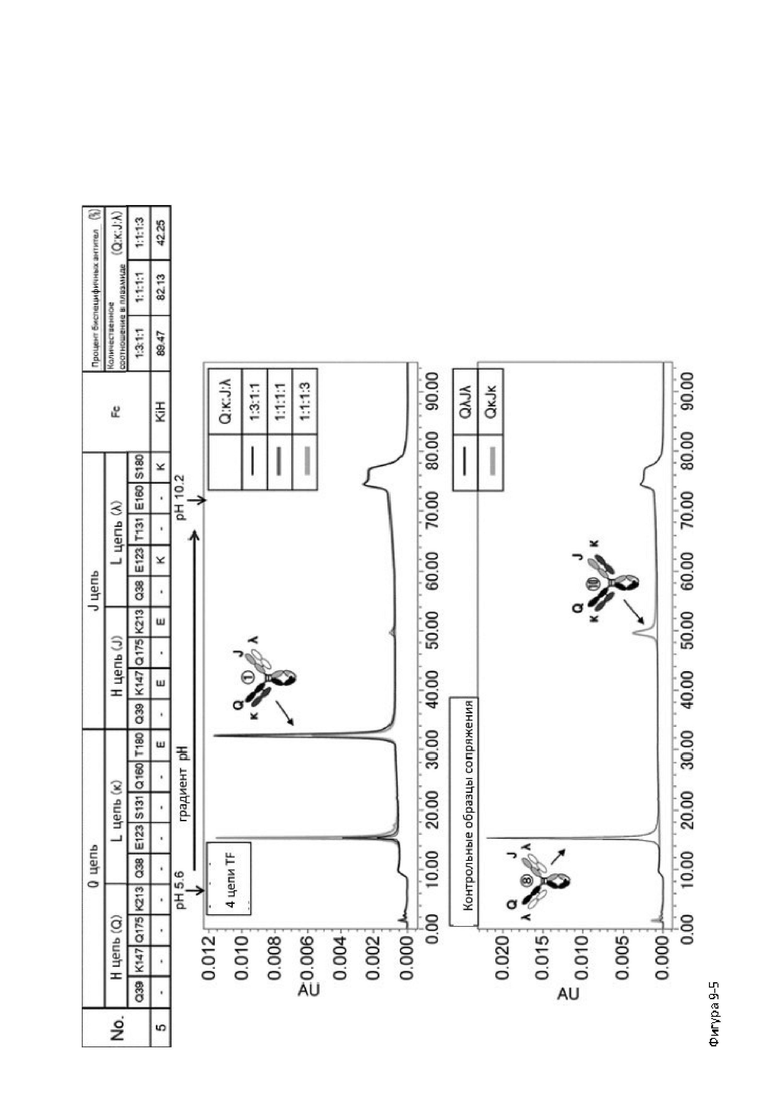
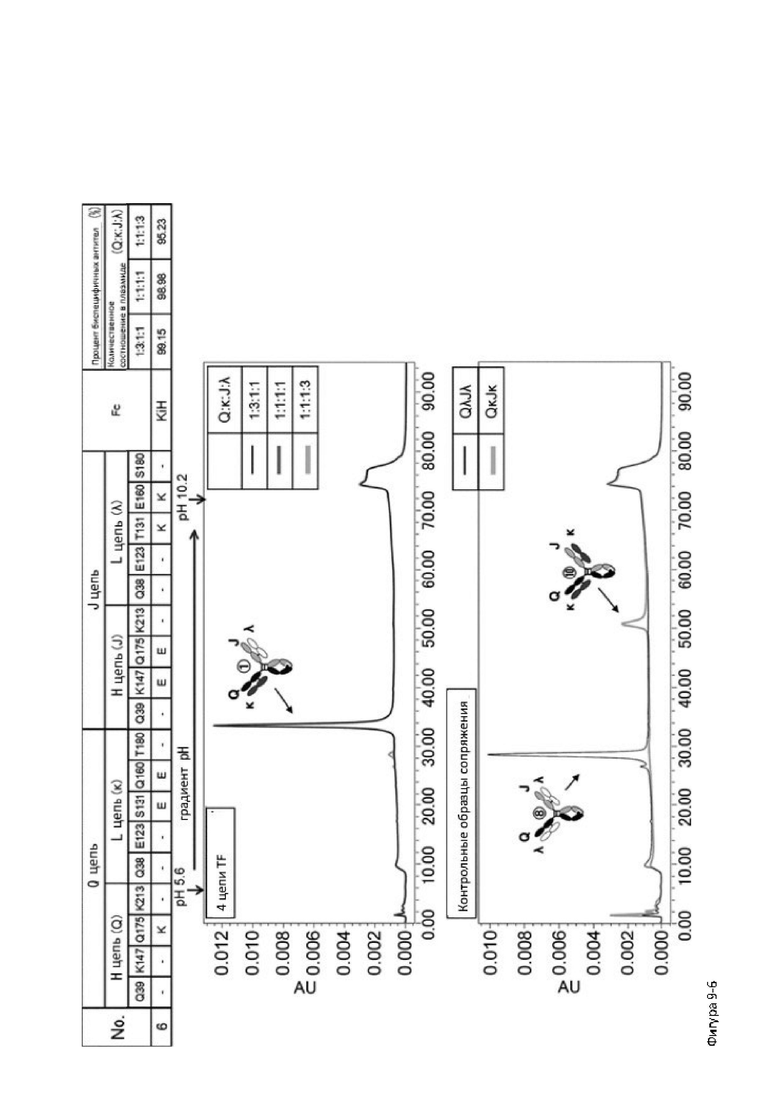
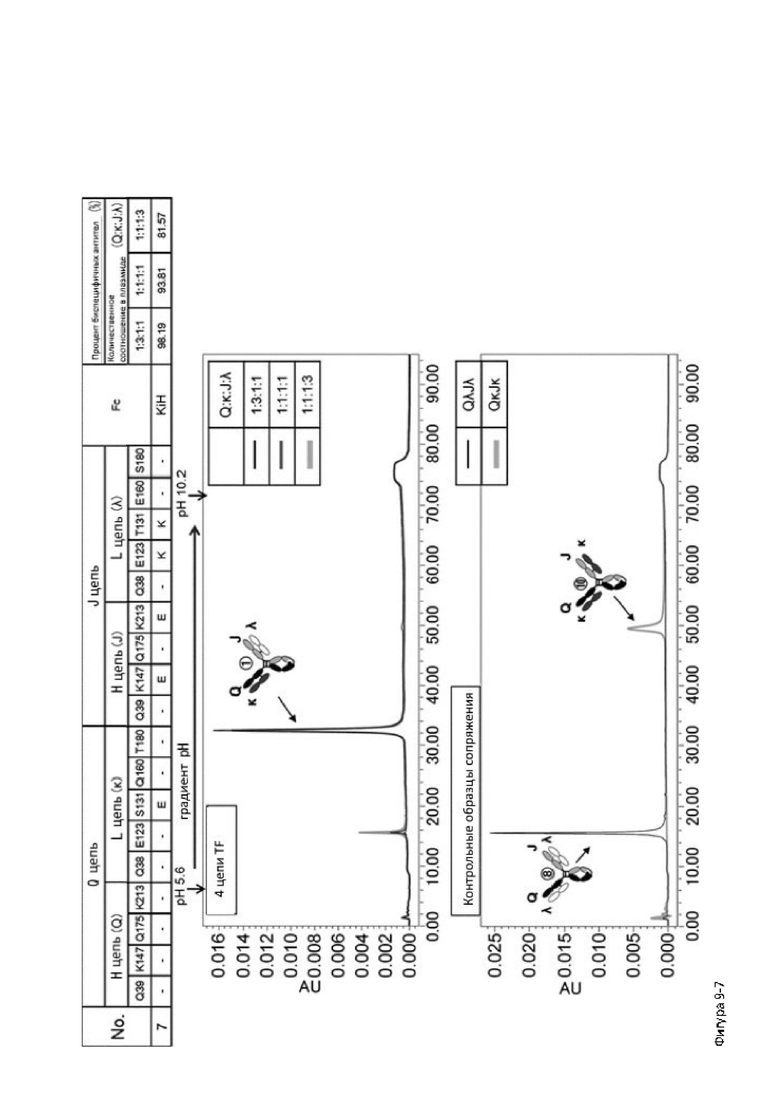
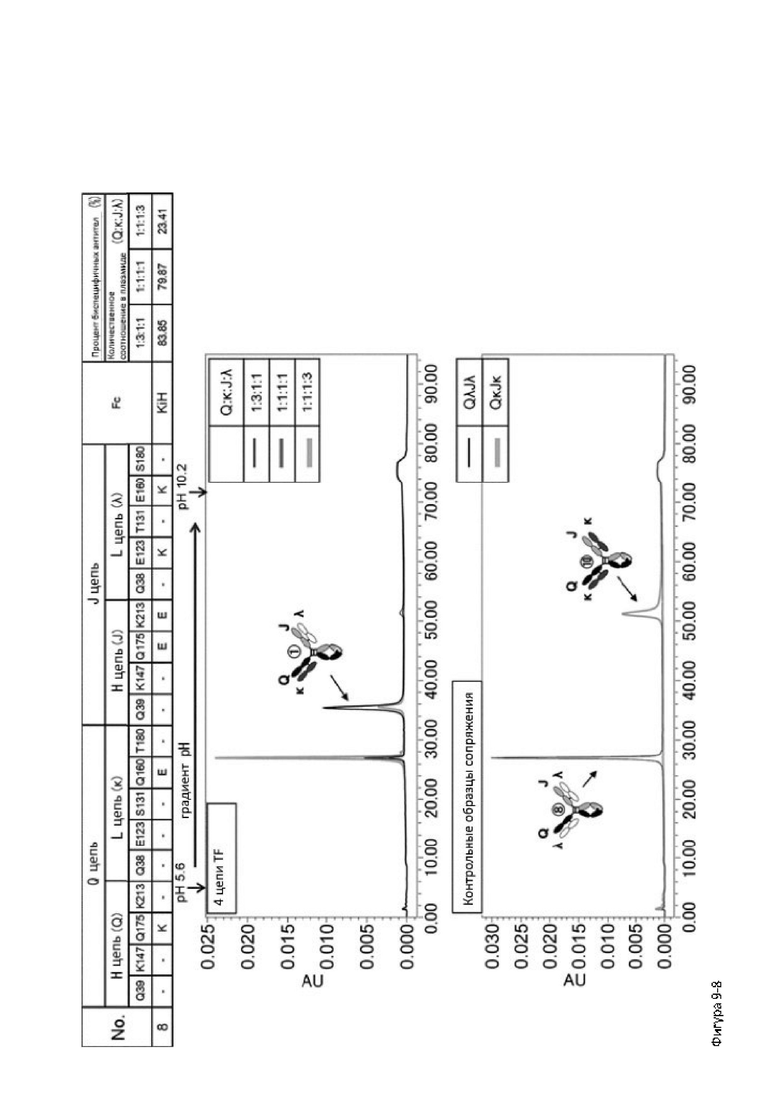
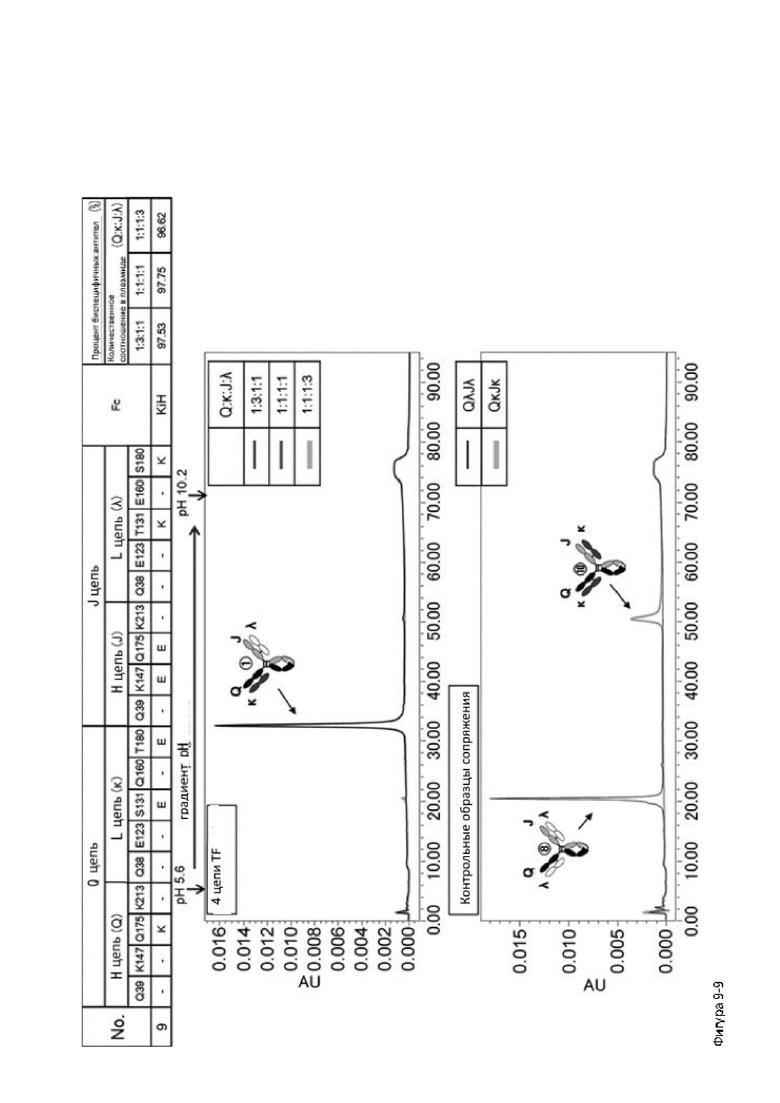
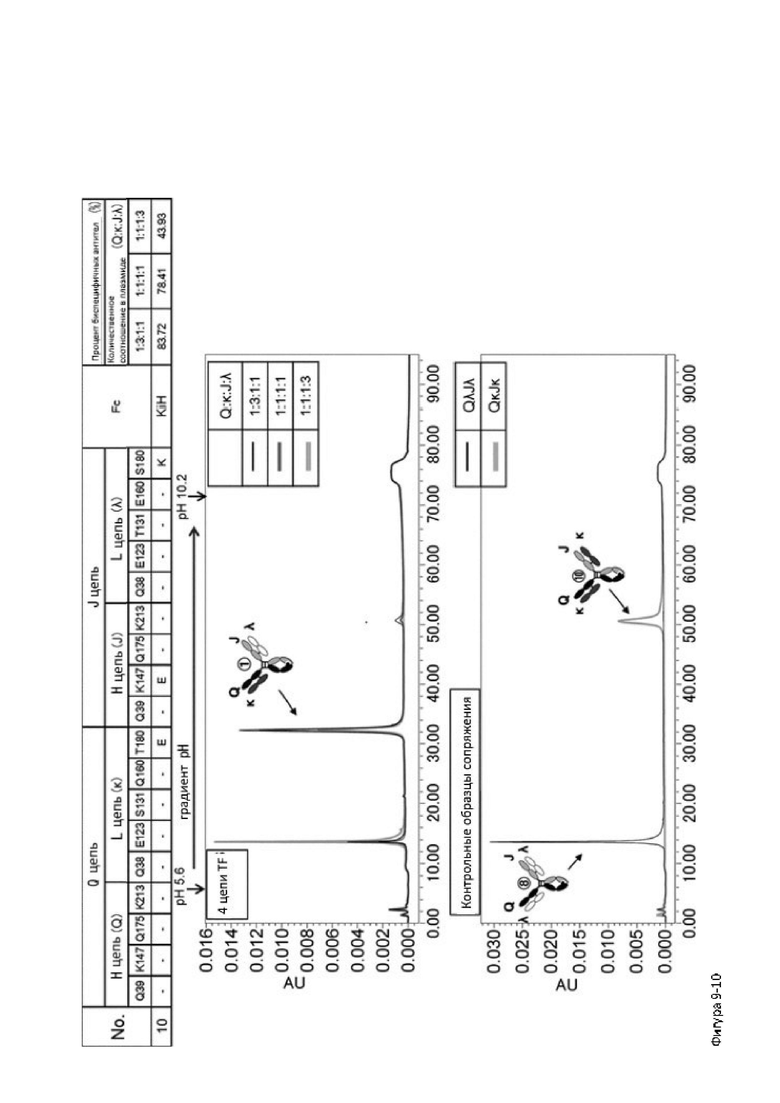
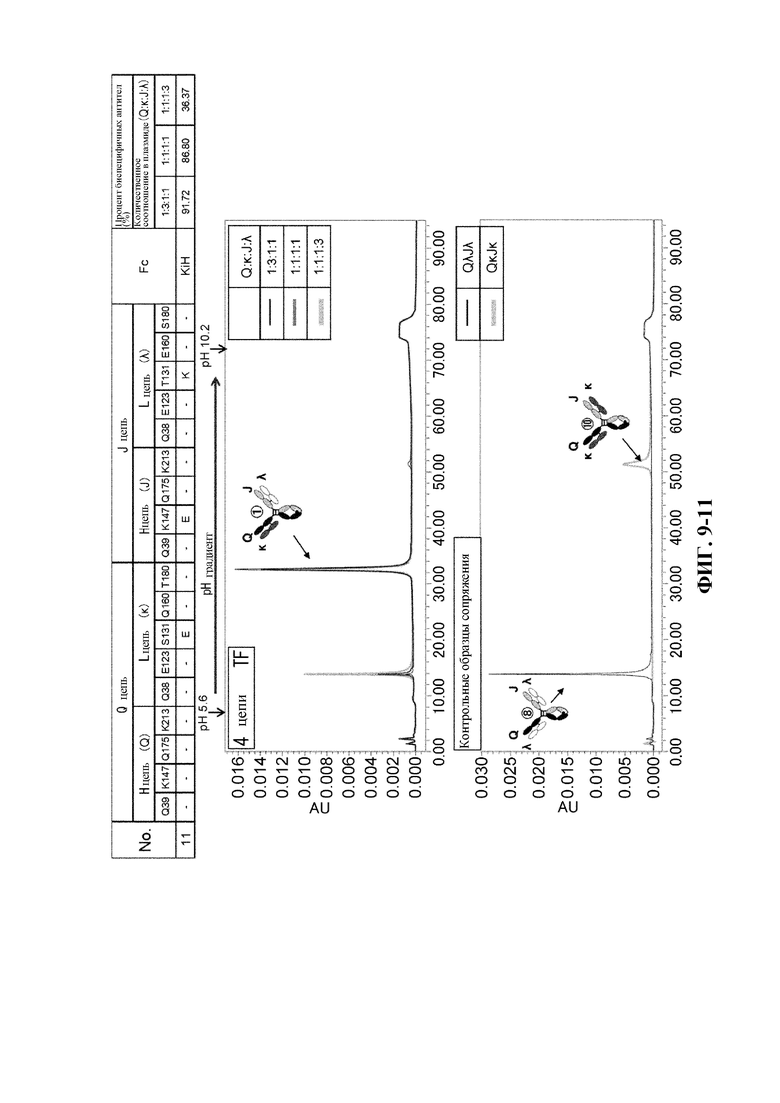
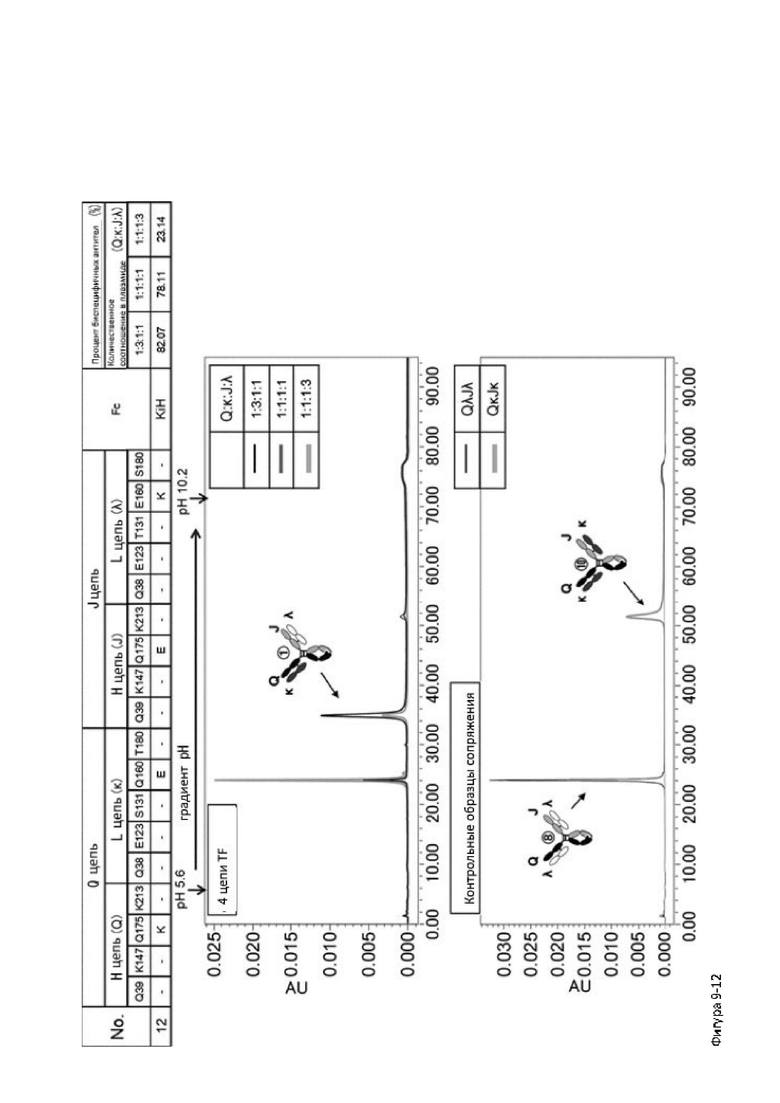
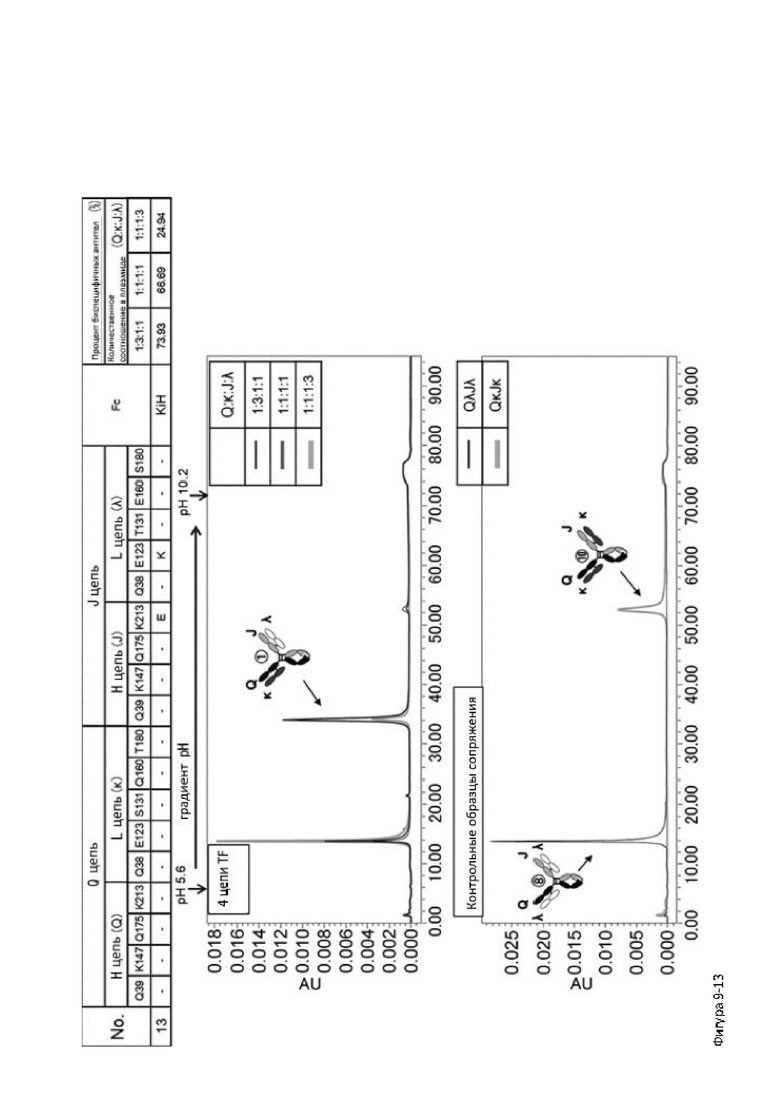
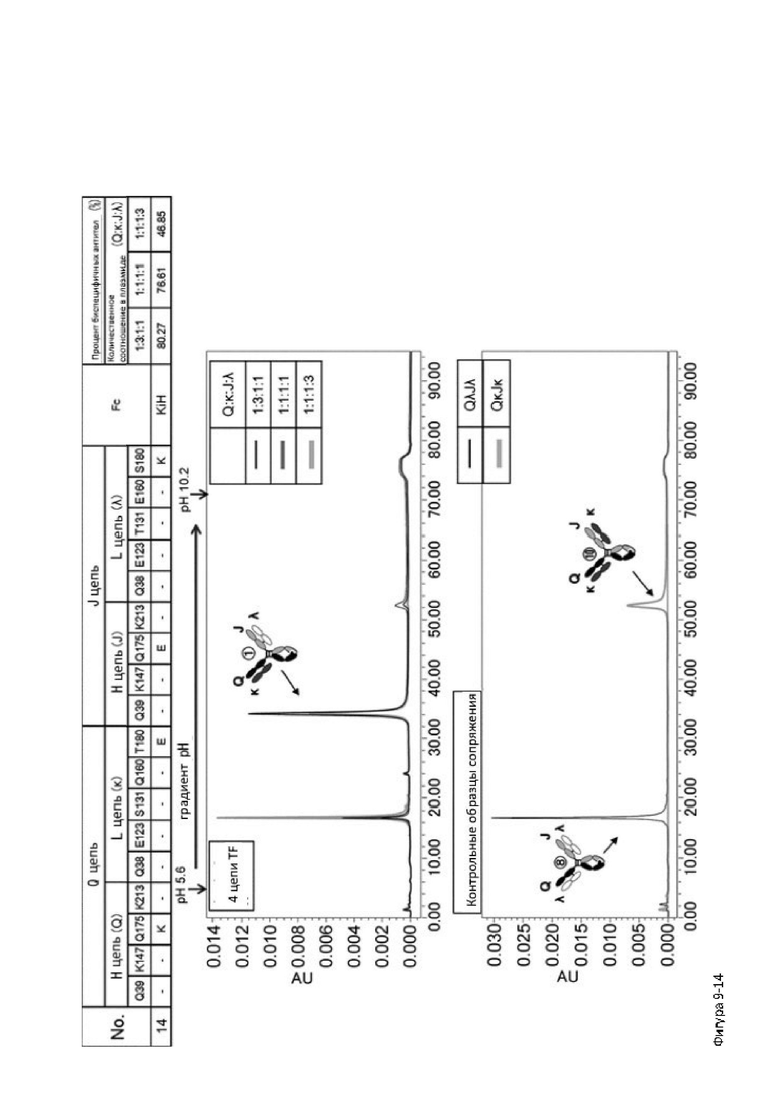
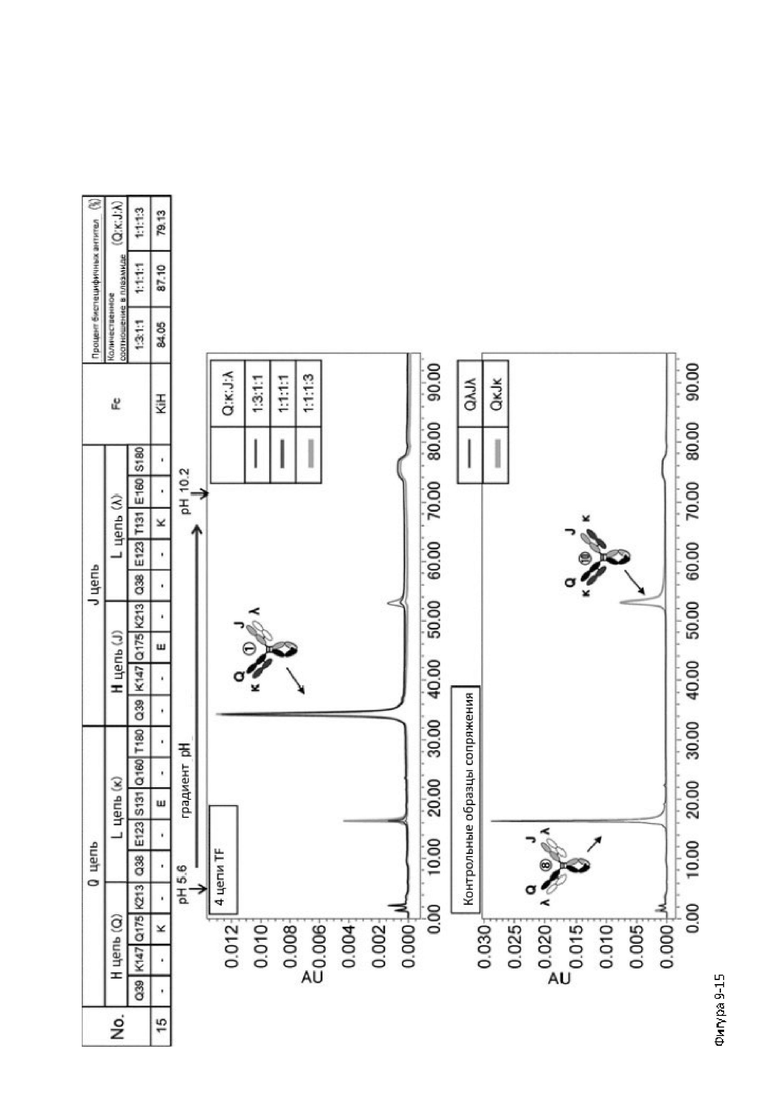
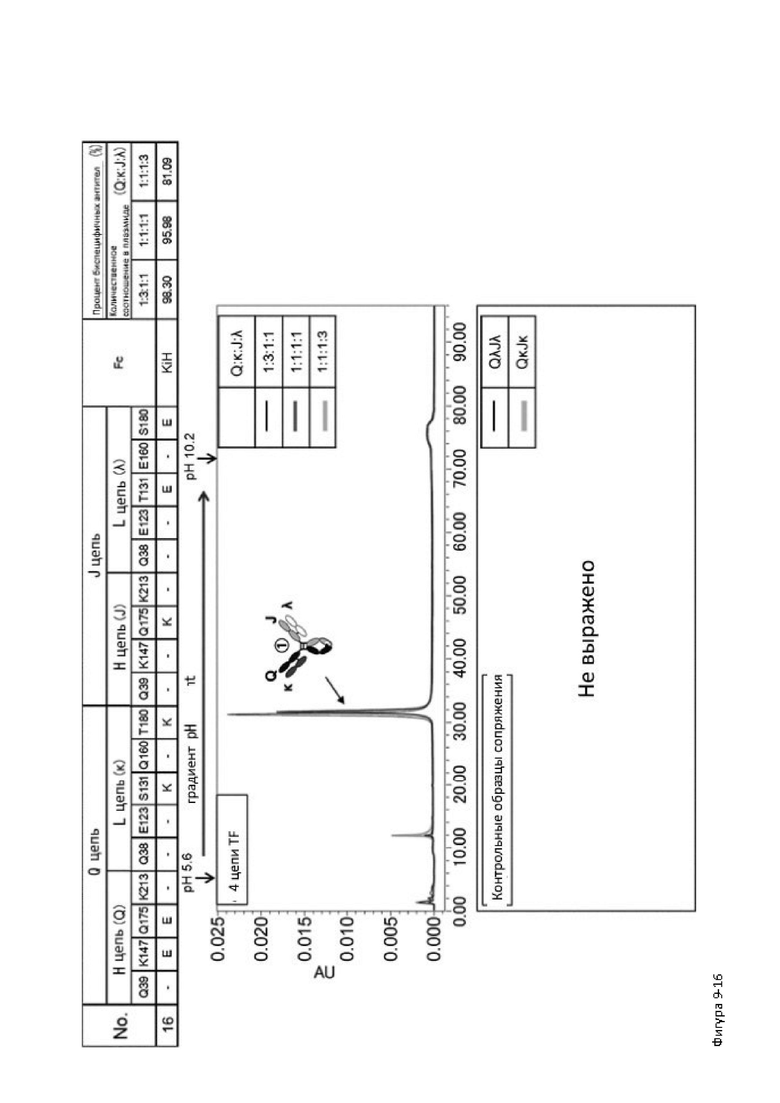
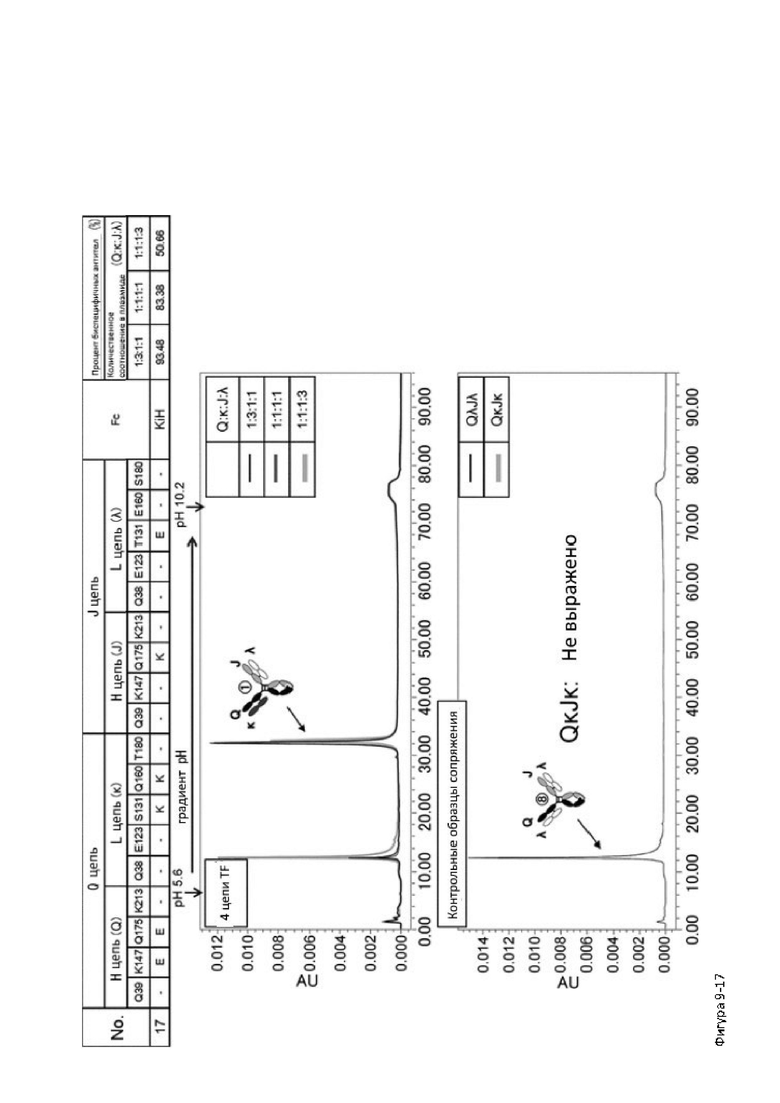
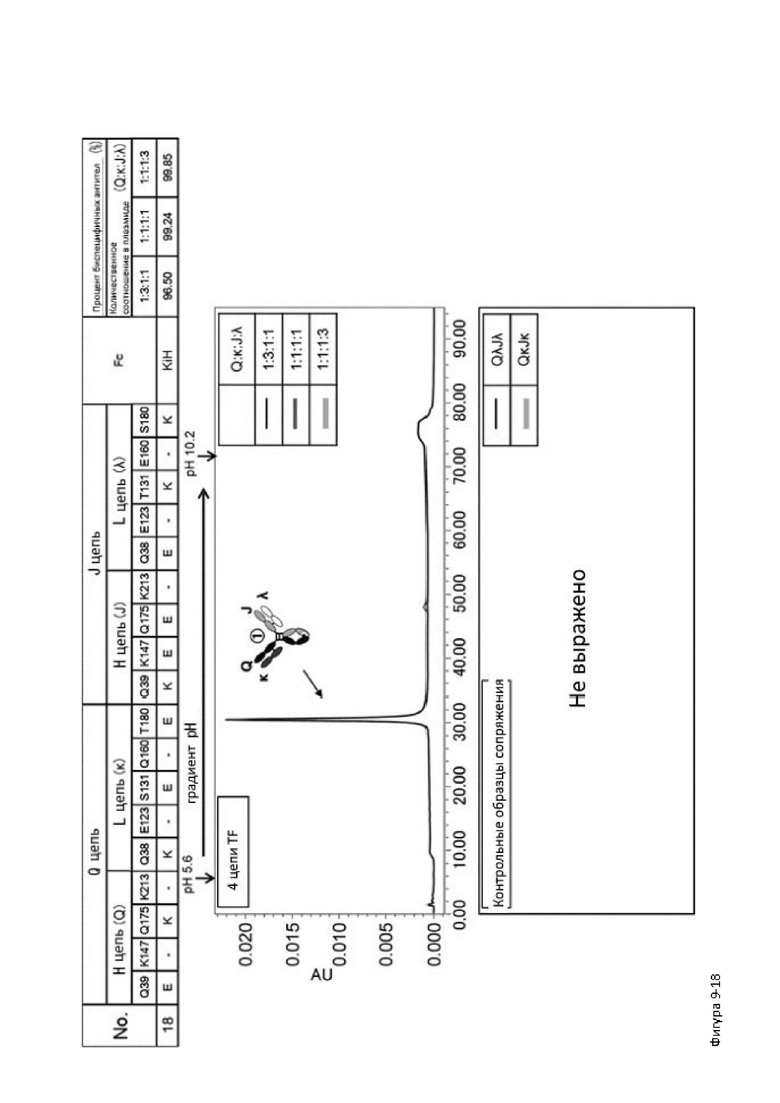
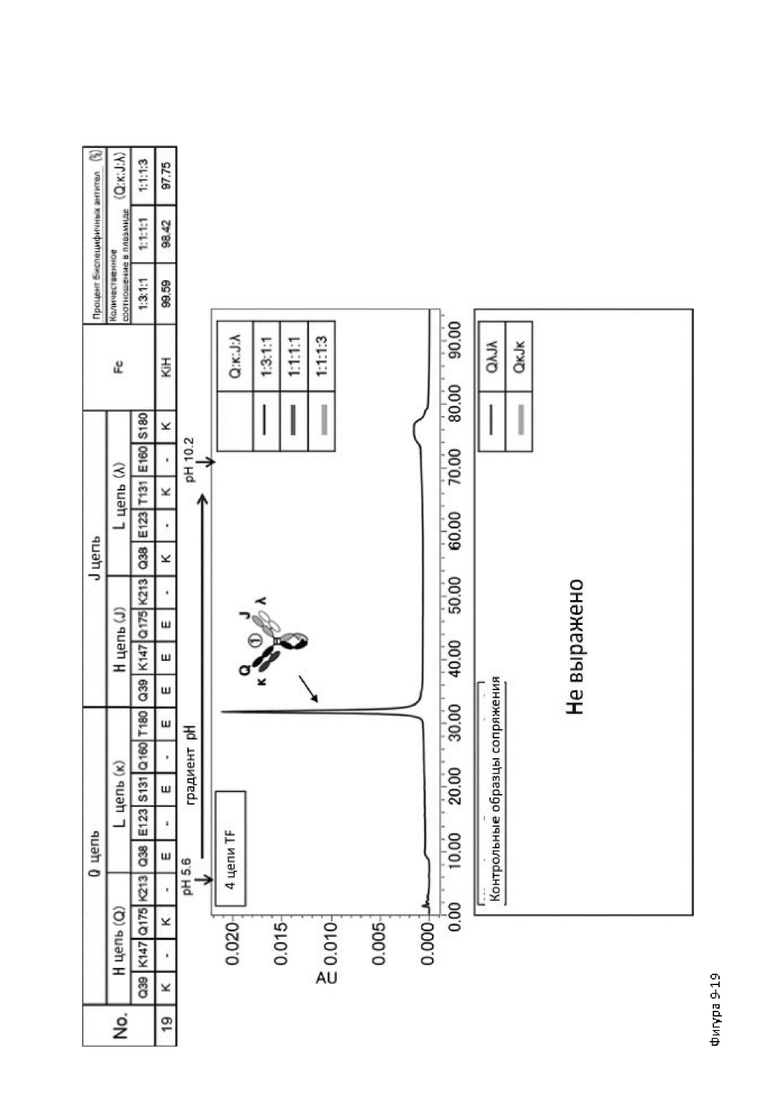
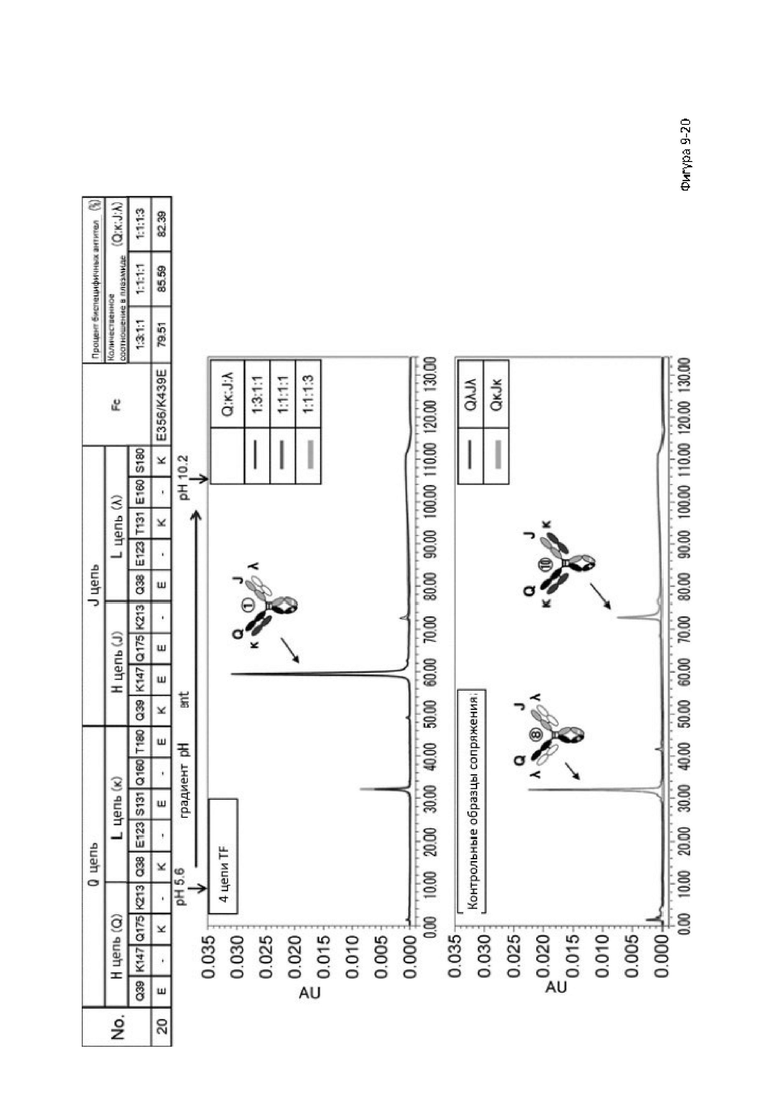

Изобретение относится к области биотехнологии, а именно к биспецифическому антителу, обладающему замещающей функциональной активностью коагулирующего фактора крови VIII, способу его получения, а также к содержащей его фармацевтической композиции. Также раскрыты выделенная нуклеиновая кислота, кодирующая вышеуказанное антитела, а также клетка-хозяин, ее содержащая. Изобретение эффективно для применения для профилактики и/или лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением. 6 н. и 9 з.п. ф-лы, 10 ил., 8 табл., 4 пр.
1. Биспецифическое антитело, обладающее замещающей функциональной активностью коагулирующего фактора крови VIII, содержащее первый антигенсвязывающий участок, который связывает коагулирующий фактор крови IX и/или активированный коагулирующий фактор крови IX, и второй антигенсвязывающий участок, который связывает коагулирующий фактор крови X, где первый антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи включает HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 1, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 2, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3, и
вариабельный домен легкой цепи включает HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 162, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 163, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 164, и
где второй антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи включает HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 4, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 5, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 6, и
вариабельный домен легкой цепи включает HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 165, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 166, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 167; причем один или более аминокислотных остатков замещены другими аминокислотными остатками, делетированы или встроены по крайней мере в одном из участков HVRs, в котором:
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 97, 98, 100, 100а, 100b и 100с в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене тяжелой цепи первого антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 53, 55, 92, 93 и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или встроен в вариабельном домене легкой цепи первого антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 51, 56, 57, 59, 61, 62, 65 и 102 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком в вариабельном домене тяжелой цепи второго антигенсвязывающего участка,
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 24, 26, 27, 29, 30, 31, 32, 50, 92, 94, 95, 95а и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене легкой цепи второго антигенсвязывающего участка.
2. Биспецифическое антитело по п. 1, в котором:
в вариабельном домене тяжелой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин или аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 26 представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене тяжелой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин или аминокислотный остаток в положении 102 представляет собой валин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин, где указанные положения соответствуют нумерации по Кабату.
3. Биспецифическое антитело по п. 1 или 2, в котором первый антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 168, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 169, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 170;
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 171, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 172, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 173;
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 174, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 175, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 176;
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 177, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 178, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 179;
5) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 180, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 181, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 182; или
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 183, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 184, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 185; и
вариабельный домен легкой цепи включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 186, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 187, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 188;
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 189, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 190, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 191;
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 192, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 193, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 194;
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 195, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 196, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 197;
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 198, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 199, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 200;
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 201, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 202, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 203;
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 204, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 205, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 206;
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 207, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 208, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 209;
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 210, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 211, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 212;
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 213, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 214, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 215;
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 216, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 217, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 218;
12) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 219, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 220, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 221; и
где второй антигенсвязывающий участок включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи,
причем вариабельный домен тяжелой цепи включает:
1) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 222, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 223, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 224;
2) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 225, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 226, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 227;
3) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 228, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 229, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 230;
4) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 231, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 232, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 233;
5) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 234, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 235, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 236;
6) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 237, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 238, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 239;
7) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 240, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 241, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 242;
8) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 243, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 244, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 245;
9) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 246, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 247, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 248;
10) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 249, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 250, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 251;
11) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 252, HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 253, и HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 254; и
вариабельный домен легкой цепи включает:
1) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 255, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 256, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 257;
2) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 258, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 259, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 260;
3) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 261, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 262, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 263;
4) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 264, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 265, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 266;
5) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 267, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 268, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 269;
6) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 270, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 271, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 272;
7) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 273, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 274, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 275;
8) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 276, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 277, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 278;
9) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 279, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 280, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 281;
10) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 282, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 283, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 284; или
11) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 285, HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 286, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 287.
4. Биспецифическое антитело по п. 1, в котором первый антигенсвязывающий участок содержит вариабельный домен тяжелой цепи SEQ ID NO: 45 и вариабельный домен легкой цепи SEQ ID NO: 13 и второй антигенсвязывающий участок содержит вариабельный домен тяжелой цепи SEQ ID NO: 46 и вариабельный домен легкой цепи SEQ ID NO: 31,
причем один или более аминокислотных остатков замещены другими аминокислотами, делетированы или встроены по крайней мере в одном из вариабельных доменов тяжелой или легкой цепи.
5. Биспецифическое антитело по п. 4, в котором
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 31, 34, 39, 97, 98, 100, 100а, 100b и 100е в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене тяжелой цепи первого антигенсвязывающего участка;
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 26, 27, 30, 31, 32, 38, 45, 53, 55, 60, 70, 76, 79, 80, 83, 85, 92, 93, 95 и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или встроен в вариабельном домене легкой цепи первого антигенсвязывающего участка;
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 28, 31, 39, 51, 56, 57, 59, 61, 62, 65, 67, 73, 82b и 102 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком в вариабельном домене тяжелой цепи второго антигенсвязывающего участка и
по крайней мере один аминокислотный остаток, выбранный из аминокислотных остатков в положениях 3, 8, 15, 24, 26, 27, 29, 30, 31, 32, 38, 48, 49, 50, 79, 92, 94, 95, 95а и 96 в соответствии с нумерацией по Кабату, замещен другим аминокислотным остатком или делетирован в вариабельном домене легкой цепи второго антигенсвязывающего участка.
6. Биспецифическое антитело по п. 4 или 5, в котором:
в вариабельном домене тяжелой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 31 представляет собой гистидин, аминокислотный остаток в положении 34 представляет собой аланин, аминокислотный остаток в положении 39 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 97 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 98 представляет собой серин, аминокислотный остаток в положении 100 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 100а представляет собой аспарагиновую кислоту или делетирован, аминокислотный остаток в положении 100b представляет собой аланин или гистидин или аминокислотный остаток в положении 100е представляет собой гистидин или изолейцин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи первого антигенсвязывающего участка аминокислотный остаток в положении 26 представляет собой треонин, аминокислотный остаток в положении 27 представляет собой аргинин, аминокислотный остаток в положении 30 представляет собой аргинин, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой аспарагиновую кислоту или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой лизин, аминокислотный остаток в положении 45 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 53 представляет собой аргинин, аминокислотный остаток в положении 55 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 60 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 70 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 76 представляет собой аспарагин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 80 представляет собой пролин или аланин, аминокислотный остаток в положении 83 представляет собой метионин или аланин, аминокислотный остаток в положении 85 представляет собой треонин, аминокислотный остаток в положении 92 представляет собой аргинин, аминокислотный остаток в положении 93 представляет собой серин или аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой пролин или аминокислотный остаток в положении 96 представляет собой глицин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене тяжелой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 28 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аспарагин, глутамин или гистидин, аминокислотный остаток в положении 39 представляет собой лизин, аминокислотный остаток в положении 51 представляет собой серин, аминокислотный остаток в положении 56 представляет собой треонин или аргинин, аминокислотный остаток в положении 57 представляет собой валин, аминокислотный остаток в положении 59 представляет собой серин, аминокислотный остаток в положении 61 представляет собой аргинин, аминокислотный остаток в положении 62 представляет собой лизин, аминокислотный остаток в положении 65 представляет собой аспарагин или глутамин, аминокислотный остаток в положении 67 представляет собой лейцин, аминокислотный остаток в положении 73 представляет собой изолейцин, аминокислотный остаток в положении 82b представляет собой глутаминовую кислоту или аминокислотный остаток в положении 102 представляет собой валин, где указанные положения соответствуют нумерации по Кабату;
в вариабельном домене легкой цепи второго антигенсвязывающего участка аминокислотный остаток в положении 3 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 8 представляет собой пролин, аминокислотный остаток в положении 15 представляет собой лейцин, аминокислотный остаток в положении 24 представляет собой треонин, аминокислотный остаток в положении 26 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 27 представляет собой глутамин, аминокислотный остаток в положении 29 представляет собой серин, аминокислотный остаток в положении 30 представляет собой глутамин, серин или глутаминовую кислоту, аминокислотный остаток в положении 31 представляет собой аргинин, аминокислотный остаток в положении 32 представляет собой глутамин или глутаминовую кислоту, аминокислотный остаток в положении 38 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 48 представляет собой изолейцин, аминокислотный остаток в положении 49 представляет собой тирозин, аминокислотный остаток в положении 50 представляет собой глутамин, аминокислотный остаток в положении 79 представляет собой глутаминовую кислоту, аминокислотный остаток в положении 92 представляет собой аланин, аминокислотный остаток в положении 94 представляет собой аспарагиновую кислоту, аминокислотный остаток в положении 95 представляет собой аспарагиновую кислоту или аланин, аминокислотный остаток в положении 95а представляет собой тирозин или делетирован или аминокислотный остаток в положении 96 представляет собой треонин, где указанные положения соответствуют нумерации по Кабату.
7. Биспецифическое антитело по любому из пп. 1-6, в котором
первый антигенсвязывающий участок содержит:
вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59 или SEQ ID NO:60, и
вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70, SEQ ID NO: 71 или SEQ ID NO: 72; и
второй антигенсвязывающий участок содержит:
вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82 или SEQ ID NO: 83, и
вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 84, SEQ ID NO: 85, SEQ ID NO: 86, SEQ ID NO: 87, SEQ ID NO: 88, SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93 или SEQ ID NO: 94.
8. Биспецифическое антитело по любому из пп. 1-7, в котором первый антигенсвязывающий участок содержит константные домены, содержащие аминокислотные последовательности (1) и (2), указанные ниже, и второй антигенсвязывающий участок содержит константные домены, содержащие аминокислотные последовательности (1) и (2), указанные ниже, которые отличаются от константных участков первого антигенсвязывающего участка:
(1) SEQ ID NO: 119 относится к константному участку тяжелой цепи и SEQ ID NO: 100 относится к константному участку легкой цепи;
(2) SEQ ID NO: 118 относится к константному участку тяжелой цепи и SEQ ID NO: 102 относится к константному участку легкой цепи.
9. Биспецифическое антитело, обладающее замещающей функциональной активностью коагулирующего фактора крови VIII, содержащее тяжелую цепь первого антитела и легкую цепь первого антитела, которые связываются с коагулирующим фактором крови IX и/или активированным коагулирующим фактором крови IX, и тяжелую цепь второго антитела и легкую цепь второго антитела, которые связываются с коагулирующим фактором крови X, причем биспецифическое антитело представляет собой любое из антител от (а) до (ф), как указано ниже:
(а) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 120, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 126, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 138, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 149;
(б) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 138, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 149;
(в) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 122, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 128, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(г) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 122, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 129, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(д) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 140, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 151;
(е) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 141, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 152;
(ж) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 121, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 127, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(з) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 130, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(и) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 131, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 139, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 150;
(к) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 131, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153;
(л) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 132, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153;
(м) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 132, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 143, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 154;
(н) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 133, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 142, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 153;
(о) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 133, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 143, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 154;
(п) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 134, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 144, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 155;
(р) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 123, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 135, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 145, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 156;
(с) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 136, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 144, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 155;
(т) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 136, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 146, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 157;
(у) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 124, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 134, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 147, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 158;
(ф) биспецифическое антитело, содержащее тяжелую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 125, легкую цепь первого антитела, содержащую аминокислотную последовательность SEQ ID NO: 137, тяжелую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 148, и легкую цепь второго антитела, содержащую аминокислотную последовательность SEQ ID NO: 159.
10. Выделенная нуклеиновая кислота, кодирующая биспецифическое антитело по любому из пп. 1-9.
11. Клетка-хозяин для получения антитела, содержащая нуклеиновую кислоту по п. 10.
12. Способ получения биспецифического антитела по любому из пп. 1-9, включающий культивирование клетки-хозяина по п. 11 в условиях, обеспечивающих продуцирование биспецифического антитела.
13. Фармацевтическая композиция, содержащая эффективное количество биспецифического антитела по любому из пп. 1-9 и фармацевтически приемлемый носитель, причем фармацевтическая композиция предназначена для применения для профилактики и/или лечения кровотечения, заболевания, в которое вовлечено кровотечение, или заболевания, вызываемого кровотечением.
14. Фармацевтическая композиция по п. 13, где кровотечение, заболевание, в которое вовлечено кровотечение, или заболевание, вызываемое кровотечением, представляет собой заболевание, развитие и/или прогрессивное течение которого обусловлено уменьшением или дефицитом активности коагулирующего фактора крови VIII и/или активированного коагулирующего фактора крови VIII.
15. Фармацевтическая композиция по п. 14, где заболевание, развитие и/или прогрессивное течение которого обусловлено уменьшением или дефицитом активности коагулирующего фактора крови VIII и/или активированного коагулирующего фактора крови VIII, представляет собой гемофилию А, заболевание, при котором непредвиденно образуется ингибитор коагулирующего фактора крови VIII и/или активированного коагулирующего фактора крови VIII, приобретенную гемофилию или болезнь Виллебранда.
| WO 2012067176 A1, 24.05.2012 | |||
| WO 2013065708 A1, 10.05.2013 | |||
| GERALD M | |||
| EDELMAN et al., THE COVALENT STRUCTURE OF AN ENTIRE γG IMMUNOGLOBULIN MOLECULE, BIOCHEMISTRY, 1969, Vol.63, N.1, pp.78-85 | |||
| LAURA M | |||
| GRIFFIN et al., Analysis of heavy and light chain sequences of conventional camelid antibodies from Camelus dromedarius and Camelus bactrianus |
