Область техники
Настоящее изобретение в целом относится к антителам, которые связываются с HLA-A2/WT1, содержащую биспецифические антигенсвязывающие молекулы, например, для активации Т-клеток. Кроме того, настоящее изобретение относится к полинуклеотидам, кодирующим такие антитела, а также векторам и клеткам-хозяевам, включающим указанные полинуклеотиды. Настоящее изобретение также относится к способам получения антител, а также к способам их применения для лечения заболевания.
Предпосылки создания изобретения
WT1 (аббревиатура от англ. Wilms Tumor 1, опухоль Вильмса 1, белок опухоли Вильмса 1) представляет собой онкогенный фактор транскрипции, участвующий в пролиферации, дифференцировке клеток, а также в клеточном апоптозе и развитии органов, при этом экспрессия этого фактора в тканях здорового взрослого человека происходит редко (Hinrichs and Restifo, Nat Biotechnol (2013) 31, 999-1008). Однако, сообщается, что WT1 гиперэкспрессируется при нескольких типах злокачественных гематологических заболеваний и широком спектре твердых опухолей (Van Driessche et al., Oncologist (2012) 17, 250-259). WT1 представляет собой ядерный белок, располагающийся внутриклеточно. Внутриклеточный белок может подвергаться деградации в протеасоме, процессироваться и презентироваться на поверхности клеток при помощи основного комплекса гистосовместимости (МНС)1 в качестве Т-клеточных эпитопов и распознаваться Т-клеточными рецепторами (TCR, аббрев. от англ. T-Cell Receptor). Соответственно, пептиды на основе WT1, такие, как WT1RMF (RMFPNAPYL) и WT1VLD (VLDFAPPGA) презентируются с участием HLA-A2 на поверхности клеток и могут служить триггером для распознавания Т-клетками.
Было предпринято несколько попыток использовать WT1 в качестве мишени для таргетной (иммуно)терапии рака, содержащую разработку вакцин на основе WT1-выделенного пептида и адоптивного Т-клеточного трансфера или WT1-специфических Т-клеток. Также были созданы TCR-подобные антитела к комплексу HLA-A2/WT1RMF, содержащую их биспецифические производные (Dao et al., Sci Transl Med (2013) 5, 176ra33; WO 2012/135854; Dao et al., Nat Biotechnol (2015) 33, 1079-1086; WO 2015/070061; WO 2017/060201).
Биспецифические антитела, которые связываются с поверхностным антигеном клеток-мишеней, а активируют Т-клеточный антиген, такой, как CD3 на поверхности Т-клеток (также обозначаемые в настоящем документе как Т-клеточные биспецифические антитела или "TCBs") представляются крайне перспективными для лечения различных раковых заболеваний. Одновременное связывание такого антитела с обеими мишенями будет усиливать временное взаимодействие между клеткой-мишенью и Т-клеткой, что будет приводить к перекрестному связыванию Т-клеточного рецептора, затем к активации какой-либо цитотоксической Т-клетки с последующим лизисом клетки-мишени. По причине высокой специфичности, способности убивать клетки-мишени, выбирать клетку-мишень самой важной задачей в применении биспецифических Т-клеточных антител является задача избежать направленной и ненаправленной токсичности. Внутриклеточные белки, такие, как WT1, являются привлекательными мишенями, но доступны лишь для антител, подобных Т-клеточным рецепторам (TCR-подобные антитела), которые связываются с молекулами основного комплекса гистосовместимости (МНС) и презентируют пептидные антигены, выделенные из внутриклеточного белка, на поверхности клеток. Характерной проблемой TCR-подобных антител является потенциальная перекрестная реактивность с молекулами МНС как таковыми, или с пептидами, презентируемыми молекулами МНС, отличными от желаемых, что может нарушать органную или тканевую селективность.
Краткое описание изобретения
Настоящее изобретение относится к новым антителам, содержащую биспецифические антитела, которые связываются с HLA-A2/WT1 и обладают особенно благоприятными свойствами для применения в терапевтических целях. Авторы настоящего изобретения разработали антитела с неожиданными, улучшенными свойствами, которые связываются с HLA-A2/WT1. В частности, антитело связывается с HLA-A2/WT1 с хорошей аффинностью и значительной специфичностью. Кроме того, авторы настоящего изобретения разработали биспецифические антигенсвязывающие молекулы, которые связываются с HLA-A2/WT1 и активирующим Т-клеточным антигеном, содержащую новое HLA-A2/WT1 антитело, и сочетают в себе высокую эффективность и продуктивность.
Согласно первому аспекту, настоящее изобретение относится к антителу, которое связывается с HLA-A2/WT1, при этом указанное антитело включает:
(i) вариабельную область тяжелой цепи (VH), содержащую область, определяющую комплементарность тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2, и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область, определяющую комплементарность легкой цепи (LCDR) 1 последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 с последовательностью SEQ ID NO: 6;
(ii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 9, HCDR 2 с последовательностью SEQ ID NO: 10, и HCDR 3 с последовательностью SEQ ID NO: 11, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 12, LCDR 2 с последовательностью SEQ ID NO: 13 и LCDR 3 с последовательностью SEQ ID NO: 14;
(iii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 17, HCDR 2 с последовательностью SEQ ID NO: 18, и HCDR 3 с последовательностью SEQ ID NO: 19, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 20, LCDR 2 с последовательностью SEQ ID NO: 21 и LCDR 3 с последовательностью SEQ ID NO: 22;
(iv) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 25, a HCDR 2 с последовательностью SEQ ID NO: 26, и HCDR 3 с последовательностью SEQ ID NO: 27, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 28, LCDR 2 с последовательностью SEQ ID NO: 29 и LCDR 3 с последовательностью SEQ ID NO: 30;
(v) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 33, и HCDR 2 с последовательностью SEQ ID NO: 34, и HCDR 3 с последовательностью SEQ ID NO: 35, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 36, LCDR 2 с последовательностью SEQ ID NO: 37 и LCDR 3 с последовательностью SEQ ID NO: 38;
(vi) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 41, HCDR 2 с последовательностью SEQ ID NO: 42, и HCDR 3 с последовательностью SEQ ID NO: 43, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 44, LCDR 2 с последовательностью SEQ ID NO: 45 и LCDR 3 с последовательностью SEQ ID NO: 46;
(vii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 49, HCDR 2 с последовательностью SEQ ID NO: 50, и HCDR 3 с последовательностью SEQ ID NO: 51, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 52, LCDR 2 с последовательностью SEQ ID NO: 53, и LCDR 3 с последовательностью SEQ ID NO: 54;
(viii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 57, HCDR 2 с последовательностью SEQ ID NO: 58, и HCDR 3 с последовательностью SEQ ID NO: 59, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 60, LCDR 2 с последовательностью SEQ ID NO: 61 и LCDR 3 с последовательностью SEQ ID NO: 62; или
(ix) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 65, HCDR 2 с последовательностью SEQ ID NO: 66, и HCDR 3 с последовательностью SEQ ID NO: 67, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 68, LCDR 2 с последовательностью SEQ ID NO: 69 и LCDR 3 с последовательностью SEQ ID NO: 70.
Согласно одному варианту осуществления изобретения, антитело включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64; или
(ix) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно одному варианту осуществления изобретения, антитело представляет собой IgG, в частности, к IgG1 антитело. Согласно одному варианту осуществления изобретения, антитело представляет собой полноразмерное антитело. Согласно другому варианту осуществления изобретения, антитело представляет собой фрагмент антитела, выбранный из группы, включающей Fv молекулу, scFv молекулу и Fab молекулу и F(ab')2 молекулу. Согласно одному варианту осуществления изобретения, антитело представляет собой мультиспецифическое антитело.
Настоящее изобретение также относится к биспецифической антигенсвязывающей молекуле, включающей:
(а) первую антигенсвязывающую молекулу, которая связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, а первая антигенсвязывающая молекула включает
(i) вариабельную область тяжелой цепи (VH), содержащую область, определяющую комплементарность тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2, и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL) содержащую область, определяющую комплементарность легкой цепи (LCDR) 1 с последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 о c последовательностью SEQ ID NO: 6;
(ii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 9, HCDR 2 с последовательностью SEQ ID NO: 10, и HCDR 3 с последовательностью SEQ ID NO: 11, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 12, LCDR 2 с последовательностью SEQ ID NO: 13 и LCDR 3 с последовательностью SEQ ID NO: 14;
(iii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 17, HCDR 2 с последовательностью SEQ ID NO: 18, и HCDR 3 с последовательностью SEQ ID NO: 19, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 20, LCDR 2 с последовательностью SEQ ID NO: 21 и LCDR 3 с последовательностью SEQ ID NO: 22;
(iv) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 25, HCDR 2 с последовательностью SEQ ID NO: 26, и HCDR 3 с последовательностью SEQ ID NO: 27, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 28, LCDR 2 с последовательностью SEQ ID NO: 29 и LCDR 3 с последовательностью SEQ ID NO: 30;
(v) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 33, HCDR 2 с последовательностью SEQ ID NO: 34, и HCDR 3 с последовательностью SEQ ID NO. 35, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO. 36, LCDR 2 с последовательностью SEQ ID NO: 37 и LCDR 3 с последовательностью SEQ ID NO: 38;
(vi) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 41, HCDR 2 с последовательностью SEQ ID NO: 42, и HCDR 3 с последовательностью SEQ ID NO: 43, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 44, LCDR 2 с последовательностью SEQ ID NO: 45 и LCDR 3 с последовательностью SEQ ID NO: 46;
(vii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 49, HCDR 2 с последовательностью SEQ ID NO: 50, и HCDR 3 с последовательностью SEQ ID NO: 51, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 52, LCDR 2 с последовательностью SEQ ID NO: 53 и LCDR 3 с последовательностью SEQ ID NO: 54;
(viii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 57, HCDR 2 с последовательностью SEQ ID NO: 58, и HCDR 3 с последовательностью SEQ ID NO: 59, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 60, LCDR 2 с последовательностью SEQ ID NO: 61 и LCDR 3 с последовательностью SEQ ID NO: 62;
(ix) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 65, HCDR 2 с последовательностью SEQ ID NO. 66, и HCDR 3 с последовательностью SEQ ID NO: 67, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 68, LCDR 2 с последовательностью SEQ ID NO: 69 и LCDR 3 с последовательностью SEQ ID NO: 70; и
(b) вторую антигенсвязывающую молекулу, которая специфически связывается со вторым антигеном.
Согласно одному варианту осуществления изобретения, первый антигенсвязывающий фрагмент включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(vii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64;
(ix) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно одному варианту осуществления изобретения, второй антиген представляет собой CD3, в частности, CD3ε. Согласно одному варианту осуществления изобретения, второй антигенсвязывающий фрагмент включает VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 115, HCDR 2 с последовательностью SEQ ID NO: 116, и HCDR 3 с последовательностью SEQ ID NO: 117, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 118, LCDR 2 с последовательностью SEQ ID NO: 119 и LCDR 3 с последовательностью SEQ ID NO: 120.
Согласно одному варианту осуществления изобретения, VH второго антигенсвязывающего фрагмента включает аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 121, a VL второго антигенсвязывающего фрагмента включает аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 122. Согласно одному варианту осуществления изобретения, первый и/или второй антигенсвязывающие фрагменты представляют собой Fab молекулы. Согласно одному варианту осуществления изобретения, второй антигенсвязывающий фрагмент представляет собой Fab молекулу, при этом вариабельные домены VL и VH или константные домены CL и СН1, в частности, вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab заменены между собой. Согласно одному варианту осуществления изобретения, первый антигенсвязывающий фрагмент представляет собой Fab молекулу, при этом в константном домене CL аминокислота в положении 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация согласно Кабату) и аминокислота в положении 123 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация согласно Кабату), а в константном домене СН1 аминокислота в положении 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно Кабату, EU индекс) и аминокислота в положении 123 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно Кабату, EU индекс). Согласно одному варианту осуществления изобретения, первый и второй антигенсвязывающие фрагменты соединены между собой, необязательно, посредством пептидного линкера. Согласно одному варианту осуществления изобретения, при этом первый и второй антигенсвязывающий фрагмент оба представляют собой Fab молекулу, при этом (а) второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи первого антигенсвязывающего фрагмента, или (b) первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи второго антигенсвязывающего фрагмента. Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает третий антигенсвязывающий фрагмент. Согласно одному варианту осуществления изобретения, при этом третий антигенсвязывающий фрагмент идентичен первому антигенсвязывающему фрагменту. Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает Fc домен, состоящий из первой и второй субъединицы. Согласно одному варианту осуществления изобретения, первый и второй и, в случае наличия, третий антигенсвязывающий фрагмент каждый представляет собой Fab молекулу; при этом (i) второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи первого антигенсвязывающего фрагмента, а первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом первой субъединицы Fc домена; или (ii) первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи вторым антигенсвязывающим фрагментом, а второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом второй субъединицы Fc домена; а третий антигенсвязывающий фрагмент, в случае его наличия, на своем С-конце Fab тяжелой цепи соединен с N-концом второй субъединицы Fc домена. Согласно одному варианту осуществления изобретения, Fc домен Fc домен представляет собой домен IgG, в частности, Fc домен IgG1. Согласно одному варианту осуществления изобретения, аминокислотный остаток в СН3 домене первой субъединицы Fc домена замещен аминокислотным остатком, имеющим больший объем боковой цепи, что создает выступ внутри СН3 домена первой субъединицы, который пространственно совпадает с впадиной внутри СН3 домена второй субъединицы, а аминокислотный остаток СН3 домена второй субъединицы Fc домена замещен аминокислотным остатком, имеющим меньший объем боковой цепи, что создает впадину внутри СН3 домена второй субъединицы, которая пространственно совпадает с выступом внутри СН3 домена первой субъединицы. Согласно одному варианту осуществления изобретения, Fc домен включает одну или более аминокислотную замену, которая снижает связывание с Fc рецептором и/или эффекторную функцию.
Согласно другому аспекту настоящего изобретения, предлагается один или более изолированный полинуклеотид(ы), кодирующий антитело или биспецифическую антигенсвязывающую молекулу, заявленную в соответствии с настоящим изобретением. Настоящее изобретение также относится к одному или более вектору(ам) экспрессии, содержащему изолированный полинуклеотид(ы), кодирующий антитело или биспецифическую антигенсвязывающую молекулу, заявленную в настоящем изобретении. Настоящее изобретение также относится к одному или более вектору экспрессии, содержащему изолированный полинуклеотид(ы), заявленный в настоящем изобретении, а также к клетке-хозяину, содержащей изолированный полинуклеотид(ы) или вектор(а) экспрессии, заявленный в соответствии с настоящим изобретением. Согласно некоторым вариантам осуществления изобретения, клетка-хозяин представляет собой эукариотическую клетку, в частности, клетку млекопитающего.
Согласно другому аспекту настоящего изобретения предлагается способ получения антитела, которое связывается с HLA-A2/WT1, содержащую этапы а) культивирования клетки-хозяина согласно изобретению в условиях, подходящих для экспрессии антитела и b) восстановление антитела. Настоящее изобретение также относится к антителу, которое связывается с HLA-A2/WT1, полученному при помощи способа, заявленного в настоящем изобретении.
Настоящее изобретение также относится к фармацевтической композиции, включающей антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению, и фармацевтически приемлемый носитель. Также настоящее изобретение включает способы применения антитела, биспецифической антигенсвязывающей молекулы и фармацевтической композиции, заявленной в настоящем изобретении. Согласно одному аспекту своего осуществления, настоящее изобретение относится к применению антитела, биспецифической антигенсвязывающей молекулы или фармацевтической композиции в качестве лекарственного средства. Согласно одному аспекту, заявленное антитело, биспецифическая антигенсвязывающая молекула или фармацевтическая композиция, согласно изобретению, применяются для лечения заболевания. Согласно специфическому варианту осуществления изобретения, заболеванием является рак.
Также предлагается применение антитела или биспецифической антигенсвязывающей молекулы согласно изобретению для создания лекарственного средства, предназначенного для лечения заболевания; а также в качестве метода лечения заболевания у индивидуума, который включает введение указанному индивидууму терапевтически эффективного количества композиции, включающей антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению в фармацевтически приемлемой форме. Согласно специфическому варианту осуществления изобретения заболеванием является рак.
Краткое описание фигур
Фигура 1. Примерные конфигурации биспецифических антигенсвязывающих молекул, заявленных в настоящем изобретении. (A, D) Изображение "1+1 CrossFab" молекулы (В, Е) Изображение "2+1 IgG Crossfab" молекулы с альтернативным порядком расположения Crossfab и Fab компонентов ("перевернуты"). (С, F) Изображение "2+1 IgG Crossfab" молекулы. (G, K) Изображение "1+1 IgG Crossfab" молекулы с альтернативным порядком расположения Crossfab и Fab компонентов ("перевернуты"). (Н, L) Изображение "1+1 IgG Crossfab" молекулы. (I, М) Изображение "2+1 IgG Crossfab" молекулы с двумя CrossFabs компонентами (J, N) Изображение "2+1 IgG Crossfab" молекулы с двумя CrossFabs компонентами и альтернативным порядком расположения Crossfab и Fab компонентов ("перевернуты"). (О, S) Изображение "Fab-Crossfab" молекулы. (Р, Т) Изображение "Crossfab-Fab" молекулы. (Q, U) Изображение "(Fab)2-Crossfab" молекулы. (R, V) Изображение "Crossfab-(Fab)2" молекулы. (W, Y) Изображение "Fab-(Crossfab)2" молекулы. (X, Z) Изображение "(Crossfab)2-Fab" молекулы. Черная точка: возможная модификация в Fc домене, способствующая гетеродимеризации. ++, --: противоположно заряженные аминокислоты опционально встроенные в СН1 и CL домены. Crossfab молекулы описываются как молекулы, имеющие замены VH и VL областей, но - в тех вариантах изобретения, когда в доменах СН1 и CL нет модификаций заряда, могут альтернативно включать замены СН1 и CL доменов.
Фигура 2. Изображение Т-клеточных биспецифических (ТСВ) молекул антител, полученных согласно Примерам. Все изучаемые ТСВ молекулы антител были получены в форме "2+1 IgG CrossFab, перевернутых" молекул с модификациями заряда (VH/VL замена CD3 связывающего компонента, модификации заряда WT1 связывающих компонентов, ЕЕ=147Е, 213Е; RK=123R, 124K).
Фигура 3. Связывание HLA-A2/WT1 IgG антител с активированными пептидами Т2 клетками. (A) 11D06 IgG, (В) 33H09-IgG, (С) 11B09-IgG, (D) 13B04-IgG, (Е) 5E11-IgG, (F) 5C01-IgG, (G) 11G06-IgG.
Фигура 4. Активация T клеток Т HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs) посредством связывания с активированными пептидами Т2 клетками (NFAT репортерный генный анализ). (A) 11D06-TCB, (В) 33Н09-ТСВ, (С) 11 В09-ТСВ, (D) 13 В04-ТСВ, (Е) ESK1-TCB, (F) 5Е11-ТСВ. (G) 5С01-ТСВ, (3) DP47GS-TCB.
Фигура 5. Уничтожение активированных пептидами Т2 клеток, опосредованное HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs). (А) 11D06-TCB, (В) 33Н09-ТСВ, (С) 13 В04-ТСВ, (D) 11 В09-ТСВ, (Е) 33F05-TCB, (F) 5С01-ТСВ.
Фигура 6. Уничтожение HLA-A2 + WT1 + клеточных опухолевых линий посредством HLA-A2/WT1 х CD3 биспецифических антител (TCBs). (А) Обзор клеточных линий. (В-Е) Уничтожение клеточных линий (В) 11D06-TCB, (С) 33Н09-ТСВ, (D) 11 В09-ТСВ, (Е) 13 В04-ТСВ. (F) Уничтожение SKM-1 клеток различными TCBs. (G) Уничтожение BJAB клеток различными TCBs.
Фигура 7. Активация Т клеток HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs) посредством связывания с HLA-A2 + WT1 + опухолевых клеточных линий. (A) SKM-1 клетки, (В) BJAB клетки.
Фигура 8. Отсутствие связывания с нецелевыми пептидами отобранных HLA-A2/WT1 х CD3 биспецифических антител (TCBs). (А-В) Связывание с активированными пептидами Т2 клетками антител (A) 11D06-TCB, (В) 33Н09-ТСВ, (С) ESK1-TCB. (D-E) Активация Т клеток посредством связывания с активированными пептидами Т клетками антителами (D) 11D06-TCB и (Е) 33Н09-ТСВ. (F) Обзор пептидов.
Фигура 9. Отсутствие связывания с дополнительными нецелевыми пептидами отобранных HLA-A2/WT1 х CD3 биспецифических антител (TCBs). (А-В) Активация Т клеток посредством связывания с активированными пептидами Т2 клетками антителами (A) 11D06-TCB и (В) 33Н09-ТСВ. Наравне с RMF пептидом были протестированы 6 установленных нецелевых пептида. (С-G) Уничтожение активированных пептидами Т2 клеток посредством антител (С, Е) 11D06-TCB, (D, F) 33Н09-ТСВ и (G) ESK1-TCB. Наравне с RMF пептидом были протестированы 6 установленных нецелевых пептида. Наравне с RMF и VLD пептидами были протестированы 19 установленных нецелевых пептида.
Фигура 10. Отсутствие уничтожения нормальных CD34 + стволовых клеток, выделенных из костного мозга, при взаимодействии с отобранными HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs). (A) 11D06-TCB, (В) 33Н09-ТСВ.
Фигура 11. Идентификация связывания аминокислотных остатков RMF пептида с отобранными HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs) посредством аланинового сканирования. (A) RMF нативный пептид, (В) RMF R1Y пептид, (С) RMF R1A пептид, (D) RMF М2А пептид, (Е) RMF F3A пептид, (F) RMF Р4А пептид, (G) RMF N5A пептид, (Н) RMF A6G пептид, (I) RMF Р7А пептид, (J) RMF Y8A пептид, (K) RMF L9A пептид. (L) Обзор пептидов. (М) Кратное изменение ЕС50 относительно ЕС50 для RMF нативного пептида. (N) Критические контактные остатки.
Фигура 12. Фармакокинетический профиль HLA-A2/WT1 х CD3 биспецифических антител (11D06-TCB и 33Н09-ТСВ) после однократного введения NSG мышам.
Фигура 13. Изучение эффективности HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") на SKM-1 ксенотрансплантате у гуманизированных мышей (А) Дизайн исследования (В) Экспериментальные группы (С) Кинетика роста опухоли (среднее) во всех экспериментальных группах. (D) Кинетика единичного опухолевого роста в контрольной группе. (Е) Кинетика единичного опухолевого роста в 11D06-TCB группе. (F) Кинетика единичного опухолевого роста в 33Н09-ТСВ группе. (G) Статистика. Расчеты основаны на данных, полученных на 38 день эксперимента (контрольная группа получала носитель). Ингибирование опухолевого роста (TGI): TGI>100 → регрессия опухоли, TGI=100 → стаз опухоли. Соотношение для групп, получавших лечение/контроль (TCR): TCR=1 → отсутствие эффекта, TCR=0 → полный регресс.
Фигура 14. Обзор кристаллической структуры комплексов HLA-A2/WT1 антитело -рМНС. Антитела (фрагменты) показаны на верху рисунка, при этом тяжелые цепи окрашены темно серым, а легкие цепи - светло-серым цветом. Кристаллизованные атомы растворителя не показаны (А) Кристаллическая структура 5С01 Fab в комплексе с HLA-A02/VLD рМНС с разрешением 1.98 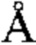 . Fab-pMHC контактная зона: ≈ 476
. Fab-pMHC контактная зона: ≈ 476 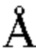 2, доля пептида: ≈ 68 2. (В) Кристаллическая структура 11D06 Fab в комплексе с HLA-A02/RMF рМНС с разрешением 2.60
2, доля пептида: ≈ 68 2. (В) Кристаллическая структура 11D06 Fab в комплексе с HLA-A02/RMF рМНС с разрешением 2.60 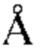 . Fab-pMHC контактная зона: ≈ 397
. Fab-pMHC контактная зона: ≈ 397 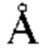 2, доля пептида: ≈ 107
2, доля пептида: ≈ 107  2. (С) Кристаллическая структура ESK1 Fab в комплексе с HLA-A02/RMF рМНС с разрешением 3.05 (опубликовано, PDB ID 4WUU). Fab-pMHC контактная зона: ≈ 505
2. (С) Кристаллическая структура ESK1 Fab в комплексе с HLA-A02/RMF рМНС с разрешением 3.05 (опубликовано, PDB ID 4WUU). Fab-pMHC контактная зона: ≈ 505  2, доля пептида: ≈ 60
2, доля пептида: ≈ 60  2
2
Фигура 15. Область связывания 5С01 Fab - HLA-A2/WT1VLD рМНС крупным планом. Основные химические взаимодействия между Fab и рМНС, выявленные при помощи программы BIOVIA Discovery Studio 4.5 ярко выделены. Атомы растворителя не показаны.
Фигура 16. Матрица взаимодействия и контакта 5С01 Fab остатков (ряды) с HLA-A2/WT1VLD рМНС остатками (колонки). N=прилегание/соседство, Н=Н-связь, Pi=Pi взаимодействия, SB=солевой мостик. Остатки, которые вступают в контакт, определены как остатки, которые подвергаются изменениям на поверхности, доступной для растворителя в присутствии/отсутствии партнера для взаимодействия.
Фигура 17. Область связывания 11D06 Fab-HLA-A2/WT1RMF рМНС крупным планом. Основные химические взаимодействия между Fab и рМНС, выявленные при помощи программы BIOVIA Discovery Studio 4.5 ярко выделены. Атомы растворителя не показаны.
Фигура 18. Матрица взаимодействия и контакта 11D06 Fab остатков (ряды) с HLA-A2/WT1RMF рМНС остатками (колонки). N=прилегание/соседство, Н=Н-связь, Pi=Pi взаимодействия, SB=солевой мостик. Остатки, которые вступают в контакт, определены как остатки, которые подвергаются изменениям на поверхности, доступной для растворителя в присутствии/отсутствии партнера для взаимодействия.
Фигура 19. Область связывания ESK1 Fab-HLA-A2/WT1RMF рМНС крупным планом (PDB ID 4WUU). Основные химические взаимодействия между Fab и рМНС, выявленные при помощи программы BIOVIA Discovery Studio 4.5 ярко выделены. Атомы растворителя не показаны.
Фигура 20. Матрица взаимодействия и контакта ESK1 Fab остатков (ряды) с HLA-A2/WT1RMF рМНС остатками (колонки). N=прилегание/соседство, Н=Н-связь, Pi=Pi взаимодействия, SB=солевой мостик. Остатки, которые вступают в контакт, определены как остатки, которые подвергаются изменениям на поверхности, доступной для растворителя в присутствии/отсутствии партнера для взаимодействия.
Фигура 21. Уничтожение HLA-A2+/WT1+SKM-1 клеток, опосредованное HLA-A2/WT1 х CD3 биспецифическими антителами с различными областями связывания CD3.
Фигура 22. Уничтожение активированных RMF пептидом Т2 клеток, опосредованное HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs).
Фигура 23. Оценка связывания нецелевых пептидов HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs) Aali-TCB (A), Daniel-TCB (В), ESK1-TCB (С) и 11D06-TCB (D).
Фигура 24. Фармакокинетический профиль HLA-A2/WT1 х CD3 биспецифических антител 11D06-TCB (V9) после однократного введения NSG мышам.
Фигура 25. Изучение эффективности HLA-A2/WT1 х CD3 биспецифического антитела 11D06-TCB (V9) на SKM-1 ксенотрансплантате у гуманизированных мышей (А) Кинетика роста опухоли (среднее) во всех экспериментальных группах. (В) Кинетика единичного опухолевого роста в контрольной группе. (С) Кинетика единичного опухолевого роста в 11D06-TCB (V9) группе. (D) Статистика. Расчеты основаны на данных, полученных на 48 день эксперимента (контрольная группа получала носитель). Ингибирование опухолевого роста (TGI): TGI>100 → регрессия опухоли, TGI=100 → стаз опухоли. Соотношение для групп, получавших лечение/контроль (TCR): TCR=1 → отсутствие эффекта, TCR=0 → полный регресс.
Фигура 26. Активация Т клеток HLA-A2/WT1 х CD3 биспецифическими антителами (TCBs) посредством связывания с СНО-K1 клетками, экспрессирующими HLA-A02/WT1RMF рМНС комплекс (NFAT репортерный генный анализ). Сплошная линия: 11D06-TCB (V9) ("2+1" формат). Пунктирная линия: аналогичные молекулы (11D06 и V9 связывающие молекулы) в "1+1 CrossMab" формате.
Детальное описание изобретения
Определения
Все термины, которые используются в настоящем документе, в целом известны из области техники и являются общепринятыми, если дополнительно не оговаривается иное.
Используемый в настоящем документе термин "антигенсвязывающая молекула" относится в широком смысле к молекуле, которая специфически связывается с антигенной детерминантой. Примерами антигенсвязывающих молекул являются иммуноглобулины и их производные, например, фрагменты.
Термин "биспецифический" означает, что антигенсвязывающая молекула способна специфически связываться по меньшей мере с двумя различными антигенными детерминантами. Обычно антигенсвязывающая молекула включает два антигенсвязывающих сайта, каждый из которых специфичен для определенной антигенной детерминанты. Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула способна одновременно связываться с двумя антигенными детерминантами, в частности, двумя антигенными детерминантами, которые экспрессируются двумя различными типами клеток.
Термин "валентный", используемый в настоящем документе, определяет наличие определенного количества антигенсвязывающих сайтов в антигенсвязывающей молекуле. Таким образом, термин "моновалентное связывание с антигеном" означает наличие одного (и не более одного) антигенспецифического сайта в составе антигенсвязывающей молекулы, специфичного для антигена.
Термин "антигенсвязывающий сайт" относится к сайту, то есть одному или более аминокислотному остатку в составе антигенсвязывающей молекулы, который обеспечивает взаимодействие с антигеном. Например, антигенсвязывающий сайт антитела включает аминокислотные остатки областей определяющих комплементарность (CDRs). Нативная молекула иммуноглобулина обычно имеет два антигенсвязывающих сайта, Fab фрагмент обычно имеет один антигенсвязывающий сайт.
Используемый в настоящем документе термин "антигенсвязывающий фрагмент" относится к полипептидному фрагменту, который специфически связывается с антигенной детерминантой. Согласно одному варианту осуществления изобретения, антигенсвязывающий фрагмент способен направлять молекулу, к которой она прикреплена (например, вторую антигенсвязывающую молекулу) к сайту-мишени, например, к специфической опухолевой клетке, несущей антигенную детерминанту. Согласно другому варианту осуществления изобретения, антигенсвязывающий фрагмент обладает способностью активировать передачу сигнала посредством таргетного антигена, например, Т-рецепторного комплексного антигена. К антигенсвязывающим фрагментам относятся антитела и их фрагменты, что раскрыто ниже в настоящем документе. Определенные антигенсвязывающие фрагменты включают антигенсвязывающий домен антитела, содержащий вариабельную область тяжелой цепи и вариабельную область легкой цепи. Согласно определенным вариантам осуществления изобретения, антигенсвязывающие фрагменты могут включать константные области антител, что дополнительно определено в настоящем документе и известно из области техники. Подходящие константные участки тяжелых цепей имеют один из пяти изотипов: α, δ, ε, γ, или μ. Подходящие константные участки легкой цепи имеют один из двух изотипов: κ и λ.
Используемый в описании термин "антигенная детерминанта" или "антиген" относится к участку на полипептидной молекуле, с которым связывается антигенсвязывающий фрагмент, образуя комплекс антигенсвязывающий фрагмент-антиген. Полезные антигенные детерминанты могут обнаруживаться, например, на поверхности опухолевых клеток, на поверхностях инфицированных вирусами клеток, на поверхности других больных клеток, на поверхности иммунных клеток, в свободной форме в сыворотке крови и/или во внеклеточном матриксе (ЕСМ аббрев. от англ. Extracellular Matrix).
Термин "эпитоп" определяет сайт антигена, как белковой, так и небелковой природы, с которым связывается антигенсвязывающий фрагмент. Эпитопы могут быть образованы как непрерывным аминокислотным участком (линейный эпитоп), так и прерывистым аминокислотным участком (конформационный эпитоп), то есть формирующим пространственное соответствие в зависимости от фолдинга антигена, то есть третичного фолдинга антигена белковой природы. Линейные эпитопы обычно все еще остаются связанными с антигенсвязывающей молекулой после воздействия денатурирующих агентов на белковый антиген, при этом конформационные эпитопы обычно разрушаются при воздействии денатурирующих агентов. Эпитоп включает, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, или 8-10 аминокислот в уникальной пространственной конформации.
"CD3" относится к любому нативному CD3 любого позвоночного животного, содержащую млекопитающих, таких, как приматы (например, люди), нечеловекообразные приматы (например, яванские макаки) и грызуны (например, мыши и крысы), если дополнительно не оговаривается иное. Термин объединяет как "полноразмерные" непроцессируемые CD3, так и любые другие формы CD3, которые образуются в результате процессинга в клетках. Термин также объединяет варианты CD3, существующие в природе, например, сплайсинговые или аллельные варианты. Согласно одному варианту осуществления изобретения CD3 представляет собой CD3 человека, в частности, субъединицу ипсилон CD3 человека (CD3ε). Аминокислотная последовательность CD3ε человека представлена в базе UniProt (www.uniprot.org) к кодом доступа Р07766 (версия доступа 189), или NCBI (www.ncbi.nlm.nih.gov/) RefSeq NP_000724.1. См. также SEQ ID NO: 107. Аминокислотная последовательность CD3 яванской макаки [Масаса fascicularis] представлена в банке NCBI GenBank под номером ВАВ71849.1. См. также SEQ ID NO: 108.
"WT1", также известный как "опухоль Вильмса 1" или "белок опухоли Вильмса", относится к любому нативному WT1 любого позвоночного животного, содержащую млекопитающих, таких, как приматы (например, люди), нечеловекообразные приматы (например, яванские макаки) и грызуны (например, мыши и крысы), если дополнительно не оговаривается иное. Термин объединяет как "полноразмерный", непроцессируемый WT1, как и любые другие формы WT1, которые образуются в результате процессинга в клетке. Термин также включает существующие в природе варианты WT1, например, сплайсинговые или аллельные варианты. Согласно одному варианту осуществления изобретения WT1 представляет собой WT1 человека, в частности белок с последовательностью SEQ ID NO: 106. Человеческий WT1 описан в базе UniProt (www.uniprot.org) под кодом доступа Р19544 (версия доступа 215), аминокислотная последовательность человеческого WT1 также показана в SEQ ID NO: 106.
Под термином "VLD", "VLD пептид" или "WTIVLD" подразумевается выделенный из WT1 пептид, имеющий аминокислотную последовательность VLDFAPPGA (SEQ ID NO: 77; положения 37-45 WT1 пептида с последовательностью SEQ ID NO: 106).
Под термином "RMF", "RMF пептид" или "WTIRMF" подразумевается выделенный из WT1 пептид, имеющий аминокислотную последовательность RMFRNAPYL (SEQ ID NO: 78; положения 126-134 WT1 пептида с последовательностью SEQ ID NO: 106).
"HLA-A2", "HLA-A*02", "HLA-A02", или "HLA-A*2" (используются взаимозаменяемо) обозначают серотипы человеческих лейкоцитарных антигенов в группе HLA-A. Белок HLA-A2 (кодируемый соответствующим HLA геном) состоит из а цепи белка соответствующего класса I МНС (основного комплекса гистосовместимости), который дополнительно включает Р2 микроглобулиновую субъединицу. Специфическим HLA=A2 белком является HLA-A201 (также обозначаемый как HLA-A0201, HLA-A02.01, или HLA-A*02:01). Согласно специфическим вариантам осуществления изобретения, HLA-A2 белок, описанный в настоящем документе, представляет собой HLA-A201.
"HLA-A2/WT1" обозначает комплекс молекулы HLA-A2 и пептида выделенного из WT1 (который в настоящем документе также обозначается как "WT1 пептид"), в частности, RMF или VLD пептида ("HLA-A2/WT1RMF" и "HLA-A2/WT1VLD", соответственно). Антитело или биспецифическая антигенсвязывающая молекула по настоящему изобретению специфически связывается как с HLA-A2/WT1RMF, так и с HLA-A2/WT1VLD комплексом.
"Специфическое связывание" подразумевает, что связывание является селективным для антигена и может быть отличимым от других нежелательных или неспецифических взаимодействий. Способность антигенсвязывающей молекулы связываться с со специфической антигенной детерминантой может измеряться как посредством иммуноэлектроблоттинга с ферментным усилением (ELISA), так и при помощи других методик, известных специалистам в данной области, например, поверхностного плазмонного резонанса (SPR) (проанализированного, например, при помощи инструмента BIAcore) (Liljeblad et al., Glyco J 17, 323-329 (2000)), и традиционных методик анализа связывания (Heeley, Endocr Res 28, 217-229 (2002)). Подходящие методы определения специфичности антитела и биспецифической антигенсвязывающей молекулы согласно настоящему изобретению описаны в настоящем документе, например, в Примерах 4,9 и 10 ниже. Согласно одному варианту осуществления изобретения, продолжительность связывания антигенсвязывающей молекулы с посторонним белком составляет менее 10% при сравнении с продолжительностью связывания антигенсвязывающей молекулы с антигеном, что можно измерить, например, с помощью SPR. Согласно определенным вариантам осуществления изобретения, антигенсвязывающий фрагмент, который связывается с антигеном, или антигенсвязывающий фрагмент, включающий такой антигенсвязывающий элемент, имеет константу диссоциации (KD) ≤1 мкм ≤100 нм, ≤10 нм, ≤1 нм, ≤0.1 нм, ≤0.01 нм, или ≤0.001 нм (например, 10-8 м или менее, например, от 10-8 м до 10-13 м, например, от 10-9 м до 10-13 м).
"Аффинность" означает силу суммарных нековалентных взаимодействий между единичным сайтом связывания молекулы (например, рецептора) и его партнера по связыванию (например, лиганда). Если не оговаривается иное, используемый в настоящем документе термин "аффинность связывания" относится к истинной аффинности связывания, которая отражает взаимодействия 1:1 между членами связывающейся пары (например, антигенсвязывающей молекулы и антигена, или рецептора и его лиганда). Аффинность молекулы X для ее партнера Y в целом может быть выражена посредством константы диссоциации (KD), которая представляет собой соотношение констант скорости диссоциации и ассоциации (koff и kon, соответственно). Таким образом, эквивалентные аффинности могут иметь различные константы скорости, при этом соотношение этих констант остается неизменным. Аффинность может измеряться при помощи способов, хорошо известных из области техники, содержащую те, что описаны в настоящем документе. Наиболее подходящим способом измерения аффинности является поверхностный плазмонный резонанс.
"Пониженное связывание", например, пониженное связывание с Fc рецептором, относится к пониженной аффинности соответствующего взаимодействия, что измеряется, например, при помощи SPR. Для большей ясности термин также включает уменьшение аффинности до нуля (или ниже определяемого уровня в случае применения метода детекции), то есть полное прекращение всех взаимодействий. Наоборот, "повышенное связывание" означает увеличение аффинности связывания для соответствующего взаимодействия.
Термин "активирующий Т-клеточный антиген", используемый в настоящем описании относится к антигенной детерминанте, экспрессируемой на поверхности Т-лимфоцита, в частности, цитотоксического Т лимфоцита, которая способна индуцировать активацию Т-клеток посредством взаимодействия с антигенсвязывающей молекулой. В особенности, взаимодействие антигенсвязывающей молекулы с активирующим Т-клеточным антигеном может индуцировать активацию Т-клеток за счет запуска сигнального каскада Т-клеточного рецепторного комплекса. Согласно определенному варианту осуществления изобретения, активирующий Т-клеточный антиген представляет собой CD3, в частности, субъединицу ипсилон CD3 (см UniProt номер Р07766 (версия 189), NCBI RefSeq номер NP_000724.1, SEQ ID NO: 107 для человеческой последовательности; или UniProt номер Q95LI5 (версия 49), NCBI GenBank номер ВАВ71849.1, SEQ ID NO: 108 для последовательности яванской макаки [Масаса fascicularis]).
Термин "активация Т клеток", используемый в настоящем документе, относится один или более клеточный ответ Т-лимфоцита, в частности, цитотоксического Т-лимфоцита, выбранный из: пролиферации, дифференцировки, секреции цитокинов, высвобождение цитотоксических эффекторных молекул, цитотоксическую активность, а также экспрессию маркеров активации. Подходящие способы измерения активации Т клеток хорошо известны из области техники и описаны в настоящем документе.
Термин "таргетный клеточный антиген", применяемый в настоящем документе, используемый в настоящем документе, относится к антигенной детерминанте, презентированной на поверхности клетки-мишени, например, опухолевой клетки, такой, как раковая клетка или другая стромальная опухолевая клетка. Согласно определенному варианту осуществления изобретения, таргетный клеточный антиген представляет собой HLA-A2/WT1, в частности HLA-A2/WT1RMF или HLA-A2/WT1VLD, в особенности HL A-A2/WT1RMF.
Используемые в настоящем описании термины "первая", "вторая" или "третья" по отношению к Fab молекулам и т.д., используются для удобства различения молекул друг от друга в том случае, когда существует более одного типа каждой молекулы. Использование указанных терминов не предназначается для указания определенного порядка или ориентации биспецифической антигенсвязывающей молекулы, если это не указано прямо.
Под термином "слитый" подразумевается, что компоненты (например, Fab молекула и субъединица Fc домена) соединены между собой пептидными связями, как напрямую, так и посредством одного или более пептидного линкера.
Термин "Fab молекула" относится к белку, включающему VH и СН1 домены тяжелой цепи ("Fab тяжелой цепи") и VL и VL домены легкой цепи ("Fab легкой цепи") иммуноглобулина.
Под термином "кроссоверная" Fab молекула (также обозначаемая как "CrossFab") понимается Fab молекула, в которой вариабельные или константные домены тяжелой или легкой Fab цепи поменяны местами (то есть, замещены друг другом), то есть кроссоверная Fab молекула включает пептидную цепь, состоящую из вариабельного домена VL легкой цепи и константного домена 1 СН1 тяжелой цепи (VL-CH1, в направлении от N-конца к С-концу), и пептидную цепь, состоящую из вариабельного домена VH тяжелой цепи и константного домена CL легкой цепи (VH-CL, в направлении от N-конца к С-концу). Для большей ясности, в кроссоверной Fab молекуле, когда вариабельные домены Fab легкой цепи и Fab тяжелой цепи поменяны местами, пептидная цепь, содержащая константный домен 1 СН1 тяжелой цепи обозначается в настоящем описании как "тяжелая цепь" (кроссоверной) Fab молекулы. И наоборот, в кроссоверной Fab молекуле, когда константные домены Fab легких цепей и Fab тяжелых цепей поменяны местами, пептидная цепь, содержащая вариабельный домен VH тяжелой цепи, обозначается в настоящем документе как "тяжелая цепь" (кроссоверной) Fab молекулы.
В противоположность этому под "традиционной" Fab молекулой понимается Fab молекула в своем естественном формате, то есть содержащая тяжелую цепь, скомпонованную из вариабельного и константного доменов тяжелой цепи (VH-CH1, в направлении от N- конца к С-концу) и легкую цепь, скомпонованную из вариабельного и константного доменов легкой цепи (VL-CL, в направлении от N-конца к С-концу).
Термин "молекула иммуноглобулина" относится к белку, имеющему структуру существующего в природе антитела. Например, иммуноглобулины класса IgG представляют собой гетеротетрамерные гликопротеины с молекулярной массой 150,000 дальтон, состоящие из двух легких цепей и двух тяжелых цепей, которые связаны между собой дисульфидными мостиками. В направлении от N-конца к С-концу каждая тяжелая цепь включает вариабельный домен (VH), также называемый вариабельный тяжелый домен или вариабельный участок тяжелой цепи, а затем три константных домена (CH1, СН2 и СН3), также называемые константными участками тяжелой цепи. Аналогично, каждая легкая цепь в направлении от N-конца к С-концу включает вариабельный домен (VL), также называемый вариабельный легкий домен или вариабельный участок легкой цепи, а затем константный домен (CL), также называемый константным участком легкой цепи. Тяжелая цепь иммуноглобулина может быть отнесена к одному из пяти типов, обозначаемых следующим образом: α (IgA), δ (IgD), ε (IgE), γ (IgG), или μ (IgM), некоторые из них могут дополнительно подразделяться на субтипы, например, γ1 (IgG1), γ2 (IgG2), γ3 (IgG3), γ4(IgG4), α1(IgA1) и α2 (LgA2). Легкая цепь иммуноглобулина может быть отнесена к одному из двух типов, обозначаемых каппа (κ) и лямбда (λ), в зависимости от аминокислотной последовательности константного домена. Иммуноглобулин, по существу, состоит из двух Fab молекул и Fc доменов, соединенных через шарнирную область иммуноглобулина.
Термин "антитело", используемый в настоящем документе, используется в широком смысле и объединяет различные антительные структуры, содержащую, без ограничений указанными, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они демонстрируют желаемую антигенсвязывающую активность.
Термин "моноклональное антитело", используемый в настоящем описании, относится к антителу, полученному из популяции практически гомогенных антител, то есть индивидуальные антитела в составе популяции являются идентичными и/или связываются с одним и тем же эпитопом, за исключением возможных вариантных антител, например, содержащих встречающиеся в природе мутации или возникающие во время получения композиции моноклональных антител, такие варианты обычно присутствуют в следовых количествах. В отличие от композиций поликлональных антител, которые обычно содержат разные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело в составе композиции моноклональных антител направлено против одной детерминанты антигена. Таким образом, термин "моноклональный" определяет именно ту характеристику антитела, что оно было получено практически из гомогенной популяции антител, а не было сконструировано при помощи какого-либо определенного метода продукции антител. Например, моноклональные антитела, которые используются в соответствии с настоящим изобретением, могут быть получены при помощи различных методик, содержащую, без ограничений указанными, метод гибридомы, методы рекомбинантной ДНК, фагово-дисплейный метод, а также методы с использованием трансгенных животных, содержащих все или часть человеческих локусов иммуноглобулина, такие способы и другие типичные способы получения моноклональных антител описаны в настоящем документе.
"Изолированное" антитело представляет собой антитело, которое было отделено от компонентов, окружающих его в естественной среде, то есть антитело вне природных условий. Никакого особого уровня очистки не требуется. Например, изолированное антитело может быть удалено из его естественного или природного окружения. Полученные при помощи рекомбинантных технологий антитела, экспрессируемые в клетках-хозяевах, считаются изолированными в целях настоящего изобретения, как и нативные или рекомбинантные антитела, которые были выделены, фракционированы или частично или полностью очищены любым подходящим способом. В таком случае, антитела и биспецифические антигенсвязывающие молекулы по настоящему изобретению являются изолированными. Согласно некоторым вариантам осуществления изобретения, антитело очищают более чем до 95% или 99% чистоты, что определяется, например, электрофоретическими (например, SDS-PAGE, изоэлектрическое фокусирование (IEF), капиллярный электрофорез) или хроматографическими (например, ионообменная HPLC или HPLC в обратной фазе) методами. Обзор способов оценки степени очистки антител представлен, например, у Flatman et al., J. Chromatogr. В 848:79-87 (2007).
Термины "полноразмерное антитело", "интактное антитело" или "целое антитело", используемые в настоящем документе взаимозаменяемо, относятся к антителу, имеющему структуру практически аналогичную структуре нативного антитела.
Термин "фрагмент антитела" относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывается с тем же антигеном, что и интактная молекула. Примерами фрагментов антител являются, без ограничений указанными, Fv, Fab, Fab', Fab'-SH, F(ab')2, диатела, линейные антитела, одноцепочечные молекулы антител (например, scFv) и однодоменные антитела. Обзор определенных фрагментов антител представлен у Hudson et al., Nat Med 9, 129-134 (2003). Для обзора scFv фрагментов, см.. например, Pliickthun, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); а также WO 93/16185; и патенты США номер. 5,571,894 и 5,587,458. Для обзора Fab и F(ab')2 молекул, содержащих эпитопы связывания рецептора реутилизации и имеющих увеличенный период полураспада in vivo, см. патент США номер 5,869,046. Диатела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть биспецифическими или бивалентными. См, например, ЕР 404,097; WO 1993/01161; Hudson et al., Nat Med 9, 129-134 (2003); and Hollinger et al., Proc Natl Acad Sci USA 90, 6444-6448 (1993). Триатела и тетратела также описаны у Hudson et al., Nat Med 9, 129-134 (2003). Однодоменные антитела представляют собой фрагменты антител, включающие все части вариабельного домена тяжелой цепи или все части вариабельного домена легкой цепи антитела. Согласно определенным вариантам осуществления изобретения, однодоменные антитела представляют собой однодоменные антитела человека (Domantis, Inc., Waltham, MA; см, например, патент США No. 6,248,516 В1). Фрагменты антител могут быть получены при помощи различных технологий, содержащую, без ограничений указанными, протеолитическое расщепление интактного антитела, а также получение с помощью рекомбинантных клеток-хозяев (например, E.coli или фага), как описано в настоящем документе.
Термин "антигенсвязывающий домен" относится к части антитела, которое включает область, которая специфически связывается с со всем антигеном или его частью и комплементарна ему. Антигенсвязывающий домен может быть представлен, например, одним или более вариабельным доменом антитела (также называемым вариабельным участком антитела). В частности, антигенсвязывающий домен включат вариабельный домен легкой цепи (VL) и вариабельный домен тяжелой цепи антитела (VH).
Термин "вариабельный участок" или "вариабельный домен" относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой и легкой цепи (VH и VL, соответственно) нативного антитела, в целом имеют идентичные структуры, при этом каждый домен включает четыре консервативных каркасных участка (FRs) и три гипервариабельных участка (HVRs). См., например, Kindt et al., Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007). Одного VH или VL домена может быть достаточно для обеспечения специфичности связывания с антигеном. В настоящем документе по отношению к последовательности вариабельных участков используется "нумерация по Кабату", система, предложенная Кабатом и соавт., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991). Также в настоящем документе все положения аминокислот во всех константных участках и доменах тяжелой или легкой цепи нумеруются согласно системе Кабата, описанной им же и соавторами, Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), в описании всегда дается отсылка на "нумерацию по Кабату" или "нумерацию согласно системе Кабата". В частности, система нумерации Кабата (см. страницы 647-660 работы Кабата и соавт.Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)) используется для константных доменов легкой цепи каппа или лямбда изотипа, а система нумерации с применением EU-индекса Кабата (см. стр. 661-723) используется для константных доменов тяжелой цепи (СН1, шарнирная область, СН2 и СН3), что в описании будет понятно из определения "нумерация согласно EU-индексу Кабата".
Термин "гипервариабельный участок" или "HVR", используемый в настоящем описании, относится к каждому из участков вариабельного домена антитела, которые имеют гипервариабельную последовательность ("области, определяющие комплементарность" или " CDRs ") и/или формируют структурно определяемые петли ("гипервариабельные петли") и/или содержат антиген-контактирующие остатки ("антигенные контакты"). В целом, антитела содержат шесть HVRs; три в VH (H1, Н2, Н3), и три в VL (L1, L2, L3). Примеры HVRs здесь включают:
(a) гипервариабельные петли, существующие на аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2), и 96-101 (Н3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987));
(b) CDRs, существующие на аминокислотных остатках 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2), и 95-102 (Н3) (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991));
(c) антигенны контакты, существующие на аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2), и 93-101 (Н3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)); и
(d) комбинации (a), (b), и/или (с), содержащую HVR аминокислотные остатки 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (H2), 93-102 (Н3), и 94-102 (Н3).
Если дополнительно не оговаривается иное, HVR остатки и другие остатки в вариабельном домене (например, FR остатки) в настоящем описании пронумерованы согласно системе Кабата, см. выше. "Каркас" или "FR" относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно располагаются в следующем порядке в VH (или VL): FR1-H1 (L1) -FR2-H2 (L2) -FR3-H3 (L3) -FR4.
Термин "гуманизированное" антитело, относится к химерному антителу, включающему аминокислотные остатки из HVRs нечеловеческой природы и аминокислотные остатки из FRs человека. Согласно определенным вариантам осуществления изобретения, гуманизированное антитело будет включать, фактически по меньшей мере один, или, как правило, два вариабельных домена, в которых все или, фактически, все HVR (например, CDR) соответствуют доменам антитела нечеловеческой природы, и все или практически все FR соответствуют таковым человеческого антитела. Такие вариабельные домены обозначаются в настоящем документе как "гуманизированные вариабельные участки". Гуманизированное антитело, необязательно, может содержать по меньшей мере участок константной области антитела, выделенный из антитела человека. Согласно некоторым вариантам осуществления изобретения, некоторые FR остатки в гуманизированном антителе замещены соответствующими остатками антитела нечеловеческой природы (например, антитела, из которого выделены HVR остатки), например, с целью восстановления или улучшения специфичности или аффинности антитела. Термин "гуманизированная форма" антитела, например нечеловеческого антитела, относится к антителу, которое подверглось гуманизации. Другие формы "гуманизированных антител", которые объединяет настоящее изобретение, представляют собой формы, в которых константные области подверглись дополнительной модификации или изменениям по сравнению с оригинальным антителом с целью получения желаемых согласно изобретению свойств, в особенности, что касается связывания Clq и/или Fc рецептора (FcR).
"Человеческое антитело" представляет собой антитело, которое имеет аминокислотную последовательность, которая соответствует последовательности антитела, синтезируемого в организме человека или клетками человека, или получена из нечеловеческого источника, который использует репертуар антител человека или другие последовательности, кодирующие антитела человека. Определение "человеческое антитело" не включает гуманизированные антитела, включающие антигенсвязывающие остатки нечеловеческой природы. Согласно определенным вариантам осуществления изобретения, человеческое антитело человека выделено из организма трансгенного животного (не человека), например, мыши, крысы или кролика. Согласно определенным вариантам осуществления изобретения, человеческое антитело получено из линии гибридомных клеток. Антитела или фрагменты антител, выделенные из библиотек человеческих антител, также считаются в настоящем документе человеческими антителами или их фрагментами.
Термин "класс" антитела или иммуноглобулина обозначает тип константного домена или константной области тяжелой цепи антитела. Существует пять основных классов антител: IgA, IgD, IgE, IgG, и IgM, a некоторые из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, LgG2, IgG3, LgG4, IgA1, и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов обозначаются α, δ, ε, γ, и μ, соответственно.
Термин "Fc домен" или "Fc область" в настоящем документе используется для обозначения С-концевого участка тяжелой цепи иммуноглобулина, который содержит по меньшей мере одну часть константного участка. Термин объединяет нативные последовательности Fc областей и варианты Fc областей. Несмотря на то, что границы Fc области тяжелой цепи IgG могут незначительно варьировать, считается, что Fc область тяжелой цепи IgG человека обычно располагается от остатка Cys226 или Pro 230, до карбоксильного конца тяжелой цепи. Однако, антитела, продуцируемые клетками-хозяевами, могут подвергаться посттрансляционному отщеплению одного или более, обычно, одного или двух, аминокислотных остатков от С-конца тяжелой цепи. Таким образом, антитело, продуцируемое клеткой-хозяином путем экспрессии специфической молекулы нуклеиновой кислоты, кодирующей полноразмерную тяжелую цепь, может включать полноразмерную тяжелую цепь, или может включать расщепленный вариант полноразмерной тяжелой цепи (также обозначаемый в настоящем документе как "расщепленный вариант тяжелой цепи"). Может быть такой вариант, когда два С-концевых аминокислотных остатка представляют собой глицин (G446) и лизин (K447, нумерация согласно EU индексу по Кабату). Таким образом, С-концевой лизин (Lys447), или С-концевой глицин (Gly446) и лизин (K447), Fc области могут как отсутствовать, так и присутствовать. Аминокислотная последовательность тяжелых цепей, содержащую Fc домены (или субъединицу Fc домена, как определено в настоящем документе) в настоящем документе обозначены как лишенные С-концевого глицин-лизинового дипептида, если дополнительно не оговаривается иное. Согласно одному варианту осуществления изобретения, тяжелая цепь, содержащую субъединицу Fc домена, как указано в описании, в составе антитела или биспецифической антигенсвязывающей молекулы согласно изобретению, содержит дополнительный С-концевой глицин-лизиновый дипептид (G446 и K447, нумерация согласно EU индексу по Кабату). Согласно одному варианту осуществления изобретения, тяжелая цепь, включающая субъединицу Fc-домена, как указано в настоящем документе, в составе антитела или биспецифической антигенсвязывающей молекулы согласно изобретению, содержит дополнительный С-концевой остаток глицина (G446, нумерация согласно EU индексу Кабата). Композиции, заявленные в соответствии с настоящим изобретением, такие, как описанные в настоящем документе фармацевтические композиции, содержат популяции антител или биспецифических антигенсвязывающих молекул согласно изобретению. Популяции антител или биспецифических антигенсвязывающих молекул могут включать молекулы, имеющие полноразмерные тяжелые цепи, и молекулы, имеющие расщепленные варианты тяжелых цепей. Популяция антител или биспецифических антигенсвязывающих молекул может состоять из смеси молекул, имеющих полноразмерную тяжелую цепь, и молекул, имеющих расщепленный вариант тяжелой цепи, где по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90% антител или биспецифических антигенсвязывающих молекул имеют расщепленный вариант тяжелой цепи. Согласно одному варианту осуществления изобретения, композиция, содержащая популяцию антител или биспецифических антигенсвязывающих молекул согласно изобретению, Согласно одному варианту осуществления изобретения, композиция, включающая популяцию антител или биспецифические антигенсвязывающие молекулы согласно изобретению, содержит антитело или биспецифическую антигенсвязывающую молекулу, включающую тяжелую цепь с субъединицей Fc домена, как предусмотрено в изобретении, с дополнительным С-концевым глицин-лизин дипептидом (G446 и K447, нумерация согласно EU индексу Кабата). Согласно одному варианту осуществления изобретения, композиция, включающая популяцию антител или биспецифические антигенсвязывающие молекулы согласно изобретению, содержит антитело или биспецифическую антигенсвязывающую молекулу, включающую тяжелую цепь с субъединицей Fc домена, как предусмотрено в изобретении, с дополнительным С-концевым остатком глицина (G446, нумерация согласно EU индексу Кабата). Согласно одному варианту осуществления изобретения, такая композиция, включающая популяцию антител или биспецифические антигенсвязывающие молекулы, состоящие из молекул, включающих тяжелую цепь с субъединицей Fc домена, как предусмотрено изобретением, с дополнительным С-концевым остатком глицина (G446, нумерация согласно EU индексу Кабата); а также молекул, включающих тяжелую цепь с субъединицей Fc домена, как предусмотрено изобретением, с дополнительным С-концевым глицин-лизин дипепидом (G446 и K447, нумерация согласно EU индексу Кабата). Если дополнительно не оговаривается иное, нумерация аминокислотных остатков в Fc области или константой области осуществляется согласно системе EU, которая также называется EU индексом, как описано у Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991 (см. выше). Термин "субъединица" Fc домена, используемый в настоящем описании, относится к одному или двум полипептидам, образующим димерный Fc домен, то есть полипептид, содержащий С-концевые константные области тяжелой цепи иммуноглобулина, способные к стабильной самоассоциации. Например, субъединица Fc домена IgG включает константные домены IgG СН2 и IgG СН3.
"Модификация, обеспечивающая ассоциацию первой и второй субъединицы Fc домена" - это манипуляция пептидного каркаса или посттрансляционные модификации субъединицы Fc домена, которые снижают или предотвращают ассоциацию полипептида, содержащего субъединицу Fc домена, с идентичным полипептидом с образованием гомодимера. Модификация, обеспечивающая ассоциацию, которая применяется в контексте настоящего изобретения, в частности, включает отдельные модификации, выполненные для каждой из двух субъединиц Fc домена, которые требуется связать (т.е. первую и вторую субъединицы Fc домена), причем модификации являются взаимодополняющими друг для друга, чтобы способствовать ассоциации двух субъединиц Fc домена. Например, модификация, обеспечивающая ассоциацию, может изменить структуру или заряд одной или обеих субъединиц Fc домена, чтобы сделать их ассоциацию стерически или электростатически более выгодной, соответственно. Таким образом, (гетеро) димеризация происходит между полипептидом, содержащим первую субъединицу Fc домена, и полипептидом, содержащим вторую субъединицу Fc домена, которые могут быть неидентичны в том смысле, что дополнительные компоненты, соединяющиеся с каждой из субъединиц (например, антигенсвязывающие молекулы) не являются одинаковыми. Согласно некоторым вариантам осуществления изобретения, модификация, обеспечивающая ассоциацию, включает аминокислотную мутацию Fc домена, а конкретно, аминокислотную замену. Согласно определенному варианту осуществления изобретения, модификация, обеспечивающая ассоциацию, включает несколько аминокислотных мутаций, а конкретно, аминокислотную замену в каждой из двух субъединиц Fc домена.
Термин "эффекторные функции" относится к тем биологическим свойствам, которые присущи Fc области антитела и отличаются в зависимости от изотипа антитела. Примерами эффекторных функций являются: связывание Clq и комплемент-зависимая цитотоксичность (CDC), связывание Fc рецептора, антителозависимая клеточноопосредованная цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP), секреция цитокинов, захват антигена антиген-презентирующими клетками, опосредованный иммунными комплексами, даун-регуляция рецепторов клеточной поверхности (например, В-клеточных рецепторов) и активация В клеток.
Используемые в настоящем документе термины "инженерия, генерировать, сгенерированный" объединяют любые манипуляции с белковым каркасом или посттрансляционные модификации существующих в природе или рекомбинантных полипептидов или их фрагментов. Инженерия включает модификации аминокислотной последовательности, паттерна гликозилирования или групп боковых цепей отдельных аминокислот, а также комбинации этих подходов.
Термин "аминокислотная мутация", используемая в настоящем документе, объединяет аминокислотные замены, делеции, вставки и модификации. Любые комбинации замен, делеций, вставок и модификаций могут осуществляться для получения финального конструкта, обеспечивая тем самым конструкту желаемые характеристики, например, ослабленное связывание с Fc рецептором, или усиленную ассоциацию с другим пептидом. Делеции и вставки аминокислотных последовательностей включают делеции и вставки амино-и/или карбокси-концевых участков аминокислот. Конкретными аминокислотными мутациями являются аминокислотные замены. С целью изменения характеристик связывания Fc области, наиболее предпочтительными являются неконсервативные аминокислотные замены, то есть замещение одной аминокислоты другой аминокислотой, имеющей другие структурные и/или химические свойства. Аминокислотные замены включают замещение аминокислотами, не существующими в природе, или производными существующих в природе двадцати стандартных аминокислот (например, 4-гидроксипролином, 3-метилгистидином, орнитином, гомосерином. 5-гидроксилизином). Аминокислотные мутации могут быть сгенерированы с использованием генетических или химических методов, хорошо известных специалистам в данной области. К генетическим методам относятся сайт-направленный мутагенез, ПЦР, синтез генов и другие. Предполагается, что методы изменения группы боковой цепи аминокислоты помимо генной инженерии, такие, как химическая модификация, также могут быть полезны. Для обозначения одной и той же аминокислотной мутации в настоящем документе могут использоваться различные обозначения. Например, замена пролина в положении 329 Fc домена на глицин может обозначаться как 329G, G329, G329, P329G, или Pro329Gly.
"Процент (%) идентичности аминокислотной последовательности" относительно контрольной полипептидной последовательности определяется как процент аминокислотных остатков в рассматриваемой последовательности, которые идентичны аминокислотным остаткам в контрольной полипептидной последовательности, после выравнивания последовательностей и введения гэпов, если необходимо, для достижения максимальной процентной идентичности и без учета любых консервативных замен как части идентичности последовательностей. Выравнивание в целях определения процента идентичности аминокислотной последовательности может быть достигнуто различными способами, хорошо известными специалистам в данной области, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, Clustal W, Megalign (DNASTAR) или пакет программ FASTA. Специалист в данной области может определить подходящие параметры для выравнивания последовательностей, содержащую любые алгоритмы, необходимые для достижения максимального выравнивания сравниваемых полноразмерных аминокислотных последовательностей. В целях настоящего изобретения, однако, величины процента (%) идентичности аминокислотных последовательностей генерируются с использованием программы ggsearch пакета FASTA версии 36.3.8 с или более поздней версии с использованием матрицы сравнения BLOSUM50. Программное обеспечение FASTA было создано W. R. Pearson and D. J. Lipman (1988), "Improved Tools for Biological Sequence Analysis", PNAS 85:2444-2448; W. R. Pearson (1996) "Effective protein sequence comparison" Meth. Enzymol. 266:227- 258; and Pearson et. al. (1997) Genomics 46:24-36, и является общедоступным на сайте http://fasta.bioch.virginia.edu/fasta_www2/fasta_down.shtml. Альтернативно, общедоступный сервер http://fasta.bioch.virginia.edu/fasta_www2/index.cgi, который может использоваться для сравнения последовательностей, используя программу ggsearch (глобально белок: белок) и параметры по умолчанию (BLOSUM50; open: -10; ext: -2; Ktup=2), чтобы обеспечить глобальное, а не локальное выравнивание. Процент аминокислотной идентичности высвечивается в заголовке окна.
Термин "полинуклеотид" относится к изолированной молекуле нуклеиновой кислоты или конструкту, например, информационной РНК (иРНК), РНК, выделенной из вируса, или плазмидной ДНК (пДНК). Полинуклеотид может включать традиционные фосфодиэфирные связи или альтернативные связи (например, амидные связи, такие, которые можно обнаружить в пептид-нуклеиновых кислотах (ПНА). Термин "молекула нуклеиновой кислоты" относится к любому одному или нескольким сегментам нуклеиновой кислоты, например, фрагментам ДНК или РНК, присутствующим в полинуклеотиде.
Под "изолированной" молекулой нуклеиновой кислоты или полинуклеотидом подразумевается молекула нуклеиновой кислоты, ДНК или РНК, которая была выделена из естественной среды. Например, рекомбинантный полинуклеотид, кодирующий полипептид, содержащийся в векторе, считается изолированным в целях настоящего изобретения. Дополнительными примерами изолированного полинуклеотида являются рекомбинантные полинуклеотиды, которые содержатся в гетерологичных клетках-хозяевах, или очищенные (частично или практически) полинуклеотиды в растворе. Изолированный полинуклеотид включает полинуклеотидную молекулу, содержащуюся в клетках, которые обычно содержат полинуклеотидную молекулу, но указанная полинуклеотидная молекула при этом присутствует вне хромосомы или в хромосоме, но локация отличается от той, которая характерна для естественных условий. Изолированные молекулы РНК включают in vivo или in vitro РНК транскрипты согласно изобретению, а также позитивные и негативные стандартные формы, а также двуцепочечные формы. Изолированные полипептиды или нуклеиновые кислоты, согласно изобретению, дополнительно включают такие молекулы, полученные синтетически. Дополнительно, полинуклеотид или нуклеиновая кислота могут представлять собой или включать регуляторный элемент, такой, как промотор, сайт связывания рибосомы или терминатор транскрипции.
"Изолированный полинуклеотид (или нуклеиновая кислота) кодирующий [например, антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению]" означает одну или более полинуклеотидную молекулу, кодирующую тяжелую или легкую цепи антитела (или их фрагменты), содержащую такие полинуклеотидные молекулы в составе единичного вектора или отдельных векторов, и такие молекулы нуклеиновых кислот, которые присутствуют в одной или нескольких локациях в клетке-хозяине.
Термин "кассета экспрессии" относится к полинуклеотиду, сгенерированному рекомбинантно или синтетически, с серией специфических элементов нуклеиновых кислот, которые обеспечивают транскрипцию определенной нуклеиновой кислоты в клетке-мишени. Рекомбинантная кассета экспрессии может быть встроена в плазмиду, хромосому, митохондриальную ДНК, пластидную ДНК, вирусный фрагмент или фрагмент нуклеиновой кислоты. Обычно часть рекомбинантной кассеты экспрессии вектора экспрессии включает, среди других последовательностей, последовательность нуклеиновой кислоты, которая транскрибируется в промотор. Согласно определенным вариантам осуществления изобретения, кассета экспрессии включает полинуклеотидные последовательности, которые кодируют антитела или биспецифические молекулы согласно изобретению или их фрагменты.
Термин "вектор" или "вектор экспрессии" относится к молекуле ДНК, которая применяется для встраивания и направления экспрессии специфического гена, с которым она функционально связана в клетке. Термин включает вектор как самореплицирующуюся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он был встроен. Вектор экспрессии в соответствии с настоящим изобретением содержит кассету экспрессии. Векторы экспрессии обеспечивают транскрипцию больших количеств стабильной иРНК. Как только вектор экспрессии оказывается внутри клетки, молекула или белок рибонуклеиновой кислоты, который кодируется геном, начинает продуцироваться клеточными механизмами транскрипции и/или трансляции. Согласно одному варианту осуществления изобретения, вектор экспрессии, согласно изобретению, содержит кассету экспрессии, которая включает полинуклеотидные последовательности, кодирующие антитела или биспецифические антигенсвязывающие молекулы согласно изобретению или их фрагменты.
Термины "клетка-хозяин", "линия клеток-хозяев," и " культура клеток-хозяев" используются взаимозаменяемо и относятся к клеткам, в которые встраиваются экзогенные молекулы нуклеиновых кислот, а также к потомству таких клеток. К клеткам-хозяевам относятся "трансформанты" и "трансформированные клетки", к которым относятся первично трансформированные клетки и их потомство, вне зависимости от количества пассажей. Потомство может быть не полностью идентично родительской клетке по своей нуклеотидной последовательности, поскольку могут содержать мутации. Мутантное потомство, которое обладает той же функциональностью или биологической активностью, которая наблюдается у оригинально трансформированных клеток, также включено в настоящее изобретение. Клетка-хозяин представляет собой любой тип клеточной системы, который может использоваться для получения антител или биспецифических антигенсвязывающих молекул по настоящему изобретению. К клеткам-хозяевам относятся культивированные клетки, например, культивированные клетки млекопитающих, такие, как HEK клетки, СНО клетки, BHK клетки, NS0 клетки, SP2/0 клетки, YO миеломные клетки, Р3Х63 мышиные миеломные клетки, PER клетки, PER.C6 клетки или гибридомные клетки, дрожжевые клетки, клетки насекомых и растений, были перечислены лишь некоторые, а также клетки трансгенных животных, трансгенных растений или культивированных тканей растений или животных.
"Активирующий Fc рецептор" представляет собой Fc рецептор, который после присоединения Fc домена антитела вызывает передачу сигнала, который стимулирует клетку, несущую рецептор, реализовывать эффекторную функцию. К активирующим Fc рецепторам человека относятся FcγRIIIa (CD 16а), FcγRI (CD64), FcγRIIa (CD32), и FcαRI (CD89).
Антителозависимая клеточная цитотоксичность (ADCC) представляет собой иммунный механизм, который приводит к лизису покрытых антителами клеток-мишеней иммунными эффекторными клетками. Клетки-мишени представляют собой клетки, с которыми специфически связываются антитела или их производные, содержащие Fc область, главным образом, посредством белковой части, которая расположена на N-конце по отношению к Fc области. Используемый в настоящем документе термин "сниженная ADCC" означает как снижение количества клеток-мишеней, которые подвергаются лизису за определенное время при заданной концентрации антитела в среде, окружающей клетки, посредством механизмов ADCC, описанных выше; так и/или увеличение концентрации антитела в окружающей клетки среде, необходимого для лизиса заданного количества клеток-мишеней за определенный период времени, посредством механизмов ADCC, описанных выше. Снижение ADCC происходит относительно ADCC, опосредованной тем же антителом, продуцируемым тем же типом клеток-хозяев, полученным с использованием тех же стандартных методов получения, очищения, формулирования и хранения (которые хорошо известны специалистам в данной области), но без использования методов генной инженерии. Например, снижение ADCC, опосредованной антителом, содержащим аминокислотную замену, приводящую к снижению ADCC, в Fc домене, происходит относительно ADCC, которая опосредуется тем же антителом, но без аминокислотной замены в Fc домене. Походящие методы анализа ADCC хорошо известны из области техники (см., например, публикацию РСТ no. WO 2006/082515 или публикацию РСТ No. WO 2012/130831).
Термин "эффективное количество" агента относится к количеству, которое является необходимым для того, чтобы вызвать физиологические изменения в клетке или ткани, в которые он вводится.
Термин "терапевтически эффективное количество" агента, например, фармацевтической композиции, относится к количеству, которое при введении в определенных дозах и с определенными временными промежутками, позволяет достичь желаемого терапевтического или профилактического результата. Терапевтически эффективное количество агента, например, устраняет, снижает, замедляет, минимизирует или препятствует негативным проявлениям заболевания.
"Индивидуум" или "субъект" представляет собой млекопитающее. К млекопитающим относятся, без ограничений указанными, одомашненные животные (например, коровы, овцы, кошки, собаки и лошади), приматы (например, люди и нечеловекообразные приматы, такие как обезьяны), кролики и грызуны (например, мыши и крысы). В частности, индивидуумом или субъектом является человек.
Термин "фармацевтическая композиция" относится к лекарственной форме, которая обеспечивает биологическую активность и эффективность активных ингредиентов в составе и не содержит дополнительных компонентов, которые являются недопустимо токсичными для субъекта, которому вводится такая композиция.
"Фармацевтически приемлемый носитель" относится к ингредиенту в составе фармацевтической композиции, отличному от активного ингредиента, который не является токсичным для субъекта. Фармацевтически доступные носители включают, без ограничений указанными, буферные агенты, эксципиенты, стабилизаторы или консерванты.
Термин "лечение", используемый в настоящем описании (и его грамматические вариации, такие, как "лечить") относится к клиническому вмешательству для изменения естественного течения заболевания у индивидуума, подлежащего лечению, которое может осуществляться также для профилактики заболевания или во время его клинического течения. К желаемым эффектам лечения относятся, без ограничений указанными, предотвращение возникновения или рецидива заболевания, смягчение симптомов, уменьшение прямых или непрямых патологических последствий заболевания, профилактика метастазирования, снижение скорости прогрессирования заболевания, смягчение или временное облегчение течения заболевания, а также ремиссия или улучшение прогноза. Согласно некоторым вариантам осуществления изобретения, антитела или биспецифические антигенсвязывающие молекулы согласно изобретению, используются для задержки развития заболевания или для замедления прогрессирования заболевания.
Термин "листок-вкладыш" используется для обозначения инструкции для пациента, которую вкладывают в промышленные упаковки лекарственных средств, содержащие информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях относительно использования таких лекарственных средств.
Детальное описание вариантов осуществления изобретения
Настоящее изобретение относится к антителам и биспецифическим антигенсвязывающим молекулам, которые связываются с HLA-A2/WT1, в особенности, с HLA-A2/WT1RMF или HLA-A2/WT1VLD, более конкретно с HLA-A2/WT1RMF, и обладают хорошей аффинностью со специфичностью, что требуется для их применения в терапевтических целях. Кроме того, заявленные молекулы также обладают благоприятными свойствами для терапевтического применения, например, с учетом их эффективности и/или безопасности, а также технологичности.
HLA-A2/WT1 антитело
Авторы настоящего изобретения разработали новые антитела, которые связываются с HLA-A2/WT1 с особенно хорошей аффинностью и специфичностью. В частности, как показано в Примерах, авторы настоящего изобретения разработали антитела, которые являются заметно более селективными по отношению к HLA-A2/WT1 (в частности, к HLA-A2/WT1RMF) и образуют более стойкие комплексы, по сравнению с комплексами HLA-A2 с другими, аналогичными по структуре пептидами.
Таким образом, согласно некоторым аспектам, настоящее изобретение относится к антителу, которое связывается с HLA-A2/WT1 и обладает любым из следующих свойств.
Согласно одному варианту осуществления изобретения, антитело обладает моновалентной аффинностью к HLA-A2/WT1 с константой диссоциации (KD) приблизительно менее, чем 100 нМ, приблизительно, менее чем 75 нМ, приблизительно, менее чем 50 нМ. Согласно одному варианту осуществления изобретения, антитело обладает бивалентной аффинностью (авидностью) по отношению к HLA-A2/WT1 с очевидной KD, равной, приблизительно, менее, 1,5 нМ, приблизительно, менее 1 нМ или, приблизительно, менее 0,75 нМ. Согласно одному варианту осуществления изобретения, бивалентная аффинность (авидность) антитела в 10 раз по меньшей мере, в 20 раз по меньшей мере, в 50 раз или по меньшей мере в 100 раз выше, чем моновалентная аффинность терминах (очевидной) KD. Согласно одному варианту осуществления изобретения, аффинность определятся при помощи Поверхностного Плазмонного Резонанса (SPR) при 25°С. Согласно одному варианту осуществления изобретения, моновалентная аффинность определяется в формате Fab молекулы антитела. Согласно одному варианту осуществления изобретения, бивалентная аффинность (авидность) определяется в формате IgG молекулы антитела. Согласно специфическому варианту осуществления изобретения, аффинность антитела определяется следующим образом:
Эксперименты осуществляют при температуре 25°С с использованием PBST в качестве электродного буфера (10 мМ PBS, рН 7.4 и 0.005% (об/об) Tween 20). Используют ProteOn XPR36 биосенсор, оборудованный GLC и GLM сенсорными чипами и связующими реагентами (10 мМ ацетата натрия рН 4.5, сульфо-N-гидроксисукцинимид [сульфо-NHS], 1-этил-3-(3-диметиламинопропил)-карбодиимид гидрохлорид [EDC] и этаноламин) поставщика BioRad Inc. (Hercules, СА). Иммобилизации осуществляют при 30 мкл/мин на чипе GLM. Античеловеческое IgG F(ab)2 специфическое антитело pAb (козла) (Jackson ImmunoResearch) соединяют в вертикальном направлении при помощи стандартной процедуры присоединения аминогрупп: все шесть лигандных каналов активируют на 5 минут смесью EDC (200 мМ) и сульфо-NHS (50 мМ). Сразу после активации всех поверхностей античеловеческое IgG F(ab)2 специфическое антитело pAb (козла) (50 мкг/мл, 10 мМ ацетат натрия, рН 5) пропускают через все шесть каналов в течение 5 минут. В конце каналы блокируют путем 5 минутного введения 1 М этаноламина--НС1 (рН 8.5). Fab варианты получают из супернатантов E.coli одновременной инъекцией по пяти горизонтальным каналам (30 мкл/мин) в течение 5 мин. Кондиционная среда вводится вдоль шестого канала для обеспечения "прямоточного" контроля для целей двойного контроля. Однократные кинетические измерения осуществляют путем введения серийных разведений HLA-WT1 (например, HLA-A2/WT1RMF или HLA-A2/WT1VLD) (100, 50, 25, 12.5, 6.25, 0 нМ, 50 мкл/мин) в течение 2 минут вдоль вертикальных каналов. Диссоциацию наблюдают в течение 3 минут. Кинетические данные анализируют при помощи программного обеспечения ProteOn Manager v. 2.1. Обработка данных о точке реакции включает применение шага между точками и шага с двумя эталонами с использованием встроенного контрольного буфера (Myszka, J Mol Recognit (1999) 12, 279-284). Данные, полученные для воспроизводимых однократных инъекций, соответствуют простой модели связывания 1: 1 Ленгмюра без переноса массы (O'Shannessy et al., Anal Biochem (1993) 212, 457-468).
Антитело, заявленное в соответствии с настоящим изобретением, специфично связывается с HLA-A2/WT1 (то есть комплексом молекулы HLA-A2 и WT1-выделенного пептида). Согласно некоторым вариантам осуществления изобретения, заявленное в изобретении антитело специфически связывается с HLA-A2/WT1RMF (то есть комплексом молекулы HLA-A2 и WT1RMF пептида). Согласно более специфическому варианту осуществления изобретения, антитело специфически связывается с HLA-A201/WT1RMF (то есть с комплексом молекулы HLA-A201 и WT1RMF пептида). К антителам, заявленным в соответствии с настоящим изобретением, которые связываются с HLA-A2/WT1RMF относятся антитела 11D06, 33Н09 и 5Е11, описанные в настоящем документе. Согласно другим вариантам осуществления изобретения, заявленное антитело специфически связывается с HLA-A2/WT1VLD (то есть с комплексом молекулы HLA-A2 и WT1VLD пептида). Согласно более специфическому варианту осуществления изобретения, антитело специфически связывается с HLA-A201/WT1VLD (то есть с комплексом HLA-A201 молекулы и WT1VLD пептида). К антителам, заявленным в соответствии с настоящим изобретением, которые связываются с HLA-A2/WT1VLD относятся антитела 11В09, 13В04 и 5С01, описанные в настоящем документе. Согласно одному варианту осуществления изобретения, специфичность связывания антитела определяют при помощи проточной цитометрии с использованием HLA-А2/пептид (например, WT1RMF или WT1VLD пептид)-экспрессирующих клеток, в частности, активированных пептидом Т2 клеток.
Согласно специфическим вариантам осуществления изобретения, специфичность связывание антитела определяют следующим образом:
Получают суспензию Т2 клеток в концентрации 106 клеток/мл в среде IMDM (Gibco by Life Technologies, Cat No. 31980-048), обогащенной 10% FBS (Gibco, Cat No. 16140-071)+1% пенициллин-стрептомицин (Gibco, Cat No. 15070-063) (полная среда), (например, WT1 VLD пептида (SEQ ID NO: 77), или RMF пептида (SEQ ID NO: 78)) при 10-2M (конечная концентрация пептида: 10-5M) в течение 2 часов при 37°С с 5% СО2. После отмывания клетки суспендируют в ледяном PBS и инкубируют с титрованными концентрациями антитела в формате IgG (например, от 10 мкг/мл до 0,00064 Клетки в общем объеме 10 мл помещают в пробирку и инкубируют с 10 мкл пептида мкг/мл) в течение 1 часа при 4°С, с последующей инкубацией с вторичным античеловеческим IgG-Fc фикоэритрин (РЕ)-конъюгированным антителом (Jackson Laboratories, Cat No. 109-116-098) в течение 30 минут. Клетки получают на FACS LSR II (BD), а данные презентируют как среднюю интенсивность флуоресценции (MFI) РЕ в Graphpad Prism.
Согласно одному варианту осуществления изобретения, антитело, заявленное в соответствии с настоящим изобретением, значимо не связывается только с HLA-A2 (то есть без присоединенного пептида) или HLA-A2 с пептидом, отличным от WT1-выделенного пептида, такого, как WT1RMF или WT1VLD.
Согласно одному варианту осуществления изобретения, антитело значимо не связывается с HLA-A2 в отсутствие WT1-выделенного пептида, в частности WT1RMF или WT1VLD. Согласно одному варианту осуществления изобретения, антитело связывается с HLA-A2/WT1 (в частности, с HLA-A2/WT1RMF или HLA-A2/WT1VLD) с ЕС50, которая по меньшей мере в 5, по меньшей мере в 10, по меньшей мере в 15, по меньшей мере в 20, по меньшей мере в 25, по меньшей мере в 50, по меньшей мере в 75 или по меньшей мере в 100 раз меньше, чем ЕС50 для связывания с HLA-A2 в отсутствие WT1-выделенного пептида (в частности, WT1RMF или WT1VLD). Согласно одному варианту осуществления изобретения, антитело связывается с HLA-A2/WT1 (в частности, HLA-A2/WT1RMF или HLA-A2/WT1VLD), но значимо не связывается с HLA-A2 с пептидом, выбранным из пептидов, представленных в Таблице 5 (пептиды SEQ ID NO: 79-105). Согласно одному варианту осуществления изобретения, антитело связывается с HLA-A2/WT1 (в частности, HLA-A2/WT1RMF или HLA-A2/WT1VLD), но значимо не связывается с HLA-A2, соединенным с любым из пептидов, представленных в Таблице 5 (пептиды SEQ ID №79-105).
Согласно одному варианту осуществления изобретения, антитело связывается с HLA-A2/WT1 (в частности, с HLA-A2/WT1RMF или HLA-A2/WT1VLD) с ЕС50, которая по меньшей мере в 5, по меньшей мере в 10, по меньшей мере15, по меньшей мере в 20, по меньшей мере в 25, по меньшей мере в 50, по меньшей мере в 75 или по меньшей мере в 100 раз ниже, чем ЕС50 для связывания HLA-A2 с любым из пептидов, представленных в Таблице 5 (пептиды SEQ ID NO: 79-105). Согласно одному варианту осуществления изобретения, антитело связывается с HLA-A2/WT1 (в частности, HLA-A2/WT1RMF или HLA-A2/WT1VLD) с ЕС50, которая по меньшей мере в 5, по меньшей мере в 10, по меньшей мере в 15, по меньшей мере в 20, по меньшей мере в 25, по меньшей мере в 50, по меньшей мере в 75 или по меньшей мере в 100 раз ниже, чем ЕС50 для связывания HLA-A2 с любым из пептидов, представленных в Таблице 5 (пептиды SEQ ID NO 79-105). Согласно одному варианту осуществления изобретения, ЕС50 определяют при помощи проточной цитометрии с использованием HLA-А2/пептид (например, WT1RMF или WT1VLD пептид)-экспрессирующих клеток, в частности, активированных пептидом Т2 клеток.
Согласно специфическому варианту осуществления изобретения, ЕС50 определяют следующим образом:
Получают суспензию Т2 клеток (АТСС, Cat. No. CRL-1992) в концентрации 106 клеток/мл в среде IMDM (Gibco by Life Technologies, Cat No. 31980-048), обогащенной 10% FBS (Gibco, Cat No. 16140-071) +1% пенициллин-стрептомицин (Gibco, Cat No. 15070-063) (полная среда). Клетки в общем объеме 10 мл помещают в пробирку и инкубируют с 10 мкл пептида (например, WT1 VLD пептида (SEQ ID NO: 77), или RMF пептида (SEQ ID NO: 78)) при 10-2 М (конечная концентрация пептида: 10-5 М) в течение 2 часов при 37°С с 5% СО2. После отмывания клетки суспендируют в ледяном PBS и инкубируют с титрованными концентрациями антитела в формате IgG (например, от 10 мкг/мл до 0,00064 мкг/мл) в течение 1 часа при 4°С, последующей инкубацией с вторичным античеловеческим IgG-Fc фикоэритрин (РЕ)-конъюгированным антителом (Jackson Laboratories, Cat No. 109-116-098) в течение 30 минут. Клетки получают на FACS LSR II (BD), а данные презентируют как среднюю интенсивность флуоресценции (MFI) РЕ в Graphpad Prism. Величины ЕС50 рассчитывают при помощи программного обеспечения Microsoft® Excel с использованием приложения XLfit® (ID Business Solutions, Guildford, UK).
Согласно одному аспекту своего осуществления, настоящее изобретение относится к антителу, которое конкурирует за связывание с HLA-A2/WT1, в частности, с HLA-A201/WT1RMF, с антителом, включающим вариабельный участок тяжелой цепи (VH) с последовательностью SEQ ID NO: 7, и вариабельный участок легкой цепи (VL) с последовательностью SEQ ID NO: 8.
Конкурентный анализ может применяться для идентификации антитела, которое конкурирует с антителом, включающим VH с последовательностью SEQ ID NO: 7 и VL с последовательностью SEQ ID NO: 8 (контрольное антитело) за связывание с HLA-A2/WT1.
Согласно определенным вариантам осуществления изобретения, такое конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), что и контрольное антитело. Детально описание примерных способов картирования эпитопов, с которыми связывается антитело, представлено, например в работе Morris (1996) "Epitope Mapping Protocols", in Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ), a также подробно описывается в настоящем документе в разделе Примеры. Согласно примерному конкурентному анализу, иммобилизованное HLA-A2/WT1 инкубировали в растворе, содержащем первое меченое антитело, которое связывается с HLA-A2/WT1 (например, антитело, включающее VH с последовательностью SEQ ID NO: 7 и VL с последовательностью SEQ ID NO: 8) и второе немеченое антитело, для которого изучают способность конкурировать с первым антителом за связывание с HLA-A2/WT1. Второе антитело может присутствовать в супернатанте гибридомы. В качестве контроля иммобилизованное HLA-A2/WT1 инкубируют в растворе, включающем первое мечено антитело без второго немеченого антитела. После инкубации в условиях, обеспечивающих связывание первого антитела с HLA-A2/WT1, удаляют избыток не связавшегося антитела и измеряют количество метки, ассоциированной с иммобилизованным HLA-A2/WT1. Если количество метки, ассоциированное с иммобилизованным HLA-A2/WT1, значительно снижается в тестируемом образце по сравнению с контрольным, это указывает на то, что второе антитело конкурирует с первым за связывание с HLA-A2/WT1. См. Harlow and Lane (1988) Antibodies: A Laboratory Manual ch. 14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
Согласно одному аспекту, настоящее изобретение относится к антителу, которое связывается с тем же эпитопом HLA-A2/WT1, в частности, HLA-A2/WT1RMF, что и антитело, включающее вариабельный участок (VH) тяжелой цепи с последовательностью SEQ ID NO: 7, и вариабельный участок легкой цепи (VL) с последовательностью SEQ ID NO: 8.
Скрининг на антитела, связывающиеся с определенным эпитопом (то есть, на те, которые связываются с одним и тем же эпитопом), может осуществляться при помощи методов, хорошо известных специалистам в данной области, например, без ограничений указанными, сканирование аланина, пептидные блоты (см. Meth. Mol. Biol. 248 (2004) 443-463), анализ пептидного расщепления, эксцизия эпитопов, экстракция эпитопов, химическая модификация антигенов (см. Prot. Sci. 9 (2000) 487-496), и эпитоп перекрестный конкурентный анализ (см "Antibodies", Harlow and Lane (Cold Spring Harbor Press, Cold Spring Harb., NY). Основанное на структуре антигена профилирование антител (ASAP), также известное, как профилирование антител с помощью модификации (MAP), позволяет сортировать множество моноклональных антител, специфически связывающихся с HLA-A2/WT1, на основании профиля связывания каждого антитела из множества с химически или ферментативно модифицированными поверхностями антигенов (см., например, патент США 2004/0101920). Антитела в каждой серии связываются с одним и тем же эпитопом, который может быть как уникальным эпитопом, так и полностью отличаться от эпитопа другой серии, либо частично перекрещиваться с ним. Также для того, чтобы понять, связывается ли антитело с тем же эпитопом HLA-A2/WT1 или конкурирует за связывание с другим антителом, может использоваться анализ конкурентного связывания с контрольным HLA-A2/WT1 антителом.
Например, "антитело, которое связывается с тем же эпитопом", как и контрольное антитело означает антитело, которое блокирует связывание контрольного антитела в конкурентном анализе на 50% или более, и, наоборот, контрольное антитело блокирует связывание антитела с антигеном в конкурентном анализе на 50% или более. Также, например, для того чтобы определить, связывается ли антитело с тем же эпитопом HLA-A2/WT1, что и контрольное антитело, позволяют контрольному антителу связаться с HLA-A2/WT1 в условиях полного насыщения. После удаления избытка контрольного антитела, оценивают способность изучаемого анти-HLA-A2/WT1 антитела связываться с HLA-A2/WT1. Если антитело способно связываться с HLA-A2/WT1 после насыщенного связывания контрольного антитела, можно сделать вывод о том, что вероятнее всего оно связывается с другим эпитопом, отличным от того, с которым связывается контрольное антитело. Но если изучаемое антитело не способно связываться с HLA-A2/WT1 после насыщенного связывания контрольного антитела, то изучаемое антитело может связываться с тем же эпитопом, что и контрольное антитело. Для подтверждения того, связывается ли указанное антитело с тем же эпитопом или связывание не происходит по стерическим причинам, можно использовать стандартные методики (например, анализ пептидной мутации и пептидного связывания с использованием ELISA, RIA, поверхностного плазмонного резонанса, проточной цитометрии или любых других количественных или качественных анализов связывания антител, доступных специалистам в данной области). Такое исследование следует проводить в два эксперимента, то есть оба антитела должны быть насыщающими. Если в обоих экспериментах только первое (насыщающее) антитело способно связываться с HLA-A2/WT1, тогда можно сделать вывод о том, что анти-HLA-A2/WT1 антитело и контрольное анти-HLA-A2/WT1 антитело конкурируют за связывание с HLA-A2/WT1. В некоторых вариантах осуществления считается, что два антитела связываются с одним и тем же или перекрывающимся эпитопом, если 1-, 5-, 10-, 20- или 100-кратный избыток одного антитела ингибирует связывание другого по меньшей мере, на 50% по меньшей мере на 75% по меньшей мере на 90% или даже на 99% или более, что измеряется в анализе конкурентного связывания (см, например, Junghans et al., Cancer Res. 50 (1990) 1495-1502). Согласно некоторым вариантам осуществления изобретения, считается, что два антитела связываются с одним и тем же эпитопом, если, по существу, все аминокислотные мутации в антигене, которые уменьшают или устраняют связывание одного антитела, также уменьшают или устраняют связывание другого. Считается, что два антитела имеют "перекрывающиеся эпитопы", если только набор аминокислотных мутаций, которые уменьшают или устраняют связывание одного антитела, уменьшают или устраняют связывание другого.
Согласно одному своему аспекту, настоящее изобретение относится к антителу, которое связывается с HLA-A2/WT1, при этом антитело связывается с эпитопом HLA-A2/WT1, в частности, HLA-A2/WT1RMF, согласно любому из представленных ниже вариантов осуществления. Эпитоп может быть точно определен при помощи анализа кристаллической структуры.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере три аминокислотных остатка WT1 пептида, в частности, WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере четыре аминокислотных остатка WT1 пептида, в частности, WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере пять аминокислотных остатков WT1 пептида, в частности, WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере шесть аминокислотных остатка WT1 пептида, в частности, WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере три аминокислотных остатка WT1 пептида, при этом указанные по меньшей мере три аминокислотных остатка выбраны из аминокислотных остатков, соответствующих аминокислотным остаткам R1, М2, Р4, N5, А6 и Y8 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере четыре аминокислотных остатка WT1 пептида, при этом указанные по меньшей мере четыре аминокислотных остатка выбраны из аминокислотных остатков, соответствующих аминокислотным остаткам R1, М2, Р4, N5, А6 и Y8 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере пять аминокислотных остатков WT1 пептида, при этом указанные по меньшей мере пять аминокислотных остатков выбраны из аминокислотных остатков, соответствующих аминокислотным остаткам R1, М2, Р4, N5, А6 и Y8 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает по меньшей мере шесть аминокислотных остатка WT1 пептида, при этом указанные по меньшей мере шесть аминокислотных остатков выбраны из аминокислотных остатков, соответствующих аминокислотным остаткам R1, М2, Р4, N5, А6 и Y8 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам R1, N5 и А6 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам R1, M2, P4, N5, A6 и Y8 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, R65, K66 и Q155 HLA-A2 последовательности, представленной в SEQ ID NO: 138.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, R65, K66 и Q155 HLA-A2 последовательности, представленной в SEQ ID NO: 138, и аминокислотные остатки, соответствующие аминокислотным остаткам R1, N5 и А6 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, R65, K66, Q155 и D61 HLA-A2 последовательности, представленной в SEQ ID NO: 138. Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, R65, K66, Q155 и А69 HLA-A2 последовательности, представленной в SEQ ID NO: 138. Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, R65, K66, Q155 и А155 HLA-А2 последовательности SEQ ID NO: 138.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, R65, K66, Q155, и одному или более остатку D61, А69 и А150 HLA-A2 последовательности, представленной в SEQ ID NO: 138, и аминокислотные остатки, соответствующие аминокислотным остаткам R1, М2, Р4, N5, А6 и/или Y8 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, D61, G62, R65, K66, А69, А150, Q155, А158, Т163, Е166, W167 и R170 HLA-А2 последовательности SEQ ID NO: 138
Согласно одному варианту осуществления изобретения, указанный эпитоп включает аминокислотные остатки, соответствующие аминокислотным остаткам Е58, D61, G62, R65, K66, А69, А150, Q155, А158, Т163, Е166, W167 и R170 HLA-А2 последовательности, представленной в SEQ ID NO: 138, и аминокислотные остатки, соответствующие аминокислотным остаткам R1, М2, Р4, N5, А6 и/или Y8 WT1RMF пептида, с последовательностью SEQ ID NO: 78.
Согласно одному варианту осуществления изобретения, указанный эпитоп не включает аминокислотные остатки, соответствующие аминокислотным остаткам G56, D106, W107, R108, Е161, G162 и/или R169 HLA-A2 последовательности SEQ ID NO: 138.
Согласно другому аспекту, настоящее изобретение относится к антителу, которое связывается с HLA-A2/WT1, при этом указанное антитело включает:
(i) вариабельную область тяжелой цепи (VH), содержащую область, определяющую комплементарность тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2, и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL) содержащую область, определяющую комплементарность легкой цепи (LCDR) 1 с последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 о с последовательностью SEQ ID NO: 6;
(ii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 9, HCDR 2 с последовательностью SEQ ID NO: 10, и HCDR 3 с последовательностью SEQ ID NO: 11, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 12, LCDR 2 с последовательностью SEQ ID NO: 13 и LCDR 3 с последовательностью SEQ ID NO: 14;
(iii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 17, HCDR 2 с последовательностью SEQ ID NO: 18, и HCDR 3 с последовательностью SEQ ID NO: 19, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 20, LCDR 2 с последовательностью SEQ ID NO: 21 и LCDR 3 с последовательностью SEQ ID NO: 22;
(iv) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 25, HCDR 2 с последовательностью SEQ ID NO: 26, и HCDR 3 с последовательностью SEQ ID NO: 27, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 28, LCDR 2 с последовательностью SEQ ID NO: 29 и LCDR 3 с последовательностью SEQ ID NO: 30;
(v) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 33, HCDR 2 с последовательностью SEQ ID NO: 34, и HCDR 3 с последовательностью SEQ ID NO: 35, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 36, LCDR 2 с последовательностью SEQ ID NO: 37 и LCDR 3 с последовательностью SEQ ID NO: 38;
(vi) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 41, HCDR 2 с последовательностью SEQ ID NO: 42, и HCDR 3 с последовательностью SEQ ID NO: 43, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 44, LCDR 2 с последовательностью SEQ ID NO: 45 и LCDR 3 с последовательностью SEQ ID NO: 46;
(vii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 49, HCDR 2 с последовательностью SEQ ID NO: 50, и HCDR 3 с последовательностью SEQ ID NO: 51, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 52, LCDR 2 с последовательностью SEQ ID NO: 53 и LCDR 3 с последовательностью SEQ ID NO: 54;
(viii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 57, HCDR 2 с последовательностью SEQ ID NO: 58, и HCDR 3 с последовательностью SEQ ID NO: 59, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 60, LCDR 2 с последовательностью SEQ ID NO: 61 и LCDR 3 с последовательностью SEQ ID NO: 62;
(ix) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 65, HCDR 2 с последовательностью SEQ ID NO: 66, и HCDR 3 с последовательностью SEQ ID NO: 67, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 68, LCDR 2 с последовательностью SEQ ID NO: 69 и LCDR 3 с последовательностью SEQ ID NO: 70.
Согласно определенному варианту осуществления изобретения, антитело включает VH, содержащую HCDR 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, и VL, содержащую LCDR 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6.
Согласно другому варианту осуществления изобретения, антитело включает VH, содержащую HCDR 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, и VL, содержащую LCDR 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14.
Согласно другому варианту осуществления изобретения, антитело включает VH, содержащую HCDR 1 с SEQ ID NO: 17, HCDR 2 с SEQ ID NO: 18, и HCDR 3 с SEQ ID NO: 19, и VL, содержащую LCDR 1 с SEQ ID NO: 20, LCDR 2 с SEQ ID NO: 21 и LCDR 3 с SEQ ID NO: 22.
Согласно еще одному варианту осуществления изобретения, антитело включает VH, содержащую HCDR 1 с SEQ ID NO: 25, HCDR 2 с SEQ ID NO: 26, и HCDR 3 с SEQ ID NO: 27, и VL, содержащую LCDR 1 с SEQ ID NO: 28, LCDR 2 с SEQ ID NO: 29 и LCDR 3 с SEQ ID NO: 30.
Согласно некоторым вариантам осуществления изобретения, антитело представляет собой человеческое антитело. Согласно одному варианту осуществления изобретения, VH представляет собой человеческий VH и/или VL представляет собой VL человека.
Согласно одному варианту осуществления изобретения, антитело включает CDRs по любому из описанных выше вариантов изобретения, и дополнительно содержит человеческий каркас, например, каркас иммуноглобулина человека.
Согласно одному варианту осуществления изобретения, антитело включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64;
(ix) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24.
Согласно дополнительному варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, антитело включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64;
(ix) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16.
Согласно другому варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24.
Согласно другому варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32.
Согласно определенным вариантам осуществления изобретения, последовательность VH или VL по меньшей мере с идентичностью 95%, 96%, 97%, 98%, или 99% содержит замены (например, консервативные замены), вставки или делеции по отношению к контрольной последовательности, но антитело, имеющее такую последовательность, сохраняет способность связывается с HLA-A2/WT1. Согласно определенным вариантам осуществления изобретения, общее количество аминокислотных замен, вставок и/или делеций в VH (SEQ ID NO: 7, 15, 23, 31, 39, 47, 55, 63 или 71) равно от 1 до 10, и/или общее количество аминокислотных замен, вставок и/или делеций в VL (SEQ ID NO: 8, 16, 24, 32, 40, 48, 56, 64 или 72) равно от 1 до 10.
Согласно определенным вариантам осуществления изобретения, замены, вставки или делеций происходят вне HVR (то есть, в FR областях).
Необязательно, антитело включает VH последовательность и/или VL последовательность, как определено выше, содержащую посттрансляционные модификации таких последовательностей.
Согласно одному варианту осуществления изобретения, антитело включает:
(i) VH, содержащую аминокислотную последовательность SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность SEQ ID NO 16;
(iii) VH, содержащую аминокислотную последовательность SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность SEQ ID NO: 55, и VL, содержащую аминокислотную последовательность SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность SEQ ID NO: 63, и VL, содержащую аминокислотную последовательность SEQ ID NO: 64;
(iv) VH, содержащую аминокислотную последовательность SEQ ID NO: 71, и VL, содержащую аминокислотную последовательность SEQ ID NO: 72.
Согласно одному варианту осуществления изобретения: (i) VH последовательность SEQ ID NO: 7, и VL последовательность SEQ ID NO: 8;
(ii) VH последовательность SEQ ID NO: 15, и VL последовательность SEQ ID NO: 16;
(iii) VH последовательность SEQ ID NO: 23, и VL последовательность SEQ ID NO: 24;
(iv) VH с последовательность SEQ ID NO: 31, и VL с последовательностью SEQ ID NO: 32;
(v) VH последовательность SEQ ID NO: 39, и VL последовательность SEQ ID NO: 40;
(vi) VH последовательность SEQ ID NO: 47, и VL последовательность SEQ ID NO: 48;
(vii) VH последовательность SEQ ID NO: 55, и VL последовательность SEQ ID NO: 56;
(viii) VH последовательность SEQ ID NO: 63, и VL последовательность SEQ ID NO: 64; или
(ix) VH последовательность SEQ ID NO: 71, и VL последовательность SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность SEQ ID NO: 7 и VL, содержащую аминокислотную последовательность SEQ ID NO: 8. Согласно другому варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность SEQ ID NO: 15 и VL, содержащую аминокислотную последовательность SEQ ID NO: 16. Согласно дополнительному варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность SEQ ID NO: 23 и VL, содержащую аминокислотную последовательность SEQ ID NO: 24. Согласно еще одному варианту осуществления изобретения, антитело включает VH, содержащую аминокислотную последовательность SEQ ID NO: 31 и VL, содержащую аминокислотную последовательность SEQ ID NO: 32.
Согласно определенному варианту осуществления изобретения, антитело включает VH последовательность SEQ ID NO: 7 и VL последовательность SEQ ID NO: 8. Согласно определенному варианту осуществления изобретения, антитело включает VH последовательность SEQ ID NO: 15 и VL последовательность SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, антитело включает VH последовательность SEQ ID NO: 23 и VL последовательность SEQ ID NO: 24.
Согласно еще одному варианту осуществления изобретения, антитело включает VH последовательность SEQ ID NO: 31 и VL последовательность SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, антитело включает константную область человека. Согласно одному варианту осуществления изобретения, антитело представляет собой молекулу иммуноглобулина, включающую константную область человека, в частности, молекулу иммуноглобулина класса IgG, включающую CH1, СН2, СН3 и/или CL домены человека. Примеры последовательностей константных доменов человека представлены в SEQ ID NOs 112 и 113 (домены CL каппа и лямбда человека, соответственно) и SEQ ID NO: 114 (константные домены СН1-СН2-СН3 тяжелых цепей IgG1 человека).
Согласно некоторым вариантам осуществления изобретения, антитело включает константную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 112 или SEQ ID NO: 113, в частности, аминокислотной последовательности SEQ ID NO: 112. Согласно некоторым вариантам осуществления изобретения, антитело включает константную область тяжелой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 114. В частности, константная область тяжелой цепи может содержать аминокислотные мутации в домене Fc, как описано в настоящем документе.
Согласно одному варианту осуществления изобретения, антитело представляет собой моноклональное антитело.
Согласно одному варианту осуществления изобретения, антитело представляет собой IgG, в частности, IgG1 антитело.
Согласно одному варианту осуществления изобретения, антитело представляет собой полноразмерное антитело.
Согласно одному варианту осуществления изобретения, антитело включает Fc домен, а именно, Fc домен IgG, более конкретно, Fc домен IgG1. Согласно данному варианту осуществления изобретения, Fc домен представляет собой Fc домен человека. Fc домен антитела может обладать любой из характеристик, как по отдельности, так и в комбинации, которые описаны в настоящем документе в контексте Fc домена биспецифической антигенсвязывающей молекулы согласно изобретению.
Согласно другому варианту осуществления изобретения, антитело представляет собой фрагмент антитела, выбранный из группы Fv молекулы, scFv молекулы, Fab молекулы и F(ab')2 молекулы; в частности, Fab молекулы.
Согласно другому варианту осуществления изобретения, фрагмент антитела представляет собой диатело, триатело или тетратело.
Согласно дополнительному аспекту осуществления изобретения, антитело по любому из представленных выше вариантов, может обладать характеристиками, как по отдельности, так и в комбинации, описанными в разделах ниже.
Варианты гликозилирования
Согласно определенным вариантам осуществления изобретения, заявленное антитело изменено для увеличения или снижения степени его гликозилирования. Добавление или удаление сайтов гликозилирования к антителу может быть легко осуществлено путем изменения аминокислотной последовательности, таким образом, что один или более сайт гликозилирования создается или удаляется.
Когда антитело содержит Fc область, присоединенный к нему олигосахарид может быть изменен. Нативные антитела, продуцируемые клетками млекопитающих, обычно включают разветвленный, биантенарный олигосахарид, который обычно прикреплен к остатку Asn297 СН2 домена Fc области посредством N-связи. См., например, Wright et al. TIBTECH 15:26-32 (1997). Олигосахариды могут включать углеводороды, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу, и сиаловую кислоту в "стержне" биантенарной олигосахаридной структуры. Согласно некоторым вариантам осуществления изобретения, модификации олигосахаридов в составе антитела согласно изобретению, могут осуществляться для создания вариантов антитела с заданными улучшенными свойствами.
Согласно одному варианту осуществления изобретения, предлагаются варианты антител, имеющие нефукозилированный олигосахарид, то есть олигосахаридную структуру, которая не имеет фукозы, прикрепленной (прямо или опосредованно) к Fc области. Такой нефукозилирванный олигосахарид (также обозначаемый термином "афукозилированный" олигосахарид), в частности, N-связанный олигосахарид, не содержит остатка фукозы, прикрепленного к первому GlcNAc в стержне биантенарной олигосахаридной структуры. Согласно одному варианту осуществления изобретения, предлагаются варианты антител, имеющие повышенное содержание нефукозилированных олигосахаридов в Fc области по сравнению с нативным или родительским антителом. Например, доля нефукозилированных олигосахаридов может составлять по меньшей мере 20% по меньшей мере 40% по меньшей мере 60% по меньшей мере 80% или даже приблизительно 100% (то есть фукозилированные олигосахариды отсутствуют). Доля нефукозилированных олигосахаридов представляет собой (среднее) количество олигосахаридов, не имеющих остатков фукозы, относительно общего количества олигосахаридов, прикрепленных к Asn 297 (например, комплексных, гибридных структур или структур с высоким содержанием маннозы), измеренное при помощи MALDI-TOF масс-спектрометрии, как описано, например, в документе WO 2006/082515. Asn297 относится к остатку аспарагина, расположенному приблизительно в положении 297 Fc области (EU нумерация остатков в Fc области); однако, Asn297 также может располагаться примерно ±3 аминокислоты выше или ниже положения 297, то есть между положениями 294 и 300, из-за незначительных вариаций последовательностей антител. Такие антитела, имеющие повышенное содержание нефукозилированных олигосахаридов в Fc области, могут иметь усиленное связывание с FcγRIIIa рецептором и/или улучшенную эффекторную функцию, конкретнее, улучшенную функцию ADCC. См. например, US 2003/0157108; US 2004/0093621.
Примерами клеточных линий, способных продуцировать антитела с пониженным фукозилированием, являются клетки линии Lec13 СНО, дефицитные по белку фукозилирования (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); US 2003/0157108; и WO 2004/056312, в частности в Примере 11), а также нокаутированные клеточные линии, такие, как СНО клеточные линии, нокаутированные по гену альфа-1,6-фукозилтрансферазы, FUT8 (см, например, Yamane-Ohnuki et al. Biotech. Bioeng. 87:614-622 (2004); Kanda, Y. et al., Biotechnol. Bioeng., 94(4):680-688 (2006); и WO 2003/085107), или клетки с пониженной или отсутствующей активностью белка, отвечающего за синтез или транспорт ГДФ-синтазы (см, например, US 2004259150, US 2005031613, US 2004132140, US 2004110282).
Согласно дополнительным вариантам осуществления изобретения, предлагаются варианты антител с бисекционным олигосахаридами, например, в которых биантенарный олигосахарид, присоединенный к области Fc антитела, делится пополам GlcNAc. Такие варианты антител обладают пониженным фукозилированием и/или улучшенной функцией ADCC, как описано выше. Примеры таких вариантов антител описаны, например, у Umana et al., Nat Biotechnol 17, 176-180 (1999); Ferrara et al., Biotechn Bioeng 93, 851-861 (2006); WO 99/54342; WO 2004/065540, WO 2003/011878. Также предлагаются варианты антител, содержащие по меньшей мере один остаток галактозы в составе олигосахарида, прикрепленного к Fc области.
Такие варианты антител могут обладать улучшенной CDC функцией. Такие варианты антител описаны, например, в документах WO 1997/30087; WO 1998/58964; и WO 1999/22764.
Цистеин-сгенерированные варианты антител
Согласно определенным вариантам осуществления изобретения, может быть желательным создать цистеин-сгенерированные антитела, например, "thioMAbs," в которых один или более аминокислотный остаток замещен остатком цистеина. Согласно определенным вариантам осуществления изобретения, замещаемые остатки располагаются в доступных участках антитела. При замещении указанных остатков цистеином в доступные участки антитела встраиваются реактивные тиоловые группы, которые могут использоваться для конъюгации антитела с другими молекулами, такими, как молекулы лекарственных средств или линкерные-лекарственные молекулы с формированием иммуноконъюгатов, как дополнительно описывается в настоящем документе. Цистеин-сгенерированные антитела могут быть получены, например, как описано в патентах США номер 7,521,541, 8,30,930, 7,855,275, 9,000,130, или публикации WO 2016040856.
Производные антител
Согласно определенным вариантам осуществления изобретения, заявленное антитело может подвергаться дополнительным модификациям, чтобы содержать дополнительные небелковые молекулы, которые хорошо известны специалистам в данной области и являются общедоступными. Молекулы, которые являются подходящими для дериватизации антител, включают, без ограничений указанными, водорастворимые полимеры. Неограничивающими примерами водорастворимых полимеров являются, без ограничений указанными, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (как гомополимеры, так и рандомные сополимеры), а также декстран или поли(n-винилпирролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерол), поливиниловый спирт, а также их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества в процессе производства благодаря своей стабильности в воде. Полимер может быть любого молекулярного веса, кроме того, он может быть как разветвленным, так и неразветвленным. Количество полимеров, прикрепленных к антителу, может отличаться, и в том случае, если прикрепляется более одного полимера, молекулы могут быть как одинаковыми, так и разными. В целом выбор количества и/или типа полимеров, которые применяются для дериватизации, может основываться на таких соображениях, как (без ограничений указанными) определенные характеристики или функции "улучшаемого" антитела, будет ли это производное антитела применяться для терапии в заданных условиях и т.д.
Согласно другому варианту осуществления изобретения, предлагаются конъюгаты антитела и небелковой молекулы могут селективно нагреваться при воздействии облучения. Согласно одному варианту осуществления изобретения, небелковая молекула представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Излучение может иметь любую длину волны, и включает при этом волны, которые не повреждают обычные клетки, но нагревают небелковую молекулу до температуры, при которой погибают клетки, приближенные к конъюгату антитело-небелковая молекула.
Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам, включающим анти-HLA-A2/WT1 антитело, как описано в настоящем документе, конъюгированное (химически связанное) с одним или более терапевтическим агентом, таким, как цитотоксический агент, химиотерапевтический агент, лекарственное средство, ингибиторы роста, токсины (например, белковые токсины, ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, а также их фрагменты), или радиоактивные изотопы.
Согласно одному варианту осуществления изобретения, иммуноконъюгат представляет собой конъюгат антитела и лекарственного средства (ADC, аббревиатура от англ. Antibody-Drug Conjugate), в котором антитело конъюгировано с одним или более терапевтическим агентом, упомянутым выше. Антитело обычно соединено с одним или более терапевтическим агентом при помощи линкеров. Обзор ADC технологий, содержащую примеры терапевтических агентов, и лекарственных средств, и линкеров представлен в источнике Pharmacol Review 68:3-19 (2016).
Согласно другому варианту осуществления изобретения, иммуноконъюгат включает антитело, как описано в настоящем документе, конъюгированное с ферментативно активным токсином или его фрагментом, содержащую, без ограничений указанными, А цепь дифтерийного токсина, необязательные активные фрагменты дифтерийного токсина, А цепь экзотоксина (выделенного из Pseudomonas aeruginosa), А цепь рицина, А цепь абрина, А цепь модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII, и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены.
Согласно другому варианту осуществления изобретения, иммуноконъюгат содержит антитело, как определено в настоящем документе, конъюгированное с радиоактивным атомом с формированием радиоконъюгата. Для получения радиоконъюгатов доступно большое разнообразие радиоактивных изотопов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 a также радиоактивные изотопы Lu. Когда радиоконъюгат используется для визуализации, он может содержать радиоактивный атом для сцинтиграфических исследований, например tc99m или I123, или спиновую метку для ядерно-магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента могут быть получены с использованием множества бифункциональных белковых связующих агентов, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил) циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl) активные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис (п-азидобензоил) гександиамин), производные бис-диазония (такие как бис- (п-диазониумбензоил)-этилендиамин) диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, рициновый иммунотоксин может быть получен, как описано у in Vitetta et al., Science 238:1098 (1987). Меченая углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен триаминопентауксусная кислота (МХ-DTPA) является примером хелатирующего агента для конъюгации радионуклида с антителом. См. публикацию WO 94/11026. Линкер может представлять собой "отщепляемый линкер", способствующий высвобождению цитотоксического лекарства в клетку. Например, может применяться кислотолабильный линкер, чувствительный к пептидазе линкер, диметиловый линкер или линкер, содержащий дисульфид (Chari et al., Cancer Res. 52:127-131 (1992); Патент США No. 5,208,020).
Иммуноконъюгаты или ADC, рассматриваемые в настоящем документе, включают конъюгаты (без ограничений указанными), полученные с использованием кросс-линкерных реагентов, таких, как (без ограничений указанными) BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC, и сульфо-SMPB, и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которая являются коммерчески доступными (например, в компании Pierce Biotechnology, Inc., Rockford, IL., U.S.A).
Мультиспецифические антитела
Согласно определенным вариантам осуществления изобретения, заявленное антитело представляет собой мультиспецифическое антитело, например, биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, которые обладают специфичностью связывания по отношению по меньшей мере к двум различным сайтам, то есть, разным эпитопам на разных антигенах или разным эпитопам на одном антигене. Согласно определенным вариантам осуществления изобретения, мультиспецифическое антитело обладает тремя специфичностями связывания или более. Одна из указанных специфичностей связывания направлена на HLA-A2/WT1, а другие (две или более) - на любые другие антигены. Согласно определенным вариантам осуществления изобретения, биспецифические антитела могут связываться с двумя (или более) различными эпитопами HLA-A2/WT1. Мультиспецифические (например, биспецифические) антитела также могут использоваться для доставки цитотоксических агентов или клеток к клеткам, экспрессирующим HLA-A2/WT1. Мультиспецифические антитела могу быть получены как в форме полноразмерных антител, так и в форме их фрагментов.
К методикам получения мультиспецифических антител относятся, без ограничений указанными, совместная рекомбинантная экспрессия двух пар тяжелых-легких цепей иммуноглобулина с различной специфичностью (см. Milstein and Cuello, Nature 305: 537 (1983)) и метод генной инженерии "выступ во впадину" (см, например., патент США No. 5,731,168, и Atwell et al., J. Mol. Biol. 270:26 (1997)). Мультиспецифические антитела также могут быть получены путем генерирования электростатических стерических эффектов для получения Fc-гетеродимерных молекул антител (см, например, WO 2009/089004); перекрестного связывания двух или более антител или их фрагментов (см, например, патент США No. 4,676,980, и Brennan et al., Science, 229: 81 (1985)); использования лейциновых застежек молний для получения биспецифических антител (см, например, Kostelny et al., J. Immunol., 148(5):1547-1553 (1992) и WO 2011/034605); применения традиционной технологии легких цепей для решения проблемы неправильного спаривания легких цепей (см, например, WO 98/50431); применения технологии "диател" для получения фрагментов биспецифических антител (см, например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993)); и применение одноцепочечных Fv (sFv) димеров (см, например, Gruber et al., J. Immunol., 152:5368 (1994)); и получение триспецифических антител, как описано, например, у Tutt et al. J. Immunol. 147: 60 (1991).
Сгенерированные антитела с тремя или более антигенсвязывающими сайтами, включают, например, "антитела-осьминоги" или DVD-Ig, которые также включены в настоящее описание (см, например WO 2001/77342 и WO 2008/024715). Другие примеры мультиспецифических антител с тремя или более антигенсвязывающими сайтами описаны в публикациях WO 2010/115589, WO 2010/112193, WO 2010/136172, WO 2010/145792, and WO 2013/026831. Биспецифическое антитело или его антигенсвязывающий фрагмент также включает "Fab с двойной активностью" или "DAF", содержащую антигенсвязывающий сайт, который связывается с HLA-A2/WT1, а также с другим отличным антигеном, или двумя различными эпитопами HLA-A2/WT1 (см, например, US 2008/0069820 и WO 2015/095539).
Мультиспецифические антитела также могут быть представлены в асимметричной форме с кроссовером доменов в одном или более связывающем плече той же антигенной специфичности, то есть путем замены VH/VL доменов (см, например, WO 2009/080252 и WO 2015/150447), CH1/CL доменов (см, например, WO 2009/080253) или полностью Fab областей (см, например., WO 2009/080251, WO 2016/016299, см также Schaefer et al, PNAS, 108 (2011) 1187-1191, and Klein at al., MAbs 8 (2016) 1010-20). Асимметричные Fab области также могут быть созданы путем встраивания заряженных или незаряженных аминокислотных мутаций в структуру доменов для коррекции спаривания Fab. См. например, WO 2016/172485. Различные другие молекулярные форматы мультиспефических антител хорошо известны специалистам в данной области и включены в настоящее описание (см, например, Mol Immunol 67 (2015) 95-106).
Определенным типом мультиспецифических антител, включенных в настоящее описание, являются биспецифические антитела, созданные для одновременного связывания с антигеном на поверхности клеток-мишеней, например, опухолевых клеток для активации инвариантного компонента Т-клеточного рецепторного (TCR) комплекса, такого, как CD3, для переориентирования Т-клеток на убийство клеток-мишеней. Следовательно, согласно определенным вариантам осуществления, заявленное антитело представляет собой мультиспецифическое антитело, в частности, биспецифическое антитело, при этом одна специфичность связывания относится к HLA-A2/WT, а другая - к CD3.
Примерами форматов биспецифических антител, которые могут быть полезны для этих целей (без ограничений указанными), являются так называемые "BiTE" молекулы (привлекающие Т-клетки биспецифические активаторы), в которых две молекулы scFv соединены между собой подвижным линкером (см, например, WO 2004/106381, WO 2005/061547, WO 2007/042261, и WO 2008/119567, Nagorsen and  Exp Cell Res 317, 1255-1260 (2011)); диатела (Holliger et al., Prot Eng 9, 299-305 (1996)) и их производные, такие, как тандемные диатела ("TandAb"; Kipriyanov et al., J Mol Biol 293, 41-56 (1999)); "DART" молекулы (переориентирующееся антитело с двойной аффинностью), которые основаны на формате диател, но имеют С-концевой дисульфидный мостик для дополнительной стабилизации (Johnson et al., J Mol Biol 399, 436-449 (2010)), а также так называемые триомабы, которые являются полными гибридными мышино/крысиными молекулами IgG (описаны Seimetz et al., Cancer Treat Rev 36, 458-467 (2010)). К определенным форматам биспецифических Т-клеточных антител, включенных в настоящее описание, являются форматы, описанные в WO 2013/026833, WO 2013/026839, WO 2016/020309; Bacac et al., Oncoimmunology 5(8) (2016) е1203498.
Exp Cell Res 317, 1255-1260 (2011)); диатела (Holliger et al., Prot Eng 9, 299-305 (1996)) и их производные, такие, как тандемные диатела ("TandAb"; Kipriyanov et al., J Mol Biol 293, 41-56 (1999)); "DART" молекулы (переориентирующееся антитело с двойной аффинностью), которые основаны на формате диател, но имеют С-концевой дисульфидный мостик для дополнительной стабилизации (Johnson et al., J Mol Biol 399, 436-449 (2010)), а также так называемые триомабы, которые являются полными гибридными мышино/крысиными молекулами IgG (описаны Seimetz et al., Cancer Treat Rev 36, 458-467 (2010)). К определенным форматам биспецифических Т-клеточных антител, включенных в настоящее описание, являются форматы, описанные в WO 2013/026833, WO 2013/026839, WO 2016/020309; Bacac et al., Oncoimmunology 5(8) (2016) е1203498.
Биспецифические антигенсвязывающие молекулы, которые связываются с HLA-A2/WT1 и вторым антигеном
Настоящее изобретение также относится к биспецифической антигенсвязывающей молекуле, то есть антигенсвязывающей молекуле, которая включает по меньшей мере два антигенсвязывающие молекулы, способные специфически связываться с двумя различными антигенными детерминантами (первым и вторым антигеном).
На основании разработанных ими же HLA-A2/WT1 антител, авторы настоящего изобретения разработали биспецифические антигенсвязывающие молекулы, которые связываются с HLA-A2/WT1 и дополнительным антигеном, в частности, антигеном, активирующим Т клетки, такие, как CD3.
Как показано в Примерах, такие биспецифические антигенсвязывающие молекулы обладают целом рядом значимых свойств, содержащую хорошую эффективность и низкую токсичность.
Таким образом, согласно определенным аспектам, настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, включающей (а) первую антигенсвязывающую молекулу, которая связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, и (b) вторую антигенсвязывающую молекулу, которая специфически связывается со вторым антигеном, при этом биспецифическая антигенсвязывающая молекула обладает любой из описанных ниже характеристик.
Биспецифическая антигенсвязывающая молекула, согласно изобретению, специфически индуцирует опосредованное Т-клетками уничтожение клеток, экспрессирующих HLA-A2/WT1 (то есть, комплекс HLA-A2 молекулы и WT1-выделенного пептида). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула, согласно изобретению, специфически индуцирует опосредованное Т-клетками уничтожение клеток, экспрессирующих HLA-A2/WT1RMF (то есть комплекс HLA-A2 молекулы и WT1RMF пептид). Согласно более специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула специфически индуцирует опосредованное Т-клетками уничтожение клеток, экспрессирующих HLA-A2.01/WT1RMF (то есть комплекс HLA-A201 молекулы и WT1RMF пептида).
Согласно одному варианту осуществления изобретения, индуцирование опосредованной Т-клетками гибели посредством биспецифической антигенсвязывающей молекул определяют с применением HLA-А2/пептид-экспрессирующих клеток (то есть, WT1RMF или WT1VLD пептид-экспрессирующих клеток), в частности, активированных пептидом Т2 клеток, а также путем измерения высвобождения лактатдегидрогеназы (ЛДГ) из указанных клеток после инкубации с биспецифической антигенсвязывающей молекулой в присутствии Т-клеток.
Согласно специфическому варианту осуществления изобретения, индуцирование опосредованной Т-клетками гибели посредством биспецифической антигенсвязывающей молекулы определяют следующим образом:
Получают суспензию Т2 клеток (АТСС, Cat. No. CRL-1992) с концентрацией 106 клеток/мл в среде IMDM (Gibco by Life Technologies, Cat No. 31980-048), обогащенной 10% FBS (Gibco, Cat No. 16140-071) +1% Пенициллином-Стрептомицином (Gibco, Cat No. 15070-063) (полная среда). Клетки в общем объеме 10 мл помещают в пробирку и инкубируют с 10 мкл пептида (например, WT1 VLD пептида (SEQ ID NO: 77), или RMF пептида (SEQ ID NO: 78)) при 10-2 M (конечная концентрация пептида: 10-5 М) в течение 2 ч при 37°С с 5% СО2. Пан CD3+ клетки изолируют от мононуклеарных клеток из лейкоцитарной пленки, при помощи градиентного центрифугирования по Ficoll (GE Healthcare, Cat. No. 17-1440-03). Общие CD3+ T клетки очищают при помощи магнитно-активированной клеточной сортировки (MACS (Miltenyi Biotec)) с использованием набора для выделения пан Т-клеток человека (Miltenyi Biotec, Cat. No. 130-096-535). Анализ цитотоксичности проводят следующим образом: активированные пептидом клетки (100 мкл) высевают на 96-луночный планшет для микротитрования с круглым дном (3×105 клеток/мл) и культивируют совместно с 50 мкл Т-клеток (6×106 клеток/мл), и с 50 мкл оттитрованной биспецифической антигенсвязывающей молекулы (например, при концентрациях от 40 мкг/мл до 0.00004 мкг/мл) в полной среде в течение 18 ч при 37°С с 5% СО2. Затем 50 мкл супернатанта переносят на новый белый планшет и в каждую лунку добавляют 25 мкл реагента CytoTox-Glo Luciferase Assay (Promega, Cat. No. G9291) и инкубируют при комнатной температуре (RT) в течение 15 минут. Люминисцентный сигнал (для измерения высвобождения ЛДГ, которое является индикатором клеточной гибели) считывают при помощи EnVision (PerkinElmer). Данные выражены в относительных единицах люминисценции (RLU).
Биспецифическая антигенсвязывающая молекула, согласно изобретению, специфически активирует Т-клетки в присутствии клеток, экспрессирующих HLA-A2/WT1 (то есть комплекса HLA-A2 молекулы и WT1-выделенного пептида). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула, согласно изобретению, специфически активирует Т клетки в присутствии клеток, экспрессирующих HLA-A2/WT1RMF (с). Согласно более специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула специфически активирует Т-клетки в присутствии клеток, экспрессирующих HLA-A201/WT1RMF (то есть комплекса HLA-A201 молекулы и WT1RMF пептида). Согласно одному варианту осуществления изобретения, активация Т клеток биспецифической антигенсвязывающей молекулой определяют путем измерения, например, с помощью проточной флуометрии, экспрессии CD25 и/или CD69 Т клетками после инкубации с биспецифической антигенсвязывающей молекулой в присутствии HLA-А2/пептид-экспрессирующих клеток (например, WT1RMF или WT1VLD пептид-экспрессирующих клеток), конкретнее, активированных пептидом Т2 клеток.
Согласно специфическому варианту осуществления изобретения, активацию Т клеток биспецифической антигенсвязывающей молекулой определяют следующим образом:
Получают суспензию Т2 клеток (АТСС, Cat. No. CRL-1992) с концентрацией 106 клеток/мл в среде IMDM (Gibco by Life Technologies, Cat No. 31980-048), обогащенной 10% FBS (Gibco, Cat No. 16140-071) +1% Пенициллином-Стрептомицином (Gibco, Cat No. 15070-063) (полная среда). Клетки в общем объеме 10 мл помещают в пробирку и инкубируют с 10 мкл пептида (например, WT1 VLD пептида (SEQ ID NO: 77), или RMF пептида (SEQ ID NO: 78)) при 10-2 М (конечная концентрация пептида: 10-5 М) в течение 2 ч при 37°С с 5% СО2. Пан CD3+ клетки изолируют от мононуклеарных клеток из лейкоцитарной пленки, при помощи градиентного центрифугирования по Ficoll (GE Healthcare, Cat. No. 17-1440-03). Общие CD3+ T клетки очищают при помощи MACS (Miltenyi Biotec) с использованием набора для выделения пан Т-клеток человека (Miltenyi Biotec, Cat. No. 130-096-535). Анализ цитотоксичности проводят следующим образом: активированные пептидом клетки (100 мкл) высевают на 96-луночный планшет для микротитрования с круглым дном (3×105 клеток/мл) и культивируют совместно с 50 мкл Т-клеток (6×106 клеток/мл), и с 50 мкл оттитрованной биспецифической антигенсвязывающей молекулы (например, при концентрациях от 40 мкг/мл до 0.00004 мкг/мл) в полной среде в течение 18 ч при 37°С с 5% СО2. Клетки собирают через 18 часов совместной инкубации и окрашивают антителами к CD3 (Biolegend Cat. No. 300321), CD25 (Biolegend Cat. No. 302606) и CD69 (Biolegend Cat. No. 310914) для измерения T-клеточной активации с помощью проточной цитометрии.
Согласно другому варианту осуществления изобретения, активацию Т клеток биспецифической антигенсвязывающей молекулой определяют с помощью HLA-А2/пептид-экспрессирующих клеток (например, WT1RMF или WT1VLD пептид-экспрессирующих клеток), в частности, активированных пептидом Т2 клеток, а также линии репортерных Т клеток, в частности, Т-клеточной линии Jurkat, которая экспрессирует люциферазный репортер, управляемый ответным элементом NFAT (ядерного фактора активированных Т клеток).
Согласно специфическому варианту осуществления изобретения, активацию Т клеток биспецифической антигенсвязывающей молекулой определяют следующим образом:
Получают суспензию Т2 клеток (АТСС, Cat. No. CRL-1992) с концентрацией 106 клеток/мл в среде IMDM (Gibco by Life Technologies, Cat No. 31980-048), обогащенной 10% FBS (Gibco, Cat No. 16140-071) +1% Пенициллином-Стрептомицином (Gibco, Cat No. 15070-063) (полная среда). Клетки в общем объеме 10 мл помещают в пробирку и инкубируют с 10 мкл пептида (например, WT1 VLD пептида (SEQ ID NO: 77), или RMF пептида (SEQ ID NO: 78)) при 10-2 М (конечная концентрация пептида: 10-5 М) в течение 2 ч при 37°С с 5% СО2.. После отмывания, 90 мкл активированных пептидом клеток в клеточной суспензии с концентрацией 2,2×105 клеток/мл высевают на 96-луночный планшет для микротитрования с круглым дном (20000 клеток на лунку, ТРР, Cat. No. 92097) и совместно культивируют с 50 мкл клеток линии Jurkat, которые экспрессирует люциферазу под действием промотора NFAT (Jurkat-NFAT; Promega, Cat. No. CS176501) (клеточная суспензия 2×106 клеток/мл), и с 10 мкл оттитрованной биспецифической антигенсвязывающей молекулы (например, в концентрации от 100 мкл/мл до 0,0064 мкг/мл в PBS) в течение 16 ч при 37°С с 5% СО2. Затем 50 мкл супернатанта удаляют и замещают реагентом для определения люциферазы Bright-Glo в количестве 100 мкл на лунку (Promega, Cat. No. Е2620) для инкубации при комнатной температуре (RT). Через пять минут, 180 мкл супернатанта переносят на новый белый планшет для измерения сигнала люминесценции с помощью EnVision (PerkinElmer). Данные представлены в относительных единицах люминесценции (RLU).
Согласно одному варианту осуществления изобретения, заявленная биспецифическая антигенсвязывающая молекула существенно не индуцирует опосредованное Т-клетками уничтожение или активацию Т-клеток в присутствии клеток, экспрессирующих только HLA-A2 (т.е. без пептида) или HLA-A2 с пептидом кроме пептида, полученного из WT1, такого как WT1RMF или WT1VLD.
Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула существенно не индуцирует опосредованное Т-клетками уничтожение или активацию Т клеток в присутствии клеток, экспрессирующих HLA-A2 без WT1-выделенного пептида, в частности, WT1RMF или WT1VLD пептида.
Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула индуцирует опосредованное Т клетками уничтожение и/или активирует Т клетки в присутствии клеток, экспрессирующих HLA-A2/WT1 (в частности, HLA-A2/WT1RMF или HLA-A2/WT1VLD), с ЕС50, которая по меньшей мере в 5, по меньшей мере в 10, по меньшей мере в 15, по меньшей мере в 20, по меньшей мере в 25,по меньшей мере в 50, по меньшей мере в 75 или по меньшей мере в 100 раз ниже, чем ЕС50 для индуцирования опосредованного Т клетками уничтожения или активации Т-клеток в присутствии клеток, экспрессирующих HLA-A2, в отсутствие пептида, выделенного из WT1 (в частности, WT1RMF или WT1). Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула индуцирует опосредованное Т-клетками уничтожение и/или активирует Т-клетки в присутствии клеток, экспрессирующих HLA-A2/WT1 (в частности, HLA-A2/WT1RMF или 6HLA-A2/WT1VLD), но не индуцирует существенно опосредованное Т клетками уничтожение, или не активирует Т клетки в присутствии клеток, экспрессирующих HLA-A2 с пептидом, выбранным из пептидов, представленных в Таблице 5 (пептиды с последовательностями SEQ ID NOs 79-105). Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула индуцирует опосредованное Т-клетками уничтожение, и/или активирует Т клетки в присутствии клеток, экспрессирующих HLA-A2/WT1 (в частности, HLA-A2/WT1RMF или HLA-A2/WT1VLD), но не индуцирует существенно опосредованное Т клетками уничтожение, или не активирует Т клетки в присутствии клеток, экспрессирующих HLA-A2 с любым из пептидов, представленных в Таблице 5 (пептиды с последовательностями SEQ ID NOs 79-105).
Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула индуцирует опосредованное Т клеточное уничтожение, и/или активирует Т клетки в присутствии клеток, экспрессирующих HLA-A2/WT1 (а частности, HLA-A2/WT1RMF или HLA-A2/WT1VLD) с ЕС50, которая по меньшей мере в 5, по меньшей мере в 10, по меньшей мере в 15, по меньшей мере в 20, по меньшей мере в 25, по меньшей мере в 50, по меньшей мере в 75 или по меньшей мере в 100 раз ниже, чем ЕС50 для индуцирования опосредованного Т клетками уничтожения или активации Т-клеток в присутствии клеток, экспрессирующих HLA-A2 с пептидом, выбранным из пептидов, представленных в Таблице 5 (пептиды с последовательностями SEQ ID NOs 79-105). Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула индуцирует опосредованное Т клеточное уничтожение, и/или активирует Т клетки в присутствии клеток, экспрессирующих HLA-A2/WT1 (в частности, HLA-A2/WT1RMF или HLA-A2/WT1VLD) с ЕС50, которая по меньшей мере в 5, по меньшей мере в 10, по меньшей мере в 15, по меньшей мере в 20, по меньшей мере в 25, по меньшей мере в 50, по меньшей мере в 75 или по меньшей мере в 100 раз ниже, чем ЕС50 для индуцирования опосредованного Т клетками уничтожения или активации Т-клеток в присутствии клеток, экспрессирующих HLA-A2 с любым из пептидов, выбранных из пептидов, представленных в Таблице 5 (пептиды с последовательностями SEQ ID NOs 79-105). Согласно определенному варианту осуществления изобретения, индуцирование опосредованного Т-клетками уничтожения и/или активации Т клеток определяют как описано выше, а ЕС50 рассчитывают в программе Microsoft® Excel с использованием приложения XLfit® (ID Business Solutions, Guildford, UK).
Согласно определенным вариантам осуществления изобретения, антигенсвязывающие элементы, содержащиеся в биспецифической антигенсвязывающей молекуле, представляют собой молекулы Fab (то есть антигенсвязывающие домены, состоящие из тяжелой и легкой цепи, каждая из которых включает вариабельный и константный домен). Согласно одному варианту осуществления изобретения, первый и/или второй антигенсвязывающий элемент представляет собой Fab молекулу. Согласно одному варианту осуществления изобретения, указанная Fab молекула является гуманизированной. В конкретном варианте осуществления указанная молекула Fab является гуманизированной. Согласно еще одному варианту осуществления изобретения, указанная Fab молекула содержит константные домены тяжелой и легкой цепи человека.
Предпочтительно по меньшей мере одна антигенсвязывающая молекула представляет собой кроссоверную Fab молекулу. Такая модификация уменьшает нарушения спаривания тяжелых и легких цепей из разных Fab молекул, что позволяет улучшить выход и степень очистки биспецифической антигенсвязывающей молекулы согласно изобретению в процессе рекомбинантного синтеза.
В наиболее полезной кроссоверной Fab молекуле в составе биспецифической антигенсвязывающей молекулы, согласно изобретению, вариабельные домены Fab легкой цепи и Fab тяжелой цепи (VL и VH, соответственно), поменяны местами. Однако даже при таком обмене доменами получение биспецифической антигенсвязывающей молекулы может сопровождаться образованием определенных побочных продуктов вследствие так называемого взаимодействия типа Бенса-Джонса между неправильно спаренными тяжелыми и легкими цепями (см. Schaefer et al, PNAS, 108 (2011) 11187-11191).
Для того чтобы дополнительно уменьшить неправильное спаривание тяжелых и легких цепей из разных молекул Fab и, таким образом, увеличить чистоту и выход желаемой биспецифической антигенсвязывающей молекулы, заряженные аминокислоты с противоположными зарядами могут быть встроены в определенные положения в доменах СН1 и CL, как в составе молекул(ы) Fab, связывающейся с первым антигеном (HLA-A2/WT1), так и в составе молекулы Fab, связывающейся со вторым антигеном (например, активирующим Т-клеточным антигеном, таким как CD3), как дополнительно описано в настоящем документе.
Модификации заряда могут производиться как в обычной молекуле(ах) Fab в составе биспецифической антигенсвязывающей молекулы (такой как показано, например, на Фиг. 1 AC, GJ), так и в VH/VL доменах кроссоверной молекулы(ах) Fab в составе биспецифической антигенсвязывающей молекулы (такой, как показано, например, на Фиг. 1 DF, KN) (но не в обоих сразу). Согласно определенным вариантам осуществления изобретения, модификации заряда осуществляются в обычной молекуле(ах) Fab, в составе биспецифической антигенсвязывающей молекулы (которая согласно определенным вариантам осуществления изобретения связывается с первым антигеном, т.е. HLA-A2/WT1).
Согласно определенному варианту осуществления изобретения, заявленная биспецифическая антигенсвязывающая молекула способна одновременно связываться с первым антигеном (то есть HLA-A2/WT1) и вторым антигеном (например, с активирующим Т-клеточным антигеном, в частности CD3). Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула способна перекрестно связывать Т-клетку и клетку-мишень путем одновременного связывания HLA-A2/WT1 и Т-клеточного активирующего антигена. Согласно дополнительному конкретному варианту осуществления изобретения, такое одновременное связывание приводит к лизису клетки-мишени, в частности опухолевой клетки, экспрессирующей HLA-A2/WT1. Согласно одному варианту осуществления изобретения, такое одновременное связывание приводит к активации Т-клеток. Согласно другому варианту осуществления изобретения, такое одновременное связывание приводит к Т-лимфоцитарному клеточному ответу, в частности, ответу цитотоксического Т-лимфоцита, при этом указанный ответ выбран из группы: пролиферации, дифференцировки, секреции цитокинов, высвобождения цитотоксической эффекторной молекулы, цитотоксической активности и экспрессии маркеров активации.
Согласно одному варианту осуществления изобретения, связывание биспецифической антигенсвязывающей молекулы с Т-клеточным активирующим антигеном, в частности CD3, без одновременного связывания с HLA-A2/WT1 не приводит к активации Т-клеток.
Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула способна перенаправлять цитотоксическую активность Т-клетки на клетку-мишень. Согласно конкретному варианту осуществления изобретения, указанное перенаправление не зависит от МНС-опосредованной презентации пептидного антигена клеткой-мишенью и/или специфичности Т-клетки.
В частности, Т-клетка в соответствии с любым из вариантов осуществления изобретения представляет собой цитотоксическую Т-клетку. В некоторых вариантах осуществления изобретения, Т-клетка представляет собой CD4+ или CD8+ Т-клетку, в частности CD8+ Т-клетку.
Первая антигенсвязывающая молекула
Биспецифическая антигенсвязывающая молекула, заявленная в соответствии с настоящим изобретением, включает по меньшей мере один антигенсвязывающий фрагмент, в частности, молекулу Fab, которая связывается с HLA-A2/WT1 (первый антиген). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула включает две антигенсвязывающие молекулы, в частности молекулы Fab, которые связываются с HLA-A2/WT1. Согласно конкретному варианту осуществления изобретения, каждая из указанных антигенсвязывающих молекул связывается с одной и той же антигенной детерминантой. Согласно такому конкретному варианту осуществления изобретения, все указанные антигенсвязывающие молекулы идентичны, то есть они имеют одинаковые аминокислотные последовательности, содержащие такие же аминокислотные замены в доменах СН1 и CL, как описано в настоящем документе (если таковые имеются). Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает не более двух антигенсвязывающих молекул, в частности молекул Fab, которые связываются с HLA-A2/WT1.
Согласно определенным вариантам осуществления изобретения, антигенсвязывающая молекула(ы), которые связываются с HLA-A2/WT1 представляют собой традиционные Fab молекулы. Согласно этому варианту осуществления изобретения, антигенсвязывающая молекула(ы), которые связываются со вторым антигеном представляют собой кроссоверную Fab молекулу, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены СН1 и CL Fab тяжелых или легких цепей поменяны местами/замещены друг другом.
Согласно альтернативным вариантам осуществления изобретения, антигенсвязывающая молекула(ы), которые связываются с HLA-A2/WT1 представляют собой кроссоверную Fab молекулу, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены СН1 и CL Fab тяжелых или легких цепей поменяны местами/замещены друг другом. Согласно этому варианту осуществления изобретения, антигенсвязывающая молекула(ы), которые связываются со вторым антигеном представляют собой традиционную Fab молекулу.
HLA-A2/WT1-связывающая молекула способна направлять биспецифическую антигенсвязывающую молекулу к месту-мишени, например, к определенному типу опухолевых клеток, которые экспрессируют HLA-A2/WT1.
Первая антигенсвязывающая молекула в составе биспецифической антигенсвязывающей молекулы может иметь любые характеристики по отдельности или в комбинации, описанные здесь в отношении антитела, связывающего HLA-A2/WT1.
Таким образом, согласно одному своему аспекту, настоящее если только это не является неразумным или невозможным с научной точки зрения. Изобретение относится к биспецифической антигенсвязывающей молекуле, включающей (а) первую антигенсвязывающую молекулу, которая связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, а первая антигенсвязывающая молекула включает:
(i) вариабельную область тяжелой цепи (VH), содержащую область, определяющую комплементарность тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2, и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область, определяющую комплементарность легкой цепи (LCDR) 1 последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 с последовательностью SEQ ID NO: 6;
(ii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 9, HCDR 2 с последовательностью SEQ ID NO: 10, и HCDR 3 с последовательностью SEQ ID NO: 11, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 12, LCDR 2 с последовательностью SEQ ID NO: 13 и LCDR 3 с последовательностью SEQ ID NO: 14;
(iii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 17, HCDR 2 с последовательностью SEQ ID NO: 18, и HCDR 3 с последовательностью SEQ ID NO: 19, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 20, LCDR 2 с последовательностью SEQ ID NO: 21 и LCDR 3 с последовательностью SEQ ID NO: 22;
(iv) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 25, a HCDR 2 с последовательностью SEQ ID NO: 26, и HCDR 3 с последовательностью SEQ ID NO: 27, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 28, LCDR 2 с последовательностью SEQ ID NO: 29 и LCDR 3 с последовательностью SEQ ID NO: 30;
(v) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 33, и HCDR 2 с последовательностью SEQ ID NO: 34, и HCDR 3 с последовательностью SEQ ID NO: 35, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 36, LCDR 2 с последовательностью SEQ ID NO: 37 и LCDR 3 с последовательностью SEQ ID NO: 38;
(vi) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 41, HCDR 2 с последовательностью SEQ ID NO: 42, и HCDR 3 с последовательностью SEQ ID NO: 43, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 44, LCDR 2 с последовательностью SEQ ID NO: 45 и LCDR 3 с последовательностью SEQ ID NO: 46;
(vii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 49, HCDR 2 с последовательностью SEQ ID NO: 50, и HCDR 3 с последовательностью SEQ ID NO: 51, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 52, LCDR 2 с последовательностью SEQ ID NO: 53, и LCDR 3 с последовательностью SEQ ID NO: 54;
(viii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 57, HCDR 2 с последовательностью SEQ ID NO: 58, и HCDR 3 с последовательностью SEQ ID NO: 59, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 60, LCDR 2 с последовательностью SEQ ID NO: 61 и LCDR 3 с последовательностью SEQ ID NO: 62; или
(ix) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 65, HCDR 2 с последовательностью SEQ ID NO: 66, и HCDR 3 с последовательностью SEQ ID NO: 67, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 68, LCDR 2 с последовательностью SEQ ID NO: 69 и LCDR 3 с последовательностью SEQ ID NO: 70.
(b) вторую антигенсвязывающую молекулу, которая связывается со вторым антигеном.
Согласно определенному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую HCDR 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, и VL, содержащую LCDR 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6.
Согласно другому варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую HCDR 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, и VL, содержащую LCDR 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14.
Согласно другому варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую HCDR 1 с SEQ ID NO: 17, HCDR 2 с SEQ ID NO: 18, и HCDR 3 с SEQ ID NO: 19, и VL, содержащую LCDR 1 с SEQ ID NO: 20, LCDR 2 с SEQ ID NO: 21 и LCDR 3 с SEQ ID NO: 22.
Согласно еще одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую HCDR 1 с SEQ ID NO: 25, HCDR 2 с SEQ ID NO: 26, и HCDR 3 с SEQ ID NO: 27, и VL, содержащую LCDR 1 с SEQ ID NO: 28, LCDR 2 с SEQ ID NO: 29 и LCDR 3 с SEQ ID NO: 30.
Согласно некоторым вариантам осуществления изобретения, первая антигенсвязывающая молекула представляет собой (выделена из) человеческое антитело. Согласно одному варианту осуществления изобретения, VH представляет собой человеческий VH и/или VL представляет собой человеческий VL. Согласно одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает CDRs согласно любому из описанных выше вариантов осуществления изобретения, и дополнительно имеет человеческий каркас, например, каркас человеческого иммуноглобулина.
Согласно одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64; или
(ix) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24.
Согласно дополнительному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает:
(i) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vii) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(viii) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64;
(ix) VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, а также VL, последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24.
Согласно дополнительному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает:
(i) VH, содержащую аминокислотную последовательность SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность по SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность по SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 24;
(vi) VH, содержащую аминокислотную последовательность по SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность по SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность по SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность по SEQ ID NO: 55, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность по SEQ ID NO: 63, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 64;
(ix) VH, содержащую аминокислотную последовательность по SEQ ID NO: 71, и VL, содержащую аминокислотную последовательность по SEQ ID NO: 72.
Согласно одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает:
(i) VH последовательность SEQ ID NO: 7, и VL последовательность SEQ ID NO: 8;
(ii) VH последовательность SEQ ID NO: 15, и VL последовательность SEQ ID NO: 16;
(iii) VH последовательность SEQ ID NO: 23, и VL последовательность SEQ ID NO: 24;
(iv) VH последовательность SEQ ID NO: 31, и VL последовательность SEQ ID NO: 32;
(v) VH последовательность SEQ ID NO: 39, и VL последовательность SEQ ID NO: 40;
(vi) VH последовательность SEQ ID NO: 47, и VL последовательность SEQ ID NO: 48;
(vii) VH последовательность SEQ ID NO: 55, и VL последовательность SEQ ID NO: 56;
(viii) VH последовательность SEQ ID NO: 63, и VL последовательность SEQ ID NO: 64; или
(ix) VH последовательность SEQ ID NO: 71, и VL последовательность SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность SEQ ID NO:7, а также VL, содержащую аминокислотную последовательность SEQ ID NO:8. Согласно другому варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность SEQ ID NO:15, а также VL, содержащую аминокислотную последовательность SEQ ID NO:16. Согласно еще одному варианту осуществления изобретения первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность SEQ ID NO:23, а также VL, содержащую аминокислотную последовательность SEQ ID NO:24. Согласно еще одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность SEQ ID NO:31, а также VL, содержащую аминокислотную последовательность SEQ ID NO:32.
Согласно определенному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность SEQ ID NO: 7, а также VL последовательность SEQ ID NO: 8. Согласно другому варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность SEQ ID NO: 15, а также VL последовательность SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность SEQ ID NO: 23, а также VL последовательность SEQ ID NO: 24. Согласно еще одному варианту осуществления изобретения, первая антигенсвязывающая молекула включает VH последовательность SEQ ID NO: 31, а также VL последовательность SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, первая антигенсвязывающая молекула содержит константную область человека. Согласно одному варианту осуществления изобретения, первая антигенсвязывающая молекула представляет собой Fab молекулу, содержащую константную область человека, а конкретно, СН1 и/или CL домен человека. Примерные последовательности константных доменов человека представлены SEQ ID NOs 112 и 113 (каппа и лямбда CL домены человека, соответственно) и SEQ ID NO: 114 (константные домены СН1-СН2-СН3 тяжелой цепи IgG1 человека). Согласно некоторым вариантам осуществления изобретения, первая антигенсвязывающая молекула включает константную область легкой цепи, имеющую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 112 или SEQ ID NO: 113, в частности, аминокислотной последовательности SEQ ID NO: 112. В частности, константная область легкой цепи может включать аминокислотные мутации, как описано в настоящем документе в разделе "модификации заряда" и/или могут включать делеции или замены одной или более (конкретнее, двух) N-концевых аминокислот, если они находятся в кроссоверной молекуле Fab. Согласно некоторым вариантам осуществления изобретения, первая антигенсвязывающая молекула включает константную область тяжелой цепи, имеющую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности СН1 домена, содержащейся в SEQ ID NO: 114. В частности, константная область тяжелой цепи (конкретно, СН1 домен) может включать аминокислотные мутации, как описано в настоящем документе в разделе "модификации заряда".
Вторая антигенсвязывающая молекула
Биспецифическая антигенсвязывающая молекула, заявленная в соответствии с настоящими изобретением, включает по меньшей мере одну антигенсвязывающую молекулу, в частности, Fab молекулу, который связывается со вторым антигеном (отличным от HLA-A2/WT1).
Согласно определенным вариантам осуществления изобретения, антигенсвязывающий фрагмент, который связывается со вторым антигеном, представляет собой кроссоверную Fab молекулу, как описано в настоящем документе, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены СН1 и CL Fab тяжелых и легких цепей поменяны местами/замещены друг другом. Согласно таким вариантам осуществления изобретения, антигенсвязывающий фрагмент(ы), который связывается с первым антигеном (то есть HLA-A2/WT1), предпочтительно, представляет собой традиционную Fab молекулу. В тех вариантах осуществления изобретения, где речь идет более чем об одном антигенсвязывающем фрагменте, конкретно, о Fab молекуле, которая связывается с HLA-A2/WT1 в составе биспецифической антигенсвязывающей молекулы, антигенсвязывающий фрагмент, который связывается со вторым антигеном, предпочтительно, представляет собой кроссоверную Fab молекулу, а антигенсвязывающие фрагменты, которые связываются с HLA-A2/WT1, представляют собой традиционные Fab молекулы.
Согласно альтернативным вариантам осуществления изобретения, антигенсвязывающий фрагмент, который связывается со вторым антигеном, представляет собой традиционную Fab молекулу. В таком варианте осуществления изобретения антигенсвязывающий фрагмент(ы), который связывается с первым антигеном (то есть, HLA-A2/WT1) представляет собой кроссоверную Fab молекулу, как описано в настоящем документе, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены СН1 и CL Fab тяжелых и легких цепей поменяны местами/замещены друг другом. В тех вариантах осуществления изобретения, где речь идет более чем об одном антигенсвязывающем фрагменте, конкретно, о Fab молекуле, которая связывается со вторым антигеном в составе биспецифической антигенсвязывающей молекулы, антигенсвязывающий фрагмент, который связывается с HLA-A2/WT1, предпочтительно, представляет собой кроссоверную Fab молекулу, а антигенсвязывающие фрагменты, которые связываются со вторым антигеном, представляют собой традиционные Fab молекулы.
Согласно некоторым вариантам осуществления изобретения, второй антиген представляет собой активирующий Т-клеточный антиген (также обозначаемый в настоящем документе как "антигенсвязывающая молекула, активирующая Т-клеточный антиген" или "Fab молекула, связывающая активирующий Т-клеточный антиген"). Согласно определенному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает не более одной антигенсвязывающей молекулы, способной специфически связываться с активирующим Т-клеточным антигеном. Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула моновалентно связывается с активирующим Т-клеточным антигеном.
Согласно определенным вариантам осуществления изобретения, второй антиген представляет собой CD3, в частности, человеческий CD3 (SEQ ID NO: 107) или CD3 яванской макаки (SEQ ID NO: 108), конкретнее, CD3 человека. Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула является перекрестно-реактивной (то есть специфически связывается) как для CD3 человека, так и для CD3 яванской макаки. Согласно некоторым вариантам осуществления изобретения, второй антиген представляет собой субъединицу ипсилон CD3 (CD3 ипсилон).
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает HCDR 1 с последовательностью SEQ ID NO: 115, а также HCDR 2 с последовательностью SEQ ID NO: 116, а также HCDR 3 с последовательностью SEQ ID NO: 117, и LCDR 1 с последовательностью SEQ ID NO: 118, а также LCDR 2 с последовательностью SEQ ID NO: 119 и LCDR 3 с последовательностью SEQ ID NO: 120.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 115, HCDR 2 с последовательностью SEQ ID NO: 116, и HCDR 3 с последовательностью SEQ ID NO: 117, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 118, LCDR 2 с последовательностью SEQ ID NO: 119. а также LCDR 3 с последовательностью SEQ ID NO: 120.
Согласно некоторым вариантам осуществления изобретения, вторая антигенсвязывающая молекула представляет собой гуманизированное антитело (или выделена из него). Согласно одному варианту осуществления изобретения, VH представляет собой гуманизированную VH и/или VL представляет собой гуманизированную VL. Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает CDR области по любому из указанных выше вариантов изобретения, а также дополнительно включает акцепторный каркас человека, например, каркас иммуноглобулина человека или консенсунсный каркас человека.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 121.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VL последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 122.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 121, а также VL последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 122.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность SEQ ID NO: 121, а также VL, содержащую аминокислотную последовательность SEQ ID NO: 122.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH последовательность SEQ ID NO: 121, и VL последовательность SEQ ID NO: 122.
Согласно определенному варианту осуществления изобретения, вторая антигенсвязывающая молекула включат HCDR1 с последовательностью SEQ ID NO: 130, HCDR 2 с последовательностью SEQ ID NO: 131, HCDR 3 с последовательностью SEQ ID NO: 132, LCDR 1 с последовательностью SEQ ID NO: 133, LCDR 2 с последовательностью SEQ ID NO: 134 и LCDR 3 с последовательностью SEQ ID NO: 135.
Согласно другому определенному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 130, HCDR 2 с последовательностью SEQ ID NO: 131, и HCDR 3 с последовательностью SEQ ID NO: 132, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 133, LCDR 2 с последовательностью SEQ ID NO: 134 и LCDR 3 с последовательностью SEQ ID NO: 135.
Согласно некоторым вариантам осуществления изобретения, вторая антигенсвязывающая молекула представляет собой гуманизированное антитело (или выделена из него). Согласно одному варианту осуществления изобретения, VH представляет собой гуманизированную VH и/или VL представляет собой гуманизированную VL. Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает CDR области по любому из указанных выше вариантов изобретения, а также дополнительно включает акцепторный каркас человека, например, каркас иммуноглобулина человека или консенсунсный каркас человека.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 136. Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VL последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 137.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 136, а также VL последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 137.
Согласно другому определенному варианту осуществления изобретения, VH второй антигенсвязывающей молекулы содержит аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 136, a VL второй антигенсвязывающей молекулы содержит аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 137.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH, содержащую аминокислотную последовательность SEQ ID NO: 136, и VL, содержащую аминокислотную последовательность SEQ ID NO: 137.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает VH последовательность SEQ ID NO: 136, и VL последовательность SEQ ID NO: 137.
Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула включает константную область человеческой природы. Согласно одному варианту осуществления изобретения, вторая антигенсвязывающая молекула представляет собой Fab молекулу, содержащую константную область человека, в частности, СН1 и/или CL домен человека. Примерные последовательности константных доменов человека представлены SEQ ID NOs 112 и 113 (каппа и лямбда CL домены, соответственно) и SEQ ID NO: 114 (константные домены СН1-СН2-СН3 тяжелой цепи IgG1 человека). Согласно некоторым вариантам осуществления изобретения, первая антигенсвязывающая молекула включает константную область легкой цепи, имеющую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 112 или SEQ ID NO: 113, в частности, аминокислотной последовательности SEQ ID NO: 112. В частности, константная область легкой цепи может включать аминокислотные мутации, как описано в настоящем документе в разделе "модификации заряда" и/или могут включать делеции или замены одной или более (конкретнее, двух) N-концевых аминокислот, если они находятся в кроссоверной молекуле Fab. Согласно некоторым вариантам осуществления изобретения, первая антигенсвязывающая молекула включает константную область тяжелой цепи, имеющую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности СН1 домена, содержащейся в SEQ ID NO: 114. В частности, константная область тяжелой цепи (конкретно, СН1 домен) может включать аминокислотные мутации, как описано в настоящем документе в разделе "модификации заряда". Согласно некоторым вариантам осуществления изобретения, вторая антигенсвязывающая молекула представляет собой Fab молекулу, при этом вариабельные домены VL и VH или константные домены CL и СН1, в частности, вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab, замещены друг другом (то есть, согласно этому варианту осуществления изобретения, вторая антигенсвязывающая молекула представляет собой кроссоверную Fab молекулу, в которой вариабельные или константные домены Fab легкой цепи и Fab тяжелой цепи поменяны местами). Согласно одному такому варианту осуществления изобретения, первая (а также третья, в случае ее наличия) антигенсвязывающая молекула представляет собой традиционную Fab молекулу. Согласно одному варианту осуществления изобретения, в биспецифической антигенсвязывающей молекуле присутствует не более одной антигенсвязывающей молекулы, которая связывается со вторым антигеном (то есть активирующим Т-клеточным антигеном, таким, как CD3), то есть биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание со вторым антигеном.
Модификации заряда
Биспецифическая антигенсвязывающая молекула, заявленная в соответствии с настоящим изобретением, может содержать аминокислотные замены в Fab молекулах, которые особенно эффективны для уменьшения количества нарушений спаривания легких цепей с обычными тяжелыми цепями (побочных продуктов Бенс-Джонса), которые могут происходить во время получения би-/мультиспецифических антигенсвязывающих молекул на основе Fab, содержащих VH/VL замены в одном (или более, в случае молекул, содержащих более двух антигенсвязывающих Fab молекул) связывающем плече (см. также публикацию РСТ номер WO 2015/150447, в частности, раздел "Примеры", которая включена в настоящее описание в качестве ссылки во всей свой полноте). Соотношение между желаемой биспецифической антигенсвязывающей молекулой и нежелательными побочными продуктами, в частности, побочными продуктами типа Бенс-Джонса, которые встречаются в составе биспецифических антигенсвязывающих молекул, содержащих замены VH/VL доменов в одном из связывающих плеч, может быть улучшено путем встраивания заряженных аминокислот с противоположными зарядами в специфические аминокислотные положения в СН1 и CL доменах (указанная модификация иногда обозначается термином "модификации заряда").
Соответственно, согласно некоторым вариантам осуществления изобретения, когда первый и второй антигенсвязывающий фрагмент в составе биспецифической антигенсвязывающей молекулы оба представляют собой Fab молекулы, при этом в одном из антигенсвязывающих фрагментов (в частности, во втором антигенсвязывающем фрагменте) вариабельные домены VL и VH Fab легкой цепи и Fab тяжелой замещены друг другом,
i) в константном домене CL первого антигенсвязывающего фрагмента аминокислота в положении 124 замещена положительно заряженной аминокислотой (нумерация по Кабату), а в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 или аминокислота в положении 213 замещена отрицательно заряженной аминокислотой (нумерация согласно EU индексу по Кабату); или
ii) в константном домене CL второго антигенсвязывающего фрагмента аминокислота в положении 124 замещена положительно заряженной аминокислотой (нумерация по Кабату), при этом в константном домене СН1 второго антигенсвязывающего фрагмента аминокислота в положении 147 или аминокислота в положении 213 замещена отрицательно заряженной аминокислотой (нумерация согласно EU индексу по Кабату).
Биспецифическая антигенсвязывающая молекула не содержит одновременно сразу две модификации, упомянутые в пунктах 1) и 2). Константные домены CL и СН1 антигенсвязывающего фрагмента, включающие VH/VL замены, не заменяются друг на друга (то есть остаются неизменными).
Согласно более специфическому варианту осуществления изобретения,
i) в константном домене CL первого антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату), а в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 или аминокислота в положении 213 независимо замещены глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу по Кабату); или
ii) в константном домене CL второго антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату), а в константном домене СН1 второго антигенсвязывающего фрагмента аминокислота в положении 147 или аминокислота в положении 213 независимо замещены глутаминовой кислотой (Е) или аспарагиновой кислотой (D) нумерация согласно EU индексу по Кабату.
Согласно одному такому варианту осуществления изобретения, в константном домене CL первого антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату), а в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 или аминокислота в положении 213 независимо замещены глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно индексу EU Кабата). Согласно дополнительному варианту осуществления изобретения, в константном домене CL первого антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату) и в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно индексу EU Кабата).
Согласно конкретному варианту осуществления изобретения, в константном домене CL первого антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату) и аминокислота в положении 123 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату), а в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно индексу EU Кабата) и аминокислота в положении 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно индексу EU Кабата).
Согласно еще более конкретному варианту осуществления изобретения, в константном домене CL первого антигенсвязывающего фрагмента аминокислота в положении 124 замещена лизином (K) (нумерация по Кабату) и аминокислота в положении 123 замещена аргинином (R) (нумерация по Кабату), а в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата) и аминокислота в положении 123 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата).
Согласно определенным вариантам осуществления изобретения, в том случае если аминокислотные замены по любому из указанных выше вариантов осуществления изобретения, производятся в константном домене CL и константном домене СН1 первого антигенсвязывающего фрагмента, константный домен CL первого антигенсвязывающего фрагмента имеет изотип каппа. Альтернативно, аминокислотные замены по любому из указанных выше вариантов осуществления изобретения, могут производиться в константном домене CL и константном домене СН1 второго антигенсвязывающего фрагмента вместо константного домена CL и константного домена СН1 второго антигенсвязывающего фрагмента. Согласно такому варианту осуществления изобретения, константный домен CL второго антигенсвязывающего фрагмента имеет изотип каппа.
Соответственно, согласно одному варианту осуществления изобретения, в константном домене CL второго антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату), а в константном домене СН1 второго антигенсвязывающего фрагмента, аминокислота в положении 147 или аминокислота в положении 213, независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата).
Согласно дополнительному варианту осуществления изобретения, в константном домене CL второго антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату), а в константном домене СН1 второго антигенсвязывающего фрагмента аминокислота в положении 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата).
Согласно еще одному варианту осуществления изобретения, в константном домене CL второго антигенсвязывающего фрагмента аминокислота в положении 124 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату) и аминокислота в положении 123 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату), а в константном домене СН1 второго антигенсвязывающего фрагмента аминокислота в положении 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата).
Согласно одному варианту осуществления изобретения, в константном домене CL второго антигенсвязывающего фрагмента аминокислота в положении 124 замещена лизином (K) (нумерация по Кабату) и аминокислота в положении 123 заменена лизином (K) (нумерация согласно Kabat), а в константном домене СН1 второго антигенсвязывающего фрагмента аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация по индексу Kabat EU) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом Kabat EU).
Согласно другому варианту осуществления изобретения, в константном домене CL второго антигенсвязывающего фрагмента аминокислота в положении 124 замещена лизином (K) (нумерация по Кабату) и аминокислота в положении 123 заменена аргинином (R) (нумерация согласно Kabat), а в константном домене СН1 второго антигенсвязывающего фрагмента аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация по индексу Kabat EU) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом Kabat EU).
Согласно определенному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула, согласно изобретению, включает:
(а) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2 / WT1, в частности HLA-A2 / WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область, определяющую комплементарность тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2 и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 с последовательностью SEQ ID NO: 6, и
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab поменяны местами/замещены друг другом; при этом в константном домене CL первого антигенсвязывающего фрагмента, аминокислота в положении 124, независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату) (в конкретном варианте осуществления изобретения независимо лизином (K)) или аргинином (R)) и аминокислота в положении 123 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату) (в конкретном варианте осуществления независимо лизином (K) или аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена независимо глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата).
Согласно другому определенному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула, согласно изобретению, включает:
(а) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2 / WT1, в частности HLA-A2 / WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащий вариабельную область тяжелой цепи (VH), содержащую область, определяющую комплементарность тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 9, HCDR 2 с последовательностью SEQ ID NO: 11 и HCDR 3 с последовательностью SEQ ID NO: 12, а также вариабельную область легкой цепи (VL), включающую область определяющую комплементарность легкой цепи (LCDR) 1 с последовательностью SEQ ID NO: 12, LCDR 2 с последовательностью SEQ ID NO: 13 и LCDR 3 с последовательностью SEQ ID NO: 14, и
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в котором вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab поменяны местами/замещены друг другом; при этом при этом в константном домене CL первого антигенсвязывающего фрагмента, аминокислота в положении 124, независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату) (в конкретном варианте осуществления изобретения независимо лизином (K)) или аргинином (R)) и аминокислота в положении 123 независимо замещена лизином (K), аргинином (R) или гистидином (Н) (нумерация по Кабату) (в конкретном варианте осуществления независимо лизином (K) или аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента аминокислота в положении 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена независимо глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно EU индексу Кабата).
Форматы биспецифической антигенсвязывающей молекулы
Компоненты биспецифической антигенсвязывающей молекулы, заявленной в соответствии с настоящим изобретением, могут быть соединены между собой в различных конфигурациях. Примерные конфигурации представлены на Фигуре 1.
Согласно определенным вариантам осуществления изобретения, антигенсвязывающие фрагменты в составе биспецифической антигенсвязывающей молекулы представляют собой Fab молекулы. Согласно таким вариантам осуществления изобретения, первый, второй, третий и т.д. антигенсвязывающий фрагмент могут обозначаться в настоящем документе как первая, вторая, третья и т.д. Fab молекула, соответственно.
Согласно одному варианту осуществления изобретения, первый и второй антигенсвязывающие фрагменты в составе биспецифической антигенсвязывающей молекулы соединены между собой, необязательно, посредством пептидного линкера. Согласно определенному варианту осуществления изобретения первый и второй антигенсвязывающие фрагменты каждый представляет собой Fab молекулу.
Согласно одному варианту осуществления изобретения, первая и вторая антигенсвязывающая молекулы соединены между собой, необязательно, посредством пептидного линкера. Согласно одному такому варианту осуществления изобретения, второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи первого антигенсвязывающего фрагмента.
Согласно другому такому варианту осуществления изобретения, при этом первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи второго антигенсвязывающего фрагмента. В вариантах осуществления изобретения, согласно которым либо (i) второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи первого антигенсвязывающего фрагмента, либо (ii) первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи второго антигенсвязывающего фрагмента, легкая цепь Fab первого антигенсвязывающего фрагмента и легкая цепь Fab второго антигенсвязывающего фрагмента могут быть соединены между собой необязательно посредством пептидного линкера.
Биспецифическая антигенсвязывающая молекула с одним антигенсвязывающим фрагментом (таким, как Fab молекула), способным специфически связываться с таргетным клеточным антигеном, таким, как HLA-A2/WT1 (например, как показано на Фигурах 1А, D, G, Н, K, L) может быть полезна в особенности в случаях, когда после высокоаффинного связывания с антигенсвязывающим фрагментом ожидается интернализация таргетного клеточного антигена. В таких случаях, наличие более чем одного антигенсвязывающего фрагмента, специфичного для таргетного клеточного антигена может усиливать интернализацию таргетного клеточного антигена, что снижает его доступность. В других случаях, однако, может оказаться преимущественным наличие биспецифической антигенсвязывающей молекулы, включающей два или более антигенсвязывающих фрагмента (например, Fab молекулы), специфических по отношению к таргетному клеточному антигену (см, примеры, представленные на Фигурах 1В, 1С, 1E, 1F, 1, 1J, 1М или 1N), например, для оптимизации направленной доставки молекулы к сайту-мишени или для обеспечения перекрестного связывания с таргетными клеточными антигенами.
Соответственно, в определенном варианте осуществления изобретения, биспецифическая антигенсвязывающая молекула, согласно изобретению, включат третий антигенсвязывающий фрагмент. Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент связывается с первым антигеном, то есть HLA-A2/WT1. Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент представляет собой Fab молекулу.
Согласно определенному варианту осуществления изобретения, третий антигенсвязывающий фрагмент идентичен первому антигенсвязывающему фрагменту. Третий антигенсвязывающий фрагмент в составе биспецифической антигенсвязывающей молекулы может обладать любыми характеристиками, отдельно или в комбинации, описанными в настоящем документе по отношению к первому антигенсвязывающему фрагменту и/или антителу, которое связывается с HLA-A2/WT1, если только это не является неразумным или невозможным с научной точки зрения.
Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает:
(i) вариабельную область тяжелой цепи (VH), содержащую область, определяющую комплементарность тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2, и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область, определяющую комплементарность легкой цепи (LCDR) 1 последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 с последовательностью SEQ ID NO: 6;
(ii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 9, HCDR 2 с последовательностью SEQ ID NO: 10, и HCDR 3 с последовательностью SEQ ID NO: 11, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 12, LCDR 2 с последовательностью SEQ ID NO: 13 и LCDR 3 с последовательностью SEQ ID NO: 14;
(iii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 17, HCDR 2 с последовательностью SEQ ID NO: 18, и HCDR 3 с последовательностью SEQ ID NO: 19, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 20, LCDR 2 с последовательностью SEQ ID NO: 21 и LCDR 3 с последовательностью SEQ ID NO: 22;
(iv) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 25, a HCDR 2 с последовательностью SEQ ID NO: 26, и HCDR 3 с последовательностью SEQ ID NO: 27, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 28, LCDR 2 с последовательностью SEQ ID NO: 29 и LCDR 3 с последовательностью SEQ ID NO: 30;
(v) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 33, и HCDR 2 с последовательностью SEQ ID NO: 34, и HCDR 3 с последовательностью SEQ ID NO: 35, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 36, LCDR 2 с последовательностью SEQ ID NO: 37 и LCDR 3 с последовательностью SEQ ID NO: 38;
(vi) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 41, HCDR 2 с последовательностью SEQ ID NO: 42, и HCDR 3 с последовательностью SEQ ID NO: 43, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 44, LCDR 2 с последовательностью SEQ ID NO: 45 и LCDR 3 с последовательностью SEQ ID NO: 46;
(vii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 49, HCDR 2 с последовательностью SEQ ID NO: 50, и HCDR 3 с последовательностью SEQ ID NO: 51, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 52, LCDR 2 с последовательностью SEQ ID NO: 53, и LCDR 3 с последовательностью SEQ ID NO: 54;
(viii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 57, HCDR 2 с последовательностью SEQ ID NO: 58, и HCDR 3 с последовательностью SEQ ID NO: 59, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 60, LCDR 2 с последовательностью SEQ ID NO: 61 и LCDR 3 с последовательностью SEQ ID NO: 62; или
(ix) VH, содержащую HCDR 1 с последовательностью SEQ ID NO. 65, HCDR 2 с последовательностью SEQ ID NO: 66, и HCDR 3 с последовательностью SEQ ID NO: 67, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 68, LCDR 2 с последовательностью SEQ ID NO: 69 и LCDR 3 с последовательностью SEQ ID NO: 70.
Согласно определенному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2, а также HCDR 3 с последовательностью SEQ ID NO: 3, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5, а также LCDR 3 с последовательностью SEQ ID NO: 6.
Согласно другому варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 9, HCDR 2 с последовательностью SEQ ID NO: 10, а также HCDR 3 с последовательностью SEQ ID NO: 11, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 12, LCDR 2 с последовательностью SEQ ID NO: 13, а также LCDR 3 с последовательностью SEQ ID NO: 14.
Согласно другому варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 17, HCDR 2 с последовательностью SEQ ID NO: 18, а также HCDR 3 с последовательностью SEQ ID NO: 19, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 20, LCDR 2 с последовательностью SEQ ID NO: 21, а также LCDR 3 с последовательностью SEQ ID NO: 22.
Согласно еще одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 25, HCDR 2 с последовательностью SEQ ID NO: 26, а также HCDR 3 с последовательностью SEQ ID NO: 27, и VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 28, LCDR 2 с последовательностью SEQ ID NO: 29, а также LCDR 3 с последовательностью SEQ ID NO: 30.
Согласно некоторым вариантам осуществления изобретения третий антигенсвязывающий фрагмент представляет собой человеческое антитело (или выделен из него). Согласно одному варианту осуществления изобретения VH представляет собой VH человека и/или VL представляет собой VL человека. Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает CDR, согласно любому из описанных выше вариантов осуществления изобретения, и дополнительно включает человеческий каркас, например, каркас иммуноглобулина человека.
Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64; или
(ix) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, и VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 5%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 5%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 5%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24.
Согласно еще одному дополнительному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность, которая по меньшей мере приблизительно, на 5%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16;
(iii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 39, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 47, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 48;
(vi) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 55, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 56;
(vii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 63, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 64;
(ix) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 71, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 15, а также VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 23, а также VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 24.
Согласно еще одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 31, а также VL последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает:
(i) VH, содержащую аминокислотную последовательность SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность SEQ ID NO: 8;
(ii) VH, содержащую аминокислотную последовательность SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность SEQ ID NO 16;
(iii) VH, содержащую аминокислотную последовательность SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность SEQ ID NO: 24;
(iv) VH, содержащую аминокислотную последовательность SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность SEQ ID NO: 32;
(v) VH, содержащую аминокислотную последовательность SEQ ID NO: 39, и VL, содержащую аминокислотную последовательность SEQ ID NO: 40;
(vi) VH, содержащую аминокислотную последовательность SEQ ID NO: 47, и VL, содержащую аминокислотную последовательность SEQ ID NO: 48;
(vii) VH, содержащую аминокислотную последовательность SEQ ID NO: 55, и VL, содержащую аминокислотную последовательность SEQ ID NO: 56;
(viii) VH, содержащую аминокислотную последовательность SEQ ID NO: 63, и VL, содержащую аминокислотную последовательность SEQ ID NO: 64;
(iv) VH, содержащую аминокислотную последовательность SEQ ID NO: 71, и VL, содержащую аминокислотную последовательность SEQ ID NO: 72.
Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает:
(i) VH последовательность SEQ ID NO: 7, и VL последовательность SEQ ID NO: 8;
(ii) VH последовательность SEQ ID NO: 15, и VL последовательность SEQ ID NO: 16;
(iii) VH последовательность SEQ ID NO: 23, и VL последовательность SEQ ID NO: 24;
(iv) VH с последовательность SEQ ID NO: 31, и VL с последовательностью SEQ ID NO: 32;
(v) VH последовательность SEQ ID NO: 39, и VL последовательность SEQ ID NO: 40;
(vi) VH последовательность SEQ ID NO: 47, и VL последовательность SEQ ID NO: 48;
(vii) VH последовательность SEQ ID NO: 55, и VL последовательность SEQ ID NO: 56;
(viii) VH последовательность SEQ ID NO: 63, и VL последовательность SEQ ID NO: 64; или
(ix) VH последовательность SEQ ID NO: 71, и VL последовательность SEQ ID NO: 72.
Согласно определенному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность SEQ ID NO: 15, и VL, содержащую аминокислотную последовательность SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность SEQ ID NO: 23, и VL, содержащую аминокислотную последовательность SEQ ID NO: 24.
Согласно еще одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность SEQ ID NO: 31, и VL, содержащую аминокислотную последовательность SEQ ID NO: 32.
Согласно определенному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность SEQ ID NO: 7, и VL, последовательность SEQ ID NO: 8.
Согласно другому варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность SEQ ID NO: 15, и VL последовательность SEQ ID NO: 16.
Согласно дополнительному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность SEQ ID NO: 23, и VL последовательность SEQ ID NO: 24.
Согласно еще одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает VH последовательность SEQ ID NO: 31, и VL последовательность SEQ ID NO: 32.
Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент включает константную область человека. Согласно одному варианту осуществления изобретения, третий антигенсвязывающий фрагмент представляет собой фрагмент Fab, содержащий человеческую константную область, в частности, человеческий домен СН1 и/или CL.
Примеры последовательностей константных доменов человека представлены в SEQ ID NOs 112 и 113 (домены CL каппа и лямбда человека, соответственно) и SEQ ID NO: 114 (константные домены СН1-СН2-СН3 тяжелых цепей IgG1 человека). Согласно некоторым вариантам осуществления изобретения, третий антигенсвязывающий фрагмент включает константную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 112 или SEQ ID NO: 113, в частности, аминокислотной последовательности SEQ ID NO: 112. В частности, константная область легкой цепи может включать аминокислотные мутации, как описано в настоящем документе в разделе "модификации заряда" и/или могут включать делеции или замены одной или более (конкретнее, двух) N-концевых аминокислот, если они находятся в кроссоверной молекуле Fab. Согласно некоторым вариантам осуществления изобретения, третий антигенсвязывающий фрагмент включает константную область тяжелой цепи, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности СН1 домена, которая включает аминокислотную последовательность SEQ ID NO: 114. В частности, константная область тяжелой цепи (в частности, СН1 домен) может содержать аминокислотные мутации как описано в настоящем документе в разделе "модификации заряда".
Согласно определенным вариантам осуществления изобретения, третий и первый антигенсвязывающий фрагмент оба представляют собой Fab молекулу, а третий антигенсвязывающий фрагмент идентичен первому антигенсвязывающему фрагменту. Таким образом, согласно данным вариантам осуществления изобретения, первый и третий антигенсвязывающие фрагменты включают одинаковые аминокислотные последовательности тяжелой и легкой цепей и имеют одинаковое расположение доменов (то есть традиционных или кроссоверных). Кроме того, согласно данным вариантам осуществления изобретения, третий антигенсвязывающий фрагмент содержит те же аминокислотные замены, если таковые имеются, что и первый антигенсвязывающий фрагмент. Например, аминокислотные замены, описанные в настоящем документе как "модификации заряда", будут сделаны в константном домене CL и константном домене СН1 каждого первого антигенсвязывающего фрагмента и третьего антигенсвязывающего фрагмента. Альтернативно, указанные аминокислотные замены могут быть сделаны в константном домене CL и константном домене СН1 второго антигенсвязывающего фрагмента (который согласно конкретным вариантам осуществления изобретения, также представляет собой Fab молекулу), но не в константном домене CL и константном домене СН1 первого антигенсвязывающего фрагмента и третьего антигенсвязывающего фрагмента. Как и первый антигенсвязывающий фрагмент, третий антигенсвязывающий фрагмент, в частности, представляет собой традиционную Fab молекулу. Однако варианты изобретения, в которых первый и третий антигенсвязывающий фрагменты представляют собой кроссоверную Fab молекулу (а второй антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу) также подразумеваются. Таким образом, согласно определенным вариантам осуществления изобретения, первый и третий антигенсвязывающий фрагменты каждый представляет собой традиционную Fab молекулу, а второй антигенсвязывающий фрагмент представляет собой кроссоверную Fab молекулу, как описано в настоящем документе, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены CL и CH1 Fab тяжелых и легких цепей поменяны местами/замещены друг другом. Согласно другим вариантам осуществления изобретения, первый и третий антигенсвязывающий фрагменты каждый представляет собой кроссоверную Fab молекулу, а второй антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу. В том случае, если третий антигенсвязывающий фрагмент присутствует, в конкретных вариантах осуществления изобретения, первый и третий фрагмент связывается с HLA-A2/WT1, а второй антигенсвязывающий фрагмент связывается со вторым антигеном, в частности, с активирующим Т-клеточным антигеном, конкретнее, с CD3, еще более конкретно, с С3 ипсилон.
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула включает Fc домен, образованный первой и второй субъединицами. Первая и вторая субъединица Fc домена способны к стабильной ассоциации.
Биспецифическая антигенсвязывающая молекула согласно изобретению, может иметь различные конфигурации, то есть первый, второй (и необязательно, третий) антигенсвязывающие фрагменты могут быть соединены друг с другом и с Fc доменом различными способами. Компоненты могут быть соединены друг с другом напрямую или, предпочтительно, посредством одного или более подходящего пептидного линкера. В том случае, если Fab молекула присоединяется к N-концу субъединицы Fc домена, это обычно происходит при помощи шарнирной области иммуноглобулина.
Согласно некоторым вариантам осуществления изобретения, первый и второй антигенсвязывающий фрагмент каждый представляет собой Fab молекулу, а второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом первой или второй субъединицы Fc домена. Согласно данному определенному варианту осуществления изобретения, указанный первый антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу, а второй антигенсвязывающий фрагмент представляет собой кроссоверную Fab молекулу, как описано в настоящем документе, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены CL и CH1 Fab тяжелых и легких цепей поменяны местами/замещены друг другом. Согласно другим таким вариантам осуществления изобретения, указанная первая Fab молекула представляет собой кроссоверную Fab молекулу, а вторая Fab молекула представляет собой традиционную Fab молекулу.
Согласно одному варианту осуществления изобретения, первый и второй антигенсвязывающий фрагмент каждый представляет собой Fab молекулу, а второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом первой или второй субъединицей Fc домена, а первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи второго антигенсвязывающего фрагмента. Согласно специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически включает первую и вторую Fab молекулу, Fc домен, образованный первой и второй субъединицей и, необязательно, одного или более пептидного линкера, при этом первая Fab молекула на своем С-конце Fab тяжелой цепи соединена с N-концом Fab тяжелой цепи второй Fab молекулы, а вторая Fab молекула на своем С-конце соединена с N-концом первой или второй субъединицы Fc домена. Такая конфигурация схематически представлена на Фигурах 1G и 1K (при этом второй антигенсвязывающий фрагмент в данных примерах представляет собой VH/VL кроссоверную Fab молекулу). Необязательно, Fab легкая цепь первой Fab молекулы и Fab легкая цепь второй Fab молекулы могут быть дополнительно соединены друг с другом.
Согласно другому варианту осуществления изобретения, первый и второй антигенсвязывающий фрагмент оба представляют собой Fab молекулу, первый и второй антигенсвязывающий фрагмент каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена. Согласно специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой и второй Fab молекул, Fc домена, образованного первой и второй субъединицами, и, необязательно, одного или более пептидного линкера, при этом первая и вторая Fab молекула каждая на своем С-конце Fab тяжелой цепи присоединена к N-концу одной из субъединиц Fc домена. Такая конфигурация схематически представлена на Фигурах 1А и 1D (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу). Первая и вторая Fab молекула может быть присоединена к Fc домену напрямую или посредством пептидного линкера. Согласно определенному варианту осуществления изобретения, первая и вторая Fab молекула оба присоединены к Fc домену посредством шарнирной области иммуноглобулина. Согласно специфическому варианту осуществления изобретения, шарнирная область иммуноглобулина представляет собой шарнирную область IgG1 человека, а конкретнее, Fc домен представляет собой Fc домен IgG1.
Согласно некоторым вариантам осуществления изобретения, первый и второй антигенсвязывающий фрагмент оба представляют собой Fab молекулу и первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи присоединен к N-концу первой или второй субъединицы Fc домена. Согласно такому варианту осуществления изобретения. Второй антигенсвязывающий фрагмент может быть на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента или (как описано выше) к N-концу любой из субъединиц Fc домена. Согласно конкретному варианту осуществления изобретения, указанный первый антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу, а второй антигенсвязывающий фрагмент представляет собой кроссоверную Fab молекулу, как описано в настоящем документе, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены CL и CH1 Fab тяжелых и легких цепей поменяны местами/замещены друг другом. Согласно другому такому варианту осуществления изобретения, указанная первая Fab молекула представляет собой кроссоверную Fab молекулу, а вторая Fab молекула представляет собой традиционную Fab молекулу.
Согласно одному варианту осуществления изобретения, первый и второй антигенсвязывающий фрагмент оба представляют собой Fab молекулу, первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи присоединен к N-концу первой или второй субъединицы Fc домена, а второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента. Согласно специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой и второй Fab молекулы, Fc домена, образованного первой и второй субъединицами, и, необязательно, одного или более пептидного линкера, при этом вторая Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи первой Fab молекулы, а первая Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу первой или второй субъединицы Fc домена. Такая конфигурация схематически представлена на Фигурах 1Н и 1L (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу). Необязательно, Fab легкая цепь первой Fab молекулы и Fab легкая цепь второй Fab молекулы могут быть дополнительно соединены друг с другом.
Согласно некоторым вариантам осуществления изобретения, третий антигенсвязывающий фрагмент, в частности, третья Fab молекула присоединена на своем С-конце Fab тяжелой цепи к N-концу первой или второй субъединицы Fc домена. Согласно такому конкретному варианту осуществления изобретения, указанная первая и третья Fab молекула обе представляют собой традиционные Fab молекулы, а вторая Fab молекула представляет собой кроссоверную Fab молекулу, как описано в настоящем документе, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены CL и CH1 Fab тяжелых и легких цепей поменяны местами/замещены друг другом. Согласно другим таким вариантам осуществления изобретения, указанная первая и третья Fab молекула обе представляют собой кроссоверные Fab молекулы, а вторая Fab молекула представляет собой традиционную Fab молекулу. Согласно определенному варианту осуществления изобретения, второй и третий антигенсвязывающие фрагменты оба на своем С-конце Fab тяжелой цепи присоединены к N-концу одной из субъединиц Fc домена, а первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второй Fab молекулы. Согласно специфическим вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой, второй и третьей Fab молекул, Fc домена, образованного первой и второй субъединицами, и, необязательно, одного или более пептидного линкера, при этом первая Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи второй Fab молекулы, а вторая Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу первой субъединицы Fc домена, при этом третья Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу второй субъединицы Fc домена. Такая конфигурация схематически представлена на Фигурах 1В и 1Е (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый и третий антигенсвязывающие фрагменты оба представляют собой традиционные Fab молекулы) и на Фигурах 1J и 1N (в этих примерах второй антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу, а первый и третий антигенсвязывающие фрагменты оба представляют собой VH/VL кроссоверные Fab молекулы). Вторая и третья Fab молекулы могут быть соединены с Fc доменом напрямую или посредством пептидного линкера.
Согласно определенному варианту осуществления изобретения, вторая и третья Fab молекулы обе присоединены к Fc домену посредством шарнирной области иммуноглобулина. Согласно специфическому варианту осуществления изобретения, шарнирная область иммуноглобулина представляет собой шарнирную область IgG1 человека, а конкретнее, Fc домен представляет собой Fc домен IgG1. Необязательно, Fab легкая цепь первой Fab молекулы и Fab легкая цепь второй Fab молекулы могут быть дополнительно соединены друг с другом.
Согласно другому такому варианту осуществления изобретения, первый и третий антигенсвязывающие фрагменты оба соединены на своих С-концах Fab тяжелых цепей с N-концом одной из субъединиц Fc домена, а второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента. Согласно специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой, второй и третьей Fab молекул, Fc домена, образованного первой и второй субъединицами, и, необязательно, одного или более пептидного линкера, при этом вторая Fab молекула на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первой Fab молекулы, а первая Fab молекула на своем С-конце своей Fab тяжелой цепи присоединена к N-концу первой субъединицы Fc домена, при этом третья Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу второй субъединицы Fc домена. Такая конфигурация схематически представлена на Фигурах 1С и 1F (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый и третий антигенсвязывающие фрагменты оба представляют собой традиционные Fab молекулы) и на Фигурах 1I и М (в этих примерах второй антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу, а первый и третий антигенсвязывающие фрагменты оба представляют собой VH/VL кроссоверные Fab молекулы). Первая и третья Fab молекулы могут быть соединены с Fc доменом напрямую или посредством пептидного линкера. Согласно определенному варианту осуществления изобретения, первая и третья Fab молекула каждая присоединена к Fc домену посредством шарнирной области иммуноглобулина. Согласно специфическому варианту осуществления изобретения, шарнирная область иммуноглобулина представляет собой шарнирную область IgG1 человека, а конкретнее, Fc домен представляет собой Fc домен IgG1. Необязательно, Fab легкая цепь первой Fab молекулы и Fab легкая цепь второй Fab молекулы могут быть дополнительно соединены друг с другом.
В тех конфигурациях биспецифической антигенсвязывающей молекулы, когда Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу каждой субъединицы Fc домена посредством шарнирных областей иммуноглобулина, обе Fab молекулы, шарнирные области и Fc домен фактически образуют молекулу иммуноглобулина. Согласно определенному варианту осуществления изобретения, молекула иммуноглобулина представляет собой молекулу иммуноглобулина IgG класса. Согласно еще более конкретному варианту осуществления изобретения, иммуноглобулин относится к подклассу IgG1 иммуноглобулинов. Согласно дополнительному конкретному варианту осуществления изобретения, иммуноглобулин представляет собой иммуноглобулин человека. Согласно другим вариантам осуществления изобретения, иммуноглобулин представляет собой химерный иммуноглобулин или гуманизированный иммуноглобулин. Согласно одному варианту осуществления изобретения, иммуноглобулин включает константную область человека, в частности, Fc область человека.
В некоторых биспецифических антигенсвязывающих молекулах согласно изобретению Fab легкая цепь первой Fab молекулы и Fab легкая цепь второй Fab молекулы соединены друг с другом, необязательно, посредством пептидного линкера. В зависимости от конфигурации первой и второй Fab молекулы, Fab легкая цепь первой Fab молекулы может быть на своем С-конце присоединена к N-концу Fab легкой цепи второй Fab молекулы, или Fab легкая цепь второй Fab молекулы может быть на своем С-конце присоединена к N-концу Fab легкой цепи первой Fab молекулы. Соединение Fab легких цепей первой и второй Fab молекул дополнительно уменьшает неправильное спаривание непарных Fab тяжелых и легких цепей, а также уменьшает количество плазмид, необходимых для экспрессии некоторых биспецифических антигенсвязывающих молекул согласно изобретению.
Антигенсвязывающие фрагменты могут быть присоединены к Fc домену или друг к другу напрямую или посредством пептидного линкера, включающего одну или более аминокислоту, обычно, приблизительно, от 2 до 20 аминокислот. Пептидные линкеры хорошо известны из области техники и описаны в настоящем документе. Подходящие неиммуногенные пептидные линкеры включают, например, (G4S)n, (SG4)n, (G4S)n или G4(SG4)n пептидные линкеры, "n" обычно представляет собой целое число от 1 до 10, обычно от 2 до 4. Согласно одному варианту осуществления изобретения, указанный пептидный линкер имеет длину по меньшей мере 5 аминокислот, согласно одному варианту осуществления изобретения, длину от 5 до 100 аминокислот, согласно дополнительному варианту осуществления изобретения, от 10 до 50 аминокислот. Согласно одному варианту осуществления изобретения, указанный пептидный линкер представляет собой (GxS)n или (GxS)nGm где G=глицин, S=серин, и (х=3, n=3, 4, 5 или 6, a m=0, 1, 2 или 3) или (х=4, n=2, 3, 4 или 5, а m=0, 1, 2 или 3), согласно одному варианту х=4 и n=2 или 3, согласно дополнительному варианту х=4 и n=2. Согласно одному варианту осуществления изобретения, указанный пептидный линкер представляет собой (G4S)2. Особенно предпочтительным пептидным линкером для соединения Fab легких цепей первой и второй Fab молекул между собой является (G4S)2. Примером пептидного линкера, подходящего для соединения Fab тяжелых цепей первой и второй Fab молекул включает последовательность (D)-(G4S)2 (SEQ ID NOs 110 и 111). Другой такой подходящий линкер включает последовательность (G4S)4. Дополнительно, линкеры могут включать шарнирную область иммуноглобулина (или ее часть). В частности, в том месте, где Fab молекула присоединена к N-концу субъединицы Fc домена, он может быть присоединена посредством шарнирной области иммуноглобулина или ее части, как с использованием пептидного линкера, так и без него.
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению, включает полипептид, в котором вариабельная область Fab легкой цепи второй молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (то есть, вторая Fab молекула включает кроссоверную Fab тяжелую цепь, при этом вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VL(2)-CH1(2)-CH2-CH3(-CH4)), и полипептид, в котором Fab тяжелая цепь первой Fab молекулы имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VH(1)-CH1(1)-CH2-CH3(-CH4)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (VH(2)-CL(2)) и полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)). Согласно определенным вариантам осуществления изобретения, полипептиды ковалентно связаны между собой, например, с помощью дисульфидной связи. Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, при этом константная область тяжелой цепи замещена константной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VH(2)-CL(2)-CH2-CH3(-CH4)), а также полипептид, в котором Fab тяжелая цепь первой Fab молекулы имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VH(1)-CH1(1)-СН2-СН3(-СН4)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (VL(2)-CH1(2)), и полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)). Согласно определенным вариантам осуществления изобретения, полипептиды ковалентно связаны между собой, например, с помощью дисульфидной связи.
Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы(то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, при этом вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепи первого Fab фрагмента, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VL(2)-CH1(2)-VH(1)-СН1(1)-СН2-СН3(-СН4)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором Fab тяжелая цепь первой Fab молекулы имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab легкой цепи второй Fab молекулы, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (то есть, вторая Fab молекула включает кроссоверную Fab тяжелую цепь, при этом вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VH(1)-CH1(1)-VL(2)-CH1(2)-CH2-CH3(-CH4)).
Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению дополнительно включает кроссоверный Fab полипептид легкой цепи второй Fab молекулы, при этом вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы(VH(2)-CL(2)) и полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)). Согласно другим вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с полипептидом Fab легкой цепи первой Fab молекулы(VH(2)-CL(2)-VL(1)-CL(1)), или полипептид, в котором полипептид Fab легкой цепи первой Fab молекулы имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab тяжелой цепи второй Fab молекулы, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (VL(1)-CL(1)-VH(2)-CL(2)), согласно установленному порядку.
Биспецифическая антигенсвязывающая молекула согласно данным вариантам осуществления изобретения, может дополнительно включать (i) полипептид субъединицы Fc домена (СН2-СН3(-СН4)), или (ii) полипептид, в котором Fab тяжелая цепь третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VH(3)-CH1(3)-CH2-CH3(-СН4)) и полипептид Fab легкой цепи третьей Fab молекулы (VL(3)-CL(3)). Согласно определенным вариантам осуществления изобретения, полипептиды ковалентно связаны между собой, например, посредством дисульфидной связи.
Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы(то есть, вторая Fab молекула включает кроссоверную Fab тяжелую цепь, при этом константная область тяжелой цепи замещена константной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепи первой Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VH(2)-CL(2)-VH(1)-CH1(1)-CH2-CH3(-CH4)). Согласно другим вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, в котором Fab тяжелая цепь первой Fab молекулы имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab тяжелой цепи второй Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (то есть, вторая Fab молекула включает кроссоверную Fab тяжелую цепь, при этом константная область тяжелой цепи замещена константной областью легкой цепи), которая, в свою очередь имеет общую карбокси-концевую пептидную связь с субъединицей Fc домена (VH(1)-CH1(1)-VH(2)-CL(2)-CH2-CH3(-CH4)).
Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает кроссоверный полипептид Fab легкой цепи второй Fab молекулы, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы(VL(2)-СН1(2)), и полипептид Fab легкой цепи первой Fab молекулы(VL(1)-CL(1)). Согласно другим таким вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с полипептидом Fab легкой цепи первой Fab молекулы (VL(2)-CH1(2)-VL(1)-CL(1)), или полипептид, в котором полипептид Fab первой Fab молекулы имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab тяжелой цепи второй Fab молекулы, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab второй Fab молекулы (VL(1)-CL(1)-VH(2)-CL(2)), согласно установленному порядку. Биспецифическая антигенсвязывающая молекула согласно указанным вариантам осуществления изобретения, может дополнительно включать (i) полипептид субъединицы Fc домена (СН2-СН3(-СН4)), или (ii) полипептид Fab тяжелой цепи третьей Fab молекулы (VL(3)-CL(3)). Согласно определенным вариантам осуществления изобретения, полипептиды ковалентно связаны между собой, например, посредством дисульфидной связи.
Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула не содержит Fc домен. Согласно некоторым таким вариантам осуществления изобретения, указанный первый, и в случае ее наличия, третья Fab молекула обе представляют собой традиционные Fab молекулы, а вторая Fab молекула представляет собой кроссоверную Fab молекулу, как описано в настоящем документе, то есть Fab молекулу, в которой вариабельные домены VH и VL или константные домены CL и CH1 Fab тяжелых и легких цепей поменяны местами/замещены друг другом. Согласно другим таким вариантам осуществления изобретения, указанная первая, и в случае ее наличия, третья Fab молекула обе представляют собой кроссоверные Fab молекулы, а вторая Fab молекула представляет собой традиционную Fab молекулу.
Согласно одному такому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой и второй Fab молекулы и, необязательно, одного или более пептидного линкера, при этом первый и второй антигенсвязывающие фрагменты оба представляют собой Fab молекулы, а первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента.
Такая конфигурация схематически представлена на Фигурах 1O и 1S (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу). Согласно другому такому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой и второй Fab молекул и, необязательно, одного или более пептидного линкера, при этом первый и второй антигенсвязывающие фрагменты оба представляют собой Fab молекулы, а второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента.
Такая конфигурация схематически представлена на Фигурах 1Р и 1Т (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый антигенсвязывающий фрагмент представляет собой традиционный Fab фрагмент).
Согласно некоторым вариантам осуществления изобретения, первая Fab молекула на своем С-конце присоединена к N-концу Fab тяжелой цепи второй Fab молекулы, а биспецифическая антигенсвязывающая молекула дополнительно включает третий антигенсвязывающий фрагмент, в частности, третья Fab молекула, при этом указанная Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи первой Fab молекулы. Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой, второй и третьей Fab молекулы, и, необязательно, одного или более пептидного линкера, при этом первая Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи второй Fab молекулы, а третья Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи первой Fab молекулы. Такая конфигурация схематически представлена на Фигурах 1Q и 1U (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый антигенсвязывающий фрагмент представляет собой традиционную Fab молекулу) или на Фигурах 1X и 1Z (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL традиционную Fab молекулу, а первый и третий антигенсвязывающие фрагменты представляют собой кроссоверные Fab молекулы).
Согласно некоторым вариантам осуществления изобретения, вторая Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи первой Fab молекулы, а биспецифическая антигенсвязывающая молекула включает третий антигенсвязывающий фрагмент, в частности, третью Fab молекулу, при этом указанная третья Fab молекула на своем N-конце Fab тяжелой цепи присоединена к С-концу Fab тяжелой цепи первой Fab молекулы. Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула фактически состоит из первой, второй и третьей Fab молекулы, и, необязательно, одного или более пептидного линкера, при этом вторая Fab молекула на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи первой Fab молекулы, а третья Fab молекула на своем N-конце Fab тяжелой цепи присоединена к С-концу Fab тяжелой цепи первой Fab молекулы. Такая конфигурация схематически представлена на Фигурах 1R и 1V (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL кроссоверную Fab молекулу, а первый и третий антигенсвязывающие фрагменты оба представляют собой традиционные Fab молекулы) или на Фигурах 1W и 1Y (в этих примерах второй антигенсвязывающий фрагмент представляет собой VH/VL традиционную Fab молекулу, а первый и третий антигенсвязывающие фрагменты представляют собой кроссоверные Fab молекулы).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором Fab тяжелая цепь первой Fab молекулы имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab легкой цепи второй Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, при этом вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи) (VH(1)-CH1(1)-VL(2)-CH1(2)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (VH(2)-CL(2)), а также полипептид Fab легкой цепи первой Fab молекулы (VL(l)-CL(1)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью первой Fab молекулы (VL(2)-CH1(2)-VH(1)-CH1(1)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (VH(2)-CL(2)), а также полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы(то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, в которой константная область тяжелой цепи замещена константной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью первой Fab молекулы (VL(2)-CL(2)-VH(1)-CH1(1)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (VL(2)-CH1(2)), а также полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы(то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью первой Fab молекулы(VL(2)-СН1(2)-VH(1)-СН1(1)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы(VH(2)-CL(2)), а также полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором Fab тяжелая цепь третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью первой Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab легкой цепи второй Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи) (VH(3)-CH1(3)-VH(1)-CH1(1)-VL(2)-CH1(2)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (VH(2)-CL(2)), а также полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид Fab легкой цепи третьей Fab молекулы (VL(3)-CL(3)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором Fab тяжелая цепь третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью первой Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab тяжелой цепи второй Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (то есть вторая Fab молекула включает кроссоверную Fab тяжелую цепь, в которой константная область тяжелой цепи замещена константной областью легкой цепи) (VH(3)-CH1(3)-VH(1)-CH1(1)-VH(2)-CL(2)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (VL(2)-CH(1)), а также полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид Fab легкой цепи третьей Fab молекулы (VL(3)-CL(3)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (то есть вторая Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью первого Fab фрагмента, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью третьей Fab молекулы (VL(2)-CH1(2)-VH(1)-CH1(1)-VH(3)-CH1(3)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (VH(2)-CL(2)) и полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)). В некоторых вариантах осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид Fab легкой цепи третьей Fab молекулы (VL(3)-CL(3)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab тяжелой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (то есть вторая Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой константная область тяжелой цепи замещена константной областью легкой цепи), который, в свою очередь имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью третьей Fab молекулы (VH(2)-CL(2)-VH(1)-CH1(1)-VH(3)-CH1(3)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи второй Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи второй Fab молекулы (VL(2)-CH1(2)) и полипептид Fab легкой цепи первой Fab молекулы (VL(1)-CL(1)). В некоторых вариантах осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид Fab легкой цепи третьей Fab молекулы (VL(3)-CL(3)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором Fab тяжелая цепь второй Fab молекулы имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab легкой цепи первого Fab фрагмента, который, в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи первой Fab молекулы (то есть первая Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab легкой цепи третьей Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи третьей Fab молекулы (то есть третья Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи) (VH(2)-CH1(2)-VL(1)-CH1(1)-VL(3)-CH1(3)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи первой Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи первой Fab молекулы (VH(1)-CL(1)) и полипептид Fab легкой цепи второй Fab молекулы (VL(2)-CL(2)). В некоторых вариантах осуществления изобретения биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи третьей Fab молекулы (VH(3)-CL(3)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором Fab тяжелая цепь второй Fab молекулы имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab тяжелой цепи первого Fab фрагмента, который, в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи первой Fab молекулы (то есть первая Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой константная область тяжелой цепи замещена константной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab тяжелой цепи третьей Fab молекулы, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи третьей Fab молекулы (то есть третья Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой константная область тяжелой цепи замещена константной областью легкой цепи) (VH(2)-CH1(2)-VL(1)-CH1(1)-VH(3)-CL(3)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи первой Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи первой Fab молекулы (VL(1)-CH1(1)) и полипептид Fab легкой цепи второй Fab молекулы (VL(2)-CL(2)). В некоторых вариантах осуществления изобретения биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи третьей Fab молекулы (VH(3)-CL(3)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab легкой цепи третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи третьей Fab молекулы (то есть третья Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab легкой цепи первой Fab молекулы (то есть первая Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), который, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепью второй Fab молекулы (VL(3)-CH1(3)-VL(1)-CH1(1)-VH(2)-CH1(2)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи первой Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи второй Fab молекулы (VL(2)-CL(2)). В некоторых вариантах осуществления изобретения биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab тяжелой цепи третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи третьей Fab молекулы (VH(3)-CL(3)).
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает полипептид, в котором вариабельная область Fab тяжелой цепи третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи третьей Fab молекулы (то есть третья Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с вариабельной областью Fab тяжелой цепи первого Fab фрагмента, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с константной областью Fab легкой цепи первой Fab молекулы (то есть первая Fab молекула содержит кроссоверную Fab тяжелую цепь, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с Fab тяжелой цепи второй Fab молекулы (VH(3)-CL(3)-VH(1)-CL(1)-VH(2)-CH1(2)). Согласно некоторым вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи первой Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи первой Fab молекулы (VL(1)-CH(1)), и полипептид Fab легкой цепи второй Fab молекулы (VL(2)-CL(2)).
В некоторых вариантах осуществления изобретения биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором вариабельная область Fab легкой цепи третьей Fab молекулы имеет общую карбокси-концевую пептидную связь с константной областью Fab тяжелой цепи третьей Fab молекулы (VL(3)-CH1(3)).
Согласно одному варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая: а) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности,
CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами.
где
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), и первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с).
Согласно определенному варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая: а) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности,
CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) третий антигенсвязывающий фрагмент, который связывается с первым антигеном и идентичен первому антигенсвязывающему фрагменту; и
d) Fc домен, образованный первой и второй субъединицами.
где
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) и третий антигенсвязывающий фрагмент по пункту с) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту d), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), а первый антигенсвязывающий фрагмент по пункту а) и третий антигенсвязывающий фрагмент по пункту с) оба на своем С-конце Fab тяжелой цепи присоединены к N-концу из субъединиц Fc домена по пункту d).
Согласно другому варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая:
а) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности, CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами.
где
(i) первый антигенсвязывающий фрагмент по пункту а) и второй антигенсвязывающий фрагмент по пункту b) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с)
Согласно одному варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности, CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами.
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), и первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с).
Согласно определенному варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности,
CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) третий антигенсвязывающий фрагмент, который связывается с первым антигеном и идентичен первому антигенсвязывающему фрагменту; и
d) Fc домен, образованный первой и второй субъединицами.
где
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) и третий антигенсвязывающий фрагмент по пункту с) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту d), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), а первый антигенсвязывающий фрагмент по пункту а) и третий антигенсвязывающий фрагмент по пункту с) оба на своем С-конце Fab тяжелой цепи присоединены к N-концу из субъединиц Fc домена по пункту d).
Согласно другому варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности, CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами.
где
(i) первый антигенсвязывающий фрагмент по пункту а) и второй антигенсвязывающий фрагмент по пункту b) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с).
Во всех различных конфигурациях биспецифической антигенсвязывающей молекулы согласно изобретению, описанные аминокислотные замены, если имеются, могут располагаться как в СН1 и CL доменах первого (и в случае его наличия) и третьего антигенсвязывающего фрагмента/Fab молекулы, так и в СН1 и CL доменах второго антигенсвязывающего фрагмента/Fab молекулы. Предпочтительно, они располагаются в СН1 и CL доменах первого и (в случае его наличия) третьего антигенсвязывающего фрагмента/Fab молекулы. В соответствии с концепцией настоящего изобретения, аминокислотные замены, как описано в настоящем документе, производятся в первом (и в случае его наличия) третьем антигенсвязывающем фрагменте/Fab молекуле, и не производятся во втором антигенсвязывающем фрагменте/Fab молекуле. В том случае, если аминокислотные замены, описанные в настоящем изобретении, производятся во втором антигенсвязывающем фрагменте/Fab молекуле, они не производятся в первом (и в случае его наличия) в третьем антигенсвязывающем фрагменте/Fab молекуле. Аминокислотные замены обычно производятся в биспецифических антигенсвязывающих молекулах, включающих Fab молекулы, в которых вариабельные домены VL и VH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом.
Согласно определенным вариантам осуществления изобретения, в биспецифической антигенсвязывающей молекуле согласно изобретению, в частности, если в ней произведены аминокислотные замены как описано в настоящем документе, в первом (и в случае его наличия, третьем) антигенсвязывающем фрагменте/Fab молекуле, константный домен CL первой (и в случае ее наличия, третьей) Fab молекулы представляет собой домен изотипа каппа. Согласно другим вариантам осуществления изобретения, в биспецифической антигенсвязывающей молекуле согласно изобретению, в частности, если в ней произведены аминокислотные замены как описано в настоящем документе, во втором антигенсвязывающем фрагменте/Fab молекуле, константный домен CL второго антигенсвязывающего фрагмента/ Fab фрагмента, представляет собой домен изотипа каппа. Согласно некоторым вариантам осуществления изобретения, константный домен CL первого (и в случае его наличия, третьего) антигенсвязывающего фрагмента/Fab молекулы и константный домен CL второго антигенсвязывающего фрагмента/Fab молекулы представляют собой домены изотипа каппа.
Согласно одному варианту своего осуществления, настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, включающей:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности, CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами;
где
в константном домене CL первого антигенсвязывающего фрагмента по пункту а) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату), а аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента по пункту а) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата), а аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата); и где
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), и первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с).
Согласно определенному варианту своего осуществления, настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, включающей:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности,
CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) третий антигенсвязывающий фрагмент, который связывается с первым антигеном и идентичен первому антигенсвязывающему фрагменту; и
d) Fc домен, образованный первой и второй субъединицами;
где
в константном домене CL первого антигенсвязывающего фрагмента по пункту а) и третьего антигенсвязывающего фрагмента по пункту с) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату), а аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента по пункту а) и третьего антигенсвязывающего фрагмента по пункту с) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата), а аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU
индексу Кабата); и
где
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) и третий антигенсвязывающий фрагмент по пункту с) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту d), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), и первый антигенсвязывающий фрагмент по пункту а) и третий антигенсвязывающий фрагмент по пункту с) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту d).
Согласно другому варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 1, HCDR 2 с SEQ ID NO: 2, и HCDR 3 с SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 4, LCDR 2 с SEQ ID NO: 5 и LCDR 3 с SEQ ID NO: 6; и
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности, 3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами; при этом в константном домене CL первого антигенсвязывающего фрагмента по пункту а) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату) и аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента по пункту а) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата); и при этом первый антигенсвязывающий фрагмент по пункту а) и второй антигенсвязывающий фрагмент по пункту б) оба на своих С-концах Fab тяжелой цепи присоединены к N-концу субъединиц Fc домена по пункту с).
Согласно одному варианту осуществления изобретения, предлагается биспецифическая антигенсвязывающая молекула, включающая:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности, CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами; при этом в константном домене CL первого антигенсвязывающего фрагмента по пункту а) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату) и аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента по пункту а) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата); и
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту с), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), а первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединены к N-концу из субъединиц Fc домена по пункту с).
Согласно определенному варианту осуществления изобретения предлагается биспецифическая антигенсвязывающая молекула, включающая:
a) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности, CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) третий антигенсвязывающий фрагмент, который связывается с первым антигеном и идентичен первому антигенсвязывающему фрагменту; и
d) Fc домен, образованный первой и второй субъединицами.
при этом в константном домене CL первого антигенсвязывающего фрагмента по пункту а) и третьего антигенсвязывающего фрагмента по пункту с) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату) и аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента по пункту а) и третьего антигенсвязывающего фрагмента по пункту с) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата); и
(i) первый антигенсвязывающий фрагмент по пункту а) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) и третий антигенсвязывающий фрагмент по пункту с) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу одной из субъединиц Fc домена по пункту d), или
(ii) второй антигенсвязывающий фрагмент по пункту b) на своем С-конце Fab тяжелой цепи присоединен к N-концу Fab тяжелой цепи первого антигенсвязывающего фрагмента по пункту а), а первый антигенсвязывающий фрагмент по пункту а) и третий антигенсвязывающий фрагмент по пункту с) каждый на своем С-конце Fab тяжелой цепи присоединены к N-концу из субъединиц Fc домена по пункту d).
Согласно другому варианту осуществления изобретения, предлагается биспецифическая антигенсвязывающая молекула, включающая:
а) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, а первый антигенсвязывающий фрагмент представляет собой Fab молекулу, содержащую вариабельную область тяжелой цепи (VH), содержащую область определяющую комплементарность тяжелой цепи (HCDR) 1 с SEQ ID NO: 9, HCDR 2 с SEQ ID NO: 10, и HCDR 3 с SEQ ID NO: 11, а также вариабельную область легкой цепи (VL), содержащую область определяющую комплементарность легкой цепи (LCDR) 1 с SEQ ID NO: 12, LCDR 2 с SEQ ID NO: 13 и LCDR 3 с SEQ ID NO: 14;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, при этом второй антиген представляет собой активирующий Т-клеточный антиген, в частности,
CD3, более конкретно CD3 ипсилон, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH или константные домены CL и CH1 Fab легкой цепи и Fab тяжелой цепи замещены друг другом;
c) Fc домен, образованный первой и второй субъединицами; при этом в константном домене CL первого антигенсвязывающего фрагмента по пункту а) и третьего антигенсвязывающего фрагмента по пункту с) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату) и аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого антигенсвязывающего фрагмента по пункту а) и третьего антигенсвязывающего фрагмента по пункту с) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата); и
первый антигенсвязывающий фрагмент по пункту а) и второй антигенсвязывающий фрагмент по пункту b) каждый на своем С-конце Fab тяжелой цепи присоединен к N-концу субъединиц Fc домена по пункту с). Согласно любому описанному выше варианту осуществления изобретения, компоненты биспецифической антигенсвязывающей молекулы (например, Fab молекулы, Fc домен) могут быть соединены между собой напрямую или посредством различных линкеров, в частности, пептидных линкеров, содержащих одну или более аминокислоту, обычно от 2 до 20 аминокислот, описанных в настоящем документе и хорошо известных из области техники. Подходящими, неиммуногенными пептидными линкерами являются, например, (G4S)n, (SG4)n, (G4S)n или G4(SG4)n пептидные линкеры, где п представляет собой целое число от 1 до 10, конкретнее, от 2 до 4.
Согласно определенному аспекту, настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, включающей:
а) первый и третий антигенсвязывающие фрагменты, которые связываются с первым антигеном; при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, при этом первый и второй антигенсвязывающие фрагменты оба представляет собой (традиционную) Fab молекулу, включающую вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO: 7 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO: 8;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном; при этом второй антиген представляет собой CD3, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH Fab легкой цепи и Fab тяжелой цепи замещены друг другом; содержащую (i) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 121, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 122, или (ii) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 136, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 137 (в частности, вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 136 и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 137);
c) Fc домен, образованный первой и второй субъединицами; при этом в константном домене CL первого и третьего антигенсвязывающих фрагментов по пункту а) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату) и аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого и третьего антигенсвязывающих фрагментов по пункту а) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата); и при этом
дополнительно первая антигенсвязывающая молекула по пункту а) на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) и третий антигенсвязывающий фрагмент по пункту а) оба на своих С-концах Fab тяжелых цепей присоединены к N-концу одной из субъединиц Fc домена по пункту с).
Согласно дополнительному аспекту, настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, включающей:
a) первый и третий антигенсвязывающий фрагменты, которые связываются с первым антигеном; при этом первый антиген представляет собой HLA-A2/WT1, в частности, HLA-A2/WT1RMF, при этом первый и второй антигенсвязывающие фрагменты оба представляет собой (традиционную) Fab молекулу, включающий вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO: 15 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO: 16;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном; при этом второй антиген представляет собой CD3, а второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH Fab легкой цепи и Fab тяжелой цепи замещены друг другом; содержащую (i) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 121, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 122, или (ii) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 136, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 137 (в частности, вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 136 и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 137);
c) Fc домен, образованный первой и второй субъединицами; при этом в константном домене CL первого и третьего антигенсвязывающих фрагментов по пункту а) аминокислота в положении 124 замещена лизином (К) (нумерация по Кабату) и аминокислота в положении 123 замещена лизином (К) или аргинином (R) (нумерация по Кабату) (более конкретно, аргинином (R)), а в константном домене СН1 первого и третьего антигенсвязывающих фрагментов по пункту а) аминокислота в положении 147 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата) и аминокислота в положении 213 замещена глутаминовой кислотой (Е) (нумерация согласно EU индексу Кабата); и при этом дополнительно
первая антигенсвязывающая молекула по пункту а) на своем С-конце Fab тяжелой цепи присоединена к N-концу Fab тяжелой цепи второго антигенсвязывающего фрагмента по пункту b), а второй антигенсвязывающий фрагмент по пункту b) и третий антигенсвязывающий фрагмент по пункту а) оба на своих С-концах Fab тяжелых цепей присоединены к N-концу одной из субъединиц Fc домена по пункту с).
Согласно одному варианту осуществления данных аспектов настоящего изобретения, в первой субъединице Fc домена остаток треонина в положении 366 замещен остатком триптофана (T366W), а во второй субъединице Fc домена остаток тирозина в положении 407 замещен остатком валина (Y407V) и, необязательно, остаток треонина в положении 366 замещен остатком серина (T366S), а остаток лейцина в положении 368 замещен остатком аланина (L368A) (нумерация согласно EU индексу Кабата).
Согласно другому варианту осуществления данных аспектов настоящего изобретения, в первой субъединице Fc домена остаток серина в положении 354 замещен остатком цистеина (S354C), или остаток глутаминовой кислоты в положении 356 замещен остатком цистеина (Е356С) (конкретнее, остаток серина в положении 354 замещен остатком цистеина),), а во второй субъединице Fc домена дополнительно остаток тирозина в положении 349 замещен остатком цистеина (Y349C) (нумерация согласно EU индексу Кабата).
Согласно еще одному другому варианту осуществления данных аспектов настоящего изобретения, в каждой первой и второй субъединице Fc домена остаток лейцина в положении 234 замещен остатком аланина (L234A), или остаток лейцина в положении 235 замещен остатком аланина (L235A), а остаток пролина в положении 329 замещен остатком глицина (P329G) (нумерация согласно EU индексу Кабата).
Согласно еще одному варианту осуществления данных аспектов настоящего изобретения, Fc домен представляет собой домен IgG1 человека.
Согласно конкретному специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 123, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 125, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 139 и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 140. Согласно дополнительному конкретному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, содержащий аминокислотную последовательность SEQ ID NO: 123, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 125, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 139 и полипептид, содержащий аминокислотную последовательность SEQ ID NO: 140.
Согласно другому специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 123, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 124, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 125 и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 129. Согласно дополнительному конкретному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, содержащий аминокислотную последовательность SEQ ID NO: 123, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 124, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 125 и полипептид, содержащий аминокислотную последовательность SEQ ID NO: 129.
Согласно другому специфическому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 126, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 127, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 128 и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, или 99% идентична последовательности SEQ ID NO: 129. Согласно дополнительному конкретному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула включает полипептид, содержащий аминокислотную последовательность SEQ ID NO: 126, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 127, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 128 и полипептид, содержащий аминокислотную последовательность SEQ ID NO: 129.
Fc домен
Согласно определенным вариантам осуществления изобретения, биспецифическая антигенсвязывающая молекула, согласно изобретению, включает Fc домен, образованный первой и второй субъединицами. Принято, что характеристики Fc домена, описанные в настоящем документе по отношению к биспецифической антигенсвязывающей молекуле, могут быть в равной степени применены к Fc домену в составе антитела согласно изобретению.
Fc домен в составе биспецифической антигенсвязывающей молекулы состоит из пары полипептидных цепей, включающих домены тяжелых цепей молекулы иммуноглобулина. Например, Fc домен иммуноглобулина G (IgG) представляет собой димер, каждая субъединица которого включает константные домены СН2 и СН3 тяжелой цепи IgG. Две субъединицы Fc домена способны к стабильной ассоциации друг с другом. Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула, согласно изобретению, содержит не более одного Fc домена.
Согласно одному варианту осуществления изобретения, Fc домен биспецифической антигенсвязывающей молекулы представляет собой Fc домен IgG. Согласно определенному варианту осуществления изобретения, Fc домен представляет собой Fc домен IgG1. Согласно другому варианту осуществления изобретения, Fc домен представляет собой Fc домен IgG4. Согласно более конкретному варианту осуществления изобретения, Fc домен представляет собой Fc домен IgG4, включающий аминокислотную замену в положении S228 (нумерация согласно EU индексу Кабата), конкретнее, аминокислотную замену в положении S228P. Аминокислотная замена приводит к уменьшению количества обменов Fab молекулами IgG4 антител in vivo (см Stubenrauch et al., Drug Metabolism and Disposition 38, 84-91 (2010)). Согласно другому конкретному варианту осуществления изобретения, Fc домен представляет собой человеческий Fc домен. Согласно еще одному конкретному варианту осуществления изобретения, Fc домен представляет собой человеческий Fc домен IgG1. Пример последовательности Fc области IgG1 человека представлен SEQ ID NO: 109.
Модификации Fc домена, усиливающие гетеродимеризацию
Биспецифическая антигенсвязывающая молекула, согласно изобретению, включает различные антигенсвязывающие фрагменты, которые могут быть соединены с одной из двух субъединиц Fc домена, таким образом, две субъединицы Fc домена обычно образованы двумя Fc домена обычно образованы двумя неидентичными полипептидными цепями. Рекомбинантная совместная экспрессия указанных полипептидов и последовательная димеризация приводит к нескольким возможным комбинациям двух полипептидов. Для того, чтобы улучить выход и степень очистки биспецифических антигенсвязывающих молекул в процессе рекомбинантного синтеза, таким образом, введение в Fc домен биспецифической антигенсвязывающей молекулы модификаций, усиливающих ассоциацию указанных полипептидов, может иметь преимущество.
Соответственно, согласно конкретным вариантам осуществления изобретения, Fc домен биспецифической антигенсвязывающей молекулы, согласно изобретению, включает модификации, усиливающие ассоциацию первой и второй субъединицы Fc домена. Сайт наиболее протяженного белок-белкового взаимодействия между двумя субъединицами Fc домена IgG человека находится в СН3 домене Fc домена. Таким образом, согласно одному варианту осуществления изобретения, указанная модификация расположена в СН3 домене Fc домена. Существует несколько подходов к осуществлению модификаций в СН3 домене Fc домена для того, чтобы усилить гетеродимеризацию, которые хорошо описаны в следующих документах WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012058768, WO 2013157954, WO 2013096291. Обычно, во всех указанных подходах СН3 домен первой субъединицы Fc домена и СН3 домен второй субъединицы Fc домена оба сгенерированы комплементарным образом так, чтобы каждый СН3 домен (или тяжелая цепь, содержащая его) не мог больше гомодимеризоваться сам с собой, но может усиленно гетеродимеризоваться с комплементарно сгенерированным другим СН3 доменом (таким образом, что первый и второй СН3 домены образуют гетеродимеры, но при этом между первыми двумя или вторыми двумя СН3 доменами гомодимеры не образуются). Эти различные подходы для улучшенной гетеродимеризации тяжелых цепей рассматриваются как различные альтернативы в сочетании с модификациями тяжелых-легких цепей (например, обмен/замещение VH и VL в одном связывающем плече или встраивание заряженных аминокислот с противоположными зарядами на границе контакта CH1 / CL) в биспецифической антигенсвязывающей молекуле для уменьшения искажений в тяжелых / легких цепях и образования побочных продуктов типа Бенс-Джонса.
Согласно специфическим вариантам осуществления изобретения, указанные модификации, усиливающие ассоциацию между первой и второй субъединицами Fc домена представляют собой так называемые модификации по типу "выступ-во-впадину", при этом модификация "выступ" располагается в одной из двух субъединиц Fc домена, а модификация "впадина" располагается в другой из двух субъединиц Fc домена. Технология "выступ-во-впадину" описана, например, в патентах US 5,731,168; US 7,695,936; и у Ridgway et al., Prot Eng 9, 617-621 (1996) и Carter, J Immunol Meth 248, 7-15 (2001). В целом, способ включает встраивание выпячивания ("выступа") на границу контакта первого полипептида и соответствующей ему полости ("впадины") на границе контакта второго полипептида, таким образом, чтобы выступ располагался во впадине, стимулировал формирование гетеродимера и препятствовал формированию гомодимера. Выпячивания конструируют путем замещения небольших боковых аминокислотных цепей на границе контакта первого полипептида большими боковыми аминокислотными цепями (например, тирозином или триптофаном). Компенсаторные впадины, размер которых идентичен или аналогичен размеру выступов, создают на границе контакта второго полипептида путем замещения больших аминокислотных боковых цепей меньшими аминокислотными боковыми цепями (например, аланином или треонином).
Соответственно, согласно определенному варианту осуществления изобретения, в СН3 домене первой субъединицы Fc домена биспецифической антигенсвязывающей молекулы аминокислотный остаток замещен аминокислотным остатком, имеющим больший размер боковой цепи, что позволяет создать выступ в структуре СН3 домена первой субъединицы, который располагается точно во впадине в структуре СН3 домена второй субъединицы, при этом в СН3 домене второй субъединицы Fc домена аминокислотный остаток замещен аминокислотным остатком с меньшим размером боковой цепи, что позволяет создать впадину в структуре СН3 домена второй субъединицы, в которой располагается выступ в структуре СН3 домена первой субъединицы.
Предпочтительно, указанный аминокислотный остаток, имеющий больший размер боковой цепи выбран из группы, включающей аргинин (R), фенилаланин (F), тирозин (Y) и триптофан (W).
Предпочтительно, указанный аминокислотный остаток, имеющий меньший размер боковой цепи выбран из группы, включающей аланин (А), серии (S), треонин (Т) и валин (V).
Выступ и впадина могут быть сгенерированы путем изменения нуклеиновой кислоты, кодирующей полипептиды, например, при помощи сайт-специфического мутагенеза, или пептидного синтеза.
Согласно специфическому варианту осуществления изобретения, в (СН3 домене) первой субъединице Fc домена (субъединице "выступа") остаток треонина в положении 366 замещен остатком триптофана (T366W), а во (СН3 домене) второй субъединицы Fc домена (субъединице "впадины") остаток тирозина в положении 407 замещен остатком валина (Y407V). Согласно одному варианту осуществления изобретения, во второй субъединице Fc домена дополнительно остаток треонина в положении 366 замещен остатком серина (T366S), а остаток лейцина в положении 368 замещен остатком аланина (L368A) (нумерация согласно EU индексу Кабата).
Согласно еще одному варианту осуществления изобретения, в первой субъединице Fc домена дополнительно остаток серина в положении 354 замещен остатком цистеина (S354C) или остаток глутаминовой кислоты в положении 366 замещен остатком цистеина (Е365С) (конкретнее, остаток серина в положении 354 замещен остатком цистеина), а во второй субъединице Fc домена дополнительно остаток тирозина в положении 349 замещен остатком цистеина (Y349C) (нумерация согласно EU индексу Кабата). Встраивание указанных двух остатков цистеина приводит к формированию дисульфидного мостика между двумя субъединицами Fc домена, что дополнительно стабилизирует димер (Carter, J Immunol Methods 248, 7-15 (2001)).
Согласно определенному варианту осуществления изобретения, первая субъединица Fc домена включает аминокислотные замены S354C и T366W, а вторая субъединица Fc домена включает аминокислотные замены Y349C, T366S, L368A и Y407V (нумерация согласно EU индексу Кабата).
Согласно определенному варианту осуществления изобретения, антигенсвязывающий фрагмент, который связывается со вторым антигеном (например, активирующим Т-клеточным антигеном) соединен (необязательно посредством первого антигенсвязывающего фрагмента, который связывается с HLA-A2/WT1, и/или пептидного линкера) с первой субъединицей Fc домена (включающей модификацию "выступ"). Без желания быть связанными какой-либо теорией, соединение антигенсвязывающего фрагмента, который связывается со вторым антигеном, таким, как активирующий Т-клеточный антиген, с содержащей "выступ" субъединицей Fc домена приведет (дополнительно) к минимальному образованию антигенсвязывающей молекулы, содержащей два антигенсвязывающих фрагмента, которые связываются с активирующим Т-клеточным антигеном (стерический конфликт двух содержащих "выступ" полипептидов).
Другие способы модификации СН3 для обеспечения гетеродимеризации рассматриваются как альтернативные в соответствии с изобретением и описаны в документах WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954, WO 2013/096291.
Согласно одному варианту осуществления изобретения, альтернативно используется подход к гетеродимеризации, описанный в документе ЕР 1870459. Этот подход основывается на встраивании заряженных аминокислот с противоположными зарядами в специфические аминокислотные положения на границе контакта СН3/СН3 доменов двух субъединиц Fc домена. Одним из предпочтительных вариантов осуществления биспецифической антигенсвязывающей молекулы, согласно изобретению, являются аминокислотные мутации R409D; K370E в одном из двух СН3 доменов (Fc домена) и аминокислотные мутации D399K; E357K в другом из двух СН3 доменов Fc домена (нумерация согласно EU индексу Кабата).
Согласно другому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает аминокислотную мутацию T366W в СН3 домене первой субъединицы Fc домена и аминокислотные мутации T366S, L368A, Y407V в СН3 домене второй субъединицы Fc домена, и дополнительные аминокислотные мутации R409D; K370E в СН3 домене первой субъединицы Fc домена и аминокислотные мутации D399K; E357K в СН3 домене второй субъединицы Fc домена (нумерация согласно EU индексу Кабата).
Согласно другому варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула согласно изобретению включает аминокислотные мутации S354C, T366W в СН3 домене первой субъединицы Fc домена и аминокислотные мутации Y349C, T366S, L368A, Y407V в СН3 домене второй субъединицы Fc домена, или указанная биспецифическая антигенсвязывающая молекула включает аминокислотные мутации Y349C, T366W в СН3 домене первой субъединицы Fc домена и аминокислотные мутации S354C, T366S, L368A, Y407V в СН3 домене второй субъединицы Fc домена и дополнительные аминокислотные мутации R409D; K370E в СН3 домене первой субъединицы Fc домена и аминокислотные мутации D399K; E357K в СН3 домене второй субъединицы Fc домена (вся нумерация согласно EU индексу Кабата).
Согласно одному варианту осуществления изобретения, альтернативно используется подход к гетеродимеризации, описанный в WO 2013/157953. Согласно одному варианту осуществления изобретения, первый СН3 домен включает аминокислотную мутацию T366K, а второй домен СН3 включает аминокислотную мутацию L351D (нумерация в соответствии с EU индексом Кабата). Согласно дополнительному варианту осуществления изобретения, первый домен СН3 включает дополнительную аминокислотную мутацию L351K. Согласно дополнительному варианту осуществления изобретения, второй домен СН3 включает дополнительную аминокислотную мутацию, выбранную из Y349E, Y349D и L368E (предпочтительно, L368E) (нумерация в соответствии с EU индексом Кабата).
Согласно одному варианту осуществления изобретения, альтернативно используется подход к гетеродимеризации, описанный в WO 2012/058768. Согласно одному варианту осуществления изобретения, первый домен СН3 включает аминокислотные мутации L351Y, Y407A, а второй домен СН3 включает аминокислотные мутации Т366А, K409F. Согласно дополнительному варианту осуществления изобретения, второй домен СН3 включает дополнительную аминокислотную мутацию в положении Т411, D399, S400, F405, N390 или K392, например, выбранном из а) Т41 IN, T411R, T411Q, T411K, Т41 ID, Т411Е или T411W, b) D399R, D399W, D399Y или D399K, с) S400E, S400D, S400R или S400K, d) F405, F405, F405, F405, F405, F555 или F405W, е) N390R, N390K или N390D, f) K392V, K392M, K392R, K392L, K392F или K392E (нумерация в соответствии с индексом EU Кабата). Согласно дополнительному варианту осуществления изобретения, первый домен СН3 включает аминокислотные мутации L351Y, Y407A, а второй домен СН3 включает аминокислотные мутации T366V, K409F. Согласно дополнительному варианту осуществления изобретения, первый домен СН3 включает аминокислотную мутацию Y407A, а второй домен СН3 включает аминокислотные мутации Т366А, K409F. Согласно дополнительному варианту осуществления изобретения, второй домен СН3 дополнительно включает аминокислотные мутации K392E, Т411Е, D399R и S400R (нумерация в соответствии с индексом EU Кабата).
Согласно одному варианту осуществления изобретения, альтернативно используется подход к гетеродимеризации, описанный в WO 2011/143545, например, с аминокислотной модификацией в положении, выбранном из группы, включающей 368 и 409 (нумерация в соответствии с индексом EU Кабата).
Согласно одному варианту осуществления изобретения, альтернативно используется подход к гетеродимеризации, описанный в WO 2011/090762, который также включает методику выступ-во-впадину, описанную выше. Согласно одному варианту осуществления изобретения, первый СН3 домен включает аминокислотную мутацию T366W, а второй СН3 домен включает аминокислотную мутацию Y407A. Согласно одному варианту осуществления изобретения, первый СН3 домен включает аминокислотную мутацию T366Y, а второй СН3 домен включает аминокислотную мутацию Y407T (нумерация в соответствии с индексом EU Кабата).
Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула или ее Fc домен относится к подклассу IgG2, при этом альтернативно применяется подход к гетеродимеризации, описанный в WO 2011/129304.
Согласно альтернативному варианту осуществления изобретения, модификация, усиливающая ассоциацию первой и второй субъединицы Fc домена, включает модификацию, которая опосредует электростатический направляющий эффект, например, как описано в заявке РСТ к WO 2009/089004. В целом, этот метод включает замещение одного или более аминокислотного остатка на границе раздела двух субъединиц Fc домена заряженными аминокислотными остатками таким образом, что образование гомодимера становится электростатически невыгодным, а гетеродимеризация, наоборот, становится электростатически выгодным. Согласно одному такому варианту осуществления изобретения, первый СН3 домен включает замену аминокислоты К392 или N392 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D), предпочтительно, K392D или N392D), а второй СН3 домен включает замену аминокислоты D399, Е356, D356, или Е357 положительно заряженной аминокислотой (например, лизином (К) или аргинином (R), предпочтительно, D399K, E356K, D356K, или E357K, и, более предпочтительно, D399K и E356K). Согласно дополнительному варианту осуществления изобретения, первый СН3 домен дополнительно включает замену аминокислоты К409 или R409 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D), предпочтительно, K409D или R409D). Согласно дополнительному варианту осуществления изобретения, первый СН3 домен дополнительно или альтернативно включает замену аминокислоты К439 и/или К370 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D)) (вся нумерация в соответствии с индексом EU Кабата).
Согласно еще одному дополнительному варианту осуществления изобретения, альтернативно используется подход к гетеродимеризации, описанный в WO 2007/147901. Согласно одному варианту осуществления изобретения, первый СН3 домен включает аминокислотные мутации K253E, D282K, и K322D, а второй СН3 домен включает аминокислотные мутации D239K, E240K и K292D (нумерация в соответствии с индексом EU Кабата).
Согласно еще одному варианту осуществления изобретения, альтернативно может использоваться подход к гетеродимеризации, описанный в WO 2007/110205. Согласно одному варианту осуществления изобретения, первый СН3 домен включает аминокислотные мутации K392D и K409D, а второй СН3 домен включает аминокислотные мутации D356K и D399K (нумерация в соответствии с индексом EU Кабата).
Модификации Fc домена, снижающие Fc рецепторное связывание и/или эффекторную функцию
Fc домен придает биспецифической антигенсвязывающей молекуле (или антителу) благоприятные фармакокинетические свойства, содержащую длительный период полувыведения, что приводит к хорошему накоплению в тканях мишенях и оптимальному распределению в крови/тканях. В то же самое время это, однако, может приводить к нежелательному направленному связыванию биспецифической антигенсвязывающей молекулы (или антитела) с клетками, экспрессирующими Fc рецепторы, а не с предпочтительными антиген-несущими клетками. Кроме того, коактивация с рецепторных сигнальных путей может приводить к высвобождению цитокинов, что в сочетании с Т-клеточными активирующими свойствами (например, в тех вариантах осуществления изобретения, согласно которым второй антигенсвязывающий фрагмент в составе биспецифической антигенсвязывающей молекулы связывается с активирующим Т-клеточным антигеном), и длительным периодом полувыведения биспецифической антигенсвязывающей молекулы приводит к избыточной активации цитокиновых рецепторов и серьезным побочным эффектам при системном введении. Активация (несущих Fc-рецептор) иммунных клеток, отличных от Т клеток, может даже снижать эффективность биспецифической антигенсвязывающей молекулы (в частности, биспецифической антигенсвязывающей молекулы, в которой второй антигенсвязывающий фрагмент связывается с активирующим Т-клеточным антигеном) за счет потенциального разрушения Т-клеток, например, NK клеток.
Соответственно, согласно определенным вариантам осуществления изобретения, Fc домен биспецифической антигенсвязывающей молекулы, согласно изобретению, обладает пониженной аффинностью связывания по отношению к Fc рецептору и/или сниженной эффекторной функцией по сравнению с Fc доменом нативного IgG1. Согласно одному такому варианту осуществления изобретения, Fc домен (или биспецифическая антигенсвязывающая молекула, содержащая указанный Fc домен) обладает аффинностью связывания к Fc рецептору, которая, предпочтительно, составляет менее 50%, предпочтительно, менее 20%, еще более предпочтительно, менее 10% и еще более предпочтительно, менее 5% от аффинности связывания Fc домена нативного IgG1 (или биспецифической антигенсвязывающей молекулы, содержащей указанный Fc домен нативного IgG1) и/или эффекторной функцией, которая составляет, менее 50%, предпочтительно, менее 20%, еще более предпочтительно, менее 10% и еще более предпочтительно, менее 5% от эффекторной функции Fc домена нативного IgG1 (или биспецифической антигенсвязывающей молекулы, содержащей указанный Fc домен нативного IgG1).
Согласно одному варианту осуществления изобретения, Fc домен (или биспецифическая антигенсвязывающая молекула, содержащая указанный Fc домен) не связывается в полной мере с Fc рецептором и/или не индуцирует эффекторную функцию. Согласно определенному варианту осуществления изобретения, Fc рецептор представляет собой Feγ рецептор. Согласно одному варианту осуществления изобретения, Fc рецептор представляет собой Fc рецептор человека. Согласно одному варианту осуществления изобретения, Fc рецептор представляет собой активирующий Fc рецептор. Согласно специфическому варианту осуществления изобретения, Fc рецептор представляет собой активирующий Feγ рецептор, более конкретно, человеческий FeγRIIIa, FeγRI или FeγRIIa рецептор, еще более конкретно, человеческий FeγRIIIa рецептор. Согласно одному варианту осуществления изобретения, эффекторная функция - это одна или более функция, выбранная из группы CDC, ADCC, ADCP и секреции цитокинов. Согласно определенному варианту осуществления изобретения эффекторная функция - это антителозависимая клеточная цитотоксичность ADCC. Согласно одному варианту осуществления изобретения, Fc домен обладает аффинностью связывания к неонатальному Fc рецептору (FcRn), которая практически полностью идентична таковой Fc домена нативного IgG1. Практически аналогичное связывание с FcRn достигается, когда Fc домен (или биспецифическая антигенсвязывающая молекула, содержащая указанный Fc домен) демонстрирует аффинность связывания с FcRn, которая составляет, приблизительно более чем 70%, конкретнее, приблизительно, более чем 80%, более конкретно, приблизительно более чем 90% аффинности связывания Fc домена нативного IgG1 (или биспецифической антигенсвязывающей молекулой, содержащей указанный Fc домен нативного IgG1). Согласно определенным вариантам осуществления изобретения Fc домен сгенерирован таким образом, чтобы обладать пониженной аффинностью связывания с Fc рецептором и/или сниженную эффекторную функцию по сравнению с доменом Fc, созданным без помощи генной инженерии. Согласно конкретным вариантам осуществления изобретения, Fc домен биспецифической антигенсвязывающей молекулы включает одну или более аминокислотную мутацию, которая уменьшает аффинность связывания Fc домена с Fc рецептором и/или его эффекторную функцию. Как правило, одна и та же или несколько аминокислотных мутаций присутствует в каждой из двух субъединиц Fc домена. Согласно одному варианту осуществления изобретения, аминокислотная мутация уменьшает аффинность связывания Fc домена с Fc рецептором. Согласно одному варианту осуществления изобретения, аминокислотная мутация уменьшает аффинность связывания Fc домена с Fc рецептором по меньшей мере в 2 раза, по меньшей мере в 5 раз или по меньшей мере в 10 раз. Согласно одному варианту осуществления изобретения, биспецифическая антигенсвязывающая молекула, содержащая сконструированный Fc-домен, имеет аффинность связывания, которая составляет менее 20%, в частности, менее 10%, более конкретно, менее 5% аффинности связывания биспецифической антигенсвязывающей молекулы, содержащей Fc домен, созданный без помощи генной инженерии. Согласно определенному варианту осуществления изобретения Fc рецептор представляет собой Feγ рецептор. Согласно некоторым вариантам осуществления изобретения, Fc рецептор представляет собой Fc рецептор человека. Согласно некоторым вариантам осуществления изобретения, Fc рецептор представляет собой активирующий Fc рецептор. Согласно специфическому варианту осуществления изобретения, Fc рецептор представляет собой человеческий активирующий Feγ рецептор, более конкретно, человеческий FeγRIIIa, FeγRI или FeγRIIa рецептор, еще более конкретно, человеческий FeγRIIIa рецептор. Предпочтительно, связывание с каждым из указанных рецепторов снижается. Согласно некоторым вариантам осуществления изобретения, аффинность связывания с компонентом системы комплемента, конкретно, аффинность связывания с C1q, также снижается. Согласно одному варианту осуществления изобретения, аффинность связывания с неонатальным Fc-рецептором (FcRn) не снижается. Почти аналогичное связывание с FcRn, то есть сохранение аффинности связывания домена Fc с указанным рецептором, достигается тогда, когда аффинность связывания Fc домена с FcRn. (или биспецифической антигенсвязывающей молекулы, содержащей указанный домен Fc) составляет, приблизительно, более 70% аффинности связывания домена Fc, полученного без использования генной инженерии (или биспецифической антигенсвязывающей молекулы, содержащей указанную несконструированную форму домена Fc). Аффинность связывания Fc домена или биспецифической антигенсвязывающей молекулы согласно изобретению, содержащей указанный Fc домен, может составлять более 80% и даже более 90% от естественной аффинности. Согласно определенным вариантам осуществления изобретения, Fc домен биспецифической антигенсвязывающей молекулы сконструирован таким образом, чтобы иметь пониженную эффекторную функцию, по сравнению с доменом, полученным без использования генной инженерии. Сниженная эффекторная функция моет включать, без ограничений указанными, одну или более из следующих функций: пониженную комплемент-зависимую цитотоксичность (CDC), пониженную антитело-зависимую опосредованную клетками цитотоксичность (ADCC), сниженный антитело-зависимый клеточный фагоцитоз (ADCP), пониженную секрецию цитокинов, сниженный опосредованный иммунными комплексами захват антигенов антиген-презентирующими клетками, пониженное связывание с NK клетками, пониженное связывание с макрофагами, пониженное связывание с моноцитами, пониженное связывание с полиморфонуклеарными клетками, снижение прямой передачи сигнала, вызывающего апоптоз, уменьшение перекрестного связывания связанных с мишенями антител, замедление процесса созревания дендритных клеток или сниженное примирование Т-клеток. Согласно одному варианту осуществления изобретения, уменьшенная эффекторная функция представляет собой одной или несколько функций, выбранных из группы: пониженной CDC, пониженной ADCC, пониженной ADCP и пониженной секреции цитокинов. Согласно конкретному варианту осуществления изобретения, пониженная эффекторная функция представляет собой пониженную ADCC. Согласно одному варианту осуществления изобретения, пониженная ADCC означает, что функция составляет менее 20% от ADCC, индуцируемой Fc доменом, полученным без применения генной инженерии (или биспецифической молекулой, содержащей такой несконструированный Fc домен). Согласно одному варианту осуществления изобретения, аминокислотная мутация, которая приводит к снижению аффинности связывания Fc домена с Fc рецептором и/или уменьшению эффекторной функции представляет собой аминокислотную замену. Согласно одному варианту осуществления изобретения, Fc домен включает аминокислотную замену в положении, выбранном из группы Е233, L234, L235, N297, Р331 и Р329 (нумерация согласно EU индексу Кабата). Согласно более специфическому варианту осуществления изобретения, Fc домен включает аминокислотную замену в положении, выбранном из группы L234, L235 и Р329 (нумерация согласно EU индексу Кабата). Согласно некоторым вариантам осуществления изобретения, Fc домен включает аминокислотные замены L234A и L235A (нумерация согласно EU индексу Кабата). Согласно одному такому варианту осуществления изобретения, Fc домен представляет собой Fc домен IgG1; в частности, Fc домен IgG1 человека. Согласно одному варианту осуществления изобретения, Fc домен включает аминокислотную замену в положении Р329. Согласно более специфическому варианту осуществления изобретения, аминокислотная замена представляет собой Р329А или P329G, в частности, P329G (нумерация согласно EU индексу Кабата). Согласно одному варианту осуществления изобретения, Fc домен включает аминокислотную замену в положении Р329 и дополнительную аминокислотную замену в положении, выбранном из Е233, L234, L235, N297 и Р331 (нумерация согласно EU индексу Кабата). Согласно более специфическому варианту осуществления изобретения, дополнительная аминокислотная замена представляет собой Е233Р, L234A, L235A, L235E, N297A, N297D или P331S. Согласно конкретному варианту осуществления изобретения, Fc домен включает аминокислотные замены в положении Р329, L234 и L235 (нумерация согласно EU индексу Кабата). Согласно более конкретному варианту осуществления изобретения, Fc домен включает аминокислотные мутации L234A, L235A и P329G ("P329G LALA", "PGLALA" или "LALAPG"). Точнее, согласно определенным вариантам осуществления изобретения, каждая субъединица Fc домена включает аминокислотные замены L234A, L235A и P329G (нумерация согласно EU индексу Кабата), то есть в каждой первой и второй субъединице Fc домена остаток лейцина в положении 234 замещен остатком аланина (L234A), остаток лейцина в положении 235 замещен остатком аланина (L235A), а остаток пролина в положении 329 замещен остатком глицина (P329G) (нумерация согласно EU индексу Кабата). Согласно одному такому варианту осуществления изобретения, Fc домен представляет собой Fc домен IgG1, в частности, Fc домен IgG1 человека. Комбинация аминокислотных замен "P329G LALA" приводит к практически полному нарушению связывания Feγ рецептора (а также молекулы комплемента) с Fc доменом IgG1 человека, как описано в публикации РСТ к WO 2012/130831, которая включена в настоящее описание во всей своей полноте. В WO 2012/130831 также описываются способы синтеза таких мутантных Fc доменов и способы определения их свойств, таких как связывание с Fc рецептором и эффекторные функции. IgG4 антитела демонстрируют пониженную аффинность связывания с Fc рецепторами и сниженные эффекторные функции по сравнению с IgG1 антителами. Следовательно, согласно некоторым вариантам осуществления изобретения, Fc домен биспецифических антигенсвязывающих молекул, согласно изобретению, представляет собой домен IgG4, в частности, Fc домен IgG4 человека. Согласно одному варианту осуществления изобретения, IgG4 Fc домен включает аминокислотные замены в положении S228, в частности, аминокислотную замену S228P (нумерация согласно EU индексу Кабата). Для того, чтобы дополнительно уменьшить аффинность связывания с Fc рецептором и/или эффекторную функцию, согласно одному варианту осуществления изобретения, IgG4 Fc домен включает аминокислотную замену в положении L235, в частности, аминокислотную замену L235E (нумерация согласно EU индексу Кабата). Согласно другому варианту осуществления изобретения, IgG4 Fc домен включает аминокислотную замену в положении Р329, в частности, аминокислотную замену P329G (нумерация согласно EU индексу Кабата). Согласно определенному варианту осуществления изобретения, IgG4 Fc домен включает аминокислотные замены в положениях S228, L235 и Р329, в частности, аминокислотные замены S228P, L235E и P329G (нумерация согласно EU индексу Кабата). Такие мутантные IgG4 Fc домены и их характеристики связывания с Feγ рецептором описаны в РСТ публикации WO 2012/130831, которая включена в настоящее описание в качестве ссылки во всей своей полноте.
Согласно определенному варианту осуществления изобретения, Fc домен, демонстрирующий пониженную аффинность связывания с Fc рецептором и/или сниженную эффекторную функцию, по сравнению с нативным IgG1 Fc доменом, представляет собой IgG1 Fc домен человека, включающий аминокислотные замены L234A, L235A и, необязательно, P329G, или IgG1 Fc домен человека, включающий аминокислотные замены S228P, L235E и, необязательно, P329G (нумерация согласно EU индексу Кабата).
Согласно определенным вариантам осуществления изобретения, было исключено N-гликозилирование Fc домена. Согласно одному такому варианту осуществления изобретения, Fc домен включает аминокислотную мутацию в положении N297, в частности, аминокислотную замену, при которой аспарагин замещен аланином (N297A) или аспарагиновой кислотой (N297D) (нумерация согласно EU индексу Кабата).
Помимо тех Fc доменов, что были описаны в настоящем документе выше или в публикации РСТ номер WO 2012/130831, к Fc доменам с пониженным связыванием с Fc рецептором и/или сниженной эффекторной функцией относятся те, которые включают замены одного или более остатка в положении 238, 265, 269, 270, 297, 327 and 329 (Патент США No. 6,737,056) (нумерация согласно EU индексу Кабата).
Такие мутантные Fc домены включают Fc домены с заменами аминокислот в одном или более положении 265, 269, 270, 297 и 327, содержащую так называемые "DANA" Fc мутантные домены с заменой остатков в положении 265 и 297 на аланин (патент США номер. 7,332,581).
Мутантные Fc домены могут быть получены путем делеции, замены, вставки или другой модификации аминокислот с использованием генетических или химических способов, хорошо известных из области техники. Генетические способы могут включать сайт-специфический мутагенез кодирующей ДНК последовательности, ПЦР, синтез генов и подобные. Правильные нуклеотидные замены можно установить, например, путем секвенирования.
Связывание с Fc рецепторами может быть легко установлено при помощи ELISA или при помощи поверхностного плазмонного резонанса (SPR) с использованием стандартного оборудования, такого как BIAcore (GE Healthcare), a Fc рецепторы при этом могут быть получены путем рекомбинантной экспрессии. Альтернативно, аффинность связывания Fc доменов или биспецифических антигенсвязывающих молекул, включающих Fc домен к Fc рецепторам, может быть оценена с использованием клеточных линий, о которых известно, что они экспрессируют определенные Fc рецепторы, таких, как NK клетки человека, экспрессирующие рецептор FeγIIIa.
Эффекторная функция Fc домена, или биспецифической антигенсвязывающей молекулы, содержащей Fc домен, может быть измерена при помощи способов, известных из области техники. Примеры исследований in vitro для оценки активности ADCC изучаемой молекулы описаны в патенте США No. 5,500,362; Hellstrom et al. Proc Natl Acad Sci USA 83, 7059-7063 (1986) и Hellstrom et al., Proc Natl Acad Sci USA 82, 1499-1502 (1985); патенте США No. 5,821,337; Bruggemann et al., J Exp Med 166, 1351-1361 (1987). Альтернативно, могут применяться нерадиоактивные способы (см, например, ACTI™ нерадиоактивный анализ цитотоксичности для проточной цитометрии (CellTechnology, Inc. Mountain View, CA); и CytoTox 96® нерадиоактивный анализ цитотоксичности (Promega, Madison, WI)). Эффекторные клетки, подходящие для таких исследований, включают мононуклеарные клетки периферической крови (РВМС) и натуральные киллеры (NK). Альтернативно или дополнительно, активность ADCC изучаемой молекулы можно оценить in vivo, например, на модельных животных, например, как описано у Clynes et al., Proc Natl Acad Sci USA 95, 652-656 (1998).
Согласно некоторым вариантам осуществления изобретения, связывание Fc домена с компонентом системы комплемента, в частности, с C1q, снижено. Соответственно, согласно тем вариантам осуществления изобретения, в которых Fc домен сконструирован при помощи генной инженерии таким образом, чтобы иметь пониженную эффекторную функцию, указанная пониженная эффекторная функция также подразумевает сниженную CDC. Анализ связывания C1q должен осуществляться для того, чтобы установить, способен ли Fc домен, или биспецифическая антигенсвязывающая молекула, содержащая Fc домен, связываться с C1q и, таким образом, обладать CDC активностью. См, например, анализ C1q и С3с связывания с помощью ELISA в WO 2006/029879 и WO 2005/100402. Для оценки активации системы комплемента может быть проведен анализ CDC (см, например, Gazzano-Santoro et al., J Immunol Methods 202, 163 (1996); Cragg et al., Blood 101, 1045-1052 (2003); and Cragg and Glennie, Blood 103, 2738-2743 (2004)).
Анализ связывания FcRn и определение клиренса/периода полувыведения in vivo также может осуществляться при помощи способов, известных из области техники (см., например, Petkova, SB. et al., Int'l. Immunol. 18(12): 1759-1769 (2006); WO 2013/120929).
Полинуклеотиды
Изобретение так же предлагает изолированные полинуклеотиды, кодирующие антитело или биспецифическую антигенсвязывающую молекулу как описано в настоящем документе или ее фрагмент. Согласно некоторым вариантам осуществления изобретения, указанный фрагмент представляет собой антигенсвязывающий фрагмент.
Полинуклеотиды, кодирующие антитела или биспецифические антигенсвязывающие молекулы, могут экспрессироваться как одиночный полинуклеотид, кодирующий полноразмерное антитело или биспецифическую антигенсвязывающую молекулу, так и в виде множества (например, двух или более) совместно экспрессируемых полинуклеотидов. Совместно экспрессируемые полипептиды, кодируемые полинуклеотидами, могут быть связаны, например, через дисульфидные связи или другими способами с формированием функционального антитела или биспецифической антигенсвязывающей молекулы.
Например, часть легкой цепи антитела или биспецифической антигенсвязывающей молекулы может кодироваться отдельным полинуклеотидом из части антитела или биспецифической антигенсвязывающей молекулы, включающей тяжелую цепь антитела или биспецифической антигенсвязывающей молекулы. При совместной экспрессии полипептиды тяжелой цепи будут связываться с полипептидами легкой цепи с формированием антитела или биспецифической антигенсвязывающей молекулы.
В другом примере часть антитела или биспецифической антигенсвязывающей молекулы, включающая одну из двух субъединиц Fc домена и, необязательно, одну или несколько Fab молекул или их часть, может быть кодирована отдельным полинуклеотидом из части антитела или биспецифической антигенсвязывающей молекулы, включающей другую из двух субъединиц Fc домена и, необязательно, Fab молекулу или ее часть. При совместной экспрессии субъединицы Fc домена будут ассоциироваться с формированием Fc домена.
Согласно некоторым вариантам осуществления изобретения, выделенный полинуклеотид кодирует полноразмерное антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению, как описано в настоящем документе. Согласно другим вариантам осуществления изобретения, выделенный полинуклеотид кодирует полипептид в составе антитела или биспецифической антигенсвязывающей молекулы согласно изобретению, как описано в настоящем документе.
Согласно определенным вариантам осуществления изобретения, полинуклеотид или нуклеиновая кислота представляет собой ДНК. Согласно другим вариантам осуществления изобретения, полинуклеотид по настоящему изобретению представляет собой РНК, например, информационную РНК (иРНК). РНК по настоящему изобретению может быть одноцепочечной или двуцепочечной.
Рекомбинантные методы
Антитела или биспецифические антигенсвязывающие молекулы, согласно изобретению, могут быть получены, например, при помощи твердофазного пептидного синтеза (например, с помощью твердофазного пептидного синтеза Меррифильда) или рекомбинантных методов. Для рекомбинантного синтеза один или несколько полинуклеотидов, кодирующих антитело или биспецифическую антигенсвязывающую молекулу (или ее фрагмент), например, как описано выше, выделяют и встраивают в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такой полинуклеотид может быть легко выделен и секвенирован с использованием стандартных методов. Согласно одному варианту осуществления изобретения, предлагается вектор, предпочтительно, вектор экспрессии, содержащий один или несколько полинуклеотидов. Методы, которые хорошо известны специалистам в данной области, могут быть использованы для конструирования векторов экспрессии, содержащих кодирующую последовательность антитела или биспецифической антигенсвязывающей молекулы (или ее фрагмента) вместе с соответствующими сигналами контроля транскрипции / трансляции. Такие методы включают технологии получения рекомбинантной ДНК in vitro, методики синтеза и in vivo рекомбинантные методики /рекомбинантные генетические методики. См., например, методы, описанные у Maniatis et al., MOLECULAR CLONING: A LABORATORY MANUAL, Cold Spring Harbor Laboratory, N.Y. (1989); и Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, Greene Publishing Associates and Wiley Interscience, N.Y (1989). Вектор экспрессии может быть частью плазмиды, вируса или может представлять собой фрагмент нуклеиновой кислоты. Вектор экспрессии включает кассету экспрессии, в которой клонируется полинуклеотид, кодирующий антитело или биспецифическую антигенсвязывающую молекулу (или ее фрагмент) (то есть кодирующую область), в оперативной связи с промотором и/или другими элементами контроля транскрипции или трансляции. Используемый в настоящем документе термин" кодирующая область" относится к части нуклеиновой кислоты, которая состоит из кодонов, транслируемых в аминокислоты. Несмотря на то что "стоп-кодон" (TAG, TGA или ТАА) не транслируется в аминокислоту, его можно рассматривать как часть кодирующей области, если он в ней присутствует, но любые фланкирующие последовательности, например промоторы, сайты связывания рибосом, терминаторы транскрипции, интроны, 5' и 3' нетранслируемые области и т.п. не являются частью кодирующей области. Две или более кодирующих области могут присутствовать в одном полинуклеотидном конструкте, например, в одном векторе или в отдельных полинуклеотидных конструктах, например, в отдельных (разных) векторах. Кроме того, любой вектор может содержать одну область кодирования или может содержать две или более области кодирования, например, вектор по данному изобретению может кодировать один или несколько полипептидов, которые после- или во время трансляции разделяются на конечные белки посредством протеолитического расщепления. Кроме того, вектор, полинуклеотид или нуклеиновая кислота согласно изобретению могут кодировать гетерологичные кодирующие области, как связанные, так и не связанные с полинуклеотидом, кодирующим антитело или биспецифическую антигенсвязывающую молекулу (или ее фрагмент) согласно изобретению или его вариант или производное. Гетерологичные кодирующие области включают без ограничений указанными, специализированные элементы или молекулы, такие как секреторный сигнальный пептид или гетерологичный функциональный домен. Оперативной связью является ситуация, когда кодирующая область генного продукта, например полипептида, связана с одной или несколькими регуляторными последовательностями таким образом, чтобы поставить экспрессию генного продукта под влияние или контроль регуляторной последовательности (последовательностей). Два фрагмента ДНК (такие как полипептидная кодирующая область и связанный с ними промотор) являются "функционально связанными", в том случае, если индукция функции промотора приводит к транскрипции иРНК, кодирующей желаемый генный продукт, а также если связь между двумя фрагментами ДНК не мешает способности регуляторных последовательностей экспрессии направлять экспрессию генного продукта или способности матрицы ДНК к транскрипции. Таким образом, промоторная область функционально связана с нуклеиновой кислотой, кодирующей полипептид, в том случае, если промотор способен влиять на транскрипцию этой нуклеиновой кислоты. Промотор может представлять собой клеточно-специфический промотор, который направляет транскрипцию ДНК только в предварительно детерминированных клетках. Другие элементы контроля транскрипции, кроме промотора, например, энхансеры, операторы, репрессоры и сигналы терминации транскрипции, могут быть функционально связаны с полинуклеотидом для направления клеточно-специфической транскрипции. Подходящие промоторы и другие области контроля транскрипции описаны в настоящем документе. Специалистам в данной области известно множество областей контроля транскрипции. Они включают, без ограничений указанными, области контроля транскрипции, которые функционируют в клетках позвоночных, такие как, без ограничений указанными, сегменты промотора и энхансера, выделенные из цитомегаловирусов (например, предранний промотор в сочетании с интроном-А), вируса обезьяньего гриппа 40 (например, ранний промотор) и ретровируса (такого как, например, вирус саркомы Рауса). Другие области контроля транскрипции включают области, выделенные из генов позвоночных, таких как актин, белок теплового шока, бычий гормон роста и β-глобин кролика, а также, другие последовательности, способные контролировать экспрессию генов в эукариотических клетках. Дополнительные подходящие области контроля транскрипции включают тканеспецифические промоторы и энхансеры, а также индуцируемые промоторы (например, индуцируемые тетрациклинами промоторы). Аналогично, специалистам в данной области техники известны различные элементы контроля трансляции. К ним относятся сайты связывания рибосом, кодоны инициации и терминации трансляции и элементы, полученные из вирусных систем (в частности, внутренний участок посадки рибосомы или IRES, также обозначаемый как CITE последовательность) и другие. Кассета экспрессии может также включать другие последовательности, такие, как точка начала репликации и/или элементы интеграции хромосом, такие как ретровирусные длинные концевые повторы (LTR) или аденоассоциированные вирусные (AAV) инвертированные концевые повторы (ITR).
Участки, кодирующие полинуклеотид и нуклеиновую кислоту, заявленные в соответствии с настоящим изобретением, могут быть связаны с дополнительными кодирующими участками, которые кодируют секреторные или сигнальные пептиды, управляющие секрецией полипептида, кодируемого полинуклеотидом по данному изобретению. Например, если желательна секреция антитела или биспецифической антигенсвязывающей молекулы, ДНК, кодирующая сигнальную последовательность, может быть размещена выше по рамке считывания, чем нуклеиновая кислота, кодирующая антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению или ее фрагмент. Согласно гипотезе сигнальной последовательности, белки, секретируемые клетками млекопитающих, имеют сигнальный пептид или секреторную лидерную последовательность, которая отщепляется от зрелого белка после инициации экспорта растущей белковой цепи через шероховатый эндоплазматический ретикулум. Специалистам в данной области известно, что полипептиды, секретируемые клетками позвоночных, обычно имеют сигнальный пептид, соединенный с N-концом полипептида, который отщепляется от транслированного полипептида с образованием секретированной или "зрелой" формы полипептида. Согласно определенным вариантам осуществления изобретения, применяется нативный сигнальный пептид, например, сигнальный пептид тяжелой или легкой цепи иммуноглобулина, или функциональное производное последовательности, которая сохраняет способность направлять секрецию полипептида, функционально связанного с ней. Альтернативно, может применяться гетерологичный сигнальный пептид млекопитающего или его функциональное производное. Например, лидерная последовательность дикого типа может быть заменена лидерной последовательностью тканевого активатора плазминогена человека (TPA) или мышиной β-глюкуронидазы.
ДНК, кодирующая короткую белковую последовательность, которая может использоваться для облегчения последующей очистки (например, от гистидиновой метки) или для облегчения присоединения метки к антителу или биспецифической антигенсвязывающей молекуле, может быть встроена внутрь или по бокам полинуклеотида, кодирующего антитело или биспецифическую антигенсвязывающую молекулу (или ее фрагмент).
Согласно другим вариантам осуществления изобретения, предлагается клетка-хозяин, содержащая один или несколько полинуклеотидов согласно изобретению. Согласно другим вариантам осуществления изобретения, предлагается клетка-хозяин, содержащая один или несколько векторов согласно изобретению. Полинуклеотиды и векторы могут иметь любые характеристики, по отдельности или в комбинации, описанные в настоящем документе в отношении полинуклеотидов и векторов, соответственно. Согласно одному такому варианту осуществления изобретения, клетка-хозяин включает (или, например, была трансформирована или трансфецирована) один или несколько векторов, содержащих один или более полинуклеотид, кодирующий антитело или биспецифическую антигенсвязывающую молекулу (или их части) согласно изобретению. Используемый в настоящем документе термин "клетка-хозяин" относится к любому типу клеточной системы, которая может быть сконструирована для получения антитела или биспецифической антигенсвязывающей молекулы согласно изобретению или ее фрагментов. Клетки-хозяева, подходящие для репликации и обеспечения экспрессии антител или биспецифических антигенсвязывающих молекул, хорошо известны из области техники. Такие клетки могут быть трансфецированы или трансдуцировани при необходимости конкретным вектором экспрессии, и большое количество клеток, содержащих вектор, может быть выращено для посева в крупные ферментеры для получения достаточных количеств антител или биспецифической антигенсвязывающей молекулы для клинического применения. Подходящие клетки-хозяева включают прокариотические микроорганизмы, такие как E.coli, или различные эукариотические клетки, такие как клетки яичника китайского хомячка (СНО), клетки насекомых и им подобные. Например, полипептиды могут продуцироваться в бактериях, особенно, когда гликозилирование не требуется. После экспрессии полипептид в растворимой фракции может быть выделен из культуры бактериальных клеток и дополнительно очищен. В дополнение к прокариотам эукариотические микробы, такие как нитчатые грибы или дрожжи, являются подходящими хозяевами для клонирования или экспрессии векторов, кодирующих полипептид; содержащую грибы и штаммы дрожжей, пути гликозилирования которых были "гуманизированы", что приводит к получению полипептида частично или полностью гликозилированного по паттерну, характерному для человека. См. Gerngross, Nat Biotech 22, 1409-1414 (2004), и Li et al., Nat Biotech 24, 210-215 (2006). Подходящие клетки-хозяева для экспрессии (гликозилированных) полипептидов также могут быть выделены из многоклеточных организмов (беспозвоночных и позвоночных). Примерами клеток беспозвоночных являются клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать совместно с клетками насекомых, особенно для трансфекции клеток Spodoptera frugiperda. Культуры растительных клеток также могут быть использованы в качестве хозяев. Смотрите, например, патенты США №5,959,177, 6,040,498, 6,420,548, 7,125,978 и 6,417,429 (описывающие технологию PLANTIBODIES™ для получения антител в трансгенных растениях). Клетки позвоночных также могут использоваться в качестве хозяев. Например, могут быть полезны клеточные линии млекопитающих, которые адаптированы для роста в суспензии. Другими примерами полезных линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированная SV40 (COS-7); человеческая эмбриональная почечная линия (клетки 293 или 293 Т, как описано, например, у Graham et al., J Gen Virol 36, 59 (1977)), клетки почки детеныша хомячка (ВНК), мышиные клетки Сертоли (клетки ТМ4, как описано, например, у Mather, Biol Reprod 23, 243-251 (1980)), клетки почки обезьяны (CV1), клетки почки африканской зеленой обезьяны (VERO-76), клетки карциномы шейки матки человека (HELA), клетки почки собаки (MDCK), клетки печени крыс буффало (BRL 3А), клетки легких человека (W138), клетки печени человека (Hep G2), клетки опухоли молочной железы мыши (ММТ 060562), клетки TRI (как описано, например, у Mather et al., Annals NY Acad Sci 383, 44-68 (1982)), клетки MRC 5 и клетки FS4. Другие полезные линии клеток-хозяев млекопитающих включают клетки яичника китайского хомячка (СНО), содержащую клетки dhfr-CHO (Urlaub et al., Proc Natl Acad Sci USA 77, 4216 (1980)); и клеточные линии миеломы, такие как YO, NS0, Р3Х63 и Sp2/0. Для обзора некоторых линий клеток-хозяев млекопитающих, подходящих для синтеза белка, см., например, Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), стр. 255-268 (2003). Клетки хозяева включают культивируемые клетки, например культивируемые клетки млекопитающих, дрожжевые клетки, клетки насекомых, бактериальные клетки и клетки растений, и это только некоторые примеры, а также клетки, содержащиеся в трансгенных животных, трансгенных растениях или культивируемых растениях или тканях животных. Согласно одному варианту осуществления изобретения, клетка-хозяин представляет собой эукариотическую клетку, предпочтительно клетку млекопитающего, такую как клетка яичника китайского хомячка (СНО), клетка эмбриональных почек человека (НЕК) или лимфоидная клетка (например, клетка YO, NSO, Sp20).
Стандартные методики экспрессии чужеродных генов в этих системах известны из области техники. Клетки, экспрессирующие полипептид, содержащий либо тяжелую, либо легкую цепь антигенсвязывающего домена, такого как антитело, могут быть сконструированы таким образом, чтобы экспрессировать также другие цепи антитела, так что продукт экспрессии будет представлять собой антитело, содержащее как тяжелую, так и легкую цепь.
Согласно одному варианту осуществления изобретения, предлагается способ получения антитела или биспецифической антигенсвязывающей молекулы согласно изобретению, при этом указанный способ включает культивирование клетки-хозяина, содержащей полинуклеотид, кодирующий антитело или биспецифическую антигенсвязывающую молекулу, как предусмотрено в настоящем документе, в условиях, подходящих для экспрессии антитела или биспецифической антигенсвязывающей молекулы, и, необязательно, восстановление антитела или биспецифической антигенсвязывающей молекулы из клетки-хозяина (или среды, в которой культивируются клетки-хозяева).
Компоненты биспецифической антигенсвязывающей молекулы (или антитела), согласно изобретению, могут быть генетически связаны друг с другом. Биспецифическая антигенсвязывающая молекула может быть сконструирована таким образом, что ее компоненты связаны между собой напрямую или посредством линкерной последовательности. Состав и длина линкера могут устанавливаться в соответствии со способами, хорошо известными из области техники, кроме того, изучается их эффективность. Примеры линкерных последовательностей между различными компонентами биспецифических антигенсвязывающих молекул представлены в настоящем документе. Дополнительные последовательности также могут быть включены для создания сайта расщепления (например, последовательности, распознаваемой эндопептидазой) для разделения отдельных компонентов, соединенных между собой, если это необходимо.
Антитело или биспецифическая антигенсвязывающая молекула, согласно изобретению, обычно включает по меньшей мере вариабельную область антитела, способную связывать антигенную детерминанту. Вариабельные области могут представлять собой часть существующих в природе или синтетических антител или их фрагментов, либо могут быть выделены из них. Способы получения поликлональных и моноклональных антител хорошо известны из области техники (см., например, Harlow and Lane, "Antibodies, a laboratory manual", Cold Spring Harbor Laboratory, 1988). He встречающиеся в природе антитела могут быть сконструированы с использованием твердофазного пептидного синтеза, могут быть получены рекомбинантными методами (например, как описано в патенте США №4186567) или могут быть получены, например, путем скрининга комбинаторных библиотек, содержащих вариабельные тяжелые цепи и вариабельные легкие цепи (см., например, патент США №5969108, выданный McCafferty).
Любое животное антитело, фрагмент такого антитела, антигенсвязывающий домен или вариабельная область могут быть использованы в составе антитела или биспецифической антигенсвязывающей молекулы согласно изобретению. Неограничивающими примерами антител, фрагментов антител, антигенсвязывающих доменов или вариабельных областей, которые могут применяться в соответствии с настоящим изобретением, могут быть получены из организма человека, примата или мыши. Если антитело или биспецифическая антигенсвязывающая молекула предназначено для применения у человека, может применяться химерная форма антитела, в которой константные области антитела имеют человеческую природу. Гуманизированная или полностью человеческая форма антитела также может быть получена в соответствии со способами, хорошо известными из области техники (см., например, патент США №5565322, Winter). Гуманизация может быть достигнута различными способами, содержащую, без ограничений указанными, (а) трансплантацию CDR областей нечеловеческого происхождения (например, донорского антитела) в каркасную область человеческого антитела (например, реципиентного антитела) и константные области с сохранением или без сохранения критичных остатков в каркасной области (например, необходимых для сохранения хорошей антигенсвязывающей аффинности или функций антител), (b) трансплантацию областей, определяющих комплементарность, только нечеловеческого происхождения (SDR или a-CDR; остатки, критичные для взаимодействия антитело-антиген) в каркасные области человеческого антитела, и константные области, или (с) трансплантацию вариабельных доменов полностью нечеловеческого происхождения, но с "маскировкой" их при помощи похожих на человеческие секциями путем замены поверхностных остатков. Гуманизированные антитела и способы их получения рассматриваются, например, у Almagro and Fransson, Front. Biosci. 13: 1619-1633 (2008) и дополнительно описаны, например, в Riechmann et al., Nature 332: 323-329 (1988); Queen et al., Proc. Nat'I Acad. Sci. США 86: 10029-10033 (1989); патенты США №5, 821, 337, 7527791, 6982321 и 7087409; Kashmiri et al, Methods 36: 25-34 (2005) (описывающий трансплантацию области, определяющей комплементарность (SDR)); Padlan, Mol. Immunol. 28: 489-498 (1991) (описание методики "обновления"); Dall'Acqua et al., Methods 36: 43-60 (2005) (описание "перестановка каркасных областей" и Osbourn et al., Methods 36: 61-68 (2005) и Klimka et al., Br. J. Cancer, 83: 252-260 (2000) (описание подхода "управляемой селекции" к перестановке каркасных областей).
Каркасные области человека, которые могут применяться для гуманизации, включают, без ограничений указанными: каркасные области, отобранные при помощи метода "наилучшего соответствия" (см., например, Sims et al. J. Immunol. 151: 2296 (1993)); каркасные области, выделенные из консенсусной последовательности вариабельных областей тяжелых и легких цепей определенной подгруппы антител человека (см., например, Carter et al. Proc. Natl. Acad. Sci. USA, 89: 4285 (1992); и Presta et al. J. Immunol., 151: 2623 (1993)); зрелые (соматически мутированные) каркасные области человека или каркасные области зародышевой линии человека (см., например, Almagro and Fransson, Front. Biosci. 13: 1619-1633 (2008)); и каркасные области, полученные при помощи скрининга FR-библиотек (см., например, Baca et al., J. Biol. Chem. 272: 10678-10684 (1997) и Rosok et al., J. Biol. Chem. 271: 22611-22618 (1996)).
Человеческие антитела могут быть получены с использованием различных методик, известных из области техники. Человеческие антитела в целом описаны у van Dijk и van de Winkel, Curr Opin Pharmacol 5, 368-74 (2001) и Lonberg, Curr Opin Immunol 20, 450-459 (2008). Человеческие антитела могут быть получены путем введения иммуногена трансгенному животному, которое было модифицировано для производства интактных человеческих антител или интактных антител с вариабельными областями человека в ответ на антигенное воздействие. Такие животные обычно содержат все или часть локусов иммуноглобулина человека, которые заменяют эндогенные локусы иммуноглобулина или которые присутствуют вне хромосом или случайным образом интегрированы в хромосомы животного. У подобных трансгенных мышей эндогенные локусы иммуноглобулина обычно инактивированы. Для обзора способов получения человеческих антител с использованием трансгенных животных, см., например, Lonberg, Nat. Biotech. 23: 1117-1125 (2005). См. также, например, патенты США №6075181 и 6150584, описывающие технологию XENOMOUSE™; патент США №5770429, описывающий технологию НиМаВ®; патент США №7044870, описывающий технологию K-М MOUSE®, и публикация заявки на патент США №US 2007/0061900, описывающая технологию VELOCIMOUSE®). Вариабельные области человека из интактных антител, генерируемых такими животными, могут быть дополнительно модифицированы, например, путем объединения с другой константной областью человека.
Человеческие антитела также могут быть получены с помощью гибридомных методов. Были описаны клеточные линии человеческой миеломы и гетеромиеломы мыши и человека для продуцирования человеческих моноклональных антител. (См., например, Kozbor J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987); and Boerner et al., J. Immunol., 147: 86 (1991).) Антитела человека, генерируемые при помощи гибридомы из В-клеток человека, также описаны у Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006). Дополнительные методы включают методы, описанные, например, в патенте США №7188926 (описывающем производство моноклональных человеческих IgM-антител из клеточной линии гибридомы) и Ni, Xiandai Mianyixue, 26 (4): 265-268 (2006) (описывающем гибридомы из разных типов клеток человека). Методика с использованием гибридомы из клеток человека (технология Trioma) также описана у Vollmers and Brandlein, Histology and Histopathology, 20(3):927-937 (2005) and Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3): 185-91 (2005).
Антитела человека также могут быть получены путем выделения из библиотек антител человека, как описано в настоящем документе.
Антитела, полезные для применения в соответствии с настоящим изобретением, могут быть выделены путем скрининга комбинаторных библиотек на антитела с желаемой активностью или активностями. Способы скрининга комбинаторных библиотек рассмотрены, например, у Lerner et al. in Nature Reviews 16:498-508 (2016). Например, из области техники известно множество способов создания фаг-дисплейных библиотек и скрининга таких библиотек на антитела, обладающие желаемыми характеристиками связывания. Такие способы раскрываются, например, у Frenzel et al. in mAbs 8:1177-1194 (2016); Bazan et al. in Human Vaccines andImmunotherapeutics 8:1817-1828 (2012) и Zhao et al. in Critical Reviews in Biotechnology 36:276-289 (2016) а так же в Hoogenboom et al. in Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001) и в Marks and Bradbury in Methods in Molecular Biology 248:161-175 (Lo, ed., Human Press, Totowa, NJ, 2003).
Согласно определенным фаг-дисплейным методам, спектры VH и VL генов клонируют по отдельности при помощи полимеразной цепной реакции (ПЦР) и случайным образом рекомбинируют в фаговых библиотеках, которые затем подвергают скринингу на антигенсвязывающий фаг, как описано у Winter et al. in Annual Review of Immunology 12: 433-455 (1994). Фаг обычно отображает фрагменты антител, как в форме одноцепочечных фрагментов Fv (scFv), так и в форме Fab фрагментов. Библиотеки из иммунизированных источников позволяют получить высокоаффинные антитела к иммуногену без необходимости конструирования гибридом. Альтернативно, наивный спектр может быть клонирован (например, от человека) для обеспечения единого источника антител к широкому спектру несобственных, а также собственных антигенов без какой-либо иммунизации, как описано у Griffiths et al. в EMBO Journal 12: 725-734 (1993). Наконец, наивные библиотеки также могут быть созданы синтетически, путем клонирования нереаранжированных сегментов V-гена из стволовых клеток и использования ПЦР-праймеров, содержащих случайную последовательность для кодирования высоко вариабельных областей CDR3 и для завершения реаранжирования in vitro, как описано у Hoogenboom and Winter in Journal of Molecular Biology 227: 381-388 (1992). Патентные публикации, описывающие фаговые библиотеки антител человека, включают, например: патенты США №5750373; 7985840; 7,785,903 и 8,679,490, а также патентные публикации США №№2005/0079574, 2007/0117126, 2007/0237764 и 2007/0292936. Дополнительные примеры методов скрининга комбинаторных библиотек на наличие антител с желаемой активностью или активностями, известные из области техники, включают рибосомный и иРНК дисплеи, а также методы антительного дисплея и селекции на бактериях, клетках млекопитающих, клетках насекомых или дрожжах. Способы отображения дрожжевого дисплея рассмотрены, например, у Scholler et al. в Methods in Molecular Biology 503: 135-56 (2012) и в Cherf et al. в Methods in Molecular biology 1319: 155-175 (2015), а также в Zhao et al. в Methods in Molecular Biology 889: 73-84 (2012). Способы рибосомного дисплея описаны, например, у Не et al. в Nucleic Acids Research 25: 5132-5134 (1997) и в Hanes et al. в PNAS 94: 4937-4942 (1997).
Антитела или биспецифичные антигенсвязывающие молекулы, полученные, как описано в настоящем документе, могут быть очищены способами, известными из области техники, такими как высокоэффективная жидкостная хроматография, ионообменная хроматография, гель-электрофорез, аффинная хроматография, эксклюзионная хроматография и тому подобными. Фактические условия, используемые для очистки конкретного белка, будут от части зависеть от таких факторов, как его суммарный заряд, гидрофобность, гидрофильность и т.д., и очевидны специалистам в данной области. Для аффинной хроматографии может использоваться антитело, лиганд, рецептор или антиген, с которым связывается антитело или биспецифическая антигенсвязывающая молекула. Например, для аффинной хроматографической очистки антител или биспецифической антигенсвязывающей молекулы, согласно изобретению, может использоваться матрица с белком А или белком G. Последовательная аффинная хроматография с белком А или G и эксклюзионная хроматография могут быть использованы для выделения антитела или биспецифической антигенсвязывающей молекулы как раскрыто в примерах. Чистота антитела или биспецифической антигенсвязывающей молекулы, может быть определена любым из множества хорошо известных аналитических методов, содержащую гель-электрофорез, жидкостную хроматографию высокого давления и им подобные.
Композиции, лекарственные формы и пути введения
Согласно дополнительному аспекту, настоящее изобретение относится к фармацевтическим композициям, содержащим любое антитело или биспецифическую антигенсвязывающую молекулу, представленную в настоящем документе, например, для применения согласно любому из перечисленных ниже терапевтических способов. Согласно одному варианту своего осуществления, фармацевтическая композиция содержит любое антитело или биспецифическую антигенсвязывающую молекулу, представленную в настоящем документе, а также фармацевтически приемлемый носитель. Согласно другому варианту осуществления изобретения, фармацевтическая композиция содержит любое антитела или биспецифическую антигенсвязывающую молекулу, представленную в настоящем документе, и по меньшей мере один дополнительный терапевтический агент, например, как описано ниже.
Также предлагается способ получения антител или биспецифической антигенсвязывающей молекулы согласно изобретению в форме, подходящей для введения in vivo, при этом указанный способ включает (а) получение антитела или биспецифической антигенсвязывающей молекулы согласно изобретению и (б) получение лекарственной формы антитела или биспецифической антигенсвязывающей молекулы по меньшей мере с одним фармацевтически приемлемым носителем в составе, в результате чего композиция антитела или биспецифической антигенсвязывающей молекулы становится пригодной для введения in vivo.
Фармацевтические композиции, заявленные в соответствии с настоящим изобретением, содержат терапевтически эффективное количество антитела или биспецифической антигенсвязывающей молекулы, растворенных или диспергированных в фармацевтически приемлемом носителе. Фразы "фармацевтический или фармакологически приемлемый" относятся к молекулярным элементам и композициям, которые в целом нетоксичны для реципиентов в используемых дозировках и концентрациях, то есть не вызывают нежелательных, аллергических или других неблагоприятных реакций при введении животному, такому как, например, человек, соответственно. Процесс получения фармацевтической композиции, содержащей антитело или биспецифическую антигенсвязывающую молекулу и, необязательно, дополнительный активный ингредиент, будет понятен специалистам в данной области в контексте настоящего изобретения, на примере, описанном в Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, данный источник включен в настоящее описание в качестве ссылки.
Кроме того, для введения животным (например, человеку) следует понимать, что лекарственные формы должны соответствовать стандартам стерильности, пирогенности, общей безопасности и чистоты, как того требует управление биологических стандартов FDA или соответствующие органы в других странах. Предпочтительными композициями являются лиофилизированные лекарственные формы или водные растворы. Используемый в настоящем описании термин "фармацевтически приемлемый носитель" включает любые или все растворители, буферы, дисперсионные среды, покрытия, сурфактанты, антиоксиданты, консерванты (например, антибактериальные агенты, противогрибковые агенты), изотонические агенты, агенты, замедляющие абсорбцию, соли, консерванты, антиоксиданты, белки, лекарственные средства, стабилизаторы лекарств, полимеры, гели, связующие вещества, эксципиенты, дезинтегрирующие агенты, любриканты, подсластители, ароматизаторы, красители, а также подобные материалы и их комбинации, что хорошо известно специалистам в данной области (см. например, Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, стр. 1289-1329; источник включен в настоящий документ в качестве ссылки). За исключением тех случаев, когда любой стандартный носитель несовместим с активным ингредиентом, его использование в терапевтических или фармацевтических композициях подразумевается.
Антитело или биспецифическая антигенсвязывающая молекула согласно изобретению (а также любой дополнительный терапевтический агент) могут вводиться любыми подходящими способами, содержащую парентеральное, внутрилегочное и интраназальное, и, если необходимо для местного лечения, внутриочаговое введение. Парентеральное введение включает внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Введение доз может осуществляться любым подходящим способом, например путем инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является введение однократным или длительным.
Парентеральные композиции включают композиции, предназначенные для инъекционного введения, например подкожного, внутрикожного, внутриочагового, внутривенного, внутриартериального, внутримышечного, интратекального или внутрибрюшинного введения. Для инъекционного введения антитела или биспецифические антигенсвязывающие молекулы, согласно изобретению, могут быть приготовлены в форме водных растворов, предпочтительно в физиологически совместимых буферах, таких как раствор Хенкса, раствор Рингера или физиологический солевой буфер. Раствор может содержать вспомогательные агенты, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. Альтернативно, антитела или биспецифические антигенсвязывающие молекулы могут быть представлены в форме порошка, который перед применением разводится подходящим носителем, например стерильной апирогенной водой. Стерильные инъекционные растворы получают путем разведения антител или биспецифических антигенсвязывающих молекул согласно изобретению в необходимом количестве в соответствующем растворителе вместе с другими различными ингредиентами, перечисленными ниже, в соответствии с инструкцией.
Стерильность может быть легко достигнута, например, путем фильтрации через стерильные фильтрационные мембраны. Обычно дисперсии получают путем разведения различных стерильных активных ингредиентов в стерильном носителе, содержащем основную дисперсионную среду и/или другие ингредиенты. В случае стерильных порошков для приготовления стерильных инъекционных растворов, суспензий или эмульсий, предпочтительными способами получения являются технологии вакуумной сушки или лиофилизации, которые позволяют получить активный ингредиент в форме порошка вместе с любым дополнительным желаемым ингредиентом, выделенным из предварительно простерилизованной фильтрацией жидкой среды. Жидкая среда при необходимости должна содержать подходящий буфер и перед введением доведена до изотонического состояния путем добавления достаточного количества солевого раствора или глюкозы. Композиция должна сохранять стабильность в условиях производства и хранения и быть защищена от загрязнения микроорганизмами, такими, как бактерии и грибы. Понятно, что контаминация эндотоксинами должна поддерживаться на минимально безопасном уровне, например, составлять менее 0,5 нг/мг белка.
Подходящие фармацевтически приемлемые носители включают следующие вещества, без ограничений указанными: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, содержащую аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; феноловый, бутиловый или бензиловый спирт; алкилпарабены, такие как метил или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (приблизительно, менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, содержащую глюкозу, маннозу или декстрины; хелатообразующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные сурфактанты, такие как полиэтиленгликоль (ПЭГ). Водные инъекционные суспензии могут содержать соединения, которые увеличивают вязкость суспензии, такие как натрийкарбоксиметилцеллюлоза, сорбит, декстран и им подобные вещества. Необязательно, суспензия также может содержать подходящие стабилизаторы или агенты, которые увеличивают растворимость соединений, что позволяет получать высококонцентрированные растворы. Кроме того, суспензии активных соединений могут быть получены в виде подходящих масляных инъекционных суспензий. Подходящие липофильные растворители или носители включают жирные масла, такие как кунжутное масло, или синтетические сложные эфиры жирных кислот, такие как триглицериды с этиловыми мостиками (ethyl cleats), или липосомы.
Активные ингредиенты могут быть заключены в микрокапсулы, полученные, например, методами коацервации или межфазной полимеризации; например, в гидроксиметилцеллюлозные капсулы или желатиновые микрокапсулы или поли-(метилметацилатные) микрокапсулы, соответственно; или в коллоидные системы доставки лекарств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методики описаны в Remington's Pharmaceutical Sciences (18-е изд. Mack Printing Company, 1990). Также могут быть получены лекарственные формы с пролонгированным высвобождением. Подходящими примерами лекарственных форм с пролонгированным высвобождением являются полупроницаемые матрицы твердых гидрофобных полимеров, содержащие полипептид, причем указанные матрицы имеют форму профилированных изделий, например пленок или микрокапсул. В конкретных вариантах пролонгированная абсорбция инъецируемой композиции может быть обусловлена применением в композициях агентов, замедляющих абсорбцию, таких как, например, моностеарат алюминия, желатин или их комбинации.
В дополнение к композициям, описанным ранее, антитела или биспецифические антигенсвязывающие молекулы также могут быть получены в форме депо-препаратов. Такие препараты пролонгированного действия можно вводить посредством имплантации (например, подкожной или внутримышечной) или внутримышечной инъекции. Таким образом, например, антитела или биспецифические антигенсвязывающие молекулы в процессе получения лекарственной формы могут быть объединены с подходящими полимерными или гидрофобными материалами (например, в форме эмульсии в подходящем масле) или ионообменными смолами, или изготовлены в форме умеренно растворимых соединений, например, умеренно растворимых солей.
Фармацевтические композиции, содержащие антитела или биспецифические антигенсвязывающие молекулы согласно изобретению, могут быть произведены при помощи стандартных процессов смешивания, растворения, эмульгирования, инкапсулирования, включения или лиофилизации. Фармацевтические композиции могут быть изготовлены традиционным способом с использованием одного или нескольких физиологически приемлемых носителей, растворителей, эксципиентов или вспомогательных веществ, которые обеспечивают получение лекарственных форм препаратов, которые могут использоваться в фармацевтических целях. Подходящая лекарственная форма определяется способом введения. Антитела или биспецифические антигенсвязывающие молекулы могут находиться в составе композиции в форме свободных кислот или оснований, либо в форме солей.
Фармацевтически приемлемые соли представляют собой соли, которые практически полностью сохраняют биологическую активность свободной кислоты или основания. К ним относятся кислотно-аддитивные соли, например соли, образованные со свободными аминогруппами белковой композиции, или соли, образованные неорганическими кислотами, такими как, например, соляная или фосфорная кислота, или такими органическими кислотами, как уксусная, щавелевая, винная или миндальная, кислота. Соли, образованные со свободными карбоксильными группами, также могут быть получены из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа; или таких органических оснований, как изопропиламин, триметиламин, гистидин или прокаин. Фармацевтические соли обычно являются более растворимыми в воде и других протонных растворителях, чем соответствующие свободные основания.
Терапевтические методы и композиции
Любое антитело или биспецифическая антигенсвязывающая молекула, завяленные в соответствии с настоящим изобретением, могут применяться в терапевтических методах. Антитела или биспецифические антигенсвязывающие молекулы, согласно изобретению, могут применяться в качестве иммунотерапевтических агентов, например, при лечении рака.
Для применения в терапевтических методах антитела или биспецифические антигенсвязывающие молекулы согласно изобретению должны быть включены в состав лекарственной формы, дозированы и введены при помощи способов, которые соответствуют надлежащей медицинской практике. В данном контексте необходимо учитывать такие факторы, как определенное заболевание, подлежащее лечению; конкретное млекопитающее животное, получающее лечение; клиническое состояние конкретного пациента; причина заболевания; место доставки лекарственного агента; способ введения, режим введения, а также другие факторы, известные практикующим врачам.
Согласно одному аспекту изобретения предлагаются антитела или биспецифические антигенсвязывающие молекулы для применения в качестве лекарственного средства. Согласно дополнительным аспектам изобретения, предлагаются антитела или биспецифические антигенсвязывающие молекулы для применения в лечении заболевания. Согласно определенным вариантам осуществления изобретения, антитела или биспецифические антигенсвязывающие молекулы согласно изобретению предназначены для применения в соответствии со способом лечения. Согласно одному варианту своего осуществления, изобретение относится к антителу или биспецифической антигенсвязывающей молекуле, как описано в настоящем документе, для применения в лечении заболевания у индивидуума, нуждающегося в этом. Согласно определенным вариантам своего осуществления, изобретение относится к антителу или биспецифической антигенсвязывающей молекуле для применения согласно способу лечения индивидуума, имеющего заболевание, при этом указанный способ включает введение индивидууму терапевтически эффективного количества антитела или биспецифической антигенсвязывающей молекулы. Согласно определенным вариантам осуществления изобретения, заболевание, которое подлежит лечению, представляет собой пролиферативное заболевание. Согласно некоторым вариантам осуществления изобретения, заболевание представляет собой рак. Согласно определенным вариантам осуществления изобретения, способ дополнительно включает введение индивидууму терапевтически эффективного количества по меньшей мере одного дополнительного терапевтического агента, например противоракового агента, если заболевание, подлежащее лечению, представляет собой рак. Согласно другим вариантам своего осуществления, изобретение относится к антителу или биспецифической антигенсвязывающей молекуле, как описано в настоящем документе, для применения для индукции лизиса клетки-мишени, в частности опухолевой клетки. Согласно определенным вариантам своего осуществления, изобретение относится к антителу или биспецифической антигенсвязывающей молекуле для применения согласно способу индукции лизиса клетки-мишени, в частности опухолевой клетки, у индивидуума, при этом указанный способ включает введение индивидууму эффективного количества антитела или биспецифической антигенсвязывающей молекулы для индукции лизиса клетки-мишени. "Индивидуум" в соответствии с любым из вышеуказанных вариантов представляет собой млекопитающее, предпочтительно человека.
Согласно другому аспекту, настоящее изобретение предусматривает применение антитела или биспецифической антигенсвязывающей молекулы согласно изобретению для производства или изготовления лекарственного средства. Согласно одному варианту осуществления изобретения, лекарственное средство предназначено для лечения заболевания у индивидуума, нуждающегося в этом. Согласно другому варианту осуществления изобретения, лекарственное средство предназначено для применения согласно способу лечения заболевания, при этом указанный способ включает введение индивидууму, страдающему от заболевания, терапевтически эффективного количества лекарственного средства. Согласно определенным вариантам осуществления изобретения, заболевание, подлежащее лечению, представляет собой пролиферативное заболевание. Согласно конкретному варианту осуществления изобретения, заболевание представляет собой рак. Согласно одному варианту осуществления изобретения, способ дополнительно включает введение индивидууму терапевтически эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, противоракового агента, если заболевание, подлежащее лечению, представляет собой рак. Согласно дополнительному варианту осуществления изобретения, лекарственное средство предназначено для индукции лизиса клетки-мишени, в частности опухолевой клетки. Согласно еще одному дополнительному варианту осуществления изобретения, лекарственное средство предназначено для применения согласно способу индукции лизиса клетки-мишени, в частности, опухолевой клетки, у индивидуума, при этом указанный способ включает введение индивидууму эффективного количества лекарственного средства для индукции лизиса клетки-мишени. "Индивидуум" в соответствии с любым из вышеуказанных вариантов осуществления изобретения, может представлять собой млекопитающее, предпочтительно, человека.
Согласно другому аспекту, настоящее изобретение относится к способу лечения заболевания. Согласно одному варианту осуществления изобретения, способ включает введение индивидууму, страдающему от такого заболевания, терапевтически эффективного количества антитела или биспецифической антигенсвязывающей молекулы согласно изобретению. Согласно другому варианту осуществления изобретения, композиция, содержащая, антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению, вводится индивидууму в фармацевтически приемлемой форме. Согласно определенным вариантам осуществления изобретения, заболевание, подлежащее лечению, представляет собой пролиферативное заболевание. Согласно конкретному варианту осуществления изобретения, заболевание представляет собой рак. Согласно некоторым вариантам осуществления изобретения, способ дополнительно включает введение индивидууму терапевтически эффективного количества по меньшей мере одного дополнительного терапевтического агента, например противоракового агента, если заболевание, подлежащее лечению, представляет собой рак. "Индивидуум" в соответствии с любым из вышеуказанных вариантов осуществления изобретения, может представлять собой млекопитающее, предпочтительно, человека.
Согласно дополнительному аспекту, изобретение относится к способу индукции лизиса клетки-мишени, в частности опухолевой клетки. Согласно одному варианту осуществления изобретения, способ включает контактирование клетки-мишени с антителом или биспецифической антигенсвязывающей молекулой согласно изобретению в присутствии Т-клетки, в частности цитотоксической Т-клетки. Согласно дополнительному варианту осуществления изобретения, предлагается способ индукции лизиса клетки-мишени, в частности опухолевой клетки, у индивидуума. Согласно одному такому варианту осуществления изобретения, способ включает введение индивидууму эффективного количества антитела или биспецифической антигенсвязывающей молекулы для индукции лизиса клетки-мишени. Согласно одному варианту осуществления изобретения, "индивидуумом" является человек.
Согласно определенным вариантам осуществления изобретения, заболевание, подлежащее лечению, представляет собой пролиферативное заболевание, в частности рак. Не имеющими ограничительного характера примерами раковых заболеваний являются гемобластоз, такой, как лейкоз, рак мочевого пузыря, рак головного мозга, рак головы и шеи, рак поджелудочной железы, рак желчевыводящих путей, рак щитовидной железы, рак легких, рак молочной железы, рак яичников, рак матки, рак шейки матки, рак эндометрия, рак пищевода, рак толстой кишки, колоректальный рак, рак прямой кишки, рак желудка, рак предстательной железы, рак кожи, плоскоклеточный рак, саркома, рак кости и рак почки. Другими примерами клеточно-пролиферативных заболеваний, которые можно лечить с применением антитела или биспецифической антигенсвязывающей молекулы по настоящему изобретению, являются, без ограничений указанными, новообразования, расположенные в: брюшной полости, костях, грудной клетке, пищеварительной системе, печени, поджелудочной железе, брюшине, эндокринных железах (надпочечники, паращитовидные железы, гипофиз, яички, яичник, тимус, щитовидная железа), глазах, голове и шее, нервной системе (центральной и периферической), лимфатической системе, тазу, кожа, мягких тканях, селезенке, грудной клетке и урогенитальной системе. Кроме того, примерами являются предраковые состояния или метастатические поражения. Согласно некоторым вариантам осуществления изобретения, раковое заболевание выбрано из группы, включающей гемобластозы (такие как лейкоз), рак почки, рак мочевого пузыря, рак кожи, рак легких, колоректальный рак, рак молочной железы, рак головного мозга, рак головы и шеи и рак простаты. Согласно одному варианту осуществления изобретения, рак представляет собой гемобластоз, в частности лейкоз, более конкретно острый лимфобластный лейкоз (ALL) или острый миелобластный лейкоз (AML). Специалисты в данной области признают, что во многих случаях антитело или биспецифическая антигенсвязывающая молекула не гарантируют полного излечения, а обеспечивают лишь временный благоприятный эффект. Согласно некоторым вариантам осуществления изобретения, физиологическое изменение, имеющее некоторый благоприятный эффект, также расценивается как положительный терапевтический ответ. Таким образом, согласно некоторым вариантам осуществления изобретения, количество антитела или биспецифической антигенсвязывающей молекулы, которое обеспечивает физиологическое изменение, считается "эффективным количеством" или "терапевтически эффективным количеством". Субъектом, пациентом или индивидуумом, нуждающимся в лечении, обычно является млекопитающее, более конкретно, человек.
Согласно некоторым вариантам, эффективное количество антитела или биспецифической антигенсвязывающей молекулы, согласно изобретению, вводят в клетку. Согласно другим вариантам осуществления изобретения, терапевтически эффективное количество антитела или биспецифической антигенсвязывающей молекулы, согласно изобретению, вводят индивидууму для лечения заболевания.
Для профилактики или лечения заболевания подходящая доза антитела или биспецифической антигенсвязывающей молекулы согласно изобретению (при использовании отдельно или в сочетании с одним или несколькими другими дополнительными терапевтическими агентами) будет зависеть от типа заболевания, подлежащего лечению, способа введения, массы тела пациента, типа антитела или биспецифической антигенсвязывающей молекулы, тяжести и течения заболевания, целей введения антитела или биспецифической антигенсвязывающей молекулы (профилактических или терапевтических), предыдущих или сопутствующих терапевтических вмешательств, анамнеза пациента, реакции на введение антитела или биспецифической антигенсвязывающей молекулы, а также собственных усмотрений лечащего врача. В любом случае, специалист, ответственный за введение, будет определять концентрацию активного ингредиента (ов) в композиции и подходящую дозу(ы) для индивидуального субъекта. Допустимы различные режимы дозирования, содержащую, без ограничений указанными: однократное или многократное введение в различные временные точки, болюсное введение и пульс-терапия. Антитело или биспецифическая антигенсвязывающая молекула соответственно вводятся пациенту однократно или на протяжении курса лечения. В зависимости от типа и тяжести заболевания начальная доза антитела или биспецифической антигенсвязывающей молекулы может составлять, приблизительно, от 1 мкг/кг до 15 мг/кг (например, от 0,1 мг/кг до 10 мг/кг) вне зависимости от того вводится она однократно или посредством многократных отдельных инъекций, или посредством непрерывной инфузии. Типичная однократная суточная доза может варьировать, приблизительно, от 1 мкг/кг до 100 мг/кг или более, в зависимости от факторов, упомянутых выше. Что касается повторных введений в течение нескольких дней или дольше, то в зависимости от состояния пациента, лечение будет продолжаться до тех пор, пока не будет достигнуто желаемое подавление симптомов заболевания. Одна примерная доза антитела или биспецифической антигенсвязывающей молекулы, находится в диапазоне, приблизительно, от 0,005 до, приблизительно, 10 мг/кг. Согласно другим не имеющим ограничительного характера примерам, доза для однократного введения также может составлять, приблизительно, от 1 микрограмм/кг массы тела, приблизительно от 5 микрограмм/кг массы тела, приблизительно, от 10 микрограмм/кг массы тела, приблизительно, от 50 микрограмм/кг массы тела, приблизительно, от 100 микрограмм/кг массы тела, приблизительно, от 200 микрограмм/кг массы тела, приблизительно, от 350 микрограмм/ кг массы тела, приблизительно, от 500 микрограмм /кг массы тела, приблизительно, от 1 миллиграмма/кг массы тела, приблизительно, от 5 миллиграмм /кг массы тела, приблизительно, от 10 миллиграмм/кг массы тела, приблизительно, от 50 миллиграмм/кг массы тела, приблизительно, от 100 миллиграмм/кг массы тела, приблизительно, от 200 миллиграмм/кг массы тела, приблизительно, от 350 миллиграмм/кг массы тела, приблизительно, от 500 миллиграмм/кг массы тела, до, приблизительно 1000 миллиграмм/кг массы тела или больше согласно любым диапазонам, представленным в настоящем документе. Согласно другим примерам, не имеющим ограничительного характера, в рамках перечисленных выше диапазонов, пациенту могут вводиться дозы, находящиеся в диапазоне, приблизительно, от 5 мг/кг массы тела до, приблизительно, 100 мг/кг массы тела, приблизительно, от 5 микрограмм/кг массы тела до приблизительно 500 миллиграмм / кг массы тела, и т.д. на основании чисел, представленных выше. Таким образом, пациенту может вводиться одна или несколько доз, составляющих, приблизительно 0,5 мг/кг, 2,0 мг/кг, 5,0 мг/кг или 10 мг/кг (или любые их комбинации). Такие дозы могут вводиться с перерывами, например, каждую неделю или каждые три недели (например, так, чтобы пациент получил, приблизительно, от двух до, приблизительно, двадцати доз; или, например, приблизительно, шесть доз антитела или биспецифической антигенсвязывающей молекулы). В начале лечения может быть назначена более высокая нагрузочная доза, после которой вводится одна или несколько более низких доз. Однако и другие режимы дозирования могут быть полезны. Эффективность терапии легко отслеживается при помощи стандартных методик и исследований.
Антитела или биспецифические антигенсвязывающие молекулы согласно изобретению, как правило, будут использоваться в количестве, эффективном для достижения намеченной цели. Для применения в целях лечения или профилактики заболевания, антитела или биспецифические антигенсвязывающие молекулы согласно изобретению или их фармацевтические композиции вводятся или применяются в терапевтически эффективном количестве. Понятие терапевтически эффективного количества понятно специалистам в данной области, особенно в свете подробного описания, представленного в настоящем документе.
Для системного введения терапевтически эффективная доза может быть первоначально определена на основании исследований in vitro, таких как исследования клеточных культур. Затем дозу определяют на модельных животных таким образом, чтобы достигался диапазон циркулирующих концентраций, который включает IC50, предварительно установленную в культуре клеток. Такая информация может быть использована для более точного определения эффективных доз для человека.
Начальные дозы также могут быть установлены при помощи данных, полученных in vivo, например, на модельных животных с использованием методик, которые хорошо известны специалистам в данной области. На основании данных, полученных на модельных животных, специалист в данной области может легко определить дозу для человека.
Дозировки и интервалы введения могут подбираться индивидуально для того, чтобы обеспечить уровни антител или биспецифических антигенсвязывающих молекул в плазме, которые являются достаточными для поддержания терапевтического эффекта. Стандартные дозы у пациентов для инъекционного введения составляют, приблизительно, от 0,1 до 50 мг/кг/сутки, обычно, приблизительно от 0,5 до 1 мг/кг/ сутки. Терапевтически эффективные уровни в плазме могут быть достигнуты путем введения нескольких доз ежедневно. Уровни в плазме можно измерить, например, при помощи высокоэффективной жидкостной хроматографии.
В случаях местного введения или селективного захвата, эффективная локальная концентрация антител или биспецифических антигенсвязывающих молекул может не коррелировать с концентрацией в плазме. Специалист в данной области сможет легко оптимизировать терапевтически эффективные локальные дозы без неоправданных экспериментов.
Антитела или биспецифические антигенсвязывающие молекулы, описанные в настоящем документе, в терапевтически эффективных дозах обычно оказывают благоприятный терапевтический эффект, не оказывая токсического действия. Токсичность и терапевтическая эффективность антител или биспецифических антигенсвязывающих молекул могут быть определены при помощи стандартных фармацевтических процедур с использованием клеточных культур или экспериментальных животных. Исследования на культуре клеток и животных можно использовать для определения LD50 (доза, летальная для 50% популяции) и ED50 (доза, терапевтически эффективная для 50% популяции). Соотношение токсической и терапевтически эффективной доз представляет собой терапевтический индекс, который может быть выражен как соотношение LD50/ED50. Предпочтительными являются антитела или биспецифические антигенсвязывающие молекулы, обладающие высокими терапевтическими индексами. Согласно одному варианту осуществления изобретения, антитело или биспецифическая антигенсвязывающая молекула демонстрирует высокий терапевтический индекс. Данные, полученные на основании исследований клеточных культур и животных, могут быть использованы для определения диапазона доз, подходящих для использования у людей. Дозировки, предпочтительно, находятся в диапазоне циркулирующих концентраций, которые включают ED50 с небольшой токсичностью или без нее. Дозировки могут варьировать в указанном диапазоне, что зависит от множества факторов, например, используемой лекарственной формы и способа введения, общего состояния субъекта и подобных факторов. Конкретная лекарственная форма, способ введения и дозировка выбираются лечащим врачом с учетом состояния пациента (см., например, Fingl et al., 1975, в: The Pharmacological Basis of Therapeutics, Ch. 1, p. 1, включен в настоящее описание в качестве ссылки во всей своей полноте).
Лечащий врач пациентов, получающих лечение антителами или биспецифическими антигенсвязывающими молекулами согласно изобретению, будет знать, как и когда прекратить, прервать или скорректировать введение лекарственных препаратов из-за возникшей цитотоксичности, дисфункции органов или подобных побочных эффектов. И наоборот, лечащий врач также будет знать, как скорректировать лечение и увеличить дозу препаратов в том случае, если клинический эффект не был адекватным (таким образом, чтобы избежать токсичности). Величина вводимой дозы для лечения заболевания, представляющего интерес, будет зависеть от тяжести этого заболевания, способа введения препаратов и подобных факторов. Тяжесть заболевания может оцениваться, например, частично при помощи стандартных методов оценки прогноза. Кроме того, доза и, возможно, частота введения также будут меняться в зависимости от возраста, массы тела и ответа на лечение конкретного пациента.
Другие агенты и способы лечения
Антитела и биспецифические антигенсвязывающие молекулы согласно изобретению в составе терапии могут вводиться в комбинации с одним или несколькими другими агентами. Например, антитело или биспецифическая антигенсвязывающая молекула, согласно изобретению, может вводиться по меньшей мере вместе с одним дополнительным терапевтическим агентом. Термин "терапевтический агент" относится к любому агенту, применяемому для лечения симптома или заболевания у индивидуума, нуждающегося в таком лечении. Такой дополнительный терапевтический агент может включать любые активные ингредиенты, подходящие для лечения конкретного состояния, подлежащего лечению, предпочтительно ингредиенты с комплементарными активностями, которые негативно не влияют друг на друга. Согласно определенным вариантам осуществления изобретения, дополнительный терапевтический агент представляет собой иммуномодулирующий агент, цитостатический агент, ингибитор клеточной адгезии, цитотоксический агент, активатор клеточного апоптоза или агент, повышающий чувствительность клеток к индукторам апоптоза. Согласно определенному варианту осуществления изобретения, дополнительный терапевтический агент представляет собой противораковое средство, например, агент, разрушающий микротрубочки, антиметаболический агент, ингибитор топоизомеразы, интеркалятор ДНК, алкилирующий агент, агент для гормональной терапии, ингибитор киназы, антагонист рецепторов, активатор апоптоза опухолевых клеток или агент, подавляющий ангиогенез.
Такие дополнительные агенты, соответственно, в составе комбинации находятся в количествах, эффективных для достижения имеющейся цели. Эффективное количество таких агентов зависит от количества используемого антитела или биспецифической антигенсвязывающей молекулы, характера заболевания и применяемого лечения, а также других факторов, описанных выше. Антитела или биспецифичные антигенсвязывающие молекулы обычно применяются в тех же дозах и вводятся теми же способами, как описано в настоящем документе, либо в дозах, которые составляют, приблизительно, от 1 до 99% от доз, указанных в настоящем документе; либо в любой дозе и посредством любого способа введения, который эмпирически/клинически был определен как подходящий.
Подходы к комбинированной терапии, описанные выше, включают как комбинированное введение композиций (когда два или более терапевтических агента вводятся в составе одной или отдельных композиций), так и раздельное введение, при котором введение антитела или биспецифической антигенсвязывающей молекулы согласно изобретению осуществляют до, одновременно и/или после введения дополнительного терапевтического агента и/или адъюванта. Антитела или биспецифичные антигенсвязывающие молекулы, согласно изобретению, также можно использовать в сочетании с лучевой терапией.
Промышленные изделия
Согласно другому аспекту, настоящее изобретение относится к промышленному изделию, включающему материалы, полезные для лечения, профилактики и/или диагностики заболеваний, описанных выше. Промышленное изделие включает контейнер и этикетку или листок-вкладыш внутри контейнера или близи него. Подходящими контейнерами являются, например, пузырьки, флаконы, шприцы, мешки для внутривенных растворов и т.д. Контейнеры могут быть сделаны из различных материалов, таких, как стекло или пластик. Контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией является эффективной для лечения, профилактики и/или диагностики заболевания; а также стерильный порт для доступа (например, контейнер может представлять собой мешок для внутривенных растворов или флакон, имеющий пробку, которую протыкают инъекционной иглой). По меньшей мере один активный агент в составе композиции представляет собой антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению. Этикетка или листок вкладыш указывают на то, что композиция применяется для лечения приоритетного заболевания. Кроме того, промышленное изделие может включать (а) первый контейнер, содержащий композицию, при этом композиция включает антитело или биспецифическую антигенсвязывающую молекулу согласно изобретению; и (b) второй контейнер, содержащий композицию, при этом указанная композиция включает дополнительный цитотоксический или другой терапевтический агент. Промышленное изделие согласно данному варианту осуществления изобретения может дополнительно содержать листок-вкладыш, указывающий на то, что композиция может применяться для лечения определенного заболевания. Альтернативно или дополнительно, промышленное изделие может дополнительно включать второй (или третий) контейнер, содержащий фармацевтически доступный буфер, такой, как вода для инъекций с бактериостатическими свойствами (BWFI), физиологический раствор с фосфатным буфером, раствор Рингера или раствор декстрозы. Оно также может дополнительно включать другие материалы в зависимости от коммерческих целей и пожеланий пользователя, содержащую другие буферные растворы, растворители, фильтры, игры и шприцы.
Способы и композиции для диагностики и обнаружения
Согласно определенным вариантам осуществления изобретения, любое из анти- HLA-A2/WT1 антител, заявленное в настоящем документе, является полезным для обнаружения присутствия HLA-A2/WT1 в биологическом образце. Термин "обнаружение", используемый в настоящем описании, относится как к количественному, так и к качественному обнаружению.
Согласно определенному варианту осуществления изобретения, биологический образец представляет собой клетку или ткань, например, ткань предстательной железы. Согласно одному варианту осуществления изобретения, предлагается анти- HLA-A2/WT1 антитело, предназначенное для применения согласно способу диагностики или обнаружения. Согласно дополнительному аспекту изобретения, предлагается способ обнаружения присутствия HLA-A2/WT1 в биологическом образце. Согласно определенным вариантам осуществления изобретения, указанный способ включает обеспечение контакта биологического образца с анти- HLA-A2/WT1 антителом, как описано в настоящем документе, в условиях, обеспечивающих связывание анти- HLA-A2/WT1 антитела с HLA-A2/WT1; а также определение формирования комплекса между анти- HLA-A2/WT1 антителом и HLA-A2/WT1. Такой способ может применяться как in vitro, так и in vivo. Согласно одному варианту осуществления изобретения, анти- HLA-A2/WT1 антитело применяется для отбора субъектов, подходящих для терапии с помощью анти- HLA-A2/WT1 антитела, то есть HLA-A2/WT1 является биомаркером для отбора пациентов.
Примерами заболеваний, которые можно диагностировать с использованием антитела, заявленного в соответствии с изобретением, является рак, в частности, рак предстательной железы.
Согласно конкретному варианту осуществления изобретения, предлагаются меченые анти- HLA-A2/WT1 антитела. Метки включают, без ограничений указанными, метки или вещества, распознаваемые напрямую (такие, как флуоресцентные, хромоформные, электронноплотные, хемилюминисцентные и радиоактивные метки), а также вещества, такие, как ферменты или лиганды, которые распознаются опосредованно, например, при помощи ферментативных реакций или молекулярных взаимодействий. Примерами меток являются, без ограничений указанными, радиоизотопы 32Р, 14С, 125I, 3Н, и 131I, флуорофоры, такие, как хелаты редкоземельных элементов или флуресцин и его производные, родамин и его производные, данзил, умбеллиферон, люцериферазы, например, люцифераза светлячков и бактериальная люцифераза (см, патент США No. 4,737,456), люциферин, 2,3-дигидрофталазинедионы, пероксидаза хрена (HRP), щелочная фосфатаза, β-галактозидаза, глюкоамилаза, лизозим, сахарид оксидазы, например, глюкозооксидаза, галактозооксидаза и глюкозо-6-фосфат дегидрогеназа, гетероциклические оксидазы, такие, как уриказа и ксантиноксидаза, объединенные с ферментом, который использует перекись водорода для окисления предшественника красителя, таким, как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и подобные вещества.
Примеры
Ниже приведены примеры способов и композиций, согласно изобретению. Следует понимать, что на основе приведенного выше описания можно осуществить и другие варианты изобретения.
Пример 1. Получение Fab, связывающихся с HLA-A2/WT1
Отбор и скрининг анти- HLA-A2/WT1 Fabs
Ahth-HLA-A2/WT1 Fabs были отобраны фагово-дисплейным методом из синтетических Fab-библиотек полностью на основе каркасных областей антител человека с разнообразными последовательностями CDR3-VL (3 различные длины) и VH-доменов (6 различных длин).
Раунды отбора ("биопэннинг") осуществляли в растворе в соответствии со следующим протоколом: 1. Предварительная очистка примерно 1012 фагмидных частиц в расчете на библиотечный пул на покрытых нейтравидином 96-луночных планшетах, обработанных 500 нМ неродственного биотинилированного HLA-A2/WT1 VLD-комплекса. 2. Инкубирование не связавшихся с HLA-A2/WT1VLD фагмидных частиц в 100 нМ биотинилированного HLA-A2/WT1RMF-комплекса в течение 5 ч в общем объеме 800 мкл. 3. Захват биотинилированного HLA-A2/WT1RMF и специфическое связывание фагов путем добавления 80 мкл покрытых стрептавидином магнитных бус в течение 20 мин в шейкере. 4. Отмывание соответствующих магнитных бус 5-10 раз 1 мл PBS/Tween 20 и 5-10 раз 1 мл PBS при использовании сепаратора магнитных бус.5. Элюирование фаговых частиц добавлением 1 мл 100 мМ триэтиламина (TEA) в течение 5-10 мин и нейтрализация добавлением 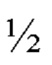 объема 1 М Трис/HCl, рН 7,4. 6. Повторное заражение клеток Е. coli TG1 в логарифмической фазе роста элюированными фаговыми частицами, заражение хелперным фагом VCSM13, инкубирование в шейкере при 30°С в течение ночи и последующая преципитация ПЭГ/NaCl фагмидных частиц, подлежащих использованию в следующем раунде отбора.
объема 1 М Трис/HCl, рН 7,4. 6. Повторное заражение клеток Е. coli TG1 в логарифмической фазе роста элюированными фаговыми частицами, заражение хелперным фагом VCSM13, инкубирование в шейкере при 30°С в течение ночи и последующая преципитация ПЭГ/NaCl фагмидных частиц, подлежащих использованию в следующем раунде отбора.
Осуществляли 3-4 раунда отбора при использовании постоянной концентрации антигена 100 нМ.
В дополнение к отбору при постоянной концентрации антигена отбор также осуществляли с понижающими концентрациями антигена 100 нм, 50 нМ, 10 нМ и 5 нМ для селекции антител с более низкой аффинностью.
Исследования связывания HLA-A2/WT1: сэндвич-ELISA для характеристики Fabs, полученных методом фагового дисплея
Индивидуальные клоны экспрессировали в бактериях в виде 1 мл-культур в 96-луночном планшете и супернатанты подвергали скринингу путем ELISA. Специфические связывающие молекулы идентифицировали как имеющие сигналы, в пять раз превышающие значения фона для HLA-A2/WT1RMF, и имеющие сигналы, в три раза меньшие, чем значения фона, для HLA-A2/WT1VLD. Более точно, нейтравидиновые 96-луночные планшеты (Thermo Fisher) покрывали 10 нМ HLA-A2/WT1RMF или 50 нМ HLA-A2/WT1VLD при 37°С в течение 30 мин и затем планшеты блокировали 2% (вес/объем) фосфатным буферным раствором с молоком (MPBS) из расчета 200 мкл/лунку в течение 1 ч при комнатной температуре. Планшеты отмывали 3 раза PBS, затем добавляли содержащие Fab бактериальные супернатанты и планшеты инкубировали при комнатной температуре в течение 1 ч. После трехкратного отмывания планшетов PBS добавляли вторые анти-FLAG-HRP антитела (Sigma, Cat. No. А8592) (разведение 1:4000) и планшеты инкубировали в шейкере в течение 1 ч. при комнатной температуре. Далее планшеты 3 раза отмывали PBS и добавляли 100 мкл/лунку ВМ Blue POD (Roche) для развития окраски. Энзиматическую реакцию останавливали добавлением 50 мкл/лунку 1М H2SO4. Оптическую плотность (OD) считывали при 450 нм (референсное значение соответствует 650 нм) для окончательного установления OD450-650. ELISA-положительные клоны подвергали экспериментальному кинетическому скринингу, как описано ниже.
Исследования связывания HLA-A2/WT1: поверхностный плазмонный резонанс для кинетической характеристики Fabs, полученных методом фагового дисплея
Специфические связывающие молекулы идентифицировали методом поверхностного плазмонного резонанса, согласно которому скринингу подвергали содержащие Fabs супернатанты бактериальных культур при использовании биосенсора ProteOn XPR36 (BioRad). Коротко говоря, после заражения клеток Е. coli TG1 в логарифмической фазе роста элюированными фаговыми частицами, отбирали единицы, формирующие одиночные колонии (cfu), и инокулировали ими среду для экспрессии в объеме 1 мл в лунках 96-луночного планшета.
Все эксперименты проводили при 25°С при использовании PBST в качестве подвижного буфера (10 мМ PBS, рН 7,4 и 0,005% (вес/объем) Tween 20). Использовали биосенсор ProteOn XPR36, оснащенный сенсорными чипами GLC и GLM, и связывающими реагентами (10 мМ ацетат натрия, рН 4,5, сульфо-N-гидроксисукцинимид [сульфо-NHS], 1-этил-3-(3-диметиламинопропил)-карбодиимид гидрохлорид [EDC] и этаноламин), от BioRad Inc. (Hercules, СА).
Иммобилизацию осуществляли при скорости 30 мкл/мин на чипе GLM. Антитела козы (pAb) против IgG человека, представляющие собой специфические анти-F(ab)2 антитела (Jackson ImmunoResearch), связывали в вертикальном направлении при использовании стандартной процедуры аминного связывания: все шесть лигандных каналов были активированы в течение 5 мин смесью EDC (200 мМ) и сульфо-NHS (50 мМ). Немедленно после активации поверхности инъецировали антитела козы (pAb) против IgG человека, представляющие собой специфические анти-F(ab)2 антитела (50 мкг/мл, 10 мМ ацетат натрия, рН 4,5), через все шесть каналов в течение 5 мин. В заключение каналы блокировали 5-минутной инъекцией 1 М этаноламином-HCl (рН 8,5). Окончательные уровни иммобилизации были схожими на всех каналах, варьируя от 11000 до 11500 RU. Fab-варианты были захвачены из супернатантов Е. coli путем одновременной инъекции вдоль пяти отдельных горизонтальных каналов (30 мл/мин) в течение 5 мин и результирующие уровни варьировали от 200 до 900 RU, в зависимости от концентрации Fab в супернатанте. Кондиционированную среду инъецировали вдоль шестого канала для обеспечения "внутрилинейного" незаполненного двойного контроля.
Одноразовые кинетические измерения проводили путем инъекции серийных разведений HLA-A2/WT1RMF или HLA-A2/WT1V1d (100, 50, 25, 12,5, 6, 25,0 нМ, 50 мкл/мин) в течение 2 мин вдоль вертикальных каналов. Осуществляли мониторинг диссоциации в течение 3 мин. Кинетические данные анализировали в биосенсоре ProteOn, версия 2.1. Обработка результатов реакции, представленных пятнами, включала применение стадии сравнения полученных пятен и двойного контроля с "внутрилинейным" незаполненным буферным контролем (Myszka, J. Mol. Recognit. (1999) 12, 279-284). Обработанные данные повторных одноразовых повторных инъекций подгоняли к простому отношению 1:1 связывающей модели Лангмура без транспортного массового переноса (O'Shannessy et. al., Anal. Biochem (1993) 212, 457-468).
Для измерения IgG из супернатантов, полученных при культивировании клеток НЕК в лунках 6-луночного планшета, варианты IgG захватывали из супернатантов НЕК293 путем одновременной инъекции вдоль пяти отдельных горизонтальных каналов (30 мл/мин) в течение 5 мин и результирующие уровни варьировали от 200 до 900 RU. Кондиционированную среду инъецировали вдоль шестого канала для обеспечения "внутрилинейного" незаполненного двойного контроля. Одноразовые кинетические измерения проводили путем инъекции серийных разведений HLA-A2/WT1RMf или HLA-A2/WT1 Vld (100, 50, 25, 12,5, 6, 25, 0 нМ, 50 мкл/мин) в течение 2 мин вдоль вертикальных каналов. Осуществляли мониторинг диссоциации в течение 3 мин. Кинетические данные анализировали как описано выше.
На основании профиля связывания и измеренной специфичности связывания с антигеном составляли шорт-лист связывающих молекул и проводили исследования связывания в клетках.
Последовательности всех отобранных связывающих молекул (11D06, 33Н09, 13 В04, 11 В09, 33F05, 5Е11, 13Е08, 5С01, 11G06) приведены в списке последовательностей, включенном в описание, и суммированы в Таблице 1, показанной ниже.
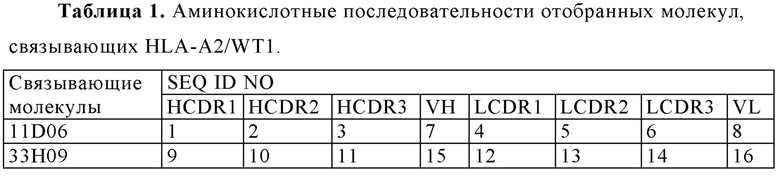
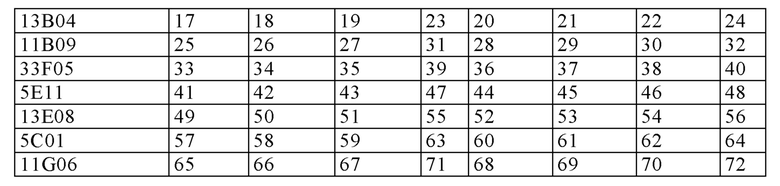
Пример 2. Получение HLA-A2/WT1 IgG и HLA-A2/WT1 × CD3 биспецифических антител
Клонирование
кДНК, кодирующие белки, были клонированы в векторной системе (Evitria) при использовании подходящих (не основанных на ПЦР) методик клонирования. Были синтезированы генные векторные плазмиды. Плазмидную ДНК получали в условиях низкого содержания эндотоксина на основе анионообменной хроматографии. Концентрацию ДНК определяли измерением абсорбции при длине волны 260 нм. Правильность последовательностей подтверждали секвенированием по Сэнгеру (до двух реакций секвенирования на плазмиду в зависимости от размера кДНК).
Продукция антител
IgG антитела и биспецифические антитела получали путем транзиторной трансфекции клеток НЕК293 EBNA (культивированных в суспензионной бессывороточной среде Excell). Клетки центрифугировали и среду замещали предварительно подогретой средой CD СНО. Векторы экспрессии смешивали в среде CD СНО, затем добавляли полиэтиленимин (PEI), раствор перемешивали в вортекс-мешалке и инкубировали в течение 10 мин при комнатной температуре. Далее клетки смешивали с раствором ДНК/PEI, переносили в шейкер и инкубировали в течение 3 ч при 37°С в термостате в атмосфере 5% СО2. После инкубации вносили среду Excell с добавками. Через один день после трансфекции вносили добавки. Клеточные супернатанты собирали через семь дней и очищали стандартными методами.
Альтернативно, для получения антител были использованы адаптированные к росту в суспензии клетки СНО K1 (полученные из АТСС и адаптированные к росту в суспензионной культуре в бессывороточной среде). Клетки выращивали в бессывороточной среде eviGrow, химически модифицированной, свободной от компонентов животного происхождения. Клетки трансфицировали реагентом трансфекции eviGrow и после трансфекции выращивали в бессывороточной среде eviMake2, свободной от компонентов животного происхождения. Супернатанты собирали центрифугированием и последующей фильтрацией (0,2-мкм фильтр).
Очистка
Белки очищали из отфильтрованных клеточных супернатантов в соответствии со стандартными протоколами. Коротко говоря, Fc-содержащие белки очищали из клеточных культуральных супернатантов аффинной хроматографией при использовании протеина А (колонка HiTrap Protein A, GE Healthcare). Элюцию осуществляли при рН 3,0 и далее образец немедленно нейтрализовали. Белок концентрировали и агрегированный белок отделяли от мономеров эксклюзионной хроматографией по размеру (колонка HiLoad Superdex 200, GE Healthcare) в 20 мМ гистидине, 140 мМ хлориде натрия, рН 6,0.
Методы анализа
Концентрацию очищенных белков определяли путем измерения оптической плотности (OD) при 280 нм при использовании молярного коэффициента экстинкции, вычисленного на основе аминокислотной последовательности согласно Расе et al. (Protein Science, 1995, 4, 2411-1423). Чистоту и молекулярный вес белков анализировали CE-SDS в присутствии и в отсутствие восстанавливающего агента с использованием системы LabChipGXII (Caliper Lifescience). Содержание агрегатов определяли HPLC-хроматографией с применением аналитической эксклюзионной колонки с разделением по размеру (TSKgel G3000 SW XL, Tosoh), уравновешенной при 25°С подвижным буфером, содержащим 25 мМ K2HPO4, 125 мМ NaCl, 200 мМ L-аргинин моногидрохлорида, рН 6,7.
CD3 биспецифические антитела также называются здесь "Т-клеточными биспецифическими антителами" или "TCBs". Схематическое изображение биспецифических антител, полученных согласно данному примеру, приведено на фиг. 2.
Примерами последовательностей TCBs являются SEQ ID Nos 123, 124, 125 и 129 (11D06-TCB) и SEQ ID Nos 126, 127, 128 и 129 (33Н09-ТСВ). Другие TCBs были сконструированы аналогичным образом при использовании последовательностей VH и VL соответствующих HLA-A2/WT1-связывающих молекул.
В качестве контроля было получено HLA-A2/WT1 × CD3 биспецифическое антитело (ТСВ), основанное на связывающих молекулах, сходных с антителом ESK1 (Dao et al., Sei. Transl. Med. (2013), 5, 176ra33; WO2012/135854), которое обозначается здесь как "ESK1-TCB" (см. SEQ ID Nos 73 и 74 для последовательностей вариабельных районов), а также, как "нетаргетное антитело ТСВ" (см. SEQ ID Nos 75 и 76 для последовательностей вариабельных районов).
Все молекулы были получены и очищены сходными методами. Качество всех полученных молекул было хорошим, практически со 100%-ным содержанием мономеров и 100%-ной степенью чистоты, подтвержденной СЕ-SDS.
Пример 3. Биохимический анализ аффинности и авидности HLA-A2 /WT1 × CD3 биспецифических антител
Для определения аффинности HLA-A2/WT1 × CD3 биспецифических антител по отношению к HLA-A2/WT1rmf осуществляли поверхностный плазмонный резонанс (SPR) при 25°С в системе Biacore Т200 с подвижным буфером HBS-EP (0,01 М HEPES, рН 7,4, 0,15 М NaCl, 0,005% сурфактант р20 (GE Healthcare). Специфическое антитело к Fc человека (GE Healthcare, номер по каталогу BR-1008-39) непосредственным образом иммобилизовали путем аминного связывания на чипе СМ5 (GE Healthcare). Биспецифические антитела захватывали в течение 30 с при 5 нМ. Серии трехкратных разведений комплекса HLA-A2/WT1rmf в HBS-EP (от 1,03 до 250 нМ) пропускали через лиганд при 30 мкл/мин в течение 120 с для регистрации фазы ассоциации. Проводили мониторинг фазы диссоциации в течение 120 с, которую запускали заменой раствора образца на HBS-EP. Поверхность чипа регенерировали после каждого цикла инъекцией 3 М MgCl2 при 10 мкл/мин в течение 30 с. Различия в показателях преломления в общем объеме корректировали вычитанием ответа, полученного на референсном потоке клеток (который содержит антитело к Fc человека, но без биспецифической конструкции, захваченной на нем). Константы аффинности выводили, исходя из констант кинетической скорости путем подгонки к отношению 1:1 связывающей модели Лангмура при использовании программного обеспечения BIAeval (GE Healthcare).
Для определения авидности и специфичности HLA-A2/WT1 × CD3 биспецифических антител по отношению к HLA-A2/WT1rmf опять осуществляли поверхностный плазмонный резонанс (SPR) при 25°С в системе Biacore Т200 с подвижным буфером HBS-EP (0,01 М HEPES, рН 7,4, 0,15 М NaCl, 0,005% сурфактант р20 (GE Healthcare). Анти-His антитело (Penta His, Qiagen, номер по каталогу 34660) непосредственным образом иммобилизовали путем аминного связывания на чипе СМ5 (GE Healthcare). HLA-A2/WT1RMf или HLA-A2/WT1Vld захватывали в течение 30 с и 10 мкл/мин при 5 или 10 нМ (для измерения биспецифического антитела или IgG, соответственно). Серии трехкратных разведений биспецифических конструкций в HBS-EP (от 1,23 до 100 нМ) пропускали через лиганд при 30 мкл/мин в течение 120 с для регистрации фазы ассоциации. Проводили мониторинг фазы диссоциации в течение 240 с, которую запускали заменой раствора образца на HBS-EP. Поверхность чипа регенерировали после каждого цикла инъекцией 10 М глицина, рН 2, в течение 60 с. Различия в показателях преломления в общем объеме корректировали вычитанием ответа, полученного на референсном потоке клеток (который содержал анти-His антитело, но без HLA-A2/WT1RMF, захваченного на нем). Даже если взаимодействие составляет 2:1 (аналит является бивалентным), константы аффинности выводят, исходя из констант кинетической скорости путем подгонки к отношению 1:1 связывающей модели Лангмура при использовании программного обеспечения BIAeval (GE Healthcare). Эти результаты, выраженные в очевидной KD, соответствуют авидности взаимодействия.
Результаты указанных экспериментов суммированы в Талице 2 ниже. Оба тестируемых HLA-A2/WT1-антитела связывают HLA-A2/WT1RMF с двузначной наномолярной (моновалентной) аффинностью/трехзначной пикомолярной (бивалентной) аффинностью (авидностью) и сохраняют ту же самую аффинность/авидность при конвертации из IgG в биспецифический формат. В то время как аффинность двух тестируемых антител различается примерно в два раза, авидность обеих молекул находится в одинаковых пределах.
Не обнаружено связывания тестируемых HLA-A2/WT1-антител с HLA-A2/WTvld (данные не приведены).
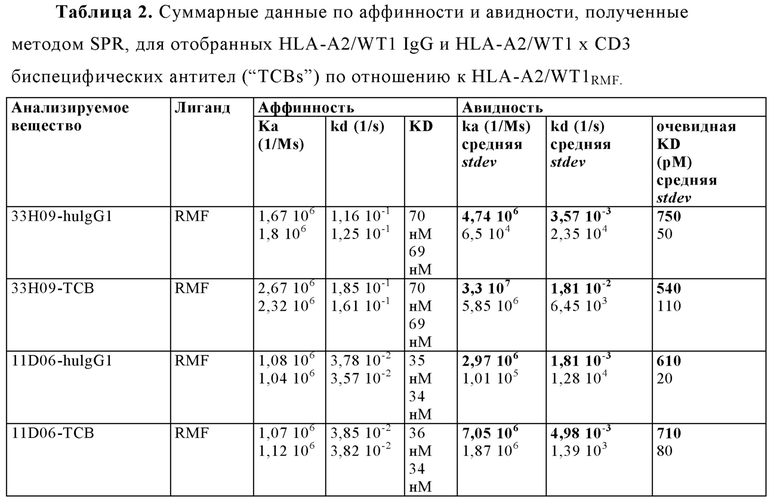
Для определения аффинности HLA-A2/WT1 × CD3 биспецифических антител по отношению к CD3 осуществляли поверхностный плазмонный резонанс (SPR) при 25°С в системе Biacore Т200 с подвижным буфером HBS-EP (0,01 М HEPES, рН 7,4, 0,15 М NaCl, 0,005% сурфактант р20 (GE Healthcare). Специфическое антитело к Fc человека (GE Healthcare, номер по каталогу 28-9583-25) непосредственным образом иммобилизовали путем аминного связывания на чипе СМ5 (GE Healthcare). Биспецифические антитела захватывали в течение 40 с при 5 нМ. Серии трехкратных разведений молекулы слияния CD3e8-Fc в HBS-EP (от 12,35 до 3000 нМ) пропускали через биспецифические антитела при 30 мкл/мин в течение 240 с для регистрации фазы ассоциации. Проводили мониторинг фазы диссоциации в течение 240 с, которую запускали заменой раствора образца на HBS-EP. Поверхность чипа регенерировали после каждого цикла двойной инъекцией 10 мМ глицина-HCl, рН 2,1, при 30 мкл/мин в течение 60 с. Различия в показателях преломления в общем объеме корректировали вычитанием ответа, полученного на референсном потоке клеток, (который содержит антитело к Fc человека, но без биспецифической конструкции, захваченной на нем). Константы аффинности выводили, исходя из констант кинетической скорости путем подгонки к отношению 1:1 связывающей модели Лангмура при использовании программного обеспечения BIAeval (GE Healthcare).
Результаты экспериментов суммированы в Таблице 3. Поскольку связывающая CD3 молекула идентична обеим тестируемым биспецифическим молекулам, как и ожидалось, их аффинность по отношению к CD3 практически одинакова.
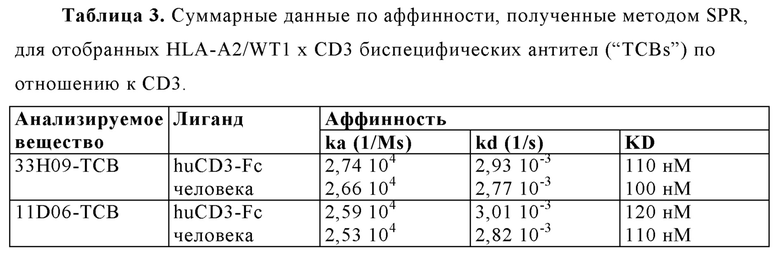
Пример 4. Отбор IgG-связывающих молекул на специфичность связывания с НЬА-А2/\¥Т1-пептидами RMF и VLD
Авторы изобретения впервые при проточной цитометрии измерили специфичность связывания обработанных пептидом клеток Т2 IgG-связывающими молекулами, полученными фагово-дисплейным методом. Коротко говоря, Т2-клетки приготавливали в виде клеточной суспензии при концентрации 106 клеток/мл в среде IMDM (Gibco от Life Technologies, номер по каталогу 31980-048) с добавлением 10% FBS (Gibco, номер по каталогу 16140-071)+1% пенициллина-стрептомицина (Gibco, номер по каталогу 15070-063) (полная среда). Клетки помещали в общем объеме 10 мл в пробирку и инкубировали с 10 мкл WT1 р37-45 VLD-пептида (SEQ ID NO: 77) или р126-134 RMF-пептида (SEQ ID NO: 78) с концентрацией 102 М (конечная концентрация пептида 10-5 М) в течение 2 ч при 37°С в атмосфере 5% СО2. После отмывания клетки суспендировали в холодном PBS и инкубировали с титруемыми концентрациями IgG-связывающих молекул (от 10 мкг/мл до 0,00064 мкг/мл) в течение 1 ч при 4°С. Далее клетки инкубировали со вторыми антителами к IgG-Fc человека, конъюгированными с фикоэритрином (РЕ) (Jackson Laboratories, номер по каталогу 109-116-098) в течение 30 мин. Клетки подвергали анализу методом FACS LSR II (BD) и данные представляли как среднее значение интенсивности флуоресценции (MIF) фикоэритрина при использовании программы Graphpad Prism.
Как показано на Фиг. 3, HD06-IgG, 33H09-IgG и 5E11-IgG все связываются с обработанными RMF-пептидом Т2-клетками, но не связываются с необработанными или обработанными VLD-пептидом Т2-клетками. В противоположность этому, HB09-IgG, 13B04-IgG и 5C01-IgG (но не HG06-IgG) все специфически связываются с VLD-пептидом, но с RMF-пептидом. На основании этих данных были отобраны связывающие молекулы для преобразования в CD3-биспецифические антитела (TCBs).
Пример 5. Активация Т-клеток в маркерном исследовании NFAT-Jurkat при определении связывании HLA-A2/WT1 × CD3 биспецифических антител ("TCBs") с обработанными пептидом Т2-клетками
Для проверки специфичности HLA-A2/WT1 × CD3 биспецифических антител ("TCBs") клетки маркерной клеточной линии Jurkat, которые экспрессируют люциферазу под контролем промотора NFAT (Jurkat-NFAT; Promega, номер по каталогу CS176501), отмывали для измерения активации Т-клеток, когда TCBs связываются с обработанными пептидом Т2-клетками (АТСС, номер по каталогу CRL-1992). Коротко говоря, Т2-клетки приготавливали в виде клеточной суспензии при концентрации 106 клеток/мл в среде IMDM (Gibco от Life Technologies, номер по каталогу 31980-048) с добавлением 10% FBS (Gibco, номер по каталогу 16140-071)+1% пенициллина-стрептомицина (Gibco, номер по каталогу 15070-063) (полная среда). Клетки помещали в общем объеме 10 мл в пробирку и инкубировали с 10 мкл WT1 р37-45 VLD-пептида или р126-134 RMF-пептида с концентрацией 10-2 М (конечная концентрация пептида 10-5 М) в течение 2 ч при 37°С в атмосфере 5% СО2. После отмывания 90 мкл обработанных пептидом клеток в клеточной суспензии с концентрацией 2,2×105 клеток/мл высевали в лунки 96-луночного планшета для микротитрования с круглым дном (20000 клеток/лунку, ТРР, номер по каталогу 92097) и культивировали совместно с 50 мкл Jurkat -NF AT (клеточная суспензия с концентрацией 2×106 клеток/мл) и с 10 мкл ТСВ в титруемых концентрациях (от 100 мкг/мл до 0,0064 мкг/мл) в течение 16 ч при 37°С в атмосфере 5% СО2. Далее удаляли 50 мкл супернатанта и замещали его реагентом для исследования люциферазы Bright-Glo (Promega, номер по каталогу Е2620) в количестве 100 мкл/лунку для инкубирования при комнатной температуре (RT). Через 5 мин 180 мкл супернатанта переносили в новый чистый планшет для измерения сигнала люминесценции с помощью En Vision (PerkinElmer). Данные представляли в относительных единицах люминесценции (RLU).
Как показано на Фиг. 4, TCBs на основе связывающих молекул 11D06 и 33Н09 (11D06-TCB и 33Н09-ТСВ, соответственно) специфически распознают комплекс HLA-A2/WT1rmf и активируют NFAT на маркерных клетках Jurkat только в присутствии Т2-клеток, обработанных RMF-пептидом. В противоположность указанному, контрольные ТСВ на основе ESK1-подобной связывающей молекулы (ESK1-TCB) не обнаруживают специфичности к RMF-пептиду. Также ТСВ на основе связывающей молекулы 5Е11 (5Е11-ТСВ) активируют NFAT контрольных Т-клеток в присутствии как Т2-клеток, обработанных RMF-пептидом, так и в присутствии как Т2-клеток, обработанных VLD-пептидом и, таким образом, эти ТСВ исключаются из следующего раунда скрининга. Кроме того, были выявлены некоторые специфичные к VLD-пептиду TCBs, идентифицированные как основанные на связывающих молекулах 11 В09 и 13 В04 (11 В09-ТСВ и 13 В04-ТСВ, соответственно). В качестве негативного контроля в данном исследовании использовались "нетаргетные" ТСВ (DP47GS-ТСВ), которые не активируют NFAT на маркерных клетках Jurkat только в присутствии Т2-клеток, обработанных как RMF-пептидом, так и VLD-пептидом. 5С01-ТСВ распознавали VLD-пептид, но перекрестно реагировали с RMF-пептидом при более высоких концентрациях.
Пример 6. Т-клеточная цитотоксичность, опосредуемая связыванием HLA-A2/WT1 × CD3 биспецифических антител ("TCBs") с обработанными пептидом Т2-клетками
Далее анализировали цитотоксичность TCBs. Клетки-мишени представляли собой обработанные пептидом Т2-клетки, как описано в Примере 5. Эффекторные клетки являлись пулом СБ3-клеток, выделенных из PBMCs, полученных из лейкоцитарной пленки путем центрифугирования в градиенте фиколла (GE Healthcare, номер по каталогу 17-1440-03). Собранные СБ3-клетки очищали методом MACS (Miltenyi Biotec) при использовании набора для выделения пула Т-клеток человека (Miltenyi Biotec, номер по каталогу 130-096-535). Исследование цитотоксичности проводили следующим образом: обработанные пептидом клетки (100 мкл) высевали в лунки 96-луночного планшета для микротитрования с круглым дном (3×105 клеток/мл) и культивировали совместно с 50 мкл Т-клеток (6х106 клеток/мл) и с 50 мкл ТСВ в титруемых концентрациях (от 40 мкг/мл до 0,00004 мкг/мл) в полной среде в течение 18 ч при 37°С в атмосфере 5% СО2. Далее 50 мкл супернатанта переносили в новый чистый планшет и добавляли реагент CytoTox-Glo для исследования люциферазы (Promega, номер по каталогу G9291) в количестве 25 мкл/лунку для инкубирования при комнатной температуре (RT) в течение 15 мин. Сигнал люминесценции (для измерения высвобождения LDH в качестве индикатора гибели клеток) считывали с помощью En Vision (PerkinElmer). Данные представляли в относительных единицах люминесценции (RLU).
На Фиг. 5 показано опосредуемое ТСВ специфическое уничтожение Т-клеток, представляющих собой клетки-мишени, экспрессирующие RMF или VLD. Было обнаружено, что как 11D06-TCB, так и 33Н09-ТСВ являются специфическими киллерами обработанных RMF-пептидом Т2-клеток. 33F05-ТСВ не опосредуют специфическое уничтожение Т2-клеток, обработанных как пептидом RMF, так и пептидом VLD. Кроме того, 13 В04-ТСВ и 11 В09-ТСВ опосредуют потенциальное уничтожение Т2-клеток, обработанных пептидом VLD. 5С01-ТСВ способны уничтожать Т2-клетки, обработанные пептидом VLD, но также могут уничтожать и Т2-клетки, обработанные пептидом RMF, при более высоких концентрациях, что согласуется с результатами маркерного исследования NFAT (Фиг. 4).
Пример 7. Т-клеточная цитотоксичность, опосредуемая связыванием HLA-A2/WT1 × CD3 биспецифических антител ("TCBs") с клеточными линиями WT1+
Для подтверждения специфического уничтожения клеток под действием TCBs осуществляли цитотоксическое исследование на опухолевых клеточных линиях WT1. Клеточные линии HLA-A2 WT1 являются SKM-1 клетками (DZMZ No. АСС 547) и клетками СНО, трансфицированными комплексом HLA-A2/WT1RMF (СНО-WT1) (in house). Отрицательным контролем служили HLA-А2+\WT1 -клетки: BJAB (DZMZ No. АСС 757), ARH-77 (DZMZ No. АСС 512) и клетки СНО, трансфицированные комплексом HLA-A2/MAGE-A4 (СНО-MAGEA4) (in house), а также HLA-A2-WT1+ клеточная линия К562 (АТСС No. CLL-243) (Фиг. 6А). Исследование цитотоксичности проводили, как описано в Примере 6. Как 11D06-TCB, так и 33Н09-ТСВ являются потенциальными киллерами клеток SKM-1 и СНО-WT1, но ни одних из HLA-A2+WT1--клеток и HLA-A2-WT1+клеток, что свидетельствует о том, что указанные два антитела TCBs обладают специфичностью по отношению к пептиду RMF WT1 (Фиг. 6 В, С). В противоположность указанному, ни одно из специфичных к VLD-пептиду TCBs не уничтожает HLA-A2WT1+клеточные линии, однако эти антитела показывают низкую способность к потенциальному уничтожению в высоких концентрациях (10 мкг/мл) WT1+ клеточных линий и ту же самую степень уничтожения в отношении WT1- клеточных линий (Фиг. 6D, Е). На основе этих функциональных данных были отобраны 11D06-TCB и 33Н09-ТСВ для дальнейших исследований.
Также было проведено сравнение отобранных TCBs с ESK1-TCB на их способность уничтожать опухолевые клетки-мишени. Интересно, что хотя ESK1-ТСВ и обладают сходным потенциалом опосредованного уничтожения SKM-1 клеток в сравнении с 11D06-TCB и 33Н09-ТСВ, они также индуцируют уничтожение HLA-A2+WT1- -клеток, указывая на то, что связывающая активность ESK1-TCB не ограничена комплексом HLA-A2/WT1RMF (Фиг. 6F, G).
Также была протестирована способность к уничтожению различных опухолевых клеточных линий, которые являются линиями HLA-A2WT1+. На основании значения ЕС50, необходимого для уничтожения опухолевых клеток, клеточные линии классифицировали на те, которые уничтожаются при ЕС50<1 мкМ (отмечены в Таблице 4 "++"), те, которые уничтожаются при ЕС50>1 мкМ и <5 мкМ (отмечены в Таблице 4 "+"), и те, которые практически не уничтожаются ((отмечены в Таблице 4 "нет").
Все отобранные TCBs 11D06-TCB и 33Н09-ТСВ опосредуют Т-клеточную цитотоксичность 6 из 13 HLA-A2+WT1- клеточных линий. Отсутствие способности к уничтожению других клеточных линий, скорее всего, обусловлена низкой экспрессией WT1, презентируемого на клеточной поверхности молекулами МНС I. В таблице 4 приведены результаты исследований для 11D06-ТСВ. Результаты исследований 33Н09-ТСВ являются сходными (причем значения ЕС50 немного выше, чем в случае 11D06-TCB).
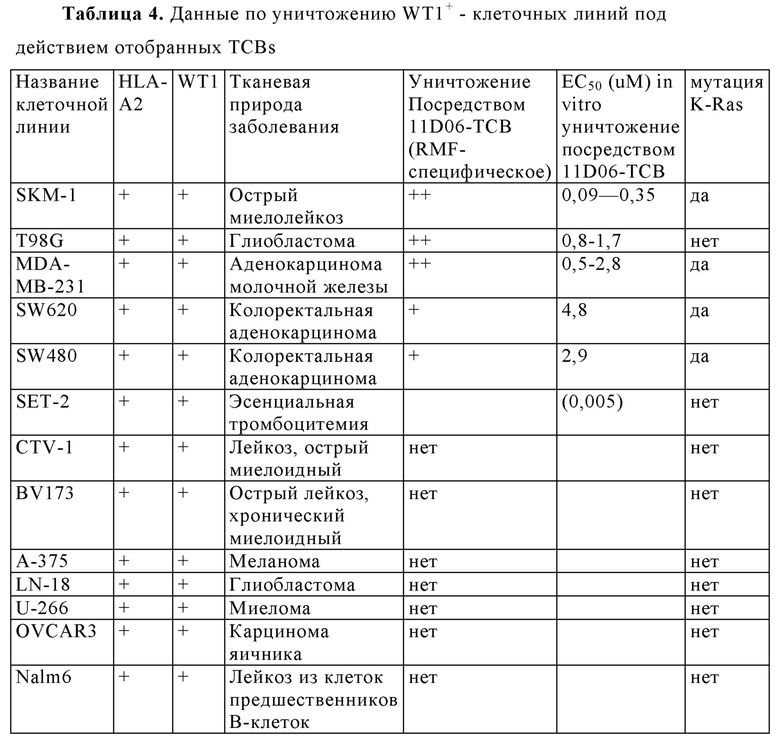
Пример 8. Активация Т-клеток, опосредуемая связыванием HLA-A2/WT1 × CD3 биспецифических антител ("TCBs") с клеточными линиями WT1+
Необходимым условием ТСВ-опосредуемой цитотоксичности по отношению к клеткам мишеням WT1+ является активация Т-клеток для приобретения ими эффекторной функции. Активационный статус Т-клеток измеряли проточной цитометрией в совместных культурах Т-клеток и HLA-А2+WTT--клеток SKM-1 или HLA-A2+WT1--клеток BJAB в присутствии двух отобранных TCBs, а именно 11D06-TCB и 33Н09-ТСВ, во время исследования по уничтожению клеток in vitro, как описано в Примере 7. Клетки собирали через 18 ч совместного инкубирования и окрашивали антителами против CD3 (Biolegend, номер по каталогу 300321) и против CD25 (Biolegend, номер по каталогу 302606) для измерения активации Т-клеток проточной цитометрией.
Как показано на Фиг. 7, оба TCBs индуцируют повышающую регуляцию CD69 и CD25 на CD3+ Т-клетках при связывании с клетками SKM-1, но не с клетками BJAB, указывая на то, что специфическое распознавание TCBs комплекса HLA-A2/WT1RMX, презентируемого SKM-1 клетками, запускает CD3-опосредуемую активацию Т-клеток, очевидным образом приводя к специфическому лизису клеток-мишеней.
Пример 9. Отсутствие связывания отобранных HLA-A2/WT1 × CD3 биспецифических антител ("TCBs") с маркерными пептидами, не являющимися целевыми мишенями
Основной потенциальный риск при получении антител, подобных Т-клеточному рецептору (TCR), заключается в перекрестной реактивности антитела с пептидами-гомологами, присутствующими среди пептидов, экспрессируемых здоровыми тканями. Как сообщалось Ataie et. al. (J. Mol. Biol. (2016) 428:194-205), ESK-1 антитело связывается пептидами, сходными по аминокислотным последовательностям с пептидами, происходящими из белков MED13L и PIGO. Поэтому авторы изобретения протестировали полученные TCBs на их перекрестную реактивность с указанными двумя пептидами. Последовательности этих пептидов приведены на Фиг. 8F и обозначены как SEQ ID Nos 79 и 80. При проточной цитометрии на основе исследования связывания, как описано в Примере 4, было обнаружено, что оба антитела 11D06-TCB и 33Н09-ТСВ не связывают пептид PIGO. Связывание пептида MED13L обеими TCBs было примерно в 100 раз меньшим по сравнению со связыванием нативного WT1 RMF-пептида (Фиг. 8А, В). Несмотря на то, что ESK-1-TCB связывается с RMF-пептидом с низкой аффинностью, оно связывает пептид PIGO сходным образом и даже более сильно связывается с MED13L (Фиг. 8С), что согласуется с опубликованными данными по связывающей активности ESK-1 IgG (Ataie et. al., J. Mol. Biol. (2016) 428:194-205)).
Также была протестирована перекрестная реактивность при использовании NFAT-маркерного исследования, как описано в Примере 5. В соответствии с данными по связыванию, величина активации NFAT на клетках Jurkat при связывании с MED13L и PIGO по меньшей мере в 100 раз слабее, чем величина активации при связывании WT1 RMF-пептида антителами 11D06-TCB и 33Н09-ТСВ (Фиг. 8D, Е).
Пример 9. Отсутствие связывания отобранных HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") с неидентифицированными пептидами, не являющимися целевыми мишенями
Перекрестная реактивность с гомологичными пептидами (в основе лежит распознавание остатков), происходящих из здоровых тканей, может вызвать серьезную и не сразу предсказуемую токсичность при терапии TCR-подобными антителами или при основанной на TCR Т-клеточной терапии (см., например, Linette et al., Blood (2013) 122:863-71; где сообщается о летальной токсичности терапии MAGE A3-TCR, обусловленной взаимодействием антител с белком, экспрессируемым сердечной тканью, который не является их мишенью). Таким образом, специфичность указанных агентов является критическим моментом. Для того, чтобы досконально установить специфичность полученных антител на основе данных по кристаллической структуре, которые подтверждают локализацию в RMF-пептиде положений аминокислотных остатков, вовлеченных в связывание с 11D06-TCB (см. Пример 15), в исследование были включены пептиды, не являющиеся возможными мишенями антител и имеющие аминокислотные последовательности, сходные с последовательностью WT1 RMF-пептида (SEQ ID No: 78), путем поиска пептидов, содержащих замаскированный участок RxxPNxxYx, в пептидомной (peptidome) базе данных (Swissprot TrEMBL). Было обнаружено дополнительно 25 пептидов, происходящих из различных белков, все из которых обладали предсказанной аффинностью к HLA-A2 (Таблица 5). Затем была протестирована перекрестная реактивность отобранных TCBs на обработанных пептидами Т2-клетках при использовании NFAT-маркерного исследования (пептиды 1-6) или Т-клеточного цитотоксического исследования (пептиды 7-25), как описано ниже.
Для первых шести протестированных пептидов не было обнаружено перекрестной реактивности ни к одному пептиду у антитела 11D06-TCB. Антитело 33Н09-ТСВ некоторым образом распознавало пептид ARHGEF11, но степень NFAT-активации была ниже более, чем в 100 раз, по сравнению с нативным RMF-пептидом (Фиг. 9Аб В). Аналогичным образом, перекрестная реактивность по отношению к остальным 19 не являющихся мишенями пептидам была по меньшей мере в 100 раз ниже, по сравнению с нативным RMF пептидом, как было измерено в исследовании прямого уничтожения Т-клеток на обработанных пептидами Т2-клетках (Фиг. 9С, D). Этот результат был подтвержден во втором эксперименте, когда были протестированы также клетки ESK1-TCB (Фиг. 9E-G).
Объединенные результаты исследований показывают, что 11D06-TCB и 33Н09-ТСВ (но не ESK1-подобная связывающая молекула) обладают специфичностью к WT1 RMF-пептиду и не имеют специфичности по отношению к не являющимся мишенями пептидам, выявленным в пептидомной базе данных.
Пример 11. Отсутствие цитотоксичности по отношению к CD34+ стволовым клеткам, опосредуемой отобранными HLA-A2/WT1 х CD3 биспецифическими антителами ("TCBs")
Сообщалось, что CD34+ гематопоэтические стволовые клетки являются WT1-положительными (Ramani и Cowell, J. Path (1996) 179:162-8), что может нанести потенциальный вред при связывании с ними TCBs. Для того, чтобы определить, способны ли СВ34-клетки эндогенно презентировать RMF-пептид, получали происходящие из HLA-A2+ костного мозга СВ34+-клетки от трех доноров от Lonza и тестировали отобранные TCBs в исследовании по уничтожению клеток, как описано выше в Примере 7. В то время как 11D06-TCB и 33Н09-ТСВ опосредуют потенциальную гибель SKM-1 клеток, они не проявляют киллерной активности по отношению к CD34+ стволовым клеткам, указывая на то, что эти стволовые клетки могут не презентировать RMF-пептид в контексте HLA-A2 (Фиг. 10).
Пример 12. Определение связывающих остатков HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") в исследовании сканирования аланина
Понятно, что полученные TCBs имеют другую степень связывания с RMF-пептидом и функциональную активность в сравнении с ESK1-TCB. Для установления потенциальных мотивов в последовательности RMF-пептида, которые связываются TCBs, проводили исследование сканирования аланина при использовании массива пептидов, полученных из нативных последовательностей путем индивидуального замещения каждой аминокислоты на аланин и измерения NFAT-маркерного сигнала Т2-клеток, обработанных указанными пептидами (Фиг. 11A-L). Данные представлены в RLU в зависимости от возрастающих концентраций ТСВ. Была построена схема кратного изменения ЕС50 относительно ЕС50 для нативного пептида, и авторы пришли к выводу, что существенным является кратное изменение ≥10, которое означает, что соответствующий аминокислотный остаток RMF-пептида может быть критическим для распознавания антителами 11D06-TCB и 33Н09-ТСВ (Фиг. 11М). Авторы заключили, что оба антитела 11D06-TCB и 33Н09-ТСВ имеют наиболее важный контакт с остатками R1, F3, N5 и А6 (Фиг. 11N), в то время как ESK1 имеют тесный контакт с остатками R1, Р4 и N5 RMF-пептида (Ataie et. al., J. Mol. Biol. (2016) 428:194-205)).
Пример 13. Фармакокинетический профиль HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") после однократной инъекции мышам NSG
Однократную дозу, составляющую 0,5 мг/кг 11D06-TCB или 33Н09-ТСВ, инъецировали мышам NSG. Все мыши были инъецированы внутривенно 200 мкл подходящего раствора. Для получения нужного количества соединений в 200 мкл вводимого раствора исходный раствор разбавляли гистидиновым буфером. У троих мышей забирали кровь в следующие временные точки: 10 мин, 1 ч, 3 ч, 6 ч, 24 ч, 48 ч, 72 ч, 96 ч, 6 дней, 8 дней, 10 дней и 12 дней. Инъецированное соединение анализировали в образцах сыворотки методом ELISA. Биотинилированные антитела к CD3-CDR человека (Roche Diagnostics, Penzberg, Germany), тестируемый образец, меченные дигоксигенином антитела к Fc человека и детектирующие антитела к дигоксигенину (связанные с пероксидазой (POD)) добавляли пошагово в лунки 96-луночного, покрытого стрептавидином планшета для микротитрования и затем инкубировали планшет после каждого шага в течение 1 ч при комнатной температуре. Планшет отмывали три раза после каждого этапа для удаления несвязавшихся субстанций. В конце исследования визуализировали связавшийся с пероксидазой комплекс путем добавления раствора субстрата ABTS (2,2’-азино-бис(3-этилбензотиазолин-6-сульфоновая кислота) для образования окрашенного продукта реакции. Интенсивность окрашивания продукта реакции, которую определяли путем фотометрии при длине волны 405 нм (в сравнении с контролем при 490 нм), пропорциональна концентрации аналита в сыворотке. Результат исследования (Фиг. 12) показывает стабильное РК-поведение для обоих тестируемых клонов, что позволяет осуществить однонедельную схему последующих исследований эффективности.
Пример 14. Исследование эффективности HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") на гуманизированных мышах с SKM-1 ксенотрансплантатом
Первое исследование эффективности HLA-A2/WT1 х CD3 биспецифических антител было направлено на сравнение эффективности двух различных WT1-связывающих молекул (11D06-TCB и 33Н09-ТСВ) на модели полностью гуманизированных мышей с ксенотрансплантатом (SKM-1; HLA-A2+, WT-1+) острого миелоидного лейкоза (AML) человека. SKM-1 клетки (AML человека) были первоначально получены из коллекции АТСС и депонированы во внутреннем клеточном банке Roche Glycart. Клетки культивировали в среде RPMI + 10% FCS + 1% глутамина в насыщенной водяным паром атмосфере в присутствии 5% СО2. Было проведено 15 пассажей in vitro для подкожной инъекции клеток с жизнеспособностью 98% в Матригеле (отношение 1:1).
Самок мышей NSG возрастом 4-5 недель на начальной стадии эксперимента (Jackson Laboratory) содержали в специальной свободной от патогенов среде с дневными циклами 12 ч свет/12 ч темнота в соответствии с утвержденными инструкциями (GV-Solas; Felasa; TierschG). Протокол экспериментальных исследований был рассмотрен и утвержден местным правительством. После получения животных выдерживали одну неделю для адаптации к новым условиям и наблюдения за ними. Постоянный мониторинг их здоровья осуществлялся на регулярной основе.
Самкам мышей NSG интраперитонеально инъецировали 15 мг/кг бисульфана через один день после внутривенной инъекции 1×105 гематопоэтических стволовых клеток человека, полученных из пуповинной крови. После 14-16 недель после введения стволовых клеток у мышей сублингвально забирали кровь, которую анализировали проточной цитометрией для оценки успешной гуманизации. Мышей с эффективно привитыми клетками в зависимости от частоты встречаемости Т-клеток человека произвольным образом разделяли на различные группы для терапии. В это время мышей подкожно инъецировали опухолевыми клетками SKM-1, как показано на Фиг. 13А, и вводили им один раз в неделю тестируемые соединения или гистидиновый буфер (носитель), когда размер опухоли достигал примерно 150 мм3 (день 10). Все мыши были инъецированы внутривенно 200 мкл соответствующего раствора. Для получения нужного количества соединений в 200 мкл вводимого раствора исходные растворы при необходимости разбавляли гистидиновым буфером. Рост опухоли отслеживали три раза в неделю при использовании кронциркуля и объем опухоли (v) рассчитывали следующим образом:
Tν: (W2/2) × L (W: ширина, L: длина)
Ингибирование роста опухоли (TGI), как и отношение опухоли к контролю, вычисляли по формулам:


Фиг. 13 показывает кинетику роста опухоли (среднее значение) во всех обработанных группах животных (Фиг. 13 С), а также кинетику роста отдельной опухоли в каждой группе (Фиг. 13D-F). Как можно видеть, оба антитела TCBs ингибируют рост опухоли, причем клон 11D06-TCB имеет наиболее сильную активность по ингибированию опухолевого роста с величиной TGI 101,3 (Фиг. 13G).
Пример 15. Кристаллическая структура комплексов HLA-A2/WT1 антитело/рМНС
Fab-фрагменты получали путем инкубации антител в течение 72 ч при 25°С в 50 мМ Трисе, рН 8,0, 150 мМ NaCl с 1,05 Ед плазмина (Roche, номер по каталогу 602361) на 1 мг. Расщепленные Fc отделяли от Fab-фрагментов при использовании 4,5 мл аффинной колонки CaptureSelect CHl (ВАС BV, номер по каталогу 191.3120), уравновешенной 50 мМ Трисом, 100 мМ глицином, 150 мМ NaCl, рН 8,0. Fab-фрагменты элюировали с колонки 50 мМ Трисом, 100 мМ глицином, 150 мМ NaCl, рН 8,0, и нейтрализовали 0,05 М фосфатом натрия, рН 8,0, до нанесения на эксклюзионную колонку S75 с разделением по размеру (GE Healthcare), уравновешенную 50 мМ Трисом, 150 мМ NaCl, рН 7,4. Качественный контроль осуществляли на эксклюзионной аналитической колонке с разделением по размеру (колонка Tosoh, TSK-Gel G3000SWXL, аналитическая система Agilent HPLC 1200) и методом CE-SDS (Caliper LabChip GXII, Perkin Elmer) в невосстанавливающих и восстанавливающих условиях.
Очищенные Fab-фрагменты замораживали в жидком азоте и хранили при -80°С.
Кристаллизация, сбор данных и определение структуры комплекса антитело 5С01/рМНС
Кристаллизация. Комплекс антитело/рМНС (Fab 5С01 HLA-A02/WT1VLD рМНС) был приготовлен путем смешивания 1,2-кратного молярного избытка HLA-A2/WT1 Fab-фрагментов связывающей молекулы 5С01 (Fab 5С01) с HLA-A2/WT1VLD пептидным комплексом (HLA-A02/WT1VLD рМНС). После инкубирования в течение 1 ч при 4°С смесь концентрировали до 10 мг/мл. Инициацию кристаллизации осуществляли в установке диффузного пара сидячей капли при 21°С. К образовавшимся в течение 1 дня каплям кристаллов добавляли 0,2 М тартрата натрия и 20% полиэтиленгликоля (ПЭГ) 3350 с 10% 2-метил-2,4-пентандиола (MPD) для подтверждения качества кристаллов. Кристаллы собирали непосредственно с планшета скрининга без дальнейшей стадии оптимизации.
Сбор данных и определение структуры. Для сбора данных кристаллы подвергали шоковой заморозке при 100К в растворе преципитанта, содержащем 15% глицерола. Дифракционные данные собирали при длине волны 1,0000  при использовании детектора PILATUS 6М на источнике синхронного излучения X10SA Swiss Light Source (Willigen, Switzerland). Данные обрабатывали с помощью XDS (Kabsch, W. Acta Cryst. D66, 133-144 (2010)) и масштабировали при использовании SADABS (BRUKER). Кристаллы комплекса принадлежали к пространственной группе P21212 с осями ячейки а=158,94
при использовании детектора PILATUS 6М на источнике синхронного излучения X10SA Swiss Light Source (Willigen, Switzerland). Данные обрабатывали с помощью XDS (Kabsch, W. Acta Cryst. D66, 133-144 (2010)) и масштабировали при использовании SADABS (BRUKER). Кристаллы комплекса принадлежали к пространственной группе P21212 с осями ячейки а=158,94  b=49,12
b=49,12  с=128,63
с=128,63  и дифракционным разрешением 1,98
и дифракционным разрешением 1,98  Структуру определяли путем молекулярного замещения с помощью PHASER (McCoy, A.J., Grosse-Kunstleve, R.W., Adams, P.D., Storoni, L.C. и Read, R.J. J. Appl. Cryst. 40, 658-674 (2007)) с применением координат кристаллической структуры из данных банка белков (PDB) со входом 4NO5 и структуры Fab, известной авторам изобретения, как поисковой модели. Использовали различия в электронной плотности для размещения пептида и замены аминокислот в соответствии с различиями в последовательностях путем фактической пространственной доработки. Структуры делали более точными с использованием программного набора ССР4 (Collaborative Computational Project, Number 4 Acta Cryst. D50, 760-763 (1994)) и BUSTER (Bricogne, G., Blanc, E., Brandl, M., Flensburg, C, Keller, P., Paciorek, W., Roversi, P., Sharff, A., Smart, O.S., Vonrhein, C, Womack, Т.О. (2011). Версия BUSTER 2.9.5. Руководства по реконструированию (Cambridge, United Kingdom: Global Phasing Ltd.) было предоставлено COOT (Emsley, P., Lohkamp, В., Scott, W.G. and Cowtan, K. Acta Cryst. D66, 133-144 (2010)).
Структуру определяли путем молекулярного замещения с помощью PHASER (McCoy, A.J., Grosse-Kunstleve, R.W., Adams, P.D., Storoni, L.C. и Read, R.J. J. Appl. Cryst. 40, 658-674 (2007)) с применением координат кристаллической структуры из данных банка белков (PDB) со входом 4NO5 и структуры Fab, известной авторам изобретения, как поисковой модели. Использовали различия в электронной плотности для размещения пептида и замены аминокислот в соответствии с различиями в последовательностях путем фактической пространственной доработки. Структуры делали более точными с использованием программного набора ССР4 (Collaborative Computational Project, Number 4 Acta Cryst. D50, 760-763 (1994)) и BUSTER (Bricogne, G., Blanc, E., Brandl, M., Flensburg, C, Keller, P., Paciorek, W., Roversi, P., Sharff, A., Smart, O.S., Vonrhein, C, Womack, Т.О. (2011). Версия BUSTER 2.9.5. Руководства по реконструированию (Cambridge, United Kingdom: Global Phasing Ltd.) было предоставлено COOT (Emsley, P., Lohkamp, В., Scott, W.G. and Cowtan, K. Acta Cryst. D66, 133-144 (2010)).
Совокупность данных и статистические показатели уточненной структуры суммированы в Таблице 6.

Для того, чтобы охарактеризовать детали взаимодействия VLD-пептида с Fab 5С01 и связывающий эпитоп и паратоп 5С01 с HLA-A02, определяли кристаллическую структуру комплекса 5С01 с HLA-A02/WT1VLD рМНС при разрешении 1,98 (Фиг. 14А). Структура показала, что основной вклад в связывание Fab 5С01 с рМНС вносят CDR1 и CDR3 легкой цепи и все CDRs тяжелой цепи. У пептида VLD (SEQ ID No: 77) остатки боковых цепей Val1, Phe4 и Pro7 находятся в непосредственном контакте с Fab. Все остальные боковые цепи пептида точечно направлены в сторону молекулы HLA-A02. Было установлено, что вклад пептида в контактирующую поверхностную область составляет около 68  в то время как общая зона контакта Fab-pMHC занимает примерно
в то время как общая зона контакта Fab-pMHC занимает примерно 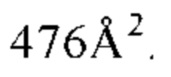 Тесная область интерфейса Fab 5С01-рМНС показана на Фиг. 15.
Тесная область интерфейса Fab 5С01-рМНС показана на Фиг. 15.
Анализ интерфейса связывания с помощью программы PISA (Е. Krissinel and Henrick (2007), J. Mol. Biol. 372, 774-797) выявил характер взаимодействия Fab 5C01 с HLA-A02 посредством 8 водородных связей, Pi-Pi-взаимодействий и Вандерваальсовых сил. Водородные связи образуются за счет боковых цепей между остатками тяжелой цепи CDR3 (Trp97) и CDR2 (Ser52, Ser53) с Glu63 и Glu166 HLA-A02. Также водородные связи устанавливаются скелетными атомами легкой цепи Tyr91 и Ile93 с Gln155. Комплекс дополнительно стабилизирован посредством Pi-Pi взаимодействий остатков легкой цепи Trp32 и Trp34 с остатками боковых цепей HLA-A02 Gln155 и His151. N-концевой валин VLD-пептида формирует водородные связи через скелетный азот с Tyrl71 HLA-А02. Его боковые цепи ориентированы по направлению к карману, образованному Glu63 и Trp167 HLA-A02 и Trp97 тяжелой цепи Fab 5С01. Более того, Phe4 пептида создает участок для тесного взаимодействия с Tyr100 тяжелой цепи. Схематическая матрица взаимодействия в комплексе Fab 5С01-рМНС, суммирующая контакты, показана на Фиг. 16.
Кристаллизация, сбор данных и определение структуры комплекса антитело 11D06/рМНС
Кристаллизация. Комплекс антитело/рМНС (Fab 11D06 HLA-A02/WT1RMF рМНС) был приготовлен путем смешивания HLA-A2/WT1 Fab-фрагментов связывающей молекулы 11D06 (Fab 11D06) с HLA-A2/WT1RMF пептидным комплексом (HLA-A02/WT1RMF рМНС) в молярном соотношении 1:1. После инкубирования в течение 4 ч при 21°С смесь концентрировали до 20 мг/мл. Инициацию кристаллизации осуществляли в установке диффузного пара сидячей капли при 21°С. Кристаллы образовывались в течение 4 дней в 0,1 М Трисе, рН 8,0, 20% полиэтиленгликоле (ПЭГ). Кристаллы собирали непосредственно с планшета скрининга без дальнейшей стадии оптимизации.
Сбор данных и определение структуры. Данные собирали, обрабатывали и масштабировали, как описано выше. Кристаллы комплекса принадлежали к пространственной группе P21 с осями ячейки а=54,11  b=67,00
b=67,00  с=139,36
с=139,36  с β и дифракционным разрешением 2,64
с β и дифракционным разрешением 2,64  Структуру определяли путем молекулярного замещения с помощью PHASER с применением координат структуры комплекса Fab (структура Fab в составе комплекса была известна авторам изобретения) и МНС как поисковой модели. Использовали различия в электронной плотности для размещения пептида и замены аминокислот в соответствии с различиями в последовательностях путем фактической пространственной доработки. Структуры делали более точными и реконструировали вручную, как описано выше.
Структуру определяли путем молекулярного замещения с помощью PHASER с применением координат структуры комплекса Fab (структура Fab в составе комплекса была известна авторам изобретения) и МНС как поисковой модели. Использовали различия в электронной плотности для размещения пептида и замены аминокислот в соответствии с различиями в последовательностях путем фактической пространственной доработки. Структуры делали более точными и реконструировали вручную, как описано выше.
Совокупность данных и статистические показатели уточненной структуры суммированы в Таблице 7.
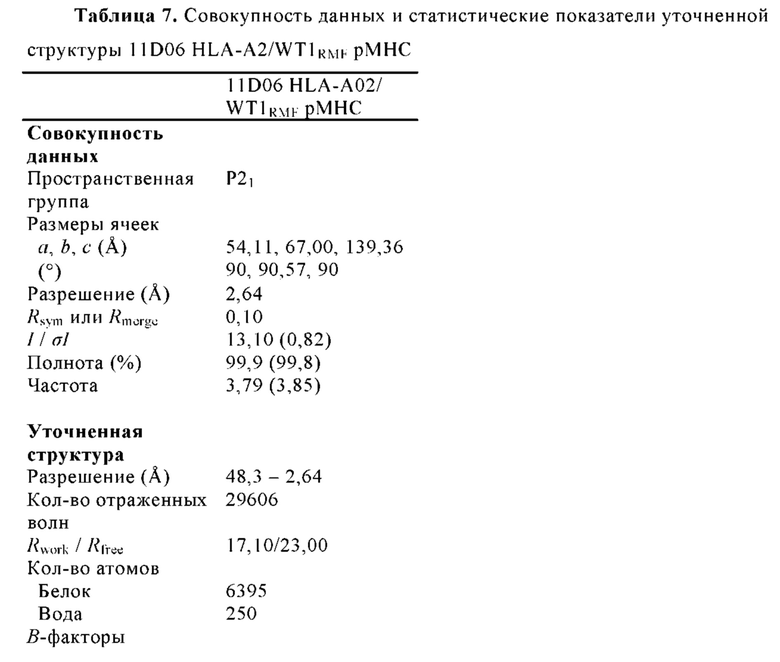

Определяли кристаллическую структуру комплекса Fab 11D06 с HLA-A02/RMF рМНС при разрешении 2, 64  (Фиг. 14 В). Структура показала, что основной вклад в связывание Fab 11D06 с рМНС вносят все CDRs. У пептида RMF (SEQ ID No: 78) остатки боковых цепей Arg1, Met2, Pro4, Asn5, Ala6 и Tyr8 находятся в непосредственном контакте с тяжелой и легкой цепью Fab. Остальные боковые цепи пептида точечно направлены в сторону молекулы HLA-А02. Вклад пептида в контактирующую поверхностную область составляет около 107
(Фиг. 14 В). Структура показала, что основной вклад в связывание Fab 11D06 с рМНС вносят все CDRs. У пептида RMF (SEQ ID No: 78) остатки боковых цепей Arg1, Met2, Pro4, Asn5, Ala6 и Tyr8 находятся в непосредственном контакте с тяжелой и легкой цепью Fab. Остальные боковые цепи пептида точечно направлены в сторону молекулы HLA-А02. Вклад пептида в контактирующую поверхностную область составляет около 107  Общая зона контакта Fab-pMHC занимает примерно 397
Общая зона контакта Fab-pMHC занимает примерно 397  Тесная область интерфейса Fab 11D06-рМНС показана на Фиг. 17.
Тесная область интерфейса Fab 11D06-рМНС показана на Фиг. 17.
Анализ интерфейса связывания с помощью программы PISA выявил характер взаимодействия Fab 11D06 с HLA-A02 посредством 4 водородных связей, многочисленных Pi-Pi-взаимодействий и Вандерваальсовых сил. Как наблюдалось, водородные связи образуются между остатками CDR1 тяжелой цепи (Ser30, Ser31) и CDR3 (Gly100A) с Glu58 и Arg65 HLA-A02. Также водородные связи устанавливаются между остатком легкой цепи Asp50 с Arg65 HLA-A02. Помимо этого, Trp100 тяжелой цепи обеспечивает Вандерваальсово и Pi-Pi взаимодействия с Arg65 и Lys66 HLA-A02 и с Pro4 RMF-пептида. N-концевой аргинин RMF-пептида вместе с его боковой цепью входит в полярный карман, сформированный Ser31, Glu97 и скелетным карбонилом Ile96 тяжелой цепи 11D06. Дополнительные полярные контакты между 11D06 и HLA-A02 обеспечиваются остатком Asn5 RMF-пептида, который является частью водородной связывающей сети с Trp32 CDR1 легкой цепи, Glu92 CDR3 легкой цепи вместе с Gln155 HLA-A02. Схематическая матрица взаимодействия Fab 11D06-рМНС, суммирующая контакты, показана на Фиг. 18.
3D-структура ESK1, полученная из открытой базы данных
Для сравнения анализировали находящуюся в открытом доступе кристаллическую структуру антитела (Fab) ESK1, связывающуюся с HLA-A02/RMF pMHC (PDB ID 4WUU; http://www.rcsb.org/pdb/explore/explore.do?structureId=4WUU), в соответствии со своими Fab-MHC контактами. На Фиг. 14С показано, что кристаллическая структура Fab ESK1 с HLA-A02/RMF рМНС формирует тот же самый угол (выравнивание по части HLA-A02), что и кристаллические структуры Fab 5С01 с HLA-A02/VLD рМНС и Fab 11D06 с HLA-A02/RMF рМНС (Фиг. 14 А и В). Тесная область интерфейса Fab ESK1-pMHC показана на Фиг. 19, а схематическая матрица взаимодействия Fab ESK1-pMHC, суммирующая контакты, показана на Фиг. 20.
Структурное сравнение выявило, что 5С01 и частично 11D06 покрывают и связываются с большей частью соответствующего WT1 пептида, чем антитело ESK1, которое образует специфические контакты исключительно с N-концевым Arg RMF-пептида, в то время как остальная часть связывающего интерфейса обеспечивается за счет HLA-A02. На основе указанных наблюдений можно заключить, что 5С01 и 11D06 должны быть в меньшей степени толерантны к пептидам, которые не являются мишенями, чем ESK1, поскольку они создают значительно большие стерические помехи для пептидов с не-VLD- или с не-RMF-подобными боковыми цепями на экспонируемой поверхности.
Все графические изображения кристаллических структур были созданы с помощью системы BIOVIA Discovery Studio 4,5, Dassault Systemes.
Остатки эпитопа и паратопа (радиус 5  )
)
Суммарно эпитопы и паратопы связывающих молекул 5С01, 11D06 ESK1 приведены ниже. Остатки эпитопов и паратопов (определение основано на соседнем радиусе 5  ) выделены жирным курсивом. Остатки CDRs выделены серыми блоками.
) выделены жирным курсивом. Остатки CDRs выделены серыми блоками.
HLA-A02
5С01
GSHSMRYFFTSVSRPGRGEPRFIAVGYVDDTQFVRFDSDAASQRMEPRAPWIEQ
EGPEYW
11D06 GSHSMRYFFTSVSRPGRGEPRFIAVGYVDDTQFVRFDSDAASQRMEPRA
PWIQEGPEYW
ESK1
GSHSMRYFFTSVSRPGRGEPRFIAVGYVDDTQFVRFDSDAASQRMEPRAPWIEQ
EGFEYW
1-60
5С01
DGETRKVKAHSQTHRVDLGTLRGYYNQSEAGSHTVQRMYGCDVGSDWRFLR
GYHQYAYDG
11D06 DGETRKVKAHSQTHRVDLGTLRGYYNQSEAGSHTVQRMYGCDVGSDW
RFLRGYHQYAYDG
ESK1
DGETRKVKAHSQTHRVDLGTLRGYYNQSEAGSHTVQRMYGCDVGSDWRFLRG
YHQYAYDG
61-120 5C01
KDYIALKEDLRSWTAADMAAQTTKHKWEAAHVAEQLRAYLEGTCVEWLRRYL
ENGKETLQ
11D06 KDYIALKEDLRSWTAADMAAQTTKHKWEAAHVAEQLRAYLEGTCVEW
LRRYLENGKETLQ
ESK1
KDYIALKEDLRSWTAADMAAQTTKHKWEAAHVAQLRAYLEGTCVEWLRRX
LENGKETLQ
121-180
5C01
RTDAPKTHMTHHAVSDHEATLRCWALSFYPAEITLTWQRDGEDQTQDTELVET
RPAGDGT
11D06 RTDAPKTHMTHHAVSDHEATLRCWALSFYPAEITLTWQRDGEDQTQDT
ELVETRPAGDGT
ESK1
RTDAPKTHMTHHAVSDHEATLRCWALSFYPAEITLTWQRDGEDQTQDTELVET
RPAGDGT
181-240
5C01 FQKWAAVVVPSGQEQRYTCHVQHEGLPKPLTLRWE
11D06 FQKWAAVVVPSGQEQRYTCHVQHEGLPKPLTLRWE
ESK1 FQKWAAVVVPSGQEQRYTCHVQHEGLPKPLTLRWE
241-275
Пептид
5С01 VLDFAPPGA
11D06 RMFPNAPYL
ESK1 RMFPNAPYL
1-9
Fab тяжелая цепь
5C01
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGS
GGSTYY
11D06 QVQLVQSGAEVKKPGSSVKVSCKASGGTFSSYAISWVRQAPGQGLEWM
GGIIPIFGTANY
ESK1
QMQLVQSGAEVKEPGESLRISCKGSGYSFTNFWSWVRQMPGKGLEWMGRVDP
GYSYSTY
1-60
5C01 ADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGSWV
SYAFDYWGQGTLVTVS
11D06 AQKFQGRVTITADKSTSTAYMELSSLRSEDTAVYYCARSIELW
WGGFDYWGQGTTVTVS
ESK1
SPSFQGHVTISADKSTSTAYLQWNSLKASDTAMYYCARVQYSGYYDWFDPWGQ
GTLVTVS
61-120
5C01
SASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFP
AVLQS
11D06 SASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSG
VHTFPAVLQS
ESK1
SASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFP
AVLQS
121-180
5C01 SGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
11D06 SGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
ESK1 SGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKS
181-224
Fab легкая цепь
5C01 DIQMTQSPSTLSASVGDRVTITCRAS Q SIS
SWLAWYQQKPGKAPKLLIYDASSLES
11D06 DIQMTQSPSTLSASVGDRVTITCRAS Q SIS
SWLAWYQQKPGKAPKLLIYDASSLES
ESK1 QAVVTQP PSASGTPGQRVTISCSG SSSNIGSN
TVNWYQQVPGTAPKLLIYSNNQRPS
1-60
5C01 GVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQQYSI
WFPYTFGQGTKVEIKRTVAAPS
11D06 GVPSRFSGSGSGTEFTLTIGSLQPDDFATYYCQQYE
DYTTFGQGTKVEIKRTVAAPS
ESK1
GVPDRFSGSKSGTSASLAISGLQSEDEADYYCAAWDDSLNGWVFGGGTKLTVL
GQPKANP
61-120
5C01
VFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDS
KDSTYS
11D06 VFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESV
TEQDSKDSTYS
ESK1
TVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADGSPVKAGVETTKPSKQ
SNNKYA
121-180
5C01 LSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
11D06 LSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
ESK1
ASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
181-220
Остатки, вовлеченные в химические взаимодействиях
Суммарно, остатки, вовлеченные в химические взаимодействия для связывающих молекул 5С01, 11D06 ESK1 приведены ниже. Остатки, вовлеченные в специфические химические взаимодействия (сравните Фиг. 16, 18 и 20) выделены жирным курсивом. Остатки CDRs выделены серыми блоками.
HLA-A02
5С01
GSHSMRYFFTSVSRPGRGEPRFIAVGYVDDTQFVRFDSDAASQRMEPRAPWIEQ
EGPEYW
11D06 GSHSMRYFFTSVSRPGRGEPRFIAVGYVDDTQFVRFDSDAASQRMEPRA
PWIEQEGPEYW
ESK1
GSHSMRYFFTSVSRPGRGEPRFIAVGYVDDTQFVRFDSDAASQRMEPRAPWIEQ
EGPEYW
1-60
5C01
DGETRKVKAHSQTHRVDLGTLRGYYNQSEAGSHTVQRMYGCDVGSDWRFLR
GYHQYAYDG
11D06 DGETRKVKAHSQTHRVDLGTLRGYYNQSEAGSHTVQRMYGCDVGSDW
RFLRGYHQYAYDG
ESK1
DGETRKVKAHSQTHRVDLGTLRGYYNQSEAGSHTVQRMYGCDVGSDWRFLR
GYHQYAYDG
61-120
5C01
KDYIALKEDLRSWTAADMAAQTTKHKWEAAHVAEQLRAYLEGTCVEWLRRY
LENGKETLQ
11D06 KDYIALKEDLRSWTAADMAAQTTKHKWEAAHVAEQLRAYLEGTCVEW
LRRYLENGKETLQ
ESK1
KDYIALKEDLRSWTAADMAAQTTKHKWEAAHVAEQLRAYLEGTCVEWLRRY
LENGKETLQ
121-180
5C01
RTDAPKTHMTHHAVSDHEATLRCWALSFYPAEITLTWQRDGEDQTQDTELVET
RPAGDGT
11D06 RTDAPKTHMTHHAVSDHEATLRCWALSFYPAEITLTWQRDGEDQTQDT
ELVETRPAGDGT
ESK1
RTDAPKTHMTHHAVSDHEATLRCWALSFYPAEITLTWQRDGEDQTQDTELVET
RPAGDGT
181-240
5C01 FQKWAAVVVPSGQEQRYTCHVQHEGLPKPLTLRWE
11D06 FQKWAAVVVPSGQEQRYTCHVQHEGLPKPLTLRWE
ESK1
FQKWAAVVVPSGQEQRYTCHVQHEGLPKPLTLRWE
241-275
Пептид
5C01 VLDFAPPGA
1D06 RMFPNAPYL
ESK1 RMFPNAPYL
1-9
Fab тяжелая цепь
5C01
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGS
GGSTYY
11D06 QVQLVQSGAEVKKPGSSVKVSCKASGGTFSSYAISWVRQAPGQGLEWM
GGIIPIFGTANY
ESK1
QMQLVQSGAEVKEPGESLRISCKGSGYSFTNFWISWVRQMPGKGLEWMGRVD
PGYSYSTY
1-60
5C01 ADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGSWV
SYAFDYWGQGTLVTVS
11D06 AQKFQGRVTITADKSTSTAYMELSSLRSEDTAVYYCARSIELW
WGGFDYWGQGTTVTVS
ESK1
SPSFQGHVTISADKSTSTAYLQWNSLKASDTAMYYCARVQYSGYYDWFDPWGQ
GTLVTVS
61-120
5C01
SASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFP
AVLQS
11D06 SASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSG
VHTFPAVLQS
ESK1
SASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFP
AVLQS
121-180
5C01 SGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
11D06 SGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
ESK1 SGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKRVEPKS
181-224
Fab легкая цепь
5C01 DIQMTQSPSTLSASVGDRVTITCRAS Q SIS
SWLAWYQQKPGKAPKLLIYDASSLES
11D06 DIQMTQSPSTLSASVGDRVTITCRAS Q SIS
SWLAWYQQKPGKAPKLLIYDASSLES
ESK1 QAVVTQP PSASGTPGQRVTISCSG SSSNIGSN
TVNWYQQVPGTAPKLLIYSNNQRPS
1-60
5C01 GVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQQYSI
WFPYTFGQGTKVEIKRTVAAPS
11D06 GVPSRFSGSGSGTEFTLTIGSLQPDDFATYYCQQYE
DYTTFGQGTKVEIKRTVAAPS
ESK1
GVPDRFSGSKSGTSASLAISGLQSEDEADYYCAAWDDSLNGWVFGGGTKLTVL
GQPKANP
61-120
5C01
VFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDS
KDSTYS
11D06 VFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESV
TEQDSKDSTYS
ESK1
TVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADGSPVKAGVETTKPSKQ
SNNKYA
121-180
5C01 LSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
11D06 LSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
ESK1 ASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
181-220
Пример 16. Сравнение HLA-A2/WT1 x CD3 биспецифических антител ("TCBs") с различными CD3 связывающими молекулами
Сравнивали потенциальную киллерную активность 11D06-TCB с активностью различных CD3 связывающих молекул при использовании клеток SKM-1 в исследовании, описанном выше (Пример 7). СН2527 представляет собой CD3 связывающую молекулу из TCBs, используемую в предварительных экспериментах (см. SEQ ID Nos 121 и 122 для последовательностей VH и VL). V9 представляет собой CD3 связывающую молекулу, описанную Rodrigues et al., Int J Cancer Suppl (1992) 7, 45-50, и в WO 1992/22653 (см. SEQ ID Nos 136 и 137 для последовательностей VH и VL), и молекула 40G5C описана в WO 2015/095392 (см. SEQ ID Nos 184 и 185 ("hu40G5C") в WO 2015/095392 для последовательностей VH и VL).
Аффинность связывающих CD3 молекул СН2527, 40G5C и V9 приведена в Таблице 8.


Пример 17. Т-клеточная цитотоксичность, опосредуемая связыванием HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") с Т-клетками, обработанными пептидом RMF
Киллерную активность 11D06-TCB (V9) (см. SEQ ID Nos 123, 125, 139, 140 для полноразмерных последовательностей) сравнивали с аналогичными TCBs различными HLA-WT1 связывающими молекулами ("Aali" и "Daniel", см. WO 2017/060201, SEQ ID Nos 5 (VH) и 6 (VL) и SEQ ID Nos 35 (VH) и 36 (VL), соответственно). ESK1-TCB и "нетаргетные" DP47-TCB также включали в исследование в качестве контроля. Эксперимент проводили, как описано в Примере 6. Клетки, обработанные RMF-пептидом (100 мкл, 2×105 клеток/мл) совместно культивировали с 50 мкл Т-клеток (2х106 клеток/мл) и серийными разведениями ТСВ (50 мкл) в течение 24 ч при 37°С в атмосфере 5% СО2.
На Фиг. 22 показана опосредуемая ТСВ специфическая киллерная активность по отношению к Т-клеткам-мишеням, экспрессирующим RMF. Было обнаружено, что 11D06-TCB и ESK1-TCB опосредуют потенциальное уничтожение Т2-клеток, обработанных RMF-пептидом, в то время как TCBs, основанные на HLA/WT1 связывающих молекулах, а именно "Aali" и "Daniel" (Aali-TCB и Daniel-TCB) показывают слабую киллерную активность по отношению к клеткам, обработанным RMF-пептидом, только при более высоких концентрациях.
Пример 18. Отсутствие связывания отобранных HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") с пептидами, не являющимися целевыми мишенями
Авторы изобретения также сравнили перекрестную реактивность 11D06-ТСВ (V9), Aali-TCB, Daniel-TCB и ESK1-TCB с пептидами, не являющимися целевыми мишенями, описанными в Примерах 9 и 10.
Для этого эксперимента использовали маркерные клетки Jurkat NFAT, экспрессирующие анти-PGLALA химерный антигенный рецептор (CAR) (исследование CAR J, см. заявку РСТ, испрашивающую приоритет по заявке на европейский патент No ЕР 17209201.7, полностью включенную в настоящее описание в качестве ссылки). Анти-PGLALA CAR распознает P329G L234A L235A ("PGLALA", система нумерации EU) мутации в Fc-области TCBs. В качестве клеток-мишеней использовали описанные выше Т2-клетки, обработанные пептидом. Принцип исследования заключается в совместном культивировании сконструированных эффекторных клеток Jurkat-NFAT с клетками-мишенями. Только при одновременном связывании TCBs с CAR (за счет мутации PGLALA) и антигеном-мишенью NFAT-промотор активируется и приводит к увеличению экспрессии люциферазы в эффекторных клетках Jurkat. При добавлении соответствующего субстрата активная люцифераза светлячков приводит к эмиссии люминесценции, которая может быть измерена как сигнал CAR-опосредуемой активации.
Коротко говоря, клетки-мишени собирали и оценивали их жизнеспособность. 20000 клеток/лунку помещали в 96-луночный планшет для микротитрования с круглым дном (Greiner bio-one, #650180) в 100 мкл среды. Необработанные пептидом Т2-клетки использовались в качестве негативного контроля. К клеткам-мишеням добавляли в разведении ТСВ в количестве 50 мкл/лунку. Далее, собирали Jurkat-NFAT репортерные клетки и оценивали их жизнеспособность. Жизнеспособные клетки суспендировали в культуральной среде и добавляли к опухолевым клеткам в количестве 100000 клеток/лунку (50 мкл/лунку) для получения конечного отношения эффектор-мишень (Е: Т) 5:1 и конечного объема 200 мкл на лунку. Клетки инкубировали в течение 20 ч при 37°С в термостате во влажной среде. После окончания времени инкубирования планшеты адаптировали при комнатной температуре (около 15 мин). Сверху из каждой лунки удаляли 125 мкл среды, добавляли 25 мкл/лунку люциферазы One-Glo (Promega #Е6120) и перемешивали. Затем 100 мкл смеси из каждой лунки переносили в 96-луночный планшет с плоским дном (Greiner bio-one, #655098) и планшеты инкубировали в течение 15 мин в темноте до выявления люминесценции с использованием Perkin Elmer.
Как показано на Фиг. 23, Aali-TCB (A), Daniel-TCB (В) и ESK1-TCB (С)обнаруживали более слабую активацию репортерной клеточной линии Jurkat-NFAT (соответствующую более слабой киллерной активности, показанной на Фиг. 22) и, более того, меньшее окно между распознаванием RMF-пептида и другими пептидами, не являющимися целевыми мишенями, по сравнению с 11D06-TCB (V9) (D).
Пример 19. Фармакокинетический профиль HLA-A2/WT1 х CD3 биспецифических антител (11D06-TCB (V9)) после однократной инъекции мышам NSG
Однократную дозу, составляющую 1 мг/кг 11D06-TCB (V9) инъецировали гуманизированным и несущим опухоли мышам NSG. Все мыши были инъецированы внутривенно 200 мкл ТСВ, разбавленного гистидиновым буфером. У троих мышей забирали кровь в следующие временные точки: 10 мин, 6 ч, 24 ч, 72 ч и 7 дней. Инъецированное соединение анализировали в образцах сыворотки методом ELISA, как описано в Примере 13.
Результат исследования (Фиг. 24) показывает стабильное РК-поведение для тестируемой молекулы ТСВ, что позволяет осуществить однонедельную схему последующих исследований эффективности.
Пример 20. Исследование эффективности HLA-A2/WT1 х CD3 биспецифических антител (11D06-TCB (V9)) на гуманизированных мышах с SKM-1 ксенотрансплантатом
Данное исследование проводили с целю оценки эффективности действия 11D06-TCB (V9) на полностью гуманизированных мышах с ксенотрансплантатом AML человека (SKM-1). Эксперимент осуществляли, как описано в Примере 14.
На Фиг. 25 показана кинетика роста опухоли (среднее значение, Фиг. 25А), а также кинетика роста отдельной опухоли в каждой группе (Фиг. 25 ВБ С). Как можно видеть, ТСВ обнаруживает ингибирование роста опухоли (TGI) 78% на 48 день исследования (Фиг. 25D).
Пример 21. Сравнение HLA-A2/WT1 х CD3 биспецифических антител ("TCBs") с различными молекулярными форматами
Сравнивали активность 11D06-TCB (V9) с аналогичными молекулами в формате "1+1 CrossMab" (как показано на Фиг. 21 А). Мониторинг активности проводили с помощью функционального исследования, которое выявляет опосредуемую ТСВ активацию маркерной клеточной линии (Jurkat-NFAT) в присутствии клеток-мишеней дозо-зависимым образом. Данное исследование, в основном, осуществляли, как описано выше в Примере 5, при использовании клеток-мишеней СНО-К1, экспрессирующих комплекс HLA-A02/RMF рМНС. Активация маркерных клеток происходит при одновременном связывании ТСВ с комплексом HLA-A02/RMF рМНС на клетках-мишенях и с доменом CD3e Т-клеточного рецептора (TCR) на маркерных клетках, что приводит к гипер-кластеризации CD3 и, таким образом, к активации TCR. Дальнейшая инициация соответствующих сигнальных путей приводит к активации транскрипционного фактора NFAT, который индуцирует экспрессию управляемого NFAT маркерного гена люциферазы. Активность маркерной люциферазы измеряется при добавлении субстрата с помощью считывания величины люминесценции.
Результаты описанного эксперимента приведены на Фиг. 26. Относительные световые единицы (RLU), отражающие зависимую от мишени активацию маркерных клеток, приведены на графике против концентрации антител. Как можно видеть на Фиг. 26, молекула формата "1+1" обнаруживает более низкую активность, чем молекула формата "2+1" в проведенном исследовании.
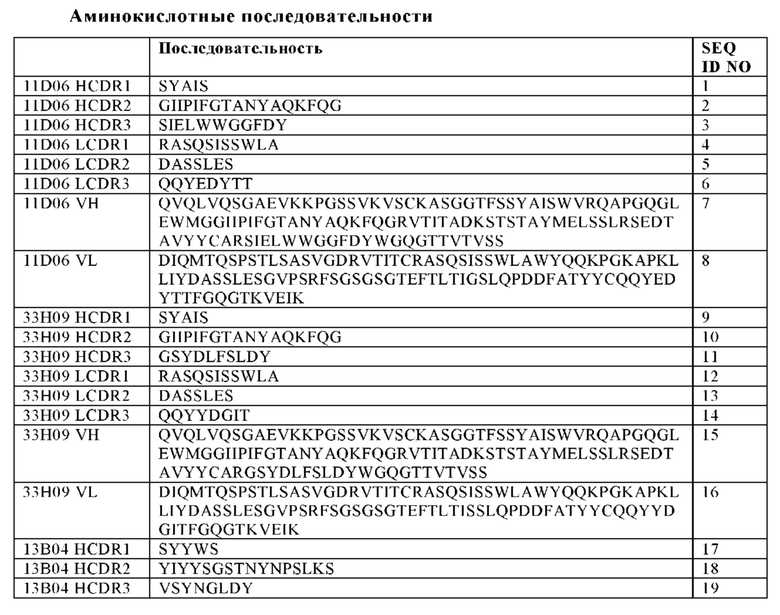
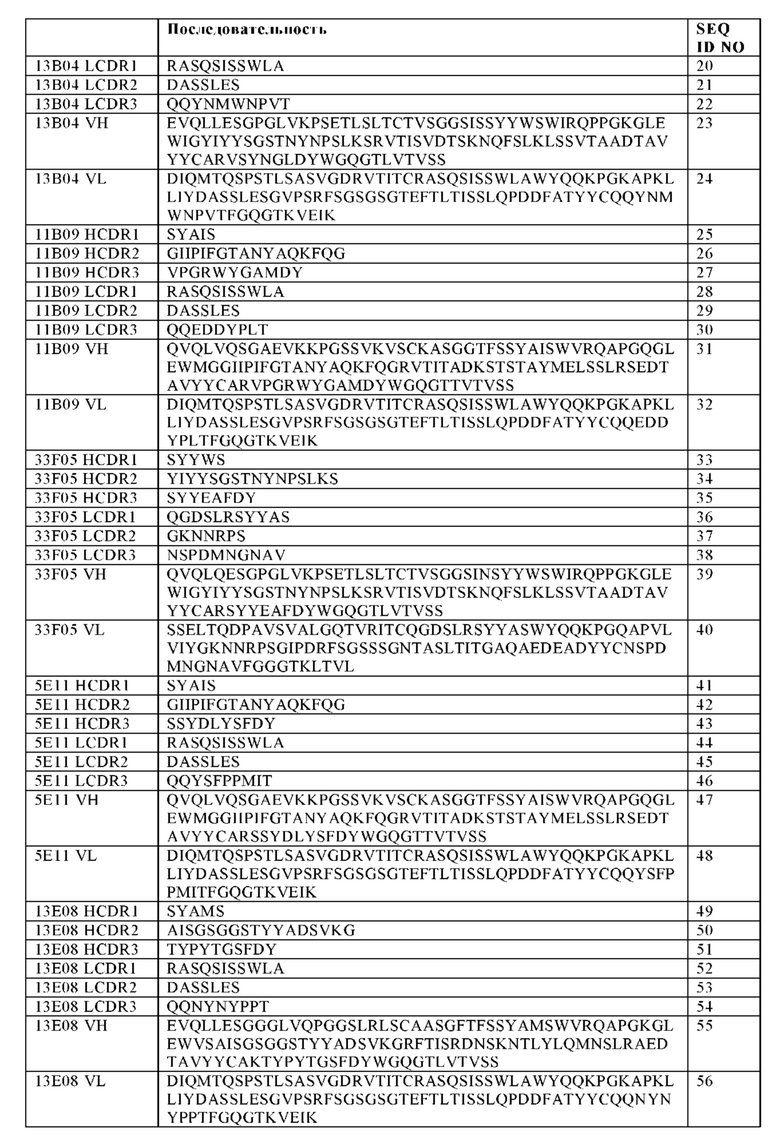
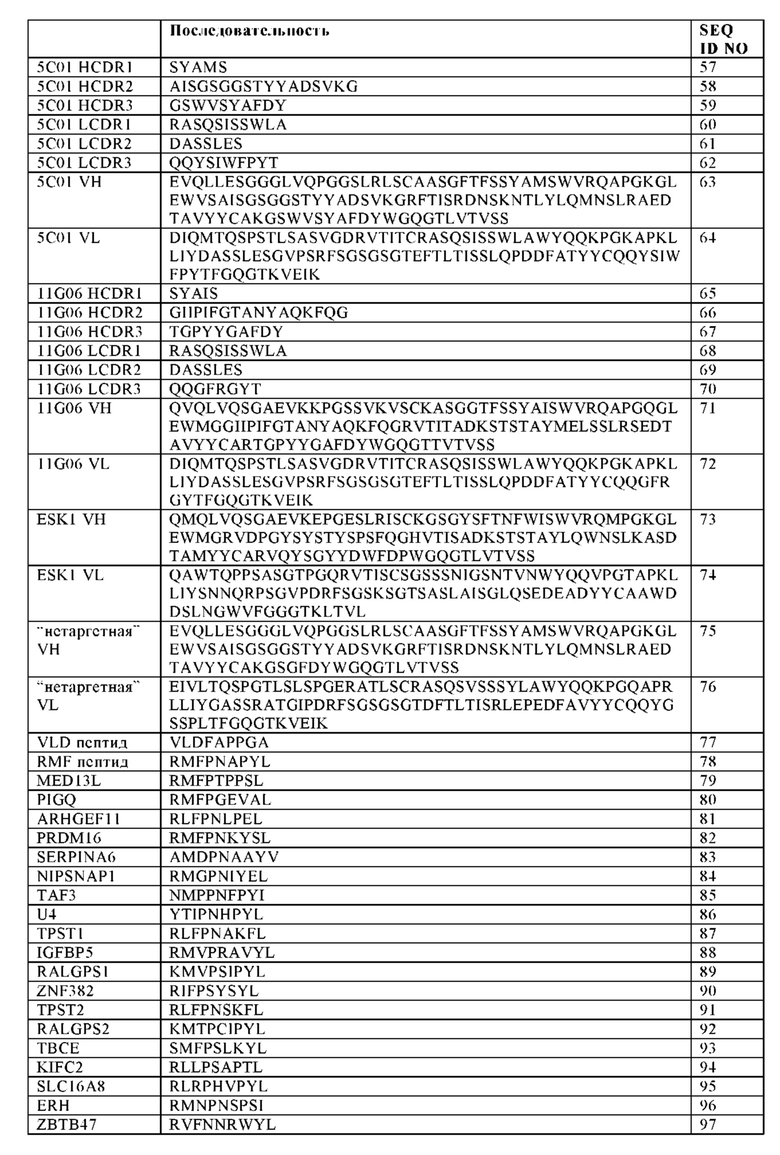
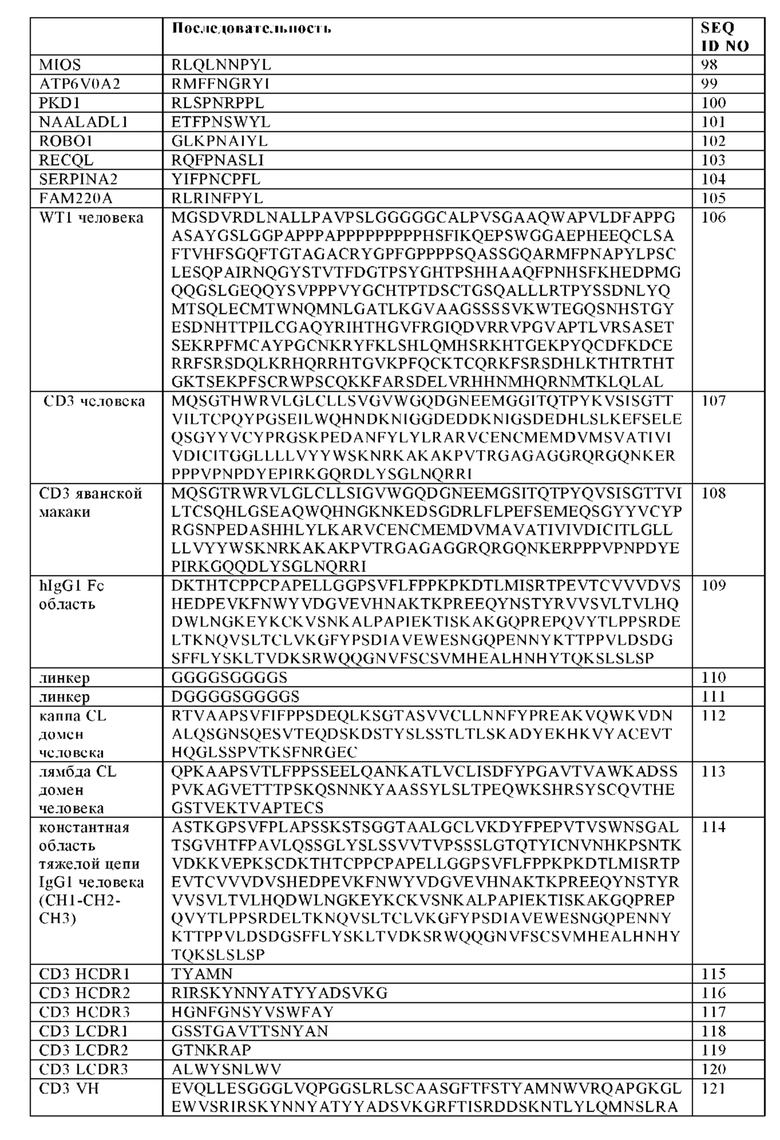
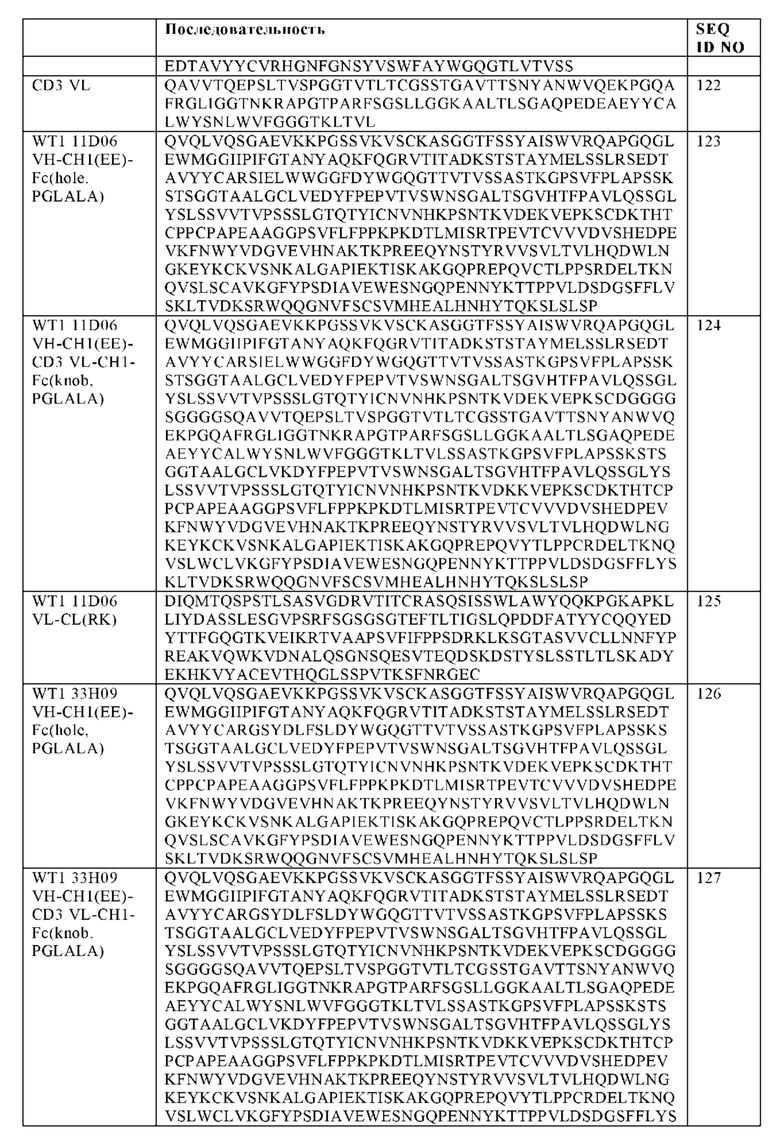

Несмотря на то что настоящее изобретение раскрыто и проиллюстрировано в определенных деталях и примерах с целью его наилучшего понимания и ясности, описание изобретения и примеры не должны пониматься, как ограничивающие объем изобретения. Раскрытие всех процитированных патентных заявок и научной литературы включено в описание изобретения исключительно в качестве ссылок.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ф.ХОФФМАНН-ЛЯ РОШ АГ
<120> АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С HLA-A2/WT1
<130> P34563
<150> EP17209205
<151> 2017-12-21
<160> 140
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 HCDR1
<400> 1
Ser Tyr Ala Ile Ser
1 5
<210> 2
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 HCDR2
<400> 2
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 3
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 HCDR3
<400> 3
Ser Ile Glu Leu Trp Trp Gly Gly Phe Asp Tyr
1 5 10
2
<210> 4
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 LCDR1
<400> 4
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 LCDR2
<400> 5
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 6
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 LCDR3
<400> 6
Gln Gln Tyr Glu Asp Tyr Thr Thr
1 5
<210> 7
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 VH
<400> 7
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
3
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ile Glu Leu Trp Trp Gly Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 8
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> 11D06 VL
<400> 8
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Gly Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Glu Asp Tyr Thr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
4
<210> 9
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 HCDR1
<400> 9
Ser Tyr Ala Ile Ser
1 5
<210> 10
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 HCDR2
<400> 10
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 11
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 HCDR3
<400> 11
Gly Ser Tyr Asp Leu Phe Ser Leu Asp Tyr
1 5 10
<210> 12
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 LCDR1
<400> 12
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 13
5
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 LCDR2
<400> 13
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 14
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 LCDR3
<400> 14
Gln Gln Tyr Tyr Asp Gly Ile Thr
1 5
<210> 15
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 VH
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
6
Ala Arg Gly Ser Tyr Asp Leu Phe Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 16
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> 33H09 VL
<400> 16
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Asp Gly Ile Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 17
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 13B04 HCDR1
<400> 17
Ser Tyr Tyr Trp Ser
1 5
<210> 18
7
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> 13B04 HCDR2
<400> 18
Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 19
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> 13B04 HCDR3
<400> 19
Val Ser Tyr Asn Gly Leu Asp Tyr
1 5
<210> 20
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 13B04 LCDR1
<400> 20
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 21
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 13B04 LCDR2
<400> 21
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 22
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
8
<223> 13B04 LCDR3
<400> 22
Gln Gln Tyr Asn Met Trp Asn Pro Val Thr
1 5 10
<210> 23
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> 13B04 VH
<400> 23
Glu Val Gln Leu Leu Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Val Ser Tyr Asn Gly Leu Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 24
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> 13B04 VL
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
9
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Met Trp Asn Pro
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 25
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 HCDR1
<400> 25
Ser Tyr Ala Ile Ser
1 5
<210> 26
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 HCDR2
<400> 26
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 27
<211> 11
10
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 HCDR3
<400> 27
Val Pro Gly Arg Trp Tyr Gly Ala Met Asp Tyr
1 5 10
<210> 28
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 LCDR1
<400> 28
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 29
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 LCDR2
<400> 29
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 30
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 LCDR3
<400> 30
Gln Gln Glu Asp Asp Tyr Pro Leu Thr
1 5
<210> 31
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 VH
11
<400> 31
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Pro Gly Arg Trp Tyr Gly Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 32
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> 11B09 VL
<400> 32
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
12
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Glu Asp Asp Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 33
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 HCDR1
<400> 33
Ser Tyr Tyr Trp Ser
1 5
<210> 34
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 HCDR2
<400> 34
Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 35
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 HCDR3
<400> 35
Ser Tyr Tyr Glu Ala Phe Asp Tyr
1 5
<210> 36
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 LCDR1
13
<400> 36
Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala Ser
1 5 10
<210> 37
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 LCDR2
<400> 37
Gly Lys Asn Asn Arg Pro Ser
1 5
<210> 38
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 LCDR3
<400> 38
Asn Ser Pro Asp Met Asn Gly Asn Ala Val
1 5 10
<210> 39
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 VH
<400> 39
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Asn Ser Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
14
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ser Tyr Tyr Glu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 40
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> 33F05 VL
<400> 40
Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Val Arg Ile Thr Cys Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Asn Ser Pro Asp Met Asn Gly Asn Ala
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 41
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 HCDR1
15
<400> 41
Ser Tyr Ala Ile Ser
1 5
<210> 42
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 HCDR2
<400> 42
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 43
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 HCDR3
<400> 43
Ser Ser Tyr Asp Leu Tyr Ser Phe Asp Tyr
1 5 10
<210> 44
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 LCDR1
<400> 44
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 45
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 LCDR2
16
<400> 45
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 46
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 LCDR3
<400> 46
Gln Gln Tyr Ser Phe Pro Pro Met Ile Thr
1 5 10
<210> 47
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 VH
<400> 47
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Tyr Asp Leu Tyr Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
17
<210> 48
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> 5E11 VL
<400> 48
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Phe Pro Pro Met
85 90 95
Ile Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 49
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 HCDR1
<400> 49
Ser Tyr Ala Met Ser
1 5
<210> 50
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 HCDR2
18
<400> 50
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 51
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 HCDR3
<400> 51
Thr Tyr Pro Tyr Thr Gly Ser Phe Asp Tyr
1 5 10
<210> 52
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 LCDR1
<400> 52
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 53
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 LCDR2
<400> 53
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 54
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 LCDR3
<400> 54
19
Gln Gln Asn Tyr Asn Tyr Pro Pro Thr
1 5
<210> 55
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 VH
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Thr Tyr Pro Tyr Thr Gly Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 56
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> 13E08 VL
<400> 56
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
20
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asn Tyr Asn Tyr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 57
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 5C01 HCDR1
<400> 57
Ser Tyr Ala Met Ser
1 5
<210> 58
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> 5C01 HCDR2
<400> 58
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 59
<211> 10
<212> PRT
<213> Artificial Sequence
21
<220>
<223> 5C01 HCDR3
<400> 59
Gly Ser Trp Val Ser Tyr Ala Phe Asp Tyr
1 5 10
<210> 60
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> 5C01 LCDR1
<400> 60
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 61
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 5C01 LCDR2
<400> 61
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 62
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> 5C01 LCDR3
<400> 62
Gln Gln Tyr Ser Ile Trp Phe Pro Tyr Thr
1 5 10
<210> 63
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> 5C01 VH
<400> 63
22
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Ser Trp Val Ser Tyr Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 64
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> 5C01 VL
<400> 64
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
23
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ile Trp Phe Pro
85 90 95
Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 65
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> 11G06 HCDR1
<400> 65
Ser Tyr Ala Ile Ser
1 5
<210> 66
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> 11G06 HCDR2
<400> 66
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 67
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> 11G06 HCDR3
<400> 67
Thr Gly Pro Tyr Tyr Gly Ala Phe Asp Tyr
1 5 10
<210> 68
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
24
<223> 11G06 LCDR1
<400> 68
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 69
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> 11G06 LCDR2
<400> 69
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 70
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> 11G06 LCDR3
<400> 70
Gln Gln Gly Phe Arg Gly Tyr Thr
1 5
<210> 71
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> 11G06 VH
<400> 71
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
25
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Pro Tyr Tyr Gly Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 72
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> 11G06 VL
<400> 72
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Phe Arg Gly Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 73
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
26
<223> ESK1 VH
<400> 73
Gln Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Glu Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Asn Phe
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Val Asp Pro Gly Tyr Ser Tyr Ser Thr Tyr Ser Pro Ser Phe
50 55 60
Gln Gly His Val Thr Ile Ser Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Asn Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Gln Tyr Ser Gly Tyr Tyr Asp Trp Phe Asp Pro Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 74
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> ESK1 VL
<400> 74
Gln Ala Trp Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln Arg
1 5 10 15
Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr
20 25 30
Val Asn Trp Tyr Gln Gln Val Pro Gly Thr Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
27
Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln Ser
65 70 75 80
Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu Asn
85 90 95
Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 75
<211> 115
<212> PRT
<213> Artificial Sequence
<220>
<223> Untargeted VH
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Ser Gly Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 76
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
28
<223> Untargeted VL
<400> 76
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 77
<211> 9
<212> PRT
<213> Homo sapiens
<400> 77
Val Leu Asp Phe Ala Pro Pro Gly Ala
1 5
<210> 78
<211> 9
<212> PRT
<213> Homo sapiens
<400> 78
Arg Met Phe Pro Asn Ala Pro Tyr Leu
1 5
<210> 79
<211> 9
<212> PRT
<213> Homo sapiens
<400> 79
29
Arg Met Phe Pro Thr Pro Pro Ser Leu
1 5
<210> 80
<211> 9
<212> PRT
<213> Homo sapiens
<400> 80
Arg Met Phe Pro Gly Glu Val Ala Leu
1 5
<210> 81
<211> 9
<212> PRT
<213> Homo sapiens
<400> 81
Arg Leu Phe Pro Asn Leu Pro Glu Leu
1 5
<210> 82
<211> 9
<212> PRT
<213> Homo sapiens
<400> 82
Arg Met Phe Pro Asn Lys Tyr Ser Leu
1 5
<210> 83
<211> 9
<212> PRT
<213> Homo sapiens
<400> 83
Ala Met Asp Pro Asn Ala Ala Tyr Val
1 5
<210> 84
<211> 9
<212> PRT
<213> Homo sapiens
<400> 84
Arg Met Gly Pro Asn Ile Tyr Glu Leu
1 5
<210> 85
<211> 9
30
<212> PRT
<213> Homo sapiens
<400> 85
Asn Met Pro Pro Asn Phe Pro Tyr Ile
1 5
<210> 86
<211> 9
<212> PRT
<213> Homo sapiens
<400> 86
Tyr Thr Ile Pro Asn His Pro Tyr Leu
1 5
<210> 87
<211> 9
<212> PRT
<213> Homo sapiens
<400> 87
Arg Leu Phe Pro Asn Ala Lys Phe Leu
1 5
<210> 88
<211> 9
<212> PRT
<213> Homo sapiens
<400> 88
Arg Met Val Pro Arg Ala Val Tyr Leu
1 5
<210> 89
<211> 9
<212> PRT
<213> Homo sapiens
<400> 89
Lys Met Val Pro Ser Ile Pro Tyr Leu
1 5
<210> 90
<211> 9
<212> PRT
<213> Homo sapiens
<400> 90
Arg Ile Phe Pro Ser Tyr Ser Tyr Leu
31
1 5
<210> 91
<211> 9
<212> PRT
<213> Homo sapiens
<400> 91
Arg Leu Phe Pro Asn Ser Lys Phe Leu
1 5
<210> 92
<211> 9
<212> PRT
<213> Homo sapiens
<400> 92
Lys Met Thr Pro Cys Ile Pro Tyr Leu
1 5
<210> 93
<211> 9
<212> PRT
<213> Homo sapiens
<400> 93
Ser Met Phe Pro Ser Leu Lys Tyr Leu
1 5
<210> 94
<211> 9
<212> PRT
<213> Homo sapiens
<400> 94
Arg Leu Leu Pro Ser Ala Pro Thr Leu
1 5
<210> 95
<211> 9
<212> PRT
<213> Homo sapiens
<400> 95
Arg Leu Arg Pro His Val Pro Tyr Leu
1 5
<210> 96
<211> 9
<212> PRT
32
<213> Homo sapiens
<400> 96
Arg Met Asn Pro Asn Ser Pro Ser Ile
1 5
<210> 97
<211> 9
<212> PRT
<213> Homo sapiens
<400> 97
Arg Val Phe Asn Asn Arg Trp Tyr Leu
1 5
<210> 98
<211> 9
<212> PRT
<213> Homo sapiens
<400> 98
Arg Leu Gln Leu Asn Asn Pro Tyr Leu
1 5
<210> 99
<211> 9
<212> PRT
<213> Homo sapiens
<400> 99
Arg Met Phe Phe Asn Gly Arg Tyr Ile
1 5
<210> 100
<211> 9
<212> PRT
<213> Homo sapiens
<400> 100
Arg Leu Ser Pro Asn Arg Pro Pro Leu
1 5
<210> 101
<211> 9
<212> PRT
<213> Homo sapiens
<400> 101
Glu Thr Phe Pro Asn Ser Trp Tyr Leu
1 5
33
<210> 102
<211> 9
<212> PRT
<213> Homo sapiens
<400> 102
Gly Leu Lys Pro Asn Ala Ile Tyr Leu
1 5
<210> 103
<211> 9
<212> PRT
<213> Homo sapiens
<400> 103
Arg Gln Phe Pro Asn Ala Ser Leu Ile
1 5
<210> 104
<211> 9
<212> PRT
<213> Homo sapiens
<400> 104
Tyr Ile Phe Pro Asn Cys Pro Phe Leu
1 5
<210> 105
<211> 9
<212> PRT
<213> Homo sapiens
<400> 105
Arg Leu Arg Ile Asn Phe Pro Tyr Leu
1 5
<210> 106
<211> 449
<212> PRT
<213> Homo sapiens
<400> 106
Met Gly Ser Asp Val Arg Asp Leu Asn Ala Leu Leu Pro Ala Val Pro
1 5 10 15
Ser Leu Gly Gly Gly Gly Gly Cys Ala Leu Pro Val Ser Gly Ala Ala
20 25 30
34
Gln Trp Ala Pro Val Leu Asp Phe Ala Pro Pro Gly Ala Ser Ala Tyr
35 40 45
Gly Ser Leu Gly Gly Pro Ala Pro Pro Pro Ala Pro Pro Pro Pro Pro
50 55 60
Pro Pro Pro Pro His Ser Phe Ile Lys Gln Glu Pro Ser Trp Gly Gly
65 70 75 80
Ala Glu Pro His Glu Glu Gln Cys Leu Ser Ala Phe Thr Val His Phe
85 90 95
Ser Gly Gln Phe Thr Gly Thr Ala Gly Ala Cys Arg Tyr Gly Pro Phe
100 105 110
Gly Pro Pro Pro Pro Ser Gln Ala Ser Ser Gly Gln Ala Arg Met Phe
115 120 125
Pro Asn Ala Pro Tyr Leu Pro Ser Cys Leu Glu Ser Gln Pro Ala Ile
130 135 140
Arg Asn Gln Gly Tyr Ser Thr Val Thr Phe Asp Gly Thr Pro Ser Tyr
145 150 155 160
Gly His Thr Pro Ser His His Ala Ala Gln Phe Pro Asn His Ser Phe
165 170 175
Lys His Glu Asp Pro Met Gly Gln Gln Gly Ser Leu Gly Glu Gln Gln
180 185 190
Tyr Ser Val Pro Pro Pro Val Tyr Gly Cys His Thr Pro Thr Asp Ser
195 200 205
Cys Thr Gly Ser Gln Ala Leu Leu Leu Arg Thr Pro Tyr Ser Ser Asp
210 215 220
Asn Leu Tyr Gln Met Thr Ser Gln Leu Glu Cys Met Thr Trp Asn Gln
225 230 235 240
Met Asn Leu Gly Ala Thr Leu Lys Gly Val Ala Ala Gly Ser Ser Ser
245 250 255
Ser Val Lys Trp Thr Glu Gly Gln Ser Asn His Ser Thr Gly Tyr Glu
260 265 270
Ser Asp Asn His Thr Thr Pro Ile Leu Cys Gly Ala Gln Tyr Arg Ile
35
275 280 285
His Thr His Gly Val Phe Arg Gly Ile Gln Asp Val Arg Arg Val Pro
290 295 300
Gly Val Ala Pro Thr Leu Val Arg Ser Ala Ser Glu Thr Ser Glu Lys
305 310 315 320
Arg Pro Phe Met Cys Ala Tyr Pro Gly Cys Asn Lys Arg Tyr Phe Lys
325 330 335
Leu Ser His Leu Gln Met His Ser Arg Lys His Thr Gly Glu Lys Pro
340 345 350
Tyr Gln Cys Asp Phe Lys Asp Cys Glu Arg Arg Phe Ser Arg Ser Asp
355 360 365
Gln Leu Lys Arg His Gln Arg Arg His Thr Gly Val Lys Pro Phe Gln
370 375 380
Cys Lys Thr Cys Gln Arg Lys Phe Ser Arg Ser Asp His Leu Lys Thr
385 390 395 400
His Thr Arg Thr His Thr Gly Lys Thr Ser Glu Lys Pro Phe Ser Cys
405 410 415
Arg Trp Pro Ser Cys Gln Lys Lys Phe Ala Arg Ser Asp Glu Leu Val
420 425 430
Arg His His Asn Met His Gln Arg Asn Met Thr Lys Leu Gln Leu Ala
435 440 445
Leu
<210> 107
<211> 207
<212> PRT
<213> Homo sapiens
<400> 107
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr
20 25 30
36
Gln Thr Pro Tyr Lys Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Pro Gln Tyr Pro Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys
50 55 60
Asn Ile Gly Gly Asp Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp
65 70 75 80
His Leu Ser Leu Lys Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr
85 90 95
Val Cys Tyr Pro Arg Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu
100 105 110
Tyr Leu Arg Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp Val Met
115 120 125
Ser Val Ala Thr Ile Val Ile Val Asp Ile Cys Ile Thr Gly Gly Leu
130 135 140
Leu Leu Leu Val Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys
145 150 155 160
Pro Val Thr Arg Gly Ala Gly Ala Gly Gly Arg Gln Arg Gly Gln Asn
165 170 175
Lys Glu Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg
180 185 190
Lys Gly Gln Arg Asp Leu Tyr Ser Gly Leu Asn Gln Arg Arg Ile
195 200 205
<210> 108
<211> 198
<212> PRT
<213> Macaca fascicularis
<400> 108
Met Gln Ser Gly Thr Arg Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Ile Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr
20 25 30
Gln Thr Pro Tyr Gln Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
37
35 40 45
Cys Ser Gln His Leu Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys
50 55 60
Asn Lys Glu Asp Ser Gly Asp Arg Leu Phe Leu Pro Glu Phe Ser Glu
65 70 75 80
Met Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro
85 90 95
Glu Asp Ala Ser His His Leu Tyr Leu Lys Ala Arg Val Cys Glu Asn
100 105 110
Cys Met Glu Met Asp Val Met Ala Val Ala Thr Ile Val Ile Val Asp
115 120 125
Ile Cys Ile Thr Leu Gly Leu Leu Leu Leu Val Tyr Tyr Trp Ser Lys
130 135 140
Asn Arg Lys Ala Lys Ala Lys Pro Val Thr Arg Gly Ala Gly Ala Gly
145 150 155 160
Gly Arg Gln Arg Gly Gln Asn Lys Glu Arg Pro Pro Pro Val Pro Asn
165 170 175
Pro Asp Tyr Glu Pro Ile Arg Lys Gly Gln Gln Asp Leu Tyr Ser Gly
180 185 190
Leu Asn Gln Arg Arg Ile
195
<210> 109
<211> 225
<212> PRT
<213> Homo sapiens
<400> 109
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
38
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro
225
<210> 110
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
39
<210> 111
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> linker
<400> 111
Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 112
<211> 107
<212> PRT
<213> Homo sapiens
<400> 112
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 113
<211> 105
<212> PRT
<213> Homo sapiens
<400> 113
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
40
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 114
<211> 328
<212> PRT
<213> Homo sapiens
<400> 114
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
41
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 115
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
42
<223> CD3 HCDR1
<400> 115
Thr Tyr Ala Met Asn
1 5
<210> 116
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 HCDR2
<400> 116
Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser
1 5 10 15
Val Lys Gly
<210> 117
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 HCDR3
<400> 117
His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe Ala Tyr
1 5 10
<210> 118
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 LCDR1
<400> 118
Gly Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1 5 10
<210> 119
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 LCDR2
43
<400> 119
Gly Thr Asn Lys Arg Ala Pro
1 5
<210> 120
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 LCDR3
<400> 120
Ala Leu Trp Tyr Ser Asn Leu Trp Val
1 5
<210> 121
<211> 125
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 VH
<400> 121
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
44
<210> 122
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 VL
<400> 122
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 123
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> WT1 11D06 VH-CH1(EE)-Fc(hole, PGLALA)
<400> 123
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ile Glu Leu Trp Trp Gly Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
46
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 124
<211> 673
<212> PRT
<213> Artificial Sequence
<220>
<223> WT1 11D06 VH-CH1(EE)-CD3 VL-CH1-Fc(knob, PGLALA)
<400> 124
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
47
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ile Glu Leu Trp Trp Gly Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr Gln
225 230 235 240
Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys
245 250 255
Gly Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp Val
260 265 270
Gln Glu Lys Pro Gly Gln Ala Phe Arg Gly Leu Ile Gly Gly Thr Asn
275 280 285
48
Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly
290 295 300
Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala Gln Pro Glu Asp Glu Ala
305 310 315 320
Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn Leu Trp Val Phe Gly Gly
325 330 335
Gly Thr Lys Leu Thr Val Leu Ser Ser Ala Ser Thr Lys Gly Pro Ser
340 345 350
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
355 360 365
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
370 375 380
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
385 390 395 400
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
405 410 415
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
420 425 430
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
435 440 445
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
450 455 460
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
465 470 475 480
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
485 490 495
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
500 505 510
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
515 520 525
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
49
530 535 540
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
545 550 555 560
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
565 570 575
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
580 585 590
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
595 600 605
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
610 615 620
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
625 630 635 640
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
645 650 655
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
660 665 670
Pro
<210> 125
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> WT1 11D06 VL-CL(RK)
<400> 125
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
50
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Gly Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Glu Asp Tyr Thr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 126
<211> 447
<212> PRT
<213> Artificial Sequence
<220>
<223> WT1 33H09 VH-CH1(EE)-Fc(hole, PGLALA)
<400> 126
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
51
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Asp Leu Phe Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
52
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser
355 360 365
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 127
<211> 672
<212> PRT
<213> Artificial Sequence
<220>
<223> WT1 33H09 VH-CH1(EE)-CD3 VL-CH1-Fc(knob, PGLALA)
<400> 127
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
53
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Asp Leu Phe Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly
210 215 220
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr Gln Glu
225 230 235 240
Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly
245 250 255
Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp Val Gln
260 265 270
54
Glu Lys Pro Gly Gln Ala Phe Arg Gly Leu Ile Gly Gly Thr Asn Lys
275 280 285
Arg Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly
290 295 300
Lys Ala Ala Leu Thr Leu Ser Gly Ala Gln Pro Glu Asp Glu Ala Glu
305 310 315 320
Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn Leu Trp Val Phe Gly Gly Gly
325 330 335
Thr Lys Leu Thr Val Leu Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
340 345 350
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
355 360 365
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
370 375 380
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
385 390 395 400
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
405 410 415
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
420 425 430
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
435 440 445
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
450 455 460
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
465 470 475 480
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
485 490 495
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
500 505 510
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
55
515 520 525
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
530 535 540
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu
545 550 555 560
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
565 570 575
Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
580 585 590
Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
595 600 605
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
610 615 620
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
625 630 635 640
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
645 650 655
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
660 665 670
<210> 128
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> WT1 33H09 VL-CL(RK)
<400> 128
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
56
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Asp Gly Ile Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 129
<211> 232
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 VH-CL
<400> 129
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
57
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val
115 120 125
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
130 135 140
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
145 150 155 160
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
165 170 175
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
180 185 190
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
195 200 205
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
210 215 220
Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 130
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 HCDR1 (V9)
58
<400> 130
Gly Tyr Thr Met Asn
1 5
<210> 131
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 HCDR2 (V9)
<400> 131
Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 132
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 HCDR3 (V9)
<400> 132
Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val
1 5 10
<210> 133
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 LCDR1 (V9)
<400> 133
Arg Ala Ser Gln Asp Ile Arg Asn Tyr Leu Asn
1 5 10
<210> 134
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 LCDR2 (V9)
<400> 134
59
Tyr Thr Ser Arg Leu Glu Ser
1 5
<210> 135
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 LCDR3 (V9)
<400> 135
Gln Gln Gly Asn Thr Leu Pro Trp Thr
1 5
<210> 136
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 VH (V9)
<400> 136
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
60
<210> 137
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 VL (V9)
<400> 137
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 138
<211> 275
<212> PRT
<213> Homo sapiens
<400> 138
Gly Ser His Ser Met Arg Tyr Phe Phe Thr Ser Val Ser Arg Pro Gly
1 5 10 15
Arg Gly Glu Pro Arg Phe Ile Ala Val Gly Tyr Val Asp Asp Thr Gln
20 25 30
Phe Val Arg Phe Asp Ser Asp Ala Ala Ser Gln Arg Met Glu Pro Arg
35 40 45
Ala Pro Trp Ile Glu Gln Glu Gly Pro Glu Tyr Trp Asp Gly Glu Thr
50 55 60
61
Arg Lys Val Lys Ala His Ser Gln Thr His Arg Val Asp Leu Gly Thr
65 70 75 80
Leu Arg Gly Tyr Tyr Asn Gln Ser Glu Ala Gly Ser His Thr Val Gln
85 90 95
Arg Met Tyr Gly Cys Asp Val Gly Ser Asp Trp Arg Phe Leu Arg Gly
100 105 110
Tyr His Gln Tyr Ala Tyr Asp Gly Lys Asp Tyr Ile Ala Leu Lys Glu
115 120 125
Asp Leu Arg Ser Trp Thr Ala Ala Asp Met Ala Ala Gln Thr Thr Lys
130 135 140
His Lys Trp Glu Ala Ala His Val Ala Glu Gln Leu Arg Ala Tyr Leu
145 150 155 160
Glu Gly Thr Cys Val Glu Trp Leu Arg Arg Tyr Leu Glu Asn Gly Lys
165 170 175
Glu Thr Leu Gln Arg Thr Asp Ala Pro Lys Thr His Met Thr His His
180 185 190
Ala Val Ser Asp His Glu Ala Thr Leu Arg Cys Trp Ala Leu Ser Phe
195 200 205
Tyr Pro Ala Glu Ile Thr Leu Thr Trp Gln Arg Asp Gly Glu Asp Gln
210 215 220
Thr Gln Asp Thr Glu Leu Val Glu Thr Arg Pro Ala Gly Asp Gly Thr
225 230 235 240
Phe Gln Lys Trp Ala Ala Val Val Val Pro Ser Gly Gln Glu Gln Arg
245 250 255
Tyr Thr Cys His Val Gln His Glu Gly Leu Pro Lys Pro Leu Thr Leu
260 265 270
Arg Trp Glu
275
<210> 139
<211> 671
<212> PRT
<213> Artificial Sequence
62
<220>
<223> WT1 11D06 VH-CH1(EE)-CD3 (V9) VL-CH1-Fc(knob, PGLALA)
<400> 139
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ile Glu Leu Trp Trp Gly Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
63
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln
225 230 235 240
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
245 250 255
Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr Leu Asn Trp Tyr Gln Gln
260 265 270
Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Tyr Thr Ser Arg Leu
275 280 285
Glu Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
290 295 300
Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr
305 310 315 320
Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp Thr Phe Gly Gln Gly Thr
325 330 335
Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
340 345 350
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
355 360 365
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
370 375 380
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
385 390 395 400
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
405 410 415
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
420 425 430
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
435 440 445
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
450 455 460
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
64
465 470 475 480
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
485 490 495
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
500 505 510
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
515 520 525
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
530 535 540
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
545 550 555 560
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
565 570 575
Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp
580 585 590
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
595 600 605
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
610 615 620
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
625 630 635 640
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
645 650 655
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
660 665 670
<210> 140
<211> 229
<212> PRT
<213> Artificial Sequence
<220>
<223> CD3 (V9) VH-CL
<400> 140
65
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro
115 120 125
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
130 135 140
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
145 150 155 160
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
165 170 175
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
180 185 190
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
195 200 205
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
210 215 220
Asn Arg Gly Glu Cys
225
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ АКТИВИРУЮЩИЕ РЕЦЕПТОРЫ И ХИМЕРНЫЕ СТИМУЛИРУЮЩИЕ РЕЦЕПТОРЫ, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2780020C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| Двухвалентное биспецифическое химерное антитело, включающее гетеродимер на основе MHC или MHC-подобных белков | 2021 |
|
RU2820683C2 |
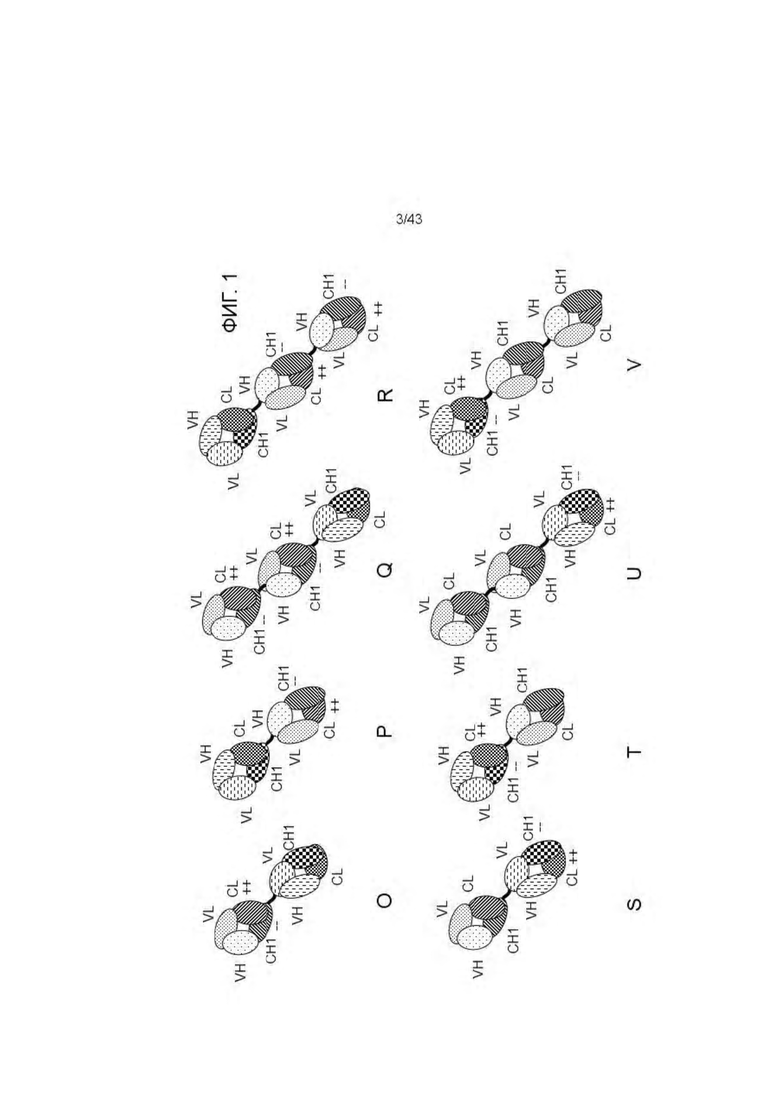

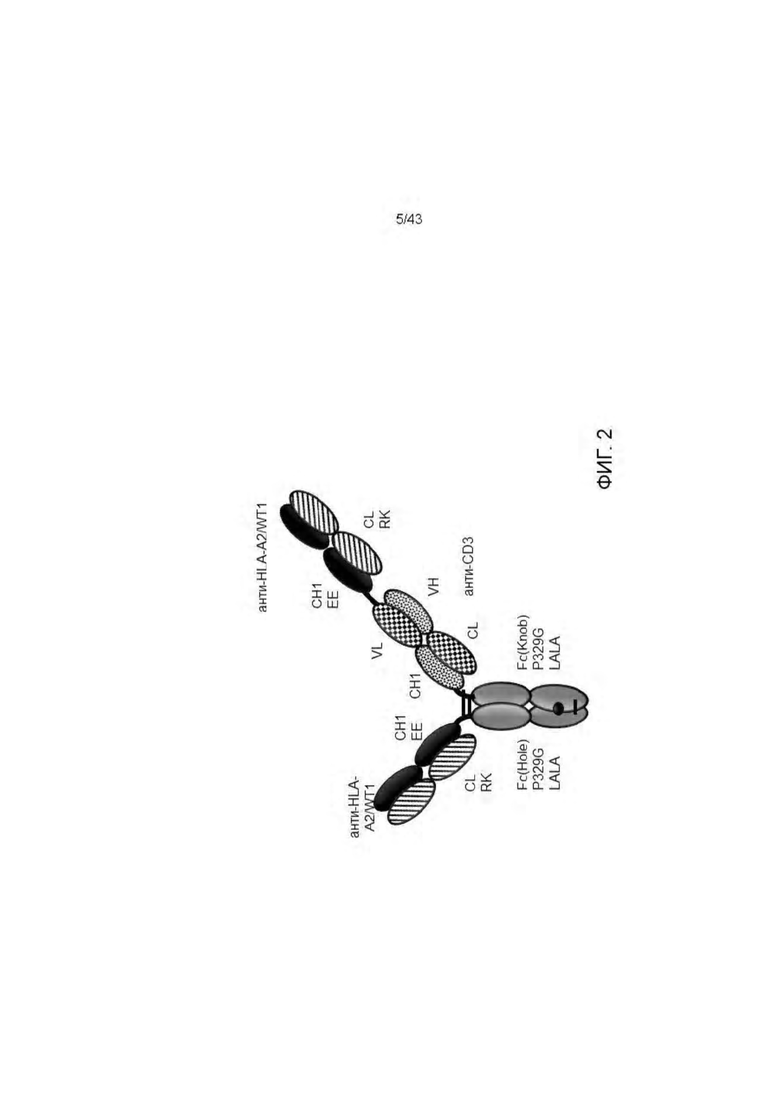
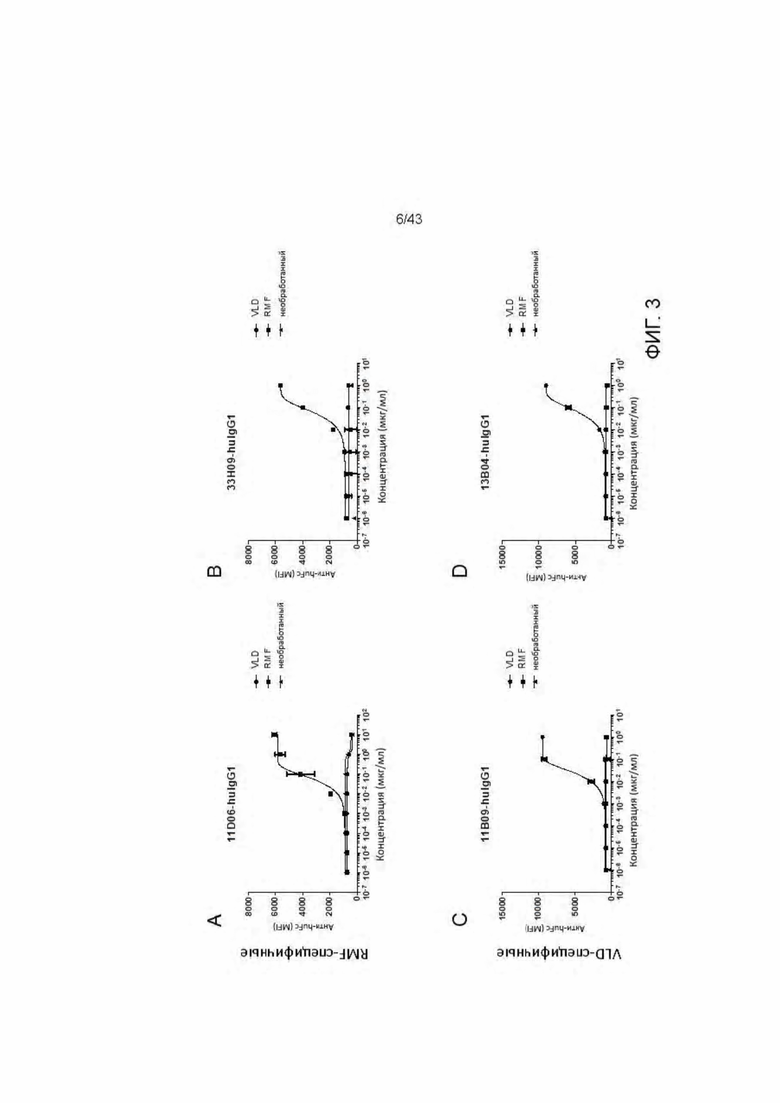
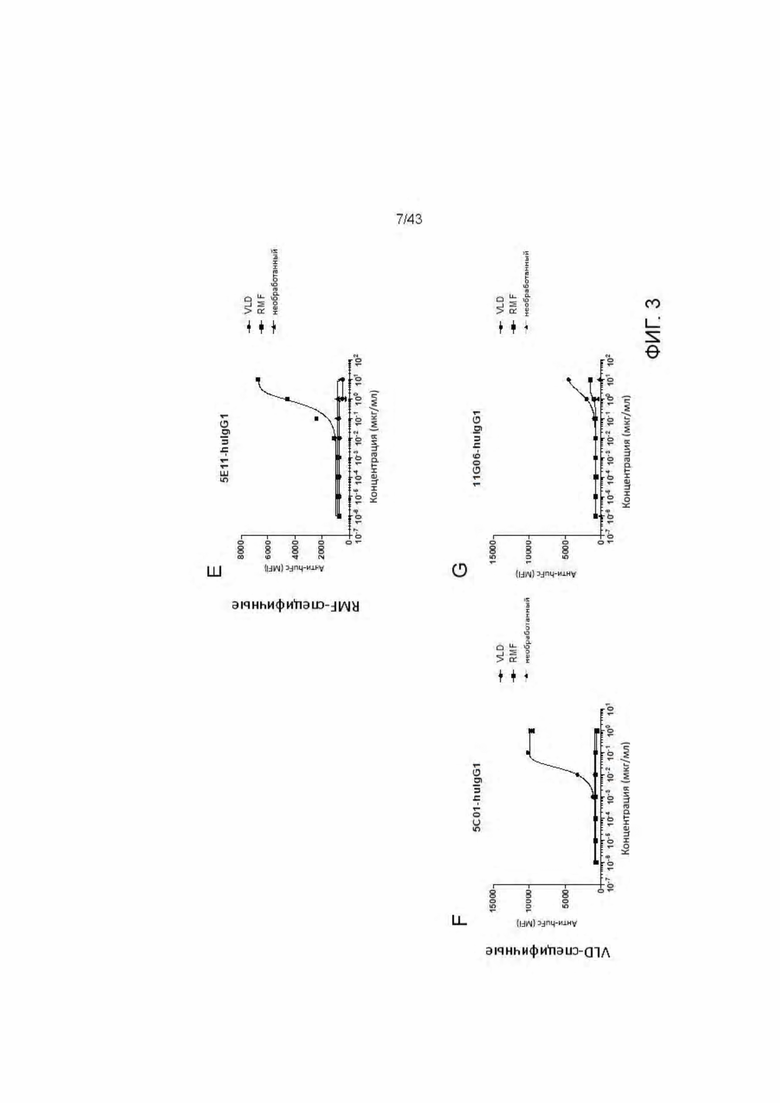
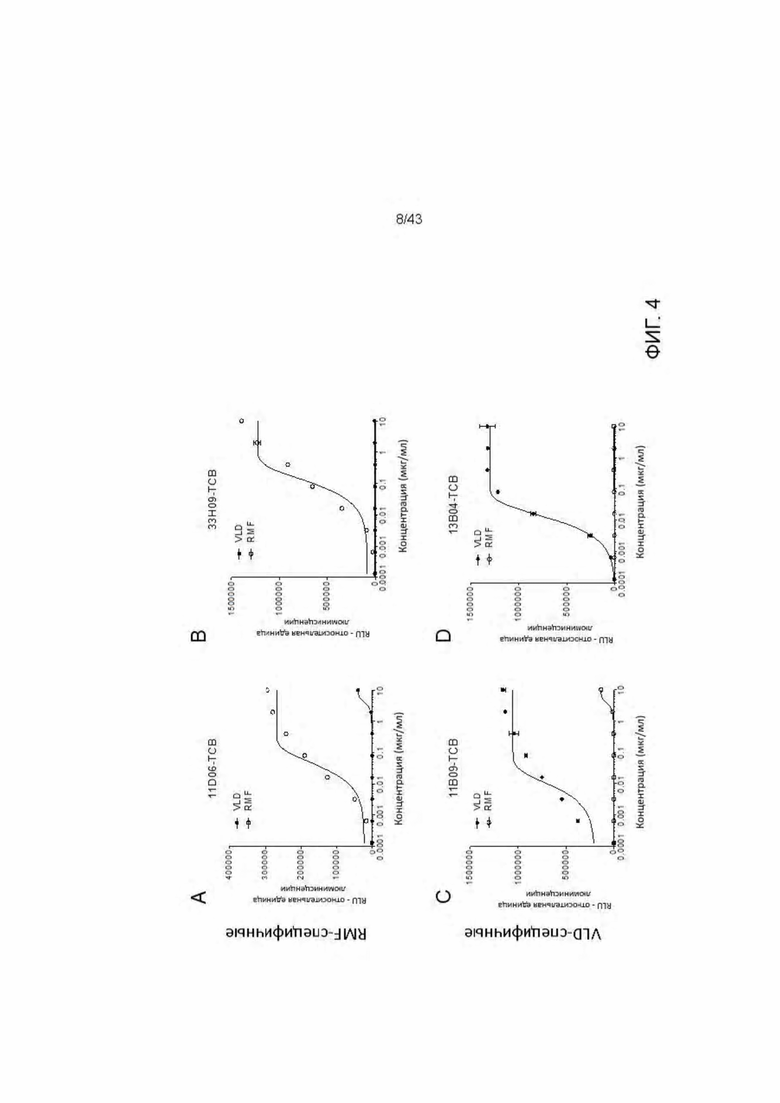
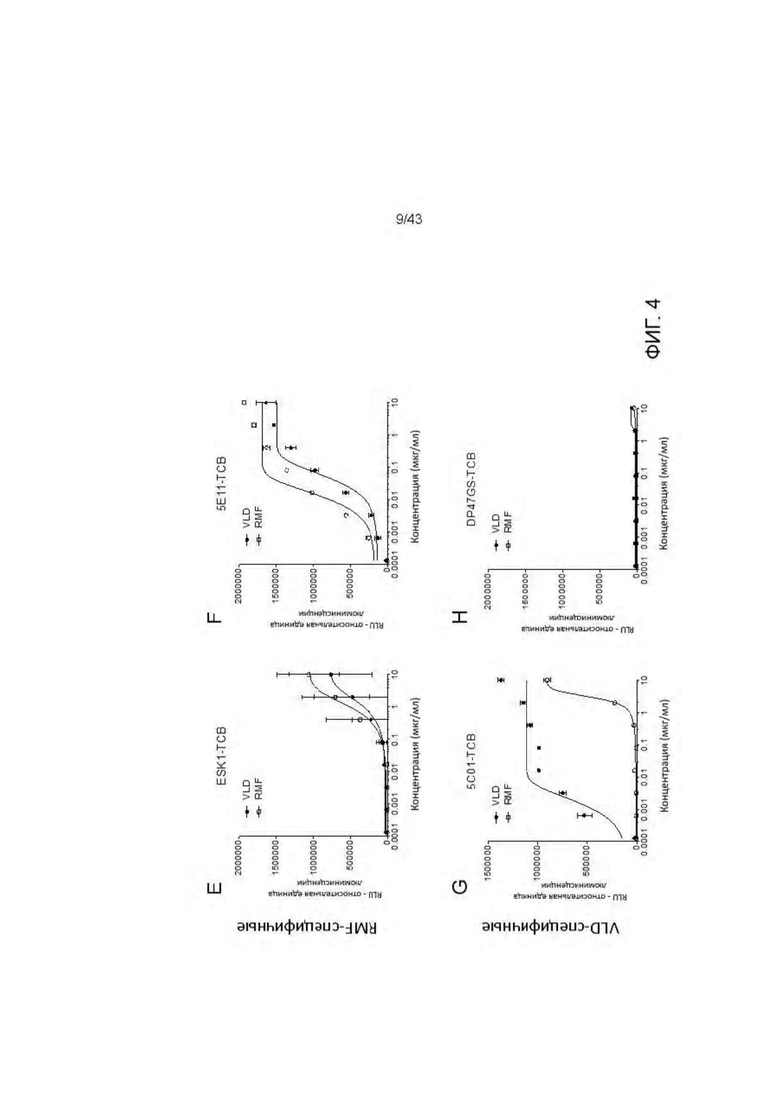
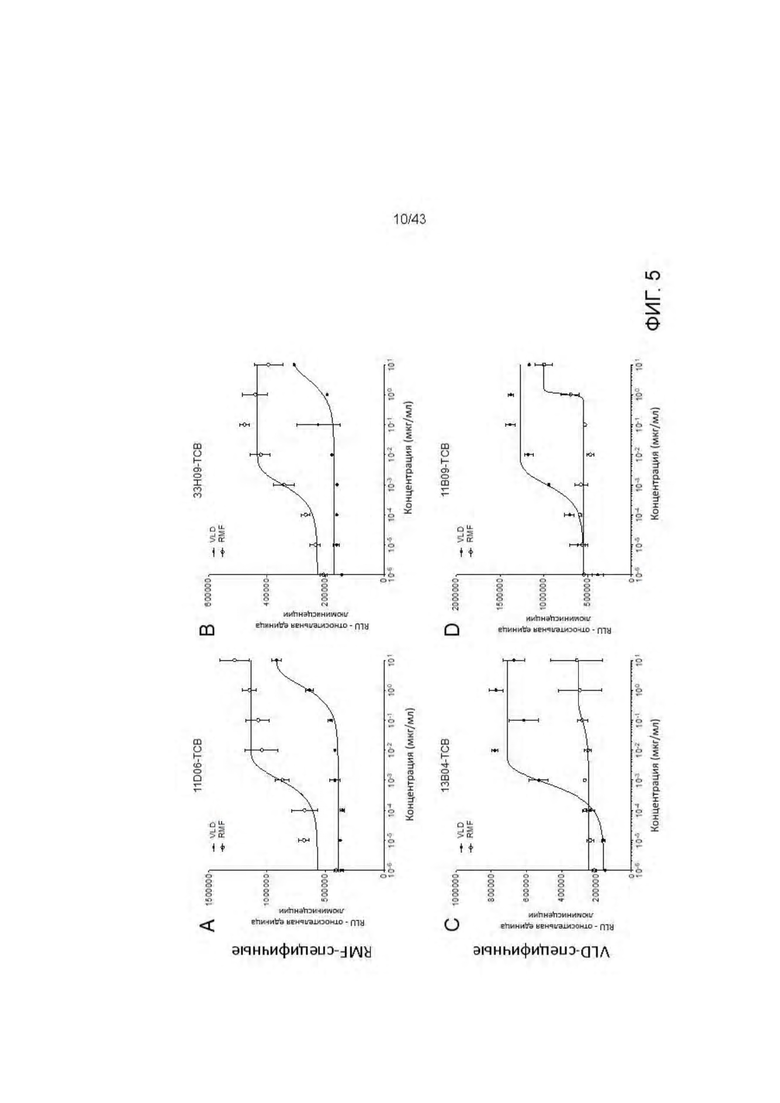
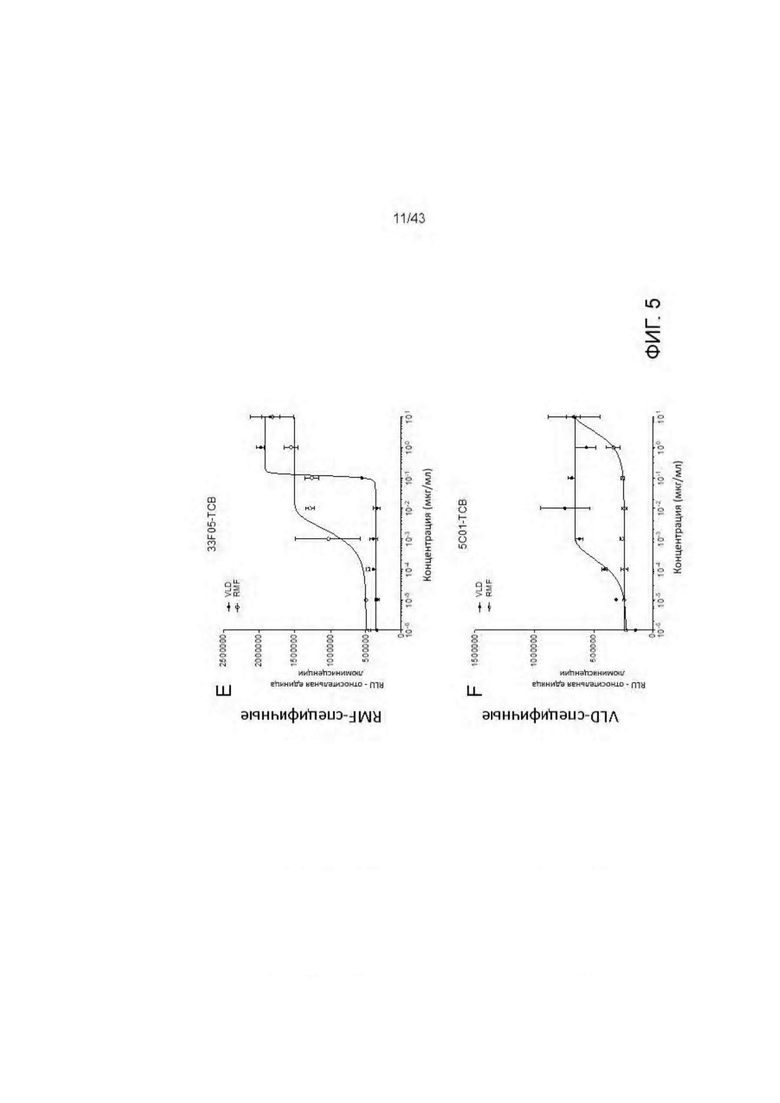
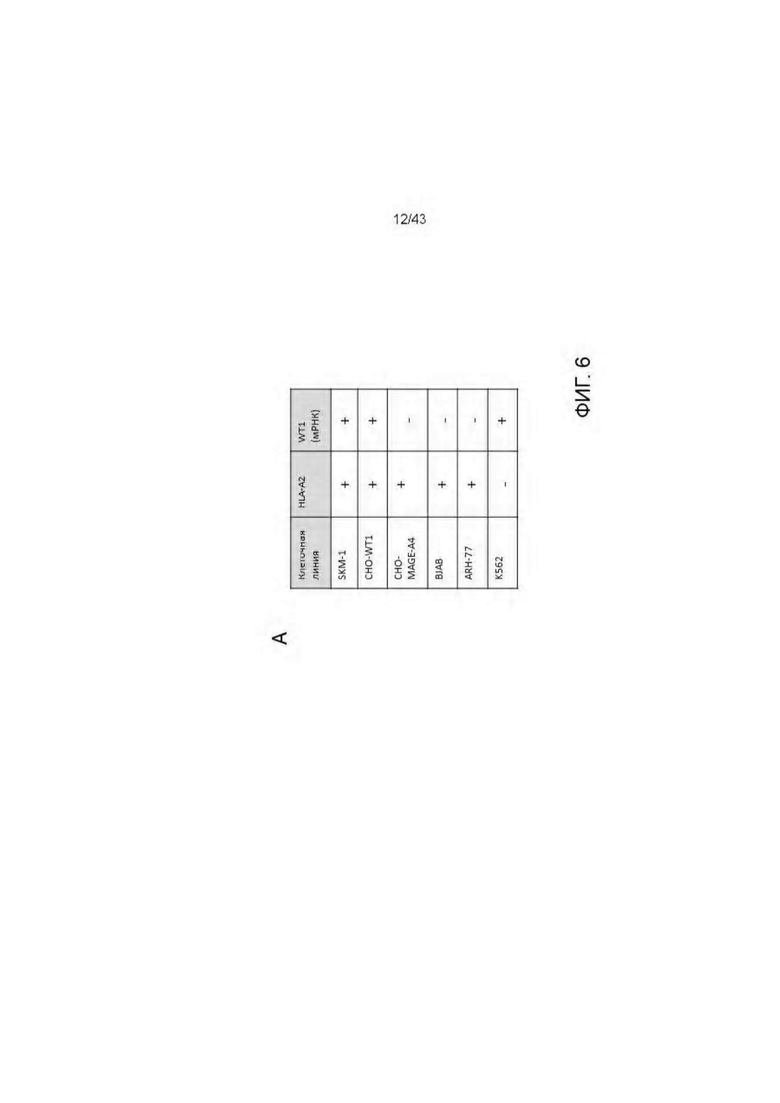
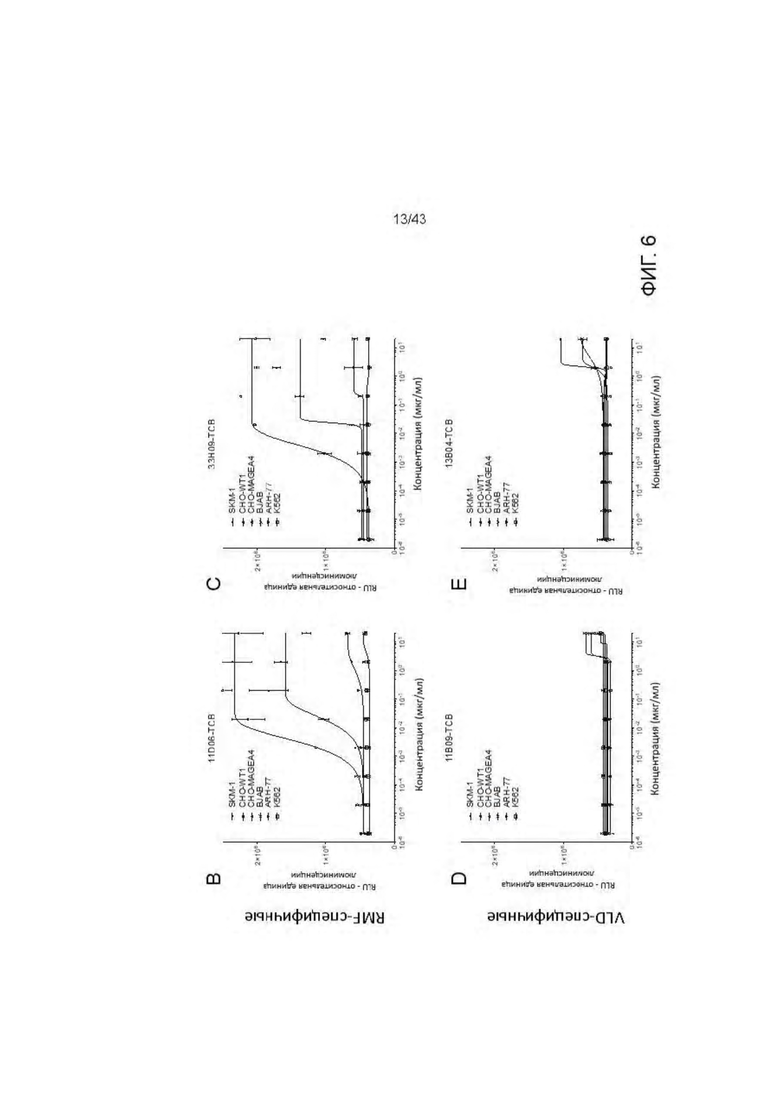
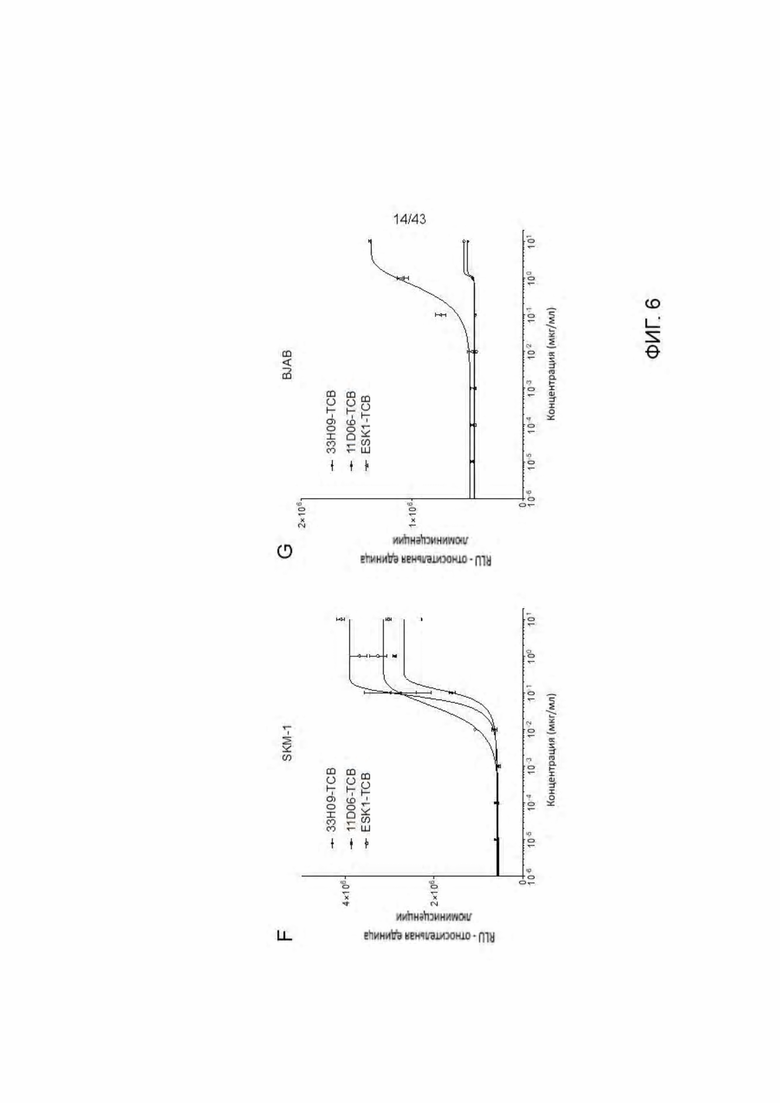
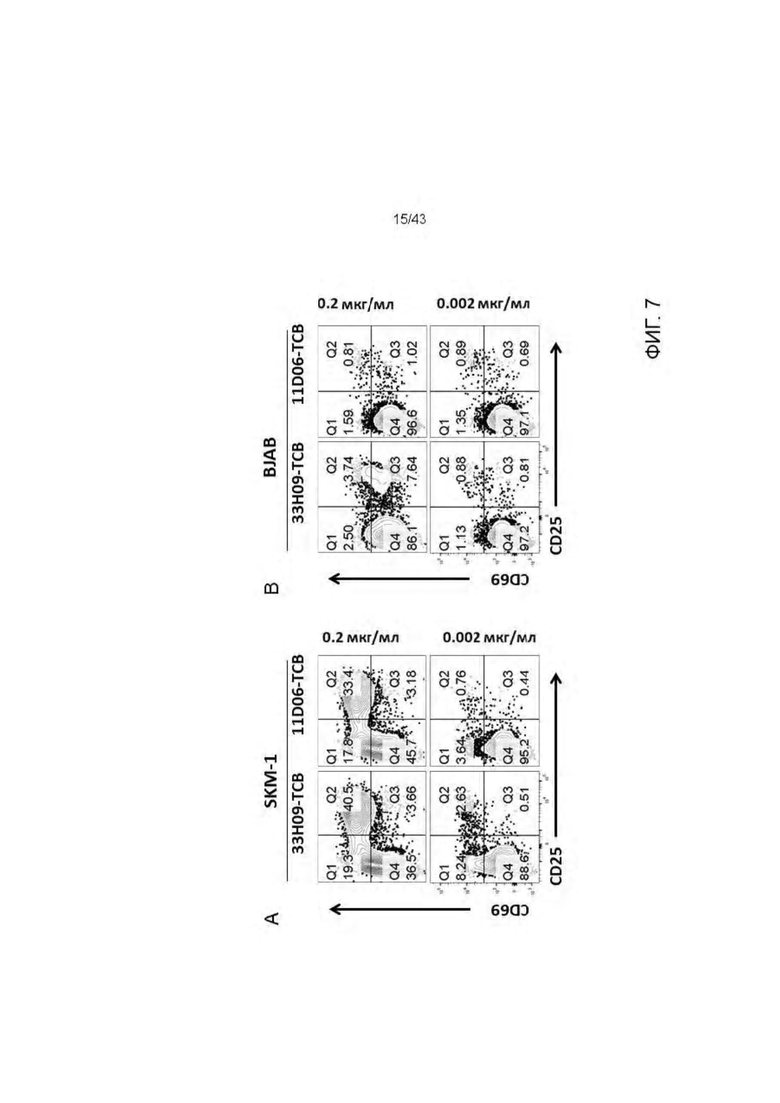
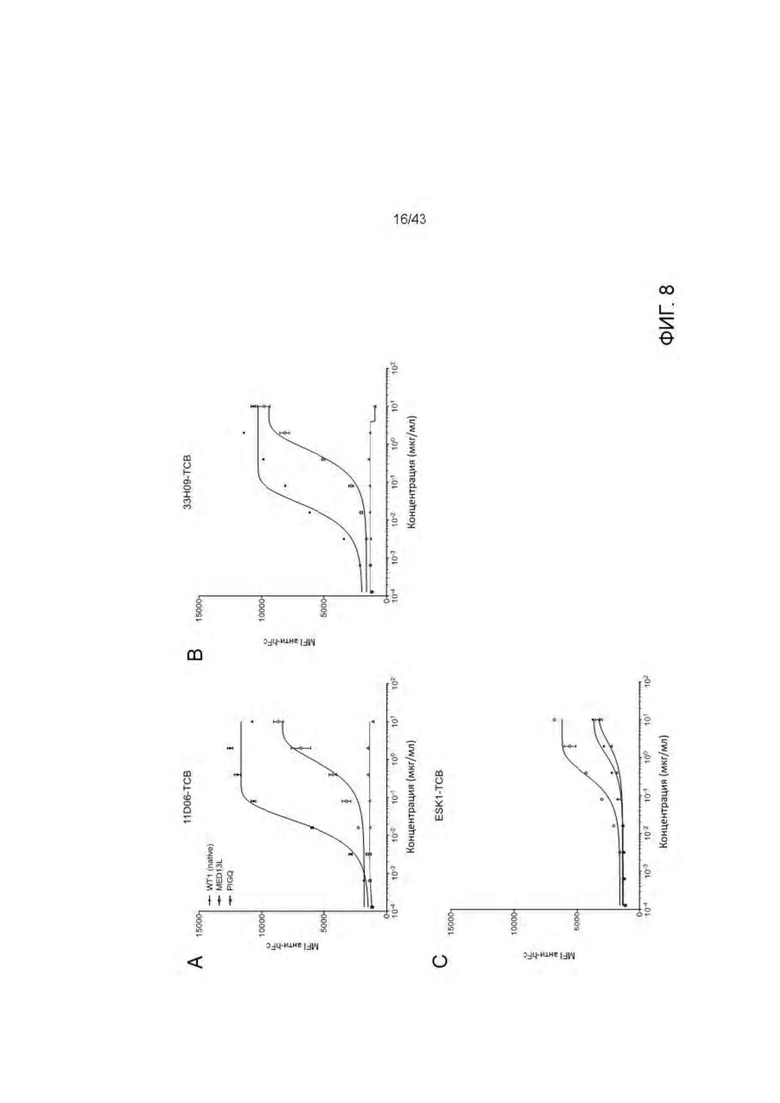
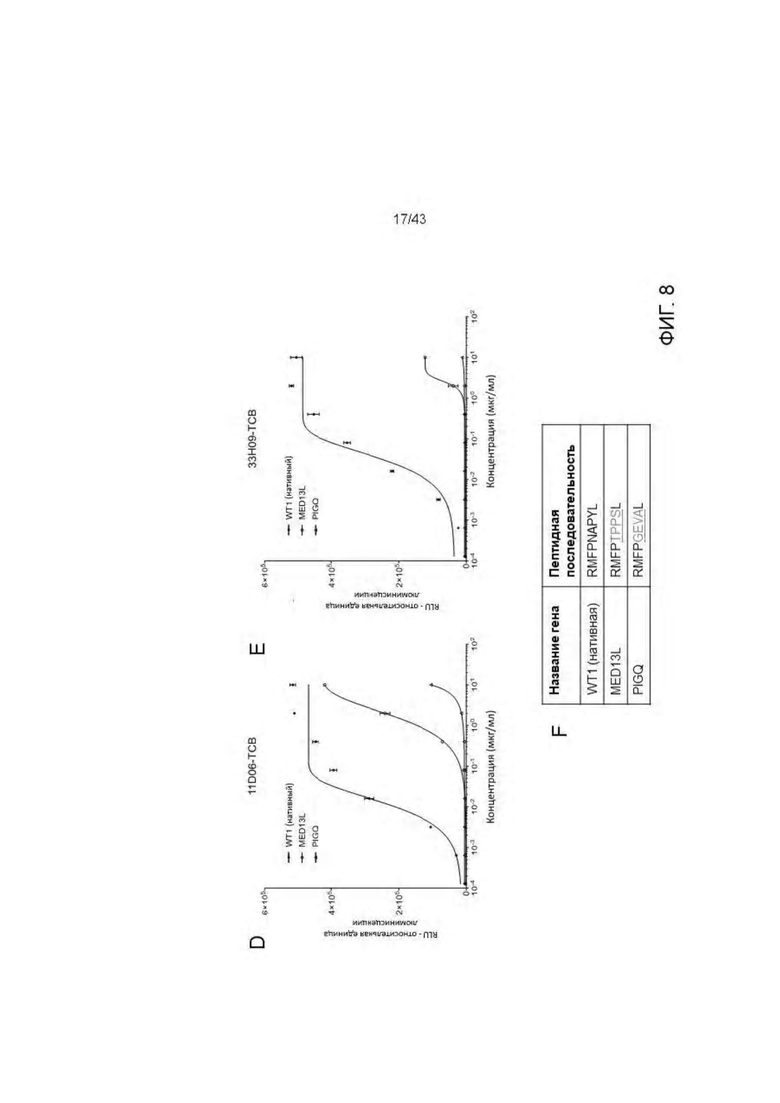
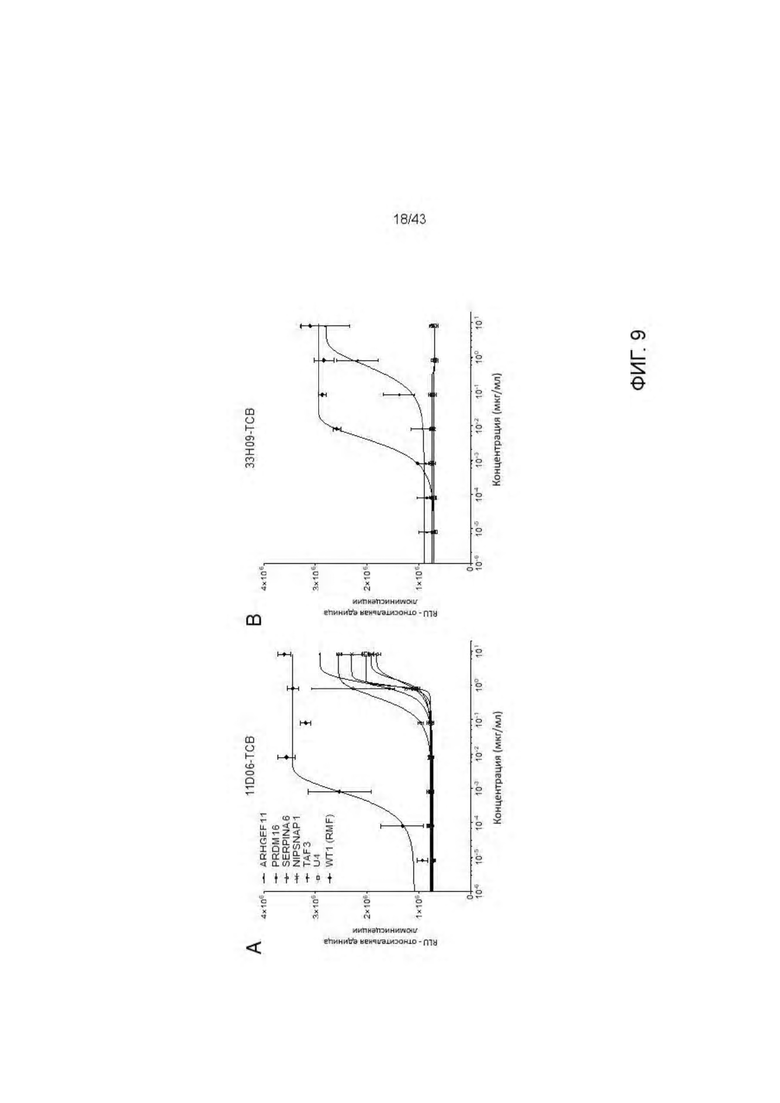
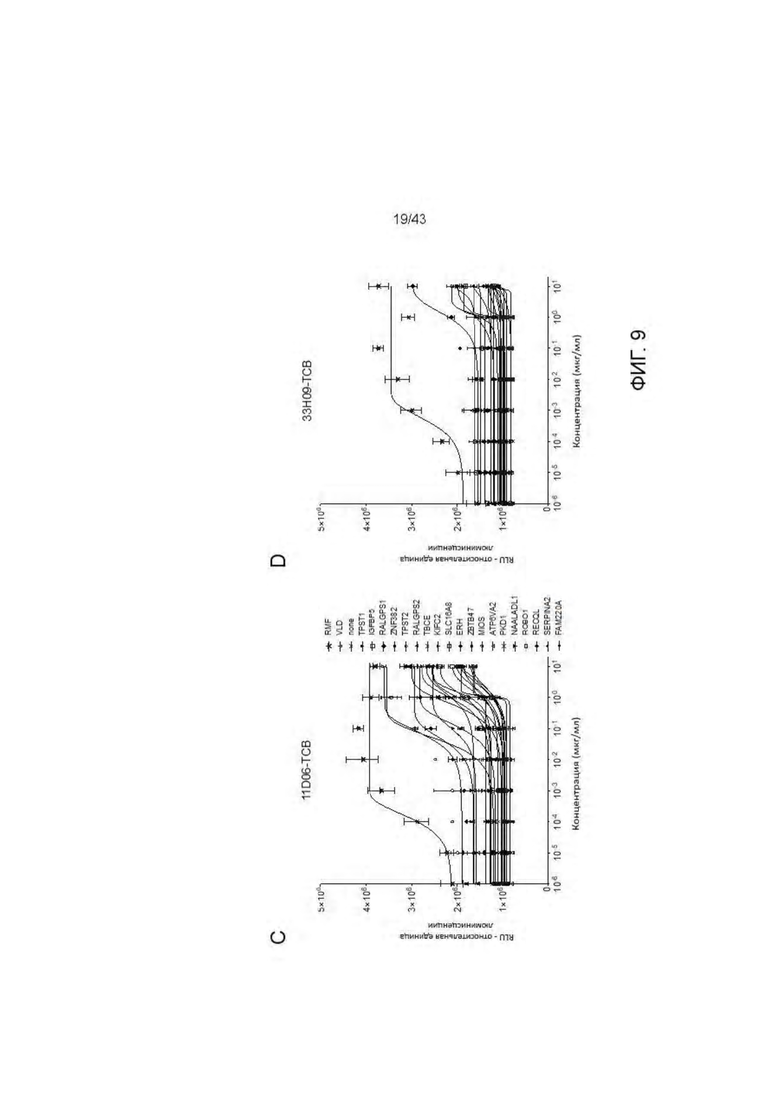
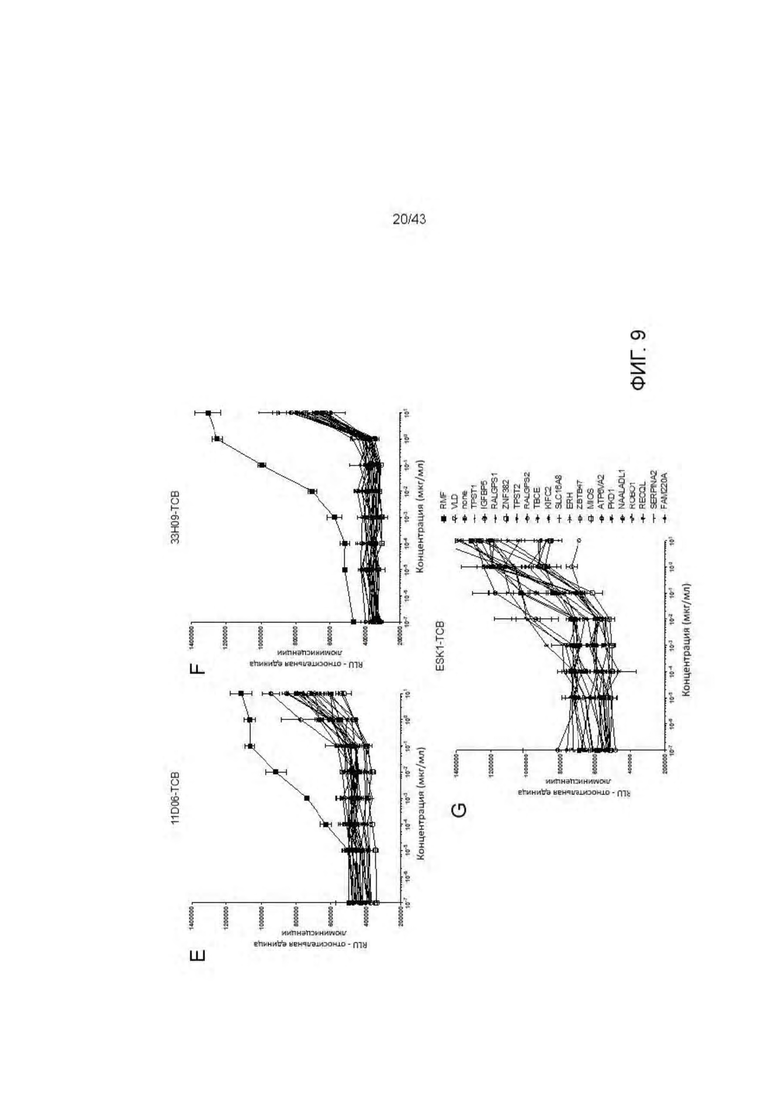
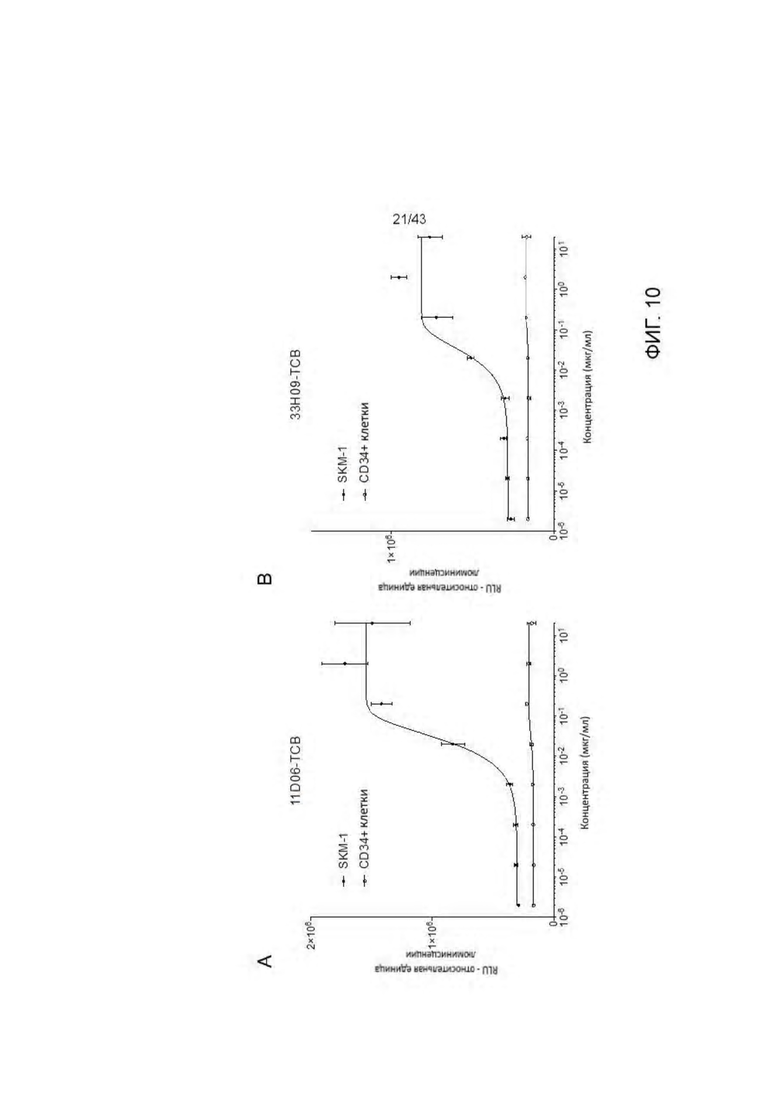
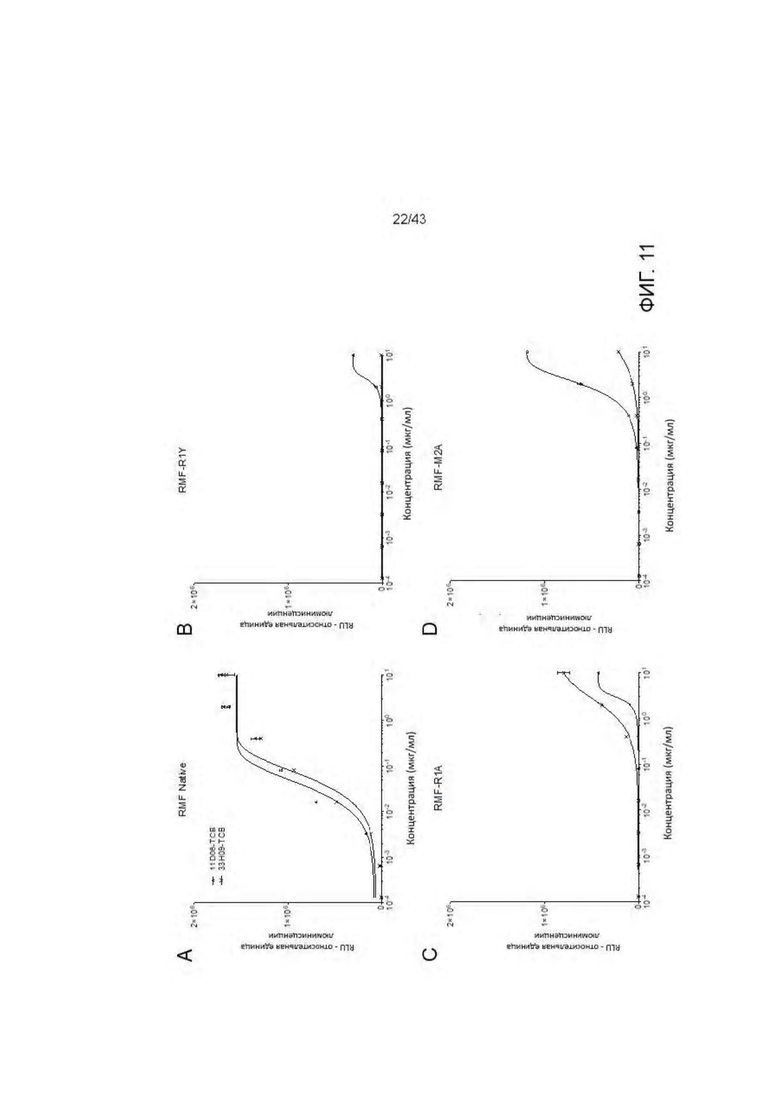
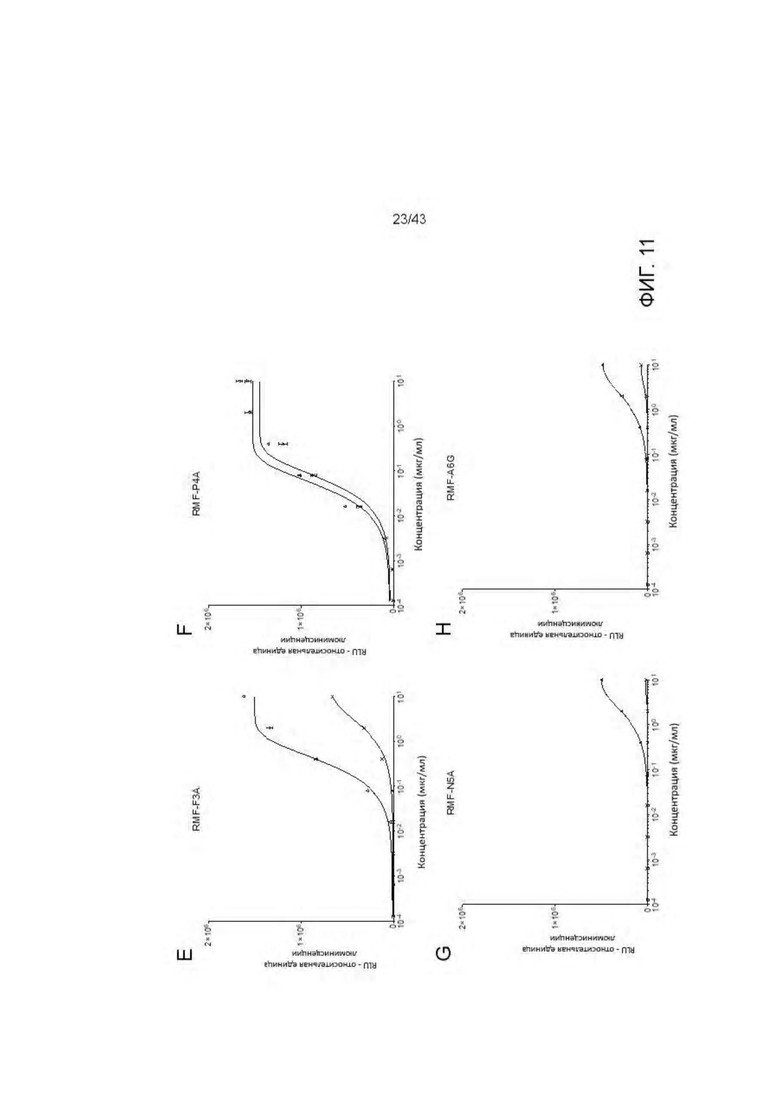
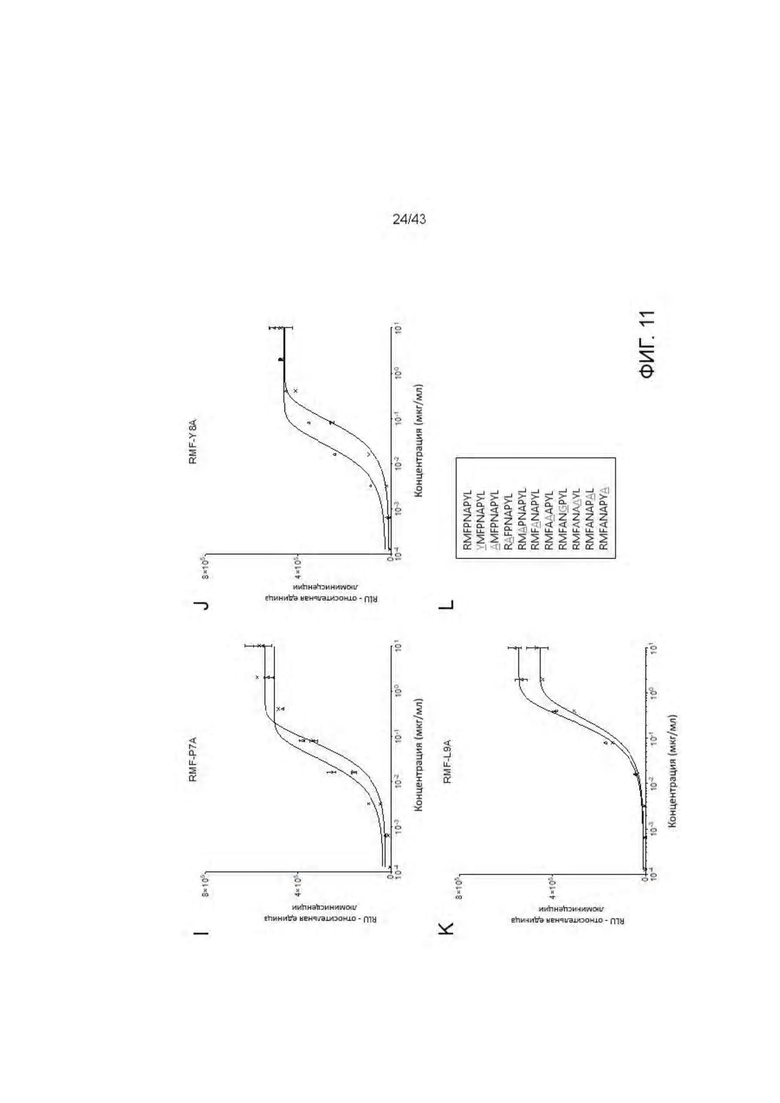
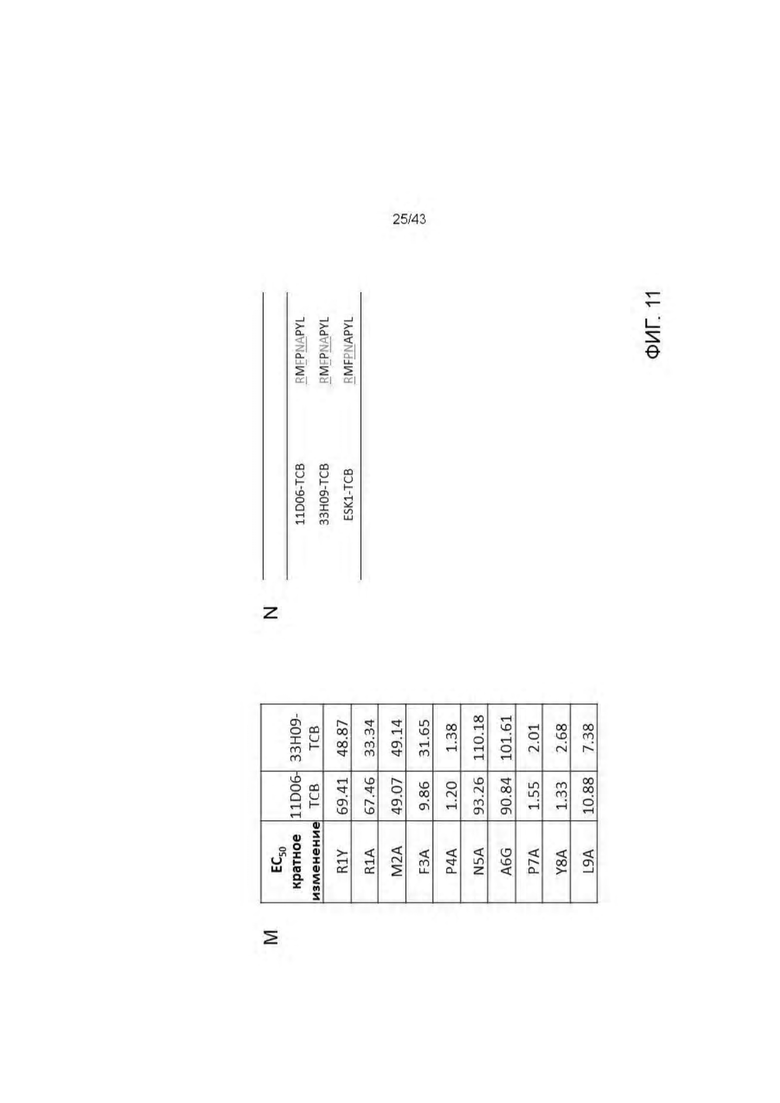
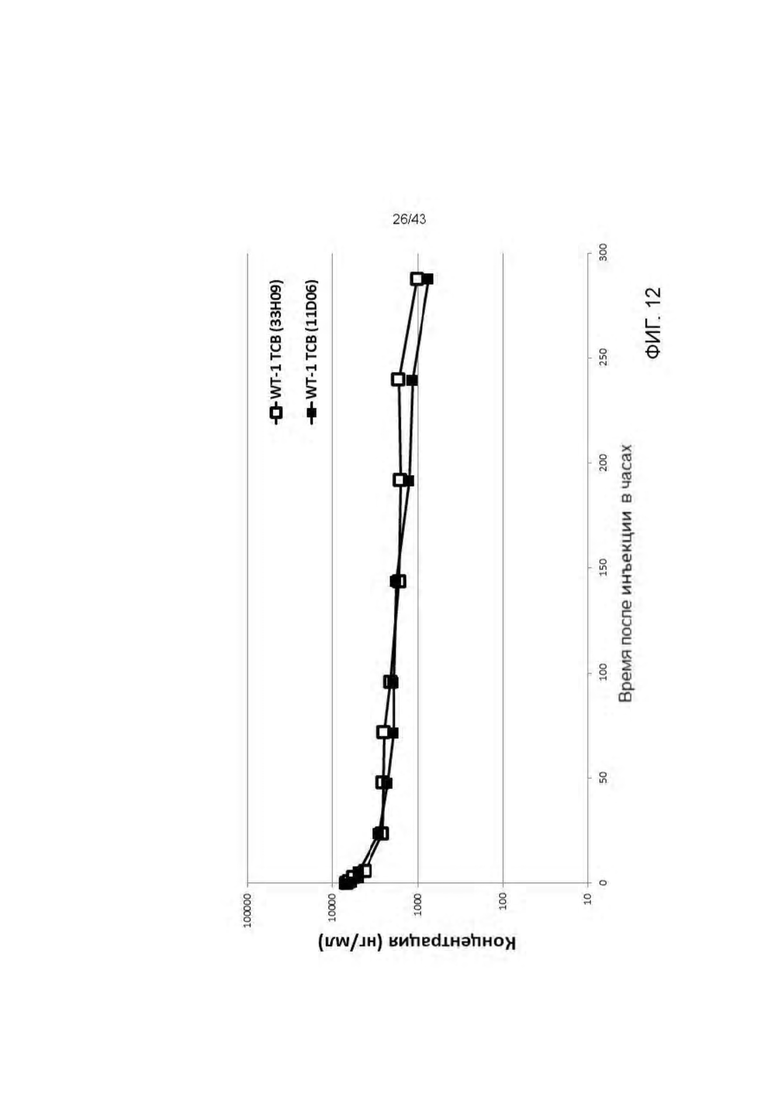
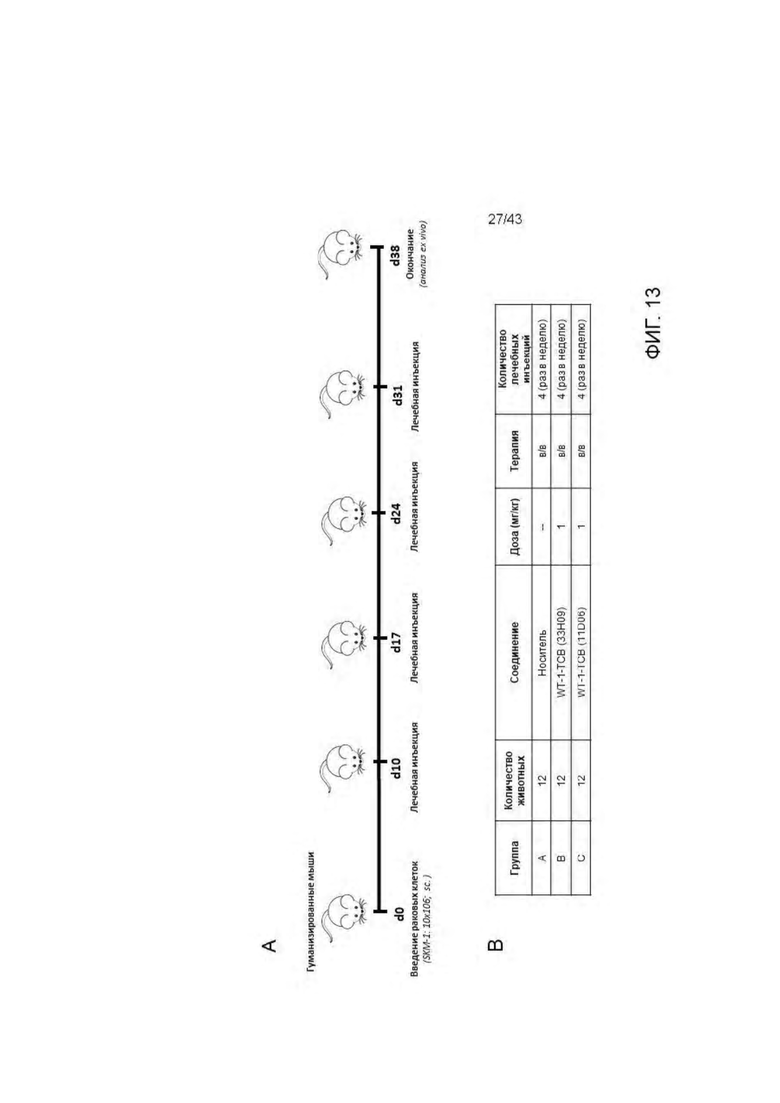
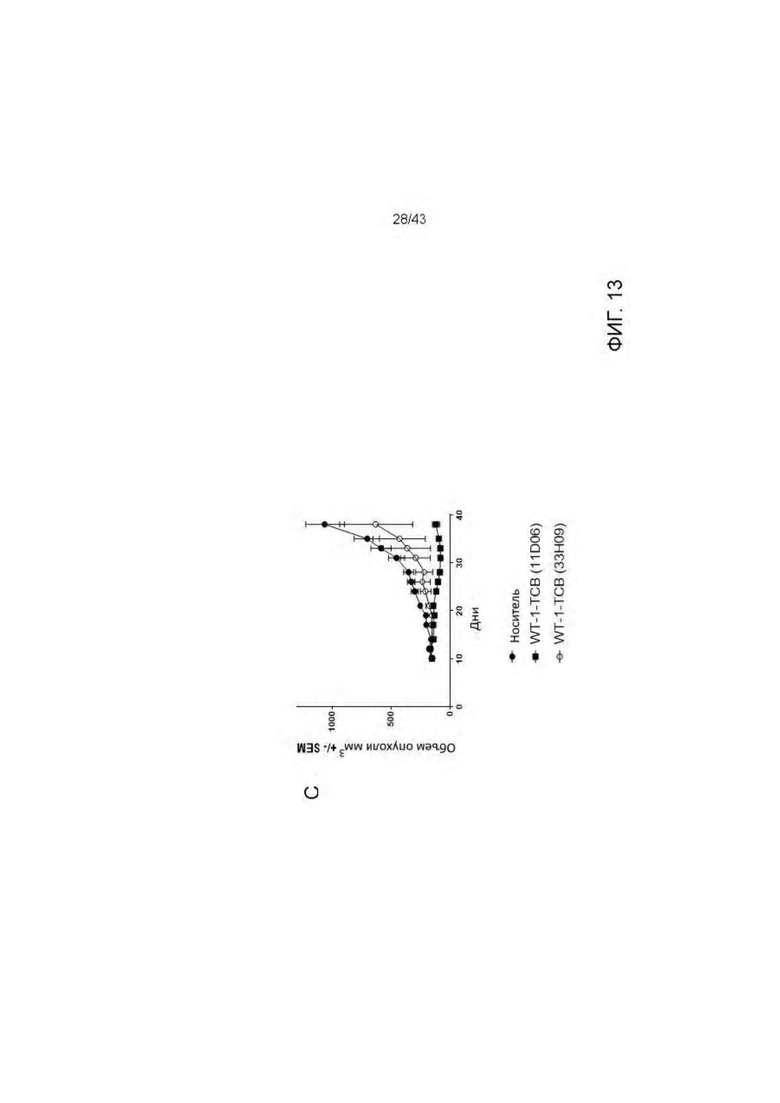
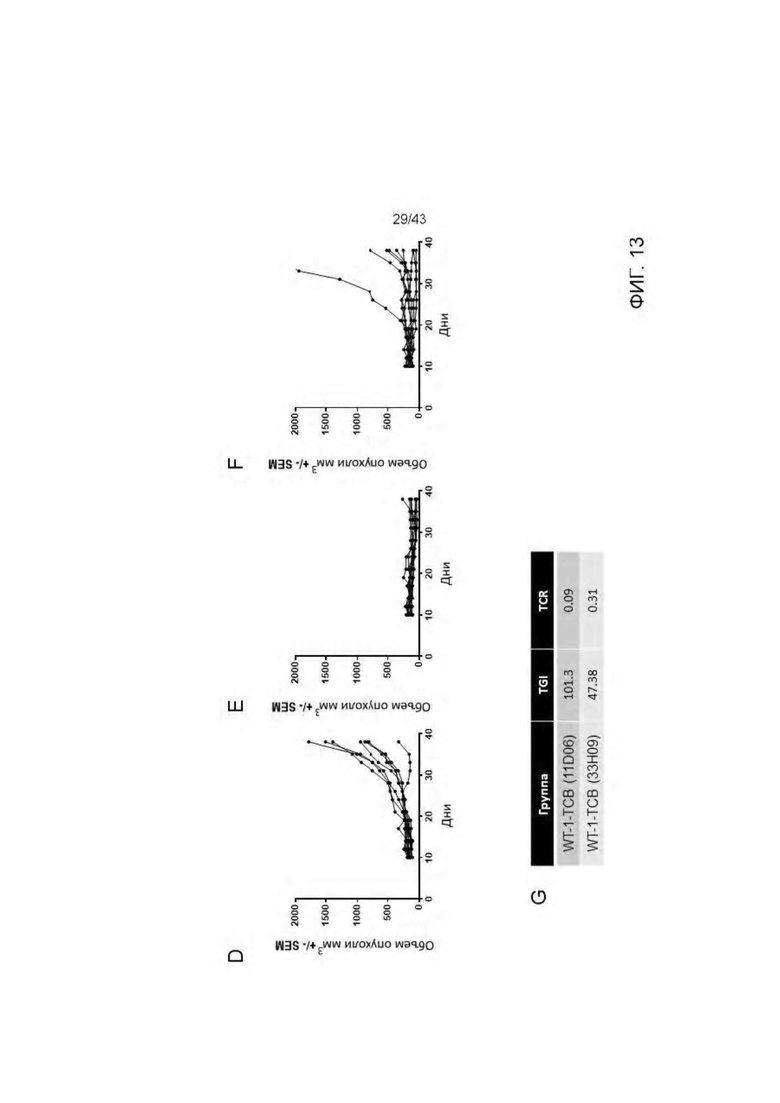
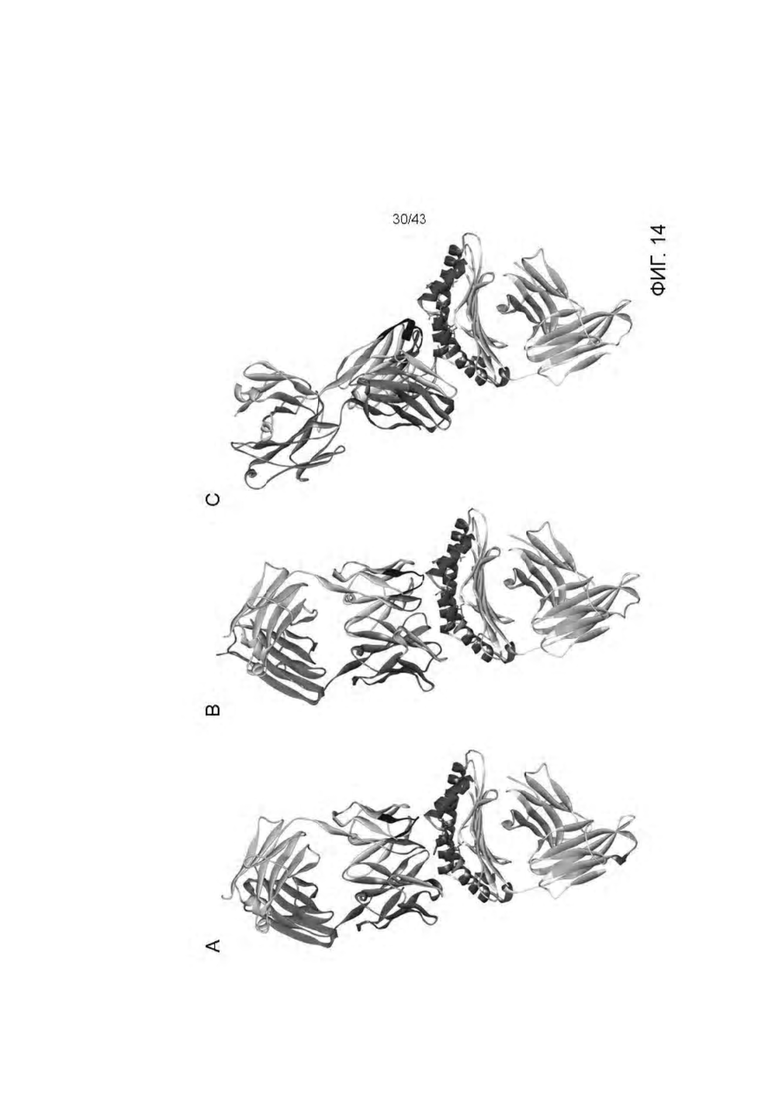


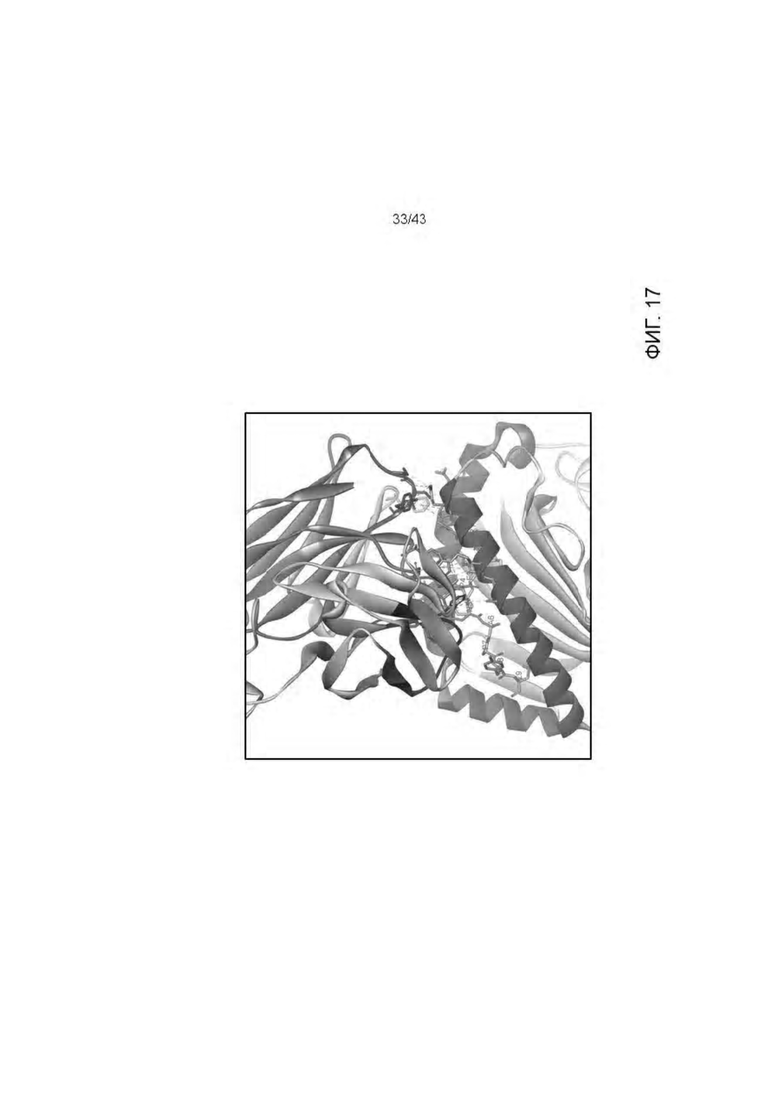
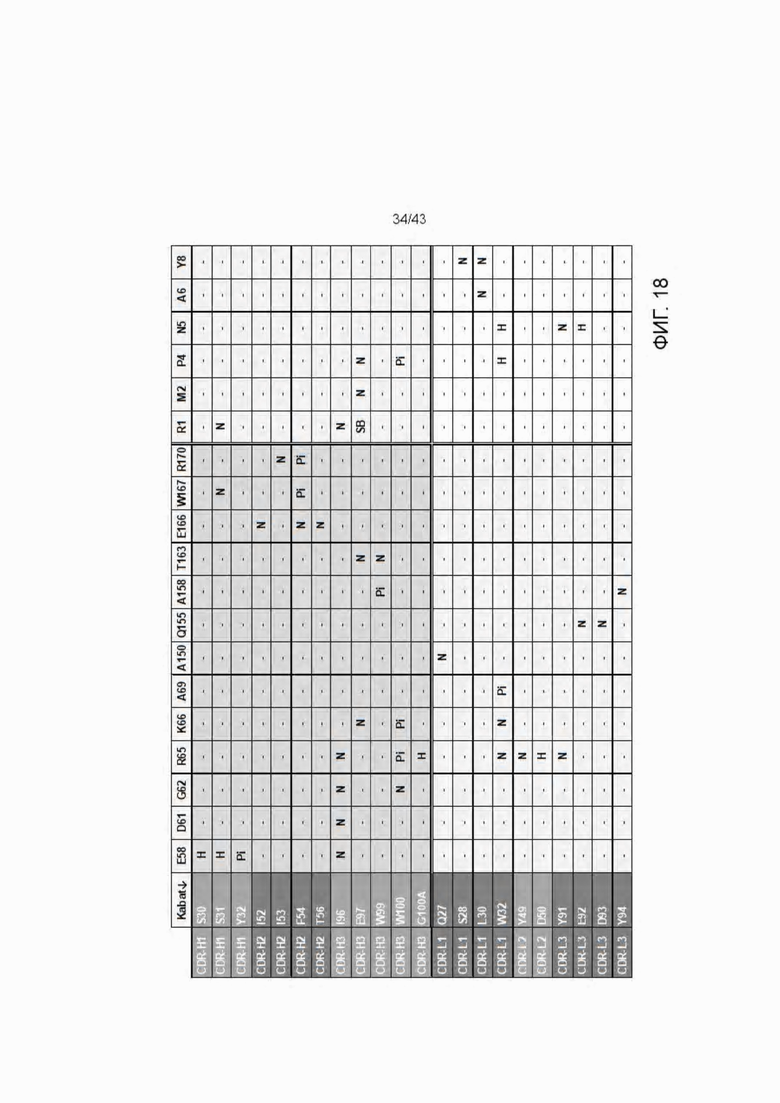


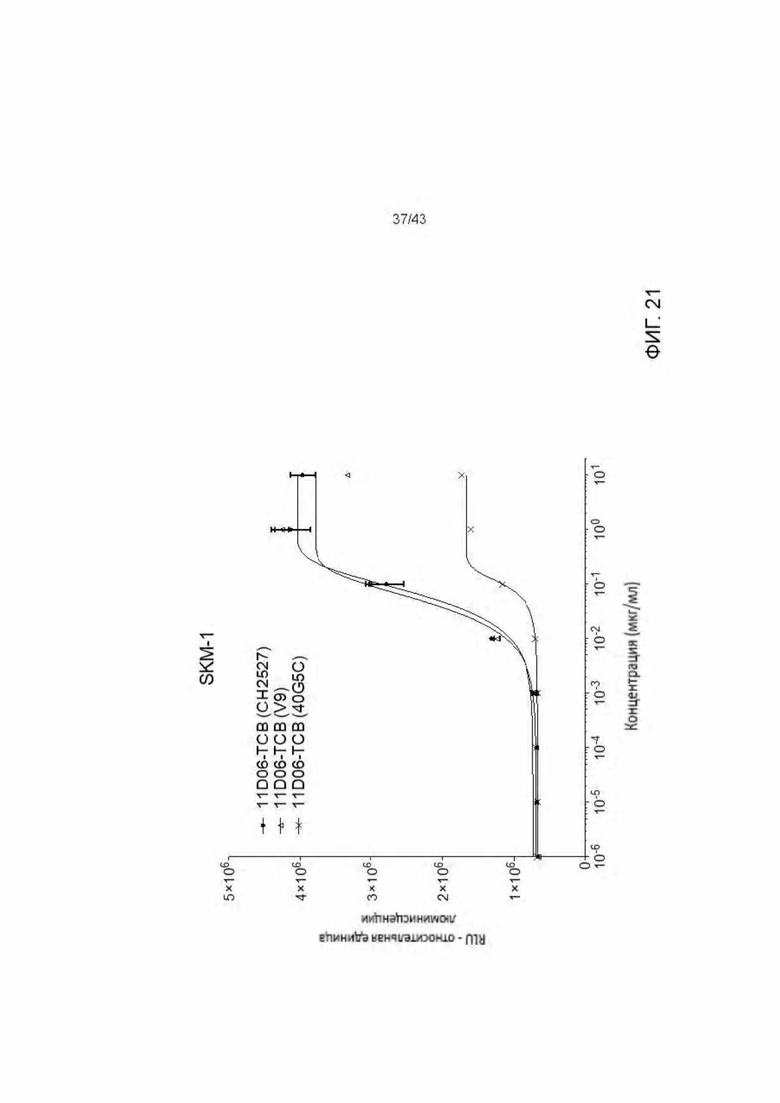
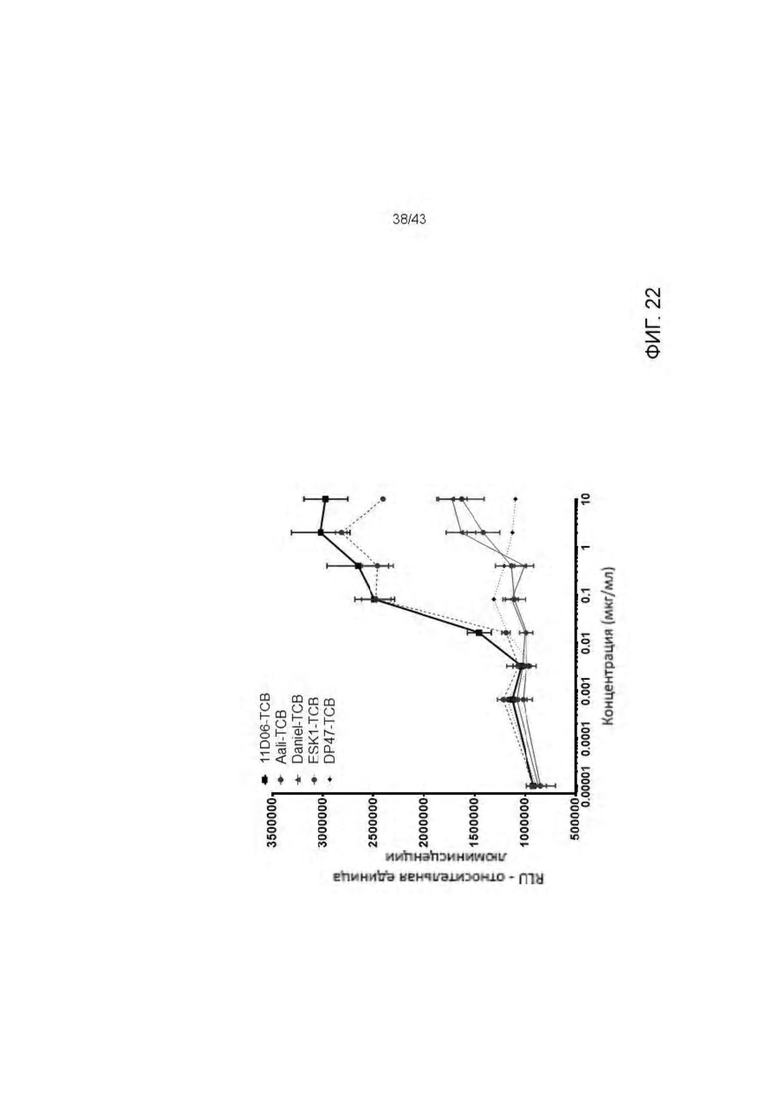
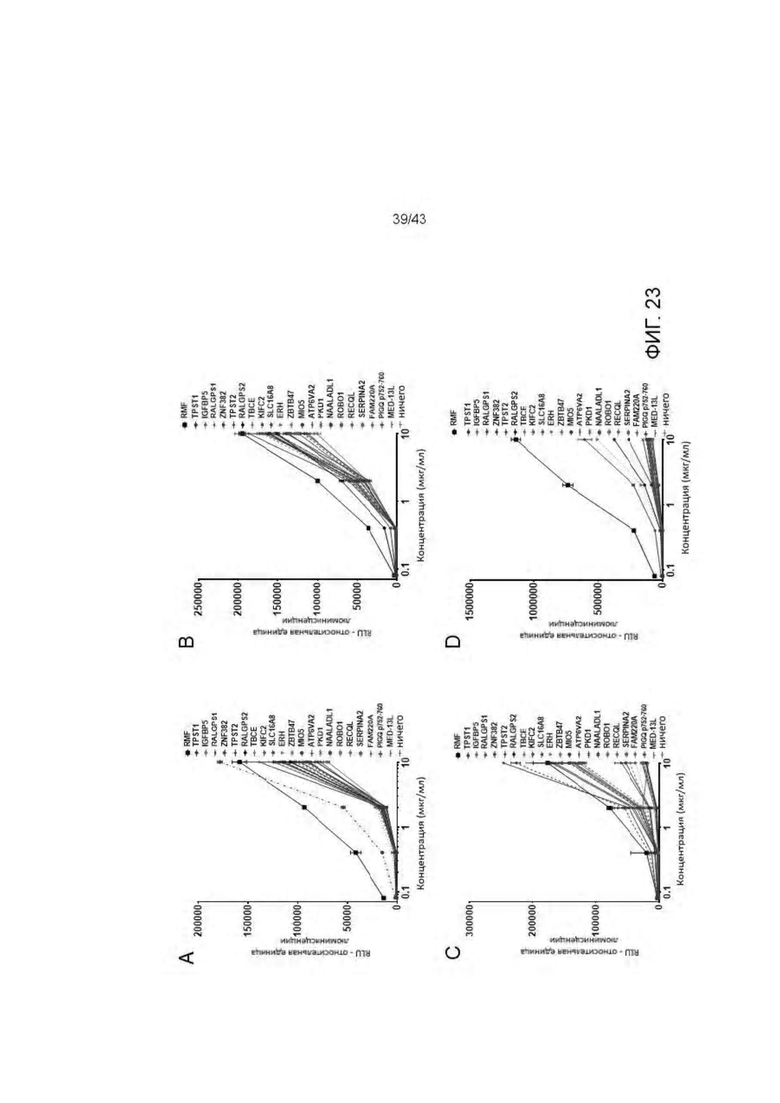
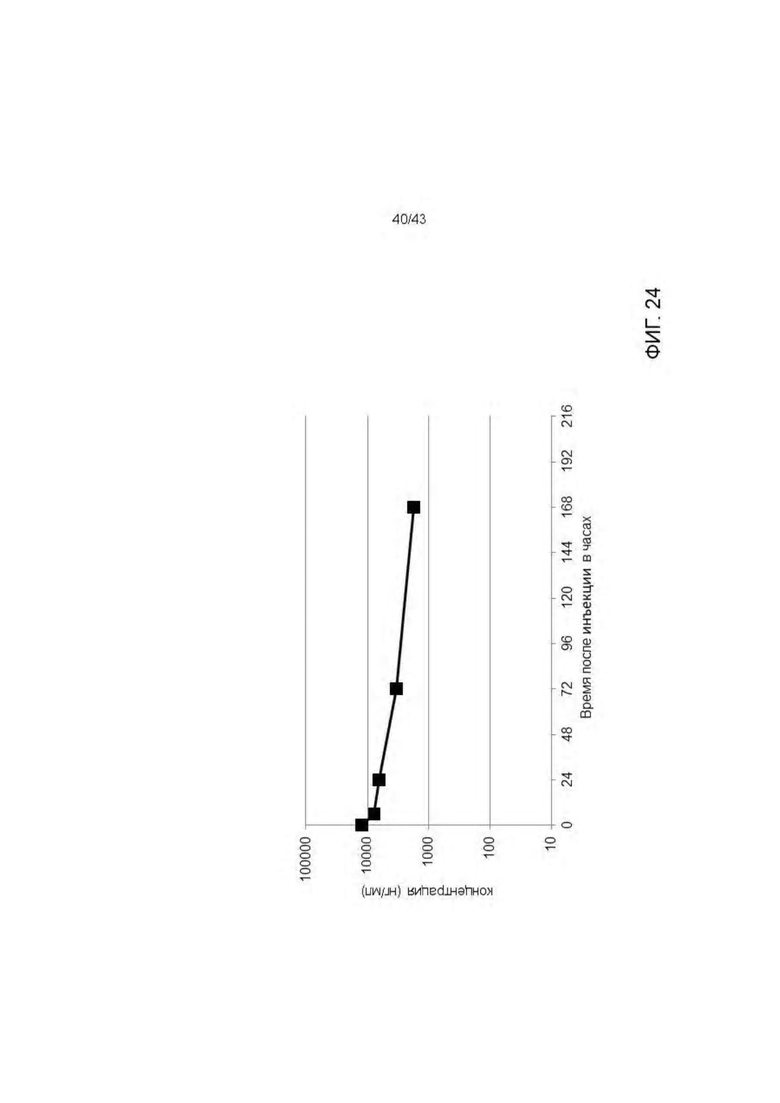
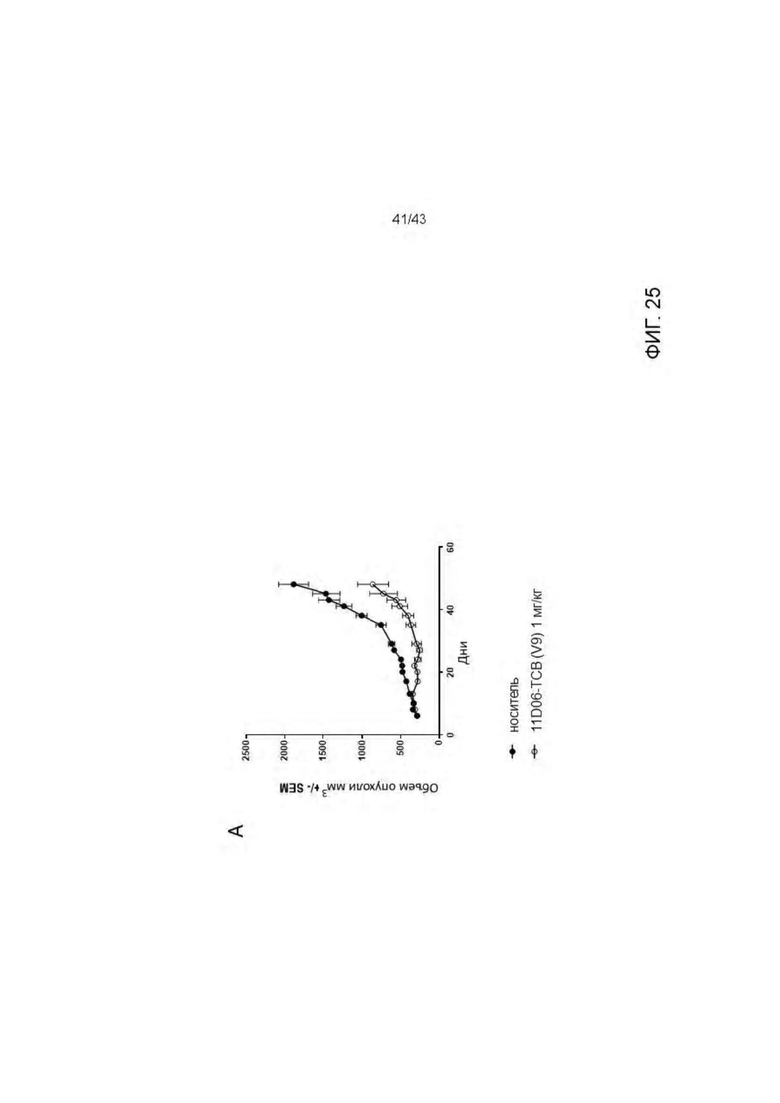

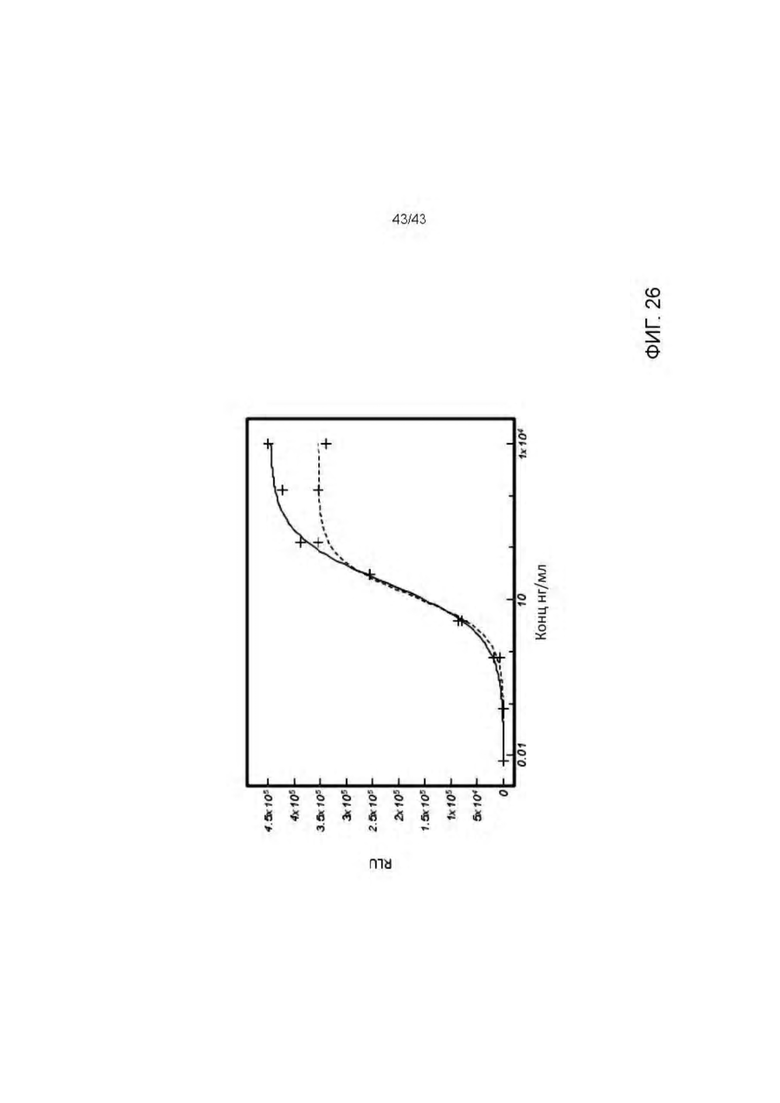
Изобретение относится к области биотехнологии. Предложены антитела, которые связываются с HLA-A2/WT1, и биспецифические антигенсвязывающие молекулы, которые связываются с HLA-A2/WT1 и CD3. Кроме того, изобретение относится к полинуклеотидам, кодирующим такие антитела или биспецифические антигенсвязывающие молекулы, а также к векторам экспрессии и клеткам-хозяевам, включающим такие полинуклеотиды. Изобретение также относится к способам получения антител и биспецифических антигенсвязывающих молекул и к способам их применения для лечения рака. Изобретение обеспечивает усиленное ингибирование опухолевого роста. 14 н. и 30 з.п. ф-лы, 43 ил., 8 табл., 21 пр.
1. Антитело, которое связывается с HLA-A2/WT1, где это антитело включает вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с последовательностью SEQID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2 и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 с последовательностью SEQ ID NO: 6.
2. Антитело по п. 1, где указанное антитело включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 8.
3. Антитело, которое связывается с HLA-A2/WT1, где это антитело включает VH, содержащую аминокислотную последовательность SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность SEQ ID NO: 8.
4. Антитело по любому из пп. 1-3, где указанное антитело представляет собой IgG, в частности антитело IgG1.
5. Антитело по любому из пп. 1-4, при этом антитело представляет собой полноразмерное антитело.
6. Антитело по любому из пп. 1-4, при этом антитело представляет собой фрагмент антитела, выбранный из группы, включающей Fv молекулы, scFv молекулы, Fab молекулы и F(ab')2 молекулы.
7. Биспецифическая антигенсвязывающая молекула, которая связывается с HLA-A2/WT1 и CD3, включающая:
(а) первый антигенсвязывающий фрагмент, который связывается с HLA-A2/WT1 и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с последовательностью SEQ ID NO: 1, HCDR 2 с последовательностью SEQ ID NO: 2 и HCDR 3 с последовательностью SEQ ID NO: 3, а также вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с последовательностью SEQ ID NO: 4, LCDR 2 с последовательностью SEQ ID NO: 5 и LCDR 3 с последовательностью SEQ ID NO: 6; и
(б) второй антигенсвязывающий фрагмент, который связывается с CD3.
8. Биспецифическая антигенсвязывающая молекула по п. 7, при этом первый антигенсвязывающий фрагмент включает VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 7, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 8.
9. Биспецифическая антигенсвязывающая молекула, которая связывается с HLA-A2/WT1 и CD3, включающая:
(а) первый антигенсвязывающий фрагмент, который связывается с HLA-A2/WT1 и содержит VH, содержащую аминокислотную последовательность SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность SEQ ID NO: 8; и
(б) второй антигенсвязывающий фрагмент, который связывается с CD3.
10. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-9, где второй антигенсвязывающий фрагмент включает:
(i) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 115, HCDR 2 с последовательностью SEQ ID NO: 116 и HCDR 3 с последовательностью SEQ ID NO: 117, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 118, LCDR 2 с последовательностью SEQ ID NO: 119 и LCDR 3 с последовательностью SEQ ID NO: 120; или
(ii) VH, содержащую HCDR 1 с последовательностью SEQ ID NO: 130, HCDR 2 с последовательностью SEQ ID NO: 131 и HCDR 3 с последовательностью SEQ ID NO: 132, а также VL, содержащую LCDR 1 с последовательностью SEQ ID NO: 133, LCDR 2 с последовательностью SEQ ID NO: 134 и LCDR 3 с последовательностью SEQ ID NO: 135.
11. Биспецифическая антигенсвязывающая молекула по п. 10, где второй антигенсвязывающий фрагмент включает:
(i) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 121, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 122, или
(ii) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 136, а также VL, содержащую аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO 137.
12. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-11, где второй антигенсвязывающий фрагмент содержит (i) VH, содержащую аминокислотную последовательность SEQ ID NO: 121, и VL, содержащую аминокислотную последовательность SEQ ID NO: 122, или (ii) VH, содержащую аминокислотную последовательность SEQ ID NO: 136, и VL, содержащую аминокислотную последовательность SEQ ID NO: 137.
13. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-12, где первый и/или второй антигенсвязывающий фрагмент представляют/ет собой Fab молекулы.
14. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-13, где второй антигенсвязывающий фрагмент представляет собой Fab молекулы, при этом вариабельные домены VL и VH или константные домены CL и СН1, в частности, вариабельные домены VL и VH, или легкая цепь Fab и тяжелая цепь Fab замещены друг другом.
15. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-14, где первый антигенсвязывающий фрагмент представляет собой Fab молекулы, при этом в константном домене CL аминокислота в положении 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация согласно Кабату) и аминокислота в положении 123 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация согласно Кабату), а в константном домене CH1 аминокислота в положении 147 независимо замещена глутаминовой кислотой (E) или аспарагиновой кислотой (D) (нумерация согласно Кабату, EU индекс) и аминокислота в положении 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно Кабату, EU индекс).
16. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-15, где первый и второй антигенсвязывающие фрагменты соединены друг с другом, необязательно при помощи пептидного линкера.
17. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-16, где первый и второй антигенсвязывающие фрагменты представляют собой Fab молекулы, при этом (i) второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи первого антигенсвязывающего фрагмента или (ii) первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи второго антигенсвязывающего фрагмента.
18. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-17, включающая третий антигенсвязывающий фрагмент.
19. Биспецифическая антигенсвязывающая молекула по п. 18, где третий антигенсвязывающий фрагмент идентичен первому антигенсвязывающему фрагменту.
20. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-19, включающая Fc домен, состоящий из первой и второй субъединицы.
21. Биспецифическая антигенсвязывающая молекула по п. 20, где первый, второй и, если присутствует, третий антигенсвязывающий фрагмент, каждый, представляет собой Fab молекулы; при этом (i) второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи первого антигенсвязывающего фрагмента, а первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом первой субъединицы Fc домена; или (ii) первый антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом Fab тяжелой цепи второго антигенсвязывающего фрагмента, а второй антигенсвязывающий фрагмент на своем С-конце Fab тяжелой цепи соединен с N-концом второй субъединицы Fc домена;
и где третий антигенсвязывающий фрагмент, если он присутствует, соединен на С-конце тяжелой цепи Fab с N-концом второй субъединицы Fc домена.
22. Биспецифическая антигенсвязывающая молекула по п. 20 или 21, где Fc домен представляет собой домен IgG, в частности Fc домен IgG1.
23. Биспецифическая антигенсвязывающая молекула по любому из пп. 20-22, где Fc домен представляет собой Fc домен человека.
24. Биспецифическая антигенсвязывающая молекула по любому из пп. 20-23, где Fc домен представляет собой Fc домен IgG1 человека.
25. Биспецифическая антигенсвязывающая молекула по любому из пп. 20-24, где аминокислотный остаток в СН3 домене первой субъединицы Fc домена замещен аминокислотным остатком, имеющим больший объем боковой цепи, что создает выступ внутри СН3 домена первой субъединицы, которая пространственно совпадает со впадиной внутри СН3 домена второй субъединицы, а аминокислотный остаток СН3 домена второй субъединицы Fc домена замещен аминокислотным остатком, имеющим меньший объем боковой цепи, что создает впадину внутри СН3 домена второй субъединицы, которая пространственно совпадает с выступом внутри СН3 домена первой субъединицы.
26. Биспецифическая антигенсвязывающая молекула по любому из пп. 20-25, где в домене CH3 первой субъединицы Fc домена остаток треонина в положении 366 заменен остатком триптофана (T366W), а в CH3 домене второй субъединицы Fc домена остаток тирозина в положении 407 заменен остатком валина (Y407V) и необязательно во второй субъединице Fc домена дополнительно остаток треонина в положении 366 заменен остатком серина (T366S) и остаток лейцина в положении 368 заменен остатком аланина (L368A) (нумерация согласно Кабату, EU индекс).
27. Биспецифическая антигенсвязывающая молекула по п. 26, где в первой субъединице Fc домена дополнительно остаток серина в положении 354 заменен остатком цистеина (S354C) или остаток глутаминовой кислоты в положении 356 заменен остатком цистеина (E356C), а во второй субъединице Fc домена дополнительно остаток тирозина в положении 349 заменен остатком цистеина (Y349C) (нумерация согласно Кабату, EU индекс).
28. Биспецифическая антигенсвязывающая молекула по любому из пп. 20-27, при этом Fc домен включает одну или более аминокислотную замену, которая снижает связывание с Fc рецептором и/или эффекторную функцию.
29. Биспецифическая антигенсвязывающая молекула по п. 28, где одна или более аминокислотных замен находятся в положении, выбранном из группы E233, L234, L235, N297, P331 и P329 (нумерация согласно Кабату, EU индекс).
30. Биспецифическая антигенсвязывающая молекула по любому из пп. 20-29, где каждая субъединица Fc домена содержит аминокислотные замены L234A, L235A и P329G (нумерация согласно Кабату, EU индекс).
31. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-30, содержащая:
а) первый и третий антигенсвязывающие фрагменты, которые связываются с первым антигеном, где первый антиген представляет собой HLA-A2/WT1 и где первый и третий антигенсвязывающие фрагменты представляют собой (традиционную) Fab молекулу, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 7, и вариабельную область легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 8;
b) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, где второй антиген представляет собой CD3 и где второй антигенсвязывающий фрагмент представляет собой Fab молекулу, в которой вариабельные домены VL и VH Fab легкой цепи и Fab тяжелой цепи заменены друг другом, включая вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 136 и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 137;
c) Fc домен IgG1 человека, состоящий из первой и второй субъединиц;
где
(i) в константном домене CL первого и третьего антигенсвязывающего фрагмента согласно а) аминокислота в положении 124 заменена лизином (К) (нумерация согласно Кабату), а аминокислота в положении 123 заменена аргинином (R) (нумерация согласно Кабату), и где в константном домене CH1 первого и третьего антигенсвязывающего фрагмента в соответствии с а) аминокислота в положении 147 заменена глутаминовой кислотой (E) (нумерация согласно Кабату, EU индекс), а аминокислота в положении 213 заменена глутаминовой кислотой (Е) (нумерация согласно Кабату, EU индекс);
(ii) первый антигенсвязывающий фрагмент в соответствии с а) соединен на С-конце Fab тяжелой цепи с N-концом Fab тяжелой цепи второго антигенсвязывающего фрагмента в соответствии с b), а второй антигенсвязывающий фрагмент в соответствии с b) и третий антигенсвязывающий фрагмент согласно а) соединен на С-конце Fab тяжелой цепи с N-концом одной из субъединиц Fc домена согласно c); и
(iii) первая субъединица Fc домена содержит аминокислотные замены S354C и T366W, а вторая субъединица Fc домена содержит аминокислотные замены Y349C, T366S, L368A и Y407V и каждая субъединица Fc домена дополнительно содержит аминокислотную замену L234A, L235A и P329G (нумерация согласно Кабату, EU индекс).
32. Биспецифическая антигенсвязывающая молекула по любому из пп. 7-31, содержащая полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 123, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 125, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 139, и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96 %, 97%, 98%, 99% или 100% идентична последовательности SEQ ID NO: 140.
33. Выделенный полинуклеотид, кодирующий антитело по любому из пп. 1-6 или биспецифическую антигенсвязывающую молекулу по любому из пп. 7-32.
34. Множество выделенных полинуклеотидов, кодирующих антитело по любому из пп. 1-6 или биспецифическую антигенсвязывающую молекулу по любому из пп. 7-32.
35. Вектор экспрессии, содержащий полинуклеотид или множество полинуклеотидов по п. 33 или 34.
36. Множество векторов экспрессии, содержащих множество полинуклеотидов по п. 34.
37. Клетка-хозяин для получения антитела, которое связывается с HLA-A2/WT1, или биспецифической антигенсвязывающей молекулы, которая связывается с HLA-A2/WT1 и CD3, содержащая полинуклеотид/полинуклеотиды по п. 33 или 34 или вектор(ы) по п. 35 или 36.
38. Способ получения антитела, которое связывается с HLA-A2/WT1, или биспецифической антигенсвязывающей молекулы, которая связывается с HLA-A2/WT1 и CD3, включающий этапы а) культивирования клетки-хозяина по п. 37 в условиях, подходящих для экспрессии антитела, и b) восстановление антитела.
39. Фармацевтическая композиция для лечения рака, включающая антитело по любому из пп. 1-6 или биспецифическую антигенсвязывающую молекулу по любому из пп. 7-32, а также фармацевтически приемлемый носитель.
40. Применение антитела по любому из пп. 1-6, биспецифической антигенсвязывающей молекулы по любому из пп. 7-32 или фармацевтической композиции по п. 39 в качестве лекарственного средства для лечения рака.
41. Применение антитела по любому из пп. 1-6, биспецифической антигенсвязывающей молекулы по любому из пп. 7-32 или фармацевтической композиции по п. 39 для лечения рака.
42. Применение по п. 41, где рак представляет собой лейкоз, в частности острый лимфобластный лейкоз (ОЛЛ) или острый миелогенный лейкоз (ОМЛ).
43. Применение антитела по любому из пп. 1-6 или биспецифической антигенсвязывающей молекулы по любому из пп. 7-32 для получения лекарственного средства, предназначенного для лечения рака.
44. Применение по п. 43, где рак представляет собой лейкоз, в частности острый лимфобластный лейкоз (ALL) или острый миелогенный лейкоз (AML).
| WO 2016199141 A2, 15.12.2016 | |||
| US 9074000 B2, 07.07.2015 | |||
| ZHAO Q | |||
| et al., Affinity maturation of T-cell receptor-like antibodies for Wilms tumor 1 peptide greatly enhances therapeutic potential, Leukemia, 2015, vol.29(11), pp.2238-2247 | |||
| КОМПОЗИЦИИ И СПОСОБЫ WT1-СПЕЦИФИЧНОЙ ИММУНОТЕРАПИИ | 1999 |
|
RU2237674C2 |

