ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0001] Настоящая заявка относится к области биомедицины, в частности, к способу получения фаговой библиотеки, экспонирующей антитела или фрагменты антител.
УРОВЕНЬ ТЕХНИКИ
[0002] В данной области существует несколько способов получения фаговой библиотеки, однако эти способы имеют некоторые недостатки. Например, емкость библиотеки является слишком малой (например, трудно получить библиотеки с емкостью более 109), и многообразие библиотеки является недостаточным для удовлетворения требованиям для скрининга высококачественных антител. В то же время, для повышения емкости библиотеки необходимо накопить множество библиотек меньшего размера. Это не только отнимает несколько лет, но также трудно гарантировать стабильность этих библиотек меньшего размера для контроля качестве полученных библиотек. Более того, при конструировании библиотеки существующими способами трудно осуществлять удобный и надежный контроль качества в ходе получения библиотеки. Обычно качество не может быть известно до тех пор, пока библиотека не получена и, таким образом, процесс имеет высокий риск и высокую вероятность неуспеха.
[0003] Более того, библиотека, сконструированная в соответствии с существующим способом, имеет высокую стоимость, требует длительного периода времени и имеет значительное снижение качества после длительного хранения, и, таким образом, трудно удовлетворить потребность промышленного массового производства.
[0004] Таким образом, существует неотложная потребность в способах получения фаговой библиотеки с большой емкостью, которая может удовлетворять потребности промышленного массового производства, имеет контролируемое качество и удобна в обращении.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0005] В рамках настоящей заявки описан способ получения фаговой библиотеки, экспонирующей антитело или фрагмент антитела, а также фаговой библиотеки, полученной с использованием способа, и способ селекции антитела или фрагмента антитела посредством фаговой библиотеки. Способ получения фаговой библиотеки, экспонирующей антитело или фрагмент антитела, описанной в рамках настоящей заявки, имеет по меньшей мере одно из следующих свойств: 1) вероятность мутаций в результате ПЦР эффективно снижается; фаговая библиотека, описанная в рамках настоящей заявки, не конструируется с использованием стратегии ПЦР с перекрыванием, используемой в общепринятом способе конструирования библиотек антител, каждый компонент, используемый для конструирования фаговой библиотеки, может быть амплифицирован способом ПЦР только один раз, а затем соединен с желаемыми компонентами для получения фаговой библиотеки Fab; 2) она может удовлетворять требованием промышленного массового производства; 3) легко проводить контроль качества; контроль качества может быть независимо и удобным образом проведен на каждой стадии, требуемой для конструирования фаговой библиотеки, описанной в рамках настоящей заявки; в то же время, после получения фаговой библиотеки, описанной в рамках настоящей заявки, ее можно просто и удобным образом подвергнуть анализу качества (например, посредством секвенирования генов или анализа экспрессии растворимых Fab); 4) процесс получения является простым и быстрым; фаговую библиотеку, описанную в рамках настоящей заявки, можно конструировать с использованием участка(ов) распознавания для специфической эндонуклеазы рестрикции, что может как обеспечить направленное соединение, так и предотвратить ошибочное соединение; в ходе соединения молекулярные количества каждого фрагмента, являющегося компонентом, можно контролировать так, чтобы они составляли 1:1, для повышения эффективности соединения и конверсии; в то же время с использованием стратегий конструирования бактериальных библиотек, таких как бактериальных библиотек с компонентами в виде легких цепей, эффективность соединения и конверсии каждого антитела или его фрагментов повышается (например, полученные продукты соединения имеют эффективность конверсии более чем приблизительно 109/мкг); посредством получения плазмид с компонентами коннексона обеспечивается наличие у полученного высвободившегося коннексона подходящего и полного адгезивного конца, так что повышается эффективность конверсии; кроме того, фаговую библиотеку, описанную в рамках настоящей заявки, можно культивировать либо посредством культивирования на чашке, либо посредством культивирования в жидкой среде; 5) полученная библиотека имеет большую емкость и высокое многообразие, способствуя быстрой и эффективной селекции антител и их антигенсвязывающих фрагментов; 6) она является в высокой степени специфической; фаговую библиотеку, описанную в рамках настоящей заявки, не конструируют с вырожденными праймерами, используемыми в общепринятых способах, тем самым избегая какой-либо ошибки, вызываемой вырожденными праймерами; в то же время, праймеры для применения в конструировании фаговой библиотеки, описанной в рамках настоящей заявки, требуют относительно высокой температуры отжига при ПЦР, что может повысить специфичность связывания между этими праймерами и их амплифицируемыми фрагментами-мишенями; и 7) она поддается расширению и ее легко сохранить; бактериальные библиотеки, включающие бактериальные библиотеки с компонентами легких цепей и бактерии, содержащие плазмиды с компонентами коннексона, могут храниться при -80°C в течение длительного времени и при необходимости могут использоваться в любой момент времени (например, для получения фаговой библиотеки, описанной в рамках настоящей заявки, требуется только двое суток), и при необходимости можно смешивать различные типы и соотношения.
[0006] В одном аспекте настоящая заявка относится к способу получения библиотеки, экспонирующей антитело или фрагмент антитела, включающему: 1) предоставление первого полинуклеотида, содержащего LC, второго полинуклеотида, содержащего коннексон, и третьего полинуклеотида, содержащего HC, где LC включает последовательность нуклеиновой кислоты, кодирующую легкую цепь или фрагмент легкой цепи антитела или фрагмента антитела, и HC включает последовательность нуклеиновой кислоты, кодирующую тяжелую цепь или фрагмент тяжелой цепи антитела или фрагмента антитела; 2) введение первого полинуклеотида в первую бактерию с получением бактериальной библиотеки с компонентом легкой цепи, введение второго полинуклеотида во вторую бактерию с получением бактерии с компонентом коннексона, и введение третьего полинуклеотида в третью бактерию с получением бактериальной библиотеки с компонентом тяжелой цепи; 3) получение плазмиды с компонентом легкой цепи, включающей LC из бактериальной библиотеки с компонентом легкой цепи, получение плазмиды с компонентом коннексона, включающей коннексон из бактерий с компонентом коннексона, и получение плазмиды с компонентом тяжелой цепи, включающей HC из бактериальной библиотеки с компонентом тяжелой цепи; 4) получение высвободившейся LC из плазмиды с компонентом легкой цепи, получение высвободившегося коннексона из плазмиды с компонентом коннексона и получение высвободившейся HC из плазмиды с компонентом тяжелой цепи; 5) предоставление вектора для дисплея и получение высвободившегося фрагмента вектора для дисплея из вектора для дисплея; 6) связывание высвободившейся LC, высвободившегося коннексона, высвободившейся HC и высвободившегося фрагмента вектора для дисплея с образованием соединенного продукта для дисплея; 7) введение соединенного продукта для дисплея в четвертую бактерию с получением библиотеки бактериального дисплея антитела или фрагмента антитела; и 8) получение фаговой библиотеки, экспонирующей антитело или фрагмент антитела, с использованием библиотеки бактериального дисплея.
[0007] В некоторых вариантах осуществления соединенный продукт для дисплея в 6) может экспрессировать легкую цепь или фрагмент легкой цепи и тяжелую цепь или фрагмент тяжелой цепи в соответствии с рамками считывания в условиях, подходящих для экспрессии антитела или фрагмента антитела.
[0008] В некоторых вариантах осуществления все из первого полинуклеотида, второго полинуклеотида и третьего полинуклеотида представляют собой линейные молекулы нуклеиновых кислот.
[0009] В некоторых вариантах осуществления последовательность нуклеиновой кислоты, кодирующая тяжелую цепь или фрагмент тяжелой цепи, и последовательность нуклеиновой кислоты, кодирующая легкую цепь или фрагмент легкой цепи, находятся в разных рамках считывания.
[0010] В некоторых вариантах осуществления коннексон включает последовательность нуклеиновой кислоты, кодирующую сигнальный пептид pelB или его фрагмент. В некоторых вариантах осуществления коннексон имеет длину от приблизительно 50 до приблизительно 200 оснований.
[0011] В некоторых вариантах осуществления в соединенном продукте для дисплея в 6) LC находится выше коннексона, и коннексон находится выше HC.
[0012] В некоторых вариантах осуществления 2) включает криоконсервацию бактериальной библиотеки с компонентом легкой цепи и/или бактериальной библиотеки с компонентом тяжелой цепи.
[0013] В некоторых вариантах осуществления 7) включает криоконсервацию библиотеку бактериального дисплея.
[0014] В некоторых вариантах осуществления 8) включает размораживание и культивирование криоконсервированной библиотеки бактериального дисплея и конструирование фаговой библиотеки с использованием культивируемой библиотеки бактериального дисплея.
[0015] В некоторых вариантах осуществления первый полинуклеотид включает структуру R1-LC-R2 в направлении от 5' к 3', второй полинуклеотид включает структуру R3-коннексон-R4 в направлении от 5' к 3', третий полинуклеотид включает структуру R5-HC-R6 в направлении от 5' к 3', и вектор для дисплея включает структуру R7-фрагмент вектора для дисплея-R8 в направлении от 5' к 3', где R1, R2, R3, R4, R5, R6, R7 и R8 представляют собой участки распознавания для эндонуклеазы рестрикции.
[0016] В некоторых вариантах осуществления конец, образующийся в результате расщепления R2 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R3 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R1, R4, R5, R6, R7 и R8 эндонуклеазой рестрикции, и наоборот.
[0017] В некоторых вариантах осуществления конец, образующийся в результате расщепления R4 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R5 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R1, R2, R3, R6, R7 и R8 эндонуклеазой рестрикции, и наоборот.
[0018] В некоторых вариантах осуществления конец, образующийся в результате расщепления R6 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R7 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R1, R2, R3, R4, R5 и R8 эндонуклеазой рестрикции, и наоборот.
[0019] В некоторых вариантах осуществления конец, образующийся в результате расщепления R8 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R1 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R2, R3, R4, R5, R6 и R7 эндонуклеазой рестрикции, и наоборот.
[0020] В некоторых вариантах осуществления конец, образующийся посредством расщепления любого из R1, R2, R3, R4, R5, R6, R7 и R8 эндонуклеазой рестрикции, представляет собой не палиндромную последовательность.
[0021] В некоторых вариантах осуществления два или более из R1, R2, R3, R4, R5, R6, R7 и R8 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции.
[0022] В некоторых вариантах осуществления R2 идентичен R3. В некоторых вариантах осуществления R4 идентичен R5. В некоторых вариантах осуществления R6 идентичен R7. В некоторых вариантах осуществления R8 идентичен R1.
[0023] В некоторых вариантах осуществления R1, R2, R3, R4, R5 и R8 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции. В некоторых вариантах осуществления R6 и R7 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции.
[0024] В некоторых вариантах осуществления эндонуклеаза рестрикции выбрана из группы, состоящей из SfiI, BsmBI и Esp3I. В некоторых вариантах осуществления R1, R2, R3, R4, R5 и R8 могут распознаваться и расщепляться посредством SfiI. В некоторых вариантах осуществления R6 и R7 могут распознаваться и расщепляться посредством BsmBI и/или Esp3I.
[0025] В некоторых вариантах осуществления R1 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 1.
[0026] В некоторых вариантах осуществления R2 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 2.
[0027] В некоторых вариантах осуществления R3 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 2.
[0028] В некоторых вариантах осуществления R4 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 3.
[0029] В некоторых вариантах осуществления R5 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 3.
[0030] В некоторых вариантах осуществления R6 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 4.
[0031] В некоторых вариантах осуществления R7 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 4.
[0032] В некоторых вариантах осуществления R8 включает последовательность нуклеиновой кислоты, представленную в SEQ ID NO: 1.
[0033] В некоторых вариантах осуществления легкая цепь или фрагмент легкой цепи включает часть вариабельной области легкой цепи и/или часть константной области легкой цепи. В некоторых вариантах осуществления тяжелая цепь или фрагмент тяжелой цепи включает часть вариабельной области тяжелой цепи и/или часть константной области тяжелой цепи. В некоторых вариантах осуществления антитело или его фрагмент включает Fab, Fab', (Fab)2 и/или (Fab')2.
[0034] В некоторых вариантах осуществления соединенный продукт для дисплея в 7) имеет эффективность соединения и конверсии по меньшей мере 108 клонов/мкг полинуклеотида.
[0035] В некоторых вариантах осуществления 2) включает расщепление первого полинуклеотида эндонуклеазой рестрикции, распознающего R1 и R2, соединение расщепленного первого полинуклеотида с фрагментом вектора для хранения легкой цепи с получением соединенного продукта для хранения легкой цепи, и введение соединенного продукта для хранения легкой цепи в первую бактерию с получением бактериальной библиотеки с компонентом легкой цепи; где вектор для хранения легкой цепи включает R1 и R2, и фрагмент вектора для хранения легкой цепи получают путем расщепления вектора для хранения легкой цепи эндонуклеазой рестрикции, распознающей R1 и R2.
[0036] В некоторых вариантах осуществления вектор для хранения легкой цепи происходит из вектора pUC. В некоторых вариантах осуществления вектор pUC представляет собой или происходит из вектора pUC19. В некоторых вариантах осуществления вектор pUC представляет собой или происходит из вектора pMD19-T.
[0037] В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R1 и R2, включает SfiI.
[0038] В некоторых вариантах осуществления 2) включает расщепление третьего полинуклеотида эндонуклеазой рестрикции, распознающей R5 и R6, соединение расщепленного третьего полинуклеотида с фрагментом вектора для хранения тяжелой цепи с получением соединенного продукта для хранения тяжелой цепи, и введение соединенного продукта для хранения тяжелой цепи в третью бактерию с получением бактериальной библиотеки с компонентом тяжелой цепи; где вектор для хранения тяжелой цепи включает R5 и R6, и фрагмент вектора для хранения тяжелой цепи получают путем расщепления вектора для хранения тяжелой цепи эндонуклеазой рестрикции, распознающей R5 и R6. В некоторых вариантах осуществления вектор для хранения тяжелой цепи происходит из вектора pUC. В некоторых вариантах осуществления вектор pUC представляет собой или происходит из вектора pUC19. В некоторых вариантах осуществления вектор pUC представляет собой или происходит из вектора pMD19-T.
[0039] В некоторых вариантах осуществления вектор для хранения тяжелой цепи включает или включает только участок распознавания BsmBI.
[0040] В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R5 и R6, включает SfiI, BsmBI и/или Esp3I.
[0041] В некоторых вариантах осуществления 2) включает соединение второго полинуклеотида с фрагментом вектора с получением вектора для хранения коннексона, и введение вектора для хранения коннексона во вторую бактерию с получением бактерии с компонентом коннексона. В некоторых вариантах осуществления вектор для хранения коннексона включает R3 и R4. В некоторых вариантах осуществления фрагмент вектора для конструирования вектора для хранения коннексона происходит из вектора pUC. В некоторых вариантах осуществления вектор pUC представляет собой или происходит из вектора pUC19. В некоторых вариантах осуществления вектор pUC представляет собой или происходит из вектора pMD19-T.
[0042] В некоторых вариантах осуществления 4) включает расщепление плазмиды с компонентом легкой цепи эндонуклеазой рестрикции, распознающей R1 и R2, с получением высвободившейся LC. В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R1 и R2, включает SfiI.
[0043] В некоторых вариантах осуществления 4) включает расщепление плазмиды с компонентом тяжелой цепи эндонуклеазой рестрикции, распознающей R5 и R6, с получением высвободившейся HC. В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R5 и R6, включает SfiI, BsmBI и/или Esp3I.
[0044] В некоторых вариантах осуществления 4) включает расщепление вектора для хранения коннексона эндонуклеазой рестрикции, распознающей R3 и R4, с получением высвободившегося коннексона. В некоторых вариантах осуществления 4) включает получение продукта амплификации вектора для хранения коннексона, где продукт амплификации включает коннексон, R3 и R4; и расщепление продукта амплификации эндонуклеазой рестрикции, распознающей R3 и R4, с получением высвободившегося коннексона. В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R3 и R4, включает SfiI.
[0045] В некоторых вариантах осуществления 5) включает расщепление вектора для дисплея эндонуклеазой рестрикции, распознающей R7 и R8, для высвобождения фрагмента вектора для дисплея. В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R7 и R8, включает SfiI, BsmBI и/или Esp3I.
[0046] В некоторых вариантах осуществления вектор для дисплея происходит из вектора pComb3x.
[0047] В некоторых вариантах осуществления получают три различных расщепленных фрагмента после расщепления вектора для дисплея эндонуклеазой рестрикции, распознающей R1, R2, R3, R4, R5 и R8. В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R1, R2, R3, R4, R5 и R8, включает SfiI. В некоторых вариантах осуществления после расщепления вектора для дисплея эндонуклеазой рестрикции, распознающей R6 и R7, вектор для дисплея линеаризуется. В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R6 и R7, включает BsmBI и/или Esp3I.
[0048] В некоторых вариантах осуществления после расщепления вектора для дисплея эндонуклеазой рестрикции, распознающей R1, R2, R3, R4, R5 R6, R7 и R8, получают четыре различных расщепленных фрагмента. В некоторых вариантах осуществления эндонуклеаза рестрикции, распознающая R1, R2, R3, R4, R5, R6, R7 и R8, включает SfiI, BsmBI и/или Esp3I.
[0049] В некоторых вариантах осуществления 2) включает проведение детекции в образцах из бактериальной библиотеки с компонентом легкой цепи, бактерий с компонентом коннексона и/или бактериальной библиотеки с компонентом тяжелой цепи для определения емкости библиотеки и/или качества бактериальной библиотеки с компонентом легкой цепи, бактерий с компонентом коннексона и/или бактериальной библиотеки с компонентом тяжелой цепи.
[0050] В некоторых вариантах осуществления бактериальная библиотека с компонентом легкой цепи включает по меньшей мере 106 различных клонов.
[0051] В некоторых вариантах осуществления бактериальная библиотека с компонентом тяжелой цепи включает по меньшей мере 107 различных клонов.
[0052] В некоторых вариантах осуществления доля эффективного клонирования в бактериальной библиотеке с компонентом легкой цепи составляет по меньшей мере приблизительно 70%.
[0053] В некоторых вариантах осуществления доля эффективного клонирования в бактериальной библиотеке с компонентом тяжелой цепи составляет по меньшей мере приблизительно 70%.
[0054] В некоторых вариантах осуществления 2) включает вычисление емкости и/или качества библиотеки бактериального дисплея на основе емкости бактериальной библиотеки с компонентом легкой цепи и/или бактериальной библиотеки с компонентом тяжелой цепи, бактерий с компонентом коннексона, и/или массы.
[0055] В некоторых вариантах осуществления библиотека бактериального дисплея включает по меньшей мере 1010 различных клонов.
[0056] В некоторых вариантах осуществления доля эффективного клонирования в библиотеке бактериального дисплея составляет по меньшей мере приблизительно 70%.
[0057] В некоторых вариантах осуществления первый полинуклеотид и третий полинуклеотид получают из материала образца. В некоторых вариантах осуществления материал образца включает материал образца, полученный из образца лимфоцитов периферической крови. В некоторых вариантах осуществления лимфоцит периферической крови представляет собой лимфоцит периферической крови человека. В некоторых вариантах осуществления материал образца включает тотальную РНК и/или мРНК, происходящую из образца.
[0058] В некоторых вариантах осуществления 8) включает приведение библиотеки бактериального дисплея в контакт с фагом-помощником с получением фаговой библиотеки.
[0059] В другом аспекте настоящая заявка относится к фаговой библиотеке, полученной описанным способом. В некоторых вариантах осуществления фаговая библиотека включает по меньшей мере 1010 различных клонов. В некоторых вариантах осуществления фаговая библиотека может экспонировать по меньшей мере 1010 типов различных антител или фрагментов антител.
[0060] В другом аспекте настоящая заявка относится к способу селекции антитела или фрагмента антитела, в том числе с использованием фаговой библиотеки.
[0061] Специалист в данной области может без труда понять другие аспекты и преимущества настоящей заявки из приведенного ниже подробного описания. Приведенное ниже подробное описание только демонстрирует и описывает иллюстративные варианты осуществления настоящей заявки. Как будет понятно специалистам в данной области, содержание настоящей заявки позволяет специалистам в данной области вносить модификации в описанные варианты осуществления без отклонения от сущности и объема изобретения в рамках настоящей заявки. Соответственно, прилагаемые чертежи и описание в рамках настоящей заявки являются только иллюстративными, а не ограничивающими.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0062] Конкретные признаки изобретения в рамках настоящей заявки приведены в прилагаемой формуле изобретения. Посредством отсылки к иллюстративным вариантам осуществления, как подробно описано ниже, и прилагаемым чертежам, признаки и преимущества изобретения, описанного в рамках настоящей заявки, могут стать лучше понятными. Краткое описание прилагаемых чертежей является следующим:
[0063] На фиг.1 представлена схематическая диаграмма вектора pUC19.
[0064] На фиг.2 представлена схематическая диаграмма модифицированного вектора pUC19, описанного в рамках настоящей заявки.
[0065] На фиг.3 представлена схематическая диаграмма вектора для хранения легкой цепи, описанного в рамках настоящей заявки.
[0066] На фиг.4 представлена схематическая диаграмма вектора для хранения коннексона, описанного в рамках настоящей заявки.
[0067] На фиг.5 представлена схематическая диаграмма вектора для хранения тяжелой цепи, описанного в рамках настоящей заявки.
[0068] На фиг.6 представлена схематическая диаграмма вектора pCom3x.
[0069] На фиг.7 представлена схематическая диаграмма вектора для дисплея, описанного в рамках настоящей заявки.
[0070] На фиг.8 представлена схематическая диаграмма плазмиды, включающей соединенный продукт для дисплея, описанной в рамках настоящей заявки.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[0071] Далее варианты осуществления изобретения заявки описаны посредством конкретных примеров. Специалисты в данной области могут без труда понять другие преимущества и эффекты изобретения, описанного в рамках настоящей заявки, из раскрытия описания.
[0072] В рамках настоящей заявки термин "фаговая библиотека", главным образом, относится к полинуклеотидной библиотеке, способной экспонировать различные рекомбинантные полипептиды на фагах. В некоторых случаях, фаговая библиотека, кроме того, может включать комбинацию фагов, трансфицированных полинуклеотидной библиотекой. В рамках настоящей заявки библиотека может относиться к коллекции или смеси диверсифицированных полинуклеотидов, включающих полинуклеотиды, кодирующие различные рекомбинантные полипептиды. Например, коллекция полинуклеотидной библиотеки может включать по меньшей мере 106, 107, 108, 109, 1010, 1011, 1012, 1013, 1014, 1015 или более типов различных полинуклеотидов. Полинуклеотиды в фаговом дисплее могут происходить из организмов одного вида (например, млекопитающие, такие как люди), или их тканей, органов и/или клеток. Библиотека может включать полинуклеотиды общего рода. Например, род может представлять собой полинуклеотиды, кодирующие определенный тип или класс полипептидов субъединиц иммуноглобулинов. Например, полинуклеотид может кодировать легкую цепь или фрагмент легкой цепи антитела. Например, полинуклеотид может кодировать тяжелую цепь или фрагмент тяжелой цепи антитела.
[0073] В рамках настоящей заявки термин "антитело", главным образом, относится к полипептидной молекуле, способной специфически распознавать и/или нейтрализовывать специфический антиген. Основным элементом четырехцепочечного антитела является гетеротетрамерный гликопротеин, состоящий из двух идентичных легких цепей и двух идентичных тяжелых цепей. В случае IgG каждая L-цепь соединена с H-цепью посредством ковалентной дисульфидной связи, в то время как две H-цепи соединены друг с другом через одну или более дисульфидных связей, где количество дисульфидных связей зависит от изотипа H-цепи. Каждая H- или L-цепь, кроме того, имеет межцепочечные дисульфидные связи с регулярными интервалами. Каждая H-цепь имеет вариабельный домен (VH) на N-конце, за которым следует три (для каждой α- или γ-цепи) или четыре (для изотипа µ или ε) константных домена (CH).
[0074] В рамках настоящей заявки термин "фрагмент антитела", главным образом, относится к части интактного антитела. Например, фрагмент антитела может включать антигенсвязывающую область и/или вариабельную область интактного антитела. Фрагмент антитела может быть получен химическим способом и/или способом генной инженерии. Например, фрагмент антитела может быть получен путем расщепления антитела протеазами, включая пепсин и папаин. В рамках настоящей заявки фрагмент антитела может представлять собой Fab.
[0075] В рамках настоящей заявки термин "Fab", главным образом, относится к двум идентичным антигенсвязывающим фрагментам, полученным посредством расщепления антитела, имеющего интактную структуру, пепсином (например, удаления Fc-области и шарнирной области). Fab может состоять из интактной легкой цепи, вариабельной области тяжелой цепи (VH) и первого константного домена тяжелой цепи (CH1). Каждый Fab может иметь один антигенсвязывающий центр.
[0076] В рамках настоящей заявки термин "коннексон", главным образом, относится к реагенту, способному соединять две или более полинуклеотидных молекул или их фрагментов. Коннексон может представлять собой полинуклеотид или его фрагмент. В рамках настоящей заявки коннексон может варьироваться по длине. Например, длина коннексона может составлять 40 п.н. или более, 50 п.н. или более, 60 п.н. или более, 70 п.н. или более, 80 п.н. или более, 90 п.н. или более, 100 п.н. или более, 150 п.н. или более, или 200 п.н. или более.
[0077] В рамках настоящей заявки термин "LC", главным образом, относится к полипептиду, включающему последовательность нуклеиновой кислоты, кодирующую легкую цепь или фрагмент легкой цепи антител или фрагментов антител. В рамках настоящей заявки легкая цепь или фрагмент легкой цепи может иметь способность к связыванию с тяжелой цепью того же или сходного антитела. В рамках настоящей заявки легкая цепь или фрагмент легкой цепи может включать вариабельную область легкой цепи (VL) и константную область легкой цепи (CL). Константная область легкой цепи может быть классифицирована как тип κ и тип λ. Легкая цепь, кроме того, включает легкую цепь из λ-вариабельной области (V-λ), соединенной с κ-константной областью (C-κ), или κ-вариабельной области (V-κ), соединенной с λ-константной областью (C-λ). Например, LC может представлять собой полипептид, включающий последовательность нуклеиновой кислоты, кодирующую легкую цепь антитела или фрагмента антитела.
[0078] В рамках настоящей заявки термин "HC", главным образом, относится к полипептиду, включающему последовательность нуклеиновой кислоты, кодирующую тяжелую цепь или фрагмент тяжелой цепи антитела или фрагмента антитела. В рамках настоящей заявки тяжелая цепь или фрагмент тяжелой цепи может иметь способность связываться с легкой цепью того же или сходного антитела. В рамках настоящей заявки тяжелая цепь или фрагмент тяжелой цепи может включать вариабельную область тяжелой цепи (VH) и константную область тяжелой цепи (CH). Константная область тяжелой цепи может включать домен CH1, шарнирную область, домен CH2 и домен CH3. В случаях IgE, IgM и IgY, константная область тяжелой цепи может включать домен CH4, но не включает шарнирную область. В рамках настоящей заявки "константная область тяжелой цепи" может представлять собой домен CH1, шарнирную область, домен CH2, CH3, CH4 или любую их комбинацию. Например, HC может представлять собой полипептид, включающий последовательность нуклеиновой кислоты, кодирующую VH и CH1 антитела или фрагмента антитела.
[0079] В рамках настоящей заявки термин "первый полинуклеотид", главным образом, относится к полинуклеотиду, включающему LC, который может представлять собой последовательность нуклеиновой кислоты, включающую легкую цепь или фрагмент легкой цепи антитела или фрагмента антитела. В рамках настоящей заявки первый полинуклеотид, кроме того, может иметь участок распознавания эндонуклеазой (например, эндонуклеаза рестрикции) на 5'-конце и/или 3'-конце. Например, первый полинуклеотид может включать, в направлении от 5' к 3', структуру R1-LC-R2, где R1, R2 могут представлять собой участки распознавания эндонуклеазой рестрикции. Например, посредством расщепления эндонуклеазой, распознающей участок распознавания эндонуклеазой в первом полинуклеотиде (например, эндонуклеаза рестрикции, распознающая R1 и R2, такая как SfiI), расщепленный первый полинуклеотид может включать LC.
[0080] В рамках настоящей заявки термин "фрагмент вектора для хранения легкой цепи", главным образом, относится к фрагментам вектора для хранения легкой цепи, полученным посредством расщепления эндонуклеазой рестрикции. В рамках настоящей заявки участок(и) распознавания эндонуклеазой рестрикции, находящийся в векторе для хранения легкой цепи, может быть соответственно таким же, как и участок(и) распознавания эндонуклеазой рестрикции, содержащийся в первом полинуклеотиде. Например, вектор для хранения легкой цепи может включать R1 и R2. В рамках настоящей заявки посредством расщепления эндонуклеазой рестрикции (например, эндонуклеаза рестрикции, распознающая R1 и R2), полученный фрагмент вектора для хранения легкой цепи может быть соединен с расщепленным первым полинуклеотидом. В рамках настоящей заявки фрагмент вектора для хранения легкой цепи может происходить из вектора pUC. Например, вектор pUC может представлять собой или может происходить из вектора pUC19; альтернативно, например, вектор pUC может представлять собой или может происходить из вектора pMD19-T.
[0081] В рамках настоящей заявки термин "соединенный продукт для хранения легкой цепи", главным образом, относится к продукту, образованному путем соединения расщепленного первого полинуклеотида с фрагментом вектора для хранения легкой цепи. В рамках настоящей заявки первый полинуклеотид и фрагмент вектора для хранения легкой цепи могут включать один и тот же участок(и) распознавания эндонуклеазой рестрикции, и могут быть соединены лигазой (например, ДНК-лигазой) с получением соединенного продукта для хранения легкой цепи.
[0082] В рамках настоящей заявки термин "первая бактерия", главным образом, относится к бактерии для введения или вмещения первого полинуклеотида. Первая бактерия может включать LC. В рамках настоящей заявки в первую бактерию может быть введен соединенный продукт для хранения легкой цепи. В рамках настоящей заявки первая бактерия может экспрессировать, реплицировать и/или хранить (например, при криоконсервации) LC, первый полинуклеотид и/или соединенный продукт для хранения легкой цепи.
[0083] В рамках настоящей заявки термин "бактериальная библиотека с компонентом легкой цепи", как правило, относится к бактериальной библиотеке, полученной путем введения первого полинуклеотида в первую бактерию. В рамках настоящей заявки бактериальная библиотека с компонентом легкой цепи может представлять собой бактериальную библиотеку, включающую последовательности нуклеиновых кислот, кодирующие легкую цепь или фрагменты легкой цепи антитела или фрагмента антитела. В рамках настоящей заявки бактериальная библиотека с компонентом легкой цепи может включать от приблизительно 105 до приблизительно 109 (например, может включать от приблизительно 105 до приблизительно 108, от приблизительно 105 до приблизительно 107, от приблизительно 106 до приблизительно 107) последовательностей нуклеиновых кислот, кодирующих различные легкие цепи или фрагменты легких цепей антител или фрагментов антитело. В рамках настоящей заявки бактериальная библиотека с компонентом легкой цепи может включать от приблизительно 107 до приблизительно 1012 (например, от приблизительно 107 до приблизительно 1011, от приблизительно 107 до приблизительно 1010, от приблизительно 107 до приблизительно 109, от приблизительно 107 до приблизительно 108) первых бактерий.
[0084] В рамках настоящей заявки термин "второй полинуклеотид", главным образом, относится к полинуклеотиду, включающему коннексон. В рамках настоящей заявки второй полинуклеотид, кроме того, может включать участок(и) распознавания эндонуклеазой (например, эндонуклеазой рестрикции), находящийся на 5'-конце и/или 3'-конце. Например, первый полинуклеотид может включать, в направлении от 5' к 3', структуру R3-коннексон-R4, где R3, R4 могут представлять собой участки распознавания эндонуклеазой рестрикции. Например, после расщепления эндонуклеазой, распознающей участок(и) распознавания эндонуклеазой, во втором полинуклеотиде (например, эндонуклеаза рестрикции, распознающая R3 и R4, такая как SfiI), расщепленный второй полинуклеотид может включать коннексон.
[0085] В рамках настоящей заявки при использовании в комбинации с вторым полинуклеотидом или коннексоном, термин "фрагмент вектора", главным образом, относится к фрагменту вектора, который способен к соединению со вторым полинуклеотидом. В рамках настоящей заявки фрагмент вектора может происходить из вектора pUC. Например, вектор pUC может представлять собой или может происходить из вектора pUC19; альтернативно, например, вектор pUC может представлять собой или происходить из вектора pMD19-T.
[0086] В рамках настоящей заявки термин "вектор для хранения коннексона", главным образом, относится к продукту, образовавшемуся путем соединения второго полинуклеотида с фрагментом вектора. В рамках настоящей заявки участок(и) распознавания эндонуклеазой рестрикции, включенный в вектор для хранения коннексона, и участок(и) распознавания эндонуклеазой рестрикции, включенный в первый полинуклеотид, могут быть соответственно одинаковыми. Например, вектор для хранения коннексона может включать R3 и R4. В рамках настоящей заявки вектор для хранения коннексона может включать только один коннексон.
[0087] В рамках настоящей заявки термин "вторая бактерия", главным образом, относится к бактерии для введения и вмещения второго полинуклеотида. Вторая бактерия может включать коннексон. В рамках настоящей заявки во вторую бактерию может быть введен вектор для хранения коннексона. В рамках настоящей заявки вторая бактерия может экспрессировать, реплицировать и/или хранить (например, при криоконсервации) коннексон и/или второй полинуклеотид.
[0088] В рамках настоящей заявки термин "бактерия с компонентом коннексона", главным образом, относится к бактерии, полученной путем введения вектора для хранения коннексона во вторую бактерию. В рамках настоящей заявки бактерии с компонентом коннексона могут представлять собой один вид бактерий. В рамках настоящей заявки один вид бактерий с компонентом коннексона может включать только один тип коннексона.
[0089] В рамках настоящей заявки термин "третий полинуклеотид", главным образом, относится к полинуклеотиду, включающему HC, который может представлять собой последовательность нуклеиновой кислоты, кодирующую тяжелую цепь или фрагмент тяжелой цепи антитела или фрагментов антитела. В рамках настоящей заявки третий полинуклеотид может дополнительно включать участок(и) распознавания эндонуклеазой (например, эндонуклеазой рестрикции) на 5'-конце и/или 3'-конце. Например, первый полинуклеотид может включать, в направлении от 5' к 3', структуру R5-HC-R6, где R5, R6 могут представлять собой участки распознавания эндонуклеазой рестрикции. Например, после расщепления эндонуклеазами, распознающими участок(и) распознавания эндонуклеазой, в третьем полинуклеотиде (например, эндонуклеаза рестрикции, распознающая R5, такая как SfiI; альтернативно, например, эндонуклеаза рестрикции, распознающая R6, такая как SfiI, BsmBI и Esp3I), расщепленный третий полинуклеотид может включать HC.
[0090] В рамках настоящей заявки термин "фрагмент вектора для хранения тяжелой цепи", главным образом, относится к фрагментам вектора для хранения тяжелой цепи, расщепленного эндонуклеазой рестрикции. В рамках настоящей заявки участок(и) распознавания эндонуклеазой рестрикции, включенный в вектор для хранения тяжелой цепи, и участок(и) распознавания эндонуклеазой рестрикции, содержащийся в третьем полинуклеотиде, могут соответственно быть одинаковыми. Например, вектор для хранения тяжелой цепи может включать R5 и R6. В рамках настоящей заявки после расщепления эндонуклеазой рестрикции (например, эндонуклеаза рестрикции, распознающая R5 и R6), полученный фрагмент вектора для хранения тяжелой цепи может быть соединен с расщепленным третьим полинуклеотидом. В рамках настоящей заявки фрагмент вектора для хранения тяжелой цепи может происходить из вектора pUC. Например, вектор pUC может представлять собой или может происходить из вектора pUC19; альтернативно, например, вектор pUC может представлять собой или может происходить из вектора pMD19-T.
[0091] В рамках настоящей заявки термин "соединенный продукт для хранения тяжелой цепи", главным образом, относится к продукту, образовавшемуся путем соединения расщепленного третьего полинуклеотида с фрагментом вектора для хранения тяжелой цепи. В рамках настоящей заявки третий полинуклеотид и фрагмент вектора для хранения тяжелой цепи могут включать один и тот же участок(и) распознавания эндонуклеазой рестрикции и могут быть соединены лигазой (например, ДНК-лигаза) с получением соединенного продукта для хранения тяжелой цепи.
[0092] В рамках настоящей заявки термин "третья бактерия", главным образом, относится к бактерии для введения и вмещения третьего полинуклеотида. Третья бактерия может включать HC. В рамках настоящей заявки в третью бактерию может быть введен соединенный продукт для хранения тяжелой цепи. В рамках настоящей заявки третья бактерия может экспрессировать, реплицировать и/или хранить (например, при криоконсервации) HC, третий полинуклеотид и/или соединенный продукт для хранения тяжелой цепи.
[0093] В рамках настоящей заявки термин "бактериальная библиотека с компонентом тяжелой цепи", главным образом, относится к бактериальной библиотеке, полученной путем введения третьего полинуклеотида в третью бактерию. В рамках настоящей заявки бактериальная библиотека с компонентом тяжелой цепи может представлять собой бактериальную библиотеку, включающую последовательности нуклеиновых кислот, кодирующие тяжелую цепь или фрагменты тяжелой цепи антител или фрагментов антител. В рамках настоящей заявки бактериальная библиотека с компонентом тяжелой цепи может включать от приблизительно 105 до приблизительно 109 (например, от приблизительно 105 до приблизительно 108, от приблизительно 105 до приблизительно 107, от приблизительно 106 до приблизительно 107) последовательностей нуклеиновых кислот, кодирующих различные тяжелые цепи или фрагменты тяжелых цепей антител или фрагментов антител. В рамках настоящей заявки бактериальная библиотека с компонентом тяжелой цепи может включать от приблизительно 107 до приблизительно 1012 (например, от приблизительно 107 до приблизительно 1011, от приблизительно 107 до приблизительно 1010, от приблизительно 107 до приблизительно 109, от приблизительно 107 до приблизительно 108) третьих бактерий.
[0094] В рамках настоящей заявки термин "плазмида с компонентом легкой цепи", главным образом, относится к плазмиде, полученной из бактерии бактериальной библиотеки с компонентом легкой цепи и содержащей LC. В рамках настоящей заявки плазмида с компонентом легкой цепи может включать LC. Например, плазмида с компонентом легкой цепи может включать первый полинуклеотид. В рамках настоящей заявки плазмида с компонентом легкой цепи может включать участок(и) распознавания эндонуклеазой рестрикции, например, R1 и R2.
[0095] В рамках настоящей заявки термин "плазмида с компонентом тяжелой цепи", главным образом, относится к плазмиде, полученной из бактерии бактериальной библиотеки с компонентом тяжелой цепи и содержащей HC. В рамках настоящей заявки плазмида с компонентом тяжелой цепи может включать HC. Например, плазмида с компонентом тяжелой цепи может включать третий полинуклеотид. В рамках настоящей заявки плазмида с компонентом тяжелой цепи может включать участок(и) распознавания эндонуклеазой рестрикции, например, R5 и R6.
[0096] В рамках настоящей заявки термин "высвободившаяся LC", главным образом, относится к LC, высвободившейся после обработки плазмиды с компонентом легкой цепи. В рамках настоящей заявки обработка может представлять собой расщепление. Например, является возможным выбор подходящей эндонуклеазы(эндонуклеаз) рестрикции (например, эндонуклеаза(ы) рестрикции, распознающая R1 и R2) для участка(ов) распознавания эндонуклеазой рестрикции в плазмиде с компонентом легкой цепи, так чтобы LC высвобождалась и отделялась от плазмиды с компонентом легкой цепи.
[0097] В рамках настоящей заявки термин "высвободившийся коннексон", главным образом, относится к коннексону, высвободившемуся после обработки плазмиды с компонентом коннексона. В рамках настоящей заявки обработка может представлять собой расщепление. Например, является возможным выбор подходящей эндонуклеазы(эндонуклеаз) рестрикции (например, эндонуклеаза(ы) рестрикции, распознающая R3 и R4) для участка(ов) распознавания эндонуклеазой рестрикции в плазмиде с компонентом коннексона, так чтобы коннексон высвобождался и отделялся от плазмиды с компонентом коннексона. Альтернативно, например, сначала плазмида с компонентом коннексона может быть взята в качестве матрицы для амплификации (например, с использованием амплификации посредством ПЦР) для включения второго полинуклеотида с последующим расщеплением полученного продукта амплификации. Например, является возможным выбор подходящей эндонуклеазы(эндонуклеаз) рестрикции (например, эндонуклеаза(ы) рестрикции, распознающая R3 и R4) для участка(ов) распознавания эндонуклеазой рестрикции в плазмиде с компонентом коннексона, так чтобы коннексон высвобождался и отделялся от плазмиды с компонентом коннексона.
[0098] В рамках настоящей заявки термин "высвободившаяся HC", главным образом, относится к HC, высвободившейся после обработки плазмидой с компонентом тяжелой цепи. В рамках настоящей заявки обработка может представлять собой расщепление. Например, является возможным выбор подходящей эндонуклеазы(эндонуклеаз) рестрикции (например, R5 и R6) для участка(ов) распознавания эндонуклеазой рестрикции в плазмиде с компонентом тяжелой цепи, так чтобы HC высвобождалась и отделялась от плазмиды с компонентом тяжелой цепи.
[0099] В рамках настоящей заявки термин "высвободившийся фрагмент вектора для дисплея", главным образом, относится к фрагментам вектора для дисплея, высвободившимся после обработки вектора для дисплея. В рамках настоящей заявки вектор для дисплея может включать, в направлении от 5' к 3', структуру R7-фрагмент вектора для дисплея-R8, где R7 и R8 могут представлять собой эндонуклеазу рестрикции. В рамках настоящей заявки обработка может представлять собой расщепление эндонуклеазой рестрикции. Например, является возможным выбор подходящей эндонуклеазы(эндонуклеаз) рестрикции (например, R7 и R8) для участка(ов) распознавания эндонуклеазой рестрикции в векторе для дисплея, так чтобы высвободившийся фрагмент вектора для дисплея высвобождался и отделялся от вектора для дисплея.
[00100] В рамках настоящей заявки термин "соединить (соединенный, соединяющий, соединение)", главным образом, относится к соединению двух или более полинуклеотидных молекул друг с другом. Например, соединения можно достигать посредством лигазы (например, ДНК-лигаза). Например, 3'-конец полинуклеотида соединяют с 5'-концом другого полинуклеотида с образованием целой полинуклеотидной молекулы.
[00101] В рамках настоящей заявки термин "соединенный продукт для дисплея", главным образом, относится к полинуклеотидной молекуле, включающей высвободившуюся LC, высвободившийся коннексон, высвободившуюся HC и высвободившийся фрагмент вектора для дисплея. В рамках настоящей заявки соединенный продукт для дисплея может быть получен путем смешения высвободившейся LC, высвободившегося коннексона, высвободившейся HC и высвободившегося фрагмента вектора для дисплея в некотором соотношении.
[00102] В рамках настоящей заявки "дисплей" может относиться к экспрессии белка (например, антитело или фрагмент антитела, такой как Fab), кодируемого соединенным продуктом для дисплея в клетке, включающей соединенный продукт для дисплея.
[00103] В рамках настоящей заявки термин "вводить (осуществляющий введение, введенный, введение)", главным образом, относится к процессу, в котором экзогенный полинуклеотид переносится или направляется в клетку. Клетка может представлять собой клетку-хозяина. Введенная клетка включает первичные клетки индивидуума и дочерние клетки. Клетка может представлять собой прокариотическую клетку, например, бактериальную клетку.
[00104] В рамках настоящей заявки термин "четвертая бактерия", главным образом, относится к бактерии для введения или включения соединенного продукта для дисплея, где соединенный продукт для дисплея включает высвободившуюся LC, высвободившийся коннексон, высвободившуюся HC и высвободившийся фрагмент вектора для дисплея. В рамках настоящей заявки в четвертую бактерию может быть введен соединенный продукт для дисплея посредством трансформации. В рамках настоящей заявки четвертая бактерия может экспрессировать легкую цепь или фрагмент легкой цепи антител или фрагментов антител, кодируемые высвободившейся LC; альтернативно, она может экспрессировать тяжелую цепь или фрагмент тяжелой цепи антител или фрагментов антител, кодируемые высвободившейся HC. В рамках настоящей заявки четвертая бактерия может экспрессировать, реплицировать и/или хранить (например, при криоконсервации) антитела или фрагменты антител.
[00105] В рамках настоящей заявки термин "библиотека бактериального дисплея", главным образом, относится к бактериальной библиотеке, включающей соединенный продукт для дисплея. В рамках настоящей заявки бактериальная библиотека для дисплея может включать четвертую бактерию. В рамках настоящей заявки библиотека бактериального дисплея может включать 1010 или более различных антител или фрагментов антител.
[00106] В рамках настоящей заявки термин "сигнальный пептид pelB", главным образом, относится к сигнальному пептиду пектинлиазы. Сигнальный пептид pelB, главным образом, используется в прокариотической экспрессирующей системе. Например, номер доступа GenBank для сигнального пептида pelB может представлять собой ABS75961.1.
[00107] В рамках настоящей заявки термин "эндонуклеаза рестрикции", главным образом, относится к ферменту, который расщепляет двухцепочечную ДНК. Эндонуклеаза рестрикции может продуцировать липкий конец с выступающей одноцепочечной ДНК, так что он может связываться ДНК-лигазой. В рамках настоящей заявки эндонуклеаза рестрикции может играть роль в распознавании и рестрикционном расщеплении. Например, участок расщепления эндонуклеазой рестрикции находится на определенном расстоянии от ее участка распознавания. Например, эндонуклеаза рестрикции может быть выбрана из группы, состоящей из SfiI, BsmBI и Esp3I.
[00108] В рамках настоящей заявки термин "линеаризовывать (линеаризованный, линеаризующийся, линеаризация)", главным образом, означает, что вектор имеет линейную форму, а не замкнут в кольцо. Например, после расщепления вектора исходная кольцевая структура вектора разрушается, так что образуется линеаризованный вектор. Например, вектор (например, вектор T) может образовывать T-конец на 3'-конце после расщепления эндонуклеазой рестрикции. В то же время, он может быть соединен с продуктом ПЦР с A-концом под действием T4-лигазы, так что продукт ПЦР соединяется с линеаризованным вектором.
[00109] В рамках настоящей заявки термин "клон", главным образом, относится к числу колоний. Например, клон может относиться к числу колоний в бактериальной библиотеке (например, бактериальная библиотека с компонентом легкой цепи, бактериальная библиотека с компонентом тяжелой цепи, библиотека бактериального дисплея и/или фаговая библиотека). В некоторых случаях клон может относиться к числу различных колоний в бактериальной библиотеке. В некоторых случаях, клон может относиться к числу дочерних популяций, образовавшихся из одного клона.
[00110] В рамках настоящей заявки термин "полинуклеотид", главным образом, относится к нуклеотидам, т.е. по меньшей мере двум нуклеотидам, соединенным вместе. Полинуклеотид может представлять собой полимер любой длины, включая, например, 200, 300, 500, 1000, 2000, 3000, 5000, 7000, 10000, 100000 и т.п. Полинуклеотид может включать фосфодиэфирную связь.
[00111] В рамках настоящей заявки термин "содержать (вмещать, включающий)", главным образом, относится к включению прямо указанного признака без исключения других элементов.
[00112] В рамках настоящей заявки термин "приблизительно", главным образом, относится к варьированию в диапазоне 0,5%-10% выше или ниже указанной величины, например, к варьированию на 0,5%, 1%, 1,5%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5%, 5%, 5,5%, 6%, 6,5%, 7%, 7,5%, 8%, 8,5%, 9%, 9,5% или 10% выше или ниже указанной величины.
[00113] В одном аспекте настоящая заявка относится к способу получения библиотеки, экспонирующей антитела или фрагменты антител, включающему: 1) предоставление первого полинуклеотида, содержащего LC, второго полинуклеотида, содержащего коннексон, и третьего полинуклеотида, содержащего HC, где LC включает последовательность нуклеиновой кислоты, кодирующую легкую цепь или фрагмент легкой цепи антитела или фрагментов антитела, и HC включает последовательность нуклеиновой кислоты, кодирующую тяжелую цепь или фрагмент тяжелой цепи антитела или фрагментов антитела; 2) введение первого полинуклеотида в первую бактерию с получением бактериальной библиотеки с компонентом легкой цепи, введение второго полинуклеотида во вторую бактерию с получением бактериальной библиотеки с компонентом коннексона, и введение третьего полинуклеотида в третью бактерию с получением бактериальной библиотеки с компонентом тяжелой цепи; 3) получение плазмиды с компонентом легкой цепи, включающей LC из бактериальной библиотеки с компонентом легкой цепи, получение плазмиды с компонентом коннексона, включающей коннексон из бактериальной библиотеки с компонентом коннексона, и получение плазмиды с компонентом тяжелой цепи, включающей HC из бактериальной библиотеки с компонентом тяжелой цепи; 4) получение высвободившейся LC из плазмиды с компонентом легкой цепи, получение высвободившегося коннексона из плазмиды с компонентом коннексона, и получение высвободившейся HC из плазмиды с компонентом тяжелой цепи; 5) предоставление вектора для дисплея и получение высвободившегося фрагмента вектора для дисплея из вектора для дисплея; 6) соединение высвободившейся LC, высвободившегося коннексона, высвободившейся HC и высвободившегося фрагмента вектора для дисплея с получением соединенного продукта для дисплея; 7) введение соединенного продукта для дисплея в четвертую бактерию с получением библиотеки бактериального дисплея антитела или фрагментов антитела; и 8) получение фаговой библиотеки, экспонирующей антитела или фрагменты антитела, с использованием библиотеки бактериального дисплея.
[00114] В рамках настоящей заявки соединенный продукт для дисплея в 6) может экспрессировать легкую цепь или фрагмент легкой цепи и тяжелую цепь или фрагмент тяжелой цепи в соответствии с рамками считывания в условиях, подходящих для экспрессии антител или фрагментов антител.
[00115] В рамках настоящей заявки все из первого полинуклеотида, второго полинуклеотида и третьего полинуклеотида могут представлять собой линейные молекулы нуклеиновых кислот.
[00116] В рамках настоящей заявки последовательность нуклеиновой кислоты, кодирующая тяжелую цепь или фрагмент тяжелой цепи, и последовательность нуклеиновой кислоты, кодирующая легкую цепь или фрагмент легкой цепи, могут находиться в различных рамках считывания. В рамках настоящей заявки рамка считывания может представлять собой непрерывную нуклеотидную последовательность, начинающуюся с инициирующего кодона и оканчивающуюся стоп-кодоном, и не включает никакого промотора или терминатора внутри нее. Например, рамка считывания может кодировать белок или полипептидный фрагмент.
[00117] В рамках настоящей заявки коннексон может включать последовательность нуклеиновой кислоты, кодирующую сигнальный пептид pelB или его фрагмент.
[00118] В рамках настоящей заявки коннексон может включать последовательность нуклеиновой кислоты, кодирующую фрагмент сигнального пептида pelB, в то время как другие последовательности нуклеиновых кислот, кодирующие фрагменты сигнального пептида pelB, могут быть дополнительно расположены в R5. Например, нуклеотид на 3'-конце последовательности нуклеиновой кислоты, кодирующей фрагмент сигнального пептида pelB, может находиться в R5. Например, когда высвободившийся коннексон соединен с высвободившейся HC, последовательность нуклеиновой кислоты, кодирующая фрагмент сигнального пептида pelB в высвободившемся коннексоне, может быть соединена с последовательностью нуклеиновой кислоты, кодирующей фрагмент сигнального пептида pelB в высвободившейся HC, с образованием последовательности нуклеиновой кислоты, кодирующей целый сигнальный пептид pelB.
[00119] В рамках настоящей заявки коннексон может иметь длину от приблизительно 50 до приблизительно 200 оснований. Например, коннексон может иметь длину от приблизительно 50 до приблизительно 200 оснований, от приблизительно 50 до приблизительно 180 оснований, от приблизительно 50 до приблизительно 160 оснований, от приблизительно 50 до приблизительно 140 оснований, от приблизительно 50 до приблизительно 120 оснований, от приблизительно 50 до приблизительно 100 оснований, от приблизительно 50 до приблизительно 90 оснований, от приблизительно 50 до приблизительно 80 оснований, от приблизительно 50 до приблизительно 75 оснований, от приблизительно 50 до приблизительно 70 оснований и от приблизительно 50 до приблизительно 60 оснований.
[00120] В рамках настоящей заявки в соединенном продукте для дисплея в 6) LC может находиться выше коннексона, и коннексон может находиться выше HC. В рамках настоящей заявки "находящийся выше" может находиться на 5'-конце нуклеотидной последовательности.
[00121] В рамках настоящей заявки 2) может включать криоконсервацию бактериальной библиотеки с компонентом легкой цепи и/или бактериальной библиотеки с компонентом тяжелой цепи. В рамках настоящей заявки 7) может включать криоконсервацию библиотеки бактериального дисплея. В рамках настоящей заявки криоконсервация может относиться к хранению при -18°C или ниже, например, к хранению при -80°C или ниже.
[00122] В рамках настоящей заявки 8) может включать размораживание и культивирование криоконсервированной библиотеки бактериального дисплея, и конструирование фаговой библиотеки с использованием культивируемой библиотеки бактериального дисплея.
[00123] В рамках настоящей заявки первый полинуклеотид может включать структуру R1-LC-R2 в направлении от 5' к 3', второй полинуклеотид включает структуру R3-коннексон-R4 в направлении от 5' к 3', третий полинуклеотид может включать структуру R5-HC-R6 в направлении от 5' к 3', и вектор для дисплея может включать структуру R7-фрагмент вектора для дисплея-R8 в направлении от 5' к 3', где R1, R2, R3, R4, R5, R6, R7 и R8 представляют собой участки распознавания эндонуклеазой рестрикции.
[00124] В рамках настоящей заявки конец, образующийся в результате расщепления R2 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R3 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R1, R4, R5, R6, R7 и R8 эндонуклеазой рестрикции, и наоборот. В рамках настоящей заявки R2 может быть идентичен R3.
[00125] В рамках настоящей заявки конец, образующийся в результате расщепления R4 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R5 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R1, R2, R3, R6, R7 и R8 эндонуклеазой рестрикции, и наоборот. В рамках настоящей заявки R4 может быть идентичен R5.
[00126] В рамках настоящей заявки конец, образующийся в результате расщепления R6 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R7 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R1, R2, R3, R4, R5 и R8 эндонуклеазой рестрикции, и наоборот. В рамках настоящей заявки R6 может быть идентичен R7.
[00127] В рамках настоящей заявки конец, образующийся в результате расщепления R8 эндонуклеазой рестрикции, может распознавать и соединяться с концом, образующимся в результате расщепления R1 эндонуклеазой рестрикции, и наоборот; и не может распознавать и соединяться с концом, образующимся в результате расщепления любого из R2, R3, R4, R5, R6 и R7 эндонуклеазой рестрикции, и наоборот. В рамках настоящей заявки R8 может быть идентичен R1.
[00128] В рамках настоящей заявки конец, образующийся в результате расщепления любого из R1, R2, R3, R4, R5, R6, R7 и R8 эндонуклеазой рестрикции, представляет собой не палиндромную последовательность.
[00129] При выборе участков распознавания эндонуклеазой (например, R1, R2, R3, R4, R5, R6, R7 и R8), является возможным выбор последовательности, которая по существу не содержится в вариабельной области антитела (например, вариабельная область легкой цепи или вариабельная область тяжелой цепи) для сохранения целостности вариабельной области антитела, чтобы предотвратить повреждение (например, деградацию) совокупности генов антитела в ходе расщепления. Более того, конец, образовавшийся посредством расщепления участка распознавания эндонуклеазой рестрикции, может представлять собой не палиндромную последовательность во избежание самосоединения, тем самым достигая уменьшения образования нецелевого соединения. Кроме того, выбор участков распознавания эндонуклеазой может сделать возможным направленное соединение множества фрагментов для повышения эффективности соединения.
[00130] В рамках настоящей заявки два или более из R1, R2, R3, R4, R5, R6, R7 и R8 (например, 2, 3, 4, 5, 6, 7 или 8) могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции.
[00131] Например, в рамках настоящей заявки R1, R2, R3, R4, R5 и R8 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции. Например, R6 и R7 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции. В некоторых случаях, эндонуклеаза рестрикции, распознающая R1, R2, R3, R4, R5 и R8, может отличаться от эндонуклеазы рестрикции, распознающей R6 и R7.
[00132] В рамках настоящей заявки эндонуклеаза рестрикции может быть выбрана из группы, состоящей из SfiI, BsmBI и Esp3I. В рамках настоящей заявки BsmBI и Esp3I могут представлять собой изоферменты, которые могут распознавать тот же участок распознавания эндонуклеазой рестрикции.
[00133] В рамках настоящей заявки, например, R1, R2, R3, R4, R5 и R8 могут распознаваться и расщепляться посредством SfiI. В рамках настоящей заявки, например, R6 и R7 могут распознаваться и расщепляться посредством BsmBI и/или Esp3I.
[00134] Например, SfiI может распознавать последовательность, состоящую из 13 оснований (от 5' к 3'): GGCCNNNN/NGGCC (SEQ ID NO:99), которая может образовывать выступающую последовательность на 3'-конце (выступающий конец, такой как одноцепочечная последовательность, содержащая 3 основания) после расщепления, где N может обозначать любое из оснований GATC. Таким образом, существует 64 различных последовательности, которые могут распознаваться посредством SfiI.
[00135] Например, BsmBI и Esp3I могут распознавать последовательность, состоящую из 12 оснований (от 5' к 3'): CGTCTCN/NNNNN (SEQ ID NO:100), которая может образовывать выступающую последовательность на 5'-конце (выступающий конец, такой как одноцепочечная последовательность, содержащая 4 основания), где N может обозначать любое из оснований GATC. Таким образом, существует 264 различных последовательности, которые могут распознаваться посредством BsmBI и Esp3I.
[00136] В рамках настоящей заявки R1 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 1. В рамках настоящей заявки R2 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 2. В рамках настоящей заявки R3 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 2. В рамках настоящей заявки R4 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 3. В рамках настоящей заявки R5 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 3. В рамках настоящей заявки R6 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 4. В рамках настоящей заявки R7 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 4. В рамках настоящей заявки R8 может включать последовательность нуклеиновой кислоты, как показано в SEQ ID NO: 1.
[00137] В рамках настоящей заявки легкая цепь или фрагмент легкой цепи может включать часть вариабельной области легкой цепи и/или часть константной области легкой цепи. В рамках настоящей заявки тяжелая цепь или фрагмент тяжелой цепи может включать часть вариабельной области тяжелой цепи и/или часть константной области тяжелой цепи.
[00138] Например, LC может включать полинуклеотид, кодирующий вариабельную область легкой цепи VL в легкой цепи или фрагменте легкой цепи. Например, первый полинуклеотид может включать полинуклеотид, кодирующий вариабельную область легкой цепи VL в легкой цепи или фрагменте легкой цепи. 5'-конец первого полинуклеотида может быть соединен с R1, и 3'-конец первого полинуклеотида может быть соединен с R2; и 5'-конец фрагмента вектора для хранения легкой цепи может быть соединен с R3, и 3'-конец фрагмента вектора для хранения легкой цепи может быть соединен с R8.
[00139] Например, HC может включать полинуклеотид, кодирующий вариабельную область тяжелой цепи VH и CH1 константной области тяжелой цепи в тяжелой цепи или фрагменте тяжелой цепи. Например, третий полинуклеотид может включать полинуклеотид, кодирующий вариабельную область тяжелой цепи VH и CH1 константной области тяжелой цепи в тяжелой цепи или фрагменте тяжелой цепи. 5'-конец третьего полинуклеотида может быть соединен с R5, и 3'-конец третьего полинуклеотида может быть соединен с R6; и 5'-конец фрагмента вектора для хранения тяжелой цепи может быть соединен с R7, и 3'-конец фрагмента вектора для хранения тяжелой цепи может быть соединен с R4.
[00140] Альтернативно, например, HC может включать полинуклеотид, кодирующий вариабельную область тяжелой цепи VH в тяжелой цепи или фрагменте тяжелой цепи. Например, третий полинуклеотид может включать полинуклеотид, кодирующий вариабельную область тяжелой цепи VH в тяжелой цепи или фрагменте тяжелой цепи. В то же время фрагмент вектора для хранения тяжелой цепи может включать полинуклеотид, кодирующий CH1 константной области тяжелой цепи в тяжелой цепи или фрагменте тяжелой цепи. 5'-конец третьего полинуклеотида может быть соединен с R5, и 3'-конец третьего полинуклеотида может быть соединен с R6; и 5'-конец фрагмента вектора для хранения тяжелой цепи может быть соединен с R7, и 3'-конец фрагмента вектора для хранения тяжелой цепи может быть соединен с R4.
[00141] В рамках настоящей заявки антитело или его фрагмент может включать Fab, Fab', (Fab)2 и/или (Fab')2.
[00142] В рамках настоящей заявки соединенный продукт для дисплея в 7) может иметь эффективность соединения и конверсии по меньшей мере 108 клонов/мкг полинуклеотида (например, по меньшей мере 109 клонов/мкг полинуклеотида, по меньшей мере 1010 клонов/мкг полинуклеотида, по меньшей мере 1011 клонов/мкг полинуклеотида и по меньшей мере 1012 клонов/мкг полинуклеотида).
[00143] В рамках настоящей заявки 2) может включать расщепление первого полинуклеотида эндонуклеазой рестрикции, распознающей R1 и R2, соединение расщепленного первого полинуклеотида с фрагментом вектора для хранения легкой цепи с образованием соединенного продукта для хранения легкой цепи, и введение соединенного продукта для хранения легкой цепи в первую бактерию с получением бактериальной библиотеки с компонентом легкой цепи; где вектор для хранения легкой цепи включает R1 и R2, и фрагмент вектора для хранения легкой цепи получают путем расщепления вектора для хранения легкой цепи эндонуклеазой рестрикции, распознающей R1 и R2.
[00144] В рамках настоящей заявки LC может иметь длину более чем приблизительно 600 п.н. Например, она может составлять более чем приблизительно 650 п.н., более чем приблизительно 680 п.н. и более чем приблизительно 700 п.н. В рамках настоящей заявки первый полинуклеотид может иметь длину более чем приблизительно 600 п.н. Например, она может составлять более чем приблизительно 650 п.н., более чем приблизительно 680 п.н. и более чем приблизительно 700 п.н. В рамках настоящей заявки фрагмент вектора для хранения легкой цепи может иметь длину более чем приблизительно 900 п.н., например, более чем приблизительно 1000 п.н., более чем приблизительно 1500 п.н., более чем приблизительно 1800 п.н. и более чем приблизительно 2100 п.н. В рамках настоящей заявки длина фрагмента вектора для хранения легкой цепи может существенно отличаться от длины LC. Например, длина фрагмента вектора для хранения легкой цепи может существенно отличаться от длины LC при использовании полиакриламидного гель-электрофореза (PAGE).
[00145] В рамках настоящей заявки вектор для хранения легкой цепи может происходить из любого вектора, такого как любой вектор, который может быть амплифицирован и/или легко хранится. В некоторых случаях, вектор, используемый в качестве вектора для хранения легкой цепи, может иметь свойства, включающие высокие количества копий, низкую молекулярную массу и т.д. Например, вектор для хранения легкой цепи может представлять собой вектор pUC. В рамках настоящей заявки вектор pUC может представлять собой pUC19 (схематическое структурное изображение которого может быть таким, как показано на фиг.1) или может происходить из вектора pUC19. В рамках настоящей заявки вектор pUC может представлять собой или может происходить из вектора pMD19-T.
[00146] В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R1 и R2, может включать SfiI.
[00147] В рамках настоящей заявки 2) может включать расщепление третьего полинуклеотида эндонуклеазой рестрикции, распознающей R5 и R6, соединение расщепленного третьего полинуклеотида с фрагментом вектора для хранения тяжелой цепи с образованием соединенного продукта для хранения тяжелой цепи, и введение соединенного продукта для хранения тяжелой цепи в третью бактерию с получением бактериальной библиотеки с компонентом тяжелой цепи; где вектор для хранения тяжелой цепи включает R5 и R6, и фрагмент вектора для хранения тяжелой цепи получают путем расщепления вектора для хранения тяжелой цепи эндонуклеазой рестрикции, распознающей R5 и R6.
[00148] В рамках настоящей заявки HC может иметь длину более чем приблизительно 320 п.н. Например, она может составлять более чем приблизительно 350 п.н., более чем приблизительно 380 п.н. и более чем приблизительно 400 п.н. В рамках настоящей заявки третий полинуклеотид может иметь длину более чем приблизительно 320 п.н. Например, она может составлять более чем приблизительно 350 п.н., более чем приблизительно 380 п.н. и более чем приблизительно 400 п.н. В рамках настоящей заявки фрагмент вектора для хранения тяжелой цепи может иметь длину более чем приблизительно 900 п.н., например, более чем приблизительно 1000 п.н., более чем приблизительно 1500 п.н., более чем приблизительно 1800 п.н. и более чем приблизительно 2100 п.н. В рамках настоящей заявки длина фрагмента вектора для хранения тяжелой цепи может существенно отличаться от длины HC. Например, длина фрагмента вектора для хранения легкой цепи может существенно отличаться от длины HC при использовании PAGE.
[00149] В рамках настоящей заявки вектор для хранения тяжелой цепи может происходить из любого вектора, такого как любой вектор, который может быть амплифицирован и/или легко хранится. В некоторых случаях, вектор, используемый в качестве вектора для хранения тяжелой цепи, может иметь свойства, такие как высокое число копий, низкая молекулярная масса и т.д. Например, вектор для хранения тяжелой цепи может представлять собой вектор pUC. В рамках настоящей заявки вектор pUC может представлять собой вектор pUC19 (схематическое структурное изображение которого может быть таким, как показано на фиг.1) или может происходить из вектора pUC19. В рамках настоящей заявки вектор pUC может представлять собой или может происходить из вектора pMD19-T.
[00150] В рамках настоящей заявки для конструирования вектора для хранения тяжелой цепи, вектор, который представляет собой или происходит из вектора pUC, может быть сконструированным/модифицированным. Например, является возможным удаление одного или более участков распознавания эндонуклеазой из вектора (например, удаление одного или более участков распознавания BsmBI из него) посредством сайт-направленного мутагенеза. В некоторых случаях, также является возможным добавление одного или более участков распознавания эндонуклеазой в вектор (например, добавление одного или более участков распознавания BsmBI в выбранных положениях) посредством сайт-направленного мутагенеза.
[00151] В одном примере является возможным удаление одного или более существующих участков распознавания эндонуклеазой BsmBI из вектора посредством сайт-направленного мутагенеза, а затем добавление одного или более дополнительных участков распознавания BsmBI в другое положение вектора с получением модифицированного вектора (например, модифицированного вектора pUC).
[00152] В рамках настоящей заявки вектор для хранения тяжелой цепи может включать и включает только один участок распознавания BsmBI.
[00153] В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R5 и R6, может включать SfiI, BsmBI и/или Esp3I. В некоторых случаях эндонуклеаза рестрикции, распознающая R5 и R6, представляет собой BsmBI и/или Esp3I.
[00154] В рамках настоящей заявки 2) включает соединение второго полинуклеотида с фрагментом вектора с образованием вектора для хранения коннексона (например, в соответствии с инструкциями продукта клона TA от Takara Inc.), и введение вектора для хранения коннексона во вторую бактерию с получением бактерии с компонентом коннексона. В рамках настоящей заявки вектор для хранения коннексона может включать участки R3 и R4.
[00155] В рамках настоящей заявки коннексон может иметь длину более чем приблизительно 70 п.н., например, приблизительно 72 п.н. или приблизительно 90 п.н. В рамках настоящей заявки второй полинуклеотид может иметь длину более чем приблизительно 80 п.н., например, более чем приблизительно 90 п.н., более чем приблизительно 100 п.н. и более чем приблизительно 120 п.н. В рамках настоящей заявки фрагмент вектора может иметь длину более чем приблизительно 800 п.н.
[00156] В рамках настоящей заявки вектор фрагмент для конструирования вектора для хранения коннексона может происходить из любого вектора, такого как любой вектор, который может быть амплифицирован и/или легко хранится. В некоторых случаях, вектор для конструирования вектора для хранения коннексона может иметь свойства, включающие высокое количество копий, низкую молекулярную массу и т.д. Например, фрагмент вектора для конструирования вектора для хранения коннексона может происходить из вектора pUC. В рамках настоящей заявки вектор pUC может представлять собой вектор pUC19 (схематическое структурное изображение которого может быть таким, как показано на фиг.1) или может происходить из вектора pUC19. В рамках настоящей заявки вектор pUC может представлять собой или может происходить из вектора pMD19-T.
[00157] В рамках настоящей заявки вектор pUC19 или вектор pMD19-T включает вектор, который происходит из него, однако является модифицированным/сконструированным.
[00158] В рамках настоящей заявки 4) может включать расщепление плазмиды с компонентом легкой цепи эндонуклеазой рестрикции, распознающей R1 и R2, с получением высвободившейся LC. В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R1 и R2, может включать SfiI.
[00159] В рамках настоящей заявки 4) может включать расщепление плазмиды с компонентом тяжелой цепи эндонуклеазой рестрикции, распознающей R5 и R6, с получением высвободившейся HC. В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R5 и R6, может включать SfiI, BsmBI и/или Esp3I.
[00160] В некоторых случаях в рамках настоящей заявки 4) может включать расщепление вектора для хранения коннексона эндонуклеазой рестрикции, распознающей R3 и R4, с получением высвободившегося коннексона.
[00161] Например, является возможным прямое расщепление вектора для хранения коннексона эндонуклеазой рестрикции, распознающей R3 и R4 (например, SfiI), с получением высвободившегося коннексона. В рамках настоящей заявки коннексон может иметь длину приблизительно 72 п.н. или более, например, приблизительно 90 п.н. или более.
[00162] В некоторых случаях в рамках настоящей заявки 4) может включать получение продукта амплификации, включающего коннексон, R3 и R4, из вектора для хранения коннексона; и расщепление продукта амплификации эндонуклеазой рестрикции, распознающей R3 и R4, с получением высвободившегося коннексона.
[00163] Например, является осуществимым получение продукта амплификации, включающего коннексон, R3 и R4, посредством ПЦР и других способов, где продукт амплификации может иметь длину приблизительно 800 п.н. или более, например, приблизительно 900 п.н. или более, приблизительно 1000 п.н. или более и приблизительно 1200 п.н. или более. В рамках настоящей заявки является возможным расщепление продукта амплификации эндонуклеазой рестрикции, распознающей R3 и R4 (например, SfiI) с получением высвободившегося коннексона. В рамках настоящей заявки коннексон может иметь длину приблизительно 72 п.н. или более и приблизительно 90 п.н. или более. В рамках настоящей заявки длина продукта амплификации может существенно отличаться от длины коннексона. Таким образом, единый коннексон может быть получен более просто и эффективно. Например, длина продукта амплификации может существенно отличаться от длины коннексона при использовании PAGE.
[00164] В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R3 и R4, может включать SfiI.
[00165] В рамках настоящей заявки 5) может включать расщепление вектора для дисплея эндонуклеазой рестрикции, распознающей R7 и R8, для высвобождения фрагмента вектора для дисплея. В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R7 и R8, может включать SfiI, BsmBI и/или Esp3I.
[00166] В рамках настоящей заявки вектор для дисплея может происходить из любого подходящего вектора, такого как вектор pComb3x. Вектор pComb3x может быть модифицирован для адаптации к цели настоящей заявки. Например, вектор pComb3x может включать один участок распознавания SfiI, находящийся на 5'-конце, и один участок распознавания SfiI, находящийся на 3'-конце. В процессе модификации участок распознавания SfiI на 3'-конце вектора pComb3x может быть удален посредством сайт-направленного мутагенеза. В некоторых случаях сайт-направленный мутагенез не может влиять на экспрессию белка рамки считывания (например, фрагменты тяжелой цепи антитела и/или фрагменты легкой цепи антитела) в векторе. Например, мутация может представлять собой не смысловую мутацию, например, мутацию, которая может изменять только последовательность оснований, но не изменяет аминокислотную последовательность.
[00167] В рамках настоящей заявки три различных расщепленных фрагмента могут быть получены после расщепления вектора для дисплея эндонуклеазой рестрикции, распознающей R1, R2, R3, R4, R5 и R8. Например, вектор для дисплея может включать 3 участка распознавания эндонуклеазой рестрикции, которые могут распознаваться SfiI. Три расщепленных фрагмента могут быть получены при расщеплении вектора для дисплея посредством SfiI.
[00168] В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R1, R2, R3, R4, R5 и R8, может включать SfiI. В рамках настоящей заявки после расщепления вектора для дисплея эндонуклеазой рестрикции, распознающей R6 и R7, вектор для дисплея может быть линеаризованным.
[00169] В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R6 и R7, может включать BsmBI и/или Esp3I.
[00170] В рамках настоящей заявки четыре различных расщепленных фрагмента могут быть получены после расщепления вектора для дисплея эндонуклеазой рестрикции, распознающей R1, R2, R3, R4, R5, R6, R7 и R8. Например, вектор для дисплея может включать 3 участка распознавания ферментом рестрикции, которые могут распознаваться SfiI, и 1 участок распознавания эндонуклеазой рестрикции, который может распознаваться BsmBI и/или Esp3I. Четыре различных расщепленных фрагмента могут быть получены, когда вектор для дисплея расщепляют посредством SfiI, BsmBI и/или Esp3I.
[00171] Например, является возможным использование эндонуклеазы рестрикции, распознающей R1 и R2, для получения высвободившейся LC; и использование эндонуклеазы рестрикции R5 и R6, для получения высвободившейся HC;
[00172] В рамках настоящей заявки эндонуклеаза рестрикции, распознающая R1, R2, R3, R4, R5, R6, R7 и R8, может включать SfiI, BsmBI и/или Esp3I.
[00173] В рамках настоящей заявки вектор (например, вектор дисплея или вектор для хранения) может включать или кодировать один или более подходящих маркеров (например, маркеры для применения для очистки/распознвания/скрининга), который может представлять собой, например, His-метку, Flag-метку, флуоресцеин, селективный антибиотик и/или авидин и т.д.
[00174] В рамках настоящей заявки 2) может включать детекцию образцов из бактериальной библиотеки с компонентом легкой цепи, бактериальной библиотеки с компонентом коннексона и/или бактериальной библиотеки с компонентом тяжелой цепи для определения емкости и/или качества бактериальной библиотеки с компонентом легкой цепи, бактериальной библиотеки с компонентом коннексона и/или бактериальной библиотеки с компонентом тяжелой цепи.
[00175] В рамках настоящей заявки бактериальная библиотека с компонентом легкой цепи может включать по меньшей мере приблизительно 106 (например, по меньшей мере приблизительно 107, по меньшей мере приблизительно 108, по меньшей мере приблизительно 109, по меньшей мере приблизительно 1010, по меньшей мере приблизительно 1011, и по меньшей мере приблизительно 1012 или более) различных клонов.
[00176] В рамках настоящей заявки бактериальная библиотека с компонентом тяжелой цепи может включать по меньшей мере приблизительно 107 (например, по меньшей мере приблизительно 108, по меньшей мере приблизительно 109, по меньшей мере приблизительно 1010, по меньшей мере приблизительно 1011, и по меньшей мере приблизительно 1012 или более) различных клонов.
[00177] В рамках настоящей заявки доля эффективного клонирования в бактериальной библиотеке с компонентом легкой цепи может по меньшей мере приблизительно 70% (например, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, и по меньшей мере приблизительно 99%).
[00178] В рамках настоящей заявки доля эффективного клонирования в бактериальной библиотеке с компонентом тяжелой цепи может составлять по меньшей мере приблизительно 70% (например, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, и по меньшей мере приблизительно 99%).
[00179] В рамках настоящей заявки 2) включает вычисление емкости и/или качества библиотеки бактериального дисплея на основе емкости бактериальной библиотеки с компонентом легкой цепи и/или бактериальной библиотеки с компонентом тяжелой цепи, бактерий с компонентом коннексона, и/или массы.
[00180] В рамках настоящей заявки библиотека бактериального дисплея может включать по меньшей мере приблизительно 1010 (например, по меньшей мере приблизительно 1011, по меньшей мере приблизительно 1012, по меньшей мере приблизительно 1013, по меньшей мере приблизительно 1014, по меньшей мере приблизительно 1015, и по меньшей мере приблизительно 1016 или более) различных клонов.
[00181] В рамках настоящей заявки доля эффективного клонирования в библиотеке бактериального дисплея может составлять по меньшей мере приблизительно 70% (например, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, и по меньшей мере приблизительно 99%).
[00182] В рамках настоящей заявки бактерия в библиотеке бактериального дисплея может подвергаться культивированию в жидкой среде. Время культивирования для культуры в жидкой среде может составлять не более чем приблизительно 8 часов, например, не более чем приблизительно 4 часа, не более чем приблизительно 5 часов, не более чем приблизительно 6 часов или не более чем приблизительно 7 часов. В рамках настоящей заявки процедура культивирования в жидкой среде является относительно простой и удобной. В некоторых случаях бактерии в библиотеке бактериального дисплея могут подвергаться культивированию на чашке с малым количеством раствора с бактериями с последующим сбором колоний. Время культивирования культуры на чашке может составлять приблизительно 12-18 часов, например, оно может составлять приблизительно 12 часов, 13 часов, 14 часов, 15 часов, 16 часов, 17 часов или 18 часов. В рамках настоящей заявки культивирование на чашке может включать селекцию колоний (например, селекцию моноклонов) для секвенирования. В рамках настоящей заявки культивирование на чашке может уменьшать количество доминантных клонов.
[00183] В рамках настоящей заявки первый полинуклеотид и третий полинуклеотид могут быть получены из материала образца. В рамках настоящей заявки материал образца может включать материал образца, полученный из образца лимфоцитов периферической крови. В рамках настоящей заявки лимфоциты периферической крови могут представлять собой лимфоциты периферической крови человека. В рамках настоящей заявки материал образца также может происходить из любых других тканей и/или клеток и не ограничивается образцом лимфоцитов периферической крови.
[00184] В рамках настоящей заявки материал образца может включать тотальную РНК и/или мРНК, происходящую из образца.
[00185] В рамках настоящей заявки 8) может включать приведение библиотеки бактериального дисплея в контакт с фагом-помощником с получением фаговой библиотеки.
[00186] В другом аспекте настоящая заявка относится к фаговой библиотеке, полученной описанным способом.
[00187] В рамках настоящей заявки фаговая библиотека может включать по меньшей мере приблизительно 1010 (например, по меньшей мере приблизительно 1011, по меньшей мере приблизительно 1012, по меньшей мере приблизительно 1013, по меньшей мере приблизительно 1014, по меньшей мере приблизительно 1015, и по меньшей мере приблизительно 1016 или более) различных клонов. В рамках настоящей заявки фаговая библиотека может экспонировать по меньшей мере приблизительно 1010 (например, по меньшей мере приблизительно 1011, по меньшей мере приблизительно 1012, по меньшей мере приблизительно 1013, по меньшей мере приблизительно 1014, по меньшей мере приблизительно 1015, и по меньшей мере приблизительно 1016 или более) различных антител или фрагментов антител.
[00188] В другом аспекте настоящая заявка относится к способу скрининга антител или фрагментов антител, в том числе с использованием фаговой библиотеки.
[00189] Например, фрагмент антитела может представлять собой Fab. Например, антитела или фрагменты антител могут специфически связываться с TNFα, IL-6, IL6R, IL17-FR, CD38, GMCSF и Siglec3.
[00190] Без связи с какой-либо теорией, приведенные ниже примеры приводятся только для иллюстрации слитых белков, способов получения и применения в рамках настоящей заявки, и не предназначены для ограничения объема изобретения настоящей заявки.
ПРИМЕРЫ
Пример 1. Конструирование библиотеки фагового дисплея поверхностных антител (Fab) с PBMC человека
1.1 Получение тотальной РНК/мРНК из иммунологических материалов
[00191] Тотальную РНК экстрагировали из лимфоцитов периферической крови человека (PBMC), и из тотальной РНК выделяли мРНК (Takara, каталожный номер # Z652N/636592, см. инструкции к продукту для конкретных методик тестирования).
1.2 Конструирование праймера для синтеза
[00192] Ссылаясь на Phage Display (A Laboratory Mannual, ISBN 0-87969-546-3), конструировали праймеры для вариабельной области тяжелой цепи VH человека, легкой цепи KLC (полноразмерная легкая цепь каппа) и легкой цепи LLC (полноразмерная легкая цепь лямбда). Среди них, в легкой цепи 5'-конец прямого праймера включает нуклеотидную последовательность R1: GGCCCAGGCGGCC (SEQ ID NO: 1), 5'-конец обратного праймера включает нуклеотидную последовательность R2: GGCCACATAGGCC (SEQ ID NO: 2); в вариабельной области тяжелой цепи 5'-конец прямого праймера включает нуклеотидную последовательность R5: GGCCCAACCGGCC (SEQ ID NO: 3), и 5'-конец обратного праймера включает нуклеотидную последовательность R6: CGTCTCCTCAGC (SEQ ID NO: 4). Праймеры были синтезированы в Genewiz, Inc.
[00193] С использованием вектора pComb3x в качестве матрицы были сконструированы и амплифицированы прямые и обратные праймеры. Среди них, 5'-конец прямого праймера включает нуклеотидную последовательность R3: GGCCACATAGGCC (SEQ ID NO: 2), и 5'-конец обратного праймера включает нуклеотидную последовательность R4: GGCCCAACCGGCC (SEQ ID NO: 3).
[00194] Для конкретных последовательностей праймеров см. таблицу 1-1 ниже.
Таблица 1-1 последовательности праймеров - 1
1.3 Получение первого полинуклеотида и третьего полинуклеотида
[00195] Генную библиотеку составных антител амплифицировали за две стадии.
[00196] На первой стадии с использованием мРНК, полученной согласно примеру 1.1, в качестве матрицы синтезировали кДНК посредством обратной транскрипции MMLV от Promega (в соответствии с инструкциями для продукта от Promega, Inc., в котором праймер представлял собой Thermo, каталожный номер # N8080127, и обратная транскриптаза представляла собой Promega, каталожный номер # M1701).
[00197] На второй стадии с использованием кДНК, полученной на первой стадии, в качестве матрицы и с использованием праймера, полученного в примере 1.2, библиотеки генов KLC, LLC и VH составных антител амплифицировали посредством ПЦР (Takara, каталожный номер # RR900A, в соответствии с инструкциями к продукту от корпорации). После очистки для выделения посредством гель-электрофореза (осуществленной с использованием набора для выделения из геля от Axygen и в соответствии с описанием в Molecular Cloning Experimental Guidelines), получали продукты ПЦР - фрагмент KLC (т.е. первый полинуклеотид настоящей заявки), фрагмент LLC (первый полинуклеотид настоящей заявки) и фрагмент VH (третий полинуклеотид настоящей заявки), соответственно.
1.4 Конструирование вектора для хранения
1.4.1 Конструирование праймеров
[00198] Праймеры конструировали с использованием методик согласно примеру 1.2.
[00199] Конструировали и синтезировали праймеры для получения векторов для хранения. Для конкретных последовательностей праймеров см. таблицу 1-2 ниже.
Таблица 1-2 Последовательности праймеров - 2
1.4. Амплификация посредством ПЦР
[00200] С использованием Fc IgG1 длиной 1 т.п.н. (SEQ ID NO: 87) в качестве матрицы проводили ПЦР с праймерами, полученными согласно примеру 1.4.1, прямого праймера и обратного праймера R1-1 т.п.н.-R2. После очистки для выделения посредством гель-электрофореза (осуществленной с использованием набора для выделения из геля от Axygen) получали продукт ПЦР - R1-1 т.п.н.-R2 (SEQ ID NO: 88).
[00201] С использованием вектора pComb3x в качестве матрицы проводили ПЦР с прямым праймером (SEQ ID NO: 83) и обратным праймером (SEQ ID NO: 84) для R3-коннексон-R4. После очистки для выделения посредством гель-электрофореза (осуществленной с использованием набора для выделения из геля от Axygen) получали продукт ПЦР - R3-коннексон-R4 (SEQ ID NO: 90), тем самым получая второй полинуклеотид в рамках настоящей заявки. Среди них, коннексон может иметь длину 72 п.н., и нуклеотидная последовательность является такой, как показано в SEQ ID NO:89.
[00202] С использованием Fc IgG1 человека длиной 1 т.п.н. (SEQ ID NO: 87) в качестве матрицы проводили ПЦР с праймерами, полученными согласно примеру 1.4.1, прямым праймером и обратным праймером для R5-1 т.п.н.-R6. После очистки для выделения посредством гель-электрофореза получали продукт ПЦР - R5-1 т.п.н.-R6 (SEQ ID NO: 91).
1.4.3 Конструирование вектора для хранения легкой цепи и вектора для хранения тяжелой цепи
[00203] Посредством способа клонирования TA (набор для клонирования TA, приобретенный от Takara Inc.), фрагмент R1-1 т.п.н.-R2, полученный согласно 1.4.2, встраивали в вектор pMD19-T с получением вектора для хранения легкой цепи DDB-R1-1 т.п.н.-R2 для встраивания в библиотеку генов полноразмерных легких цепей. Карта вектора является такой, как показано на фиг.3.
[00204] Посредством способа клонирования TA (набор для клонирования TA, приобретенный от Takara Inc.) фрагмент R5-1 т.п.н.-R6, полученный согласно 1.4.2, встраивали в вектор pMD19-T с получением вектора, включающего фрагмент R5-1 т.п.н.-R6. Далее, с использованием этого вектора в качестве матрицы исходный участок расщепления BsmBI удаляли из этого вектора посредством праймера для внесения мутации с получением вектора для хранения тяжелой цепи DDB-R5-1 т.п.н.-R6 для встраивания в библиотеку генов VH. Карта вектора является такой, как показано на фиг.5.
[00205] Для конкретных последовательностей праймеров см. таблицу 1-3 ниже.
Таблица 1-3 Последовательности праймеров - 3
1.4.4 Конструирование вектора для хранения коннексона и получение бактерий с компонентом коннексона
[00206] С использованием способа клонирования TA (набор для клонирования TA, приобретенный от Takara Inc.), R3-коннексон-R4, полученный согласно примеру 1.4.2, встраивали в вектор pMD19-T с получением вектора для хранения коннексона DDB-R3-коннексон-R4. Карта вектора является такой, как показано на фиг.4. Вектор для хранения коннексона можно использовать для трансформации компетентных бактерий TG1 (Lucigen, Inc.), с последующим культивированием на чашке при 37°C в течение ночи. Колонии секвенировали, а затем собирали с получением бактерий с компонентом коннексона. Бактерии с компонентом коннексона можно криоконсервировать для применения.
1.5 Получение бактериальной библиотеки с компонентами
1.5.1 Получение бактериальной библиотеки с компонентом легкой цепи
[00207] Первый полинуклеотид (включая KLC и LLC), полученный согласно примеру 1.3, расщепляли эндонуклеазой рестрикции, распознающей R1 и R2, с получением целевого фрагмента легкой цепи (приблизительно 0,65 т.п.н.).
[00208] Вектор для хранения легкой цепи DDB-R1-1 т.п.н.-R2, полученный согласно примеру 1.4, расщепляли эндонуклеазой рестрикции R1 и R2. Получали фрагмент вектора для хранения легкой цепи (приблизительно 2,7 т.п.н.).
[00209] Полученный целевой фрагмент легкой цепи и фрагмент вектора для хранения легкой цепи смешивали и соединяли с использованием ДНК-лигазы T4 (приобретенной от NEB, Thermo) с получением соединенного продукта для хранения легкой цепи. Затем соединенный продукт для хранения легкой цепи использовали для трансформации компетентных бактерий TG1 (Lucigen, каталожный номер # 60502-2, использованный в соответствии с инструкциями изготовителя) путем культивирования на чашке с ампициллином (Thermo, каталожный номер #240845) при 37°C в течение ночи. Колонии отбирали для секвенирования, а затем колонии собирали с получением бактериальной библиотеки с компонентом легкой цепи. Бактериальная библиотека с компонентом легкой цепи может быть протестирована в отношении ее качества и/или бактериальная библиотека с компонентом легкой цепи может быть криоконсервирована для применения.
1.5.2 Получение бактериальной библиотеки с компонентом тяжелой цепи
[00210] Третий полинуклеотид, полученный согласно примеру 1.3, расщепляли эндонуклеазой рестрикции, распознающей R5 и R6, с получением целевого фрагмента вариабельной области тяжелой цепи (приблизительно 0,35 т.п.н.).
[00211] Вектор для хранения тяжелой цепи DDB-R5-1 т.п.н.-R6, полученный согласно примеру 1.4, расщепляли эндонуклеазой рестрикции, распознающей R5 и R6. Получали фрагмент вектора для хранения тяжелой цепи (приблизительно 2,7 т.п.н.).
[00212] Полученный целевой фрагмент вариабельной области тяжелой цепи и фрагмент вектора для хранения тяжелой цепи смешивали и соединяли посредством ДНК-лигазы T4 (приобретенной от NEB, Thermo) с получением соединенного продукта для хранения тяжелой цепи. Затем соединенный продукт для хранения тяжелой цепи использовали для трансформации компетентных бактерий TG1 (Lucigen, каталожный номер # 60502-2, использованный в соответствии с инструкциями производителя) путем культивирования в чашке с ампициллином (Thermo, каталожный номер #240845) при 37°C в течение ночи. Колонии отбирали для секвенирования, а затем все колонии собирали с получением бактериальной библиотеки с компонентом тяжелой цепи. Бактериальная библиотека с компонентом тяжелой цепи может быть протестирована в отношении ее качества и/или бактериальная библиотека с компонентом тяжелой цепи может быть криоконсервирована для применения.
1.6 Получение плазмиды с компонентом легкой цепи, плазмиды с компонентом тяжелой цепи и фрагмента коннексона
[00213] Плазмиды в бактериальной библиотеке с компонентом легкой цепи, полученной согласно примеру 1.5.1, и бактериальной библиотеке с компонентом тяжелой цепи, полученной согласно примеру 1.5.2, экстрагировали с использованием набора для экстракции плазмид (приобретенного от Axygen) с получением плазмиды с компонентом легкой цепи и плазмиды с компонентом тяжелой цепи, соответственно.
[00214] Плазмиду с компонентом легкой цепи, полученную согласно примеру 1.5.1, расщепляли эндонуклеазой рестрикции, распознающей R1 и R2, и очищали и выделяли посредством гель-электрофореза с получением фрагмента со вставкой легкой цепи LC.
[00215] Плазмиду с компонентом тяжелой цепи, полученную согласно примеру 1.5.2, расщепляли эндонуклеазой рестрикции, распознающей R5 и R6, и очищали и выделяли посредством гель-электрофореза с получением фрагмента с вставкой тяжелой цепи HC.
[00216] Плазмиды в бактериях с компонентом коннексона, полученные согласно примеру 1.4.4, экстрагировали с использованием набора для экстракции плазмид (приобретенного от Axygen) с получением плазмиды с компонентом коннексона. С использованием плазмиды с компонентом коннексона или вектора для хранения коннексона согласно примеру 1.4.4 в качестве матрицы амплифицировали содержащий коннексон фрагмент размером 0,8 т.п.н. с использованием прямого праймера (SEQ ID NO: 79) и обратного праймера (SEQ ID NO: 80) для коннексона, а затем продукт ПЦР размером 0,8 т.п.н. расщепляли эндонуклеазой рестрикции, распознающей R3 и R4, и очищали для выделения посредством гель-электрофореза (с выделением из геля малых фрагментов, LifeFeng Biotech, каталожный номер # DK402) с получением фрагмента коннексона размером 72 п.н.
1.7 Получение вектора для дисплея
[00217] Приобретали вектор pComb3x путем покупки. Карта вектора является такой, как показано на фиг.6, где карта модифицированного вектора pComb3x-fab для дисплея Fab антител является такой, как показано на фиг.7.
[00218] Участок расщепления SfiI на 3'-конце гена Fab в векторе pComb3x-fab удаляли посредством не смысловой мутации.
[00219] Затем участок расщепления для эндонуклеазы рестрикции, распознающей R2, добавляли ниже стоп-кодона легкой цепи в векторе с не смысловой мутацией, вносили участок расщепления эндонуклеазой рестрикции, распознающей R5, на конец сигнального пептида вариабельной области тяжелой цепи посредством не смысловой мутации, с получением модифицированного вектора для дисплея DDB-R1R2R5R6. Его карта является такой, как показано на фиг.8.
1.8 Получение библиотеки бактериального дисплея
[00220] Вектор для дисплея DDB-R1R2R5R6, полученный согласно примеру 1.7, расщепляли эндонуклеазой рестрикции, распознающей R7, и эндонуклеазой рестрикции, распознающей R8, с получением фрагмента вектора для дисплея размером 3,6 т.п.н.
[00221] Фрагмент вставки легкой цепи LC (0,65 т.п.н.), фрагмент вставки тяжелой цепи HC (0,35 т.п.н.), фрагмент коннексона (72 п.н.), и фрагмент вектора для дисплея (3,6 т.п.н.), полученный согласно примеру 1.6, смешивали в молекулярном соотношении 1:1:1:1 и соединяли посредством ДНК-лигазы T4 при 20°C в течение 20 часов или более с получением соединенного продукта для дисплея.
[00222] Соединенный продукт очищали с использованием PCR-Clean-up, трансформировали им компетентные бактерии TG1 (Lucigen, каталожный номер #60502-2, использованный в соответствии с инструкциями изготовителя), культивировали при встряхивании в свободной от антибиотиков культуральной среде 2YT при 37°C при 250 об/мин в течение 60 минут, и культивировали на чашке с ампициллином (Thermo, каталожный номер #240845) при 37°C для выращивания в течение ночи. Колонии отбирали для секвенирования. Все колонии, выращенные на чашке, собирали с получением библиотеки бактериального дисплея. Библиотеку бактериального дисплея можно хранить для применения.
1.9 Получение библиотеки фагового дисплея антител
[00223] Подходящее количество раствора с бактериями отбирали из библиотеки бактериального дисплея, полученной согласно примеру 1,8, и культивировали в культуральной среде 2YT (содержавшей 100 мкг/мл ампициллина и 2% глюкозу) при 37°C до достижения OD600 0,5. Затем в раствор с бактериями добавляли вспомогательный фаг M13KO7 (приобретенный от NEB, каталожный номер # N0315S, с MOI приблизительно 10-20), хорошо перемешивали и оставляли стоять при 37°C в течение 30 минут. Затем смесь встряхивали при 37°C, 250 об/мин в течение 30 минут и центрифугировали. Супернатант, содержавший вспомогательный фаг M13KO7, выбрасывали. Клетки ресуспендировали в 4x первоначальном объеме раствора с бактериями (содержавшего ампициллин и канамицин), и встряхивали при 30°C, 250 об/мин в течение ночи. На следующие сутки фаг собирали посредством преципитации с ПЭГ, титровали до требуемой концентрации фага и подвергали субупаковыванию с получением фаговой библиотеки для дисплея антител.
Пример 2 Анализ качества бактериальной библиотеки компонентов
[00224] Бактериальную библиотеку компонентов, полученную согласно примеру 1.5, анализировали в отношении ее качества. Результаты представлены в таблице 2.
[00225] При конструировании библиотеки бактериального дисплея соотношение бактериальной библиотеки с компонентом легкой цепи (KLC) и бактериальной библиотеки с компонентом легкой цепи (LLC) составляет 2:1. В соответствии с результатами качества, как описано выше, может быть оценено, что доля эффективного клонирования библиотеки бактериального дисплея достигнет 84% или более. Емкость библиотеки, полученной путем случайного комбинирования бактериальной библиотеки с компонентом легкой цепи и бактериальная библиотеки с компонентом тяжелой цепи теоретически, может достигнуть 1015.
Пример 3. Анализ качества библиотеки бактериального дисплея
[00226] 197 мкг выделенного и очищенного соединенного продукта для дисплея, полученного согласно примеру 1.8, использовали для трансформации 398 компетентных бактерий TG1 (Lucigen, каталожный номер # 60502-2, в соответствии с инструкциями производителя). Их культивировали при встряхивании в свободной от антибиотиков культуральной среде 2YT при 37°C при 250 об/мин в течение 60 минут, а затем культивировали в 197 чашках с ампициллином (Thermo, каталожный номер #240845) при 37°C в течение ночи. Все колонии собирали и подвергали субупаковыванию для хранения (с получением библиотеки бактериального дисплея, полученной согласно примеру 1.8). Емкость библиотеки бактериального дисплея составляет 1011, и фоновая доля клонирования составляет 2%.
[00227] Библиотеку бактериального дисплея анализировали в отношении ее качества, и результаты представлены в таблице 3:
[00228] Пригодные результаты секвенирования (всего 158 в двух партиях) в таблице 3 анализировали, из которых 131 клон имел правильные рамки считывания (включая клоны, включающие как правильные вариабельные области тяжелой цепи, так и легкие цепи). Можно видеть, что доля эффективного клонирования библиотеки бактериального дисплея достигает 83%, что согласуется с оцененными результатами примера 2.
Пример 4. Конструирование фаговой библиотеки дисплея поверхностных антител (Fab) с PBMC человека
4.1 Экстрагирование тотальной РНК из иммунологических материалов
[00229] Тотальную РНК экстрагировали из лимфоцитов периферической крови (приобретенных от MT Bio, каталожный номер # PB100C) (с использованием набора RNA mini kit от Qiagen, каталожный номер # 74101, см. инструкции набора для конкретных методик тестирования).
4.2 Конструирование праймера для синтеза
[00230] Ссылаясь на Phage Display (A Laboratory Mannual, ISBN 0-87969-546-3), конструировали праймеры для вариабельной области тяжелой цепи VH человека, легкой цепи KLC (полноразмерная легкая цепь каппа) и легкой цепи LLC (полноразмерная легкая цепь лямбда). Среди них, в легкой цепи 5'-конец прямого праймера включает нуклеотидную последовательность R1: GGCCCAGGCGGCC (SEQ ID NO: 1), 5'-конец обратного праймера включает нуклеотидную последовательность R2: GGCCACATAGGCC (SEQ ID NO: 2); в вариабельной области тяжелой цепи 5'-конец прямого праймера включает нуклеотидную последовательность R5: GGCCCAACCGGCC (SEQ ID NO: 3), 5'-конец обратного праймера включает нуклеотидную последовательность R6: CGTCTCCTCAGC (SEQ ID NO: 4). Праймеры были синтезированы в Genewiz, Inc.
[00231] С использованием вектора pComb3x в качестве матрицы были сконструированы и амплифицированы прямые и обратные праймеры для коннексона. Среди них, 5'-конец прямого праймера включает нуклеотидную последовательность R3: GGCCACATAGGCC (SEQ ID NO: 2), и 5'-конец обратного праймера включает нуклеотидную последовательность R4: GGCCCAACCGGCC (SEQ ID NO: 3).
[00232] Для конкретных последовательностей праймеров см. таблицу 4-1 ниже.
4.3 Получение первого полинуклеотида и третьего полинуклеотида
[00233] Генную библиотеку составных антител амплифицировали за две стадии.
[00234] На первой стадии с использованием мРНК, полученной согласно примеру 4.1, в качестве матрицы синтезировали кДНК посредством обратной транскрипции MMLV от Promega (в соответствии с инструкциями для продукта от Promega, Inc., в котором праймер представлял собой Thermo, каталожный номер # N8080127, и обратная транскриптаза представляла собой Promega, каталожный номер # M1701).
[00235] На второй стадии с использованием кДНК, полученной на первой стадии, в качестве матрицы и с использованием праймера, полученного в примере 4.2, библиотеки генов KLC, LLC и VH составных антител амплифицировали посредством ПЦР (Takara, каталожный номер # RR900A, в соответствии с инструкциями к продукту от корпорации). После очистки для выделения посредством гель-электрофореза (осуществленной с использованием набора для выделения из геля от Axygen и в соответствии с описанием в Molecular Cloning Experimental Guidelines), получали продукты ПЦР - фрагмент KLC (т.е. первый полинуклеотид настоящей заявки), фрагмент LLC (первый полинуклеотид настоящей заявки) и фрагмент VH (третий полинуклеотид настоящей заявки), соответственно.
4.4 Конструирование вектора для хранения
4.4.1 Конструирование праймеров
[00236] Конструировали и синтезировали праймеры для получения векторов для хранения. Для конкретных последовательностей праймеров см. таблицу 4-2 ниже.
4.4.2 Амплификация посредством ПЦР
[00237] С использованием Fc IgG1 длиной 1 т.п.н. (SEQ ID NO: 87) в качестве матрицы проводили ПЦР с праймерами, полученными согласно примеру 4.4.1, прямого праймера и обратного праймера для R1-1 т.п.н.-R2. После очистки для выделения посредством гель-электрофореза (осуществленной с использованием набора для выделения из геля от Axygen) получали продукт ПЦР - R1-1 т.п.н.-R2 (SEQ ID NO: 88).
[00238] С использованием вектора pComb3x в качестве матрицы проводили ПЦР с прямым праймером для R3-коннексон-R4 (SEQ ID NO: 83) и обратным праймером для R3-коннексон-R4 (SEQ ID NO: 84). После очистки для выделения посредством гель-электрофореза (осуществленной с использованием набора для выделения из геля от Axygen) получали продукт ПЦР - R3-коннексон-R4 (SEQ ID NO: 95), тем самым получая второй полинуклеотид в рамках настоящей заявки. Среди них, коннексон может иметь длину 90 п.н., и нуклеотидная последовательность является такой, как показано в SEQ ID NO:89.
[00239] С использованием Fc IgG1 человека длиной 1 т.п.н. (SEQ ID NO: 87) в качестве матрицы проводили ПЦР с праймерами, полученными согласно примеру 4.4.1, прямым праймером и обратным праймером для R5-1 т.п.н.-R6. После очистки для выделения посредством гель-электрофореза получали продукт ПЦР - R5-1 т.п.н.-R6 (SEQ ID NO: 91).
4.4.3 Конструирование вектора для хранения легкой цепи и вектора для хранения тяжелой цепи
[00240] Посредством способа клонирования TA (набор для клонирования TA, приобретенный от Takara Inc.), фрагмент R1-1 т.п.н.-R2, полученный согласно 4.4.2, встраивали в вектор pMD19-T с получением вектора для хранения легкой цепи DDB-R1-1 т.п.н.-R2 для встраивания в библиотеку генов полноразмерных легких цепей. Карта вектора является такой, как показано на фиг.3.
[00241] Посредством способа клонирования TA (набор для клонирования TA, приобретенный от Takara Inc.) фрагмент R5-1 т.п.н.-R6, полученный согласно 4.4.2, встраивали в вектор pMD19-T с получением вектора, включающего фрагмент R5-1 т.п.н.-R6. Далее, с использованием этого вектора в качестве матрицы исходный участок расщепления BsmBI удаляли из этого вектора посредством праймера для внесения мутации с получением вектора для хранения тяжелой цепи DDB-R5-1 т.п.н.-R6 для встраивания в библиотеку генов VH. Карта вектора является такой, как показано на фиг.5.
[00242] Для конкретных последовательностей праймеров см. таблицу 4-3 ниже.
4.4.4 Конструирование вектора для хранения коннексона и получение бактерий с компонентом коннексона
[00243] С использованием способа клонирования TA (набор для клонирования TA, приобретенный от Takara Inc.), R3-коннексон-R4, полученный согласно примеру 4.4.2, встраивали в вектор pMD19-T с получением вектора для хранения коннексона DDB-R3-коннексон-R4. Карта вектора является такой, как показано на фиг.4. Вектор для хранения коннексона можно использовать для трансформации компетентных бактерий TG1 (Lucigen, Inc.), с последующим культивированием на чашке при 37°C в течение ночи. Колонии секвенировали, а затем собирали с получением бактерий с компонентом коннексона. Бактерии с компонентом коннексона можно криоконсервировать для применения.
4.5 Получение бактериальной библиотеки с компонентами
4.5.1 Получение бактериальной библиотеки с компонентом легкой цепи
[00244] Первый полинуклеотид (включая KLC и LLC), полученный согласно примеру 4.3, расщепляли эндонуклеазой рестрикции, распознающей R1 и R2, с получением целевого фрагмента легкой цепи (приблизительно 0,65 т.п.н.).
[00245] Вектор для хранения легкой цепи DDB-R1-1 т.п.н.-R2, полученный согласно примеру 4.4, расщепляли эндонуклеазой рестрикции R1 и R2. Получали фрагмент вектора для хранения легкой цепи (приблизительно 2,7 т.п.н.).
[00246] Полученный целевой фрагмент легкой цепи и фрагмент вектора для хранения легкой цепи смешивали и соединяли с использованием ДНК-лигазы T4 (приобретенной от NEB, Thermo) с получением соединенного продукта для хранения легкой цепи. Затем соединенный продукт для хранения легкой цепи использовали для трансформации компетентных бактерий TG1 (Lucigen, каталожный номер # 60502-2, использованный в соответствии с инструкциями изготовителя) путем культивирования на чашке с ампициллином (Thermo, каталожный номер #240845) при 37°C в течение ночи. Колонии отбирали для секвенирования, а затем колонии собирали с получением бактериальной библиотеки с компонентом легкой цепи. Бактериальная библиотека с компонентом легкой цепи может быть протестирована в отношении ее качества и/или бактериальная библиотека с компонентом легкой цепи может быть криоконсервирована для применения.
4.5.2 Получение бактериальной библиотеки с компонентом тяжелой цепи
[00247] Третий полинуклеотид, полученный согласно примеру 4.3, расщепляли эндонуклеазой рестрикции, распознающей R5 и R6, с получением целевого фрагмента вариабельной области тяжелой цепи (приблизительно 0,35 т.п.н.).
[00248] Вектор для хранения тяжелой цепи DDB-R5-1 т.п.н.-R6, полученный согласно примеру 4.4, расщепляли эндонуклеазой рестрикции, распознающей R5 и R6. Получали фрагмент вектора для хранения тяжелой цепи (приблизительно 2,7 т.п.н.).
[00249] Полученный целевой фрагмент вариабельной области тяжелой цепи и фрагмент вектора для хранения тяжелой цепи смешивали и соединяли посредством ДНК-лигазы T4 (приобретенной от NEB, Thermo) с получением соединенного продукта для хранения тяжелой цепи, а затем соединенный продукт для хранения тяжелой цепи использовали для трансформации компетентных бактерий TG1 (Lucigen, каталожный номер # 60502-2, использованный в соответствии с инструкциями производителя). Смесь культивировали при встряхивании в свободной от антибиотиков культуральной среде 2YT при 37°C, 250 об/мин в течение 60 минут, а затем культивировали на чашке с ампициллином (Thermo, каталожный номер #240845) при 37°C в течение ночи. Колонии отбирали для секвенирования, а затем все колонии собирали с получением бактериальной библиотеки с компонентом тяжелой цепи. Бактериальная библиотека с компонентом тяжелой цепи может быть протестирована в отношении ее качества и/или бактериальная библиотека с компонентом тяжелой цепи может быть криоконсервирована для применения.
4.6 Получение плазмиды с компонентом легкой цепи, плазмиды с компонентом тяжелой цепи и фрагмента коннексона
[00250] Плазмиды в бактериальной библиотеке с компонентом легкой цепи, полученной согласно примеру 4.5.1, и бактериальной библиотеке с компонентом тяжелой цепи, полученной согласно примеру 4.5.2, экстрагировали с использованием набора для экстракции плазмид (приобретенного от Axygen) с получением плазмиды с компонентом легкой цепи и плазмиды с компонентом тяжелой цепи, соответственно.
[00251] Плазмиду с компонентом легкой цепи, полученную согласно примеру 4.5.1, расщепляли эндонуклеазой рестрикции, распознающей R1 и R2, и очищали и выделяли посредством гель-электрофореза с получением фрагмента со вставкой легкой цепи LC.
[00252] Плазмиду с компонентом тяжелой цепи, полученную согласно примеру 4.5.2, расщепляли эндонуклеазой рестрикции, распознающей R5 и R6, и очищали и выделяли посредством гель-электрофореза с получением фрагмента с вставкой тяжелой цепи HC.
[00253] Плазмиды в бактериях с компонентом коннексона, полученные согласно примеру 4.4.4, экстрагировали с использованием набора для экстракции плазмид (приобретенного от Axygen) с получением плазмиды с компонентом коннексона. С использованием плазмиды с компонентом коннексона или вектора для хранения коннексона согласно примеру 4.4.4 в качестве матрицы амплифицировали содержащий коннексон фрагмент размером 0,8 т.п.н. с использованием прямого праймера (SEQ ID NO.96 или SEQ ID NO.97) и обратного праймера (SEQ ID NO: 98) для коннексона, а затем продукт ПЦР размером 0,8 т.п.н. расщепляли эндонуклеазой рестрикции, распознающей R3 и R4, и очищали для выделения посредством гель-электрофореза (с выделением из геля малых фрагментов, LifeFeng Biotech, каталожный номер # DK402) с получением фрагмента коннексона размером 90 п.н.
4.7 Получение вектора для дисплея
[00254] Приобретали вектор pComb3x путем покупки. Карта вектора является такой, как показано на фиг.6, где карта модифицированного вектора pComb3x-fab для дисплея Fab антител является такой, как показано на фиг.7.
[00255] Участок расщепления SfiI на 3'-конце гена Fab в векторе pComb3x-fab удаляли посредством не смысловой мутации.
[00256] Затем участок расщепления для эндонуклеазы рестрикции, распознающей R2, добавляли ниже стоп-кодона легкой цепи в векторе с не смысловой мутацией, вносили участок расщепления эндонуклеазой рестрикции, распознающей R5, на конец сигнального пептида вариабельной области тяжелой цепи посредством не смысловой мутации, с получением модифицированного вектора для дисплея DDB-R1R2R5R6. Его карта является такой, как показано на фиг.8.
4.8 Получение библиотеки бактериального дисплея
[00257] Вектор для дисплея DDB-R1R2R5R6, полученный согласно примеру 4.7, расщепляли эндонуклеазой рестрикции, распознающей R7, и эндонуклеазой рестрикции, распознающей R8, с получением фрагмента вектора для дисплея размером 3,6 т.п.н.
[00258] Фрагмент вставки легкой цепи LC (0,65 т.п.н.), фрагмент вставки тяжелой цепи HC (0,35 т.п.н.), фрагмент коннексона (90 п.н.) и фрагмент вектора для дисплея (3,6 т.п.н.), полученный согласно примеру 4.6, смешивали в молекулярном соотношении 1:1:1:1 и соединяли посредством ДНК-лигазы T4 при 20°C в течение 20 часов или более с получением соединенного продукта для дисплея.
[00259] Соединенный продукт очищали с использованием PCR-Clean-up, трансформировали им компетентные бактерии TG1 (Lucigen, каталожный номер #60502-2, использованный в соответствии с инструкциями изготовителя), культивировали при встряхивании в свободной от антибиотиков культуральной среде 2YT при 37°C при 250 об/мин в течение 60 минут, а затем подвергали экспансии в соотношении 1:20 в содержавшей ампициллин культуральной среде 2YT при 37°C, 250 об/мин в течение 4,5 часов. Бактерии собирали с использованием центрифуги для криоконсервации, т.е. библиотеки бактериального дисплея. Перед экспансией культуры с ампициллином небольшое количество раствора с бактериями отбирали в культуру в чашке при 37°C в течение ночи. Колонии отбирали для секвенирования. Библиотеку бактериального дисплея можно хранить для применения.
4.9 Получение библиотеки фагового дисплея антител
[00260] Подходящее количество раствора с бактериями отбирали из библиотеки бактериального дисплея, полученной согласно примеру 4,8, и культивировали в культуральной среде 2YT (содержавшей 100 мкг/мл ампициллина и 2% глюкозу) при 37°C до достижения OD600 0,5. Затем в раствор с бактериями добавляли вспомогательный фаг M13KO7 (приобретенный от NEB, каталожный номер # N0315S, с MOI приблизительно 10-20), хорошо перемешивали и оставляли стоять при 37°C в течение 30 минут. Затем смесь встряхивали при 37°C, 250 об/мин в течение 30 минут и центрифугировали. Супернатант, содержавший вспомогательный фаг M13KO7, выбрасывали. Клетки ресуспендировали в 4x первоначальном объеме раствора с бактериями (содержавшего ампициллин и канамицин) и встряхивали при 30°C, 250 об/мин в течение ночи. На следующие сутки фаг собирали посредством преципитации с ПЭГ, титровали до требуемой концентрации фага и подвергали субупаковыванию с получением фаговой библиотеки для дисплея антител.
Пример 5 Анализ качества бактериальной библиотеки компонентов
[00261] Бактериальную библиотеку компонентов, полученную согласно примеру 4.5, анализировали в отношении ее качества. Результаты представлены в таблице 5.
[00262] При конструировании библиотеки бактериального дисплея соотношение бактериальной библиотеки с компонентом легкой цепи (KLC) и бактериальной библиотеки с компонентом легкой цепи (LLC) составляет 2:1. В соответствии с приведенными выше результатами качества, может быть оценено, что доля эффективного клонирования библиотеки бактериального дисплея достигнет 82%. Емкость библиотеки, полученной путем случайного комбинирования бактериальной библиотеки с компонентом легкой цепи и бактериальной библиотеки с компонентом тяжелой цепи, теоретически может достигнуть 1018.
Пример 6. Анализ качества библиотеки бактериального дисплея
[00263] 16 мкг выделенного и очищенного соединенного продукта для дисплея, полученного согласно примеру 4.8, использовали для трансформации 25 компетентных бактерий TG1. После амплификации культуры бактерии собирали посредством центрифуги с получением всего 2,4×1012 бактерий, которые подвергали субупаковыванию для хранения (с получением библиотеки бактериального дисплея, полученной согласно примеру 4.8). Емкость библиотеки бактериального дисплея составляет 4×1010, доля фонового клонирования составляет 3%, и эффективность конверсии на мкг соединенного продукта достигает 2,5×109.
[00264] Качество библиотеки бактериального дисплея анализировали двумя способами:
[00265] Первый способ: 90 колоний отбирали случайным образом для секвенирования. Результаты показывают, что 56 колоний имели правильные рамки считывания и не имели повторяющихся последовательностей. Доля клонирования правильных рамок считывания составляла 62,2%.
[00266] Второй способ: чашку покрывали антителом против Fab человека и определяли экспрессию Fab в выше чем 90 клонах посредством ELISA. Результаты показывают, что результаты для двух отрицательных контролей составляли 0,07 и 0,082, соответственно. Всего был 71 клон с результатами выше 0,1, которые были определены как экспрессирующие Fab. Доля положительных клонов составляет 79%.
[00267] 34 клона с ошибками последовательности анализировали, где 18 имели экспрессию Fab, что составляет 20% от 90 клонов. Клоны с правильным секвенированием и клоны с экспрессией, но неправильным секвенированием, суммировали (56+18), и доля эффективных клонов достигала 82,2% (74/90), что соответствовало оцененным результатам в примере 5.
Пример 7 Анализ качества библиотеки фагового дисплея антител
[00268] Фаговую библиотеку дисплея антител согласно примеру 4.9 преципитировали с ПЭГ. После преципитации посредством центрифугирования клетки ресуспендировали в 280 мл PBS. Добавляли 60 мл глицерина. Смесь хорошо перемешивали и подвергали субупаковыванию для криоконсервации. Криоконсервированную библиотеку фагового дисплея антител анализировали в отношении титра фага библиотеки. Результаты показывают, что титр фаговой библиотеки составляет 2×1013/мл, общее количество фагов в библиотеке составляет вплоть до 6,8×1015, и 100 мкл включают количество фагов, соответствующее 50-кратной емкости библиотеки.
[00269] Для анализа того, можно ли проводить скрининг антиген-специфических антител из сконструированной фаговой библиотеки, выбирали три секретируемых цитокина (IL6, IL17A/F, GMCSF) и три белка клеточной мембраны (IL6R, EGFR, Siglec3), и для каждого антигена проводили скрининг с использованием 100 мкл фаговой библиотеки.
[00270] Антиген-специфический Fab подвергали скринингу способом скрининга в жидкой фазе с использованием адсорбции на связанные с антигеном меченные авидином магнитные гранулы. Вышеупомянутые шесть типов меченных биотином антигенов приобретали от AcroBiosystems (последовательные каталожные номера продуктов представляют собой # IL6-H8218, ILF-H82W1, GMF-H8214, CD6-H82E8, EGR-H82E3 и CD3-H82E7), и меченные авидином магнитные гранулы приобретали от Thermo (каталожный номер #11206D). Концентрации антигена в трех раундах скрининга в жидкой фазе составляли 5 мкг/мл, 3 мкг/мл и 1 мкг/мл, соответственно. Фаги, собранные посредством элюирования в первом раунде скрининга, амплифицировали для применения во втором раунде скрининга. Фаги, собранные посредством элюирования во втором раунде скрининга, прямо использовали в третьем раунде скрининга без амплификации.
[00271] Фаги собирали посредством элюирования в третьем раунде скрининга инфицированных бактерий TG1, и проводили культивирование на покрытых ампициллином чашках при 37°C в течение ночи. На следующие сутки 96 моноклонов отбирали из каждой антиген-специфической чашки для культивирования и амплификации (культуральная среда в чашке включала 2YT, 100 мкг/мл ампициллина и 0,2% глюкозу) до достижения OD600 0,6, а затем добавляли IPTG (до конечной концентрации 1 мМ) для индукции в течение ночи. На 3 сутки антиген-специфический Fab в культуральной среде анализировали посредством ELISA. Результаты представлены в таблице 6.
[00272] В таблице 6 клоны, чей результат ELISA более чем в два раза превышает минимальный результат ELISA, приведены в качестве положительных клонов.
[00273] Результаты в таблице 6 указывают на то, что является возможным скрининг и получение Fab, специфичных к IL-6, IL6R, IL17-FR, GMCSF, EGFR и Siglec3, из фаговой библиотеки антител, и доля положительных клонов может достигать приблизительно 25-87%.
[00274] Из каждой группы антигенспецифические положительные клоны с наилучшими 12 результатами ELISA отбирали для анализа с использованием секвенирования для сопоставления аминокислотных последовательностей 12 клонов в каждой группе. Результаты представлены в таблице 7. Результаты в таблице 7 указывают на то, что каждая группа имеет множество уникальных аминокислотных последовательностей.
[00275] Вышеуказанное подробное описание приведено в качестве иллюстрации и примера, и подразумевается, что оно не ограничивает объем прилагаемой формулы изобретения. Различные модификации вариантов осуществления, приведенные в рамках настоящей заявки, являются очевидными специалистам в данной области и охватываются объемом прилагаемых пунктов формулы изобретения и их эквивалентов.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> DDBio.Co,Ltd.,(Shang Hai)
<120> СПОСОБ ПОЛУЧЕНИЯ ФАГОВОЙ БИБЛИОТЕКИ
<130> 0112-PA-004RU
<160> 100
<170> PatentIn version 3.5
<210> 1
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R1/R8
<400> 1
ggcccaggcg gcc 13
<210> 2
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R2/R3
<400> 2
ggccacatag gcc 13
<210> 3
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R4/R5
<400> 3
ggcccaaccg gcc 13
<210> 4
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R6/R7
<400> 4
ggccctcagc ggcc 14
<210> 5
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 5
tcgctaccgt ggcccaggcg gccgacatcc agatgaccca gtctcc 46
<210> 6
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 6
tcgctaccgt ggcccaggcg gccgatattg tgatgaccca gactccac 48
<210> 7
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 7
tcgctaccgt ggcccaggcg gccgccatcc agttgaccca gtctcc 46
<210> 8
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 8
tcgctaccgt ggcccaggcg gccgccatcc ggatgaccca gtctcc 46
<210> 9
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 9
tcgctaccgt ggcccaggcg gccgatgttg tgatgactca gtctccac 48
<210> 10
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 10
tcgctaccgt ggcccaggcg gccgatattg tgatgactca gtctccac 48
<210> 11
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 11
tcgctaccgt ggcccaggcg gccgaaattg tgttgacgca gtctccag 48
<210> 12
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 12
tcgctaccgt ggcccaggcg gccgaaatag tgatgacgca gtctccag 48
<210> 13
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 13
tcgctaccgt ggcccaggcg gccgaaattg tgttgacaca gtctccag 48
<210> 14
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 14
tcgctaccgt ggcccaggcg gccgaaattg tgctgactca gtctcc 46
<210> 15
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 15
tcgctaccgt ggcccaggcg gccaacatcc agatgaccca gtctcc 46
<210> 16
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 16
tcgctaccgt ggcccaggcg gccgacatcc agttgaccca gtctcc 46
<210> 17
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 17
tcgctaccgt ggcccaggcg gccgtcatct ggatgaccca gtctcc 46
<210> 18
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 18
tcgctaccgt ggcccaggcg gccgccatcc agatgaccca gtctcc 46
<210> 19
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 19
tcgctaccgt ggcccaggcg gccgaaattg taatgacaca gtctccagc 49
<210> 20
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 20
tcgctaccgt ggcccaggcg gccgacatcg tgatgaccca gtctcc 46
<210> 21
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 21
tcgctaccgt ggcccaggcg gccgaaacga cactcacgca gtctcc 46
<210> 22
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 22
tcgctaccgt ggcccaggcg gccgatgttg tgatgacaca gtctccag 48
<210> 23
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для легкой цепи KLC
<400> 23
taaattcctc ggcctatgtg gcctattaac actctcccct gttgaagctc t 51
<210> 24
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 24
tcgctaccgt ggcccaggcg gcccagtctg tgctgactca gccacc 46
<210> 25
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 25
tcgctaccgt ggcccaggcg gcccagtctg ccctgactca gcctc 45
<210> 26
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 26
tcgctaccgt ggcccaggcg gcccagtctg ccctgactca gcctg 45
<210> 27
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 27
tcgctaccgt ggcccaggcg gcctcctatg agctgacaca gccacc 46
<210> 28
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 28
tcgctaccgt ggcccaggcg gcctcctatg agctgacaca gccatc 46
<210> 29
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 29
tcgctaccgt ggcccaggcg gcctcctatg agctgactca gccacc 46
<210> 30
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 30
tcgctaccgt ggcccaggcg gcctcctatg agctgactca gccactc 47
<210> 31
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 31
tcgctaccgt ggcccaggcg gcccagtctg tgctgacgca gccgcc 46
<210> 32
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 32
tcgctaccgt ggcccaggcg gcccagtctg tgttgacgca gccgc 45
<210> 33
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 33
tcgctaccgt ggcccaggcg gcctcttctg agctgactca ggaccc 46
<210> 34
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 34
tcgctaccgt ggcccaggcg gcctcctatg tgctgactca gccacc 46
<210> 35
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 35
tcgctaccgt ggcccaggcg gcctcctatg agctgacaca gctacc 46
<210> 36
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 36
tcgctaccgt ggcccaggcg gcctcctatg agctgatgca gccacc 46
<210> 37
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 37
tcgctaccgt ggcccaggcg gccctgcctg tgctgactca gcccc 45
<210> 38
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 38
tcgctaccgt ggcccaggcg gcccagcctg tgctgactca atcatcc 47
<210> 39
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 39
tcgctaccgt ggcccaggcg gcccagcttg tgctgactca atcgcc 46
<210> 40
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 40
tcgctaccgt ggcccaggcg gcccagcctg tgctgactca gccacc 46
<210> 41
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 41
tcgctaccgt ggcccaggcg gcccaggctg tgctgactca gccgg 45
<210> 42
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 42
Thr Cys Gly Cys Thr Ala Cys Cys Gly Thr Gly Gly Cys Cys Cys Ala
1 5 10 15
Gly Gly Cys Gly Gly Cys Cys Cys Ala Gly Cys Cys Thr Gly Thr Gly
20 25 30
Cys Thr Gly Ala Cys Thr Cys Ala Gly Cys Cys Ala Thr Cys
35 40 45
<210> 43
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 43
tcgctaccgt ggcccaggcg gccaatttta tgctgactca gccccac 47
<210> 44
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 44
tcgctaccgt ggcccaggcg gcccagactg tggtgactca ggagcc 46
<210> 45
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 45
tcgctaccgt ggcccaggcg gcccaggctg tggtgactca ggagc 45
<210> 46
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 46
tcgctaccgt ggcccaggcg gcccagactg tggtgaccca ggagc 45
<210> 47
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 47
tcgctaccgt ggcccaggcg gcccagcctg tgctgactca gccac 45
<210> 48
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 48
tcgctaccgt ggcccaggcg gcccaggcag ggctgactca gccac 45
<210> 49
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для легкой цепи LLC
<400> 49
taaattcctc ggcctatgtg gcctattatg aacattctgt aggggccact g 51
<210> 50
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 50
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagtct g 51
<210> 51
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 51
tattacttgc ggcccaaccg gccatggccc aggtgcagct gcaggagtcg g 51
<210> 52
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 52
tattacttgc ggcccaaccg gccatggccc aggtgcagct ggtgcagtct g 51
<210> 53
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 53
tattacttgc ggcccaaccg gccatggccc aggtgcagct ggtggagtct g 51
<210> 54
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 54
tattacttgc ggcccaaccg gccatggccc aggtcacctt gaaggagtct gg 52
<210> 55
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 55
tattacttgc ggcccaaccg gccatggccg aagtgcagct ggtggagtct gg 52
<210> 56
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 56
tattacttgc ggcccaaccg gccatggccc aggtccagct tgtgcagtct gg 52
<210> 57
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 57
tattacttgc ggcccaaccg gccatggccc aggttcagct ggtgcagtct gg 52
<210> 58
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 58
tattacttgc ggcccaaccg gccatggccc aggtccagct ggtacagtct gg 52
<210> 59
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 59
tattacttgc ggcccaaccg gccatggccc agatgcagct ggtgcagtct gg 52
<210> 60
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 60
tattacttgc ggcccaaccg gccatggccc aaatgcagct ggtgcagtct gg 52
<210> 61
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 61
tattacttgc ggcccaaccg gccatggccg aggtccagct ggtacagtct gg 52
<210> 62
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 62
tattacttgc ggcccaaccg gccatggccc agatcacctt gaaggagtct gg 52
<210> 63
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 63
tattacttgc ggcccaaccg gccatggccg aggtgcagct gttggagtct gg 52
<210> 64
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 64
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagact g 51
<210> 65
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 65
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagtcc gg 52
<210> 66
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 66
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagtct cg 52
<210> 67
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 67
tattacttgc ggcccaaccg gccatggccc agctgcagct gcaggagtcc gg 52
<210> 68
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 68
tattacttgc ggcccaaccg gccatggccc aggtgcagct acagcagtgg gg 52
<210> 69
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 69
tattacttgc ggcccaaccg gccatggccc agctgcagct gcaggagtcg gg 52
<210> 70
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 70
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtgcagtct gg 52
<210> 71
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 71
tattacttgc ggcccaaccg gccatggccg aagtgcagct ggtgcagtct gg 52
<210> 72
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 72
tattacttgc ggcccaaccg gccatggccc aggtacagct gcagcagtca gg 52
<210> 73
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 73
tattacttgc ggcccaaccg gccatggccc aggtgcagct ggtgcaatct gg 52
<210> 74
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 74
cttggtggag gctgaggaga cggtgaccag ggtgccctgg cc 42
<210> 75
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 75
cttggtggag gctgaggaga cggtgaccag ggtgccacgg cc 42
<210> 76
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 76
cttggtggag gctgaggaga cggtgaccat tgtcccttgg cc 42
<210> 77
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 77
cttggtggag gctgaggaga cggtgaccag ggttccctgg cc 42
<210> 78
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 78
cttggtggag gctgaggaga cggtgaccgt ggtcccttgg cc 42
<210> 79
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для коннексон
<400> 79
acagcttgtc tgtaagcgga tg 22
<210> 80
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для коннексон
<400> 80
tgtggataac cgtattaccg cc 22
<210> 81
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для R1-1 т.п.н.-R2
<400> 81
ttagcgaatt ccagacatct aatggcccag gcggcctcag ctagcaccaa gggcccatc 59
<210> 82
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для R1-1 т.п.н.-R2
<400> 82
cgggtgatca ccagtcttct attggcctat gtggccttat catttacccg gagacaggg 59
<210> 83
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для R3-коннексон-R4
<400> 83
taggccacat aggccgagga atttaaaatg aaatacctat tgcctacggc agccg 55
<210> 84
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для R3-коннексон-R4
<400> 84
atggccggtt gggccgcaag taataacaat ccagcggctg ccgtaggcaa tagg 54
<210> 85
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для R5-1 т.п.н.-R6
<400> 85
ttagcgaatt ccagacatct aatggcccaa ccggcctcag ctagcaccaa gggcccatc 59
<210> 86
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для R5-1 т.п.н.-R6
<400> 86
tgatcaccag tcttctattg gctgaggaga cgttatcatt tacccggaga c 51
<210> 87
<211> 987
<212> ДНК
<213> Homo sapiens
<400> 87
tcagctagca ccaagggccc atcggtcttc cccctggcgc cctgctccag gagcacctcc 60
gagagcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg 120
tcgtggaact caggcgctct gaccagcggc gtgcacacct tcccagctgt cctacagtcc 180
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcaactt cggcacccag 240
acctacacct gcaacgtaga tcacaagccc agcaacacca aggtggacaa gacagttgag 300
cgcaaatgtt gtgtcgagtg cccaccgtgc ccagcaccac ctgtggcagg accgtcagtc 360
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcacg 420
tgcgtggtgg tggacgtgag ccacgaagac cccgaggtcc agttcaactg gtacgtggac 480
ggcgtggagg tgcataatgc caagacaaag ccacgggagg agcagttcaa cagcacgttc 540
cgtgtggtca gcgtcctcac cgttgtgcac caggactggc tgaacggcaa ggagtacaag 600
tgcaaggtct ccaacaaagg cctcccagcc cccatcgaga aaaccatctc caaaaccaaa 660
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggagga gatgaccaag 720
aaccaggtca gcctgacctg cctggtcaaa ggcttctacc ccagcgacat cgccgtggag 780
tgggagagca atgggcagcc ggagaacaac tacaagacca cacctcccat gctggactcc 840
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg 900
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc 960
ctctccctgt ctccgggtaa atgataa 987
<210> 88
<211> 1013
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R1-1 т.п.н.-R2
<400> 88
ggcccaggcg gcctcagcta gcaccaaggg cccatcggtc ttccccctgg cgccctgctc 60
caggagcacc tccgagagca cagcggccct gggctgcctg gtcaaggact acttccccga 120
accggtgacg gtgtcgtgga actcaggcgc tctgaccagc ggcgtgcaca ccttcccagc 180
tgtcctacag tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcaa 240
cttcggcacc cagacctaca cctgcaacgt agatcacaag cccagcaaca ccaaggtgga 300
caagacagtt gagcgcaaat gttgtgtcga gtgcccaccg tgcccagcac cacctgtggc 360
aggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac 420
ccctgaggtc acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tccagttcaa 480
ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccacggg aggagcagtt 540
caacagcacg ttccgtgtgg tcagcgtcct caccgttgtg caccaggact ggctgaacgg 600
caaggagtac aagtgcaagg tctccaacaa aggcctccca gcccccatcg agaaaaccat 660
ctccaaaacc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga 720
ggagatgacc aagaaccagg tcagcctgac ctgcctggtc aaaggcttct accccagcga 780
catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacacctcc 840
catgctggac tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag 900
gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta 960
cacgcagaag agcctctccc tgtctccggg taaatgataa ggccacatag gcc 1013
<210> 89
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Коннексон длиной 72 п.н.
<400> 89
aggccgagga atttaaaatg aaatacctat tgcctacggc agccgctgga ttgttattac 60
ttgcggccca ac 72
<210> 90
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R3-коннексон-R4 (72 п.н.)
<400> 90
ggccacatag gccgaggaat ttaaaatgaa atacctattg cctacggcag ccgctggatt 60
gttattactt gcggcccaac cggcc 85
<210> 91
<211> 1012
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R5-1 т.п.н.-R6
<400> 91
ggcccaaccg gcctcagcta gcaccaaggg cccatcggtc ttccccctgg cgccctgctc 60
caggagcacc tccgagagca cagcggccct gggctgcctg gtcaaggact acttccccga 120
accggtgacg gtgtcgtgga actcaggcgc tctgaccagc ggcgtgcaca ccttcccagc 180
tgtcctacag tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcaa 240
cttcggcacc cagacctaca cctgcaacgt agatcacaag cccagcaaca ccaaggtgga 300
caagacagtt gagcgcaaat gttgtgtcga gtgcccaccg tgcccagcac cacctgtggc 360
aggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac 420
ccctgaggtc acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tccagttcaa 480
ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccacggg aggagcagtt 540
caacagcacg ttccgtgtgg tcagcgtcct caccgttgtg caccaggact ggctgaacgg 600
caaggagtac aagtgcaagg tctccaacaa aggcctccca gcccccatcg agaaaaccat 660
ctccaaaacc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga 720
ggagatgacc aagaaccagg tcagcctgac ctgcctggtc aaaggcttct accccagcga 780
catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacacctcc 840
catgctggac tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag 900
gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta 960
cacgcagaag agcctctccc tgtctccggg taaatgataa cgtctcctca gc 1012
<210> 92
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для внесения мутации в вектор pUC19
<400> 92
atgacggtgt taacctctga cacatgcagc tcccggagaa ggtcacagct tgtctgtaag 60
c 61
<210> 93
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для внесения мутации в вектор pUC19
<400> 93
gtcagaggtt aacaccgtca tcaccgaaac gcgcgacacg aaagggcctc gtgatac 57
<210> 94
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Коннексон длиной 90 п.н.
<400> 94
aggccttcta gataattaat taggaggaat ttaaaatgaa atacctattg cctacggcag 60
ccgctggatt gttattactt gcggcccaac 90
<210> 95
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R3-коннексон-R4 (90 п.н.)
<400> 95
ggccacatag gccttctaga taattaatta ggaggaattt aaaatgaaat acctattgcc 60
tacggcagcc gctggattgt tattacttgc ggcccaaccg gcc 103
<210> 96
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для 90 п.н. коннексона
<400> 96
agtgttaata ggccacatag gccttctaga taattaatta ggag 44
<210> 97
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для 90 п.н. коннексона
<400> 97
ccttctagat aattaattag gaggaattta aaatgaaata cc 42
<210> 98
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для 90 п.н. коннексона
<400> 98
acctcggcca tggccggttg ggccgcaag 29
<210> 99
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность распознавания для SfiI
<220>
<221> дополнительный признак
<222> (5)..(9)
<223> n представляет собой a, c, g или t
<400> 99
ggccnnnnng gcc 13
<210> 100
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность распознавания для BsmBI и Esp3I
<220>
<221> дополнительный признак
<222> (7)..(12)
<223> n представляет собой a, c, g или t
<400> 100
cgtctcnnnn nn 12
<---
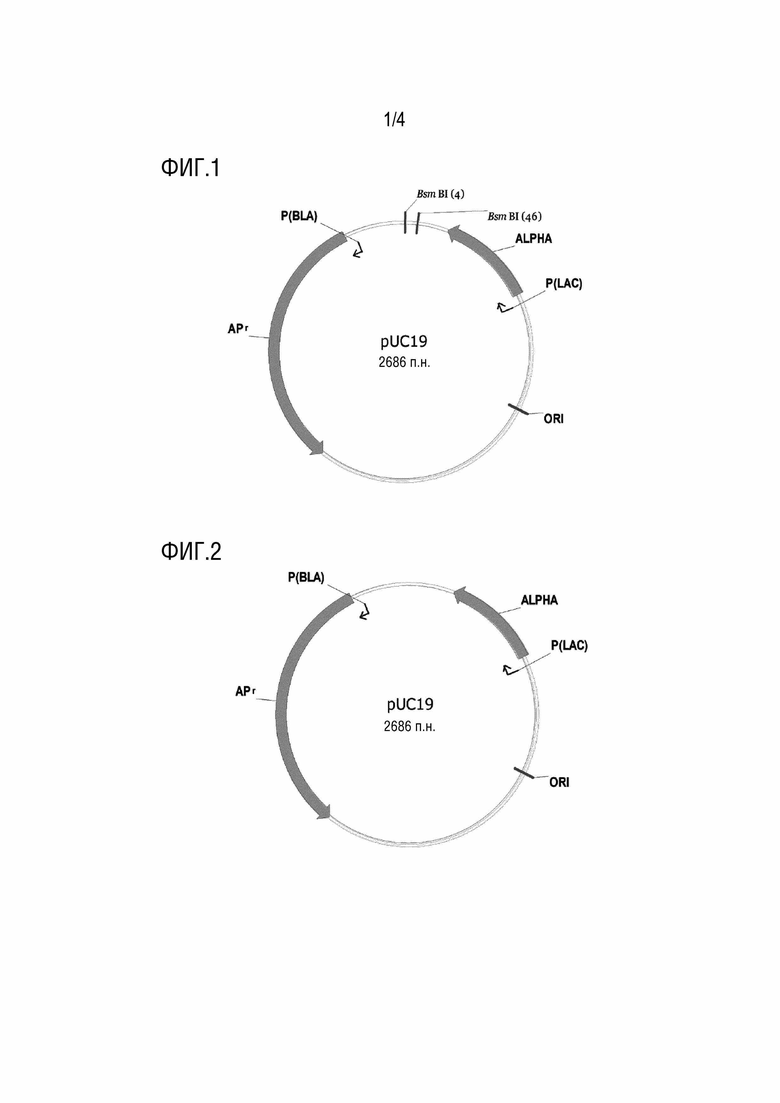
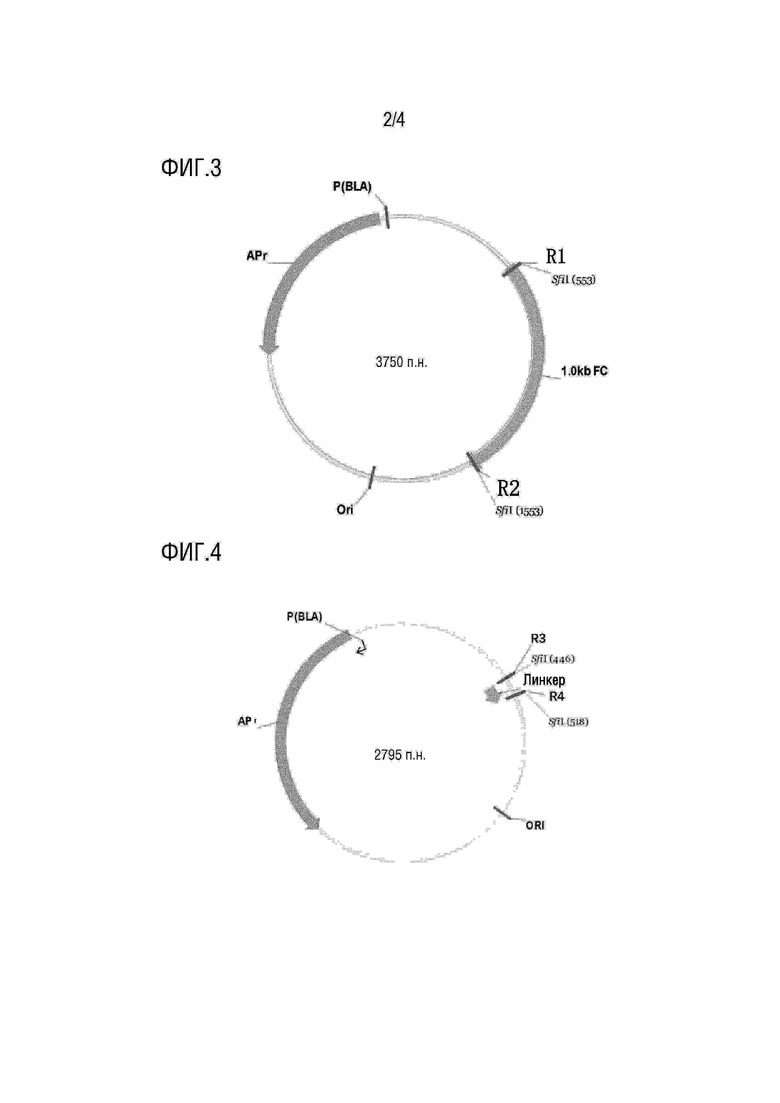
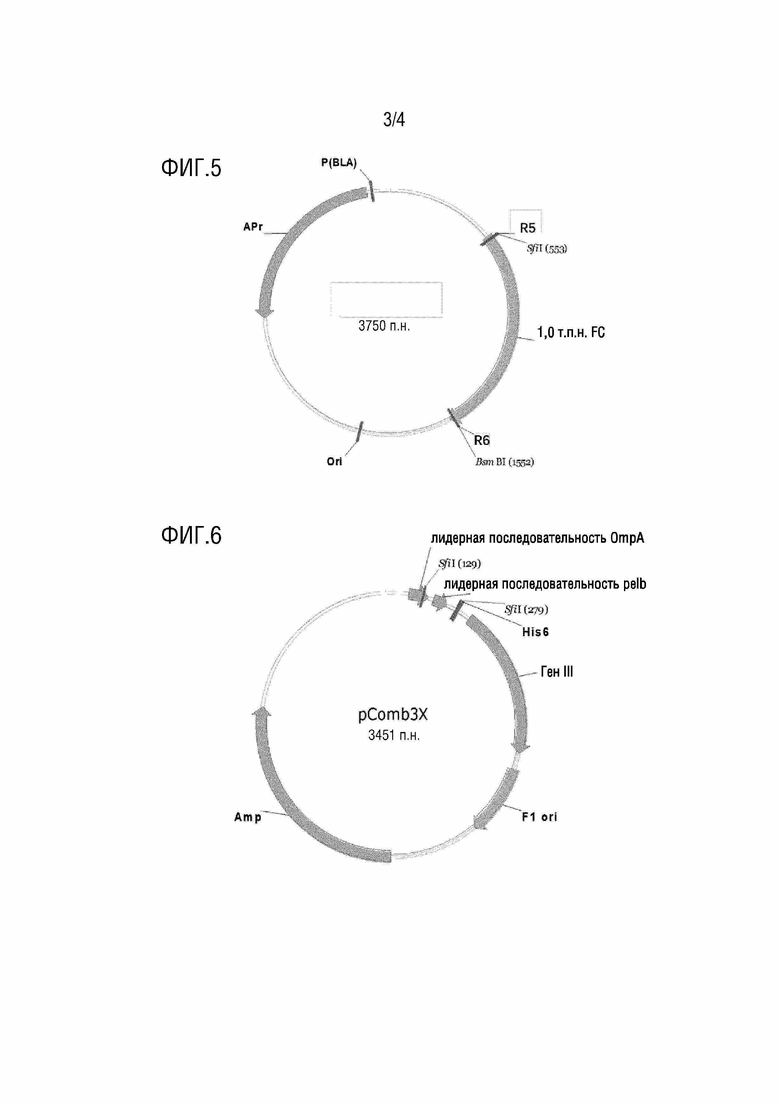
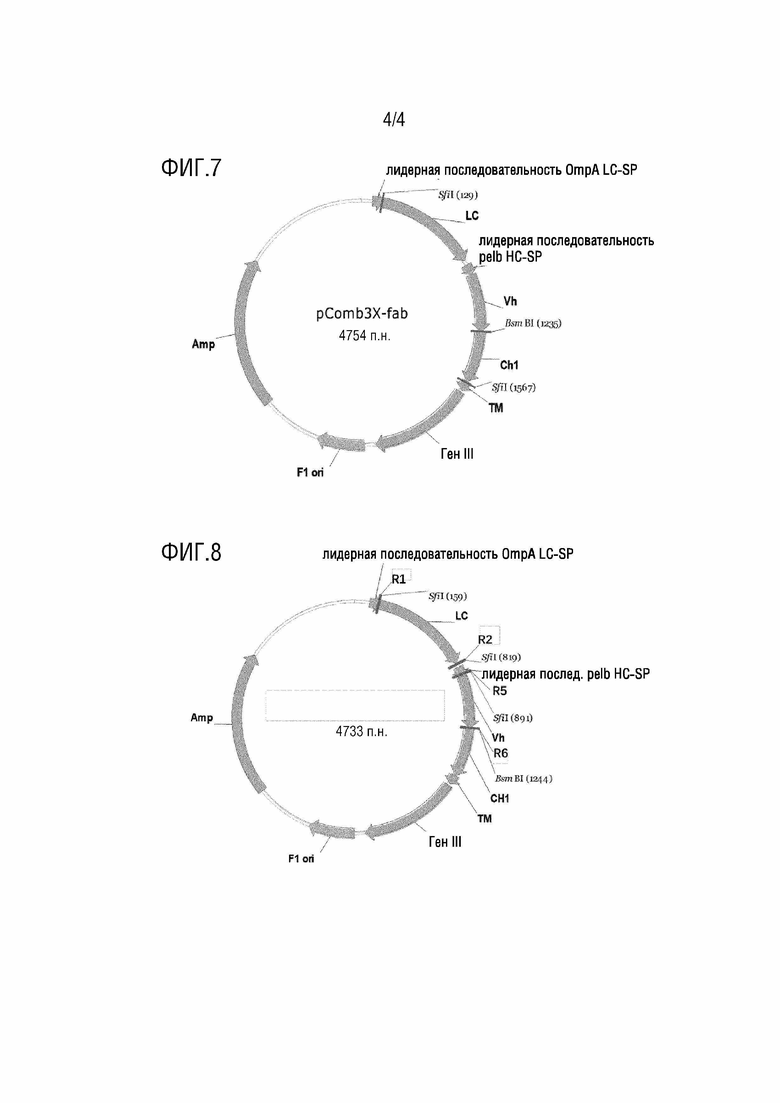
Изобретение относится к области биотехнологии. Представлен способ получения фаговой библиотеки, экспонирующей антитела или фрагменты антител. Способ включает предоставление первого, второго и третьего полинуклеотидов, содержащих LC, коннексон и HC соответственно, введение указанных полинуклеотидов в первую, вторую и третью бактерию с получением бактериальной библиотеки с компонентом легкой цепи, бактериальной библиотеки с компонентом коннексона и бактериальной библиотеки с компонентом тяжелой цепи, получение плазмид с компонентом легкой цепи, компонентом коннексона и компонентом тяжелой цепи из библиотек, получение высвободившихся LC, коннексона и HC из плазмид, соединение высвободившихся сегментов вектора для дисплея с образованием соединенного продукта для дисплея, введение третьей бактерии с получением библиотеки бактериального дисплея и использование библиотеки бактериального дисплея для получения фаговой библиотеки для дисплея антител или фрагментов антител. Изобретение позволяет получить фаговую библиотеку с большой емкостью, которая может удовлетворять потребности промышленного массового производства. 24 з.п. ф-лы, 8 ил., 11 табл., 7 пр.
1. Способ получения фаговой библиотеки, экспонирующей антитело или фрагмент антитела, включающий:
1) предоставление первого полинуклеотида, второго полинуклеотида и третьего полинуклеотида, причем указанный первый полинуклеотид содержит LC, указанный второй полинуклеотид содержит коннексон, и указанный третий полинуклеотид содержит HC, где указанная LC содержит последовательность нуклеиновой кислоты, кодирующую легкую цепь или фрагмент легкой цепи антитела или фрагмента антитела, и указанная HC содержит последовательность нуклеиновой кислоты, кодирующую тяжелую цепь или фрагмент тяжелой цепи антитела или фрагмента антитела;
2) введение указанного первого полинуклеотида в первую бактерию с получением бактериальной библиотеки с компонентом легкой цепи, введение указанного второго полинуклеотида в указанную вторую бактерию с получением бактериальной библиотеки с компонентом коннексона, и введение указанного третьего полинуклеотида в третью бактерию с получением бактериальной библиотеки с компонентом тяжелой цепи;
3) получение плазмиды с компонентом легкой цепи, содержащей указанную LC из указанной бактериальной библиотеки с компонентом легкой цепи, получение плазмиды с компонентом коннексона, содержащей указанный коннексон из указанной бактерии с компонентом коннексона, и получение плазмиды с компонентом тяжелой цепи, содержащей указанную HC из указанной бактериальной библиотеки с компонентом тяжелой цепи;
4) получение высвободившейся LC из указанной плазмиды с компонентом легкой цепи, получение высвободившегося коннексона из указанной плазмиды с компонентом коннексона, и получение высвободившейся HC из указанной плазмиды с компонентом тяжелой цепи;
5) предоставление вектора для дисплея и получение высвободившегося фрагмента вектора для дисплея из указанного вектора для дисплея;
6) соединение высвободившейся LC, высвободившегося коннексона, высвободившейся HC и высвободившегося фрагмента вектора для дисплея с образованием соединенного продукта для дисплея;
7) введение указанного соединенного продукта для дисплея в четвертую бактерию с получением библиотеки бактериального дисплея антитела или фрагмента антитела; и
8) получение указанной фаговой библиотеки, экспонирующей антитело или фрагмент антитела, с использованием указанной библиотеки бактериального дисплея.
2. Способ по п.1, где указанный соединенный продукт для дисплея в 6) может экспрессировать легкую цепь или фрагмент легкой цепи и тяжелую цепь или фрагмент тяжелой цепи в соответствии с рамкой считывания в условиях, подходящих для экспрессии антитела или фрагмента антитела.
3. Способ по любому из пп.1, 2, где все из указанного первого полинуклеотида, указанного второго полинуклеотида и указанного третьего полинуклеотида представляют собой линейные молекулы нуклеиновой кислоты.
4. Способ по любому из пп.1-3, где указанный коннексон включает последовательность нуклеиновой кислоты, кодирующую сигнальный пептид pelB или его фрагмент.
5. Способ по любому из пп.1-4, где указанный первый полинуклеотид содержит структуру R1-LC-R2 в направлении от 5' к 3', указанный второй полинуклеотид содержит структуру R3-коннексон-R4 в направлении от 5' к 3', указанный третий полинуклеотид содержит структуру R5-HC-R6 в направлении от 5' к 3', и указанный вектор для дисплея содержит структуру R7-фрагмент вектора для дисплея-R8 в направлении от 5' к 3', где R1, R2, R3, R4, R5, R6, R7 и R8 представляют собой участки распознавания для эндонуклеазы рестрикции, где конец, образовавшийся посредством расщепления указанного R2 эндонуклеазой рестрикции, может распознавать и может быть соединен с концом, образовавшимся посредством расщепления указанного R3 эндонуклеазой рестрикции, и наоборот, и не может распознавать или быть соединен с концом, образовавшимся посредством расщепления любого из указанных R1, R4, R5, R6, R7 и R8 эндонуклеазой рестрикции, и наоборот; конец, образовавшийся посредством расщепления указанного R4 эндонуклеазой рестрикции, может распознавать и быть соединен с концом, образовавшимся посредством расщепления указанного R5 эндонуклеазой рестрикции, и наоборот; и не может распознавать или быть соединен с концом, образовавшимся посредством расщепления любого из указанных R1, R2, R3, R6, R7 и R8 эндонуклеазой рестрикции, и наоборот; конец, образовавшийся посредством расщепления указанного R6 эндонуклеазой рестрикции, может распознавать и быть соединен с концом, образовавшимся посредством расщепления указанного R7 эндонуклеазой рестрикции, и наоборот; и не может распознавать или быть соединен с концом, образовавшимся посредством расщепления любого из указанных R1, R2, R3, R4, R5 и R8 эндонуклеазой рестрикции, и наоборот; конец, образовавшийся посредством расщепления указанного R8 эндонуклеазой рестрикции, может распознавать и быть соединен с концом, образовавшимся посредством расщепления указанного R1 эндонуклеазой рестрикции, и наоборот; и не может распознавать или быть соединен с концом, образовавшимся посредством расщепления любого из указанных R2, R3, R4, R5, R6 и R7 эндонуклеазой рестрикции, и наоборот.
6. Способ по п.5, где конец, образовавшийся посредством расщепления любого из указанных R1, R2, R3, R4, R5, R6, R7 и R8 эндонуклеазой рестрикции, представляет собой не палиндромную последовательность.
7. Способ по любому из пп.5-6, где два или более из указанных R1, R2, R3, R4, R5, R6, R7 и R8 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции.
8. Способ по любому из пп.5-7, где указанные R1, R2, R3, R4, R5 и R8 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции.
9. Способ по любому из пп.5-8, где указанные R6 и R7 могут распознаваться и расщепляться одной и той же эндонуклеазой рестрикции.
10. Способ по любому из пп.1-9, где указанное антитело или его фрагмент содержит Fab, Fab', (Fab)2 и/или (Fab')2.
11. Способ по любому из пп.1-10, где 2) включает расщепление указанного первого полинуклеотида эндонуклеазой рестрикции, распознающей указанные R1 и R2, соединение расщепленного первого полинуклеотида с фрагментом вектора для хранения легкой цепи с образованием соединенного продукта для хранения легкой цепи, и введение указанного соединенного продукта для хранения легкой цепи в указанную первую бактерию с получением указанной бактериальной библиотеки с компонентом легкой цепи; где указанный вектор для хранения легкой цепи содержит указанные R1 и R2, и указанный фрагмент вектора для хранения легкой цепи получают путем расщепления указанного вектора для хранения легкой цепи эндонуклеазой рестрикции, распознающей указанные R1 и R2.
12. Способ по п.11, где 2) включает расщепление указанного третьего полинуклеотида эндонуклеазой рестрикции, распознающей указанные R5 и R6, соединение расщепленного третьего полинуклеотида с фрагментом вектора для хранения тяжелой цепи с образованием соединенного продукта для хранения тяжелой цепи, и введение указанного соединенного продукта для хранения тяжелой цепи в указанную третью бактерию с получением указанной бактериальной библиотеки с компонентом тяжелой цепи; где указанный вектор для хранения тяжелой цепи содержит указанные R5 и R6, и указанный фрагмент вектора для хранения тяжелой цепи получают путем расщепления указанного вектора для хранения тяжелой цепи эндонуклеазой рестрикции, распознающей указанные R5 и R6.
13. Способ по п.12, где указанный вектор для хранения тяжелой цепи содержит или содержит только участок распознавания для BsmBI.
14. Способ по любому из пп.1-13, где указанный 2) включает соединение указанного второго полинуклеотида в фрагментом вектора с образованием вектора для хранения коннексона, и введение указанного вектора для хранения коннексона в указанную вторую бактерию с получением указанной бактерии с компонентом коннексона.
15. Способ по п.14, где указанный вектор для хранения коннексона содержит указанные R3 и R4.
16. Способ по любому из пп.1-15, где 4) включает расщепление указанной плазмиды с компонентом легкой цепи эндонуклеазой рестрикции, распознающей указанные R1 и R2, с получением указанной высвободившейся LC.
17. Способ по любому из пп.1-16, где 4) включает расщепление указанной плазмиды с компонентом тяжелой цепи эндонуклеазой рестрикции, распознающей указанные R5 и R6, с получением указанной высвободившейся HC.
18. Способ по любому из пп.1-17, где 4) включает расщепление указанного вектора для хранения коннексона эндонуклеазой рестрикции, распознающей указанные R3 и R4, с получением указанного высвободившегося коннексона.
19. Способ по любому из пп.1-18, где 4) включает получение продукта амплификации из указанного вектора для хранения коннексона, указанный продукт амплификации содержит указанный коннексон, указанный R3 и указанный R4; и указанный продукт амплификации расщепляют с использованием эндонуклеазы рестрикции, распознающей указанные R3 и R4, с получением указанного высвободившегося коннексона.
20. Способ по любому из пп.1-19, где 5) включает расщепление указанного вектора для дисплея эндонуклеазой рестрикции, распознающей указанные R7 и R8, высвобождая указанный фрагмент вектора для дисплея.
21. Способ по любому из пп.1-20, где указанный вектор для дисплея происходит из вектора pComb3x.
22. Способ по любому из пп.5-21, где три различных расщепленных фрагмента получают после расщепления указанного вектора для дисплея эндонуклеазой рестрикции, распознающей указанные R1, R2, R3, R4, R5 и R8.
23. Способ по любому из пп.5-22, где указанный вектор для дисплея является линеаризованным после расщепления указанного вектора для дисплея эндонуклеазой рестрикции, распознающей указанные R6 и R7.
24. Способ по любому из пп.5-23, где четыре различных расщепленных фрагмента получают после расщепления указанного вектора для дисплея эндонуклеазой рестрикции, распознающей указанные R1, R2, R3, R4, R5, R6, R7 и R8.
25. Способ по любому из пп.1-24, где 2) включает проведение детекции в образцах из указанной бактериальной библиотеки с компонентом легкой цепи, указанной бактерии с компонентом коннексона и/или указанной бактериальной библиотеки с компонентом тяжелой цепи для определения емкости и/или качества указанной бактериальной библиотеки с компонентом легкой цепи, указанной бактерии с компонентом коннексона и/или указанной бактериальной библиотеки с компонентом тяжелой цепи.
| WO03029456 A1, 10.04.2003 | |||
| CN 102732974 A, 17.10.2012 | |||
| CEN X | |||
| et al., Construction of a Large Phage Display Antibody Library by In Vitro Package and In Vivo Recombination, Appl Microbiol Biotechnol, 2006, vol | |||
| Контрольный стрелочный замок | 1920 |
|
SU71A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| CRAMERI A | |||
| et al., Construction and evolution of antibody-phage libraries by DMA shuffling, Nature Medicine, 1996, | |||

