Область техники, к которой относится изобретение
Настоящее изобретение относится к области биомедицины, и конкретно, к способу конструирования вектора дисплея генов антигенспецифических связывающих полипептидов, который можно использовать для скрининга антигенспецифических связывающих полипептидов.
Уровень техники
В настоящее время существует две главных категории способов обнаружения антител, общеупотребительных на предшествующем уровне техники: технология гибридомы и технология дисплея антител. Технология гибридомы дополнительно включает два типа мышиной гибридомы и гибридомы трансгенной мыши. Существует в основном три типа технологии дисплея антител: фаговый дисплей, дрожжевой дисплей и дисплей клеток млекопитающих, соответственно. Каждая технология обнаружения антител имеет очень явные преимущества, но также очень значительные недостатки и ограничения. Например, отсутствие контроля качества в процессе конструирования исходного банка лекарственных средств на основе антител приводит к плохому качеству, малой емкости библиотеки, низкому разнообразию, низкой доле эффективных клонов библиотеки антител, таким образом, делая сложным скрининг лидирующих антител высокого качества; альтернативно, технология скрининга библиотеки антител не имеет количественного скрининга, и производительность скрининга мала, эффект скрининга слабый, и скрининг требует больших затрат времени.
Таким образом, необходима инновационная технология обнаружения антител для улучшения качества, количества и разнообразия лидирующих молекул антител для скрининга инновационных лекарственных средств на основе антител, ускорения разработки лекарственных средств на основе антител и улучшения показателя успешности разработки.
Сущность изобретения
Настоящее изобретение относится к способу конструирования вектора дисплея генов антигенспецифических связывающих полипептидов. Вектор дисплея генов антигенспецифических связывающих полипептидов состоит из четырех фрагментов, в которых 5’- и 3’-концы четырех фрагментов представлены с липкими концами специфических последовательностей посредством конструирования библиотек компонентов и векторов дисплея таким образом, чтобы они направленным образом циклизовались с формированием вектора дисплея генов антигенспецифических связывающих полипептидов. Способ конструирования вектора дисплея генов антигенспецифических связывающих полипептидов по настоящему изобретению и способ скрининга антигенспецифического связывающего полипептида посредством использованием вектора дисплея генов антигенспецифических связывающих полипептидов по настоящему изобретения имеет по меньшей мере одно из следующих свойств: 1) специфические участки узнавания для рестрикционной эндонуклеазы можно использовать в конструировании вектора дисплея генов антигенспецифических связывающих полипептидов по настоящему изобретению, что не только обеспечивает направленное лигирование, но также предотвращает неправильное лигирование, и количество молекул фрагмента каждого компонента можно контролировать до 1:1 во время лигирования, таким образом, улучшая эффективность лигирования и трансформации; в то же время, способы конструирования библиотеки компонента VH и библиотеки компонента LC используют для улучшения эффективности лигирования и трансформации каждого фрагмента; 2) способ комбинаторной ПЦР, используемый в общепринятых способах конструирования библиотек антител, не используют, эффективно уменьшая вероятность введения мутаций, вызванного ПЦР; 3) проще осуществлять контроль качества, и он может удовлетворять нуждам промышленной массовой продукции; 4) можно осуществлять прямой скрининг вектора дисплея посредством экспериментов анализа биологической активности после введения в клетку, эффективно укорачивая время от конструирования вектора дисплея генов антигенспецифических связывающих полипептидов до скрининга антигенспецифического полипептида уникальной последовательности. Например, время от конструирования вектора дисплея до скрининга положительного клона уникальной последовательности может составлять по меньшей мере приблизительно 1 неделю (по меньшей мере приблизительно 10 суток, по меньшей мере приблизительно 2 недели, по меньшей мере приблизительно 3 недели, по меньшей мере приблизительно 4 недели, по меньшей мере приблизительно 5 недель, по меньшей мере приблизительно 6 недель, по меньшей мере приблизительно 7 недель, по меньшей мере приблизительно 8 недель); 5) разнообразие клонов в бактериальной библиотеке вектора дисплея является большим, и эффективность скрининга является высокой. В некоторых случаях, в библиотеке, содержащей векторы дисплея генов антигенспецифических связывающих полипептидов, доля эффективных клонов может составлять вплоть до более чем приблизительно 50% (например, более чем приблизительно 55%, более чем приблизительно 60%, более чем приблизительно 65%, более чем приблизительно 70%, более чем приблизительно 75%, более чем приблизительно 80%, более чем приблизительно 85%, более чем приблизительно 90%, более чем приблизительно 95% или выше), так что эффективность трансформации является высокой, и частота успешного конструирования библиотеки является высокой.
В одном аспекте настоящее изобретение относится к способу конструирования вектора дисплея генов антигенспецифических связывающих полипептидов. Способ включает: a) получение первого полинуклеотида вектора дисплея, содержащего B2-дисплей VH-B3 в направлении от 5' до 3'; b) получение второго полинуклеотида вектора дисплея, содержащего S5-дисплей LC-S6 в направлении от 5' до 3'; c) получение третьего полинуклеотида вектора дисплея, содержащего B3-фрагмент I вектора дисплея-S5 в направлении от 5' до 3'; d) получение четвертого полинуклеотида вектора дисплея, содержащего S6-фрагмент II вектора дисплея-B2 в направлении от 5' до 3'; e) специфическое расщепление первого полинуклеотида вектора дисплея, второго полинуклеотида вектора дисплея, третьего полинуклеотида вектора дисплея и четвертого полинуклеотида вектора дисплея с использованием рестрикционной эндонуклеазы для получения расщепленного первого полинуклеотида вектора дисплея, расщепленного второго полинуклеотида вектора дисплея, расщепленного третьего полинуклеотида вектора дисплея и расщепленного четвертого полинуклеотида вектора дисплея; где рестрикционная эндонуклеаза специфически узнает B2, B3, S5 и S6, соответственно; f) смешивание расщепленного первого полинуклеотида вектора дисплея, расщепленного второго полинуклеотида вектора дисплея, расщепленного третьего полинуклеотида вектора дисплея и расщепленного четвертого полинуклеотида вектора дисплея таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования вектора дисплея генов антигенспецифических связывающих полипептидов; где дисплей VH кодирует вариабельную область тяжелой цепи антигенспецифического связывающего полипептида, дисплей LC кодирует легкую цепь антигенспецифического связывающего полипептида; где каждый из B2, B3, S5 и S6 независимо представляют собой участки узнавания для рестрикционной эндонуклеазы.
В некоторых вариантах осуществления, конец, образованный в результате специфического расщепления B2 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B3, S5 и S6 посредством соответствующей рестрикционной эндонуклеазы.
В некоторых вариантах осуществления, конец, образованный в результате специфического расщепления B3 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B2, S5 и S6 посредством соответствующей рестрикционной эндонуклеазы.
В некоторых вариантах осуществления, конец, образованный в результате специфического расщепления S5 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B2, B3 и S6 посредством соответствующей рестрикционной эндонуклеазы.
В некоторых вариантах осуществления, конец, образованный в результате специфического расщепления S6 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B2, B3 и S5 посредством соответствующей рестрикционной эндонуклеазы.
В некоторых вариантах осуществления, рестрикционная эндонуклеаза выбрана из SfiI, Esp3I и BsmBI.
В некоторых вариантах осуществления, B2 и B3 могут быть специфически узнаны и расщеплены посредством фермента, выбранного из группы, состоящей из: BsmBI и Esp3I.
В некоторых вариантах осуществления, S5 и S6 могут быть специфически узнаны и расщеплены посредством Sfil.
В некоторых вариантах осуществления, B2 включает последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 8.
В некоторых вариантах осуществления, B3 включает последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 9.
В некоторых вариантах осуществления, S5 включает последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 10.
В некоторых вариантах осуществления, S6 включает последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 11.
В некоторых вариантах осуществления, способ дополнительно включает введение первого полинуклеотида вектора дисплея в первую бактерию дисплея для получения бактериальной библиотеки компонента дисплея VH.
В некоторых вариантах осуществления, способ включает вставку первого полинуклеотида вектора дисплея в вектор компонента дисплея для формирования продукта лигирования для хранения дисплея VH и введение продукта лигирования для хранения дисплея VH в первую бактерию дисплея для получения бактериальной библиотеки компонента дисплея VH.
В некоторых вариантах осуществления, способ дополнительно включает введение второго полинуклеотида вектора дисплея во вторую бактерию дисплея для получения бактериальной библиотеки компонента дисплея LC.
В некоторых вариантах осуществления, способ включает вставку второго полинуклеотида вектора дисплея в вектор компонента дисплея для формирования продукта лигирования для хранения дисплея LC и введение продукта лигирования для хранения дисплея LC во вторую бактерию дисплея для получения бактериальной библиотеки компонента дисплея LC.
В некоторых вариантах осуществления, способ дополнительно включает введение третьего полинуклеотида вектора дисплея в третью бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея I.
В некоторых вариантах осуществления, способ включает вставку третьего полинуклеотида вектора дисплея в вектор компонента дисплея для формирования вектора дисплея продукта лигирования для хранения фрагмента I и введение продукта лигирования для хранения в третью бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея I.
В некоторых вариантах осуществления, способ дополнительно включает введение четвертого полинуклеотида вектора дисплея в четвертую бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея II.
В некоторых вариантах осуществления, способ включает вставку четвертого полинуклеотида вектора дисплея в вектор компонента дисплея для формирования вектора дисплея продукта лигирования для хранения фрагмента II и введение продукта лигирования для хранения в четвертую бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея II.
В некоторых вариантах осуществления, вектор компонента дисплея происходит из вектора pUC.
В некоторых вариантах осуществления, вектор pUC представляет собой вектор pUC19 или происходит из вектора pUC19.
В некоторых вариантах осуществления, способ дополнительно включает получение плазмиды компонента дисплея VH, содержащей первый полинуклеотид вектора дисплея, из бактериальной библиотеки компонента дисплея VH, и получение расщепленного первого полинуклеотида вектора дисплея из плазмиды компонента дисплея VH.
В некоторых вариантах осуществления, способ включает расщепление плазмиды компонента дисплея VH с использованием рестрикционной эндонуклеазы, специфически узнающей B2 и B3, таким образом, получение расщепленного первого полинуклеотида вектора дисплея.
В некоторых вариантах осуществления, способ дополнительно включает получение плазмиды компонента дисплея LC, содержащей второй полинуклеотид вектора дисплея, из бактериальной библиотеки компонента дисплея LC; и получение расщепленного второго полинуклеотида вектора дисплея из плазмиды компонента дисплея LC.
В некоторых вариантах осуществления, способ включает расщепление плазмиды компонента дисплея LC с использованием рестрикционной эндонуклеазы, специфически узнающей S5 и S6, таким образом, получение расщепленного второго полинуклеотида вектора дисплея.
В некоторых вариантах осуществления, способ дополнительно включает получение плазмиды фрагмента I компонента дисплея, содержащей третий полинуклеотид вектора дисплея, из бактериальной библиотеки экспрессирующего вектора компонента I; и получение расщепленного третьего полинуклеотида вектора дисплея из плазмиды фрагмента I компонента дисплея.
В некоторых вариантах осуществления, способ включает расщепление плазмиды фрагмента I компонента дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей B3 и S5, таким образом, получение расщепленного третьего полинуклеотида вектора дисплея.
В некоторых вариантах осуществления, способ дополнительно включает получение плазмиды фрагмента II компонента дисплея, содержащей четвертый полинуклеотид вектора дисплея, из бактериальной библиотеки экспрессирующего вектора компонента II, и получение расщепленного четвертого полинуклеотида вектора дисплея из плазмиды фрагмента II компонента дисплея.
В некоторых вариантах осуществления, способ включает расщепление плазмиды фрагмента II компонента дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей S6 и B2, таким образом, получение расщепленного четвертого полинуклеотида вектора дисплея.
В некоторых вариантах осуществления, способ включает:
a) получение пятого полинуклеотида, содержащего B-антигенспецифическую VH-B в направлении от 5' до 3';
b) получение вектора компонента VH, где вектор компонента VH включает шестой полинуклеотид, содержащий B3-фрагмент лигирования вектора компонента VH-B2 в направлении от 5' до 3';
c) расщепление пятого полинуклеотида и вектора компонента VH с использованием рестрикционной эндонуклеазы для получения расщепленного пятого полинуклеотида и высвобожденного шестого полинуклеотида;
d) смешивание расщепленного пятого полинуклеотида и высвобожденного шестого полинуклеотида таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования библиотеки компонента антигенспецифической VH;
где B представляет собой участок узнавания для рестрикционной эндонуклеазы, которая может специфически узнавать B2 и/или B3, и антигенспецифическая VH кодирует вариабельную область тяжелой цепи антигенспецифического связывающего полипептида.
В некоторых вариантах осуществления, способ включает:
a) получение седьмого полинуклеотида, содержащего S-антигенспецифическую LC-S в направлении от 5' до 3';
b) получение вектора компонента LC, где вектор компонента LC включает восьмой полинуклеотид, содержащий S6-фрагмент лигирования вектора компонента LC-S5 в направлении от 5' до 3';
c) расщепление седьмого полинуклеотида и вектора компонента LC с использованием рестрикционной эндонуклеазы для получения расщепленного седьмого полинуклеотида и высвобожденного восьмого полинуклеотида;
d) смешивание расщепленного седьмого полинуклеотида и высвобожденного восьмого полинуклеотида таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования библиотеки компонента антигенспецифической LC,
где S представляет собой участок узнавания для рестрикционной эндонуклеазы, которая может специфически узнавать S5 и/или S5, и антигенспецифическая LC кодирует легкую цепь антигенспецифического связывающего полипептида.
В некоторых вариантах осуществления, способ включает:
a) получение девятого полинуклеотида, содержащего B2-рабочий фрагмент вектора компонента VH-B3 в направлении от 5' до 3';
b) вставку девятого полинуклеотида в экспрессирующий вектор компонента для получения вектора компонента VH.
В некоторых вариантах осуществления, способ включает:
a) получение десятого полинуклеотида, содержащего S5-рабочий фрагмент вектора компонента LC-S6 в направлении от 5' до 3';
b) вставку десятого полинуклеотида в экспрессирующий вектор компонента для получения вектора компонента LC.
В некоторых вариантах осуществления, экспрессирующий вектор компонента происходит из вектора pMD.
В некоторых вариантах осуществления, вектор pMD представляет собой вектор pMD19 или происходит из вектора pMD19.
В некоторых вариантах осуществления, способ включает стадии:
a) введения вектора компонента VH в девятую бактерию для получения бактериальной библиотеки вектора для хранения компонента VH;
b) получения плазмиды вектора для хранения компонента VH из бактериальной библиотеки вектора для хранения компонента VH;
c) получения высвобожденного шестого полинуклеотида из плазмиды вектора для хранения компонента VH.
В некоторых вариантах осуществления, способ включает расщепление плазмиды вектора для хранения компонента VH с использованием рестрикционной эндонуклеазы, специфически узнающей B2 и B3, таким образом, получение высвобожденного шестого полинуклеотида.
В некоторых вариантах осуществления, способ включает стадии:
a) введения вектора компонента LC в десятую бактерию для получения бактериальной библиотеки вектора для хранения компонента LC;
b) получения плазмиды вектора для хранения компонента LC из бактериальной библиотеки вектора для хранения компонента LC;
c) получения высвобожденного восьмого полинуклеотида из плазмиды вектора для хранения компонента LC.
В некоторых вариантах осуществления, способ включает расщепление плазмиды вектора для хранения компонента LC с использованием рестрикционной эндонуклеазы, специфически узнающей S5 и S6, таким образом, получение высвобожденного восьмого полинуклеотида.
В некоторых вариантах осуществления, способ включает:
a) получение библиотеки компонента антигенспецифической VH, где библиотека компонента антигенспецифической VH включает первый полинуклеотид, содержащий B2-антигенспецифическую VH-B3 в направлении от 5' до 3';
b) получение библиотеки компонента антигенспецифической LC, где библиотека компонента антигенспецифической LC включает второй полинуклеотид, содержащий S5-антигенспецифическую LC-S6 в направлении от 5' до 3';
c) получение вектора дисплея, включающего третий полинуклеотид, содержащий B3-фрагмент I вектора дисплея-S5 в направлении от 5' до 3', и четвертый полинуклеотид, содержащий S6-фрагмент II вектора дисплея-B2 в направлении от 5' до 3';
d) специфическое расщепление библиотеки компонента антигенспецифической VH, библиотеки компонента антигенспецифической LC и вектора дисплея с использованием рестрикционной эндонуклеазы для получения высвобожденного первого полинуклеотида, высвобожденного второго полинуклеотида, высвобожденного третьего полинуклеотида и высвобожденного четвертого полинуклеотида; где рестрикционная эндонуклеаза специфически узнает B2, B3, S5 и S6, соответственно;
e) смешивание высвобожденного первого полинуклеотида, высвобожденного второго полинуклеотида, высвобожденного третьего полинуклеотида и высвобожденного четвертого полинуклеотида таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования вектора дисплея генов антигенспецифических связывающих полипептидов;
где антигенспецифическая LC кодирует легкую цепь антигенспецифического связывающего полипептида, и антигенспецифическая VH кодирует вариабельную область тяжелой цепи антигенспецифического связывающего полипептида;
где каждый из B2, B3, S5 и S6 независимо представляют собой участки узнавания для рестрикционной эндонуклеазы.
В некоторых вариантах осуществления, способ включает расщепление библиотеки компонента антигенспецифической VH с использованием рестрикционной эндонуклеазы, специфически узнающей B2 и B3, таким образом, получение высвобожденного первого полинуклеотида.
В некоторых вариантах осуществления, способ включает расщепление библиотеки компонента антигенспецифической LC с использованием рестрикционной эндонуклеазы, специфически узнающей S5 и S6, таким образом, получение высвобожденного второго полинуклеотида.
В некоторых вариантах осуществления, способ включает расщепление вектора дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей B3, и рестрикционной эндонуклеазы, специфически узнающей S5, таким образом, получение высвобожденного третьего полинуклеотида.
В некоторых вариантах осуществления, способ включает расщепление вектора дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей S6, и рестрикционной эндонуклеазы, специфически узнающей B2, таким образом, получение высвобожденного четвертого полинуклеотида.
В некоторых вариантах осуществления, пятый полинуклеотид, седьмой полинуклеотид, девятый полинуклеотид, десятый полинуклеотид, первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и/или четвертый полинуклеотид вектора дисплея получают из материалов образцов.
В некоторых вариантах осуществления, материалы образцов включают антитела, нацеленные на специфические антигены, или их антигенсвязывающие фрагменты и/или IgG.
В некоторых вариантах осуществления, антитела или их антигенсвязывающие фрагменты нацелены на ROR1, PD-1 и/или PD-L1.
В некоторых вариантах осуществления, IgG представляет собой IgG человека.
В некоторых вариантах осуществления, IgG человека представляет собой IgG1 человека или IgG2 человека.
В некоторых вариантах осуществления, направленное лигирование включает использование лигазы.
В некоторых вариантах осуществления, лигаза включает ДНК-лигазу T4.
В некоторых вариантах осуществления, способ включает введение вектора дисплея генов антигенспецифических связывающих полипептидов в клетку и получение антигенспецифического связывающего полипептида из клетки.
В некоторых вариантах осуществления, способ включает:
a) введение вектора дисплея генов антигенспецифических связывающих полипептидов в первую бактерию для получения бактериальной библиотеки дисплея генов антигенспецифических связывающих полипептидов;
b) получение библиотеки дисплея генов антигенспецифических связывающих полипептидов из бактериальной библиотеки дисплея генов антигенспецифических связывающих полипептидов;
c) получение ДНК экспрессирующего вектора антигенспецифического связывающего полипептида из библиотеки дисплея генов антигенспецифических связывающих полипептидов;
d) введение ДНК экспрессирующего вектора антигенспецифического связывающего полипептида в клетку;
e) получение антигенспецифического связывающего полипептида из клетки.
В некоторых вариантах осуществления, способ включает криоконсервирование бактериальной библиотеки дисплея генов антигенспецифических связывающих полипептидов, бактериальной библиотеки вектора для хранения компонента VH, бактериальной библиотеки вектора для хранения компонента LC, бактериальной библиотеки компонента дисплея VH, бактериальной библиотеки компонента дисплея LC, бактериальной библиотеки вектора компонента дисплея I и бактериальной библиотеки вектора компонента дисплея II.
В некоторых вариантах осуществления, бактериальная библиотека вектора для хранения компонента VH включает по меньшей мере 10 различных клонов.
В некоторых вариантах осуществления, бактериальная библиотека вектора для хранения компонента LC включает по меньшей мере 10 различных клонов.
В некоторых вариантах осуществления, бактериальная библиотека компонента дисплея VH включает по меньшей мере 10 различных клонов.
В некоторых вариантах осуществления, бактериальная библиотека компонента дисплея LC включает по меньшей мере 10 различных клонов.
В некоторых вариантах осуществления, бактериальная библиотека вектора компонента дисплея I включает по меньшей мере 10 идентичных клонов.
В некоторых вариантах осуществления, бактериальная библиотека вектора компонента дисплея II включает по меньшей мере 10 идентичных клонов.
В некоторых вариантах осуществления, доля эффективных клонов в бактериальной библиотеке дисплея генов антигенспецифических связывающих полипептидов составляет по меньшей мере приблизительно 10%.
В некоторых вариантах осуществления, клетка представляет собой клетку млекопитающего.
В другом аспекте, настоящее изобретение относится к способу скрининга антигенспецифического связывающего полипептида или его фрагментов, включающему использование вектора дисплея генов антигенспецифических связывающих полипептидов.
В другом аспекте, настоящее изобретение относится к вектору дисплея генов антигенспецифических связывающих полипептидов, полученному посредством этого способа.
В другом аспекте, настоящее изобретение относится к бактериальной библиотеке дисплея генов антигенспецифических связывающих полипептидов, полученной посредством этого способа.
Другие аспекты и преимущества настоящего изобретения может легко осознать специалист в данной области из следующего подробного описания. В следующем подробном описании, только иллюстративные варианты осуществления настоящего изобретения показаны и описаны. Как будет понятно специалисту в данной области, содержание настоящего изобретения позволяет специалисту в данной области вносить изменения в описанные конкретные варианты осуществления без отклонения от содержания и объема изобретения, включенного в настоящую заявку. Соответственно, чертежи и раскрытие в описании настоящего изобретения являются просто иллюстративными, а не ограничивающими.
Краткое описание чертежей
Конкретные признаки изобретения, включенного в настоящую заявку, являются такими, как показано в прилагаемой формуле изобретения. Характеристики и преимущества изобретения, включенного в настоящую заявку, можно лучше понять со ссылкой на иллюстративные варианты осуществления, подробно описанные ниже, и сопровождающие чертежи. Краткое описание чертежей является таким, как ниже:
На ФИГ. 1 показана структура вектора дисплея по настоящему изобретению;
На ФИГ. 2 показан пример вектора дисплея по настоящему изобретению;
На ФИГ. 3 показана структура вектора компонента VH по настоящему изобретению;
На ФИГ. 4 показана структура вектора компонента LC по настоящему изобретению;
На ФИГ. 5A показана аминокислотная последовательность антигенспецифической LC по настоящему изобретению, и на ФИГ. 5B показана аминокислотная последовательность антигенспецифической VH по настоящему изобретению;
На ФИГ. 6 показана экспрессия специфического для антигена ROR1 связывающего полипептида на поверхности клеток CHO, как анализировано посредством FACS;
На ФИГ. 7 показан анализ электрофореза SDS-PAGE в денатурирующем восстанавливающем геле 8 иллюстративных антител;
На ФИГ. 8 показаны результаты анализа FACS положительных антител, отобранных посредством скрининга способом по настоящему изобретению;
На ФИГ. 9 показана схематическая диаграмма вектора для хранения легкой цепи в способе конструирования фаговой библиотеки;
На ФИГ. 10 показана схематическая диаграмма вектора для хранения тяжелой цепи в способе конструирования фаговой библиотеки;
На ФИГ. 11 показана схематическая диаграмма вектора для хранения линкера в способе конструирования фаговой библиотеки;
На ФИГ. 12 показана схематическая диаграмма вектора pCom3x в способе конструирования фаговой библиотеки;
На ФИГ. 13 показана схематическая диаграмма вектора фагового дисплея в способе конструирования фаговой библиотеки;
На ФИГ. 14 показана схематическая диаграмма плазмиды для продукта лигирования для дисплея в способе конструирования фаговой библиотеки.
Подробное описание вариантов осуществления
Осуществление настоящего изобретения будет проиллюстрировано ниже посредством конкретных примеров, и другие преимущества и эффекты настоящего изобретению может легко узнать специалист в данной области из содержания, раскрытого в описании.
Определение терминов
В настоящей заявке термин «антигенсвязывающий полипептид», в общем, относится к молекуле полипептида, способной к специфическому узнаванию и/или нейтрализации специфических антигенов. Этот термин может включать антитело, или его антигенсвязывающую группу или антигенсвязывающую область, и/или вариабельную область антитела из интактного антитела. Основная 4-цепочечная единица антитела представляет собой гетеротетрамерный гликопротеин, состоящий из двух идентичных легких цепей и двух идентичных тяжелых цепей. В случае IgG, каждая цепь L связана с цепью H посредством ковалентной дисульфидной связи, в то время как две цепи H связаны друг с другом посредством одной или нескольких дисульфидных связей, количество которых зависит от изотипа цепи H. Каждая цепь H и L также имеет расположенные с регулярными интервалами внутрицепочечные дисульфидные связи. Каждая цепь H имеет на N-конце вариабельный домен (VH), за которым следуют три (для каждой из цепей α и γ) или четыре (для изотипов μ и ε) константных домена (CH). Антигенсвязывающий полипептид можно получать посредством химических способов и/или способов генной инженерии. Например, антитела можно расщеплять посредством использования протеазы, включая пепсин и папаин, для получения антигенсвязывающих фрагментов. В настоящей заявке, фрагменты антитела могут представлять собой Fab.
В настоящей заявке, термин «Fab», в общем, относится к двум идентичным антигенсвязывающим фрагментам, полученным посредством расщепления антитела с интактной структурой (например, с удаленными Fc и шарнирными областями) посредством папаина. Fab может состоять из интактной легкой цепи, вариабельной области тяжелой цепи (VH) и первого константного домена тяжелой цепи (CH1). Каждый Fab может иметь один антигенсвязывающий участок.
В настоящей заявке, термин «первый полинуклеотид», в общем, относится к полинуклеотиду, содержащему антигенспецифическую VH, который может иметь участки узнавания для эндонуклеаз (например, рестрикционных эндонуклеаз) на 5'-конце и/или 3'-конце. Например, первый полинуклеотид может включать B2-антигенспецифическую VH-B3 в направлении от 5' до 3', где B2, B3 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Например, после расщепления с использованием эндонуклеазы, узнающей участок узнавания эндонуклеазы в первом полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей B2 и B3, такой как BsmBI), высвобожденный первый полинуклеотид может содержать антигенспецифическую VH, и два конца антигенспецифической VH могут также иметь липкие концы специфической последовательности после расщепления.
В настоящей заявке, термин «второй полинуклеотид», в общем, относится к полинуклеотиду, содержащему антигенспецифическую LC, который может иметь участки узнавания для эндонуклеаз (например, рестрикционных эндонуклеаз) на 5'-конце и/или 3'-конце. Например, первый полинуклеотид может включать S5-антигенспецифическую LC-S6 в направлении от 5' до 3', где S5, S6 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Например, после расщепления с использованием эндонуклеазы, узнающей участок узнавания эндонуклеазы во втором полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей S5 и S6, такой как SfiI), высвобожденный второй полинуклеотид может включать антигенспецифическую LC, которая может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «антигенспецифическая VH», в общем, относится к нуклеотиду, кодирующему вариабельную область тяжелой цепи антитела, способного специфически связываться с антигеном, и термин «антигенспецифическая LC», в общем, относится к нуклеотиду, кодирующему легкую цепь антитела, способного специфически связываться с антигеном. Последовательности антигенспецифической VH и антигенспецифической LC можно получать посредством любых способов, известных в данной области, включая, но без ограничения, технологию фагового дисплея, технологию дисплея на поверхности дрожжей, технологию рибосомного дисплея, технологию дисплея мРНК и/или технологию гибридомы. Например, их можно получать посредством способа библиотеки фагового дисплея.
В настоящей заявке, термин «третий полинуклеотид», в общем, относится к полинуклеотиду, содержащему фрагмент I вектора дисплея, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, третий полинуклеотид может включать B3-фрагмент I вектора дисплея-S5 в направлении от 5' до 3', где B3, S5 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Третий полинуклеотид может быть включен в вектор дисплея. После расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в третьем полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей B3 и S5, такой как SfiI, BsmBI и/или Esp3I), третий полинуклеотид можно высвобождать. Высвобожденный третий полинуклеотид может включать фрагмент I вектора дисплея, который может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «высвобожденный третий полинуклеотид», в общем, относится к фрагменту третьего полинуклеотида, высвобожденному после обработки с использованием вектора дисплея. В настоящей заявке, обработка может представлять собой расщепление с использованием рестрикционной эндонуклеазы. Например, подходящие рестрикционные эндонуклеазы (например, SfiI, BsmBI и/или Esp3I) можно выбирать для участков узнавания для рестрикционной эндонуклеазы на векторе дисплея, таким образом, чтобы высвобожденный третий полинуклеотид можно было высвобождать из вектора дисплея и выделять.
В настоящей заявке, термин «четвертый полинуклеотид», в общем, относится к полинуклеотиду, содержащему фрагмент II вектора дисплея, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, четвертый полинуклеотид может включать S6-фрагмент II вектора дисплея-B2 в направлении от 5' до 3', где S6, B2 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Четвертый полинуклеотид может быть включен в вектор дисплея. После расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в четвертом полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей S6 и B2, такой как SfiI, BsmBI и/или Esp3I), четвертый полинуклеотид можно высвобождать. Высвобожденный четвертый полинуклеотид может включать фрагмент II вектора дисплея, который может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «высвобожденный четвертый полинуклеотид», в общем, относится к фрагменту четвертого полинуклеотида, высвобожденному после обработки с использованием вектора дисплея. В настоящей заявке, обработка может представлять собой расщепление с использованием рестрикционной эндонуклеазы. Например, подходящие рестрикционные эндонуклеазы (например, SfiI, BsmBI и/или Esp3I) можно выбирать для участков узнавания для рестрикционной эндонуклеазы на векторе дисплея, таким образом, чтобы высвобожденный четвертый полинуклеотид можно было высвобождать из вектора дисплея и выделять.
В настоящей заявке, термин «пятый полинуклеотид», в общем, относится к полинуклеотиду, содержащему антигенспецифическую VH, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, пятый полинуклеотид может включать B-антигенспецифическую VH-B в направлении от 5' до 3', где B может представлять собой участок узнавания для рестрикционной эндонуклеазы, и рестрикционная эндонуклеаза может представлять собой рестрикционную эндонуклеазу, способную узнавать B2 и/или B3. После расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в пятом полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей B, такой как BsmBI и/или Esp3I), расщепленный пятый полинуклеотид может включать антигенспецифическую VH.
В настоящей заявке, термин «седьмой полинуклеотид», в общем, относится к полинуклеотиду, содержащему антигенспецифическую LC, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, седьмой полинуклеотид может включать S-антигенспецифическую LC-S в направлении от 5' до 3', где S может представлять собой участок узнавания для рестрикционной эндонуклеазы, и рестрикционная эндонуклеаза может представлять собой рестрикционную эндонуклеазу, способную узнавать S5 и/или S6. После расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в седьмом полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей S, такой как SfiI), расщепленный седьмой полинуклеотид может включать антигенспецифическую LC.
В настоящей заявке, термин «вектор компонента VH», в общем, относится к кольцевому полинуклеотиду, содержащему шестой полинуклеотид и/или рабочий фрагмент вектора компонента VH.
В настоящей заявке, термин «вектор компонента LC», в общем, относится к кольцевому полинуклеотиду, содержащему восьмой полинуклеотид и/или рабочий фрагмент вектора компонента LC.
В настоящей заявке, термин «вектор дисплея», в общем, относится к кольцевому полинуклеотиду, содержащему фрагмент I вектора дисплея и фрагмент II вектора дисплея, который может дополнительно включать дисплей VH и дисплей LC. После обработки, вектор дисплея может высвобождать первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и/или четвертый полинуклеотид вектора дисплея.
В настоящей заявке, термин «первый полинуклеотид вектора дисплея», в общем, относится к полинуклеотиду, содержащему дисплей VH, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, первый полинуклеотид вектора дисплея может включать B2-дисплей VH-B3 в направлении от 5' до 3', где B2, B3 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Например, после расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в первом полинуклеотиде вектора дисплея (например, рестрикционной эндонуклеазы, узнающей B2 и B3, такой как BsmBI), расщепленный первый полинуклеотид вектора дисплея может включать дисплей VH, который может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «второй полинуклеотид вектора дисплея», в общем, относится к полинуклеотиду, содержащему дисплей LC, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, второй полинуклеотид вектора дисплея может включать S5-дисплей LC-S6 в направлении от 5' до 3', где S5, S6 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Например, после расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы во втором полинуклеотиде вектора дисплея (например, рестрикционной эндонуклеазы, узнающей S5 и S6, такой как SfiI), расщепленный второй полинуклеотид вектора дисплея может включать дисплей LC, который может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «дисплей VH», в общем, относится к нуклеотиду, кодирующему вариабельную область тяжелой цепи антигенсвязывающего полипептида, и термин «дисплей LC», в общем, относится к нуклеотиду, кодирующему легкую цепь антигенсвязывающего полипептида. «Дисплей VH» и «антигенспецифическая VH» в настоящей заявке могут представлять собой нуклеотиды, кодирующие вариабельные области тяжелой цепи, происходящие из связывающих полипептидов, нацеленных против одного и того же антигена, и могут также представлять собой нуклеотиды, кодирующие вариабельные области тяжелой цепи, происходящие из связывающих полипептидов, нацеленных против различных антигенов. «Дисплей LC» и «антигенспецифическая LC» в настоящей заявке могут представлять собой нуклеотиды, кодирующие легкие цепи, происходящие из связывающих полипептидов, нацеленных против одного и того же антигена, и могут также представлять собой нуклеотиды, кодирующие легкие цепи, происходящие из связывающих полипептидов, нацеленных против различных антигенов.
В настоящей заявке, термин «фрагменты вектора дисплея», в общем, относится к фрагментам, полученным посредством расщепления вектора дисплея с использованием рестрикционных эндонуклеаз (например, BsmBI и/или SfiI), таким как фрагмент I вектора дисплея и фрагмент II вектора дисплея. 5'-конец и 3'-конец фрагментов вектора дисплея могут включать участки узнавания для рестрикционных эндонуклеаз.
В настоящей заявке, термин «третий полинуклеотид вектора дисплея», в общем, относится к полинуклеотиду, содержащему фрагмент I вектора дисплея, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, третий полинуклеотид вектора дисплея может включать B3-фрагмент I вектора дисплея-S5 в направлении от 5' до 3', где B3, S5 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Например, после расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в третьем полинуклеотиде вектора дисплея (например, рестрикционной эндонуклеазы, узнающей S5, такой как SfiI, или рестрикционной эндонуклеазы, узнающей B3, такой как BsmBI и/или Esp3I), расщепленный третий полинуклеотид вектора дисплея может включать фрагмент I вектора дисплея, который может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «четвертый полинуклеотид вектора дисплея», в общем, относится к полинуклеотиду, содержащему фрагмент II вектора дисплея, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, четвертый полинуклеотид вектора дисплея может включать S6-фрагмент II вектора дисплея-B2 в направлении от 5' до 3', где S6, B2 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Например, после расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в четвертом полинуклеотиде вектора дисплея (например, рестрикционной эндонуклеазы, узнающей S6, такой как SfiI, или рестрикционной эндонуклеазы, узнающей B2, такой как BsmBI и/или Esp3I), расщепленный четвертый полинуклеотид вектора дисплея может включать фрагмент II вектора дисплея, который может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «вектор компонента VH», в общем, относится к кольцевому полинуклеотиду, полученному посредством вставки девятого полинуклеотида в экспрессирующий вектор компонента.
В настоящей заявке, термин «вектор компонента LC», в общем, относится к кольцевому полинуклеотиду, полученному посредством вставки десятого полинуклеотида в экспрессирующий вектор компонента.
В настоящей заявке, термин «экспрессирующий вектор компонента», в общем, относится к вектору, в который можно вставлять полинуклеотиды (например, девятый полинуклеотид и/или десятый полинуклеотид). Экспрессирующий вектор компонента может происходить из вектора pMD. Например, экспрессирующий вектор компонента может представлять собой вектор pMD19 или происходить из вектора pMD19.
В настоящей заявке, термин «девятый полинуклеотид», в общем, относится к полинуклеотиду, содержащему рабочий фрагмент вектора компонента VH, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, девятый полинуклеотид может включать B2-рабочий фрагмент вектора компонента VH-B3 в направлении от 5' до 3', где B2, B3 могут представлять собой участки узнавания для рестрикционной эндонуклеазы.
В настоящей заявке, термин «десятый полинуклеотид», в общем, относится к полинуклеотиду, содержащему рабочий фрагмент вектора компонента LC, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, десятый полинуклеотид может включать S5-рабочий фрагмент вектора компонента LC-S6 в направлении от 5' до 3', где S5, S6 могут представлять собой участки узнавания для рестрикционной эндонуклеазы.
В настоящей заявке, термин «рабочий фрагмент вектора компонента», в общем, относится к любому полинуклеотиду, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце, но никаких участков узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) внутри него. Длина рабочего фрагмента вектора компонента обычно отличается от длины антигенспецифической VH и антигенспецифической LC. В некоторых случаях, длина рабочего фрагмента вектора компонента может составлять приблизительно 1 т.п.о. В некоторых случаях, рабочий фрагмент вектора компонента может происходить из области Fc IgG. Например, рабочий фрагмент вектора компонента может происходить из области Fc, выбранной из группы, состоящей из IgG1 человека и IgG2 человека. Например, участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) могут представлять собой B2 и B3. В качестве другого примера, участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) могут представлять собой S5 и S6.
В настоящей заявке, термин «вектор компонента», в общем, относится к кольцевому полинуклеотиду, полученному посредством вставки девятого полинуклеотида и/или десятого полинуклеотида в экспрессирующий вектор компонента.
В настоящей заявке, термин «девятая бактерия», в общем, относится к бактерии для введения или содержания девятого нуклеотида. Девятая бактерия может включать вектор компонента VH. В настоящей заявке, девятая бактерия может экспрессировать, реплицировать и/или хранить (например, посредством криоконсервирования) девятый нуклеотид и/или вектор компонента VH. В настоящей заявке, плазмиду вектора для хранения компонента VH, содержащую вектор компонента VH, можно получать из девятой бактерии.
В настоящей заявке, термин «десятая бактерия», в общем, относится к бактерии для введения или содержания десятого нуклеотида. Десятая бактерия может включать вектор компонента LC. В настоящей заявке, десятая бактерия может экспрессировать, реплицировать и/или хранить (например, посредством криоконсервирования) десятый нуклеотид и/или вектор компонента LC. В настоящей заявке, плазмиду вектора для хранения компонента LC, содержащую вектор компонента LC, можно получать из десятой бактерии.
В настоящей заявке, термин «шестой полинуклеотид», в общем, относится к полинуклеотиду, содержащему фрагмент лигирования вектора компонента VH, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, шестой полинуклеотид может включать B3-фрагмент лигирования вектора компонента VH-B2 в направлении от 5' до 3', где B3, B2 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Шестой полинуклеотид может быть включен в вектор компонента VH. После расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в шестом полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей B3 и B2, такой как BsmBI и/или Esp3I), шестой полинуклеотид можно высвобождать. Высвобожденный шестой полинуклеотид может включать антигенспецифическую VH, которая может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «высвобожденный шестой полинуклеотид», в общем, относится к фрагменту шестого полинуклеотида, высвобожденному после обработки с использованием вектора компонента VH. В настоящей заявке, обработка может представлять собой расщепление с использованием рестрикционной эндонуклеазы. Например, подходящие рестрикционные эндонуклеазы (например, BsmBI и/или Esp3I) можно выбирать для участков узнавания для рестрикционной эндонуклеазы на векторе компонента VH, таким образом, чтобы высвобожденный шестой полинуклеотид можно было высвобождать из вектора компонента VH и выделять.
В настоящей заявке, термин «восьмой полинуклеотид», в общем, относится к полинуклеотиду, содержащему фрагмент лигирования вектора компонента LC, который может иметь участки узнавания для эндонуклеазы (например, рестрикционной эндонуклеазы) на 5'-конце и/или 3'-конце. Например, восьмой полинуклеотид может включать S6-фрагмент лигирования вектора компонента LC-S5 в направлении от 5' до 3', где S6, S5 могут представлять собой участки узнавания для рестрикционной эндонуклеазы. Восьмой полинуклеотид может быть включен в вектор компонента LC. После расщепления с использованием эндонуклеазы, узнающей участки узнавания эндонуклеазы в восьмом полинуклеотиде (например, рестрикционной эндонуклеазы, узнающей S6 и S5, такой как SfiI), восьмой полинуклеотид можно высвобождать. Высвобожденный восьмой полинуклеотид может включать антигенспецифическую LC, которая может также иметь липкие концы специфической последовательности после расщепления на двух своих концах.
В настоящей заявке, термин «высвобожденный восьмой полинуклеотид», в общем, относится к фрагменту восьмого полинуклеотида, высвобожденного после обработки с использованием вектора компонента LC. В настоящей заявке, обработка может представлять собой расщепление с использованием рестрикционной эндонуклеазы. Например, подходящие рестрикционные эндонуклеазы (например, SfiI) можно выбирать для участков узнавания для рестрикционной эндонуклеазы на векторе компонента LC, таким образом, чтобы высвобожденный восьмой полинуклеотид можно было высвобождать из вектора компонента LC и выделять.
В настоящей заявке, термин «рестрикционная эндонуклеаза», в общем, относится к ферменту, который расщепляет двухцепочечную ДНК. Рестрикционная эндонуклеаза может образовывать липкие концы с выступающей одноцепочечной ДНК, которые могут связываться с ДНК-лигазой. В настоящей заявке, рестрикционная эндонуклеаза может оказывать эффекты узнавания и рестрикционного расщепления. Например, участок расщепления для рестрикционной эндонуклеазы находится на определенном расстоянии от ее участка узнавания. Например, рестрикционная эндонуклеаза может быть выбрана из SfiI, BsmBI и Esp3I.
В настоящей заявке, термин «первая бактерия», в общем, относится к бактерии для введения или содержания вектора дисплея генов антигенспецифических связывающих полипептидов. Первая бактерия может включать антигенспецифическую VH, антигенспецифическую LC, фрагмент I вектора дисплея и фрагмент II вектора дисплея. В настоящей заявке, первая бактерия может экспрессировать, реплицировать и/или хранить (например, посредством криоконсервирования) антигенспецифическую VH, антигенспецифическую LC, фрагмент I вектора дисплея и фрагмент II вектора дисплея, или ДНК экспрессирующего вектора антигенспецифического связывающего полипептида.
В настоящей заявке, термин «бактериальная библиотека дисплея генов антигенспецифических связывающих полипептидов», в общем, относится к бактериальной библиотеке, полученной посредством введения вектора дисплея генов антигенспецифических связывающих полипептидов в первую бактерию. В настоящей заявке, бактериальная библиотека дисплея генов антигенспецифических связывающих полипептидов может представлять собой бактериальную библиотеку, содержащую последовательность нуклеиновой кислоты, кодирующую легкую цепь антигенспецифического связывающего полипептида или вариабельную область тяжелой цепи антигенспецифического полипептида. В настоящей заявке, бактериальная библиотека дисплея генов антигенспецифических связывающих полипептидов может включать от приблизительно 105 до приблизительно 109 (например, она может включать от приблизительно 105 до приблизительно 108, от приблизительно 105 до приблизительно 107, от приблизительно 106 до приблизительно 107) последовательностей нуклеиновой кислоты, кодирующих антигенспецифический связывающий полипептид. В настоящей заявке, бактериальная библиотека дисплея генов антигенспецифических связывающих полипептидов может включать от приблизительно 107 до приблизительно 1012 (например, она может включать от приблизительно 107 до приблизительно 1011, от приблизительно 107 до приблизительно 1010, от приблизительно 107 до приблизительно 109, от приблизительно 107 до приблизительно 108) первых бактерий.
В настоящей заявке, термин «первая бактерия дисплея», в общем, относится к бактерии для введения или содержания первого полинуклеотида вектора дисплея. Первая бактерия может включать дисплей VH. В настоящей заявке, первая бактерия дисплея может экспрессировать, реплицировать и/или хранить (например, посредством криоконсервирования) дисплей VH и/или первый полинуклеотид вектора дисплея.
В настоящей заявке, термин «бактериальная библиотека компонента дисплея VH», в общем, относится к бактериальной библиотеке, полученной посредством введения первого полинуклеотида вектора дисплея в первую бактерию дисплея. В настоящей заявке, бактериальная библиотека компонента дисплея VH может представлять собой бактериальную библиотеку, содержащую последовательность нуклеиновой кислоты, кодирующую вариабельную область тяжелой цепи антигенспецифического полипептида. В настоящей заявке, бактериальная библиотека компонента дисплея VH может включать от приблизительно 105 до приблизительно 109 (например, она может включать от приблизительно 105 до приблизительно 108, от приблизительно 105 до приблизительно 107, от приблизительно 106 до приблизительно 107) последовательностей нуклеиновой кислоты, кодирующих дисплей VH. В настоящей заявке, бактериальная библиотека компонента дисплея VH может включать от приблизительно 107 до приблизительно 1012 (например, она может включать от приблизительно 107 до приблизительно 1011, от приблизительно 107 до приблизительно 1010, от приблизительно 107 до приблизительно 109, от приблизительно 107 до приблизительно 108) первых бактерий дисплея.
В настоящей заявке, термин «вторая бактерия дисплея», в общем, относится к бактерии для введения или содержания второго полинуклеотида вектора дисплея. Вторая бактерия может включать дисплей LC. В настоящей заявке, вторая бактерия дисплея может экспрессировать, реплицировать и/или хранить (например, посредством криоконсервирования) дисплей LC, и/или второй полинуклеотид вектора дисплея.
В настоящей заявке, термин «бактериальная библиотека компонента дисплея LC», в общем, относится к бактериальной библиотеке, полученной посредством введения второго полинуклеотида вектора дисплея во вторую бактерию дисплея. В настоящей заявке, бактериальная библиотека компонента дисплея LC может представлять собой бактериальную библиотеку, содержащую последовательность нуклеиновой кислоты, кодирующую легкую цепь антигенспецифического полипептида. В настоящей заявке, бактериальная библиотека компонента дисплея LC может включать от приблизительно 105 до приблизительно 109 (например, она может включать от приблизительно 105 до приблизительно 108, от приблизительно 105 до приблизительно 107, от приблизительно 106 до приблизительно 107) последовательностей нуклеиновой кислоты, кодирующих дисплей LC. В настоящей заявке, бактериальная библиотека компонента дисплея LC может включать от приблизительно 107 до приблизительно 1012 (например, она может включать от приблизительно 107 до приблизительно 1011, от приблизительно 107 до приблизительно 1010, от приблизительно 107 до приблизительно 109, от приблизительно 107 до приблизительно 108) вторых бактерий дисплей.
В настоящей заявке, термин «третья бактерия дисплея», в общем, относится к бактерии для введения или содержания третьего полинуклеотида вектора дисплея. Третья бактерия может включать фрагмент I вектора дисплея. В настоящей заявке, третья бактерия дисплея может экспрессировать, реплицировать и/или хранить (например, посредством криоконсервирования) фрагмент I вектора дисплея, и/или третий полинуклеотид вектора дисплея.
В настоящей заявке, термин «бактериальная библиотека вектора компонента дисплея I», в общем, относится к бактериальной библиотеке, полученной посредством введения третьего полинуклеотида вектора дисплея в третью бактерию дисплея. В настоящей заявке, бактериальная библиотека вектора компонента дисплея I может представлять собой бактериальную библиотеку, содержащую последовательность нуклеиновой кислоты, кодирующую вектор компонента дисплея I. В настоящей заявке, бактериальная библиотека вектора компонента дисплея I может включать от приблизительно 105 до приблизительно 109 (например, она может включать от приблизительно 105 до приблизительно 108, от приблизительно 105 до приблизительно 107, от приблизительно 106 до приблизительно 107) последовательностей нуклеиновой кислоты, кодирующих дисплей LC. В настоящей заявке, бактериальная библиотека вектора компонента дисплея I может включать от приблизительно 107 до приблизительно 1012 (например, она может включать от приблизительно 107 до приблизительно 1011, от приблизительно 107 до приблизительно 1010, от приблизительно 107 до приблизительно 109, от приблизительно 107 до приблизительно 108) третьих бактерий дисплея.
В настоящей заявке, термин «четвертая бактерия дисплея», в общем, относится к бактерии для введения или содержания четвертого полинуклеотида вектора дисплея. Четвертая бактерия может включать фрагмент II вектора дисплея. В настоящей заявке, четвертая бактерия дисплея может экспрессировать, реплицировать и/или хранить (например, посредством криоконсервирования) фрагмент II вектора дисплея и/или четвертый полинуклеотид вектора дисплея.
В настоящей заявке, термин «бактериальная библиотека вектора компонента дисплея II», в общем, относится к бактериальной библиотеке, полученной посредством введения четвертого полинуклеотида вектора дисплея в четвертую бактерию дисплея. В настоящей заявке, бактериальная библиотека вектора компонента дисплея II может представлять собой бактериальную библиотеку, содержащую последовательность нуклеиновой кислоты, кодирующую вектор компонента дисплея II. В настоящей заявке, бактериальная библиотека вектора компонента дисплея II может включать от приблизительно 105 до приблизительно 109 (например, она может включать от приблизительно 105 до приблизительно 108, от приблизительно 105 до приблизительно 107, от приблизительно 106 до приблизительно 107) последовательностей нуклеиновой кислоты, кодирующих дисплей LC. В настоящей заявке, бактериальная библиотека вектора компонента дисплея II может включать от приблизительно 107 до приблизительно 1012 (например, она может включать от приблизительно 107 до приблизительно 1011, от приблизительно 107 до приблизительно 1010, от приблизительно 107 до приблизительно 109, от приблизительно 107 до приблизительно 108) четвертых бактерий дисплея.
В настоящей заявке, термин «введение», в общем, относится к способу переноса или введения экзогенного полинуклеотида в клетку. Клетка может представлять собой клетку-хозяина. Введенная клетка включает первичные клетки субъекта и их потомство. Клетка может представлять собой прокариотическую клетку, например, она может представлять собой бактериальную клетку.
В настоящей заявке, термин «лигирование», в общем, относится к лигированию двух или более молекул полинуклеотидов вместе. Например, лигирование можно осуществлять посредством лигазы (например, ДНК-лигазы). Например, 3'-конец одного полинуклеотида лигируют с 5'-концом другого полинуклеотида для формирования интактной молекулы полинуклеотида.
В настоящей заявке, термин «клоны», в общем, относится к количеству колоний. Например, клоны могут представлять собой количество колоний в бактериальной библиотеке (например, в бактериальной библиотеке компонента легкой цепи, бактериальной библиотеке компонента тяжелой цепи, бактериальной библиотеке дисплея и/или фаговой библиотеке). В некоторых случаях, клоны могут представлять собой количество различных колоний в бактериальной библиотеке. В некоторых случаях, клоны могут представлять собой количество популяций потомства, образованных отдельным клоном.
В настоящей заявке, термины «полинуклеотид», «нуклеотид», «нуклеотидная последовательность», «нуклеиновая кислота» и «олигонуклеотид» могут быть использованы взаимозаменяемо, и, в общем, относятся к полимерной форме нуклеотидов любой длины, таких как дезоксирибонуклеотид или рибонуклеотид, или их аналоги, включая, например, 200, 300, 500, 1000, 2000, 3000, 5000, 7000, 10000, 100000 и т.д. Полинуклеотид может содержать фосфодиэфирные связи.
В настоящей заявке, термин «и/или» следует понимать как обозначающий любую или обе из альтернатив.
В настоящей заявке, термин «содержать», в общем, относится к включению явно указанных признаков, но не к исключению других элементов.
В настоящей заявке, термин «приблизительно», в общем, относится к изменчивости в рамках диапазона 0,5%-10% выше или ниже указанного значения, например, к изменчивости в рамках диапазона 0,5%, 1%, 1,5%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5%, 5%, 5,5%, 6%, 6,5%, 7%, 7,5%, 8%, 8,5%, 9%, 9,5% или 10% выше или ниже указанного значения.
Подробное описание
В одном аспекте, настоящее изобретение относится к способу конструирования вектора дисплея генов антигенспецифических связывающих полипептидов.
Вектор дисплея генов антигенспецифических связывающих полипептидов по настоящему изобретению может состоять из четырех фрагментов, связанных посредством направленной циклизации, которые могут, соответственно, включать антигенспецифическую VH, антигенспецифическую LC, фрагмент I вектора дисплея и фрагмент II вектора дисплея.
Вектор компонента
Способ по настоящему изобретению может включать конструирование вектора компонента, который может включать получение полинуклеотидов, например, девятого полинуклеотида и десятого полинуклеотида. Девятый полинуклеотид может включать B2-рабочий фрагмент вектора компонента VH-B3 в направлении от 5' до 3', и десятый полинуклеотид включает S5-рабочий фрагмент вектора компонента LC-S6 в направлении от 5' до 3'. Рабочий фрагмент вектора может происходить из фрагмента Fc IgG. Например, фрагмента Fc IgG1 человека. В качестве другого примера, фрагмент Fc IgG2 человека. В некоторых случаях, девятый нуклеотид можно получать посредством амплификации с использованием DDB214 и DDB215 в качестве праймеров, и фрагмента Fc IgG1 человека в качестве матрицы, где DDB214 может включать аминокислотную последовательность, как указано в SEQ ID NO: 1, и DDB215 может включать аминокислотную последовательность, как указано в SEQ ID NO: 2. В некоторых случаях, десятый нуклеотид можно получать посредством амплификации с использованием DDB216 и DDB217 в качестве праймеров, и фрагмента Fc IgG1 человека в качестве матрицы, где DDB216 может включать аминокислотную последовательность, как указано в SEQ ID NO: 3, и DDB217 может включать аминокислотную последовательность, как указано в SEQ ID NO: 4. Девятый нуклеотид может включать участки узнавания B2 и B3 рестрикционной эндонуклеазы на двух своих концах, и десятый нуклеотид может включать участки узнавания S5 и S6 рестрикционной эндонуклеазы на двух своих концах.
Способ может включать вставку полинуклеотидов (например, девятого полинуклеотида и десятого полинуклеотида) в экспрессирующий вектор компонента для получения вектора компонента (например, вектора компонента VH и вектора компонента LC).
Экспрессирующий вектор компонента может происходить из любого вектора, например, любого вектора, который можно амплифицировать и/или легко сохранять. В некоторых случаях, вектор, используемый в качестве экспрессирующего вектора компонента, может иметь такие свойства, как высокое количество копий, небольшая молекулярная масса и т.д. В некоторых случаях, экспрессирующий вектор компонента может происходить из вектора pMD. Например, экспрессирующий вектор компонента может представлять собой вектор pMD19 или происходить из вектора pMD19.
В настоящей заявке, для цели конструирования экспрессирующего вектора компонента, вектор pMD или вектор, происходящий из pMD, можно конструировать/модифицировать. Например, один или несколько участков узнавания для эндонуклеазы в векторе можно удалять посредством сайт-направленного мутагенеза (например, удаления одного или нескольких участков узнавания для BsmBI и/или SfiI в нем). В некоторых случаях, один или несколько участков узнавания для эндонуклеазы можно также добавлять в вектор посредством сайт-направленного мутагенеза (например, добавления одного или нескольких участков узнавания для BsmBI и/или SfiI в выбранных локализациях).
Например, посредством сайт-направленного мутагенеза, один или несколько участков узнавания для BsmBI, исходно содержащиеся в векторе, можно удалять, и один или несколько дополнительных участков узнавания для BsmBI можно затем добавлять в другой локализации в векторе, для получения таким образом модифицированного вектора (например, модифицированного вектора pMD).
В настоящей заявке, экспрессирующий вектор компонента (например, экспрессирующий вектор компонента из вектора компонента VH) может включать участок узнавания для BsmBI. В некоторых случаях, экспрессирующий вектор компонента может включать два участка узнавания для BsmBI.
В качестве другого примера, посредством сайт-направленного мутагенеза, один или несколько участков узнавания для SfiI, исходно содержащиеся в векторе, можно удалять, и один или несколько дополнительных участков узнавания для SfiI можно затем добавлять в другой локализации в векторе, для получения таким образом модифицированного вектора (например, модифицированного вектора pMD).
В настоящей заявке, экспрессирующий вектор компонента (например, экспрессирующий вектор компонента из вектора компонента LC) может включать участок узнавания для SfiI. В некоторых случаях, экспрессирующий вектор компонента может включать два участка узнавания для SfiI.
В некоторых случаях, способ может дополнительно включать вставку полинуклеотидов (например, девятого полинуклеотида и десятого полинуклеотида) в экспрессирующий вектор компонента для получения плазмиды вектора для хранения компонента (например, плазмиды вектора для хранения компонента VH и плазмиды вектора для хранения компонента LC), и затем введение плазмиды вектора для хранения компонента в бактерии (например, девятую бактерию и десятую бактерию) для получения бактериальной библиотеки вектора для хранения компонента (например, бактериальной библиотеки вектора для хранения компонента VH и бактериальной библиотеки вектора для хранения компонента LC).
Например, на ФИГ. 3 показан вектор компонента VH по настоящему изобретению, который можно получать посредством лигирования, посредством вставки девятого полинуклеотида в экспрессирующий вектор компонента. Вектор компонента VH включает шестой полинуклеотид, который может включать B3-фрагмент лигирования вектора компонента VH-B2 в направлении от 5' до 3', где B2 и B3 могут быть специфически узнаны и расщеплены посредством BsmBI и/или Esp3I, соответственно. Например, B2 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 8, и B3 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 9.
После обработки (например, расщепления), вектор компонента VH может образовывать высвобожденный шестой полинуклеотид, который может иметь липкие концы специфической последовательности, полученные после расщепления, на его 5'- и 3'-концах.
Например, на ФИГ. 4 показан вектор компонента LC по настоящему изобретению, который можно получать посредством лигирования, посредством вставки десятого полинуклеотида в экспрессирующий вектор компонента. Вектор компонента LC включает восьмой полинуклеотид, который может включать S6-фрагмент лигирования вектора компонента LC-S5 в направлении от 5' до 3', где S6 и S5 могут быть специфически узнаны и расщеплены посредством SfiI, соответственно. Например, S6 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 11, и S5 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 10.
После обработки (например, расщепления), вектор компонента LC может образовывать высвобожденный восьмой полинуклеотид, который может иметь липкие концы специфической последовательности, полученные после расщепления, на его 5'- и 3'-концах.
Антигенспецифическая VH и антигенспецифическая LC
Способ по настоящему изобретению включает получение пятого полинуклеотида, содержащего B-антигенспецифическую VH-B в направлении от 5' до 3', где B представляет собой участок узнавания для рестрикционной эндонуклеазы, которая может специфически узнавать B2 и/или B3. Например, антигенспецифическую VH можно амплифицировать с использованием антигенспецифического фрагмента VH в качестве матрицы, таким образом, чтобы присоединять 5'- и 3’-концы антигенспецифической VH к участкам узнавания для рестрикционной эндонуклеазы (например, BsmBI и/или Esp3I).
Способ по настоящему изобретению включает получение седьмого полинуклеотида, содержащего S-антигенспецифическую LC-S в направлении от 5' до 3', где S представляет собой участок узнавания для рестрикционной эндонуклеазы, которая может специфически узнавать S5 и/или S6. Например, антигенспецифическую LC можно амплифицировать с использованием антигенспецифического фрагмента LC в качестве матрицы, таким образом, чтобы присоединять 5' и 3'-концы антигенспецифической LC к участкам узнавания для рестрикционной эндонуклеазы (например, SfiI).
В некоторых случаях, антигенспецифический фрагмент VH и антигенспецифический фрагмент LC можно получать посредством способов из предшествующей области техники. Например, их можно получать от животных, иммунизированных с использованием антигенов, и можно также получать из библиотеки антител, включая комбинаторную библиотеку антител, библиотеку фагового дисплея, библиотеку дисплея на поверхности дрожжей, библиотеку рибосомного дисплея, библиотеку дисплея мРНК.
Библиотека компонента
Способ по настоящему изобретению может включать расщепление пятого полинуклеотида и вектора компонента VH с использованием рестрикционной эндонуклеазы для получения расщепленного пятого полинуклеотида и высвобожденного шестого полинуклеотида, и затем смешивание расщепленного пятого полинуклеотида и высвобожденного шестого полинуклеотида, таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования библиотеки компонента антигенспецифической VH.
Библиотека компонента антигенспецифической VH может включать антигенспецифическую VH. После расщепления библиотеки компонента антигенспецифической VH с использованием рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, узнающей B2 и B3), можно получать высвобожденную антигенспецифическую VH, 5'- и 3'-концы которой могут иметь липкие концы специфической последовательности.
Способ по настоящему изобретению может включать расщепление седьмого полинуклеотида и вектора компонента LC с использованием рестрикционной эндонуклеазы для получения расщепленного седьмого полинуклеотида и высвобожденного восьмого полинуклеотида, и затем смешивание расщепленного седьмого полинуклеотида и высвобожденного восьмого полинуклеотида, таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования библиотеки компонента антигенспецифической LC.
Библиотека компонента антигенспецифической LC может включать антигенспецифическую LC. После расщепления библиотеки компонента антигенспецифической LC с использованием рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, узнающей S5 и S6), можно получать высвобожденную антигенспецифическую LC, 5'- и 3'-концы которой могут иметь липкие концы специфической последовательности.
Вектор дисплея
Способ по настоящему изобретению может также включать конструирование вектора дисплея, который может состоять из четырех полинуклеотидов вектора дисплея (например, первого полинуклеотида вектора дисплея, второго полинуклеотида вектора дисплея, третьего полинуклеотида вектора дисплея и четвертого полинуклеотида вектора дисплея).
Полинуклеотиды вектора дисплея по настоящему изобретению (первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и четвертый полинуклеотид вектора дисплея) могут включать антигенсвязывающие полипептиды или их фрагменты, такие как дисплей LC и/или дисплей VH. В настоящей заявке, дисплей LC может кодировать легкую цепь антигенсвязывающего полипептида, дисплей VH может кодировать вариабельную область тяжелой цепи антигенсвязывающего полипептида, и легкая цепь может связываться с вариабельной областью тяжелой цепи для формирования Fab, узнающего мишень. В некоторых случаях, мишень может представлять собой антиген. Например, мишень представляет собой PD-1.
Полинуклеотиды вектора дисплея по настоящему изобретению (например, первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и четвертый полинуклеотид вектора дисплея) могут включать фрагменты вектора дисплея, такие как фрагмент I вектора дисплея и фрагмент II вектора дисплея. Желательную длину или тип фрагмента I вектора дисплея и фрагмента II вектора дисплея можно выбирать, соответственно, в соответствии с длиной или характером антигенсвязывающего полипептида или его фрагментов, подлежащих экспрессии, и длиной или характером участков рестрикции.
В некоторых случаях, фрагмент I вектора дисплея и фрагмент II вектора дисплея могут происходить из любого фрагмента вектора, способного экспрессировать ген-мишень. Например, фрагмент I экспрессирующего вектора и фрагмент II экспрессирующего вектора могут происходить из фрагментов вектора дисплея pDGB4 (применительно к pDGB4, см. Ivan Zhou, et al., «Four-way ligation for construction of a mammalian cell-based full-length antibody display library», Acta Biochim Biophys Sin 2011, 43: 232-238).
Фрагменты вектора дисплея по настоящему изобретению (например, фрагмент I вектора дисплея и фрагмент II вектора дисплея) могут включать нуклеотидные последовательности со специфическими функциями, включая, но без ограничения, промоторы, энхансеры, сигнальные пептиды, маркеры для скрининга (например, они могут включать участки узнавания ферментов, гены устойчивости, репортерные гены и гены для скрининга), которые может корректировать в фрагментах вектора дисплея специалист в данной области, в соответствии с желательной функцией (вставляя/заменяя и/или делетируя вышеуказанные нуклеотидные последовательности со специфическими функциями). В некоторых случаях, фрагменты вектора дисплея можно корректировать в различных случаях для получения различных нуклеотидных последовательностей.
В настоящей заявке, первый полинуклеотид вектора дисплея может включать B2-дисплей VH-B3 в направлении от 5' до 3', где каждый из B2 и B3 могут независимо представлять собой участки узнавания для рестрикционной эндонуклеазы, и дисплей VH может кодировать вариабельную область тяжелой цепи антигенсвязывающего полипептида. В некоторых случаях, B2 и B3 могут быть специфически узнаны и расщеплены посредством BsmBI, соответственно. Например, B2 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 8, и B3 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 9.
Второй полинуклеотид вектора дисплея может включать S5-дисплей LC-S6 в направлении от 5' до 3', где каждый из S5 и S6 могут независимо представлять собой участки узнавания для рестрикционной эндонуклеазы, и дисплей LC может кодировать легкую цепь антигенсвязывающего полипептида. В некоторых случаях, S5 и S6 могут быть специфически узнаны и расщеплены посредством SfiI, соответственно. Например, S5 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 10, и S6 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 11.
Третий полинуклеотид вектора дисплея может включать B3-фрагмент I вектора дисплея-S5 в направлении от 5' до 3', где каждый из B3 и S5 могут независимо представлять собой участки узнавания для рестрикционной эндонуклеазы. В некоторых случаях, S5 может быть специфически узнан и расщеплен посредством Sfil, и B3 может быть специфически узнан и расщеплен посредством BsmBI и/или Esp3I. Например, B3 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 9, и S5 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 10.
Четвертый полинуклеотид вектора дисплея может включать S6-фрагмент II вектора дисплея-B2 в направлении от 5' до 3', где каждый из S6 и B2 могут независимо представлять собой участки узнавания для рестрикционной эндонуклеазы. В некоторых случаях, S6 может быть специфически узнан и расщеплен посредством Sfil, и B2 может быть специфически узнан и расщеплен посредством BsmBI и/или Esp3I. Например, B2 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 8, и S6 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 11.
Первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и/или четвертый полинуклеотид вектора дисплея по настоящему изобретению можно получать из материалов образцов. В некоторых случаях, материалы образцов могут включать нацеленные на антигены антитела или их антигенсвязывающие фрагменты. Антигены могут представлять собой любые иммуногенные фрагменты или детерминанты, включая, но без ограничения, PD-1, PD-L1, LAG-3, CD47, CD3. Например, антитела или их антигенсвязывающие фрагменты нацелены на PD-1.
Для скрининга положительных бактерий, в которые введены полинуклеотиды вектора дисплея, полинуклеотиды вектора дисплея (например, первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и четвертый полинуклеотид вектора дисплея) могут также включать последовательности нуклеиновой кислоты, кодирующие сигнальные пептиды, например, сигнальные пептиды, экспрессирующие природные гены устойчивости. В одном примере, 3'-конец последовательности нуклеиновой кислоты, кодирующей сигнальный пептид, может связываться с участком рестрикции на 5'-конце полинуклеотида. В некоторых случаях, для введения подходящего участка рестрикции в 3’-концевую часть последовательности нуклеиновой кислоты, кодирующей сигнальный пептид, ее последовательность оснований может быть изменена посредством непреднамеренной мутации, однако, аминокислотная последовательность сигнального пептида остается неизменной. Например, последовательность нуклеиновой кислоты, кодирующая сигнальный пептид, может включать последовательность нуклеиновой кислоты, как указано в любой, выбранной из SEQ ID NO: 12 и SEQ ID NO: 14; альтернативно, сигнальный пептид может включать аминокислотную последовательность, как указано в любой, выбранной из SEQ ID NO: 13 и SEQ ID NO: 15.
Полинуклеотиды можно получать посредством общепринятых в данной области способов, которые могут включать, но без ограничения: стандартную ПЦР, протяженную ПЦР, ПЦР с горячим стартом, qПЦР, RT-ПЦР и изотермическую амплификацию. В некоторых случаях, праймеры можно конструировать, в соответствии с последовательностями целевых фрагментов (например, дисплея LC, дисплея VH, фрагмента I вектора дисплея и фрагмента II вектора дисплея), соответственно, которые в таком случае используют в качестве матриц для амплификации, для получения полинуклеотидов. Например, праймеры для амплификации дисплея LC могут включать нуклеотидные последовательности, как указано в SEQ ID NO: 20 и SEQ ID NO: 21. Например, праймеры для амплификации дисплея VH могут включать нуклеотидные последовательности, как указано в SEQ ID NO: 22 и SEQ ID NO: 23. Например, праймеры для амплификации фрагмента I вектора дисплея могут включать нуклеотидные последовательности, как указано в SEQ ID NO: 18 и SEQ ID NO: 19. Например, праймеры для амплификации фрагмента II вектора дисплея могут включать нуклеотидные последовательности, как указано в SEQ ID NO: 16 и SEQ ID NO: 17.
После получения полинуклеотидов вектора дисплея, их можно по отдельности вводить в бактерию для получения бактериальной библиотеки. Таким образом, способ по настоящему изобретению может дополнительно включать следующие стадии: введения первого полинуклеотида вектора дисплея в первую бактерию дисплея для получения бактериальной библиотеки компонента дисплея VH; введения второго полинуклеотида вектора дисплея во вторую бактерию дисплея для получения бактериальной библиотеки компонента дисплея LC; введения третьего полинуклеотида вектора дисплея в третью бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея I; и введения четвертого полинуклеотида вектора дисплея в четвертую бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея II.
В настоящей заявке, первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и четвертый полинуклеотид вектора дисплея все могут представлять собой линейные молекулы нуклеиновой кислоты.
В некоторых случаях, полинуклеотиды вектора дисплея можно вставлять в вектор компонента дисплея для получения продуктов лигирования для хранения. В некоторых случаях, полинуклеотиды можно вставлять в векторы компонентов посредством использования клонирования продуктов ПЦР. Векторы компонентов могут включать плазмидные векторы (например, pBR322, векторы pUC), фаговые векторы (например, вектор M13, вектор λ), происходящие из фагов плазмиды (например, фагмиду, космиду) и бактериальную искусственную хромосому (BAC). В некоторых вариантах осуществления, вектор компонента может происходить из вектора pUC. Например, вектор компонента может представлять собой вектор pUC19 или происходить из вектора pUC19.
Затем продукты лигирования для хранения можно вводить в бактерию для получения бактериальной библиотеки дисплея.
В настоящей заявке, бактериальная библиотека дисплея (например, бактериальная библиотека компонента дисплея VH и бактериальная библиотека компонента дисплея LC) может включать приблизительно по меньшей мере 10 (например, приблизительно по меньшей мере 100, по меньшей мере 200, по меньшей мере 300, по меньшей мере 400, по меньшей мере 500, по меньшей мере 600, по меньшей мере 800, приблизительно по меньшей мере 1000, приблизительно по меньшей мере 10000 или более) различных клонов.
В настоящей заявке, бактериальная библиотека дисплея (например, бактериальная библиотека вектора компонента дисплея I и бактериальная библиотека вектора компонента дисплея II) может включать приблизительно по меньшей мере 10 (например, приблизительно по меньшей мере 100, по меньшей мере 200, по меньшей мере 300, по меньшей мере 400, по меньшей мере 500, по меньшей мере 600, по меньшей мере 800, приблизительно по меньшей мере 1000, приблизительно по меньшей мере 10000 или более) идентичных клонов.
В настоящей заявке, доля эффективных клонов в бактериальной библиотеке дисплея (например, бактериальной библиотеке компонента дисплея VH, бактериальной библиотеке компонента дисплея LC, бактериальной библиотеке вектора компонента дисплея I и бактериальной библиотеке вектора компонента дисплея II) может составлять по меньшей мере приблизительно 50% (например, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 99%).
В настоящей заявке, бактерии в бактериальной библиотеке дисплея (например, бактериальной библиотеке компонента дисплея VH, бактериальной библиотеке компонента дисплея LC, бактериальной библиотеке вектора компонента дисплея I и бактериальной библиотеке вектора компонента дисплея II) можно культивировать в жидкости. Время для жидкой культуры может составлять не более чем приблизительно 8 часов, например, оно может составлять не более чем приблизительно 4 часа, не более чем приблизительно 5 часов, не более чем приблизительно 6 часов или не более чем приблизительно 7 часов. В настоящей заявке, операции с жидкой культурой являются простыми. В некоторых случаях, бактерии в бактериальной библиотеке дисплея можно культивировать в небольшом количестве бактериального бульона с распределением по чашке, и затем колонии сортируют. Время для культуры на чашках может составлять приблизительно 12-18 часов, например, оно может составлять приблизительно 12 часов, 13 часов, 14 часов, 15 часов, 16 часов, 17 часов или 18 часов. В настоящей заявке, культура на чашках позволяет отбор колоний (например, отбор моноклонов) до анализа секвенирования.
Вектор дисплея генов антигенспецифических связывающих полипептидов
Способ по настоящему изобретению может дополнительно включать специфическое расщепление библиотеки компонента VH, библиотеки компонента LC и вектора дисплея с использованием рестрикционных эндонуклеаз (например, рестрикционных эндонуклеаз, специфически узнающих S5, S6, B2 и B3) для получения высвобожденного первого полинуклеотида, высвобожденного второго полинуклеотида, высвобожденного третьего полинуклеотида и высвобожденного четвертого полинуклеотида.
5’-конец высвобожденного первого полинуклеотида может иметь липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей B2, например, BsmBI и/или Esp3I), и 3'-конец может иметь липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей B3, например, BsmBI и/или Esp3I).
5'-конец высвобожденного второго полинуклеотида имеет липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей S5, например, SfiI), и 3'-конец имеет липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей S6, например, SfiI).
5'-конец высвобожденного третьего полинуклеотида имеет липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей B3, например, BsmBI и/или Esp3I), и 3'-конец имеет липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей S5, например, SfiI).
5'-конец высвобожденного четвертого полинуклеотида имеет липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей S6, например, SfiI), и 3'-конец имеет липкие концы после расщепления посредством рестрикционной эндонуклеазы (например, рестрикционной эндонуклеазы, специфически узнающей B2, например, BsmBI и/или Esp3I).
В настоящей заявке, первый полинуклеотид, второй полинуклеотид, третий полинуклеотид и четвертый полинуклеотид все могут представлять собой линейные молекулы нуклеиновой кислоты.
Способ по настоящему изобретению может дополнительно включать смешивание высвобожденного первого полинуклеотида, высвобожденного второго полинуклеотида, высвобожденного третьего полинуклеотида и высвобожденного четвертого полинуклеотида таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования вектора дисплея генов антигенспецифических связывающих полипептидов. В некоторых случаях, направленное лигирование может включать использование лигазы, например, ДНК-лигазы T4.
В некоторых случаях, вектор дисплея генов антигенспецифических связывающих полипептидов можно вводить в бактерию для получения бактериальной библиотеки дисплея генов антигенспецифических связывающих полипептидов.
В настоящей заявке, бактериальная библиотека дисплея генов антигенспецифических связывающих полипептидов может включать приблизительно по меньшей мере 10 (например, приблизительно по меньшей мере 100, приблизительно по меньшей мере 200, приблизительно по меньшей мере 300, приблизительно по меньшей мере 400, приблизительно по меньшей мере 500, приблизительно по меньшей мере 1000, приблизительно по меньшей мере 10000 или более) клонов.
В настоящей заявке, доля эффективных клонов в бактериальной библиотеке дисплея генов антигенспецифических связывающих полипептидов может составлять по меньшей мере приблизительно 50% (например, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 99%).
Время, необходимое от конструирования вектора дисплея до скрининга антигенспецифического связывающего полипептида уникальной последовательности посредством использования способа по настоящему изобретению, может составлять по меньшей мере приблизительно 1 неделю (например, по меньшей мере приблизительно 10 суток, по меньшей мере приблизительно 2 недели, по меньшей мере приблизительно 3 недели, по меньшей мере приблизительно 4 недели).
В настоящей заявке, бактерии в бактериальной библиотеке дисплея генов антигенспецифических связывающих полипептидов можно культивировать в жидкости. Время для жидкой культуры может составлять не более чем приблизительно 24 часа, например, оно может составлять не более чем приблизительно 5 часов, может составлять не более чем приблизительно 10 часов, может составлять не более чем приблизительно 15 часов, не более чем приблизительно 20 часов, не более чем приблизительно 22 часа или не более чем приблизительно 24 часа. В настоящей заявке, операции с жидкой культурой являются простыми. В некоторых случаях, бактерии в бактериальной библиотеке можно культивировать в небольшом количестве бактериального бульона с распределением по чашке, и затем колонии отбирают. Время для культуры на чашках может составлять приблизительно 12-18 часов, например, оно может составлять приблизительно 12 часов, 13 часов, 14 часов, 15 часов, 16 часов, 17 часов или 18 часов. В настоящей заявке, культура на чашках позволяет отбор колоний (например, отбор моноклонов) до анализа секвенирования.
Способ может дополнительно включать введение вектора дисплея генов антигенспецифических связывающих полипептидов в клетку и получение антигенспецифического связывающего полипептида из клетки. Например, клетка может представлять собой клетку млекопитающего. Антигенспецифический связывающий полипептид можно затем получать из клетки.
Участок рестрикции
В настоящей заявке, последовательность участка узнавания для рестрикционной эндонуклеазы сконструирована как не включенная в полинуклеотид, кодирующий антигенсвязывающий полипептид или его фрагменты. Рестрикционные эндонуклеазы по настоящему изобретению могут специфически узнавать B2, B3, S5 и S6, соответственно. При этом, каждый из B2, B3, S5 и S6 могут независимо представлять собой участки узнавания для рестрикционной эндонуклеазы.
Участки узнавания для рестрикционных эндонуклеаз в настоящей заявке могут подвергаться специфическому узнаванию посредством 1, 2, 3, 4 или более рестрикционных эндонуклеаз, соответственно. В некоторых случаях, рестрикционные эндонуклеазы могут быть выбраны из SfiI, BsmBI и/или Esp3I. В других случаях, могут быть выбраны также другие целесообразные рестрикционные эндонуклеазы.
В настоящей заявке, рестрикционная эндонуклеаза может быть выбрана из SfiI, BsmBI и Esp3I. В настоящей заявке, BsmBI и Esp3I могут представлять собой изоферменты, которые могут узнавать одинаковые участки узнавания для рестрикционной эндонуклеазы.
В настоящей заявке, например, S5 и S6 могут быть узнаны и расщеплены посредством SfiI. В настоящей заявке, например, B2 и B3 могут быть узнаны и расщеплены посредством BsmBI и/или Esp3I.
Например, SfiI может узнавать последовательность, состоящую из 13 оснований (от 5' до 3') GGCCNNNN/NGGCC, которая может формировать выступающую последовательность (например, одноцепочечную последовательность, содержащую 3 основания) на 3'-конце после ферментного расщепления, где N может представлять собой любое из четырех оснований GATC. Таким образом, существуют 4^5 различных последовательностей, которые могут быть узнаны посредством SfiI.
Например, BsmBI и Esp3I могут узнавать последовательность, состоящую из 12 оснований (от 5' до 3') CGTCTCN/NNNNN, которая может формировать выступающую последовательность (например, одноцепочечную последовательность, содержащую 4 основания) на 5'-конце после ферментного расщепления, где N может представлять собой любое из четырех оснований GATC. Таким образом, существуют 4^6 различных последовательностей, которые могут быть узнаны посредством BsmBI и Esp3I.
В некоторых случаях, участки узнавания для рестрикционных эндонуклеаз могут представлять собой участки, специфически узнаваемые и расщепляемые посредством SfiI. Например, они могут быть обозначены как S5 и S6, соответственно. Например, S5 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 10. В качестве другого примера, S6 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 11.
Участки узнавания для рестрикционных эндонуклеаз могут представлять собой участки, специфически узнаваемые и расщепляемые посредством BsmBI и/или Esp3I. Например, они могут быть обозначены как B2 и B3, соответственно. Например, B2 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 8. В качестве другого примера, B3 может включать последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 9.
Следует отметить, что участки узнавания для рестрикционных эндонуклеаз в настоящей заявке включают, но без ограничения, участки узнавания, перечисленные в настоящем описании, и могут также включать участки узнавания для других рестрикционных эндонуклеаз, не перечисленные, так же как другие участки узнавания для рестрикционных эндонуклеаз, при условии, что они не вызывают нежелательного узнавания или расщепления целевой последовательности (например, полинуклеотида, кодирующего антигенсвязывающий полипептид или его фрагменты).
В другом аспекте, настоящее изобретение, кроме того, относится к бактериальной библиотеке дисплея генов антигенспецифических связывающих полипептидов.
В другом аспекте, настоящее изобретение, кроме того, относится к вектору дисплея, полученному посредством этого способа.
Без намерения быть ограниченными какой-либо теорией, следующие примеры представлены только для иллюстрации слитого белка, способа его получения и применения по настоящему изобретению, и не использованы для ограничения изобретательского объема настоящего изобретения.
Примеры
Пример 1 Конструирование библиотеки дисплея антител (Fab) на поверхности фага из PBMC человека
1.1 Получение материалов тотальной РНК/мРНК после иммунизации
Тотальную РНК выделяли из лимфоцитов периферической крови человека, и мРНК далее выделяли из тотальной РНК (Takara, кат. # Z652N/636592, конкретные стадии тестирования см. в инструкции к продукту).
1.2 Конструирование и синтез праймеров
Руководствуясь Phage Display (A Laboratory Manual, ISBN 0-87969-546-3), конструировали праймеры для вариабельной области тяжелой цепи VH, легкой цепи KLC (полноразмерной легкой цепи каппа) и легкой цепи LLC (полноразмерной легкой цепи лямбда) человека. При этом, 5'-конец прямого праймера для легкой цепи содержит нуклеотидную последовательность GGCCCAGGCGGCC (SEQ ID NO: 78) из R1, и 5'-конец обратного праймера содержит нуклеотидную последовательность GGCCACATAGGCC (SEQ ID NO: 79) из R2; 5'-конец прямого праймера для вариабельной области тяжелой цепи содержит нуклеотидную последовательность GGCCCAACCGGCC (SEQ ID NO: 80) из R5, и 5'-конец обратного праймера содержит нуклеотидную последовательность GGCCCTCAGCGGCC (SEQ ID NO: 81) из R6. Праймеры были синтезированы в GENEWIZ.
Прямой и обратный праймеры для амплификации линкеров конструировали с использованием вектора pComb3x в качестве матрицы. При этом, 5'-конец прямого праймера содержит нуклеотидную последовательность GGCCACATAGGCC (SEQ ID NO: 79) из R3, и 5'-конец обратного праймера содержит нуклеотидную последовательность GGCCCAACCGGCC (SEQ ID NO: 80) из R4.
Конкретные последовательности праймеров можно увидеть в таблице 1-1 ниже:
1.3 Получение первого полинуклеотида и третьего полинуклеотида
Компонент библиотеки генов антител амплифицировали посредством двухстадийного способа.
Стадия 1. С использованием мРНК, полученной в примере 1.1, в качестве матрицы, кДНК синтезировали посредством обратной транскрипции с использованием MMLV от Promega (в соответствии с инструкцией к продукту от Promega Co., где праймер представлял собой Thermo, кат. # N8080127, и обратная транскриптаза представляла собой Promega, кат. # M1701).
Стадия 2. С использованием кДНК, полученной на стадии 1, в качестве матрицы, и с использованием праймеров, полученных в примере 1.2, библиотеки генов компонентов антител KLC, LLC и VH амплифицировали посредством ПЦР (Takara, кат. # RR900A, в соответствии с инструкцией к продукту от компании). После очистки и выделения посредством электрофореза в геле (с использованием набора для экстракции из геля Axygen, и действий в соответствии с инструкцией в «Molecular Cloning: A Laboratory Manual»), получали продукты ПЦР, представляющие собой фрагменты KLC, фрагменты LLC и фрагменты VH, соответственно.
1.4 Конструирование векторов для хранения
1.4.1 Конструирование праймеров
Праймеры конструировали, руководствуясь содержанием примера 1.2.
Праймеры для получения векторов для хранения конструировали и синтезировали. Конкретные последовательности праймеров можно увидеть в таблице 1-2 ниже:
Таблица 1-2 Последовательности праймеров-2
1.4.2 Амплификация ПЦР
С использованием Fc IgG1 человека (SEQ ID NO: 164) длиной 1 т.п.о. в качестве матрицы, ПЦР проводили с использованием праймеров, полученных в примере 1.4.1, представляющих собой прямой праймер для R1-1,0 т.п.о.-R2 и обратный праймер для R1-1,0 т.п.о.-R2. После очистки и выделения посредством электрофореза в геле (с использованием набора для экстракции из геля Axygen), получали продукт ПЦР, R1-1,0 т.п.о.-R2 (SEQ ID NO: 165).
С использованием вектор pComb3x в качестве матрицы, ПЦР проводили с использованием прямого праймера для R3-линкера-R4 (SEQ ID NO: 160) и обратного праймера для R3-линкера-R4 (SEQ ID NO: 161). После очистки и выделения посредством электрофореза в геле (с использованием набора для экстракции из геля Axygen), получали продукт ПЦР R3-линкер-R4 (SEQ ID NO: 167). При этом, линкер может иметь длину 72 п.о., и его нуклеотидная последовательность является такой, как указано в SEQ ID NO:166.
С использованием Fc IgG1 человека (SEQ ID NO: 164) длиной 1 т.п.о. в качестве матрицы, ПЦР проводили с использованием праймеров, полученных в примере 1.4.1, представляющих собой прямой праймер для R5-1,0 т.п.о.-R6 и обратный праймер для R5-1,0 т.п.о.-R6. После очистки и выделения посредством электрофореза в геле, получали продукт ПЦР R5-1,0 т.п.о.-R6 (SEQ ID NO: 168).
1.4.3 Конструирование векторов для хранения легкой цепи и векторов для хранения тяжелой цепи
Посредством способа клонирования TA (набора для клонирования TA, закупленного из Takara Co.), фрагмент R1-1,0 т.п.о.-R2, полученный в 1.4.2, вставляли в вектор pMD19-T для получения вектора для хранения легкой цепи DDB-R1-1,0 т.п.о.-R2 для вставки в библиотеку генов полноразмерных легких цепей, и карта вектора для нее показана на ФИГ. 9.
Посредством способа клонирования TA (набора для клонирования TA, закупленного из Takara Co.), фрагмент R5-1,0 т.п.о.-R6, полученный в 1.4.2, вставляли в вектор pMD19-T для получения вектора, содержащего фрагмент R5-1,0 т.п.о.-R6, который затем использовали в качестве матрицы для удаления исходного участка рестрикции BsmBI в этом векторе посредством мутации праймера, для получения таким образом вектора для хранения тяжелой цепи DDB-R5-1,0 т.п.о.-R6, для вставки в библиотеку генов VH, и карта вектора для нее показана на ФИГ. 10.
Конкретные последовательности праймеров можно увидеть в таблице 1-3 ниже:
1.4.4 Конструирование векторов для хранения линкеров и получение бактерий для компонента линкера
Посредством способа клонирования TA (набора для клонирования TA, закупленного из Takara Co.), R3-линкер-R4, поученный в примере 1.4.2, вставляли в вектор pMD19-T для получения вектора для хранения линкера DDB-R3-линкер-R4, и карта вектора для него показана на ФИГ. 11. Компетентные бактерии TG1 (Lucigen Co.) трансформировали с использованием вектора для хранения линкера, и культивировали в течение ночи при 37°C на чашке. Колонии посылали на секвенирование, и затем колонии собирали для получения бактерий для компонента линкера, которые можно криоконсервировать для позднейшего использования.
1.5 Получение бактериальной библиотеки компонента
1.5.1 Получение бактериальной библиотеки компонента легкой цепи
Полинуклеотиды, включающие KLC и LLC, полученные в примере 1.3, расщепляли с использованием рестрикционных эндонуклеаз R1 и R2 для получения целевых фрагментов легкой цепи (приблизительно 0,65 т.п.о.).
Вектор для хранения легкой цепи DDB-R1-1,0 т.п.о.-R2, полученный в примере 1.4, расщепляли с использованием рестрикционных эндонуклеаз R1 и R2 для получения фрагментов вектора для хранения легкой цепи (приблизительно 2,7 т.п.о.).
Полученные в результате целевые фрагменты легкой цепи смешивали с фрагментами вектора для хранения легкой цепи, затем лигировали с использованием ДНК-лигазы T4 (закупленной из NEB, Thermo) для получения продукта лигирования для хранения легкой цепи. Компетентные бактерии TG1 (Lucigen, кат. # 60502-2, используемые в соответствии с инструкциями производителя) трансформировали с использованием продукта лигирования для хранения легкой цепи, распределяли на чашках для анализа устойчивости к ампициллину (Thermo, кат. #240845) и культивировали в течение ночи при 37°C. Колонии посылали на секвенирование, и затем все колонии собирали для получения бактериальной библиотеки компонента легкой цепи, качество которой можно детектировать и/или которую можно криоконсервировать для позднейшего использования.
1.5.2 Получение бактериальной библиотеки компонента тяжелой цепи
Полинуклеотиды, полученные в примере 1.3, расщепляли с использованием рестрикционных эндонуклеаз R5 и R6 для получения целевых фрагментов вариабельной области тяжелой цепи (приблизительно 0,35 т.п.о.).
Вектор для хранения тяжелой цепи DDB-R5-1,0 т.п.о.-R6, полученный в примере 1.4, расщепляли с использованием рестрикционных эндонуклеаз R5 и R6 для получения фрагментов вектора для хранения тяжелой цепи (приблизительно 2,7 т.п.о.).
Полученные в результате целевые фрагменты вариабельной области тяжелой цепи смешивали с фрагментами вектора для хранения тяжелой цепи, затем лигировали с использованием ДНК-лигазы T4 (закупленной из NEB, Thermo) для получения продукта лигирования для хранения тяжелой цепи. Компетентные бактерии TG1 (Lucigen, кат. # 60502-2, используемые в соответствии с инструкциями производителя) трансформировали с использованием продукта лигирования для хранения тяжелой цепи, распределяли на чашках для анализа устойчивости к ампициллину (Thermo, кат. #240845) и культивировали в течение ночи при 37°C. Колонии посылали на секвенирование, и затем все колонии собирали для получения бактериальной библиотеки компонента тяжелой цепи, качество которой можно детектировать и/или которую можно криоконсервировать для позднейшего использования.
1.6 Получение плазмиды компонента легкой цепи, плазмиды компонента тяжелой цепи и фрагментов линкеров
С использованием набора для выделения плазмид (закупленного из Axygen), плазмиды из бактериальной библиотеки компонента легкой цепи, полученной в примере 1.5.1, и из бактериальной библиотеки компонента тяжелой цепи, полученной в примере 1.5.2, по отдельности выделяли для получения плазмиды компонента легкой цепи и плазмиды компонента тяжелой цепи, соответственно.
Плазмиду компонента легкой цепи, полученную в примере 1.5.1, расщепляли с использованием рестрикционных эндонуклеаз R1 и R2, затем очищали и выделяли посредством электрофореза в геле для получения вставляемых фрагментов легкой цепи LC.
Плазмиду компонента тяжелой цепи, полученную в примере 1.5.2, расщепляли с использованием рестрикционных эндонуклеаз R5 и R6, затем очищали и выделяли посредством электрофореза в геле для получения вставляемых фрагментов тяжелой цепи HC.
С использованием набора для выделения плазмид (закупленного из Axygen), плазмиду из бактерий с компонентом линкера, полученных в примере 1.4.4, выделяли для получения плазмиды компонента линкера. С использованием плазмиды компонента линкера или вектора для хранения линкера в примере 1.4.4 в качестве матрицы, фрагмент 0,8 т.п.о., содержащий линкер, амплифицировали с использованием прямого праймера для линкера (SEQ ID NO: 156) и обратного праймера для линкера (SEQ ID NO: 157), и затем продукт ПЦР 0,8 т.п.о. расщепляли с использованием рестрикционных эндонуклеаз R3 и R4, затем очищали и выделяли посредством электрофореза в геле (с использованием набора для экстракции небольших фрагментов из геля, закупленного из Lifefeng Biotech, кат. # DK402) для получения фрагментов линкеров 72 п.о.
1.7 Получение векторов дисплея
Вектор pComb3x был закуплен, и карта вектора для него показана на ФИГ. 12, и карта модифицированного вектора pComb3x-fab, используемого для дисплея Fab антитела, показана на ФИГ. 13.
Участок рестрикции SfiI на 3'-конце гена Fab в векторе pComb3x-fab удаляли посредством нонсенс-мутации.
После этого, участок рестрикции рестрикционной эндонуклеазы R2 добавляли ниже стоп-кодона легкой цепи в векторе после нонсенс-мутации, и участок рестрикции рестрикционной эндонуклеазы R5 вводили на конце сигнального пептида вариабельной области тяжелой цепи посредством нонсенс-мутации, для получения модифицированного вектора фагового дисплея DDB-R1R2R5R6, карта которого показана на ФИГ. 14.
1.8 Получение бактериальной библиотеки дисплея
Вектор дисплея DDB-R1R2R5R6, полученный в примере 1.7, расщепляли с использованием рестрикционной эндонуклеазы R7 и рестрикционной эндонуклеазы R8 для получения фрагментов вектора дисплея 3,6 т.п.о.
Вставляемый фрагмент легкой цепи LC (0,65 т.п.о.), вставляемый фрагмент тяжелой цепи HC (0,35 т.п.о.), фрагмент линкера (72 п.о.) и фрагмент вектора фагового дисплея (3,6 т.п.о.), полученные в примере 1.6, смешивали в молекулярном соотношении 1:1:1:1, и лигировали с использованием ДНК-лигазы T4 при 20°C в течение более чем 20 часов для получения продукта лигирования для дисплея.
Продукт лигирования очищали посредством очистки продуктов ПЦР, переносили в компетентные бактерии TG1 (Lucigen, кат. # 60502-2, используемые в соответствии с инструкциями производителя), и культивировали в среде 2YT без антибиотиков при 37°C, с встряхиванием в то же время при 250 об./мин, в течение 60 минут, затем распределяли на чашках для анализа устойчивости к ампициллину (Thermo, кат. #240845) и выращивали в течение ночи при 37°C. Колонии сортировали для секвенирования, и собирали все колонии, выросшие на чашках, то есть, бактериальную библиотеку фагового дисплея, которую можно сохранять для позднейшего использования.
1.9 Получение библиотеки фагового дисплея антител
Соответствующее количество бактериальной жидкости отбирали из бактериальной библиотеки дисплея, полученной в примере 1.8, и культивировали в среде 2YT (содержащей 100 мкг/мл ампициллина и 2% глюкозы) при 37°C до достижения OD600 0,5. Затем фаг-помощник M13KO7 (закупленный из NEB, кат. # N0315S, MOI приблизительно 10-20) добавляли к бактериальному раствору и перемешивали до однородности. После перемешивания, ему позволяли стоять при 37°C в течение 30 минут, и встряхивали при 37°C и 250 об./мин в течение 30 минут. После центрифугирования, часть супернатанта из культуральной среды, содержащую фаг-помощник M13KO7, отбрасывали. Бактерии ресуспендировали в культуральной среде (содержащей ампициллин и канамицин), в 4 раза больше исходного объема бактериального раствора, и встряхивали в течение ночи при 30°C и 250 об./мин. На следующие сутки, фаги собирали посредством преципитации с использованием PEG. Концентрацию фагов титровали, и фаги хранили в аликвотах для получения библиотеки фагового дисплея антител.
Пример 2 Конструирование векторов дисплея
2.1 Получение материалов образцов
Для конструирования векторов дисплея, как показано на ФИГ. 1, нацеленное на PD-1 антитело пембролизумаб, так же как вектор pDGB4, выбраны в качестве примеров. Нуклеотидная последовательность легкой цепи пембролизумаба представляла собой: SEQ ID NO: 5, нуклеотидная последовательность вариабельной области тяжелой цепи пембролизумаба представляла собой: SEQ ID NO: 6, и нуклеотидная последовательность вектора pDGB4 представляла собой: SEQ ID NO: 7.
2.2 Конструирование участков рестрикции
Рестрикционные эндонуклеазы BsmBI и SfiI были выбраны для конструирования последовательностей 2 участков узнавания BsmBI (B2 и B3) и последовательностей 2 участков узнавания SfiI (S5 и S6), где нуклеотидная последовательность B2 являлась такой, как указано в SEQ ID NO: 8, нуклеотидная последовательность B3 являлась такой, как указано в SEQ ID NO: 9, нуклеотидная последовательность S5 являлась такой, как указано в SEQ ID NO: 10, и нуклеотидная последовательность S6 являлась такой, как указано в SEQ ID NO: 11.
2.3 Выбор сигнальных пептидов
Выбраны два сигнальных пептида, экспрессирующих нативные гены антител: SP1 и SP2. Для введения подходящих участков рестрикции в 3'-концевую часть сигнального пептида, последовательности оснований двух сигнальных пептидов были изменены посредством непреднамеренной мутации, однако, аминокислотные последовательности сигнальных пептидов оставались неизменными. SP1 экспрессировал дисплей VH, нуклеотидная последовательность которого являлась такой, как указано в SEQ ID NO: 12, и аминокислотная последовательность которого являлась такой, как указано в SEQ ID NO: 13; SP2 экспрессировал дисплей LC, нуклеотидная последовательность которого являлась такой, как указано в SEQ ID NO: 14, и аминокислотная последовательность которого являлась такой, как указано в SEQ ID NO: 15.
2.4 Получение полинуклеотидов вектора дисплея
Праймеры для дисплея VH, дисплея LC, так же как фрагмента I вектора дисплея и фрагмента II вектора дисплея, конструировали соответственно, и экспрессией их всех управлял промотор CMV. Синтетические праймеры амплифицировали посредством ПЦР с последовательностями в 2.1 в качестве матрицы. Последовательности перечислены в таблице 2.
Четыре полинуклеотида вектора дисплея амплифицировали посредством ПЦР (LA Taq, Takara Co., проведенной в соответствии с инструкцией к продукту от компании), и использованные последовательности матриц и праймеров показаны в таблице 2. Продукты ПЦР получали, соответственно, после очистки и выделения посредством электрофореза в геле (действуя в соответствии с инструкцией в «Molecular Cloning: A Laboratory Manual»). Посредством способа клонирования TA (набора для клонирования TA, закупленного из Takara Co.), продукты ПЦР вставляли в плазмидный вектор pUC19 для получения продукта лигирования для хранения. Компетентные бактерии DH5a (Takara Co.) трансформировали с использованием вектора для хранения продукта и культивировали в течение ночи при 37°C на чашке. Колонии посылали на секвенирование, и затем получали бактерии, представляющие собой дисплей полинуклеотидов вектора, содержащих желательные последовательности, представляющих собой первый полинуклеотид вектора дисплея, содержащий дисплей VH, второй полинуклеотид вектора дисплея, содержащий дисплей LC, третий полинуклеотид вектора дисплея, содержащий фрагмент I вектора дисплея, и четвертый полинуклеотид вектора дисплея, содержащий фрагмент II экспрессирующего вектора. Бактерии можно криоконсервировать в форме бактериальной библиотеки для позднейшего использования.
2.5 Расщепление
Плазмиды из бактерий в бактериальной библиотеке из примера 2.4, соответственно, выделяли посредством использования набора для выделения плазмид (закупленного из Axygen). Затем плазмидные векторы расщепляли с использованием рестрикционных эндонуклеаз BsmBI и SfiI, выделяли и очищали посредством электрофореза для получения четырех расщепленных полинуклеотидов вектора дисплея.
2.6 Получение векторов дисплея посредством лигирования
Четыре расщепленных полинуклеотида вектора дисплея, полученных в примере 2.5, смешивали в равных молекулярных пропорциях, к которым добавляли лигазу таким образом, чтобы они подвергались направленному лигированию и циклизации для формирования экспрессирующего вектора. Экспрессирующий вектор переносили в компетентные бактерии DH5a (Takara, используемые в соответствии с инструкциями производителя), и культивировали в среде 2YT без антибиотиков при 37°C, с встряхиванием в то же время при 250 об./мин в течение 60 минут, затем распределяли на чашках для анализа устойчивости к ампициллину (Thermo, кат. #240845) и выращивали в течение ночи при 37°C. Колонии сортировали для секвенирования, и получали векторы дисплея, содержащие корректные последовательности. Конкретные структуры векторов дисплея показаны на ФИГ. 2.
Пример 3 Конструирование вектора компонента VH и вектора компонента LC
3.1 Вектор компонента VH
С использованием IgG1 человека Fc длиной 1 т.п.о. в качестве матрицы, ПЦР проводили с использованием праймеров DDB214 и DDB215. После очистки и выделения посредством электрофореза в геле, получали продукт ПЦР B2-т.п.о.-B3, где нуклеотидная последовательность DDB214 являлась такой, как указано в SEQ ID NO: 1, и нуклеотидная последовательность DDB215 являлась такой, как указано в SEQ ID NO: 2. B2-т.п.о.-B3 вставляли в экспрессирующий вектор компонента, вектор pMD19, для получения плазмидного вектора для хранения компонента VH. Компетентные бактерии TG1 (Lucigen Co.) трансформировали с использованием плазмидного вектора для хранения компонента VH, и культивировали в течение ночи при 37°C на чашке. Колонии сортировали для секвенирования, для определения вектора компонента VH, содержащего корректную последовательность. Структура вектора компонента VH показана на ФИГ. 3.
3.2 Вектор компонента LC
С использованием IgG1 человека Fc длиной 1 т.п.о. в качестве матрицы, проводили ПЦР с использованием праймеров DDB216 и DDB217. После очистки и выделения посредством электрофореза в геле, получали продукт ПЦР S5-т.п.о.-S6, где нуклеотидная последовательность DDB216 являлась такой, как указано в SEQ ID NO: 3, и нуклеотидная последовательность DDB217 являлась такой, как указано в SEQ ID NO: 4. S5-т.п.о.-S6 вставляли в экспрессирующий вектор компонента, вектор pMD19, для получения плазмидного вектора для хранения компонента LC. Компетентные бактерии TG1 (Lucigen Co.) трансформировали с использованием плазмидного вектора для хранения компонента LC, и культивировали в течение ночи при 37°C на чашке. Колонии сортировали для секвенирования, для определения вектора компонента LC, содержащего корректную последовательность. Структура вектора компонента LC показана на ФИГ. 4.
Пример 4 Получение антигенспецифической VH и антигенспецифической LC
4.1 Первый цикл скрининга
Отбирали 500 мкл фаговой библиотеки, сконструированной в примере 1 (библиотека Fab, исходная емкость библиотеки составляла 4×1010, эффективные клоны составляли более чем 80%, и полученная фаговая библиотека составляла 2×1013/мл). Меченный биотином антиген ROR1 (Acro Biosystems, кат. # RO1-H82E6) смешивали с фаговой библиотекой (с концентрацией антигена 10 мкг/мл), и встряхивали при комнатной температуре в течение 2 часов, чтобы позволить фагу, экспонирующему антигенспецифический Fab, связаться с меченным биотином антигеном. Затем 80 мкл магнитных бусин (закупленных из Invitrogen) смешивали с фаговой библиотекой-антигеном, встряхивали при комнатной температуре в течение 20 минут, и антигенспецифический фаг захватывали посредством связывания авидина и биотина на поверхности магнитных бусин с формированием продукта перекрестного сшивания магнитной бусины-авидина-биотина-антигена-фрагмента Fab антитела. Затем образованный продукт перекрестного сшивания, несущий специфический для антигена ROR1 Fab, собирали посредством магнитного штатива, и фаг, экспонирующий специфический для антигена ROR1 Fab, элюировали с использованием раствора глицина, pH 2,2, и нейтрализовали с использованием Трис-буфера, pH 8,0, до pH 7,0, наконец получая 550 мкл раствора фага.
4.2 Второй цикл скрининга
250 мкл раствора фага, полученного из первого цикла скрининга, смешивали с равным количеством 4% молока-PBS до конечного объема 0,5 мл. Затем 4 мкг меченного биотином антигена смешивали с раствором фага до конечной концентрации антигена 8 мкг/мл, и встряхивали при комнатной температуре в течение 3 часов, чтобы позволить фагу, экспонирующему антигенспецифический Fab, связаться с меченным биотином антигеном. Затем 40 мкл магнитных бусин смешивали с раствором фага-антигеном, встряхивали при комнатной температуре в течение 20 минут, и антигенспецифический фаг захватывали посредством связывания авидина и биотина на поверхности магнитных бусин с формированием продукта перекрестного сшивания магнитной бусины-авидина-биотина-антигена-фрагмента Fab антитела. Образованный продукт перекрестного сшивания, несущий специфический для антигена ROR1 Fab, собирали посредством магнитного штатива. Затем магнитные бусины промывали с использованием 1×PBST 4 раза, и затем промывали с использованием 1×PBS 4 раза. Наконец, фаг, экспонирующий специфический для антигена ROR1 Fab, элюировали с использованием 50 мкл раствора глицина, pH 2,2, и нейтрализовали с использованием 20 мкл Трис-буфера, pH 8,0, до pH 7,0, наконец получая 75 мкл раствора фага.
4.3 Инфекция бактерий TG1
75 мкл раствора фага, полученного из второго цикла скрининга в примере 4.2, смешивали с 500 мкл бактерий TG1 в логарифмической фазе роста, с выстаиванием при 37°C в течение 30 минут. Затем раствор инфицированных бактерий TG1 распределяли на чашках для анализа устойчивости к Amp и культивировали в течение ночи при 37°C.
4.4 Скрининг положительных клонов посредством ELISA
Колонии, растущие на чашке, подсчитывали, инокулировали в два фрагмента 96-луночных планшетов с глубокими лунками, где каждая лунка содержит 400 мкл культуральной среды (2YT+Amp+0,2% глюкоза), и культивировали при 37°C с встряхиванием в течение 6 часов. В каждую лунку добавляли 400 мкл культуральной среды, содержащей IPTG (2YT+Amp+2 мМ IPTG), с конечной концентрацией IPTG 1 мМ, и культивировали в течение ночи при 30°C с встряхиванием в то же время при 250 об./мин. Два фрагмента 96-луночных планшетов для ELISA покрывали антигеном ROR1 без мечения биотином при 100 нг/100 мкл/лунку, при 4°C в течение ночи.
После промывки и блокирования 96-луночных планшетов для ELISA, которые покрывали антигеном в течение ночи, 100 мкл раствора культивированных в течение ночи бактерий добавляли в каждую лунку, инкубировали при 37°C в течение 1 часа, и снова промывали; и вторичные антитела (меченные HRP антитела против IgG-Fab человека) затем добавляли и инкубировали при 37°C в течение 40 минут. После промывки, добавляли проявляющий раствор и сохраняли в темноте в течение 30 минут. Значения OD600 считывали посредством считывателя для микропланшетов, с результатами, показанными в таблицах 3-1 и 3-2 ниже.
Таблица 3-1 Предварительные результаты скрининга положительных клонов-1
Всего 35 клонов со считываниями более чем 0,25 посланы на секвенирование. Анализ последовательности показал, что присутствовало 34 уникальных VH и 28 уникальных LC. Результаты сравнения аминокислотных последовательностей легкой цепи показаны на ФИГ. 5A, и результаты сравнения аминокислотных последовательностей вариабельной области тяжелой цепи показаны на ФИГ. 5B.
Пример 5 Конструирование библиотеки компонента VH и библиотеки компонента LC
5.1 Амплификация антигенспецифической VH и антигенспецифической LC
Конструировали праймеры, содержащие участки узнавания для рестрикционных эндонуклеаз (Esp3I и SfiI). Существовало 29 праймеров для амплификации антигенспецифической VH, включая 24 прямых праймера и 5 обратных праймеров; 19 праймеров для амплификации антигенспецифической KLC (легкой цепи каппа), включая 18 прямых праймеров и 1 обратный праймер; 26 праймеров для амплификации антигенспецифической LLC (легкая цепь лямбда), включая 25 прямых праймеров и 1 обратный праймер. Каждый праймер в каждой группе прямых праймеров смешивали в равных пропорциях, обратные праймеры для VH также смешивали в равных пропорциях, и затем прямые и обратные праймеры смешивали в равных пропорциях для получения трех групп праймеров, которые использовали для амплификации VH, KLC и LLC соответственно. В качестве приведенной для примера иллюстрации, KLC взята в качестве примера в этом варианте осуществления, где прямые праймеры для VH являлись такими, как указано в SEQ ID NO: 30-53, обратные праймеры для VH являлись такими, как указано в SEQ ID NO: 54-58, прямые праймеры для KLC являлись такими, как указано в SEQ ID NO: 59-76, и обратный праймер для KLC являлся таким, как указано в SEQ ID NO 77.
Минипрепараты ДНК для 35 положительных клонов, полученных посредством скрининга в примере 4, смешивали в равных количествах, и амплифицировали с использованием трех групп праймеров, описанных выше, соответственно. Очищенные антигенспецифическую VH и антигенспецифическую LC (с KLC в качестве примера) с участком узнавания, полученным посредством ПЦР, анализировали посредством электрофореза.
5.2 Расщепление и лигирование
Антигенспецифическую VH, полученную в примере 5.1, расщепляли с использованием Esp3I, и очищенную антигенспецифическую VH после расщепления анализировали посредством электрофореза. Вектор компонента VH, полученный в примере 3.1, расщепляли с использованием Esp3I, и очищенный фрагмент 2,8 т.п.о. вектора компонента после расщепления анализировали посредством электрофореза. Очищенные антигенспецифическую VH и фрагмент 2,8 т.п.о. вектора компонента лигировали для получения библиотеки компонента специфического для ROR1 VH.
Антигенспецифическую LC, полученную в примере 5,1, расщепляли с использованием SfiI, и очищенную антигенспецифическую LC после расщепления анализировали посредством электрофореза. Вектор компонента LC, полученный в примере 3,2, расщепляли с использованием SfiI, и очищенный фрагмент 2,8 т.п.о. вектора компонента после расщепления анализировали посредством электрофореза. Очищенные антигенспецифическую LC и фрагмент 2,8 т.п.о. вектора компонента лигировали для получения библиотеки компонента специфической для ROR1 LC.
Пример 6 Конструирование библиотеки дисплея антигенспецифических связывающих полипептидов
Библиотеку компонента VH расщепляли с использованием Esp3I, и очищенные 0,35 т.п.о. из антигенспецифической VH (т.е., высвобожденный первый полинуклеотид) с липкими концами после расщепления анализировали посредством электрофореза. Библиотеку компонента KLC расщепляли с использованием SfiI, и очищенные 0,65 т.п.о. из антигенспецифической LC (т.е., высвобожденный второй полинуклеотид) с липкими концами после расщепления анализировали посредством электрофореза. Вектор дисплея, полученный в примере 1, подвергали двойному расщеплению с использованием Esp3I и SfiI, и очищали для получения фрагмента I 3 т.п.о. вектора дисплея (т.е., высвобожденного третьего полинуклеотида) с липкими концами и фрагмента II 5 т.п.о. вектора дисплея (т.е., высвобожденного четвертого полинуклеотида) с липкими концами. Вышеуказанные четыре расщепленных фрагмента смешивали в равных молекулярных пропорциях и лигировали при 20°C в течение 4 часов в 10 мкл системы лигирования, содержащих суммарное количество 25 нг фрагментов, для получения вектора дисплея генов антигенспецифических связывающих полипептидов.
Продукт лигирования очищали посредством очистки продуктов ПЦР, и собирали с использованием 10 мкл ddH2O. 4 мкл очищенного продукта лигирования брали для электропорации (Takara DH5a, подвергнутые электропорации компетентные бактерии), распределяли на чашке и культивировали в течение ночи при 37°C. Колонии подсчитывали, и когда емкость библиотеки достигала 2,3×105, получали бактериальную библиотеку дисплея генов антигенспецифических связывающих полипептидов. Собирали все колонии, из которых ДНК вектора выделяли для получения библиотеки дисплея антигенспецифических связывающих полипептидов.
Пример 7 Скрининг моноклональных антител
ДНК экспрессирующего вектора специфического для ROR1 связывающего полипептида получали из библиотеки дисплея антигенспецифического связывающего полипептида, полученного в примере 6, и 40 мкг ДНК трансформировали в клетки FCHO. Через 60 часов после трансформации, экспрессию полноразмерных антител на поверхности клетки, и специфичность для антигена ROR1 антител анализировали посредством FACS. Результаты из ФИГ. 6 показали, что присутствовали полноразмерные антитела против ROR1, экспрессированные на поверхности клетки, и экспрессированные антитела могли специфически связываться с меченным FITC ROR1 (полученным посредством мечения с использованием набора для мечения FITC). Антигены ROR1 без биотиновой метки получены из Acro Biosystems (кат. # RO1-H5250-1 мг). На ФИГ. 6 показаны клетки после двойного окрашивания с использованием меченного PE антитела мыши против легкой цепи каппа человека и меченного FITC антигена ROR1, так же как флуоресцентные сигналы на поверхности клетки, как анализировано посредством FACS, где A обозначает отрицательный контроль; B обозначает клеточную библиотеку, экспрессирующую специфическое для ROR1 антитело.
Стабильно трансформированную клеточную библиотеку FCHO подвергали скринингу с использованием воздействия гигромицина (при концентрации гигромицина 500 мкг/мл). Через 10 суток культивирования под воздействием гигромицина, получали стабильно трансформированную клеточную библиотеку. Клеточную библиотеку подвергали двойному окрашиванию с использованием меченного PE антитела мыши против легкой цепи каппа человека (BD) и меченного FITC антигена ROR1, и двойные положительные по PE и FITC клетки сортировали посредством FACS. Клоны отдельных клеток добавляли в 96-луночные планшеты, с одной клеткой на лунку, и культивировали с использованием воздействия гигромицина.
Через 14 суток после культивирования с использованием воздействия гигромицина, получили 92 стабильно трансформированных клона отдельных клеток. Клетки расщепляли с использованием 0,5 мМ буфера ЭДТА-PBS. 92 клона отдельных клеток подвергали двойному окрашиванию с использованием меченного PE антитела мыши против легкой цепи каппа человека и меченного FITC антигена ROR1 (концентрация антигена составляла 0,15 нг/50 мкл).
Посредством анализа FACS, получили 71 двойной положительный по флуоресценции PE и FITC клон клеток, и частота положительных составляла 77% (71/92=77%).
Пример 8 Получение последовательностей положительных клонов
В соответствии с локализацией положительной популяции клеток на графике анализа FACS, всего 30 клонов клеток, локализованных в различных положениях (представляющих различную аффинность), отбирали для амплификации ПЦР генов антител. Положительные клоны клеток, соответственно, собирали посредством центрифугирования, супернатант отбрасывали, и 20 мкл раствора для экстракции клеточного генома (Quick Extraction Buffer, Lucigen) использовали для экстракции клеточной геномной ДНК, в соответствии с инструкцией к реагенту. 2 мкл экстракта клеточной геномной ДНК отбирали из каждого клона, и VH и LC из каждого клона амплифицировали посредством ПЦР. Прямой праймер для амплификации фрагментов VH представлял собой TGGGCTCTGCTCCTCCTGACC (SEQ ID NO: 24), обратный праймер для амплификации фрагментов VH представлял собой AGTTCCACGACACCGTCACCGGTTC (SEQ ID NO: 25), прямой праймер для амплификации фрагментов LC представлял собой GGACCTGGAGGATCCTCTTCTTGG (SEQ ID NO: 26), и обратный праймер для амплификации фрагментов LC представлял собой TAAATTCCTCGGCCGTGCAGGCCTTATCAACACTCTCCCCTGTTGAAGCTCT (SEQ ID NO: 27).
Фрагменты VH и LC, амплифицированные посредством ПЦР, выделяли и очищали посредством электрофореза. Очищенные фрагменты VH и LC анализировали посредством секвенирования, определив 14 уникальных VH и 13 уникальных LC, которые можно было комбинировать для получения 17 положительных клонов с уникальными последовательностями (6 CDR из легких и тяжелых цепей, отличающиеся по меньшей мере одной аминокислотой). В качестве примера, уникальные последовательности одной пары VH и LC перечислены в настоящем описании, где аминокислотная последовательность VH уникальной последовательности являлась такой, как указано в SEQ ID NO: 28, и аминокислотная последовательность LC каппа уникальной последовательности являлась такой, как указано в SEQ ID NO: 29.
Пример 9 Анализ аффинности антител
Положительные фрагменты VH с уникальными последовательностями, полученные в примере 8, расщепляли с использованием Esp3I, и положительные фрагменты LC с уникальными последовательностями, полученные в примере 8, расщепляли с использованием SfiI. Расщепленные фрагменты VH и LC очищали посредством очистки продуктов ПЦР. Фрагменты VH и фрагменты LC по отдельности вставляли в экспрессирующие векторы растворимой тяжелой цепи, и колонии определяли посредством секвенирования. Выделяли ДНК экспрессирующих векторов VH и LC, определенных посредством секвенирования.
Клетки 293 EXP амплифицировали посредством суспензионной культуры, и всего 17 пар экспрессирующих векторов легких и тяжелых цепей антитела формировали, в соответствии со спариванием легких и тяжелых цепей, как определено в примере 8. Каждую пару смешивали в соотношении 18 мкг экспрессирующего вектора легкой цепи и 12 мкг экспрессирующего вектора тяжелой цепи, и использовали для трансформации 30 мл суспендированных клеток 293EXP (1,2×106/мл). На сутки 6 трансформации, культуральный супернатант собирали, и антитело очищали с использованием магнитных бусин от GenScript (кат. # L00695), в соответствии с инструкциями к продукту, уравновешивали посредством диализа против раствора PBS-антитела, и хранили при -80°C. Анализ проводили с использованием денатурирующего электрофореза в геле SDS-PAGE (ФИГ. 7), с результатами, показывающими, что чистота антитела достигла более 90%. Аффинность связывания очищенных антител с антигенами анализировали посредством ELISA, и EC50 8 иллюстративных антител перечислены в настоящем описании для иллюстрации, с результатами, показанными в таблице 4 ниже.
Пример 10 Быстрый скрининг антигенспецифического полипептида
В соответствии с данными ELISA положительных клонов, полученных посредством скрининга из фаговой библиотеки в примере 4, 15 клонов со считываниями ELISA более чем 0,8, выбраны из таблиц 3-1 и 3-2. После смешивания с равным количеством бактериальной жидкости, проводили экстракцию векторной ДНК в мини-масштабе. В соответствии со способами в примере 5, антигенспецифическую VH и антигенспецифическую LC из 15 клонов, соответственно, амплифицировали посредством ПЦР, и очищали посредством расщепления. В соответствии со способами в примере 6, конструировали библиотеку дисплея антигенспецифических связывающих полипептидов. Затем, 48 колоний сортировали случайным образом и посылали на секвенирование. После анализа результатов секвенирования, сортировали 36 клонов с корректными VH и LC, из которых экстрагировали векторную ДНК в мини-масштабе и временно трансфицировали в клетки CHO. Через 60 часов, клетки расщепляли с использованием 0,5 мМ буфера ЭДТА-PBS. 36 популяций клеток подвергали двойному окрашиванию с использованием меченного PE антитела мыши против легкой цепи каппа человека и меченного FITC антигена ROR1 (концентрация антигена составляла 0,15 нг/50 мкл). Результаты анализа FACS показаны на ФИГ. 8. Получили шесть двойных положительных по PE и FITC клонов клеток, и частота положительных составляла 17% (6/36). На ФИГ. 8 показаны клетки, подвергнутые двойному окрашиванию с использованием меченного PE антитела мыши против легкой цепи каппа человека и меченного FITC антигена ROR1, так же как флуоресцентные сигналы на поверхности клетки, как анализировано посредством FACS. A обозначает отрицательный контроль; B обозначает клоны клеток, экспрессирующие не специфические для ROR1 антитела; C-H обозначает 6 иллюстративных положительных клонов клеток, экспрессирующих специфические для ROR1 антитела.
--->
Список последовательностей
<110> DDBio.Co,Ltd.,(Shang Hai)
<120> СПОСОБ КОНСТРУИРОВАНИЯ И ПРИМЕНЕНИЕ ВЕКТОРА ДИСПЛЕЯ ГЕНОВ
АНТИГЕНСПЕЦИФИЧЕСКИХ СВЯЗЫВАЮЩИХ ПОЛИПЕПТИДОВ
<130> 0112-PA-006RU
<160> 175
<170> PatentIn версии 3.5
<210> 1
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DDB214
<400> 1
tccagacatc taatggcgtc tcagtggttc agctagcacc aagggcccat 50
<210> 2
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DDB215
<400> 2
accagtcttc tattggctga tgagacgtta tcatttaccc ggagacaggg 50
<210> 3
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DDB216
<400> 3
atggggccac aggggcctca gctagcacca agggcccatc 40
<210> 4
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> DDB217
<400> 4
ttgggccgtg caggccttat catttacccg gagacaggg 39
<210> 5
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PD1-LC
<400> 5
gaaattgtgc tgacccagag ccctgccaca ctcagcctga gccctggaga gagggccacc 60
ctgtcctgca gagccagcaa gggcgtgagc accagcggct acagctacct gcactggtac 120
cagcagaagc ccggccaggc tcccagactg ctgatctacc tggcttctta tttagaaagc 180
ggcgtgcctg ctagattcag cggctccggc tccggcaccg actttaccct gaccatcagc 240
tccctggagc ccgaggactt cgccgtgtac tactgccagc acagcaggga cctgcccctg 300
acctttggcg gaggcaccaa ggtggagatc aagaggaccg tggccgctcc cagcgtgttc 360
atcttccccc ccagcgacga gcagctgaag agcggaaccg ccagcgtggt gtgcctgctg 420
aacaacttct accccaggga ggccaaggtg cagtggaagg tcgacaacgc cctgcagagc 480
ggcaatagcc aggagtccgt gaccgagcag gacagcaagg acagcaccta cagcctgagc 540
agcaccctca ccctgtccaa ggccgactac gagaagcata aggtgtacgc ctgtgaggtg 600
acccaccagg gcctgtccag ccctgtgacc aaaagcttta accgcggcga atgctga 657
<210> 6
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PD1-VH
<400> 6
caggtgcagc tggtgcagag cggcgtggag gtgaaaaagc ccggcgccag cgtgaaggtg 60
tcctgtaagg cctccggcta cacattcacc aactactaca tgtactgggt gaggcaagcc 120
cctggccagg gactggaatg gatgggcgga atcaacccct ccaatggagg caccaacttt 180
aacgagaagt tcaagaacag ggtgacactg acaaccgaca gcagcacaac aaccgcttac 240
atggagctga agagcctgca gttcgatgac accgccgtgt actactgcgc caggagggac 300
tacaggttcg acatgggctt tgattactgg ggccagggca ccaccgtgac cgtctcatca 360
<210> 7
<211> 9109
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pDGB4
<400> 7
tctggctaac tagagaaccc actgcttact ggcttatcga aattaatacg actcactata 60
gggagaccca agctggctag gccacatagg cctgaaccac catggtgttg cagacccagg 120
tcttcatttc tctgttgctc tggatctctg tgattacagg tgccgacggg gacatcgtga 180
tgacccagtc tccagactcc ctggctgtgt ctctgggcga gagggccacc atcaactgca 240
agtccagcca gagtgtttta tacagcccca acaatgagaa cttcttagct tggtaccaac 300
agaagccagg acagtctcct aagttgctca tttactgggc gtctacccgg gaatccgggg 360
tccctgaccg attcagtggc agcgggtctg ggacagattt cactctcacc atcagcaacc 420
tgcaggctga agatgtggca gtttactact gtcaacaata ttatagtgct cctatcactt 480
tcggcggagg gaccaaggtg gagatcaaac gaactgtggc tgcaccatct gtcttcatct 540
tcccgccatc tgatgagcag ttgaaatctg gaactgcctc tgttgtgtgc ctgctgaata 600
acttctatcc cagagaggcc aaagtacagt ggaaggtgga taacgccctc caatcgggta 660
actcccagga gagtgtcaca gagcaggaca gcaaggacag cacctacagc ctcagcagca 720
ccctgacgct gagcaaagca gactacgaga aacacaaact ctacgcctgc gaagtcaccc 780
atcagggcct gagctcgccc gtcacaaaga gcttcaacag gggagagtct tgataaggcc 840
tgcacggccc tcgagtctag agggcccgtt taaacccgct gatcagcctc gactgtgcct 900
tctagttgcc agccatctgt tgtttgcccc tcccccgtgc cttccttgac cctggaaggt 960
gccactccca ctgtcctttc ctaataaaat gaggaaattg catcgcattg tctgagtagg 1020
tgtcattcta ttctgggggg tggggtgggg caggacagca agggggagga ttgggaagac 1080
aatagcaggc atgctgggga tgcggtgggc tctatggctt ctgaggcgga aagaaccagc 1140
tggggctcta gggggtatcc ccacgcgccc tgtagcggcg cattaagcgc ggcgggtgtg 1200
gtggttacgc gcagcgtgac cgctacactt gccagcgccc tagcgcccgc tcctttcgct 1260
ttcttccctt cctttctcgc cacgttcgcc ggctttcccc gtcaagctct aaatcggggg 1320
ctccctttag ggttccgatt tagtgcttta cggcacctcg accccaaaaa acttgattag 1380
ggtgatggtt cacgtaccta gaagttccta ttccgaagtt cctattctct agaaagtata 1440
ggaacttcct tggccaaaaa gcctgaactc accgcgacgt ctgtcgagaa gtttctgatc 1500
gaaaagttcg acagcgtgtc cgacctgatg cagctctcgg agggcgaaga atctcgtgct 1560
ttcagcttcg atgtaggagg gcgtggatat gtcctgcggg taaatagctg cgccgatggt 1620
ttctacaaag atcgttatgt ttatcggcac tttgcatcgg ccgcgctccc gattccggaa 1680
gtgcttgaca ttggggaatt cagcgagagc ctgacctatt gcatctcccg ccgtgcacag 1740
ggtgtcacgt tgcaagacct gcctgaaacc gaactgcccg ctgttctgca gccggtcgcg 1800
gaggccatgg atgcgatcgc tgcggccgat cttagccaga cgagcgggtt cggcccattc 1860
ggaccgcaag gaatcggtca atacactaca tggcgtgatt tcatatgcgc gattgctgat 1920
ccccatgtgt atcactggca aactgtgatg gacgacaccg tcagtgcgtc cgtcgcgcag 1980
gctctcgatg agctgatgct ttgggccgag gactgccccg aagtccggca cctcgtgcac 2040
gcggatttcg gctccaacaa tgtcctgacg gacaatggcc gcataacagc ggtcattgac 2100
tggagcgagg cgatgttcgg ggattcccaa tacgaggtcg ccaacatctt cttctggagg 2160
ccgtggttgg cttgtatgga gcagcagacg cgctacttcg agcggaggca tccggagctt 2220
gcaggatcgc cgcggctccg ggcgtatatg ctccgcattg gtcttgacca actctatcag 2280
agcttggttg acggcaattt cgatgatgca gcttgggcgc agggtcgatg cgacgcaatc 2340
gtccgatccg gagccgggac tgtcgggcgt acacaaatcg cccgcagaag cgcggccgtc 2400
tggaccgatg gctgtgtaga agtactcgcc gatagtggaa accgacgccc cagcactcgt 2460
ccgagggcaa aggaatagca cgtactacga gatttcgatt ccaccgccgc cttctatgaa 2520
aggttgggct tcggaatcgt tttccgggac gccggctgga tgatcctcca gcgcggggat 2580
ctcatgctgg agttcttcgc ccaccccaac ttgtttattg cagcttataa tggttacaaa 2640
taaagcaata gcatcacaaa tttcacaaat aaagcatttt tttcactgca ttctagttgt 2700
ggtttgtcca aactcatcaa tgtatcttat catgtctgta taccgtcgac ctctagctag 2760
agcttggcgt aatcatggtc atagctgttt cctgtgtgaa attgttatcc gctcacaatt 2820
ccacacaaca tacgagccgg aagcataaag tgtaaagcct ggggtgccta atgagtgagc 2880
taactcacat taattgcgtt gcgctcactg cccgctttcc agtcgggaaa cctgtcgtgc 2940
cagctgcatt aatgaatcgg ccaacgcgcg gggagaggcg gtttgcgtat tgggcgctct 3000
tccgcttcct cgctcactga ctcgctgcgc tcggtcgttc ggctgcggcg agcggtatca 3060
gctcactcaa aggcggtaat acggttatcc acagaatcag gggataacgc aggaaagaac 3120
atgtgagcaa aaggccagca aaaggccagg aaccgtaaaa aggccgcgtt gctggcgttt 3180
ttccataggc tccgcccccc tgacgagcat cacaaaaatc gacgctcaag tcagaggtgg 3240
cgaaacccga caggactata aagataccag gcgtttcccc ctggaagctc cctcgtgcgc 3300
tctcctgttc cgaccctgcc gcttaccgga tacctgtccg cctttctccc ttcgggaagc 3360
gtggcgcttt ctcatagctc acgctgtagg tatctcagtt cggtgtaggt cgttcgctcc 3420
aagctgggct gtgtgcacga accccccgtt cagcccgacc gctgcgcctt atccggtaac 3480
tatcgtcttg agtccaaccc ggtaagacac gacttatcgc cactggcagc agccactggt 3540
aacaggatta gcagagcgag gtatgtaggc ggtgctacag agttcttgaa gtggtggcct 3600
aactacggct acactagaag aacagtattt ggtatctgcg ctctgctgaa gccagttacc 3660
ttcggaaaaa gagttggtag ctcttgatcc ggcaaacaaa ccaccgctgg tagcggtggt 3720
ttttttgttt gcaagcagca gattacgcgc agaaaaaaag gatctcaaga agatcctttg 3780
atcttttcta cggggtctga cgctcagtgg aacgaaaact cacgttaagg gattttggtc 3840
atgagattat caaaaaggat cttcacctag atccttttaa attaaaaatg aagttttaaa 3900
tcaatctaaa gtatatatga gtaaacttgg tctgacagtt accaatgctt aatcagtgag 3960
gcacctatct cagcgatctg tctatttcgt tcatccatag ttgcctgact ccccgtcgtg 4020
tagataacta cgatacggga gggcttacca tctggcccca gtgctgcaat gataccgcga 4080
gacccacgct caccggctcc agatttatca gcaataaacc agccagccgg aagggccgag 4140
cgcagaagtg gtcctgcaac tttatccgcc tccatccagt ctattaattg ttgccgggaa 4200
gctagagtaa gtagttcgcc agttaatagt ttgcgcaacg ttgttgccat tgctacaggc 4260
atcgtggtgt cacgctcgtc gtttggtatg gcttcattca gctccggttc ccaacgatca 4320
aggcgagtta catgatcccc catgttgtgc aaaaaagcgg ttagctcctt cggtcctccg 4380
atcgttgtca gaagtaagtt ggccgcagtg ttatcactca tggttatggc agcactgcat 4440
aattctctta ctgtcatgcc atccgtaaga tgcttttctg tgactggtga gtactcaacc 4500
aagtcattct gagaatagtg tatgcggcga ccgagttgct cttgcccggc gtcaatacgg 4560
gataataccg cgccacatag cagaacttta aaagtgctca tcattggaaa acgttcttcg 4620
gggcgaaaac tctcaaggat cttaccgctg ttgagatcca gttcgatgta acccactcgt 4680
gcacccaact gatcttcagc atcttttact ttcaccagcg tttctgggtg agcaaaaaca 4740
ggaaggcaaa atgccgcaaa aaagggaata agggcgacac ggaaatgttg aatactcata 4800
ctcttccttt ttcaatatta ttgaagcatt tatcagggtt attgtctcat gagcggatac 4860
atatttgaat gtatttagaa aaataaacaa ataggggttc cgcgcacatt tccccgaaaa 4920
gtgccacctg acgtcgacgg atcgggagat ctcccgatcc cctatggtgc actctcagta 4980
caatctgctc tgatgccgca tagttaagcc agtatctgct ccctgcttgt gtgttggagg 5040
tcgctgagta gtgcgcgagc aaaatttaag ctacaacaag gcaaggcttg accgacaatt 5100
gcatgaagaa tctgcttagg gttaggcgtt ttgcgctgct tcgcgatgta cgggccagat 5160
atacgcgttg acattgatta ttgactagtt attaatagta atcaattacg gggtcattag 5220
ttcatagccc atatatggag ttccgcgtta cataacttac ggtaaatggc ccgcctggct 5280
gaccgcccaa cgacccccgc ccattgacgt caataatgac gtatgttccc atagtaacgc 5340
caatagggac tttccattga cgtcaatggg tggagtattt acggtaaact gcccacttgg 5400
cagtacatca agtgtatcat atgccaagta cgccccctat tgacgtcaat gacggtaaat 5460
ggcccgcctg gcattatgcc cagtacatga ccttatggga ctttcctact tggcagtaca 5520
tctacgtatt agtcatcgct attaccatgg tgatgcggtt ttggcagtac atcaatgggc 5580
gtggatagcg gtttgactca cggggatttc caagtctcca ccccattgac gtcaatggga 5640
gtttgttttg gcaccaaaat caacgggact ttccaaaatg tcgtaacaac tccgccccat 5700
tgacgcaaat gggcggtagg cgtgtacggt gggaggtcta tataagcaga gctctctggc 5760
taactagaga acccactgct tactggctta tcgaaattaa tacgactcac tatagggaga 5820
gggagaccca agctggctag ccacctgatt ggcgtctcta gtccaccatg gactttgggc 5880
tgagctggct ttttcttgtg gctattttaa aaggtgtcca gtgtgaggtg cagctgttgg 5940
agtctggggg aggcttggta cagcctgggg ggtccctgag actctcctgt gcagcctctg 6000
gattcacctt tagcagctat gccatgagct gggtccgcca ggctccaggg aaggggctgg 6060
agtgggtctc aggtattact gggagtggtg gtagtacata ctacgcagac tccgtgaagg 6120
gccggttcac catctccaga gacaattcca agaacacgct gtatctgcaa atgaacagcc 6180
tgagagccga ggacacggcc gtatattact gtgcgaaaga tccagggact acggtgatta 6240
tgagttggtt cgacccctgg ggccagggaa ccctggtcac cgtctcctca gctagcacca 6300
agggcccatc ggtcttcccc ctggcgccct gctccaggag cacctccgag agcacagcgg 6360
ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg tggaactcag 6420
gcgctctgac cagcggcgtg cacaccttcc cagctgtcct acagtcctca ggactctact 6480
ccctcagcag cgtggtgacc gtgccctcca gcaacttcgg cacccagacc tacacctgca 6540
acgtagatca caagcccagc aacaccaagg tggacaagac agttgagcgc aaatgttgtg 6600
tcgagtgccc accgtgccca gcaccacctg tggcaggacc gtcagtcttc ctcttccccc 6660
caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacgtgc gtggtggtgg 6720
acgtgagcca cgaagacccc gaggtccagt tcaactggta cgtggacggc gtggaggtgc 6780
ataatgccaa gacaaagcca cgggaggagc agttcaacag cacgttccgt gtggtcagcg 6840
tcctcaccgt tgtgcaccag gactggctga acggcaagga gtacaagtgc aaggtctcca 6900
acaaaggcct cccagccccc atcgagaaaa ccatctccaa aaccaaaggg cagccccgag 6960
aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac caggtcagcc 7020
tgacctgcct ggtcaaaggc ttctacccca gcgacatcgc cgtggagtgg gagagcaatg 7080
ggcagccgga gaacaactac aagaccacac ctcccatgct ggactccgac ggctccttct 7140
tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac gtcttctcat 7200
gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc tccctgtctc 7260
cgggtaaaaa tgctgtgggc caggacacgc aggaggtcat cgtggtgcca cactccttgc 7320
cctttaaggt ggtggtgatc tcagccatcc tggccctggt ggtgctcacc atcatctccc 7380
ttatcatcct catcatgctt tggcagaaga agccacgttg ataaccatga ctctggacca 7440
tgggaaatgt cagagtggag aaccacaccg agtgccactg cagcacttgt tattatcaca 7500
aatcctaata gtttgcagtg ggccttgctg atgatggctg acttgctcaa aaggaaaatt 7560
aatttgtcca gtgtctatgg ctttgtgaga taaaaccctc cttttccttg ccataccatt 7620
tttaacctgc tttgagaata tactgcagct ttattgcttt tctccttatc ctacaatata 7680
atcagtagtc ttgatctttt catttggaat gaaatatggc atttagcatg accataaaaa 7740
gctgattcca ctggaaataa agtcttttaa atcatcactc tatcactgaa ttctaatttt 7800
ttctgaaaag tttcaagcca gttacttttg ataggattaa cggaagggag tgagccagtg 7860
ggtgaggtgg gttcccatgt agtcaatggc ctaatactgg agaatcttat tctaaccaag 7920
ccttccagag caagctgtga gcccctcaga cagtgggcta ctcatgagac agtccattgg 7980
ggtaaaggaa gaaaatataa cttctatttc tattcatttg cacattgtct ttagatgccc 8040
atttgggtga gttttataga agtacagcta cattaaaaaa tagaactgat aatagataag 8100
gctttaaaaa aacttcattc atcaccagtt tgtcaagatt ccatttcaaa gtgaaaaacc 8160
aatttctaac gggttggtaa acacagcaga tggcagggtg aaaaattaaa gtgagtgcat 8220
gtacctttaa agaaacactg aaatgcacac acattactta acctgctcat tcatttattt 8280
acatatagtc ttgggtgtac aaaatttaga aataaataca tagatctccc gatcccctat 8340
ggtgcactct cagtacaatc tgctctgatg ccgcatagtt aagccagtat ctgctccctg 8400
cttgtgtgtt ggagcaaaat ttaagctaca acaaggcaag gcttgaccga caattgcatg 8460
aagaatctgc ttagggttag gcgttttgcg ctgcttcgcg atgtacgggc cagatatacg 8520
cgttgacatt gattattgac tagttattaa tagtaatcaa ttacggggtc attagttcat 8580
agcccatata tggagttccg cgttacataa cttacggtaa atggcccgcc tggctgaccg 8640
cccaacgacc cccgcccatt gacgtcaata atgacgtatg ttcccatagt aacgccaata 8700
gggactttcc attgacgtca atgggtggag tatttacggt aaactgccca cttggcagta 8760
catcaagtgt atcatatgcc aagtacgccc cctattgacg tcaatgacgg taaatggccc 8820
gcctggcatt atgcccagta catgacctta tgggactttc ctacttggca gtacatctac 8880
gtattagtca tcgctattac catggtgatg cggttttggc agtacatcaa tgggcgtgga 8940
tagcggtttg actcacgggg atttccaagt ctccacccca ttgacgtcaa tgggagtttg 9000
ttttggcacc aaaatcaacg ggactttcca aaatgtcgta acaactccgc cccattgacg 9060
caaatgggcg gtaggcgtgt acggtgggag gtctatataa gcagagctc 9109
<210> 8
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> B2
<400> 8
cgtctcagtg gt 12
<210> 9
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> B3
<400> 9
cgtctcatca gc 12
<210> 10
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S5
<400> 10
ggccacaggg gcc 13
<210> 11
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> S6
<400> 11
ggcctgcacg gcc 13
<210> 12
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> SP1
<400> 12
atgccctggg ctctgctcct cctgaccctc ctcactcact ctgccgtctc agtggtc 57
<210> 13
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> SP1
<400> 13
Met Pro Trp Ala Leu Leu Leu Leu Thr Leu Leu Thr His Ser Ala Val
1 5 10 15
Ser Val Val
<210> 14
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> SP2
<400> 14
atggactgga cctggaggat cctcttcttg gtggcagcgg ccacaggggc ccactcc 57
<210> 15
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> SP2
<400> 15
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Ala His Ser
<210> 16
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P1
<400> 16
aaggcctgca cggccctcga gtctagaggg cccgttt 37
<210> 17
<211> 86
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P2
<400> 17
gggagaccca agctggctag tagtccacca tgccctgggc tctgctcctc ctgaccctcc 60
tcactcactc tgccgtctca gtggtc 86
<210> 18
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P3
<400> 18
caccgtgacc gtctcatcag ctagcaccaa gggcccatcg gtc 43
<210> 19
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P4
<400> 19
tgggcccctg tggccgctgc caccaagaag aggatcctcc aggtccagtc catggtggtt 60
catagccagc ttgggtctcc cta 83
<210> 20
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P5
<400> 20
tggcagcggc cacaggggcc cactccgaaa ttgtgctgac ccagagccc 49
<210> 21
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P6
<400> 21
tagactcgag ggccgtgcag gccttatcag cattcgccgc ggttaaagct tttg 54
<210> 22
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P7
<400> 22
ctcactctgc cgtctcagtg gtccaggtgc agctggtgca gagcg 45
<210> 23
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P8
<400> 23
ccttggtgct agctgatgag acggtcacgg tggtgccctg gc 42
<210> 24
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> фрагмент VH, прямой праймер
<400> 24
tgggctctgc tcctcctgac c 21
<210> 25
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH fragment reverse primer
<400> 25
agttccacga caccgtcacc ggttc 25
<210> 26
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> фрагмент LC, прямой праймерs
<400> 26
ggacctggag gatcctcttc ttgg 24
<210> 27
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> фрагмент LC, обратный праймер
<400> 27
taaattcctc ggccgtgcag gccttatcaa cactctcccc tgttgaagct ct 52
<210> 28
<211> 122
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH антигенсвязывающего пептида
<400> 28
Gln Met Gln Leu Val Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gly
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Gly Ser Ile Ser Gly Ser
20 25 30
Asn Trp Trp Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Glu Ile Phe His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Val Asp Lys Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Asn Leu Asn Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Gly Gly Tyr Tyr Asp Ser Ser Gly Tyr Ser His Trp
100 105 110
Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 29
<211> 214
<212> PRT
<213> Искусственная последовательность
<220>
<223> KLC антигенсвязывающего пептида
<400> 29
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Gly Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Arg Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 30
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 1-й прямой праймер
<400> 30
ctcactctgc cgtctcagtg gtcgaggtgc agctggtgga gtctg 45
<210> 31
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 2-й прямой праймер
<400> 31
ctcactctgc cgtctcagtg gtccaggtgc agctgcagga gtcgg 45
<210> 32
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 3-й прямой праймер
<400> 32
ctcactctgc cgtctcagtg gtccaggtgc agctggtgca gtctg 45
<210> 33
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 4-й прямой праймер
<400> 33
ctcactctgc cgtctcagtg gtccaggtgc agctggtgga gtctg 45
<210> 34
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 5-й прямой праймер
<400> 34
ctcactctgc cgtctcagtg gtccaggtca ccttgaagga gtctgg 46
<210> 35
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 6-й прямой праймер
<400> 35
ctcactctgc cgtctcagtg gtcgaagtgc agctggtgga gtctgg 46
<210> 36
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 7-й прямой праймер
<400> 36
ctcactctgc cgtctcagtg gtccaggtcc agcttgtgca gtctgg 46
<210> 37
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 8-й прямой праймер
<400> 37
ctcactctgc cgtctcagtg gtccaggttc agctggtgca gtctgg 46
<210> 38
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 9-й прямой праймер
<400> 38
ctcactctgc cgtctcagtg gtccaggtcc agctggtaca gtctgg 46
<210> 39
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 10-й прямой праймер
<400> 39
ctcactctgc cgtctcagtg gtccagatgc agctggtgca gtctgg 46
<210> 40
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 11-й прямой праймер
<400> 40
ctcactctgc cgtctcagtg gtccaaatgc agctggtgca gtctgg 46
<210> 41
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 12-й прямой праймер
<400> 41
ctcactctgc cgtctcagtg gtcgaggtcc agctggtaca gtctgg 46
<210> 42
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 13-й прямой праймер
<400> 42
ctcactctgc cgtctcagtg gtccagatca ccttgaagga gtctgg 46
<210> 43
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 14-й прямой праймер
<400> 43
ctcactctgc cgtctcagtg gtcgaggtgc agctgttgga gtctgg 46
<210> 44
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 15-й прямой праймер
<400> 44
ctcactctgc cgtctcagtg gtcgaggtgc agctggtgga gactg 45
<210> 45
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 16-й прямой праймер
<400> 45
ctcactctgc cgtctcagtg gtcgaggtgc agctggtgga gtccgg 46
<210> 46
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 17-й прямой праймер
<400> 46
ctcactctgc cgtctcagtg gtcgaggtgc agctggtgga gtctcg 46
<210> 47
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 18-й прямой праймер
<400> 47
ctcactctgc cgtctcagtg gtccagctgc agctgcagga gtccgg 46
<210> 48
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 19-й прямой праймер
<400> 48
ctcactctgc cgtctcagtg gtccaggtgc agctacagca gtgggg 46
<210> 49
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 20-й прямой праймер
<400> 49
ctcactctgc cgtctcagtg gtccagctgc agctgcagga gtcggg 46
<210> 50
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 21-й прямой праймер
<400> 50
ctcactctgc cgtctcagtg gtcgaggtgc agctggtgca gtctgg 46
<210> 51
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 22-й прямой праймер
<400> 51
ctcactctgc cgtctcagtg gtcgaagtgc agctggtgca gtctgg 46
<210> 52
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 23-й прямой праймер
<400> 52
ctcactctgc cgtctcagtg gtccaggtac agctgcagca gtcagg 46
<210> 53
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 24-й прямой праймер
<400> 53
ctcactctgc cgtctcagtg gtccaggtgc agctggtgca atctgg 46
<210> 54
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 1-й обратный праймер
<400> 54
cttggtggag gctgatgaga cggtgaccag ggtgccctgg cc 42
<210> 55
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 2-й обратный праймер
<400> 55
cttggtggag gctgatgaga cggtgaccag ggtgccacgg cc 42
<210> 56
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 3-й обратный праймер
<400> 56
cttggtggag gctgatgaga cggtgaccat tgtcccttgg cc 42
<210> 57
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 4-й обратный праймер
<400> 57
cttggtggag gctgatgaga cggtgaccag ggttccctgg cc 42
<210> 58
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH, 5-й обратный праймер
<400> 58
cttggtggag gctgatgaga cggtgaccgt ggtcccttgg cc 42
<210> 59
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 1-й прямой праймер
<400> 59
tggtggcagc ggccacaggg gcccactccg acatccagat gacccagtct cc 52
<210> 60
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 2-й прямой праймер
<400> 60
tggtggcagc ggccacaggg gcccactccg atattgtgat gacccagact ccac 54
<210> 61
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 3-й прямой праймер
<400> 61
tggtggcagc ggccacaggg gcccactccg ccatccagtt gacccagtct cc 52
<210> 62
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 4-й прямой праймер
<400> 62
tggtggcagc ggccacaggg gcccactccg ccatccggat gacccagtct cc 52
<210> 63
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 5-й прямой праймер
<400> 63
tggtggcagc ggccacaggg gcccactccg atgttgtgat gactcagtct ccac 54
<210> 64
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 6-й прямой праймер
<400> 64
tggtggcagc ggccacaggg gcccactccg atattgtgat gactcagtct ccac 54
<210> 65
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 7-й прямой праймер
<400> 65
tggtggcagc ggccacaggg gcccactccg aaattgtgtt gacgcagtct ccag 54
<210> 66
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 8-й прямой праймер
<400> 66
tggtggcagc ggccacaggg gcccactccg aaatagtgat gacgcagtct ccag 54
<210> 67
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 9-й прямой праймер
<400> 67
tggtggcagc ggccacaggg gcccactccg aaattgtgtt gacacagtct ccag 54
<210> 68
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 10-й прямой праймер
<400> 68
tggtggcagc ggccacaggg gcccactccg aaattgtgct gactcagtct cc 52
<210> 69
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 11-й прямой праймер
<400> 69
tggtggcagc ggccacaggg gcccactcca acatccagat gacccagtct cc 52
<210> 70
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 12-й прямой праймер
<400> 70
tggtggcagc ggccacaggg gcccactccg acatccagtt gacccagtct cc 52
<210> 71
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 13-й прямой праймер
<400> 71
tggtggcagc ggccacaggg gcccactccg tcatctggat gacccagtct cc 52
<210> 72
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 14-й прямой праймер
<400> 72
tggtggcagc ggccacaggg gcccactccg ccatccagat gacccagtct cc 52
<210> 73
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 15-й прямой праймер
<400> 73
tggtggcagc ggccacaggg gcccactccg aaattgtaat gacacagtct ccagc 55
<210> 74
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 16-й прямой праймер
<400> 74
tggtggcagc ggccacaggg gcccactccg acatcgtgat gacccagtct cc 52
<210> 75
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 17-й прямой праймер
<400> 75
tggtggcagc ggccacaggg gcccactccg aaacgacact cacgcagtct cc 52
<210> 76
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, 18-й прямой праймер
<400> 76
tggtggcagc ggccacaggg gcccactccg atgttgtgat gacacagtct ccag 54
<210> 77
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LC, обратный праймер
<400> 77
taaattcctc ggccgtgcag gccttatcaa cactctcccc tgttgaagct ct 52
<210> 78
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R1 или R8
<400> 78
ggcccaggcg gcc 13
<210> 79
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R2 или R3
<400> 79
ggccacatag gcc 13
<210> 80
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R4 или R5
<400> 80
ggcccaaccg gcc 13
<210> 81
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R6 или R7
<400> 81
cgtctcctca gc 12
<210> 82
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 82
tcgctaccgt ggcccaggcg gccgacatcc agatgaccca gtctcc 46
<210> 83
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 83
tcgctaccgt ggcccaggcg gccgatattg tgatgaccca gactccac 48
<210> 84
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 84
tcgctaccgt ggcccaggcg gccgccatcc agttgaccca gtctcc 46
<210> 85
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 85
tcgctaccgt ggcccaggcg gccgccatcc ggatgaccca gtctcc 46
<210> 86
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 86
tcgctaccgt ggcccaggcg gccgatgttg tgatgactca gtctccac 48
<210> 87
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 87
tcgctaccgt ggcccaggcg gccgatattg tgatgactca gtctccac 48
<210> 88
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 88
tcgctaccgt ggcccaggcg gccgaaattg tgttgacgca gtctccag 48
<210> 89
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 89
tcgctaccgt ggcccaggcg gccgaaatag tgatgacgca gtctccag 48
<210> 90
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 90
tcgctaccgt ggcccaggcg gccgaaattg tgttgacaca gtctccag 48
<210> 91
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 91
tcgctaccgt ggcccaggcg gccgaaattg tgctgactca gtctcc 46
<210> 92
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 92
tcgctaccgt ggcccaggcg gccaacatcc agatgaccca gtctcc 46
<210> 93
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 93
tcgctaccgt ggcccaggcg gccgacatcc agttgaccca gtctcc 46
<210> 94
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 94
tcgctaccgt ggcccaggcg gccgtcatct ggatgaccca gtctcc 46
<210> 95
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 95
tcgctaccgt ggcccaggcg gccgccatcc agatgaccca gtctcc 46
<210> 96
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 96
tcgctaccgt ggcccaggcg gccgaaattg taatgacaca gtctccagc 49
<210> 97
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 97
tcgctaccgt ggcccaggcg gccgacatcg tgatgaccca gtctcc 46
<210> 98
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 98
tcgctaccgt ggcccaggcg gccgaaacga cactcacgca gtctcc 46
<210> 99
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи KLC
<400> 99
tcgctaccgt ggcccaggcg gccgatgttg tgatgacaca gtctccag 48
<210> 100
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для легкой цепи KLC
<400> 100
taaattcctc ggcctatgtg gcctattaac actctcccct gttgaagctc t 51
<210> 101
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 101
tcgctaccgt ggcccaggcg gcccagtctg tgctgactca gccacc 46
<210> 102
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 102
tcgctaccgt ggcccaggcg gcccagtctg ccctgactca gcctc 45
<210> 103
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 103
tcgctaccgt ggcccaggcg gcccagtctg ccctgactca gcctg 45
<210> 104
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 104
tcgctaccgt ggcccaggcg gcctcctatg agctgacaca gccacc 46
<210> 105
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 105
tcgctaccgt ggcccaggcg gcctcctatg agctgacaca gccatc 46
<210> 106
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 106
tcgctaccgt ggcccaggcg gcctcctatg agctgactca gccacc 46
<210> 107
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 107
tcgctaccgt ggcccaggcg gcctcctatg agctgactca gccactc 47
<210> 108
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 108
tcgctaccgt ggcccaggcg gcccagtctg tgctgacgca gccgcc 46
<210> 109
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 109
tcgctaccgt ggcccaggcg gcccagtctg tgttgacgca gccgc 45
<210> 110
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 110
tcgctaccgt ggcccaggcg gcctcttctg agctgactca ggaccc 46
<210> 111
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 111
tcgctaccgt ggcccaggcg gcctcctatg tgctgactca gccacc 46
<210> 112
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 112
tcgctaccgt ggcccaggcg gcctcctatg agctgacaca gctacc 46
<210> 113
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 113
tcgctaccgt ggcccaggcg gcctcctatg agctgatgca gccacc 46
<210> 114
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 114
tcgctaccgt ggcccaggcg gccctgcctg tgctgactca gcccc 45
<210> 115
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 115
tcgctaccgt ggcccaggcg gcccagcctg tgctgactca atcatcc 47
<210> 116
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 116
tcgctaccgt ggcccaggcg gcccagcttg tgctgactca atcgcc 46
<210> 117
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 117
tcgctaccgt ggcccaggcg gcccagcctg tgctgactca gccacc 46
<210> 118
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 118
tcgctaccgt ggcccaggcg gcccaggctg tgctgactca gccgg 45
<210> 119
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 119
Thr Cys Gly Cys Thr Ala Cys Cys Gly Thr Gly Gly Cys Cys Cys Ala
1 5 10 15
Gly Gly Cys Gly Gly Cys Cys Cys Ala Gly Cys Cys Thr Gly Thr Gly
20 25 30
Cys Thr Gly Ala Cys Thr Cys Ala Gly Cys Cys Ala Thr Cys
35 40 45
<210> 120
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 120
tcgctaccgt ggcccaggcg gccaatttta tgctgactca gccccac 47
<210> 121
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 121
tcgctaccgt ggcccaggcg gcccagactg tggtgactca ggagcc 46
<210> 122
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 122
tcgctaccgt ggcccaggcg gcccaggctg tggtgactca ggagc 45
<210> 123
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 123
tcgctaccgt ggcccaggcg gcccagactg tggtgaccca ggagc 45
<210> 124
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 124
tcgctaccgt ggcccaggcg gcccagcctg tgctgactca gccac 45
<210> 125
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для легкой цепи LLC
<400> 125
tcgctaccgt ggcccaggcg gcccaggcag ggctgactca gccac 45
<210> 126
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для легкой цепи LLC
<400> 126
taaattcctc ggcctatgtg gcctattatg aacattctgt aggggccact g 51
<210> 127
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 127
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagtct g 51
<210> 128
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 128
tattacttgc ggcccaaccg gccatggccc aggtgcagct gcaggagtcg g 51
<210> 129
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 129
tattacttgc ggcccaaccg gccatggccc aggtgcagct ggtgcagtct g 51
<210> 130
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 130
tattacttgc ggcccaaccg gccatggccc aggtgcagct ggtggagtct g 51
<210> 131
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 131
tattacttgc ggcccaaccg gccatggccc aggtcacctt gaaggagtct gg 52
<210> 132
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 132
tattacttgc ggcccaaccg gccatggccg aagtgcagct ggtggagtct gg 52
<210> 133
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 133
tattacttgc ggcccaaccg gccatggccc aggtccagct tgtgcagtct gg 52
<210> 134
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 134
tattacttgc ggcccaaccg gccatggccc aggttcagct ggtgcagtct gg 52
<210> 135
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 135
tattacttgc ggcccaaccg gccatggccc aggtccagct ggtacagtct gg 52
<210> 136
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 136
tattacttgc ggcccaaccg gccatggccc agatgcagct ggtgcagtct gg 52
<210> 137
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 137
tattacttgc ggcccaaccg gccatggccc aaatgcagct ggtgcagtct gg 52
<210> 138
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 138
tattacttgc ggcccaaccg gccatggccg aggtccagct ggtacagtct gg 52
<210> 139
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 139
tattacttgc ggcccaaccg gccatggccc agatcacctt gaaggagtct gg 52
<210> 140
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 140
tattacttgc ggcccaaccg gccatggccg aggtgcagct gttggagtct gg 52
<210> 141
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 141
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagact g 51
<210> 142
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 142
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagtcc gg 52
<210> 143
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 143
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtggagtct cg 52
<210> 144
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 144
tattacttgc ggcccaaccg gccatggccc agctgcagct gcaggagtcc gg 52
<210> 145
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 145
tattacttgc ggcccaaccg gccatggccc aggtgcagct acagcagtgg gg 52
<210> 146
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 146
tattacttgc ggcccaaccg gccatggccc agctgcagct gcaggagtcg gg 52
<210> 147
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 147
tattacttgc ggcccaaccg gccatggccg aggtgcagct ggtgcagtct gg 52
<210> 148
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 148
tattacttgc ggcccaaccg gccatggccg aagtgcagct ggtgcagtct gg 52
<210> 149
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 149
tattacttgc ggcccaaccg gccatggccc aggtacagct gcagcagtca gg 52
<210> 150
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для VH
<400> 150
tattacttgc ggcccaaccg gccatggccc aggtgcagct ggtgcaatct gg 52
<210> 151
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 151
cttggtggag gctgaggaga cggtgaccag ggtgccctgg cc 42
<210> 152
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 152
cttggtggag gctgaggaga cggtgaccag ggtgccacgg cc 42
<210> 153
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 153
cttggtggag gctgaggaga cggtgaccat tgtcccttgg cc 42
<210> 154
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 154
cttggtggag gctgaggaga cggtgaccag ggttccctgg cc 42
<210> 155
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для VH
<400> 155
cttggtggag gctgaggaga cggtgaccgt ggtcccttgg cc 42
<210> 156
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для линкера
<400> 156
acagcttgtc tgtaagcgga tg 22
<210> 157
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для линкера
<400> 157
tgtggataac cgtattaccg cc 22
<210> 158
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для R1-1,0 т.п.о.-R2
<400> 158
ttagcgaatt ccagacatct aatggcccag gcggcctcag ctagcaccaa gggcccatc 59
<210> 159
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для R1-1,0 т.п.о.-R2
<400> 159
cgggtgatca ccagtcttct attggcctat gtggccttat catttacccg gagacaggg 59
<210> 160
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для R3-линкер-R4
<400> 160
taggccacat aggccgagga atttaaaatg aaatacctat tgcctacggc agccg 55
<210> 161
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для R3-линкер-R4
<400> 161
atggccggtt gggccgcaag taataacaat ccagcggctg ccgtaggcaa tagg 54
<210> 162
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для R5-1,0 т.п.о.-R6
<400> 162
ttagcgaatt ccagacatct aatggcccaa ccggcctcag ctagcaccaa gggcccatc 59
<210> 163
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для R5-1,0 т.п.о.-R6
<400> 163
tgatcaccag tcttctattg gctgaggaga cgttatcatt tacccggaga c 51
<210> 164
<211> 987
<212> ДНК
<213> Homo sapiens
<400> 164
tcagctagca ccaagggccc atcggtcttc cccctggcgc cctgctccag gagcacctcc 60
gagagcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg 120
tcgtggaact caggcgctct gaccagcggc gtgcacacct tcccagctgt cctacagtcc 180
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcaactt cggcacccag 240
acctacacct gcaacgtaga tcacaagccc agcaacacca aggtggacaa gacagttgag 300
cgcaaatgtt gtgtcgagtg cccaccgtgc ccagcaccac ctgtggcagg accgtcagtc 360
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcacg 420
tgcgtggtgg tggacgtgag ccacgaagac cccgaggtcc agttcaactg gtacgtggac 480
ggcgtggagg tgcataatgc caagacaaag ccacgggagg agcagttcaa cagcacgttc 540
cgtgtggtca gcgtcctcac cgttgtgcac caggactggc tgaacggcaa ggagtacaag 600
tgcaaggtct ccaacaaagg cctcccagcc cccatcgaga aaaccatctc caaaaccaaa 660
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggagga gatgaccaag 720
aaccaggtca gcctgacctg cctggtcaaa ggcttctacc ccagcgacat cgccgtggag 780
tgggagagca atgggcagcc ggagaacaac tacaagacca cacctcccat gctggactcc 840
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg 900
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc 960
ctctccctgt ctccgggtaa atgataa 987
<210> 165
<211> 1013
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R1-1,0 т.п.о.-R2
<400> 165
ggcccaggcg gcctcagcta gcaccaaggg cccatcggtc ttccccctgg cgccctgctc 60
caggagcacc tccgagagca cagcggccct gggctgcctg gtcaaggact acttccccga 120
accggtgacg gtgtcgtgga actcaggcgc tctgaccagc ggcgtgcaca ccttcccagc 180
tgtcctacag tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcaa 240
cttcggcacc cagacctaca cctgcaacgt agatcacaag cccagcaaca ccaaggtgga 300
caagacagtt gagcgcaaat gttgtgtcga gtgcccaccg tgcccagcac cacctgtggc 360
aggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac 420
ccctgaggtc acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tccagttcaa 480
ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccacggg aggagcagtt 540
caacagcacg ttccgtgtgg tcagcgtcct caccgttgtg caccaggact ggctgaacgg 600
caaggagtac aagtgcaagg tctccaacaa aggcctccca gcccccatcg agaaaaccat 660
ctccaaaacc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga 720
ggagatgacc aagaaccagg tcagcctgac ctgcctggtc aaaggcttct accccagcga 780
catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacacctcc 840
catgctggac tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag 900
gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta 960
cacgcagaag agcctctccc tgtctccggg taaatgataa ggccacatag gcc 1013
<210> 166
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 72 п.о.- длинный линкер
<400> 166
aggccgagga atttaaaatg aaatacctat tgcctacggc agccgctgga ttgttattac 60
ttgcggccca ac 72
<210> 167
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R3-линкер(72 п.о.)-R4
<400> 167
ggccacatag gccgaggaat ttaaaatgaa atacctattg cctacggcag ccgctggatt 60
gttattactt gcggcccaac cggcc 85
<210> 168
<211> 1012
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R5-1,0 т.п.о.-R6
<400> 168
ggcccaaccg gcctcagcta gcaccaaggg cccatcggtc ttccccctgg cgccctgctc 60
caggagcacc tccgagagca cagcggccct gggctgcctg gtcaaggact acttccccga 120
accggtgacg gtgtcgtgga actcaggcgc tctgaccagc ggcgtgcaca ccttcccagc 180
tgtcctacag tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcaa 240
cttcggcacc cagacctaca cctgcaacgt agatcacaag cccagcaaca ccaaggtgga 300
caagacagtt gagcgcaaat gttgtgtcga gtgcccaccg tgcccagcac cacctgtggc 360
aggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac 420
ccctgaggtc acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tccagttcaa 480
ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccacggg aggagcagtt 540
caacagcacg ttccgtgtgg tcagcgtcct caccgttgtg caccaggact ggctgaacgg 600
caaggagtac aagtgcaagg tctccaacaa aggcctccca gcccccatcg agaaaaccat 660
ctccaaaacc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga 720
ggagatgacc aagaaccagg tcagcctgac ctgcctggtc aaaggcttct accccagcga 780
catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacacctcc 840
catgctggac tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag 900
gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta 960
cacgcagaag agcctctccc tgtctccggg taaatgataa cgtctcctca gc 1012
<210> 169
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для мутации вектора pUC19
<400> 169
atgacggtgt taacctctga cacatgcagc tcccggagaa ggtcacagct tgtctgtaag 60
c 61
<210> 170
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для мутации вектора pUC19
<400> 170
gtcagaggtt aacaccgtca tcaccgaaac gcgcgacacg aaagggcctc gtgatac 57
<210> 171
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 90 п.о. длинный линкер
<400> 171
aggccttcta gataattaat taggaggaat ttaaaatgaa atacctattg cctacggcag 60
ccgctggatt gttattactt gcggcccaac 90
<210> 172
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> R3-линкер(90 п.о.)-R4
<400> 172
ggccacatag gccttctaga taattaatta ggaggaattt aaaatgaaat acctattgcc 60
tacggcagcc gctggattgt tattacttgc ggcccaaccg gcc 103
<210> 173
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для линкера 90 п.о.
<400> 173
agtgttaata ggccacatag gccttctaga taattaatta ggag 44
<210> 174
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для линкера 90 п.о.
<400> 174
ccttctagat aattaattag gaggaattta aaatgaaata cc 42
<210> 175
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для линкера 90 п.о.
<400> 175
acctcggcca tggccggttg ggccgcaag 29
<---
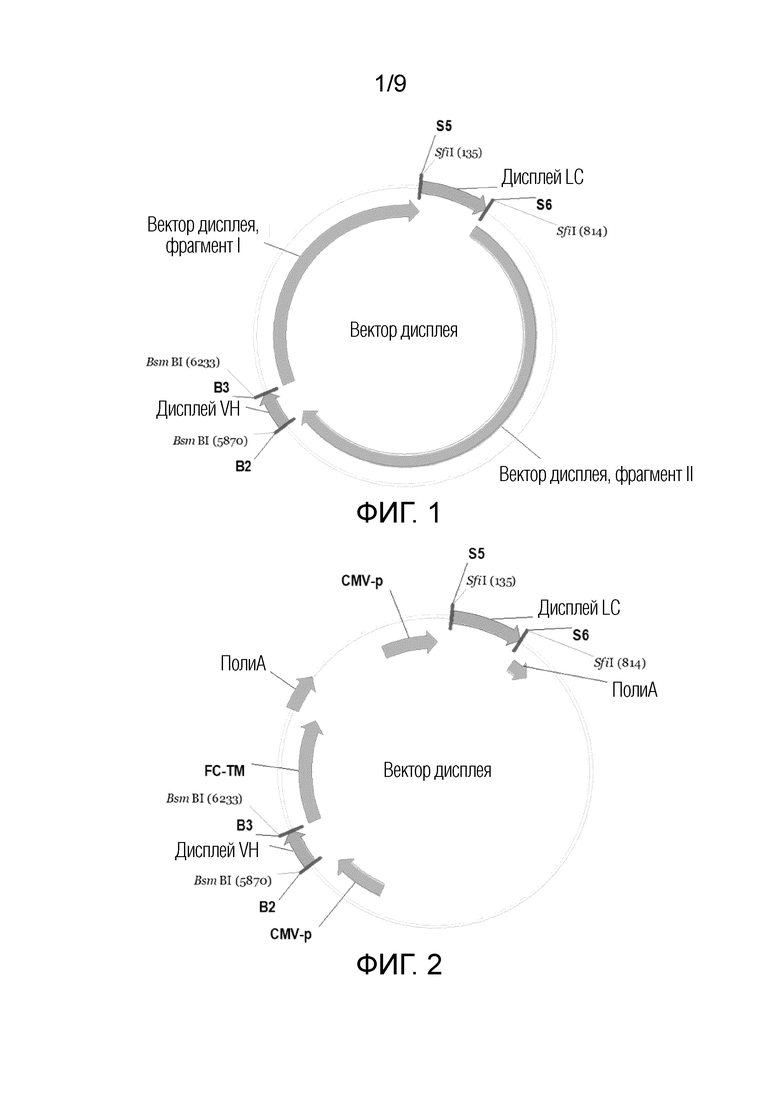
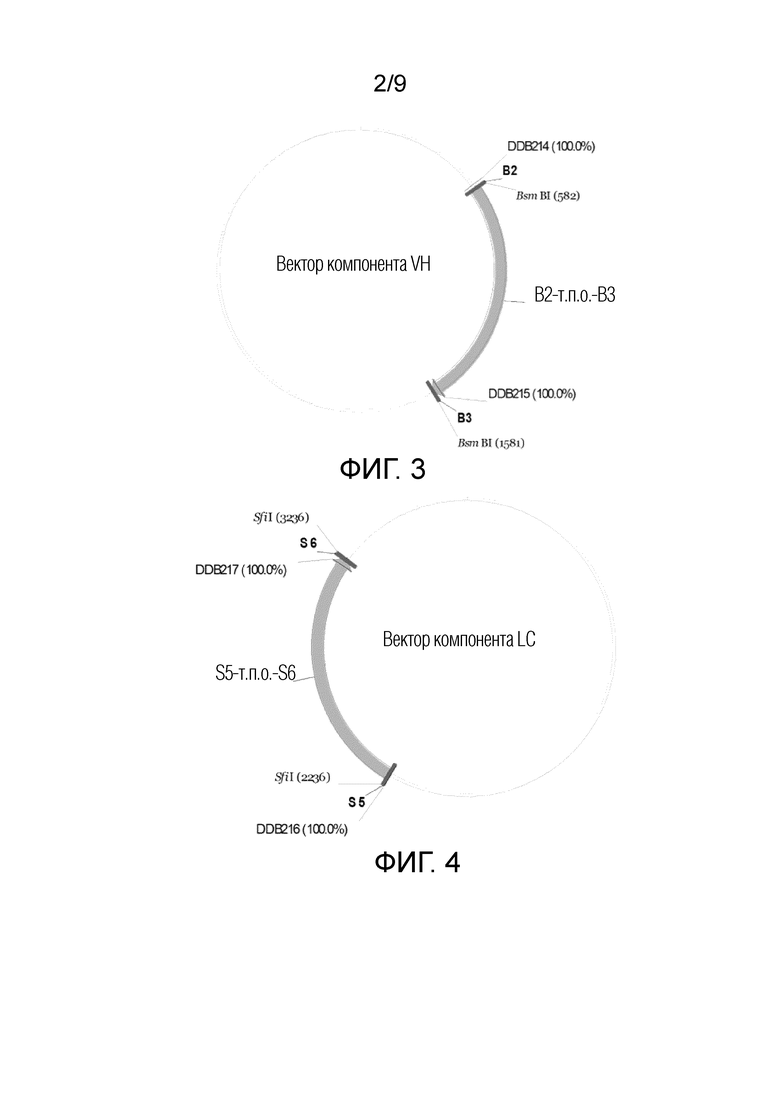


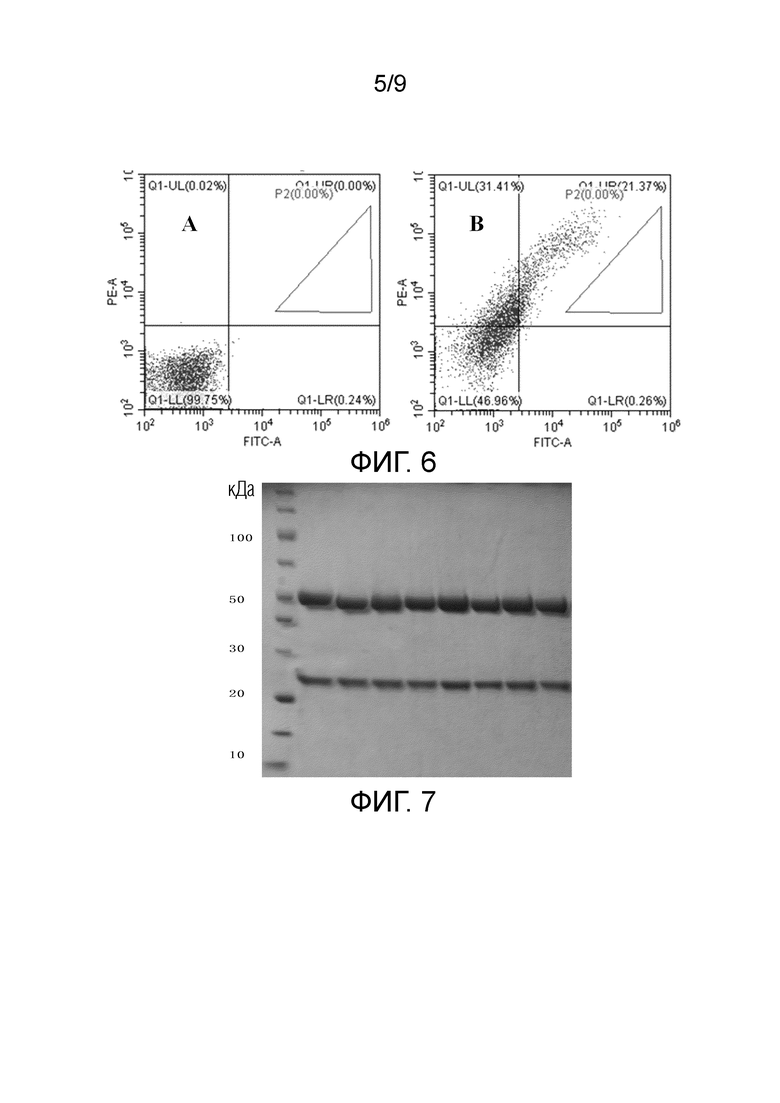
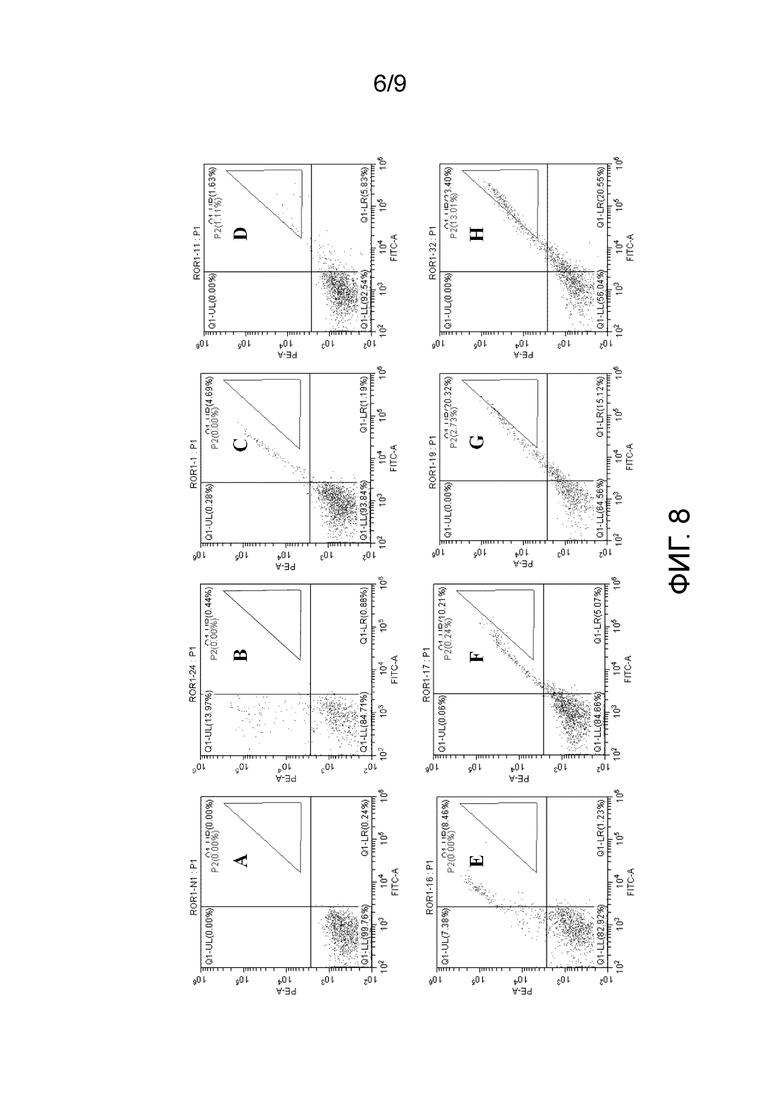
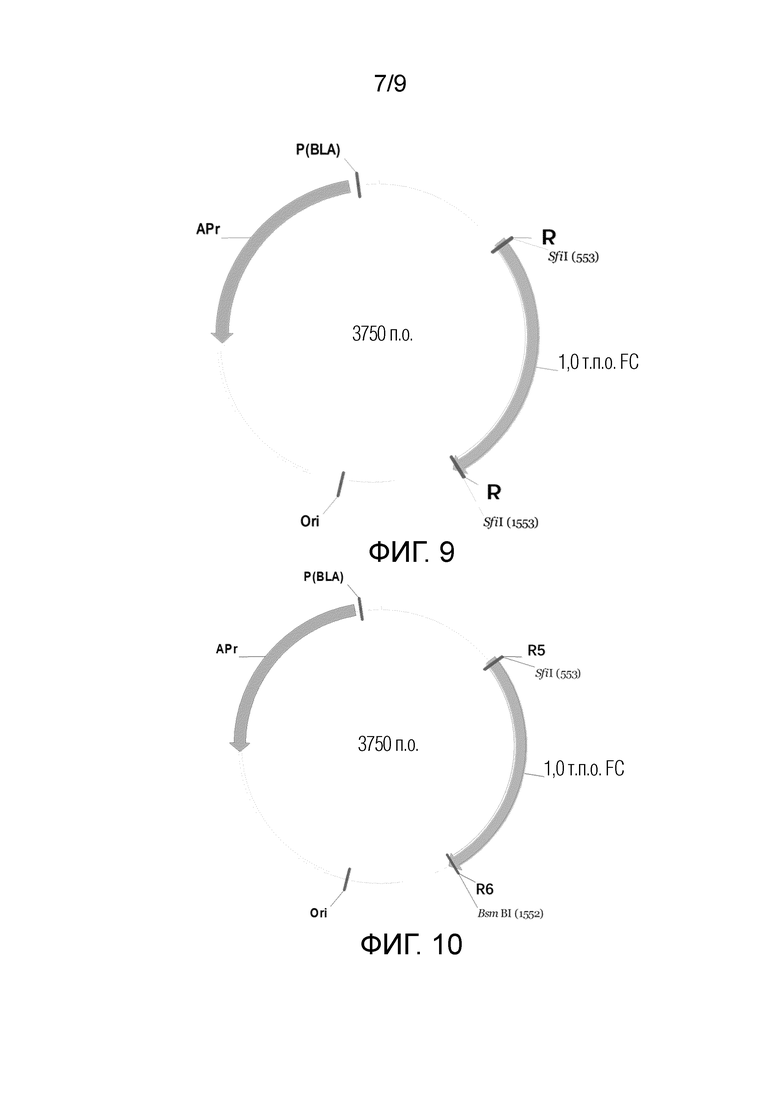
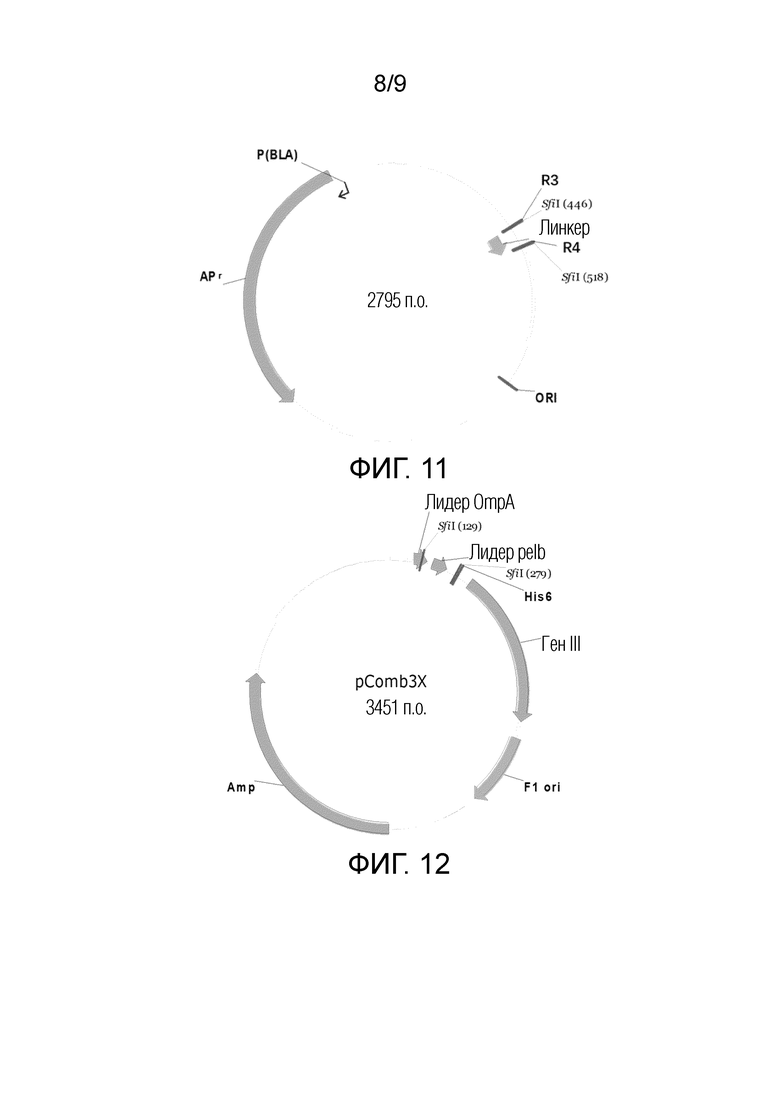
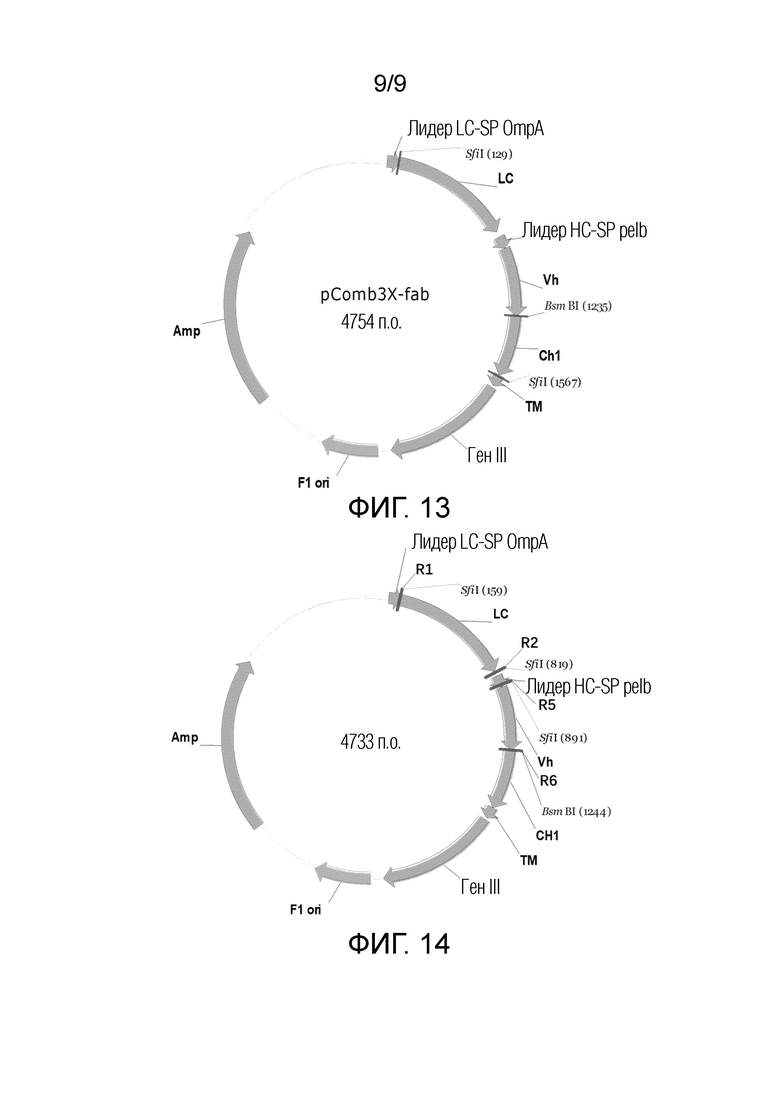
Изобретение относится к биотехнологии, в частности к способу конструирования вектора дисплея генов антигенспецифических связывающих полипептидов. Способ включает обработку посредством использования рестрикционной эндонуклеазы, специфически узнающей участок рестрикции, для получения четырех фрагментов нуклеиновой кислоты, имеющих специфические липкие концы, и затем обеспечение возможности направленного лигирования фрагментов нуклеиновой кислоты. Способ можно использовать для эффективного скрининга антигенспецифических антигенсвязывающих полипептидов или их фрагментов. 23 з.п. ф-лы, 14 ил., 4 табл., 10 пр.
1. Способ конструирования вектора дисплея генов антигенспецифических связывающих полипептидов, включающий:
a) получение первого полинуклеотида вектора дисплея, содержащего B2-дисплей VH-B3 в направлении от 5' до 3';
b) получение второго полинуклеотида вектора дисплея, содержащего S5-дисплей LC-S6 в направлении от 5' до 3';
c) получение третьего полинуклеотида вектора дисплея, содержащего B3-фрагмент I вектора дисплея-S5 в направлении от 5' до 3';
d) получение четвертого полинуклеотида вектора дисплея, содержащего S6-фрагмент II вектора дисплея-B2 в направлении от 5' до 3';
e) специфическое расщепление первого полинуклеотида вектора дисплея, второго полинуклеотида вектора дисплея, третьего полинуклеотида вектора дисплея и четвертого полинуклеотида вектора дисплея с использованием рестрикционной эндонуклеазы для получения расщепленного первого полинуклеотида вектора дисплея, расщепленного второго полинуклеотида вектора дисплея, расщепленного третьего полинуклеотида вектора дисплея и расщепленного четвертого полинуклеотида вектора дисплея; где рестрикционная эндонуклеаза специфически узнает B2, B3, S5 и S6, соответственно;
f) смешивание расщепленного первого полинуклеотида вектора дисплея, расщепленного второго полинуклеотида вектора дисплея, расщепленного третьего полинуклеотида вектора дисплея и расщепленного четвертого полинуклеотида вектора дисплея таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования вектора дисплея генов антигенспецифических связывающих полипептидов;
где дисплей VH кодирует вариабельную область тяжелой цепи антигенспецифического связывающего полипептида, и дисплей LC кодирует легкую цепь антигенспецифического связывающего полипептида;
где каждый из B2, B3, S5 и S6 независимо представляют собой участки узнавания для рестрикционной эндонуклеазы.
2. Способ по п. 1, где конец, образованный в результате специфического расщепления B2 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B3, S5 и S6 посредством соответствующей рестрикционной эндонуклеазы;
конец, образованный в результате специфического расщепления B3 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B2, S5 и S6 посредством соответствующей рестрикционной эндонуклеазы;
конец, образованный в результате специфического расщепления S5 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B2, B3 и S6 посредством соответствующей рестрикционной эндонуклеазы; и/или
конец, образованный в результате специфического расщепления S6 посредством рестрикционной эндонуклеазы, специфически узнающей его, не узнает конец или не связывается с концом, образованным в результате специфического расщепления любого из B2, B3 и S5 посредством соответствующей рестрикционной эндонуклеазы.
3. Способ по любому из пп. 1, 2, где рестрикционная эндонуклеаза выбрана из SfiI, Esp3I и BsmBI.
4. Способ по любому из пп. 1-3, где B2 и B3 могут быть специфически узнаны и расщеплены посредством фермента, выбранного из группы, состоящей из: BsmBI и Esp3I; S5 и S6 могут быть специфически узнаны и расщеплены посредством Sfil.
5. Способ по любому из пп. 1-4, где B2 содержит последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 8; B3 содержит последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 9; S5 содержит последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 10; и/или S6 содержит последовательность нуклеиновой кислоты, как указано в SEQ ID NO: 11.
6. Способ по п. 1, дополнительно включающий введение первого полинуклеотида вектора дисплея в первую бактерию дисплея для получения бактериальной библиотеки компонента дисплея VH; введение второго полинуклеотида вектора дисплея во вторую бактерию дисплея для получения бактериальной библиотеки компонента дисплея LC; введение третьего полинуклеотида вектора дисплея в третью бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея I; и/или введение четвертого полинуклеотида вектора дисплея в четвертую бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея II.
7. Способ по п. 6, включающий вставку первого полинуклеотида вектора дисплея в вектор компонента дисплея для формирования продукта лигирования для хранения дисплея VH и введение продукта лигирования для хранения дисплея VH в первую бактерию дисплея для получения бактериальной библиотеки компонента дисплея VH; вставку второго полинуклеотида вектора дисплея в вектор компонента дисплея для формирования продукта лигирования для хранения дисплея LC и введение продукта лигирования для хранения дисплея LC во вторую бактерию дисплея для получения бактериальной библиотеки компонента дисплея LC; вставку третьего полинуклеотида вектора дисплея в вектор компонента дисплея для формирования вектора дисплея продукта лигирования для хранения фрагмента I и введение продукта лигирования для хранения в третью бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея I; и вставку четвертого полинуклеотида вектора дисплея в вектор компонента дисплея для формирования вектора дисплея продукта лигирования для хранения фрагмента II и введение продукта лигирования для хранения в четвертую бактерию дисплея для получения бактериальной библиотеки вектора компонента дисплея II.
8. Способ по п. 7, где вектор компонента дисплея происходит из вектора pUC.
9. Способ по любому из пп. 6-8, дополнительно включающий получение плазмиды компонента дисплея VH, содержащей первый полинуклеотид вектора дисплея, из бактериальной библиотеки компонента дисплея VH, и получение расщепленного первого полинуклеотида вектора дисплея из плазмиды компонента дисплея VH;
получение плазмиды компонента дисплея LC, содержащей второй полинуклеотид вектора дисплея, из бактериальной библиотеки компонента дисплея LC, и получение расщепленного второго полинуклеотида вектора дисплея из плазмиды компонента дисплея LC;
получение плазмиды фрагмента I компонента дисплея, содержащей третий полинуклеотид вектора дисплея, из бактериальной библиотеки экспрессирующего вектора компонента I, и получение расщепленного третьего полинуклеотида вектора дисплея из плазмиды фрагмента I компонента дисплея;
и/или получение плазмиды фрагмента II компонента дисплея, содержащей четвертый полинуклеотид вектора дисплея, из бактериальной библиотеки экспрессирующего вектора компонента II, и получение расщепленного четвертого полинуклеотида вектора дисплея из плазмиды фрагмента II компонента дисплея.
10. Способ по любому из пп. 6-9, включающий расщепление плазмиды компонента дисплея VH с использованием рестрикционной эндонуклеазы, специфически узнающей B2 и B3, таким образом, получение расщепленного первого полинуклеотида вектора дисплея;
расщепление плазмиды компонента дисплея LC с использованием рестрикционной эндонуклеазы, специфически узнающей S5 и S6, таким образом, получение расщепленного второго полинуклеотида вектора дисплея;
расщепление плазмиды фрагмента I компонента дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей B3 и S5, таким образом, получение расщепленного третьего полинуклеотида вектора дисплея; и/или
расщепление плазмиды фрагмента II компонента дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей S6 и B2, таким образом, получение расщепленного четвертого полинуклеотида вектора дисплея.
11. Способ по любому из пп. 1-10, включающий:
a) получение пятого полинуклеотида, содержащего B-антигенспецифическую VH-B в направлении от 5' до 3';
b) получение вектора компонента VH, где вектор компонента VH содержит шестой полинуклеотид, содержащий B3-фрагмент лигирования вектора компонента VH-B2 в направлении от 5' до 3';
c) расщепление пятого полинуклеотида и вектора компонента VH с использованием рестрикционной эндонуклеазы для получения расщепленного пятого полинуклеотида и высвобожденного шестого полинуклеотида;
d) смешивание расщепленного пятого полинуклеотида и высвобожденного шестого полинуклеотида таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования библиотеки компонента антигенспецифической VH;
где B представляет собой участок узнавания для рестрикционной эндонуклеазы, которая может специфически узнавать B2 и/или B3, и антигенспецифическая VH кодирует вариабельную область тяжелой цепи антигенспецифического связывающего полипептида.
12. Способ по любому из пп. 1-11, включающий:
a) получение седьмого полинуклеотида, содержащего S-антигенспецифическую LC-S в направлении от 5' до 3';
b) получение вектора компонента LC, где вектор компонента LC содержит восьмой полинуклеотид, содержащий S6-фрагмент лигирования вектора компонента LC-S5 в направлении от 5' до 3';
c) расщепление седьмого полинуклеотида и вектора компонента LC с использованием рестрикционной эндонуклеазы для получения расщепленного седьмого полинуклеотида и высвобожденного восьмого полинуклеотида;
d) смешивание расщепленного седьмого полинуклеотида и высвобожденного восьмого полинуклеотида таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования библиотеки компонента антигенспецифической LC,
где S представляет собой участок узнавания для рестрикционной эндонуклеазы, которая может специфически узнавать S5 и/или S5, и антигенспецифическая LC кодирует легкую цепь антигенспецифического связывающего полипептида.
13. Способ по любому из пп. 1-12, включающий
a) получение девятого полинуклеотида, содержащего B2-рабочий фрагмент вектора компонента VH-B3 в направлении от 5' до 3';
b) вставку девятого полинуклеотида в экспрессирующий вектор компонента для получения вектора компонента VH.
14. Способ по любому из пп. 1-13, включающий
a) получение десятого полинуклеотида, содержащего S5-рабочий фрагмент вектора компонента LC-S6 в направлении от 5' до 3';
b) вставку десятого полинуклеотида в экспрессирующий вектор компонента для получения вектора компонента LC.
15. Способ по любому из пп. 13, 14, где экспрессирующий вектор компонента происходит из вектора pMD.
16. Способ по любому из пп. 11-15, включающий стадии:
введения вектора компонента VH в девятую бактерию для получения бактериальной библиотеки вектора для хранения компонента VH; получения плазмиды вектора для хранения компонента VH из бактериальной библиотеки вектора для хранения компонента VH; получения высвобожденного шестого полинуклеотида из плазмиды вектора для хранения компонента VH; и/или
введения вектора компонента LC в десятую бактерию для получения бактериальной библиотеки вектора для хранения компонента LC; получения плазмиды вектора для хранения компонента LC из бактериальной библиотеки вектора для хранения компонента LC; получения высвобожденного восьмого полинуклеотида из плазмиды вектора для хранения компонента LC.
17. Способ по п. 16, включающий расщепление плазмиды вектора для хранения компонента VH с использованием рестрикционной эндонуклеазы, специфически узнающей B2 и B3, таким образом, получение высвобожденного шестого полинуклеотида; и/или расщепление плазмиды вектора для хранения компонента LC с использованием рестрикционной эндонуклеазы, специфически узнающей S5 и S6, таким образом, получение высвобожденного восьмого полинуклеотида.
18. Способ по любому из пп. 11-17, включающий:
a) получение библиотеки компонента антигенспецифической VH, где библиотека компонента антигенспецифической VH содержит первый полинуклеотид, содержащий B2-антигенспецифическую VH-B3 в направлении от 5' до 3';
b) получение библиотеки компонента антигенспецифической LC, где библиотека компонента антигенспецифической LC содержит второй полинуклеотид, содержащий S5-антигенспецифическую LC-S6 в направлении от 5' до 3';
c) получение вектора дисплея, содержащего третий полинуклеотид, содержащий B3-фрагмент I вектора дисплея-S5 в направлении от 5' до 3', и четвертый полинуклеотид, содержащий S6-фрагмент II вектора дисплея-B2 в направлении от 5' до 3';
d) специфическое расщепление библиотеки компонента антигенспецифической VH, библиотеки компонента антигенспецифической LC и вектора дисплея с использованием рестрикционной эндонуклеазы для получения высвобожденного первого полинуклеотида, высвобожденного второго полинуклеотида, высвобожденного третьего полинуклеотида и высвобожденного четвертого полинуклеотида; где рестрикционная эндонуклеаза специфически узнает B2, B3, S5 и S6, соответственно;
e) смешивание высвобожденного первого полинуклеотида, высвобожденного второго полинуклеотида, высвобожденного третьего полинуклеотида и высвобожденного четвертого полинуклеотида, таким образом, чтобы их можно было направленно лигировать и циклизовать для формирования вектора дисплея генов антигенспецифических связывающих полипептидов;
где антигенспецифическая LC кодирует легкую цепь антигенспецифического связывающего полипептида, и антигенспецифическая VH кодирует вариабельную область тяжелой цепи антигенспецифического связывающего полипептида;
где каждый из B2, B3, S5 и S6 независимо представляют собой участки узнавания для рестрикционной эндонуклеазы.
19. Способ по п. 18, включающий расщепление библиотеки компонента антигенспецифической VH с использованием рестрикционной эндонуклеазы, специфически узнающей B2 и B3, таким образом, получение высвобожденного первого полинуклеотида; расщепление библиотеки компонента антигенспецифической LC с использованием рестрикционной эндонуклеазы, специфически узнающей S5 и S6, таким образом, получение высвобожденного второго полинуклеотида; расщепление вектора дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей B3, и рестрикционной эндонуклеазы, специфически узнающей S5, таким образом, получение высвобожденного третьего полинуклеотида; расщепление вектора дисплея с использованием рестрикционной эндонуклеазы, специфически узнающей S6, и рестрикционной эндонуклеазы, специфически узнающей B2, таким образом, получение высвобожденного четвертого полинуклеотида.
20. Способ по любому из пп. 18, 19, где пятый полинуклеотид, седьмой полинуклеотид, девятый полинуклеотид, десятый полинуклеотид, первый полинуклеотид вектора дисплея, второй полинуклеотид вектора дисплея, третий полинуклеотид вектора дисплея и/или четвертый полинуклеотид вектора дисплея получают из материалов образцов.
21. Способ по п. 20, где материалы образцов содержат антитела, нацеленные на специфические антигены, или их антигенсвязывающие фрагменты и/или IgG.
22. Способ по любому из пп. 1-21, где направленное лигирование включает применение лигазы.
23. Способ по любому из пп. 1-22, включающий введение вектора дисплея генов антигенспецифических связывающих полипептидов в клетку и получение антигенспецифического связывающего полипептида из клетки.
24. Способ по любому из пп. 1-23, включающий:
a) введение вектора дисплея генов антигенспецифических связывающих полипептидов в первую бактерию для получения бактериальной библиотеки дисплея генов антигенспецифических связывающих полипептидов;
b) получение библиотеки дисплея генов антигенспецифических связывающих полипептидов из бактериальной библиотеки дисплея генов антигенспецифических связывающих полипептидов;
c) получение ДНК экспрессирующего вектора антигенспецифического связывающего полипептида из библиотеки дисплея генов антигенспецифических связывающих полипептидов;
d) введение ДНК экспрессирующего вектора антигенспецифического связывающего полипептида в клетку;
e) получение антигенспецифического связывающего полипептида из клетки.
| SOLFOROSI L | |||
| et al., A phage display vector optimized for the generation of human antibody combinatorial libraries and the molecular cloning of monoclonal antibody fragments, New Microbiol, 2012, vol | |||
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| РЕЛЬСОВАЯ ПЕДАЛЬ | 1920 |
|
SU289A1 |
| LI T | |||
| et al., A neutralization scFv antibody against IL-1β isolated from a NIPA-based bacterial display library, Curr Pharm | |||

