ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к антителу, которое связывается протеинтирозинфосфатазой σ рецепторного типа человека. Далее “протеинтирозинфосфатазу” сокращенно обозначают как PTP, “протеинтирозинфосфатазу рецепторного типа” сокращенно обозначают как RPTP или PTPR, “протеинтирозинфосфатазу σ рецепторного типа” иногда сокращенно обозначают как RPTP-σ, PTP-σ или PTPRS, и “человека” и “мышь” иногда обозначают с помощью приставок h и m, соответственно.
УРОВЕНЬ ТЕХНИКИ
Интерфероны (далее “интерферон” иногда сокращенно обозначается как IFN) являются наиболее важными цитокинами при противовирусном иммунном ответе. Интерферон-продуцирующая клетка (IPC: IPC представляет собой недифференцированную лимфоцитарную дендритную клетку, которая определяется как клетка-предшественник дендритной клетки (DC). IPC также иногда называют плазмацитоидной дендритной клеткой или подобной плазматической клетке дендритной клеткой (плазмацитоидная дендритная клетка: pDC). Далее в настоящем описании считается, что IPC и pDC имеют одинаковое значение, и далее они стандартно обозначаются как pDC в качестве общего правила.) в крови человека экспрессирует белок главного комплекса гистосовместимости класса II вместе с CD4. Однако поскольку количество таких клеток является небольшим и клетки быстро вызывают апоптоз и лишены маркера линии дифференцировки, эти клетки не были выделены или детально охарактеризованы до настоящего времени. Показано, что pDC представляет собой клетку-предшественник дендритной клетки типа CD4+CD11c-2, и что она продуцирует в 200-1000 раз больше IFN, чем другие клетки крови после стимуляции микроорганизмом. Таким образом, pDC2 является решающей эффекторной клеткой иммунной системы при противовирусном и противоопухолевом иммунных ответах.
IFNα и IFNβ известны как IFN типа I, имеющие противовирусную активность или противоопухолевую активность. С другой стороны, было установлено, что IFNα связан с аутоиммунными заболеваниями. Например, аномальная продукция IFNα была описана у пациентов, страдающих указанными ниже аутоиммунными заболеваниями. Более того, было сделано предположение о возможности смягчения аутоиммунного состояния путем нейтрализации IFNα.
Были описаны системный эритематоз (Shiozawa et al., Arthr. & Rheum. 35, 412, 1992) и хронический ревматоидный артрит (Hopkins et al., Clin. Exp. Immunol 73, 88, 1988), и, более того, примеры, в которых состояние аутоиммунного заболевания проявлялось или ухудшалось путем введения рекомбинантного IFNα2 или IFN (Wada et al., Am. J. Gastroenterol. 90, 136, 1995; Perez et al., Am. J. Hematol. 49, 365, 1995; Wilson LE et al., Semin Arthritis, Rheum. 32, 163-173, 2002).
Более того, также было установлено, что IFNα индуцирует дифференцировку дендритных клеток (DC). Поскольку дендритная клетка также является антигенпредставляющей клеткой, считается, что индукция дифференцировки дендритной клетки представляет собой важный механизм аутоиммунных заболеваний. В действительности, было предположено, что индукция дифференцировки дендритных клеток посредством IFNα тесно связана с возникновением системного эритематоза (Blanco et al., Science, 16: 294, 1540-1543, 2001). Таким образом, была отмечена противоопухолевая активность и тесная связь IFNα с аутоиммунными заболеваниями. Более того, IFNα также тесно связан с возникновением псориаза (Nestle FO et al., J. Exp. Med. 202, 135-143, 2005).
В крови присутствует только небольшое количество pDC. Считается, что доля pDC среди лимфоцитов периферической крови составляет 1% или менее. Однако pDC обладают чрезвычайно высокой способностью продуцировать IFN. Способность pDC продуцировать IFN достигает, например, 3000 пг/мл/104 клеток. Следовательно, можно считать, что, хотя количество этих клеток является небольшим, при проникновении вирусной инфекции основная часть IFNα или IFNβ в крови продуцируется pDC.
pDC дифференцируется в дендритную клетку при вирусной стимуляции, что индуцирует продукцию IFN-γ и IL-10 T-клеткой. Более того, pDC также дифференцируется в дендритную клетку при стимуляции IL-3. Дендритная клетка, дифференцированная посредством стимуляции IL-3, индуцирует продукцию Th2-цитокинов (IL-4, IL-5, IL-10) T-клеткой. Таким образом, pDC имеет характеристику, состоящую в том, что она дифференцируется в различные дендритные клетки, в зависимости от различий в стимуляции.
Таким образом, pDC представляет собой клетку, имеющую два аспекта: один из них представляет собой аспект в качестве IFN-продуцирующей клетки, а другой представляет собой аспект в качестве клетки-предшественника для дендритной клетки. Оба из них играют важные роли в иммунной системе. Это означает, что pDC является одной из важных клеток, которая поддерживает иммунную систему с различных аспектов.
Для контроля активности гуморального фактора, такого как IFN, является эффективным введение антитела, которое распознает этот фактор. Например, попытка лечить аутоиммунное заболевание антителом против интерлейкина (IL)-1 или IL-4 нашла практическое применение (Guler et al., Arthritis Rheum, 44, S307, 2001). Более того, считается, что нейтрализованное антитело может стать терапевтическим средством от аутоиммунных заболеваний также в случае интерферонов (IFN) (Stewart, TA. Cytokine Growth Factor Rev. 14; 139-154, 2003). Можно ожидать, что сходный подход может быть эффективным для IFN, продуцируемых pDC. Однако такой подход основан на ингибировании действия продуцированного гуморального фактора. Если бы продуцирование рассматриваемого гуморального фактора можно было контролировать прямо, то можно было бы достигнуть более существенного терапевтического эффекта.
Было описано антитело, которое распознает pDC человека. Например, моноклональное антитело против BDCA-2 представляет собой моноклональное антитело, которое является специфичным к pDC человека (Dzionek A. et al., J. Immunol 165: 6037-6046, 2000). Было установлено, что моноклональное антитело против BDCA-2 обладает действием подавления продукции IFN из pDC человека (J. Exp. Med. 194: 1823-1834, 2001). Более того, также было описано, что моноклональное антитело, которое распознает интерферон-продуцирующую клетку мыши, подавляет продукцию интерферонов (Blood 2004 Jun 1; 103/11: 4201-4206. Epub 2003 Dec). Было описано, что моноклональное антитело против pDC мыши снижало количество дендритных клеток (J. Immunol. 2003, 171: 6466-6477).
Было бы полезным, если бы было предоставлено антитело, которое аналогичным образом распознает pDC человека и может контролировать их активность. Например, авторы настоящего изобретения уже установили, что антитело, которое распознает Ly49Q, специфично связывается с pDC мыши. Однако антитело против Ly49Q не препятствует активности pDC мыши (Blood, 1 Apri1 2005, vol. 105, No. 7, pp. 2787-2792: WO2004/13325A1).
Протеинфосфатазы представляют собой дефосфорилированные ферменты, которые были выявлены в исследованиях метаболизма гликогена. Помимо протеинтирозинфосфатазы (PTP), были открыты сериновая/треониновая протеинфосфатаза, фосфолипид-специфическая фосфатаза и т.п., и они составляют суперсемейство протеинфосфатаз. Среди них, протеинтирозинфосфатаза представляет собой фермент, который ответственен за фосфорилирование в обратимых модификациях фосфорилированием, которое наблюдается в остатках тирозина белков. С другой стороны, протеинтирозинкиназа (PTK) является примером фермента, который ответственен за фосфорилирование в обратимых модификациях фосфорилированием, которое наблюдается в остатках тирозина белков.
Протеинтирозинфосфатаза (PTP) преобразует информацию о связывании лиганда в его внеклеточном домене с активностью фосфатазы внутриклеточного домена, и считается, что протеинтирозинкиназа (PTK) активируется путем связывания лиганда, в то время как протеинтирозинфосфатаза (PTP), главным образом, инактивируется связыванием лиганда. Таким образом, в случае как протеинтирозинфосфатазы (PTP), так и протеинтирозинкиназы (PTK), стимуляция лиганда приводит к увеличению уровня фосфорилирования, в то время как в свойствах сигнала ожидается существенное отличие. В случае протеинтирозинкиназы (PTK) осуществляется контроль с положительной обратной связью, при котором рецепторы фосфорилируются друг другом и активируются, и локальная активация молекул протеинтирозинкиназы (PTK) передается другим молекулам протеинтирозинкиназы (PTK) на клеточной мембране, таким образом, фосфорилирование увеличивается в широком диапазоне. С другой стороны, в случае протеинтирозинфосфатазы (PTP) инактивируются только молекулы, с которыми связаны лиганды, и фосфорилирование субстрата увеличивается только местно. Протеинтирозинфосфатаза (PTP), которая вовлечена во множество физиологических функций и клеточных функций, привлекает значительное внимание в широких областях нейробиохимии головного мозга, иммунологии, злокачественных опухолей, сахарного диабета и т.п. (копия домашней страницы Division of Molecular Neurobiology, National Institute for Basic Biology, http://niwww3.nibb.ac.jp/RPTP.pdf).
Семейство протеинтирозинфосфатаз может быть подразделено на рецепторный тип, имеющий область, проходящую через мембрану клетки, и нерецепторный тип. Существует 21 молекула протеинтирозинфосфатаз рецепторного типа (также сокращенно обозначаемых как RPTP или PTPR) у млекопитающих, которые подразделяются на восемь подсемейств, и каждое подсемейство имеет присущую ему внеклеточную структуру, в которой выявлены иммуноглобулин-подобный домен, домен, подобный фибронектину типа III, домен, подобный карбонатдегидратазе, домен MAM и т.п. (Nat Rev Mol Cell Biol., Vol. 7, 833-846, 2006).
Протеинтирозинфосфатаза σ рецепторного типа человека (сокращенно обозначаемая как hRPTP-σ, hPTP-σ или hPTPRS, и в настоящем описании в основном используется сокращение hPTPRS) принадлежит семейству R2A вместе с LAR (связанная с лейкоцитарным антигеном протеинтирозинфосфатаза) и протеинтирозинфосфатазой δ рецепторного типа (PTP-δ). Ферменты семейства PTPR экспрессируются в различных тканях, включая нервную систему с начала формирования до времени после созревания животных, однако установлено мало их физиологических функций, поскольку идентификация молекул лигандов и молекул субстратов является непростой.
Дендритные клетки (DC) представляют собой основные антигенпредставляющие клетки в живом организме, которые присутствуют в крови, лимфоидных тканях и т.п. и их приближенно классифицируют на миелоидные дендритные клетки (mDC) и плазмацитоидные дендритные клетки (pDC). pDC селективно экспрессирует TLR7 и TLR9 в качестве Toll-подобных рецепторов на ее клеточной поверхности и продуцирует интерфероны α и β типа I, особенно интерферон α.
Недавние испытания установили различные молекулы лигандов, которые действуют на дендритные клетки, контролируя их созревание и активацию, и внутриклеточные механизмы передачи сигнала их рецепторов становятся понятными. Однако существует множество неясных моментов относительно механизмов модификации и контроля функций дендритных клеток. Аналогично тому, что установлено для многих других клеток, считается, что фосфорилирование белков играет важную роль также и в дендритных клетках для контроля передачи сигнала от рецепторов, движения/миграции клеток и т.п.
Протеинфосфатазы, которые представляют собой факторы отрицательного контроля фосфорилирования белков, являются преобладающими кандидатами в качестве факторов поддержания надлежащей интенсивности и длительности сигналов, чтобы модулировать активацию и функции дендритных клеток. (Nobuhiro Tanuma (Institute for Genetic Medicine, Hokkaido University), “Functional Analysis Of Tyrosine Phosphatase Induced in Maturing of Dendritic Cells” in the homepage of the Northern Advancement Center for Science & Technology (abbreviation: NOASTEC), http://www.noastec.jp/kinouindex/data2005/pdf/01/01_20.pdf).
В международной публикации № WO95/9656A1 описана RPTP-σ (PTPRS) и нуклеиновая кислота, кодирующая ее; однако описанная аминокислотная последовательность представляет собой последовательность крысы, и в публикации не упоминается антитело, специфичное к PTPRS. В международной публикации № WO95/9656A1 также не описано антитело против PTPRS человека.
Международная публикация № WO2007/41317A1 относится к выделенному антителу, которое специфично связывается по меньшей мере с RPTP-σ или RPTP-δ, подавляя иммунный ответ иммунной клетки, или к его антигенсвязывающему фрагменту. В документе описано, что связывание полипептида поксвируса посредством A41L и RPTP конкурентно ингибируется с использованием антитела, которое специфично связывается с RPTP, посредством чего достигается подавление иммунного ответа иммунной клетки. Однако в этом документе не описано, что антитело, которое специфично связывается с RPTP-σ (PTPRS), было в действительности получено, и, при рассмотрении описания примеров, в примерах только подтверждается, что RPTP, экспрессируемый в иммунной клетке, которая связывается с A41L, является частью RPTP-σ, RPTP-δ и LAR, которые принадлежат к одному и тому же подтипу R2A, и получен слитый белок иммуноглобулин-подобного домена LAR и Fc (слитый белок LAR (Ig домен)-Fc). Трудно сказать, что в международной публикации № WO2007/41317A1 описано антитело, специфичное только к RPTP-σ, и его получение.
До настоящего времени не было получено антитело, которое связывается только с RPTP-σ, т.е. конкретным участком PTPRS, описанным в настоящей заявке, и антитело, которое может специфично связываться с RPTP-σ (PTPRS), но не с RPTP-δ и LAR, которые принадлежат тому же подтипу R2A. PTPRS человека представляет собой молекулу, для которой наблюдается специфическая экспрессия в pDC, однако до настоящего времени не было получено какого-либо антитела против PTPRS человека.
Список ссылок
Патентная литература
PTL 1: WO2004/13325A1
PTL 2: WO95/9656A1
PTL 3: WO2007/41317A1
Непатентная литература
NPL 1: Shiozawa et al., Arthr. & Rheum. 35, 412, 1992
NPL 2: Hopkins et al., Clin. Exp. Immunol. 73, 88, 1988
NPL 3: Wada et al., Am.J. Gastroenterol. 90, 136, 1995
NPL 4: Perez et al., Am. J. Hematol. 49, 365, 1995
NPL 5: Wilson LE et al, Semin Arthritis. Rheum. 32, 163-173, 2002
NPL 6: Blanco et al., Science, 16:294, 1540-1543,2001
NPL 7: Nestle FO et al., J. Exp. Med. 202, 135-143, 2005
NPL 8: Guler et al., Arthritis Rheum., 44. S307, 2001
NPL 9: Stewart, TA. Cytokine Growth Factor Rev. 14; 139-154, 2003
NPL 10: Dzionek, A. et al. J.Immunol. 165: 6037-6046, 2000
NPL 11: J. Exp. Med. 194:1823-1834, 2001
NPL 12: Blood 2004 Jun 1; 103/11:4201-4206. Epub 2003 Dec
NPL 13: J. Immunol. 2003, 171:6466-6477
NPL 14: Blood, 1 April 2005, Vol. 105, No. 7, pp. 2787-2792
NPL 15: http://niwww3.nibb.ac.jp/RPTP.pdf
NPL 16: Nat Rev Mol Cell Biol., Vol. 7, 833-846, 2006
NPL 17: http://www.noastec.jp/kinouindex/data2005/pdf/01/01_20.pdf
Сущность изобретения
Техническая проблема
Задачей настоящего изобретения является предоставление антитела, которое связывается с протеинтирозинфосфатазой σ рецепторного типа человека (PTPRS человека, hRPTP-σ), и выявление, идентификация или выделение pDC. Более того, задачей настоящего изобретения является модулирование активности pDC.
Авторы настоящего изобретения подтвердили посредством исследований, касающихся pDC человека, что экспрессия PTPRS в pDC специфически усиливается. Таким образом, авторы настоящего изобретения предприняли попытку получить антитело против PTPRS и проверить его действие.
Для получения антитела, которое распознает следовое количество белка, происходящего из живого организма, в качестве иммуногена обычно используют белок, полученный технологией рекомбинации генов. Авторы настоящего изобретения предприняли попытку экспрессировать PTPRS человека на основе последовательности оснований кДНК PTPRS человека, которая уже установлена, и информации об аминокислотной последовательности, кодируемой ей (номер доступа GenBank № NM_002856.3).
Для получения антитела против белка в качестве иммуногена часто пытаются использовать неполную аминокислотную последовательность природного белка. Однако, чтобы антитело распознавало молекулу на клеточной поверхности, следует выбирать область, которая составляет часть, которая распознается антителом в качестве эпитопа на клеточной поверхности. Таким образом, считалось, что получение антитела, которое является специфичным для PTPRS человека, с использованием аминокислотной последовательности фрагмента в качестве иммуногена произойдет еще нескоро.
Решение этой проблемы
В такой ситуации авторы настоящего изобретения установили, что антитело, которое связывается с pDC, можно получать с использованием определенного иммуногена. Более того, они также подтвердили, что полученное таким образом антитело специфически распознает pDC человека и обладает действием модулирования его активности, и осуществили настоящее изобретение.
Таким образом, настоящее изобретение относится к следующему антителу против PTPRS человека, способу его получения и его применениям.
Настоящее изобретение состоит в следующем.
(1) Моноклональное антитело, которое связывается с внеклеточным доменом протеинтирозинфосфатазы σ рецепторного типа человека (PTPRS человека), или фрагмент, включающий его антигенсвязывающую область.
(2) Моноклональное антитело или фрагмент, включающий его антигенсвязывающую область согласно упомянутому выше (1), которые связываются с плазмацитоидной дендритной клеткой.
(3) Моноклональное антитело, продуцируемое гибридомой 9H5-4, которая была депонирована в качестве номера доступа № FERM ABP-11356, гибридомой 10F7-38, которая была депонирована в качестве номера доступа № FERM ABP-11357, гибридомой 13G5-52, которая была депонирована в качестве номера доступа № FERM ABP-11358, гибридомой 13G5-57, которая была депонирована в качестве номера доступа № FERM ABP-11359, гибридомой 14A8-85, которая была депонирована в качестве номера доступа № FERM ABP-11360, гибридомой 22H8-84, которая была депонирована в качестве номера доступа № FERM ABP-11361, гибридомой 49F2-30, которая была депонирована в качестве номера доступа № FERM ABP-11362, или гибридомой 55E7-79, которая была депонирована в качестве номера доступа № FERM ABP-11363, или фрагмент, включающий его антигенсвязывающую область.
(4) Гибридома, которая продуцирует любое из моноклональных антител согласно упомянутым выше (1) или (2).
(5) Моноклональное антитело, продуцируемое гибридомой 9H5-4, которая была депонирована в качестве номера доступа № FERM ABP-11356, гибридомой 10F7-38, которая была депонирована в качестве номера доступа № FERM ABP-11357, гибридомой 13G5-52, которая была депонирована в качестве номера доступа № FERM ABP-11358, гибридомой 13G5-57, которая была депонирована в качестве номера доступа № FERM ABP-11359, гибридомой 14A8-85, которая была депонирована в качестве номера доступа № FERM ABP-11360, гибридомой 22H8-84, которая была депонирована в качестве номера доступа № FERM ABP-11361, гибридомой 49F2-30, которая была депонирована в качестве номера доступа № FERM ABP-11362, или гибридомой 55E7-79, которая была депонирована в качестве номера доступа № FERM ABP-11363, или фрагмент, включающий его антигенсвязывающую область.
(6) Способ получения моноклонального антитела, который включает культивирование гибридомы согласно упомянутому выше (5), и сбор моноклонального антитела из культуры.
(7) Способ получения клетки, которая продуцирует моноклональное антитело, которое связывается с PTPRS человека, который включает:
1) введение клетки, которая экспрессирует экзогенный белок, включающий внеклеточный домен PTPRS человека, для иммунизации животного, и
2) выбор антителопродуцирующей клетки, которая продуцирует антитело, которое связывается с PTPRS человека, из антителопродуцирующих клеток иммунизированного животного.
(8) Способ согласно упомянутому выше (7), где клетка, которая экспрессирует PTPRS человека, представляет собой клетку, которая содержит экспрессируемый экзогенный полинуклеотид, который кодирует аминокислотную последовательность, включающую внеклеточный домен PTPRS человека.
(9) Способ согласно упомянутому выше (8), где клетка представляет собой клетку животного.
(10) Способ согласно упомянутому выше (9), где клетка представляет собой клетку, происходящую из мыши.
(11) Способ согласно упомянутому выше (10), где происходящая из человека клетка представляет собой клетку D2SC.
(12) Способ согласно любому из упомянутых выше (7)-(11), который дополнительно включает клонирование полученной антителопродуцирующей клетки.
(13) Способ получения моноклонального антитела, которое связывается с внеклеточным доменом PTPRS человека, который включает культивирование антителопродуцирующей клетки, полученной способом согласно упомянутому выше (9), и сбор моноклонального антитела из культуры.
(14) Моноклональное антитело, которое распознает PTPRS человека, которое получают с помощью следующих стадий, или фрагмент, включающий его антигенсвязывающую область:
1) введение клетки животному для его иммунизации, которая экзогенным путем экспрессирует белок, включающий внеклеточный домен PTPRS человека;
2) выбор антителопродуцирующей клетки, которая продуцирует антитело, которое связывается с PTPRS человека, из антителопродуцирующих клеток иммунизированного животного; и
3) культивирование антителопродуцирующей клетки, выбранной в (2), и сбор антитела, которое распознает PTPRS человека, из культуры.
(15) (a) Иммуноген для получения антитела, которое связывается с PTPRS человека, который включает клетку животного, которая содержит экзогенный и экспрессируемый полинуклеотид, который кодирует аминокислотную последовательность, включающую внеклеточный домен PTPRS человека, или ее фракцию клеточной мембраны.
(16) Иммуноген согласно упомянутому выше (15), где клетка животного представляет собой происходящую из мыши клетку.
(17) Способ выявления плазмацитоидной дендритной клетки, который включает контактирование моноклонального антитела, которое связывается с внеклеточным доменом PTPRS человека, или фрагмента, включающего его антигенсвязывающую область, с рассматриваемой клеткой, и выявление моноклонального антитела, которое связалось с клеткой, или фрагмента, включающего его антигенсвязывающую область.
(18) Средство для выявления плазмацитоидной дендритной клетки, которое включает моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека, или фрагмент, включающий его антигенсвязывающую область.
(19) Способ подавления активности плазмацитоидной дендритной клетки, который включает контактирование любого из следующих компонентов с плазмацитоидной дендритной клеткой:
(a) моноклональное антитело, которое связывается с PTPRS человека, подавляя активность плазмацитоидной дендритной клетки, или фрагмент, включающий его антигенсвязывающую область, и
(b) иммуноглобулин, в который пересажена определяющая комплементарность область моноклонального антитела согласно (a), или фрагмент, включающий его антигенсвязывающую область.
(20) Способ подавления активности плазмацитоидной дендритной клетки в живом организме, который включает введение любого из следующих компонентов в живой организм:
(a) моноклональное антитело, которое связывается с PTPRS человека, подавляя активность плазмацитоидной дендритной клетки, или фрагмент, включающий его антигенсвязывающую область, и
(b) иммуноглобулин, в который пересажена определяющая комплементарность область моноклонального антитела согласно (a), или фрагмент, включающий его антигенсвязывающую область.
(21) Способ согласно упомянутому выше (19) или (20), где активность плазмацитоидной дендритной клетки представляет собой одну или обе из активности продукции интерферона и выживания интерферон-продуцирующей клетки.
(22) Средство для подавления активности плазмацитоидной дендритной клетки, которое включает любой из следующих компонентов в качестве активного ингредиента:
(a) моноклональное антитело, которое связывается с PTPRS человека, подавляя активность плазмацитоидной дендритной клетки, или фрагмент, включающий его антигенсвязывающую область,
(b) иммуноглобулин, в который пересажена определяющая комплементарность область моноклонального антитела согласно (a), или фрагмент, включающий его антигенсвязывающую область.
(23) Средство для подавления активности интерферон-продуцирующей клетки согласно упомянутому выше (22), где активность плазмацитоидной дендритной клетки представляет собой одну или обе из активности продукции интерферона и выживания интерферон-продуцирующей клетки.
Преимущественные эффекты изобретения
Настоящее изобретение относится к антителу, которое специфически распознает PTPRS человека, к иммуногену, который пригоден для получения антитела, и к способу получения антитела против PTPRS человека с использованием иммуногена. PTPRS человека представляет собой мембранный белок, который принадлежит семейству RPTP. Авторы настоящего изобретения установили, что может быть без труда получено антитело, которое специфически распознает PTPRS человека. Антитело против PTPRS человека, которое может быть получено в рамках настоящего изобретения, представляет собой антитело, имеющее высокую специфичность, которая отличает pDC человека от клеток, которые экспрессируют другие белки семейства RPTP.
В предпочтительном варианте осуществления антитело против PTPRS человека, предусматриваемое настоящим изобретением, связывается с pDC человека. Более того, антитело по настоящему изобретению специфически распознает pDC человека. Таким образом, оно пригодно для выявления и выделения pDC. pDC представляет собой клетку, которая продуцирует наибольшую часть IFN 1 типа. Таким образом, ее выявление и выделение является важным в диагностике и исследовании заболеваний, в которые вовлечены pDC, таких как аутоиммунные заболевания.
Более того, в предпочтительном варианте осуществления антитело против PTPRS человека, предусматриваемое настоящим изобретением, обладает действием модулирования активности pDC человека. Таким образом, антитело против PTPRS человека по настоящему изобретению можно использовать для подавления активности pDC. Таким образом, при использовании подавления активности pDC с использованием антитела по настоящему изобретению, терапевтический эффект можно ожидать даже у пациента с аутоиммунным заболеванием, у которого усилена экспрессия IFNα.
pDC продуцируют большое количество IFN при малом количестве клеток. Для нейтрализации IFN необходимо антитело, соответствующее молекулярному количеству IFN. Однако в рамках настоящего изобретения активность продуцирующей клетки подавляется прямо. В результате, можно ожидать более высокого эффекта подавления IFN с помощью меньшего количества антитела по сравнению с нейтрализации антителом против IFN. Более того, в случае, когда IFN продуцируется постоянно, можно ожидать, что нейтрализация IFN антителом подавляется только временно, в то время как в рамках настоящего изобретения происходит подавление pDC и, таким образом, можно ожидать эффекта подавления продуцирования IFN в течение длительного времени.
Краткое описание чертежей
На фиг.1 представлена аминокислотная последовательность PTPRS (SEQ ID NO: 1). PTPRS представляет собой однократно трансмембранный белок, имеющий иммуноглобулин-подобный домен (Ig-подобный домен) и домен, подобный фибронектину типа III, во внеклеточной области. Более того, он имеет две области протеинтирозинфосфатазы (PTP-домены) во внутриклеточной области;
На фиг.2 представлен график, на котором показаны относительные уровни экспрессии PTPRS в различных иммунных клетках. Было показано, что PTPRS экспрессируется специфическим для pDC образом;
На фиг.3 представлен график, на котором показано сравнение экспрессии гена PTPRS между тканями. мРНК PTPRS демонстрирует относительно высокую экспрессию в селезенке и яичниках, а также широко экспрессируется в других тканях;
На фиг.4 показан отбор клеток, экспрессирующих PTPRS человека (hPTPRS), с помощью сортировки FACS;
На фиг.5 показан скрининг способом FACS гибридом с использованием иммунизированной клетки hPTPRS/D2SC/1. Было получено тринадцать гибридом, которые продуцируют антитело против hPTPRS;
На фиг.6 показан скрининг FACS с использованием клетки CAL-1;
На фиг.7 показан скрининг FACS с использованием pDC периферической крови человека;
На фиг.8 представлен график, на котором показана гомология hPTPRS с другими PTPR. PTPRS принадлежит семейству PTPR, в котором аминокислотные последовательности нескольких молекул семейства имеют высокую гомологию с аминокислотной последовательностью PTPRS;
На фиг.9 представлен результат испытания, на котором показано, связываются ли специфически супернатанты десяти типов гибридом (2G6, 4B2, 2G2, 9H5, 10F7, 22H8, 49F2, 14A8, 55E7, 13G5), которые распознают PTPRS и продуцируют антитело, которое специфично связывается с pDC человека, только с PTPRS (hPTPRE не экспрессировалась на клеточной поверхности). В результате для 2G6 была показана перекрестная реактивность с PTPRF (фиг.9, D), и для 4B2 была показана перекрестная реактивность с PTPRD (фиг.9, C). Другие 9 типов антител показали специфическое связывание с PTPRS (фиг.9, A-D);
На фиг.10 представлен результат испытания перекрестной реактивности антител против PTPRS в отношении обезьяны. Все супернатанты клеточных культур гибридом специфически связывались с группой клеток pDC (линия-CD123+ HLA-DR+) яванского макака;
На фиг.11 показана сигнализационная сортировка гибридом. Сортировку единичных клеток проводили с использованием FACS Aria (BD), и клетки D2SC/1 и клетки hPTPRS/D2SC/1 (A и B), клетки CAL-1 (C) и pDC человека (D) окрашивали с использованием клеточных культуральных супернатантов гибридом, и отбирали единичные гибридомы;
На фиг.12 показаны концентрации эндотоксина в очищенных антителах, полученных из культуральных супернатантов гибридом. Все концентрации представляли собой стандартную величину 0,3 ЕЭ/мг Ab или менее;
На фиг.13 представлен результат испытания способности очищенных антител связываться с PTPRS человека на клеточной поверхности. Можно было подтвердить, что все из антител сохраняли их способность связываться;
На фиг.14 представлено специфическое связывание очищенных антител с группами клеток pDC (BDCA2+) периферической крови человека;
На фиг.15 представлен результат исследования того, связываются ли антитела против PTPRS человека также с PTPRS мыши. 49F2-30, 13G5-52, 13G5-57 и 22H8-84 связываются с mPTPRS/CHO;
На фиг.16 показана комплемент-зависимая цитотоксическая активность антител против PTPRS в отношении клетки, экспрессирующей hPTPRS. Определяли комплемент-зависимую цитотоксическую активность антител против PTPRS в отношении PTPRS человека/CHO (фиг.16A) и PTPRS мыши/CHO (фиг.16B). В результате 13G5-52 и 13G5-57 продемонстрировали приблизительно 20% активности CDC против мишени в виде PTPRS человека/CHO (A), между тем как 13G5-52 и 13G5-57 продемонстрировали приблизительно 100% активности CDC против мишени в виде PTPRS мыши/CHO (B);
На фиг.17 показано, что химерные антитела против hPTPRS ch49F2-30 (abu17A), ch9H5-4, ch13G5-57 и ch22H8-84 (фиг.17B) повреждают клетку-мишень hPTPRS/CHO зависимым от количества эффекторных клеток образом; и
На фиг.18 показано, что продукция IFNα полностью ингибируется обработкой химерным антителом против PTPRS ch49F2-30, ch9H5-4, ch13G5-57 и ch22H8-84 (фиг.18A), и было установлено, что популяция pDC снижалась в большей степени, чем при обработке контрольным антителом Synagis (фиг.18B и 18C).
Описание вариантов осуществления
PTPRS человека представляет собой молекулу, специфическую экспрессию которой наблюдают в подобной плазматической клетке дендритной клетке pDC. Однако еще не разработан какой-либо способ продуцирования антитела, которое распознает PTPRS человека.
Известны четыре изоформы PTPRS человека, которые включают изоформу 1, которая состоит из 1948 аминокислотных остатков, изоформу 2, которая состоит из 1910 аминокислотных остатков, изоформу 3, которая состоит из 1501 аминокислотного остатка, и изоформу 4, которая состоит из 1505 аминокислотных остатков. В их структурах наблюдают три иммуноглобулин-подобных домена (первый Ig-домен, второй Ig-домен и третий Ig-домен), домен, подобный фибронектину типа III, трансмембранный домен (трансмембранный домен, TM-область) в качестве внеклеточных структур, и два фосфатазных домена (домены D1 и D2) в качестве внутриклеточных структур. Только домен D1, который расположен вблизи клеточной мембраны, обладает активностью протеинтирозинфосфатазы (PTP). На фиг.1, сигнальные пептиды и типичные домены обозначены в аминокислотной последовательности.
Изоформа 3 PTPRS человека представляет собой проходящий через мембрану белок, включающий аминокислоты с 831 по 851 SEQ ID NO: 1 (фиг.1) в качестве трансмембранного домена. Среди 1501 аминокислотного остатка, включая N-конец, 29 аминокислотных остатков (1-29 в SEQ ID NO: 1) представляют собой сигнальную последовательность, и остатки 30-830 представляют собой внеклеточный домен. С другой стороны, C-концевая сторона является внутриклеточным доменом. Считается, что лиганды во внеклеточной среде контролируют активность PTPRS.
Авторы настоящего изобретения подтвердили с помощью анализа экспрессии генов, что PTPRS специфически экспрессируется в pDC человека. Они заключили, что, если бы можно было получить антитело, которое может отличить PTPRS человека от других молекул, это было бы полезным для исследования pDC. Однако в семействе PTP, включающем PTPRS человека, имеется множество молекул, имеющих сходные структуры. В частности, молекулы, такие как PTPRS, которая представляет собой RPTP-σ, и PTPRA (RPTP-α), PTPRD (RPTP-δ), PTPRE (RPTP-ε, PTPRF (RPTP-ζ), включают аминокислотные последовательности, имеющие высокую гомологию (фиг.8). Таким образом, считается, что трудно получить антитело, которое может различать эти молекулы между собой с использованием доменного пептида, в котором в качестве иммуногена используется неполная последовательность аминокислотной последовательности, которая составляет внеклеточный домен. Таким образом, авторы настоящего изобретения предприняли попытку получить антитело против PTPRS человека с использованием клетки, которая экспрессирует PTPRS человека в качестве иммуногена.
Авторы настоящего изобретения провели обширные испытания для получения антитела, которое распознает PTPRS человека, и установили, что рассматриваемое антитело можно получать с использованием специфически трансформированной клетки в качестве иммуногена, и осуществили настоящее изобретение. Это означает, что настоящее изобретение относится к моноклональному антителу, которое связывается с внеклеточным доменом PTPRS человека, или к фрагменту, включающему его антигенсвязывающую область.
В рамках настоящего изобретения PTPRS человека может быть определена как природная молекула, которая экспрессируется в pDC человека, или молекула, которая иммунологически эквивалентна PTPRS человека, которая экспрессируется в pDC человека. В рамках настоящего изобретения то, что антитело связывается с PTPRS человека, можно подтверждать, например, следующим образом.
- Подтверждение на основе реактивности в отношении клеток человека:
В соответствии с данными, полученными авторами настоящего изобретения, можно считать, что PTPRS человека можно использовать в качестве маркера для pDC, поскольку наблюдается экспрессия, специфическая для pDC человека.
Исходя из такого профиля экспрессии PTPRS человека, прежде всего, активность в отношении связывания по меньшей мере с частью подгруппы pDC является одной из важных характеристик антитела, которое связывается с PTPRS человека, в рамках настоящего изобретения. То, что определенная клетка представляет собой pDC, можно подтвердить с помощью маркера клеточной поверхности, который является присущим каждой группе клеток. Например, связывание с рассматриваемой клеткой подтверждают с помощью двойного окрашивания антителом, которое связывается с маркером клеточной поверхности, и антителом, активность связывания которого предстоит подтвердить. Это означает, что pDC в рамках настоящего изобретения включает, например, клетку, которая экспрессирует BDCA2.
- Подтверждение на основе реактивности в отношении трансформированной клетки, которая экспрессирует ген PTPRS человека:
Авторы настоящего изобретения подтвердили, что, когда ген PTPRS человека экспрессируется в определенных условиях, иммунологические характеристики PTPRS человека, экспрессированной в pDC человека, воспроизводятся. Таким образом, реактивность в отношении PTPRS человека можно подтверждать, исходя из реактивности антитела против клетки, в которую искусственно введен ген, который кодирует PTPRS человека. Это означает, что настоящее изобретение относится к моноклональному антителу, которое связывается с молекулой, включающей аминокислотную последовательность, которая составляет внеклеточный домен PTPRS человека в качестве внеклеточного домена, или фрагменту, включающему его антигенсвязывающую область. Между тем, внеклеточный домен составляет аминокислотная последовательность, соответствующая аминокислотам 30-830 в SEQ ID NO: 1 (фиг.1) с N-конца аминокислотной последовательности, представленной в SEQ ID NO: 1.
Например, в клетке, которая трансформирована экспрессирующим вектором, включающим ДНК, которая кодирует PTPRS человека, иммунологические характеристики PTPRS, которая экспрессируется в pDC человека, сохраняются. Таким образом, трансформированная клетка, которая экспрессирует PTPRS человека, является предпочтительной в качестве клетки для подтверждения связывающих свойств антитела в отношении внеклеточного домена PTPRS человека по настоящему изобретению. Когда реактивность антитела подтверждают с помощью трансформированной клетки по настоящему изобретению, в качестве контроля желательно использовать клетку, которая не была трансформирована.
Далее, антитело, которое связывается с PTPRS человека, по настоящему изобретению может представлять собой антитело, у которого наблюдается или не наблюдается перекрестная реактивность в отношении группы клеток, о которой известно, что она экспрессирует представителя семейства PTP, отличного от PTPRS. Антитело, у которого не наблюдают перекрестную реактивность, является предпочтительным в качестве антитела, которое связывается с PTPRS человека, в рамках настоящего изобретения. В частности, в рамках настоящего изобретения антитело, связывание которого с группой клеток, о которой известно, что она экспрессирует представителя семейства PTP, отличного от PTPRS человека, в тех же условиях, что и условия, в которых подтверждено связывание с pDC, является предпочтительным в качестве антитела, которое связывается с PTPRS человека.
Это означает, что в рамках настоящего изобретения моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека, предпочтительно включает моноклональное антитело, имеющее следующие иммунологические характеристики.
a) Оно связывается с pDC человека.
b) В условиях, в которых оно связывается с pDC человека, его связывание с одним типом или множеством типов клеток, выбранных из группы, состоящей из моноцита, макрофага, B-клетки и CD34-положительной клетки, и дендритными клетками, происходящими из этих клеток, не может быть подтверждено.
В частности, антитело, связывание которого с одним типом или несколькими типами клеток, выбранными из группы, состоящей из моноцита, макрофага, B-клетки и CD34-положительной клетки, и дендритными клетками, происходящими из этих клеток, не может быть подтверждено в условиях, в которых антитело связывается с pDC человека, является предпочтительным в качестве моноклонального антитела по настоящему изобретению.
Альтернативно моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека, в рамках настоящего изобретения предпочтительно включает моноклональное антитело, имеющее следующие иммунологические характеристики.
c) Оно связывается с трансформированной клеткой, которая трансформирована экспрессирующим вектором, который сохраняет экспрессирующуюся ДНК, которая кодирует PTPRS человека.
d) В условиях связывания с трансформированной клеткой согласно c), связывание с клеткой-хозяином до трансформации согласно c) не может быть подтверждено.
В рамках настоящего изобретения то, что моноклональное антитело против PTPRS человека не реагирует перекрестно с другими молекулами семейства PTP, может быть подтверждено с использованием клетки, в которой каждый представитель семейства PTP принудительно экспрессируется. Это означает, что кДНК, которая кодирует аминокислотную последовательность каждого представителя семейства PTP, экспрессируется принудительно путем введения в подходящую клетку-хозяина. Моноклональное антитело против PTPRS человека, перекрестную реактивность которого требуется подтвердить, контактируют с полученной трансформированной клеткой. Затем, если связывание с клеткой, которая экспрессирует другую молекулу семейства PTP, отличную от PTPRS, не наблюдается, то это может подтвердить, что антитело может иммунологически отличать PTPRS человека от другой молекулы семейства PTP. Например, в примерах, упомянутых ниже, было подтверждено, что большинство из моноклональных антител против PTPRS человека, полученных в рамках настоящего изобретения, не реагировали перекрестно с PTPRA, PTPRD и PTPRF, которые, в частности, имели высокую гомологию с PTPRS. Таким образом, моноклональное антитело, которое связывается с PTPRS человека и связывание которого с PTPRA, PTPRD и PTPRF в тех же условиях не выявляется, является предпочтительным моноклональным антителом в рамках настоящего изобретения. Если использовать антитело, которое может иммунологически отличать эти молекулы семейства PTP от PTPRS, можно выявить специфическое изменение экспрессии PTPRS. Кроме того, было показано, что, среди молекул, имеющих высокую гомологию с PTPRS, экспрессия PTPRE может быть подтверждена в клетке, но PTPRE не экспрессируется вне клетки. Таким образом, антитело не связывается с PTPRE.
Связывание между моноклональным антителом, связывающую активность которого требуется подтвердить, и различных клеток можно подтверждать, например, по принципу проточной цитометрии. Для подтверждения реактивность антитела по принципу проточной цитометрии, является преимущественным предварительное мечение антитела молекулой или атомной группой, которая генерирует поддающийся детекции сигнал. Как правило, используют флуоресцентную метку или испускающую свет метку. Для анализа связывания между флуоресцентно меченным антителом и клеткой по принципу проточной цитометрии можно использовать активируемый флуоресценцией клеточный сортер (FACS). С использованием FACS можно эффективно подтвердить связывание между множеством антител и клеток.
В частности, например, антитело A, для которого заранее было установлено, что оно способно идентифицировать pDC, и антитело B, чье свойство связывания с pDC анализируют, одновременно подвергают реакции с группой клеток, включающей pDC. Антитело A и антитело B предварительно метят флуоресцентными сигналами, которые можно отличить друг от друга. Если два сигнала выявляют в одной и той же группе клеток, то можно подтвердить, что эти антитела связываются с той же группой клеток. Это означает, что может быть выявлено, что антитело A и антитело B имеют одинаковое свойство связывания. Если они связываются с различными группами клеток, то понятно, что их свойства связывания отличаются.
Примеры предпочтительного моноклонального антитела в рамках настоящего изобретения могут включать моноклональное антитело, продуцируемое гибридомами 9H5-4, 10F7-38, 13G5-52, 13G5-57, 14A8-85, 22H8-84, 49F2-30 или 55E7-79.
Гибридомы 9H5-4, 10F7-38, 13G5-52, 13G5-57, 14A8-85, 22H8-84, 49F2-30 и 55E7-79 были депонированы под номерами доступа № FERM ABP-11356, FERM ABP-11357, FERM ABP-11358, FERM ABP-11359, FERM ABP-11360, FERM ABP-11361, FERM ABP-11362 и FERM ABP-11363, соответственно, в International Patent Organism Depositary (IPOD), National Institute of Advanced Industrial Science and Technology (NAIST) 1 апреля 2011 года. Далее описано содержимое указанного депозита.
(a) Название организации депозитария: International Patent Organism Depositary (IPOD), National Institute of Advanced Industrial Science and Technology (NAIST)Адрес: Tsukuba Central 6. 1-1-1 Higashi, Tsukuba, Ibaraki 305-8566, Япония
(b) Дата депонирования: 1 апреля 2011 года
(c) Номер доступа № FERM ABP-11356 (гибридома 9H5-4)
(c) Номер доступа № FERM: ABP-11357 (гибридома 10F7-38)
(c) Номер доступа № FERM ABP-11358 (гибридома 13G5-52)
(c) Номер доступа № FERM ABP-11359 (гибридома 13G5-57)
(c) Номер доступа № FERM ABP-11360 (гибридома 14A8-85)
(c) Номер доступа № FERM ABP-11361 (гибридома 22H8-84)
(c) Номер доступа № FERM ABP-11362 (гибридома 49F2-30)
(c) Номер доступа № FERM ABP-11363 (гибридома 55E7-79)
Моноклональное антитело по настоящему изобретению может представлять собой фрагмент, включающий его антигенсвязывающую область. Например, фрагмент антитела, который включает антигенсвязывающий центр, полученный ферментативным расщеплением IgG, также можно использовать в качестве антитела в рамках настоящего изобретения. В частности, фрагмент антитела, такой как Fab или F(ab’)2, можно получать путем расщепления папаином или пепсином. Хорошо известно, что эти фрагменты антител можно использовать в качестве молекул антител, обладающих аффинностью связывания с антителом. Альтернативно, при условии сохранения требуемой активности связывания антигена, можно использовать антитело, сконструированное путем рекомбинации генов. Примеры антитела, сконструированного путем рекомбинации генов, могут включать химерные антитела, антитела с пересаженными CDR, одноцепочечные Fv, диантитела, линейные антитела, полиспецифические антитела, которые образованы из фрагментов антител, и т.п. Способы получения этих антител на основе моноклональных антител или антитело-продуцирующих клеток, которые продуцируют моноклональные антитела, известны.
Моноклональное антитело по настоящему изобретению можно получать с использованием определенной трансформированной клетки в качестве иммуногена. Это означает, что настоящее изобретение относится к способу получения клетки, которая продуцирует моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека, который включает:
(1) введение клетки, которая экспрессирует экзогенный белок, включающий внеклеточный домен PTPRS человека, для иммунизации животного, и
(2) отбор антителопродуцирующей клетки, которая связывается с PTPRS человека, из антителопродуцирующих клеток иммунизированного животного.
Путем культивирования полученной таким образом антителопродуцирующей клетки или антителопродуцирующей клетки, которую подвергли иммортализации, из культуры можно собирать рассматриваемое моноклональное антитело. Известны различные способы иммортализации антителопродуцирующей клетки.
Трансформированную клетку, которую используют в качестве иммуногена в рамках настоящего изобретения, можно получать, например, путем получения следующей клетки, которая сохраняет экспрессию экзогенного полинуклеотида (a), который кодирует аминокислотную последовательность, включающую внеклеточный домен PTPRS человека.
В рамках настоящего изобретения экзогенный полинуклеотид относится к полинуклеотиду, который искусственным образом введен в клетку-хозяина. В случае, когда в качестве клетки используют клетку человека, ген человека вводят в клетку человека. Также в такой комбинации искусственно введенный полинуклеотид называют экзогенным полинуклеотидом. Таким образом, экспрессия экзогенного полинуклеотида охватывает эктопическую экспрессию PTPRS человека.
В рамках настоящего изобретения внеклеточный домен PTPRS человека относится к аминокислотной последовательности в положениях 30-830, которая соответствует внеклеточному домену аминокислотной последовательности, описанной в SEQ ID NO: 1. Например, аминокислотная последовательность, которая включает соответствующие области в порядке со стороны N-конца, упомянутом ниже, является предпочтительной аминокислотной последовательностью, включающей внеклеточный домен PTPRS человека в рамках настоящего изобретения.
[Сигнальная последовательность + внеклеточный домен + трансмембранный домен+ внеклеточная область]
Альтернативно аминокислотная последовательность, которая частично лишена внутриклеточных областей, как описано ниже, также охватывается аминокислотной последовательностью, включающей внеклеточный домен PTPRS человека, в рамках настоящего изобретения.
[Сигнальная последовательность + внеклеточный домен + трансмембранный домен + часть внутриклеточной области]
Более того, структура, которая лишена внутриклеточной области, как указано ниже, также охватывается аминокислотной последовательностью, включающей внеклеточный домен PTPRS человека, в рамках настоящего изобретения.
[Сигнальная последовательность+внеклеточный домен+трансмембранный домен]
В упомянутых выше структурах области, отличные от внеклеточного домена, могут иметь последовательность, выбранную из аминокислотной последовательности, представленной в SEQ ID NO: 1, или могут включать другую гомологичную аминокислотную последовательность в комбинации. Например, аминокислотные последовательности, которые составляют сигнальную последовательность, трансмембранный домен и внутриклеточную область, могут представлять собой аминокислотные последовательности молекул семейства PTP, отличных от PTPRS человека. Альтернативно можно вносить в комбинации аминокислотную последовательность молекулы семейства PTP вида, отличного от человека. Более того, аминокислотные последовательности, которые составляют области, отличные от внеклеточного домена, могут включать мутацию до такой степени, чтобы могли сохраняться функции соответствующих областей. Более того, между соответствующими областями может быть встроена другая область. Например, между сигнальной последовательностью и внеклеточным доменом может быть встроена эпитопная метка, такая как FLAG. В частности, сигнальная последовательность представляет собой область, которая транслируется в белок, процессируется на стадии переноса на поверхность клеточной мембраны, и удаляется. Таким образом, любую аминокислотную последовательность, которая индуцирует прохождение транслируемого белка через клеточную мембрану, можно использовать в качестве сигнальной последовательности. Более конкретно, аминокислотная последовательность PTPRS человека (SEQ ID NO: 1) является предпочтительной в качестве аминокислотной последовательности, включающей внеклеточный домен PTPRS человека.
Таким образом, для полинуклеотида, который составляет упомянутый выше (a) в рамках настоящего изобретения, можно использовать любую последовательность оснований, которая кодирует аминокислотную последовательность, которая составляет упомянутую выше структуру [сигнальная последовательность+внеклеточный домен+трансмембранный домен+внутриклеточная область]. Например, аминокислотная последовательность SEQ ID NO: 1 кодируется последовательностью оснований, описанной в SEQ ID NO: 2.
В рамках настоящего изобретения для получения трансформированной клетки, предназначенной для применения в качестве иммуногена, необходимо только ввести экспрессирующий вектор, в котором сохраняется экспрессия упомянутого выше полинуклеотида (a), в подходящую клетку-хозяина.
Клетка-хозяина в рамках настоящего изобретения предпочтительно представляет собой клетку млекопитающего. В частности, в качестве клетки-хозяина можно использовать клетку, происходящую из человека, обезьяны, мыши или крысы. В частности, происходящая из человека клетка является предпочтительной в качестве клетки-хозяина. Например, клетка HEK-293T является предпочтительной клеточной линией почки, происходящей из эмбриона человека, которую можно использовать в качестве клетки-хозяина в рамках настоящего изобретения. Клетка HEK-293T доступна в качестве ATCC CRL-11268. Также в качестве клеток-хозяев можно использовать другие клетки, происходящие из иммунизированных животных. Когда клетку, происходящую из иммунизированного животного, используют в качестве иммуногена, иммунный ответ против клетки-хозяина является небольшим. Таким образом, можно эффективно получать антитело против внеклеточного домена PTPRS человека, которое экспрессируется эндогенно. Таким образом, например, когда мышь используют в качестве иммунизированного животного, в качестве клетки-хозяина можно использовать происходящую из мыши клетку.
Упомянутым выше полинуклеотидом можно трансформировать клетку путем помещения полинуклеотида в вектор, который может индуцировать экспрессию в клетке-хозяине. Можно использовать коммерчески доступный вектор, который может индуцировать экспрессию в клетке млекопитающего. В рамках настоящего изобретения можно использовать экспрессирующие векторы, такие как вектор pCMV-Script (R), вектор pSG5 (производимый Stratagene) и pcDNA3.1 (изготавливаемый Invitrogen).
Полученную таким образом трансформированную клетку вводят для иммунизации животного вместе с дополнительными компонентами, такими как адъювант, при необходимости. В качестве адъюванта можно использовать полный адъювант Фрейнда и т.п. В случае, когда в качестве иммунизированного животного используют мышь, трансформированную клетку можно вводить в количестве от 104 до 109 клеток, более конкретно, от 104 до 106 клеток. Как правило, иммуноген вводят несколько раз с некоторыми интервалами до тех пор, пока титр антитела не возрастет. Например, в случае кратковременного процесса иммунизации, трансформированную клетку можно вводить с интервалами от 2 до 4 суток, в частности 3 суток, и антителопродуцирующие клетки можно собирать после введения от 2 до 3 раз. Альтернативно антителопродуцирующие клетки можно собирать после введения от 5 до 6 раз с интервалами приблизительно один раз в неделю.
В рамках настоящего изобретения собранную антителопродуцирующую клетку клонируют для получения моноклонального антитела. Для клонирования является предпочтительным иммортализовать антителопродуцирующую клетку. Например, в качестве способа иммортализации антителопродуцирующей клетки можно использовать способ слияния клеток, такой как способ гибридом, или трансформацию вирусом Эпштейна-Барр (EBV).
В антителопродуцирующих клетках одна клетка продуцирует один тип антитела. Таким образом, если можно получить (т.е. клонировать) группу клеток, происходящую из одной клетки, можно получить моноклональное антитело. Способ гибридом относится к способу, в котором антителопродуцирующую клетку подвергают слиянию с подходящим клеточным штаммом, иммортализуют и клонируют. Иммортализованную антителопродуцирующую клетку можно клонировать способом, таким как способ лимитирующих разведений. Известно множество штаммов клеток, пригодных для способа гибридом. Эти клеточные штаммы имеют различные генетические маркеры, которые являются превосходными с точки зрения эффективности иммортализации лимфоцитарной клетки и необходимы для селекции клетки, в которой слияние клеток было успешным. Более того, в случае, когда намереваются получить антителопродуцирующую клетку, также можно использовать клеточный штамм, который лишен способности продуцировать антитело.
Например, миелому мыши P3×63Ag8.653 (ATCC CRL-1580) и P3×63Ag8U.1 (ATCC CRL-1597) широко используют в качестве клеточных штаммов, которые пригодны в способах слияния клеток у мышей и крыс. Как правило, гибридому получают путем слияния гомологичных клеток, однако моноклональное антитело можно получать из близко родственных гетерологичных гетерогибридом.
Конкретный протокол слияния клеток известен. Это означает, что антителопродуцирующую клетку иммунизированного животного смешивают с подходящим партнером по слияния для достижения слияния клеток. Для антителопродуцирующей клетки используют клетку селезенки, лимфоцитарную клетку, взятую из лимфатического узла, B-клетку периферической крови и т.п. В качестве партнера по слиянию можно использовать различные штаммы клеток, упомянутые выше. Для слияния клеток используют способ с полиэтиленгликолем или способ электрического слияния.
Далее клетку, в которой успешно произошло слияние клеток, отбирают на основе селективного маркера, которым обладает слитая клетка. Например, в случае, когда для слияния клеток используют чувствительный к HAT штамм клеток, клетку, в которой успешно произошло слияние леток, отбирают путем селекции клеток, которые растут в среде HAT. Более того, подтверждают, что антитело, продуцированное отобранной клеткой, обладает предполагаемой реактивностью.
Каждую гибридому подвергают скринингу в отношении реактивности антитела. Это означает, что гибридому, которая продуцирует антитело, которое связывается с PTPRS человека, отбирают способом, упомянутым выше. Предпочтительно, отобранную гибридому субклонируют и в случае, когда продуцирование рассматриваемого антитела окончательно подтверждено, ее отбирают в качестве гибридомы, которая продуцирует моноклональное антитело по настоящему изобретению.
В частности, рассматриваемую гибридому можно отбирать, исходя из реактивности в отношении клеток человека или реактивности в отношении трансформированной клетки, которая экспрессирует ген PTPRS человека. Антитело, которое связывается с клеткой, можно выявлять по принципу иммуноанализа. Например, для выявления рассматриваемого антитела можно использовать ELISA с использованием клетки в качестве антигена. В частности, культуральный супернатант гибридомы контактируют с подложкой, на которой фиксированы pDC человека или трансформированная клетка, используемые в качестве иммуногена. В случае, когда культуральный супернатант включает рассматриваемое антитело, антитело улавливается клеткой, фиксированной на подложке. Затем, твердую фазу отделяют от культурального супернатанта и промывают при необходимости, тем самым можно выявлять антитело, уловленное на твердой фазе. Для выявления антитела можно использовать антитело, которое распознает антитело. Например, антитело мыши можно выявлять с помощью антитела против иммуноглобулина мыши. Если антитело, которое распознает антитело, является предварительно меченым, его выявление является легким. В качестве метки можно использовать фермент, флуоресцентный пигмент, испускающий свет пигмент и т.п.
С другой стороны, в качестве подложки для фиксации клеток можно использовать частицы или внутреннюю стенку микропланшета для титрования. Клетку можно фиксировать путем физической адсорбции на поверхности частиц или контейнера, изготовленного из пластмассы. Например, в качестве подложки для фиксации клетки можно использовать гранулы или реакционный контейнер, изготовленный из полистирола.
При выборе гибридомы в некоторых случаях можно ожидать продуцирования антитела не против PTPRS человека, а против клеток хозяина трансформированной клетки, использованной в качестве иммуногена. Например, как показано в примерах, когда клетку человека используют в качестве иммуногена и мышь используют в качестве иммунизированного животного, клетка человека распознается в качестве чужеродного агента и можно ожидать продукции антитела, которое связывается с ней. Настоящее изобретение нацелено на получение антитела, которое распознает PTPRS человека. Таким образом, не является необходимым получение антитела, которое распознает антиген клетки человека, отличный от PTPRS. Для исключения гибридомы, которая продуцирует такое антитело посредством скрининга, непредусмотренное антитело можно абсорбировать заранее до подтверждения реактивности антитела.
Непредусмотренное антитело можно абсорбировать с помощью антигена, с которым связывается антителом, присутствие которого ожидается. В частности, например, антитела против антигенов клеток человека, отличные от PTPRS человека, можно абсорбировать с помощью клеток, в которых экспрессия PTPRS человека не может быть выявлена. В рамках настоящего изобретения, клетка-хозяин, используемая в качестве иммуногена, является предпочтительной в качестве антигена для абсорбции непредусмотренного антитела.
Когда необходимо, подтверждают истинный эффект на активность pDC моноклонального антитела, для которого активность связывания с антигеном подтверждена. Эффект на pDC можно подтверждать, например, способом, упомянутым ниже.
Моноклональное антитело по настоящему изобретению можно собирать из культуры, полученной путем культивирования гибридомы, которая продуцирует моноклональное антитело. Гибридому можно культивировать in vitro или in vivo. Гибридому можно культивировать in vitro с использованием известной среды, такой как RPMI1640. В культуральном супернатанте накапливается иммуноглобулин, секретируемый гибридомой. Таким образом, моноклональное антитело по настоящему изобретению можно получать путем сбора культурального супернатанта и очистки при необходимости. Очистка иммуноглобулина является более простой в случае, когда в среду не добавляют сыворотку. Однако для целей более быстрой пролиферации гибридомы и ускорения продуцирования антитела, в среду можно добавлять приблизительно 10% эмбриональной телячьей сыворотки.
Гибридому также можно культивировать in vivo. В частности, путем инокуляции гибридомы в брюшную полость мышей nude, гибридому можно культивировать в брюшной полости. Моноклональное антитело накапливается в асцитной жидкости. Таким образом, желаемое моноклональное антитело можно получать путем сбора асцитной жидкости и очистки при необходимости. Полученное моноклональное антитело можно пригодным образом модифицировать или обрабатывать в зависимости от назначения.
Моноклональное антитело по настоящему изобретению можно экспрессировать путем получения кДНК, которая кодирует антигенсвязывающую область антитела из гибридомы, и встраивания ее в подходящий экспрессирующий вектор. Способ получения кДНК, которая кодирует вариабельную область антитела, и экспрессии в подходящей клетке-хозяине, известен. Более того, также известен способ связывания вариабельной области, включающей антигенсвязывающую область, с константной областью с получением химерного антитела.
Например, в качестве предпочтительного моноклонального антитела в рамках настоящего изобретения может быть указано моноклональное антитело, продуцируемое гибридомой 9H5-4, которая была депонирована в качестве номера доступа № FERM ABP-11356, гибридомой 10F7-38, которая была депонирована в качестве номера доступа № FERM ABP-11357, гибридомой 13G5-52, которая была депонирована в качестве номера доступа № FERM ABP-11358, гибридомой 13G5-57, которая была депонирована в качестве номера доступа № FERM ABP-11359, гибридомой 14A8-85, которая была депонирована в качестве номера доступа № FERM ABP-11360, гибридомой 22H8-84, которая была депонирована в качестве номера доступа № FERM ABP-11361, гибридомой 49F2-30, которая была депонирована в качестве номера доступа № FERM ABP-11362, или гибридомой 55E7-79, которая была депонирована в качестве номера доступа № FERM ABP-11363, и т.п.
В качестве химерного антитела, включающего вариабельную область, или гуманизированного антитела, в который пересажена CDR, которая составляет вариабельную область, охватывается антитело, имеющее константную область, происходящую из IgG или IgM, в качестве предпочтительного антитела в рамках настоящего изобретения. Авторы настоящего изобретения подтвердили, что моноклональное антитело против PTPRS обладает действием CDC против экспрессирующей PTPRS клетки. Таким образом, антитело, имеющее константную область, происходящую из IgG или IgM, обладает цитотоксическим действием против экспрессирующей PTPRS клетки посредством действия CDC. Такое антитело является пригодным для подавления ряда экспрессирующих PTPRS клеток, таких как pDC.
Химерное антитело, которое распознает PTPRS человека, или гуманизированное антитело можно получать способами генной инженерии с использованием полинуклеотида, который кодирует антитело.
Приблизительно четыре года уже прошло с момента установления структуры PTPRS человека, как описано в WO2007/041317 (JP2009-510102A); однако еще не получено антитело, которое специфически распознает PTPRS человека. Антитело, которое распознает PTPRS человека, было впервые предоставлено с помощью иммуногена по настоящему изобретению. Это означает, что настоящее изобретение предусматривает антитело, которое распознает PTPRS человека, которое можно получать следующим способом:
(1) введение животному для его иммунизации белка, включающего внеклеточный домен PTPRS человека;
(2) отбор антителопродуцирующей клетки, которая продуцирует антитело, которое связывается с PTPRS человека, из антителопродуцирующих клеток иммунизированного животного; и
(3) культивирование антителопродуцирующей клетки, отобранной согласно (2), и сбор антитела, которое распознает PTPRS человека, из культуры.
Было установлено, что PTPRS человека специфически экспрессируется в pDC человека. Специфическую экспрессию в pDC человека авторы настоящего изобретения также подтвердили в анализе экспрессии генов SAGE. Однако в предшествующих сообщениях уровень экспрессии PTPRS человека во всех случаях анализировали на основе мРНК. Поскольку не было предоставлено антитело, с помощью которого возможно выявление PTPRS человека, белок в экспрессированном состоянии в прошлом не анализировали. Антитело, которое связывается с внеклеточным доменом PTPRS человека, которое предоставлено в рамках настоящего изобретения, позволило осуществить анализ белка PTPRS человека.
В соответствии с фактическим подтверждением, осуществленным авторами настоящего изобретения, моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека на основе настоящего изобретения, специфически выявляло pDC человека. Это означает, что настоящее изобретение относится к способу выявления плазмацитоидной дендритной клетки, который включает контактирование моноклонального антитела, которое связывается с внеклеточным доменом PTPRS человека или фрагментом, включающим его антигенсвязывающую область, с рассматриваемой клеткой, и выявление моноклонального антитела или фрагмента, включающего его антигенсвязывающую область, которое связалось с клеткой.
Посредством выявления PTPRS человека на основе настоящего изобретения можно подтвердить, является или не является ли определенная клетка pDC. Это означает, что настоящее изобретение относится к способу идентификации pDC с использованием PTPRS человека в качестве показателя. Альтернативно pDC человека можно выделять путем выделения клетки, в которой выявлен PTPRS человека в соответствии с настоящим изобретением. Это означает, что настоящее изобретение относится к способу выделения pDC с использованием PTPRS человека в качестве показателя.
В рамках настоящего изобретения моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека или фрагментом, включающим его антигенсвязывающую область, можно метить заранее. Например, антитело можно без труда выявлять путем мечения испускающим свет пигментом или флуоресцентным пигментом. Более конкретно, меченное флуоресцентным пигментом антитело контактируют с агрегатом клеток, который возможно включает pDC, тем самым можно выявлять клетку, с которой связано антитело по настоящему изобретению, с использованием флуоресцентного пигмента в качестве показателя. Более того, если выделить клетки, в которых выявлен флуоресцентный пигмент, можно выделить pDC. Этот набор процессов можно без труда проводить по принципу FACS.
Альтернативно антитело по настоящему изобретению можно предварительно связывать с твердофазной подложкой, такой как магнитные частицы. Антитело, связанное с твердофазной подложкой, распознает PTPRS человека, и pDC улавливаются на твердофазной подложке. В результате можно выявлять и выделять pDC.
Антитело, требуемое для выявления pDC на основе настоящего изобретения, можно предоставлять в качестве средства для выявления pDC. Это означает, что настоящее изобретение относится средству для выявления pDC, включающему моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека, или фрагмент, включающий его антигенсвязывающую область. В средство для выявления pDC по настоящему изобретению, помимо антитела можно добавлять положительный контроль или отрицательный контроль. Например, в качестве положительного контроля можно использовать трансформированную клетку, которая экспрессирует внеклеточный домен PTPRS человека, которую использовали в качестве иммуногена, pDC, взятую от человека и т.п. Как правило, только очень мало pDC человека можно получить из периферической крови. Таким образом, трансформированная клетка является особенно предпочтительной в качестве положительного контроля в средстве по настоящему изобретению. С другой стороны, в качестве отрицательного контроля можно использовать любую клетку, которая не экспрессирует PTPRS человека.
Это означает, что настоящее изобретение относится набору для выявления pDC человека, который включает моноклональное антитело, которое связывается с внеклеточным доменом PTPRS человека, или фрагменту, включающему его антигенсвязывающую область.
Более того, авторы настоящего изобретения проанализировали эффект антитела, которое связывается с внеклеточным доменом PTPRS человека, на pDC. В результате, они подтвердили, что антитело, которое связывается с внеклеточным доменом PTPRS человека, подавляет активность pDC. Это означает, что настоящее изобретение относится к способу подавления активности интерферон-продуцирующей клетки, который включает контактирование любого из следующих компонентов с pDC:
(a) моноклональное антитело, которое связывается с PTPRS человека, подавляя активность pDC, или фрагмент, включающий его антигенсвязывающую область, и
(b) иммуноглобулин, в который пересажена определяющая комплементарность область моноклонального антитела согласно (a), или фрагмент, включающий его антигенсвязывающую область.
Альтернативно настоящее изобретение относится к способу подавления активности pDC в живом организме, который включает введение любого из следующих компонентов в живой организм:
(a) моноклональное антитело, которое связывается с PTPRS человека, подавляя активность pDC, или фрагмент, включающий его антигенсвязывающую область,
(b) иммуноглобулин, в который пересажена определяющая комплементарность область моноклонального антитела согласно (a), или фрагмент, включающий его антигенсвязывающую область, и
(c) полинуклеотид, который кодирует компонент, описанный в (a) или (b).
В рамках настоящего изобретения pDC относится к клетке, которая обладает способностью продуцировать IFN, и экспрессирует PTPRS человека на клеточной поверхности. Далее, если нет иных указаний, pDC охватывает не только клетку, которая является клеткой-предшественником дендритной клетки, но также клетку, которая обладает способностью продуцировать IFN и экспрессировать PTPRS человека на клеточной поверхности. Способ идентификации таких pDC известен. Например, pDC можно отличать от других клеток крови с использованием нескольких маркеров в качестве показателей. В частности, профиль маркеров клеточной поверхности pDC человека является таким, как представлено ниже (Shortman, K. and Liu, YJ, Nature Reviews 2: 151-161, 2002). Также в последние годы было описано, что положительные по BDCA-2 клетки определяются как pDC (Dzionek, A. et al. J. Immunol. 165: 6037-6046, 2000).
[Профиль антигенов клеточной поверхности pDC человека]
CD4-положительные, CD123-положительные, с негативной линией дифференцировки (CD3, CD14, CD16, CD19, CD20, CD56), CD11c-отрицательные
Таким образом, клетка, имеющая профиль экспрессии этих известных маркеров и также обладающая способностью продуцировать IFN, также может быть обозначена как pDC. Более того, даже в случае группы клеток, имеющих профиль, который отличается от характера экспрессии профиля экспрессии этих маркеров в клетке в живом организме, способных продуцировать IFN, эти клетки охватываются pDC.
Более того, в качестве характеристик, которые часто наблюдаются в pDC человека, могут быть показаны следующие характеристики.
[Характеристики формы клеток]
- она напоминает плазматическую клетку.
- она представляет собой круглую клетку, имеющую гладкую поверхность клетки.
- она имеет относительно большое ядро.
[Функциональные характеристики клетки]
- она продуцирует большое количество IFN типа I за короткий период в ходе вирусной инфекции.
- она дифференцируется в дендритную клетку после вирусной инфекции.
В рамках настоящего изобретения подавление активности pDC относится к подавлению по меньшей мере одной функции, которой обладают pDC. В качестве функций pDC могут быть показаны продукция IFN и выживание клеток. Иными словами выживанием клеток можно назвать количество клеток. Таким образом, подавление одной или обеих из этих функций относится к подавлению активности pDC. Было установлено, что IFN типа I, продуцируемый pDC, вызывает различные заболевания. Таким образом, является полезным подавление количества клеток pDC и продуцирования IFN в качестве терапевтической стратегии при этих заболеваниях.
Например, была отмечена взаимосвязь между патологическими состояниями при аутоиммунных заболеваниях и IFNα. Наибольшая часть IFNα продуцируется pDC. Таким образом, если их продукция подавляется, могут смягчаться патологические состояния, вызванные IFNα. Между тем, в рамках настоящего изобретения подавление продукции IFN посредством pDC относится к подавлению продукции по меньшей мере одного типа IFN среди IFN, продуцируемых pDC. IFN типа I являются предпочтительными IFN в рамках настоящего изобретения. Среди них важным является IFNα.
Это означает, что настоящее изобретение относится к средству для подавления продуцирования IFN, которое включает антитело, которое связывается с внеклеточным доменом PTPRS человека в качестве активного ингредиента. Альтернативно настоящее изобретение относится к способу подавления продуцирования IFN, который включает введение антитела, который связывается с внеклеточным доменом PTPRS человека. Более того, настоящее изобретение относится к применению антитела, которое связывается с внеклеточным доменом PTPRS человека, для получения фармацевтической композиции для подавления продуцирования IFN.
pDC включает клетку, которая продуцирует большое количество IFN при небольшом количестве клеток. Например, клетка-предшественник дендритной клетки, стимулированная вирусом и т.п., продуцирует наибольшее количество IFN, продуцируемого в живом организме. Следовательно, снижение количества клеток pDC, которые продуцируют большое количество IFN, приводит к снижению продуцируемого количества IFN. Таким образом, патологические состояния, вызываемые IFNα, также можно смягчать путем снижения количества клеток pDC.
В предпочтительном варианте осуществления настоящего изобретения было подтверждено, что моноклональное антитело против PTPRS человека связывается с клеткой, экспрессирующей PTPRS человека, и обеспечивает цитотоксическое действие посредством CDC (комплементзависимая цитотоксичность). Действие CDC является одним из важных механизмов действия лекарственных средств на основе антител. Моноклональное антитело против PTPRS человека по настоящему изобретению также обладает сильным цитотоксическим действием против клеток, экспрессирующих PTPRS человека, таких как pDC, посредством их действия путем CDC. Это означает, что в предпочтительном варианте осуществления эффект подавления продуцирования IFN можно ожидать также посредством цитотоксического действия против pDC, помимо механизма подавления продукции IFN.
Антитело, которое распознает внеклеточный домен PTPRS человека, используемый в рамках настоящего изобретения, можно получать на основе способа, упомянутого выше. Антитело в рамках настоящего изобретения может представлять собой антитело любого класса. Более того, также не ограничен вид организма, из которого происходит антитело. Более того, в качестве антитела можно использовать фрагмент, включающий антигенсвязывающую область антитела. Например, фрагмент антитела, который включает антигенсвязывающий центр, полученный ферментативным расщеплением IgG, также можно использовать в качестве антитела в рамках настоящего изобретения. В частности, фрагмент антитела, такой как Fab или F(ab’)2, можно получать путем расщепления папаином или пепсином. Хорошо известно, что эти фрагменты антител можно использовать в качестве молекул антител, обладающих аффинностью связывания с антигенами. Альтернативно также можно использовать антитело, сконструированное путем рекомбинации генов, при условии, что оно сохраняет необходимую активность связывания антигена. Примеры антитела, сконструированного путем рекомбинации генов, могут включать химерные антитела, антитела с пересаженными CDR, одноцепочечные Fv, диантитела и линейные антитела, и полиспецифические антитела, образованные из фрагментов антител, и т.п. Способы получения этих антител на основе моноклональных антител известны.
В рамках настоящего изобретения антитело можно модифицировать при необходимости. В соответствии с настоящим изобретением, антитело, которое распознает внеклеточный домен PTPRS человека, обладает действием подавления активности pDC. Это означает, что рассматривалась возможность того, что само по себе антитело обладает цитотоксическим действием против pDC. Подкласс антител, проявляющий высокое эффекторное действие, известен. Альтернативно, путем модификации антитела цитотоксическим веществом (цитотоксическим агентом), можно далее усиливать эффект подавления активности pDC. Примеры цитотоксического вещества могут включать следующие вещества.
Токсины: эндотоксин Pseudomonas (PE), дифтеротоксин
Лизин
Радиоизотопные элементы: Tc99m, Sr89, I131, Y90
Средства против злокачественной опухоли: калихеамицин, митомицин, паклитаксел
Токсины, состоящие из белка, могут быть связаны с антителом или его фрагментом и т.п. с помощью бифункционального агента. Альтернативно ген, который кодирует токсины, можно связывать с геном, который кодирует антитело, с получением слитого белка двух генов. Способ связывания радиоизотопного элемента с антителом также известен. Например, известен способ мечения антитела радиоизотопным элементом с использованием хелатирующего агента. Более того, средство против злокачественной опухоли может быть связано с антителом с использованием цепи сахаров или бифункционального агента.
В рамках настоящего изобретения, в качестве активного ингредиента также можно использовать антитело, структура которого модифицирована искусственным путем. Например, известны различные способы модификации для смягчения цитотоксического действия и стабильности антитела. В частности, известен иммуноглобулин, в котором модифицирована цепь сахаров тяжелой цепи (Shinkawa, T. et al., J. Biol. Chem 278:3466-3473. 2003.). Путем модификации цепи сахаров усиливали активность ADCC (антителозависимая клеточно-опосредуемая цитотоксичность) иммуноглобулина.
Когда антитело, которое связывается с внеклеточным доменом PTPRS человека, контактируют с pDC, его активность снижается. Таким образом, эти антитела можно использовать в качестве средства или способа подавления активности pDC. Это означает, что настоящее изобретение относится к средству для подавления активности pDC, которое включает по меньшей мере один тип компонента, выбранный из группы, состоящей из следующих (a)-(c), в качестве активного ингредиента. Альтернативно настоящее изобретение относится к способу подавления активности pDC, который включает введение по меньшей мере одного типа компонента, выбранного из группы, состоящей из следующих (a)-(c). Более того, настоящее изобретение относится к применению компонента, выбранного из группы, состоящей из следующих (a)-(c), для получения средства для подавления активности pDC.
(a) Антитело, которое связывается с внеклеточным доменом PTPRS человека, или фрагмент, включающий его антигенсвязывающую область, и
(b) Иммуноглобулин, в который пересажена определяющая комплементарность область моноклонального антитела согласно (a), или фрагмент, включающий его антигенсвязывающую область.
В рамках настоящего изобретения в качестве моноклонального антитела, которое подавляет активность pDC, можно использовать моноклональное антитело, которое распознает внеклеточный домен PTPRS человека. В рамках настоящего изобретения можно использовать один тип или несколько типов моноклональных антител. Например, в настоящее изобретение может быть включено и может использоваться несколько типов моноклональных антител, которые распознают внеклеточный домен PTPRS человека.
То, что антитело обладает действием подавления активности продуцирования IFN из pDC можно подтверждать следующим образом. pDC продуцирует большое количество IFN при стимуляции вирусом. Путем предоставления антитела против pDC до или после стимуляции вирусом, или одновременно со стимуляцией в вирусом, и с использованием pDC, против которых антитело не предоставляется, в качестве контроля, сравнивают способность продуцировать IFN. Способность продуцирования IFN можно оценивать путем измерения IFN-α и IFN-β, включенных в культуральный супернатант pDC. В результате сравнения, когда количество IFN в супернатанте значительно снижается при добавлении антитела, это может подтвердить, что исследуемое антитело обладает действием подавления способности продуцировать IFN. Способ измерения этих IFN известен. pDC представляет собой клетку, которая продуцирует большинство IFN в живом организме. Таким образом, путем подавления способности pDC продуцировать IFN, можно модулировать состояние продуцирования IFN в живом организме.
В рамках настоящего изобретения активность pDC включает поддержание количества клеток pDC. Таким образом, подавление активности pDC в рамках настоящего изобретения включает снижение количества клеток pDC. Если подтверждают то, что количество клеток pDC снижается в присутствии антитела, то устанавливают, что антитело подавляет активность pDC. В качестве контроля для сравнения продуцирования IFN можно использовать инертный иммуноглобулин, происходящий из того же вида животных, что и антитело, активность которого требуется подтвердить. Количество клеток pDC можно сравнивать количественно путем подсчета количества клеток. Количество клеток можно подсчитывать с помощью FACS или микроскопа.
Более того, также считается, что pDC дифференцируется в клетку, которая индуцирует Th2, называемую DC2 (дендритная клетка 2) в результате инфекции вирусом и т.п. Если продуцирование IFN посредством pDC при стимуляции вирусом может подавляться, то также возможно, что может подавляться дифференцировка в Th2. Таким образом, для моноклонального антитела по настоящему изобретению, которое подавляет продуцирование IFN, можно ожидать терапевтических эффектов в отношении различных аллергических заболеваний.
В случае, когда антитело, которое распознает внеклеточный домен PTPRS человека, вводят хозяину, отличающемуся от вида организма, из которого происходит антитело, является желательным преобразовать антитело в форму, которая практически не распознается в качестве чужеродного вещества для хозяина. Например, путем преобразования следующих молекул может стать трудным распознавание иммуноглобулина в качестве чужеродного вещества. Известен следующий способ преобразования молекулы иммуноглобулина.
- Фрагмент, включающий антигенсвязывающую область, которая лишена константной области (Monoclonal Antibodies: Principles and Practice, third edition, Academic Press Limited. 1995; Antibody Engineering, A Practical Approach, IRL PRESS, 1996)
- Химерное антитело, которое образовано антигенсвязывающей областью моноклонального антитела и константной областью иммуноглобулина хозяина (Experimental Manual for Gene Expression, Kodansha Ltd., 1994 (edited by Isao Ishida and Tamie Ando))
- Антитело с замененной CDR, полученное путем замены определяющей комплементарность области (CDR) в иммуноглобулине хозяина на CDR моноклонального антитела (Experimental Manual for Gene Expression, Kodansha Ltd., 1994 (edited by Isao Ishida and Tamie Ando)).
Альтернативно ген вариабельной области иммуноглобулина человека может быть получен способом фагового дисплея (McCafferty J. et al., Nature 348: 552-554, 1990; Kretzschmar T et al., Curr Opin Biotechnol 2002 Dec: 13 (6): 598-602.). В способе фагового дисплея ген, который кодирует вариабельную область иммуноглобулина человека, встраивают в ген фага. Библиотеку фагов можно получать с использованием различных генов иммуноглобулинов в качестве источников. Фаг экспрессирует вариабельную область в качестве слитого белка с белком, который является компонентом самого фага. Вариабельная область на поверхности фага, которая экспрессируется фагом, сохраняет активность связывания с антигеном. Таким образом, путем выбора фага, который связывается с клеткой, которая экспрессирует антиген, или антигеном и т.п., можно проводить скрининг фага, который экспрессирует вариабельную область, имеющую предполагаемую активность связывания, из библиотеки. Более того, ген, который кодирует вариабельную область, имеющую намеченную активность связывания, сохраняется в фаговых частицах, выбранных подобным образом. Это означает, что в способе фагового дисплея ген, который кодирует вариабельную область, имеющую намеченную активность связывания, можно получать с использованием активности связывания вариабельной области в качестве показателя.
Средство или способ подавления активности pDC в соответствии с настоящим изобретением, антитело, которое распознает внеклеточный домен PTPRS человека, или фрагмент антитела, включающий по меньшей мере его антигенсвязывающую область, можно вводить в качестве белка или полинуклеотида, который кодирует белок. Для введения полинуклеотида желательно использовать вектор, в который помещены нуклеотиды, которые кодируют намеченный белок, под контролем подходящего промотора, так чтобы намеченный белок мог экспрессироваться. В вектор также может быть помещен энхансер или терминатор. Вектор, в котором сохранены гены тяжелой цепи и легкой цепи, которые составляют иммуноглобулин, и который может экспрессировать молекулу иммуноглобулина, известен. Вектор, который может экспрессировать иммуноглобулин, можно вводить путем введения в клетку. При введении в живой организм, вектор, который может быть перенесен в клетку путем введения в живой организм, можно вводить как есть. Альтернативно вектор можно вводить в лимфоцит, который был когда-то отделен от живого организма и после этого возвращен в живой организм (ex vivo).
В средстве или способе подавления активности pDC в соответствии с настоящим изобретением количество моноклонального антитела, подлежащего введению в живой организм в качестве иммуноглобулина, как правило, составляет от 0,5 мг до 100 мг, например, от 1 мг до 50 мг, предпочтительно от 2 мг до 10 мг, на 1 кг массы тела. Интервалы между введением антитела в живой организм можно пригодным образом модулировать так, чтобы могла поддерживаться эффективная концентрация иммуноглобулина в живом организме в ходе периода лечения. В частности, антитело можно вводить с интервалами от 1 до 2 недель. Путь введения является произвольным. Специалист в данной области может пригодным образом выбирать эффективный путь введения для лечения. В частности, введение можно проводить перорально или парентерально. Например, антитело можно вводить системно или местно путем внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции или подкожной инъекции и т.п. Примеры составов, которые пригодны для парентерального введения в рамках настоящего изобретения, могут включать инъецируемое средство, суппозиторий, аэрозоль и т.п. Более того, когда антитело предоставляют клетке, иммуноглобулин обычно предоставляют в количестве 1 мкг/мл, предпочтительно 10 мкг/мл или более, более предпочтительно 50 мкг/мл или более, более предпочтительно 0,5 мг/мл или более.
В средстве или способе подавления активности pDC в соответствии с настоящим изобретением, моноклональное антитело можно вводить в живой организм любым способом. Как правило, моноклональное антитело составляют с фармацевтически приемлемым носителем. Когда это необходимо, в моноклональное антитело можно включать добавки, такие как загуститель, стабилизатор, антисептик и солюбилизатор. Примеры такого носителя или добавки могут включать лактозу, лимонную кислоту, стеариновую кислоту, стеарат магния, сахарозу, крахмал, тальк, желатин, агар, растительные масла, этиленгликоль и т.п. Термин “фармацевтически приемлемый” относится к одобренному контролирующим органом правительства каждой страны или приведенный в фармакопее каждой страны или общепризнанной фармакопее в отношении применения у животных, млекопитающих и, в частности, у человека. Средство для подавления активности pDC по настоящему изобретению может быть предоставлено в форме лиофилизированного порошка или таблетки, включающей одну дозу или несколько доз. Лиофилизированный порошок или таблетку можно далее комбинировать с инъецируемой стерилизованной водой, физиологическим раствором или буфером для растворения композиции, так чтобы получить желаемую концентрацию перед введением.
Более того, при введении в форме вектора, который экспрессирует иммуноглобулин, каждую плазмиду можно вводить в количестве от 0,1 до 10 мг, например, от 1 до 5 мг на 1 кг массы тела, учитывая, что тяжелая цепь и легкая цепь котрансфицированы в качестве отдельных плазмид. Более того, для введения в клетку in vitro, используют вектор в количестве от 1 до 5 мкг/106 клеток.
Далее настоящее изобретение пояснено более конкретно с помощью примеров.
Все из документов уровня техники, цитированных в настоящем описании, включены в настоящее описание в качестве ссылок.
Далее настоящее изобретение будет объяснено более конкретно с помощью примеров, однако настоящее изобретение не следует истолковывать как ограничивающееся этими примерами.
ПРИМЕРЫ
Пример 1
A. Анализ экспрессии PTPRS
A-1) Анализ с использованием библиотеки SAGE
Экспрессию гена в моноцитах человека, pDC человека и pDC человека, обработанных вирусом простого герпеса (HSV), сравнивали и анализировали способом SAGETM (серийный анализ экспрессии генов). Способ анализа является следующим.
Моноциты выделяли в качестве CD14-положительной клетки и pDC выделяли в качестве положительной по BDCA-4 клетки из мононуклеарных клеток периферической крови человека с помощью клеточного сортера. Более того, pDC культивировали в присутствии HSV в течение 12 часов для получения активированных pDC. Из соответствующих клеток получали РНК и получали библиотеку SAGE с использованием набора I-SAGETM (Invitrogen). Полученные данные о последовательности информации с приблизительно 100000 маркеров анализировали с помощью программного обеспечения SAGE Analysis Software (Invitrogen). В результате в качестве гена, имеющего оценочную величину для моноцитов/pDC/pDC+HSV, составляющую 0/7/0, т.е. гена, который проявляет pDC-специфическую экспрессию, был выявлен известный ген: PTPRS (GenBank Acc#NM_002856.3). PTPRS кодируется последовательностью оснований, представленной в SEQ ID NO: 2. Более того, он имеет единый трансмембранный домен, имеющий иммуноглобулин-подобный домен (Ig-подобный домен) и домен, подобный фибронектину типа III, во внеклеточной области. Кроме того, он имеет две области протеинтирозинфосфатазы (PTP-домена) во внутриклеточной области (фиг.1).
A-2) Анализ экспрессии мРНК PTPRS в различных иммунокомпетентных клетках человека с помощью количественной ОТ-ПЦР
Экспрессию PTPRS в иммунных клетках анализировали более детально. Каждую клетку выделяли из периферической крови человека с помощью клеточного сортера. РНК экстрагировали из каждой популяции клеток и синтезировали кДНК. С использованием полученной кДНК в качестве матрицы, проводили количественную ОТ-ПЦР в соответствии с общим способом для анализа уровня экспрессии мРНК PTPRS. Путем нормализации с уровнем экспрессии гена GAPDH (глицеральдегид-3-фосфат дегидрогеназа), о котором известно, что он экспрессируется постоянно, сравнивали экспрессию гена PTPRS между иммунными клетками.
Последовательности оснований используемых праймеров и условия ПЦР являются следующими.
Прямой праймер для PTPRS: 5’ CAC GGC CTA TGA CCT CCA 3’ (SEQ ID NO: 3)
Обратный праймер для PTPRS: 5’ AAG TTC TTG GGC GAG ACT TG 3’ (SEQ ID NO: 4)
Прямой праймер для GAPDH: 5’ CCA CCC ATG GCA AAT TCC 3’ (SEQ ID NO: 5)
Обратный праймер для GAPDH: 5’ TGG GAT TTC CAT TGA TGA CAA G 3’ (SEQ ID NO: 6)
1 цикл при 50°C в течение 2 минут,
1 цикл при 95°C в течение 10 минут, и
50 циклов [при 95°C в течение 15 секунд и при 60°C в течение 60 секунд].
Моноциты, pDC, pDC, стимулированные HSV, B-клетки (CD19+ клетки), T-клетки (CD3+ клетки), активированные T-клетки, стимулированные PMA (форбол 12-миристат 13-ацетат), и NK-клетки (CD56+ клетки) анализировали, и было показано, что PTPRS экспресировалась специфическим для pDC образом. Более того, было открыто в качестве характерного признака, что экспрессия PTPRS снижается в pDC, стимулированных HSV (фиг.2).
A-3) Анализ экспрессии мРНК PTPRS в ткани человека с помощью количественной ОТ-ПЦР
Более того, экспрессию в тканях исследовали с помощью количественной ПЦР с использованием ABI PRISM 7000 (Applied Biosystem). В качестве панелей кДНК, использовали панель ДНК множества тканей BD™ MTC (Human I; каталожный номер № 636742, Human immune; каталожный номер № 636748, Human blood fractions; каталожный номер № 636750; все от Becton Dickinson). Использованные последовательности оснований праймеров представлены ниже.
Прямой праймер для PTPRS: 5’ ACT CAC CCA CAC CCT ACA AGA 3’(SEQ ID NO: 7)
Обратный праймер для PTPRS: 5’ CTT GGT GGT ACG GCC ATC 3’(SEQ ID NO: 8)
Прямой праймер для GAPDH: 5’ CCA CCC ATG GCA AAT TCC 3’(SEQ ID NO: 5)
Обратный праймер для GAPDH: 5’ TGG GAT TTC CAT TGA TGA CAA G 3’(SEQ ID NO: 6)
С использованием набора SYBR green PCR master mix kit (Applied Biosystem) проводили ПЦР с помощью ABI PRISM 7000, доступного от той же компании. Для анализа использовали программное обеспечение Sequence Detection System Software от той же компании. Условия реакции являются следующими.
Стадия 1: 1 цикл при 50°C в течение 2 минут
Стадия 2: 1 цикл при 95°C в течение 10 минут
Стадия 3: 40 циклов при 95°C в течение 15 секунд и при 60°C в течение 1 минуты
Путем нормализации по уровню экспрессии гена GAPDH (глицеральдегид-3-фосфатдегидрогеназа), о котором известно, что экспрессируется постоянно, экспрессию гена PTPRS сравнивали между тканями. В результате, мРНК PTPRS широко экспрессировалась в тканях (фиг.3).
B. Получение экспрессирующего вектора PTPRS
Для экспрессии белка PTPRS проводили получение экспрессирующего вектора для гена PTPRS. Из клона кДНК PTPRS, который был включен в клонирующий вектор pCR4-TOPO (Open Biosystem cc# MHS1010-98052887), брали только ген PTPRS и включали его в экспрессирующий вектор pcDNA3.1 (PTPRS/pcDNA3.1). С использованием полученной плазмиды PTPRS/pcDNA3.1 в качестве матрицы, ген PTPRS амплифицировали с праймером, включающим EcoRI, NotI и последовательность Kozak (GCC GCC ACC) (информация о праймерах представлена ниже). Продукт ПЦР клонировали в ретровирусный вектор pMX-IP в участки EcoRI и NotI (PTPRS/pMX-IP). Для реакции ПЦР использовали одну единицу ДНК-полимеразы KOD Plus (TOYOBO), и условия реакции представляли собой 1 цикл при 94°C в течение 2 минут и 25 циклов при 94°C в течение 15 секунд и при 68°C в течение 4 минут и 30 секунд].
Прямой праймер (SEQ ID NO: 9): 5’ aaa GAA TTC gcc gcc acc ATG GCG CCC ACC TGG GGC CCT3’
Обратный праймер (SEQ ID NO: 10): 5’ aaa gcg gcc gcT TAG GTT GCA TAG TGG TCA AAG C3’
В упомянутых выше последовательностях оснований прописные буквы соответствуют участкам расщепления ферментом рестрикции EcoRI или участкам NotI. aaa на 5’-конце представляют собой дополнительные основания для ферментативного расщепления.
C. Получение клетки, экспрессирующей PTPRS человека (hPTPRS)
Для получения ретровируса, содержащего ген PTPRS, клетку HEK-293T, которая представляет собой линию клеток почки эмбриона человека, временно трансфицировали PTPRS/pMX-IP и ретровирусным упаковывающим вектором PCL-ECO из набора FuGENE (Roche). Через двое суток собирали супернатант клеточной культуры, в котором присутствовал вирус, включающий ген hPTPRS, и инфицировали клетки D2SC/1, которые представляют собой дендритные клетки, происходящие из селезенки мыши BALB/c (их получали, основываясь на Paglia et al., J. Exp. Med., 178, 1893-1901 (1993)). Поскольку ретровирусный вектор pMX-IP включает ген устойчивости к пуромицину, при культивировании инфицированных клеток D2SC/1 с пуромицином может выжить только клетка, которая экспрессирует hPTPRS, тем самым, становится возможной селекция. Клетки D2SC/1, экспрессирующие hPTPRS, отбирали посредством сортировки FACS и культивировали. Для подтверждения экспрессии hPTPRS, к отобранным клеткам hPTPRS/D2SC/1 добавляли 10 мкг/мл IgG козы (SantaCruz) и коммерчески доступного поликлонального антитела против hPTPRS (pAb; R&D) по 100 мкл каждого, и смесь инкубировали при 4°C в течение 30 минут. Клетки промывали PBS, а затем добавляли меченное FITC антитело против IgG козы (SantaCruz), разбавленное в 100 раз, в количестве 50 мкл, и смесь инкубировали при 4°C в течение 30 минут. После промывания PBS, импортировали данные с помощью FACSCalibur (BD) (фиг.4).
Пример 2
A. Получение моноклонального антитела против PTPRS человека
A-1) Иммунизация
В качестве клетки, используемой в качестве иммуногена, использовали упомянутые выше клетки hPTPRS/D2SC/I. Мышей BALB/c подвергали анестезии и эмульсию полного адъюванта Фрейнда (CFA) инъецировали подкожно в подушечки лап по 50 мкл на каждую ступню. Всего вводили 100 мкл/мышь. На следующие сутки получали эмульсию с использованием клеток hPTPRS/D2SC/1, полученных в качестве иммуногена, и неполного адъюванта Фрейнда (IFA) и инъецировали подкожно в подушечки лап (50 мкл/ступня, всего 100 мкл/мышь). Иммунизацию проводили раз в двое суток всего три раза, и дренирующие лимфатические узлы извлекали через 3 суток после последней иммунизации.
A-2) Слияние клеток
Клетки дренирующих лимфатических узлов собирали из обеих ступней иммунизированной мыши и смешивали с клетками миеломы мыши P3-X63-Ag8.563, которые культивировали в среде RPMI1640 (SIGMA), включающей 10% FBS, так чтобы соотношение клеток лимфатических узлов и клеток миеломы стало 5:4, и клетки собирали центрифугированием. К смешанным клеткам добавляли PEG1500 (Roche) для слияния клеток. Слитые клетки (гибридомы) промывали и культивировали в 10% эмбриональной телячьей сыворотке (FBS), включающей добавку для роста клеток + среда HAT (Sigma)-RPMI1640 (включающая 2 мМ L-глутамин, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина, 10 мМ HEPES, 1 мМ пируват натрия, 50 мкМ 2-ME).
A-3) Скрининг с помощью FACS гибридом с использованием иммунизированных клеток hPTPRS/D2SC/1
Антитело против CD16/32 (2.4G2), приготовленное в концентрации 2,5 мкг/мл, добавляли в количестве 50 мкл к клеткам D2SC/1 или клеткам hPTPRS/D2SC/1 в количестве 3×105/лунка для блокирования FC-рецептора. После промывания PBS добавляли IgG козы, приготовленное в концентрации 10 мкг/мл, коммерчески доступное pAb против hPTPRS (R&D), IgG2ak мыши (BioLegend) и культуральный супернатант культивируемой гибридомы в количестве 60 мкл каждого, и смесь инкубировали при 4°C в течение 60 минут. После промывания PBS к клеткам добавляли разбавленное в 50 раз меченное FITC антитело против IgG козы и разбавленное в 100 раз меченное PE антитело против IgG мыши (BD) по 50 мкл каждого, и смесь инкубировали при 4°C в течение 30 минут в защищенном от света месте. После промывания PBS клетки суспендировали в 200 мкл PBS. Данные получали с помощью FACS Calibur (BD). Полученные данные получали с помощью точечных диаграмм FSC и SSC для ограничения области живых клеток. Данные собирали до тех пор, пока данные для клеток в этой области не достигали количества 2000. В результате смогли получить 13 гибридом, которые продуцируют антитело против hPTPRS (2G6, 28G10, 4B2, 2G2, 9H5, 10F7, 22H8, 49F2, 9D2, 14A8, 55E7, 13G5, 16H2) (фиг.5).
A-4) Скрининг с помощью FACS с использованием клеток CAL-1
3×105 клеток pDC-подобной линии клеток CAL-1 окрашивали в 50 мкл культурального супернатанта каждой из упомянутых выше гибридом в течение 15 минут 4°C. Клетки промывали один раз буфером FACS (1% FBS+PBS), а затем центрифугировали для удаления супернатанта. Затем 2 мкг/мл меченного PE антитела против IgG мыши подвергали реакции при 4°C в течение 20 минут. Клетки промывали один раз буфером FACS и центрифугировали. Клеточный осадок ресуспендировали в буфере FACS и анализировали с помощью Calibur. В результате 2G6, 4B2, 2G2, 9H5, 10F7, 22H8, 49F2, 14A8, 55E7, 13G5 и 16H2 в культуральном супернатанте гибридом хорошо реагировали с CAL-1. С другой стороны, 28G10 и 9D2 реагировали слабо (фиг.6).
A-5) Скрининг с помощью FACS с использованием pDC периферической крови человека
[Выделение PBMC человека]
Проводили взятие 20 мл периферической крови от здорового человека, и выделяли мононуклеарные клетки периферической крови (PBMC) с помощью гравитационного центрифугирования с использованием HISTOPAQUE-1077 (SIGMA). 1×106 PBMC окрашивали с помощью каждого образца. Клетки промывали буфером FACS, добавляли реагент Fc-block reagent (Miltenyi) по 25 мкл, разведенный в 5 раз, и реакцию проводили при 4°C в течение 15 минут. После промывания буфером FACS, добавляли 50 мкл супернатанта клеточной культуры каждой гибридомы, 10 мкг/мл IgG козы, pAb против hPTPRS и IgG2a,κ мыши, и реакцию проводили при 4°C в течение 20 минут. После промывания буфером FACS, добавляли 8 мкг/мл меченного FITC антитела против IgG козы или 2 мкг/мл меченного PE антитела против IgG мыши, и реакцию проводили при 4°C в течение 20 минут. После промывания буфером FACS 50 мкл меченного APC антитела против BDCA2 в 10-кратном разведении подвергали реакции при 4°C в течение 20 минут. После промывания буфером FACS клетки ресуспендировали в 300 мкл буфера FACS и анализировали с помощью FACS calibur. В результате 2G6, 28G10, 4B2, 2G2, 9H5, 10F7, 22H8, 49F2, 14A8, 55E7 и 13G5 продемонстрировали реакцию связывания, специфичную к популяции клеток pDC. 9D2 продемонстрировало связывание с pDC, и также продемонстрировало реакции с группой клеток, отличных от pDC (BDCA2-). 16H2 не показало реакции с PBMC (фиг.7).
Испытание специфичности антитела против PTPRS
PTPRS принадлежит семейству PTPR, и аминокислотные последовательности нескольких молекул этого семейства обладают высокой гомологией с аминокислотной последовательностью PTPRS (фиг.8).
A-6) Исследовали то, связываются или не связываются ли 10 типов культуральных супернатантов клеток гибридом, которые продуцируют антитело, которое распознает PTPRS и специфично связывается с pDC человека (2G6, 4B2, 2G2, 9H5, 10F7, 22H8, 49F2, 14A8, 55E7, 13G5) специфично только с PTPRS. Трансфицированные клетки PTPRA (40%), PTPRD (76%) и PTPRF (67%), которые имели особенно высокую гомологию с PTPRS, получали путем экспрессии FLAG-метки на N-конце молекулы, и окрашивали. Экспрессию hPTPRE в трансфицированных клетках подтверждали вестерн-блоттингом, однако экспрессию на клеточной поверхности не могли подтвердить. Таким образом, hPTPRE не экспрессировался на клеточной поверхности. В результате 4B2 реагировало с hPTPRD (фиг.9C), и 2G6 показало перекрестную реактивность с hPTPRF (фиг.9D). Другие 8 типов антител продемонстрировали специфическое связывание с PTPRS (фиг.9A-D).
A-7) Перекрестная реактивность антитела против PTPRS в отношении обезьяны
PBMC яванского макака выделяли из периферической крови (10 мл; Shin-Nippon Biomedical Laboratories, Ltd.) гравитационным центрифугированием с использованием HISTOPAQUE-1077 (SIGMA). Для FACS использовали 5×105 клеток на один образец. Клетки промывали буфером FACS и к ним добавляли 10 мкл 10% сыворотки яванского макака, разбавленной буфером FACS, и реакцию проводили при 4°C в течение 20 минут. После промывания буфером FACS добавляли 100 мкл каждого культурального супернатанта каждой гибридомы и 10 мкг/мл IgG2a,κ мыши или IgG1,κ мыши (BioLegend) и реакцию проводили при 4°C в течение 15 минут. После промывания буфером FACS добавляли 1 мкг/мл меченного APC антитела против IgG мыши (BD), и реакцию проводили при 4°C в течение 20 минут. После промывания буфером FACS меченное FITC антитело против Lineagel (BD), меченное PE антитело против CD123 (BD) и меченное PerCP7Cy5.5 антитело против HLA-DR (BD) в количестве 25 мкл в 10-кратном разведении подвергали реакции при 4°C в течение 15 минут. После промывания буфером FACS клетки ресуспендировали в 300 мкл буфера FACS и анализировали с помощью FACS calibur. В качестве использованных культуральных супернатантов гибридомы отобрали 7 типов: 49F2, 55E7, 14A8, 13G5, 10F7, 22H8 и 9H5, которые являются специфичными к PTPRS и хорошо связываются с клеткой CAL-1 и pDC человека. В результате все супернатанты клеточной культуры гибридом специфично связывались с группой популяции pDC (линия-CD123+HLA-DR+) яванского макака (фиг.10).
A-8) Сигнализация гибридом
Каждый из упомянутых выше 7 типов гибридом (49F2, 55E7, 14A8, 13G5, 10F7, 22H8 и 9H5) собирали и суспендировали в буфере для сортировки (1% FBS/PBS) до 1×105 клеток/мл. С использованием FACS Aria (BD) проводили сортировку единичных клеток. Данные получали, и полученные данные преобразовывали в двухмерную точечную диаграмму с осью X: FSC и осью Y: SSC. Ограничивали живые клетки, дающие сигнал выше порогового значения, на точечном графике. Проводили ограничение по пороговому значению для удаления дублетов каждой клетки в ограниченной области живых клеток, и популяцию клеток распределяли в 96-луночный планшет с плоским дном до 1 клетки/лунка. Клетки, подвергнутые сортировке отдельных клеток, культивировали в среде HAT (RPMI1640+2 мМ L-глутамин, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина, 10 мМ HEPES, l мМ пируват натрия, и 50 мкМ 2-ME)+ добавка для роста гибридом HFCS (Roche). После этого клетки D2SC и клетки hPTPRS/D2SC (фиг.11A и B), клетки CAL-1 (фиг.11C) и pDC человека (фиг.11D) окрашивали с использованием клеточного культурального супернатанта гибридомы, и отбирали единичные гибридомы.
Пример 3
Очистка антитела
Восемь типов очищенных антител (9H5-4, 10F7-38, 13G5-52, 13G5-57, 14A8-85, 22H8-84, 49F2-30 и 55E7-79) получали из культурального супернатанта гибридом посредством очистки с использованием Protein G Sepharose FastFlow (GE Healthcare). С использованием набора для определения изотипов Pierce rapid ELISA mouse mAb Isotyping Kit (Thermo Fisher Scientific) определяли изотипы. В результате 13G5-52 и 13G5-57 представляли собой IgG2b, κ мыши, 55E7-79 имело как IgG2b, κ мыши, так и IgG1, κ мыши, а другие представляли собой IgG1, κ мыши. Если очищенное антитело включает эндотоксин, это может повлиять на результат испытания по определению свойств. Таким образом, измеряли концентрацию эндотоксина. Использованные наборы представляли собой набор Endospecy ES-50M, набор Toxicolor DIA-MP и набор стандартного продукта эндотоксина CSE-L (все от Seikagaku Biobusiness Corporation). В результате, все очищенные антитела имели концентрацию эндотоксина, равную или меньшую стандартное величины 0,3 ЕЭ/мг Ab (фиг.12).
Исследование реактивности очищенного антитела
Способность связывания очищенных антител подтверждали с помощью клеток лини pDC-подобных клеток человека CAL-1 (фиг.13). Кроме того, все антитела сохраняли способность связываться с популяцией pDC человека периферической крови человека (BDCA2+) (фиг.14).
Гомология аминокислотной последовательности PTPRS человека с PTPRS мыши (mPTPRS) составляет приблизительно 96%. Поскольку они являются достаточно сходными друг с другом, исследовали то, связывается или не связывается ли полученное антитело против PTPRS человека с PTPRS мыши. Клетку CHO, в которой принудительно экспрессировался ген mPTPRS (клетка яичника китайского хомяка; далее обозначаемая как mPTPRS/CHO) окрашивали посредством 10 мкг/мл каждого антитела против PTPRS. Количество клеток составляло 2×105 на один образец. После промывания буфером FACS, PE-меченное антитело против IgG мыши разбавляли в 50 раз и проводили окрашивание количеством 25 мкл. В результате, 49F2-30, 13G5-52, 13G5-57 и 22H8-84 связывались с mPTPRS/CHO (фиг.15).
Пример 4
Комплемент-зависимая клеточная цитотоксичность антитела против PTPRS в отношении экспрессирующей hPTPRS клетки
С использованием комплемента детенышей кролика измеряли комплементзависимую клеточную цитотоксичность (далее обозначаемую как активность CDC) антитела против PTPRS в отношении клеток CHO, которые экспрессируют PTPRS человека (далее обозначаемых как hPTPRS/CHO) и клеток PTPRS/CHO мыши (далее обозначаемых как mPTPRS/CHO). Активность получали с использованием клеточной токсичности, которую вычисляли из измеренной величины уровня лактатдегидрогеназы (LDH), высвобожденной из клеток, в качестве показателя. Каждую клетку распределяли в 96-луночный планшет с U-образным дном в количестве 2×104 клеток/50 мкл/лунка. 18% комплемент (CEDARLANE) приготавливали в среде для CDC (RPMI1640+0,1%BSA+10 мМ HEPES+2 мМ L-глутамин+100 единиц/мл пенициллина +100 мкг/мл стрептомицина). Два типа: 3,3 мкг/ мл и 30 мкг/мл приготавливали для контрольного антитела (IgG1, κ мыши или IgG2b, κ мыши) и антитела против PTPRS. Анализ проводили с использованием набора CytoTox 96 Non-Radioactive Cytotoxicity Assay (Promega). В результате 13G5-52 и 13G5-57 показали приблизительно 20% активность CDC против мишени hPTPRS/CHO (фиг.16A). С другой стороны, 13G5-52 и 13G5-57 показали приблизительно 100% активность CDC в отношении mPTPRS/CHO (фиг.16B).
Пример 5
Получение химеризованного антитела
В качестве гибридомы для получения антитела против PTPRS мыши, использовали следующие.
Гибридома 9H5-4 (номер доступа №: FERM ABP-11356)
Гибридома 10F7-38 (номер доступа №: FERM ABP-11357)
Гибридома 13G5-52 (номер доступа №: FERM ABP-11358)
Гибридома 13G5-57 (номер доступа №: FERM ABP-11359)
Гибридома 14A8-85 (номер доступа №: FERM ABP-11360)
Гибридома 22H8-84 (номер доступа №: FERM ABP-11361)
Гибридома 49F2-30 (номер доступа №: FERM ABP-11362)
1. Подтверждение изотипа константной области
Подтверждали изотип константной области каждого антитела мыши, продуцированного семью гибридомами (9H5-4,10F7-38,13G5-52, 13G5-57, 14A8-85, 22H8-84 и 49F2-30).
Для подтверждения использовали набор для изотипирования моноклональных антител мыши (каталожный номер №: MMT1; Serotec Product; Oxford, UK) или набор Pierce Rapid ELISA mouse mAb Isotyping Kit (Thermo Fisher Scientific) и культуральный супернатант гибридом 9H5-4,10F7-38,13G5-52, 13G5-57, 14A8-85, 22H8-84 и 49F2-30 в качестве образца.
В результате изотип антитела, продуцированный гибридомами 13G5-52 и 13G5-57, представлял собой изотип, включающий IgG2b мыши в качестве тяжелой цепи и κ в качестве легкой цепи. С другой стороны, изотип антител, продуцируемых гибридомами 9H5-4, 10F7-38, 14A8-85, 22H8-84 и 49F2-30, представлял собой изотип, включающий IgG1 мыши в качестве тяжелой цепи и κ в качестве легкой цепи.
2. Клонирование кДНК, которая кодирует вариабельную область антитела против PTPRS мыши
2-1) Выделение тотальной РНК
С использованием коммерчески доступного набора “RNeasy Mini Kit” (Qiagen, каталожный номер №: 74106), выделяли тотальную РНК из семи гибридом в соответствии с инструкцией, прилагаемой к набору. Получали приблизительно 30 мкг тотальной РНК из линии клеток гибридомы в количестве 5×106 клеток.
2-2) Амплификация и фрагментация кДНК, которая кодирует вариабельную область тяжелой цепи мыши
С использованием 5 мкг тотальной РНК, выделенной согласно 2-1), кДНК, которая кодирует вариабельную область тяжелой цепи мыши, амплифицировали способом 5’ RACE ПЦР. В амплификации использовали коммерчески доступный набор “5’ RACE System for Rapid Amplification of cDNA ENDs, Version 2.0 Kit” (Invitrogen, каталожный номер №: 18374-058). Детали являются следующими. Сначала синтезировали первую цепь кДНК с тотальной РНК, полученной согласно 2-1) с помощью обратной транскриптазы. В этом случае использовали антисмысловой праймер (GSP1), представленный ниже.
Праймер GSP1, использованный для амплификации кДНК, используют в зависимости от изотипа каждой тяжелой цепи мыши. Например, следующие антисмысловые праймеры используют для клонирования вариабельной области тяжелой цепи гибридом 9H5-4, 10F7-38, 14A8-85, 22H8-84 и 49F2-30, включающих IgG1 мыши в качестве тяжелой цепи.
Праймер GSP1: mu IgG1 VH-GSP1
Последовательность: 5’-CCA GGA GAG TGG GAG AGG CTC TTC TCA GTA TGG TGG-3’ (36-мер) (SEQ ID NO: 39)
Праймер GSP2: mu IgG1 VH-GSP2
Последовательность: 5’-GGC TCA GGG AAA TAG CCC TTG ACC AGG CAT CC-3’ (32-мер) (SEQ ID NO: 40)
Также, например, следующие антисмысловые праймеры можно использовать для клонирования вариабельной области тяжелой цепи гибридом 9H5-4, 10F7-38, 14A8-85, 22H8-84 и 49F2-30, включающих IgG1 мыши в качестве тяжелой цепи.
Праймер GSP1: mu IgGHγ1-GSP1
Последовательность: 5’-TCC AGA GTT CCA GGT CAC TGT CAC-3’ (24-мер) (SEQ ID NO: 11)
Праймер: mu IgG Hγ1-GSP2
Последовательность: 5’-AGG GGC CAG TGG ATA GAC AGA TGG-3’ (32-мер) (SEQ ID NO: 13)
Далее, следующие антисмысловые праймеры используют для клонирования вариабельной области тяжелой цепи гибридом 13G5-52 и 13G5-57, включающих IgG2b мыши в качестве тяжелой цепи.
Праймер GSP1: mu IgGHγ 2B-GSP1
Последовательность: 5’-TCC AGA GTT CCA AGT CAC AGT CAC-3’ (24-мер) (SEQ ID NO: 41)
Праймер GSP2: mu IgG Hγ 2B-GSP2
Последовательность: 5’-AGG GGC CAG TGG ATA GAC TGA TGG-3’ (24-мер) (SEQ ID NO: 42)
Более того, с использованием терминальной дезоксинуклеотидилтрансферазы (TdT) на 3’-конце кДНК первой цепи, добавляли нуклеотидный гомополимер dC. Более того, с использованием якорного праймера, имеющего нуклеотидный полимер, который коплементарен dC (якорная последовательность) (SEQ ID NO: 12), и антисмыслового праймера (GSP2), кДНК амплифицировали способом ПЦР. Более того, с использованием полученного продукта ПЦР в качестве матрицы и с использованием праймера AUAP (SEQ ID NO: 14) и антисмыслового праймера (GSP2), кДНК амплифицировали способом гнездовой ПЦР. Более того, этот продукт ПЦР очищали способом с 1,5% агарозой с низкой температурой плавления.
Якорный праймер для 5’RACE (SEQ ID NO: 12): 5’-GGC CAC GCG TCG ACT AGT ACG GGI IGG GII GGG IIG-3’ (36-мер)
Праймер AUAP для 5’RACE (SEQ ID NO: 14): 5’-GGC CAC GCG TCG ACT AGT AC-3’ (20-мер)
2-3) Амплификация и фрагментация кДНК, которая кодирует вариабельную область легкой цепи мыши
Из тотальной РНК, выделенной согласно 2-1), кДНК, которая кодирует вариабельную область легкой цепи мыши, амплифицировали аналогично тому, как в 2-2).
Поскольку эти семь антител включают легкую цепь κ Ig мыши, следующие антисмысловые праймеры используют для клонирования легкой цепи.
Праймер GSP1: Mu IgVL5RACE-GSP1
Последовательность: 5’-TTC ACT GCC ATC AAT CTT CCA CTT-3’ (24-мер) (SEQ ID NO: 15)
Праймер GSP2: Mu IgVL5RACE-GSP2
Последовательность: 5’-GAT GGA TAC AGT TGG TGC AGC-3’ (21-мер) (SEQ ID NO: 16)
Полученный продукт ПЦР очищали способом с 1,5% агарозой с низкой температурой плавления.
2-4) Подтверждение последовательности оснований кДНК и определение области CDR
Каждый из фрагментов кДНК вариабельной области тяжелой цепи, полученные согласно 2-2), и вариабельной области легкой цепи, полученные согласно 2-3) клонировали в вектор pCR4Blunt-TOPO с использованием коммерчески доступного набора “Zero Blunt TOPO PCR Cloning Kit” (Invitrogen, каталожный номер №: 1325137), в соответствии с инструкцией, прилагаемой к набору, и вводили в компетентную клетку E. coli с получением трансформанта E. coli. Из этого трансформанта получали плазмиду и образец плазмидной ДНК отсылали в Operon Biotechnology Co. Ltd (Tokyo) для анализа последовательности для подтверждения последовательности оснований кДНК в плазмиде. Для анализа последовательностей использовали программное обеспечение “Sequencher DNA sequence assembly and analysis software version 4.2.2 (Gene Codes Corporation)” и “GENETYX-MAC Version 11.1.1” (GENETYX CORPORATION).
Трансформанты, которые приводили к неактивной РНК вследствие сдвига рамки считывания РНК, нонсенс-мутации и т.п., появившиеся рядом с определяющей комплементарность областью (далее обозначаемой как “область CDR”) исключали, и выделяли трансформанты, имеющие правильные последовательности. Более того, проводили подтверждение в Immunoglobulin Database(IgBLAST, URL:www.ncbi.nlm.nih.gov/igblast/) и подтверждали гомологию для последовательности оснований кДНК, включенных в плазмиду, для определения последовательностей области CDR (CDR; CDR1, CDR2, CDR3) в каждой вариабельной области, каркасной области, и последовательность вариабельной области определяли в соответствии со способом анализа с использованием системы нумерации Kabat (Kabat et al., 1991, Sequences of Proteins of Immunological Interest, National Institutes of Health Publication No. 91-3242, 5th ed., United States Department of Health and Human Services, Bethesda, MD).
Последовательность нуклеиновой кислоты вариабельной области тяжелой цепи полученного антитела мыши против PTPRS 9H5-4 представляла собой SEQ ID NO: 43, и аминокислотная последовательность представляла собой SEQ ID NO: 44. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области тяжелой цепи антитела мыши 9H5-4 представляли собой SEQ ID NO: 45, SEQ ID NO: 46 и SEQ ID NO: 47, соответственно.
Последовательность нуклеиновой кислоты вариабельной области легкой цепи полученного антитела мыши против PTPRS 9H5-4 представляет собой SEQ ID NO: 48, и аминокислотная последовательность представляет собой SEQ ID NO: 49. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области легкой цепи антитела мыши 9H5-4 представляют собой SEQ ID NO: 50, SEQ ID NO: 51 и SEQ ID NO: 52, соответственно.
Далее, последовательности нуклеиновых кислот вариабельной области тяжелой цепи и вариабельной области легкой цепи полученного антитела мыши против PTPRS 10F7-38 и антитела 14A8-85 были такими же, как и у антитела 9H5-4, включая последовательности CDR1, CDR2 и CDR3.
Последовательность нуклеиновой кислоты вариабельной области тяжелой цепи полученного антитела мыши против PTPRS 13G5-57 представляла собой SEQ ID NO: 53, и аминокислотная последовательность представляла собой SEQ ID NO: 54. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области тяжелой цепи антитела мыши 13G5-57 представляли собой SEQ ID NO: 55, SEQ ID NO: 56 и SEQ ID NO: 57, соответственно.
Последовательность нуклеиновой кислоты вариабельной области легкой цепи полученного антитела мыши против PTPRS 13G5-57 представляет собой SEQ ID NO: 58, и аминокислотная последовательность представляет собой SEQ ID NO: 59. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области легкой цепи антитела мыши 13G5-57 представляют собой SEQ ID NO: 60, SEQ ID NO: 61 и SEQ ID NO: 62, соответственно.
Далее, последовательности нуклеиновых кислот вариабельной области тяжелой цепи и вариабельной области легкой цепи полученного антитела мыши против 13G5-52 были такими же, как и у антитела 13G5-57, включая последовательности CDR1, CDR2 и CDR3.
Последовательность нуклеиновой кислоты вариабельной области тяжелой цепи полученного антитела мыши против PTPRS 22H8-84 представляла собой SEQ ID NO: 63, и аминокислотная последовательность представляла собой SEQ ID NO: 64. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области тяжелой цепи антитела мыши 22H8-84 представляли собой SEQ ID NO: 65, SEQ ID NO: 66 и SEQ ID NO: 67, соответственно.
Последовательность нуклеиновой кислоты вариабельной области легкой цепи полученного антитела мыши против PTPRS 22H8-84 представляет собой SEQ ID NO: 68, и аминокислотная последовательность представляет собой SEQ ID NO: 69. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области легкой цепи антитела 22H8-84 мыши представляют собой SEQ ID NO: 70, SEQ ID NO: 71 и SEQ ID NO: 72, соответственно.
Последовательность нуклеиновой кислоты вариабельной области тяжелой цепи полученного антитела мыши против PTPRS 49F2-30 представляла собой SEQ ID NO: 25, и аминокислотная последовательность представляла собой SEQ ID NO: 26. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области тяжелой цепи антитела мыши 49F2-30 представляют собой SEQ ID NO: 27, SEQ ID NO: 28 и SEQ ID NO: 29, соответственно.
Последовательность нуклеиновой кислоты вариабельной области легкой цепи полученного антитела мыши против PTPRS 49F2-30 представляет собой SEQ ID NO: 30, и аминокислотная последовательность представляет собой SEQ ID NO: 31. Аминокислотные последовательности CDR1, CDR2 и CDR3 в вариабельной области легкой цепи антитела мыши 49F2-30 представляют собой SEQ ID NO: 32, SEQ ID NO: 33 и SEQ ID NO: 34, соответственно.
Последовательность нуклеиновой кислоты (471 п.н.) вариабельной области тяжелой цепи полученного антитела мыши против PTPRS 9H5-4 представлена ниже (SEQ ID NO: 43). Заглавными буквами показана вариабельная область VH 9H5-4 мыши и строчными буквами показана константная область тяжелой цепи IgG1 мыши.
ATGGAGTTGGGACTGAGCTGGGTATTTCTTGTGGCTCTTTTGAATGGTGTCCAGTGTCAGGTGCAGCTTGTAGAGACCGGGGGAGGCTTGGTGAGGCCTGGAAATTCTCTGAAACTCTCCTGTGTTACCTCGGGATTCACTTTCAGTAACTACCGGATGCACTGGCTTCGCCAGCCTCCAGGGAAGAGGCTGGAGTGGATTGCTGTAATTACAGTCAAATCTGATAATTATGGAGCAAATTATGCAGAGTCTGTGAAAGGCAGATTCACTATTTCAAGAGATGATTCAAAAAGCAGTGTCTACCTGCAGATGAACAGATTAAGAGAGGAAGACACTGCCACTTATTATTGTAGTAGATCGGTCTACTATGGTTACGTCCTAGCCTTTGACTACTGGGGCCAAGGCACCACTCTCACAGTCTCCTCAgccaaaacgacacccccatctgtctatccactggcccctaagggc
Аминокислотная последовательность (157 а.к.) вариабельной области тяжелой цепи антитела мыши 9H5-4 представлена ниже (SEQ ID NO: 44). Заглавными буквами показана последовательность вариабельной области VH и строчными буквами показана константная область тяжелой цепи IgG1 мыши. Подчеркнутая часть означает сигнальную последовательность и часть с двойным подчеркиванием означает область CDR (CDR1, CDR2, CDR3).
MELGLSWVFLVALLNGVQCQVQLVETGGGLVRPGNSLKLSCVTSGFTFSNYRMH
WLRQPPGKRLEWIAVITVKSDNYGANYAESVKGRFTISRDDSKSSVYLQMNRLR
EEDTATYYCSRSVYYGYVLAFDYWGQGTTLTVSSakttppsvyplapkg
CDR1 вариабельной области тяжелой цепи антитела 9H5-4 представляет собой NYRMH (SEQ ID NO: 45), CDR2 вариабельной области тяжелой цепи антитела 9H5-4 представляет собой VITVKSDNYGANYAESVKG (SEQ ID NO: 46) и CDR3 вариабельной области тяжелой цепи антитела 9H5-4 представляет собой SVYYGYVLAFDY (SEQ ID NO: 47).
Последовательность нуклеиновой кислоты (402 п.н.) вариабельной области легкой цепи полученного антитела мыши против PTPRS 9H5-4 представлена ниже (SEQ ID NO: 48). Заглавными буквами показана вариабельная область VH 9H5-4 мыши и строчными буквами показана константная область легкой цепи κ Ig.
ATGATGTCCTCTGCTCAGTTCCTTGGTCTCCTGTTGCTCTGTTTTCAAGGTACCAGATGTGATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCAAGTCAGGACATTAGCAATTATTTAAACTGGTATCAGCAGAAACCAGATGGAACTGTTAAACTCCTGATCTACTACACATCAAGATTACACTCAGGAGTCCCATCAAGGTTCAGTGGCAGTGGGTCTGGAACAGATTATTCTCTCACCATTAGCAACCTGGAGCAAGAAGATATTGCCACTTACTTTTGCCAACAGGGTAATACGCTTCCGTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAAcgggctgatgctgcaccaact
Аминокислотная последовательность (134 а.к.) вариабельной области легкой цепи антитела мыши 9H5-4 представлена ниже (SEQ ID NO: 49). Заглавными буквами показана последовательность вариабельной области VH 9H5-4 мыши и строчными буквами показана константная область легкой цепи κ Ig. Подчеркнутая часть означает сигнальную последовательность и часть с двойным подчеркиванием означает область CDR (CDR1, CDR2, CDR3).
MMSSAQFLGLLLLCFQGTRCDIQMTQTTSSLSASLGDRVTISCRASQDISNYLNW
YQQKPDGTVKLLIYYTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFC
QQGNTLPWTFGGGTKLEIKradaapt
CDR1 вариабельной области легкой цепи антитела 9H5-4 представляет собой RASQDISNYLN (SEQ ID NO: 50), CDR2 вариабельной области легкой цепи антитела 9H5-4 представляет собой YTSRLHS (SEQ ID NO: 51), и CDR3 вариабельной области легкой цепи антитела 9H5-4 представляет собой QQGNTLP (SEQ ID NO: 52).
Последовательность нуклеиновой кислоты последовательность (465 п.н.) вариабельной области тяжелой цепи антитела мыши против PTPRS 13G5-57 представлена ниже (SEQ ID NO: 53). Заглавными буквами показана вариабельная область VH 13G5-57 мыши и строчными буквами показана константная область тяжелой цепи IgG2b мыши.
ATGAACTTGGGGCTCAGCTTGATTTTCCTTGTCCTTGTTTTAAAAGGTGTCCAGTGTGAAGTGAAGCTGGTGGAGTCTGGGGGAGGCTTAGTGCAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAACCTCTGGATTCACTTTCAGTGACTATTACATGTATTGGGTTCGCCAGACTCCAGAGAAGAGGCTGGAGTGGGTCGCATACATTAGTAATGGTGGTGGTAGCACCTATTATCCAGACACTGTAAAGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAACACCCTGTACCTGCAAATGAGCCGTCTGAAGTCTGAGGACACAGCCATGTATTACTGTGCAAGACATGTTTACTACGGGAGGAACTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCAgccaaaacaacacccccatcagtctatccactggcccctaagggc
Аминокислотная последовательность (155 а.к.) вариабельной области тяжелой цепи антитела мыши 13G5-57 представлена ниже (SEQ ID NO: 54). Заглавными буквами показана последовательность вариабельной области VH и строчными буквами показана константная область тяжелой цепи IgG2b мыши. Подчеркнутая часть означает сигнальную последовательность и часть с двойным подчеркиванием означает область CDR (CDR1, CDR2, CDR3).
MNLGLSLIFLVLVLKGVQCEVKLVESGGGLVQPGGSLKLSCATSGFTFSDYYMY
WVRQTPEKRLEWVAYISNGGGSTYYPDTVKGRFTISRДНКKNTLYLQMSRLKSE
DTAMYYCARHVYYGRNYAMDYWGQGTSVTVSSakttppsvyplapkg
CDR1 вариабельной области тяжелой цепи антитела 13G5-57 представляет собой DYYMY (SEQ ID NO: 55), CDR2 вариабельной области тяжелой цепи антитела 13G5-57 представляет собой YISNGGGSTYYPDTVKG (SEQ ID NO: 56), и CDR3 вариабельной области тяжелой цепи антитела 13G5-57 представляет собой HVYYGRNYAMDY (SEQ ID NO: 57).
Последовательность нуклеиновой кислоты (465 п.н.) вариабельной области легкой цепи полученного антитела мыши против PTPRS 13G5-57 представлена ниже (SEQ ID NO: 58). Заглавными буквами показана вариабельная область VH 13G5-57 мыши и строчными буквами показана константная область легкой цепи κ Ig мыши.
ATGAACTTGGGGCTCAGCTTGATTTTCCTTGTCCTTGTTTTAAAAGGTGTCCAGTGTGAAGTGAAGCTGGTGGAGTCTGGGGGAGGCTTAGTGCAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAACCTCTGGATTCACTTTCAGTGACTATTACATGTATTGGGTTCGCCAGACTCCAGAGAAGAGGCTGGAGTGGGTCGCATACATTAGTAATGGTGGTGGTAGCACCTATTATCCAGACACTGTAAAGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAACACCCTGTACCTGCAAATGAGCCGTCTGAAGTCTGAGGACACAGCCATGTATTACTGTGCAAGACATGTTTACTACGGGAGGAACTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCAgccaaaacaacacccccatcagtctatccactggcccctaagggc
Аминокислотная последовательность (155 а.к.) вариабельной области легкой цепи антитела мыши 13G5-57 представлена ниже (SEQ ID NO: 59). Заглавными буквами показана последовательность вариабельной области VH 13G5-57 мыши и строчными буквами показана константная область константная область легкой цепи κ Ig мыши. Подчеркнутая часть означает сигнальную последовательность и часть с двойным подчеркиванием означает область CDR (CDR1, CDR2, CDR3).
MMSSAQFLGLLLLCFQGTRCDIQMTQTTSSLSASLGDRVTISCRASQDISNYLNW
YQQKPDGTVKLLIYYTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFC
QQGNTLPYTFGGGTKLEIKradaaptvsifppsseqltsggasvvcf
CDR1 вариабельной области легкой цепи антитела 13G5-57 представляет собой RASQDISNYLN (SEQ ID NO: 60), CDR2 вариабельной области легкой цепи антитела 13G5-57 представляет собой YTSRLHS (SEQ ID NO: 61), и CDR3 вариабельной области легкой цепи антитела 13G5-57 представляет собой QQGNTLPY (SEQ ID NO: 62).
Последовательность нуклеиновой кислоты (458 п.н.) вариабельной области тяжелой цепи антитела мыши против PTPRS 22H8-84 представлена ниже (SEQ ID NO: 63). Заглавными буквами показана вариабельная область VH 22H8-84 мыши и строчными буквами показана константная область тяжелой цепи IgG1 мыши.
ATGGAATGTAACTGGATACTTCCTTTTATTCTGTCAGTAACTTCAGGTGTCTACTCACAGGTTCAGCTCCAGCAGTCTGGGGCTGAGCTGGCAAGACCTGGGGCTTCAGTGAAGTTGTCCTGCAAGGCTTCTGGCTACACCTTTACTAGCTACTGGATGCAGTGGGTAAAACAGAGGCCTGGACAGGGTCTGGAATGGATTGGGGCTATTTATCCTGGAGATGGTGATACTAGGTACACTCAGAAGTTCAAGGGCAAGGCCACATTGACTGCAGATAAATCCTCCAGCACAGCCTACATGCAACTCAGCAGCTTGGCATCTGAGGACTCTGCGGTCTATTACTGTGCAAGAAGGATTTACTACGGCTATTACTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCagccaaaacgacacccccatctgtctatccactggcccc
Аминокислотная последовательность (152 а.к.) вариабельной области тяжелой цепи антитела мыши 22H8-84 представлена ниже (SEQ ID NO: 64). Заглавными буквами показана последовательность вариабельной области VH и строчными буквами показана константная область тяжелой цепи IgG1 мыши. Подчеркнутая часть означает сигнальную последовательность и часть с двойным подчеркиванием означает область CDR (CDR1, CDR2, CDR3).
MECNWILPFILSVTSGVYSQVQLQQSGAELARPGASVKLSCKASGYTFTSYWMQ
WVKQRPGQGLEWIGAIYPGDGDTRYTQKFKGKATLTADKSSSTAYMQLSSLASE
DSAVYYCARRIYYGYYYAMDYWGQGTSVTVSSakttppsvypla
CDR1 вариабельной области тяжелой цепи антитела 22H8-84 представляет собой SYWMQ (SEQ ID NO: 65), CDR2 вариабельной области тяжелой цепи антитела 22H8-84 представляет собой AIYPGDGDTRYTQKFKG (SEQ ID NO: 66), и CDR3 вариабельной области тяжелой цепи антитела 22H8-84 представляет собой RIYYGYYYAMDY (SEQ ID NO: 67).
Последовательность нуклеиновой кислоты (430 п.н.) вариабельной области легкой цепи полученного антитела мыши против PTPRS 22H8-84 представлена ниже (SEQ ID NO: 68). Заглавными буквами показана вариабельная область VH 22H8-84 мыши и строчными буквами показана константная область легкой цепи κ Ig мыши.
ATGGAGACAGACACAATCCTGCTATGGGTGCTGCTGCTCTGGGTTCCAGGCTCCACTGGTGACATTGTGCTGACCCAATCTCCAGCTTCTTTGGCTGTGTCTCTAGGGCAGAGGGCCACCATCTCCTGCAAGGCCAGCCAAAGTGTTGATTATGATGGTGATAGTTATATGAACTGGTACCAACAGAAACCAGGACAGCCACCCAAACTCCTCATCTATGCTGCATCCAATCTAGAATCTGGGATCCCAGCCAGGTTTAGTGGCAGTGGGTCTGGGACAGACTTCACCCTCAACATCCATCCTGTGGAGGAGGAGGATGCTGCAACCTATTACTGTCAGCAAAGTAATGAGGATCCTCTCACGTTCGGTGCTGGGACCAAGCTGGAGCTGAAAcgggctgatgctgcaccaactgtatccatcaagggcg
Аминокислотная последовательность (143 а.к.) вариабельной области легкой цепи антитела 22H8-84 мыши представлена ниже (SEQ ID NO: 69). Заглавными буквами показана последовательность вариабельной области VH 22H8-84 мыши и строчными буквами показана константная область легкой цепи κ Ig мыши. Подчеркнутая часть означает сигнальную последовательность и часть с двойным подчеркиванием означает область CDR (CDR1, CDR2, CDR3).
METDTILLWVLLLWVPGSTGDIVLTQSPASLAVSLGQRATISCKASQSVDYDGDS
YMNWYQQKPGQPPKLLIYAASNLESGIPARFSGSGSGTDFTLNIHPVEEEDAATY
YCQQSNEDPLTFGAGTKLELKradaaptvsikg
CDR1 вариабельной области легкой цепи антитела 22H8-84 представляет собой KASQSVDYDGDSYMN (SEQ ID NO: 70), CDR2 вариабельной области легкой цепи антитела 22H8-84 представляет собой AASNLES (SEQ ID NO: 71), и CDR3 вариабельной области легкой цепи антитела 22H8-84 представляет собой QQSNEDPL (SEQ ID NO: 72).
Последовательность нуклеиновой кислоты вариабельной области тяжелой цепи антитела мыши против PTPRS 49F2-30 (469 п.н.) представлена ниже (SEQ ID NO: 25). Заглавными буквами показана вариабельная область VH 49F2-30 мыши и строчными буквами показана константная область тяжелой цепи IgG1 мыши.
ATGAACTTCGGGCTCAGGTTGATTTTCCTTGCCCTCATTTTAAAAGGTGTCCAGTGTGAGGTGCAGCTGGTGGAGTCTGGGGGAGACTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCATTTTCAGTAGCTATGGCATGTCTTGGGTTCGCCAGACTCCAGACAAGAGGCTGGAGTGGGTCGCAACCATTAGTAGTGGTGGTAGTGACACCTATTATCCAGACAGTGTGAAGGGGCGATTCACCATCTCCAGAGACAATGCCAACAACACCCTGTACCTGCAAATGAGCAGTCTGAAGTCTGAGGACACAGCCATGTATTACTGTGCAAGACAGGTCTACTATGGTCTTTACTGGTATTTCGATGTCTGGGGCGCAGGGACCACGGTCACCGTCTCCTCAgccaaaacgacacccccatctgtctatccactggcccctaagggcgaat
Аминокислотная последовательность вариабельной области тяжелой цепи антитела 49F2-30 мыши (156 а.к.) представлена ниже (SEQ ID NO: 26). Заглавными буквами показана вариабельная область VH и строчными буквами показана константная область тяжелой цепи IgG1 мыши. Подчеркнутые последовательности указывают на сигнальные последовательности, и последовательности с двойным подчеркиванием указывают на области CDR (CDR1, CDR2, CDR3).
CDR1 вариабельной области тяжелой цепи антитела 49F2-30 представляет собой SYGMS (SEQ ID NO: 27), CDR2 вариабельной области тяжелой цепи антитела 49F2-30 представляет собой TISSGGSDTYYPDSVKG (SEQ ID NO: 28), и CDR3 вариабельной области тяжелой цепи антитела 49F2-30 представляет собой QVYYGLYWYFDV (SEQ ID NO: 29).
Последовательность нуклеиновой кислоты вариабельной области легкой цепи полученного антитела мыши против PTPRS 49F2-30 (413 п.н.) представлена ниже (SEQ ID NO: 30). Заглавными буквами показана вариабельная область VL 49F2-30 мыши и строчными буквами показана константная область легкой цепи κ Ig мыши.
ATGGAGTCACAGATTCAGGTCTTTGTATTCGTGTTTCTCTGGTTGTCTGGTGTTGACGGAGACATTGTGATGACCCAGTCTCACAAATTCATGTCCACATCAGTAGGAGACAGGGTCAGCATCATTTGTAAGGCCAGTCAGGATGTGAATACTGCTGTAGCCTGGTATCAACAGAAACCAGGACAATCTCCTAAATTACTGATTTACTCGGCATCCTACCGGTACACTGGAGTCCCTGATCGCTTCACTGGCAGTGGATCTGGGACGGATTTCACTTTCACCATCAGCAGTGTGCAGGCTGAAGACCTGGCAATTTATTACTGTCAGCAACATTATAGTACTCCGTACACGTTCGGAGGGGGGACCAAGCTGGAAATAAAAcgggctgatgctgcaccaactgtatccatcaa
Аминокислотная последовательность вариабельной области легкой цепи антитела мыши 49F2-30 (137 а.к.) представлена ниже (SEQ ID NO: 31). Заглавными буквами показана вариабельная область VL 49F2-30 мыши и строчными буквами показана константная область легкой цепи κ Ig мыши. Подчеркнутые последовательности указывают на сигнальные последовательности, и последовательности с двойным подчеркиванием указывают на области CDR (CDR1, CDR2, CDR3).

CDR1 вариабельной области легкой цепи антитела 49F2-30 представляет собой KASQDVNTAVA (SEQ ID NO: 32), CDR2 вариабельной области легкой цепи антитела 49F2-30 представляет собой SASYRYT (SEQ ID NO: 33), и CDR3 вариабельной области легкой цепи антитела 49F2-30 представляет собой QQHYSTP (SEQ ID NO: 34).
3. Получение экспрессирующего вектора для химеризованного антитела
3-1) Клонирование кДНК, которая кодирует константную область Ig человека
кДНК константной области тяжелой цепи IgG1 человека и константной области легкой цепи Ig κ человека клонировали из тотальной РНК PBMC человека, и каждую из них клонировали в вектор pCR4Blunt-TOPO и вводили в компетентные клетки E. coli с использованием коммерчески доступного набора “Zero Blunt TOPO PCR Cloning Kit” (Invitrogen, Catalog No.: 1325137) в соответствии с инструкциями, прилагаемыми к набору, с получением трансформанта E. coli. Из этого трансформанта получали упомянутую выше плазмиду и образец плазмидной ДНК отсылали в Operon Biotechnology Co., Ltd.(Tokyo) для анализа последовательности для подтверждения последовательности оснований кДНК в плазмиде.
3-2) Получение экспрессирующего вектора для химеризованного антитела 9H5-4 (10F7-38, 14A8-85)
Для получения кДНК, которая кодирует тяжелую цепь химеризованного антитела против PTPRS, вариабельную область тяжелой цепи антитела мыши 9H5-4, полученную согласно 2-2, и вектор pEE6.4 (Lonza Biologics, Slough, Великобритания), в который была встроена константная область тяжелой цепи IgG человека подвергали слиянию, и вариабельную область тяжелой цепи антитела мыши 9H5-4 амплифицировали способом ПЦР и получали продукт ПЦР, который имел длину приблизительно 450 оснований. В этом случае праймеры были такими, как показано ниже. Полученный продукт ПЦР очищали способом с 1,5% агарозой с низкой температурой плавления.
Праймер для экспрессии тяжелой цепи в химерном антителе 9H5-4
1) прямой праймер: chi10F7VH-IF(Hind3)
Последовательность: 5' ttt AAG CTT gcc gcc acc ATG GAG TTG GGA CTG AGC TGG 3' (39-мер) (SEQ ID NO: 73)
2) обратный праймер: chi10F7VH-462R(ApaI)
Последовательность: 5' cga tgg gcc ctt ggt gct agc TGA GGA GAC TGT GAG AGT GGT 3' (42-мер) (SEQ ID NO: 74)
“Продукт ПЦР, который кодировал вариабельную область тяжелой цепи 9H5-4” получали из вариабельной области тяжелой цепи антитела мыши 9H5-4, полученной согласно 2-2, способом ПЦР. Продукт ПЦР, который кодировал вариабельную область тяжелой цепи 9H5-4, расщепляли ферментом рестрикции Hind III и an Apa I и очищали способом с 1,5% агарозным гелем. Его растворяли ddH2O с получением раствора фрагмента кДНК, который кодировал вариабельную область тяжелой цепи.
Кодирующую область VH 9H5-4 в полученной кДНК амплифицировали способом ПЦР из клона плазмиды pCR4Blunt-TOPO, включающего кодирующую область VH 9H5-4, с использованием праймеров chi10F7VH-IF (Hind3) и chi10F7VH-462R (ApaI), в которые были внесены предпочтительные участки рестрикции для клонирования в вектор pEE6.4 (Lonza Biologics, Slough, Великобритания) (HindIII и ApaI) и идеальная последовательность Kozak (GCCGCCACC) с использованием HindIII и ApaI в качестве участков клонирования. Вектор chi9H5-4VH-pEE6.4 включает константную область тяжелой цепи IgG1 человека. Фрагмент ПЦР VH встраивали в вектор pEE6.4 в рамке считывания с использованием HindIII и ApaI. Конструкцию исследовали анализом последовательности оснований кДНК и образец плазмидной ДНК отсылали в Operon Biotechnology Co., Ltd. (Tokyo) для анализа последовательности для подтверждения последовательности оснований кДНК в плазмиде.
Для получения кДНК, которая кодирует легкую цепь химеризованного антитела 9H5-4, амплифицировали продукт ПЦР длиной приблизительно 730 оснований способом на основе перекрывающейся ПЦР с удлинением из фрагмента ПЦР, в котором были слиты вариабельная область легкой цепи антитела мыши 9H5-4, полученная согласно 2-3, и константная область легкой цепи κ Ig человека, полученная согласно 3-2.
Продукт ПЦР, который кодировал вариабельную область легкой цепи 9H5-4, расщепляли ферментом рестрикции HindIII и EcoRI, и очищали способом с 1,5% агарозным гелем. Его растворяли в ddH2O с получением раствора фрагмента кДНК, который кодирует вариабельную область легкой цепи.
Полученную кДНК, кодирующую VL 9H5-4, амплифицировали способом ПЦР из плазмидного клона pCR4Blunt-TOPO, включающего VL-область 9H5-4 с использованием праймеров chi11G9VL-IF (Hind) и chi11G9VL-726R (RI), в которые были встроены предпочтительные участки рестрикции (HindIII и EcoRI) для клонирования в вектор pEE14.4 (Lonza Biologics) и идеальная последовательность Kozak. Вектор Chi9H5-4VL-pEE14.4 включает константную область легкой цепи каппа. ПЦР-фрагмент VL встраивали в вектор pEE14.4 в рамке считывания с использованием HindIII и EcoRI. Конструкцию исследовали с помощью анализа последовательности оснований кДНК.
Праймер для экспрессии легкой цепи в химерном антителе 9H5-4
1) Прямой праймер: chi11G9VL-IF(Hind)
Последовательность: 5' acc AAG CTT gcc gcc acc ATG ATG TCC TCT GCT CAG TTC 3' (39-мер) (SEQ ID NO: 75)
2) обратный праймер: chi11G9VL-408R
Последовательность: 5’ agc cac agt tcg TTT GAT TTC CAG CTT GGT GCC 3' (33-мер) (SEQ ID NO: 76)
3) Прямой праймер: chi11G9VL-385F
Последовательность: 5’ CTG GAA ATC AAA cga act gtg gct gca cca tct 3' (33-мер) (SEQ ID NO: 77)
4) обратный праймер: chi11G9VL-726R(RI)
Последовательность: 5' aaa GAA TTC cta gca ctc tcc cct gtt gaa 3' (30-мер) (SEQ ID NO: 78)
3-2) Получение экспрессирующего вектора для химеризованного антитела 13G5-57(13G5-52)
Для получения кДНК, которая кодирует тяжелую цепь химеризованного антитела PTPRS, проводили слияние вариабельной области тяжелой цепи антитела мыши 13G5-57, полученного согласно 2-2, и вектора pEE6.4 (Lonza Biologics, Slough, UK), в который была встроена константная область тяжелой цепи IgG человека. В способе, сходном со способом для антитела 9H5-4, получали продукт ПЦР и очищали его. В этом случае праймеры были следующими.
Праймер для экспрессии тяжелой цепи в химерном антителе 13G5-57
1) прямой праймер: chi13G5.57VH-IF(Hind3)
Последовательность: 5' ttt AAG CTT gcc gcc acc ATG AAC TTG GGG CTC AGC TTG 3' (39-мер) (SEQ ID NO: 79)
2) обратный праймер: chi13G5.57VH-456R(ApaI)
Последовательность: 5' cga tgg gcc ctt ggt gct agc TGA GGA GAC GGT GAC TGA GGT 3' (42-мер) (SEQ ID NO: 80)
Для получения кДНК, которая кодирует легкую цепь химеризованного антитела 13G5-57 амплифицировали продукт ПЦР длиной приблизительно 730 оснований способом на основе перекрывающейся ПЦР с удлинением из фрагмента ПЦР, в котором были слиты вариабельная область легкой цепи антитела мыши 13G5-57, полученная согласно 2-3, и константная область легкой цепи κ Ig человека, полученная согласно 3-2.
Праймер для экспрессии легкой цепи в химерном антителе 13G5-57
1) Прямой праймер: chi11G9VL-IF(Hind)
Последовательность: 5' acc AAG CTT gcc gcc acc ATG ATG TCC TCT GCT CAG TTC 3' (39-мер) (SEQ ID NO: 81)
2) обратный праймер: chi11G9VL-408R
Последовательность: 5’ agc cac agt tcg TTT GAT TTC CAG CTT GGT GCC 3' (33-мер) (SEQ ID NO: 82)
3) Прямой праймер: chi11G9VL-385F
Последовательность: 5’ CTG GAA ATC AAA cga act gtg gct gca cca tct 3' (33-мер) (SEQ ID NO: 83)
4) обратный праймер: chi11G9VL-726R(RI)
Последовательность: 5' aaa GAA TTC cta gca ctc tcc cct gtt gaa 3' (30-мер) (SEQ ID NO: 84)
Аналогично получению экспрессирующих векторов химеризованного антитела 9H5-4, получали экспрессирующие векторы для такой тяжелой цепи и легкой цепи химеризованного антитела 13G5-57.
3-2) Получение экспрессирующего вектора для химеризованного антитела 22H8-84
Для получения кДНК, которая кодирует тяжелую цепь химеризованного антитела PTPRS, проводили слияние вариабельной области тяжелой цепи антитела мыши 22H8-84, полученную согласно 2-2, и вектор pEE6.4 (Lonza Biologics, Slough, Великобритания), в который была встроена константная область тяжелой цепи IgG человека. В способе, сходном со способом для антитела 9H5-4, получали продукт ПЦР и очищали его. В этом случае праймеры были следующими.
Праймер для экспрессии тяжелой цепи химерного антитела 22H8-84
1) прямой праймер: chi22H8VH-IF(Hind3)
Последовательность: 5' ttt AAG CTT gcc gcc acc ATG GAA TGT AAC TGG ATA CTT 3' (39-мер) (SEQ ID NO: 85)
2) обратный праймер: chi22H8VH-456R(ApaI)
Последовательность: 5' cga tgg gcc ctt ggt gct agc TGA GGA GAC GGT GAC TGA GGT 3' (42-мер) (SEQ ID NO: 86)
Для получения кДНК, которая кодирует легкую цепь химеризованного антитела 22H8-84, амплифицировали продукт ПЦР длиной приблизительно 730 оснований способом на основе перекрывающейся ПЦР с удлинением из фрагмента ПЦР, в котором были слиты вариабельная область легкой цепи антитела мыши 22H8-84, полученная согласно 2-3, и константная область легкой цепи κ Ig человека, полученная согласно 3-2.
Праймер, экспрессирующий легкую цепь химерного антитела 22H8-84
1) Прямой праймер: chi22H8VL-IF(Hind)
Последовательность: 5' acc AAG CTT gcc gcc acc ATG GAG ACA GAC ACA ATC CTG 3' (39-мер) (SEQ ID NO: 87)
2) обратный праймер: chi22H8VL-420R
Последовательность: 5’ agc cac agt tcg TTT CAG CTC CAG CTT GGT CCC 3' (33-мер) (SEQ ID NO: 88)
3) Прямой праймер: chi22H8VL-397F
Последовательность: 5’ CTG GAG CTG AAA cga act gtg gct gca cca tct 3' (33-мер) (SEQ ID NO: 89)
4) обратный праймер: chi49F2VL-726R(RI)
Последовательность: 5' aaa GAA TTC cta gca ctc tcc cct gtt gaa 3' (30-мер) (SEQ ID NO: 90)
Аналогично получению экспрессирующих векторов химеризованного антитела 9H5-4, получали экспрессирующие векторы для такой тяжелой цепи и легкой цепи химеризованного антитела 22H8-84.
3-2) Получение кДНК, которая кодирует тяжелую цепь химеризованного антитела PTPRS
Для получения кДНК, которая кодирует тяжелую цепь химеризованного антитела PTPRS, два фрагмента ПЦР изменяли по методике на основе способа перекрывающейся ПЦР с удлинением во фрагменте ПЦР, в котором были слиты вариабельная область тяжелой цепи антитела мыши 49F2-30, полученная согласно 2-2, и константная область тяжелой цепи IgG человека, полученная согласно 3-1, и амплифицировали продукт ПЦР длиной 1434 оснований, который позволяет частичное образование двунитевой молекулы в результате гибридного взаимодействия. В этом случае праймеры (SEQ ID NO: 17-24) представляли собой праймеры, представленные в таблице 1. Полученный продукт ПЦР очищали способом с 1,5% агарозой с низкой температурой плавления.


Существует область, в которой кДНК перекрывает вариабельную область тяжелой цепи антитела мыши 49F2-30, полученную согласно 2-2, и константную область тяжелой цепи IgG1 человека, полученную согласно 3-1. Таким образом, с использованием этой области получали “продукт ПЦР, который кодирует вариабельную область тяжелой цепи 49F2-30” амплифицировали способом перекрывающейся ПЦР с удлинением. Продукт ПЦР, который кодирует вариабельную область тяжелой цепи 49F2-30, расщепляли ферментом рестрикции Hind III и EcoR I и очищали способом с 1,5% агарозным гелем. Его растворяли с помощью ddH2O с получением раствора фрагмента кДНК, который кодирует вариабельную область тяжелой цепи.
Кодирующую область VH 49F2-30 в полученной кДНК амплифицировали способом ПЦР из клона плазмиды pCR4Blunt-TOPO, включающего кодирующую область VH 49F2-30, с использованием праймеров chi49F2VH-IF (Hind3) и chi49F2VH-1434R (RI), в которые встроены предпочтительные участки рестрикции для клонирования в вектор pEE6.4 (Lonza Biologics, Slough, Великобритания) (Hind III и EcoRI) и идеальная последовательность Kozak (GCCGCCACC), с использованием Hind III и EcoRI в качестве участков клонирования. Вектор chi49F2VH-pEE6.4 включает константную область тяжелой цепи IgG1 человека. ПЦР-фрагмент VH встраивали в вектор pEE6.4 в рамке считывания с использованием HindIII и EcoRI. Конструкцию исследовали с помощью анализа последовательности оснований кДНК.
3-3) Получение кДНК, которая кодирует легкую цепь химеризованного антитела против PTPRS
Для получения кДНК, которая кодирует легкую цепь химеризованного антитела против PTPRS, амплифицировали продукт ПЦР длиной 726 оснований способом на основе перекрывающейся ПЦР с удлинением из фрагмента ПЦР, в котором были слиты вариабельная область легкой цепи антитела мыши 49F2-30, полученная согласно 2-3, и константная область легкой цепи κ Ig человека, полученная согласно 3-2.
Продукт ПЦР, который кодирует вариабельную область легкой цепи 49F2-30, расщепляли ферментом рестрикции HindIII и EcoRI, и очищали способом с 1,5% агарозным гелем. Его растворяли в ddH2O с получением раствора фрагмента кДНК, который кодирует вариабельную область легкой цепи.
Полученную кДНК, кодирующую VL 49F2-30, амплифицировали способом ПЦР из клона плазмиды pCR4Blunt-TOPO, включающего область VL 49F2-30, с использованием праймеров chi49F2VL-IF (Hind) и chi 49F2VL-726R (RI), в которые были встроены предпочтительные участки рестрикции (HindIII и EcoRI) для клонирования в вектор pEE14.4 (Lonza Biologics) и идеальная последовательность Kozak. Вектор Chi49F2VL-pEE14.4 включает константную область легкой цепи каппа. ПЦР-фрагмент VL встраивали в вектор pEE14.4 в рамке считывания с использованием HindIII и EcoRI. Конструкцию исследовали с помощью анализа последовательности оснований кДНК.
3-4) Конструирование экспрессирующего вектора Lonza с двумя генами для химеризованного антитела против PTPRS
Экспрессирующий вектор для химеризованного антитела против PTPRS (с двумя генами) Lonza, в котором были скомбинированы экспрессирующий вектор тяжелой цепи химеризованного антитела против PTPRS и экспрессирующий вектор легкой цепи химеризованного антитела PTPRS, в одном векторе с двумя геном, конструировали с помощью стандартной технологии клонирования.
4. Временная экспрессия в клетках HEK-293F
Использовали следующие ДНК векторов для временной экспрессии (80 мкг).
1) ДНК вектора chi9H5-4VH/VL DG Lonza
2) ДНК вектора chi13G5-57VH/VL DG Lonza
3) ДНК вектора chi22H8-84VH/VL DG Lonza
4) ДНК вектора chi49F2-30VH/VL DG Lonza
В день, предшествующий трансфекции, клетки 293F доводили до 80 мл с количеством 8×105 клеток/мл в 250-мл колбе Эрленмейера (Corning#431144), и культивировали при качании в условиях 37°C и концентрации CO2 8% в течение 7 суток.
После культивирования в течение 7 суток культуральную жидкость клеток 293F, которые подвергли трансфекции, собирали в 50-мл пробирку и центрифугировали в условиях 2070 g и 4°C в течение 5 минут. Супернатант фильтровали с помощью шприцевого фильтра (каталожный номер № 431220; CORNING), имеющего размер пор 0,45 мкм, и культуральные супернатанты объединяли.
5. Очистка химеризованного антитела против PTPRS
Химеризованное антитело 9H5-4.13G5-57.22H8-84 и 49F2-30 очищали аффинной хроматографией с белком A. Каждую неочищенную жидкость с антителом, полученную согласно 4, очищали на аффинной колонке с белком A (rProtein A Sepharose Fast Flow (каталожный №17-1279-01; партия 311272; GE Healthcare). Условия колонки являются такими, как представлено ниже. Аффинную очистку проводили с использованием буфера для связывания (20 мМ фосфат натрия, 0,15 M NaCl, pH 7,4) и буфера для элюирования (0,1 M глицин-HCl, pH 2,7). pH элюированной фракции доводили приблизительно до 7,2 добавлением буфера для нейтрализации (1 M Tris-HCl pH 9,5). Для замены буфера с очищенным антителом на PBS, буфер заменяли с использованием установки для диализа Slide-A-Lyzer MINI Dialysis unit 10kMWCO.
Концентрацию очищенного антитела вычисляли путем измерения поглощения при 280 нм и определения 1 мг/л как 1,38 OD.
Очищенное химеризованное антитело против PTPRS (ch9H5-4Ab, ch13G5-57Ab, ch22H8-84Ab и ch49F2-30Ab) анализировали способом SDS-PAGE и способом проточной цитометрии.
Последовательности нуклеиновых кислот и аминокислотные последовательности тяжелой цепи и легкой цепи полученного химерного антитела 9H5-4 представлены, соответственно, с помощью следующих номеров последовательностей.
(последовательность нуклеиновой кислоты)
(последовательность нуклеиновой кислоты)
(аминокислотная последовательность)
(аминокислотная последовательность)
Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 9H5-4 (1419 п.н.) представлена ниже (SEQ ID NO: 91). Заглавными буквами показана вариабельная область VH химерного 9H5-4, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
ATGGAGTTGGGACTGAGCTGGGTATTTCTTGTGGCTCTTTTGAATGGTGTCCAGTGTCAGGTGCAGCTTGTAGAGACCGGGGGAGGCTTGGTGAGGCCTGGAAATTCTCTGAAACTCTCCTGTGTTACCTCGGGATTCACTTTCAGTAACTACCGGATGCACTGGCTTCGCCAGCCTCCAGGGAAGAGGCTGGAGTGGATTGCTGTAATTACAGTCAAATCTGATAATTATGGAGCAAATTATGCAGAGTCTGTGAAAGGCAGATTCACTATTTCAAGAGATGATTCAAAAAGCAGTGTCTACCTGCAGATGAACAGATTAAGAGAGGAAGACACTGCCACTTATTATTGTAGTAGATCGGTCTACTATGGTTACGTCCTAGCCTTTGACTACTGGGGCCAAGGCACCACTCTCACAGTCTCCTCAgctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga
Аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 9H5-4 (472 а.к.) представлена ниже (SEQ ID NO: 92). Заглавными буквами показана вариабельная область VH химерного 9H5-4, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
MELGLSWVFLVALLNGVQCQVQLVETGGGLVRPGNSLKLSCVTSGFTFSNYRMHWLRQPPGKRLEWIAVITVKSDNYGANYAESVKGRFTISRDDSKSSVYLQMNRLREEDTATYYCSRSVYYGYVLAFDYWGQGTTLTVSSastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
Последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 9H5-4 (705 п.н.) представлена ниже (SEQ ID NO: 93). Заглавными буквами показана вариабельная область VL химерного 9H5-4, и строчными буквами показана константная область легкой цепи κ Ig человека.
ATGATGTCCTCTGCTCAGTTCCTTGGTCTCCTGTTGCTCTGTTTTCAAGGTACCAGATGTGATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCAAGTCAGGACATTAGCAATTATTTAAACTGGTATCAGCAGAAACCAGATGGAACTGTTAAACTCCTGATCTACTACACATCAAGATTACACTCAGGAGTCCCATCAAGGTTCAGTGGCAGTGGGTCTGGAACAGATTATTCTCTCACCATTAGCAACCTGGAGCAAGAAGATATTGCCACTTACTTTTGCCAACAGGGTAATACGCTTCCGTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAAcgaactgtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgctag
Аминокислотная последовательность легкой цепи химерного антитела против PTPRS 9H5-4 (234 а.к.) представлена ниже (SEQ ID NO: 94). Заглавными буквами показана вариабельная область VL химерного 9H5-4, и строчными буквами показана константная область легкой цепи κ Ig человека.
MMSSAQFLGLLLLCFQGTRCDIQMTQTTSSLSASLGDRVTISCRASQDISNYLNWYQQKPDGTVKLLIYYTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPWTFGGGTKLEIKrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
<13G5-57>
Последовательности нуклеиновых кислот и аминокислотные последовательности тяжелой цепи и легкой цепи полученного химерного антитела 13G5-57 представлены, соответственно, с помощью следующих номеров последовательностей.
(последовательность нуклеиновой кислоты)
(последовательность нуклеиновой кислоты)
(аминокислотная последовательность)
(аминокислотная последовательность)
Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 13G5-57 (1413 п.н.) представлена ниже (SEQ ID NO: 95). Заглавными буквами показана вариабельная область VH химерного 13G5-57, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
ATGAACTTGGGGCTCAGCTTGATTTTCCTTGTCCTTGTTTTAAAAGGTGTCCAGTGTGAAGTGAAGCTGGTGGAGTCTGGGGGAGGCTTAGTGCAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAACCTCTGGATTCACTTTCAGTGACTATTACATGTATTGGGTTCGCCAGACTCCAGAGAAGAGGCTGGAGTGGGTCGCATACATTAGTAATGGTGGTGGTAGCACCTATTATCCAGACACTGTAAAGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAACACCCTGTACCTGCAAATGAGCCGTCTGAAGTCTGAGGACACAGCCATGTATTACTGTGCAAGACATGTTTACTACGGGAGGAACTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCAgctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga
Аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 13G5-57 (470 а.к.) представлена ниже (SEQ ID NO: 96). Заглавными буквами показана вариабельная область VH 13G5-57, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
MNLGLSLIFLVLVLKGVQCEVKLVESGGGLVQPGGSLKLSCATSGFTFSDYYMYWVRQTPEKRLEWVAYISNGGGSTYYPDTVKGRFTISRДНКKNTLYLQMSRLKSEDTAMYYCARHVYYGRNYAMDYWGQGTSVTVSSastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
Последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 13G5-57 (705 п.н.) представлена ниже (SEQ ID NO: 97). Заглавными буквами показана вариабельная область VL 13G5-57, и строчными буквами показана константная область легкой цепи κ Ig человека.
ATGATGTCCTCTGCTCAGTTCCTTGGTCTCCTGTTGCTCTGTTTTCAAGGTACCAGATGTGATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCAAGTCAGGACATTAGCAATTATTTAAACTGGTATCAGCAGAAACCAGATGGAACTGTTAAACTCCTGATCTACTACACATCAAGATTACACTCAGGAGTCCCATCAAGGTTCAGTGGCAGTGGGTCTGGAACAGATTATTCTCTCACCATTAGCAACCTGGAGCAAGAAGATATTGCCACTTACTTTTGCCAACAGGGTAATACGCTTCCGTACACGTTCGGAGGGGGGACCAAGCTGGAAATAAAAcgaactgtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgctag
Аминокислотная последовательность легкой цепи химерного антитела против PTPRS 13G5-57 (234 а.к.) представлена ниже (SEQ ID NO: 98). Заглавными буквами показана вариабельная область VL химерного 13G5-57 вариабельная область, и строчными буквами показана константная область легкой цепи κ Ig человека.
MMSSAQFLGLLLLCFQGTRCDIQMTQTTSSLSASLGDRVTISCRASQDISNYLNWYQQKPDGTVKLLIYYTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEIKrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
Последовательности нуклеиновых кислот и аминокислотные последовательности тяжелой цепи и легкой цепи полученного химерного антитела 22H8-84 представлены, соответственно, с помощью следующих номеров последовательностей.
(последовательность нуклеиновой кислоты)
(последовательность нуклеиновой кислоты)
(аминокислотная последовательность)
(аминокислотная последовательность)
Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 22H8-84 (1413 п.н.) представлена ниже (SEQ ID NO: 99). Заглавными буквами показана вариабельная область VH химерного 22H8-84, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
ATGGAATGTAACTGGATACTTCCTTTTATTCTGTCAGTAACTTCAGGTGTCTACTCACAGGTTCAGCTCCAGCAGTCTGGGGCTGAGCTGGCAAGACCTGGGGCTTCAGTGAAGTTGTCCTGCAAGGCTTCTGGCTACACCTTTACTAGCTACTGGATGCAGTGGGTAAAACAGAGGCCTGGACAGGGTCTGGAATGGATTGGGGCTATTTATCCTGGAGATGGTGATACTAGGTACACTCAGAAGTTCAAGGGCAAGGCCACATTGACTGCAGATAAATCCTCCAGCACAGCCTACATGCAACTCAGCAGCTTGGCATCTGAGGACTCTGCGGTCTATTACTGTGCAAGAAGGATTTACTACGGCTATTACTATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCAgctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga
Аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 22H8-84 (470 а.к.) представлена ниже (SEQ ID NO: 100). Заглавными буквами показана вариабельная область VH химерного 22H8-84, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
MECNWILPFILSVTSGVYSQVQLQQSGAELARPGASVKLSCKASGYTFTSYWMQWVKQRPGQGLEWIGAIYPGDGDTRYTQKFKGKATLTADKSSSTAYMQLSSLASEDSAVYYCARRIYYGYYYAMDYWGQGTSVTVSSastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
Последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 22H8-84 (717 п.н.) представлена ниже (SEQ ID NO: 101). Заглавными буквами показана вариабельная область VL химерного 22H8-84, и строчными буквами показана константная область легкой цепи κ Ig человека.
ATGGAGACAGACACAATCCTGCTATGGGTGCTGCTGCTCTGGGTTCCAGGCTCCACTGGTGACATTGTGCTGACCCAATCTCCAGCTTCTTTGGCTGTGTCTCTAGGGCAGAGGGCCACCATCTCCTGCAAGGCCAGCCAAAGTGTTGATTATGATGGTGATAGTTATATGAACTGGTACCAACAGAAACCAGGACAGCCACCCAAACTCCTCATCTATGCTGCATCCAATCTAGAATCTGGGATCCCAGCCAGGTTTAGTGGCAGTGGGTCTGGGACAGACTTCACCCTCAACATCCATCCTGTGGAGGAGGAGGATGCTGCAACCTATTACTGTCAGCAAAGTAATGAGGATCCTCTCACGTTCGGTGCTGGGACCAAGCTGGAGCTGAAAcgaactgtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgctag
Аминокислотная последовательность легкой цепи химерного антитела против PTPRS 22H8-84 (238 а.к.) представлена ниже (SEQ ID NO: 102). Заглавными буквами показана вариабельная область VL химерного 22H8-84, и строчными буквами показана константная область легкой цепи κ Ig человека.
METDTILLWVLLLWVPGSTGDIVLTQSPASLAVSLGQRATISCKASQSVDYDGDSYMNWYQQKPGQPPKLLIYAASNLESGIPARFSGSGSGTDFTLNIHPVEEEDAATYYCQQSNEDPLTFGAGTKLELKrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
Последовательности нуклеиновых кислот и аминокислотные последовательности тяжелой цепи и легкой цепи полученного химерного антитела 49F2-30 представлены, соответственно, с помощью следующих номеров последовательностей.
(последовательность нуклеиновой кислоты)
(последовательность нуклеиновой кислоты)
(аминокислотная последовательность)
(аминокислотная последовательность)
Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 49F2-30 (1413 п.н.) представлена ниже (SEQ ID NO: 35). Заглавными буквами показана вариабельная область VH химерного 49F2-30, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
ATGAACTTCGGGCTCAGGTTGATTTTCCTTGCCCTCATTTTAAAAGGTGTCCAGTGTGAGGTGCAGCTGGTGGAGTCTGGGGGAGACTTAGTGAAGCCTGGAGGGTCCCTGAAACTCTCCTGTGCAGCCTCTGGATTCATTTTCAGTAGCTATGGCATGTCTTGGGTTCGCCAGACTCCAGACAAGAGGCTGGAGTGGGTCGCAACCATTAGTAGTGGTGGTAGTGACACCTATTATCCAGACAGTGTGAAGGGGCGATTCACCATCTCCAGAGACAATGCCAACAACACCCTGTACCTGCAAATGAGCAGTCTGAAGTCTGAGGACACAGCCATGTATTACTGTGCAAGACAGGTCTACTATGGTCTTTACTGGTATTTCGATGTCTGGGGCGCAGGGACCACGGTCACCGTCTCCTCAgctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga
Аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 49F2-30 (470 а.к.) представлена ниже (SEQ ID NO: 36). Заглавными буквами показана вариабельная область VH химерного 49F2-30, и строчными буквами показана константная область тяжелой цепи IgG1 человека.
MNFGLRLIFLALILKGVQCEVQLVESGGDLVKPGGSLKLSCAASGFIFSSYGMSWVRQTPDKRLEWVATISSGGSDTYYPDSVKGRFTISRДНКNNTLYLQMSSLKSEDTAMYYCARQVYYGLYWYFDVWGAGTTVTVSSastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
Последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 49F2-30 (705 п.н.) представлена ниже (SEQ ID NO: 37). Заглавными буквами показана вариабельная область VL химерного 49F2-30, и строчными буквами показана константная область легкой цепи κ Ig человека.
ATGGAGTCACAGATTCAGGTCTTTGTATTCGTGTTTCTCTGGTTGTCTGGTGTTGACGGAGACATTGTGATGACCCAGTCTCACAAATTCATGTCCACATCAGTAGGAGACAGGGTCAGCATCATTTGTAAGGCCAGTCAGGATGTGAATACTGCTGTAGCCTGGTATCAACAGAAACCAGGACAATCTCCTAAATTACTGATTTACTCGGCATCCTACCGGTACACTGGAGTCCCTGATCGCTTCACTGGCAGTGGATCTGGGACGGATTTCACTTTCACCATCAGCAGTGTGCAGGCTGAAGACCTGGCAATTTATTACTGTCAGCAACATTATAGTACTCCGTACACGTTCGGAGGGGGGACCAAGCTGGAAATAAAAcgaactgtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgctag
Аминокислотная последовательность легкой цепи химерного антитела против PTPRS 49F2-30 (234 а.к.) представлена ниже (SEQ ID NO: 38). Заглавными буквами показана вариабельная область VL химерного 49F2-30, и строчными буквами показана константная область легкой цепи κ Ig человека.
MESQIQVFVFVFLWLSGVDGDIVMTQSHKFMSTSVGDRVSIICKASQDVNTAVAWYQQKPGQSPKLLIYSASYRYTGVPDRFTGSGSGTDFTFTISSVQAEDLAIYYCQQHYSTPYTFGGGTKLEIKrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
Пример 6
Антителозависимая клеточная цитотоксичность полученного химерного антитела против PTPRS (ch49F2-30, ch9H5-4, ch13G5-57 и ch22H8-84)
Измеряли антителозависимую клеточную цитотоксичность (активность ADCC). Активность получали с использованием клеточной цитотоксичности, вычисленного уровня высвобожденной лактатдегидрогеназы (LDH) в качестве показателя. Мононуклеарные клетки периферической крови человека, предназначенные для того, чтобы быть эффекторными клетками, очищали гравитационным центрифугированием с использованием HISTOPAQUE-1077. В качестве клетки, предназначенной для того, чтобы быть мишенью, использовали клетку с вынужденной трансформацией гена hPTPRS с использованием CHO (линия клеток яичника китайского хомяка) (2×104/лунка). Эффекторные клетки и клетки-мишени смешивали так, чтобы их соотношение становилось равным 10:1, 20:1, 40:1 и 80:1, добавляли 10 мкг/мл полученного химерного антитела против PTPRS человека (ch49F2-30, ch9H5-4, ch13G5-57 и ch22H8-84) или контрольного антитела Synagis, и смесь культивировали в течение 4 часов при 37°C для оценки эффекта клеточной цитотоксичности антитела. В результате химерные антитела против hPTPRS h49F2-30, ch9H5-4, ch13G5-57 и ch22H8-84 осуществляли лизис клеток-мишеней hPTPRS/CHO зависимым от количества эффекторных клеток образом (фиг.17A и фиг.17B). Этот результат показал, что полученное химерное антитело против PTPRS селективно продемонстрировано цитотоксичность в отношении клеток, экспрессирующих PTPRS.
Исследовали эффект антитела против PTPRS на pDC. PBMC выделяли из периферической крови, смешивали с 10 мкг/мл химерного антитела против PTPRS человека и культивировали в течение 24 часов. После этого проводили стимуляцию в течение 24 часов с помощью CpG2216, который является лигандом Toll-подобного рецептора 9, экспрессируемого в pDC, для индукции продуцирования IFNα. Через 24 ч после стимуляции CpG исследовали продуцированное количество IFNα. В результате продукция IFNα полностью ингибировалась обработкой полученных химерным антителом против PTPRS человека (ch49F2-30, ch9H5-4, ch13G5-57 и ch22H8-84) (фиг.18A). Более того, когда клетки собирали через 6 часов после обработки ch49F2-30, ch9H5-4, ch13G5-57 и ch22H8-84 и pDC l подтверждали посредством двойного окрашивания антителом против BDCA2 и антителом против BDCA4, было открыто, что популяция pDC снижалась в большей степени, чем при обработке контрольным антителом Synagis (фиг.18B и фиг.18C).
Промышленная применимость
Настоящее изобретение относится к антителу, которое специфически распознает PTPRS человека - иммуноген, который пригоден для продуцирования антитела, и к способу получения антитела против PTPRS человека с использованием этого иммуногена.
Номера доступа
FERM ABP-11356
FERM ABP-11357
FERM ABP-11358
FERM ABP-11359
FERM ABP-11360
FERM ABP-11361
FERM ABP-11362
FERM ABP-11363
Свободный текст списка последовательностей
SEQ ID NO: 3: прямой праймер
SEQ ID NO: 4: обратный праймер
SEQ ID NO: 5: прямой праймер
SEQ ID NO: 6: обратный праймер
SEQ ID NO: 7: прямой праймер
SEQ ID NO: 8: обратный праймер
SEQ ID NO: 9: прямой праймер
SEQ ID NO: 10: обратный праймер
SEQ ID NO: 11: антисмысловой праймер
SEQ ID NO: 12: якорный праймер
SEQ ID NO: 12: n представляет собой дезоксиинозин.
SEQ ID NO: 13: антисмысловой праймер
SEQ ID NO: 14: праймер AUAP
SEQ ID NO: 15: антисмысловой праймер
SEQ ID NO: 16: антисмысловой праймер
SEQ ID NO: 17: праймер
SEQ ID NO: 18: праймер
SEQ ID NO: 19: праймер
SEQ ID NO: 20: праймер
SEQ ID NO: 21: праймер
SEQ ID NO: 22: праймер
SEQ ID NO: 23: праймер
SEQ ID NO: 24: праймер
SEQ ID NO: 35: последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 49F2-30
SEQ ID NO: 36: аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 49F2-30
SEQ ID NO: 37: последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 49F2-30
SEQ ID NO: 38: аминокислотная последовательность легкой цепи химерного антитела против PTPRS 49F2-30
SEQ ID NO: 39: праймер
SEQ ID NO: 40: праймер
SEQ ID NO: 41: праймер
SEQ ID NO: 42: праймер
SEQ ID NO: 73: прямой праймер
SEQ ID NO: 74: обратный праймер
SEQ ID NO: 75: прямой праймер
SEQ ID NO: 76: обратный праймер
SEQ ID NO: 77: прямой праймер
SEQ ID NO: 78: обратный праймер
SEQ ID NO: 79: прямой праймер
SEQ ID NO: 80: обратный праймер
SEQ ID NO: 81: прямой праймер
SEQ ID NO: 82: обратный праймер
SEQ ID NO: 83: прямой праймер
SEQ ID NO: 84: обратный праймер
SEQ ID NO: 85: прямой праймер
SEQ ID NO: 86: обратный праймер
SEQ ID NO: 87: прямой праймер
SEQ ID NO: 88: обратный праймер
SEQ ID NO: 89: прямой праймер
SEQ ID NO: 90: обратный праймер
SEQ ID NO: 91: последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 9H5-4
SEQ ID NO: 92: аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 9H5-4
SEQ ID NO: 93: последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 9H5-4
SEQ ID NO: 94: аминокислотная последовательность легкой цепи химерного антитела против PTPRS 9H5-4
SEQ ID NO: 95: последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 13G5-57
SEQ ID NO: 96: аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 13G5-57
SEQ ID NO: 97: последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 13G5-57
SEQ ID NO: 98: аминокислотная последовательность легкой цепи химерного антитела против PTPRS 13G5-57
SEQ ID NO: 99: последовательность нуклеиновой кислоты тяжелой цепи химерного антитела против PTPRS 22H8-84
SEQ ID NO: 100: аминокислотная последовательность тяжелой цепи химерного антитела против PTPRS 22H8-84
SEQ ID NO: 101: последовательность нуклеиновой кислоты легкой цепи химерного антитела против PTPRS 22H8-84
SEQ ID NO: 102: аминокислотная последовательность легкой цепи химерного антитела против PTPRS 22H8-84.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SBI Biotech Co., Ltd.
<120> АНТИТЕЛО ПРОТИВ ПРОТЕИНТИРОЗИНФОСФАТАЗЫ σ РЕЦЕПТОРНОГО ТИПА ЧЕЛОВЕКА
<130> W6373-000000
<150> JP 2011-101752
<151> 2011-04-28
<160> 102
<170> PatentIn version 3.4
<210> 1
<211> 1501
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> ПЕПТИД
<222> (1)..(1501)
<400> 1
Met Ala Pro Thr Trp Gly Pro Gly Met Val Ser Val Val Gly Pro Met
1 5 10 15
Gly Leu Leu Val Val Leu Leu Val Gly Gly Cys Ala Ala Glu Glu Pro
20 25 30
Pro Arg Phe Ile Lys Glu Pro Lys Asp Gln Ile Gly Val Ser Gly Gly
35 40 45
Val Ala Ser Phe Val Cys Gln Ala Thr Gly Asp Pro Lys Pro Arg Val
50 55 60
Thr Trp Asn Lys Lys Gly Lys Lys Val Asn Ser Gln Arg Phe Glu Thr
65 70 75 80
Ile Glu Phe Asp Glu Ser Ala Gly Ala Val Leu Arg Ile Gln Pro Leu
85 90 95
Arg Thr Pro Arg Asp Glu Asn Val Tyr Glu Cys Val Ala Gln Asn Ser
100 105 110
Val Gly Glu Ile Thr Val His Ala Lys Leu Thr Val Leu Arg Glu Asp
115 120 125
Gln Leu Pro Ser Gly Phe Pro Asn Ile Asp Met Gly Pro Gln Leu Lys
130 135 140
Val Val Glu Arg Thr Arg Thr Ala Thr Met Leu Cys Ala Ala Ser Gly
145 150 155 160
Asn Pro Asp Pro Glu Ile Thr Trp Phe Lys Asp Phe Leu Pro Val Asp
165 170 175
Pro Ser Ala Ser Asn Gly Arg Ile Lys Gln Leu Arg Ser Gly Ala Leu
180 185 190
Gln Ile Glu Ser Ser Glu Glu Thr Asp Gln Gly Lys Tyr Glu Cys Val
195 200 205
Ala Thr Asn Ser Ala Gly Val Arg Tyr Ser Ser Pro Ala Asn Leu Tyr
210 215 220
Val Arg Val Arg Arg Val Ala Pro Arg Phe Ser Ile Leu Pro Met Ser
225 230 235 240
His Glu Ile Met Pro Gly Gly Asn Val Asn Ile Thr Cys Val Ala Val
245 250 255
Gly Ser Pro Met Pro Tyr Val Lys Trp Met Gln Gly Ala Glu Asp Leu
260 265 270
Thr Pro Glu Asp Asp Met Pro Val Gly Arg Asn Val Leu Glu Leu Thr
275 280 285
Asp Val Lys Asp Ser Ala Asn Tyr Thr Cys Val Ala Met Ser Ser Leu
290 295 300
Gly Val Ile Glu Ala Val Ala Gln Ile Thr Val Lys Ser Leu Pro Lys
305 310 315 320
Ala Pro Gly Thr Pro Met Val Thr Glu Asn Thr Ala Thr Ser Ile Thr
325 330 335
Ile Thr Trp Asp Ser Gly Asn Pro Asp Pro Val Ser Tyr Tyr Val Ile
340 345 350
Glu Tyr Lys Ser Lys Ser Gln Asp Gly Pro Tyr Gln Ile Lys Glu Asp
355 360 365
Ile Thr Thr Thr Arg Tyr Ser Ile Gly Gly Leu Ser Pro Asn Ser Glu
370 375 380
Tyr Glu Ile Trp Val Ser Ala Val Asn Ser Ile Gly Gln Gly Pro Pro
385 390 395 400
Ser Glu Ser Val Val Thr Arg Thr Gly Glu Gln Ala Pro Ala Ser Ala
405 410 415
Pro Arg Asn Val Gln Ala Arg Met Leu Ser Ala Thr Thr Met Ile Val
420 425 430
Gln Trp Glu Glu Pro Val Glu Pro Asn Gly Leu Ile Arg Gly Tyr Arg
435 440 445
Val Tyr Tyr Thr Met Glu Pro Glu His Pro Val Gly Asn Trp Gln Lys
450 455 460
His Asn Val Asp Asp Ser Leu Leu Thr Thr Val Gly Ser Leu Leu Glu
465 470 475 480
Asp Glu Thr Tyr Thr Val Arg Val Leu Ala Phe Thr Ser Val Gly Asp
485 490 495
Gly Pro Leu Ser Asp Pro Ile Gln Val Lys Thr Gln Gln Gly Val Pro
500 505 510
Gly Gln Pro Met Asn Leu Arg Ala Glu Ala Arg Ser Glu Thr Ser Ile
515 520 525
Thr Leu Ser Trp Ser Pro Pro Arg Gln Glu Ser Ile Ile Lys Tyr Glu
530 535 540
Leu Leu Phe Arg Glu Gly Asp His Gly Arg Glu Val Gly Arg Thr Phe
545 550 555 560
Asp Pro Thr Thr Ser Tyr Val Val Glu Asp Leu Lys Pro Asn Thr Glu
565 570 575
Tyr Ala Phe Arg Leu Ala Ala Arg Ser Pro Gln Gly Leu Gly Ala Phe
580 585 590
Thr Pro Val Val Arg Gln Arg Thr Leu Gln Ser Ile Ser Pro Lys Asn
595 600 605
Phe Lys Val Lys Met Ile Met Lys Thr Ser Val Leu Leu Ser Trp Glu
610 615 620
Phe Pro Asp Asn Tyr Asn Ser Pro Thr Pro Tyr Lys Ile Gln Tyr Asn
625 630 635 640
Gly Leu Thr Leu Asp Val Asp Gly Arg Thr Thr Lys Lys Leu Ile Thr
645 650 655
His Leu Lys Pro His Thr Phe Tyr Asn Phe Val Leu Thr Asn Arg Gly
660 665 670
Ser Ser Leu Gly Gly Leu Gln Gln Thr Val Thr Ala Trp Thr Ala Phe
675 680 685
Asn Leu Leu Asn Gly Lys Pro Ser Val Ala Pro Lys Pro Asp Ala Asp
690 695 700
Gly Phe Ile Met Val Tyr Leu Pro Asp Gly Gln Ser Pro Val Pro Val
705 710 715 720
Gln Ser Tyr Phe Ile Val Met Val Pro Leu Arg Lys Ser Arg Gly Gly
725 730 735
Gln Phe Leu Thr Pro Leu Gly Ser Pro Glu Asp Met Asp Leu Glu Glu
740 745 750
Leu Ile Gln Asp Ile Ser Arg Leu Gln Arg Arg Ser Leu Arg His Ser
755 760 765
Arg Gln Leu Glu Val Pro Arg Pro Tyr Ile Ala Ala Arg Phe Ser Val
770 775 780
Leu Pro Pro Thr Phe His Pro Gly Asp Gln Lys Gln Tyr Gly Gly Phe
785 790 795 800
Asp Asn Arg Gly Leu Glu Pro Gly His Arg Tyr Val Leu Phe Val Leu
805 810 815
Ala Val Leu Gln Lys Ser Glu Pro Thr Phe Ala Ala Ser Pro Phe Ser
820 825 830
Asp Pro Phe Gln Leu Asp Asn Pro Asp Pro Gln Pro Ile Val Asp Gly
835 840 845
Glu Glu Gly Leu Ile Trp Val Ile Gly Pro Val Leu Ala Val Val Phe
850 855 860
Ile Ile Cys Ile Val Ile Ala Ile Leu Leu Tyr Lys Asn Lys Pro Asp
865 870 875 880
Ser Lys Arg Lys Asp Ser Glu Pro Arg Thr Lys Cys Leu Leu Asn Asn
885 890 895
Ala Asp Leu Ala Pro His His Pro Lys Asp Pro Val Glu Met Arg Arg
900 905 910
Ile Asn Phe Gln Thr Pro Gly Met Leu Ser His Pro Pro Ile Pro Ile
915 920 925
Ala Asp Met Ala Glu His Thr Glu Arg Leu Lys Ala Asn Asp Ser Leu
930 935 940
Lys Leu Ser Gln Glu Tyr Glu Ser Ile Asp Pro Gly Gln Gln Phe Thr
945 950 955 960
Trp Glu His Ser Asn Leu Glu Val Asn Lys Pro Lys Asn Arg Tyr Ala
965 970 975
Asn Val Ile Ala Tyr Asp His Ser Arg Val Ile Leu Gln Pro Ile Glu
980 985 990
Gly Ile Met Gly Ser Asp Tyr Ile Asn Ala Asn Tyr Val Asp Gly Tyr
995 1000 1005
Arg Arg Gln Asn Ala Tyr Ile Ala Thr Gln Gly Pro Leu Pro Glu
1010 1015 1020
Thr Phe Gly Asp Phe Trp Arg Met Val Trp Glu Gln Arg Ser Ala
1025 1030 1035
Thr Ile Val Met Met Thr Arg Leu Glu Glu Lys Ser Arg Ile Lys
1040 1045 1050
Cys Asp Gln Tyr Trp Pro Asn Arg Gly Thr Glu Thr Tyr Gly Phe
1055 1060 1065
Ile Gln Val Thr Leu Leu Asp Thr Ile Glu Leu Ala Thr Phe Cys
1070 1075 1080
Val Arg Thr Phe Ser Leu His Lys Asn Gly Ser Ser Glu Lys Arg
1085 1090 1095
Glu Val Arg Gln Phe Gln Phe Thr Ala Trp Pro Asp His Gly Val
1100 1105 1110
Pro Glu Tyr Pro Thr Pro Phe Leu Ala Phe Leu Arg Arg Val Lys
1115 1120 1125
Thr Cys Asn Pro Pro Asp Ala Gly Pro Ile Val Val His Cys Ser
1130 1135 1140
Ala Gly Val Gly Arg Thr Gly Cys Phe Ile Val Ile Asp Ala Met
1145 1150 1155
Leu Glu Arg Ile Lys Pro Glu Lys Thr Val Asp Val Tyr Gly His
1160 1165 1170
Val Thr Leu Met Arg Ser Gln Arg Asn Tyr Met Val Gln Thr Glu
1175 1180 1185
Asp Gln Tyr Ser Phe Ile His Glu Ala Leu Leu Glu Ala Val Gly
1190 1195 1200
Cys Gly Asn Thr Glu Val Pro Ala Arg Ser Leu Tyr Ala Tyr Ile
1205 1210 1215
Gln Lys Leu Ala Gln Val Glu Pro Gly Glu His Val Thr Gly Met
1220 1225 1230
Glu Leu Glu Phe Lys Arg Leu Ala Asn Ser Lys Ala His Thr Ser
1235 1240 1245
Arg Phe Ile Ser Ala Asn Leu Pro Cys Asn Lys Phe Lys Asn Arg
1250 1255 1260
Leu Val Asn Ile Met Pro Tyr Glu Ser Thr Arg Val Cys Leu Gln
1265 1270 1275
Pro Ile Arg Gly Val Glu Gly Ser Asp Tyr Ile Asn Ala Ser Phe
1280 1285 1290
Ile Asp Gly Tyr Arg Gln Gln Lys Ala Tyr Ile Ala Thr Gln Gly
1295 1300 1305
Pro Leu Ala Glu Thr Thr Glu Asp Phe Trp Arg Met Leu Trp Glu
1310 1315 1320
Asn Asn Ser Thr Ile Val Val Met Leu Thr Lys Leu Arg Glu Met
1325 1330 1335
Gly Arg Glu Lys Cys His Gln Tyr Trp Pro Ala Glu Arg Ser Ala
1340 1345 1350
Arg Tyr Gln Tyr Phe Val Val Asp Pro Met Ala Glu Tyr Asn Met
1355 1360 1365
Pro Gln Tyr Ile Leu Arg Glu Phe Lys Val Thr Asp Ala Arg Asp
1370 1375 1380
Gly Gln Ser Arg Thr Val Arg Gln Phe Gln Phe Thr Asp Trp Pro
1385 1390 1395
Glu Gln Gly Val Pro Lys Ser Gly Glu Gly Phe Ile Asp Phe Ile
1400 1405 1410
Gly Gln Val His Lys Thr Lys Glu Gln Phe Gly Gln Asp Gly Pro
1415 1420 1425
Ile Ser Val His Cys Ser Ala Gly Val Gly Arg Thr Gly Val Phe
1430 1435 1440
Ile Thr Leu Ser Ile Val Leu Glu Arg Met Arg Tyr Glu Gly Val
1445 1450 1455
Val Asp Ile Phe Gln Thr Val Lys Met Leu Arg Thr Gln Arg Pro
1460 1465 1470
Ala Met Val Gln Thr Glu Asp Glu Tyr Gln Phe Cys Tyr Gln Ala
1475 1480 1485
Ala Leu Glu Tyr Leu Gly Ser Phe Asp His Tyr Ala Thr
1490 1495 1500
<210> 2
<211> 4665
<212> ДНК
<213> Homo sapiens
<220>
<221> gene
<222> (1)..(4665)
<300>
<301> Strausberg t al.
<302> Generation and initial analysis of more than 15,000 full-length
human and mouse cDNA sequences
<303> Proc. Natl. Acad. Sci. U.S.A.
<304> 99
<305> 26
<306> 16899-16903
<307> 2002
<400> 2
gggctcgagg cctctctgtg agggaccggg gggccatccc cctccagggc ggagatcgga 60
ggtcgctgcc aagcatggcg cccacctggg gccctggcat ggtgtctgtg gttggtccca 120
tgggcctcct tgtggtcctg ctcgttggag gctgtgcagc agaagagccc cccaggttta 180
tcaaagaacc caaggaccag atcggcgtgt cggggggtgt ggcctctttc gtgtgtcagg 240
ccacgggtga ccccaagcca cgagtgacct ggaacaagaa gggcaagaag gtcaactctc 300
agcgctttga gacgattgag tttgatgaga gtgcaggggc agtgctgagg atccagccgc 360
tgaggacacc gcgggatgaa aacgtgtacg agtgtgtggc ccagaactcg gttggggaga 420
tcacagtcca tgccaagctt actgtcctcc gagaggacca gctgccctct ggcttcccca 480
acatcgacat gggcccacag ttgaaggtgg tggagcggac acggacagcc accatgctct 540
gtgcagccag cggcaaccct gaccctgaga tcacctggtt caaggacttc ctgcctgtgg 600
atcctagtgc cagcaatgga cgcatcaaac agctgcgatc aggagccctg cagattgaaa 660
gcagtgagga aaccgaccag ggcaaatatg agtgtgtggc caccaacagc gccggcgtgc 720
gctactcctc acctgccaac ctctacgtgc gagtccgccg cgtggccccg cgcttctcca 780
tcctgcccat gagccacgag atcatgccag ggggcaacgt gaacatcacc tgcgtggccg 840
tgggctcgcc catgccatac gtgaagtgga tgcagggggc cgaggacctg acccccgagg 900
atgacatgcc cgtgggtcgg aacgtgctgg aactcacaga tgtcaaggac tcggccaact 960
acacctgcgt ggccatgtcc agcctgggcg tcattgaggc ggttgctcag atcacggtga 1020
aatctctccc caaagctccc gggactccca tggtgactga gaacacagcc accagcatca 1080
ccatcacgtg ggactcgggc aacccagatc ctgtgtccta ttacgtcatc gaatataaat 1140
ccaagagcca agacgggccg tatcagatta aagaggacat caccaccaca cgttacagca 1200
tcggcggcct gagccccaac tcggagtacg agatctgggt gtcggccgtc aactccatcg 1260
gccaggggcc ccccagcgag tccgtggtca cccgcacagg cgagcaggcc ccggccagcg 1320
cgccgcggaa cgtgcaagcc cggatgctca gcgcgaccac catgattgtg cagtgggagg 1380
agccggtgga gcccaacggc ctgatccgcg gctaccgcgt ctactacacc atggaaccgg 1440
agcaccccgt gggcaactgg cagaagcaca acgtggacga cagcctgctg accaccgtgg 1500
gcagcctgct ggaggacgag acctacaccg tgcgggtgct cgccttcacc tccgtcggcg 1560
acgggcccct ctcggacccc atccaggtca agacgcagca gggagtgccg ggccagccca 1620
tgaacctgcg ggccgaggcc aggtcggaga ccagcatcac gctgtcctgg agccccccgc 1680
ggcaggagag tatcatcaag tacgagctcc tcttccggga aggcgaccat ggccgggagg 1740
tgggaaggac cttcgacccg acgacttcct acgtggtgga ggacctgaag cccaacacgg 1800
agtacgcctt ccgcctggcg gcccgctcgc cgcagggcct gggcgccttc acccccgtgg 1860
tgcggcagcg cacgctgcag tccatctcgc ccaagaactt caaggtgaaa atgatcatga 1920
agacatcagt tctgctcagc tgggagttcc ctgacaacta caactcaccc acaccctaca 1980
agatccagta caatgggctc acactggatg tggatggccg taccaccaag aagctcatca 2040
cgcacctcaa gccccacacc ttctacaact ttgtgctgac caatcgcggc agcagcctgg 2100
gcggcctcca gcagacggtc accgcctgga ctgccttcaa cctgctcaac ggcaagccca 2160
gcgtcgcccc caagcctgat gctgacggct tcatcatggt gtatcttcct gacggccaga 2220
gccccgtgcc tgtccagagc tatttcattg tgatggtgcc actgcgcaag tctcgtggag 2280
gccaattcct gaccccgctg ggtagcccag aggacatgga tctggaagag ctcatccagg 2340
acatctcacg gctacagagg cgcagcctgc ggcactcgcg tcagctggag gtgccccggc 2400
cctatattgc agctcgcttc tctgtgctgc cacccacgtt ccatcccggc gaccagaagc 2460
agtatggcgg cttcgataac cggggcctgg agcccggcca ccgctatgtc ctcttcgtgc 2520
ttgccgtgct tcagaagagc gagcctacct ttgcagccag tcccttctca gaccccttcc 2580
agctggataa cccggacccc cagcccatcg tggatggcga ggaggggctt atctgggtga 2640
tcgggcctgt gctggccgtg gtcttcataa tctgcattgt cattgctatc ctgctctaca 2700
agaacaaacc cgacagtaaa cgcaaggact cagaaccccg caccaaatgc ctcctgaaca 2760
atgccgacct cgcccctcac caccccaagg accctgtgga aatgagacgc attaacttcc 2820
agactccagg catgcttagc cacccgccaa ttcccatcgc agacatggcg gagcacacgg 2880
agcggctcaa ggccaacgac agcctcaagc tctcccagga gtatgagtcc atcgaccctg 2940
gacagcagtt cacatgggaa cattccaacc tggaagtgaa caagccgaag aaccgctatg 3000
ccaacgtcat cgcctatgac cactcccgtg tcatcctcca gcccattgaa ggcatcatgg 3060
gcagtgatta catcaatgcc aactacgtgg acggctaccg gcgtcagaac gcgtacattg 3120
ccacgcaggg gccgctgcct gagacctttg gggacttctg gcgtatggtg tgggagcagc 3180
ggtcggcgac catcgtcatg atgacgcggc tggaggagaa gtcacggatc aagtgtgatc 3240
agtattggcc caacagaggc acggagacct acggcttcat ccaggtcacg ttgctagata 3300
ccatcgagct ggccacattc tgcgtcagga cattctctct gcacaagaat ggctccagtg 3360
agaaacgcga ggtccgccag ttccagttta cggcgtggcc ggaccatggc gtgcccgaat 3420
acccaacgcc cttcctggct ttcctgcgga gagtcaagac ctgcaacccg ccagatgccg 3480
gccccatcgt ggttcactgc agtgccggtg tgggccgcac aggctgcttt atcgtcatcg 3540
acgccatgct tgagcggatc aagccagaga agacagtcga tgtctatggc cacgtgacgc 3600
tcatgaggtc ccagcgcaac tacatggtgc agacggagga ccagtacagc ttcatccacg 3660
aggccctgct ggaggccgtg ggctgtggca acacagaagt gcccgcacgc agcctctatg 3720
cctacatcca gaagctggcc caggtggagc ctggcgaaca cgtcactggc atggaactcg 3780
agttcaagcg gctggctaac tccaaggccc acacgtcacg cttcatcagt gccaatctgc 3840
cttgtaacaa gttcaagaac cgcctggtga acatcatgcc ctatgagagc acacgggtct 3900
gtctgcaacc catccggggt gtggagggct ctgactacat caacgccagc ttcattgatg 3960
gctacaggca gcagaaggcc tacatcgcga cacaggggcc gctggcggag accacggaag 4020
acttctggcg catgctgtgg gagaacaatt cgacgatcgt ggtgatgctg accaagctgc 4080
gggagatggg ccgggagaag tgtcaccagt actggccggc cgagcgctct gcccgctacc 4140
agtactttgt ggtagatccg atggcagaat acaacatgcc tcagtatatc ctgcgagagt 4200
tcaaggtcac agatgcccgg gatggccagt cccggactgt ccggcagttc cagttcacag 4260
actggccgga acagggtgtg ccaaagtcgg gggagggctt catcgacttc attggccaag 4320
tgcataagac taaggagcag tttggccagg acggccccat ctctgtccac tgcagtgccg 4380
gcgtgggcag gacgggcgtc ttcatcacgc ttagcatcgt gctggagcgg atgcggtatg 4440
aaggcgtggt ggacatcttt cagacggtga agatgctacg aacccagcgg ccggccatgg 4500
tgcagacaga ggatgagtac cagttctgtt accaggcggc actggagtac ctcggaagct 4560
ttgaccacta tgcaacctaa agccatggtt ccccccaggc ccgacaccac tggccccgga 4620
tgcctctgcc cctcccgggc ggacctcctg aggcctggac cccca 4665
<210> 3
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер
<400> 3
cacggcctat gacctcca 18
<210> 4
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер
<400> 4
aagttcttgg gcgagacttg 20
<210> 5
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер
<400> 5
ccacccatgg caaattcc 18
<210> 6
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер
<400> 6
tgggatttcc attgatgaca ag 22
<210> 7
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер
<400> 7
actcacccac accctacaag a 21
<210> 8
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер
<400> 8
cttggtggta cggccatc 18
<210> 9
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер
<400> 9
aaagaattcg ccgccaccat ggcgcccacc tggggccct 39
<210> 10
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер
<400> 10
aaagcggccg cttaggttgc atagtggtca aagc 34
<210> 11
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой праймер
<400> 11
tccagagttc caggtcactg tcac 24
<210> 12
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Якорный праймер
<220>
<221> модифицированное основание
<222> (24)..(25)
<223> n представляет собой дезоксиинозин
<220>
<221> модифицированное основание
<222> (29)..(30)
<223> n представляет собой дезоксиинозин
<220>
<221> модифицированное основание
<222> (34)..(35)
<223> n представляет собой дезоксиинозин
<400> 12
ggccacgcgt cgactagtac gggnngggnn gggnng 36
<210> 13
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой праймер
<400> 13
aggggccagt ggatagacag atgg 24
<210> 14
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> AUAP праймер
<400> 14
ggccacgcgt cgactagtac 20
<210> 15
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой праймер
<400> 15
ttcactgcca tcaatcttcc actt 24
<210> 16
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Антисмысловой праймер
<400> 16
gatggataca gttggtgcag c 21
<210> 17
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 17
accaagcttg ccgccaccat gaacttcggg ctcaggttg 39
<210> 18
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 18
cttggtgcta gctgaggaga cggtgaccgt ggt 33
<210> 19
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 19
accgtctcct cagctagcac caagggccca tcg 33
<210> 20
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 20
tttgaattct catttacccg gagacaggga 30
<210> 21
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 21
accaagcttg ccgccaccat ggagtcacag attcaggtc 39
<210> 22
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 22
agccacagtt cgttttattt ccagcttggt ccc 33
<210> 23
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 23
ctggaaataa aacgaactgt ggctgcacca tct 33
<210> 24
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 24
aaagaattcc tagcactctc ccctgttgaa 30
<210> 25
<211> 469
<212> ДНК
<213> Mus musculus
<400> 25
atgaacttcg ggctcaggtt gattttcctt gccctcattt taaaaggtgt ccagtgtgag 60
gtgcagctgg tggagtctgg gggagactta gtgaagcctg gagggtccct gaaactctcc 120
tgtgcagcct ctggattcat tttcagtagc tatggcatgt cttgggttcg ccagactcca 180
gacaagaggc tggagtgggt cgcaaccatt agtagtggtg gtagtgacac ctattatcca 240
gacagtgtga aggggcgatt caccatctcc agagacaatg ccaacaacac cctgtacctg 300
caaatgagca gtctgaagtc tgaggacaca gccatgtatt actgtgcaag acaggtctac 360
tatggtcttt actggtattt cgatgtctgg ggcgcaggga ccacggtcac cgtctcctca 420
gccaaaacga cacccccatc tgtctatcca ctggccccta agggcgaat 469
<210> 26
<211> 156
<212> БЕЛОК
<213> Mus musculus
<400> 26
Met Asn Phe Gly Leu Arg Leu Ile Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Ser Gly Gly Ser Asp Thr Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Asn Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Gln Val Tyr Tyr Gly Leu Tyr Trp Tyr Phe Asp
115 120 125
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Ala Lys Thr Thr
130 135 140
Pro Pro Ser Val Tyr Pro Leu Ala Pro Lys Gly Glu
145 150 155
<210> 27
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 27
Ser Tyr Gly Met Ser
1 5
<210> 28
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 28
Thr Ile Ser Ser Gly Gly Ser Asp Thr Tyr Tyr Pro Asp Ser Val Lys
1 5 10 15
Gly
<210> 29
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 29
Gln Val Tyr Tyr Gly Leu Tyr Trp Tyr Phe Asp Val
1 5 10
<210> 30
<211> 413
<212> ДНК
<213> Mus musculus
<400> 30
atggagtcac agattcaggt ctttgtattc gtgtttctct ggttgtctgg tgttgacgga 60
gacattgtga tgacccagtc tcacaaattc atgtccacat cagtaggaga cagggtcagc 120
atcatttgta aggccagtca ggatgtgaat actgctgtag cctggtatca acagaaacca 180
ggacaatctc ctaaattact gatttactcg gcatcctacc ggtacactgg agtccctgat 240
cgcttcactg gcagtggatc tgggacggat ttcactttca ccatcagcag tgtgcaggct 300
gaagacctgg caatttatta ctgtcagcaa cattatagta ctccgtacac gttcggaggg 360
gggaccaagc tggaaataaa acgggctgat gctgcaccaa ctgtatccat caa 413
<210> 31
<211> 137
<212> БЕЛОК
<213> Mus musculus
<400> 31
Met Glu Ser Gln Ile Gln Val Phe Val Phe Val Phe Leu Trp Leu Ser
1 5 10 15
Gly Val Asp Gly Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser
20 25 30
Thr Ser Val Gly Asp Arg Val Ser Ile Ile Cys Lys Ala Ser Gln Asp
35 40 45
Val Asn Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro
50 55 60
Lys Leu Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Asp
65 70 75 80
Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser
85 90 95
Ser Val Gln Ala Glu Asp Leu Ala Ile Tyr Tyr Cys Gln Gln His Tyr
100 105 110
Ser Thr Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Ala Asp Ala Ala Pro Thr Val Ser Ile
130 135
<210> 32
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 32
Lys Ala Ser Gln Asp Val Asn Thr Ala Val Ala
1 5 10
<210> 33
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 33
Ser Ala Ser Tyr Arg Tyr Thr
1 5
<210> 34
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 34
Gln Gln His Tyr Ser Thr Pro
1 5
<210> 35
<211> 1413
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела
против PTPRS 49F2-30
<400> 35
atgaacttcg ggctcaggtt gattttcctt gccctcattt taaaaggtgt ccagtgtgag 60
gtgcagctgg tggagtctgg gggagactta gtgaagcctg gagggtccct gaaactctcc 120
tgtgcagcct ctggattcat tttcagtagc tatggcatgt cttgggttcg ccagactcca 180
gacaagaggc tggagtgggt cgcaaccatt agtagtggtg gtagtgacac ctattatcca 240
gacagtgtga aggggcgatt caccatctcc agagacaatg ccaacaacac cctgtacctg 300
caaatgagca gtctgaagtc tgaggacaca gccatgtatt actgtgcaag acaggtctac 360
tatggtcttt actggtattt cgatgtctgg ggcgcaggga ccacggtcac cgtctcctca 420
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 480
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 540
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 600
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 660
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 720
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 780
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 840
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 900
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 960
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 1020
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1080
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 1140
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1200
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1260
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 1320
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1380
cagaagagcc tctccctgtc tccgggtaaa tga 1413
<210> 36
<211> 470
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи химерного антитела против
PTPRS 49F2-30
<400> 36
Met Asn Phe Gly Leu Arg Leu Ile Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Ser Gly Gly Ser Asp Thr Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Asn Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Gln Val Tyr Tyr Gly Leu Tyr Trp Tyr Phe Asp
115 120 125
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 37
<211> 705
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеиновой кислоты легкой цепи химерного антитела
против PTPRS 49F2-30
<400> 37
atggagtcac agattcaggt ctttgtattc gtgtttctct ggttgtctgg tgttgacgga 60
gacattgtga tgacccagtc tcacaaattc atgtccacat cagtaggaga cagggtcagc 120
atcatttgta aggccagtca ggatgtgaat actgctgtag cctggtatca acagaaacca 180
ggacaatctc ctaaattact gatttactcg gcatcctacc ggtacactgg agtccctgat 240
cgcttcactg gcagtggatc tgggacggat ttcactttca ccatcagcag tgtgcaggct 300
gaagacctgg caatttatta ctgtcagcaa cattatagta ctccgtacac gttcggaggg 360
gggaccaagc tggaaataaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 420
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 480
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 540
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 600
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 660
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gctag 705
<210> 38
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность легкой цепи химерного антитела против
PTPRS 49F2-30
<400> 38
Met Glu Ser Gln Ile Gln Val Phe Val Phe Val Phe Leu Trp Leu Ser
1 5 10 15
Gly Val Asp Gly Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser
20 25 30
Thr Ser Val Gly Asp Arg Val Ser Ile Ile Cys Lys Ala Ser Gln Asp
35 40 45
Val Asn Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro
50 55 60
Lys Leu Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Asp
65 70 75 80
Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser
85 90 95
Ser Val Gln Ala Glu Asp Leu Ala Ile Tyr Tyr Cys Gln Gln His Tyr
100 105 110
Ser Thr Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 39
<211> 36
<212> ДНК
<213> Искусственная
<220>
<223> праймер
<400> 39
ccaggagagt gggagaggct cttctcagta tggtgg 36
<210> 40
<211> 32
<212> ДНК
<213> Искусственная
<220>
<223> праймер
<400> 40
ggctcaggga aatagccctt gaccaggcat cc 32
<210> 41
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> праймер
<400> 41
tccagagttc caagtcacag tcac 24
<210> 42
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> праймер
<400> 42
aggggccagt ggatagactg atgg 24
<210> 43
<211> 471
<212> ДНК
<213> Mus musculus
<400> 43
atggagttgg gactgagctg ggtatttctt gtggctcttt tgaatggtgt ccagtgtcag 60
gtgcagcttg tagagaccgg gggaggcttg gtgaggcctg gaaattctct gaaactctcc 120
tgtgttacct cgggattcac tttcagtaac taccggatgc actggcttcg ccagcctcca 180
gggaagaggc tggagtggat tgctgtaatt acagtcaaat ctgataatta tggagcaaat 240
tatgcagagt ctgtgaaagg cagattcact atttcaagag atgattcaaa aagcagtgtc 300
tacctgcaga tgaacagatt aagagaggaa gacactgcca cttattattg tagtagatcg 360
gtctactatg gttacgtcct agcctttgac tactggggcc aaggcaccac tctcacagtc 420
tcctcagcca aaacgacacc cccatctgtc tatccactgg cccctaaggg c 471
<210> 44
<211> 157
<212> БЕЛОК
<213> Mus musculus
<400> 44
Met Glu Leu Gly Leu Ser Trp Val Phe Leu Val Ala Leu Leu Asn Gly
1 5 10 15
Val Gln Cys Gln Val Gln Leu Val Glu Thr Gly Gly Gly Leu Val Arg
20 25 30
Pro Gly Asn Ser Leu Lys Leu Ser Cys Val Thr Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Arg Met His Trp Leu Arg Gln Pro Pro Gly Lys Arg Leu
50 55 60
Glu Trp Ile Ala Val Ile Thr Val Lys Ser Asp Asn Tyr Gly Ala Asn
65 70 75 80
Tyr Ala Glu Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser
85 90 95
Lys Ser Ser Val Tyr Leu Gln Met Asn Arg Leu Arg Glu Glu Asp Thr
100 105 110
Ala Thr Tyr Tyr Cys Ser Arg Ser Val Tyr Tyr Gly Tyr Val Leu Ala
115 120 125
Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Lys
130 135 140
Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Lys Gly
145 150 155
<210> 45
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 45
Asn Tyr Arg Met His
1 5
<210> 46
<211> 19
<212> БЕЛОК
<213> Mus musculus
<400> 46
Val Ile Thr Val Lys Ser Asp Asn Tyr Gly Ala Asn Tyr Ala Glu Ser
1 5 10 15
Val Lys Gly
<210> 47
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 47
Ser Val Tyr Tyr Gly Tyr Val Leu Ala Phe Asp Tyr
1 5 10
<210> 48
<211> 402
<212> ДНК
<213> Mus musculus
<400> 48
atgatgtcct ctgctcagtt ccttggtctc ctgttgctct gttttcaagg taccagatgt 60
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagagtcacc 120
atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca 180
gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca 240
aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa 300
gaagatattg ccacttactt ttgccaacag ggtaatacgc ttccgtggac gttcggtgga 360
ggcaccaagc tggaaatcaa acgggctgat gctgcaccaa ct 402
<210> 49
<211> 134
<212> БЕЛОК
<213> Mus musculus
<400> 49
Met Met Ser Ser Ala Gln Phe Leu Gly Leu Leu Leu Leu Cys Phe Gln
1 5 10 15
Gly Thr Arg Cys Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val
50 55 60
Lys Leu Leu Ile Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser
85 90 95
Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn
100 105 110
Thr Leu Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Ala Asp Ala Ala Pro Thr
130
<210> 50
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 50
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 51
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 51
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 52
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 52
Gln Gln Gly Asn Thr Leu Pro
1 5
<210> 53
<211> 465
<212> ДНК
<213> Mus musculus
<400> 53
atgaacttgg ggctcagctt gattttcctt gtccttgttt taaaaggtgt ccagtgtgaa 60
gtgaagctgg tggagtctgg gggaggctta gtgcagcctg gagggtccct gaaactctcc 120
tgtgcaacct ctggattcac tttcagtgac tattacatgt attgggttcg ccagactcca 180
gagaagaggc tggagtgggt cgcatacatt agtaatggtg gtggtagcac ctattatcca 240
gacactgtaa agggccgatt caccatctcc agagacaatg ccaagaacac cctgtacctg 300
caaatgagcc gtctgaagtc tgaggacaca gccatgtatt actgtgcaag acatgtttac 360
tacgggagga actatgctat ggactactgg ggtcaaggaa cctcagtcac cgtctcctca 420
gccaaaacaa cacccccatc agtctatcca ctggccccta agggc 465
<210> 54
<211> 155
<212> БЕЛОК
<213> Mus musculus
<400> 54
Met Asn Leu Gly Leu Ser Leu Ile Phe Leu Val Leu Val Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Tyr Met Tyr Trp Val Arg Gln Thr Pro Glu Lys Arg Leu
50 55 60
Glu Trp Val Ala Tyr Ile Ser Asn Gly Gly Gly Ser Thr Tyr Tyr Pro
65 70 75 80
Asp Thr Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Arg Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg His Val Tyr Tyr Gly Arg Asn Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr
130 135 140
Pro Pro Ser Val Tyr Pro Leu Ala Pro Lys Gly
145 150 155
<210> 55
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 55
Asp Tyr Tyr Met Tyr
1 5
<210> 56
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 56
Tyr Ile Ser Asn Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Thr Val Lys
1 5 10 15
Gly
<210> 57
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 57
His Val Tyr Tyr Gly Arg Asn Tyr Ala Met Asp Tyr
1 5 10
<210> 58
<211> 465
<212> ДНК
<213> Mus musculus
<400> 58
atgaacttgg ggctcagctt gattttcctt gtccttgttt taaaaggtgt ccagtgtgaa 60
gtgaagctgg tggagtctgg gggaggctta gtgcagcctg gagggtccct gaaactctcc 120
tgtgcaacct ctggattcac tttcagtgac tattacatgt attgggttcg ccagactcca 180
gagaagaggc tggagtgggt cgcatacatt agtaatggtg gtggtagcac ctattatcca 240
gacactgtaa agggccgatt caccatctcc agagacaatg ccaagaacac cctgtacctg 300
caaatgagcc gtctgaagtc tgaggacaca gccatgtatt actgtgcaag acatgtttac 360
tacgggagga actatgctat ggactactgg ggtcaaggaa cctcagtcac cgtctcctca 420
gccaaaacaa cacccccatc agtctatcca ctggccccta agggc 465
<210> 59
<211> 155
<212> БЕЛОК
<213> Mus musculus
<400> 59
Met Met Ser Ser Ala Gln Phe Leu Gly Leu Leu Leu Leu Cys Phe Gln
1 5 10 15
Gly Thr Arg Cys Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val
50 55 60
Lys Leu Leu Ile Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser
85 90 95
Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn
100 105 110
Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
130 135 140
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe
145 150 155
<210> 60
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 60
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 61
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 61
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 62
<211> 8
<212> БЕЛОК
<213> Mus musculus
<400> 62
Gln Gln Gly Asn Thr Leu Pro Tyr
1 5
<210> 63
<211> 458
<212> ДНК
<213> Mus musculus
<400> 63
atggaatgta actggatact tccttttatt ctgtcagtaa cttcaggtgt ctactcacag 60
gttcagctcc agcagtctgg ggctgagctg gcaagacctg gggcttcagt gaagttgtcc 120
tgcaaggctt ctggctacac ctttactagc tactggatgc agtgggtaaa acagaggcct 180
ggacagggtc tggaatggat tggggctatt tatcctggag atggtgatac taggtacact 240
cagaagttca agggcaaggc cacattgact gcagataaat cctccagcac agcctacatg 300
caactcagca gcttggcatc tgaggactct gcggtctatt actgtgcaag aaggatttac 360
tacggctatt actatgctat ggactactgg ggtcaaggaa cctcagtcac cgtctcctca 420
gccaaaacga cacccccatc tgtctatcca ctggcccc 458
<210> 64
<211> 152
<212> БЕЛОК
<213> Mus musculus
<400> 64
Met Glu Cys Asn Trp Ile Leu Pro Phe Ile Leu Ser Val Thr Ser Gly
1 5 10 15
Val Tyr Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Met Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asp Gly Asp Thr Arg Tyr Thr
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Arg Ile Tyr Tyr Gly Tyr Tyr Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr
130 135 140
Pro Pro Ser Val Tyr Pro Leu Ala
145 150
<210> 65
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 65
Ser Tyr Trp Met Gln
1 5
<210> 66
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 66
Ala Ile Tyr Pro Gly Asp Gly Asp Thr Arg Tyr Thr Gln Lys Phe Lys
1 5 10 15
Gly
<210> 67
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 67
Arg Ile Tyr Tyr Gly Tyr Tyr Tyr Ala Met Asp Tyr
1 5 10
<210> 68
<211> 430
<212> ДНК
<213> Mus musculus
<400> 68
atggagacag acacaatcct gctatgggtg ctgctgctct gggttccagg ctccactggt 60
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 120
atctcctgca aggccagcca aagtgttgat tatgatggtg atagttatat gaactggtac 180
caacagaaac caggacagcc acccaaactc ctcatctatg ctgcatccaa tctagaatct 240
gggatcccag ccaggtttag tggcagtggg tctgggacag acttcaccct caacatccat 300
cctgtggagg aggaggatgc tgcaacctat tactgtcagc aaagtaatga ggatcctctc 360
acgttcggtg ctgggaccaa gctggagctg aaacgggctg atgctgcacc aactgtatcc 420
atcaagggcg 430
<210> 69
<211> 143
<212> БЕЛОК
<213> Mus musculus
<400> 69
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Asn Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu
115 120 125
Glu Leu Lys Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Lys Gly
130 135 140
<210> 70
<211> 15
<212> БЕЛОК
<213> Mus musculus
<400> 70
Lys Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr Met Asn
1 5 10 15
<210> 71
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 71
Ala Ala Ser Asn Leu Glu Ser
1 5
<210> 72
<211> 8
<212> БЕЛОК
<213> Mus musculus
<400> 72
Gln Gln Ser Asn Glu Asp Pro Leu
1 5
<210> 73
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 73
tttaagcttg ccgccaccat ggagttggga ctgagctgg 39
<210> 74
<211> 42
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 74
cgatgggccc ttggtgctag ctgaggagac tgtgagagtg gt 42
<210> 75
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 75
accaagcttg ccgccaccat gatgtcctct gctcagttc 39
<210> 76
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 76
agccacagtt cgtttgattt ccagcttggt gcc 33
<210> 77
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 77
ctggaaatca aacgaactgt ggctgcacca tct 33
<210> 78
<211> 30
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 78
aaagaattcc tagcactctc ccctgttgaa 30
<210> 79
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 79
tttaagcttg ccgccaccat gaacttgggg ctcagcttg 39
<210> 80
<211> 42
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 80
cgatgggccc ttggtgctag ctgaggagac ggtgactgag gt 42
<210> 81
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 81
accaagcttg ccgccaccat gatgtcctct gctcagttc 39
<210> 82
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 82
agccacagtt cgtttgattt ccagcttggt gcc 33
<210> 83
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 83
ctggaaatca aacgaactgt ggctgcacca tct 33
<210> 84
<211> 30
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 84
aaagaattcc tagcactctc ccctgttgaa 30
<210> 85
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 85
tttaagcttg ccgccaccat ggaatgtaac tggatactt 39
<210> 86
<211> 42
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 86
cgatgggccc ttggtgctag ctgaggagac ggtgactgag gt 42
<210> 87
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 87
accaagcttg ccgccaccat ggagacagac acaatcctg 39
<210> 88
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 88
agccacagtt cgtttcagct ccagcttggt ccc 33
<210> 89
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер
<400> 89
ctggagctga aacgaactgt ggctgcacca tct 33
<210> 90
<211> 30
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер
<400> 90
aaagaattcc tagcactctc ccctgttgaa 30
<210> 91
<211> 1419
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела
против PTPRS 9H5-4
<400> 91
atggagttgg gactgagctg ggtatttctt gtggctcttt tgaatggtgt ccagtgtcag 60
gtgcagcttg tagagaccgg gggaggcttg gtgaggcctg gaaattctct gaaactctcc 120
tgtgttacct cgggattcac tttcagtaac taccggatgc actggcttcg ccagcctcca 180
gggaagaggc tggagtggat tgctgtaatt acagtcaaat ctgataatta tggagcaaat 240
tatgcagagt ctgtgaaagg cagattcact atttcaagag atgattcaaa aagcagtgtc 300
tacctgcaga tgaacagatt aagagaggaa gacactgcca cttattattg tagtagatcg 360
gtctactatg gttacgtcct agcctttgac tactggggcc aaggcaccac tctcacagtc 420
tcctcagcta gcaccaaggg cccatcggtc ttccccctgg caccctcctc caagagcacc 480
tctgggggca cagcggccct gggctgcctg gtcaaggact acttccccga accggtgacg 540
gtgtcgtgga actcaggcgc cctgaccagc ggcgtgcaca ccttcccggc tgtcctacag 600
tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcag cttgggcacc 660
cagacctaca tctgcaacgt gaatcacaag cccagcaaca ccaaggtgga caagaaagtt 720
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 780
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 840
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 900
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 960
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 1020
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 1080
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 1140
gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 1200
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1260
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1320
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1380
tacacgcaga agagcctctc cctgtctccg ggtaaatga 1419
<210> 92
<211> 472
<212> БЕЛОК
<213> Искусственная
<220>
<223> Аминокислотная последовательность тяжелой цепи химерного антитела
против PTPRS 9H5-4
<400> 92
Met Glu Leu Gly Leu Ser Trp Val Phe Leu Val Ala Leu Leu Asn Gly
1 5 10 15
Val Gln Cys Gln Val Gln Leu Val Glu Thr Gly Gly Gly Leu Val Arg
20 25 30
Pro Gly Asn Ser Leu Lys Leu Ser Cys Val Thr Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Arg Met His Trp Leu Arg Gln Pro Pro Gly Lys Arg Leu
50 55 60
Glu Trp Ile Ala Val Ile Thr Val Lys Ser Asp Asn Tyr Gly Ala Asn
65 70 75 80
Tyr Ala Glu Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser
85 90 95
Lys Ser Ser Val Tyr Leu Gln Met Asn Arg Leu Arg Glu Glu Asp Thr
100 105 110
Ala Thr Tyr Tyr Cys Ser Arg Ser Val Tyr Tyr Gly Tyr Val Leu Ala
115 120 125
Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Ser
130 135 140
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
145 150 155 160
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
165 170 175
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
180 185 190
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
195 200 205
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
210 215 220
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
225 230 235 240
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
245 250 255
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
260 265 270
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
275 280 285
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
290 295 300
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
305 310 315 320
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
325 330 335
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
340 345 350
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
355 360 365
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
370 375 380
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
385 390 395 400
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
405 410 415
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
420 425 430
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
435 440 445
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
450 455 460
Ser Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 93
<211> 705
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность нуклеиновой кислоты легкой цепи химерного антитела
против PTPRS 9H5-4
<400> 93
atgatgtcct ctgctcagtt ccttggtctc ctgttgctct gttttcaagg taccagatgt 60
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagagtcacc 120
atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca 180
gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca 240
aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa 300
gaagatattg ccacttactt ttgccaacag ggtaatacgc ttccgtggac gttcggtgga 360
ggcaccaagc tggaaatcaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 420
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 480
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 540
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 600
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 660
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gctag 705
<210> 94
<211> 234
<212> БЕЛОК
<213> Искусственная
<220>
<223> Аминокислотная последовательность легкой цепи химерного антитела против
PTPRS 9H5-4
<400> 94
Met Met Ser Ser Ala Gln Phe Leu Gly Leu Leu Leu Leu Cys Phe Gln
1 5 10 15
Gly Thr Arg Cys Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val
50 55 60
Lys Leu Leu Ile Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser
85 90 95
Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn
100 105 110
Thr Leu Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 95
<211> 1413
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела
против PTPRS 13G5-57
<400> 95
atgaacttgg ggctcagctt gattttcctt gtccttgttt taaaaggtgt ccagtgtgaa 60
gtgaagctgg tggagtctgg gggaggctta gtgcagcctg gagggtccct gaaactctcc 120
tgtgcaacct ctggattcac tttcagtgac tattacatgt attgggttcg ccagactcca 180
gagaagaggc tggagtgggt cgcatacatt agtaatggtg gtggtagcac ctattatcca 240
gacactgtaa agggccgatt caccatctcc agagacaatg ccaagaacac cctgtacctg 300
caaatgagcc gtctgaagtc tgaggacaca gccatgtatt actgtgcaag acatgtttac 360
tacgggagga actatgctat ggactactgg ggtcaaggaa cctcagtcac cgtctcctca 420
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 480
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 540
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 600
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 660
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 720
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 780
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 840
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 900
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 960
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 1020
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1080
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 1140
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1200
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1260
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 1320
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1380
cagaagagcc tctccctgtc tccgggtaaa tga 1413
<210> 96
<211> 470
<212> БЕЛОК
<213> Искусственная
<220>
<223> Аминокислотная последовательность тяжелой цепи химерного антитела
против PTPRS 13G5-57
<400> 96
Met Asn Leu Gly Leu Ser Leu Ile Phe Leu Val Leu Val Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Tyr Met Tyr Trp Val Arg Gln Thr Pro Glu Lys Arg Leu
50 55 60
Glu Trp Val Ala Tyr Ile Ser Asn Gly Gly Gly Ser Thr Tyr Tyr Pro
65 70 75 80
Asp Thr Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Arg Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg His Val Tyr Tyr Gly Arg Asn Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 97
<211> 705
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность нуклеиновой кислоты легкой цепи химерного антитела
против PTPRS 13G5-57
<400> 97
atgatgtcct ctgctcagtt ccttggtctc ctgttgctct gttttcaagg taccagatgt 60
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagagtcacc 120
atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca 180
gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca 240
aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa 300
gaagatattg ccacttactt ttgccaacag ggtaatacgc ttccgtacac gttcggaggg 360
gggaccaagc tggaaataaa acgaactgtg gctgcaccat ctgtcttcat cttcccgcca 420
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 480
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 540
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 600
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 660
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gctag 705
<210> 98
<211> 234
<212> БЕЛОК
<213> Искусственная
<220>
<223> Аминокислотная последовательность легкой цепи химерного антитела против
PTPRS 13G5-57
<400> 98
Met Met Ser Ser Ala Gln Phe Leu Gly Leu Leu Leu Leu Cys Phe Gln
1 5 10 15
Gly Thr Arg Cys Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val
50 55 60
Lys Leu Leu Ile Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser
85 90 95
Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn
100 105 110
Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 99
<211> 1413
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность нуклеиновой кислоты тяжелой цепи химерного антитела
против PTPRS 22H8-84
<400> 99
atggaatgta actggatact tccttttatt ctgtcagtaa cttcaggtgt ctactcacag 60
gttcagctcc agcagtctgg ggctgagctg gcaagacctg gggcttcagt gaagttgtcc 120
tgcaaggctt ctggctacac ctttactagc tactggatgc agtgggtaaa acagaggcct 180
ggacagggtc tggaatggat tggggctatt tatcctggag atggtgatac taggtacact 240
cagaagttca agggcaaggc cacattgact gcagataaat cctccagcac agcctacatg 300
caactcagca gcttggcatc tgaggactct gcggtctatt actgtgcaag aaggatttac 360
tacggctatt actatgctat ggactactgg ggtcaaggaa cctcagtcac cgtctcctca 420
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 480
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 540
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 600
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 660
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 720
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 780
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 840
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 900
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 960
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 1020
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1080
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 1140
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1200
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1260
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 1320
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1380
cagaagagcc tctccctgtc tccgggtaaa tga 1413
<210> 100
<211> 470
<212> БЕЛОК
<213> Искусственная
<220>
<223> Аминокислотная последовательность тяжелой цепи химерного антитела
против PTPRS 22H8-84
<400> 100
Met Glu Cys Asn Trp Ile Leu Pro Phe Ile Leu Ser Val Thr Ser Gly
1 5 10 15
Val Tyr Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Met Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Ala Ile Tyr Pro Gly Asp Gly Asp Thr Arg Tyr Thr
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Arg Ile Tyr Tyr Gly Tyr Tyr Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 101
<211> 717
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность нуклеиновой кислоты легкой цепи химерного антитела
против PTPRS 22H8-84
<400> 101
atggagacag acacaatcct gctatgggtg ctgctgctct gggttccagg ctccactggt 60
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 120
atctcctgca aggccagcca aagtgttgat tatgatggtg atagttatat gaactggtac 180
caacagaaac caggacagcc acccaaactc ctcatctatg ctgcatccaa tctagaatct 240
gggatcccag ccaggtttag tggcagtggg tctgggacag acttcaccct caacatccat 300
cctgtggagg aggaggatgc tgcaacctat tactgtcagc aaagtaatga ggatcctctc 360
acgttcggtg ctgggaccaa gctggagctg aaacgaactg tggctgcacc atctgtcttc 420
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 480
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 540
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 600
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 660
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgctag 717
<210> 102
<211> 238
<212> БЕЛОК
<213> Искусственная
<220>
<223> Аминокислотная последовательность легкой цепи химерного антитела
против PTPRS 22H8-84
<400> 102
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Asn Glu Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu
115 120 125
Glu Leu Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| СВЯЗЫВАЮЩИЕ ILT7 МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2756109C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| АНТИТЕЛО ПРОТИВ SIRPα | 2019 |
|
RU2791002C2 |
| АНТИТЕЛО К CD40, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2779128C2 |
| АНТИТЕЛО К B7-H4, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2792748C2 |
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| АНТИТЕЛО К PD-L1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2778085C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |

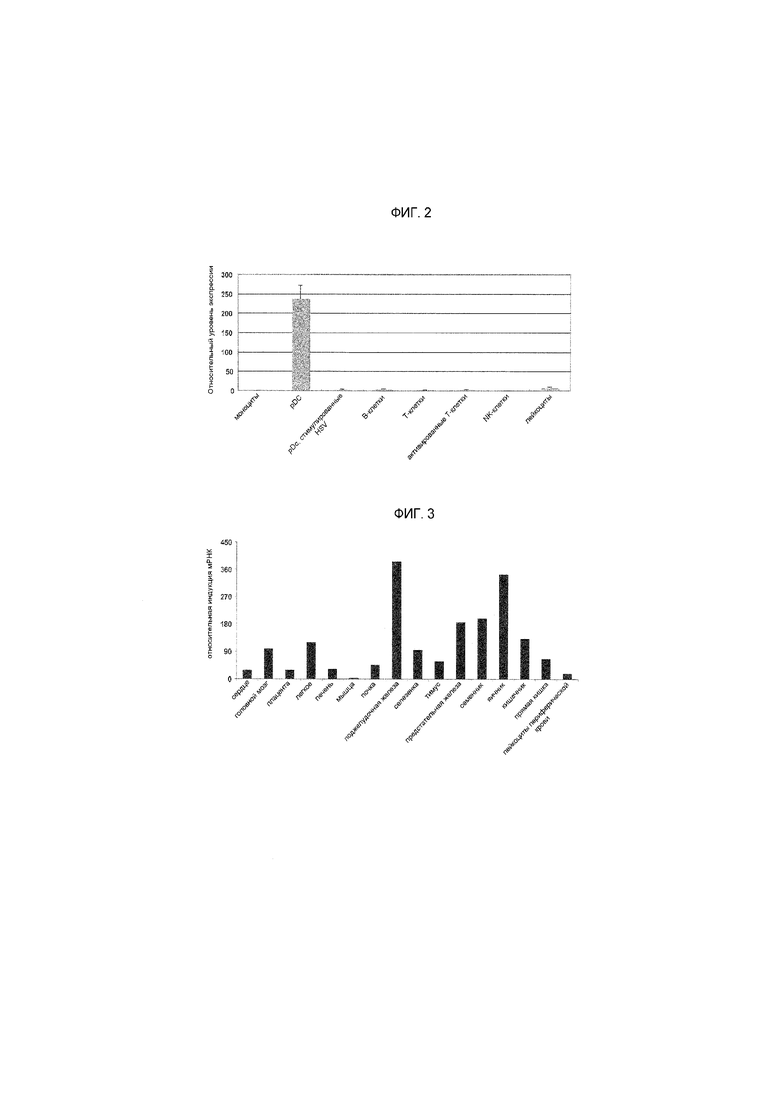
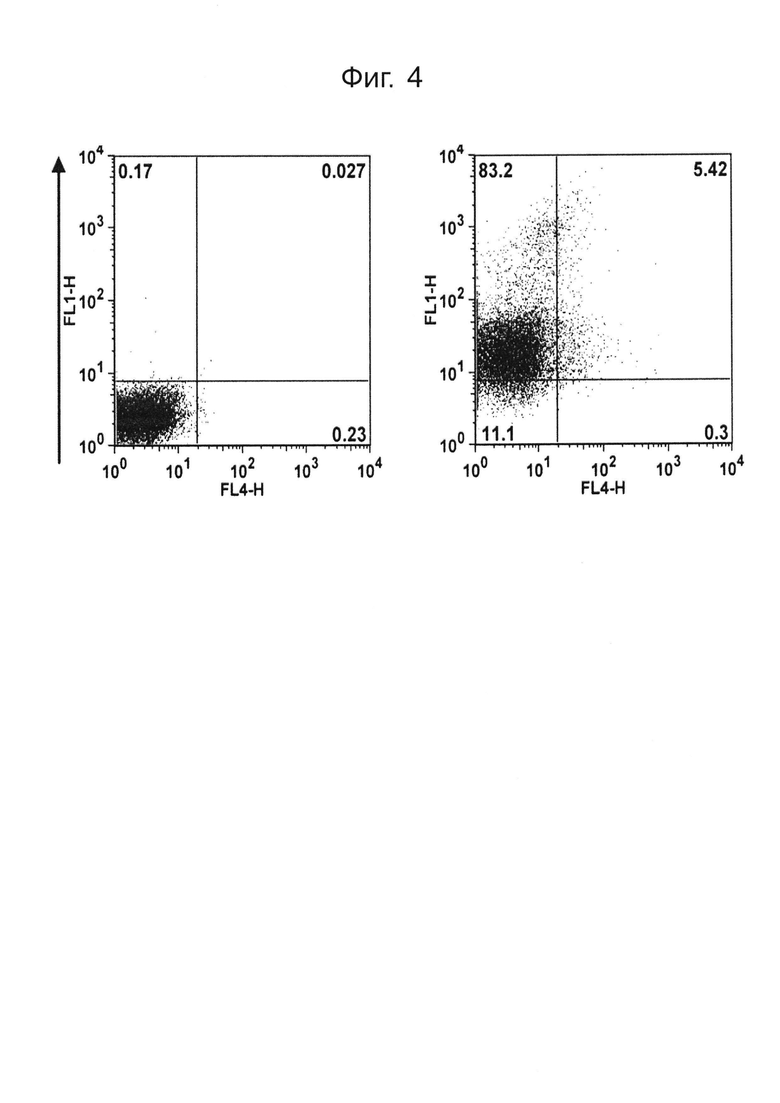


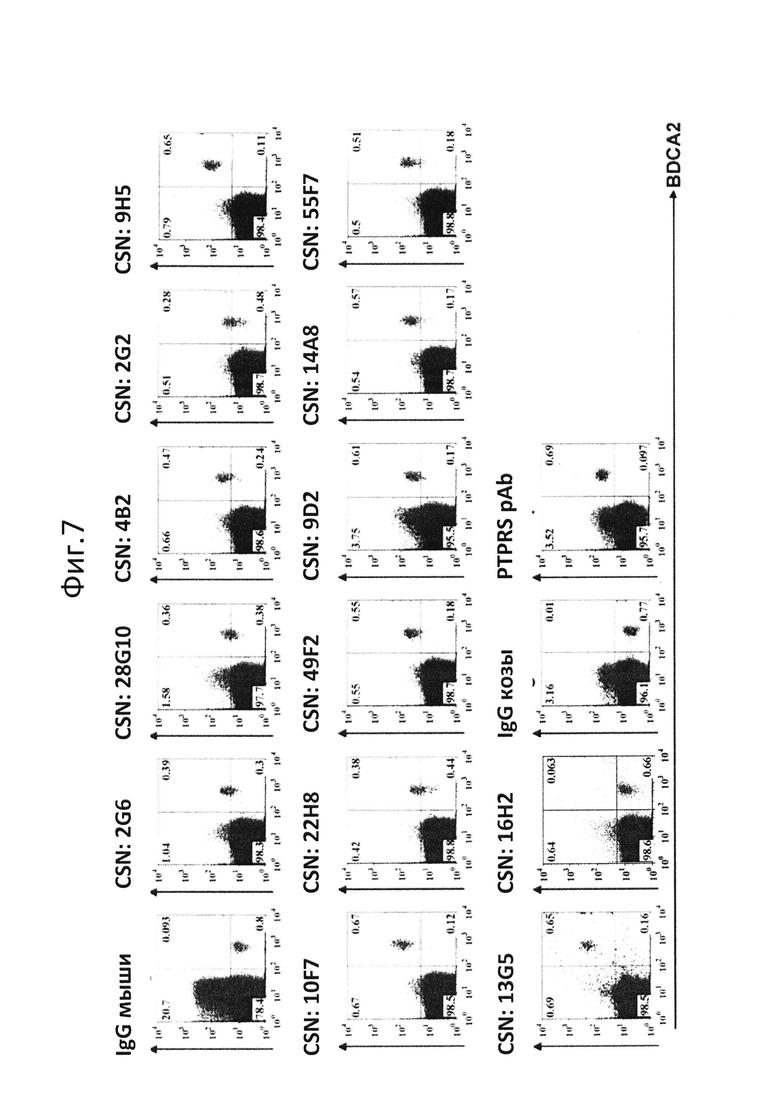

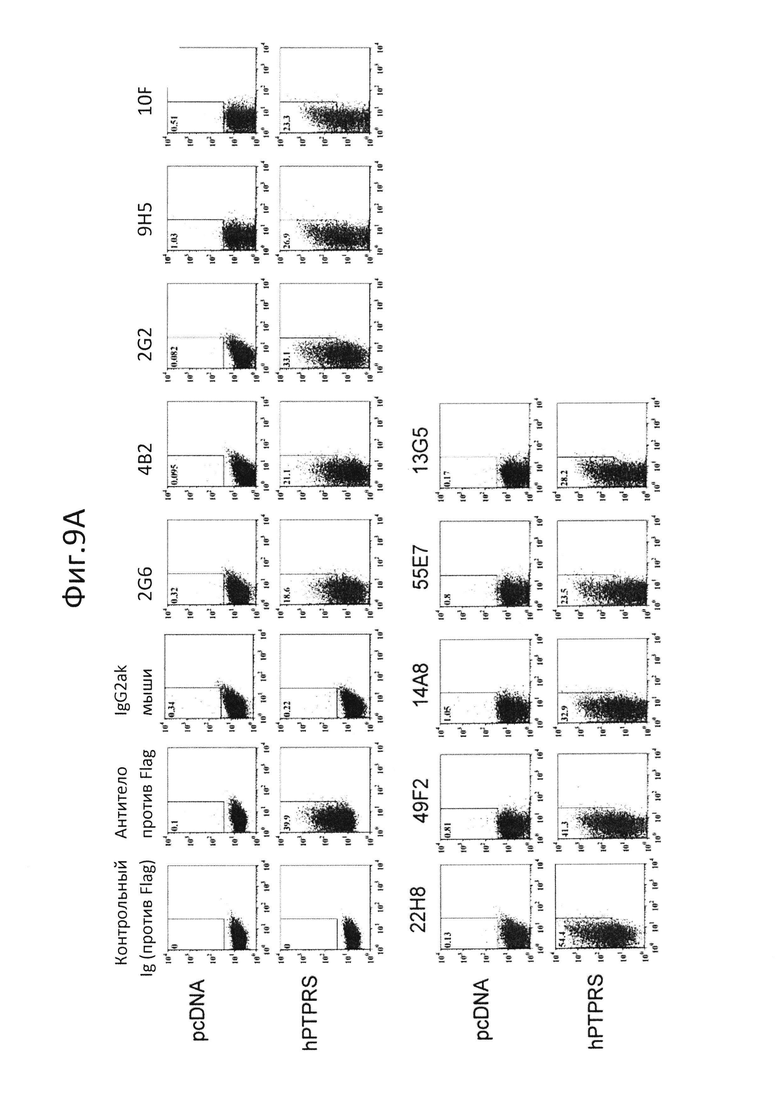


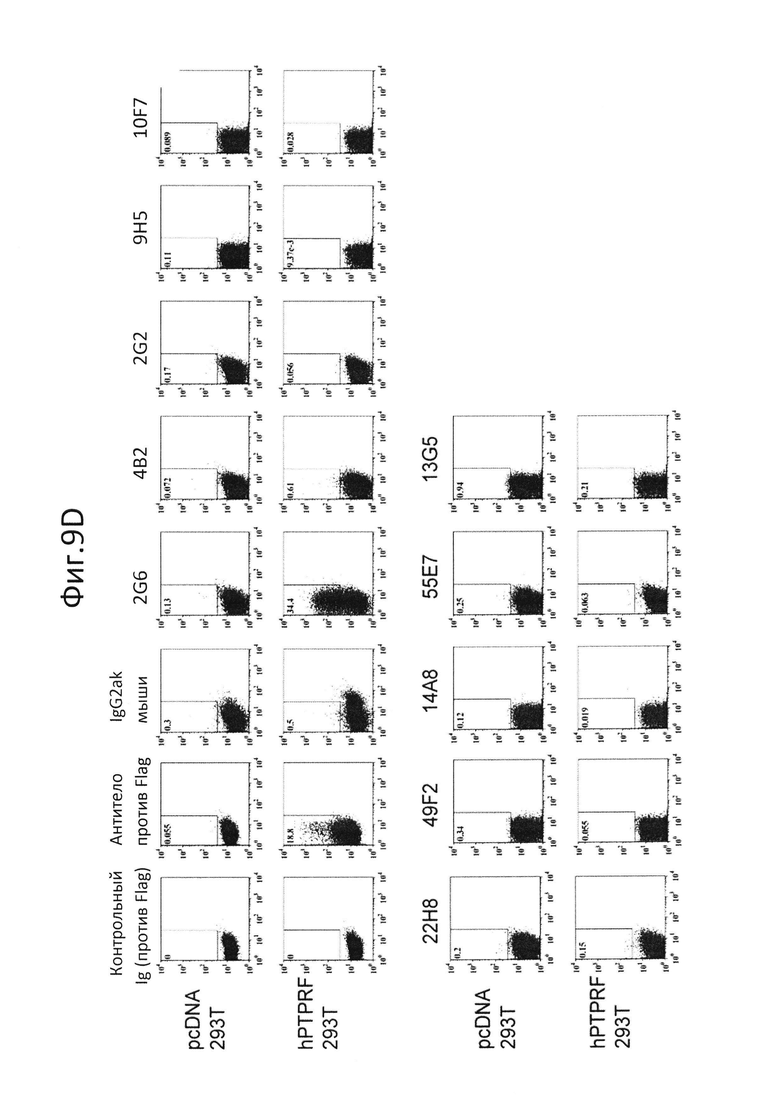

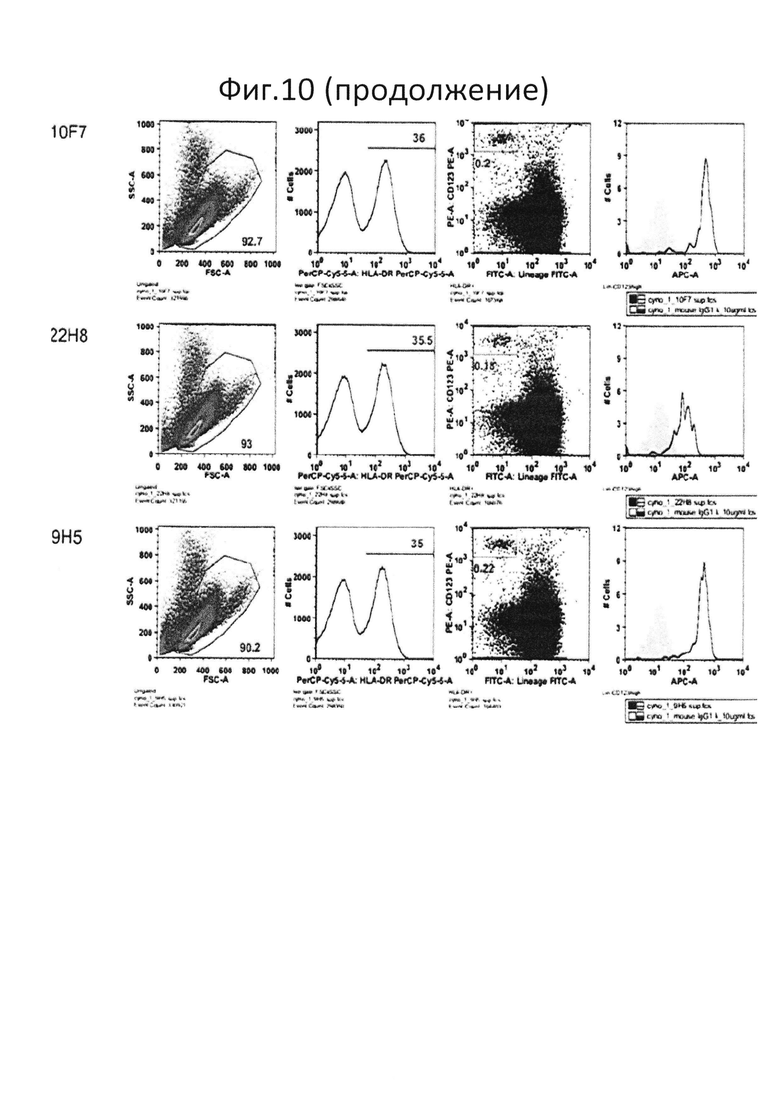
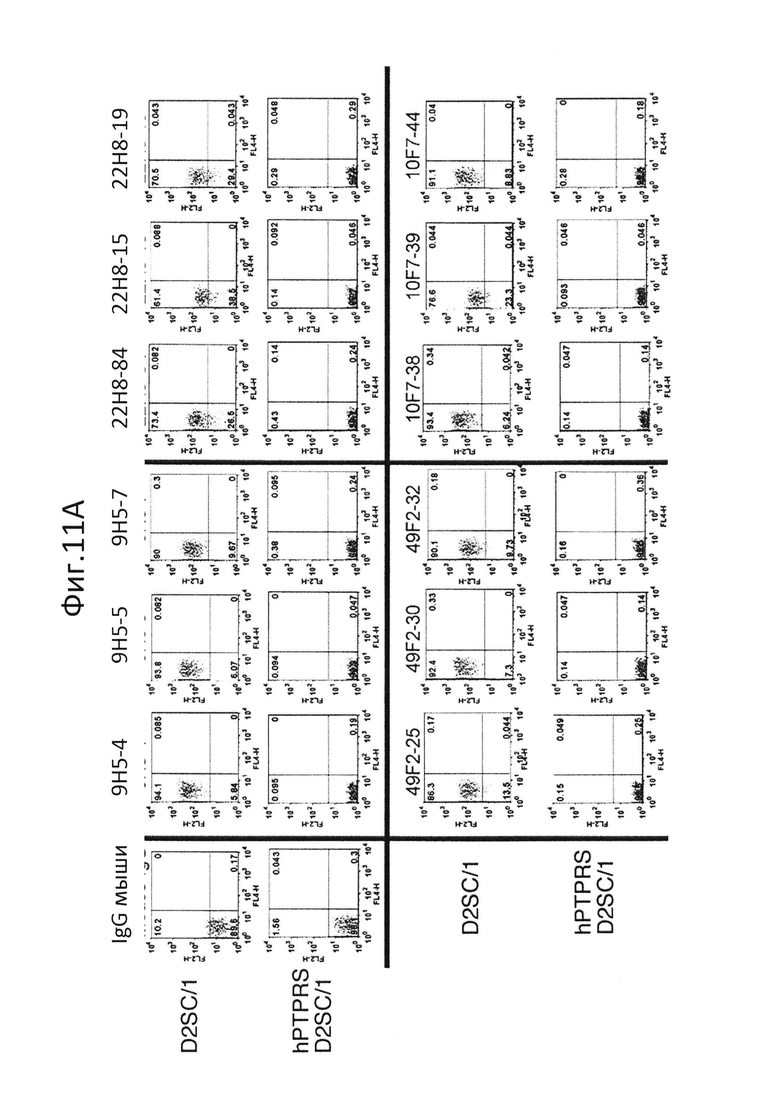
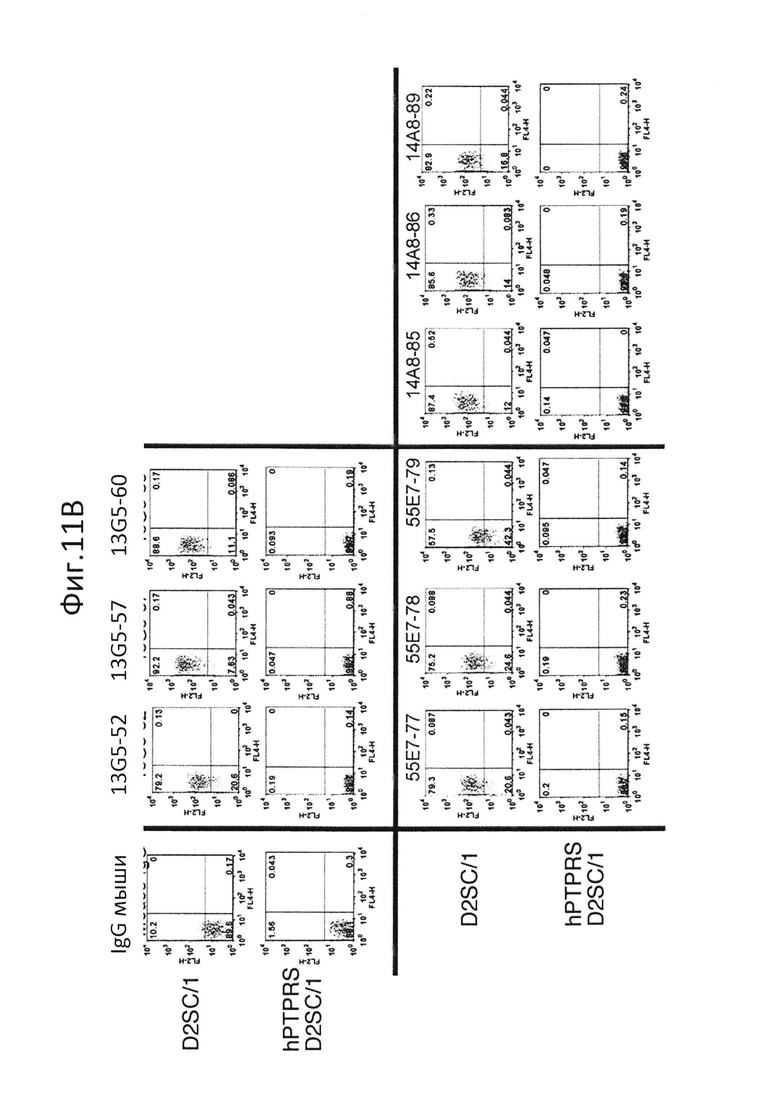
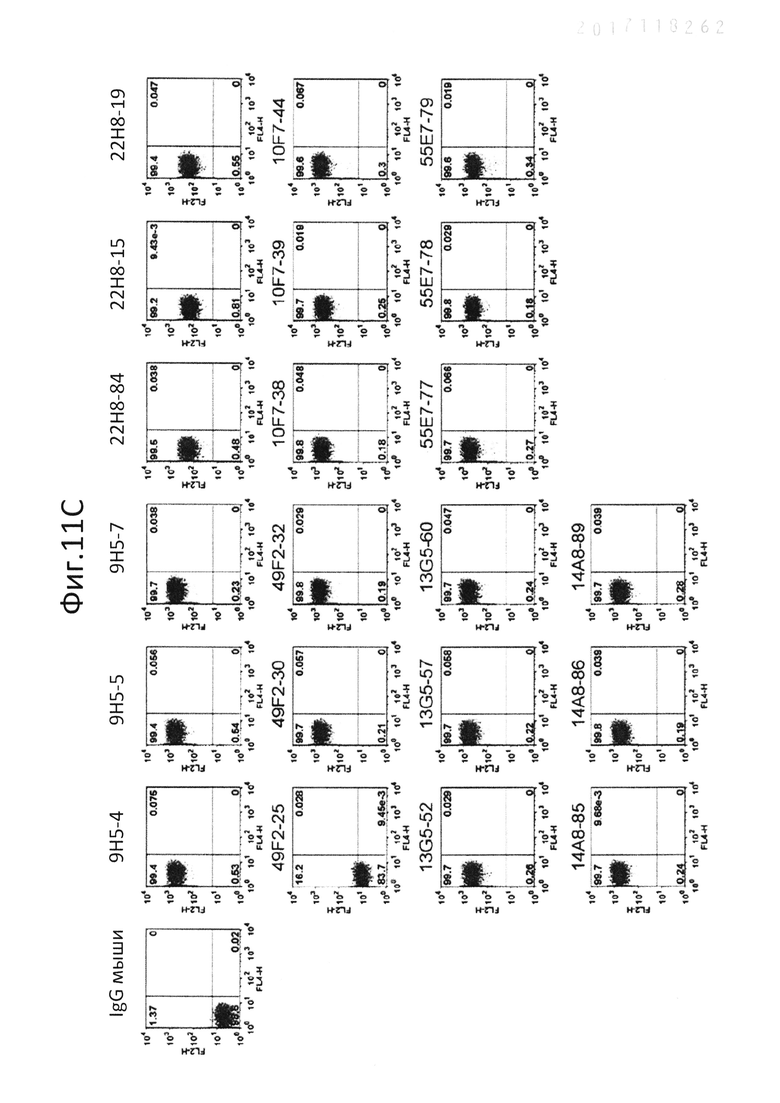
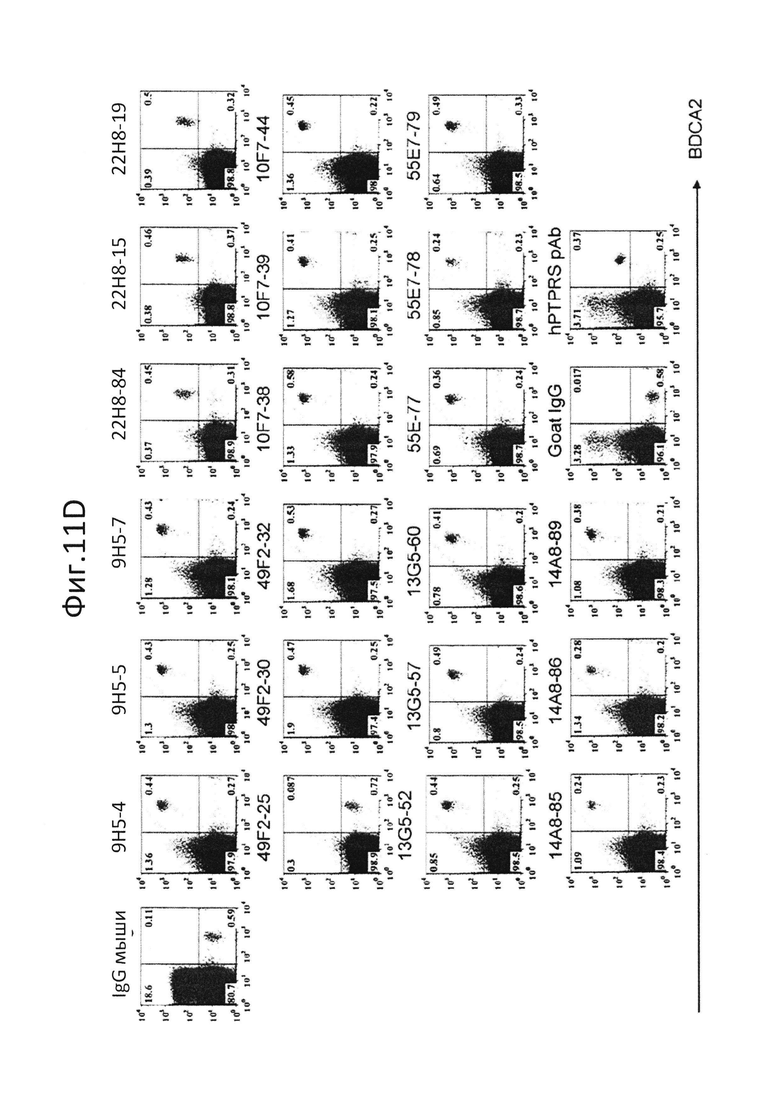
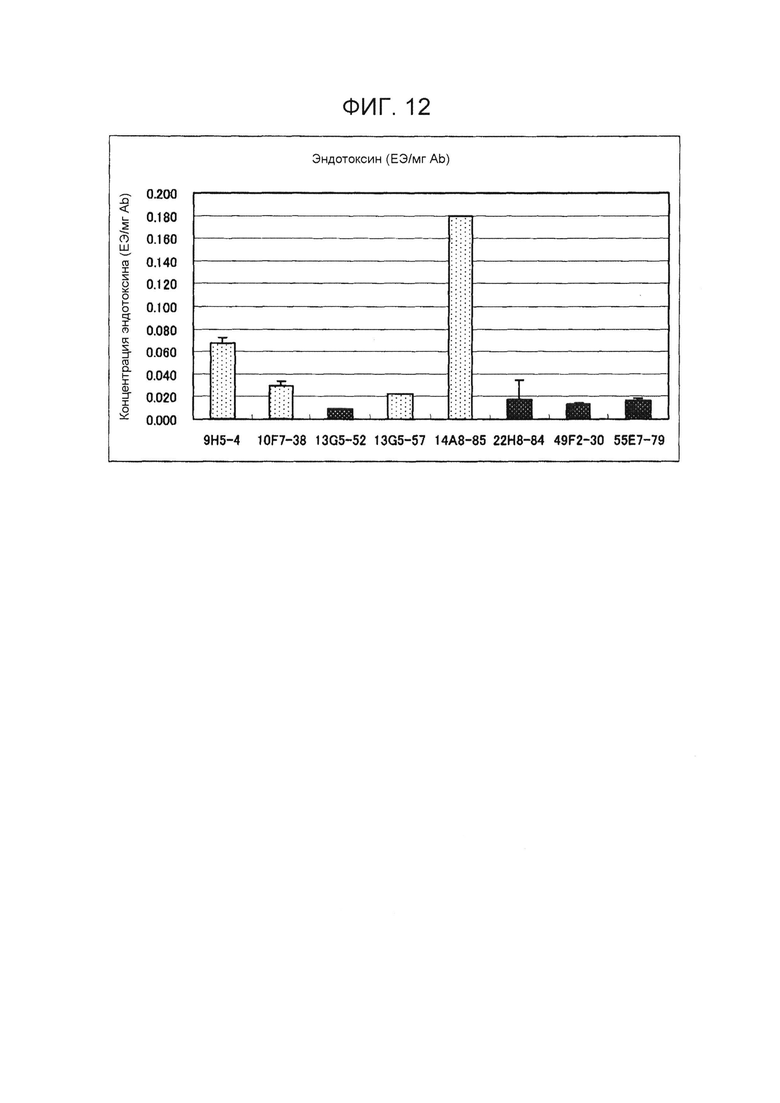
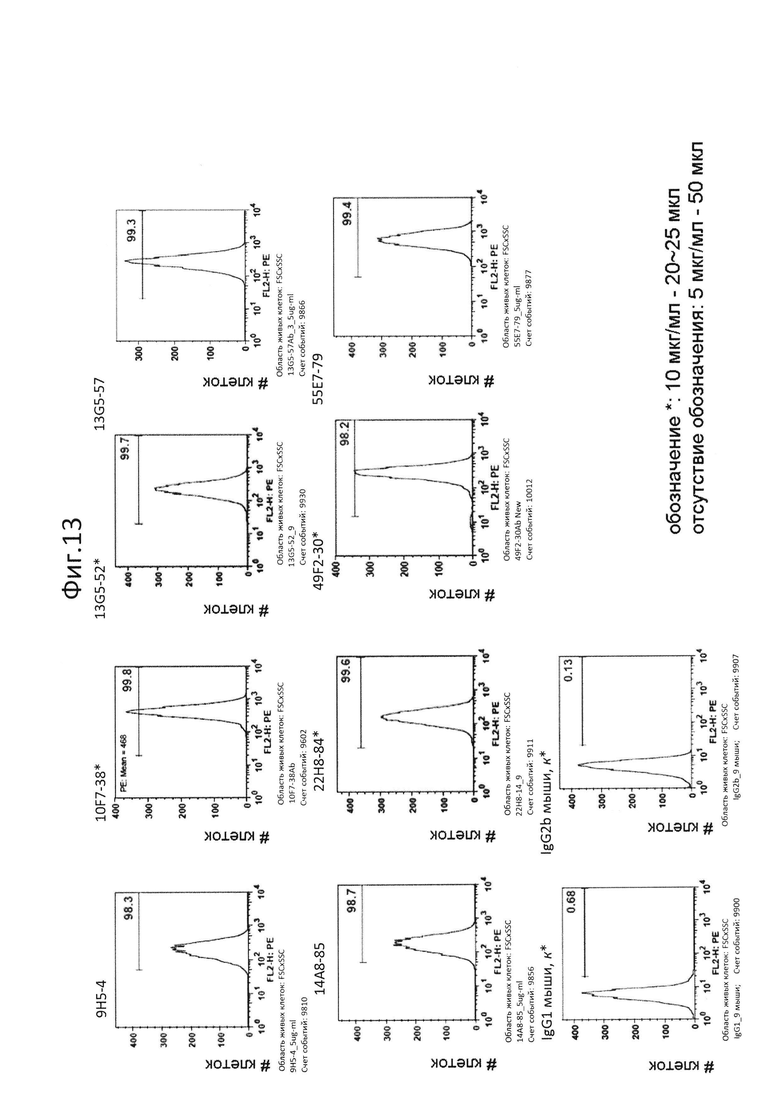
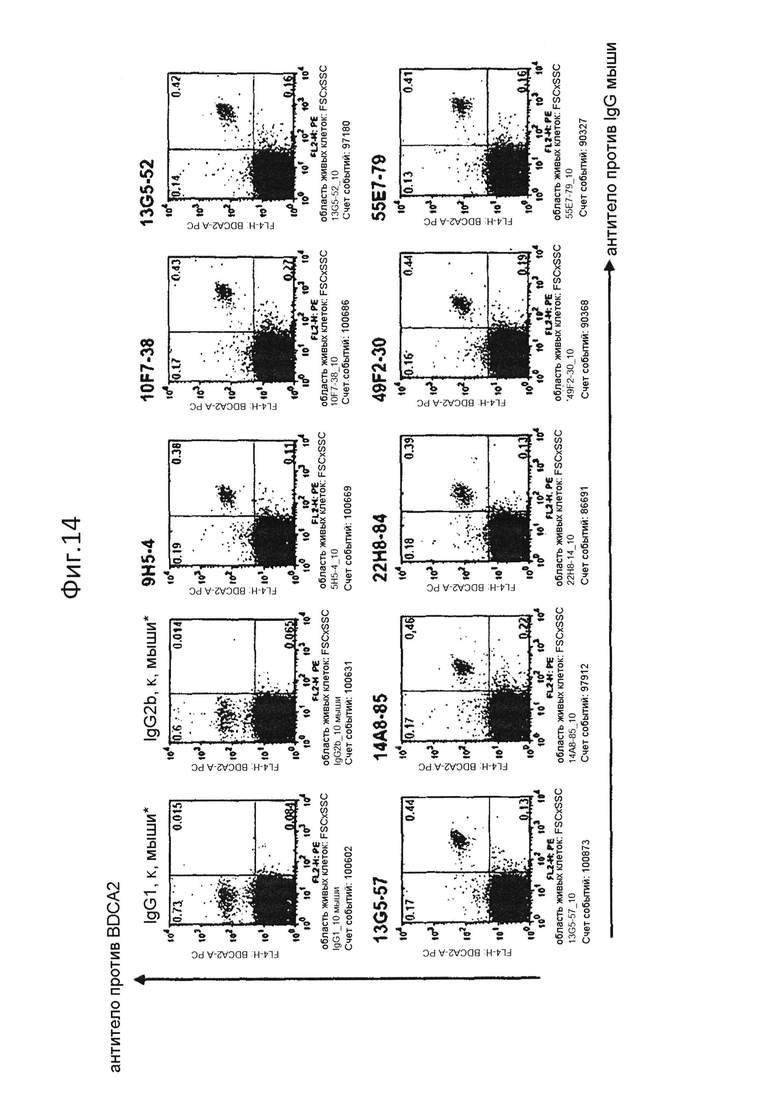
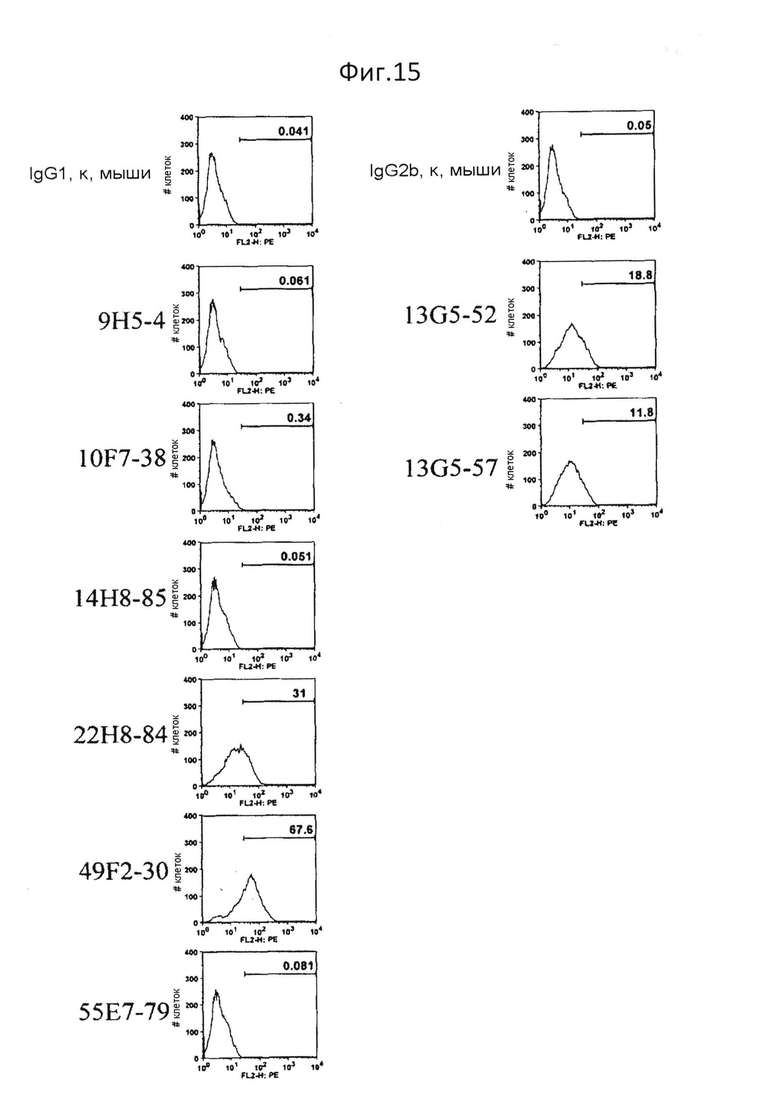

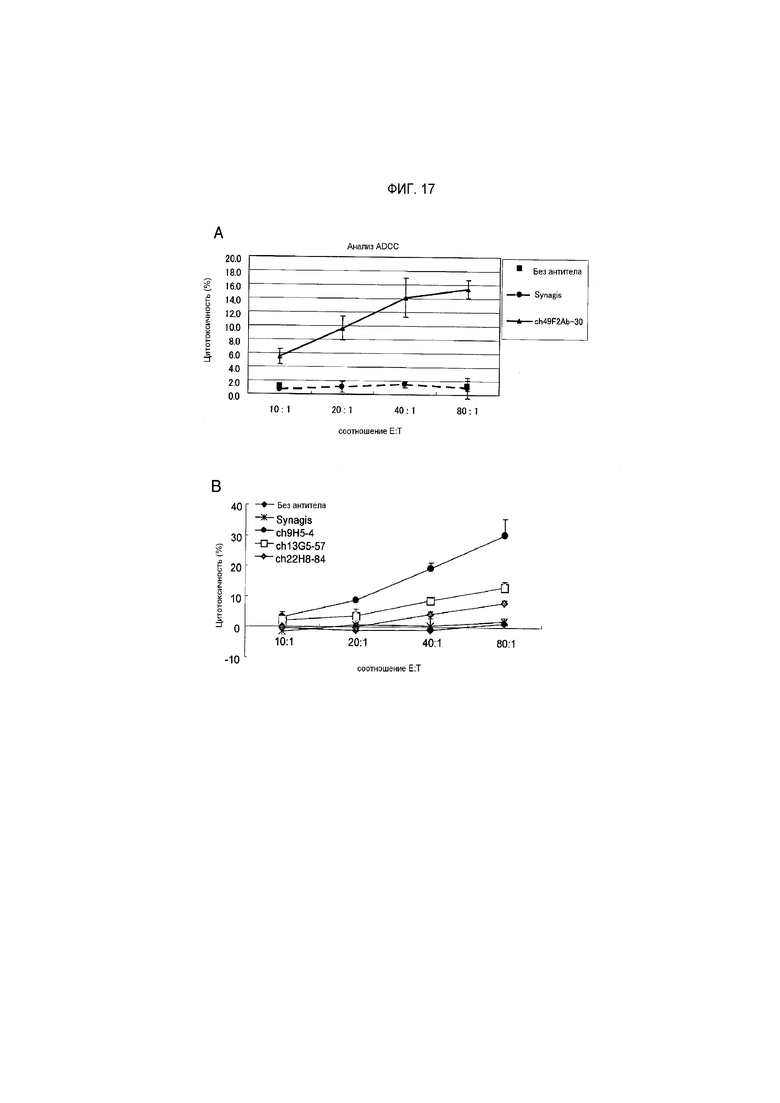
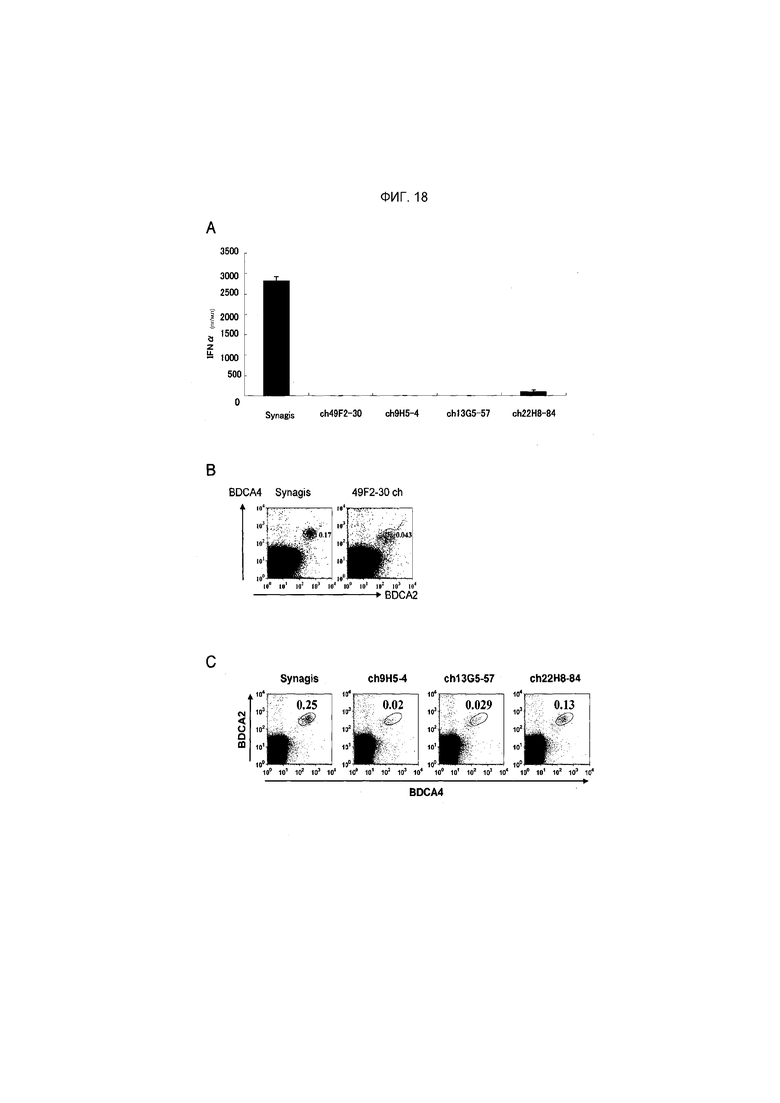
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая моноклональное антитело или его антиген-связывающий фрагмент, которое связывается с внеклеточным доменом протеинтирозинфосфатазы σ рецепторного типа человека (PTPRS человека) на плазмоцитоидной дендритной клетке (варианты), гибридому для получения вышеуказанного антитела, способ получения клетки, которая продуцирует антитело, способ получения антитела, распознающего PTPRS человека, применение клетки животного, которая сохраняет способность экспрессировать эндогенный полинуклеотид, который кодирует аминокислотную последовательность, включающую внеклеточный домен PTPRS человека, или фракцию мембран этой клетки для получения моноклонального антитела, способ детекции плазмацитоидной дендритной клетки, применение антитела или его фрагмента для детекции плазмоцитоидной дендритной клетки, способ подавления активности плазмацитоидной дендритной клетки ex vivo, применение антитела или его фрагмента для лечения заболевания, связанного с рDC и для лечения аутоиммунного заболевания, связанного с IFNальфа. Изобретение расширяет арсенал антител, связывающихся с PTPRS человека. 13 н. и 3 з.п. ф-лы, 18 ил., 1 табл., 6 пр.
1. Моноклональное антитело или его антиген-связывающий фрагмент, которое связывается с внеклеточным доменом протеинтирозинфосфатазы σ рецепторного типа человека (PTPRS человека) на плазмоцитоидной дендритной клетке, полученное способом с помощью белка PTPRS с последовательностью SEQ ID NO:1, причем белок PTPRS не денатурирован фиксаторами.
2. Моноклональное антитело или его антиген-связывающий фрагмент, которое связывается с внеклеточным доменом протеинтирозинфосфатазы σ рецепторного типа человека (PTPRS человека) на плазмацитоидной дендритной клетке и подавляет продукцию IFN клетки,
где моноклональное антитело, или его фрагмент, получено способом, включающим следующие стадии:
1) введение животному клетки, которая экзогенно экспрессирует белок, содержащий внеклеточный домен PTPRS человека, для иммунизации животного этой клеткой;
2) отбор антитело-продуцирующих клеток, которые продуцируют антитело, связывающееся с PTPRS человека, из антитело-продуцирующих клеток иммунизированного животного; и
3) культивирование антитело-продуцирующих клеток, отобранных на стадии (2), и сбор из культуры антитела, которое распознает PTPRS человека, связывается с плазмоцитоидной дендритной клеткой и подавляет продукцию IFN клеткой.
3. Моноклональное антитело или его антиген-связывающий фрагмент, которое связывается с внеклеточным доменом протеинтирозинфосфатазы σ рецепторного типа человека (PTPRS человека) на плазмацитоидной дендритной клетке и подавляет выживание клетки,
где моноклональное антитело, или его фрагмент, получено способом, включающим следующие стадии:
1) введение животному клетки, которая экзогенно экспрессирует белок, содержащий внеклеточный домен PTPRS человека, для иммунизации животного этой клеткой;
2) отбор антитело-продуцирующих клеток, которые продуцируют антитело, связывающееся с PTPRS человека, из антитело-продуцирующих клеток иммунизированного животного; и
3) культивирование антитело-продуцирующих клеток, отобранных на стадии (2), и сбор из культуры антитела, которое распознает PTPRS человека, связывается с плазмоцитоидной дендритной клеткой и подавляет выживание клетки.
4. Моноклональное антитело или его антиген-связывающий фрагмент, которое связывается с внеклеточным доменом протеинтирозинфосфатазы σ рецепторного типа человека (PTPRS человека) на плазмацитоидной дендритной клетке, содержащее:
(А) CDR1 тяжелой цепи с SEQ ID NO:45,
CDR2 тяжелой цепи с SEQ ID NO:46,
CDR3 тяжелой цепи с SEQ ID NO:47,
CDR1 легкой цепи с SEQ ID NO:50,
CDR2 легкой цепи с SEQ ID NO:51 и
CDR3 легкой цепи с SEQ ID NO:52;
(В) CDR1 тяжелой цепи с SEQ ID NO:55,
CDR2 тяжелой цепи с SEQ ID NO:56,
CDR3 тяжелой цепи с SEQ ID NO:57,
CDR1 легкой цепи с SEQ ID NO:60,
CDR2 легкой цепи с SEQ ID NO:61 и
CDR3 легкой цепи с SEQ ID NO:62;
(С) CDR1 тяжелой цепи с SEQ ID NO:65,
CDR2 тяжелой цепи с SEQ ID NO:66,
CDR3 тяжелой цепи с SEQ ID NO:67,
CDR1 легкой цепи с SEQ ID NO:70,
CDR2 легкой цепи с SEQ ID NO:71 и
CDR3 легкой цепи с SEQ ID NO:72;
(D) CDR1 тяжелой цепи с SEQ ID NO:27,
CDR2 тяжелой цепи с SEQ ID NO:28,
CDR3 тяжелой цепи с SEQ ID NO:29,
CDR1 легкой цепи с SEQ ID NO:32,
CDR2 легкой цепи с SEQ ID NO:33 и
CDR3 легкой цепи с SEQ ID NO:34;
(E) вариабельную область тяжелой цепи с аминокислотами в положении 20-142 последовательности SEQ ID NO:44 и
вариабельную область легкой цепи с аминокислотами в положении 21-127 последовательности SEQ ID NO:49;
(F) вариабельную область тяжелой цепи с аминокислотами в положении 20-140 последовательности SEQ ID NO:54 и
вариабельную область легкой цепи с аминокислотами в положении 21-127 последовательности SEQ ID NO:59;
(G) вариабельную область тяжелой цепи с аминокислотами в положении 20-140 последовательности SEQ ID NO:64 и
вариабельную область легкой цепи с аминокислотами в положении 21-131 последовательности SEQ ID NO:69; или
(H) вариабельную область тяжелой цепи с аминокислотами в положении 20-140 последовательности SEQ ID NO:26 и
вариабельную область легкой цепи с аминокислотами в положении 21-127 последовательности SEQ ID NO:31.
5. Моноклональное антитело или его фрагмент по п.4, которое является химерным.
6. Моноклональное антитело или его фрагмент по п.4, содержащее:
(А) тяжелую цепь с последовательностью SEQ ID NO:92 и
легкую цепь с последовательностью SEQ ID NO:94;
(В) тяжелую цепь с последовательностью SEQ ID NO:96 и
легкую цепь с последовательностью SEQ ID NO:98;
(С) тяжелую цепь с последовательностью SEQ ID NO:100 и
легкую цепь с последовательностью SEQ ID NO:102; или
(D) тяжелую цепь с последовательностью SEQ ID NO:36 и
легкую цепь с последовательностью SEQ ID NO:38.
7. Гибридома для получения антитела по любому из пп.1-3, которая выбрана из группы, состоящей из:
гибридомы 9H5-4, которая депонирована под номером доступа FERM BP-11356,
гибридомы 10F7-38, которая депонирована под номером доступа FERM BP-11357,
гибридомы 13G5-52, которая депонирована под номером доступа FERM BP-11358,
гибридомы 13G5-57, которая депонирована под номером доступа FERM BP-11359,
гибридомы 14А8-85, которая депонирована под номером доступа FERM BP-11360,
гибридомы 22H8-84, которая депонирована под номером доступа FERM BP-11361,
гибридомы 49F2-30, которая депонирована под номером доступа FERM BP-11362, или
гибридомы 55Е7-79, которая депонирована под номером доступа FERM BP-11363,
где указанные гибридомы депонированы в International Patent Organism Depositary (IPOD), National Institute of Advanced Industrial Science and Technology (AIST), Япония.
8. Способ получения клетки, которая продуцирует моноклональное антитело по любому из пп.1-6, включающий:
1) введение животному клетки, которая экзогенно экспрессирует экзогенный белок, содержащий внеклеточный домен PTPRS человека, для иммунизации животного этой клеткой, и
2) отбор антитело-продуцирующих клеток, продуцирующих антитело по любому из пп.1-6, из иммунизированного животного.
9. Способ получения моноклонального антитела, распознающего PTPRS человека по любому из пп.1-6, включающий:
1) введение к животному клетки, которая экзогенно экспрессирует белок, содержащий внеклеточный домен PTPRS человека, для иммунизации животного клеткой;
2) отбор антитело-продуцирующих клеток, продуцирующих антитело, которое связывается с PTPRS человека, из антитело-продуцирующих клеток иммунизированного животного; и
3) культивирование антитело-продуцирующих клеток, отобранных в (2), и сбор антитела, которое распознает PTPRS человека, из культуры.
10. Применение клетки животного, которая сохраняет способность экспрессировать эндогенный полинуклеотид, который кодирует аминокислотную последовательность, включающую внеклеточный домен PTPRS человека, или фракцию мембран этой клетки для получения моноклонального антитела по любому из пп.1-6.
11. Применение по п.10, при котором клетка животного является мышиной клеткой.
12. Способ детекции плазмацитоидной дендритной клетки, который включает:
приведение моноклонального антитела или его антиген-связывающего фрагмента, которое связывается с внеклеточным доменом PTPRS человека, по любому из пп.1-6, в контакт с клеткой, подвергающейся воздействию,
детекцию указанного моноклонального антитела или его фрагмента, которое связано с клеткой, таким образом детектирование плазмоцитоидной дендритной клетки.
13. Применение моноклонального антитела или его фрагмента по любому из пп.1-6 для детекции плазмоцитоидной дендритной клетки.
14. Способ подавления активности плазмацитоидной дендритной клетки ex vivo, который включает приведение моноклонального антитела или его фрагмента по любому из пп.1-11 в контакт с плазмацитоидной дендритной клеткой,
где указанной активностью плазмоцитоидной дендритной клетки является продукция IFN и/или клеточное выживание, таким образом, подавляя активность плазмацитоидной дендритной клетки.
15. Применение моноклонального антитела или его фрагмента по любому из пп.1-6 для лечения заболевания, связанного с рDC.
16. Применение моноклонального антитела или фрагмента по любому из пп.1-6 для лечения аутоиммунного заболевания, связанного с IFNальфа.
| WO 2010056332 A1 A1 20.05.2010 | |||
| WO 1995009656 A1 13.04.1995 | |||
| WO 2011097327 A2 11.08.2011 | |||
| WO 2007041317 A2 12.04.2007 | |||
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО И ПРОДУЦИРУЮЩАЯ ЕГО ГИБРИДОМА | 2004 |
|
RU2312109C2 |

