Область техники
Настоящее изобретение относится к соединению, а также к способам его получения и применениям, и, в частности, относится к классу конъюгатов цистеин-модифицированное антитело-токсин (TDC) и к способам их получения и к их применениям.
Уровень техники:
Конъюгаты антитело-лекарственное средство (ADC) в настоящее время представляют большой интерес в области направленной терапии, причем два лекарственных средства уже одобрено в США: адцетрис и кадсила, которые демонстрируют высокую клиническую эффективность, и более 50 ADC в настоящее время исследуют в качестве части испытаний клинической фазы. Новый конъюгат цистеин-модифицированное антитело-токсин (TDC), описанный в этом патенте, имеет преимущество, состоящее в лучшей однородности лекарственного средства и меньших побочных эффектах по сравнению с ADC с присоединением не в фиксированной точке, и в доклинических испытаниях было показано, что он имеет значительно лучшие характеристики, чем ADC с присоединением не в фиксированной точке.
Сущность изобретения:
Для конъюгата цистеин-модифицированное антитело-токсин (TDC) по настоящему изобретению соответствующие участки встраивания цистеина включают один или несколько участков, выбранных из следующих 15 участков встраивания: положение 110 легкой цепи (номер согласно Kabat с аминокислотной последовательностью EIKRTCVAAPS), положение 111 легкой цепи (номер согласно Kabat с аминокислотной последовательностью IKRTVCAAPSV), положение 142 легкой цепи (номер согласно Kabat с аминокислотной последовательностью NNFYPCREAKV), положение 254 тяжелой цепи (c аминокислотной последовательностью ISRTPCEVTCV), положение 255 тяжелой цепи (с аминокислотной последовательностью SRTPECVTCVV), положение 258 тяжелой цепи (с аминокислотной последовательностью PEVTCCVVVDV), положение 259 тяжелой цепи (с аминокислотной последовательностью EVTCVCVVDVS), положение 354 тяжелой цепи (с аминокислотной последовательностью PSRDECLTKNQ), положение 355 тяжелой цепи (с аминокислотной последовательностью SRDELCTKNQV), положение 357 тяжелой цепи (с аминокислотной последовательностью DELTKCNQVSL), положение 378 тяжелой цепи (с аминокислотной последовательностью IAVEWCESNGQ), положение 379 тяжелой цепи (с аминокислотной последовательностью AVEWECSNGQP), положение 386 тяжелой цепи (с аминокислотной последовательностью GQPENCNYKTT), положение 387 тяжелой цепи (с аминокислотной последовательностью QPENNCYKTTP) и положение 410 тяжелой цепи (с аминокислотной последовательностью KLTVDCKSRWQ). Антитела, которые содержат одну или несколько мутаций-инсерций цистеина, описанных выше, сохраняют способность их исходного антитела связываться с соответствующими антигенами (аффинность). Путем проведения связывания конъюгата антитело-токсин (TDC) в фиксированной точке между сульфгидрильной группой цистеина, встроенного в легкую цепь, и/или сульфгидрильной группой цистеина, встроенной в тяжелую цепь, может быть достигнуто соотношение токсина и антитела (DAR) от 1,6 до 2,0 или от 3,2 до 4,0.
Примеры
Пример 1: Синтез mc
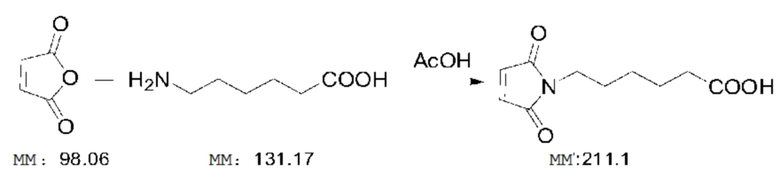
3,9 г (0,03 моль) 6-аминокапроновой кислоты и 3,5 г (0,036 моль) 1,2 экв. малеинового ангидрида добавляли к 30 мл ледяной уксусной кислоты. Реакционный раствор перемешивали при 120°C в течение от 4 до 6 часов. После завершения реакции нагревание прекращали и раствору позволяли остыть до комнатной температуры естественным образом. Затем раствор концентрировали при пониженном давлении при 60°C для удаления большей части уксусной кислоты. Полученную таким образом коричневато-желтую вязкую жидкость выливали в воду, проводили экстракцию этилацетатом три раза (20 мл каждый раз) и органические слои объединяли. Органический слой последовательно промывали водой и насыщенным рассолом, сушили безводным сульфатом натрия и фильтровали, и фильтрат концентрировали при пониженном давлении с получением коричневато-желтого маслянистого вещества, после чего добавляли 50 мл воды и проводили перемешивание, что приводило к преципитации не совсем белого твердого вещества, которое фильтровали и сушили при 50°C при пониженном давлении с получением 5,08 г заданного продукта с выходом 80%. mp: от 89 до 92°C. m/z: 212,2 [M+H]+. 1H-ЯМР (400 МГц, DMSO): 13,21 (ушир., 1H, COOH), 6,75 (с, 2H, COCH=CHCO), 3,63 (т, 2H, J=7,2 Гц, NCH2CH2), 2,42(т, 2H, J=7,4 Гц, CH2COOH), 1,52-1,68 (м, 4H, NCH2CH2CH2CH2), 1,30-1,42 (м, 2H, NCH2CH2CH2CH2).
Пример 2: Синтез Mc-OSu
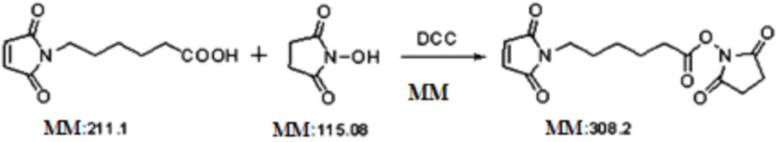
4,7 г (22 ммоль) MC и 25 г (22 ммоль) HOSu добавляли к 50 мл ацетонитрила в атмосфере газообразазного азота. 4,5 г (22 ммоль) DCC отдельно растворяли в 25 мл ацетонитрила при поддержании внутренней температуры приблизительно 0°C, и этот раствор медленно капельно добавляли к описанному выше реакционному раствору. Реакционному раствору позволяли реагировать при 0°C в течение двух часов, а затем доводили до комнатной температуры, после чего реакции позволяли продолжаться в течение ночи. После фильтрации полученный остаток на фильтре промывали ацетонитрилом три раза (по 10 мл каждый раз) и фильтрат концентрировали при пониженном давлении до сухого состояния. Полученное таким образом масло сушили при пониженном давлении при комнатной температуре в течение шести часов с получением 6,4 г светло-коричневого твердого вещества с выходом 95%. (Затем его использовали непосредственно в следующей реакции без дальнейшей очистки) m/z: 309,2 [M+H]+. 1H-ЯМР (400 МГц, CDCl3): 1-2 (м, 6H, CCH2CH2CH2C), 2,68 (т, 2H, CH2CO), 2,95 (с, 4H, COCH2CH2CO), 3,68 (т, 2H, CH2N), 6,81 (с, 2H, CH=CH).
Пример 3: Синтез Fmoc-Val-OSu
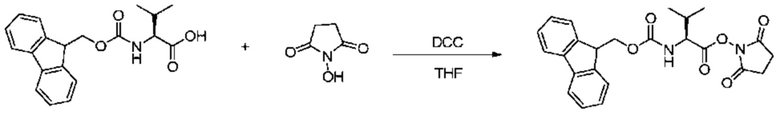
10 г Fmoc-Val и 3,4 г HOSu добавляли к 100 мл THF. 6 г DCC отдельно растворяли в 50 мл ацетонитрила при поддержании внутренней температуры приблизительно 0°C, и раствор медленно капельно добавляли к описанному выше реакционному раствору. Реакционный раствор перемешивали при комнатной температуре в течение 24 часов. После фильтрации остаток на фильтре промывали THF, и фильтрат концентрировали при пониженном давлении с получением прозрачного масла. Маслянистое вещество использовали непосредственно в следующей реакции без дальнейшей очистки. m/z: 437,4 [M+H]+.
Пример 4: Синтез Fmoc-vc
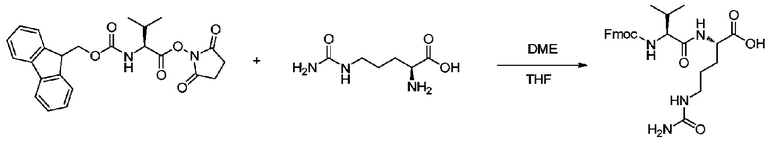
4,0 г (1,05 экв.) Cit и 60 мл водного раствора бикарбоната натрия (NaHCO3, 2 г, 1,05 экв.) добавляли к 20 мл THF. 22,35 ммоль Fmoc-Val-OSu отдельно растворяли в 60 мл DME, и полученный раствор добавляли к реакционному раствору. Реакционный раствор перемешивали при комнатной температуре в течение 24 часов. После завершения реакции к системе добавляли 110 мл 15% водного раствора лимонной кислоты и проводили экстракцию два раза посредством EA, и органические слои объединяли и концентрировали при пониженном давлении с получением белого твердого вещества. К белому твердому веществу добавляли 100 мл метил трет-бутилового эфира и проводили перемешивание с последующей фильтрацией; остаток на фильтре подвергали сушке при пониженном давлении при 40°C в течение 4 ч с получением 4,83 г продукта с выходом 65%. m/z: 497,6 (M+H)+. 1H-ЯМР (400 МГц, DMSO): 0,92 (6H, м), 1,35-1,65 (4H, м), 2,10 (1H, м), 3,01 (2H, кв), 3,99 (1H, т), 4,01-4,45 (2H, м), 4,45 (2H, т), 5,46 (2H, ушир.), 6,03 (1H, т), 7,20-8,02 (8H, м), 8,25 (1H, д).
Пример 5: Синтез Fmoc-vc-PABOH
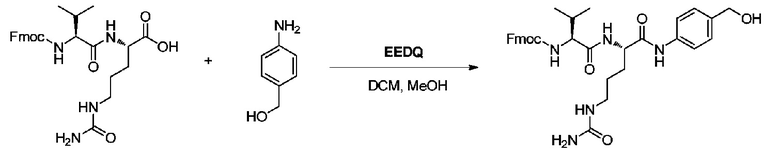
60 мл смешанного растворителя DCM/MeOH=2/1 добавляли в реакционную колбу, а затем добавляли 2 г (4,2 ммоль) Fmoc-vc и 1,04 г (2 экв.) PABOH, после чего смесь перемешивали до частичного растворения и добавляли 2,0 г (2 экв.) EEDQ. Реакционной системе позволяли продолжать реагировать при перемешивании в течение 2,0 суток в темноте при комнатной температуре. После завершения реакции реакционный раствор концентрировали при пониженном давлении при 40°C с получением белого твердого вещества. Белое твердое вещество собирали, добавляли 100 мл метил трет-бутилового эфира и проводили перемешивание, а затем фильтрацию; остаток на фильтре промывали метил трет-бутиловым эфиром и полученное белое твердое вещество сушили при пониженном давлении при 40°C с получением 2,2 г продукта с выходом приблизительно 88%. m/z: 602,6 (M+H)+. 1H-ЯМР (400 МГц, DMSO): 0,95 (6H, м), 1,45-1,69 (4H, м), 2,10 (1H, м), 3,11 (2H, м), 3,99 (1H, м), 4,30 (2H, д), 4,05-4,66 (2H, м), 4,55 (2H, д), 5,21 (1H, т), 5,51 (2H, ушир.), 6,11 (1H, т), 7,09-8,10 (12H, м), 8,21 (1H, д), 10,51 (1H, ушир.).
Пример 6: Синтез vc-PABOH
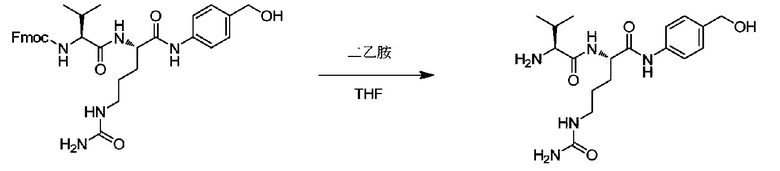
490 мг (0,815 ммоль) Fmoc-vc-PABOH добавляли к 10 мл NMP и смесь перемешивали до растворения, после чего добавляли 2 мл диэтиламина. Затем реакции позволяли протекать при перемешивании при комнатной температуре в течение 24 часов. После завершения реакции раствор концентрировали при пониженном давлении при 40°C, к полученному таким образом маслянистому веществу добавляли 20 мл DCM и перемешивание продолжали до получения кристаллов, которые отфильтровывали; затем остаток на фильтре промывали DCM, и полученное таким образом твердое вещество сушили при пониженном давлении с получением 277 мг продукта с выходом 90%. m/z: 380,2 (M+H)+. 1H-ЯМР (400 МГц, DMSO): 0,89 (6H, м), 1,31-1,61 (4H, м), 1,82 (1H, м), 2,86 (1H, м), 2,89 (2H, д), 4,38 (2H, д), 4,44 (1H, м), 5,01 (1H, ушир.), 5,35 (2H, ушир.), 5,84 (1H, ушир.), 7,14 (2H, д), 7,42 (2H, д), 8,08 (1H, ушир.), 9,88 (1H, ушир.).
Пример 7: Синтез mc-vc-PABOH
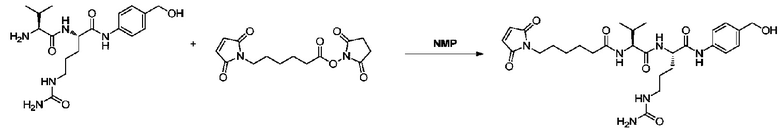
205 мг (0,54 ммоль) vc-PABOH и 184 мг (1,1 экв.) MC-OSu добавляли к 10 мл NMP, и после завершения добавления реакции позволяли протекать при перемешивании при комнатной температуре в течение 24 часов. После завершения реакции реакционный раствор концентрировали при пониженном давлении при 40°C, к полученному таким образом маслянистому веществу добавляли 20 мл метил трет-бутилового эфира и проводили перемешивание до получения кристаллов. Затем кристаллы отфильтровывали и остаток на фильтре промывали метил трет-бутиловым эфиром с получением 310 мг продукта с выходом 100%. m/z: 573,3 (M+H)+. 1H-ЯМР (400 МГц, DMSO): 0,89 (6H, м), 1,15-1,99 (10H, м), 2,11 (1H, м), 2,31 (2H, т), 3,21 (2H, м), 3,53 (2H, т), 4,32 (1H, t), 4,51 (1H, м), 4,59 (2H, ушир.), 5,24 (1H, ушир.), 5,56 (2H, ушир.), 6,20 (1H, ушир.), 7,12 (2H, с), 7,23 (2H, д), 7,58 (2H, д), 7,94 (1H, д), 8,17 (1H, д), 10,21 (1H, ушир.).
Пример 8: Синтез mc-vc-PAB-PNP
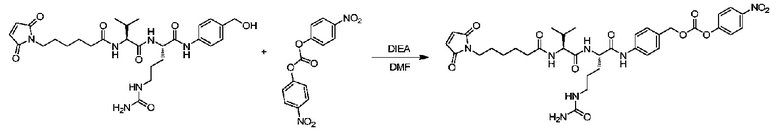
168,6 мг (0,294 ммоль) mc-vc-PABOH растворяли в 5 мл безводного пиридина в атмосфере газообразного азота, и реакционную систему охлаждали приблизительно до 0°C. 179 мг (3 экв.) PNP отдельно растворяли в 5 мл DCM, и полученный раствор медленно добавляли в реакционную систему. После поддержания температуры при 0°C в течение 10 мин ледяную баню удаляли и реакционный раствор перемешивали при комнатной температуре в течение трех часов. После завершения реакции добавляли 70 мл EA и 100 мл 15% водного раствора лимонной кислоты, и органический слой отделяли. Органический слой последовательно промывали лимонной кислотой, водой и насыщенным рассолом, а затем сушили над безводным сульфатом натрия и фильтровали, и фильтрат концентрировали при пониженном давлении до сухого состояния с получением светло-желтого масла, после чего добавляли метил трет-бутиловый эфир для индукции кристаллизации и получали 86 мг не совсем белого твердого вещества с выходом 40%. m/z: 738 (M+H)+. 1H-ЯМР (400 МГц, CDCl3/CD3OD): 0,84 (6H, м), 1,11-1,84 (10H, м), 2,05 (1H, м), 2,15 (2H, т), 3,09 (2H, м), 3,32 (2H, т), 4,12(1H, м), 4,38 (1H, м), 5,15 (2H, с), 6,61 (2H, с), 6,84 (1H, д), 7,61 (1H, д), 7,21 (2H, д), 7,50 (2H, д), 7,61 (2H, д), 8,18 (2H, д), 9,59 (1H, ушир.).
Пример 9: Синтез mc-vc-PAB-MMAE
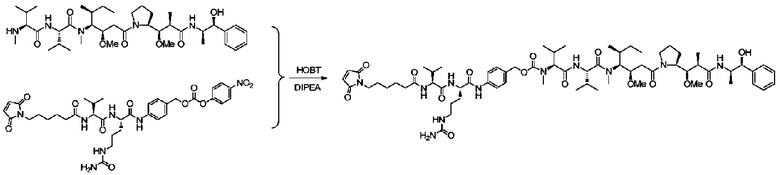
20 мг mc-vc-PAB-PNP (1,5 экв.) и 3 мг HOBT добавляли к 2 мл DMF. После кратковременного перемешивания смеси при комнатной температуре добавляли 13 мг MMAE, 0,5 мл пиридина и 25 мкл DIEA. Реакционный раствор перемешивали при комнатной температуре в течение 2 суток. После завершения реакции реакционный раствор прямо очищали с использованием препаративной колонки и требуемые компоненты собирали, концентрировали и лиофилизировали с получением приблизительно 10 мг продукта с выходом приблизительно 42%. m/z: 1317,1 (M+H)+.
Пример 10: Синтез mc-vc-PAB-MMAF
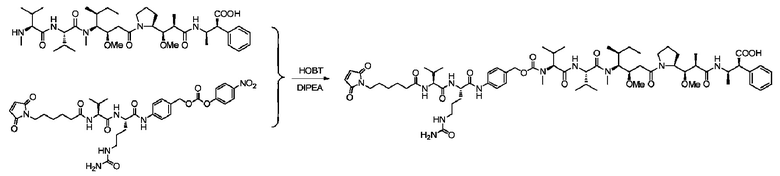
С использованием той же методики, что и в примере 9, получали приблизительно 12,5 мг mc-vc-PAB-MMAF с выходом 45,2%; m/z: 1345,7 (M+H)+.
Пример 11 Синтез mc-vc-PAB-PBD
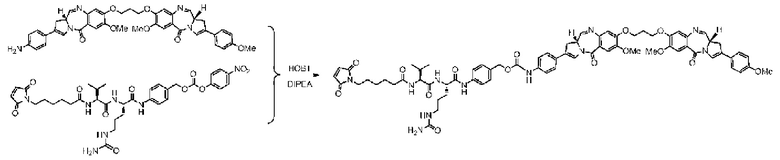
С использованием той же методики, что и в примере 9, получали приблизительно 9,5 мг mc-vc-PAB-PBD с выходом 32,5%; m/z: 1325,4 (M+H)+.
Пример 12 Синтез mc-vc-PAB-DOX
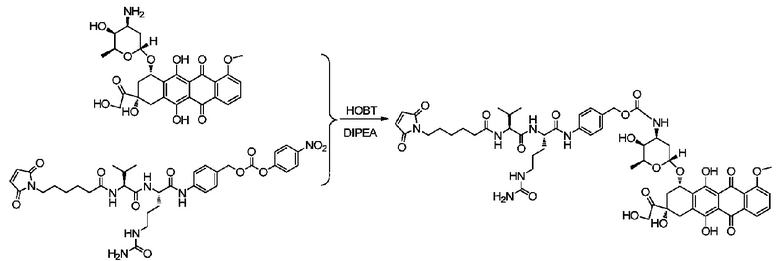
С использованием той же методики, что и в примере 9, получали приблизительно 11,2 мг mc-vc-PAB-DOX с выходом 38,9%; m/z: 1143,2 (M+H)+.
Пример 13: Синтез mc-vc-PAB-SN-38
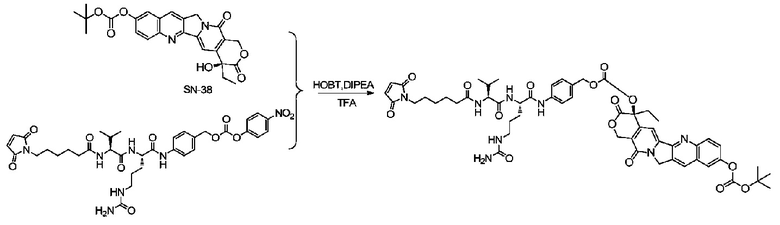
После растворения 100 мг коммерчески приобретенного 10-0-Boc-SN-38 в 10 мл сухого дихлорметана добавляли 25,6 мг (1 экв.) DMAP, и капельно добавляли раствор трифосгена в дихлорметане (62 мг трифосгена, растворенные в 2 мл дихлорметана) при 0°C, после чего реакции позволяли протекать при 0°C в течение 12 ч; затем дихлорметан удаляли при пониженном давлении и полученное вещество растворяли в 10 мл сухого DMF, после чего добавляли 144 мг mc-vc-PABOH и полученную смесь перемешивали в течение 24 часов при комнатной температуре и проводили препаративную ВЭЖХ с получением 41 мг mc-vc-PAB-SN-38 с общим выходом за две стадии 19,7%, m/z: 1063,2 (M+H)+.
Пример 14: Экспрессия и очистка антитела 2A1-HC-Cys254ins
Для экспрессии антитела 2A1-HC-Cys254ins использовали суспензию клеток FreestyleTM 293-F (Invitrogen). За одни сутки до трансфекции клетки высевали с плотностью 6×105 клеток/мл во вращающийся флакон объемом 1 л, содержавший 300 мл полной среды F17 (среда для экспрессии FreestyleTM F17, Gibco), и инкубировали в течение ночи при 37°C и 5% CO2 на устройстве для встряхивания клеток при 120 об/мин. На следующий день проводили трансфекцию экспрессирующей антитело плазмидой с использованием PEI, где соотношение плазмиды и PEI составляло 2:1. Через одни сутки после трансфекции добавляли питательную среду TN1 в количестве 2,5% (об./об.), культивирование проводили в течение дополнительных четырех суток и супернатант собирали центрифугированием.
Полученный супернатант для экспрессии клеток собирали и элюировали 0,1 M лимонной кислотой (pH 3,0) с использованием колонки для аффинной хроматографии с белком A (Mabselect Sure LX, GE), и уловленные антитела доводили до pH 7,0 с использованием 1 M Tris-HCl (pH 9,0) в количестве 1/10 (об./об.), а затем пропускали через хроматографическую колонку для гель-фильтрации SEC (Superdex 200, GE) для удаления примесей, таких как агрегаты и эндотоксины, в то время как буфер для антитела заменяли на PBS (pH 7,4), и заданные образцы пика при УФ 280 нм собирали и концентрировали до 2 мг/мл с использованием центрифужных пробирок для ультрафильтрации (30KD, Pall).
Полученные таким образом антитела 2A1-HC-Cys254ins продемонстрировали концентрацию 2 мг/мл с долей заданного мономера антитела (POI%) более 90%, и их использовали в последующих экспериментах.
Пример 15: Получение образцов TDC 2A1-HC-Cys357ins-mc-vc-PAB-MMAE посредством связывания антитела 2A1-HC-Cys357ins и mc-vc-PAB-MMAE
Экспрессированные клетками антитела 2A1-HC-Cys357ins очищали с использованием Mabselect Sure и после элюирования при pH сразу добавляли раствор Tris для достижения нейтрализации и раствор заменяли на буфер Tris-HCl при pH 7,5. Соединение mc-vc-PAB-MMAE в виде белого порошка и растворяли в DMA для последующего применения. Антитело сначала необходимо было восстановить для устранения барьера на мутантном остатке цистеина. К раствору антитела 2A1-HC-Cys357ins добавляли 1 M водный раствор DTT в молярном количестве 40 экв. и полученную смесь хорошо перемешивали и позволяли ей реагировать при 20°C в течение двух часов. После этого pH образца доводили до 5,0 и DTT и барьер в образце устраняли посредством катионообменной хроматографии с SP Sepharose FF. Далее к образцу добавляли раствор DHAA в молекулярном соотношении 20x и реакции позволяли протекать в течение четырех часов при 25°C в темноте для восстановления дисульфидных связей между цепями антител. После этого добавляли раствор mc-vc-PAB-MMAE для индукции связывания между mc-vc-PAB-MMAE и антителом с мутацией на цистеин, после чего проводили тщательное перемешивание и реакции позволяли протекать при 25°C в течение двух часов. После завершения реакции использовали катионообменную хроматографию с SP Sepharose FF для удаления mc-vc-PAB-MMAE, который не был связан с молекулой антитела, и получали образцы TDC 2A1-HC-Cys357ins-mc-vc-PAB-MMAE.
Пример 16: Измерение посредством HIC-ВЭЖХ соотношения токсина и антитела (DAR)
Образцы TDC анализировали посредством гидрофобной высокоэффективной жидкостной хроматографии, и DAR вычисляли, исходя их соответствующей площади пика. Конкретный использованный способ был следующим:
Колонка: Proteimix® HICBu-NP5 (5 мкм, 4,6×35 мм);
Подвижные фазы: A: 2 M сульфат аммония, 0,025M, фосфатный буфер с pH 7; B: 0,025M, фосфатный буфер с pH 7; C: 100% изопропанол;
Буфер A уравновешивали, буфер B и буфер C использовали для проведения градиентного элюирования, и детекцию проводили при 25°C и 214 нм.
На фиг.1A-1D показаны: Детекция посредством HIC-ВЭЖХ TDC DAR 2A1-HC-Cys357ins-mc-vc-PAB-MMAE, 2A1-HC-Cys378ins-mc-vc-PAB-MMAE, 2A1-HC-Cys387ins-mc-vc-PAB-MMAE и 2A1-HC- Cys410ins-mc-vc-PAB-MMAE, соответственно.
Данные, приведенные на фиг.1A-1D, использовали для вычисления DAR для присоединения в фиксированной точке, которые находились в диапазоне от 1,6 до 1,7, и была продемонстрирована высокая однородность соединения.
В прилагаемой таблице 1: TDC 2A1-LC-Cys110ins-mc-vc-PAB-MMAE,
TDC 2A1-LC-Cys111ins-mc-vc-PAB-MMAE, TDC 2A1-LC-Cys142ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys254ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys255ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys258ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys259ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys354ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys355ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys357ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys378ins-mc-vc-PAB-MMAE TDC, TDC 2A1-HC-Cys379ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys386ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys387ins-mc-vc-PAB-MMAE и 2A1-HC-Cys410ins-mc-vc-PAB-MMAE
Прилагаемая таблица 1: Таблица, демонстрирующая эффективность присоединения в TDC (DAR)


В прилагаемой таблице 1 показано, что эффективность присоединения в соединениях TDC после присоединения в фиксированной точке посредством модификации путем мутации-инсерции цистеина является относительно высокой (теоретическая максимальная величина составляет 2,0), и DAR составляло ≥ 1,6.
Пример 17: Определение посредством SEC-ВЭЖХ стабильности основной цепи антитела TDC и агрегации TDC
Образцы антитела TDC хранили при 37°C, и агрегацию анализировали посредством SEC-ВЭЖХ на 0, 7, 14 и 21 сутки. Конкретная методология была следующей:
Колонка: TSKgel SuperSW mAb HR (7,8 мм × 30 см)
Подвижная фаза: 0,1 M сульфат натрия, 0,1 M фосфатный буфер, pH 6,7.
Детекцию проводили при 25°C и 280 нм.
Прилагаемая таблица 2
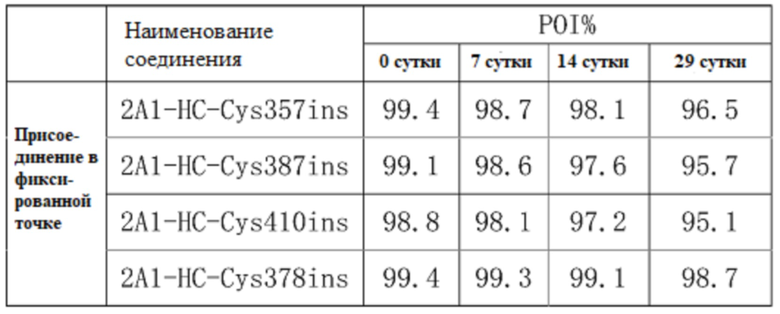
На фиг.2A-2D показаны: 2A1-HC-Cys378ins Результаты SEC на 0, 7, 14 и 21 сутки. Данные показывают, что образуц был в высокой степени стабильным при хранении при 37°C в течение трех недель.
Прилагаемая таблица 2: Определение посредством SEC-ВЭЖХ агрегации антительных каркасов TDC 2A1-HC-Cys357ins, 2A1-HC-Cys387ins, 2A1-HC-Cys378ins, и 2A1-HC-Cys410ins; образцы хранили при 37°C в течение трех недель практически без изменения содержания мономера POI;
TDC 2A1-LC-Cys110ins-mc-vc-PAB-MMAE, TDC 2A1-LC-Cys111ins-mc-vc-PAB-MMAE, TDC 2A1-LC-Cys142ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys254ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys255ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys258ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys259ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys354ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys355ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys357ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys378ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys379ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys386ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys387ins-mc-vc-PAB-MMAE и 2A1-HC-Cys410ins-mc-vc-PAB-MMAE
Прилагаемая таблица 3: Содержание заданного мономера TDC
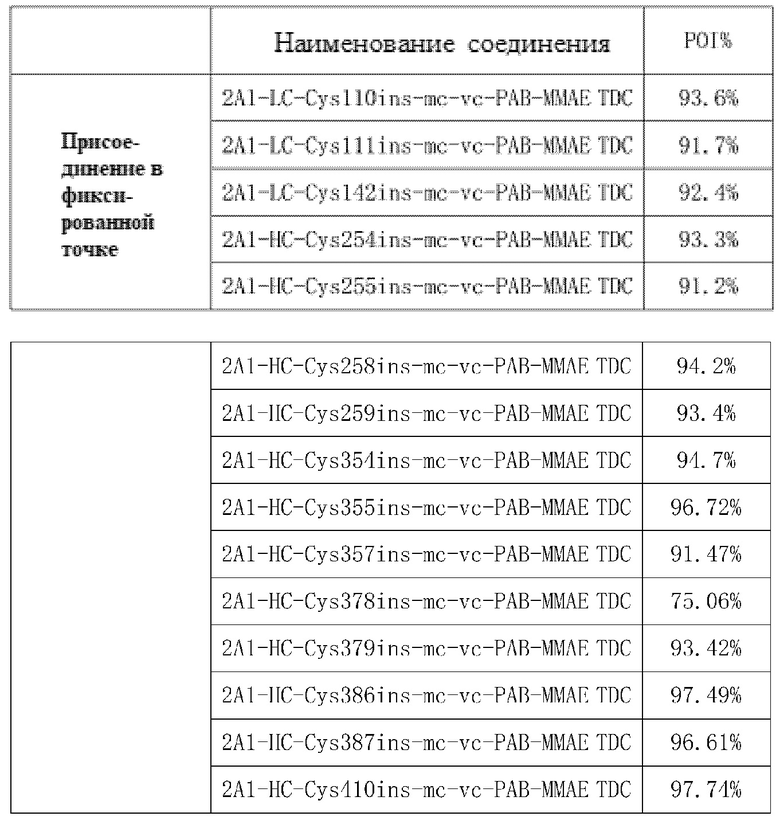
В прилагаемой таблице 3 показано, что содержание заданного мономера в соединениях TDC с присоединением цистеина в фиксированной точке составляло более 90%.
Пример 18: Аффинность антительного каркаса после модификации посредством мутации-инсерции цистеина и исходного антитела 2A1 в отношении EGFRvIII
ELISA использовали для сравнения относительной аффинности TDC 2A1-LC-Cys110ins-mc-vc-PAB-MMAE, TDC 2A1-LC-Cys111ins-mc-vc-PAB-MMAE, TDC 2A1-LC-Cys142ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys254ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys255ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys258ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys259ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys354ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys355ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys357ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys378ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys379ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys386ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys387ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys410ins-mc-vc-PAB-MMAE и TDC 2A1 в отношении EGFRvIII. Конкретный использованный способ был следующим:
Рекомбинантный антиген EGFRvIII-His*6 использовали для нанесения на планшеты; в качестве заливки использовали желатин рыбьей кожи; разведение каждого мутантного антитела с инсерцией проводили по отдельности вплоть до максимальной концентрации 10 мкг/мл и 4-кратное градиентное разведение проводили всего для 11 концентраций; для инкубации использовали меченные HRP вторичные антитела; для проявки использовали TMB и детекцию поглощения проводили при 450 нм. Результаты анализа наносили на график в качестве парных концентраций A450, и, как показано на фиг.3A-3E и в таблице 3, антитела с мутацией посредством инсерции цистеина сохраняли сходную аффинность в отношении 2A1 с величинами EC50, очень близкими друг к другу, что указывает на то, что мутации с инсерциями в легкой или тяжелой цепях на 2A1 не влияют на аффинность в отношении антигена EGFRvIII.
Прилагаемая таблица 4: Определение аффинности мутантного антитела

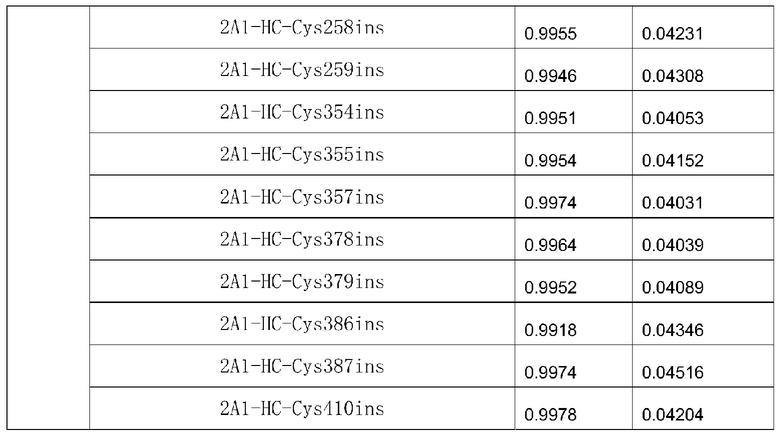
На фиг.3 и в прилагаемой таблице 4: Антитела 2A1-LC-Cys110ins, 2A1-LC-Cys111ins, 2A1-LC-Cys142ins, 2A1-HC-Cys254ins, 2A1-HC-Cys255ins, 2A1-HC-Cys258ins, 2A1-HC-Cys259ins, 2A1 -HC-Cys354ins, 2A1-HC-Cys355ins, 2A1-HC-Cys357ins, 2A1-HC-Cys378ins, 2A1-HC-Cys379ins, 2A1-HC-Cys386ins, 2A1-HC-Cys387ins и 2A1-HC-Cys410ins сохраняли аффинность 2A1 в отношении антигена EGFRvIII.
Пример 19: Тестирование цитотоксичности
Цитотоксическую активность TDC определяли по следующей экспериментальной методике: TDC добавляли в сверхэкспрессирующую EGFR или экспрессирующую EGFRVIII культуральную среду клеток опухоли человека и определяли жизнеспособность клеток после культивирования клеток в течение 72 часов. Для определения жизнеспособности клеток, цитотоксичности и запрограммированной клеточной смерти, индуцированных TDC по настоящему изобретению, использовали клеточные эксперименты in vitro.
Эффективность in vitro конъюгатов антитело-токсин определяли посредством анализа клеточной пролиферации. Анализ CellTiter96® AqueousOne Solution Cell Proliferation Assay приобретали коммерчески от Promega Corp. (Madison, WI). Анализ CellTiter96® Aqueous One Solution Cell Proliferation Assay (a) представляет собой колориметрический способ, используемый для определения количества живых клеток в экспериментах по определению пролиферации и цитотоксичности клеток. Указанный выше реагент содержит новое соединение тетразола [3-(4,5-диметилтиазол-2-ил)-5-(3-карбоксиметоксифенил)-2-(4-сульфофенил)-2H-тетразолий, внутренняя соль; MTS) а также агент электронного присоединения (феназин этосульфат; PES). PES демонстрирует повышенную химическую стабильность, которая позволяет смешение его с MTS с получением стабильного раствора. Эта удобная модель "одного раствора" является усовершенствованием относительно первого поколения анализов CellTiter96® Aqueous Assay, в котором агент электронного присоединения PMS предоставлялся отдельно от раствора MTS. MTS (раегент Оуэна) восстанавливается клеточными механизмами в окрашенный продукт формазан, который может прямо растворяться в культуральной среде, как показано на схеме ниже. Это конвертирование вероятно облегчается посредством NADPH или NADH, продуцируемых дегидрогеназой, присутствующей в метаболически активных клетках. Для анализа небольшое количество реагента CellTiter96® Aqueous One Solution Reagent добавляют прямо в среду, находящуюся в лунках планшета для культивирования, после чего проводят инкубацию в течение от 1 до 4 часов, с последующим считыванием величины поглощения при 490 нм с использованием устройства для считывания микропланшетов.

Структура MTS, соли тетразолия, и его продукта формазана.
Количество продукта формазана, выявленное при 490 нм, пропорционально количеству жизнеспособных клеток в культуре. Поскольку продукт MTS формазан является растворимым в среде для культивирования анализ CellTiter96® Aqueous One Solution Assay имеет меньше стадий, чем способы с MTT или INT.
A431 (клетки, сверхэкспрессирующие EGFR) и U87-EGFRVIII (стабильная клеточная линия с мутантным EGFRVIII) использовали в рамках настоящего изобретения в качестве исследуемой системы для проведения анализов эффективности in vitro. В 96-луночных планшетах клетки высевали в количестве 6000 клеток/лунка и через 24 часа проводили добавление антитела. Дозируемые концентрации для A431 и U87-EGFRvIII находились в диапазоне от 200 нМ до 10 пМ при четырехкратном разведении, и концентрация U87-EGFRVIII находилась в диапазоне от 500 нМ до 30 пМ. Жизнеспособность клеток анализировали с использованием MTS через 72 часа после обработки.
На фиг.4A и 4B показаны: Результаты определения IC50 для 2A1-HC-Cys357ins-mc-vc-PAB-MMAE, 2A1-HC-Cys378ins-mc-vc-PAB-MMAE, 2A1-HC-Cys387ins-mc-vc-PAB-MMAE и 2A1-HC-Cys410ins-mc-vc-PAB-MMAE на сверхэкспрессирующей EGFRwt клеточной линии плоскоклеточной карциномы кожи человека A431 и сверхэкспрессирующей EFGRvIII клеточной линии глиомы человека U87-EGFRvIII.
Прилагаемая таблица 5. Результаты определения IC50, демонстрирующие цитотоксичность TDC в отношении клеточной линии A431, сверхэкспрессирующей EGFRwt, и клеточной линии U87-EGFRvIII, сверхэкспрессирующей EFGRvIII
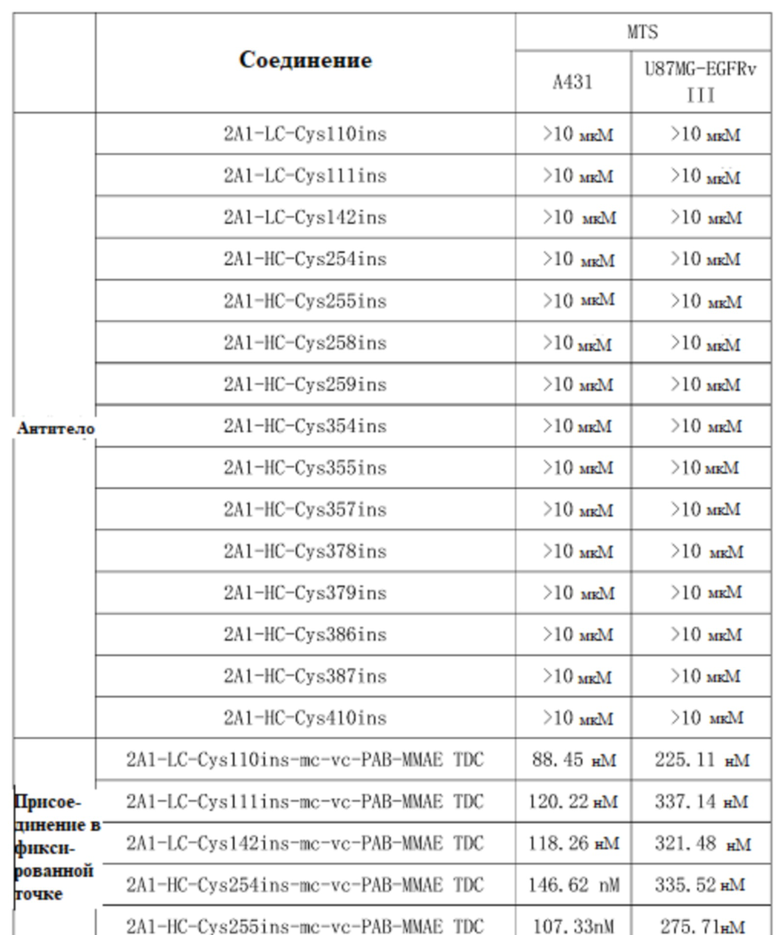
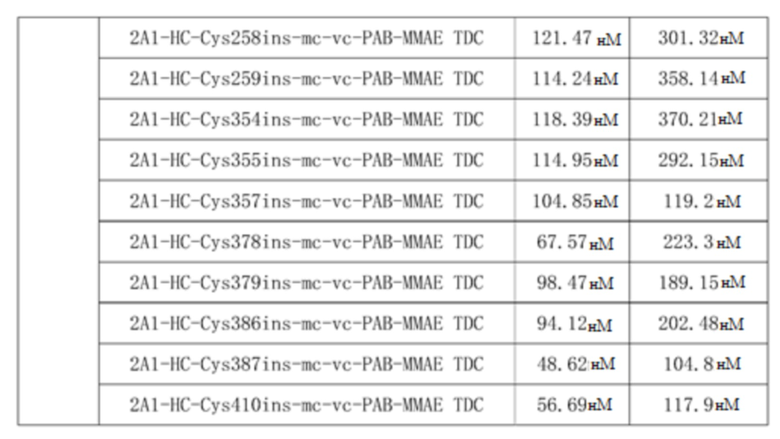
Результаты, приведенные в таблице 5, показывают, что TDC 2A1-LC-Cys110ins-mc-vc-PAB-MMAE, TDC 2A1-LC-Cys111ins-mc-vc-PAB-MMAE, TDC 2A1-LC-Cys142ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys254ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys255ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys258ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys259ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys354ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys355ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys357ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys378ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys379ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys386ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys387ins-mc-vc-PAB-MMAE и 2A1-HC-Cys410ins-mc-vc-PAB-MMAE, демонстрируют значительную цитотоксическую активность против клеточной линии A431, сверхэкспрессирующей EGFRwt, и клеточной линии U87-EGFRvIII, сверхэкспрессирующей EFGRvIII.
Объем настоящего изобретения не ограничивается вариантами осуществления, раскрытыми в примерах; указанные примеры используются для иллюстрации нескольких аспектов изобретения, и любые варианты осуществления, которые являются функционально эквивалентными, входят в объем настоящего изобретения. Действительно, в дополнение к примерам, показанным и описанных в настоящем описании, дополнительные варианты изобретения также станут понятными специалисту в данной области и входят в объем формулы изобретения, прилагаемой к ним.
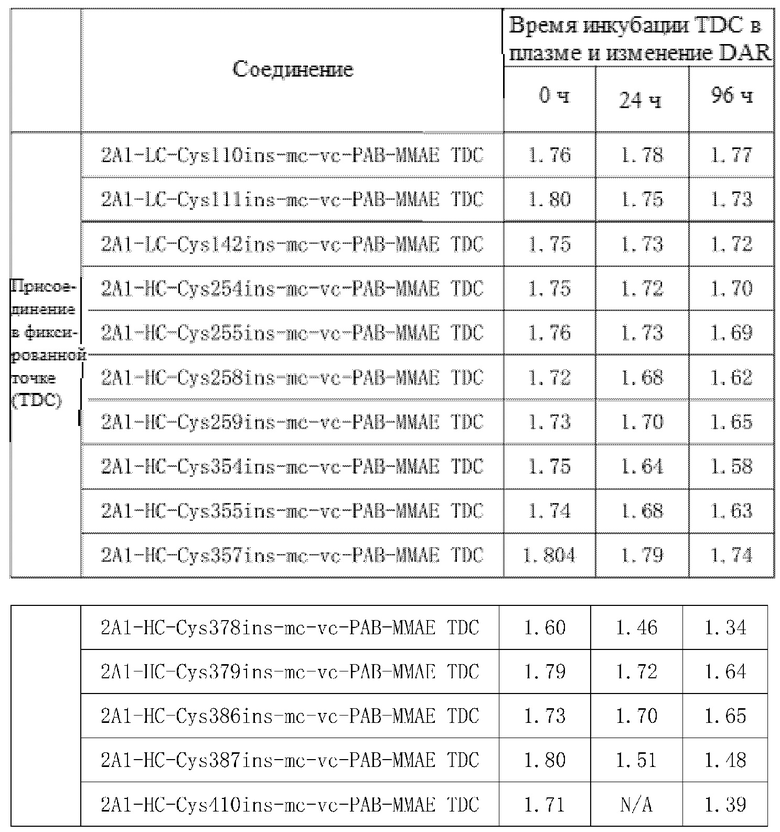
Пример 20. Тестирование стабильности в плазме
Фиксированное количество образца TDC добавляли к плазме человека, из которой IgG человека уже был удален, причем каждую пробирку с TDC подготавливали в трех экземплярах; далее проводили инкубацию на водяной бане при 37°C в течение 0 часов, 24 часов и 72 часов, после чего образец TDC извлекали, а затем в каждую пробирку добавляли 100 мкл смолы с белком A (MabSelect SuReTM LX, партия:#10221479GE, промытая PBS); затем позволяли происходить адсорбции при встряхивании на вертикальном смесителе в течение двух часов, а затем происходило промывание и элюирование с получением инкубированного TDC. Тестирование HIC-ВЭЖХ и RP-ВЭЖХ проводили для образцов TDC, инкубированных в течение определенного периода времени, для оценки стабильности образцов в плазме.
На фиг.5A-5D показаны: Результаты испытания стабильности TDC 2A1-HC-Cys357ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys410ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys387ins-mc-vc-PAB-MMAE, TDC 2A1-HC-Cys378ins-mc-vc-PAB-MMAE в плазме посредством HIC-ВЭЖХ. Результаты, представленные на фиг.5A-5D, демонстрируют, что эти три молекулы демонстрируют хорошую стабильность в плазме.
Прилагаемая таблица 6. Результаты испытания стабильности TDC в плазме
Пример 21: Тестирование эффективности лекарственного средства у мышей, имеющих опухоль
В рамках настоящего изобретения авторы настоящего изобретения разработали модель с подкожной трансплантации опухоли клеточной линии рака поджелудочной железы человека BxPC3 у мышей nude для оценки эффективности in vivo конъюгатов TDC, таких как 2A1-HC-Cys357ins-mc-vc-PAB-MMAE. 5×106 клеток A431 инъецировали подкожно в бока мышей BALB/c nude в возрасте от 4 до 6 недель, и, когда средний размер опухоли достигал приблизительно 150-200 мм3, мышей случайным образом распределяли на группы по шесть мышей в каждой. На 1 сутки вводили 2A1-HC-Cys357ins-mc-vc-PAB-MMAE (2A1-HC-Cys357ins TDC), 2A1-HC-Cys410ins-mc-vc-PAB-MMAE (2A1-HC-Cys410ins TDC), 2A1-HC-Cys387ins-mc-vc-PAB-MMAE (2A1-HC-Cys387ins TDC) и 2A1-HC-Cys378ins-mc-vc-PAB-MMAE (2A1-HC-Cys378ins TDC) посредством однократной инъекции в хвостовую вену в дозе 5 мг/кг (фиг.6) и получали данные в форме среднего объема опухоли ± SE в момент измерения.
На фиг.6: 2A1-HC-Cys357ins-mc-vc-PAB-MMAE (2A1-HC-Cys357ins TDC), 2A1-HC-Cys410ins-mc-vc-PAB-MMAE (2A1-HC-Cys410ins TDC), 2A1-HC-Cys387ins-mc-vc-PAB-MMAE (2A1-HC-Cys387ins TDC) и 2A1-HC-Cys378ins-mc-vc-PAB-MMAE (2A1-HC-Cys378ins TDC) вводили посредством однократной инъекции в хвостовую вену в дозе 5 мг/кг, достигая значительного противоопухолевого эффекта.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Sichuan Baili Pharmaceutical Co. Ltd
<120> CКРИНИНГ УЧАСТКОВ ПРИСОЕДИНЕНИЯ С ФИКСИРОВАННОЙ ТОЧКОЙ В КОНЪЮГАТЕ ЦИСТЕИН-МОДИФИЦИРОВАННОЕ АНТИТЕЛО-ТОКСИН (TDC)
<130> BACN1905US
<160> 59
<170> PatentIn version 3.5
<210> 1
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 1
gatgtgcagc ttcaggagtc gggacctagc ctggtgaaac cttctcagtc tctgtccctc 60
acctgcactg tcactggcta ctcaatcacc agtgattttg cctggaactg gattcggcag 120
tttccaggaa acaagctgga gtggatgggc tacataagtt atagtggtaa cactaggtac 180
aacccatctc tcaaaagtcg aatctctatc actcgcgaca catccaagaa ccaattcttc 240
ctgcagttga actctgtgac tattgaggac acagccacat attactgtgt aacggcggga 300
cgcgggtttc cttattgggg ccaagggact ctggtcactg tctctgca 348
<210> 2
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 2
Asp Val Gln Leu Gln Glu Ser Gly Pro Ser Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Asp
20 25 30
Phe Ala Trp Asn Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Gly Tyr Ile Ser Tyr Ser Gly Asn Thr Arg Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Ile Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Val Thr Ala Gly Arg Gly Phe Pro Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ala
115
<210> 3
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 3
Ser Asp Phe Ala Trp Asn
1 5
<210> 4
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 4
Tyr Ile Ser Tyr Ser Gly Asn Thr Arg Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 5
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 5
Val Thr Ala Gly Arg Gly Phe Pro Tyr
1 5
<210> 6
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 6
gacatcctga tgacccaatc tccatcctcc atgtctgtat ctctgggaga cacagtcagc 60
atcacttgcc attcaagtca ggacattaac agtaatatag ggtggttgca gcagagacca 120
gggaaatcat ttaagggcct gatctatcat ggaaccaact tggacgatga agttccatca 180
aggttcagtg gcagtggatc tggagccgat tattctctca ccatcagcag cctggaatct 240
gaagattttg cagactatta ctgtgtacag tatgctcagt ttccgtggac gttcggtgga 300
ggcaccaagc tggaaatcaa acgt 324
<210> 7
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 7
Asp Ile Leu Met Thr Gln Ser Pro Ser Ser Met Ser Val Ser Leu Gly
1 5 10 15
Asp Thr Val Ser Ile Thr Cys His Ser Ser Gln Asp Ile Asn Ser Asn
20 25 30
Ile Gly Trp Leu Gln Gln Arg Pro Gly Lys Ser Phe Lys Gly Leu Ile
35 40 45
Tyr His Gly Thr Asn Leu Asp Asp Glu Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Ala Asp Tyr Tyr Cys Val Gln Tyr Ala Gln Phe Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 8
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 8
His Ser Ser Gln Asp Ile Asn Ser Asn Ile Gly
1 5 10
<210> 9
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 9
His Gly Thr Asn Leu Asp Asp Glu
1 5
<210> 10
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 10
Val Gln Tyr Ala Gln Phe Pro Trp Thr
1 5
<210> 11
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 11
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccgggtaaa 990
<210> 12
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 12
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 13
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 13
acggtggctg caccatctgt cttcatcttc ccgccatctg atgagcagtt gaaatctgga 60
actgcctctg ttgtgtgcct gctgaataac ttctatccca gagaggccaa agtacagtgg 120
aaggtggata acgccctcca atcgggtaac tcccaggaga gtgtcacaga gcaggacagc 180
aaggacagca cctacagcct cagcagcacc ctgacgctga gcaaagcaga ctacgagaaa 240
cacaaagtct acgcctgcga agtcacccat cagggcctga gctcgcccgt cacaaagagc 300
ttcaacaggg gagagtgtta g 321
<210> 14
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 14
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 15
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 15
gaaatcaaac gtacgtgtgt ggctgcacca tctgtcttca tcttcccgcc atctgatgag 60
cagttgaaat ctggaactgc ctctgttgtg tgcctgctga ataacttcta tcccagagag 120
gccaaagtac agtggaaggt ggataacgcc ctccaatcgg gtaactccca ggagagtgtc 180
acagagcagg acagcaagga cagcacctac agcctcagca gcaccctgac gctgagcaaa 240
gcagactacg agaaacacaa agtctacgcc tgcgaagtca cccatcaggg cctgagctcg 300
cccgtcacaa agagcttcaa caggggagag tgttag 336
<210> 16
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 16
Glu Ile Lys Arg Thr Cys Val Ala Ala Pro Ser Val Phe Ile Phe Pro
1 5 10 15
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
20 25 30
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
35 40 45
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
50 55 60
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
65 70 75 80
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
85 90 95
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
100 105 110
<210> 17
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 17
gaaatcaaac gtacggtgtg tgctgcacca tctgtcttca tcttcccgcc atctgatgag 60
cagttgaaat ctggaactgc ctctgttgtg tgcctgctga ataacttcta tcccagagag 120
gccaaagtac agtggaaggt ggataacgcc ctccaatcgg gtaactccca ggagagtgtc 180
acagagcagg acagcaagga cagcacctac agcctcagca gcaccctgac gctgagcaaa 240
gcagactacg agaaacacaa agtctacgcc tgcgaagtca cccatcaggg cctgagctcg 300
cccgtcacaa agagcttcaa caggggagag tgttag 336
<210> 18
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 18
Glu Ile Lys Arg Thr Val Cys Ala Ala Pro Ser Val Phe Ile Phe Pro
1 5 10 15
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
20 25 30
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
35 40 45
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
50 55 60
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
65 70 75 80
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
85 90 95
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105 110
<210> 19
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 19
gaaatcaaac gtacggtggc tgcaccatct gtcttcatct tcccgccatc tgatgagcag 60
<210> 20
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 20
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
1 5 10 15
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
20 25 30
Asn Asn Phe Tyr Pro Cys Arg Glu Ala Lys Val Gln Trp Lys Val Asp
35 40 45
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
50 55 60
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
65 70 75 80
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
85 90 95
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105 110
<210> 21
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 21
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
tgtgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 480
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 540
aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 600
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 660
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 720
gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 780
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 22
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 22
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Cys Glu Val Thr
130 135 140
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
145 150 155 160
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
165 170 175
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
180 185 190
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
195 200 205
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
210 215 220
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
225 230 235 240
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 23
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 23
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gagtgtgtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 480
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 540
aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 600
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 660
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 720
gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 780
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 24
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 24
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Cys Val Thr
130 135 140
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
145 150 155 160
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
165 170 175
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
180 185 190
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
195 200 205
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
210 215 220
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
225 230 235 240
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 25
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 25
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gctgtgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 480
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 540
aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 600
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 660
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 720
gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 780
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 26
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 26
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
145 150 155 160
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
165 170 175
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
180 185 190
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
195 200 205
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
210 215 220
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
225 230 235 240
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 27
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 27
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtgtgtgt ggtggacgtg agccacgaag accctgaggt caagttcaac 480
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 540
aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 600
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 660
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 720
gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 780
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 28
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 28
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Cys Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
145 150 155 160
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
165 170 175
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
180 185 190
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
195 200 205
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
210 215 220
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
225 230 235 240
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 29
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 29
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
tgtctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 780
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 30
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 30
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Cys Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 31
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 31
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgtgtacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 780
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 32
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 32
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Cys Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 33
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 33
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaagt gtaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 780
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 34
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 34
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Cys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 35
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 35
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt ggtgtgagag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 36
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 36
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Cys Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 37
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 37
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagtgtag caatgggcag ccggagaaca actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 38
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 38
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Cys Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 39
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 39
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaactgta actacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 40
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 40
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Cys Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 41
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 41
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact gttacaagac cacgcctccc 840
gtgctggact ccgacggctc cttcttcctc tatagcaagc tcaccgtgga caagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 42
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 42
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
ln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Cys Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 43
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 43
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaatt cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggactg taagagcagg 900
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 960
acgcagaaga gcctctccct gtctccgggt taa 993
<210> 44
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 44
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Cys Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
325 330
<210> 45
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 45
Glu Ile Lys Arg Thr Cys Val Ala Ala Pro Ser
1 5 10
<210> 46
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 46
Ile Lys Arg Thr Val Cys Ala Ala Pro Ser Val
1 5 10
<210> 47
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 47
Asn Asn Phe Tyr Pro Cys Arg Glu Ala Lys Val
1 5 10
<210> 48
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 48
Ile Ser Arg Thr Pro Cys Glu Val Thr Cys Val
1 5 10
<210> 49
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 49
Ser Arg Thr Pro Glu Cys Val Thr Cys Val Val
1 5 10
<210> 50
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 50
Pro Glu Val Thr Cys Cys Val Val Val Asp Val
1 5 10
<210> 51
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 51
Glu Val Thr Cys Val Cys Val Val Asp Val Ser
1 5 10
<210> 52
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 52
Pro Ser Arg Asp Glu Cys Leu Thr Lys Asn Gln
1 5 10
<210> 53
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 53
Ser Arg Asp Glu Leu Cys Thr Lys Asn Gln Val
1 5 10
<210> 54
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 54
Asp Glu Leu Thr Lys Cys Asn Gln Val Ser Leu
1 5 10
<210> 55
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 55
Ile Ala Val Glu Trp Cys Glu Ser Asn Gly Gln
1 5 10
<210> 56
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 56
Ala Val Glu Trp Glu Cys Ser Asn Gly Gln Pro
1 5 10
<210> 57
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 57
Gly Gln Pro Glu Asn Cys Asn Tyr Lys Thr Thr
1 5 10
<210> 58
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 58
Gln Pro Glu Asn Asn Cys Tyr Lys Thr Thr Pro
1 5 10
<210> 59
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 59
Lys Leu Thr Val Asp Cys Lys Ser Arg Trp Gln
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНЪЮГАТ "ЦИСТЕИН-МОДИФИЦИРОВАННОЕ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО" И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2017 |
|
RU2762594C2 |
| НАЦЕЛЕННЫЕ НА uPARAP КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2740311C2 |
| САЙТ-СПЕЦИФИЧНАЯ КОНЪЮГАЦИЯ ЛИНКЕРНЫХ ЛЕКАРСТВЕННЫХ ПРЕПАРАТОВ С АНТИТЕЛАМИ И ПОЛУЧАЕМЫЕ В РЕЗУЛЬТАТЕ ADC | 2015 |
|
RU2773536C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2791984C2 |
| КОНЪЮГАТ АНТИТЕЛО ПРОТИВ C-MET-ЛЕКАРСТВЕННОЕ СРЕДСТВО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2822497C1 |
| АНТИ-МЕЗОТЕЛИН АНТИТЕЛО И ЕГО КОНЪЮГАТ С ЛЕКАРСТВЕННЫМИ СРЕДСТВАМИ | 2019 |
|
RU2747995C1 |
| САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПОСРЕДСТВОМ ГЛИКОИНЖЕНЕРИИ | 2014 |
|
RU2831409C2 |
| АНТИТЕЛО ПРОТИВ КЛАУДИНА 18.2 И ЕГО КОНЪЮГАТ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2022 |
|
RU2814164C2 |
| КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО НА ОСНОВЕ ЭРИБУЛИНА И СПОСОБЫ ПРИМЕНЕНИЯ | 2017 |
|
RU2754369C2 |
| ПОЛУЧЕННЫЕ С ПОМОЩЬЮ ГЕННОЙ ИНЖЕНЕРИИ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ И ИХ КОНЪЮГАТЫ | 2011 |
|
RU2755066C2 |
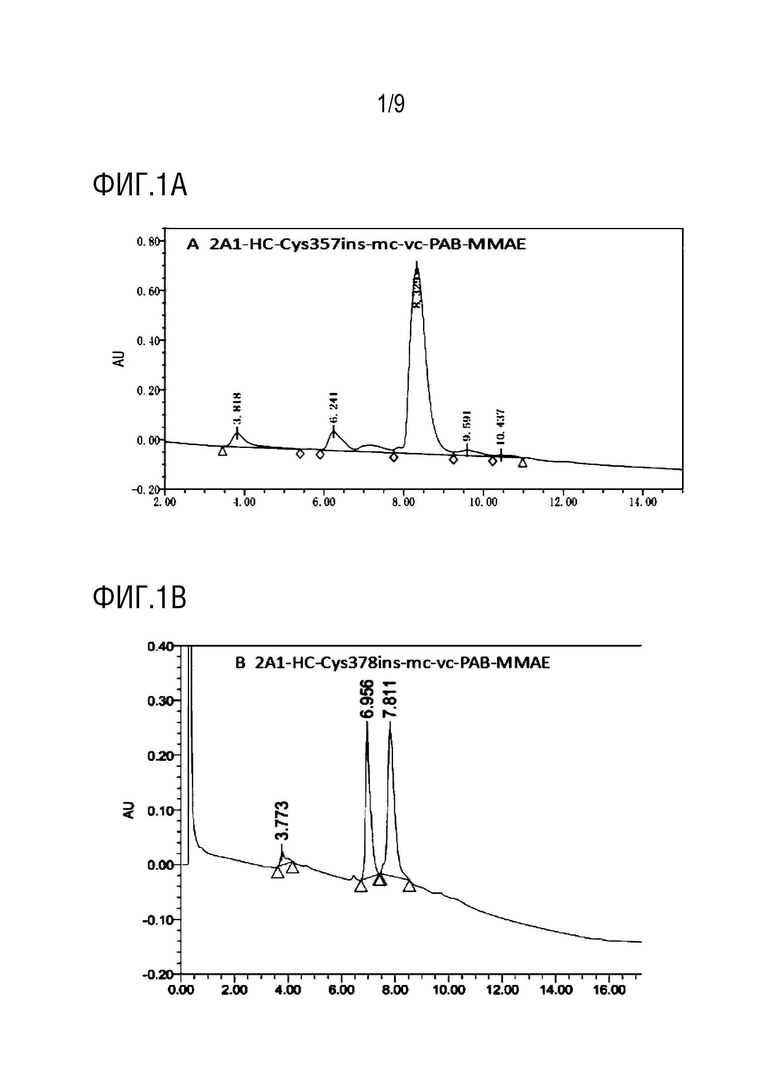

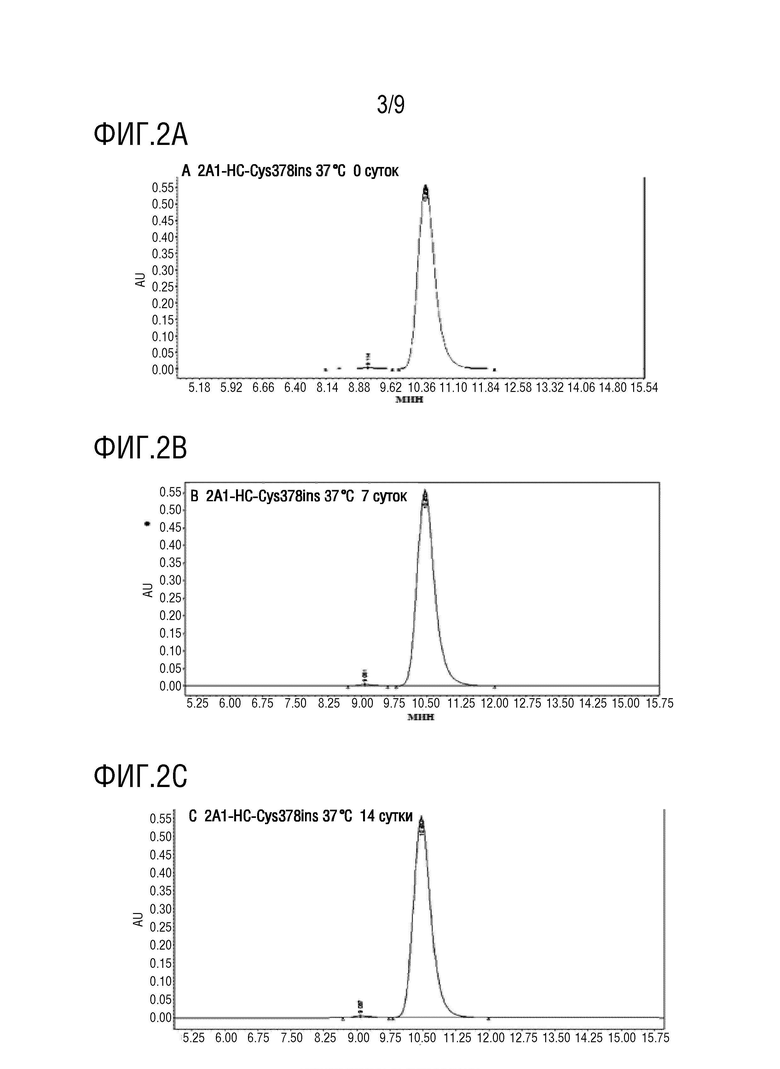
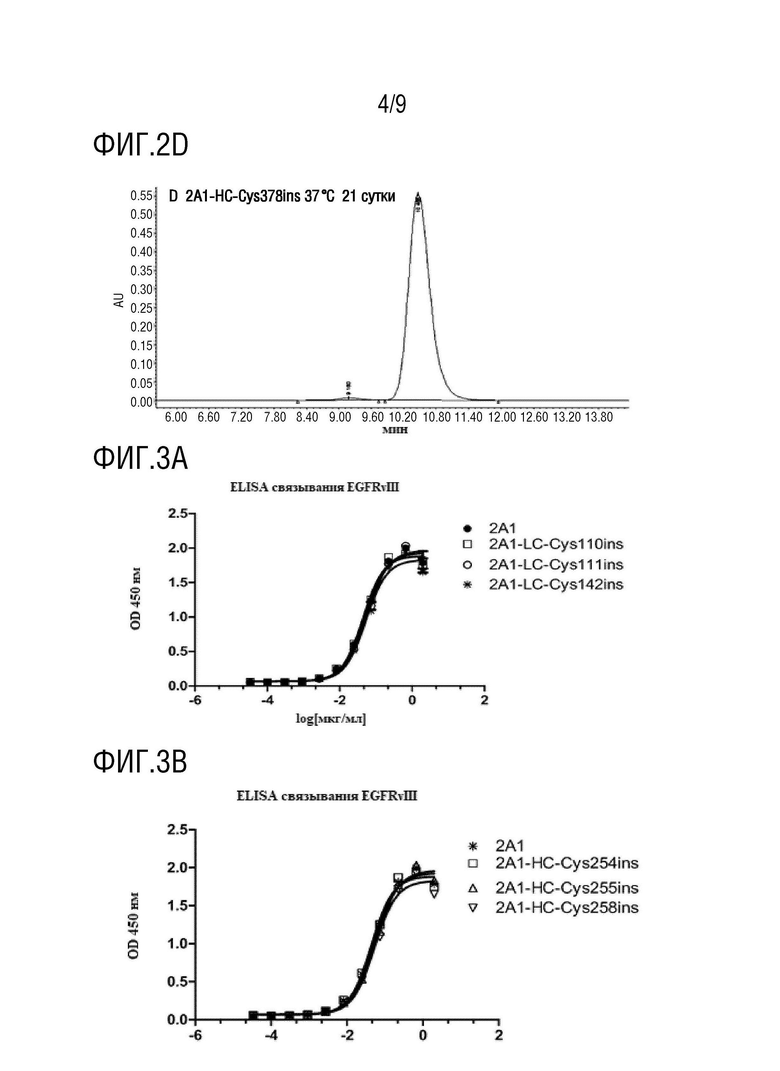
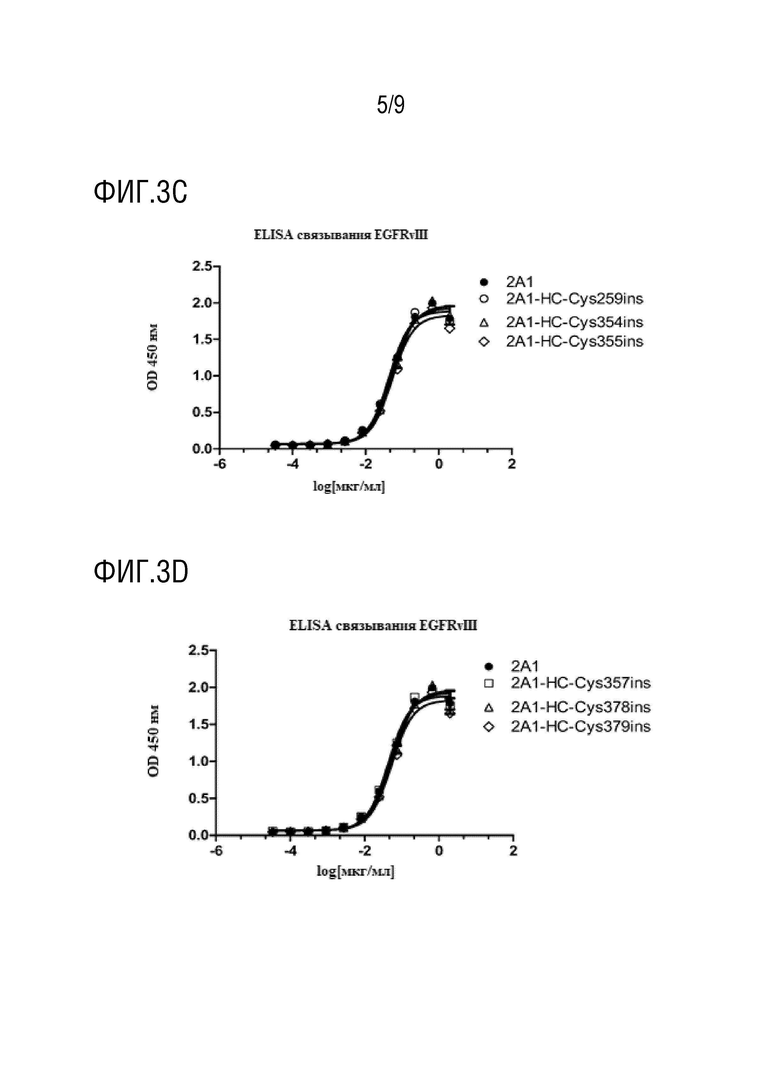
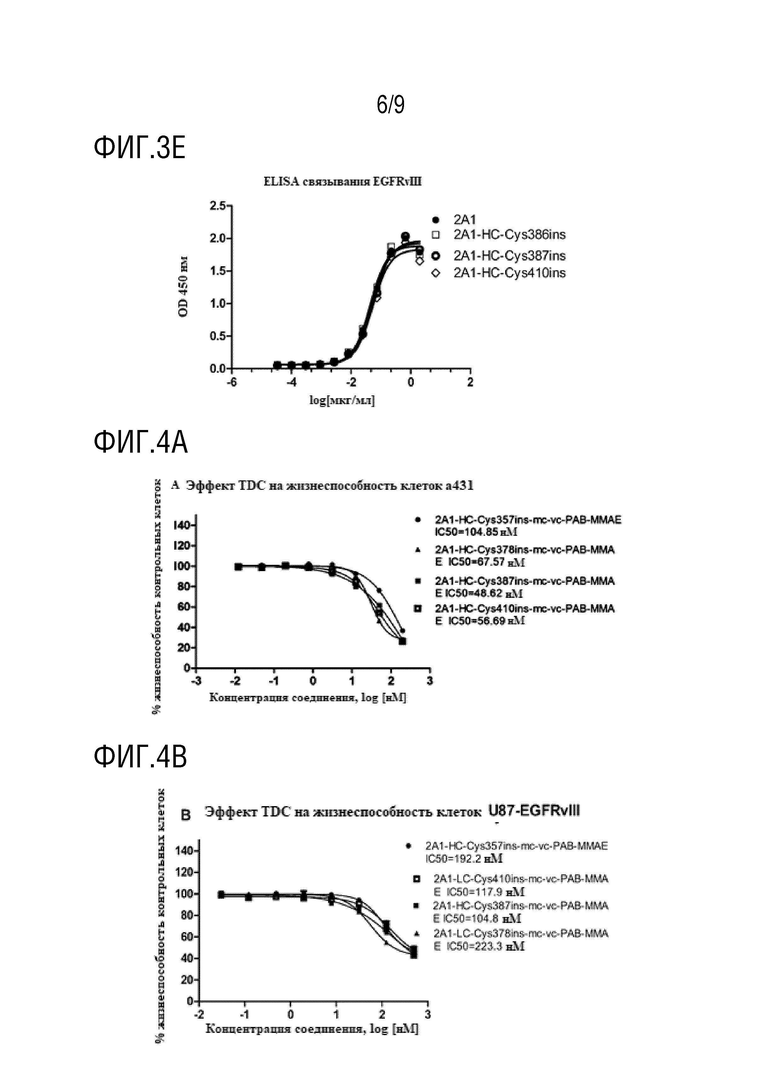
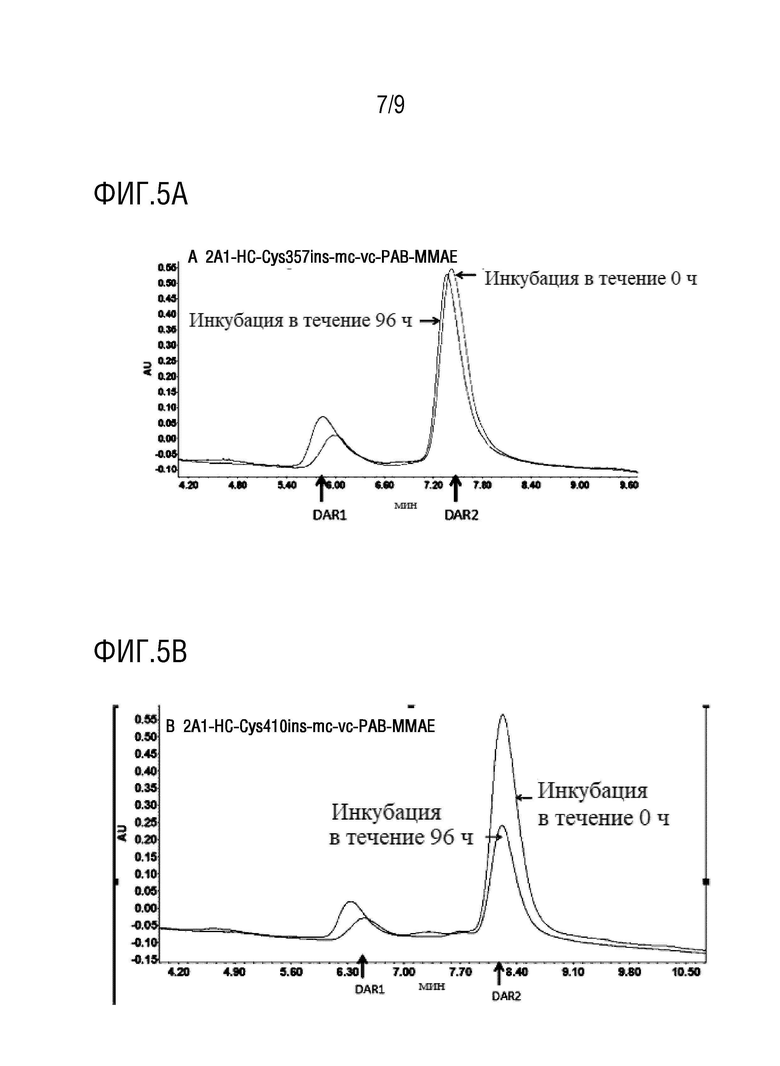

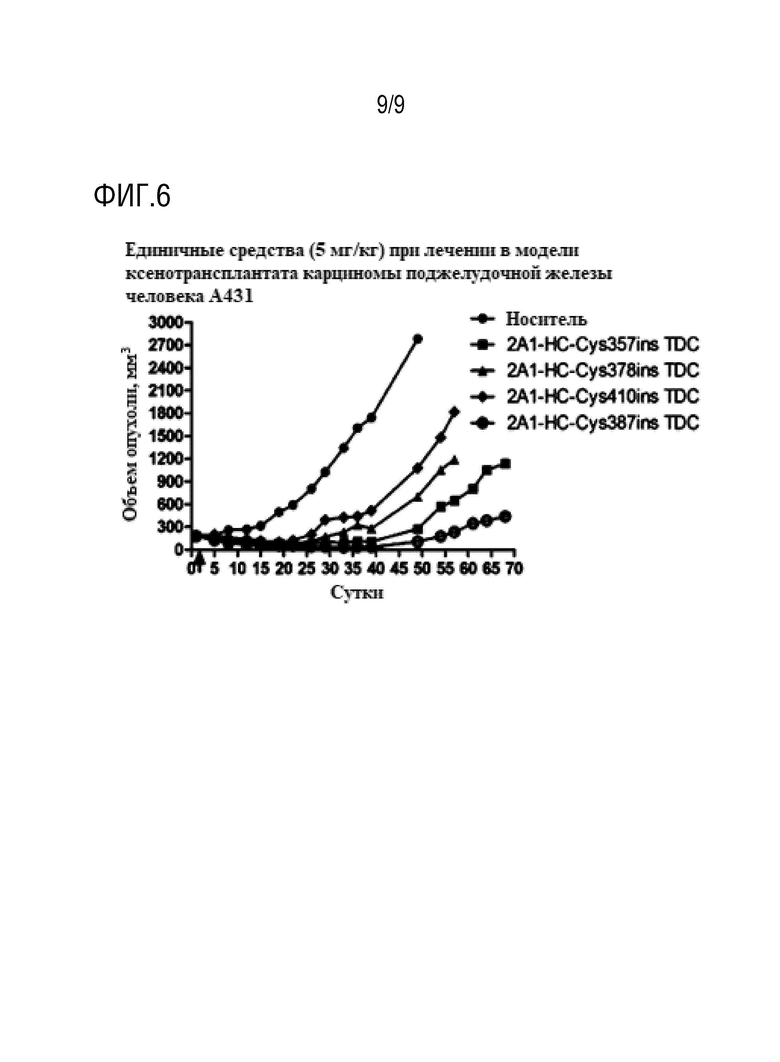
Изобретение относится к иммунологии и биотехнологии. Предложен конъюгат цистеин-модифицированное антитело-токсин, обладающий цитотоксической активностью в отношении опухолевых клеток. Антитело содержит встроенный цистеин в одном или нескольких участках встраивания цистеина, который включает один или несколько из положений константной области легкой цепи каппа/λ 110, 111, 142, положений константной области тяжелой цепи IgG-антитела 254, 255, 258, 259, 354, 355, 357, 378, 379, 386, 387 или 410 (номер согласно Kabat). Высокоактивный цитотоксин выбран из MMAE, MMAF, PBD, SN-38, Dox. Изобретение позволяет получить со связанными в фиксированной точке антителом и токсином и достичь соотношения токсина и антитела (DAR) от 1,6 до 2,0 или от 3,2 до 4,0. 7 з.п. ф-лы, 20 ил., 6 табл., 21 пр.
1. Конъюгат цистеин-модифицированное антитело-токсин, обладающий цитотоксической активностью в отношении опухолевых клеток, содержащий антитело и высокоактивный цитотоксин, где антитело содержит встроенный цистеин в одном или нескольких участках встраивания цистеина, где участок встраивания цистеина включает один или несколько из положений константной области легкой цепи каппа/λ 110, 111, 142, положений константной области тяжелой цепи IgG-антитела 254, 255, 258, 259, 354, 355, 357, 378, 379, 386, 387 или 410 (номер согласно Kabat), где аминокислотная последовательность указанного участка встраивания цистеина включает:
LC-110ins: EIKRTCVAAPS (SEQ ID NO: 45);
LC-111ins: IKRTVCAAPSV (SEQ ID NO: 46);
LC-142ins: NNFYPCREAKV (SEQ ID NO: 47);
HC-254ins: ISRTPCEVTCV (SEQ ID NO: 48);
HC-255ins: SRTPECVTCVV (SEQ ID NO: 49);
HC-258ins: PEVTCCVVVDV (SEQ ID NO: 50);
HC-259ins: EVTCVCVVDVS (SEQ ID NO: 51);
HC-354ins: PSRDECLTKNQ (SEQ ID NO: 52);
HC-355ins: SRDELCTKNQV (SEQ ID NO: 53);
HC-357ins: DELTKCNQVSL (SEQ ID NO: 54);
HC-378ins: IAVEWCESNGQ (SEQ ID NO: 55);
HC-379ins: AVEWECSNGQP (SEQ ID NO: 56);
HC-386ins: GQPENCNYKTT (SEQ ID NO: 57);
HC-387ins: QPENNCYKTTP (SEQ ID NO: 58);
HC-410ins: KLTVDCKSRWQ (SEQ ID NO: 59), и
где указанный высокоактивный цитотоксин выбран из группы, состоящей из MMAE, MMAF, PBD, SN-38, Dox, имеющих следующие молекулярные формулы, и их производных:
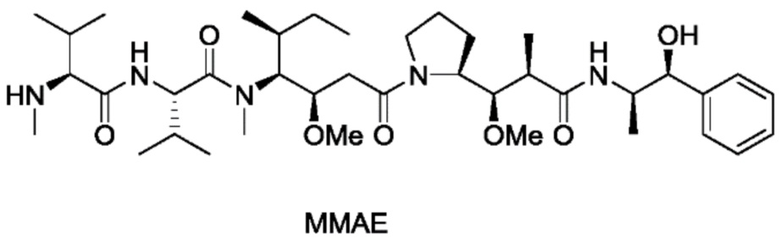
2. Конъюгат цистеин-модифицированное антитело-токсин по п.1, где высокоактивный цитотоксин конъюгирован с встроенным цистеином путем присоединения через его сульфгидрильную группу посредством линкера, и где линкер и легкая цепь антитела имеют аминокислотную последовательность, соответствующую: EIKRTCVAAPS, IKRTVCAAPSV и NNFYPCREAKV, где линкер и тяжелая цепь антитела имеют аминокислотную последовательность, соответствующую: ISRTPCEVTCV, SRTPECVTCVV, PEVTCCVVVDV, EVTCVCVVDVS, PSRDECLTKNQ, SRDELCTKNQV, DELTKCNQVSL, IAVEWCESNGQ, AVEWECSNGQP, GQPENCNYKTT, QPENNCYKTTP или KLTVDCKSRWQ, и где C соответствует встроенному цистеину в легкой или тяжелой цепи цистеин-модифицированного антитела.
3. Конъюгат цистеин-модифицированное антитело-токсин по п.1, где указанная легкая цепь антитела включает изотипы каппа или лямбда.
4. Конъюгат цистеин-модифицированное антитело-токсин по п.1, где указанная тяжелая цепь антитела включает изотипы IgG1, IgG2, IgG3 или IgG4.
5. Конъюгат цистеин-модифицированное антитело-токсин по п.1, где указанный встроенный цистеин содержит сульфгидрильную группу, связанную с высокоактивным цитотоксином.
6. Конъюгат цистеин-модифицированное антитело-токсин по п.5, где указанная сульфгидрильная группа способна к химическому присоединению.
7. Конъюгат цистеин-модифицированное антитело-токсин по п.1, где соотношение указанного высокоактивного цитотоксина и антитела находится в диапазоне от 1,6 до 2,0 или от 3,2 до 4,0.
8. Конъюгат цистеин-модифицированное антитело-токсин по п.1, полученный путем
восстановления цистеин-модифицированного антитела, содержащего защищенный цистеин, имеющий защищенную сульфгидрильную группу и защитную группу,
использования восстановителя для удаления защитной группы из защищенного цистеина с получением свободной защитной группы и восстановленного антитела, содержащего цистеин с удаленной защитной группой, имеющий сульфгидрильную группу,
удаления свободной защитной группы и какого-либо избытка восстановителя с использованием способа, включающего катионообменную хроматографию или ультрафильтрацию,
окисления восстановленного антитела с использованием окислителя для восстановления межцепочечных дисульфидных связей антитела,
присоединения высокоактивного цитотоксина к сульфгидрильной группе цистеина с удаленной защитной группой с получением конъюгата цистеин-модифицированное антитело-токсин, и
удаления какого-либо несвязанного высокоактивного цитотоксина с использованием способа, включающего катионообменную хроматографию или ультрафильтрацию.
| WO 2015157595 A1, 15.10.2015 | |||
| WO 2014124316 A2, 14.08.2014 | |||
| JUNUTULA, J | |||
| R., et al | |||
| Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index | |||
| Nature Biotechnology, 2008, 26(8), p.925-932 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| CN106467575 A, 01.03.2017 | |||
| Способ подготовки кинолент перед обработкой их жидкостями | 1926 |
|
SU5404A1 |
| МОДИФИЦИРОВАННЫЕ АНТИТЕЛА, С КОТОРЫМИ СВЯЗАН МОТИВ, ВКЛЮЧАЮЩИЙ ОСТАТОК ЦИСТЕИНА, КОНЪЮГАТЫ ЭТИХ МОДИФИЦИРОВАННЫХ АНТИТЕЛ С ЛЕКАРСТВЕННЫМИ ВЕЩЕСТВАМИ И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2013 |
|
RU2582259C2 |

