[0001] Данная заявка заявляет преимущество приоритета предварительной патентной заявки США №62/302,562, поданной 2-го марта 2016 года, полное содержание которой включено в данный документ посредством ссылки.
[0002] Данное изобретение относится к конъюгатам антитело-лекарственное средство (ADC), которые связывают онкологические антигенные цели (мишени) человека, такие как рецептор фолиевой кислоты альфа и/или обеспечивают активность антитубулинового лекарственного средства. Данное изобретение дополнительно относится к способам и композициям, полезным для лечения и диагностики злокачественных новообразований, которые экспрессируют рецептор фолиевой кислоты альфа и/или поддаются лечению путем разрушения тубулина.
[0003] Рак является одной из ведущих причин заболеваемости и смертности во всем мире, с, приблизительно, 14 миллионов новых случаев заболевания и 8,2 миллиона смертей связанных с раком в 2012 году. Наиболее распространенной причиной смерти от рака является рак: легких (1,59 миллиона смертей); печени (745000 смертей); желудка (723000 смертей); колоректальный рак (694000 смертей); груди (521000 смертей); и пищевода (400000 смертей). Ожидается, что в течение следующих двух десятилетий число новых случаев заболеваемости раком возрастет на около 70%, до около 22 миллионов новых случаев заболеваемости раком в год (World Cancer Report 2014 год).
[0004] Микротрубочки представляют собой динамические нитевидные цитоскелетные белки, которые участвуют в различных клеточных функциях, включая внутриклеточную миграцию и транспорт, клеточную сигнализацию и поддержание формы клеток. Микротрубочки также играют важную роль в делении митотических клеток, образуя митотическое веретено, необходимое для сегрегации хромосом на две дочерних клетки. Биологические функции микротрубочек во всех клетках в значительной степени регулируются их динамикой полимеризации, которая происходит путем обратимого нековалентного добавления димеров α и β тубулина на обоих концах микротрубочек. Такое динамическое поведение и получающийся в результате контроль над длиной микротрубочек имеют жизненно важное значение для правильного функционирования митотического веретена. Даже незначительное изменение динамики микротрубочек может запускать контрольную точку сборки веретена деления, блокировать прогрессию клеточного цикла при митозе и впоследствии привести к гибели клеток (Muhtar и соавт. (2014 год) Mol. Cancer Ther. 13:275-84). Из-за быстрого деления клеток раковые клетки обычно более чувствительны к соединениям, которые связываются с тубулином и нарушают его нормальную функцию по сравнению с нормальными клетками. По этой причине ингибиторы тубулина и другие агенты, нацеленные на микротрубочки, стали многообещающим классом лекарственных средств для лечения рака (Dumontet and Jordan (2010 год) Nat. Rev. Drug Discov. 9:790-803).
[0005] Рецептор фолиевой кислоты альфа (FRA) представляет собой гликофосфатидилинозитол (ГФИ)-связанный мембранный белок, который связывает фолат. В то время как роль FRA в биологии нормальной и раковой ткани еще не полностью понята, он сверхэкспрессируется в большом количестве злокачественных новообразований яичников эпителиального происхождения (O'Shannessy и соавт. (2013 год) Int. J. Gynecol. Pathol. 32(3):258-68), а также в определенном проценте немелкоклеточных карцином легких (Christoph и соавт. (2014 год) Clin. Lung Cancer 15(5):320-30). FRA также обладает ограниченной экспрессией в нормальных тканях. Эти свойства делают FRA привлекательной целью для иммунотерапии рака.
[0006] Протоонкоген рецептора человеческого эпидермального фактора роста 2 типа (HER2) кодирует трансмембранный рецептор тирозинкиназы, который принадлежит к семейству рецепторов человеческого эпидермального фактора роста (EGFR) (King и соавт. (1985 год) Science 229:974-6). Сверхэкспрессия HER2 обеспечивает конститутивную активацию сигнальных путей фактора роста, таких как путь PI3K-AKT-mTOR, и, таким образом, служит онкогенным фактором при нескольких видах рака, включая приблизительно 20% инвазивных карцином молочной железы. (Slamon и соавт. (1989 год) Science 244:707-12; Gajria and Chandarlapaty (2011 год) Expert Rev. Anticancer Ther. 11:263-75). Учитывая, что амплификация HER2 опосредует трансформированный фенотип, HER2 является еще одной перспективной целью для лечения рака.
[0007] Данное изобретение, в частности, обеспечивает новые соединения с биологической активностью против опухолевых клеток. Соединения могут ингибировать рост опухоли у млекопитающих и могут быть полезны для лечения пациентов, страдающих от рака.
[0008] Данное изобретение более конкретно относится к веществам конъюгат антитело-лекарственное средство, которые способны связывать, поглощать и убивать опухолевые клетки (например, опухолевые клетки, экспрессирующие FRA). Описаны конъюгаты антитело-лекарственное средство, содержащие линкер, который прикрепляет лекарственный фрагмент к фрагменту антитела. Конъюгаты антитело-лекарственное средство (ADC) могут быть представлены Формулой I:
Ab-(L-D)p (I)
в которой Ab представляет собой интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевую клетку;
D представляет собой эрибулин;
L представляет собой расщепляемый линкер, который ковалентно присоединяет Ab к D; а также
p представляет собой целое число от 1 до 20.
[0009] В некоторых вариантах реализации изобретения линкер стабилен вне клетки, так что ADC остается интактным, в случае если он присутствует во внеклеточных условиях, но способен расщепляться при интернализации в клетке, например, раковой клетке. В некоторых вариантах реализации изобретения фрагмент лекарственного средства эрибулина отщепляется от фрагмента антитела, в случае если ADC входит в клетку, которая экспрессирует антиген, специфичный к фрагменту антитела ADC, и расщепление высвобождает немодифицированную форму эрибулина. В некоторых вариантах реализации изобретения линкер содержит расщепляемый фрагмент, который расположен так, что часть линкера или фрагмента антитела не остается связанной с эрибулиновым фрагментом лекарственного средства при расщеплении.
[0010] В некоторых вариантах реализации изобретения расщепляемый фрагмент в линкере представляет собой расщепляемый пептидный фрагмент. В некоторых вариантах реализации изобретения ADC, который содержит расщепляемый пептидный фрагмент, демонстрирует более низкие уровни агрегации, улучшенное соотношение антитело:лекарственное средство, увеличение целевого уничтожения раковых клеток, уменьшение нецелевого уничтожения не раковых клеток и/или более высокое содержание лекарственного средства (p) относительно ADC, который содержит альтернативный расщепляемый фрагмент. В некоторых вариантах реализации изобретения добавление расщепляемого фрагмента увеличивает цитотоксичность и/или активность по сравнению с нерасщепляемым линкером. В некоторых вариантах реализации изобретения повышенная активность и/или цитотоксичность присутствует в раке, экспрессирующем умеренные уровни антигена, являющегося целью фрагмента антитела ADC (например, умеренная экспрессия FRA). В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент расщепляется ферментом, а линкер представляет собой расщепляемый ферментом линкер. В некоторых вариантах реализации изобретения фермент представляет собой катепсин, а линкер представляет собой расщепляемый катепсином линкер. В некоторых вариантах реализации изобретения расщепляемый ферментом линкер (например, расщепляемый катепсином линкер) проявляет одно или более улучшенных свойств, упомянутых выше, по сравнению с альтернативным механизмом расщепления.
[0011] В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент в линкере содержит аминокислотную единицу. В некоторых вариантах реализации изобретения аминокислотная единица содержит валин-цитруллин (Val-Cit). В некоторых вариантах реализации изобретения ADC, который включает в себя Val-Cit, демонстрирует повышенную стабильность, уменьшение нецелевого уничтожения клеток, увеличение целевого уничтожения клеток, снижение уровней агрегации и/или более высокое содержание лекарственного средства по сравнению с ADC, который содержит альтернативную аминокислотную единицу или альтернативный расщепляемый фрагмент.
[0012] В некоторых вариантах реализации изобретения линкер содержит, по меньшей мере, одну спейсерную единицу, соединяющую фрагмент антитела с отщепляемым фрагментом. В некоторых вариантах реализации изобретения спейсерная единица в линкере может содержать по меньшей мере один фрагмент полиэтиленгликоля (ПЭГ). ПЭГ-фрагмент может, например, содержать -(ПЭГ)m-, где m представляет собой целое число от 1 до 10. В некоторых вариантах реализации изобретения спейсерная единица в линкере содержит (ПЭГ)2. В некоторых вариантах реализации изобретения ADC, который содержит более короткую спейсерную единицу (например, (ПЭГ)2), демонстрирует более низкие уровни агрегации и/или более высокое содержание лекарственного средства по сравнению с ADC, который содержит более длинную спейсерную единицу (например, (ПЭГ)8), несмотря на более короткую длину линкера.
[0013] В некоторых вариантах реализации изобретения спейсерная единица в линкере присоединяется к фрагменту антитела ADC через малеимидный фрагмент (Mal). В некоторых вариантах реализации изобретения ADC, который содержит линкер, присоединенный к фрагменту антитела через Mal, демонстрирует более высокое содержание лекарственного средства по сравнению с ADC, который содержит линкер, присоединенный к фрагменту антитела через альтернативный фрагмент. В некоторых вариантах реализации изобретения Mal в линкере реагирует с остатком цистеина на фрагменте антитела. В некоторых вариантах реализации изобретения Mal в линкере соединен с фрагментом антитела через остаток цистеина. В некоторых вариантах реализации изобретения Mal-спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)m, например, Mal-(ПЭГ)2. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2. В некоторых вариантах реализации изобретения Mal-спейсерная единица прикрепляет фрагмент антитела к отщепляемому фрагменту в линкере. В некоторых вариантах реализации изобретения расщепляемый фрагмент в линкере представляет собой расщепляемый пептидный фрагмент, например, аминокислотную единицу. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2-Val-Cit.
[0014] В некоторых вариантах реализации изобретения расщепляемый фрагмент в линкере непосредственно соединяется с эрибулиновым фрагментом лекарственного средства ADC, и расщепляемый фрагмент либо непосредственно связан с фрагментом антитела, либо связан через спейсерную единицу. В некоторых вариантах реализации изобретения спейсерная единица также прикрепляет расщепляемый фрагмент в линкере к эрибулиновому фрагменту лекарственного средства. В некоторых вариантах реализации изобретения спейсерная единица, которая прикрепляет расщепляемый фрагмент в линкере к эрибулиновому фрагменту лекарственного средства, является саморасщепляющейся. В некоторых вариантах реализации изобретения саморасщепляющийся спейсер способен высвобождать немодифицированный эрибулин в целевой клетке. В некоторых вариантах реализации изобретения саморасщепляющаяся единица содержит п-аминобензиловый спирт. В некоторых вариантах реализации изобретения саморасщепляющаяся спейсерная единица содержит п-аминобензилоксикарбонил (pAB). pAB в линкере, в некоторых вариантах реализации изобретения, прикрепляет расщепляемый фрагмент к эрибулиновому фрагменту лекарственного средства. В некоторых вариантах реализации изобретения расщепляемый фрагмент представляет собой расщепляемый пептидный фрагмент, например, аминокислотную единицу. В некоторых вариантах реализации изобретения линкер содержит Val-Cit-pAB. В некоторых вариантах реализации изобретения линкер содержит Val-Cit-pAB и спейсерную единицу ПЭГ, соединяющую линкер с фрагментом антитела через Mal.
[0015] В некоторых вариантах реализации изобретения p представляет собой целое число от 1 до 6, от 2 до 5 или, предпочтительно, от 3 до 4. В некоторых вариантах реализации изобретения p представляет собой 4. В некоторых вариантах реализации изобретения предоставляется пул ADC, а средний p в пуле составляет около 4 (например, 3,5-4,5, например, около 3,8). В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2-Val-Cit-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2-Val-Cit-pAB, а p представляет собой 4. В некоторых вариантах реализации изобретения предоставляется пул ADC, при этом каждый ADC содержит линкер Mal-(ПЭГ)2-Val-Cit-pAB, а средний p в пуле составляет около 4 (например, 3,5-4,5, например, около 3,8).
[0016] В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент (фрагмент Ab или Ab) ADC представляет собой антитело к рецептору фолиевой кислоты альфа (FRA) или интернализирующийся фрагмент антитела и может связывать опухолевые клетки, экспрессирующие FRA (то есть, ADC нацеливается на клетки, экспрессирующие FRA). В некоторых вариантах реализации изобретения ADC, содержащий фрагмент Ab к FRA и расщепляемый пептидный фрагмент, демонстрирует более низкие уровни агрегации, улучшенное соотношение антитело:лекарственное средство, увеличение целевого уничтожения раковых клеток, уменьшение нецелевого уничтожения не раковых клеток, более высокое содержание лекарственного средства (p), повышенную цитотоксичность и/или активность по отношению к нерасщепляемому линкеру или альтернативному механизму расщепления. В некоторых вариантах реализации изобретения повышенная активность и/или цитотоксичность присутствует в раке, экспрессирующем умеренные уровни антигена, являющегося целью фрагмента антитела ADC (например, умеренная экспрессия FRA). В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент расщепляется ферментом, а линкер представляет собой расщепляемый ферментом линкер. В некоторых вариантах реализации изобретения фермент представляет собой катепсин, а линкер представляет собой расщепляемый катепсином линкер. В некоторых вариантах реализации изобретения расщепляемый ферментом линкер (например, расщепляемый катепсином линкер) проявляет одно или более улучшенных свойств, упомянутых выше, по сравнению с альтернативным механизмом расщепления. В некоторых вариантах реализации изобретения линкер представляет собой Mal-(ПЭГ)m-Val-Cit-pAB.
[0017] В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент связывается с рецептором фолиевой кислоты альфа (FRA) и нацеливается на опухолевые клетки, экспрессирующие FRA. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит три области, определяющие комплементарность (CDR) тяжелой цепи и три CDR легкой цепи, при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 2, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 3 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 4; и три CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 7, CDR2 легкой цепи, состоящий из SEQ ID NO: 8 и CDR3 легкой цепи, состоящий из SEQ ID NO: 9, как определено системой нумерации Кабата; или при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 13, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 14 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 15; и CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 16, CDR2 легкой цепи, состоящий из SEQ ID NO: 17 и CDR3 легкой цепи, состоящий из SEQ ID NO: 18, как определено системой нумерации IMGT. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит каркасные последовательности человека. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит вариабельный домен тяжелой цепи из SEQ ID NO: 23 и вариабельный домен легкой цепи из SEQ ID NO: 24. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент конкурирует за связывание и/или связывает тот же эпитоп, что и антитело, содержащее вариабельный домен тяжелой цепи из SEQ ID NO: 23 и вариабельный домен легкой цепи из SEQ ID NO: 24. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент связывается с эпитопом, содержащим аланин-гистидин-лизин-аспарагиновую кислоту (AHKD) (SEQ ID NO: 365) (O'Shannessy и соавт., (2011 год) Oncotarget 2:1227-43). В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент связывается с эпитопом, содержащим NTSQEAHKDVSYL (SEQ ID NO: 366).
[0018] В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент является интернализирующимся антителом к FRA или интернализирующимся антигенсвязывающим фрагментом. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи, при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 2, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 3 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 4; и три CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 7, CDR2 легкой цепи, состоящий из SEQ ID NO: 8 и CDR3 легкой цепи, состоящий из SEQ ID NO: 9, как определено системой нумерации Кабата; или при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 13, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 14 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 15; и CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 16, CDR2 легкой цепи, состоящий из SEQ ID NO: 17 и CDR3 легкой цепи, состоящий из SEQ ID NO: 18, как определено системой нумерации IMGT; линкер содержит Mal-(ПЭГ)2-Val-Cit-pAB; и p представляет собой 4. В некоторых вариантах реализации изобретения предоставляется пул таких ADC b p составляет около 4 (например, около 3,8). В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит вариабельный домен тяжелой цепи из SEQ ID NO: 23 и вариабельный домен легкой цепи из SEQ ID NO: 24. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент конкурирует за связывание и/или связывает тот же эпитоп, что и антитело, содержащее вариабельный домен тяжелой цепи из SEQ ID NO: 23 и вариабельный домен легкой цепи из SEQ ID NO: 24. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент связывается с эпитопом, содержащим SEQ ID NO: 365. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент связывается с эпитопом, содержащим SEQ ID NO: 366.
[0019] В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент связывается с рецептором человеческого эпидермального фактора роста 2 типа (her2) и нацеливается на клетки опухоли, экспрессирующие her2. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит три области, определяющие комплементарность (CDR) тяжелой цепи и три CDR легкой цепи, при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 71, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 72 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 73; и три CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 74, CDR2 легкой цепи, состоящий из SEQ ID NO: 75 и CDR3 легкой цепи, состоящий из SEQ ID NO: 76, как определено системой нумерации Кабата; или при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 191, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 192 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 193; и CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 194, CDR2 легкой цепи, состоящий из SEQ ID NO: 195 и CDR3 легкой цепи, состоящий из SEQ ID NO: 196, как определено системой нумерации IMGT. В некоторых вариантах реализации изобретения антитело или интернализирующийся антигенсвязывающий фрагмент содержит каркасные последовательности человека. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит вариабельный домен тяжелой цепи из SEQ ID NO: 27 и вариабельный домен легкой цепи из SEQ ID NO: 28. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующееся связывание антигена конкурирует за связывание и/или связывает тот же эпитоп, что и антитело, содержащее вариабельный домен тяжелой цепи из SEQ ID NO: 27 и вариабельный домен легкой цепи из SEQ ID NO: 28.
[0020] В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент является интернализирующимся антителом к her2 или интернализирующимся антигенсвязывающим фрагментом. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи, при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 71, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 72 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 73; и три CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 74, CDR2 легкой цепи, состоящий из SEQ ID NO: 75 и CDR3 легкой цепи, состоящий из SEQ ID NO: 76, как определено системой нумерации Кабата; или при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 191, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 192 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 193; и CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 194, CDR2 легкой цепи, состоящий из SEQ ID NO: 195 и CDR3 легкой цепи, состоящий из SEQ ID NO: 196, как определено системой нумерации IMGT; линкер содержит Mal-(ПЭГ)2-Val-Cit-pAB; и p представляет собой 4. В некоторых вариантах реализации изобретения предоставляется пул таких ADC b p составляет около 4 (например, около 3,8). В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит вариабельный домен тяжелой цепи из SEQ ID NO: 27 и вариабельный домен легкой цепи из SEQ ID NO: 28. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующееся связывание антигена конкурирует за связывание и/или связывает тот же эпитоп, что и антитело, содержащее вариабельный домен тяжелой цепи из SEQ ID NO: 27 и вариабельный домен легкой цепи из SEQ ID NO: 28.
[0021] В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент связывается с мезотелином (MSLN) и нацеливается на опухолевые клетки, экспрессирующие MSLN. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит три области, определяющие комплементарность (CDR) тяжелой цепи и три CDR легкой цепи, при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 65, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 66 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 67; и три CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 68, CDR2 легкой цепи, состоящий из SEQ ID NO: 69 и CDR3 легкой цепи, состоящий из SEQ ID NO: 70, как определено системой нумерации Кабата; или при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 185, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 186 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 187; и CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 188, CDR2 легкой цепи, состоящий из SEQ ID NO: 189 и CDR3 легкой цепи, состоящий из SEQ ID NO: 190, как определено системой нумерации IMGT. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит вариабельный домен тяжелой цепи из SEQ ID NO: 25 и вариабельный домен легкой цепи из SEQ ID NO: 26. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующееся связывание антигена конкурирует за связывание и/или связывает тот же эпитоп, что и антитело, содержащее вариабельный домен тяжелой цепи из SEQ ID NO: 25 и вариабельный домен легкой цепи из SEQ ID NO: 26.
[0022] В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент является интернализирующимся антителом к MSLN или интернализирующимся антигенсвязывающим фрагментом. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи, при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 65, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 66 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 67; и три CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 68, CDR2 легкой цепи, состоящий из SEQ ID NO: 69 и CDR3 легкой цепи, состоящий из SEQ ID NO: 70, как определено системой нумерации Кабата; или при этом CDR тяжелой цепи содержат CDR1 тяжелой цепи, состоящий из SEQ ID NO: 185, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 186 и CDR3 тяжелой цепи, состоящий из SEQ ID NO: 187; и CDR легкой цепи содержат CDR1 легкой цепи, состоящий из SEQ ID NO: 188, CDR2 легкой цепи, состоящий из SEQ ID NO: 189 и CDR3 легкой цепи, состоящий из SEQ ID NO: 190, как определено системой нумерации IMGT; линкер содержит Mal-(ПЭГ)2-Val-Cit-pAB; и p представляет собой 4. В некоторых вариантах реализации изобретения предоставляется пул таких ADC b p составляет около 4 (например, около 3,8). В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит вариабельный домен тяжелой цепи из SEQ ID NO: 25 и вариабельный домен легкой цепи из SEQ ID NO: 26. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующийся антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig. В некоторых вариантах реализации изобретения интернализирующееся антитело или интернализирующееся связывание антигена конкурирует за связывание и/или связывает тот же эпитоп, что и антитело, содержащее вариабельный домен тяжелой цепи из SEQ ID NO: 25 и вариабельный домен легкой цепи из SEQ ID NO: 26.
[0023] Также в данном документе представлены композиции, содержащие множественные копии любого из описанных ADC, при этом среднее содержание лекарственного средства (среднее p) ADC в композиции составляет от около 3 до 4 или от около 3,5 до около 4,5, или около 4. В некоторых вариантах реализации изобретения среднее p находится между около 3,2 и 3,8. В некоторых вариантах реализации изобретения среднее p находится между около 3,6 и 4,4.
[0024] Также в данном документе представлены композиции, содержащие -L-D, при этом D представляет собой эрибулин; а L представляет собой расщепляемый линкер, ковалентно прикрепленный к D. В некоторых вариантах реализации изобретения расщепляемый линкер ковалентно присоединяется к амину C-35 на эрибулине. В некоторых вариантах реализации изобретения расщепляемый линкер содержит Val-Cit. В некоторых вариантах реализации изобретения расщепляемый линкер содержит спейсерную единицу ПЭГ. В некоторых вариантах реализации изобретения расщепляемый линкер содержит Mal-(ПЭГ)2-Val-Cit-pAB.
[0025] Далее в данном документе представлены фармацевтические композиции, содержащие ADC и фармацевтически приемлемый разбавитель, носитель и/или вспомогательное вещество.
[0026] Другой аспект данного изобретения включает терапевтическое и диагностическое применение описанных соединений и композиций ADC, например, при лечении рака. Другой аспект включает способы лечения рака, который экспрессирует антиген, являющийся целью фрагмента антитела ADC, такой как FRA. В различных вариантах реализации изобретения предлагаются способы уничтожения или ингибирования пролиферации опухолевых клеток или раковых клеток путем введения терапевтически эффективного количества и/или применения курса лечения любого из описанных ADC. Другой аспект включает способы обнаружения опухолевых клеток или раковых клеток, которые экспрессируют FRA с применением описанных ADC, и способы скрининга пациентов, страдающих на рак, которые будут реагировать на лечение описанными ADC. В некоторых вариантах реализации изобретения рак представляет собой рак желудка, серозный рак яичников, светлоклеточный рак яичника, немелкоклеточный рак легкого, колоректальный рак, трижды негативный рак молочной железы, рак эндометрия, серозную карциному эндометрия, карциноид легких или остеосаркому. Способы получения описанных ADC также раскрыты.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0027] На Фиг. 1 изображена одна из методик, применяемых для получения ADC MORAb-003, как описано в некоторых вариантах реализации изобретения. В этом подходе неспаренные цистеины образуются путем частичного восстановления с ограниченными молярными эквивалентами восстанавливающего агента TCEP, не содержащего тиол. Этот подход предпочтительно уменьшает межцепочечные дисульфидные связи, связывающие легкую цепь и тяжелую цепь (по одной паре на спаривание H-L) и две тяжелые цепи в шарнирной области (две пары на спаривание H-H в случае IgG1 человека), оставляя внутрицепочечные дисульфидные связи интактными.
[0028] На Фиг. 2 изображен способ синтеза малеимид-(ПЭГ)2-Val-Cit-pAB-эрибулина (mal-(ПЭГ)2-VCP-эрибулина), как описано в некоторых вариантах реализации изобретения.
[0029] На Фиг. 3 изображен анализ условий восстановления для MORAb-003 с применением ДСН-ПААГ-электрофореза. Дорожки указаны справа от фигуры. Дорожка M соответствует стандарту белка; дорожка 1 соответствует необработанному MORAb-003; дорожка 2 соответствует 5,3 мг/мл, восстановленному в 70,6 мкМ TCEP; дорожка 3 соответствует MORAb-003 5,3 мг/мл, восстановленному в 141,2 мкМ TCEP; дорожка 4 соответствует MORAb-003 1,5 мг/мл, восстановленному в 20 мкМ TCEP; и дорожка 5 соответствует MORAb-003 1,5 мг/мл, восстановленному в 40 мкМ TCEP. Идентичности каждой полосы указаны на нижнем правом геле. "Н" обозначает тяжелую цепь. "L" обозначает легкую цепь.
[0030] На Фиг. 4 изображен анализ условий восстановления для MORAb-003 с применением ДСН-ПААГ-электрофореза. Дорожка 1 соответствует стандарту белка; дорожка 2 соответствует необработанному MORAb-003; дорожка 3 соответствует MORAb-003, обработанному в соотношении MORAb-003:TCEP 1:1; дорожка 4 соответствует MORAb-003, обработанному в соотношении MORAb-003:TCEP 1:2; дорожка 5 соответствует MORAb-003, обработанному в отношении MORAb-003:TCEP 1:3; и дорожка 6 соответствует MORAb-003, обработанному в соотношении MORAb-003:TCEP 1:4.
[0031] На Фиг. 5 изображен анализ в невосстанавливающем ДСН-ПААГ электрофорезе выбранных ADC MORAb-003, включая M-MMAE (дорожка 2), M-DM1 (дорожка 3), M-0026 (дорожка 4), M-0260 (дорожка 5), M-0267 (дорожка 6), M-0272 (дорожка 7), M-0285 (дорожка 8), M-0292 (дорожка 9), M-027-0381 (дорожка 10) и M-0284 (дорожка 11).
[0032] На Фиг. 6A изображены результаты независимого исследования анализа цитотоксичности MORAb-003-малеимидо-ПЭГ2-Val-Cit-pAB-эрибулина (M3-VCP-эрибулина или "MORAb-202"). На Фиг. 6B изображены результаты независимого исследования анализа цитотоксичности MORAb-003-малеимидо-(CH2)5-Val-Cit-pAB-ER-001150828 (M3-ER-61318). На Фиг. 6C изображены результаты независимого исследования анализа цитотоксичности MORAb-003-ПЭГ-pAB-дуостатина 3 (M3-027-0285). Информация, изображенная в легендах соответствующей фигуры, обеспечивает тестируемые клеточную линию:агент (клеточная линия/культивированные клеточные линии, плотность посева 1ой/2ой клеточной линии).
[0033] На Фиг. 7A и 7B изображено распределение соотношения лекарственного средства-к-антителу (DAR - drug-to-antibody) для ADC MORAb-003-VCP-эрибулин (Фиг. 7A) и MORAb-003-0285 (Фиг. 7B) по отношению к неконъюгированному MORAb-003, как описано в некоторых вариантах реализации изобретения. Числа над каждым пиком указывают на DAR отдельных видов.
[0034] -На Фиг. 8 изображены результаты анализа цитотоксичности - конкуренция MORAb-003-VCP-эрибулина с неконъюгированным MORAb-003 (2 мкМ) в клетках IGROV1 или SJSA-1.
[0035] На Фиг. 9 изображена кинетика массы тела для каждой группы мышей CD-1 (среднее по группе и СОС), получавших одну внутривенную дозу носителя (ФСБ) или MORAb-202 в количестве 10, 20, 40 или 80 мг/кг.
[0036] На Фиг. 10 изображена кинетика массы тела для каждой группы мышей CD-1 (среднее по группе и СОС), получавших внутривенно ФСБ или эрибулин в количестве 0,4, 0,8, 1,6 или 3,2 мг/кг в соответствии с режимом дозирования q4dx3 (дозы вводились один раз каждые четыре дня, всего 3 дозы).
[0037] На Фиг. 11 изображена кинетика роста опухоли для каждой группы мышей CB17-SCID, имплантированных клетками НМРЛ человека NCI-H2110 (среднее по группе и СОС), и получавших одну внутривенную дозу ФСБ, MORAb-003-VCP-эрибулин (MORAb-202) при 1, 2,5 или 5 мг/кг, или MORAb-003-0285 при 5 мг /кг.
[0038] На Фиг. 12 изображены объемы опухолей отдельных мышей CB17-SCID, имплантированных клетками НМРЛ человека NCI-H2110, а также среднее по группам и СОС на 17-й день. Группы получали одну дозу ФСБ внутривенно, MORAb-003-VCP-эрибулина (MORAb-202) при 1, 2,5 или 5 мг/кг или MORAb-003-0285 при 5 мг/кг.
[0039] На Фиг. 13 изображена кинетика массы тела для каждой группы мышей CB17-SCID, имплантированных NCI-H2110 (среднее по группе и СОС), получавших одну внутривенную дозу ФСБ, MORAb-003-VCP-эрибулин (MORAb-202) при 1, 2,5 или 5 мг/кг, или MORAb-003-0285 при 5 мг /кг.
[0040] На Фиг. 14 изображена кинетика роста опухоли для каждой группы мышей CB17-SCID, имплантированных NCI-H2110 (среднее по группе и СОС), получавших внутривенно носитель (ФСБ) или эрибулин при 0,5, 0,2, 0,8 или 1,6 мг/кг, в соответствии с режимом дозирования q4dx3.
[0041] На Фиг. 15 изображены объемы опухолей отдельных мышей CB17-SCID, имплантированных NCI-H2110, а также среднее по группам и СОС на 24-й день. Группы получали внутривенно носитель (ФСБ) или эрибулин при 0,5, 0,2, 0,8 или 1,6 мг/кг в соответствии с режимом дозирования q4dx3.
[0042] На Фиг. 16 изображена кинетика изменения массы тела для каждой группы мышей CB17-SCID, имплантированных NCI-H2110 (среднее по группе и СОС), получавших внутривенно носитель (ФСБ) или эрибулин при 0,5, 0,2, 0,8 или 1,6 мг/кг, в соответствии с режимом дозирования q4dx3.
[0043] На Фиг. 17 изображена активность MORAb-003-VCP-эрибулина (MORAb-202) на клетках IGROV1, OVCAR3, NCI-H2110, A431-A3 и SJSA-1, как измерено анализом цитотоксичности с кристаллическим фиолетовым.
[0044] На Фиг. 18 изображена кинетика роста опухоли для каждой группы мышей CB17-SCID, имплантированных NCI-H2110 (среднее по группе и СОС), получавших одну внутривенную дозу ФСБ или MORAb-003-VCP-эрибулин (MORAb-202) при 1, 2,5 или 5 мг/кг.
[0045] На Фиг. 19A и 19B изображена кинетика роста опухоли (Фиг. 19A) и кинетика изменения массы тела (Фиг. 19B) для каждой группы PDx НМРЛ (LXFA-737) мышей-опухоленосителей (среднее по группе и СОС), получавших одну внутривенную дозу носителя (ФСБ), MORAb-003 в количестве 5 мг/кг или MORAb-003-VCP-эрибулин (MORAb-202) в количестве 5 мг/кг.
[0046] На Фиг. 20A и 20B изображены отдельные соотношения объемов опухолей (Фиг. 20A) и кинетика изменения массы тела (Фиг. 20B) для каждой группы мышей-опухоленосителей рака эндометрия PDx (Endo-12961) (среднее по группе и СОС), получавших одну внутривенную дозу PBS, эрибулина в количестве 0,1 или 3,2 мг/кг или MORAb-003-VCP-эрибулин (MORAb-202) в количестве 5 мг/кг. На Фиг. 20C и 20D изображена кинетика роста опухоли (Фиг. 20C) и кинетика изменения массы тела (Фиг. 20D) для каждой группы мышей-опухоленосителей рака эндометрия PDx (Endo-10590) (среднее по группе и СОС), получавших одну внутривенную дозу PBS, эрибулина в количестве 0,1 или 3,2 мг/кг или MORAb-003-VCP-эрибулин (MORAb-202) в количестве 5 мг/кг.
[0047] На Фиг. 21A изображено иммуногистохимическое (ИГХ) окрашивание опухолевой ткани мышей-опухоленосителей PDx ТНРМЖ (OD-BRE-0631) с антителом к человеческому IgG. Опухолевые ткани из мышей, получавших одну внутривенную дозу носителя (справа) или MORAb-003-VCP-эрибулин (MORAb-202) в количестве 5 мг/кг (слева), собирали и окрашивали через 5 дней после лечения. На Фиг. 21B изображено IHC окрашивание опухолевой ткани мышей-опухоленосителей PDx ТНРМЖ (OD-BRE-0631) с антителом α-гладкомышечного актина (ГМА)-FITC. Опухолевые ткани у мышей, не получавших лечение собирали за 2 дня до лечения (слева), тогда как опухолевые ткани у мышей, получавших одну внутривенную дозу MORAb-003-VCP-эрибулина (MORAb-202) в количестве 5 мг/кг, собирали через 5 дней после лечения (справа). На Фиг. 21C изображена кинетика роста опухоли для каждой группы мышей-опухоленосителей PDx ТНРМЖ (OD-BRE-0631) (среднее по группе и СОС), получавших одну внутривенную дозу носителя (ФСБ) или MORAb-003-VCP-эрибулина (MORAb-202) в количестве 5 мг/кг.
[0048] На Фиг. 22 изображено дифференцирование человеческих мезенхимальных стволовых клеток костного мозга (МСК-КМ) в культуре с клетками MKN-74 после введения носителя (ФСБ или этанола), эрибулина, MORAb-003 или MORAb-003-VCP-эрибулина (MORAb-202), как измерено методом проточной цитометрии. Stro-1+/CD105+, CD34+/CD31- и NG2+ являются маркерами MSC, адипоцитов и перицитов, соответственно.
[0049] На Фиг. 23 изображен анализ временной динамики опухолевых тканей из мышей CB17-SCID, имплантированных NCI-H2110, получавших одну внутривенную дозу носителя (ФСБ) или MORAb-003-VCP-эрибулина (MORAb-202) в количестве 5 мг/кг, окрашенных с применением антитела α-гладкомышечного актина (ГМА)-FITC. Опухолевые ткани собирали и окрашивали в день 0 и в дни 3, 5, 7 и 9 после лечения. Ось Y:%=[подсчитаны окрашенные клетки/подсчитано общее количество клеток] * 100. Ось X: день (подсчитано общее количество клеток).
ПОДРОБНОЕ ОПИСАНИЕ ИЛЛЮСТРАТИВНЫХ ВАРИАНТОВ РЕАЛИЗАЦИИ ИЗОБРЕТЕНИЯ
[0050] Описанные композиции и способы могут быть поняты более легко со ссылкой на следующее подробное описание, взятое в связи с прилагаемыми фигурами, которые составляют часть данного описания. Следует понимать, что описанные композиции и способы не ограничены конкретными композициями и способами, описанными и/или показанными в данном документе, и что применяемая в данном документе терминология предназначена для описания конкретных вариантов реализации изобретения только в качестве примера и не предназначена для ограничения заявленных композиций и способов.
[0051] В этом тексте описания относятся к композициям и способам применения указанных композиций. В тех случаях, когда данное описание описывает или претендует на признак или вариант реализации изобретения, связанные с композицией, такой признак или вариант реализации изобретения в равной степени применимы к способам применения указанной композиции. Аналогичным образом, если данное описание описывает или заявляет признак, или вариант реализации изобретения, связанный со способом применения композиции, такой признак или вариант реализации изобретения в равной степени применимы к композиции.
[0052] В случае если выражен диапазон значений, он включает в себя варианты реализации изобретения, применяющие любое конкретное значение в пределах диапазона. Дополнительно, ссылка на значения, указанные в диапазонах, включает каждое значение в этом диапазоне. Все диапазоны включают в себя их конечные точки и являются комбинируемыми. В случае если значения выражены в виде приближений при помощи предшествующего термина "около", то следует понимать, что конкретное значение образует другой вариант реализации изобретения. Ссылка на конкретное числовое значение включает в себя, по меньшей мере, это конкретное значение, если из контекста явно не следует иное. Применение "или" означает "и/или", если конкретный контекст его применения не указывает иное. Все источники в данном описании включены посредством ссылки для любых целей. Если ссылка и спецификация вступают в конфликт, спецификация будет преобладать.
[0053] Следует принимать во внимание, что некоторые особенности описанных композиций и способов, которые для ясности, описанные в данном документе в контексте отдельных вариантов реализации изобретения, также могут быть представлены в комбинации в одном варианте реализации изобретения. И наоборот, различные признаки описанных композиций и способов, которые для краткости описаны в контексте одного варианта реализации изобретения, также могут быть представлены по отдельности или в любой подкомбинации.
Определения
[0054] Различные термины, относящиеся к аспектам данного описания, применяются в описании и формуле изобретения. Такие термины должны иметь свое обычное значение в данной области техники, если не указано иное. Другие конкретно определенные термины должны толковаться в соответствии с определениями, представленными в данном документе.
[0055] Как применяются в данном документе формы единственного числа включают в себя формы множественного числа, если из контекста явно не следует иное.
[0056] Термины "около" или "приблизительно" в контексте числовых значений и диапазонов относятся к значениям или диапазонам, которые приближены или близки к указанным значениям или диапазонам, таким образом, что вариант реализации изобретения может выполняться так, как предполагалось, например, имеющий нужное количество нуклеиновых кислот или полипептидов в реакционной смеси, как это очевидно специалисту в данной области техники из приведенных в данном документе принципов. Это связано, по меньшей мере частично, с различными свойствами вариаций композиций нуклеиновых кислот, возраста, рас, пола, анатомии и физиологии и неточностями биологических систем. Таким образом, эти термины охватывают значения, выходящие за рамки систематических ошибок.
[0057] В данном документе термины "конъюгат антитело-лекарственное средство", "конъюгат антитела", "конъюгат", "иммуноконъюгат" и "ADC" применяются взаимозаменяемо и относятся к соединению или его производному, которое связано с антителом (например, антителом к FRA) и определяется общей формулой:Ab-(L-D)p (Формула I), где Ab=фрагмент антитела (то есть, антитело или антигенсвязывающий фрагмент), L=линкерный фрагмент, D=фрагмент лекарственного средства и p=количество фрагментов лекарственного средства на фрагмент антитела.
[0058] Термин "антитело" применяется в самом широком смысле, чтобы ссылаться на молекулу иммуноглобулина, которая распознает и специфически связывается с целью, такой как белок, полипептид, углевод, полинуклеотид, липид или комбинации вышеизложенного через, по меньшей мере, один сайт распознавания антигена в вариабельной области молекулы иммуноглобулина. Тяжелая цепь антитела состоит из вариабельного домена тяжелой цепи (VH) и константной области тяжелой цепи (CH). Легкая цепь состоит из вариабельного домена легкой цепи (VL) и константной области легкой цепи (CL). Для целей данной заявки вариабельные домены зрелой тяжелой цепи и легкой цепи содержат три области, определяющие комплементарность (CDR1, CDR2 и CDR3) в четырех каркасных областях (FR1, FR2, FR3 и FR4), расположенных от N-конца до C-конца:FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. "Антитело" может быть естественного происхождения или искусственным, таким как моноклональные антитела, продуцируемые традиционной гибридомной технологией. Термин "антитело" включает полноразмерные моноклональные антитела и полноразмерные поликлональные антитела, а также фрагменты антител, такие как Fab, Fab', F(ab')2, Fv и одноцепочечные антитела. Антитело может быть любым из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM или их подклассов (например, изотипы IgG1, IgG2, IgG3, IgG4). Термин дополнительно охватывает человеческие антитела, химерные антитела, гуманизированные антитела и любую модифицированную молекулу иммуноглобулина, содержащую сайт распознавания антигена, при условии, что он проявляет желаемую биологическую активность.
[0059] Термин "моноклональное антитело" в данном документе относится к антителу, полученному из популяции в значительной степени однородных антител, то есть отдельные антитела в составе популяции являются идентичными, за исключением мутаций, происходящих по естественным причинам, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высокоспецифичными, поскольку они направлены против одного антигенного эпитопа. В противоположность этому, обычные (поликлональные) препараты антител обычно включают множество антител, направленных против (или специфических к) разных эпитопов. Определение "моноклональное" указывает на то, что антитело получено из практически однородной популяции антител; его не следует интерпретировать как требование продукции антитела посредством какого-либо конкретного способа. Например, моноклональные антитела, которые должны применяться в соответствии с данным изобретением, могут быть получены с помощью гибридомного способа, впервые описанного Kohler и соавт. (1975 год) Nature 256:495 или могут быть получены с помощью способов рекомбинантной ДНК (см., например, патент США №4,816,567). Моноклональные антитела также могут быть выделены из библиотек фаговых антител, применяя методы, описанные в Clackson и соавт. (1991 год) Nature 352:624-8, и Marks и соавт. (1991 год) J. Mol. Biol. 222:581-97, например.
[0060] Моноклональные антитела описанные в данном документе конкретно включают "химерные" антитела, в которых участок тяжелой и/или легкой цепи идентичен или гомологичен соответствующим последовательностям антител, полученных от конкретного вида или относящихся к конкретному классу или подклассу антител, тогда как остаток цепи(ей) идентичен или гомологичен соответствующим последовательностям антител, полученным от другого вида или относящихся к другому классу или подклассу антител, а также фрагменты таких антител при условии, что они специфически связывают целевой антиген и/или проявляют желаемую биологическую активность.
[0061] В данном контексте термин "человеческое антитело" означает антитело, продуцируемое человеком или антитело, имеющее аминокислотную последовательность антитела, продуцируемого человеком.
[0062] В данном контексте термин "химерное антитело" относится к антителам, в которых аминокислотная последовательность молекулы иммуноглобулина получена из двух или более видов. В некоторых случаях вариабельные области как тяжелых, так и легких цепей соответствуют вариабельным областям антител, полученных от одного вида с желаемой специфичностью, аффинностью и активностью, тогда как константные области гомологичны антителам, полученным из другого вида (например, человека) для минимизации иммунного ответа у последнего вида.
[0063] В данном контексте термин "гуманизированное антитело" относится к формам антител, которые содержат последовательности из нечеловеческих (например, мышиных) антител, а также человеческих антител. Такие антитела представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, нечеловеческого происхождения. В общем, гуманизированное антитело будет содержать практически все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или практически все гипервариабельные петли соответствуют таким участкам иммуноглобулина нечеловеческого происхождения, а все или практически все шарнирные (FR) области соответствуют таким участкам последовательности иммуноглобулина человека. Гуманизированное антитело также не обязательно будет содержать по меньшей мере часть константной области иммуноглобулина (Fc), обычно происходящей из иммуноглобулина человека. Гуманизированное антитело может быть дополнительно модифицировано путем замещения остатков либо в каркасной области Fv, и/или в замещенных остатках нечеловеческого происхождения, для уточнения и оптимизации специфичности, аффинности и/или активности антител.
[0064] В данном документе термин "антигенсвязывающий фрагмент" или "антигенсвязывающая часть" антитела, относится к одному или более фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, FRA). Антигенсвязывающие фрагменты предпочтительно также сохраняют способность к интернализации в клетку, экспрессирующую антиген. В некоторых вариантах реализации изобретения антигенсвязывающие фрагменты также сохраняют активность иммунного эффекторна. Было показано, что фрагменты полноразмерного антитела могут выполнять антигенсвязывающую функцию полноразмерного антитела. Примеры связывающих фрагментов, охватываемых термином "антигенсвязывающий фрагмент" или "антигенсвязывающая часть" антитела, включают (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) фрагмент F(ab')2, двухвалентный фрагмент, содержащий два фрагмента Fab, соединенных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и CH1; (iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; (v) фрагмент dAb, который содержит один вариабельный домен, например домен VH (см., например, Ward и соавт. (1989 год) Nature 341:544-6; и Winter и соавт., WO 90/05144); и (vi) выделенную область, определяющую комплементарность (CDR). Кроме того, хотя два домена фрагмента Fv, VL и VH, кодируются отдельными генами, они могут соединяться, с применением рекомбинантных способов, синтетическим линкером, который позволяет им производиться в виде одной белковой цепи, в которой области VL и VH спариваются с образованием одновалентных молекул (известные как одноцепочечные Fv (scFv)). См., например, Bird и соавт. (1988 год) Science 242:423-6; и Huston и соавт. (1988 год) Proc. Natl. Acad. Sci. USA 85:5879-83. Такие одноцепочечные антитела также охватываются термином "антигенсвязывающий фрагмент" или "антигенсвязывающая часть" антитела и известны в данной области техники как типовый тип связывающего фрагмента, который может интернализоваться в клетки при связывании. См., например, Zhu и соавт. (2010 год) 9:2131-41; He и соавт. (2010 год) J. Nucl. Med. 51:427-32; и Fitting и соавт. (2015 год) MAbs 7:390-402. В некоторых вариантах реализации изобретения молекулы scFv могут быть включены в слитый белок. Также охватываются другие формы одноцепочечных антител, такие как диатела. Диатела представляют собой двухвалентные, биспецифические антитела, в которых домены VH и VL экспрессируются в одной полипептидной цепи, но с применением линкера, который является слишком коротким, чтобы обеспечить спаривание между двумя доменами в одной и той же цепи, тем самым заставляя домены спариваться с комплементарными доменами другой цепи и создавая два сайта связывания антигена (см., например, Holliger и соавт. (1993 год) Proc. Natl. Acad. Sci. USA 90:6444-8; и Poljak и соавт. (1994 год) Structure 2:1121-3). Антигенсвязывающие фрагменты получают с применением обычных методов, известных специалистам в данной области техники, а связывающие фрагменты подвергают скринингу на полезность (например, аффинность связывания, интернализацию) таким же образом, как и интактные антитела. Антигенсвязывающие фрагменты могут быть получены расщеплением интактного белка, например, протеазой или химическим расщеплением.
[0065] В данном контексте термин "интернализирующийся" в отношении антитела или антигенсвязывающего фрагмента, относится к антителу или антигенсвязывающему фрагменту, который способен проходить через липидную бислойную мембрану клетки во внутренний компартмент (то есть "интернализированный") при связывании с клеткой, предпочтительно в деградирующий компартмент в клетке. Например, интернализирующееся антитело к FRA представляет собой антитело, которое может быть принято в клетку после связывания с FRA на клеточной мембране.
[0066] В данном контексте термин "рецептор фолиевой кислоты альфа" или "FRA" относится к любой нативной форме человеческого FRA. Данный термин охватывает полноразмерный FRA (например, NCBI Reference Sequence:NP_000793; SEQ ID NO: 19), а также любую форму человеческого FRA, которая является результатом процессинга в клетке. Данный термин также включает варианты FRA естественного происхождения, включая, но не ограничиваясь ими, сплайсинговые варианты, аллельные варианты и изоформы. FRA может быть выделен из человека или может быть получен рекомбинантно или с помощью синтетических способов.
[0067] Термин "антитело к FRA" или "антитело, которое специфически связывает FRA" относится к любой форме антитела или его фрагмента, которая специфически связывает FRA и охватывает моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела и биологически функциональные фрагменты антител, при условии, что они специфически связывают FRA. Предпочтительно антитело к FRA, применяемое в описанных в данном документе ADC, является интернализирующимся антителом или интернализирующимся фрагментом антитела. MORAb-003 представляет собой типовое интернализирующееся антитело к FRA человека. В данном контексте термин "специфический", "специфически связывается" и "связывается специфически" относится к селективному связыванию антитела с эпитопом целевого антигена. Антитела могут быть протестированы на специфичность связывания путем сравнения связывания с соответствующим антигеном со связыванием с нерелевантным антигеном или антигенной смесью при заданном наборе условий. Если антитело связывается с соответствующим антигеном с по меньшей мере в 2, 5, 7 и, предпочтительно, 10 раз большей аффинностью, чем с нерелевантным антигеном или антигенной смесью, то это считается специфическим связыванием. В одном варианте реализации изобретения специфическое антитело представляет собой антитело, которое связывает только антиген FRA, но не связывает (или проявляет минимальное связывание) с другими антигенами.
[0068] В данном контексте термин "рецептор человеческого эпидермального фактора роста 2 типа", "her2" или "her2/neu" относится к любой нативной форме her2 человека. Данный термин охватывает полноразмерный her2 (например, NCBI Reference Sequence:NP_004439. 2; SEQ ID NO: 21), а также любую форму человеческого her2, которая является результатом процессинга в клетке. Данный термин также включает варианты her2 естественного происхождения, включая, но не ограничиваясь ими, сплайсинговые варианты, аллельные варианты и изоформы. Her2 может быть выделен из человека или может быть получен рекомбинантно или с помощью синтетических способов.
[0069] Термин "антитело к her2" или "антитело, которое специфически связывает her2" относится к любой форме антитела или его фрагмента, которая специфически связывает her2 и охватывает моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела и биологически функциональные фрагменты антител, при условии, что они специфически связывают her2. В патенте США №5,821,337 (включенном в данный документ в качестве ссылки) представлены типовые последовательности связывания her2, включая типовые последовательности антитела к her2. Предпочтительно антитело к her2, применяемое в описанных в данном документе ADC, является интернализирующимся антителом или интернализирующимся фрагментом антитела. Трастузумаб представляет собой типовое интернализирующееся антитело к her2 человека.
[0070] Термин "эпитоп" относится к части антигена, которая может быть распознана и специфически связана антителом. Когда антиген является полипептидом, эпитопы могут быть образованы из смежных аминокислот или несмежных аминокислот, сопоставленных третичной укладкой полипептида. Эпитоп, связанный антителом, может быть идентифицирован с применением любого метода картирования эпитопов, известного в данной области техники, включая рентгеновскую кристаллографию для идентификации эпитопа путем прямой визуализации комплекса антиген-антитело, а также контроля связывания антитела с фрагментами или мутированными вариантами антигена или контроля доступности для растворителя различных частей антитела и антигена. Типовые стратегии, применяемые для картирования эпитопов антител, включают, но не ограничиваются ими, олигопептидное сканирование на основе матрицы, ограниченный протеолиз, сайт-направленный мутагенез, мутагенезное картирование с высокой пропускной способностью, водород-дейтериевый обмен и масс-спектрометрию (см., например, Gershoni и соавт. (2007 год) 21:145-56, и Hager-Braun and Tomer (2005 год) Expert Rev. Proteomics 2:745-56).
[0071] Конкурентное связывание и связывание эпитопа также могут быть применены для определения антител, имеющих идентичные или перекрывающиеся эпитопы. Конкурентное связывание может быть оценено с применением эпитоп-перекрестного конкурентного анализа, такого как анализ, описанный в "Antibodies, A Laboratory Manual," Cold Spring Harbor Laboratory, Harlow and Lane (1е издание 1988 год, 2е издание 2014 год). В некоторых вариантах реализации изобретения конкурентное связывание идентифицируют, когда тестируемое антитело или связывающий белок уменьшает связывание эталонного антитела или связывающего белка с целевым антигеном, таким как FRA или her2 (например, связывающий белок, содержащий CDR и/или вариабельные домены, выбранные из тех, которые указаны в Таблицах 2, 4 и 6), по меньшей мере, на 50% в эпитоп-перекрестном конкурентном анализе (например, на 50%, 60%, 70%, 80%, 90%, 95%, 99%, 99,5% или более или любой процент между ними) и/или наоборот. В некоторых вариантах реализации изобретения конкурентное связывание может быть связано с общими или сходными (например, частично перекрывающимися) эпитопами или из-за стерического затруднения, в случае если антитела или связывающие белки связываются в соседних эпитопах. См., например, Tzartos, Methods in Molecular Biology (Morris, ред. (1998 год) том 66, стр. 55-66). В некоторых вариантах реализации изобретения конкурентное связывание может быть применено для сортировки групп связывающих белков, которые имеют сходные эпитопы, например, те, которые конкурируют за связывание, могут быть "сгруппированы" в виде группы связывающих белков, которые имеют перекрывающиеся или близлежащие эпитопы, тогда как те, которые не конкурируют помещают в отдельную группу связывающих белков, которые не имеют перекрывающихся или близлежащих эпитопов.
[0072] Термин "kon" или "ka" относится к константе скорости для ассоциации антитела с антигеном с образованием комплекса антитело/антиген. Скорость может быть определена с применением стандартных анализов, таких как анализ Biacore или ИФА.
[0073] Термин "koff" или "kd" относится к константе скорости диссоциации антитела из комплекса антитело/антиген. Скорость может быть определена с применением стандартных анализов, таких как анализ Biacore или ИФА.
[0074] Термин "KD" относится к равновесной константе диссоциации конкретного взаимодействия антитело-антиген. KD вычисляется по ka/kd. Скорость может быть определена с применением стандартных анализов, таких как анализ Biacore или ИФА.
[0075] Термин "p" или "соотношение антитело: лекарственное средство" или "соотношение лекарственного средства-к-антителу" или "DAR" относится к числу фрагментов лекарственного средства на фрагмент антитела, то есть, содержанию лекарственного средства или количеству фрагментов L-D на антитело или антигенсвязывающий фрагмент (Ab) в ADC Формулы I. В композициях, содержащих несколько копий ADC Формулы I, "p" относится к среднему числу фрагментов L-D на антитело или антигенсвязывающий фрагмент, также называемый средним содержанием лекарственного средства.
[0076] "Линкер" или "линкерный фрагмент" представляет собой любой химический фрагмент, который способен ковалентно соединяться с соединением, обычно фрагментом лекарственного средства, таким как химиотерапевтический агент, с другим фрагментом, таким как фрагмент антитела. Линкеры могут быть подвержены или практически устойчивыми к вызванному кислотой расщеплению, расщеплению, вызванному пептидазой, индуцированному светом расщеплению, расщеплению, вызванному эстеразой и/или расщеплению дисульфидной связи в условиях, когда соединение или антитело остаются активными.
[0077] В данном контексте термин "агент" применяется для обозначения химического соединения, смеси химических соединений, биологической макромолекулы или экстракта, полученного из биологических материалов. Термин "терапевтический агент", "лекарственное средство" или "фрагмент лекарственного средства" относится к агенту, который способен модулировать биологический процесс и/или обладает биологической активностью.
[0078] В данном контексте термин "химиотерапевтический агент" или "противораковый агент" применяется для обозначения всех химических соединений, которые эффективны при лечении рака независимо от механизма действия. Ингибирование метастазов или ангиогенеза часто является свойством химиотерапевтического агента. Неограничивающие примеры химиотерапевтических агентов включают алкилирующие агенты, например, азотистые иприты, этилениминовые соединения и алкилсульфонаты; антиметаболиты, например фолиевую кислоту, пуриновые или пиримидиновые антагонисты; антимитотические агенты, например антитубулиновые агенты, такие как эрибулин или мезилат эрибулина (Halaven™) или их производные, алкалоиды барвинка и ауристатины; цитотоксические антибиотики; соединения, которые повреждают или мешают экспрессии или репликации ДНК, например, белки, связывающиеся с малой бороздой ДНК; и антагонисты рецепторов фактора роста. Кроме того, химиотерапевтические агенты включают в себя антитела, биологические молекулы и малые молекулы. Химиотерапевтический агент может быть цитотоксическим или цитостатическим агентом. Термин "цитостатический агент" относится к агенту, который ингибирует или подавляет рост клеток и/или размножение клеток.
[0079] Термин "цитотоксический агент" относится к веществу, которое вызывает гибель клеток, главным образом, путем вмешательства в экспрессионную активность клетки и/или ее функционирование. Примеры цитотоксических агентов включают в себя, но не ограничиваются ими, антимитотические агенты, такие как эрибулин, ауристатин (например, монометил ауристатин E (MMAE), монометил ауристатин F (MMAF)), майтанзиноиды (например, майтансин), доластатины, дуостатины, криптофицины, алкалоиды барвинка (например, винкристин, винбластин), таксаны, таксолы и колхицины; антрациклины (например, даунорубицин, доксорубицин, дигидроксиантрациндион); цитотоксические антибиотики (например, митомицины, актиномицины, дуокармицины (например, CC-1065), ауромицины, дуомицины, калихеамицины, эндомицины, феномицины); алкилирующие агенты (например, цисплатин); интеркалирующие агенты (например, бромид этидия); ингибиторы топоизомеразы (например, этопозид, тенопозид); радиоизотопы, такие как At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212 или 213, P32, и радиоактивные изотопы лютеция (например, Lu177); а также токсины бактериального, грибкового, растительного или животного происхождения (например, рицин (например, A-цепь рицина), дифтерийный токсин, экзотоксин A Pseudomonas (например, PE40), эндотоксин, митогеллин, комбрестатин, рестриктоцин, гелонин, альфа-сарцин, абрин (например, А-цепь абрина), модецин (например, A-цепь модецина), курицин, кротин, ингибитор Sapaonaria officinalis, глюкокортикоид).
[0080] В данном контексте термин "эрибулин" относится к синтетическому аналогу галихондрина В, макроциклического соединения, которое первоначально было выделено из морской губки Halichondria okadais. Термин "фрагмент лекарственного средства эрибулина" относится к компоненту ADC, который имеет структуру эрибулина, и присоединен к линкеру ADC через его амин С-35. Эрибулин является ингибитором динамики микротрубочек, который, как считается, связывает тубулин и индуцирует остановку клеточного цикла в фазе G2/M путем ингибирования сборки митотического веретена. Термин "мезилат эрибулина" относится к мезилатной соли эрибулина, которая продается под торговым названием Halaven™.
[0081] Термин "гомолог" относится к молекуле, которая проявляет гомологию с другой молекулой, имея, например, последовательности химических остатков, которые являются одинаковыми или похожими в соответствующих положениях.
[0082] В данном контексте термин "ингибировать" или "ингибирование" означает уменьшение на измеримое количество и может включать, но не требует полного предотвращения или ингибирования.
[0083] Термин "отрицательный на наличие цели" или "отрицательный на наличие целевого антигена" относится к отсутствию экспрессии целевого антигена клеткой или тканью. Термин "положительный на наличие цели" или "положительный на наличие целевого антигена" относится к присутствию экспрессии целевого антигена. Например, клетка или клеточная линия, которая не экспрессирует целевой антиген, могут быть описаны как отрицательные на наличие цели, тогда как клетка или клеточная линия, которая экспрессирует целевой антиген, может быть описана как положительные на наличие цели.
[0084] Термин "неспецифический цитолиз" или "эффект "свидетеля"" относится к убийству клеток, отрицательных на наличие цели в присутствии клеток, положительных на наличие цели, при этом убийство клеток, отрицательных на наличие цели не наблюдается в отсутствие клеток, положительных на наличие цели. Клеточный контакт или, по меньшей мере, близость между клетками, положительными на наличие цели и отрицательными на наличие цели делает возможным неспецифический цитолиз. Этот тип уничтожения отличается от "нецелевого уничтожения", которое относится к неизбирательному убийству клеток, отрицательных на наличие цели. "Нецелевое уничтожение" может наблюдаться в отсутствие клеток, положительных на наличие цели.
[0085] Термин "рак" относится к физиологическому состоянию у млекопитающих, в которых популяция клеток характеризуется нерегулируемым ростом клеток. Примеры рака включают, но не ограничиваются ими, карциному, лимфому, бластому, саркому и лейкемию. Более конкретные примеры таких злокачественных новообразований включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточную карциному легкого, рак органов брюшной полости, гепатоцеллюлярный рак, рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак молочной железы (например, трижды негативный рак молочной железы), остеосаркому, меланому, рак толстой кишки, колоректальный рак, рак эндометрия (например, серозный) или рак матки, карциному слюнных желез, рак почки, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, карцинома печени и различные типы рака головы и шеи. Трижды негативный рак молочной железы относится к раку молочной железы, который является отрицательным на экспрессию генов рецептора эстрогена (ER), рецептора прогестерона (PR) или Her2/neu.
[0086] Термины "опухоль" и "новообразование" относятся к любой массе ткани, которая является результатом чрезмерного роста или пролиферации клеток, доброкачественных или злокачественных, включая предраковые поражения.
[0087] Термины "раковая клетка" и "опухолевая клетка" относятся к отдельным клеткам или общей популяции клеток, полученных из опухоли, включая как неопухолегенные клетки, так и раковые стволовые клетки. Применяемый в данном документе термин "опухолевая клетка" будет модифицироваться термином "неопухолегенный", когда он относится исключительно к тем опухолевым клеткам, у которых нет способности обновляться и дифференцироваться, чтобы отличать эти опухолевые клетки от раковых стволовых клеток.
[0088] Термины "субъект" и "пациент" применяются в данном документе взаимозаменяемо для обозначения любого животного, такого как любое млекопитающее, включая, но не ограничиваясь ими, людей, приматов, отличных от человека, грызунов и тому подобное. В некоторых вариантах реализации изобретения млекопитающее представляет собой мышь. В некоторых вариантах осуществления млекопитающее представляет собой человека.
[0089] Термин "совместное введение" или введение "в сочетании с" одним или более терапевтическими агентами включает одновременное и последовательное введение в любом порядке.
[0090] "Фармацевтическая композиция" относится к препарату, который находится в такой форме, которая позволяет вводить, а затем обеспечивает предполагаемую биологическую активность активного ингредиента(ов) и/или достигать терапевтического эффекта, и которая не содержит дополнительных компонентов, которые неприемлемо токсичны для субъекта, которому вводили препарат. Фармацевтическая композиция может быть стерильной.
[0091] "Фармацевтический наполнитель" включает материал, такой как адъювант, носитель, рН-регулирующие и буферные агенты, агенты для регулирования тоничности, смачивающие агенты, консервант и тому подобное.
[0092] "Фармацевтически приемлемые" средства одобряются или утверждаются регулирующим органом федерального правительства или правительства штата, или внесены в в Фармакопею США или другую общепризнанную фармакопею для применения на животных и, более конкретно, на людях.
[0093] "Эффективное количество" ADC, как описано в данном документе, представляет собой количество, достаточное для выполнения конкретно заявленной цели, например, для получения терапевтического эффекта после введения, такого как уменьшение скорости роста опухоли или объема опухоли, уменьшения симптома рака или некоторых других признаков эффективности лечения. Эффективное количество может быть определено в обычном порядке в отношении заявленной цели. Термин "терапевтически эффективное количество" относится к количеству ADC, эффективного для лечения заболевания или расстройства у субъекта. В случае рака терапевтически эффективное количество ADC может уменьшать количество раковых клеток, уменьшать размер опухоли, ингибировать (например, замедлять или останавливать) метастазы опухолей, ингибировать (например, замедлять или останавливать) рост опухоли и/или облегчать один или более симптомов. "Профилактически эффективное количество" относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для достижения желаемого профилактического результата. Как правило, поскольку профилактическая доза применяется у субъектов до или на более ранней стадии заболевания, профилактически эффективное количество будет меньше терапевтически эффективного количества.
[0094] Применяемый в данном документе термин "лечить" или "терапевтический" и грамматически связанные термины относится к любому улучшению любых последствий заболевания, таких как пролонгированная выживаемость, меньшая заболеваемость и/или уменьшение побочных эффектов, которые являются побочными продуктами альтернативного терапевтического воздействия. Как легко понять в данной области техники, полное искоренение болезни является предпочтительным, но не обязательным для лечения. В данном контексте "лечение" или "лечить" относится к введению описанного ADC субъекту, например, пациенту. Лечение может заключаться в терапии, излечении, облегчении, уменьшении, изменении, залечивании, устранении, ослаблении, улучшении или влиянии на расстройство, симптомы расстройства или предрасположенность к расстройству, например, раку.
[0095] В некоторых вариантах реализации изобретения применяется меченный ADC. Подходящие "метки" включают радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные фрагменты, хемилюминесцентные фрагменты, магнитные частицы и тому подобное.
[0096] Под "белком", как применяется в данном документе, подразумевается по меньшей мере две ковалентно присоединенные аминокислоты. Данный термин охватывает полипептиды, олигопептиды и пептиды. В некоторых вариантах реализации изобретения две или более ковалентно присоединенные аминокислоты присоединены с помощью пептидной связи. Белок может состоять из аминокислот естественного происхождения и пептидных связей, например, в случае если белок получают рекомбинантно с применением экспрессионных систем и клеток-хозяев. В альтернативном варианте, белок может включать в себя синтетические аминокислоты (например, гомофенилаланин, цитруллин, орнитин и норлейцин) или пептидомиметические структуры, то есть "пептидные или белковые аналоги", такие как пептоиды. Пептоиды представляют собой типовый класс пептидомиметиков, боковые цепи которых присоединены к атому азота пептидного остова, а не к α-атомам углерода (так как они находятся в аминокислотах) и имеют различные водородные связи и конформационные характеристики по сравнению с пептидами (см., например, Simon и соавт. (1992 год) Proc. Natl. Acad. Sci. USA 89:9367). Как таковые, пептоиды могут быть устойчивыми к протеолизу или другим физиологическим условиям или условиям хранения и эффективны при проникновении через клеточные мембраны. Такие синтетические аминокислоты могут быть включены, в частности, в случае если антитело синтезировано in vitro общепринятыми способами, хорошо известными в данной области техники. Кроме того, можно применять любую комбинацию пептидомиметических, синтетических и встречающихся в природе остатков/структур. Термин "аминокислота" также включает аминокислотные остатки, такие как пролин и гидроксипролин. Аминокислотная "группа R" или "боковая цепь" может быть либо в (L)-, либо в (S)-конфигурации. В конкретном варианте реализации изобретения аминокислоты находятся в (L)- или (S)-конфигурации.
[0097] "Рекомбинантный белок" представляет собой белок, полученный с применением рекомбинантных методов с применением любых методов и способов, известных в данной области техники, то есть посредством экспрессии рекомбинантной нуклеиновой кислоты. Способы и методы получения рекомбинантных белков хорошо известны в данной области техники.
[0098] "Выделенный" белок не сопровождается, по меньшей мере, некоторым продуктом, с которым он обычно связан в его естественном состоянии, например, составляющим по меньшей мере около 5% или по меньшей мере около 50% от общего веса белка в данном образце. Понятно, что выделенный белок может составлять от 5 до 99,9% массы общего содержания белка в зависимости от обстоятельств. Например, белок может быть получен в значительно более высокой концентрации за счет применения индуцибельного промотора или промотора повышенной экспрессии, так что белок получают в повышенных концентрациях. Определение включает продуцирование антитела в самых разных организмах и/или клетках-хозяевах, известных в данной области техники.
[0099] Для аминокислотных последовательностей идентичность последовательности и/или сходство могут быть определены с применением стандартных методов, известных в данной области техники, включая, но без ограничений, алгоритм идентификации локальной последовательности по Smith and Waterman (1981 год) Adv. Appl. Math. 2:482, алгоритм выравнивания идентичности последовательности по Needleman and Wunsch (1970 год) J. Mol. Biol. 48:443, способ поиска сходства по Pearson and Lipman (1988 год) Proc. Nat. Acad. Sci. USA 85:2444, компьютерные реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Мэдисон, штат Висконсин), программу последовательности Best Fit, описанную Devereux и соавт. (1984 год) Nucl. Acid Res. 12:387-95, предпочтительно применяя настройки по умолчанию или непосредственно проверкой. Предпочтительно, процентная идентичность вычисляется с помощью FastDB на основе следующих параметров: штраф несоответствия 1; штраф за пропуск в последовательности 1; размер штрафа за пропуск в последовательности 0,33; и штраф соединения 30 ("Current Methods in Sequence Comparison and Analysis," Macromolecule Sequencing and Synthesis, Selected Methods and Applications, стр. 127-149 (1988 год), Alan R. Liss, Inc).
[00100] Примером полезного алгоритма является PILEUP. PILEUP создает множественное выравнивание последовательностей из группы подобных последовательностей с применением прогрессивных попарных выравниваний. Он также может отображать схему, показывающую отношения кластеризации, применяемые для создания выравнивания. PILEUP применяет упрощение способа прогрессивного выравнивания по Feng & Doolittle (1987 год) J. Mol. Evol. 35:351-60; способ аналогичен описанному Higgins and Sharp (1989 год) CABIOS 5:151-3. Полезные параметры PILEUP включают штраф за открытие разрыва по умолчанию 3,00, штраф за продолжение разрыва по умолчанию 0,10 и оцениваемые концевые разрывы.
[00101] Другим примером полезного алгоритма является алгоритм BLAST, описанный в: Altschul и соавт. (1990 год) J. Mol. Biol. 215:403-10; Altschul и соавт. (1997 год) Nucleic Acids Res. 25:3389-402; и Karin и соавт. (1993 год) Proc. Natl. Acad. Sci. USA 90:5873-87. Особенно полезной программой BLAST является программа WU-BLAST-2, полученная от Altschul и соавт. (1996 год) Methods in Enzymology 266:460-80. WU-BLAST-2 применяет несколько параметров поиска, большинство из которых имеют значения по умолчанию. Регулируемые параметры задаются со следующими значениями: длина перекрывания=l, доля перекрытия=0,125, пороговая длина слова (T)=II. Параметры HSP S и HSP S2 являются динамическими значениями и определяются самой программой в зависимости от состава конкретной последовательности и состава конкретной базы данных, с которой осуществляется поиск последовательности, представляющей интерес; однако значения могут быть скорректированы для повышения точности.
[00102] Дополнительным полезным алгоритмом является gapped BLAST, как сообщается в Altschul и соавт. (1993 год) Nucl. Acids Res. 25:3389-402. Gapped BLAST применяет очки замещения BLOSUM-62; пороговый параметр T установлен на 9; метод двойного удара применяется для запуска расширений ungapped, цена длины разрыва k составляет 10+k; Xu установлен на 16, а Xg установлен на 40 для этапа поиска в базе данных и на 67 для этапа вывода алгоритмов. Выравнивания gapped запускаются с помощью оценки, соответствующей около 22 битам.
[00103] Как правило, гомология аминокислот, сходство или идентичность между белками, описанными в данном документе, и их вариантами, включая варианты FRA, варианты her2, варианты последовательностей тубулина и варианты вариабельных доменов антитела (включая отдельные варианты CDR), составляют по меньшей мере 80% по отношению к последовательностям, представленным в данном документе, и как правило с предпочтительно возрастающими гомологиями или идентичностями по меньшей мере 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и почти 100% или 100%.
[00104] Аналогичным образом, "процент (%) идентичности аминокислотной последовательности" по отношению к последовательности нуклеиновой кислоты антител и других белков, определенных в данном документе, определяют, как процентное содержание нуклеотидных остатков в последовательности-кандидате, которые идентичны остаткам нуклеотидов в кодирующей последовательности антигенсвязывающего белка. В конкретном способе используется модуль BLASTN WU-BLAST-2 с параметрами по умолчанию, с длиной перекрывания и долей перекрывания, установленным на 1 и 0,125, соответственно.
[00105] Хотя сайт или область для введения вариации аминокислотной последовательности предопределены, мутация сама по себе не обязательно должна быть предопределена. Например, с целью оптимизации производительности мутации на данном сайте, случайный мутагенез может быть проведен в целевом кодоне или области, а экспрессированные варианты CDR антигенсвязывающего белка, скринированы на оптимальное сочетание желаемой активности. Методы получения замещающих мутаций на заранее определенных участках ДНК, имеющих известную последовательность, являются хорошо известными, например, мутагенез с применением праймера MI3 и мутагенез с применением ПЦР.
Конъюгаты антитело-лекарственное средство
[00106] Соединения по данному изобретению включают соединения с противораковой активностью. В частности, соединения включают в себя фрагмент антитела (включая его антигенсвязывающий фрагмент), конъюгированный (то есть ковалентно связанный линкером) с фрагментом лекарственного средства, при этом фрагмент лекарственного средства, в случае если он не конъюгирован с фрагментом антитела, имеет цитотоксический или цитостатический эффект. В различных вариантах реализации изобретения фрагмент лекарственного средства проявляет пониженную цитотоксичность или ее отсутствие в случае если связан в конъюгате, но возобновляет цитотоксичность после расщепления из линкера и фрагмента антитела. В различных вариантах реализации изобретения фрагмент лекарственного средства демонстрирует уменьшение или отсутствие неспецифического цитолиза в случае если связан в конъюгате (например, с применением нерасщепляемого линкера), но проявляет повышенный неспецифический цитолиз после отщепления от конъюгата (например, конъюгата, имеющего расщепляемый фрагмент Val-Cit).
[00107] Разработка и производство ADC для применения в качестве терапевтического агента для человека, например, в качестве онкологического агента, может потребовать больше, чем идентификация антитела, способного связываться с желаемой целью или целями и прикрепляться к лекарственному средству, применяемому самостоятельно для лечения рака. Связывание антитела с лекарственным средством может оказывать значительное и непредсказуемое воздействие на активность одного или обоих антител и лекарственного средства, воздействия, которые будут варьироваться в зависимости от типа линкера и/или выбранного лекарственного средства. Поэтому в некоторых вариантах реализации изобретения компоненты ADC выбирают так, чтобы (i) сохранять одно или более терапевтических свойств, проявляемых антителом и фрагментами лекарственного средства при выделении, (ii) поддерживать специфические связывающие свойства фрагмента антитела; (iii) оптимизировать содержание лекарственного средства и соотношения лекарственного средства-к-антителу; (iv) позволить доставку, например, внутриклеточную доставку фрагмента лекарственного средства посредством стабильного присоединения к фрагменту антитела; (v) сохранять стабильность ADC в виде интактного конъюгата до транспортировки или доставки к целевому сайту; (vi) минимизировать агрегацию ADC до или после введения; (vii) позволить терапевтический эффект, например цитотоксический эффект, фрагмента лекарственного средства после расщепления в клеточной среде; (viii) демонстрировать эффективность противораковой терапии in vivo сравнимую или превосходящую эффективность антитела и фрагментов лекарственного средства при выделении; (ix) свести к минимуму нецелевое уничтожение с помощью фрагмента лекарственного средства; и/или (х) демонстрировать желаемые фармакокинетические и фармакодинамические свойства, формулируемость и токсикологические/иммунологические профили. Скрининг каждого из этих свойств может потребоваться для идентификации улучшенного ADC с целью терапевтического применения (Ab и соавт. (2015 год) Mol. Cancer The 14:1605-13).
[00108] В различных вариантах реализации изобретения описанные в данном документе ADC демонстрируют неожиданно благоприятные свойства в некоторых или каждой из перечисленных выше категорий. Например, в некоторых вариантах реализации изобретения конструкции ADC, содержащие присоединение Mal к антителу, спейсерную единицу ПЭГ (предпочтительно короткую спейсерную единицу ПЭГ) и/или пептидный расщепляемый линкер (например, линкер Val-Cit) демонстрируют удивительно благоприятное содержание лекарственного средства, агрегацию и/или профили стабильности, и/или сохраняют функцию связывания антител, активность лекарственного средства, и/или улучшенный неспецифический цитолиз, в то же время уменьшая нецелевое уничтожение, по сравнению с ADC, которые применяют другие расщепляемые или нерасщепляемые линкерные структуры.
[00109] В некоторых вариантах реализации ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с антителом (например, антителом к FRA, таким как MORAb-003), проявляет особенно благоприятные свойства по перечисленным категориям по сравнению с другими расщепляемыми или нерасщепляемыми линкерами, соединяющими эрибулин с фрагментом антитела. В некоторых вариантах реализации изобретения ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с антителом (например, антителом к FRA, таким как MORAb-003), проявляет особенно благоприятные свойства неспецифического цитолиза по сравнению с нерасщепляемым ADC. В некоторых вариантах реализации изобретения ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с антителом (например, антителом к FRA, таким как MORAb-003), проявляет особенно благоприятные свойства неспецифического цитолиза по сравнению с ADC, который применяет альтернативные расщепляемые линкерные структуры.
[00110] В некоторых вариантах реализации изобретения ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с MORAb-003, обладает более высоким и более желательным соотношением лекарственное средство:антитело (то есть, соотношение около 3-4) по отношению к ADC, например, содержащим линкер, присоединенный к антителу через альтернативный фрагмент (например, сукцинимидный фрагмент). В некоторых вариантах реализации изобретения ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с MORAb-003, обладает более высоким и более желательным соотношением лекарственное средство:антитело и/или более низкими уровнями агрегации по отношению к ADC, например, содержащим более длинную спейсерную единицу (например, (ПЭГ)8). В некоторых вариантах реализации изобретения ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с MORAb-003, демонстрирует более высокое и более желательное соотношение лекарственное средство:антитело, более низкие уровни агрегации, повышенное целевое уничтожение и/или уменьшенное нецелевое уничтожение по сравнению с ADC, например, содержащим альтернативный расщепляемый фрагмент (то есть непептидный расщепляемый фрагмент, такой как расщепляемый дисульфид или сульфонамид). В некоторых вариантах реализации изобретения ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с MORAb-003, демонстрирует повышенную стабильность, увеличенное целевое уничтожение, уменьшенное нецелевое уничтожение, более низкие уровни агрегации и/или более высокое и более желательное соотношение лекарственное средство:антитело по отношению к ADC, например, содержащим альтернативную аминокислотную единицу (например, Ala-Ala-Asn) или альтернативный расщепляемый фрагмент (например, расщепляемый дисульфид или сульфонамид).
[00111] В некоторых вариантах реализации изобретения ADC, содержащий линкер Mal-(ПЭГ)2-Val-Cit-pAB, соединяющий эрибулин с MORAb-003, может наблюдаться с ADC, содержащими линкерный токсин Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин, конъюгированный с антителом к Her2, таким как трастузумаб, или антителом к мезотелину.
[00112] Соединения ADC по данному изобретению могут избирательно доставлять эффективную дозу цитотоксического или цитостатического агента в раковые клетки или в опухолевую ткань. Было обнаружено, что описанные ADC обладают сильной цитотоксической и/или цитостатической активностью против клеток, экспрессирующих соответствующий целевой антиген (например, FRA или her2). В некоторых вариантах реализации изобретения цитотоксическая и/или цитостатическая активность ADC зависит от уровня экспрессии целевого антигена в клетке. В некоторых вариантах реализации изобретения, описанные ADC особенно эффективны при убийстве раковых клеток, экспрессирующих целевой антиген на высоком уровне, по сравнению с раковыми клетками, экспрессирующими один и тот же антиген на низком уровне. В некоторых вариантах реализации изобретения, описанные ADC особенно эффективны при убийстве раковых клеток, экспрессирующих целевой антиген на умеренном уровне, по сравнению с раковыми клетками, экспрессирующими тот же самый антиген на низком уровне. Типовые раки с высоким уровнем экспрессии FRA включают в себя, но без ограничений, рак яичников (например, серозный рак яичников, светлоклеточный рак яичников), карциноид легких, трижды негативный рак молочной железы, рак эндометрия и немелкоклеточный рак легкого (например, аденокарцинома). Типовые раки с умеренным уровнем экспрессии FRA включают в себя, но без ограничений, рак желудка и колоректальный рак. Типовые раки с низким уровнем экспрессии FRA включают в себя, но без ограничений, меланому и лимфому. Типовые раки с высоким уровнем экспрессии her2 включают в себя, но без ограничений, рак молочной железы, рак желудка, рак пищевода, рак яичников и рак эндометрия. Типовые раки с умеренным уровнем экспрессии her2 включают в себя, но без ограничений, рак легких и рак мочевого пузыря.
[00113] В некоторых вариантах реализации изобретения расщепление ADC высвобождает эрибулин из фрагмента антитела и линкера. В некоторых вариантах реализации изобретения расщепление и высвобождение эрибулина улучшает цитотоксичность ADC. В некоторых вариантах реализации изобретения ADC, содержащий расщепляемый линкер, особенно эффективен при убийстве раковых клеток, включая неспецифический цитолиз, по сравнению с сопоставимым лечением с ADC, содержащим нерасщепляемый линкер. В некоторых вариантах реализации изобретения ADC, содержащий расщепляемый линкер (например, линкер Val-Cit), демонстрирует повышенное целевое уничтожение клеток и/или уменьшенное нецелевое уничтожение клеток по сравнению с ADC, содержащим нерасщепляемый линкер, (например, нерасщепляемый линкер (ПЭГ)2 или (ПЭГ)4), при этом в частности клетки и/или рак, получавшие ADC, не экспрессируют высокие уровни целевого антигена.
[00114] В некоторых вариантах реализации изобретения, описанные ADC также демонстрируют активность неспецифического цитолиза, но низкую нецелевую цитотоксичность. Не ограничиваясь теорией, активность неспецифического цитолиза ADC может быть особенно полезной, в случае если его проникновение в солидную опухоль ограничено и/или экспрессия целевого антигена среди опухолевых клеток является гетерогенной. В некоторых вариантах реализации изобретения ADC, содержащий расщепляемый линкер, особенно эффективен при неспецифическом цитолизе и/или демонстрирует улучшенную активность неспецифического цитолиза, по сравнению с сопоставимым лечением с ADC, содержащим нерасщепляемый линкер.
[00115] В данном изобретении представлены соединения ADC, содержащие антитело или его антигенсвязывающий фрагмент (Ab), который нацелен на опухолевую клетку, фрагмент лекарственного средства (D) и линкерный фрагмент (L), который ковалентно присоединяет Ab к D. В некоторых аспектах антитело или антигенсвязывающий фрагмент могут связываться с опухолеассоциированным антигеном (например, FRA или her2) с высокой специфичностью и высокой аффинностью. В некоторых вариантах реализации изобретения антитело или антигенсвязывающий фрагмент интернализируются в целевую клетку при связывании, например, в деградирующий компартмент в клетке. Предпочтительными ADC являются, таким образом, те, которые интернализируются при связывании с целевой клеткой, подвергаются деградации и высвобождают фрагмент лекарственного средства для уничтожения раковых клеток. Фрагмент лекарственного средства может быть высвобожден из антитела и/или линкерного фрагмента ADC путем ферментативного действия, гидролиза, окисления или любого другого механизма.
[00116] Типовый ADC соответствует Формуле I:
Ab-(L-D)p (I)
где Ab=фрагмент антитела (то есть, антитело или антигенсвязывающий фрагмент), L=линкерный фрагмент, D=фрагмент лекарственного средства и p=количество фрагментов лекарственного средства на фрагмент антитела.
Антитела
[00117] Фрагмент антитела (Ab) Формулы I включает в свой объем любое антитело или антигенсвязывающий фрагмент, который специфически связывается с целевым антигеном в раковой клетке. Антитело или антигенсвязывающий фрагмент может связываться с целевым антигеном с константой диссоциации (KD) ≤1 мМ, ≤100 нМ или ≤10 нМ или любым значением между ними, как измерено, например, с помощью анализа BIAcore®. В определенных вариантах реализации изобретения KD составляет от 1 пМ до 500 пМ. В некоторых вариантах реализации изобретения KD составляет от 500 пМ до 1 мкМ, от 1 мкМ до 100 нМ или от 100 мМ до 10 нМ.
[00118] В некоторых вариантах реализации изобретения фрагмент антитела представляет собой четырехцепочечное антитело (также называемое иммуноглобулином), содержащее две тяжелые цепи и две легкие цепи. В некоторых вариантах реализации изобретения фрагмент антитела представляет собой двухцепочечное тело (одну легкую цепь и одну тяжелую цепь) или антигенсвязывающий фрагмент иммуноглобулина.
[00119] В некоторых вариантах реализации изобретения фрагмент антитела является интернализирующимся антителом или его интернализирующимся антигенсвязывающим фрагментом. В некоторых вариантах реализации изобретения интернализирующееся антитело связывается с целевым раковым антигеном, экспрессируемым на поверхности клетки, и проникает в клетку при связывании. В некоторых вариантах реализации изобретения фрагмент лекарственного средства ADC высвобождается из фрагмента антитела ADC после того, как ADC проникает в клетку, и присутствует в клетке, экспрессирующей целевой раковый антиген (то есть после того, как ADC был интернализирован).
[00120] Аминокислоты и последовательности нуклеиновых кислот типовых антител по данному изобретению приведены в Таблицах 1-9.
Таблица 1. Антитела
Сокращения: xi - химерное; zu - гуманизированное.
Таблица 2. Аминокислотные последовательности вариабельных областей мАт
Таблица 3. Последовательности нуклеиновой кислоты, кодирующие вариабельные области мАт
Таблица 4. Аминокислотные последовательности CDR мАт по Кабату
Таблица 5. Последовательности нуклеиновой кислоты, кодирующие CDR мАт по Кабату
Таблица 6. Аминокислотные последовательности CDR мАт IMGT
Таблица 7. Последовательности нуклеиновой кислоты, кодирующие CDR мАт IMGT
Таблица 8. Аминокислотные последовательности полноразмерных цепей мАт Ig
Таблица 9. Последовательности нуклеиновой кислоты, кодирующие полноразмерные цепи мАт Ig+
+ Представленные последовательности нуклеиновых кислот не включают лидерные последовательности.
[00121] В различных вариантах реализации изобретения, описанный в данном документе ADC может содержать любой набор вариабельных доменов тяжелой и легкой цепей, перечисленных в приведенных выше таблицах (например, вариабельные домены тяжелой и легкой цепи MORAb-003 или вариабельные домены тяжелой и легкой цепи трастузумаба) или набор из шести последовательностей CDR из набора тяжелых и легких цепей. В некоторых вариантах реализации изобретения ADC дополнительно содержит константные домены тяжелой и легкой цепи человека или их фрагменты. Например, ADC может содержать константный домен тяжелой цепи IgG человека (такой как IgG1) и константный домен легкой цепи каппа или лямбда человека. В различных вариантах реализации изобретения фрагмент антитела описанных ADC содержит константный домен тяжелой цепи подтипа 1 иммуноглобулина G (IgG1) человека с константным доменом легкой цепи Ig каппа человека.
[00122] В различных вариантах реализации изобретения целевой раковый антиген для ADC представляет собой рецептор фолиевой кислоты альфа ("FRA").
[00123] В различных вариантах реализации изобретения антитело к FRA или его антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи следующим образом: CDR1 тяжелой цепи (HCDR1), состоящий из SEQ ID NO: 2, CDR2 тяжелой цепи (HCDR2), состоящий из SEQ ID NO: 3, CDR3 тяжелой цепи (HCDR3), состоящий из SEQ ID NO: 4; CDR1 легкой цепи (LCDR1), состоящий из SEQ ID NO: 7, CDR2 легкой цепи (LCDR2), состоящий из SEQ ID NO: 8 и CDR3 легкой цепи (LCDR3), состоящий из SEQ ID NO: 9, как определено системой нумерации Кабат (Kabat, Sequences of Proteins of Immunological Interest (National Institutes of Health, Бетесда, штат Мэриленд (1987 год и 1991 год))).
[00124] В некоторых вариантах реализации изобретения антитело к FRA или его антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи следующим образом: CDR1 тяжелой цепи, состоящий из SEQ ID NO: 13, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 14, CDR3 тяжелой цепи, состоящий из SEQ ID NO: 15; CDR1 легкой цепи, состоящий из SEQ ID NO: 16, CDR2 легкой цепи, состоящий из SEQ ID NO: 17 и CDR3 легкой цепи, состоящий из SEQ ID NO: 18, как определено системой нумерации IMGT (International ImMunoGeneTics Information System (IMGT®)).
[00125] В различных вариантах реализации изобретения антитело к FRA или его антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 23, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 24. В некоторых вариантах реализации изобретения антитело к FRA или его антигенсвязывающий фрагмент содержит аминокислотную последовательность вариабельной области тяжелой цепи из SEQ ID NO: 23 и аминокислотную последовательность вариабельной области легкой цепи из SEQ ID NO: 24 или последовательности, которые по меньшей мере на 95% идентичны вышеупомянутым последовательностям. В некоторых вариантах реализации изобретения антитело к FRA или его антигенсвязывающий фрагмент имеет аминокислотную последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична SEQ ID NO: 23 и аминокислотную последовательность вариабельной области легкой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична SEQ ID NO: 24.
[00126] В различных вариантах реализации изобретения антитело к FRA содержит константный домен тяжелой цепи IgG1 человека с константным доменом легкой цепи Ig каппа человека.
[00127] В различных вариантах реализации изобретения антитело к FRA содержит аминокислотную последовательность тяжелой цепи из SEQ ID NO: 1 или последовательность, которая по меньшей мере на 95% идентична последовательности SEQ ID NO: 1, и аминокислотную последовательность легкой цепи SEQ ID NO: 6 или последовательность, которая по меньшей мере на 95% идентична последовательности SEQ ID NO: 6. В конкретных вариантах реализации изобретения антитело содержит аминокислотную последовательность тяжелой цепи из SEQ ID NO: 1 и аминокислотную последовательность легкой цепи из SEQ ID NO: 6 или последовательности, которые по меньшей мере на 95% идентичны вышеупомянутым последовательностям. В некоторых вариантах реализации изобретения антитело к FRA имеет аминокислотную последовательность тяжелой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична последовательности SEQ ID NO: 1 и/или аминокислотную последовательность легкой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична последовательности SEQ ID NO: 6. В некоторых вариантах реализации изобретения антитело к FRA содержит тяжелую цепь, кодируемую нуклеотидной последовательностью SEQ ID NO: 11 (с нуклеотидами, кодирующими лидерную последовательность) или SEQ ID NO: 345 (без нуклеотидов, кодирующих лидерную последовательность); и легкую цепь, кодируемую нуклеотидной последовательностью SEQ ID NO: 12 (с нуклеотидами, кодирующими лидерную последовательность) или SEQ ID NO: 346 (без нуклеотидов, кодирующих лидерную последовательность). В некоторых вариантах реализации изобретения аминокислотная последовательность тяжелой цепи не имеет С-концевого лизина. В различных вариантах реализации изобретения антитело к FRA имеет аминокислотную последовательность антитела, продуцируемого клеточной линией, депонированной на условиях в соответствии с Будапештским договором с Американской коллекцией типовых культур (ATCC, 10801 University Blvd., Манассас, штат Виргиния20110-2209) 24 апреля 2006 года под № доступа PTA -7552 или такие последовательности, лишенные C-концевого лизина тяжелой цепи. В различных вариантах реализации изобретения антитело к FRA представляет собой MORAb-003 (название USAN: фарлетузумаб) (Ebel и соавт. (2007 год) Cancer Immunity 7:6) или его антигенсвязывающий фрагмент.
[00128] В различных других вариантах реализации изобретения целевой раковый антиген для ADC представляет собой рецептор человеческого эпидермального фактора роста 2 типа ("her2").
[00129] В различных вариантах реализации изобретения антитело к her2 или его антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи следующим образом: CDR1 тяжелой цепи (HCDR1), состоящий из SEQ ID NO: 71, CDR2 тяжелой цепи (HCDR2), состоящий из SEQ ID NO: 72, CDR3 тяжелой цепи (HCDR3), состоящий из SEQ ID NO: 73; CDR1 легкой цепи (LCDR1), состоящий из SEQ ID NO: 74, CDR2 легкой цепи (LCDR2), состоящий из SEQ ID NO: 75 и CDR3 легкой цепи (LCDR3), состоящий из SEQ ID NO: 76, как определено системой нумерации Кабат.
[00130] В некоторых вариантах реализации изобретения антитело к her2 или его антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи следующим образом: CDR1 тяжелой цепи, состоящий из SEQ ID NO: 191, CDR2 тяжелой цепи, состоящий из SEQ ID NO: 192, CDR3 тяжелой цепи, состоящий из SEQ ID NO: 193; CDR1 легкой цепи, состоящий из SEQ ID NO: 194, CDR2 легкой цепи, состоящий из SEQ ID NO: 195 и CDR3 легкой цепи, состоящий из SEQ ID NO: 196, как определено системой нумерации IMGT.
[00131] В различных вариантах реализации изобретения антитело к her2 или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 27, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 28. В некоторых вариантах реализации изобретения антитело к her2 или его антигенсвязывающий фрагмент содержит аминокислотную последовательность вариабельной области тяжелой цепи из SEQ ID NO: 27 и аминокислотную последовательность вариабельной области легкой цепи из SEQ ID NO: 28 или последовательности, которые по меньшей мере на 95% идентичны вышеупомянутым последовательностям. В некоторых вариантах реализации изобретения антитело к her2 или его антигенсвязывающий фрагмент имеет аминокислотную последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична SEQ ID NO: 27 и/или аминокислотную последовательность вариабельной области легкой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична SEQ ID NO: 28.
[00132] В различных вариантах реализации изобретения антитело к her2 содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи Ig каппа человека.
[00133] В различных вариантах реализации изобретения антитело к her2 содержит аминокислотную последовательность тяжелой цепи из SEQ ID NO: 327 или последовательность, которая по меньшей мере на 95% идентична последовательности SEQ ID NO: 327, и аминокислотную последовательность легкой цепи из SEQ ID NO: 328 или последовательность, которая по меньшей мере на 95% идентична последовательности SEQ ID NO: 328. В конкретных вариантах реализации изобретения антитело содержит аминокислотную последовательность тяжелой цепи из SEQ ID NO: 327 и аминокислотную последовательность легкой цепи из SEQ ID NO: 328 или последовательности, которые по меньшей мере на 95% идентичны вышеупомянутым последовательностям. В некоторых вариантах реализации изобретения антитело к her2 имеет аминокислотную последовательность тяжелой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична последовательности SEQ ID NO: 327 и аминокислотную последовательность легкой цепи, которая по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична последовательности SEQ ID NO: 328. В различных вариантах реализации изобретения антитело к her2 представляет собой трастузумаб или его антигенсвязывающий фрагмент.
[00134] В различных вариантах реализации изобретения антитело к FRA или его антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи MORAb-003 или при этом CDR включают не более одного, двух, трех, четырех, пяти или шести аминокислотных добавлений, делеций или замещений в HCDR1 (SEQ ID NO: 2 в соответствии с Кабатом или SEQ ID NO: 13 в соответствии с IMGT), HCDR2 (SEQ ID NO: 3 в соответствии с Кабатом или SEQ ID NO: 14 в соответствии с IMGT), HCDR3 (SEQ ID NO: 4 в соответствии с Кабатом, или SEQ ID NO: 15 в соответствии с IMGT); LCDR1 (SEQ ID NO: 7 в соответствии с Кабатом или SEQ ID NO: 16 в соответствии с IMGT), LCDR2 (SEQ ID NO: 8 в соответствии с Кабатом или SEQ ID NO: 17 в соответствии с IMGT) и LCDR3 (SEQ ID NO: 9 в соответствии с Кабатом, или SEQ ID NO: 18 в соответствии с IMGT).
[00135] В различных других вариантах реализации изобретения антитело к her2 или его антигенсвязывающий фрагмент содержит три CDR тяжелой цепи и три CDR легкой цепи трастузумаба или при этом CDR включают не более одного, двух, трех, четырех, пяти или шести аминокислотных добавлений, делеций или замещений в HCDR1 (SEQ ID NO: 71 в соответствии с Кабатом или SEQ ID NO: 191 в соответствии с IMGT), HCDR2 (SEQ ID NO: 72 в соответствии с Кабатом или SEQ ID NO: 192 в соответствии с IMGT), HCDR3 (SEQ ID NO: 73 в соответствии с Кабатом, или SEQ ID NO: 193 в соответствии с IMGT); LCDR1 (SEQ ID NO: 74 в соответствии с Кабатом или SEQ ID NO: 194 в соответствии с IMGT), LCDR2 (SEQ ID NO: 75 в соответствии с Кабатом или SEQ ID NO: 195 в соответствии с IMGT) и LCDR3 (SEQ ID NO: 76 в соответствии с Кабатом, или SEQ ID NO: 196 в соответствии с IMGT).
[00136] В различных вариантах реализации изобретения аминокислотные замены представляют собой замены одного остатка. Вставки обычно будут иметь порядок от около 1 до около 20 аминокислотных остатков, хотя значительно более крупные вставки могут быть допустимы, пока сохраняется биологическая функция (например, связывание с FRA или her2). Делеции обычно варьируются от около 1 до около 20 аминокислотных остатков, хотя в некоторых случаях делеции могут иметь значительно больший размер. Замены, делеции, вставки или любая их комбинация могут применяться для получения окончательного производного или варианта. Как правило, эти изменения производятся на нескольких аминокислотах, чтобы свести к минимуму изменение молекулы, особенно иммуногенность и специфичность антигенсвязывающего белка. Тем не менее, большие изменения могут допускаться при определенных обстоятельствах. Консервативные замены обычно выполняются в соответствии со следующей таблицей, изображенной как Таблица 10.

[00137] Существенные изменения функции или иммунологической идентичности производятся путем отбора замен, которые являются менее консервативными, чем те, которые показаны в Таблице 10. Например, могут быть сделаны замены, которые более существенно влияют на: структуру полипептидного остова в области изменения, например, альфа-спиральную или бета-складчатую структуру; заряд или гидрофобность молекулы на целевом сайте; или объемную часть боковой цепи. Ожидается, что замены, которые в общем будут давать наибольшие изменения в свойствах полипептида, представляют собой такие, в которых (а) гидрофильный остаток, например, серил или треонил, замещен (или замещается) гидрофобным остатком, например, лейцилом, изолейцилом, фенилаланилом, валилом или аланилом; (b) цистеин или пролин замещен (или замещается) любым другим остатком; (c) остаток, имеющий электроположительную боковую цепь, например лизил, аргинил или гистидил, замещен (или замещается) электроотрицательным остатком, например, глутамилом или аспартилом; или (d) остаток, имеющий объемную боковую цепь, например фенилаланин, замещен (или замещается) одним остатком, не имеющим боковой цепи, например глицином.
[00138] В различных вариантах реализации изобретения, где варианты последовательностей антитела применяются в ADC, варианты обычно проявляют такую же качественную биологическую активность и будут вызывать тот же иммунный ответ, хотя варианты могут также быть выбраны для модификации характеристик антигенсвязывающих белков по мере необходимости. В альтернативном варианте, вариант может быть сконструирован таким образом, что биологическая активность антигенсвязывающего белка изменяется. Например, сайты гликозилирования могут быть изменены или удалены, как обсуждалось в данном документе.
[00139] Различные антитела могут применяться с применяемым в данном документе ADC для нацеливания на раковые клетки. Как показано ниже, линкер-токсины в ADC, описанных в данном документе, являются неожиданно эффективными с различными антиген-нацеливающими антителами против опухоли. В данной области техники известны подходящие антигены, экспрессирующиеся на опухолевых клетках, но не на здоровых клетках, или экспрессирующиеся на опухолевых клетках на более высоком уровне, чем на здоровых клетках, а также антитела, направленные против них. Эти антитела можно применять с линкерами и токсином (например, эрибулином), описанным в данном документе. В некоторых вариантах реализации изобретения фрагмент антитела нацелен на FRA. В некоторых вариантах реализации изобретения фрагмент антитела, нацеленного на FRA, представляет собой MORAb-003. В некоторых вариантах реализации изобретения, в то время как описанные линкеры и токсин (эрибулин) являются неожиданно эффективными с несколькими различными антителами, нацеленными на опухоль, фрагменты антител, нацеленные на FRA, такие как MORAb-003, обеспечивают особенно улучшенные соотношение лекарственное средство:антитело, нацеливание на опухоль, неспецифический цитолиз, эффективность лечения, а также уменьшенное нецелевое уничтожение. Повышенная эффективность лечения может быть измерена in vitro или in vivo, и может включать снижение скорости роста опухоли и/или уменьшение объема опухоли.
[00140] В некоторых вариантах реализации изобретения применяют антитела к другим целям антигена и обеспечивают, по меньшей мере, некоторые из благоприятных функциональных свойств ADC, содержащего фрагмент антитела, нацеленного на FRA, такой как MORAb-003 (например, улучшенное соотношение лекарственное средство:антитело, улучшенная эффективность лечения, снижение нецелевого уничтожения и тому подобное). В некоторых вариантах реализации изобретения некоторые или все из этих благоприятных функциональных свойств наблюдаются, в случае если описанные линкеры и токсин (эрибулин) конъюгированы с фрагментом антитела, нацеленного на her2, таким как трастузумаб. В некоторых вариантах реализации изобретения фрагмент антитела нацелен на her2. В некоторых вариантах реализации изобретения фрагмент антитела, нацеленного на her2, представляет собой трастузумаб. В некоторых вариантах реализации изобретения некоторые или все из этих благоприятных функциональных свойств наблюдаются, в случае если описанные линкеры и токсин (эрибулин) конъюгированы с фрагментом антитела, нацеленного на MSLN, таким как MORAb-009. В некоторых вариантах реализации изобретения фрагмент антитела нацелен на MSLN. В некоторых вариантах реализации изобретения фрагмент антитела, нацеленного на MSLN, представляет собой MORAb-009.
Линкеры
[00141] В различных вариантах реализации изобретения линкер в ADC является внеклеточно стабильным достаточным образом, чтобы быть терапевтически эффективным. В некоторых вариантах реализации изобретения линкер стабилен вне клетки, так что ADC остается интактным, в случае если он присутствует во внеклеточных условиях (например, перед транспортировкой или доставкой в клетку). Термин "интактный", применяемый в контексте ADC, означает, что фрагмент антитела остается присоединенным к фрагменту лекарственного средства. В данном контексте термин "стабильный" в контексте линкера или ADC, содержащего линкер, означает, что не более чем 20%, не более чем около 15%, не более чем около 10%, не более чем около 5%, не более чем около 3% или не более чем около 1% линкеров (или любой процент между ними) в образце ADC расщепляются (или в случае общего ADC, в противном случае не интактного), в случае если ADC находится во внеклеточных условиях.
[00142] Стабилен ли линкер внеклеточно, можно определить, например, путем включения ADC в плазму крови в течение заданного периода времени (например, 2, 4, 6, 8, 16 или 24 часов), а затем количественного определения количества свободного фрагмента лекарственного средства присутствующего в плазме крови. Стабильность может предоставить время для ADC с целью локализации целевых опухолевых клеток и предотвратить преждевременное высвобождение лекарственного средства, что может снизить терапевтический индекс ADC, из-за повреждения как нормальных, так и опухолевых тканей. В некоторых вариантах реализации изобретения линкер стабилен вне целевой клетки и высвобождает часть лекарственного средства из ADC один раз внутри клетки, таким образом, что фрагмент лекарственного средства может связываться со своей целью (например, с микротрубочками). Таким образом, эффективный линкер будет:(i) поддерживать специфические связывающие свойства фрагмента антитела; (ii) позволять доставку, например, внутриклеточную доставку фрагмента лекарственного средства посредством стабильного присоединения к фрагменту антитела; (iii) оставаться стабильными и интактным до тех пор, пока ADC не будет транспортирован или доставлен к его целевому сайту; и (iv) позволять терапевтический эффект, например цитотоксический эффект, фрагмента лекарственного средства после расщепления.
[00143] Линкеры могут влиять на физико-химические свойства ADC. Поскольку многие цитотоксические агенты являются гидрофобными по своей природе, связывание их с антителом с дополнительным гидрофобным фрагментом может привести к агрегации. Агрегаты ADC нерастворимы и часто ограничивают достижимое содержание лекарственного средства на антителе, что может отрицательно влиять на эффективность ADC. Протеиновые агрегаты биологических препаратов, в общем, также были связаны с повышенной иммуногенностью. Как показано ниже, линкеры, описанные в данном документе, приводят к ADC с низким уровнем агрегации и желательным уровнем содержания лекарственного средства.
[00144] Линкер может быть "расщепляемым" или "нерасщепляемым" (Ducry and Stump, Bioconjugate Chem. (2010 год) 21:5-13). Расщепляемые линкеры предназначены для высвобождения лекарственного средства при воздействии определенных факторов окружающей среды, например, при интернализации в целевую клетку, тогда как нерасщепляемые линкеры обычно полагаются на деградацию самого фрагмента антитела.
[00145] В некоторых вариантах реализации изобретения линкером является нерасщепляемый линкер. В некоторых вариантах реализации изобретения фрагмент лекарственного средства ADC высвобождается путем деградации фрагмента антитела. Нерасщепляемые линкеры, как правило, остаются ковалентно связанными с, по меньшей мере, одной аминокислотой антитела и лекарственным средством при интернализации и деградации в целевой клетке. Нерасщепляемые линкеры обычно включают в себя тиоэфирную связь, которую получают путем конъюгации тиольной группы на лекарственном средстве или антителе с малеимидной или галоацетамидной группой на антителе или лекарственном средстве, соответственно (Goldmacher и соавт., In Cancer Drug Discovery and Development: Antibody-Drug Conjugates and Immunotoxins (G. L. Phillips ред., Springer, 2013 год)). Типовый нерасщепляемый линкер содержит тиоэфир, циклогексил, N-сукцинимидил-4-(N-малеимидометил) циклогексан-1 карбоксилат (SMCC), N-гидроксисукцинимид (NHS) или один или более фрагментов полиэтиленгликоля (ПЭГ), например, 1, 2, 3, 4, 5 или 6 ПЭГ. В некоторых вариантах реализации изобретения нерасщепляемый линкер содержит (ПЭГ)2. В других вариантах реализации изобретения нерасщепляемый линкер содержит (ПЭГ)4.
[00146] В некоторых вариантах реализации изобретения линкером является расщепляемый линкер. Расщепляемый линкер относится к любому линкеру, который содержит расщепляемый фрагмент. В данном контексте термин "расщепляемый фрагмент" относится к любой химической связи, которая может быть расщеплена. Подходящие расщепляемые химические связи хорошо известны в данной области техники и включают, но без ограничений, кислотолабильные связи, протеазо/пептидазолабильные связи, фотолабильные связи, дисульфидные связи и эстеразолабильные связи. Линкеры, содержащие расщепляемый фрагмент, могут позволить высвобождение фрагмента лекарственного средства из ADC через расщепление на определенном участке в линкере. В различных вариантах реализации изобретения расщепление антитела от связанного токсина активирует или увеличивает активность токсина. В некоторых вариантах реализации изобретения ADC, содержащий расщепляемый линкер (например, линкер Val-Cit), демонстрирует повышенное целевое уничтожение клеток и/или уменьшенное нецелевое уничтожение клеток по сравнению с ADC, содержащим нерасщепляемый линкер, (например, нерасщепляемый линкер (ПЭГ)2 или (ПЭГ)4). В некоторых вариантах реализации изобретения ADC, содержащий расщепляемый линкер, демонстрирует улучшенную эффективность лечения по отношению к ADC, содержащим нерасщепляемый линкер, в случае если клетки и/или рак, получавшие лечение в виде ADC, не экспрессируют целевой антиген на высоких уровнях (например, FRA или her2). В некоторых вариантах реализации изобретения расщепление антитела от связанного токсина требуется для достижения улучшенной эффективности лечения ADC, как измерено in vitro и/или in vivo.
[00147] В некоторых вариантах реализации изобретения линкер расщепляется при внутриклеточных условиях, так что расщепление линкера в достаточной степени высвобождает фрагмент лекарственного средства от фрагмента антитела во внутриклеточной среде для активации лекарственного средства и/или делает лекарственное средство терапевтически эффективным. В некоторых вариантах реализации изобретения фрагмент лекарственного средства не расщепляется от фрагмента антитела до тех пор, пока ADC не попадет в клетку, которая экспрессирует антиген, специфичный для фрагмента антитела ADC, а фрагмент лекарственного средства отщепляется от фрагмента антитела после входа в клетку. В некоторых вариантах реализации изобретения линкер содержит расщепляемый фрагмент, который расположен так, что часть линкера или фрагмента антитела не остается связанной с фрагментом лекарственного средства при расщеплении. Примеры расщепляемых линкеров включают кислотолабильные линкеры, линкеры, чувствительные к протеазе/пептидазе, фотолабильные линкеры, диметил-, дисульфид- или сульфонамидсодержащие линкеры.
[00148] В некоторых вариантах реализации изобретения линкером является линкер, чувствительный к рН и чувствительный к гидролизу при определенных значениях рН. Как правило, линкер, чувствительный к рН расщепляется в кислых условиях. Эта стратегия расщепления обычно использует более низкие значения рН в эндосомальных (pH ~ 5-6) и лизосомальных (pH ~ 4,8) внутриклеточных компартментах по сравнению с цитозолем (pH ~ 7,4) для запуска гидролиза кислотолабильной группы в линкере, таком как гидразон (Jain и соавт. (2015 год) Pharm Res 32:3526-40). В некоторых вариантах реализации изобретения линкер является кислотолабильным и/или гидролизуемым линкером. Например, может быть применен кислотолабильный линкер, который гидролизуется в лизосоме, и содержит кислотолабильную группу (например, гидразон, семикарбазон, тиосемикарбазон, цис-аконитовый амид, ортоэфир, ацеталь, кеталь или тому подобное). См., например, патентыСША №5,122,368; 5,824,805; 5,622,929; Dubowchik and Walker (1999 год) Pharm. Therapeutics 83:67-123; Neville и соавт. (1989 год) Biol. Chem. 264:14653-61. Такие линкеры относительно стабильны при нейтральных условиях рН, например, в крови, но нестабильны при рН ниже 5,5 или 5,0, приблизительном значении рН лизосомы. В некоторых вариантах реализации изобретения гидролизуемый линкер представляет собой тиоэфирный линкер (такой как, например, тиоэфир, присоединенный к терапевтическому агенту через ацилгидразоновую связь). См., например, патент США №5,622,929.
[00149] В некоторых вариантах реализации изобретения линкер расщепляется в восстановительных условиях. В некоторых вариантах реализации изобретения линкер расщепляется в присутствии восстанавливающего агента, такого как глутатион или дитиотреитол. В некоторых вариантах реализации изобретения линкер представляет собой расщепляемый дисульфидный линкер или расщепляемый сульфонамидный линкер.
[00150] В некоторых вариантах реализации изобретения линкер представляет собой расщепляемый дисульфидный линкер. В данной области техники известны различные дисульфидные линкеры, включая, например, те, которые могут быть получены с применением SATA (N-сукцинимидил-5-ацетилтиоацетата), SPDP (N-сукцинимидил-3-(2-пиридилдитио)пропионата), SPDB (N-сукцинимидил-3-(2-пиридилдитио)бутирата) и SMPT (N-сукцинимидилоксикарбонил-альфа-метил-альфа-(2-пиридил-дитио)толуола), SPDB и SMPT. См., например, Thorpe и соавт. (1987 год) Cancer Res. 47:5924-31; Wawrzynczak и соавт., In Immunoconjugates:Antibody Conjugates in Radioimagery and Therapy of Cancer (C. W. Vogel ред., Oxford U. Press, 1987 год). См., также патент США №4,880,935. Дисульфидные линкеры обычно применяют с целью использования распространенных внутриклеточных тиолов, которые могут способствовать расщеплению их дисульфидных связей. Внутриклеточные концентрации наиболее распространенного внутриклеточного тиола, восстановленного глутатиона, обычно находятся в диапазоне 1-10 нМ, что в около 1000 раз выше, чем у наиболее распространенного низкомолекулярного тиола в крови (то есть, цистеина) около 5 мкМ (Goldmacher и соавт., In Cancer Drug Discovery and Development:Antibody-Drug Conjugates and Immunotoxins (G. L. Phillips ред., Springer, 2013 год)). Внутриклеточные ферменты семейства белков дисульфид-изомеразы могут также способствовать внутриклеточному расщеплению дисульфидного линкера. Как применяется в данном документе, расщепляемый дисульфидный линкер относится к любому линкеру, который содержит расщепляемый дисульфидный фрагмент. Термин "расщепляемый дисульфидный фрагмент" относится к дисульфидной связи, которая может быть расщеплена и/или восстановлена, например, тиолом или ферментом. В некоторых вариантах реализации изобретения расщепляемый дисульфидный фрагмент представляет собой дисульфидил-диметил.
[00151] В некоторых вариантах реализации изобретения линкером является расщепляемый сульфонамидный линкер. Как применяется в данном документе, расщепляемый сульфонамидный линкер относится к любому линкеру, который содержит расщепляемый сульфонамидный фрагмент. Термин "расщепляемый сульфонамидный фрагмент" относится к сульфонамидной группе, то есть, к сульфонильной группе, присоединенной к аминогруппе, при этом серо-азотная связь может быть расщеплена.
[00152] В некоторых вариантах реализации изобретения линкер может быть линкером дендритного типа для ковалентного присоединения более чем одного фрагмента лекарственного средства к фрагменту антитела через разветвляющийся, многофункциональный фрагмент линкера. См., например, Sun и соавт. (2002 год) Bioorg. Med. Chem. Lett. 12:2213-5; Sun и соавт. (2003 год) Bioorg. Med. Chem. 11:1761-8. Дендритные линкеры могут увеличивать молярное соотношение лекарственного средства по отношению к антителу, то есть, содержание лекарственного средства, что связано с эффективностью ADC. Таким образом, если фрагмент антитела несет только одну реакционноспособную цистеиновую тиольную группу, например, через дендритный линкер можно присоединить множество фрагментов лекарственного средства. В некоторых вариантах реализации изобретения линкерный фрагмент или фрагмент линкер-лекарственное средство могут быть присоединены к антителу посредством химии восстановленных дисульфидных мостиков или ограниченной технологии ограниченной утилизации лизина. См., например, Международные публикации № WO 2013173391 и WO 2013173393.
[00153] В некоторых вариантах реализации изобретения линкер расщепляется расщепляющим агентом, например, ферментом, который присутствует во внутриклеточной среде (например, в лизосоме или эндосоме, или кавеоле). Линкером может быть, например, пептидный линкер, который расщепляется внутриклеточной пептидазой или ферментом протеазы, включая, но без ограничений, лизосомальную или эндосомальную протеазу. В некоторых вариантах реализации изобретения линкером является расщепляемый пептидный линкер. Как применяется в данном документе, расщепляемый пептидный линкер относится к любому линкеру, который содержит расщепляемый пептидный фрагмент. Термин "расщепляемый пептидный фрагмент" относится к любой химической связи, связывающей аминокислоты (натуральные или синтетические производные аминокислот), которые могут быть расщеплены агентом, который присутствует во внутриклеточной среде. Например, линкер может содержать последовательность аланин-аланин-аспарагин (Ala-Ala-Asn) или валин-цитруллин (Val-Cit), которая расщепляется пептидазой, такой как катепсин, например, катепсин B.
[00154] В некоторых вариантах реализации изобретения линкер представляет собой расщепляемый ферментом линкер и расщепляемый пептидный фрагмент в линкере расщепляется ферментом. В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент расщепляется лизосомальным ферментом, например, катепсином. В некоторых вариантах реализации изобретения линкер представляет собой расщепляемый катепсином линкер. В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент в линкере расщепляется лизосомальным цистеиновым катепсином, таким как катепсин B, C, F, H, K, L, O, S, V, X или W. В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент расщепляется катепсином B. Типовым дипептидом, который может быть расщеплен катепсином В, является валин-цитруллин (Val-Cit) (Dubowchik и соавт., 2002 год) Bioconjugate Chem. 13:855-69). В некоторых вариантах реализации ADC, который содержит расщепляемый пептидный фрагмент, демонстрирует более низкие уровни агрегации и/или более высокое содержание лекарственного средства (p) по сравнению с ADC, который содержит альтернативный расщепляемый фрагмент (например, расщепляемый дисульфидный фрагмент или расщепляемый сульфонамидный фрагмент).
[00155] В некоторых вариантах реализации изобретения линкер или расщепляемый пептидный фрагмент в линкере содержит аминокислотную единицу. В некоторых вариантах реализации изобретения аминокислотная единица позволяет расщеплять линкер с помощью протеазы, тем самым облегчая высвобождение фрагмента лекарственного средства из ADC при воздействии одной или более внутриклеточных протеаз, таких как один или более лизосомальных ферментов (Doronina и соавт. (2003 год) Nat. Biotechnol. 21:778-84; Dubowchik and Walker (1999 год) Pharm. Therapeutics 83:67-123). Типовые аминокислотные единицы включают в себя, но без ограничений, дипептиды, трипептиды, тетрапептиды и пентапептиды. Типовые дипептиды включают в себя, но без ограничений, валин-цитруллин (Val-Cit), аланин-аспарагин (Ala-Asn), аланин-фенилаланин (Ala-Phe), фенилаланин-лизин (Phe-Lys), аланин-лизин (Ala-Lys), аланин-валин (Ala-Val), валин-аланин (Val-Ala), валин-лизин (Val-Lys), лизин-лизин (Lys-Lys), фенилаланин-цитруллин (Phe-Cit), лейцин-цитруллин (Leu-Cit), изолейцин-цитруллин (Ile-Cit), триптофан-цитруллин (Trp-Cit) и фенилаланин-аланин (Phe-Ala). Типовые трипептиды включают в себя, но без ограничений, аланин-аланин-аспарагин (Ala-Ala-Asn), глицин-валин-цитруллин (Gly-Val-Cit), глицин-глицин-глицин (Gly-Gly-Gly), фенилаланин-фенилаланин-лизин (Phe-Phe-Lys) и глицин-фенилаланин-лизин (Gly-Phe-Lys). Другие типовые аминокислотные единицы включают, но без ограничений, Gly-Phe-Leu-Gly, Ala-Leu-Ala-Leu, Phe-N9-тозил-Arg и Phe-N9-Нитро-Arg, как описано, например, в патентеСША №6,214,345. В некоторых вариантах реализации изобретения аминокислотная единица в линкере содержит Val-Cit. В некоторых вариантах реализации изобретения аминокислотная единица в линкере содержит Ala-Ala-Asn. В некоторых вариантах реализации изобретения ADC, который включает в себя Val-Cit, демонстрирует уменьшение нецелевого уничтожения клеток, увеличение целевого уничтожения клеток, снижение уровней агрегации и/или более высокое содержание лекарственного средства (p) по сравнению с ADC, который содержит альтернативную аминокислотную единицу или альтернативный расщепляемый фрагмент. Аминокислотная единица может содержать аминокислотные остатки, которые встречаются в природе и/или второстепенные аминокислоты, и/или не встречающиеся в природе аналоги аминокислот, такие как цитруллин. Аминокислотные единицы могут быть сконструированы и оптимизированы для ферментативного расщепления конкретным ферментом, например, опухолеассоциированной протеазой, лизосомальной протеазой, такой как катепсин В, С, D или S, или плазминовой протеазой.
[00156] В некоторых вариантах реализации изобретения линкер в любом из описанных в данном документе ADC может содержать по меньшей мере одну спейсерную единицу, соединяющую фрагмент антитела с фрагментом лекарственного средства. В некоторых вариантах реализации изобретения спейсерная единица соединяет сайт расщепления (например, расщепляемый пептидный фрагмент) в линкере с фрагментом антитела. В некоторых вариантах реализации изобретения линкер и/или спейсерная единица в линкере являются по существу гидрофильными. Гидрофильный линкер может применяться для уменьшения степени, в которой лекарственное средство может выкачиваться из устойчивых раковых клеток из-за множественной устойчивости к лекарственным средствам (MDR) или функционально аналогичных транспортеров. В некоторых аспектах линкер включает один или более фрагментов полиэтиленгликоля (ПЭГ), например, 1, 2, 3, 4, 5 или 6 фрагментов ПЭГ. В некоторых вариантах реализации изобретения линкер является более коротким линкером ПЭГ и обеспечивает улучшенную стабильность и уменьшенную агрегацию по сравнению с более длинными линкерами ПЭГ.
[00157] В некоторых вариантах реализации изобретения спейсерная единица в линкере содержит один или более фрагментов ПЭГ. В некоторых вариантах реализации изобретения спейсерная единица содержит -(ПЭГ)m- и m представляет собой целое число от 1 до 10В некоторых вариантах реализации изобретения m находится в диапазоне от 1 до 10; от 2 до 8; от 2 до 6; от 2 до 5; от 2 до 4; или от 2 до 3. В некоторых вариантах реализации изобретения m равно 8. В некоторых вариантах реализации изобретения m равно 4. В некоторых вариантах реализации изобретения m равно 3. В некоторых вариантах реализации изобретения m равно 2. В некоторых вариантах реализации изобретения спейсерная единица содержит (ПЭГ)2, (ПЭГ)4, (ПЭГ)8, (ПЭГ)9, (ПЭГ)3-триазол-(ПЭГ)3, (ПЭГ)4-триазол-(ПЭГ)3, или дибензилциклооктен-триазол-(ПЭГ)3. В некоторых предпочтительных вариантах реализации изобретения спейсерная единица содержит (ПЭГ)2. В некоторых вариантах реализации изобретения ADC, который содержит более короткую спейсерную единицу (например, (ПЭГ2), демонстрирует более низкие уровни агрегации и/или более высокое содержание лекарственного средства (p) по сравнению с ADC, который содержит более длинную спейсерную единицу (например, (ПЭГ)8).
[00158] В некоторых вариантах реализации изобретения спейсерная единица в линкере содержит алкильный фрагмент. В некоторых вариантах реализации изобретения спейсерная единица содержит -(CH2)n-, и n представляет собой целое число от 1 до 10 (то есть, n может быть 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10). В некоторых вариантах реализации изобретения n равно 5. В некоторых вариантах реализации изобретения ADC, который содержит более короткую спейсерную единицу (например, (CH2)5), демонстрирует более низкие уровни агрегации и/или более высокое содержание лекарственного средства (p) по сравнению с ADC, который содержит более длинную спейсерную единицу (например, (ПЭГ)8).
[00159] Спейсерная единица может быть применена, например, для связывания фрагмента антитела с фрагментом лекарственного средства, прямо или косвенно. В некоторых вариантах реализации изобретения спейсерная единица непосредственно связывает фрагмент антитела с фрагментом лекарственного средства. В некоторых вариантах реализации изобретения фрагмент антитела и фрагмент лекарственного средства присоединяются через спейсерную единицу, содержащую одну или более фрагментов ПЭГ (например, (ПЭГ)2 или (ПЭГ)4). В некоторых вариантах реализации изобретения спейсерная единица опосредованно связывает фрагмент антитела с фрагментом лекарственного средства. В некоторых вариантах реализации изобретения спейсерная единица связывает фрагмент антитела с фрагментом лекарственного средства опосредованно через расщепляемый фрагмент (например, расщепляемый пептид, расщепляемый дисульфид или расщепляемый сульфонамид) и/или присоединенный фрагмент для присоединения спейсерной единицы к фрагменту антитела, например, малеимидному фрагменту.
[00160] Спейсерная единица в различных вариантах реализации изобретения прикрепляется к фрагменту антитела (то есть к антителу или антигенсвязывающему фрагменту) через малеимидный фрагмент (Mal). В некоторых вариантах реализации изобретения ADC, который содержит линкер, присоединенный к фрагменту антитела через малеимидный фрагмент, демонстрирует более высокое содержание лекарственного средства (p) по сравнению с ADC, который содержит линкер, присоединенный к фрагменту антитела через альтернативный присоединенный фрагмент, такой как сукцинимидный фрагмент.
[00161] Спейсерная единица, которая прикрепляется к антителу или антигенсвязывающему фрагменту через Mal, упоминается в данном документе как "Mal-спейсерная единица". В данном контексте термин "малеимидный фрагмент" означает соединение, которое содержит малеимидную группу, и которое реагирует с сульфгидрильной группой, например, сульфгидрильной группой остатка цистеина на фрагменте антитела. Другие функциональные группы, которые реагируют с сульфгидрильными группами (тиолами), включают иодацетамид, бромацетамид, винилпиридин, дисульфид, пиридилдисульфид, изоцианат и изотиоцианат, но не ограничиваются ими. В некоторых вариантах реализации изобретения Mal-спейсерная единица реагирует с остатком цистеина на антителе или антигенсвязывающем фрагменте. В некоторых вариантах реализации изобретения Mal-спейсерная единица соединяется с антителом или антигенсвязывающим фрагментом через остаток цистеина. В некоторых вариантах реализации изобретения Mal-спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения Mal-спейсерная единица содержит алкильный фрагмент.
[00162] В определенных вариантах реализации изобретения линкер содержит Mal-спейсерная единица и расщепляемый пептидный фрагмент. В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент содержит аминокислотную единицу. В некоторых вариантах реализации изобретения аминокислотная единица содержит Val-Cit. В некоторых вариантах реализации изобретения аминокислотная единица содержит Ala-Ala-Asn. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица и Val-Cit. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2 и Val-Cit. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)m и Val-Cit, где m составляет от 2 до 8 или от 2 до 5, или 2, 3, 4 или 5. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)8 и Val-Cit. В определенных вариантах реализации изобретения линкер содержит Mal-(CH2)5 и Val-Cit. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица и Ala-Ala-Asn. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2 и Ala-Ala-Asn.
[00163] В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица и расщепляемый дисульфидный фрагмент. В некоторых вариантах реализации изобретения расщепляемый дисульфидный фрагмент представляет собой дисульфидил-диметил. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица и дисульфидил-диметил. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)4-триазол-(ПЭГ)3 и дисульфидил-диметил.
[00164] В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица и расщепляемый сульфонамидный фрагмент. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)4-триазол-(ПЭГ)3 и сульфонамид.
[00165] В различных вариантах реализации изобретения спейсерная единица прикрепляется к антителу или антигенсвязывающему фрагменту через сукцинимидный фрагмент (OSu). Спейсерная единица, которая прикрепляется к антителу или антигенсвязывающему фрагменту через OSu, упоминается в данном документе как "OSu-спейсерная единица". В данном контексте термин "сукцинимидный фрагмент" означает соединение, которое содержит сукцинимидное соединение, которое реагирует с аминогруппой, например, аминогруппой лизинового остатка на фрагменте антитела. Типовый сукцинимидный фрагмент представляет собой N-гидроксисукцинимид (NHS). В некоторых вариантах реализации изобретения OSu-спейсерная единица реагирует с остатком лизина на антителе или антигенсвязывающем фрагменте. В некоторых вариантах реализации изобретения OSu-спейсерная единица соединяется с антителом или антигенсвязывающим фрагментом через остаток лизина. В некоторых вариантах реализации изобретения OSu-спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения OSu-спейсерная единица содержит алкильный фрагмент.
[00166] В определенных вариантах реализации изобретения линкер содержит OSu-спейсерная единица и расщепляемый пептидный фрагмент. В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент содержит аминокислотную единицу. В некоторых вариантах реализации изобретения аминокислотная единица содержит Val-Cit. В некоторых вариантах реализации изобретения аминокислотная единица содержит Ala-Ala-Asn. В некоторых вариантах реализации изобретения линкер содержит OSu-спейсерная единица и Val-Cit. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)2 и Val-Cit. В других вариантах реализации изобретения линкер содержит OSu-(ПЭГ)9 и Val-Cit. В других вариантах реализации изобретения линкер содержит OSu-(CH2)5 и Val-Cit. В определенных вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3 и Val-Cit. В некоторых вариантах реализации изобретения линкер содержит OSu-спейсерная единица и Ala-Ala-Asn. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)2 и Ala-Ala-Asn.
[00167] В некоторых вариантах реализации изобретения линкер содержит OSu-спейсерная единица и расщепляемый дисульфидный фрагмент. В некоторых вариантах реализации изобретения расщепляемый дисульфидный фрагмент представляет собой дисульфидил-диметил. В некоторых вариантах реализации изобретения линкер содержит OSu-спейсерная единица и дисульфидил-диметил. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3 и дисульфидил-диметил. В других вариантах реализации изобретения линкер содержит OSu-дибензилциклооктен-триазол-(ПЭГ)3 и дисульфидил-диметил.
[00168] В некоторых вариантах реализации изобретения линкер содержит OSu-спейсерная единица и расщепляемый сульфонамидный фрагмент. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3 и сульфонамид. В других вариантах реализации изобретения линкер содержит OSu-дибензилциклооктен-триазол-(ПЭГ)3 и сульфонамид.
[00169] В некоторых вариантах реализации изобретения Mal-спейсерная единица или OSu-спейсерная единица присоединяют фрагмент антитела (то есть антитело или антигенсвязывающий фрагмент) к расщепляемому фрагменту в линкере. В некоторых вариантах реализации изобретения Mal-спейсерная единица или OSu-спейсерная единица присоединяют антитело или антигенсвязывающий фрагмент к расщепляемому пептидному фрагменту. В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент содержит аминокислотную единицу. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица-аминокислотная единица или OSu-спейсерная единица-аминокислотная единица. В некоторых вариантах реализации изобретения Mal-спейсерная единица или OSu-спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения Mal-спейсерная единица или OSu-спейсерная единица содержит алкильный фрагмент. В некоторых вариантах реализации изобретения аминокислотная единица содержит Val-Cit. В других вариантах реализации изобретения аминокислотная единица содержит Ala-Ala-Asn.
[00170] В некоторых вариантах реализации линкер содержит структуру: Mal-спейсерная единица-Val-Cit. В некоторых вариантах реализации линкер содержит структуру: Mal-(ПЭГ)2-Val-Cit. В некоторых вариантах реализации линкер содержит структуру: Mal-(ПЭГ)2-Val-Cit-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)8-Val-Cit. В определенных вариантах реализации изобретения линкер содержит Mal-(CH2)5-Val-Cit. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица-Ala-Ala-Asn. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2-Ala-Ala-Asn.
[00171] В некоторых вариантах реализации изобретения линкер содержит OSu-спейсерная единица-Val-Cit. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)2-Val-Cit. В других вариантах реализации изобретения линкер содержит OSu-(ПЭГ)9-Val-Cit. В других вариантах реализации изобретения линкер содержит OSu-(CH2)5-Val-Cit. В других вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-Val-Cit. В некоторых вариантах реализации изобретения линкер содержит OSu-спейсерная единица-Ala-Ala-Asn. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)2-Ala-Ala-Asn.
[00172] В различных вариантах реализации изобретения Mal-спейсерная единица или OSu-спейсерная единица присоединяют антитело или антигенсвязывающий фрагмент к расщепляемому дисульфидному фрагменту. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица-дисульфид или OSu-спейсерная единица-дисульфид. В некоторых вариантах реализации изобретения дисульфид представляет собой дисульфидил-диметил. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица-дисульфидил-диметил. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)4-триазол-(ПЭГ)3-дисульфидил-диметил. В других вариантах реализации изобретения линкер содержит OSu-спейсерная единица-дисульфидил-диметил. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-дисульфидил-диметил. В других вариантах реализации изобретения линкер содержит OSu-дибензилциклооктен-триазол-(ПЭГ)3-дисульфидил-диметил.
[00173] В определенных вариантах реализации изобретения Mal-спейсерная единица или OSu-спейсерная единица присоединяют антитело или антигенсвязывающий фрагмент к расщепляемому сульфонамидному фрагменту. В некоторых вариантах реализации изобретения линкер содержит Mal-спейсерная единица-сульфонамид или OSu-спейсерная единица-сульфонамид. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)4-триазол-(ПЭГ)3-сульфонамид. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-сульфонамид. В других вариантах реализации изобретения линкер содержит OSu-дибензилциклооктен-триазол-(ПЭГ)3-сульфонамид.
[00174] В различных вариантах реализации изобретения расщепляемый фрагмент в линкере соединяется непосредственно с фрагментом лекарственного средства. В других вариантах реализации изобретения другая спейсерная единица применяется для прикрепления расщепляемого фрагмента в линкере к фрагменту лекарственного средства. В различных вариантах реализации изобретения фрагмент лекарственного средства представляет собой эрибулин. В различных вариантах реализации изобретения эрибулин присоединен к расщепляемому фрагменту в линкере с помощью спейсерной единицы. В некоторых вариантах реализации изобретения эрибулин присоединен к расщепляемому фрагменту в линкере с помощью саморасщепляющейся спейсерной единицы. В некоторых вариантах реализации изобретения эрибулин присоединен к расщепляемому фрагменту в линкере с помощью саморасщепляющейся спейсерной единицы, расщепляемый фрагмент содержит Val-Cit, а дополнительная спейсерная единица, содержащая ПЭГ, соединяет расщепляемый фрагмент с фрагментом антитела. В определенных вариантах реализации изобретения эрибулин соединяется с антителом к FRA через Mal-спейсерная единица в линкере, соединенном с расщепляемым фрагментом Val-Cit и саморасщепляющейся спейсерной единицей pAB. В определенных других вариантах реализации изобретения эрибулин соединяется с антителом к her2 через Mal-спейсерная единица в линкере, соединенном с расщепляемым фрагментом Val-Cit и саморасщепляющейся спейсерной единицей pAB.
[00175] Спейсерная единица может быть "саморасщепляющейся" или "не саморасщепляющейся". "Не саморасщепляющаяся" спейсерная единица, представляет собой спейсерную единицу, полностью или частично остающуюся связанной с фрагментом лекарственного средства при расщеплении линкера. Примеры не саморасщепляющихся спейсерных единиц включают глициновую спейсерную единицу и глицин-глициновую спейсерную единицу, но не ограничиваются ими. Не саморасщепляющиеся спейсерные единицы могут со временем деградировать, но не могут легко в полной мере высвобождать связанное нативное лекарственное средство в клеточных условиях. "Саморасщепляющаяся" спейсерная единица обеспечивает высвобождение фрагмента нативного лекарственного средства во внутриклеточных условиях. "Нативное лекарственное средство" представляет собой лекарственное средство, в котором после расщепления/деградации спейсерной единицы не остается части спейсерной единицы или другой химической модификации.
[00176] Химия саморасщепления известна в данной области техники и может быть легко выбрана для описанных ADC. В различных вариантах реализации изобретения спейсерная единица, прикрепляющая расщепляемый фрагмент в линкере к фрагменту лекарственного средства (например, эрибулина), является саморасщепляющейся и подвергается саморасщеплению одновременно с или незадолго до/после расщепления расщепляемого фрагмента при внутриклеточных условиях.
[00177] В некоторых вариантах реализации изобретения саморасщепляющаяся спейсерная единица в линкере содержит п-аминобензильную группу. В некоторых вариантах реализации изобретения, п-аминобензиловый спирт (pABOH) присоединен к аминокислотной единице или другому расщепляемому фрагменту в линкере посредством амидной связи, и между pABOH и фрагментом лекарственного средства образуется карбамат, метилкарбамат или карбонат (Hamann и соавт. (2005 год) Expert Opin. Ther. Patents 15:1087-103). В некоторых вариантах реализации изобретения саморасщепляющаяся спейсерная единица представляет собой или содержит п-аминобензилоксикарбонил (pAB). Не привязываясь ктеории, считается, что саморасщепляемый pAB включает спонтанную реакцию 1,6-элиминации (Jain и соавт. (2015 год) Pharm Res 32:3526-40).
[00178] В различных вариантах реализации изобретения структура п-аминобензилоксикарбонила (pAB), применяемого в описанных ADC, показана ниже:
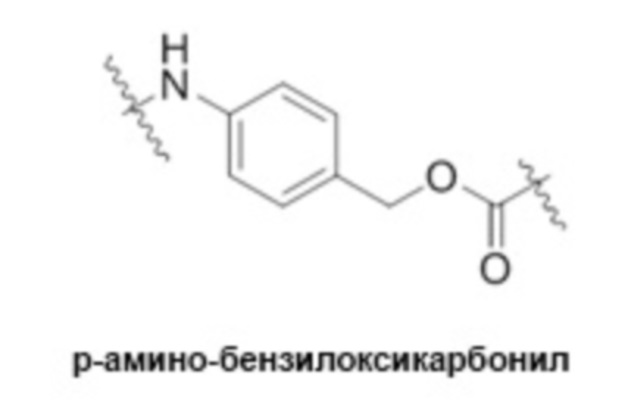
[00179] В различных вариантах реализации изобретения саморасщепляющаяся спейсерная единица прикрепляет расщепляемый фрагмент в линкере к амину C-35 на эрибулине. В некоторых вариантах реализации изобретения саморасщепляющаяся спейсерная единица представляет собой pAB. В некоторых вариантах реализации изобретения pAB прикрепляет расщепляемый фрагмент в линкере к амину C-35 на эрибулине. В некоторых вариантах реализации изобретения pAB подвергается саморасщеплению при расщеплении расщепляемого фрагмента, и эрибулин высвобождается из ADC в его нативной, активной форме. В некоторых вариантах реализации изобретения антитело к FRA (например, MORAb-003) соединяется с амином C-35 эрибулина с помощью линкера, содержащего Mal-(ПЭГ)2-Val-Cit-pAB. В других вариантах реализации изобретения антитело к her2 (например, трастузумаб) соединяется с амином C-35 эрибулина с помощью линкера, содержащего Mal-(ПЭГ)2-Val-Cit-pAB.
[00180] В некоторых вариантах реализации изобретения pAB подвергается саморасщеплению при расщеплении расщепляемого пептидного фрагмента в линкере. В некоторых вариантах реализации изобретения расщепляемый пептидный фрагмент содержит аминокислотную единицу. В некоторых вариантах реализации изобретения линкер содержит аминокислотную единицу-pAB. В некоторых вариантах реализации изобретения аминокислотная единица представляет собой Val-Cit. В некоторых вариантах реализации изобретения линкер содержит Val-Cit-pAB (VCP). В определенных вариантах реализации изобретения аминокислотная единица представляет собой Ala-Ala-Asn. В некоторых вариантах реализации изобретения линкер содержит Ala-Ala-Asn-pAB.
[00181] В некоторых вариантах реализации изобретения pAB подвергается саморасщеплению при расщеплении расщепляемого дисульфидного фрагмента в линкере. В некоторых вариантах реализации изобретения линкер содержит дисульфид-pAB. В некоторых вариантах реализации изобретения линкер содержит дисульфидил-диметил-pAB.
[00182] В некоторых вариантах реализации изобретения pAB подвергается саморасщеплению при расщеплении расщепляемого сульфонамидного фрагмента в линкере. В некоторых вариантах реализации изобретения линкер содержит сульфонамид-pAB.
[00183] В различных аспектах фрагмент антитела ADC конъюгирует с фрагментом лекарственного средства посредством линкера, при этом линкер содержит Mal-спейсерная единица, расщепляемую аминокислотную единицу и pAB. В некоторых вариантах реализации изобретения спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения спейсерная единица содержит алкильный фрагмент. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2-Val-Cit-pAB. В других вариантах реализации изобретения линкер содержит Mal-(ПЭГ)2-Ala-Ala-Asn-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)8-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)8-Val-Cit-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(CH2)5-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(CH2)5-Val-Cit-pAB.
[00184] В различных вариантах реализации изобретения фрагмент антитела ADC конъюгирует с фрагментом лекарственного средства посредством линкера, при этом линкер содержит Mal-спейсерная единица-дисульфид-pAB. В некоторых вариантах реализации изобретения спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)4-триазол-(ПЭГ)3-дисульфид-pAB. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)4-триазол-(ПЭГ)3-дисульфидил-диметил-pAB.
[00185] В некоторых вариантах реализации изобретения фрагмент антитела ADC конъюгирует с фрагментом лекарственного средства посредством линкера, при этом линкер содержит Mal-спейсерная единица-сульфонамид-pAB. В некоторых вариантах реализации изобретения спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения линкер содержит Mal-(ПЭГ)4-триазол-(ПЭГ)3-сульфонамид-pAB.
[00186] В некоторых аспектах фрагмент антитела ADC конъюгирует с фрагментом лекарственного средства посредством линкера, при этом линкер содержит OSu-спейсерная единица-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения спейсерная единица содержит алкильный фрагмент. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)2-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)2-Val-Cit-pAB. В других вариантах реализации изобретения линкер содержит OSu-(ПЭГ)2-Ala-Ala-Asn-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)9-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)9-Val-Cit-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(CH2)5-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(CH2)5-Val-Cit-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-аминокислотная единица-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-Val-Cit-pAB.
[00187] В некоторых вариантах реализации изобретения фрагмент антитела ADC конъюгирует с фрагментом лекарственного средства посредством линкера, при этом линкер содержит OSu-спейсерная единица-дисульфид-pAB. В некоторых вариантах реализации изобретения спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-дисульфид-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-дисульфидил-диметил-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-дибензилциклооктен-триазол-(ПЭГ)3-дисульфид-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-дибензилциклооктен-триазол-(ПЭГ)3-дисульфидил-диметил-pAB.
[00188] В некоторых вариантах реализации изобретения фрагмент антитела ADC конъюгирует с фрагментом лекарственного средства посредством линкера, при этом линкер содержит OSu-спейсерная единица-сульфонамид-pAB. В некоторых вариантах реализации изобретения спейсерная единица содержит ПЭГ-фрагмент. В некоторых вариантах реализации изобретения линкер содержит OSu-(ПЭГ)3-триазол-(ПЭГ)3-сульфонамид-pAB. В некоторых вариантах реализации изобретения линкер содержит OSu-дибензилциклооктен-триазол-(ПЭГ)3-сульфонамид-pAB.
[00189] В различных вариантах реализации изобретения линкер предназначен для облегчения неспецифичного цитолиза (убийства соседних клеток) посредством расщепления после клеточной интернализации и диффузии фрагмента линкер-лекарственное средство и/или фрагмента лекарственного средства отдельно к соседним клеткам. В некоторых вариантах реализации изобретения линкер способствует клеточной интернализации. В некоторых вариантах реализации изобретения линкер предназначен для сведения к минимуму расщепления во внеклеточной среде и, таким образом, для снижения токсичности по отношению к нецелевой ткани (например, не раковой ткани), при сохранении связывания ADC с целевой клеткой и неспецифическим цитолизом раковой ткани, которая не экспрессирует антиген, нацеленный на фрагмент антитела ADC, но окружает целевую раковую ткань, экспрессирующую этот антиген. В некоторых вариантах реализации изобретения линкер, содержащий малеимидный фрагмент (Mal), фрагмент полиэтиленгликоля (ПЭГ), валин-цитруллин (Val-Cit или "vc") и pAB, обеспечивает эти функциональные признаки. В некоторых вариантах реализации изобретения линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB особенно эффективен для обеспечения этих функциональных признаков, например, при соединении фрагмента антитела к FRA, такого как MORAb-003, и фрагмента лекарственного средства, такого как эрибулин. В некоторых вариантах реализации изобретения, по меньшей мере, некоторые из этих функциональных признаков также могут наблюдаться без фрагмента антитела к FRA и/или без MORAb-003. Например, в некоторых вариантах реализации изобретения линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB эффективен для обеспечения всех или некоторых из этих функциональных признаков, например, при соединении фрагмента антитела к her2, такого как трастузумаб, и фрагмента лекарственного средства, такого как эрибулин.
[00190] В некоторых вариантах реализации изобретения фрагмент антитела конъюгирован с фрагментом лекарственного средства посредством линкера, содержащего малеимидный фрагмент (Mal), фрагмент полиэтиленгликоля (ПЭГ), валин цитруллин (Val-Cit или "vc") и pAB. В этих вариантах реализации изобретения малеимидный фрагмент ковалентно присоединяет фрагмент линкера-лекарственного средства к фрагменту антитела, а pAB действует как саморасщепляющаяся спейсерная единица. Такой линкер может упоминаться как линкер "m-vc-pAB", линкер "Mal-VCP", линкер "Mal-(ПЭГ)2-VCP", или линкер "Mal-(ПЭГ)2-Val-Cit-pAB". В некоторых вариантах реализации изобретения фрагмент лекарственного средства представляет собой эрибулин. Структура Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин приведена в Таблице 46. pAB линкера Mal-(ПЭГ)2-Val-Cit-pAB присоединен к амину C-35 на эрибулине.
[00191] Было обнаружено, что ADC, содержащие Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин, демонстрируют конкретную комбинацию желательных свойств, особенно в случае если спарены с антителом к FRA, таким как MORAb-003, или его антигенсвязывающим фрагментом. Эти свойства включают, но не ограничиваются ими, эффективные уровни содержания лекарственного средства (p ~ 4), низкие уровни агрегации, стабильность в условиях хранения или при циркуляции в организме (например, стабильность в сыворотке крови), сохранения аффинности к клеткам, экспрессирующим цель по сравнению с неконъюгированным антителом, сильной цитотоксичностью против клеток, экспрессирующих цель, низкие уровни нецелевого уничтожения клеток, высокие уровни неспецифического цитолиза и/или эффективную противораковою активность in vivo, все по сравнению с ADC с применением других линкер-токсинов и/или антител. В то время как многочисленные варианты линкера и комбинации спейсеров и сайтов расщепления были известны в данной области техники и могут обеспечить определенные преимущества в одной или более из этих функциональных категорий, конкретная комбинация Mal-(ПЭГ)2-Val-Cit-pAB, соединяющая эрибулин с фрагментом антитела, таким как антитело к FRA (например, MORAb-003), может обеспечивать хорошие или превосходные свойства по всему спектру желаемых функциональных свойств для терапевтического ADC. В некоторых вариантах реализации изобретения хорошие или превосходные функциональные свойства, обеспечиваемые конкретной комбинацией линкера Mal-(ПЭГ)2-Val-Cit-pAB, соединяющего эрибулин с фрагментом антитела, могут наблюдаться с этим линкер-токсином, конъюгированным с, например, антителом к her2, таким как трастузумаб.
[00192] В некоторых вариантах реализации изобретения ADC содержит Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин и фрагмент антитела, содержащий интернализирующееся антитело или его антигенсвязывающий фрагмент, который сохраняет способность к нацеливанию и интернализации в опухолевой клетке. В некоторых вариантах реализации изобретения ADC содержит Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин и интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие FRA. В некоторых вариантах реализации изобретения интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевую клетку, экспрессирующую FRA, содержит три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 2 (HCDR1), SEQ ID NO: 3 (HCDR2) и SEQ ID NO: 4 (HCDR3); три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 7 (LCDR1), SEQ ID NO: 8 (LCDR2) и SEQ ID NO: 9 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 13 (HCDR1), SEQ ID NO: 14 (HCDR2) и SEQ ID NO: 15 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 16 (LCDR1), SEQ ID NO: 17 (LCDR2) и SEQ ID NO: 18 (LCDR3), как определено системой нумерации IMGT. В некоторых вариантах реализации изобретения интернализирующееся антитело его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие FRA, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 23, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 24. В некоторых вариантах реализации изобретения интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие FRA, содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig.
[00193] В некоторых вариантах реализации изобретения ADC имеет Формулу I:
Ab-(L-D)p (I)
в которой:
(i) Ab представляет собой интернализирующееся антитело к рецептору фолиевой кислоты альфа (FRA) или его интернализирующийся антигенсвязывающий фрагмент, содержащий три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 2 (HCDR1), SEQ ID NO: 3 (HCDR2) и SEQ ID NO: 4 (HCDR3); три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 7 (LCDR1), SEQ ID NO: 8 (LCDR2) и SEQ ID NO: 9 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 13 (HCDR1), SEQ ID NO: 14 (HCDR2) и SEQ ID NO: 15 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 16 (LCDR1), SEQ ID NO: 17 (LCDR2) и SEQ ID NO: 18 (LCDR3), как определено системой нумерации IMGT;
(ii) D представляет собой эрибулин;
(iii) L представляет собой расщепляемый линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB; и
(iv) p представляет собой целое число от 1 до 20.
[00194] В некоторых вариантах реализации изобретения интернализирующееся антитело его интернализирующийся антигенсвязывающий фрагмент, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 23, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 24. В некоторых вариантах реализации изобретения интернализирующееся антитело представляет собой MORAb-003. В некоторых вариантах реализации изобретения p составляет от 1 до 8 или от 1 до 6. В некоторых вариантах реализации изобретения p составляет от 2 до 8 или от 2 до 5. В некоторых вариантах реализации изобретения p составляет от 3 до 4. В некоторых вариантах реализации изобретения p представляет собой 4.
[00195] В других вариантах реализации изобретения ADC содержит Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин и интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие her2. В некоторых вариантах реализации изобретения интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевую клетку, экспрессирующую her2, содержит три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 71 (HCDR1), SEQ ID NO: 72 (HCDR2) и SEQ ID NO: 73 (HCDR3); три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 74 (LCDR1), SEQ ID NO: 75 (LCDR2) и SEQ ID NO: 76 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 191 (HCDR1), SEQ ID NO: 192 (HCDR2) и SEQ ID NO: 193 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 194 (LCDR1), SEQ ID NO: 195 (LCDR2) и SEQ ID NO: 196 (LCDR3), как определено системой нумерации IMGT. В некоторых вариантах реализации изобретения интернализирующееся антитело его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие her2, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 27, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 28. В некоторых вариантах реализации изобретения интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие her2, содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig.
[00196] В некоторых вариантах реализации изобретения ADC имеет Формулу I:
Ab-(L-D)p (I)
в которой:
(i) Ab представляет собой интернализирующееся антитело к рецептору человеческого эпидермального фактора роста 2 типа (her2) или его интернализирующийся антигенсвязывающий фрагмент, содержащий три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 71 (HCDR1), SEQ ID NO: 72 (HCDR2) и SEQ ID NO: 73 (HCDR3); три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 74 (LCDR1), SEQ ID NO: 75 (LCDR2) и SEQ ID NO: 76 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 191 (HCDR1), SEQ ID NO: 192 (HCDR2) и SEQ ID NO: 193 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 194 (LCDR1), SEQ ID NO: 195 (LCDR2) и SEQ ID NO: 196 (LCDR3), как определено системой нумерации IMGT;
(ii) D представляет собой эрибулин;
(iii) L представляет собой расщепляемый линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB; и
(iv) p представляет собой целое число от 1 до 20.
[00197] В некоторых вариантах реализации изобретения интернализирующееся антитело его интернализирующийся антигенсвязывающий фрагмент, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 27, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 28. В некоторых вариантах реализации изобретения интернализирующееся антитело представляет собой трастузумаб. В некоторых вариантах реализации изобретения p составляет от 1 до 8 или от 1 до 6. В некоторых вариантах реализации изобретения p составляет от 2 до 8 или от 2 до 5. В некоторых вариантах реализации изобретения p составляет от 3 до 4. В некоторых вариантах реализации изобретения p представляет собой 4.
[00198] В других вариантах реализации изобретения ADC содержит Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин и интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие мезотелин (MSLN). В некоторых вариантах реализации изобретения интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевую клетку, экспрессирующую MSLN, содержит три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 65 (HCDR1), SEQ ID NO: 66 (HCDR2) и SEQ ID NO: 67 (HCDR3); три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 68 (LCDR1), SEQ ID NO: 69 (LCDR2) и SEQ ID NO: 70 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 185 (HCDR1), SEQ ID NO: 186 (HCDR2) и SEQ ID NO: 187 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 188 (LCDR1), SEQ ID NO: 189 (LCDR2) и SEQ ID NO: 190 (LCDR3), как определено системой нумерации IMGT. В некоторых вариантах реализации изобретения интернализирующееся антитело его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие MSLN, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 25, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 26. В некоторых вариантах реализации изобретения интернализирующееся антитело или его интернализирующийся антигенсвязывающий фрагмент, который нацелен на опухолевые клетки, экспрессирующие MSLN, содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи каппа Ig.
[00199] В некоторых вариантах реализации изобретения ADC имеет Формулу I:
Ab-(L-D)p (I)
в которой:
(i) Ab представляет собой интернализирующееся антитело к мезотелину или его интернализирующийся антигенсвязывающий фрагмент, содержащий три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 65 (HCDR1), SEQ ID NO: 66 (HCDR2) и SEQ ID NO: 67 (HCDR3); три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 68 (LCDR1), SEQ ID NO: 69 (LCDR2) и SEQ ID NO: 70 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 185 (HCDR1), SEQ ID NO: 186 (HCDR2) и SEQ ID NO: 187 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 188 (LCDR1), SEQ ID NO: 189 (LCDR2) и SEQ ID NO: 190 (LCDR3), как определено системой нумерации IMGT;
(ii) D представляет собой эрибулин;
(iii) L представляет собой расщепляемый линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB; и
(iv) p представляет собой целое число от 1 до 20.
[00200] В некоторых вариантах реализации изобретения интернализирующееся антитело его интернализирующийся антигенсвязывающий фрагмент, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 25, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 26. В некоторых вариантах реализации изобретения интернализирующееся антитело представляет собой MORAb-003, MORAb-009 или трастузумаб. В некоторых вариантах реализации изобретения p составляет от 1 до 8 или от 1 до 6. В некоторых вариантах реализации изобретения p составляет от 2 до 8 или от 2 до 5. В некоторых вариантах реализации изобретения p составляет от 3 до 4. В некоторых вариантах реализации изобретения p представляет собой 4.
Фрагменты лекарственных средств
[00201] Фрагмент лекарственного средства (D) ADC, описанного в данном документе, может быть любым химиотерапевтическим агентом. Полезные классы химиотерапевтических агентов включают, например, антитубулиновые агенты. В определенных вариантах реализации изобретения фрагмент лекарственного средства представляет собой противотубулиновый агент. Примеры антитубулиновых агентов включают в себя криптофицин и эрибулин. Предпочтительным фрагментом лекарственного средства для применения в описанных ADC является эрибулин.
[00202] В различных вариантах реализации изобретения фрагмент лекарственного средства представляет собой эрибулин. В этих вариантах реализации изобретения линкер ADC присоединен через амин C-35 на эрибулине.
[00203] В различных вариантах реализации изобретения естественная форма эрибулина применяется для соединения с линкером и фрагментом антитела, показана ниже:
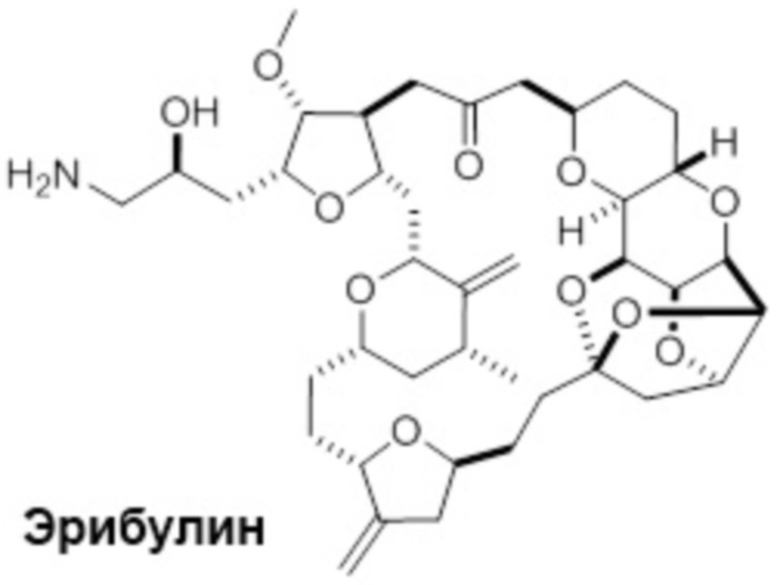
[00204] В некоторых вариантах реализации изобретения промежуточное соединение, которое является предшественником линкера, подвергают взаимодействию с фрагментом лекарственного средства в соответствующих условиях. В определенных вариантах реализации изобретения реактивные группы применяют на лекарственном средстве и/или в промежуточном соединении или линкере. Продукт реакции между лекарственным средством и промежуточным соединением или дериватизированным лекарственным средством затем подвергают взаимодействию с антителом или антигенсвязывающим фрагментом в соответствующих условиях. В альтернативном варианте, линкер или промежуточное соединение могут сначала взаимодействовать с антителом или дериватизированным антителом, а затем реагировать с лекарственным средством или дериватизированным лекарственным средством.
[00205] Существует множество различных реакций для ковалентного присоединения лекарственных средств и/или линкеров к фрагменту антитела. Это часто достигается реакцией одного или более аминокислотных остатков молекулы антитела, включая аминогруппы лизина, свободные карбоксильные группы глутаминовой кислоты и аспарагиновой кислоты, сульфгидрильные группы цистеина и различные фрагменты ароматических аминокислот. Например, неспецифическое ковалентное присоединение может быть осуществлено с применением карбодиимидной реакции для связывания карбокси (или амино) группы на соединении с амино или (карбокси) группой на фрагменте антитела. Кроме того, бифункциональные агенты, такие как диальдегиды или имидоэфиры, также могут быть применены для связывания аминогруппы на соединении с аминогруппой на фрагменте антитела. Также для прикрепления лекарственного средства к связующим агентам доступна реакция основания Шиффа. Этот способ включает окисление периодата лекарственного средства, которое содержит гликолевые или гидроксильные группы, образуя таким образом альдегид, который затем подвергают взаимодействию со связывающим агентом. Присоединение происходит путем образования основания Шиффа с аминогруппами связывающего агента. Изотиоцианаты также могут быть применены в качестве сопрягающих агентов для ковалентного прикрепления лекарственных средств к связывающими агентам. Другие методы известны специалисту в данной области техники и находятся в рамках данного описания.
Содержание лекарственного средства
[00206] Содержание лекарственного средства представлено p, а также упоминается в данном документе как соотношение лекарственного средства к антителу (DAR). Содержание лекарственного средства может составлять от 1 до 20 фрагментов лекарственного средства на фрагмент антитела. В некоторых вариантах реализации изобретения p представляет собой целое число от 1 до 20. В некоторых вариантах реализации изобретения p представляет собой целое число от 1 до 10, от 1 до 9, от 1 до 8, от 1 до 7, от 1 до 6, от 1 до 5, от 1 до 4, от 1 до 3 или от 1 до 2. В некоторых вариантах реализации изобретения p представляет собой целое число от 2 до 10, от 2 до 9, от 2 до 8, от 2 до 7, от 2 до 6, от 2 до 5, от 2 до 4 или от 2 до 3. В некоторых вариантах реализации изобретения p представляет собой целое число от 3 до 4. В других вариантах реализации изобретения p составляет 1, 2, 3, 4, 5 или 6, предпочтительно 3 или 4.
[00207] Содержание лекарственного средства может быть ограничено количеством сайтов присоединения на фрагменте антитела. В некоторых вариантах реализации изобретения фрагмент линкера (L) ADC прикрепляется к фрагменту антитела через химически активную группу на одном или более аминокислотных остатках на фрагменте антитела. Например, линкер может быть присоединен к фрагменту антитела через свободную амино, имино, гидроксильную, тиольную или карбоксильную группу (например, к N- или С-концу, к эпсилон-аминогруппе одного или более остатков лизина, к свободной карбоксильной группе одного или более остатков глутаминовой кислоты или аспарагиновой кислоты или к сульфгидрильной группе одного или более остатков цистеина). Сайт, к которому присоединен линкер, может быть остатком естественного происхождения в аминокислотной последовательности фрагмента антитела или может быть введен в фрагмент антитела, например, с помощью рекомбинантной технологии ДНК (например, путем введения остатка цистеина в аминокислотную последовательность) или биохимии белка (например, путем восстановления, регулированием рН или гидролизом).
[00208] В некоторых вариантах реализации количество молекул лекарственного вещества, которые можно конъюгировать с фрагментом антитела, ограничено количеством свободных остатков цистеина. Например, если присоединяемая группа представляет собой тиольную группу цистеина, антитело может содержать только одну или несколько тиольных групп цистеина, или может содержать только одну или несколько достаточно реакционноспособных тиольных групп, к которым можно присоединить линкер. Как правило, антитела не содержат многих свободных и реакционноспособных тиольных групп цистеина, которые могут быть связаны с фрагментом лекарственного средства. Действительно, большинство тиольных остатков в антителах существуют в виде дисульфидных мостиков. Чрезмерное присоединение линкер-токсина к антителу может дестабилизировать антитело путем уменьшения остатков цистеина, доступных для образования дисульфидных мостиков. Следовательно, оптимальное соотношение лекарственное средство:антитело должно увеличивать активность ADC (путем увеличения количества присоединенных фрагментов лекарственного средства на антитело) без дестабилизации фрагмента антитела. В некоторых вариантах реализации изобретения оптимальное соотношение может составлять около 3-4.
[00209] В некоторых вариантах реализации изобретения линкер, присоединенный к фрагменту антитела через фрагмент Mal, дает отношение около 3-4. В некоторых вариантах реализации изобретения линкер, присоединенный к фрагменту антитела через альтернативный фрагмент (например, фрагмент OSu), может обеспечивать менее оптимальное соотношение (например, более низкое соотношение, такое как около 0-3). В некоторых вариантах реализации изобретения линкер, содержащий короткую спейсерную единицу (например, короткую спейсерную единицу ПЭГ, такую как (ПЭГ)2 или (ПЭГ)4, или короткую алкильную спейсерную единицу, такую как (CH2)5) обеспечивает соотношение около 3-4. В некоторых вариантах реализации изобретения линкер, который содержит более длинную спейсерную единицу (например, (ПЭГ)8), может обеспечить менее оптимальное соотношение (например, более низкое соотношение, например, около 0-3). В некоторых вариантах реализации изобретения линкер, содержащий пептидный расщепляемый фрагмент, обеспечивает соотношение около 3-4. В некоторых вариантах реализации изобретения линкер, который содержит альтернативный расщепляемый фрагмент (например, расщепляемый дисульфид или расщепляемый сульфонамид), может обеспечить менее оптимальное соотношение (например, меньшее соотношение, например, около 0-3). В некоторых вариантах реализации изобретения ADC, содержащий Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин, присоединенный к антителу, такому как антитело к FRA (например, MORAb-003) имеет соотношение около 3-4. В некоторых вариантах реализации изобретения соотношение около 3-4 наблюдается с ADC, содержащим Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин, присоединенный к другому антителу, такому как антитело к her2 (например, трастузумабу). В некоторых вариантах реализации изобретения оптимальное соотношение, наблюдаемое с ADC, содержащим линкер-токсин Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин, является антитело-независимым.
[00210] В некоторых вариантах реализации изобретения фрагмент антитела подвергают воздействию восстановительных условий до конъюгации с целью получения одного или более свободных остатков цистеина. Антитело, в некоторых вариантах реализации изобретения, можно восстановить с помощью восстанавливающего агента, например, дитиотрейтола (ДТТ) или трис(2-карбоксиэтил)фосфина (TCEP) при частично или полностью восстановительных условиях с целью получения реакционноспособных тиольных групп цистеина. Неспаренные цистеины могут быть получены путем частичного восстановления с ограниченными молярными эквивалентами TCEP, что предпочтительно уменьшает межцепочечные дисульфидные связи, которые связывают легкую цепь и тяжелую цепь (по одной паре на спаривание H-L) и две тяжелые цепи в шарнирной области (две пары на спаривание H-H в случае IgG1 человека), оставляя внутрицепочечные дисульфидные связи интактными (Stefano и соавт. (2013 год) Methods Mol. Biol., 1045:145-71). В вариантах реализации изобретения дисульфидные связи внутри антител восстанавливаются электрохимически, например, с использованием рабочего электрода, который создает переменное восстановительное и окисляющее напряжение. Этот подход может обеспечить связывание восстановления дисульфидной связи в режиме реального времени с аналитическим устройством (например, электрохимическим детекторным устройством, ЯМР-спектрометром или масс-спектрометром) или устройством химического разделения (например, жидкостным хроматографом (например, ВЭЖХ) или электрофорезом (см., например, публикациюСША №20140069822)). В некоторых вариантах реализации изобретения антитело подвергают воздействию денатурирующих условий с целью выявления реакционноспособных нуклеофильных групп на аминокислотных остатках, например, лизине или цистеине.
[00211] Содержание лекарственного средства ADC может контролироваться по-разному, например, с помощью:(i) ограничения молярного избытка промежуточного вещества лекарственное средство-линкер или линкерного реагента по отношению к антителу; (ii) ограничения времени или температуры реакции конъюгации; (iii) частичных или ограничивающих восстановительных условий для модификации тиола цистеина; и/или (iv) конструирования с помощью рекомбинантных методов аминокислотной последовательности антитела, так что количество и положение остатков цистеина модифицировано для контроля количества и/или положения присоединений линкер-лекарственное средство.
[00212] В некоторых вариантах реализации изобретения свободные остатки цистеина вводят в аминокислотную последовательность фрагмента антитела. Например, могут быть получены антитела, сконструированные с помощью цистеина, при этом одна или более аминокислот исходного антитела замещены аминокислотой цистеина. Любая форма антитела может быть сконструирована таким образом, то есть мутирована. Например, фрагмент Fab исходного антитела может быть сконструирован для образования сконструированного с помощью цистеина Fab, называемого "ThioFab". Аналогично, исходное моноклональное антитело может быть сконструировано с образованием "ThioMab". Мутация в одном сайте дает один искусственно введенный цистеиновый остаток в ThioFab, тогда как мутация в одном сайте дает два искусственно введенных цистеиновых остатка в ThioMab из-за димерной природы антитела IgG. ДНК, кодирующая вариант аминокислотной последовательности исходного полипептида, может быть получена различными способами, известными в данной области техники (см., например, способы, описанные в WO 2006/034488). Эти способы включают, но без ограничений, приготовление с помощью сайт-направленного (или опосредованного олигонуклеотидами) мутагенеза, ПЦР-мутагенеза и кассетного мутагенеза ранее приготовленной ДНК, кодирующей полипептид. Варианты рекомбинантных антител также могут быть сконструированы также с помощью манипуляции с рестрикционным фрагментом или с помощью ПЦР с перекрывающимися синтетическими олигонуклеотидами. ADC формулы I содержат, но без ограничений, антитела, которые имеют 1, 2, 3, или 4 искусственно введенных аминокислотных остатков цистеина (Lyon и соавт. (2012 год) Methods in Enzymol. 502:123-38). В некоторых вариантах реализации один или более свободных остатков цистеина уже присутствуют в фрагменте антитела без применения инженерии, и в этом случае существующие свободные остатки цистеина можно применять для конъюгирования фрагмента антитела с фрагментом лекарственного средства.
[00213] В некоторых вариантах реализации высокое содержание лекарственного средства (например, p > 5), может привести к агрегации, нерастворимости, токсичности или потере способности некоторых конъюгатов антитело-лекарственное средство проникать в клетку. Более высокое содержание лекарственного средства также может отрицательно влиять на фармакокинетику (например, клиренс) определенных ADC. В некоторых вариантах реализации изобретения более низкое содержание лекарственного средства (например, p < 3) может снизить активность некоторых ADC против клеток, экспрессирующих цель и/или случайных клеток. В некоторых вариантах реализации изобретения содержание лекарственного средства для ADC по данному изобретению варьируется от 1 до около 8; от около 2 до около 6; от около 2 до около 5; от около 3 до около 5; или от около 3 до около 4.
[00214] Когда более чем одна нуклеофильная группа реагирует с промежуточным соединением лекарственного средства-линкера или реагентом фрагмента линкера с последующим реагентом фрагмента лекарственного средства, в реакционной смеси, содержащей множественные копии фрагмента антитела и фрагмента линкера, затем полученный продукт может представлять собой смесь соединений ADC с распределением одного или более фрагментов лекарственного средства, прикрепленных к каждой копии фрагмента антитела в смеси. В некоторых вариантах реализации изобретения содержание лекарственного средства в смеси ADC, образующихся в результате реакции конъюгации, варьирует от 1 до 20 фрагментов лекарственного средства, прикрепленных на фрагмент антитела. Среднее количество фрагментов лекарственного средства на фрагмент антитела (то есть среднее содержание лекарственного средства или среднее значение p) может быть вычислено любым общепринятым способом, известным в данной области техники, например, с помощью масс-спектрометрии (например, обращенно-фазовой ЖХ-МС) и/или высокоэффективной жидкостной хроматографии (например, HIC-ВЭЖХ). В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела определяют с помощью хроматографии с гидрофобным взаимодействием-высокоэффективной жидкостной хроматографией (HIC-ВЭЖХ). В некоторых вариантах реализации изобретения среднее число фрагментов лекарственного средства на фрагмент антитела определяют обращенно-фазовой жидкостной хроматографией-масс-спектрометрией (ЖХ-МС). В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от около 3 до около 4; от около 3,1 до около 3,9; от около 3,2 до около 3,8; от около 3,2 до около 3,7; от около 3,2 до около 3,6; от около 3,3 до около 3,8; или от около 3,3 до около 3,7. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от около 3,2 до около 3,8. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет около 3,8. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от 3 до 4; от 3,1 до 3,9; от 3,2 до 3,8; от 3,2 до 3,7; от 3,2 до 3,6; от 3,3 до 3,8; или от 3,3 до 3,7. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от 3,2 до 3,8. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет 3,8.
[00215] В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от около 3,5 до около 4,5; от около 3,6 до около 4,4; от около 3,7 до около 4,3; от около 3,7 до около 4,2; или от около 3,8 до около 4,2. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от около 3,6 до около 4,4. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет около 4,0. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от 3,5 до 4,5; от 3,6 до 4,4; от 3,7 до 4,3; от 3,7 до 4,2; или от 3,8 до 4,2. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет от 3,6 до 4,4. В некоторых вариантах реализации изобретения среднее количество фрагментов лекарственного средства на фрагмент антитела составляет 4,0.
[00216] В различных вариантах реализации изобретения термин "около", применяемый в отношении среднего количества фрагментов лекарственного средства на фрагмент антитела, составляет +/- 10%.
[00217] Отдельные соединения ADC или "виды" могут быть идентифицированы в смеси с помощью масс-спектроскопии и разделены с помощью СВЭЖХ или ВЭЖХ, например, хроматографии с гидрофобным взаимодействием (HIC-ВЭЖХ). В некоторых вариантах реализации изобретения из конъюгационной смеси можно выделить однородный или почти однородный ADC с одинаковым значением содержания, например, посредством электрофореза или хроматографии.
[00218] В некоторых вариантах реализации изобретения содержание лекарственного средства и/или среднее содержание лекарственного средства около 4 обеспечивает полезные свойства. В некоторых вариантах реализации изобретения содержание лекарственного средства и/или среднее содержание лекарственного средства менее чем около 4 может приводить к неприемлемо высокому уровню неконъюгированных видов антител, которые могут конкурировать с ADC за связывание с целевым антигеном и/или обеспечивать снижение эффективности лечения. В некоторых вариантах реализации изобретения содержание лекарственного средства и/или среднее содержание лекарственного средства более чем около 4 может приводить к неприемлемо высокому уровню неоднородности продукта и/или агрегации ADC. Содержание лекарственного средства и/или среднее содержание лекарственного средства более чем около 4 может также влиять на стабильность ADC из-за потери одной или более химических связей, необходимых для стабилизации фрагмента антитела.
[00219] В некоторых вариантах реализации изобретения ADC имеет Формулу I:
Ab-(L-D)p (I)
в которой:
(i) Ab представляет собой интернализирующееся антитело к рецептору фолиевой кислоты альфа или его антигенсвязывающий фрагмент, содержащий вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 23, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 24;
(ii) D представляет собой эрибулин;
(iii) L представляет собой расщепляемый линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB; и
(iv) p представляет собой целое число от 3 до 4.
[00220] В других вариантах реализации изобретения ADC имеет Формулу I:
Ab-(L-D)p (I)
где:
(i) Ab представляет собой интернализирующееся антитело к рецептору человеческого эпидермального фактора роста 2 типа или его антигенсвязывающий фрагмент, содержащий вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 27, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 28;
(ii) D представляет собой эрибулин;
(iii) L представляет собой расщепляемый линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB; и
(iv) p представляет собой целое число от 3 до 4.
[00221] В некоторых вариантах реализации изобретения p представляет собой 4.
[00222] Данное изобретение включает в себя способы получения описанных ADC. Вкратце, ADC содержат антитело или антигенсвязывающий фрагмент в качестве фрагмента антитела, фрагмента лекарственного средства и линкера, который соединяет фрагмент лекарственного средства и фрагмент антитела. В некоторых вариантах реализации изобретения ADC можно получить с применением линкера, содержащего реакционноспособные функциональные группы для ковалентного присоединения к фрагменту лекарственного средства и фрагменту антитела. Например, в некоторых вариантах реализации изобретения цистеиновый тиол фрагмента антитела может образовывать связь с реакционноспособной функциональной группой линкера или промежуточного соединения лекарственное средство-линкер (например, малеимидный фрагмент) с образованием ADC. Получение ADC может быть выполнено любым способом, известным специалисту в данной области техники.
[00223] В некоторых вариантах реализации изобретения ADC получают путем контактирования фрагмента антитела с линкером и фрагментом лекарственного средства последовательным образом, таким образом что фрагмент антитела сначала ковалентно связывается с линкером, а затем предварительно сформированное промежуточное соединение антитело-линкер реагирует с фрагментом лекарственного средства. Промежуточное соединение антитело-линкер может подвергаться этапу очистки или не может подвергаться этапу очистки перед контактированием с фрагментом лекарственного средства. В других вариантах реализации изобретения ADC получают путем контактирования фрагмента антитела с соединением линкер-лекарственное средство, предварительно сформированным путем взаимодействия линкера с фрагментом лекарственного средства. Предварительно сформированное соединение линкер-лекарственное средство может или не может подвергаться этапу очистки перед контактированием с фрагментом антитела. В других вариантах реализации изобретения фрагмент антитела контактирует с линкером и фрагментом лекарственного средства в одной реакционной смеси, позволяя одновременное образование ковалентных связей между фрагментом антитела и линкером и между линкером и фрагментом лекарственного средства. Этот способ получения ADC может включать реакцию, в которой фрагмент антитела контактирует с фрагментом антитела перед добавлением линкера к реакционной смеси и наоборот. В некоторых вариантах реализации изобретения ADC получают путем взаимодействия фрагмента антитела с линкером, присоединенным к фрагменту лекарственного средства, такому как Mal-(ПЭГ)2-Val-Cit-pAB-эрибулин, в условиях, допускающих конъюгирование.
[00224] ADC, полученные в соответствии со способами, описанными выше, могут подвергаться этапу очистки. Этап очистки может включать любые биохимические способы, известные в данной области техники для очистки белков, или любую комбинацию их способов. Они включают, но без ограничения, тангенциальную поточную фильтрацию (ТПФ), аффинную хроматографию, ионообменную хроматографию, любую хроматографию на основе заряда или изоэлектрической точки, хроматографию со смешанным режимом, например, CHT (керамический гидроксиапатит), хроматографию с гидрофобным взаимодействием, эксклюзионную хроматографию, диализ, фильтрацию, селективное осаждение или любую их комбинацию.
Терапевтические применения и композиции
[00225] В данном документе описаны способы применения описанных ADC для лечения субъекта, имеющего расстройство, например, онкологическое расстройство. ADC можно вводить отдельно или в сочетании со вторым терапевтическим агентом и могут вводиться в любой фармацевтически приемлемой композиции, дозировке и режиме дозирования. Эффективность лечения ADC может быть оценена как по токсичности, так и по показателям эффективности и соответствующим образом скорректирована. Измерения эффективности включают, но не ограничиваются ими, наблюдаемый in vitro или in vivo цитостатический и/или цитотоксический эффект, уменьшенный объем опухоли, ингибирование роста опухоли и/или пролонгированное выживание.
[00226] Известны способы определения того, оказывает ли ADC цитостатический и/или цитотоксический эффект на клетку. Например, цитотоксическую или цитостатическую активность ADC можно измерить путем: обработки клеток млекопитающих, экспрессирующих целевой белок ADC в клеточной культуральной среде; культивирования клеток в течение периода от около 6 часов до около 5 дней; и измерение жизнеспособности клеток. Исследования in vitro на клетках также могут быть применены для измерения жизнеспособности (пролиферации), цитотоксичности и индукции апоптоза (активации каспазы) ADC.
[00227] Для определения того, оказывает ли конъюгат антитело-лекарственное средство цитостатический эффект, может быть применен анализ включения тимидина. Например, раковые клетки, экспрессирующие целевой антиген с плотностью 5000 клеток/лунка 96-луночного планшета, могут быть культивированы в течение 72-часового периода и подвергаются воздействию 0,5 мкКи 3H-тимидина в течение последних 8 часов 72-часового периода. Включение 3H-тимидина в клетки культуры измеряется в присутствии и отсутствии ADC.
[00228] Для определения цитотоксичности могут быть измерены некроз или апоптоз (запрограммированная гибель клеток). Некроз обычно сопровождается повышенной проницаемостью плазматической мембраны; клеточным отеком и разрывом плазматической мембраны. Апоптоз обычно характеризуется пузырением мембраны, конденсацией цитоплазмы и активацией эндогенных эндонуклеаз. Определение любого из этих эффектов на раковые клетки указывает на то, что ADC полезен при лечении рака.
[00229] Жизнеспособность клеток может быть измерена, например, путем определения в клетке поглощения красителя, такого как нейтральный красный, трипановый синий, кристаллический фиолетовый или ALAMAR™ синий (см., например, Page и соавт. (1993 год) Intl. J. Oncology 3:473-6). В таком анализе клетки инкубируют в средах, содержащих краситель, клетки промывают, а оставшийся краситель, отражающий клеточное поглощение красителя, измеряют спектрофотометрически. В некоторых вариантах реализации изобретения in vitro активность полученных ADC оценивают, применяя анализ с кристаллическим фиолетовым. Кристаллический фиолетовый представляет собой триарилметановый краситель, который накапливается в ядре жизнеспособных клеток. В этом анализе клетки подвергаются воздействию ADC или контрольных агентов в течение определенного периода времени, после чего клетки окрашивают кристаллическим фиолетовым, тщательно промывают водой, затем солюбилизируют 1% SDS и считывают спектрофотометрически. Белоксвязывающий краситель сульфородамин B (SRB) также может быть применен для измерения цитотоксичности (Skehan и соавт., 1990 год) J. Natl. Cancer Inst. 82:1107-12).
[00230] Апоптоз можно количественно определить, например, путем измерения фрагментации ДНК. Доступны коммерческие фотометрические способы количественного определения фрагментации ДНК in vitro. Примеры таких анализов, включая TUNEL (который обнаруживает включение меченых нуклеотидов в фрагментированной ДНК) и анализы на основе ИФА, описаны в Biochemica (1999 год) №2, стр. 34-37 (Roche Molecular Biochemicals).
[00231] Апоптоз можно также определить путем измерения морфологических изменений в клетке. Например, как и при некрозе, потеря целостности плазматической мембраны может быть определена путем измерения поглощения определенных красителей (например, флуоресцентного красителя, такого как, например, акридиновый оранжевый или бромид этидия). Способ измерения числа апоптотических клеток был описан Duke and Cohen, Current Protocols in Immunology (Coligan и соавт., ред. (1992 год) стр. 3. 17. 1-3. 17. 16). Клетки также могут быть помечены красителем ДНК (например, акридиновым оранжевым, бромидом этидия или иодидом пропидия) и клетками, которые наблюдаются на наличие конденсации и скопления хроматина вдоль внутренней ядерной мембраны. Другие морфологические изменения, которые можно измерить для определения апоптоза, включают, например, цитоплазматическую конденсацию, повышенное пузырение мембраны и уменьшение размера клеток.
[00232] Описанные ADC также могут быть оценены на активность неспецифического цитолиза. Активность неспецифического цитолиза может быть определена, например, анализом с применением двух клеточных линий, одной содержащей целевой антиген и одной не содержащей целевой антиген. Чтобы различать клеточные линии их предпочтительно метят. Например, клетки IGROV1 (FRA+), помеченные Nuclight™ Green (NLG) и HL-60 (FRA-), помеченные Nuclight™ Red (NLR), могут быть совместно культивированы, обработаны ADC к FRA с последующим контролем цитотоксичности. Убийство клеток, не содержащих целевой антиген при их смешивании с клетками, содержащими целевой антиген свидетельствует о неспецифическом цитолизе, тогда как убийство клеток, не содержащих целевой антиген в отсутствие клеток, содержащих целевой антиген свидетельствует об нецелевом уничтожении.
[00233] В некоторых аспектах данное описание включает способ убийства, ингибирования или модулирования роста, или нарушения метаболизма раковой клетки или ткани путем разрушения тубулина. Этот способ может применяться с любым субъектом, где нарушение тубулина обеспечивает терапевтическое преимущество. Субъекты, которые могут получать пользу от разрушения тубулина, включают, но без ограничения, те, которые имеют или могут иметь рак желудка, рак яичников (например, серозный рак яичников), рак легкого (например, немелкоклеточный рак легких), рак груди (например, трижды негативный рак молочной железы), рак эндометрия (например, серозную карциному эндометрия), остеосаркому, саркому Капоши, рак половых клеток яичка, лейкемию, лимфому (например, болезнь Ходжкина, неходжкинскую лимфома), миелому, рак головы и шеи, рак пищевода, рак поджелудочной железы, рак предстательной железы, рак мозга (например, глиобластому), рак щитовидной железы, колоректальный рак и/или рак кожи (например, меланому) или любые метастазы (Dumontet and Jordan, 2010) Nat. Rev. Drug Discov. 9:790-803). В различных вариантах реализации изобретения описанные ADC можно вводить в любой клетке или ткани, которая экспрессирует FRA, такой как раковая клетка или ткань, экспрессирующая FRA. Типовый вариант реализации изобретения включает в себя способ ингибирования передачи сигналов, опосредуемых FRA, или способ убийства клетки. Этот способ можно применять на любой клетке или ткани, которые экспрессирует FRA, таких как раковая клетка или метастатическое поражение. Неограничивающие примеры раков, экспрессирующих FRA, включают в себя рак желудка, серозный рак яичников, светлоклеточный рак яичника, немелкоклеточный рак легкого, колоректальный рак, трижды негативный рак молочной железы, рак эндометрия, серозную карциному эндометрия, карциноид легких и остеосаркому. Неограничивающие примеры клеток, экспрессирующих FRA, включают клетки карциномы яичника IGROV1 и OVCAR3 человека, клетки немелкоклеточной карциномы легких NC-H2110 человека и клетки, содержащие рекомбинантную нуклеиновую кислоту, кодирующую FRA или ее часть.
[00234] В различных других вариантах реализации изобретения описанные ADC можно вводить в любой клетке или ткани, которая экспрессирует her2, такой как раковая клетка или ткань, экспрессирующая her2. Типовый вариант реализации изобретения включает в себя способ ингибирования передачи сигналов, опосредуемых her2, или способ убийства клетки. Этот способ можно применять на любой клетке или ткани, которые экспрессирует her2, таких как раковая клетка или метастатическое поражение. Неограничивающие примеры раков, экспрессирующих her2, опухолей включают рак молочной железы, рак желудка, рак мочевого пузыря, карциному уротелиальных клеток, рак пищевода, рак легких, рак шейки матки, рак эндометрия и рак яичников (English и соавт. (2013 год) Mol. Diagn. Ther. 17:85-99). Неограничивающие примеры клеток, экспрессирующих her2, включают клетки желудочной карциномы NCI-N87-luc человека, клетки проточной карциномы молочной железы ZR75 и BT-474 человека и клетки, содержащие рекомбинантную нуклеиновую кислоту, кодирующую her2 или ее часть.
[00235] В различных других вариантах реализации изобретения описанные ADC можно вводить любую клетку или ткань, которая экспрессирует мезотелин (MSLN), такую как раковая клетка или ткань, экспрессирующая MSLN. Типовый вариант реализации изобретения включает в себя способ ингибирования передачи сигналов, опосредуемых MSLN, или способ убийства клетки. Этот способ можно применять на любой клетке или ткани, которые экспрессирует MSLN, таких как раковая клетка или метастатическое поражение. Неограничивающие примеры раков, экспрессирующих MSLN, включают мезотелиому, рак поджелудочной железы (например, аденокарциному поджелудочной железы), рак яичников и рак легких (например, аденокарциному легких) (Wang и соавт. (2012 год) PLoS ONE 7:e33214). Неограничивающие примеры клеток, экспрессирующих MSLN включают клетки карциномы яичника OVCAR3 человека, эндометроидные клетки HEC-251 человека, плоские клетки мезотелиомы легких H226 человека и клетки, содержащие рекомбинантную нуклеиновую кислоту, кодирующую MSLN или ее часть.
[00236] Типовые способы включают в себя этапы контактирования клетки с ADC, как описано в данном документе, в эффективном количестве, то есть количестве, достаточном для уничтожения клетки. Способ может быть применен на клетках в культуре, например, in vitro, in vivo, ex vivo, или in situ. Например, клетки, которые экспрессируют FRA, her2 и/или MSLN (например, клетки, собранные путем биопсии опухолевого или метастатического поражения, клетки из верифицированной линии раковых клеток; или рекомбинантные клетки), могут быть культивированы in vitro в культуральной среде, а этап контактирования может быть осуществлен путем добавления ADC к культуральной среде. Указанный способ приведет к уничтожению клеток, экспрессирующих FRA, her2 и/или MSLN, включая, в частности, опухолевые клетки, экспрессирующие FRA, her2 и/или MSLN. В альтернативном варианте, чтобы иметь эффект in vivo ADC можно вводить субъекту любым подходящим способом введения (например, внутривенным, подкожным или прямым контактом с опухолевой тканью).
[00237] Эффект in vivo описанной терапевтической композиции ADC может быть оценен на подходящей животной модели. Например, могут быть применены модели ксеногенного рака, при этом экспланты рака или перевиваемые ксенотрансплантатные ткани вводятся в животных с ослабленным иммунитетом, таких как безтимусные мыши или мыши SCID (Klein и соавт., 1997 год) Nature Med., 3:402-8). Эффективность может быть предсказана с применением анализов, которые измеряют ингибирование образования опухолей, регрессии опухолей или метастазов и тому подобного.
[00238] Также могут быть применены анализы in vivo, которые оценивают продвижение апоптоза. В одном варианте реализации изобретения ксенотрансплантаты от мышей-опухоленосителей, получавших терапевтическую композицию, можно исследовать на наличие апоптотических очагов и сравнить с контрольными мышами, не получавшими лечение, переносящими ксенотрансплантат. Степень, в которой апоптотические очаги обнаруживаются в опухолях мышей, получавших лечение, указывает на терапевтическую эффективность композиции.
[00239] Далее в данном документе описаны способы лечения рака. ADC, описанные в данном документе, могут быть введены млекопитающему, отличному от человека, или человеку в терапевтических целях. Терапевтические способы включают введение млекопитающему, имеющему опухоль, биологически эффективного количества ADC, содержащего выбранный химиотерапевтический агент (например, эрибулин), связанный с нацеливающим антителом, которое связывается с экспрессируемым антигеном, который доступен для связывания или локализован на поверхности раковой клетки. Типовый вариант реализации изобретения представляет собой способ доставки химиотерапевтического агента в клетку, экспрессирующую FRA, включающий конъюгирование химиотерапевтического агента с антителом, которое иммуноспецифически связывается с эпитопом FRA и подвержение клетки воздействию ADC. Типовые опухолевые клетки, которые экспрессируют FRA, для которых указаны ADC по данному изобретению, включают клетки рака желудка, серозного рака яичников, немелкоклеточного рака легких, колоректального рака, рака молочной железы (например, трижды негативного рака молочной железы), карциноида легких, остеосаркомы, рака эндометрия и карциномы эндометрия с серозной гистологией.
[00240] Другой типовый вариант реализации изобретения представляет собой способ доставки химиотерапевтического агента в клетку, экспрессирующую her2, включающий конъюгирование химиотерапевтического агента с антителом, которое иммуноспецифически связывается с эпитопом her2 и подвержение клетки воздействию ADC. Типовые опухолевые клетки, которые экспрессируют her2, для которых указаны ADC по данному изобретению, включают клетки рака молочной железы, рака желудка, рака мочевого пузыря, карциномы уротелиальных клеток, рака пищевода, рака легких, рака шейки матки, рака эндометрия и рака яичников.
[00241] Другой типовый вариант реализации изобретения представляет собой способ доставки химиотерапевтического агента в клетку, экспрессирующую MSLN, включающий конъюгирование химиотерапевтического агента с антителом, которое иммуноспецифически связывается с эпитопом MSLN и подвержение клетки воздействию ADC. Типовые опухолевые клетки, которые экспрессируют MSLN, для которых указаны ADC по данному изобретению, включают клетки из мезотелиомы, рака поджелудочной железы (например, аденокарциномы поджелудочной железы), рака яичников и рака легкого (например, аденокарциномы легкого).
[00242] Другим типовым вариантом реализации изобретения является способ лечения пациента, имеющего рак или подверженного риску возникновения рака, экспрессирующего целевой антиген для фрагмента антитела ADC, такой как FRA, her2 или MSLN, содержащий введение пациенту терапевтически эффективного количества ADC по данному изобретению. В некоторых вариантах реализации изобретения пациент не реагирует или слабо реагирует на лечение антителом к FRA при введении в монотерапии и/или на лечение фрагментом лекарственного средства (например, эрибулином) при введении в монотерапии. В других вариантах реализации изобретения пациент не реагирует или слабо реагирует на лечение антителом к her2 при введении в монотерапии и/или на лечение фрагментом лекарственного средства (например, эрибулином) при введении в монотерапии. В других вариантах реализации изобретения пациент не реагирует или слабо реагирует на лечение антителом к MSLN при введении в монотерапии и/или на лечение фрагментом лекарственного средства (например, эрибулином) при введении в монотерапии. В других вариантах реализации изобретения пациент имеет непереносимость к лечению фрагментом лекарственного средства (например, эрибулином) при введении в монотерапии. Например, пациент может иметь необходимость в дозах эрибулина для лечения рака, которые приводят к системной токсичности, которую преодолевают путем целенаправленной доставки в рак, экспрессирующий целевой антиген для фрагмента антитела ADC, такой как FRA, her2 или MSLN, тем самым уменьшая нецелевое уничтожение.
[00243] Другим типовым вариантом реализации изобретения является способ уменьшения или ингибирования роста опухоли, экспрессирующей целевой антиген (например, опухоли, экспрессирующей FRA, опухоли, экспрессирующей her2 или опухоли, экспрессирующей MSLN), включающий введение терапевтически эффективного количества ADC. В некоторых вариантах реализации изобретения лечение является достаточным для уменьшения или ингибирования роста опухоли пациента, уменьшения количества или размера метастатических поражений, снижения опухолевой массы, снижения массы первичной опухоли, снижения инвазивности, продления времени выживания и/или поддержания или улучшения качества жизни. В некоторых вариантах реализации изобретения опухоль устойчива или резистентна к лечению с помощью антитела к FRA при введении в монотерапии и/или на лечение фрагментом лекарственного средства (например, эрибулином) при введении в монотерапии. В других вариантах реализации изобретения опухоль устойчива или резистентна к лечению с помощью антитела к her2 при введении в монотерапии и/или на лечение фрагментом лекарственного средства (например, эрибулином) при введении в монотерапии. В некоторых вариантах реализации изобретения опухоль устойчива или резистентна к лечению с помощью антитела к MSLN при введении в монотерапии и/или на лечение фрагментом лекарственного средства (например, эрибулином) при введении в монотерапии.
[00244] Более того, антитела по данному изобретению могут быть введены млекопитающему, отличному от человека, которое экспрессирует антиген, с которым ADC способен связываться для ветеринарных целей или как животная модель человеческого заболевания. Что касается последнего, такие животные модели могут быть полезны для оценки терапевтической эффективности описанных ADC (например, тестирование дозировок и временной динамики введения).
[00245] Далее в данном документе описано терапевтическое применение описанных ADC. Типовым вариантом реализации изобретения является применение ADC для лечения рака, экспрессирующего целевой антиген (например, рака, экспрессирующего FRA, рака, экспрессирующего her2 или рака, экспрессирующего MSLN). Также описаны ADC для применения в лечении рака, экспрессирующего целевой антиген (например, рака, экспрессирующего FRA, рака, экспрессирующего her2 или рака, экспрессирующего MSLN). Способы идентификации субъектов, имеющих раки, которые экспрессируют FRA, her2 и/или MSLN, известны в данной области техники и могут быть применены для идентификации подходящих пациентов с целью лечения описанным ADC.
[00246] Другим вариантом реализации изобретения является применение ADC в способе изготовления медицинского препарата для лечения рака, экспрессирующего целевой антиген (например, рака, экспрессирующего FRA, рака, экспрессирующего her2 или рака, экспрессирующего MSLN).
[00247] Терапевтические композиции, применяемые в практике вышеприведенных способов, могут быть составлены в виде фармацевтических композиций, содержащих фармацевтически приемлемый носитель, подходящий для желаемого способа доставки. Типовый вариант реализации изобретения представляет собой фармацевтическую композицию, содержащую ADC по данному изобретению и фармацевтически приемлемый носитель. Подходящие носители включают любой материал, который в сочетании с терапевтической композицией сохраняет противоопухолевую функцию терапевтической композиции и, как правило, не вступает в реакции с иммунной системой пациента. Фармацевтически приемлемые носители включают любые и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и замедляющие всасывание агенты и тому подобные, которые физиологически совместимы. Примеры фармацевтически приемлемых носителей включают одно или более из воды, физиологического раствора, фосфатного буферного раствора, декстрозы, глицерола, этанола, мезилатной соли и тому подобного, а также их комбинаций. Во многих случаях будет предпочтительным включать в композицию изотонические агенты, например, сахара, полиспирты, такие как маннит, сорбит или натрия хлорид. Фармацевтически приемлемые носители могут дополнительно содержать незначительные количества вспомогательных веществ, таких как увлажняющие или эмульсифицирующие агенты, консерванты или буферы, которые увеличивают срок хранения или эффективность ADC.
[00248] Терапевтические составы могут быть солюбилизированы и введены любым способом, способным доставлять терапевтическую композицию в расположение опухоли. Потенциально эффективные пути введения включают, но без ограничений, внутривенное, парентеральное, внутрибрюшинное, внутримышечное, внутриопухолевое, внутрикожное, внутриоргановое, ортотопическое и тому подобное. Терапевтические белковые препараты можно лиофилизировать и хранить в виде стерильных порошков, предпочтительно под вакуумом, а затем ресуспендировать в бактериостатической воде (содержащей, например, консервант бензилового спирта) или в стерильной воде перед инъекцией. Терапевтические составы могут содержать ADC или его фармацевтически приемлемую соль, например мезилатную соль.
[00249] Описанные в данном документе ADC могут вводиться в дозе от 0,2 мг/кг до около 10 мг/кг пациенту, нуждающемуся в этом. В некоторых вариантах реализации изобретения ADC вводится пациенту ежедневно, раз в два месяца или в любой промежуток времени между ними. Дозировки и протоколы введения для лечения злокачественных новообразований с применением вышеуказанных способов будут варьироваться в зависимости от способа и целевого рака и будут в целом зависеть от ряда других факторов, признанных в данной области техники.
[00250] Известны различные системы доставки и могут применяться для введения одного или более ADC по данному изобретению. Способы введения ADC включают, но без ограничения, парентеральное введение (например, внутрикожное внутримышечное, внутрибрюшинное, внутривенное и подкожное), эпидуральное введение, внутриопухолевое введение и мукозальное введение (например, интраназальный и оральный пути). Кроме того, введение пульмональным способом может быть осуществлено, например, с применением ингалятора или распылителя и состава с аэрозольным агентом. См., например, композиции и способы для пульмонального введения, описанные в патентах США №6,019,968, 5,985,320, 5,985,309, 5,934,272, 5,874,064, 5,855,913, 5,290,540 и 4,880,078; и публикациях PCT № WO 92/19244, WO 97/32572, WO 97/44013, WO 98/31346 и WO 99/66903. ADC можно быть введена любым удобным путем, например, путем инфузии или болюсной инъекции или путем абсорбции через эпителиальные или кожно-слизистые выстилающие ткани (например, слизистую оболочку полости рта, прямой кишки и слизистой оболочки кишечника и тому подобное). Введение может быть, как системным, так и местным.
[00251] Описанные в данном документе терапевтические композиции могут быть стерильными и стабильными в условиях производства и хранения. В некоторых вариантах реализации изобретения один или более ADC, или фармацевтических композиций поставляются в виде сухого стерилизованного лиофилизированного порошка или безводного концентрата в герметично закрытом контейнере и могут быть ресуспендированы (например, водой или физиологическим раствором) до соответствующей концентрации для введения субъекту. Предпочтительно один или более профилактических, или терапевтических агентов или фармацевтических композиций поставляются в виде сухого стерильного лиофилизированного порошка в герметично закрытом контейнере при единичной дозе по меньшей мере 5 мг, по меньшей мере 10 мг, по меньшей мере 15 мг, по меньшей мере 25 мг, по меньшей мере 35 мг, по меньшей мере 45 мг, по меньшей мере 50 мг, по меньшей мере 75 мг или по меньшей мере 100 мг, или любое количество между данными значениями. В некоторых вариантах реализации изобретения лиофилизированные ADC или фармацевтические композиции хранят при температуре от 2°С до 8°С в оригинальной упаковке. В некоторых вариантах реализации изобретения один или более ADC или фармацевтических композиций, описанных в данном документе, поставляются в жидкой форме в герметично закрытом контейнере, например, в контейнере, указывающем количество и концентрацию агента. В некоторых вариантах реализации изобретения жидкая форма вводимой композиции поставляется в герметично закрытом контейнере по меньшей мере 0,25 мг/мл, по меньшей мере 0,5 мг/мл, по меньшей мере 1 мг/мл, по меньшей мере 2,5 мг/мл, по меньшей мере 5 мг/мл, по меньшей мере 8 мг/мл, по меньшей мере 10 мг/мл, по меньшей мере 15 мг/мл, по меньшей мере 25 мг/мл, по меньшей мере 50 мг/мл, по меньшей мере 75 мг/мл или по меньшей мере 100 мг/мл ADC. Жидкая форма может храниться при температуре от 2°С до 8°С в оригинальной упаковке.
[00252] В некоторых вариантах реализации изобретения, описанные ADC могут быть включены в фармацевтическую композицию, подходящую для парентерального введения. Раствор для инъекций может состоять из жидкой или лиофилизированной лекарственной формы в стеклянном прозрачном флаконе или флаконе из темного (оранжевого) стекла, ампуле или предварительно заполненном шприце или другом известном устройстве доставки или хранения.
[00253] Композиции, описанные в данном документе, могут быть в различных формах. К ним относятся, например, жидкие, полутвердые и твердые лекарственные формы, такие как жидкие растворы (например, инъекционные и инфузионные растворы), дисперсии или суспензии, таблетки, пилюли, порошки, липосомы и суппозитории. Предпочтительная форма зависит от предполагаемого способа введения и терапевтического применения.
[00254] В различных вариантах реализации изобретения лечение включает единичный болюс или повторное введение препарата ADC через приемлемый путь введения.
[00255] Пациенты могут быть оценены на уровни целевого антигена в данном образце (например, на уровни клеток, экспрессирующих целевой антиген), чтобы помочь в определении наиболее эффективного режима дозирования и тому подобного. Типовый вариант реализации изобретения представляет собой способ определения, будет ли пациент реагировать на лечение ADC по данному изобретению, включая предоставление биологического образца от пациента и контактирование биологического образца с ADC. Типовые биологические образцы включают в себя ткань или биологическую жидкость, такую как воспалительный экссудат, кровь, сыворотку, кишечную жидкость, пробу фекалий или биопсию опухоли (например, биопсию опухоли, полученную от пациента, имеющего или подверженного риску заболевания на рак, экспрессирующий целевой антиген, например, рак, экспрессирующий FRA, рак, экспрессирующий her2 или рак, экспрессирующий MSLN). В некоторых вариантах реализации изобретения образец (например, ткань и/или биологическая жидкость) может быть получен от субъекта, и подходящий иммунологический способ может быть применен для выявления и/или измерения экспрессии белка целевого антигена (например, FRA, her2 или MSLN). Такие оценки также применяются с целью мониторинга во время терапии и полезны для оценки терапевтического успеха в сочетании с оценкой других параметров.
[00256] В некоторых вариантах реализации изобретения эффективность ADC может быть оценена путем контактирования образца опухоли из субъекта с ADC и оценки скорости роста или объема опухоли. В некоторых вариантах реализации изобретения, в случае если определено, что ADC является эффективным, его можно вводить субъекту.
[00257] Вышеуказанные терапевтические подходы могут быть объединены с любым из множества разнообразных дополнительных хирургических, химиотерапевтических или лучевых курсами лечения.
[00258] Также в данном документе описано применение одного или более описанных ADC при изготовлении лекарственного препарата для лечения рака, например, в соответствии со способами, описанными выше. В некоторых вариантах реализации изобретения, описанные в данном документе ADC применяются для лечения рака, например, в соответствии со способами, описанными выше.
[00259] В различных вариантах реализации изобретения наборы для применения в лабораторных и терапевтических применениях, описанные в данном документе, входят в объем данного изобретения. Такие наборы могут содержать носитель, упаковку или контейнер, которые разделены на части чтобы вмещать один или более контейнеров, таких как флаконы, трубки и тому подобное, причем каждый из контейнеров содержит один из отдельных элементов, которые должны применяться в описанном в данном документе способе, наряду с этикеткой или вкладышем, содержащей инструкции для применения, такие как применение, описанное в данном документе. Наборы могут содержать контейнер, содержащий фрагмент лекарственного средства. Данное описание также обеспечивает один или более ADC, или их фармацевтических композиций, упакованных в герметично закрытый контейнер, такой как ампула или фильтр-пакет, с указанием количества агента.
[00260] Наборы могут содержать контейнер, описанный выше, и один или более других контейнеров, связанных с ним, которые содержат материалы, желательные с коммерческой и пользовательской точек зрения, включая буферы, разбавители, фильтры, иглы, шприцы; носители, упаковки, контейнеры, флаконы и/или этикетки с надписью, и/или инструкции для применения, а также листки-вкладыши с инструкциями по применению.
[00261] Этикетка может присутствовать на контейнере или в нем для указания, что композиция применяется для конкретной терапии или нетерапевтического применения, такого как прогностическое, профилактическое, диагностическое или лабораторное применение. Этикетка может также указывать направления для применения in vivo или in vitro, такие как те, что описаны в данном документеУказания и/или другая информация также могут быть указаны на вкладыше(шах) или этикетке(ках), которые включены в набор или находится на наборе. Этикетка может быть включена в контейнер или связана с контейнером. Этикетка может находиться на контейнере, когда буквы, цифры или другие символы, образующие этикетку, прессуются или вытравливаются на самом контейнере. Этикетка может быть связана с контейнером, в случае если она присутствует в емкости или носителе, которая также удерживает контейнер, например, в виде листа-вкладыша. Этикетка может указывать на то, что композиция применяется для диагностики или лечения состояния, такого как рак, описанный в данном документе.
[00262] Специалистам в данной области техники будет очевидно, что другие подходящие модификации и приспособления, описанных в данном документе способов изобретения очевидны, и могут быть сделаны с применением подходящих эквивалентов, не отступая от объема изобретения или описанных в данном документе вариантов реализации изобретения. Опираясь на подробное описание изобретения, оно будет более понятным со ссылкой на следующие примеры, которые включены исключительно в целях иллюстрации и не предназначены для ограничения.
ПРИМЕР 1
1. Материалы и способы
[00263] MORAb-003, применяемый для приготовления ADC, был из партии #AA0312.
1.1 Цитотоксины
[00264] Структуры конъюгируемых цитотоксинов приведены в Таблице 11.
Таблица 11. Конъюгируемые цитотоксины
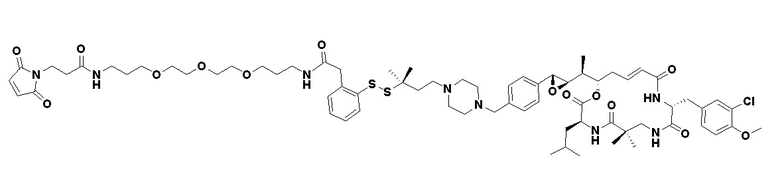
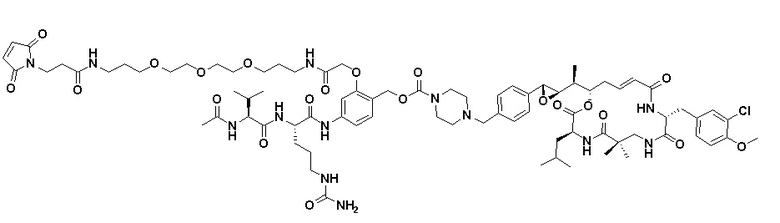
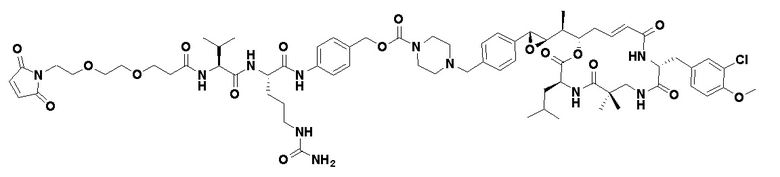
(ER-001159569)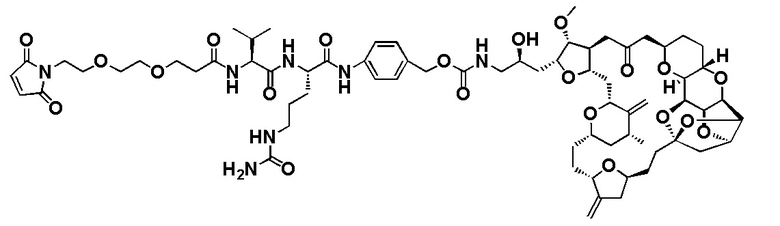
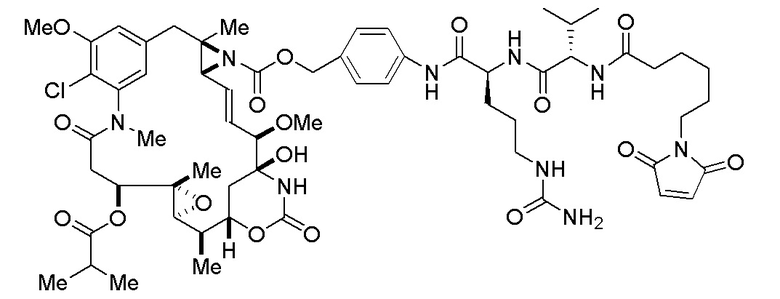
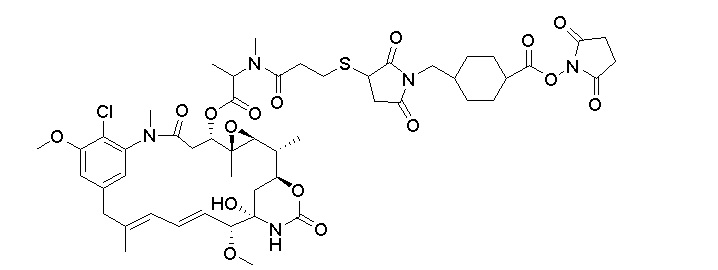
Сокращения: Ala, аланин; Asn, аспарагин; Cit, цитруллин; NHS, N-гидроксисукцинимид; pAB, п-аминобензилоксикарбонил; ПЭГ, полиэтиленгликоль; SMCC, сукцинимидил-4-(N-малеимидометил) циклогексан-1-карбоксилат; Val, валин; VCP, Val-Cit-pAB.
1.2 Конъюгация антитела-лекарственного средства
1.2.1 Частичное восстановление с помощью TCEP
[00265] Частично восстанавливающие условия для MORAb-003 были установлены путем изменения концентрации восстанавливающего агента трис (2-карбоксиэтил) фосфина (TCEP), не содержащего тиол, изменения концентрации антител и времени восстановления. Для MORAb-003 заменяли буфер на фосфатно-солевой буфер Дульбекко (DPBS), содержащий 1 мМ этилендиаминтетрауксусной кислоты (ЭДТК), затем концентрировали до 10 мг/мл, применяя центрифужную концентрацию с центрифужными фильтрами с границей отсечки 10 кДа по молекулярному весу задерживаемых компонентов (MWCO - molecular weight cut-off). Антитела разводили до соответствующей концентрации и добавляли TCEP в указанной конечной концентрацией и легко перемешивали в течение 1 часа при комнатной температуре. TCEP удаляли путем обессоливания, применяя 5- или 10-мииллилитровые вращающиеся колонки для обессоливания Zeba™ с DPBS/1 мМ ЭДТК в качестве буфера (Thermo Fisher, 40 кДа MWCO) в соответствии с протоколом производителя. Образцы анализировали на содержание свободных тиолов с помощью набора флуорометрического количественного определения тиола (Abcam) в соответствии с протоколом производителя. Анализ ДСН-ПААГ-электрофореза в невосстанавливающих условиях проводили для определения степени и места разрыва дисульфидной связи, как описано в разделе 1. 3. 3. В некоторых случаях обессоленные мАт доводили до 1-2 мг/мл путем разбавления в DPBS и подвергали биотинилированию для определения способности конъюгировать и соотношения лекарственного средства-к-антителу (DAR). 10 мМ малеимидо-ПЭГ2-биотина (Thermo Fisher) в диметилсульфоксиде (ДМСО) добавляли к антителу (мАт) при молярном соотношении 10:1 и инкубировали при комнатной температуре в течение 4 часов при легком перемешивании. После конъюгации непрореагировавшее соединение удаляли путем обессоливания с применением вращающихся колонок для обессоливания Zeba™ (Thermo Fisher). Затем образцы анализировали с помощью ЖХ-МС для определения DAR, как описано в разделе 1. 3. 4.
1.2.2 Конъюгация цитотоксинов
[00266] Частично восстановленное антитело доводили до 2,5 мг/мл в 0,5X DPBS, 0,5 мМ ЭДТК и тщательно перемешивали. Затем, если их применяли, добавляли органические сорастворители и тщательно перемешивали. В качестве сорастворителей использовали пропиленгликоль (конечная концентрация 20% и 50%), диметилсульфоксид (ДМСО) (10%), N,N-диметилформамид (20%), N,N-диметилацетамид (20%) и N,N-диметилпропионамид (20%). Малеимид-модифицированный цитотоксин (6 мМ исходный раствор в ДМСО) добавляли к антителам при молярном соотношении 1:6 (мАт:соединение) и тщательно перемешивали. Конъюгация продолжалась при комнатной температуре в течение 3,5 часов при легком перемешивании. 50% пропиленгликоль при 50% был выбран в качестве конечного органического модификатора и применялся во всех последующих реакциях конъюгации.
1.2.3 Очистка
[00267] Конъюгированное антитело очищали с помощью колонки(ок) для обессоливания 26/10 HiTrap® (GE Healthcare) с хроматографией, выполненной на жидкостной экспресс-хроматографии белков (ЖЭХБ) (GE Healthcare), для удаления непрореагировавшего малеимидо-цитотоксина и пропиленгликоля. ADC MORAb-003, включая MORAb-003-mal-VCP-эрибулин (MORAb-202), были приготовлены в DPBS (буфер для составов применяли в качестве подвижного буфера во время ЖЭХБ-хроматографии).
1.3 Биофизическая характеристика
1.3.1 Анализ БХК
[00268] Готовый реагент (200 мкл) бицинхониновой кислоты (БХК) добавляли к 25 мкл серийно разведенных ADC или к 2 мг/мл стандарта бычьего гамма-глобина (Thermo Fisher), и образцы тщательно перемешивали. Образцы инкубировали при 37°С в течение 20 мин. Планшеты считывали при 595 нм на планшет-ридере SpectraMax® M5 (Molecular Devices). Данные анализировали с помощью SoftMax® Pro (версия 3. 2) с моделью 4-параметрической подгонки.
1.3.2 Анализ эксклюзионной ВЭЖХ
[00269] Агрегацию антител анализировали с помощью эксклюзионной высокоэффективной жидкостной хроматографии (эксклюзионной ВЭЖХ) с применением Agilent 1100. мАт разбавляли до 1 мг/мл в DPBS. Антитело (20 мкл) вводили в защитную колонку TSKgel® SuperSW (4,6 мм×3,5 см, размер пор 4 мкм, Tosoh Bioscience), а затем в колонку TSKgel® SuperSW3000 (4,6 мм×30 см, размер пор 4 мкм), элюировали из колонки 0,1 М фосфатом натрия, содержащим 0,15 М NaCl и 0,05% NaN3, при рН 7,4 при скорости потока 0,3 мл/мин в течение 20 мин. Все данные были проанализированы с применением программного обеспечения Agilent ChemStation. Процент агрегации рассчитывался как [PAагрегата/PAобщий]*100, где PA=интегрированная площадь пика.
1.3.3 Анализ ДСН-ПААГ-электрофореза
[00270] Образцы белка (0,1-10 мкг) доводили до 1X буфером для образцов, содержащим додецилсульфата лития (ДСЛ). Для невосстановленных образцов инкубацию проводили при комнатной температуре в течение 10 мин перед электрофорезом. Для восстановленных образцов добавляли дитиотреитол (ДТТ) до конечной концентрации 20 мМ и образцы нагревали до 95°С в течение 10 мин и помещали на лед перед электрофорезом. Образцы загружали в 10-, 12- или 15-луночные гели Бис-Трис для ДСН-ПААГ-электрофореза (Thermo Fisher) с 1X МОПС или 1X МЭС в качестве электродного буфера. -Электрофорез проводили при 185 В (постоянное напряжение) в течение 1 часа. Гели окрашивали красящим раствором InstantBlue (Expedeon) и обесцвечивали в воде. Документацию выполняли в системе документирования гелей UltraLum с применением оранжевых фильтров с длиной волны 600 нм.
1.3.4 Анализ СВЭЖХ/ЭРИ-МС соотношения лекарственного средства-к-антителу (DAR)
[00271] ADC были дегликозилированы, применяя PNGазы F (New England BioLabs). Буфер G7 (10 мкл) и PNGаза F (2 мкл) добавляли к мАт (90 мкл, 1 мг/мл в DPBS). Реакцию инкубировали в микроволновой печи (СЕМ) Discover в течении двух циклов (1) микроволновая мощность 10 Вт, 37°С, 10 мин, далее следует 5-минутная пауза; (2) микроволновая мощность 2 Вт, 37°С, 10 мин. Часть образца восстанавливали с помощью добавления ДТТ до конечной концентрации 20 мМ с последующей инкубацией при 60°С в течение 3 мин. Затем образцы анализировали, применяя сверхвысокоэффективную жидкостную хроматографию (СВЭЖХ) Waters Acquity и квадрупольную время-пролетную (Q-Tof - quadrupole time of flight) масс-спектрометрию Premier. Образцы (0,5-2 мкг каждого) вводили в микроколонку для обессоливания MassPrep™ при 65°С, элюировали из колонки с 5-минутным уравновешиванием в 95% подвижной фазы А, 10-минутным градиентом (5-90% В), и 10-минутным повторным уравновешиванием в 95% подвижной фазы А при 0,05 мл/мин. Подвижная фаза А составляла 0,1% муравьиной кислоты в воде. Подвижная фаза B составляла 0,1% муравьиной кислоты в ацетонитриле. Масс-спектрометр Q-Tof работал в V-режиме определения положительных ионов с детектированием в диапазоне 500-4000 м/з. Параметры источника были следующими:капиллярное напряжение, 2,25 кВ (интактное антитело)-2,50 кВ (восстановленное антитело); напряжение пробоотборного конуса, 65,0 В (интактное антитело) или 50,0 В (восстановленное антитело); температура источника, 100°C; температура десольватации, 250°С; поток газа десольватации, 550 л/час. Пик белка деконволюционировали с применением функции MassLynx® MaxEnt 1. Относительные интенсивности масс каждой неконъюгированной, однократно-конъюгированной и многократно-конъюгированной тяжелой и легкой цепи объединяли для расчета общего DAR по формуле:
2[[ILC+1+2(ILC+2)+3(ILC+3)+…n(ILC+n)]/ΣILCобщ]+2[[IHC+1+2(IHC+2)+3(IHC+3)+…n(IHC+n)]/ΣIHCобщ]
где ILC+1 является массовой интенсивностью легкой цепи, конъюгированной с одним цитотоксином, ILC+2 является массовой интенсивностью легкой цепи, конъюгированной с двумя цитотоксинами, и так далее. IHC представляют собой интенсивности соответствующих конъюгированных тяжелых цепей и ΣILCобщ и ΣIHCобщ представляют собой объединенные интенсивности всех неконъюгированных и конъюгированных легких цепей и тяжелых цепей соответственно.
1.3.5 HIC-ВЭЖХ анализ DAR
[00272] В дополнение к анализу DAR с помощью анализа СВЭЖХ/электрораспылительной ионизации (ЭРИ)-МС, DAR MORAb-003-vcp-эрибулин и DAR MORAb-003-0285 также анализировали с применением ВЭЖХ с гидрофобным взаимодействием (HIC-ВЭЖХ). Образцы вводили в TSKgel® Ether-5 PW, 7,5 мм ID x 7,5 см, размер пор 10 мкм, и элюировали из колонки с 3-минутным уравновешиванием в 100% подвижной фазы А, 15-минутным градиентом (0-100% B), 5-минутным выдерживанием в 100% В, 1-минутной заменой на 100% А и 5-минутным повторным уравновешиванием в 100% подвижной фазы А при 0,7 мл/мин. Подвижная фаза А составляла 25 мМ фосфата натрия, 1,5 М сульфата аммония, рН 7,0. Подвижная фаза В составляла 25 мМ фосфата натрия, 25% изопропанола, рН 7,0. Детекцию проводили при 280 нм (контроль 320 нм). DAR определяли по формуле:
[AUC+1+2(AUC+2)+3(AUC+3)+…n(AUC+n)]/ΣAUCобщ]
где AUC+1 является площадью под кривой для пика мАт, соответствующего ADC, конъюгированному с одним цитотоксином, AUC+2 является площадью под кривой для пика мАт, соответствующего ADC, конъюгированному с двумя цитотоксинами, и так далее. ΣAUCобщ представляет собой объединенную площадь под кривой для всех пиков.
1.4 Анализ цитотоксичности
1.4.1 Анализ окрашивания кристаллическим фиолетовым
[00273] Клетки IGROV1 (FRhi) и SJSA-1 (FRneg) субкультивировали и высевали при 10000 клеток/лунку в готовую питательную среду в 96-луночных планшетах для культивирования тканей, инкубировали при 37°C, 5% CO2 в течение ночи (16 часов). Как правило, реагенты для испытания серийно разводили 1:4 в планшетах для разведения с 2-миллилитровыми глубокими лунками, начиная с 1 мкМ (всего 10 разведений). 100 мкл разбавленных образцов добавляли к клеточным планшетам (при начальной концентрации образцов для испытания 500 нМ). Планшеты инкубировали при 37°С, 5% CO2 в течение дополнительных 48 часов. Среду удаляли, планшеты промывали один раз 200 мкл DPBS, окрашивали 50 мкл 0,2% раствора кристаллического фиолетового при комнатной температуре в течение 15 мин и затем тщательно промывали водопроводной водой. Планшеты сушили на воздухе, а кристаллический фиолетовый растворяли с 200 мкл 1% раствора ДСН. Планшеты считывали при 570 нм. Данные анализировали с применением GraphPad Prism 6. Анализы проводили, применяя плотности посева 1000 клеток на лунку, а воздействие соединения составляло в общей сложности 5 дней. Когда требовалось более короткое воздействие, через 4 часа удаляли среду, содержащую цитотоксические агенты, и заменяли свежей питательной средой перед 5-дневной инкубацией. Для OVCAR3, CaOV3 и NCI-H2110, клетки высевали при 3000 клеток/лунку и инкубировали в течение 5 дней с ADC. Для конкурентных экспериментов титруемые ADC предварительно инкубировали с 2 мкМ (конечная) неконъюгированного MORAb-003 перед инкубацией с клетками.
1.4.2 Анализ неспецифического цитолиза
[00274] За день до начала исследования клетки IGROV1 Nuclight™ Green (NLG) высевали при 5000 клеток/лунку в 96-луночные круглодонные планшеты с последующим центрифугированием при 1000 об/мин в течение 3 мин при комнатной температуре для обеспечения образования клеточного осадка. Планшет помещали в резервуар в Incucyte Zoom® (EssenBio science) и инкубировали при 37° C/5% CO2 в течение ночи. Программа была настроена каждые два часа собирать изображения роста клеток и определять общее количество клеток с окрашенными в зеленый цвет ядрами и окрашенными в красный цвет ядрами, а также фазу и конфлюентность клеток. В день эксперимента ADC MORAb-003 или чистое лекарственное средство разбавляли в полной среде RPMI и серийно разводили, начиная с 400 нМ. 50 мкл раствора цитотоксина добавляли к клеткам NLG-IGROV1 и инкубировали в течение 30 мин. В течение инкубационного периода клетки HL-60 (FRneg) Nuclight™ Red (NLR) разбавляли свежей средой до 2×105, 1×105 или 5×104 клеток/мл. В лунки с NLG-IGROV1 добавляли 50 мкл суспензии клеток NLR-HL60 или отдельно среды с последующим центрифугированием при 1000 об/мин в течение 3 мин при комнатной температуре для обеспечения повторного образования клеточного осадка. Планшет помещали обратно в резервуар в Incucyte Zoom (EssenBio science) и инкубировали при 37°C/5% CO2 в течение 5 дней. Относительный рост клеток NLG-IGROV1 определяли, применяя подсчет зеленых клеток, путем сравнения с образцами без ADC или образцами, к которым добавили отдельно чистое лекарственное средство. Относительный рост клеток HL60 определяли аналогичным образом, за исключением того, что определяли количество красных клеток. Определение значений ИК50 для NLG-IGROV1 и NLR-HL-60 определялось с помощью Prism (GraphPad).
1.4.3 Анализ стабильности сыворотки
[00275] 20 мкл ADC MORAb-003 тщательно смешивали с 80 мкл DPBS, нормальной объединенной сывороткой человека (Bioreclamation, партия BRH552911) или нормальной объединенной сывороткой мыши (Bioreclamation, партия MSE152591) и инкубировали при 37°C в течение 0, 4, 24 и 48 часов. После инкубации образцы замораживали и хранили при -20°C до оценивания в анализах цитотоксичности и связывания. Для анализа цитотоксичности образцы оценивали на клетках IGROV1 и SJSA-1, как описано в разделе 1. 4. 1. Для оценки связывания образцы оценивали, применяя анализ MSD ECL на основе раствора. Образцы инкубировали с биотинилированным рецептором фолиевой кислоты альфа и сульфо-меткой к MORAb-003 перед улавливанием на стрептавидин в планшете и детектировали с применением электрохемилюминесценции с помощью устройства MSD Sector Imager 2400.
2. Результаты
2. 1 Приготовление ADC MORAb-003
[00276] Чтобы выбрать лучшую комбинацию линкера и цитотоксина для конъюгации с MORAb-003, ADC были подготовлены с помощью трех методик. В соответствии со стратегией конъюгации, изображенной на Фиг. 1, неспаренные цистеины образуются путем частичного восстановления с ограниченными молярными эквивалентами восстанавливающего агента TCEP, не содержащего тиол. Эта стратегия предпочтительно уменьшает межцепочечные дисульфидные связи, которые связывают легкую цепь и тяжелую цепь (по одной паре на спаривание H-L) и две тяжелые цепи в шарнирной области (две пары на спаривание H-H в случае IgG1 человека), оставляя внутрицепочечные дисульфидные связи интактными.
[00277] Вторая стратегия конъюгации для получения ADC MORAb-003 использовала химию восстановленных дисульфидных мостиков. Химия восстановленных дисульфидных мостиков перестраивает мостики свободных тиолов цистеиновых остатков, освобождаемых в процессе частичного восстановления, имитируя роль дисульфидной связи и тем самым сохраняя стабильность и функцию ADC.
[00278] В третьей стратегии конъюгации для получения ADC MORAb-003 применяли ограниченное использование лизина. Ограниченное использование лизина приводит к конъюгации очень ограниченного количества вычисленных 70+ гридрофильных лизинов, доступных на типичной молекуле человеческого IgG и потенциально может давать смеси продуктов ADC с более низкой гомогенностью по сравнению со стратегиями, включающими модификацию цистеина.
2.1.1 Приготовление VCP-эрибулина для ADC MORAb-003
[00279] Эрибулин (1) (10 мг, 14 мкмоль) (Фиг. 2) растворяли в N,N-диметилформамиде (ДМФА) (1мл) и хорошо перемешивали. Добавляли N,N-диизопропилэтиламин (основание Хунига или iPr2NEt) (3,6 мкл, 21 мкмоль) и Фмок-Val-Cit-пара-аминобензил-пара-нитрофенол (Фмок-VCP-PNP) (2) (16 мг, 21 мкмоль, Concortis Biosystems, cat# VC1003). Реакционную смесь перемешивали при комнатной температуре в течение 4-16 часов, отслеживали с помощью набора для нингидриновой реакции (Anaspec, cat# 25241) до тех пор, пока реакция не была завершена. Затем в реакционную смесь добавляли диэтиламин (Et2NH) (0,014 мл, 0,14 мкмоль), перемешивали в течение 2 часов при 18-25°C для удаления защитной группы Фмок. Реакцию отслеживали с помощью набора для нингидриновой реакции. По окончании, растворитель выпаривали под вакуумом чтобы получить неочищенный VCP-эрибулин (3) (16 мг), очищали с применением колонки ZOBAX SB-C18 (размер пор 5 мкм, 9,4×150 мм) в системе ВЭЖХ Waters Alliance e2695 в подвижной фазе H2O-CH3CN, содержащей 0,1% муравьиной кислоты, через градиент В 15-70%. VCP-эрибулин (3) (16 мг) растворяли в ДМФА (1 мл). Основание Хунига (7,2 мкл, 41мкмоль) и малеимид-ПЭГ2-NHS (4) (9,7 мг, 27 мкмоль) добавляли. Реакционную смесь перемешивали при 18-25°C в течение 3 часов. Реакционную смесь очищали с помощью ВЭЖХ (H2O-CH3CN), содержащей 0,1% муравьиной кислоты) через градиент В 15-70%. Растворитель удаляли путем лиофилизации, чтобы получить mal-(ПЭГ)2-Val-Cit-п-аминобензилоксикарбонил (pAB)-эрибулин (mal-(ПЭГ)2-VCP-эрибулин) (5).
2. 1. 2 Оптимизация условий восстановления
[00280] ADC MORAb-003 получали путем образования неспаренных цистеинов вследствие частичного восстановления с ограниченными молярными эквивалентами восстанавливающего агента трис(2-карбоксиэтил)фосфина (TCEP). Первоначальное исследование было выполнено на MORAb-003, в котором варьировались концентрация антитела, концентрация TCEP и время инкубации, с целью создания в среднем 4 конъюгируемых сайтов на молекулу антитела. Количество сайтов свободных тиолов определяли с помощью анализа флуорометрического количественного определения тиола. Результаты этого анализа приведены в таблице 12. Степень разрушения связей H-H и H-L после 10 мин, 30 мин, 60 мин или 120 мин инкубации также анализировали с помощью ДСН-ПААГ-электрофореза (Фиг. 3). Для этого анализа невосстановленные и восстановленные образцы загружали на гель для ДСН-ПААГ-электрофореза, и электрофорез проводили при 185 В в течение 1 часа. На Фиг. 3 дорожка М соответствует стандартному белку. Дорожка 1 соответствует необработанному, невосстановленному MORAb-003. Дорожка 2 соответствует MORAb-003 (5,3 мг/мл), восстановленному в 70,6 мкМ TCEP. Дорожка 3 соответствует MORAb-003 (5,3 мг/мл), восстановленному в 141,2 мкМ TCEP. Дорожка 4 соответствует MORAb-003 (1,5 мг/мл), восстановленному в 20 мкМ TCEP. Дорожка 5 соответствует MORAb-003 (1,5 мг/мл), восстановленному в 40 мкМ TCEP. Идентичности каждой полосы указаны на нижнем правом геле. "Н" обозначает тяжелую цепь, тогда как "L" обозначает легкую цепь.
Таблица 12. Оптимизация условий восстановления MORAb-003
мкМ (мг/мл)
мкМ
мкМ
мкМ
мкМ
мкМ
[00281] Анализ ДСН-ПААГ-электрофореза и содержания тиола показал, что 60-минутная инкубация 5,3 мг/мл мАт при 4-кратном молярном соотношении TCEP к мАт обеспечивала удовлетворительное исходное положение, поскольку, как представляется, присутствовало ограниченное восстановление внутримолекулярных дисульфидов (что определено по содержанию свободного тиола) и оставалось очень мало невосстановленных мАт (невосстановленное мАт будет действовать как конкурентный ингибитор в исследованиях in vitro и in vivo при применении приготовленных ADC). Дальнейшие исследования проводились с MORAb-003 при начальных концентрациях 5,0 мг/мл для подтверждения даного оптимизированного молярного соотношения TCEP к мАт с применением анализа ДСН-ПААГ-электрофореза (Фиг. 4). На Фиг. 4 дорожка 1 соответствует стандартному белку. Дорожка 2 соответствует необработанному, невосстановленному MORAb-003. Дорожка 3 соответствует MORAb-003, обработанному в соотношении 1:1 MORAb-003:TCEP. Дорожка 4 соответствует MORAb-003, обработанному в соотношении 1:2 MORAb-003:TCEP. Дорожка 5 соответствует MORAb-003, обработанному в соотношении 1:3 MORAb-003:TCEP. Дорожка 6 соответствует MORAb-003, обработанному в соотношении 1:4 MORAb-003:TCEP. Также следом за восстановлением и удалением ТСЕР проводили конъюгацию с применением малеимидо-ПЭГ2-биотина для того, чтобы стимулировать конъюгацию цитотоксина для приготовления ADC. Анализ DAR проводили, применяя ЖХ-МС. Результаты этих исследований приведены в таблице 13.
Таблица 13. Оптимизация условий восстановления MORAb-003 - уровни конъюгации с малеимидо-ПЭГ2-биотином
LC, уровень биотина легкой цепи; HC, уровень биотина тяжелой цепи; DAR, биотин на мАт [DAR=2(LC)+2(HC)].
[00282] После конъюгации биотина анализ свободных тиолов показал, что в MORAb-003-биотине нет свободных тиолов. Это указывало на то, что после восстановления дисульфидных связей конъюгация обычно происходила как при образовании тиолов, так и в результате того, что любые неконъюгированные, восстановленные дисульфиды подвергались повторному окислению для переобразования дисульфидных связей. Конечными условиями, выбранными для восстановления для генерации ADC, были концентрация антитела 5,0 мг/мл, концентрация ТСЕР 110 мкМ и время инкубации 60 мин. Это приводит к мАт с DAR 4 после конъюгации.
2.1.3 Оптимизация конъюгации ADC
[00283] В качестве первого цитотоксина, применяемого для приготовления ADC, применяли криптофицин, который представляет собой гидрофобное соединение, эксперименты по оптимизации начальной конъюгации проводили с помощью "суррогатного" антитела к мезотелину человека, имеющего два непарных цистеина, доступных для конъюгации (по одному на легкую цепь) в определенных положениях. Это значительно облегчает анализ эффективности конъюгации с помощью масс-спектрометрии, поскольку нужно анализировать только легкую цепь. Затем проводили титрование пропиленгликоля во время конъюгации малеимидо-LL3-криптофицина с суррогатным антителом с последующим анализом эффективности конъюгации легкой цепи с помощью ЖХ-МС (таблица 14).
Таблица 14. Оптимизация концентрации пропиленгликоля в реакции конъюгации
Массы LC: неконъюгированная, 23536 Да; конъюгированная, 24367 Да.
[00284] 50% пропиленгликоль приводил к полному заполнению доступных сайтов и был выбран в качестве конечной концентрации для применения. После конъюгации не наблюдалось потери связывания мАт (данные не показаны), что указывает на то, что пропиленгликоль не оказывал вредного воздействия на антитело. Таким образом, выбранные конечные условия реакции конъюгации составляли 2,5 мг/мл конечной концентрации мАт, 6:1 молярное соотношение малеимидо-линкер-цитотоксин:мАт в 0,5Х DPBS (конечная концентрация после добавления пропиленгликоля), 0,5 мМ ЭДТК, 50% пропиленгликоля, pH 7,2, в течение 3,5-4 часов при комнатной температуре. В этих реакциях перед добавлением малеимидо-линкер-цитотоксина добавляют пропиленгликоль.
2.1.4 Приготовление ADC и биофизическая характеристика
[00285] Установленные условия восстановления и конъюгации, описанные в разделе 2. 1. 2, были применены для приготовления первых 10 ADC MORAb-003, перечисленных в таблице 15. Остальные ADC, за исключением M-MMAE и M-DM1, были приготовлены с помощью либо восстановленных дисульфидных мостиков, либо ограниченного использования лизина. M-MMAE и M-DM1 были приготовлены компанией Concortis Biosystems, Inc. и получены в конъюгированной форме.
[00286] Химия восстановленных дисульфидных мостиков строит мостики по свободным тиолам, образующимся в процессе частичного восстановления, давая один цитотоксин на восстановленный дисульфид. Согласно теории, антитело с DAR=4 будет иметь перестроенные дисульфидные мостики в H-L и шарнире, придавая ADC повышенную стабильность и однородность по сравнению с традиционными подходами конъюгации. Ограниченное использование лизина приводит к конъюгации очень ограниченного количества вычисленных 70+ гридрофильных лизинов, доступных на типичной молекуле человеческого IgG. Конъюгаты MORAb-003, приготовленные с применением данного способа, приводили к 2,0 DAR, что указывало на то, что на пару H-L был использован один лизин.
[00287] Все ADC очищали путем хроматографии с обессоливанием HiPrep 26/10 и составляли в DPBS. Анализ DAR проводили на всех приготовленных ADC с помощью ЖХ-МС и уровни агрегации определяли с помощью эксклюзионной ВЭЖХ. Результаты данных анализов DAR и агрегации перечислены в таблице 15 рядом с соответствующим ADC.
Таблица 15. Биофизический анализ ADC MORAb-003
[00288] Значения DAR для всех ADC находились в предварительно определенном диапазоне (DAR между 3 и 4). Уровни агрегатов для ADC на основе криптофицина были значительно выше желаемого (>10%), тогда как малеимидо-линкер-цитотоксины на основе эрибулина (VCP-эрибулин) и на основе майтансина (ER-001161318, ER-001161319 и M-MMAE) демонстрировали приемлемые уровни агрегатов. Исследование других органических сорастворителей проводили на реакциях конъюгации MORAb-003 с применением VCP-криптофицина. Испытанными сорастворителями были ДМСО (10%), N,N-диметилформамид (20%), N,N-диметилацетамид (20%) и N,N-диметилпропионамид (20%). Уровни агрегатов после конъюгации с применением этих сорастворителей были равными или превышали уровни для 50% пропиленгликоля.
[00289] Анализ невосстанавливающего ДСН-ПААГ-электрофореза был выполнен на подмножестве ADC (Фиг. 5). Поскольку DAR для всех этих ADC было определено как 4, считалось, что эти ADC должны мигрировать как интактный IgG ~ 160 кДа, так как дисульфидные мостики должны быть перестроены в обоих H-L и обоих шарнирах. Это подмножество ADC включает в себя M-MMAE (дорожка 2), M-DM1 (дорожка 3), M-0026 (дорожка 4), M-0260 (дорожка 5), M-0267 (дорожка 6), M-0272 (дорожка 7), M-0285 (дорожка 8), M-292 (дорожка 9), M-027-0381 (дорожка 10) и M-0384 (дорожка 11) (Фиг. 5). На Фиг. 5 дорожка 1 соответствует стандартному белку.
[00290] Из этого анализа ясно, что, для ADC, созданных способом химии восстановленных дисульфидных мостиков (дорожки 4-9, 11), в дополнение к интактному ADC, имеются значительные моновалентные виды H-L (80 кДа). Это указывает на то, что, в дополнение к образованию межцепочечных мостиков в шарнире, имеется значительное образование внутрицепочечных дисульфидных мостиков в шарнире. Анализ эксклюзионной ВЭЖХ показывает, что ADC мигрируют как один интактный IgG, что указывает на то, что для этих ADC с образованными внутрицепочечными H-H мостиками тяжелые цепи ассоциированы нековалентно в конечном ADC.
2.2 Анализ in vitro активности ADC MORAb-003
2.2.1 Цитотоксичность на клетках IGROV1 и SJSA-1
[00291] Активностьприготовленных ADC in vitro оценивали с помощью анализа окрашивания кристаллическим фиолетовым, как подробно описано в разделе 1. 4. 1.
[00292] Первоначальный скрининг всех ADC MORAb-003 выполняли на клетках IGROV1 (FRhi(+++)) и SJSA-1 (FRneg(-)). Клетки IGROV1 имеют происхождение эпителиальной карциномы яичников человека и экспрессируют высокие уровни рецептора фолиевой кислоты альфа (FR), целевого антигена MORAb-003. Клетки SJSA-1 представляют собой линию опухолевых клеток остеосаркомы человека, у которых отсутствует рецептор фолиевой кислоты альфа. Скрининг выбранных ADC также выполнялся на клетках CaOV3 (карцинома яичника человека, FRmed(++)), NCI-H2110 (немелкоклеточная карцинома легких человека, FRmed(++)) и/или OVCAR3 (карцинома яичника человека, FRmed(++)). Результаты этого скрининга приведены в таблице 16.
Таблица 16. Скрининг цитотоксичности (ИК50) ADC MORAb-003 на различных линиях опухолевых клеток
Все значения представлены ИК50 в нМ и являются средними значениями выполненных повторных экспериментов.
[00293] ADC VCP-эрибулин был эффективным (54 пМ) на клетках IGROV1 и оказывал низкое уничтожение клеток SJSA-1. Для этих клеточных линий ADC VCP-эрибулин демонстрировал более высокую активность и специфичность по сравнению с ADC с эквивалентными значениями DAR, такими как M-MMAE и M-DM1. ADC VCP-эрибулин также демонстрировал высокую цитотоксичность на дополнительных линиях опухолевых клеток, экспрессирующих FR, происходящих из яичника (CaOV3 и OVCAR3) и немелкоклеточной карциномы легких (NC-H2110).
[00294] ADC VCP-эрибулина, LL2-криптофицина, LL3-криптофицина, VCP-криптофицина, ER-001161318, ER-001161319 и ER-001159200 показали специфическую цитотоксичность (> 2-log специфичности) на клетках CaOV3 (FRmed(++)). Множество этих ADC проявили субнаномолярную активность. Конъюгаты криптофицина также демонстрировали высокие уровни активности (23 пМ - 86 пМ) на клетках IGROV1, но, за исключением VCP-криптофицина, также демонстрировали заметную цитотоксичность на клетках SJSA-1. Расщепляемые конъюгаты майтансина ER-001161318 и ER-001161319 проявляли промежуточную активность на клетках IGROV1 (0,26 нМ и 0,49 нМ) и незначительное нецелевое уничтожение клеток SJSA-1.
[00295] Все конъюгаты, полученные путем ограниченного использования лизина, не демонстрировали никакой специфичности и далее не оценивались. Все расщепляемые конъюгаты, приготовленные с помощью технологии восстановленных дисульфидных мостиков дуостаза-3 (M-0285), дуостатина-5 (M-0115) и дуостатина-14 (M-292 и M-0026) демонстрировали специфическую цитотоксичность на клеточной линии IGROV1, незначительную цитотоксичность на клеточной линии SJSA-1. Конъюгаты дуостатина-3 и дуостатина-5, производные ауристатина, имели немного более высокую активность, чем конъюгаты дуостатина-14, который является производным майтансина. Эффективности и специфичности были сопоставимы с контрольным конъюгатом M-MMAE, который применяет линкер Val-Cit-pAB (VCP), присоединенный к монометилу E. Все нерасщепляемые конъюгаты, приготовленные с помощью химии восстановленных дисульфидов, не обладали достаточной активностью или специфичностью, и далее не анализировались.
2.2.2. Цитотоксичность на клеточной линии рака яичника человека CaOV3, экспрессирующей рецептор фолиевой кислоты
[00296] Активность выбранных ADC MORAb-003 также определяли на линиях опухолевых клеток яичника человека OVCAR3 и CaOV3, а также на клеточной линии НМРЛ человека NCI-H2110 (таблица 16). На клеточной линии яичника человека CaOV3 конъюгаты криптофицина демонстрировали заметно более высокую активность, чем конъюгат VCP-эрибулин, в отличие от наблюдаемых на клетках IGROV1. Это может быть связано с более низким уровнем экспрессии рецептора фолиевой кислоты альфа в клетках CaOV3 по сравнению с IGROV1 или с более высокой активностью криптофицина в этих клетках по сравнению с эрибулином. Конъюгаты на основе майтансина ER-001161318, ER-001161319 и ER-001159200 все показывали эффективности подобные или более низкие, чем VCP-эрибулин.
2.3 Неспецифический цитолиз VCP-эрибулина, ER-001161318 и M-0285
[00297] Чтобы оценить активность неспецифического цитолиза, анализ был настроен с применением двух меченных клеточных линий. В этом анализе клетки IGROV1 (FRhi), меченные Nuclight™ Green, и HL-60 (FRneg), меченные Nuclight™ Red, совместно культивировали в разных соотношениях количества клеток и обрабатывали титрами ADC MORAb-003 VCP-эрибулином, ER-001161318 или M-0285. VCP-эрибулин представляет собой ADC, приготовленный на основе эрибулина, содержащий расщепляемый линкер малеимидо-ПЭГ2-Val-Cit-pAB, тогда как ER-001161318 представляет собой ADC, приготовленный на основе майтансина, содержащий расщепляемый линкер малеимидо-(CH2)5-Val-Cit-pAB и M-0285 представляет собой ADC, приготовленный на основе дуостатина, содержащий расщепляемый линкер ПЭГ-pAB. Цитотоксичность отслеживали с помощью системы визуализации клеток Incucyte Zoom®. Результаты данного анализа неспецифической цитотоксичности приведены в таблице 17 и на Фиг. 6A-C.
Таблица 17. Активность неспецифического цитолиза VCP-эрибулина на со-культуре FR-положительных и FR-отрицательных клеточных линий
(со-культура с IGROV-1)
(эрибулин)
[00298] Когда клетки HL-60 (FRneg) культивировали при соотношении 2:1 к клеткам IGROV1 (FRhi), обработка MORAb003-VCP-эрибулином приводила к 2-log увеличению уничтожения клеток HL-60 по сравнению с клетками HL-60 отдельно (таблица 17 и Фиг. 6A). Эти данные свидетельствуют о том, что клетки, отрицательные на наличие целевого рецептора фолиевой кислоты альфа (FR), более эффективно уничтожаются MORAb003-VCP-эрибулином при совместном культивировании с клетками, положительными на наличие целевого FR, упоминаются в данном документе как неспецифический цитолиз. Неспецифический цитолиз отличается от нецелевого уничтожения, которое определяется как самостоятельное уничтожение клеток, отрицательных на наличие цели, в отсутствие и независимо от совместного культивирования с клетками, положительными на наличие цели. Наблюдаемое увеличение неспецифического цитолиза также было почти идентично увеличению, наблюдаемому после обработки клеток HL-60 свободным эрибулином, что указывает на потенциальный механизм неспецифического эффекта. Не желая быть связанными соответствием какой-либо теории, авторы полагают, что MORAb003-VCP-эрибулин может быть расщеплен в или вблизи FR-позитивных клеток IGROV1, которые также подвергаются апоптозу и высвобождают свободный эрибулин в культуру. Высвобожденный цитотоксин может уничтожить FR-негативные клетки HL-60.
[00299] Напротив, для MORAb003-ER-001161318 наблюдался лишь незначительный сдвиг (Фиг. 6B) и не наблюдалось никакого сдвига для MORAb003-0285 (Фиг. 6C). Когда соотношение HL-60:IGROV1 было снижено с 2:1 до 1:2, для MORAb003-ER-001161318 наблюдалось заметное уничтожение клеток HL-60 по сравнению с клетками HL-60 отдельно, в то время как для MORAb003-0285 неспецифический эффект все еще оставался низким, хотя и детектируемым. Эти данные показывают, что на предмет неспецифического цитолиза оцененные ADC MORAb-003 могут ранжироваться как VCP-эрибулин > ER-001161318 > M-0285.
2.4 Анализ стабильности сыворотки
[00300] Учитывая длительный период полужизни ADC в кровотоке in vivo и потенциал токсичности, если цитотоксины высвобождаются в кровоток, ADC должны демонстрировать стабильность в сыворотке. ADC MORAb-003 VCP-эрибулин, ER-001161319 и M-0285 были предварительно инкубированы в сыворотке человека или мыши при 37°C в течение 48 часов, а затем оценены в анализе цитотоксичности на клетках IGROV1 и SJSA-1. ER-001161319 представляет собой ADC, созданный на основе майтансина, содержащий тот же расщепляемый линкер, что и VCP-эрибулин, малеимидо-ПЭГ2-Val-Cit-pAB. Контроли ФСБ и сыворотка были включены для коррекции любых эффектов сыворотки на выполнение анализа. Результаты этого исследования приведены в таблице 18.
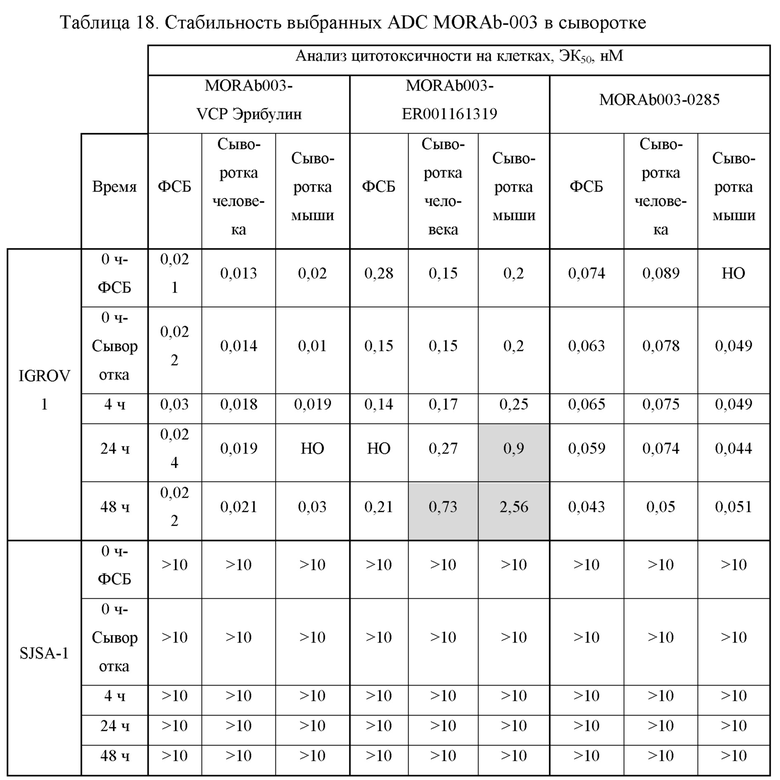
Заштрихованные ящики указывают на значительное снижение активности сравнительно образца Т=0.
В то время, как VCP-эрибулин и M-0285 были стабильными в течение по меньшей мере 48 часов в обеих сыворотках, ER-001161319 демонстрировал значительное снижение активности через 48 часов. Это может быть связано с азиридин-карбаматной связью с майтансином, которая ранее не была описана в литературе. Форма высвобождаемого соединения может не обладать высокой эффективностью, поскольку не наблюдалось увеличения цитотоксичности на клетках SJSA-1.
2.5 Исследования in vitro с MORAb003-VCP-эрибулином
2.5.1 HIC-ВЭЖХ анализ DAR и гетерогенности продукта
[00301] MORAb003-VCP-эрибулин и MORAb003-0285 анализировали с помощью HIC-ВЭЖХ с целью оценки DAR альтернативным способом и изучения гетерогенности продукта, а также содержания неконъюгированного антитела (конкурента). Показано, что MORAb003-VCP-эрибулин имеет виды DAR 0, 2, 4 и 6, что предусмотрено способом, применяемым для восстановления и конъюгации (Фиг. 7A). Наблюдалось очень небольшое количество видов с DAR=0. Общий DAR, основанный на расчетах AUC, составил 3,80, что соответствует значениям, определенным ЖХ-МС. На HIC-ВЭЖХ MORAb003-0285 мигрировал как единственный пик, что указывает на один вид DAR (Фиг. 7B). Это было определено как DAR 4. 0.
2.5.2 Специфичность по конкурентному анализу
[00302] Антигенная специфичность цитотоксичности MORAb-003-VCP-эрибулина была продемонстрирована для конъюгата VCP-эрибулин с помощью формата конкурентного анализа (Фиг. 8). В этом эксперименте титры MORAb-003-VCP-эрибулина (начальная концентрация 100 нМ) совместно инкубировали с 2 мкМ неконъюгированного MORAb-003. Неконъюгированный MORAb-003 обеспечивал 2-log сдвиг активности на клетках IGROV1, аналогичный результатам, полученным с IMGN853, ADC к человеческому рецептору фолиевой кислоты альфа-майтансину от Immunogen, находящемуся в данный момент на II-й фазе клинических испытаний, на клетках KB (Moore и соавт., 2015 Американское общество клинической онкологии (ASCO - American Society of Clinical Oncology) ежегодное собрание, Реферат 5518).
2.5.3 Цитотоксичность на клетках НМРЛ NCI-H2110
[00303] Цитотоксичность на клеточной линии НМРЛ человека NCI-H2110, как для MORAb003-VCP-эрибулина, так и для MORAb003-0285, определяли с помощью анализа окрашивания кристаллическим фиолетовым. Результаты этого анализа приведены в таблице 16. MORAb003-VCP-эрибулин имел ИК50 0,73 нМ, тогда как MORAb003-0285 имел ИК50 14 нМ.
2.6 In vivo исследования
2.6.1 Максимальная переносимая доза (МПД) MORAb-003-VCP-эрибулина (MORAb-202) на линии мышей CD-1
[00304] Мышам CD-1, ранее не получавшим лечения, вводили внутривенно 200 мкл MORAb-202 в соответствии со схемой из таблицы 19. Вес тела измеряли в день дозирования до ввода дозы, через 24 часа после дозы и три раза в неделю в последующее время. Животных наблюдали для клинического хорошего самочувствия на протяжении всей продолжительности исследования. Через две недели после дозирования измеряли и регистрировали конечную массу тела. В конце исследования мыши, подверженные эвтаназии, (а также если какая-либо мышь была подвергнута эвтаназии или найдена мертвой во время исследования) были обработаны для вскрытия. Органы были обследованы на признаки повреждения тканей.
Таблица 19. План исследования
[00305] По сравнению с контрольной группой, получавшей ФСБ, ни в одной из групп лечения не наблюдалось значимой потери массы тела, или же любых результатов клинических исследований, свидетельствующих о токсичности во время лечения. Масса тела отдельных мышей приведена в таблице 20, а среднее по группе и СОС приведены в таблице 21. Кинетика изменения массы тела для каждой группы (среднее значение по группе и СОС) изображены на Фиг. 9. MORAb-202 при дозах до 80 мг/кг через внутривенное введение болюса не вызывало токсичности. Следовательно, МПД выше 80 мг/кг.
Таблица 20
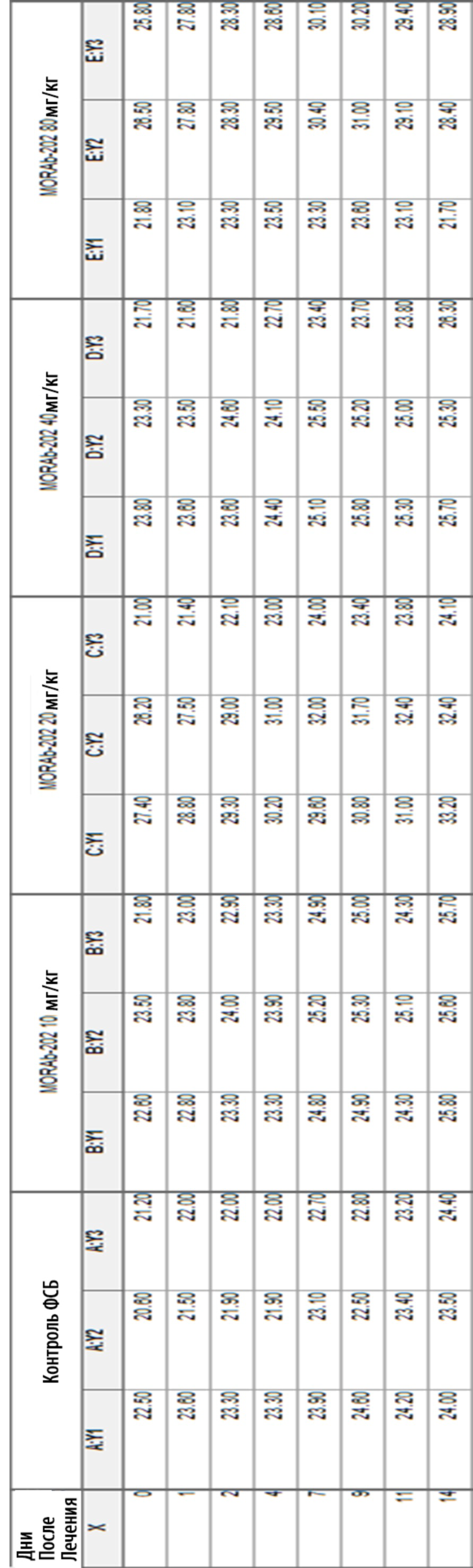
Таблица 21
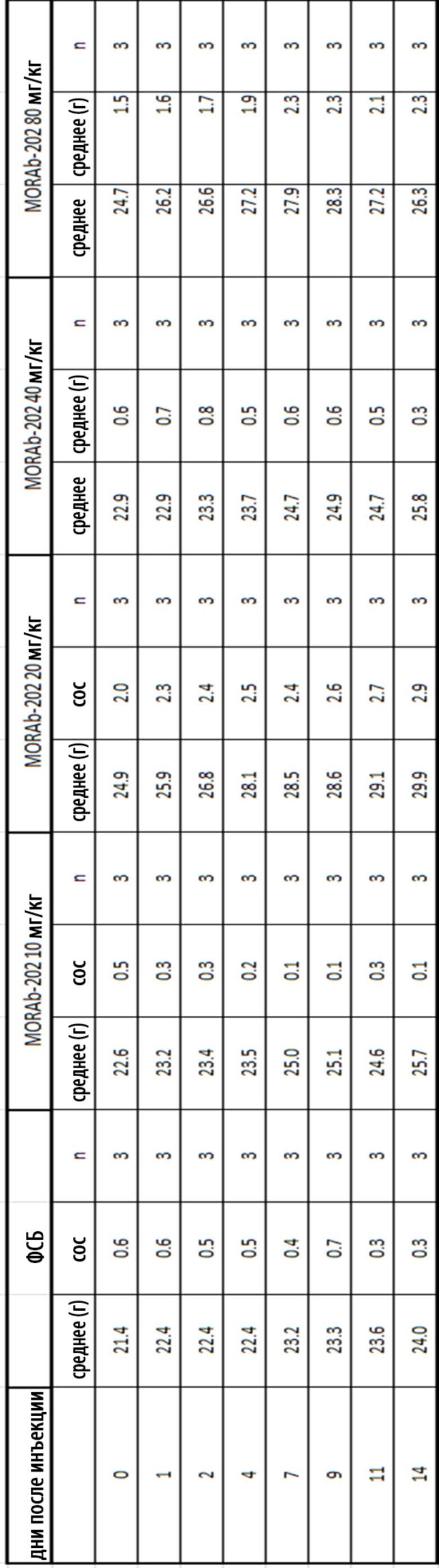
2.6.2 Максимальная переносимая доза эрибулина у мышей CD-1
[00306] Мышам CD-1, ранее не получавшим лечения, вводили внутривенно 200 мкл эрибулина в соответствии со схемой из таблицы 22. Вес тела измеряли три раза в неделю, включая измерение перед введением дозы в каждый день дозирования и через 24 часа после каждой дозы. Животных наблюдали для клинического хорошего самочувствия на протяжении всей продолжительности исследования (две недели после последней дозы). Измеряли и регистрировали конечную массу тела. В конце исследования мыши, подверженные эвтаназии, (а также если какая-либо мышь была подвергнута эвтаназии или найдена мертвой во время исследования) были обработаны для вскрытия. Органы были обследованы на признаки повреждения тканей.
Таблица 22. План исследования
[00307] У животных, которым вводили эрибулин в дозах до 1,6 мг/кг, с применением режима дозирования q4dx3 (один раз каждые четыре дня, всего 3 дозы), не наблюдали никакой значимой потери веса тела или результатов клинических исследований, свидетельствующих о токсичности. Введение 3,2 мг/кг по той же схеме вызывало у всех трех мышей пилоэрекцию после второй дозы. Наблюдалась серьезная потеря веса (потеря 23% у одной мыши, #552, после второй дозы, 17% и 8% у остальных, #551 и #552, после третьей дозы) по сравнению с контролем ФСБ. Макроскопических изменений в органах мышей при вскрытии не наблюдалось. Масса тела отдельных мышей приведена в таблице 23, а среднее по группе и СОС приведены в таблице 24. Кинетика изменения массы тела для каждой группы (среднее по группе и СОС) изображены на Фиг. 10.
[00308] Эрибулин при дозах до 1,6 мг/кг с применением режима дозирования q4dx3 не вызывал токсичности, тогда как 3,2 мг/кг вызывало серьезную потерю веса. Таким образом, МПД эрибулина в этом исследовании составляет 1,6 мг/кг, q4dx3.
Таблица 23

Каждый столбец отображает отдельное животное.
*9003: подвергали эвтаназии при потере веса > 20%.
Таблица 24
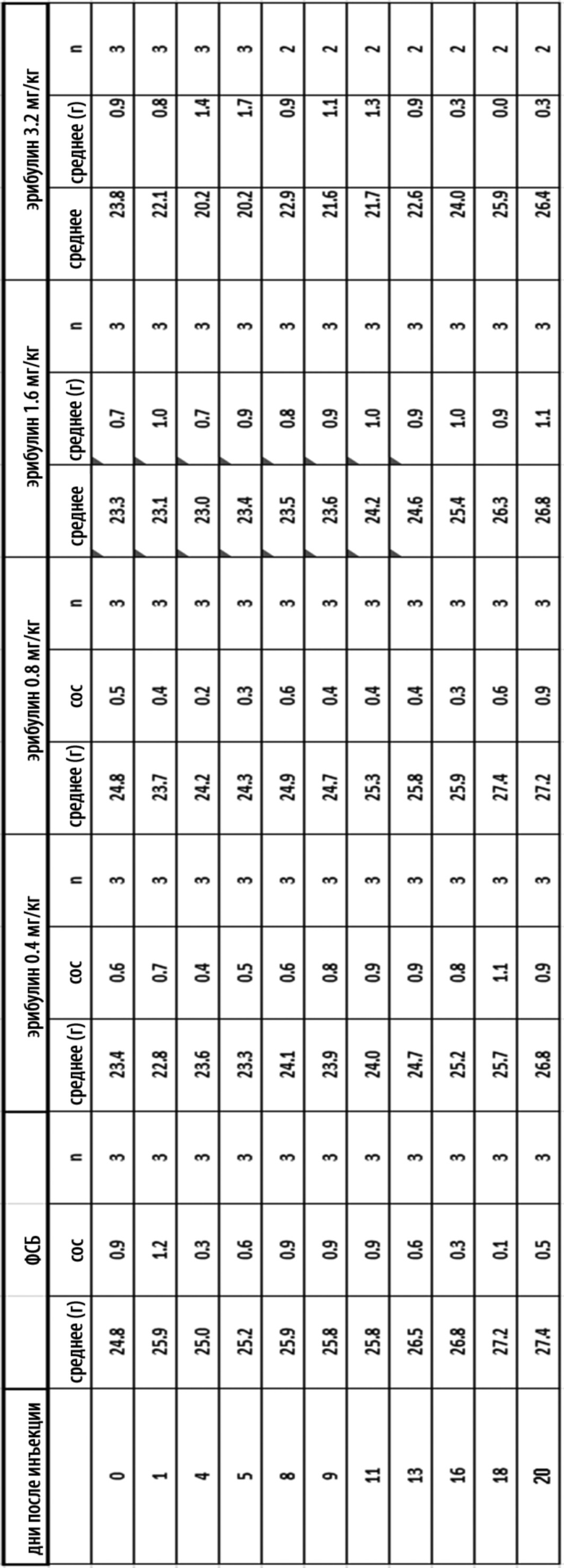
2.6.3 Оценка минимальной эффективной дозы MORAb003-VCP-эрибулина
(MORAb-202) на модели НМРЛ человека NCI-H2110 у мышей CB17-SCID
[00309] НМРЛ человека, клетки NCI-H2110, пассаж 47, имплантировали подкожно 30 мышам CB17 SCID (самка, в возрасте от 5 до 6 недель, весом 20 грамм). Через 14 дней после имплантации мышей распределяли случайным образом на пять групп. Средний объем опухоли в каждой группе в день лечения (день 0) варьировал от 154 до 175 мм3 (таблица 27). Мышей, включенных в исследование, лечили MORAb003-VCP-эрибулином (MORAb-202) (партия# NB2900-87E 10/07/15) при 1, 2,5 или 5 мг/кг, MORAb-003-0285 (партия# 042-150-002) в качестве контроля при 5 мг/кг, или ФСБ, в соответствии с планом исследования (таблица 25). Каждая группа была исключена из исследования, когда объем опухоли у любого животного в группе составлял >2000 мм3. Последняя группа была прервана на 61-й день.
Таблица 25. План исследования
1 Четыре мыши в этой группе. Одна мышь была исключена из этой группы из-за ошибки введения препарата, которая была подтверждена отсутствием соединения в сыворотке животного на основе данных электрохемилюминесцентного иммуноанализа (ЭХЛА).
[00310] Объемы опухолей отдельных мышей приведены в таблице 26, а среднее по группе и СОС приведены в таблице 27. Кинетика роста опухоли для каждой группы (среднее по группе и стандартная ошибка среднего, СОС) приведены на Фиг. 11, а объемы опухолей у отдельных мышей, а также среднее по группе и СОС приведены на Фиг. 12. Основываясь на объемах опухолей на 17-й день (когда наблюдался первый объем опухоли >2000 мм3), MORAb-202 вызывал ингибирование роста опухоли (TGI - tumor growth inhibition) на 47% при 1 мг/кг (p=0,002 по сравнению с физиологическим раствором), 96% TGI при 2,5 мг/кг (p < 0,0001 по сравнению с физиологическим раствором). Однако, регрессированные опухоли восстанавливаются через одну-две недели после окончания лечения. У мышей, обработанных 5 мг/кг MORAb-202, опухоль не обнаружена. Эти мыши оставались без опухоли свыше 60-ти дней после лечения однократной дозой. MORAb-003-0285 вызывало 89,7% TGI при 5 мг/кг (p < 0,0001 по сравнению с физиологическим раствором).
[00311] Масса тела отдельных мышей приведена в таблице 28, а среднее по группе и СОС приведены в таблице 29. Кинетика изменения массы тела для каждой группы (среднее по группе и СОС) изображены на Фиг. 13.
[00312] По сравнению с контролем, ни в одной из групп лечения не наблюдалось значимой потери массы тела.
[00313] MORAb-202 оказал значимый эффект на рост опухоли NCI-H2110. Регрессия опухоли была достигнута путем болюсного лечения при 2,5 мг/кг с 94% TGI (по сравнению с ФСБ). Следовательно, минимальная эффективная доза MORAb-202, испытанная на этой модели, составляет 2,5 мг/кг. Полное уничтожение опухоли было достигнуто однократной дозой 5 мг/кг. Рост опухоли не наблюдался более 60-ти дней.
Таблица 26. Объемы опухолей
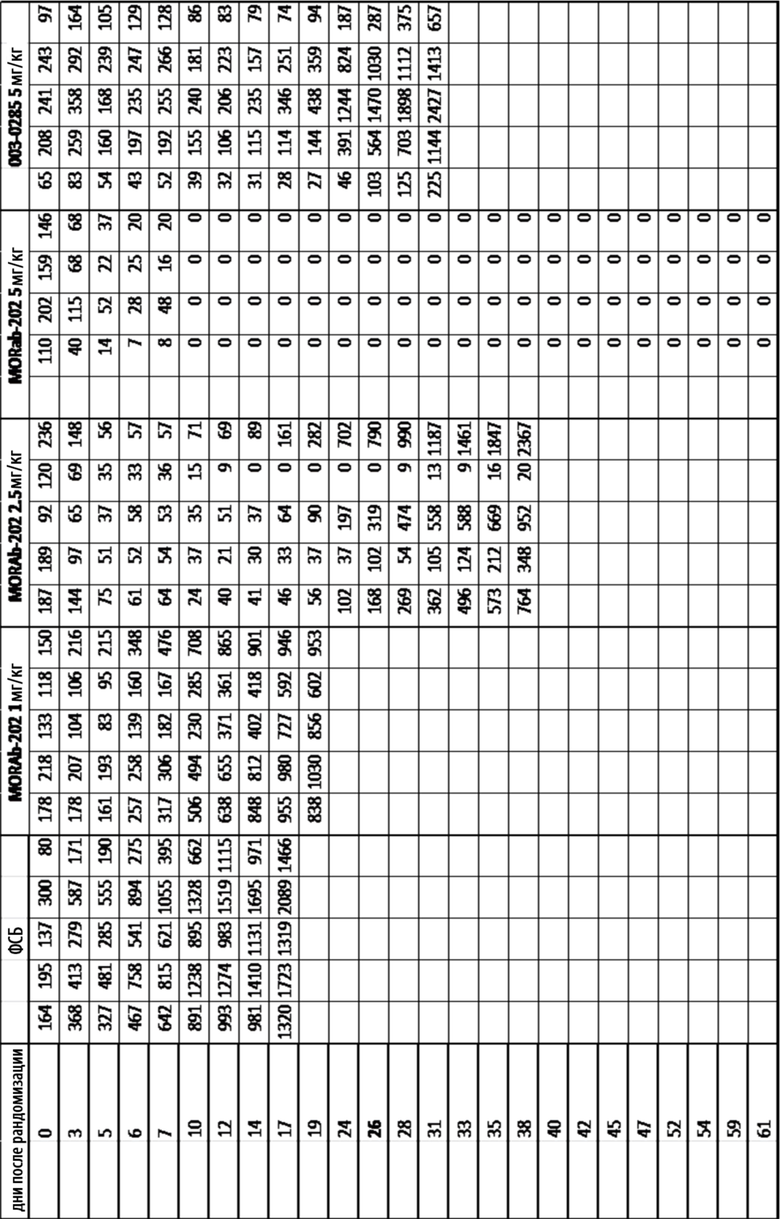
Каждый столбец отображает отдельное животное.
Таблица 27
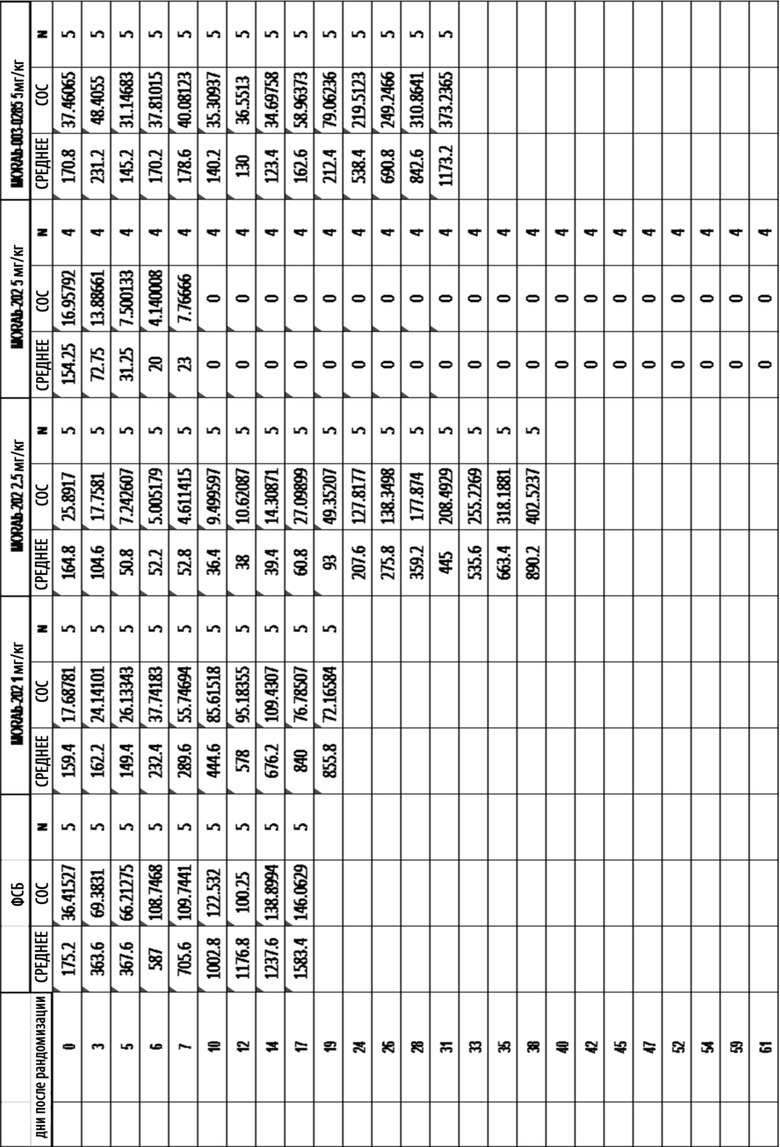
Таблица 28
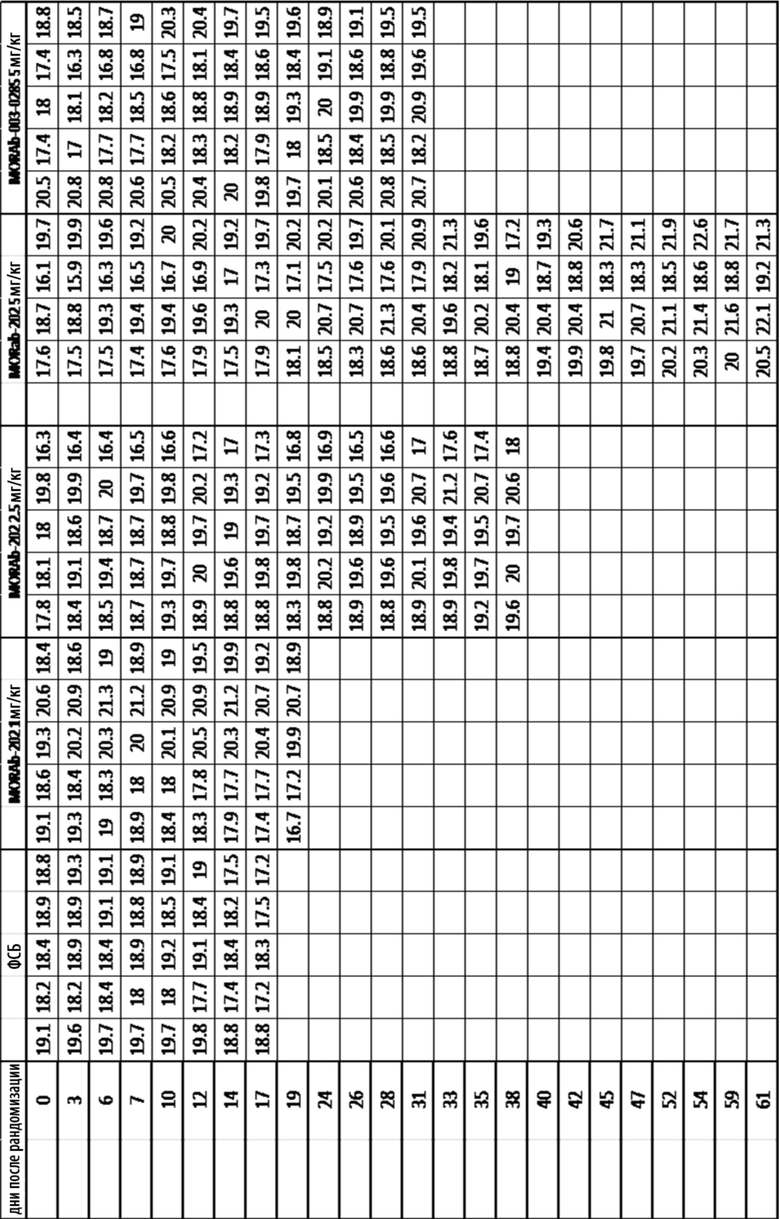
Таблица 29
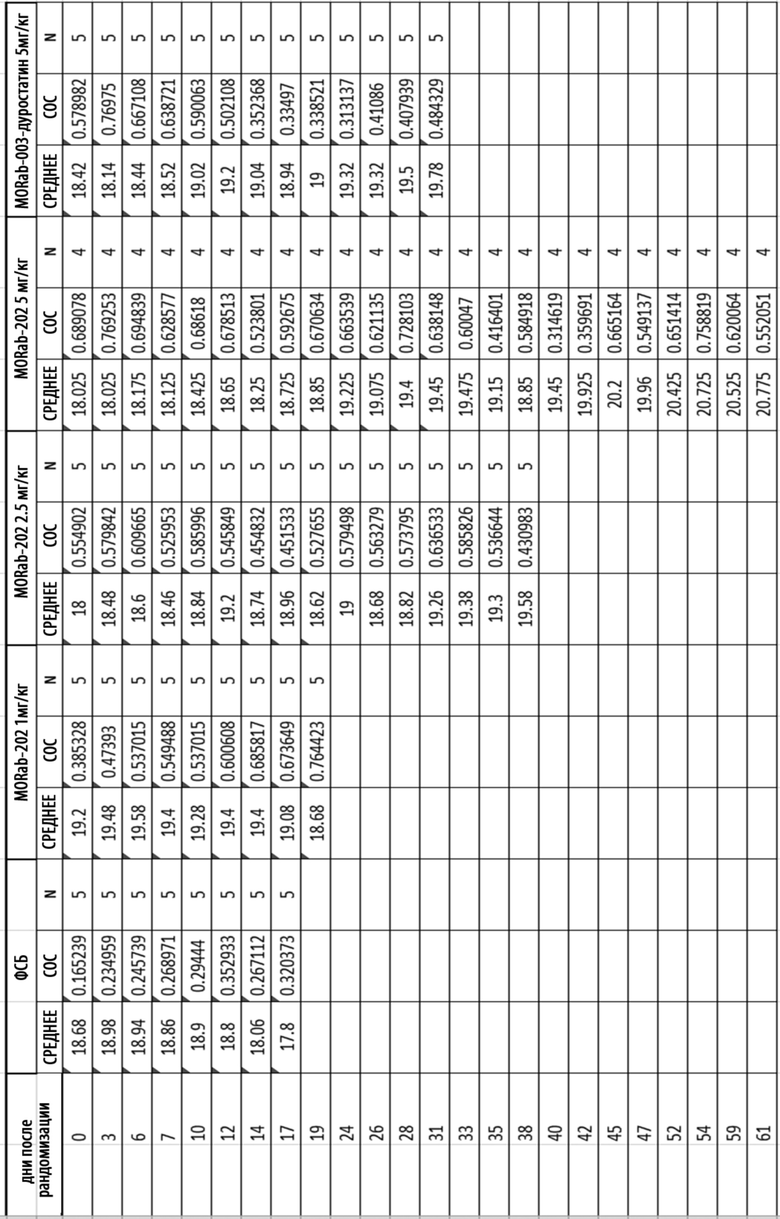
2.6.4 Оценка минимальной эффективной дозы эрибулина на модели НМРЛ человека NCI-H2110 у мышей CB17-SCID
[00314] НМРЛ человека, клетки H2110, пассаж 46, имплантировали подкожно 30-ти мышам CB17 SCID (самка, в возрасте от 5 до 6 недель, весом 20 грамм). Через 11 дней после имплантации мышей распределяли случайным образом на пять групп. Пять животных с объемами опухолей, наиболее отклоняющимися от среднего, были исключены. Средний объем опухоли в каждой группе в день лечения (день 0) варьировал от 87,6 до 89,4 мм3 (таблица 32). Мышей, включенных в исследование, лечили эрибулином (партия# N1201193) при 0,05, 0,2, 0,8 или 1,6 мг/кг, либо ФСБ, в соответствии с планом исследования (таблица 30). Каждая группа была исключена из исследования, соответственно, когда внутри группы впервые наблюдали объем опухоли >2000 мм3. Исследование было прекращено на 38-й день (30 дней после последней дозы).
Таблица 30 План исследования
[00315] Объемы опухолей отдельных мышей приведены в таблице 31, а среднее по группе и СОС приведены в таблице 32. Кинетика роста опухоли для каждой группы (среднее по группе и СОС) изображена на Фиг. 14, а объемы опухолей у отдельных мышей, а также среднее по группе и СОС на 24-й день (когда у мышей, обработанных ФСБ, наблюдали объем опухоли >2000 мм3), изображены на Фиг. 15. Эрибулин вызывал 50,5% TGI (без наблюдаемой регрессии опухоли) при 0,05 мг/кг (p=0,0026 по сравнению с физиологическим раствором); ~ 99% TGI при 0,2, 0,8 или 1. 6 мг/кг (p значение составляло < 0,0001 для всех 3 групп по сравнению с физиологическим раствором). Минимальная эффективная доза, которая вызывала регрессию опухоли, составляет 0,2 мг/кг. Однако большинство регрессированных опухолей у этих мышей (3/5 в группе 0,2 мг/кг, 4/5 в группе 0,8 мг/кг и 2/5 в группе 1,6 мг/кг) вновь выросли или оставались измеримыми на протяжении всего исследования (30 дней после последней дозы).
[00316] Масса тела отдельных мышей приведена в таблице 33, а среднее по группе и СОС приведены в таблице 34. Кинетика изменения массы тела для каждой группы (среднее по группе и СОС) изображены на Фиг. 16.
[00317] По сравнению с контрольной группой, получавшей физиологический раствор, ни в одной из групп лечения не наблюдалось значимой потери массы тела. Результаты клинических исследований, свидетельствующие о токсичности во время лечения, не наблюдались.
[00318] Эрибулин оказал значимый эффект на рост опухоли H2110 при введении в/в q4dX3 при 0,2 мг/кг. Регрессия опухоли была достигнута. Регрессия опухоли не была достигнута, когда вводили более низкую дозу (при 0,05 мг/кг). Следовательно, минимальная эффективная доза, испытанная в этом исследовании, составляет 0,2 мг/кг.
Таблица 31

Таблица 32
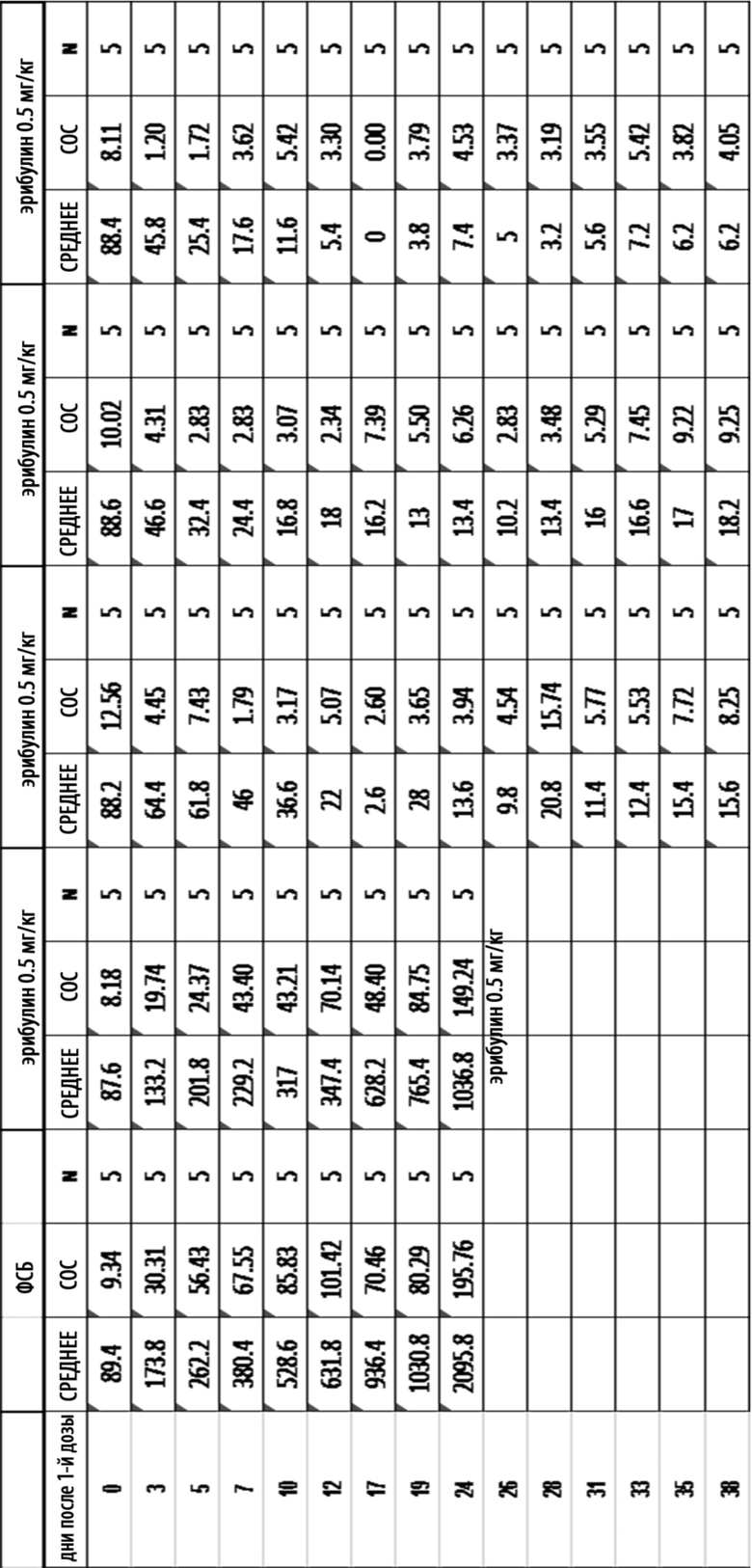
Таблица 33

Каждый столбец отображает отдельное животное.
Таблица 34
ПРИМЕР 2
1. Материалы и способы
[00319] MORAb003-VCP-эрибулин (MORAb-202) синтезировали путем конъюгации MORAb-003 (гуманизированного антитела к рецептору фолиевой кислоты альфа человека) с соединением MAL-ПЭГ2-Val-Cit-PAB-эрибулин (ER-001159569), описанным в разделе 1. 1 Примера 3. Способ конъюгации описан в разделе 1. 4. 1 Примера 4.
1.1 Опухолевые модели
[00320] Линии опухолевых клеток человека, применяемые в дополнительной оценке MORAb-202 in vitro, включают IGROV1 (карциному яичника человека, FRhi (+++)), OVCAR3 (карциному яичника человека, FRmed(++)), NCI-H2110 (немелкоклеточную карциному легких человека, FRmed(++)), A431-A3 (родительскую клеточную линияю A431, стабильно трансфицированную мезотелином человека, FRlo(+/-)), SJSA-1 (остеосаркому человека, FRneg(-)) и HL-60 (лейкоз человека, FRneg(-)). Все эти клеточные линии были получены непосредственно из Американской коллекции типовых культур (ATCC). Для исследований in vivo ксенотрансплантатные модели опухоли у мышей, полученные из пациентов с немелкоклеточным раком легкого, трижды негативным раком молочной железы и раком эндометрия, были созданы и поддерживались в Oncotest GmbH (Фрайбург, Германия), Oncodesign (Дижон, Франция) и EPO Berlin-Buch GmbH (Берлин, Германия), соответственно.
1.2 Анализы цитотоксичности in vitro
1.2.1 Анализ окрашивания кристаллическим фиолетовым
[00321] Клетки IGROV1 (FRhi (+++)), A431-A3 (FRlo(+/-)) и SJSA-1 (FRneg(-)) субкультивировали и высевали при 10000 клеток/лунку в готовую питательную среду в 96-луночных планшетах для культивирования тканей, инкубировали при 37°C, 5% CO2 в течение ночи (16 часов). Как правило, реагенты для испытания серийно разводили 1:4 в планшетах для разведения с 2-миллилитровыми глубокими лунками, начиная с 1 мкМ (всего 10 разведений). 100 мкл разбавленных образцов добавляли к клеточным планшетам (при начальной концентрации образцов для испытания 100 нМ). Планшеты инкубировали при 37°С, 5% CO2 в течение дополнительных 48 часов. Среду удаляли, планшеты промывали один раз 200 мкл DPBS, окрашивали 50 мкл 0,2% раствора кристаллического фиолетового при комнатной температуре в течение 15 мин и затем тщательно промывали водопроводной водой. Планшеты сушили на воздухе, а кристаллический фиолетовый растворяли с 200 мкл 1% раствора ДСН. Планшеты считывали при 570 нм. Данные анализировали с применением GraphPad Prism 6. Для OVCAR3 (FRmed(++)) и NCI-H2110 (FRmed(++)), клетки высевали при 3000 клеток/лунку и инкубировали в течение 5 дней с MORAb-202.
1.3 Исследования in vivo
1.3.1 Ксенотрансплантатная модель NCI-H2110
[00322] Подготовка животных: Мышей CB17 SCID (самка, в возрасте 6 недель) размещали по 5 мышей на вентилируемую клетку. Стерилизованные пищевые гранулы и бутылка с водой были доступны животным ad lib. Животных адаптировали в течение 5-7 дней до имплантации опухоли.
[00323] Культура клеток: Человеческие клетки NCI-H2110 размораживали из замороженной исходной культуры (NB2813-65) и культивировали в среде RPMI-1640, дополненной 10% фетальной бычьей сывороткой (ФБС) в 5% CO2 при 37°C. После двух пассажей при достижении примерно 70% конфлюэнтности клетки собирали с помощью раствора для диссоциации клеток, дважды промывали бессывороточной средой и подсчитывали.
[00324] Имплантация опухоли: Клеточную суспензию в бессывороточной среде смешивали 1:1 (об/об) с ледяным матригелем до конечной концентрации 1,0×108 клеток/мл. Каждой мыши вводили подкожно 100 мкл смеси при 1,0×107 клеток/мышь. Для всех инъекций применяли иглу 27G. Мышей наблюдали для клинического хорошего самочувствия и опухоли измеряли цифровым штангенциркулем три раза в неделю, начиная на 3-й день после имплантации. Объем опухоли (мм3) был рассчитан по формуле: Ш (мм) × Д (мм) × Г (мм) × π/6. Когда опухоли достигали ~100 мм3 (в среднем от >70 до ~130 мм3), животных распределяли случайным образом по 4-5 на группу. 5 животных с объемами опухолей, наиболее отклоняющимися от среднего, были исключены.
[00325] План исследования: Включенным в исследование подопытным мышам вводили внутривенно 200 мкл носителя или MORAb-202 при 1,0, 2,5 и 5 мг/кг в соответствии с планом исследования (таблица 35) в день случайного распределения. Вес тела измеряли до ввода дозы и два раза в неделю во время исследования. В конце исследования измеряли и регистрировали конечную массу тела. Животных подвергали эвтаназии, когда отдельный объем опухоли превышал 2000 мм3. Критерии досрочного прекращения исследования до достижения максимально допустимого объема опухоли:(1) язва опухоли более 50% опухоли (об/об); (2) паралич; (3) >20% потери массы тела; и (4) 50% животных в группе поддались исключению. Любую мышь, которая подвергалась эвтаназии или была найдена мертвой во время исследования, обрабатывали в соответствии с описанной выше процедурой исключения.
Таблица 35. План исследования
1.3.2 Ксенотрансплантатные модели, полученные из пациентов (PDx - patient-derived xenograft)
1.3.2.1 PDx модель немелкоклеточного рака легкого (НМРЛ): LXFA-737 (Oncotest)
[00326] Имплантация опухоли: Фрагменты опухоли НМРЛ были получены из опухолевых ксенотрансплантатов LXFA-737, серийно перевиваемые голым мышам. После удаления из донорных мышей опухоли разрезали на фрагменты (длина кромок 3-4 мм) и помещали в фосфатно-солевой буфер (ФСБ), содержащий 10% пенициллина/стрептомицина. Животных-реципиентов анестезировали путем ингаляции изофлурана и подкожно имплантировали полученную одностороннюю или двустороннюю опухоль в бок. Опухолевые ксенотрансплантаты были имплантированы одной или двумя опухолями на мышь с частотой приживания < 65%. В случае двустороннего взятия одна из этих опухолей была эксплантирована до случайного распределения. Животных и опухолевые имплантаты ежедневно наблюдали, пока рост солидной опухоли не был обнаружен у достаточного количества животных. При случайном распределении был определен объем растущих опухолей. Животные, отвечающие критериям случайного распределения (то есть имеющие опухоли 50-250 мм3, предпочтительно 80-200 мм3), были распределены на экспериментальные группы, состоящие из 5-6 животных на группу, направленные на сопоставимые медианные и средние объемы опухоли группы приблизительно 100-120 мм3. Животные, не применяемые для экспериментов, подвергались эвтаназии. День случайного распределения был обозначен как 0-вой день эксперимента.
[00327] План исследования: Включенным в исследование подопытным мышам вводили внутривенно носитель, MORAb-003 при 5 мг/кг или MORAb-202 при 5 мг/кг, в соответствии с планом исследования (таблица 36) в день случайного распределения. Вес тела измеряли в каждый день дозирования до ввода дозы и два раза в неделю во время исследования. В конце исследования измеряли и регистрировали конечную массу тела. Животных подвергали эвтаназии, когда отдельный объем опухоли превышал 2000 мм3.
Таблица 36. План исследования
1.3.2.2 PDx модель трижды негативного рака молочной железы (ТНРМЖ): OD-BRE-0631 (Oncodesign)
[00328] Имплантация опухоли: Девяти самкам мышей SWISS nude вводили подкожно в правый бок фрагменты ТНРМЖ опухоли, полученные из пациентов. Мышей-опухоленосителей подвергали эвтаназии, когда объем опухоли достигал 500-1000 мм3, а опухоли вырезали хирургическим путем. Фрагменты опухолей (30-50 мг) ортотопически имплантировали в область жировой ткани молочной железы у 34 самок мышей SWISS nude 24 до 72 часа после облучения всего тела гамма-источником (2 Гр, 60Co, BioMEP, Франция). Когда опухоли достигли среднего объема 200-300 мм3, 24 из 34 животных были распределяли случайным образом на две группы (n=12 животных) в соответствии с их индивидуальным объемом опухоли, применяя программное обеспечение Vivo Manager® (Biosystemes, Кутернон, Франция). Был проведен статистический тест (анализ дисперсии) для оценки однородности между группами. День случайного распределения был обозначен как 0-вой день эксперимента.
[00329] План исследования: На 1-й день отбирали (через один день после случайного распределения и за два дня до лечения) 3 мыши из каждой из двух групп не получавших лечение. Оставшимся подопытным мышам на 3-й день вводили внутривенно носитель или MORAb-202 при 5 мг/кг, в соответствии с планом исследования (Таблица 37). На 8-й день (через пять дней после лечения) 3 мыши из каждой из двух обработанных групп были отобраны. Сразу же после окончания опухолевую ткань собирали и фиксировали в 4% забуференном нейтральном формалине от 24 до 48 часов, а затем заключали в парафин (Histosec®, Merck, Дармштадт, Германия). Заключенный в парафин образец хранили при комнатной температуре для последующего иммуногистохимического анализа.
Таблица 37. План исследования
[00330] Иммуногистохимический анализ (ИГХ): Было проведено ИГХ окрашивание фиксированных в формалине, заключенных в парафин опухолевых тканей для оценки как расположения MORAb-202, так и экспрессии ассоциированных с раком фибробластов. Перед окрашиванием слайды депарафинизировали и антиген извлекали в Lab Vision™ PT Module (Thermo Scientific) в цитратном буфере (pH 6,0), предварительно нагретом до 85°C, применяя следующую программу:нагреть до 97°C; инкубировать при 97°С в течение 30 мин; и охладить до 60°C. Затем слайды переносили в бидистиллированную воду при комнатной температуре на 5 мин. Окрашивание проводили в Lab Vision™ Autostainer 360 (Thermo Scientific). Вкратце, слайды дважды промывали в 1Х Трис-буферизированном физрастворе/Твин-20 (TBST - Tris-buffered saline/Tween-20) в течение 6 мин/промывка. Затем тканевые срезы инкубировали в блокирующем буфере (300 мкл) (10% козьей сыворотки (Jackson Immunoresearch Laboratory Inc., Cat No. 005-000-121), разбавленной в 3% бычьем сывороточном альбумине (БСА) - фосфатно-солевом буфере (ФСБ)) в течение 1 часа, инкубировали с конъюгированным антителом (200 мкл) (таблица 38) в течение 1 часа и промывали пять раз в 1X TBST в течение 6 мин/промывка. Слайды докрашивали DAPI в заливочных средах и накрытые покровным стеклом слайды оставляли затвердеть на 30 мин. Слайды обрабатывались на сканере Panoramic MIDI (3DHISTECH), а ИГХ изображения анализировали с помощью программного обеспечения Halo (Indica Labs). Антитела, применяемые в этом анализе, нацелены на α-гладкомышечный актин (ГМА), который является специфическим маркером для ассоциированных с раком фибробластов, и человеческий IgG, который может выявлять присутствие MORAb-202.
Таблица 38. ИГХ антитела
1.3.2.3 PDx модели рака эндометрия: Endo-12961 и Endo-10590 (EPO Berlin)
[00331] Имплантация опухоли: Фрагмены опухоли рака эндометрия получали из серийно перевиваемых опухолевых ксенотрансплантатов Endo-12961 и Endo-10590 и хранили как исходную культуру в жидком азоте. Фрагменты опухоли имплантировали подкожно в левый бок 40 самок мышей NMRI nu/nu и наблюдали объем опухоли. Мышей с объемом опухоли 100-160 мм3 распределяли случайным образом в одну из четырех групп (группы A-D, Таблица 39). Спутниковые мыши для случайного распределения были включены в пятую группу (группа E, Таблица 39). Каждая группа состояла из 8 животных. День случайного распределения был обозначен как 0-вой день эксперимента.
[00332] План исследования: Включенным в исследование подопытным мышам вводили внутривенно ФСБ, эрибулин при 3,2 мг/кг или 0,1 мг/кг, или MORAb-202 при 5 мг/кг, в соответствии с планом исследования (Таблица 39) в день случайного распределения. Рост опухоли оценивали путем измерения двух перпендикулярных диаметров два раза в неделю, и рассчитывали объем опухоли (TV - tumor volume), относительный объем опухоли (RTV - relative tumor volume) и значения лечение/контроль (T/C - treated/control). Вес тела также оценивали два раза в неделю в качестве параметра на токсичность, с расчетом массы тела на группу и изменениями массы тела (BWC - body weight change) по сравнению с началом лечения. Животных умерщвляли, когда отдельный объем опухоли превышал 1 см3, или в конце исследования.
Таблица 39. План исследования
1.4 Механизм действия
1.4.1 Трехмерная (3D) система совместной культуры в zPredicta
[00333] Все эксперименты с 3D со-культурой, содержащей мезенхимальные стволовые клетки (МСК), проводились в zPredicta с применением органоспецифических 3D-систем внеклеточного матрикса, таких как rStomach™. Мезенхимальные стволовые клетки костного мозга (МСК КМ) в rStomach™ были совместно культивированы с клеточной линией рака желудка Nuc Red Light MKN-74 в четырех повторностях в 48-луночном формате в течение 12 дней. Ранее было показано, что клетки MKN-74 экспрессируют достаточное количество рецептора фолиевой кислоты альфа (FR) в ответ на обработку MORAb-202, чтобы индуцировать клеточный апоптоз. Перед культивированием, МСК КМ оценивали на экспрессию целевого антигена и на маркеры дифференциации МСК (Таблица 40) с помощью проточной цитометрии.
Таблица 40. Маркеры дифференциации МСК
[00334] Культуры rStomach™ обрабатывали либо MORAb-202, неконъюгированным антителом MORAb-003, эрибулином, или контролем, как приведено в Таблице 41. Контроль включал необработанные и обработанные носителем (ФСБ и ДМСО) культуры. Дифференциацию МСК наблюдали с помощью световой микроскопии. Как только наблюдалась видимая дифференциация, образцы собирали для окрашивания и анализа проточной цитометрией.
Таблица 41. Лечения со-культур
1.4.2 Анализ временной динамики воздействия MORAb-202 на ассоциированные с раком фибробласты
[00335] Мышей-опухоленосителей, с подкожным ксенотрансплантатом H2110, получали, как описано в разделе 1. 3. 1. Образцы опухолей собирали в дни 0, 3, 5, 7 и 9 после введения носителя или MORAb-202 при 5 мг/кг. Собранные образцы опухолей обрабатывали на слайдах, а экспрессию ассоциированных с раком фибробластов анализировали путем ИГХ, как описано в разделе 1. 3. 2. 2.
2. Результаты
2.1 Анализы цитотоксичности in vitro
2.1.1 Цитотоксичность MORAb-202
[00336] Активность MORAb-202 in vitro оценивали с помощью анализа окрашивания кристаллическим фиолетовым, как описано в разделе 1. 2. 1. Скрининг выполняли на клетках IGROV1 (FRhi (+++)), OVCAR3 (FRmed(++)), NCI-H2110 (FRmed(++)), A431-A3 (FRlo(+/-)) и SJSA-1 (FRneg(-)). Результаты этого скрининга представлены на Фиг. 17 и в Таблице 42.
Таблица 42. Скрининг цитотоксичности (ЭК50) MORAb-202 на различных линиях опухолевых клеток
(FR+++)
(FR++)
(FR++)
(FR+/-)
(FR-)
[00337] MORAb-202 проявлял цитотоксичность к линиям опухолевых клеток, зависящую от экспрессии рецептора фолиевой кислоты альфа, и низкие уровни нецелевого уничтожения. MORAb-202 продемонстрировал наивысший уровень активности (0,01 нМ) на клетках IGROV1 с небольшой цитотоксичностью (> 100 нМ) на клетках SJSA-1, не содержащих рецептор фолиевой кислоты альфа. Промежуточная активность наблюдалась на клетках OVCAR3 и NCI-H2110 (0,16 нМ и 0,74 нМ).
2.2 Исследования in vivo
2.2.1 Эффективность MORAb-202 на ксенотрансплантатной модели NC1-H2110
[00338] Мышам-опухоленосителям, с подкожным H2110, внутривенно вводили носитель или MORAb-202 при 1, 2,5 и 5 мг/кг. Значимая регрессия опухоли наблюдалась после однократной дозы MORAb-202 при 5 мг/кг (Фиг. 18 и Таблица 43). Показано, что применяя эту ксенотрансплантатную модель с высоким уровнем экспрессии рецептора фолиевой кислоты альфа и введениями однократной дозы, терапевтический диапазон для MORAb-202 составляет 1 мг/кг для задержки роста опухоли (со стабильным заболеванием) и ≥ 2,5 мг/кг для регрессии опухоли. В этом исследовании MORAb-202 в дозе 2,5 мг/кг приводил к частичному ответу, а MORAb-202 в дозе 5 мг/кг приводил к полному ответу.
Таблица 43. Противоопухолевая активность MORAb-202 на ксенотрансплантатной модели NC1-H2110
2.2.2 Эффективность MORAb-202 на PDx модели НМРЛ:LXFA-737
[00339] Мышам-опухоленосителям, с подкожным PDx НМРЛ, внутривенно вводили носитель, MORAb-003 при 5 мг/кг или MORAb-202 при 5 мг/ кг. Однократная доза MORAb-202 (5 мг/кг) приводила к значимой регрессии опухоли на этой модели, в отличие от однократной дозы неконъюгированного антитела MORAb-003 (5 мг/кг), которая не демонстрировала значимую противоопухолевую активность (Фиг. 19А). Пять из всех шести мышей, получавших MORAb-202, считались не имеющими опухолей на 32-й день исследования (Таблица 44), и четыре оставались без опухолей до 74-го дня (прекращение исследования). Дополнительно, в группе лечения не наблюдалась значимая потеря массы тела по сравнению с контрольной группой, получавшей носитель, что указывало на отсутствие токсичности во время лечения (Фиг. 19B).
Таблица 44. Противоопухолевая активность MORAb-202 на PDx модели НМРЛ
5 мг/кг, однократная доза (n=6)
5 мг/кг, однократная доза (n=6)
(4/6 без опухоли)
2.2.3 Относительная эффективность MORAb-202 и эрибулина на PDx моделях рака эндометрия: Endo-12961 и Endo-10590
[00340] Ксенотрансплантаты Endo-12961 и Endo-10590 экспрессируют высокие уровни рецептора фолиевой кислоты альфа. Мышам-опухоленосителям, с подкожным PDx раком эндометрия, вводили внутривенно ФСБ, эрибулин при 3,2 мг/кг или 0,1 мг/кг или MORAb-202 при 5 мг/кг. Максимальная переносимая доза (МПД) эрибулина на этой модели составляет 3,2 мг/кг, тогда как 0,1 мг/кг является эквивалентной дозе эрибулина, предоставленной MORAb-202, вводимым при 5 мг/кг. На протяжении начала исследования значимая противоопухолевая активность наблюдалась после лечения MORAb-202 (5 мг/кг) и дозой МПД эрибулина (3,2 мг/кг) на обеих животных моделях, в то время как никакой значимой противоопухолевой активности не наблюдалось после лечения эрибулином при 0,1 мг/кг (Фиг. 20А и 20С). Однако, регрессированные опухоли у мышей, получавших эрибулин при 3,2 мг/кг, начали вновь расти во время исследования, тогда как у мышей, получавших MORAb-202, не наблюдалось значимого повторного роста опухоли. В этом исследовании MORAb-202 оказался значительно более эффективным, чем эрибулин. Лечение эрибулином также временно влияло на массу тела в течение первой недели после лечения (Фиг. 20B и 20D). Напротив, у животных, получавших MORAb-202, не наблюдалась потеря массы тела.
2.3 Механизм действия MORAb-202
2.3.1 ИГХ и эффективность MORAb-202 на PDx модели ТНРМЖ: OD-BRE-0631
[00341] Мышам-опухоленосителям, с подкожным PDx ТНРМЖ, вводили внутривенно носитель или MORAb-202 при 5 мг/кг. У мышей в каждой группе собирали опухолевую ткань до начала лечения (День 1) и после лечения (День 8). Анализы ИГХ собранных опухолевых тканей показали, что, после введения на 3-й день однократной дозы (5 мг/кг), MORAb-202 располагается в опухолевых клетках, экспрессирующих рецептор фолиевой кислоты альфа, через пять дней после лечения (День 8). Расположение в клетках оценивали с применением антитела к IgG человека (Фиг. 21A). Было также показано, что лечение MORAb-202 приводит к уменьшению структуры ассоциированных с раком фибробластов, что показано путем ИГХ окрашивания с антителом к α-гладкомышечному актину (ГМА)-FITC (Фиг. 21B). С точки зрения эффективности, лечение MORAb-202 приводило к максимальной регрессии опухоли через 11 дней после лечения, измеренной по относительному объему опухоли (RTV) 0,62 (Фиг. 21C).
2.3.2 Влияние MORAb-202, MORAb-003 и эрибулина на систему 3D со-культуры
[00342] Мезенхимальные стволовые клетки костного мозга (МСК КМ) в rStomach™ (zPredicta) были совместно культивированы с линией клеток рака желудка MKN-74 в течение 12 дней. Перед культивированием, МСК КМ оценивали на экспрессию рецептора фолиевой кислоты альфа и на маркеры дифференциации МСК с помощью проточной цитометрии. Культуры rStomach™ обрабатывали либо MORAb-202, неконъюгированным антителом MORAb-003, эрибулином или контролем. Как только при помощи световой микроскопии наблюдалась видимая дифференциация МСК, образцы собирали для окрашивания и анализа проточной цитометрией. Результаты этих анализов изображены на Фиг. 22.
[00343] Общая продолжительность обработки 7 дней с дополнением обработки в течение этого периода была достаточной для того, чтобы произвести измеряемый эффект на дифференциацию МСК КМ человека в культуре с клетками MKN-74. Сравнительно к контролю носителя, обработка MORAb-202 (10 нМ) приводила к увеличению в популяциях МСК и адипоцитов и уменьшению в популяциях перицитов (Таблица 45). Эти данные показывают, что MORAb-202 может оказывать значительное влияние на микроокружение опухоли.
Таблица 45. Влияние MORAb-202, MORAb-003 и эрибулина на систему 3D со-культуры
2.3.3 Анализ временной динамики воздействия MORAb-202 на ассоциированные с раком фибробласты
[00344] Образцы опухолей собирали у мышей-опухоленосителей, с подкожным ксенотрансплантатом H2110, в дни 0, 3, 5, 7 и 9 после введения носителя или MORAb-202 при 5 мг/кг. Собранные образцы опухолей обрабатывали на слайдах, а экспрессию ассоциированных с раком фибробластов (CAF) анализировали путем ИГХ. Структура сети CAF, как оценивали и количественно определяли путем окрашивания с антителом к α-гладкомышечному актину (ГМА)-FITC, была видна на 3-й день и 5-й день после введения однократной дозы MORAb-202 при 5 мг/кг (Фиг. 23). Однако, к 7-му дню большая часть этой структуры значительно уменьшилась.
ПРИМЕР 3
1. Материалы и способы
[00345] Конъюгируемые соединения эрибулина имеющие структуры, приведенные в Таблице 46, синтезировали в соответствии со следующими процедурами и применяли в приготовлении ADC (Пример 4).
[00346] Все растворители, применяемые в реакциях синтеза, были в безводном виде (EMD Millipore). Все растворители, применяемые для клинического обследования или очистки, были степени чистоты для высокоэффективной жидкостной хроматографии (ВЭЖХ) (EMD Millipore). Все химические вещества были коммерчески доступными, если не указано иное. Колоночную хроматографию проводили с применением Biotage® SP4. Удаление растворителя проводили с применением либо роторного испарителя (Büchi Labortechik AG), либо центробежного испарителя (Genevac, SP scientific). Подготовительную жидкостную хроматографию-масс-спектрометрию (ЖХ-МС) проводили с применением Waters AutoPurification System и колонки XTerra MS C18 (5 мкм, 19 мм × 100 мм) в условиях кислой подвижной фазы. Спектры ядерного магнитного резонанса (ЯМР) были получены с применением дейтерированного хлороформа (CDCl3), если не указано иное, и зарегистрированы при 400 или 500 МГц с применением инструмента Varian (Agilent Technologies). Масс-спектры получали с применением сверхвысокоэффективной ЖХ-МС Waters Acquity. Применяемый в данном документе термин "инертный" относится к замещению воздуха в реакторе (например, реакционном резервуаре, колбе, стеклянном реакторе) с по существу не содержащим влаги инертным газом, таким как азот или аргон. Мультиплетности указаны с применением следующих сокращений: с=синглет, д=дублет, т=триплет, кв=квартет, пентет=квинтет, секстет=секстет, м=мультиплет, дд=дублет-дублетов, ддд=дублет-дублетов-дублетов, дт=дублет-триплетов, ш. с=широкий синглет.
Таблица 46. Конъюгируемые соединения эрибулина
MAL-ПЭГ2-Val-Cit-PAB-эрибулин
(ER-001159569)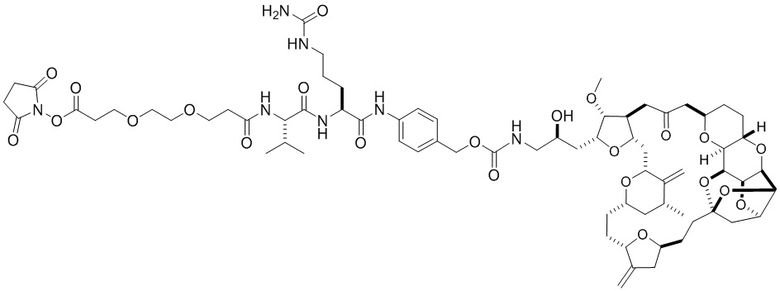
NHS-ПЭГ2-Val-Cit-PAB-эрибулин
(ER-001236940)
NHS-(CH2)5-Val-Cit-PAB-эрибулин
(ER-001236941)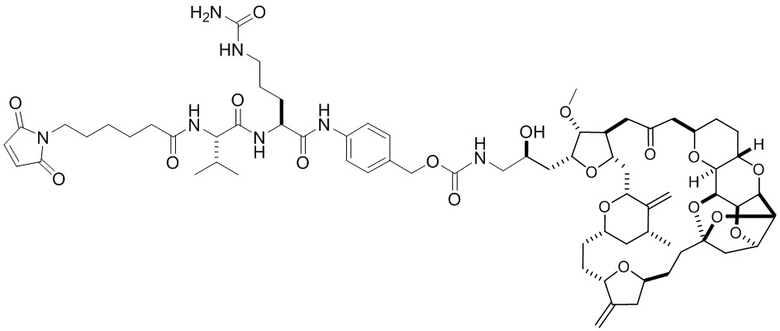
Mal-(CH2)5-Val-Cit-PAB-эрибулин
(ER-001235638)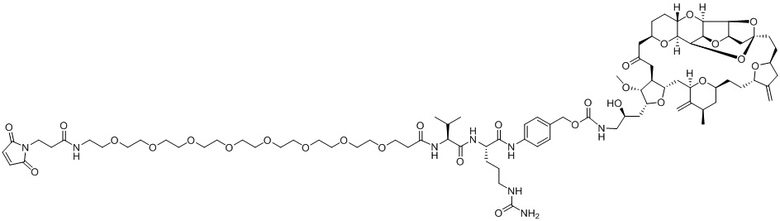
Mal-ПЭГ8-Val-Cit-PAB-эрибулин
(ER-001242287)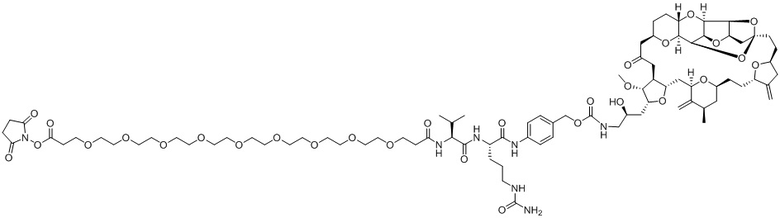
NHS-ПЭГ9-Val-Cit-PAB-эрибулин
(ER-001242288)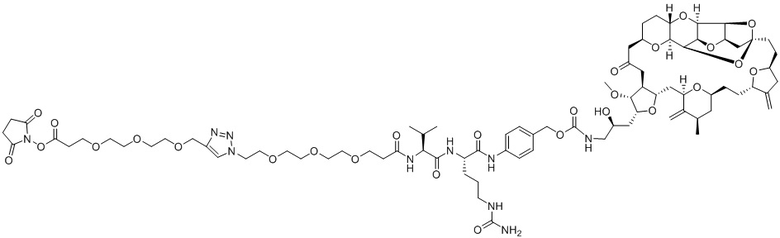
NHS-ПЭГ3-триазол-ПЭГ3-Val-Cit-PAB-эрибулин
(ER-001243700)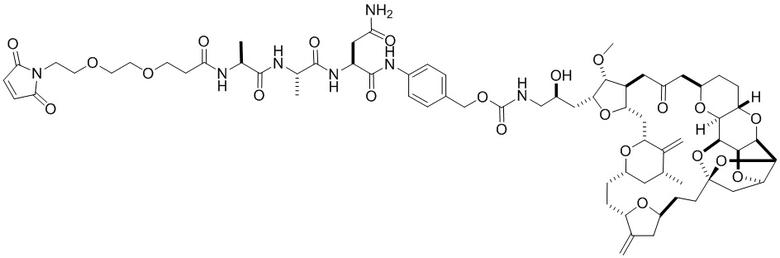
Mal-ПЭГ2-Ala-Ala-Asn-PAB-эрибулин
(ER-001231679)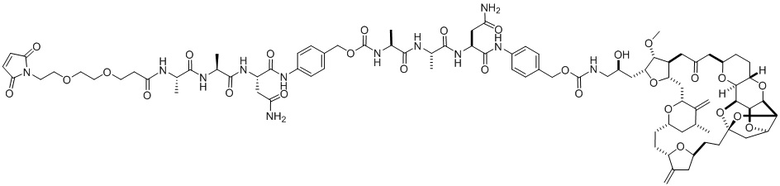
Mal-ПЭГ2-(Ala-Ala-Asn-PAB)2-эрибулин
(ER-001231690)
NHS-ПЭГ2-Ala-Ala-Asn-PAB-эрибулин
(ER-001231691)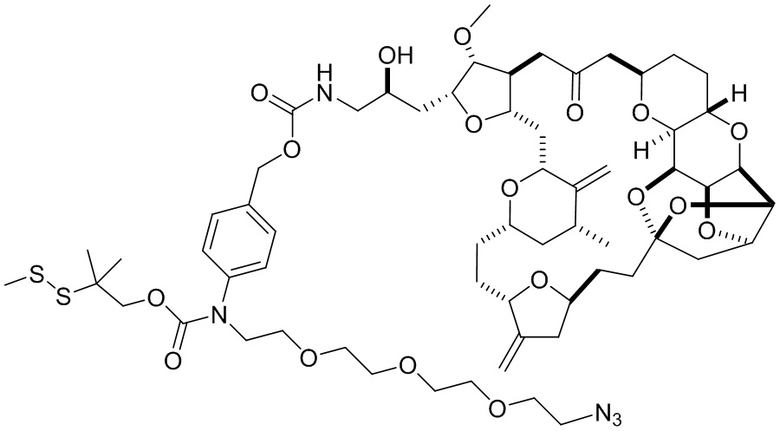
Азид-ПЭГ3-дисульфид-PAB-эрибулин
(ER-001237508)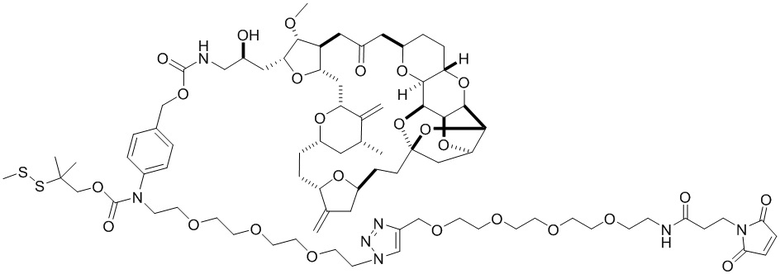
Mal-ПЭГ4-триазол-ПЭГ3-дисульфид-PAB-эрибулин
(ER-001237504)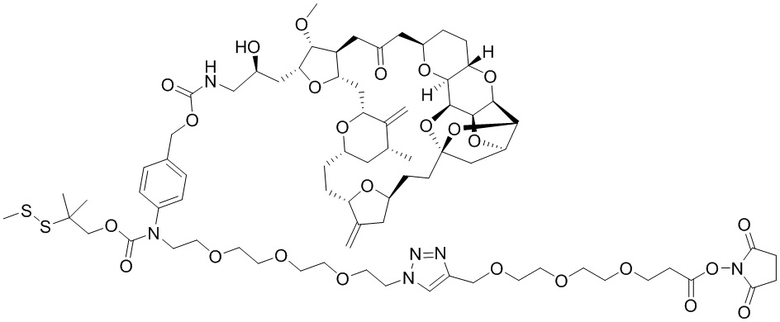
NHS-ПЭГ3-триазол-ПЭГ3-дисульфид-PAB-эрибулин
(ER-001244129)
Азид-ПЭГ3-сульфонамид-PAB-эрибулин
(ER-001138856)
Mal-ПЭГ4-триазол-ПЭГ3-сульфонамид-PAB-эрибулин
(ER-001237505)
NHS-ПЭГ3-триазол-ПЭГ3-сульфонамид-PAB-эрибулин
(ER-001244623)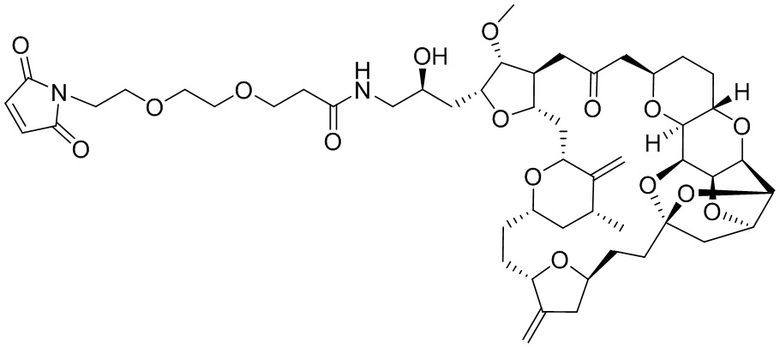
Mal-ПЭГ2-эрибулин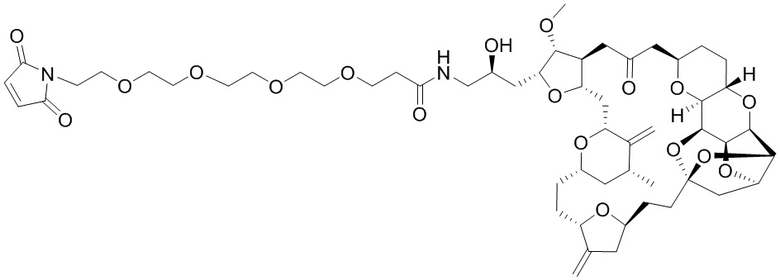
Mal-ПЭГ4-эрибулин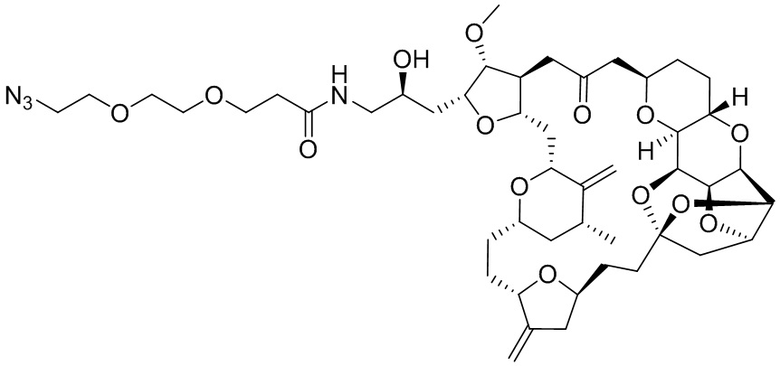
Азидо-ПЭГ2-эрибулин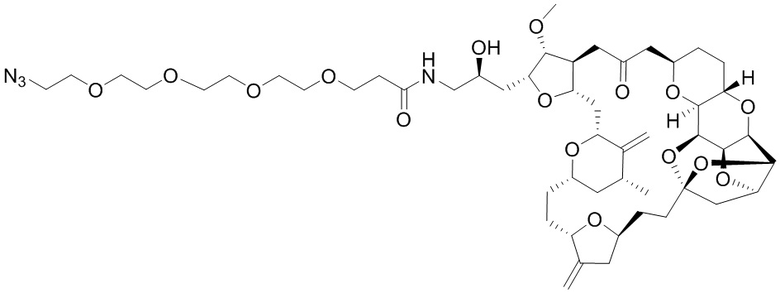
Азидо-ПЭГ4-эрибулин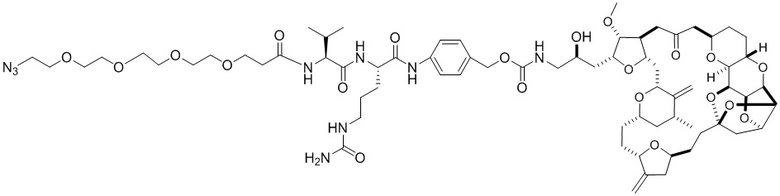 Азидо-ПЭГ4-Val-Cit-PAB-эрибулин
Азидо-ПЭГ4-Val-Cit-PAB-эрибулин
1. 1 Приготовление MAL-ПЭГ2-Val-Cit-PAB-эрибулина (ER-001159569)
[00347] Эрибулин (ER-000086526) (61,5 мг, 0,074 ммоль) расстворяли в N,N-диметилформамиде (ДМФА) (6,0 мл) и затем смешивали с основанием Хунига (0,027 мл, 0,156 ммоль) и Фмок-Val-Cit-PAB-PNP (86 мг, 0,112 ммоль). Реакционную смесь перемешивали при комнатной температуре в течение 18 часов, до тех пор, пока связывание не было завершено, что было определено с помощью анализа высокоэффективной жидкостной хроматографии (ВЭЖХ). К смеси добавляли диэтиламин (0,078 мл, 0,745 ммоль) и смесь перемешивали в течение дополнительных 2 часов до завершения реакции. Растворитель удаляли выпариванием и остаток очищали с помощью флэш-хроматографии для получения Val-Cit-PAB-эрибулина (ER-001228950) в виде белого твердого вещества (60 мг, выход 71%). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,56 (д, J=8,4 Гц, 2H), 7,32 (д, J=8,4 Гц, 2H), 5,14 (с, 1H), 5,06 (д, J=12,4 Гц, 1H), 5,03 (с, 1H), 5,01 (d, J=12,4 Гц, 1H), 4,87 (с, 1H), 4,83 (с, 1H), 4,71 (т, J=4,4 Гц, 1H), 4,62 (т, J=4,4 Гц, 1H), 4,57 (дд, J=4,8, 8,8 Гц, 1H), 4,47 (д, J=10,8 Гц, 1H), 4,32-4,27 (м, 2H), 4,18 (дд, J=4,8, 6,4 Гц, 1H), 4,13-4,07 (м, 2H), 3,98 (т, J=10,4 Гц, 1H), 3,88-3,82 (м, 3H), 3,76-3,70 (м, 4H), 3,60 (д, J=6,0 Гц, 1H), 3,38 (с, 3H), 3,26-3,10 (м, 3H), 2,93 (дд, J=2,0, 11,2 Гц, 1H), 2,91-2,84 (м, 1H), 2,75-2,64 (м, 2H), 2,44-2,29 (м, 5H), 2,21-1,97 (м, 8H), 1,93-1,83 (м, 3H), 1,79-1,72 (м, 5H), 1,68-1,29 (м, 8H), 1,11 (д, J=6,8 Гц, 3H), 1,07-1,01 (м, 1H), 1,06 (д, J=7,2 Гц, 3H), 1,02 (д, J=7,2 Гц, 3H). ЖХ-МС (M+H) =1135,7.
[00348] Val-Cit-PAB-эрибулин (ER-001228950) (16 мг, 14 мкмоль) растворяли в ДМФА (1 мл). N,N-диизопропилэтиламин (7,2 мкл, 41 мкмоль) и Mal-ПЭГ2-NHS (9,7 мг, 27 мкмоль) добавляли к этому раствору при комнатной температуре и реакционную смесь перемешивали при комнатной температуре в течение 1 часа. По завершении реакции неочищенную смесь очищали с помощью обращенно-фазовой ВЭЖХ с применением подвижной фазы ацетонитрил-вода, содержащей 0,1% муравьиной кислоты. Собранные фракции концентрировали в вакууме при комнатной температуре на охлаждаемой водяной бане для получения Mal-ПЭГ2-Val-Cit-PAB-эрибулина (ER-001159569) (7,1 мг, 5,2 мкмоль, выход 38%). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,59 (д, J=8,4 Гц, 2H), 7,31 (д, J=8,4 Гц, 2H), 6,81 (с, 2H), 5,13 (с, 1H), 5,06 (д, J=12,4 Гц, 1H), 5,02 (с, 1H), 5,01 (д, J=12,4 Гц, 1H), 4,87 (с, 1H), 4,82 (с, 1H), 4,71 (т, J=4,0 Гц, 1H), 4,61 (т, J=4,4 Гц, 1H), 4,50 (дд, J=5,2, 9,2 Гц, 1H), 4,47 (д, J=10,8 Гц, 1H), 4,32-4,27 (м, 2H), 4,19 (дд, J=6,8, 11,6 Гц, 1H), 4,13-4,07 (м, 2H), 3,98 (т, J=10,4 Гц, 1H), 3,88-3,82 (м, 3H), 3,76-3,64 (м, 6H), 3,62-3,51 (м, 6H), 3,38 (с, 3H), 3,22-3,08 (м, 4H), 2,93 (дд, J=2,4, 9,6 Гц, 1H), 2,92-2,84 (м, 1H), 2,76-2,63 (м, 2H), 2,52 (т, J=6,0 Гц, 2H), 2,44-2,29 (м, 5H), 2,21-1,97 (м, 8H), 1,93-1,83 (м, 3H), 1,80-1,66 (м, 5H), 1,66-1,28 (м, 10H), 1,11 (д, J=6,4 Гц, 3H), 1,07-1,01 (м, 1H), 0,99 (д, J=6,8 Гц, 3H), 0,97 (д, J=6,4 Гц, 3H). ЖХ-МС (M+H) =1374,9.
1. 2 Приготовление NHS-ПЭГ2-Val-Cit-PAB-эрибулина (ER-001236940)

[00349] Val-Cit-PAB-эрибулин (ER-001228950) (45 мг, 0,04 ммоль) и бис(2,5-диоксопирролидин-1-ил) 3,3'-(этан-1,2-диилбис(окси))дипропаноат (79 мг, 0,198 ммоль) смешивали в ДМФА (1,5 мл) и затем добавляли Et3N (44,2 мкл, 0,317 ммоль). Смесь перемешивали в течение 18 часов до завершения реакции, что было определено с помощью анализа ВЭЖХ. Растворитель выпаривали и остаток очищали с помощью флэш-хроматографии для получения NHS-ПЭГ2-Val-Cit-PAB-эрибулина (ER-001236940) в виде белого твердого вещества (38 мг, выход 68%). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,58 (д, J=8,4 Гц, 2H), 7,33 (д, J=8,4 Гц, 2H), 5,14 (с, 1H), 5,05 (д, J=12,4 Гц, 1H), 5,03 (с, 1H), 5,01 (д, J=12,4 Гц, 1H), 4,87 (с, 1H), 4,83 (с, 1H), 4,71 (т, J=4,4 Гц, 1H), 4,62 (т, J=4,4 Гц, 1H), 4,51 (дд, J=4,8, 8,8 Гц, 1H), 4,50-4,47 (м, 1H), 4,32-4,27 (м, 2H), 4,21 (дд, J=4,8, 6,4 Гц, 1H), 4,14-4,08 (м, 2H), 3,99 (т, J=10,4 Гц, 1H), 3,88-3,82 (м, 3H), 3,78-3,70 (м, 4H), 3,62 (с, 2H), 3,62-3,58 (м, 1H), 3,50-3,46 (м, 2H), 3,39 (с, 4H), 3,36 (с, 3H), 3,22-3,08 (м, 3H), 2,93 (дд, J=2,0, 11,2 Гц, 1H), 2,91-2,87 (м, 1H), 2,84 (с, 2H), 2,80 (с, 2H), 2,75-2,64 (м, 2H), 2,59-2,52 (м, 2H), 2,44-2,29 (м, 5H), 2,21-1,97 (м, 10H), 1,93-1,83 (м, 3H), 1,79-1,72 (м, 5H), 1,68-1,29 (м, 8H), 1,11 (д, J=6,8 Гц, 3H), 1,08-0,98 (м, 1H), 1,00 (д, J=7,2 Гц, 3H), 0,98 (д, J=7,2 Гц, 3H). ЖХ-МС (M+H) =1421,0.
1. 3 Приготовление NHS-(CH2)5-Val-Cit-PAB-эрибулина (ER-001236941)
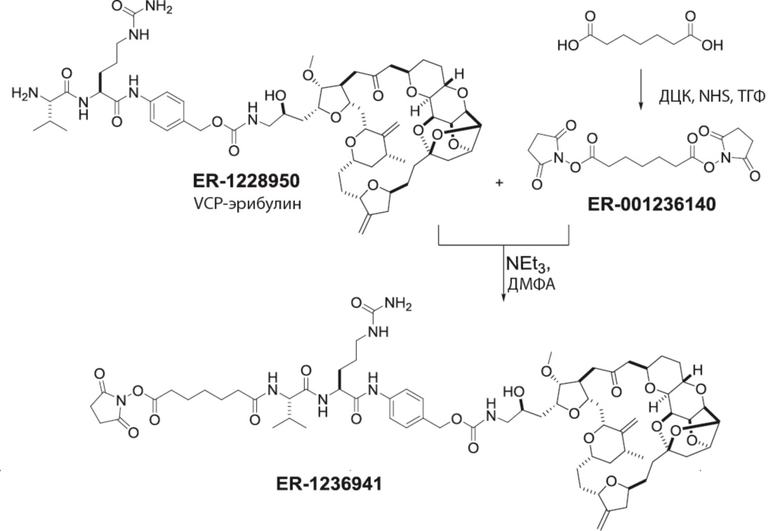
[00350] Пимелиновую кислоту (1,6 г, 9,99 ммоль) растворяли в тетрагидрофуране (ТГФ) (100 мл) и затем добавляли 1-гидроксипирролидин-2,5-дион (2,299 г, 19,98 ммоль) с последующим добавлением ДЦК (4,12 г, 19,98 ммоль). Смесь перемешивали при комнатной температуре в течение 18 часов, пока анализ ВЭЖХ не показал завершение реакции. Твердое вещество удаляли с помощью фильтрования через целитовую прокладку и промывали ТГФ (3 × 2 мл). Объединенный фильтрат концентрировали и очищали с помощью флэш-хроматографии для получения бис(2,5-диоксопирролидин-1-ил) гептанедиоата (ER-001236140) в виде белого твердого вещества (2,5 г, выход 71%). 1H ЯМР (400 МГц) δ м. д. 2,83 (с, 8H), 2,64 (т, J=7,6 Гц, 4H), 1,80 (дт, J=7,6 Гц, 4H), 1,59-1,51 (м, 2H). ЖХ-МС (M+H) =355,2.
[00351] NHS-(CH2)5-Val-Cit-PAB-эрибулин (ER-001236941) был получен (8,5 мг, выход 47%) из VCP-эрибулина (ER-001228950) и бис(2,5-диоксопирролидин-1-ил) гептанедиоата (ER-001236140) с применением той же процедуры, что описана выше для приготовления NHS-ПЭГ2-Val-Cit-PAB-эрибулина (ER-001236940). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,56 (д, J=8,4 Гц, 2H), 7,30 (д, J=8,4 Гц, 2H), 5,13 (с, 1H), 5,04 (д, J=12,0 Гц, 1H), 5,01 (с, 1H), 5,00 (д, J=12,4 Гц, 1H), 4,86 (с, 1H), 4,82 (с, 1H), 4,70 (т, J=4,4 Гц, 1H), 4,60 (т, J=4,4 Гц, 1H), 4,50 (дд, J=4,8, 8,8 Гц, 1H), 4,46 (д, J=10,8 Гц, 1H), 4,36-4,25 (м, 2H), 4,17 (дд, J=4,8, 6,4 Гц, 1H), 4,13-4,06 (м, 2H), 3,97 (т, J=10,4 Гц, 1H), 3,87-3,80 (м, 3H), 3,74-3,68 (м, 2H), 3,37 (с, 3H), 3,20-3,06 (м, 4H), 2,94 (дд, J=2,0, 11,2 Гц, 1H), 2,90-2,82 (м, 1H), 2,82 (с, 4H), 2,74-2,65 (м, 2H), 2,61 (т, J=8,0 Гц, 2H), 2,46-2,26 (м, 7H), 2,24-1,81 (м, 13H), 1,78-1,28 (м, 19H), 1,10 (д, J=6,8 Гц, 3H), 1,06-0,96 (м, 1H), 0,97 (д, J=7,2 Гц, 3H), 0,95 (д, J=7,2 Гц, 3H). ЖХ-МС (M+H) =1375,1.
1. 4 Приготовление Mal-(CH2)5-Val-Cit-PAB-эрибулина (ER-001235638)
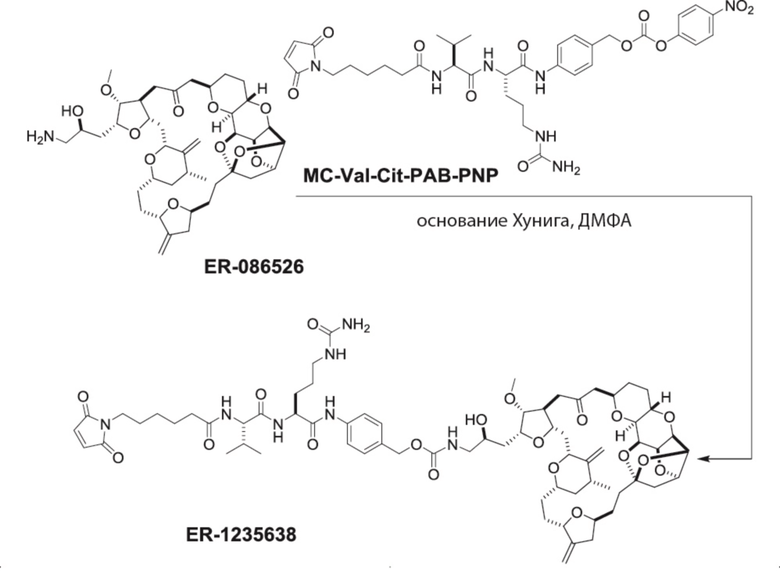
[00352] Эрибулин (ER-000086526) (10 мг, 0,012 ммоль) расстворяли в ДМФА (1 мл) и смешивали с MC-Val-Cit-PAB-PNP (9,02 мг, 0,012 ммоль) и основанием Хунига (4,44 мкл, 0,025 ммоль). Затем смесь перемешивали при комнатной температуре в течение 12 часов, пока анализ ВЭЖХ не показал завершение реакции. Реакционную смесь концентрировали и очищали с помощью флеш-хроматографии для получения Mal-(CH2)5-Val-Cit-PAB-эрибулина (ER-001235638) в виде белого твердого вещества (11,3 мг, выход 63%). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,57 (д, J=8,4 Гц, 2H), 7,31 (д, J=8,4 Гц, 2H), 6,79 (с, 2H), 5,13 (с, 1H), 5,05 (д, J=12,4 Гц, 1H), 5,02 (с, 1H), 5,00 (д, J=12,4 Гц, 1H), 4,87 (с, 1H), 4,83 (с, 1H), 4,71 (т, J=4,4 Гц, 1H), 4,61 (т, J=4,4 Гц, 1H), 4,56-4,46 (м, 3H), 4,35-4,27 (м, 2H), 4,20-4,07 (м, 4H), 3,98 (т, J=10,8 Гц, 1H), 3,87-3,83 (м, 3H), 3,73-3,70 (м, 2H), 3,48 (т, J=7,6 Гц, 2H), 3,38 (с, 3H), 3,20-3,08 (м, 4H), 2,93 (дд, J=1,6, 9,6 Гц, 1H), 2,89-2,85 (м, 1H), 2,69 (дт, J=11,2, 16,8 Гц, 2H), 2,44-2,33 (м, 5H), 2,27-1,83 (м, 13H), 1,78-1,68 (м, 5H), 1,66-1,27 (м, 14H), 1,11 (д, J=7,2 Гц, 3H), 1,07-0,98 (м, 1H), 0,98 (д, J=7,2 Гц, 3H), 0,96 (д, J=7,2 Гц, 3H). ЖХ-МС (M+H)=1328,9.
1. 5 Приготовление Mal-ПЭГ8-Val-Cit-PAB-эрибулина (ER-001242287)
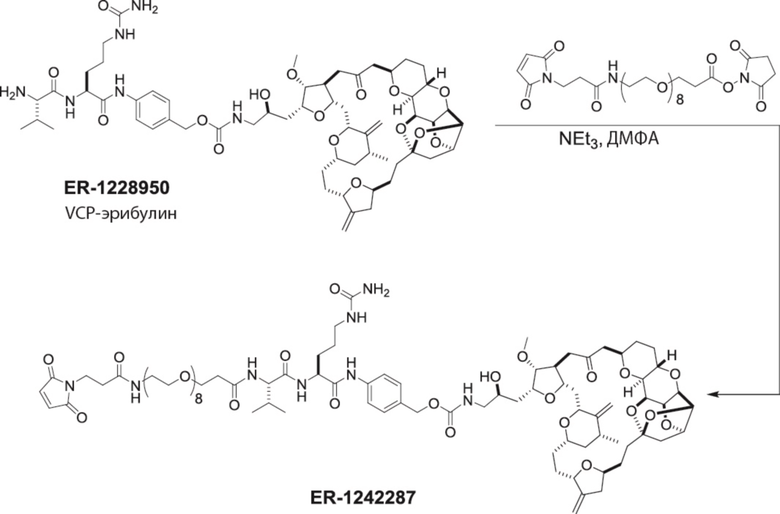
[00353] VCP-эрибулин (ER-001228950) (10 мг, 8,808 мкмоль) и 2,5-диоксопирролидин-1-ил 1-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)-3-оксо-7,10,13,16,19,22,25,28-октаокса-4-азагентриаконтан-31-оат (6,07 мг, 8,808 мкмоль) смешивали с ДМФА (1 мл) с последующим добавлением Et3N (9,82 мкл, 0,07 ммоль). Реакционную смесь перемешивали при комнатной температуре в течение 18 часов, пока анализ ВЭЖХ не показал завершение реакции. Растворитель удаляли выпариванием и остаток очищали с помощью флэш-хроматографии для получения Mal-ПЭГ8-Val-Cit-PAB-эрибулина (ER-001242287) в виде белого твердого вещества (3,0 мг, выход 20%). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,58 (д, J=8,4 Гц, 2H), 7,29 (д, J=8,4 Гц, 2H), 6,80 (с, 2H), 5,12 (с, 1H), 5,04 (д, J=12,4 Гц, 1H), 5,01 (с, 1H), 4,99 (д, J=12,4 Гц, 1H), 4,85 (с, 1H), 4,80 (с, 1H), 4,69 (т, J=4,4 Гц, 1H), 4,59 (т, J=4,4 Гц, 1H), 4,50-4,42 (м, 2H), 4,32-4,24 (м, 2H), 4,20-4,14 (м, 2H), 4,12-4,04 (м, 3H), 3,96 (т, J=10,4 Гц, 1H), 3,86-3,80 (м, 3H), 3,76-3,57 (м, 4H), 3,48 (т, J=6,0 Гц, 1H), 3,36 (с, 3H), 3,20-3,08 (м, 3H), 2,91 (дд, J=2,0, 11,2 Гц, 1H), 2,90-2,82 (м, 1H), 2,74-2,60 (м, 2H), 2,44-2,29 (м, 5H), 2,21-1,97 (м, 10H), 1,93-1,83 (м, 3H), 1,79-1,20 (м, 19H), 1,09 (д, J=6,8 Гц, 3H), 1,04-0,98 (м, 1H), 0,99 (д, J=7,2 Гц, 3H), 0,97 (д, J=7,2 Гц, 3H). ЖХ-МС (M+H) =1711,6.
1. 6 Приготовление NHS-ПЭГ9-Val-Cit-PAB-эрибулина (ER-001242288)
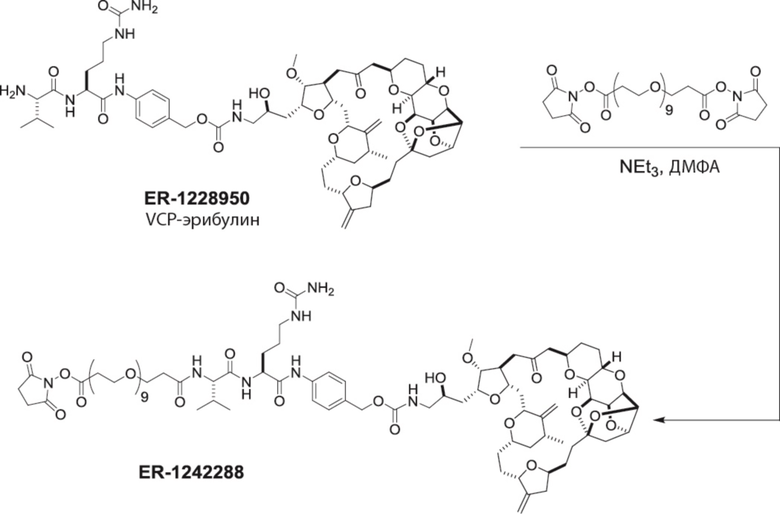
[00354] NHS-ПЭГ9-Val-Cit-PAB-эрибулин (ER-001242288) был получен (13 мг, выход 85%) из VCP-эрибулина (ER-001228950) и БисNHS-ПЭГ9 с применением той же процедуры, что описана выше для приготовления NHS-ПЭГ2-Val-Cit-PAB-эрибулина (ER-001236940). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,61 (д, J=8,4 Гц, 2H), 7,32 (д, J=8,4 Гц, 2H), 5,16 (с, 1H), 5,06 (д, J=12,4 Гц, 1H), 5,01 (с, 1H), 5,00 (д, J=12,4 Гц, 1H), 4,87 (с, 1H), 4,82 (с, 1H), 4,71 (т, J=4,4 Гц, 1H), 4,61 (т, J=4,4 Гц, 1H), 4,52-4,45 (м, 2H), 4,34-4,26 (м, 2H), 4,20-4,19 (м, 1H), 4,14-4,06 (м, 2H), 3,98 (т, J=10,4 Гц, 1H), 3,88-3,80 (м, 3H), 3,76-3,70 (м, 4H), 3,66-3,58 (м, 37H), 3,38 (с, 3H), 3,24-3,10 (м, 3H), 2,93 (дд, J=2,0, 11,2 Гц, 1H), 2,91-2,84 (м, 1H), 2,84 (с, 4H), 2,76-2,64 (м, 2H), 2,58-2,50 (м, 4H), 2,46-2,28 (м, 5H), 2,22-1,96 (м, 8H), 1,91-1,82 (м, 3H), 1,79-1,68 (м, 5H), 1,64-1,24 (м, 8H), 1,11 (д, J=6,8 Гц, 3H), 1,08-0,96 (м, 1H), 0,99 (д, J=7,2 Гц, 3H), 0,97 (д, J=7,2 Гц, 3H). ЖХ-МС (M+H) =1729,7.
1. 7 Приготовление NHS-ПЭГ3-триазол-ПЭГ3-Val-Cit-PAB-эрибулина (ER-001243700)
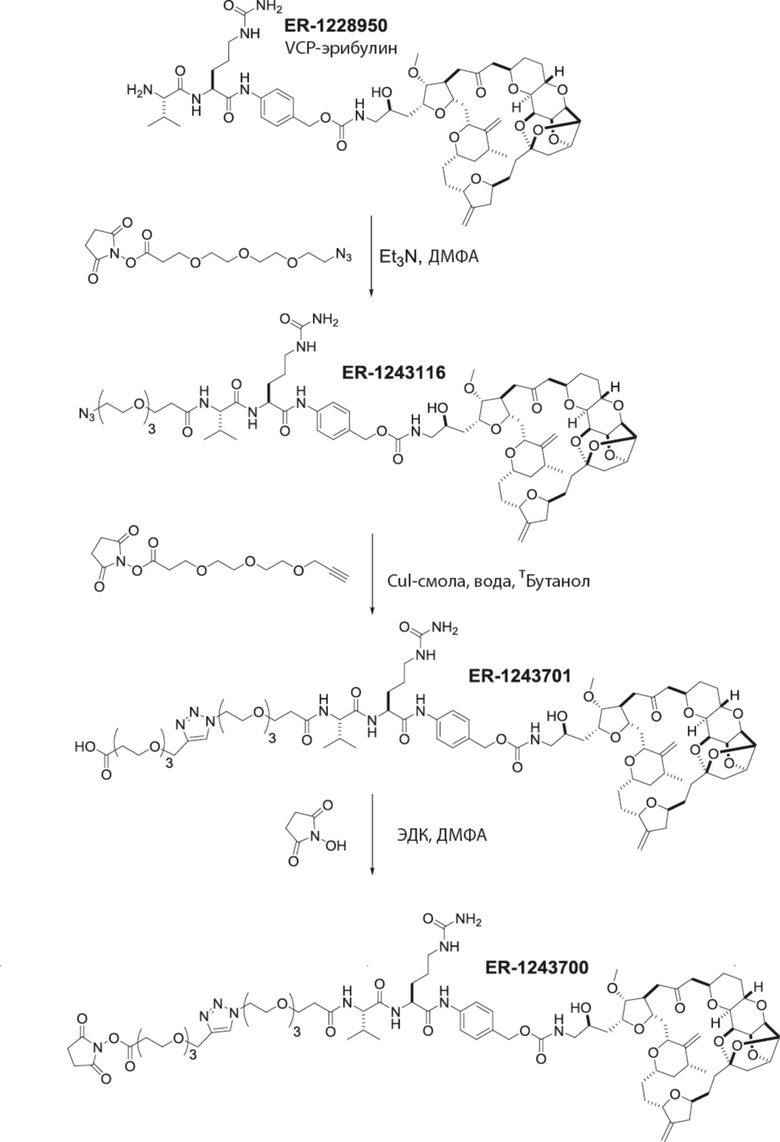
[00355] VCP-эрибулин (ER-001228950) (25 мг, 0,022 ммоль) расстворяли в ДМФА (2,5 мл) и затем смешивали с Et3N (24,55 мкл, 0,176 ммоль) и азид-ПЭГ3-NHS (8,34 мг, 0,024 ммоль). Смесь перемешивали при комнатной температуре в течение 18 часов, пока анализ ВЭЖХ не показал завершение реакции. Смесь концентрировали в вакууме и остаток очищали препаративной ВЭЖХ (MeCN и вода с 0,1% муравьиной кислотой). Фракции, содержащие азид-ПЭГ3-Val-Cit-PAB-эрибулин, экстрагировали дихлорметаном (CH2Cl2) (3×20 мл), а CH2Cl2 выпаривали для получения азид-ПЭГ3-Val-Cit-PAB-эрибулина (ER-001243116) в виде белого твердого вещества (18,9 мг, выход 63%). 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,58 (д, J=8,4 Гц, 2H), 7,30 (д, J=8,4 Гц, 2H), 5,14 (с, 1H), 5,04 (д, J=12,4 Гц, 1H), 5,03 (с, 1H), 5,01 (д, J=12,4 Гц, 1H), 4,85 (с, 1H), 4,81 (с, 1H), 4,70 (т, J=4,4 Гц, 1H), 4,61 (т, J=4,4 Гц, 1H), 4,52-4,48 (м, 2H), 4,31-4,25 (м, 2H), 4,20-4,15 (м, 1H), 4,13-4,07 (м, 2H), 3,99 (м, J=10,4 Гц, 1H), 3,84-3,79 (м, 3H), 3,77-3,65 (м, 4H), 3,64-3,56 (м, 13H), 3,38 (с, 3H), 3,20-3,05 (м, 3H), 2,95-2,80 (м, 2H), 2,75-2,60 (м, 2H), 2,55-2,50 (м, 2H), 2,43-2,25 (м, 5H), 2,21-1,97 (м, 8H), 1,93-1,83 (м, 3H), 1,79-1,72 (м, 5H), 1,68-1,29 (м, 10H), 1,08 (д, J=6,8 Гц, 3H), 1,05-0,95 (м, 1H), 0,98 (д, J=7,2 Гц, 3H), 0,95 (д, J=7,2 Гц, 3H). ЖХ-МС (M+H) =1365,1.
[00356] Азид-ПЭГ3-VCP-эрибулин (ER-001243116) (9,6 мг, 7,035 мкмоль) и 2,5-диоксопирролидин-1-ил 3-(2-(2-(проп-2-ин-1-илокси)этокси)этокси)пропаноат (6,61 мг, 0,021 ммоль) смешивали в воде (0,6 мл) и т-бутаноле (1,8 мл). Смесь барботировали с N2 в течение 45 мин. К смеси добавляли йодид меди на амберлист-21 (1,23 ммоль/г, 10 мг) и через смесь барботировали N2 в течение следующих 30 мин. Затем реакционную смесь перемешивали при комнатной температуре в течение 72 часов до полного потребления исходного материала. С помощью ЖХ-МС не было обнаружено никакого желаемого продукта сложного эфира NHS, а только гидролизованную карбоновую кислоту. Смесь фильтровали через короткую целитовую прокладку для удаления смолы CuI. Фильтрат концентрировали в вакууме и полученный остаток очищали препаративной тонкослойной хроматографией (преп-ТСХ) (20% MeOH/CH2Cl2) для получения кислоты-ПЭГ3-триазол-ПЭГ3-Val-Cit-PAB-эрибулина (ER-001243701) в виде белого твердого вещества (3,7 мг, выход 33%). ЖХ-МС (ES) (M+H)=1581,2.
[00357] Кислота-ПЭГ3-триазол-PEG3-Val-Cit-PAB-эрибулин (ER-001243701) (3,0 мг, 1,898 мкмоль) расстворяли в ДМФА (200 мкл) и добавляли 1-гидроксипирролидин-2,5-дион (0,437 мг, 3,796 мкмоль) с последующим добавлением 1-этил-3-(3-диметиламинопропил)карбодиимид (ЭДК) (0,728 мг, 3,796 мкмоль). После перемешивания при комнатной температуре в течение 18 часов реакция прошла примерно на 50%. Добавляли ЭДК (1,46 мг, 7,8 мкмоль) и смесь перемешивали в течение следующих 18 часов, пока анализ ВЭЖХ не показал >95% конверсии в NHS-ПЭГ3-триазол-ПЭГ3-Val-Cit-PAB-эрибулин. Смесь концентрировали в вакууме и остаток очищали преп-ТСХ (15% MeOH/CH2Cl2) для получения NHS-ПЭГ3-триазол-ПЭГ3-Val-Cit-PAB-эрибулина (ER-001243700) в виде белого твердого вещества (2,2 мг, выход 69%). 1H ЯМР (400 МГц, CD3OD) δ м. д. 8,00 (с, 1H), 7,59 (д, J=8,0 Гц, 2H), 7,31 (д, J=8,4 Гц, 2H), 5,13 (с, 1H), 5,04 (д, J=12,4 Гц, 1H), 5,02 (с, 1H), 5,00 (д, J=12,4 Гц, 1H), 4,87 (с, 1H), 4,83 (с, 1H), 4,71 (т, J=4,0 Гц, 1H), 4,63 (с, 2H), 4,61 (т, J=4,4 Гц, 1H), 4,57-4,55 (м, 2H), 4,51-4,45 (м, 1H), 4,32-4,28 (м, 2H), 4,21-4,17 (м, 2H), 4,13-4,10 (м, 2H), 3,98 (т, J=10,8 Гц, 1H), 3,88-3,80 (м, 5H), 3,75-3,70 (м, 4H), 3,68-3,55 (м, 18H), 3,45-3,40 (м, 2H), 3,38 (с, 3H), 3,20-3,08 (м, 4H), 2,93-2,80 (м, 2H), 2,75-2,50 (м, 2H), 2,68 (с, 4H), 2,48-2,30 (м, 7H), 2,28-1,92 (м, 10H), 1,90-1,68 (м, 8H), 1,65-1,27 (м, 8H), 1,11 (д, J=6,8 Гц, 3H), 1,05-0,95 (м, 1H), 0,99 (д, J=7,2 Гц, 3H), 0,97 (д, J=6,8 Гц, 3H). ЖХ-МС (M+H) =1678,3.
1. 8 Приготовление Mal-ПЭГ2-Ala-Ala-Asn-PAB-эрибулина (ER-001231679) и Mal-ПЭГ2-(Ala-Ala-Asn-PAB)2-эрибулина (ER-001231690)
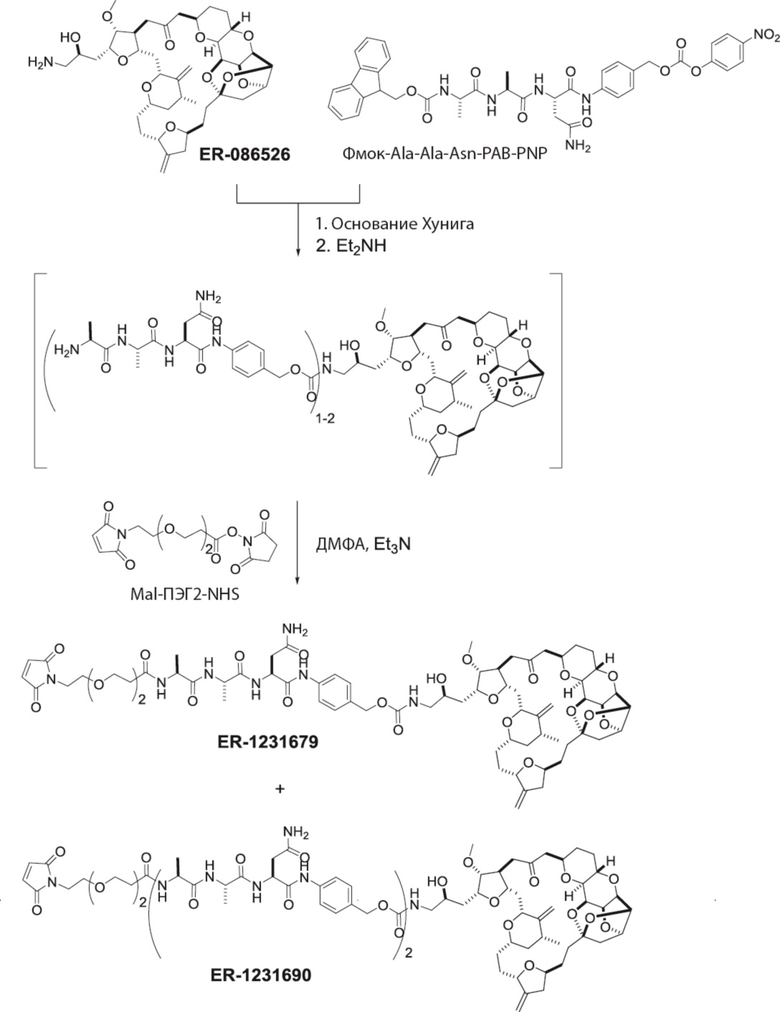
[00358] Эрибулин (ER-000086526) (10 мг, 0,014 ммоль) расстворяли в ДМФА (0,5 мл) и смешивали с основанием Хунига (3,59 мкл, 0,021 ммоль). Затем добавляли (9H-флуорен-9-ил)метил ((S)-1-(((S)-1-(((S)-4-амино-1-((4-((((4-нитрофенокси)карбонил)окси)метил)фенил)амино)-1,4-диоксобутан-2-ил)амино)-1-оксопропан-2-ил)амино)-1-оксопропан-2-ил)карбамат (15,76 мг, 0,021 ммоль) и полученный желтый раствор перемешивали при комнатной температуре в течение 3 дней, пока анализ ВЭЖХ не показал полного потребления исходного материала. К реакционной смеси добавляли диэтиламин (14,23 мкл, 0,137 ммоль), который затем перемешивали при комнатной температуре в течение дополнительных 2 часов до тех пор, пока не было 100% расщепления Фмок защиты. Реакционную смесь концентрировали для удаления диэтиламина и остаток повторно растворяли в ДМФА (1,5 мл). Et3N (0,015 мл, 0,11 ммоль) добавляли при комнатной температуре с последующим добавлением 2,5-диоксопирролидин-1-ил 3-(2-(2-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)этокси)этокси)пропаноата (9,71 мг, 0,027 ммоль). Реакционную смесь перемешивали при комнатной температуре в течение 16 часов до завершения реакции, что было определено с помощью анализа ЖХ-МС. Смесь концентрировали под глубоким вакуумом и очищали с помощью флэш-хроматографии для получения Mal-ПЭГ2-Ala-Ala-Asn-PAB-эрибулина (ER-001231679) (9,2 мг, выход 49%) и Mal-ПЭГ2-(Ala-Ala -An-PAB)2-эрибулина (ER-001231690) (6,0 мг, выход 18%) в виде бесцветных масел.
[00359] Mal-ПЭГ2-Ala-Ala-Asn-PAB-эрибулин (ER-001231679): 1H ЯМР (400 МГц) δ м. д. 9,23 (с, 1H), 8,00 (д, J=7,6 Гц, 1H), 7,61 (д, J=8,4 Гц, 2H), 7,38 (д, J=6,8 Гц, 1H), 7,24 (д, J=8,4 Гц, 2H), 7,13 (д, J=7,2 Гц, 1H), 6,68 (с, 2H), 6,30 (ш. с, 1H), 6,04-6,00 (м, 1H), 5,77 (ш. с, 1H), 5,42 (ш. с, 1H), 5,07 (с, 1H), 5,06-4,98 (м, 2H), 4,93 (с, 1H), 4,88 (с, 1H), 4,90-4,82 (м, 1H), 4,80 (с, 1H), 4,69 (т, J=4,0 Гц, 1H), 4,60 (т, J=4,0 Гц, 1H), 4,49-4,42 (м, 1H), 4,38-4,25 (м, 4H), 4,19 (т, J=4,8 Гц, 1H), 4,15-4,08 (м, 1H), 4,03 (т, J=4,8 Гц, 1H), 3,97-3,85 (м, 3H), 3,83-3,50 (м, 12H), 3,41 (с, 3H), 3,50-3,10 (м, 3H), 3,02-2,64 (м, 6H), 2,52-2,30 (м, 7H), 2,30-1,65 (м, 14H), 1,65-1,20 (м, 12H), 1,10 (д, J=6,8 Гц, 3H), 1,13-1,05 (м, 1H). ЖХ-МС (M+Na) =1396,6.
[00360] Mal-ПЭГ2-(Ala-Ala-Asn-PAB)2-эрибулин (ER-001231690): 1H ЯМР (400 МГц, CD3OD) δ м. д. 7,65 (д, J=8,4 Гц, 2H), 7,60 (д, J=8,4 Гц, 2H), 7,28 (д, J=8,8 Гц, 2H), 7,23 (д, J=8,4 Гц, 2H), 6,79 (с, 2H), 5,13 (с, 1H), 5,02 (с, 1H), 5,06-4,98 (м, 4H), 4,87 (с, 1H), 4,82 (с, 1H), 4,85-4,72 (м, 2H), 4,71 (т, J=4,8 Гц, 1H), 4,61 (т, J=4,4 Гц, 1H), 4,47 (д, J=11,2 Гц, 1H), 4,30-4,06 (м, 9H), 3,97 (т, J=4,8 Гц, 1H), 3,89-3,80 (м, 3H), 3,75-3,48 (м, 12H), 3,38 (с, 3H), 3,17 (д, J=6,8 Hz, 2H), 2,94-2,62 (м, 8H), 2,50-2,28 (м, 7H), 2,22-1,65 (м, 14H), 1,58-1,30 (м, 18H), 1,10 (д, J=6,8 Гц, 3H), 1,06-0,97 (м, 1H). ЖХ-МС (M+Na) =1802,8.
1.9 Приготовление NHS-ПЭГ2-Ala-Ala-Asn-PAB-эрибулина (ER-001231691)
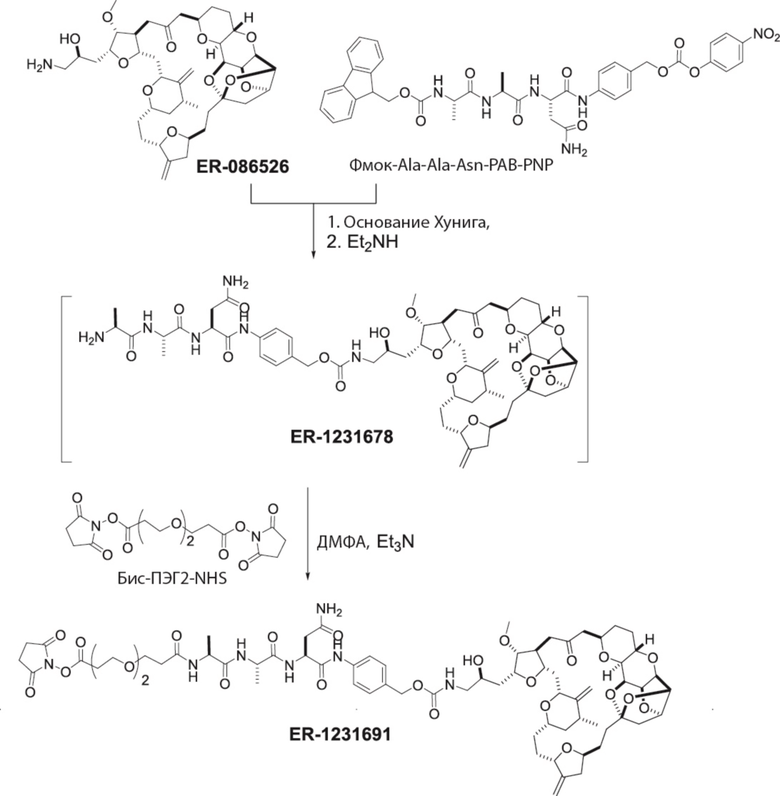
[00361] Ala-Ala-Asn-PAB-эрибулин (ER-001231678) были приготовлен (15 мг, количественный выход) из эрибулина (ER-000086526) и Фмок-Ala-Ala-Asn-PAB-PNP с применением той же процедуры, что описана выше для приготовления Val-Cit-PAB-эрибулина (ER-001228950). ЖХ-МС (M+H) =1135,5.
[00362] NHS-ПЭГ2-Ala-Ala-Asn-PAB-эрибулин (ER-001231691) был получен (12,4 мг, выход 64%) из Ala-Ala-Asn-PAB-эрибулина (ER-001231678) и БисNHS-ПЭГ2 с применением той же процедуры, что описана выше для приготовления NHS-ПЭГ2-Val-Cit-PAB-эрибулина (ER-001236940). 1H ЯМР (400 МГц) δ м. д. 9,21 (с, 1H), 7,95 (д, J=8,0 Гц, 1H), 7,62 (д, J=8,8 Гц, 2H), 7,58-7,52 (м, 1H), 7,28 (ш. с, 1H), 7,24 (д, J=8,4 Гц, 2H), 7,10 (ш. с, 1H), 6,29 (д, J=12,4 Гц, 1H), 5,83 (ш. с, 1H), 5,38 (ш. с, 1H), 5,07 (с, 1H), 5,05-4,95 (м, 2H), 4,93 (с, 1H), 4,88 (с, 1H), 4,90-4,83 (м, 1H), 4,81 (с, 1H), 4,69 (т, J=4,4 Гц, 1H), 4,60 (т, J=4,4 Гц, 1H), 4,46-4,41 (м, 1H), 4,36-4,25 (м, 4H), 4,19 (дд, J=4,8, 6,0 Гц, 1H), 4,15-4,09 (м, 1H), 4,03 (дд, J=4,8, 6,0 Гц, 1H), 3,99-3,89 (м, 3H), 3,85-3,50 (м, 10H), 3,41 (с, 3H), 3,40-3,10 (м, 3H), 3,01-2,60 (м, 10H), 2,60-2,35 (м, 7H), 2,35-1,65 (м, 14H), 1,65-1,20 (м, 14H), 1,10 (д, J=6,8 Гц, 3H), 1,15-1,03 (м, 1H). ЖХ-МС (ES) (M+H) =1442,7.
1. 10 Приготовление азид-ПЭГ3-дисульфид-PAB-эрибулина (ER-001237508) 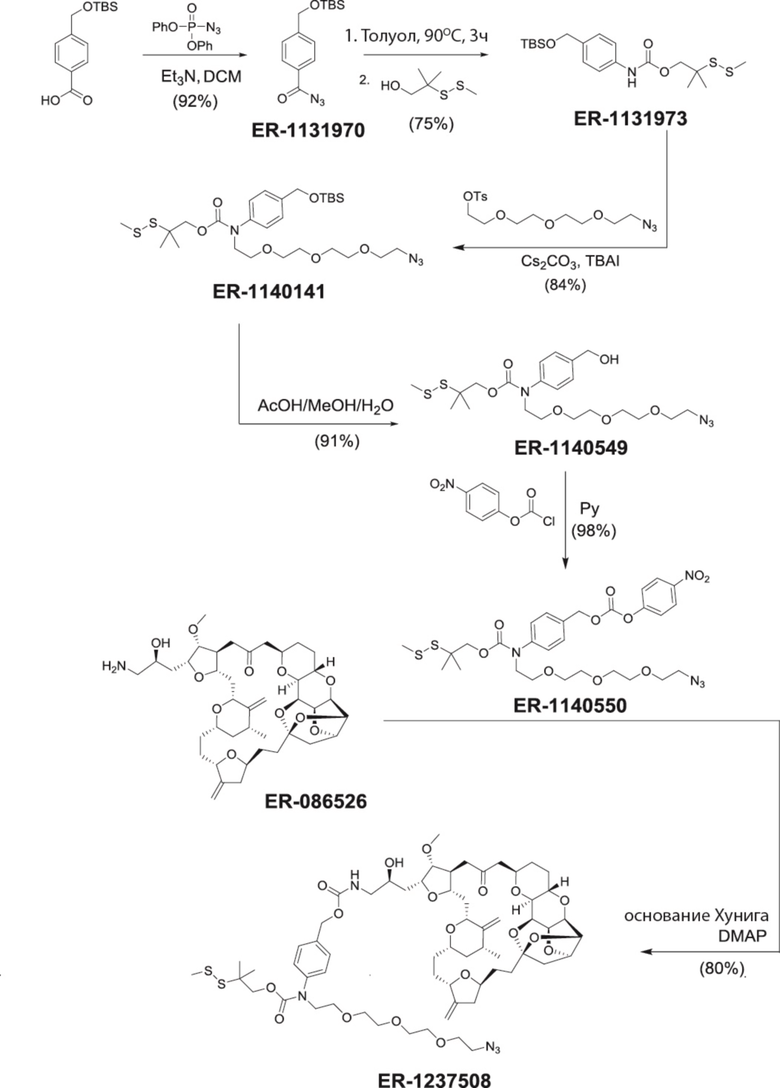
[00363] 4-(((трет-бутилдиметилсилил)окси)метил)бензойную кислоту (1,0 г, 3,754 ммоль) растворяли в дихлорметане (ДХМ) (25 мл), охлаждали до 0°С. Затем добавляли триэтиламин (0,549 мл, 3,941 ммоль), а затем -дифенилфосфоразидат (1,085 мг, 3,941 ммоль). Реакционную смесь медленно нагревали до комнатной температуры и перемешивали в течение 14 часов. Неочищенную смесь разбавляли с этилацетатом (EtOAc)/Hep (1:1, 100 мл) и пропускали через короткую кремниевую пробку, элюируя EtOAc/Hep (50%). Растворитель удаляли под вакуумом для получения 1,10 г 4-(((трет-бутилдиметилсилил)окси)метил)бензойной кислоты (ER-001131970). 1H ЯМР (400 МГц) δ м. д. 7,98 (д, 2 H, J=8,0 Гц), 7,40 (д, 2 H, J=8,0 Гц), 4,79 (с, 2 H), 0,94 (с, 9 H), 0,10 (с, 6 H).
[00364] 4-(((трет-бутилдиметилсилил)окси)метил)бензойную кислоту (ER-001131970) (1,1 г, 3,775 ммоль) растворяли в толуоле (20 мл), нагревали при 110°С в течение 3 часов. Несмотря на то, что продукт не проявился как одно пятно, анализ тонкослойной хроматографии (ТСХ) показал, что исходный материал был израсходован. Затем реакционную смесь охлаждали до комнатной температуры и переносили в виалу, герметизировали в токе азота, и хранили в виде раствора в толуоле (1 мл=32,6 мг) при -20°С.
[00365] К раствору трет-бутил((4-изоцианатобензил)окси)диметилсилана (165 мг, 0,626 ммоль) в толуоле (5 мл) добавляли триэтиламин (0,099 мл, 0,709 ммоль), затем спирт (90,0 мг, 0,591 ммоль), и реакционную смесь перемешивали в течение 6 часов при 36°С. Ход реакции проверяли с помощью СВЭЖХ/МС. Затем добавляли насыщенный раствор гидрокарбоната натрия (NaHCO3) (10 мл) экстрагировали EtOAc/Hep (1:1, 60 мл), промывали соляным раствором, сушили сульфатом натрия и концентрировали. Неочищенный материал очищали с помощью флэш-хроматографии (EtOAc/Hep 10% - 40%) для получения 215 мг 2-метил-2-(метилдисульфанил)пропил(4-(((трет-бутилдиметилсилил)окси)метил)фенил)карбамата (ER-001131973). 1H ЯМР (400 МГц) δ м. д. 7,34 (д, 2 H, J=8,4 Гц), 7,26 (д, 2 H, J=7,6 Гц), 6,63 (ш. с, 1 H), 4,69 (с, 2 H), 4,17 (с, 2 H), 2,42 (с, 3 H), 1,35 (с, 6 H), 0,93 (с, 9 H), 0,08 (с, 6 H).
[00366] 2-метил-2-(метилдисульфанил)пропил (4-(((трет-бутилдиметилсилил)окси)метил)фенил)карбамат (ER-001131973) (198 мг, 0,476 ммоль) и 2-(2-(2-(2-азидоэтокси)этокси)этокси)этил 4-метилбензолсульфонат (325 мг, 0,87 ммоль) растворяли в ДМФА (6,6 мл). Затем добавляли карбонат цезия (621 мг, 1,905 ммоль), а затем тетрабутиламмониййодид (45 мг, 0,122 ммоль) и реакционную смесь перемешивали в течение 15 часов при 36°С. Ход реакции проверяли с помощью СВЭЖХ/МС. Затем добавляли насыщенный раствор NH4Cl (30 мл), экстрагировали EtOAc/Hep (2:1, 150 мл), промывали соляным раствором (10 мл), сушили сульфатом натрия и концентрировали под вакуумом. Неочищенный материал очищали с помощью флэш-хроматографии (EtOAc/Hep 20% - 50%) для получения 248 мг 2-метил-2-(метилдисульфанил)пропил (2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)(4-(((трет-бутилдиметилсилил)окси)метил)фенил)карбамата (ER-001140141). 1H ЯМР (400 МГц) δ м. д. 7,28 (д, 2 H, J=8,4 Гц), 7,20 (д, 2 H, J=8,0 Гц), 4,73 (с, 2 H), 4,06 (ш. с, 2 H), 3,83 (дд, 2 H, J=6,4, 5,6 Гц), 3,68- 3,56 (м, 12 H), 3,37 (дд, 2 H, J=5,6, 5,2 Гц), 2,33 (с, 3 H), 1,14 (ш. с, 6 H), 0,93 (с, 9 H), 0,09 (с, 6 H).
[00367] 2-метил-2-(метилдисульфанил)пропил (2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)(4-(((трет-бутилдиметилсилил)окси)метил)фенил)карбамат (ER-001140141) (81 мг, 0,131 ммоль) растворяли в смеси метанола (5 мл) и воды (0,5 мл). Затем к реакционной смеси добавляли уксусную кислоту (0,5 мл, 8,734 ммоль) и перемешивали в течение 14 часов при 38°С. Реакционную смесь охлаждали до комнатной температуры и растворитель удаляли под вакуумом. Остаток разбавляли EtOAc (30 мл), промывали водой (2×5 мл), NaHCO3 и соляным раствором (3 мл), сушили сульфатом натрия и концентрировали под вакуумом. Неочищенный материал очищали с помощью флэш-хроматографии (EtOAc/Hep 30% - 90%) для получения 61,0 мг 2-метил-2-(метилдисульфанил)пропил (2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)(4-(гидроксиметил)фенил)карбамата (ER-001140549). 1H ЯМР (400 МГц) δ м. д. 7,34 (д, 2 H, J=8,8 Гц), 7,26 (д, 2 H, J=8,0 Гц), 4,69 (д, 2 H, J=4,4 Гц), 4,06 (ш. с, 2 H), 3,84 (дд, 2 H, J=6,2, 6,2 Гц), 3,66-3,56 (м, 12 H), 3,37 (дд, 2 H, J=5,2, 5,2 Гц), 2,33 (с, 3 H), 1,74 (ш. с, 1 H), 1,14 (ш. с, 6 H).
[00368] 2-метил-2-(метилдисульфанил)пропил (2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)(4-(гидроксиметил)фенил)карбамат (ER-001140549) (60 мг, 0,119 ммоль) растворяли в ДХМ (2 мл) и Py (0,019 мл, 0,239 ммоль) охлаждали до 0°C. Затем добавляли 4-нитрофенилкарбонохлоридат (38,5 мг, 0,191 ммоль) в ДХМ (2 мл) и диметиламинопиридин (ДМАП) (2,9 мг, 0,024 ммоль), и реакционную смесь перемешивали в течение 30 мин при 0°С. Реакционную смесь медленно нагревали до комнатной температуры и перемешивали до тех пор, пока исходный материал не был потреблен (приблизительно 2,5 часа). Растворитель удаляли под вакуумом и остаток очищали путем флэш-хроматографии (EtOAc/Hep 10% - 35%) для получения 78 мг 2-метил-2-(метилдисульфанил)пропил (2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)(4-((((4нитрофенокси)карбонил)окси)метил)фенил)карбамата (ER-001140550). 1H ЯМР (400 МГц) δ м. д. 8,27 (дд, 2 H, J=6,8, 2,4 Гц), 7,41 (д, 2 H, J=8,8 Гц), 7,37 (дд, 2 H, J=7,2, 2,4 Гц), 7,33(д, 2 H, J=8,8 Гц), 5,27 (с, 2 H), 4,08 (ш. с, 2 H), 3,85 (дд, 2 H, J=5,8, 5,8 Гц), 3,66- 3,57 (м, 12 H), 3,36 (дд, 2 H, J=5,2, 5,2 Гц), 2,33 (ш. с, 3 H), 1,19 (ш. с, 6 H).
[00369] 2-метил-2-(метилдисульфанил)пропил (2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)(4-((((4-нитрофенокси)карбонил)окси)метил)фенил)карбамат (ER-001140550) (30 мг, 0,045 ммоль) в ДХМ (3 мл, 46,625 ммоль) помещали в 25-миллилитровую колбу в токе азота и охлаждали до 0°С. Добавляли амин (40,8 мг, 0,049 ммоль) в ДХМ (2 мл) и основание Хунига (0,024 мл, 0,135 ммоль), а затем ДМАП (1,4 мг, 0,011 ммоль). Затем реакционную смесь медленно нагревали до комнатной температуры, перемешивали в течение 3 часов, концентрировали под вакуумом и очищали с помощью флэш-хроматографии (EtOAc/Hep 50% - 100%, с последующим MeOH/EtOAc 3% - 8%) для получения 45,0 мг чистого азид-ПЭГ3-дисульфид-PAB-эрибулина (ER-001237508). 1H ЯМР (400 МГц) δ м. д. 7,32 (д, 2 H, J=8,0 Hz), 7,25 (д, 2 H, J=7,2 Гц), 5,28 (дд, 1 H, J=5,6, 5,6 Гц), 5,11-5,04 (м, 3 H), 4,93 (с, 1 H), 4,88 (с, 1 H), 4,81 (с, 1 H), 4,69 (дд, 1 H, J=4,4, 4,4 Гц), 4,60 (дд, 1 H, J=4,2, 4,2 Гц), 4,36 (ш. с, 1 H), 4,33(дд, 1 H, J=4,0, 2,0), 4,29 (ддд, 1 H, J=9,6, 4,4, 4,4 Гц), 4,18 (дд, 1 H, J=6,4, 4,4 Гц), 4,14-4,04 (м, 3 H), 4,03 (дд, 1 H, J=6,4, 4,4 Гц), 3,97-3,89 (м, 3 H), 3,84-3,78 (м, 3 H), 3,67-3,56 (м, 14 H), 3,42 (с, 3 H), 3,40-3,35 (м, 1 H), 3,37 (дд, 2 H, J=5,2, 5,2 Гц), 3,27 (д, 1 H, J=3,2 Гц), 3,20 (ддд, 1 H, J=12,8, 6,0, 6,0 Гц), 2,91-2,83 (м, 2 H), 2,70 (дд, 1 H, J=16,0, 10,0 Гц), 2,52-2,40 (м, 3 H), 2,35-2,13 (м, 9 H), 2,10-2,06 (м, 1 H), 2,01-1,89 (м, 4 H), 1,78-1,64 (м, 4 H), 1,60-1,52 (м, 4 H), 1,49-1,28 (м, 5 H), 1,22-1,07 (м, 6 H), 1,09 (д, 3 H, J=6,0 Гц).
1. 11 Приготовление Mal-ПЭГ4-триазол-ПЭГ3-дисульфид-PAB-эрибулина (ER-001237504)
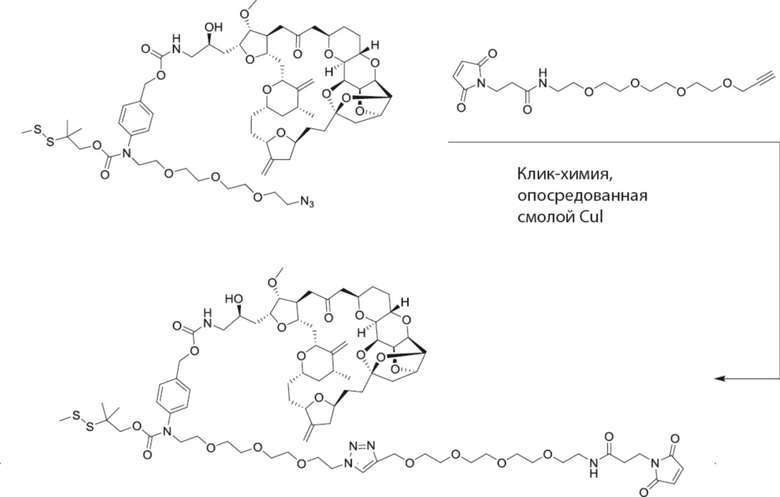
[00370] Смесь азида (9,0 мг, 7,151 мкмоль) и 3-(2,5-диоксо-2,5-дигодро-1H-пиррол-1-ил)-N-(3,6,9,12-тетраоксапентадек-14-ин-1-ил)пропанамида (6,8 мг, 0,018 ммоль) в трет-бутаноле (1,5 мл) и воде (0,5 мл) дегазировали в течение 45 мин. Затем добавляли йодид меди на амберлист-21 (1,23 ммоль/г, 10 мг) и дегазировали в течение дополнительных 30 мин. Реакционную смесь перемешивали при комнатной температуре в течение 18 часов и проверяли с помощью СВЭЖХ/МС. Большая часть исходного материала была потреблена и желаемый продукт проявился как основной пик. Смесь затем отделяли от смолы и очищали на ВЭЖХ (ацетонитрил/вода с 0,05% муравьиной кислотой) для получения 1,5 мг Mal-ПЭГ4-триазол-ПЭГ3-дисульфид-PAB-эрибулина (ER-001237504). 1H ЯМР(400 МГц) δ м. д. 7,74 (с, 1 H), 7,32 (д, 2 H, J=8,4 Гц), 7,27-7,25 (м, 2 H), 6,69 (ш. с, 2 H), 5,43 (дд, 1 H, J=5,6, 5,6 Гц), 5,14-5,06 (м, 3 H), 4,95 (с, 1 H), 4,89 (с, 1 H), 4,82 (с, 1 H), 4,70 (дд, 1 H, J=4,4, 4,4 Гц), 4,66 (с, 2 H), 4,62 (дд, 1 H, J=4,4, 4,4 Гц), 4,52(дд, 1 H, J=5,2, 5,2 Гц), 4,38-4,31 (м, 2 H), 4,30 (ддд, 1 H, J=10,4, 4,0, 4,0 Гц), 4,20 (дд, 1 H, J=6,4, 4,4 Гц), 4,16-4,05 (м, 3 H), 4,04 (дд, 1 H, J=6,4, 4,4 Гц), 3,99-3,91 (м, 3 H), 3,87-3,80 (м, 6 H), 3,70-3,59 (м, 22 H), 3,53 (дд, 2 H, J=5,2, 5,2 Гц), 3,44 (с, 3 H), 3,43-3,36 (м, 3 H), 3,29 (д, 1 H, J=2,8 Гц), 3,18 (ддд, 1 H, J=12,9, 6,2, 6,2 Гц), 2,92-2,84 (м, 2 H), 2,72 (дд, 1 H, J=16,0, 10,0 Гц), 2,54-2,42 (м, 5 H), 2,37-1,90 (м, 19 H), 178-1,52 (м, 3 H), 1,50-1,14 (м, 16 H), 1,10 (д, 3 H, J=6,0 Гц). ЖХ-МС (M+H) =1642,1.
1. 12 Приготовление NHS-ПЭГ3-триазол-ПЭГ3-дисульфид-PAB-эрибулина (ER-001244129)
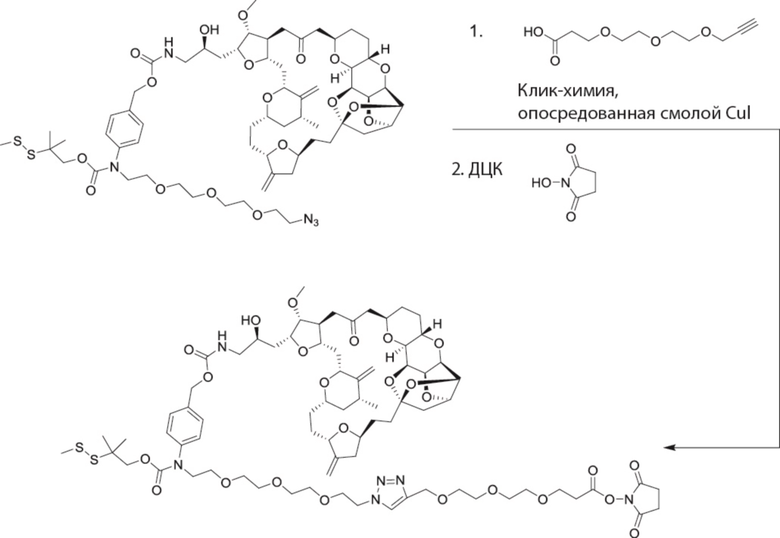
[00371] Смесь азида (9 мг, 7,151 мкмоль) и 2,5-диоксопирролидин-1-ил 3-(2-(2-(проп-2-ин-1-илокси)этокси)этокси)пропаноата (4,5 мг, 14,30 мкмоль) в трет-бутаноле (1 мл) и воде (0,5 мл) дегазировали в течение 45 мин. Затем добавляли йодид меди на амберлист-21 (1,23 ммоль/г, 10 мг, 7,151 мкмоль) и дегазировали в течение дополнительных 30 мин. Реакционную смесь перемешивали при комнатной температуре в течение 18 часов и проверяли с помощью СВЭЖХ/МС. Большая часть исходного материала была потреблена и желаемый продукт проявился как основной пик. Затем смесь отделяли от смолы фильтрованием, экстрагировали с ДХМ (15 мл), промывали соляным раствором (3 × 3 мл), сушили сульфатом натрия и концентрировали под вакуумом. Остаток (5 мг, 3,39 мкмоль) подвергали азеотропной перегонке с толуолом, растворяли в ТГФ (1 мл) и охлаждали до 0 °С. Добавляли ДЦК (4,2 мг, 0,02 ммоль) с последующим добавлением 1-гидроксипирролидин-2,5-диона (2,2 мг, 0,019 ммоль) и реакционную смесь перемешивали при комнатной температуре в течение 18 часов. Большая часть исходного материала была потреблена и желаемый продукт проявился как основной пик, что определено СВЭЖХ/МС. Затем реакционную смесь концентрировали и очищали на препаративной ТСХ (ДХМ/i-пропанол, 8%) для получения 2,5 мг NHS-ПЭГ3-триазол-ПЭГ3-дисульфид-PAB-эрибулина (ER-001244129) в виде бесцветного масла. 1H ЯМР (400 МГц, CD2Cl2) δ м. д. 7,72 (с, 1 H), 7,32 (д, 2 H, J=8,8 Гц), 7,25 (д, 2 H, J=8,8 Гц), 5,08-5,04 (м, 3 H), 4,93 (с, 1 H), 4,85 (с, 1 H), 4,78 (с, 1 H), 4,64 (дд, 1 H, J=4,4, 4,4 Гц), 4,58 (с, 2 H), 4,55 (дд, 1 H, J=4,4, 4,4 Гц), 4,48 (дд, 2 H, J=5,0, 5,0 Гц), 4,32 (д, 1 H, J=6,6 Гц), 4,27-4,22 (м, 2 H), 4,14 (дд, 1 H, J=6,6, 4,8 Гц), 4,10-4,01 (м, 3 H), 4,00 (дд, 1 H, J=6,8, 4,4 Гц), 3,92-3,78 (м, 9 H), 3,65-3,53 (м, 19 H), 3,44-3,39 (м, 4 H), 3,37 (с, 3 H), 3,26 (д, 1 H, J=3,2 Гц), 3,13 (ддд, 1 H, J=12,4, 6,0, 6,0 Гц), 2,91-2,73 (м, 11 H), 2,70-2,64 (м, 2 H), 2,54-2,41 (м, 3 H), 2,38-1,80 (м, 16 H), 1,74-1,52 (м, 3 H), 1,41-1,13 (м, 10 H), 1,07 (д, 3 H, J=6,4 Гц). ЖХ-МС (M+H) =1572,3.
1. 13 Приготовление азид-ПЭГ3-сульфонамид-PAB-эрибулина (ER-001138856)
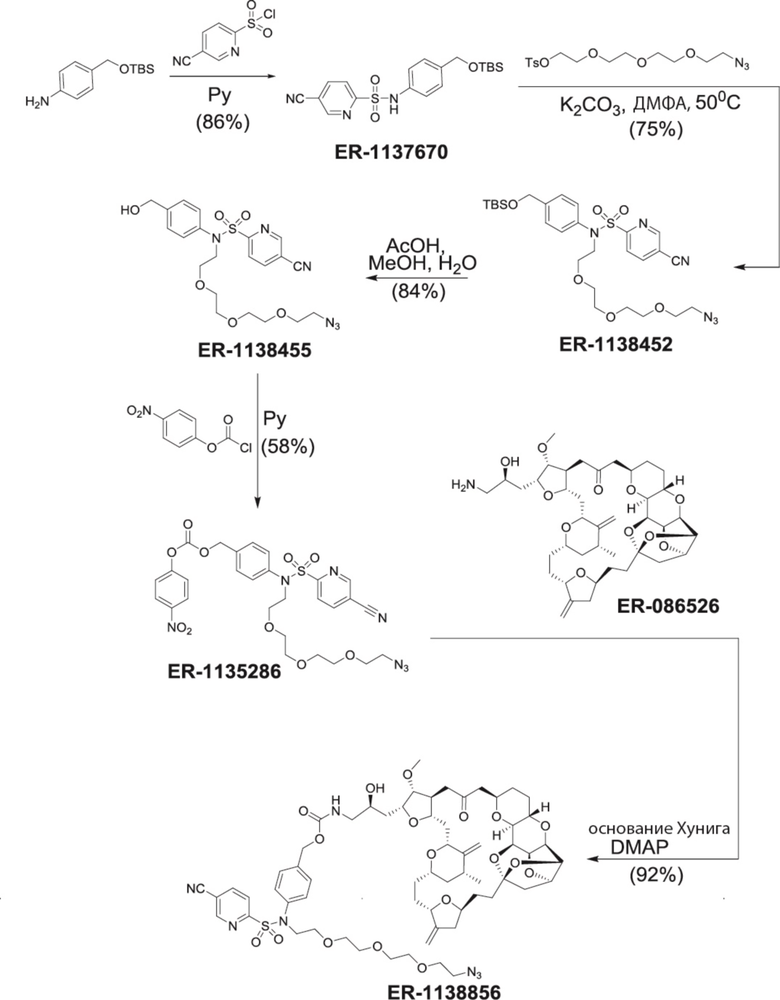
[00372] 4-(((трет-бутилдиметилсилил)окси)метил)анилин (315 г, 1,327 ммоль) растворяли в ДХМ (10 мл), охлаждали до 0°С. Затем добавляли пиридин (0,268 мл, 3,317 ммоль) с последующим добавлением 5-цианопиридин-2-сульфонилхлорида (365 мг, 1,801 ммоль) в ДХМ (10 мл) в течение 15 мин. Реакционную смесь медленно нагревали до комнатной температуры в течение 1 часа и перемешивали в течение 2 часов. Реакционную смесь разбавляли EtOAc (50 мл), промывали соляным раствором, сушили сульфатом натрия и концентрировали под вакуумом для получения 610 мг (103%) N-(4-(((трет-бутилдиметилсилил)окси)метил)фенил)-5-цианопиридин-2-сульфонамида (ER-001137670). Неочищенный продукт был достаточно чистым, хотя и окрашенным. 1H ЯМР (400 МГц) δ м. д. 8,94 (дд, 1 H, J=1,8, 0,6 Гц), 8,10 (дд, 1 H, J=8,4, 2,0 Гц), 7,99 (дд, 1 H, J=8,0, 0,8 Гц), 7,18 (д, 2 H, J=8,2 Гц), 7,15 (ш. с, 1 H), 7,11 (дд, 2 H, J=6,8, 0,8 Гц), 4,64 (с, 2 H), 0,90 (с, 9 H), 0,05 (с, 6 H).
[00373] N-(4-(((трет-бутилдиметилсилил)окси)метил)фенил)-5-цианопиридин-2-сульфонамид (ER-001137670) (105,0 мг, 0,26 ммоль) и 2-(2-(2-(2-азидоэтокси)этокси)этокси)этил 4-метилбензолсульфонат (143 мг, 0,383 ммоль) растворяли в ДМФА (4 мл). Затем добавляли карбонат калия (K2CO3) (144 мг, 1,041 ммоль), а затем тетрабутиламмониййодид (19,2 мг, 0,052 ммоль) и реакционную смесь перемешивали в течение 36 часов при 50°С. Ход реакции проверяли с помощью СВЭЖХ/MS. Добавляли насыщенный раствор NH4Cl (10 мл), экстрагировали EtOAc/Hep (2:1, 30 мл), промывали соляным раствором, сушили сульфатом натрия и концентрировали. Неочищенный материал очищали с помощью флэш-хроматографии (EtOAc/Hep 25% - 80%) для получения 118,0 мг N-(2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)-N-(4-(((трет-бутилдиметилсилил)окси)метил)фенил)-5-цианопиридин-2-сульфонамида (ER-001138452) (75%). 1H ЯМР (400 МГц) δ м. д. 8,99 (дд, 1 H, J=1,8, 0,6 Гц), 8,08 (дд, 1 H, J=8,2, 2,2 Гц), 7,86 (дд, 1 H, J=8,0, 0,8 Гц), 7,24 (д, 2 H, J=10 Гц), 7,09 (д, 2 H, J=8,8 Гц), 4,69 (с, 2 H), 4,06 (дд, 2 H, J=6,0, 6,0 Гц), 3,67 (дд, 2 H, J=5,2, 5,2 Гц), 3,65-3,62 (м, 4 H), 3,58 (дд, 2 H, J=6,2, 6,2 Гц), 3,56-3,53 (м, 4 H), 3,38 (дд, 2 H, J=5,2, 5,2 Гц), 0,93 (с, 9 H), 0,08 (с, 6 H).
[00374] N-(2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)-N-(4-(((трет-бутилдиметилсилил)окси)метил)фенил)-5-цианопиридин-2-сульфонамид ( ER-001138452) (150 мг, 0,248 ммоль) растворяли в метаноле (6 мл). Затем добавляли воду (0,60 мл) с последующей уксусной кислотой (AcOH) (0,60 мл, 10,481 ммоль). Реакционную смесь медленно нагревали до 38°C и перемешивали в течение 14 часов. Большую часть растворителя удаляли под вакуумом. Остаток разбавляли EtOAc (30 мл), промывали водой (2×5 мл), NaHCO3 и соляным раствором, сушили сульфатом натрия и концентрировали под вакуумом. Неочищенный материал очищали с помощью флэш-хроматографии (EtOAc/Hep 35% - 90%) для получения 105,0 мг N-(2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)-5-циано-N-(4-гидроксиметил)фенил)пиридин-2-сульфонамида (ER-001138455) (84%). 1H ЯМР (400 МГц) δ м. д. 8,99 (д, 1 H, J=1,2 Гц), 8,09 (дд, 1 H, J=8,4, 2,0 Гц), 7,88 (дд, 1 H, J=8,4, 0,8 Гц), 7,30 (д, 2 H, J=8,8 Гц), 7,15 (д, 2 H, J=8,4 Гц), 4,67 (с, 2 H), 4,06 (дд, 2 H, J=6,2, 6,2 Гц), 3,66 (дд, 2 H, J=5,0, 5,0 Гц), 3,65-3,58 (м, 6 H), 3,55-3,51 (м, 4 H), 3,38 (дд, 2 H, J=5,2, 5,2 Гц.
[00375] N-(2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)-5-циано-N-(4-гидроксиметил)фенил)пиридин-2-сульфонамид ( ER-001138455) (45 мг, 0,092 ммоль) растворяли в ДХМ (3 мл) и охлаждали до 0°С после добавления пиридина (0,015 мл, 0,183 ммоль). Затем добавляли 4-нитрофенилкарбонохлоридат (20,3 мг, 0,101 ммоль) в ДХМ (2 мл) и ДМАП (2,3 мг, 0,018 ммоль). Реакционную смесь медленно нагревали до комнатной температуры и перемешивали в течение 2 часов. СВЭЖХ/МС показала, что осталось немного исходного материала. Затем реакционную смесь концентрировали под вакуумом и очищали с помощью флэш-хроматографии (EtOAc/Hep 12% - 40%) для получения 35 мг 4-((N-(2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)-5-цианопиридин)-2-сульфонамидо)бензил (4-нитрофенил)карбоната (ER-001235286) (58%) и 20 мг исходного материала. 1H ЯМР (400 МГц) δ м. д. 8,99 (д, 1 H, J=0,8 Гц), 8,27 (дд, 2 H, J=9,2, 2,0 Гц), 8,12 (дд, 1 H, J=7,6, 2,0 Гц), 7,92 (д, 1 H, J=8,4 Гц), 7,38 (д, 4 H, J=9,6 Гц), 7,26 (д, 2 H, J=8,8 Гц), 5,45 (с, 2 H), 4,06 (дд, 2 H, J=5,8, 5,8 Гц), 3,67-3,58 (м, 8 H), 3,58-3,50 (м, 4 H), 3,38 (дд, 2 H, J=6,1, 6,1 Гц).
[00376] 4-(N-(2-(2-(2-(2-азидоэтокси)этокси)этокси)этил)-5-цианопиридин-2-сульфонамидо)бензил (4-нитрофенил)карбонат (ER-001235286) (35,0 мг, 0,053 ммоль) помещали в 25-миллилитровую в токе азота и охлаждали до 0°С. Затем добавляли амин (48,5 мг, 0,059 ммоль) в ДХМ (3 мл, 46,625 ммоль) и основание Хунига (0,037 мл, 0,214 ммоль), а затем ДМАП (2,61 мг, 0,021 ммоль). Реакционную смесь перемешивали в течение 30 мин при 0°С и затем перемешивали в течение дополнительных 6 часов при комнатной температуре. Затем реакционную смесь концентрировали под вакуумом и очищали с помощью флэш-хроматографии (EtOAc/Hep 50% - 100%, с последующим MeOH/EtOAc 3% - 8%) для получения 61,0 мг чистого азид-ПЭГ3-сульфонамид-PAB-эрибулина (ER-001138856). 1H ЯМР (400 МГц) δ м. д. 8,98 (д, 1 H, J=1,2 Гц), 8,10 (дд, 1 H, J=8,2, 1,8 Гц), 7,87 (д, 1 H, J=8,0 Гц), 7,26 (д, 2 H, J=6,8 Гц), 7,13 (д, 2 H, J=8,4 Гц), 5,29 (дд, 1 H, J=5,6, 5,6 Гц), 5,08-5,00 (м, 3 H), 4,92 (с, 1 H), 4,87 (с, 1 H), 4,80 (с, 1 H), 4,68 (дд, 1 H, J=4,6, 4,6 Гц), 4,59 (дд, 1 H, J=4,6, 4,6 Гц), 4,38-4,30 (м, 2 H), 4,28 (ддд, 1 H, J=10,4, 4,0, 4,0 Гц), 4,17 (дд, 1 H, J=6,2, 4,6 Гц), 4,13-4,01 (м, 4 H), 3,97-3,88 (м, 3 H), 3,82-3,78 (м, 1 H), 3,67-3,50 (м, 15 H), 3,41 (с, 3 H), 3,40-3,33 (м, 1 H), 3,37 (дд, 2 H, J=4,8, 4,8 Гц), 3,27 (д, 1 H, J=3,2 Гц), 3,15 (ддд, 1 H, J=12,8, 6,4, 6,4 Гц), 2,90-2,82 (м, 2 H), 2,70 (дд, 1 H, J=16,0, 10,0 Гц), 2,51-2,40 (м, 3 H), 2,34-2,13 (м, 7 H), 2,10-2,05 (м, 1 H), 1,99-1,88 (м, 4 H), 1,78-1,64 (м, 5 H), 1,62-1,52 (м, 2 H), 1,50-1,29 (м, 4 H), 1,08 (д, 3 H, J=6,8 Гц).
1.14 Приготовление Mal-ПЭГ4-триазол-ПЭГ3-сульфонамид-PAB-эрибулина (ER-001237505)
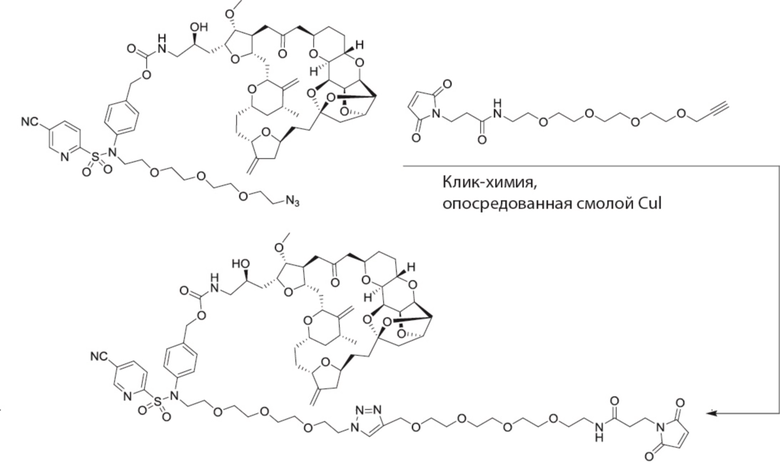
[00377] Смесь азида (10 мг, 8,023 мкмоль) и 3-(2,5-диоксо-2,5-дигодро-1H-пиррол-1-ил)-N-(3,6,9,12-тетраоксапентадек-14-ин-1-ил)пропанамида (9,20 мг, 0,024 ммоль) в трет-бутаноле (2,1 мл) и воде (0,7 мл) дегазировали в течение 45 мин. Затем добавляли йодид меди на амберлист-21 (1,23 ммоль/г, 15 мг) и дегазировали в течение дополнительных 30 мин. Реакционную смесь перемешивали при комнатной температуре в течение 18 часов и проверяли с помощью СВЭЖХ/МС. Большая часть исходного материала была потреблена и желаемый продукт проявился как основной пик. Затем реакционную смесь отделяют от смолы и очищают на препаративной ТСХ (ДХМ/метанол, 7%) для получения 5,5 мг Mal-ПЭГ4-триазол-ПЭГ3-сульфонамид-PAB-эрибулина (ER-001237505). 1H ЯМР (400 МГц, CD2Cl2) δ м. д. 9,01 (с, 1 H), 8,15 (дд, 1 H, J=8,0, 1,8 Гц), 7,87 (д, 1 H, J=8,0 Гц), 7,75 (с, 1 H), 7,28 (д, 2 H, J=8,0 Гц), 7,14 (д, 2 H, J=8,4 Гц), 6,68 (с, 2 H), 6,47 (ш. с, 1 H), 5,44 (ш. с, 1 H), 5,10-5,02 (м, 3 H), 4,94 (с, 1 H), 4,86 (с, 1 H), 4,80 (с, 1 H), 4,68 (дд, 1 H, J=4,4, 4,4 Гц), 4,59 (с, 2 H), 4,56 (дд, 1 H, J=4,4, 4,4 Гц), 4,51(дд, 2 H, J=5,2, 5,2, Гц), 4,34(д, 1 H, J=7,6, Гц), 4,30-4,23 (м, 2 H), 4,19-4,14 (м, 2 H), 4,08 (дд, 1 H, J=4,0, 4,0 Гц), 4,03-3,98 (м, 2 H), 3,94-3,72 (м, 8 H), 3,68-3,46 (м, 28 H), 3,38 (с, 3 H), 3,38-3,33 (м, 3 H), 3,27 (д, 1 H, J=3,2 Гц), 3,16-3,02 (м, 2 H), 2,90-2,81 (м, 2 H), 2,68 (дд, 1 H, J=16,2, 9,8 Гц), 2,54-2,40 (м, 7H), 2,40-1,8 (м, 11 H), 1,80-1,50 (м, 3 H), 1,48-1,25 (м, 3 H), 1,09 (д, 3 H, J=6,4 Гц). ЖХ-МС (M+H) =1630,0.
1.15 Приготовление NHS-ПЭГ3-триазол-ПЭГ3-сульфонамид-PAB-эрибулина (ER-001244623)
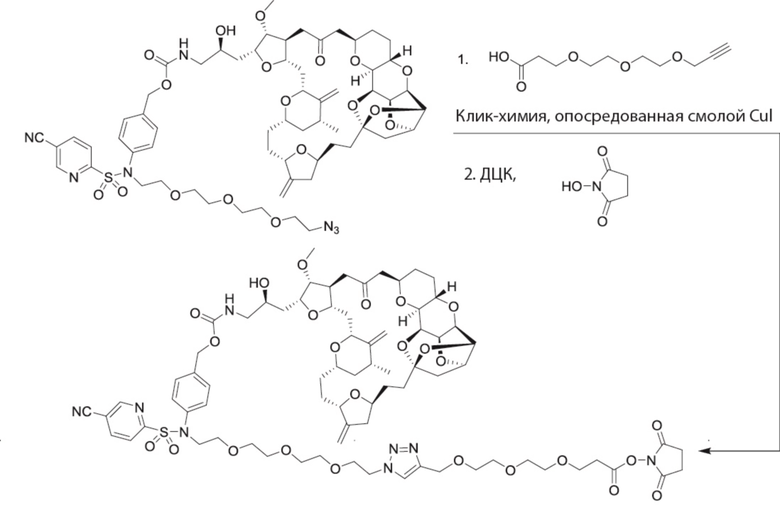
[00378] Смесь азида (14 мг, 0. 011 ммоль) и 2,5-диоксопирролидин-1-ил 3-(2-(2-(проп-2-ин-1-илокси)этокси)этокси)пропаноата (8,80 мг, 0,028 мкмоль) в трет-бутаноле (2 мл) и воде (1 мл) дегазировали в течение 45 мин. Затем добавляли йодид меди на амберлист-21 (1,23 ммоль/г, 20 мг) и дегазировали в течение дополнительных 30 мин. Реакционную смесь перемешивали при комнатной температуре в течение 18 часов и проверяли с помощью СВЭЖХ/МС. Большая часть исходного материала была потреблена и желаемый продукт проявился как основной пик. Реакционную смесь затем отделяли от смолы путем экстракции с ДХМ (2 × 10 мл). Слой ДХМ промывали соляным раствором (4 × 5 мл), сушили сульфатом натрия и концентрировали до желаемого продукта (который применяли на следующем этапе без дополнительной очистки).
[00379] Неочищенную кислоту (15,0 мг, 10,255 мкмоль) расстворяли в ТГФ (1,5 мл) и охлаждали до 0°C. Затем добавляли ДЦК (15,2 мг, 0,074 ммоль), а затем 1-гидроксипирролидин-2,5-дион (8,3 мг, 0,072 ммоль). Реакционную смесь перемешивали при комнатной температуре в течение 18 часов. СВЭЖХ/МС показала, что большая часть исходного материала была потреблена и желаемый продукт проявился как основной пик. Затем реакционную смесь концентрировали и очищали на препаративной ТСХ (ДХМ/i-пропанол, 8%) для получения 2,5 мг NHS-ПЭГ3-триазол-ПЭГ3-сульфонамид-PAB-эрибулина (ER-001244623). 1H ЯМР (400 МГц, CD2Cl2) δ м. д. 9,00 (с, 1 H), 8,12 (д, 1 H, J=8,4 Гц), 8,00 (д, 1 H, J=8,0 Гц), 7,72 (с, 1 H), 7,26 (д, 2 H, J=8,0 Гц), 7,12 (д, 2 H, J=8,0 Гц), 5,37 (ш. с, 1 H), 5,08-5,02 (м, 3 H), 4,93 (с, 1 H), 4,85 (с, 1 H), 4,78 (с, 1 H), 4,66-4,62 (м, 1 H), 4,58-4,56 (м, 4 H), 4,33 (д, 1 H, J=10,8 Гц), 4,29-4,21 (м, 2 H), 4,10-3,96 (м, 4 H), 3,93-3,76 (м, 6 H), 3,74-3,44 (м, 27 H), 3,36 (с, 3 H), 3,34-3,24 (м, 2 H), 3,15-3,06 (м, 1 H), 2,97 (ш. с, 1 H), 2,90-2,78 (м, 8 H), 2,74-2,08 (м, 13 H), 2,05 -1,78 (м, 5 H), 1,73-1,50 (м, 2 H), 1,41-1,25 (м, 4 H), 1,07 (д, 3 H, J=6,0 Гц). ЖХ-МС (M+H) =1560,0.
1.16 Приготовление Mal-ПЭГ2-эрибулина
[00380] Эрибулин (5мг, 7 ммоль) расстворяли в ДМФА (0,5 мл) и смешивали с малеимидо-ПЭГ2-NHS (5 мг, 14 мкмоль; Broadpharm, Cat No. BP-21680) и основанием Хунига (2,4 мкл, 14 мкмоль). Реакционную смесь перемешивали при комнатной температуре в течение 2 часов. Затем реакционную смесь очищали с помощью ВЭЖХ (вода-градиент ацетонитрила 30-70%, содержащие 0,1% муравьиной кислоты). Элюент собирали по массе и лиофилизировали досуха. Конечный выход составил 3,7 мг (3,8 мкмоль, 54%). Прогнозируемая точная масса составляла 968,5 Да. Измеренная масса составляла 969,6 Да [М+Н].
1.17 Приготовление Mal-ПЭГ4-эрибулина
[00381] Эрибулин (5мг, 7 ммоль) расстворяли в ДМФА (0,5 мл) и смешивали с малеимид-ПЭГ4-NHS (6,2 мг, 14 мкмоль; Broadpharm, Cat No. BP-20554) и основанием Хунига (2,4 мкл, 14 мкмоль). Реакционную смесь перемешивали при комнатной температуре в течение 2 часов. Затем реакционную смесь очищали с помощью ВЭЖХ (вода-градиент ацетонитрила 30-70%, содержащие 0,1% муравьиной кислоты). Элюент собирали по массе и лиофилизировали досуха. Конечный выход составил 3,7 мг (3,5 мкмоль, 50%). Прогнозируемая точная масса составляла 1056,5 Да. Измеренная масса составляла 1057,7 Да [М+Н].
1.18 Приготовление азидо-ПЭГ2-эрибулина
[00382] Эрибулин (5мг, 7 ммоль) расстворяли в ДМФА (0,5 мл) и смешивали с азидо-ПЭГ2-NHS (4,2 мг, 14 мкмоль; Broadpharm, Cat No. BP-20524) и основанием Хунига (2,4 мкл, 14 мкмоль). Реакционную смесь перемешивали при комнатной температуре в течение 2 часов. Затем реакционную смесь очищали с помощью ВЭЖХ (вода-градиент ацетонитрила 30-70%, содержащие 0,1% муравьиной кислоты). Элюент собирали по массе и лиофилизировали досуха. Конечный выход составил 2,2 мг (2,4 мкмоль, 34%). Прогнозируемая точная масса составляла 914,5 Да. Измеренная масса составляла 915,7 Да [М+Н].
1.19 Приготовление азидо-ПЭГ4-эрибулина
[00383] Эрибулин (5 мг, 7 ммоль) расстворяли в ДМФА (0,5 мл) и смешивали с азидо-ПЭГ4-NHS (5,5 мг, 14 мкмоль; Broadpharm, Cat No. BP-20518) и основанием Хунига (2,4 мкл, 14 мкмоль). Реакционную смесь перемешивали при комнатной температуре в течение 2 часов. Затем реакционную смесь очищали с помощью ВЭЖХ (вода-градиент ацетонитрила 30-70%, содержащие 0,1% муравьиной кислоты). Элюент собирали по массе и лиофилизировали досуха. Конечный выход составил 3,0 мг (3,0 мкмоль, 43%). Прогнозируемая точная масса составляла 1002,5 Да. Измеренная масса составляла 1003,7 Да [М+Н].
1.20 Приготовление азидо-ПЭГ4-Val-Cit-PAB-эрибулна
[00384] Эрибулин (15 мг, 21 ммоль) расстворяли в ДМФА (1,5 мл) и хорошо перемешивали. Затем добавляли основание Хунига (5,5 мкл, 32 мкмоль) и Фмок-VCP-PNP (24 мг, 22 мкмоль, Levena Biopharma, Cat No. VC1003). Реакционную смесь перемешивали при комнатной температуре в течение ночи (16 часов). По завершении реакции к реакционной смеси добавляли диэтиламин (20 мкл, 0,21 ммоль) и перемешивали в течение 2 часов при комнатной температуре для удаления защитной группы Фмок. Реакцию удаления защитных групп отслеживали с применением масс-спектрометра Waters SQD. По завершении реакции реакционную смесь переносили в предварительно взвешенную 1,5 мл микроцентрифужную пробирку. Растворитель выпаривали под вакуумом с применением охлажденного концентратора Centrivap с температурой, установленной на 30°С. Выход составлял 16 мг (14 мкмоль) неочищенного NH2-Val-Cit-pAB-эрибулина (точная масса 1134,6 Да, выход 67%).
[00385] NH2-Val-Cit-pAB-эрибулин (16 мг, 14,1 мкмоль) расстворяли в ДМФА (1,5 мл). Затем добавляли основание Хунига (7,2 мкл, 41 мкмоль) и азидо-ПЭГ4-NHS (11 мг, 28,2 мкмоль). Реакционную смесь перемешивали при комнатной температуре в течение 3 часов. Затем реакционную смесь очищали с помощью ВЭЖХ (вода-градиент ацетонитрила 48-72%, содержащие 0,1% муравьиной кислоты). Элюент собирали при 1409 м/з и лиофилизировали с получением азидо-ПЭГ4-Val-Cit-PAB-эрибулина (точная масса 1407,7 Да). Получали 13 мг (9,2 мкмоль) азидо-ПЭГ4-Val-Cit-PAB-эрибулина (выход на этапе 65%, 44% в целом).
Пример 4
1. Материалы и способы
[00386] Все примененные реагенты были получены от коммерческих поставщиков, высокой степени чистоты или выше, если не указано иное.
1.1 Антитела
[00387] MORAb-003 (гуманизированное антитело к рецептору фолиевой кислоты альфа человека, 25 мг/мл) и MORAb-009 (химерное антитело мыши/человека к мезотелину человека, 25 мг/мл), применяемые в следующих исследованиях, были из партии #NB02962-19 и партии #030A14, соответственно. Трастузумаб был приобретен коммерчески (Clingen) и был из партии #503345.
[00388] Химерные антитела кролика/человека и гуманизированные антитела к мезотелину человека, имеющие неспаренный цистеин в LCcys80 (Таблица 1), транзиентно экспрессировали в клетках 293F или как стабильно выбранные пулы. Кондиционированную среду очищали и децистеинилировали, как описано в разделе 1.4.1.2.1.
1.2 Цитотоксины
[00389] Конъюгируемые соединения эрибулина синтезировали, как описано в Примере 3 (таблица 46). Исходный раствор (10 мМ) готовили в ДМСО и хранили при -20°С до применения.
1.3 Линии опухолевых клеток
[00390] Линии опухолевых клеток человека, применяемые в анализах ADC MORAb-003, MORAb-009 и Трастузумаба, приготовленных с соединениями малеимидо/сукцинимида (OSu)/азидо-линкер-эрибулина (таблица 46), включают IGROV1 (карциному яичника человека, FRhi, MSLNneg), NCI-H2110 (немелкоклеточную карциному легкого человека, FRmed, MSLNmed), A431 (FRneg, MSLNneg), NCI-N87-luc (карциному желудка человека, FRlo, MSLNmed, her2hi), NUGC3 (аденокарциному желудка человекаа, FRneg, MSLNneg, her2neg), ZR75 (протоковую карциному молочной железы человека, FRneg, MSLNneg, her2med) и BT-474 (протоковую карциному молочной железы человека, FRneg, MSLNneg, her2hi). Линии опухолевых клеток человека, применяемые в анализах химерных антител кролика/человека и гуманизированных антител к LCcys80 мезотелина человека, конъюгированных с MAL-ПЭГ2-Val-Cit-PAB-эрибулином (ER-001159569), представляли собой A3 (A431, стабильно трансфицированную мезотелином человека, MSLNhi), OVCAR3 (карциному яичника человека, MSLNhi), HEC-251 (эндометроид человека, MSLNmed), H226 (плоскоклеточную мезотелиому легких человека, MSLNlo) и исходную A431(MSLNneg). Все применяемые клеточные линии были получены непосредственно из Американской коллекции типовых культур (ATCC), за исключением IGROV1 (получено с разрешения от Национального института рака) и A3 (созданную в Morphotek из исходной A431).
1.4 Конъюгация антитела-лекарственного средства
1.4.1 Конъюгация на основе цистеина с применением малеимидов
1.4.1.1 Конъюгация межцепочечных дисульфидов
1.4.1.1.1 Частичное восстановление
[00391] Для MORAb-003 и MORAb-009 заменяли буфер на фосфатно-солевой буфер Дульбекко (DPBS) и затем концентрировали до 20 мг/мл, применяя центрифужную концентрацию. Добавляли равный объем 270 мкМ трис(2-карбоксиэтил)фосфина (TCEP) в 1X DPBS с 2 мМ ЭДТК и восстановление проводили путем осторожного перемешивания в течение 80 мин при комнатной температуре. Аналогичным образом частично восстанавливали трастузумаб, за исключением того, что восстановление проводили путем осторожного перемешивания в течение 40 мин при комнатной температуре.
1.4.1.1.2 Конъюгация
[00392] Соединение малеимидо-линкер-эрибулин (в ДМСО) конъюгировали с частично восстановленными антителами при молярном соотношении 1:6 (мАт:соединение). Соединение добавляли к 50% пропиленгликолю в DPBS и хорошо перемешивали. Затем добавляли равный объем частично восстановленного антитела и осторожно перемешивали (конечная концентрация пропиленгликоля 25%). Конъюгация продолжалась от 3,5 до 4 часов при комнатной температуре.
1.4.1.2 Конъюгация с LCcys80
1.4.1.2.1 Децистеинилирование
[00393] Применяя ÄKTA Explorer (GE Healthcare), колонку с белком А (GE Healthcare) уравновешивали 10-тью объемами колонки (CV - column volume) 20 мМ фосфата натрия, 10 мМ ЭДТК, pH 7,2 (уравновешивающий буфер). Затем загружали кондиционированную среду, а затем промывали несвязанный материал 10-тью CV уравновешивающего буфера. Колонку промывали 16-тью CV 20 мМ фосфата натрия, 10 мМ ЭДТК, 5 мМ цистеина, рН 7,2, при 0,5 мл/мин в течение 16 часов для удаления кэпирующей группы. Затем колонку промывали 60-тью CV 20 мМ Триса, pH 7,5, при 0,5 мл/мин в течение 60 часов. Децистеинилированное антитело элюировали с применением 5-ти CV 0,1 M глицина, pH 2,9 и тотчас нейтрализовали, применяя 5% объема 2 M Триса, pH 9,0. Фракции, содержащие антитела, объединяли и диализовали в DPBS с применением 20K MWCO Slide-A-Lyzer (Thermo Fisher).
1.4.1.2.2 Конъюгация
[00394] Децистеинилированное антитело доводили до 5,0 мг/мл в DPBS, 1 мМ ЭДТК и 50% пропиленгликоль готовили в DPBS, 1 мМ ЭДТК. MAL-ПЭГ2-Val-Cit-PAB-эрибулин(ER-001159569) (12 мМ в ДМСО) добавляли к 50% пропиленгликолю и тщательно перемешивали. Затем добавляли равный объем децистеинилированного антитела в молярном соотношении 1:4 (мАт:соединение) и осторожно перемешивали. Конъюгация продолжалась от 3,5 до 4 часов при комнатной температуре.
1.4.2 Конъюгация на основе амина с применением сукцинимидов
1.4.2.1 Конъюгация
[00395] Антитело (MORAb-003 или MORAb-009, не восстановленное) доводили до 10,0 мг/мл в 0,1 М бикарбонате натрия, рН 8,3. 50% пропиленгликоль готовили в 0,1 М бикарбонате натрия, рН 8,3. Сукцинимид (OSu)-линкер-эрибулин (в ДМСО) добавляли к 50% пропиленгликолю и тщательно перемешивали. Затем добавляли равный объем антитела в молярном соотношении 1:4 (мАт:соединение) и тщательно перемешивали. Конъюгация продолжалась 1 час при комнатной температуре. Реакцию конъюгации останавливали добавлением 1:20 объема 1 М Триса, рН 8,0, и ADC очищали, как описано в разделе 1.4.4.
1.4.3 Двухстадийная конъюгация на основе амина с применением химии азид-алкинового циклоприсоединения, промотируемого напряжением цикла (SPAAC)
1.4.3.1 Дериватизация дибензилциклооктина (ДБЦО)
[00396] Антитело (MORAb-003 или MORAb-009, не восстановленное) доводили до 10,0 мг/мл в 0,1 М бикарбонате натрия, рН 8,3. 50% пропиленгликоль готовили в 0,1 М бикарбонате натрия, рН 8,3. NHS-ПЭГ4-ДБЦО (Click Chemistry Tools, 50 мМ в ДМСО) добавляли к 50% пропиленгликолю и тщательно перемешивали. Затем добавляли равный объем антитела в молярном соотношении 1:4 (мАт:соединение) и тщательно перемешивали. Конъюгация продолжалась 1 час при комнатной температуре. Непрореагировавший NHS-ПЭГ4-ДБЦО удаляли, как описано в разделе 1.4.4.
1.4.3.2 Конъюгация
[00397] 50% пропиленгликоль готовили в DPBS. Азидо-линкер-эрибулин добавляли к 50% пропиленгликолю и тщательно перемешивают. Затем в смесь добавляли равный объем ДБЦО-модифицированного MORAb-003 или MORAb-009 в молярном соотношении 1:4 (мАт:соединение) и тщательно перемешивали. SPAAC конъюгацию оставляли на ночь при комнатной температуре. Непрореагировавший NHS-ПЭГ4-ДБЦО удаляли, как описано в разделе 1.4.4.
1.4.4 Очистка
[00398] Конъюгированное антитело очищали с помощью колонки(ок) для обессоливания HiTrap (GE Healthcare). Хроматографию проводили на быстрой белковой жидкостной хроматографии (ББЖХ) (GE Healthcare), применяя 1X DPBS в качестве подвижного буфера, чтобы удалить малеимидо/OSu/азидо-линкер-эрибулин и пропиленгликоль. Конечное содержание белка определяли с помощью анализа БХК, как описано в разделе 1.3.1 Примера 1.
1.5 Биофизическая характеристика
1.5.1 Анализ эксклюзионной ВЭЖХ
[00399] Агрегацию ADC анализировали с помощью эксклюзионной высокоэффективной жидкостной хроматографии (эксклюзионной ВЭЖХ) с применением ВЭЖХ Agilent 1260. ADC разбавляли до 1 мг/мл в DPBS. Затем ADC (10 мкл) вводили в защитную колонку Advanced SEC 300A (4,6 мм×3,5 см, размер пор 2,7 мкм, Agilent), а затем в колонку AdvancedBio 300A (4.6 мм×30 см, размер пор 2,7 мкм). ADC элюировали из колонки 0,1 М фосфатом натрия, содержащим 0,15 М NaCl и 5% ИПС, pH 7,4 при скорости потока 0,25 мл/мин в течение 28 мин. Все данные были проанализированы с помощью программного обеспечения Agilent ChemStation. Процент агрегации рассчитывался как [PAагрегата/PAобщий]*100, где PA=интегрированная площадь пика.
1.5.2 Анализ HIC-ВЭЖХ соотношения лекарственного средства-к-антителу (DAR)
[00400] DAR анализировали с применением ВЭЖХ с гидрофобным взаимодействием (HIC-ВЭЖХ). Образцы вводили в колонку TSKgel® Butyl-NP5, 4,6 мм ID x 3,5 см, непристый размер 2,5 мкм (Tosoh Bioscience), и элюировали с 3-минутным уравновешиванием в 100% подвижной фазы А, 15-минутным градиентом (0-100% B), 5-минутным выдерживанием в 100% В, 1-минутной заменой на 100% А и 5-минутным повторным уравновешиванием в 100% подвижной фазы А при 0,7 мл/мин. Подвижная фаза А составляла 25 мМ фосфата натрия, 1,5 М сульфата аммония, рН 7,0. Подвижная фаза В составляла 25 мМ фосфата натрия, 25% изопропанола, рН 7,0. Детекцию проводили при 280 нм (контроль 320 нм). DAR определяли по формуле:
[AUC+1+2(AUC+2)+3(AUC+3)+…n(AUC+n)]/ΣAUCобщ]
где AUC+1 является площадью под кривой для пика антитела, соответствующего ADC, конъюгированному с одним цитотоксином, AUC+2 является площадью под кривой для пика антитела, соответствующего ADC, конъюгированному с двумя цитотоксинами, и так далее. ΣAUCобщ представляет собой объединенную площадь под кривой для всех пиков.
1.5.3 ЖХ-МС анализ DAR
[00401] DAR также анализировали с применением способа ЖХ-МС с ВЭЖХ Waters Alliance с SQD/PDA детекцией. Образцы вводили в колонку Proteomix RP-1000 (5 мкМ, 1000А, 4,6 мм×15 см, Sepax) при 65°С и элюировали с 3-минутным уравновешиванием в 25% В, 27-минутным линейным градиентом 25% - 55% В, 5-минутным выдерживанием в 55% В, 1-минутной заменой на 90% В, 5-минутным выдерживанием в 90% В, 1-минутной заменой на 25% В и 5-минутным повторным уравновешиванием в 25% В. Подвижная фаза A представляла собой 0,1% ТФУ в воде и подвижная фаза В представляла собой 0,1% ТФУ в ацетонитриле. Элюат затем разделяли (10:1) в детекторы PDA и SQD. Детектор SQD был настроен в режиме позитивных ионов (ES positive), капиллярное напряжение 3,2 кВ, напряжение конуса 40 В, экстрактор 3 В и радиочастотная линза 0,2 В, температура источника 150°С и температура десольватации 250°С. Данные масс были получены при 200-2000 м/з в течение 40 минут, непрерывном режиме, времени сканирования 1 секунда. Данные анализировали и деконволюционировали в автономном режиме с применением MassLynx и MaxEnt1. DAR рассчитывали, применяя формулу:
2[[AUCLC+1+2(AUCLC+2)+3(AUCLC+3)+…n(AUCLC+n)]/ΣILCобщ]+
2[[AUCHC+1+2(AUCHC+2)+3(AUCHC+3)+…n(AUCHC+n)]/ΣAUCHCобщ]
где AUCLC+1 представляет собой площадь под кривой пика легкой цепи, конъюгированной с одним цитотоксином, AUCLC+2 представляет собой площадь под кривой пика легкой цепи, конъюгированной с двумя цитотоксинами, и так далее. AUCHC представляет собой площадь под кривой соответствующих тяжелых цепей и ΣAUCLCобщ и ΣAUCHCобщ представляют собой объединенную площадь под кривой всех неконъюгированных и конъюгированных легких цепей и тяжелых цепей, соответственно.
1.5.4 Анализ СВЭЖХ/ЭРИ-МС DAR ADC LCcys80
[00402] ADC (1 мг/мл) восстанавливали добавлением ДТТ до конечной концентрации 20 мМ с последующей инкубацией при 60°С в течение 3 мин. Затем образцы анализировали с применением сверхвысокоэффективной жидкостной хроматографии Waters Acquity и масс-спектрометра Q-Tof Premier. Образцы (0,5-2 мкг каждого) вводили в микроколонку для обессоливания MassPrep при 65°С, элюировали из колонки с 5-минутным уравновешиванием в 95% подвижной фазы А, 10-минутным градиентом (5-90% В), и 10-минутным повторным уравновешиванием в 95% подвижной фазы А при 0,05 мл/мин. Подвижная фаза А составляла 0,1% муравьиной кислоты в воде. Подвижная фаза B составляла 0,1% муравьиной кислоты в ацетонитриле. Масс-спектрометр Q-Tof работал в V-режиме определения положительных ионов с детектированием в диапазоне 500-4000 м/з. Параметры источника были следующими: капиллярное напряжение, 2,25 кВ (интактное антитело)-2,50 кВ (восстановленное антитело); напряжение пробоотборного конуса, 65,0 В (интактное антитело) или 50,0 В (восстановленное антитело); температура источника, 105°C; температура десольватации, 250°С; поток газа десольватации, 550 л/час. Пик белка легкой цепи деконволюционировали с применением функции MassLynx MaxEnt 1. Относительные интенсивности масс неконъюгированной и однократно-конъюгированной легкой цепи применяли для расчета общего DAR по формуле:
2[LC+1/ΣLCобщ]
где LC+1 является массовой интенсивностью легкой цепи, конъюгированной с одним цитотоксином и ΣLCобщ представляет собой объединенные интенсивности неконъюгированных и конъюгированных легких цепей.
1.6 Характеристика связывания
1.6.1 BIAcore
[00403] Концентрации антител доводили до 2 мкг/мл в буфере HBS-P+ (GE Healthcare). Немодифицированные антитела или ADC вводили через сенсор для антител к IgG человека на BIAcore T100 (GE Healthcare) в течение 1 мин при скорости потока 10 мкл/мин. Для регистрации ассоциации антигенов с захваченным антителом вводили серии возрастающих концентраций антигена в течение 300 с при скорости потока 30 мкл/мин. Для антител к мезотелину диапазон концентраций составлял 10 нМ - 0,041 нМ. Для ADC MORAb-003 и MORAb-009 диапазон концентраций составлял 100 нМ - 0,41 нМ. Диссоциацию антигена проверяли в течение 30 мин при той же скорости потока. Поверхность сенсора восстанавливали с помощью введения 3 М MgCl2 в течение 2×30 с при скорости потока 30 мкл/мин. Сенсограммы анализировали с помощью программного обеспечения для оценки Biacore T100, применяя модель связывания Ленгмюра 1:1.
1.6.2 ИФА - Рецептор фолиевой кислоты альфа
[00404] Рекомбинантный рецептор фолиевой кислоты альфа человека разбавляли до 115 нг/мл в буфере для покрытия (50 мМ карбонат-бикарбонатного буфера, рН 9,6) и наносили в черные 96-луночные планшеты Maxisorp (Thermo, Cat No. 43711, 100 мкл/лунку) при 4°С, на ночь. Раствор для покрытия выбрасывали и планшеты промывали три раза, применяя 1X ФСБ с 0,05% буфером Твин-20 (ФСБ-Твин). Планшеты блокировали в 300 мкл блокирующего буфера (1% БСА в ФСБ-Твин) при комнатной температуре в течение 2 часов на орбитальном шейкере. ADC MORAb-003 и MORAb-003 разводили до 1000 нг/мл в блокирующем буфере, затем 2-кратно серийно разводили для получения диапазона от 1000 нг/мл до 0,98 нг/мл. Блокирующий буфер выбрасывали и в планшеты добавляли 100 мкл/лунку разбавленного антитела. Планшеты инкубировали при комнатной температуре в течение 2 часов на орбитальном шейкере. Раствор антитела выбрасывали и планшеты промывали три раза, применяя ФСБ-Твин. К планшетам добавляли 100 мкл/лунку раствора антитела козы к человеческому IgG (H+L)-HRP (разведение 1:10000 в блокирующем буфере) и планшеты инкубировали при комнатной температуре в течение 1 часа на орбитальном шейкере. Раствор вторичных антител выбрасывали и планшеты промывали три раза, применяя ФСБ-Твин. К планшетам добавляли 100 мкл/лунку рабочего раствора флуорогенного субстрата пероксидазы QuantaBlu (Thermo, Cat No. 15169) и планшеты инкубировали при комнатной температуре в течение 30 мин. Флуоресценцию считывали при возбуждении 325 нм/эмиссии 420 нм, применяя SpectraMax M5 (Molecular Devices). Данные анализировали с применением программного обеспечения SoftMaxPro 5. 4. 2 с 4-параметрической подгонкой.
1.6.3 ИФА - Мезотелин
[00405] Рекомбинантный мезотелин человека разбавляли до 1 мкг/мл в буфере для покрытия (50 мМ карбонат-бикарбонатного буфера, рН 9,6) и наносили в черные 96-луночные планшеты Maxisorp (Thermo, Cat No. 43711, 100 мкл/лунку) при 4°С, на ночь. Раствор для покрытия выбрасывали и планшеты промывали три раза, применяя 1X ФСБ с 0,05% буфером Твин-20 (ФСБ-Твин). Планшеты блокировали в 300 мкл блокирующего буфера (1% БСА в ФСБ-Твин) при комнатной температуре в течение 2 часов на орбитальном шейкере. ADC MORAb-009 и MORAb-009 разводили до 1000 нг/мл в блокирующем буфере, затем 2,5-кратно серийно разводили для получения диапазона от 1000 нг/мл до 0,105 нг/мл. Блокирующий буфер выбрасывали и в планшеты добавляли 100 мкл/лунку разбавленного антитела. Планшеты инкубировали при комнатной температуре в течение 2 часов на орбитальном шейкере. Раствор антитела выбрасывали и планшеты промывали три раза, применяя ФСБ-Твин. К планшетам добавляли 100 мкл/лунку раствора антитела козы к человеческому IgG (H+L)-HRP (разведение 1:10000 в блокирующем буфере) и планшеты инкубировали при комнатной температуре в течение 1 часа на орбитальном шейкере. Раствор вторичных антител выбрасывали и планшеты промывали три раза, применяя ФСБ-Твин. К планшетам добавляли 100 мкл/лунку рабочего раствора флуорогенного субстрата пероксидазы QuantaBlu (Thermo, Cat No. 15169) и планшеты инкубировали при комнатной температуре в течение 30 мин. Флуоресценцию считывали при возбуждении 325 нм/эмиссии 420 нм, применяя SpectraMax M5 (Molecular Devices). Данные анализировали с применением программного обеспечения SoftMaxPro 5. 4. 2 с 4-параметрической подгонкой.
1.7 Анализ цитотоксичности
1.7.1 Анализ окрашивания кристаллическим фиолетовым
[00406] Клетки IGROV1 (FRhi, MSLNneg), NCI-H2110 (FRmed, MSLNmed) и A431 (FRneg, MSLNneg) субкультивировали и высевали при 5000 клеток/лунку в готовую питательную среду в 96-луночных планшетах для культивирования тканей, инкубировали при 37°C, 5% CO2 в течение ночи (16 часов). Реагенты для испытания серийно разводили 1:3 в планшетах для разведения с 2-миллилитровыми глубокими лунками, начиная с 200 нМ (всего 10 разведений). Разбавленные образцы (100 мкл) добавляли к клеточным планшетам (при начальной концентрации образцов для испытания 100 нМ). Планшеты инкубировали при 37°С, 5% CO2 в течение дополнительных 5 дней. Затем среду выбрасывали. Планшеты промывали один раз с 200 мкл DPBS, окрашивали с 50 мкл 0,2% раствора кристаллического фиолетового при комнатной температуре в течение 15 мин и затем тщательно промывали водопроводной водой. Планшеты сушили на воздухе, а кристаллический фиолетовый растворяли с 200 мкл 1% раствора ДСН. Планшеты считывали при 570 нм. Данные анализировали с применением GraphPad Prism 6.
2. Результаты
2.1 Биофизическая характеристика ADC MORAb-003, MORAb-009 и трастузумаба
[00407] ADC MORAb-003 (гуманизированного антитела к рецептору фолиевой кислоты альфа человека), MORAb-009 (химерного антитела мыши/человека к мезотелину человека) и трастузумаба (гуманизированного антитела к her2) были приготовлены с применением конъюгируемых соединений эрибулина, перечисленных в таблице 46, в соответствии с одним из трех способов конъюгации, включая:(1) частичное восстановление межцепочечных дисульфидов антитела с применением восстанавливающего агента TCEP, не содержащего тиол, с последующей конъюгацией с применением тиол-реактивных конструкций малеимидо-спейсер-линкер-эрибулина; (2) прямая конъюгация с остатками лизина антитела с применением конструкций сукцинимид (OSu)-спейсер-линкер-эрибулина; и (3) конъюгация с остатками лизина антитела с применением двухстадийного подхода, в результате чего сначала конъюгировали OSu-ПЭГ4-дибензилциклооктин с остатками лизина, затем проводили ортогональную конъюгацию конструкций азидо-спейсер-линкер-эрибулина с применением SPAAC.
[00408] После очистки уровни агрегации для всех ADC MORAb-003, MORAb-009 и трастузумаба определяли с помощью эксклюзионной ВЭЖХ и соотношение лекарственного средства-к-антителу (DAR) анализировали с применением обращенно-фазовой ЖХ-МС и/или HIC-ВЭЖХ. DAR для всех ADC, созданных на основе малеимида, анализировали с применением как обращенно-фазовой ЖХ-МС, так и HIC-ВЭЖХ. Между этими двумя способами обычно наблюдалась разница значений DAR менее, чем 0,3. Напротив, DAR для всех ADC, полученных путем конъюгации через остатки лизина, анализировали только с помощью ЖХ-МС, поскольку высокая степень гетерогенности этих ADC препятствует разделению отдельных видов DAR путем HIC-ВЭЖХ. Для ADC MORAb-003 и MORAb-009 связывание с целевым антигеном также анализировали с помощью ИФА. Результаты анализов агрегации и DAR приведены в таблице 47 рядом с соответствующим ADC.
Биофизический анализ ADC MORAb-003, MORAb-009 и трастузумаба
Фраг.
нМ
клик
клик
клик
клик
клик
клик
клик
клик
клик
клик
Сокращения:% Агр., % агрегации; % Фраг, % фрагментации.
2.1.1 ADC MORAb-003, MORAb-009 и трастузумаба
[00409] Никаких значимых различий между MORAb-003, MORAb-009 и трастузумабом не наблюдалось с точки зрения, как эффективности конъюгации, так и биофизических параметров. Все ADC демонстрировали подобные значения DAR и уровни образования агрегатов.
2.1.2 ADC на основе малеимида
[00410] Для ADC на основе малеимида, как спейсеры пентил и ПЭГ2, спаренные с сайтом расщепления val-cit-pAB, так и спейсер ПЭГ2, спаренный с сайтом расщепления ala-ala-asn-pAB, давали значения DAR между 3,5 и 4,0 по обращенно-фазной ЖХ-МС и HIC-ВЭЖХ, в дополнение к низким (<5%) уровням агрегатов. Однако, когда спейсер для ПЭГ8 (спаренному с сайтом расщепления val-cit-pAB) удлиняли, уровни агрегатов возрастали (11-18%), а эффективность конъюгации снижалась, что приводило к значениям DAR между 1,1 и 2,3. См., например, процент агрегации и значения DAR для MORAb003/MORAb009-ER-001159569 (короткий ПЭГ-линкер) и MORAb003/MORAb009-1242287 (длинный ПЭГ-линкер) в таблице 47.
[00411] Для ADC, приготовленных с сайтом расщепления дисульфидил-pAB, наблюдали низкие значения DAR (1,0-1,6) вместе с относительно высокими уровнями агрегатов (10-14%). Значительно более низкие значения DAR наблюдались, когда эти ADC анализировали с помощью ЖХ-МС, чем с помощью HIC-ВЭЖХ (см., например, ЖХ-МС/HIC-ВЭЖХ значения DAR для MORAb003/MORAb009-ER1237504 и MORAb003/MORAb009-ER1237505 в таблице 47). Этот результат предполагает, что линкерный сайт расщепления показывает связанную с рН нестабильность, поскольку подвижная фаза анализа ЖХ-МС имеет приблизительно 3,0, тогда как подвижная фаза анализа HIC-ВЭЖХ является нейтральной.
[00412] Для ADC, приготовленных с сульфонамидным сайтом расщепления, наблюдали низкие (<5%) уровни агрегатов. Подобно к ADC дисульфидил-pAB, наблюдали более низкие значения DAR при анализе с помощью ЖХ-МС (1,8-2,3), чем при HIC-ВЭЖХ (3,9), что еще раз указывает на то, что линкерный сайт расщепления показывает связанную с рН нестабильность.
[00413] Для нерасщепляемых линкеров ПЭГ2 и ПЭГ4 наблюдали эффективную конъюгацию, приводящую к значениям DAR от 4,0 до 4,7. ADC MORAb-009 с этими нерасщепляемыми линкерами также демонстрировали низкие уровни агрегации (<2%), в то время как наблюдались несколько более высокие уровни агрегации для соответствующих ADC MORAb-003 (4% и 10% для ПЭГ2 и ПЭГ4, соответственно).
2.1.3 ADC на основе сукцинимида
[00414] Все ADC, приготовленные с применением сукцинимида, соединенного со спейсер-линкер-эрибулином, давали значения DAR < 1,0. Чтобы подтвердить, что данная сниженная эффективность конъюгации (по отношению к малеимидам) не была следствием самой процедуры конъюгации, эти ADC были переделаны с применением более высокого соотношения соединение:антитело и повторно проанализированы с применением тех же способов анализа DAR. Были получены аналогичные результаты, что, без привязки к теории, означает, что более низкие значения DAR являются постоянным свойством комбинации сукцинимида и эрибулина и что малеимиды могут быть более эффективно конъюгированы. Эффективность конъюгации сукцинимида повышали с помощью применения двухстадийного способа, в соответствии с чем сначала добавляли ДБЦО к антителу применяя NHS-DBCO с последующим добавлением азидосоединений. Этот подход приводит к более высоким значениям DAR, что измерено с помощью анализа обращенно-фазной ВЭЖХ, по сравнению с конъюгацией непосредственно с остатками лизина антитела. Для ADC на основе сукцинимида, имещих линкеры сульфонамид (расщепляемый), val-cit-PAB (расщепляемый) или ПЭГ2/ПЭГ4 (наресщепляемый), значения DAR, полученные в результате двухстадийной конъюгации, были аналогичны значениям, определенным для ADC на основе малеимида, имеющих сульфонамидный сайт расщепления. Не привязываясь к теории, этот результат снова указывает на то, что более низкие значения DAR для реакций конъюгации сукцинимид-спейсер-линкер-эрибулина являются постоянным свойством комбинации сукцинимида и эрибулина.
2.2 Характеристика связывания ADC MORAb-003 и MORAb-009
[00415] Для ADC MORAb-003 в отношении связывания целевого антигена не наблюдалось значимых различий между ADC с нерасщепляемым, созданным на основе малеимида линкером-эрибулином, и исходным MORAb-003. Для других ADC MORAb-003, с созданным на основе малеимида линекром-эрибулином, обычно, с помощью ИФА, наблюдали 2-3-кратную потерю связывания целевого антигена по сравнению с исходным MORAb-003. Однако не наблюдалось никакой корреляции между длиной линкера или составом линкера и более низкими значениями ЭК50. Аналогично, для ADC MORAb-003, с созданным на основе сукцинимида линекром-эрибулином, обычно наблюдали 0-3-кратную потерю связывания целевого антигена по сравнению с неконьюгированным MORAb-003. Опять же, никакой корреляции между длиной линкера или составом линкера и более низкими значениями ЭК50 не было. Для ADC MORAb-009 все ADC имели менее чем 2-кратное снижение значений ЭК50 относительно исходного MORAb-009.
2.3 Анализы цитотоксичности ADC MORAb-003, MORAb-009 и трастузумаба in vitro
[00416] Активность приготовленных ADC MORAb-003, MORAb-009 и трастузумаба in vitro оценивали, применяя анализ цитотоксичности на клетках на основе окрашивания кристаллическим фиолетовым. Клеточные линии, выбранные для скрининга ADC MORAb-003 и MORAb-009, представляли собой IGROV1, NCI-H2110 и A431. Клетки IGROV1 имеют происхождение эпителиальной карциномы яичников человека и экспрессируют высокие уровни рецептора фолиевой кислоты альфа, но не имеют мезотелин (то есть, MORAb-003-реактивный). Клетки NCI-H2110 имеют происхождение немелкоклеточной карциномы легких человека и экспрессируют умеренные уровни как рецептора фолиевой кислоты альфа, так и мезотелина (то есть, MORAb-003- и MORAb-009-реактивные). Контрольные клетки A431 имеют происхождение эпидермальной карциномы человека и не экспрессируют никакой целевой антиген. Результаты этого скрининга приведены в таблице 48. ADC MORAb-003, MORAb-009 и трастузумаба, содержащие линкер-токсин малеимидо-ПЭГ2-val-cit-pAB-эрибулин (VCP-эрибулин), также оценивали на дополнительных линиях клеток рака желудка и рака молочной железы, включая NCI-N87 (FRlo, MSLNmed, her2hi), BT-474 (FRneg, MSLNneg, her2hi), ZR-75 (FRneg, MSLNneg, her2med) и NUGC3 (FRneg, MSLNneg, her2neg). Результаты этого скрининга приведены в таблице 49.
(FRhi, MSLNneg)
(FRmed, MSLNmed)
(FRneg, MSLNneg)
(Партия NB3073-88L)
(Партия NB3073-88F)
(Партия NB3142-62A)
(Партия NB3142-62D)
клик
клик
клик
клик
VCP эрибулин
клик
VCP эрибулин
клик
клик
клик
клик
клик
Все значения ИК50 представлены в нМ и представляют собой средние значения повторных экспериментов. СО - стандартное отклонение.
(FRlo, MSLNmed,
her2hi)
(FRneg, MSLNneg, her2hi)
(FRneg, MSLNneg, her2med)
(Партия NB3073-88L)
(Партия NB3073-88F)
(Партия NB3142-62A)
(Партия NB3142-62D)
Все значения ИК50 представлены в нМ и представляют собой средние значения повторных экспериментов. СО - стандартное отклонение.
2.1.1
2.3.1 Цитотоксичность ADC на основе малеимида
[00417] Все ADC MORAb-003 и MORAb-009, созданные на основе малеимида, проявляли специфическую цитотоксичность на клетках IGROV1 с наблюдаемой между антителами разницей по эффективности в 2-3 порядка. ADC MORAb-003 val-cit-pAB-эрибулин продемонстрировали более высокую активность на клеточной линии IGROV1, чем нерасщепляемые ADC MORAb-003 ПЭГ2 или ПЭГ4, однако уровень специфичности не изменился. Аналогичные тенденции наблюдались для ADC MORAb-009, с нерасщепляемыми ADC MORAb-009, демонстрирующими более низкую цитотоксичность на клетках IGROV1, чем ADC MORAb-009 val-cit-pAB-эрибулин.
[00418] ADC MORAb-009 на основе малеимида с линкерами на основе дисульфидила и сульфонамида продемонстрировали более высокую активность на клеточной линии NCI-H2110, чем на клеточной линии IGROV1. Это может быть связано с потенциальной нестабильностью линкеров в культуре, как указано ниже. Также наблюдалась высокая цитотоксичность с соответствующими ADC MORAb-003. Напротив, ADC MORAb-003 и MORAb-009 на основе малеимида с нерасщепляемыми линкерами демонстрировали относительно низкую активность на клетках NCI-H2110. Не привязываясь к теории, этот результат указывает на то, что, при более низкой целевой экспрессии, эффективное расщепление и высвобождение полезной нагрузки может увеличить цитотоксичность.
[00419] ADC с расщепляемым ферментом линкером val-cit-pAB или нерасщепляемым линкером продемонстрировали низкие уровни нецелевого уничтожения на контрольных клетках A431 (ИК50 > 100 нМ), тогда как ADC с расщепляемым ферментом линкером ala-ala-asn-pAB проявили слабое, но обнаруживаемое уничтожение данных контрольных клеток. Это указывает на то, что расщепляемые ферментом линкеры val-cit-pAB могут быть более стабильными в культуре, чем расщепляемые ферментом линкеры ала-ala-asn-pAB. Кроме того, ADC MORAb-009 с более коротким спейсером ПЭГ2 продемонстрировали более высокую цитотоксичность на клетках IGROV1, чем соответствующие ADC с более длинным спейсером ПЭГ8. Аналогичная тенденция наблюдалась на клетках NCI-H2110 как для ADC MORAb-003, так и для ADC MORAb-009 с более короткой длиной спейсера, приводящей к более высокой цитотоксичности.
[00420] ADC с линкерами на основе сульфонамида обычно демонстрировали более высокие значения DAR и более низкие уровни агрегатов по сравнению с соответствующими ADC с линкерами на основе дисульфидила. Однако в обеих этих категориях ADC наблюдалось уничтожение контрольных клеток A431 на наномолярном уровне, указывая на то, что линкеры на основе дисульфидила и сульфонамида были менее стабильными в культуре, чем расщепляемые ферментом линкеры в проверенных условиях анализа.
[00421] Конкретный линкер-токсин малеимидо-ПЭГ2-val-cit-pAB-эрибулин (VCP-эрибулин) дополнительно исследовали на предмет специфичности и активности на различных линиях клеток рака желудка и рака молочной железы. VCP-эрибулин конъюгировали с MORAb-003 и MORAb-009, в дополнение к антителу к her2 человека трастузумабу. MORAb-003-VCP-эрибулин демонстрировал слабое, но специфическое уничтожение клеток NCI-N87, которые экспрессируют низкие уровни рецептора фолиевой кислоты альфа (FR), и незначительное уничтожение на оставшихся трех FR-негативных клеточных линиях. MORAb-009-VCP-эрибулин также демонстрировал высокую цитотоксичность на клетках NCI-N87, которые экспрессируют умеренные уровни мезотелина. Трастузумаб-VCP-эрибулин был очень сильнодействующим (3-6 пМ, ИК50) на клетки NCI-N87 и BT-474, две клеточные линии, которые экспрессируют высокие уровни her2, а также сильнодействующим на клетки рака молочной железы ZR-75, которые лишь умеренно экспрессируют her2. Все ADC MORAb-003, MORAb-009 и трастузумаба VCP-эрибулина продемонстрировали низкую цитотоксичность на клетках NUGC3, которая не экспрессирует FR, мезотелин или her2, соответствующие целевые антигены.
2.3.2 Цитотоксичность ADC на основе сукцинимида
[00422] Тенденции цитотоксичности ADC на основе сукцинимида были сходны с ADC на основе малеимида для клеток IGROV1, с ADC со спейсером ПЭГ8, демонстрирующими низкую цитотоксичность в дополнение к низким значениям DAR. Более низкая цитотоксичность как на клетках IGROV1, так и на NCI-H2110 обычно наблюдалась для ADC на основе сукцинимида с расщепляемыми ферментом линкерами по сравнению с соответствующими ADC на основе малеимида, что, скорее всего, связано с их более низкими значениями DAR. Нецелевое уничтожение клеток A431 наблюдалось также с линкерами на основе дисульфидила и сульфонамида, аналогичными соответствующим ADC на основе малеимида. Это указывает на повышенную нестабильность, скорее потенциально возникающую из-за сайта расщепления, чем из-за химии конъюгации.
[00423] Когда выполняли двухстадийную конъюгацию, наблюдали более высокие значения DAR относительно тех, которые наблюдались при подходе прямой конъюгаци сукцинимида. Эти более высокие значения DAR коррелировали с более высокой эффективностью. Для ADC MORAb-003 VCP-эрибулина наблюдали высокую цитотоксичность как на клетках IGROV1, так и на NCI-H2110. Несмотря на то, что нерасщепляемые ADC MORAb-003 продемонстрировали эффективность на клетках IGROV1 (1-4 нМ), они все еще были менее эффективными, чем ADC MORAb-003 VCP-эрибулин, приготовленные с помощью этого способа (38 пМ), хотя значения DAR были сопоставимыми. Кроме того, нерасщепляемые ADC MORAb-003, приготовленные с применением двухстадийного способа, были немного менее эффективными на клеточной линии IGROV1, чем соответствующие ADC на основе малеимида, что может быть связано с их более низкими значениями DAR. Подобно их аналогам на основе малеимида, нерасщепляемые ADC, приготовленные с применением двухстадийного способа, также утратили почти всю цитотоксичность на клетках NCI-2110.
2.4 Биофизическая характеристика ADC антитела к мезотелину человека (LCcys80)
[00424] MAL-ПЭГ2-Val-Cit-PAB-эрибулин (ER-001159569) конъюгировали с восемью различными антителами к мезотелину человека (таблица 1). Аффинности связывания исходных антител определяли путем анализа BIAcore, как описано выше в разделе 1. 6. 1. Уровни агрегации для всех ADC антител к мезотелину человека определяли с помощью эксклюзионной ВЭЖХ и DAR анализировали с применением HIC-ВЭЖХ. Активность in vitro оценивали, применяя анализ цитотоксичности на клетках на основе окрашивания кристаллическим фиолетовым, на клетках A3 (A431, стабильно трансфицированная мезотелином человека (MSLN), MSLNhi), OVCAR3 (яичник человека, MSLNhi), HEC-251 (эндометроид человека, MSLNmed), H226 (плоскоклеточная мезотелиома легких человека, MSLNlo) и исходной A431 (MSLNneg). Результаты анализа DAR, агрегации и цитотоксичности приведены в таблице 50.
Таблица 50. Биофизическая характеристика ADC антитела к мезотелину человека (LCcys80)
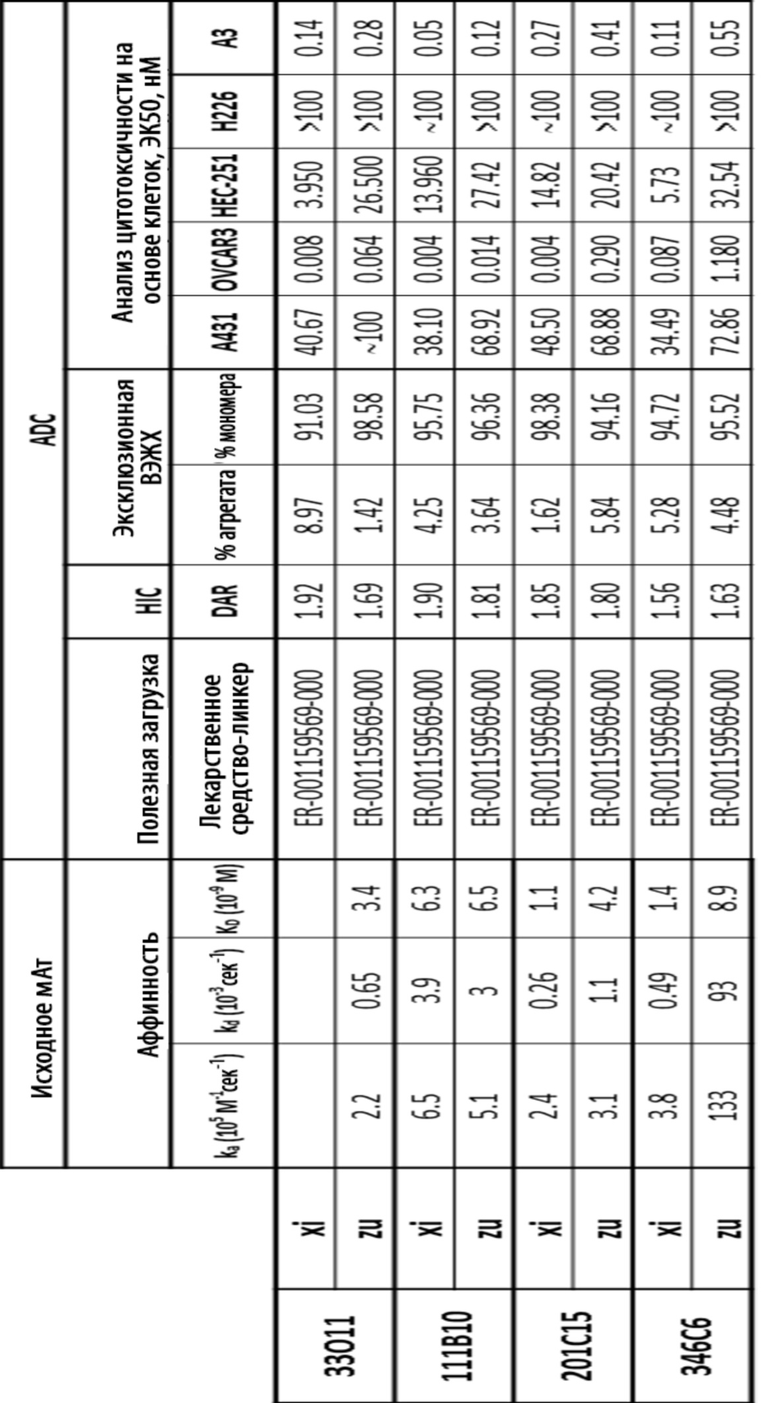
Сокращения: xi - химерное; zu - гуманизированное.
[00425] Все ADC антитела к мезотелину человека сохраняли низкие уровни агрегации (<10% агрегата) и демонстрировали высокую активность на целевых клеточных линиях. Высокая активность наблюдалась на A3 и OVCAR3, тогда как клетки HEC-251 и H226 были относительно устойчивы к цитотоксичности ADC.
Выбранные последовательности:
SEQ ID NO: 1 (тяжелая цепь (HC) MORAb-003)
1 EVQLVESGGG VVQPGRSLRL SCSASGFTFS GYGLSWVRQA PGKGLEWVAM
51 ISSGGSYTYY ADSVKGRFAI SRDNAKNTLF LQMDSLRPED TGVYFCARHG
101 DDPAWFAYWG QGTPVTVSSA STKGPSVFPL APSSKSTSGG TAALGCLVKD
151 YFPEPVTVSW NSGALTSGVH TFPAVLQSSG LYSLSSVVTV PSSSLGTQTY
201 ICNVNHKPSN TKVDKKVEPK SCDKTHTCPP CPAPELLGGP SVFLFPPKPK
251 DTLMISRTPE VTCVVVDVSH EDPEVKFNWY VDGVEVHNAK TKPREEQYNS
301 TYRVVSVLTV LHQDWLNGKE YKCKVSNKAL PAPIEKTISK AKGQPREPQV
351 YTLPPSRDEL TKNQVSLTCL VKGFYPSDIA VEWESNGQPE NNYKTTPPVL
401 DSDGSFFLYS KLTVDKSRWQ QGNVFSCSVM HEALHNHYTQ KSLSLSPGK
SEQ ID NO: 2 (CDR1 HC MORAb-003; по Кабату): GYGLS
SEQ ID NO: 3 (CDR2 HC MORAb-003; по Кабату): MISSGGSYTYYADSVKG
SEQ ID NO: 4 (CDR3 HC MORAb-003; по Кабату): HGDDPAWFAY
SEQ ID NO:5 (аминокислотная последовательность полноразмерного белка-предшественника тяжелой цепи MORAb-003; лидерная последовательность подчеркнута)
1 MGWSCIILFL VATATGVHSE VQLVESGGGV VQPGRSLRLS CSASGFTFSG
51 YGLSWVRQAP GKGLEWVAMI SSGGSYTYYA DSVKGRFAIS RDNAKNTLFL
101 QMDSLRPEDT GVYFCARHGD DPAWFAYWGQ GTPVTVSSAS TKGPSVFPLA
151 PSSKSTSGGT AALGCLVKDY FPEPVTVSWN SGALTSGVHT FPAVLQSSGL
201 YSLSSVVTVP SSSLGTQTYI CNVNHKPSNT KVDKKVEPKS CDKTHTCPPC
251 PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV
301 DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP
351 APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV KGFYPSDIAV
401 EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ GNVFSCSVMH
451 EALHNHYTQK SLSLSPGK
SEQ ID NO: 6 (легкая цепь (LC) MORAb-003)
1 DIQLTQSPSS LSASVGDRVT ITCSVSSSIS SNNLHWYQQK PGKAPKPWIY
51 GTSNLASGVP SRFSGSGSGT DYTFTISSLQ PEDIATYYCQ QWSSYPYMYT
101 FGQGTKVEIK RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ
151 WKVDNALQSG NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT
201 HQGLSSPVTK SFNRGEC
SEQ ID NO: 7 (CDR1 LC MORAb-003; по Кабату): SVSSSISSNNLH
SEQ ID NO: 8 (CDR2 LC MORAb-003: по Кабату): GTSNLAS
SEQ ID NO: 9 (CDR3 LC MORAb-003; по Кабату): QQWSSYPYMYT
SEQ ID NO: 10 аминокислотная последовательность полноразмерного белка-предшественника легкой цепи MORAb-003 (лидерная последовательность подчеркнута)
1 MGWSCIILFL VATATGVHSD IQLTQSPSSL SASVGDRVTI TCSVSSSISS
51 NNLHWYQQKP GKAPKPWIYG TSNLASGVPS RFSGSGSGTD YTFTISSLQP
101 EDIATYYCQQ WSSYPYMYTF GQGTKVEIKR TVAAPSVFIF PPSDEQLKSG
151 TASVVCLLNN FYPREAKVQW KVDNALQSGN SQESVTEQDS KDSTYSLSST
201 LTLSKADYEK HKVYACEVTH QGLSSPVTKS FNRGEC
SEQ ID NO:11 (нт HC MORAb-003)
1 ATGGGATGGA GCTGTATCAT CCTCTTCTTG GTAGCAACAG CTACAGGTGT
51 CCACTCCGAG GTCCAACTGG TGGAGAGCGG TGGAGGTGTT GTGCAACCTG
101 GCCGGTCCCT GCGCCTGTCC TGCTCCGCAT CTGGCTTCAC CTTCAGCGGC
151 TATGGGTTGT CTTGGGTGAG ACAGGCACCT GGAAAAGGTC TTGAGTGGGT
201 TGCAATGATT AGTAGTGGTG GTAGTTATAC CTACTATGCA GACAGTGTGA
251 AGGGTAGATT TGCAATATCG CGAGACAACG CCAAGAACAC ATTGTTCCTG
301 CAAATGGACA GCCTGAGACC CGAAGACACC GGGGTCTATT TTTGTGCAAG
351 ACATGGGGAC GATCCCGCCT GGTTCGCTTA TTGGGGCCAA GGGACCCCGG
401 TCACCGTCTC CTCAGCCTCC ACCAAGGGCC CATCGGTCTT CCCCCTGGCA
451 CCCTCCTCCA AGAGCACCTC TGGGGGCACA GCGGCCCTGG GCTGCCTGGT
501 CAAGGACTAC TTCCCCGAAC CGGTGACGGT GTCGTGGAAC TCAGGCGCCC
551 TGACCAGCGG CGTGCACACC TTCCCGGCTG TCCTACAGTC CTCAGGACTC
601 TACTCCCTCA GCAGCGTGGT GACCGTGCCC TCCAGCAGCT TGGGCACCCA
651 GACCTACATC TGCAACGTGA ATCACAAGCC CAGCAACACC AAGGTGGACA
701 AGAAAGTTGA GCCCAAATCT TGTGACAAAA CTCACACATG CCCACCGTGC
751 CCAGCACCTG AACTCCTGGG GGGACCGTCA GTCTTCCTCT TCCCCCCAAA
801 ACCCAAGGAC ACCCTCATGA TCTCCCGGAC CCCTGAGGTC ACATGCGTGG
851 TGGTGGACGT GAGCCACGAA GACCCTGAGG TCAAGTTCAA CTGGTACGTG
901 GACGGCGTGG AGGTGCATAA TGCCAAGACA AAGCCGCGGG AGGAGCAGTA
951 CAACAGCACG TACCGTGTGG TCAGCGTCCT CACCGTCCTG CACCAGGACT
1001 GGCTGAATGG CAAGGAGTAC AAGTGCAAGG TCTCCAACAA AGCCCTCCCA
1051 GCCCCCATCG AGAAAACCAT CTCCAAAGCC AAAGGGCAGC CCCGAGAACC
1101 ACAGGTGTAC ACCCTGCCCC CATCCCGGGA TGAGCTGACC AAGAACCAGG
1151 TCAGCCTGAC CTGCCTGGTC AAAGGCTTCT ATCCCAGCGA CATCGCCGTG
1201 GAGTGGGAGA GCAATGGGCA GCCGGAGAAC AACTACAAGA CCACGCCTCC
1251 CGTGCTGGAC TCCGACGGCT CCTTCTTCTT ATATTCAAAG CTCACCGTGG
1301 ACAAGAGCAG GTGGCAGCAG GGGAACGTCT TCTCATGCTC CGTGATGCAT
1351 GAGGCTCTGC ACAACCACTA CACGCAGAAG AGCCTCTCCC TGTCTCCCGG
1401 GAAATGA
SEQ ID NO: 12 (нт LC MORAb-003)
1 ATGGGATGGA GCTGTATCAT CCTCTTCTTG GTAGCAACAG CTACAGGTGT
51 CCACTCCGAC ATCCAGCTGA CCCAGAGCCC AAGCAGCCTG AGCGCCAGCG
101 TGGGTGACAG AGTGACCATC ACCTGTAGTG TCAGCTCAAG TATAAGTTCC
151 AACAACTTGC ACTGGTACCA GCAGAAGCCA GGTAAGGCTC CAAAGCCATG
201 GATCTACGGC ACATCCAACC TGGCTTCTGG TGTGCCAAGC AGATTCAGCG
251 GTAGCGGTAG CGGTACCGAC TACACCTTCA CCATCAGCAG CCTCCAGCCA
301 GAGGACATCG CCACCTACTA CTGCCAACAG TGGAGTAGTT ACCCGTACAT
351 GTACACGTTC GGCCAAGGGA CCAAGGTGGA AATCAAACGA ACTGTGGCTG
401 CACCATCTGT CTTCATCTTC CCGCCATCTG ATGAGCAGTT GAAATCTGGA
451 ACTGCCTCTG TTGTGTGCCT GCTGAATAAC TTCTATCCCA GAGAGGCCAA
501 AGTACAGTGG AAGGTGGATA ACGCCCTCCA ATCGGGTAAC TCCCAGGAGA
551 GTGTCACAGA GCAGGACAGC AAGGACAGCA CCTACAGCCT CAGCAGCACC
601 CTGACGCTGA GCAAAGCAGA CTACGAGAAA CACAAAGTCT ACGCCTGCGA
651 AGTCACCCAT CAGGGCCTGA GCTCGCCCGT CACAAAGAGC TTCAACAGGG
701 GAGAGTGTTA A
SEQ ID NO: 13 (CDR1 HC MORAb-003; IMGT): GFTFSGYG
SEQ ID NO: 14 (CDR2 HC MORAb-003; IMGT): ISSGGSYT
SEQ ID NO: 15 (CDR3 HC MORAb-003; IMGT): ARHGDDPAWFAY
SEQ ID NO: 16 (CDR1 LC MORAb-003; IMGT): SSISSNN
SEQ ID NO: 17 (CDR2 LC MORAb-003; IMGT): GTS
SEQ ID NO: 18 (CDR3 LC MORAb-003; IMGT): QQWSSYPYMYT
SEQ ID NO: 19 (FRA человека)
1 maqrmttqll lllvwvavvg eaqtriawar tellnvcmna khhkekpgpe dklheqcrpw
61 rknaccstnt sqeahkdvsy lyrfnwnhcg emapackrhf iqdtclyecs pnlgpwiqqv
121 dqswrkervl nvplckedce qwwedcrtsy tcksnwhkgw nwtsgfnkca vgaacqpfhf
181 yfptptvlcn eiwthsykvs nysrgsgrci qmwfdpaqgn pneevarfya aamsgagpwa
241 awpfllslal mllwlls
SEQ ID NO: 20 (нуклеотид FRA человека)
1 cattccttgg tgccactgac cacagctctt tcttcaggga cagacatggc tcagcggatg
61 acaacacagc tgctgctcct tctagtgtgg gtggctgtag taggggaggc tcagacaagg
121 attgcatggg ccaggactga gcttctcaat gtctgcatga acgccaagca ccacaaggaa
181 aagccaggcc ccgaggacaa gttgcatgag cagtgtcgac cctggaggaa gaatgcctgc
241 tgttctacca acaccagcca ggaagcccat aaggatgttt cctacctata tagattcaac
301 tggaaccact gtggagagat ggcacctgcc tgcaaacggc atttcatcca ggacacctgc
361 ctctacgagt gctcccccaa cttggggccc tggatccagc aggtggatca gagctggcgc
421 aaagagcggg tactgaacgt gcccctgtgc aaagaggact gtgagcaatg gtgggaagat
481 tgtcgcacct cctacacctg caagagcaac tggcacaagg gctggaactg gacttcaggg
541 tttaacaagt gcgcagtggg agctgcctgc caacctttcc atttctactt ccccacaccc
601 actgttctgt gcaatgaaat ctggactcac tcctacaagg tcagcaacta cagccgaggg
661 agtggccgct gcatccagat gtggttcgac ccagcccagg gcaaccccaa tgaggaggtg
721 gcgaggttct atgctgcagc catgagtggg gctgggccct gggcagcctg gcctttcctg
781 cttagcctgg ccctaatgct gctgtggctg ctcagctgac ctccttttac cttctgatac
841 ctggaaatcc ctgccctgtt cagccccaca gctcccaact atttggttcc tgctccatgg
901 tcgggcctct gacagccact ttgaataaac cagacaccgc acatgtgtct tgagaattat
961 ttggaaaaaa aaaaaaaaaa aa
SEQ ID NO: 21 (her2 человека)
1 melaalcrwg lllallppga astqvctgtd mklrlpaspe thldmlrhly qgcqvvqgnl
61 eltylptnas lsflqdiqev qgyvliahnq vrqvplqrlr ivrgtqlfed nyalavldng
121 dplnnttpvt gaspgglrel qlrslteilk ggvliqrnpq lcyqdtilwk difhknnqla
181 ltlidtnrsr achpcspmck gsrcwgesse dcqsltrtvc aggcarckgp lptdccheqc
241 aagctgpkhs dclaclhfnh sgicelhcpa lvtyntdtfe smpnpegryt fgascvtacp
301 ynylstdvgs ctlvcplhnq evtaedgtqr cekcskpcar vcyglgmehl revravtsan
361 iqefagckki fgslaflpes fdgdpasnta plqpeqlqvf etleeitgyl yisawpdslp
421 dlsvfqnlqv irgrilhnga ysltlqglgi swlglrslre lgsglalihh nthlcfvhtv
481 pwdqlfrnph qallhtanrp edecvgegla chqlcarghc wgpgptqcvn csqflrgqec
541 veecrvlqgl preyvnarhc lpchpecqpq ngsvtcfgpe adqcvacahy kdppfcvarc
601 psgvkpdlsy mpiwkfpdee gacqpcpinc thscvdlddk gcpaeqrasp ltsiisavvg
661 illvvvlgvv fgilikrrqq kirkytmrrl lqetelvepl tpsgampnqa qmrilketel
721 rkvkvlgsga fgtvykgiwi pdgenvkipv aikvlrents pkankeilde ayvmagvgsp
781 yvsrllgicl tstvqlvtql mpygclldhv renrgrlgsq dllnwcmqia kgmsyledvr
841 lvhrdlaarn vlvkspnhvk itdfglarll dideteyhad ggkvpikwma lesilrrrft
901 hqsdvwsygv tvwelmtfga kpydgipare ipdllekger lpqppictid vymimvkcwm
961 idsecrprfr elvsefsrma rdpqrfvviq nedlgpaspl dstfyrslle dddmgdlvda
1021 eeylvpqqgf fcpdpapgag gmvhhrhrss strsgggdlt lglepseeea prsplapseg
1081 agsdvfdgdl gmgaakglqs lpthdpsplq rysedptvpl psetdgyvap ltcspqpeyv
1141 nqpdvrpqpp spregplpaa rpagatlerp ktlspgkngv vkdvfafgga venpeyltpq
1201 ggaapqphpp pafspafdnl yywdqdpper gappstfkgt ptaenpeylg ldvpv
SEQ ID NO: 22 (нуклеотид her2 человека)
1 ATGGAGCTGG CGGCCTTGTG CCGCTGGGGG CTCCTCCTCG CCCTCTTGCC CCCCGGAGCC
61 GCGAGCACCC AAGTGTGCAC CGGCACAGAC ATGAAGCTGC GGCTCCCTGC CAGTCCCGAG
121 ACCCACCTGG ACATGCTCCG CCACCTCTAC CAGGGCTGCC AGGTGGTGCA GGGAAACCTG
181 GAACTCACCT ACCTGCCCAC CAATGCCAGC CTGTCCTTCC TGCAGGATAT CCAGGAGGTG
241 CAGGGCTACG TGCTCATCGC TCACAACCAA GTGAGGCAGG TCCCACTGCA GAGGCTGCGG
301 ATTGTGCGAG GCACCCAGCT CTTTGAGGAC AACTATGCCC TGGCCGTGCT AGACAATGGA
361 GACCCGCTGA ACAATACCAC CCCTGTCACA GGGGCCTCCC CAGGAGGCCT GCGGGAGCTG
421 CAGCTTCGAA GCCTCACAGA GATCTTGAAA GGAGGGGTCT TGATCCAGCG GAACCCCCAG
481 CTCTGCTACC AGGACACGAT TTTGTGGAAG GACATCTTCC ACAAGAACAA CCAGCTGGCT
541 CTCACACTGA TAGACACCAA CCGCTCTCGG GCCTGCCACC CCTGTTCTCC GATGTGTAAG
601 GGCTCCCGCT GCTGGGGAGA GAGTTCTGAG GATTGTCAGA GCCTGACGCG CACTGTCTGT
661 GCCGGTGGCT GTGCCCGCTG CAAGGGGCCA CTGCCCACTG ACTGCTGCCA TGAGCAGTGT
721 GCTGCCGGCT GCACGGGCCC CAAGCACTCT GACTGCCTGG CCTGCCTCCA CTTCAACCAC
781 AGTGGCATCT GTGAGCTGCA CTGCCCAGCC CTGGTCACCT ACAACACAGA CACGTTTGAG
841 TCCATGCCCA ATCCCGAGGG CCGGTATACA TTCGGCGCCA GCTGTGTGAC TGCCTGTCCC
901 TACAACTACC TTTCTACGGA CGTGGGATCC TGCACCCTCG TCTGCCCCCT GCACAACCAA
961 GAGGTGACAG CAGAGGATGG AACACAGCGG TGTGAGAAGT GCAGCAAGCC CTGTGCCCGA
1021 GTGTGCTATG GTCTGGGCAT GGAGCACTTG CGAGAGGTGA GGGCAGTTAC CAGTGCCAAT
1081 ATCCAGGAGT TTGCTGGCTG CAAGAAGATC TTTGGGAGCC TGGCATTTCT GCCGGAGAGC
1141 TTTGATGGGG ACCCAGCCTC CAACACTGCC CCGCTCCAGC CAGAGCAGCT CCAAGTGTTT
1201 GAGACTCTGG AAGAGATCAC AGGTTACCTA TACATCTCAG CATGGCCGGA CAGCCTGCCT
1261 GACCTCAGCG TCTTCCAGAA CCTGCAAGTA ATCCGGGGAC GAATTCTGCA CAATGGCGCC
1321 TACTCGCTGA CCCTGCAAGG GCTGGGCATC AGCTGGCTGG GGCTGCGCTC ACTGAGGGAA
1381 CTGGGCAGTG GACTGGCCCT CATCCACCAT AACACCCACC TCTGCTTCGT GCACACGGTG
1441 CCCTGGGACC AGCTCTTTCG GAACCCGCAC CAAGCTCTGC TCCACACTGC CAACCGGCCA
1501 GAGGACGAGT GTGTGGGCGA GGGCCTGGCC TGCCACCAGC TGTGCGCCCG AGGGCACTGC
1561 TGGGGTCCAG GGCCCACCCA GTGTGTCAAC TGCAGCCAGT TCCTTCGGGG CCAGGAGTGC
1621 GTGGAGGAAT GCCGAGTACT GCAGGGGCTC CCCAGGGAGT ATGTGAATGC CAGGCACTGT
1681 TTGCCGTGCC ACCCTGAGTG TCAGCCCCAG AATGGCTCAG TGACCTGTTT TGGACCGGAG
1741 GCTGACCAGT GTGTGGCCTG TGCCCACTAT AAGGACCCTC CCTTCTGCGT GGCCCGCTGC
1801 CCCAGCGGTG TGAAACCTGA CCTCTCCTAC ATGCCCATCT GGAAGTTTCC AGATGAGGAG
1861 GGCGCATGCC AGCCTTGCCC CATCAACTGC ACCCACTCCT GTGTGGACCT GGATGACAAG
1921 GGCTGCCCCG CCGAGCAGAG AGCCAGCCCT CTGACGTCCA TCATCTCTGC GGTGGTTGGC
1981 ATTCTGCTGG TCGTGGTCTT GGGGGTGGTC TTTGGGATCC TCATCAAGCG ACGGCAGCAG
2041 AAGATCCGGA AGTACACGAT GCGGAGACTG CTGCAGGAAA CGGAGCTGGT GGAGCCGCTG
2101 ACACCTAGCG GAGCGATGCC CAACCAGGCG CAGATGCGGA TCCTGAAAGA GACGGAGCTG
2161 AGGAAGGTGA AGGTGCTTGG ATCTGGCGCT TTTGGCACAG TCTACAAGGG CATCTGGATC
2221 CCTGATGGGG AGAATGTGAA AATTCCAGTG GCCATCAAAG TGTTGAGGGA AAACACATCC
2281 CCCAAAGCCA ACAAAGAAAT CTTAGACGAA GCATACGTGA TGGCTGGTGT GGGCTCCCCA
2341 TATGTCTCCC GCCTTCTGGG CATCTGCCTG ACATCCACGG TGCAGCTGGT GACACAGCTT
2401 ATGCCCTATG GCTGCCTCTT AGACCATGTC CGGGAAAACC GCGGACGCCT GGGCTCCCAG
2461 GACCTGCTGA ACTGGTGTAT GCAGATTGCC AAGGGGATGA GCTACCTGGA GGATGTGCGG
2521 CTCGTACACA GGGACTTGGC CGCTCGGAAC GTGCTGGTCA AGAGTCCCAA CCATGTCAAA
2581 ATTACAGACT TCGGGCTGGC TCGGCTGCTG GACATTGACG AGACAGAGTA CCATGCAGAT
2641 GGGGGCAAGG TGCCCATCAA GTGGATGGCG CTGGAGTCCA TTCTCCGCCG GCGGTTCACC
2701 CACCAGAGTG ATGTGTGGAG TTATGGTGTG ACTGTGTGGG AGCTGATGAC TTTTGGGGCC
2761 AAACCTTACG ATGGGATCCC AGCCCGGGAG ATCCCTGACC TGCTGGAAAA GGGGGAGCGG
2821 CTGCCCCAGC CCCCCATCTG CACCATTGAT GTCTACATGA TCATGGTCAA ATGTTGGATG
2881 ATTGACTCTG AATGTCGGCC AAGATTCCGG GAGTTGGTGT CTGAATTCTC CCGCATGGCC
2941 AGGGACCCCC AGCGCTTTGT GGTCATCCAG AATGAGGACT TGGGCCCAGC CAGTCCCTTG
3001 GACAGCACCT TCTACCGCTC ACTGCTGGAG GACGATGACA TGGGGGACCT GGTGGATGCT
3061 GAGGAGTATC TGGTACCCCA GCAGGGCTTC TTCTGTCCAG ACCCTGCCCC GGGCGCTGGG
3121 GGCATGGTCC ACCACAGGCA CCGCAGCTCA TCTACCAGGA GTGGCGGTGG GGACCTGACA
3181 CTAGGGCTGG AGCCCTCTGA AGAGGAGGCC CCCAGGTCTC CACTGGCACC CTCCGAAGGG
3241 GCTGGCTCCG ATGTATTTGA TGGTGACCTG GGAATGGGGG CAGCCAAGGG GCTGCAAAGC
3301 CTCCCCACAC ATGACCCCAG CCCTCTACAG CGGTACAGTG AGGACCCCAC AGTACCCCTG
3361 CCCTCTGAGA CTGATGGCTA CGTTGCCCCC CTGACCTGCA GCCCCCAGCC TGAATATGTG
3421 AACCAGCCAG ATGTTCGGCC CCAGCCCCCT TCGCCCCGAG AGGGCCCTCT GCCTGCTGCC
3481 CGACCTGCTG GTGCCACTCT GGAAAGGCCC AAGACTCTCT CCCCAGGGAA GAATGGGGTC
3541 GTCAAAGACG TTTTTGCCTT TGGGGGTGCC GTGGAGAACC CCGAGTACTT GACACCCCAG
3601 GGAGGAGCTG CCCCTCAGCC CCACCCTCCT CCTGCCTTCA GCCCAGCCTT CGACAACCTC
3661 TATTACTGGG ACCAGGACCC ACCAGAGCGG GGGGCTCCAC CCAGCACCTT CAAAGGGACA
3721 CCTACGGCAG AGAACCCAGA GTACCTGGGT CTGGACGTGC CAGTGTGA
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> EISAI CO., LTD.
EISAI INC.
MORPHOTEK, INC.
<120> КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО НА ОСНОВЕ ЭРИБУЛИНА
И СПОСОБЫ ПРИМЕНЕНИЯ
<130> 08061.0024-00304
<140> PCT/США2017/020529
<141> 2017-03-02
<150> 62/302,562
<151> 2016-03-02
<160> 368
<170> PatentIn версия 3,5
<210> 1
<211> 449
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид"
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Gly Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Met Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Arg His Gly Asp Asp Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 2
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 2
Gly Tyr Gly Leu Ser
1 5
<210> 3
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 3
Met Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 4
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 4
His Gly Asp Asp Pro Ala Trp Phe Ala Tyr
1 5 10
<210> 5
<211> 468
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 5
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Thr Phe
35 40 45
Ser Gly Tyr Gly Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Met Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Ala Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Phe Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val
100 105 110
Tyr Phe Cys Ala Arg His Gly Asp Asp Pro Ala Trp Phe Ala Tyr Trp
115 120 125
Gly Gln Gly Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Lys
465
<210> 6
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 6
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Val Ser Ser Ser Ile Ser Ser Asn
20 25 30
Asn Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Trp
35 40 45
Ile Tyr Gly Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Tyr Met Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 7
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 7
Ser Val Ser Ser Ser Ile Ser Ser Asn Asn Leu His
1 5 10
<210> 8
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 8
Gly Thr Ser Asn Leu Ala Ser
1 5
<210> 9
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 9
Gln Gln Trp Ser Ser Tyr Pro Tyr Met Tyr Thr
1 5 10
<210> 10
<211> 236
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 10
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Val Ser Ser Ser Ile
35 40 45
Ser Ser Asn Asn Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Pro Trp Ile Tyr Gly Thr Ser Asn Leu Ala Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser
100 105 110
Ser Tyr Pro Tyr Met Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 11
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 11
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctacaggtgt ccactccgag 60
gtccaactgg tggagagcgg tggaggtgtt gtgcaacctg gccggtccct gcgcctgtcc 120
tgctccgcat ctggcttcac cttcagcggc tatgggttgt cttgggtgag acaggcacct 180
ggaaaaggtc ttgagtgggt tgcaatgatt agtagtggtg gtagttatac ctactatgca 240
gacagtgtga agggtagatt tgcaatatcg cgagacaacg ccaagaacac attgttcctg 300
caaatggaca gcctgagacc cgaagacacc ggggtctatt tttgtgcaag acatggggac 360
gatcccgcct ggttcgctta ttggggccaa gggaccccgg tcaccgtctc ctcagcctcc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttctt atattcaaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctcccgg gaaatga 1407
<210> 12
<211> 711
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 12
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctacaggtgt ccactccgac 60
atccagctga cccagagccc aagcagcctg agcgccagcg tgggtgacag agtgaccatc 120
acctgtagtg tcagctcaag tataagttcc aacaacttgc actggtacca gcagaagcca 180
ggtaaggctc caaagccatg gatctacggc acatccaacc tggcttctgg tgtgccaagc 240
agattcagcg gtagcggtag cggtaccgac tacaccttca ccatcagcag cctccagcca 300
gaggacatcg ccacctacta ctgccaacag tggagtagtt acccgtacat gtacacgttc 360
ggccaaggga ccaaggtgga aatcaaacga actgtggctg caccatctgt cttcatcttc 420
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac 480
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccctcca atcgggtaac 540
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc 600
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat 660
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgtta a 711
<210> 13
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 13
Gly Phe Thr Phe Ser Gly Tyr Gly
1 5
<210> 14
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 14
Ile Ser Ser Gly Gly Ser Tyr Thr
1 5
<210> 15
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 15
Ala Arg His Gly Asp Asp Pro Ala Trp Phe Ala Tyr
1 5 10
<210> 16
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 16
Ser Ser Ile Ser Ser Asn Asn
1 5
<210> 17
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 17
Gly Thr Ser
1
<210> 18
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 18
Gln Gln Trp Ser Ser Tyr Pro Tyr Met Tyr Thr
1 5 10
<210> 19
<211> 257
<212> ПРТ
<213> Homo sapiens
<400> 19
Met Ala Gln Arg Met Thr Thr Gln Leu Leu Leu Leu Leu Val Trp Val
1 5 10 15
Ala Val Val Gly Glu Ala Gln Thr Arg Ile Ala Trp Ala Arg Thr Glu
20 25 30
Leu Leu Asn Val Cys Met Asn Ala Lys His His Lys Glu Lys Pro Gly
35 40 45
Pro Glu Asp Lys Leu His Glu Gln Cys Arg Pro Trp Arg Lys Asn Ala
50 55 60
Cys Cys Ser Thr Asn Thr Ser Gln Glu Ala His Lys Asp Val Ser Tyr
65 70 75 80
Leu Tyr Arg Phe Asn Trp Asn His Cys Gly Glu Met Ala Pro Ala Cys
85 90 95
Lys Arg His Phe Ile Gln Asp Thr Cys Leu Tyr Glu Cys Ser Pro Asn
100 105 110
Leu Gly Pro Trp Ile Gln Gln Val Asp Gln Ser Trp Arg Lys Glu Arg
115 120 125
Val Leu Asn Val Pro Leu Cys Lys Glu Asp Cys Glu Gln Trp Trp Glu
130 135 140
Asp Cys Arg Thr Ser Tyr Thr Cys Lys Ser Asn Trp His Lys Gly Trp
145 150 155 160
Asn Trp Thr Ser Gly Phe Asn Lys Cys Ala Val Gly Ala Ala Cys Gln
165 170 175
Pro Phe His Phe Tyr Phe Pro Thr Pro Thr Val Leu Cys Asn Glu Ile
180 185 190
Trp Thr His Ser Tyr Lys Val Ser Asn Tyr Ser Arg Gly Ser Gly Arg
195 200 205
Cys Ile Gln Met Trp Phe Asp Pro Ala Gln Gly Asn Pro Asn Glu Glu
210 215 220
Val Ala Arg Phe Tyr Ala Ala Ala Met Ser Gly Ala Gly Pro Trp Ala
225 230 235 240
Ala Trp Pro Phe Leu Leu Ser Leu Ala Leu Met Leu Leu Trp Leu Leu
245 250 255
Ser
<210> 20
<211> 982
<212> ДНК
<213> Homo sapiens
<400> 20
cattccttgg tgccactgac cacagctctt tcttcaggga cagacatggc tcagcggatg 60
acaacacagc tgctgctcct tctagtgtgg gtggctgtag taggggaggc tcagacaagg 120
attgcatggg ccaggactga gcttctcaat gtctgcatga acgccaagca ccacaaggaa 180
aagccaggcc ccgaggacaa gttgcatgag cagtgtcgac cctggaggaa gaatgcctgc 240
tgttctacca acaccagcca ggaagcccat aaggatgttt cctacctata tagattcaac 300
tggaaccact gtggagagat ggcacctgcc tgcaaacggc atttcatcca ggacacctgc 360
ctctacgagt gctcccccaa cttggggccc tggatccagc aggtggatca gagctggcgc 420
aaagagcggg tactgaacgt gcccctgtgc aaagaggact gtgagcaatg gtgggaagat 480
tgtcgcacct cctacacctg caagagcaac tggcacaagg gctggaactg gacttcaggg 540
tttaacaagt gcgcagtggg agctgcctgc caacctttcc atttctactt ccccacaccc 600
actgttctgt gcaatgaaat ctggactcac tcctacaagg tcagcaacta cagccgaggg 660
agtggccgct gcatccagat gtggttcgac ccagcccagg gcaaccccaa tgaggaggtg 720
gcgaggttct atgctgcagc catgagtggg gctgggccct gggcagcctg gcctttcctg 780
cttagcctgg ccctaatgct gctgtggctg ctcagctgac ctccttttac cttctgatac 840
ctggaaatcc ctgccctgtt cagccccaca gctcccaact atttggttcc tgctccatgg 900
tcgggcctct gacagccact ttgaataaac cagacaccgc acatgtgtct tgagaattat 960
ttggaaaaaa aaaaaaaaaa aa 982
<210> 21
<211> 1255
<212> ПРТ
<213> Homo sapiens
<400> 21
Met Glu Leu Ala Ala Leu Cys Arg Trp Gly Leu Leu Leu Ala Leu Leu
1 5 10 15
Pro Pro Gly Ala Ala Ser Thr Gln Val Cys Thr Gly Thr Asp Met Lys
20 25 30
Leu Arg Leu Pro Ala Ser Pro Glu Thr His Leu Asp Met Leu Arg His
35 40 45
Leu Tyr Gln Gly Cys Gln Val Val Gln Gly Asn Leu Glu Leu Thr Tyr
50 55 60
Leu Pro Thr Asn Ala Ser Leu Ser Phe Leu Gln Asp Ile Gln Glu Val
65 70 75 80
Gln Gly Tyr Val Leu Ile Ala His Asn Gln Val Arg Gln Val Pro Leu
85 90 95
Gln Arg Leu Arg Ile Val Arg Gly Thr Gln Leu Phe Glu Asp Asn Tyr
100 105 110
Ala Leu Ala Val Leu Asp Asn Gly Asp Pro Leu Asn Asn Thr Thr Pro
115 120 125
Val Thr Gly Ala Ser Pro Gly Gly Leu Arg Glu Leu Gln Leu Arg Ser
130 135 140
Leu Thr Glu Ile Leu Lys Gly Gly Val Leu Ile Gln Arg Asn Pro Gln
145 150 155 160
Leu Cys Tyr Gln Asp Thr Ile Leu Trp Lys Asp Ile Phe His Lys Asn
165 170 175
Asn Gln Leu Ala Leu Thr Leu Ile Asp Thr Asn Arg Ser Arg Ala Cys
180 185 190
His Pro Cys Ser Pro Met Cys Lys Gly Ser Arg Cys Trp Gly Glu Ser
195 200 205
Ser Glu Asp Cys Gln Ser Leu Thr Arg Thr Val Cys Ala Gly Gly Cys
210 215 220
Ala Arg Cys Lys Gly Pro Leu Pro Thr Asp Cys Cys His Glu Gln Cys
225 230 235 240
Ala Ala Gly Cys Thr Gly Pro Lys His Ser Asp Cys Leu Ala Cys Leu
245 250 255
His Phe Asn His Ser Gly Ile Cys Glu Leu His Cys Pro Ala Leu Val
260 265 270
Thr Tyr Asn Thr Asp Thr Phe Glu Ser Met Pro Asn Pro Glu Gly Arg
275 280 285
Tyr Thr Phe Gly Ala Ser Cys Val Thr Ala Cys Pro Tyr Asn Tyr Leu
290 295 300
Ser Thr Asp Val Gly Ser Cys Thr Leu Val Cys Pro Leu His Asn Gln
305 310 315 320
Glu Val Thr Ala Glu Asp Gly Thr Gln Arg Cys Glu Lys Cys Ser Lys
325 330 335
Pro Cys Ala Arg Val Cys Tyr Gly Leu Gly Met Glu His Leu Arg Glu
340 345 350
Val Arg Ala Val Thr Ser Ala Asn Ile Gln Glu Phe Ala Gly Cys Lys
355 360 365
Lys Ile Phe Gly Ser Leu Ala Phe Leu Pro Glu Ser Phe Asp Gly Asp
370 375 380
Pro Ala Ser Asn Thr Ala Pro Leu Gln Pro Glu Gln Leu Gln Val Phe
385 390 395 400
Glu Thr Leu Glu Glu Ile Thr Gly Tyr Leu Tyr Ile Ser Ala Trp Pro
405 410 415
Asp Ser Leu Pro Asp Leu Ser Val Phe Gln Asn Leu Gln Val Ile Arg
420 425 430
Gly Arg Ile Leu His Asn Gly Ala Tyr Ser Leu Thr Leu Gln Gly Leu
435 440 445
Gly Ile Ser Trp Leu Gly Leu Arg Ser Leu Arg Glu Leu Gly Ser Gly
450 455 460
Leu Ala Leu Ile His His Asn Thr His Leu Cys Phe Val His Thr Val
465 470 475 480
Pro Trp Asp Gln Leu Phe Arg Asn Pro His Gln Ala Leu Leu His Thr
485 490 495
Ala Asn Arg Pro Glu Asp Glu Cys Val Gly Glu Gly Leu Ala Cys His
500 505 510
Gln Leu Cys Ala Arg Gly His Cys Trp Gly Pro Gly Pro Thr Gln Cys
515 520 525
Val Asn Cys Ser Gln Phe Leu Arg Gly Gln Glu Cys Val Glu Glu Cys
530 535 540
Arg Val Leu Gln Gly Leu Pro Arg Glu Tyr Val Asn Ala Arg His Cys
545 550 555 560
Leu Pro Cys His Pro Glu Cys Gln Pro Gln Asn Gly Ser Val Thr Cys
565 570 575
Phe Gly Pro Glu Ala Asp Gln Cys Val Ala Cys Ala His Tyr Lys Asp
580 585 590
Pro Pro Phe Cys Val Ala Arg Cys Pro Ser Gly Val Lys Pro Asp Leu
595 600 605
Ser Tyr Met Pro Ile Trp Lys Phe Pro Asp Glu Glu Gly Ala Cys Gln
610 615 620
Pro Cys Pro Ile Asn Cys Thr His Ser Cys Val Asp Leu Asp Asp Lys
625 630 635 640
Gly Cys Pro Ala Glu Gln Arg Ala Ser Pro Leu Thr Ser Ile Ile Ser
645 650 655
Ala Val Val Gly Ile Leu Leu Val Val Val Leu Gly Val Val Phe Gly
660 665 670
Ile Leu Ile Lys Arg Arg Gln Gln Lys Ile Arg Lys Tyr Thr Met Arg
675 680 685
Arg Leu Leu Gln Glu Thr Glu Leu Val Glu Pro Leu Thr Pro Ser Gly
690 695 700
Ala Met Pro Asn Gln Ala Gln Met Arg Ile Leu Lys Glu Thr Glu Leu
705 710 715 720
Arg Lys Val Lys Val Leu Gly Ser Gly Ala Phe Gly Thr Val Tyr Lys
725 730 735
Gly Ile Trp Ile Pro Asp Gly Glu Asn Val Lys Ile Pro Val Ala Ile
740 745 750
Lys Val Leu Arg Glu Asn Thr Ser Pro Lys Ala Asn Lys Glu Ile Leu
755 760 765
Asp Glu Ala Tyr Val Met Ala Gly Val Gly Ser Pro Tyr Val Ser Arg
770 775 780
Leu Leu Gly Ile Cys Leu Thr Ser Thr Val Gln Leu Val Thr Gln Leu
785 790 795 800
Met Pro Tyr Gly Cys Leu Leu Asp His Val Arg Glu Asn Arg Gly Arg
805 810 815
Leu Gly Ser Gln Asp Leu Leu Asn Trp Cys Met Gln Ile Ala Lys Gly
820 825 830
Met Ser Tyr Leu Glu Asp Val Arg Leu Val His Arg Asp Leu Ala Ala
835 840 845
Arg Asn Val Leu Val Lys Ser Pro Asn His Val Lys Ile Thr Asp Phe
850 855 860
Gly Leu Ala Arg Leu Leu Asp Ile Asp Glu Thr Glu Tyr His Ala Asp
865 870 875 880
Gly Gly Lys Val Pro Ile Lys Trp Met Ala Leu Glu Ser Ile Leu Arg
885 890 895
Arg Arg Phe Thr His Gln Ser Asp Val Trp Ser Tyr Gly Val Thr Val
900 905 910
Trp Glu Leu Met Thr Phe Gly Ala Lys Pro Tyr Asp Gly Ile Pro Ala
915 920 925
Arg Glu Ile Pro Asp Leu Leu Glu Lys Gly Glu Arg Leu Pro Gln Pro
930 935 940
Pro Ile Cys Thr Ile Asp Val Tyr Met Ile Met Val Lys Cys Trp Met
945 950 955 960
Ile Asp Ser Glu Cys Arg Pro Arg Phe Arg Glu Leu Val Ser Glu Phe
965 970 975
Ser Arg Met Ala Arg Asp Pro Gln Arg Phe Val Val Ile Gln Asn Glu
980 985 990
Asp Leu Gly Pro Ala Ser Pro Leu Asp Ser Thr Phe Tyr Arg Ser Leu
995 1000 1005
Leu Glu Asp Asp Asp Met Gly Asp Leu Val Asp Ala Glu Glu Tyr
1010 1015 1020
Leu Val Pro Gln Gln Gly Phe Phe Cys Pro Asp Pro Ala Pro Gly
1025 1030 1035
Ala Gly Gly Met Val His His Arg His Arg Ser Ser Ser Thr Arg
1040 1045 1050
Ser Gly Gly Gly Asp Leu Thr Leu Gly Leu Glu Pro Ser Glu Glu
1055 1060 1065
Glu Ala Pro Arg Ser Pro Leu Ala Pro Ser Glu Gly Ala Gly Ser
1070 1075 1080
Asp Val Phe Asp Gly Asp Leu Gly Met Gly Ala Ala Lys Gly Leu
1085 1090 1095
Gln Ser Leu Pro Thr His Asp Pro Ser Pro Leu Gln Arg Tyr Ser
1100 1105 1110
Glu Asp Pro Thr Val Pro Leu Pro Ser Glu Thr Asp Gly Tyr Val
1115 1120 1125
Ala Pro Leu Thr Cys Ser Pro Gln Pro Glu Tyr Val Asn Gln Pro
1130 1135 1140
Asp Val Arg Pro Gln Pro Pro Ser Pro Arg Glu Gly Pro Leu Pro
1145 1150 1155
Ala Ala Arg Pro Ala Gly Ala Thr Leu Glu Arg Pro Lys Thr Leu
1160 1165 1170
Ser Pro Gly Lys Asn Gly Val Val Lys Asp Val Phe Ala Phe Gly
1175 1180 1185
Gly Ala Val Glu Asn Pro Glu Tyr Leu Thr Pro Gln Gly Gly Ala
1190 1195 1200
Ala Pro Gln Pro His Pro Pro Pro Ala Phe Ser Pro Ala Phe Asp
1205 1210 1215
Asn Leu Tyr Tyr Trp Asp Gln Asp Pro Pro Glu Arg Gly Ala Pro
1220 1225 1230
Pro Ser Thr Phe Lys Gly Thr Pro Thr Ala Glu Asn Pro Glu Tyr
1235 1240 1245
Leu Gly Leu Asp Val Pro Val
1250 1255
<210> 22
<211> 3768
<212> ДНК
<213> Homo sapiens
<400> 22
atggagctgg cggccttgtg ccgctggggg ctcctcctcg ccctcttgcc ccccggagcc 60
gcgagcaccc aagtgtgcac cggcacagac atgaagctgc ggctccctgc cagtcccgag 120
acccacctgg acatgctccg ccacctctac cagggctgcc aggtggtgca gggaaacctg 180
gaactcacct acctgcccac caatgccagc ctgtccttcc tgcaggatat ccaggaggtg 240
cagggctacg tgctcatcgc tcacaaccaa gtgaggcagg tcccactgca gaggctgcgg 300
attgtgcgag gcacccagct ctttgaggac aactatgccc tggccgtgct agacaatgga 360
gacccgctga acaataccac ccctgtcaca ggggcctccc caggaggcct gcgggagctg 420
cagcttcgaa gcctcacaga gatcttgaaa ggaggggtct tgatccagcg gaacccccag 480
ctctgctacc aggacacgat tttgtggaag gacatcttcc acaagaacaa ccagctggct 540
ctcacactga tagacaccaa ccgctctcgg gcctgccacc cctgttctcc gatgtgtaag 600
ggctcccgct gctggggaga gagttctgag gattgtcaga gcctgacgcg cactgtctgt 660
gccggtggct gtgcccgctg caaggggcca ctgcccactg actgctgcca tgagcagtgt 720
gctgccggct gcacgggccc caagcactct gactgcctgg cctgcctcca cttcaaccac 780
agtggcatct gtgagctgca ctgcccagcc ctggtcacct acaacacaga cacgtttgag 840
tccatgccca atcccgaggg ccggtataca ttcggcgcca gctgtgtgac tgcctgtccc 900
tacaactacc tttctacgga cgtgggatcc tgcaccctcg tctgccccct gcacaaccaa 960
gaggtgacag cagaggatgg aacacagcgg tgtgagaagt gcagcaagcc ctgtgcccga 1020
gtgtgctatg gtctgggcat ggagcacttg cgagaggtga gggcagttac cagtgccaat 1080
atccaggagt ttgctggctg caagaagatc tttgggagcc tggcatttct gccggagagc 1140
tttgatgggg acccagcctc caacactgcc ccgctccagc cagagcagct ccaagtgttt 1200
gagactctgg aagagatcac aggttaccta tacatctcag catggccgga cagcctgcct 1260
gacctcagcg tcttccagaa cctgcaagta atccggggac gaattctgca caatggcgcc 1320
tactcgctga ccctgcaagg gctgggcatc agctggctgg ggctgcgctc actgagggaa 1380
ctgggcagtg gactggccct catccaccat aacacccacc tctgcttcgt gcacacggtg 1440
ccctgggacc agctctttcg gaacccgcac caagctctgc tccacactgc caaccggcca 1500
gaggacgagt gtgtgggcga gggcctggcc tgccaccagc tgtgcgcccg agggcactgc 1560
tggggtccag ggcccaccca gtgtgtcaac tgcagccagt tccttcgggg ccaggagtgc 1620
gtggaggaat gccgagtact gcaggggctc cccagggagt atgtgaatgc caggcactgt 1680
ttgccgtgcc accctgagtg tcagccccag aatggctcag tgacctgttt tggaccggag 1740
gctgaccagt gtgtggcctg tgcccactat aaggaccctc ccttctgcgt ggcccgctgc 1800
cccagcggtg tgaaacctga cctctcctac atgcccatct ggaagtttcc agatgaggag 1860
ggcgcatgcc agccttgccc catcaactgc acccactcct gtgtggacct ggatgacaag 1920
ggctgccccg ccgagcagag agccagccct ctgacgtcca tcatctctgc ggtggttggc 1980
attctgctgg tcgtggtctt gggggtggtc tttgggatcc tcatcaagcg acggcagcag 2040
aagatccgga agtacacgat gcggagactg ctgcaggaaa cggagctggt ggagccgctg 2100
acacctagcg gagcgatgcc caaccaggcg cagatgcgga tcctgaaaga gacggagctg 2160
aggaaggtga aggtgcttgg atctggcgct tttggcacag tctacaaggg catctggatc 2220
cctgatgggg agaatgtgaa aattccagtg gccatcaaag tgttgaggga aaacacatcc 2280
cccaaagcca acaaagaaat cttagacgaa gcatacgtga tggctggtgt gggctcccca 2340
tatgtctccc gccttctggg catctgcctg acatccacgg tgcagctggt gacacagctt 2400
atgccctatg gctgcctctt agaccatgtc cgggaaaacc gcggacgcct gggctcccag 2460
gacctgctga actggtgtat gcagattgcc aaggggatga gctacctgga ggatgtgcgg 2520
ctcgtacaca gggacttggc cgctcggaac gtgctggtca agagtcccaa ccatgtcaaa 2580
attacagact tcgggctggc tcggctgctg gacattgacg agacagagta ccatgcagat 2640
gggggcaagg tgcccatcaa gtggatggcg ctggagtcca ttctccgccg gcggttcacc 2700
caccagagtg atgtgtggag ttatggtgtg actgtgtggg agctgatgac ttttggggcc 2760
aaaccttacg atgggatccc agcccgggag atccctgacc tgctggaaaa gggggagcgg 2820
ctgccccagc cccccatctg caccattgat gtctacatga tcatggtcaa atgttggatg 2880
attgactctg aatgtcggcc aagattccgg gagttggtgt ctgaattctc ccgcatggcc 2940
agggaccccc agcgctttgt ggtcatccag aatgaggact tgggcccagc cagtcccttg 3000
gacagcacct tctaccgctc actgctggag gacgatgaca tgggggacct ggtggatgct 3060
gaggagtatc tggtacccca gcagggcttc ttctgtccag accctgcccc gggcgctggg 3120
ggcatggtcc accacaggca ccgcagctca tctaccagga gtggcggtgg ggacctgaca 3180
ctagggctgg agccctctga agaggaggcc cccaggtctc cactggcacc ctccgaaggg 3240
gctggctccg atgtatttga tggtgacctg ggaatggggg cagccaaggg gctgcaaagc 3300
ctccccacac atgaccccag ccctctacag cggtacagtg aggaccccac agtacccctg 3360
ccctctgaga ctgatggcta cgttgccccc ctgacctgca gcccccagcc tgaatatgtg 3420
aaccagccag atgttcggcc ccagccccct tcgccccgag agggccctct gcctgctgcc 3480
cgacctgctg gtgccactct ggaaaggccc aagactctct ccccagggaa gaatggggtc 3540
gtcaaagacg tttttgcctt tgggggtgcc gtggagaacc ccgagtactt gacaccccag 3600
ggaggagctg cccctcagcc ccaccctcct cctgccttca gcccagcctt cgacaacctc 3660
tattactggg accaggaccc accagagcgg ggggctccac ccagcacctt caaagggaca 3720
cctacggcag agaacccaga gtacctgggt ctggacgtgc cagtgtga 3768
<210> 23
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 23
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Gly Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Met Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Arg His Gly Asp Asp Pro Ala Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser
115
<210> 24
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 24
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Val Ser Ser Ser Ile Ser Ser Asn
20 25 30
Asn Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Trp
35 40 45
Ile Tyr Gly Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro
85 90 95
Tyr Met Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 25
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 25
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Glu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Leu Ile Thr Pro Tyr Asn Gly Ala Ser Ser Tyr Asn Gln Lys Phe
50 55 60
Arg Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Asp Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Tyr Asp Gly Arg Gly Phe Asp Tyr Trp Gly Ser Gly
100 105 110
Thr Pro Val Thr Val Ser Ser
115
<210> 26
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 26
Asp Ile Glu Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Gly Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Asn Ser Tyr Ser Leu Thr Ile Ser Ser Val Glu Ala Glu
65 70 75 80
Asp Asp Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Lys His Pro Leu Thr
85 90 95
Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105
<210> 27
<211> 120
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 28
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 28
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 29
<211> 123
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 29
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Ser Leu Ser Ser Asp Ala
20 25 30
Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Ile Ile Asn Gly Gly Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Ile
85 90 95
Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr Gly Met Asp Leu
100 105 110
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 30
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 30
Glu Val Leu Met Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Asp Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Ser Ser Val
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Thr Asn Tyr Gly Thr Ser Ser
85 90 95
Ser Asn Tyr Gly Phe Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 31
<211> 126
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 31
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Ser Leu Ser Ser Asp
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ile Ile Asn Gly Gly Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg His Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Ile Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr Gly
100 105 110
Met Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 32
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 32
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Val
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Cys
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Thr Asn Tyr Gly Thr Ser Ser
85 90 95
Ser Asn Tyr Gly Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 33
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 33
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Asn Asn Tyr Ala
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ser Ile Ser Thr Gly Gly Leu Ala Phe Tyr Ala Asn Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Gly Arg Asn Gly
85 90 95
Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 34
<211> 108
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 34
Ala Phe Glu Leu Thr Gln Thr Pro Ser Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Ile Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Phe Cys Gln Ser Tyr Tyr Asp Ile Gly Thr
85 90 95
Ser Thr Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105
<210> 35
<211> 121
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 35
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Asn Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Ser Thr Gly Gly Leu Ala Phe Tyr Ala Asn Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asn Gly Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 36
<211> 108
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 36
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Cys
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Ser Tyr Tyr Asp Ile Gly Thr
85 90 95
Ser Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 37
<211> 114
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 37
Gln Ser Val Lys Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr Ala
20 25 30
Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Thr Ile Asn Ile Gly Gly Arg Val Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Ala Pro
65 70 75 80
Ser Leu Thr Ala Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Tyr Tyr
85 90 95
Asn Gly Gly Ser Tyr Asp Ile Trp Gly Pro Gly Thr Leu Val Thr Val
100 105 110
Ser Leu
<210> 38
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 38
Asp Val Val Met Thr Gln Thr Pro Ala Ser Ala Ser Glu Pro Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Glu Ser Ile Tyr Arg Val
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Thr Ser Thr Leu Ala Ser Gly Ala Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Tyr Gly Thr Glu Phe Thr Leu Thr Ile Ser Gly Val Gln Cys
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Tyr Tyr Ala Asp Ser
85 90 95
Tyr Gly Ile Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 39
<211> 117
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 39
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Thr Ile Asn Ile Gly Gly Arg Val Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Tyr Tyr Asn Gly Gly Ser Tyr Asp Ile Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 40
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Tyr Arg Val
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Thr Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Cys
65 70 75 80
Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Tyr Tyr Ala Asp Ser
85 90 95
Tyr Gly Ile Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 41
<211> 122
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 41
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Lys Pro Asp Glu Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr Ala
20 25 30
Met Ile Trp Val Arg Gln Ala Pro Gly Glu Gly Leu Glu Trp Ile Gly
35 40 45
Thr Ile Ser Thr Gly Gly Ile Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Gly
85 90 95
Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr Phe Asp Leu Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 42
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 42
Ala Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Asn Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Cys Asp Asp
85 90 95
Asp Ala Asp Thr Phe Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 43
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 43
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Ala Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Ser Thr Gly Gly Ile Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Gly Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr Phe
100 105 110
Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 44
<211> 112
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 44
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu
35 40 45
Leu Ile Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Cys Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Cys Asp Asp
85 90 95
Asp Ala Asp Thr Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 45
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 45
gaggtccaac tggtggagag cggtggaggt gttgtgcaac ctggccggtc cctgcgcctg 60
tcctgctccg catctggctt caccttcagc ggctatgggt tgtcttgggt gagacaggca 120
cctggaaaag gtcttgagtg ggttgcaatg attagtagtg gtggtagtta tacctactat 180
gcagacagtg tgaagggtag atttgcaata tcgcgagaca acgccaagaa cacattgttc 240
ctgcaaatgg acagcctgag acccgaagac accggggtct atttttgtgc aagacatggg 300
gacgatcccg cctggttcgc ttattggggc caagggaccc cggtcaccgt ctcctca 357
<210> 46
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 46
gacatccagc tgacccagag cccaagcagc ctgagcgcca gcgtgggtga cagagtgacc 60
atcacctgta gtgtcagctc aagtataagt tccaacaact tgcactggta ccagcagaag 120
ccaggtaagg ctccaaagcc atggatctac ggcacatcca acctggcttc tggtgtgcca 180
agcagattca gcggtagcgg tagcggtacc gactacacct tcaccatcag cagcctccag 240
ccagaggaca tcgccaccta ctactgccaa cagtggagta gttacccgta catgtacacg 300
ttcggccaag ggaccaaggt ggaaatcaaa 330
<210> 47
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 47
caggtacaac tgcagcagtc tgggcctgag ctggagaagc ctggcgcttc agtgaagata 60
tcctgcaagg cttctggtta ctcattcact ggctacacca tgaactgggt gaagcagagc 120
catggaaaga gccttgagtg gattggactt attactcctt acaatggtgc ttctagctac 180
aaccagaagt tcaggggcaa ggccacatta actgtagaca agtcatccag cacagcctac 240
atggacctcc tcagtctgac atctgaagac tctgcagtct atttctgtgc aagggggggt 300
tacgacggga ggggttttga ctactgggga tccgggaccc cggtcaccgt ctcctca 357
<210> 48
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 48
gacatcgagc tcactcagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccagctc aagtgtaagt tacatgcact ggtaccagca gaagtcaggc 120
acctccccca aaagatggat ttatgacaca tccaaactgg cttctggagt cccaggtcgc 180
ttcagtggca gtgggtctgg aaactcttac tctctcacaa tcagcagcgt ggaggctgaa 240
gatgatgcaa cttattactg ccagcagtgg agtaagcacc ctctcacgtt cggatccggg 300
accaaggtgg aaatcaaa 318
<210> 49
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 49
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcaccgtct ctggaatctc cctcagtagc gatgcaataa gctgggtccg ccaggctcca 120
gggaaggggc tcgaatacat cggaatcatt aatggtggtg gtaacacata ctacgcgagc 180
tgggcgaaag gccgattcac catctccaaa acctcgacca cggtggatct gaaaatcacc 240
agtccgacaa ccgaggacac ggccacctat ttctgtgcca gaggcattca acatggtggt 300
ggtaatagtg attattatta ttacggcatg gacctctggg gcccaggcac cctggtcact 360
gtctcttca 369
<210> 50
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 50
gaagtgttga tgacccagac tccatcctcc gtgtctgcag ctgtgggaga cacagtcacc 60
atcaagtgcc aggccagtca gagcattagt agtgtcttgt cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctatctg gcatccactc tggcatctgg ggtcccatcg 180
cggttcagcg gcagtagatc tgggacagag ttcactctca ccatcagcga cctggagtgt 240
gacgatgctg ccacttacta ctgtcaaacc aattatggta ctagtagtag taattatggt 300
tttgctttcg gcggagggac cgaggtggtc gtcaaa 336
<210> 51
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 51
gaagtccaac tggtggaaag cgggggagga ctggtgcagc cgggcggatc cctccggctg 60
tcatgtgctg catcgggaat ttccctctcc tccgacgcga ttagctgggt cagacaggcc 120
cccggaaagg ggctggagta catcggtatc atcaacggcg gcggaaacac ctactacgcc 180
tcctgggcca agggccgctt caccatctcg cggcataatt ccaagaacac tctgtacttg 240
caaatgaact ccctgagggc cgaggacacc gccgtgtact actgcgcgcg cggcatccag 300
cacggtggtg gaaacagcga ctactactac tatgggatgg atctgtgggg ccagggaact 360
cttgtgaccg tgtcgtca 378
<210> 52
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 52
gacattcaga tgacccagtc cccaagctcg ctgtccgcct ccgtgggcga ccgcgtgacc 60
atcacgtgcc aggcgtccca gtcaattagc agcgtgctct cctggtacca acagaagccg 120
gggaaagcac ccaagctgct gatctacttg gcctccactc tggcctcggg agtgccttca 180
cggttctccg gatcgggatc tggtactgat ttcaccctca ccatctcgag ccttcagtgc 240
gaggacatcg ctacttacta ttgtcaaacc aactacggaa cctccagctc caactacggc 300
tttgccttcg gtggcgggac caaggtcgaa atcaaa 336
<210> 53
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 53
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcacagtct ctggattctc cctcaataac tatgcaatga gctgggtccg ccaggctcca 120
gggaaggggc tggaatggat cggatccatt agtactggtg gtctcgcatt ctacgcgaac 180
tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaaatgacc 240
agtctgacaa ccgaggacac ggccacctat ttctgtggca gaaatggtgg tggtagttat 300
attttctatt attttgactt gtggggccaa ggcaccctcg tcactgtctc ttca 354
<210> 54
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 54
gcattcgaat tgacccagac tccatcctcc gtggaggcag ctgtgggagg cacaatcacc 60
atcaagtgcc aggccagtca gagcattagt agttacttat cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctattct gcatccactc tggcatctgg ggtctcatcg 180
cggttcaaag gcagtggatc tgggacagag tacactctca ccatcagcga cctggagtgt 240
gccgatgctg ccacttactt ctgtcaaagc tattatgata ttggtactag tactttcggc 300
ggagggaccg aggtggtcgt caaa 324
<210> 55
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 55
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
tcttgtgccg cctccggctt ctccctgaac aactacgcca tgtcctgggt gcgacaggcc 120
cctggcaaag gcctggaatg gatcggctcc atcagcacag gcggcctggc cttctacgcc 180
aattgggcca agggccggtt caccatcagc cgggacaact ccaagaacac cctgtacctc 240
cagatgaact ccctgcgggc cgaggacacc gccgtgtact actgtgccag aaacggcgga 300
ggctcctaca tcttctacta cttcgacctg tggggccagg gcaccctcgt gacagtgtca 360
tct 363
<210> 56
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 56
gatattcaga tgacccagtc cccctccagc ctgtccgctt ctgtgggcga cagagtgacc 60
atcacctgtc aggcctccca gtccatctcc tcctacctgt cctggtatca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactct gcctccacac tggcctccgg cgtgccctct 180
agattctccg gctctggctc tggcaccgac tttaccctga ccatcagctc cctccagtgc 240
gaggatgccg ccacctacta ctgccagtcc tactacgaca tcggcacctc caccttcggc 300
ggaggcacca aggtggaaat caaa 324
<210> 57
<211> 342
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 57
cagtcagtga aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcacagtct ctggaatcga cctcagtagc tatgcaatgg gctggttccg ccaggctcca 120
gggaaggggc tggaatacat cggaaccatt aatattggtg gtcgcgtata ttacgcgagc 180
tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaagcgccc 240
agtctgacag ccgaggacac ggccacctat ttctgtgcca gatattataa tggtggtagt 300
tatgacatct ggggcccagg caccctggtc accgtctctt ta 342
<210> 58
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 58
gatgttgtga tgacccagac tccagcctcc gcgtctgaac ctgtgggagg cacagtcacc 60
atcaagtgcc aggccagtga gagcatttat cgcgtattgg cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctatgat acatccactc tggcatctgg ggccccatcg 180
cggttcaaag gcagtggata tgggacagag ttcactctca ccatcagcgg cgtgcagtgt 240
gaagatgctg ccacttacta ctgtcaaggc ggttattatg ctgatagtta tggtattgct 300
ttcggcggag ggaccgaggt ggtggtcaaa 330
<210> 59
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 59
caggtgcagc tggtggaatc tggcggagga ctggtgcagc ctggcggctc tctgagactg 60
tcctgttccg cctccggaat cgacctgtcc tcctacgcta tgggctgggt gcgacaggct 120
cctggcaagg gcctggagta catcggcacc atcaacatcg gcggcagagt gtactacgcc 180
tcctgggcca agggccggtt caccatctcc agagacaact ccaagaacac cctgtacctc 240
cagatgaact ccctgcgggc cgaggacacc gccgtgtact actgcgcccg gtactacaac 300
ggcggctcct acgatatctg gggccagggc acactcgtga ccgtgtcctc t 351
<210> 60
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 60
gatatccaga tgacccagtc cccctccacc ctgtctgcct ctgtgggcga cagagtgacc 60
atcacctgtc aggcctccga gtccatctac cgggtgctgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacgac accagcacac tggcctccgg cgtgccctct 180
agattctccg gctctggctc tggcaccgag tttaccctga ccatctccag cctccagtgc 240
gacgacgccg ccacctacta ttgtcagggc ggctactacg ccgactccta cggaatcgct 300
ttcggcggag gcaccaaggt ggaaatcaaa 330
<210> 61
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 61
cagtcggtgg aggagtccgg cggtcgcctg gtaaagcctg acgaatccct gacactcacc 60
tgcacagcct ctggattctc cctcagtagt tatgcaatga tctgggtccg ccaggctcca 120
ggggaggggc tggaatggat cggaaccatt agtactggtg gtatcacata ctacgcgagc 180
tgggcgaaag gccgattcac catctccaaa acctcgacca cggtggatct gaaaatcacc 240
agtccgacaa ccgaggacac ggccacctat ttctgtgcca gagggggata tgctgctagt 300
agtgcttatt atctcccgta ctactttgac ttgtggggcc aagggaccct ggtcaccgtc 360
tcctca 366
<210> 62
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 62
gcagccgtgc tgacccagac accatcaccc gtgtctgcag ctgtgggagg cacagtcacc 60
atcagttgcc agtccagtca gagtgtttat aataataaca acttagcctg gtttcagcag 120
aaacccgggc agcctcccaa gcttctgatc tatctggcat ccactctggc atctggggtc 180
ccatcacggt tcagcggcag tggatctggg acacagttca ctctcaccat cagcggcgtg 240
cagtgtgacg atgctgccac ttattactgt ctaggtggtt gtgatgatga tgctgatact 300
tttgctttcg gcggagggac tgaggtggtg gtcaaa 336
<210> 63
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 63
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
tcttgtgccg cctccggctt ctccctgtcc tcctacgcta tgatctgggt gcgacaggcc 120
cctggcaagg gcctggaatg gatcggcacc atctctaccg gcggaattac ctactacgcc 180
tcctgggcca agggccggtt caccatctcc agagacaact ccaagaacac cctgtacctc 240
cagatgaact ccctgcgggc cgaggacacc gccgtgtact attgtgctag aggcggctac 300
gccgccagct ccgcttacta cctgccctac tacttcgacc tgtggggcca gggcaccctc 360
gtgacagtgt catct 375
<210> 64
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 64
gatattcaga tgacccagtc cccctccagc ctgtccgctt ctgtgggcga cagagtgacc 60
atcacctgtc agtcctccca gtccgtgtat aacaacaaca acctggcctg gtatcagcag 120
aaacccggca aggtgcccaa gctgctgatc tacctggcct ccacactggc ctctggcgtg 180
ccctctagat tctccggctc tggctctggc accgacttta ccctgaccat cagctccctc 240
cagtgcgagg atgccgccac ctactattgc ctgggcggct gcgacgacga cgccgatacc 300
tttgcttttg gcggaggcac caaggtggaa atcaaa 336
<210> 65
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 65
Gly Tyr Thr Met Asn
1 5
<210> 66
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 66
Leu Ile Thr Pro Tyr Asn Gly Ala Ser Ser Tyr Asn Gln Lys Phe Arg
1 5 10 15
Gly
<210> 67
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 67
Gly Gly Tyr Asp Gly Arg Gly Phe Asp Tyr
1 5 10
<210> 68
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 68
Ser Ala Ser Ser Ser Val Ser Tyr Met His
1 5 10
<210> 69
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 69
Asp Thr Ser Lys Leu Ala Ser
1 5
<210> 70
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 70
Gln Gln Trp Ser Lys His Pro Leu Thr
1 5
<210> 71
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 71
Asp Thr Tyr Ile His
1 5
<210> 72
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 72
Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 73
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 73
Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 74
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 74
Arg Ala Ser Gln Asp Val Asn Thr Ala Val Ala
1 5 10
<210> 75
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 75
Ser Ala Ser Phe Leu Tyr Ser
1 5
<210> 76
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 76
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 77
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 77
Ser Asp Ala Ile Ser
1 5
<210> 78
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 78
Ile Ile Asn Gly Gly Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 79
<211> 18
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 79
Gly Ile Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr Gly Met
1 5 10 15
Asp Leu
<210> 80
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 80
Gln Ala Ser Gln Ser Ile Ser Ser Val Leu Ser
1 5 10
<210> 81
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 81
Leu Ala Ser Thr Leu Ala Ser
1 5
<210> 82
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 82
Gln Thr Asn Tyr Gly Thr Ser Ser Ser Asn Tyr Gly Phe Ala
1 5 10
<210> 83
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 83
Ser Asp Ala Ile Ser
1 5
<210> 84
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 84
Ile Ile Asn Gly Gly Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 85
<211> 18
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 85
Gly Ile Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr Gly Met
1 5 10 15
Asp Leu
<210> 86
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 86
Gln Ala Ser Gln Ser Ile Ser Ser Val Leu Ser
1 5 10
<210> 87
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 87
Leu Ala Ser Thr Leu Ala Ser
1 5
<210> 88
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 88
Gln Thr Asn Tyr Gly Thr Ser Ser Ser Asn Tyr Gly Phe Ala
1 5 10
<210> 89
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 89
Asn Tyr Ala Met Ser
1 5
<210> 90
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 90
Ser Ile Ser Thr Gly Gly Leu Ala Phe Tyr Ala Asn Trp Ala Lys Gly
1 5 10 15
<210> 91
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 91
Asn Gly Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu
1 5 10
<210> 92
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 92
Gln Ala Ser Gln Ser Ile Ser Ser Tyr Leu Ser
1 5 10
<210> 93
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 93
Ser Ala Ser Thr Leu Ala Ser
1 5
<210> 94
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 94
Gln Ser Tyr Tyr Asp Ile Gly Thr Ser Thr
1 5 10
<210> 95
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 95
Asn Tyr Ala Met Ser
1 5
<210> 96
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 96
Ser Ile Ser Thr Gly Gly Leu Ala Phe Tyr Ala Asn Trp Ala Lys Gly
1 5 10 15
<210> 97
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 97
Asn Gly Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu
1 5 10
<210> 98
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 98
Gln Ala Ser Gln Ser Ile Ser Ser Tyr Leu Ser
1 5 10
<210> 99
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 99
Ser Ala Ser Thr Leu Ala Ser
1 5
<210> 100
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 100
Gln Ser Tyr Tyr Asp Ile Gly Thr Ser Thr
1 5 10
<210> 101
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 101
Ser Tyr Ala Met Gly
1 5
<210> 102
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 102
Thr Ile Asn Ile Gly Gly Arg Val Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 103
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 103
Tyr Tyr Asn Gly Gly Ser Tyr Asp Ile
1 5
<210> 104
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 104
Gln Ala Ser Glu Ser Ile Tyr Arg Val Leu Ala
1 5 10
<210> 105
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 105
Asp Thr Ser Thr Leu Ala Ser
1 5
<210> 106
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 106
Gln Gly Gly Tyr Tyr Ala Asp Ser Tyr Gly Ile Ala
1 5 10
<210> 107
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 107
Ser Tyr Ala Met Gly
1 5
<210> 108
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 108
Thr Ile Asn Ile Gly Gly Arg Val Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 109
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 109
Tyr Tyr Asn Gly Gly Ser Tyr Asp Ile
1 5
<210> 110
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 110
Gln Ala Ser Glu Ser Ile Tyr Arg Val Leu Ala
1 5 10
<210> 111
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 111
Asp Thr Ser Thr Leu Ala Ser
1 5
<210> 112
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 112
Gln Gly Gly Tyr Tyr Ala Asp Ser Tyr Gly Ile Ala
1 5 10
<210> 113
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 113
Ser Tyr Ala Met Ile
1 5
<210> 114
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 114
Thr Ile Ser Thr Gly Gly Ile Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 115
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 115
Gly Gly Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr Phe Asp
1 5 10 15
Leu
<210> 116
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 116
Gln Ser Ser Gln Ser Val Tyr Asn Asn Asn Asn Leu Ala
1 5 10
<210> 117
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 117
Leu Ala Ser Thr Leu Ala Ser
1 5
<210> 118
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 118
Leu Gly Gly Cys Asp Asp Asp Ala Asp Thr Phe Ala
1 5 10
<210> 119
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 119
Ser Tyr Ala Met Ile
1 5
<210> 120
<211> 16
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 120
Thr Ile Ser Thr Gly Gly Ile Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 121
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 121
Gly Gly Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr Phe Asp
1 5 10 15
Leu
<210> 122
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 122
Gln Ser Ser Gln Ser Val Tyr Asn Asn Asn Asn Leu Ala
1 5 10
<210> 123
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 123
Leu Ala Ser Thr Leu Ala Ser
1 5
<210> 124
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 124
Leu Gly Gly Cys Asp Asp Asp Ala Asp Thr Phe Ala
1 5 10
<210> 125
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 125
ggctatgggt tgtct 15
<210> 126
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 126
atgattagta gtggtggtag ttatacctac tatgcagaca gtgtgaaggg t 51
<210> 127
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 127
catggggacg atcccgcctg gttcgcttat 30
<210> 128
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 128
agtgtcagct caagtataag ttccaacaac ttgcac 36
<210> 129
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 129
ggcacatcca acctggcttc t 21
<210> 130
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 130
caacagtgga gtagttaccc gtacatgtac acg 33
<210> 131
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 131
ggctacacca tgaac 15
<210> 132
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 132
cttattactc cttacaatgg tgcttctagc tacaaccaga agttcagggg c 51
<210> 133
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 133
gggggttacg acgggagggg ttttgactac 30
<210> 134
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 134
agtgccagct caagtgtaag ttacatgcac 30
<210> 135
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 135
gacacatcca aactggcttc t 21
<210> 136
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 136
cagcagtgga gtaagcaccc tctcacg 27
<210> 137
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 137
agcgatgcaa taagc 15
<210> 138
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 138
atcattaatg gtggtggtaa cacatactac gcgagctggg cgaaaggc 48
<210> 139
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 139
ggcattcaac atggtggtgg taatagtgat tattattatt acggcatgga cctc 54
<210> 140
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 140
caggccagtc agagcattag tagtgtcttg tcc 33
<210> 141
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 141
ctggcatcca ctctggcatc t 21
<210> 142
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 142
caaaccaatt atggtactag tagtagtaat tatggttttg ct 42
<210> 143
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 143
tccgacgcga ttagc 15
<210> 144
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 144
atcatcaacg gcggcggaaa cacctactac gcctcctggg ccaagggc 48
<210> 145
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 145
ggcatccagc acggtggtgg aaacagcgac tactactact atgggatgga tctg 54
<210> 146
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 146
caggcgtccc agtcaattag cagcgtgctc tcc 33
<210> 147
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 147
ttggcctcca ctctggcctc g 21
<210> 148
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 148
caaaccaact acggaacctc cagctccaac tacggctttg cc 42
<210> 149
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 149
aactatgcaa tgagc 15
<210> 150
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 150
tccattagta ctggtggtct cgcattctac gcgaactggg caaaaggc 48
<210> 151
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 151
aatggtggtg gtagttatat tttctattat tttgacttg 39
<210> 152
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 152
caggccagtc agagcattag tagttactta tcc 33
<210> 153
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 153
tctgcatcca ctctggcatc t 21
<210> 154
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 154
caaagctatt atgatattgg tactagtact 30
<210> 155
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 155
aactacgcca tgtcc 15
<210> 156
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 156
tccatcagca caggcggcct ggccttctac gccaattggg ccaagggc 48
<210> 157
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 157
aacggcggag gctcctacat cttctactac ttcgacctg 39
<210> 158
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 158
caggcctccc agtccatctc ctcctacctg tcc 33
<210> 159
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 159
tctgcctcca cactggcctc c 21
<210> 160
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 160
cagtcctact acgacatcgg cacctccacc 30
<210> 161
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 161
agctatgcaa tgggc 15
<210> 162
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 162
accattaata ttggtggtcg cgtatattac gcgagctggg caaaaggc 48
<210> 163
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 163
tattataatg gtggtagtta tgacatc 27
<210> 164
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 164
caggccagtg agagcattta tcgcgtattg gcc 33
<210> 165
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 165
gatacatcca ctctggcatc t 21
<210> 166
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 166
caaggcggtt attatgctga tagttatggt attgct 36
<210> 167
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 167
tcctacgcta tgggc 15
<210> 168
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 168
accatcaaca tcggcggcag agtgtactac gcctcctggg ccaagggc 48
<210> 169
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 169
tactacaacg gcggctccta cgatatc 27
<210> 170
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 170
caggcctccg agtccatcta ccgggtgctg gcc 33
<210> 171
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 171
gacaccagca cactggcctc c 21
<210> 172
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 172
cagggcggct actacgccga ctcctacgga atcgct 36
<210> 173
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 173
agttatgcaa tgatc 15
<210> 174
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 174
accattagta ctggtggtat cacatactac gcgagctggg cgaaaggc 48
<210> 175
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 175
gggggatatg ctgctagtag tgcttattat ctcccgtact actttgactt g 51
<210> 176
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 176
cagtcctccc agtccgtgta taacaacaac aacctggcc 39
<210> 177
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 177
ctggcatcca ctctggcatc t 21
<210> 178
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 178
ctaggtggtt gtgatgatga tgctgatact tttgct 36
<210> 179
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 179
tcctacgcta tgatc 15
<210> 180
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 180
accatctcta ccggcggaat tacctactac gcctcctggg ccaagggc 48
<210> 181
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 181
ggcggctacg ccgccagctc cgcttactac ctgccctact acttcgacct g 51
<210> 182
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 182
cagtcctccc agtccgtgta taacaacaac aacctggcc 39
<210> 183
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 183
ctggcctcca cactggcctc t 21
<210> 184
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 184
ctgggcggct gcgacgacga cgccgatacc tttgct 36
<210> 185
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 185
Gly Tyr Ser Phe Thr Gly Tyr Thr
1 5
<210> 186
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 186
Ile Thr Pro Tyr Asn Gly Ala Ser
1 5
<210> 187
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 187
Ala Arg Gly Gly Tyr Asp Gly Arg Gly Phe Asp Tyr
1 5 10
<210> 188
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 188
Ser Ser Val Ser Tyr
1 5
<210> 189
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 189
Asp Thr Ser
1
<210> 190
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 190
Gln Gln Trp Ser Lys His Pro Leu Thr
1 5
<210> 191
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 191
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 192
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 192
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 193
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 193
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 194
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 194
Gln Asp Val Asn Thr Ala
1 5
<210> 195
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 195
Ser Ala Ser
1
<210> 196
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 196
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 197
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 197
Gly Ile Ser Leu Ser Ser Asp Ala
1 5
<210> 198
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 198
Ile Asn Gly Gly Gly Asn Thr
1 5
<210> 199
<211> 20
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 199
Ala Arg Gly Ile Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Leu
20
<210> 200
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 200
Gln Ser Ile Ser Ser Val
1 5
<210> 201
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 201
Leu Ala Ser
1
<210> 202
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 202
Gln Thr Asn Tyr Gly Thr Ser Ser Ser Asn Tyr Gly Phe Ala
1 5 10
<210> 203
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 203
Gly Ile Ser Leu Ser Ser Asp Ala
1 5
<210> 204
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 204
Ile Asn Gly Gly Gly Asn Thr
1 5
<210> 205
<211> 20
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 205
Ala Arg Gly Ile Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Leu
20
<210> 206
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 206
Gln Ser Ile Ser Ser Val
1 5
<210> 207
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 207
Leu Ala Ser
1
<210> 208
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 208
Gln Thr Asn Tyr Gly Thr Ser Ser Ser Asn Tyr Gly Phe Ala
1 5 10
<210> 209
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 209
Gly Phe Ser Leu Asn Asn Tyr Ala
1 5
<210> 210
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 210
Ile Ser Thr Gly Gly Leu Ala
1 5
<210> 211
<211> 15
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 211
Gly Arg Asn Gly Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu
1 5 10 15
<210> 212
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 212
Gln Ser Ile Ser Ser Tyr
1 5
<210> 213
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 213
Ser Ala Ser
1
<210> 214
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 214
Gln Ser Tyr Tyr Asp Ile Gly Thr Ser Thr
1 5 10
<210> 215
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 215
Gly Phe Ser Leu Asn Asn Tyr Ala
1 5
<210> 216
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 216
Ile Ser Thr Gly Gly Leu Ala
1 5
<210> 217
<211> 15
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 217
Ala Arg Asn Gly Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu
1 5 10 15
<210> 218
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 218
Gln Ser Ile Ser Ser Tyr
1 5
<210> 219
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 219
Ser Ala Ser
1
<210> 220
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 220
Gln Ser Tyr Tyr Asp Ile Gly Thr Ser Thr
1 5 10
<210> 221
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 221
Gly Ile Asp Leu Ser Ser Tyr Ala
1 5
<210> 222
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 222
Ile Asn Ile Gly Gly Arg Val
1 5
<210> 223
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 223
Ala Arg Tyr Tyr Asn Gly Gly Ser Tyr Asp Ile
1 5 10
<210> 224
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 224
Glu Ser Ile Tyr Arg Val
1 5
<210> 225
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 225
Asp Thr Ser
1
<210> 226
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 226
Gln Gly Gly Tyr Tyr Ala Asp Ser Tyr Gly Ile Ala
1 5 10
<210> 227
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 227
Gly Ile Asp Leu Ser Ser Tyr Ala
1 5
<210> 228
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 228
Ile Asn Ile Gly Gly Arg Val
1 5
<210> 229
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 229
Ala Arg Tyr Tyr Asn Gly Gly Ser Tyr Asp Ile
1 5 10
<210> 230
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 230
Glu Ser Ile Tyr Arg Val
1 5
<210> 231
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 231
Asp Thr Ser
1
<210> 232
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 232
Gln Gly Gly Tyr Tyr Ala Asp Ser Tyr Gly Ile Ala
1 5 10
<210> 233
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 233
Gly Phe Ser Leu Ser Ser Tyr Ala
1 5
<210> 234
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 234
Ile Ser Thr Gly Gly Ile Thr
1 5
<210> 235
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 235
Ala Arg Gly Gly Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr
1 5 10 15
Phe Asp Leu
<210> 236
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 236
Gln Ser Val Tyr Asn Asn Asn Asn
1 5
<210> 237
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 237
Leu Ala Ser
1
<210> 238
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 238
Leu Gly Gly Cys Asp Asp Asp Ala Asp Thr Phe Ala
1 5 10
<210> 239
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 239
Gly Phe Ser Leu Ser Ser Tyr Ala
1 5
<210> 240
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 240
Ile Ser Thr Gly Gly Ile Thr
1 5
<210> 241
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 241
Ala Arg Gly Gly Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr
1 5 10 15
Phe Asp Leu
<210> 242
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 242
Gln Ser Val Tyr Asn Asn Asn Asn
1 5
<210> 243
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 243
Leu Ala Ser
1
<210> 244
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 244
Leu Gly Gly Cys Asp Asp Asp Ala Asp Thr Phe Ala
1 5 10
<210> 245
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 245
ggcttcacct tcagcggcta tggg 24
<210> 246
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 246
attagtagtg gtggtagtta tacc 24
<210> 247
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 247
gcaagacatg gggacgatcc cgcctggttc gcttat 36
<210> 248
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 248
tcaagtataa gttccaacaa c 21
<210> 249
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 249
ggcacatcc 9
<210> 250
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 250
caacagtgga gtagttaccc gtacatgtac acg 33
<210> 251
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 251
ggttactcat tcactggcta cacc 24
<210> 252
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 252
attactcctt acaatggtgc ttct 24
<210> 253
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 253
gcaagggggg gttacgacgg gaggggtttt gactac 36
<210> 254
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 254
tcaagtgtaa gttac 15
<210> 255
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 255
gacacatcc 9
<210> 256
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 256
cagcagtgga gtaagcaccc tctcacg 27
<210> 257
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 257
ggaatctccc tcagtagcga tgca 24
<210> 258
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 258
attaatggtg gtggtaacac a 21
<210> 259
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 259
gccagaggca ttcaacatgg tggtggtaat agtgattatt attattacgg catggacctc 60
<210> 260
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 260
cagagcatta gtagtgtc 18
<210> 261
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 261
ctggcatct 9
<210> 262
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 262
caaaccaatt atggtactag tagtagtaat tatggttttg ct 42
<210> 263
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 263
ggaatttccc tctcctccga cgcg 24
<210> 264
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 264
atcaacggcg gcggaaacac c 21
<210> 265
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 265
gcgcgcggca tccagcacgg tggtggaaac agcgactact actactatgg gatggatctg 60
<210> 266
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 266
cagtcaatta gcagcgtg 18
<210> 267
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 267
ttggcctcc 9
<210> 268
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 268
caaaccaact acggaacctc cagctccaac tacggctttg cc 42
<210> 269
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 269
ggattctccc tcaataacta tgca 24
<210> 270
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 270
attagtactg gtggtctcgc a 21
<210> 271
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 271
ggcagaaatg gtggtggtag ttatattttc tattattttg acttg 45
<210> 272
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 272
cagagcatta gtagttac 18
<210> 273
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 273
tctgcatcc 9
<210> 274
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 274
caaagctatt atgatattgg tactagtact 30
<210> 275
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 275
ggcttctccc tgaacaacta cgcc 24
<210> 276
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 276
atcagcacag gcggcctggc c 21
<210> 277
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 277
gccagaaacg gcggaggctc ctacatcttc tactacttcg acctg 45
<210> 278
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 278
cagtccatct cctcctac 18
<210> 279
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 279
tctgcctcc 9
<210> 280
<400> 280
000
<210> 281
<400> 281
000
<210> 282
<400> 282
000
<210> 283
<400> 283
000
<210> 284
<400> 284
000
<210> 285
<400> 285
000
<210> 286
<400> 286
000
<210> 287
<400> 287
000
<210> 288
<400> 288
000
<210> 289
<400> 289
000
<210> 290
<400> 290
000
<210> 291
<400> 291
000
<210> 292
<400> 292
000
<210> 293
<400> 293
000
<210> 294
<400> 294
000
<210> 295
<400> 295
000
<210> 296
<400> 296
000
<210> 297
<400> 297
000
<210> 298
<400> 298
000
<210> 299
<400> 299
000
<210> 300
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 300
cagtcctact acgacatcgg cacctccacc 30
<210> 301
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 301
ggaatcgacc tcagtagcta tgca 24
<210> 302
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 302
attaatattg gtggtcgcgt a 21
<210> 303
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 303
gccagatatt ataatggtgg tagttatgac atc 33
<210> 304
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 304
gagagcattt atcgcgta 18
<210> 305
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 305
gatacatcc 9
<210> 306
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 306
caaggcggtt attatgctga tagttatggt attgct 36
<210> 307
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 307
ggaatcgacc tgtcctccta cgct 24
<210> 308
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 308
atcaacatcg gcggcagagt g 21
<210> 309
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 309
gcccggtact acaacggcgg ctcctacgat atc 33
<210> 310
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 310
gagtccatct accgggtg 18
<210> 311
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 311
gacaccagc 9
<210> 312
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 312
cagggcggct actacgccga ctcctacgga atcgct 36
<210> 313
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 313
ggattctccc tcagtagtta tgca 24
<210> 314
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 314
attagtactg gtggtatcac a 21
<210> 315
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 315
gccagagggg gatatgctgc tagtagtgct tattatctcc cgtactactt tgacttg 57
<210> 316
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 316
cagagtgttt ataataataa caac 24
<210> 317
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 317
ctggcatcc 9
<210> 318
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 318
ctaggtggtt gtgatgatga tgctgatact tttgct 36
<210> 319
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 319
ggcttctccc tgtcctccta cgct 24
<210> 320
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 320
atctctaccg gcggaattac c 21
<210> 321
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 321
gctagaggcg gctacgccgc cagctccgct tactacctgc cctactactt cgacctg 57
<210> 322
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 322
cagtccgtgt ataacaacaa caac 24
<210> 323
<211> 9
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 323
ctggcctcc 9
<210> 324
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
олигонуклеотид”
<400> 324
ctgggcggct gcgacgacga cgccgatacc tttgct 36
<210> 325
<211> 449
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 325
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Glu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Leu Ile Thr Pro Tyr Asn Gly Ala Ser Ser Tyr Asn Gln Lys Phe
50 55 60
Arg Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Asp Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Tyr Asp Gly Arg Gly Phe Asp Tyr Trp Gly Ser Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 326
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 326
Asp Ile Glu Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Gly Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Asn Ser Tyr Ser Leu Thr Ile Ser Ser Val Glu Ala Glu
65 70 75 80
Asp Asp Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Lys His Pro Leu Thr
85 90 95
Phe Gly Ser Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 327
<211> 451
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 327
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 328
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 328
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 329
<211> 453
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 329
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Ser Leu Ser Ser Asp Ala
20 25 30
Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Ile Ile Asn Gly Gly Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Ile
85 90 95
Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr Gly Met Asp Leu
100 105 110
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 330
<211> 219
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 330
Glu Val Leu Met Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Asp Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Ser Ser Val
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Thr Asn Tyr Gly Thr Ser Ser
85 90 95
Ser Asn Tyr Gly Phe Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 331
<211> 456
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 331
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Ser Leu Ser Ser Asp
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ile Ile Asn Gly Gly Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg His Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Ile Gln His Gly Gly Gly Asn Ser Asp Tyr Tyr Tyr Tyr Gly
100 105 110
Met Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 332
<211> 219
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 332
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Val
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Cys
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Thr Asn Tyr Gly Thr Ser Ser
85 90 95
Ser Asn Tyr Gly Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 333
<211> 448
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 333
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Asn Asn Tyr Ala
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ser Ile Ser Thr Gly Gly Leu Ala Phe Tyr Ala Asn Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Gly Arg Asn Gly
85 90 95
Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 334
<211> 215
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 334
Ala Phe Glu Leu Thr Gln Thr Pro Ser Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Ile Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Phe Cys Gln Ser Tyr Tyr Asp Ile Gly Thr
85 90 95
Ser Thr Phe Gly Gly Gly Thr Glu Val Val Val Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 335
<211> 451
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 335
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Asn Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Ser Thr Gly Gly Leu Ala Phe Tyr Ala Asn Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asn Gly Gly Gly Ser Tyr Ile Phe Tyr Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 336
<211> 215
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 336
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Cys
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Ser Tyr Tyr Asp Ile Gly Thr
85 90 95
Ser Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 337
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 337
Gln Ser Val Lys Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr Ala
20 25 30
Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Thr Ile Asn Ile Gly Gly Arg Val Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Ala Pro
65 70 75 80
Ser Leu Thr Ala Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Tyr Tyr
85 90 95
Asn Gly Gly Ser Tyr Asp Ile Trp Gly Pro Gly Thr Leu Val Thr Val
100 105 110
Ser Leu Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 338
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 338
Asp Val Val Met Thr Gln Thr Pro Ala Ser Ala Ser Glu Pro Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Glu Ser Ile Tyr Arg Val
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Thr Ser Thr Leu Ala Ser Gly Ala Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Tyr Gly Thr Glu Phe Thr Leu Thr Ile Ser Gly Val Gln Cys
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Tyr Tyr Ala Asp Ser
85 90 95
Tyr Gly Ile Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 339
<211> 447
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 339
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Thr Ile Asn Ile Gly Gly Arg Val Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Tyr Tyr Asn Gly Gly Ser Tyr Asp Ile Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 340
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 340
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ser Ile Tyr Arg Val
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Thr Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Cys
65 70 75 80
Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Tyr Tyr Ala Asp Ser
85 90 95
Tyr Gly Ile Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 341
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 341
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Lys Pro Asp Glu Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr Ala
20 25 30
Met Ile Trp Val Arg Gln Ala Pro Gly Glu Gly Leu Glu Trp Ile Gly
35 40 45
Thr Ile Ser Thr Gly Gly Ile Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Gly
85 90 95
Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr Phe Asp Leu Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 342
<211> 219
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 342
Ala Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Asn Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Cys Asp Asp
85 90 95
Asp Ala Asp Thr Phe Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 343
<211> 455
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 343
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Ala Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Ser Thr Gly Gly Ile Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Gly Tyr Ala Ala Ser Ser Ala Tyr Tyr Leu Pro Tyr Tyr Phe
100 105 110
Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 344
<211> 219
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полипептид”
<400> 344
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu
35 40 45
Leu Ile Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Cys Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Cys Asp Asp
85 90 95
Asp Ala Asp Thr Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 345
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 345
gaggtccaac tggtggagag cggtggaggt gttgtgcaac ctggccggtc cctgcgcctg 60
tcctgctccg catctggctt caccttcagc ggctatgggt tgtcttgggt gagacaggca 120
cctggaaaag gtcttgagtg ggttgcaatg attagtagtg gtggtagtta tacctactat 180
gcagacagtg tgaagggtag atttgcaata tcgcgagaca acgccaagaa cacattgttc 240
ctgcaaatgg acagcctgag acccgaagac accggggtct atttttgtgc aagacatggg 300
gacgatcccg cctggttcgc ttattggggc caagggaccc cggtcaccgt ctcctcagcc 360
tccaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cttatattca aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc cgggaaatga 1350
<210> 346
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 346
gacatccagc tgacccagag cccaagcagc ctgagcgcca gcgtgggtga cagagtgacc 60
atcacctgta gtgtcagctc aagtataagt tccaacaact tgcactggta ccagcagaag 120
ccaggtaagg ctccaaagcc atggatctac ggcacatcca acctggcttc tggtgtgcca 180
agcagattca gcggtagcgg tagcggtacc gactacacct tcaccatcag cagcctccag 240
ccagaggaca tcgccaccta ctactgccaa cagtggagta gttacccgta catgtacacg 300
ttcggccaag ggaccaaggt ggaaatcaaa cgaactgtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg ttaa 654
<210> 347
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 347
caggtacaac tgcagcagtc tgggcctgag ctggagaagc ctggcgcttc agtgaagata 60
tcctgcaagg cttctggtta ctcattcact ggctacacca tgaactgggt gaagcagagc 120
catggaaaga gccttgagtg gattggactt attactcctt acaatggtgc ttctagctac 180
aaccagaagt tcaggggcaa ggccacatta actgtagaca agtcatccag cacagcctac 240
atggacctcc tcagtctgac atctgaagac tctgcagtct atttctgtgc aagggggggt 300
tacgacggga ggggttttga ctactgggga tccgggaccc cggtcaccgt ctcctcagcc 360
tccaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc cgggaaatga 1350
<210> 348
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 348
gacatcgagc tcactcagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccagctc aagtgtaagt tacatgcact ggtaccagca gaagtcaggc 120
acctccccca aaagatggat ttatgacaca tccaaactgg cttctggagt cccaggtcgc 180
ttcagtggca gtgggtctgg aaactcttac tctctcacaa tcagcagcgt ggaggctgaa 240
gatgatgcaa cttattactg ccagcagtgg agtaagcacc ctctcacgtt cggatccggg 300
accaaggtgg aaatcaaacg aactgtggct gcaccatctg tcttcatctt cccgccatct 360
gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc 420
agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag 480
agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg 540
agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg 600
agctcgcccg tcacaaagag cttcaacagg ggagagtgtt aa 642
<210> 349
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 349
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcaccgtct ctggaatctc cctcagtagc gatgcaataa gctgggtccg ccaggctcca 120
gggaaggggc tcgaatacat cggaatcatt aatggtggtg gtaacacata ctacgcgagc 180
tgggcgaaag gccgattcac catctccaaa acctcgacca cggtggatct gaaaatcacc 240
agtccgacaa ccgaggacac ggccacctat ttctgtgcca gaggcattca acatggtggt 300
ggtaatagtg attattatta ttacggcatg gacctctggg gcccaggcac cctggtcact 360
gtctcttcag catccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag 900
cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggatgagc tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcttatatt caaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct cccgggaaat ga 1362
<210> 350
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 350
gaagtgttga tgacccagac tccatcctcc gtgtctgcag ctgtgggaga cacagtcacc 60
atcaagtgcc aggccagtca gagcattagt agtgtcttgt cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctatctg gcatccactc tggcatctgg ggtcccatcg 180
cggttcagcg gcagtagatc tgggacagag ttcactctca ccatcagcga cctggagtgt 240
gacgatgctg ccacttacta ctgtcaaacc aattatggta ctagtagtag taattatggt 300
tttgctttcg gcggagggac cgaggtggtc gtcaaacgaa ctgtggctgc accatctgtc 360
ttcatcttcc cgccatctga tgagcagttg aaatctggaa ctgcctctgt tgtgtgcctg 420
ctgaataact tctatcccag agaggccaaa gtacagtgga aggtggataa cgccctccaa 480
tcgggtaact cccaggagag tgtcacagag caggacagca aggacagcac ctacagcctc 540
agcagcaccc tgacgctgag caaagcagac tacgagaaac acaaagtcta cgcctgcgaa 600
gtcacccatc agggcctgag ctcgcccgtc acaaagagct tcaacagggg agagtgttga 660
<210> 351
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 351
gaagtccaac tggtggaaag cgggggagga ctggtgcagc cgggcggatc cctccggctg 60
tcatgtgctg catcgggaat ttccctctcc tccgacgcga ttagctgggt cagacaggcc 120
cccggaaagg ggctggagta catcggtatc atcaacggcg gcggaaacac ctactacgcc 180
tcctgggcca agggccgctt caccatctcg cggcataatt ccaagaacac tctgtacttg 240
caaatgaact ccctgagggc cgaggacacc gccgtgtact actgcgcgcg cggcatccag 300
cacggtggtg gaaacagcga ctactactac tatgggatgg atctgtgggg ccagggaact 360
cttgtgaccg tgtcgtcagc atccaccaag ggcccatcgg tcttccccct ggcaccctcc 420
tccaagagca cctctggggg cacagcggcc ctgggctgcc tggtcaagga ctacttcccc 480
gaaccggtga cggtgtcgtg gaactcaggc gccctgacca gcggcgtgca caccttcccg 540
gctgtcctac agtcctcagg actctactcc ctcagcagcg tggtgaccgt gccctccagc 600
agcttgggca cccagaccta catctgcaac gtgaatcaca agcccagcaa caccaaggtg 660
gacaagaaag ttgagcccaa atcttgtgac aaaactcaca catgcccacc gtgcccagca 720
cctgaactcc tggggggacc gtcagtcttc ctcttccccc caaaacccaa ggacaccctc 780
atgatctccc ggacccctga ggtcacatgc gtggtggtgg acgtgagcca cgaagaccct 840
gaggtcaagt tcaactggta cgtggacggc gtggaggtgc ataatgccaa gacaaagccg 900
cgggaggagc agtacaacag cacgtaccgt gtggtcagcg tcctcaccgt cctgcaccag 960
gactggctga atggcaagga gtacaagtgc aaggtctcca acaaagccct cccagccccc 1020
atcgagaaaa ccatctccaa agccaaaggg cagccccgag aaccacaggt gtacaccctg 1080
cccccatccc gggatgagct gaccaagaac caggtcagcc tgacctgcct ggtcaaaggc 1140
ttctatccca gcgacatcgc cgtggagtgg gagagcaatg ggcagccgga gaacaactac 1200
aagaccacgc ctcccgtgct ggactccgac ggctccttct tcttatattc aaagctcacc 1260
gtggacaaga gcaggtggca gcaggggaac gtcttctcat gctccgtgat gcatgaggct 1320
ctgcacaacc actacacgca gaagagcctc tccctgtctc ccgggaaatg a 1371
<210> 352
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 352
gacattcaga tgacccagtc cccaagctcg ctgtccgcct ccgtgggcga ccgcgtgacc 60
atcacgtgcc aggcgtccca gtcaattagc agcgtgctct cctggtacca acagaagccg 120
gggaaagcac ccaagctgct gatctacttg gcctccactc tggcctcggg agtgccttca 180
cggttctccg gatcgggatc tggtactgat ttcaccctca ccatctcgag ccttcagtgc 240
gaggacatcg ctacttacta ttgtcaaacc aactacggaa cctccagctc caactacggc 300
tttgccttcg gtggcgggac caaggtcgaa atcaaacgaa ctgtggctgc accatctgtc 360
ttcatcttcc cgccatctga tgagcagttg aaatctggaa ctgcctctgt tgtgtgcctg 420
ctgaataact tctatcccag agaggccaaa gtacagtgga aggtggataa cgccctccaa 480
tcgggtaact cccaggagag tgtcacagag caggacagca aggacagcac ctacagcctc 540
agcagcaccc tgacgctgag caaagcagac tacgagaaac acaaagtcta cgcctgcgaa 600
gtcacccatc agggcctgag ctcgcccgtc acaaagagct tcaacagggg agagtgttga 660
<210> 353
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 353
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcacagtct ctggattctc cctcaataac tatgcaatga gctgggtccg ccaggctcca 120
gggaaggggc tggaatggat cggatccatt agtactggtg gtctcgcatt ctacgcgaac 180
tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaaatgacc 240
agtctgacaa ccgaggacac ggccacctat ttctgtggca gaaatggtgg tggtagttat 300
attttctatt attttgactt gtggggccaa ggcaccctcg tcactgtctc ttcagcatcc 360
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 420
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 480
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 540
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 600
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 660
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 720
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 780
acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 840
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 900
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 960
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1020
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1080
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1140
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1200
tccgacggct ccttcttctt atattcaaag ctcaccgtgg acaagagcag gtggcagcag 1260
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1320
agcctctccc tgtctcccgg gaaatga 1347
<210> 354
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 354
gcattcgaat tgacccagac tccatcctcc gtggaggcag ctgtgggagg cacaatcacc 60
atcaagtgcc aggccagtca gagcattagt agttacttat cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctattct gcatccactc tggcatctgg ggtctcatcg 180
cggttcaaag gcagtggatc tgggacagag tacactctca ccatcagcga cctggagtgt 240
gccgatgctg ccacttactt ctgtcaaagc tattatgata ttggtactag tactttcggc 300
ggagggaccg aggtggtcgt caaacgaact gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttga 648
<210> 355
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 355
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
tcttgtgccg cctccggctt ctccctgaac aactacgcca tgtcctgggt gcgacaggcc 120
cctggcaaag gcctggaatg gatcggctcc atcagcacag gcggcctggc cttctacgcc 180
aattgggcca agggccggtt caccatcagc cgggacaact ccaagaacac cctgtacctc 240
cagatgaact ccctgcgggc cgaggacacc gccgtgtact actgtgccag aaacggcgga 300
ggctcctaca tcttctacta cttcgacctg tggggccagg gcaccctcgt gacagtgtca 360
tctgcatcca ccaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct 420
gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg 480
tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc 540
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag 600
acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gaaagttgag 660
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 720
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 780
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 840
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 900
aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 960
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 1020
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 1080
gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 1140
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 1200
gtgctggact ccgacggctc cttcttctta tattcaaagc tcaccgtgga caagagcagg 1260
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1320
acgcagaaga gcctctccct gtctcccggg aaatga 1356
<210> 356
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 356
gatattcaga tgacccagtc cccctccagc ctgtccgctt ctgtgggcga cagagtgacc 60
atcacctgtc aggcctccca gtccatctcc tcctacctgt cctggtatca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactct gcctccacac tggcctccgg cgtgccctct 180
agattctccg gctctggctc tggcaccgac tttaccctga ccatcagctc cctccagtgc 240
gaggatgccg ccacctacta ctgccagtcc tactacgaca tcggcacctc caccttcggc 300
ggaggcacca aggtggaaat caaacgaact gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttga 648
<210> 357
<211> 1335
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 357
cagtcagtga aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcacagtct ctggaatcga cctcagtagc tatgcaatgg gctggttccg ccaggctcca 120
gggaaggggc tggaatacat cggaaccatt aatattggtg gtcgcgtata ttacgcgagc 180
tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaagcgccc 240
agtctgacag ccgaggacac ggccacctat ttctgtgcca gatattataa tggtggtagt 300
tatgacatct ggggcccagg caccctggtc accgtctctt tagcatccac caagggccca 360
tcggtcttcc ccctggcacc ctcctccaag agcacctctg ggggcacagc ggccctgggc 420
tgcctggtca aggactactt ccccgaaccg gtgacggtgt cgtggaactc aggcgccctg 480
accagcggcg tgcacacctt cccggctgtc ctacagtcct caggactcta ctccctcagc 540
agcgtggtga ccgtgccctc cagcagcttg ggcacccaga cctacatctg caacgtgaat 600
cacaagccca gcaacaccaa ggtggacaag aaagttgagc ccaaatcttg tgacaaaact 660
cacacatgcc caccgtgccc agcacctgaa ctcctggggg gaccgtcagt cttcctcttc 720
cccccaaaac ccaaggacac cctcatgatc tcccggaccc ctgaggtcac atgcgtggtg 780
gtggacgtga gccacgaaga ccctgaggtc aagttcaact ggtacgtgga cggcgtggag 840
gtgcataatg ccaagacaaa gccgcgggag gagcagtaca acagcacgta ccgtgtggtc 900
agcgtcctca ccgtcctgca ccaggactgg ctgaatggca aggagtacaa gtgcaaggtc 960
tccaacaaag ccctcccagc ccccatcgag aaaaccatct ccaaagccaa agggcagccc 1020
cgagaaccac aggtgtacac cctgccccca tcccgggatg agctgaccaa gaaccaggtc 1080
agcctgacct gcctggtcaa aggcttctat cccagcgaca tcgccgtgga gtgggagagc 1140
aatgggcagc cggagaacaa ctacaagacc acgcctcccg tgctggactc cgacggctcc 1200
ttcttcttat attcaaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc 1260
tcatgctccg tgatgcatga ggctctgcac aaccactaca cgcagaagag cctctccctg 1320
tctcccggga aatga 1335
<210> 358
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 358
gatgttgtga tgacccagac tccagcctcc gcgtctgaac ctgtgggagg cacagtcacc 60
atcaagtgcc aggccagtga gagcatttat cgcgtattgg cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctatgat acatccactc tggcatctgg ggccccatcg 180
cggttcaaag gcagtggata tgggacagag ttcactctca ccatcagcgg cgtgcagtgt 240
gaagatgctg ccacttacta ctgtcaaggc ggttattatg ctgatagtta tggtattgct 300
ttcggcggag ggaccgaggt ggtggtcaaa cgaactgtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg ttga 654
<210> 359
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 359
caggtgcagc tggtggaatc tggcggagga ctggtgcagc ctggcggctc tctgagactg 60
tcctgttccg cctccggaat cgacctgtcc tcctacgcta tgggctgggt gcgacaggct 120
cctggcaagg gcctggagta catcggcacc atcaacatcg gcggcagagt gtactacgcc 180
tcctgggcca agggccggtt caccatctcc agagacaact ccaagaacac cctgtacctc 240
cagatgaact ccctgcgggc cgaggacacc gccgtgtact actgcgcccg gtactacaac 300
ggcggctcct acgatatctg gggccagggc acactcgtga ccgtgtcctc tgcatccacc 360
aagggcccat cggtcttccc cctggcaccc tcctccaaga gcacctctgg gggcacagcg 420
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca 480
ggcgccctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac 540
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc 600
aacgtgaatc acaagcccag caacaccaag gtggacaaga aagttgagcc caaatcttgt 660
gacaaaactc acacatgccc accgtgccca gcacctgaac tcctgggggg accgtcagtc 720
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca 780
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac 840
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac 900
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag 960
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa 1020
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaccaag 1080
aaccaggtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 1140
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 1200
gacggctcct tcttcttata ttcaaagctc accgtggaca agagcaggtg gcagcagggg 1260
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc 1320
ctctccctgt ctcccgggaa atga 1344
<210> 360
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 360
gatatccaga tgacccagtc cccctccacc ctgtctgcct ctgtgggcga cagagtgacc 60
atcacctgtc aggcctccga gtccatctac cgggtgctgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacgac accagcacac tggcctccgg cgtgccctct 180
agattctccg gctctggctc tggcaccgag tttaccctga ccatctccag cctccagtgc 240
gacgacgccg ccacctacta ttgtcagggc ggctactacg ccgactccta cggaatcgct 300
ttcggcggag gcaccaaggt ggaaatcaaa cgaactgtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg ttga 654
<210> 361
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 361
cagtcggtgg aggagtccgg cggtcgcctg gtaaagcctg acgaatccct gacactcacc 60
tgcacagcct ctggattctc cctcagtagt tatgcaatga tctgggtccg ccaggctcca 120
ggggaggggc tggaatggat cggaaccatt agtactggtg gtatcacata ctacgcgagc 180
tgggcgaaag gccgattcac catctccaaa acctcgacca cggtggatct gaaaatcacc 240
agtccgacaa ccgaggacac ggccacctat ttctgtgcca gagggggata tgctgctagt 300
agtgcttatt atctcccgta ctactttgac ttgtggggcc aagggaccct ggtcaccgtc 360
tcctcagcat ccaccaaggg cccatcggtc ttccccctgg caccctcctc caagagcacc 420
tctgggggca cagcggccct gggctgcctg gtcaaggact acttccccga accggtgacg 480
gtgtcgtgga actcaggcgc cctgaccagc ggcgtgcaca ccttcccggc tgtcctacag 540
tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcag cttgggcacc 600
cagacctaca tctgcaacgt gaatcacaag cccagcaaca ccaaggtgga caagaaagtt 660
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 720
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 780
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 840
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 900
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 960
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 1020
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 1080
gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 1140
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1200
cccgtgctgg actccgacgg ctccttcttc ttatattcaa agctcaccgt ggacaagagc 1260
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1320
tacacgcaga agagcctctc cctgtctccc gggaaatga 1359
<210> 362
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 362
gcagccgtgc tgacccagac accatcaccc gtgtctgcag ctgtgggagg cacagtcacc 60
atcagttgcc agtccagtca gagtgtttat aataataaca acttagcctg gtttcagcag 120
aaacccgggc agcctcccaa gcttctgatc tatctggcat ccactctggc atctggggtc 180
ccatcacggt tcagcggcag tggatctggg acacagttca ctctcaccat cagcggcgtg 240
cagtgtgacg atgctgccac ttattactgt ctaggtggtt gtgatgatga tgctgatact 300
tttgctttcg gcggagggac tgaggtggtg gtcaaacgaa ctgtggctgc accatctgtc 360
ttcatcttcc cgccatctga tgagcagttg aaatctggaa ctgcctctgt tgtgtgcctg 420
ctgaataact tctatcccag agaggccaaa gtacagtgga aggtggataa cgccctccaa 480
tcgggtaact cccaggagag tgtcacagag caggacagca aggacagcac ctacagcctc 540
agcagcaccc tgacgctgag caaagcagac tacgagaaac acaaagtcta cgcctgcgaa 600
gtcacccatc agggcctgag ctcgcccgtc acaaagagct tcaacagggg agagtgttga 660
<210> 363
<211> 1368
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 363
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
tcttgtgccg cctccggctt ctccctgtcc tcctacgcta tgatctgggt gcgacaggcc 120
cctggcaagg gcctggaatg gatcggcacc atctctaccg gcggaattac ctactacgcc 180
tcctgggcca agggccggtt caccatctcc agagacaact ccaagaacac cctgtacctc 240
cagatgaact ccctgcgggc cgaggacacc gccgtgtact attgtgctag aggcggctac 300
gccgccagct ccgcttacta cctgccctac tacttcgacc tgtggggcca gggcaccctc 360
gtgacagtgt catctgcatc caccaagggc ccatcggtct tccccctggc accctcctcc 420
aagagcacct ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa 480
ccggtgacgg tgtcgtggaa ctcaggcgcc ctgaccagcg gcgtgcacac cttcccggct 540
gtcctacagt cctcaggact ctactccctc agcagcgtgg tgaccgtgcc ctccagcagc 600
ttgggcaccc agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtggac 660
aagaaagttg agcccaaatc ttgtgacaaa actcacacat gcccaccgtg cccagcacct 720
gaactcctgg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 780
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 840
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 900
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 960
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 1020
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 1080
ccatcccggg atgagctgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 1140
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 1200
accacgcctc ccgtgctgga ctccgacggc tccttcttct tatattcaaa gctcaccgtg 1260
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 1320
cacaaccact acacgcagaa gagcctctcc ctgtctcccg ggaaatga 1368
<210> 364
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
полинуклеотид”
<400> 364
gatattcaga tgacccagtc cccctccagc ctgtccgctt ctgtgggcga cagagtgacc 60
atcacctgtc agtcctccca gtccgtgtat aacaacaaca acctggcctg gtatcagcag 120
aaacccggca aggtgcccaa gctgctgatc tacctggcct ccacactggc ctctggcgtg 180
ccctctagat tctccggctc tggctctggc accgacttta ccctgaccat cagctccctc 240
cagtgcgagg atgccgccac ctactattgc ctgggcggct gcgacgacga cgccgatacc 300
tttgcttttg gcggaggcac caaggtggaa atcaaacgaa ctgtggctgc accatctgtc 360
ttcatcttcc cgccatctga tgagcagttg aaatctggaa ctgcctctgt tgtgtgcctg 420
ctgaataact tctatcccag agaggccaaa gtacagtgga aggtggataa cgccctccaa 480
tcgggtaact cccaggagag tgtcacagag caggacagca aggacagcac ctacagcctc 540
agcagcaccc tgacgctgag caaagcagac tacgagaaac acaaagtcta cgcctgcgaa 600
gtcacccatc agggcctgag ctcgcccgtc acaaagagct tcaacagggg agagtgttga 660
<210> 365
<211> 4
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 365
Ala His Lys Asp
1
<210> 366
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 366
Asn Thr Ser Gln Glu Ala His Lys Asp Val Ser Tyr Leu
1 5 10
<210> 367
<211> 4
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 367
Gly Phe Leu Gly
1
<210> 368
<211> 4
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетический
пептид”
<400> 368
Ala Leu Ala Leu
1
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ КОНЪЮГАТА АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО НА ОСНОВЕ ЭРИБУЛИНА | 2020 |
|
RU2840013C1 |
| КОНЪЮГАТЫ АНТИТЕЛА И ЛЕКАРСТВЕННОГО СРЕДСТВА, СОДЕРЖАЩИЕ МОДУЛЯТОР СПЛАЙСИНГА НА ОСНОВЕ ГЕРБОКСИДИЕНА, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2820607C2 |
| АМАТОКСИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛА С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826004C2 |
| НАЦЕЛЕННЫЕ НА uPARAP КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2740311C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ROR2 | 2018 |
|
RU2784586C2 |
| КОНЪЮГАТ АНТИТЕЛО ПРОТИВ C-MET-ЛЕКАРСТВЕННОЕ СРЕДСТВО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2822497C1 |
| АНТИТЕЛО ПРОТИВ КЛАУДИНА 18.2 И ЕГО КОНЪЮГАТ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2022 |
|
RU2814164C2 |
| СКРИНИНГ УЧАСТКОВ ПРИСОЕДИНЕНИЯ С ФИКСИРОВАННОЙ ТОЧКОЙ В КОНЪЮГАТЕ ЦИСТЕИН-МОДИФИЦИРОВАННОЕ АНТИТЕЛО-ТОКСИН (TDC) | 2018 |
|
RU2816510C2 |
| ЛИНКЕРЫ НА ОСНОВЕ СУЛЬФОМАЛЕИМИДА И СООТВЕТСТВУЮЩИЕ КОНЪЮГАТЫ | 2019 |
|
RU2815199C2 |
| КОНЪЮГАТЫ АНТИ-HER2 БИПАРАТОПНЫХ АНТИТЕЛ-ЛЕКАРСТВЕННЫХ ВЕЩЕСТВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2806049C2 |
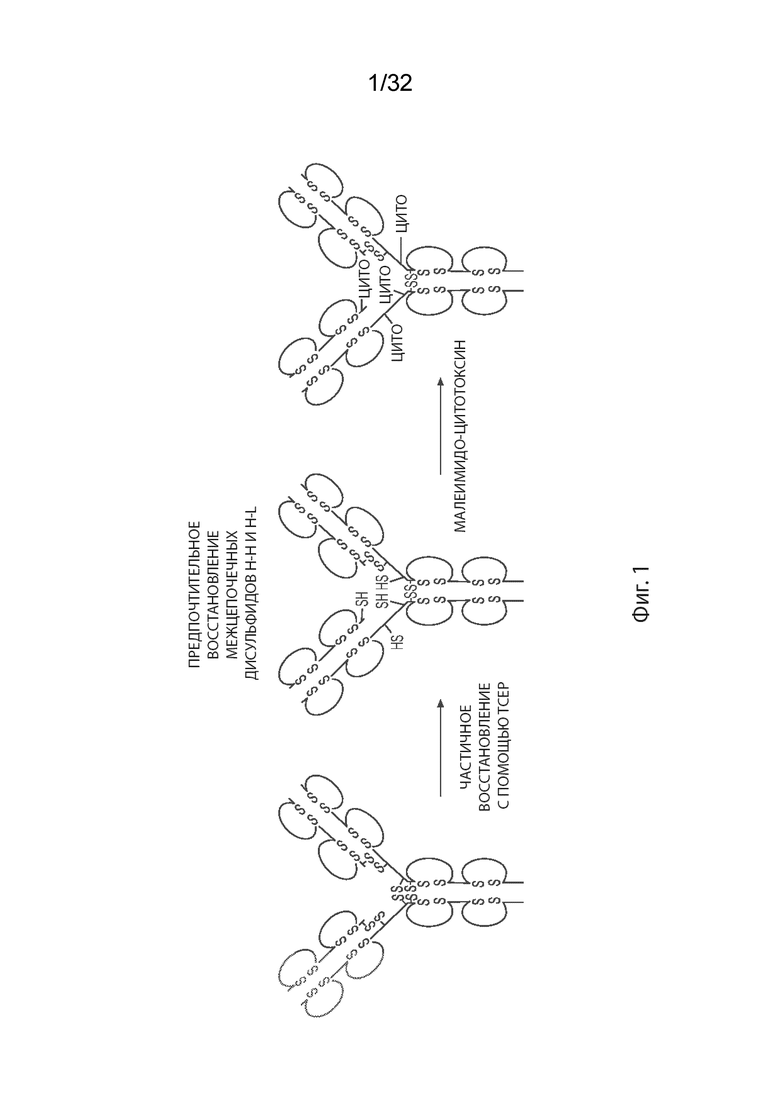


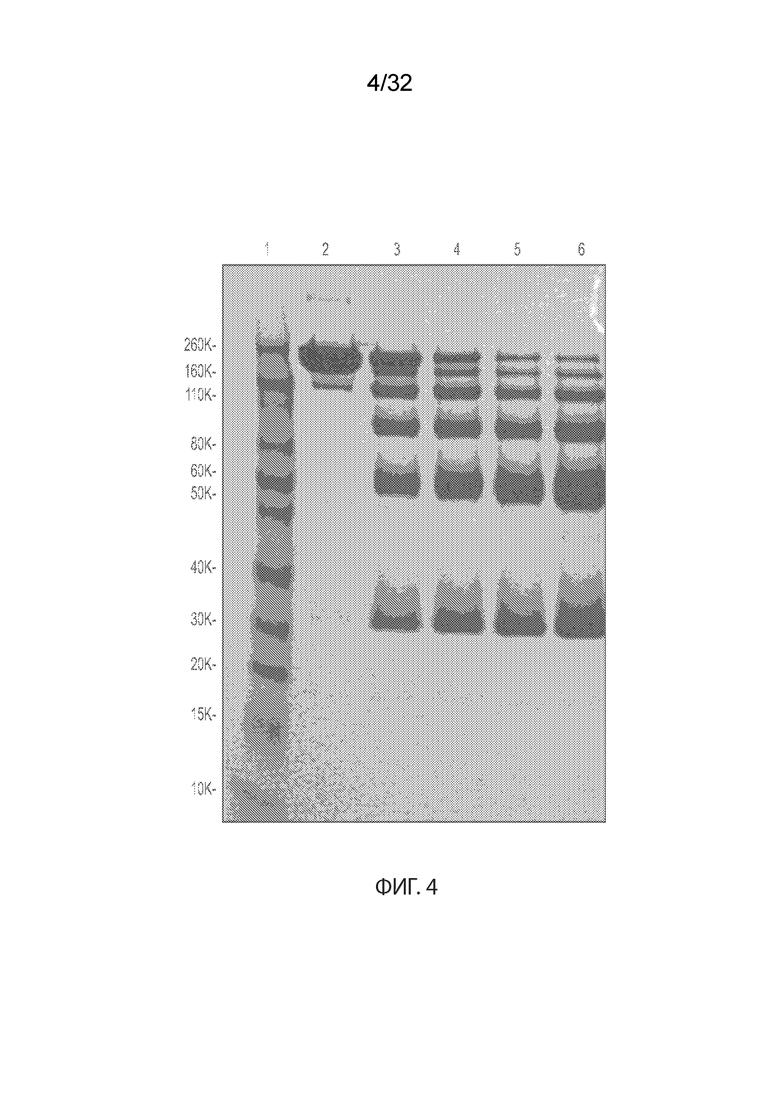

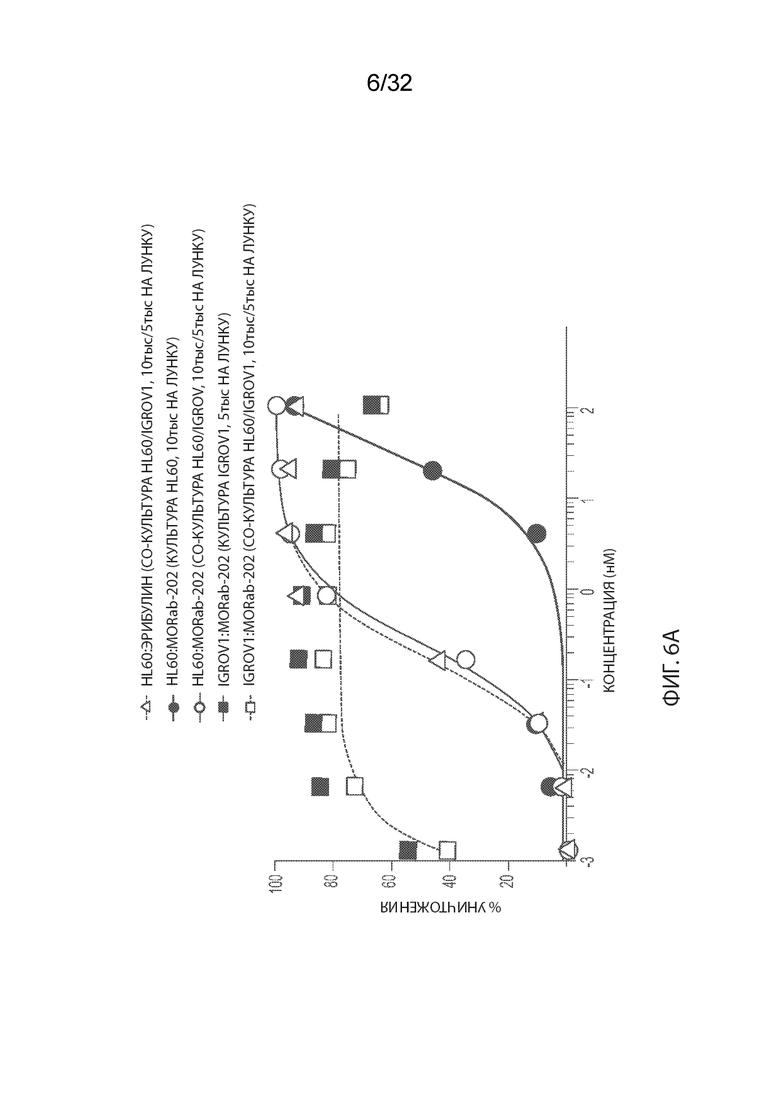
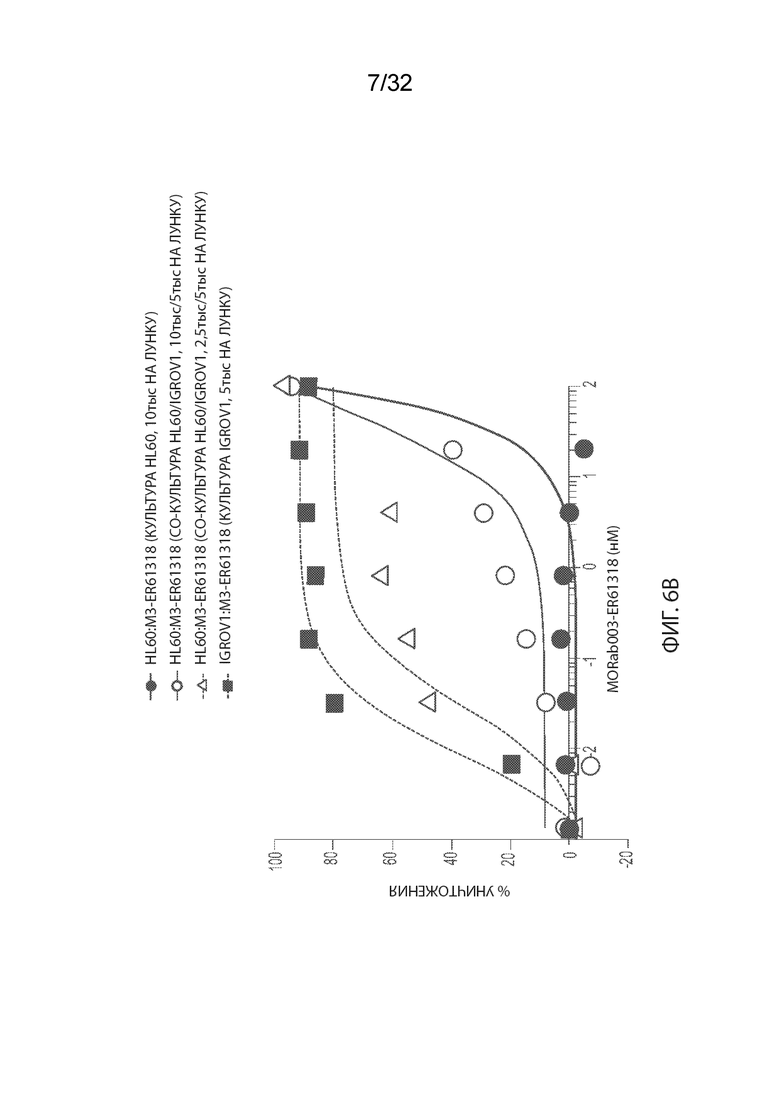

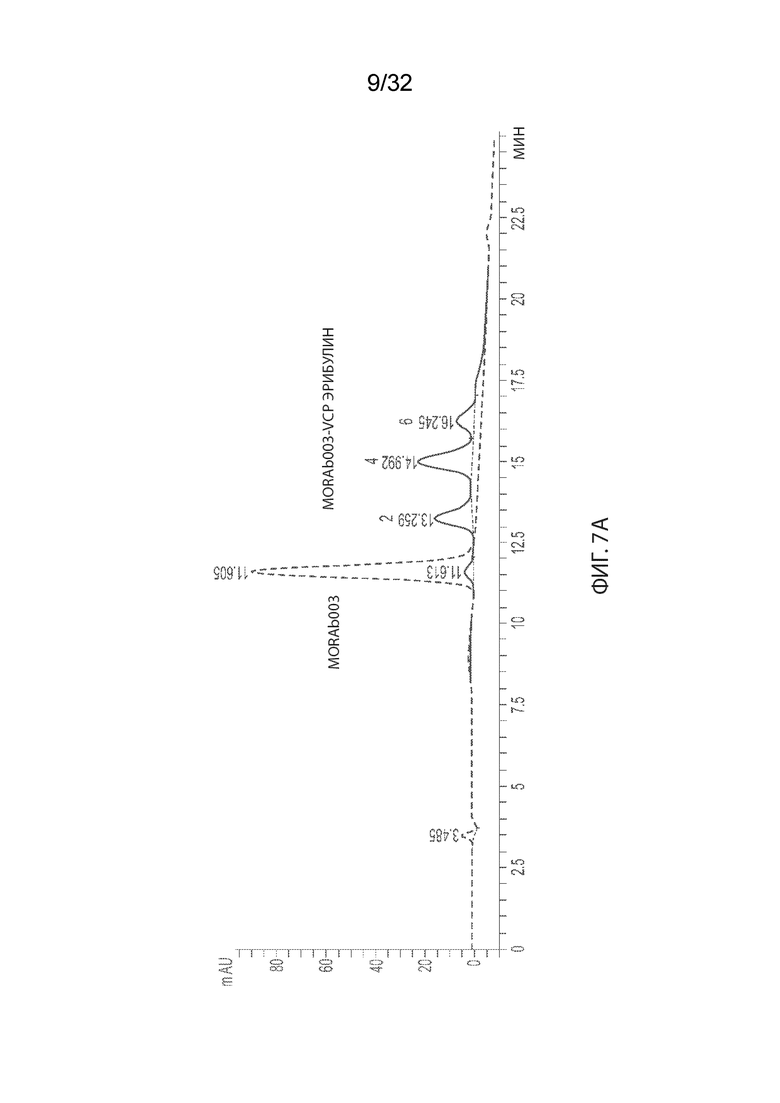
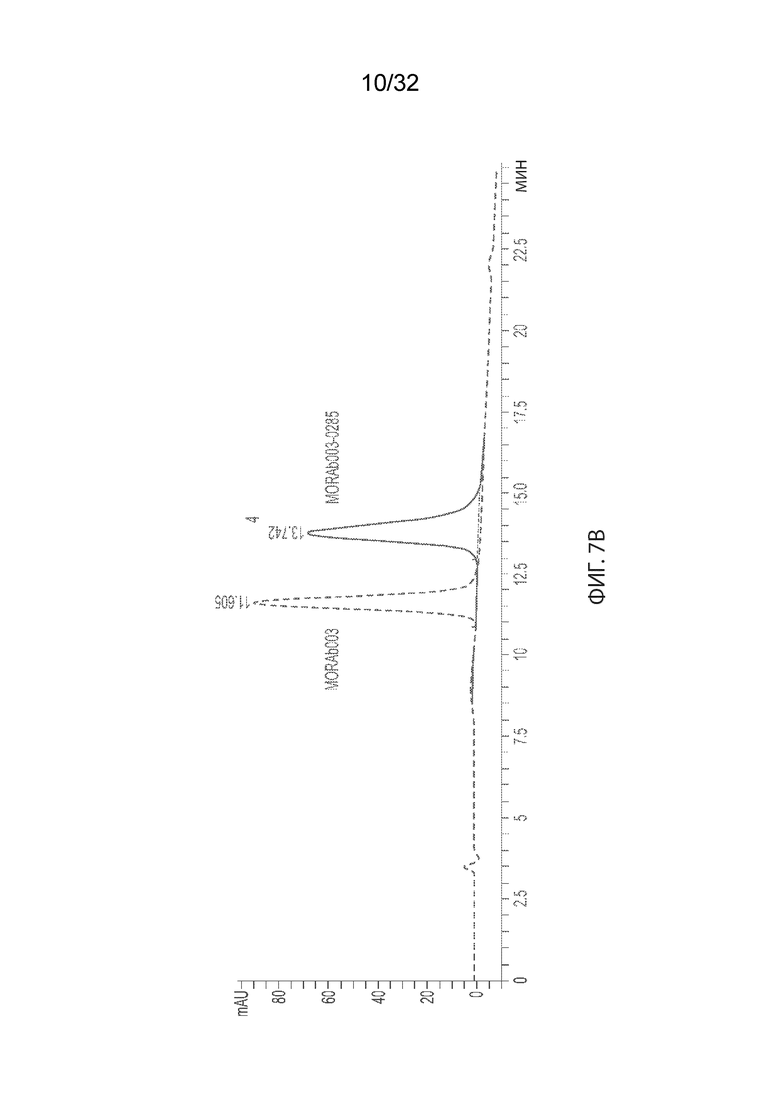
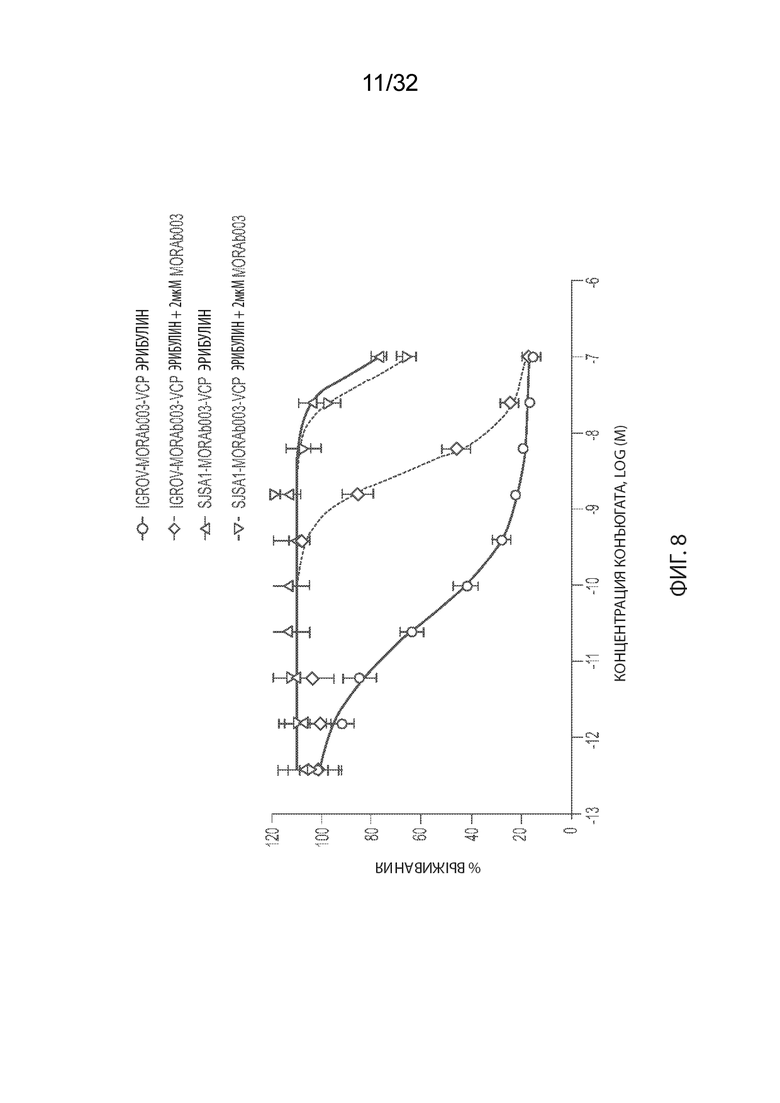
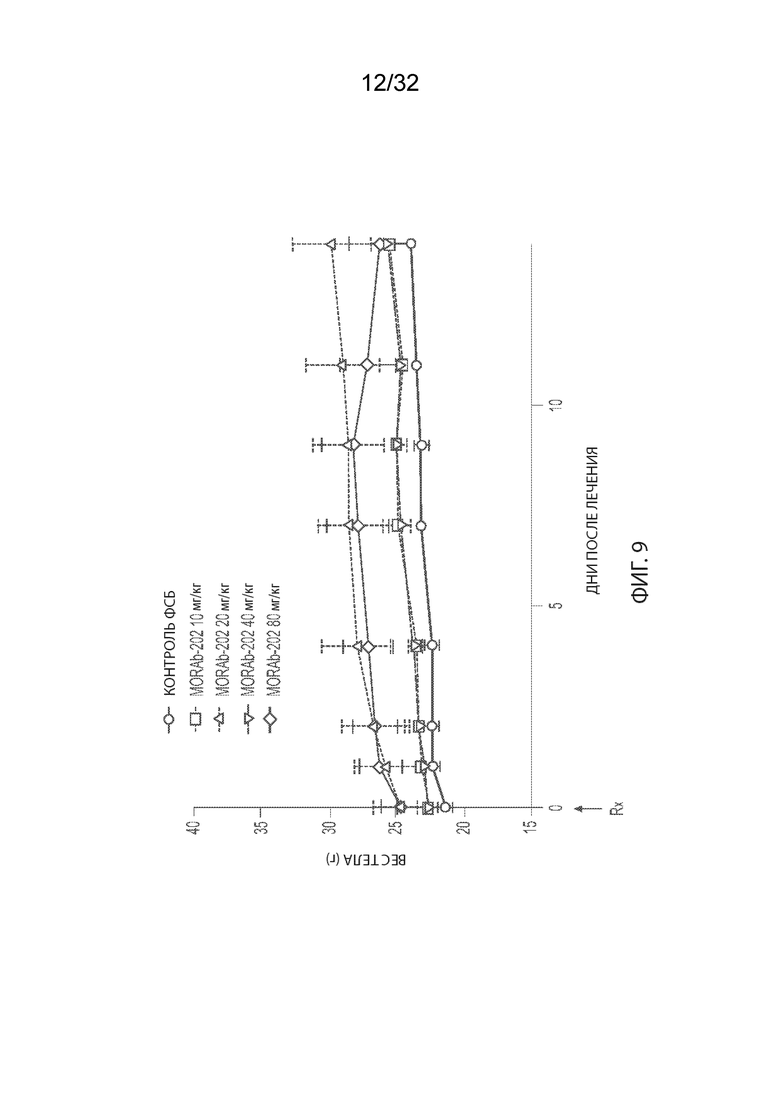
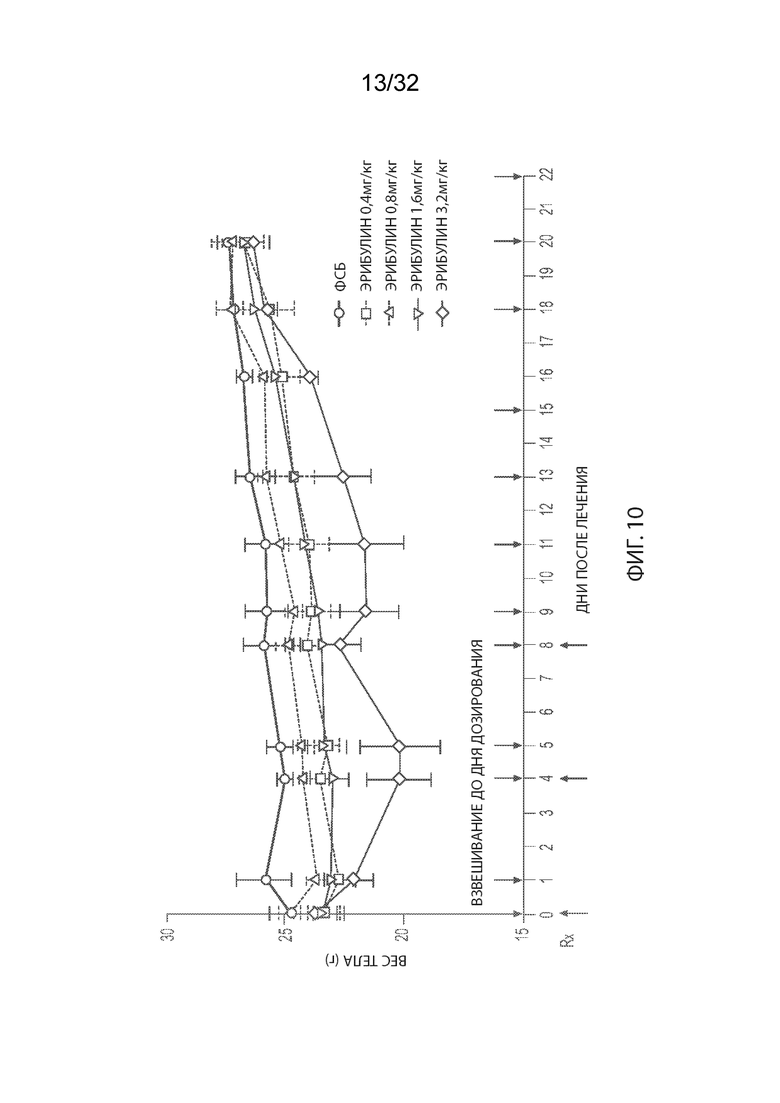
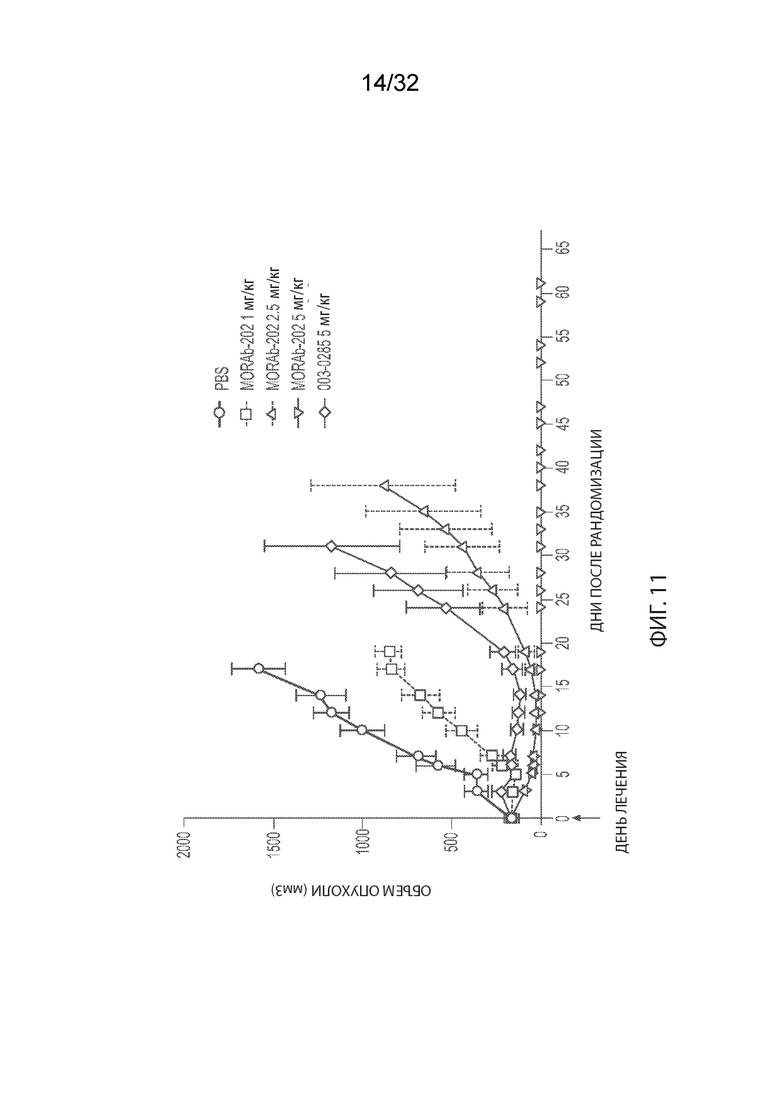
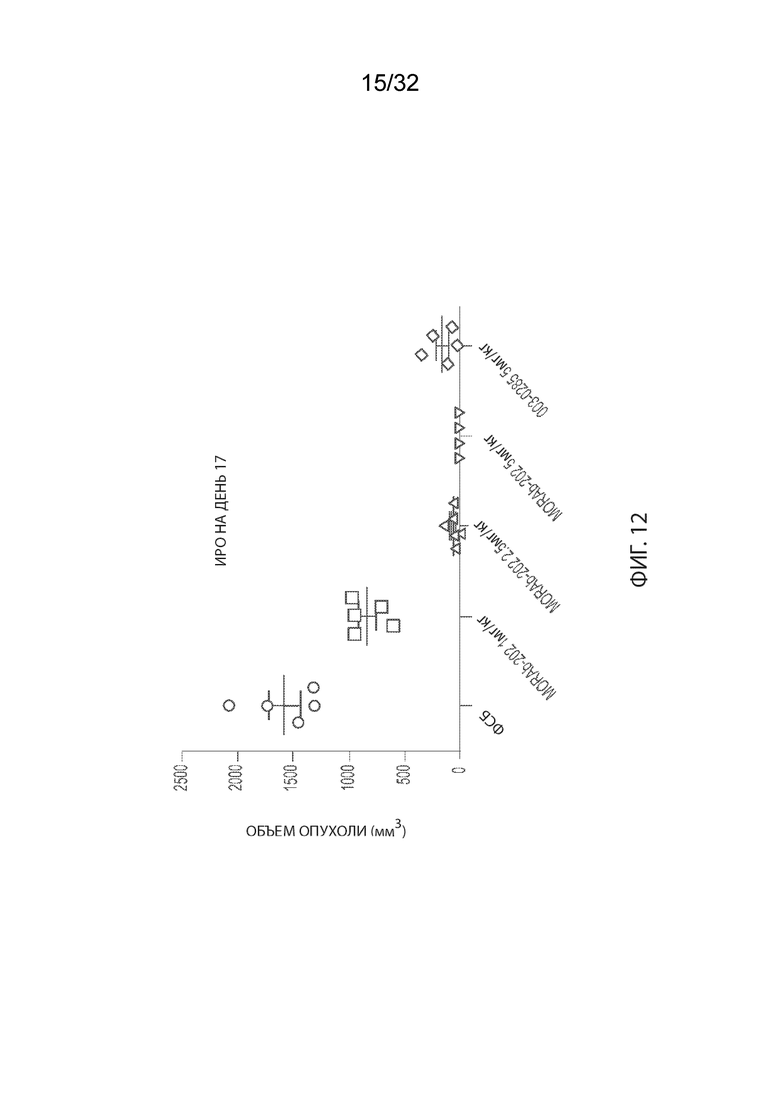
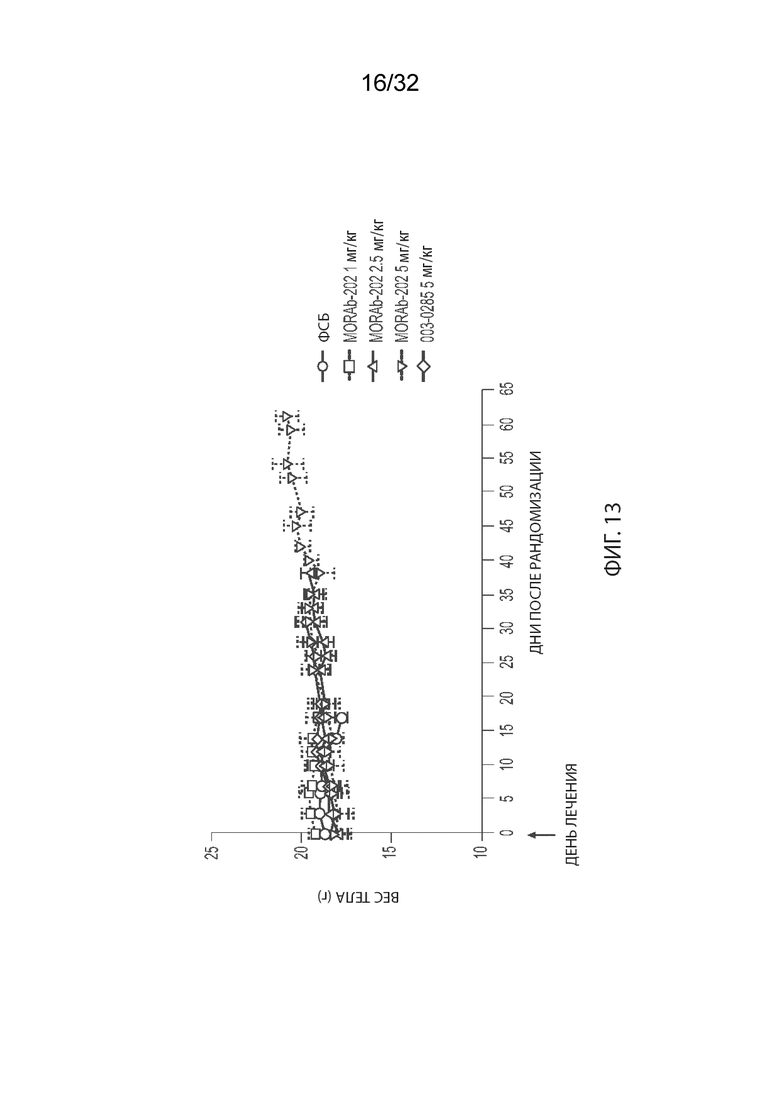

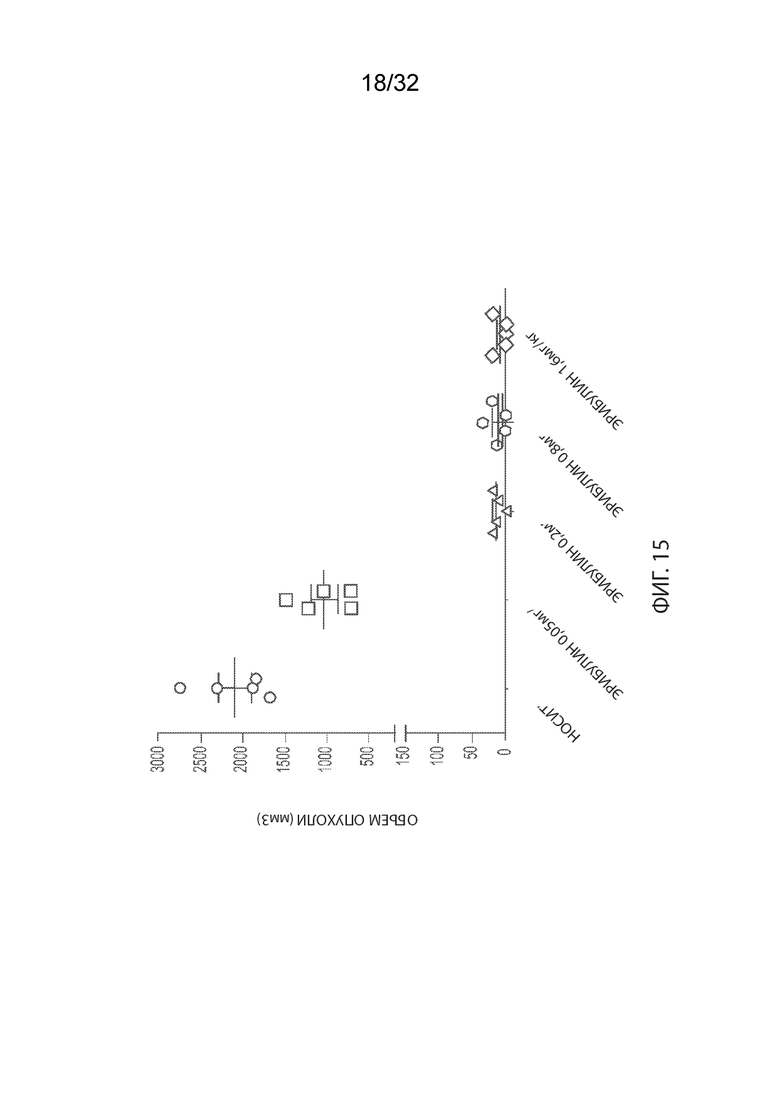
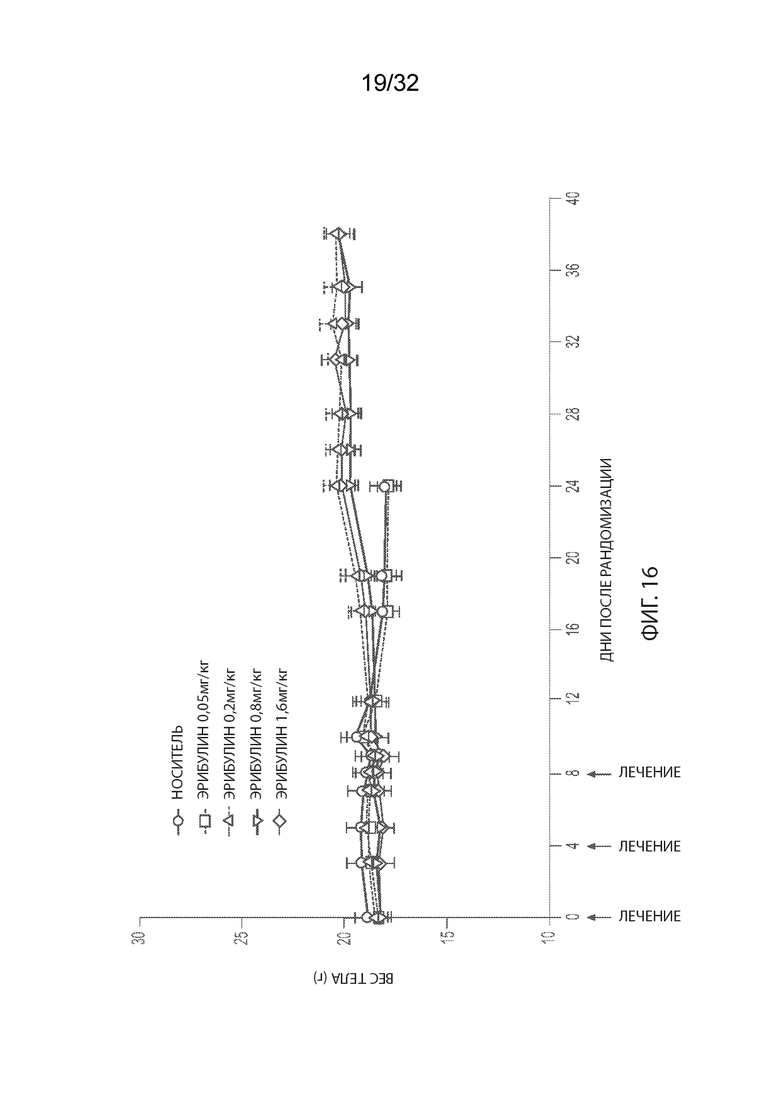
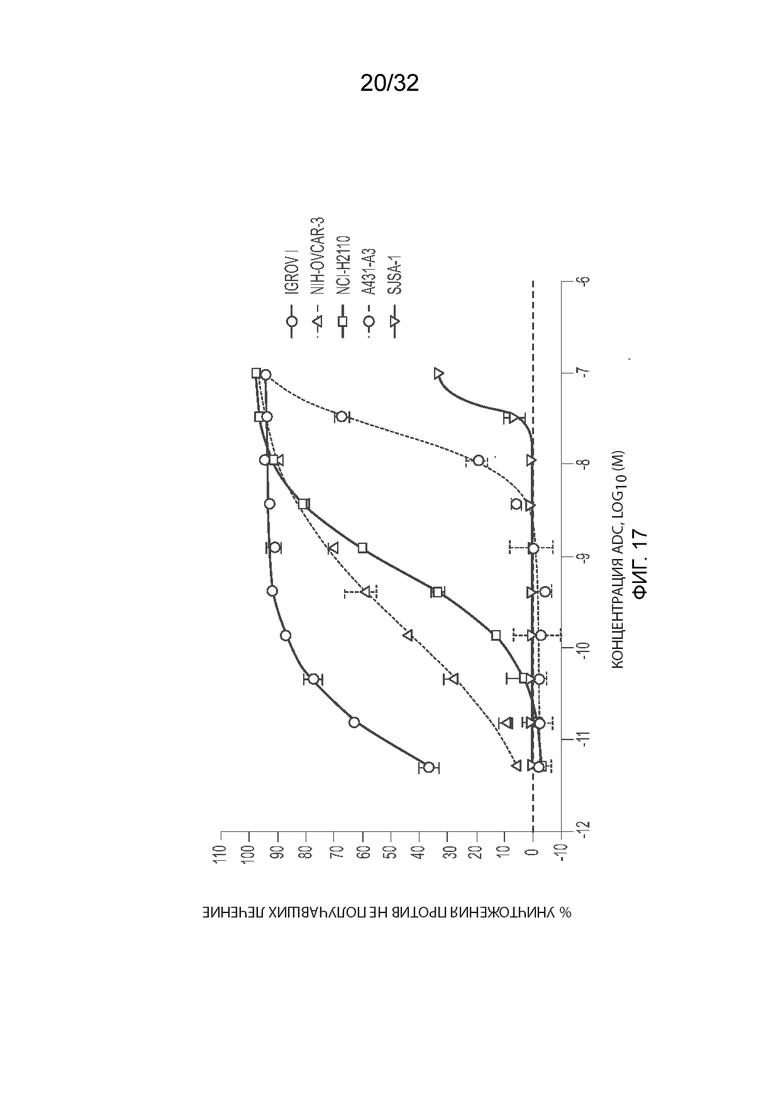
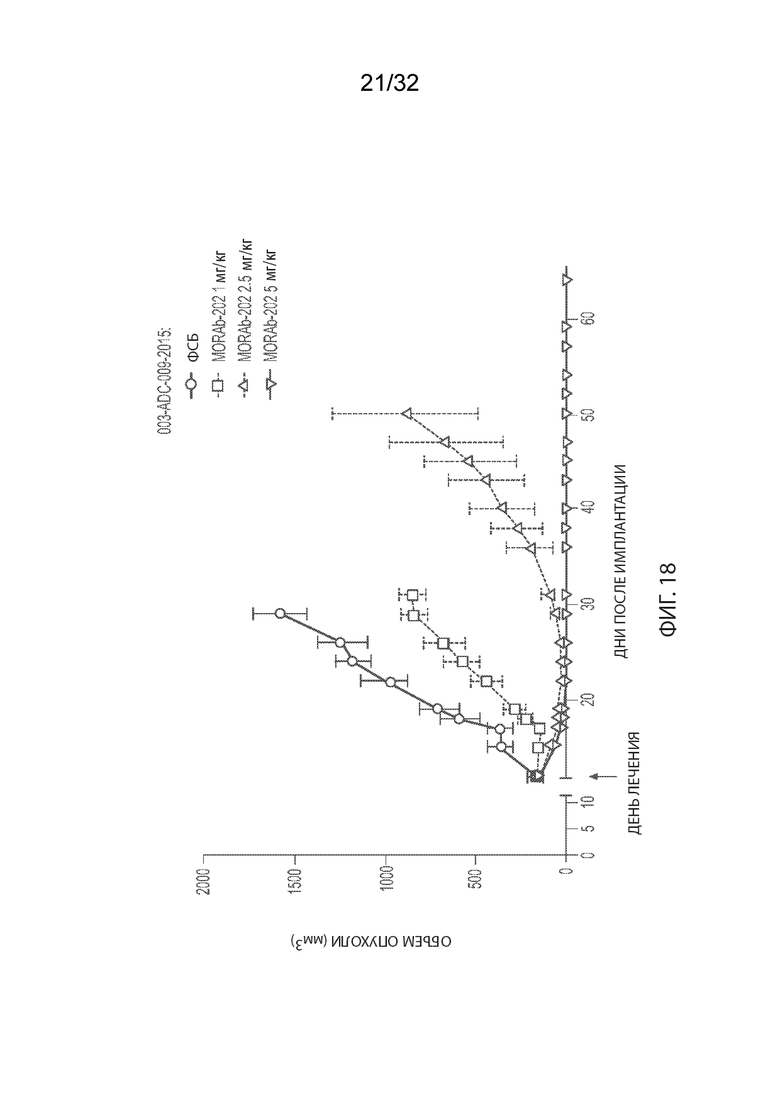

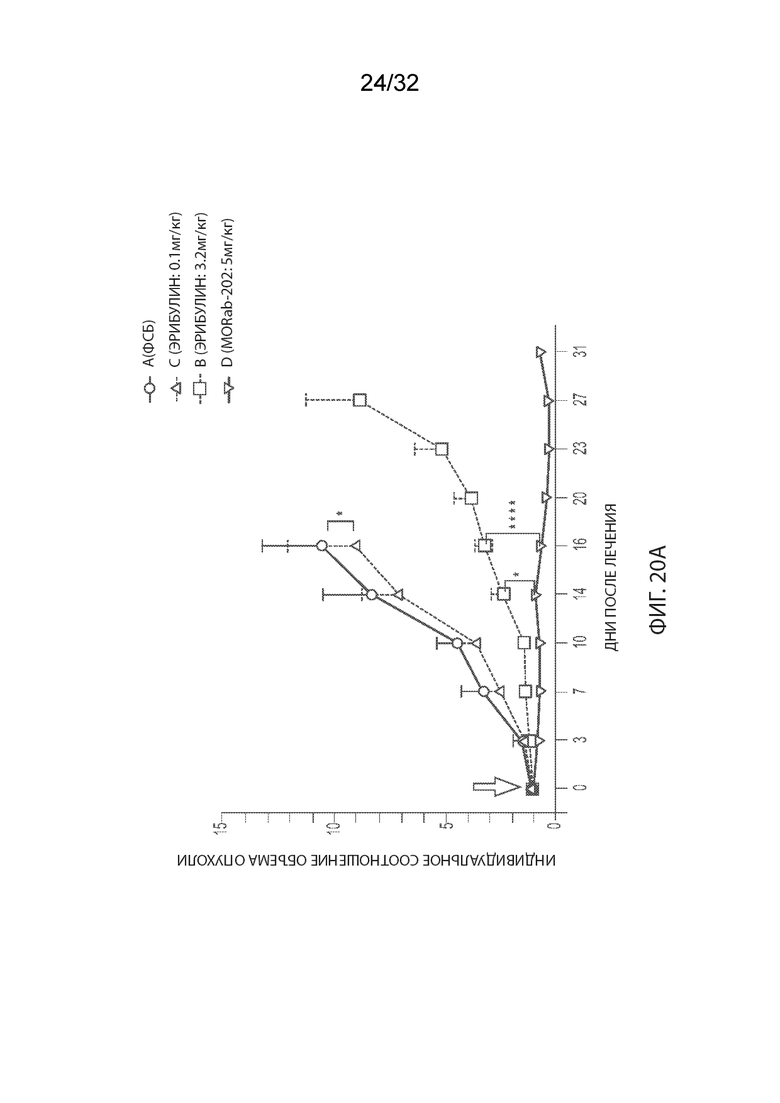
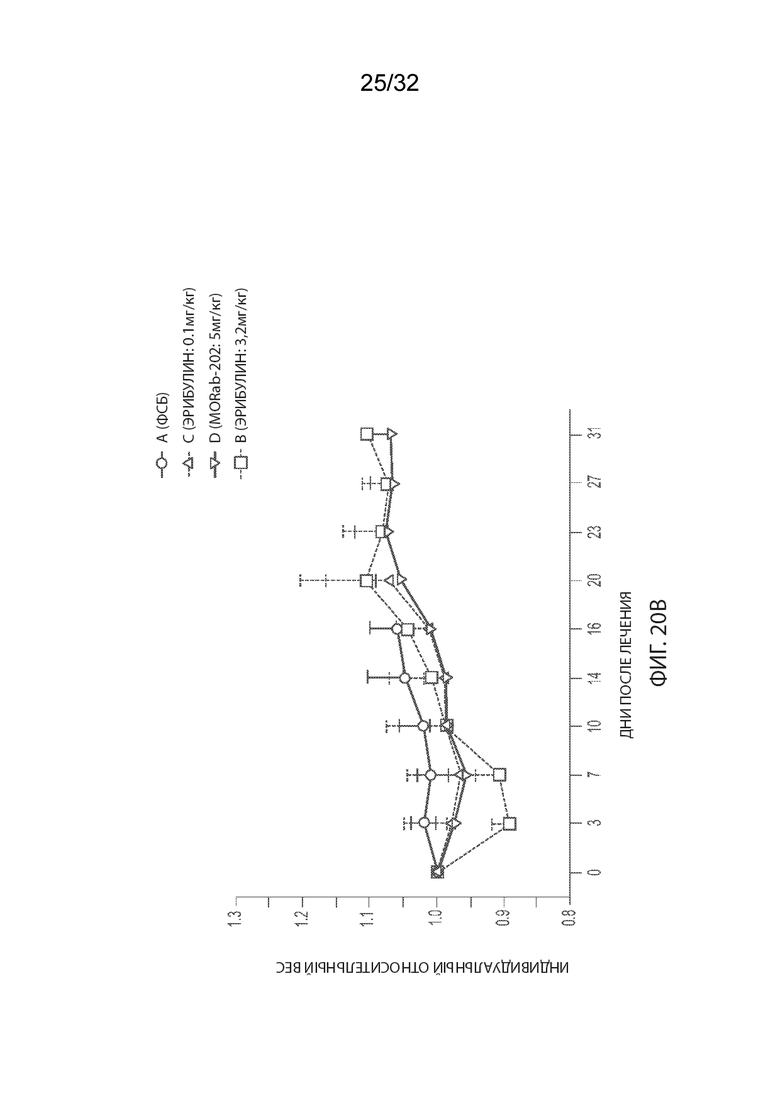
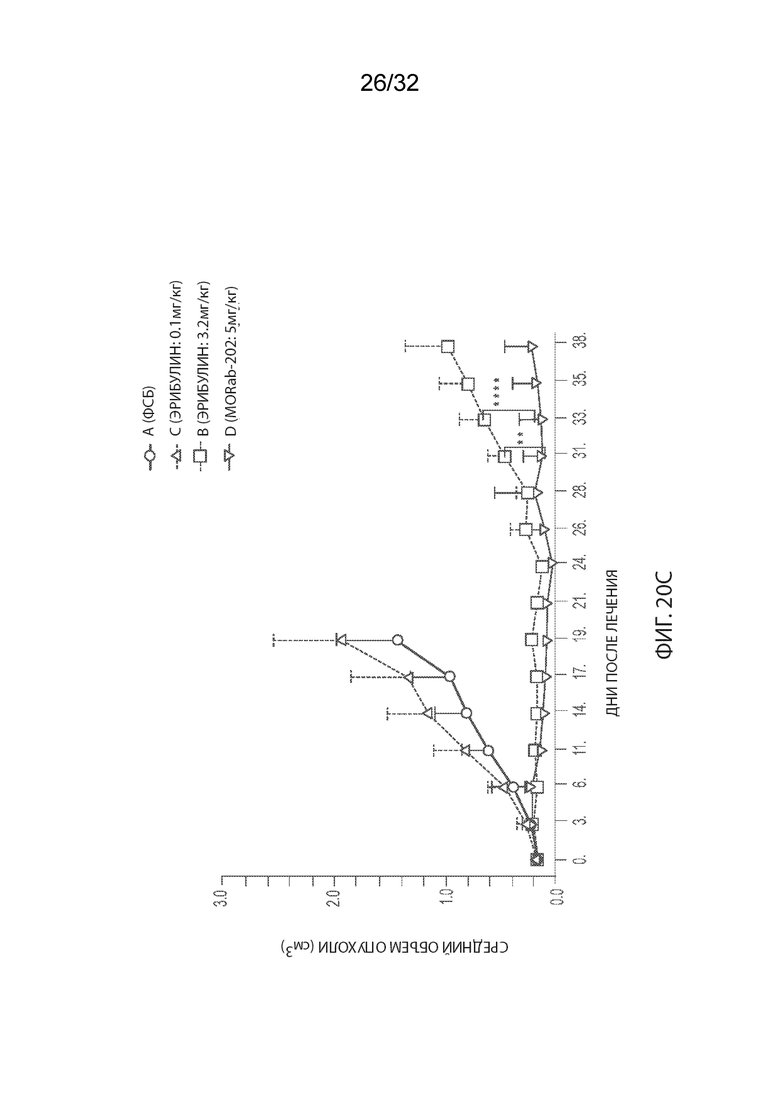
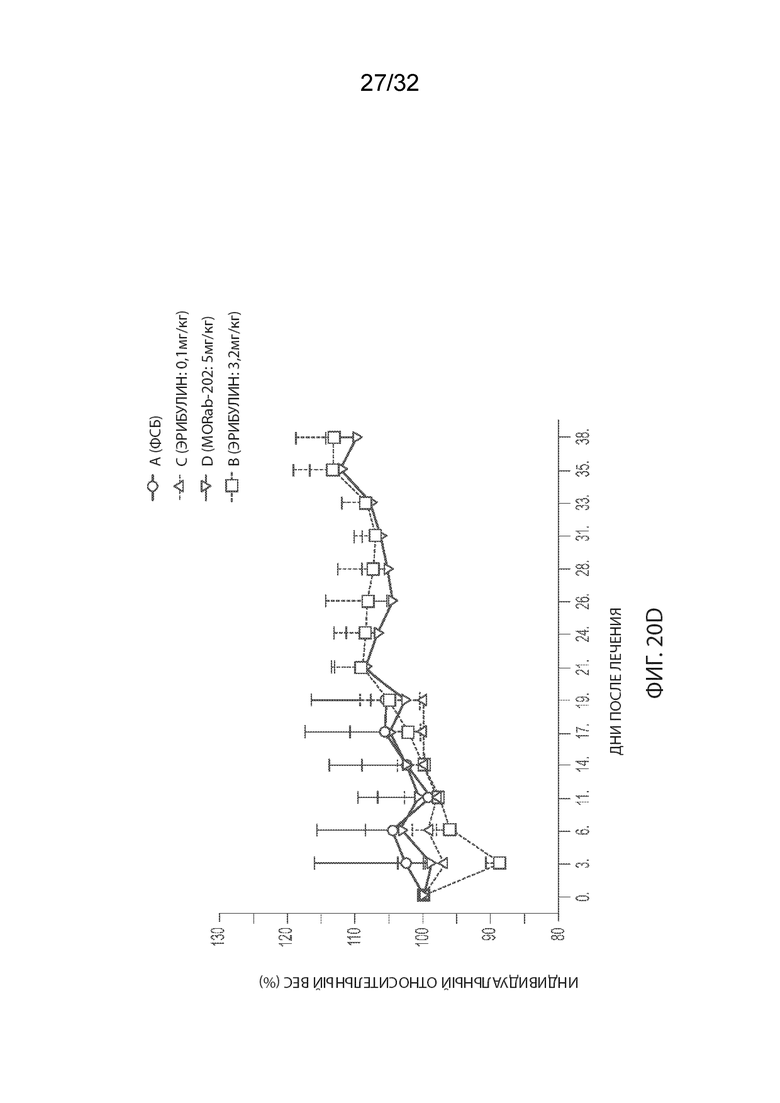
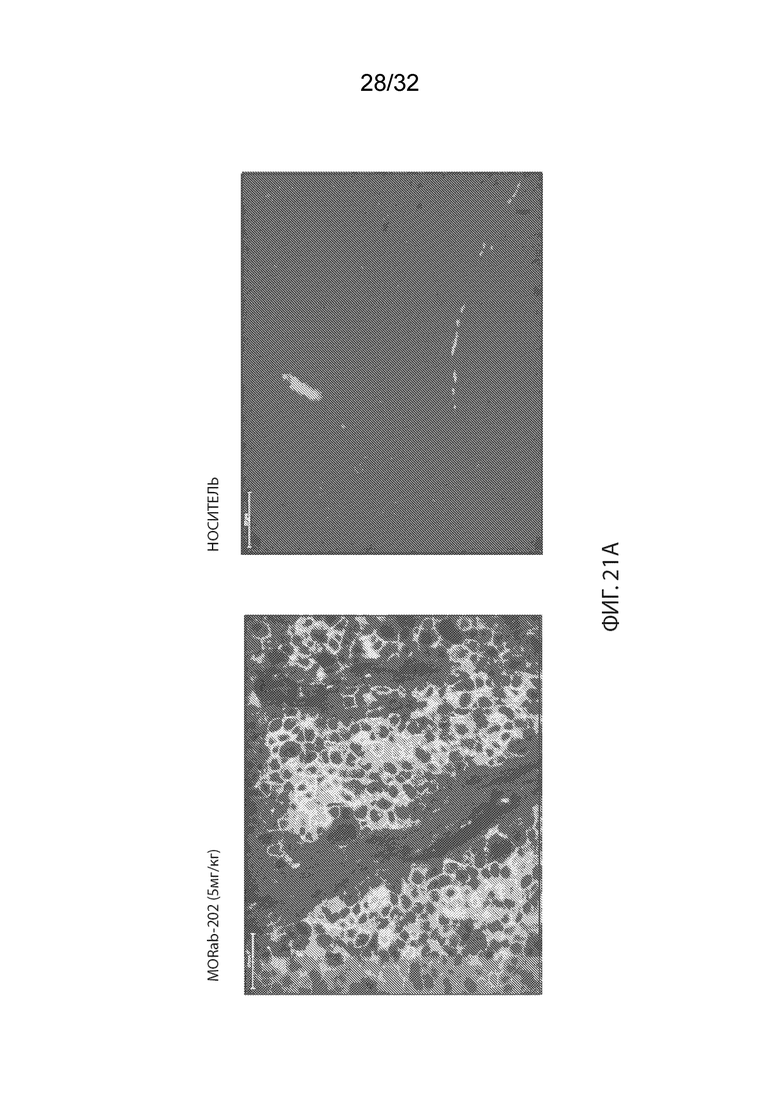
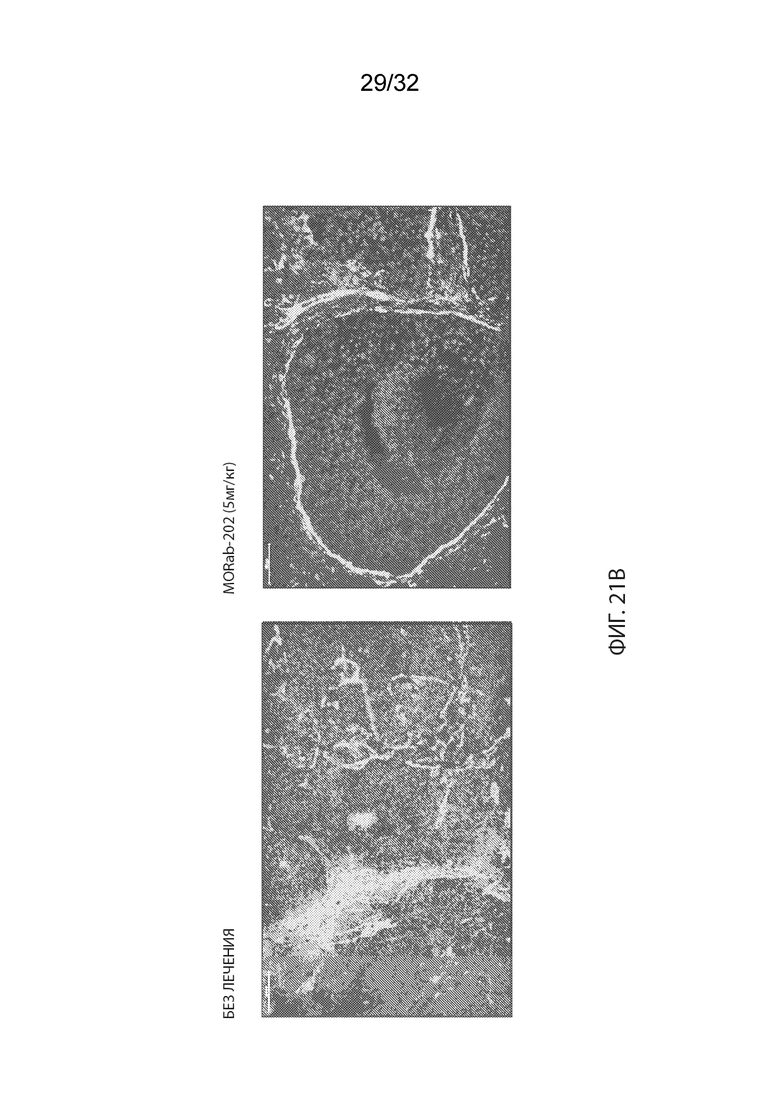
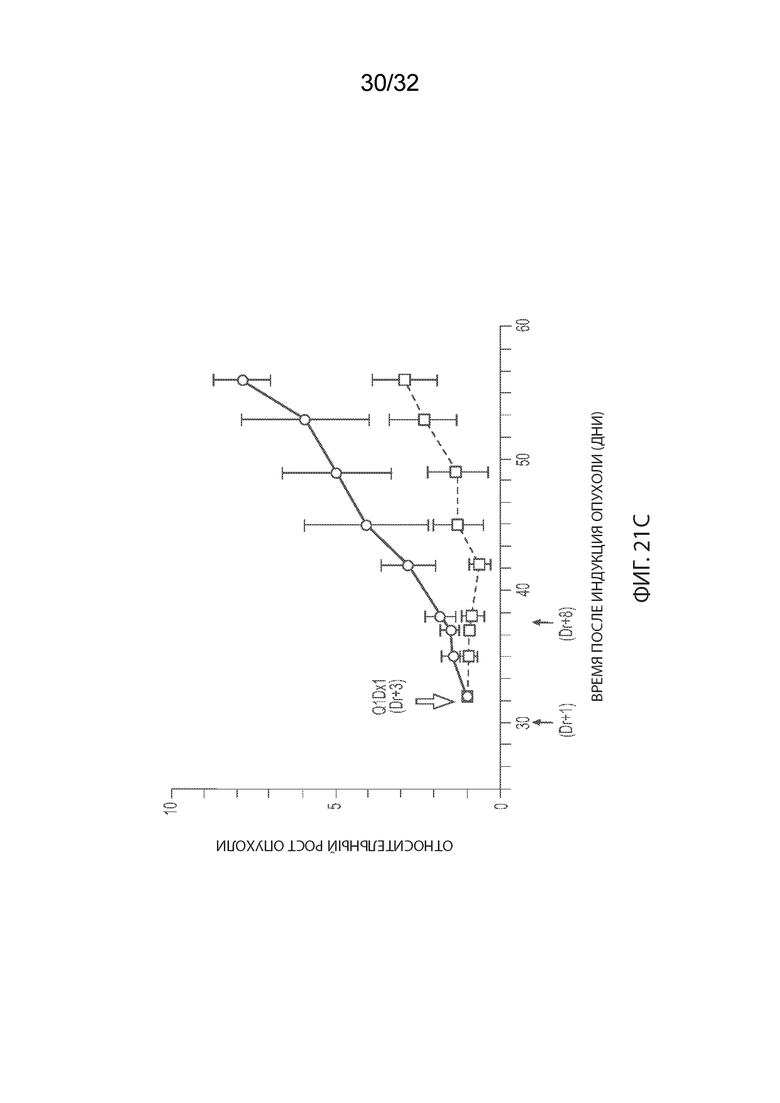
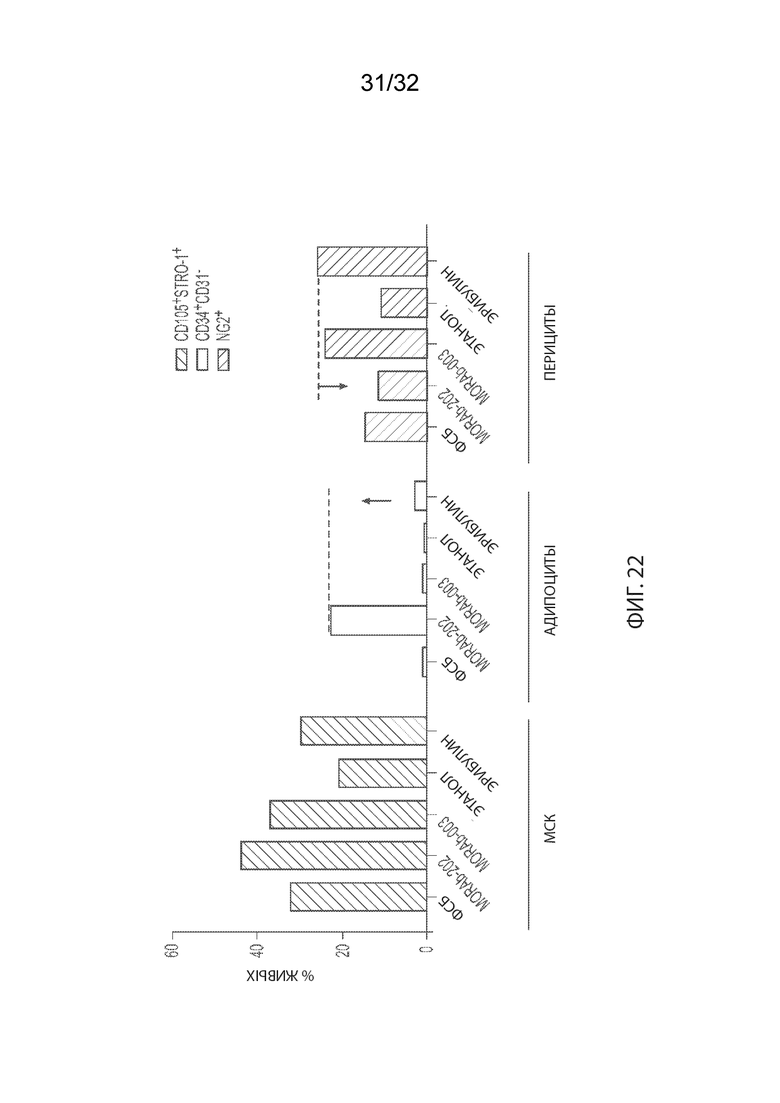
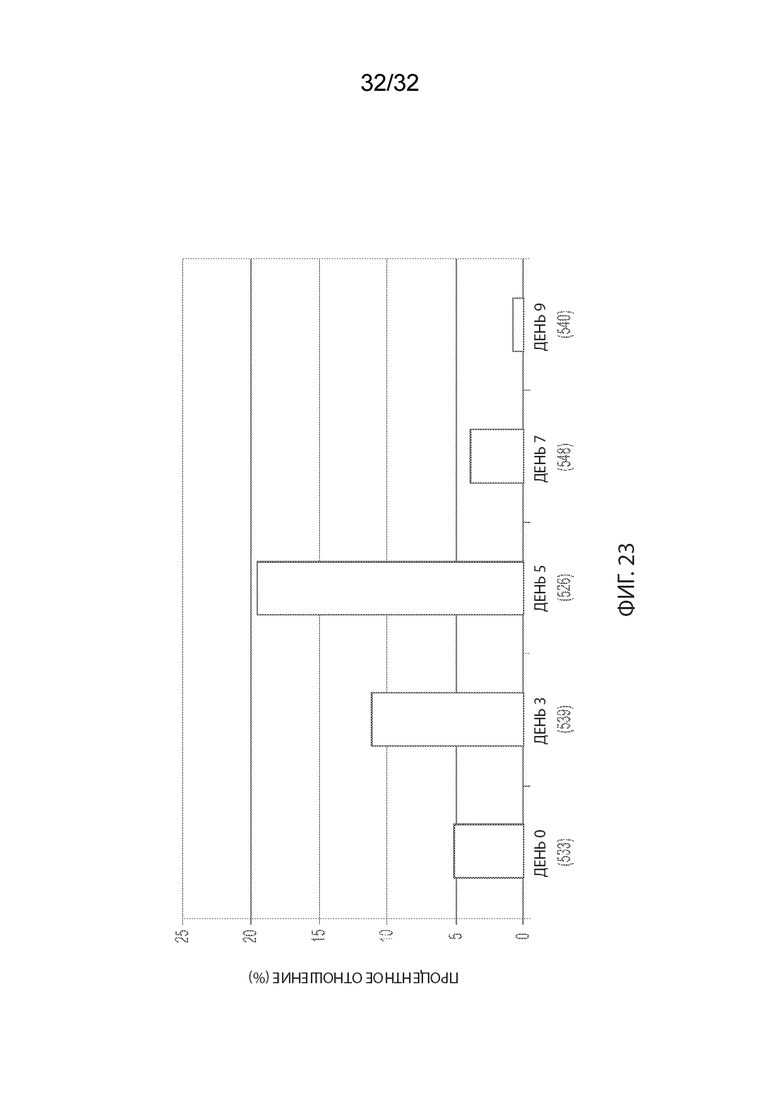
Настоящее изобретение относится к области биотехнологии, а именно к конъюгату типа антитело-лекарственное средство, и может быть использовано в медицине. Изобретение позволяет получить конъюгат антитела (или его Fab-фрагмента) к рецептору фолиевой кислоты альфа с эрибулином — противоопухолевым агентом, ингибирующим фазу роста микротрубочек в ходе митотического деления клеток. Фрагмент антитела в составе конъюгата по настоящему изобретению способствует направленной доставке терапевтического агента (эрибулина) к клеткам опухоли-мишени в организме пациента. Изобретение может быть применимо в качестве лекарственного средства в терапии злокачественных новообразований, клетки которых сверхэкспрессируют рецептор фолиевой кислоты, например, при лечении рака яичников или опухолей эпителиального происхождения. 8 н. и 36 з.п. ф-лы, 23 ил., 50 табл., 4 пр.
1. Конъюгат антитело-лекарственное средство Формулы (I) для лечения рака, который экспрессирует рецептор фолиевой кислоты альфа:
Ab-(L-D)p , (I)
в которой Ab представляет собой интернализирующееся антитело к рецептору фолиевой кислоты альфа или его интернализирующийся антигенсвязывающий фрагмент, содержащие три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности SEQ ID NO: 2 (HCDR1), SEQ ID NO: 3 (HCDR2) и SEQ ID NO: 4 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности SEQ ID NO: 7 (LCDR1), SEQ ID NO: 8 (LCDR2) и SEQ ID NO: 9 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности SEQ ID NO: 13 (HCDR1), SEQ ID NO: 14 (HCDR2) и SEQ ID NO: 15 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности SEQ ID NO: 16 (LCDR1), SEQ ID NO: 17 (LCDR2) и SEQ ID NO: 18 (LCDR3), как определено системой нумерации IMGT;
D представляет собой эрибулин;
L представляет собой расщепляемый линкер, содержащий группу присоединения антитела, спейсерную единицу и расщепляемую единицу, где группа присоединения антитела присоединена к Ab, и расщепляемая единица прямо или опосредованно присоединена к D; а также
p представляет собой целое число от 1 до 8.
2. Конъюгат антитело-лекарственное средство по п. 1, отличающийся тем, что p равно 3 или 4.
3. Конъюгат антитело-лекарственное средство по п. 1 или 2, где группа присоединения антитела прямо или опосредованно присоединена к спейсерной единице.
4. Конъюгат антитело-лекарственное средство по любому из пп. 1-3, где спейсерная единица прямо или опосредованно присоединена к расщепляемой единице.
5. Конъюгат антитело-лекарственное средство по любому из пп. 1-4, отличающийся тем, что расщепляемая единица расположена так, что никакая часть линкера, или антитела, или антигенсвязывающего фрагмента не остается связанной с эрибулином после расщепления.
6. Конъюгат антитело-лекарственное средство по любому из пп. 1-3, отличающийся тем, что расщепляемая единица содержит расщепляемый пептидный фрагмент.
7. Конъюгат антитело-лекарственное средство по любому из пп. 1-6, отличающийся тем, что расщепляемый пептидный фрагмент или расщепляемая единица содержит аминокислотную единицу.
8. Конъюгат антитело-лекарственное средство по п. 7, отличающийся тем, что аминокислотная единица содержит валин-цитруллин (Val-Cit).
9. Конъюгат антитело-лекарственное средство по любому из пп. 1-8, отличающийся тем, что спейсерная единица содержит полиэтиленгликолевый (ПЭГ) фрагмент.
10. Конъюгат антитело-лекарственное средство по п. 9, отличающийся тем, что ПЭГ-фрагмент содержит -(ПЭГ)m- и m представляет собой целое число от 1 до 10
11. Конъюгат антитело-лекарственное средство по п. 10, отличающийся тем, что m равно 2.
12. Конъюгат антитело-лекарственное средство по любому из пп. 1-11, отличающийся тем, что спейсерная единица прикрепляется к антителу или антигенсвязывающему фрагменту через группу присоединения к антителу.
13. Конъюгат антитело-лекарственное средство по п. 12, где группа присоединения антитела содержит малеимидный (Mal) фрагмент (Mal-спейсерная единица).
14. Конъюгат антитело-лекарственное средство по п. 13, отличающийся тем, что Mal-спейсерная единица соединена с антителом или антигенсвязывающим фрагментом через остаток цистеина на антителе или антигенсвязывающем фрагменте.
15. Конъюгат антитело-лекарственное средство по п. 13 или 14, отличающийся тем, что расщепляемый линкер содержит Mal-спейсерную единицу и расщепляемый пептидный фрагмент.
16. Конъюгат антитело-лекарственное средство по п. 15, отличающийся тем, что расщепляемый пептидный фрагмент содержит Val-Cit.
17. Конъюгат антитело-лекарственное средство по любому из пп. 13-16, отличающийся тем, что Mal-спейсерная единица прикрепляет антитело или антигенсвязывающий фрагмент к расщепляемой единице в линкере.
18. Конъюгат антитело-лекарственное средство по п. 17, отличающийся тем, что расщепляемая единица в линкере содержит расщепляемый пептидный фрагмент.
19. Конъюгат антитело-лекарственное средство по п. 18, отличающийся тем, что расщепляемый пептидный фрагмент содержит Val-Cit.
20. Конъюгат антитело-лекарственное средство по любому из пп. 17-19, отличающийся тем, что расщепляемый линкер содержит Mal-(ПЭГ)2-Val-Cit.
21. Конъюгат антитело-лекарственное средство по любому из пп. 1-20, отличающийся тем, что расщепляемая единица в линкере соединена с эрибулином прямо или опосредованно через необязательную дополнительную спейсерную единицу.
22. Конъюгат антитело-лекарственное средство по п. 21, отличающийся тем, что указанная дополнительная спейсерная единица содержит п-аминобензилоксикарбонил (pAB).
23. Конъюгат антитело-лекарственное средство по п. 21 или 22, отличающийся тем, что расщепляемая единица в линкере содержит расщепляемый пептидный фрагмент.
24. Конъюгат антитело-лекарственное средство по п. 23, отличающийся тем, что расщепляемый пептидный фрагмент содержит Val-Cit.
25. Конъюгат антитело-лекарственное средство по любому из пп. 21-24, отличающийся тем, что расщепляемый линкер содержит Val-Cit-pAB.
26. Конъюгат антитело-лекарственное средство по любому из пп. 21-25, отличающийся тем, что расщепляемый линкер содержит Mal-(PEG)2-Val-Cit-pAB.
27. Конъюгат антитело-лекарственное средство по любому из пп. 21-26, отличающийся тем, что расщепляемый линкер ковалентно присоединяется к эрибулину через амин C-35.
28. Конъюгат антитело-лекарственное средство по любому из пп. 1-27, отличающийся тем, что антитело или антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 23, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 24.
29. Конъюгат антитело-лекарственное средство по любому из пп. 1-28, отличающийся тем, что антитело или антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи Ig каппа человека.
30. Конъюгат антитело-лекарственное средство по п. 28 или 29, отличающийся тем, что p представляет собой 3 или 4.
31. Конъюгат антитело-лекарственное средство Формулы (I) для лечения рака, который экспрессирует рецептор фолиевой кислоты альфа:
Ab-(L-D)p , (I)
в которой Ab представляет собой интернализирующееся антитело к рецептору фолиевой кислоты альфа или его интернализирующийся антигенсвязывающий фрагмент, содержащий три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 2 (HCDR1), SEQ ID NO: 3 (HCDR2) и SEQ ID NO: 4 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 7 (LCDR1), SEQ ID NO: 8 (LCDR2) и SEQ ID NO: 9 (LCDR3), как определено системой нумерации Кабата; или три определяющие комплементарность области тяжелой цепи (HCDR), содержащие аминокислотные последовательности из SEQ ID NO: 13 (HCDR1), SEQ ID NO: 14 (HCDR2) и SEQ ID NO: 15 (HCDR3); и три определяющие комплементарность области легкой цепи (LCDR), содержащие аминокислотные последовательности из SEQ ID NO: 16 (LCDR1), SEQ ID NO: 17 (LCDR2) и SEQ ID NO: 18 (LCDR3), как определено системой нумерации IMGT;
D представляет собой эрибулин;
L представляет собой расщепляемый линкер, содержащий Mal-(ПЭГ)2-Val-Cit-pAB, где Mal присоединен к Ab, и pAb присоединен к D; и
p представляет собой целое число от 1 до 8.
32. Конъюгат антитело-лекарственное средство по п. 31, отличающийся тем, что антитело или антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 23, и вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO: 24.
33. Конъюгат антитело-лекарственное средство по п. 31 или 32, отличающийся тем, что антитело или антигенсвязывающий фрагмент содержит константный домен тяжелой цепи IgG1 человека и константный домен легкой цепи Ig каппа человека.
34. Конъюгат антитело-лекарственное средство по любому из пп. 31-33, отличающийся тем, что p представляет собой 3 или 4.
35. Композиция для лечения рака, который экспрессирует рецептор фолиевой кислоты альфа, отличающаяся тем, что композиция содержит множественные копии конъюгата антитело-лекарственное средство по любому из пп. 1-34, и тем, что среднее p конъюгатов антитело-лекарственное средство в указанной композиции составляет от около 3,2 до около 4,4.
36. Способ лечения пациента, имеющего рак или имеющего риск возникновения рака, который экспрессирует рецептор фолиевой кислоты альфа, включающий введение пациенту терапевтически эффективного количества конъюгата антитело-лекарственное средство по любому из пп. 1-34 или композиции по п. 35.
37. Способ по п. 36, согласно которому рак представляет собой рак желудка, серозный рак яичников, светлоклеточный рак яичника, немелкоклеточный рак легкого, колоректальный рак, трижды негативный рак молочной железы, рак эндометрия, карциноид легких, остеосаркому, рак мочевого пузыря или карциному уротелиальных клеток.
38. Способ снижения или ингибирования роста опухоли, которая экспрессирует рецептор фолиевой кислоты альфа, включающий введение терапевтически эффективного количества конъюгата антитело-лекарственное средство по любому из пп. 1-34 или композиции по п. 35.
39. Способ по п. 38, согласно которому указанная опухоль представляет собой рак желудка, серозный рак яичников, светлоклеточный рак яичника, немелкоклеточный рак легкого, колоректальный рак, трижды негативный рак молочной железы, рак эндометрия, карциноид легких, остеосаркому, рак мочевого пузыря или карциному уротелиальных клеток.
40. Применение конъюгата антитело-лекарственное средство по п. 1-34 или композиции по п. 35 в лечении рака, который экспрессирует рецептор фолиевой кислоты альфа.
41. Применение по п. 40, отличающееся тем, что рак представляет собой рак желудка, серозный рак яичников, светлоклеточный рак яичника, немелкоклеточный рак легкого, колоректальный рак, трижды негативный рак молочной железы, рак эндометрия, карциноид легких, остеосаркому, рак мочевого пузыря или карциному уротелиальных клеток.
42. Применение конъюгата антитело-лекарственное средство по любому из пп. 1-34 или композиции по п. 35 в способе изготовления медицинского препарата для лечения рака, который экспрессирует рецептор фолиевой кислоты альфа.
43. Применение по п. 42, отличающееся тем, что рак представляет собой рак желудка, серозный рак яичников, светлоклеточный рак яичника, немелкоклеточный рак легкого, колоректальный рак, трижды негативный рак молочной железы, рак эндометрия, карциноид легких, остеосаркому, рак мочевого пузыря или карциному уротелиальных клеток.
44. Фармацевтическая композиция для лечения рака, который экспрессирует рецептор фолиевой кислоты альфа, содержащая конъюгат антитело-лекарственное средство по любому из пп. 1-34 или композицию по п. 35 и фармацевтически приемлемый носитель.
| WO 2014089335 A2, 12.06.2014 | |||
| ZHENG W | |||
| et al | |||
| Macrocyclic ketone analogues of halichondrin B, BIOORG | |||
| & MED | |||
| CHEM | |||
| LETTERS, 2004, v | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| Локомотив, работающий под давлением продуктов горения и паром | 1925 |
|
SU5551A1 |
| NARAYAN S | |||
| et al | |||
| Novel second generation analogs of eribulin | |||
| Part II: Orally available and active against resistant tumors in vivo, BIOORG | |||
| & MED | |||
| CHEM | |||
| LETTERS, 2011, v | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |

