Ссылка на перечень последовательностей, поданный в электронном виде
Перечень последовательностей настоящей заявки подан в электронном виде через EFS-Web в виде ASCII отформатированного перечня последовательностей с названием файла “24608WOPCT-SEQLIST-02JUl2019.txt”, имеющего дату создания 2 июля 2019 г. и размер 80,5 кБ. Этот перечень последовательностей, поданный через EFS-Web, является частью описания изобретения и включен в настоящую заявку посредством ссылки во всей полноте.
Предпосылки создания изобретения
4’-Этинил-2’-дезоксинуклеозидные аналоги известны как обладающие активностью против ВИЧ, СПИДа и связанных с ними заболеваний.
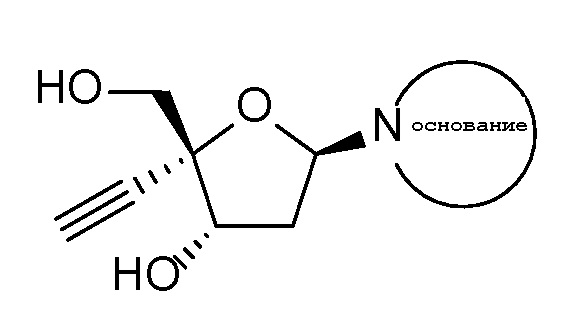
Одним примером 4’-этинилнуклеозидного аналога является 4’-этинил-2-фтор-2’-дезоксиаденозин (EFdA, также известный как MK-8591), который является нуклеозидным ингибитором транслокации обратной транскриптазы, который блокирует репликацию вирусов ВИЧ-1 и SIV in vitro (Kawamoto, A., Kodama, E., Sarafianos S. F. et al, Int. J. Biochem. Cell Biol.; 40(11):2410-20 [2008]; Ohrui, H., Kohgo, S., Hayakawa, H. et al, Nucleosides, Nucleotides & Nucleic Acids, 26, 1543-1546 [2007]) и in vivo (Hattori, S., Ide, K., Nakata, H. et al. Antimicrobial. Agents and Chemotherapy, 53, 3887-3893 [2009]). EFdA заявлен в Патенте США № 7339053 (указан в ‘053 патенте как 2’-дезокси-4’-C-этинил-2-фтораденозин). EFdA имеет следующую химическую структуру:
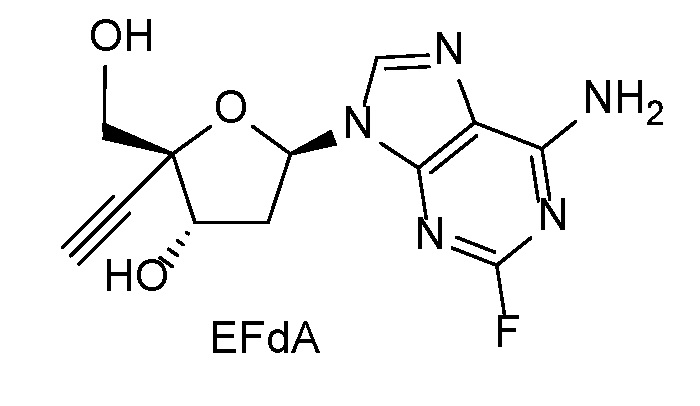
EFdA метаболизируется в клетках до своего активного трифосфатного анаболита, который ингибирует обратную транскриптазу ВИЧ. В отличие от нуклеозидных ингибиторов обратной транскриптазы (NsRTI) и нуклеотидных ингибиторов обратной транскриптазы (NtRTI), доступных в настоящее время для лечения ВИЧ-инфекции, которые отсутствует 3'-ОН группа для блокирования включения входящего нуклеотида, EFdA сохраняет 3'-ОН группу и действует в качестве терминатора цепи, предотвращая транслокацию праймера:матрицы в активном сайте обратной транскриптазы (RT) и предотвращая связывание входящих дезоксирибонуклеотидтрифосфатов (dNTP). Кроме того, предполагается, что сморщивание модифицированного рибозного кольца EFdA вносит вклад в ингибирование обратной транскриптазы, помещая 3'-OH в вектор, в котором фосфоперенос от поступающего нуклеотида неэффективен. (Michailidis E, et al., Mechanism of inhibition of HIV-1 reverse transcriptase by 4’-ethynyl-2-fluoro-2’-deoxyadenosine triphosphate, J Biol Chem 284:35681-35691 [2009]; Michailidis E, et al., 4’-Ethynyl-2-fluoro-2’-deoxyadenosine (EFdA) inhibits HIV-1 reverse transcriptase with multiple mechanisms, J Biol Chem 289:24533-24548 [2014]).
В анализах репликации ВИЧ in vitro EFdA является мощным антиретровирусным препаратом и проявляет сопоставимую противовирусную активность против клинических изолятов всех подтипов, которые были оценены. Он быстро анаболизируется как в линиях лимфоидных клеток, так и в мононуклеарных клетках периферической крови до активного трифосфата in vitro, а внутриклеточный период полужизни EFdA-трифосфата (EFdA-TP) превышает 72 часа. (Stoddart, C. A., Galkina, et al., Oral Administration of the Nucleoside EFdA (4’-Ethynyl-2-Fluoro-2’-Deoxyadenosine) Provides Rapid Suppression of HIV Viremia in Humanized Mice and Favorable Pharmacokinetic Properties in Mice and the Rhesus Macaque, Antimicrob Agents Chemother, 2015 Jul; 59(7): 4190-4198, Published online 2015 May 4).
Было показано, что EFdA обладает эффективностью на животных моделях ВИЧ-инфекции, включая гуманизированные мышиные модели и модель макак-резус, инфицированных SIV. Фармакокинетические исследования перорального введения EFdA на мышах и макаках-резус продемонстрировали быструю абсорбцию и высокие концентрации в плазме. Длительный внутриклеточный период полужизни был продемонстрирован тем фактом, что выделенные мононуклеарные клетки периферической крови макака-резуса были невосприимчивы к инфекции SIV через 24 часа после введения лекарства. (Там же.)
Предыдущие способы синтеза 4'-этинилнуклеозидных аналогов, включая EFdA, страдают умеренной стереоселективностью при образовании C-N связи между этинилдезоксирибозным сахаром и 2-фторадениновым (также называемым 2-фтор-9H-пурин-6-амином) нуклеооснованием. Предыдущие способы синтеза также требуют защитных групп для осуществления реакции гликозилирования, что снижает эффективность синтеза.
Синтез, описанный в Kei Fukuyama, et al., Synthesis EFdA via the Diastereoselective Aldol Reaction of a Protected 3-Keto Furanose, Organic Letters 2015, 17 (4), pp. 828-831; DOI: 10.1021/ol5036535) представляет собой 14-ступенчатый синтез из D-глюкоза диацетонида, в котором используются диастереоселективные реакции для установки трех стереоцентров. Стереохимию аномерного центра контролируют при помощи 2'-ацетокси направляющей группы, которую впоследствии удаляют посредством гидролиза и деоксигенирования. Для этого способа требуются 4 хроматографических очистки и стехиометрическое использование токсичного оловоорганического реагента для поздней стадии деоксигенирования.
В другом пути (см. Mark McLaughlin, et al., Enantioselective Synthesis of 4′-Ethynyl-2-fluoro-2′-deoxyadenosine (EFdA) via Enzymatic Desymmetrization, Organic Letters 2017, 19 (4), pp. 926-929) полностью замещенный 4'-карбинол получают стереоселективно с ферментативной десимметризацией. 3'-стереоцентр устанавливают путем каталитического гидрирования с асимметричным переносом, а аномерную 1'-связь устанавливают с умеренной стереоселективностью с использованием контроля субстрата, с повышением стереохимической чистоты, достигаемым кристаллизацией промежуточного продукта. Этот способ требует 15 стадий, требует использования нескольких защитных групп и образует гликозильную связь между нуклеооснованием и фрагментами сахара с низкой стереоселективностью (1,8:1)
12-стадийный синтез для получения EFdA из R-глицеральдегид ацетонида описан в Kageyama, M., et al., Concise Synthesis of the Anti-HIV Nucleoside EFdA, Biosci. Biotechnol. Biochem, 2012, 76, pp. 1219 - 1225; and Enantioselective Total Synthesis of the Potent Anti-HIV Nucleoside EFdA, Masayuki Kageyama, et al., Organic Letters 2011 13 (19), pp. 5264-5266 [DOI: 10.1021/ol202116k]. В этих способах синтеза используют хиральное исходное вещество для установки 3'-стереоцентра с умеренной диастереоселективностью. После хроматографического разделения стереоизомеров новый стереоцентр используется для направления диастереоселективного присоединения алкина для установки полностью замещенного 4'-стереоцентра. Аномерное 1'-положение устанавливается с низким стереоконтролем, и требуется хроматография для разделения аномеров. Этот путь требует хроматографического разделения диастереоизомеров на двух разных стадиях и начинается с дорогостоящего хирального исходного вещества.
Kohgo, S., et al., Design, Efficient Synthesis, and Anti-HIV Activity of 4′-C-Cyano- and 4′-C-Ethynyl-2′-deoxy Purine Nucleosides, Nucleosides, Nucleotides and Nucleic Acids, 2004, 23, pp. 671-690 [DOI: 10.1081/NCN-120037508] описывает путь синтеза, который начинается с существующего нуклеозида и модифицирует как сахарные части, так и нуклеооснования. Это 18-стадийный синтез, исходящий из 2-амино-2'-дезоксиаденозина, с низким общим выходом 2,5%.
Известно, что ферменты, такие как пурин-нуклеозидфосфорилаза (PNP, EC 2.4.2.1), могут образовывать гликозильную связь в нуклеозидах и нуклеозидных аналогах с высокой стереоселективностью и без использования защитных групп. См., например, обзор: New Trends in Nucleoside Biotechnology, Mikhailopulo, I.A., Miroshnikov, A.I,. Acta Naturae 2010, 2, pp. 36-58. Однако существующий объем сахарных фрагментов, которые могут подвергаться реакции, катализируемой PNP, ограничен α-1-фосфатами природной рибозы и дезоксирибозы, а также небольшим количеством аналогов с небольшими H, NH2 или F заместителями в положениях C2' и C3' и заменами C5' OH группы. Не было никаких сообщений об успешном гликозилировании, катализируемом PNP, с использованием сахаров с углеродными заместителями в кольце или любого замещения в положении C4’.
Доступ к рибозным и дезоксирибоза α-1-фосфатным субстратам для катализируемого PNP гликозилирования был продемонстрирован путем транслокации фосфатной группы из положения 5'-гидроксила в 1'-гидроксил с использованием фермента фосфопентомутазы (PPM, EC 5.4.2.7) (см. Mikhailopulo, I.A., et al. выше). Однако диапазон сахаров, для которых PPM способен катализировать эту реакцию, ограничен рибозой, арабинозой, 2-дезоксирибозой и 2,3-дидезоксирибозой. Не сообщалось о примерах успешной реакции с сахарными фосфатами, содержащими какие-либо дополнительные заместители.
Известно, что ферменты дезоксирибозофосфатальдолазы (DERA, EC 4.1.2.4) катализируют альдольное присоединение ацетальдегида к другим короткоцепочечным альдегидам (см. обзор: Stephen M. Dean, et al., Recent Advances in Aldolase-Catalyzed Asymmetric Synthesis, Adv. Synth. Catal. 2007, 349, pp. 1308-1320; DOI: 10.1002/adsc.200700115). Однако не сообщалось ни о каких примерах с альдегидами, несущими полностью замещенный углерод α относительно альдегида.
Патент США № 7229797 описывает образование дезоксирибонуклеозидов из природного незамещенного дезоксирибозо-1-фосфата с использованием пурин-нуклеозидфосфорилазы (PNP) и дополнительным использованием ферментов, таких как сахарозофосфорилаза, для удаления неорганического фосфатного побочного продукта и поддержания равновесия. Он не раскрывает ферментную инженерию для создания PNP ферментов, которые могут генерировать нуклеозиды из неприродного 4-этинил-D-2-дезоксирибозо-1-фосфата, а также то, что путем конструирования ферментов PPM и DERA для воздействия на неприродные субстраты можно получить 4-этинил-D-2-дезоксирибозо-1-фосфат.
С учетом сложных и длительных вариантов синтеза, разработанных на сегодняшний день для получения 4'-этинилнуклеозидных аналогов, было бы желательно разработать улучшенный ферментативный синтез 4'-этинилнуклеозидных аналогов, таких как EFdA, который сокращает количество стадий процесса, сводит к минимуму использование защитных групп, улучшает стереоселективность гликозилирования и позволяет избежать использования токсичных материалов.
Неожиданно было обнаружено, что PPM ферменты обладают некоторой активностью с 3-атомным этинильным заместителем в 4’ положении рибозы, и что активность PPM фермента может быть улучшена путем введения мутаций в ферменты для успешного развития реакции изомеризации 4-этинил-D-2-дезоксирибозо-5-фосфата (6) до 4-этинил-D-2-дезоксирибозо-1-фосфата (6.5), катализируемой PPM, что предполагает возможность более эффективного способа получения 4'-этинил-2'-дезоксинуклеозидов.
Кроме того, было обнаружено, что PNP ферменты обладают некоторой активностью с 3-атомным этинильным заместителем в положении 4 дезоксирибозы, и что ферментативная активность PNP может быть улучшена путем введения мутаций в ферменты для успешного развития реакции гликозилирования, катализируемой PNP, что предполагает возможность более эффективного способа получения 4'-этинил-2'-дезоксинуклеозидов.
Дальнейшее улучшение общего способа синтеза исходило из открытия, что DERA ферменты, в частности DERA из Shewanella halifaxensis, обладают активностью для альдольной реакции с 2-этинилглицеральдегид-3-фосфатом, который имеет полностью замещенный α-углерод. Это открытие позволило эффективно синтезировать 4-этинил-D-2-дезоксирибозо-5-фосфат, предшественник 4’-этинил-2’-дезоксинуклеозидных аналогов, например, включая EFdA.
Сущность изобретения
Настоящее изобретение включает использование сконструированных ферментов в новом ферментативном синтезе 4’-этинил-2’-дезоксинуклеозидных аналогов, включая EFdA, который исключает использование защитных групп на промежуточных соединениях, улучшает стереоселективность гликозилирования и значительно уменьшает количество стадий процесса, необходимых для получения указанных соединений, по сравнению с предшествующими способами, среди прочих технологических усовершествований. Изобретение также относится к новым промежуточным соединениям, которые являются неотъемлемой частью ферментативного способа.
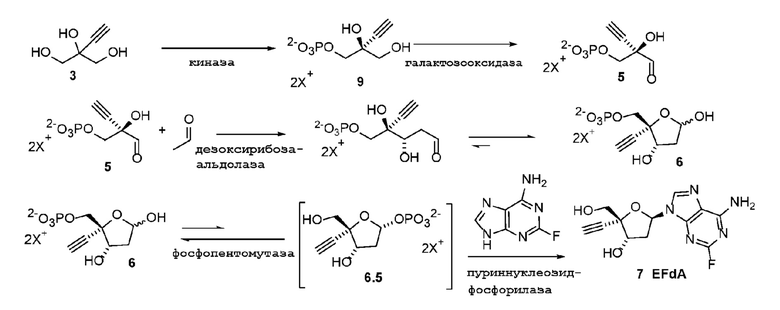
Общий способ представлен ниже на Схеме 1 и Схеме 2; последняя схема представляет альтернативный способ для получения соединения 5:
Схема 1
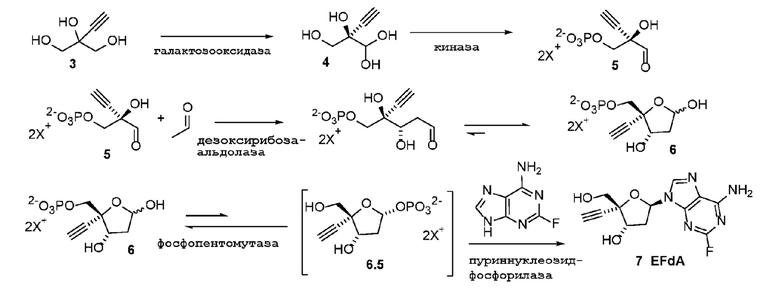
Схема 1A
В способе, описанном в настоящей заявке, можно использовать кислотную форму или соли фосфатных промежуточных соединений, и они не ограничиваются конкретными кислотными или солевыми формами, представленными в иллюстративных примерах стадий способа в настоящей заявке. Для всех фосфатных промежуточных соединений, описанных в настоящей заявке, 2X+ представляет собой любую комбинацию двух протонов, одного протона с одним другим одновалентным катионом, двух одновалентных катионов (одинаковых или отличных друг от друга) или один двухвалентный катион. Фосфатные промежуточные соединения, показанные как -HO3PO-, также могут иметь любую комбинацию двух протонов, одного протона с одним другим одновалентным катионом, двух одновалентных катионов (одинаковых или отличных друг от друга) или один двухвалентный катион, связанную с фосфатной группой. Примеры включают, но не ограничиваются этим, соли кальция, магния или цинка; моно- или динатриевые соли, моно- или дикалиевые соли, моно- или дилитиевые соли; моно- или диаммониевые соли; или одно- или двухвалентные соли с первичными, вторичными или третичными аминами.
Как хорошо известно в данной области техники, промежуточные соединения, показанные или названные на стадиях синтеза как альдегид или гидрат, могут существовать в любой форме или смеси таких форм в реакциях, описанных в настоящей заявке. Например, соединения (4) и (5) показаны на Схеме 1 как гидрат и альдегид, соответственно, но каждый может существовать в форме гидрата или альдегида или их смеси на стадиях реакции, где каждый присутствует. Каждая такая форма охватывается посредством ссылки на номера соединений (4) или (5) в рамках стадий способа, описанных в настоящей заявке:
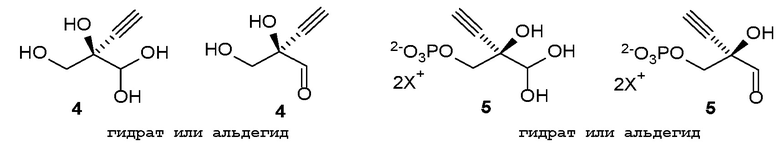 .
.
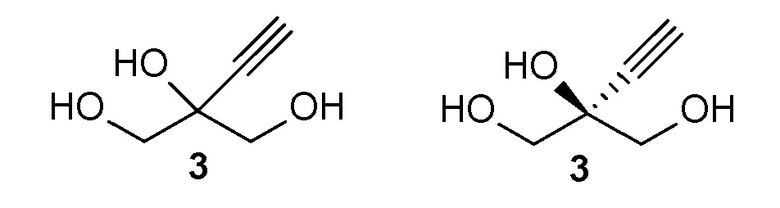
Соединение (3) является ахиральным и может быть показано как любое из следующих:
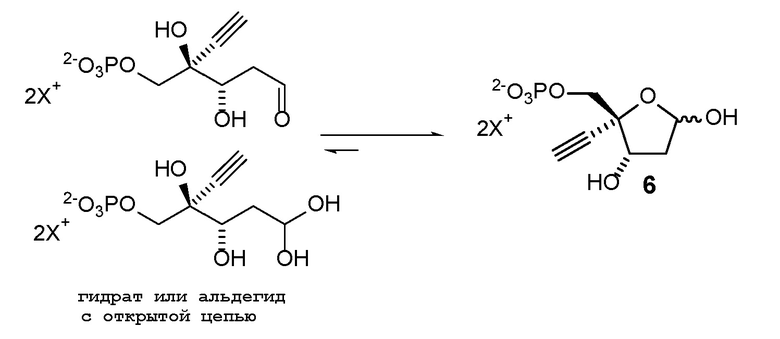
Соединение (6) может существовать в его кольцевой форме или в виде альдегида или гидрата с открытой цепью, каждое в виде его кислотно или его солевой формы, на стадиях реакции где оно присутствует:
Подробное описание изобретения
4’-Этинил-2’-дезоксинуклеозиды и их аналоги
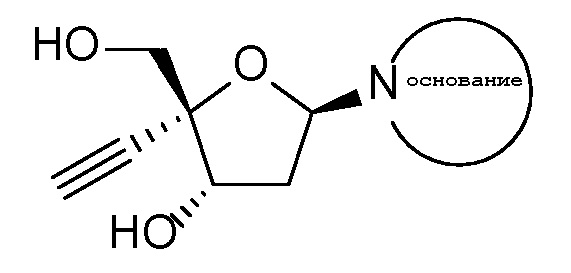
имеющие аномерную C-N связь, были исследованы на активность против ВИЧ, СПИДа и связанных с ними заболеваний. 4’-Этинил-2’-дезоксинуклеозиды и их аналоги включают 4’-этинил-2’-дезоксирибозу, присоединенную через аномерную C-N связь к пуриновому или пиримидиновому нуклеооснованию (аденин, гуанин, цитозин, тимин или урацил) или модифицированному пуриновому или пиримидиновому нуклеооснованию.
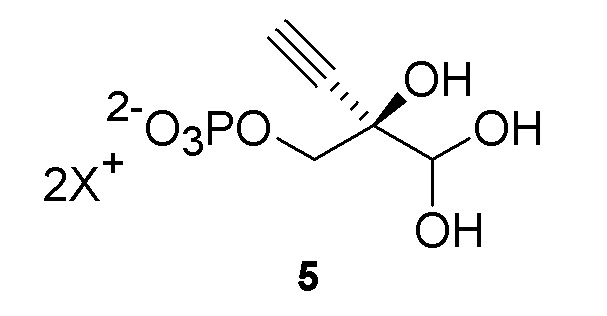
Было обнаружено, что 4’-этинил-2’-дезоксинуклеозидные аналоги, такие как EfdA, можно синтезировать с использованием способа, осуществляемого в одном реакционном сосуде, на конечной стадии путем объединения 4-этинил-D-2-дезоксирибозо-5-фосфата (6) с двумя ферментами фосфопентомутазой (PPM) [например, но не ограничиваясь этим, SEQ ID NO.: 8] и пуриннуклеозид-фосфорилазой (PNP) [например, но не ограничиваясь этим, SEQ ID NO.: 9, SEQ ID NO.: 15], как показано на Схеме 2.
Схема 2
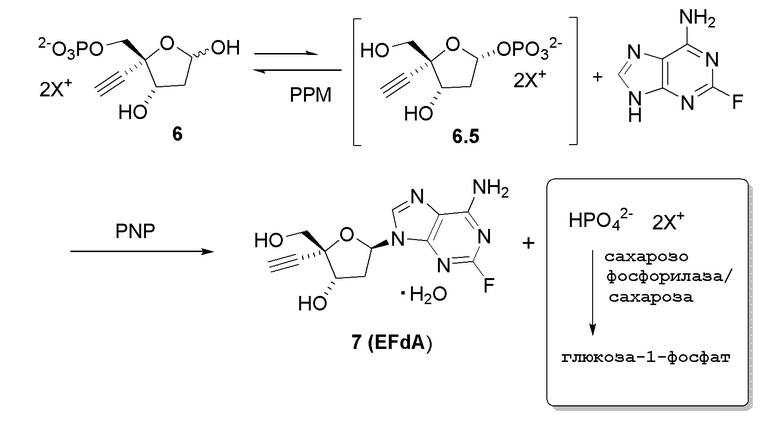
Схема 2A
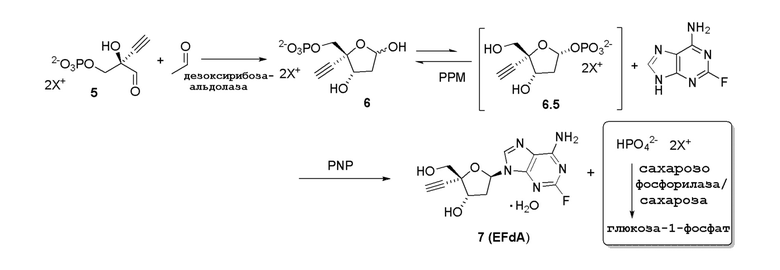
Как показано на Схеме 2, на конечной стадии синтеза используют 2-ферментную реакцию с необязательным 3-м ферментом для направления равновесия реакции на желаемый конечный продукт. Конечная стадия начинается с соединения (6) или его соли, где (6) представляет собой 4-этинил-2-дезоксирибозо-5-фосфат в кольцевой форме, как показано выше, или в форме его альдегида или гидрата с открытой цепью.
Соединение (6) объединяют с фосфопентомутазой (PPM), пуриннуклеозид-фосфорилазой (PNP), сахарозо-фосфорилазой, сахарозой и нуклеооснованием, например незамещенным или замещенным аденином, в буферном растворе, содержащем соль марганца (II), и доводят, по мере необходимости, до pH в диапазоне около 6,5-8,0 или более, конкретно около 7,0-7,5. Молярное соотношение сахароза:соединение (6) может составлять, но не ограничивается этим, от около 1:1 до 4:1. Компоненты этой реакции, осуществляемой в одном реакционном сосуде, можно объединять в любом порядке.
Реакционную смесь перемешивают в температурных пределах, которые не приводят к денатурации ферментов, например около 30- 45°C, и более конкретно около 35-45°C. До определенного момента более низкие температуры могут работать, но это будет замедлять скорость реакции.
Любой буфер с подходящим pH и содержащий соль марганца (II), можно использовать в реакции. Примеры таких буферов включают, но не ограничиваются этим, следующие: триэтаноламин; PIPES, например пиперазин-N, N′-бис(2-этансульфоновая кислота); MOPS, например 3-(N-морфолино)пропансульфоновая кислота или 3-морфолинопропан-1-сульфоновая кислота; HEPES, например 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота или 2-[4-(2-гидроксиэтил)пиперазин-1-ил]этансульфоновая кислота; TRIS, например, трис(гидроксиметил)аминометан или 2-амино-2-(гидроксиметил)пропан-1,3-диол; и BIS-TRIS метан, например 2-[бис(2-гидроксиэтил)амино]-2-(гидроксиметил)пропан-1,3-диол. Более конкретно буфер представляет собой триэтаноламин. Соль марганца (II) в буфере может представлять собой, например, хлорид марганца, хлорид марганца гидрат, бромид марганца, иодид марганца, нитрат марганца и/или сульфат марганца. Концентрация марганца в буфере может быть в диапазоне от около 0,05 мМ до около 10 мМ и, в частности, составляет около 5 мМ.
Равновесие реакции можно направить на высокую конверсию конечного продукта путем расходования неорганической фосфатной соли, являющейся побочным продуктом, путем фосфоролиза сахарозы в D-фруктозу и α-D-глюкоза-1-фосфат, катализируемого сахарозо-фосфорилазой (EC 2.4.1.7), добавленной к реакционной смеси. Однако можно использовать любые другие способы для удаления фосфата в процессе реакции, например, добавление кальция, магния или марганца к реакционной смеси для осаждения фосфатной соли вместо использования сахарозо-фосфорилазы и сахарозы. Этот высокоэффективный и экологически безопасный способ обладает преимуществом образования аномерной связи между сахаром и нуклеооснованием с очень высокой стереоселективностью без использования защитных групп или органических растворителей, и его можно осуществить как реакцию, осуществляемую в одном реакционном сосуде.
После завершения реакции конечный продукт можно выделить с использованием стандартных процедур, известных специалистам в данной области, таких как, но не ограничиваясь этим, выделение путем кристаллизации конечного продукта и его сбор фильтрованием, или экстрагирование в подходящий растворитель с последующей кристаллизацией.
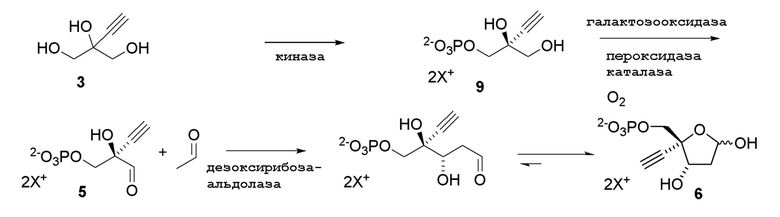
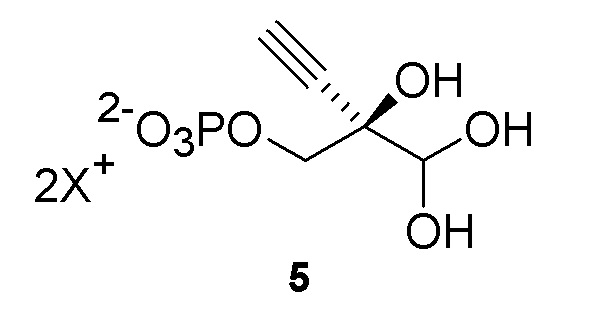
Как показано на Схеме 2A, на конечной стадии синтеза можно альтернативно использовать 3-ферментную реакцию с необязательным 4-м ферментом для направления равновесия реакции на желаемый конечный продукт. Конечная стадия начинается с соединения (5) или его соли, где (5) представляет собой (R)-2-этинил-глицеральдегид 3-фосфат или его гидрат.
Соединение (5) объединяют с дезоксирибозо-фосфат-альдолазой (DERA), ацетальдегидом, фосфопентомутазой (PPM), пуриннуклеозид-фосфорилазой (PNP), сахарозо-фосфорилазой, сахарозой и нуклеооснованием или его аналогом, например незамещенным или замещенным аденином, в буферном растворе, содержащем соль марганца (II), и доводят, по мере необходимости, до pH в диапазоне около 4-10, или конкретно около 6,5-8,0, или более конкретно около 7,0-7,5. Молярное соотношение сахароза:соединение (5) может составлять, но не ограничивается этим, от около 1:1 до 4:1. Компоненты этой реакции, осуществляемой в одном реакционном сосуде, можно объединять в любом порядке.
Реакцию осуществляют в диапазоне температур, который не приводят к денатурации ферментов, например около 30-45°C или конкретно около 35-45°C. До определенного момента более низкие температуры могут работать, но это будет замедлять скорость реакции.
Ацетальдегид добавляют в виде раствора, и более конкретно в виде 40% масс. раствора в изопропиловом спирте. В реакции можно использовать любой подходящий раствор ацетальдегида или чистый ацетальдегид. Примеры таких растворов включают, но не ограничиваются этим: раствор ацетальдегида в изопропаноле, раствор ацетальдегида в этаноле, раствор ацетальдегида в воде, раствор ацетальдегида в THF. Молярное соотношение альдегид:соединение (5) может составлять, но не ограничивается этим, от около 0,5:1 до 4:1, и более конкретно 1,5:1.
Любой буфер с подходящим pH и содержащий соль марганца (II) можно использовать в реакции. Примеры таких буферов включают, но не ограничиваются этим, следующие: триэтаноламин; PIPES, например пиперазин-N, N′-бис(2-этансульфоновая кислота); MOPS, например, 3-(N-морфолино)пропансульфоновая кислота или 3-морфолинопропан-1-сульфоновая кислота; HEPES, например 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота или 2-[4-(2-гидроксиэтил)пиперазин-1-ил]этансульфоновая кислота; TRIS, например, трис(гидроксиметил)аминометан или 2-амино-2-(гидроксиметил)пропан-1,3-диол; и BIS-TRIS метан, например 2-[бис(2-гидроксиэтил)амино]-2-(гидроксиметил)пропан-1,3-диол. Более конкретно буфер представляет собой триэтаноламин. Соль марганца (II) в буфере может представлять собой, например, хлорид марганца, хлорид марганца гидрат, бромид марганца, иодид марганца, нитрат марганца и/или сульфат марганца. Концентрация марганца в буфере может быть в диапазоне от около 0,05 мМ до около 10 мМ и, в частности, составляет около 5 мМ.
Равновесие реакции можно направить на высокую конверсию конечного продукта путем расходования неорганической фосфатной соли, являющейся побочным продуктом, путем фосфоролиза сахарозы в D-фруктозу и α-D-глюкоза-1-фосфат, катализируемого сахарозо-фосфорилазой (EC 2.4.1.7), добавленной к реакционной смеси. Однако можно использовать любые другие способы для удаления фосфата в процессе реакции, например, добавление кальция, магния или марганца к реакционной смеси для осаждения фосфатной соли вместо использования сахарозо-фосфорилазы и сахарозы. Этот высокоэффективный и экологически безопасный способ обладает преимуществом образования аномерной связи между сахаром и нуклеооснованием с очень высокой стереоселективностью без использования защитных групп или органических растворителей, и его можно осуществить как реакцию, осуществляемую в одном реакционном сосуде.
После завершения реакции конечный продукт можно выделить с использованием стандартных процедур, известных специалистам в данной области, таких как, но не ограничиваясь этим, выделение путем кристаллизации конечного продукта и его сбор фильтрованием, или экстрагирование в подходящий растворитель с последующей кристаллизацией.
Некоторые предшествующие промежуточные соединения, используемые в представленном способе для синтеза конечного продукта 4’-этинил-2’-дезоксинуклеозидов и их аналогов, также получают способами с использованием ферментативных реакций, как показано на Схеме 3; Схеме 3A и Схеме 3B
Схема 3
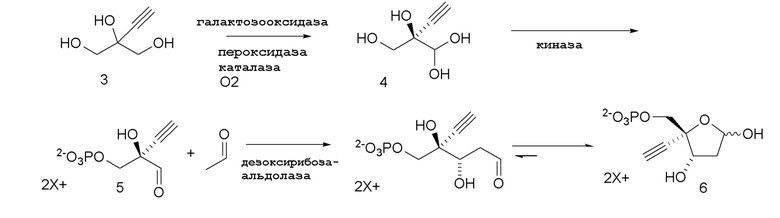
Схема 3A
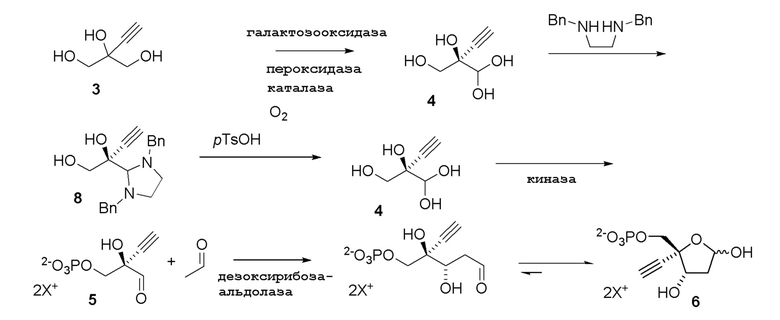
Схема 3B
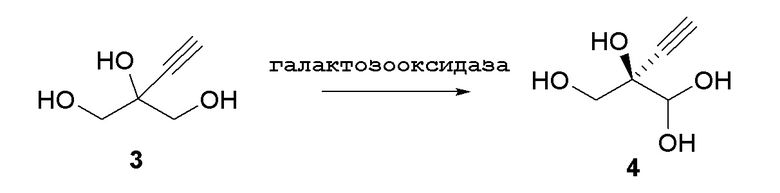
Соединение 4: Оксидазная реакция
Как показано на Схеме 3, (R)-2-этинил-глицеральдегид (4) получают путем взаимодействия галактозооксидазы с 2-этинил-пропан-1,2,3-триолом (3) в буферном растворе, доведенном, по мере необходимости, до pH в диапазоне около 3-10 или более конкретно около 6-8. Можно использовать любой буфер, имеющий подходящий диапазон pH, например, но не ограничиваясь этим, фосфат натрия; ацетат натрия; PIPES, например пиперазин-N, N′-бис(2-этансульфоновая кислота); MOPS, например 3-(N-морфолино)пропансульфоновая кислота или 3-морфолинопропан-1-сульфоновая кислота; HEPES, например 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота или 2-[4-(2-гидроксиэтил)пиперазин-1-ил]этансульфоновая кислота; TRIS, например, трис(гидроксиметил)аминометан или 2-амино-2-(гидроксиметил)пропан-1,3-диол; и BIS-TRIS метан, например 2-[бис(2-гидроксиэтил)амино]-2-(гидроксиметил)пропан-1,3-диол; борат; CAPS, например N-циклогексил-3-аминопропансульфоновая кислота; MES, например 2-(N-морфолино)этансульфоновая кислота; CHES, например N-циклогексил-2-аминоэтансульфоновая кислота; глицин; или бицин (N, N-бис(2-гидроксиэтил)глицин); при этом фосфат натрия является предпочтительным.
В реакции для активации галактозооксидазы (GOase) используют как медь, так и пероксидазу. Медь можно вводить в реакционную смесь путем добавления CuSO4, Cu(OAc)2, CuCl2 или других солей Cu(II) или Cu(I). Пероксидаза может представлять собой пероксидазу хрена или пероксидазу, полученную из других организмов, или ее можно заменить окислителем, таким как феррицианид, иридат, соли марганца (III), персульфатные соли и другие одноэлектронные или двухэлектронные окислители, или неорганические или органические окислители. Предпочтительно пероксидаза представляет собой пероксидазу хрена. Каталазу также добавляют для предотвращения дезактивации GOase. Каталаза может происходить от млекопитающих (бык) или из бактериального или грибкового источника, такого как Corynebacterium, Aspergillus или другие организмы, известные в данной области для этой цели.
Реакция протекает в присутствии кислорода. Один из удобных способов включает продувку реакционной смеси воздухом. Альтернативно, можно использовать другие системы для генерации кислорода, такие как пероксид водорода/каталаза, супероксид, или использовать другие методы, известные в данной области техники, для этой цели.
Реакцию можно осуществить с концентрацией субстрата около 10-180 г/л и, в частности, 20-50 г/л. Реакцию можно осуществлять при температуре от около 0-40°C и, в частности, около 10-30°C.
Соединение 8: Образование аминаля
Как проиллюстрировано на Схеме 3A, (R)-2-этинил-глицеральдегид (4) можно выделить в форме его аминаля (например, соединение 8) путем его взаимодействия с любым амином, диамином или аминоспиртом, который образует стабильный N, N-ацеталь или N, O-ацеталь, таким как, но не ограничиваясь этим, N, N’-дибензилэтан-1,2-диамин, N, N’-диметилэтан-1,2-диамин, N, N’-дифенилэтан-1,2-диамин и N-бензилэтаноламин; при этом N, N’-дибензилэтан-1,2-диамин является предпочтительным. Реакцию осуществляют в органическом растворителе при температуре около 50°C или ниже, предпочтительно 20-30°C, чтобы избежать разложения аминаля. Можно использовать любой растворитель, не смешивающийся с водой, например, но не ограничиваясь этим, MTBE, 2-MeTHF, CPME, диэтиловый эфир, диизопропиловый эфир, этилацетат, изопропилацетат, толуол, DCM или их смесь, при этом MTBE является предпочтительным. Реакцию можно осуществить с концентрацией субстрата около 10-100 г/л и, в частности, 20-50 г/л.
Необязательно, аминаль затем можно очистить кристаллизацией из органического растворителя, такого как, но не ограничиваясь этим, MTBE, 2-MeTHF, CPME, диэтиловый эфир, диизопропиловый эфир, этилацетат, изопропилацетат, толуол, DCM или их смесь, при этом MTBE является предпочтительным. Кристаллизацию осуществляют при 50°C или ниже, например при около 40°C, чтобы избежать разложения аминаля.
Реакция протекает в отсутствие кислорода. Один удобный способ включает продувку реакционной смеси при помощи N2. Альтернативно, можно использовать другие системы для исключения кислорода, такие как аргон, гелий или использование других способов, известных в данной области, для этих целей.
Соединение 4: Восстановление альдегида 4 из аминаля 8
(R)-2-Этинил-глицеральдегид (4) может быть восстановлен из его соответствующего аминаля путем его взаимодействия с органической или неорганической кислотой в присутствии не смешиваемого с водой органического растворителя при температуре при 50°C или ниже, например около 0-15°C, чтобы избежать разложения аминаля. Можно использовать любую органическую или неорганическую кислоту, такую как, но не ограничиваясь этим, п-толуолсульфоновая кислотп, метансульфоновая кислотп, камфорсульфоновая кислота, уксусная кислота, хлористоводородная кислота, фосфорная кислота, серная кислота. п-Толуолсульфоновая кислота является предпочтительной в реакции с аминалем 8 из-за низкой растворимости N, N’-дибензилэтан-1,2-диамин бис п-толуолсульфонатной соли в воде. Можно использовать любой растворитель, не смешивающийся с водой, например, но не ограничиваясь этим, MTBE, 2-MeTHF, CPME, диэтиловый эфир, диизопропиловый эфир, этилацетат, изопропилацетат, толуол, DCM или их смесь; при этом MTBE и 2-MeTHF являются предпочтительными. Реакцию можно осуществить с концентрацией субстрата около 5-100 г/л и, в частности, 20-50 г/л.
Необязательно, раствор альдегида 4 можно затем обработать смолой для удаления избытка органической или неорганической кислоты. Обработку смолой можно осуществить с использованием основных смол, таких как DOWEX™ MARATHON™ A смола (гидроксидная форма) и AMBERLYST® 15 смола (водородная форма) или их смесь, предпочтительно смесь DOWEX™ MARATHON™ A смолы (гидроксидная форма) и AMBERLYST® 15 смолы.
Необязательно, затем можно осуществить упаривание в вакууме раствора альдегида 4 или продувку газом для удаления избытка органического растворителя.
Соединение 5: Киназная реакция
Как показано на Схеме 3 и Схеме 3A, (R)-2-этинил-глицеральдегид 3-фосфат гидрат (5) получают путем взаимодействия пантотенаткиназы (PanK) дикого типа из E. coli или ее варианта с соединением (4) в буферном растворе, доведенном, по мере необходимости, до pH в диапазоне около 4-10, или конкретно около 6,5-8,5, или более конкретно 5,5-8,5. Можно использовать любой буфер, имеющий подходящий диапазон pH, например, но не ограничиваясь этим, фосфат натрия, PIPES, например пиперазин-N, N′-бис(2-этансульфоновая кислота); BIS-TRIS метан, например 2-[бис(2-гидроксиэтил)амино]-2-(гидроксиметил)пропан-1,3-диол; борат; HEPES, например 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота или 2-[4-(2-гидроксиэтил)пиперазин-1-ил]этансульфоновая кислота; триэтаноламин и TRIS, например TRIS, например трис(гидроксиметил)аминометан или 2-амино-2-(гидроксиметил)пропан-1,3-диол; при этом фосфат натрия является предпочтительным. Реакцию можно осуществить в присутствии любой подходящей соли двухвалетного металла, например, но не ограничиваясь этим, соли магния, например хлорида магния, и солей кобальта, марганца, цинка или кальция.
В этой реакции используют аденозин 5’-дифосфат (АДФ) в качестве источника фосфата, который требует восстановления до 5’-трифосфата (АТФ). АТФ может быть образован in situ и дополнительно восстановлен любым способом, известным в данной области, из АДФ, аденозин 5’-монофосфата (АМФ) или аденозина. Например, можно использовать комбинацию ацетилфосфата вместе с ацетаткиназой для восстановления АДФ до АТФ. Например, в присутствии пирувата, фосфата и кислорода комбинация пируватоксидазы и каталазы генерирует ацетилфосфат, и поэтому в присутствии ацетаткиназы может использоваться для восстановления АДФ до АТФ.
Реакцию можно осуществить с концентрацией субстрата около 10-100 г/л и, в частности, около 20-40 г/л. Реакцию можно осуществлять при температуре около 0-40°C и, в частности, при около 10-25°C.
Реакцию также можно осуществить с пантотенаткиназой (PanK), иммобилизованной на смоле, или с PanK и ацетаткиназой, иммобилизованными на смоле. Можно использовать любой подходящий способ иммобилизации фермента, известный в данной области, например, но не ограничиваясь этим, смолу для аффинной хроматографии на иммобилизованных ионах металла (IMAC) или иммобилизацию на аффинной смоле с использованием других биологических меток, ковалентную иммобилизацию, иммобилизацию на ионных смолах, иммобилизацию путем адсорбции, инкапсуляцию и/или сшитые ферменты. Например, можно использовать смолу для аффинной хроматографии на иммобилизованных ионах металла (IMAC), или можно использовать любую подходящую комбинацию IMAC смолы и двухвалентного катиона, где катион может представлять собой, например, но не ограничиваясь этим, никель, кобальт, медь, цинк, железо и/или алюминий. Конкретно, можно использовать IMAC смолу, нагруженную никелем. Предпочтительно, когда и ацетаткиназа и пантотенаткиназа (PanK) иммобилизованы на смоле.
Соединение 9: Киназная реакция
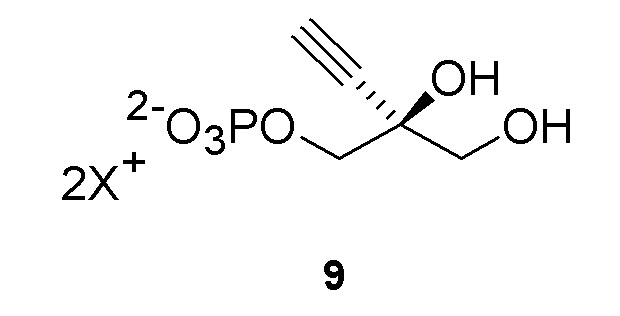
Как показано на Схеме 3B, (S)-2-этинил-пропан-1,2,3-триол 1-фосфат (9) получают путем взаимодействия пантотенаткиназы (PanK) дикого типа из E. coli или ее варианта с соединением (3) в буферном растворе, доведенном, по мере необходимости, до pH в диапазоне от около 4-10, или конкретно около 6,5-8,5, или более конкретно 5,5-8,5. Можно использовать любой буфер, имеющий подходящий диапазон pH, например, но не ограничиваясь этим, фосфат натрия, PIPES, например пиперазин-N, N′-бис(2-этансульфоновая кислота); BIS-TRIS метан, например 2-[бис(2-гидроксиэтил)амино]-2-(гидроксиметил)пропан-1,3-диол; борат; HEPES, например 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота или 2-[4-(2-гидроксиэтил)пиперазин-1-ил]этансульфоновая кислота;, триэтаноламин и TRIS, например трис(гидроксиметил)аминометан или 2-амино-2-(гидроксиметил)пропан-1,3-диол, при этом фосфат натрия является предпочтительным. Реакцию можно осуществить в присутствии любой подходящей соли двухвалетного металла, например, но не ограничиваясь этим, соли магния, например хлорида магния, и солей кобальта, марганца, цинка или кальция.
В этой реакции используют аденозин 5’-дифосфат (АДФ) в качестве источника фосфата, который требует восстановления до 5’-трифосфата (АТФ). АТФ может быть образован in situ и дополнительно восстановлен любым способом, известным в данной области, из АДФ, аденозин 5’-монофосфата (АМФ) или аденозина. Например, можно использовать комбинацию ацетилфосфата вместе с ацетаткиназой для восстановления АДФ до АТФ. Альтернативно, (a) комбинацию пируватоксидаза, каталазы и ацетаткиназы в присутствии пирувата, фосфата и кислорода можно использовать для восстановления АДФ до АТФ, или (b) комбинацию пируватоксидазы, каталазы и ацетаткиназы в присутствии пирувата, фосфата и кислорода в комбинации с ацетилфосфатом и ацетаткиназой можно использовать для восстановления АТФ из АДФ.
Реакцию можно осуществить с концентрацией субстрата около 10-100 г/л и, в частности, около 20-40 г/л. Реакцию можно осуществлять при температуре от около 0-40°C и, в частности, при около 10-25°C.
Реакцию также можно осуществить с пантотенаткиназой (PanK), иммобилизованной на смоле, или с PanK и ацетаткиназой, иммобилизованными на смоле. Можно использовать любой подходящий способ иммобилизации фермента, известный в данной области, например, но не ограничиваясь этим, смолу для аффинной хроматографии на иммобилизованных ионах металла (IMAC) или иммобилизацию на аффинной смоле с использованием других биологических меток, ковалентную иммобилизацию, иммобилизацию на ионных смолах, иммобилизацию путем адсорбции, инкапсуляцию и/или сшитые ферменты. Например, можно использовать смолу для аффинной хроматографии на иммобилизованных ионах металла (IMAC), или можно использовать любую подходящую комбинацию IMAC смолы и двухвалентного катиона, где катион может представлять собой, например, но не ограничиваясь этим, никель, кобальт, медь, цинк, железо и/или алюминий. Конкретно, можно использовать IMAC смолу, нагруженную никелем. Предпочтительно, когда и ацетаткиназа и пантотенаткиназа (PanK) иммобилизованы на смоле.
Соединение 5: Оксидазная реакция
Как показано на Схеме 3B, (R)-2-этинил-глицеральдегид гидрат 3-фосфат (5) получают путем взаимодействия галактозооксидазы с (S)-2-этинил-пропан-1,2,3-триол 1-фосфатом (9) в буферном растворе, доведенном, по мере необходимости, до pH в диапазоне около 3-10 или более конкретно около 6-8. Можно использовать любой буфер, имеющий подходящий диапазон pH, такой как, но не ограничиваясь этим, фосфат натрия; ацетат натрия; PIPES, например пиперазин-N, N′-бис(2-этансульфоновая кислота); MOPS, например, 3-(N-морфолино)пропансульфоновая кислота или 3-морфолинопропан-1-сульфоновая кислота; HEPES, например 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота или 2-[4-(2-гидроксиэтил)пиперазин-1-ил]этансульфоновая кислота; TRIS, например трис(гидроксиметил)аминометан или 2-амино-2-(гидроксиметил)пропан-1,3-диол; и BIS-TRIS метан, например 2-[бис(2-гидроксиэтил)амино]-2-(гидроксиметил)пропан-1,3-диол; борат; CAPS, например N-циклогексил-3-аминопропансульфоновая кислота; MES, например 2-(N-морфолино)этансульфоновая кислота; CHES, например N-циклогексил-2-аминоэтансульфоновая кислота; глицин; или бицин (N, N-бис(2-гидроксиэтил)глицин); при этом фосфат натрия является предпочтительным.
В реакции для активации галактозооксидазы (GOase) используют как медь, так и пероксидазу. Медь можно вводить в реакционную смесь путем добавления CuSO4, Cu(OAc)2, CuCl2 или других солей Cu(II) или Cu(I). Пероксидаза может представлять собой пероксидазу хрена или пероксидазу, полученную из других организмов, или ее можно заменить окислителем, таким как феррицианид, иридат, соли марганца (III), персульфатные соли и другие одноэлектронные или двухэлектронные окислители, или неорганические или органические окислители. Предпочтительно пероксидаза представляет собой пероксидазу хрена. Каталазу также добавляют для предотвращения дезактивации GOase. Каталаза может происходить от млекопитающих (бычья) или из бактериального или грибкового источника, такого как Corynebacterium, Aspergillus или другие организмы, известные в данной области для этой цели.
Реакция протекает в присутствии кислорода. Один из удобных способов включает продувку реакционной смеси воздухом. Альтернативно, можно использовать другие системы для генерации кислорода, такие как пероксид водорода/каталаза, супероксид, или использовать другие методы, известные в данной области техники, для этой цели.
Реакцию можно осуществить с концентрацией субстрата около 10-180 г/л и, в частности, 20-50 г/л. Реакцию можно осуществлять при температуре около 0-40°C и, в частности, около 10-30°C.
Соединение 6: Дезоксирибозо-фосфат-альдолазная (DERA) реакция
Важным преимуществом этого нового пути для получения соединения (6) по сравнению с предлшествующими известными способами является то, что он создает сахарный каркас при правильном состоянии окисления без использования защитных групп.
4-Этинил-D-2-дезоксирибозо-5-фосфат (6) получают путем взаимодействия дезоксирибозо-фосфат-альдолазы (DERA) с (R)-2-этинил-глицеральдегид 3-фосфатом (5) в кислотной или солевой форме и ацетальдегидом в водном растворе, доведенном, по мере необходимости, до pH в диапазоне около 5-9 или более конкретно около 6-8. Примеры солей (5) включают, но не ограничиваются этим, кальциевые, магниевые, цинковые, моно- или ди-Na соли, моно- или ди-K соли, или моно- или ди-Li соли; моно- или ди-аммониевые соли; или одновалентные или двухвалентные соли с первичными, вторичными или третичными аминами. Реакцию можно осуществить в открытом сосуде, или ее предпочтительно осуществляют в герметично закрытом сосуде для предотвращения испарения ацетальдегида.
Реакцию можно осуществить с концентрацией субстрата около 10-100 г/л, конкретно около 30-60 г/л. Ее можно осуществить при температуре около 0-40°C и, в частности, около 25-35°C.
Реакцию можно осуществлять без каких-либо буферов. Альтернативно, можно использовать буферы, например, но не ограничиваясь этим, триэтаноламин; фосфат; MOPS, например 3-(N-морфолино)пропансульфоновую кислоту или 3-морфолинопропан-1-сульфоновую кислоту; HEPES, например 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновую кислоту или 2-[4-(2-гидроксиэтил)пиперазин-1-ил]этансульфоновую кислоту; BIS-TRIS метан, например 2-[бис(2-гидроксиэтил)амино]-2-(гидроксиметил)пропан-1,3-диол; борат; PIPES, например пиперазин-N, N′-бис(2-этансульфоновая кислота); MES, например 2-(N-морфолино)этансульфоновую кислоту; и борат; или другие буферы, имеющие подходящий диапазон pH, которые не содержат никаких первичных амино групп.
Каждую стадию и процедуру способов, описанных в настоящей заявке, которые включают использование одного или нескольких ферментов, осуществляют при температуре, которая не вызывает денатурацию указанных одного или нескольких ферментов. Каждую стадию и процедуру способов, описанных в настоящей заявке, которые включают использование одного или нескольких ферментов, можно осуществить при pH в диапазоне около 3-10 или около 4-10.
“Нуклеооснование” (или “азотистое основание” или “основание”) представляет собой пиримидиновый или пуриновый гетероцикл нуклеиновых кислот, таких как ДНК и РНК. В контексте настоящей заявки нуклеооснование включает аденин, гуанин, цитозин, тимин или урацил, а также нуклеооснования с неприродными модификациями, например, где основание имеет один или несколько неприродных заместителей, или модификацией, влияющей на гетероатом(гетероатомы) в основании, исключая любое изменение аномерной C-N связи.
4’-этинил-2’-дезоксинуклеозид содержит нуклеооснование. В контексте настоящей заявки аналог 4’-этинил-2’-дезоксинуклеозида означает неприродную модификацию основания нуклеозида, например, где основание имеет один или несколько неприродных заместителей, или модификацию, влияющую на гетероатом(гетероатомы) в основании, исключая любое изменение аномерной C-N связи.
В контексте настоящей заявки ферменты “фосфопентомутазы” (“PPM”) (например EC 5.4.2.7) представляют собой ферменты, которые катализируют обратимую изомеризацию рибозо-1-фосфата до рибозо-5-фосфата и родственных соединений, таких как дезоксирибозофосфат и аналоги рибозофосфата и дезоксирибозофосфата.
В контексте настоящей заявки ферменты “пуриннуклеозид-фосфорилазы” (“PNP”) (EC 2.4.2.2) представляют собой ферменты, которые катализируют обратимый фосфоролиз пуриновых рибонуклеозидов и родственных соединений (например, дезоксирибонуклеозидов и аналогов рибонуклеозидов и дезоксирибонуклеозидов) в свободное пуриновое основание и рибозо-1-фосфат (и их аналоги).
В контексте настоящей заявки ферменты “сахарозо-фосфорилазы” (“SP”) (EC 2.4.1.7) представляют собой ферменты, которые катализируют обратимый фосфоролиз сахарозы в D-фруктозное основание и глюкоза-1-фосфат (и их аналоги). Сахарозо-фосфорилазу (SP) в комбинации с сахарозой используют в комбинации с пуриннуклеозид-фосфорилазой (PNP) и фосфомутазой (PPM) для удаления свободных фосфат-ионов из реакции, где комбинация ферментов катализирует образование нуклеозида MK-8591 (EFdA), тогда как в некоторых вариантах осуществления это можно заменить другими способами, известными в данной области.
В контексте настоящей заявки “дезоксирибозо-фосфат-альдолаза” (“DERA”) (например, EC 4.1.2.4) относится к ферменту семейства лиаз, которые обратимо расщепляют или создают углерод-углеродные связи. Дезоксирибозо-фосфат-альдолазы, в контексте настоящей заявки, включают природную (дикого типа) дезоксирибозо-фосфат-альдолазу, а также неприродные сконструированные полипептиды, получаемые путем манипулирования, осуществляемого человеком. Дезоксирибозо-фосфат-альдолаза дикого типа катализирует обратимую реакцию 2-дезокси-D-рибозо-5-фосфата с образованием D-глицеральдегид 3-фосфата и ацетальдегида.
В контексте настоящей заявки “пантотенаткиназа” (“PanK”) относится к ферментам (EC 2.7.1.33), которые в природе фосфорилируют пантотенат с образованием 4’-фосфопантотената. Варианты ферментов, полученные из таких ферментов PanK, могут проявлять улучшенную активность и стереоселективность в отношении 3’OH-группы D-этинилглицеральдегида независимо от того, сохраняют ли такие варианты свою природную функцию в отношении пантотената.
В контексте настоящей заявки ферменты “галактозоксидазы” (“GOase”; EC 1.1.3.9) представляют собой медь-зависимые ферменты, которые в присутствии бимолекулярного кислорода катализируют окисление первичных спиртов до соответствующих альдегидов. Они действуют как регио-, так и энантиоспецифическим образом, делая возможными синтетические подходы, которые не требуют или требуют в незначительной степени защиты функциональных групп и дают желаемый стереоизомер. Способ окисления является мягким и контролируемым, так что активность не приводит к чрезмерному окислению спирта до его соответствующей карбоновой кислоты.
В контексте настоящей заявки термин “пероксидаза хрена” (HRP, EC 1.11.1.7) означает железо-зависимый фермент, который активирует и поддерживает каталитическую активность GOase, окисляя неактивное окислительно-восстановительное состояние активного центра, которое возникает во время нормального каталитического цикла GOase. HRP типа I используется каталитическим образом в примерах, включенных в настоящую заявку, однако он не предназначен исключительно для этой роли, так как существуют другие ферменты, переносящие электрон, которые принадлежат к этому и другим классам ферментов, а также химические реагенты, которые могут выполнять эту роль.
В контексте настоящей заявки термин “каталаза” относится к гем-зависимому ферменту (EC 1.11.1.6), который действует на пероксид водорода, побочный продукт реакций галактозооксидазы или пируватоксидазы, который может сделать ферменты неактивными при содержании перекиси водорода выше определенных уровней. Каталаза используется в качестве каталитического поддерживающего фермента в приведенных примерах для превращения пероксида водорода в воду и кислород, хотя в некоторых вариантах осуществления это можно заменить другими методами, такими как электрохимическое разложение пероксида водорода. Гем-зависимая каталаза используется каталитическим образом в примерах, включенных в настоящую заявку, однако она не предназначена исключительно для этой роли, поскольку существуют другие ферменты, принадлежащие к этому классу, которые могут выполнять эту роль.
В контексте настоящей заявки термин “ацетаткиназа” (“AcK”) относится к ферменту (EC 2.7.2.1), который катализирует образование ацетилфосфата из ацетата и аденозинтрифосфата (АТФ). Он также может катализировать обратную реакцию, когда он фосфорилирует аденозин-5’-дифосфат (АДФ) до аденозин-5’-трифосфата (АТФ) в присутствии ацетилфосфата. Ацетаткиназа используется для рециркуляции АТФ, необходимого для пантотенаткиназы (PanK) в приведенных примерах, тогда как в некоторых вариантах осуществления комбинация рециркуляции ацетилфосфат-ацетаткиназа может быть заменена другими методами, известными в данной области.
В контексте настоящей заявки термин “пируватоксидаза” (“PO”) относится к ферменту (EC 1.2.3.3), зависящему от флавинадениндинуклеотида (FAD) и тиаминдифосфата. Пируватоксидаза представляет собой фермент, принадлежащий к семейству оксидоредуктаз, в частности тех, которые действуют на альдегидную или оксогруппу донора с кислородом в качестве акцептора, и он катализирует химическую реакцию между пируватом, фосфат-ионом и бимолекулярным кислородом с образованием ацетилфосфата, диоксида углерода и пероксид водорода. Пируватоксидаза (PO) используется в комбинации с ацетаткиназой (AcK) и каталазой в качестве каталитической регенерирующей АТФ комбинации в приведенных примерах, где комбинация ферментов катализирует образование АТФ из АДФ в присутствии кислорода, пирувата и фосфат-ионов, хотя в некоторых вариантах осуществления они могут быть заменены другими методами, известными в данной области.
В контексте настоящей заявки термины фермент “дикого типа” и “природный” фермент относятся к форме, встречающейся в природе. Например, полипептидная последовательность дикого типа представляет собой последовательность, присутствующую в организме, которая может быть выделена из природного источника и которая не была намеренно модифицирована осуществляемыми человеком манипуляциями.
В контексте настоящей заявки термины “сконструированный”, “вариант”, “мутант” и “неприродный” при использовании в отношении фермента, включающего полипептид, относятся к материалу или материалу, соответствующему природной или нативной форме материала, который был модифицирован так, как в ином случае он не существовал бы в природе. В некоторых вариантах осуществления полипептид идентичен встречающемуся в природе полипептиду, но его получают или производят из синтетических материалов и/или путем манипуляции с использованием рекомбинантных методов.
В контексте настоящей заявки “процент идентичности последовательностей”, “процент идентичности” и “идентичность в процентах” по отношению к ферментам используются для обозначения сравнений между полинуклеотидными последовательностями или полипептидными последовательностями и определяются путем сравнения двух оптимально выровненных последовательностей в окне сравнения, где часть полинуклеотидной или полипептидной последовательности в окне сравнения может содержать добавления или делеции (то есть гэпы) по сравнению с эталонной последовательностью для оптимального выравнивания двух последовательностей. Процент рассчитывают путем определения количества положений, в которых либо идентичное основание нуклеиновой кислоты, либо аминокислотный остаток встречается в обеих последовательностях, либо основание нуклеиновой кислоты или аминокислотный остаток выравнивают с гэпом для получения количества совпадающих положений, деля количество совпадающих положений на общее количество положений в окне сравнения и умножая результат на 100 с получением процента идентичности последовательностей. Определение оптимального выравнивания и процента идентичности последовательностей осуществляют с использованием алгоритмов BLAST и BLAST 2.0 (см., например, Altschul et al., 1990, J. Mol. Biol. 215: 403-410 и Altschul et al., 1977, Nucleic Acids Res. 3389-3402). Программное обеспечение для осуществления анализов BLAST общедоступно на веб-сайте Национального центра биотехнологической информации.
Вкратце, анализ BLAST включает сначала идентификацию пар последовательностей с высокой оценкой (HSP) путем определения коротких слов длины W в искомой последовательности, которые либо совпадают, либо удовлетворяют некоторой положительно оцениваемой пороговой оценке T при выравнивании со словом такой же длины в последовательности из базы данных. T обозначается как порог оценки соседнего слова (Altschul. et al., см. выше). Эти начальные совпадения соседних слов действуют как семена для инициирования поиска, чтобы найти более длинные HSP, содержащие их. Затем совпадения слов расширяются в обоих направлениях вдоль каждой последовательности до тех пор, пока может быть увеличена совокупная оценка выравнивания. Совокупные баллы рассчитываются с использованием, для нуклеотидных последовательностей, параметров M (поощрительный балл за пару совпадающих остатков; всегда >0) и N (штрафной балл за несовпадающие остатки; всегда <0). Для аминокислотных последовательностей используется оценочная матрица для расчета совокупного балла. Расширение совпадений слов в каждом направлении прекращается, когда: совокупная оценка выравнивания падает на величину X от максимального достигнутого значения; совокупная оценка становится равной нулю или ниже из-за накопления одного или нескольких отрицательно оцениваемых выравниваний остатков; либо достигнут конец любой последовательности. Параметры алгоритма BLAST W, T и X определяют чувствительность и скорость выравнивания. Программа BLASTN (для нуклеотидных последовательностей) использует по умолчанию длину слова (W), равную 11, ожидание €, равное 10, M=5, N= -4 и сравнение обеих цепей. Для аминокислотных последовательностей программа BLASTP использует по умолчанию длину слова (W), равную 3, ожидание €(E), равное 10, и матрицу оценки BLOSUM62 (см. Henikoff and Henikoff, 1989, Proc Natl Acad Sci USA 89: 10915).
Доступно множество других алгоритмов, которые работают аналогично BLAST, обеспечивая процент идентичности для двух последовательностей. Оптимальное выравнивание последовательностей для сравнения можно осуществить, например, с использованием алгоритма локальной гомологии Smith and Waterman, 1981, Adv. Appl. Math. 2:482, алгоритма выравнивания гомологии Needleman and Wunsch, 1970, J. Mol. Biol. 48:443, метода поиска сходства Pearson and Lipman, 1988, Proc. Natl. Acad. Sci. USA 85:2444, компьютеризированных реализаций этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения GCG Wisconsin) или путем визуального осмотра (см. Current Protocols в Molecular Biology, F. M. Ausubel. et al., eds., Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (1995 Supplement) (Ausubel)). Кроме того, для определения выравнивания последовательностей и процента идентичности последовательностей можно использовать программы BESTFIT или GAP в пакете программ GCG Wisconsin (Accelerys, Madison WI) с использованием параметров по умолчанию.
“Существенная идентичность” относится к полинуклеотидной или полипептидной последовательности, которая имеет идентичность последовательности по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более по сравнению с эталонной последовательностью в пределах окна сравнения, состоящего по меньшей мере из 20 положений остатков, часто в диапазоне по меньшей мере 30-50 остатков, где процент идентичности последовательностей определяется путем сравнения эталонной последовательности с последовательностью, которая включает делеции или добавления, которые составляют 20 или менее процентов эталонной последовательности в окне сравнения. В конкретных вариантах осуществления, применяемых к полипептидам, термин “существенная идентичность” означает, что две полипептидные последовательности при оптимальном выравнивании, например, при помощи программ GAP или BESTFIT с использованием штрафов за открытие гэпа по умолчанию, имеют по меньшей мере 80-процентную идентичность последовательностей, предпочтительно по меньшей мере 89-процентную идентичность последовательностей, более предпочтительно по меньшей мере 95-процентную идентичность последовательностей или более (например, идентичность последовательностей 99%). Предпочтительно, положения остатков, которые не являются неидентичными, отличаются консервативными аминокислотными заменами.
“Стереоселективность” относится к преимущественному образованию в химической или ферментативной реакции одного стереоизомера по сравнению с другим. Стереоселективность может быть частичной, когда образование одного стереоизомера предпочтительнее другого, или может быть полной, когда образуется только один стереоизомер. Когда стереоизомеры являются энантиомерами, стереоселективность указывается как энантиоселективность, доля (обычно указываемая в процентах) одного энантиомера в сумме обоих. В данной области ее обычно альтернативно указывают (обычно в процентах) как энантиомерный избыток (э.и.), рассчитанный в соответствии с формулой [основной энантиомер - второстепенный энантиомер]/[основной энантиомер+второстепенный энантиомер]. Если стереоизомеры представляют собой диастереоизомеры, стереоселективность указывается как диастереоселективность, доля (обычно указывается в процентах) одного диастереомера в смеси двух диастереомеров, обычно также указывается как диастереомерный избыток (д.и.). Энантиомерный избыток и диастереомерный избыток являются типами стереомерного избытка.
Фраза “подходящие реакционные условия” относится к тем условиям в реакционном растворе для ферментативного превращения (например, диапазоны загрузки фермента, загрузки субстрата, температуры, pH, буферы, сорастворители и т.д.), при которых каждый полипептид, используемый в настоящем изобретении, способен превращать субстрат в желаемое соединение, являющееся продуктом. В настоящей заявке представлены некоторые иллюстративные подходящие реакционные условия.
Используемый в настоящей заявке термин “субстрат” в контексте процесса реакции ферментативного превращения относится к соединению или молекуле, на которые действуют сконструированные ферменты, используемые в настоящей заявке.
Используемый в настоящей заявке термин “продукт” в контексте процесса ферментативного превращения относится к соединению или молекуле, полученным в результате действия ферментативного полипептида на субстрат.
В контексте настоящей заявки термин “увеличение” выхода продукта (например, аналога 4'-этинил-2'-дезоксирибозы фосфата или аналога 4'-этинил-2'-дезоксинуклеозида) реакции происходит, когда конкретный компонент, присутствующий во время реакции (например, фермент), вызывает образование большего количества продукта по сравнению с реакцией, осуществляемой в тех же условиях с тем же субстратом, но в отсутствие представляющего интерес компонента.
В контексте настоящей заявки термины “уравновешивание” или “равновесие” относятся к процессу, приводящему к устойчивой концентрации химических веществ в химической или ферментативной реакции (например, взаимопревращение двух видов A и B), включая взаимопревращение стереоизомеров, что определяется константой скорости прямой и константой скорости обратной химической или ферментативной реакции.
“Энантиомерный избыток” (э.и.) является показателем чистоты, используемым для хиральных веществ. Он отражает степень, в которой образец содержит один энантиомер в большем количестве, чем другой. Например, рацемическая смесь имеет э.и. 0%, в то время как один полностью чистый энантиомер имеет э.и. 100%; и образец с 70% одного энантиомера и 30% другого имеет э.и. 40% (70% - 30%). Диастереомерный избыток (д.и.) рассчитывается так же, как и э.и., когда в смеси присутствуют только два диастереоизомера.
"Белок", “фермент”, "полипептид" и "пептид" используются взаимозаменяемо для обозначения полимера по меньшей мере из двух аминокислот, ковалентно связанных амидной связью, независимо от длины или посттрансляционной модификации (например, гликозилирования, фосфорилирования, липидирования, миристилирования, 10 убиквитинирования и т.д.). В это определение включены D- и L-аминокислоты, а также смеси D- и L-аминокислот.
В контексте настоящей заявки термин “около” означает допустимую ошибку для конкретного значения. В некоторых случаях “около” означает в пределах 0,05%, 0,5%, 1,0% или 2,0% от нижнего и верхнего предела заданного диапазона значений. Что касается pH, “около” означает плюс или минус 0,5.
В контексте настоящей заявки термин “по существу чистый” полипептид или “очищенный” белок относится к композиции, в которой вид полипептида является преобладающим присутствующим видом (т.е. в молярном или массовом отношении он преобладает по сравнению с любыми другими отдельными макромолекулярными видами в композиции), и обычно представляет собой по существу очищенную композицию, когда целевой вид составляет по меньшей мере около 50 процентов от присутствующих макромолекулярных частиц в молях или % масс. Однако в некоторых вариантах осуществления композиция, содержащая полипептид, включает полипептид, который имеет чистоту менее 50% (например, около 10%, около 20%, около 30%, около 40% или около 50%). Как правило, по существу чистая полипептидная композиция включает около 60% или более, около 70% или более, около 80% или более, около 90% или более, около 95% или более и около 98% или более всех макромолекулярных видов, в молях или % масс., присутствующих в композиции. В некоторых вариантах осуществления полипептид очищают до существенной гомогенности (т.е. загрязняющие примеси не могут быть обнаружены в композиции обычными методами детекции), при этом композиция состоит по существу из одного макромолекулярного вида. Растворители, малые молекулы (<500 Дальтон) и элементарные ионы не считаются макромолекулярными видами. В некоторых вариантах осуществления выделенные полипептиды представляют собой по существу чистые полипептидные композиции.
В контексте настоящей заявки термин “улучшенное свойство” фермента относится по меньшей мере к одному улучшенному свойству фермента. В некоторых вариантах осуществления в настоящем изобретеним используются сконструированные полипептиды PPM, PNP, DERA, PanK, AcK, SP и/или GOase, которые демонстрируют улучшение любого ферментного свойства по сравнению с эталонным PPM, PNP, DERA, PanK, AcK, SP или GOase полипептидом, соответственно, и/или PPM, PNP, DERA, PanK, AcK, SP или GOase полипептидом дикого типа, соответственно, и/или другим сконструированным PPM, PNP, DERA, PanK, AcK, SP или GOase полипептидом, соответственно. Таким образом, уровень “улучшения” можно определить и сравнить между различными полипептидами, включая полипептиды дикого типа, а также сконструированные полипептиды. Улучшенные свойства включают, но не ограничиваются этим, такие свойства, как повышенная экспрессия белка, повышенное продуцирование целевого продукта, повышенная специфичность или сродство к субстрату (т.е. повышенная активность в отношении субстрата), повышенная термоактивность, повышенная термостабильность, повышенная pH активность, повышенная стабильность, повышенная ферментативная активность, повышенная специфическая активность, повышенная резистентность к ингибированию субстрата или конечного продукта, повышенная химическая стабильность, улучшенная хемоселективность, улучшенная устойчивость к растворителю, повышенная устойчивость к кислому pH, повышенная устойчивость к протеолитической активности (т.е. пониженная чувствительность к протеолизу), уменьшенная агрегация, повышенная растворимость и измененный температурный профиль. В дополнительных вариантах осуществления этот термин используется в отношении по меньшей мере одного улучшенного свойства PPM, PNP, DERA, PanK, AcK, SP и/или GOase ферментов. В некоторых вариантах осуществления в настоящем изобретении используются сконструированные PPM, PNP, DERA, PanK, AcK, SP и/или GOase полипептиды, которые демонстрируют улучшение любого ферментного свойства по сравнению с эталонным PPM, PNP, DERA, PanK, AcK, SP и/или GOase полипептидом, соответственно; и/или полипептидом дикого типа, и/или другим сконструированным PPM, PNP, DERA, PanK, AcK, SP и/или GOase полипептидом, соответственно. Таким образом, уровень “улучшения” можно определить и сравнить между различными полипептидами, включая полипептиды дикого типа, а также сконструированные полипептиды.
В контексте настоящей заявки термин “конверсия” (“конв” или “конв.”) относится к ферментативному преобразованию (или биотрансформации) субстрата(субстратов) в соответствующий продукт(продукты). “Процент” конверсии относится к проценту субстрата, который превращается в продукт за определенный период времени при определенных условиях. Таким образом, “ферментативная активность” или “активность” полипептида может быть выражена как процент конверсии субстрата в продукт за определенный период времени.
В контексте настоящей заявки термин “стереоселективность” относится к преимущественному образованию в химической или ферментативной реакции одного стереоизомера по сравнению с другим. Стереоселективность может быть частичной, когда образование одного стереоизомера предпочтительнее другого, или может быть полной, когда образуется только один стереоизомер. Когда стереоизомеры являются энантиомерами, стереоселективность указывается как энантиоселективность, доля (обычно указываемая в процентах) одного энантиомера в сумме обоих. В данной области ее обычно альтернативно указывают (обычно в процентах) как энантиомерный избыток (э.и.), рассчитанный в соответствии с формулой [основной энантиомер - второстепенный энантиомер]/[основной энантиомер+второстепенный энантиомер]. Если стереоизомеры представляют собой диастереоизомеры, стереоселективность указывается как диастереоселективность, доля (обычно указывается в процентах) одного диастереомера в смеси двух диастереомеров, обычно также указывается как диастереомерный избыток (д.и.). Энантиомерный избыток и диастереомерный избыток являются типами стереомерного избытка.
Настоящее изобретение включает использование сконструированных PPM, PNP, DERA, PanK, AcK, SP и GOase полипептидов, в частности полипептидов, имеющих SEQ ID NO: 1-21, и указанные последовательности, которые включают одну или несколько консервативных аминокислотных замен, которые могут называться консервативно модифицированными вариантами каждой из SEQ ID NO. 1-21.
В контексте настоящей заявки термин “консервативная” аминокислотная замена относится к заменам аминокислот в белке другими аминокислотами, имеющими аналогичные характеристики (например, кислотные, основные, положительно или отрицательно заряженные, полярные или неполярные, размер боковой цепи, гидрофобность/гидрофильность, конформация и жесткость скелета и т.д.) так, чтобы изменения можно было часто осуществлять без изменения биологической активности белка. Это включает одну или несколько замен аминокислоты в полипептиде другой аминокислотой в пределах того же или подобного определенного класса аминокислот. Специалисты в данной области должно быть понятно, что, как правило, замены отдельных аминокислот в несущественных областях полипептида существенно не изменяют биологическую активность (см., например, Watson et al. (1987) Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th Ed.)). Кроме того, замены структурно или функционально схожих аминокислот с меньшей вероятностью нарушат биологическую активность. В качестве примера, а не ограничения, в некоторых вариантах осуществления аминокислота с алифатической боковой цепью заменена другой алифатической аминокислотой (например, аланин, валин, лейцин и изолейцин); аминокислота с гидроксильной боковой цепью заменена другой аминокислотой с гидроксильной боковой цепью (например, серин и треонин); аминокислота, имеющая ароматические боковые цепи, заменена другой аминокислотой, имеющей ароматическую боковую цепь (например, фенилаланин, тирозин, триптофан и гистидин); аминокислота с основной боковой цепью заменена другой аминокислотой с основной боковой цепью (например, лизин и аргинин); аминокислота с кислотной боковой цепью заменена другой аминокислотой с кислотной боковой цепью (например, аспарагиновая кислота или глутаминовая кислота); и/или гидрофобная или гидрофильная аминокислота заменена другой гидрофобной или гидрофильной аминокислотой, соответственно. Дополнительные иллюстративные консервативные аминокислотные замены представлены в Таблице 1.
Иллюстративные консервативные аминокислотные замены
Термин "набор аминокислотных замен" или "набор замен" относится к группе аминокислотных замен в полипептидной последовательности по сравнению с эталонной последовательностью. Набор замен может включать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или больше аминокислотных замен.
“Функциональный фрагмент” относится к полипептиду, который имеет амино-концевую и/или карбокси-концевую делецию(делеции) и/или внутренние делеции, но где оставшаяся аминокислотная последовательность идентична соответствующим положениям в последовательности, с которой ее сравнивают (например, полноразмерный сконструированный фермент PPM, PNP, DERA, PanK, AcK, SP или GOase, используемый в настоящем изобретении), и которая сохраняет практически всю активность полноразмерного полипептида.
В контексте настоящей заявки термин “делеция” относится к модификации полипептида путем удаления одной или нескольких аминокислот из эталонного полипептида. Делеции могут включать удаление 1 или более аминокислот, 2 или более аминокислот, 5 или более аминокислот, 10 или более аминокислот, 15 или более аминокислот или 20 или более аминокислот, до 10% от общего количества аминокислот или до 20% от общего количества аминокислот, составляющих эталонный фермент, при сохранении ферментативной активности и/или сохранении улучшенных свойств сконструированного PPM, PNP, DERA, PanK, AcK, SP или GOase фермента. Делеции могут быть направлены на внутренние части и/или концевые части полипептида. В различных вариантах осуществления делеция может включать непрерывный сегмент или может быть прерывистой. Делеции обычно обозначаются как “-” в аминокислотных последовательностях.
В контексте настоящей заявки термин “вставка” относится к модификации полипептида путем добавления одной или нескольких аминокислот из эталонного полипептида. Вставки могут иметь место во внутренних частях полипептида или на карбокси- или амино-конце. Используемые вставки включают слитые белки, как известно в данной области. Вставка может представлять собой непрерывный сегмент аминокислот или может быть разделена одной или несколькими аминокислотами в природном полипептиде.
Дополнительные акронимы и аббревиатуры, используемые в настоящей заявке, являются следующими:
хроматография
Экспериментальные процедуры
Получение 2-этинил-2-гидроксипропан-1,3-диил диацетата (2)
Способ A:
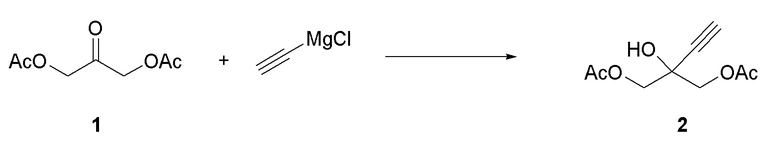
К -35°C раствору диацетоксиацетона (1) (159 г, 914,0 ммоль) в THF (1000 мл) добавляли 1600 мл 0,5 M раствора этинилмагний хлорида в THF, поддерживая температуру ниже -20°C. Когда реакция завершалась, добавляли по каплям уксусную кислоту (78 мл) в 400 мл метил трет-бутилового эфира (MTBE), поддерживая температуру ниже -20°C. Затем добавляли MTBE (800 мл) и смесь нагревали до комнатной температуры. Добавляли насыщенный раствор NaCl в воде (1000 мл) с последующим добавлением насыщенного раствора NH4Cl в воде (1050 мл). Органический слой отделяли, сушили над Na2SO4 и упаривали с получением соединения (2) в виде масла (160 г, 88%). 1H ЯМР (CDCl3, 500 МГц): δ 4,26 (дд, 4 H), 2,55 (с, 1H), 2,14 (с, 6H).
Получение 2-этинил-пропан-1,2,3-триола (3)
Способ B:
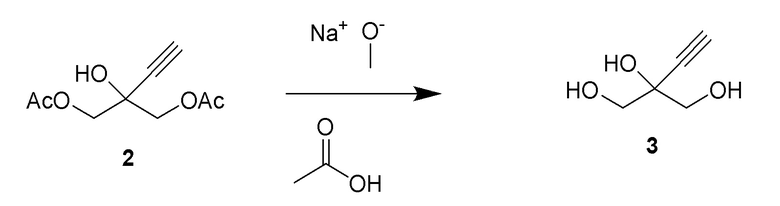
К раствору 2-этинил-2-гидроксипропан-1,3-диил диацетата (2) (70 г, 350 ммоль) в этаноле добавляли 0,5M раствор метоксилата натрия в метаноле (69,9 мл, 35,0 ммоль) при комнатной температуре (комн.темп). Реакционную смесь перемешивали при комнатной температуре в течение 2 часов (ч) для достижения завершения реакции. Растворители выпаривали и остаток снова растворяли в 100 мл воды и экстрагировали при помощи 3 × 50 мл MTBE. Водный слой продували азотом для удаления остаточных растворителей с получением 40,9% раствора 2-этинил-пропан-1,2,3-триола (3) (108 г, 100% выход), как определено методом ядерного магнитного резонанса(ЯМР) (малеиновая кислота в качестве внутреннего стандарта). 1H ЯМР (D2O, 500 МГц): δ 3,60 (дд, 4 H), 2,85 (с, 1H).
Альтернативные получения (R)-2-этинил-глицеральдегида (4)
Способ C1:
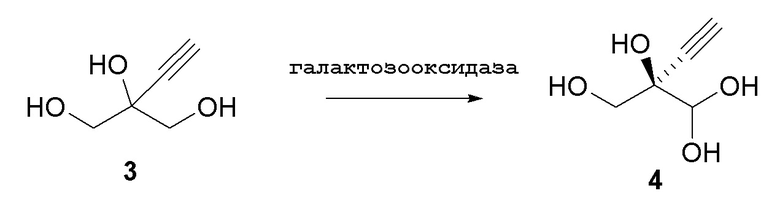
В перемешиваемом реакторе 2-этинил-пропан-1,2,3-триол (3) (1,1 г, 9,47 ммоль) в натрий-фосфатном буфере (30 мл, 100 мМ, pH 7,0), содержащем пеногаситель 204 (Sigma A6426, 1 капля ~ 20 мкл), нагревали до 30°C с продувкой воздухом при 12,5 ст.см3/мин. Галактозооксидазу (GOase, SEQ ID NO.: 1) (250 мг), пероксидазу хрена* (Тип I, 5 мг) и бычью каталазу** (5 мг), растворенные в натрий-фосфатном буфере (5 мл 100 мМ, pH 7,0), добавляли в реактор, с последующим добавлением водного раствора CuSO4 (100 мМ, 150 мл). Реакционную смесь перемешивали при 600 об/мин с продувкой воздухом в течение 47ч с получением (R)-2-этинил-глицеральдегида (4) при 47% конверсии (по данным ЯМР) и 72% э.и. (Продукт не выделяли). 1H ЯМР (D2O, 500 МГц): δ 4,29 (с, 1H), 3,65 (дд, 2H), 2,83 (с, 1H).
* Пероксидаза хрена: дикого типа пероксидаза из хрена Типа I, коммерчески доступная от SIGMA (P8125), выделена из корней хрена (Amoracia rusticana).
** Бычья каталаза: гем-зависимая каталаза из бычьего источника, коммерчески доступная от Sigma (C1345)
Способ C2:
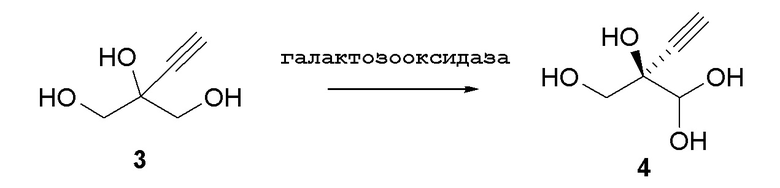
В перемешиваемый 100-л снабженный кожухом реактор, загруженный деионизированной водой (56,2 кг), добавляли дигидрофосфат натрия (1,212 кг, 10 моль). pH доводили до 7,02 с использованием 10 N раствор гидроксида натрия (852,6 г) при 25°C. В реактор загружали пеногаситель 204 (A6426, 10 мл), затем CuSO4•5H2O (6,5 г). Добавляли галактозооксидазу (451,2 г) (SEQ ID NO.: 10) и перемешивали в течение 15 минут при продувке воздухом. Добавляли пероксидазу хрена* (200,2 г) и каталазу** (502,6 г) и реактор промывали водой (2,0 кг). Затем добавляли раствор 2-этинил-пропан-1,2,3-триола (3) в воде (9,48%, 30,34 кг, 24,72 моль) с последующим добавлением дополнительной порции пеногасителя 204 (A6426, 10 мл). Реакционную смесь продували воздухом и перемешивали в течение ночи с получением 94,0 кг (R)-2-этинил-глицеральдегида (4) при 66% конверсии (по данным ЯМР) и 84% э.и. Выход 60%: 1H ЯМР (D2O, 500 МГц): δ 4,29 (с, 1H), 3,65 (дд, 2H), 2,83 (с, 1H).
* Пероксидаза хрена: дикого типа пероксидаза из очищенного хрена, коммерчески доступная от Toyobo (PEO-301), выделенная из корней хрена (Amoracia rusticana).
** Бычья каталаза: гем-зависимая каталаза из бычьего источника, коммерчески доступная от Sigma (C1345).
Описанную выше реакцию также осуществляли с использованием галактозооксидазы(SEQ ID NO.: 11) и продукт (4) получали при 67% конверсии (по данным ЯМР) и 88% э.и. с выходом 59%: 1H ЯМР (D2O, 500 МГц): δ 4,29 (с, 1H), 3,65 (дд, 2H), 2,83 (с, 1H).
Способ C3:
В 100-мл EasyMax сосуд, снабженный барботером и регулятором потока, загружали воду (82 мл) и PIPES калиевый буфер (5 мл, 0,5 M). pH доводили до 7,5 с использованием 5 M KOH раствора при 25°C. Добавляли пеногаситель 204 (200 мкл) с последующим добавлением модифицированной галактозооксидазы (SEQ ID NO.: 17, 450 мг порошкообразного фермента) и сульфата меди(II) пентагидрата (100 мкл, 100 мМ). Реакционную смесь продували воздухом при 125 стандартных кубических сантиметров в минуту (ст.см3/мин) в течение 15 мин. Загружали бычью каталазу (C1345, Sigma-Aldrich, 150 мг, 2000-5000 Ед./мг, 0,75 МЕд.) с последующим добавлением пероксидазы хрена (HRP, Toyobo PEO-301, 100 мг, 130 Ед./мг, 1,3 кЕд.) и водный раствор 2-этинил-пропан-1,2,3-триола (3) (25% масс., 12 мл, 25,8 ммоль). Реакционную смесь перемешивали при 30°C с продувкой воздухом при 125 ст.см3/мин и осуществляли отбор проб с использованием EasySampler через 20ч, достигая 70% конверсии и образования соединения (4) ((R)-2-этинил-глицеральдегид) с 58% выходом и 99% э.и. 1H ЯМР (D2O, 500 МГц): δ 4,29 (с, 1H), 3,65 (дд, 2H), 2,83 (с, 1H). Неочищенный реакционный поток непосредственно использовали на последующей стадии фосфорилирования.
Способ C4: Окисление иммобилизованной галактозооксидазой
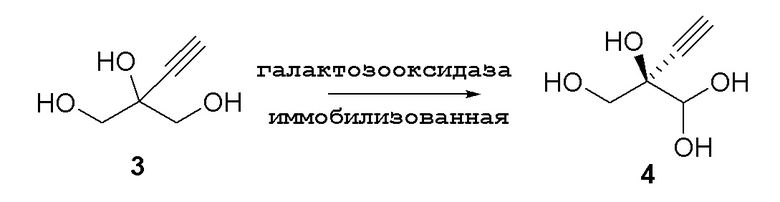
Процедура иммобилизации фермента:
Nuvia IMAC Ni-нагруженную смолу (16 мл в расчете на насыпной объем) добавляли в фильтр-воронку и промывали связывающим буфером (10 колоночных объемов, 160 мл; 500 мМ хлорида натрия, 50 мМ фосфата натрия, 15 мМ имидазола, pH 8,0) для удаления исходного содержащего смолу раствора. В сосуде лиофилизированные порошки модифицированной галактозооксидазы (SEQ ID NO.: 17, 2,00 г) ресуспендировали в растворе сульфата меди (II) (100 мкМ; 5,00 мл), с последующим добавлением связывающего буфера (50 мл) и смолы. Раствор смешивали с использованием вращающегося смесителя при 20°C в течение 5ч. Смолу фильтровали и промывали связывающим буфером (10 колоночных объемов, 160 мл) и калиевым PIPES буфером (10 колоночных объемов, 160 мл; 50 мМ, pH 7,5), и ее непосредственно использовали в реакции.
Процедура реакции:
В 100-мл EasyMax сосуд, снабженный барботером и регулятором потока, загружали воду (82 мл) и PIPES калиевый буфер (5 мл, 1 M). pH доводили до 7,5 с использованием 5 M KOH раствора при 25°C. Добавляли пеногаситель 204 (200 мкл) с последующим добавлением модифицированной галактозооксидазы, иммобилизованной на смоле (SEQ ID NO.: 17, 750 мг порошкообразного фермента на 6 мл смолы), и сульфата меди(II) пентагидрата (100 мкл, 100 мМ). Реакционную смесь продували воздухом при 125 стандартных кубических сантиметров в минуту (ст.см3/мин) в течение 15 мин. Добавляли бычью каталазу (C1345, Sigma-Aldrich, 210 мг, 2000-5000 Ед./мг, 1,05 МЕд.) с последующим добавлением пероксидазы хрена (HRP, Toyobo PEO-301, 100 мг, 130 Ед./мг, 1,3 кЕд.) и водного раствора 2-этинил-пропан-1,2,3-триола (3) (25% масс., 13 мл, 29,4 ммоль). Реакционную смесь перемешивали при 25°C с продувкой воздухом при 125 ст.см3/мин. Через 22ч реакция достигала 91% конверсии, с получением 200 мМ раствора (R)-2-этинил-глицеральдегида (4) (100 мл, 68% выход, 97% э.и. 1H ЯМР (D2O, 500 МГц): δ 4,29 (с, 1H), 3,65 (дд, 2H), 2,83 (с, 1H). Неочищенный реакционный поток непосредственно использовали на последующей стадии фосфорилирования.
Способ C5: Необязательное выделение альдегида через образование аминаля (8)
Стадия 1: Получение (S)-2-(1,3-дибензилимидазолидин-2-ил)бут-3-ин-1,2-диола
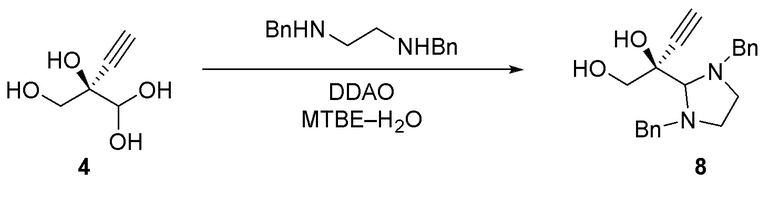
В 100-л снабженный кожухом цилиндрический сосуд снабженный устройством для барботирования азота, механической мешалкой и термопарой, загружали неочищенный оксидазный реакционный поток, содержащий (R)-2-этинил-глицеральдегид ((4), 26,0 кг, 1,85% масс. альдегида, 3,64 моль) и создавали инертную атмосферу N2. Водный раствор нагревали до 20°C и добавляли N, N-диметилдодекан-1-амин оксид (DDAO) (30% масс. в воде, 798 г, 0,96 моль) с последующим добавлением MTBE (55,3 кг, 76 л) и N, N’-дибензилэтан-1,2-диамина (1,55 кг, 6,43 моль). Коричневую бифазную смесь перемешивали в течение ночи при 20°C в атмосфере азота. Через 17 часов перемешивание прекращали и органическую фазу удаляли и отбрасывали. Получали светло-коричневый MTBE раствор (S)-2-(1,3-дибензилимидазолидин-2-ил)бут-3-ин-1,2-диола (56,5 кг, 2,02% масс. аминаля, 3,39 ммоль, 93% выход).
Шесть аналогичных MTBE растворов обрабатывали вместе на одной стадии дистилляции и кристаллизации (в общей сложности 374,4 кг раствора, содержащего 7,91 кг аминаля).
В 50-л снабженный кожухом цилиндрический сосуд, снабженный механической мешалкой, дистилляционной насадкой (холодильник при -20°C) и термопарой, загружали раствор аминаля (45 л). В сосуде создавали вакуум (65-95 тор) и температуру кожуха устанавливали на 40°C. Растворитель удаляли дистилляцией до достижения объема 35 л. В этот момент внутренняя температура была 6,1°C, и начинало кристаллизоваться не совсем белое твердое вещество. медленно добавляли оставшийся MTBE раствор, поддерживая постоянный объем 35-40 л и внутреннюю температуру 0-10°C. Когда весь MTBE раствор был добавлен, объем уменьшался до 25 л. Дистилляцию останавливали, в сосуде создавали инертную атмосферу азота и температуру кожуха снижали до 10°C. Полученную бледно-желтую суспензию подвергали старению при этой температуре в течение 2 часов и твердые частицы собирали фильтрованием. Фильтровальную лепешку промывали холодным (-2°C) MTBE (12,7 кг) и затем сушили под потоком азота в течение 7 часов. (S)-2-(1,3-дибензилимидазолидин-2-ил)-бут-3-ин-1,2-диол получали в виде не совсем белого кристаллического твердого вещества (5,75 кг). 1H ЯМР (500 МГц, DMSO-d6) δ 7,42-7,35 (м, 4H), 7,32 (тд, J=7,5, 1,6 Гц, 4H), 7,27-7,21 (м, 2H), 5,10 (т, J=5,6 Гц, 1H), 5,03 (с, 1H), 4,28 (д, J=13,3Гц, 1H), 4,16 (д, J=13,3 Гц, 1H), 3,76 (с, 1H), 3,70-3,58 (м, 4H), 3,21 (д, J=0,9 Гц, 1H), 2,90-2,80 (м, 2H), 2,60-2,51 (м, 2H), 13C ЯМР (126 МГц, DMSO-d6) δ 140,0, 140,0, 128,5, 128,3, 128,2, 128,1, 126,8, 126,8, 88,6, 86,9, 75,0, 74,0, 66,4, 60,7, 60,5, 50,4, 50,3, 39,5. HR-MS (ESI) Аминаль (M+H+) C21H25N2O2+ рассчитано 337,1911; найдено 337,1922.
Стадия 2: Получение (R)-2-этинил-глицеральдегида (4) из аминаля (8)
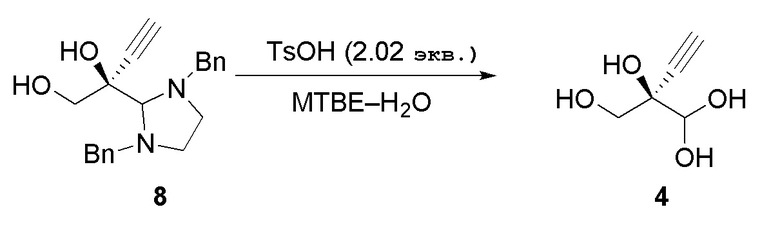
В 4-л снабженный кожухом цилиндрический сосуд, снабженный устройством для барботирования азота и механической мешалкой, загружали TsOH•H2O (12,0 г, 63,1 ммоль), воду (60 мл), (S)-2-(1,3-дибензилимидазолидин-2-ил)бут-3-ин-1,2-диол (110 г, 327 ммоль) и MTBE (1700 мл). Бифазную смесь помещали в атмосферу азота и температуру кожуха устанавливали на 15°C. Добавляли раствор TsOH•H2O (114 г, 599,3 ммоль) в воде (600 мл) по каплям в течение 1,5 часа при перемешивании верхнеприводной мешалкой (200 об/мин). Когда добавление было завершено, температуру кожуха понижали до 5°C и полученную суспензию подвергали старению в течение 1 часа. Твердые частицы удаляли фильтрованием и промывали холодной водой (270 мл). Бифазный раствор переносили в делительную воронку и органическую фазу удаляли и отбрасывали. Водную фазу обрабатывали DOWEX™ MARATHON™ A смолой (гидроксидная форма, 11,0 г) и AMBERLYST® 15 смолой (водородная форма, 11,0 г) при продувке N2 при скорости 200 ст.см3/мин в течение 24 часов для удаления остаточного MTBE. Смолы удаляли фильтрованием с получением бесцветного водного раствора (R)-2-гидрокси-2-(гидроксиметил)бут-3-иналя (774 г, 4,6% масс. альдегида, 82% выход). 1H ЯМР (500 МГц, D2O) δ 5,01 (с, 1H), 3,77 (д, J=11,7 Гц, 1H), 3,73 (д, J=11,7 Гц, 1H), 2,92 (с, 1H). 13C ЯМР (126 МГц, D2O) δ 129,4, 125,4, 90,3, 81,0, 76,0, 73,9, 65,3. HRMS (ESI) Альдегидный димер (2M+Na+) C10H12NaO6+ рассчитано 251,0526; найдено 251,0530.
Альтернативные получения (R)-2-этинил-глицеральдегид 3-фосфата (5):
Способ D1: Ацетаткиназа: система АТФ-восстановления

В перемешиваемом реакторе к раствору динатриевой соли аденозиндифосфата (40 мг, 0,087 ммоль) и хлорида магния (38 мг, 0,400 ммоль) в HEPES буфере (66 мМ, pH 7,5, 30 мл) добавляли (R)-2-этинил-глицеральдегид (4) (1,9 мл, 210 г/л раствор в воде, 3,51 ммоль) с последующим добавлением ацетаткиназы (SEQ ID NO.: 3) (40 мг) и пантотенаткиназы (SEQ ID NO.: 2) (120 мг). Реакционную смесь нагревали до 25°C и добавляли по каплям раствор ацетилфосфат литиево-калиевой соли (1,3 г, 7,01 ммоль) в HEPES буфере (50 мМ, pH 7,5, 10 мл) в течение 4 часов, поддерживая pH при 7,5 с использованием 5M гидроксида натрия. Реакционную смесь перемешивали в течение 18 часов с получением (R)-2-этинил-глицеральдегид 3-фосфата (5) при 85% конверсии (по данным ВЭЖХ) (Продукт не выделяли). 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H). ЖХ-МС: (ES, m/z): рассчитано для C5H7O6P (M-H): 193,1; найдено 193,0.
Способ D2: Пируватоксидаза: система АТФ-восстановления
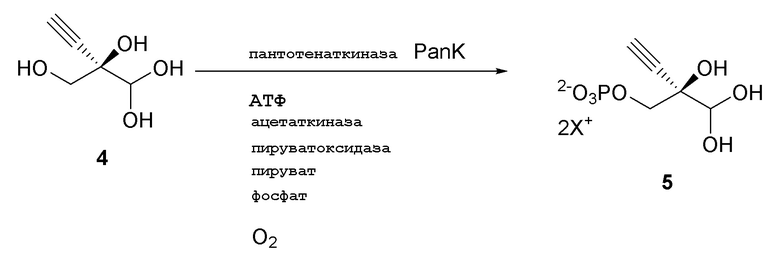
В перемешиваемом реакторе в раствор пирувата натрия (3,11 г, 28 ммоль) и фосфорной кислоты (0,523 мл, 7,71 ммоль) в 76 мл воды pH 7,5 загружали (R)-2-этинил-глицеральдегид (4) (3,8 мл, 210 г/л раствор в воде, 7,01 ммоль), динатриевую соль аденозиндифосфата (80 мг, 0,174 ммоль), тиаминпирофосфат (40 мг, 0,086 ммоль), гидрат динатриевой соли флавинадениндинуклеотида (64 мг, 0,077 ммоль) и хлорид магния (400 мкл, 1 M раствор в воде, 0,4 ммоль). pH снова доводили до 7,5 водным раствором 5M гидроксида натрия и объем реакции снова доводили до 80 мл при помощи воды. Добавляли ацетаткиназу (SEQ ID NO.: 3) (80 мг), пируватоксидазу (SEQ ID NO.: 4) (80 мг, лиофилизированный не содержащий клетки экстракт), пантотенаткиназу (SEQ ID NO.: 2) (400 мг) и каталазу (800 мкл, суспензия сульфата аммония CAT-101, Biocatalytics). Реакционную смесь перемешивали при 500 об/мин и 30°C с продувкой воздухом в течение 72 часов с получением (R)-2-этинил-глицеральдегид 3-фосфата 5 при 95% конверсии (по данным ВЭЖХ) (Продукт не выделяли). 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H). ЖХ-МС: (ES, m/z): рассчитано для C5H7O6P (M-H): 193,1; найдено 193,0.
Описанную выше реакцию также осуществляли с использованием пантотенаткиназы (SEQ ID NO.: 13) и продукт 5 получали при 66% конверсии. (Продукт не выделяли). 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H).
Способ D3: Ацетаткиназа: система АТФ-восстановления с использованием иммобилизованных ферментов
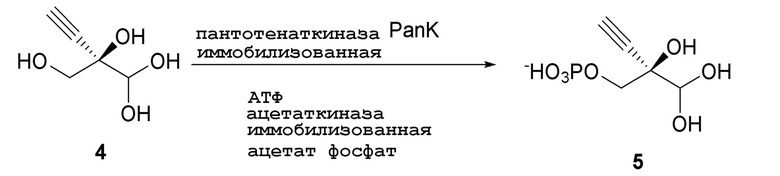
Процедура иммобилизации ферментов:
NUVIA™ никель-нагруженную смолу для аффинной хроматографии на иммобилизованных ионах металла (IMAC) (168 мл в расчете на насыпной объем) добавляли в фильтр-воронку и промывали связывающим буфером (1,6 л; 500 мМ хлорида натрия, 50 мМ фосфата натрия, pH 8,0). В сосуде пантотенаткиназу (8,4 г) (SEQ ID NO.: 12) и ацетаткиназу (2,8 г) (SEQ ID NO.: 3) растворяли в связывающем буфере (500 мл). Промытую смолу загружали в сосуд и раствор перемешивали в течение 4 часов при 20°C. Смолу фильтровали и промывали сначала связывающим буфером (1,6 л), затем пиперазин-N, N′-бис(2-этансульфоновая кислота) (PIPES) буфером (840 мл; 50 мМ, pH 6,5). Промытую смолу использовали непосредственно на следующей стадии.
Процедура реакции:
В 1-л реактор загружали раствор (R)-2-этинил-глицеральдегида (4) в воде (608,7 г, 4,6% масс., 212 ммоль) и охлаждали до 5°C. К охлажденному раствору добавляли пиперазин-N, N′-бис(2-этансульфоновая кислота) (PIPES) буфер (32,7 мл, 1 M, pH 6,5, 32,7 ммоль), хлорид магния (9,33 мл, 1 M, 9,33 ммоль), ацетилфосфат диаммониевую соль (51,8 г, 265 ммоль), гидрат динатриевой соли аденозиндифосфата (1,17 г, 2,12 ммоль) и воду (192 мл). Раствор оставляли для перемешивания и pH доводили до 6,4 с использованием 5 N KOH. Реакционную смесь нагревали до 20°C и добавляли 168 мл смолы с ко-иммобилизованными пантотенаткиназой (SEQ ID NO.: 12) и ацетаткиназой (SEQ ID NO.: 3). Реакционную смесь перемешивали в течение 10 часов, используя 5 N KOH для поддержания pH=6,4, с получением (R)-2-этинил-глицеральдегид 3-фосфата (5) при 92% конверсии (по данным ВЭЖХ) и с 91% выходом (по данным 31P ЯМР с тетрафенилфосфоний хлоридом в качестве внутреннего стандарта) (продукт не выделяли). 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H). ЖХ-МС: (ES, m/z): рассчитано для C5H7O6P (M-H): 193,1; найдено 193,0.
Получение 4-этинил-D-2-дезоксирибозо-5-фосфата (6)
Способ E:

К раствору (R)-2-этинил-глицеральдегид 3-фосфата (5) (5, 20 мл, 5,3 ммоль) в воде добавляли раствор ацетальдегида в воде (40% масс., 2,02 мл, 15,9 ммоль) при комнатной температуре с последующим добавлением дезоксирибозо-фосфат-альдолазы (DERA) (SEQ ID NO.: 6), 25 мг раствора в триэтаноламин гидрохлоридном буфере (1 мл, 1 M, pH 7,0). Реактор герметично закрывали и смесь перемешивали в течение ночи при 30°C и 600 об/мин с получением 4-этинил-D-2-дезоксирибозо-5-фосфата (6) при 99% конв. и 99% э.и., 99% д.и. в виде 1:1 смеси аномеров (Продукт не выделяли). α-аномер: 1H ЯМР (D2O, 600 МГц) δ 5,31 (т, 1H), 4,13 (т, 1H), 3,81-3,72 (м, 2H), 2,89 (с, 1H), 2,42-2,34 (м, 1H), 1,87-1,79 (м, 1H); 13C ЯМР (D2O, 151 МГц) δ 97,7 (с), 81,4 (д), 79,4 (с), 78,9 (с), 71,1 (с), 67,7 (д), 39,6 (с). β-аномер: 1H ЯМР (D2O, 600 МГц) δ 5,40 (дд, 1H), 4,28 (т, 1H), 3,88-3,80 (м, 2H), 2,87 (с, 1H), 2,13-2,06 (м, 1H), 2,04-1,97 (м, 1H); 13C ЯМР (D2O, 151 МГц) δ 97,3 (с), 82,2 (д), 78,7 (с), 78,5 (с), 71,3 (с), 68,4 (д), 39,6 (с). ЖХ-МС: (ES, m/z): рассчитано для C7H10O7P (M-H): 237,0; найдено 237,0
Альтернативные получения (2R,3S,5R)-5-(6-амино-2-фтор-9H-пурин-9-ил)-2-этинил-2-(гидроксиметил)тетрагидрофуран-3-ола моногидрата (7) [альтернативное название 4’-этинил-2-фтор-2’-дезоксиаденозин или EFdA]
Способ F1:
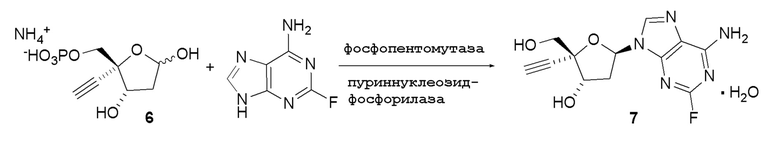
Аммоний ((2R,3S)-2-этинил-3,5-дигидрокситетрагидрофуран-2-ил)метил гидрофосфат (1,00 г, 3,91 ммоль) растворяли в 10 мл pH 7,5 буфера (100 мМ триэтаноламин HCl, содержащий 5 мМ MnCl2). pH раствора доводили до 7,3 при помощи 5 N NaOH. К раствору добавляли 2-фтораденин (0,599 г, 3,91 ммоль) и сахарозу (2,68 г, 7,82 ммоль). Раствор ферментов получали путем растворения фосфопентомутазы (SEQ ID NO.: 8) (100 мг), пуриннуклеозид-фосфорилазы (SEQ ID NO.: 9) (50 мг) и сахарозо-фосфорилазы (SEQ ID NO.: 7) (10 мг) в 10 мл pH 7,5 буфера. Раствор ферментов добавляли к смеси реагентов и полученную суспензию встряхивали при 40°C. Через 20 ч суспензию охлаждали до 0°C и фильтровали, промывая холодной водой. Твердое вещество сушили в вакууме с получением указанного в заголовке соединения (1,12 г, 92%) в виде единственного изомера.
1H ЯМР: (300 МГц, DMSO-d6, м.д.): δ 7,68 (шир.с, 2H), 7,32 (д, J=2,0 Гц, 1H), 6,44 (т, J=5,8 Гц, 1H), 5,52 (д, J=5,6 Гц, 1H), 5,27 (т, J=6,0 Гц, 1H), 4,44 (кв., J=6,4 Гц, 1H), 3,60 (кв., J=6,0 Гц, 1H), 3,53 (кв., J=6,4 Гц, 1H), 3,48 (с, 1H), 2,48-2,41 (м, 1H), 2,37-2,30 (м, 1H). 13C ЯМР (150,92 МГц, DMSO-d6, м.д.) δ 158,5 (д, JCF=203,5), 157,6 (д, JCF=21,2), 150,2 (д, JCF=20,2), 139,7 (д, JCF=2,4), 117,4 (д, JCF=4,0), 85,1, 82,0, 81,4, 78,7, 70,1, 64,2, 38,1. ЖХ-МС: (ES, m/z): рассчитано для C12H12FN5O3 (M+Na): 316,0822; найдено 316,0818.
PPM и PNP ферменты, использумые на этой стадии, каждый был получен в результате мутаций, исходя из ферментов из E. coli (Escherichia coli). Сахарозо-фосфорилазу (SP), использумую на этой стадии, получали из Alloscardovia omnicolens; SP, выделенную из других организмов, также можно использовать.
Способ F2:
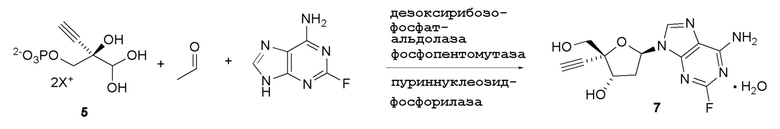
К водному раствору (R)-2-этинил-глицеральдегид 3-фосфата (5) (950 мл, 157 ммоль), содержащему пиперазин-N, N′-бис(2-этансульфоновая кислота) (PIPES) буфер при pH около 5,5-6,0, добавляли триэтаноламин (7,09 г, 47,5 ммоль). pH раствора доводили от 7,1 до 7,6 с использованием гидроксида калия (8 мл, 8M). Добавляли хлорид марганца(II) гидрат (0,592 г, 4,70 ммоль) с последующим добавлением сахарозы (161 г, 470 ммоль), получая pH=7,5. К раствору добавляли следующие ферменты: дезоксирибозо-фосфат-альдолазу (SEQ ID NO.: 14) (461 мг), сахарозо-фосфорилазу (SEQ ID NO.: 7) (494 мг), фосфопентомутазу (SEQ ID NO.: 8)(2,63 г) и пуриннуклеозид-фосфорилазу (SEQ ID NO.: 15) (659 мг). После растворения ферментов добавляли 2-фтораденин (19,80 г, 125 ммоль). Реакционную смесь нагревали до 35°C и добавляли ацетальдегид (40% масс. в изопропиловом спирте, 29,8 мл, 235 ммоль). После взаимодействия в течение 2ч в смесь вносили в качестве затравки EFdA кристаллический продукт (0,96 г, 2 моль%). После взаимодействия в течение 26 ч при 35°C суспензию охлаждали до 0°C и твердые частицы собирали фильтрованием, промывая водой два раза (40 мл каждый раз). Твердые частицы сушили при продувке азотом. Выход 43,2 г, 92% масс., 96,2% скорректированный. 1H ЯМР: (300 МГц, DMSO-d6, м.д.): δ 7,68 (шир.с, 2H), 7,32 (д, J=2,0 Гц, 1H), 6,44 (т, J=5,8 Гц, 1H), 5,52 (д, J=5,6 Гц, 1H), 5,27 (т, J=6,0 Гц, 1H), 4,44 (кв., J=6,4 Гц, 1H), 3,60 (кв., J=6,0 Гц, 1H), 3,53 (кв., J=6,4 Гц, 1H), 3,48 (с, 1H), 2,48-2,41 (м, 1H), 2,37-2,30 (м, 1H). 13C ЯМР (150,92 МГц, DMSO-d6, м.д.) δ 158,5 (д, JCF=203,5), 157,6 (д, JCF=21,2), 150,2 (д, JCF=20,2), 139,7 (д, JCF=2,4), 117,4 (д, JCF=4,0), 85,1, 82,0, 81,4, 78,7, 70,1, 64,2, 38,1. ЖХ-МС: (ES, m/z): рассчитано для C12H12FN5O3 (M+Na): 316,0822; найдено 316,0818.
Альтернативные получения (S)-2-этинил-пропан-1,2,3-триол l 1-фосфата (9) :
Способ G1: Ацетаткиназа: система АТФ-восстановления с использованием ферментов SEQ. ID No.: 2 и SEQ. ID No.: 3

В 50-мл реактор загружали раствор 2-этинил-пропан-1,2,3-триола (3) в воде (9,29 г, 9,46% масс., 7,57 ммоль), калиевый PIPES буфер (1,02 мл, 1 M, pH 6,5, 1,02 ммоль), хлорид магния (292 мкл, 1 M, 0,292 ммоль), ацетилфосфат диаммониевую соль (1,851 г, 89% масс., 9,46 ммоль), аденозиндифосфат динатриевую соль гидрат (АДФ , 42 мг, 0,076 ммоль, 0,01 экв.) и воду (28 мл). pH доводили до 6,4 с использованием 5 M KOH, раствор нагревали до 20°C и добавляли модифицированную пантотенаткиназу PanK SEQ. ID No.: 2 (264 мг) и ацетаткиназу AcK SEQ. ID No.: 3 (88 мг). Реакционную смесь перемешивали в течение 16 часов, поддерживая pH при 6,4 с использованием 5 N KOH. Конечная реакционная смесь содержала (S)-2-этинил-пропан-1,2,3-триол 1-фосфат (9) при >95% э.и. и 99% конверсии (по данным 31P ЯМР). Продукт не выделяли. 1H ЯМР (D2O, 500 МГц) δ 3,89 (м, 2H), 3,72 (д, J=11,6 Гц, 1 H), 3,65 (д, J=11,6 Гц, 1H), 2,93 (с, 1H). 13C ЯМР (D2O, 126 МГц) δ 82,9 (с), 75,1 (с), 71,0 (д, J=6,9 Гц), 67,0 (д, J=4,5 Гц), 64,7 (с). 31P ЯМР (D2O, 202 МГц) δ 3,39. HRMS: (ESI, m/z): рассчитано для [M-1]- C5H8O6P: 195,0058; Найдено 195,0068 [M-H]-: 195,0058.
Способ G2: Ацетаткиназа: система АТФ-восстановления с использованием фермента SEQ. ID No.: 20 и фермента SEQ. ID No.: 21

В снабженный кожухом реактор загружали водный раствор 2-этинил-пропан-1,2,3-триола (3) (11,47 кг, 8,7% масс., 8,61 моль) и воду (7,5 кг) с последующим добавлением 1M BIS-TRIS метанового буфера pH 6,5 (1 л) и хлорида магния (41,4 г). Добавляли АТФ (48 г, 0,086 моль, 0,01 эквивалента) и диаммоний ацетилфосфат (2,021 кг, 89%, 10,33 ммоль), раствор нагревали до 20°C и pH раствора снова доводили до 6,8 с использованием KOH (270,4 г). Затем загружали модифицированную пантотенаткиназу SEQ. ID No.: 20 (20,4 г) и модифицированную ацетаткиназу SEQ. ID No.: 21 (3 г) в виде твердых веществ. Реакционную смесь перемешивали при 20°C в течение 16ч, за это время pH снижался до 5,5. Получали количественную конверсию 2-этинил-пропан-1,2,3-триола (3), судя по данным 1H и 31P ЯМР. Такой полученный раствор (S)-2-этинил-пропан-1,2,3-триол 1-фосфата (9) (397 мМ, 22,5 кг, 98% выход) использовали на последующей стадии окисления без какой-либо дополнительной очистки. 1H ЯМР (D2O, 500 МГц) δ 3,89 (м, 2H), 3,72 (д, J=11,6 Гц, 1 H), 3,65 (д, J=11,6 Гц, 1H), 2,93 (с, 1H).
Способ G3: Ацетаткиназа: система АТФ-восстановления с использованием фермента SEQ. ID No.: 20 и фермента SEQ. ID No.: 21 с дейтерированным соединением (3) для определения абсолютной стереохимии и демонстрации десимметризирующего фосфорилирования.
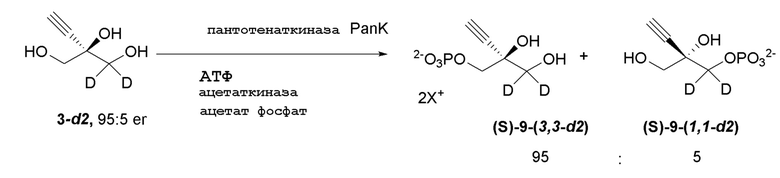
Модифицированную пантотенаткиназу SEQ. ID No.: 20 (100 мкл 10 г/л раствора в воде) и модифицированную ацетаткиназу SEQ. ID No.: 21 (100 мкл 2 г/л раствора в воде) добавляли к раствору, содержащему диаммоний ацетилфосфат (41 мг), 2-этинил-пропан-1,1-d2-1,2,3-триол ((R)-3-d2, 20 мг, 170 мкмоль), хлорид магния (10 мкл 1 M раствора в воде), АДФ (10 мкл 100 г/л раствора в воде) и натрийфосфатный буфер (10 мкл 1 M раствора в воде) в воде (800 мкл) при pH 6,5. Реакционную смесь инкубировали в течение 24ч при комнатной температуре с получением дейтерированных 2-этинил-пропан-1,2,3-триол 1-фосфатных аналогов (S)-9-(3,3-d2) и (S)-9-(1,1-d2) в соотношении 95:5 и с общим выходом 99%. Соотношение фосфорилированных соединений, определенное методом 31P ЯМР, составляло ~95:5, подтверждая стереоселективное фосфорилирование 2-этинил-пропан-1,2,3-триола (3) по про-(S) гидроксильной группе (т.е. десимметризирующее фосфорилирование). 1H ЯМР (D2O, 500 МГц) δ 3,89 (м, 2H), 3,72 (д, J=11,6 Гц, 1 H), 3,65 (д, J=11,6 Гц, 1H), 2,93 (с, 1H). 13C ЯМР (D2O, 126 МГц) δ 82,9 (с), 75,1 (с), 71,0 (д, J=6,9 Гц), 67,0 (д, J=4,5 Гц), 64,7 (с).
Способ G4: Ацетаткиназа: система АТФ-восстановления с использованием иммобилизованных ферментов SEQ. ID No.: 20 и SEQ. ID No.: 21

Процедура иммобилизации ферментов:
Nuvia IMAC Ni-нагруженную смолу (75 мл в расчете на насыпной объем) добавляли в фильтр-воронку и промывали водой (9 колоночных объемов, 3 × 225 мл) и связывающим буфером (1 колоночный объем, 75 мл; 500 мМ хлорида натрия, 50 мМ фосфата натрия, 15 мМ имидазола, pH 8,0). В сосуде пантотенаткиназу (SEQ ID NO.: 20, 6,0 г) в виде лиофилизированного порошка ресуспендировали в связывающем буфере (200 мл) и добавляли промытую смолу. Раствор смешивали с использованием вращающегося смесителя при 25°C в течение 6ч. Смолу фильтровали и промывали связывающим буфером (6 колоночных объемов, 6×225 мл) и BIS-TRIS буфером (8 колоночных объемов, 600 мл; 50 мМ, pH 6,2).
Процедура реакции:
Водный раствор 2-этинил-пропан-1,2,3-триола (3) (574 г, 8,7% масс., 0,430 моль) и воды (350 мл) загружали в снабженный кожухом реактор с последующим добавлением 1M BIS-TRIS метанового буфера pH 6,5 (50 мл) и хлорида магния (2,033 г, 0,01 моль). Добавляли АТФ (2,37 г, 0,0043 моль, 0,01 эквивалент) и диаммоний ацетилфосфат (101 г, 89%, 0,530 ммоль, 1,2 экв.), раствор нагревали до 20°C и pH раствора снова доводили до 6,8 с использованием 5M KOH. Затем загружали смолу с иммобилизованной пантотенаткиназой SEQ. ID No.: 20 (25 мл) и модифицированной ацетаткиназой SEQ. ID No.: 21 (0,15 г) в виде твердых веществ. Реакционную смесь перемешивали при 20°C в течение 16ч, за это время pH снижался до 5,5. Получали количественную конверсию 2-этинил-пропан-1,2,3-триола (3) в (S)-2-этинил-пропан-1,2,3-триол 1-фосфат (9), судя по данным 1H и 31P ЯМР. 1H ЯМР (D2O, 500 МГц) δ 3,89 (м, 2H), 3,72 (д, J=11,6 Гц, 1 H), 3,65 (д, J=11,6 Гц, 1H), 2,93 (с, 1H).
Альтернативные получения (R)-2-этинил-глицеральдегид 3-фосфата (5):
Способ H1: Иммобилизованные галактозооксидазы SEQ ID No.: 16
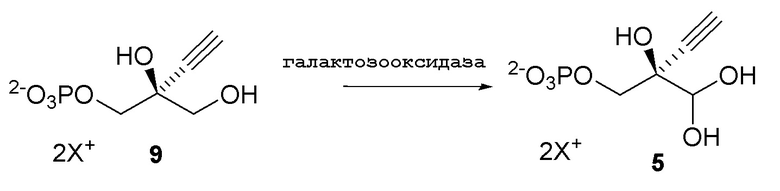
Процедура иммобилизации ферментов:
Nuvia IMAC Ni-нагруженную смолу (10 мл в расчете на насыпной объем) добавляли в фильтр-воронку и промывали связывающим буфером (10 колоночных объемов, 100 мл; 500 мМ хлорида натрия, 50 мМ фосфата натрия, 15 мМ имидазола, pH 8,0) для удаления исходного содержащего смолу раствора и получения 16 г промытой смолы. В сосуде лиофилизированные порошки модифицированной галактозооксидазы (SEQ ID NO.: 16, 750 мг) ресуспендировали в растворе сульфата меди (II) (100 мкМ; 5,00 мл) с последующим добавлением связывающего буфера (20 мл) и промытой смолы (3,0 г). Раствор смешивали с использованием вращающегося смесителя при 20°C в течение 5ч. Смолу фильтровали и промывали связывающим буфером (10 колоночных объемов, 100 мл) и BIS-TRIS буфером (10 колоночных объемов, 100 мл; 50 мМ, pH 7,5), и ее использовали непосредственно в реакции гликозилирования.
Процедура реакции:
Смолу с иммобилизованной галактозооксидазой SEQ ID NO.: 16 (3,0 г) добавляли к раствору (S)-2-этинил-пропан-1,2,3-триол 1-фосфата (9, 5,4 ммоль, 270 мМ, 20 мл) в BIS-TRIS метановом буфере (35 мМ, pH доводили до 7,2), с последующим добавлением раствора сульфата меди (II) в воде (30 мкл, 100 мМ) и пероксидазы хрена (PEO-301, 18 мг) и бычьей каталазы (C1345, 120 мг), ресуспендированных в воде (600 мкл). Реакционную смесь герметично закрывали газопроницаемой мембраной и интенсивно встряхивали при 22°C в течение 4 дней для достижения конечной конверсии 77% и получения (R)-2-этинил-глицеральдегид 3-фосфата (5) при 95% э.и. Несущую ферменты смолу отфильтровывали и раствор (R)-2-этинил-глицеральдегид 3-фосфата (5) использовали непосредственно в реакции гликозилирования. 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H). ЖХ-МС: (ES, m/z): рассчитано для C5H7O6P (M-H): 193,1; найдено 193,0.
Способ H2: Иммобилизованные галактозооксидазы SEQ ID No.: 17

Процедура иммобилизации ферментов:
Nuvia IMAC Ni-нагруженную смолу (10 мл в расчете на насыпной объем) добавляли в фильтр-воронку и промывали связывающим буфером (10 колоночных объемов, 100 мл; 500 мМ хлорида натрия, 50 мМ фосфата натрия, 15 мМ имидазола, pH 8,0) для удаления исходного содержащего смолу раствора и получения 16 г промытой смоле. В сосуде лиофилизированные порошки модифицированной галактозооксидазы (SEQ ID NO.: 16, 750 мг) ресуспендировали в растворе сульфата меди (II) (100 мкМ; 5,00 мл) с последующим добавлением связывающего буфера (20 мл) и промытой смолы (3,0 г). Раствор смешивали с использованием вращающегося смесителя при 20°C в течение 5ч. Смолу фильтровали и промывали связывающим буфером (10 колоночных объемов, 100 мл) и BIS-TRIS метановым буфером (10 колоночных объемов, 100 мл; 50 мМ, pH 7,5) и ее непосредственно использовали в реакции.
Процедура реакции:
Смолу с иммобилизованной модифицированной галактозооксидазой SEQ ID NO.: 17 (3,0 г) добавляли к раствору (S)-2-этинил-пропан-1,2,3-триол 1-фосфата (9, 5,4 ммоль, 270 мМ, 20 мл) в BIS-TRIS метановом буфере (35 мМ, pH доводили до 7,2), с последующим добавлением раствора сульфата меди (II) в воде (30 мкл, 100 мМ) и пероксидазы хрена (PEO-301, 18 мг) и бычьей каталазы (C1345, 120 мг), ресуспендированных в воде (600 мкл). Реакционную смесь герметично закрывали газопроницаемой мембраной и интенсивно встряхивали при 22°C в течение 4 дней до достижения конечной конверсии 77% и получения (R)-2-этинил-глицеральдегид 3-фосфата (5) при 95% э.и. Несущую ферменты смолу отфильтровывали и раствор (R)-2-этинил-глицеральдегид 3-фосфата (5) использовали непосредственно в реакции гликозилирования. 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H). ЖХ-МС: (ES, m/z): рассчитано для C5H7O6P (M-H): 193,1; найдено 193,0.
Способ H3: Иммобилизованные галактозооксидазы SEQ ID No.: 18
Процедура иммобилизации ферментов:
Nuvia IMAC Ni-нагруженную смолу (3 мл в расчете на насыпной объем) добавляли в фильтр-воронку и промывали связывающим буфером (10 колоночных объемов, 30 мл; 500 мМ хлорида натрия, 50 мМ фосфата натрия, 15 мМ имидазола, pH 8,0) для удаления исходного содержащего смолу раствора и получения 2,4 г промытой смолы. В сосуде лиофилизированные порошки модифицированной галактозооксидазы (SEQ ID NO.: 18, 75 мг) ресуспендировали в растворе сульфата меди (II) (100 мкМ; 1,00 мл) с последующим добавлением связывающего буфера (5 мл) и промытой смолы (400 мг). Раствор смешивали с использованием вращающегося смесителя при 20°C в течение 5ч. Смолу фильтровали и промывали связывающим буфером (10 колоночных объемов, 4 мл) и BIS-TRIS метановым буфером (10 колоночных объемов, 4 мл; 50 мМ, pH 7,5) и использовали непосредственно в реакции.
Процедура реакции:
Иммобилизованную модифицированную GOase SEQ ID NO.: 18 добавляли (400 мг) к раствору (S)-2-этинил-пропан-1,2,3-триол 1-фосфата ((9), 5,4 ммоль, 270 мМ, 1 мл) в BIS-TRIS метановом буфере (35 мМ, pH доводили до 7,2), с последующим добавлением пероксидазы хрена (PEO-301, 1 мг) и каталазы из Corynebacterium glutamicum (Roche, лиофилизат, #11650645103, 3 мг) ресуспендировали в воде (100 мкл). Реакционную смесь герметично закрывали газопроницаемой мембраной и интенсивно встряхивали при 30°C в течение 48ч. Конечная конверсия через 2 дня достигала 90%, и (R)-2-этинил-глицеральдегид 3-фосфат (5) получали с >99% э.и. Несущую ферменты смолу отфильтровывали и раствор (R)-2-этинил-глицеральдегид 3-фосфата (5) использовали непосредственно без дополнительной очистки. 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H). ЖХ-МС: (ES, m/z): рассчитано для C5H7O6P (M-H): 193,1; найдено 193,0.
Способ H4: Иммобилизованные галактозооксидазы SEQ ID No.: 19

Процедура иммобилизации ферментов:
Nuvia IMAC Ni-нагруженную смолу (3 мл в расчете на насыпной объем) добавляли в фильтр-воронку и промывали связывающим буфером (10 колоночных объемов, 30 мл; 500 мМ хлорида натрия, 50 мМ фосфата натрия, 15 мМ имидазола, pH 8,0) для удаления исходного содержащего смолу раствора и получения 2,4 г промытой смолы. В сосуде лиофилизированные порошки модифицированной галактозооксидазы (SEQ ID NO.: 19, 75 мг) ресуспендировали в растворе сульфата меди (II) (100 мкМ; 1,00 мл), с последующим добавлением связывающего буфера (5 мл) и промытой смолы (400 мг). Раствор смешивали с использованием вращающегося смесителя при 20°C в течение 5ч. Смолу фильтровали и промывали связывающим буфером (10 колоночных объемов, 4 мл) и BIS-TRIS метановым буфером (10 колоночных объемов, 4 мл; 50 мМ, pH 7,5) и использовали непосредственно в реакции.
Процедура реакции:
Иммобилизованную модифицированную GOase SEQ ID NO.: 18 добавляли (400 мг) к раствору (S)-2-этинил-пропан-1,2,3-триол 1-фосфата (9, 5,4 ммоль, 270 мМ, 1 мл) в BIS-TRIS метановом буфере (35 мМ, pH доводили до 7,2) с последующим добавлением пероксидазы хрена (PEO-301, 1 мг) и каталазы из Corynebacterium glutamicum (Roche, лиофилизат, #11650645103, 3 мг), ресуспендированных в воде (100 мкл). Реакционную смесь герметично закрывали газопроницаемой мембраной и интенсивно встряхивали при 30°C в течение 48ч. Конечная конверсия через 2 дня достигала 100%, и (R)-2-этинил-глицеральдегид 3-фосфат (5) получали при >99% э.и. Несущую ферменты смолу отфильтровывали и раствор (R)-2-этинил-глицеральдегид 3-фосфата (5) использовали непосредственно без дополнительной очистки. 1H ЯМР (D2O, 400 МГц): δ 5,02 (с, 1H), 4,00 (дкв., 2 H), 2,88 (с, 1H). ЖХ-МС: (ES, m/z): рассчитано для C5H7O6P (M-H): 193,1; найдено 193,0.
“Аминокислоты” обозначаются их общеизвестными однобуквенными символами, которые рекомендованы Комиссией по биохимической номенклатуре IUPAC-IUB. Для целей настоящего описания коды, используемые для генетически кодируемых аминокислот для ферментов, используемых в способах, описанных в настоящей заявке, показаны в Таблице 2:
Номера ID последовательностей для ферментов, используемых или которые можно использовать в способах синтеза EfdA, описанных в настоящей заявке, и в проиллюстрированных стадиях способа в экспериментальных процедурах, описанных в настоящей заявке, представлены, но не ограничиваются этим, в Таблице 3.
MASAPIGSAIPRNNWAVTCDSAQSGNECNKAIDGNKDTFWHTFYGANGDPKPPHTYTIDMKTTQNVNGLSVLPRQDGNQNGWIGRHEVYLSSDGTNWGSPVASGSWFADSTTKYSNFETRPARYVRLVAITEANGQPWTSIAEINVFQASSYTAPQPGLGRWGPTIDLPIVPAAAAIEPTSGRVLMWSSYRNDAFEGSPGGITLTSSWDPSTGIVSDRTSTVTKHDMFCPGISMDGNGQIVVDETATGGNDAKKTSLYDSSSDSWIPGPDMQVARGYQSSATMSDGRVFTIGGSFSGGRVEKNGEVYSPSSKTWTSLPNAKVNPMLTADKQGLYRSDNHAWLFGWKKGSVFQAGPSTAMNWYYTSGSGDVKSAGKRQSNRGVAPDAMCGNAVMYDAVKGKILTFGGSPDYEDSDATTNAHIITLGEPGTSPNTVFASNGLYFARTFHTSVVLPDGSTFITGGQRRGIPTEDSTPVFTPEIYVPEQDTFYKQNPNSIVRAYHSISLLLPDGRVFNGGGGLCGDCTTNHFDAQIFTPNYLYDSNGNLATRPKITRTSTQSVKVGGRITISTDSSISKASLIRYGTATHTVNTDQRRIPLTLTNNGGNSYSFQVPSDSGVALPGYWMLFVMNSAGVPSVASTIRVTQGGGGSWSHPQFEK
MSIKEQTLMTPYLQFDRNQWAALRDSVPMTLSEDEIARLKGINEDLSLEEVAEIYLPLSRLLNFYISSNLRRQAVLEQFLGTNGQRIPYIISIAGSVAVGKSTTARVLQALLSRWPEHRRVELITTDGFLHPNQVLKERGLMKKKGFPESYDMHRLVKFVSDLKSGVPNVTAPVYSHLIYDVIPDGDKTVVQPDILILEGLNVLQSGMDYPHDPHHVFVSDFVDFSIYVDAPEDLLQTWYINRFLKFREGAFTDPDSYFHNYAKLTKEEAIKTAMTIWKEMNWLNLKQNILPTRERASLILTKSANHAVEEVRLRK
MGSHHHHHHGSRVLVINSGSSSIKYQLIEMEGEKVLCKGIAERIGIEGSRLVHRVGDEKHVIERELPDHEEALKLILNTLVDEKLGVIKDLKEIDAVGHRVVHGGERFKESVLVDEEVLKAIEEVSPLAPLHNPANLMGIKAAMKLLPGVPNVAVFDTAFHQTIPQKAYLYAIPYEYYEKYKIRRYGFHGTSHRYVSKRAAEILGKKLEELKIITCHIGNGASVAAVKYGKCVDTSMGFTPLEGLVMGTRSGDLDPAIPFFIMEKEGISPQEMYDILNKKSGVYGLSKGFSSDMRDIEEAALKGDEWCKLVLEIYDYRIAKYIGAYAAAMNGVDAIVFTAGVGENSPITREDVCSYLEFLGVKLDKQKNEETIRGKEGIISTPDSRVKVLVVPTNEELMIARDTKEIVEKIGR
MGSSHHHHHHSSGLVPRGSHMTVGKTKVSTASLKVLAGWGIDTIYGIPSGTLAPLMEALGEQEETDIKFLQVKHEEVGAMAAVMQWKFGGKLGVCVGSGGPGASHLINGLYDAAMDNTPVLAILGSPPQRELNMDAFQELNQNPMYDHIAVYNRRVAYAEQLPKLIDDAIRTAISKRGVAVLEVPGDFGYKEIANDAFYSSGHSYRDYVSSAINEVDIDAAVEVLNKSKRAVIYAGIGTMGHGPAVQELSRKIKAPIITTAKNFETFDYDFEGLTGSTYRVGWKPANEAVKEADTVLFVGSNFPFAEVEGTFSNVENFIQIDNNPTMLGKRHNADVAILGDAGEAVQMLLEKVAPVEESAWWNANLKNIQNWRDYMTKLETKENGPLQLYQVYNAINKYADEDAIYSIDVGNTTQTSIRHLHMTPKNMWRTSPLFASMGIALPGGIGAKNVYPERQVFNLMGDGAFSMNYQDIVTNVRYNMPVINVVFTNTEYGFIKNKYEDTNTNTFGTEFTDVDYAMIGEAQGAVGFTVSRIEDMDQVMAAAVKANKEGKTVVIDAKITKDRPIPVETLKLDPALYSEEEIKAYKERYEAEELVPFSEFLKAEGLESKVAK
MSDLKKAAQQAISLMDLTTLNDDDTDQKVIELCHKAKTPAGDTAAICIYPRFIPIARKTLNEIGGDDIKIATVTNFPHGNDDIAIAVLETRAAVAYGADEVDVVFPYRALMEGNETVGFELVKACKEACGEDTILKVIIESGVLADPALIRKASELSIDAGADFIKTSTGKVAVNATLEAAEIMMTVISEKNPKVGFKPAGGVKDAAAAAEFLGVAARLLGDDWATPATFRFGASSLLTNLLHTLELADAPQGAQGY
MCDLKKAAQRAISLMDLTTLNDDDTDQKVIELCHKAKTPAGDTAAIVIYPRFIPIARKTLNEIGGLDIKIVTVTNFPHGNDDIAIAVLETRAAVAYGADEVDVVFPYRALMEGNETVGFELVKACKEACGEDTILKVIIESGVLKDPALIRKASEISIDAGADFIKTSTGKVAVNATLEAAEIIMTVISEKNPKVGFKPAGGIKDAAAAAEFLGVAARLLGDDWATPATFRFGATDLLTNLLHTLELADAPQGAQGY
MKNKVQLITYADRLGDGTLKSMTETLRKHFEGVYEGVHILPFFTPFDGADAGFDPVDHTKVDPRLGSWDDVAELSTTHDIMVDTIVNHMSWESEQFQDVMAKGEDSEYYPMFLTMSSIFPDGVTEEDLTAIYRPRPGLPFTHYNWGGKTRLVWTTFTPQQVDIDTDSEMGWNYLLSILDQLSQSHVSQIRLDAVGYGAKEKNSSCFMTPKTFKLIERIKAEGEKRGLETLIEVHSYYKKQVEIASKVDRVYDFAIPGLLLHALEFGKTDALAQWIDVRPNNAVNVLDTHDGIGVIDIGSDQMDRSLAGLVPDEEVDALVESIHRNSKGESQEATGAAASNLDLYQVNCTYYAALGSDDQKYIAARAVQFFMPGVPQVYYVGALAGSNDMDLLKRTNVGRDINRHYYSAAEVASEVERPVVQALNALGRFRNTLSAFDGEFSYSNADGVLTMTWADDATRATLTFAPKANSNGASVARLEWTDAAGEHATDDLIANPPVVA
MKRAFIMVLDSFGIGATEDAERFGDVGADTLGHIAEACAKGEADNGRKGPLNLPNLTRLGLAKAHEGSTGFIPAGMDGNAEVIGAYAWAHEMSSGKDSVSGHWEIAGVPVLFEWGYFSDHENSFPQELLDKLVERANLPGYLGNCRSSGTVILDQLGEEHMKTGKPIFYTSAASVFQIACHEETFGLDKLYELCEIAREELTNGGYNIGRVIARPFIGDKAGNFQRTGNRRDLAVEPPAPTVLQKLVDEKHGQVVSVGKIADIYANCGITKKVKATGLDALFDATIKEMKEAGDNTIVFTNFVDFDSSWGHRRDVAGYAAGLELFDRRLPELMSLLRDDDILILTADHGCDPTWTGTDHTREHIPVLVYGPKVKPGSLGHRETFADIGQTLAKYFGTSDMEYGKAMF
MATPHINAEMGDFADVVLMPGDPLRAKYIAETFLEDAREVNNVRGMLGFTGTYKGRKISVMGHGAGIPSCSIYTKELITDFGVKKIIRVGSCGAVLPHVKLRDVVIGMGACTDSKVNRIRFKDHDFAAIADFDMVRNAVDAAKALGIDARVGNLFSADLFYSPDGEMFDVMEKYGILGVEMEAAGIYGVAAEFGAKALTICTVSDHIRTHEQTTAAERQTTFNDMIKIALESVLLGDKE
MASAPIGVAIPRNNWAVTCDSAQSGNECNKAIDGNKDTFWHTQYGVNGDPKPPHTITIDMKTVQNVNGLSVLPRQDGNQNGWIGRHEVYLSSDGVNWGSPVASGSWFADSTTKYSNFETRPARYVRLVAITEANGQPWTSIAEINVFQASSYTAPQPGLGRWGPTIDLPIVPSAAAIEPTSGRVLMWSSYRQDAFEGSPGGITLTSSWDPSTGIVSDRTSTVTKHDMFCPGISMDGNGQIVVSGGNDAKKTSLYDSSSDSWIPGPDMQVARGYQSSATMSDGRVFTIGGSFSGGQVEKNGEVYSPSSKTWTSLPNAKVNPMLTADKQGLYRSDNHAWLFGWKKGSVFQAGPSTAMNWYYTSGSGDVKSAGKRQSNRGVAPDAMCGNAVMYDAVKGKILTFGGSPDYEDSDATTNAHIITLGEPGTSPNTVFASNGLYFARTFHTSVVLPDGSTFITGGQQRGIPTEDSTPVFTPEIYVPEQDTFYKQNPNSIVRAYHSISLLLPDGRVFNGGGGLCGDCTTNHFDAQIFTPNYLYDSNGNLATRPKITRTSTQSVVVGGWITIWTDMSISAASLIRYGTATHTVNTDQRRIPLTLTNNGGNSYSFQVPSDSGVALPGYWMLFVMNSAGVPSVASTIRVTQGQTGHHHHHH
MASAPIGVAIPRNNWAVTCDSAQSGNECNKAIDGNKDTFWHTQYGVNGDPKPPHTITIDMKTVQNVNGLSVLPRQDGNQNGWIGRHEVYLSSDGVNWGSPVASGSWFADSTTKYSNFETRPARYVRLVAITEANGQPWTSIAEINVFQASSYTAPQPGLGRWGPTIDLPIVPSAAAIEPTSGRVLMWSSYRQDAFEGSPGGITLTSSWDPSTGIVSDRTSTVTGHDMFCPGISMDGNGQIVVSGGNDAKKTSLYDSSSDSWIPGPDMQVARGYNSSATMSDGRVFTIGGSFSGGQVEKNGEVYSPSSKTWTSLPNAKVNPMLTADKQGLYRSDNHAWLFGWKKGSVFQAGPSTAMNWYYTSGSGDVKSAGKRQSNRGVAPDAMCGNAVMYDAVKGKILTFGGSPDYQDSDATTNAHIITLGEPGTSPNTVFASNGLLFARTFHTSVVLPDGSTFITGGQQRGIPTEDSTPVFTPEIYVPEQDTFYKQNPNSIVRAYHSISLLLPDGRVFNGGGGLCGDCETNHFDAQIFTPNYLYDSNGNLATRPKITRTSTQSVVVGGWITIWTDMSISAASLIRYGTATHTVNTDQRRIPLTLTNNGGNSYSFQVPSDSGVALPGYWMLFVMNSAGVPSVASTINVTQGQTGHHHHHH
MSIKEQTLMTPYLQLDRNQWAALRDSNPMTLSEDEIARLKGINEDLSLEEVAEVYLPLSRLLNFYISSNLRRQAVLEQFLGTNGQRIPYIISIAGSVAVGKSTTARVLQALLSRWPEHRRVELITTDGFLHPNQVLKERGLMKKKGFPESYDMHRLMKFVKDLKSGVPNVTAPVYSHLIYDVIPDGDKTVVQPDILILEGLNVLQSGMDYPHDPHHVFVSDFVDFSIYVDAPEDLLQTWYINRFLKFREGAFTDPDSYFHGYAKLTKEEAIKTAMTIWKEMNHLNLKQNILPTRERASLILTKSANHIVEEVRLRK
MHHHHHHGGMSIKEQTLMTPYLQLDRNQWAALRDSNPMTLSEDEIARLKGINEDLSLEEVAEVYLPLSRLLNFYISSNLRRQAVLEQFLGTNGQRIPYIISIAGSVAVGKSTTARVLQALLSRWPEHRRVEHITTDGFLHPNQVLKERGLMGKKGFPESYDMHRLMKFVKDLKSGVPNVTAPVYSHLIYDVIPDGDKTVVQPDILILEGLNVLQSGMDYPHDPHHVFVSDFVDFSIYVDAPEDLLQTWYINRFLKFREGAFTDPDSYFHGYAKLTKEEAIKTAMTIWKEMNHLNLKQNILPTRERASLILTKSANHIVEEVRLRK
MHHHHHHCDLKKAAQRAISLMDLTTLNDDDTDQKVIELCHKAKTPAGDTAAIVIYPRFIPIARKTLNEIGGLDIKIVTVTNFPHGNDDIAIAVLETRAAVAYGADEVDVVFPYRALMEGNETVGFELVKACKEACGEDTILKVIIESGVLKDPALIRKASEISIDAGADFIKTSTGKVAVNATLEAAEIIMTVISEKNPKVGFKPAGGIKDAAAAAEFLGVAARLLGDDWATPATFRFGATDLLTNLLHTLELADAPQGAQGY
MATPHINAEMGDFADVVLMPGDPLRAKYIAETFLEDAREVNNVRGMLGFTGTYKGRKISVMGHGMGIPSCSIYTKELITDFGVKKIIRVGSCGAVLPHVKLRDVVIGMGACTDSKVNRIRFKDHDFAAIADFDMVRNAVDAAKALGIDARVGNLFSADLFYSPDGEMFDVMEKYGILGVEMEAAGIYGVAAEFGAKALTICTVSDHIRTHEQTTAAERQTTFNDMIKIALESVLLGDKE
MASAPIGVAIPRNNWAVTCDSAQSGNECNKAIDGNKDTFWHTQYGVNGDPKPPHTITIDMKTVQNVNGLSVLPRQDGNQNGWIGRHEVYLSSDGVNWGSPVASGSWFADSTTKYSNFETRPARYVRLVAITEANGQPWTSIAEINVFQASSYTAPQPGLGRWGPTIDLPIVPSAAAIEPTSGRVLMWSSYRQDAFEPSPGGITLTSSWDPSTGIVSDRTSTVTGHDMFCPGISMDGNGQIVVSGGNDAKKTSLYDSSSDSWIPGPDMQVARGYNSSATMSDGRVFTIGGSYSGGQVEKNGEVYSPSSKTWTSLPNAKVNPMLTADKQGLYRSDNHAWLFGWKKGSVFQAGPSTAMNWYYTSGSGDVKSAGKRQSDRGVAPDAMCGNAVMYDAVKGKILTFGGSPDYQDSDATTNAHIITLGEPGTSPNTVFASNGLLFARTFHTSVVLPDGSVFITGGQQRGVPLEDSTPVFTPEIYVPEQDTFYKQNPNSIVRAYHSISLLLPDGRVFNGGGGLCGDCETNHFDAQIFTPNYLYDSNGNLATRPKITRTSTQSVVVGGWITIWTDMSISAASLIRYGTATHTVNTDQRRIGLTLTNNGGNSYSFQVPSDSGVALPGYWMLFVMNSAGVPSVASTINVTQGQTGHHHHHH
MASAPIGVAIPRNNWAVTCDSAQSGNECIKAIDGNKDTFWHTQYGVNGDPKPPHTITIDMKTVQNVNGLSVLPRQDGNQNGWIGRHEVYLSSDGVNWGSPVASGSWFADSTTKYSNFETRPARYVRLVAITEANGQPWTSIAEINVFQASSYTAPQPGLGRWGPTIDLPIVPSAAAIEPTSGRVLMWSSYRQDAFEDSPGGITLTSSWDPSTGIVSDRTSTVTGHDMFCPGISMDGNGQIVVSGGNDAKKTSLYDSSSDSWIPGPDMQVARGYNSSATMSDGRVFTIGGSYSGGQVEKNGEVYSPSSKTWTSLPNAKVNPMLTADKQGLYRSDNHAWLFGWKKGSVFQAGPSTAMNWYYTSGSGDVKSAGKRQSDRGVAPDAMCGNAVMYDAVKGKILTFGGSPDYQDSDATTNAHIITLGEPGTSPNTVFASNGLLFARTFHTSVVLPDGSVFITGGQQRGVPLEDSTPVFTPEIYVPEQDTFYKQNPNSIVRAYHSISLLLPDGRVFNGGGGLCGDCETNHFDAQIFTPNYLYDSNGNLATRPKITRTSTQSVVVGGWITIWTDMSISAASLIRYGTATHTVNTDQRRIGLTLTNNGGNSYSFQVPSDSGVALPGYWMLFVMNSAGVPSVASTINVTQGQTGHHHHHH
MASAPIGVAIPRNNWAVTCDSAQSGNECIKAIDGNKDTFWHTQYGVNGDPKPPHTITIDMKTVQNVNGLSVLPRQDGNQNGWIGRHEVYLSSDGVNWGSPVASGSWFADSTTKYSNFETRPARYVRLVAITEANGQPWTSIAEINVFQASSYTAPQPGLGRWGPTIDLPIVPSAAAIEPTSGRVLMWSSYRQDAFEDSPGGITLTSSWDPSTGIVSDRTSTVTGHDMFCPGISMDGNGQIVVSGGNDAKKTSLYDSSSDSWIPGPDMQVARGYNSSATMSDGRVFTIGGSYSGGQVEKNGEVYSPSSKTWTSLPNAKVNPMLTADKRGLYRSDNHAWLFGWKKGSVFQAGPSTAMNWYYTSGSGDVKSAGKRQSDRGVAPDAMCGNAVMYDAVKGKILTFGGSPDYQDSDATTNAHIITLGEPGTSPNTVFASNGLLFARTFHTSVVLPDGSVFITGGQQRGVPLEDSTPVFTPEIYVPEQDTFYKQNPNSIVRAYHSISLLLPDGRVFNGGGGLCGDCETNHFDAQIFTPNYLYDSNGNLATRPKITRTSTQSVVVGGWITIWTDMSISAASLIRYGTATHTVNTDQRRIGLTLTNNGGNSYSFQVPSDSGVALPGYWMLFVMNSAGVPSVASTINVTQGQTGHHHHHH
MASAPIGVAIPRNNWAVTCDSAQSGNECIKAIDGNKDTFWHTQYGVNGDPKPPHTITIDMKTVQNVNGLSVLPRQDGNQNGWIGRHEVYLSSDGVNWGSPVASGSWFADSTTKYSNFETRPARYVRLVAITEANGQPWTSIAEINVFQASSYTAPQPGLGRWGPTIDLPIVPSAAAIEPTSGRVLMWSSYRQDAFRDSPGGITLTSSWDPSTGIVSDRTSTVTGHDMFCPGISMDGNGQIVVSGGNDAKKTSLYDSSSDSWIPGPDMQVARGYNSSATMSDGRVFTIGGSYSGGQVEKNGEVYSPSSKTWTSLPNAKVNPMLTADKQGLYRSDNHAWLFGWKKGSVFQAGPSTAMNWYYTSGSGDVKSAGKRQSDRGVAPDAMCGNAVMYDAVKGKILTFGGSPDYQDSDATTNAHIITLGEPGTSPNTVFASNGLLFARTFHTSVVLPDGSVFITGGQQRGVPLEDSTPVFTPEIYVPEQDTFYKQNPNSIVRAYHSISLLLPDGRVFNGGGGLCGDCETNHFDAQIFTPNYLYDSNGNLATRPKITRTSTQSVVVGGWITIWTDMSISAASLIRYGTATHTVNTDQRRIGLTLTNNGGNSYSFQVPSDSGVALPGYWMLFVMNSAGVPSVASTINVTQGQTGHHHHHH
MHHHHHHGGSGSIKEQTLMTPYLQLDRNQWAALRDSNPMTLSEDEIARLKGINEDLSLEEVAEVYLPLSRLLNFYISSNLRRQAQLEQFLGTNGQRIPYIISIAGSVAVGKSTFARVLQALLSRWPEHRRVEHITTDGFLHPNQVLKERGLMGKKGFPESYDMHRLMKFVKDLKSGVPNVTAPVYSHLIYDVIPDGDKTVVQPDILILEGLNVLQSGMDYPHDPHHVFVSDFVDFSIYVDAPEDLLQTWYINRFLKFREGAFTDPDSYFHGYAKLTKEEAIKTAMTIWKEMNHVNLKQNILPTRERASLILTKSANHIVEEVRLRK
MGSHHHHHHGSRVLNINSGSSSIKYQLIEMEGEKVLCKGIAERIGIEGSRLVHRVGDEKHVIERELPDHEEALKLILNTLVDEKLGVIKDLKEIDAVGHRVVHGGERFKESVLVDEEVLKAIEEVSPLAPLHNPANLMGIKAAMKLLPGVPNVQVFDTAFHQTIPQKAYLYAIPYEYYEKYKIRRYGFHGISHRYVSKRAAEILGKKLEELKIITCHIGNGASVAAVKYGKCVDTSMGFTPLEGLVMGTRSGDLDPAIPFFIMEKEGISPQEMYDILNKKSGVYGLSKGFSSDMRDNLEAALKGDEWCKLVLEIYDYRIAKYIGAYAAAMNGVDAIVFTAGVGENSPITREDVCKYLEFLGVKLDKQKNEETIRGKEGIISTPDSRVKVLVVPTNEELMIARDTKEIVEKIGR
Пероксидаза хрена: пероксидаза дикого типа из хрена Типа I, коммерчески доступная от SIGMA (P8125), выделена из корней хрена (Amoracia rusticana).
Каталаза: (1) каталаза дикого типа из бычьей печени коммерчески доступа от SIGMA (C1345); или (2) CAT-101, Biocatalytics; или (3) из Corynebacterium glutamicum (Roche, #11650645103).
Дополнительные варианты осуществления настоящего изобретения включают, но не ограничиваются этим, использование следующих ферментов на стадиях способа синтеза, описанного в настоящей заявке, для получения 4’-этинил 2’-дезоксинуклеозида или его аналога, например EFdA.
А. Пуриннуклеозид-фосфорилаза.
1A. Сконструированная пуриннуклеозид-фосфорилаза, включающая полипептидную последовательность, имеющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с SEQ ID NO.: 9 или SEQ ID NO.: 15, или ее функциональный фрагмент, где полипептидная последовательность указанной сконструированной пуриннуклеозид-фосфорилазы включает по меньшей мере одну аминокислотную замену или набор аминокислотных замен по сравнению с SEQ ID NO: 9 или SEQ ID NO.: 15.
2A. Сконструированная пуриннуклеозид-фосфорилаза по п. 1A, где указанная сконструированная пуриннуклеозид-фосфорилаза включает полипептидную последовательность, которая по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична SEQ ID NO: 9 или SEQ ID NO.: 15.
3A. Сконструированная пуриннуклеозид-фосфорилаза, которая состоит из полипептидной последовательности, представленной в SEQ ID NO: 9 или SEQ ID NO.: 15.
4А. Сконструированная пуриннуклеозид-фосфорилаза по любому из пп. 1A-3A, которая включает по меньшей мере одно улучшенное свойство по сравнению с пуриннуклеозид-фосфорилазой дикого типа E. coli.
5A. Сконструированная пуриннуклеозид-фосфорилаза по п. 4A, где указанное улучшенное свойство включает улучшенную активность в отношении субстрата-соединения 6,5 (в его кольцевой форме или в виде альдегида или гидрата с открытой цепью, или соли любого из вышеуказанных) по сравнению с пуриннуклеозид-фосфорилазой дикого типа E. coli.
6A. Сконструированная пуриннуклеозид-фосфорилаза по п. 4A, где указанное улучшенное свойство включает повышенную продукцию EFdA (соединение 7) по сравнению с пуриннуклеозид-фосфорилазой дикого типа E. coli.
7A. Сконструированная пуриннуклеозид-фосфорилаза по любому из пп. A1-6A, где указанная сконструированная пуриннуклеозид-фосфорилаза является очищенной.
8A. Сконструированная пуриннуклеозид-фосфорилаза по любому из пп. 1A-7A, где по меньшей мере одна аминокислотная замена (т.е. одна или несколько аминокислотных замен) представляет собой консервативную аминокислотную замену(замены).
B. Фосфопентомутаза.
1B. Сконструированная фосфопентомутаза, включающая полипептидную последовательность, имеющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с SEQ ID NO.: 8, или ее функциональный фрагмент, где полипептидная последовательность указанной сконструированной фосфопентомутазы включает по меньшей мере одну аминокислотную замену или набор аминокислотных замен по сравнению с SEQ ID NO: 8.
2B. Сконструированная фосфопентомутаза по п. 1B, где указанная сконструированная фосфопентомутаза включает полипептидную последовательность, которая по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична SEQ ID NO.: 8.
3B. Сконструированная фосфопентомутаза, которая состоит из полипептидной последовательности, представленной в SEQ ID NO.: 8.
4B. Сконструированная фосфопентомутаза по любому из пп. 1B-3B, которая включает по меньшей мере одно улучшенное свойство по сравнению с фосфопентомутазой дикого типа E. coli.
5B. Сконструированная фосфопентомутаза по п. 4B, где указанное улучшенное свойство включает улучшенную активность в отношении субстрата-соединения 6 (в его кольцевой форме или в виде альдегида или гидрата с открытой цепью, или соли любого из вышеуказанных) по сравнению с фосфопентомутазой дикого типа E. coli.
6B. Сконструированная фосфопентомутаза по п. 4B, где указанное улучшенное свойство включает повышенную продукцию соединения 6,5 или соединения 7 (EFdA) по сравнению с фосфопентомутазой дикого типа E. coli.
7B. Сконструированная фосфопентомутаза по любому из пп. 1B-6B, где указанная сконструированная фосфопентомутаза является очищенной.
8B. Сконструированная фосфопентомутаза по любому из пп. 1B-7B, где по меньшей мере одна аминокислотная замена (т.е. одна или несколько аминокислотных замен) представляет собой консервативную аминокислотную замену(замены).
C. Дезоксирибозо-фосфат-альдолаза.
1C. Дезоксирибозо-фосфат-альдолаза, которая состоит из полипептидной последовательности дикого типа из Shewanella halifaxensis, представленной в SEQ ID NO.: 5.
2C. Сконструированная дезоксирибозо-фосфат-альдолаза, которая состоит из полипептидной последовательности, представленной в SEQ ID NO.: 6 или SEQ ID NO.: 14.
3C. Сконструированная дезоксирибозо-фосфат-альдолаза, где указанная сконструированная дезоксирибозо-фосфат-альдолаза включает полипептидную последовательность, которая по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична SEQ ID NO.: 5, SEQ ID NO.: 6 или SEQ ID NO.: 14.
4C. Сконструированная дезоксирибозо-фосфат-альдолаза, включающая полипептидную последовательность, имеющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с SEQ ID NO.: 5, SEQ ID NO.: 6 или SEQ ID NO.: 14, или ее функциональный фрагмент, где полипептидная последовательность указанной сконструированной дезоксирибозо-фосфат-альдолазы включает по меньшей мере одну аминокислотную замену или набор аминокислотных замен по сравнению с SEQ ID NO.: 5, SEQ ID NO.: 6 или SEQ ID NO.: 14.
5C. Дезоксирибозо-фосфат-альдолаза по любому из пп. 1C-4C, которая обладает активностью в отношении субстрата-соединения 5 ((R)-2-этинил-глицеральдегид 3-фосфата, его гидрата или соли любого из вышеуказанных).
6C. Дезоксирибозо-фосфат-альдолаза по любому из пп. 1C-5C, которая включает способность к образованию соединения 6 (4-этинил-D-2-дезоксирибозо-5-фосфата или формы его альдегида или гидрата с открытой цепью, или соли любого из вышеуказанных) без необходимости в защитных группах на субстрате-соединении 5 ((R)-2-этинил-глицеральдегид 3-фосфате, его гидрате или соли любого из вышеуказанных) в процессе реакции.
7C. Сконструированная дезоксирибозо-фосфат-альдолаза по любому из пп. 2C-6C, где дезоксирибозо-фосфат-альдолаза обладает улучшенным свойством, которое включает повышенную продукцию соединения 6 (4-этинил-D-2-дезоксирибозо-5-фосфата, или формы его альдегида или гидрата с открытой цепью, или соли любого из вышеуказанных), по сравнению с дезоксирибозо-фосфат-альдолазой дикого типа Shewanella halifaxensis.
8C. Дезоксирибозо-фосфат-альдолаза по любому из пп. 1C-7C, где указанная дезоксирибозо-фосфат-альдолаза является очищенной.
9C. Сконструированная дезоксирибозо-фосфат-альдолаза по любому из пп. 2C-7C, где по меньшей мере одна аминокислотная замена (т.е. одна или несколько аминокислотных замен) представляет собой консервативную аминокислотную замену(замены).
D. Пантотенаткиназа.
1D. Сконструированная пантотенаткиназа, включающая полипептидную последовательность, имеющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с SEQ ID NO: 2, SEQ ID NO.: 12, SEQ ID NO.: 13 или SEQ ID NO.: 20, или ее функциональный фрагмент, где полипептидная последовательность указанной сконструированной пантотенаткиназы включает по меньшей мере одну аминокислотную замену или набор аминокислотных замен по сравнению с SEQ ID NO: 2, SEQ ID NO.: 12, SEQ ID NO.: 13 или SEQ ID NO.: 20.
2D. Сконструированная пантотенаткиназа по п. 1D, где указанная сконструированная пантотенаткиназа включает полипептидную последовательность, которая по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична SEQ ID NO.: 2, SEQ ID NO.: 12, SEQ ID NO.: 13 или SEQ ID NO.: 20.
3D. Сконструированная пантотенаткиназа, которая состоит из полипептидной последовательности, представленной в SEQ ID NO.: 2, SEQ ID NO.: 12, SEQ ID NO.: 13 или SEQ ID NO.: 20.
4D. Сконструированная пантотенаткиназа по любому из пп. 1D-3D, которая включает по меньшей мере одно улучшенное свойство по сравнению с пантотенаткиназой дикого типа E. coli.
5D. Сконструированная пантотенаткиназа по п. 4D, где указанное улучшенное свойство включает улучшенную активность в отношении субстрата-соединения 4 ((R)-2-этинил-глицеральдегида или его гидратированной формы) по сравнению с пантотенаткиназой дикого типа e. coli.
6D. Сконструированная пантотенаткиназа по п. 5D, где указанное улучшенное свойство включает повышенную продукцию соединения 5 ((R)-2-этинил-глицеральдегид 3-фосфата) по сравнению с пантотенаткиназой дикого типа.
7D. Сконструированная пантотенаткиназа по п. 4D, где указанное улучшенное свойство включает улучшенную активность в отношении субстрата-соединения 3 (2-этинил-пропан-1,2,3-триола) по сравнению с пантотенаткиназой дикого типа e. coli.
8D. Сконструированная пантотенаткиназа по п. 7D, где указанное улучшенное свойство включает повышенную продукцию соединения 9 ((S)-2-этинил-пропан-1,2,3-триол 1-фосфата) по сравнению с пантотенаткиназой дикого типа.
9D. Сконструированная пантотенаткиназа по любому из пп. 1D-8D, где указанная пантотенаткиназа является очищенной.
10D. Сконструированная пантотенаткиназа по любому из пп. 1D-9D, где по меньшей мере одна аминокислотная замена (т.е. одна или несколько аминокислотных замен) представляет собой консервативную аминокислотную замену(замены).
E. Галактозооксидаза.
1E.Сконструированная галактозооксидаза, включающая полипептидную последовательность, имеющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с SEQ ID NO.: 1, 10, 11, 16, 17, 18 или 19, или ее функциональный фрагмент, где полипептидная последовательность указанной сконструированной галактозооксидазы включает по меньшей мере одну аминокислотную замену или набор аминокислотных замен по сравнению с SEQ ID NOs.: 1, 10, 11, 16, 17, 18 или 19.
2E. Сконструированная галактозооксидаза по п. 1E, где указанная сконструированная галактозооксидаза включает полипептидную последовательность, которая по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична SEQ ID NOs.: 1, 10, 11, 16, 17, 18 или 19.
3E. Сконструированная галактозооксидаза, которая состоит из полипептидной последовательности, представленной в SEQ ID NOs.: 1, 10, 11, 16, 17, 18 или 19.
4E. Сконструированная галактозооксидаза по любому из пп. 1E-3E, которая включает по меньшей мере одно улучшенное свойство по сравнению с галактозооксидазой дикого типа F. graminearum.
5E. Сконструированная галактозооксидаза по п. 4E, где указанное улучшенное свойство включает улучшенную активность в отношении субстрата, который представляет собой первичный спирт, по сравнению с галактозооксидазой дикого типа F. graminearum.
6E. Сконструированная галактозооксидаза по п. 4E, где указанное улучшенное свойство включает улучшенную активность в отношении субстрата-соединения 3 (2-этинил-пропан-1,2,3-триол) по сравнению с галактозооксидазой дикого типа F. graminearum.
7E. Сконструированная галактозооксидаза по п. 6E, где указанное улучшенное свойство включает повышенную продукцию соединения 4 ((R)-2-этинил-глицеральдегида или его гидратированной формы) по сравнению с галактозооксидазой дикого типа F. graminearum.
8E. Сконструированная галактозооксидаза по п. 4E, где указанное улучшенное свойство включает улучшенную активность в отношении субстрата-соединения 9 ((S)-2-этинил-пропан-1,2,3-триол 1-фосфат) по сравнению с галактозооксидазой дикого типа F. graminearum.
9E. Сконструированная галактозооксидаза по п. 8E, где указанное улучшенное свойство включает повышенную продукцию соединения 5 ((R)-2-этинил-глицеральдегид 3-фосфата или его гидратированной формы) по сравнению с галактозооксидазой дикого типа F. graminearum.
10E. Сконструированная галактозооксидаза по любому из пп. 1E-9E, где указанная галактозооксидаза является очищенной.
11E. Сконструированная галактозооксидаза по любому из пп. 1E-10E, где по меньшей мере одна аминокислотная замена (т.е. одна или несколько аминокислотных замен) представляет собой консервативную аминокислотную замену(замены).
F. Ацетаткиназа.
1F. Ацетаткиназа, которая состоит из полипептидной последовательности дикого типа из Thermotoga maritima, представленной в SEQ ID NO.: 3 или SEQ ID NO.: 21.
2F. Сконструированная ацетаткиназа, где указанная сконструированная ацетаткиназа включает полипептидную последовательность, которая по меньшей мере на 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична SEQ ID NO.: 3 или SEQ ID NO.: 21.
3F. Сконструированная ацетаткиназа, включающая полипептидную последовательность, имеющую по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с SEQ ID NO.: 3 или SEQ ID NO.: 21, или ее функциональный фрагмент, где полипептидная последовательность указанной сконструированной ацетаткиназы включает по меньшей мере одну аминокислотную замену или набор аминокислотных замен по сравнению с SEQ ID NO.: 3 или SEQ ID NO: 21.
4F. Ацетаткиназа по п. 2F или 3F, которая включает по меньшей мере одно улучшенное свойство по сравнению с ацетаткиназой дикого типа T. maritima.
5F. Ацетаткиназа по п. 4F, где указанное улучшенное свойство включает улучшенную активность для рецикла АТФ-кофактора в реакции фосфорилирования в отношении субстрата-соединения 4 ((R)-2-этинил-глицеральдегида или его гидратированной формы) по сравнению с ацетаткиназой дикого типа Thermotoga maritima.
6F. Ацетаткиназа по п. 5F, где указанное улучшенное свойство включает повышенную продукцию соединения 5 ((R)-2-этинил-глицеральдегид 3-фосфата или его гидрата, или соли любого из вышеуказанных) по сравнению с ацетаткиназой дикого типа Thermotoga maritima.
7F. Ацетаткиназа по п. 4F, где указанное улучшенное свойство включает улучшенную активность для рецикла АТФ-кофактора в реакции фосфорилирования в отношении субстрата-соединения 3 (2-этинил-пропан-1,2,3-триол) по сравнению с ацетаткиназой дикого типа Thermotoga maritima.
8F. Ацетаткиназа по п. 7F, где указанное улучшенное свойство включает повышенную продукцию соединения 9 ((S)-2- этинил-пропан-1,2,3-триол 1-фосфата или соли любого из вышеуказанных) по сравнению с ацетаткиназой дикого типа Thermotoga maritima.
9F. Ацетаткиназа по любому из пп. 1F-8F, где указанная ацетаткиназа является очищенной.
10F. Сконструированная ацетаткиназа по любому из пп. 2F-7F, где по меньшей мере одна аминокислотная замена (т.е. одна или несколько аминокислотных замен) представляет собой консервативную аминокислотную замену(замены).
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНАЦИЯ ТЕРМОСТАБИЛЬНЫХ БИОКАТАЛИЗАТОРОВ ДЛЯ СИНТЕЗА НУКЛЕОЗИДОВ | 2010 |
|
RU2569110C2 |
| НОВАЯ АЛЬДОЛАЗА И СПОСОБ ПОЛУЧЕНИЯ ОПТИЧЕСКИ АКТИВНЫХ IHOG И МОНАТИНА | 2005 |
|
RU2313579C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПРОИЗВОДНОГО ГЛЮТАМАТА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МОНАТИНА | 2002 |
|
RU2280078C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДОЛ-3-ПИРОВИНОГРАДНОЙ КИСЛОТЫ И ЕЕ ПРОИЗВОДНЫХ | 2006 |
|
RU2325442C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ПУРИННУКЛЕОЗИД-ФОСФОРИЛАЗЫ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pERPUPHO1 И ШТАММ ESCHERICHIA COLI BL21(DE3)/pERPUPHO1 ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2000 |
|
RU2179188C2 |
| Новая термостабильная фруктозо-6-фосфат-3-эпимераза и способ получения аллюлозы с ее использованием | 2017 |
|
RU2733427C2 |
| СПОСОБ ПОЛУЧЕНИЯ 1-β-D-РИБОФУРАНОЗИЛ-1,2,4-ТРИАЗОЛ-3-КАРБОКСИАМИДА (РИБАВИРИНА) | 2002 |
|
RU2230118C2 |
| НОВАЯ АЛЬДОЛАЗА И СПОСОБ ПОЛУЧЕНИЯ ЗАМЕЩЕННЫХ α-КЕТОКИСЛОТ | 2003 |
|
RU2307871C2 |
| ПРОИЗВОДНЫЕ ПАНТОТЕНАТА ДЛЯ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2653497C2 |
| ПИТЬЕВАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ МОНАТИН, И СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ | 2004 |
|
RU2380989C2 |
Изобретение относится к способу синтеза 4’-этинил-2’-дезоксинуклеозида, включающему объединение соединения 6.5
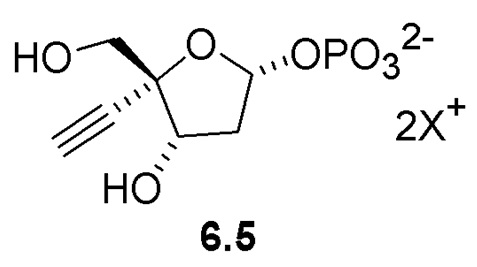 с пуриннуклеозид-фосфорилазой и нуклеооснованием в буферном растворе, содержащем соль марганца (II), где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион, и где 4’-этинил-2’-дезоксинуклеозид представляет собой
с пуриннуклеозид-фосфорилазой и нуклеооснованием в буферном растворе, содержащем соль марганца (II), где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион, и где 4’-этинил-2’-дезоксинуклеозид представляет собой
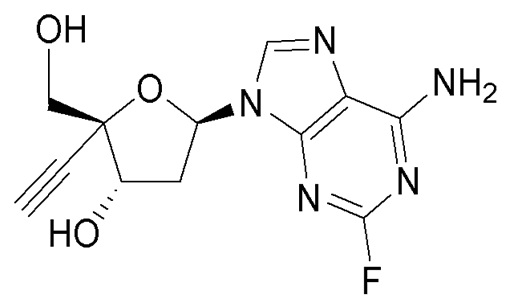 . Изобретение также относится к новым промежуточным соединениям и способам их получения. Технический результат – разработан новый способ получения 4’-этинил-2’-дезоксинуклеозида, который исключает использование защитных групп на промежуточных соединениях, улучшает стереоселективность гликозилирования и уменьшает количество стадий процесса. 13 н. и 19 з.п. ф-лы, 3 табл.
. Изобретение также относится к новым промежуточным соединениям и способам их получения. Технический результат – разработан новый способ получения 4’-этинил-2’-дезоксинуклеозида, который исключает использование защитных групп на промежуточных соединениях, улучшает стереоселективность гликозилирования и уменьшает количество стадий процесса. 13 н. и 19 з.п. ф-лы, 3 табл.
1. Способ синтеза 4’-этинил-2’-дезоксинуклеозида, включающий объединение соединения 6.5:
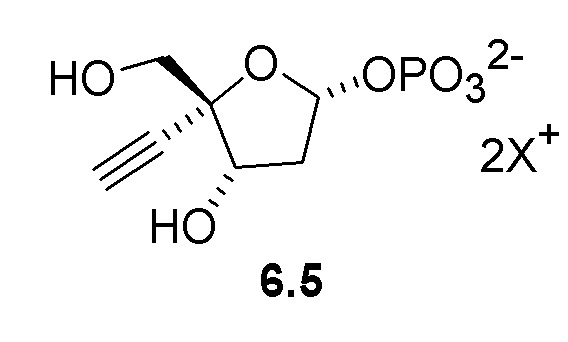 ,
,
с пуриннуклеозид-фосфорилазой и нуклеооснованием в буферном растворе, содержащем соль марганца (II),
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион, и где 4’-этинил-2’-дезоксинуклеозид представляет собой
 .
.
2. Способ по п. 1, дополнительно включающий выделение
 .
.
3. Способ по п. 1 для синтеза 4’-этинил-2’-дезоксинуклеозида, дополнительно включающий объединение соединения 6
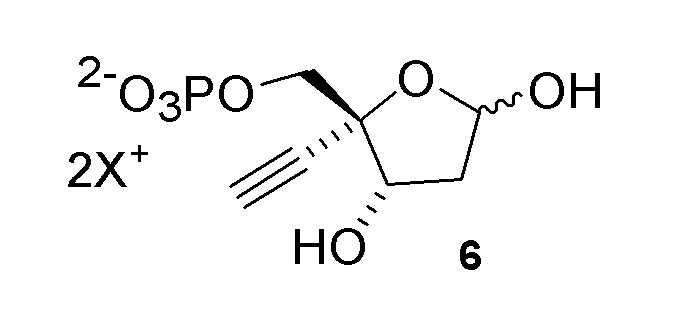
и фосфопентомутазы с пуриннуклеозид-фосфорилазой и нуклеооснованием в буферном растворе, содержащем соль марганца (II).
4. Способ по п. 3, дополнительно включающий удаление неорганического фосфатного побочного продукта из реакционной смеси.
5. Способ по п. 4, включающий удаление неорганического фосфатного побочного продукта из реакционного раствора путем (a) добавления сахарозо-фосфорилазы и сахарозы к реакционной смеси или (b) добавления кальция, магния или марганца к реакционной смеси.
6. Способ по любому из пп. 3-5, дополнительно включающий выделение 4’-этинил-2’-дезоксинуклеозида.
7. Способ по п. 3, дополнительно включающий стадию синтеза соединения 6, где синтез включает объединение соединения 5
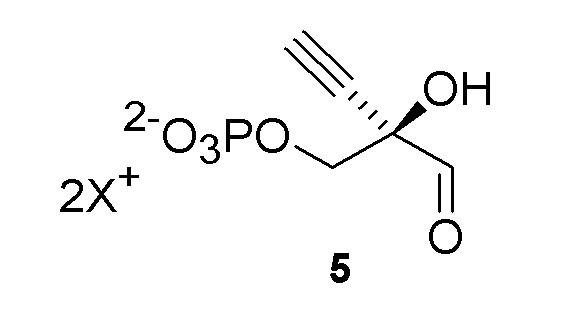
с ацетальдегидом и дезоксирибозо-фосфат-альдолазой в водном растворе с получением соединения 6;
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
8. Способ по п. 7, где реакцию осуществляют в герметично закрытом сосуде.
9. Способ по п. 7 или 8, дополнительно включающий стадию синтеза соединения 5, где синтез включает объединение соединения 4
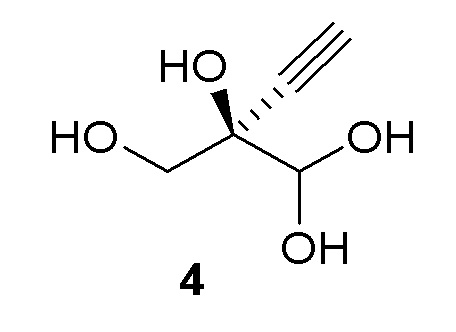
с пантотенаткиназой в буферном растворе в присутствии соли двухвалетного металла с АТФ в качестве источника фосфата, где АТФ восстанавливают in situ,
с получением соединения 5.
10. Способ по п. 9, где АТФ восстанавливают in situ с использованием (a) ацетилфосфата и ацетаткиназы, или (b) пируватоксидазы, каталазы и ацетаткиназы в присутствии пирувата, фосфата и кислорода, или (c) комбинации вышеуказанных.
11. Способ по п. 10, где (a) пантотенаткиназа является иммобилизованной или (b) пантотенаткиназа и ацетаткиназа являются иммобилизованными.
12. Способ по п. 9, дополнительно включающий стадию синтеза соединения 4, где синтез включает объединение соединения 3
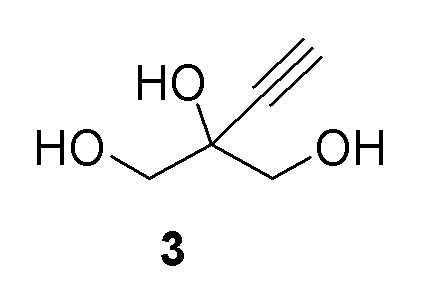
с (a)галактозооксидазой, медью, каталазой и (b)пероксидазой или окислителем;
в присутствии кислорода в буферном растворе с получением соединения 4.
13. Способ по п. 12, где галактозооксидаза является иммобилизованной.
14. Способ синтеза 4’-этинил-2’-дезоксинуклеозида, включающий объединение соединения 5
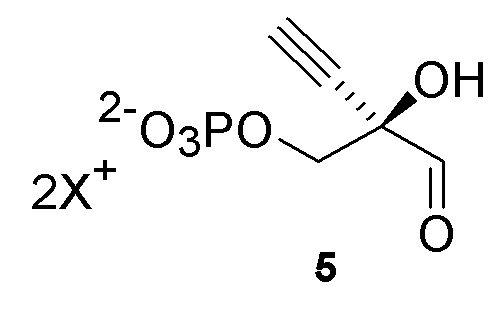 , ацетальдегида и нуклеооснования с дезоксирибозо-фосфат-альдолазой, фосфопентомутазой и пуриннуклеозид-фосфорилазой в буферном растворе, содержащем соль марганца (II),
, ацетальдегида и нуклеооснования с дезоксирибозо-фосфат-альдолазой, фосфопентомутазой и пуриннуклеозид-фосфорилазой в буферном растворе, содержащем соль марганца (II),
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион, и
где 4’-этинил-2’-дезоксинуклеозид представляет собой
 .
.
15. Способ по п. 14, дополнительно включающий удаление неорганического фосфатного побочного продукта из реакционной смеси.
16. Способ по п. 15, включающий удаление неорганического фосфатного побочного продукта из реакционной смеси путем (a) добавления сахарозо-фосфорилазы и сахарозы к реакционной смеси или (b) добавления кальция, магния или марганца к реакционной смеси.
17. Способ по любому из пп. 14-16, дополнительно включающий выделение 4’-этинил-2’-дезоксинуклеозида.
18. Способ по п. 17, дополнительно включающий выделение
 .
.
19. Способ синтеза соединения 6.5
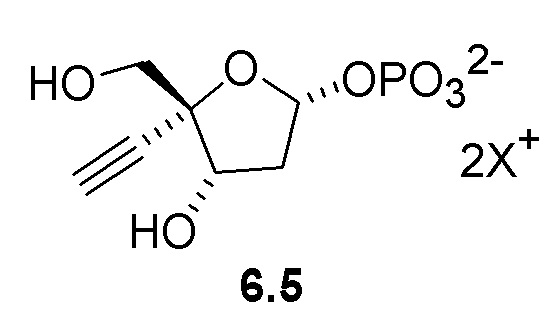 ,
,
включающий объединение соединения 6
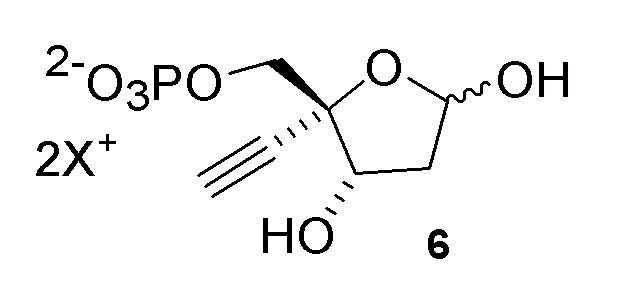
с фосфопентомутазой в буферном растворе, содержащем соль марганца (II),
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
20. Способ синтеза соединения 6
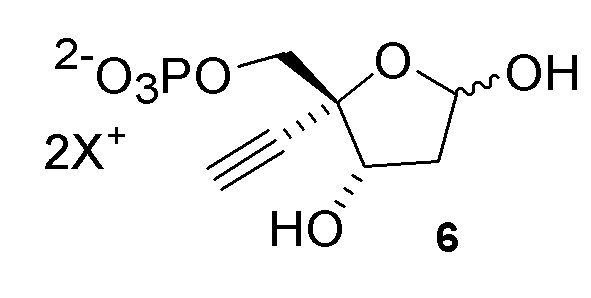 ,
,
включающий объединение соединения 5
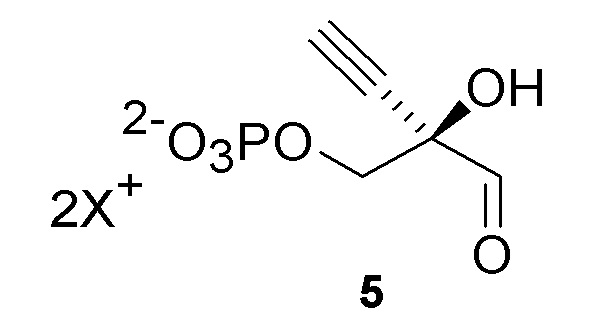
с ацетальдегидом и дезоксирибозо-фосфат-альдолазой в водном растворе с получением соединения 6;
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
21. Способ синтеза соединения 5
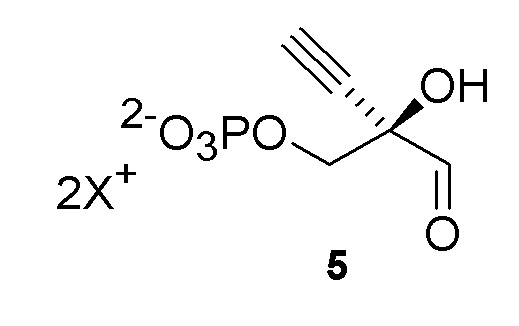 ,
,
включающий объединение соединения 4
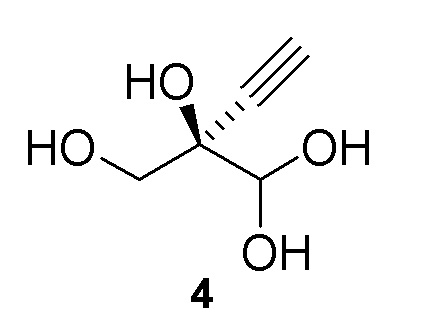
с пантотенаткиназой в буферном растворе в присутствии соли двухвалетного металла с АТФ в качестве источника фосфата, где АТФ восстанавливают in situ, и
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
22. Способ по п. 21, где АТФ восстанавливают in situ с использованием (a) ацетилфосфата и ацетаткиназы, или (b) пируватоксидазы, каталазы и ацетаткиназы в присутствии пирувата, фосфата и кислорода, или (c) комбинации вышеуказанных.
23. Способ по п. 22, где (a) пантотенаткиназа является иммобилизованной или (b) пантотенаткиназа и ацетаткиназа являются иммобилизованными.
24. Способ синтеза соединения 4
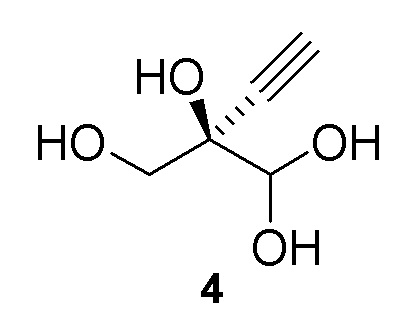 ,
,
включающий объединение соединения 3
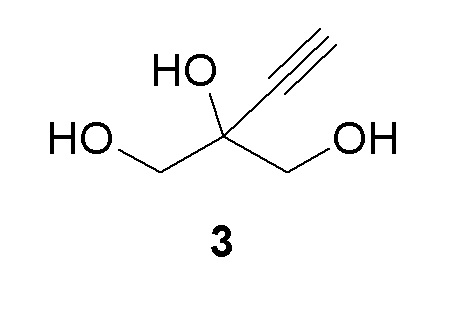
с (а) галактозооксидазой, медью, каталазой и (b) пероксидазой или окислителем;
в присутствии кислорода, в буферном растворе с получением соединения 4.
25. Способ по п. 24, где галактозооксидаза является иммобилизованной.
26. Способ выделения соединения 4
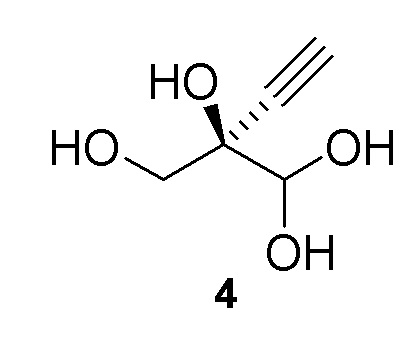 ,
,
включающий
(1) взаимодействие соединения 4 с амином, диамином или аминоспиртом, который образует стабильный N, N-ацеталь или N, O-ацеталь, в органическом растворителе, который не смешивается с водой,
в отсутствие кислорода с образованием аминаля; и
(2) взаимодействие аминаля с органической или неорганической кислотой в присутствии не смешиваемого с водой органического растворителя для восстановления соединения 4.
27. Способ синтеза соединения 5
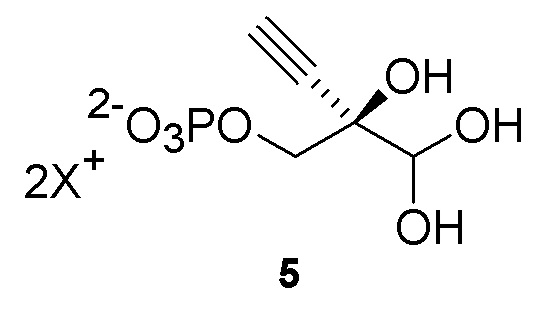 ,
,
включающий объединение соединения 9
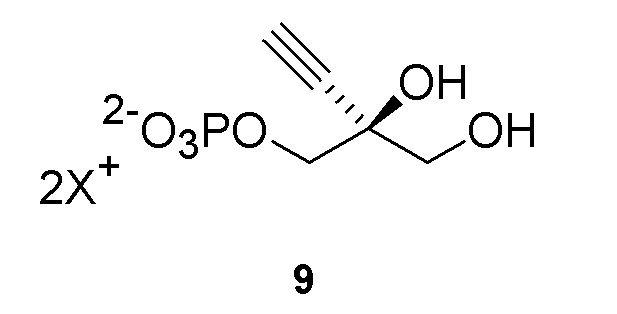
с галактозооксидазой в буферном растворе в присутствии кислорода, каталазы и либо пероксидазы, либо химического окислителя,
с получением соединения 5,
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый указанный катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
28. Способ синтеза соединения 9
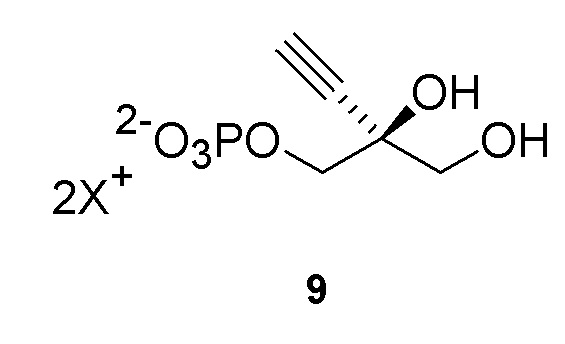 ,
,
включающий объединение соединения 3
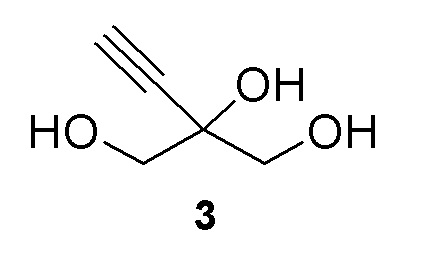
с пантотенаткиназой в буферном растворе в присутствии соли двухвалетного металла с АТФ в качестве источника фосфата, где АТФ восстанавливают in situ,
с получением соединения (9),
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый указанный катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
29. Соединение
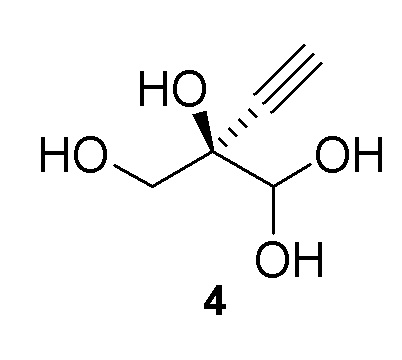 .
.
30. Соединение
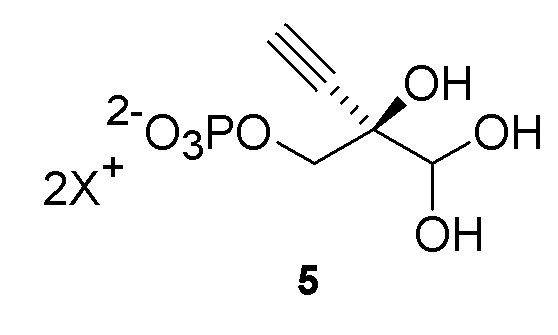 ,
,
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый указанный катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
31. Соединение
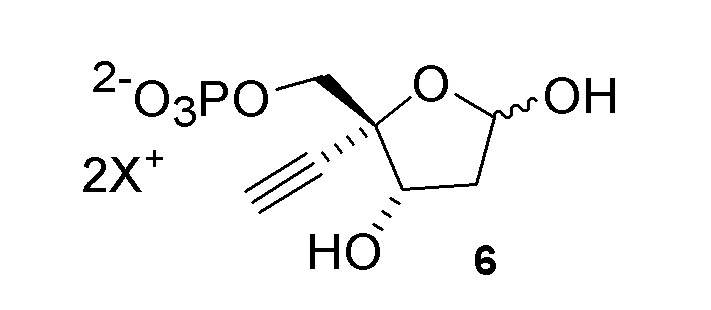 ,
,
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый указанный катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
32. Соединение
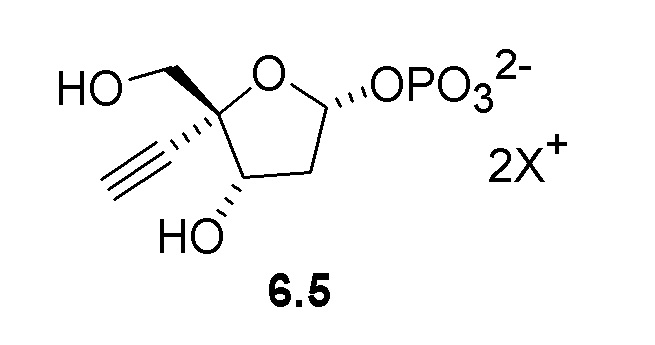 ,
,
где 2X+ представляет собой (a) два протона, (b) один протон и один одновалентный катион, (c) два одновалентных катиона, где каждый указанный катион является одинаковым или они отличны друг от друга, или (d) один двухвалентный катион.
| KAGEYAMA | |||
| et al | |||
| Concise Synthesis of the Anti-HIV Nucleoside EFdA | |||
| Bioscience, Biotechnology, and Biochemistry, 2012, vol.76, pp | |||
| Комнатная печь | 1917 |
|
SU1219A1 |
| McLaughlin M | |||
| et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Organic Letters, 2017, vol.19(4), pp | |||
| ЭЛЕКТРИЧЕСКИЙ ТРЕХФАЗНЫЙ КАБЕЛЬ | 1924 |
|
SU926A1 |
| Способ и промежуточные соединения для получения прегабалина | 2014 |
|
RU2628298C2 |
| US | |||

