Данная заявка имеет приоритет заявки США Сер. №60/497627 от 25 августа 2003 г., полное содержание которой включено в данное описание путем ссылки.
Настоящее изобретение относится к новым питьевым композициям, содержащим монатин, а также к способам получения таких композиций. Настоящее изобретение также относится к композициям напитков, содержащим определенные стереоизомеры монатина, определенные смеси стереоизомеров монатина и/или монатин, полученный путем биосинтеза in vivo (например, в клетках) или in vitro.
Применение некалорийных высокоинтенсивных подсластителей увеличивается вследствие беспокойства, вызываемого такими заболеваниями, как детское ожирение, диабет типа II и родственные заболевания. Следовательно, существует потребность в подсластителях, которые имеют значительно более высокую сладость, чем традиционные подсластители, такие как гранулированный сахар (сахароза). Многие высокоинтенсивные подсластители имеют неприятный посторонний привкус и/или неожиданные и менее желательные вкусовые профили сладости. В промышленности проводятся многочисленные исследования по ингибиторам горечи, методам маскировки неприятных привкусов и смесям подсластителей, направленные на преодоление указанных проблем и достижение вкусовой характеристики сладости, подобной вкусовой характеристике сладости сахарозы.
Монатин (2-гидрокси-2-(индол-3-илметил)-4-аминоглутаровая кислота) представляет собой природный, высокоинтенсивный подсластитель, выделенный из растения Sclerochiton ilicifolius, обнаруженного в области Трансвааль Южной Африки. Монатин не содержит углевода или сахара и почти не содержит калорий в отличие от сахарозы или других пищевых подсластителей при равном уровне сладости.
Настоящее изобретение относится к композициям напитков, содержащим монатин (2-гидрокси-2-(индол-3-илметил)-4-аминоглутаровая кислота, также известная как 4-амино-2-гидрокси-2-(1H-индол-3-илметил)пентандиовая кислота, или, по альтернативной системе нумерации, 4-гидрокси-4-(3-индолилметил)глутаминовая кислота), соединение, имеющее формулу:
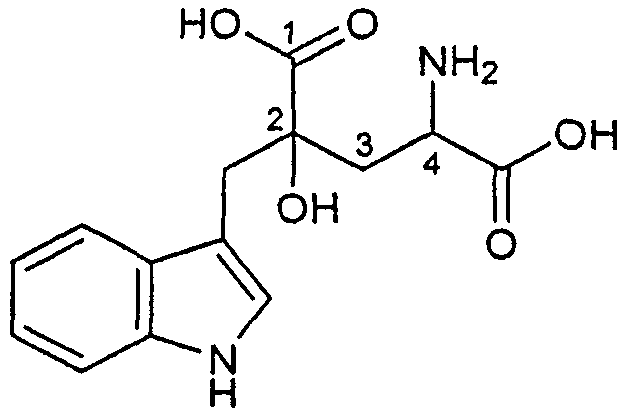
Монатин представляет собой природный высокоинтенсивный подсластитель. Монатин имеет четыре стереоизомерные формы: 2R,4R ("R,R-стереоизомер" или "R,R-монатин"), 2S,4S ("S,S-стереоизомер" или "S,S-монатин"), 2R,4S ("R,S-стереоизомер" или "R,S-монатин") и 2S,4R ("S,R-стереоизомер" или "S,R-монатин"). В данном описании, если не указано иначе, термин "монатин" относится ко всем четырем стереоизомерам монатина, а также к любым смесям любых сочетаний стереоизомеров монатина (например, к смеси R,R- и S,S,-стереоизомеров монатина).
Монатин обладает сладостью превосходного качества. Вкусовой профиль монатина является таким же чистым или даже чище, чем вкусовые профили других известных высокоинтенсивных подсластителей. Полученная для монатина кривая "доза-ответ" является более линейной и, следовательно, она более близка к соответствующей кривой сахарозы, чем кривые других высокоинтенсивных подсластителей, таких как сахарин. Превосходный вкусовой профиль сладости монатина делает его пригодным для применения в столовых подсластителях, пищевых продуктах, напитках и других продуктах.
В производстве подсластителей можно использовать разные стереоизомеры монатина, в том числе R,R- и S,S-стереоизомеры, как в виде отдельных ингредиентов, так и в виде смесей. Монатин имеет желательный вкусовой профиль сам по себе или в смеси с углеводами. Предполагается, что монатин и смеси стереоизомеров монатина с другими подсластителями, такими как углеводы, обладают превосходными вкусовыми профилями и/или физическими свойствами по сравнению с другими высокоинтенсивными подсластителями. Например, монатин более стабилен, чем аспартам (также известный, как "APM"), имеет более выраженный вкус, чем сахарин, а один стереоизомер (R,R-монатин) является более сладким, чем сукралоза. Подобным образом, монатиновые подсластители не имеют горького послевкусия, присущего сахарину, или металлического, кислого, вяжущего или раздражающего горло послевкусия некоторых других высокоэффективных подсластителей. Кроме того, монатиновые подсластители не оставляют лакричного послевкусия, свойственного некоторым природным подсластителям, таким как стевиозид и глицирризин.
Кроме того, в отличие от аспартамовых подсластителей, монатиновые подсластители не требуют ограничения по фенилаланину у пациентов, страдающих фенилкетонурией. Также предполагается, что монатин не относится к кариесогенным веществам (т.е. не вызывает разрушения зубов), поскольку он не содержит поддающихся ферментации углеводов. Кроме того, результаты тестирования рН показали, что при смешивании со слюной монатин не вызывает снижения рН ниже 5,7 (которое может быть вредным для зубов). Из-за интенсивного сладкого вкуса R,R-стереоизомер является особенно конкурентоспособным с экономической точки зрения по сравнению с другими высокоинтенсивными подсластителями.
Один объект настоящего изобретения касается питьевой композиции, содержащей монатин или его соль, например, R,R-, S,S-, R,S- или S,R-монатин, или смесь разных стереоизомеров. В данном описании термин "питьевая композиция" относится к композиции, которая пригодна для питья как есть (т.е. не требует разбавления или "готова к употреблению"), или представляет собой концентрат, который можно разбавить или смешать с пригодной для питья жидкостью с получением готового к употреблению напитка. Например, питьевая композиция может представлять собой сухую смесь для получения напитка (например, смесь для получения шоколадного напитка, смесь для получения фруктового напитка, смесь для получения солодового напитка или лимонадная смесь), которую можно смешать, например, с водой или молоком с получением готового к употреблению напитка. Другим примером питьевой композиции может служить сироп, который можно разбавить, например, газированной водой с получением газированного безалкогольного напитка. В другом примере сироп или смесь можно разбавить водой со льдом и добавить один или несколько других ингредиентов (например, текилу) с получением алкогольного напитка (например, "маргариты"). Как указано в данном описании, монатином можно заменять другие традиционные подсластители-наполнители без существенного изменения вкуса. Газированные напитки, содержащие монатин, имеют улучшенные вкусовые профили по сравнению с газированными безалкогольными напитками типа кола, подслащенными аспартамом. В условиях кислого безалкогольного напитка монатин более стабилен, чем аспартам, и предполагается, что монатин имеет более длительный срок годности. В данном описании термин "газированный" означает, что напиток содержит как растворенный, так и диспергированный диоксид углерода.
В некоторых вариантах питьевые композиции содержат смесь монатина и подсластителя (например, сахарозы или высокофруктозного кукурузного сиропа). В других вариантах питьевые композиции, содержащие монатин, включают ароматизатор, кофеин и/или подсластитель-наполнитель. Подсластителями-наполнителями могут быть, например, подсластители, содержащие сахар, подсластители, не содержащие сахар, и низкогликемические углеводы (т.е. углеводы с более низким гликемическим индексом, чем у глюкозы). В других вариантах монатин-содержащие питьевые композиции включают высокоинтенсивный подсластитель и/или низкогликемический углевод. В следующих вариантах монатин-содержащие питьевые композиции включают усилители сладкого вкуса.
В некоторых вариантах питьевые композиции содержат монатин, который в основном состоит из S,S- или R,R-монатина. В других вариантах композиции содержат преимущественно S,S- или R,R-монатин. Термин "преимущественно" означает, что среди стереоизомеров монатина доля конкретного стереоизомера составляет более 90%. В некоторых вариантах композиции практически не содержат S,S- или R,R-монатина. Термин "практически не содержит" означает, что среди стереоизомеров монатина, присутствующих в композиции, доля конкретного стереоизомера составляет менее 2%. В добавление к вышесказанному, или альтернативно, термин "практически не содержит" в применении к монатину, получаемому биосинтетическим способом, включает количество стереоизомера (например, S,S-монатина), являющегося побочным продуктом биосинтетического способа, в котором участвуют стереоспецифичные ферменты (например, дегидрогеназы D-аминокислот или аминотрансферазы D-аминокислот) и/или стереоспецифичные субстраты (например, у которых атом углерода имеет R-конфигурацию) и образуется другой стереоизомер (например, R,R-монатин).
Другой объект настоящего изобретения касается питьевой композиции, содержащей обогащенную конкретным изомером смесь монатинов, полученную биосинтетическим способом. Термин "обогащенная конкретным стереоизомером смесь монатинов" означает, что смесь содержит более одного стереоизомера монатина и по меньшей мере 60% стереоизомеров монатина в смеси составляет конкретный изомер, например, R,R, S,S, S,R или R,S. В других вариантах смесь содержит более 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99% конкретного стереоизомера монатина. В другом варианте питьевая композиция содержит обогащенный стереоизомером R,R- или S,S-монатин. Термин "обогащенный стереоизомером" R,R-монатин означает, что монатин содержит по меньшей мере 60% R,R-монатина. Термин "обогащенный стереоизомером" S,S-монатин означает, что монатин содержит по меньшей мере 60% S,S-монатина. В других вариантах "обогащенный стереоизомером" монатин содержит более 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99% R,R- или S,S-монатина.
Монатин можно выделить из коры корней растения Sclerochiton ilicifolius. Например, кору можно измельчить, экстрагировать водой, отфильтровать и высушить из замороженного состояния с получением темно-коричневой аморфной массы. Массу можно снова растворить в воде и подвергнуть взаимодействию с катионообменной смолой в кислой форме, например, "Biorad" AG50W ×8 в HCl-форме (Bio-Rad Laboratories, Richmond, CA). Смолу можно промыть водой и связанные со смолой соединения элюировать водным раствором аммиака. Элюат можно высушить из замороженного состояния и подвергнуть водной гель-фильтрации. См., например, патент США №5128164. Альтернативно, монатин можно синтезировать химически. См., например, способы, описанные в Holzapfel and Olivier, Synth. Commun. 23: 2511 (1993); Holzapfel et al., Synth. Commun. 38: 7025 (1994); патенте США №5128164; патенте США №4975298; и патенте США №5994559. Монатин также можно получить рекомбинантными способами.
В одном из аспектов настоящее изобретение предлагает способ получения питьевой композиции, содержащей монатин. Данный способ предусматривает биосинтетическое получение монатина либо in vivo, либо in vitro. "Биосинтетический путь" предусматривает по меньшей мере одну стадию биологического превращения. В некоторых вариантах биосинтетический путь представляет собой многостадийный процесс, в котором по меньшей мере одна стадия представляет собой стадию биологического превращения. В других вариантах биосинтетический путь представляет собой многостадийный процесс, включающий стадии и биологического, и химического превращения. В некоторых вариантах получаемый монатин представляет собой обогащенную конкретным стереоизомером смесь монатинов.
В другом аспекте настоящего изобретения предоставляется питьевая композиция, содержащая биосинтетический монатин. Хотя монатин можно синтезировать химически, при применении в напитках биосинтетический монатин может иметь преимущество по сравнению с монатином, синтезированным химическими методами, поскольку монатин, синтезированный химическими методами, может содержать нежелательные примеси.
В другом аспекте настоящего изобретения предлагается несколько биосинтетических способов получения монатина из субстратов, выбранных из глюкозы, триптофана, индол-3-молочной кислоты, а также индол-3-пирувата и 2-гидрокси 2-(индол-3-илметил)-4-кетоглутаровой кислоты (также известной как "предшественник монатина", "MP", или альфа-кетоформа монатина). Примеры биосинтетических способов производства или получения монатина или его промежуточных соединений иллюстрируются на фиг.1-3 и 11-13, причем промежуточные и конечные продукты представлены в рамках. В данных способах происходит превращение одного продукта в другой, например, глюкозы в триптофан, триптофана в индол-3-пируват, индол-3-пирувата в MP, MP в монатин или индол-3-молочной кислоты (индоллактата) в индол-3-пируват.
Указанные превращения в биосинтетических путях можно осуществлять химическими и/или биологическими способами. Термин "превращать" относится к изменению первого промежуточного соединения с получением второго промежуточного соединения с использованием любых химических средств или по меньшей мере одного полипептида. Превращения могут происходить in vivo или in vitro. Термин "химическое превращение" относится к реакции, в которой полипептиды не принимают активного участия. Термин "биологическое превращение" относится к реакции, в которой активно участвуют один или несколько полипептидов. Для проведения биологических превращений полипептиды и/или клетки можно иммобилизовать на подложке, например, путем химического присоединения к полимерной подложке. Превращение можно проводить в любом реакторе, известном рядовому специалисту в данной области, например, в реакторе периодического действия или в реакторе непрерывного действия.
Примеры полипептидов и последовательностей, кодирующих данные полипептиды, которые могут использоваться для проведения биологических превращений, представлены на фиг.1-3 и 11-13. Для получения монатина можно использовать полипептиды, несущие одну или несколько точечных мутаций, которые изменяют субстратную специфичность и/или активность полипептидов. Для получения монатина можно использовать выделенные рекомбинантные клетки, экспрессирующие такие полипептиды. Такими клетками могут быть любые клетки, например, растительные, животные, бактериальные, дрожжевые клетки, клетки водорослей, архебактерий или грибов.
Например, монатин-продуцирующие клетки могут содержать один или несколько (например, два или более, или три или более) из нижеследующих ферментов: триптофан-аминотрансфераза (EC 2.6.1.27), тирозин (ароматический)-аминотрансфераза (EC 2.6.1.5), триптофан-дегидрогеназа (EC 1.4.1.19), глутамат-дегидрогеназа (EC 1.4.1.2, 1.4.1.3, 1.4.1.4), фенилаланин-дегидрогеназа (EC 1.4.1.20), триптофанфенилпируват-трансаминаза (EC 2.6.1.28), аминотрансфераза, использующая разные субстраты (EC 2.6.1.-), аспартат-аминотрансфераза (EC 2.6.1.1), оксидаза L-аминокислот (EC 1.4.3.2), триптофан-оксидаза (номер EC отсутствует, Hadar et al., J Bacteriol 125: 1096-1104, 1976, и Furuya et al., Biosci Biotechnol Biochem 64: 1486-93, 2000), D-триптофан-аминотрансфераза (Kohiba and Mito, Proceedings of the 8th International Symposium on Vitamin B6 and Carbonyl Catalysis, Osaka, Japan 1990), дегидрогеназа D-аминокислот (EC 1.4.99.1), оксидаза D-аминокислот (EC 1.4.3.3), D-аланин-аминотрансфераза (EC 2.6.1.21), синтаза/лиаза (EC 4.1.3.-), такая как 4-гидрокси-2-оксоглутарат-альдолаза (EC 4.1.3.16) или 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (EC 4.1.3.17) и/или синтаза/лиаза (4.1.2.-).
В другом примере клетки могут содержать один или несколько (например, два или более, или три или более) из нижеследующих ферментов: индоллактат-дегидрогеназа (EC 1.1.1.110), R-4-гидроксифениллактат-дегидрогеназа (EC 1.1.1.222), 3-(4)-гидроксифенилпируват-редуктаза (EC 1.1.1.237), лактат-дегидрогеназа (EC 1.1.1.27, 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил)лактат-дегидрогеназа (EC 1.1.1.111), лактат-оксидаза (EC 1.1.3.-), синтаза/лиаза (4.1.3.-), такая как 4-гидрокси-2-оксоглутарат-альдолаза (EC 4.1.3.16) или 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (EC 4.1.3.17), синтаза/лиаза (4.1.2.-), триптофан-аминотрансфераза (EC 2.6.1.27), тирозин (ароматический)-аминотрансфераза (EC 2.6.1.5), триптофан-дегидрогеназа (EC 1.4.1. 19), глутамат-дегидрогеназа (EC 1.4.1.2, 1.4.1.3, 1.4.1.4), фенилаланин-дегидрогеназа (EC 1.4.1.20), триптофанфенилпируват-трансаминаза (EC 2.6.1.28), аминотрансфераза, использующая разные субстраты (EC 2.6.1.-), аспартат-аминотрансфераза (EC 2.6.1.1), D-триптофан-аминотрансфераза, дегидрогеназа D-аминокислот (EC 1.4.99.1) и/или D-аланин-аминотрансфераза (EC 2.6.1.21).
Кроме того, клетки могут содержать один или несколько (например, два или более, или три или более) из нижеследующих ферментов: триптофан-аминотрансфераза (EC 2.6.1.27), тирозин (ароматический)-аминотрансфераза (EC 2.6.1.5), триптофан-дегидрогеназа (EC 1.4.1.19), глутамат-дегидрогеназа (EC 1.4.1.2, 1.4.1.3, 1.4.1.4), фенилаланин-дегидрогеназа (EC 1.4.1.20), триптофанфенилпируват-трансаминаза (EC 2.6.1.28), аминотрансфераза, использующая разные субстраты (EC 2.6.1.-), аспартат-аминотрансфераза (EC 2.6.1.1), оксидаза L-аминокислот (EC 1.4.3.2), триптофан-оксидаза, D-триптофан-аминотрансфераза, дегидрогеназа D-аминокислот (EC 1.4.99.1), оксидаза D-аминокислот (EC 1.4.3.3), D-аланин-аминотрансфераза (EC 2.6.1.21), индоллактат-дегидрогеназа (EC 1.1.1.110), R-4-гидроксифениллактат-дегидрогеназа (EC 1.1.1.222), 3-(4)-гидроксифенилпируват-редуктаза (EC 1.1.1.237), лактат-дегидрогеназа (EC 1.1.1.27, 1.1.1.28, 1.1.2.3), (3-имидазол-5-ил)лактат-дегидрогеназа (EC 1.1.1.111), лактат-оксидаза (EC 1.1.3.-), синтаза/лиаза (EC 4.1.3.-), такая как 4-гидрокси-2-оксоглутарат-альдолаза (EC 4.1.3.16) или 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (EC 4.1.3.17) и/или синтаза/лиаза (4.1.2.-).
В другом примере клетки могут содержать одну или несколько из нижеследующих альдолаз: KHG-альдолаза, ProA-альдолаза, KDPG-альдолаза и/или родственные полипептиды (KDPH), транскарбоксибензальпируват-гидратаза-альдолаза, 4-(2-карбоксифенил)-2-оксобут-3-еноат-альдолаза, транс-O-гидроксибензилиденпируват-гидратаза-альдолаза, 3-гидроксиаспартат-альдолаза, бензоин-альдолаза, дигидронеоптерин-альдолаза, L-трео-3-фенилсерин-бензальдегид-лиаза (фенилсерин-альдолаза), 4-гидрокси-2-оксовалерат-альдолаза, 1,2-дигидроксибензилпируват-альдолаза и/или 2-гидроксибензальпируват-альдолаза.
Монатин можно получить способами, которые включают контактирование триптофана и/или индол-3-молочной кислоты с первым полипептидом, причем первый полипептид превращает триптофан и/или индол-3-молочную кислоту в индол-3-пируват (в качестве субстрата, превращаемого в индол-3-пируват, можно использовать как D-, так и L-форму триптофана или индол-3-молочной кислоты; специалистам в данной области известно, что для данной стадии следует выбирать полипептиды, обладающие соответствующей специфичностью), контактирование полученного индол-3-пирувата со вторым полипептидом, причем второй полипептид превращает индол-3-пируват в 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту (MP), и контактирование MP с третьим полипептидом, причем третий полипептид превращает MP в монатин. Примеры полипептидов, которые можно использовать для данных превращений, представлены на фиг.2 и 3.
Получение монатина биосинтетическим способом, включающим одно или несколько биологических превращений, имеет определенные преимущества. Например, используя в биосинтетических способах конкретные полипептиды и/или определенные субстраты, можно получить смесь, обогащенную конкретным стереоизомером, и/или получить смесь монатинов, практически не содержащую одного или нескольких стереоизомеров.
Монатиновая композиция может содержать примеси, природа которых зависит от используемого способа синтеза монатина. Монатиновые композиции, получаемые исключительно синтетическими способами (т.е. не включающими ни одного биологического превращения), содержат примеси, отличающиеся от примесей, содержащихся в монатиновых композициях, получаемых биосинтетическим способом. Например, в зависимости от используемых сырьевых веществ монатиновые композиции, получаемые исключительно синтетическими способами, могут содержать нефтехимические, токсичные и/или другие вредные примеси, не подходящие для потребления человеком. Примерами таких примесей являются вредные химические вещества, такие как LDA, водород-Pd/C, диазометан, KCN, реактив Гриньяра и Na/Hg. С другой стороны, монатиновая композиция, получаемая биосинтетическим способом, может содержать пищевые или питьевые примеси, но не содержит нефтехимических, токсичных и/или других вредных примесей.
Предположительно, биосинтетический способ получения монатина, включающий одно или несколько биологических превращений, дает меньше токсичных или вредных примесей и/или позволяет получать более высокий процент конкретного стереоизомера, по сравнению с исключительно синтетическим способом. Например, при получении монатина с использованием дегидрогеназ D-аминокислот, D-аланин(аспартат)-аминотрансфераз, D-ароматических аминотрансфераз или D-метионин-аминотрансфераз можно получить по меньшей мере 60% R,R-монатина и менее 40% S,S-, S,R- и/или R,S-монатина. Также можно ожидать, например, что при получении монатина с использованием вышеуказанных D-ферментов, а также по меньшей мере одного субстрата (например, предшественника монатина), содержащего атом углерода в R-стереоконфигурации, можно получить по меньшей мере 95% R,R-монатина и менее 5% S,S-, S,R- и/или R,S-монатина. Наоборот, при получении монатина исключительно синтетическими способами можно получить 25%-50% целевого стереоизомера.
В одном варианте биосинтетический способ получения монатина, например, включающий одно или несколько биологических превращений, позволяет избежать образования нефтехимических, токсичных или вредных примесей. Термин "нефтехимические, токсичные или вредные примеси" относится к любому нефтехимическому, токсичному, вредному и/или иному веществу, не подходящему для потребления человеком, в том числе, к примесям, представляющим собой сырьевое вещество или образовавшимся в процессе получения монатина исключительно синтетическими способами. В другом варианте при биосинтетическом способе получения монатина, например, включающем одно или несколько биологических превращений, образуются только пищевые или питьевые вещества. Термин "пищевое или питьевое вещество" относится к одному или нескольким соединениям или веществам, пригодным для еды или питья, или для иного безопасного потребления человеком. Примеры пищевого или питьевого вещества включают монатин, триптофан, пируват, глутамат, другие аминокислоты, а также другие природные соединения или вещества, присутствующие в организме.
В одном варианте питьевая композиция, включающая монатин или его соль, при сравнимом уровне сладости содержит меньше калорий и углеводов, чем такое же количество питьевой композиции, содержащей вместо монатина или его соли сахарозу или высокофруктозный кукурузный сироп.Термин "сравнимая сладость" или "сравнимый уровень сладости" означает, что эксперт по органолептической оценке определяет, что в среднем сладость первой композиции составляет от 80% до 120% от сладости второй композиции.
В других вариантах питьевая композиция, включающая монатин или его соль, дополнительно содержит цитрусовый ароматизатор, причем монатин или его соль присутствуют в количестве, усиливающем аромат цитрусового ароматизатора. В другом варианте питьевая композиция также содержит цитрусовый аромат и углевод, причем монатин или его соль и углевод присутствуют в количестве, усиливающем аромат цитрусового ароматизатора. Углевод, без ограничения, можно выбрать из эритрита, мальтодекстрина, сахарозы и их комбинации.
В одном варианте газированный напиток содержит композицию сиропа в количестве, составляющем приблизительно от 15% до 25% от веса газированного напитка, причем композиция сиропа содержит монатин или его соль.
В другом варианте питьевая композиция содержит приблизительно от 3 до 10000 м.д. монатина или его соли. В других вариантах питьевая композиция содержит приблизительно от 3 до 30 м.д. монатина, или от более чем 2500 до приблизительно 10000 м.д. монатина. В другом варианте питьевая композиция представляет собой сироп или сухую смесь для приготовления напитка, причем композиция содержит приблизительно от 10 до 10000 м.д. монатина или его соли. Например, питьевая композиция может быть сиропом, представляющим собой концентрат, который можно разбавлять напитком в соотношении приблизительно от 1 части сиропа: 3 части напитка до 1 части сиропа: 5,5 частей напитка. В одном варианте сироп содержит приблизительно от 600 до 10000 м.д. S,S-монатина или его соли. В другом варианте сироп содержит приблизительно от 18 до 300 м.д. R,R-монатина или его соли. Альтернативно, сироп содержит приблизительно от 0 до 10000 м.д. S,S-монатина или его соли и от 0 до приблизительно 300 м.д. R,R-монатина или его соли.
В другом варианте питьевая композиция представляет собой сухую смесь для приготовления напитка, содержащую приблизительно от 10 до 10000 м.д. монатина или его соли. В одном варианте сухая смесь для приготовления напитка содержит приблизительно от 600 до 10000 м.д. S,S монатина или его соли. В другом варианте сухая смесь для приготовления напитка содержит приблизительно от 10 до 450 м.д. R,R монатина или его соли. Альтернативно, сухая смесь для приготовления напитка содержит приблизительно от 0 до 10000 м.д. S,S-монатина или его соли и приблизительно от 0 до 450 м.д. R,R монатина или его соли.
В других вариантах питьевая композиция содержит приблизительно от 3 до 10000 м.д. монатина или его соли и практически не содержит R,R-монатина или его соли, или практически не содержит S,S-монатина или его соли. В другом варианте питьевая композиция содержит приблизительно от 3 до 450 м.д. R,R-монатина или его соли (например, приблизительно от 6 до 225 м.д. R,R-монатина или его соли). В другом варианте питьевая композиция содержит приблизительно от 3 до 10000 м.д. S,S-монатина или его соли (например, приблизительно от 60 до 4600 м.д. S,S-монатина или его соли). В другом варианте питьевая композиция содержит приблизительно от 0 до 10000 м.д. S,S-монатина или его соли и приблизительно от 0 до 450 м.д. R,R-монатина или его соли.
В одном варианте питьевая композиция представляет собой готовую к употреблению композицию, содержащую приблизительно от 3 до 2000 м.д. монатина или его соли. В других вариантах готовая к употреблению композиция содержит приблизительно от 5 до 50 м.д. R,R-монатина или его соли, или приблизительно от 60 до 2000 м.д. S,S-монатина или его соли.
В другом варианте питьевая композиция содержит приблизительно 450 или менее м.д. R,R-монатина или его соли и практически не содержит S,S-, S,R- или R,S-монатина или его соли. Альтернативно, питьевая композиция содержит приблизительно 10000 или менее м.д. S,S-монатина или его соли и практически не содержит R,R-, S,R- или R,S-монатина или его соли. В некоторых вариантах монатин или его соль в питьевой композиции в основном состоит из R,R-монатина или его соли, или в основном состоит из S,S-монатина или его соли. В других вариантах монатин или его соль в питьевой композиции представляет собой смесь, обогащенную R,R-монатином или его солью, или представляет собой смесь, обогащенную S,S-монатином или его солью. В других вариантах монатин или его соль в питьевой композиции содержит по меньшей мере 95% R,R-монатина или его соли, или по меньшей мере 95% S,S-монатина или его соли.
В одном варианте питьевая композиция содержит монатин или его соль, полученные биосинтетическим способом. В другом варианте питьевая композиция содержит смесь монатинов, обогащенную конкретным изомером, причем смесь монатинов получают биосинтетическим способом. В одном варианте биосинтетический способ представляет собой многостадийный способ, в котором по меньшей мере одна стадия представляет собой химическое превращение. В других вариантах смесь монатинов, полученная биосинтетическим способом, преимущественно состоит из R,R-монатина или его соли, или она преимущественно состоит из S,S-монатина или его соли.
В одном варианте питьевая композиция содержит монатиновую композицию, полученную биосинтетическим способом, причем монатиновая композиция не содержит нефтехимических, токсичных или вредных примесей. В другом варианте питьевая композиция содержит монатин или его соль, причем монатин или его соль получают биосинтетическим способом и выделяют из рекомбинантной клетки, причем рекомбинантная клетка не содержит нефтехимических, токсичных или вредных примесей.
В одном варианте питьевая композиция, содержащая монатин или его соль, является не кариесогенной. В других вариантах питьевая композиция, содержащая монатин или его соль, дополнительно содержит эритрит, трегалозу, цикламат, D-тагатозу или их комбинацию.
В других вариантах питьевая композиция, содержащая монатин или его соль, дополнительно содержит подсластитель-наполнитель, высокоинтенсивный подсластитель, углевод с низким гликемическим индексом, ароматизатор, антиоксидант, кофеин, усилитель сладости или их комбинацию. Например, ароматизатор можно выбрать из ароматизатора кола, цитрусового ароматизатора и их комбинации. Например, подсластитель-наполнитель можно выбрать из кукурузных подсластителей, сахарозы, декстрозы, инвертного сахара, мальтозы, декстрина, мальтодекстрина, фруктозы, левулозы, высокофруктозного кукурузного сиропа, сухого кукурузного сиропа, левулозы, галактозы, трегалозы, изомальтулозы, фрукто-олигосахаридов и их комбинаций. Например, высокоинтенсивный подсластитель можно выбрать из сукралозы, аспартама, сахарина, ацесульфама K, алитама, тауматина, дигидрохалконов, неотама, цикламатов, стевиозида, могрозида, глицирризина, филлодульцина, монеллина, мабинлина, бразеина, циркулина, пентадина и их комбинаций. Например, углевод с низким гликемическим индексом можно выбрать из D-тагатозы, сорбита, маннита, ксилита, лактита, эритрита, мальтита, гидрированных гидролизатов крахмала, изомальта, D-псикозы, 1,5-ангидро-D-фруктозы и их комбинаций. Например, усилитель сладости можно выбрать из куркулина, миракулина, цинарина, хлорогеновой кислоты, кофейной кислоты, строгинов, арабиногалактана, мальтола, дигидроксибензойных кислот и их комбинаций.
В другом варианте питьевая композиция содержит монатин, который представляет собой смесь R,R- и S,S-изомеров, или его соль. Кроме того, питьевая композиция может содержать смесь монатина или его соли и отличного от монатина подсластителя. Отличный от монатина подсластитель можно выбрать, например, из сахарозы и высокофруктозного кукурузного сиропа.
В некоторых вариантах способы получения питьевой композиции, содержащей монатин или его соль, включают получение монатина или его соли по меньшей мере из одного субстрата, выбранного из глюкозы, триптофана, индол-3-молочной кислоты, индол-3-пирувата и предшественника монатина. Данные способы также могут включать объединение монатина или его соли по меньшей мере с одним другим ингредиентом, отличным от монатина или его соли (например, с эритритом, трегалозой, цикламатом, D-тагатозой, мальтодекстрином или их комбинацией). В некоторых вариантах другой ингредиент можно выбрать, например, из наполнителей, подсластителей-наполнителей, жидких подсластителей, углеводов с низким гликемическим индексом, высокоинтенсивных подсластителей, загустителей, жиров, масел, эмульгаторов, антиоксидантов, усилителей сладости, красителей, ароматизаторов, кофеина, кислот, порошков, средств, повышающих текучесть, буферов, белковых источников, усилителей вкуса, стабилизаторов вкуса и их комбинаций. Подсластители-наполнители можно выбрать, например, из содержащих сахар подсластителей, не содержащих сахар подсластителей, углеводов с низким гликемическим индексом и их комбинаций. В других вариантах питьевые композиции, полученные с помощью указанных способов, содержат приблизительно от 0 до 10000 м.д. S,S-монатина или его соли и приблизительно от 0 до 450 м.д. R,R-монатина или его соли.
В других вариантах способы получения питьевой композиции, содержащей монатин или его соль, включают получение монатина или его соли биосинтетическим способом. В некоторых вариантах способы получения питьевой композиции, содержащей монатин или его соль, включают получение монатина или его соли с использованием по меньшей мере одного биологического превращения, или с использованием только биологических превращений. В другом варианте способ получения питьевой композиции, содержащей монатиновую композицию, предусматривает: (a) получение монатина или его соли биосинтетическим способом в рекомбинантной клетке; (b) выделение монатиновой композиции из рекомбинантной клетки, причем монатиновая композиция состоит из монатина или его соли и другого пищевого или питьевого вещества.
В других вариантах способ получения питьевой композиции, содержащей монатиновую композицию, предусматривает получение монатиновой композиции биосинтетическим способом, причем монатиновая композиция не содержит нефтехимических, токсичных или вредных примесей. В других вариантах способ получения питьевой композиции, содержащей монатиновую композицию, предусматривает получение монатиновой композиции из субстрата многостадийным способом, причем одна или несколько стадий многостадийного способа представляют собой биологическое превращение, причем монатиновая композиция не содержит нефтехимических, токсичных или вредных примесей.
В другом варианте способ получения питьевой композиции, содержащей монатиновую композицию, предусматривает получение монатиновой композиции биосинтетическим способом, причем монатиновая композиция состоит из монатина или его соли и другого пищевого или питьевого вещества. В другом варианте способ получения питьевой композиции, содержащей монатиновую композицию, предусматривает получение монатиновой композиции из субстрата многостадийным способом, причем одна или несколько стадий многостадийного способа представляют собой биологическое превращение, причем монатиновая композиция состоит из монатина или его соли и другого пищевого или питьевого вещества.
Если не указано иначе, все технические и научные термины, используемые в данном описании, имеют значения, традиционно используемые специалистами в области, к которой относится данное изобретение. Для осуществления или испытания настоящего изобретения можно использовать способы и вещества, подобные или эквивалентные способам, описанным ниже в данном изобретении. Все публикации, патентные заявки, патенты и другие ссылки, упоминающиеся в данном описании, включены в него путем ссылки во всей их полноте. В случае конфликта настоящее описание, включая определения, будет проверяться. Кроме того, вещества, способы и примеры являются только иллюстративными и не предназначаются для ограничения.
Из представленных здесь объяснений специалисту в данной области должно быть понятно, что конкретные варианты настоящего изобретения могут быть направлены на один или несколько из указанных выше аспектов, а также на другие аспекты. Другие признаки и преимущества данного изобретения станут понятны из нижеследующего подробного описания.
КРАТКОЕ ОПИСАНИЕ ИЛЛЮСТРАЦИЙ
На фиг.1 показаны биосинтетические способы получения монатина и/или индол-3-пирувата. В одном способе индол-3-пируват получают из триптофана, а в другом способе индол-3-пируват получают из индол-3-молочной кислоты. Затем через промежуточное соединение MP получают монатин.
Представленные в рамках соединения представляют собой субстраты и продукты, получаемые биосинтетическими способами. Рядом со стрелками указаны кофакторы или реагенты, используемые в процессе превращения субстрата в продукт. Тип используемого кофактора или реагента зависит от типа полипептида, используемого на конкретной стадии биосинтетического способа. Кофактор PLP (пиридоксаль 5'-фосфат) может катализировать реакции независимо от полипептида, и, следовательно, лишь участие PLP может обеспечить превращение субстрата в продукт.
На фиг.2 представлена более подробная схема биосинтетического способа, в котором используется промежуточное соединение MP. Субстраты, используемые на каждой стадии данного способа, представлены в рамках. Полипептиды, осуществляющие превращение субстратов, указаны рядом со стрелками, находящимися между субстратами. Для каждого фермента представлено тривиальное название и номер в соответствии с классификацией ферментов (EC).
На фиг.3 представлена более подробная схема биосинтетического способа превращения индол-3-молочной кислоты в индол-3-пируват. Субстраты представлены в рамках, а полипептиды, осуществляющие превращение субстратов, указаны рядом со стрелками, находящимися между субстратами. Для каждого фермента представлено тривиальное название и номер EC.
На фиг.4 представлена химическая реакция, с помощью которой можно получить MP.
На фиг.5A и 5B представлены хроматограммы, демонстрирующие ЖХ/МС идентификацию монатина, полученного ферментативным способом.
На фиг.6 представлен анализ полученного ферментативным способом монатина методом масс-спектрометрии с электрораспылением.
На фиг.7A и 7B представлены хроматограммы анализа полученного ферментативным способом монатина методом ЖХ/МС/МС дочерних ионов.
На фиг.8 представлена хроматограмма высокоразрешающего измерения массы полученного ферментативным способом монатина.
На фиг.9A-9C представлены хроматограммы хирального разделения (A) R-триптофана, (B) S-триптофана и (C) полученного ферментативным способом монатина.
На фиг.10 представлена гистограмма, демонстрирующая относительное количество монатина, полученного в бактериальных клетках после индукции IPTG. (-) Указывает на отсутствие субстрата (триптофан или пируват не добавляются).
На фиг.11-12 представлены схематические диаграммы, демонстрирующие способы, использующиеся для увеличения выхода монатина, получаемого из триптофана или индол-3-пирувата.
На фиг.13 представлена схематическая диаграмма, демонстрирующая способ, который может использоваться для увеличения выхода монатина, получаемого из триптофана или индол-3-пирувата.
На фиг.14 представлена кривая зависимости "доза-эффект", полученная для R,R-стереоизомера монатина.
На фиг.15 представлена кривая зависимости "доза-эффект", полученная для смеси R,R/S,S-стереоизомеров монатина.
На фиг.16 проводится сравнение кривой зависимости "доза-эффект", полученной для смеси R,R/S,S-стереоизомеров монатина, с кривой дозовой зависимости, полученной для сахарина.
На фиг.17 представлены результаты хроматографии на обращенной фазе стандартных образцов синтетического монатина.
На фиг.18 представлены результаты хиральной хроматографии стандартных образцов монатина.
ПОДРОБНОЕ ОПИСАНИЕ
Обзор биосинтетических способов получения монатина
Нижеследующие определения терминов и методов представлены для того, чтобы лучше разъяснить настоящее изобретение, а также в качестве руководства по реализации настоящего изобретения для специалистов в данной области. В данном описании термин "включающий" означает "содержащий". Кроме того, если в контексте не указано иначе, единственное число включает также и множественное число.
Термин "приблизительно" охватывает диапазон экспериментальной ошибки, которая встречается во всех экспериментах. Если не указано иначе, предполагается, что все экспериментальные данные представлены в приблизительном значении, даже если перед ними не употребляется слово "приблизительно". Термин "% мас./об." или "% вес./об." означает отношение массы к объему, где 100% вес./об. составляет 1 г/мл. Так, например, 1 г/100 мл соответствует 1% вес./об. (в жидких композициях). Термин "м.д." относится к частям на миллион. Восемьдесят м.д. монатина, например, соответствует 80 граммам (г) монатина в миллионе граммов. Подобным образом, 1 м.д.=0,0001% вес./вес.или, в водных растворах,=1 мг/л=1 мкг/мл=0,0001% вес./об.
Как показано на фиг.1-3 и 11-13, многие биосинтетические способы можно использовать для получения монатина или его промежуточных соединений, таких как индол-3-пируват или MP. Для превращения конкретного субстрата (например, глюкозы, триптофана, индол-3-молочной кислоты, индол-3-пирувата и MP) в продукт (например, триптофан, индол-3-пируват, MP и монатин), можно использовать несколько разных полипептидов. Более того, данные реакции можно проводить in vivo, in vitro или можно использовать сочетание реакций in vivo и in vitro, таких как in vitro реакции, которые включают неферментативные химические реакции. Таким образом, на фиг.1-3 и 11-13 представлено несколько разных способов получения целевых продуктов.
Превращение глюкозы в триптофан
Многие организмы могут синтезировать триптофан из глюкозы. В таких организмах можно клонировать конструкт(ы), содержащие ген(ы), необходимый для получения монатина, MP и/или индол-3-пирувата из глюкозы и/или триптофана. В данном документе показано, что триптофан можно превратить в монатин.
В других примерах с использованием известных полипептидов можно создать организм, осуществляющий продукцию или сверхпродукцию триптофана. Например, в патенте США №4371614 описан штамм E.coli, трансформированный плазмидой, содержащей оперон триптофана дикого типа.
Максимальный титр триптофана в патенте США №4371614 составляет приблизительно 230 м.д. Подобным образом, в WO 8701130 описывается полученный методом генной инженерии штамм E.coli, продуцирующий триптофан, и обсуждается повышенная продукция L-триптофана. Специалистам в данной области известно, что организмы, способные синтезировать триптофан из глюкозы, также могут утилизировать другие углеродные и энергетические источники, превращая их в глюкозу или фруктозу-6-фосфат с подобным результатом. Примеры углеродных и энергетических источников включают, без ограничения, сахарозу, фруктозу, крахмал, целлюлозу или глицерин.
Превращение триптофана в индол-3-пируват
Для превращения триптофана в индол-3-пируват можно использовать разные полипептиды. Примеры полипептидов включают, без ограничения, ферменты классов (EC) 2.6.1.27, 1.4.1.19, 1.4.99.1, 2.6.1.28, 1.4.3.2, 1.4.3.3, 2.6.1.5, 2.6.1.-, 2.6.1.1 и 2.6.1.21. Данные классы включают, без ограничения, такие полипептиды, как триптофан-аминотрансфераза (также называемая L-фенилаланин-2-оксоглутарат-аминотрансфераза, триптофан-трансаминаза, 5-гидрокситриптофан-кетоглутаровой кислоты трансаминаза, гидрокситриптофан-аминотрансфераза, L-триптофан-аминотрансфераза, L-триптофан-трансаминаза и L-триптофан:2-оксоглутарат-аминотрансфераза), которая превращает L-триптофан и 2-оксоглутарат в индол-3-пируват и L-глутамат; D-триптофан-аминотрансфераза, которая превращает D-триптофан и 2-оксокислоту в индол-3-пируват и аминокислоту; триптофан-дегидрогеназа (также называемая NAD(P)-L-триптофан-дегидрогеназа, L-триптофан-дегидрогеназа, L-Trp-дегидрогеназа, TDH и L-триптофан:NAD(P)-оксидоредуктаза (дезаминирующая)), которая превращает L-триптофан и NAD(P) в индол-3-пируват, NH3 и NAD(P)H; дегидрогеназа D-аминокислот, которая превращает D-аминокислоты и FAD в индол-3-пируват, NH3 и FADH2; триптофанфенилпируват-трансаминаза (также называемая L-триптофан-α-кетоизокапроат-аминотрансфераза и L-триптофан:фенилпируват-аминотрансфераза), которая превращает L-триптофан и фенилпируват в индол-3-пируват и L-фенилаланин; оксидаза L-аминокислот (также называемая оксидаза офио-аминокислот и L-аминокислоты:кислород-оксидоредуктаза (дезаминирующая)), которая превращает L-аминокислоты, H2О и O2 в 2-оксокислоты, NH3 и H2О2; оксидаза D-аминокислот (также называемая оксидаза офио-аминокислот и D-аминокислоты:кислород-оксидоредуктаза (дезаминирующая)), которая превращает D-аминокислоты, H2О и O2 в 2-оксокислоты, NH3 и H2О2; и триптофан-оксидаза, которая превращает L-триптофан, H2О и О2 в индол-3-пируват, NH3 и H2О2. Данные классы также включают тирозин (ароматический)-аминотрансферазу, аспартат-аминотрансферазу, аминотрансферазу D-аминокислот (или D-аланина) и аминотрансферазу широкого спектра действия (имеющую несколько субстратов), которая обладает несколькими аминотрансферазными активностями, некоторые из них могут превращать триптофан и 2-оксокислоту в индол-3-пируват и аминокислоту.
Одиннадцать членов класса аминотрансфераз, обладающих такой активностью, описаны ниже в примере 1, в том числе, новая аминотрансфераза с последовательностями SEQ ID NO: 11 и 12. Таким образом, в данном описании представлены нуклеотидная и аминокислотная последовательности, идентичные по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98% или даже по меньшей мере на 99% последовательностям, описанным в SEQ ID NO: 11 и 12, соответственно. Данное описание также охватывает фрагменты и гибриды последовательностей, описанных в SEQ ID NO: 11 и 12, которые кодируют полипептид, обладающий аминотрансферазной активностью или сохраняющий аминотрансферазную активность. Примеры фрагментов включают, не ограничиваясь ими, по меньшей мере 10, 12, 15, 20, 25, 50, 100, 200, 500 или 1000 последовательных нуклеотидов SEQ ID NO: 11 или по меньшей мере 6, 10, 15, 20, 25, 50, 75, 100, 200, 300 или 350 последовательных аминокислот SEQ ID NO: 12. Описанные последовательности (а также их варианты, фрагменты и гибриды) могут составлять часть вектора. Такой вектор можно использовать для трансформации клеток-хозяев с получением рекомбинантных клеток, которые могут синтезировать индол-3-пируват из триптофана, и в некоторых примерах также могут продуцировать MP и/или монатин.
Оксидазы L-аминокислот (1.4.3.2) известны, их последовательности можно выделить из нескольких разных источников, таких как Vipera lebetine (sp P81375), Ophiophagus hannah (sp P81383), Agkistrodon rhodostoma (spP81382), Crotalus atrox (sp P56742), Burkholderia cepacia, Arabidopsis thaliana, Caulobacter cresentus, Chlamydomonas reinhardtii, Mus musculus, Pseudomonas syringae и Rhodococcus str. Кроме того, в литературе описаны оксидазы триптофана, которые можно выделить, например, из Coprinus sp.SF-1, китайской капусты с клубеньковой болезнью корней, Arabidopsis thaliana и печени млекопитающих. В представленном ниже примере 3 описан один член класса оксидаз L-аминокислот, которые могут превращать триптофан в индол-3-пируват, а также альтернативные источники для молекулярного клонирования. Для молекулярного клонирования можно использовать гены многочисленных оксидаз D-аминокислот, имеющиеся в базах данных.
Триптофан-дегидрогеназы известны, их можно выделить, например, из шпината, Pisum sativum, Prosopis juliflora, гороха, мескитового дерева, пшеницы, маиса, томатов, табака, Chromobacterium violaceum и Lactobacilli. Известны последовательности генов многих дегидрогеназ D-аминокислот.
Как показано на фиг.11-13, если для превращения триптофана в индол-3-пируват используется оксидаза аминокислот, например, оксидаза триптофана, для уменьшения уровня пероксида водорода или устранения его можно добавить каталазу.
Превращение индол-3-лактата в индол-3-пируват
Реакция конверсии индол-3-лактата в индол-3-пируват может катализироваться рядом полипептидов, например, полипептидами классов 1.1.1.110, 1.1.1.27, 1.1.1.28, 1.1.2.3, 1.1.1.222, 1.1.1.237, 1.1.3.- или 1.1.1.111. Класс полипептидов 1.1.1.110 включает индоллактат-дегидрогеназы (также называемые оксидоредуктазы индолмолочной кислоты: NAD+). Классы 1.1.1.27, 1.1.1.28 и 1.1.2.3 включают лактат-дегидрогеназы (также называемые дегидрогеназы молочной кислоты, лактат:NAD+-оксидоредуктаза). Класс 1.1.1.222 включает (R)-4-гидроксифениллактат-дегидрогеназу (также называемую D-ароматическая лактат-дегидрогеназа, R-ароматическая лактат-дегидрогеназа и R-3-(4-гидроксифенил)лактат:NAD(P)+-2-оксидоредуктаза), а класс 1.1.1.237 включает 3-(4-гидроксифенилпируват)редуктазу (также называемую гидроксифенилпируват-редуктаза и 4-гидроксифениллактат:NAD+-оксидоредуктаза). Класс 1.1.3.- включает лактат-оксидазы, а класс 1.1.1.111 включает (3-имидазол-5-ил)лактат-дегидрогеназы (также называемые (S)-3-(имидазол-5-ил)лактат:NAD(P)+-оксидоредуктазы). Вероятно, некоторые полипептиды данных классов катализируют превращение индол-3-молочной кислоты в индол-3-пируват. Примеры осуществления данного превращения представлены в примере 2.
Для превращения индол-3-молочной кислоты в индол-3-пируват можно также использовать химические реакции. Такие химические реакции включают стадию окисления, которую можно проводить несколькими способами, например: путем окисления воздухом с использованием катализатора B2 (China Chemical Reporter, vol. 13, no. 28, pg. 18(1), 2002), путем разбавления перманганатом и перхлоратом, или пероксидом водорода, в присутствии металлических катализаторов.
Превращение индол-3-пирувата в 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту (MP)
Для превращения индол-3-пирувата в MP можно использовать несколько известных полипептидов. Примеры классов полипептидов включают 4.1.3.-, 4.1.3.16, 4.1.3.17 и 4.1.2.-. Данные классы включают углерод-углеродные синтазы/лиазы, такие как альдолазы, катализирующие конденсацию двух карбоновых кислот. Класс полипептидов EC 4.1.3.- включает синтазы/лиазы, которые образуют углерод-углеродные связи, используя в качестве электрофилов оксо-кислоты (например, индол-3-пируват), тогда как EC 4.1.2.- включает синтазы/лиазы, которые образуют углерод-углеродные связи, используя в качестве электрофилов альдегиды (такие как бензальдегид).
Например, полипептид, описанный в EP 1045-029 (EC 4.1.3.16, 4-гидрокси-2-оксоглутаратглиоксилат-лиаза, также называемая 4-гидрокси-2-оксоглутарат-альдолаза, 2-оксо-4-гидроксиглутарат-альдолаза или KHG-альдолаза), превращает глиоксиловую кислоту и пируват в 4-гидрокси-2-кетоглутаровую кислоту, а полипептид 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (EC 4.1.3.17, также называемый 4-гидрокси-4-метил-2-оксоглутаратпируват-лиаза или ProA-альдолаза), катализирует конденсацию двух кето-кислот, например, двух пируватов, с образованием 4-гидрокси-4-метил-2-оксоглутарата. Реакции, проводимые с использованием указанных лиаз, описаны в данном документе.
На фиг.1-2 и 11-13 представлены схематические диаграммы данных реакций, в которых 3-углеродную (C3) молекулу соединяют с индол-3-пируватом. Многие члены EC 4.1.2.- и 4.1.3.-, особенно полипептиды, использующие PLP, в качестве C3-молекул могут использовать аминокислоты, такие как серин, цистеин и аланин, или их производные. В альдольных конденсациях, катализируемых членами классов EC 4.1.2.- и 4.1.3.-, трехуглеродная молекула должна представлять собой пируват или производное пирувата. Однако в качестве С3-источника можно использовать другие соединения, которые могут быть превращены в пируват. Пируват можно получить путем трансаминирования аланина под действием многих трансаминаз, использующих PLP, в том числе, многих из указанных выше. Пируват и аммиак можно получить путем бета-элиминирования (например, катализируемых триптофаназой или β-тирозиназой) L-серина, L-цистеина и производных серина и цистеина, имеющих подходящие уходящие группы, такие как О-метил-L-серин, О-бензил-L-серин, S-метилцистеин, S-бензилцистеин, S-алкил-L-цистеин, О-ацил-L-серин и 3-хлор-L-аланин. Аспартат может служить источником пирувата в PLP-опосредованных бета-лиазных реакциях, например, катализируемых триптофаназой (EC 4.1.99.1) и/или β-тирозиназой (EC 4.1.99.2, также называемой тирозинфенол-лиаза). Скорость бета-лиазных реакций можно увеличить путем сайт-направленного мутагенеза (4.1.99.1-2) полипептидов, как описано в Mouratou et al. (J. Biol. Chem 274: 1320-5, 1999) и в примере 8. Данные модификации придают полипетидам способность акцептировать в качестве субстратов дикарбоновые аминокислоты. Лактат также может служить источником пирувата, его окисление до пирувата проводят путем добавления лактат-дегидрогеназы и окисленного кофактора, или лактат-оксидазы и кислорода. Примеры данных реакций описаны ниже. Например, как показано на фиг.2 и фиг.11-13, ProA-альдолазу можно привести в контакт с индол-3-пируватом, если в качестве С3-молекулы используется пируват.
MP также можно получить с помощью химических реакций, таких как альдольные конденсации, представленные в примере 5.
Превращение MP в монатин
Превращение MP в монатин можно катализировать одним или несколькими из следующих полипептидов: триптофан-аминотрансфераза (2.6.1.27), триптофан-дегидрогеназа (1.4.1.19), дегидрогеназа D-аминокислот (1.4.99.1), глутамат-дегидрогеназа (1.4.1.2-4), фенилаланин-дегидрогеназа (EC 1.4.1.20), триптофан-фенилпируват-трансаминаза (2.6.1.28), или чаще членами семейства аминотрансфераз (2.6.1.-) такими как аспартат-аминотрансфераза (EC 2.6.1.1), тирозин (ароматический)-аминотрансфераза (2.6.1.5), D-триптофан-аминотрансфераза или D-аланин (2.6.1.21)-аминотрансфераза (фиг.2). Одиннадцать членов класса аминотрансфераз описаны ниже (пример 1), в том числе, новый член класса с последовательностями SEQ ID NO: 11 и 12, а реакции, демонстрирующие активность ферментов аминотрансфераз и дегидрогеназ, представлены в примере 7.
Данное превращение также можно проводить посредством химических реакций. Аминирование кетокислоты (MP) проводят путем восстановительного аминирования с использованием аммиака и цианоборгидрида натрия.
На фиг.11-13 показаны другие полипептиды, которые могут превращать MP в монатин, а также способствуют увеличению выхода при получении монатина из индол-3-пирувата или триптофана. Например, если в качестве донора аминогруппы используется аспартат, для превращения аспартата в оксалоацетат можно использовать аспартат-аминотрансферазу (фиг.11). Превращение оксалоацетата в пируват и диоксид углерода катализируется декарбоксилазой, такой как оксалоацетат-декарбоксилаза (фиг.11). Кроме того, если в качестве донора аминогруппы используется лизин, для превращения лизина в аллизин можно использовать лизин-эпсилон-аминотрансферазу (фиг.12). Аллизин спонтанно превращается в 1-пиперидин-6-карбоксилат (фиг.12). Если для превращения МР в монатин используется полипептид, способный катализировать реакции восстановительного аминирования (например, глутамат-дегидрогеназа), можно использовать полипептид, утилизирующий NAD(P)H и/или продуцирующий летучий продукт (фиг.13), например, формиат-дегидрогеназу.
Другие реакции конденсации биосинтетических способов
Чтобы увеличить выход продукта, к продуцирующим клеткам можно добавить кофакторы, субстраты и/или другие полипептиды, в зависимости от типа полипептидов, используемых для получения индол-3-пирувата, MP и/или монатина. Кроме того, чтобы увеличить выход продуктов, например, индол-3-пирувата, MP и/или монатина, можно провести генетические модификации. Можно также провести оптимизацию клетки-хозяина, используемой для получения монатина.
Удаление пероксида водорода
Пероксид водорода (H2О2) представляет собой продукт, который, в случае образования, может повреждать продуцирующие клетки, полипептиды или получаемые продукты (например, промежуточные соединения). Описанная выше оксидаза L-аминокислот генерирует H2О2 в качестве продукта. Следовательно, чтобы уменьшить вероятность повреждения клетки или продукта, при использовании оксидазы L-аминокислот образующийся H2О2 нужно удалять или снижать его уровень.
Для уменьшения уровня H2О2 в клетке можно использовать каталазы (фиг.11-13). Продуцирующая клетка может экспрессировать генную последовательность или последовательность кДНК, кодирующую каталазу (EC 1.11.1.6), которая катализирует разложение пероксида водорода на воду и газообразный кислород. Например, экспрессия каталазы может происходить из вектора, трансфицированного в продуцирующую клетку. Примеры пригодных для использования каталаз включают, без ограничения: tr|Q9EV50 (Staphylococcus xylosus), tr|Q9KBE8 (Bacillus halodurans), tr|Q9URJ7 (Candida albicans), tr|P77948 (Streptomyces coelicolor), tr|Q9RBJ5 (Xanthomonas campestris) (SwissProt Accession Nos.). Биокаталитические реакторы, в которых используется оксидаза L-аминокислот, оксидаза D-аминокислот или оксидаза триптофана, также могут содержать каталазный полипептид.
Модуляция доступности пиридоксаль-5'-фосфата (PLP)
Как показано на фиг.1, PLP можно использовать в одной или нескольких из описанных здесь биосинтетических стадий. Концентрацию PLP можно повышать так, чтобы PLP не стал фактором, ограничивающим общую эффективность реакции.
Биосинтетический путь витамина B6 (предшественника PLP) тщательно изучался в E.coli и некоторые белки были выделены в кристаллическом виде (Laber et al., FEBS Letters, 449: 45-8, 1999). Для других метаболических путей требуется два гена (epd или gapB и serС), тогда как для биосинтеза пиридоксальфосфата нужны три гена (pdxA, pdxB и pdxJ). Одним из исходных веществ пути в E.coli является 1-дезокси-D-ксилулоза-5-фосфат (DXP). Синтез данного предшественника из обычных 2- и 3-углеродных основных метаболитов катализируется полипептидом 1-дезокси-D-ксилулоза-5-фосфат-синтазой (DXS). Другим предшественником является производное треонина, полученное из 4-углеродного сахара, D-эритроза-4-фосфата. Для превращения в фосфо-4-гидроксил-L-треонин (HTP) нужны гены epd, pdxB и serC. Последняя реакция в пути синтеза PLP представляет собой сложную внутримолекулярную конденсацию и реакцию замыкания цикла между DXP и HTP, катализируемую продуктами генов pdxA и pdxJ.
Если PLP становится ограничивающим питательным ингредиентом в процессе ферментации с получением монатина, для увеличения выхода монатина можно увеличить экспрессию одного или нескольких генов пути в продуцирующей клетке-хозяине. Организм-хозяин может содержать несколько копий нативных генов пути либо в геном организма могут быть введены копии ненативных генов пути. Кроме того, в организме хозяина можно клонировать несколько копий генов "реутилизационного" пути.
В одном реутилизационном пути, консервативном для всех организмов, различные производные витамина В6 метаболизируются до активной формы PLP. В данном пути участвуют такие полипептиды, как киназа pdxK, оксидаза pdxH и киназа pdxY. Сверхэкспрессия одного или нескольких из указанных генов может увеличивать доступность PLP.
Уровни витамина B6 можно увеличить путем устранения или подавления метаболической регуляции нативных генов биосинтетического пути в организме хозяина. PLP подавляет активность полипептидов, участвующих в биосинтезе предшественника треонина, у штамма 238-7 бактерии Flavobacterium sp. Данный бактериальный штамм, у которого отсутствует метаболический контроль, осуществляет сверхпродукцию пиридоксальных производных и может экскретировать до 20 мг/л PLP. С организмом хозяина, продуцирующим монатин, можно провести генетические манипуляции, позволяющие увеличить продукцию PLP без сверхэкспрессии генов биосинтетического пути.
Использование аммиака
Реакции с участием триптофаназы можно направить на синтез (получение триптофана из индола), увеличив доступность аммиака, или путем удаления воды. Реакции восстановительного аминирования, например, катализируемые глутамат-дегидрогеназой, также могут стимулироваться избытком аммиака.
Аммиак может быть доступным в виде карбоната аммония или фосфата аммония в карбонатной или фосфатной буферной системе. Аммиак также может быть введен в систему в виде пирувата аммония или формиата аммония. Альтернативно, аммиак может появляться, если реакция сочетается с реакцией, генерирующей аммиак, такой как реакция с участием глутамат-дегидрогеназы или триптофан-дегидрогеназы. Аммиак может образоваться в результате добавления природных субстратов EC 4.1.99.- (тирозина или триптофана), которые гидролизуются до фенола или индола, пирувата и NH3. Это также приводит к увеличению выхода синтетического продукта по сравнению с нормальным равновесным количеством в результате гидролиза ферментом предпочтительного субстрата.
Удаление продуктов и побочных продуктов
При превращении триптофана в индол-3-пируват под действием триптофан-аминотрансферазы скорость образования индол-3-пирувата может снижаться, поскольку в данной реакции образуется глутамат и требуется присутствие второго субстрата 2-оксоглутарата (α-кетоглутарата). Глутамат может ингибировать аминотрансферазу, а для проведения реакции может потребоваться большое количество второго субстрата. Кроме того, высокая концентрация глутамата может оказать неблагоприятное влияние на протекание последующих процессов разделения.
Полипептид глутамат-дегидрогеназа (GLDH) превращает глутамат в 2-оксоглутарат, который используется в качестве второго субстрата в реакции, катализируемой триптофан-аминотрансферазой. GLDH также может генерировать восстанавливающие компоненты (NADH или NADPH), которые могут использоваться как источники энергии в клетке (ATP) в аэробных условиях. Использование глутамата GLDH также снижает образование побочного продукта. Кроме того, в данной реакции образуется аммиак, который может служить источником азота в клетке и субстратом в реакции восстановительного аминирования на последней стадии, представленной на фиг.1. Следовательно, продуцирующую клетку со сверхэкспрессией полипептида GLDH можно использовать для увеличения выхода и снижения стоимости среды и/или процессов выделения.
В способе синтеза монатина из триптофана донор аминогруппы третьей стадии (например, глутамат или аспартат) можно снова превратить в акцептор аминогруппы, необходимый на стадии 1 (например, 2-оксоглутарат или оксалоацетат), если используется аминотрансфераза соответствующего класса. Применение двух разных трансаминаз может увеличить эффективность данного способа, если субстрат одной трансаминазы не ингибирует конкурентным образом активность другой трансаминазы.
Многие реакции в описанных способах являются обратимыми, и, следовательно, можно достичь равновесия между субстратами и продуктами. Выход в данном способе можно увеличить путем непрерывного удаления продуктов из сферы действия полипептидов. Например, секреция монатина в ферментационную среду с использованием пермеазы или другого транспортного белка, или избирательная кристаллизация монатина из потока биокаталитического реактора, сопровождающаяся возвращением в процесс субстратов, увеличивает выход реакции.
Удаление побочных продуктов путем дополнительных ферментативных реакций или путем замены аминодонорных групп является другим способом увеличения выхода реакции. Некоторые примеры описаны в примере 13 и показаны на фиг.11-13. Например, можно получить побочный продукт, который не может участвовать в обратной реакции либо по причине фазового перехода (упаривания), либо по причине спонтанного превращения в нереакционноспособный конечный продукт, такой как диоксид углерода.
Модуляция субстратных пулов
Индольный пул можно модулировать путем увеличения продукции предшественников триптофана и/или изменения катаболических путей, включающих индол-3-пируват и/или триптофан. Например, образование индол-3-уксусной кислоты из индол-3-пирувата можно уменьшить или прекратить путем функциональной делеции гена, кодирующего EC 4.1.1.74 в клетке-хозяине. Образование индола из триптофана можно уменьшить или прекратить путем функциональной делеции гена, кодирующего EC 4.1.99.1 в клетке-хозяине. Альтернативно, избыток индола можно использовать в качестве субстрата в in vitro или in vivo процессах в сочетании с повышенным количеством гена, кодирующего EC 4.1.99.1 (Kawasaki et al., R Ferm. and Bioeng., 82: 604-6, 1996). Кроме того, уровень таких промежуточных соединений, как D-эритроза-4-фосфат и хоризмат, можно увеличить путем генетических модификаций.
Продукция триптофана регулируется в большинстве организмов. Одним из механизмов является ингибирование некоторых ферментов данного пути по типу обратной связи: по мере увеличения уровней триптофана скорость образования триптофана уменьшается. Так, если клетка-хозяин сконструирована для получения монатина через триптофан как промежуточное соединение, можно использовать организм, не чувствительный к концентрациям триптофана. Например, штамм Catharanthus roseus, устойчивый к ингибированию роста под действием разных аналогов триптофана, выбирают, подвергая его повторяющемуся воздействию высоких концентраций 5-метилтриптофана (Schallenberg and Berlin, Z Naturforsch 34: 541-5, 1979). Конечная активность триптофан-синтазы данного штамма менее подвержена ингибированию продуктом, а также она меньше зависит от мутаций в гене. Подобным образом, можно оптимизировать клетку-хозяина, используемую для получения монатина.
Продукцию триптофана можно оптимизировать путем применения направленной эволюции с отбором полипептидов, которые менее чувствительны к ингибированию продуктом. Например, скрининг можно проводить на чашках, среда которых не содержит триптофана, но содержит высокие уровни неметаболизируемых аналогов триптофана. В патентах США №5756345; 4742007 и 4,371614 описаны способы, используемые для увеличения продукции триптофана в ферментируемом организме. Последней стадией биосинтеза триптофана является добавление серина к индолу; следовательно, для увеличения продукции триптофана можно увеличить доступность серина.
Количество монатина, продуцируемого ферментируемым организмом, можно увеличить путем увеличения количества пирувата, продуцируемого организмом-хозяином. В способах, описанных в данном документе, можно использовать некоторые дрожжи, например, Trichosporon cutaneum (Wang et al., Lett. Appl. Microbiol. 35: 338-42, 2002) и Torulopsis glabrata (Li et al., Appl Microbiol. Biotechnol. 57: 451-9, 2001), которые осуществляют сверхпродукцию пирувата. Кроме того, организмы могут быть подвергнуты генетическим модификациям, целью которых является продукция пировиноградной кислоты, таким как модификации, осуществленные в штамме E.coli W14851ip2 (Kawasaki et al., J. Ferm. and Bioeng. 82: 604-6, 1996).
Регуляция хиральности
Вкусовой профиль монатина можно изменить путем регуляции его стереохимии (хиральности). Например, для разных пищевых систем могут потребоваться разные соотношения стереоизомеров монатина. Хиральность можно регулировать, варьируя рН и тип полипептидов.

Рацемизацию по положению C-4 монатина (нумерация молекулы представлена выше) можно проводить путем депротонирования и репротонирования альфа-углерода в результате изменения рН или путем взаимодействия с кофактором PLP, связанным с ферментом, таким как рацемаза, или находящимся в растворе в свободном состоянии. В микроорганизме вряд ли можно изменить рН в достаточной степени, чтобы вызвать рацемизацию, а PLP присутствует в избытке. Способы регуляции хиральности с использованием полипептидов выбирают в зависимости от биосинтетического способа, используемого для получения монатина.
Если монатин получают по способу, описанному на фиг.2, можно использовать следующий способ регуляции хиральности. В биокаталитической реакции хиральность углерода 2 можно определить с помощью фермента, превращающего индол-3-пируват в MP. Многие ферменты (например, классов EC 4.1.2.-, 4.1.3.-) могут превращать индол-3-пируват в MP, следовательно, можно выбрать фермент, который дает желательный стереоизомер. Альтернативно, энантиоспецифичность фермента, превращающего индол-3-пируват в MP, можно модифицировать путем направленной эволюции, либо можно создать каталитические антитела, катализирующие желательную реакцию. После получения MP (ферментативного или путем химической конденсации) аминогруппу можно ввести стереоспецифически с помощью трансаминазы, например, выбранной из описанных в данном документе. Может образовываться либо R-, либо S-конфигурация, в зависимости от того, используется ли аминотрансфераза D- или L-ароматических кислот. Большинство аминотрансфераз специфичны к L-стереоизомеру; однако, в некоторых растениях присутствуют D-триптофан-аминотрансферазы (Kohiba and Mito, Proceedings of the 8th International Symposium on Vitamin B6 and Carbonyl Catalysis, Osaka, Japan 1990). Кроме того, были идентифицированы D-аланин-аминотрансферазы (2.6.1.21), D-метионинпируват-аминотрансферазы (2.6.1.41) и как (R)-3-амино-2-метилпропаноат-аминотрансфераза (2.6.1.61), так и (S)-3-амино-2-метилпропаноат-аминотрансфераза (2.6.1.22). Некоторые аминотрансферазы могут только акцептировать субстрат для данной реакции с конкретной конфигурацией по углероду C2. Таким образом, даже если образование MP не является стереоспецифичным, стереохимию конечного продукта можно регулировать путем выбора соответствующей трансаминазы. Поскольку реакции являются обратимыми, непрореагировавший MP (нежелательный стереоизомер) можно снова использовать с получением рацемической смеси MP.
Активация субстратов
В описанных здесь реакциях можно использовать фосфорилированные субстраты, такие как фосфоенолпируват (PEP). Фосфорилированные субстраты могут быть энергетически выгодными, и, следовательно, их можно использовать для увеличения скорости реакции и/или выхода. В реакциях альдольной конденсации добавление фосфатной группы стабилизирует енольный таутомер нуклеофильного субстрата, увеличивая его реакционную способность. В других реакциях фосфорилированный субстрат может предоставлять более хорошую уходящую группу. Подобным образом, субстраты можно активировать путем превращения в производные CoA или производные пирофосфата.
Применение монатина в питьевой композиции
S,S-Стереоизомер монатина по весу приблизительно в 50-200 раз слаще, чем сахароза. R,R-Стереоизомер монатина по весу приблизительно в 2000-2400 раз слаще, чем сахароза. Сладость монатина рассчитывают по результатам, полученным с помощью экспертов по органолептической оценке в процедуре сравнения сладости, при которой сладость тестируемого раствора подсластителя сравнивают со сладостью одного из нескольких стандартных растворов. Растворы можно получить, например, с использованием буфера, содержащего 0,16% (вес./об.) лимонной кислоты и 0,02% (вес./об.) цитрата натрия при pH ~3,0.
Конкретно, сладость подсластителя по сравнению с сахарозой может определяться группой экспертов по органолептической оценке в процедуре оценки сладости. Все образцы (в одинаковых буферах) используют с двойными повторами при температуре 22°C±1°C. Растворы образцов можно получить, например, с использованием буфера, содержащего 0,16% (вес./об.) лимонной кислоты и 0,02% (вес./об.) цитрата натрия при pH ~3,0. Тестируемые растворы, маркированные 3-значными случайными номерными кодами, индивидуально выдают экспертам по органолептической оценке в случайном порядке. Также выдаются стандартные растворы сахарозы с концентрацией в интервале 2,0-10,0% (вес./об.) с шагом 0,5% (вес./об.). Экспертов просят оценить сладость путем сравнения сладости тестируемого раствора со сладостью стандартных растворов сахарозы. Чтобы оценить сладость, делают последовательно 3 глотка тестируемого раствора, глоток воды, 3 глотка стандартного раствора сахарозы, глоток воды и т.д. Эксперты оценивают сладость до первого десятичного знака, например, 6,8, 8,5. Разные тестируемые растворы оценивают с перерывом в пять минут.Экспертов также просят хорошо сполоснуть рот и съесть крекер, чтобы уменьшить любые возможные эффекты наложения вкуса.
Строят график зависимости значения сахарозного эквивалента (SEV) (например, % сахарозы), определяемого группой экспертов по органолептической оценке, от концентрации монатина и получают кривую зависимости "доза-эффект". К кривой зависимости "доза-эффект" применяют полиномиальную подгонку кривой и для конкретной точки, например, 8% SEV, рассчитывают интенсивность или мощность сладости, например, путем деления значения сахарозного эквивалента (SEV) на концентрацию монатина (например, % монатин). См., например, фиг.15 (кривая зависимости "доза-эффект" R,R/S,S-монатина); фиг.14 (кривая зависимости "доза-эффект" R,R-монатина). Вышеуказанная интенсивность сладости S,S- и R,R-монатина (т.е. приблизительно в 50-200 раз слаще и приблизительно в 2000-2400 раз слаще по весу, чем сахароза, соответственно) определена приблизительно при 8% SEV.
Монатин растворяется в водных растворах в концентрациях, подходящих для употребления. Различные смеси стереоизомеров монатина могут обладать более хорошим качеством в определенных растворах или в смеси с другими подсластителями. Смеси монатина с другими подсластителями можно использовать для достижения максимальной интенсивности вкуса, и/или максимально благоприятной вкусовой характеристики, а также для минимизации стоимости. Монатин можно использовать в сочетании с другими подсластителями и/или другими ингредиентами для получения временной вкусовой характеристики, подобной вкусовой характеристике сахарозы, или для других целей.
Например, монатин можно смешивать с другими пищевыми и непищевыми подсластителями для получения конкретных вкусовых характеристик или определенного количества калорий. Так, композиции подсластителя могут содержать сочетания монатина с одним или несколькими из подсластителей следующих типов: (1) сахарных спиртов (таких как эритрит, сорбит, мальтит, маннит, лактит, ксилит, изомальт, сиропы с низким гликемическим индексом и т.д.); (2) других высокоинтенсивных подсластителей (таких как аспартам, сукралоза, сахарин, ацесульфам-K, стевиозид, цикламат, неотам, тауматин, алитам, дигидрохалкон, монеллин, глицирризин, могрозид, филлодульцин, мабинлин, бразеин, циркулин, пентадин и т.д.) и (3) пищевых подсластителей (таких как сахароза, D-тагатоза, инвертный сахар, фруктоза, кукурузный сироп, высокофруктозный кукурузный сироп (HFCS), глюкоза/декстроза, трегалоза, изомальтулоза и т.д.). Монатин можно использовать в таких смесях, как модификатор вкуса для подавления послевкусия, усиления других ароматов, таких как лимон, или улучшения временной вкусовой характеристики. Данные также указывают, что монатин проявляет качественный синергизм с цикламатами (которые не используются в Европе), но никакого заметного качественного синергизма не было отмечено с аспартамом, сахарином, ацесульфамом-K, сукралозой или углеводными подсластителями.
Поскольку монатин не является углеводом, его можно использовать для снижения содержания углеводов в композициях напитков. В одном варианте некоторое количество питьевой композиции, содержащей монатин, содержит меньше калорий и углеводов, чем такое же количество питьевой композиции, содержащей сахар (например, сахарозу и/или высокофруктозный кукурузный сироп) вместо монатина. В других вариантах питьевые композиции, содержащие монатин (например, содержащие монатин и один или несколько углеводов), оставляют с течением времени ощущение во рту, вкус и сладость, сравнимые с аналогичными характеристиками подобных композиций напитков, содержащих в качестве подсластителей только углеводы.
Монатин является стабильным в сухом виде и обладает желательным вкусовым профилем сам по себе или в смеси с углеводами. Он не подвергается необратимому распаду, но при низких значениях рН иногда образует лактоны и/или лактамы (в водных буферах), которые находятся в равновесии с основной формой. С течением времени он может медленно подвергаться рацемизации по 4 положению, однако, чаще всего это происходит при высоких значениях pH. Как правило, стабильность монатина сравнима со стабильностью аспартама или превосходит ее, а вкусовой профиль монатина сравним с вкусовым профилем качественно других подсластителей, таких как аспартам, алитам и сукралоза, или превосходит его. Монатин не имеет нежелательного послевкусия, присущего некоторым другим высокоинтенсивным подсластителям, таким как сахарин и стевиозид.
В некоторых вариантах питьевые композиции, содержащие монатин, также содержат один или несколько из следующих ингредиентов: буферы, наполнители, загустители, жиры, ароматизаторы, окрашивающие средства (также называемые красителями или пигментами), подсластители и средства, повышающие текучесть. Питьевые композиции с определенным профилем сладости можно получить, например, путем варьирования в напитке количества монатина или других подсластителей, или путем варьирования в композиции количества или типа других добавок, в том числе ароматизаторов или кислот. В других вариантах все ингредиенты, присутствующие в композициях напитков, имеют качество пищевых марок и, как правило, являются безопасными.
В некоторых вариантах питьевые композиции, содержащие монатин, также содержат антиоксиданты пищевых марок. Примеры таких антиоксидантов включают витамин C (например, аскорбиновую кислоту, аскорбилфосфат магния), эриторбат (изоаскорбиновую кислоту), каротиноиды, такие как лютеин, ликопен и бета-каротин, токоферолы (например, α-токоферол (природный витамин E), γ-токоферол, δ-токоферол), гидроксициннаматы (например, неохлорогеновая кислота и хлорогеновая кислота), глутатион, фенольные смолы (например, фенолы какао, фенолы красного вина, фенольные смолы чернослива), бутилированный гидроксианизол (BHA), бутилированный гидрокситолуол (BHT), четвертичный бутилгидрохинон (TBHQ), пропилгаллат, низин, экстракт зеленого чая и экстракт розмарина. В других вариантах питьевые композиции, содержащие монатин, также содержат известные консерванты, такие как бензоат натрия и/или сорбат калия.
В других вариантах питьевые композиции, содержащие монатин, также содержат один или несколько ингредиентов, предотвращающих неферментативные реакции, приводящие к потемнению (например, потемнению в результате реакций Майяра). Такие ингредиенты могут включать, не ограничиваясь ими, сульфиты и сульфитирующие агенты (например, диоксид серы, сульфит натрия, бисульфит калия или натрия, метабисульфиты, сульфгидрил-содержащие аминокислоты), хлорид кальция и другие неорганические галогениды, антиоксиданты и соединения, которые воздействуют на активность воды (такие как глицерин, сорбит и трегалоза).
В некоторых вариантах монатин-содержащие концентрации напитков, такие как сухая смесь для приготовления напитков, можно легко диспергировать с получением шоколадных напитков, фруктовых напитков, солодовых напитков или лимонада. В других вариантах концентрат напитка представляет собой сироп напитка, который можно использовать для получения газированных безалкогольных напитков. Газированный напиток можно приготовить, например, путем разбавления сиропа напитка, содержащего воду, монатин и ароматизаторы, газированной водой. В некоторых вариантах сироп напитка также содержит другие подсластители и/или добавки. Сиропы напитков можно получить, например, путем смешивания всех ингредиентов с последующим нагреванием до их растворения. Сиропы напитков могут содержать, например, по меньшей мере 80% воды (например, по меньшей мере 85%, 90% или 95% воды).
В некоторых вариантах монатин присутствует в количестве, составляющем приблизительно от 0,0003 до 1% от веса питьевой композиции (т.е. приблизительно от 3 до 10000 м.д.) (например, приблизительно от 0,0005 до 0,2%), включая любое конкретное значение в указанном интервале (например, 0,0003%, 0,005%, 0,06% или 0,2% от веса питьевой композиции). Например, питьевая композиция может содержать от 0,0005 до 0,005% (например, от 0,001 до 0,0045%) R,R-монатина, или от 0,005 до 0,2% (например, от 0,01 до 0,175%) S,S-монатина.
Специалисту в данной области известно, что для получения питьевой композиции с желательным вкусом и подходящим количеством калорий можно использовать сочетание подсластителей. Так, количество подсластителя в питьевой композиции зависит от выбора подсластителей и желаемой интенсивности сладости. Подсластители можно получить, например, от Cargill Inc. (Wayzata, MN) и McNeil Specialty (Fort Washington, PA). В одном варианте питьевая композиция включает смесь монатина и подсластителя (например, сахарозы или высокофруктозного кукурузного сиропа). Например, питьевая композиция может включать монатин и подсластитель-наполнитель.
Подсластители-наполнители можно выбрать, например, из подсластителей, содержащих сахар, подсластителей, не содержащих сахар, углеводов с низким гликемическим индексом и их комбинаций. Подсластители, содержащие сахар, могут включать, например, кукурузный подсластитель, сахарозу, декстрозу (например, декстрозу Cerelose), мальтозу, декстрин, мальтодекстрин, инвертный сахар, фруктозу, высокофруктозный кукурузный сироп, левулозу, галактозу, сухой кукурузный сироп, галактозу, трегалозу, изомальтулозу, фруктоолигосахариды (такие как кестоза или нистоза), высокомолекулярные фруктоолигосахариды или их комбинации. Высокофруктозный кукурузный сироп (HFCS) и другие кукурузные подсластители представляют собой, например, сочетания декстрозы (глюкозы) и фруктозы. Кроме того, сахарсодержащие подсластители включают фруктовые сахара, кленовый сироп и мед или их комбинации. В одном варианте питьевая композиция может содержать от 0,0003 до 0,15% монатина (например, от 0,0006 до 0,004% R,R-монатина) и от 2 до 10% (например, от 3 до 10% или от 4 до 6%) сахарозы или высокофруктозного кукурузного сиропа.
В другом варианте питьевая композиция содержит не содержащий сахара подсластитель и/или углевод с низким гликемическим индексом (т.е. гликемический индекс углевода ниже, чем у глюкозы). Не содержащие сахара подсластители или углеводы с низким гликемическим индексом включают, не ограничиваясь ими, D-тагатозу, сорбит (в том числе, аморфный и кристаллический сорбит), маннит, ксилит, лактит, эритрит, мальтит, гидрированные гидролизаты крахмала, изомальт, D-псикозу, 1,5-ангидро-D-фруктозу или их комбинации.
В некоторых вариантах питьевые композиции, содержащие монатин, также содержат высокоинтенсивные подсластители. В некоторых вариантах высокоинтенсивные подсластители по меньшей мере в 20 раз слаще, чем сахароза (т.е. 20X сахарозы). Такие высокоинтенсивные подсластители включают, не ограничиваясь ими, сукралозу, аспартам, сахарин и его соли, соли ацесульфама (например, ацесульфама K), алитам, тауматин, дигидрохалконы (например, неогесперидин дигидрохалкон), неотам, цикламовую кислоту и ее соли (т.е. цикламаты), стевиозид (экстрагированный из листьев Stevia rebaudiana), могрозид (экстрагированный из фруктов Lo Han Guo), глицирризин, филлодульцин (экстрагированный из листьев Hydrangea macrophylla, приблизительно 400-600X сахарозы), монеллин, мабинлин, бразеин, циркулин, пентадин, по отдельности или в сочетании друг с другом.
Питьевая композиция также может содержать усилители сладости, которые обладают сладким вкусом только в присутствии других соединений, таких как кислоты. Неограничивающие примеры усилителей сладости (также известных, как потенцирующие факторы сладости) включают куркулин, миракулин, цинарин, хлорогеновую кислоту, кофейную кислоту, строгины, арабиногалактан, мальтол и дигидроксибензойные кислоты. В некоторых вариантах питьевые композиции, содержащие монатин, также содержат усилители вкуса или стабилизаторы, такие как Sucramask™ или трегалоза.
Питьевые композиции могут необязательно содержать природные или искусственные красители пищевых сортов. Данные красители могут быть выбраны из широко известных и доступных в данной области красителей, включающих синтетические красители (например, азокрасители, трифенилметаны, ксантены, хинины и индигоидные красители), жженый сахар, диоксид титана, красный #3, красный #40, синий #1 и желтый #5. Также можно использовать, например, природные красители, такие как свекловичный сок (свекольный красный), кармин, куркумин, лютеин, морковный сок, ягодные соки, пряные экстракты (куркума, аннатто и/или паприка) и каротиноиды. Тип и количество выбранного красителя зависят от конечного продукта и потребительского предпочтения.
В некоторых вариантах питьевые композиции также содержат один или несколько природных или синтетических ароматизаторов. Подходящие ароматизаторы включают цитрусовые и не цитрусовые фруктовые ароматизаторы; пряности; травы; растительное сырье; шоколад, какао или шоколадный ликер; кофе; ароматизаторы, полученные из бобов ванили; ореховые экстракты; ликеры и ликерные экстракты; дистилляты фруктового бренди; ароматические химикаты, имитаторы ароматов; а также их концентраты, экстракты или эссенции. Цитрусовые ароматы включают, например, лимон, лайм, апельсин, мандарин, грейпфрут, цитрон или кумкват. Многие ароматизаторы можно получить, например, от Rhodia USA (Cranbury, NJ); IFF (South Brunswick, NJ); Wild Flavors, Inc. (Erlanger, KY); Silesia Flavors, Inc. (Hoffman Estates, IL), Chr. Hansen (Milkwaukee, WI) и Firmenisch (Princeton, NJ).
Например, сироп напитка для получения газированного безалкогольного напитка может содержать природный ароматизатор колы (например, из орехового экстракта колы), который можно использовать для придания напитку аромата колы. В некоторых вариантах ароматизаторы могут находиться в виде эмульсии, которую затем диспергируют в сиропе напитка. Удельная плотность капелек эмульсии меньше, чем у воды, следовательно, они могут образовывать отдельную фазу. Для стабилизации капелек ароматизирующей эмульсии можно использовать утяжелители, эмульгаторы и стабилизаторы эмульсий. Примеры таких эмульгаторов и стабилизаторов эмульсий включают смолы, пектины, целлюлозу, полисорбаты, сложные эфиры сорбитана и альгинаты пропиленгликоля. В некоторых вариантах эмульсии ароматизатора колы составляют от 0,8 до 1,5% от веса сиропа напитка. В других вариантах дополнительные ароматизаторы, которые можно использовать для усиления аромата колы, включают цитрусовые ароматы, такие как лимон, лайм, апельсин, мандарин, грейпфрут, цитрон или кумкват, и ароматы пряностей, такие как гвоздика и ваниль. В других вариантах цитрусовые ароматы (например, природный аромат лимона или лайма) составляют приблизительно от 0,03 до 0,06% от веса сиропа напитка, а ароматы пряностей (например, ваниль) составляют от 0,5 до 1,5% от веса сиропа напитка.
pH сиропа напитка можно регулировать добавлением кислот (например, неорганических или органических кислот). Как правило, pH сиропа напитка варьирует от 2,5 до 5 (например, от 2,5 до 4,0). Из неорганических кислот наиболее часто используется фосфорная кислота, которая может присутствовать в недиссоциированной форме или в виде соли щелочного металла (например, в виде гидрофосфата калия или натрия, или дигидрофосфата калия или натрия). Неограничивающие примеры подходящих органических кислот включают лимонную кислоту, яблочную кислоту, фумаровую кислоту, адипиновую кислоту, глюконовую кислоту, глюкуронолактон, гидроксилимонную кислоту, винную кислоту, аскорбиновую кислоту, уксусную кислоту или их смеси. Данные кислоты могут присутствовать в недиссоциированной форме или в виде соответствующих солей.
В некоторых вариантах сироп напитка дополнительно содержит кофеин (например, из природного аромата колы). Кофеин также можно добавлять отдельно.
В одном варианте газированный напиток можно приготовить путем разбавления сиропа напитка газированной водой так, чтобы полученный напиток содержал от 15 до 25% сиропа и от 75 до 85% воды. Альтернативно, при приготовлении напитка для разбавления сиропа можно использовать негазированную воду и затем в напиток можно ввести углекислый газ. В другом варианте газированный напиток обычно помещают в контейнер, такой как бутылка или банка, и затем закупоривают. Для получения газированного напитка данного изобретения можно использовать любой традиционный метод насыщения углекислым газом.
В некоторых вариантах питьевые композиции могут представлять собой сухие смеси напитков. Следует отметить, что "сухое" вещество может содержать остаточные уровни жидкости. Например, смесь напитка может представлять собой солодовую смесь напитка, смесь напитка с шоколадным ароматом или порошкообразную смесь фруктового напитка, такую как Kool-Aid® или Crystal Light®. В одном варианте сухие смеси напитков можно получить путем влажного смешивания жидких ингредиентов в растворе, вакуумной сушки ингредиентов с получением сухого кека и последующего распыления его в базовый порошок. Для внесения других сухих ингредиентов, например, для добавления порошка какао к базовому порошку, можно использовать такие ингредиенты, как масло, эмульгаторы и вода.
В другом варианте базовый порошок напитка, который обычно не содержит подсластителя, такой как лимонадный пакет, и который, как правило, объединяется с сахарозой потребителем, можно смешать с высокоинтенсивным подсластителем, таким как монатин. Смешивание можно облегчить, например, путем применения разбавителя или наполнителя, такого как мальтодекстрин, гидролизованный крахмал, декстроза, полидекстрин и инулин.
В других вариантах солодовые смеси напитков включают сухие ингредиенты напитков, такие как, например, порошкообразный источник белков, такой как сухое молоко, сухое обезжиренное молоко, порошок яичного белка, растительные или зерновые белковые изоляты, такие как изоляты соевого белка, солодовые порошки, гидролизованные зерновые порошки, порошки крахмала, порошки других углеводов, витамины, минералы, порошки какао и порошкообразные ароматизаторы, или любое сочетание указанных ингредиентов. Жидкие ингредиенты солодового напитка могут включать, например, один или несколько из жиров и масел, жидких солодовых экстрактов, жидких подсластителей, таких как мед и сироп глюкозы, и жидких источников белков, таких как концентраты растительных белков, или любые их сочетания. Подходящие жиры включают, без ограничения, частично или полностью гидрированные растительные масла, такие как хлопковое масло, соевое масло, кукурузное масло, подсолнечное масло, пальмовое масло, масло канолы, косточковое пальмовое масло, арахисовое масло, рисовое масло, сафлоровое масло, кокосовое масло, рапсовое масло и их заменители со средним или высоким содержанием масла; или любые их сочетания. Также можно использовать животные жиры, такие как жир коровьего масла. Количество каждого ингредиента солодового напитка может варьировать в зависимости от желаемого состава. В некоторых вариантах монатин можно объединить с подсластителем-наполнителем, как указано выше.
В некоторых вариантах заранее приготовленные смеси фруктовых напитков содержат лимонную кислоту (например, от 60 до 70%), ароматизаторы (например, от 2 до 4%), красители (например, от 0,001 до 1%), монатин, фосфат кальция (например, от 0 до 25%), средство, вызывающее помутнение (например, от 0 до 5%), и аскорбиновую кислоту (например, от 0 до 2%). Например, смесь фруктового напитка может содержать 64,9% лимонной кислоты, 20,5% фосфата кальция, 3,9% средства, вызывающего помутнение, 0,78 аскорбиновой кислоты, 2,7% ароматизаторов, 0,1% красителей и монатин. В некоторых вариантах монатин можно объединить с подсластителем-наполнителем, как указано выше. В другом варианте для получения фруктового напитка заранее приготовленную смесь можно разбавить водой так, чтобы полученный напиток содержал приблизительно от 0,5 до 1,5% (например, 0,75%) смеси.
В одном варианте сухая композиция шоколадного напитка может содержать сухое обезжиренное молоко (например, приблизительно от 20 до 30%), сухую сыворотку (например, от 35 до 45%), забеливатель для кофе (например, от 10 до 15%), обезжиренный порошок какао (например, от 15 до 20%), бикарбонат калия (например, от 0,1 до 10%), гуаровую смолу (например, от 0,06 до 2%), каррагенан (например, от 0,05 до 5%), ароматизаторы (например, шоколад и/или ваниль) и монатин. Например, сухая композиция шоколадного напитка может содержать 26% сухого обезжиренного молока, 40% сухой сыворотки, 12% забеливателя для кофе, 18% обезжиренного порошка какао, 1% бикарбоната калия, 0,6% гуаровой смолы, 0,5% каррагенана, шоколадный ароматизатор, ванильный ароматизатор и монатин. В другом варианте для получения шоколадного напитка заранее приготовленную смесь можно разбавить водой или молоком так, чтобы полученный напиток содержал приблизительно от 0,5 до 1,5% (например, 0,8%) смеси.
В некоторых вариантах смеси сухих ингредиентов, используемые для получения питьевой композиции, смеси влажных ингредиентов, используемые с той же целью, или жидкие смеси (дисперсии) сухих и влажных ингредиентов предоставляются как композиции. Такие композиции могут предоставляться в виде готовых изделий, которые могут быть упакованы в контейнеры (например, мешочки, ведерки, картонные пакеты), подходящие для транспортировки к пунктам продажи и приготовления, а также для удобства наливания и/или смешивания. Готовое изделие может содержать необязательные элементы, такие как посуда, контейнеры для смешивания и др. Готовое изделие может также содержать инструкции для получения композиций напитков.
Ожидается, что содержащийся в напитках монатин по сравнению с другими подсластителями напитков будет иметь более длительный срок годности, повышенную устойчивость к воздействию нагревания и кислот, а также улучшенные вкусовые профили и маркетинговые преимущества. Далее данное изобретение будет описано более подробно с помощью нижеследующих примеров, которые не ограничивают объем описываемого изобретения.
ПРИМЕРЫ
ПРИМЕР 1
Клонирование и экспрессия триптофан-аминотрансфераз
В данном примере описываются способы клонирования триптофан-аминотрансфераз, используемых для превращения триптофана в индол-3-пируват.
Экспериментальная часть
Одиннадцать генов, кодирующих аминотрансферазы, клонируют в E.coli. К данным аминотрансферазам относятся следующие: D-аланин-аминотрансфераза Bacillus subtilis (dat, №по каталогу Genbank Y14082.1, п.о. 28622-29470, и №по каталогу Genbank NP_388848.1, нуклеотидная последовательность и аминокислотная последовательность, соответственно), тирозин-аминотрансфераза Sinorhizobium meliloti (также называемых Rhizobium meliloti) (tatA, SEQ ID NO: 1 и 2, нуклеотидная последовательность и аминокислотная последовательность, соответственно), тирозин-аминотрансфераза Rhodobacter sphaeroides, штамм 2.4.1 (tatA утверждена по гомологии, SEQ ID NO: 3 и 4, нуклеотидная последовательность и аминокислотная последовательность, соответственно), тирозин-аминотрансфераза R. sphaeroides 35053 (утверждена по гомологии, SEQ ID NO: 5 и 6, нуклеотидная последовательность и аминокислотная последовательность, соответственно), многосубстратная аминотрансфераза Leishmania major (bsat, утверждена по гомологии с пептидными фрагментами из L. mexicana, SEQ ID NO: 7 и 8, нуклеотидная последовательность и аминокислотная последовательность, соответственно), ароматическая аминотрансфераза Bacillus subtilis (araT, утверждена по гомологии, SEQ ID NO: 9 и 10, нуклеотидная последовательность и аминокислотная последовательность, соответственно), ароматическая аминотрансфераза Lactobacillus amylovorus (araT утверждена по гомологии, SEQ ID NO: 11 и 12, нуклеотидная последовательность и аминокислотная последовательность, соответственно), многосубстратная аминотрансфераза R. sphaeroides 35053 (утверждена по гомологии, SEQ ID NO: 13 и 14, нуклеотидная последовательность и аминокислотная последовательность, соответственно), многосубстратная аминотрансфераза Rhodobacter sphaeroides, штамм 2.4.1 (msa утверждена по гомологии, №по каталогу Genbank AAAE01000093.1, п.о. 14743-16155, и №по каталогу Genbank ZP00005082.1, нуклеотидная последовательность и аминокислотная последовательность, соответственно), аспартат-аминотрансфераза Escherichia coli (aspC, №по каталогу Genbank AE000195.1, п.о. 2755-1565, и №по каталогу Genbank AAC74014.1, нуклеотидная последовательность и аминокислотная последовательность, соответственно) и тирозин-аминотрансфераза E.coli (tyrB, SEQ ID NO: 31 и 32, нуклеотидная последовательность и аминокислотная последовательность, соответственно).
Гены клонируют, экспрессируют и анализируют способность кодируемых ими ферментов превращать триптофан в индол-3-пируват, наряду с коммерчески доступными ферментами. Все одиннадцать клонов обладали активностью.
Идентификация бактериальных штаммов, содержащих полипептиды с целевой активностью
В базе данных NCBI (Национальный центр информации по биотехнологии - National Center for Biotechnology Information) не зарегистрированы гены триптофан-аминотрансферазы. Однако идентифицированы организмы, обладающие данной ферментативной активностью. Активность L-триптофан-аминотрансферазы (TAT) обнаружена в клеточных экстрактах или очищенных белках из следующих источников: изолят клубеньковых бактерий из Festuca octoflora, митохондрии и цитозоль гороха, клетки корневого рака подсолнечника, Rhizobium leguminosarum biovar trifoli, Erwinia herbicola pv gypsophilae, Pseudomonassyringae pv. savastanoi, Agrobacterium tumefaciens, Azospirillum lipferum & brasilense, Enterobacter cloacae, Enterobacter agglonzeras, Bradyrhizobium elkanii, Cadida maltosa, Azotobacter vinelandii, мозг крысы, печень крысы, Sinorhizobium meliloti, Pseudomonas fluorescens CHA0, Lactococcuslactis, Lactobacillus casei, Lactobacillus helveticus, сеянцы пшеницы, ячмень, Phaseolus aureus (порченные бобы), Saccharomyces uvarum (carlsbergensis), Leishmania sp., кукуруза, побеги томатов, растения гороха, табак, свиньи, Clostridium sporogenes и Streptomyces griseus.
ПРИМЕР 2
Превращение индол-3-лактата в индол-3-пируват
Как показано на фиг.1 и 3, для получения индол-3-пирувата можно использовать индол-3-молочную кислоту. Превращение молочной кислоты в пируват является обратимой реакцией, как и превращение индол-3-пирувата в индол-3-лактат. Окисление индоллактата обычно обнаруживают по высокому уровню фона при 340 нм, обусловленному присутствием индол-3-пирувата.
Стандартная аналитическая смесь объемом 0,1 мл содержит 100 мМ фосфата калия, pH 8,0, 0,3 мМ NAD+, 7 единиц лактат-дегидрогеназы (LDH) (Sigma-L2395, St. Louis, MO) и 2 мМ субстрата. Анализ проводят с двойными повторами в титрационном микропланшете, прозрачном для УФ-света, на спектрофотометре для прочтения планшетов Molecular Devices SpectraMax Plus. Полипептид и буфер смешивают и с помощью пипетки вносят в лунки, содержащие индол-3-молочную кислоту и NAD+, затем, после краткого перемешивания, в каждой лунке измеряют поглощение при 340 нм с интервалами 9 секунд. Реакционную смесь инкубируют при 25°C в течение 5 минут. Увеличение поглощения при 340 нм показывает, что происходит образование NADH из NAD+. Отдельные отрицательные контроли не содержат NAD+и субстрата. D-LDH из Leuconostoc mesenteroides (номер по каталогу Sigma L2395) является более активной по отношению к индольным субстратам, чем L-LDH из Bacillus stearothermophilus (номер по каталогу Sigma L5275).
Подобные способы применяются для D-молочной кислоты и NAD+, или NADH и пирувата, природных субстратов полипептидов D-LDH. Vmax реакции восстановления пирувата в 100-1000 раз выше, чем Vmax реакции окисления лактата. Vmax реакции окисления индол-3-молочной кислоты, катализируемой D-LDH, составляет приблизительно одну пятую от Vmax реакции окисления молочной кислоты. Присутствие индол-3-пирувата также определяют по изменению поглощения при 327 нм (енолборатное производное) с использованием 50 мМ натрий-боратного буфера, содержащего 0,5 мМ EDTA и 0,5 мМ арсената натрия. Как для L-, так и для D-LDH полипептидов наблюдают небольшие, но воспроизводимые изменения поглощения по сравнению с отрицательными контролями.
Кроме того, лактат-дегидрогеназы широкой специфичности (ферменты с активностью, связанной с EC 1.1.1.27, EC 1.1.1.28 и/или EC 1.1.2.3) можно клонировать и использовать для получения индол-3-пирувата из индол-3-молочной кислоты. Источники дегидрогеназ широкой специфичности включают E.coli, Neisseria gonorrhoeae и Lactobacillus plantarum.
Альтернативно, индол-3-пируват можно получить путем контактирования индол-3-лактата с клеточными экстрактами Clostridium sporogenes, содержащими индоллактат-дегидрогеназу (EC 1.1.1.110); или с клеточными экстрактами Trypanosoma cruzi epimastigotes, содержащими п-гидроксифениллактат-дегидрогеназу (EC 1.1.1.222), которая, как известно, активна по отношению к индол-3-пирувату; или с клеточными экстрактами Pseudomonas acidovorans или E.coli, содержащими имидазол-5-иллактат-дегидрогеназу (EC 1.1.1.111); или с Coleus blumei, содержащими гидроксифенилпируват-редуктазу (EC 1.1.1.237); или с Candida maltosa, содержащими ароматический D-лактат-дегидрогеназу (EC 1.1.1.222). Ссылки, в которых описаны указанные ферменты, включают Nowicki et al. (FEMS Microbiol Lett 71: 119-24, 1992), Jean and DeMoss (Canadian J Microbiol. 14, 1968, Coote and Hassall (Biochem. J 111: 237-9, 1969), Cortese et al. (C. R. Seances Soc. Biol. Fil. 162390-5, 1968), Petersen and Alfermann (Z. Naturforsch. C: Biosci. 43 501-4, 1988) и Bhatnagar et al. (J. Gen Microbiol 135: 353-60, 1989). Кроме того, для окисления индол-3-молочной кислоты до индол-3-пирувата можно использовать лактат-оксидазу, например, из Pseudomonas sp.(Gu et al. J. Mol. Catalysis B: Enzymatic: 18: 299-305, 2002).
ПРИМЕР 3
Превращение L-триптофана в индол-3-пируват с использованием оксидазы L-аминокислот
В данном примере описаны способы превращения триптофана в индол-3-пируват с помощью оксидазы (EC 1.4.3.2), как альтернатива применению триптофан-аминотрансферазы, описанному в примере 1. Оксидазу L-аминокислот выделяют из Crotalus durissus (Sigma, St. Louis, MO, номер по каталогу A-2805). Номера по каталогу оксидаз L-аминокислот для молекулярного клонирования включают: CAD21325.1, AAL14831, NP_490275, BAB78253, A38314, CAB71136, JE0266, T08202, S48644, CAC00499, P56742, P81383, О93364, P81382, P81375, S62692, P23623, AAD45200, AAC32267, CAA88452, AP003600 и Z48565.
Реакции проводят в микроцентрифужных пробирках в общем объеме 1 мл, инкубируют 10 минут при встряхивании при 37°C. Реакционная смесь содержит 5 мМ L-триптофана, 100 мМ натрий-фосфатного буфера pH 6,6, 0,5 мМ арсената натрия, 0,5 мМ EDTA, 25 мМ тетрабората натрия, 0,016 мг каталазы (83 ед., Sigma C-3515), 0,008 мг FAD (Sigma) и 0,005-0,125 единиц оксидазы L-аминокислот. Отрицательные контроли содержат все компоненты, кроме триптофана, а пустые контроли содержат все компоненты, кроме оксидазы. Каталазу используют для удаления пероксида водорода, образующегося в процессе окислительного дезаминирования. Тетраборат и арсенат натрия используют для стабилизации енолборатной формы индол-3-пирувата, которая обладает максимальным поглощением при 327 нм. Получают стандартные растворы индол-3-пирувата с концентрацией 0,1-1 мМ в реакционной смеси.
Получаемая оксидаза L-аминокислот имеет удельную активность 540 мкг индол-3-пирувата в минуту на мг белка. Значение удельной активности ферментов триптофан-аминотрансфераз имеет такой же порядок.
ПРИМЕР 4
Превращение индол-3-пирувата в 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровую кислоту с использованием альдолазы
В данном примере описаны способы превращения индол-3-пирувата в MP с помощью альдолазы (лиазы) (фиг.2). Альдольные конденсации представляют собой реакции образования углерод-углеродных связей между β-углеродом альдегида или кетона и углеродом карбонильной группы другого альдегида или кетона. На атоме углерода, находящемся рядом с карбонильной группой субстрата, образуется карбанион, который служит нуклеофилом, атакующим карбонильный атом углерода второго субстрата (электрофильный атом углерода). Обычно электрофильным субстратом является альдегид, поэтому большинство альдолаз относится к категории EC 4.1.2. Чаще всего электрофильным субстратом является пируват. Реже альдолазы катализируют конденсацию двух кетокислот или двух альдегидов.
Однако идентифицированы альдолазы, катализирующие конденсацию двух карбоновых кислот. Например, в EP 1045-029 описано получение L-4-гидрокси-2-кетоглутаровой кислоты из глиоксиловой кислоты и пирувата с использованием культуры Pseudomonas (EC 4.1.3.16). Кроме того, 4-гидрокси-4-метил-2-оксоглутарат-альдолаза (4-гидрокси-4-метил-2оксоглутаратпируват-лиаза, EC 4.1.3.17) может катализировать конденсацию двух кетокислот. Следовательно, подобные альдолазные полипептиды можно использовать для катализа конденсации индол-3-пирувата с пируватом.
Клонирование
4-Гидрокси-4-метил-2-оксоглутаратпируват-лиаза (ProA-альдолаза, EC 4.1.3.17) и 4-гидрокси-2-оксоглутаратглиоксилат-лиаза (KHG-альдолаза, EC 4.1.3.16) катализируют реакции, подобные альдолазной реакции, представленной на фиг.2. Синтезируют праймеры, содержащие липкие концы, совместимые с вектором pET30Xa/LIC (Novagen, Madison, WI).
Результаты измерения активности продуктов гена proA
Генные конструкты C. testosteroni proA и S. meliloti SMc00502 имеют высокие уровни экспрессии при индуцировании IPTG. Рекомбинантные белки обладают высокой растворимостью, как определено при анализе SDS-PAGE общего белка и образцов клеточных экстрактов. Чистота продукта гена C. testosteroni составляет>95%. Поскольку продукт гена S. meliloti имеет очень низкий выход после аффинной хроматографии с использованием картриджа His-Bind, для ферментативных анализов используют клеточный экстракт.
Обе рекомбинантные альдолазы катализируют образование MP из индол-3-пирувата и пирувата. Для ферментативной активности требуется присутствие как двухвалентного магния, так и фосфата калия. Образование продукта не наблюдается, если отсутствует индол-3-пируват, пируват или фосфат калия. Небольшое количество продукта образуется при отсутствии фермента (обычно на порядок меньше, чем в присутствии фермента).
Продукт элюируется с колонки с обращенной фазой C18 несколько позже, чем стандарт индол-3-пируват, масс-спектр пика продукта показывает наличие родительского иона, индуцированного столкновением ([M+H]+) 292,1, родительский ион относят к продукту MP. Основные дочерние фрагменты, присутствующие в масс-спектре, включают фрагменты, имеющие m/z: 158 (ион 1H-индол-3-карбальдегидкарбония), 168 (ион 3-бута-1,3-диенил-1H-индолкарбония), 274 (292-H2O), 256 (292-2H2О), 238 (292-3 H2О), 228 (292-CH4О3) и 204 (отщепление пирувата). Продукт также имеет УФ-спектр, характерный для других индолсодержащих соединений, таких как триптофан, с λmax 279-280 и небольшим плечом приблизительно при 290 нм.
Количество MP, продуцируемое альдолазой C. testosteroni, увеличивается при увеличении температуры реакции от комнатной до 37°C, количества субстрата и количества магния. Синтетическая активность фермента уменьшается при увеличении pH, причем максимальное количество продукта образуется при pH 7. С помощью триптофановых стандартов обнаружено, что количество MP, продуцируемое в стандартном анализе с использованием 20 мкг очищенного белка, составляет приблизительно 10-40 мкг на один мл реакционной смеси.
Поскольку последовательности S. meliloti и C. testosteroni ProA, кодирующие альдолазы, в высокой степени гомологичны другим генам, описанным выше, предполагается, что все продукты рекомбинантных генов могут катализировать данную реакцию. Более того, предполагается, что альдолазы, содержащие треонин (T) в положениях 59 и 87, аргинин (R) в положении 119, аспартат (D) в положении 120 и гистидин (H) в положениях 31 и 71 (по системе нумерации C. testosteroni), будут обладать подобной активностью.
Результаты измерения активности продуктов гена khg
Генные конструкты khg как B. subtilis, так и E.coli, имеют высокие уровни экспрессии белка при индуцировании IPTG, тогда как S. meliloti khg имеет низкий уровень экспрессии. Рекомбинантные белки обладают высокой растворимостью, как определено при анализе SDS-PAGE общих белков и клеточных экстрактов. Чистота продуктов генов B. subtilis и E.coli khg составляет>95%; продукт гена S. meliloti имеет не такой высокий выход после аффинной хроматографии с использованием картриджа His-Bind.
Не существует данных, свидетельствующих о том, что для активности указанного фермента необходимо присутствие магния и фосфата. Однако в литературе описано, что данные анализы проводят в натрий-фосфатном буфере, и что указанный фермент является бифункциональным и обладает активностью по отношению к фосфорилированным субстратам, таким как 2-кето-3-дезокси-6-фосфоглюконат (KDPG). Ферментативные анализы проводят, как описано выше, и в некоторых случаях фосфат не используют. Полученные результаты показывают, что рекомбинантные альдолазы KHG продуцируют MP, но не обладают активностью альдолаз ProA. В некоторых случаях уровень MP, продуцируемый в присутствии KHG, практически идентичен количеству, получаемому в присутствии только магния и фосфата. Фосфат не повышает активность KHG. Фермент Bacillus имеет самую высокую активность, приблизительно на 20-25% выше, чем активность в присутствии только магния и фосфата, как определено методом SRM (см. пример 10). Фермент Sinorhizobium имеет самую низкую активность, что может быть обусловлено свертыванием и проблемами с растворимостью при экспрессии. Все три фермента содержат глутамат в активном центре (положение 43 по системе нумерации B. subtilis), кроме того, для образования основания Шиффа с пируватом необходим лизин (положение 130); однако, фермент B. subtilis в положении 47, в качестве остатка активного центра, чаще содержит треонин, чем аргинин. KHG B. subtilis меньше и имеет кластерные отличия от ферментов S. meliloti и E.coli, причем другие ферменты содержат треонин в активном центре. Различия в активном центре могут быть причиной повышенной активности фермента B. subtilis.
Повышение активности альдолазы
Каталитические антитела могут обладать такой же активностью, как и природные альдолазы, распознавая широкий диапазон субстратов, и могут использоваться для катализа реакции, представленной на фиг.2.
Альдолазы можно усовершенствовать путем направленной эволюции, например, как ранее описано для альдолазы KDPG (в высокой степени гомологичной описанной выше KHG), отбирая путем перетасовки ДНК и допускающей ошибки ПЦР образцы, не требующие присутствия фосфата и с инвертированной энантиоселективностью. Альдолазные полипептиды KDPG используются в биохимических реакциях, поскольку они являются высокоспецифичными по отношению к донорному субстрату (в данном описании пирувату), но допускают некоторые вариации в отношении акцепторного субстрата (т.е. индол-3-пирувата) (Koeller & Wong, Nature 409: 232-9, 2001). Альдолаза KHG катализирует конденсацию пирувата с рядом карбоновых кислот. Считается, что версии альдолазы KHG млекопитающих обладают более широкой специфичностью, чем бактериальные версии, в том числе, они обладают более высокой активностью по отношению к 4-гидрокси-4-метил-2-оксоглутарату и воспринимают оба стереоизомера 4-гидрокси-2-кетоглутарата. Для бактериальных источников R-стереоизомер предпочтительнее в 10 раз. В геномных базах данных существует около 100 гомологов KHG, их активность продемонстрирована на Pseudomonas, Paracoccus, Providencia, Sinorhizobium, Morganella, E.coli и тканях млекопитающих. Данные ферменты можно использовать в качестве отправных веществ при подборе энантиоспецифичности, необходимой для получения монатина.
Альдолазы, использующие пируват и другой субстрат, который представляет собой кетокислоту и/или имеет массивную гидрофобную группу, как индол, могут участвовать в отборе полипептидов по специфичности, скорости и селективности. Кроме упомянутых здесь альдолаз KHG и ProA, примеры данных ферментов включают, не ограничиваясь ими: альдолазу KDPG и родственные полипептиды (KDPH); транс-карбоксибензальпируват-гидратазу-альдолазу из Nocardioides st; 4-(2-карбоксифенил)-2-оксобут-3-еноат-альдолазу (2'-карбоксибензальпируват-альдолазу), которая катализирует конденсацию пирувата и 2-карбоксибензальдегида (субстрата, содержащего ароматический цикл); транс-O-гидроксибензилиденпируват-гидратазу-альдолазу из Pseudomonas putida и Sphingomonas aromaticivorans, которая также использует в качестве субстратов пируват и альдегид, содержащий ароматический цикл; 3-гидроксиаспартат-альдолазу (эритро-3-гидрокси-L-аспартатглиоксилат-лиазу), которая использует в качестве субстратов 2-оксокислоту и, как полагают, присутствует в организме Micrococcus denitrificans; бензоин-альдолазу (бензальдегид-лиазу), которая использует субстраты, содержащие бензильные группы; дигидронеоптерин-альдолазу; L-трео-3-фенилсеринбензальдегид-лиазу (фенилсерин-альдолазу), которая катализирует конденсацию глицина с бензальдегидом; 4-гидрокси-2-оксовалерат-альдолазу; 1,2-дигидроксибензилпируват-альдолазу; и 2-гидроксибензальпируват-альдолазу.
Полипептид, обладающий желательной активностью, можно выбрать путем скрининга представляющих интерес клонов с использованием нижеследующих методов. Триптофановые ауксотрофы трансформируют векторами, несущими представляющие интерес клоны в кассетах экспрессии, и выращивают на среде, содержащей небольшое количество монатина или MP. Поскольку реакции аминотрансфераз и альдолаз являются обратимыми, клетки могут продуцировать триптофан из рацемической смеси монатина. Подобным образом, организмы (как рекомбинантные, так и дикого типа) можно подвергнуть скринингу по способности использовать MP или монатин в качестве источника углерода или энергии. Источником целевых альдолаз могут служить экспрессионные библиотеки различных штаммов Pseudomonas и клубеньковых бактерий. Pseudomonads имеют много необычных катаболических путей деградации ароматических молекул, и, кроме того, они содержат много альдолаз; тогда как клубеньковые бактерии, которые, как известно, растут в ризосфере растений, содержат альдолазы и много генов, описанных для конструирования биосинтетического пути монатина.
ПРИМЕР 5
Химический синтез предшественника монатина
В примере 4 описан способ применения альдолазы для превращения индол-3-пирувата в MP. Данный пример описывает альтернативный способ химического синтеза MP. MP можно получить путем обычной альдольной конденсации (фиг.4). Коротко говоря, типичная альдольная реакция включает образование карбаниона сложного эфира пирувата с использованием сильного основания, такого как LDA (диизопропиламид лития), литийгексаметилдисилазан или бутиллитий. Полученный карбанион взаимодействует с индол-пируватом с образованием конденсированного продукта.
Защитные группы, которые можно использовать для защиты индольного азота, включают, не ограничиваясь ими: трет-бутилоксикарбонил (Boc) и бензилоксикарбонил (Cbz). Блокирующие группы для карбоновых кислот включают, не ограничиваясь ими, алкиловые сложные эфиры (например, метиловые, этиловые, бензиловые сложные эфиры). При использовании таких защитных групп невозможно контролировать стереохимию образующегося продукта. Однако если R2 и/или R3 представляют собой хиральные защитные группы (фиг.4), такие как (S)-2-бутанол, ментол или хиральный амин, можно достичь преимущественного образования одного энантиомера MP.
ПРИМЕР 6
Превращение триптофана или индол-3-пирувата в монатин
Способ in vitro с использованием двух ферментов, аминотрансферазы и альдолазы, позволяет получать монатин из триптофана и пирувата. На первой стадии альфа-кетоглутарат принимает аминогруппу триптофана в реакции трансаминирования с образованием индол-3-пирувата и глутамата. Альдолаза катализирует вторую реакцию, в которой пируват взаимодействует с индол-3-пируватом в присутствии Mg2+и фосфата с образованием альфа-кето-производного монатина (MP), 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровой кислоты. Перенос аминогруппы от глутамата, образованного в первой реакции, дает целевой продукт, монатин. Идентификация продукта после очистки показывает, что получен S,S-стереоизомер монатина. Описываются альтернативные субстраты, ферменты и условия, а также усовершенствования, которые можно применить к данному способу.
Ферменты
Альдолазу, 4-гидрокси-4-метил-2-оксоглутаратпируват-лиазу, (альдолаза ProA, ген proA) (EC 4.1.3.17) из Comamonas testosteroni клонируют, экспрессируют и очищают, как описано в примере 4. 4-Гидрокси-2-оксоглутаратглиоксилат-лиазы (KHG альдолазы) (EC 4.1.3.16) из B. subtilis, E.coli и S. meliloti клонируют, экспрессируют и очищают, как описано в примере 4.
В сочетании с альдолазами для получения монатина используют следующие аминотрансферазы: L-аспартат-аминотрансферазу, кодируемую геном aspC E.coli, тирозин-аминотрансферазу, кодируемую геном E.coli, фермент TatA S. meliloti, многосубстратную аминотрансферазу, кодируемую геном bsat L. major, или трансаминазу глутаминовой-щавелево-уксусной кислот из сердца свиньи (тип IIa). Клонирование, экспрессия и очистка белков организмов, отличных от млекопитающих, описаны в примере 1. Трансаминазу глутаминовой-щавелево-уксусной кислот из сердца свиньи (тип IIa) получают от Sigma (# G7005).
Способ с использованием альдолазы ProA и L-аспартат-аминотрансферазы
Реакционная смесь содержит в одном литре 50 мМ ацетата аммония, pH 8,0, 4 мМ MgCl2, 3 мМ фосфата калия, 0,05 мМ пиридоксальфосфата, 100 мМ пирувата аммония, 50 мМ триптофана, 10 мМ альфа-кетоглутарата, 160 мг рекомбинантной альдолазы ProA C. testosteroni (неочищенный клеточный экстракт, ~30% альдолазы), 233 мг рекомбинантной L-аспартат-аминотрансферазы E.coli (неочищенный клеточный экстракт, ~40% аминотрансферазы). Все компоненты за исключением ферментов смешивают вместе и инкубируют при 30°C до растворения триптофана. Затем добавляют ферменты и реакционную смесь инкубируют при 30°C при осторожном перемешивании (100 об/мин) в течение 3,5 часов. Через 0,5 и 1 час после добавления ферментов к реакционной смеси добавляют аликвоты твердого триптофана (по 50 ммоль каждая). Весь добавленный триптофан не растворяется, но концентрация поддерживается при 50 мМ или выше. Через 3,5 часа твердый триптофан отфильтровывают. Анализ реакционной смеси методом ЖХ/МС с использованием определенного количества триптофана в качестве стандарта показывает, что концентрация триптофана в растворе составляет 60,5 мМ, а концентрация монатина составляет 5,81 мМ (1,05 г).
Для очистки конечного продукта используются нижеследующие способы. Девяноста процентов прозрачного раствора наносят на колонку BioRad со смолой AG50W-X8 (225 мл; связывающая емкость 1,7 мэкв./мл). Колонку промывают водой, собирая фракции по 300 мл, пока поглощение при 280 нм не будет составлять <5% от поглощения первой проточной фракции. Затем колонку элюируют 1M ацетатом аммония, pH 8,4 и собирают 4 фракции по 300 мл. Все 4 фракции содержат монатин, их упаривают до объема 105 мл, используя роторный испаритель с умеренно теплой водяной баней. В процессе упаривания осадок, образующийся по мере уменьшения объема, отфильтровывают.
Анализ элюированных с колонки фракций методом ЖХ/МС показывает, что 99% триптофана и монатина связаны с колонкой. Осадок, образовавшийся при упаривании, содержит >97% триптофана и <2% монатина. Соотношение триптофана к продукту в супернатанте составляет приблизительно 2:1.
Супернатант (7 мл) наносят на колонку Fast Flow с DEAE-сефарозой (Amersham Biosciences) объемом 100 мл, заранее переведенную в ацетатную форму путем промывания 0,5 л 1M NaOH, 0,2 л воды, 1,0 л 1,0M ацетата аммония, pH 8,4, и 0,5 л воды. Супернатант загружают со скоростью <2 мл/мин, после чего колонку промывают водой со скоростью 3-4 мл/мин до тех пор, пока поглощение при 280 нм не станет приблизительно равно 0. Монатин элюируют 100 мМ ацетатом аммония, pH 8,4, собирая 4 фракции по 100 мл.
Анализ фракций показывает, что соотношение триптофана к монатину в промывочных фракциях составляет приблизительно 85:15, а соотношение в элюируемых фракциях составляет 7:93. Расчеты, проведенные на основе допущения, что коэффициенты экстинкции триптофана и монатина при 280 нм примерно равны, показывают, что элюируемые фракции содержат 0,146 ммоль продукта. Экстраполяция к общему объему реакционной смеси 1 л дает количество монатина ~2,4 ммоль (~710 мг) при извлечении 68%.
Фракции, элюируемые с колонки с DEAE-сефарозой, упаривают до объема <20 мл. Для характеристики монатина в аналитическом масштабе часть продукта подвергают дополнительной очистке путем нанесения на препаративную колонку с обращенной фазой С8, используя такие же условия хроматографии, как в примере 10. Для запуска автоматического сбора фракций монатина на основе детекции иона с m/z=293 используют программное обеспечение Waters Fractionlynx™. Фракцию, содержащую соответствующий протонированный молекулярный ион монатина, собирают с колонки С8, упаривают досуха и затем растворяют в небольшом количестве воды. Данную фракцию используют для идентификации продукта.
Полученный продукт характеризуют с помощью нижеследующих методов.
Спектроскопия в УФ/видимой области. Спектры полученного ферментативно монатина в УФ/видимой области снимают с помощью спектрофотометра УФ/видимой части спектра Cary 100 Bio. Очищенный продукт, растворенный в воде, имеет максимум поглощения при 280 нм с плечом при 288 нм, что характерно для индол-содержащих соединений.
Анализ ЖХ/МС. Анализы смесей монатина, полученного с помощью биохимических реакций in vitro, проводят, как описано в примере 10. Типичный анализ методом ЖХ/МС монатина в полученной in vitro ферментативной синтетической смеси представлен на фиг.5. В нижней части фиг.5 представлена хроматограмма выбранного иона для протонированного молекулярного иона монатина при m/z=293. Данная идентификация монатина в смеси подтверждается масс-спектром, представленным на фиг.6. Анализ очищенного продукта методом ЖХ/МС показывает наличие одного пика с молекулярным ионом 293 и поглощением при 280 нм. Масс-спектр идентичен представленному на фиг.6.
Анализ МС/МС. Эксперименты ЖХ/МС/МС с дочерними ионами, описанные в примере 10, также проводят для монатина. Масс-спектр дочернего иона монатина представлен на фиг.7. Осуществляют ориентировочные структурные отнесения всех фрагментарных ионов, отмеченных на фиг.7. Данные ионы включают фрагментарные ионы с m/z=275 (293-H2O), 257 (293-(2×H2O)), 230 (275-COOH), 212 (257-COOH), 168 (ион 3-бута-1,3-диенил-1H-индолкарбония), 158 (ион 1H-индол-3-карбальдегидкарбония), 144 (ион 3-этил-1H-индолкарбония), 130 (ион 3-метилeн-1H-индолкарбония) и 118 (ион индолкарбония). Многие из них идентичны ионам MP (пример 4), полученным при распаде индольного фрагмента молекулы. Некоторые из них имеют массу на 1 единицу выше, чем аналогичные ионы MP, из-за того, что вместо кетогруппы присутствует аминогруппа.
Точное определение массы монатина. На фиг.8 представлен масс-спектр, полученный для очищенного монатина с помощью гибридного квадрупольного/времяпролетного масс-спектрометра Applied Biosystems-Perkin Elmer Q-Star. Масса протонированного монатина, измеренная с использованием триптофана в качестве внутреннего стандарта для определения массы, составляет 293,1144. Масса протонированного монатина, рассчитанная на основе элементного состава C14H17N2O5, составляет 293,1137. Ошибка при измерении массы составляет менее 2 частей на миллион (м.д.), что является убедительным подтверждением элементного состава монатина, полученного ферментативным способом.
ЯМР-спектроскопия. ЯМР-эксперименты проводят на приборе Varian Inova, 500 МГц. Образец монатина (~3 мг) растворяют в 0,5 мл D2О. Вначале растворитель (D2О) используют в качестве внутреннего стандарта при 4,78 м.д. Поскольку вода имеет большой пик, 1H-ЯМР проводят с подавлением пика воды. Затем, из-за широты пика воды, в качестве стандартного пика используют C-2 протон монатина и устанавливают на опубликованном в литературе значении 7,192 м.д.
При проведении 13C-ЯМР первые несколько сотен сканов показывают, что образец является слишком разбавленным для получения адекватного 13C-спектра в выделенное время. Затем проводят эксперимент по гетероядерному множественному квантовому согласованию (HMQC), который делает возможным проведение корреляции между атомами водорода и атомами углерода, к которым они присоединены, а также предоставляет информацию по химическим сдвигам атомов углерода.
Суммарные результаты 1H-ЯМР и HMQC представлены в таблицах 1 и 2. Сравнение данных ЯМР с опубликованными значениями показывает, что полученный ферментативным способом монатин представляет собой один из (S,S)- и (R,R)-изомеров, или смесь обоих изомеров.
Хиральный анализ ЖХ/МС. Чтобы установить, что полученный in vitro монатин представляет собой один стереоизомер, а не смесь (R,R)- и (S,S)-энантиомеров, проводят хиральный анализ ЖХ/МС с помощью прибора, описанного в примере 10.
Хиральные ЖХ разделения проводят при комнатной температуре, используя хиральную хроматографическую колонку Chirobiotic T (Advanced Separations Technology). Разделение и детекцию оптимизируют для R- (D) и S- (L) стереоизомеров триптофана, следуя инструкциям поставщика. Подвижная фаза ЖХ состоит из A) воды, содержащей 0,05% (об./об.) трифторуксусной кислоты; B) метанола, содержащего 0,05% (об./об.) трифторуксусной кислоты. Проводят изократическое элюирование, используя смесь 70% A и 30% B. Скорость потока составляет 1,0 мл/мин, а поглощение PDA регистрируют в диапазоне от 200 нм до 400 нм. Инструментальные параметры, используемые для хирального ЖХ/МС анализа триптофана и монатина, идентичны параметрам ЖХ/МС анализа, описанным в примере 10. Используют набор масс-спектров для участка m/z 150-400. Хроматограммы выбранных ионов для протонированных молекулярных ионов ([M+H]+=205 для R- и S-триптофана и [M+H]+=293 для монатина) позволяют непосредственно идентифицировать данные вещества в смесях.
Хроматограммы R- и S-изомеров триптофана и монатина, разделенных методом хиральной хроматографии с регистрацией МС, представлены на фиг.9. Единственный пик в хроматограмме монатина указывает, что соединение представляет собой один стереоизомер, причем время удерживания данного пика почти идентично времени удерживания S-триптофана.
Данные 1H-ЯМР
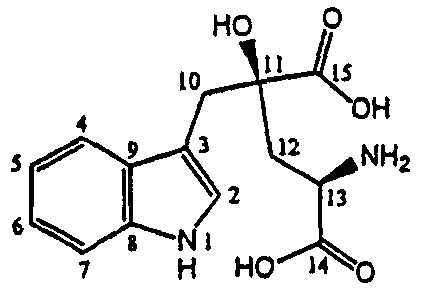
2Takeshi and Shusuke (JP2002060382, 2002-02-26).
Данные 13С-ЯМР (из спектра HMQC)
Поляриметрия. Оптическое вращение измеряют на поляриметре Rudolph Autopol III. Получают раствор монатина в воде с концентрацией 14,6 мг/мл. Ожидаемое удельное вращение ([a]D 20) S,S-монатина (в виде соли) составляет -49,6 для раствора в воде с концентрацией 1 г/мл (Vleggaar et al.). Для очищенного, полученного ферментативным способом монатина наблюдаемое значение [a]D 20 составляет -28,1, указывая, что монатин представляет собой S,S-стереоизомер.
Усовершенствования
Оптимизируют условия реакций, в том числе, концентрации реагентов и ферментов, и получают выход 5-10 мг/мл при использовании следующей смеси реагентов: 50 мМ ацетат аммония pH 8,3, 2 мМ MgCl2, 200 мМ пируват (натриевая или аммониевая соль), 5 мМ альфа-кетоглутарат (натриевая соль), 0,05 мМ пиридоксальфосфат, деаэрированная вода до получения конечного объема 1 мл после добавления ферментов, 3 мМ фосфат калия, 50 мкг/мл рекомбинантной альдолазы ProA (клеточный экстракт; концентрация общего белка 167 мкг/мл), 1000, мкг/мл L-аспартат-аминотрансферазы, кодируемой геном aspC E.coli (клеточный экстракт; концентрация общего белка 2500 мкг/мл), и твердый триптофан, обеспечивающий концентрацию в растворе >60 мМ (насыщение; некоторое количество триптофана остается нерастворенным на протяжении всей реакции). Смесь инкубируют при 30°C в течение 4 часов при осторожном взбалтывании или перемешивании.
Замены
Концентрацию альфа-кетоглутарата можно уменьшить до 1 мМ и добавить 9 мМ аспартата, при этом выход монатина останется прежним. На первой стадии можно использовать альтернативные акцепторы аминокислот, такие как оксалоацетат.
При использовании рекомбинантной многосубстратной аминотрансферазы L. major вместо L-аспартат-аминотрансферазы E.coli выход монатина остается на прежнем уровне. Однако с помощью анализа ЖХ-МС обнаруживают второй не идентифицированный продукт (3-10% от основного продукта) с молекулярной массой 292. При использовании в качестве аминотрансфераз фермента, кодируемого геном tyrb E.coli, фермента, кодируемого геном tat A S. meliloti, или трансаминазы глутаминовой-щавелево-уксусной кислот из сердца свиньи (тип IIa) получают концентрации монатина 0,1-0,5 мг/мл. Если в качестве исходного вещества используют индол-3-пируват, восстановительное аминирование на последней стадии можно проводить с использованием глутамат-дегидрогеназы и NADH (как описано в примере 7).
Для ферментативного получения монатина в сочетании с L-аспартат-аминотрансферазой E.coli используют альдолазы KHG из B. subtilis, E.coli и S. meliloti. Используют следующие условия реакции: 50 мМ NH4-OAc pH 8,3, 2 мМ MgCl2, 200 мМ пируват, 5 мМ глутамат, 0,05 мМ пиридоксальфосфат, деаэрированная вода до получения конечного объема 0,5 мл после добавления ферментов, 3 мМ фосфат калия, 20 мкг/мл рекомбинантной альдолазы KHG B. subtilis (очищенной), приблизительно 400 мкг/мл неочищенной L-аспартат-аминотрансферазы (AspC) E.coli из клеточного экстракта и 12 мМ индол-3-пируват. Реакции инкубируют при 30°C в течение 30 минут при встряхивании. Количество монатина, полученного при использовании фермента B. subtilis, составляет 80 нг/мл и увеличивается при повышении количества альдолазы. Если индол-3-пируват и глутамат заменить насыщающими количествами триптофана и 5 мМ альфа-кетоглутаратом, выход монатина увеличится до 360 нг/мл. Реакции повторяют, используя 30 мкг/мл каждого из трех ферментов KHG в 50 мМ Tris pH 8,3 и триптофан в насыщающей концентрации и оставляют протекать в течение часа, чтобы увеличить уровень детектируемых веществ. Фермент Bacillus имеет наивысшую активность, как показано в примере 4, продуцируя приблизительно 4000 нг/мл монатина. KHG E.coli продуцирует 3000 нг/мл монатина, а фермент S. meliloti продуцирует 2300 нг/мл.
ПРИМЕР 7
Взаимопревращение MP и монатина
Аминирование MP с образованием монатина можно катализировать аминотрансферазами, например, описанными в примерах 1 и 6, или дегидрогеназами, которые требуют присутствия восстанавливающего кофактора, такого как NADH или NADPH. Данные реакции являются обратимыми и могут определяться в любом направлении. При использовании фермента дегидрогеназы направление в основном контролируют по концентрации солей аммония.
Дегидрогеназная активность. Окислительное дезаминирование монатина регистрируют по увеличению поглощения при 340 нм при превращении NAD(P)+ в более хромофорный NAD(P)H. Ферментативное получение и очистку монатина проводят по способу примера 6.
Типичная аналитическая смесь содержит 50 мМ Tris-HCl, pH 8,0-8,9, 0,33 мМ NAD+или NADP+, от 2 до 22 единиц глутамат-дегидрогеназы (Sigma) и 10-15 мМ субстрата в 0,2 мл. Анализ проводят с двойными повторами в титрационном микропланшете, прозрачном для УФ-света, на спектрофотометре для прочтения планшетов Molecular Devices SpectraMax Plus. Смесь фермента, буфера и NADP+с помощью пипетки вносят в лунки, содержащие субстрат, и затем, после краткого перемешивания, в каждой лунке регистрируют увеличение поглощения при 340 нм с интервалами 10 секунд. Реакционную смесь инкубируют при 25°C в течение 10 минут. Отрицательные контроли не содержат субстрата, а в качестве положительного контроля используют глутамат. Глутамат-дегидрогеназа типа III из бычьей печени (Sigma #G-7882) катализирует превращение монатина в предшественник монатина, причем скорость данного превращения составляет приблизительно одну сотую от скорости превращения глутамата в альфа-кетоглутарат.
Трансаминирующая активность. Анализы монатин-аминотрансферазы проводят с использованием аспартат-аминотрансферазы (AspC) из E.coli, тирозин-аминотрансферазы (TyrB) из E.coli, многосубстратной аминотрансферазы (BSAT) из L. major и двух коммерчески доступных свиных глутаматоксалоацетат-аминотрансфераз, описанных в примере 1. И оксалоацетат и альфа-кетоглутарат тестируют в качестве акцепторов аминогруппы. Аналитическая смесь содержит (в 0,5 мл) 50 мМ Tris-HCl, pH 8,0, 0,05 мМ PLP, 5 мМ акцептора аминогруппы, 5 мМ монатина и 25 мкг аминотрансферазы. Аналитические смеси инкубируют 30 минут при 30°C, после чего реакцию останавливают добавлением 0,5 мл изопропилового спирта. Убывание монатина регистрируют с помощью ЖХ/МС (пример 10). Наивысшая активность наблюдается при использовании BSAT L. major и оксалоацетата в качестве акцептора аминогруппы и затем при использовании того же фермента и альфа-кетоглутарата в качестве акцептора аминогруппы. При использовании оксалоацетата активность изменяется следующим образом: BSAT>AspC>свиная аминотрансфераза типа IIa>свиная аминотрансфераза типа I=TyrB. При использовании альфа-кетоглутарата активность изменяется следующим образом: BSAT>AspC>свиная аминотрансфераза типа I>свиная аминотрансфераза типа IIa>TyrB.
ПРИМЕР 8
Получение монатина из триптофана и C3-источников, отличных от пирувата
Как описано выше в примере 6, индол-3-пируват или триптофан можно превратить в монатин, используя пируват в качестве C3-молекулы. Однако в некоторых случаях нежелательно использовать пируват в качестве исходного вещества. Например, пируват может быть более дорогим, чем другие C3-источники, или он может оказывать неблагоприятное действие на процессы ферментации при добавлении в среду. Аланин может подвергаться аминированию под действием многих ферментов PLP с получением пирувата. Триптофаназа-подобные ферменты осуществляют реакции бета-элиминирования с более высокими скоростями, чем другие ферменты PLP, такие как аминотрансферазы. Ферменты данного класса (4.1.99.-) могут продуцировать аммиак и пируват из аминокислот, таких как L-серин, L-цистеин и производные серина и цистеина, содержащих подходящие уходящие группы, например, O-метил-L-серин, О-бензил-L-серин, S-метилцистеин, S-бензилцистеин, S-алкил-L-цистеин, О-ацил-L-серин, 3-хлор-L-аланин.
Способы получения монатина с использованием полипептидов EC 4.1.99.- можно улучшить путем введения мутаций в β-тирозиназу (TPL) или триптофаназу по способу Mouratou et al. (J. Biol. Chem 274: 1320-5, 1999). Mouratou et al. сообщают, что β-тирозиназу можно превратить в β-лиазу дикарбоновых аминокислот, которая, по имеющимся данным, в природе не встречается. Изменение специфичности происходит в результате замены валина (V) 283 на аргинин (R) и аргинина (R) 100 на треонин (T). Указанные аминокислотные замены обусловливают способность лиазы акцептировать дикарбоновые аминокислоты (такие как аспартат) и осуществлять реакцию гидролитического дезаминирования. Таким образом, аспартат можно использовать в качестве источника пирувата, используемого в последующих реакциях альдольной конденсации.
Кроме того, к клеткам или в ферментативные реакторы можно добавить лактат и фермент, превращающий лактат в пируват. Примеры ферментов, способных катализировать данную реакцию, включают лактат-дегидрогеназу и лактат-оксидазу.
Реакционная смесь содержит 50 мМ Tris-Cl pH 8,3, 2 мМ MgCl2, 200 мМ C3-источник, 5 мМ альфа-кетоглутарат, натриевую соль, 0,05 мМ пиридоксальфосфат, деаэрированную воду до получения конечного объема 0,5 мл после добавления ферментов, 3 мМ фосфат калия pH 7,5, 25 мкг неочищенной рекомбинантной альдолазы ProA C. testosterone, полученной по способу примера 4, 500 мкг неочищенной L-аспартат-аминотрансферазы (AspC), полученной по способу примера 1, и твердый триптофан, обеспечивающий концентрацию в растворе >60 мМ (насыщение; некоторое количество триптофана остается нерастворенным на протяжении всей реакции). Смесь инкубируют при перемешивании в течение 30 минут при 30°C. В качестве 3-углеродных источников добавляют серин, аланин и аспартат. Анализы проводят в присутствии или в отсутствие второго фермента PLP (очищенного), способного осуществлять бета-элиминирование и бета-лиазные реакции (триптофаназа (TNA), β-триптофаназа с двойной мутацией, тирозиназа (TPL)). Результаты представлены в таблице 3:
Получение монатина с использованием альтернативных C3-источников
С помощью сканирующего анализа дочерних ионов ЖХ/МС/МС подтверждают, что монатин, полученный из аланина и серина, используемых в качестве 3-углеродных источников, идентичен идентифицированному монатину, полученному в примере 6. Аланин, трансаминирование которого катализируется ферментом AspC, является наилучшим альтернативным вариантом из тестируемых. Количество продуцируемого монатина увеличивается при добавлении триптофаназы, у которой способность к трансаминированию является второй активностью. Количество монатина, получаемого с использованием серина в качестве углеродного источника, почти удваивается при добавлении триптофаназных ферментов, даже если количество добавляемой триптофаназы составляет только одну пятую от количества аминотрансферазы. AspC способна одна проявлять такой же уровень бета-элиминирующей активности. Результаты, полученные с использованием аспартата, показывают, что триптофаназная активность в отношении аспартата не повышается при введении таких же сайт-направленных мутаций, как и в случае β-тирозиназы. Предполагается, что в отношении получения монатина мутантная β-тирозиназа является более активной.
ПРИМЕР 9
Химический синтез монатина
Монатин можно получить путем добавления аланина к индол-3-пировиноградной кислоте, и данную реакцию проводят с использованием реактива Гриньяра или органолитиевого реагента.
Например, к 3-хлор- или 3-бромаланину с подходящим образом блокированными карбоксильной и аминогруппой добавляют магний в безводных условиях. Затем добавляют индол-3-пируват (блокированный подходящим образом), что приводит к образованию продукта конденсации, и после удаления защитных групп получают монатин. Особенно пригодные защитные группы включают THP (простой эфир тетрагидропиранила), который можно легко присоединять и удалять.
ПРИМЕР 10
Детекция триптофана, монатина и MP
В данном примере описаны способы обнаружения монатина или его предшественника 2-гидрокси-2-(индол-3-илметил)-4-кетоглутаровой кислоты.
Анализ ЖХ/МС
Анализ смесей монатина, MP и/или триптофана, полученных посредством биохимических реакций in vitro или in vivo, проводят на оборудовании Waters/Micromass для анализа методом жидкостной хроматографии-тандемной масс-спектрометрии (ЖХ/МС/МС), которое включает жидкостной хроматограф Waters 2690 с монитором поглощения с фотодиодной матрицей Waters 996 (PDA), находящимся между хроматографом и трехполюсным квадрупольным масс-спектрометром Micromass Quattro Ultima. ЖХ разделения проводят на колонке с обращенной фазой C18 Supelco Discovery, 2,1 мм (150 мм или на колонке с обращенной фазой C8 Xterra MS, 2,1 мм (250 мм, при комнатной температуре. Подвижная фаза ЖХ состоит из A) воды, содержащей 0,05% (об./об.) трифторуксусной кислоты, и B) метанола, содержащего 0,05% (об./об.) трифторуксусной кислоты.
Для элюирования используют следующие градиенты: линейный от 5% B до 35% B, 0-9 мин, линейный от 35% B до 90% B, 9-16 мин, изократический 90% B, 16-20 мин, линейный от 90% B до 5% B, 20-22 мин, с 10-минутным периодом уравновешивания между прогонами. Скорость потока составляет 0,25 мл/мин, а PDA поглощение регистрируют в интервале от 200 нм до 400 нм. Все параметры ESI-MS оптимизируют и выбирают по генерации протонированных молекулярных ионов ([M+H]+) представляющих интерес анализируемых веществ и по продукции характеристических фрагментарных ионов.
Для ЖХ/МС анализа монатина используют следующие инструментальные параметры: капилляр: 3,5 кВ; конус: 40 V; гекс.1: 20 В; апертура: 0 В; гекс.2: 0 В; температура источника: 100°C; температура десольватации: 350°C; газ десольватации: 500 л/ч; газ конуса: 50 л/ч; низкое массовое разрешение (Q1): 15,0; высокое массовое разрешение (Q1): 15,0; энергия ионов: 0,2; вход: 50 В; энергия столкновения: 2; выход: 50 В; низкое массовое разрешение (Q2): 15; высокое массовое разрешение (Q2): 15; энергия ионов (Q2): 3,5; умножитель: 650. Погрешности опубликованных значений отношения масса/заряд (m/z) и молекулярной массы составляют ±0,01%. Начальную детекцию альфа-кетокислотной формы монатина (MP) и монатина в смесях проводят путем ЖХ/МС мониторинга со сбором масс-спектров в диапазоне m/z 150-400. Хроматограммы выбранных ионов для протонированных молекулярных ионов ([M+H]+=292 для MP, [M+H]+=293 для монатина) позволяют осуществлять непосредственную идентификацию указанных анализируемых веществ в смесях.
Анализ МС/МС
ЖХ/МС/МС анализ монатина с использованием дочерних ионов проводят следующим образом. Анализ с использованием дочерних ионов включает передачу представляющего интерес дочернего иона (например, с m/z=293 для монатина) из первого масс-анализатора (Q1) в ячейку столкновений масс-спектрометра, куда вводят аргон и где происходит химическая диссоциация родительского иона на фрагментарные (дочерние) ионы. Затем данные фрагментарные ионы детектируют с помощью второго масс-анализатора (Q2) и используют для подтверждения структуры родительского иона. Триптофан характеризуют и определяют его количество таким же способом посредством трансмиссии и фрагментации иона с m/z=205.
Для ЖХ/МС/МС анализа монатина используют следующие инструментальные параметры: капилляр: 3,5 кВ; конус: 40 V; гекс.1: 20 В; апертура: 0 В; гекс.2: 0 В; температура источника: 100°C; температура десольватации: 350°C; газ десольватации: 500 л/ч; газ конуса: 50 л/ч; низкое массовое разрешение (Q1): 13,0; высокое массовое разрешение (Q1): 13,0; энергия ионов: 0,2; вход: 5 В; энергия столкновения: 14; выход: 1 В; низкое массовое разрешение (Q2): 15; высокое массовое разрешение (Q2): 15; энергия ионов (Q2): 3,5; умножитель: 650.
Анализ монатина и триптофана с высокой разрешающей способностью
Анализы с высокой разрешающей способностью (<5 мин/образец) смесей монатина и триптофана, полученных посредством реакций in vitro или in vivo, проводят с использованием описанного выше приборного обеспечения и параметров, описанных для ЖХ/МС/МС. ЖХ разделение проводят на колонке 4,6 мм (50 мм Advanced Separation Technologies Chirobiotic T при комнатной температуре. Подвижная фаза ЖХ состоит из A) воды, содержащей 0,25% уксусной кислоты; B) метанола, содержащего 0,25% уксусной кислоты. Изократическое элюирование проводят с использованием смеси, содержащей 50% B, в течение 0-5 мин. Скорость потока составляет 0,6 мл/мин. Все параметры системы ESI-МС/МС оптимизируют и выбирают на основе оптимальной генерации в источнике протонированного молекулярного иона триптофана с использованием внутреннего стандарта 2H5-триптофана, а также индуцированной столкновением генерации характерных для аминокислот фрагментарных ионов в экспериментах с мониторингом нескольких реакций (MRM). Для ЖХ/МС/МС анализа монатина и триптофана в режиме мониторинга нескольких реакций (MRM) с использованием положительных ионов используют следующие инструментальные параметры: капилляр: 3,5 кВ; конус: 20 V; гекс.1: 15 В; апертура: 1 В; гекс.2: 0 В; температура источника: 100°C; температура десольватации: 350°C; газ десольватации: 500 л/ч; газ конуса: 40 л/ч; низкое массовое разрешение (Q1): 12,0; высокое массовое разрешение (Q1): 12,0; энергия ионов: 0,2; вход: 5 В; энергия столкновения: 14; выход: 1 В; низкое массовое разрешение (Q2): 15; высокое массовое разрешение (Q2): 15; энергия ионов (Q2): 0,5; умножитель: 650. Параметры MRM: запаздывание внутри канала: 0,03 с; запаздывание между сканами: 0,03 с; пауза: 0,05 с.
Точное определение массы монатина
МС анализ высокого разрешения проводят на гибридном квадрупольном/времяпролетном масс-спектрометре Applied Biosystems-Perkin Elmer Q-Star. Массу протонированного монатина измеряют, используя триптофан в качестве внутреннего стандарта определения массы. Масса протонированного монатина, рассчитанная на основе элементного состава C14H17N2O5, составляет 293,1137. Измеренная масса монатина, полученного биокаталитическим способом, описанным в примере A, составляет 293,1144. Ошибка при измерении массы составляет менее 2 частей на миллион (м.д.), что является подтверждением элементного состава монатина, полученного ферментативным способом.
ПРИМЕР 11
Получение монатина с помощью бактерий
В данном примере описаны способы получения монатина в клетках E.coli. Специалистам в данной области известно, что подобные методы можно использовать для получения монатина в других бактериальных клетках. Кроме того, можно использовать векторы, содержащие другие гены из синтетического пути монатина (фиг.2).
Среду Trp-1+глюкоза, минимальную среду, которую используют для увеличения продукции триптофана в клетках E.coli (Zeman et al. Folia Microbiol. 35: 200-4, 1990), получают следующим образом. К 700 мл чистой на наноуровне воды добавляют следующие реагенты: 2 г (NH4)2SO4, 13,6 г KH2PО4, 0,2 г MgSО4·7H2О, 0,01 г CaCl2·2H2О и 0,5 мг FeSО4·7H2О. pH доводят до 7,0, объем увеличивают до 850 мл и проводят автоклавирование среды. Отдельно получают 50% раствор глюкозы, который стерилизуют фильтрацией. Сорок мл полученного раствора добавляют к основной среде (850 мл) до конечного объема 1 л.
Готовят раствор L-триптофана, 10 г/л, в 0,1M фосфате натрия, pH 7 и стерилизуют его фильтрацией. Одну десятую часть объема обычно добавляют к культурам, как описано ниже. Также получают 10% раствор пирувата натрия, который стерилизуют фильтрацией. На литр культуры обычно добавляют 10 мл данного раствора. Получают исходные растворы ампициллина (100 мг/мл), канамицина (25 мг/мл) и IPTG (840 мМ), стерилизуют их фильтрацией и перед применением хранят при -20°C. Твин 20 (полиоксиэтилен-20-сорбитанмонолаурат) используют в конечной концентрации 0,2% (об./об.). Ампициллин используют в нелетальных концентрациях, как правило, его конечная концентрация составляет 1-10 мкг/мл.
Готовят свежие плашки, содержащие E.coli BL21 (DE3):C. testosteroni proA/pET 30 Xa/LIC (описанные в примере 4) в среде LB, содержащей 50 мкг/мл канамицина. После культивирования в течение ночи культуры (5 мл) от одной колонии высевают и выращивают при 30°C в среде LB, содержащей канамицин. Как правило, для индукции в среде trp-1+глюкоза используют от 1 до 50 инокулята. Добавляют свежий антибиотик с конечной концентрацией 50 мг/л. Перед индукцией колбы инкубируют при 37°C при встряхивании.
Каждый час отбирают образцы клеточной суспензии и регистрируют значение оптической плотности, пока не будет получено значение OD600 0,35-0,8. Затем клетки индуцируют 0,1 мМ IPTG и температуру уменьшают до 34°C. Перед индукцией отбирают образцы (1 мл) (нулевой момент времени) и центрифугируют их при 5000×g. Супернатант замораживают при -20°C для последующего анализа ЖХ/МС. Через четыре часа после индукции снова отбирают образец объемом 1 мл и центрифугируют его, чтобы отделить клетки от среды. Триптофан, пируват натрия, ампициллин и твин добавляют, как описано выше.
Клетки выращивают в течение 48 часов после индукции, снова отбирают образец объемом 1 мл и обрабатывают его, как описано выше. Через 48 часов добавляют еще аликвоту триптофана и пирувата. Весь объем культуры центрифугируют приблизительно через 70 часов (после индукции), в течение 20 минут при 4°C и 3500 об/мин. Супернатант декантируют, среду и клетки замораживают при -80°C. Фракции среды отфильтровывают и анализируют методом ЖХ/МС. Высоту и площадь пиков [M+H]+=293 регистрируют, как описано в примере 10. Фоновое значение среды вычитают. Результаты клеточного роста также нормализуют путем нанесения на график высоты пика [M+H]+=293, разделенной на оптическую плотность культуры при 600 нм.
При добавлении пирувата, ампициллина и твина через 4 часа после индукции получают более высокие уровни монатина, чем в случае добавления указанных реагентов во время индукции. Другие добавки, такие как PLP, дополнительное количество фосфата или дополнительное количество MgCl2, не увеличивают продукцию монатина. Более высокие титры монатина получают, если вместо индол-3-пирувата используют триптофан и если триптофан добавляют после индукции, а не во время инокуляции или индукции. Перед индукцией и через 4 часа после индукции (во время добавления субстрата) монатин не детектируется ни в ферментационной среде, ни в клеточных экстрактах. Для отрицательных контролей используют клетки, содержащие только вектор pET30a, а также культуры, не содержащие триптофана и пирувата. МС анализ родительского иона показывает, что соединение с (m+1)/z=293 не является продуктом распада более крупных молекул, а анализ дочерних ионов (проводимый как в примере 10) показывает сходство с монатином, полученным in vitro.
Влияние твина исследуют, используя твин-20 с конечными концентрациями 0, 0,2% (об./об.) и 0,6%. Наибольшее количество монатина получают во встряхиваемых колбах при концентрации твина 0,2%. Концентрация ампициллина варьирует от 0 до 10 мкг/мл. Количество монатина в клеточном бульоне быстро увеличивается (2,5X) от 0 до 1 мкг/мл, и увеличение составляет 1,3X, если концентрация ампициллина повышается от 1 до 10 мкг/мл.
Типичные результаты экспериментов по исследованию зависимости от времени представлены на фиг.10. Количество монатина, секретируемого в клеточную среду, увеличивается, даже если значения нормализуются по клеточному росту. Используя коэффициент молярной экстинкции триптофана, определяют количество монатина в среде, которое составляет менее 10 мкг/мл. Такой же эксперимент проводят с клетками, содержащими вектор, не имеющий вставки proA. Многие значения являются отрицательными, указывая на то, что высота пика m/z=293 в данных культурах меньше, чем просто в среде (фиг.10). В отсутствие триптофана и пирувата значения стабильно ниже, это указывает на то, что продукция монатина является результатом ферментативной реакции, катализируемой ферментом альдолазой.
Получение монатина in vivo в бактериальных клетках проводят во встряхиваемых колбах объемом 800 мл и в ферментаторах. Образец монатина объемом 250 мл (в среде, не содержащей клеток) очищают методом анионообменной хроматографии и препаративной жидкостной хроматографии на обращенной фазе. Полученный образец упаривают и анализируют методом МС высокого разрешения (описанным в примере 6). МС высокого разрешения показывает, что полученный метаболит представляет собой монатин.
Исследования in vitro показывают, что аминотрансфераза должна присутствовать на более высоком уровне, чем альдолаза (см. пример 6), следовательно, сверхэкспрессия аспартат-аминотрансферазы из E.coli в сочетании с геном альдолазы приводит к увеличению количества продуцируемого монатина. Для введения proA C. testosteroni в оперон, содержащий aspC/pET30 Xa/LIC, получают следующие праймеры: 5'-праймер: ACTCGGATCCGAAGGAGATATACATATGTACGAACTGGGACT (SEQ ID NO: 67) и 3'-праймер: CGGCTGTCGACCGTTAGTCAATATATTTCAGGC (SEQ ID NO: 68). 5'-Праймер содержит сайт BamHI, 3'-праймер содержит сайт SalI. ПЦР проводят по способу примера 4, очистку осуществляют методом гель-хроматографии. Конструкт aspC/pET30 Xa/LIC получают в результате расщепления BamHI и SalI, как и продукт ПЦР. Продукты расщепления очищают с помощью вращающейся колонки Qiagen. Продукт ПЦР proA встраивают в вектор с использованием набора для лигирования Roche Rapid DNA (Indianapolis, IN) в соответствии с инструкциями производителя. Химические превращения проводят, используя Novablues Singles (Novagen), как описано в примере 1. Колонии выращивают в среде LB, содержащей 50 мг/л канамицина, а ДНК-плазмиду выделяют, используя мининабор Qiagen. Скрининг клонов проводят по результатам рестрикционного расщепления, последовательность подтверждают с помощью Seqwright (Houston, TX). Конструкты субклонируют в BLR(DE3), BLR(DE3)pLysS, BL21(DE3) и BL21(DE3)pLysS (Novagen). Конструкт proA/pET30 Xa/LIC также трансформируют в BL21(DE3)pLysS.
Начальное сравнение образцов BLR(DE3) из встряхиваемых колб в стандартных условиях, описанных выше, показало, что добавление второго гена (aspС) увеличивает количество продуцируемого монатина в семь раз. Для ускорения роста используют штаммы-хозяева, полученные из BL21(DE3). Экспрессию клонов proA и клонов двух генных оперонов индуцируют в среде Trp-1, как описано выше, в случае хозяев pLysS к среде также добавляют хлорамфеникол (34 мг/л). Эксперименты во встряхиваемых колбах проводят в присутствии и в отсутствие 0,2% твин-20 и 1 мг/л ампициллина. Количество монатина в среде рассчитывают, используя в качестве стандарта очищенный монатин, полученный in vitro. Анализы SRM проводят по способу, описанному в примере 10. Образцы клеток берут после выращивания в течение ноля часов, 4 часов, 24 часов, 48 часов, 72 часов и 96 часов.
В таблице 4 представлены результаты, соответствующие максимальным количествам, продуцируемым в культуральных средах. В большинстве случаев конструкт, содержащий два гена, дает более высокие значения, чем конструкт, содержащий только proA. Штаммы pLysS, оболочка которых обладает большей пропускающей способностью, секретируют более высокие уровни монатина, несмотря на то, что обычно они растут медленнее. Добавление твина и ампициллина оказывает благоприятное действие.
Количество монатина, продуцируемого бактериями E.coli
ПРИМЕР 12
Получение монатина в дрожжах
В данном примере описываются способы, используемые для получения монатина в эукариотических клетках. Специалистам в данной области будет понятно, что подобные способы можно использовать для получения монатина в любой клетке, представляющей интерес. Кроме того, в сочетании с генами, описанными в данном примере, или вместо них, можно использовать другие гены (например, перечисленные на фиг.2).
Для клонирования и экспрессии генов aspC E.coli и proA C. testosteroni в Saccharomyces cerevisiae используют векторную систему pESC Yeast Epitope Tagging (Stratagene, La Jolla, CA). Векторы pESC содержат как промотор GAL1, так и промотор GAL10 на противоположных цепях, а также два разных сайта множественного клонирования, обеспечивающих экспрессию двух генов в одно время. Вектор pESC-His также содержит ген His3 для комплементации ауксотрофии гистидина в хозяине (YPH500). Промоторы GAL1 и GAL10 подавляются глюкозой и индуцируются галактозой; для достижения оптимальной экспрессии в дрожжах используют последовательность Козака. Плазмиды pESC представляют собой "челночные" векторы, позволяющие получать исходный конструкт в E.coli (содержащий ген bla для облегчения селекции); однако, в сайтах множественного клонирования отсутствуют участки, связывающие бактериальные рибосомы.
Для клонирования в pESC-His синтезируют следующие праймеры (сайты рестрикции подчеркнуты, последовательность Козака выделена жирным шрифтом): aspC (BamHI/SalI), GAL1: 5'-CGCGGATCC ATAATGGTTGAGAACATTACCG-3' (SEQ ID NO: 69) и 5'-ACGCGTCGACTTACAGCACTGCCACAATCG-3' (SEQ ID NO: 70). proA (EcoRI/NotI), GAL10: 5'-CCGGAATTC ATAATGGTCGAACTGGGAGTTGT-3' (SEQ ID NO: 71) и 5'-GAATGCGGCCGCTTAGTCAATATATTTCAGGCC-3' (SEQ ID NO: 72).
Вследствие введения последовательности Козака во втором кодоне обоих зрелых белков ароматическая аминокислота заменяется валином. Представляющие интерес гены амплифицируют с помощью мининабора для получения ДНК pET30 Xa/LIC, а в качестве матрицы используют клоны, описанные в примерах 1 и 4. ПЦР проводят, используя градиентный термоблок Eppendorf Master cycler и следующую схему реакции для объема 50 мкл: 1,0 мкл матрицы, 1,0 мкМ каждого праймера, 0,4 мМ каждого dNTP, 3,5 ед. полимеразы Expand, обеспечивающей высокую точность воспроизведения (Roche, Indianapolis, IN), и 1X буфер Expand™, содержащий Mg. Для термоблока используется программа, включающая горячий старт при 94°C в течение 5 минут, затем 29 повторов следующих стадий: 94°C в течение 30 секунд, 50°C в течение 1 минуты 45 секунд и 72°C в течение 2 минут 15 секунд. После 29 повторов образец держат 10 минут при 72°C и затем хранят при 4°C. Продукты ПЦР очищают разделением на 1% TAE-агарозном геле и затем извлекают, используя набор для экстракции в геле QIAquick (Qiagen, Valencia, CA).
Вектор ДНК pESC-His (2,7 мкг) расщепляют BamHI/SalI и очищают в геле, как описано выше. Продукт ПЦР aspC расщепляют BamHI/SalI и очищают на колонке для ПЦР-очистки QIAquick. Лигирование проводят с помощью набора для лигирования Roche Rapid DNA, следуя инструкциям производителя. Обессоленные продукты лигирования вводят в компетентные клетки Electromax DH10B (Invitrogen), 40 мкл, методом электропорации в одноразовой кювете Biorad, 0,2 см, используя генератор импульсов Biorad Gene Pulser II с импульсным контроллером, в соответствии с инструкциями производителя. После восстановления в течение 1 часа в 1 мл среды SOC трансформированные клетки помещают в плашки в среде LB, содержащей 100 мкг/мл ампициллина. Препараты плазмидной ДНК для клонирования получают с помощью наборов QIAprep Spin Miniprep. Плазмидную ДНК подвергают скринингу, используя рестрикционное расщепление, и последовательность подтверждают путем секвенирования (Seqwright) с использованием праймеров, синтезированных для вектора.
Клон aspC/pESC-His расщепляют EcoRI и NotI, как и продукт ПЦР proA. ДНК очищают и лигируют, как описано выше. Конструкт, содержащий два гена, вводят в клетки DH10B и подвергают скринингу путем рестрикционного расщепления и ДНК-секвенированию.
Конструкт вводят в штамм S. cerevisiae YPH500 с помощью набора для трансформации S.c. EasyComp((Invitrogen). Реакции трансформации проводят в минимальной среде SC-His (Invitrogen pYES2 manual), содержащей 2% глюкозы. Проводят скрининг отдельных колоний дрожжей по присутствию генов proA и aspC путем ПЦР колоний с использованием указанных выше ПЦР-праймеров. Клеточные осадки (2 мкл) суспендируют в 20 мкл буфера Y-Lysis (Zymo Research), содержащего 1 мкл зимолазы, и нагревают при 37°C в течение 10 минут. Затем четыре мкл данной суспензии вносят в реакционную смесь ПЦР объемом 50 мкл, причем используют реакционную смесь и программу, описанные выше.
Суспензии культур объемом пять мл выращивают в течение ночи на среде SC-His+глюкоза при 30°C и 225 об/мин. Клетки постепенно адаптируют к росту на рафинозе, чтобы минимизировать лаг-период перед индукцией галактозой. Приблизительно после 12 часов роста измеряют поглощение при 600 нм, соответствующий объем суспензии центрифугируют и клеточный осадок ресуспендируют в свежей среде SC-His до получения OD 0,4. Для индукции последовательно используют следующие источники углерода: 1% рафинозы+1% глюкозы, 0,5% глюкозы+1,5% рафинозы, 2% рафинозы и, наконец, 1% рафинозы+2% галактозы.
Приблизительно после 16 часов выращивания в индукционной среде 50 мл суспензий культур разделяют на дубликаты по 25 мл и к одному из дубликатов добавляют следующие ингредиенты: (указаны конечные концентрации) 1 г/л L-триптофана, 5 мМ фосфата натрия pH 7,1, 1 г/л пирувата натрия, 1 мМ MgCl2. Образцы бульонов и клеточных осадков после выращивания в неиндукционной среде, а также после 16-часового культивирования перед добавлением субстратов пути синтеза монатина сохраняют в качестве отрицательных контролей. Кроме того, в качестве дополнительного отрицательного контроля используют конструкты, содержащие только функциональный ген aspC (и усеченный ген proA). После индукции клетки оставляют расти в течение 69 часов. Иногда дрожжевые клетки индуцируют при более низких значениях OD и выращивают перед добавлением триптофана и пирувата только 4 часа. Однако поскольку данные монатиновые субстраты ингибируют рост, эффективнее добавлять их при более высоком значении OD.
Клеточные осадки культур лизируют, используя 5 мл YeastBuster(+50 мкл THP (Novagen) на грамм (влажной массы) клеток в соответствии с инструкциями производителя, с добавлением ингибиторов протеаз и бензоназы нуклеазы, как описано в предыдущих примерах. Культуральную среду и клеточные экстракты фильтруют и анализируют методом SRM, как описано в примере 10. С помощью данного метода монатин в среде не обнаруживается, это указывает на то, что в используемых условиях клетки не секретируют монатин. В данных условиях протонная тяга может быть недостаточной, либо общие транспортеры аминокислот могут быть насыщены триптофаном. Экспрессия белка находится на уровне, который не позволяет детектировать изменения с помощью SDS-PAGE.
Транзиентную экспрессию монатина детектируют (приблизительно 60 нг/мл) в клеточных экстрактах культуры, содержащей два функциональных гена, если в среду добавляют триптофан и пируват. В клеточных экстрактах отрицательных контролей монатин не обнаруживается. Анализы монатина in vitro проводят в дубликатах, содержащих 4,4 мг/мл общего белка (приблизительно в два раза больше, чем в случае клеточных экстрактов E.coli), с использованием оптимизированного способа, описанного в примере 6. Другие анализы проводят с добавлением либо 32 мкг/мл альдолазы ProA C. testosterone, либо 400 мкг/мл аминотрансферазы AspC, чтобы определить, какой фермент является лимитирующим для клеточных экстрактов. В отрицательные контроли фермент не добавляют, либо добавляют только аминотрансферазу AspC (альдольная конденсация может происходить в некоторой степени в отсутствие фермента). Положительные контроли содержат частично чистые ферменты (30-40%), 16 мкг/мл альдолазы и 400 мкг/мл аминотрансферазы.
Результаты экспериментов in vitro анализируют методом SRM. Анализ клеточных экстрактов показал, что, если триптофан добавляют в среду после индукции, он эффективно проникает в клетки, поэтому его уровень на два порядка выше, чем в том случае, когда дополнительный триптофан не добавляют. Результаты анализа монатина in vitro представлены в таблице 5 (в нг/мл).
Продукция монатина в экстрактах дрожжевых клеток
Положительные результаты получают для экстрактов клеток, содержащих двухгенный конструкт, при добавлении субстрата и без добавления субстрата к питательной среде. Данные результаты при сравнении с положительными контролями показывают, что ферменты экспрессируются на уровне, близком к 1% от общего содержания белка в дрожжах. Количество продуцируемого монатина в экстрактах клеток, содержащих конструкт aspC (с усеченным proA), при анализе в присутствии альдолазы значительно выше, чем при анализе просто клеточных экстрактов, и указывает, что рекомбинантная аминотрансфераза AspC составляет приблизительно 1-2% от общего содержания белка в дрожжах. В клеточных экстрактах неиндуцированных культур в присутствии альдолазы наблюдается небольшой уровень активности вследствие наличия в клетках нативной аминотрансферазы. В присутствии аминотрансферазы AspC активность в экстрактах неиндуцированных клеток увеличивается до количества монатина, продуцируемого отрицательным контролем, содержащим AspC (приблизительно 200 нг/мл). Наоборот, в экстрактах клеток, содержащих двухгенный конструкт, при добавлении аминотрансферазы активность выше, чем при добавлении альдолазы. Поскольку оба гена экспрессируются на одном уровне, данный факт указывается на то, что количество продуцируемого монатина достигает максимума, если уровень аминотрансферазы выше уровня альдолазы, что находится в соответствии с результатами, полученными в примере 6.
Добавление пирувата и триптофана не только ингибирует клеточный рост, но также, по-видимому, ингибирует экспрессию белка. Для коррекции ауксотрофии триптофана в клетках-хозяевах YPH500 можно добавить плазмиду pESC-Trp, чтобы снизить влияние триптофана на рост, экспрессию и секрецию.
ПРИМЕР 13
Усовершенствование ферментативных процессов с использованием сопряженных реакций
Теоретически, при отсутствии побочных реакций или распада субстратов или промежуточных соединений, максимальное количество продукта ферментативной реакции, представленной на фиг.1, прямо пропорционально константам равновесия каждой реакции и концентрациям триптофана и пирувата. Триптофан является малорастворимым субстратом, а концентрации пирувата, превышающие 200 мМ, оказывают негативное действие на выход (см. пример 6).
В идеале, концентрацию монатина максимизируют по отношению к субстратам, чтобы уменьшить стоимость разделения. Физическое разделение можно проводить, удаляя монатин из реакционной смеси, что препятствует протеканию обратных реакций. Исходные вещества и катализаторы можно впоследствии регенерировать. Из-за того, что по размеру, заряду и гидрофобности монатин похож на некоторые реагенты и промежуточные соединения, физическое разделение затруднено, если не использовать большого количества вещества, обладающего сродством к монатину (как, например, в методе аффинной хроматографии). Однако реакции, приводящие к получению монатина, могут быть сопряжены с другими реакциями так, чтобы равновесие в системе сдвигалось в сторону образования монатина. Далее представлены примеры способов улучшения выхода монатина, получаемого из триптофана или индол-3-пирувата.
Сопряженные реакции с использованием оксалоацетат-декарбоксилазы (EC 4.1.1.3)
На фиг.11 представлена схема реакции. Чтобы направить реакцию в сторону образования индол-3-пирувата, используют триптофан-оксидазу и каталазу. Каталазу используют в избытке, чтобы пероксид водорода не участвовал в обратной реакции или не разрушал ферменты или промежуточные соединения. В процессе каталазной реакции кислород регенерируется. Альтернативно, в качестве субстрата можно использовать индол-3-пируват.
Аспартат используют в качестве донора аминогруппы для аминирования MP, катализируемого аспартат-аминотрансферазой. В идеале, используют аминотрансферазу, которая обладает низкой специфичностью по отношению к реакции триптофан/индол-3-пируват по сравнению с реакцией превращения MP в монатин, чтобы аспартат не использовался для повторного аминирования индол-3-пирувата. Для превращения оксалоацетата в пируват и диоксид углерода можно добавить оксалоацетат-декарбоксилазу (из Pseudomonas sp.). Поскольку CО2 является летучим, он не вступает в реакцию с ферментами, в результате уменьшается вероятность протекания обратных реакций или предотвращается их протекание. Пируват, образующийся на данной стадии, также можно использовать в реакции альдольной конденсации. Можно использовать другие декарбоксилазы, известно, что гомологичные ферменты присутствуют в Actinobacillus actinomycetemcomitans, Aquifex aeolicus, Archaeoglobus fulgidus, Azotobacter vinelandii, Bacteroides fragilis, некоторых видах Bordetella, Campylobacter jejuni, Chlorobium tepidum, Chloroflexus aurantiacus, Enterococcus faecalis, Fusobacterium nucleatum, Klebsiella pneumoniae, Legionella pneumophila, Magnetococcus MC-1, Mannheimia haemolytica, Methylobacillus flagellatus KT, Pasteurella multocida Pm70, Petrotoga miotherma, Porphyromonas gingivalis, некоторых видах Pseudomonas, некоторых видах Pyrococcus, Rhodococcus, некоторых видах Salmonella, некоторых видах Streptococcus, Thermochromatium tepidum, Thermotoga maritima, Treponema pallidum и некоторых видах Vibrio.
Анализы триптофан-аминотрансферазы проводят с использованием аспартат-аминотрансферазы (AspC) из E. Coli, тирозин-аминотрансферазы (TyrB) из E. Coli, полисубстратной аминотрасферазы (BSAT) из L. major и двух коммерчески доступных свиных глутамат-оксалоацетат-аминотрансфераз, как описано в примере 1. И оксалоацетат, и альфа-кетоглутарат анализируют в качестве акцепторов аминогруппы. Чтобы определить, какой фермент обладает наивысшей специфичностью в отношении монатин-аминотрансферазной реакции, сравнивают активность, наблюдающуюся при использовании монатина, с активностью, наблюдающейся при использовании триптофана. Результаты показывают, что наивысшей специфичностью по отношению к монатиновой реакции по сравнению с триптофановой реакцией обладает свиная глутамат-оксалоацетат-аминотрансфераза типа II, GOAT (Sigma G7005). Данная специфичность не зависит от типа используемого аминоакцептора. Следовательно, указанный фермент используют в реакциях, сопряженных с оксалоацетат-декарбоксилазными реакциями.
Типичная реакционная смесь с индол-3-пируватом в качестве исходного вещества включает (указаны конечные концентрации) 50 мМ Tris-HCl pH 7,3, 6 мМ индол-3-пируват, 6 мМ пируват натрия, 6 мМ аспартат, 0,05 мМ PLP, 3 мМ фосфат калия, 3 мМ MgCl2, 25 мкг/мл аминотрансферазы, 50 мкг/мл альдолазы ProA C. tectosteroni и 3 ед./мл декарбоксилазы (Sigma O4878). Реакцию оставляют протекать в течение 1 часа при 26°C. В некоторых случаях декарбоксилазу исключают или аспартат заменяют на альфа-кетоглутарат (в случае отрицательных контролей). Чтобы подтвердить ранее проводимые эксперименты по специфичности, вместо GOAT также анализируют описанные выше аминотрансферазы. Образцы фильтруют и анализируют методом ЖХ/МС, как описано в примере 10. Результаты показывают, что фермент GOAT продуцирует максимальное количество монатина на мг белка, и при этом образуется минимальное количество триптофана как побочного продукта. Кроме того, применение GOAT дает 2-3-кратное улучшение по сравнению с применением фермента декарбоксилазы. Фермент AspC E.coli также продуцирует большие количества монатина по сравнению с другими аминотрансферазами.
Продукция монатина увеличивается в результате: 1) периодического добавления 2 мМ индолпирувата, пирувата и аспартата (с интервалом от получаса до часа), 2) проведения реакций в анаэробной среде или с дегазацией буферов, 3) проведения реакции в течение ночи и 4) использования свежеполученной декарбоксилазы, которая не подвергалась несколько раз замораживанию-оттаиванию. Пируват ингибирует декарбоксилазу в концентрациях, превышающих 12 мМ. При концентрациях индол-3-пирувата более 4 мМ ускоряются побочные реакции с участием индол-3-пирувата. Количество индол-3-пирувата, используемое в реакции, можно увеличить, если увеличивается количество альдолазы. Обнаружено, что высокие уровни фосфата (50 мМ) и аспартата (50 мМ) ингибируют фермент декарбоксилазу. В случае реакции, проводимой в течение одного часа, добавляемое количество фермента декарбоксилазы можно уменьшить до 0,5 ед./мл без снижения продукции монатина. Количество продуцируемого монатина возрастает при увеличении температуры от 26°C до 30°C и от 30°C до 37°C; однако, при 37°C ускоряются побочные реакции с участием индол-3-пирувата. Количество продуцируемого монатина возрастает при увеличении pH от 7 до 7,3 и является относительно стабильным в интервале pH 7,3-8,3.
Типичная реакционная смесь с триптофаном в качестве исходного вещества включает (указаны конечные концентрации) 50 мМ Tris-HCl pH 7,3, 20 мМ триптофан, 6 мМ аспартат, 6 мМ пируват натрия, 0,05 мМ PLP, 3 мМ фосфат калия, 3 мМ MgCl2, 25 мкг/мл аминотрансферазы, 50 мкг/мл альдолазы ProA C. tectosteroni, 4 ед./мл декарбоксилазы, 5-200 мЕд./мл оксидазы L-аминокислот (Sigma A-2805), 168 ед./мл каталазы (Sigma C-3515) и 0,008 мг FAD. Реакции проводят в течение 30 минут при 30°C. Улучшение наблюдается при добавлении декарбоксилазы. Максимальное количество монатина продуцируется при использовании 50 мЕд./мл оксидазы. Улучшения достигаются так же, как и при использовании индол-3-пирувата в качестве субстрата. Кроме того, количество продуцируемого монатина возрастает, если 1) используется низкий уровень триптофана (т.е. если данный уровень ниже Km фермента аминотрансферазы и, следовательно, триптофан не способен конкурировать с MP за связывание с активным центром), и 2) отношение оксидазы к альдолазе и количество аминотрансферазы поддерживаются на таком уровне, что индол-3-пируват не накапливается.
Если в качестве исходных веществ используют либо индол-3-пируват, либо триптофан, количество монатина, продуцируемого в реакции со временем инкубации 1-2 часа, возрастает, если количества всех ферментов увеличиваются в 2-4 раза при сохранении прежнего отношения ферментов. При использовании любого субстрата получают концентрацию монатина, составляющую приблизительно 1 мг/мл. Если в качестве исходного вещества используют индол-пируват, количество образующегося триптофана обычно составляет менее 20% от количества продукта, что является благоприятным результатом применения сопряженных реакций. Производительность и выход можно существенно увеличить путем дополнительной оптимизации и регуляции концентраций промежуточных соединений и побочных реакций.
Сопряженные реакции с использованием лизин-эпсилон-аминотрансферазы (EC 2.6.1.36)
Лизин-эпсилон-аминотрансфераза (L-лизин-6-трансаминаза) обнаружена в некоторых организмах, в том числе, Rhodococcus, Mycobacterium, Streptomyces, Nocardia, Flavobacterium, Candida utilis и Streptomyces. Она используется организмами на первой стадии получения некоторых бета-лактамных антибиотиков (Rius and Demain, J. Microbiol. Biotech., 7: 95-100, 1997). Данный фермент превращает лизин в L-2-аминоадипат 6-семиальдегид (аллизин) путем PLP-опосредованного трансаминирования лизина по C-6 с использованием альфа-кетоглутарата в качестве аминоакцептора. Аллизин нестабилен и спонтанно подвергается внутримолекулярной дегидратации с образованием циклической молекулы 1-пиперидин-6-карбоксилата. Это приводит к эффективному ингибированию любых побочных реакций. Схема реакции изображена на фиг.12. Также можно использовать альтернативный фермент, лизин-пируват-6-трансаминазу (EC 2.6.1.71).
Типичная реакционная смесь содержит в 1 мл: 50 мМ Tris-HCl pH 7,3, 20 мМ индол-3-пируват, 0,05 мМ PLP, 6 мМ фосфат калия pH 8, 2-50 мМ пируват натрия, 1,5 мМ MgCl2, 50 мМ лизин, 100 мкг аминотрансферазы (лизин-эпсилон-аминотрансфераза LAT-101, BioCatalytics Pasadena, CA) и 200 мкг альдолазы ProA C. testosteroni. Количество продуцируемого монатина возрастает при увеличении концентрации пирувата. В данных условиях реакции (при концентрации пирувата 50 мМ) максимальное количество в 10 раз меньше, чем в случае применения сопряженных реакций с использованием оксалоацетат-декарбоксилазы (приблизительно 0,1 мг/мл).
Время элюирования пика, соответствующего [M+H]+=293, совпадает со временем элюирования монатина, а масс-спектр содержит несколько таких же фрагментов, как и в случае других ферментативных процессов. Второй пик с соответствующим соотношением массы к заряду (293) элюируется немного раньше, что характерно для S,S-монатина, полученного в примере 6, и может указывать на присутствие другого стереоизомера монатина. Данный фермент продуцирует очень мало триптофана. Однако имеет место некоторая активность в отношении пирувата (образуется аланин в качестве побочного продукта). Кроме того, известно, что данный фермент нестабилен. С помощью экспериментов по направленной эволюции можно добиться таких улучшений, как увеличение стабильности, уменьшение активности в отношении пирувата и увеличение активности в отношении MP. Данные реакции могут сочетаться с реакциями, катализируемыми оксидазой/каталазой L-аминокислот, как описано выше.
Другие сопряженные реакции
Другие сопряженные реакции, которые могут улучшить выход монатина, получаемого из триптофана или индолпирувата, представлены на фиг.13. Формиат-дегидрогеназа (EC 1.2.1.2 или 1.2.1.43) является обычным ферментом. Для функционирования одних формиат-дегидрогеназ требуется присутствие NADH, тогда как другие используют NADPH. Глутамат-дегидрогеназа катализирует описанное в предыдущих примерах взаимопревращение предшественника монатина и монатина, которое проводят в буферах на основе аммония. Присутствие формиата аммония и формиат-дегидрогеназы обеспечивает эффективную регенерацию кофакторов, а образование диоксида углерода эффективно снижает скорость обратных реакций (Bommarius et al., Biocatalysis 10: 37, 1994 and Galkin et al. Appl. Environ. Microbiol. 63: 4651-6, 1997). Кроме того, в буфере для реакции можно растворить большие количества формиата аммония. Выход монатина, получаемого с помощью реакций, катализируемых глутамат-дегидрогеназой (или подобных реакций восстановительного аминирования), можно улучшить путем добавления формиат-дегидрогеназы и формиата аммония.
Для сдвига равновесия в сторону образования монатина можно использовать другие способы. Например, если аминопропан используется в качестве донора аминокислоты в реакции превращения MP в монатин с использованием аминотрансферазы омега-аминокислот (EC 2.6.1.18), такой как описанная в патентах США 5360724 и 5300437, то одним из продуктов является ацетон, более летучий продукт, чем субстрат аминопропан. Чтобы удалить ацетон и таким образом сдвинуть равновесие, можно периодически повышать температуру в течение коротких промежутков времени. Температура кипения ацетона составляет 47°C, данная температура при использовании в течение коротких промежутков времени, по всей вероятности, не вызывает разложения промежуточных соединений. Большинство аминотрансфераз, обладающих активностью по отношению к альфа-кетоглутарату, также обладают активностью по отношению к предшественнику монатина. Подобным образом, если аминотрансфераза глиоксилата/ароматических кислот (EC 2.6.1.60) используется вместе с глицином в качестве аминодонора, то образуется глиоксилат, который является относительно нестабильным и имеет гораздо более низкую температуру кипения, чем глицин.
ПРИМЕР 14
Кривая "доза-ответ"
Растворы монатина (смесь, содержащая приблизительно 96% 2R,4R/2S,4S-энантиомерной пары и 4% 2R,4S/2S,4R-энантиомерной пары, также называемая "рацемическая смесь" монатина) с концентрацией 15, 30, 45, 60, 75 и 90 м.д. получают в модельной системе безалкогольного напитка, pH 3,2, содержащей 0,14% (вес./об.) лимонной кислоты и 0,04% (вес./об.) цитрата натрия. Сладость монатина по сравнению с сахарозой определяют с помощью описанного ниже метода оценки сладости. Все определения проводятся с использованием дубликатов группой экспертов (n=6-8), обладающих опытом по определению сладости данным методом. Все образцы анализируют при температуре 20°C±1°C.
Растворы монатина маркируют и индивидуально передают экспертам в случайном порядке. Также предоставляются стандартные растворы сахарозы, имеющие концентрацию сахарозы в интервале 2,0-11,0% (вес./об.) с шагом 0,5% (вес./об.). Экспертов просят оценить сладость путем сравнения сладости тестируемого раствора со сладостью стандартных растворов сахарозы. Чтобы провести оценку, делают последовательно 3 глотка тестируемого раствора, глоток воды, 3 глотка стандартного раствора сахарозы, глоток воды и т.д. Эксперты оценивают сладость до первого десятичного знака, например, 6,8, 8,5. Разные тестируемые растворы оценивают с перерывом в пять минут. Экспертов также просят хорошо сполоснуть рот и съесть крекер, чтобы уменьшить любые возможные эффекты наложения вкуса. Значения сахарозного эквивалента (SEV) и стандартные отклонения представлены в таблице 6.
Установлено, что смеси характеризуются быстрым появлением сладкого вкуса и развитием сладости до максимальной интенсивности. Ослабление сладости тоже происходит быстро. Установлено, что большинство смесей имеет меньше фруктового аромата, чем сахароза, за исключением смеси монатин/глюкоза. Отмечено, что остается незначительное сладкое послевкусие и очень слабые горькие/металлические оттенки. Лакричного или охлаждающего послевкусия не обнаружено.
Данные для кривой "доза-ответ" монатина
ПРИМЕР 15
Смешивание монатина с углеводными подсластителями
Получают смеси монатина (как описано в примере 14) с сахарозой, HFCS (55% фруктозы) и сиропом глюкозы (63 эквивалента декстрозы, DE), эквивалентные по сладости 10,0% (вес./об.) раствору сахарозы. Для каждого углеводного подсластителя отношение монатин:подсластитель подбирают так, чтобы вклад монатина в общую сладость составлял 25, 50 и 75%. Соответствие сладости 10,0% (вес./об.) раствору сахарозы определяют с помощью метода оценки сладости, описанного в примере 14. Как и в примере 14, все анализы проводятся на модельной системе безалкогольного напитка, pH 3,2, группой из 6-8 экспертов, с использованием дубликатов каждой пробы. Результаты представлены в таблицах 7-9. Монатин подобен сукралозе, с легкой задержкой появления сладости.
Сравнение сладости смесей монатина и растворов сахарозы
(%; вес./об.)
Сравнение сладости смесей монатина и HFCS
Сравнение сладости смесей монатина и сиропа глюкозы
Затем качество эквивалентной сладости смесей монатин/углевод (50:50) оценивается по сравнению с сахарозой небольшой группой опытных экспертов. Данную оценку проводят "двойным слепым" методом. Подслащенную сахарозой систему идентифицируют как контроль, а другие продукты маркируют случайным образом. Экспертов просят оценить маркированный случайным образом образец по сравнению с контролем по следующим параметрам: характеристика сладости: появление, развитие и ослабление; характеристика аромата: кислотность, горькость и другие свойства; ощущение во рту; и послевкусие. Экспертов также просят оценить качество системы подсластителя по шкале (1; плохо - 5; хорошо). Итоговые комментарии и оценки представлены в таблице 10.
Вкусовые профили смесей монатин/углевод
ПРИМЕР 16
Временной профиль интенсивности монатина в системе безалкогольного напитка
Получают растворы, содержащие 80 м.д. монатина (рацемическая смесь монатина, описанная в примере 14), 10,0% (вес./об.) сахарозы и 200 м.д. сукралозы в модельной системе безалкогольного напитка, pH 3,2, описанной в примере 14. Затем для полученных растворов определяют временной профиль интенсивности с помощью нижеследующего метода. В исследовании участвуют шесть экспертов. Данных экспертов отбирают по общей остроте ощущений, а также по чувствительности к интенсивности сладости и к различиям в качестве сладости. Все эксперты обладают опытом тестирования подсластителей и прошли специальную подготовку по проведению оценки временной интенсивности. Сеансы обучения проводят вначале, чтобы ознакомить эксперта с методом анализа и оценки образцов с течением времени с помощью компьютеризованной системы ввода данных.
Образцы каждого раствора (13 мл) маркируют и индивидуально передают экспертам в случайном порядке. Для каждого эксперта сразу после проглатывания компьютер регистрирует изменение интенсивности во времени по шкале 0-100 каждую секунду, в течение 60 секунд. Каждый раствор оценивают с дубликатами. Результаты оценки временной интенсивности представлены в таблице 11.
Результаты исследования временной интенсивности
Полученные результаты показывают, что временные характеристики вкуса монатина сравнимы с соответствующими характеристиками сахарозы, что указывает на высокое качество подсластителя. Кроме того, монатин выигрывает по сравнению с сукралозой, широко распространенным подсластителем с высокой интенсивностью.
ПРИМЕР 17
Получение напитков кола и лимон/лайм, содержащих монатин
Напитки кола и лимон/лайм получают, используя нижеследующие композиции, и подслащивают их сахарозой, HFCS (55% фруктозы), аспартамом, сукралозой, монатином (рацемическая смесь, описанная в примере 14), монатин/сахарозой или монатин/HFCS. Одну часть сиропа смешивают с 5,5 частями газированной воды и анализируют.
Композиция сиропа лимон/лайм:
Композиция сиропа колы:
Концентрации подсластителей в газированном напитке лимон/лайм или кола:
Оценки проводятся "двойным слепым" методом группой опытных дегустаторов. Продукт, подслащенный сахарозой, идентифицируют как контроль, а все другие продукты маркируют случайным образом. Экспертов просят оценить маркированный случайным образом образец по сравнению с контролем по следующим параметрам:
горькость
другие свойства
развитие
интенсивность
ослабление
Послевкусие
Экспертов также просят оценить качество системы подсластителя по шкале (1; плохо - 5; хорошо). Итоговые комментарии, сделанные вместе с присвоением оценки, представлены в таблицах 12 и 13 для газированных напитков лимон/лайм и кола, соответственно. В аромате лимон/лайм монатин сравним по вкусу с аспартамом. Смеси монатин/углевод оценены выше. В коле монатин похож на аспартам.
Вкусовые профили газированных напитков лимон/лайм
Вкусовые профили газированных напитков лимон/лайм
Вкусовые профили газированных напитков кола
Вкусовые профили газированных напитков кола
сахароза
Обсуждение
Использующийся в данном примере монатин обладает чистым, сладким вкусовым профилем, практически не содержащим горькости, охлаждающих и лакричных ароматов, зачастую свойственных природным высокоинтенсивным подсластителям. Используемая в данном примере смесь стереоизомеров монатина позволяет получать сглаженную правильную кривую "доза-ответ" с относительной интенсивностью сладости 1250 (по сравнению с раствором сахарозы с концентрацией 10,0% (вес./об.) SEV.
Результаты исследования по изменению интенсивности во времени показывают, что временной профиль сладости монатина ориентировочно совпадает с профилями сахарозы и сукралозы. По сравнению с сахарозой у монатина немного дольше достигается максимальная интенсивность и медленнее происходит ослабление вкуса при более сильном ощущении сладости в конце анализа (60 с). Однако наблюдаемые различия не являются статистически значимыми.
При смешивании с углеводными подсластителями монатин обеспечивает интенсивность сладости, превосходящую интенсивность сладости сахарозы в 1500-2000 раз. Полученные смеси имеют очень высокое качество сладости и хорошие характеристики вкуса. Небольшое замедление появления сладости наблюдается только при низком уровне детектируемой задержки сладости. Смеси монатина и углеводных подсластителей можно использовать, например, для получения напитков со средним содержанием калорий.
Анализ монатина дает хорошие результаты как при его отдельном использовании, так и в смеси с углеводными подсластителями. В случае газированных напитков лимон/лайм продукты, подслащенные только монатином, имеют такой же вкусовой профиль, как и напитки, подслащенные аспартамом и сукралозой. Напиток, подслащенный смесью монатин/сахароза, обладает особенно хорошими характеристиками и реально оценивается как более приемлемый, чем контрольный продукт, подслащенный сахарозой. Предполагается, что монатин в смеси с другими углеводными подсластителями усиливает лимонный/лаймовый аромат. В случае напитков кола при использовании смеси монатина с HFCS получают продукт, приемлемый в такой же степени, как и контроль, полученный с использованием HFCS.
ПРИМЕР 18
Органолептическая стабильность монатина в воде
Органолептическую стабильность монатина (рацемическая смесь, описанная в примере 14) в воде (8% SEV) исследуют после хранения при комнатной температуре в течение 0-6 часов. Значения SEV регистрируют (как описано выше в примере 14) либо через 0-1 часов, либо через 5-6 часов после получения раствора монатина. После 6 часов хранения при комнатной температуре уменьшение значения SEV монатина не наблюдается; полученные результаты подтверждаются аналитическими исследованиями с использованием метода ЖХ/МС (например, по отсутствию лактонизации).
ПРИМЕР 19
Получение предварительной смеси солодового напитка
Предварительную смесь солодового напитка получают, используя ингредиенты, перечисленные в таблице 14.
ПРИМЕР 20
Получение предварительной смеси шоколадного напитка
Предварительную смесь шоколадного напитка получают, используя ингредиенты, перечисленные в таблице 15. Немолочные сливки могут включать растительное масло, загуститель, лецитин, белок, витамины, минералы, эмульгаторы (такие как лецитин, DATEM и моно- и диглицериды) и наполнители (например, сухой кукурузный сироп, низкокалорийные наполнители).
ПРИМЕР 21
Получение предварительной смеси апельсинового напитка
Предварительную смесь апельсинового напитка получают, используя ингредиенты, перечисленные в таблице 16.
Апельсиновый напиток можно получить путем смешивания приблизительно 1 унции сухой смеси с 8 унциями воды, с последующим перемешиванием и встряхиванием до полной гидратации. Таким образом, полученный готовый к употреблению напиток содержит приблизительно от 66 до 440 м.д. S,S-монатина, приблизительно от 6 до 13 м.д. R,R-монатина, или их смесь.
ПРИМЕР 22
Получение лимонада с использованием монатина в качестве подсластителя
Можно получить удобные одноразовые пакетики с подсластителем, содержащим монатин, причем подсластитель обеспечивает сладость, эквивалентную 2 чайным ложкам (~8 грамм) гранулированного сахара. Поскольку S,S-монатин в 50-200 раз слаще, чем сахароза, 40-160 мг S,S-монатина обеспечивают сладость, эквивалентную 8 граммам гранулированного сахара. Так, например, с учетом оптимизации сладости+/-25%, одноразовый пакетик, содержащий 1 грамм композиции монатина, может содержать приблизительно 40-200 мг S,S-монатина.
Подобным образом, поскольку R,R-монатин в 2000-2400 раз слаще, чем сахароза, 3,3-4,0 мг R,R-монатина обеспечивают сладость, эквивалентную 8 граммам сахара. Так, в другом варианте, с учетом оптимизации сладости+/-25%, одноразовый пакетик, содержащий 1 грамм композиции монатина, может содержать приблизительно 3,3-5,0 мг R,R-монатина. В другом варианте пакетик с композицией может содержать 40-200 мг S,S-монатина, 3,3-5,0 мг R,R-монатина или их комбинацию в таких же или меньших количествах на грамм общей массы, чтобы обеспечить сладость, эквивалентную 2 чайным ложкам гранулированного сахара.
Чтобы получить лимонад, в фужере смешивают 2 столовые ложки лимонного сока и 3 пакетика (3 г) композиции монатина с 3/4 чашки воды до растворения. Добавляют лед. Лимонад, подслащенный монатином, почти эквивалентен по сладости и в равной степени предпочтителен лимонаду, подслащенному 6 чайными ложками (24 г) сахарозы, но имеет значительно меньше калорий (приблизительно 0 калорий по сравнению с 96 калориями).
ПРИМЕР 23
Оценка подсластителей, содержащих R,R-монатин, в кофе и чае со льдом
Монатин-содержащие композиции подсластителей, содержащие R,R-монатин или сочетания R,R-монатин/эритрит, оценивают по сравнению с другими известными подсластителями (аспартамом и сукралозой) в кофе и чае со льдом. Основные оцениваемые органолептические параметры включают качество сладости, послевкусие, горький вкус и его послевкусие. Проводят качественную оценку.
Композиции продуктов
(i) Кофе
Для оценки характеристик подсластителя используют стандартный кофе (таблица 17).
Композиция кофе
Подсластители добавляют в кофе в следующих концентрациях:
(ii) Чай со льдом
Для оценки характеристик подсластителя получают композицию чая со льдом (таблица 18).
Композиция чая со льдом
Подсластители добавляют в чай в следующих концентрациях:
Органолептическая оценка
Оценка полученных напитков чая и кофе проводится группой экспертов (n=6), имеющих опыт органолептических оценок, которые анализируют кофейные продукты во время одной дегустации, а чайные продукты во время другой дегустации. Результаты проведенных анализов представлены в таблице 19.
Органолептическая оценка кофе и чая (размер порции 200 мл)
концентрация
Обсуждение
В композициях подсластителей монатин обеспечивает неожиданное улучшение характеристик, в том числе, отчетливо улучшает органолептические свойства. При добавлении монатина в кофе отчетливо ощущается увеличение уровня кофейного аромата. Данное улучшение дополнительно усиливается при добавлении низких концентраций эритрита, который способен уравновешивать и сглаживать аромат, а также ускорять появление сладости. В чае со льдом, особенно в подкисленном чае, монатин усиливает оттенки лимонного аромата. Опять же, эритрит в смеси с монатином способствует дополнительному улучшению аромата.
Монатин обеспечивает улучшение органолептических свойств (например, уменьшает послевкусие, уменьшает посторонний привкус, не маскирует вкус) широко употребляемых напитков, таких как чай и кофе. Подслащенный монатином кофе содержит около 0 калорий, в отличие от кофе, подслащенного 2 чайными ложками (~8 г) сахарозы, который содержит 32 калории.
Ожидается, что в композициях напитков монатин будет усиливать все цитрусовые ароматы, а также обеспечивать более благоприятный временной профиль интенсивности сладости по сравнению с аспартамом или сукралозой. Также ожидается, что в композициях напитков смесь монатина и эритрита будет дополнительно усиливать цитрусовые ароматы и обеспечивать более благоприятные профили сладости по сравнению с аспартамом или сукралозой. Ожидается, что смеси монатина и эритрита будут обеспечивать данные улучшения в любых композициях напитков, таких как безалкогольные напитки, газированные напитки, сиропы, сухие смеси для приготовления напитков и концентрированные напитки, хранящиеся при низких температурах.
ПРИМЕР 24
Оценка R,R-монатина в напитках
Получают напитки (кола, лимон-лайм и апельсин) и подслащивают их аспартамом, сукралозой или R,R-монатином. Проводят качественную оценку.
Композиции продуктов
Получение и оценка композиций безалкогольных напитков представлены в таблице 20. Термин "приводить" относится к разбавлению водой. Например, представление "1+4" означает смешивание 1 части композиции концентрата с 4 частями воды. Так, если композиция концентрата содержит 0,021% вес./об. (т.е. 210 м.д.) R,R-монатина, например, представление 1+4 означает получение раствора, содержащего 42 м.д. (210 м.д./5) R,R-монатина.
Композиции безалкогольных напитков (концентраты)
(%; вес./об.)
(ii) Сукралоза 0,100
(iii) R,R-монатин 0,021
(ii) Сукралоза 0,1430
(iii) R,R-монатин 0,0293
(ii) Сукралоза 0,110
(iii) R,R-монатин 0,0225
Полученные готовые к употреблению напитки (после представления) содержат подсластители в следующих концентрациях:
Органолептическая оценка напитков
Оценка полученных безалкогольных напитков проводится группой экспертов (n=6), которые оценивают каждый набор напитков на отдельном сеансе дегустации. Результаты оценок представлены в таблице 21.
Органолептическая оценка безалкогольных напитков
концентрация
Обсуждение
Напиткам лимон/лайм, оранжад и кола монатин придает сладкий вкус, по качеству сходный со вкусом аспартама и немного превосходящий вкус сукралозы, оба из которых являются высококачественными подсластителями. В случае напитка лимон/лайм, композиция, содержащая монатин, оставляет меньшее послевкусие, чем композиция, содержащая аспартам. Кроме того, эффективность R,R-монатина выше, чем эффективность аспартама и сукралозы.
ПРИМЕР 25
Кривая "доза-ответ" для сладости монатина и сахарина
Сладость монатина и сахарина оценивается 20 опытными дегустаторами, делающими оценки в дубликатах. Тестируемые и контрольные растворы получают в буфере лимонная кислота/цитрат при pH 3,2. См. фиг.16. Более линейный ответ R,R/S,S-монатина по сравнению с сахарином согласуется с более сахароподобным профилем вкуса. Плато выше 10% SEV указывает на отсутствие/низкие уровни "смесь-подавляющих" привкусов и остаточных вкусов. Монатин имеет такую же форму кривой "доза-ответ", как и аспартам, сукралоза и алитам, которые являются "качественными" подсластителями.
Если в модельной системе (pH 3,2) R,R/S,S-монатин является единственным подсластителем, система имеет следующие характеристики: (1) небольшая задержка появления сладкого вкуса; (2) достаточно быстрое ослабление сладкого вкуса; (3) легкий "аспартамо-подобное" послевкусие, слегка сладкое послевкусие, отсутствие горькости в послевкусии; и (4) остаточное холодящее ощущение в неароматизированных системах.
ПРИМЕР 26
Стабильность монатина при pH 3 и повышении температуры
Образец синтетического монатина подвергают воздействию pH 3 при температуре 25°C, 50°C и 100°C. При комнатной температуре и pH 3 наблюдается уменьшение количества монатина на 14% в течение 48 часов. Данное уменьшение обусловлено образованием лактона. При 50°C и pH 3 наблюдается уменьшение количества монатина на 23% в течение 48 часов. Данное уменьшение обусловлено образованием лактона и появлением неизвестного соединения приблизительно через 15,5 минут. При 100°C и pH 3 через 24 часа исчезает почти весь монатин. Основным детектируемым компонентом является неизвестное соединение, образующееся через 15,5 минут.
ПРИМЕР 27
Органолептическая стабильность монатина и аспартама при pH 2,5, 3,0, 4,0 и 40°C
Органолептическую стабильность растворов монатина, полученных при pH 2,5, 3,0 и 4,0 и хранящихся при 40°C, отслеживают в течение 100 дней. Утрату сладости в данных растворах сравнивают с утратой сладости в растворах аспартама, полученных и хранящихся в таких же условиях.
Органолептическую стабильность монатина (8% SEV, ~55 м.д., синтетическая смесь, содержащая приблизительно 96% энантиомерной пары 2R,4R/2S,4S и 4% энантиомерной пары 2R,4S/2S,4R) в фосфат/цитратных буферах, имеющих pH 2,5, 3,0 и 4,0, определяют после хранения при 40°C. Стабильность монатина сравнивают со стабильностью аспартама (400 м.д.) в таких же буферах. Готовят три контрольных раствора сахарозы в таких же фосфат/цитратных буферах, что и растворы монатина и аспартама. Все полученные растворы хранят в темноте.
Буферные композиции:
Сладость каждого подсластителя по сравнению с сахарозой оценивается в дубликатах группой дегустаторов (n=8), имеющих опыт по оценке сладости. Все образцы (в одинаковых буферах) анализируют с дубликатами при температуре 22°C±1°C. Растворы монатина (тестируемые), маркированные 3-значными случайными номерными кодами, индивидуально выдают экспертам по органолептической оценке в случайном порядке. Также выдаются стандартные растворы сахарозы с концентрацией в интервале 4,0-10,0% (вес./об.) с шагом 0,5% (вес./об.). Экспертов просят оценить сладость путем сравнения сладости тестируемого раствора со сладостью стандартных растворов сахарозы. Чтобы оценить сладость, делают последовательно 3 глотка тестируемого раствора, глоток воды, 3 глотка стандартного раствора сахарозы, глоток воды и т.д. Эксперты оценивают сладость до первого десятичного знака, например, 6,8, 8,5. Разные тестируемые растворы оценивают с перерывом в пять минут. Экспертов также просят хорошо сполоснуть рот и съесть крекер, чтобы уменьшить любые возможные эффекты наложения вкуса.
В таблицах 22 и 23 представлены результаты анализа стабильности в фосфат-цитратных буферах. При каждом значении pH после 100 дней хранения при 40°C в темноте процент сохранения сладости монатина превышает процент сохранения сладости аспартама. При рН 4,0 утрата сладости раствора монатина почти стабилизируется, поскольку зарегистрировано очень незначительное изменение интенсивности сладости между 17 и 100 днем, тогда как раствор аспартама продолжал терять сладость.
Органолептическая стабильность монатина: сладость после 100 дней хранения при 40°C
А.
Стабильность: Количество сладости, оставшееся после 100 дней хранения при определенном значении pH и 40°C
Соответствующие буферы эффективно поддерживают pH, как видно из таблицы 24:
Если принять, что реакция разложения имеет псевдопервый порядок, график зависимости logn процента сохранения от времени (logn%RTN v. t) позволяет рассчитать период полужизни (t1/2) и константу скорости (κ) утраты сладости в любых заданных условиях. Рассчитанную таким образом кинетику утраты сладости монатина и аспартама можно представить, как в таблице 25.
(κ; день-1)
При каждом значении pH после 100 дней хранения при 40°C процент сохранения сладости монатина превышает процент сохранения сладости аспартама. При рН 4,0 утрата сладости раствора монатина почти стабилизируется, поскольку зарегистрировано очень незначительное изменение интенсивности сладости между 17 и 100 днем, тогда как раствор аспартама продолжал терять сладость.
Определение периода полужизни монатина и аспартама показывает, что сладость монатина уменьшается медленнее, чем сладость аспартама. Период полужизни сладости монатина при pH 2,5, 3,0 и 4,0 составляет 65 дней, 115 дней и 230 дней, соответственно. Период полужизни сладости аспартама в тех же условиях составляет 55 дней, 75 дней и 140 дней.
Таким образом, в кислых условиях, после хранения при 40°C, монатин обеспечивает более стабильную сладость, чем аспартам. Стабильность монатина выше стабильности аспартама в напитках кола и в других напитках, имеющих более низкое значение pH, а также при более высокой температуре. Поскольку монатин обладает более высокой стабильностью, чем аспартам, а также достигает равновесия и не подвергается необратимому распаду при pH 3, ожидается, что монатин будет обеспечивать длительную стабильную сладость в напитках, имеющих низкое значение pH, таких как напитки кола.
Кроме того, было обнаружено (данные не показаны), что при воздействии ультрафиолетового (УФ) света монатин в фосфорном/цитратном буфере при pH 3,0 (при температуре окружающей среды) тоже является стабильным и немного более стабилен, чем аспартам. УФ-нестабильность может увеличиваться в присутствии некоторых ароматизирующих систем. УФ-поглощающий упаковочный материал, красители и/или антиоксиданты могут защищать ароматизаторы от индуцированных УФ-светом взаимодействий в монатин-содержащих напитках.
ПРИМЕР 28
Хроматография стереоизомеров монатина
Приготовление образцов - Приблизительно 50-75 мкг лиофилизированного вещества помещают в микроцентрифужную пробирку. Добавляют 1,0 мл метанола категории чистый для ВЭЖХ. Раствор встряхивают 30 минут, центрифугируют и аликвоту супернатанта берут для анализа.
ВЭЖХ на обращенной фазе - Хроматографию двух разных диастереомерных пиков (R,R/S,S и R,S/S,R) проводят, используя ВЭЖХ колонку 2,1×250 мм Xterra(MS C8 5 мкм (Waters Corporation). Детекцию проводят, используя трехполюсный квадрупольный масс-спектрометр Ultima(от Micromass. В качестве подвижной фазы используют следующий градиент:
Хиральная ВЭЖХ - Хроматографию двух разных стереоизомеров монатина (R,R и S,S) проводят, используя ВЭЖХ колонку 250×4,6 мм Chirobiotic T (Advanced Separations Technologies, Inc.). Детекцию проводят, используя трехполюсный квадрупольный масс-спектрометр Ultima™ от Micromass. В качестве фазы используют метанол, содержащий 0,2% уксусной кислоты и 0,05% гидроксида аммония.
Масс-спектрометрия (МС/МС) - Присутствие монатина детектируют, используя метод контроля селективных реакций (SRM). Протонированный молекулярный ион монатина ([M+H]+) имеет m/z=293,3. Фрагментация данного молекулярного иона дает характеристический ион с m/z=257,3 образующийся в результате множественной дегидратации молекулярного иона. Показано, что данный переход является очень специфичным для монатина, поэтому его выбирают как переход (от 293,3 к 257,3) для мониторинга на протяжении эксперимента SRM. Данный способ детекции используют как для обращеннофазового, так и хирального разделения монатина.
Результаты - Стандартные образцы R,S/S,R и S,S/R,R анализируют методом ВЭЖХ на обращенной фазе. Образцы получают путем дериватизации и ферментативного разложения. Хроматограммы стандартных растворов представлены на фиг.17. После анализа на обращенной фазе проводят хиральную хроматографию, чтобы определить, какие именно стереоизомеры присутствуют в образцах. Результаты хиральной хроматографии стандартных растворов S,S- и R,R-монатина представлены на фиг.18.
ПРИМЕР 29
Стабильность монатина при высокой температуре (80°C) и нейтральном значении pH
В качестве исходного раствора используют 100 миллилитров раствора, содержащего 75 м.д. монатина при pH 7. Образец синтетического монатина содержит приблизительно 96% энантиомерной пары 2R,4R/2S,4S и 4% энантиомерной пары 2R,4S/2S,4R. Образцы инкубируют при 80°C и pH 7 в течение всего эксперимента, пробы отбирают через 0, 1, 2, 3, 4 часа и через 1, 2, 4, 7, 14, 21 и 35 дней. Все экспериментальные условия повторяют в дубликатах.
Разделение и количественное определение методом ЖХ-МС с использованием хроматографии на обращенной фазе - Строят кривую ответа для пиков обоих детектируемых диастереомеров синтетического монатина. Используя в качестве стандарта синтетический монатин, растворенный в DI воде, устанавливают интервал 5-150 м.д. Разделение двух диастереомеров проводят, используя ВЭЖХ колонку 3,9×150 мм Novapak C18 (Waters Corporation). Для детекции и количественного определения используют линейно подсоединенные детектор в ультрафиолетовой и видимой области спектра (УФ) и масс-спектрометр (MS). Монатин и его лактон имеют УФmax при 279 нм, что способствует точной детекции. Количественное определение проводят, используя метод контроля селективных реакций (SIM), по ионам с m/z 293,3 и m/z 275,3 в режиме электрораспыления с использованием положительных ионов.
Результаты - Обнаружено, что степень разрушения монатина при нейтральном рН является незначительной, даже через 7-35 дней. Исчезновение монатина со временем сильно зависит от pH, поскольку первичные побочные продукты подвергаются циклизации и, возможно, в очень небольшой степени, рацемизации. В процессе эксперимента при 80°C и pH 7 изменений в концентрации рацемического RR/SS-монатина или его лактонов не обнаружено в пределах точности количественного определения методом ЖХ-МС.
Поскольку монатин является термически стабильным при нейтральных значениях pH, ожидается, что монатин будет иметь подходящую стабильность в напитках с нейтральными значениями pH (таких как молочные или порошкообразные питьевые композиции). Также ожидается, что монатин будет иметь более длительный срок хранения в указанных композициях напитков, чем другие высокоинтенсивные подсластители (например, аспартам). Кроме того, ожидается, что монатин будет более стабилен в условиях обработки, такой как горячая заливка.
ПРИМЕР 30
Другие безалкогольные композиции (концентраты)
Представление 1+4. Разбавленный готовый к употреблению напиток содержит 1980 м.д. S,S-монатина.
Представление 1+4. Разбавленный готовый к употреблению напиток содержит 80 м.д. рацемической смеси монатина.
Представление 1+4. Разбавленный готовый к употреблению напиток содержит 550 м.д. S,S-монатина и 32 м.д. R,R-монатина.
С точки зрения многих возможных модификаций настоящего изобретения следует понимать, что показанные конкретные варианты представляют собой только конкретные примеры, не ограничивающие объем изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВАЯ АЛЬДОЛАЗА И СПОСОБ ПОЛУЧЕНИЯ ОПТИЧЕСКИ АКТИВНЫХ IHOG И МОНАТИНА | 2005 |
|
RU2313579C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПРОИЗВОДНОГО ГЛЮТАМАТА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МОНАТИНА | 2002 |
|
RU2280078C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДОЛ-3-ПИРОВИНОГРАДНОЙ КИСЛОТЫ И ЕЕ ПРОИЗВОДНЫХ | 2006 |
|
RU2325442C2 |
| ФЕРМЕНТАТИВНЫЙ СИНТЕЗ 4’-ЭТИНИЛНУКЛЕОЗИДНЫХ АНАЛОГОВ | 2019 |
|
RU2816846C2 |
| РЕБАУДИОЗИД Е И ПИЩЕВЫЕ ПРОДУКТЫ, ПОДСЛАЩЕННЫЕ РЕБАУДИОЗИДОМ Е | 2014 |
|
RU2658361C2 |
| СПОСОБ ПОЛУЧЕНИЯ МОНАТИНА | 2011 |
|
RU2505606C2 |
| БИОЛОГИЧЕСКИЙ СИНТЕЗ ДИФУНКЦИОНАЛЬНЫХ АЛКАНОВ ИЗ АЛЬФА-КЕТОКИСЛОТ | 2009 |
|
RU2496880C2 |
| КОМПОЗИЦИЯ ПОДСЛАСТИТЕЛЯ | 2005 |
|
RU2392834C2 |
| ПИТЬЕВЫЕ ПРОДУКТЫ С НЕКАЛОРИЙНЫМ ПОДСЛАСТИТЕЛЕМ И ГОРЬКИМ АГЕНТОМ | 2010 |
|
RU2536134C1 |
| ПОДСЛАСТИТЕЛИ НА ОСНОВЕ РЕБАУДИОЗИДА D И ПИЩЕВЫЕ ПРОДУКТЫ, ПОДСЛАЩЕННЫЕ РЕБАУДИОЗИДОМ D | 2012 |
|
RU2575042C2 |



































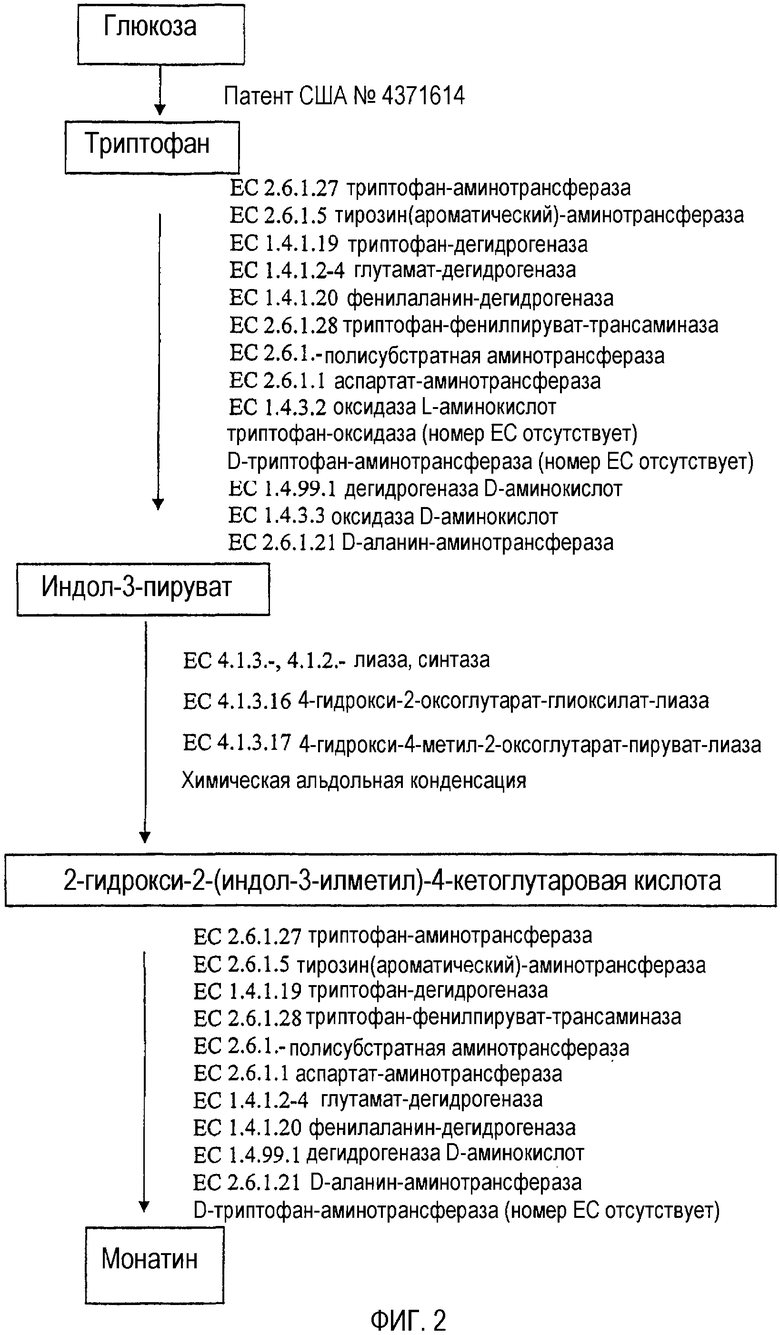
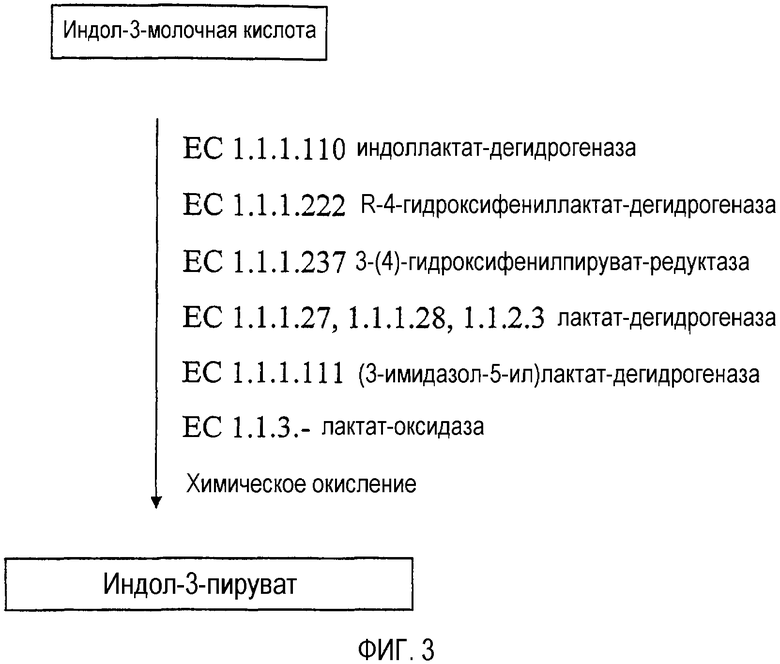
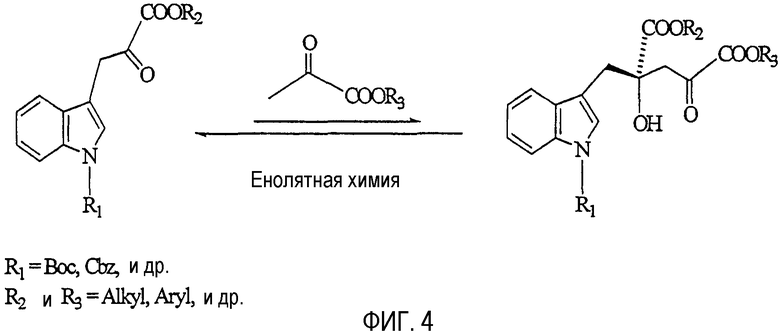



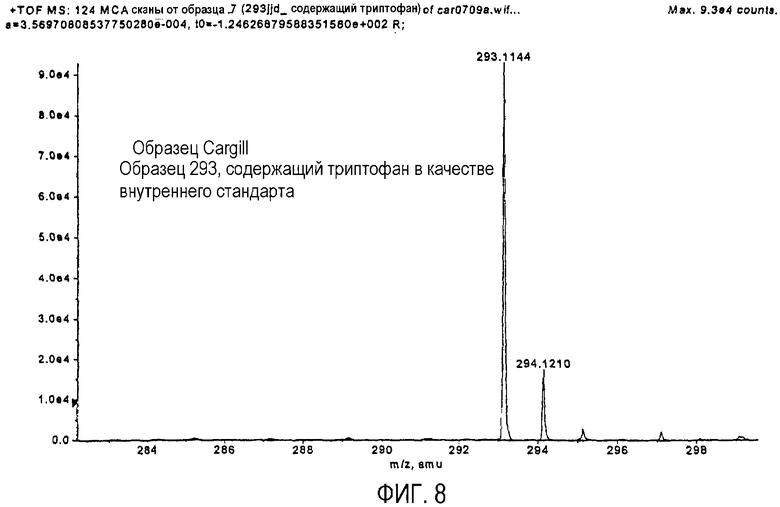
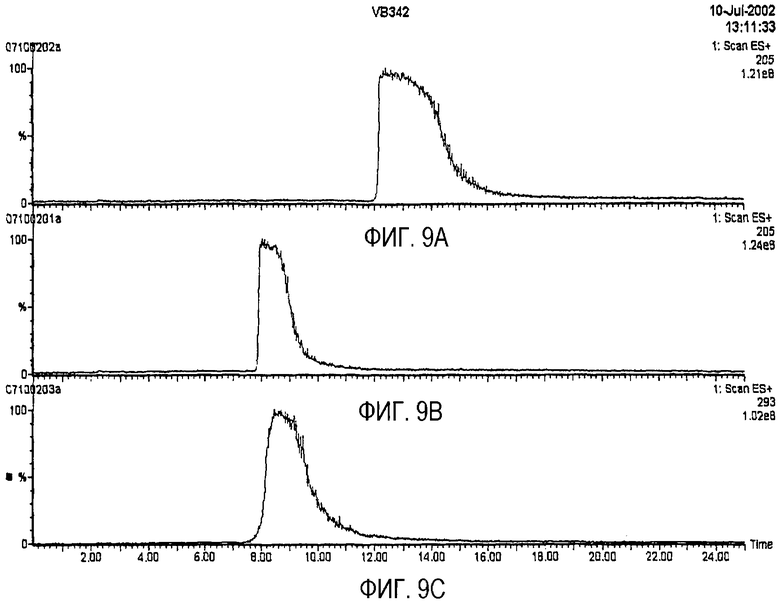

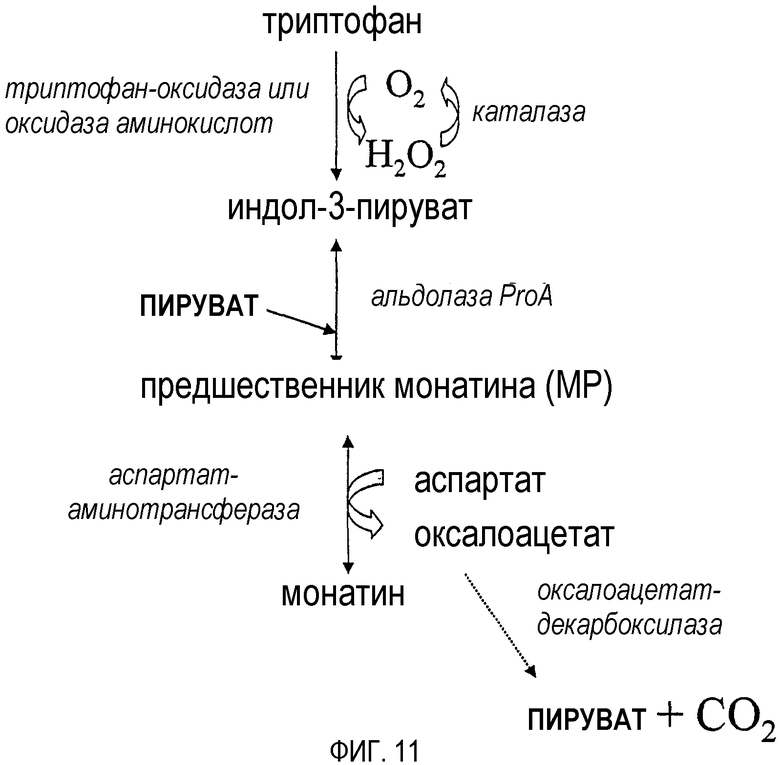
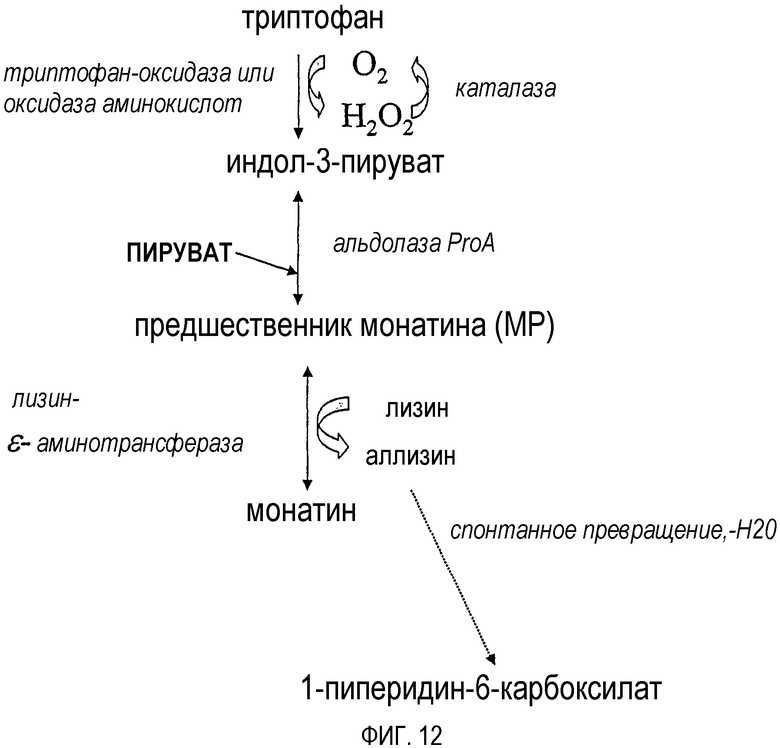

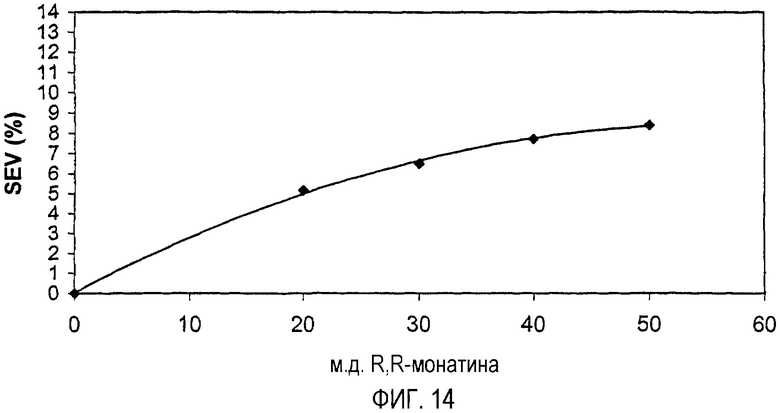
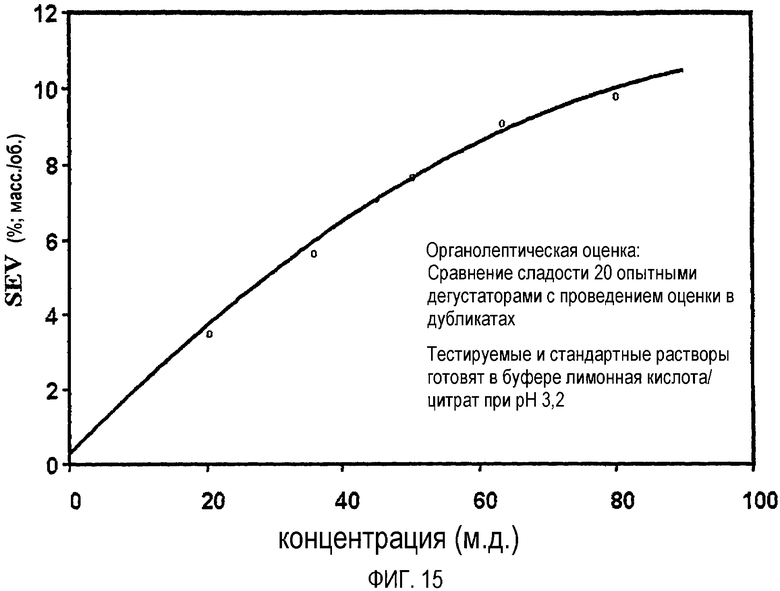
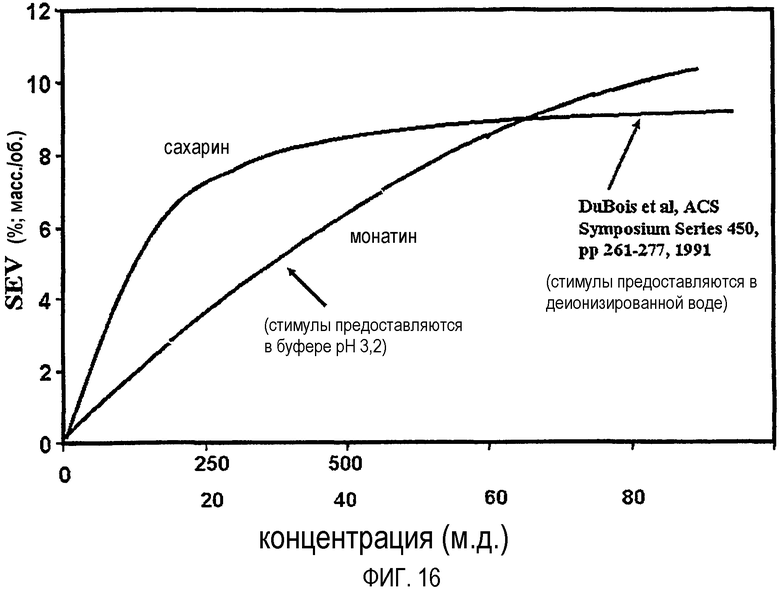

Изобретение относиться к композициям напитков, содержащим монатин, обогащенный его R,R-стереоизомерами, или его соль. Также к способам получения таких композиций биосинтетическим способом и не содержащих нефтехимических, токсичных или вредных примесей относится способ получения композиции напитка путем объединения монатина, обогащенного R,R-стереоизомерами или его соль по меньшей мере с одним другим ингредиентом. Изобретение относится к газированному напитку, содержащему сироп в количестве, варьирующем приблизительно от 15% до 25% по отношению к массе газированного напитка, причем сироп содержит монатин, обогащенный R,R-стереоизомером, или его соль. Изобретение позволяет получить монатин, который будет иметь более длительный срок хранения в композициях напитков, чем другие подсластители. Также более стабилен в условиях обработки, такой как горячая заливка. 8 н. и 34 з.п.ф-лы, 18 ил., 25 табл.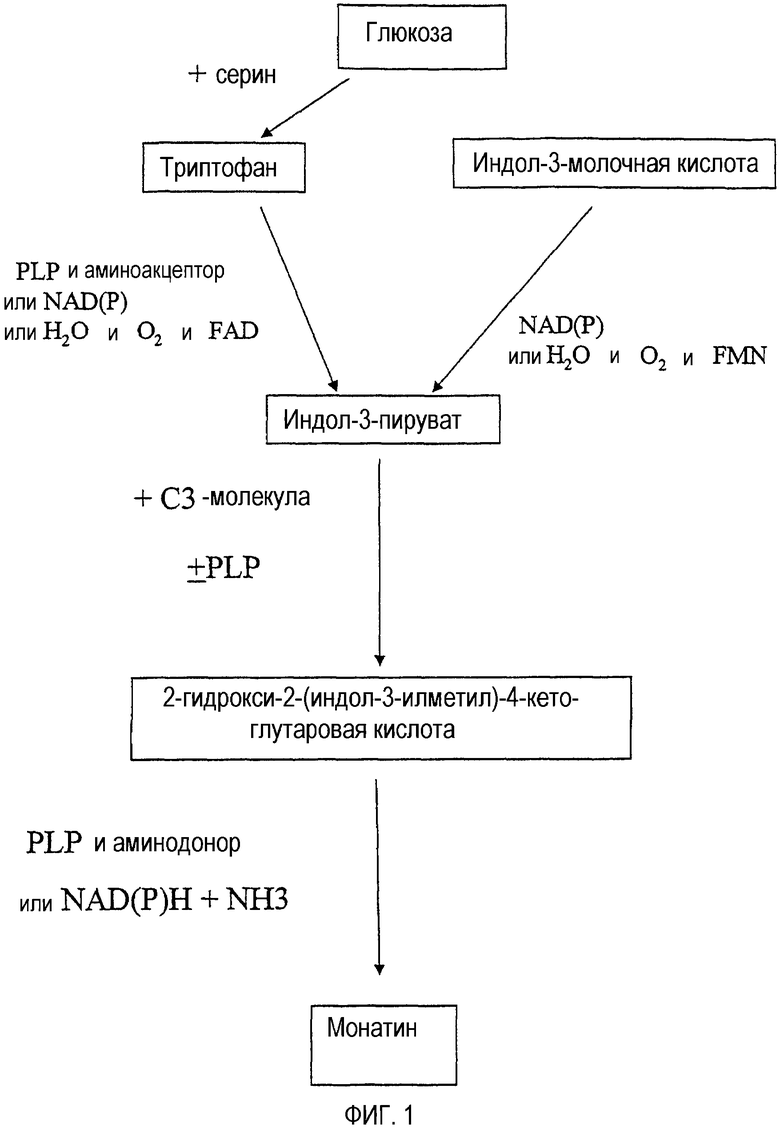
1. Композиция напитка, содержащая монатин, обогащенный его R,R-стереоизомером, или его соль.
2. Композиция напитка по п.1, дополнительно содержащая цитрусовый ароматизатор, причем монатин или его соль присутствуют в количестве, усиливающем аромат цитрусового ароматизатора.
3. Композиция напитка по п.1, дополнительно содержащая цитрусовый ароматизатор и углевод, причем монатин или его соль и углевод присутствуют в количестве, усиливающем аромат цитрусового ароматизатора.
4. Композиция напитка по п.1, в которой углевод выбран из эритрита, мальтодекстрина, сахарозы и их комбинации.
5. Композиция напитка по п.1, содержащая приблизительно от 3 до 10000 м.д. (миллионных долей) монатина или его соли.
6. Композиция напитка по п.1, которая представляет собой сироп или сухую смесь для приготовления готового напитка, причем композиция содержит приблизительно от 10 до 10000 м.д. монатина или его соли.
7. Композиция напитка по п.6, которая представляет собой сироп, причем сироп представляет собой концентрат, пригодный для разбавления напитком в интервале приблизительно от 1 часть сиропа: 3 части напитка до 1 часть сиропа: 5,5 частей напитка.
8. Композиция напитка по п.7, в которой сироп содержит приблизительно от 18 до 300 м.д. R,R-монатина или его соли.
9. Композиция напитка по п.1, которая представляет собой сухую смесь для приготовления напитка, содержащую приблизительно от 10 до 10000 м.д. монатина или его соли.
10. Композиция напитка по п.9, содержащая приблизительно от 10 до 450 м.д. R,R-монатина или его соли.
11. Питьевая композиция по п.1, по существу не содержащая S,S-монатина или его соли.
12. Композиция напитка по п.1, в которой композиция содержит приблизительно от 3 до 450 м.д. R,R-монатина или его соли.
13. Композиция напитка по п.12, содержащая приблизительно от 6 до 225 м.д. R,R-монатина или его соли.
14. Композиция напитка по п.1, представляющая собой композицию готового к употреблению напитка, содержащую приблизительно от 3 до 2000 м.д. монатина или его соли.
15. Композиция напитка по п.14, содержащая приблизительно от 5 до 50 м.д. R,R-монатина или его соли.
16. Композиция напитка по п.1, содержащая приблизительно 450 или менее м.д. R,R-монатина или его соли, причем монатин или его соль практически не содержат
S,S-, S,R- или R,S-монатина или его соли.
17. Композиция напитка по п.1, в которой монатин или его соль состоят в основном из R,R-монатина или его соли.
18. Композиция напитка по п.1, в которой монатин или его соль содержат по меньшей мере 95% R,R-монатина или его соли.
19. Композиция напитка по п.1, в которой монатин или его соль получен биосинтетическим способом.
20. Композиция напитка по п.1, дополнительно содержащая эритрит, трегалозу, цикламат, D-тагатозу или их комбинацию.
21. Композиция напитка по п.1, которая является некариесогенной.
22. Композиция напитка по п.19, в которой биосинтетический способ представляет собой многостадийный путь и по меньшей мере одна стадия данного многостадийного пути представляет собой химическое превращение.
23. Композиция напитка по п.1, в которой монатин или его соль представляют собой смесь R,R- и S,S-монатина или их солей.
24. Композиция напитка по п.1, дополнительно содержащая подсластитель-наполнитель, высокоинтенсивный подсластитель, углевод с низким гликемическим индексом, ароматизатор, антиоксидант, кофеин, усилитель сладости или их комбинацию.
25. Композиция напитка по п.24, в которой ароматизатор выбран из ароматизатора колы, цитрусового ароматизатора и их комбинации, подсластитель-наполнитель выбран из кукурузных подсластителей, сахарозы, декстрозы, инвертного сахара, мальтозы, декстрина, мальтодекстрина, фруктозы, левулозы, высокофруктозного кукурузного сиропа, сухого кукурузного сиропа, левулозы, галактозы, трегалозы, изомальтулозы, фруктовых олигосахаридов и их комбинации, высокоинтенсивный подсластитель выбран из сукралозы, аспартама, сахарина, ацесульфама К, алитама, тауматина, дигидрохалконов, неотама, цикламатов, стевиозида, могрозида, глицирризина, филлодульцина, монеллина, мабинлина, бразеина, циркулина, пентадина и их комбинации, углевод с низким гликемическим индексом выбран из D-тагатозы, сорбита, маннита, ксилита, лактита, эритрита, мальтита, гидролизатов гидрированного крахмала, изомальта, D-псикозы, 1,5-ангидро-D-фруктозы и их комбинации, и усилитель сладости выбран из куркулина, миракулина, цинарина, хлорогеновой кислоты, кофейной кислоты, строгинов, арабиногалактана, мальтола, дигидроксибензойных кислот и их комбинаций.
26. Композиция напитка по п.1, содержащая смесь монатина или его соли с отличающимся от монатина подсластителем.
27. Композиция напитка по п.26, в которой указанный отличающийся от монатина подсластитель выбран из сахарозы и высокофруктозного кукурузного сиропа.
28. Композиция напитка, содержащая монатин, полученный биосинтетическим способом, причем монатин не содержит нефтехимических, токсичных или вредных примесей.
29. Композиция напитка по п.28, в которой указанный монатин выделен из рекомбинантной клетки, не содержащей нефтехимических, токсичных или вредных примесей.
30. Газированный напиток, содержащий сироп в количестве, варьирующем приблизительно от 15 до 25% по отношению к массе газированного напитка, причем сироп содержит монатин, обогащенный R,R-стереоизомером, или его соль.
31. Способ получения композиции напитка по п.1, включающий получение монатина, обогащенного R,R-стереоизомером, или его соли биосинтетическим способом; и объединение монатина или его соли с по меньшей мере одним другим ингредиентом для получения монатин-содержащей композиции напитка.
32. Способ по п.31, в котором монатин или его соль получены из по меньшей мере одного субстрата, выбранного из глюкозы, триптофана, индол-3-молочной кислоты, индол-3-пирувата и предшественника монатина.
33. Способ по пп.31 и 32, в котором монатин или его соль, получены с использованием по меньшей мере одного биологического превращения.
34. Способ по п.33, в котором монатин или его соль, получены с использованием только биологических превращений.
35. Способ по п.31, в котором монатин или его соль объединяют с ингредиентами, выбранными из группы, состоящей из наполняющих агентов, подсластителей-наполнителей, жидких подсластителей, углеводов с низким гликемическим индексом, высокоинтенсивных подсластителей, загустителей, жиров, масел, эмульгаторов, антиоксидантов, усилителей сладости, красителей, ароматизаторов, кофеина, кислот, порошков, средств, повышающих текучесть, буферов, источников белков, усилителей вкуса, стабилизаторов вкуса и их комбинаций.
36. Способ по п.35, в котором монатин или его соль объединяют с эритритом, трегалозой, цикламатом, D-тагатозой, мальтодекстрином или их комбинацией.
37. Способ по п.31, в котором указанный биосинтетический способ включает стадии: (а) получение монатина или его соли биосинтетическим способом в рекомбинантной клетке; (b) выделение монатиновой композиции из рекомбинантной клетки, причем выделенная монатиновая композиция состоит из монатина или его соли и другого пищевого или питьевого вещества.
| US 5128164 А, 07.07.1992 | |||
| СЛАБОАЛКОГОЛЬНЫЙ НАПИТОК | 1999 |
|
RU2154098C2 |
| RU 99100470 A, 20.10.2000 | |||
| СУХАЯ, СВОБОДНО ТЕКУЧАЯ КОМПОЗИЦИЯ ДЛЯ НАПИТКА | 1996 |
|
RU2197873C2 |
| VLEGGAAR R., LOUIS G.J | |||
| ACKERMAN - Structure Elucidation of Monatin, a High-intensity Sweetener Isolated from the Plant Schlerochiton ilicifolius, CHEM | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| I, 1992. | |||

