Перекрестные ссылки на родственные заявки
Эта заявка испрашивает приоритет в соответствии с 35 U.S.C. § 119 (e) на основании временных заявок США No. 62/874 287 от 15 июля 2019 г., No. 62/926 047, от 25 октября 2019 г.и No. 63/048 873, от 7 июля 2020 г., содержание заявок США No. 62/926 047 и No. 63/048873 включено в настоящее описание в качестве ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к антителам к белкам, участвующим в костимулирующих или коингибиторных сигнальных путях, включающих CTLA-4. Более конкретно, настоящее изобретение также относится к канинизированным антителам к собачьему CTLA-4, которые имеют специфические последовательности и высокую аффинность связывания с собачьим CTLA-4. Настоящее изобретение также относится к применению антител по настоящему изобретению для лечения онкологических заболеваний у собак.
УРОВЕНЬ ТЕХНИКИ
Инициирование или прекращение иммунных ответов опосредуется сигнальными путями, которые активируются сложными взаимодействиями между набором белков, экспрессируемых на поверхности многих иммунных клеток, в первую очередь Т-лимфоцитов и антигенпрезентирующих клеток (APC). Костимулирующие сигнальные пути приводят к развитию иммунных ответов и, как было показано, опосредуются, в первую очередь, посредством взаимодействия CD28 на поверхности Т-клеток и B7.1 (также известного как CD80) и B7.2 (также известного как CD86) на поверхности APC. Считается, что B7.1 и B7.2 выполняют аналогичные функции.
Напротив, коингибирующие пути приводят к ингибированию или прекращению иммунных ответов и, как было показано, опосредуются посредством взаимодействия между белком 4, ассоциированным с цитотоксическими T-лимфоцитами, (CTLA-4) на Т-клетках и белками CD80/CD86 на APC. Дополнительные коингибирующие сигнальные пути, как было показано, опосредуются посредством взаимодействия между рецептором 1 запрограммированной клеточной смерти (PD-1) на Т-клетках и белками лигандов 1 или 2 рецептора запрограммированной клеточной смерти (PD-L1/PD-L2) на APC. Кроме того, также было показано, что взаимодействие между PD-L1 и CD80 также может приводить к ингибирующим сигналам в Т-клетках.
CD80 и CD86 являются членами суперсемейства иммуноглобулинов (Ig) [Sharpe and Freeman, Nature Reviews, 2: 116-126 (2002)]. CD80 экспрессируется на активированных В-клетках, активированных Т-клетках, а также на макрофагах и дендритных клетках [Swanson and Hall, Eur J. Immunol., 23:295-298 (1993); Razi-Wolfe et al., PNAS, 89:4210-4214 (1992)]. CD86 конститутивно экспрессируется на дендритных клетках, клетках Лангерганса и В-клетках. Кроме того, CD86 экспрессируется на моноцитах и активируется после стимуляции IFN-гамма [Larsen et al., Immunol., 152: 5208-5219 (1994); Inaba, J. Exp.Med. 180: 1849-1860 (1994)].
CD80 и CD86 связываются с CD28 и CTLA-4 с различными функциональными последствиями [Linsley et al., PNAS, 87: 5031-5035 (1990); Linsley et al., J. Exp.Med., 173: 721-730 (1991); Azuma et al., Nature 366: 76-79 (1993); Freeman et al., Science 262: 909-912 (1993)]. Связывание CD80 и CD86 с CTLA-4 имеет гораздо более высокую аффинность, чем связывание CD80/CD86 с CD28 [van der Merwe, J. Exp.Med. 185: 393-402 (1997)].
CD28 представляет собой гомодимерный гликопротеин, который является членом суперсемейства Ig [Aruffo and Seed, PNAS, 84: 8573-8577 (1987)]. Зрелый белок имеет единственный внеклеточный вариабельный домен из 134 аминокислотных остатков, содержащий гексапептидный мотив MYPPPY, который необходим для связывания противорецептора [Riley and June, Blood, 105: 13-21 (2005)]. Цитоплазматический домен CD28 из 41 аминокислоты содержит четыре остатка тирозина, которые могут фосфорилироваться при активации [Sharpe and Freeman, Nat. Rev. Immunol., 2: 116-126 (2002)]. CD28 экспрессируется на большинстве CD4+Т-клеток и примерно на 50% CD8+Т-клеток [Gross et al., J. Immunol., 149: 380388 (1992); Riley and June, Blood,105:13-21 (2005)]. После лигирования Т-клеточного рецептора (TCR) связывание B7.1/B7.2 с CD28 обеспечивает критический костимулирующий сигнал для Т-клетки, позволяющий активировать Т-клетки и последующее развитие иммунного ответа [Reiser et al., PNAS, 89: 271-275 (1992); Jenkins et al., J. Immunol., 147: 2461-2466 (1991)]. Было показано, что в отсутствие сигнала CD28 Т-клетки подвергаются апоптозу или переходят в состояние невосприимчивости [Jenkins et al., J. Exp.Med. 165: 302-319 (1987); Jenkins et al., PNAS, 84: 5409-5413 (1987); Schwartz, Science, 248: 1349-1356 (1990)]. Связывание CD28-B7.1/B7.2 может изменять пороговый уровень лигирования TCR (например, количество комплекса антиген-MHC), необходимый для активации, сокращать время, необходимое для стимуляции наивных клеток, и увеличивать величину ответа Т-клеток. [Soskic et al., Advances in Immunology,124:96-123 (2014)].
CTLA-4 (CD152) также является членом суперсемейства Ig и состоит из единственного внеклеточного домена, трансмембранного домена и короткого цитоплазматического хвоста [Swanson, Immunology; 1010: 169-177 (2000)]. Кроме того, CTLA-4 имеет примерно 30% аминокислотной идентичности с CD28. CTLA-4 не экспрессируется конститутивно на наивных Т-клетках, хотя он быстро активируется вскоре после лигирования CD28 и активации Т-клеток с пиковым уровнем экспрессии CTLA-4 примерно через 48-96 часов после начальной активации Т-клеток [Alegre et al., J. Immunol., 157: 4762-4770 (1996); Freeman et al., J. Immunol., 149: 3795-3801 (1992)]. CTLA-4 связывается как с B7.1, так и с B7.2 с гораздо более высокой аффинностью, чем CD28 [van der Merwe et al., J. Exp.Med., 185: 393-402 (1997)]. Однако, в отличие от стимулирующих эффектов связывания CD28 B7.1 или B7.2, CTLA-4 действует как ингибирующий рецептор, который жизненно важен для подавления иммунного ответа [Walnus et al., Immunity, 1: 405- 413 (1994); Walnus, J.Exp.Med., 183: 2541-2550 (1996); Krummeland Allison, J. Exp.Med., 183: 2533-2540 (1996)]. Механизм, с помощью которого CTLA-4 опосредует свои иммунные ингибирующие функции, связан с его способностью действовать как конкурентный ингибитор взаимодействия между CD28 и CD80/CD86 [обзор в Swanson, Immunology, 1010: 169-177 (2000)]. Критическая роль CTLA-4 в подавлении иммунитета продемонстрирована на мышах с дефицитом CTLA-4, которые умирают в возрасте 3-5 недель из-за развития лимфопролиферативного заболевания, характеризующегося инфильтрацией Т-лимфоцитами множества органов [Tivol et al., Immunity, 3: 541-5417 (1995); Waterhouse et al., Science, 270: 985-988 (1995)]. Было также продемонстрировано, что последствия нокаута CTLA-4 зависят от взаимодействия CD28 с его лигандами CD80 и CD86, что демонстрируется отсутствием заболевания у мышей с тройным нокаутом CTLA-4/CD80/CD86 [Mandelbrot et al., J. Exp.Med., 189: 435-440 (1999)]. Это также подтверждается защитой от лимфопролиферации, обеспечиваемой повторным введением CTLA-4 Ig мышам с нокаутом CTLA-4 [Tivol et al., J. Immunol., 158: 5091-5094 (1997)].
Кроме того, было показано, что блокирование эффекта CTLA-4 с помощью антител усиливает ответы Т-клеток in vitro и in vivo и усиливает противоопухолевые иммунные ответы [Leach et al., Science, 271: 1734-1736 (1996)]. На основании этих результатов была предпринята разработка блокаторов CTLA-4, таких как моноклональные антитела, для обеспечения терапевтических возможностей лечения онкологических заболеваний [Hodi et al., PNAS, 100 (8): 4712-4717 (2003); Phan GQ et al., PNAS, 100 (14): 8372-8377 (2003); Attia, Journal of Clinical Oncology, 23(25):6043-6053 (2005); Comin-Anduix et al., Journal of Translational Medicine, 6: 22-22 (2008); WO2000037504 A2; США 8 017 114 В2; WO2010097597A1; WO2012120125 A1; и Boutros et al., Nat Rev Clin Oncol., 13 (8): 473-486 (2016)].
PD-1 является членом семейства иммуномодулирующих рецепторов CD28/CTLA-4. PD-1 также является членом суперсемейства Ig и содержит внеклеточный вариабельный домен, который связывает его лиганды, и цитоплазматический хвост, который связывает сигнальные молекулы [обзор в Zak et al., Cell Structure, 25: 1163-1174 (2017)]. Цитоплазматический хвост PD-1 содержит два сигнальных мотива на основе тирозина [Zhang et al., Immunity 20: 337-347 (2004)]. Экспрессия PD-1 не обнаруживается на нестимулированных Т-клетках, В-клетках или миелоидных клетках. Однако экспрессия PD1 в этих клетках повышается после активации [Chemnitz et al., J. Immunol., 173: 945-954 (2004); Petrvas et al., J. Exp.Med., 203: 2281-2292 (2006)]. PD-1 наиболее близок к CTLA-4, имея с ним приблизительно 24% аминокислотной идентичности [Jin et al., Current Topics in Microbiology and Immunology, 350: 17-37 (2010)]. PD-1 ослабляет активацию Т-клеток при связывании с PD-L1 и PD-L2, которые экспрессируются на поверхности APC. Связывание любого из этих лигандов с PD-1 негативно регулирует передачу сигналов антигена через Т-клеточный рецептор (TCR). На сегодняшний день обнаружено, что только PD-L1 и PD-L2 действуют как лиганды для PD-1. Как и CTLA-4, лигирование PD-1, по-видимому, передает отрицательный иммуномодулирующий сигнал. Лигирование PD-1 с помощью PD-L1 или PD-L2 приводит к ингибированию TCR-опосредованной пролиферации и продукции цитокинов [Jin et al., Current Topics in Microbiology and Immunology, 350: 17-37 (2010)]. В отличие от животных с дефицитом CTLA-4, мыши с дефицитом PD-1 умирают намного позже и демонстрируют признаки аутоиммунитета, хотя тяжесть наблюдаемых эффектов не так велика, как у животных с дефицитом CTLA-4 [Nishimura et al., Immunity, 11 (2): 141-151 (1999); Nishimura et al., Science, 291 (5502): 319-322 (2001)]. Хотя сигнальные пути PD-1 в настоящее время интенсивно исследуются, на сегодняшний день исследования показывают, что взаимодействия PD-L1/PD-L2/PD-1 участвуют в негативной регуляции некоторых иммунных ответов из-за ослабления сигналов ниже по сигнальному пути от стимуляции TCR, приводящего к снижению секреции цитокинов и нарушению пролиферации Т-клеток и снижению продукции цитотоксических молекул Т-клетками [Freeman et al., J. Exp.Med., 192 (7): 1027-1034 (2000)].
PD-L1 (CD274) представляет собой мембранный белок типа 1 и состоит из IgV-подобных и IgC-подобных внеклеточных доменов, гидрофобного трансмембранного домена и состоящего из 30 аминокислот короткого цитоплазматического хвоста с неизвестными свойствами сигнальной трансдукции. PD-L1 признан членом семейства B7 и имеет приблизительно 20% аминокислотной идентичности с членами семейства B7. PDL1 связывается со своим рецептором PD-1, обнаруженным на активированных Т-клетках, В-клетках и миелоидных клетках. PD-L1 также связывается с костимулирующей молекулой CD80, но не с CD86 [Butte et al., Immunology, 45 (13): 3567-3572 (2008)]. Аффинность CD80 к PD-L1 является промежуточной между его аффинностью к CD28 и CTLA-4. Родственная молекула PD-L2 не имеет аффинности ни к CD80, ни к CD86, но связана с PD-1 в качестве рецептора. Взаимодействие PD-L1 с его рецептором PD-1 на Т-клетках доставляет сигнал, который ингибирует TCR-опосредованную продукцию IL-2 и пролиферацию Т-клеток. Связывание PD-L1 с PD-1 также способствует понижающей модуляции TCR, индуцированной лигандом, во время презентации антигена наивным Т-клеткам. Кроме того, связывание PD-L1 с CD80 на Т-клетках приводит к апоптозу Т-клеток. Роль PD1 и PD-L1 как ингибиторов активации Т-клеток была продемонстрирована во многих исследованиях. На основании этих результатов была предпринята разработка блокаторов PD-1 и PD-L1, таких как моноклональные антитела, для обеспечения терапевтических возможностей для лечения онкологических и инфекционных заболеваний.
Гуманизированные моноклональные антитела, которые блокируют связывание и активность собачьих PD-1, PDL1 и CTLA-4, были разработаны и в настоящее время доступны для использования при лечении людей, у которых диагностирован один из нескольких различных типов онкологических заболеваний. Аналогичным образом также сообщалось о канинизированных моноклональных антителах, которые блокируют связывание и активность собачьих PD-1 и PDL1 [US 9944704 B2, US 10106607 B2 и US.2018/0237535A1, содержание которых включено в настоящее описание в качестве ссылки в полном объеме]. Однако до сих пор не было сообщений о канинизированных моноклональных антителах, которые блокируют связывание и активность собачьего CTLA-4.
Цитирование любой ссылки настоящего описания не следует рассматривать как допущение того, что такая ссылка доступна в качестве «предшествующего уровня техники» по отношению к настоящей заявке.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам против собачьего белка 4, ассоциированного с цитотоксическими Т-лимфоцитами, (CTLA-4), которые связываются с собачьим CTLA-4. В конкретных вариантах осуществления антитела к собачьему CTLA-4 специфически связываются с собачьим CTLA-4. В более конкретных вариантах осуществления антитела к собачьему CTLA-4 также обладают способностью блокировать связывание собачьего CTLA-4 с собачьим CD80. В других конкретных вариантах осуществления антитела к собачьему CTLA-4 также обладают способностью блокировать связывание собачьего CTLA-4 с собачьим CD86. В других конкретных вариантах осуществления антитела к собачьему CTLA-4 обладают способностью как блокировать связывание собачьего CTLA-4 с собачьим CD80, так и блокировать связывание собачьего CTLA-4 с собачьим CD86.
Кроме того, настоящее изобретение относится к определяющим комплементарность областям (CDR), содержащимся в этих антителах, и к комбинации этих CDR (например, полученных из мышиных антител против собачьего CTLA-4) в каркасах собаки с образованием канинизированных антител против собачьего CTLA-4. Настоящее изобретение также относится к применению таких антител при лечении таких состояний, как онкологическое заболевание.
Соответственно, настоящее изобретение относится к уникальным наборам CDR из шести (6) иллюстративных мышиных антител против собачьего CTLA-4. Шесть иллюстративных мышиных антител против собачьего CTLA-4 имеют уникальные наборы CDR, то есть три CDR легкой цепи: CDR легкой цепи 1 (CDRL1), CDR легкой цепи 2 (CDRL2) и CDR легкой цепи 3 (CDRL3), и три CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3). Как подробно описано ниже, имеется существенная гомология последовательностей в каждой группе CDR и даже некоторая избыточность (например, см. набор VL CDR-3 ниже в Таблице 1). Следовательно, настоящее изобретение относится не только к аминокислотным последовательностям шести CDR из шести иллюстративных мышиных антител против собачьего CTLA-4, но также относится к консервативно модифицированным вариантам этих CDR, а также вариантам, которые включают (например, имеют общую) одинаковую каноническую структура и/или связываются с одним или более (например, 1, 2, 3, 4 или более) аминокислотными остатками собачьего CTLA-4, которые содержатся в эпитопе собачьего CTLA-4.
Один аспект настоящего изобретения относится к антителам млекопитающего, которые связываются с собачьим белком 4, ассоциированным с цитотоксическими T-лимфоцитами, (CTLA-4). В конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент по настоящему изобретению представляет собой мышиное антитело. В предпочтительных вариантах осуществления антитела млекопитающих по настоящему изобретению, включающие мышиные антитела по настоящему изобретению, или их антигенсвязывающие фрагменты представляют собой канинизированные антитела или их канинизированные антигенсвязывающие фрагменты.
В конкретных вариантах осуществления антитела млекопитающих специфически связываются с собачьим CTLA-4. В более конкретных вариантах осуществления антитела млекопитающего к собачьему CTLA-4 также обладают способностью блокировать связывание собачьего CTLA-4 с собачьим CD80. В других конкретных вариантах осуществления антитела млекопитающих к собачьему CTLA-4 также обладают способностью блокировать связывание собачьего CTLA-4 с собачьим CD86. В других конкретных вариантах осуществления антитела млекопитающих к собачьему CTLA-4 обладают способностью как блокировать связывание собачьего CTLA-4 с собачьим CD80, так и блокировать связывание собачьего CTLA-4 с собачьим CD86.
В некоторых вариантах осуществления антитела млекопитающих, которые связываются с собачьим CTLA-4, представляют собой выделенные антитела. Настоящее изобретение также относится к антигенсвязывающим фрагментам любого из этих антител млекопитающих, которые связываются с собачьим CTLA-4. В конкретных вариантах осуществления антитела содержат три определяющие комплементарность области легкой цепи (CDR): CDR легкой цепи 1 (CDRL1), CDR легкой цепи 2 (CDRL2) и CDR легкой цепи 3 (CDRL3); и три CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3).
В конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент содержит CDRH3, которая содержит аминокислотную последовательность SEQ ID NO: 90, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 90 или вариант SEQ ID NO: 90, который включает каноническую структуру класса 7. В более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRH2, которая содержит аминокислотную последовательность SEQ ID NO: 88, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 88 или вариант SEQ ID NO: 88, который включает каноническую структуру класса 2A. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRH1, которая содержит аминокислотную последовательность SEQ ID NO: 86, CDRH1, которая содержит консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 86 или вариант SEQ ID NO: 86, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL3, которая содержит аминокислотную последовательность SEQ ID NO: 96, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 96 или вариант SEQ ID NO: 96, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRL2, которая содержит аминокислотную последовательность SEQ ID NO: 94, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 94 или вариант SEQ ID NO: 94, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL1, которая содержит аминокислотную последовательность SEQ ID NO: 92, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 92, или Вариант SEQ ID NO: 92, который включает каноническую структуру класса 4.
В альтернативных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент содержит CDRH3, которая содержит аминокислотную последовательность SEQ ID NO: 102, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 102 или вариант SEQ ID NO: 102, который включает каноническую структуру класса 9. В более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRH2, которая содержит аминокислотную последовательность SEQ ID NO: 100, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 100 или вариант SEQ ID NO: 100, который включает каноническую структуру класса 4. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRH1, которая содержит аминокислотную последовательность SEQ ID NO: 98, CDRH1, которая содержит консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 98 или вариант SEQ ID NO: 98, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL3, которая содержит аминокислотную последовательность SEQ ID NO: 108, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 108 или вариант SEQ ID NO: 108, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL2, которая содержит аминокислотную последовательность SEQ ID NO: 106, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 106 или вариант SEQ ID NO: 106, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL1, которая содержит аминокислотную последовательность SEQ ID NO: 104, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 104, или Вариант SEQ ID NO: 104, который включает каноническую структуру класса 1.
В других альтернативных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент содержит CDRH3, которая содержит аминокислотную последовательность SEQ ID NO: 113, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 113 или вариант SEQ ID NO: 113, который включает каноническую структуру класса 7. В более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRH2, которая содержит аминокислотную последовательность SEQ ID NO: 88, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 88 или вариант SEQ ID NO: 88, который включает каноническую структуру класса 2A. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRH1, которая содержит аминокислотную последовательность SEQ ID NO: 86, CDRH1, которая содержит консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 86 или вариант SEQ ID NO: 86, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL3, которая содержит аминокислотную последовательность SEQ ID NO: 96, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 96 или вариант SEQ ID NO: 96, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL2, которая содержит аминокислотную последовательность SEQ ID NO: 94, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 94 или вариант SEQ ID NO: 94, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL1, которая содержит аминокислотную последовательность SEQ ID NO: 117, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 117, или вариант SEQ ID NO: 117, который включает каноническую структуру класса 4.
Еще в других альтернативных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент содержит CDRH3, которая содержит аминокислотную последовательность SEQ ID NO: 115, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 115 или вариант SEQ ID NO: 115, который включает каноническую структуру класса 7. В более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRH2, которая содержит аминокислотную последовательность SEQ ID NO: 88, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 88 или вариант SEQ ID NO: 88, который включает каноническую структуру класса 2A. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRH1, которая содержит аминокислотную последовательность SEQ ID NO: 86, CDRH1, которая содержит консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 86 или вариант SEQ ID NO: 86, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL3, которая содержит аминокислотную последовательность SEQ ID NO: 96, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 96 или вариант SEQ ID NO: 96, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL2, которая содержит аминокислотную последовательность SEQ ID NO: 122, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 122 или вариант SEQ ID NO: 122, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL1, которая содержит аминокислотную последовательность SEQ ID NO: 119, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 119, или вариант SEQ ID NO: 119, который включает каноническую структуру класса 4.
В других альтернативных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент содержит CDRH3, которая содержит аминокислотную последовательность SEQ ID NO: 114, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 114 или вариант SEQ ID NO: 114, который включает каноническую структуру класса 7. В более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRH2, которая содержит аминокислотную последовательность SEQ ID NO: 111, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 111 или вариант SEQ ID NO: 111, который включает каноническую структуру класса 2A. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRH1, которая содержит аминокислотную последовательность SEQ ID NO: 109, CDRH1, которая содержит консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 109 или вариант SEQ ID NO: 109, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL3, которая содержит аминокислотную последовательность SEQ ID NO: 96, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 96 или вариант SEQ ID NO: 96, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRL2, которая содержит аминокислотную последовательность SEQ ID NO: 121, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 121 или вариант SEQ ID NO: 121, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL1, которая содержит аминокислотную последовательность SEQ ID NO: 118, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 118, или вариант SEQ ID NO: 118, который включает каноническую структуру класса 4.
Еще в других альтернативных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент содержит CDRH3, которая содержит аминокислотную последовательность SEQ ID NO: 116, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 116 или вариант SEQ ID NO: 116, который включает каноническую структуру класса 12. В более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент дополнительно содержит CDRH2, которая содержит аминокислотную последовательность SEQ ID NO: 112, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 112 или вариант SEQ ID NO: 112, который включает каноническую структуру класса 2A. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRH1, которая содержит аминокислотную последовательность SEQ ID NO: 110, CDRH1, которая содержит консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 110 или вариант SEQ ID NO: 110, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL3, которая содержит аминокислотную последовательность SEQ ID NO: 124, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 124 или вариант SEQ ID NO: 124, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL2, которая содержит аминокислотную последовательность SEQ ID NO: 123, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 123 или вариант SEQ ID NO: 123, который включает каноническую структуру класса 1. В еще более конкретных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент также дополнительно содержит CDRL1, которая содержит аминокислотную последовательность SEQ ID NO: 120, консервативно модифицированный вариант аминокислотной последовательности SEQ ID NO: 120, или вариант SEQ ID NO: 120, который включает каноническую структуру класса 2.
Как указано выше, канинизированные антитела к собачьему CTLA-4 или их канинизированные антигенсвязывающие фрагменты являются важным аспектом настоящего изобретения, и настоящее изобретение обеспечивает канинизированные антитела млекопитающих, включая канинизированные мышиные антитела, из всех таких антител млекопитающих. Соответственно, настоящее изобретение также относится к выделенному канинизированному антителу или его антигенсвязывающему фрагменту, которое специфически связывается с CTLA-4, и содержит тяжелую цепь собачьего IgG и собачью легкую цепь каппа или лямбда. В конкретных вариантах осуществления этого типа собачья легкая цепь каппа или лямбда содержит три определяющих комплементарность области легкой цепи (CDR): CDR легкой цепи 1 (CDRL1), CDR легкой цепи 2 (CDRL2) и CDR легкой цепи 3 (CDRL3); и тяжелая цепь собачьего IgG содержит три CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3), которые получены из мышиных антител против собачьего CTLA-4. Конкретные варианты осуществления канинизированных антител и их антигенсвязывающих фрагментов по настоящему изобретению связываются с собачьим CTLA-4 и/или блокируют связывание собачьего CTLA-4 с собачьим CD80 и/или собачьим CD86.
Канинизированное антитело по настоящему изобретению или его канинизированный антигенсвязывающий фрагмент может содержать IgGD, который содержит шарнирную область, которая содержит аминокислотную последовательность SEQ ID NO: 128. В родственном варианте осуществления шарнирная область содержит аминокислотную последовательность SEQ ID NO: 129. Еще в одном родственном варианте осуществления шарнирная область содержит аминокислотную последовательность SEQ ID NO: 130. Еще в одном родственном варианте осуществления шарнирная область содержит аминокислотную последовательность SEQ ID NO: 131.
В альтернативных вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 62. В конкретных вариантах осуществления этого типа тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 61. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 64. В конкретных вариантах осуществления этого типа тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 63. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 66. В конкретных вариантах осуществления этого типа тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 65. В более конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 50. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 49. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 52. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 51. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 54. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 53.
В альтернативных вариантах осуществления канинизированное антитело содержит модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 74. В конкретном варианте этого типа модифицированная тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 73. В других вариантах осуществления канинизированное антитело содержит модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 76. В конкретном варианте этого типа модифицированная тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 75. В других вариантах осуществления канинизированное антитело содержит модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 78. В конкретных вариантах осуществления этого типа модифицированная тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 77. В более конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 50. В конкретном варианте этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 49. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 52. В конкретном варианте этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 51. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 54. В конкретном варианте осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 53.
В конкретных вариантах осуществления канинизированные антитела содержат тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 66, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 52. В других вариантах осуществления канинизированные антитела содержат тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 66, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 54.
В альтернативных вариантах осуществления канинизированные антитела содержат модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 78, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 52. В других вариантах осуществления канинизированные антитела содержат модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 78, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 54.
В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 68. В конкретных вариантах осуществления этого типа тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 67. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 70. В конкретных вариантах осуществления этого типа тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 69. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 72. В конкретных вариантах осуществления этого типа тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 71. В более конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 56. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 55. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 58. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 57. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 60. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 59.
В альтернативных вариантах осуществления канинизированное антитело содержит модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 80. В конкретном варианте осуществления этого типа модифицированная тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 79. В других вариантах осуществления канинизированное антитело содержит модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 82. В конкретном варианте этого типа модифицированная тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 81. Еще в других вариантах осуществления канинизированное антитело содержит модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 84. В конкретных вариантах осуществления этого типа модифицированная тяжелая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 83. В более конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 56. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 55. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 58. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 57. В других конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 60. В конкретных вариантах осуществления этого типа легкая цепь кодируется нуклеотидной последовательностью SEQ ID NO: 59.
В конкретных вариантах осуществления канинизированные антитела содержат модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 72, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 58. В других вариантах осуществления канинизированные антитела содержат тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 72, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 60.
В альтернативных вариантах осуществления канинизированные антитела содержат модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 84, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 58. В других вариантах осуществления канинизированные антитела содержат модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 84, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 60.
Настоящее изобретение также относится к антителам млекопитающих или их антигенсвязывающим фрагментам, которые связываются с собачьим CTLA-4 с константой диссоциации (Kd), которая ниже 1×10-12 M (например, 5×10-13 M или ниже). В других вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 с константой диссоциации от 1×10-5 M до 1×10-12 М. В более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 с константой диссоциации от 1×10-7 М до 1×10-11 М. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 с константой диссоциации от 1×10-8 М до 1×10-11 М. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 с константой диссоциации от 1×10-8 M до 1×10-10 M.
Настоящее изобретение также относится к антителам млекопитающих или их антигенсвязывающим фрагментам, которые связываются с собачьим CTLA-4 со скоростью ассоциации (kon) выше чем 1×107 M-1 с-1. В других вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 со скоростью ассоциации от 1×102 M-1 с-1 до 1×107 M-1 с-1. В более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 со скоростью ассоциации от 1×103 M-1 с-1 до 1×106 M-1 с-1. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 со скоростью ассоциации от 1×103 M-1 с-1 до 1×105 M-1 с-1. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 со скоростью ассоциации от 1×104 M-1 с-1 до 1×105 M-1 с-1.
Настоящее изобретение дополнительно относится к антителам млекопитающих или их антигенсвязывающим фрагментам, которые связываются с собачьим CTLA-4 со скоростью диссоциации (koff) ниже, чем 1×10-7 с-1. В других вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 со скоростью диссоциации от 1×10-3 с-1 до 1×10-8 с-1. В более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 со скоростью диссоциации от 1×10-4 с-1 до 1×10-7 с-1. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим CTLA-4 со скоростью диссоциации от 1×10-5 с-1 до 1×10-7 с-1.
В конкретных вариантах осуществления антитело млекопитающего по настоящему изобретению (включая химерные антитела) блокирует связывание собачьего CD80 и/или CD86 с собачьим CTLA-4. В более конкретных вариантах осуществления антитело блокирует связывание собачьего CD80 и/или CD86 с собачьим CTLA-4 с минимальной ЕС50 от 1×10-8 M до 1×10-9 M или даже в более низкой концентрации. В еще более конкретных вариантах осуществления ЕС50 составляет от 5×10-9 M до 5×10-13 М. В еще более конкретных вариантах осуществления ЕС50 составляет от 5×10-9 M до 5×10-11 М.
Соответственно, в конкретных вариантах осуществления антитела по настоящему изобретению могут проявлять одно, два, три, четыре или все из этих свойств, то есть вышеупомянутые константы диссоциации с собачьим CTLA-4, указанные выше скорости связывания с собачьим CTLA-4, указанные выше скорости диссоциации от связывающего комплекса антитело-собачий CTLA-4 или эффективное лечение онкологического заболевания у больного животного.
Настоящее изобретение также относится к канинизированным антителам млекопитающих и антигенсвязывающим фрагментам, которые перекрестно конкурируют с антителами млекопитающих, описанными в настоящем описании. В конкретных вариантах осуществления канинизированные антитела млекопитающих перекрестно конкурируют с антителом, содержащим 6 CDR 45A9 [см. Таблицу 1 ниже]. В родственных вариантах осуществления канинизированные антитела млекопитающих перекрестно конкурируют с антителом, содержащим 6 CDR 27G12 [см. Таблицу 1 ниже]. Еще в других родственных вариантах осуществления канинизированные антитела млекопитающих перекрестно конкурируют с антителом, содержащим 6 CDR 22A11. [см. Таблицу 1 ниже]. Еще в других родственных вариантах осуществления канинизированные антитела млекопитающих перекрестно конкурируют с антителом, содержащим 6 CDR 110E3 [см. Таблицу 1 ниже]. В конкретных вариантах осуществления канинизированные антитела млекопитающих перекрестно конкурируют с антителом, содержащим 6 CDR 12B3 [см. Таблицы 1 и 3 ниже]. В других конкретных вариантах осуществления канинизированные антитела млекопитающих перекрестно конкурируют с антителом, содержащим 6 CDR 39A11 [см. Таблицы 1 и 3 ниже]. В конкретных вариантах осуществления анализ представляет собой стандартный анализ связывания. В одном таком варианте осуществления стандартный анализ связывания проводят с помощью BIACore®. В другом таком варианте осуществления стандартный анализ связывания проводят с помощью ИФА. Еще в одном таком варианте осуществления стандартный анализ связывания проводят с помощью проточной цитометрии.
Как указано выше, антитела (и их антигенсвязывающие фрагменты) по настоящему изобретению, включая вышеупомянутые антитела (и их антигенсвязывающие фрагменты), могут быть моноклональными антителами (и их антигенсвязывающими фрагментами), антителами млекопитающих (и их антигенсвязывающими фрагментами), например, мышиными (мышиные) антителами (и их антигенсвязывающими фрагментами), канинизированными антителами (и их антигенсвязывающими фрагментами), включая канинизированные мышиные антитела (и их антигенсвязывающие фрагменты). В некоторых вариантах осуществления антитела (и их антигенсвязывающие фрагменты) являются выделенными.
В предпочтительных вариантах осуществления канинизированное антитело по настоящему изобретению или его антигенный фрагмент связывается с эпитопом аминокислотной последовательности собачьего CTLA-4. В конкретном варианте осуществления канинизированное антитело взаимодействует с одним или более аминокислотными остатками в положениях T35, R38, T51, T53, Y90, K93, Y98 и Y102 аминокислотной последовательности SEQ ID NO: 138. В другом варианте осуществления канинизированное антитело взаимодействует с одним или более аминокислотными остатками в положениях 35T, R38, S42, K93 и Y102 аминокислотной последовательности SEQ ID NO: 138.
Настоящее изобретение также относится к канинизированным антителам, которые связываются с одним или более эпитопами или их частями аминокислотных последовательностей SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136 и SEQ ID NO: 137. В конкретных вариантах осуществления канинизированное антитело по настоящему изобретению или его антигенный фрагмент связывается с эпитопом или его частью, состоящим из аминокислотной последовательности SEQ ID NO: 132. В более конкретном варианте осуществления этого типа эпитоп или его часть состоит из аминокислотной последовательности SEQ ID NO: 134. В другом варианте осуществления этого типа эпитоп или его часть состоит из аминокислотной последовательности SEQ ID NO: 135. В некоторых вариантах осуществления эпитоп или его часть состоит из аминокислотной последовательности SEQ ID NO: 133. В более конкретном варианте осуществления этого типа эпитоп или его часть состоит из аминокислотной последовательности SEQ ID NO: 136. В родственных вариантах осуществления канинизированные антитела связываются с одним или более эпитопами или их частями, которые состоят из аминокислотных последовательностей SEQ ID NO: 134 и/или SEQ ID NO: 136 и/или SEQ ID NO: 135.
Настоящее изобретение также относится к нуклеиновым кислотам (включая выделенные и/или рекомбинантные нуклеиновые кислоты), которые кодируют любую из легких цепей канинизированного антитела по настоящему изобретению. Точно так же настоящее изобретение относится к выделенным нуклеиновым кислотам (включая выделенные и/или рекомбинантные нуклеиновые кислоты), которые кодируют любую из тяжелых цепей канинизированного антитела по настоящему изобретению.
Настоящее изобретение также относится к экспрессирующим векторам, которые содержат одну или более нуклеиновых кислот (включая выделенные нуклеиновые кислоты) по настоящему изобретению. Настоящее изобретение также относится к клеткам-хозяевам, которые содержат один или более экспрессирующих векторов по настоящему изобретению.
В конкретных вариантах осуществления, антитело представляет собой рекомбинантное антитело или его антигенсвязывающий фрагмент. В родственных вариантах осуществления вариабельный домен тяжелой цепи и вариабельный домен легкой цепи соединены гибким линкером с образованием одноцепочечного антитела. В конкретных вариантах осуществления антитело или антигенсвязывающий фрагмент представляет собой Fab-фрагмент. В других вариантах осуществления, антитело или антигенсвязывающий фрагмент представляет собой Fab'-фрагмент. Еще в других вариантах осуществления антитело или антигенсвязывающий фрагмент представляет собой фрагмент (Fab')2. Еще в других вариантах осуществления, антитело или антигенсвязывающий фрагмент представляет собой диантитело. В конкретном варианте осуществления, антитело или антигенсвязывающий фрагмент представляет собой домен-содержащее антитело. В конкретных вариантах осуществления антитело или антигенсвязывающий фрагмент представляет собой однодоменное антитело.
В конкретных вариантах осуществления канинизированное мышиное антитело против собачьего CTLA-4 или антигенсвязывающий фрагмент связывается с CTLA-4 у животного (например, собаки), которое подвергают лечению от онкологического заболевания. В более конкретных вариантах осуществления введение канинизированного мышиного антитела против собачьего CTLA-4 или антигенсвязывающего фрагмента по настоящему изобретению служит для облегчения одного или более симптомов онкологического заболевания у животного (например, собаки), которое подвергают лечению.
Настоящее изобретение также относится к выделенным нуклеиновым кислотам, кодирующим мышиные антитела против собачьего CTLA-4 или их части. В родственных вариантах осуществления, такие антитела или антигенсвязывающие фрагменты могут использоваться для получения лекарственного средства для лечения онкологического заболевания у собаки. Альтернативно или совместно, в настоящем изобретении предложено применение любого из антител или фрагментов антител по настоящему изобретению для диагностических целей. Еще в дополнительных вариантах осуществления предложен набор, содержащий любое из канинизированных антител или антигенсвязывающих фрагментов, раскрытых в настоящем описании.
Настоящее изобретение также относится к выделенным пептидам, которые связываются с канинизированным антителом по настоящему изобретению, которые содержат от 5 до 25 аминокислотных остатков и которые на 90% или более идентичны аминокислотной последовательности SEQ ID NO: 132. В конкретных вариантах осуществления выделенные пептиды идентичны аминокислотной последовательности SEQ ID NO: 132. В более конкретных вариантах осуществления выделенные пептиды содержат от 10 до 20 аминокислотных остатков. В родственных вариантах осуществления выделенные пептиды связываются с канинизированным антителом по настоящему изобретению, содержат от 5 до 25 аминокислотных остатков и на 90% или более идентичны аминокислотной последовательности SEQ ID NO: 133. В конкретных вариантах осуществления выделенные пептиды идентичны аминокислотной последовательности SEQ ID NO: 133. В более конкретных вариантах осуществления этого типа выделенные пептиды содержат от 10 до 20 аминокислотных остатков.
В других вариантах осуществления выделенные пептиды, которые связываются с канинизированным антителом по настоящему изобретению, содержат аминокислотные последовательности, которые на 90% или более идентичны аминокислотной последовательности SEQ ID NO: 134. В других вариантах осуществления выделенные пептиды содержат аминокислотные последовательности, которые идентичны аминокислотной последовательности SEQ ID NO: 134. В других вариантах осуществления выделенные пептиды, которые связываются с канинизированным антителом по настоящему изобретению, содержат аминокислотные последовательности, которые на 90% или более идентичны аминокислотной последовательности SEQ ID NO: 135. В других вариантах осуществления выделенные пептиды содержат аминокислотные последовательности, которые идентичны аминокислотной последовательности SEQ ID NO: 135. В других вариантах осуществления выделенные пептиды, которые связываются с канинизированным антителом по настоящему изобретению, содержат аминокислотные последовательности, которые на 90% или более идентичны аминокислотной последовательности SEQ ID NO: 136. В других вариантах осуществления выделенные пептиды содержат аминокислотные последовательности, которые идентичны аминокислотной последовательности SEQ ID NO: 136.
Кроме того, настоящее изобретение относится к слитым белкам, которые содержат такие выделенные пептиды, которые связываются с канинизированным антителом по настоящему изобретению. Настоящее изобретение дополнительно относится к слитым белкам, которые содержат любой из вышеупомянутых пептидов. В конкретном варианте осуществления слитый белок содержит такой антигенный пептид и Fc-область антитела IgG млекопитающего, отличного от собаки. В более конкретном варианте осуществления слитый белок содержит Fc-область антитела IgG млекопитающего, отличного от собаки. В некоторых вариантах осуществления антитело IgG млекопитающего, отличного от собаки, представляет собой мышиный IgG. В альтернативных вариантах осуществления антитело IgG млекопитающего, отличного от собаки, представляет собой IgG человека. В других вариантах осуществления антитело IgG млекопитающего, отличного от собаки, представляет собой IgG лошади. В других вариантах осуществления антитело IgG млекопитающего, отличного от собак, представляет собой IgG свиньи. Еще в других вариантах осуществления антитело IgG млекопитающего, отличного от собаки, представляет собой IgG крупного рогатого скота.
В конкретных вариантах осуществления антитело IgG млекопитающего, отличного от собаки, представляет собой IgG1. В других вариантах осуществления антитело IgG млекопитающего, отличного от собаки, представляет собой IgG2a. В других вариантах осуществления антитело IgG млекопитающего, отличного от собаки, представляет собой IgG3. Еще в других вариантах осуществления антитело IgG млекопитающего, отличного от собак, представляет собой IgG4. В других вариантах осуществления слитый белок включает любой из вышеупомянутых антигенных пептидов и мальтоза-связывающий белок. Еще в других вариантах осуществления слитый белок содержит любой из вышеупомянутых антигенных пептидов и бета-галактозидазу. В других вариантах осуществления слитый белок содержит любой из вышеуказанных антигенных пептидов и глутатионстрансферазу. Еще в других вариантах осуществления слитый белок содержит любой из вышеупомянутых антигенных пептидов и тиоредоксин. В других вариантах осуществления слитый белок содержит любой из вышеуказанных антигенных пептидов и Gro EL. Еще в других вариантах осуществления слитый белок содержит любой из вышеупомянутых антигенных пептидов и NusA.
Настоящее изобретение также относится к нуклеиновым кислотам (включая выделенные и/или рекомбинантные нуклеиновые кислоты), которые кодируют один или более выделенных иммуногенных и/или антигенных пептидов и/или слитых белков по настоящему изобретению. Настоящее изобретение также относится к экспрессирующим векторам, содержащим такие выделенные нуклеиновые кислоты, а также к клеткам-хозяевам, которые содержат один или более экспрессирующих векторов по настоящему изобретению.
Фармацевтические композиции могут также содержать антигенные пептиды (включая выделенные антигенные пептиды) собачьего CTLA-4, слитые белки, содержащие антигенные пептиды собачьего CTLA-4 по настоящему изобретению, нуклеиновые кислоты (включая выделенные нуклеиновые кислоты), кодирующие антигенные фрагменты, и/или слитые белки по настоящему изобретению, экспрессирующие векторы, содержащие такие нуклеиновые кислоты или любую их комбинацию, и фармацевтически приемлемый носитель или разбавитель. Кроме того, настоящее изобретение включает фармацевтические композиции, содержащие антитела против собачьего CTLA-4 (включая канинизированные мышиные антитела против собачьего CTLA-4) или их антигенсвязывающие фрагменты по настоящему изобретению. Такие фармацевтические композиции можно использовать для лечения онкологического заболевания, инфекции или инфекционного заболевания, использовать в качестве вакцинного адъюванта и/или в способе повышения активности иммунной клетки, включающем введение пациенту терапевтически эффективного количества фармацевтической композиции.
В конкретных вариантах осуществления такие фармацевтические композиции дополнительно содержат антитело против собачьего PD-1 (включая канинизированное мышиное антитело против собачьего PD-1) или его антигенсвязывающий фрагмент. В более конкретных вариантах осуществления антитело против собачьего PD-1 представляет собой канинизированное мышиное антитело против собачьего PD-1 или антигенсвязывающий фрагмент канинизированного мышиного антитела против собачьего PD-1.
В родственных вариантах осуществления такие фармацевтические композиции дополнительно содержат антитело против собачьего PDL1 (включая канинизированное мышиное антитело против собачьего PD-L1) или его антигенсвязывающий фрагмент. В конкретных вариантах осуществления антитело против собачьего PD-L1 представляет собой канинизированное мышиное антитело против собачьего PD-1 или антигенсвязывающий фрагмент канинизированного мышиного антитела против собачьего антитела против PD-1.
Соответственно, настоящее изобретение относится к фармацевтическим композициям, которые содержат один, два, три или более из следующих элементов: антитело против собачьего PD-L1, антитело против собачьего PD-1, антитело против собачьего CTLA-4, антигенсвязывающий фрагмент антитела против собачьего PD-L1, антигенсвязывающий фрагмент антитела против собачьего PD-1 или антигенсвязывающий фрагмент антитела против собачьего CTLA-4. В конкретных вариантах осуществления такие антитела против собачьего белка (т.е. против собачьего PD-L1, PD-1 или CTLA-4) или их антигенсвязывающие фрагменты представляют собой мышиные антитела против собачьего белка. В других вариантах осуществления такие антитела против собачьего белка или их антигенсвязывающие фрагменты представляют собой канинизированные антитела против собачьего белка. В более конкретных вариантах осуществления антитела против собачьего белка или их антигенсвязывающие фрагменты представляют собой канинизированные мышиные антитела против собачьего белка.
Кроме того, настоящее изобретение относится к способам повышения активности иммунной клетки, включающие введение пациенту терапевтически эффективного количества фармацевтической композиции по настоящему изобретению. В некоторых вариантах осуществления способ используют при лечении онкологического заболевания. В других вариантах осуществления, способ используют при лечении инфекции или инфекционного заболевания. Еще в других вариантах осуществления, канинизированное антитело по настоящему изобретению или его антигенсвязывающий фрагмент применяют в качестве вакцинного адъюванта. В конкретных вариантах осуществления фармацевтическая композиция, содержащая канинизированное мышиное антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент, может быть введена до, после или одновременно с канинизированным мышиным антителом против собачьего PD-1 или его антигенсвязывающим фрагментом и/или канинизированным мышиным антителом против собачьего PD-L1 или его антигенсвязывающим фрагментом.
Эти и другие аспекты настоящего изобретения будут лучше понятны при ссылке на следующее краткое описание чертежей и на подробное описание.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фигуре 1 показана активность связывания шести антител с собачьим CTLA-4 (cCTLA-4). Соответственно, на Фигуре 1 изображен график количества индивидуальных собачьих антител против CTLA-4 в нг/мл (Ab Log), добавленных к собачьему CTLA-4 в ИФА, демонстрирующий связывающую активность антител с cCTLA-4. Индивидуальные антитела к собачьему CTLA-4 обозначены как 27G12, 110E3, 12B3, 45A9, 39A11 и 22A11.
На Фигуре 2 изображены антитела, блокирующие взаимодействие собачьего CD86 с CTLA-4. На Фигуре показан график количества индивидуальных собачьих антител против CTLA-4 в нг/мл (Ab Log), добавленных к cCTLA-4, чтобы препятствовать связыванию собачьих CTLA-4 с CD86. Индивидуальные антитела к собачьему CTLA-4 обозначены как 39A11, 27G12, 45A9, 12B3, 110E3 и 22A11. Как видно, антитела могут блокировать взаимодействие собачьего CD86 с CTLA-4.
На Фигуре 3 показаны антитела, блокирующие взаимодействие собачьего CD80 с CTLA-4. На фигуре показан график количества индивидуальных собачьих антител против CTLA-4 в нг/мл (Ab Log), добавленных к cCTLA-4, чтобы препятствовать связыванию собачьих CTLA-4 с CD80. Индивидуальные антитела к собачьему CTLA-4 обозначены как 39A11, 27G12, 45A9,12B3, 110E3 и 22A11. Как видно, антитела также могут блокировать взаимодействие собачьего CD80 с CTLA-4.
На Фигурах 4A-4G изображены антитела, связывающиеся с клетками CHO, которые экспрессируют собачий CTLA-4. Фиг.4A представляет собой Iso-контроль, Фиг.4B - 39A11, Фиг.4C - 27G12, Фиг.4D - 12B3, Фиг.4E - 45A9, Фиг.4F - 110E3, а Фиг.4G - 22A11. Как видно, антитела могут связываться с клетками CHO, экспрессирующими cCTLA-4.
На Фигуре 5 изображена гистограмма, которая количественно определяет три уменьшающихся концентрации индивидуальных собачьих антител CTLA-4, добавленных в 25 мкг/мл, 50 мкг/мл или 100 мкг/мл (Ab), которые активируют собачьи клетки PBMC в присутствии конканавалина A (CoA) для продуцирования IFNγ. Тестируемые антитела нанесены на абсциссу и помечены как моноклональные антитела CTLA-4 (mAb xCTLA-4). Как можно видеть, антитела могут активировать собачьи клетки PBMC для продуцирования IFNγ.
На Фигуре 6 изображен график количества моноклональных антител CTLA-4 (xCTLA-4; Ab Log нг/мл), которые обладают такой же реактивностью по отношению к собачьим CTLA-4, как и родительские антитела. Результаты ИФА показывают, что как 12B3, так и 39A11 были успешно канинизированы. Канинизированные c12B3L3H2 и L3H3 обладают такой же реактивностью в отношении cCTLA-4, что и родительское 12B3, а канинизированное c39A11L3H3 обладает такой же реактивностью в отношении cCTLA-4, что и родительское 39A11.
На Фигуре 7A-7B представлены связывающие эпитопы на cCTLA-4 для c12B3 (Фигура 7A) и c39A11 (Фигура 7B). Изображены две области собачьего белка CTLA-4, которые имеют аминокислотные последовательности SEQ ID NO: 132 и SEQ ID NO: 133, соответственно (см. Таблицу 8 ниже). Оба антитела связываются с аминокислотной последовательностью SEQ ID NO: 136, которая содержит мотив MYPPPY (SEQ ID NO: 137), и с аминокислотной последовательностью SEQ ID NO: 134. c12B3 также связывается с аминокислотной последовательностью SEQ ID NO: 135.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
СОКРАЩЕНИЯ
В тексте подробного описания и в примерах изобретения используются следующие сокращения:
ADCC Антитело-зависимая клеточная цитотоксичность
CDC Комплемент-зависимая циотоксичность
CDR Определяющая комплементарность область в вариабельных областях иммуноглобулина, определенная с использованием системы нумерации Kabat
CHO клетки Яичники китайского хомячка
EC50 концентрация, приводящая к 50% эффективности или связывания
ИФА твердофазный Иммуноферментный Анализ
FR Каркасная область антитела: вариабельные области иммуноглобулина за исключением области CDR.
HRP Пероксидаза хрена
IFN интерферон
IC50 концентрация, приводящая к 50% ингибирования
IgG Иммуноглобулин G
Kabat Система выравнивания и нумерации иммуноглобулинов, впервые разработанная Elvin A. Kabat [Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)]
mAb Моноклональное антитело (также Mab или MAb)
MES 2-(N-морфолин)этансульфокислота
MOA Механизм действия
NHS Сыворотка здорового человека
ПЦР Полимеразная цепная реакция
PK Фармакокинетика
SEB Энтеротоксин B стафилококка
TT Столбнячный анатоксин
V область Сегмент цепей IgG, последовательность которого является вариабельной между разными антителами. Он простирается до остатка Kabat 109 в легкой цепи и 113 в тяжелой цепи.
VH Вариабельная область тяжелой цепи иммуноглобулина
VL Вариабельная область легкой цепи иммуноглобулина
VK Вариабельная область легкой цепи каппа иммуноглобулина
ОПРЕДЕЛЕНИЯ
Для того чтобы легче понять изобретение, ниже специально приведены определения некоторых технических и научных терминов. До тех пор пока специально не определено иное где-либо в настоящем описании, все остальные технические и научные термины, используемые в настоящем описании, имеют общепринятые значения, понятные специалисту в области, к которой относится изобретение.
При использовании в настоящем описании, включая прилагаемую формулу изобретения, формы единственного числа включает их соответствующие множественные значения до тех пор, пока контекст ясно не указывает иное.
«CTLA-4» представляет собой сокращение от «белка 4, ассоциированного с цитотоксическими Т-лимфоцитами», также известного как CD152 (кластер дифференцировки 152), который представляет собой рецептор белка, который функционирует как иммунная контрольная точка и подавляет иммунные ответы. Аминокислотная последовательность собачьего CTLA-4 представляет собой SEQ ID NO: 126. Настоящее изобретение также относится к канинизированным мышиным антителам к собачьему CTLA-4.
«Активация» при применении к клеткам или к рецепторам относится к активации или к обработке клетки или рецептора с помощью лиганда до тех пор, пока не указано иное с помощью контекста или в прямой форме. «Активация» может относиться к активации клеток, регулируемой внутренними механизмами, а также внешними факторами или факторами окружающей среды.
«Лиганд» охватывает природные и синтетические лиганды, например, цитокины, варианты цитокинов, аналоги, мутантные белки и связывающие соединение, выделенные из антител. «Лиганд» также охватывает низкомолекулярные соединения, например, пептидомиметики цитокинов и пептидомиметики антител.
«Активность» молекулы может описывать или относиться к связыванию молекулы с лигандом или с рецептором, к каталитической активности; к способности стимулировать экспрессию генов и клеточную сигнальную систему, к дифференцировке или созреванию; к антигенной активности, к модулированию активностей других молекул и тому подобное. «Активность» молекулы также может относиться к активности в модулировании или поддержании межклеточных взаимодействий, например, адгезии, или активности в поддержании структуры клетки, например, клеточных мембран или цитоскелета. «Активность» также может означать специфическую активность, например, [каталитическая активность]/[мг белка] или [иммунологическая активность]/[мг белка], концентрация в биологическом компартменте и т.п. «Активность» может относиться к модулированию компонентов врожденного или приобретенного иммунитета.
«Введение» и «лечение» применительно к животному, например собаке, клетке, ткани, органу или биологической жидкости, относятся к контакту экзогенного фармацевтического, терапевтического, диагностического агента или композиции с животным, например, собакой, клеткой, тканью, органом или биологической жидкостью. Обработка клетки охватывает контакт реагента с клеткой, а также контакт реагента с жидкостью, где жидкость находится в контакте с клеткой.
«Введение» и «лечение» также означают лечение in vitro и ex vivo, например, клетки, реагентом, диагностическим, связывающим соединением или другой клеткой.
Термин «объект» включает любой организм, предпочтительно животное, более предпочтительно млекопитающее (например, собаку, кошку или человека) и наиболее предпочтительно собаку.
«Лечить» или «лечение» означает введение терапевтического агента, такого как композиция, содержащая любое из антител или антигенсвязывающих фрагментов по настоящему изобретению, внутрь или экстракорпорально, например, собаке или пациенту, имеющему один или более симптомов заболевания, или подозрение на заболевание, в отношении которого агент обладает терапевтической активностью.
Обычно агент вводят в количестве, эффективном для облегчения и/или облегчения одного или более симптомов заболевания у подвергающегося лечению субъекта или популяции, либо путем индукции регресса или ингибирования прогрессирования такого симптома(симптомов) в любой клинически измеримой степени. Количество терапевтического агента, которое эффективно для облегчения какого-либо конкретного симптома заболевания (также называемое «терапевтически эффективное количество»), может варьироваться в зависимости от таких факторов, как состояние болезни, возраст и масса пациента (например, собаки), и от способности фармацевтической композиции вызывать целевой ответ у пациента. Облегчение или улучшение симптома заболевания можно оценить с помощью любого клинического измерения, как правило, используемого ветеринарами или другими медицинскими работниками, для оценки состояния тяжести или прогрессии данного симптома. В то время как вариант осуществления настоящего изобретения (например, способ лечения или готовое изделие) может быть не эффективным в облегчении симптомов целевого заболевания у каждого пациента, он должен облегчать симптомы целевого заболевания у статистически значимого количества пациентов, как определено с помощью статистического теста, известного в данной области, такого как тест Стъюдента, критерий хиквадрат, U-тест Манна и Уитни, критерий Крускала-Уоллиса (H-тест), Критерий Джонкхиера-Терпстра и критерий Уилкоксона.
«Лечение», применительно к человеку, ветеринарии (например, к собаке) или объекту исследования, относится к терапевтическому лечению, а также к исследованиям и диагностическим применениям. «Лечение» применительно к человеку, ветеринарии (например, собаке) или объекту исследования, клетке, ткани или органу включает контакт антител или антигенсвязывающих фрагментов по настоящему изобретению, например, с собакой или другим животным, клеткой, тканью, физиологическим компартментом или физиологической жидкостью.
Используемый в настоящем описании термин «собака» включает всех домашних собак, семейство волчьих или семейство псовых до тех пор, пока не указано иное.
Используемый в настоящем описании термин «кошка» относится к любому члену семейства кошачьих. Члены этого семейства включают диких, зоопарковских и домашних представителей, включая домашних кошек, чистопородных и/или беспородных домашних кошек, выставочных кошек, лабораторных кошек, клонированных кошек и диких или одичавших кошек.
Используемый в настоящем описании термин «собачий каркас» относится к аминокислотной последовательности тяжелой цепи и легкой цепи собачьего антитела, кроме остатков гипервариабельной области, определенных здесь как остатки CDR. Что касается канинизированного антитела, в большинстве вариантов осуществления аминокислотные последовательности нативных собачьих CDR заменены соответствующими чужеродными CDR (например, из мышиного антитела) в обеих цепях. Необязательно тяжелая и/или легкая цепи собачьего антитела могут содержать некоторые чужеродные остатки, не относящиеся к CDR, например, для сохранения конформации чужеродных CDR внутри собачьего антитела и/или для модификации функции Fc, как проиллюстрировано ниже.
Было обнаружено, что собачий CTLA-4 содержит аминокислотную последовательность SEQ ID NO: 126 (включая сигнальную последовательность). В конкретном варианте осуществления собачий CTLA-4 кодируется нуклеиновой кислотой, которая содержит нуклеотидную последовательность SEQ ID NO: 125. Последовательности собачьего CTLA-4 могут отличаться, например, наличием консервативных вариаций в неконсервативных областях, но собачий CTLA-4 будет иметь по существу ту же биологическую функцию, что и собачий CTLA-4, содержащий аминокислотную последовательность SEQ ID NO: 126.
Используемый в настоящем описании термин «замена аминокислотного остатка» другим аминокислотным остатком в аминокислотной последовательности антитела, например, эквивалентен «замене аминокислотного остатка» другим аминокислотным остатком и означает, что конкретный аминокислотный остаток в конкретном положении в аминокислотной последовательности был заменен (или заменен) другим аминокислотным остатком. Такие замены могут быть специально разработаны, т.е. целенаправленные замены аланина серином в конкретном положении в аминокислотной последовательности, например, с помощью технологии рекомбинантной ДНК. Альтернативно, конкретный аминокислотный остаток или цепочка аминокислотных остатков антитела могут быть заменены одним или более аминокислотными остатками с помощью более естественных процессов отбора, например, на основе способности антитела, продуцируемого клеткой, связываться с заданной областью на этом антигене, например антигене, содержащем эпитоп или его часть, и/или антитело, содержащее конкретную CDR, которая сохраняет ту же каноническую структуру, что и заменяемая CDR. Такие замены/замены могут привести к получению «вариантов» CDR и/или вариантов антител.
Костимулирующие сигнальные пути приводят к развитию иммунных ответов и, как было показано, опосредуются взаимодействием CD28 на поверхности Т-клеток и CD80 (также известного как B7.1) и CD86 (также известного как B7.2). CTLA-4 связывается как с CD80, так и с CD86 с гораздо более высокой аффинностью, чем CD28, и, таким образом, действует как ингибирующий рецептор, который жизненно важен для подавления иммунного ответа. Действительно, механизм, с помощью которого CTLA-4 опосредует свои иммунные ингибирующие функции, связан с его способностью действовать в качестве конкурентного ингибитора взаимодействия CD28 с CD80 и CD86. Соответственно, в настоящем изобретении описано получение и характеристика моноклональных антител, которые блокируют связывание собачьего CD80 и собачьего CD86 с CTLA-4 и, таким образом, разрешает костимулирующую передачу сигналов за счет связывания собачьего CD28 с собачьими CD80 и CD86. Таким образом, эти антитела могут применяться для лечения онкологического заболевания, а также других заболеваний у домашних животных, как описано в настоящем описании.
Конкретная аминокислотная последовательность собачьего CTLA-4 обычно по меньшей мере на 90% идентична собачьему CTLA-4, содержащему аминокислотную последовательность SEQ ID NO: 126, за исключением сигнальной последовательности. В некоторых случаях собачий CTLA-4 может быть по меньшей мере на 95% или даже по меньшей мере на 96%, 97%, 98% или на 99% идентичен собачьему CTLA-4, содержащему аминокислотную последовательность SEQ ID NO: 126, за исключением сигнальной последовательности. В некоторых вариантах осуществления аминокислотная последовательность собачьего CTLA-4 будет демонстрировать не более чем 10 аминокислотных отличий от собачьего CTLA-4, содержащего аминокислотную последовательность SEQ ID NO: 126, за исключением сигнальной последовательности. В некоторых вариантах осуществления аминокислотная последовательность собачьего CTLA-4 может отображать не более чем 5 или даже не более чем 4, 3, 2 или 1 аминокислотных отличий от собачьего CTLA-4, содержащего аминокислотную последовательность SEQ ID NO: 126, за исключением сигнальной последовательности. Процент идентичности может определяться, как описано в настоящем описании ниже.
Термин «иммунный ответ» относится к действию, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитарных клеток, гранулоцитов и растворимых макромолекул, продуцируемых указанными выше клетками или печенью (включая антитела, цитокины и комплемент), что приводит к селективному повреждению, разрушению или удалению из организма млекопитающего (например, тела собаки) опухолевых клеток, клеток или тканей, инфицированных патогенами, или инвазивных патогенов.
Антитела против собачьего CTLA-4
Настоящее изобретение относится к выделенным антителам (в частности, мышиным антителам против собачьего CTLA-4 и их канинизированным антителам) или их антигенсвязывающим фрагментам, которые связываются с собачьим CTLA-4, и к применениям таких антител или их фрагментов. В конкретных вариантах осуществления предложены CDR из мышиных антител против собачьих CTLA-4, которые, как было показано, как и связываются с собачьим CTLA-4, так и блокируют связывание собачьего CTLA-4 с одним или обоими его лигандами, собачьим CD86 или CD80. Эти CDR могут быть вставлены в модифицированный собачий каркас собачьего антитела для создания канинизированного мышиного антитела против собачьего CTLA-4.
Используемый в настоящем описании термин «антитело против собачьего CTLA-4» относится к антителу, которое было индуцировано против собачьего CTLA-4 (например, у млекопитающего, такого как мышь или кролик), и которое специфически связывается с собачьим CTLA-4. Антитело, которое «специфически связывается с собачьим CTLA-4» и, в частности, с собачьим CTLA-4, или антитело, которое «специфически связывается с полипептидом, содержащим аминокислотную последовательность собачьего CTLA-4», представляет собой антитело, которое проявляет преимущественное связывание с собачьим CTLA-4 по сравнению с другие собачьи антигенами, но эта специфичность не требует абсолютной специфичности связывания. Антитело против собачьего CTLA-4 считается «специфическим» для собачьего CTLA-4, если его связывание является определяющим для присутствия собачьего CTLA-4 в образце, который ограничен собачьими белками, или если оно способно изменять активность собачьего CTLA-4 без чрезмерного вмешательства в активность других молекул в образце собаки, например без получения нежелательных результатов, таких как ложные положительные результаты в диагностическом контексте или побочные эффекты в терапевтическом контексте. Степень специфичности, необходимая для антитела против собачьего CTLA-4, может зависеть от предполагаемого использования антитела и, в любом случае, определяется его пригодностью для использования по назначению. Антитело или связывающее соединение, полученное из антигенсвязывающего сайта антитела, рассматриваемого способа связывается со своим антигеном или его вариантом или мутеином с аффинностью, которая по меньшей мере в два раза больше, предпочтительно по меньшей мере в десять раз больше, более предпочтительно, по меньшей мере, в 20 раз больше, а наиболее предпочтительно, по меньшей мере, в 100 раз больше, чем аффинность к любому другому собачьему антигену.
В данном контексте считается, что антитело специфически связывается с полипептидом, содержащим данную антигенную последовательность (в данном случае часть аминокислотной последовательности собачьего CTLA-4), если оно связывается с полипептидами, составляющими часть аминокислотной последовательности собачьего CTLA-4, но не связывается с другими собачьими белками, лишенными этой части последовательности собачьего CTLA-4. Например, антитело, которое специфически связывается с полипептидом, содержащим собачий CTLA-4, может связываться с меченной FLAG® формой собачьего CTLA-4, но не будет связываться с другими собачьими белками, меченными FLAG®. Антитело или связывающее соединение, происходящее из антигенсвязывающего сайта антитела, связывается со своим собачьим антигеном или его вариантом или мутеином «со специфичностью», когда оно имеет аффинность к этому собачьему антигену или его варианту или мутеину, которая по меньшей мере в десять раз больше, более предпочтительно по меньшей мере в 20 раз больше и даже более предпочтительно по меньшей мере в 100 раз больше, чем его аффинность к любому другому протестированному собачьему антигену.
Используемый в настоящем описании термин «антитело» относится к любой форме антитела, которая проявляет желаемую биологическую активность. Таким образом, он используется в самом широком смысле и конкретно охватывает, но не ограничивается этим, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, полиспецифические антитела (например, биспецифические антитела), канинизированные антитела, полностью собачьи антитела, химерные антитела и камелизированные однодоменные антитела. «Родительские антитела» представляют собой антитела, полученные путем воздействия на иммунную систему антигена перед модификацией антител для предполагаемого использования, такого как канинизация антитела для использования в качестве терапевтического антитела для собак.
В контексте настоящего описания, если не указано иное, «фрагмент антитела» или «антигенсвязывающий фрагмент» относится к антигенсвязывающим фрагментам антител, то есть фрагментам антител, которые сохраняют способность специфически связываться с антигеном, связанным с полноразмерным антителом, например фрагменты, которые сохраняют одну или более областей CDR. Примеры антигенсвязывающих фрагментов включают в частности Fab-, Fab'-, F(ab')2-, и Fv-фрагменты; диантитела; линейные антитела; молекулы одноцепочечных антител, например, sc-Fv; наноантитела и полиспецифичные антитела, образованные из фрагментов антител.
«Fab-фрагмент» состоит из одной легкой цепи, а также CH1 и вариабельной области одной тяжелой цепи. Тяжелая цепь молекулы Fab не может образовывать дисульфидную связь с другой молекулой тяжелой цепи. «Fab-фрагмент» может быть продуктом расщепления антитела папаином.
Область «кристаллизующегося фрагмента» («Fc») содержит два фрагмента тяжелой цепи, содержащие домены CH3 и CH2 антитела. Два фрагмента тяжелой цепи удерживаются вместе двумя или более дисульфидными связями и гидрофобными взаимодействиями доменов CH3.
«Фрагмент Fab'» содержит одну легкую цепь и часть или фрагмент одной тяжелой цепи, которая содержит домен VH и домен CH1, а также область между доменами CH1 и CH2, так что межцепочечная дисульфидная связь может быть образована между две тяжелые цепи двух фрагментов Fab' с образованием молекулы F(ab')2.
«Фрагмент F(ab')2» содержит две легкие цепи и две тяжелые цепи, содержащие часть константной области между доменами CH1 и CH2, так что между двумя тяжелыми цепями образуется межцепочечная дисульфидная связь. Таким образом, фрагмент F(ab')2 состоит из двух фрагментов Fab', которые удерживаются вместе дисульфидной связью между двумя тяжелыми цепями. «Фрагмент F(ab')2» может быть продуктом пепсинового расщепления антитела.
«Область Fv» содержит вариабельные области как тяжелой, так и легкой цепей, но не имеет константных областей.
Термин «одноцепочечное Fv» или «scFv» антитело относится к фрагментам антитела, содержащим домены VH и VL антитела, где эти домены присутствуют в одной полипептидной цепи. Как правило, полипептид Fv дополнительно содержит полипептидный линкер между доменами VH и VL, который позволяет scFv образовывать желаемую структуру для связывания антигена. [См. Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113 Rosenburg and Moore eds., Springer-Verlag, New York, pp.269-315 (1994); WO 88/01649; и US 4946778 и US 5260203].
В данном контексте, антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент, которое «блокирует», «является блокирующим» или «блокирует связывание» собачьего CTLA-4 с его партнером по связыванию (лигандом), например собачьим CD80 или собачьим CD 86, представляет собой антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент, который блокирует (частично или полностью) связывание собачьего CTLA-4 с собачьим CD86 и/или CD80, как определено в стандартных анализах связывания (например, BIACore®, ИФА или проточной цитометрии). Такое «блокирование» проиллюстрировано в Примере 4 ниже с использованием анализа блокирования на основе ИФА.
Используемый в настоящем описании термин «каноническая структура» относится к локальной конформации, которая может быть принята каждой из гипервариабельных областей тяжелой и легкой цепи антитела в пределах каркасной области, в которой они находятся. Для каждой гипервариабельной области существует небольшое количество канонических структур (обычно обозначаемых простыми целыми числами, такими как 1 или 2 и т.д.), которые можно предсказать с большой точностью по аминокислотным последовательностям соответствующей гипервариабельной области [особенно в контексте аминокислотной последовательности его каркаса для соответствующих вариабельных доменов против собачьего CTLA-4]. Эти канонические структуры могут быть определяющими в отношении того, приведет ли модификация аминокислотной последовательности данной CDR к сохранению или потере способности связываться с его антигенным партнером по связыванию [См. Chothia and Lesk, Canonical Structures for the hypervariable regions of immunoglobulins, J. Mol. Biol. 196:901-917(1987); Chothia et al., Conformation of immunoglobulin hypervaribale regions, Nature, 34:877-883(1989); and Al-Lazikani et al., Standard Conformations for the canonical structures of immunoglobulins, J. Mol. Biol. 273: 927-948 (1997)].
«Доменное антитело» представляет собой иммунологически функциональный фрагмент иммуноглобулина, содержащий только вариабельную область тяжелой цепи или вариабельную область легкой цепи. В некоторых случаях две или более областей VH ковалентно соединяются с пептидным линкером для создания двухвалентного доменного антитела. Две области VH двухвалентного доменного антитела могут нацеливаться на один и тот же или разные антигены.
«Двухвалентное антитело» включает два антигенсвязывающих сайта. В некоторых случаях два сайта связывания имеют одинаковую антигенную специфичность. Однако двухвалентные антитела могут быть биспецифичными (см. ниже).
В некоторых вариантах осуществления настоящего изобретения моноклональные антитела также включают камелизированные однодоменные антитела. [См., например, Muyldermans et al., Trends Biochem. Sci. 26:230 (2001); Reichmann et al.,J. Immunol. Methods 231:25 (1999); WO 94/04678; WO 94/25591; U.S. 6,005,079]. В одном варианте осуществления настоящее изобретение относится к однодоменным антителам, содержащим два домена VH с такими модификациями, что образуются однодоменные антитела.
Используемый в настоящем описании термин «диантитела» относится к небольшим фрагментам антител с двумя антигенсвязывающими сайтами, причем эти фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в той же полипептидной цепи (VH-VL или VL-VH). При использовании линкера, который является слишком коротким чтобы позволить спаривание между двумя вариабельными доменами на одной и той же цепи, домены диантитела вынуждены спариваться с комплементарными доменами связывания другой цепи и создают два антигенсвязывающих сайта. [См. EP 0404097 B1; WO 93/11161; и Holliger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993)]. Для обзора сконструированных вариантов антител [обычно см. Holliger and Hudson Nat. Biotechnol. 23: 1126-1136 (2005)].
Обычно антитело или антигенсвязывающий фрагмент по изобретению сохраняет по меньшей мере 10% своей активности связывания с собачьим CTLA-4 (по сравнению с исходным антителом), когда эта активность выражается на молярной основе. Предпочтительно антитело или антигенсвязывающий фрагмент по изобретению сохраняет по меньшей мере 20%, 50%, 70%, 80%, 90%, 95% или 100% или более аффинности связывания собачьего CTLA-4 в качестве родительского антитела. Также предполагается, что антитело или антигенсвязывающий фрагмент по изобретению может включать консервативные или неконсервативные аминокислотные замены (называемые «консервативными вариантами» или «функционально консервативными вариантами» антитела), которые существенно не изменяют его биологическую активность.
«Выделенное антитело» относится к статусу очистки и в таком контексте означает молекулу, которая по существу свободна от других биологических молекул, таких как нуклеиновые кислоты, белки, липиды, углеводы или другой материал, такой как клеточный дебрис и ростовая среда. Как правило, термин «выделенный» не предназначен для обозначения полного отсутствия такого материала или отсутствия воды, буферов или солей, если только они не присутствуют в количествах, которые существенно мешают экспериментальному или терапевтическому применению связывающего соединения, как описано здесь.
В данном контексте «химерное антитело» представляет собой антитело, имеющее вариабельный домен из первого антитела и константный домен из второго антитела, где первое и второе антитело относятся к разным видам. [U.S. 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)]. Обычно вариабельные домены получают из антитела от экспериментального животного («родительское антитело»), такого как грызун, а последовательности константных доменов получают из антител животного-объекта, например человека или собаки, так что полученное химерное антитело будет с меньшей вероятностью вызвать неблагоприятный иммунный ответ у человека или собаки, соответственно, чем родительское (например, грызуна) антитело.
Используемый в настоящем описании термин «канинизированное антитело» относится к формам антител, которые содержат последовательности как собачьих, так и не собачьих (например, мышиных) антител. В общем, канинизированное антитело будет включать практически все из, по меньшей мере, одного или более обычно двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют таковым у иммуноглобулина, отличного от собачьего (например, содержащего 6 CDR мышиного антитела против собачьего CTLA-4, как проиллюстрировано ниже), и все или по существу все каркасные области (FR) (и обычно весь или по существу весь оставшийся каркас) являются таковыми из последовательности собачьего иммуноглобулина. Как проиллюстрировано здесь, канинизированное антитело включает как три CDR тяжелой цепи, так и три CDR легкой цепи из мышиного антитела против собачьего CTLA-4 вместе с собачьим каркасом или модифицированным собачьим каркасом. Модифицированный собачий каркас включает одну или более замен аминокислот, как проиллюстрировано здесь, которые дополнительно оптимизируют эффективность канинизированного антитела, например, для увеличения его связывания с собачьим CTLA-4 и/или его способности блокировать связывание собачьего CTLA-4 с собачьими CD86 и/или CD80.
Термин «полностью собачье антитело» относится к антителу, которое содержит белковые последовательности только собачьего иммуноглобулина. Полностью собачьи антитела могут содержать мышиные углеводные цепи, если они продуцируются у мыши, в клетке мыши или в гибридоме, полученной из клетки мыши. Точно так же «мышиное антитело» относится к антителу, которое содержит только последовательности иммуноглобулина мыши. Альтернативно, полностью собачье антитело может содержать крысиные углеводные цепи, если оно продуцируется крысой, крысиной клеткой или гибридомой, полученной из крысиной клетки. Аналогичным образом, «крысиное антитело» относится к антителу, которое содержит только последовательности иммуноглобулина крысы.
Существует четыре известных подтипа тяжелой цепи IgG собачьего IgG, и они называются IgG-A, IgG-B, IgG-C и IgG-D. Два известных подтипа легкой цепи называются лямбда и каппа.
Вариабельные области каждой пары легкой/тяжелой цепи образуют сайт связывания антитела. Таким образом, вообще интактное антитело содержит два сайта связывания. За исключением бифункциональных или биспецифических антител два сайта связывания, в основном, одинаковые.
Как правило, вариабельные домены обеих, тяжелой и легкой, цепей содержат три гипервариабельные области, также называемые гипервариабельными областями (CDR), локализованными внутри относительно консервативных каркасных областей (FR). CDR обычно выравниваются по каркасным областям, что позволяет связываться со специфическим эпитопом. Вообще, от N-конца к C-концу вариабельные домены обеих цепей, легкой и тяжелой, содержат FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Распределение аминокислот в каждом домене, как правило, согласуется с определениями Sequences of Proteins of Immunological Interest, Kabat, et al.; National Institutes of Health, Bethesda, Md.; 5th ed.; NIH Publ. No. 91-3242 (1991); Kabat, Adv. Prot. Chem. 32:1-75 (1978); Kabat, et al., J. Biol. Chem. 252:6609-6616 (1977); Chothia, et al., J. Mol. Biol. 196: 901-917 (1987) или Chothia, et al., Nature 342: 878-883 (1989)].
Используемый в настоящем описании термин «гипервариабельная область» относится к аминокислотным остаткам антитела, которые отвечают за антигенное связывание. Гипервариабельная область содержит аминокислотные остатки из «определяющей комплементарность области» или «CDR» (т.е. CDRL1, CDRL2 и CDRL3 в вариабельном домене легкой цепи и CDRH1, CDRH2 и CDRH3 в вариабельном домене тяжелой цепи). [См. Kabat et al. Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), для определения областей CDR антитела по последовательности; см. также Chothia and Lesk, J. Mol. Biol. 196: 901-917 (1987), определение областей CDR антитела по структуре]. При использовании в настоящем описании, термин «каркасные» или «FR» остатки относится к таким остаткам вариабельных доменов, отличным от остатков гипервариабельных областей, определенных в настоящем описании как CDR-остатки.
В конкретных вариантах осуществления изобретения, помимо связывания и активации собачьих иммунных клеток, собачье или канинизированное антитело против CTLA-4 оптимально имеет два атрибута:
1. Оно лишено эффекторных функций, таких как антитело-зависимая цитотоксичность (ADCC) и комплемент-зависимая цитотоксичность (CDC), и
2. легко очищается в больших масштабах с использованием стандартных промышленных технологий, таких как хроматография на протеине А.
Ни один из природных изотипов собачьего IgG не удовлетворяет обоим критериям. Например, IgG-B может очищаться с использованием протеина A, но при этом оно имеет высокий уровень ADCC-активности. С другой стороны, IgG-A слабо связывается с протеином A, но также проявляет активность ADCC. Более того, ни IgG-C, ни IgG-D нельзя очистить на колонках с протеином А, хотя IgG-D не проявляет активности ADCC. (IgG-C имеет значительную ADCC-активность). Одним из способов решения этих проблем в настоящем изобретении является предложение модифицированных собачьих антител IgG-B, специфичных к CTLA-4, которые лишены эффекторных функций, таких как ADCC, и которые могут быть легко очищены с использованием стандартной хроматографии на основе протеина A.
В альтернативных вариантах осуществления настоящего изобретения собачьи антитела IgG-B или IgG-C, специфичные к CTLA-4, намеренно не модифицируются для удаления/существенного уменьшения эффекторных функций, таких как ADCC, и, следовательно, сохраняют эффекторные функции, такие как ADCC.
«Гомология» относится к сходству между двумя полинуклеотидными последовательностями или между двумя полипептидными последовательностями, когда они оптимально выравнены. Когда положение в обеих из двух сравниваемых последовательностей занято одним и тем же основанием или субъединицей мономера аминокислоты, например, если положение в каждой из двух молекул ДНК занято аденином, то молекулы гомологичны в этом положении. Процент гомологии представляет собой количество гомологичных положений, общих для двух последовательностей, деленное на общее количество сравниваемых положений × 100. Например, если 6 из 10 положений в двух последовательностях совпадают или гомологичны, когда последовательности оптимально выровнены, то две последовательности гомологичны на 60%. Как правило, сравнение осуществляют, когда две последовательности выровнены с получением максимального процента гомологии.
«Выделенная молекула нуклеиновой кислоты» означает ДНК или РНК геномного происхождения, мРНК-, кДНК- или синтетического происхождения, или некоторую их комбинацию, которая не связана со всем или частью полинуклеотида, в котором выделенный полинуклеотид встречается в природе или связан с полинуклеотидом, с которым он не связан в природе. Для целей настоящего описания, следует понимать, что «молекула нуклеиновой кислоты, содержащая» конкретную нуклеотидную пследовательность, не охватывает интактные хромосомы. Выделенные молекулы нуклеиновой кислоты, «содержащие» конкретные последовательности нуклеиновых кислот, могут включать дополнительно к конкретным последовательностям кодирующие последовательности до десяти или даже до двадцати или более других белков или их частей, или их фрагментов, или могут включать функционально связанные регуляторные последовательности, которые контролируют экспрессию кодирующей области упомянутых последовательностей нуклеиновых кислот, и/или могут включать векторные последовательности.
Фраза «контрольные последовательности» относится к ДНК-последовательностям, необходимым для экспрессии функционально связанной кодирующей области в конкретном организме-хозяине. Контрольные последовательности, которые подходят для прокариот, например, включают промотор, необязательно последовательность оператора и сайт связывания рибосом. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Последовательность нуклеиновой кислоты является «функционально связанной», если она находится в функциональном взаимодействии с другой последовательностью нуклеиновой кислоты. Например, ДНК для пред-последовательности или секреторного лидера функционально связана с ДНК для полипептида, если она экспрессируется как пред-белок, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы облегчить трансляцию. Обычно «функционально связанный» означает, что связываемые последовательности ДНК являются смежными, а в случае секреторного лидера - смежными и находятся в рамке считывания. Однако энхансеры не должны быть непрерывными. Связывание осуществляется лигированием по подходящим сайтам рестрикции. Если таких сайтов не существует, синтетические олигонуклеотидные адаптеры или линкеры используют в соответствии с обычной практикой.
При использовании в настоящем описании выражения «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо и все такие обозначения включают потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают первичную рассматриваемую клетку и полученные из нее культуры без учета количества переносов. Также понятно, что не все потомство будет иметь точно идентичное содержание ДНК, что связано с преднамеренными или со спонтанными мутациями. Включено мутантное потомство, которое обладает такой же функцией или биологической активностью, которые выявляются при скрининге в исходно трансформированной клетке. В случае, где подразумеваются различные обозначения, это будет понятно из контекста.
Используемый в настоящем описании термин «зародышевая последовательность» относится к нереаранжированным ДНК-последовательностям иммуноглобулинов. Можно использовать любой подходящий источник нереаранжированных последовательностей иммуноглобулинов. Последовательности зародышевой линии человека могут быть получены, например, из баз данных зародышевой линии JOINSOLVER® на веб-сайте Национального института артрита и скелетно-мышечных и кожных заболеваний Национального института здравоохранения США. Последовательности зародышевой линии мыши могут быть получены, например, как описано в Giudicelli et al. [Nucleic Acids Res. 33: D256-D261 (2005)].
Свойства мышиных антител против собачьего CTLA-4 и
канинизированных мышиных антител против собачьего CTLA-4
Настоящее изобретение относится к выделенным мышиным антителам против собачьего CTLA-4 и их канинизированным антителам, способам применения этих антител или их антигенсвязывающих фрагментов при лечении заболевания, например, лечения онкологического заболевания у собак. У собак имеется четыре тяжелые цепи IgG, обозначаемые как A, B, C и D. Эти тяжелые цепи представляют четыре различных подкласса IgG собаки, которые обозначаются как IgGA, IgGB, IgGC и IgGD. Каждая из двух тяжелых цепей состоит из одного вариабельного домена (VH) и трех константных доменов, называемых СН-1, СН-2 и СН-3. Домен CH-1 соединен с доменом CH-2 через аминокислотную последовательность, называемую «шарнир» или, альтернативно, «шарнирная область».
ДНК и аминокислотные последовательности этих четырех тяжелых цепей были впервые идентифицированы Tang et al. [Vet. Immunol. Immunopathol. 80: 259-270 (2001)]. Последовательности аминокислот и ДНК для этих тяжелых цепей также доступны в базах данных GenBank. Например, аминокислотная последовательность тяжелой цепи IgGA имеет номер доступа AAL35301.1, IgGB имеет номер доступа AAL35302.1, IgGC имеет номер доступа AAL35303.1, а IgGD имеет номер доступа (AAL35304.1). Собачьи антитела также содержат два типа легких цепей, каппа и лямбда. ДНК и аминокислотную последовательность этих легких цепей можно получить из баз данных GenBank. Например, аминокислотная последовательность легкой цепи каппа имеет номер доступа ABY 57289.1, а легкая цепь лямбда имеет номер доступа ABY 55569.1.
В настоящем изобретении аминокислотная последовательность для каждого из четырех Fc-фрагментов собачьего IgG основана на идентифицированной границе доменов CH1 и CH2, как определено Tang et al, выше. Канизированные мышиные антитела против собачьего CTLA-4, которые связываются с собачьим CTLA-4, включают, но не ограничиваются этим: антитела, которые содержат тяжелые цепи собачьих IgG-A, IgG-B, IgG-C и IgG-D и/или собачьи легкие цепи каппа вместе с CDR мышиных антител против собачьего CTLA-4. Соответственно, в настоящем изобретении предложены выделенные мышиные антитела против собачьего CTLA-4 и/или канинизированные мышиные антитела против собачьего CTLA-4 или их антигенсвязывающие фрагменты, которые связываются с собачьим CTLA-4 и блокируют связывание собачьего CTLA-4 с собачьим CD86 и/или собачьим CD80.
Настоящее изобретение также относится к полноразмерным собачьим тяжелым цепям, которые могут быть сопоставлены с соответствующими легкими цепями для получения канинизированного антитела. Соответственно, настоящее изобретение дополнительно относится к канинизированным мышиным антителам против собачьего антигена (включая выделенные мышиные антитела против собачьего CTLA-4) и к способам применения этих антител или их антигенсвязывающих фрагментов при лечении заболевания, например, лечения онкологического заболевания у собак.
Настоящее изобретение также относится к канинизированным мышиным антителам против собачьего CTLA-4, которые содержат кристаллизующуюся область собачьего фрагмента (cFc-область), в которой cFc был генетически модифицирован для усиления, уменьшения или устранения одной или более эффекторных функций. В одном из аспектов настоящего изобретения генетически модифицированная cFc снижает или устраняет одну или более эффекторных функций. В другом аспекте изобретения генетически модифицированная cFc усиливает одну или более эффекторных функций. В некоторых вариантах осуществления генетически модифицированная cFc-область представляет собой генетически модифицированную Fc-область собачьего IgGB. В другом таком варианте осуществления генетически модифицированная cFc-область представляет собой генетически модифицированную Fc-область собачьего IgGC. В конкретном варианте осуществления эффекторная функция представляет собой антителозависимую цитотоксичность (ADCC), которая усиливается, снижается или устраняется. В другом варианте осуществления эффекторная функция представляет собой комплемент-зависимую цитотоксичность (CDC), которая усиливается, снижается или устраняется. Еще в одном варианте осуществления cFc-область была генетически модифицирована для усиления, уменьшения или устранения как ADCC, так и CDC.
Для создания вариантов собачьего IgG, лишенных эффекторных функций, был создан ряд мутантных тяжелых цепей собачьего IgG. Эти варианты могут включать одну или более из следующих одиночных или комбинированных замен в Fc-части аминокислотной последовательности тяжелой цепи: P4A, D31A, N63A, G64P, T65A, A93G и P95A. Варианты тяжелых цепей (т.е. содержащие такие аминокислотные замены) клонировали в экспрессирующие плазмиды и трансфецировали в клетки НЕК 293 плазмиду, содержащую ген, кодирующий легкую цепь. Интактные антитела, экспрессированные и очищенные из клеток HEK 293, оценивали на связывание с FcγRI и C1q, чтобы оценить их потенциал для опосредования иммунных эффекторных функций. [См. Патент США 10106607 B2, содержание которого включено в настоящее описание в качестве ссылки в полном объеме].
Настоящее изобретение также относится к модифицированным собачьим IgGD, которые вместо его естественного шарнирного участка IgGD содержат шарнирную область из:
IgGA: fnecrctdtppcpvpep, SEQ ID NO: 128;
IgGB: pkrengrvprppdcpkcpapem, SEQ ID NO: 129; или
IgGC: akececkcncnncpcpgcgl, SEQ ID NO: 130.
Альтернативно, шарнирная область IgGD может быть генетически модифицирована путем замены остатка серина остатком пролина, то есть pkestckciPpcpvpes, SEQ ID NO: 131 (с остатком пролина (P) подчеркнутым и выделенным жирным шрифтом вместо встречающегося в природе остатка серина). Такие модификации могут привести к тому, что собачий IgGD лишится обмена Fab-плечей. Модифицированные собачьи IgGD могут быть сконструированы с использованием стандартных методов технологии рекомбинантной ДНК [например, Maniatis et al., Molecular Cloning, A Laboratory Manual (1982)]. Чтобы сконструировать эти варианты, нуклеиновые кислоты, кодирующие аминокислотную последовательность собачьего IgGD, можно модифицировать так, чтобы она кодировала модифицированные IgGD. Затем модифицированные последовательности нуклеиновых кислот клонируют в экспрессирующие плазмиды для экспрессии белка.
Антитело или его антигенсвязывающий фрагмент, который связывается с собачьим CTLA-4, может содержать три, четыре, пять или шесть определяющих комплементарность областей (CDR) мышиного антитела против собачьего белка, как описано в настоящем описании. Три, четыре, пять или шесть CDR могут быть независимо выбраны из последовательностей CDR, представленных ниже. В дополнительном варианте осуществления выделенное антитело или его антигенсвязывающий фрагмент, который связывается с собачьим CTLA-4, содержит легкую цепь каппа или лямбда собачьего антитела, содержащую CDR-1, CDR-2 и/или CDR3 мышиной легкой цепи, и IgG тяжелой цепи собачьего антитела, содержащий CDR-1, CDR-2 и/или CDR3 мышиной тяжелой цепи.
В других вариантах осуществления изобретение относится к антителам или их антигенсвязывающим фрагментам, которые специфически связываются с собачьим CTLA-4 и имеют легкие цепи каппа или лямбда собачьего антитела, содержащие заданный набор из трех CDR, имеющих по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотными последовательностями SEQ ID NO: 92, 94 и 96 для VLCDR-1, VLCDR-2 и VLCDR-3, соответственно, и IgG тяжелой цепи собачьего антитела, содержащие данный набор из трех CDR, имеющих по меньшей мере 80% 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотными последовательностями SEQ ID NO: 86, 88 и 90 для VHCDR-1, VHCDR-2 и VHCDR-3, соответственно; или легкие цепи каппа или лямбда собачьего антитела, содержащие заданный набор из трех CDR, имеющих по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотными последовательностями SEQ ID NO: 104, 106, и 108, для VLCDR-1, VLCDR-2 и VLCDR-3, соответственно, и IgG тяжелой цепи собачьего антитела, содержащего набор различных CDR, имеющих по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотными последовательностями SEQ ID NO: 98, 100 и 102 для VHCDR-1, VHCDR-2 и VHCDR-3, соответственно; или легкие цепи каппа или лямбда собачьего антитела, содержащие заданный набор из трех CDR, содержащих по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотными последовательностями SEQ ID NO: 117, 94 и 96, для VLCDR-1, VLCDR-2 и VLCDR-3, соответственно, и IgG тяжелой цепи собачьего антитела, содержащего набор различных CDR, имеющих по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотными последовательностями SEQ ID NO: 86, 88 и 113 для VHCDR-1, VHCDR-2 и VHCDR-3, соответственно; легкие цепи каппа или лямбда собачьего антитела, содержащие заданный набор из трех CDR, имеющих по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотными последовательностями SEQ ID NO: 119, 122 и 96 для VLCDR-1, VLCDR-2 и VLCDR-3, соответственно, и IgG тяжелой цепи собачьего антитела, содержащего набор различных CDR, имеющих по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности с аминокислотные последовательности SEQ ID NO: 86, 88 и 115 для VHCDR-1, VHCDR-2 и VHCDR-3, соответственно; проявляя при этом целевые связывающие и функциональные свойства. В другом варианте осуществления антитело или антигенсвязывающий фрагмент по настоящему изобретению содержит собачий каркас, содержащий комбинацию последовательности тяжелой цепи IgG с легкой цепью каппа или лямбда, имеющей один или более из вышеупомянутых наборов из трех CDR легкой цепи и трех CDR тяжелой цепи, содержащих 0, 1, 2, 3, 4 или 5 консервативных или неконсервативных аминокислотных замен, все еще демонстрируя целевые связывающие и функциональные свойства.
Идентичность последовательности относится к степени, в которой аминокислоты двух полипептидов являются аналогичными в эквивалентных положениях, при оптимальном выравнивании двух последовательностей. В данном контексте одна аминокислотная последовательность на 100% «идентична» второй аминокислотной последовательности, если аминокислотные остатки обеих последовательностей идентичны. Соответственно, аминокислотная последовательность на 50% «идентична» второй аминокислотной последовательности, когда 50% аминокислотных остатков двух аминокислотных последовательностей идентичны. Сравнение последовательностей выполняется по непрерывному блоку аминокислотных остатков, содержащимся в данном белке, например, белке или части сравниваемого полипептида. В конкретных вариантах осуществления учитываются выбранные делеции или вставки, которые в противном случае могли бы изменить соответствие между двумя аминокислотными последовательностями.
Сходство последовательностей включает идентичные остатки и неидентичные биохимически родственные аминокислоты. Обсуждаются биохимически родственные аминокислоты, которые обладают схожими свойствами и могут быть взаимозаменяемыми.
«Консервативно модифицированные варианты» или «консервативная замена» относится к заменам аминокислот в белке другими аминокислотами, имеющими аналогичные характеристики (например, заряд, размер боковой цепи, гидрофобность/гидрофильность, конформацию и жесткость основной цепи и т.д.), таким образом, что изменения часто могут быть сделаны без изменения биологической активности белка. Специалисты в данной области признают, что, как правило, замены отдельных аминокислот в несущественных областях полипептида существенно не изменяют биологическую активность [см., например, Watson et al., Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p.224 (4th Ed.; 1987)]. Кроме того, замены структурно или функционально сходных аминокислот с меньшей вероятностью нарушают биологическую активность. Типичные консервативные замены представлены непосредственно в таблице A ниже.

Функционально-консервативные варианты антител по изобретению также рассматриваются настоящим изобретением. «Функционально-консервативные варианты» в контексте настоящего описания относятся к антителам или фрагментам, в которых один или более аминокислотных остатков был изменен без изменения целевого свойства, такого как аффинность к антигену и/или специфичность. Такие варианты включают, но не ограничиваются ими, замену аминокислоты на аминокислоту, имеющую аналогичные свойства, такие как консервативные замены аминокислот в Таблице А выше.
Нуклеиновые Кислоты
Настоящее изобретение дополнительно включает нуклеиновые кислоты, кодирующие иммуноглобулиновые цепи мышиных антител против собачьего CTLA-4 и/или канинизированных мышиных антител против собачьего CTLA-4 и их антигенсвязывающие фрагменты, раскрытые в настоящем описании (см., например, примеры ниже).
В настоящее изобретение также включены нуклеиновые кислоты, которые кодируют полипептиды иммуноглобулинов, содержащие аминокислотные последовательности, которые идентичны по меньшей мере примерно на 70%, предпочтительно идентичны по меньшей мере примерно на 80%, более предпочтительно идентичны по меньшей мере примерно на 90% и наиболее предпочтительно идентичны по меньшей мере примерно на 95% (например, 95%, 96%, 97%, 98%, 99%, 100%) аминокислотным последовательностям канинизированных антител, представленных в настоящем описании, когда сравнение выполняется с помощью алгоритма BLAST, в котором параметры алгоритма выбраны, чтобы получить наибольшее совпадение между соответствующими последовательностями по всей длине соответствующих контрольных последовательностей. Настоящее изобретение также относится к нуклеиновым кислотам, которые кодируют полипептиды иммуноглобулинов, содержащие аминокислотные последовательности, которые схожи по меньшей мере примерно на 70%, предпочтительно схожи по меньшей мере примерно на 80%, более предпочтительно по схожи меньшей мере примерно на 90% и наиболее предпочтительно схожи по меньшей мере примерно на 95% (например, 95%, 96%, 97%, 98%, 99%, 100%) с любой из эталонных аминокислотных последовательностей, когда сравнение выполняется с помощью алгоритма BLAST, где параметры алгоритма выбираются так, чтобы дать наибольшее соответствие между соответствующими последовательностями по всей длине соответствующих контрольных последовательностей.
В данном контексте идентичность нуклеотидной и аминокислотной последовательностей в процентах может быть определена с использованием C, MacVector (MacVector, Inc. Cary, NC 27519), Vector NTI (Informax, Inc. MD), Oxford Molecular Group PLC (1996) и алгоритма Clustal W с параметрами выравнивания по умолчанию и параметрами по умолчанию для идентификации. Эти коммерчески доступные программы также можно использовать для определения сходства последовательностей с использованием тех же или аналогичных параметров по умолчанию. В качестве альтернативы можно использовать расширенный поиск Blast в условиях фильтрации по умолчанию, например, с помощью программы pileup GCG (Genetics Computer Group, Руководство по программе для пакета GCG, версия 7, Мэдисон, Висконсин) с параметрами по умолчанию.
Следующие ссылки относятся к алгоритмам BLAST, часто используемым для анализа последовательностей: АЛГОРИТМЫ BLAST: Altschul, S.F., et al., J. Mol. Biol. 215:403-410 (1990); Gish, W., et al., Nature Genet. 3:266-272 (1993); Madden, T.L., et al., Meth. Enzymol. 266:131-141(1996); Altschul, S.F., et al., Nucleic Acids Res. 25: 3389-3402 (1997); Zhang, J., et al., Genome Res. 7:649-656 (1997); Wootton, J.C., et al., Comput. Chem. 17:149-163 (1993); Hancock, J.M. et al., Comput. Appl. Biosci. 10: 67-70 (1994); СИСТЕМЫ ОЦЕНКИ ВЫРАВНИВАНИЯ: Dayhoff, M.O., et al., «A model of evolutionary change in proteins» in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3. M.O. Dayhoff (ed.), pp.345-352, (1978); Natl. Biomed. Res. Found., Washington, DC; Schwartz, R.M., et al., «Matrices for detecting distant relationships.» in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3. (1978), М.О. Dayhoff (ed.), pp.353-358 (1978), Natl. Biomed. Res. Found., Washington, DC; Altschul, S.F., J. Mol. Biol. 219: 555-565 (1991); States, D.J., et al., Methods 3: 66-70 (1991); Henikoff, S., et al., Proc. Natl. Acad. Sci. USA 89: 10915-10919 (1992); Altschul, S.F., et al., J. Mol. Evol. 36:290-300 (1993); СТАТИСТИКА ВЫРАВНИВАНИЯ: Karlin, S., et al., Proc. Natl. Acad. Sci. USA 87: 2264-2268 (1990); Karlin, S., et al., Proc. Natl. Acad. Sci.USA 90: 5873-5877 (1993); Dembo, A., et al., Ann. Prob. 22:2022-2039 (1994); и Altschul, S.F. «Evaluating the statistical significance of multiple distinct local alignments». in Theoretical and Computational Methods in Genome Research (S. Suhai, ed.), pp.1-14, Plenum, New York (1997).
В настоящем изобретении также предложены экспрессирующие векторы, содержащие нуклеиновые кислоты по изобретению, где нуклеиновая кислота функционально связана с контрольными последовательностями, которые распознаются клеткой-хозяином, когда она трансфецирована вектором. Также предложены клетки-хозяева, содержащие экспрессирующий вектор по настоящему изобретению, и способы получения антитела или его антигенсвязывающего фрагмента, раскрытые в настоящем описании, включающие культивирование клетки-хозяина, несущей экспрессирующий вектор, кодирующий антитело или антигенсвязывающий фрагмент, в культуральной среде и выделение антигена или его антигенсвязывающий фрагмент из клетки-хозяина или культуральной среды.
Канинизированное мышиное антитело против собачьего CTLA-4 может быть получено рекомбинантно способами, известными в данной области. Клеточные линии млекопитающих, доступные в качестве хозяев для экспрессии антител или фрагментов, раскрытых в настоящем описании, хорошо известны в данной области и включают множество иммортализованных клеточных линий, доступных из Американской Типированной Коллекции Клеточных Культур (ATCC). К ним относятся, среди прочего, клетки яичника китайского хомячка (СНО), клетки NSO, SP2, клетки HeLa, клетки почек детеныша хомячка (BHK), клетки почек обезьяны (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), клетки A549, клетки 3T3, клетки HEK-293 и ряд других клеточных линий. Клетки-хозяева млекопитающих включают клетки человека, мыши, крысы, собаки, обезьяны, свиньи, козы, коровы, лошади и хомяка. Особенно предпочтительные клеточные линии выбирают посредством определения того, в какой клеточной линии будет высокий уровень экспрессии. Другие клеточные линии, которые могут использоваться, представляют собой клеточные линии насекомых, такие как клетки Sf9, клетки амфибий, бактериальные клетки, растительные клетки и грибные клетки. Когда рекомбинантные экспрессирующие векторы, кодирующие тяжелую цепь или антигенсвязывающую часть или его фрагмент, легкую цепь и/или ее антигенсвязывающий фрагмент вводят в клетки-хозяева млекопитающих, антитела получают путем культивирования клеток-хозяев в течение периода времени, достаточного для того, чтобы дать возможность экспрессии антитела в клетках-хозяевах или более предпочтительно секреции антитела в культуральную среду, в которой растят клетки-хозяева.
Антитела могут быть извлечены из культуральной среды с использованием стандартных методов очистки белка. Кроме того, экспрессия антител по изобретению (или других их компонентов) из продуцирующих клеточных линий может повышаться с использованием ряда известных методов. Например, система экспрессии гена глутаминсинтетазы (система GS) является распространенным подходом для повышения экспрессии при определенных условиях. Система GS обсуждается в целом или частично в связи с Европейскими патентами No. 0216846 0256055 и 0323997 и Европейской патентной заявкой No. 89303964.4.
В общем, гликопротеины, продуцируемые в конкретной клеточной линии или трансгенном животном, будут иметь профиль гликозилирования, который характерен для гликопротеинов, продуцируемых в клеточной линии или трансгенном животном. Таким образом, конкретный профиль гликозилирования антитела будет зависеть от конкретной клеточной линии или трансгенного животного, используемых для продуцирования антитела. Однако настоящее изобретение включает все антитела, кодируемые молекулами нуклеиновых кислот, представленных в настоящем описании, независимо от профиля гликозилирования, который могут иметь антитела. Аналогичным образом, в конкретных вариантах осуществления могут быть полезными антитела с профилем гликозилирования, включающим только нефукозилированные N-гликаны, поскольку было показано, что эти антитела обычно проявляют более высокую эффективность, чем их фукозилированные аналоги как in vitro, так и in vivo [см., например, Shinkawa et al., J. Biol. Chem. 278: 3466-3473 (2003); Пат.США №6946292 и 7214775].
Настоящее изобретение также включает фрагменты антител мышиных антител против собачьего CTLA-4, описанных в настоящем документе. Фрагменты антител включают фрагменты F(ab)2, которые могут быть получены ферментативным расщеплением IgG, например, пепсином. Fab-фрагменты могут быть получены, например, восстановлением F(ab)2 дитиотреитолом или меркаптоэтиламином. Fab-фрагмент представляет собой цепь VL-CL, присоединенную к цепи VH-CH1 дисульфидным мостиком. Фрагмент F(ab)2 представляет собой два Fab-фрагмента, которые, в свою очередь, соединены двумя дисульфидными мостиками. Fab-часть молекулы F(ab)2 включает часть Fc-области, между которой расположены дисульфидные мостики. Фрагмент FV представляет собой область VL или VH.
В одном варианте осуществления антитело или антигенсвязывающий фрагмент содержит константную область тяжелой цепи, например, константную область собаки, такую как константная область тяжелой цепи собачьего IgGA, IgGB, IgGC и IgGD или ее вариант.В другом варианте осуществления, антитело или антигенсвязывающий фрагмент содержит константную область легкой цепи, например, константную область собачьей легкой цепи, как например, константную область собачьей легкой цепи лямбда или каппа или ее вариант.В качестве примера, но не ограничения, константная область тяжелой цепи собаки может происходить из IgG-B, а константная область легкой цепи собаки может происходить из каппа-цепи.
Конструирование антитела
Канинизированные мышиные антитела против собачьего CTLA-4 по настоящему изобретению могут быть сконструированы для включения модификаций собачьего каркаса и/или остатков собачьего каркаса в вариабельных доменах родительского (т.е. собачьего) моноклонального антитела, например для улучшения свойств антитела.
Связывание эпитопа и аффинность связывания
Настоящее изобретение также относится к антителам или их антигенсвязывающим фрагментам, которые связываются с аминокислотными остатками того же эпитопа собачьего CTLA-4, что и раскрытые в настоящем описании мышиные антитела против собачьего CTLA-4. В конкретных вариантах осуществления мышиные антитела против собачьего CTLA-4 или их антигенсвязывающие фрагменты также способны ингибировать/блокировать связывание собачьего CTLA-4 с собачьим CD86 и/или CD80. В родственных вариантах осуществления канинизированные мышиные антитела против собачьего CTLA-4 или их антигенсвязывающие фрагменты также способны ингибировать/блокировать связывание собачьего CTLA-4 с собачьим CD86 и/или CD80.
Экспериментальные и диагностические применения
Мышиные антитела против собачьего CTLA-4 и/или канинизированные антитела против собачьего CTLA-4 или их антигенсвязывающие фрагменты по настоящему изобретению также могут быть полезны в диагностических анализах собачьего белка CTLA-4, например, для определения его экспрессии в комбинации с и/или по отношению к раку.
Например, такой способ включает следующие стадии:
(а) покрывают субстрат (например, поверхность лунки микропланшета, например, пластикового планшета) мышиным антителом против собачьего CTLA-4 или его антигенсвязывающим фрагментом;
(b) наносят образец для тестирования на присутствие собачьего CTLA-4 на субстрат;
(c) промывают планшет, чтобы удалить несвязанный материал в образце;
(d) наносят детектируемые меченые антитела (например, ферментно-связанные антитела), которые также специфичны к антигену CTLA-4;
(e) промывают субстрат, чтобы удалить несвязанные меченые антитела;
(f) если меченые антитела связаны с ферментом, применяют химическое вещество, которое ферментом преобразуется во флуоресцентный сигнал; и
(g) детектируют присутствие меченого антитела.
В другом варианте осуществления меченое антитело метят пероксидазой, которая взаимодействует с ABTS [например, 2,2'-азино-бис (3-этилбензтиазолин-6-сульфоновая кислота)] или 3,3',5,5'-тетраметилбензидином (TMB) для изменения цвета, которое можно детектировать. Альтернативно, меченое антитело метят детектируемым радиоизотопом (например, 3H), который может быть обнаружен сцинтилляционным счетчиком в присутствии сцинтиллятора. Мышиные антитела против собачьих CTLA-4 по изобретению можно использовать в процедуре вестерн-блоттинга или иммуноблоттинга.
Такая процедура образует часть настоящего изобретения и включает, например:
(i) контакт мембраны или другого твердого субстрата, подлежащего тестированию на наличие связанного собачьего CTLA-4 или его фрагмента, с канинизированным мышиным антителом против собачьего CTLA-4 или его антигенсвязывающим фрагментом по настоящему изобретению. Такая мембрана может иметь форму мембраны на основе нитроцеллюлозы или винила [например, поливинилиденфторида (PVDF)], на которую переносили белки, подлежащие тестированию на присутствие собачьего CTLA-4 в неденатурирующем геле PAGE (электрофорез в полиакриламидном геле) или геле SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), (например, после электрофоретического разделения в геле). Перед контактом мембраны с канинизированным мышиным антителом против собачьего CTLA-4 или его антигенсвязывающим фрагментом мембрану необязательно блокируют, например, обезжиренным сухим молоком или подобным, чтобы связать неспецифические сайты связывания белка на мембране.
(ii) промывание мембраны один или более раз для удаления несвязавшегося канинизированного мышиного антитела против собачьего CTLA-4 или его антигенсвязывающего фрагмента и других несвязанных веществ; и
(iii) детектирование связанного канинизированного мышиного антитела против собачьего CTLA-4 или его антигенсвязывающего фрагмента.
Детектирование связанного антитела или антигенсвязывающего фрагмента может осуществляться путем связывания антитела или антигенсвязывающего фрагмента с вторичным антителом (антитело против иммуноглобулина), которое помечено с возможностью детектирования, и затем путем детектирования присутствия вторичного антитела.
Мышиные антитела против собачьего CTLA-4, канинизированные мышиные антитела против собачьего CTLA-4 и/или их антигенсвязывающие фрагменты, раскрытые в настоящем описании, также могут быть использованы для иммуногистохимии. Такой способ составляет часть настоящего изобретения и включает, например, (1) контакт клетки, подлежащей тестированию на присутствие собачьего CTLA-4, например, с мышиным антителом против собачьего CTLA-4 или его антигенсвязывающим фрагментом по настоящему изобретению; и (2) детектирование антитела или фрагмента на клетке или внутри нее. Если антитело или антигенсвязывающий фрагмент само по себе помечено детектируемой меткой, то оно может быть детектировано напрямую. Альтернативно, антитело или антигенсвязывающий фрагмент может связываться со вторичным антителом, меченным детектируемой меткой, которое детектируют.
Методы визуализации включают визуализацию SPECT (однофотонная эмиссионная компьютерная томография) или PET (позитрон-эмиссионная томография). Метки включают, например, йод-123 (123I) и технеций-99m (99mTc), например, в комбинации с визуализацией SPECT или 11C, 13N, 15O или 18F, например, в комбинации с визуализацией PET или Индий-111 [см., например, Gordon и др., International Rev. Neurobiol. 67: 385-440 (2005)].
Перекрестно-блокирующие антитела
Кроме того, антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент по настоящему изобретению включает любое антитело или его антигенсвязывающий фрагмент, которое связывается с тем же эпитопом собачьего CTLA-4, с которым связываются обсуждаемые в настоящем описании антитела и фрагменты, и любое антитело или антигенсвязывающий фрагмент, которое перекрестно блокирует (частично или полностью) или перекрестно блокируется (частично или полностью) антителом или фрагментом, обсуждаемым в настоящем описании, для связывания собачьего CTLA-4; а также любой его вариант.
Перекрестно-блокирующие антитела и их антигенсвязывающие фрагменты, обсуждаемые в настоящем описании, могут быть идентифицированы на основе их способности перекрестно конкурировать с антителами, раскрытыми в настоящем описании (на основе CDR, как представлено ниже в Примере 5), то есть 45A9, 27G12, 22A11, 110E3; и более конкретно, 12B3 и/или 39A11 в стандартных анализах связывания (например, BIACore®, ИФА, как проиллюстрировано ниже, или проточная цитометрия). Например, можно использовать стандартные анализы ИФА, в которых рекомбинантный собачий белок CTLA-4 иммобилизован на планшете, одно из антител помечено флуоресцентной меткой, и оценивается способность немеченых антител конкурировать за связывание меченого антитела. Дополнительно или альтернативно, анализ BIAcore® можно использовать для оценки способности антител к перекрестной конкуренции. Способность тестируемого антитела ингибировать связывание, например, 27G12, 45A9, 110E3 и/или 22A11; и даже более конкретно 12B3 и/или 39A11, с собачьим CTLA-4 демонстрирует, что тестируемое антитело может конкурировать с 27G12, 45A9, 110E3 и/или 22A11, и/или 12B3, и/или 39A11 за связывание с собачьим CTLA-4 и, таким образом, может в некоторых случаях связываются с одним и тем же эпитопом на собачьем CTLA-4 как 27G12, 45A9, 110E3 и/или 22A11, и/или 12B3, и/или 39A11. Как указано выше, антитела и фрагменты, которые связываются с тем же эпитопом, что и любое из антител против собачьего CTLA-4 или их фрагментов по настоящему изобретению, также составляют часть настоящего изобретения.
Фармацевтические композиции и Введение
Для приготовления фармацевтических или стерильных композиций канинизированного мышиного антитела против собачьего CTLA-4 или его антигенсвязывающего фрагмента его можно смешать с фармацевтически приемлемым носителем или вспомогательным веществом. [См., например, Remington's Pharmaceutical Sciences и Фармакопея США: Национальный формуляр, Mack Publishing Company, Истон, Пенсильвания (1984)].
Составы терапевтических и диагностических агентов могут быть приготовлены путем смешивания с приемлемыми носителями, вспомогательными веществами или стабилизаторами в форме, например, лиофилизированных порошков, суспензий, водных растворов или суспензий [см., например, Hardman, et al. (2001) Goodman and Gilman’s The Pharmacological Basis of Therapeutics, McGraw-Hill, New York, NY; Gennaro (2000) Remington: The Science and Practice of Pharmacy, Lippincott, Williams, and Wilkins, New York, NY; Avis, et al. (eds.) (1993) Pharmaceutical Dosage Forms: Parenteral Medications, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Tablets, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Disperse Systems, Marcel Dekker, NY; Weiner и Kotkoskie (2000) Excipient Toxicity and Safety, Marcel Dekker, Inc., New York, NY]. В одном варианте осуществления антитела против CTLA-4 по настоящему изобретению разбавляют до подходящей концентрации в растворе ацетата натрия с pH 5-6, и для придания тоничности добавляют NaCl или сахарозу. Дополнительные агенты, такие как полисорбат 20 или полисорбат 80, могут быть добавлены для повышения стабильности.
Токсичность и терапевтическая эффективность композиций антител, вводимых отдельно или в комбинации с другим агентом, можно определить стандартными фармацевтическими процедурами на культурах клеток или экспериментальных животных, например, для определения LD50 (доза, летальная для 50% популяции) и ED50 (доза, терапевтически эффективная для 50% популяции). Соотношение доз между токсическим и терапевтическим эффектами представляет собой терапевтический индекс (LD50/ED50). В конкретных аспектах, целевыми являются антитела, демонстрирующие высокие терапевтические индексы. Данные, полученные из таких анализов клеточных культур и исследований на животных, могут использоваться при составлении диапазона дозировки для применения у собак. Дозировка таких соединений предпочтительно находится в диапазоне циркулирующих концентраций, который включает ED50 с небольшой токсичностью или без нее. Дозировка может варьироваться внутри данного диапазона в зависимости от применяемой дозировки и пути введения.
Способ введения может варьироваться. Подходящие пути введения включают пероральный, ректальный, чрезслизистый, интестинальный, парентеральный; внутримышечный, подкожный, внутрикожный, костномозговой, внутриоболочечный, прямой внутрижелудочковый, внутривенный, внутрибрюшинный, интраназальный, внутриглазной, ингаляционный, инсуфляционный, местный, кожный, чрескожный или внутриартериальный. В конкретных вариантах осуществления канинизированное мышиное антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент можно вводить инвазивным путем, например путем инъекции. В дополнительных вариантах осуществления изобретения канинизированное мышиное антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент или его фармацевтическая композиция вводится внутривенно, подкожно, внутримышечно, внутриартериально или путем ингаляции, с доставкой в виде аэрозоля. Введение неинвазивными путями (например, перорально; например, в пилюле, капсуле или таблетке) также входит в объем настоящего изобретения.
Композиции могут вводить с использованием медицинских устройств, известных в данной области. Например, фармацевтическая композиция по изобретению может вводиться путем инъекции с помощью иглы для подкожных инъекций, включая, например, предварительно заполненный шприц или автоинжектор. Фармацевтические композиции, раскрытые в настоящем описании, также можно вводить с использованием безыгольного гиподермического инъекционного устройства; такого как устройства, раскрытые в Патентах No.: 6620135; 6096002; 5399163; 5383851; 5312335; 5064413; 4941880; 4790824 или 4596556.
Фармацевтические композиции, раскрытые в настоящем описании, также можно вводить с помощью инфузии. Примеры хорошо известных имплантатов и модулей для введения фармацевтических композиций включают: Патент США No. 4487603, в котором раскрыт имплантируемый насос для микроинфузий для дозирования препарата котролируемым путем; Патент США No. 4447233, в котором раскрыт насос для инфузии препарата для доставки препарата с точной скоростью инфузии; Патент США No. 4447224, в котором раскрыто имплантируемое устройство для инфузии с изменяемым потоком для непрерывной доставки лекарственного средства; Патент США No. 4439196, в котором раскрыта осмотическая система доставки лекарственного средства, содержащая многокамерные компартменты. Специалисту в данной области хорошо известно множество таких имплантов, систем доставки и модулей.
Альтернативно, можно вводить мышиное антитело против собачьего CTLA-4 или канинизированное мышиное антитело против собачьего CTLA-4 местным, а не системным образом, например, путем инъекции антитела непосредственно в сустав, пораженный артритом, или в вызванное патогеном поражение, характеризующееся иммунопатологией, часто в составе депо или с замедленным высвобождением. Кроме того, можно вводить антитело в системе направленной доставки лекарственного средства, например, в липосоме, покрытой тканеспецифическим антителом, нацеленным, например, на артритный сустав или вызванное патогеном поражение, характеризующееся иммунопатологией. Липосомы будут направлены в пораженную ткань и селективно поглощены там.
Схема введения зависит от нескольких факторов, включая метаболизм терапевтического антитела в сыворотке или ткани, уровень симптомов, иммуногенность терапевтического антитела и доступность клеток-мишеней в биологической матрице. Предпочтительно, схема введения доставляет достаточное количество терапевтического антитела для воздействия на улучшение целевого болезненного состояния при одновременной минимизации нежелательных побочных эффектов. Соответственно, количество доставляемого биологического препарата частично зависит от конкретного терапевтического антитела и тяжести состояния, которое подвергается лечению. Доступны инструкции по выбору подходящих доз терапевтических антител [см., например, Wawrzynczak Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK (1996); Kresina (ed.) Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, NY (1991); Bach (ed.) Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, NY (1993); Baert, et al. New Engl. J. Med. 348: 601-608 (2003); Milgrom et al. New Engl. J. Med. 341:1966-1973 (1999); Slamon et al. New Engl. J. Med. 344:783-792 (2001); Beniaminovitz et al. New Engl. J. Med. 342: 613-619 (2000); Ghosh et al. New Engl. J. Med. 348: 24-32 (2003); Lipsky et al. New Engl. J. Med. 343: 1594-1602 (2000)].
Подходящую дозу определяет ветеринар, например, с использованием параметров или факторов, известных или предполагаемых в данной области как влияющих на лечение. Как правило, доза начинается с количества, чуть меньшего, чем оптимальная доза, и повышается маленькими шагами до тех пор, пока не будет достигнут целевой или оптимальный эффект относительно любого негативного побочного эффекта. Важные диагностические меры включают симптомы, например, размер опухоли.
Антитела или их антигенсвязывающие фрагменты, раскрытые в настоящем описании, могут быть обеспечены путем непрерывной инфузии или с помощью доз, вводимых, например, ежедневно, 1-7 раз в неделю, еженедельно, раз в две недели, ежемесячно, раз в два месяца, ежеквартально, раз в полгода, ежегодно и т.д. Дозы могут вводиться, например, внутривенно, подкожно, местно, перорально, назально, ректально, внутримышечно, интрацеребрально, внутриспинально или путем ингаляции. Общая недельная доза обычно составляет по меньшей мере 0,05 мкг/кг массы тела, в более общем случае по меньшей мере 0,2 мкг/кг, 0,5 мкг/кг, 1 мкг/кг, 10 мкг/кг, 100 мкг/кг, 0,25 мг/кг, 1 мг/кг, 2 мг/кг, 5 мг/мл, 10 мг/кг, 25 мг/кг, 50 мг/кг или более [см., например, Yang, et al. New Engl. J. Med. 349:427-434 (2003); Herold, et al.New Engl. J. Med. 346:1692-1698 (2002); Liu, et al.J.Neurol. Neurosurg. Psych. 67: 451-456 (1999); Portielji, et al. Cancer Immunol. Immunother. 52: 133-144 (2003)]. Также могут быть предусмотрены дозы для достижения заранее определенной целевой концентрации канинизированного мышиного антитела против CTLA-4 в сыворотке пациента, например 0,1, 0,3, 1, 3, 10, 30, 100, 300 мкг/мл или более. В других вариантах осуществления канинизированное мышиное антитело против собачьего CTLA-4 по настоящему изобретению вводят подкожно или внутривенно, еженедельно, раз в две недели, «каждые 4 недели», ежемесячно, раз в два месяца или ежеквартально в количестве 10, 20, 50, 80, 100, 200, 500, 1000 или 2500 мг/на объект.
Антигенные пептиды (например, пептиды, содержащие эпитопы или их части из CTLA-4), которые распознаются антителами mAb против собачьего CTLA-4, также можно использовать в качестве вакцин для выработки антител, которые блокируют связывание собачьего CTLA-4 с собачьими CD80 и/или CD86. Такие вакцины могут быть полезны в качестве терапевтических вакцин от таких заболеваний, как онкологическое заболевание. Чтобы использовать эти антигенные пептиды в качестве вакцин, один или более из этих пептидов могут быть связаны химически или с помощью технологий рекомбинантной ДНК с другим белком-носителем, чтобы повысить иммуногенность этих пептидов и вызвать выработку пептид-специфических антител. Способы соединения пептидов с белками-носителями известны специалистам в данной области. Пептидные вакцины можно использовать для вакцинации животных внутримышечно, подкожно, перорально, с помощью спрея или in ovo. Пептидные вакцины могут использоваться в виде субъединичных белков, экспрессируемых из бактериальных, вирусных, дрожжевых или бакуловирусных систем. Альтернативно такие пептидные вакцины могут быть доставлены после введения множества вирусных или бактериальных векторов, которые экспрессируют такие пептидные вакцины, что может быть осуществлено методами, известными специалистам в данной области. Пептидные вакцины могут вводиться в дозах от 1 до 1000 мкг и необязательно могут содержать адъювант и приемлемый фармацевтический носитель.
Используемый в настоящем описании термин «ингибировать», или «лечить», или «лечение» включает отсрочку развития симптомов, связанных с расстройством, и/или уменьшение тяжести симптомов такого расстройства. Термины дополнительно включают улучшение существующих неконтролируемых или нежелательных симптомов, предотвращение дополнительных симптомов и улучшение или предотвращение причин, обуславливающих такие симптомы. Таким образом, термины обозначают, что положительный результат был получен у позвоночного объекта (например, собаки) с расстройством, заболеванием или симптомом или с возможностью развития такого расстройства, заболевания или симптома.
Используемые в настоящем описании термины «терапевтически эффективное количество», «терапевтически эффективная доза» и «эффективное количество» относятся к количеству канинизированного мышиного антитела против собачьего CTLA-4 или его антигенсвязывающего фрагмента по настоящему изобретению, которое при введении отдельно или в комбинации с дополнительным терапевтическим агентом в клетку, ткань или объекту, эффективно, чтобы вызвать измеримое улучшение одного или более симптомов заболевания или состояния или прогрессирования такого заболевания или состояния. Терапевтически эффективная доза также относится к тому количеству связывающего соединения, которое достаточно для того, чтобы привести, по меньшей мере, к частичному облегчению симптомов, например, к лечению, исцелению, профилактике или облегчению соответствующего медицинского состояния или к увеличению скорости лечения, излечения, предотвращения или улучшения таких состояний. При применении к индивидуальному активному ингредиенту, который вводят индивидуально, терапевтически эффективная доза относится только к такому ингредиенту. При применении к комбинации, терапевтически эффективная доза относится к объединенным количествам активных ингредиентов, которые приводят к получению терапевтического эффекта при введении либо в комбинации, последовательно, либо одновременно. Эффективное количество терапевтического средства приведет к улучшению диагностического показателя или параметра по меньшей мере на 10%; обычно по меньшей мере на 20%; предпочтительно по меньшей мере примерно на 30%; более предпочтительно по меньшей мере на 40% и наиболее предпочтительно по меньшей мере на 50%. Эффективное количество также может приводить к улучшению объективного показателя в случаях, где объективные показатели используют для оценки тяжести заболевания.
Другие комбинированные терапии
Как описано ранее, канинизированное мышиное антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент и/или антигенный пептид по настоящему изобретению можно вводить совместно с одним или более другими терапевтическими агентами (такими как ингибитор, как обсуждается в следующем абзаце) и/или с канинизированным мышиным антителом против собачьего PD-1 [см., например, US 9944704 B2 и US 10106107 B2, содержание каждого из которых включено в настоящее описание в качестве ссылки в полном объеме] и/или с канинизированным мышиным антителом против собачьего PD-L1 [см., например, US 20180237535 A1, содержание которого включено в настоящее описание в качестве ссылки в полном объеме]. Антитело (антитела) может быть связано с агентом (в виде иммунокомплекса) и/или может вводиться отдельно от агента или другого антитела. В последнем случае (раздельное введение) антитела можно вводить до, после или одновременно с агентом или можно вводить совместно с другими известными методами лечения.
Наборы
Кроме того, предложены наборы, содержащие один или более компонентов, которые включают, но не ограничиваются ими, антитело или антигенсвязывающий фрагмент, как обсуждается в настоящем описании, которое специфически связывается с CTLA-4 (например, канинизированное мышиное антитело против собачьего CTLA-4 или его антигенсвязывающий фрагмент) в комбинации с одним или более дополнительными компонентами, включая канинизированное мышиное антитело против собачьего PD-1 и/или канинизированное мышиное антитело против собачьего PD-L1. Связывающие композиции, описанные непосредственно выше, могут быть приготовлены в виде чистой композиции или в комбинации с фармацевтически приемлемым носителем в фармацевтической композиции.
В одном варианте осуществления набор включает связывающую композицию по настоящему изобретению (например, канинизированное мышиное антитело против собачьего CTLA-4 или его фармацевтическую композицию в одном контейнере (например, в стерильном стеклянном или пластиковом флаконе), канинизированное мышиное антитело против собачьего PD-1 и/или канинизированное мышиное антитело против собачьего PD-L1 или их фармацевтическую композицию (композиции) в другом контейнере (например, в стерильном стеклянном или пластиковом флаконе).
Если набор включает фармацевтическую композицию для парентерального введения пациенту, набор также может включать устройство для выполнения такого введения. Например, набор может включать одну или более гиподермических игл или других устройств для инъекции, как обсуждалось выше. Набор также может включать лист-вкладыш, включающий информацию, касающуюся фармацевтических композиций и лекарственных форм в наборе. Как правило, такая информация помогает владельцам домашних животных и ветеринарам эффективно и безопасно использовать прилагаемые фармацевтические композиции и лекарственные формы. Например, следующая информация касательно комбинации по изобретению может быть представлена во вкладыше: фармакокинетика, фармакодинамика, клинические исследования, параметры эффективности, показания к применению, противопоказания, предупреждения, предостережения, побочные реакции, передозировки, корректные дозировки и введение, как поставляется, корректные условия хранения, ссылки, информация производителя/поставщика и патентная информация.
Для удобства антитело или специфический связывающий агент, раскрытое в настоящем описании, может быть предоставлено в наборе, то есть в виде упакованной комбинации реагентов в заранее определенных количествах с инструкциями по выполнению диагностического или детектирующего анализа. Если антитело или антитела помечены ферментом, набор будет включать субстраты и кофакторы, необходимые для фермента (например, предшественник субстрата, который обеспечивает детектируемый хромофор или флуорофор). Кроме того, могут быть добавлены другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или лизирующий буфер) и т.п.Относительные количества различных реагентов могут широко варьироваться для обеспечения концентраций реагентов в растворе, которые по существу оптимизируют чувствительность анализа. Конкретно, реагенты могут предлагаться в виде сухих порошков, обычно лиофилизированных, включающих вспомогательные вещества, которые при растворении обеспечивают раствор реагентов, имеющих подходящую концентрацию.
ПРИМЕРЫ
ПРИМЕР 1
Получение мышиных моноклональных антител к собачьему CTLA-4 и соответствующих химерных антител мышь-собака
Моноклональные антитела мыши были получены с использованием технологии мышиных гибридом с собачьим рекомбинантным белком CTLA-4 (cCTLA-4) в качестве иммуногена. Положительные клоны гибридомы отбирали на основе реактивности антитела с cCTLA-4 и блокирования взаимодействия собачьего CD86 или CD80 с cCTLA-4 (блокирующая активность) с помощью анализов ИФА и FACS. Отобранные гибридомные клоны секвенировали быстрой амплификацией концов кДНК (RACE) для фрагментов антител с последовательностями VH и VL. Шесть отобранных моноклональных антител обозначены как: 12B3, 27G12, 39A11, 45A9, 110E3 и 22A11, соответственно. Аминокислотные последовательности шести антител представляют собой SEQ ID NO: 2, 4, 6, 8, 10 и 12 для вариабельной области тяжелой цепи, соответственно, и SEQ ID NO: 14, 16, 18, 20, 22 и 24 для вариабельная область легкой цепи, соответственно. CDR подчеркнуты в последовательностях, представленных ниже [см. также Таблицу 1 ниже]. Соответствующие нуклеотидные последовательности, которые кодируют идентифицированные выше аминокислотные последовательности соответствуют SEQ ID NO: 1, 3, 5, 7, 9 и 11 для вариабельной области тяжелой цепи, соответственно, и SEQ ID NO: 13, 15, 17, 19, 21 и 23 для вариабельной области легкой цепи, соответственно. Нуклеотидные последовательности вариабельных областей тяжелой цепи были слиты с нуклеотидной последовательностью модифицированной константной области тяжелой цепи собаки (CH1-шарнир-CH2-CH3), соответственно, для получения химерной «мышь-собака» нуклеотидной последовательности тяжелой цепи, SEQ ID NO: 25, 27, 29, 31, 33 и 35. Жирным шрифтом выделены вариабельные области. Нуклеотидные последовательности вариабельной области легкой цепи были слиты с нуклеотидной последовательностью константного домена легкой цепи каппа собаки, соответственно, для получения химерной «мышь-собака» нуклеотидной последовательности легкой цепи, SEQ ID NO: 37, 39, 41, 43, 45 и 47. Вариабельные области выделены жирным шрифтом. Аминокислотные последовательности, кодируемые химерными «мышь-собака» нуклеотидными последовательностями тяжелой цепи, соответствуют SEQ ID NO: 26, 28, 30, 32, 34 и 36. Аминокислотные последовательности, кодируемые химерными «мышь-собака» нуклеотидными последовательностями легкой цепи, соответствуют SEQ ID NO: 38, 40, 42, 44, 46 и 48. Вариабельные области выделены жирным шрифтом, а CDR подчеркнуты. Химерные «человек-собака» тяжелые и легкие цепи клонировали в отдельные экспрессирующие плазмиды с использованием стандартных методов молекулярной биологии. Плазмиды, содержащие гены тяжелой и легкой цепей, трансфецировали в клетки НЕК 293, и экспрессированные антитела очищали из супернатанта клеток НЕК 293 с использованием Протеина А.
ПРИМЕР 2
АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ CDR МЫШИ
CDR мышиных моноклональных антител против собачьего CTLA-4 перечислены в Таблице 1 ниже.
LysGly
Назначение индивидуальных канонических структур для шести CDR каждого из шести антител представлено в Таблице 2 ниже.
ПРИМЕР 3
Реактивность химерных антител с собачьим CTLA-4
Химерное антитело обычно обладает той же реактивностью, что и его родительское мышиное антитело. Чтобы подтвердить реактивность шести антител с cCTLA-4, были продуцированы химерные антитела «мышь-собака» и протестированы на их реактивность с cCTLA-4 с помощью ИФА следующим образом:
1. Наносили покрытие в виде 200 нг/лунку cCTLA-4 на иммунопланшете и инкубировали планшет при 4°C в течение ночи.
2. Планшет промывали 3 раза PBS с 0,05% Tween 20 (PBST).
3. Блокировали планшет 0,5% BSA в PBS в течение 45-60 мин при комнатной температуре.
4. Планшет промывали 3 раза PBST.
5. Трехкратно разводили антитела в каждом столбце или ряду планшета для разведения.
6. Переносили разведенные антитела в каждый столбец или ряд планшета и инкубировали планшет в течение 45-60 мин при комнатной температуре.
7. Планшет промывали 3 раза PBST.
8. В каждую лунку планшета добавляли разбавленный 1:2000 меченный пероксидазой хрена Fc против собачьего IgG и инкубировали планшет в течение 45-60 мин при комнатной температуре.
9. Планшет промывали 3 раза PBST.
10. Добавляли субстрат TMB в каждую лунку планшета и инкубировали планшет в течение 10-15 минут при комнатной температуре, чтобы была возможность проявки цвета.
11. Добавляли 100 мкл 1,5 М фосфорной кислоты в каждую лунку для остановки реакции.
12. Считывали планшет при 450 нм с эталонной длиной волны 540 нм.
Результаты ИФА показывают, что химерные антитела могут связываться с cCTLA-4 [см. Фигуру 1].
ПРИМЕР 4
БЛОКИРУЮЩАЯ АКТИВНОСТЬ ХИМЕРНЫХ АНТИТЕЛ В ОТНОШЕНИИ
ВЗАИМОДЕЙСТВИЯ СОБАЧЬЕГО CD86 ИЛИ CD80 С СОБАЧНЫМ CTLA-4
Для исследования блокирующей активности химерных антител был проведен анализ блокирования на основе ИФА следующим образом:
1. Наносили покрытие в виде 200 нг/на лунку cCTLA-4 в иммунопланшете и инкубировали планшет при 4°C в течение ночи.
2. Промывали планшет 3 раза PBS с 0,05% Tween 20 (PBST).
3. Блокировали планшет 0,5% BSA в PBS на 45-60 мин при комнатной температуре.
4. Планшет промывали 3 раза PBST.
5. Трехкратно разбавляли антитела в каждом столбце или ряду планшета для разведения, а затем добавляли 100 нг/на лунку биотинилированного CD86 или CD80. Далее смешивали с антителами.
6. Переносили смесь в каждый столбец или ряд иммунопланшета и инкубировали планшет в течение 45-60 мин при комнатной температуре.
7. Планшет промывали 3 раза PBST.
8. В каждую лунку планшета добавляли разбавленный 1:2000 стрептавидин, конъюгированный с пероксидазой хрена, и инкубировали планшет в течение 45-60 мин при комнатной температуре.
9. Планшет промывали 3 раза PBST.
10. Добавляли субстрат TMB в каждую лунку планшета и инкубировали планшет в течение 10-15 минут при комнатной температуре для проявки цвета.
11. Добавляли 100 мкл 1,5 М фосфорной кислоты в каждую лунку для остановки реакции.
12. Считывали планшет при 450 нм с эталонной длиной волны 540 нм.
Было обнаружено, что химерные антитела блокируют взаимодействие cCTLA-4 с CD86 [Фигура 2] и с CD80 [Фигура 3].
ПРИМЕР 5
АНАЛИЗ FACS ДЛЯ ТЕСТИРОВАНИЯ СВЯЗЫВАЮЩЕЙ АКТИВНОСТИ
ХИМЕРНЫХ АНТИТЕЛ НА CHO-cCTLA-4
Была создана клеточная линия CHO-K1, стабильно экспрессирующая cCTLA-4. Клетки использовали для тестирования связывающей и блокирующей активности антител в проточном анализе FACS. Чтобы проверить cCTLA-4-связывающую активность химерных антител, FACS-анализ проводили следующим образом:
1. Выращивали клетки CHO-K1-cCTLA-4 в культуральной среде в колбе Т-75. Клетки пассировали, когда конфлюэнтность клеток достигала 90%.
Питательная среда: F12K (Gibco, номер в каталоге 21127-022), 10% FBS (Gibco, номер в каталоге 10099-141) и пуромицин 4 мкг/мл (Gibco, номер в каталоге A1113803).
2. Клетки открепляли раствором трипсин-ЭДТА, ресуспендировали клетки в культуральной среде и подсчитывали жизнеспособные клетки с жизнеспособностью более 95%.
3. Клетки центрифугировали, супернатант аспирировали, затем ресуспендировали клетки в буфере FACS (Thermo Fisher Scientific, номер в каталоге BDB554656) до 1×107 клеток/мл.
4. Добавляли антитело в 100 мкл клеток, инкубировали при комнатной температуре в течение 30 мин при осторожном встряхивании.
5. Клетки промывали 3×250 мкл буфера FACS и ресуспендировали клетки в 100 мкл буфера FACS.
6. Окрашивали клетки FITC-конъюгированным антителом против собачьего IgG, инкубировали при комнатной температуре в течение 30 мин при осторожном встряхивании.
7. Клетки промывали 3×250 мкл буфера FACS и ресуспендировали клетки в 500 мкл буфера FACS.
8. Считывали 10000 клеток с помощью проточной цитометрии.
Результаты FACS показывают, что химерное антитело может связываться с клетками CHO-cCTLA-4 [см. Фигуры 4A-4G].
ПРИМЕР 6
ГЕНЕРИРОВАНИЕ ИНТЕРФЕРОН ГАММА (IFNγ) СОБАЧЬИХ PBMC, АКТИВИРОВАННЫХ ХИМЕРНЫМИ АНТИТЕЛАМИ
Выделение мононуклеарных клеток периферической крови собак
1. Отбирали ~ 20 мл цельной крови в пробирку с ЭДТА или гепарином натрия.
2. Переносили кровь в полистирольную пробирку на 50 мл и разбавляли в соотношении 50:50 HBSS (Thermo Fisher Scientific, номер в каталоге 21022CM).
3. Добавляли 15 мл Ficoll-Plaque Plus в четыре пробирки SepMate™ по 50 мл (STEMCELL Technologies, номер в каталоге 15460). Затем медленно добавляли ~ 10 мл разведенной крови 50:50 на стенку каждой пробирки SepMate™, содержащей фиколл.
4. Пробирки центрифугировали при 1200xg в течение 20 минут.
5. Клетки собирали из границы раздела градиента и переносили клетки в полипропиленовую пробирку на 50 мл. Добавляли HBSS до отметки 40-45 мл и центрифугировали клетки при 800xg в течение 10 минут.
6. Отбрасывали супернатант, ресуспендировали клетки в 40-45 мл HBSS и снова центрифугировали пробирку при 800xg в течение 10 минут.
7. Отбрасывали супернатант и ресуспендировали клетки из каждой пробирки с 2 мл Среды для лимфоцитов собак (среда RPMI, Lonza, номер в каталоге 12-167Q). Клетки объединяли от одного и того же животного.
8. Брали небольшие аликвоты клеточной суспензии, смешивали с 0,04% трипановым синим и подсчитывали количество клеток.
9. Клеточную суспензию хранили при 2-7°C до использования, но не более чем за 24 часа до использования.
Анализ пролиферации клеток для мононуклеаров периферической крови собак
1. Разводили антитела в среде для лимфоцитов собак для достижения конечной концентрации 40 мкг/мл (готовили 160 мкг/мл) и стерилизовали с использованием шприцевого фильтра 0,2 мкм. Двукратно разводили антитела в стерильном планшете для разведения и откладывали.
2. Разводили клетки до 2,5×106 клеток/мл в среде для лимфоцитов собак и распределяли по 100 мкл на лунку всего 96-луночного культурального планшета.
3. Разводили Con A в среде для лимфоцитов собак до достижения конечной концентрации 250 нг/мл (готовили 1000 нг/мл), стерилизовали с использованием шприцевого фильтра 0,2 мкм и добавляли 50 мкл во все лунки. (Не добавляли Con A в один столбец из восьми лунок для контроля только с клетками и в лунки, предназначенные для контроля только «клетки+mAb».)
4. Добавляли 100 мкл среды для лимфоцитов собак на лунку в лунки только для клеток и 50 мкл среды в колонку, содержащую контрольные лунки с Con A (Con A+клетки без обработки mAb).
5. В дублированные лунки добавляли 50 мкл разведенных mAb.
6. Инкубировали планшеты при 36±2°C, 4-6% CO2 в увлажненном инкубаторе в течение 68-124 часов.
IFNγ ИФА
1. Через 68-124 часов после инкубации планшет центрифугировали при 800xg в течение 10 минут.
2. Собирали супернатант из каждой лунки и пул дублировали. Эти образцы можно заморозить при температуре ≤ -50°C для дальнейшего использования или немедленного тестирования.
3. При необходимости разводили образцы супернатанта соответствующим образом и выполняли IFN-гамма-ИФА в соответствии с инструкциями к набору для IFN-гамма-ИФА Quantikine для собак [R&D Systems номер в каталоге CAIF00].
Результаты демонстрируют, что отобранные антитела, включая 12B3, могут активировать собачьи Т-клетки для продуцирования IFNγ [см. Фигуру 5 ниже].
ПРИМЕР 7
КОНСТРУКЦИЯ КАНИНИЗИРОВАННОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА 2B3 И 39A11 ПРОТИВ cCTLA-4
Обладая сильной аффинностью связывания с cCTLA-4 и их блокирующей активностью в отношении cCTLA-4 с его лигандами, CD86 и CD80, мышиные антитела 12B3 и 39A11 были отобраны для создания исходных канинизированных антител. Для выполнения процесса канинизации была определена последовательность ДНК, кодирующая тяжелую и легкую цепи собачьего IgG. ДНК- и белковая последовательность собачьей тяжелой и легкой цепи известны в данной области и могут быть получены путем поиска в генной и белковой базах данных NCBI. Существует четыре известных IgG-подтипа IgG собаки, и они называются IgGA, IgGB, IgGC и IgGD. Подобно человеческому IgG1, собачий IgGB обладает сильной эффекторной функцией. Чтобы нокаутировать эффекторную функцию IgGB, был сконструирован модифицированный IgGB (IgGBm), удаляющий нативные функции ADCC и CDC [см. US 10106107 B2, полностью включено в настоящее описание в качестве ссылки в полном объеме]. В собачьих антителах есть два типа легких цепей, которые называются каппа и лямбда. Не ограничиваясь каким-либо конкретным подходом, общий процесс производства канинизированных тяжелых и легких цепей, которые можно смешивать в различных комбинациях для получения канинизированных mAb против собачьего CTLA-4, может включать следующий протокол:
i) Идентифицировали CDR H и L цепей отобранных антител. Обратно транслировали аминокислотные последовательности CDR в подходящую последовательность ДНК.
ii) Идентифицировали подходящую последовательность ДНК для H и L цепи собачьего IgG (например, тяжелой цепи IgGB и легкой каппа-цепи).
iii) Идентифицированы последовательности ДНК, кодирующие эндогенные CDR H- и L-цепей собачьего IgG, ДНК указанной выше последовательности.
iv) Заменяли последовательность ДНК, кодирующую эндогенные CDR H и L цепей собак, на последовательности ДНК, кодирующие CDR отобранных антител. Кроме того, необязательно заменяли ДНК, кодирующую некоторые аминокислотные остатки каркасной области собаки, на ДНК, кодирующую отобранные аминокислотные остатки из отобранных каркасных областей антитела.
v) Синтезировали ДНК из стадии (iv) и клонировали ее в подходящую экспрессирующую плазмиду.
vi) Трансфецировали синтезированные плазмиды в клетки НЕК 293.
vii) Очищали экспрессированное канинизированное антитело из супернатанта НЕК 293.
viii) Тестировали очищенное канинизированное антитело на связывание с собачьим CTLA-4.
Нуклеотидные и аминокислотные последовательности CDR 12B3 и 39A11 представлены в Таблице 3 ниже.
Конструировали набор канинизированных последовательностей легкой и тяжелой цепей. Их идентификационные номера указаны в Таблицах 4-6 ниже.
(Аминокислота)1
(ДНК)2
(Аминокислота)1
(ДНК)2
(Аминокислота)1
(Нуклеиновая кислота)
(Аминокислота)
Настоящее изобретение относится к канинизированным антителам 12B3 и 39A11, образованным комбинацией канинизированных тяжелых и легких цепей каждого антитела, представленного в таблицах выше; такие антитела демонстрируют особенно прочное связывание с cCTLA-4. Как показано на Фигуре 6, результаты ИФА демонстрируют, что оба, 12B3 и 39A11, успешно канинизированы. Канинизированные c12B3L3H2 и L3H3 обладают такой же реактивностью с cCTLA-4, что и родительское 12B3; канинизированное c39A11L3H3 обладает такой же реактивностью с cCTLA-4, что и родительское 39A11. Химеры 12B3 и 39A11 представляют собой их родительские антитела.
НУКЛЕОТИДНЫЕ (NA) и АМИНОКИСЛОТНЫЕ (AA) ПОСЛЕДОВАТЕЛЬНОСТИ
SEQ ID NO: 1: NA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 12B3
cagatccagttggtgcagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaaactatggaatgaactgggtgaagcaggctccaggaaagggtttaaagtggatgggctggataaacacctacactggagagccaacatatgctgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctatttgcagatcaacaacctcaaaaatgaggacatggctacatatttctgtgcaagacggtcaatttattacccgtactggggccaaggcaccactctcacagtctcctca
SEQ ID NO: 2: AA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 12B3
QIQLVQSGPELKKPGETVKISCKASGYTFTNYGMNWVKQAPGKGLKWMGWINTYTG
EPTYADDFKGRFAFSLETSASTAYLQINNLKNEDMATYFCARRSIYYPYWGQGTTLTVSS
SEQ ID NO: 3: NA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 27G12
cagatccagttggtacagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaacctatggagtgagctgggtgaaacaggctccaggaaagggtttaaggtggatgggctggataaacacctactctggaatgccaacatatgttgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctttttgcagatcaacaacctcaaaaatgaggacacggctatatatttctgtgcaagacggggtatctcctttgactactggggccaaggcaccactctcacagtctcctca
SEQ ID NO: 4: AA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 27G12

SEQ ID NO: 5: NA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 39A11
gaggtgaagctggtggagtctggaggaggcttggtacagcctgggggttccctgagtctctcctgtgcaacttctggattcaccttcagtgattactacatgagctgggtccgccagtctccggggaaggcacttgagtggatgggttttattagaaacaaagctaatggttacacaacagagtacagcgcatctctgaagggtcggttcaccatctccagagataattcccaaagcatcctctatcttcaaatgaatgtcctgagagctgaggacagtgccacttattactgtgtaagatttgggttaatgtactactttgactactggggccaaggcaccactctcacagtctcctca
SEQ ID NO: 6: AA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 39A11
EVKLVESGGGLVQPGGSLSLSCATSGFTFSDYYMSWVRQSPGKALEWMGFIRNKANG
YTTEYSASLKGRFTISRDNSQSILYLQMNVLRAEDSATYYCVRFGLMYYFDYWGQGTTLTVSS
SEQ ID NO: 7: NA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 45A9
cagatccagttggtgcagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaaactatggaatgaactgggtgaagcaggctccaggaaagggtttaaagtggatgggctggataaacacctacactggagagccaacatatgctgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctatttgcagatcaacaacctcaaaaatgaggacacggctacatatttctgtgcaagaagggggacctactataggccctggggccaaggcaccactctcacagtctcctca
SEQ ID NO: 8: AA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 45A9
QIQLVQSGPELKKPGETVKISCKASGYTFTNYGMNWVKQAPGKGLKWMGWINTYTG
EPTYADDFKGRFAFSLETSASTAYLQINNLKNEDTATYFCARRGTYYRPWGQGTTLTVSS
SEQ ID NO: 9: NA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 110E3
cagatccagttggtgcagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctggatataccttcacaaactatggaatgaactgggtgaagcaggctccaggaaagggtttaaagtggatgggctggataaacacctacactggagagccaacatatgctgatgacttcaagggacgggttgccttctctttggaaacctctgccagcactgcctttttgcagatcaacaacctcaaaaatgaggacacggctacatatttctgtgcaaggcggggggtacgactggactactggggccaaggcaccactctcacagtctcctca
SEQ ID NO: 10: AA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 110E3
QIQLVQSGPELKKPGETVKISCKASGYTFTNYGMNWVKQAPGKGLKWMGWINTYTG
EPTYADDFKGRVAFSLETSASTAFLQINNLKNEDTATYFCARRGVRLDYWGQGTTLTVSS
SEQ ID NO: 11: NA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 22A11
caggtccaactgcagcagcctgggactgaactggtgaagcctggggcttcagtgaagctgtcctgcaaggcctctggctataccttcaccagctactggatgcactgggtgaagcagaggcctggacaaggccttgagtggattggaaatatcaatcctagcaatggtggtactaggttcaatgagaagttcaagaacaaggccacactgactgaagacaaatcctccagcacagcctacatgcagctcagtagcctgacatctgaggactctgcggtctattattgtgcaagatcgaactacggtagtggctgggcctggtttgcttactggggccaagggactctggtcactgtctctgca
SEQ ID NO: 12: AA-последовательность вариабельной области тяжелой цепи мышиного моноклонального антитела 22A11
QVQLQQPGTELVKPGASVKLSCKASGYTFTSYWMHWVKQRPGQGLEWIGNINPSNG
GTRFNEKFKNKATLTEDKSSSTAYMQLSSLTSEDSAVYYCARSNYGSGWAWFAYWG
QGTLVTVSA
SEQ ID NO: 13: NA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 12B3
gatgttttgatgacccaaactccactctccctgcctgtcagtcttggagatcaagcctccatctcttgcagatctagtcagagcattgtatatagtaatggaaacacctatttagaatggtacctgcagaaaccaggccagtctccaaagctcctgatctacaaagtttccaaccgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaa
SEQ ID NO: 14: AA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 12B3
DVLMTQTPLSLPVSLGDQASISCRSSQSIVYSNGNTYLEWYLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYYCFQGSHVPWTFGGGTKLEIK
SEQ ID NO: 15: NA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 27G12
Gatgttttgatgacccagactccactctccctgcctgtcagtcttggagatcacgcctccatctcttgcaaatctagtcagagcattgtatatattaatggaaacacctatttagaatggtacctgcagaagccaggccagtctccaaagctcctgatctacaaagtttccaaacgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaa
SEQ ID NO: 16: AA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 27G12

SEQ ID NO: 17: NA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 39A11
gaaaatgtgctcatccagtctccagcaatcatgtctgcttctccaggggaaaaggtcaccatgacctgcagggccagctcaagtgtaagttccagttacttgcactggtaccagcagaagtcaggtgcctcccccaaactctggatttttagcacatccaacttggcttctggagtccctgctcgcttcagtggcagtgggtctgggacctcttattctctcacaatcaacagtgtggaggctgaagatgctgccacttattactgccagcagtacagtggtctcccactcacgttcggaggggggaccaagctggaaataaaa
SEQ ID NO: 18: AA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 39A11
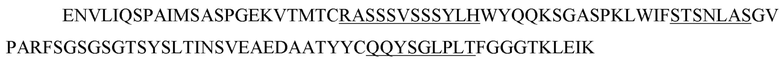
SEQ ID NO: 19: NA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 45A9
gatgttttgatgacccaaactccactctccctgcctgtcagtcttggagatcaagcctccatctcttgcagatctagtcagagtattgtatatagtcatggaaacacctatttagaatggtacctgcagaaaccaggccagtctccaaaggtcctgatctacaaagtttccaaccgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaa
SEQ ID NO: 20: AA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 45A9

SEQ ID NO: 21: NA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 110E3
gatgttttgatgacccaaactccactctccctgcctgtcagtcttggagatcaagcctccatctcttgcagatctagtcagagcattgtatatattagtggaagcacctatttagaatggtatctgcagaaaccaggccagtctccaaagctcctgatctacaaagtttccagtcgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaa
SEQ ID NO: 22: AA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 110E3
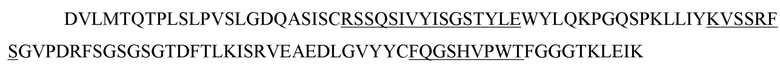
SEQ ID NO: 23: NA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 22A11
gacatccagatgaaccagtctccatccagtctgtctgcatcccttggagacacaattaccatcacttgccatgccagtcagaacattaatgtttggttaagctggtaccagcagaaaccaggaaatattcctaaacttttgatctataagtcttccaacttgcacacaggcgtcccatcaaggtttagtggcagtggatctggaacaggtttcacattaaccatcagcagcctgcagcctgaagacattgccacttactactgtcaacagggtcaaagttatccgtggacgttcggtggaggcaccaagctggaaatcaaa
SEQ ID NO: 24: AA-последовательность вариабельной области легкой цепи мышиного моноклонального антитела 22A11

SEQ ID NO: 25: NA-последовательность тяжелой цепи химерного антитела мышь-собака 12B3
cagatccagttggtgcagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaaactatggaatgaactgggtgaagcaggctccaggaaagggtttaaagtggatgggctggataaacacctacactggagagccaacatatgctgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctatttgcagatcaacaacctcaaaaatgaggacatggctacatatttctgtgcaagacggtcaatttattacccgtactggggccaaggcaccactctcacagtctcctcagcgagcaccaccgcgccgagcgtgtttccgctggcgccgagctgcggcagcaccagcggcagcaccgtggcgctggcgtgcctggtgagcggctattttccggaaccggtgaccgtgagctggaacagcggcagcctgaccagcggcgtgcatacctttccgagcgtgctgcagagcagcggcctgtatagcctgagcagcatggtgaccgtgccgagcagccgctggccgagcgaaacctttacctgcaacgtggcgcatccggcgagcaaaaccaaagtggataaaccggtgccgaaacgcgaaaacggccgcgtgccgcgcccgccggattgcccgaaatgcccggcgccggaaatgctgggcggcccgagcgtgtttatttttccgccgaaaccgaaagataccctgctgattgcgcgcaccccggaagtgacctgcgtggtggtggatctggatccggaagatccggaagtgcagattagctggtttgtggatggcaaacagatgcagaccgcgaaaacccagccgcgcgaagaacagtttaacggcacctatcgcgtggtgagcgtgctgccgattggccatcaggattggctgaaaggcaaacagtttacctgcaaagtgaacaacaaagcgctgccgagcccgattgaacgcaccattagcaaagcgcgcggccaggcgcatcagccgagcgtgtatgtgctgccgccgagccgcgaagaactgagcaaaaacaccgtgagcctgacctgcctgattaaagatttttttccgccggatattgatgtggaatggcagagcaacggccagcaggaaccggaaagcaaatatcgcaccaccccgccgcagctggatgaagatggcagctattttctgtatagcaaactgagcgtggataaaagccgctggcagcgcggcgatacctttatttgcgcggtgatgcatgaagcgctgcataaccattatacccaggaaagcctgagccatagcccgggcaaa
SEQ ID NO: 26: AA-последовательность тяжелой цепи химерного антитела мышь-собака 12B3

SEQ ID NO: 27: NA-последовательность тяжелой цепи химерного антитела мышь-собака 27G12
cagatccagttggtacagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaacctatggagtgagctgggtgaaacaggctccaggaaagggtttaaggtggatgggctggataaacacctactctggaatgccaacatatgttgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctttttgcagatcaacaacctcaaaaatgaggacacggctatatatttctgtgcaagacggggtatctcctttgactactggggccaaggcaccactctcacagtctcctcagcgagcaccaccgcgccgagcgtgtttccgctggcgccgagctgcggcagcaccagcggcagcaccgtggcgctggcgtgcctggtgagcggctattttccggaaccggtgaccgtgagctggaacagcggcagcctgaccagcggcgtgcatacctttccgagcgtgctgcagagcagcggcctgtatagcctgagcagcatggtgaccgtgccgagcagccgctggccgagcgaaacctttacctgcaacgtggcgcatccggcgagcaaaaccaaagtggataaaccggtgccgaaacgcgaaaacggccgcgtgccgcgcccgccggattgcccgaaatgcccggcgccggaaatgctgggcggcccgagcgtgtttatttttccgccgaaaccgaaagataccctgctgattgcgcgcaccccggaagtgacctgcgtggtggtggatctggatccggaagatccggaagtgcagattagctggtttgtggatggcaaacagatgcagaccgcgaaaacccagccgcgcgaagaacagtttaacggcacctatcgcgtggtgagcgtgctgccgattggccatcaggattggctgaaaggcaaacagtttacctgcaaagtgaacaacaaagcgctgccgagcccgattgaacgcaccattagcaaagcgcgcggccaggcgcatcagccgagcgtgtatgtgctgccgccgagccgcgaagaactgagcaaaaacaccgtgagcctgacctgcctgattaaagatttttttccgccggatattgatgtggaatggcagagcaacggccagcaggaaccggaaagcaaatatcgcaccaccccgccgcagctggatgaagatggcagctattttctgtatagcaaactgagcgtggataaaagccgctggcagcgcggcgatacctttatttgcgcggtgatgcatgaagcgctgcataaccattatacccaggaaagcctgagccatagcccgggcaaa
SEQ ID NO: 28: AA-последовательность тяжелой цепи химерного антитела мышь-собака 27G12

SEQ ID NO: 29: NA-последовательность тяжелой цепи химерного антитела мышь-собака 39A11
gaggtgaagctggtggagtctggaggaggcttggtacagcctgggggttccctgagtctctcctgtgcaacttctggattcaccttcagtgattactacatgagctgggtccgccagtctccggggaaggcacttgagtggatgggttttattagaaacaaagctaatggttacacaacagagtacagcgcatctctgaagggtcggttcaccatctccagagataattcccaaagcatcctctatcttcaaatgaatgtcctgagagctgaggacagtgccacttattactgtgtaagatttgggttaatgtactactttgactactggggccaaggcaccactctcacagtctcctcagcgagcaccaccgcgccgagcgtgtttccgctggcgccgagctgcggcagcaccagcggcagcaccgtggcgctggcgtgcctggtgagcggctattttccggaaccggtgaccgtgagctggaacagcggcagcctgaccagcggcgtgcatacctttccgagcgtgctgcagagcagcggcctgtatagcctgagcagcatggtgaccgtgccgagcagccgctggccgagcgaaacctttacctgcaacgtggcgcatccggcgagcaaaaccaaagtggataaaccggtgccgaaacgcgaaaacggccgcgtgccgcgcccgccggattgcccgaaatgcccggcgccggaaatgctgggcggcccgagcgtgtttatttttccgccgaaaccgaaagataccctgctgattgcgcgcaccccggaagtgacctgcgtggtggtggatctggatccggaagatccggaagtgcagattagctggtttgtggatggcaaacagatgcagaccgcgaaaacccagccgcgcgaagaacagtttaacggcacctatcgcgtggtgagcgtgctgccgattggccatcaggattggctgaaaggcaaacagtttacctgcaaagtgaacaacaaagcgctgccgagcccgattgaacgcaccattagcaaagcgcgcggccaggcgcatcagccgagcgtgtatgtgctgccgccgagccgcgaagaactgagcaaaaacaccgtgagcctgacctgcctgattaaagatttttttccgccggatattgatgtggaatggcagagcaacggccagcaggaaccggaaagcaaatatcgcaccaccccgccgcagctggatgaagatggcagctattttctgtatagcaaactgagcgtggataaaagccgctggcagcgcggcgatacctttatttgcgcggtgatgcatgaagcgctgcataaccattatacccaggaaagcctgagccatagcccgggcaaa
SEQ ID NO: 30: AA-последовательность тяжелой цепи химерного антитела мышь-собака 39A11

SEQ ID NO: 31: NA-последовательность тяжелой цепи химерного антитела мышь-собака 45A9
cagatccagttggtgcagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaaactatggaatgaactgggtgaagcaggctccaggaaagggtttaaagtggatgggctggataaacacctacactggagagccaacatatgctgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctatttgcagatcaacaacctcaaaaatgaggacacggctacatatttctgtgcaagaagggggacctactataggccctggggccaaggcaccactctcacagtctcctcagcgagcaccaccgcgccgagcgtgtttccgctggcgccgagctgcggcagcaccagcggcagcaccgtggcgctggcgtgcctggtgagcggctattttccggaaccggtgaccgtgagctggaacagcggcagcctgaccagcggcgtgcatacctttccgagcgtgctgcagagcagcggcctgtatagcctgagcagcatggtgaccgtgccgagcagccgctggccgagcgaaacctttacctgcaacgtggcgcatccggcgagcaaaaccaaagtggataaaccggtgccgaaacgcgaaaacggccgcgtgccgcgcccgccggattgcccgaaatgcccggcgccggaaatgctgggcggcccgagcgtgtttatttttccgccgaaaccgaaagataccctgctgattgcgcgcaccccggaagtgacctgcgtggtggtggatctggatccggaagatccggaagtgcagattagctggtttgtggatggcaaacagatgcagaccgcgaaaacccagccgcgcgaagaacagtttaacggcacctatcgcgtggtgagcgtgctgccgattggccatcaggattggctgaaaggcaaacagtttacctgcaaagtgaacaacaaagcgctgccgagcccgattgaacgcaccattagcaaagcgcgcggccaggcgcatcagccgagcgtgtatgtgctgccgccgagccgcgaagaactgagcaaaaacaccgtgagcctgacctgcctgattaaagatttttttccgccggatattgatgtggaatggcagagcaacggccagcaggaaccggaaagcaaatatcgcaccaccccgccgcagctggatgaagatggcagctattttctgtatagcaaactgagcgtggataaaagccgctggcagcgcggcgatacctttatttgcgcggtgatgcatgaagcgctgcataaccattatacccaggaaagcctgagccatagcccgggcaaa
SEQ ID NO: 32: AA-последовательность тяжелой цепи химерного антитела мышь-собака 45A9

SEQ ID NO: 33: NA-последовательность тяжелой цепи химерного антитела мышь-собака 110E3
cagatccagttggtgcagtctggacctgagctgaagaagcctggagagacagtcaagatctcctgcaaggcttctggatataccttcacaaactatggaatgaactgggtgaagcaggctccaggaaagggtttaaagtggatgggctggataaacacctacactggagagccaacatatgctgatgacttcaagggacgggttgccttctctttggaaacctctgccagcactgcctttttgcagatcaacaacctcaaaaatgaggacacggctacatatttctgtgcaaggcggggggtacgactggactactggggccaaggcaccactctcacagtctcctcagcgagcaccaccgcgccgagcgtgtttccgctggcgccgagctgcggcagcaccagcggcagcaccgtggcgctggcgtgcctggtgagcggctattttccggaaccggtgaccgtgagctggaacagcggcagcctgaccagcggcgtgcatacctttccgagcgtgctgcagagcagcggcctgtatagcctgagcagcatggtgaccgtgccgagcagccgctggccgagcgaaacctttacctgcaacgtggcgcatccggcgagcaaaaccaaagtggataaaccggtgccgaaacgcgaaaacggccgcgtgccgcgcccgccggattgcccgaaatgcccggcgccggaaatgctgggcggcccgagcgtgtttatttttccgccgaaaccgaaagataccctgctgattgcgcgcaccccggaagtgacctgcgtggtggtggatctggatccggaagatccggaagtgcagattagctggtttgtggatggcaaacagatgcagaccgcgaaaacccagccgcgcgaagaacagtttaacggcacctatcgcgtggtgagcgtgctgccgattggccatcaggattggctgaaaggcaaacagtttacctgcaaagtgaacaacaaagcgctgccgagcccgattgaacgcaccattagcaaagcgcgcggccaggcgcatcagccgagcgtgtatgtgctgccgccgagccgcgaagaactgagcaaaaacaccgtgagcctgacctgcctgattaaagatttttttccgccggatattgatgtggaatggcagagcaacggccagcaggaaccggaaagcaaatatcgcaccaccccgccgcagctggatgaagatggcagctattttctgtatagcaaactgagcgtggataaaagccgctggcagcgcggcgatacctttatttgcgcggtgatgcatgaagcgctgcataaccattatacccaggaaagcctgagccatagcccgggcaaa
SEQ ID NO: 34: AA-последовательность тяжелой цепи химерного антитела мышь-собака 110E3

SEQ ID NO: 35: NA-последовательность тяжелой цепи химерного антитела мышь-собака 22A11
caggtccaactgcagcagcctgggactgaactggtgaagcctggggcttcagtgaagctgtcctgcaaggcctctggctataccttcaccagctactggatgcactgggtgaagcagaggcctggacaaggccttgagtggattggaaatatcaatcctagcaatggtggtactaggttcaatgagaagttcaagaacaaggccacactgactgaagacaaatcctccagcacagcctacatgcagctcagtagcctgacatctgaggactctgcggtctattattgtgcaagatcgaactacggtagtggctgggcctggtttgcttactggggccaagggactctggtcactgtctctgcagcgagcaccaccgcgccgagcgtgtttccgctggcgccgagctgcggcagcaccagcggcagcaccgtggcgctggcgtgcctggtgagcggctattttccggaaccggtgaccgtgagctggaacagcggcagcctgaccagcggcgtgcatacctttccgagcgtgctgcagagcagcggcctgtatagcctgagcagcatggtgaccgtgccgagcagccgctggccgagcgaaacctttacctgcaacgtggcgcatccggcgagcaaaaccaaagtggataaaccggtgccgaaacgcgaaaacggccgcgtgccgcgcccgccggattgcccgaaatgcccggcgccggaaatgctgggcggcccgagcgtgtttatttttccgccgaaaccgaaagataccctgctgattgcgcgcaccccggaagtgacctgcgtggtggtggatctggatccggaagatccggaagtgcagattagctggtttgtggatggcaaacagatgcagaccgcgaaaacccagccgcgcgaagaacagtttaacggcacctatcgcgtggtgagcgtgctgccgattggccatcaggattggctgaaaggcaaacagtttacctgcaaagtgaacaacaaagcgctgccgagcccgattgaacgcaccattagcaaagcgcgcggccaggcgcatcagccgagcgtgtatgtgctgccgccgagccgcgaagaactgagcaaaaacaccgtgagcctgacctgcctgattaaagatttttttccgccggatattgatgtggaatggcagagcaacggccagcaggaaccggaaagcaaatatcgcaccaccccgccgcagctggatgaagatggcagctattttctgtatagcaaactgagcgtggataaaagccgctggcagcgcggcgatacctttatttgcgcggtgatgcatgaagcgctgcataaccattatacccaggaaagcctgagccatagcccgggcaaa
SEQ ID NO: 36: AA-последовательность тяжелой цепи химерного антитела мышь-собака 22A11

SEQ ID NO: 37: NA-последовательность легкой цепи химерного антитела мышь-собака 12B3
gatgttttgatgacccaaactccactctccctgcctgtcagtcttggagatcaagcctccatctcttgcagatctagtcagagcattgtatatagtaatggaaacacctatttagaatggtacctgcagaaaccaggccagtctccaaagctcctgatctacaaagtttccaaccgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaacgcaacgatgcgcagccggcggtgtatctgtttcagccgagcccggatcagctgcataccggcagcgcgagcgtggtgtgcctgctgaacagcttttatccgaaagatattaacgtgaaatggaaagtggatggcgtgattcaggataccggcattcaggaaagcgtgaccgaacaggatagcaaagatagcacctatagcctgagcagcaccctgaccatgagcagcaccgaatatctgagccatgaactgtatagctgcgaaattacccataaaagcctgccgagcaccctgattaaaagctttcagcgcagcgaatgccagcgcgtggat
SEQ ID NO: 38: AA-последовательность легкой цепи химерного антитела мышь-собака 12B3

SEQ ID NO: 39: NA-последовательность легкой цепи химерного антитела мышь-собака 27G12
gatgttttgatgacccagactccactctccctgcctgtcagtcttggagatcacgcctccatctcttgcaaatctagtcagagcattgtatatattaatggaaacacctatttagaatggtacctgcagaagccaggccagtctccaaagctcctgatctacaaagtttccaaacgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaacgcaacgatgcgcagccggcggtgtatctgtttcagccgagcccggatcagctgcataccggcagcgcgagcgtggtgtgcctgctgaacagcttttatccgaaagatattaacgtgaaatggaaagtggatggcgtgattcaggataccggcattcaggaaagcgtgaccgaacaggatagcaaagatagcacctatagcctgagcagcaccctgaccatgagcagcaccgaatatctgagccatgaactgtatagctgcgaaattacccataaaagcctgccgagcaccctgattaaaagctttcagcgcagcgaatgccagcgcgtggat
SEQ ID NO: 40: AA-последовательность легкой цепи химерного антитела мышь-собака 27G12

SEQ ID NO: 41: NA-последовательность легкой цепи химерного антитела мышь-собака 39A11
gaaaatgtgctcatccagtctccagcaatcatgtctgcttctccaggggaaaaggtcaccatgacctgcagggccagctcaagtgtaagttccagttacttgcactggtaccagcagaagtcaggtgcctcccccaaactctggatttttagcacatccaacttggcttctggagtccctgctcgcttcagtggcagtgggtctgggacctcttattctctcacaatcaacagtgtggaggctgaagatgctgccacttattactgccagcagtacagtggtctcccactcacgttcggaggggggaccaagctggaaataaaacgcaacgatgcgcagccggcggtgtatctgtttcagccgagcccggatcagctgcataccggcagcgcgagcgtggtgtgcctgctgaacagcttttatccgaaagatattaacgtgaaatggaaagtggatggcgtgattcaggataccggcattcaggaaagcgtgaccgaacaggatagcaaagatagcacctatagcctgagcagcaccctgaccatgagcagcaccgaatatctgagccatgaactgtatagctgcgaaattacccataaaagcctgccgagcaccctgattaaaagctttcagcgcagcgaatgccagcgcgtggat
SEQ ID NO: 42: AA-последовательность легкой цепи химерного антитела мышь-собака 39A11

SEQ ID NO: 43: NA-последовательность легкой цепи химерного антитела мышь-собака 45A9
gatgttttgatgacccaaactccactctccctgcctgtcagtcttggagatcaagcctccatctcttgcagatctagtcagagtattgtatatagtcatggaaacacctatttagaatggtacctgcagaaaccaggccagtctccaaaggtcctgatctacaaagtttccaaccgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaacgcaacgatgcgcagccggcggtgtatctgtttcagccgagcccggatcagctgcataccggcagcgcgagcgtggtgtgcctgctgaacagcttttatccgaaagatattaacgtgaaatggaaagtggatggcgtgattcaggataccggcattcaggaaagcgtgaccgaacaggatagcaaagatagcacctatagcctgagcagcaccctgaccatgagcagcaccgaatatctgagccatgaactgtatagctgcgaaattacccataaaagcctgccgagcaccctgattaaaagctttcagcgcagcgaatgccagcgcgtggat
SEQ ID NO: 44: AA-последовательность легкой цепи химерного антитела мышь-собака 45A9
DVLMTQTPLSLPVSLGDQASISCRSSQSIVYSHGNTYLEWYLQKPGQSPKVLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYYCFQGSHVPWTFGGGTKLEIKRNDAQPAVYLFQPSPDQLHTGSASVVCLLNSFYPKDINVKWKVDGVIQDTGIQESVTEQDSKDSTYSLSSTLTMSSTEYLSHELYSCEITHKSLPSTLIKSFQRSECQRVD
SEQ ID NO: 45: NA-последовательность легкой цепи химерного антитела мышь-собака 110E3
gatgttttgatgacccaaactccactctccctgcctgtcagtcttggagatcaagcctccatctcttgcagatctagtcagagcattgtatatattagtggaagcacctatttagaatggtatctgcagaaaccaggccagtctccaaagctcctgatctacaaagtttccagtcgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtggacgttcggtggaggcaccaagctggaaatcaaacgcaacgatgcgcagccggcggtgtatctgtttcagccgagcccggatcagctgcataccggcagcgcgagcgtggtgtgcctgctgaacagcttttatccgaaagatattaacgtgaaatggaaagtggatggcgtgattcaggataccggcattcaggaaagcgtgaccgaacaggatagcaaagatagcacctatagcctgagcagcaccctgaccatgagcagcaccgaatatctgagccatgaactgtatagctgcgaaattacccataaaagcctgccgagcaccctgattaaaagctttcagcgcagcgaatgccagcgcgtggat
SEQ ID NO: 46: AA-последовательность легкой цепи химерного антитела мышь-собака 110E3
DVLMTQTPLSLPVSLGDQASISCRSSQSIVYISGSTYLEWYLQKPGQSPKLLIYKVSSRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYYCFQGSHVPWTFGGGTKLEIKRNDAQPAVYLFQPSPDQLHTGSASVVCLLNSFYPKDINVKWKVDGVIQDTGIQESVTEQDSKDSTYSLSSTLTMSSTEYLSHELYSCEITHKSLPSTLIKSFQRSECQRVD
SEQ ID NO: 47: NA-последовательность легкой цепи химерного моноклонального антитела мышь-собака 22A11
gacatccagatgaaccagtctccatccagtctgtctgcatcccttggagacacaattaccatcacttgccatgccagtcagaacattaatgtttggttaagctggtaccagcagaaaccaggaaatattcctaaacttttgatctataagtcttccaacttgcacacaggcgtcccatcaaggtttagtggcagtggatctggaacaggtttcacattaaccatcagcagcctgcagcctgaagacattgccacttactactgtcaacagggtcaaagttatccgtggacgttcggtggaggcaccaagctggaaatcaaacgcaacgatgcgcagccggcggtgtatctgtttcagccgagcccggatcagctgcataccggcagcgcgagcgtggtgtgcctgctgaacagcttttatccgaaagatattaacgtgaaatggaaagtggatggcgtgattcaggataccggcattcaggaaagcgtgaccgaacaggatagcaaagatagcacctatagcctgagcagcaccctgaccatgagcagcaccgaatatctgagccatgaactgtatagctgcgaaattacccataaaagcctgccgagcaccctgattaaaagctttcagcgcagcgaatgccagcgcgtggat
SEQ ID NO: 48: AA-последовательность легкой цепи химерного антитела мышь-собака 22A11

SEQ ID NO: 49: NA-последовательность легкой цепи канинизированного 12B3 (VL1)
gatattgtgatgacccagaccccgctgagcctgagcgtgagcccgggcgaaccggcgagcattagctgccgcagcagccagagcattgtgtatagcaacggcaacacctatctggaatggtttcagcagaaaccgggccagagcccgcagcgcctgatttataaagtgagcaaccgctttagcggcgtgccggatcgctttagcggcagcggcagcggcaccgattttaccctgcgcattagccgcgtggaagcggatgatgcgggcgtgtattattgctttcagggcagccatgtgccgtggacctttggcggcggcaccaaactggaaattaaaaggaacgacgctcagccagccgtgtacctcttccagccttcgccggaccagcttcatacggggtcagcgtcggtggtgtgcctgttgaactcgttttaccccaaggacattaacgtgaagtggaaggtagacggggtaattcaagacactggcattcaagagtccgtcacggaacaagactcaaaagactcaacgtattcactgtcgtcaaccttgacgatgtcaagcaccgagtatcttagccatgagctgtattcgtgcgagatcacccacaagtccctcccctccactcttatcaaatcctttcagcggtcggaatgtcagcgggtcgat
SEQ ID NO: 50: AA-последовательность легкой цепи канинизированного 12B3 (VL1)

SEQ ID NO: 51: NA-последовательность легкой цепи канинизированного 12B3 (VL2)
gatattgtgatgacccagaccccgctgagcctgagcgtgagcccgggcgaaccggcgagcattagctgccgcagcagccagagcattgtgtatagcaacggcaacacctatctggaatggtatcagcagaaaccgggccagagcccgaaactgctgatttataaagtgagcaaccgctttagcggcgtgccggatcgctttagcggcagcggcagcggcaccgattttaccctgcgcattagccgcgtggaagcggatgatgcgggcgtgtattattgctttcagggcagccatgtgccgtggacctttggcggcggcaccaaactggaaattaaaaggaacgacgctcagccagccgtgtacctcttccagccttcgccggaccagcttcatacggggtcagcgtcggtggtgtgcctgttgaactcgttttaccccaaggacattaacgtgaagtggaaggtagacggggtaattcaagacactggcattcaagagtccgtcacggaacaagactcaaaagactcaacgtattcactgtcgtcaaccttgacgatgtcaagcaccgagtatcttagccatgagctgtattcgtgcgagatcacccacaagtccctcccctccactcttatcaaatcctttcagcggtcggaatgtcagcgggtcgat
SEQ ID NO: 52: AA-последовательность легкой цепи канинизированного 12B3 (VL2)

SEQ ID NO: 53: NA-последовательность легкой цепи канинизированного 12B3 (VL3)
gatgtgctgatgacccagaccccgctgagcctgagcgtgagcccgggcgaaccggcgagcattagctgccgcagcagccagagcattgtgtatagcaacggcaacacctatctggaatggtatctgcagaaaccgggccagagcccgaaactgctgatttataaagtgagcaaccgctttagcggcgtgccggatcgctttagcggcagcggcagcggcaccgattttaccctgcgcattagccgcgtggaagcggatgatgcgggcgtgtattattgctttcagggcagccatgtgccgtggacctttggcggcggcaccaaactggaactgaaaaggaacgacgctcagccagccgtgtacctcttccagccttcgccggaccagcttcatacggggtcagcgtcggtggtgtgcctgttgaactcgttttaccccaaggacattaacgtgaagtggaaggtagacggggtaattcaagacactggcattcaagagtccgtcacggaacaagactcaaaagactcaacgtattcactgtcgtcaaccttgacgatgtcaagcaccgagtatcttagccatgagctgtattcgtgcgagatcacccacaagtccctcccctccactcttatcaaatcctttcagcggtcggaatgtcagcgggtcgat
SEQ ID NO: 54: AA-последовательность легкой цепи канинизированного 12B3 (VL3)

SEQ ID NO: 55: NA-последовательность легкой цепи канинизированного 39A11 (VL1)
gaaattgtgatgacccagagcccggcgagcctgagcctgagccaggaagaaaaagtgaccattacctgccgcgcgagcagcagcgtgagcagcagctatctgcattggtatcagcagaaaccgggccaggcgccgaaactgctgatttatagcaccagcaacctggcgagcggcgtgccgagccgctttagcggcagcggcagcggcaccgattttagctttaccattagcagcctggaaccggaagatgtggcggtgtattattgccagcagtatagcggcctgccgctgacctttggcggcggcaccaaactggaaattaaaaggaacgacgctcagccagccgtgtacctcttccagccttcgccggaccagcttcatacggggtcagcgtcggtggtgtgcctgttgaactcgttttaccccaaggacattaacgtgaagtggaaggtagacggggtaattcaagacactggcattcaagagtccgtcacggaacaagactcaaaagactcaacgtattcactgtcgtcaaccttgacgatgtcaagcaccgagtatcttagccatgagctgtattcgtgcgagatcacccacaagtccctcccctccactcttatcaaatcctttcagcggtcggaatgtcagcgggtcgat
SEQ ID NO: 56: AA-последовательность легкой цепи канинизированного 39A11 (VL1)

SEQ ID NO: 57: NA-последовательность легкой цепи канинизированного 39A11 (VL2)
gaaaacgtgctgacccagagcccggcgagcctgagcctgagccaggaagaaaaagtgaccattacctgccgcgcgagcagcagcgtgagcagcagctatctgcattggtatcagcagaaaccgggccaggcgccgaaactgtggatttttagcaccagcaacctggcgagcggcgtgccgagccgctttagcggcagcggcagcggcaccgattatagctttaccattagcagcctggaaccggaagatgtggcggtgtattattgccagcagtatagcggcctgccgctgacctttggcggcggcaccaaactggaactgaaaaggaacgacgctcagccagccgtgtacctcttccagccttcgccggaccagcttcatacggggtcagcgtcggtggtgtgcctgttgaactcgttttaccccaaggacattaacgtgaagtggaaggtagacggggtaattcaagacactggcattcaagagtccgtcacggaacaagactcaaaagactcaacgtattcactgtcgtcaaccttgacgatgtcaagcaccgagtatcttagccatgagctgtattcgtgcgagatcacccacaagtccctcccctccactcttatcaaatcctttcagcggtcggaatgtcagcgggtcgat
SEQ ID NO: 58: AA-последовательность легкой цепи канинизированного 39A11 (VL2)

SEQ ID NO: 59: NA-последовательность легкой цепи канинизированного 39A11 (VL3)
gaaaacgtgctgacccagagcccggcgagcctgagcctgagcccgggcgaaaaagtgaccattacctgccgcgcgagcagcagcgtgagcagcagctatctgcattggtatcagcagaaaccgggccagagcccgaaactgtggatttttagcaccagcaacctggcgagcggcgtgccgagccgctttagcggcagcggcagcggcaccagctatagctttaccattagcagcctggaaccggaagatgtggcggtgtattattgccagcagtatagcggcctgccgctgacctttggcggcggcaccaaactggaactgaaaaggaacgacgctcagccagccgtgtacctcttccagccttcgccggaccagcttcatacggggtcagcgtcggtggtgtgcctgttgaactcgttttaccccaaggacattaacgtgaagtggaaggtagacggggtaattcaagacactggcattcaagagtccgtcacggaacaagactcaaaagactcaacgtattcactgtcgtcaaccttgacgatgtcaagcaccgagtatcttagccatgagctgtattcgtgcgagatcacccacaagtccctcccctccactcttatcaaatcctttcagcggtcggaatgtcagcgggtcgat
SEQ ID NO: 60: AA-последовательность легкой цепи канинизированного 39A11 (VL3)

SEQ ID NO: 61: NA-последовательность тяжелой цепи канинизированного 12B3 (VH1) с IgGB
gaagtgcagctggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgtggcgagcggctatacctttaccaactatggcatgaactgggtgcgccaggcgccgggcaaaggcctgcagtgggtggcgtggattaacacctataccggcgaaccgacctatgcggatgattttaaaggccgctttaccattagccgcgataacgcgaaaaacaccctgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgcgcgccgcagcatttattatccgtattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggaccttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcaacggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 62: AA-последовательность тяжелой цепи канинизированного 12B3 (VH1) с IgGB

SEQ ID NO: 63: NA-последовательность тяжелой цепи канинизированного 12B3 (VH2) с IgGB
gaaattcagctggtgcagagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcaaagcgagcggctatacctttaccaactatggcatgaactgggtgcgccaggcgccgggcaaaggcctgcagtggatgggctggattaacacctataccggcgaaccgacctatgcggatgattttaaaggccgctttacctttagcctggataacgcgaaaaacaccctgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtatttttgcgcgcgccgcagcatttattatccgtattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggaccttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcaacggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 64: AA-последовательность тяжелой цепи канинизированного 12B3 (VH2) с IgGB

SEQ ID NO: 65: NA-последовательность тяжелой цепи канинизированного 12B3 (VH3) с IgGB
gaaattcagctggtgcagagcggcggcgatctggtgaaaccgggcggcagcgtgcgcctgagctgcaaagcgagcggctatacctttaccaactatggcatgaactgggtgaaacaggcgccgggcaaaggcctgcagtggatgggctggattaacacctataccggcgaaccgacctatgcggatgattttaaaggccgctttacctttagcctggataacgcgaaaaacaccgcgtatctgcagattaacagcctgcgcgcggaagataccgcggtgtatttttgcgcgcgccgcagcatttattatccgtattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggaccttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcaacggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 66: AA-последовательность тяжелой цепи канинизированного 12B3 (VH3) с IgGB

SEQ ID NO: 67: NA-последовательность тяжелой цепи канинизированного 39A11 (VH1) с IgGB
gaagtgcagctggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgtggcgagcggctttacctttagcgattattatatgagctgggtgcgccaggcgccgggcaaaggcctggaatgggtggcgtttattcgcaacaaagcgaacggctataccaccgaatatagcgcgagcctgaaaggccgctttaccattagccgcgataacgcgaaaaacatggcgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgcgagctttggcctgatgtattattttgattattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggaccttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcaacggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 68: AA-последовательность тяжелой цепи канинизированного 39A11 (VH1) с IgGB

SEQ ID NO: 69: NA-последовательность тяжелой цепи канинизированного 39A11 (VH2) с IgGB
gaagtgcagctggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgcgaccagcggctttacctttagcgattattatatgagctgggtgcgccaggcgccgggcaaaggcctggaatggatgggctttattcgcaacaaagcgaacggctataccaccgaatatagcgcgagcctgaaaggccgctttaccattagccgcgataacgcgaaaaacatggcgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgtgcgctttggcctgatgtattattttgattattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggaccttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcaacggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 70: AA-последовательность тяжелой цепи канинизированного 39A11 (VH2) с IgGB

SEQ ID NO: 71: NA-последовательность тяжелой цепи канинизированного 39A11 (VH3) с IgGB
gaagtgaaactggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgcgaccagcggctttacctttagcgattattatatgagctgggtgcgccaggcgccgggcaaagcgctggaatggatgggctttattcgcaacaaagcgaacggctataccaccgaatatagcgcgagcctgaaaggccgctttaccattagccgcgataacgcgaaaaacatgctgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgtgcgctttggcctgatgtattattttgattattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggaccttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcaacggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 72: AA-последовательность тяжелой цепи канинизированного 39A11 (VH3) с IgGB

SEQ ID NO: 73: NA-последовательность тяжелой цепи канинизированного 12B3 (VH1) с IgGBm
gaagtgcagctggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgtggcgagcggctatacctttaccaactatggcatgaactgggtgcgccaggcgccgggcaaaggcctgcagtgggtggcgtggattaacacctataccggcgaaccgacctatgcggatgattttaaaggccgctttaccattagccgcgataacgcgaaaaacaccctgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgcgcgccgcagcatttattatccgtattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggctcttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcgccggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 74: AA-последовательность тяжелой цепи канинизированного 12B3 (VH1) с IgGBm

SEQ ID NO: 75: NA-последовательность тяжелой цепи канинизированного 12B3 (VH2) с IgGBm
gaaattcagctggtgcagagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcaaagcgagcggctatacctttaccaactatggcatgaactgggtgcgccaggcgccgggcaaaggcctgcagtggatgggctggattaacacctataccggcgaaccgacctatgcggatgattttaaaggccgctttacctttagcctggataacgcgaaaaacaccctgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtatttttgcgcgcgccgcagcatttattatccgtattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggctcttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcgccggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 76: AA-последовательность тяжелой цепи канинизированного 12B3 (VH2) с IgGBm

SEQ ID NO: 77. NA-последовательность тяжелой цепи канинизированного 12B3 (VH3) с IgGBm
gaaattcagctggtgcagagcggcggcgatctggtgaaaccgggcggcagcgtgcgcctgagctgcaaagcgagcggctatacctttaccaactatggcatgaactgggtgaaacaggcgccgggcaaaggcctgcagtggatgggctggattaacacctataccggcgaaccgacctatgcggatgattttaaaggccgctttacctttagcctggataacgcgaaaaacaccgcgtatctgcagattaacagcctgcgcgcggaagataccgcggtgtatttttgcgcgcgccgcagcatttattatccgtattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggctcttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcgccggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 78: AA-последовательность тяжелой цепи канинизированного 12B3 (VH3) с IgGBm

SEQ ID NO: 79: NA-последовательность тяжелой цепи канинизированного 39A11 (VH1) с IgGBm
gaagtgcagctggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgtggcgagcggctttacctttagcgattattatatgagctgggtgcgccaggcgccgggcaaaggcctggaatgggtggcgtttattcgcaacaaagcgaacggctataccaccgaatatagcgcgagcctgaaaggccgctttaccattagccgcgataacgcgaaaaacatggcgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgcgagctttggcctgatgtattattttgattattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggctcttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcgccggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 80: AA-последовательность тяжелой цепи канинизированного 39A11 (VH1) с IgGBm

SEQ ID NO: 81: NA-последовательность тяжелой цепи канинизированного 39A11 (VH2) с IgGBm
gaagtgcagctggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgcgaccagcggctttacctttagcgattattatatgagctgggtgcgccaggcgccgggcaaaggcctggaatggatgggctttattcgcaacaaagcgaacggctataccaccgaatatagcgcgagcctgaaaggccgctttaccattagccgcgataacgcgaaaaacatggcgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgtgcgctttggcctgatgtattattttgattattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggctcttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcgccggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 82: AA-последовательность тяжелой цепи канинизированного 39A11 (VH2) с IgGBm

SEQ ID NO: 83: NA-последовательность тяжелой цепи канинизированного 39A11 (VH3) с IgGBm
gaagtgaaactggtggaaagcggcggcgatctggtgaaaccgggcggcagcctgcgcctgagctgcgcgaccagcggctttacctttagcgattattatatgagctgggtgcgccaggcgccgggcaaagcgctggaatggatgggctttattcgcaacaaagcgaacggctataccaccgaatatagcgcgagcctgaaaggccgctttaccattagccgcgataacgcgaaaaacatgctgtatctgcagatgaacagcctgcgcgcggaagataccgcggtgtattattgcgtgcgctttggcctgatgtattattttgattattggggccagggcaccaccctgaccgtgagcagcgcttccacaaccgcgccatcagtctttccgttggccccatcatgcgggtcgacgagcggatcgactgtggccctggcgtgcttggtgtcgggatactttcccgaacccgtcacggtcagctggaactccggatcgcttacgagcggtgtgcatacgttcccctcggtcttgcaatcatcagggctctactcgctgtcgagcatggtaacggtgccctcatcgaggtggccctccgaaacgttcacatgtaacgtagcacatccagcctccaaaaccaaggtggataaacccgtgccgaaaagagagaatgggcgggtgcctcgaccccctgattgccccaagtgtccggctccggaaatgctcggtggaccctcagtgtttatcttccctccgaagcccaaggacactctgctgatcgcgcgcactccagaagtaacatgtgtagtggtggctcttgatcccgaggaccccgaagtccagatctcctggtttgtagatgggaaacagatgcagaccgcaaaaactcaacccagagaggagcagttcgccggaacataccgagtggtatccgtccttccgattggccaccaggactggttgaaagggaagcagtttacgtgtaaagtcaacaataaggggttgcctagccctattgagcggacgatttcgaaagctaggggacaggcccaccagccatcggtctatgtccttccgccttcccgcgaggagctctcgaagaatacagtgagccttacatgcctcattaaggatttcttcccgcctgatatcgacgtagagtggcaatcaaacggtcaacaggagccggaatccaagtatagaaccactccgccccagcttgacgaggacggatcatactttttgtattcaaaactgtcggtggataagagccggtggcagagaggtgacaccttcatctgtgcggtgatgcacgaagcactccataatcactacacccaagagagcctctcgcattcccccggaaag
SEQ ID NO: 84: AA-последовательность тяжелой цепи канинизированного 39A11 (VH3) с IgGBm

SEQ ID NO: 125: NA-последовательность собачьего CTLA-4 (NCBI Эталонная последовательность: NP_001003106).
atggcgggctttggctttcgccgccatggcgcgcagccggatctggcgagccgcacctggccgtgcaccgcgctgtttagcctgctgtttattccggtgtttagcaaaggcatgcatgtggcgcagccggcggtggtgctggcgagcagccgcggcgtggcgagctttgtgtgcgaatatggcagcagcggcaacgcggcggaagtgcgcgtgaccgtgctgcgccaggcgggcagccagatgaccgaagtgtgcgcggcgacctataccgtggaagatgaactggcgtttctggatgatagcacctgcaccggcaccagcagcggcaacaaagtgaacctgaccattcagggcctgcgcgcgatggataccggcctgtatatttgcaaagtggaactgatgtatccgccgccgtattatgtgggcatgggcaacggcacccagatttatgtgattgatccggaaccgtgcccggatagcgattttctgctgtggattctggcggcggtgagcagcggcctgtttttttatagctttctgattaccgcggtgagcctgagcaaaatgctgaaaaaacgcagcccgctgaccaccggcgtgtatgtgaaaatgccgccgaccgaaccggaatgcgaaaaacagtttcagccgtattttattccgattaac
SEQ ID NO: 126: AA-последовательность собачьего CTLA-4 (NCBI Эталонная последовательность: NP_001003106).SEQ ID NO: 138, зрелая последовательность (т.е., без сигнальной последовательности) выделено жирным шрифтом:

SEQ ID NO: 127: Генетически модифицированная cFc-область собачьего IgG B (из U.S. 10106107 B2)

ПРИМЕР 8
КАРТИРОВАНИЕ ЭПИТОПА КАНИНИЗИРОВАННОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА 2B3 И 39A11 ПРОТИВ cCTLA-4
Взаимодействие антител с их родственными белковыми антигенами опосредуется связыванием определенных аминокислот антител (паратопов) со специфическими аминокислотами (эпитопами) антигенов-мишеней. Эпитоп представляет собой антигенную детерминанту, которая вызывает специфическую реакцию иммуноглобулина. Эпитоп состоит из группы аминокислот на поверхности антигена. Интересующий белок может содержать несколько эпитопов, которые распознаются разными антителами. Эпитопы, распознаваемые антителами, классифицируются как линейные или конформационные эпитопы. Линейные эпитопы образуются отрезком непрерывной последовательности аминокислот в белке, в то время как конформационные эпитопы состоят из аминокислот, которые прерываются (например, располагаются далеко друг от друга) в первичной аминокислотной последовательности, но объединяются в трехмерной белковой структуре.
Картирование эпитопов относится к процессу идентификации аминокислотных последовательностей (т.е. эпитопов), которые распознаются антителами на своих антигенах-мишенях. Идентификация эпитопов, распознаваемых моноклональными антителами (mAb) на антигенах-мишенях, имеет важные применения. Например, это может помочь в разработке новых терапевтических средств, диагностических средств и вакцин. Картирование эпитопов также может помочь в выборе оптимизированных терапевтических mAb и помочь выяснить механизмы их действия. Информация об эпитопах собачьего CTLA-4 может также выявить уникальные эпитопы и определить защитные или патогенные эффекты вакцин. Идентификация эпитопа также может привести к разработке субъединичных вакцин на основе химического или генетического связывания идентифицированного пептидного эпитопа с белком-носителем или другими иммуностимулирующими агентами.
Картирование эпитопа может быть выполнено с использованием поликлональных или моноклональных антител, и несколько методов используются для идентификации эпитопа в зависимости от предполагаемой природы эпитопа (т.е. линейной или конформационной). Картирование линейных эпитопов является более эффективным и его относительно проще осуществить. Для этой цели в коммерческих службах картирования линейных эпитопов часто используется сканирование пептидов. В этом случае перекрывающийся набор коротких пептидных последовательностей целевого белка химически синтезируется и тестируется на их способность связываться с интересующими антителами. Стратегия является быстрой, высокопроизводительной и относительно недорогой в исполнении. С другой стороны, картирование прерывистого эпитопа является более сложным с технической точки зрения и требует более специализированных методов, таких как совместная рентгеновская кристаллография моноклонального антитела вместе с его белком-мишенью, водородно-дейтериевый обмен (H/D), масс-спектрометрия в комбинации с ферментативным расщеплением, а также несколько других методов, известных специалистам в данной области.
Картирование эпитопов собачьего рецептора альфа CTLA-4 с помощью масс-спектроскопии:
Чтобы определить эпитоп для канинизированного 12B3 (например, 12B3L2H3) и 39A11 (например, 39A11L3H3) на собачьем CTLA-4, каждый из комплексов cCTLA-4/c12B3L2H3 и cCTLA-4/c39A11L2H3 инкубировали с дейтерированными кросс-линкерами и подвергали мультиферментативному расщеплению. После обогащения перекрестно связанных пептидов образцы анализировали масс-спектрометрией высокого разрешения (nLC-LTQ-Orbitrap MS), а полученные данные анализировали с использованием программного обеспечения XQuest и Stavrox.
Анализ показывает, что c12B3L2H3 взаимодействует с аминокислотными остатками в положениях 35, 38, 51, 53, 90, 93, 98 и 102 на cCTLA-4, содержащем аминокислотную последовательность SEQ ID NO: 138 (Фиг.7A); c39A11L2H3 взаимодействует с аминокислотными остатками в положениях 35, 38, 42, 93 и 102 на cCTLA-4, содержащем аминокислотную последовательность SEQ ID NO: 138 (Фиг.7B). Две специфические области собачьего белка CTLA-4 изображены на Фиг.7A и 7B: аминокислотные последовательности SEQ ID NO: 132 и SEQ ID NO: 133, соответственно, (см. Таблицу 8 ниже). Примечательно, что оба антитела связываются с SEQ ID NO: 134 и SEQ ID NO: 136, которые содержат мотив MYPPPY (SEQ ID NO: 137), на cCTLA-4. Мотив MYPPPY формирует петлю связывания с CD80 и CD86, которые являются консервативным мотивом для CTLA-4 у всех видов. c12B3 также, по-видимому, связывается с одной дополнительной областью собачьего CTLA-4, содержащей аминокислотную последовательность SEQ ID NO: 135. В комбинации с результатами примера 4 результаты картирования эпитопа дополнительно подтверждают, что как c12B3, так и c39A11 являются функциональными антителами со способностью блокировать взаимодействие собачьего CTLA-4 с его лигандом CD80 и CD86. Более того, канинизированные антитела, которые связываются с эпитопами в SEQ ID NO: 134 и SEQ ID NO: 136, также являются частью настоящего изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>ИНТЕРВЕТ ИНТЕРНЭШНЛ Б.В.
<120>Антитела против собачьего CTLA-4
<130>24814-US-PCT
<150>US 62/874,287
<151>2019-07-15
<150>US 63/048,873
<151>2020-07-07
<150>US 62/926,047
<151>2019-10-25
<160>138
<170>PatentIn version 3.5
<210>1
<211>348
<212>ДНК
<213>Mus musculus
<400>1
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca aactatggaa tgaactgggt gaagcaggct 120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct acactggaga gccaacatat 180
gctgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgcctat 240
ttgcagatca acaacctcaa aaatgaggac atggctacat atttctgtgc aagacggtca 300
atttattacc cgtactgggg ccaaggcacc actctcacag tctcctca 348
<210>2
<211>116
<212>Белок
<213>Mus musculus
<400>2
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Met Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210>3
<211>348
<212>ДНК
<213>Mus musculus
<400>3
cagatccagt tggtacagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca acctatggag tgagctgggt gaaacaggct 120
ccaggaaagg gtttaaggtg gatgggctgg ataaacacct actctggaat gccaacatat 180
gttgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgccttt 240
ttgcagatca acaacctcaa aaatgaggac acggctatat atttctgtgc aagacggggt 300
atctcctttg actactgggg ccaaggcacc actctcacag tctcctca 348
<210>4
<211>116
<212>Белок
<213>Mus musculus
<400>4
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Val Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Arg Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Ser Gly Met Pro Thr Tyr Val Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Phe
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Ile Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Ile Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210>5
<211>360
<212>ДНК
<213>Mus musculus
<400>5
gaggtgaagc tggtggagtc tggaggaggc ttggtacagc ctgggggttc cctgagtctc 60
tcctgtgcaa cttctggatt caccttcagt gattactaca tgagctgggt ccgccagtct 120
ccggggaagg cacttgagtg gatgggtttt attagaaaca aagctaatgg ttacacaaca 180
gagtacagcg catctctgaa gggtcggttc accatctcca gagataattc ccaaagcatc 240
ctctatcttc aaatgaatgt cctgagagct gaggacagtg ccacttatta ctgtgtaaga 300
tttgggttaa tgtactactt tgactactgg ggccaaggca ccactctcac agtctcctca 360
<210>6
<211>120
<212>Белок
<213>Mus musculus
<400>6
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Ser Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ser Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Val Leu Arg Ala Glu Asp Ser Ala Thr Tyr
85 90 95
Tyr Cys Val Arg Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser
115 120
<210>7
<211>348
<212>ДНК
<213>Mus musculus
<400>7
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca aactatggaa tgaactgggt gaagcaggct 120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct acactggaga gccaacatat 180
gctgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgcctat 240
ttgcagatca acaacctcaa aaatgaggac acggctacat atttctgtgc aagaaggggg 300
acctactata ggccctgggg ccaaggcacc actctcacag tctcctca 348
<210>8
<211>116
<212>Белок
<213>Mus musculus
<400>8
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Thr Tyr Tyr Arg Pro Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210>9
<211>348
<212>ДНК
<213>Mus musculus
<400>9
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctggata taccttcaca aactatggaa tgaactgggt gaagcaggct 120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct acactggaga gccaacatat 180
gctgatgact tcaagggacg ggttgccttc tctttggaaa cctctgccag cactgccttt 240
ttgcagatca acaacctcaa aaatgaggac acggctacat atttctgtgc aaggcggggg 300
gtacgactgg actactgggg ccaaggcacc actctcacag tctcctca 348
<210>10
<211>116
<212>Белок
<213>Mus musculus
<400>10
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Val Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Phe
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Val Arg Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210>11
<211>363
<212>ДНК
<213>Mus musculus
<400>11
caggtccaac tgcagcagcc tgggactgaa ctggtgaagc ctggggcttc agtgaagctg 60
tcctgcaagg cctctggcta taccttcacc agctactgga tgcactgggt gaagcagagg 120
cctggacaag gccttgagtg gattggaaat atcaatccta gcaatggtgg tactaggttc 180
aatgagaagt tcaagaacaa ggccacactg actgaagaca aatcctccag cacagcctac 240
atgcagctca gtagcctgac atctgaggac tctgcggtct attattgtgc aagatcgaac 300
tacggtagtg gctgggcctg gtttgcttac tggggccaag ggactctggt cactgtctct 360
gca 363
<210>12
<211>121
<212>Белок
<213>Mus musculus
<400>12
Gln Val Gln Leu Gln Gln Pro Gly Thr Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Ser Asn Gly Gly Thr Arg Phe Asn Glu Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Glu Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Asn Tyr Gly Ser Gly Trp Ala Trp Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ala
115 120
<210>13
<211>336
<212>ДНК
<213>Mus musculus
<400>13
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gagcattgta tatagtaatg gaaacaccta tttagaatgg
120
tacctgcaga aaccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt
180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc
240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg
300
tggacgttcg gtggaggcac caagctggaa atcaaa 336
<210>14
<211>112
<212>Белок
<213>Mus musculus
<400>14
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210>15
<211>336
<212>ДНК
<213>Mus musculus
<400>15
gatgttttga tgacccagac tccactctcc ctgcctgtca gtcttggaga tcacgcctcc 60
atctcttgca aatctagtca gagcattgta tatattaatg gaaacaccta tttagaatgg 120
tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaagtttc caaacgattt 180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc 240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg 300
tggacgttcg gtggaggcac caagctggaa atcaaa 336
<210>16
<211>112
<212>Белок
<213>Mus musculus
<400>16
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp His Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val Tyr Ile
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210>17
<211>324
<212>ДНК
<213>Mus musculus
<400>17
gaaaatgtgc tcatccagtc tccagcaatc atgtctgctt ctccagggga aaaggtcacc 60
atgacctgca gggccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag 120
tcaggtgcct cccccaaact ctggattttt agcacatcca acttggcttc tggagtccct 180
gctcgcttca gtggcagtgg gtctgggacc tcttattctc tcacaatcaa cagtgtggag 240
gctgaagatg ctgccactta ttactgccag cagtacagtg gtctcccact cacgttcgga 300
ggggggacca agctggaaat aaaa 324
<210>18
<211>108
<212>Белок
<213>Mus musculus
<400>18
Glu Asn Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Ser Gly Ala Ser Pro Lys Leu Trp
35 40 45
Ile Phe Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Asn Ser Val Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Gly Leu Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210>19
<211>336
<212>ДНК
<213>Mus musculus
<400>19
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gagtattgta tatagtcatg gaaacaccta tttagaatgg 120
tacctgcaga aaccaggcca gtctccaaag gtcctgatct acaaagtttc caaccgattt 180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc 240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg 300
tggacgttcg gtggaggcac caagctggaa atcaaa 336
<210>20
<211>112
<212>Белок
<213>Mus musculus
<400>20
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ser
20 25 30
His Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Val Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210>21
<211>336
<212>ДНК
<213>Mus musculus
<400>21
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gagcattgta tatattagtg gaagcaccta tttagaatgg 120
tatctgcaga aaccaggcca gtctccaaag ctcctgatct acaaagtttc cagtcgattt 180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc 240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg 300
tggacgttcg gtggaggcac caagctggaa atcaaa 336
<210>22
<211>112
<212>Белок
<213>Mus musculus
<400>22
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ile
20 25 30
Ser Gly Ser Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Ser Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210>23
<211>321
<212>ДНК
<213>Mus musculus
<400>23
gacatccaga tgaaccagtc tccatccagt ctgtctgcat cccttggaga cacaattacc 60
atcacttgcc atgccagtca gaacattaat gtttggttaa gctggtacca gcagaaacca 120
ggaaatattc ctaaactttt gatctataag tcttccaact tgcacacagg cgtcccatca 180
aggtttagtg gcagtggatc tggaacaggt ttcacattaa ccatcagcag cctgcagcct 240
gaagacattg ccacttacta ctgtcaacag ggtcaaagtt atccgtggac gttcggtgga 300
ggcaccaagc tggaaatcaa a 321
<210>24
<211>107
<212>Белок
<213>Mus musculus
<400>24
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ser Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gly Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Ser Tyr Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210>25
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>25
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca aactatggaa tgaactgggt gaagcaggct
120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct acactggaga gccaacatat
180
gctgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgcctat
240
ttgcagatca acaacctcaa aaatgaggac atggctacat atttctgtgc aagacggtca
300
atttattacc cgtactgggg ccaaggcacc actctcacag tctcctcagc gagcaccacc
360
gcgccgagcg tgtttccgct ggcgccgagc tgcggcagca ccagcggcag caccgtggcg
420
ctggcgtgcc tggtgagcgg ctattttccg gaaccggtga ccgtgagctg gaacagcggc
480
agcctgacca gcggcgtgca tacctttccg agcgtgctgc agagcagcgg cctgtatagc
540
ctgagcagca tggtgaccgt gccgagcagc cgctggccga gcgaaacctt tacctgcaac
600
gtggcgcatc cggcgagcaa aaccaaagtg gataaaccgg tgccgaaacg cgaaaacggc
660
cgcgtgccgc gcccgccgga ttgcccgaaa tgcccggcgc cggaaatgct gggcggcccg
720
agcgtgttta tttttccgcc gaaaccgaaa gataccctgc tgattgcgcg caccccggaa
780
gtgacctgcg tggtggtgga tctggatccg gaagatccgg aagtgcagat tagctggttt
840
gtggatggca aacagatgca gaccgcgaaa acccagccgc gcgaagaaca gtttaacggc
900
acctatcgcg tggtgagcgt gctgccgatt ggccatcagg attggctgaa aggcaaacag
960
tttacctgca aagtgaacaa caaagcgctg ccgagcccga ttgaacgcac cattagcaaa
1020
gcgcgcggcc aggcgcatca gccgagcgtg tatgtgctgc cgccgagccg cgaagaactg
1080
agcaaaaaca ccgtgagcct gacctgcctg attaaagatt tttttccgcc ggatattgat
1140
gtggaatggc agagcaacgg ccagcaggaa ccggaaagca aatatcgcac caccccgccg
1200
cagctggatg aagatggcag ctattttctg tatagcaaac tgagcgtgga taaaagccgc
1260
tggcagcgcg gcgatacctt tatttgcgcg gtgatgcatg aagcgctgca taaccattat
1320
acccaggaaa gcctgagcca tagcccgggc aaa 1353
<210>26
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>26
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Met Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>27
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>27
cagatccagt tggtacagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca acctatggag tgagctgggt gaaacaggct
120
ccaggaaagg gtttaaggtg gatgggctgg ataaacacct actctggaat gccaacatat
180
gttgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgccttt
240
ttgcagatca acaacctcaa aaatgaggac acggctatat atttctgtgc aagacggggt
300
atctcctttg actactgggg ccaaggcacc actctcacag tctcctcagc gagcaccacc
360
gcgccgagcg tgtttccgct ggcgccgagc tgcggcagca ccagcggcag caccgtggcg
420
ctggcgtgcc tggtgagcgg ctattttccg gaaccggtga ccgtgagctg gaacagcggc
480
agcctgacca gcggcgtgca tacctttccg agcgtgctgc agagcagcgg cctgtatagc
540
ctgagcagca tggtgaccgt gccgagcagc cgctggccga gcgaaacctt tacctgcaac
600
gtggcgcatc cggcgagcaa aaccaaagtg gataaaccgg tgccgaaacg cgaaaacggc
660
cgcgtgccgc gcccgccgga ttgcccgaaa tgcccggcgc cggaaatgct gggcggcccg
720
agcgtgttta tttttccgcc gaaaccgaaa gataccctgc tgattgcgcg caccccggaa
780
gtgacctgcg tggtggtgga tctggatccg gaagatccgg aagtgcagat tagctggttt
840
gtggatggca aacagatgca gaccgcgaaa acccagccgc gcgaagaaca gtttaacggc
900
acctatcgcg tggtgagcgt gctgccgatt ggccatcagg attggctgaa aggcaaacag
960
tttacctgca aagtgaacaa caaagcgctg ccgagcccga ttgaacgcac cattagcaaa
1020
gcgcgcggcc aggcgcatca gccgagcgtg tatgtgctgc cgccgagccg cgaagaactg
1080
agcaaaaaca ccgtgagcct gacctgcctg attaaagatt tttttccgcc ggatattgat
1140
gtggaatggc agagcaacgg ccagcaggaa ccggaaagca aatatcgcac caccccgccg
1200
cagctggatg aagatggcag ctattttctg tatagcaaac tgagcgtgga taaaagccgc
1260
tggcagcgcg gcgatacctt tatttgcgcg gtgatgcatg aagcgctgca taaccattat
1320
acccaggaaa gcctgagcca tagcccgggc aaa 1353
<210>28
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>28
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Val Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Arg Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Ser Gly Met Pro Thr Tyr Val Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Phe
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Ile Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Ile Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>29
<211>1365
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>29
gaggtgaagc tggtggagtc tggaggaggc ttggtacagc ctgggggttc cctgagtctc 60
tcctgtgcaa cttctggatt caccttcagt gattactaca tgagctgggt ccgccagtct
120
ccggggaagg cacttgagtg gatgggtttt attagaaaca aagctaatgg ttacacaaca
180
gagtacagcg catctctgaa gggtcggttc accatctcca gagataattc ccaaagcatc
240
ctctatcttc aaatgaatgt cctgagagct gaggacagtg ccacttatta ctgtgtaaga
300
tttgggttaa tgtactactt tgactactgg ggccaaggca ccactctcac agtctcctca
360
gcgagcacca ccgcgccgag cgtgtttccg ctggcgccga gctgcggcag caccagcggc
420
agcaccgtgg cgctggcgtg cctggtgagc ggctattttc cggaaccggt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg catacctttc cgagcgtgct gcagagcagc
540
ggcctgtata gcctgagcag catggtgacc gtgccgagca gccgctggcc gagcgaaacc
600
tttacctgca acgtggcgca tccggcgagc aaaaccaaag tggataaacc ggtgccgaaa
660
cgcgaaaacg gccgcgtgcc gcgcccgccg gattgcccga aatgcccggc gccggaaatg
720
ctgggcggcc cgagcgtgtt tatttttccg ccgaaaccga aagataccct gctgattgcg
780
cgcaccccgg aagtgacctg cgtggtggtg gatctggatc cggaagatcc ggaagtgcag
840
attagctggt ttgtggatgg caaacagatg cagaccgcga aaacccagcc gcgcgaagaa
900
cagtttaacg gcacctatcg cgtggtgagc gtgctgccga ttggccatca ggattggctg
960
aaaggcaaac agtttacctg caaagtgaac aacaaagcgc tgccgagccc gattgaacgc
1020
accattagca aagcgcgcgg ccaggcgcat cagccgagcg tgtatgtgct gccgccgagc
1080
cgcgaagaac tgagcaaaaa caccgtgagc ctgacctgcc tgattaaaga tttttttccg
1140
ccggatattg atgtggaatg gcagagcaac ggccagcagg aaccggaaag caaatatcgc
1200
accaccccgc cgcagctgga tgaagatggc agctattttc tgtatagcaa actgagcgtg
1260
gataaaagcc gctggcagcg cggcgatacc tttatttgcg cggtgatgca tgaagcgctg
1320
cataaccatt atacccagga aagcctgagc catagcccgg gcaaa 1365
<210>30
<211>455
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>30
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Ser Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ser Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Val Leu Arg Ala Glu Asp Ser Ala Thr Tyr
85 90 95
Tyr Cys Val Arg Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210>31
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>31
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca aactatggaa tgaactgggt gaagcaggct
120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct acactggaga gccaacatat
180
gctgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgcctat
240
ttgcagatca acaacctcaa aaatgaggac acggctacat atttctgtgc aagaaggggg
300
acctactata ggccctgggg ccaaggcacc actctcacag tctcctcagc gagcaccacc
360
gcgccgagcg tgtttccgct ggcgccgagc tgcggcagca ccagcggcag caccgtggcg
420
ctggcgtgcc tggtgagcgg ctattttccg gaaccggtga ccgtgagctg gaacagcggc
480
agcctgacca gcggcgtgca tacctttccg agcgtgctgc agagcagcgg cctgtatagc
540
ctgagcagca tggtgaccgt gccgagcagc cgctggccga gcgaaacctt tacctgcaac
600
gtggcgcatc cggcgagcaa aaccaaagtg gataaaccgg tgccgaaacg cgaaaacggc
660
cgcgtgccgc gcccgccgga ttgcccgaaa tgcccggcgc cggaaatgct gggcggcccg
720
agcgtgttta tttttccgcc gaaaccgaaa gataccctgc tgattgcgcg caccccggaa
780
gtgacctgcg tggtggtgga tctggatccg gaagatccgg aagtgcagat tagctggttt
840
gtggatggca aacagatgca gaccgcgaaa acccagccgc gcgaagaaca gtttaacggc
900
acctatcgcg tggtgagcgt gctgccgatt ggccatcagg attggctgaa aggcaaacag
960
tttacctgca aagtgaacaa caaagcgctg ccgagcccga ttgaacgcac cattagcaaa
1020
gcgcgcggcc aggcgcatca gccgagcgtg tatgtgctgc cgccgagccg cgaagaactg
1080
agcaaaaaca ccgtgagcct gacctgcctg attaaagatt tttttccgcc ggatattgat
1140
gtggaatggc agagcaacgg ccagcaggaa ccggaaagca aatatcgcac caccccgccg
1200
cagctggatg aagatggcag ctattttctg tatagcaaac tgagcgtgga taaaagccgc
1260
tggcagcgcg gcgatacctt tatttgcgcg gtgatgcatg aagcgctgca taaccattat
1320
acccaggaaa gcctgagcca tagcccgggc aaa 1353
<210>32
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>32
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Thr Tyr Tyr Arg Pro Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>33
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>33
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctggata taccttcaca aactatggaa tgaactgggt gaagcaggct
120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct acactggaga gccaacatat
180
gctgatgact tcaagggacg ggttgccttc tctttggaaa cctctgccag cactgccttt
240
ttgcagatca acaacctcaa aaatgaggac acggctacat atttctgtgc aaggcggggg
300
gtacgactgg actactgggg ccaaggcacc actctcacag tctcctcagc gagcaccacc
360
gcgccgagcg tgtttccgct ggcgccgagc tgcggcagca ccagcggcag caccgtggcg
420
ctggcgtgcc tggtgagcgg ctattttccg gaaccggtga ccgtgagctg gaacagcggc
480
agcctgacca gcggcgtgca tacctttccg agcgtgctgc agagcagcgg cctgtatagc
540
ctgagcagca tggtgaccgt gccgagcagc cgctggccga gcgaaacctt tacctgcaac
600
gtggcgcatc cggcgagcaa aaccaaagtg gataaaccgg tgccgaaacg cgaaaacggc
660
cgcgtgccgc gcccgccgga ttgcccgaaa tgcccggcgc cggaaatgct gggcggcccg
720
agcgtgttta tttttccgcc gaaaccgaaa gataccctgc tgattgcgcg caccccggaa
780
gtgacctgcg tggtggtgga tctggatccg gaagatccgg aagtgcagat tagctggttt
840
gtggatggca aacagatgca gaccgcgaaa acccagccgc gcgaagaaca gtttaacggc
900
acctatcgcg tggtgagcgt gctgccgatt ggccatcagg attggctgaa aggcaaacag
960
tttacctgca aagtgaacaa caaagcgctg ccgagcccga ttgaacgcac cattagcaaa
1020
gcgcgcggcc aggcgcatca gccgagcgtg tatgtgctgc cgccgagccg cgaagaactg
1080
agcaaaaaca ccgtgagcct gacctgcctg attaaagatt tttttccgcc ggatattgat
1140
gtggaatggc agagcaacgg ccagcaggaa ccggaaagca aatatcgcac caccccgccg
1200
cagctggatg aagatggcag ctattttctg tatagcaaac tgagcgtgga taaaagccgc
1260
tggcagcgcg gcgatacctt tatttgcgcg gtgatgcatg aagcgctgca taaccattat
1320
acccaggaaa gcctgagcca tagcccgggc aaa 1353
<210>34
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>34
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Val Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Phe
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Arg Gly Val Arg Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>35
<211>1368
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>35
caggtccaac tgcagcagcc tgggactgaa ctggtgaagc ctggggcttc agtgaagctg 60
tcctgcaagg cctctggcta taccttcacc agctactgga tgcactgggt gaagcagagg
120
cctggacaag gccttgagtg gattggaaat atcaatccta gcaatggtgg tactaggttc
180
aatgagaagt tcaagaacaa ggccacactg actgaagaca aatcctccag cacagcctac
240
atgcagctca gtagcctgac atctgaggac tctgcggtct attattgtgc aagatcgaac
300
tacggtagtg gctgggcctg gtttgcttac tggggccaag ggactctggt cactgtctct
360
gcagcgagca ccaccgcgcc gagcgtgttt ccgctggcgc cgagctgcgg cagcaccagc
420
ggcagcaccg tggcgctggc gtgcctggtg agcggctatt ttccggaacc ggtgaccgtg
480
agctggaaca gcggcagcct gaccagcggc gtgcatacct ttccgagcgt gctgcagagc
540
agcggcctgt atagcctgag cagcatggtg accgtgccga gcagccgctg gccgagcgaa
600
acctttacct gcaacgtggc gcatccggcg agcaaaacca aagtggataa accggtgccg
660
aaacgcgaaa acggccgcgt gccgcgcccg ccggattgcc cgaaatgccc ggcgccggaa
720
atgctgggcg gcccgagcgt gtttattttt ccgccgaaac cgaaagatac cctgctgatt
780
gcgcgcaccc cggaagtgac ctgcgtggtg gtggatctgg atccggaaga tccggaagtg
840
cagattagct ggtttgtgga tggcaaacag atgcagaccg cgaaaaccca gccgcgcgaa
900
gaacagttta acggcaccta tcgcgtggtg agcgtgctgc cgattggcca tcaggattgg
960
ctgaaaggca aacagtttac ctgcaaagtg aacaacaaag cgctgccgag cccgattgaa
1020
cgcaccatta gcaaagcgcg cggccaggcg catcagccga gcgtgtatgt gctgccgccg
1080
agccgcgaag aactgagcaa aaacaccgtg agcctgacct gcctgattaa agattttttt
1140
ccgccggata ttgatgtgga atggcagagc aacggccagc aggaaccgga aagcaaatat
1200
cgcaccaccc cgccgcagct ggatgaagat ggcagctatt ttctgtatag caaactgagc
1260
gtggataaaa gccgctggca gcgcggcgat acctttattt gcgcggtgat gcatgaagcg
1320
ctgcataacc attataccca ggaaagcctg agccatagcc cgggcaaa 1368
<210>36
<211>456
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>36
Gln Val Gln Leu Gln Gln Pro Gly Thr Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Ser Asn Gly Gly Thr Arg Phe Asn Glu Lys Phe
50 55 60
Lys Asn Lys Ala Thr Leu Thr Glu Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Asn Tyr Gly Ser Gly Trp Ala Trp Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ala Ala Ser Thr Thr Ala Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val
130 135 140
Ala Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val
180 185 190
Pro Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His
195 200 205
Pro Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn
210 215 220
Gly Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu
225 230 235 240
Met Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Leu Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly
275 280 285
Lys Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn
290 295 300
Gly Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp
305 310 315 320
Leu Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro
325 330 335
Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln
340 345 350
Pro Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn
355 360 365
Thr Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile
370 375 380
Asp Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr
385 390 395 400
Arg Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr
405 410 415
Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe
420 425 430
Ile Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu
435 440 445
Ser Leu Ser His Ser Pro Gly Lys
450 455
<210>37
<211>669
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>37
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gagcattgta tatagtaatg gaaacaccta tttagaatgg
120
tacctgcaga aaccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt
180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc
240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg
300
tggacgttcg gtggaggcac caagctggaa atcaaacgca acgatgcgca gccggcggtg
360
tatctgtttc agccgagccc ggatcagctg cataccggca gcgcgagcgt ggtgtgcctg
420
ctgaacagct tttatccgaa agatattaac gtgaaatgga aagtggatgg cgtgattcag
480
gataccggca ttcaggaaag cgtgaccgaa caggatagca aagatagcac ctatagcctg
540
agcagcaccc tgaccatgag cagcaccgaa tatctgagcc atgaactgta tagctgcgaa
600
attacccata aaagcctgcc gagcaccctg attaaaagct ttcagcgcag cgaatgccag
660
cgcgtggat 669
<210>38
<211>223
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>38
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
115 120 125
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
145 150 155 160
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu
180 185 190
Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser
195 200 205
Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210>39
<211>669
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>39
gatgttttga tgacccagac tccactctcc ctgcctgtca gtcttggaga tcacgcctcc 60
atctcttgca aatctagtca gagcattgta tatattaatg gaaacaccta tttagaatgg
120
tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaagtttc caaacgattt
180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc
240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg
300
tggacgttcg gtggaggcac caagctggaa atcaaacgca acgatgcgca gccggcggtg
360
tatctgtttc agccgagccc ggatcagctg cataccggca gcgcgagcgt ggtgtgcctg
420
ctgaacagct tttatccgaa agatattaac gtgaaatgga aagtggatgg cgtgattcag
480
gataccggca ttcaggaaag cgtgaccgaa caggatagca aagatagcac ctatagcctg
540
agcagcaccc tgaccatgag cagcaccgaa tatctgagcc atgaactgta tagctgcgaa
600
attacccata aaagcctgcc gagcaccctg attaaaagct ttcagcgcag cgaatgccag
660
cgcgtggat 669
<210>40
<211>223
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>40
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp His Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Ile Val Tyr Ile
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
115 120 125
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
145 150 155 160
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu
180 185 190
Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser
195 200 205
Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210>41
<211>657
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>41
gaaaatgtgc tcatccagtc tccagcaatc atgtctgctt ctccagggga aaaggtcacc 60
atgacctgca gggccagctc aagtgtaagt tccagttact tgcactggta ccagcagaag
120
tcaggtgcct cccccaaact ctggattttt agcacatcca acttggcttc tggagtccct
180
gctcgcttca gtggcagtgg gtctgggacc tcttattctc tcacaatcaa cagtgtggag
240
gctgaagatg ctgccactta ttactgccag cagtacagtg gtctcccact cacgttcgga
300
ggggggacca agctggaaat aaaacgcaac gatgcgcagc cggcggtgta tctgtttcag
360
ccgagcccgg atcagctgca taccggcagc gcgagcgtgg tgtgcctgct gaacagcttt
420
tatccgaaag atattaacgt gaaatggaaa gtggatggcg tgattcagga taccggcatt
480
caggaaagcg tgaccgaaca ggatagcaaa gatagcacct atagcctgag cagcaccctg
540
accatgagca gcaccgaata tctgagccat gaactgtata gctgcgaaat tacccataaa
600
agcctgccga gcaccctgat taaaagcttt cagcgcagcg aatgccagcg cgtggat 657
<210>42
<211>219
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>42
Glu Asn Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Ser Gly Ala Ser Pro Lys Leu Trp
35 40 45
Ile Phe Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Asn Ser Val Glu
65 70 75 80
Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Gly Leu Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Asn Asp Ala
100 105 110
Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr
115 120 125
Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu
180 185 190
Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys
195 200 205
Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210>43
<211>669
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>43
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gagtattgta tatagtcatg gaaacaccta tttagaatgg
120
tacctgcaga aaccaggcca gtctccaaag gtcctgatct acaaagtttc caaccgattt
180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc
240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg
300
tggacgttcg gtggaggcac caagctggaa atcaaacgca acgatgcgca gccggcggtg
360
tatctgtttc agccgagccc ggatcagctg cataccggca gcgcgagcgt ggtgtgcctg
420
ctgaacagct tttatccgaa agatattaac gtgaaatgga aagtggatgg cgtgattcag
480
gataccggca ttcaggaaag cgtgaccgaa caggatagca aagatagcac ctatagcctg
540
agcagcaccc tgaccatgag cagcaccgaa tatctgagcc atgaactgta tagctgcgaa
600
attacccata aaagcctgcc gagcaccctg attaaaagct ttcagcgcag cgaatgccag
660
cgcgtggat 669
<210>44
<211>223
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>44
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ser
20 25 30
His Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Val Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
115 120 125
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
145 150 155 160
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu
180 185 190
Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser
195 200 205
Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210>45
<211>669
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>45
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gagcattgta tatattagtg gaagcaccta tttagaatgg
120
tatctgcaga aaccaggcca gtctccaaag ctcctgatct acaaagtttc cagtcgattt
180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc
240
agcagagtgg aggctgagga tctgggagtt tattactgct ttcaaggttc acatgttccg
300
tggacgttcg gtggaggcac caagctggaa atcaaacgca acgatgcgca gccggcggtg
360
tatctgtttc agccgagccc ggatcagctg cataccggca gcgcgagcgt ggtgtgcctg
420
ctgaacagct tttatccgaa agatattaac gtgaaatgga aagtggatgg cgtgattcag
480
gataccggca ttcaggaaag cgtgaccgaa caggatagca aagatagcac ctatagcctg
540
agcagcaccc tgaccatgag cagcaccgaa tatctgagcc atgaactgta tagctgcgaa
600
attacccata aaagcctgcc gagcaccctg attaaaagct ttcagcgcag cgaatgccag
660
cgcgtggat 669
<210>46
<211>223
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>46
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ile
20 25 30
Ser Gly Ser Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Ser Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
115 120 125
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
145 150 155 160
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu
180 185 190
Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser
195 200 205
Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210>47
<211>654
<212>ДНК
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>47
gacatccaga tgaaccagtc tccatccagt ctgtctgcat cccttggaga cacaattacc 60
atcacttgcc atgccagtca gaacattaat gtttggttaa gctggtacca gcagaaacca
120
ggaaatattc ctaaactttt gatctataag tcttccaact tgcacacagg cgtcccatca
180
aggtttagtg gcagtggatc tggaacaggt ttcacattaa ccatcagcag cctgcagcct
240
gaagacattg ccacttacta ctgtcaacag ggtcaaagtt atccgtggac gttcggtgga
300
ggcaccaagc tggaaatcaa acgcaacgat gcgcagccgg cggtgtatct gtttcagccg
360
agcccggatc agctgcatac cggcagcgcg agcgtggtgt gcctgctgaa cagcttttat
420
ccgaaagata ttaacgtgaa atggaaagtg gatggcgtga ttcaggatac cggcattcag
480
gaaagcgtga ccgaacagga tagcaaagat agcacctata gcctgagcag caccctgacc
540
atgagcagca ccgaatatct gagccatgaa ctgtatagct gcgaaattac ccataaaagc
600
ctgccgagca ccctgattaa aagctttcag cgcagcgaat gccagcgcgt ggat 654
<210>48
<211>218
<212>Белок
<213>Искусственная последовательность
<220>
<223>mus musculus-canis familiaris, химерная
<400>48
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ser Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gly Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Ser Tyr Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Asn Asp Ala Gln
100 105 110
Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr Gly
115 120 125
Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu Tyr
180 185 190
Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys Ser
195 200 205
Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210>49
<211>669
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>49
gatattgtga tgacccagac cccgctgagc ctgagcgtga gcccgggcga accggcgagc 60
attagctgcc gcagcagcca gagcattgtg tatagcaacg gcaacaccta tctggaatgg
120
tttcagcaga aaccgggcca gagcccgcag cgcctgattt ataaagtgag caaccgcttt
180
agcggcgtgc cggatcgctt tagcggcagc ggcagcggca ccgattttac cctgcgcatt
240
agccgcgtgg aagcggatga tgcgggcgtg tattattgct ttcagggcag ccatgtgccg
300
tggacctttg gcggcggcac caaactggaa attaaaagga acgacgctca gccagccgtg
360
tacctcttcc agccttcgcc ggaccagctt catacggggt cagcgtcggt ggtgtgcctg
420
ttgaactcgt tttaccccaa ggacattaac gtgaagtgga aggtagacgg ggtaattcaa
480
gacactggca ttcaagagtc cgtcacggaa caagactcaa aagactcaac gtattcactg
540
tcgtcaacct tgacgatgtc aagcaccgag tatcttagcc atgagctgta ttcgtgcgag
600
atcacccaca agtccctccc ctccactctt atcaaatcct ttcagcggtc ggaatgtcag
660
cgggtcgat 669
<210>50
<211>223
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>50
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Phe Gln Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Arg Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
115 120 125
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
145 150 155 160
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu
180 185 190
Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser
195 200 205
Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210>51
<211>669
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>51
gatattgtga tgacccagac cccgctgagc ctgagcgtga gcccgggcga accggcgagc 60
attagctgcc gcagcagcca gagcattgtg tatagcaacg gcaacaccta tctggaatgg
120
tatcagcaga aaccgggcca gagcccgaaa ctgctgattt ataaagtgag caaccgcttt
180
agcggcgtgc cggatcgctt tagcggcagc ggcagcggca ccgattttac cctgcgcatt
240
agccgcgtgg aagcggatga tgcgggcgtg tattattgct ttcagggcag ccatgtgccg
300
tggacctttg gcggcggcac caaactggaa attaaaagga acgacgctca gccagccgtg
360
tacctcttcc agccttcgcc ggaccagctt catacggggt cagcgtcggt ggtgtgcctg
420
ttgaactcgt tttaccccaa ggacattaac gtgaagtgga aggtagacgg ggtaattcaa
480
gacactggca ttcaagagtc cgtcacggaa caagactcaa aagactcaac gtattcactg
540
tcgtcaacct tgacgatgtc aagcaccgag tatcttagcc atgagctgta ttcgtgcgag
600
atcacccaca agtccctccc ctccactctt atcaaatcct ttcagcggtc ggaatgtcag
660
cgggtcgat 669
<210>52
<211>223
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>52
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Gln Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
115 120 125
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
145 150 155 160
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu
180 185 190
Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser
195 200 205
Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210>53
<211>669
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>53
gatgtgctga tgacccagac cccgctgagc ctgagcgtga gcccgggcga accggcgagc 60
attagctgcc gcagcagcca gagcattgtg tatagcaacg gcaacaccta tctggaatgg
120
tatctgcaga aaccgggcca gagcccgaaa ctgctgattt ataaagtgag caaccgcttt
180
agcggcgtgc cggatcgctt tagcggcagc ggcagcggca ccgattttac cctgcgcatt
240
agccgcgtgg aagcggatga tgcgggcgtg tattattgct ttcagggcag ccatgtgccg
300
tggacctttg gcggcggcac caaactggaa ctgaaaagga acgacgctca gccagccgtg
360
tacctcttcc agccttcgcc ggaccagctt catacggggt cagcgtcggt ggtgtgcctg
420
ttgaactcgt tttaccccaa ggacattaac gtgaagtgga aggtagacgg ggtaattcaa
480
gacactggca ttcaagagtc cgtcacggaa caagactcaa aagactcaac gtattcactg
540
tcgtcaacct tgacgatgtc aagcaccgag tatcttagcc atgagctgta ttcgtgcgag
600
atcacccaca agtccctccc ctccactctt atcaaatcct ttcagcggtc ggaatgtcag
660
cgggtcgat 669
<210>54
<211>223
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>54
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val Tyr Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105 110
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
115 120 125
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
145 150 155 160
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu
180 185 190
Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser
195 200 205
Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210>55
<211>657
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>55
gaaattgtga tgacccagag cccggcgagc ctgagcctga gccaggaaga aaaagtgacc 60
attacctgcc gcgcgagcag cagcgtgagc agcagctatc tgcattggta tcagcagaaa
120
ccgggccagg cgccgaaact gctgatttat agcaccagca acctggcgag cggcgtgccg
180
agccgcttta gcggcagcgg cagcggcacc gattttagct ttaccattag cagcctggaa
240
ccggaagatg tggcggtgta ttattgccag cagtatagcg gcctgccgct gacctttggc
300
ggcggcacca aactggaaat taaaaggaac gacgctcagc cagccgtgta cctcttccag
360
ccttcgccgg accagcttca tacggggtca gcgtcggtgg tgtgcctgtt gaactcgttt
420
taccccaagg acattaacgt gaagtggaag gtagacgggg taattcaaga cactggcatt
480
caagagtccg tcacggaaca agactcaaaa gactcaacgt attcactgtc gtcaaccttg
540
acgatgtcaa gcaccgagta tcttagccat gagctgtatt cgtgcgagat cacccacaag
600
tccctcccct ccactcttat caaatccttt cagcggtcgg aatgtcagcg ggtcgat 657
<210>56
<211>219
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>56
Glu Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ser Leu Ser Gln Glu
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Ser Phe Thr Ile Ser Ser Leu Glu
65 70 75 80
Pro Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Tyr Ser Gly Leu Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Asn Asp Ala
100 105 110
Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr
115 120 125
Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu
180 185 190
Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys
195 200 205
Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210>57
<211>657
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>57
gaaaacgtgc tgacccagag cccggcgagc ctgagcctga gccaggaaga aaaagtgacc 60
attacctgcc gcgcgagcag cagcgtgagc agcagctatc tgcattggta tcagcagaaa
120
ccgggccagg cgccgaaact gtggattttt agcaccagca acctggcgag cggcgtgccg
180
agccgcttta gcggcagcgg cagcggcacc gattatagct ttaccattag cagcctggaa
240
ccggaagatg tggcggtgta ttattgccag cagtatagcg gcctgccgct gacctttggc
300
ggcggcacca aactggaact gaaaaggaac gacgctcagc cagccgtgta cctcttccag
360
ccttcgccgg accagcttca tacggggtca gcgtcggtgg tgtgcctgtt gaactcgttt
420
taccccaagg acattaacgt gaagtggaag gtagacgggg taattcaaga cactggcatt
480
caagagtccg tcacggaaca agactcaaaa gactcaacgt attcactgtc gtcaaccttg
540
acgatgtcaa gcaccgagta tcttagccat gagctgtatt cgtgcgagat cacccacaag
600
tccctcccct ccactcttat caaatccttt cagcggtcgg aatgtcagcg ggtcgat 657
<210>58
<211>219
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>58
Glu Asn Val Leu Thr Gln Ser Pro Ala Ser Leu Ser Leu Ser Gln Glu
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Leu Trp
35 40 45
Ile Phe Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Tyr Ser Phe Thr Ile Ser Ser Leu Glu
65 70 75 80
Pro Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Tyr Ser Gly Leu Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys Arg Asn Asp Ala
100 105 110
Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr
115 120 125
Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu
180 185 190
Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys
195 200 205
Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210>59
<211>657
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>59
gaaaacgtgc tgacccagag cccggcgagc ctgagcctga gcccgggcga aaaagtgacc 60
attacctgcc gcgcgagcag cagcgtgagc agcagctatc tgcattggta tcagcagaaa
120
ccgggccaga gcccgaaact gtggattttt agcaccagca acctggcgag cggcgtgccg
180
agccgcttta gcggcagcgg cagcggcacc agctatagct ttaccattag cagcctggaa
240
ccggaagatg tggcggtgta ttattgccag cagtatagcg gcctgccgct gacctttggc
300
ggcggcacca aactggaact gaaaaggaac gacgctcagc cagccgtgta cctcttccag
360
ccttcgccgg accagcttca tacggggtca gcgtcggtgg tgtgcctgtt gaactcgttt
420
taccccaagg acattaacgt gaagtggaag gtagacgggg taattcaaga cactggcatt
480
caagagtccg tcacggaaca agactcaaaa gactcaacgt attcactgtc gtcaaccttg
540
acgatgtcaa gcaccgagta tcttagccat gagctgtatt cgtgcgagat cacccacaag
600
tccctcccct ccactcttat caaatccttt cagcggtcgg aatgtcagcg ggtcgat 657
<210>60
<211>219
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>60
Glu Asn Val Leu Thr Gln Ser Pro Ala Ser Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Ser Ser
20 25 30
Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Trp
35 40 45
Ile Phe Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Ser Tyr Ser Phe Thr Ile Ser Ser Leu Glu
65 70 75 80
Pro Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Tyr Ser Gly Leu Pro
85 90 95
Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys Arg Asn Asp Ala
100 105 110
Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr
115 120 125
Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp
130 135 140
Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu
180 185 190
Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys
195 200 205
Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210>61
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>61
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgtgg cgagcggcta tacctttacc aactatggca tgaactgggt gcgccaggcg
120
ccgggcaaag gcctgcagtg ggtggcgtgg attaacacct ataccggcga accgacctat
180
gcggatgatt ttaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gcgccgcagc
300
atttattatc cgtattgggg ccagggcacc accctgaccg tgagcagcgc ttccacaacc
360
gcgccatcag tctttccgtt ggccccatca tgcgggtcga cgagcggatc gactgtggcc
420
ctggcgtgct tggtgtcggg atactttccc gaacccgtca cggtcagctg gaactccgga
480
tcgcttacga gcggtgtgca tacgttcccc tcggtcttgc aatcatcagg gctctactcg
540
ctgtcgagca tggtaacggt gccctcatcg aggtggccct ccgaaacgtt cacatgtaac
600
gtagcacatc cagcctccaa aaccaaggtg gataaacccg tgccgaaaag agagaatggg
660
cgggtgcctc gaccccctga ttgccccaag tgtccggctc cggaaatgct cggtggaccc
720
tcagtgttta tcttccctcc gaagcccaag gacactctgc tgatcgcgcg cactccagaa
780
gtaacatgtg tagtggtgga ccttgatccc gaggaccccg aagtccagat ctcctggttt
840
gtagatggga aacagatgca gaccgcaaaa actcaaccca gagaggagca gttcaacgga
900
acataccgag tggtatccgt ccttccgatt ggccaccagg actggttgaa agggaagcag
960
tttacgtgta aagtcaacaa taaggggttg cctagcccta ttgagcggac gatttcgaaa
1020
gctaggggac aggcccacca gccatcggtc tatgtccttc cgccttcccg cgaggagctc
1080
tcgaagaata cagtgagcct tacatgcctc attaaggatt tcttcccgcc tgatatcgac
1140
gtagagtggc aatcaaacgg tcaacaggag ccggaatcca agtatagaac cactccgccc
1200
cagcttgacg aggacggatc atactttttg tattcaaaac tgtcggtgga taagagccgg
1260
tggcagagag gtgacacctt catctgtgcg gtgatgcacg aagcactcca taatcactac
1320
acccaagaga gcctctcgca ttcccccgga aag 1353
<210>62
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>62
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>63
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>63
gaaattcagc tggtgcagag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcaaag cgagcggcta tacctttacc aactatggca tgaactgggt gcgccaggcg
120
ccgggcaaag gcctgcagtg gatgggctgg attaacacct ataccggcga accgacctat
180
gcggatgatt ttaaaggccg ctttaccttt agcctggata acgcgaaaaa caccctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt atttttgcgc gcgccgcagc
300
atttattatc cgtattgggg ccagggcacc accctgaccg tgagcagcgc ttccacaacc
360
gcgccatcag tctttccgtt ggccccatca tgcgggtcga cgagcggatc gactgtggcc
420
ctggcgtgct tggtgtcggg atactttccc gaacccgtca cggtcagctg gaactccgga
480
tcgcttacga gcggtgtgca tacgttcccc tcggtcttgc aatcatcagg gctctactcg
540
ctgtcgagca tggtaacggt gccctcatcg aggtggccct ccgaaacgtt cacatgtaac
600
gtagcacatc cagcctccaa aaccaaggtg gataaacccg tgccgaaaag agagaatggg
660
cgggtgcctc gaccccctga ttgccccaag tgtccggctc cggaaatgct cggtggaccc
720
tcagtgttta tcttccctcc gaagcccaag gacactctgc tgatcgcgcg cactccagaa
780
gtaacatgtg tagtggtgga ccttgatccc gaggaccccg aagtccagat ctcctggttt
840
gtagatggga aacagatgca gaccgcaaaa actcaaccca gagaggagca gttcaacgga
900
acataccgag tggtatccgt ccttccgatt ggccaccagg actggttgaa agggaagcag
960
tttacgtgta aagtcaacaa taaggggttg cctagcccta ttgagcggac gatttcgaaa
1020
gctaggggac aggcccacca gccatcggtc tatgtccttc cgccttcccg cgaggagctc
1080
tcgaagaata cagtgagcct tacatgcctc attaaggatt tcttcccgcc tgatatcgac
1140
gtagagtggc aatcaaacgg tcaacaggag ccggaatcca agtatagaac cactccgccc
1200
cagcttgacg aggacggatc atactttttg tattcaaaac tgtcggtgga taagagccgg
1260
tggcagagag gtgacacctt catctgtgcg gtgatgcacg aagcactcca taatcactac
1320
acccaagaga gcctctcgca ttcccccgga aag 1353
<210>64
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>64
Glu Ile Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>65
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>65
gaaattcagc tggtgcagag cggcggcgat ctggtgaaac cgggcggcag cgtgcgcctg 60
agctgcaaag cgagcggcta tacctttacc aactatggca tgaactgggt gaaacaggcg
120
ccgggcaaag gcctgcagtg gatgggctgg attaacacct ataccggcga accgacctat
180
gcggatgatt ttaaaggccg ctttaccttt agcctggata acgcgaaaaa caccgcgtat
240
ctgcagatta acagcctgcg cgcggaagat accgcggtgt atttttgcgc gcgccgcagc
300
atttattatc cgtattgggg ccagggcacc accctgaccg tgagcagcgc ttccacaacc
360
gcgccatcag tctttccgtt ggccccatca tgcgggtcga cgagcggatc gactgtggcc
420
ctggcgtgct tggtgtcggg atactttccc gaacccgtca cggtcagctg gaactccgga
480
tcgcttacga gcggtgtgca tacgttcccc tcggtcttgc aatcatcagg gctctactcg
540
ctgtcgagca tggtaacggt gccctcatcg aggtggccct ccgaaacgtt cacatgtaac
600
gtagcacatc cagcctccaa aaccaaggtg gataaacccg tgccgaaaag agagaatggg
660
cgggtgcctc gaccccctga ttgccccaag tgtccggctc cggaaatgct cggtggaccc
720
tcagtgttta tcttccctcc gaagcccaag gacactctgc tgatcgcgcg cactccagaa
780
gtaacatgtg tagtggtgga ccttgatccc gaggaccccg aagtccagat ctcctggttt
840
gtagatggga aacagatgca gaccgcaaaa actcaaccca gagaggagca gttcaacgga
900
acataccgag tggtatccgt ccttccgatt ggccaccagg actggttgaa agggaagcag
960
tttacgtgta aagtcaacaa taaggggttg cctagcccta ttgagcggac gatttcgaaa
1020
gctaggggac aggcccacca gccatcggtc tatgtccttc cgccttcccg cgaggagctc
1080
tcgaagaata cagtgagcct tacatgcctc attaaggatt tcttcccgcc tgatatcgac
1140
gtagagtggc aatcaaacgg tcaacaggag ccggaatcca agtatagaac cactccgccc
1200
cagcttgacg aggacggatc atactttttg tattcaaaac tgtcggtgga taagagccgg
1260
tggcagagag gtgacacctt catctgtgcg gtgatgcacg aagcactcca taatcactac
1320
acccaagaga gcctctcgca ttcccccgga aag 1353
<210>66
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>66
Glu Ile Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Asn Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>67
<211>1365
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>67
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgtgg cgagcggctt tacctttagc gattattata tgagctgggt gcgccaggcg
120
ccgggcaaag gcctggaatg ggtggcgttt attcgcaaca aagcgaacgg ctataccacc
180
gaatatagcg cgagcctgaa aggccgcttt accattagcc gcgataacgc gaaaaacatg
240
gcgtatctgc agatgaacag cctgcgcgcg gaagataccg cggtgtatta ttgcgcgagc
300
tttggcctga tgtattattt tgattattgg ggccagggca ccaccctgac cgtgagcagc
360
gcttccacaa ccgcgccatc agtctttccg ttggccccat catgcgggtc gacgagcgga
420
tcgactgtgg ccctggcgtg cttggtgtcg ggatactttc ccgaacccgt cacggtcagc
480
tggaactccg gatcgcttac gagcggtgtg catacgttcc cctcggtctt gcaatcatca
540
gggctctact cgctgtcgag catggtaacg gtgccctcat cgaggtggcc ctccgaaacg
600
ttcacatgta acgtagcaca tccagcctcc aaaaccaagg tggataaacc cgtgccgaaa
660
agagagaatg ggcgggtgcc tcgaccccct gattgcccca agtgtccggc tccggaaatg
720
ctcggtggac cctcagtgtt tatcttccct ccgaagccca aggacactct gctgatcgcg
780
cgcactccag aagtaacatg tgtagtggtg gaccttgatc ccgaggaccc cgaagtccag
840
atctcctggt ttgtagatgg gaaacagatg cagaccgcaa aaactcaacc cagagaggag
900
cagttcaacg gaacataccg agtggtatcc gtccttccga ttggccacca ggactggttg
960
aaagggaagc agtttacgtg taaagtcaac aataaggggt tgcctagccc tattgagcgg
1020
acgatttcga aagctagggg acaggcccac cagccatcgg tctatgtcct tccgccttcc
1080
cgcgaggagc tctcgaagaa tacagtgagc cttacatgcc tcattaagga tttcttcccg
1140
cctgatatcg acgtagagtg gcaatcaaac ggtcaacagg agccggaatc caagtataga
1200
accactccgc cccagcttga cgaggacgga tcatactttt tgtattcaaa actgtcggtg
1260
gataagagcc ggtggcagag aggtgacacc ttcatctgtg cggtgatgca cgaagcactc
1320
cataatcact acacccaaga gagcctctcg cattcccccg gaaag 1365
<210>68
<211>455
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>68
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Ser Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210>69
<211>1365
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>69
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgcga ccagcggctt tacctttagc gattattata tgagctgggt gcgccaggcg
120
ccgggcaaag gcctggaatg gatgggcttt attcgcaaca aagcgaacgg ctataccacc
180
gaatatagcg cgagcctgaa aggccgcttt accattagcc gcgataacgc gaaaaacatg
240
gcgtatctgc agatgaacag cctgcgcgcg gaagataccg cggtgtatta ttgcgtgcgc
300
tttggcctga tgtattattt tgattattgg ggccagggca ccaccctgac cgtgagcagc
360
gcttccacaa ccgcgccatc agtctttccg ttggccccat catgcgggtc gacgagcgga
420
tcgactgtgg ccctggcgtg cttggtgtcg ggatactttc ccgaacccgt cacggtcagc
480
tggaactccg gatcgcttac gagcggtgtg catacgttcc cctcggtctt gcaatcatca
540
gggctctact cgctgtcgag catggtaacg gtgccctcat cgaggtggcc ctccgaaacg
600
ttcacatgta acgtagcaca tccagcctcc aaaaccaagg tggataaacc cgtgccgaaa
660
agagagaatg ggcgggtgcc tcgaccccct gattgcccca agtgtccggc tccggaaatg
720
ctcggtggac cctcagtgtt tatcttccct ccgaagccca aggacactct gctgatcgcg
780
cgcactccag aagtaacatg tgtagtggtg gaccttgatc ccgaggaccc cgaagtccag
840
atctcctggt ttgtagatgg gaaacagatg cagaccgcaa aaactcaacc cagagaggag
900
cagttcaacg gaacataccg agtggtatcc gtccttccga ttggccacca ggactggttg
960
aaagggaagc agtttacgtg taaagtcaac aataaggggt tgcctagccc tattgagcgg
1020
acgatttcga aagctagggg acaggcccac cagccatcgg tctatgtcct tccgccttcc
1080
cgcgaggagc tctcgaagaa tacagtgagc cttacatgcc tcattaagga tttcttcccg
1140
cctgatatcg acgtagagtg gcaatcaaac ggtcaacagg agccggaatc caagtataga
1200
accactccgc cccagcttga cgaggacgga tcatactttt tgtattcaaa actgtcggtg
1260
gataagagcc ggtggcagag aggtgacacc ttcatctgtg cggtgatgca cgaagcactc
1320
cataatcact acacccaaga gagcctctcg cattcccccg gaaag 1365
<210>70
<211>455
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>70
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210>71
<211>1365
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>71
gaagtgaaac tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgcga ccagcggctt tacctttagc gattattata tgagctgggt gcgccaggcg
120
ccgggcaaag cgctggaatg gatgggcttt attcgcaaca aagcgaacgg ctataccacc
180
gaatatagcg cgagcctgaa aggccgcttt accattagcc gcgataacgc gaaaaacatg
240
ctgtatctgc agatgaacag cctgcgcgcg gaagataccg cggtgtatta ttgcgtgcgc
300
tttggcctga tgtattattt tgattattgg ggccagggca ccaccctgac cgtgagcagc
360
gcttccacaa ccgcgccatc agtctttccg ttggccccat catgcgggtc gacgagcgga
420
tcgactgtgg ccctggcgtg cttggtgtcg ggatactttc ccgaacccgt cacggtcagc
480
tggaactccg gatcgcttac gagcggtgtg catacgttcc cctcggtctt gcaatcatca
540
gggctctact cgctgtcgag catggtaacg gtgccctcat cgaggtggcc ctccgaaacg
600
ttcacatgta acgtagcaca tccagcctcc aaaaccaagg tggataaacc cgtgccgaaa
660
agagagaatg ggcgggtgcc tcgaccccct gattgcccca agtgtccggc tccggaaatg
720
ctcggtggac cctcagtgtt tatcttccct ccgaagccca aggacactct gctgatcgcg
780
cgcactccag aagtaacatg tgtagtggtg gaccttgatc ccgaggaccc cgaagtccag
840
atctcctggt ttgtagatgg gaaacagatg cagaccgcaa aaactcaacc cagagaggag
900
cagttcaacg gaacataccg agtggtatcc gtccttccga ttggccacca ggactggttg
960
aaagggaagc agtttacgtg taaagtcaac aataaggggt tgcctagccc tattgagcgg
1020
acgatttcga aagctagggg acaggcccac cagccatcgg tctatgtcct tccgccttcc
1080
cgcgaggagc tctcgaagaa tacagtgagc cttacatgcc tcattaagga tttcttcccg
1140
cctgatatcg acgtagagtg gcaatcaaac ggtcaacagg agccggaatc caagtataga
1200
accactccgc cccagcttga cgaggacgga tcatactttt tgtattcaaa actgtcggtg
1260
gataagagcc ggtggcagag aggtgacacc ttcatctgtg cggtgatgca cgaagcactc
1320
cataatcact acacccaaga gagcctctcg cattcccccg gaaag 1365
<210>72
<211>455
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>72
Glu Val Lys Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210>73
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>73
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgtgg cgagcggcta tacctttacc aactatggca tgaactgggt gcgccaggcg
120
ccgggcaaag gcctgcagtg ggtggcgtgg attaacacct ataccggcga accgacctat
180
gcggatgatt ttaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gcgccgcagc
300
atttattatc cgtattgggg ccagggcacc accctgaccg tgagcagcgc ttccacaacc
360
gcgccatcag tctttccgtt ggccccatca tgcgggtcga cgagcggatc gactgtggcc
420
ctggcgtgct tggtgtcggg atactttccc gaacccgtca cggtcagctg gaactccgga
480
tcgcttacga gcggtgtgca tacgttcccc tcggtcttgc aatcatcagg gctctactcg
540
ctgtcgagca tggtaacggt gccctcatcg aggtggccct ccgaaacgtt cacatgtaac
600
gtagcacatc cagcctccaa aaccaaggtg gataaacccg tgccgaaaag agagaatggg
660
cgggtgcctc gaccccctga ttgccccaag tgtccggctc cggaaatgct cggtggaccc
720
tcagtgttta tcttccctcc gaagcccaag gacactctgc tgatcgcgcg cactccagaa
780
gtaacatgtg tagtggtggc tcttgatccc gaggaccccg aagtccagat ctcctggttt
840
gtagatggga aacagatgca gaccgcaaaa actcaaccca gagaggagca gttcgccgga
900
acataccgag tggtatccgt ccttccgatt ggccaccagg actggttgaa agggaagcag
960
tttacgtgta aagtcaacaa taaggggttg cctagcccta ttgagcggac gatttcgaaa
1020
gctaggggac aggcccacca gccatcggtc tatgtccttc cgccttcccg cgaggagctc
1080
tcgaagaata cagtgagcct tacatgcctc attaaggatt tcttcccgcc tgatatcgac
1140
gtagagtggc aatcaaacgg tcaacaggag ccggaatcca agtatagaac cactccgccc
1200
cagcttgacg aggacggatc atactttttg tattcaaaac tgtcggtgga taagagccgg
1260
tggcagagag gtgacacctt catctgtgcg gtgatgcacg aagcactcca taatcactac
1320
acccaagaga gcctctcgca ttcccccgga aag 1353
<210>74
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>74
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>75
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>75
gaaattcagc tggtgcagag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcaaag cgagcggcta tacctttacc aactatggca tgaactgggt gcgccaggcg
120
ccgggcaaag gcctgcagtg gatgggctgg attaacacct ataccggcga accgacctat
180
gcggatgatt ttaaaggccg ctttaccttt agcctggata acgcgaaaaa caccctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt atttttgcgc gcgccgcagc
300
atttattatc cgtattgggg ccagggcacc accctgaccg tgagcagcgc ttccacaacc
360
gcgccatcag tctttccgtt ggccccatca tgcgggtcga cgagcggatc gactgtggcc
420
ctggcgtgct tggtgtcggg atactttccc gaacccgtca cggtcagctg gaactccgga
480
tcgcttacga gcggtgtgca tacgttcccc tcggtcttgc aatcatcagg gctctactcg
540
ctgtcgagca tggtaacggt gccctcatcg aggtggccct ccgaaacgtt cacatgtaac
600
gtagcacatc cagcctccaa aaccaaggtg gataaacccg tgccgaaaag agagaatggg
660
cgggtgcctc gaccccctga ttgccccaag tgtccggctc cggaaatgct cggtggaccc
720
tcagtgttta tcttccctcc gaagcccaag gacactctgc tgatcgcgcg cactccagaa
780
gtaacatgtg tagtggtggc tcttgatccc gaggaccccg aagtccagat ctcctggttt
840
gtagatggga aacagatgca gaccgcaaaa actcaaccca gagaggagca gttcgccgga
900
acataccgag tggtatccgt ccttccgatt ggccaccagg actggttgaa agggaagcag
960
tttacgtgta aagtcaacaa taaggggttg cctagcccta ttgagcggac gatttcgaaa
1020
gctaggggac aggcccacca gccatcggtc tatgtccttc cgccttcccg cgaggagctc
1080
tcgaagaata cagtgagcct tacatgcctc attaaggatt tcttcccgcc tgatatcgac
1140
gtagagtggc aatcaaacgg tcaacaggag ccggaatcca agtatagaac cactccgccc
1200
cagcttgacg aggacggatc atactttttg tattcaaaac tgtcggtgga taagagccgg
1260
tggcagagag gtgacacctt catctgtgcg gtgatgcacg aagcactcca taatcactac
1320
acccaagaga gcctctcgca ttcccccgga aag 1353
<210>76
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>76
Glu Ile Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>77
<211>1353
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>77
gaaattcagc tggtgcagag cggcggcgat ctggtgaaac cgggcggcag cgtgcgcctg 60
agctgcaaag cgagcggcta tacctttacc aactatggca tgaactgggt gaaacaggcg
120
ccgggcaaag gcctgcagtg gatgggctgg attaacacct ataccggcga accgacctat
180
gcggatgatt ttaaaggccg ctttaccttt agcctggata acgcgaaaaa caccgcgtat
240
ctgcagatta acagcctgcg cgcggaagat accgcggtgt atttttgcgc gcgccgcagc
300
atttattatc cgtattgggg ccagggcacc accctgaccg tgagcagcgc ttccacaacc
360
gcgccatcag tctttccgtt ggccccatca tgcgggtcga cgagcggatc gactgtggcc
420
ctggcgtgct tggtgtcggg atactttccc gaacccgtca cggtcagctg gaactccgga
480
tcgcttacga gcggtgtgca tacgttcccc tcggtcttgc aatcatcagg gctctactcg
540
ctgtcgagca tggtaacggt gccctcatcg aggtggccct ccgaaacgtt cacatgtaac
600
gtagcacatc cagcctccaa aaccaaggtg gataaacccg tgccgaaaag agagaatggg
660
cgggtgcctc gaccccctga ttgccccaag tgtccggctc cggaaatgct cggtggaccc
720
tcagtgttta tcttccctcc gaagcccaag gacactctgc tgatcgcgcg cactccagaa
780
gtaacatgtg tagtggtggc tcttgatccc gaggaccccg aagtccagat ctcctggttt
840
gtagatggga aacagatgca gaccgcaaaa actcaaccca gagaggagca gttcgccgga
900
acataccgag tggtatccgt ccttccgatt ggccaccagg actggttgaa agggaagcag
960
tttacgtgta aagtcaacaa taaggggttg cctagcccta ttgagcggac gatttcgaaa
1020
gctaggggac aggcccacca gccatcggtc tatgtccttc cgccttcccg cgaggagctc
1080
tcgaagaata cagtgagcct tacatgcctc attaaggatt tcttcccgcc tgatatcgac
1140
gtagagtggc aatcaaacgg tcaacaggag ccggaatcca agtatagaac cactccgccc
1200
cagcttgacg aggacggatc atactttttg tattcaaaac tgtcggtgga taagagccgg
1260
tggcagagag gtgacacctt catctgtgcg gtgatgcacg aagcactcca taatcactac
1320
acccaagaga gcctctcgca ttcccccgga aag 1353
<210>78
<211>451
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>78
Glu Ile Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Asn Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Arg Ser Ile Tyr Tyr Pro Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu
130 135 140
Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp
180 185 190
Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr
195 200 205
Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg
210 215 220
Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp Pro Glu Asp
260 265 270
Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr
275 280 285
Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln
305 310 315 320
Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg
325 330 335
Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val
340 345 350
Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210>79
<211>1365
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>79
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgtgg cgagcggctt tacctttagc gattattata tgagctgggt gcgccaggcg
120
ccgggcaaag gcctggaatg ggtggcgttt attcgcaaca aagcgaacgg ctataccacc
180
gaatatagcg cgagcctgaa aggccgcttt accattagcc gcgataacgc gaaaaacatg
240
gcgtatctgc agatgaacag cctgcgcgcg gaagataccg cggtgtatta ttgcgcgagc
300
tttggcctga tgtattattt tgattattgg ggccagggca ccaccctgac cgtgagcagc
360
gcttccacaa ccgcgccatc agtctttccg ttggccccat catgcgggtc gacgagcgga
420
tcgactgtgg ccctggcgtg cttggtgtcg ggatactttc ccgaacccgt cacggtcagc
480
tggaactccg gatcgcttac gagcggtgtg catacgttcc cctcggtctt gcaatcatca
540
gggctctact cgctgtcgag catggtaacg gtgccctcat cgaggtggcc ctccgaaacg
600
ttcacatgta acgtagcaca tccagcctcc aaaaccaagg tggataaacc cgtgccgaaa
660
agagagaatg ggcgggtgcc tcgaccccct gattgcccca agtgtccggc tccggaaatg
720
ctcggtggac cctcagtgtt tatcttccct ccgaagccca aggacactct gctgatcgcg
780
cgcactccag aagtaacatg tgtagtggtg gctcttgatc ccgaggaccc cgaagtccag
840
atctcctggt ttgtagatgg gaaacagatg cagaccgcaa aaactcaacc cagagaggag
900
cagttcgccg gaacataccg agtggtatcc gtccttccga ttggccacca ggactggttg
960
aaagggaagc agtttacgtg taaagtcaac aataaggggt tgcctagccc tattgagcgg
1020
acgatttcga aagctagggg acaggcccac cagccatcgg tctatgtcct tccgccttcc
1080
cgcgaggagc tctcgaagaa tacagtgagc cttacatgcc tcattaagga tttcttcccg
1140
cctgatatcg acgtagagtg gcaatcaaac ggtcaacagg agccggaatc caagtataga
1200
accactccgc cccagcttga cgaggacgga tcatactttt tgtattcaaa actgtcggtg
1260
gataagagcc ggtggcagag aggtgacacc ttcatctgtg cggtgatgca cgaagcactc
1320
cataatcact acacccaaga gagcctctcg cattcccccg gaaag 1365
<210>80
<211>455
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>80
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Ser Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210>81
<211>1365
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>81
gaagtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgcga ccagcggctt tacctttagc gattattata tgagctgggt gcgccaggcg
120
ccgggcaaag gcctggaatg gatgggcttt attcgcaaca aagcgaacgg ctataccacc
180
gaatatagcg cgagcctgaa aggccgcttt accattagcc gcgataacgc gaaaaacatg
240
gcgtatctgc agatgaacag cctgcgcgcg gaagataccg cggtgtatta ttgcgtgcgc
300
tttggcctga tgtattattt tgattattgg ggccagggca ccaccctgac cgtgagcagc
360
gcttccacaa ccgcgccatc agtctttccg ttggccccat catgcgggtc gacgagcgga
420
tcgactgtgg ccctggcgtg cttggtgtcg ggatactttc ccgaacccgt cacggtcagc
480
tggaactccg gatcgcttac gagcggtgtg catacgttcc cctcggtctt gcaatcatca
540
gggctctact cgctgtcgag catggtaacg gtgccctcat cgaggtggcc ctccgaaacg
600
ttcacatgta acgtagcaca tccagcctcc aaaaccaagg tggataaacc cgtgccgaaa
660
agagagaatg ggcgggtgcc tcgaccccct gattgcccca agtgtccggc tccggaaatg
720
ctcggtggac cctcagtgtt tatcttccct ccgaagccca aggacactct gctgatcgcg
780
cgcactccag aagtaacatg tgtagtggtg gctcttgatc ccgaggaccc cgaagtccag
840
atctcctggt ttgtagatgg gaaacagatg cagaccgcaa aaactcaacc cagagaggag
900
cagttcgccg gaacataccg agtggtatcc gtccttccga ttggccacca ggactggttg
960
aaagggaagc agtttacgtg taaagtcaac aataaggggt tgcctagccc tattgagcgg
1020
acgatttcga aagctagggg acaggcccac cagccatcgg tctatgtcct tccgccttcc
1080
cgcgaggagc tctcgaagaa tacagtgagc cttacatgcc tcattaagga tttcttcccg
1140
cctgatatcg acgtagagtg gcaatcaaac ggtcaacagg agccggaatc caagtataga
1200
accactccgc cccagcttga cgaggacgga tcatactttt tgtattcaaa actgtcggtg
1260
gataagagcc ggtggcagag aggtgacacc ttcatctgtg cggtgatgca cgaagcactc
1320
cataatcact acacccaaga gagcctctcg cattcccccg gaaag 1365
<210>82
<211>455
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>82
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210>83
<211>1365
<212>ДНК
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>83
gaagtgaaac tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
agctgcgcga ccagcggctt tacctttagc gattattata tgagctgggt gcgccaggcg
120
ccgggcaaag cgctggaatg gatgggcttt attcgcaaca aagcgaacgg ctataccacc
180
gaatatagcg cgagcctgaa aggccgcttt accattagcc gcgataacgc gaaaaacatg
240
ctgtatctgc agatgaacag cctgcgcgcg gaagataccg cggtgtatta ttgcgtgcgc
300
tttggcctga tgtattattt tgattattgg ggccagggca ccaccctgac cgtgagcagc
360
gcttccacaa ccgcgccatc agtctttccg ttggccccat catgcgggtc gacgagcgga
420
tcgactgtgg ccctggcgtg cttggtgtcg ggatactttc ccgaacccgt cacggtcagc
480
tggaactccg gatcgcttac gagcggtgtg catacgttcc cctcggtctt gcaatcatca
540
gggctctact cgctgtcgag catggtaacg gtgccctcat cgaggtggcc ctccgaaacg
600
ttcacatgta acgtagcaca tccagcctcc aaaaccaagg tggataaacc cgtgccgaaa
660
agagagaatg ggcgggtgcc tcgaccccct gattgcccca agtgtccggc tccggaaatg
720
ctcggtggac cctcagtgtt tatcttccct ccgaagccca aggacactct gctgatcgcg
780
cgcactccag aagtaacatg tgtagtggtg gctcttgatc ccgaggaccc cgaagtccag
840
atctcctggt ttgtagatgg gaaacagatg cagaccgcaa aaactcaacc cagagaggag
900
cagttcgccg gaacataccg agtggtatcc gtccttccga ttggccacca ggactggttg
960
aaagggaagc agtttacgtg taaagtcaac aataaggggt tgcctagccc tattgagcgg
1020
acgatttcga aagctagggg acaggcccac cagccatcgg tctatgtcct tccgccttcc
1080
cgcgaggagc tctcgaagaa tacagtgagc cttacatgcc tcattaagga tttcttcccg
1140
cctgatatcg acgtagagtg gcaatcaaac ggtcaacagg agccggaatc caagtataga
1200
accactccgc cccagcttga cgaggacgga tcatactttt tgtattcaaa actgtcggtg
1260
gataagagcc ggtggcagag aggtgacacc ttcatctgtg cggtgatgca cgaagcactc
1320
cataatcact acacccaaga gagcctctcg cattcccccg gaaag 1365
<210>84
<211>455
<212>Белок
<213>Искусственная последовательность
<220>
<223>канинизированная mus musculus
<400>84
Glu Val Lys Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Ala Leu Glu Trp Met
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Phe Gly Leu Met Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210>85
<211>15
<212>ДНК
<213>Mus musculus
<400>85
aactatggaa tgaac 15
<210>86
<211>5
<212>Белок
<213>Mus musculus
<400>86
Asn Tyr Gly Met Asn
1 5
<210>87
<211>51
<212>ДНК
<213>Mus musculus
<400>87
tggataaaca cctacactgg agagccaaca tatgctgatg acttcaaggg a 51
<210>88
<211>17
<212>Белок
<213>Mus musculus
<400>88
Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210>89
<211>21
<212>ДНК
<213>Mus musculus
<400>89
cggtcaattt attacccgta c 21
<210>90
<211>7
<212>Белок
<213>Mus musculus
<400>90
Arg Ser Ile Tyr Tyr Pro Tyr
1 5
<210>91
<211>48
<212>ДНК
<213>Mus musculus
<400>91
agatctagtc agagcattgt atatagtaat ggaaacacct atttagaa 48
<210>92
<211>16
<212>Белок
<213>Mus musculus
<400>92
Arg Ser Ser Gln Ser Ile Val Tyr Ser Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210>93
<211>21
<212>ДНК
<213>Mus musculus
<400>93
aaagtttcca accgattttc t 21
<210>94
<211>7
<212>Белок
<213>Mus musculus
<400>94
Lys Val Ser Asn Arg Phe Ser
1 5
<210>95
<211>27
<212>ДНК
<213>Mus musculus
<400>95
tttcaaggtt cacatgttcc gtggacg 27
<210>96
<211>9
<212>Белок
<213>Mus musculus
<400>96
Phe Gln Gly Ser His Val Pro Trp Thr
1 5
<210>97
<211>15
<212>ДНК
<213>Mus musculus
<400>97
gattactaca tgagc 15
<210>98
<211>5
<212>Белок
<213>Mus musculus
<400>98
Asp Tyr Tyr Met Ser
1 5
<210>99
<211>57
<212>ДНК
<213>Mus musculus
<400>99
tttattagaa acaaagctaa tggttacaca acagagtaca gcgcatctct gaagggt 57
<210>100
<211>19
<212>Белок
<213>Mus musculus
<400>100
Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala Ser
1 5 10 15
Leu Lys Gly
<210>101
<211>27
<212>ДНК
<213>Mus musculus
<400>101
tttgggttaa tgtactactt tgactac 27
<210>102
<211>9
<212>Белок
<213>Mus musculus
<400>102
Phe Gly Leu Met Tyr Tyr Phe Asp Tyr
1 5
<210>103
<211>36
<212>ДНК
<213>Mus musculus
<400>103
agggccagct caagtgtaag ttccagttac ttgcac 36
<210>104
<211>12
<212>Белок
<213>Mus musculus
<400>104
Arg Ala Ser Ser Ser Val Ser Ser Ser Tyr Leu His
1 5 10
<210>105
<211>21
<212>ДНК
<213>Mus musculus
<400>105
agcacatcca acttggcttc t 21
<210>106
<211>7
<212>Белок
<213>Mus musculus
<400>106
Ser Thr Ser Asn Leu Ala Ser
1 5
<210>107
<211>27
<212>ДНК
<213>Mus musculus
<400>107
cagcagtaca gtggtctccc actcacg 27
<210>108
<211>9
<212>Белок
<213>Mus musculus
<400>108
Gln Gln Tyr Ser Gly Leu Pro Leu Thr
1 5
<210>109
<211>5
<212>Белок
<213>Mus musculus
<400>109
Thr Tyr Gly Val Ser
1 5
<210>110
<211>5
<212>Белок
<213>Mus musculus
<400>110
Ser Tyr Trp Met His
1 5
<210>111
<211>17
<212>Белок
<213>Mus musculus
<400>111
Trp Ile Asn Thr Tyr Ser Gly Met Pro Thr Tyr Val Asp Asp Phe Lys
1 5 10 15
Gly
<210>112
<211>17
<212>Белок
<213>Mus musculus
<400>112
Asn Ile Asn Pro Ser Asn Gly Gly Thr Arg Phe Asn Glu Lys Phe Lys
1 5 10 15
Asn
<210>113
<211>7
<212>Белок
<213>Mus musculus
<400>113
Arg Gly Thr Tyr Tyr Arg Pro
1 5
<210>114
<211>7
<212>Белок
<213>Mus musculus
<400>114
Arg Gly Ile Ser Phe Asp Tyr
1 5
<210>115
<211>7
<212>Белок
<213>Mus musculus
<400>115
Arg Gly Val Arg Leu Asp Tyr
1 5
<210>116
<211>12
<212>Белок
<213>Mus musculus
<400>116
Ser Asn Tyr Gly Ser Gly Trp Ala Trp Phe Ala Tyr
1 5 10
<210>117
<211>16
<212>Белок
<213>Mus musculus
<400>117
Arg Ser Ser Gln Ser Ile Val Tyr Ser His Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210>118
<211>16
<212>Белок
<213>Mus musculus
<400>118
Lys Ser Ser Gln Ser Ile Val Tyr Ile Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210>119
<211>16
<212>Белок
<213>Mus musculus
<400>119
Arg Ser Ser Gln Ser Ile Val Tyr Ile Ser Gly Ser Thr Tyr Leu Glu
1 5 10 15
<210>120
<211>11
<212>Белок
<213>Mus musculus
<400>120
His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser
1 5 10
<210>121
<211>7
<212>Белок
<213>Mus musculus
<400>121
Lys Val Ser Lys Arg Phe Ser
1 5
<210>122
<211>7
<212>Белок
<213>Mus musculus
<400>122
Lys Val Ser Ser Arg Phe Ser
1 5
<210>123
<211>7
<212>Белок
<213>Mus musculus
<400>123
Lys Ser Ser Asn Leu His Thr
1 5
<210>124
<211>9
<212>Белок
<213>Mus musculus
<400>124
Gln Gln Gly Gln Ser Tyr Pro Trp Thr
1 5
<210>125
<211>669
<212>ДНК
<213>Canis familiaris
<400>125
atggcgggct ttggctttcg ccgccatggc gcgcagccgg atctggcgag ccgcacctgg 60
ccgtgcaccg cgctgtttag cctgctgttt attccggtgt ttagcaaagg catgcatgtg
120
gcgcagccgg cggtggtgct ggcgagcagc cgcggcgtgg cgagctttgt gtgcgaatat
180
ggcagcagcg gcaacgcggc ggaagtgcgc gtgaccgtgc tgcgccaggc gggcagccag
240
atgaccgaag tgtgcgcggc gacctatacc gtggaagatg aactggcgtt tctggatgat
300
agcacctgca ccggcaccag cagcggcaac aaagtgaacc tgaccattca gggcctgcgc
360
gcgatggata ccggcctgta tatttgcaaa gtggaactga tgtatccgcc gccgtattat
420
gtgggcatgg gcaacggcac ccagatttat gtgattgatc cggaaccgtg cccggatagc
480
gattttctgc tgtggattct ggcggcggtg agcagcggcc tgttttttta tagctttctg
540
attaccgcgg tgagcctgag caaaatgctg aaaaaacgca gcccgctgac caccggcgtg
600
tatgtgaaaa tgccgccgac cgaaccggaa tgcgaaaaac agtttcagcc gtattttatt
660
ccgattaac 669
<210>126
<211>223
<212>Белок
<213>Canis familiaris
<400>126
Met Ala Gly Phe Gly Phe Arg Arg His Gly Ala Gln Pro Asp Leu Ala
1 5 10 15
Ser Arg Thr Trp Pro Cys Thr Ala Leu Phe Ser Leu Leu Phe Ile Pro
20 25 30
Val Phe Ser Lys Gly Met His Val Ala Gln Pro Ala Val Val Leu Ala
35 40 45
Ser Ser Arg Gly Val Ala Ser Phe Val Cys Glu Tyr Gly Ser Ser Gly
50 55 60
Asn Ala Ala Glu Val Arg Val Thr Val Leu Arg Gln Ala Gly Ser Gln
65 70 75 80
Met Thr Glu Val Cys Ala Ala Thr Tyr Thr Val Glu Asp Glu Leu Ala
85 90 95
Phe Leu Asp Asp Ser Thr Cys Thr Gly Thr Ser Ser Gly Asn Lys Val
100 105 110
Asn Leu Thr Ile Gln Gly Leu Arg Ala Met Asp Thr Gly Leu Tyr Ile
115 120 125
Cys Lys Val Glu Leu Met Tyr Pro Pro Pro Tyr Tyr Val Gly Met Gly
130 135 140
Asn Gly Thr Gln Ile Tyr Val Ile Asp Pro Glu Pro Cys Pro Asp Ser
145 150 155 160
Asp Phe Leu Leu Trp Ile Leu Ala Ala Val Ser Ser Gly Leu Phe Phe
165 170 175
Tyr Ser Phe Leu Ile Thr Ala Val Ser Leu Ser Lys Met Leu Lys Lys
180 185 190
Arg Ser Pro Leu Thr Thr Gly Val Tyr Val Lys Met Pro Pro Thr Glu
195 200 205
Pro Glu Cys Glu Lys Gln Phe Gln Pro Tyr Phe Ile Pro Ile Asn
210 215 220
<210>127
<211>215
<212>Белок
<213>Искусственная последовательность
<220>
<223>генетически модифицированная canis familiaris
<400>127
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
1 5 10 15
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu
20 25 30
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
35 40 45
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly
50 55 60
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
65 70 75 80
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
85 90 95
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
100 105 110
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
115 120 125
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
130 135 140
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
145 150 155 160
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
165 170 175
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
180 185 190
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
195 200 205
Leu Ser His Ser Pro Gly Lys
210 215
<210>128
<211>17
<212>Белок
<213>Canis familiaris
<400>128
Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Pro
<210>129
<211>22
<212>Белок
<213>Canis familiaris
<400>129
Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys Pro Lys
1 5 10 15
Cys Pro Ala Pro Glu Met
20
<210>130
<211>20
<212>Белок
<213>Canis familiaris
<400>130
Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn Asn Cys Pro Cys Pro
1 5 10 15
Gly Cys Gly Leu
20
<210>131
<211>17
<212>Белок
<213>Искусственная последовательность
<220>
<223>генетически модифицированная canis familiarus
<400>131
Pro Lys Glu Ser Thr Cys Lys Cys Ile Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Ser
<210>132
<211>31
<212>Белок
<213>Canis familiaris
<400>132
Ala Glu Val Arg Val Thr Val Leu Arg Gln Ala Gly Ser Gln Met Thr
1 5 10 15
Glu Val Cys Ala Ala Thr Tyr Thr Val Glu Asp Glu Leu Ala Phe
20 25 30
<210>133
<211>21
<212>Белок
<213>Canis familiaris
<400>133
Tyr Ile Cys Lys Val Glu Leu Met Tyr Pro Pro Pro Tyr Tyr Val Gly
1 5 10 15
Met Gly Asn Gly Thr
20
<210>134
<211>8
<212>Белок
<213>Canis familiaris
<400>134
Thr Val Leu Arg Gln Ala Gly Ser
1 5
<210>135
<211>5
<212>Белок
<213>Canis familiaris
<400>135
Ala Thr Tyr Thr Val
1 5
<210>136
<211>13
<212>Белок
<213>Canis familiaris
<400>136
Tyr Ile Cys Lys Val Glu Leu Met Tyr Pro Pro Pro Tyr
1 5 10
<210>137
<211>6
<212>Белок
<213>Canis familiaris
<400>137
Met Tyr Pro Pro Pro Tyr
1 5
<210>138
<211>186
<212>Белок
<213>Canis familiaris
<400>138
Met His Val Ala Gln Pro Ala Val Val Leu Ala Ser Ser Arg Gly Val
1 5 10 15
Ala Ser Phe Val Cys Glu Tyr Gly Ser Ser Gly Asn Ala Ala Glu Val
20 25 30
Arg Val Thr Val Leu Arg Gln Ala Gly Ser Gln Met Thr Glu Val Cys
35 40 45
Ala Ala Thr Tyr Thr Val Glu Asp Glu Leu Ala Phe Leu Asp Asp Ser
50 55 60
Thr Cys Thr Gly Thr Ser Ser Gly Asn Lys Val Asn Leu Thr Ile Gln
65 70 75 80
Gly Leu Arg Ala Met Asp Thr Gly Leu Tyr Ile Cys Lys Val Glu Leu
85 90 95
Met Tyr Pro Pro Pro Tyr Tyr Val Gly Met Gly Asn Gly Thr Gln Ile
100 105 110
Tyr Val Ile Asp Pro Glu Pro Cys Pro Asp Ser Asp Phe Leu Leu Trp
115 120 125
Ile Leu Ala Ala Val Ser Ser Gly Leu Phe Phe Tyr Ser Phe Leu Ile
130 135 140
Thr Ala Val Ser Leu Ser Lys Met Leu Lys Lys Arg Ser Pro Leu Thr
145 150 155 160
Thr Gly Val Tyr Val Lys Met Pro Pro Thr Glu Pro Glu Cys Glu Lys
165 170 175
Gln Phe Gln Pro Tyr Phe Ile Pro Ile Asn
180 185
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КАНИНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CTLA-4 | 2020 |
|
RU2822460C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2801209C2 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2840044C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2838943C1 |
| АНТИТЕЛА К PD-L1, СВЯЗЫВАЮЩИЕ PD-L1 СОБАКИ | 2015 |
|
RU2722562C2 |
| АНТИТЕЛА К PD-L1, СВЯЗЫВАЮЩИЕ PD-L1 СОБАКИ | 2015 |
|
RU2832918C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2732604C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2795485C2 |

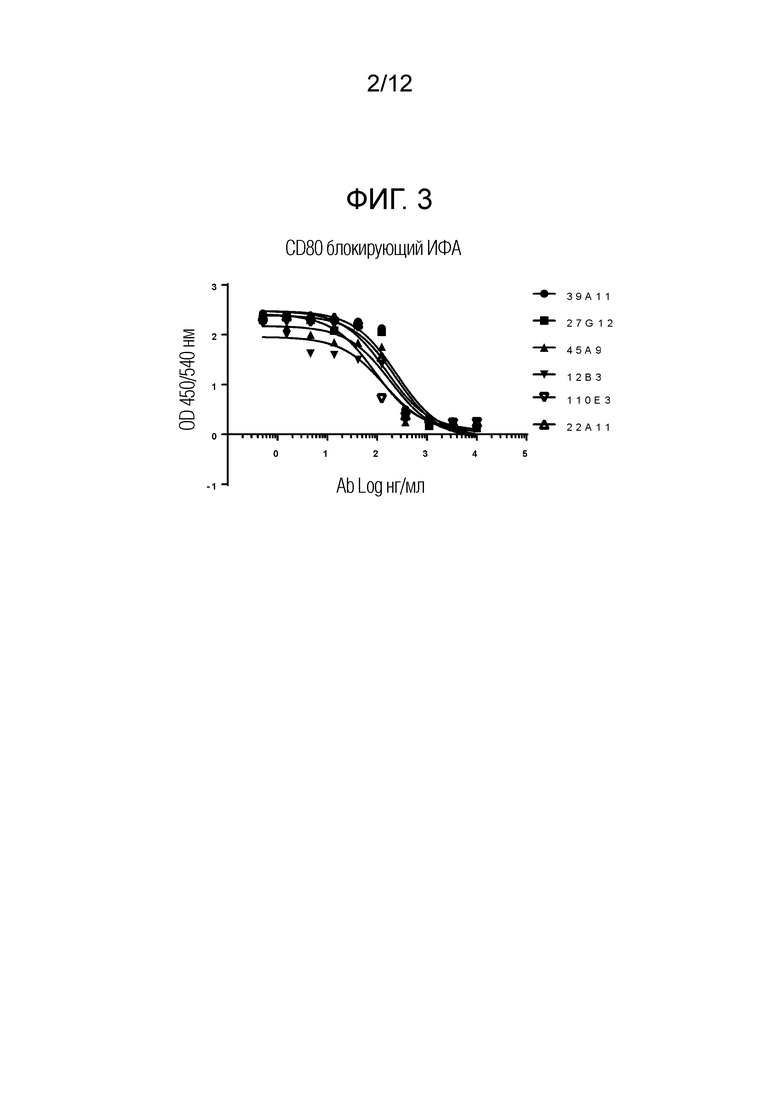


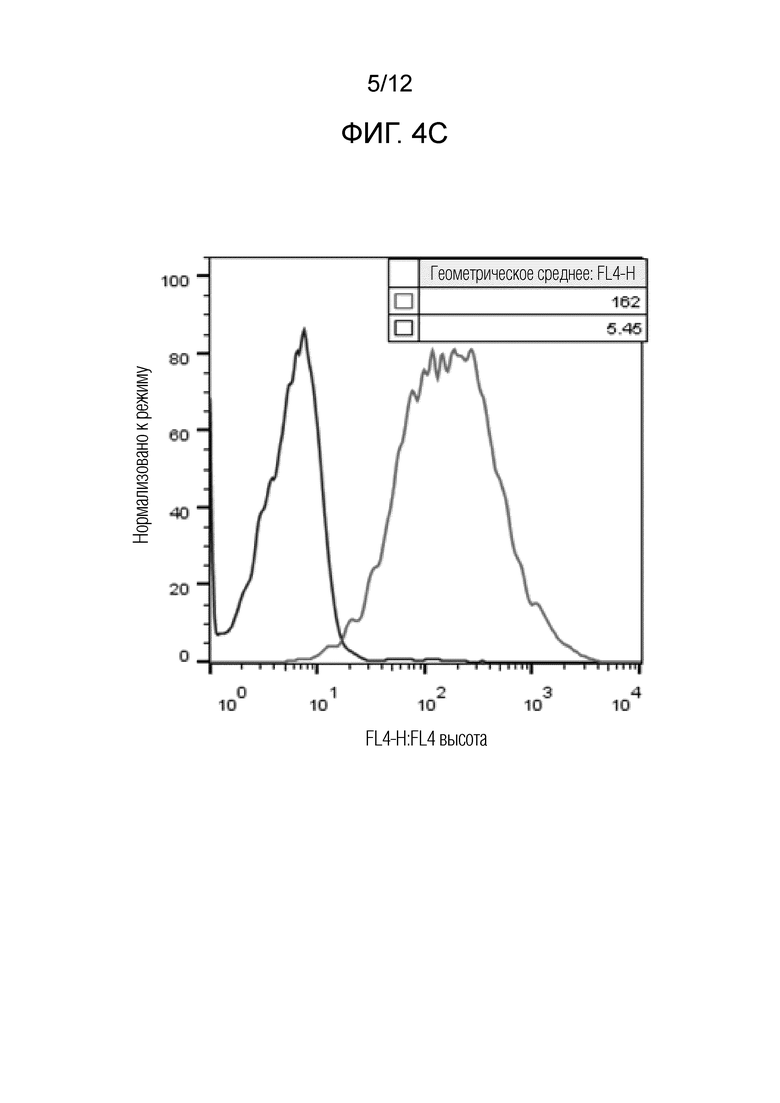





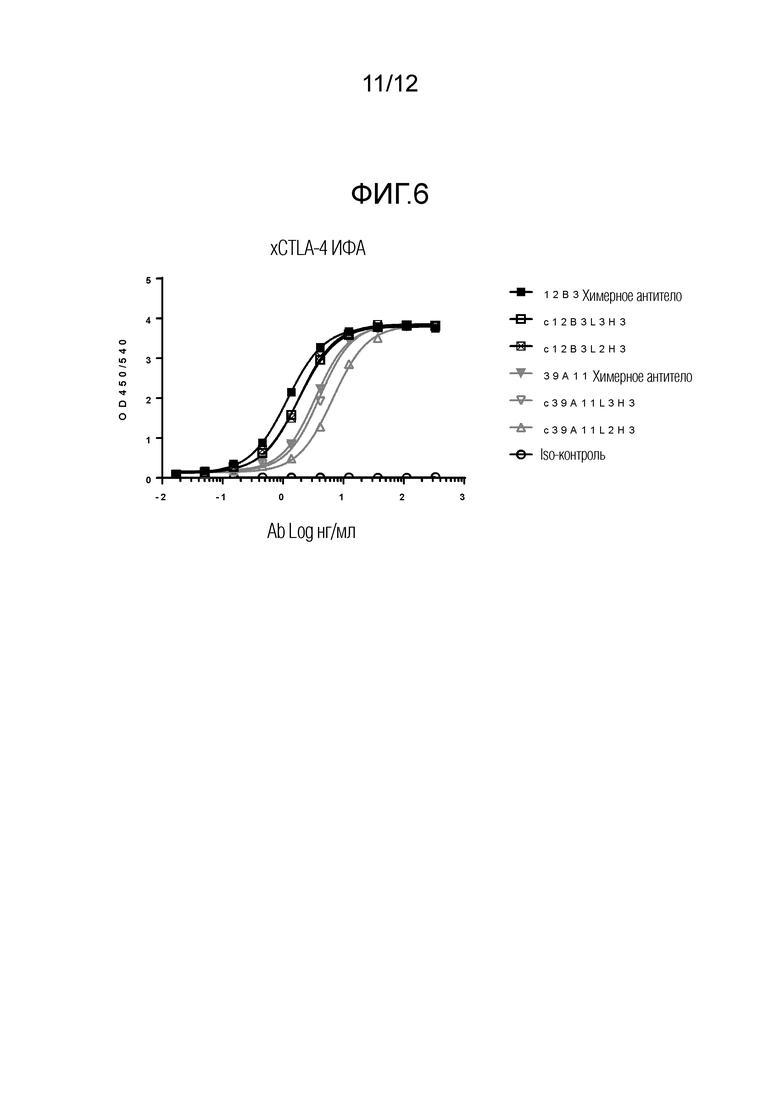
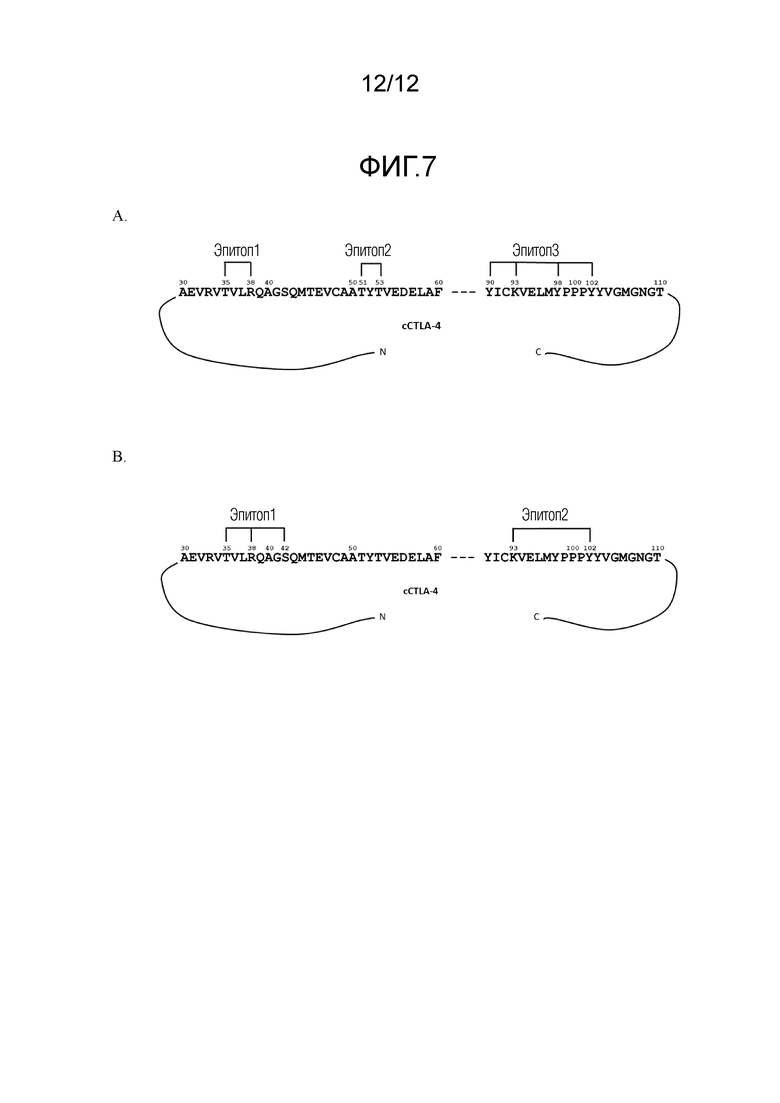
Изобретение относится к области биотехнологии, в частности к выделенному антителу млекопитающего или его антигенсвязывающему фрагменту, которое связывается с собачьим белком 4, ассоциированным с цитотоксическими T-лимфоцитами, (CTLA-4), а также к композиции, его содержащей. Изобретение эффективно для повышения активности иммунной клетки собаки. 3 н. и 12 з.п. ф-лы, 7 ил., 8 табл., 8 пр.
1. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент, которое связывается с собачьим белком 4, ассоциированным с цитотоксическими T-лимфоцитами, (CTLA-4), и блокирует связывание собачьего CTLA-4 с собачьим CD80, блокирует связывание собачьего CTLA-4 с собачьим CD86 или блокирует как связывание собачьего CTLA-4 с собачьим CD80, так и связывание собачьего CTLA-4 с собачьим CD86;
где указанное антитело содержит набор из шести областей, определяющих комплементарность, (CDR), три из которых представляют собой CDR легкой цепи: CDR легкой цепи 1 (CDRL1), CDR легкой цепи 2 (CDRL2) и CDR легкой цепи 3 (CDRL3); и три из которых представляют собой CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3);
где набор из шести CDR выбран из группы наборов, состоящих из (i), (ii), (iii), (iv), (v) и (vi);
где для набора (i):
CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 92, консервативно модифицированного варианта SEQ ID NO: 92;
CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 94, консервативно модифицированного варианта SEQ ID NO: 94;
CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 96, консервативно модифицированного варианта SEQ ID NO: 96;
CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 86, консервативно модифицированного варианта SEQ ID NO: 86;
CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 88, консервативно модифицированного варианта SEQ ID NO: 88; и
CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 90, консервативно модифицированного варианта SEQ ID NO: 90;
где для набора (ii):
CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 104, консервативно модифицированного варианта SEQ ID NO: 104;
CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 106, консервативно модифицированного варианта SEQ ID NO: 106;
CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 108, консервативно модифицированного варианта SEQ ID NO: 108;
CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, консервативно модифицированного варианта SEQ ID NO: 98;
CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 100, консервативно модифицированного варианта SEQ ID NO: 100; и
CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 102, консервативно модифицированного варианта SEQ ID NO: 102;
где для набора (iii):
CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 117, консервативно модифицированного варианта SEQ ID NO: 117;
CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 94, консервативно модифицированного варианта SEQ ID NO: 94;
CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 96, консервативно модифицированного варианта SEQ ID NO: 96;
CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 86, консервативно модифицированного варианта SEQ ID NO: 86;
CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 88, консервативно модифицированного варианта SEQ ID NO: 88; и
CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 113, консервативно модифицированного варианта SEQ ID NO: 113;
где для набора (iv):
CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 119, консервативно модифицированного варианта SEQ ID NO: 119;
CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 122, консервативно модифицированного варианта SEQ ID NO: 122;
CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 96, консервативно модифицированного варианта SEQ ID NO: 96;
CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 86, консервативно модифицированного варианта SEQ ID NO: 86;
CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 88, консервативно модифицированного варианта SEQ ID NO: 88; и
CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 115, консервативно модифицированного варианта SEQ ID NO: 115;
где для набора (v):
CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 118, консервативно модифицированного варианта SEQ ID NO: 118;
CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 121, консервативно модифицированного варианта SEQ ID NO: 121;
CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 96, консервативно модифицированного варианта SEQ ID NO: 96;
CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 109, консервативно модифицированного варианта SEQ ID NO: 109;
CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 111, консервативно модифицированного варианта SEQ ID NO: 111; и
CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 114, консервативно модифицированного варианта SEQ ID NO: 114; и
где для набора (vi):
CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 120, консервативно модифицированного варианта SEQ ID NO: 120;
CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 123, консервативно модифицированного варианта SEQ ID NO: 123;
CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 124, консервативно модифицированного варианта SEQ ID NO: 124;
CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 110, консервативно модифицированного варианта SEQ ID NO: 110;
CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 112, консервативно модифицированного варианта SEQ ID NO: 112; и
CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 116, консервативно модифицированного варианта SEQ ID NO: 116.
2. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 1, которое представляет собой канинизированное антитело или его канинизированный антигенсвязывающий фрагмент.
3. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 2, которое содержит шарнирную область, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130 и SEQ ID NO: 131.
4. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 2 или 3, где
(a) CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 92, консервативно модифицированного варианта SEQ ID NO: 92;
(b) CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 94, консервативно модифицированного варианта SEQ ID NO: 94;
(c) CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 96, консервативно модифицированного варианта SEQ ID NO: 96;
(d) CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 86, консервативно модифицированного варианта SEQ ID NO: 86;
(e) CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 88, консервативно модифицированного варианта SEQ ID NO: 88; и
(f) CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 90, консервативно модифицированного варианта SEQ ID NO: 90.
5. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 4, содержащее тяжелую цепь, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 62, SEQ ID NO: 64 и SEQ ID NO: 66; или
модифицированную тяжелую цепь, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 74, SEQ ID NO: 76 и SEQ ID NO: 78; или
легкую цепь, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 50, SEQ ID NO: 52 и SEQ ID NO: 54, или
комбинацию указанной тяжелой цепи или указанной модифицированной тяжелой цепи с указанной легкой цепью.
6. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 5, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 66, или модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 78.
7. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 6, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 52 или аминокислотную последовательность SEQ ID NO: 54.
8. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 2 или 3, где
(a) CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 104, консервативно модифицированного варианта SEQ ID NO: 104;
(b) CDRL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 106, консервативно модифицированного варианта SEQ ID NO: 106;
(c) CDRL3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 108, консервативно модифицированного варианта SEQ ID NO: 108;
(d) CDRH1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 98, консервативно модифицированного варианта SEQ ID NO: 98;
(e) CDRH2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 100, консервативно модифицированного варианта SEQ ID NO: 100; и
(f) CDRH3 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 102, консервативно модифицированного варианта SEQ ID NO: 102.
9. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 8, содержащее тяжелую цепь, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 68, SEQ ID NO: 70 и SEQ ID NO: 72; или
модифицированную тяжелую цепь, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 80, SEQ ID NO: 82 и SEQ ID NO: 84; или
легкую цепь, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 56, SEQ ID NO: 58 и SEQ ID NO: 60, или
комбинацию указанной тяжелой цепи или указанной модифицированной тяжелой цепи с указанной легкой цепью.
10. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 9, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 72, или модифицированную тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 84.
11. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по п. 10, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 58 или аминокислотную последовательность SEQ ID NO: 60.
12. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по любому из пп. 2-11, где указанное антитело или его антигенсвязывающий фрагмент проявляют одно, два, три, четыре или все пять из следующих свойств:
(i) связывание с собачьим CTLA-4 с константой диссоциации (Kd) 1х10-5 M - 1х10-12 M;
(ii) связывание с собачьим CTLA-4 с константой скорости ассоциации (kon) 1х102 M-1⋅с-1-1х107 М-1⋅с-1;
(iii) связывание с собачьим CTLA-4 с константой скорости диссоциации (koff) 1х10-3 с-1-1х10-8 с-1;
(iv) блокирование связывания собачьего CTLA-4 с собачьим CD80; и
(v) блокирование связывания собачьего CTLA-4 с собачьим CD86.
13. Выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по любому из пп. 2-12, где антитело или его антигенсвязывающий фрагмент связывается с любой одной или более аминокислотными последовательностями, выбранными из группы, состоящей из SEQ ID NO: 132, SEQ ID NO: 133, SEQ ID NO: 134, SEQ ID NO: 135, SEQ ID NO: 136 и SEQ ID NO: 137.
14. Фармацевтическая композиция для повышения активности иммунной клетки собаки, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 2-13 и фармацевтически приемлемый носитель или разбавитель.
15. Способ повышения активности иммунной клетки собаки, включающий введение пациенту терапевтически эффективного количества фармацевтической композиции по п. 14.
| WO 2016183469 A1, 17.11.2016 | |||
| MICHIHITO TAGAWA et al., Evaluation of costimulatory molecules in dogs with B cell high grade lymphoma, PLoS ONE, 2018, 13(7):e0201222 | |||
| LISA M | |||
| BERGERON et al., Comparative functional characterization of canine IgG subclasses, Veterinary Immunology and Immunopathology, 2014, Volume 157, Issues 1-2, pp | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| WO | |||

