ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По этой заявке испрашивается приоритет на основании временной заявки США с серийным номером 61/918946, поданной 20 декабря 2013 года, временной заявке США с серийным номером 61/918847, поданной 20 декабря 2013 года, и временной заявки США с серийным номером 62/030812, поданной 30 июля 2014 года, содержание которых полностью включено в настоящее описание в качестве ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к мышиным антителам к PD-1 собак, которые содержат специфические последовательности и обладают высоким сродством связывания по отношению к PD-1 собак. Изобретение также относится к применению антител согласно настоящему изобретению в способе лечения рака у собак.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Иммуноингибиторный рецептор, который в основном экспрессируется на активированных Т и В-клетках, рецептор программируемой гибели клеток 1, также упоминаемый как рецептор программируемой смерти 1 (PD-1), является членом суперсемейства иммуноглобулинов, связанного с CD28 и CTLA-4. PD-1 и подобные ему члены семейства относятся к трансмембранным гликопротеинам типа I, которые содержат внеклеточный домен, относящийся к типу вариабельных доменов Ig (V-тип) и способный связывать лиганды, и цитоплазматический хвост, который связывает сигнальные молекулы. Цитоплазматический хвост PD-1 содержит два тирозин-содержащих сигнальных мотива, ITIM (иммунорецепторный тирозин-содержащий ингибиторный мотив) и ITSM (иммунорецепторный тирозин-содержащий переключающий мотив).
PD-1 ослабляет Т-клеточный ответ при связывании с лигандом программируемой гибели клеток 1, также упоминаемым как лиганд программируемой смерти 1 (PD-L1), и/или лигандом программируемой гибели клеток 2, также упоминаемым как лиганд программируемой смерти 2 (PD-L2). Связывание любого из этих лигандов с PD-1 оказывает негативное влияние на антиген-рецепторный сигнальный путь. Блокирование связывания PD-L1 с PD-1 повышает опухоль-специфичный иммунитет, опосредованный Т-клетками CD8+, способствуя уничтожению опухолевых клеток иммунной системой. Трехмерная структура мышиного PD-1, а также со-кристаллическая структура комплекса мышиного PD-1 с человеческим PD-L1 описаны в [Zhang et al., Immunity 20: 337-347 (2004); Lin et al., Proc. Natl. Acad. Sci. USA 105: 3011-3016 (2008)].
PD-L1 и PD-L2 представляют собой трансмембранные лиганды типа I, которые содержат как IgV-, так и IgC-подобные внеклеточные домены наряду с короткими цитоплазматическими участками, не содержащими каких-либо известных сигнальных мотивов. И PD-L1, и PD-L2, либо конститутивно экспрессируются, либо индуцируются в клетках разных типах, в том числе в не кроветворных тканях, а также в опухолях разных типов. PD-L1 экспрессируется не только на B-, T-, миелоидных и дендритных клетках (DC), но и на периферических клетках, таких как клетки эндотелия микрососудов и клетки не лимфоидных органов, например, клетки сердца или легких. PD-L2 же обнаружен только на макрофагах и DC. Характер экспрессии лигандов PD-1 позволяет предположить, что PD-1 играет важную роль в поддержании периферической резистентности и, кроме того, может участвовать в регуляции аутореактивных Т- и В-клеточных ответов в периферических тканях.
В любом случае, теперь совершенно ясно, что PD-1 играет важную роль в развитии, по меньшей мере, некоторых злокачественных опухолей человека, предположительно, путем опосредования механизма ускользания от иммунного надзора. В соответствии с приведенными выше данными показано, что PD-L1 экспрессируется на клетках ряда мышиных и человеческих опухолей и индуцируется IFN-гамма в большинстве линий PD-L1-отрицательных опухолевых клеток [Iwai et al., Proc. Natl. Acad. Sci. U.S.A. 99: 12293-12297 (2002); Strome et al., Cancer Res., 63: 6501-6505 (2003)]. Кроме того, экспрессия PD-1 на опухоль-инфильтрующих лимфоцитах, и/или PD-L1 на опухолевых клетках идентифицирована в ряде биопсийных образцов первичных опухолей человека. Такие опухоли включают в себя рак легкого, печени, яичников, шейки матки, кожи, толстой кишки, глиомы, мочевого пузыря, молочной железы, почек, пищевода, желудка, плоскоклеточный рак полости рта, рак уротелиальных клеток и поджелудочной железы, а также опухоли головы и шеи [Brown et al., J. Immunol. 170: 1257-1266 (2003); Dong et al., Nat. Med. 8: 793-800 (2002); Wintterle et al., Cancer Res. 63: 7462-7467 (2003); Strome et al., Cancer Res., 63: 6501-6505 (2003); Thompson et al., Cancer Res. 66: 3381-5 (2006); Thompson et al., Clin. Cancer Res. 13: 1757-1761 (2007); Nomi et al., Clin.Cancer Res. 13: 2151-2157. (2007)]. Более неожиданным является то, что экспрессия лиганда PD на опухолевых клетках коррелирует с плохим прогнозом у больных раком людей при разных видах опухолей [обзор на эту тему представлен Okazaki and Honjo, Int. Immunol. 19: 813-824 (2007)].
Кроме того, Nomi et al. [Clin. Cancer Res. 13: 2151-2157 (2007)] демонстрируют терапевтическую эффективность блокирования связывания PD-L1 с PD-1 у мышиной модели агрессивного рака поджелудочной железы путем введения антитела, направленного на PD-1 или на PD-L1. Указанные антитела эффективно стимулируют инфильтрацию в опухоль опухольреактивных Т-клеток CD8+, обеспечивая повышающую регуляцию противоопухолевых эффекторов, таких как IFN-гамма, гранзим B и перфорин. Подобным образом, блокирование связывания PD-L1 и PD-1 под действием антител приводит к значительному ингибированию роста опухоли у мышиной модели плоскоклеточной карциномы [Tsushima et al., Oral Oncol. 42: 268-274 (2006)].
В других исследованиях обнаружено, что трансфекция линии мышиной мастоцитомы, содержащей PD-L1, приводит к снижению лизиса опухолевых клеток при совместном культивировании с опухоль-специфичным клоном CTL. После добавления моноклонального антитела против PD-L1 лизис восстанавливается [Iwai et al., Proc. Natl. Acad. Sci. U.S.A. 99: 12293-12297 (2002)]. Показано, что блокада взаимодействия PD1/PD-L1 in vivo увеличивает эффективность адоптивной терапии с переносом T-клеток мышиной модели опухоли [Strome et al., Cancer Res. 63: 6501-6505 (2003)]. Другим доказательством возможности применения PD-1 в способе лечения рака являются результаты экспериментов, проведенных с использованием мышей, нокаутных по PD-1, где клетки миеломы, экспрессирующие PD-L1, растут только у животных дикого типа (приводя к росту опухоли и связанной с ним гибели животных), но не у мышей с дефицитом PD-1 [Iwai Y. et al., Proc. Natl. Акад. Sci. USA, 99: 12293-12297 (2002)]. Совсем недавно продемонстрировано успешное, по меньшей мере на первоначальном этапе, применение антител к PD-1 (включающих гуманизированные мышиные моноклональные антитела против человеческого PD-1) для лечения рака у человека [см., например, US 8354509 B2, US 8008449 B2 и US 7595048 B2].
Антитела к PD-1 также можно использовать при хронической вирусной инфекции. Т-клетки памяти CD8+, образующиеся после острой вирусной инфекции, являются в высокой степени функциональными и представляют собой важный компонент защитного иммунитета. И наоборот, хронические инфекции часто характеризуются разной степенью функциональных нарушений (истощения) вирус-специфических Т-клеточных ответов, причем указанные нарушения являются основной причиной неспособности организма-хозяина устранить присутствующий патоген. Хотя на ранних стадиях инфекции образуются функциональные эффекторные Т-клетки, по мере течения хронической инфекции они постепенно утрачивают способность к функционированию. Barber et al. [Nature 439: 682-687 (2006)] демонстрируют, что у мышей, инфицированных лабораторным штаммом LCMV, развивается хроническая инфекция, приводящая к высокому уровню вируса в крови и других тканях. У этих мышей вначале развивается надежный Т-клеточный ответ, однако в конечном счете они поддаются инфекции в результате истощения Т-клеток. Barber et al. обнаружили, что число и функции эффекторных Т-клеток у хронически инфицированных мышей можно восстановить путем инъекции антитела, блокирующего взаимодействие между PD-1 и PD-L1.
Цитирование любой ссылки в настоящем описании не должно истолковываться как допущение того, что такая ссылка относится к "уровню техники, предшествующему" настоящей заявке.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам к PD-1 собак, которые обладают высоким сродством связывания в отношении PD-1 собак и способностью блокировать связывание PD-1 собак с PD-L1 собак. В конкретных вариантах осуществления такие антитела к PD-1 собак представляют собой мышиные антитела к PD-1 собак. В конкретных вариантах осуществления антитела к PD-1 собак обладают высоким сродством связывания в отношении PD-1 собак и способностью блокировать связывание PD-1 собак с PD-L2 собак.
Кроме того, настоящее изобретение относится к гипервариабельным участкам (CDR), входящим в состав указанных антител, и комбинации таких CDR (например, полученных из мышиных антител к PD-1 собак) с собачьими каркасными участками с получением канинизированного (caninized) антитела к PD-1 собак. Настоящее изобретение также относится к применению указанных антител для лечения таких заболеваний, как рак, и/или заболеваний, возникающих в результате инфекций.
Соответственно, настоящее изобретение предлагает уникальные наборы CDR из семи приведенных в качестве примера мышиных антител к PD-1 собак. Хотя каждое из семи приведенных в качестве примера мышиных антител к PD-1 собак имеет уникальный набор CDR, а именно, три CDR легкой цепи: CDR легкой цепи 1 (CDRL1), CDR легкой цепи 2 (CDRL2) и CDR легкой цепи 3 (CDRL3), и три CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3), как описано ниже, существует значительная гомология последовательностей в пределах каждой группы CDR, например, в совокупности CDRL1. Таким образом, настоящее изобретение предлагает не только аминокислотные последовательности шести CDR из семи приведенных в качестве примера мышиных антител к PD-1 собак, но и консервативно модифицированные варианты этих CDR, а также варианты, которые содержат такую же каноническую структуру (например, обладают такой же канонической структурой), и/или связываются с одним или несколькими (например, с 1-4 или со всеми) аминокислотными остатками PD-1 собак, которые входят в состав эпитопа PD-1 собак.
Таким образом, настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, способным специфически связываться с собачьим рецептором программируемой смерти 1 (PD-1 собак) и содержащим гипервариабельный участок легкой цепи 1 (VL CDR1), который содержит аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, и/или гипервариабельный участок легкой цепи 2 (VL CDR2), который содержит аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 21, и/или гипервариабельный участок легкой цепи 3 (VL CDR3), который содержит аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26, и/или гипервариабельный участок тяжелой цепи 1 (VH CDR1), где CDRH1 содержит аминокислотную последовательность SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30, и/или гипервариабельный участок тяжелой цепи 2 (VH CDR2), который содержит аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, и/или гипервариабельный участок тяжелой цепи 3 (VH CDR3), который содержит аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 114. В конкретных вариантах осуществления изобретения антитело представляет собой антитело млекопитающего. В более конкретных вариантах осуществления антитело представляет собой канинизированное антитело.
Соответственно, канинизированное антитело согласно настоящему изобретению, или его антигенсвязывающий фрагмент, содержит один или несколько из гипервариабельных участков тяжелой цепи 1 (VH CDR1), имеющих аминокислотные последовательности SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30. В другом варианте осуществления гипервариабельный участок тяжелой цепи 2 (VH CDR2) содержит аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35. В следующем варианте осуществления гипервариабельный участок тяжелой цепи 3 (VH CDR3) содержит аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 114. В конкретном варианте осуществления данного типа канинизированное антитело, или его антигенсвязывающий фрагмент, содержит как VH CDR1, содержащий аминокислотную последовательность SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30, так и VH CDR2, содержащий аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35. В другом таком варианте осуществления канинизированное антитело, или его антигенсвязывающий фрагмент, содержит как VH CDR1, содержащий аминокислотную последовательность SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30, так и VH CDR3, содержащий аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 114. В следующем таком варианте осуществления канинизированное антитело, или его антигенсвязывающий фрагмент, содержит как VH CDR2, содержащий аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34, или SEQ ID NO: 35, так и VH CDR3, содержащий аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 114. В другом таком варианте осуществления канинизированное антитело, или его антигенсвязывающий фрагмент, содержит VH CDR1, содержащий аминокислотную последовательность SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30, VH CDR2, содержащий аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, и VH CDR3, содержащий аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 114.
В конкретных вариантах осуществления канинизированное антитело, или его антигенсвязывающий фрагмент, также содержит гипервариабельный участок легкой цепи 1 (VL CDR1), содержащий аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15. В родственных вариантах осуществления гипервариабельный участок легкой цепи 2 (VL CDR2) содержит аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 21. В другом варианте осуществления гипервариабельный участок легкой цепи 3 (VL CDR3) содержит аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26. В конкретном варианте осуществления данного типа канинизированное антитело, или его антигенсвязывающий фрагмент, содержит как VL CDR1, содержащий аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, так и VL CDR2, содержащий аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 21.
В других таких вариантах осуществления канинизированное антитело, или его антигенсвязывающий фрагмент, содержит как VL CDR1, содержащий аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, так и VL CDR3, содержащий аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26. В других таких вариантах осуществления канинизированное антитело, или его антигенсвязывающий фрагмент, содержит как VL CDR2, содержащий аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 21, так и VL CDR3, содержащий аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26. В других таких вариантах осуществления канинизированное антитело, или его антигенсвязывающий фрагмент, содержит VL CDR1, содержащий аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, VL CDR2, содержащий аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 21, и VL CDR3, содержащий аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26.
В конкретных вариантах осуществления канинизированное антитело к PD-1 собак также содержит гипервариабельные участки (CDR), которые имеют следующие канонические структуры: H1-1, H2-1 и H3-6, соответственно для CDR1, CDR2 и CDR3 тяжелой цепи, т.е. CDR1 тяжелой цепи имеет каноническую структуру класса 1, CDR2 тяжелой цепи имеет каноническую структуру класса 1, а CDR3 тяжелой цепи имеет каноническую структуру класса 6. В более конкретных вариантах осуществления CDR соответствующих легких цепей имеют следующие канонические структуры: L1-3, L2-1 и L3-1, соответственно для CDR1, CDR2 и CDR3 легкой цепи. В других вариантах осуществления канинизированное антитело к PD-1 собак также содержит гипервариабельные участки (CDR), которые имеют следующие канонические структуры: H1-1, H2-1 и H3-11, соответственно для CDR1, CDR2 и CDR3 тяжелой цепи. В еще более конкретных вариантах осуществления данного типа CDR соответствующих легких цепей имеют следующие канонические структуры: L1-2A, L2-1 и L3-1, соответственно для CDR1, CDR2 и CDR3 легкой цепи. В других вариантах осуществления канинизированное антитело к PD-1 собак также содержит гипервариабельные участки (CDR), которые имеют следующие канонические структуры: H1-1, H2-2A и H3-11, соответственно для CDR1, CDR2, и CDR3 тяжелой цепи. В еще более конкретных вариантах осуществления данного типа CDR соответствующих легких цепей имеют канонические структуры: L1-2A, L2-1 и L3-1, соответственно для CDR1, CDR2 и CDR3 легкой цепи. В других вариантах осуществления канинизированное антитело к PD-1 собак дополнительно содержит гипервариабельные участки (CDR), которые имеют следующие канонические структуры: H1-1, H2-2A и H3-13, соответственно для CDR1, CDR2, и CDR3 тяжелой цепи. В еще более конкретных вариантах осуществления данного типа CDR соответствующих легких цепей имеют следующие канонические структуры: L1-4, L2-1 и L3-1, соответственно для CDR1, CDR2 и CDR3 легкой цепи.
Кроме того, настоящее изобретение относится к антителам к PD-1 собак, например, моноклональным антителам, которые содержат варианты CDR согласно настоящему изобретению, имеющие соответствующие канонические структуры, описанные в настоящем описании, и способны связываться с аминокислотной последовательностью SEQ ID NO: 103. В конкретных вариантах осуществления этого типа константа диссоциации (Kd) комплекса антитело-PD-1 собак находится в диапазоне от 1×10-5 до 1×10-12 M. В более конкретных вариантах осуществления антитела к PD-1 собак содержат варианты CDR настоящего изобретения, которые имеют соответствующие канонические структуры, описанные в настоящем описании, и связываются с аминокислотной последовательностью SEQ ID NO: 104.
Настоящее изобретение также относится к выделенному канинизированному антителу или его антигенсвязывающему фрагменту, способным специфически связывать рецептор программируемой смерти 1 (PD-1) и содержащим тяжелую цепь собачьего IgG и собачью легкую цепь каппа или лямбда. В конкретных вариантах осуществления этого типа собачью легкую цепь каппа или лямбда, которая содержит три гипервариабельных участка (CDR) легкой цепи: CDR легкой цепи 1 (CDRL1), CDR легкой цепи 2 (CDRL2) и CDR легкой цепи 3 (CDRL3); и тяжелую цепь собачьего IgG, которая содержит три CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3), получают из мышиных антител к PD-1 собак. Конкретные варианты осуществления канинизированных антител согласно настоящему изобретению и их антигенсвязывающих фрагментов связывают PD-1 собак и/или блокируют связывание PD-1 собак с собачьим лигандом программируемой смерти 1 (PD-L1).
В конкретных вариантах осуществления настоящее изобретение относится к выделенному антителу млекопитающего или его антигенсвязывающему фрагменту, которые обладают способностью специфически связывать собачий рецептор программируемой смерти 1 (PD-1 собак) и содержат три гипервариабельныех участка легкой цепи (CDR): CDR-1 легкой цепи (CDRL1), CDR легкой цепи 2 (CDRL2), и CDR легкой цепи 3 (CDRL3); и три CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3). В некоторых вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 13, вариант SEQ ID NO: 13, консервативно модифицированный вариант SEQ ID NO: 13, вариант SEQ ID NO: 13, который содержит каноническую структуру класса 3, SEQ ID NO: 15, вариант SEQ ID NO: 15, консервативно модифицированный вариант SEQ ID NO: 15, или вариант SEQ ID NO: 15, который содержит каноническую структуру класса 2А; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 16, вариант SEQ ID NO: 16, консервативно модифицированный вариант SEQ ID NO: 16, вариант SEQ ID NO: 16, который содержит каноническую структуру класса 1, SEQ ID NO: 18, вариант SEQ ID NO: 18, консервативно модифицированный вариант SEQ ID NO: 18, вариант SEQ ID NO: 18, который содержит каноническую структуру класса 1, SEQ ID NO: 19, вариант SEQ ID NO: 19, консервативно модифицированный вариант SEQ ID NO: 19, вариант SEQ ID NO: 19, который содержит каноническую структуру класса 1, SEQ ID NO: 20, вариант SEQ ID NO: 20, консервативно модифицированный вариант SEQ ID NO: 20, вариант SEQ ID NO: 20, который содержит каноническую структуру класса 1, SEQ ID NO: 21, вариант SEQ ID NO: 21, консервативно модифицированный вариант SEQ ID NO: 21, или вариант SEQ ID NO: 21, который содержит каноническую структуру класса 1, CDRL3 содержит аминокислотную последовательность SEQ ID NO: 22, вариант SEQ ID NO: 22, консервативно модифицированный вариант последовательности SEQ ID NO: 22, или вариант SEQ ID NO: 22, который содержит каноническую структуру класса 1, SEQ ID NO: 24, вариант SEQ ID NO: 24, консервативно модифицированный вариант SEQ ID NO: 24, вариант SEQ ID NO: 24, который содержит каноническую структуру класса 1, SEQ ID NO: 25, вариант SEQ ID NO: 25, консервативно модифицированный вариант SEQ ID NO: 25, вариант SEQ ID NO: 25, который содержит каноническую структуру класса 1, SEQ ID NO: 26, вариант SEQ ID NO: 26, консервативно модифицированный вариант SEQ ID NO: 26, или вариант SEQ ID NO: 26, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 27, вариант SEQ ID NO: 27, консервативно модифицированный вариант SEQ ID NO: 27, вариант SEQ ID NO: 27, который содержит каноническую структуру класса 1, SEQ ID NO: 29, вариант SEQ ID NO: 29, консервативно модифицированный вариант SEQ ID NO: 29, вариант SEQ ID NO: 29, который содержит каноническую структуру класса 1, SEQ ID NO: 30, вариант SEQ ID NO: 30, консервативно модифицированный вариант SEQ ID NO: 30, или вариант SEQ ID NO: 30, который содержит каноническую структуру класса 1, CDRH2 содержит аминокислотную последовательность SEQ ID NO: 31, вариант SEQ ID NO: 31, консервативно модифицированный вариант SEQ ID NO: 31, или вариант SEQ ID NO: 31, который содержит каноническую структуру класса 1, SEQ ID NO: 33, вариант SEQ ID NO: 33, консервативно модифицированный вариант SEQ ID NO: 33, вариант SEQ ID NO: 33, который содержит каноническую структуру класса 2А, SEQ ID NO: 34, вариант SEQ ID NO: 34, консервативно модифицированный вариант SEQ ID NO: 34, вариант SEQ ID NO: 34, который содержит каноническую структуру класса 1, SEQ ID NO: 35, вариант SEQ ID NO: 35, консервативно модифицированный вариант SEQ ID NO: 35, или вариант SEQ ID NO: 35, который содержит каноническую структуру класса 1, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 36, вариант SEQ ID NO: 36, консервативно модифицированный вариант SEQ ID NO: 36, вариант SEQ ID NO: 35, который содержит каноническую структуру класса 6, SEQ ID NO: 38, вариант SEQ ID NO: 38, консервативно модифицированный вариант SEQ ID NO: 38, или вариант последовательности SEQ ID NO: 38, который содержит каноническую структуру класса 11, SEQ ID NO: 114, вариант SEQ ID NO: 114, консервативно модифицированный вариант SEQ ID NO: 114, или вариант SEQ ID NO: 114, который содержит каноническую структуру класса 11. В конкретных вариантах осуществления изобретения антитело и антигенсвязывающий фрагмент связывают PD-1 собак и блокируют связывание PD-1 собак с собачьим лигандом программируемой смерти 1 (PD-L1). В родственных вариантах осуществления изобретения антитело также блокирует связывание PD-1 собак с собачьим лигандом программируемой смерти 2 (PD-L2). В конкретных вариантах осуществления выделенное антитело млекопитающего представляет собой канинизированное антитело. В более конкретных вариантах осуществления при связывании с собачьим PD-1 антитело или его антигенсвязывающий фрагмент связываются с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, из SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104.
В других вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 13, вариант SEQ ID NO: 13, консервативно модифицированный вариант SEQ ID NO: 13, или вариант SEQ ID NO: 13, который содержит каноническую структуру класса 3; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 16, вариант SEQ ID NO: 16, консервативно модифицированный вариант SEQ ID NO: 16, или вариант SEQ ID NO: 16, который содержит каноническую структуру класса 1; CDRL3 содержит аминокислотную последовательность SEQ ID NO: 22, вариант SEQ ID NO: 22, консервативно модифицированный вариант SEQ ID NO: 22, или вариант SEQ ID NO: 22, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 27, вариант SEQ ID NO: 27, консервативно модифицированный вариант SEQ ID NO: 27, или вариант SEQ ID NO: 27, который содержит каноническую структуру класса 1; CDRH2 содержит аминокислотную последовательность SEQ ID NO: 31, вариант SEQ ID NO: 31, консервативно модифицированный вариант SEQ ID NO: 31, и вариант SEQ ID NO: 31, который содержит каноническую структуру класса 1, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 36, вариант SEQ ID NO: 36, консервативно модифицированный вариант SEQ ID NO: 36, или вариант SEQ ID NO: 36, который содержит каноническую структуру класса 6. В конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104. В более конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав последовательности SEQ ID NO: 102.
В других вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 13, вариант SEQ ID NO: 13, консервативно модифицированный вариант SEQ ID NO: 13, или вариант SEQ ID NO: 13, который содержит каноническую структуру класса 3; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 19, вариант SEQ ID NO: 19, консервативно модифицированный вариант SEQ ID NO: 19, или вариант SEQ ID NO: 19, который содержит каноническую структуру класса 1; CDRL3 содержит аминокислотную последовательность SEQ ID NO: 25, вариант SEQ ID NO: 25, консервативно модифицированный вариант SEQ ID NO: 25, или вариант SEQ ID NO: 25, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 27, вариант SEQ ID NO: 27, консервативно модифицированный вариант SEQ ID NO: 27, или вариант SEQ ID NO: 27, который содержит каноническую структуру класса 1; CDRH2 содержит аминокислотную последовательность SEQ ID NO: 31, вариант SEQ ID NO: 31, консервативно модифицированный вариант SEQ ID NO: 31, и вариант SEQ ID NO: 31, который содержит каноническую структуру класса 1, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 36, вариант SEQ ID NO: 36, консервативно модифицированный вариант SEQ ID NO: 36, или вариант SEQ ID NO: 36, который содержит каноническую структуру класса 6. В конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104. В более конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с одним из аминокислотных остатков R75 и R90, входящих в состав SEQ ID NO: 2, или с обоими указанными аминокислотными остатками.
В других вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 13, вариант SEQ ID NO: 13, консервативно модифицированный вариант SEQ ID NO: 13, или вариант SEQ ID NO: 13, который содержит каноническую структуру класса 3; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 20, вариант SEQ ID NO: 20, консервативно модифицированный вариант SEQ ID NO: 20, или вариант SEQ ID NO: 20, который содержит каноническую структуру класса 1; CDRL3 содержит аминокислотную последовательность SEQ ID NO: 25, вариант SEQ ID NO: 25, консервативно модифицированный вариант SEQ ID NO: 25, или вариант SEQ ID NO: 25, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 27, вариант SEQ ID NO: 27, консервативно модифицированный вариант SEQ ID NO: 27, или вариант SEQ ID NO: 27, который содержит каноническую структуру класса 1; CDRH2 содержит аминокислотную последовательность SEQ ID NO: 34, вариант SEQ ID NO: 34, консервативно модифицированный вариант SEQ ID NO: 34, и вариант SEQ ID NO: 34, который содержит каноническую структуру класса 1, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 36, вариант SEQ ID NO: 36, консервативно модифицированный вариант SEQ ID NO: 36, или вариант SEQ ID NO: 36, который содержит каноническую структуру класса 6. В конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104.
В других вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 13, вариант SEQ ID NO: 13, консервативно модифицированный вариант SEQ ID NO: 13, или вариант SEQ ID NO: 13, который содержит каноническую структуру класса 3; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 16, вариант SEQ ID NO: 16, консервативно модифицированный вариант SEQ ID NO: 16, или вариант SEQ ID NO: 16, который содержит каноническую структуру класса 1; CDRL3 содержит аминокислотную последовательность SEQ ID NO: 22, вариант SEQ ID NO: 22, консервативно модифицированный вариант SEQ ID NO: 22, или вариант SEQ ID NO: 22, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 30, вариант SEQ ID NO: 30, консервативно модифицированный вариант SEQ ID NO: 30, или вариант SEQ ID NO: 30, который содержит каноническую структуру класса 1; CDRH2 содержит аминокислотную последовательность SEQ ID NO: 31, вариант SEQ ID NO: 31, консервативно модифицированный вариант SEQ ID NO: 31, и вариант SEQ ID NO: 31, который содержит каноническую структуру класса 1, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 36, вариант SEQ ID NO: 36, консервативно модифицированный вариант SEQ ID NO: 36, или вариант SEQ ID NO: 36, который содержит каноническую структуру класса 6. В конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104.
В других вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 15, вариант SEQ ID NO: 15, консервативно модифицированный вариант SEQ ID NO: 15, или вариант SEQ ID NO: 15, который содержит каноническую структуру класса 2А; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 18, вариант SEQ ID NO: 18, консервативно модифицированный вариант SEQ ID NO: 18, или вариант SEQ ID NO: 18, который содержит каноническую структуру класса 1; CDRL3 содержит аминокислотную последовательность SEQ ID NO: 24, вариант SEQ ID NO: 24, консервативно модифицированный вариант SEQ ID NO: 24, или вариант SEQ ID NO: 24, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 29, вариант SEQ ID NO: 29, консервативно модифицированный вариант SEQ ID NO: 29, или вариант SEQ ID NO: 29, который содержит каноническую структуру класса 1; CDRH2 содержит аминокислотную последовательность SEQ ID NO: 33, вариант SEQ ID NO: 33, консервативно модифицированный вариант SEQ ID NO: 33 и вариант SEQ ID NO: 33, который содержит каноническую структуру класса 1, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 38, вариант SEQ ID NO: 38, консервативно модифицированный вариант SEQ ID NO: 38, или вариант SEQ ID NO: 38, который содержит каноническую структуру класса 11. В конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, из SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104. В более конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав SEQ ID NO: 84.
В следующих вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 15, вариант SEQ ID NO: 15, консервативно модифицированный вариант SEQ ID NO: 15, или вариант SEQ ID NO: 15, который содержит каноническую структуру класса 2А; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 21, вариант SEQ ID NO: 21, консервативно модифицированный вариант SEQ ID NO: 21, или вариант SEQ ID NO: 21, который содержит каноническую структуру класса 1; CDRL3 содержит аминокислотную последовательность SEQ ID NO: 26, вариант SEQ ID NO: 26, консервативно модифицированный вариант SEQ ID NO: 26, или вариант SEQ ID NO: 26, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 29, вариант SEQ ID NO: 29, консервативно модифицированный вариант SEQ ID NO: 29, или вариант SEQ ID NO: 29, который содержит каноническую структуру класса 1; CDRH2 содержит аминокислотную последовательность SEQ ID NO: 35, вариант SEQ ID NO: 35, консервативно модифицированный вариант SEQ ID NO: 35, и вариант SEQ ID NO: 35, который содержит каноническую структуру класса 1, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 114, вариант SEQ ID NO: 114, консервативно модифицированный вариант SEQ ID NO: 114 или вариант SEQ ID NO: 114, который содержит каноническую структуру класса 11. В конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104.
В других вариантах осуществления CDRL1 содержит аминокислотную последовательность SEQ ID NO: 14, вариант SEQ ID NO: 14, консервативно модифицированный вариант SEQ ID NO: 14, или вариант SEQ ID NO: 14, который содержит каноническую структуру класса 4; CDRL2 содержит аминокислотную последовательность SEQ ID NO: 17, вариант SEQ ID NO: 17, консервативно модифицированный вариант SEQ ID NO: 17, или вариант SEQ ID NO: 17, который содержит каноническую структуру класса 1; CDRL3 содержит аминокислотную последовательность SEQ ID NO: 23, вариант SEQ ID NO: 23, консервативно модифицированный вариант SEQ ID NO: 23, или вариант SEQ ID NO: 23, который содержит каноническую структуру класса 1, CDRH1 содержит аминокислотную последовательность SEQ ID NO: 28, вариант SEQ ID NO: 28, консервативно модифицированный вариант SEQ ID NO: 28, или вариант SEQ ID NO: 28, который содержит каноническую структуру класса 1; CDRH2 содержит аминокислотную последовательность SEQ ID NO: 32, вариант SEQ ID NO: 32, консервативно модифицированный вариант SEQ ID NO: 32, и вариант SEQ ID NO: 32, который содержит каноническую структуру класса 2А, CDRH3 содержит аминокислотную последовательность SEQ ID NO: 37, вариант SEQ ID NO: 37, консервативно модифицированный вариант SEQ ID NO: 37, или вариант SEQ ID NO: 37, который содержит каноническую структуру класса 13. В конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, из SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и/или SEQ ID NO: 104. В более конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним аминокислотным остатком, входящим в состав SEQ ID NO: 83, SEQ ID NO: 84 и/или SEQ ID NO: 100. В более конкретных вариантах осуществления при связывании с собачьим PD-1 антитело, или его антигенсвязывающий фрагмент, связывается с по меньшей мере одним или несколькими из следующих аминокислотных остатков аргинина: R62, R69, R72 и R75, входящих в состав SEQ ID NO: 2.
Настоящее изобретение относится к антителам и антигенсвязывающим фрагментам, способным специфически связывать собачий рецептор программируемой смерти 1 (PD-1 собак), так, что при этом антитело связывается с по меньшей мере одним аминокислотным остатком в последовательности SEQ ID NO: 103. В конкретных вариантах осуществления данного типа антитела и антигенсвязывающие фрагменты связывают PD-1 собак и блокируют связывание PD-1 собак с собачьим лигандом программируемой смерти 1 (PD-L1). В более конкретных вариантах осуществления антитела и антигенсвязывающие фрагменты связывают PD-1 собак, а также блокируют связывание PD-1 собак с собачьим лигандом программируемой смерти 2 (PD-L2).
Соответственно, в конкретных вариантах осуществления при связывании с собачьим PD-1 антитело (такое как антитело, содержащее один или несколько вариантов CDR, например, вариант, включающий в себя консервативно модифицированный вариант и/или вариант, который содержит каноническую структуру определенного класса) связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или нескольких из следующих аминокислотных последовательностей: SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102 и/или SEQ ID NO: 104. В еще более конкретных вариантах осуществления при связывании с собачьим PD-1 антитела или их антигенсвязывающие фрагменты связываются с одним или несколькими из следующих остатков аргинина: R62, R69, R72, R75 и R90, входящих в состав SEQ ID NO: 2. В еще более конкретных вариантах осуществления при связывании с собачьим PD-1 антитела или их антигенсвязывающие фрагменты связываются, по меньшей мере, с одним аминокислотным остатком в последовательности SEQ ID NO: 104. В более конкретных вариантах осуществления при связывании с собачьим PD-1 антитела или их антигенсвязывающие фрагменты связываются с одним или несколькими из следующих остатков аргинина: R62, R69, R72 и R75, входящих в состав SEQ ID NO: 2. В еще более конкретных вариантах осуществления при связывании с собачьим PD-1 антитела или их антигенсвязывающие фрагменты связываются с R75, входящим в состав SEQ ID NO: 2.
Настоящее изобретение также относится к антителам млекопитающих или их антигенсвязывающим фрагментам, способным связываться с собачьим PD-1 с константой диссоциации (Kd) ниже 1×10-12М (например, 1×10-13 М, или ниже). В конкретных вариантах осуществления антитела млекопитающих или антигенсвязывающие фрагменты связываются с собачьим PD-1 с константой диссоциации от 1×10-5М до 1×10-12 М. В более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим PD-1 с константой диссоциации от 1×10-7М до 1×10-11 М. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим PD-1 с константой диссоциации от 1×10-8М до 1×10-11 М. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим PD-1 с константой диссоциации от 1×10-8М до 1×10-10 М.
Настоящее изобретение также относится к антителам млекопитающих или их антигенсвязывающим фрагментам, которые связываются с собачьим PD-1 со скоростью (kon), превышающей 1×107 М-1с-1. В конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим PD-1 со скоростью, варьирующей от 1×102 М-1с-1 до 1×107 М-1с-1. В более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим PD-1 со скоростью, варьирующей от 1×103 М-1с-1 до 1×106 М-1с-1. В еще более конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты связываются с собачьим PD-1 со скоростью, варьирующей от 1×103 М-1с-1 до 1×105 М-1с-1. В еще более конкретных вариантах осуществления антитела млекопитающих или ихантигенсвязывающие фрагменты связываются с собачьим PD-1 со скоростью, варьирующей от 1×104 М-1с-1 до 1×105 М-1с-1.
Настоящее изобретение также относится к антителам млекопитающих или их антигенсвязывающим фрагментам, которые связываются с собачьим PD-1, образуя комплексы, характеризующиеся скоростью диссоциации (koff) ниже 1×10-7 с-1. В конкретных вариантах осуществления скорость диссоциации комплексов антител млекопитающих или их антигенсвязывающих фрагментов с собачьим PD-1 находится в диапазоне от 1×10-3 с-1 до 1×10-8 с-1. В более конкретных вариантах осуществления скорость диссоциации комплексов антител млекопитающих или их антигенсвязывающих фрагментов с собачьим PD-1 находится в диапазоне от 1×10-4 с-1 до 1×10-7 с-1. В еще более конкретных вариантах осуществления скорость диссоциации комплексов антител млекопитающих или их антигенсвязывающих фрагментов с собачьим PD-1 находится в диапазоне от 1×10-5 с-1 до 1×10-7 с-1.
В родственных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты, стимулируют ответы на опухоль или патоген, характеризующиеся антиген-специфической памятью. В конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты стимулируют гуморальный ответ in vivo. В других конкретных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты стимулируют иммунный ответ у животного. В более конкретных вариантах осуществления животное является собакой. В родственном варианте осуществления животное является кошкой.
Соответственно, любое из антител согласно настоящему изобретению может обладать одним, двумя, тремя, четырьмя, пятью, или всеми указанными свойствами, включающими в себя вышеуказанные константы диссоциации комплексов с собачьим PD-1, вышеуказанные скорости образования комплексов с собачьим PD-1, вышеуказанные скорости диссоциации комплексов антитела с собачьим PD-1, стимуляцию ответов на опухоль или патоген, характеризующихся антиген-специфической памятью, стимуляцию гуморального ответа in vivo, и/или стимуляцию иммунного ответа у животного.
Как указано выше, антитела (и их антигенсвязывающие фрагменты) согласно настоящему изобретению, в том числе вышеуказанные антитела (и их антигенсвязывающие фрагменты), могут представлять собой моноклональные антитела (и их антигенсвязывающие фрагменты), антитела млекопитающих (и их антигенсвязывающие фрагменты), например, мышиные антитела (антитела мыши) (и их антигенсвязывающие фрагменты), канинизированные антитела (и их антигенсвязывающие фрагменты), включающие в себя канинизированные мышиные антитела (и их антигенсвязывающие фрагменты), причем в некоторых вариантах осуществления антитела (и их антигенсвязывающие фрагменты) являются выделенными.
Настоящее изобретение также относится к нуклеиновым кислотам (в том числе выделенным нуклеиновым кислотам), которые кодируют одну из легких цепей канинизированного антитела согласно настоящему изобретению. Подобным образом, настоящее изобретение относится к выделенным нуклеиновым кислотам, которые кодируют одну из тяжелых цепей канинизированного антитела согласно настоящему изобретению. Примеры конкретных нуклеотидных последовательностей приведены в настоящем описании.
Настоящее изобретение также относится к векторам экспрессии, которые содержат одну или несколько из нуклеиновых кислот (включающих выделенные нуклеиновые кислоты) согласно настоящему изобретению. Настоящее изобретение также относится к клеткам-хозяевам, которые содержат один или несколько векторов экспрессии согласно настоящему изобретению.
В конкретных вариантах осуществления изобретения антитело представляет собой рекомбинантное антитело или его антигенсвязывающий фрагмент. В родственных вариантах осуществления вариабельный домен тяжелой цепи и вариабельный домен легкой цепи соединены между собой гибким линкером с образованием одноцепочечного антитела.
В конкретных вариантах осуществления изобретения антитело или его антигенсвязывающий фрагмент представляет собой фрагмент Fab.
В других вариантах осуществления изобретения антитело или его антигенсвязывающий фрагмент представляет собой фрагмент Fab'. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент представляет собой фрагмент (Fab')2. В следующих вариантах осуществления антитело или его антигенсвязывающий фрагмент представляет собой диатело. В конкретных вариантах осуществления антитело или его антигенсвязывающий фрагмент представляет собой доменное антитело. В конкретных вариантах осуществления антитело или его антигенсвязывающий фрагмент представляет собой камелизированное (camelized) однодоменное антитело.
В конкретных вариантах осуществления канинизированное мышиное антитело к PD-1 собак, или его антигенсвязывающий фрагмент, усиливает иммунный ответ у собаки, подлежащей лечению.
Настоящее изобретение также относится к выделенным нуклеиновым кислотам, которые кодируют канинизированные мышиные антитела к PD-1 собак, или их фрагменты. В родственных вариантах осуществления такие антитела или их антигенсвязывающие фрагменты можно использовать в способе получения лекарственного средства для лечения рака у собак. В качестве альтернативы, или помимо вышеуказанного, настоящее изобретение предлагает применение любого из антител или фрагментов антител согласно настоящему изобретению для диагностики. В других вариантах осуществления изобретение относится к набору, содержащему одно из раскрытых здесь канинизированных антител или их антигенсвязывающих фрагментов.
В других вариантах осуществления изобретение относится к вектору экспрессии, содержащему выделенную нуклеиновую кислоту, кодирующую одно из канинизированное мышиных антител к PD-1 собак или их антигенсвязывающих фрагментов согласно настоящему изобретению. Изобретение также относится к клетке-хозяину, содержащей один из описанных в настоящем описании векторов экспрессии. В конкретных вариантах осуществления указанные нуклеиновые кислоты, векторы экспрессии или полипептиды согласно настоящему изобретению можно использовать в способах получения антител.
Настоящее изобретение также относится к антигенным пептидам (в том числе выделенным антигенным пептидам), которые содержат 80 или менее аминокислотных остатков, составляющих аминокислотную последовательность SEQ ID NO: 103, и/или SEQ ID NO: 83, и/или SEQ ID NO: 84, и/или SEQ ID NO: 99, и/или SEQ ID NO: 100, и/или SEQ ID NO: 101, и/или SEQ ID NO: 102, и/или SEQ ID NO: 104. В родственных вариантах осуществления антигенные пептиды (в том числе выделенные пептиды) содержат 60 или менее аминокислотных остатков, составляющих аминокислотную последовательность SEQ ID NO: 103, и/или SEQ ID NO: 83, и/или SEQ ID NO: 84 и/или SEQ ID NO: 99, и/или SEQ ID NO: 100, и/или SEQ ID NO: 101, и/или SEQ ID NO: 102, и/или SEQ ID NO: 104. В других вариантах осуществления изобретения антигенные пептиды содержат от 10 до 44 аминокислотных остатков, входящих в состав аминокислотной последовательности SEQ ID NO: 103. В других вариантах осуществления изобретения пептиды содержат от 15 до 45 аминокислотных остатков, входящих в состав аминокислотной последовательности SEQ ID NO: 103.
Настоящее изобретение также относится к антигенным пептидам (в том числе выделенным пептидам), которые содержат 80 или менее аминокислотных остатков, составляющих аминокислотную последовательность, которая на 80%, 85%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 103 и/или SEQ ID NO: 83, и/или SEQ ID NO: 84, и/или SEQ ID NO: 99, и/или SEQ ID NO: 100, и/или SEQ ID NO: 101, и/или SEQ ID NO: 102, и/или SEQ ID NO: 104, и способна связываться с выделенным антителом млекопитающего, или антигенсвязывающим фрагментом настоящего изобретения. В родственных вариантах осуществления антигенные пептиды (в том числе выделенные антигенные пептиды) содержат 60 или менее аминокислотных остатков, составляющих аминокислотную последовательность, которая на 80%, 85%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 103, и/или SEQ ID NO: 83, и/или SEQ ID NO: 84, и/или SEQ ID NO: 9, и/или SEQ ID NO: 100, и/или SEQ ID NO: 101, и/или SEQ ID NO: NO: 102, и/или SEQ ID NO: 104, и способна связываться с выделенным антителом млекопитающего или его антигенсвязывающим фрагментом. В других вариантах осуществления пептиды содержат от 10 до 44 аминокислотных остатков аминокислотной последовательности, которая на 80%, 85%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 103, и/или SEQ ID NO: 83, и/или SEQ ID NO: 84, и/или SEQ ID NO: 99, и/или SEQ ID NO: 100, и/или SEQ ID NO: 101, и/или SEQ ID NO: 102, и/или SEQ ID NO: NO: 104, и способна связываться с выделенным антителом млекопитающего или его антигенсвязывающим фрагментом. В конкретных вариантах осуществления изобретения антитело представляет собой IB5. В других вариантах осуществления антитело представляет собой 3B6. В других конкретных вариантах осуществления изобретения антитело представляет собой 2H9. В следующих вариантах осуществления антитело представляет собой 2G9. В других вариантах осуществления изобретения антитело представляет собой 1Al. В других вариантах осуществления антитело представляет собой 1E4.
Настоящее изобретение также относится к гибридным белкам, которые содержат один из вышеупомянутых антигенных пептидов. В конкретном варианте осуществления гибридный белок содержит такой антигенный пептид и Fc-участок антитела IgG отличного от собаки млекопитающего. В более конкретном варианте осуществления гибридный белок содержит Fc-участок антитела IgG отличного от собаки млекопитающего. В некоторых вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой мышиный IgG. В альтернативных вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой человеческий IgG. В других вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой лошадиный IgG. В других вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой свиной IgG. В других вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой бычий IgG.
В конкретных вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой IgG1. В других вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой IgG2a. В других вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой IgG3. В следующих вариантах осуществления антитело IgG отличного от собаки млекопитающего представляет собой IgG4.
В других вариантах осуществления гибридный белок содержит один из вышеупомянутых антигенных пептидов и мальтоза-связывающий белок. В других вариантах осуществления настоящего изобретения гибридный белок содержит один из указанных выше антигенных пептидов и бета-галактозидазу. В других вариантах осуществления гибридный белок содержит один из вышеупомянутых антигенных пептидов и глутатион-S-трансферазу. В других вариантах осуществления гибридный белок содержит один из указанных выше антигенных пептидов и тиоредоксин. В следующих вариантах осуществления гибридный белок содержит один из вышеупомянутых антигенных пептидов и Gro EL. В других вариантах осуществления гибридный белок содержит один из вышеупомянутых антигенных пептидов и NusA.
Настоящее изобретение также относится к нуклеиновым кислотам (в том числе выделенным нуклеиновым кислотам), которые кодируют антигенные пептиды и соответствующие гибридные белки согласно настоящему изобретению. Настоящее изобретение также относится к векторам экспрессии, которые содержат указанные нуклеиновые кислоты.
Кроме того, настоящее изобретение относится к фармацевтическим композициям, содержащим антитела к PD-1 собак или их антигенсвязывающие фрагменты согласно настоящему изобретению, антигенные пептиды (в том числе выделенные антигенные пептиды) из PD-1 собак, гибридные белки, содержащие антигенные пептиды из PD-1 собак согласно настоящему изобретению, нуклеиновые кислоты (в том числе выделенные нуклеиновые кислоты), кодирующие антигенные фрагменты и/или гибридные белки согласно настоящему изобретению, векторы экспрессии, содержащие такие нуклеиновые кислоты, или любую их комбинацию, а также фармацевтически приемлемый носитель или разбавитель.
Кроме того, настоящее изобретение относится к способам повышения активности иммунной клетки, включающим введение пациенту, терапевтически эффективного количества таких фармацевтических композиций. В некоторых вариантах осуществления способ используют для лечения рака. В других вариантах осуществления способ используют для лечения инфекции или инфекционного заболевания. В других вариантах осуществления канинизированное антитело согласно настоящему изобретению или его антигенсвязывающий фрагмент используют в качестве адъювантной вакцины.
Нижеследующие краткое описание чертежей и подробное описание изобретения приведены для облегчения понимания указанных и других аспектов настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 показана реакционная способность мышиных mAb против внеклеточного домена PD-1 собак. Разные мышиные mAb тестируют на связывание с внеклеточным доменом PD-1 собак методом ELISA. Тестируемые mAb обозначаются как ♦ 3B6, ■ 5F3, - 5G5, × 4D12, * 2H9, • 2C12, + 2G9, ▲ неспецифическое mAb.
На фигуре 2 показана реакционная способность мышиных mAb в отношении экспрессируемого на клеточной поверхности PD-1 собак. Разные мышиные mAb тестируют на способность связываться с собачьим PD-1, экспрессируемым на клетках СНО, методом CELISA. Антитела обозначают следующим образом: 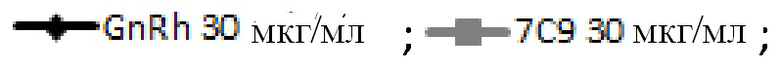
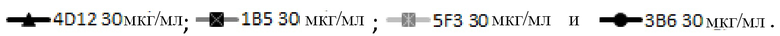
На фигуре 3А показана блокада лиганда мышиными mAb к PD-1 собак. Разные мышиные mAb тестируют на способность ингибировать связывание PD-1, экспрессируемого на клетках CHO, с PD-L1. Антитела обозначают следующим образом: 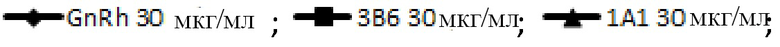
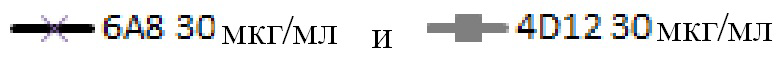 .
.
На фигуре 3В показана блокада лиганда мышиными mAb к PD-1 собак. Разные мышиные mAb тестируют на способность ингибировать связывание PD-1, экспрессируемого на клетках CHO, с PD-L1. Антитела обозначают следующим образом: 
 .
.
На фигуре 3С показана блокада лиганда мышиными mAb к PD-1 собак. Разные мышиные mAb тестируют на способность ингибировать связывание PD-1, экспрессируемого на клетках CHO, с PD-L1. Антитела обозначают следующим образом: 
 .
.
На фигуре 4 показано связывание мышиных mAb с собачьим PD-1 на Т-клетках CD+ в РВМС здоровых собак. Разные мышиные mAb тестируют на способность связываться с собачьим PD-1, экспрессируемым на Т-клетках CD+ в РВМС здоровых собак. Антитела тестируют, используя 2-кратные разведения, начиная с диапазона 0,156-20 мкг/мл.
На фигуре 5 показано связывание мышиных mAb с собачьим PD-1 на Т-клетках CD+ в РВМС собак, страдающих от рака. Указанные мышиные mAb тестируют на способность связываться с собачьим PD-1, экспрессируемым на Т-клетках CD8+ собак, страдающих от рака (саркома). Антитела тестируют в концентрации 2,5 и 5 мкг/мл.
На фигуре 6 показана секреция цитокинов, индуцированная мышиными mAb к PD-1 собак. Разные мышиные mAb тестируют на способность индуцировать секрецию цитокинов из РВМС здоровых собак.
На фигуре 7 показана секреция цитокинов, индуцированная мышиными mAb к PD-1 собак. Разные мышиные mAb тестируют на способность индуцировать секрецию цитокинов из РВМС собак, страдающих от рака (гемангиосаркомы).
На фигуре 8 приведены результаты выравнивания константных участков тяжелых цепей (CH) собачьих IgGB, утративших функцию ADCC. Изображены собачий IgB дикого типа [cIgGB дикого типа], собачий IgGB(+)A-шарнир [cIgGB(+)A-шарнир], собачий IgGB(+)D-шарнир [cIgGB(+)D-шарнир] и собачий IgGB(-)ADCC [cIgGB(-) ADCC]. (+) A-шарнир получают в результате замены на шарнирный участок IgG-A, содержащий аминокислотные замены на лизин и аспарагин, как показано; (+) D-шарнир получают в результате замены на шарнирный участок IgG-D, содержащий аминокислотные замены на лизин и аспарагин, как показано. (-)ADCC означает наличие аминокислотной замены на лизин и аспарагин.
На фигуре 9A изображена поверхность взаимодействия PD-1 собак и канинизированного антитела 2G9. Аминокислотные положения соответствуют положениям в аминокислотной последовательности PD-1, не содержащей сигнальной последовательности, т.е. SEQ ID NO: 2. Определение проводят методом химической сшивки, масс-спектрометрии MALDI для частиц с большой массой и масс-спектрометрии nLC-Orbitrap.
На фигуре 9В изображена поверхность взаимодействия PD-1 собак и канинизированного антитела 3B6. Аминокислотные положения соответствуют положениям в аминокислотной последовательности PD-1, не содержащей сигнальной последовательности, т.е. SEQ ID NO: 2. Определение проводят методом химической сшивки, масс-спектрометрии MALDI для частиц с большой массой и масс-спектрометрии nLC-Orbitrap.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
СОКРАЩЕНИЯ
В подробном описании и примерах настоящего изобретения используют следующие сокращения:
ADCC - антитело-зависимая клеточная цитотоксичность
CDC - комплемент-зависимая цитотоксичность
CDR - гипервариабельный участок вариабельного домена иммуноглобулина, определяемый с использованием системы нумерации Kabat
CHO - клетки яичника китайского хомячка
ЕС50 - концентрация, обеспечиваящая 50% эффективность или 50% связывание
ELISA - иммуноферментный анализ
FR - каркасной участок антитела: вариабельный домен иммуноглобулина за исключением участков CDR
HRP - пероксидаза хрена
IFN - интерферон
IC50 - концентрация, обеспечивающая 50% ингибирование
IgG - иммуноглобулин G
Kabat - система выравнивания и нумерации иммуноглобулинов, впервые разработанная Elvin A. Kabat [Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)]
mAb - моноклональное антитело (также обозначаемое Mab или MAb)
MES - 2-(N-морфолино)этансульфоновая кислота
MOA - механизм действия
NHS - нормальная человеческая сыворотка
ПЦР - полимеразная цепная реакция
PK - фармакокинетика
SEB - стафилококковый энетротоксин B
TT - столбнячный анатоксин
V-участок - сегмент цепи IgG, последовательность которого варьирует среди разных антител. Он простирается от остатка 109 легкой цепи до остатка 113 тяжелой цепи, в соответствии с нумерацией Kabat
VH - вариабельный участок тяжелой цепи иммуноглобулина
VK - вариабельной участок легкой цепи каппа иммуноглобулина
ОПРЕДЕЛЕНИЯ
Для облегчения понимания настоящего изобретения ниже приведены конкретные определения некоторых технических и научных терминов. Если в настоящем описании специально не указано иначе, все другие используемые здесь технические и научные термины имеют традиционные значения, известные рядовым специалистам в области, к которой принадлежит настоящее изобретение.
В данном описании, если из контекста явно не следует иное, термины, встречающиеся в единственном числе, включают в себя соответствующие ссылки на множественное число.
Термин "активация", используемый в применении к клеткам или рецепторам, относится к активации клетки или рецептора, или к обработке клеток или рецепторов лигандом, если контекст в явном виде не указывает иное. Термин "лиганд" охватывает природные и синтетические лиганды, например, цитокины, варианты, аналоги, мутеины цитокинов и связывающие соединения, полученные из антител. Термин "лиганд" также охватывает небольшие молекулы, например, пептидные миметики цитокинов и пептидные миметики антител. Термин "активация" может относиться к активации клеток под действием внутренних механизмов, а также внешних факторов или факторов окружающей среды.
Термин "активность" в применении к молекуле может относиться к связыванию молекулы с лигандом или с рецептором, к каталитической активности; к способности стимулировать экспрессию гена или клеточный сигнальный путь, дифференцировку или созревание клеток; к антигенной активности, к модуляции активности других молекул и т.п. Термин "активность" в применении к молекуле также может относиться к способности модулировать или поддерживать межклеточные взаимодействия, такие как адгезия, или к способности поддерживать структуру клетки, например, клеточные мембраны или цитоскелет. "Активность" также может включать в себя удельную активность, такую как [каталитическая активность]/[мг белка] или [иммунологическая активность]/[мг белка], накопление в биологическом отсеке и т.п. "Активность" может включать в себя способность модулировать компоненты врожденной или адаптивной иммунной системы.
Термины "введение" и "обработка", используемые в применении к животному, такому как подопытная собака, а также к клеткам, тканям, органам или биологическим жидкостям собаки, включает в себя приведение экзогенных фармацевтических, терапевтических, диагностических средств или композиций в контакт с животным, например, с организмом, клеткой, тканью, органом или биологической жидкостью собаки. Обработка клетки включает в себя приведение реагента в контакт с клеткой, а также приведение реагента в контакт с жидкостью, если жидкость находится в контакте с клеткой. Термины "введение" и "обработка" также включают в себя обработку in vitro и ex vivo, например, клетки, реагентом, диагностическим средством, связывающим соединением или другой клеткой. Термин "индивидуум" включает в себя любой организм, предпочтительно животное, более предпочтительно млекопитающее (такое как собака, кошка или человек), и наиболее предпочтительно собаку.
В данном описании термин "замена аминокислотного остатка" другим аминокислотным остатком в аминокислотной последовательности антитела, например, эквивалентен термину "замещение аминокислотного остатка" другим аминокислотным остатком, и означает, что конкретный аминокислотный остаток в определенном положении аминокислотной последовательности заменяют (или замещают) другим аминокислотным остатком. Такие замены можно специально спланировать, например, можно целенаправленно заменить аланин на серин в определенном положении аминокислотной последовательности, с помощью, например, технологии рекомбинантных ДНК. Альтернативно, конкретный аминокислотный остаток или последовательность аминокислотных остатков антитела могут быть заменены одним или несколькими аминокислотными остатками, в результате более естественных процессов отбора, например, обеспечивающих способность антитела, продуцируемого клеткой, связываться с заданным участком на конкретном антигене, например, на антигене, содержащем эпитоп, или его часть, и/или обеспечивающих продукцию антитела, содержащего конкретный CDR, который имеет такую же каноническую структуру, как и замещаемый CDR. Такие замены/замещения могут привести к образованию "вариантных" CDR и/или вариантных антител.
Термин "лечить" или "лечение" относится к внутреннему или наружнему введению терапевтического средства, такого как композиция, содержащая антитело или антигенсвязывающий фрагмент настоящего изобретения, собаке или пациенту, имеющим один или несколько симптомов заболевания, или предрасположенным к заболеванию, против которого направлена терапевтическая активность указанного средства.
Как правило, средство вводят в количестве, эффективном для облегчения и/или улучшения одного или нескольких симптомов заболевания у подлежащего лечению субъекта, или подлежащей лечению популяции, в результате индукции регрессии или ингибирования развития такого симптома (симптомов) в клинически значимой степени. Количество терапевтического средства, эффективное для облегчения какого-либо конкретного симптома заболевания (также называемое "терапевтически эффективное количество"), может варьировать в зависимости от таких факторов, как болезненное состояние, возраст и масса пациента (например, собаки), а также способность фармацевтической композиции вызывать желательный ответ у индивидуума. Облегчение или улучшение симптома заболевания можно оценить с помощью любого клинического анализа, обычно используемого ветеринарами или другими специалистами в области медицины для оценки тяжести или степени развития данного симптома. Хотя вариант осуществления настоящего изобретения (например, способ лечения или изделие промышленного производства) может не обеспечивать эффективного облегчения целевого симптома (симптомов) заболевания у каждого субъекта, оно должно облегчать целевой симптом (симптомы) заболевания у статистически значимого числа индивидуумов, определенного с помощью любого статистического теста, известного в данной области, такого как t-тест Стьюдента, тест хи2, U-тест Манна и Уитни, тест Крускала-Уоллиса (H-тест), тест Джонкхиера-Терпстра и тест Вилкоксона.
Термин "лечение", используемый в применении к человеку, животному (такому как собака) или подопытному субъекту, относится к терапевтическому лечению, а также к исследовательскому и диагностическому применению. Термин "лечение", используемый в применении к человеку, животному (такому как собака) или подопытному субъекту, либо к клетке, ткани или органу, включает в себя приведение антител или антигенсвязывающих фрагментов настоящего изобретения в контакт с организмом собаки или другого животного, с клеткой, тканью, физиологическим компартментом, или физиологической жидкостью.
Если не указано иное, используемый в данном описании термин "собака" включает в себя домашних собак, Canis lupus familiaris или Canis Familiaris.
Используемый в данном описании термин "кошка" относится к любому члену семейства кошачьих. Члены данного семейства включают в себя диких, живущих в зоопарке и домашних животных, например, членов подсемейств Felinae, таких как кошки, львы, тигры, пумы, ягуары, леопарды, снежные барсы, пантеры, североамериканские горные львы, гепарды, рыси, рыси рыжие, каракалы или любые их помеси. К кошкам также относятся домашние кошки, чистопородные и/или беспородные кошки-компаньоны, выставочные кошки, лабораторные кошки, клонированные кошки и дикик или бездомные кошки.
Используемый в данном описании термин "собачий каркасный участок" относится к аминокислотной последовательности тяжелой цепи и легкой цепи собачьего антитела, за исключением остатков гипервариабельных участков, определенных в данном описании как остатки CDR. Что касается канинизированного антитела, в большинстве вариантов осуществления аминокислотные последовательности нативных собачьих CDR в обеих цепях заменены соответствующими чужеродными CDR (например, полученными из мышиного антитела). При необходимости тяжелые и/или легкие цепи собачьего антитела могут содержать некоторые чужеродные остатки, отличные от CDR, например, позволяющие сохранить конформацию чужеродных CDR в собачьем антителе, и/или модифицировать функции Fc, как описано ниже.
Установлено, что PD-1 собак содержит аминокислотную последовательность SEQ ID NO: 2. В конкретном варианте осуществления PD-1 собак кодируется нуклеиновой кислотой, которая содержит нуклеотидную последовательность SEQ ID NO: 1. Последовательности PD-1 собак могут отличаться, например, они могут содержать консервативные различия в неконсервативных участках, однако полученный в результате PD-1 собак должен выполнять по существу такую же биологическую функцию, как и PD-1 собак, содержащий аминокислотную последовательность SEQ ID NO: 2. Например, биологическая функция PD-1 включает в себя ослабление Т-клеточного ответа в результате связывании с PD-L1 и/или PD-L2. То есть, PD-1 можно считать отрицательным регулятором. Следует отметить, что цитоплазматический хвост PD-1 содержит два сигнальных мотива на основе тирозина, ITIM (иммунорецепторный тирозин-содержащий ингибиторный мотив) и ITSM (иммунорецепторный тирозин-содержащий переключающий мотив). Кроме того, биологическая функция PD-1 собак может включать в себя, например, специфическое связывание антитела согласно настоящему изобретению посредством эпитопа во внеклеточном домене.
Установлено, что собачий PD-L1 содержит аминокислотную последовательность SEQ ID NO: 8. В конкретном варианте осуществления собачий PD-L1 кодируется нуклеотидной последовательностью, содержащей SEQ ID NO: 7. Последовательности собачьего PD-L1 могут отличаться, например, они могут содержать консервативные различия в неконсервативных участках, однако полученный в результате собачий PD-L1 должен выполнять по существу такую же биологическую функцию, как и собачий PD-L1, содержащий аминокислотную последовательность SEQ ID NO: 8. Например, одна из биологических функций PD-L1 включает в себя ослабление Т-клеточного ответа в результате связывании с PD-1.
Конкретная аминокислотная последовательность PD-1 собак или PD-L1, как правило, по меньшей мере, на 90% идентична аминокислотной последовательности SEQ ID NO: 2 PD-1 собак, или аминокислотной последовательности SEQ ID NO: 8 собачьего PD-L1, соответственно. В некоторых случаях PD-1 собак или PD-L1 может быть идентичен соответственно PD-1 собак, содержащему аминокислотную последовательность SEQ ID NO: 2, или собачему PD-L1, содержащему аминокислотную последовательность SEQ ID NO: 8, по меньшей мере, на 95%, или даже, по меньшей мере, на 96%, 97%, 98% или 99%. В некоторых вариантах осуществления аминокислотная последовательность PD-1 собак или PD-L1 может содержать не более 10 различий по сравнению с собачьим PD-1, содержащим аминокислотную последовательность SEQ ID NO: 2, или PD-L1 собак, содержащим аминокислотную последовательность SEQ ID NO: 8, соответственно. В некоторых вариантах осуществления аминокислотная последовательность PD-1 собак или PD-L1 может содержать не более 5 различий, или даже не более 4, 3, 2 или 1 различия по сравнению с собачьим PD-1, содержащим аминокислотную последовательность SEQ ID NO: 2, или PD-L1 собак, содержащим аминокислотную последовательность SEQ ID NO: 8, соответственно. Процент идентичности можно определить с помощью описанного ниже способа.
Термин "иммунный ответ" относится к функционированию, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитарных клеток, гранулоцитов и растворимых макромолекул, продуцирцемых указанными клетками или печенью (таких как антитела, цитокины, и комплемент), приводящему к селективному повреждению, разрушению или удалению из организма млекопитающего (например, из организма собаки) раковых клеток, клеток или тканей, инфицированных патогенами, или инвазивных патогенов.
Антитела к PD-1 собак
Настоящее изобретение предлагает выделенные антитела (такие как мышиные антитела к PD-1 собак и соответствующие им канинизированные антитела) или их антигенсвязывающие фрагменты, способные связыватьтся с собачьим PD-1, а также применение таких антител или их фрагментов. В конкретных вариантах осуществления изобретение предлагает мышиные CDR к PD-1 собак, полученные из мышиных антител к PD-1 собак, которые, как показано, способны связываться с собачьим PD-1 и блокировать связывание PD-1 собак с его лигандом, PD-L1 собак. Указанные CDR можно вставить в модифицированный собачий каркасный участок собачьего антитела с получением канинизированного мышиного антитела к PD-1 собак.
Используемый здесь термин "антитело к PD-1 собак" относится к антителу, вырабатываемому к PD-1 собак (например, в организме млекопитающего, такого как мышь или кролик), и способному специфически связываться с собачьим PD-1. Антитело, способное "специфически связываться с собачьим PD-1", в частности, с собачьим PD-1, или антитело, способное "специфически связываться с полипептидом, содержащим аминокислотную последовательность PD-1 собак", представляет собой антитело, способное избирательно связывать PD-1 собак среди других антигенов, однако данная специфичность не является абсолютной специфичностью связывания. Антитело к PD-1 собак считается "специфическим" в отношении PD-1 собак, если с его помощью можно определить присутствие PD-1 собак в образце, или, если оно способно изменить активность PD-1 собак, не препятствуя проявлению активности других молекул в собачьем образце, например, при отсутствии нежелательных результатов, таких как ложноположительные результаты в диагностическом контексте, или побочные эффекты в терапевтическом контексте. Степень специфичности, необходимая для антитела к PD-1 собак, может зависеть от предполагаемого применения антитела, и, в любом случае, определяется его пригодностью для использования по прямому назначению. Антитело или связывающее соединение, полученное из антигенсвязывающего участка антитела, предназначенное для применения в предполагаемом способе, связывается со своим антигеном, или его вариантом или мутеином, со сродством, которое превышает сродство к любому другому антигену, по меньшей мере, в два раза, предпочтительно, по меньшей мере, в десять раз, более предпочтительно, по меньшей мере, в 20 раз, и, наиболее предпочтительно, по меньшей мере, в 100 раз.
В соответствии с данным описанием считают, что антитело специфически связывается с полипептидом, содержащим конкретную антигенную последовательность (в данном случае фрагмент аминокислотной последовательности PD-1 собак), если оно связывается с полипептидами, содержащими фрагмент аминокислотной последовательности PD-1 собак, но не связывается с другими собачьими белками, не содержащими указанный фрагмент последовательности PD-1 собак. Например, антитело, способное специфически связыватьтся с полипептидом, содержащим PD-1 собак, может связываться с FLAG®-меченной формой PD-1 собак, но не с другими FLAG®-меченными собачьими белками. Антитело или связывающее соединение, полученное из антигенсвязывающего участка антитела, способно "специфически" связываться с собачьим антигеном, или его вариантом или мутеином, если его сродство к указанному собачьему антигену, или его варианту или мутеину, превышает сродство к любому другому тестируемому собачьему антигену, по меньшей мере, в десять раз, более предпочтительно, по меньшей мере, в 20 раз, и еще более предпочтительно, по меньшей мере, в 100 раз.
В данном описании термин "антитело" относится к любой форме антитела, которая проявляет желательную биологическую активность. Таким образом, данный термин используется в самом широком смысле и конкретно включает в себя, без ограничения, моноклональные антитела (в том числе полноразмерные моноклональные антитела), поликлональные антитела, полиспецифические антитела (например, биспецифические антитела), канинизированные антитела, полностью собачьи антитела, химерные антитела и камелизированные однодоменные антитела. "Исходные антитела" представляют собой антитела, полученные в результате воздействия антигена на иммунную систему, до модификации антител в целях предполагаемого применения, такой как канинизация антитела с целью его применения в качестве собачьего терапевтического антитела.
Если не указано иное, в данном описании термин "фрагмент антитела", или "антигенсвязывающий фрагмент", включает в себя антигенсвязывающие фрагменты антител, то есть, фрагменты антител, которые сохраняют способность специфически связываться с антигеном, связываемым полноразмерным антителом, например, фрагменты, которые сохраняют один или несколько участков CDR. Примеры антигенсвязывающих фрагментов включают в себя, без ограничения, Fab, Fab', F(аb')2 и Fv-фрагменты; диатела; линейные антитела; молекулы одноцепочечных антител, например, SC-Fv; нанотела и полиспецифические антитела, полученные из фрагментов антител.
"Фрагмент Fab" содержит одну легкую цепь и CH1, а также вариабельные участки одной тяжелой цепи. Тяжелая цепь молекулы Fab не может образовывать дисульфидные связи с тяжелой цепью другой молекулы. "Фрагмент Fab" может представлять собой продукт расщепления антитела папаином.
"Кристаллизующийся фрагмент" ("Fc") представляет собой участок, который содержит два фрагмента тяжелых цепей, содержащие домены антитела CH1 и СН2. Два фрагмента тяжелых цепей удерживаются вместе двумя или более дисульфидными связями и гидрофобными взаимодействиями доменов CH3.
"Фрагмент Fab'" содержит одну легкую цепь и часть или фрагмент одной тяжелой цепи, содержащий домен VH и домен СH1, а также участок между доменами CH1 и CH2, так, что между двумя тяжелыми цепями двух фрагментов Fab' может образовываться дисульфидная связь с получением молекулы F(аb')2.
"Фрагмент F(аb')2" содержит две легкие цепи и две тяжелые цепи, содержащие часть константного участка между доменами CH1 и CH2, так, что между двумя тяжелыми цепями образуется межцепочная дисульфидная связь. Таким образом, фрагмент F(аb')2 состоит из двух фрагментов Fab', которые удерживаются вместе дисульфидной связью, образованной между двумя тяжелыми цепями. "Фрагмент F(аb')2" может являться продуктом расщепления антитела пепсином.
"Участок Fv" содержит вариабельные участки тяжелой и легкой цепей, но не содержит константные участки.
Термин "одноцепочечный Fv" или антитело "scFv" относится к фрагментам антител, включающим в себя домены VH и VL антитела, присутствующие в одной полипептидной цепи. Как правило, полипептид Fv дополнительно содержит полипептидный линкер между доменами VH и VL, который позволяет ScFv формировать структуру, необходимую для связывания антигена. [см. Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113 Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); WO 88/01649; US 4946778 и US 5260203].
Используемый в данном описании термин "каноническая структура" относится к локальной конформации, которую может приобретать каждый из гипервариабельных участков тяжелой и легкой цепей антитела в пределах каркасного участка, в котором они находятся. Для каждого гипервариабельного участка, существует небольшое число канонических структур (как правило, обозначаемыми простыми целыми числами, такими как 1, 2 и т.д.), которые можно предсказать с высокой степенью точности на основании аминокислотных последовательностей соответствующего гипервариабельного участка (в частности, с учетом аминокислотной последовательности его каркасного участка, как описано ниже для соответствующих вариабельных доменов к PD-1 собак). С использованием указанных канонических структур можно определить, приводит ли модификация аминокислотной последовательности данного CDR к сохранению или потере способности связываться с антигеном как партнером по связыванию [см. Chothia and Lesk, Canonical Structures for the hypervariable regions of immunoglobulins, J. Mol. Biol. 196: 901-917(1987); Chothia et al., Conformation of immunoglobulin hypervaribale regions, Nature, 34: 877-883(1989); и Al-Lazikani et al., Standard Conformations for the canonical structures of immunoglobulins, J. Mol. Biol. 273: 927-948 (1997)].
"Доменное антитело" представляет собой иммунологически функциональный фрагмент иммуноглобулина, содержащий только вариабельный участок тяжелой цепи, или вариабельной участок легкой цепи. В некоторых случаях два или более участка VH ковалентно соединяют пептидным линкером с получением двухвалентного доменного антитела. Два участка VH двухвалентного доменного антитела могут быть специфичными к одному и тому же антигену, или к разным антигенам.
"Двухвалентное антитело" содержит два антигенсвязывающих участка. В некоторых случаях два связывающих участка обладают одинаковой антигенной спефичностью. Однако двухвалентные антитела могут быть биспецифическими (см. ниже).
В некоторых вариантах осуществления моноклональные антитела согласно настоящему изобретению также включают в себя камелизированные однодоменные антитела. [см., например, Muyldermans et al., Trends Biochem. Sci. 26: 230 (2001); Reichmann et al., J. Immunol. Methods 231: 25 (1999); WO 94/04678; WO 94/25591; US 6005079]. В одном варианте осуществления настоящее изобретение предлагает однодоменные антитела, содержащие два домена VH, модифицированные таким образом, чтобы образовались однодоменные антитела.
Используемый в настоящем описании термин "диатела" относится к небольшим фрагментам антител, содержащим два антигенсвязывающих участка, где фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH-VL или VL-VH). При использовании линкера, который является слишком коротким для того, чтобы обеспечить спаривание двух доменов, находящихся в одной цепи, домены вынуждены спариваться с комплементарными доменами другой цепи, образуя два антигенсвязывающих участка. [См., EP 0404097B1; WO 93/11161; и Holliger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993)]. Обзор по рекомбинантным вариантам антител можно найти в [Holliger and Hudson Nat. Biotechnol. 23:1126-1136 (2005)].
Как правило, антитело или антигенсвязывающий фрагмент настоящего изобретения сохраняет, по меньшей мере, 10% активности, направленной на связывание PD-1 собак (по сравнению с исходным антителом), если эта активность выражается в молях. Предпочтительно сродство связывания антитела или антигенсвязывающего фрагмента настоящего изобретения в отношении PD-1 собак составляет, по меньшей мере, 20%, 50%, 70%, 80%, 90%, 95%, 100% или более от сродства связывания исходного антитела в отношении PD-1 собак. Также подразумевается, что антитело или антигенсвязывающий фрагмент настоящего изобретения может содержать консервативные или неконсервативные аминокислотные замены (так называемые "консервативные варианты" или "функционально консервативные варианты" антитела), которые практически не влияют на биологическую активность.
Термин "выделенное антитело" относится к степени очистки и в таком контексте означает, что молекула практически не содержит других биологических молекул, таких как нуклеиновые кислоты, белки, липиды, углеводы, или других материалов, таких как клеточный дебрис и питательная среда. Как правило, термин "выделенный" не означает полного отсутствия таких материалов, или отсутствия воды, буферов или солей, если они не присутствуют в количествах, которые в значительной степени препятствуют экспериментальному или терапевтическому применению описанного здесь связывающего соединения.
В данном описании "химерное антитело" представляет собой антитело, содержащее вариабельный домен первого антитела и константный домен второго антитела, где первое и второе антитела получены от разных видов. [US 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)]. Как правило, вариабельный домен получают из антитела ("исходное антитело") экспериментального животного, такого как грызун, а последовательности константного домена получают из антитела животного-пациента, например, человека или собаки, чтобы полученное в результате химерное антитело с меньшей вероятностью вызывало неблагоприятные иммунные ответы у человека или собаки, соответственно, чем исходное антитело (например, антитело грызуна).
Используемый в данном описании термин "канинизированное антитело" относится к формам антител, которые содержат последовательности как собачьих, так и несобачьих (например, мышиных) антител. Как правило, канинизированное антитело содержит практически все из, по меньшей мере, одного или, чаще, двух вариабельных доменов, в которых все, или почти все гипервариабельные петли получены из несобачьего иммуноглобулина (например, содержащего 6 мышиных CDR к PD-1 собак, как описано ниже), а все или почти все каркасные участки (FR) (и обычно весь или почти весь оставшийся каркас) получены из последовательности собачьего иммуноглобулина. Как описано в настоящем описании, канинизированное антитело содержит три CDR тяжелой цепи и три CDR легкой цепи, полученные из мышиного антитела к PD-1 собак, а также собачий каркас или модифицированный собачий каркас. Модифицированный собачий каркас содержит одну или несколько аминокислотных замен, примеры которых описаны в настоящем описании, которые позволяют дополнительно оптимизировать эффективность канинизированного антитела, например, повысить его способность связывать PD-1 собак и/или блокировать связывание PD-1 собак с PD-L1 собак.
Термин "полностью собачье антитело" относится к антителу, которое содержит белковые последовательности только собачьих иммуноглобулинов. Полностью собачье антитело может содержать мышиные углеводные цепи, если оно продуцируется в организме мыши, мышиной клеткой, или гибридомой, полученной из мышиной клетки. Подобным образом, "мышиное антитело" представляет собой антитело, которое содержит последовательности только мышиных иммуноглобулинов. Альтернативно полностью собачье антитело может содержать крысиные углеводные цепи, если оно продуцируется в организме крысы, крысиной клеткой, или гибридомой, полученной из крысиной клетки. Подобным образом, "крысиное антитело" представляет собой антитело, которое содержит последовательности только крысиных иммуноглобулинов.
Существуют четыре известных подтипа тяжелых цепей IgG собаки, которые обозначают IgG-А, IgG-B, IgG-C и IgG-D. Два известных подтипа легких цепей называют лямбда и каппа.
Вариабельные участки каждой пары легкая цепь/тяжелая цепь образуют связывающий участок антитела. Таким образом, интактное антитело обычно содержит два связывающих участка. За исключением бифункциональных или биспецифических антител, два связывающих участка являются, как правило, одинаковыми.
Как правило, вариабельные домены как тяжелых, так и легких цепей содержат три гипервариабельных участка, называемых также участки, определяющие комплементарность (CDR), которые находятся в относительно консервативных каркасных участках (FR). Каркасные участки отвечают за правильную ориентацию CDR, обеспечивающую связывание с конкретным эпитопом. Как правило, в направлении от N-конца к С-концу вариабельные домены как легких, так и тяжелых цепей содержат FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Отнесение аминокислот к каждому домену обычно осуществляют в соответствии с правилами, описанными в Sequences of Proteins of Immunological Interest, Kabat, et al.; National Institutes of Health, Bethesda, Md.; 5th ed.; NIH Publ. No. 91-3242 (1991); Kabat, Adv. Prot. Chem. 32: 1-75 (1978); Kabat, et al., J. Biol. Chem. 252: 6609-6616 (1977); Chothia, et al., J. Mol. Biol. 196: 901-917 (1987) или Chothia, et al., Nature 342: 878-883 (1989)].
Используемый в данном описании термин "гипервариабельный участок" относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельный участок содержит аминокислотные остатки из "участка, определяющего комплементарность" или "CDR" (т.е. CDRL1, CDRL2 и CDRL3 в вариабельном домене легкой цепи и CDRH1, CDRH2 и CDRH3 в вариабельном домене тяжелой цепи). [Определение участков CDR антитела по последовательности описано в Kabat et al. Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991). Определение участков CDR антитела по структуре также можно найти в Chothia and Lesk, J. Mol. Biol. 196: 901-917 (1987)]. Используемый в данном описании термин остатки "каркасного участка" или "FR" относится к остаткам, отличным от остатков гипервариабельных участков, определенных в настоящем документе как остатки CDR.
Помимо связывания и способности активировать собачьи иммунные клетки, собачье или канинизированное антитело против PD-1 в идеале должно обладать следующими двумя характеристиками:
1. отсутствие эффекторных функций, таких как антитело-зависимая цитотоксичность (ADCC) и комплемент-зависимая цитотоксичность (CDC), и
2. способность легко подвергаться очистке в крупных масштабах с помощью стандартных промышленных технологий, таких, как хроматография с использованием белка A.
Ни один из встречающихся в природе изотипов собачьего IgG не удовлетворяет обоим критериям. Например, IgG-В можно очистить с использованием белка A, но он характеризуется высоким уровнем активности ADCC. IgG-А, наоборот, слабо связывается с белком А, но проявляет нежелательную активность ADCC. Кроме того, ни IgG-С, ни IgG-D нельзя очистить на колонках с белком А, хотя IgG-D не проявляет активности ADCC. (IgG-С обладает значительной активностью ADCC). Настоящее изобретение позволяет преодолеть данную проблему путем предоставления мутантных собачьих антител IgG-B, специфичных к PD-1; такие антитела не обладают эффекторными функциями, такими как ADCC, и могут легко подвергаться очистке стандартным методом промышленной хроматографии с использованием белка A.
Термин "гомология" относится к подобию двух полинуклеотидных последовательностей или двух полипептидных последовательностей, определяемому при их оптимальном выравнивании. Если в некотором положении обеих сравниваемых последовательностей находятся одинаковые основания или аминокислотные мономерные субъединицы, например, если в конкретном положении каждой из двух молекул ДНК находится аденина, то молекулы являются гомологичными по данному положению. Процент гомологии рассчитывают как число гомологичных положений в двух последовательностях, деленное на общее число положений и ×100. Например, если 6 из 10 положений в двух последовательностях являются совпадающими или гомологичными при оптимальном выравнивании последовательностей, то две указанные последовательности являются гомологичными на 60%. Как правило, сравнение осуществляют при выравнивании двух последовательностей с получением максимального процента гомологии.
Термин "выделенная молекула нуклеиновой кислоты" относится к молекуле ДНК или РНК геномного или синтетического происхождения, мРНК, кДНК, или некоторым их сочетаниям, которая не связана со всем полинуклеотидом, в котором выделенная молекула полинуклеотида встречается в природе, или с его частью, или связана с полинуклеотидом, с которым она не связана в природе. Следует понимать, что в целях данного описания термин "молекула нуклеиновой кислоты, содержащая" конкретную нуклеотидную последовательность, не охватывает интактные хромосомы. Выделенные молекулы нуклеиновых кислот, "содержащие" указанные нуклеотидные последовательности, могут включать в себя, в дополнение к указанным последовательностям, последовательности, кодирующие до десяти или даже до двадцати или более других белков, или их частей или фрагментов, или они могут включать в себя функционально связанные регуляторные последовательности, контролирующие экспрессию кодирующего участка указанных нуклеотидных последовательностей, и/или они могут включать в себя векторные последовательности.
Выражение "контролирующие последовательности" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Контролирующие последовательности, подходящие для прокариотов, например, включают в себя промотор, необязательно последовательность оператора и участок связывания рибосом. Эукариотические клетки, как известно, используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является "функционально связанной", если она находится в функциональной зависимости от другой нуклеотидной последовательности. Например, ДНК, кодирующая предпоследовательность или секреторную лидерную последовательность, функционально связана с ДНК, кодирующей полипептид, если она экспрессируется в виде белка-предшественника, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или участок связывания рибосом функционально связан с кодирующей последовательностью, если его расположение обеспечивает проведение трансляции. Как правило, термин "функционально связанный" означает, что функционально связанные последовательности ДНК являются смежными, а в случае секреторной лидерной последовательности они являются смежными и находятся в одной рамке считывания. Однако энхансеры не обязательно находятся в смежном положении. Связывание осуществляют путем лигирования по подходящим участкам рестрикции. Если такие участки отсутствуют, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с традиционной практикой.
В данном описании выражения "клетка", "клеточная линия" и "клеточная культура" используются как взаимозаменяемые, и все они включают в себя потомство. Так, термины "трансформанты" и "трансформированные клетки" включают в себя первичную исследуемую клетку и полученные из нее культуры, независимо от числа пересеваний. Следует также понимать, что не все потомство будет иметь совершенно одинаковый состав ДНК вследствие преднамеренных или случайных мутаций. Мутантное потомство, которое имеет ту же функцию или биологическую активность, по которой проводится скрининг исходно трансформированных клеток, входит в объем настоящего изобретения. При наличии разных обозначений контекст определяет конкретное применение.
Используемый в данном описании термин "зародышевая последовательность" относится к нереаранжированной последовательности ДНК иммуноглобулина. Можно использовать любой подходящий источник нереаранжированных последовательностей иммуноглобулина. Человеческие зародышевые последовательности можно получить, например, из зародышевой базы данных JOINSOLVER® на веб-сайте Национального института артрита, костно-мышечных и кожных заболеваний Национальных институтов здравоохранения США. Мышиные зародышевые последовательности можно получить, например, по способу, описанному в Giudicelli et al. [Nucleic Acids Res. 33:D256-D261 (2005)].
Характеристики мышиных антител к PD-1 собак и канинизированных мышиных антител к PD-1 собак
Настоящее изобретение предлагает выделенные мышиные антитела к PD-1 собак и полученные из них канинизированные антитела, а также способы применения таких антител или их антигенсвязывающих фрагментов для лечения заболевания, например, для лечения рака у собак. У собак обнаружены четыре тяжелые цепи IgG, обозначаемые А, В, С и D. Эти тяжелые цепи соответствуют четырем разным подклассам собачьих IgG, обозначаемым IgGA, IgGB, IgGC и IgGD. Аминокислотные последовательности указанных четырех тяжелых цепей и кодирующие их последовательности ДНК были впервые идентифицированы Tang et al. [Vet. Immunol. Immunopathol. 80: 259-270 (2001)]. Аминокислотные последовательности указанных тяжелых цепей и кодирующие их последовательности ДНК также можно получить из базы данных GenBank. Например, аминокислотная последовательность тяжелой цепи IgGA имеет номер доступа AAL35301.1, IgGB имеет номер доступа AAL35302.1, IgGC имеет номер доступа AAL35303.1 и IgGD имеет номер доступа (AAL35304.1). Собачьи антитела также содержат два типа легких цепей, каппа и лямбда. Аминокислотные последовательности указанных легких цепей и кодирующие их последовательности ДНК можно получить из базы данных GenBank. Например, аминокислотная последовательность легкой цепи каппа имеет номер доступа ABY 57289.1, а последовательность легкой цепи лямбда имеет номер доступа ABY 55569.1. В настоящем изобретении аминокислотные последовательности Fc-фрагментов всех четырех собачьих IgG определяют с использованием идентифицированной границы между доменами CH1 и СН2, как описано в Tang et al, выше. Канинизированные мышиные антитела к PD-1 собак, способные связывать PD-1 собак, включают в себя, без ограничения: антитела, которые содержат тяжелые цепи собачьих IgG-A, IgG-B и IgG-D, и/или собачьи легкие цепи каппа наряду с мышиными CDR к PD-1 собак. Соответственно, настоящее изобретение предлагает выделенные мышиные антитела к PD-1 собак, и/или канинизированные мышиные антитела к PD-1 собак, или их антигенсвязывающие фрагменты, способные связываться с собачьим PD-1 и блокировать связывание PD-1 собак с PD-L1 собак.
Настоящее изобретение также предлагает полноразмерные собачьи тяжелые цепи, которые могут спариваться с соответствующими легкими цепями, с образованием канинизированного антитела. Соответственно, настоящее изобретение также предлагает канинизированные мышиные антитела против собачьего антигена (в том числе выделенные канинизированные мышиные антитела к PD-1 собак), а также способы применения таких антител, или их антигенсвязывающих фрагментов, для лечения заболеваний, например, для лечения рака у собак.
Выделенное антитело, или его антигенсвязывающий фрагмент, способные связывать PD-1 собак, могу содержать один, два, три, четыре, пять или шесть гипервариабельных участков (CDR) мышиных антител против собачьего антигена, как описано в настоящем документе. Один, два, три, четыре, пять или шесть CDR могут быть независимо выбраны из последовательностей CDR, приведенных ниже. В другом варианте осуществления выделенное антитело, или его антигенсвязывающий фрагмент, способные связывать PD-1 собак, содержат легкую цепь каппа собачьего антитела, содержащую CDR-1, CDR-2 и/или CDR-3 мышиной легкой цепи, и тяжелую цепь собачьего антитела IgG, содержащую CDR-1, CDR-2 и/или CDR-3 мышиной тяжелой цепи.
В других вариантах осуществления изобретение предлагает антитела, или их антигенсвязывающие фрагменты, способные специфически связывать PD-1, которые содержат легкие цепи каппа собачьего антитела, содержащие от одного до шести разных CDR, по меньшей мере, на 80%, 85%, 90%, 95%, 98% или 99% идентичных аминокислотным последовательностям SEQ ID NO: 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 и/или 26, и тяжелые цепи собачьего антитела IgG, содержащие от одного до шести разных CDR, по меньшей мере, на 80%, 85%, 90%, 95%, 98% или 99% идентичных аминокислотным последовательностям SEQ ID NO: 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38 и/или 114, и при этом обладающих желательными характеристиками связывания и функциональными свойствами. В другом варианте осуществления антитело или антигенсвязывающий фрагмент настоящего изобретения содержит собачий каркас, содержащий сочетание последовательности тяжелой цепи IgG с легкой цепью каппа, содержащей одну или несколько из вышеупомянутых аминокислотных последовательностей CDR, отличающихся на 0, 1, 2, 3, 4 или 5 консервативных или неконсервативных аминокислотных замен, и при этом обладающих желательными характеристиками связывания и функциональными свойствами.
Термин "идентичность последовательностей" относится к степени совпадения аминокислот двух полипептидов, находящихся в эквивалентных положениях при оптимальном выравнивании последовательностей. В соответствии с данным описанием одна аминокислотная последовательность на 100% "идентична" второй аминокислотной последовательности, если аминокислотные остатки обеих последовательностей идентичны. Соответственно, аминокислотная последовательность на 50% "идентична" второй аминокислотной последовательности, если 50% аминокислотных остатков двух аминокислотных последовательностей являются идентичными. Сравнение последовательностей осуществляют на протяжении непрерывного блока аминокислотных остатков, входящих в состав конкретного белка, например, подлежащего сравнению белка, или фрагмента полипептида. В конкретном варианте осуществления учитывают отдельные делеции или инсерции, которые в противном случае могли бы привести к изменению соответствия двух аминокислотных последовательностей.
Термин "подобие последовательностей" относится к присутствию идентичных остатков, а также неидентичных остатков биохимически родственных аминокислот. Учитываются биохимически родственные аминокислоты, которые обладают подобными свойствами и могут использоваться как взаимозаменяемые.
Термин "консервативно модифицированные варианты" или "консервативное замещение" относится к заменам аминокислот в белке на другие аминокислоты, имеющие аналогичные характеристики (например заряд, размер боковой цепи, гидрофобность/гидрофильность, конформация скелета, жесткость и т.д.), которые зачастую не приводят к изменению биологической активности белка. Специалистам в данной области техники известно, что одиночные аминокислотные замены в участках полипептида, не играющих важной роли в поддержании его функции, не оказывают существенного влияния на биологическую активность [см., например, Watson et al., Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th Ed.; 1987)]. Кроме того, замены на структурно или функционально подобные аминокислоты с меньшей вероятностью нарушают биологическую активность. Примеры консервативных замен приведены ниже в таблице 3.
Примеры консервативных аминокислотных замен
Функционально консервативные варианты антител согласно настоящему изобретению также входят в объем настоящего изобретения. В данном описании термин "функционально консервативные варианты" относится к антителам или фрагментам, которые содержат замены одного или нескольких аминокислотных остатков, не приводящие к изменению желательного свойства, такого как сродство и/или специфичность к антигену. Такие варианты включают в себя, без ограничения, молекулы, полученные в результате замены аминокислоты на аминокислоту, обладающую сходными свойствами, например, в результате консервативных аминокислотных замен, приведенных выше в таблице 3.
Нуклеиновые кислоты
Настоящее изобретение также предлагает нуклеиновые кислоты, кодирующие иммуноглобулиновые цепи мышиных антител к PD-1 собак, и/или канинизированных мышиных антител к PD-1 собак, и их антигенсвязывающие фрагменты, описанные в настоящем описании (см. раздел Примеры ниже).
Кроме того, настоящее изобретениие охватывает нуклеиновые кислоты, которые кодируют иммуноглобулиновые полипептиды, содержащие аминокислотные последовательности, которые, по меньшей мере, примерно на 70%, предпочтительно, по меньшей мере, примерно на 80%, более предпочтительно, по меньшей мере, примерно на 90%, и, наиболее предпочтительно, по меньшей мере, примерно на 95% (например, на 95%, 96%, 97%, 98%, 99%, 100%) идентичны описанным здесь аминокислотным последовательностям CDR и антителам, где сравнение проводят с использованием алгоритма BLAST, параметры которого выбирают так, чтобы получить наибольшее совпадение соответствующих последовательностей на протяжении всей длины соответствующих стандартных последовательностей. Настоящее изобретение также предлагает нуклеиновые кислоты, которые кодируют иммуноглобулиновые полипептиды, содержащие аминокислотные последовательности, которые, по меньшей мере, примерно на 70%, предпочтительно, по меньшей мере, примерно на 80%, более предпочтительно, по меньшей мере, примерно на 90%, и, наиболее предпочтительно, по меньшей мере, примерно на 95% (например, на 95%, 96%, 97%, 98%, 99%, 100%) подобны любой из стандартных аминокислотных последовательностей, где сравнение проводят с использованием алгоритма BLAST, параметры которого выбирают так, чтобы получить наибольшее совпадение соответствующих последовательностей на протяжении всей длины соответствующих стандартных последовательностей.
В соответствии с данным описанием, процент идентичности нуклеотидных и аминокислотных последовательностей можно определить с помощью алгоритмов C, MacVector (MacVector, Inc. Cary, NC 27519), Vector NTI (InforMax, Inc. MD), Oxford Molecular Group PLC (1996) и Clustal W, используя параметры по умолчанию для выравнивания и определения идентичности. С помощью указанных коммерчески доступных программ также можно определять подобие последовательностей, используя такие же или аналогичные параметры по умолчанию. В качестве альтернативы можно использовать расширенный поиск Blast в условиях фильтрования по умолчанию, например, с помощью программы для наложения GCG (Genetics Computer Group, руководство по программированию для пакета GCG, версия 7, Madison, Wisconsin), используя параметры по умолчанию.
Следующие ссылки, относящиеся к алгоритмам BLAST, часто используют для анализа последовательностей: BLAST ALGORITHMS: Altschul, S.F., et al., J. Mol. Biol. 215: 403-410 (1990); Gish, W., et al., Nature Genet. 3: 266-272 (1993); Madden, T.L., et al., Meth. Enzymol. 266: 131-141 (1996); Altschul, S.F., et al., Nucleic Acids Res. 25: 3389-3402 (1997); Zhang, J., et al., Genome Res. 7: 649-656 (1997); Wootton, J.C., et al., Comput. Chem. 17: 149-163 (1993); Hancock, J.M. et al., Comput. Appl. Biosci. 10: 67-70 (1994); ALIGNMENT SCORING SYSTEMS: Dayhoff, M.O., et al., "A model of evolutionary change in proteins" in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3. M.O. Dayhoff (ed.), pp. 345-352, (1978); Natl. Biomed. Res. Found., Washington, DC; Schwartz, R.M., et al., "Matrices for detecting distant relationships" in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3." (1978), M.O. Dayhoff (ed.), pp. 353-358 (1978), Natl. Biomed. Res. Found., Washington, DC; Altschul, S.F., J. Mol. Biol. 219: 555-565 (1991); States, D.J., et al., Methods 3: 66-70(1991); Henikoff, S., et al., Proc. Natl. Acad. Sci. USA 89: 10915-10919 (1992); Altschul, S.F., et al., J. Mol. Evol. 36: 290-300 (1993); ALIGNMENT STATISTICS: Karlin, S., et al., Proc. Natl. Acad. Sci. USA 87: 2264-2268 (1990); Karlin, S., et al., Proc. Natl. Acad. Sci. USA 90: 5873-5877 (1993); Dembo, A., et al., Ann. Prob. 22: 2022-2039 (1994); и Altschul, S.F. "Evaluating the statistical significance of multiple distinct local alignments" in Theoretical and Computational Methods in Genome Research (S. Suhai, ed.), pp. 1-14, Plenum, New York (1997).
Настоящее изобретение также предлагает векторы экспрессии, содержащие выделенные нуклеиновые кислоты настоящего изобретения, в которых нуклеиновая кислота функционально связана с регуляторными последовательностями, которые распознаются клеткой-хозяином, трансфицированной вектором. Изобретение также предлагает клетки-хозяева, содержащие вектор экспрессии настоящего изобретения, и способы получения раскрытого здесь антитела или его антигенсвязывающего фрагмента, включающие в себя культивирование клетки-хозяина, несущей вектор экспрессии, кодирующий антитело или его антигенсвязывающий фрагмент, в культуральной среде, и выделение антигена или его антигенсвязывающего фрагмента из клетки-хозяина или культуральной среды.
Связывание эпитопа и сродство связывания
Настоящее изобретение также предлагает антитела или их антигенсвязывающие фрагменты, способные связываться с аминокислотными остатками эпитопа PD-1 собак, такие как мышиные антитела к PD-1 собак, раскрытые в настоящем описании. В конкретных вариантах осуществления мышиные антитела к PD-1 собак, или их антигенсвязывающие фрагменты, также способны ингибировать/блокировать связывание PD-1 собак с PD-L1 собак.
Канинизированное мышиное антитело к PD-1 собак можно получить с помощью рекомбинантных методов, известных в данной области. Линии клеток млекопитающих, которые можно использовать в качестве хозяев для экспрессии раскрытых здесь антител или их фрагментов, хорошо известны в данной области и включают в себя многочисленные иммортализованные клеточные линии, которые можно получить из Американской коллекции типовых культур (АТСС). Такие клетки включают в себя, в числе прочего, клетки яичника китайского хомячка (СНО), NSO, клетки SP2, клетки HeLa, клетки почки детеныша хомячка (ВНК), клетки почки обезьяны (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), клетки А549 , клетки 3Т3, клетки НЕК-293 и ряд других клеточных линий. Клетки млекопитающих, которые можно использовать в качестве хозяев, включают в себя клетки человека, мыши, крысы, собаки, обезьяны, свиньи, козы, крупного рогатого скота, лошади и хомяка. Особенно предпочтительными являются клеточные линии, характеризующиеся высоким уровнем экспрессии. Другие подходящие клеточные линии включают в себя линии клеток насекомых, таких как клетки Sf9, клеток амфибий, бактериальных клеток, клеток растений и клеток грибов. Если рекомбинантные векторы экспрессии, кодирующие тяжелую цепь или ее антигенсвязывающий участок или фрагмент, легкую цепь и/или ее антигенсвязывающий фрагмент, вводят в клетки-хозяева млекопитающих, антитела получают путем культивирования клеток-хозяев в течение периода времени, достаточного для обеспечения экспрессии антитела в клетках-хозяевах, или, более предпочтительно, секреции антитела в культуральную среду, в которой выращивают клетки-хозяева.
Антитела можно выделить из культуральной среды с помощью стандартных способов очистки белков. Кроме того, экспрессию антител согласно настоящему изобретению (или их фрагментов) продуцирующими клеточными линиями можно повысить с помощью ряда известных методов. Например, система экспрессии гена глутаминсинтетазы (система GS) является общим способом повышения экспрессии в определенных условиях. Система GS описана полностью или частично в европейских патентах №№ 0216846, 0256055 и 0323997 и заявке на европейский патент 89303964.4.
Как правило, гликопротеины, продуцирующиеся в конкретной клеточной линии или в конкретном трансгенном животном, определяют характер гликозилирования, специфичный для гликопротеинов, продуцирующихся в клеточной линии или в трансгенном животном. Таким образом, конкретный характер гликозилирования антитела зависит от конкретной клеточной линии или трансгенного животного, используемого для получения антитела. Однако все антитела, кодируемые описанными здесь молекулами нуклеиновых кислот, или содержащие описанные здесь аминокислотные последовательности, входят в объем настоящего изобретения независимо от характера гликозилирования антител. Подобным образом, в конкретных вариантах осуществления антитела с характером гликозилирования, включающим в себя только нефукозилированные N-гликаны, являются предпочтительными, поскольку показано, что данные антитела обычно являются более эффективными, чем их фукозилированные аналоги, как in vitro, так и in vivo [см., например, Shinkawa et al., J. Biol. Chem. 278: 3466-3473 (2003); патенты США №№ 6946292 и 7214775].
Настоящее изобретение также предлагает фрагменты мышиных антител к PD-1 собак, раскрытых в настоящем описании. Фрагменты антител включают в себя фрагменты F(аb)2, которые можно получить путем ферментативного расщепления IgG, например, пепсином. Фрагменты Fab можно получить, например, путем восстановления F(аb)2 дитиотреитолом или меркаптоэтиламином. Фрагмент Fab представляет собой цепь VL-CL, присоединенную к цепи VH-CH1 посредством дисульфидного мостика. Фрагмент F(аb)2 состоит из двух фрагментов Fab, которые, в свою очередь, соединяются двумя дисульфидными мостиками. Фрагменты Fab молекулы F(аb)2 содержат фрагменты Fc-участков, между которыми образуются дисульфидные мостики. Фрагмент Fv представляет собой участок VL или VH.
В одном варианте осуществления антитело или его антигенсвязывающий фрагмент содержит константный участок тяжелой цепи, например, собачий константный участок, такой как константной участок тяжелой цепи собачьего IgG-А, IgG-B, IgG-C и IgG-D, или его вариант. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент содержит константный участок легкой цепи, например, константный участок собачей легкой цепи, такой как участок собачей легкой цепи лямбда или каппа, или его вариант. В качестве неограничивающего примера константный участок собачей тяжелой цепи можно получить из IgG-B, а константный участок собачей легкой цепи можно получить из легкой цепи каппа.
Конструирование антител рекомбинантными методами
С помощью рекомбинантных методов можно получить канинизированные мышиные антитела к PD-1 собак настоящего изобретения, которые содержат модификации в собачьем каркасном участке и/или среди собачьих каркасных остатков вариабельных доменов исходного (т.е. собачьего) моноклонального антитела, например, с целью улучшения свойств антитела.
Экспериментальные и диагностические способы применения
Мышиные антитела к PD-1 собак, и/или канинизированные мышиные антитела к PD-1 собак настоящего изобретения, или их антигенсвязывающие фрагменты также можно использовать в диагностических анализах на собачий белок PD-1, например, путем детекции его экспрессии в конкретных опухолевых клетках, тканях, или в сыворотке. Такие диагностические методы можно использовать для диагностики разных заболеваний, особенно некоторых видов рака у собак.
Например, такой способ включает в себя следующие стадии:
(a) покрытие подложки (такой как поверхность лунки планшета для микротитрования, например, пластикового планшета) мышиным антителом к PD-1 собак, или его антигенсвязывающим фрагментом;
(b) нанесение на подложку образца, тестируемого на наличие PD-1 собак;
(c) промывание планшета, чтобы удалить несвязанные вещества;
(d) нанесение антител, содержащих детектируемую метку (например, связанных с ферментом антител) и специфичных в отношении антигена PD-1;
(e) промывание подложки, чтобы удалить несвязанные меченые антитела;
(f) если меченые антитела связаны с ферментом, добавление химического вещества, способного к превращению под действием фермента с генерированием флуоресцентного сигнала; и
(g) детекция присутствия меченого антитела.
В другом варианте осуществления антитело метят пероксидазой, которая взаимодействует с ABTS [2,2'-азино-бис(3-этилбензтиазолин-6-сульфоновая кислота)] или 3,3',5,5'-тетраметилбензидином, вызывая изменение цвета, которое можно детектировать. Альтернативно антитело метят детектируемым радиоактивным изотопом (например, 3H), который можно обнаружить путем подсчета сцинтилляций с помощью сцинтиллятора. Мышиные антитела к PD-1 собак настоящего изобретения можно использовать при проведении вестерн-блоттинга или процедуры иммуноанализа белков методом блоттинга.
Такая процедура является частью настоящего изобретения и включает в себя, например:
(i) приведение мембраны или другой твердой подложки, тестируемой на наличие связанного PD-1 собак или его фрагмента, в контакт с мышиным антителом к PD-1 собак или антигенсвязывающего фрагмента настоящего изобретения. Такая мембрана может представлять собой нитроцеллюлозную или виниловую [например, из поливинилиденфторида (PVDF)] мембрану, на которую переносят (например, после электрофоретического разделения в геле) белки, подлежащие тестированию на наличие PD-1 собак методом неденатурирующего PAGE (электрофорез в полиакриламидном геле) или SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия). Перед приведением в контакт с мышиным антителом к PD-1 собак или его антигенсвязывающим фрагментом, мембрану необязательно блокируют, например, обезжиренным сухим молоком или подобным средством, чтобы блокировать участки неспецифического связывания белков на мембране;
(ii) промывание мембраны один или несколько раз, чтобы удалить несвязанное мышиное антитело к PD-1 собак или его антигенсвязывающего фрагмента, и другие несвязанные вещества; и
(iii) детекцию связанного мышиного антитела к PD-1 собак или его антигенсвязывающего фрагмента.
Детекцию связанного антитела или его антигенсвязывающего фрагмента можно осуществлять путем связывания антитела или его антигенсвязывающего фрагмента со вторичным антителом (антителом против иммуноглобулина), содержащим детектируемую метку, с последующей детекцией вторичного антитела.
Описанные здесь мышиные антитела к PD-1 собак и их антигенсвязывающие фрагменты также можно использовать для иммуногистохимического анализа. Такой анализ входит в объем настоящего изобретения и включает в себя, например, (1) приведение клетки, подлежащей тестированию на присутствие PD-1 собак, в контакт с мышиным антителом к PD-1 собак или антигенсвязывающим фрагментом настоящего изобретения; и (2) детекцию антитела или его фрагмента на клетке, или в клетке. Если антитело или его антигенсвязывающий фрагмент содержит детектируемую метку, его можно обнаружить непосредственно. Альтернативно антитело, или его антигенсвязывающий фрагмент, можно подвергнуть связыванию со вторичным антителом, содержащим детектируемую метку, с его последующей детекцией.
Некоторые описанные здесь мышиные антитела к PD-1 собак и их антигенсвязывающие фрагменты можно использовать для визуализации опухоли in vivo. Такой способ может включать в себя введение меченных радиоактивным изотопом мышиных антител к PD-1 собак, или их антигенсвязывающих фрагментов, в организм собаки, подлежащей тестированию на наличие опухоли, связанной с экспрессией PD-1 собак, с последующей ядерной визуализацией организма пациента с целью детекции присутствия меченого антитела или его антигенсвязывающего фрагмента, например, в локусах, содержащих высокую концентрацию антитела или его антигенсвязывающего фрагмента, связанных с опухолью.
Методы визуализации включают в себя томографию SPECT (однофотонная эмиссионная компьютерная томография) или PET (позитронно-эмиссионная томография). В качестве меток можно использовать, например, йод-123 (123I) и технеций-99m (99mTc), например, в сочетании с томографией SPECT, или 11C, 13N, 15O или 18F, например, в сочетании с PET, или индий-111 [см., например, Gordon et al., International Rev. Neurobiol. 67:385-440 (2005)].
Перекрестно-блокирующие антитела
Кроме того, антитело к PD-1 собак или антигенсвязывающий фрагмент настоящего изобретения включают в себя любое антитело или его антигенсвязывающий фрагмент, способные связываться с тем же эпитопом PD-1 собак, с которым связываются описанные здесь антитела и фрагменты, и любое антитело или его антигенсвязывающий фрагмент, способные перекрестно блокировать (частично или полностью) связывание описанного здесь антитела или его фрагмента с собачьим PD-1, или связывание которых с собачьим PD-1 перекрестно блокируется (частично или полностью) описанным здесь антителом или его фрагментом; а также любые их варианты.
Описанные здесь поперечно блокирующие антитела и их антигенсвязывающие фрагменты можно идентифицировать по их способности перекрестно конкурировать с любым из IB5, 3B6, 4D12, 7C9, 2H9, 5G5 и/или 2G9 в стандартных анализах связывания (таких как BIACore®, ELISA, примеры которых приведены ниже, или проточная цитометрия). Например, можно использовать стандартные анализы ELISA, включающие в себя иммобилизацию рекомбинантного собачьего белка PD-1 на планшете, присоединение флуоресцентной метки к одному из антител и определение способности немеченого антитела конкурировать с меченным антителом за связывание. Дополнительно или альтернативно, анализ BIAcore® можно использовать для оценки способности антител к перекрестной конкуренции. Способность тестируемого антитела ингибировать связывание, например, IB5, 3B6, 4D12, 7C9, 2H9, 5G5 и/или 2G9, с собачьим PD-1 свидетельствует о том, что тестируемое антитело может конкурировать с IB5, 3B6, 4D12 , 7C9, 2H9, 5G5 и/или 2G9 за связывание с собачьим PD-1 и, следовательно, в некоторых случаях может связываться с тем же эпитопом PD-1 собак, что и IB5, 3B6, 4D12, 7C9, 2H9, 5G5 и/или 2G9. Как указано выше, антитела и фрагменты, способные связываться с тем же эпитопом, что и любое из описанных в настоящем изобретении антител к PD-1 собак, или их фрагментов, также составляют часть настоящего изобретения.
Фармацевтические композиции и способы введения
Фармацевтические или стерильные композиции канинизированного мышиного антитела к PD-1 собак или его антигенсвязывающего фрагмента можно получить путем смешивания их с фармацевтически приемлемым носителем или наполнителем. [см., например, Remington's Pharmaceutical Sciences and U.S. Pharmacopeia: National Formulary, Mack Publishing Company, Easton, PA (1984)].
Композиции терапевтических и диагностических средств можно получить путем их смешивания с приемлемыми носителями, наполнителями или стабилизаторами, например, в виде лиофилизированных порошков, взвесей, водных растворов или суспензий [см., например, Hardman, et al. (2001) Goodman and Gilman's The Pharmacological Basis of Therapeutics, McGraw-Hill, New York, NY; Gennaro (2000) Remington: The Science and Practice of Pharmacy, Lippincott, Williams, and Wilkins, New York, NY; Avis, et al. (eds.) (1993) Pharmaceutical Dosage Forms: Parenteral Medications, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Tablets, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Disperse Systems, Marcel Dekker, NY; Weiner and Kotkoskie (2000) Excipient Toxicity and Safety, Marcel Dekker, Inc., New York, NY]. В одном из вариантов осуществления антитела против PD-1 настоящего изобретения разбавляют до подходящей концентрации раствором ацетата натрия, рН 5-6, и для поддержания тоничности добавляют NaCl или сахарозу. Для повышения стабильности можно добавить вспомогательные средства, такие как полисорбат 20 или полисорбат 80.
Токсичность и терапевтическую эффективность композиций антител, вводимых отдельно или в сочетании с другим средством, можно определить с помощью стандартных фармацевтических процедур, проводимых на клеточных культурах или на экспериментальных животных, например, путем измерения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной для 50% популяции). Отношение токсического и терапевтического эффектов дозы называют терапевтическим индексом (LD50/ED50). В конкретных аспектах желательными являются антитела с высокими терапевтическими индексами. Результаты, полученные с помощью указанных анализов клеточных культур и исследований на животных, можно использовать для определения диапазона доз, подходящего для собак. Доза таких соединений предпочтительно находится в диапазоне, обеспечивающем концентрации в кровотоке, включающие в себя ED50 при низкой токсичности, или при отсутствии токсичности. Доза может варьировать в пределах указанного диапазона в зависимости от используемой лекарственной формы и способа введения.
Можно использовать разные способы введения. Подходящие способы введения включают в себя пероральное введение, ректальное введение, введение через слизистую оболочку, парентеральное введение; внутримышечное введение, подкожное введение, внутрикожное введение, интрамедуллярное введение, интратекальное введение, прямое внутрижелудочковое введение, внутривенное введение, внутрибрюшинное введение, интраназальное введение, внутриглазное введение, введение путем ингаляции, путем вдувания, местное введение, кожное введение, трансдермальное введение или внутриартериальное введение.
В конкретных вариантах осуществления мышиное антитело к PD-1 собак, или его антигенсвязывающий фрагмент, можно вводить инвазивным способом, например путем инъекции. В других вариантах осуществления настоящего изобретения мышиное антитело к PD-1 собак, или его антигенсвязывающий фрагмент, или его фармацевтическую композицию вводят внутривенно, подкожно, внутримышечно, внутриартериально, внутрь опухоли, или путем ингаляции, аэрозольной доставки. Введение неинвазивными способами (например, пероральное введение композиции в виде пилюль, капсул или таблеток) также входит в объем настоящего изобретения.
Композиции можно вводить с помощью медицинских устройств, известных в данной области техники. Например, фармацевтическую композицию настоящего изобретения можно вводить с помощью иглы для подкожных инъекций, в том числе, например, с использовнием предварительно заполненного шприца или автоинъектора. Описанные здесь фармацевтические композиции также можно вводить с помощью безыгольного устройства для подкожных инъекций; включающего в себя устройства, раскрытые в патентах США №№ 6620135; 6096002; 5399163; 5383851; 5312335; 5064413; 4941880; 4790824 или 4596556.
Описанные здесь фармацевтические композиции также можно вводить путем инфузии. Примеры введения фармацевтических композиций с использованием хорошо известных имплантатов и модульных форм описаны, например, в патенте США № 4487603, который раскрывает имплантируемый микроинфузионный насос для дозирования лекарства с контролируемой скоростью; патенте США № 4447233, который раскрывает инфузионный насос для доставки лекарственного средства с точной скоростью инфузии; патенте США № 4447224, который раскрывает имплантируемое инфузионное устройство с переменным потоком для непрерывной доставки лекарственного средства; патенте США № 4439196, который раскрывает осмотическую многокамерную систему доставки лекарственного средства. Специалистам в данной области техники хорошо известны многие другие подобные имплантаты, системы доставки и модули.
Альтернативно мышиное антитело к PD-1 собак или канинизированное мышиное антитело к PD-1 собак можно вводить местно, а не системно, например, путем инъекции антитела непосредственно в артритический сустав или участок индуцированного патогеном повреждения, характеризующегося иммунопатологией, часто в виде депо или композиции с замедленным высвобождением. Кроме того, антитело можно вводить с использованием системы направленной доставки лекарственного средства, такой как липосомы, покрытые тканесецифическим антителом, для доставки, например, в артритический сустав или участок индуцированного патогеном повреждения, характеризующегося иммунопатологией. Такие липосомы селективно направляются к пораженной ткани и поглощаются ею.
Режим введения зависит от нескольких факторов, включающих скорость превращения терапевтического антитела в сыворотке крови или ткани, тяжесть симптомов, иммуногенность терапевтического антитела и доступность клеток-мишеней в биологической среде. Предпочтительно режим введения обеспечивает количество терапевтического антитела, достаточное для улучшения состояния целевого заболевания при одновременной минимизации нежелательных побочных эффектов. Соответственно, количество доставляемого биологического средства отчасти зависит от конкретного терапевтического антитела и тяжести состояния, подлежащего лечению. Существует руководство по выбору подходящих доз терапевтических антител [см., например, Wawrzynczak Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK (1996); Kresina (ed.) Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, NY (1991); Bach (ed.) Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, NY (1993); Baert, et al. New Engl. J. Med. 348:601-608 (2003); Milgrom et al. New Engl. J. Med. 341:1966-1973 (1999); Slamon et al. New Engl. J. Med. 344:783-792 (2001); Beniaminovitz et al. New Engl. J. Med. 342:613-619 (2000); Ghosh et al. New Engl. J. Med. 348:24-32 (2003); Lipsky et al. New Engl. J. Med. 343:1594-1602 (2000)].
Определение подходящей дозы осуществляет ветеринар, например, с учетом параметров или факторов, которые, как известно или предполагается в данной области техники, могут влиять на лечение. Как правило, начинают с дозы, которая несколько меньше, чем оптимальная, и затем увеличивают ее с небольшим шагом до тех пор, пока не будет достигнут желательный или оптимальный эффект с учетом каких-либо негативных побочных эффектов. Важные диагностические показатели включают в себя такие симптомы, как, например, воспаление или уровень продуцируемых воспалительных цитокинов.
Описанные здесь антитела или их антигенсвязывающие фрагменты моно доставлять путем непрерывной инфузии, или путем введения доз, например, ежедневно, 1-7 раз в неделю, раз в неделю, раз в две недели, раз в месяц, два раза в месяц, раз в квартал, раз в полгода, раз в год и т.д. Дозы можно вводить, например, внутривенно, подкожно, местно, перорально, интраназально, ректально, внутримышечно, интрацеребрально, интраспинально, или путем ингаляции. Общая недельная доза предпочтительно составляет, по меньшей мере, 0,05 мкг/кг массы тела, более предпочтительно, по меньшей мере, 0,2 мкг/кг, 0,5 мг/кг, 1 мкг/кг, 10 мкг кг, 100 мкг/кг, 0,25 мг/кг, 1,0 мг/кг, 2,0 мг/кг, 5,0 мг/мл, 10 мг/кг, 25 мг/кг, 50 мг/кг или более [см., например, Yang, et al. New Engl. J. Med. 349:427-434 (2003); Herold, et al. New Engl. J. Med. 346:1692-1698 (2002); Liu, et al. J. Neurol. Neurosurg. Psych. 67:451-456 (1999); Portielji, et al. Cancer Immunol. Immunother. 52:133-144 (2003)]. Также можно использовать дозы, позволяющие достичь заранее определенной концентрации канинизированного мышиного антитела к PD-1 собак в сыворотке индивидуума, такой как 0,1, 0,3, 1, 3, 10, 30, 100, 300 мкг/мл или более. В других вариантах осуществления канинизированное мышиное антитело к PD-1 собак настоящего изобретения вводят подкожно или внутривенно, раз в неделю, раз в две недели, "каждые 4 недели," раз в месяц, два раза в месяц, или ежеквартально в дозе 10, 20, 50, 80, 100, 200, 500, 1000 или 2500 мг/индивидуума.
Антигенные пептиды, распознаваемые mAb против собачьих PD-1 и PDL-1, также можно использовать в качестве вакцин, вызывающих образование антител, которые блокируют связывание PD-1 с PDL-1, приводя к активации Т-клеток и усилению иммунного ответа. Данные вакцины можно использовать в качестве терапевтических вакцин против таких заболеваний, как рак, или для усиления иммунного ответа на другие вакцины. Чтобы использовать указанные антигенные пептиды в качестве вакцин, один или несколько таких пептидов можно присоединить химически или с помощью методов рекомбинантных ДНК к другому белку-носителю с целью усиления иммуногенности этих пептидов и индукции пептид-специфичных антител. Способы присоединения пептидов к белкам-носителям известны специалистам в данной области техники. Пептидные вакцины можно использовать для вакцинации животных посредством в.м., п/к, перорального, аэрозольного или in ovo способов. Пептидные вакцины можно использовать в виде субъединичных белков, экспрессированных бактериальными, вирусными, дрожжевыми или бакуловирусными системами. Альтернативно такие пептидные вакцины можно доставлять путем введения ряда вирусных или бактериальных векторов, экспрессирующих такие пептидные вакцины, с использованием способов, известных специалистам в данной области техники. Пептидные вакцины, которые можно вводить в дозах от 1-1000 мкг, могут необязательно содержать адъювант и приемлемый фармацевтический носитель.
Используемый здесь термин "ингибировать", или "лечить", или "лечение" включает в себя отсрочку развития симптомов, связанных с расстройством, и/или уменьшение тяжести симптомов такого расстройства. Данные термины также включают в себя облегчение существующих неконтролируемых или нежелательных симптомов, предотвращение появления других симптомов, а также ослабление или предотвращение причин появления таких симптомов. Таким образом, указанные термины означают достижение положительного результата у позвоночного, страдающего от расстройства, заболевания или симптома, или имеющего риск развития такого расстройства, заболевания или симптома.
В данном описании термин "терапевтически эффективное количество", "терапевтически эффективная доза" и "эффективное количество" относятся к количеству канинизированного мышиного антитела к PD-1 собак настоящего изобретения, или его антигенсвязывающего фрагмента, которое при введении указанного антитела, или его фрагмента, отдельно или в сочетании с другим терапевтическим средством, в клетку, ткань или организм индивидуума, способно эффективно вызывать измеримое улучшение одного или нескольких симптомов заболевания или состояния, или предотвращать развитие такого заболевания или состояния. Терапевтически эффективная доза также относится к количеству связывающего соединения, достаточному для достижения, по меньшей мере, частичного облегчения симптомов, например, для лечения, исцеления, предотвращения или улучшения соответствующего медицинского состояния, или увеличения скорости лечения, заживления, профилактики или облегчения таких состояний. При применении к отдельному активному ингредиенту в чистом виде терапевтически эффективная доза относится только к этому ингредиенту. При применении к сочетанию, терапевтически эффективная доза относится к суммарному количеству активных ингредиентов, которое обеспечивает достижение терапевтического эффекта, независимо от того, вводят ли сочетание последовательно или одновременно. Эффективное количество терапевтического средства приводит к улучшению диагностического показателя или параметра, по меньшей мере, на 10%; как правило, по меньшей мере, на 20%; предпочтительно, по меньшей мере, примерно на 30%; более предпочтительно, по меньшей мере, на 40%, и наиболее предпочтительно, по меньшей мере, на 50%. Эффективное количество также может приводить к улучшению субъективного показателя, если для оценки тяжести заболевания используют субъективные показатели.
Другие сочетанные способы лечения
Как указано ранее, канинизированное мышиное антитело к PD-1 собак, или его антигенсвязывающий фрагмент, и/или антигенный пептид настоящего изобретения, можно вводить в сочетании с одним или несколькими другими терапевтическими средствами (такими как химиотерапевтическое средство). Антитело может быть связано с таким средством (с образованием иммунокомплекса), или его можно вводить отдельно от указанного средства. В последнем случае (раздельное введение) антитело можно вводить до, после или одновременно с указанным средством, или его можно вводить совместно с другими известными терапевтическими средствами.
Наборы
Кроме того, изобретение предлагает наборы, содержащие один или несколько компонентов, которые включают в себя, без ограничения, описанные здесь антитело или его антигенсвязывающий фрагмент, способные специфически связываться с PD-1 (например, канинизированное мышиное антитело к PD-1 собак или его антигенсвязывающий фрагмент), наряду с одним или несколькими другими компонентами, включающими в себя, без ограничения, фармацевтически приемлемый носитель и/или химиотерапевтическое средство, как указано в настоящем описании. Связывающую композицию и/или химиотерапевтическое средство можно использовать в виде чистого средства, или в сочетании с фармацевтически приемлемым носителем в составе фармацевтической композиции.
В одном варианте осуществления набор содержит связывающую композицию настоящего изобретения (например, канинизированное мышиное антитело к PD-1 собак, или его фармацевтическую композицию, в одном контейнере (например, в стерильном стакане или пластиковом флаконе)) и ее фармацевтическую композицию и/или химиотерапевтическое средство в другом контейнере (например, в стерильном стеклянном или пластиковом флаконе).
Если набор содержит фармацевтическую композицию для парентерального введения индивидууму, он также может содержать устройство для осуществления такого введения. Например, набор может содержать одну или несколько игл для подкожных инъекций или других инъекционных устройств, описанных выше. Набор также может включать в себя вкладыш в упаковку, содержащий информацию, касающуюся фармацевтических композиций и лекарственных форм, присутствующих в наборе. Как правило, такая информация помогает владельцам домашних животных и ветеринарам эффективно и безопасно использовать входящие в состав набора фармацевтические композиции и лекарственные формы. Например, вкладыш может содержать следующую информацию о сочетании настоящего изобретения: фармакокинетика, фармакодинамика, результаты клинических исследований, параметры эффективности, показания и способы применения, противопоказания, предупреждения, меры предосторожности, побочные реакции, передозировка, подходящие дозы и способы введения, формы выпуска, надлежащие условия хранения, ссылки, информация о производителе/дистрибьюторе и патентная информация.
Для удобства набор может содержать описанное здесь антитело или специфическое связывающее средство, т.е. упакованное сочетание реагентов в заранее определенных количествах с инструкциями по проведению диагностического или детекционного анализа. В случае применения меченного ферментом антитела набор может содержать субстраты и кофакторы, необходимые для функционирования фермента (например, субстратный предшественник, который предоставляет детектируемый хромофор или флуорофор). Кроме того, состав набора могут входить другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или буфер для лизиса) и т.п. Относительные количества разных реагентов, которые могут широко варьировать, обеспечивают концентрации реагентов в растворе, способные существенно оптимизировать чувствительность анализа. В частности, реагенты, включающие в себя вспомогательные вещества, могут предоставляться в виде сухих порошков, обычно лиофилизированных, которые можно растворить с получением раствора реагента, имеющего подходящую концентрацию.
ПРИМЕРЫ
ПРИМЕР 1
PD-1 СОБАК и PD-L1
Идентификация и клонирование PD-1 собак:
Нуклеиновую кислоту, кодирующую полноразмерный PD-1 собак (cPD-1), идентифицируют посредством поиска в базе данных банка генов NCBI (номер доступа XM_543338.4, SEQ ID NO: 1). Транслируемую аминокислотную последовательность SEQ ID NO: 2 (номер доступа XP-543338.3), которая соответствует предполагаемому собачьему белку PD-1, также идентифицируют путем поиска в белковой базе данных банка генов (NCBI) с последующим выравниванием идентифицированной аминокислотной последовательности с аминокислотными последовательностями мышиного, кошачьего и человеческого PD-1. Синтезируют последовательность ДНК, соответствующую полноразмерному гену PD-1 собак и оптимизированную по кодонам для клеток СНО, после чего ее клонируют в плазмиде, обозначаемой p96793. Сравнение последовательности ДНК и белковой последовательности предполагаемого PD-1 собак с известными последовательностями ДНК и белковыми последовательностями PD-1 позволяет идентифицировать последовательности ДНК, кодирующие внеклеточный домен (ECD) PD-1 собак (SEQ ID NO: 3), и аминокислотную последовательность ECD PD-1 собак (SEQ ID NO: 4).
Синтезируют последовательность ДНК, кодирующую ECD PD-1 собак, наряду с линкером GT и 8 остатками гистидина, и затем клонируют ее в плазмиде, обозначаемой LPD2726. Нуклеотидную последовательность (SEQ ID NO: 5), соответствующую ECD PD-1 собак, содержащему линкер GT и Fc-участок гена Fc человеческого IgG1, синтезируют с помощью химических способов и клонируют в плазмиде, обозначаемой LPD2727. ECD PD-1 собак и Fc-участок человеческого IgG1 содержат аминокислотную последовательность SEQ ID NO: 6.
Идентификация и клонирование собачьего PD-L1:
Нуклеиновую кислоту, кодирующую полноразмерный собачий PD-L1, идентифицируют посредством поиска в базе данных банка генов NCBI (номер доступа XM_541302.4, SEQ ID NO: 7). Транслируемую аминокислотную последовательность (номер доступа XP-541302.4; SEQ ID NO: 8), которая соответствует предполагаемому собачьему белку PD-L1, идентифицируют путем поиска в белковой базе данных банка генов (NCBI) с последующим выравниванием идентифицированной аминокислотной последовательности с известными мышиными и человеческими последовательностями PD-L1.
Сравнение ДНК, кодирующей собачий PD-L1, с известными последовательностями PD-L1 позволяет идентифицировать последовательность ДНК, соответствующую домену ECD собачьего PD-L1 (SEQ ID NO: 9; кодон-оптимизированная для клеток СНО). Предсказанная аминокислотная последовательность ECD собачьего PD-L1 описана в SEQ ID NO: 10. Синтезируют ДНК, кодирующую ECD PD-L1, наряду с линкером GT и 8 остатками гистидина, и затем клонируют ее в плазмиде, обозначаемой LPD2695.
Последовательность ДНК, кодирующую аминокислотную последовательность ECD собачьего PD-L1, содержащую линкер GT и Fc-участок человеческого IgG1 (SEQ ID NO: 11), синтезируют с помощью химических способов и клонируют в плазмиде, обозначаемой LPD2697. ECD собачьего PD-L1 плюс линкер GT и Fc-участок человеческого IgG1 содержат аминокислотную последовательность SEQ ID NO: 12. В таблице 4 приведено описание упомянутых выше плазмид экспрессии.
Плазмиды, содержащие ДНК, кодирующую PD-1 или PD-L1
Экспрессия белков PD-1 и PD-L1:
Экспрессионные плазмиды, кодирующие белки PD-1ECD-HIS, PD-1ECD-Fc, PDL-1ECD-HIS и PD-L1ECD-Fc, трансфицируют в клетки НЕК 293, после чего белки выделяют из супернатанта трансфицированных клеток методом хроматографии на колонке, содержащей белок A в случае Fc-гибридных белков, или никель (Ni) в случае HIS-меченых белков. Очищенные белки используют для ELISA или анализов связывания, описанных ниже. Экспрессированные белки анализируют методом SDS-PAGE.
Последовательность ДНК полноразмерного PD-1 собак: сигнальная последовательность подчеркнута и выделена жирным шрифтом
В SEQ ID NO: 1 сигнальная последовательность отсутствует; в SEQ ID NO: 105 сигнальная последовательность присутствует.
atggggagccggcgggggccctggccgctcgtctgggccgtgctgcagctgggctggtggccaggatggctcctagactcccctgacaggccctggagcccgctcaccttctccccggcgcagctcacggtgcaggagggagagaacgccacgttcacctgcagcctggccgacatccccgacagcttcgtgctcaactggtaccgcctgagcccccgcaaccagacggacaagctggccgccttccaggaggaccgcatcgagccgggccgggacaggcgcttccgcgtcatgcggctgcccaacgggcgggacttccacatgagcatcgtcgctgcgcgcctcaacgacagcggcatctacctgtgcggggccatctacctgccccccaacacacagatcaacgagagtccccgcgcagagctctccgtgacggagagaaccctggagccccccacacagagccccagccccccacccagactcagcggccagttgcaggggctggtcatcggcgtcacgagcgtgctggtgggtgtcctgctactgctgctgctgacctgggtcctggccgctgtcttccccagggccacccgaggtgcctgtgtgtgcgggagcgaggacgagcctctgaaggagggccccgatgcagcgcccgtcttcaccctggactacggggagctggacttccagtggcgagagaagacgccggagcccccggcgccctgtgccccggagcagaccgagtatgccaccatcgtcttcccgggcaggccggcgtccccgggccgcagggcctcggccagcagcctgcagggagcccagcctccgagccccgaggacggacccggcctgtggcccctctga
Аминокислотная последовательность полноразмерного PD-1 собак: сигнальная последовательность подчеркнута и выделена жирным шрифтом
В SEQ ID NO: 2 сигнальная последовательность отсутствует; в SEQ ID NO: 106 сигнальная последовательность присутствует.
MGSRRGPWPLVWAVLQLGWWPGWLLDSPDRPWSPLTFSPAQLTVQEGENATFTCSLADIPDSFVLNWYRLSPRNQTDKLAAFQEDRIEPGRDRRFRVMRLPNGRDFHMSIVAARLNDSGIYLCGAIYLPPNTQINESPRAELSVTERTLEPPTQSPSPPPRLSGQLQGLVIGVTSVLVGVLLLLLLTWVLAAVFPRATRGACVCGSEDEPLKEGPDAAPVFTLDYGELDFQWREKTPEPPAPCAPEQTEYATIVFPGRPASPGRRASASSLQGAQPPSPEDGPGLWPL
Последовательность ДНК внеклеточного домена сбачьего PD-1: SEQ ID NO: 3 (кодон-оптимизированная для экспрессии в клетках СНО)
ctggattcccccgacagaccctggagccctctcaccttctcccctgcccagctgaccgtccaggaaggcgagaatgccaccttcacctgcagcctcgccgacatccccgacagcttcgtgctgaactggtacagactgagccccaggaaccagaccgacaagctggccgctttccaggaggacaggatcgaacccggcagggacaggaggtttagggtcatgaggctgcccaacggcagggacttccacatgtccatcgtggccgccagactgaacgactccggcatctacctgtgcggcgctatctacctgccccccaacacccagatcaacgagagccccagggccgaactgagcgtgacagagagaaccctggaacctcccacccagagcccttcccctcctcctagactgagcggacagctgcagggcctggtg
Внеклеточный домен PD-1 собак: SEQ ID NO: 4:
LDSPDRPWSPLTFSPAQLTVQEGENATFTCSLADIPDSFVLNWYRLSPRNQTDKLAAFQEDRIEPGRDRRFRVMRLPNGRDFHMSIVAARLNDSGIYLCGAIYLPPNTQINESPRAELSVTERTLEPPTQSPSPPPRLSGQLQGLV
Последовательность ДНК внеклеточного домена сбачьего PD-1-Fc человеческого IgG1: SEQ ID NO: 5 (кодон-оптимизированная для экспрессии в клетках НЕК-293)
ctggattcccccgacagaccctggagccctctcaccttctcccctgcccagctgaccgtccaggaaggcgagaatgccaccttcacctgcagcctcgccgacatccccgacagcttcgtgctgaactggtacagactgagccccaggaaccagaccgacaagctggccgctttccaggaggacaggatcgaacccggcagggacaggaggtttagggtcatgaggctgcccaacggcagggacttccacatgtccatcgtggccgccagactgaacgactccggcatctacctgtgcggcgctatctacctgccccccaacacccagatcaacgagagccccagggccgaactgagcgtgacagagagaaccctggaacctcccacccagagcccttcccctcctcctagactgagcggacagctgcagggcctggtgggtaccgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga
Гибридный белок, содержащий внеклеточный домен PD-1 собак и Fc человеческого IgG1: сигнальная последовательность подчеркнута и выделена жирным шрифтом:
В SEQ ID NO: 6 сигнальная последовательность отсутствует; в SEQ ID NO: 113 сигнальная последовательность присутствует.
MNFLLSWVHWSLALLLYLHHAKWSQALDSPDRPWSPLTFSPAQLTVQEGENATFTCSLADIPDSFVLNWYRLSPRNQTDKLAAFQEDRIEPGRDRRFRVMRLPNGRDFHMSIVAARLNDSGIYLCGAIYLPPNTQINESPRAELSVTERTLEPPTQSPSPPPRLSGQLQGLVGTDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Последовательность ДНК полноразмерного собачьего PD-L1: сигнальная последовательность подчеркнута и выделена жирным шрифтом
В SEQ ID NO: 7 сигнальная последовательность отсутствует; в SEQ ID NO: 107 сигнальная последовательность присутствует.
atgagaatgtttagtgtctttacattcatggcctactgccatttgctaaaagcatttacgatcacagtttctaaggacctgtatgtggtagagtatggtggcaatgtgacaatggaatgcaaattcccggtggaaaaacagttaaacttgtttgcactaatcgtctactgggaaatggaggataaaaaaattatacaatttgtgaatggaaaggaagacctgaaagttcagcacagcagctacagccagagggctcagctattgaaggaccagctcttcttggggaaggctgcgcttcagatcacagatgtgagattgcaggatgcaggggtttactgctgcttgatcggctatggcggtgctgactacaagcggattactttgaaagttcatgccccgtaccgcaacatcagccaaagaatttctgtggatcctgtcacctctgaacatgaactaatgtgtcaggctgagggttaccctgaggctgaagtcatctggacaagcagtgaccaccgagtcctgagtggcaaaaccaccatcactaattccaatagggaagagaagcttttcaatgtgaccagcacgctgaacatcaatgcaacagctaatgagattttctactgcacttttcaaagatcaggtcctgaggaaaacaatactgccgagttggtcatcccagaacgactgcccgttccagcaagtgagaggactcatttcatgattctgggacctttcctgttgcttcttggtgtagtcctggcagtcactttctgtctaaaaaaacatgggagaatgatggatgtggaaaaatgttgcacccgagataggaactcaaagaaacgaaatgatatacaatttgaagagacataa
Полноразмерный собачий PD-L1: сигнальная последовательность подчеркнута и выделена жирным шрифтом
В SEQ ID NO: 8 сигнальная последовательность отсутствует; в SEQ ID NO: 108 сигнальная последовательность присутствует.
MRMFSVFTFMAYCHLLKAFTITVSKDLYVVEYGGNVTMECKFPVEKQLNLFALIVYWEMEDKKIIQFVNGKEDLKVQHSSYSQRAQLLKDQLFLGKAALQITDVRLQDAGVYCCLIGYGGADYKRITLKVHAPYRNISQRISVDPVTSEHELMCQAEGYPEAEVIWTSSDHRVLSGKTTITNSNREEKLFNVTSTLNINATANEIFYCTFQRSGPEENNTAELVIPERLPVPASERTHFMILGPFLLLLGVVLAVTFCLKKHGRMMDVEKCCTRDRNSKKRNDIQFEET
Последовательность ДНК внеклеточного домена собачьего PD-L1: SEQ ID NO: 9 (кодон-оптимизированная для экспрессии в клетках СНО)
tttaccatcaccgtgtccaaggacctgtacgtggtcgagtacggcggcaatgtgaccatggagtgcaagttccccgtggagaagcagctgaacctgttcgccctcatcgtgtactgggagatggaggacaagaagatcatccagttcgtgaacggcaaggaggacctgaaggtgcagcactccagctactcccagagagcccagctgctgaaggaccagctgttcctgggcaaggccgccctgcagatcaccgacgtgagactgcaggacgccggcgtgtattgctgcctgatcggctacggaggcgccgactacaagaggatcaccctgaaggtgcatgcaccctacaggaacatcagccagaggatcagcgtcgatcccgtgaccagcgagcacgagctgatgtgccaagccgagggctatcccgaggccgaagtgatctggaccagcagcgaccacagggtcctgagcggcaagaccaccatcaccaacagcaacagggaggagaagctgttcaacgtgaccagcaccctcaacatcaacgccaccgccaacgagatcttctactgcaccttccagaggagcggccccgaagagaacaacaccgccgagctggtgatccccgagagactgcctgtgcctgccagcgagaggacccac
Белковая последовательность внеклеточного домена собачьего PD-L1: SEQ ID NO: 10
FTITVSKDLYVVEYGGNVTMECKFPVEKQLNLFALIVYWEMEDKKIIQFVNGKEDLKVQHSSYSQRAQLLKDQLFLGKAALQITDVRLQDAGVYCCLIGYGGADYKRITLKVHAPYRNISQRISVDPVTSEHELMCQAEGYPEAEVIWTSSDHRVLSGKTTITNSNREEKLFNVTSTLNINATANEIFYCTFQRSGPEENNTAELVIPERLPVPASERTH
Последовательность ДНК внеклеточного домена собачьего PD-L1-Fc человеческого IgG1: SEQ ID NO: 11 (кодон-оптимизированная для экспрессии в клетках НЕК-293)
tttaccatcaccgtgtccaaggacctgtacgtggtcgagtacggcggcaatgtgaccatggagtgcaagttccccgtggagaagcagctgaacctgttcgccctcatcgtgtactgggagatggaggacaagaagatcatccagttcgtgaacggcaaggaggacctgaaggtgcagcactccagctactcccagagagcccagctgctgaaggaccagctgttcctgggcaaggccgccctgcagatcaccgacgtgagactgcaggacgccggcgtgtattgctgcctgatcggctacggaggcgccgactacaagaggatcaccctgaaggtgcatgcaccctacaggaacatcagccagaggatcagcgtcgatcccgtgaccagcgagcacgagctgatgtgccaagccgagggctatcccgaggccgaagtgatctggaccagcagcgaccacagggtcctgagcggcaagaccaccatcaccaacagcaacagggaggagaagctgttcaacgtgaccagcaccctcaacatcaacgccaccgccaacgagatcttctactgcaccttccagaggagcggccccgaagagaacaacaccgccgagctggtgatccccgagagactgcctgtgcctgccagcgagaggacccacggtaccgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga
Гибридный белок, содержащий внеклеточный домен собачьего PD-L1 и Fc человеческого IgG1: SEQ ID NO: 12
FTITVSKDLYVVEYGGNVTMECKFPVEKQLNLFALIVYWEMEDKKIIQFVNGKEDLKVQHSSYSQRAQLLKDQLFLGKAALQITDVRLQDAGVYCCLIGYGGADYKRITLKVHAPYRNISQRISVDPVTSEHELMCQAEGYPEAEVIWTSSDHRVLSGKTTITNSNREEKLFNVTSTLNINATANEIFYCTFQRSGPEENNTAELVIPERLPVPASERTHGTDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
ПРИМЕР 2
АНТИТЕЛА к PD-1 собак
Получение моноклональных антител против собачьего PD1:
В общей сложности трех мышей Balb/c иммунизируют несколько раз (10 мкг каждый раз) в течение 17 дней. В качестве иммунизирующего антигена используют гибридный белок ECD PD-1 собак-Fc. После иммунизации собирают сыворотку от каждой мыши и анализируют ее на реакционноспособность в отношении HIS-меченного белка ECD PD-1 собак. Клетки селезенки мыши с самым высоким уровнем титра сыворотки против PD-1 ECD-HIS гибридизуют с миеломной клеточной линией P3X63Ag8.653. Примерно через 2 недели после гибридизации супернатант предполагаемых гибридомных клеток анализируют методом ELISA на реакционноспособность в отношении меченного белка PD-1 ECD-HIS. Гибридомы, продуцирующие интенсивные положительные сигналы в анализе ELISA, субклонируют методом серийных разведений и повторно анализируют на реакционноспособность в отношении меченного белка PD-1 собак ECD-HIS.
Подтверждение реакционноспособности моноклональных мышиных антител в отношении PD-1 собак:
Реакционную способность антител, секретируемых гибридомами в отношении ECD PD-1 собак подтверждают методом ELISA. Гибридомные клетки культивируют с использованием биореакторов CELLine (Integra-biosciences) в течение 10-30 дней. Вначале клетки культивируют в среде DMEM, содержащей 4 мМ L-глутамин и 10% фетальной бычьей сыворотки (FBS) со сверхнизким содержанием IgG, Gibco. Гибридомные клетки высевают в клеточные камеры биореактора CELLine при плотности клеток примерно 2×106 клеток/мл в 15 мл той же среды с концентрацией FBS, повышенной до 20%. Внешнюю камеру заполняют 1 л питательной среды (DMEM, содержащая 4 мМ L-глутамин и 2% стандартной FBS). Клетки гибридомы в клеточной камере выращивают до плотности примерно 2,5×107 клеток/мл в течение 3-7 дней. Затем из клеточной камеры собирают 10 мл суспензии клеток и заменяют свежей средой, чтобы обеспечить новый рост клеток и последующий сбор. Данную процедуру повторяют, по мере необходимости, нужное число раз чтобы получить достаточное количество mAb от каждого гибридомного клона. Собранные клеточные суспензии центрифугируют и супернатанты фильтруют через фильтрующие мембраны 0,2 мкм. Чтобы выделить антитела, супернатант каждого клона очищают с использованием быстропроточной колонки объемом 5 мл, содержащей сефарозу 4-белок G (GE Healthcare), с гравитационным течением. После промывания буфером трис-ЭДТА (ТЕ), рН 8,0, связанные антитела элюируют буфером, содержащим 0,1М глицин, рН 2,7, после чего рН нейтрализуют с помощью 1М трис, рН 8,0. Антитела концентрируют и буфер заменяют на забуференный фосфатом физиологический раствор (PBS) с использованием центрифужных фильтров Centriprep YM-10,10 кДа NMWL (Millipore). Концентрацию антител количественно определяют методом спектрофотометрии.
Очищенные mAb к PD-1 собак тестируют на реакционноспособность в отношении HIS-меченного домена ECD PD-1 собак методом ELISA следующим образом: HIS-меченный белок ECD PD-1 собак разводят до 10 мкг/мл буфером для нанесения покрытия (карбонат/бикарбонатный, рН 9,0) и вносят по 100 мкл/лунку в 96-луночные плоскодонные планшеты ELISA (Nunc). Планшеты инкубируют при 4°С в течение ночи. Затем планшеты трижды промывают фосфатно-буферным раствором, содержащим 0,05% Tween-20 (PBST). Затем в каждую лунку добавляют 200 мкл блокирующего буфера (5% обезжиренное молоко в PBST) и планшеты инкубировали при 37°С в течение 60 минут. Затем планшеты промывают три раза PBST. В первые лунки соответствующих столбцов добавляют 100 мкл тестируемых mAb, разведенных блокирующим буфером. Тестируемые mAb разводят в два раза в соответствующих положениях планшета. После инкубации при температуре 37°С в течение 60 минут планшеты промывают PBST три раза. Затем в планшеты добавляют 100 мкл на лунку пероксидазы хрена, конъюгированной с козьим антителом против мышиного IgG (KPL), в разведении 1:2000, и инкубируют при 37°С в течение 60 минут. Планшеты промывают три раза PBST и добавляют 100 мкл/лунку 3,3',5,5'-тетраметилбензидина (ТМВ), используемого в качестве субстрата (KPL). Цветной реакции дают развиваться в течение 5-20 минут при 37°C и затем измеряют оптическую плотность при 650 нм.
Клетки СНО, экспрессирующие собачий белок PD-1:
Ген полноразмерного PD-1 собак клонируют в плазмиде p96793. В данной плазмиде экспрессию белка PD-1 регулирует промотор HCMV. Клетки CHO DXB11 (dhfr-) поддерживают в среде MEM-альфа (Gibco), содержащей 10% фетальной бычьей сыворотки. Трансфекцию клеток СНО плазмидой p96793 проводят в колбах объемом 75 см2, содержащих примерно 6×106 клеток, путем опосредованной липосомами доставки генов с использованием липофектамина (Invitrogen). Через 48 часов клетки пересаживают в среду МЕМ-альфа, не содержащую нуклеозидов, дополненную 10% FBS и 400 мкг/мл гигромицина В (селективная среда). Клонирование методом серийных разведений проводят на пуле dhfr+, устойчивых к гигромицину клеток. Клоны тестируют на экспрессию PD-1 собак путем анализа иммунофлуоресценции. Коротко говоря, монослои клеток фиксируют в 96-луночных планшетах 80% ацетоном. Затем фиксированные и высушенные клеточные монослои инкубируют в течение 1 часа с поликлональным козьим антителом против человеческого PD-1 (R&D Systems). Планшеты промывают PBS и затем инкубируют в течение 1 часа с меченным флуоресцеином кроличьим антителом против козьего IgG (KPL). Планшеты промывали PBS. Клоны, генерирующие флуоресценцию, культивируют и создают запасы клеток.
Реакционная способность мышиных mAb в отношении собачьих белков PD-1, экспрессируемых на клетках СНО:
Реакционную способность мышиных mAb к PD-1 собак в отношении PD-1 собак на клетках СНО определяют с помощью клеточного анализа, используя клетки СНО, которые экспрессируют PD-1. Коротко говоря, клетки СНО, экспрессирующие PD-1 собак, культивируют до 80-100% слияния в 50 мкл среды (DMEM/HAM's F12, 10% FBS). Затем добавляют 50 мкл среды, содержащей разные концентрации очищенных mAb, и инкубируют в течение 1 часа при 37°С. После трех промываний PBS-Tween добавляют 100 мкл раствора козьего антитела против мышиных иммуноглобулинов, конъюгированного с пероксидазой хрена (HRP), разведенного 1:1000 культуральной средой, и инкубируют в течение одного часа при 37°С. Промывают еще три раза PBS-Tween и затем связанные mAb визуализируют с помощью субстрата пероксидазы (ТМВ). Увеличение поглощения света, связанное с активностью пероксидазы, измеряют при 450 нм с помощью ридера для микропланшетов.
Анализ связывания мышиных mAb к PD-1 собак и канинизированных мышиных mAb к PD-1 собак с собачьим PD-1
Примерно 70 резонансных единиц (RU) собачьего антигена PD-1 иммобилизуют непосредственно путем аминного сочетания. Измерение сродства проводят методом поверхностного плазмонного резонанса в отсутствии меток (например, Biacore® T200) с временем ассоциации 300 секунд, временем диссоциации 1200 секунд, при концентрациях 50, 100, 200 (×2) 400 и 800 наномолей (нмоль). Используют соответствующую модель связывания 1:1. Антиген (PD-1 собак) иммобилизуют на сенсорном чипе путем аминного сочетания и затем четыре антитела, указанные ниже в таблице 5, используемые в качестве анализируемых веществ, пропускают над поверхностью, содержащей антиген. Результаты демонстрируют, что антитела согласно настоящему изобретению к PD-1 собак обладают высоким сродством к собачьему антигену PD-1, которое характеризуется константой диссоциации (Kd) в наномолярном и даже субнаномолярном диапазоне. Кроме того, мышиное моноклональное антитело к PD-1 собак и соответствующее канинизированное мышиное моноклональное антитело к PD-1 собак из того же клона дают поразительно сходные значения Kd (см. приведенную ниже таблицу 5).
Определение константы связывания
M-1с-1
с-1
M
(RU2)
Блокада лиганда мышиными mAb к PD-1 собак:
Клеточный анализ ELISA (CELISA), проводимый на клеточной линии СНО, экспрессирующей PD-1 собак, используют для идентификации мышиных mAb, способных взаимодействовать с собачьим PD-1 (cPD-1). Блокаду лиганда подтверждают с помощью указанного анализа, используя биотинилированный белок cPD-L1/Fc. Коротко говоря, клетки cPD-1 CHO высевают в 96-луночные планшеты при плотности 4×104 клеток на лунку и инкубируют при 37°С в течение 18-24 часов, до достижения 95-100% слияния. Среду для культивирования клеток отсасывают и планшеты промывали 3 раза PBS+0,05% Tween20 и 1× средой CHO. Готовят трехкратные серийные разведения mAb против cPD1 в среде CHO, начиная с 30 мкг/мл, и каждое разведение антитела вносят в планшет в объеме 50 мкл/лунку. Инкубацию проводили при 37°C, 5% CO2, непрерывно встряхивая, в течение 30 мин. Добавляют 50 мкл/лунку cPD-L1-Fc-биотина (2 мкг/мл в исходной среде CHO) и инкубации при 37°C, 5% CO2, продолжая встряхивать, в течение 45 мин. Планшеты 6 раз промывают PBS+0,05% Tween 20. Добавляют 100 мкл/лунку стрептавидин-HRP (1:2000) в среде CHO и инкубируют в течение 30-60 мин при 37°C, 5% CO2. Планшеты промывают 5 раз PBS+0,05% Tween 20 и затем добавляют 100 мкл/лунку ТМВ, развивающего окраску субстрата. Развитие окрашивания останавливают путем добавления 50 мкл/лунку 1М фосфорной кислоты. Измеряют оптическую плотность (OD) при A450-A620 с помощью планшет-ридера ELISA.
Реакционная способность мышиных mAb в отношении PD-1, экспрессированного на РВМС, полученных от здоровых и больных раком собак:
РВМС получают из образцов крови, обработанных EDTA, полученных от здоровых и больных раком собак, путем разделения на фиколле. РВМС, ресуспендированные в буфере FACS (PBS, 1% FBS и 0,1% азида натрия), добавляют в концентрации 2,5×105 клеток на лунку и инкубируют с тестируемыми моноклональными антителами (mAb) в разных концентрациях. Клетки инкубируют в течение 30 мин при комнатной температуре и затем промывают два раза. После этого клетки ресуспендируют и инкубируют с ослиным антителом против мышиных IgG (цепи H+L), конъюгированным с Alexa-488, в течение 30 мин при комнатной температуре и затем промывают два раза. Затем клетки инкубируют с PB- и PE-конъюгированными антителами против собачьих CD4 и CD8 в течение 30 мин и промывают. Промытые клетки ресуспендируют в буфере FACS и анализируют с помощью проточной цитометрии, чтобы определить процент Т-клеток CD4 или CD8, связывающихся с моноклональными антителами против PD-1. В качестве контролей используют клетки, инкубированные только со вторичными антителами, или с mAb, обладающими другой специфичностью, но относящимися к тому же изотипу.
Высвобождение цитокинов из РВМС, полученных от здоровых и больных раком собак:
РВМС получают из образцов крови, обработанных EDTA, полученных от здоровых и больных раком собак, путем разделения на фиколле. Клетки промывают 3 раза, ресуспендируют в полной среде для культивирования тканей при концентрации 2,5×105 клеток на лунку и вносят в 96-луночные планшеты с тройными повторами. Клетки активируют конканавалином А, 1 мкг/мл. Тестируемые антитела добавляют в разных концентрациях и культуры инкубируют в течение 96 часов. В качестве контролей используют клетки, инкубированные с conA в отсутствии антитела или с conA и антителом, обладающим другой специфичностью, но относящимся к тому же изотипу. После культивирования в течение 96 часов супернатанты собирают и анализируют на высвобождение IFN-гамма, используя коммерческий набор для определения собачьего IFN-гамма методом ELISA (R&D Systems).
Клонирование и идентификация последовательностей ДНК, соответствующих вариабельным участкам мышиных mAb:
ДНК-последовательность мышиных цепей VH и VL и ДНК-последовательности, кодирующие их CDR, идентифицируют после выделения мРНК из каждой гибридомы с помощью стандартных методов молекулярной биологии. SEQ ID NO предсказанных аминокислотных последовательностей CDR, полученных из данных гибридом перечислены ниже:
Следует отметить, что существуют значительная степень гомологии между аминокислотными последовательностями CDR каждого из семи мышиных антител к PD-1 собак, примеры которых приведены ниже.
Аминокислотные последовательности CDR


Канонические структуры (классы) CDR VH-цепи:

Канонические структуры (классы) CDR VL-цепи:

ПРИМЕР 3
МУТАНТНЫЕ СОБАЧЬИ АНТИТЕЛА IgG-B, СПЕЦИФИЧНЫЕ К PD-1
Существуют четыре известных подтипа тяжелой цепи IgG собаки, обозначаемые IgG-А, IgG-B, IgG-C и IgG-D. Два известных подтипа легкой цепи называют лямбда и каппа. Однако помимо связывания и способности активировать собачьи иммунные клетки, собачье или канинизированное антитело против PD-1 в идеале должно обладать следующими двумя характеристиками:
1. отсутствие эффекторных функций, таких как антитело-зависимая цитотоксичность (ADCC) и комплемент-зависимая цитотоксичность (CDC), и
2. способность легко подвергаться очистке в крупных масштабах с помощью стандартных промышленных технологий, таких, как хроматография с использованием белка A.
Ни один из встречающихся в природе изотипов собачьего IgG не удовлетворяет обоим критериям. Например, IgG-В можно очистить с использованием белка A, но он характеризуется высоким уровнем активности ADCC. IgG-C также обладает значительной активностью ADCC. IgG-А, наоборот, слабо связывается с белком А, но проявляет нежелательную активность ADCC. Кроме того, ни IgG-С, ни IgG-D нельзя очистить на колонках с белком А, хотя IgG-D не проявляет активности ADCC. Настоящее изобретение позволяет преодолеть данную проблему путем предоставления мутантных собачьих антител IgG-B, специфичных к PD-1; такие антитела не обладают эффекторными функциями, такими как ADCC, и могут легко подвергаться очистке стандартным методом промышленной хроматографии с использованием белка A. Конкретные модификации показаны на фигуре 8.
Описанные варианты IgG-B с пониженными эффекторными функциями включают в себя первый вариант IgG-B, который получают путем замены каждого из остатка лизина (D 277) и остатка аспарагина (N 325) на остаток аланина [cIgGB(-)ADCC], второй вариант, который получают путем замены шарнирного участка IgG-B на шарнирный участок IgG-D [cIgGB(+)D-шарнир], третий вариант, который получают путем замены шарнирного участка IgG-B на шарнирный участок IgG-A [cIgGB(+)A-шарнир]. Кроме того, второй и третий варианты также содержат замену тех же остатков лизина и аспарагина, что и в первом варианте, на остаток аланина. Нумерация остатков лизина и аспарагина, заменяемых в данном изобретении, соответствует схеме нумерации, описанной для тяжелых цепей собачьих IgG в Tang et al., [Vet Immunol and Immunopathol, 80:259-270 (2001)].
Собачий IgGB дикого типа
sasttapsvfplapscgstsgstvalaclvsgyfpepvtvswnsgsltsgvhtfpsvlqssglyslssmvtvpssrwpsetftcnvahpasktkvdkpvpkrengrvprppdcpkcpapemlggpsvfifppkpkdtlliartpevtcvvvdldpedpevqiswfvdgkqmqtaktqpreeqfngtyrvvsvlpighqdwlkgkqftckvnnkalpspiertiskargqahqpsvyvlppsreelskntvsltclikdffppdidvewqsngqqepeskyrttppqldedgsyflysklsvdksrwqrgdtficavmhealhnhytqeslshspgkSEQ ID NO:39
Собачий IgGB(+)A-шарнир
sasttapsvfplapscgstsgstvalaclvsgyfpepvtvswnsgsltsgvhtfpsvlqssglyslssmvtvpssrwpsetftcnvahpasktkvdkpvfnecrctdtppcpapemlggpsvfifppkpkatlliartpevtcvvvdldpedpevqiswfvdgkqmqtaktqpreeqfagtyrvvsvlpighqdwlkgkqftckvnnkalpspiertiskargqahqpsvyvlppsreelskntvsltclikdffppdidvewqsngqqepeskyrttppqldedgsyflysklsvdksrwqrgdtficavmhealhnhytqeslshspgkSEQ ID NO:40
Собачий IgGB(+)D-шарнир
sasttapsvfplapscgstsgstvalaclvsgyfpepvtvswnsgsltsgvhtfpsvlqssglyslssmvtvpssrwpsetftcnvahpasktkvdkpvpkestckcispcpapemlggpsvfifppkpkatlliartpevtcvvvdldpedpevqiswfvdgkqmqtaktqpreeqfagtyrvvsvlpighqdwlkgkqftckvnnkalpspiertiskargqahqpsvyvlppsreelskntvsltclikdffppdidvewqsngqqepeskyrttppqldedgsyflysklsvdksrwqrgdtficavmhealhnhytqeslshspgkSEQ ID NO: 41
Собачий IgGB(-)ADCC
sasttapsvfplapscgstsgstvalaclvsgyfpepvtvswnsgsltsgvhtfpsvlqssglyslssmvtvpssrwpsetftcnvahpasktkvdkpvpkrengrvprppdcpkcpapemlggpsvfifppkpkatlliartpevtcvvvdldpedpevqiswfvdgkqmqtaktqpreeqfagtyrvvsvlpighqdwlkgkqftckvnnkalpspiertiskargqahqpsvyvlppsreelskntvsltclikdffppdidvewqsngqqepeskyrttppqldedgsyflysklsvdksrwqrgdtficavmhealhnhytqeslshspgkSEQ ID NO: 42
ПРИМЕР 4
ИНФОРМАЦИЯ О ПОСЛЕДОВАТЕЛЬНОСТЯХ АНТИТЕЛ
(из приведенного выше ПРИМЕРА 2)
Лидерная последовательность подчеркнута; последовательности CDR выделены жирным шрифтом; а последовательность каркасного участка не подчеркнута и не выделена жирным шрифтом.
mAb 1B5: Вариабельный участок тяжелой цепи, ДНК (SEQ ID NO: 43)
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
cagatccagttggtacagtctggacctgaactgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaacctatggaatgagctgggtgaaacaggctccaggaaagggtttaaagtggatgggctggattaatatctactctggaatcccaacatatgctgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctatttgcagatcgacaacctcaaaaatgaggacacggctacatatttctgtgcaagatttgatggtcccgactactggggccaaggcaccactctcaccgtctcccca
mAb 1B5: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 44)
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
QIQLVQSGPELKKPGETVKISCKASGYTFTTYGMSWVKQAPGKGLKWMGWINIYSGIPTYADDFKGRFAFSLETSASTAYLQIDNLKNEDTATYFCARFDGPDYWGQGTTLTVSP
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asp Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Pro
mAb 1B5: вариабельную участок легкой цепи, ДНК (SEQ ID NO: 45)
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
gacattgtgatgacacagtctccatcctccctggctatgtcagtaggacagaaggtcactatgagctgcaagtccagtcagagccttttaaatagtgtcaatcaaaagaactatttggcctggtaccagcagaaaccaggacagtctcctaaagttctggtatactttgcatccactagggtatctggggtccctgatcgcttcataggcagtggatctgggacagatttcactcttaccatcaccagtgtgcaggctgaagacctgacaacttacctctgtcagcaatattttagcactcctctcacgttcggtgctgggaccaagctggaaataaaa
mAb 1B5: вариабельный участок легкой цепи, белок (SEQ ID NO: 46)
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
DIVMTQSPSSLAMSVGQKVTMSCKSSQSLLNSVNQKNYLAWYQQKPGQSPKVLVYFASTRVSGVPDRFIGSGSGTDFTLTITSVQAEDLTTYLCQQYFSTPLTFGAGTKLEIK
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Met Ser Val Gly Gln Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Thr Arg Val Ser Gly Val Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Thr Ser Val Gln Ala Glu Asp Leu Thr Thr Tyr Leu Cys Gln Gln Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
mAb 2G9: вариабельный участок тяжелой цепи, ДНК (SEQ ID NO: 47)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgggattcagcaggatctttctcttcctcctgtcagtaactacaggtgtccactcccaggcttatctacagcagtctggggctgagctggtgaggcctggggcctcagtgaagatctcctgcaaggcttctggctacacatttaccagatacaatatgcactgggtaaagcagacacctagacagggcctggaatggattggaactatttatcccggatatggtgatacttcttacaatcagaaattcaagggcaaggccacactgactgtagacatatcctccagcacagcctacatgcagctcaccagcctgacatctgaggactctgcggtctatttctgttcaagggagtttgccgatgattaccccattcccccctttgactactggggccaaggcaccactctcacagtctcctca
mAb 2G9: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 48)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MGFSRIFLFLLSVTTGVHSQAYLQQSGAELVRPGASVKISCKASGYTFTRYNMHWVKQTPRQGLEWIGTIYPGYGDTSYNQKFKGKATLTVDISSSTAYMQLTSLTSEDSAVYFCSREFADDYPIPPFDYWGQGTTLTVSS
Met Gly Phe Ser Arg Ile Phe Leu Phe Leu Leu Ser Val Thr Thr Gly Val His Ser Gln Ala Tyr Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Asn Met His Trp Val Lys Gln Thr Pro Arg Gln Gly Leu Glu Trp Ile Gly Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Ile Ser Ser Ser Thr Ala Tyr Met Gln Leu Thr Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys Ser Arg Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
mAb 2G9: вариабельный участок легкой цепи, ДНК (SEQ ID NO: 49)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgaagttgcctgttaggctgttggtgctgatgttctggattcctgcttccagcagtgatgttttgatgacccaaactccactctccctgcctgtcagtcttggagatcaagcctccatctcttgtagatctagtcagaacattgtacatagtaatggaaacacctacttagaatggtacctgcagaaaccaggccagtctccaaagctcctgatctacaaagtttccaaccgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggaatttattactgctttcaaggttcacatgttccgtacacgttcggaggggggaccaagctggaaataaaa
mAb 2G9: вариабельный участок легкой цепи, белок (SEQ ID NO: 50)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MKLPVRLLVLMFWIPASSSDVLMTQTPLSLPVSLGDQASISCRSSQNIVHSNGNTYLEWYLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGIYYCFQGSHVPYTFGGGTKLEIK
Met Lys Leu Pro Val Arg Leu Leu Val Leu Met Phe Trp Ile Pro Ala Ser Ser Ser Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Asn Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Phe Gln Gly Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
mAb 2H9: вариабельный участок тяжелой цепи, ДНК (SEQ ID NO: 51)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgaaattcagctgggtcatcttcttcctgatggcagtggttacaggggtcaattcagaggttcagctgcagcagtctgtggcagagcttgtgaggccaggggcctcagtcaagttgtcctgcacagcttctggcttcaacattaaaaacacctatatgcactggataaaacagaggcctgaacagggcctggagtggattggaaggattgctcctgcgaatgttgatactaaatatgccccgaagttccagggcaaggccactataactgcagacacatcctccaacacagcctacatgcagctcagcaccctgacatcggaggacactgccatctattactgtgtcctgatctactatgattacgacggggacatcgatgtctggggcacagggaccacggtcaccgtctcctca
mAb 2H9: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 52)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MKFSWVIFFLMAVVTGVNSEVQLQQSVAELVRPGASVKLSCTASGFNIKNTYMHWIKQRPEQGLEWIGRIAPANVDTKYAPKFQGKATITADTSSNTAYMQLSTLTSEDTAIYYCVLIYYDYDGDIDVWGTGTTVTVSS
Met Lys Phe Ser Trp Val Ile Phe Phe Leu Met Ala Val Val Thr Gly Val Asn Ser Glu Val Gln Leu Gln Gln Ser Val Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys Asn Thr Tyr Met His Trp Ile Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile Gly Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe Gln Gly Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr Met Gln Leu Ser Thr Leu Thr Ser Glu Asp Thr Ala Ile Tyr Tyr Cys Val Leu Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Thr Gly Thr Thr Val Thr Val Ser Ser
mAb 2H9: вариабельный участок легкой цепи, ДНК (SEQ ID NO: 53)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgagggtccttgctgagctcctggggctgctgctgttctgctttttaggtgtgagatgtgacatccagatgaaccagtctccatccagtctgtctgcatcccttggagacacaattaccatcacttgccatgccagtcagaacattaatgtttggttaagttggtaccagcagagaccaggaaatattcctaaactattgatctataaggcttctcacttacacacaggcgtcccatcaaggtttagtggcagtggatctggaacaggtttcacattaaccatcagcagcctgcagcctgaagacattgccacttactactgtcaacagggtcaaagttggccgctcacgttcggtgctgggaccaaactggagctgaaa
mAb 2H9: вариабельный участок легкой цепи, белок (SEQ ID NO: 54)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MRVLAELLGLLLFCFLGVRCDIQMNQSPSSLSASLGDTITITCHASQNINVWLSWYQQRPGNIPKLLIYKASHLHTGVPSRFSGSGSGTGFTLTISSLQPEDIATYYCQQGQSWPLTFGAGTKLELK
Met Arg Val Leu Ala Glu Leu Leu Gly Leu Leu Leu Phe Cys Phe Leu Gly Val Arg Cys Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly Asp Thr Ile Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser Trp Tyr Gln Gln Arg Pro Gly Asn Ile Pro Lys Leu Leu Ile Tyr Lys Ala Ser His Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Gly Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Ser Trp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
mAb 3B6: вариабельный участок тяжелой цепи, ДНК (SEQ ID NO: 55)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgggttggctgtggaacttgctattcctgatggcagctgcccaaagtgcccaaacacagatccagttggtacagtctggacctgaactgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaacctatggaatgagctgggtgaaacaggctccaggaaagggtttaaagtggatgggctggattaatatctactctggaatcccaacatatgctgatgacttcaagggacgatttgccttctctttggaaacctctgccagcactgcctatttgcagatcgacaacctcaaaaatgaggacacggctacatatttctgtgcaagatttgatggtcccgactactggggccaaggcaccactctcacagtctcctca
mAb 3B6: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 56)
MGWLWNLLFLMAAAQSAQTQIQLVQSGPELKKPGETVKISCKASGYTFTTYGMSWVKQAPGKGLKWMGWINIYSGIPTYADDFKGRFAFSLETSASTAYLQIDNLKNEDTATYFCARFDGPDYWGQGTTLTVSS
Met Gly Trp Leu Trp Asn Leu Leu Phe Leu Met Ala Ala Ala Gln Ser Ala Gln Thr Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asp Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
mAb 3B6: вариабельный участок легкой цепи, ДНК (SEQ ID NO: 57)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atggaatcacagacccaggtcctcatgtttcttctgctctgggtatctggtgcctgtgcagacattgtgatgacacagtctccatcctccctggctgtgtcagtaggacggaaggtcactatgagctgcaagtccagtcagagccttttaaatagtgtcaatcaaaagaactatttggcctggtaccagcagaaaccaggacagtctcctaaagttctggtatactttgcatccgctagggtatctggggtccctgatcgcttcataggcagtggatctgggacagatttcactcttgccatcagcagtgtgcaggctgaagacctgacaacttacttctgtcagcaatattttagcactcctctcacgttcggtgctgggaccaagctggaactgaaa
mAb 3B6: вариабельный участок легкой цепи, белок (SEQ ID NO: 58)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MESQTQVLMFLLLWVSGACADIVMTQSPSSLAVSVGRKVTMSCKSSQSLLNSVNQKNYLAWYQQKPGQSPKVLVYFASARVSGVPDRFIGSGSGTDFTLAISSVQAEDLTTYFCQQYFSTPLTFGAGTKLELK
Met Glu Ser Gln Thr Gln Val Leu Met Phe Leu Leu Leu Trp Val Ser Gly Ala Cys Ala Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly Arg Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Ala Arg Val Ser Gly Val Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ala Ile Ser Ser Val Gln Ala Glu Asp Leu Thr Thr Tyr Phe Cys Gln Gln Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
mAb 4D12: вариабельный участок тяжелой цепи, ДНК (SEQ ID NO: 59)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgggttggctgtggaacttgctattcctgatggcagctgcccaaagtgcccaagcacagatccagttggtacagtctggacctgaactgaagaagcctggagagacagtcaagatctcctgcaaggcttctgggtataccttcacaacctatggaatgagctgggtgaaacaggcgccaggaaagggtttaaagtggatgggctggataaatatctactctggaatgccaacatatgctgatgacttcaagggacggtttgccttctctttggaaacctctgtcagcactgcctatttgcagatcaacaacctcaaaaatgaggacacggctacatatttctgtgcaagatttgatggtcccgactactggggccaaggcaccactctcacagtctcctca
mAb 4D12: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 60)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MGWLWNLLFLMAAAQSAQAQIQLVQSGPELKKPGETVKISCKASGYTFTTYGMSWVKQAPGKGLKWMGWINIYSGMPTYADDFKGRFAFSLETSVSTAYLQINNLKNEDTATYFCARFDGPDYWGQGTTLTVSS
Met Gly Trp Leu Trp Asn Leu Leu Phe Leu Met Ala Ala Ala Gln Ser Ala Gln Ala Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Val Ser Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
mAb 4D12: вариабельный участок легкой цепи, ДНК (SEQ ID NO: 61)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atggaatcacagacccaggtcctcatgtttcttctgctctgggtatctggtgcctgtgcagacattgtgatgacacagtctccatcctccctggctatgtcagtaggacagaaggtcactatgagctgcaagtccagtcagagccttttaaatagtgtcaatcaaaagaactatttggcctggtaccagcagaaaccaggacagtctcctaaagttctggtatactttgcatccactaggatatctggggtccctgatcgcttcataggcagtggatctgggacagatttcactcttaccatcagcagtgtgcaggctgaagacctggcagattacttctgtcagcaatattttagcactcctctcacgttcggtgctgggaccaagctggagctgaaa
mAb 4D12: вариабельный участок легкой цепи, белок (SEQ ID NO: 62)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MESQTQVLMFLLLWVSGACADIVMTQSPSSLAMSVGQKVTMSCKSSQSLLNSVNQKNYLAWYQQKPGQSPKVLVYFASTRISGVPDRFIGSGSGTDFTLTISSVQAEDLADYFCQQYFSTPLTFGAGTKLELK
Met Glu Ser Gln Thr Gln Val Leu Met Phe Leu Leu Leu Trp Val Ser Gly Ala Cys Ala Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Met Ser Val Gly Gln Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Thr Arg Ile Ser Gly Val Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
mAb 5G5: вариабельный участок тяжелой цепи, ДНК (SEQ ID NO 63)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgaaattcagctgggtcatcttcttcctgatggcagtggttacaggggtcaattcagaggttcagctgcagcagtctgtggcagagcttgtgaggccaggggcctcagtcaagttgtcctgcacagtttctggcttcaacattaaaaacacctatatgcactgggtgaagcagaggcctgaacagggcctggagtggattggaagaattgatcctgcgaatgttaatactaaatatgccccgaagttccagggcaaggccactataactacagacacatcctccaacacagcctacatgcagctcagcagcctgacatcggaggacactgccatctattactgtgtcctgattttctatgattacgacggggacatcgatgtctggggcacagggaccaaggtcaccgtctcctca
mAb 5G5: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 64)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MKFSWVIFFLMAVVTGVNSEVQLQQSVAELVRPGASVKLSCTVSGFNIKNTYMHWVKQRPEQGLEWIGRIDPANVNTKYAPKFQGKATITTDTSSNTAYMQLSSLTSEDTAIYYCVLIFYDYDGDIDVWGTGTKVTVSS
Met Lys Phe Ser Trp Val Ile Phe Phe Leu Met Ala Val Val Thr Gly Val Asn Ser Glu Val Gln Leu Gln Gln Ser Val Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Val Ser Gly Phe Asn Ile Lys Asn Thr Tyr Met His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile Gly Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe Gln Gly Lys Ala Thr Ile Thr Thr Asp Thr Ser Ser Asn Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Ile Tyr Tyr Cys Val Leu Ile Phe Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Thr Gly Thr Lys Val Thr Val Ser Ser
mAb 5G5: вариабельный участок легкой цепи, ДНК (SEQ ID NO: 65)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgagggtccttgctgagctcctggggctgctgctgttctgctttttaggtgtgagatgtgacatccagatgaaccagtctccatccagtctgtctgcatcccttggagacacaattaccatcacttgccatgccagtcagaacattaatgtttggttaagctggtaccagcagaaaccaggaaatattcctaaactattgatctataaggcttccaacttacacacaggcgtcccatcaaggtttagtggcagtggatctggaacagatttcacattaaccatcagcagcctgcagcctgaagacattgccacttactactgtcaacagggtcaaagttatccgctcacgttcggtgctgggaccaagctggagctgaaa
mAb 5G5: вариабельный участок легкой цепи, белок (SEQ ID NO: 66)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MRVLAELLGLLLFCFLGVRCDIQMNQSPSSLSASLGDTITITCHASQNINVWLSWYQQKPGNIPKLLIYKASNLHTGVPSRFSGSGSGTDFTLTISSLQPEDIATYYCQQGQSYPLTFGAGTKLELK
Met Arg Val Leu Ala Glu Leu Leu Gly Leu Leu Leu Phe Cys Phe Leu Gly Val Arg Cys Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly Asp Thr Ile Thr Ile Thr Cys His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
mAb 7C9: вариабельный участок тяжелой цепи, ДНК (SEQ ID NO: 67)
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
caggtgcagctgaagcagtcaggacctggcctagtgcagccctcacagagcctgtccataacctgcacagtctctggtttctcattaactagctatggtgtacactgggttcgccagtctccaggaaagggtttaaagtggatgggctggattaatatctactctggaatcccaacatatgctgatgacttcaagggacggtttgccttctctttggaaacctctgccagcactgcctatttgcagatcgacaacctcaaaaatgaggacacggctacatatttctgtgcaagatttgatggtcccgactactggggccaaggcatcactctcactgtctccgca
mAb 7C9: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 68)
QVQLKQSGPGLVQPSQSLSITCTVSGFSLTSYGVHWVRQSPGKGLKWMGWINIYSGIPTYADDFKGRFAFSLETSASTAYLQIDNLKNEDTATYFCARFDGPDYWGQGITLTVSA
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asp Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Ile Thr Leu Thr Val Ser Ala
mAb 7C9: вариабельный участок легкой цепи, ДНК (SEQ ID NO: 69)
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
gacattgtgatgacacagtctccatcctccctggctatgtcagtaggacagaaggtcactatgagctgcaagtccagtcagagccttttaaatagtgtcaatcaaaagaactatttggcctggtaccagcagaaaccaggacagtctcctaaagttctggtatactttgcatccactagggtatctggggtccctgatcgcttcataggcagtggatctgggacagatttcactcttaccatcaccagtgtgcaggctgaagacctgacaacttacttctgtcagcaatattttagcactcctctcacgttcggtgctgggaccaagctggaactgaaa
mAb 7C9: вариабельный участок легкой цепи, белок (SEQ ID NO: 70)
FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
DIVMTQSPSSLAMSVGQKVTMSCKSSQSLLNSVNQKNYLAWYQQKPGQSPKVLVYFASTRVSGVPDRFIGSGSGTDFTLTITSVQAEDLTTYFCQQYFSTPLTFGAGTKLELK
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Met Ser Val Gly Gln Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Thr Arg Val Ser Gly Val Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Thr Ser Val Gln Ala Glu Asp Leu Thr Thr Tyr Phe Cys Gln Gln Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
mAb 1E4: вариабельный участок тяжелой цепи, ДНК (SEQ ID NO: 109)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgggattcagcaggatctttctcttcctcctgtcagtaactacaggtgtccactcccaggcttatctacagcagtctggggctgagctggtgaggcctggggcctcagtgaagatgtcctgcaaggcttttggctacacatttaccagttacaatatgcactgggtgaagcagacacctagacagggcctggaatggattggaaccatttatccaggagatggtgacgcttcctacaatcagaaattccaggacaaggccacactgactgttgacaaatcctccagcacagcctacatgcagctcagcagcctgacatctgaagactctgcggtctatttctgttcaagggagtttgccgatgcttaccccattcccccctttgactactggggccaaggcaccactctcacagtctcctca
mAb 1E4: вариабельный участок тяжелой цепи, белок (SEQ ID NO: 110)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MGFSRIFLFLLSVTTGVHSQAYLQQSGAELVRPGASVKMSCKAFGYTFTSYNMHWVKQTPRQGLEWIGTIYPGDGDASYNQKFQDKATLTVDKSSSTAYMQLSSLTSEDSAVYFCSREFADAYPIPPFDYWGQGTTLTVSS
mAb 1E4: вариабельный участок легкой цепи, ДНК (SEQ ID NO: 111)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
atgaagttgcctgttaggctgttggtgctgattttctggattcctgcttccagtagtgatgttttgatgacccaaactccactctccctggttgtcagtcttggagatcaggcctccatctcttgcagatctagtcagagcattgtatatagtaatggaaacacctatttagaatggtacctgcaaaaaccaggccagtctccaaagctcctgatttacaaagtttccaaccgattttctggggtcccagacaggttcagtggcagtggatcagggacagatttcacactcaagatcagcagagtggaggctgaggatctgggagtttattactgctttcaaggttcacatgttccgtacacgttcggaggggggaccaagctggaaataaaa
mAb 1E4: вариабельный участок легкой цепи, белок (SEQ ID NO: 112)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
MKLPVRLLVLIFWIPASSSDVLMTQTPLSLVVSLGDQASISCRSSQSIVYSNGNTYLEWYLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYYCFQGSHVPYTFGGGTKLEIK
ПРИМЕР 5
ЭПИТОПНОЕ КАРТИРОВАНИЕ АНТИТЕЛ к PD-1 собак
Введение
Взаимодействие антител с распознаваемыми ими белковыми антигенами осуществляется посредством связывания специфических аминокислот (паратопов) антитела с определенными аминокислотами (эпитопами) антигенов-мишеней. Эпитоп представляет собой антигенную детерминанту, способную вступать в специфическое взаимодействие с иммуноглобулином. Он состоит из группы аминокислот на поверхности антигена.
Представляющий интерес белок может содержать несколько эпитопов, которые распознаются разными антителами. Эпитопы, распознаваемые антителами, классифицируются как линейные или конформационные эпитопы. Линейные эпитопы образованы непрерывной цепочкой аминокислот, входящих в состав белка, тогда как конформационные эпитопы состоят из аминокислот, которые не являются смежными (например, находятся далеко друг от друга) в первичной аминокислотной последовательности, но сближаются в результате трехмерной укладки белка.
Картирование эпитопов представляет собой процесс идентификации аминокислотных последовательностей (т.е. эпитопов), которые распознаются антителами на антигенах-мишенях. Идентификация эпитопов, распознаваемых моноклональными антителами (mAb) на антигенах-мишенях, имеет важное значение. Например, она может способствовать разработке новых терапевтических средств, диагностических средств и вакцин. Картирование эпитопов также можно использовать для выбора оптимизированных терапевтических моноклональных антител и выяснения механизмов их действия. Информацию по эпитопам также можно использовать для идентификации уникальных эпитопов рака и определения защитных или патогенных эффектов вакцин.
Картирование эпитопов можно проводить с помощью поликлональных или моноклональных антител, используя разные методы для идентификации эпитопов, в зависимости от предполагаемой природы эпитопа (т.е. линейной или конформационной). Картирование линейных эпитопов является более простым и относительно легко выполняется. Для картирования линейных эпитопов коммерческие службы часто используют сканирование пептида. В данном случае путем химического синтеза получают множество перекрывающихся коротких пептидных последовательностей белка-мишени и тестируют их на способность связывать представляющие интерес антитела. Данная стратегия является быстрой, относительно недорогой и характеризуется высокой пропускной способностью. С другой стороны, картирование прерывистого эпитопа является технически более сложным и требует более специализированных методов, таких как рентгеновская совместная кристаллография моноклонального антитела вместе с белком-мишенью, водород-дейтериевый (H/D) обмен, и/или масс-спектроскопия в сочетании с ферментативным расщеплением.
Картирование эпитопов PD-1 с использованием микрочипа ProImmune®:
Чтобы идентифицировать аминокислоты, которые образуют эпитопы для mAb против PD1, химически синтезируют в общей сложности 28 пептидов, которые содержат 15 аминокислот в длину и перекрываются по 10 аминокислотам. Такая библиотека перекрывающихся пептидов охватывает всю длину собачьего белка PD-1. Последовательности указанных пептидов приведены в таблице 6 ниже. Определение связывания пептид-антитело проводят путем присоединения образцов антитела к пептидному микрочипу ProArray Ultra®, с последующей инкубацией с вторичным антителом, содержащим флуоресцентную метку. Все пептиды синтезируют отдельно и затем присоединяют к рабочей поверхности ProArray Ultra® наряду с мышиными IgG ProImmune®, используемыми в качестве контроля. Данный оптимизированный способ гарантирует, что пептиды, присутствующие на чипе, будут точно имитировать свойства соответствующего участка белка, позволяет избежать физикохимических вариаций, присущих свободным пептидам, и получить совместимую, объединенную пептидную и белковую матричную платформу. Тестируемые анализируемые вещества (пептиды) распределяют на пластинах ProArray Ultra® в отдельных точках и затем соответствующие gal-файлы обеспечивают точное совпадение полученных параметров чипа с нанесенными анализируемыми веществами. Пластины ProArray Ultra® блокируют проверенным блокирующим буфером, чтобы уменьшить неспецифическое связывание mAb. После этого их инкубируют с образцами mAb и затем со специфическим флуоресцентно-меченным вторичным антителом. После нескольких стадий промывки чипы ProArray Ultra® сушат и сканируют с помощью системы для сканирования флуоресцентных микрочипов с высоким разрешением. После сканирования флуоресцентно-меченных пластин ProArray Ultra®, сканер регистрирует изображение, которое анализируют с помощью программного обеспечения для анализа изображений, позволяющего интерпретировать и количественно оценивать уровни интенсивности флуоресценции в каждом флуоресцентном пятне на сканируемой пластине микрочипа. Результаты данного эксперимента свидетельствуют о том, что некоторые пептиды PD-1 собак распознаются отдельными анализируемыми mAb. Конкретные mAb и распознаваемые ими аминокислотные последовательности приведены в таблице 7. Результаты данного исследования демонстрируют, что mAb 2H9 распознает эпитоп, расположенный во внеклеточном домене PD-1 собак и содержащий аминокислотную последовательность, описанную в SEQ ID NO: 84, а mAb 1A1 распознает эпитоп, содержащий аминокислотную последовательность, описанную в SEQ ID NO: 84, и перекрывающуюся аминокислотную последовательность, описанную в SEQ ID NO: 83.
Картирование эпитопов PD-1 методом масс-спектрометрии:
Чтобы идентифицировать потенциально прерывающиеся эпитопы, распознаваемые антителом к PD-1 собак, используют способ, включающий в себя химическую сшивку и детекцию методом масс-спектрометрии (CovalX® Instrument Incorporated). Применение данной технологии для картирования эпитопов PD-1 собак позволило идентифицировать, по меньшей мере, часть эпитопов, распознаваемых mAb, перечисленными в таблице 8. Как видно из таблицы 8, mAb 3B6 распознает, по меньшей мере, часть эпитопа, расположенного во внеклеточном домене PD-1 собак, в аминокислотной последовательности SEQ ID NO: 99, а mAb 2G9 распознает, по меньшей мере, часть эпитопа, расположенного в аминокислотной последовательности SEQ ID NO: 100. С другой стороны, mAb 1E4 и mAb 1B5 распознают, по меньшей мере, часть эпитопа в аминокислотной последовательности SEQ ID NO: 101 и в аминокислотной последовательности SEQ ID NO: 102, соответственно.
Как показано на фигуре 9а, результаты исследования, проводимого с использованием химической сшивки, масс-спектрометрии MALDI для анализа частиц с большой массой и масс-спектрометрии nLC-Orbitrap, показывают, что эпитоп на собачьем PD-1, распознаваемый канинизированным антителом 2G9, включает в себя R62, R69, R72 и R75 из последовательности SEQ ID NO: 2. Результаты аналогичного анализа демонстрируют, что эпитоп на собачьем PD-1, распознаваемый канинизированным антителом 3B6, включает в себя R75 и R90 из последовательности SEQ ID NO: 2. Соответственно, R75 представляет собой особенно важный остаток аминокислоты, входящий в состав одного или нескольких эпитопов PD-1 собак. Интересно отметить, что после проведения указанных анализов было обнаружено, что аминокислотные последовательности CDR 1A1 идентичны аминокислотным последовательностям CDR 2G9. Совпадающие результаты, полученные с помощью двух указанных очень разных методов для участка PD-1, с которым связывается 2G9, и для участка, с которым связывается 1A1, свидетельствуют о том, что данный участок содержит аминокислотные остатки, входящие в состав эпитопа PD-1, который распознается канинизированными антителами против PD-1 (см. таблицы 7 и 8 ниже).
Кроме того, участок аминокислотной последовательности PD-1, который распознается тестируемыми блокирующими антителами настоящего изобретения, находится во внеклеточном домене PD-1 собак. Распознаваемый участок входит в состав приведенного ниже пептида (см. таблицы 7 и 8 ниже).
NQTDKLAAFQEDRIEPGRDRRFRVM*RLPNGRDFHMSIVAARLNDS (SEQ ID NO: 103)
В пределах данного пептида существует короткий пептид, который выделен жирным шрифтом. Указанный короткий пептид распознается микрочипом ProImmune® (см. таблицу 7).
DRIEPGRDRRFRVM*RLPNGR (SEQ ID NO:104)
Следует отметить, что все R62, R69, R72 и R75 из последовательности SEQ ID NO: 2 входят в состав как более длинного (SEQ ID NO: 103), так и более короткого пептида (SEQ ID NO: 104), тогда как R90 из последовательности SEQ ID NO: 2 входит в состав как более длинного пептида. Пять указанных остатков аргинина, по-видимому, являются важными аминокислотными остатками одного или нескольких эпитопов PD-1 собак. Как указано в таблицах 6-8, отмеченный звездочкой остаток метионина (*) также описывается как остаток треонина.
ПЕПТИДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ КАРТИРОВАНИЯ ЭПИТОПОВ С ПОМОЩЬЮ МИКРОЧИПА ProImmune®
ЭПИТОПЫ PD-1, РАСПОЗНАВАЕМЫЕ mAb к PD-1 собак, ИДЕНТИФИЦИРОВАННЫЕ С ПОМОЩЬЮ МИКРОЧИПА ProImmune®
# CDR антитела 1A1 идентичны CDR антитела 2G9.
ЭПИТОПЫ PD-1, РАСПОЗНАВАЕМЫЕ mAb к PD-1 собак, ИДЕНТИФИЦИРОВАННЫЕ МЕТОДОМ МАСС-СПЕКТРОМЕТРИИ
ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ PD-1 СОБАК и СОБАЧЬЕГО PD-L1
сигнальная последовательность
ОПИСАНИЕ МОДИФИКАЦИЙ ПОСЛЕДОВАТЕЛЬНОСТИ СОБАЧЬЕГО IgGB
Все ссылки, приведенные в данном описании, включены в настоящее описание в качестве ссылки в такой степени, как если бы каждая отдельная ссылка, включающая в себя публикацию, компонент базы данных (такой как последовательность GenBank или значение GeneID), заявку на патент или патент, была бы конкретно и отдельно указана как включенная в качестве ссылки. Данное заявление о включении путем ссылки приведено заявителями в соответствии с 37 C.F.R. §1.57(b)(1) в отношении ко всем и каждой из отдельных ссылок, включающих публикацию, компонент базы данных (такой как последовательность GenBank или значение GeneID), заявку на патент или патент, каждая из которых четко определена в соответствии с 37 C.F.R. §1.57(b)(2), даже если такая цитата не находится в непосредственной близости к указанному заявлению о включении путем ссылки. Упоминание указанных заявлений о включении путем ссылки, если таковые имеются, в пределах описания не ослабляет каким-либо образом общее заявление о включении путем ссылки. Цитирование ссылок в настоящем документе не следует рассматривать как допущение, что ссылка относится к предшествующему уровню техники, а также как допущение, касающееся содержания или даты публикации указанных документов.
Объем настоящего изобретения не должен ограничиваться конкретными вариантами осуществления, описанными в настоящем описании. В действительности специалисты в данной области смогут осуществить разные модификации изобретения, помимо описанных здесь, на основании приведенного выше описания и прилагаемых чертежей. Такие модификации также входят в объем приложенной формулы изобретения.
Приведенное выше описание считается достаточным для того, чтобы специалист в данной области техники смог осуществить настоящее изобретение. На основании приведенного выше описания специалисты в данной области могут осуществить разные модификации изобретения, помимо показанных и описанных здесь, которые входят в объем прилагаемой формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Intervet International B.V.
Morsey, Mohamad
Zhang, Yuanzheng
Bartels-Morozov, Denise
Erskine, Jason
Tarpey, Ian
<120> Антитела против собачьего PD-1
<130> 23705
<150> 61/918946
<151> 2013-12-20
<150> 61/918847
<151> 2013-12-20
<150> 62/030812
<151> 2014-07-30
<160> 114
<170> PatentIn version 3.5
<210> 1
<211> 795
<212> ДНК
<213> Canis familiaris
<400> 1
ctagactccc ctgacaggcc ctggagcccg ctcaccttct ccccggcgca gctcacggtg 60
caggagggag agaacgccac gttcacctgc agcctggccg acatccccga cagcttcgtg 120
ctcaactggt accgcctgag cccccgcaac cagacggaca agctggccgc cttccaggag 180
gaccgcatcg agccgggccg ggacaggcgc ttccgcgtca tgcggctgcc caacgggcgg 240
gacttccaca tgagcatcgt cgctgcgcgc ctcaacgaca gcggcatcta cctgtgcggg 300
gccatctacc tgccccccaa cacacagatc aacgagagtc cccgcgcaga gctctccgtg 360
acggagagaa ccctggagcc ccccacacag agccccagcc ccccacccag actcagcggc 420
cagttgcagg ggctggtcat cggcgtcacg agcgtgctgg tgggtgtcct gctactgctg 480
ctgctgacct gggtcctggc cgctgtcttc cccagggcca cccgaggtgc ctgtgtgtgc 540
gggagcgagg acgagcctct gaaggagggc cccgatgcag cgcccgtctt caccctggac 600
tacggggagc tggacttcca gtggcgagag aagacgccgg agcccccggc gccctgtgcc 660
ccggagcaga ccgagtatgc caccatcgtc ttcccgggca ggccggcgtc cccgggccgc 720
agggcctcgg ccagcagcct gcagggagcc cagcctccga gccccgagga cggacccggc 780
ctgtggcccc tctga 795
<210> 2
<211> 264
<212> Белок
<213> Canis familiaris
<400> 2
Leu Asp Ser Pro Asp Arg Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala
1 5 10 15
Gln Leu Thr Val Gln Glu Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu
20 25 30
Ala Asp Ile Pro Asp Ser Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro
35 40 45
Arg Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu
50 55 60
Pro Gly Arg Asp Arg Arg Phe Arg Val Met Arg Leu Pro Asn Gly Arg
65 70 75 80
Asp Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser Gly Ile
85 90 95
Tyr Leu Cys Gly Ala Ile Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu
100 105 110
Ser Pro Arg Ala Glu Leu Ser Val Thr Glu Arg Thr Leu Glu Pro Pro
115 120 125
Thr Gln Ser Pro Ser Pro Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly
130 135 140
Leu Val Ile Gly Val Thr Ser Val Leu Val Gly Val Leu Leu Leu Leu
145 150 155 160
Leu Leu Thr Trp Val Leu Ala Ala Val Phe Pro Arg Ala Thr Arg Gly
165 170 175
Ala Cys Val Cys Gly Ser Glu Asp Glu Pro Leu Lys Glu Gly Pro Asp
180 185 190
Ala Ala Pro Val Phe Thr Leu Asp Tyr Gly Glu Leu Asp Phe Gln Trp
195 200 205
Arg Glu Lys Thr Pro Glu Pro Pro Ala Pro Cys Ala Pro Glu Gln Thr
210 215 220
Glu Tyr Ala Thr Ile Val Phe Pro Gly Arg Pro Ala Ser Pro Gly Arg
225 230 235 240
Arg Ala Ser Ala Ser Ser Leu Gln Gly Ala Gln Pro Pro Ser Pro Glu
245 250 255
Asp Gly Pro Gly Leu Trp Pro Leu
260
<210> 3
<211> 438
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированная
<400> 3
ctggattccc ccgacagacc ctggagccct ctcaccttct cccctgccca gctgaccgtc 60
caggaaggcg agaatgccac cttcacctgc agcctcgccg acatccccga cagcttcgtg 120
ctgaactggt acagactgag ccccaggaac cagaccgaca agctggccgc tttccaggag 180
gacaggatcg aacccggcag ggacaggagg tttagggtca tgaggctgcc caacggcagg 240
gacttccaca tgtccatcgt ggccgccaga ctgaacgact ccggcatcta cctgtgcggc 300
gctatctacc tgccccccaa cacccagatc aacgagagcc ccagggccga actgagcgtg 360
acagagagaa ccctggaacc tcccacccag agcccttccc ctcctcctag actgagcgga 420
cagctgcagg gcctggtg 438
<210> 4
<211> 438
<212> Белок
<213> Canis familiaris
<400> 4
Cys Thr Gly Gly Ala Thr Thr Cys Cys Cys Cys Cys Gly Ala Cys Ala
1 5 10 15
Gly Ala Cys Cys Cys Thr Gly Gly Ala Gly Cys Cys Cys Thr Cys Thr
20 25 30
Cys Ala Cys Cys Thr Thr Cys Thr Cys Cys Cys Cys Thr Gly Cys Cys
35 40 45
Cys Ala Gly Cys Thr Gly Ala Cys Cys Gly Thr Cys Cys Ala Gly Gly
50 55 60
Ala Ala Gly Gly Cys Gly Ala Gly Ala Ala Thr Gly Cys Cys Ala Cys
65 70 75 80
Cys Thr Thr Cys Ala Cys Cys Thr Gly Cys Ala Gly Cys Cys Thr Cys
85 90 95
Gly Cys Cys Gly Ala Cys Ala Thr Cys Cys Cys Cys Gly Ala Cys Ala
100 105 110
Gly Cys Thr Thr Cys Gly Thr Gly Cys Thr Gly Ala Ala Cys Thr Gly
115 120 125
Gly Thr Ala Cys Ala Gly Ala Cys Thr Gly Ala Gly Cys Cys Cys Cys
130 135 140
Ala Gly Gly Ala Ala Cys Cys Ala Gly Ala Cys Cys Gly Ala Cys Ala
145 150 155 160
Ala Gly Cys Thr Gly Gly Cys Cys Gly Cys Thr Thr Thr Cys Cys Ala
165 170 175
Gly Gly Ala Gly Gly Ala Cys Ala Gly Gly Ala Thr Cys Gly Ala Ala
180 185 190
Cys Cys Cys Gly Gly Cys Ala Gly Gly Gly Ala Cys Ala Gly Gly Ala
195 200 205
Gly Gly Thr Thr Thr Ala Gly Gly Gly Thr Cys Ala Thr Gly Ala Gly
210 215 220
Gly Cys Thr Gly Cys Cys Cys Ala Ala Cys Gly Gly Cys Ala Gly Gly
225 230 235 240
Gly Ala Cys Thr Thr Cys Cys Ala Cys Ala Thr Gly Thr Cys Cys Ala
245 250 255
Thr Cys Gly Thr Gly Gly Cys Cys Gly Cys Cys Ala Gly Ala Cys Thr
260 265 270
Gly Ala Ala Cys Gly Ala Cys Thr Cys Cys Gly Gly Cys Ala Thr Cys
275 280 285
Thr Ala Cys Cys Thr Gly Thr Gly Cys Gly Gly Cys Gly Cys Thr Ala
290 295 300
Thr Cys Thr Ala Cys Cys Thr Gly Cys Cys Cys Cys Cys Cys Ala Ala
305 310 315 320
Cys Ala Cys Cys Cys Ala Gly Ala Thr Cys Ala Ala Cys Gly Ala Gly
325 330 335
Ala Gly Cys Cys Cys Cys Ala Gly Gly Gly Cys Cys Gly Ala Ala Cys
340 345 350
Thr Gly Ala Gly Cys Gly Thr Gly Ala Cys Ala Gly Ala Gly Ala Gly
355 360 365
Ala Ala Cys Cys Cys Thr Gly Gly Ala Ala Cys Cys Thr Cys Cys Cys
370 375 380
Ala Cys Cys Cys Ala Gly Ala Gly Cys Cys Cys Thr Thr Cys Cys Cys
385 390 395 400
Cys Thr Cys Cys Thr Cys Cys Thr Ala Gly Ala Cys Thr Gly Ala Gly
405 410 415
Cys Gly Gly Ala Cys Ala Gly Cys Thr Gly Cys Ala Gly Gly Gly Cys
420 425 430
Cys Thr Gly Gly Thr Gly
435
<210> 5
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная по собачьим и человеческим кодонам
<400> 5
ctggattccc ccgacagacc ctggagccct ctcaccttct cccctgccca gctgaccgtc 60
caggaaggcg agaatgccac cttcacctgc agcctcgccg acatccccga cagcttcgtg 120
ctgaactggt acagactgag ccccaggaac cagaccgaca agctggccgc tttccaggag 180
gacaggatcg aacccggcag ggacaggagg tttagggtca tgaggctgcc caacggcagg 240
gacttccaca tgtccatcgt ggccgccaga ctgaacgact ccggcatcta cctgtgcggc 300
gctatctacc tgccccccaa cacccagatc aacgagagcc ccagggccga actgagcgtg 360
acagagagaa ccctggaacc tcccacccag agcccttccc ctcctcctag actgagcgga 420
cagctgcagg gcctggtggg taccgacaaa actcacacat gcccaccgtg cccagcacct 480
gaactcctgg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 540
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 600
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 660
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 720
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 780
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 840
ccatcccggg atgagctgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 900
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 960
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 1020
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 1080
cacaaccact acacgcagaa gagcctctcc ctgtctccgg gtaaatga 1128
<210> 6
<211> 375
<212> Белок
<213> Искусственная последовательность
<220>
<223> собачья и человеческая
<400> 6
Leu Asp Ser Pro Asp Arg Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala
1 5 10 15
Gln Leu Thr Val Gln Glu Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu
20 25 30
Ala Asp Ile Pro Asp Ser Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro
35 40 45
Arg Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu
50 55 60
Pro Gly Arg Asp Arg Arg Phe Arg Val Met Arg Leu Pro Asn Gly Arg
65 70 75 80
Asp Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser Gly Ile
85 90 95
Tyr Leu Cys Gly Ala Ile Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu
100 105 110
Ser Pro Arg Ala Glu Leu Ser Val Thr Glu Arg Thr Leu Glu Pro Pro
115 120 125
Thr Gln Ser Pro Ser Pro Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly
130 135 140
Leu Val Gly Thr Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
145 150 155 160
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
165 170 175
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
180 185 190
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
195 200 205
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
210 215 220
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
225 230 235 240
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
245 250 255
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
260 265 270
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
275 280 285
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
290 295 300
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
305 310 315 320
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
325 330 335
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
340 345 350
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
355 360 365
Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 7
<211> 816
<212> ДНК
<213> Canis familiaris
<400> 7
tttacgatca cagtttctaa ggacctgtat gtggtagagt atggtggcaa tgtgacaatg 60
gaatgcaaat tcccggtgga aaaacagtta aacttgtttg cactaatcgt ctactgggaa 120
atggaggata aaaaaattat acaatttgtg aatggaaagg aagacctgaa agttcagcac 180
agcagctaca gccagagggc tcagctattg aaggaccagc tcttcttggg gaaggctgcg 240
cttcagatca cagatgtgag attgcaggat gcaggggttt actgctgctt gatcggctat 300
ggcggtgctg actacaagcg gattactttg aaagttcatg ccccgtaccg caacatcagc 360
caaagaattt ctgtggatcc tgtcacctct gaacatgaac taatgtgtca ggctgagggt 420
taccctgagg ctgaagtcat ctggacaagc agtgaccacc gagtcctgag tggcaaaacc 480
accatcacta attccaatag ggaagagaag cttttcaatg tgaccagcac gctgaacatc 540
aatgcaacag ctaatgagat tttctactgc acttttcaaa gatcaggtcc tgaggaaaac 600
aatactgccg agttggtcat cccagaacga ctgcccgttc cagcaagtga gaggactcat 660
ttcatgattc tgggaccttt cctgttgctt cttggtgtag tcctggcagt cactttctgt 720
ctaaaaaaac atgggagaat gatggatgtg gaaaaatgtt gcacccgaga taggaactca 780
aagaaacgaa atgatataca atttgaagag acataa 816
<210> 8
<211> 271
<212> Белок
<213> Canis familiaris
<400> 8
Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr Gly Gly
1 5 10 15
Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asn Leu
20 25 30
Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile Ile Gln
35 40 45
Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser Tyr Ser
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp Pro Val
115 120 125
Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly Lys Thr
145 150 155 160
Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val Thr Ser
165 170 175
Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys Thr Phe
180 185 190
Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val Ile Pro
195 200 205
Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His Phe Met Ile Leu
210 215 220
Gly Pro Phe Leu Leu Leu Leu Gly Val Val Leu Ala Val Thr Phe Cys
225 230 235 240
Leu Lys Lys His Gly Arg Met Met Asp Val Glu Lys Cys Cys Thr Arg
245 250 255
Asp Arg Asn Ser Lys Lys Arg Asn Asp Ile Gln Phe Glu Glu Thr
260 265 270
<210> 9
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная по собачьим кодонам
<400> 9
tttaccatca ccgtgtccaa ggacctgtac gtggtcgagt acggcggcaa tgtgaccatg 60
gagtgcaagt tccccgtgga gaagcagctg aacctgttcg ccctcatcgt gtactgggag 120
atggaggaca agaagatcat ccagttcgtg aacggcaagg aggacctgaa ggtgcagcac 180
tccagctact cccagagagc ccagctgctg aaggaccagc tgttcctggg caaggccgcc 240
ctgcagatca ccgacgtgag actgcaggac gccggcgtgt attgctgcct gatcggctac 300
ggaggcgccg actacaagag gatcaccctg aaggtgcatg caccctacag gaacatcagc 360
cagaggatca gcgtcgatcc cgtgaccagc gagcacgagc tgatgtgcca agccgagggc 420
tatcccgagg ccgaagtgat ctggaccagc agcgaccaca gggtcctgag cggcaagacc 480
accatcacca acagcaacag ggaggagaag ctgttcaacg tgaccagcac cctcaacatc 540
aacgccaccg ccaacgagat cttctactgc accttccaga ggagcggccc cgaagagaac 600
aacaccgccg agctggtgat ccccgagaga ctgcctgtgc ctgccagcga gaggacccac 660
<210> 10
<211> 220
<212> Белок
<213> Canis familiaris
<400> 10
Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr Gly Gly
1 5 10 15
Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asn Leu
20 25 30
Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile Ile Gln
35 40 45
Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser Tyr Ser
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp Pro Val
115 120 125
Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly Lys Thr
145 150 155 160
Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val Thr Ser
165 170 175
Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys Thr Phe
180 185 190
Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val Ile Pro
195 200 205
Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His
210 215 220
<210> 11
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная по собачьим и человеческим кодонам
<400> 11
tttaccatca ccgtgtccaa ggacctgtac gtggtcgagt acggcggcaa tgtgaccatg 60
gagtgcaagt tccccgtgga gaagcagctg aacctgttcg ccctcatcgt gtactgggag 120
atggaggaca agaagatcat ccagttcgtg aacggcaagg aggacctgaa ggtgcagcac 180
tccagctact cccagagagc ccagctgctg aaggaccagc tgttcctggg caaggccgcc 240
ctgcagatca ccgacgtgag actgcaggac gccggcgtgt attgctgcct gatcggctac 300
ggaggcgccg actacaagag gatcaccctg aaggtgcatg caccctacag gaacatcagc 360
cagaggatca gcgtcgatcc cgtgaccagc gagcacgagc tgatgtgcca agccgagggc 420
tatcccgagg ccgaagtgat ctggaccagc agcgaccaca gggtcctgag cggcaagacc 480
accatcacca acagcaacag ggaggagaag ctgttcaacg tgaccagcac cctcaacatc 540
aacgccaccg ccaacgagat cttctactgc accttccaga ggagcggccc cgaagagaac 600
aacaccgccg agctggtgat ccccgagaga ctgcctgtgc ctgccagcga gaggacccac 660
ggtaccgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtctcc gggtaaatga 1350
<210> 12
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> собачья и человеческая
<400> 12
Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr Gly Gly
1 5 10 15
Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asn Leu
20 25 30
Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile Ile Gln
35 40 45
Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser Tyr Ser
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp Pro Val
115 120 125
Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly Lys Thr
145 150 155 160
Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val Thr Ser
165 170 175
Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys Thr Phe
180 185 190
Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val Ile Pro
195 200 205
Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His Gly Thr Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 13
<211> 17
<212> Белок
<213> Mus musculus
<400> 13
Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 14
<211> 16
<212> Белок
<213> Mus musculus
<400> 14
Arg Ser Ser Gln Asn Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 15
<211> 11
<212> Белок
<213> Mus musculus
<400> 15
His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser
1 5 10
<210> 16
<211> 7
<212> Белок
<213> Mus musculus
<400> 16
Phe Ala Ser Thr Arg Val Ser
1 5
<210> 17
<211> 7
<212> Белок
<213> Mus musculus
<400> 17
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 18
<211> 7
<212> Белок
<213> Mus musculus
<400> 18
Lys Ala Ser His Leu His Thr
1 5
<210> 19
<211> 7
<212> Белок
<213> Mus musculus
<400> 19
Phe Ala Ser Ala Arg Val Ser
1 5
<210> 20
<211> 7
<212> Белок
<213> Mus musculus
<400> 20
Phe Ala Ser Thr Arg Ile Ser
1 5
<210> 21
<211> 7
<212> Белок
<213> Mus musculus
<400> 21
Lys Ala Ser Asn Leu His Thr
1 5
<210> 22
<211> 9
<212> Белок
<213> Mus musculus
<400> 22
Gln Gln Tyr Phe Ser Thr Pro Leu Thr
1 5
<210> 23
<211> 9
<212> Белок
<213> Mus musculus
<400> 23
Phe Gln Gly Ser His Val Pro Tyr Thr
1 5
<210> 24
<211> 9
<212> Белок
<213> Mus musculus
<400> 24
Gln Gln Gly Gln Ser Trp Pro Leu Thr
1 5
<210> 25
<211> 9
<212> Белок
<213> Mus musculus
<400> 25
Gln Gln Tyr Phe Ser Thr Pro Leu Thr
1 5
<210> 26
<211> 9
<212> Белок
<213> Mus musculus
<400> 26
Gln Gln Gly Gln Ser Tyr Pro Leu Thr
1 5
<210> 27
<211> 10
<212> Белок
<213> Mus musculus
<400> 27
Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser
1 5 10
<210> 28
<211> 10
<212> Белок
<213> Mus musculus
<400> 28
Gly Tyr Thr Phe Thr Arg Tyr Asn Met His
1 5 10
<210> 29
<211> 10
<212> Белок
<213> Mus musculus
<400> 29
Gly Phe Asn Ile Lys Asn Thr Tyr Met His
1 5 10
<210> 30
<211> 10
<212> Белок
<213> Mus musculus
<400> 30
Gly Phe Ser Leu Thr Ser Tyr Gly Val His
1 5 10
<210> 31
<211> 17
<212> Белок
<213> Mus musculus
<400> 31
Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 32
<211> 17
<212> Белок
<213> Mus musculus
<400> 32
Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 33
<211> 17
<212> Белок
<213> Mus musculus
<400> 33
Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe Gln
1 5 10 15
Gly
<210> 34
<211> 17
<212> Белок
<213> Mus musculus
<400> 34
Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 35
<211> 17
<212> Белок
<213> Mus musculus
<400> 35
Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe Gln
1 5 10 15
Gly
<210> 36
<211> 6
<212> Белок
<213> Mus musculus
<400> 36
Phe Asp Gly Pro Asp Tyr
1 5
<210> 37
<211> 13
<212> Белок
<213> Mus musculus
<400> 37
Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr
1 5 10
<210> 38
<211> 11
<212> Белок
<213> Mus musculus
<400> 38
Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val
1 5 10
<210> 39
<211> 336
<212> Белок
<213> Canis familiaris
<400> 39
Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys
1 5 10 15
Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly
20 25 30
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr
35 40 45
Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr
50 55 60
Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu
65 70 75 80
Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp
85 90 95
Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp
100 105 110
Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe
115 120 125
Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg Thr Pro
130 135 140
Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val
145 150 155 160
Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr
165 170 175
Gln Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val
180 185 190
Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys
195 200 205
Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser
210 215 220
Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro
225 230 235 240
Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile
245 250 255
Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly
260 265 270
Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp
275 280 285
Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser
290 295 300
Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala
305 310 315 320
Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330 335
<210> 40
<211> 331
<212> Белок
<213> Искусственная последовательность
<220>
<223> модифицированная собачья
<400> 40
Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys
1 5 10 15
Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly
20 25 30
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr
35 40 45
Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr
50 55 60
Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu
65 70 75 80
Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp
85 90 95
Lys Pro Val Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro Cys Pro
100 105 110
Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys
115 120 125
Pro Lys Ala Thr Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val
130 135 140
Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe
145 150 155 160
Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu
165 170 175
Gln Phe Ala Gly Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His
180 185 190
Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys
195 200 205
Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln
210 215 220
Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu
225 230 235 240
Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro
245 250 255
Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu
260 265 270
Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr
275 280 285
Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly
290 295 300
Asp Thr Phe Ile Cys Ala Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330
<210> 41
<211> 331
<212> Белок
<213> Искусственная последовательность
<220>
<223> модифицированная собачья
<400> 41
Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys
1 5 10 15
Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly
20 25 30
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr
35 40 45
Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr
50 55 60
Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu
65 70 75 80
Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp
85 90 95
Lys Pro Val Pro Lys Glu Ser Thr Cys Lys Cys Ile Ser Pro Cys Pro
100 105 110
Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys
115 120 125
Pro Lys Ala Thr Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val
130 135 140
Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe
145 150 155 160
Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu
165 170 175
Gln Phe Ala Gly Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His
180 185 190
Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys
195 200 205
Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln
210 215 220
Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu
225 230 235 240
Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro
245 250 255
Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu
260 265 270
Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr
275 280 285
Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly
290 295 300
Asp Thr Phe Ile Cys Ala Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330
<210> 42
<211> 336
<212> Белок
<213> Искусственная последовательность
<220>
<223> модифицированная собачья
<400> 42
Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys
1 5 10 15
Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly
20 25 30
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr
35 40 45
Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr
50 55 60
Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu
65 70 75 80
Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp
85 90 95
Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp
100 105 110
Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe
115 120 125
Ile Phe Pro Pro Lys Pro Lys Ala Thr Leu Leu Ile Ala Arg Thr Pro
130 135 140
Glu Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val
145 150 155 160
Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr
165 170 175
Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr Tyr Arg Val Val Ser Val
180 185 190
Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys
195 200 205
Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser
210 215 220
Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro
225 230 235 240
Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile
245 250 255
Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly
260 265 270
Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp
275 280 285
Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser
290 295 300
Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala
305 310 315 320
Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330 335
<210> 43
<211> 345
<212> ДНК
<213> Mus musculus
<400> 43
cagatccagt tggtacagtc tggacctgaa ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca acctatggaa tgagctgggt gaaacaggct 120
ccaggaaagg gtttaaagtg gatgggctgg attaatatct actctggaat cccaacatat 180
gctgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgcctat 240
ttgcagatcg acaacctcaa aaatgaggac acggctacat atttctgtgc aagatttgat 300
ggtcccgact actggggcca aggcaccact ctcaccgtct cccca 345
<210> 44
<211> 115
<212> Белок
<213> Mus musculus
<400> 44
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asp Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
100 105 110
Val Ser Pro
115
<210> 45
<211> 339
<212> ДНК
<213> Mus musculus
<400> 45
gacattgtga tgacacagtc tccatcctcc ctggctatgt cagtaggaca gaaggtcact 60
atgagctgca agtccagtca gagcctttta aatagtgtca atcaaaagaa ctatttggcc 120
tggtaccagc agaaaccagg acagtctcct aaagttctgg tatactttgc atccactagg 180
gtatctgggg tccctgatcg cttcataggc agtggatctg ggacagattt cactcttacc 240
atcaccagtg tgcaggctga agacctgaca acttacctct gtcagcaata ttttagcact 300
cctctcacgt tcggtgctgg gaccaagctg gaaataaaa 339
<210> 46
<211> 113
<212> Белок
<213> Mus musculus
<400> 46
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Met Ser Val Gly
1 5 10 15
Gln Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Thr Arg Val Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Thr Ser Val Gln Ala Glu Asp Leu Thr Thr Tyr Leu Cys Gln Gln
85 90 95
Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 47
<211> 423
<212> ДНК
<213> Mus musculus
<400> 47
atgggattca gcaggatctt tctcttcctc ctgtcagtaa ctacaggtgt ccactcccag 60
gcttatctac agcagtctgg ggctgagctg gtgaggcctg gggcctcagt gaagatctcc 120
tgcaaggctt ctggctacac atttaccaga tacaatatgc actgggtaaa gcagacacct 180
agacagggcc tggaatggat tggaactatt tatcccggat atggtgatac ttcttacaat 240
cagaaattca agggcaaggc cacactgact gtagacatat cctccagcac agcctacatg 300
cagctcacca gcctgacatc tgaggactct gcggtctatt tctgttcaag ggagtttgcc 360
gatgattacc ccattccccc ctttgactac tggggccaag gcaccactct cacagtctcc 420
tca 423
<210> 48
<211> 141
<212> Белок
<213> Mus musculus
<400> 48
Met Gly Phe Ser Arg Ile Phe Leu Phe Leu Leu Ser Val Thr Thr Gly
1 5 10 15
Val His Ser Gln Ala Tyr Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Asn Met His Trp Val Lys Gln Thr Pro Arg Gln Gly Leu
50 55 60
Glu Trp Ile Gly Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Ile Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Thr Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ser Arg Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
130 135 140
<210> 49
<211> 393
<212> ДНК
<213> Mus musculus
<400> 49
atgaagttgc ctgttaggct gttggtgctg atgttctgga ttcctgcttc cagcagtgat 60
gttttgatga cccaaactcc actctccctg cctgtcagtc ttggagatca agcctccatc 120
tcttgtagat ctagtcagaa cattgtacat agtaatggaa acacctactt agaatggtac 180
ctgcagaaac caggccagtc tccaaagctc ctgatctaca aagtttccaa ccgattttct 240
ggggtcccag acaggttcag tggcagtgga tcagggacag atttcacact caagatcagc 300
agagtggagg ctgaggatct gggaatttat tactgctttc aaggttcaca tgttccgtac 360
acgttcggag gggggaccaa gctggaaata aaa 393
<210> 50
<211> 131
<212> Белок
<213> Mus musculus
<400> 50
Met Lys Leu Pro Val Arg Leu Leu Val Leu Met Phe Trp Ile Pro Ala
1 5 10 15
Ser Ser Ser Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val
20 25 30
Ser Leu Gly Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Asn Ile
35 40 45
Val His Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro
50 55 60
Gly Gln Ser Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser
65 70 75 80
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys
100 105 110
Phe Gln Gly Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu
115 120 125
Glu Ile Lys
130
<210> 51
<211> 417
<212> ДНК
<213> Mus musculus
<400> 51
atgaaattca gctgggtcat cttcttcctg atggcagtgg ttacaggggt caattcagag 60
gttcagctgc agcagtctgt ggcagagctt gtgaggccag gggcctcagt caagttgtcc 120
tgcacagctt ctggcttcaa cattaaaaac acctatatgc actggataaa acagaggcct 180
gaacagggcc tggagtggat tggaaggatt gctcctgcga atgttgatac taaatatgcc 240
ccgaagttcc agggcaaggc cactataact gcagacacat cctccaacac agcctacatg 300
cagctcagca ccctgacatc ggaggacact gccatctatt actgtgtcct gatctactat 360
gattacgacg gggacatcga tgtctggggc acagggacca cggtcaccgt ctcctca 417
<210> 52
<211> 139
<212> Белок
<213> Mus musculus
<400> 52
Met Lys Phe Ser Trp Val Ile Phe Phe Leu Met Ala Val Val Thr Gly
1 5 10 15
Val Asn Ser Glu Val Gln Leu Gln Gln Ser Val Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile
35 40 45
Lys Asn Thr Tyr Met His Trp Ile Lys Gln Arg Pro Glu Gln Gly Leu
50 55 60
Glu Trp Ile Gly Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala
65 70 75 80
Pro Lys Phe Gln Gly Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn
85 90 95
Thr Ala Tyr Met Gln Leu Ser Thr Leu Thr Ser Glu Asp Thr Ala Ile
100 105 110
Tyr Tyr Cys Val Leu Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val
115 120 125
Trp Gly Thr Gly Thr Thr Val Thr Val Ser Ser
130 135
<210> 53
<211> 381
<212> ДНК
<213> Mus musculus
<400> 53
atgagggtcc ttgctgagct cctggggctg ctgctgttct gctttttagg tgtgagatgt 60
gacatccaga tgaaccagtc tccatccagt ctgtctgcat cccttggaga cacaattacc 120
atcacttgcc atgccagtca gaacattaat gtttggttaa gttggtacca gcagagacca 180
ggaaatattc ctaaactatt gatctataag gcttctcact tacacacagg cgtcccatca 240
aggtttagtg gcagtggatc tggaacaggt ttcacattaa ccatcagcag cctgcagcct 300
gaagacattg ccacttacta ctgtcaacag ggtcaaagtt ggccgctcac gttcggtgct 360
gggaccaaac tggagctgaa a 381
<210> 54
<211> 127
<212> Белок
<213> Mus musculus
<400> 54
Met Arg Val Leu Ala Glu Leu Leu Gly Leu Leu Leu Phe Cys Phe Leu
1 5 10 15
Gly Val Arg Cys Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Asp Thr Ile Thr Ile Thr Cys His Ala Ser Gln Asn
35 40 45
Ile Asn Val Trp Leu Ser Trp Tyr Gln Gln Arg Pro Gly Asn Ile Pro
50 55 60
Lys Leu Leu Ile Tyr Lys Ala Ser His Leu His Thr Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Gly Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln
100 105 110
Ser Trp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
115 120 125
<210> 55
<211> 402
<212> ДНК
<213> Mus musculus
<400> 55
atgggttggc tgtggaactt gctattcctg atggcagctg cccaaagtgc ccaaacacag 60
atccagttgg tacagtctgg acctgaactg aagaagcctg gagagacagt caagatctcc 120
tgcaaggctt ctgggtatac cttcacaacc tatggaatga gctgggtgaa acaggctcca 180
ggaaagggtt taaagtggat gggctggatt aatatctact ctggaatccc aacatatgct 240
gatgacttca agggacgatt tgccttctct ttggaaacct ctgccagcac tgcctatttg 300
cagatcgaca acctcaaaaa tgaggacacg gctacatatt tctgtgcaag atttgatggt 360
cccgactact ggggccaagg caccactctc acagtctcct ca 402
<210> 56
<211> 134
<212> Белок
<213> Mus musculus
<400> 56
Met Gly Trp Leu Trp Asn Leu Leu Phe Leu Met Ala Ala Ala Gln Ser
1 5 10 15
Ala Gln Thr Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Gly Met Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala
65 70 75 80
Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asp Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr
115 120 125
Thr Leu Thr Val Ser Ser
130
<210> 57
<211> 399
<212> ДНК
<213> Mus musculus
<400> 57
atggaatcac agacccaggt cctcatgttt cttctgctct gggtatctgg tgcctgtgca 60
gacattgtga tgacacagtc tccatcctcc ctggctgtgt cagtaggacg gaaggtcact 120
atgagctgca agtccagtca gagcctttta aatagtgtca atcaaaagaa ctatttggcc 180
tggtaccagc agaaaccagg acagtctcct aaagttctgg tatactttgc atccgctagg 240
gtatctgggg tccctgatcg cttcataggc agtggatctg ggacagattt cactcttgcc 300
atcagcagtg tgcaggctga agacctgaca acttacttct gtcagcaata ttttagcact 360
cctctcacgt tcggtgctgg gaccaagctg gaactgaaa 399
<210> 58
<211> 133
<212> Белок
<213> Mus musculus
<400> 58
Met Glu Ser Gln Thr Gln Val Leu Met Phe Leu Leu Leu Trp Val Ser
1 5 10 15
Gly Ala Cys Ala Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala
20 25 30
Val Ser Val Gly Arg Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser
35 40 45
Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln
50 55 60
Lys Pro Gly Gln Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Ala Arg
65 70 75 80
Val Ser Gly Val Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Ala Ile Ser Ser Val Gln Ala Glu Asp Leu Thr Thr Tyr
100 105 110
Phe Cys Gln Gln Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr
115 120 125
Lys Leu Glu Leu Lys
130
<210> 59
<211> 402
<212> ДНК
<213> Mus musculus
<400> 59
atgggttggc tgtggaactt gctattcctg atggcagctg cccaaagtgc ccaagcacag 60
atccagttgg tacagtctgg acctgaactg aagaagcctg gagagacagt caagatctcc 120
tgcaaggctt ctgggtatac cttcacaacc tatggaatga gctgggtgaa acaggcgcca 180
ggaaagggtt taaagtggat gggctggata aatatctact ctggaatgcc aacatatgct 240
gatgacttca agggacggtt tgccttctct ttggaaacct ctgtcagcac tgcctatttg 300
cagatcaaca acctcaaaaa tgaggacacg gctacatatt tctgtgcaag atttgatggt 360
cccgactact ggggccaagg caccactctc acagtctcct ca 402
<210> 60
<211> 134
<212> Белок
<213> Mus musculus
<400> 60
Met Gly Trp Leu Trp Asn Leu Leu Phe Leu Met Ala Ala Ala Gln Ser
1 5 10 15
Ala Gln Ala Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Thr Tyr Gly Met Ser Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala
65 70 75 80
Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Val Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr
115 120 125
Thr Leu Thr Val Ser Ser
130
<210> 61
<211> 399
<212> ДНК
<213> Mus musculus
<400> 61
atggaatcac agacccaggt cctcatgttt cttctgctct gggtatctgg tgcctgtgca 60
gacattgtga tgacacagtc tccatcctcc ctggctatgt cagtaggaca gaaggtcact 120
atgagctgca agtccagtca gagcctttta aatagtgtca atcaaaagaa ctatttggcc 180
tggtaccagc agaaaccagg acagtctcct aaagttctgg tatactttgc atccactagg 240
atatctgggg tccctgatcg cttcataggc agtggatctg ggacagattt cactcttacc 300
atcagcagtg tgcaggctga agacctggca gattacttct gtcagcaata ttttagcact 360
cctctcacgt tcggtgctgg gaccaagctg gagctgaaa 399
<210> 62
<211> 133
<212> Белок
<213> Mus musculus
<400> 62
Met Glu Ser Gln Thr Gln Val Leu Met Phe Leu Leu Leu Trp Val Ser
1 5 10 15
Gly Ala Cys Ala Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala
20 25 30
Met Ser Val Gly Gln Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser
35 40 45
Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln
50 55 60
Lys Pro Gly Gln Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Thr Arg
65 70 75 80
Ile Ser Gly Val Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Asp Tyr
100 105 110
Phe Cys Gln Gln Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr
115 120 125
Lys Leu Glu Leu Lys
130
<210> 63
<211> 417
<212> ДНК
<213> Mus musculus
<400> 63
atgaaattca gctgggtcat cttcttcctg atggcagtgg ttacaggggt caattcagag 60
gttcagctgc agcagtctgt ggcagagctt gtgaggccag gggcctcagt caagttgtcc 120
tgcacagttt ctggcttcaa cattaaaaac acctatatgc actgggtgaa gcagaggcct 180
gaacagggcc tggagtggat tggaagaatt gatcctgcga atgttaatac taaatatgcc 240
ccgaagttcc agggcaaggc cactataact acagacacat cctccaacac agcctacatg 300
cagctcagca gcctgacatc ggaggacact gccatctatt actgtgtcct gattttctat 360
gattacgacg gggacatcga tgtctggggc acagggacca aggtcaccgt ctcctca 417
<210> 64
<211> 139
<212> Белок
<213> Mus musculus
<400> 64
Met Lys Phe Ser Trp Val Ile Phe Phe Leu Met Ala Val Val Thr Gly
1 5 10 15
Val Asn Ser Glu Val Gln Leu Gln Gln Ser Val Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Val Ser Gly Phe Asn Ile
35 40 45
Lys Asn Thr Tyr Met His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu
50 55 60
Glu Trp Ile Gly Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala
65 70 75 80
Pro Lys Phe Gln Gly Lys Ala Thr Ile Thr Thr Asp Thr Ser Ser Asn
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Ile
100 105 110
Tyr Tyr Cys Val Leu Ile Phe Tyr Asp Tyr Asp Gly Asp Ile Asp Val
115 120 125
Trp Gly Thr Gly Thr Lys Val Thr Val Ser Ser
130 135
<210> 65
<211> 381
<212> ДНК
<213> Mus musculus
<400> 65
atgagggtcc ttgctgagct cctggggctg ctgctgttct gctttttagg tgtgagatgt 60
gacatccaga tgaaccagtc tccatccagt ctgtctgcat cccttggaga cacaattacc 120
atcacttgcc atgccagtca gaacattaat gtttggttaa gctggtacca gcagaaacca 180
ggaaatattc ctaaactatt gatctataag gcttccaact tacacacagg cgtcccatca 240
aggtttagtg gcagtggatc tggaacagat ttcacattaa ccatcagcag cctgcagcct 300
gaagacattg ccacttacta ctgtcaacag ggtcaaagtt atccgctcac gttcggtgct 360
gggaccaagc tggagctgaa a 381
<210> 66
<211> 127
<212> Белок
<213> Mus musculus
<400> 66
Met Arg Val Leu Ala Glu Leu Leu Gly Leu Leu Leu Phe Cys Phe Leu
1 5 10 15
Gly Val Arg Cys Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Asp Thr Ile Thr Ile Thr Cys His Ala Ser Gln Asn
35 40 45
Ile Asn Val Trp Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro
50 55 60
Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln
100 105 110
Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
115 120 125
<210> 67
<211> 345
<212> ДНК
<213> Mus musculus
<400> 67
caggtgcagc tgaagcagtc aggacctggc ctagtgcagc cctcacagag cctgtccata 60
acctgcacag tctctggttt ctcattaact agctatggtg tacactgggt tcgccagtct 120
ccaggaaagg gtttaaagtg gatgggctgg attaatatct actctggaat cccaacatat 180
gctgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgcctat 240
ttgcagatcg acaacctcaa aaatgaggac acggctacat atttctgtgc aagatttgat 300
ggtcccgact actggggcca aggcatcact ctcactgtct ccgca 345
<210> 68
<211> 115
<212> Белок
<213> Mus musculus
<400> 68
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asp Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Ile Thr Leu Thr
100 105 110
Val Ser Ala
115
<210> 69
<211> 339
<212> ДНК
<213> Mus musculus
<400> 69
gacattgtga tgacacagtc tccatcctcc ctggctatgt cagtaggaca gaaggtcact 60
atgagctgca agtccagtca gagcctttta aatagtgtca atcaaaagaa ctatttggcc 120
tggtaccagc agaaaccagg acagtctcct aaagttctgg tatactttgc atccactagg 180
gtatctgggg tccctgatcg cttcataggc agtggatctg ggacagattt cactcttacc 240
atcaccagtg tgcaggctga agacctgaca acttacttct gtcagcaata ttttagcact 300
cctctcacgt tcggtgctgg gaccaagctg gaactgaaa 339
<210> 70
<211> 113
<212> Белок
<213> Mus musculus
<400> 70
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Met Ser Val Gly
1 5 10 15
Gln Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Val Leu Val Tyr Phe Ala Ser Thr Arg Val Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Thr Ser Val Gln Ala Glu Asp Leu Thr Thr Tyr Phe Cys Gln Gln
85 90 95
Tyr Phe Ser Thr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
100 105 110
Lys
<210> 71
<211> 15
<212> Белок
<213> Canis familiaris
<400> 71
Leu Asp Ser Pro Asp Arg Pro Trp Ser Pro Leu Thr Phe Ser Pro
1 5 10 15
<210> 72
<211> 15
<212> Белок
<213> Canis familiaris
<400> 72
Arg Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala Gln Leu Thr Val
1 5 10 15
<210> 73
<211> 15
<212> Белок
<213> Canis familiaris
<400> 73
Leu Thr Phe Ser Pro Ala Gln Leu Thr Val Gln Glu Gly Glu Asn
1 5 10 15
<210> 74
<211> 15
<212> Белок
<213> Canis familiaris
<400> 74
Ala Gln Leu Thr Val Gln Glu Gly Glu Asn Ala Thr Phe Thr Cys
1 5 10 15
<210> 75
<211> 15
<212> Белок
<213> Canis familiaris
<400> 75
Gln Glu Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu Ala Asp Ile
1 5 10 15
<210> 76
<211> 15
<212> Белок
<213> Canis familiaris
<400> 76
Ala Thr Phe Thr Cys Ser Leu Ala Asp Ile Pro Asp Ser Phe Val
1 5 10 15
<210> 77
<211> 15
<212> Белок
<213> Canis familiaris
<400> 77
Ser Leu Ala Asp Ile Pro Asp Ser Phe Val Leu Asn Trp Tyr Arg
1 5 10 15
<210> 78
<211> 15
<212> Белок
<213> Canis familiaris
<400> 78
Pro Asp Ser Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro Arg Asn
1 5 10 15
<210> 79
<211> 15
<212> Белок
<213> Canis familiaris
<400> 79
Leu Asn Trp Tyr Arg Leu Ser Pro Arg Asn Gln Thr Asp Lys Leu
1 5 10 15
<210> 80
<211> 15
<212> Белок
<213> Canis familiaris
<400> 80
Leu Ser Pro Arg Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu
1 5 10 15
<210> 81
<211> 15
<212> Белок
<213> Canis familiaris
<400> 81
Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro
1 5 10 15
<210> 82
<211> 15
<212> Белок
<213> Canis familiaris
<400> 82
Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg
1 5 10 15
<210> 83
<211> 15
<212> Белок
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa может обозначать Met или Thr
<400> 83
Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg
1 5 10 15
<210> 84
<211> 15
<212> Белок
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa может обозначать Met или Thr
<400> 84
Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg
1 5 10 15
<210> 85
<211> 15
<212> Белок
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa может обозначать Met или Thr
<400> 85
Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg Asp Phe His Met Ser
1 5 10 15
<210> 86
<211> 15
<212> Белок
<213> Canis familiaris
<400> 86
Leu Pro Asn Gly Arg Asp Phe His Met Ser Ile Val Ala Ala Arg
1 5 10 15
<210> 87
<211> 15
<212> Белок
<213> Canis familiaris
<400> 87
Asp Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser Gly
1 5 10 15
<210> 88
<211> 15
<212> Белок
<213> Canis familiaris
<400> 88
Ile Val Ala Ala Arg Leu Asn Asp Ser Gly Ile Tyr Leu Cys Gly
1 5 10 15
<210> 89
<211> 15
<212> Белок
<213> Canis familiaris
<400> 89
Leu Asn Asp Ser Gly Ile Tyr Leu Cys Gly Ala Ile Tyr Leu Pro
1 5 10 15
<210> 90
<211> 15
<212> Белок
<213> Canis familiaris
<400> 90
Ile Tyr Leu Cys Gly Ala Ile Tyr Leu Pro Pro Asn Thr Gln Ile
1 5 10 15
<210> 91
<211> 15
<212> Белок
<213> Canis familiaris
<400> 91
Ala Ile Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu Ser Pro Arg
1 5 10 15
<210> 92
<211> 15
<212> Белок
<213> Canis familiaris
<400> 92
Pro Asn Thr Gln Ile Asn Glu Ser Pro Arg Ala Glu Leu Ser Val
1 5 10 15
<210> 93
<211> 15
<212> Белок
<213> Canis familiaris
<400> 93
Asn Glu Ser Pro Arg Ala Glu Leu Ser Val Thr Glu Arg Thr Leu
1 5 10 15
<210> 94
<211> 15
<212> Белок
<213> Canis familiaris
<400> 94
Ala Glu Leu Ser Val Thr Glu Arg Thr Leu Glu Pro Pro Thr Gln
1 5 10 15
<210> 95
<211> 15
<212> Белок
<213> Canis familiaris
<400> 95
Thr Glu Arg Thr Leu Glu Pro Pro Thr Gln Ser Pro Ser Pro Pro
1 5 10 15
<210> 96
<211> 15
<212> Белок
<213> Canis familiaris
<400> 96
Glu Pro Pro Thr Gln Ser Pro Ser Pro Pro Pro Arg Leu Ser Gly
1 5 10 15
<210> 97
<211> 15
<212> Белок
<213> Canis familiaris
<400> 97
Ser Pro Ser Pro Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly Leu
1 5 10 15
<210> 98
<211> 15
<212> Белок
<213> Canis familiaris
<400> 98
Pro Ser Pro Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly Leu Val
1 5 10 15
<210> 99
<211> 25
<212> Белок
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa может обозначать Met или Thr
<400> 99
Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg Asp Phe His Met Ser
1 5 10 15
Ile Val Ala Ala Arg Leu Asn Asp Ser
20 25
<210> 100
<211> 26
<212> Белок
<213> Canis familiaris
<220>
<221> misc_feature
<222> (20)..(20)
<223> Xaa может обозначать любую природную аминокислоту
<400> 100
Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg
1 5 10 15
Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg
20 25
<210> 101
<211> 31
<212> Белок
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (15)..(15)
<223> Xaa может обозначать Met или Thr
<400> 101
Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg
1 5 10 15
Leu Pro Asn Gly Arg Asp Phe His Met Ser Ile Val Ala Ala Arg
20 25 30
<210> 102
<211> 31
<212> Белок
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (25)..(25)
<223> Xaa может обозначать Met или Thr
<400> 102
Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro
1 5 10 15
Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg
20 25 30
<210> 103
<211> 45
<212> Белок
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (25)..(25)
<223> Xaa может обозначать Met или Thr
<400> 103
Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro
1 5 10 15
Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg Asp
20 25 30
Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser
35 40 45
<210> 104
<211> 20
<212> Белок
<213> Canis familiaris
<220>
<221> misc_feature
<222> (14)..(14)
<223> Xaa может обозначать Met или Thr
<400> 104
Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu
1 5 10 15
Pro Asn Gly Arg
20
<210> 105
<211> 867
<212> ДНК
<213> Canis familiaris
<400> 105
atggggagcc ggcgggggcc ctggccgctc gtctgggccg tgctgcagct gggctggtgg 60
ccaggatggc tcctagactc ccctgacagg ccctggagcc cgctcacctt ctccccggcg 120
cagctcacgg tgcaggaggg agagaacgcc acgttcacct gcagcctggc cgacatcccc 180
gacagcttcg tgctcaactg gtaccgcctg agcccccgca accagacgga caagctggcc 240
gccttccagg aggaccgcat cgagccgggc cgggacaggc gcttccgcgt catgcggctg 300
cccaacgggc gggacttcca catgagcatc gtcgctgcgc gcctcaacga cagcggcatc 360
tacctgtgcg gggccatcta cctgcccccc aacacacaga tcaacgagag tccccgcgca 420
gagctctccg tgacggagag aaccctggag ccccccacac agagccccag ccccccaccc 480
agactcagcg gccagttgca ggggctggtc atcggcgtca cgagcgtgct ggtgggtgtc 540
ctgctactgc tgctgctgac ctgggtcctg gccgctgtct tccccagggc cacccgaggt 600
gcctgtgtgt gcgggagcga ggacgagcct ctgaaggagg gccccgatgc agcgcccgtc 660
ttcaccctgg actacgggga gctggacttc cagtggcgag agaagacgcc ggagcccccg 720
gcgccctgtg ccccggagca gaccgagtat gccaccatcg tcttcccggg caggccggcg 780
tccccgggcc gcagggcctc ggccagcagc ctgcagggag cccagcctcc gagccccgag 840
gacggacccg gcctgtggcc cctctga 867
<210> 106
<211> 288
<212> Белок
<213> Canis familiaris
<400> 106
Met Gly Ser Arg Arg Gly Pro Trp Pro Leu Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Trp Pro Gly Trp Leu Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Ser Pro Leu Thr Phe Ser Pro Ala Gln Leu Thr Val Gln Glu Gly Glu
35 40 45
Asn Ala Thr Phe Thr Cys Ser Leu Ala Asp Ile Pro Asp Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Leu Ser Pro Arg Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Gln Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg
85 90 95
Val Met Arg Leu Pro Asn Gly Arg Asp Phe His Met Ser Ile Val Ala
100 105 110
Ala Arg Leu Asn Asp Ser Gly Ile Tyr Leu Cys Gly Ala Ile Tyr Leu
115 120 125
Pro Pro Asn Thr Gln Ile Asn Glu Ser Pro Arg Ala Glu Leu Ser Val
130 135 140
Thr Glu Arg Thr Leu Glu Pro Pro Thr Gln Ser Pro Ser Pro Pro Pro
145 150 155 160
Arg Leu Ser Gly Gln Leu Gln Gly Leu Val Ile Gly Val Thr Ser Val
165 170 175
Leu Val Gly Val Leu Leu Leu Leu Leu Leu Thr Trp Val Leu Ala Ala
180 185 190
Val Phe Pro Arg Ala Thr Arg Gly Ala Cys Val Cys Gly Ser Glu Asp
195 200 205
Glu Pro Leu Lys Glu Gly Pro Asp Ala Ala Pro Val Phe Thr Leu Asp
210 215 220
Tyr Gly Glu Leu Asp Phe Gln Trp Arg Glu Lys Thr Pro Glu Pro Pro
225 230 235 240
Ala Pro Cys Ala Pro Glu Gln Thr Glu Tyr Ala Thr Ile Val Phe Pro
245 250 255
Gly Arg Pro Ala Ser Pro Gly Arg Arg Ala Ser Ala Ser Ser Leu Gln
260 265 270
Gly Ala Gln Pro Pro Ser Pro Glu Asp Gly Pro Gly Leu Trp Pro Leu
275 280 285
<210> 107
<211> 870
<212> ДНК
<213> Canis familiaris
<400> 107
atgagaatgt ttagtgtctt tacattcatg gcctactgcc atttgctaaa agcatttacg 60
atcacagttt ctaaggacct gtatgtggta gagtatggtg gcaatgtgac aatggaatgc 120
aaattcccgg tggaaaaaca gttaaacttg tttgcactaa tcgtctactg ggaaatggag 180
gataaaaaaa ttatacaatt tgtgaatgga aaggaagacc tgaaagttca gcacagcagc 240
tacagccaga gggctcagct attgaaggac cagctcttct tggggaaggc tgcgcttcag 300
atcacagatg tgagattgca ggatgcaggg gtttactgct gcttgatcgg ctatggcggt 360
gctgactaca agcggattac tttgaaagtt catgccccgt accgcaacat cagccaaaga 420
atttctgtgg atcctgtcac ctctgaacat gaactaatgt gtcaggctga gggttaccct 480
gaggctgaag tcatctggac aagcagtgac caccgagtcc tgagtggcaa aaccaccatc 540
actaattcca atagggaaga gaagcttttc aatgtgacca gcacgctgaa catcaatgca 600
acagctaatg agattttcta ctgcactttt caaagatcag gtcctgagga aaacaatact 660
gccgagttgg tcatcccaga acgactgccc gttccagcaa gtgagaggac tcatttcatg 720
attctgggac ctttcctgtt gcttcttggt gtagtcctgg cagtcacttt ctgtctaaaa 780
aaacatggga gaatgatgga tgtggaaaaa tgttgcaccc gagataggaa ctcaaagaaa 840
cgaaatgata tacaatttga agagacataa 870
<210> 108
<211> 289
<212> Белок
<213> Canis familiaris
<400> 108
Met Arg Met Phe Ser Val Phe Thr Phe Met Ala Tyr Cys His Leu Leu
1 5 10 15
Lys Ala Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Gly Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu
35 40 45
Asn Leu Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile
50 55 60
Ile Gln Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser
65 70 75 80
Tyr Ser Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr
100 105 110
Cys Cys Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu
115 120 125
Lys Val His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp
130 135 140
Pro Val Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro
145 150 155 160
Glu Ala Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly
165 170 175
Lys Thr Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val
180 185 190
Thr Ser Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys
195 200 205
Thr Phe Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val
210 215 220
Ile Pro Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His Phe Met
225 230 235 240
Ile Leu Gly Pro Phe Leu Leu Leu Leu Gly Val Val Leu Ala Val Thr
245 250 255
Phe Cys Leu Lys Lys His Gly Arg Met Met Asp Val Glu Lys Cys Cys
260 265 270
Thr Arg Asp Arg Asn Ser Lys Lys Arg Asn Asp Ile Gln Phe Glu Glu
275 280 285
Thr
<210> 109
<211> 423
<212> ДНК
<213> Mus musculus
<400> 109
atgggattca gcaggatctt tctcttcctc ctgtcagtaa ctacaggtgt ccactcccag 60
gcttatctac agcagtctgg ggctgagctg gtgaggcctg gggcctcagt gaagatgtcc 120
tgcaaggctt ttggctacac atttaccagt tacaatatgc actgggtgaa gcagacacct 180
agacagggcc tggaatggat tggaaccatt tatccaggag atggtgacgc ttcctacaat 240
cagaaattcc aggacaaggc cacactgact gttgacaaat cctccagcac agcctacatg 300
cagctcagca gcctgacatc tgaagactct gcggtctatt tctgttcaag ggagtttgcc 360
gatgcttacc ccattccccc ctttgactac tggggccaag gcaccactct cacagtctcc 420
tca 423
<210> 110
<211> 141
<212> Белок
<213> Mus musculus
<400> 110
Met Gly Phe Ser Arg Ile Phe Leu Phe Leu Leu Ser Val Thr Thr Gly
1 5 10 15
Val His Ser Gln Ala Tyr Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Phe Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Asn Met His Trp Val Lys Gln Thr Pro Arg Gln Gly Leu
50 55 60
Glu Trp Ile Gly Thr Ile Tyr Pro Gly Asp Gly Asp Ala Ser Tyr Asn
65 70 75 80
Gln Lys Phe Gln Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ser Arg Glu Phe Ala Asp Ala Tyr Pro Ile Pro Pro Phe
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
130 135 140
<210> 111
<211> 393
<212> ДНК
<213> Mus musculus
<400> 111
atgaagttgc ctgttaggct gttggtgctg attttctgga ttcctgcttc cagtagtgat 60
gttttgatga cccaaactcc actctccctg gttgtcagtc ttggagatca ggcctccatc 120
tcttgcagat ctagtcagag cattgtatat agtaatggaa acacctattt agaatggtac 180
ctgcaaaaac caggccagtc tccaaagctc ctgatttaca aagtttccaa ccgattttct 240
ggggtcccag acaggttcag tggcagtgga tcagggacag atttcacact caagatcagc 300
agagtggagg ctgaggatct gggagtttat tactgctttc aaggttcaca tgttccgtac 360
acgttcggag gggggaccaa gctggaaata aaa 393
<210> 112
<211> 131
<212> Белок
<213> Mus musculus
<400> 112
Met Lys Leu Pro Val Arg Leu Leu Val Leu Ile Phe Trp Ile Pro Ala
1 5 10 15
Ser Ser Ser Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Val Val
20 25 30
Ser Leu Gly Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile
35 40 45
Val Tyr Ser Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro
50 55 60
Gly Gln Ser Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser
65 70 75 80
Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys
100 105 110
Phe Gln Gly Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu
115 120 125
Glu Ile Lys
130
<210> 113
<211> 401
<212> Белок
<213> Искусственная последовательность
<220>
<223> собачья и человеческая
<400> 113
Met Asn Phe Leu Leu Ser Trp Val His Trp Ser Leu Ala Leu Leu Leu
1 5 10 15
Tyr Leu His His Ala Lys Trp Ser Gln Ala Leu Asp Ser Pro Asp Arg
20 25 30
Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala Gln Leu Thr Val Gln Glu
35 40 45
Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu Ala Asp Ile Pro Asp Ser
50 55 60
Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro Arg Asn Gln Thr Asp Lys
65 70 75 80
Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg
85 90 95
Phe Arg Val Met Arg Leu Pro Asn Gly Arg Asp Phe His Met Ser Ile
100 105 110
Val Ala Ala Arg Leu Asn Asp Ser Gly Ile Tyr Leu Cys Gly Ala Ile
115 120 125
Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu Ser Pro Arg Ala Glu Leu
130 135 140
Ser Val Thr Glu Arg Thr Leu Glu Pro Pro Thr Gln Ser Pro Ser Pro
145 150 155 160
Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly Leu Val Gly Thr Asp Lys
165 170 175
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
180 185 190
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
195 200 205
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
210 215 220
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
225 230 235 240
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
245 250 255
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
260 265 270
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
275 280 285
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
290 295 300
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
305 310 315 320
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
325 330 335
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
340 345 350
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
355 360 365
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
370 375 380
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
385 390 395 400
Lys
<210> 114
<211> 11
<212> Белок
<213> Mus musculus
<400> 114
Ile Phe Tyr Asp Tyr Asp Gly Asp Ile Asp Val
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО CTLA-4 | 2020 |
|
RU2818586C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2801209C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| АНТИТЕЛА К PD-L1, СВЯЗЫВАЮЩИЕ PD-L1 СОБАКИ | 2015 |
|
RU2832918C2 |
| АНТИТЕЛА К PD-L1, СВЯЗЫВАЮЩИЕ PD-L1 СОБАКИ | 2015 |
|
RU2722562C2 |
| КАНИНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CTLA-4 | 2020 |
|
RU2822460C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИТЕЛА К ПАРВОВИРУСУ ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2830441C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2795485C2 |
| АНТИ-PD-L1 АНТИТЕЛО ДЛЯ ДЕТЕКЦИИ PD-L1 | 2018 |
|
RU2758723C2 |
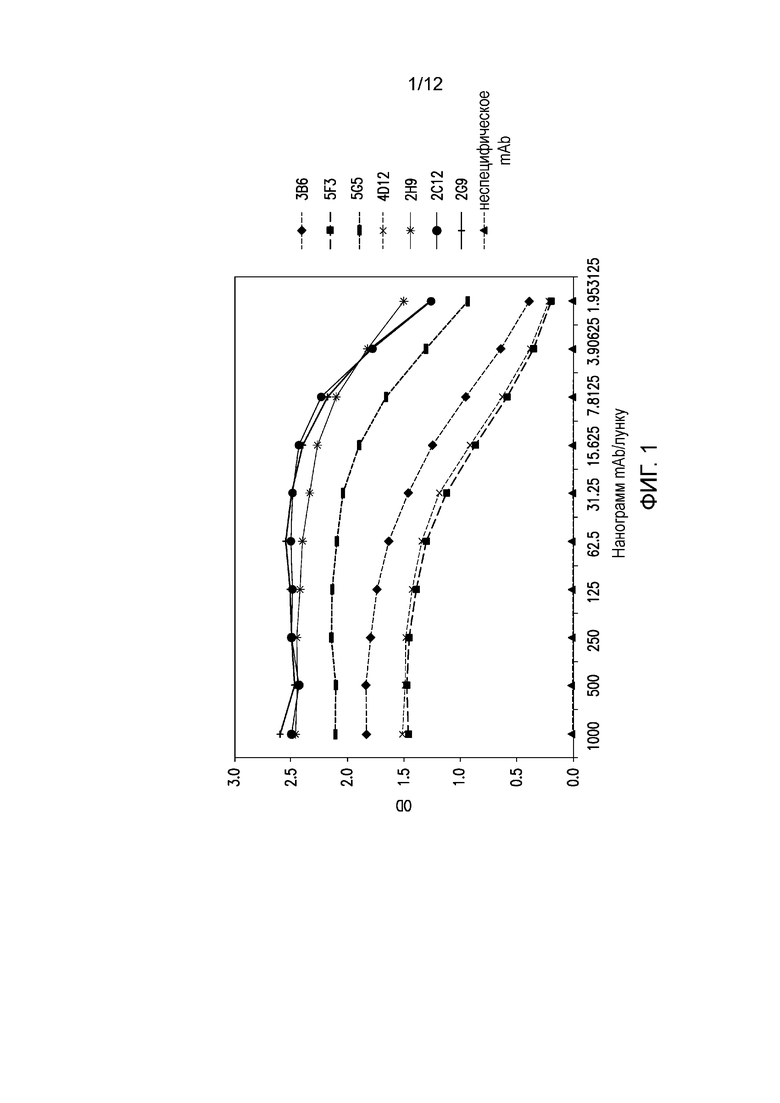
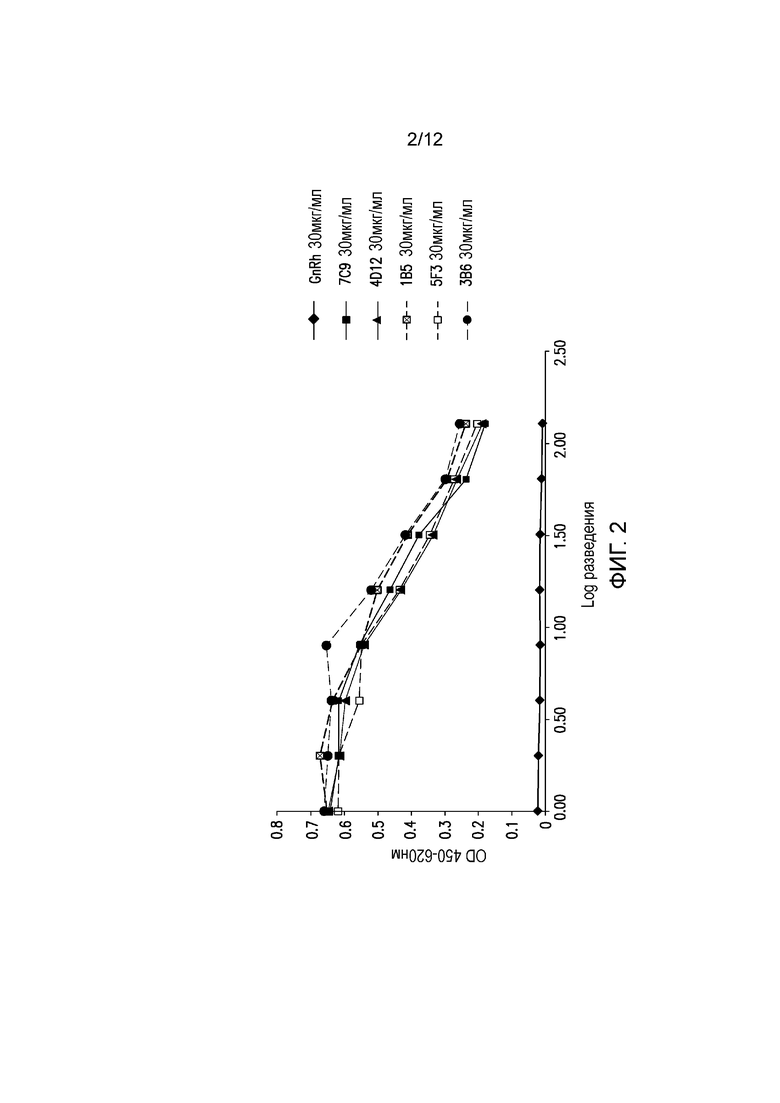
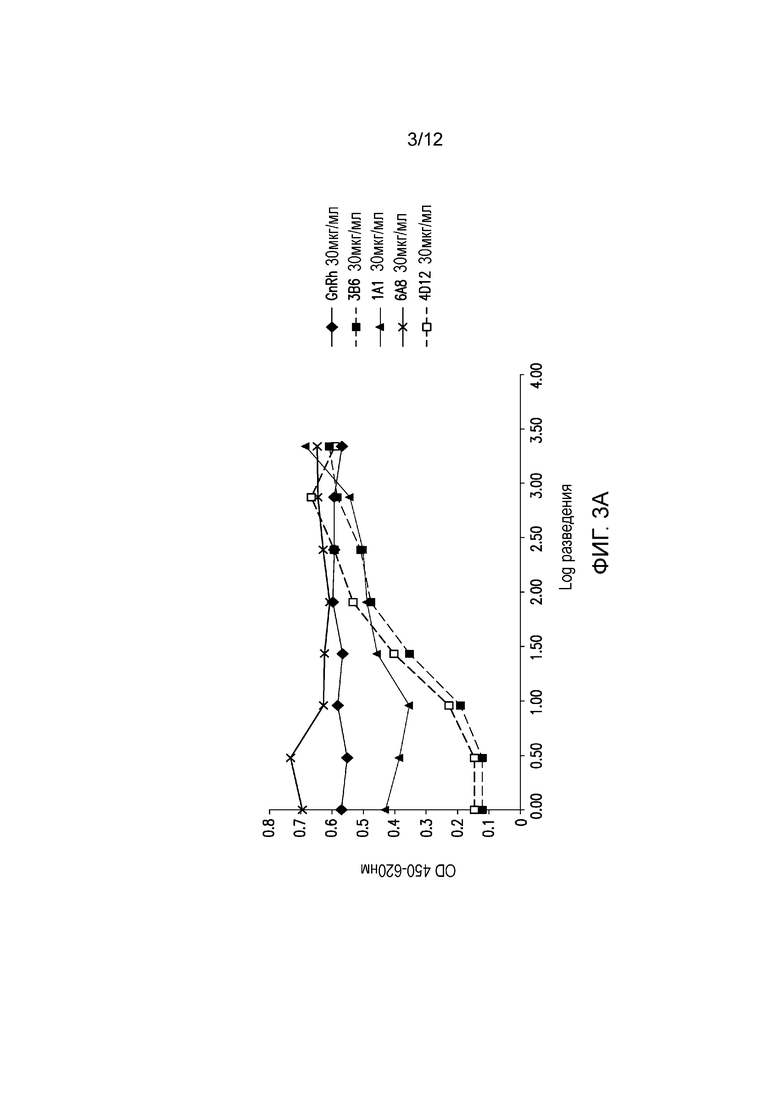
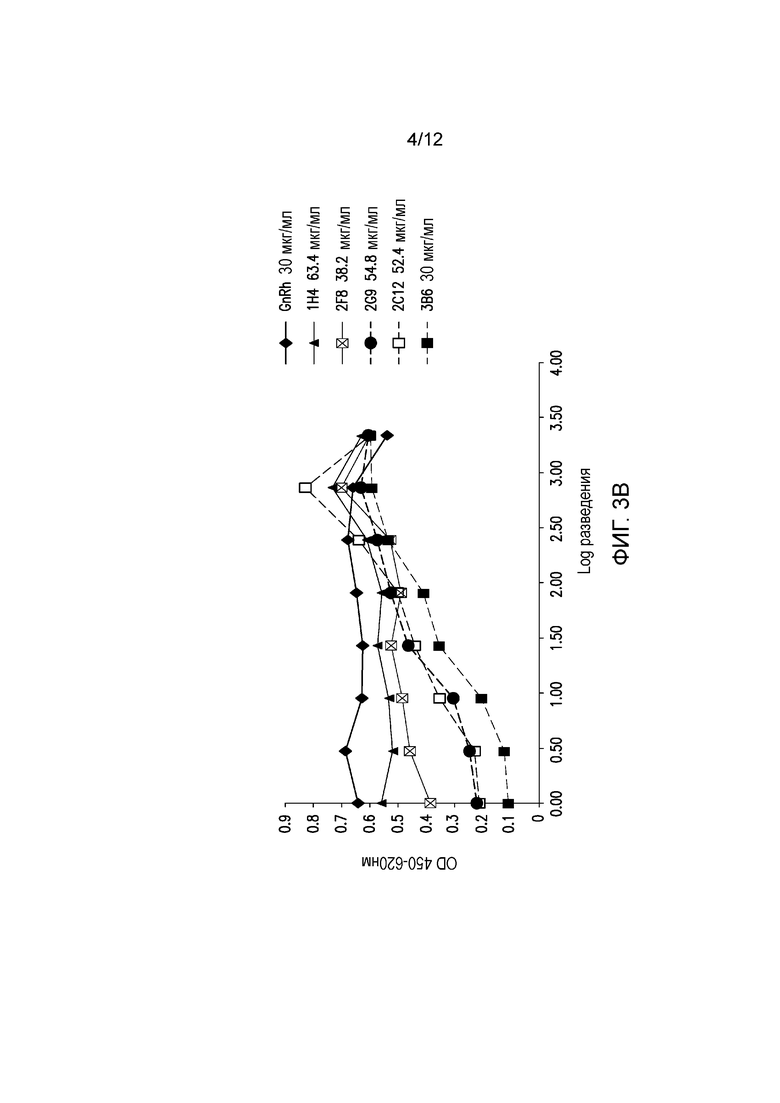
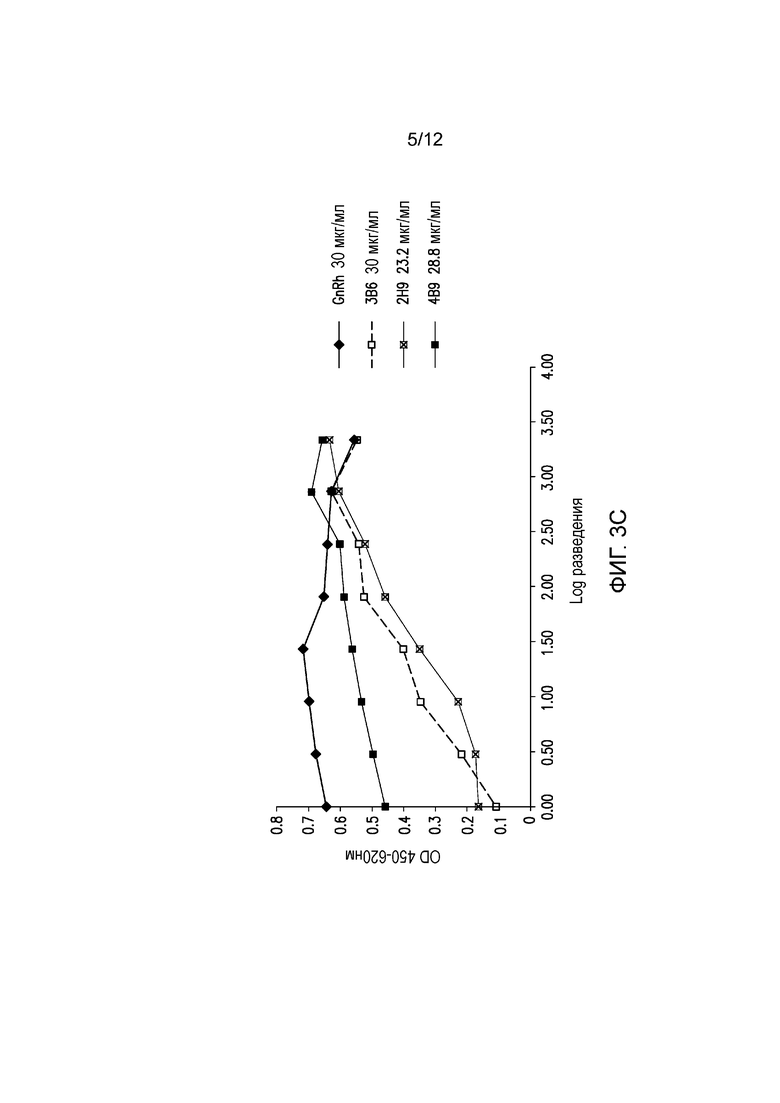
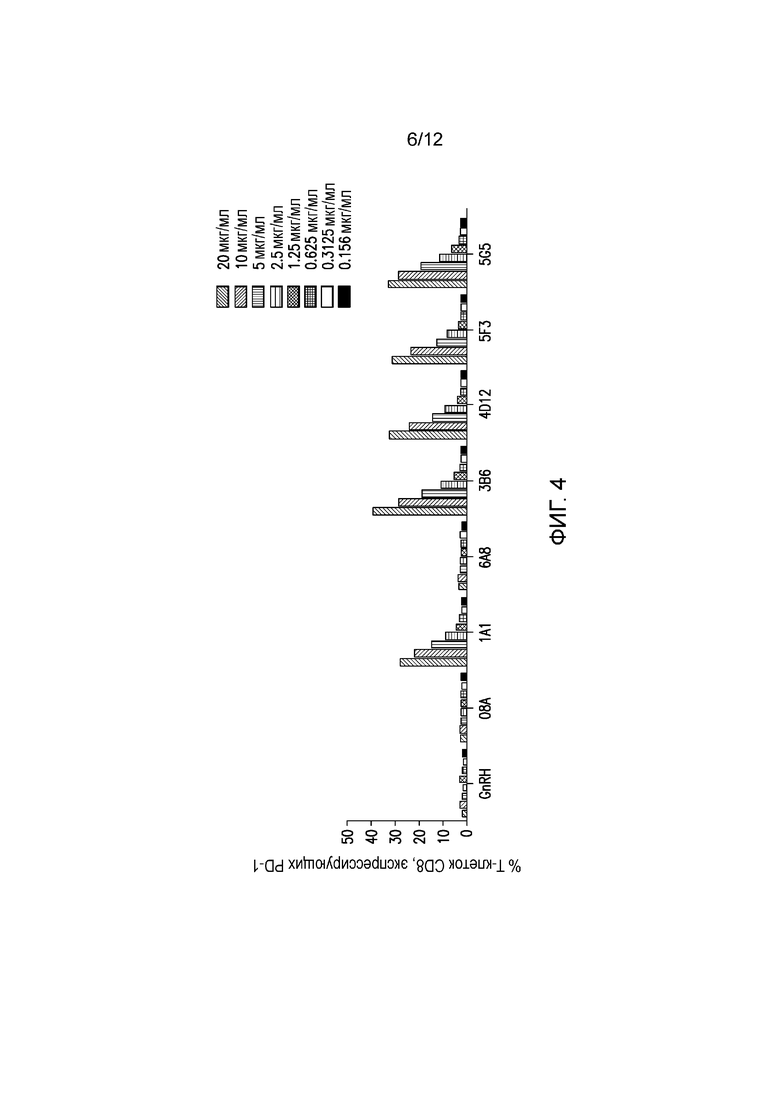
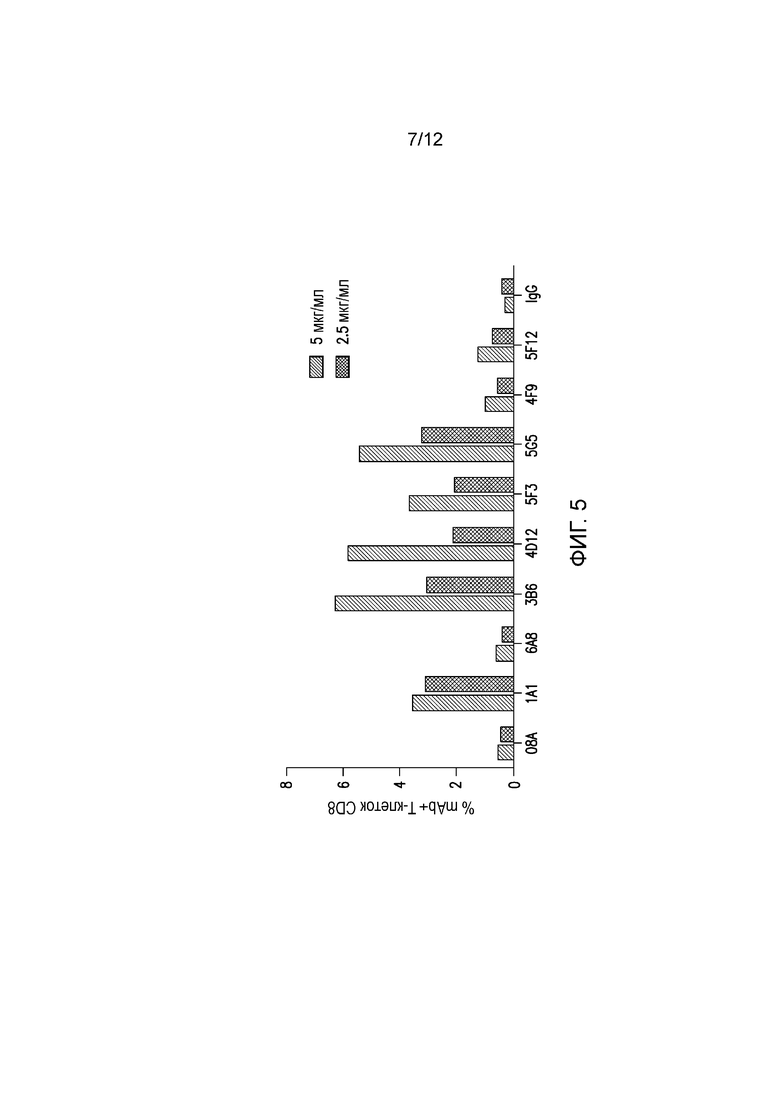
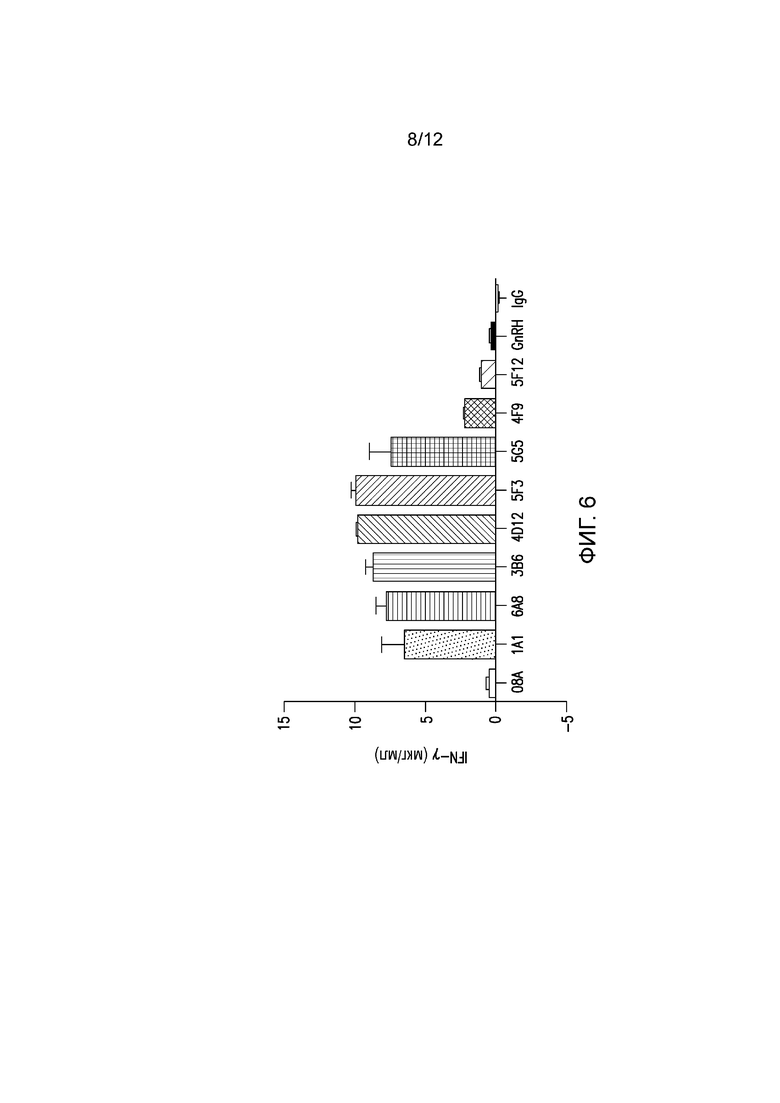
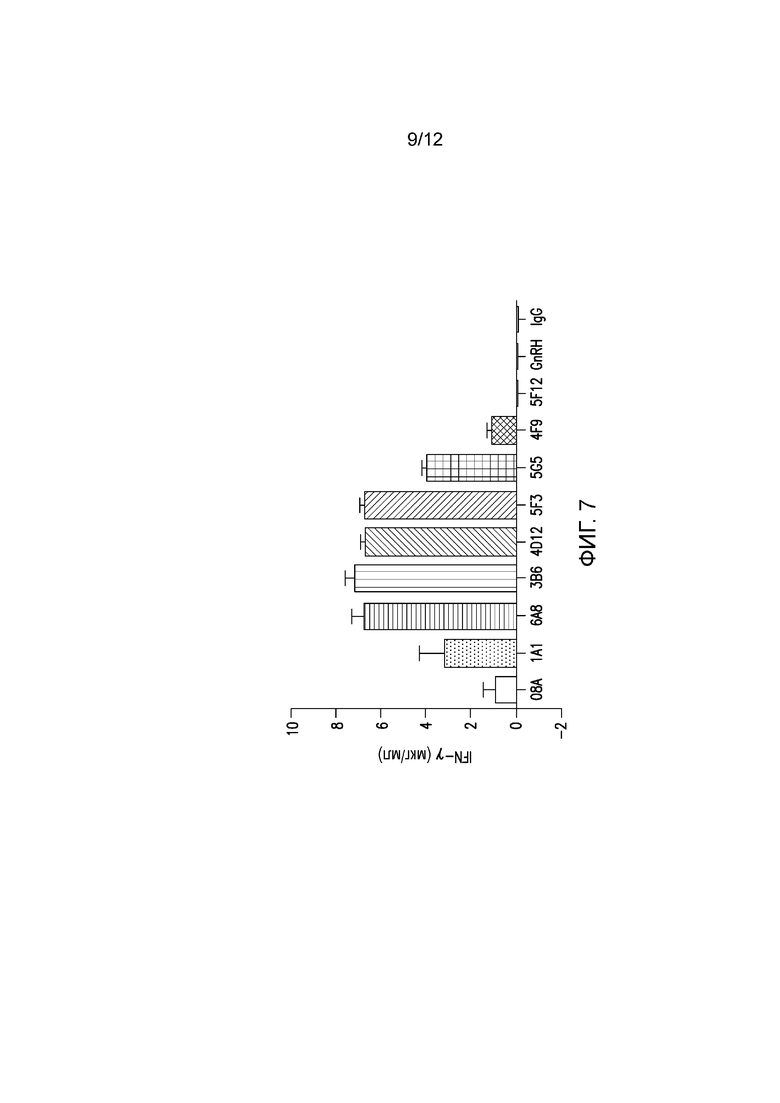
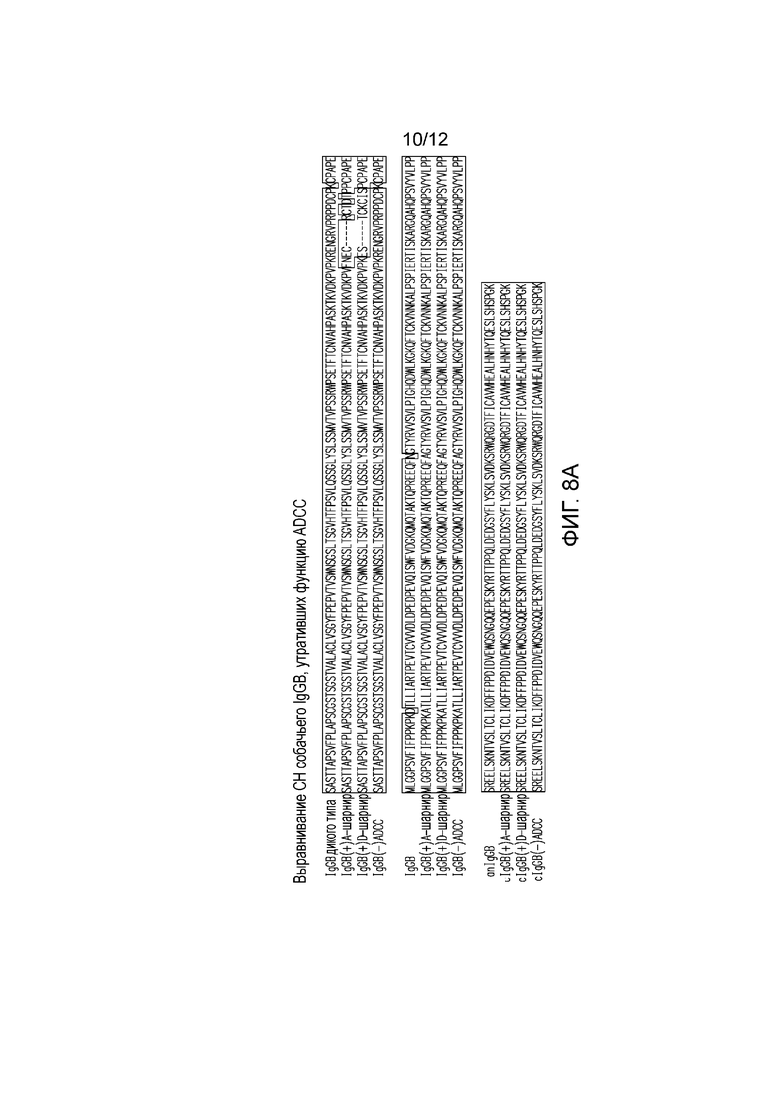


Изобретение относится к области биохимии, в частности к выделенному канинизированному антителу для лечения рака, ассоциированного с экспрессией PD-L1, собак. Также раскрыты нуклеиновая кислота, кодирующая указанное антитело, вектор экспрессии, содержащий указанную нуклеиновую кислоту, фармацевтическая композиция, содержащая указанное антитело. Изобретение позволяет эффективно лечить заболевания, ассоциированные с экспрессией PD-L1, собак. 5 н. и 5 з.п. ф-лы, 8 табл., 9 ил., 5 пр.
1. Выделенное антитело млекопитающего для лечения рака, ассоциированного с экспрессией PD-L1, причем указанное антитело млекопитающего специфически связывается с собачьим рецептором программируемой смерти 1 (PD-1 собак), где указанное антитело содержит набор из шести участков, определяющих комплементарность (CDR), которые включают три определяющих комплементарность участка (CDR) легкой цепи: CDR легкой цепи 1 (CDRL1), CDR легкой цепи 2 (CDRL2) и CDR легкой цепи 3 (CDRL3); и три CDR тяжелой цепи: CDR тяжелой цепи 1 (CDRH1), CDR тяжелой цепи 2 (CDRH2) и CDR тяжелой цепи 3 (CDRH3); причем указанный набор из шести CDR выбран из группы, состоящей из (a)-(f) или (g)-(l):
(a) где CDRL1 содержит аминокислотную последовательность SEQ ID NO: 15;
(b) где CDRL2 содержит аминокислотную последовательность SEQ ID NO: 18;
(c) где CDRL3 содержит аминокислотную последовательность SEQ ID NO: 24;
(d) где CDRH1 содержит аминокислотную последовательность SEQ ID NO: 29;
(e) где CDRH2 содержит аминокислотную последовательность SEQ ID NO: 33; и
(f) где CDRH3 содержит аминокислотную последовательность SEQ ID NO: 38;
или
(g) где CDRL1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13;
(h) где CDRL2 содержит аминокислотную последовательность SEQ ID NO: 19;
(i) где CDRL3 содержит аминокислотную последовательность SEQ ID NO: 25;
(j) где CDRH1 содержит аминокислотную последовательность SEQ ID NO: 27;
(k) где CDRH2 содержит аминокислотную последовательность SEQ ID NO: 31;
(l) где CDRH3 содержит аминокислотную последовательность SEQ ID NO: 36;
где антитело связывает PD-1 собак и блокируют связывание PD-1 собак с собачьим лигандом программируемой смерти 1 (PD-L1).
2. Выделенное антитело млекопитающего по п. 1, в котором вариабельный участок тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 52, и вариабельный участок легкой цепи содержит аминокислотную последовательность SEQ ID NO: 54.
3. Выделенное антитело млекопитающего по п. 1, в котором вариабельный участок тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 56, и вариабельный участок легкой цепи содержит аминокислотную последовательность SEQ ID NO: 58.
4. Выделенное антитело млекопитающего по пп. 1, 2 или 3, где при связывании с собачьим PD-1 указанное антитело связывается с по меньшей мере одним аминокислотным остатком, входящим в состав одной или более последовательностей, выбранных из группы, состоящей из SEQ ID NO: 83, SEQ ID NO: 84, SEQ ID NO: 99, SEQ ID NO: 100, SEQ ID NO: 101, SEQ ID NO: 102, SEQ ID NO: 103 и SEQ ID NO: 104; причем антитело связывает PD-1 собак и блокирует связывание PD-1 собак с собачьим лигандом программируемой смерти 1 (PD-L1).
5. Выделенное антитело млекопитающего по п. 4, где при связывании с собачьим PD-1 указанное антитело связывается с по меньшей мере одним аминокислотным остатком, входящим в состав последовательности SEQ ID NO: 104.
6. Выделенное антитело млекопитающего по п. 5, где при связывании с собачьим PD-1 указанное антитело связывается с одним или несколькими аминокислотными остатками, выбранными из группы, включающей R62, R69, R72, R75 и R90 из последовательности SEQ ID NO: 2.
7. Выделенная нуклеиновая кислота, которая кодирует тяжелую цепь антитела млекопитающего по пп. 1-6.
8 Выделенная нуклеиновая кислота, которая кодирует легкую цепь антитела млекопитающего по пп. 1-6.
9. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 7 или 8.
10. Фармацевтическая композиция для лечения рака, ассоциированного с экспрессией PD-L1, содержащая эффективное количество антитела млекопитающего по пп. 1-5 или 6 или любой их комбинации и фармацевтически приемлемый носитель или разбавитель.
| RU 2007145419 A, 20.06.2009 | |||
| US 20120237522 A1, 20.09.2012 | |||
| US 20100203056 A1, 12.08.2010 | |||
| FOLKL A | |||
| AND ET.AL., Feline programmed death and its ligand: Characterization and changes with feline immunodeficiency virus infection, Veterinary Immunology and Immunopathology, 2010, n.134 pp.107-114. |

