ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей патентной заявке испрашивается приоритет по патентной заявке США серийный № 62/030,812, поданной 30 июля 2914 года, по патентной заявке США серийный № 61/918,847, поданной 20 декабря 2013 года, и по патентной заявке США серийный № 61/918,946, поданной 20 декабря 2014 года, содержание каждой из которых введено здесь ссылкой в полном объеме.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к канинизированным антителам со специфическими свойствами. Также настоящее изобретение относится к канинизированным антителам против собачьего PD-1, которые имеют специфические последовательности и высокую аффинность связывания с собачьим PD-1. Дополнительно настоящее изобретение относится к применению антитела по настоящему изобретению для лечения собак, включая лечение рака.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Собачьи антитела (также указываемые как иммуноглобулин G или IgG) представляют крупные тетрамерные белки около 150 кДа. Каждый белок IgG состоит из двух идентичных легких цепей около 25 кДа каждая и двух идентичных тяжелых цепей около 50 кДа каждая. Существует четыре известных подкласса тяжелых цепей собачьего IgG, и они указываются как IgGA, IgGB, IgGC и IgGD. Существует два типа легких цепей: каппа и лямбда цепи. Каждая из каппа или лямбда легких цепей состоит из одного вариабельного домена (VL) и одного константного домена (CL). Каждая из двух тяжелых цепей состоит из одного вариабельного домена (VH) и трех константных доменов, указываемых как CH-1, CH-2 и CH-3. Домен CH-1 соединен с доменом CH-2 аминокислотной последовательностью, указанной как «шарнир», или в качестве альтернативы, как «шарнирная область». У людей IgG присутствует в виде одного из четырех подклассов, указываемых как IgG1, IgG2, IgG3 и IgG4. Подкласс IgG главным образом определяется последовательностью шарнирной области, которая отличается у каждого из четырех подклассов IgG. Две тяжелые цепи соединены друг с другом дисульфидными связями, и каждая тяжелая цепь также связана с одной из легких цепей через дисульфидную связь.
Расщепление антитела IgG ферментом папаином разрушает молекулу антитела в шарнирной области и в результате приводит к образованию трех фрагментов. Два из этих фрагментов являются идентичными и каждый состоит из легкой цепи, удерживающей вместе домены VH и CH1 тяжелой цепи. Эти фрагменты называются «Fab» фрагментами и они содержат антигенсвязывающие участки антитела. Третий фрагмент, образовавшийся в результате расщепления папаином, называется «Fc» и он содержит остаток двух тяжелый цепей, удерживаемых вместе дисульфидными связями. Следовательно, Fc содержит димер, состоящий из CH2 и CH3 домена, каждый из которых состоит из двух тяжелых цепей. Fab позволяет антителу связываться с его родственным эпитопом, Fc позволяет антителу опосредовать функции иммунного эффектора, такие как антитителозависимая клеточная цитотоксичность (ADCC), антителозависимый фагоцитоз (ADCP) и комплиментзависимая цитотоксичность (CDC).
Из предшествующего уровня техники хорошо известно, что антитела IgG опосредуют эффекторные функции, такие как ADCC и ADCP через связывание их Fc фрагментом с семейством белков, известных как Fcᵧ рецепторы, в свою очередь CDC опосредовано связыванием через Fc с первым компонентом комплимента C1q. Также из предшествующего уровня техники хорошо известно, что различные подклассы IgG отличаются по своей способности опосредовать эти эффекторные функции. Например, человеческий IgG1 демонстрирует сильную ADCC и CDC, в то время как IgG4 демонстрирует от слабой ADCC и CDC до отсутствия ADCC и CDC. Дополнительно, из предшествующего уровня техники хорошо известны способы определения, какой из подклассов IgG демонстрирует или у него отсутствуют эффекторные функции.
Подходы, основывающиеся на применении моноклональных антител для терапевтических целей, требуют конструкции, соответствующей целевому назначению антитела или фрагментов антитела для достижения заданного терапевтического ответа. Например, некоторые терапевтические подходы для рака требуют терапевтического антитела, обладающего усиленными эффекторными функциями, в то время как другие требуют эффекторные функции, которые будут значительно ослаблены или совсем элиминированы. Усиление или элиминирование эффекторных функций может быть достигнуто через введение одной или более мутации аминокислоты (замещения) в Fc части антитела, таким образом, чтобы усилить или ослабить связывание с рецепторами Fcᵧ и первым компонентом комплимента. В предшествующем уровне техники имеет место множество сообщений, в которых описываются замещения, которые могут быть введены в молекулу антитела для модулирования ее эффекторых функций. Например, Shields et al., [J. of Biol. Chem., 276 (9): 6591-6604 (2001)] описано замещение аспарагина на аланин (N297A), которое в результате позволяет получить не гликозилированное антитело, значительно ослабленное связывание антитела с отдельными Fcᵧ рецепторами. Дополнительно Shields et al. описано, что замещение аспарагиновой кислоты на аланин (D265A) также значительно снижает связывание антитела с рецепторами Fcᵧ. Каждое из замещений N297A и D265A также демонстрирует значительное ухудшение CDC. Существуют и другие аналогичные сообщения, описывающие потенциальные замещения для ослабления или элиминирования эффекторной функции антитела [например, Sazinsky et al., Proc.Nat.Acad.Sci.,105:20167-20172 (2008), Alegre et al., Transplantation, 57:1537-1543 (1994), Hutchins et al., Proc.Nat.Acad.Sci. 92:11980-11984 (1994), McEarchem et al., Blood, 109:1185-1192 (2007)].
Иммуноингибирующий рецептор, который главным образом экспрессируется на активированных T и B клетках, рецептор запрограммированной гибели клеток 1, также указываемый, как рецептор запрограммированной смерти 1 (PD-1), является членом суперсемейства иммуноглобулина, относящегося к CD28 и CTLA-4. PD-1, как и члены семейства, представляют трансмембранные гликобелки типа I, содержащие внеклеточный вариабельный (V-тип) домен Ig, который связывается своими лигандами и цитоплазматический концевой сегмент, который связывается с сигнальными молекулами. Цитоплазматический концевой сегмент PD-1 содержит два сигнальных мотива на основе тирозина, ITIM (тирозинсодержащий ингибирующий мотив иммунорецептора) и ITSM (тирозинсодержащий активирующий мотив иммунорецептора).
PD-1 ослабляет T-клеточные ответы, когда связан с лигандом запрограммированной гибели клеток 1, также указываемым, как лиганд запрограммированной смерти 1 (PD-L1), и/или лигандом запрограммированной гибели клеток 2, также указываемым как лиганд запрограммированной смерти 2 (PD-L2). Связывание любого из этих лигандов с PD-1 негативно регулирует передачу сигнала через рецептор антигена. Блокирование связывания PD-L1 с PD-1 усиливает опухоль-специфический CD8+ T-клеточный иммунитет, при этом способствуя клиренсу опухолевых клеток иммунной системой. О пространственной структуре мышиного PD-1, наряду с сокристаллической структурой мышиного PD-1 с человеческим PD-L1 сообщалось в [Zhang et al., Immunity 20: 337-347 (2004); Lin et al., Proc. Natl. Acad. Sci. USA 105: 3011-3016 (2008)].
PD-L1 и PD-L2 представляют трансмембранные лиганды типа I, которые содержат оба IgV- и IgC-подобные домены во внеклеточной области вместе с короткими цитоплазматическими участками с неизвестными сигнальными мотивами. Оба и PD-L1 и PD-L2 либо постоянно экспрессируются либо могут быть индуцированы в различных типах клеток, включая не кроветворные ткани наряду с различными типами опухолей. PD-L1 не только экспрессируется на B, T, миелоидных и дендритных клетках (DCs), но также на периферических клетках, таких как эндотелиальные клетки микрососудов, и в не лимфоидных органах, например, сердце или легкие. В противоположность, PD-L2 обнаруживается только на макрофагах и DCs. Паттерн экспрессии лигандов PD-1 предполагает, что PD-1 играет роль в поддержании периферической толерантности и может дополнительно служить для регуляции аутореактивных T- и B-клеточных ответов в периферии.
В любом случае, совершенно ясно, что PD-1 играет решающую роль по меньшей мере в определенных видах рака человека, предположительно опосредуемых ускользанием от механизмов иммунологического надзора. Соответственно, PD-L1 продемонстрировал способность экспрессироваться на множестве мышиных и человеческих опухолей и индуцируется IFN- γ в большинстве PD-L1 негативных линий опухолевых клеток [Iwai et al., Proc. Natl. Acad. Sci. U.S.A. 99: 12293-12297 (2002); Strome et al., Cancer Res., 63: 6501-6505 (2003)]. Дополнительно была определена экспрессия PD-1 на проникающих в опухоль лимфоцитах и/или PD-L1 на опухолевых клетках во множестве биопсий первичных опухолей человека. Такие опухолевые ткани включают рак легкого, печени, яичников, шейки матки, кожи, толстой кишки, глиомы, рака мочевого пузыря, груди, почек, пищевода, желудка, сквамозные клетки ротовой полости, уротелиальные клетки и поджелудочной железы, наряду с опухолями головы и шеи [Brown et al., J. Immunol. 170: 1257-1266 (2003); Dong et al., Nat. Med. 8: 793-800 (2002); Wintterle et al., Cancer Res. 63: 7462-7467 (2003); Strome et al., Cancer Res., 63: 6501-6505 (2003); Thompson et al., Cancer Res. 66: 3381-5 (2006); Thompson et al., Clin. Cancer Res. 13: 1757-1761 (2007); Nomi et al., Clin.Cancer Res. 13: 2151-2157. (2007)]. Очень удивительно, что экспрессия PD-лигандов на опухолевых клетках коррелирует с плохим прогнозом у пациентов с раком у человека со множеством различных типов опухолей [reviewed in Okazaki and Honjo, Int. Immunol. 19: 813-824 (2007)].
Дополнительно в Nomi et al. [Clin. Cancer Res. 13: 2151-2157 (2007)] продемонстрирован терапевтический эффект блокирования связывания PD-L1 с PD-1 на мышиной модели агрессивного рака поджелудочной железы посредством введения PD-1 или PD-L1 специфического антитела. Эти антитела эффективно промотируют опухолереактивную CD8+ T клеточную инфильтрацию в опухоли с позитивной регуляцией противоопухолевых эффекторов, включая IFN-γ, гранзим B, и перфорин. Аналогично применение антител для блокировки связывания PD-L1 и PD-1 значительно ингибирует рост опухоли на мышиной модели карциномы сквамозных клеток [Tsushima et al., Oral Oncol. 42: 268-274 (2006)].
В других исследованиях трансфекция линии мышиной мастоциомы PD-L1 привела к снижению лизиса опухолевых клеток при сокультивировании с опухоль-специфическим CTL клоном. Лизис был восстановлен введение анти -PD-L1 моноклонального антитела [Iwai et al., Proc. Natl. Acad. Sci. U.S.A. 99: 12293-12297 (2002)]. In vivo блокирование взаимодействия PD1/PD-L1 показало повышение эффективности терапии переносом адоптивных T-клеток на мышиной опухолевой модели [Strome et al., Cancer Res. 63: 6501-6505 (2003)]. Дополнительное свидетельство роли PD-1 в лечении рака получено в ходе экспериментов, проведенных при участии мышей с выключенным PD-1, у которых PD-L1 экспрессирующие клетки миеломы выросли только у животных дикого типа (с ростом опухоли и связанной с этим смерти животного), но не у мышей дефицитных по PD-1 [Iwai Y. et al., Proc. Natl. Acad. Sci. U.S.A. 99: 12293-12297 (2002)]. Совсем недавно антитела против PD-1 (включая гуманизированное мышиное моноклональное антитело против человеческого PD 1) показали по меньшей мере успех на начальном этапе в терапии рака у людей [смотрите, например, US 8,354,509 B2, US 8,008,449 B2, и US 7,595,048 B2].
Анти-PD-1 антитело также может быть использовано при хронической вирусной инфекции. CD8+ T -клетки памяти, генерированные после острой вирусной инфекции, представляют высоко функциональные и являются важным компонентом защитного иммунитета. В противоположность, хронические инфекции часто характеризуются различными степенями функциональных нарушений (истощение) вирус-специфических T-клеточных ответов, и этот дефект является основной причиной неспособности хозяина избавиться от персистентного патогена. Хотя функциональные эффекторные T клетки сначала генерируются во время ранних стадий инфекции, их функция постепенно истощается во время течения хронической инфекции. Barber et al. [Nature 439: 682-687 (2006)] показали, что у мышей, инфицированных лабораторным штаммом LCMV, развилась хроническая инфекция с высокими уровнями вируса в крови и других тканях. У этих мышей сначала развился активный T клеточный ответ, но со временем прекратилось сопротивление инфекции при истощении T клеток. Barber et al. обнаружили, что снижение числа и функции эффекторных T клеток у хронически инфицированных мышей может быть обращено вспять инъекцией антитела, блокирующего взаимодействие между PD-1 и PD-L1.
Цитирование любой ссылки в описании настоящей патентной заявки не следует понимать, как признание того, что такая ссылка делает настоящую патентную заявку частью «предшествующего уровня техники».
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение обеспечивает кристаллизуемую область фрагмента собачьего (cFc область) антитела, где cFc был генетически модифицирован для усиления, ослабления или элиминирования одной или более эффекторной функции. В одном аспекте настоящего изобретения генетически модифицированный cFc снижает или элиминирует одну или более эффекторную функцию. В другом аспекте настоящего изобретения генетически модифицированный cFc усиливает одну или более эффекторную функцию.
В некоторых вариантах воплощения настоящего изобретения генетически модифицированная cFc область представляет генетически модифицированную Fc область собачьего IgGB. В другом таком варианте воплощения настоящего изобретения генетически модифицированная cFc область представляет генетически модифицированную Fc область собачьего IgGC. В конкретном варианте воплощения настоящего изобретения эффекторная функция представляет антителозависимую цитотоксичность (ADCC), которая усилена, снижена или элиминирована. В другом варианте воплощения настоящего изобретения эффекторная функция представляет комплиментзависимую цитотоксичность (CDC), которая усилена, снижена или элиминирована. В другом варианте воплощения настоящего изобретения cFc область была генетически модифицирована для усиления, снижения или элиминирования обоих, и ADCC, и CDC.
Дополнительно, настоящее изобретение обеспечивает собачьи каркасы и/или полноразмерные тяжелые цепи, которые содержат генетически модифицированные cFc области. Соответственно, настоящее изобретение обеспечивает полноразмерные тяжелые цепи антитела, в котором полноразмерные тяжелые цепи содержат генетически модифицированные cFc области по настоящему изобретению. Такие полноразмерные тяжелые цепи также могут быть скомбинированы с соответствующими собачьими легкими (каппа или лямбда) цепями с образованием полного антитела. В конкретных вариантах воплощения для такого типа по настоящему изобретению полученное в результате антитело специфически связывается с конкретным собачьим антигеном. В таких конкретных вариантах воплощения настоящего изобретения собачий антиген представляет собачий PD-1. В других вариантах воплощения настоящего изобретения собачий антиген представляет собачий PD-L1. В других вариантах воплощения настоящего изобретения собачий антиген представляет альфа цепь рецептора IL-4. В других вариантах воплощения настоящего изобретения собачий антиген представляет собачий тимусовый стромальный лимфопоэтиновый белок (cTSLP) [смотрите, США 7,718,772 B2, содержимое которого введено здесь ссылкой в полном объеме].
В некоторых вариантах воплощения настоящего изобретения генетически модифицированная cFc область содержит аминокислотную последовательность SEQ ID NO: 130 (или SEQ ID NO: 132), в которой от одного до семи следующих остатков замещены другим аминокислотным остатком в указанных позициях: P4, D31, N63, G64, T65, A93 или P95. Аминокислотные замещения для P4, D31, N63, G64, T65, A93 и/или P95 индивидуально выбирают из одной из других 19 стандартных природных аминокислот, приведенных в Таблице 1 ниже. Дополнительно, настоящее изобретение обеспечивает варианты генетически модифицированных cFc областей, которые содержат аминокислотные последовательности, имеющие 90%, 95%, 98% или 99% идентичность аминокислотной последовательности таких генетически модифицированных cFc областей и сохраняют по меньшей мере 50%, 75%, 90%, 95% или более усиления, снижения или элиминирования ADCC и/или CDC как генетически модифицированные cFc области, содержащие аминокислотную последовательность SEQ ID NO: 130 (или SEQ ID NO: 132), в которой один или более из следующих аминокислотных остатков были замещены: то есть в P4, D31, N63, G64, T65, A93 или P95.
В других вариантах воплощения настоящего изобретения от двух до пяти следующих аминокислотных остатков замещены другим аминокислотным остатком в указанных позициях: P4, D31, N63, G64, T65, A93 или P95. В конкретных вариантах воплощения для такого типа по настоящему изобретению генетически модифицированная cFc область содержит аминокислотную последовательность SEQ ID NO: 130 или SEQ ID NO: 132 со следующими замещениями: P4A, D31A, N63A, A93G и P95A. В близких вариантах воплощения настоящего изобретения генетически модифицированная cFc область содержит аминокислотную последовательность SEQ ID NO: 130 или SEQ ID NO: 132 со следующими замещениями: P4A, D31A, N63A и P95A. В других вариантах воплощения настоящего изобретения генетически модифицированная cFc область содержит аминокислотную последовательность SEQ ID NO: 130 или SEQ ID NO: 132 с замещениями в D31 и N63. В конкретных вариантах воплощения для такого типа по настоящему изобретению остаток аспарагиновой кислоты в позиции 31 замещен остатком глутаминовой кислоты, остатком аспарагина или остатком аланина, при этом остаток аспарагина в позиции 63 замещен остатком глутамина, остатком гистидина или остатком аланина. В более конкретном варианте воплощения настоящего изобретения генетически модифицированная cFc область содержит аминокислотную последовательность SEQ ID NO: 130 или SEQ ID NO: 132 со следующими замещениями: D31A и N63A. В конкретных вариантах воплощения настоящего изобретения генетически модифицированная cFc область кодируется нуклеотидной последовательностью SEQ ID NO: 129 или SEQ ID NO: 131, содержащей нуклеотидные цепи, которые соответствуют аминокислотным последовательностям, которые они кодируют.
В другом варианте воплощения настоящего изобретения генетически модифицированная cFc область содержит аминокислотную последовательность SEQ ID NO: 130 или SEQ ID NO: 132 с замещением в A93. В конкретном варианте воплощения для такого типа по настоящему изобретению замещение представляет A93G. В соответствующем варианте воплощения настоящего изобретения замещение представляет A93S. Как показано ниже в Примере 4, замещение A93G приводит к усилению связывания C1q комплемента, что указывает на повышение активности CDC.
В близких вариантах воплощения настоящего изобретения генетически модифицированная cFc область дополнительно содержит шарнирную область, которая сдержит аминокислотную последовательность SEQ ID NO: 109. В других вариантах воплощения настоящего изобретения генетически модифицированная Fc область дополнительно содержит шарнирную область, которая содержит аминокислотную последовательность SEQ ID NO: 110. В других вариантах воплощения настоящего изобретения генетически модифицированная Fc область дополнительно содержит шарнирную область, которая содержит аминокислотную последовательность SEQ ID NO: 111. В других вариантах воплощения настоящего изобретения генетически модифицированная Fc область дополнительно содержит генетически модифицированную шарнирную область, которая содержит аминокислотную последовательность SEQ ID NO: 112.
В альтернативных вариантах воплощения настоящее изобретение обеспечивает Fc область собачьего IgGD с генетически модифицированной шарнирной областью из собачьего IgGD антитела, шарнирную область из собачьего IgGA антитела, шарнирную область из собачьего IgGB антитела или шарнирную область из собачьего IgGC антитела. Дополнительно настоящее изобретение обеспечивает полноразмерные тяжелые цепи антител, в которых полноразмерные тяжелые цепи содержат Fc область собачьего IgGD по настоящему изобретению с генетически модифицированной шарнирной областью из собачьего IgGD антитела, шарнирную область из собачьего IgGA антитела, шарнирную область из собачьего IgGB антитела или шарнирную область из собачьего IgGC антитела. Такие полноразмерные тяжелые цепи также могут быть скомбинированы с соответствующими собачьими легкими (каппа или лямбда) цепями с получением полного антитела.
Соответственно настоящее изобретение обеспечивает Fc область собачьего IgGD, которая дополнительно содержит генетически модифицированную шарнирную область из собачьего IgGD антитела. В конкретных вариантах воплощения для такого типа по настоящему изобретению Fc область собачьего IgGD и генетически модифицированная шарнирная область содержат аминокислотную последовательность SEQ ID NO: 6 или аминокислотную последовательность, которая на 90%, 95%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 6, которая содержит остаток пролина в позиции 10 (P10). В более конкретном варианте воплощения настоящего изобретения Fc область собачьего IgGD и генетически модифицированная шарнирная область кодируется нуклеотидной последовательностью SEQ ID NO: 5. В других вариантах воплощения настоящего изобретения Fc область собачьего IgGD дополнительно содержит шарнирную область из собачьего IgGA антитела. В конкретных вариантах воплощения для такого типа по настоящему изобретению Fc область собачьего IgGD и шарнирная область содержат аминокислотную последовательность SEQ ID NO: 8 или аминокислотную последовательность, которая на 90%, 95%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 8. В более конкретном варианте воплощения настоящего изобретения Fc область собачьего IgGD и шарнирная область кодируется нуклеотидной последовательностью SEQ ID NO: 7. В других вариантах воплощения настоящего изобретения Fc область собачьего IgGD дополнительно содержит шарнирную область из собачьего IgGB антитела. В конкретных вариантах воплощения для такого типа по настоящему изобретению Fc область собачьего IgGD и шарнирная область содержит аминокислотную последовательность SEQ ID NO: 10 или аминокислотную последовательность, которая на 90%, 95%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 10. В более конкретном варианте воплощения настоящего изобретения Fc область собачьего IgGD и шарнирная область кодируется нуклеотидной последовательностью SEQ ID NO: 9. В других вариантах воплощения настоящего изобретения Fc область собачьего IgGD дополнительно содержит шарнирную область из собачьего IgGC антитела. В конкретных вариантах воплощения для такого типа по настоящему изобретению cFc область собачьего IgGD и шарнирная область содержат аминокислотную последовательность SEQ ID NO: 12 или аминокислотную последовательность, которая на 90%, 95%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 12. В более конкретном варианте воплощения настоящего изобретения cFc область собачьего IgGD и шарнирная область кодируется нуклеотидной последовательностью SEQ ID NO: 11. Дополнительно, настоящее изобретение обеспечивает канинизированные антитела, которые содержат Fc области собачьего IgGD и шарнирные области. В конкретном варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент специфически связывает собачий рецептор запрограммированной смерти 1 (собачий PD-1).
Следовательно, настоящее изобретение обеспечивает канинизированные антитела против собачьего PD-1, специфически связывающиеся и/или имеющие высокую аффинность связывания с собачьим PD 1. В конкретных вариантах воплощения настоящего изобретения канинизированные антитела против собачьего PD-1 также имеют способность блокировать связывание собачьего PD 1 с собачим PD L1. В конкретных вариантах воплощения настоящего изобретения канинизированные антитела против собачьего PD-1 имеют высокую аффинность связывания с собачим PD 1, наряду со способностью также блокировать связывание собачьего PD 1 с собачим PD L2. Канинизированные антитела или их антигенсвязывающие фрагменты, которые специфично связываются с собачьим PD-1, могут содержать тяжелую цепь собачьего IgG по настоящему изобретению и легкую каппа или лямбда цепь собачьего IgG. В конкретных вариантах воплощения настоящего изобретения канинизированные антитела против собачьего PD-1 представляют канинизированные мышиные антитела против собачьего PD-1. Также настоящее изобретение относится к применению таких канинизированных антител в лечении таких заболеваний, как рак, и/или вызванных инфекцией.
В конкретных вариантах воплощения настоящего изобретения канинизированное антитело против собачьего PD-1 содержит генетически модифицированную cFc область по настоящему изобретению. В альтернативных вариантах воплощения настоящего изобретения канинизированное антитело против собачьего PD-1 содержит Fc область собачьего IgGD с генетически модифицированной шарнирной областью из собачьего IgGD антитела, шарнирную область из собачьего IgGA антитела, шарнирную область из собачьего IgGB антитела или шарнирную область из собачьего IgGC антитела. Дополнительно, настоящее изобретение обеспечивает такие канинизированные антитела против собачьего PD-1, содержащие собачьи каркасы по настоящему изобретению в комбинации с CDRs, полученными из мышиных антител против собачьего PD-1, то есть три CDRs легкой цепи: CDR 1 из легкой (CDRL1), CDR 2 из легкой (CDRL2) и CDR 3 из легкой (CDRL3), и три CDRs тяжелой цепи: CDR 1 из тяжелой (CDRH1), CDR 2 из тяжелой (CDRH2) и CDR 3 из тяжелой (CDRH3).
В конкретных вариантах воплощения настоящее изобретение относится к канинизированному мышиным против собачьего PD-1 антителам, содержащим генетически модифицированную cFc область IgGB или IgGC по настоящему изобретению или в качестве альтернативы, Fc область собачьего IgGD вместе с генетически модифицированной шарнирной областью собачьего IgGD антитела, шарнирную область собачьего IgGA антитела, шарнирную область собачьего IgGB антитела, или шарнирную область собачьего IgGC антитела в комбинации с CDRs, полученными из мышиных антител против собачьего PD-1. Кроме того, настоящее изобретение не только обеспечивает канинизированные мышиные антитела против собачьего PD-1 со специфическими CDRs, как приведено в описании настоящей патентной заявки, но дополнительно обеспечивает канинизированные мышиные антитела против собачьего PD-1, содержащие консервативно модифицированные варианты таких CDRs наряду с вариантами, которые содержат (например, часть) идентичные канонические структуры.
Соответственно, в конкретных вариантах воплощения настоящего изобретения канинизированное антитело против собачьего PD-1 дополнительно содержит определяющие комплиментарность области (CDRs), в которых CDRs имеют каноническую структуру из: H1-1, H2-1, и H3-6, соответственно для CDR1, CDR2, и CDR3 тяжелой цепи, то есть CDR1 тяжелой цепи имеет каноническую структуру класса 1, CDR2 тяжелой цепи имеет каноническую структуру класса 1, и CDR3 тяжелой цепи имеет каноническую структуру класса 6. В более конкретных вариантах воплощения настоящего изобретения CDRs для соответствующих легких цепей имеют канонические структуры: L1-3, L2-1 и L3-1, соответственно для CDR1, CDR2 и CDR3 легкой цепи. В других вариантах воплощения настоящего изобретения канинизированное антитело против собачьего PD-1 дополнительно содержит определяющие комплементарность области (CDRs), в которых CDRs имеют канонические структуры: H1-1, H2-1 и H3-11, соответственно, для CDR1, CDR2 и CDR3 тяжелой цепи. В еще более конкретных вариантах воплощения для таких типов по настоящему изобретению CDRs для соответствующих легких цепей имеют канонические структуры: L1-2A, L2-1 и L3-1, соответственно, для CDR1, CDR2 и CDR3 легкой цепи. В других вариантах воплощения настоящего изобретения канинизированное антитело против собачьего PD-1 дополнительно содержит определяющие комплементарность области (CDRs), в которых CDRs имеют канонические структуры: H1-1, H2-2A и H3-11, соответственно, для CDR1, CDR2 и CDR3 тяжелой цепи. В еще более конкретных вариантах воплощения для таких типов по настоящему изобретению CDRs для соответствующих легких цепей имеют канонические структуры L1-2A, L2-1 и L3-1, соответственно, для CDR1, CDR2 и CDR3 легкой цепи. В других вариантах воплощения настоящего изобретения канинизированное антитело против собачьего PD-1 дополнительно содержит определяющие комплементарность области (CDRs), в которой CDRs имеют канонические структуры: H1-1, H2-2A и H3-13, соответственно, для CDR1, CDR2 и CDR3 тяжелой цепи. В еще более конкретных вариантах воплощения для таких типов по настоящему изобретению CDRs для соответствующих легких цепей имеют канонические структуры: L1-4, L2-1 и L3-1, соответственно, для CDR1, CDR2 и CDR3 легкой цепи.
В более конкретных вариантах воплощения настоящего изобретения канинизированное антитело по настоящему изобретению или его антигенсвязывающий фрагмент содержит одну или более определяющую комплементарность область 1 (VH CDR1) тяжелой цепи с аминокислотной последовательностью SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30. В другом варианте воплощения настоящего изобретения определяющая комплементарность область 2 (VH CDR2) тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35. В другом варианте воплощения настоящего изобретения определяющая комплементарность область 3 (VH CDR3) тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 146. В конкретном варианте воплощения для такого типа по настоящему изобретению канинизированное антитело или антигенсвязывающий фрагмент содержит обе, и VH CDR1, содержащую аминокислотную последовательность SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30, и VH CDR2, содержащую аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35. В другом таком варианте воплощения настоящего изобретения канинизированное антитело или антигенсвязывающий фрагмент содержит обе, и VH CDR1, содержащую аминокислотную последовательность SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30 и VH CDR3, содержащую аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 146. В другом таком варианте воплощения настоящего изобретения канинизированное антитело или антигенсвязывающий фрагмент содержит обе, и VH CDR2, содержащую аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, и VH CDR3, содержащую аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 146. В другом таком варианте воплощения настоящего изобретения канинизированное антитело или антигенсвязывающий фрагмент содержит VH CDR1, содержащую аминокислотную последовательность SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30, VH CDR2, содержащую аминокислотную последовательность SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33, SEQ ID NO: 34 или SEQ ID NO: 35, и VH CDR3, содержащую аминокислотную последовательность SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38 или SEQ ID NO: 146.
В конкретных вариантах воплощения настоящего изобретения канинизированное антитело или антигенсвязывающий фрагмент также содержит определяющую комплементарность область 1 (VL CDR1) легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15. В близких вариантах воплощения настоящего изобретения определяющая комплементарность область 2 (VL CDR2) легкой цепи содержит аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 21. В другом варианте воплощения настоящего изобретения определяющая комплементарность область 3 (VL CDR3) легкой цепи содержит аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26. В конкретном варианте воплощения для такого типа по настоящему изобретению канинизированное антитело или антигенсвязывающий фрагмент содержит обе, и VL CDR1, содержащую аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, и VL CDR2 содержащую аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 21.
В другом таком варианте воплощения настоящего изобретения канинизированное антитело или антигенсвязывающий фрагмент содержит обе, и VL CDR1, содержащую аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, и VL CDR3 содержащую аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26. В другом таком варианте воплощения настоящего изобретения канинизированное антитело или антигенсвязывающий фрагмент содержит обе, и VL CDR2, содержащую аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, или SEQ ID NO: 21, и VL CDR3 содержащую аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26. В другом таком варианте воплощения настоящего изобретения канинизированное антитело или антигенсвязывающий фрагмент содержит VL CDR1, содержащую аминокислотную последовательность SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, VL CDR2, содержащую аминокислотную последовательность SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, или SEQ ID NO: 21, и VL CDR3, содержащую аминокислотную последовательность SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25 или SEQ ID NO: 26.
Дополнительно, настоящее изобретение обеспечивает канинизированные антитела, которые содержат аминокислотную последовательность SEQ ID NO: 40 или таковую на 90%, 95%, 98%, или 99% идентичную аминокислотной последовательности SEQ ID NO: 40; SEQ ID NO: 42 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 42; SEQ ID NO: 44 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 44; SEQ ID NO: 46 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 46; SEQ ID NO: 48 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 48; SEQ ID NO: 50 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 50; SEQ ID NO: 52 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 52; SEQ ID NO: 54 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 54; SEQ ID NO: 56 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 56; SEQ ID NO: 58 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 58; SEQ ID NO: 60 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 60; SEQ ID NO: 62 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 62; SEQ ID NO: 64 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 64; или SEQ ID NO: 66 или таковую на 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO: 66, или антигенсвязывающие фрагменты таких канинизированных антител.
В конкретных вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 40, 52, 56, или 64), которая содержит (i) P, A, G или S в позиции 239, (ii) A, G или S в позиции 266, (iii) A, G или S в позиции 298, (iv) G, P или A в позиции 299, (v) T, A, G или S в позиции 300, (vi) A, G или S в позиции 328, и (vii) P, A, G или S в позиции 330. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 42, 54, 58, или 66), которая содержит (i) P, A, G или S в позиции 237, (ii) A, G или S в позиции 264, (iii) A, G или S в позиции 296, (iv) G, P или A в позиции 297, (v) T, A, G или S в позиции 298, (vi) A, G или S в позиции 326, и (vii) P, A, G или S в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 44, 50 или 60), которая содержит (i) P, A, G или S в позиции 244, (ii) A, G или S в позиции 271, (iii) A, G или S в позиции 303, (iv) G, P или A в позиции 304, (v) T, A, G или S в позиции 305, (vi) A, G или S в позиции 333, и (vii) P, A, G или S в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) P, A, G или S в позиции 242, (ii) A, G или S в позиции 269, (iii) A, G или S в позиции 301, (iv) G, P или A в позиции 302, (v) T, A, G или S в позиции 303, (vi) A, G или S в позиции 331, и (vii) P, A, G или S в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 48) которая содержит (i) P, A, G или S в позиции 246, (ii) A, G или S в позиции 273, (iii) A, G или S в позиции 305, (iv) G, P или A в позиции 306, (v) T, A, G или S в позиции 307, (vi) A, G или S в позиции 335, и (vii) P, A, G или S в позиции 337.
В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 40, 52, 56 или 64), которая содержит (i) P, A, G или S в позиции 239, (ii) A в позиции 266, (iii) A в позиции 298, (iv) G, P или A в позиции 299, (v) T, A, G или S в позиции 300, (vi) A, G или S в позиции 328, и (vii) P, A, G или S в позиции 330. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 42, 54, 58 или 66), которая содержит (i) P, A, G или S в позиции 237, (ii) A в позиции 264, (iii) A в позиции 296, (iv) G, P или A в позиции 297, (v) T, A, G, или S в позиции 298, (vi) A, G, или S в позиции 326, и (vii) P, A, G, или S в позиции 328. В других вариантах воплощения настоящего изобретения, тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50, или 60 (или 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 44, 50, или 60) которая содержит (i) P, A, G, или S в позиции 244, (ii) A в позиции 271, (iii) A в позиции 303, (iv) G, P или A в позиции 304, (v) T, A, G, или S в позиции 305, (vi) A, G или S в позиции 333, и (vii) P, A, G или S в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) P, A, G или S в позиции 242, (ii) A в позиции 269, (iii) A в позиции 301, (iv) G, P или A в позиции 302, (v) T, A, G или S в позиции 303, (vi) A, G или S в позиции 331, и (vii) P, A, G или S в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 48), которая содержит (i) P, A, G или S в позиции 246, (ii) A в позиции 273, (iii) A в позиции 305, (iv) G, P или A в позиции 306, (v) T, A, G или S в позиции 307, (vi) A, G или S в позиции 335, и (vii) P, A, G или S в позиции 337.
В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 40, 52, 56 или 64), которая содержит (i) A в позиции 239, (ii) A в позиции 266, (iii) A в позиции 298, (iv) P в позиции 299, (v) A в позиции 300, (vi) G, в позиции 328, и (vii) A в позиции 330. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 42, 54, 58, или 66), которая содержит (i) A в позиции 237, (ii) A в позиции 264, (iii) A в позиции 296, (iv) P в позиции 297, (v) A в позиции 298, (vi) G в позиции 326, и (vii) A в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 44, 50 или 60), которая содержит (i) A в позиции 244, (ii) A в позиции 271, (iii) A в позиции 303, (iv) P в позиции 304, (v) A в позиции 305, (vi) G в позиции 333, и (vii) A в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) A в позиции 242, (ii) A в позиции 269, (iii) A в позиции 301, (iv) P в позиции 302, (v) A в позиции 303, (vi) G в позиции 331, и (vii) A в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 48), которая содержит (i) A в позиции 246, (ii) A в позиции 273, (iii) A в позиции 305, (iv) P в позиции 306, (v) A в позиции 307, (vi) G в позиции 335, и (vii) A в позиции 337.
В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 40, 52, 56 или 64), которая содержит (i) P в позиции 239, (ii) A, G или S в позиции 266, (iii) A, G или S в позиции 298, (iv) G в позиции 299, (v) T в позиции 300, (vi) A в позиции 328, и (vii) P в позиции 330. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 42, 54, 58 или 66), которая содержит (i) P в позиции 237, (ii) A, G или S в позиции 264, (iii) A, G или S в позиции 296, (iv) G в позиции 297, (v) T в позиции 298, (vi) A в позиции 326, и (vii) P в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 44, 50 или 60), которая содержит (i) P в позиции 244, (ii) A, G или S в позиции 271, (iii) A, G или S в позиции 303, (iv) G в позиции 304, (v) T в позиции 305, (vi) A в позиции 333, и (vii) P в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) P в позиции 242, (ii) A, G или S в позиции 269, (iii) A, G или S в позиции 301, (iv) G в позиции 302, (v) T в позиции 303, (vi) A в позиции 331, и (vii) P в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или 90%, 95%, 98% или 99% идентичную SEQ ID NO: 48), которая содержит (i) P в позиции 246, (ii) A, G или S в позиции 273, (iii) A, G или S в позиции 305, (iv) G в позиции 306, (v) T в позиции 307, (vi) A в позиции 335, и (vii) P в позиции 337.
В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 40, 52, 56 или 64), которая содержит (i) P в позиции 239, (ii) A в позиции 266, (iii) A в позиции 298, (iv) G в позиции 299, (v) T в позиции 300, (vi) A в позиции 328, и (vii) P в позиции 330. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 42, 54, 58 или 66), которая содержит (i) P в позиции 237, (ii) A в позиции 264, (iii) A в позиции 296, (iv) G в позиции 297, (v) T в позиции 298, (vi) A в позиции 326, и (vii) P в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60 (или 90%, 95%, 98% или 99% идентичную SEQ ID NO: 44, 50 или 60), которая содержит (i) P в позиции 244, (ii) A в позиции 271, (iii) A в позиции 303, (iv) G в позиции 304, (v) T в позиции 305, (vi) A в позиции 333, и (vii) P в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) P в позиции 242, (ii) A в позиции 269, (iii) A в позиции 301, (iv) G в позиции 302, (v) T в позиции 303, (vi) A в позиции 331, и (vii) P в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 48), которая содержит (i) P в позиции 246, (ii) A в позиции 273, (iii) A в позиции 305, (iv) G в позиции 306, (v) T в позиции 307, (vi) A в позиции 335, и (vii) P в позиции 337.
В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 40, 52, 56 или 64), которая содержит (i) P, A, G или S в позиции 239, (ii) A, G или S в позиции 266, (iii) A, G или S в позиции 298, (iv) G в позиции 299, (v) T в позиции 300, (vi) A, G или S в позиции 328, и (vii) P, A, G или S в позиции 330. В других таких вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 42, 54, 58 или 66), которая содержит (i) P, A, G или S в позиции 237, (ii) A, G или S в позиции 264, (iii) A, G или S в позиции 296, (iv) G в позиции 297, (v) T в позиции 298, (vi) A, G или S в позиции 326, and (vii) P, A, G или S в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 44, 50 или 60), которая содержит (i) P, A, G или S в позиции 244, (ii) A, G или S в позиции 271, (iii) A, G или S в позиции 303, (iv) G в позиции 304, (v) T в позиции 305, (vi) A, G, или S в позиции 333, и (vii) P, A, G или S в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) P, A, G или S в позиции 242, (ii) A, G или S в позиции 269, (iii) A, G или S в позиции 301, (iv) G в позиции 302, (v) T в позиции 303, (vi) A, G или S в позиции 331, и (vii) P, A, G или S в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 48), которая содержит (i) P, A, G или S в позиции 246, (ii) A, G или S в позиции 273, (iii) A, G или S в позиции 305, (iv) G в позиции 306, (v) T в позиции 307, (vi) A, G или S в позиции 335, и (vii) P, A, G или S в позиции 337.
В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 40, 52, 56 или 64), которая содержит (i) P, A, G или S в позиции 239, (ii) A в позиции 266, (iii) A в позиции 298, (iv) G в позиции 299, (v) T в позиции 300, (vi) A, G или S в позиции 328, и (vii) P, A, G или S в позиции 330. В других таких вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 42, 54, 58 или 66), которая содержит (i) P, A, G или S в позиции 237, (ii) A в позиции 264, (iii) A в позиции 296, (iv) G в позиции 297, (v) T в позиции 298, (vi) A, G или S в позиции 326, и (vii) P, A, G или S в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 44, 50 или 60), которая содержит (i) P, A, G или S в позиции 244, (ii) A в позиции 271, (iii) A в позиции 303, (iv) G в позиции 304, (v) T в позиции 305, (vi) A, G или S в позиции 333, и (vii) P, A, G или S в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98%, или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) P, A, G или S в позиции 242, (ii) A в позиции 269, (iii) A в позиции 301, (iv) G в позиции 302, (v) T в позиции 303, (vi) A, G или S в позиции 331, и (vii) P, A, G или S в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 48), которая содержит (i) P, A, G или S в позиции 246, (ii) A в позиции 273, (iii) A в позиции 305, (iv) G в позиции 306, (v) T в позиции 307, (vi) A, G или S в позиции 335, и (vii) P, A, G или S в позиции 337.
В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56 или 64 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 40, 52, 56 или 64), которая содержит (i) A в позиции 239, (ii) A в позиции 266, (iii) A в позиции 298, (iv) G в позиции 299, (v) T в позиции 300, (vi) G в позиции 328, and (vii) A в позиции 330. В других таких вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 42, 54, 58 или 66), которая содержит (i) A в позиции 237, (ii) A в позиции 264, (iii) A в позиции 296, (iv) G в позиции 297, (v) T в позиции 298, (vi) G в позиции 326, и (vii) A в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 44, 50 или 60), которая содержит (i) A в позиции 244, (ii) A в позиции 271, (iii) A в позиции 303, (iv) G в позиции 304, (v) T в позиции 305, (vi) G в позиции 333, и (vii) A в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 46 или 62), которая содержит (i) A в позиции 242, (ii) A в позиции 269, (iii) A в позиции 301, (iv) G в позиции 302, (v) T в позиции 303, (vi) G в позиции 331, и (vii) A в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48 (или на 90%, 95%, 98% или 99% идентичную SEQ ID NO: 48), которая содержит (i) A в позиции 246, (ii) A в позиции 273, (iii) A в позиции 305, (iv) G в позиции 306, (v) T в позиции 307, (vi) G в позиции 335, и (vii) A в позиции 337.
Дополнительно, настоящее изобретение обеспечивает канинизированное антитело или его антигенсвязывающий фрагмент, которое дополнительно содержит собачью легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 72, SEQ ID NO: 78, SEQ ID NO: 84, SEQ ID NO: 90, SEQ ID NO: 96, SEQ ID NO: 102 или SEQ ID NO: 108.
Соответственно, дополнительно настоящее изобретение обеспечивает канинизированное антитело или его антигенсвязывающий фрагмент, которое содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 68, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 72. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 70, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 72. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 74, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 78. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 76, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 78. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 80, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 84. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 82, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 84. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 86, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 90. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 88, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 90. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 92, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 96. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 94, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 96. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 98, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 102. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 100, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 102. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 104, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 108. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 106, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 108.
Дополнительно настоящее изобретение обеспечивает канинизированное антитело или его антигенсвязывающий фрагмент, которое содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 40, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 72. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 42, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 72. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 44, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 78. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 46, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 78. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 48, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 84. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 50, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 84. В другом варианте воплощения настоящего изобретения, канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 52, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 90. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 54, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 90. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 56, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 96. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 58, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 96.
В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 60, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 102. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 62, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 102. В другом варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 64, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 108. В близком варианте воплощения настоящего изобретения канинизированное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 66, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 108.
Дополнительно, настоящее изобретение обеспечивает нуклеиновые кислоты, которые кодируют любую из аминокислотных последовательностей по настоящему изобретению, включая CDRs, cFc области, cFc области с шарнирными областями, и тяжелые цепи, и легкие цепи канинизированных антител по настоящему изобретению. Дополнительно, настоящее изобретение обеспечивает векторы экспрессии, которые содержат одну или более из нуклеиновых кислот по настоящему изобретению. Дополнительно, настоящее изобретение обеспечивает клетки-хозяева, которые содержат один или более векторы экспрессии по настоящему изобретению, и способы экспрессирования CDRs, и/или cFc областей, и/или cFc областей с шарнирными областями, и/или тяжелых цепей, и/или легких цепей канинизированных антител по настоящему изобретению при использовании таких клеток-хозяев. Настоящее изобретение также обеспечивает клетки-хозяева, которые подверглись воздействию генной инженерии для экспрессии CDRs, и/или cFc областей, и/или cFc областей с шарнирными областями, и/или тяжелых цепей, и/или легких цепей канинизированных антител по настоящему изобретению при отсутствии таких векторов. В конкретных вариантах воплощения настоящего изобретения эти нуклеиновые кислоты, векторы экспрессии, полипептиды или клетки-хозяева по настоящему изобретению используют в способах получения антител.
В конкретных вариантах воплощения настоящего изобретения антитело представляет рекомбинантное антитело или его антигенсвязывающий фрагмент. В близких вариантах воплощения настоящего изобретения вариабельный домен тяжелой цепи и вариабельный домен легкой цепи соединены гибким линкером с образованием одноцепопчечного антитела.
В конкретных вариантах воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент представляет Fab фрагмент.
В других вариантах воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент представляет Fab' фрагмент. В других вариантах воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент представляет (Fab')2 фрагмент. В других вариантах воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент представляет диатело. В конкретных вариантах воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент представляет домен антитела. В конкретных вариантах воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент представляет однодоменное камелизованное антитело.
В конкретных вариантах воплощения настоящего изобретения канинизированное мышиное антитело против собачьего PD-1 или антигенсвязывающий фрагмент повышает иммунный ответ у лечимой им собаки.
В конкретных вариантах воплощения настоящего изобретения, когда канинизированное антитело или его антигенсвязывающий фрагмент связаны с собачьим PD-1, то связываются по меньшей мере с одним аминокислотным остатком в одной или более аминокислотной последовательности из следующих: SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144, и/или SEQ ID NO: 145.
Дополнительно, настоящее изобретение обеспечивает канинизированные антитела к собачьему PD-1, которые содержат варианты CDRs по настоящему изобретению, которые имеют соответствующие канонические структуры, обеспеченные в описании настоящей патентной заявки, и/или которые связываются с аминокислотной последовательностью SEQ ID NO: 144. В конкретных вариантах воплощения для такого типа по настоящему изобретению константа диссоциации (Kd) для связывания канинизированное антитело-собачий PD-1 составляет от 1×10-5 до 1×10-12 M. В более конкретных вариантах воплощения настоящего изобретения канинизированные антитела к собачьему PD-1 содержат варианты CDRs по настоящему изобретению, которые имеют соответствующие канонические структуры, обеспеченные в описании настоящей патентной заявки, и связывающиеся с аминокислотной последовательностью SEQ ID NO: 145. Следовательно, настоящее изобретение включает канинизированные антитела и их антигенсвязывающие фрагменты, которые специфически связываются с собачьим PD-1, и которые, когда они связаны с собачьим PD-1, то антитело связывается по меньшей мере с одним аминокислотным остатком в SEQ ID NO: 144. В конкретных вариантах воплощения для такого типа по настоящему изобретению антитела и их антигенсвязывающие фрагменты связывают собачий PD-1 и блокируют связывание собачьего PD 1 с собачьим лигандом запрограммированной смерти 1 (PD-L1).
Соответственно, в конкретных вариантах воплощения настоящего изобретения, когда канинизированное антитело (включая антитела с одним или более вариантом CDR, например, вариант, включающий консервативно модифицированный вариант и/или вариант, который содержит определенный класс канонической структуры) связано с собачьим PD-1, то оно связывается по меньшей мере с одним аминокислотным остатком в одной или более аминокислотной последовательности из следующих: SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143 и/или SEQ ID NO: 145. В еще более конкретных вариантах воплощения настоящего изобретения, когда канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1, то они связываются с одним или более аминокислотным остатком из следующих остатков аргинина: R62, R69, R72, R75 и R90 SEQ ID NO: 114. В конкретных вариантах воплощения настоящего изобретения, когда канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1, то они связываются по меньшей мере с одним аминокислотным остатком в SEQ ID NO: 145. В конкретных вариантах воплощения настоящего изобретения, когда антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1, то они связываются с одним или более аминокислотным остатком из следующих остатков аргинина: R62, R69, R72 и R75 SEQ ID NO: 114. В более конкретных вариантах воплощения настоящего изобретения, когда антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1, то они связываются с R75 SEQ ID NO: 114.
Дополнительно, настоящее изобретение обеспечивает канинизированные антитела или их антигенсвязывающие фрагменты, которые связываются с собачьим PD-1 с константой диссоциации (Kd), которая составляет менее (например, 1×10-13 M, или ниже), чем 1×10-12 M. В конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты к собачьему PD-1 имеют константу диссоциации от 1×10-5 M до 1×10-12 M. В более конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 с константой диссоциации от1×10-7 M до 1×10-11 M. В более конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 с константой диссоциации от 1×10-8 M до 1×10-11 M. В других конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 с константой диссоциации от 1×10-8 M до 1×10-10 M.
Настоящее изобретение также обеспечивает канинизированные антитела или их антигенсвязывающие фрагменты, которые связаны с собачьим PD-1 со скоростью ассоциации (kon), составляющей более чем 1×107 M-1s-1. В конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты thereof связаны с собачьим PD-1 со скоростью ассоциации от 1×102 M-1s-1 до 1×107 M-1s-1. В более конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 со скоростью ассоциации от 1×103 M-1s-1 до 1×106 M-1s-1. В еще более конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 со скоростью ассоциации от 1×103 M-1s-1 до 1×105 M-1s-1. В еще более конкретных вариантах воплощения настоящего изобретения, канинизированные антитела или антигенсвязывающие фрагменты связаны с собачьим PD-1 со скоростью ассоциации от 1×104 M-1s-1 до 1×105 M-1s-1.
Дополнительно, настоящее изобретение обеспечивает канинизированные антитела или их антигенсвязывающие фрагменты, которые связаны с собачьим PD-1 со скоростью диссоциации (koff) медленнее, чем 1×10-7с-1. В конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 со скоростью диссоциации от 1×10-3с-1 до 1×10-8с-1. В более конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 со скоростью диссоциации от 1×10-4с-1 до 1×10-7с-1. В еще более конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты связаны с собачьим PD-1 со скоростью диссоциации от 1×10-5с-1 до 1×10-7с-1.
В близких вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты стимулируют память антиген-специфических ответов на опухоль или патоген. В конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты стимулируют гуморальный иммунный ответ in vivo. В других конкретных вариантах воплощения настоящего изобретения канинизированные антитела или их антигенсвязывающие фрагменты стимулируют иммунный ответ у животного-субъекта. В более конкретных вариантах воплощения настоящего изобретения животное-субъект представляет представителя собачьих. В близком варианте воплощения настоящего изобретения животное-субъект представляет представителя кошачьих.
Соответственно, любое канинизированное антитело по настоящему изобретению может демонстрировать одно, два, три, четыре, пять или все эти свойства, то есть указанные выше константы диссоциации с собачьим PD-1, указанные выше скорости связывания с собачьим PD-1, указанные выше скорости диссоциации для диссоциирования из комплекса связывания канинизированное антитело-собачий PD-1, стимуляцию ответов антигенспецифической памяти на опухоль или патоген, стимуляцию гуморального иммунного ответа in vivo, и/или стимуляцию иммунного ответа у животного-субъекта.
В более конкретных вариантах воплощения настоящего изобретения канинизированные антитела и их антигенсвязывающие фрагменты по настоящему изобретению связываются с собачьим PD-1 и также блокируют связывание собачьего PD 1 с PD-L1. В еще более конкретных вариантах воплощения настоящего изобретения канинизированные антитела и их антигенсвязывающие фрагменты по настоящему изобретению связываются с собачьим PD-1, блокируют связывание собачьего PD 1 с PD-L1, и также блокируют связывание собачьего PD-1 с PD-L2.
Дополнительно, настоящее изобретение обеспечивает нуклеиновые кислоты, которые кодируют канинизированное мышиное антитело против собачьего PD-1 или его части по настоящему изобретению. В близких вариантах воплощения настоящего изобретения такие антитела или их антигенсвязывающие фрагменты могут быть использованы для получения лекарственного средства для лечения рака у субъекта-животного. В качестве альтернативы или в совокупности, настоящее изобретение обеспечивает использование одного или более антитела или фрагментов антитела по настоящему изобретению для диагностики. В дополнительных вариантах воплощения настоящего изобретения обеспечивается набор, содержащий любое из канинизированных антител или его антигенсвязывающих фрагментов, как приведено в описании настоящей патентной заявки.
Дополнительно настоящее изобретение включает фармацевтические композиции, содержащие антитело против собачьего антигена или его связывающие фрагменты (например, антитело против собачьего PD -1 или его антигенсвязывающий фрагмент) вместе с фармацевтически приемлемым носителей или разбавителем. Настоящее изобретение также обеспечивает способ повышения активности иммунной клетки, включающий введение субъекту (например, собаке), нуждающемуся в нем, терапевтически эффективного количества фармацевтической композиции по настоящему изобретению. В конкретных вариантах воплощения настоящего изобретения способ используют для лечения рака. В других вариантах воплощения настоящего изобретения способ используют для лечения инфекции или инфекционных заболеваний. В других вариантах воплощения настоящего изобретения канинизированное антитело по настоящему изобретению или его антигенсвязывающий фрагмент используют в качестве адъюванта вакцины. В другом варианте воплощения настоящего изобретения канинизированное анти-TSLP антитело вводят собаке для лечения атопического дерматита.
Эти другие аспекты настоящего изобретения будут более понятны понятны из следующего краткого описания чертежей и подробного описания, на которые делается ссылка.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 - реактивность канинизированных моноклональных антител (mAbs) против внеклеточного домена собачьего PD-1, как функции OD 650/490 от log mAb (nM). Протестировали различные канинизированные mAbs на их связывание с внеклеточным доменом собачьего PD-1 при использовании ELISA. Четыре протестированных mAbs указаны, как: 2H9 VH4 IgGB/VL3, 3B6 VH3 IgGB/ VL3, 2H9 VH4 IgGB (YZZ1062)/VL3 и 2H9 VH4 IgGB (YZZ1068)/VL3.
Фигура 2 - реактивность канинизированных mAbs против собачьего PD-1, экспрессированного на поверхности клеток. Протестировали различные мышиные mAbs на их связывание с собачьим PD-1, экспрессированным на CHO клетках при использовании CELISA как функцию OD 450/540 от log mAb (nM). Шесть протестированных mAbs были указаны, как: 3B6 VH3/VL4, 3B6 VH3/VL1, 3B6 VH3/VL3, 3B6 VH3/VL2, 3B6 VH1/VL1 и 3B6 m-c Chimera.
Фигура 3 - блокирование лиганда канинизированными mAbs против собачьего PD-1. Протестировали различные канинизированные mAbs на способность ингибировать связывание PD-1, экспрессированного на CHO клетках с PD- L1 как функции OD 450/540 от log mAb (nM). Протестировали шесть mAbs, обозначенные как: 3B6 VH3/VL4, 3B6 VH3/VL1, 3B6 VH3/VL3, 3B6 VH3/VL2, 3B6 VH1/VL1 и 3B6 m-c Chimera.
Фигура 4 - секреция цитокина, индуцированная канинизированными mAbs против собачьего PD-1. Протестировали различные канинизированные mAbs и их варианты на способность индуцировать секрецию цитокина PBMC у здоровых собак.
Фигура 5A и 5B - связывание канинизированных mAbs и их вариантов (при начальной 1 μг/мл) с FcᵧRI. Протестировали различные mAbs на их способность связываться с FcRI. Антитела обозначили как: can 2H9 ADCC (1062) VH4/VL3, can 2H9 ADCC mut 1 VH4/VL3, can 2H9 ADCC mut 2 VH4/VL3, can 2H9 IgGD VH4/VL3, can 2H9 VH4/VL3 и can 3B6 VH4/VL4 на Фигуре 5A; и can 2H9 ADCC (1059) VH4/VL3, can 2H9 ADCC (1060) VH4/VL3, can 2H9 ADCC (1061) VH4/VL3, can 2H9 IgGB ADCC (1068) VH4/VL3, can 2H9 VH4/VL3 и can 3B6 VH4/VL4 на Фигуре 5B.
Фигура 6A и 6B - связывание канинизированных mAbs и их вариантов (при начальной 1 μг/мл) с C1Q. Протестировали различные mAbs на их способность связываться с C1Q. Антитела обозначили как: can 2H9 VH4 IgGB ADCC (1062) /VL3, can 2H9 VH4 IgGB ADCC (mut 1)/VL3, can 2H9 VH4 IgGB ADCC (mut 2)/VL3, can 2H9 VH4 IgGD/VL3, can 2H9 VH4/VL3 и can 3B6 VH4/VL4 IgGB на Фигуре 6A; и can 2H9 VH4 IgGB ADCC (1059) /VL3, can 2H9 VH4 IgGB ADCC (1060)/VL3, can 2H9 VH4 IgGB ADCC (1061)/VL3, can 2H9 VH4 IgGB ADCC (1068)/VL3, can 2H9 VH4/VL3 IgGB и can 3B6 VH4/VL4 IgGB на Фигуре 6B.
Фигура 7A - характеристика области контакта собачьего PD-1 и канинизированного антитела 2G9. Положение аминокислот приведено относительно аминокислотной последовательности PD-1 без сигнальной последовательности, то есть SEQ ID NO: 114. Определение провели при использовании химического перекрестного сшивания, матрично-активированной лазерной десорбции-ионизации в комбинации с детектором частиц с высокой массой и в сочетании с масс-спектрометрией (High-Mass MALDI mass spectrometry) и не линейной хроматографии с орбитальной ионной ловушкой в сочетании с масс- спектрометрией (nLC-Orbitrap mass spectrometry).
Фигура 7B - характеристика области контакта собачьего PD-1 и канинизированного антитела 3B6. Положение аминокислот приведено относительно аминокислотной последовательности PD-1 без сигнальной последовательности, то есть SEQ ID NO: 114. Определение провели при использовании химического перекрестного сшивания, матрично-активированной лазерной десорбции-ионизации в комбинации с детектором частиц с высокой массой и в сочетании с масс-спектрометрией (High-Mass MALDI mass spectrometry) и не линейной хроматографии с орбитальной ионной ловушкой в сочетании с масс-спектрометрией (nLC-Orbitrap mass spectrometry).
ПОДРОБНОЕ ОПИСАНИЕ
Сокращения
В подробном описании и примерах настоящей патентной заявки используются следующие сокращения:
ADCC Антитело-зависимая клеточная цитотоксичность
CDC Комплимент-зависимая цитотоксичность
CDR Определяющая комплементарность область в вариабельных областях иммуноглобулина, определенных для человеческих антител при использовании номенклатуры Кэбота
CHO Яичник китайского хомячка
EC50 концентрация, обеспечивающая 50% эффективность или связывание
ELISA Иммуносорбентный анализ с ферментной меткой
FR Каркасный участок антитела: вариабельные области иммуноглобулина, исключая CDR области.
HRP Пероксидаза хрена
IFN Интерферон
IC50 концентрация, обеспечивающая 50% ингибирование
IgG Иммуноглобулин G
Kabat Выравнивание иммуноглобулина и номенклатура человеческих антител, введенные Elvin A. Kabat [Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)]
mAb Моноклональное антитело (также Mab или MAb)
MES 2-(N-морфолино)этансульфоновая кислота
MOA Механизм действия
NHS Сыворотка здорового человека
PCR Полимеразная цепная реакция
PK Фармакокинетика
SEB Энтеротоксин B стафилококка
TT Столбнячный токсин
V область Сегмент цепей человеческого IgG, последовательность которого у различных антител вариабельны. Он располагается до 109 остатка по Кэботу в легкой цепи и до 113 остатка в тяжелой цепи.
VH Вариабельный участок тяжелой цепи иммуноглобулина
VK Вариабельный участок каппа легкой цепи иммуноглобулина
ОПРЕДЕЛЕНИЯ
Для лучшего понимания настоящего изобретения ниже приведены некоторые технические и научные термины. Если в этом документе ясно не указано иное, все другие технические и научные термины, используемые в описании настоящей патентной заявки, имеют значение общепринятое в области техники, к которой относится настоящее изобретение.
Используемые в описании настоящей патентной заявки формы единственного числа включают и множественное число, если в контексте ясно не просматривается иное.
«Активация» в отношении клеток или рецепторов относится к активации или воздействию лигандов на клетку или рецептор, если из контекста не просматривается иное, или ясно не указано иное. «Лиганд» включает в объем понятия натуральные и синтетические лиганды, например, цитокины, варианты цитокинов, аналоги, мутантные белки и связывающие соединения, полученные из антител. «Лиганд» также включает в объем понятия малые молекулы, например, пептидные миметики цитокинов и пептидные миметики антител. «Активация» может относиться к клеточной активации, как регулируемой внутренними механизмами, наряду с внешними факторами или факторами окружающей среды.
«Активность» молекулы может описываться или относиться к связыванию молекулы с лигандом или рецептором, каталитической активности; к способности стимулировать генную экспрессию или сигнальной системе клетки, дифференцировке или созреванию; к антигенной активности, модуляции активностей других молекул и аналогичному им. «Активность» молекулы может также относиться к активности в модулируемых или поддерживаемым взаимодействиям клетка-клетка, например, адгезия, или активности в сохранении структуры клетки, клеточных мембран или цитоскелета. «Активность» также может означать специфическую активность, например, [каталитическая активность]/[мг белка], или [иммунологическая активность]/[мг белка], концентрация в биологическом компартменте, или аналогичное им. «Активность» может относиться к модуляции компонентов врожденной или адаптивной иммунных систем.
«Введение» или «лечебное воздействие» в отношении животного, например, подопытного, относящегося к собачьим, клетки, ткани, органа или биологической жидкости относится к контакту экзогенного фармацевтического, терапевтического, диагностического агента или композиции с животным, например, субъектом, относящимся к собачьим, клеткой, тканями, органом или биологической жидкостью. Воздействие на клетку включает контакт реагента с клеткой, наряду с контактом реагента с жидкостью, где жидкость находится в контакте с клеткой. «Введение» и «лечебное воздействие» также означает in vitro и ex vivo воздействие, например, на клетку реагентом, диагностическим соединением, связывающим соединением или другой клеткой.
Используемый в описании настоящей патентной заявки термин «субъект» включает в объем понятия любой организм, предпочтительно животное, более предпочтительно млекопитающее (например, относящееся к собачьим, кошачьим или человеку) и наиболее предпочтительно относящийся к собачьим.
Используемый в описании настоящей патентной заявки термин «относящийся к кошачьим» относится к любому члену семейства кошачьих. Члены этого семейства включают диких, содержащихся в зоопарках и домашних членов, таких как любой член из подсемейства кошачьих, например, кошки, львы, тигры, пумы, ягуары, леопарды, снежные леопарды, пантеры, североамериканские горные львы, гепарды, рыси, рыжие рыси, каракалы и любые их гибриды. Кошки также включают домашних кошек, чистопородных и/или беспородных кошек-компаньонов, выставочных кошек, лабораторных кошек, клонированных кошек и диких или бродячих кошек.
Используемый в описании настоящей патентной заявки термин «замещение аминокислотного остатка» другим аминокислотным остатком в аминокислотной последовательности является эквивалентом «замена аминокислотного остатка» другим аминокислотным остатком и означает, что конкретный аминокислотный остаток в специфической позиции в аминокислотной последовательности замещается (или заменяется) на отличающуюся аминокислоту. Например, одно такое замещение (замена) обозначается как P4A Fc области аминокислотной последовательности IgGB или IgGC, в таком случае остаток пролина в позиции 4 аминокислотной последовательности Fc области IgGB или Fc области IgGC замещен на (заменен) остатком аланина.
Соответственно, такие аминокислотные замещения могут быть разработаны специально, то есть целенаправленное замещение аланина серином в специфической позиции в аминокислотной последовательности, например, при использовании технологии рекомбинантной ДНК. В качестве альтернативы, конкретный аминокислотный остаток или ряд аминокислотных остатков антитела могут быть замещены одним или более аминокислотным остатком при использовании более естественных процессов отбора, например, на основе способности антитела, продуцируемого клеткой, связываться с заданной областью такого антигена, например, таковой, содержащей эпитоп или его часть, и/или для включения в антитело конкретной CDR, которая сохраняет такую же каноническую структуру, как и замещаемая CDR. Такие замещения/замены могут привести к «варианту» CDRs и/или антител.
«Лечение» или «лечебное воздействие» означает введение терапевтического агента, такого как композиция, содержащая любое антитело или его антигенсвязывающие фрагменты по настоящему изобретению, внутренне или внешне субъекту, относящемуся к собачьим или пациенту, имеющему один или более симптом заболевания или предрасположенному к возникновению заболевания, для которого терапевтический агент имеет терапевтическую активность.
Как правило, агент вводят в количестве, эффективном для облегчения и/или улучшения одного или более симптома заболевания у лечимого субъекта или популяции, либо за счет индуцирования регрессии, либо за счет ингибирования прогрессирования такого симптома(ов) в любой клинически измеримой степени. Количество терапевтического агента, которое является эффективным для облегчения любого конкретного симптома заболевания (также указанное как «терапевтически эффективное количество»), может варьировать согласно факторам, таким как стадия заболевания, возраст и масса субъекта (например, относящегося к собачьим), и способности фармацевтической композиции вызвать заданный ответ у субъекта. Облегчение или улучшение симптомов заболевания может быть оценено при использовании любого клинического измерения, как правило используемого ветеринарами или другими медицинскими работниками, способными оценить тяжесть состояния или прогресс состояния такого симптома. Хотя вариант воплощения настоящего изобретения (например, способ лечения или изделие промышленного производства) может не быть эффективным в облегчении целевого симптома(ов) заболевания у каждого субъекта, это должно быть облегчение целевого симптома(ов) у статистически значимого числа субъектов, как определяется при использовании любого статистического теста, известного из предшествующего уровня техники, такого как параметрический t-критерий Стьюдента, критерий хи2, U-критерий Манна-Уитни, критерий Крускала-Уоллиса (H-критерий), критерий Джонкхиера- Терпстра и критерий Уилкоксона.
«Лечение» в отношении человека, ветеринарного субъекта (например, относящегося к собачьим) или субъекта, участвующего в исследованиях, относится к терапевтическому лечению, наряду с исследовательскими применениями и диагностическими применениями. «Лечение» в отношении человека, ветеринарного субъекта (например, относящегося к собачьим) или субъекта, участвующего в исследованиях или клетки, ткани, или органа включает контакт канинизированного антитела или антигенсвязывающих фрагментов по настоящему изобретению с относящемся к собачьим или другим субъектом-животным, клеткой, тканями, физиологическим компартментом или физиологической жидкостью.
Было обнаружено, что собачий PD- 1 содержит аминокислотную последовательность SEQ ID NO: 114 [Предварительная заявка на патент США № 61/918,946, поданная 20 декабря 2013, содержание которой введено здесь ссылкой в полном объеме]. В конкретном варианте воплощения настоящего изобретения собачий PD- 1 кодируется нуклеиновой кислотой, которая содержит нуклеотидную последовательность SEQ ID NO: 113.
Было обнаружено, что собачий PD- 1 содержит аминокислотную последовательность SEQ ID NO: 120 [Предварительная заявка на патент США № 61/918,946, поданная 20 декабря 2013, как указано выше]. В конкретном варианте воплощения настоящего изобретения собачий PD- L1 кодируется нуклеиновой кислотой, которая содержит нуклеотидную последовательность SEQ ID NO: 119.
Используемый в описании настоящей патентной заявки термин «иммунный ответ» относится к воздействию, например, лимфоцитов, антиген-презентирующих клеток, фагоцитов, гранулоцитов и растворимых макромолекул, продуцированных указанными выше клетками или печенью (включая антитела, цитокины, и комплемент), которые приводят к селективному повреждению, разрушению или элиминированию в организме млекопитающего (например, в организме относящегося к собачьим) раковых клеток, клеток или тканей, инфицированных патогенами или инвазированных патогенами.
Канинизированные антитела против собачьего антигена
Используемый в описании настоящей патентной заявки термин «относящийся к собачьим» включает в объем понятия всех домашних собак, животных семейства волчьих или семейства собачьих, если не указано иное.
Как известно антитело специфически связывается с полипептидом, содержащим заданную антигенную последовательность (в этом случае часть аминокислотной последовательности собачьего антигена, например, собачий PD -1), если оно связывается с полипептидом, содержащим эту часть аминокислотной последовательности собачьего антигена, например, собачий PD-1, то не связывается с другими собачьими белками, у которых отсутствует часть последовательности собачьего антигена, например, собачьего PD-1. Например, антитело, которое специфически связывается с полипептидом, содержащим собачий PD-1 может связываться с FLAG®-меченной формой собачьего PD-1, но не связывается специфически с другими FLAG®-меченными собачьими белками. Антитело или связывающее соединение, полученное из антигенсвязывающего участка антитела, «специфически» связывается с собачьим антигеном или его вариантом, или его мутантными белками, когда оно имеет сродство с таковым собачьим антигеном или его вариантом, или его мутантными белками, которое по меньшей мере в десять раз больше, более предпочтительно по меньшей мере в 20 раз больше, и еще более предпочтительно по меньшей мере в 100 раз больше, чем его сродство с любым другим тестируемым собачьим антигеном.
Используемый в описании настоящей патентной заявки термин «антитело» относится к любой форме антитела, которая демонстрирует заданную биологическую активность. Следовательно, он используется в самом широком смысле и конкретно включает в объем понятия без ограничения моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела), канинизированные антитела, полностью собачьи антитела, химерные антитела и однодоменные камелизованные антитела. «Родительские антитела» представляют антитела, полученные воздействием иммунной системы на антиген перед модификацией антитела для предполагаемого использования, такого как канинизация антитела для применения в качестве собачьего терапевтического антитела.
Используемый в описании настоящей патентной заявки термин, если не указано иное, «фрагмент антитела» или «антигенсвязывающий фрагмент» относится к антигенсвязывающим фрагментам антитела, то есть фрагментам антитела, которые сохранили способность специфически связываться с антигеном, связанным полноразмерным антителом, например фрагменты, которые сохранили одну или более CDR область. Примеры антигенсвязывающих фрагментов включают без ограничения Fab, Fab', F(ab')2 и Fv фрагменты; диатела; линейные антитела; молекулами одноцепочечных антител, например, sc-Fv; нанотела и мультиспецифические антитела, образованные из фрагментов антител.
«Fab фрагмент» состоит из одной легкой цепи и CH1, и вариабельных участков одной тяжелой цепи. Молекула Fab тяжелой цепи не может образовывать дисульфидную связь с другой молекулой тяжелой цепи. «Fab фрагмент» может представлять продукт расщепления антитела папаином.
«Кристаллизуемый фрагмент» («Fc») области содержит два фрагмента тяжелой цепи (то есть два идентичных полипептида), содержащих CH2 и CH3 домены антитела. Два фрагмента тяжелой цепи удерживаются вместе двумя или более дисульфидными связями и гидрофобными взаимодействиями доменов CH3. В настоящем изобретении аминокислотная последовательность каждого из четырех Fc фрагментов собачьего IgG располагается на определенной границе доменов CH1 и CH2, как определено Tang et al. [Vet. Immunol. Immunopathol. 80: 259-270 (2001)].
«Fab' фрагмент» содержит одну легкую цепь и часть или фрагмент одной тяжелой цепи, которая содержит VH домен и CH1 домен, также область между CH1 и CH2 доменами, таким образом, что между двумя Fab' фрагментами тяжелой цепи может быть образована межцепьевая дисульфидная связь с образованием молекулы F(ab')2.
«F(ab')2 фрагмент» содержит две легких цепи и две тяжелых цепи, содержащие часть константной области между CH1 и CH2 доменами, таким образом, что между двумя тяжелыми цепями образуется межцепьевая дисульфидная связь. Следовательно, F(ab')2 фрагмент состоит из двух фрагментов Fab', которые удерживаются вместе дисульфидной связью между двумя тяжелыми цепями. «F(ab')2 фрагмент» может представлять продукт расщепления пепсином антитела.
«Fv область» содержит вариабельные области обеих, и тяжелой и легко цепи, но не содержат константные области.
Используемый в описании настоящей патентной заявки термин «одноцепочечный Fv» или «scFv» антитела относится к фрагментам антител, содержащим VH и VL домены антител, где эти домены присутствуют в единственной полипептидной цепи. Как правило, полипептидный Fv дополнительно содержит полипептидный линкер между VH и VL доменами, которые позволяют scFv образовывать заданную структуру для антиген-связывания. [Смотрите, Pluckthun, THE PHARMACOLOGY OF MONOCLONAL ANTIBODIES, vol. 113 Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994); WO 88/01649; и США 4,946,778 и США 5,260,203.]
Используемый в описании настоящей патентной заявки термин «каноническая структура» относится к локальной конформации, которая может быть адаптирована для каждой гипервариабельной области тяжелой и легкой цепи антитела в каркасе, в котором они находятся. Для каждой гипервариабельной области существует небольшое число канонических структур (как правило, обозначаются простыми числами, таким как 1 или 2, и тому подобными), которое может быть спрогнозировано с высокой точностью по аминокислотным последовательностям соответствующей гипервариабельной области (в частности в контексте аминокислотной последовательности этого каркаса, как приведено ниже для соответствующих вариабельных доменов канонизированного мышиного антитела против собачьего PD-1). Эти канонические структуры могут быть определяющими относительно того, приведет ли модификация аминокислотной последовательности заданной CDR в результате к сохранению или к потере способности связываться с его антиген-связывающим партнером [Смотрите, Chothia and Lesk, Canonical Structures for the hypervariable regions of immunoglobulins, J. Mol. Biol. 196:901-917(1987); Chothia et al., Conformation of immunoglobulin hypervaribale regions, Nature, 34:877-883(1989); and Al-Lazikani et al., Standard Conformations for the canonical structures of immunoglobulins, J. Mol. Biol. 273:927-948 (1997)].
«Домен антитела» представляет иммунологически функциональный фрагмент иммуноглобулина, содержащий только вариабельную область тяжелой цепи или вариабельную область легкой цепи. В некоторых случаях, две или более VH области ковалентно связаны с пептидным линкером с образованием домена бивалентного антитела. Две VH области домена бивалетного антитела могут быть нацелены на идентичные или различные антигены.
«Бивалентное антитело» содержит два антигенсвязывающих участка. В некоторых случаях два сайта связывания обладают идентичной антиген-специфичностью. Однако бивалентные антитела могут быть биспецифическими (смотрите, ниже).
В некоторых вариантах воплощения настоящего изобретения моноклональные антитела по настоящему изобретению также включают однодоменные камелизованные антитела. [Смотрите, например, Muyldermans et al., Trends Biochem. Sci. 26:230 (2001); Reichmann et al., J. Immunol. Methods 231:25 (1999); WO 94/04678; WO 94/25591; U.S. 6,005,079]. В одном варианте воплощения настоящее изобретение обеспечивает однодоменные антитела, содержащие два VH домена с модификациями, таким образом, что образуются однодоменные антитела.
Используемый в описании настоящей патентной заявки термин «диатела» относится к малым фрагментам антител с двумя участками связывания антигенов, такие фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в той же самой полипептидной цепи (VH-VL или VL-VH). Использование линкера, который является слишком коротким, позволяет объединить два домена одной и той же цепи, домены вынуждены соединяться с комплементарными доменами другой цепи и создают два участка связывания антигенов. [Смотрите, EP 0 404 097 B1; WO 93/11161; and Holliger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993)]. Для обзора сконструированных вариантов антител [смотрите, Holliger and Hudson Nat. Biotechnol. 23:1126-1136 (2005)].
Как правило, антитело или антигенсвязывающий агент по настоящему изобретению сохраняет по меньшей мере 10% своей связывающей активности собачьего PD-1 (по сравнению с родительским антителом), когда активность выражается в молях. Предпочтительно антитело или антигенсвязывающий агент по настоящему изобретению сохраняет по меньшей мере 20%, 50%, 70%, 80%, 90%, 95% или 100% или более аффинности связывания собачьего антигена, (например, PD-1) по сравнению с родительским антителом. Также предполагается, что канинизированное антитело или антиген-связывающий фрагмент по настоящему изобретению могут включать консервативные или не консервативные аминокислотные замещения (указанные, как «консервативные варианты» или «варианты консервативных функций» антитела), которые по существу не изменяют их биологическую активность.
«Выделенное антитело» относится к статусу очистки и в таком контексте означает молекулу по существу свободную от других биологических молекул, таких как нуклеиновые кислоты, белки, липиды, углеводы или другой материал, такой как продукт распада клеток и ростовая среда. Как правило, используемый в описании настоящей патентной заявки термин «выделенный» относится к полному отсутствию такого материала или к отсутствию воды, буферов или солей, если они не присутствуют в количествах по существу препятствующим экспериментальному или терапевтическому применению приведенного в описании настоящей патентной заявки связывающего соединения.
Используемый в описании настоящей патентной заявки термин «химерное антитело» относится к антителу, имеющему вариабельный домен от первого антитела и константный домен от второго антитела, где первое и второе антитела относятся к различным видам. [США 4,816,567; and Morrison et al., Proc. Natl. Acad. Sci. USA 81: 6851-6855 (1984)]. Как правило, вариабельные домены получают из антител от экспериментального животного («родительское антитело»), такого как грызун, а последовательности константного домена получают из антитела животного-субъекта, например, относящегося к собачьим, таким образом, полученное в результате химерное антитело будет с меньшей вероятностью вызывать нежелательный иммунный ответ у субъекта, относящегося к собачьим, по сравнению с родительским (например, грызуна) антителом.
Используемый в описании настоящей патентной заявки термин «канинизированное антитело» относится к формам антитела, которые содержат последовательности от обоих, и от относящегося к собачьим и от не относящегося к собачьим (например, относящиеся к мышиным) антителам. Традиционно канинизированное антитело содержит по существу все из, по меньшей мере один, и как правило, два вариабельных домена, в котором все или по существу все гипервариабельные петли, соответствуют таковым не собачьего иммуноглобулина (например, содержащего 6 CDRs мышиного антитела против собачьего PD-1, как приведено в качестве примера ниже), все или по существу все из каркаса, относящегося к собачьим.
Используемый в описании настоящей патентной заявки термин «полностью собачье антитело» относится к антителу, которое содержит только последовательности белка иммуноглобулина, относящегося к собачьим. Полностью собачье антитело может содержать углеводные цепи, в случае, когда они получены в мыши, в клетке мыши или в гибридоме, полученной из мышиной клетки. Аналогично «мышиное антитело» относится к антителу, которое содержит только последовательности мышиного иммуноглобулина. В качестве альтернативы, полностью собачье антитело может содержать крысиные углеводные цепи, в случае, когда они получены в крысе, в клетке крысы или в гибридоме, полученной из крысиной клетки. Аналогично «крысиное антитело» относится к антителу, которое содержит только последовательности крысиного иммуноглобулина.
Вариабельные области каждой пары легкой/тяжелой цепи образуют сайт связывания антитела. Следовательно, как правило, интактное антитело имеет два сайта связывания. За исключением бифункционального или биспецифического антитела, где два сайта связывания являются, как правило, идентичными.
Как правило, вариабельные домены обеих, и тяжелой и легкой цепи содержат три гипервариабельные области, которые также называются областями, определяющими комплементарность (CDRs), расположенными в относительно консервативных областях каркаса (FR). CDRs, как правило, фланкированы областями каркаса, позволяющими связываться со специфическим эпитопом. Как правило, от N-конца до C-конца вариабельные домены обеих, и легкой и тяжелой цепи содержат FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Соотнесение аминокислот к каждому домену для человеческого антитела, как правило, находится в соответствии с определениями Sequences of Proteins of Immunological Interest, Kabat, et al.; National Institutes of Health, Bethesda, Md. ; 5th ed.; NIH Publ. No. 91-3242 (1991); Kabat, Adv. Prot. Chem. 32:1-75 (1978); Kabat, et al., J. Biol. Chem. 252:6609-6616 (1977); Chothia, et al., J. Mol. Biol. 196:901-917 (1987) or Chothia, et al., Nature 342:878-883 (1989)].
Используемый в описании настоящей патентной заявки термин «гипервариабельная область» относится к аминокислотным остаткам антитела, которые ответственны за антиген-связывание. Гипервариабельная область содержит аминокислотные остатки из «области, определяющей комплиментарность» или «CDR» (то есть, CDRL1, CDRL2 и CDRL3, в вариабельном домене легкой цепи и CDRH1, CDRH2 и CDRH3 в вариабельном домене тяжелой цепи). [Смотрите, Kabat et al. Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), definining the CDR regions of a human antibody by sequence; see also Chothia and Lesk, J. Mol. Biol. 196: 901-917 (1987) defining the CDR regions of an antibody by structure]. Используемый в описании настоящей патентной заявки термин «каркасные» или «FR» остатки относится к таковым остаткам вариабельного домена иным, чем остатки гипервариабельной области, определенные в описании настоящей патентной заявки, как CDR остатки.
Используемый в описании настоящей патентной заявки термин «собачий каркас» относится к аминокислотной последовательности тяжелой цепи и легкой цепи собачьего антитела иной, чем остатки гипервариабельной области, определенные в описании настоящей патентной заявки, как CDR остатки. В обеих цепях аминокислотные последовательности нативных собачьих CDRs замещены соответствующими привнесенными CDRs (например, таковыми из мышиного антитела). Необязательно тяжелые и/или легкие цепи собачьего антитела могут содержать некоторые привнесенные не CDR остатки, например, сохраняя, таким образом, конформацию привнесенных в собачье антитело CDRs, и/или модифицируя Fc функцию, как приведено в качестве примера ниже.
Используемый в описании настоящей патентной заявки термин «антитело против собачьего PD-1» относится к антителу, которое было создано против собачьего PD-1 (у млекопитающих, таких как мышь или крыса) и которое специфически связывается с собачьим PD-1. Антитело, которое «специфически связывается с собачьим PD-1», или антитело, которое «специфически связывается с полипептидом, содержащим аминокислотную последовательность SEQ ID NO: 114», представляет антитело, которое демонстрирует преферентивное связывание с собачьим PD-1 по сравнению с другими антигенами, например, «специфическое» связывание с собачьим PD-1. Связывание не требует абсолютной специфичности связывания. Антитело против собачьего PD-1 считается «специфичным» к собачьему PD-1, если его связывание является определено присутствием в образце собачьего PD-1, если оно способно изменять активность собачьего PD-1 без излишнего препятствования активности других молекул в образце, относящемся к собачьим, например, без приведения к нежелательным результатам, таким как ложноположительный результат в диагностическом контексте или побочные эффекты в терапевтическом контексте. Степень специфичности, необходимая для антитела против собачьего PD-1, может зависеть от предполагаемого использования антитела, и по меньшей мере определяется его пригодностью для использования в предполагаемых целях.
Соответственно, настоящее изобретение обеспечивает канинизированное антитело против собачьего PD-1 или его антигенсвязывающие фрагменты (включая в выделенной форме), которые связываются с собачьим PD-1 (например, специфически), и применение такого антитела или его фрагментов. В конкретных вариантах воплощения настоящее изобретение обеспечивает CDRs мышиного антитела против собачьего PD-1 из мышиных антител против собачьего PD-1, которые продемонстрировали оба, и связывание с собачьим PD-1, и блокирование связывания собачьего PD-1 по меньшей мере с одним его лигандом, например, собачьим PD-L1. Эти CDRs могут быть вставлены в модифицированный собачий каркас по настоящему изобретению с получением канинизированного мышиного антитела против собачьего PD-1, как приведено в качестве примера ниже в описании настоящей патентной заявки.
В частности «канинизированное мышиное антитело против PD-1» по настоящему изобретению относится к антителу, которое содержит три CDRs тяжелой цепи и три CDRs легкой цепи с образованием мышиного антитела против собачьего PD-1 вместе с собачьим каркасом или модифицированным собачьим каркасом. Модифицированный собачий каркас содержит одно или более изменение аминокислоты, как приведено в качестве примера ниже в описании настоящей патентной заявки, что дополнительно оптимизирует эффективность канинизированного антитела, например, усилены, снижены или элиминированы эффекторные свойства антитела для усиления его связывания с собачьим антигеном, например, собачьим PD-1, и/или усиления его способности блокировать связывание собачьего антигена, например, собачьего PD-1 с его природным партнером связывания (например, собачий PD-L1 в случае, когда антиген представляет собачий PD-1).
«Гомология» относится к сходству последовательности между двумя полинуклеотидными последовательностями или между двумя полипептидными последовательностями, когда они оптимально выровнены. Когда позиция в обоих из двух, сравниваемых последовательностей, занята идентичными основаниями или аминокислотной мономерной субъединицей, например, в случае, когда позиция в каждой из двух молекул ДНК занята аденином, то молекулы являются гомологичными в этой позиции. Процент гомологии является числом гомологичных позиций, разделенным на две последовательности, деленным на общее число сравниваемых позиций ×100. Например, в случае, когда 6 из 10 позиций в двух последовательностях совпадают или гомологичны при оптимальном выравнивании последовательностей, то две последовательности гомологичны на 60%. Как правило, сравнение проводят, когда две последовательности выравнены с достижением максимального процента гомологии.
«Выделенная молекула нуклеиновой кислоты» относится к геномной ДНК или РНК, mРНК, cДНК или синтетического происхождения или некой их комбинации, которая не связана со всеми или с частью полинуклеотида, в котором выделенный полинуклеотид находится в природе, или связан с полинуклеотидом, с которым он не связан в природе. Для целей настоящего изобретения следует понимать, что «молекула нуклеиновой кислоты, содержащая» конкретную нуклеотидную последовательность, не входит в интактные хромосомы. Выделенные молекулы нуклеиновой кислоты, «содержащие» специфические последовательности нуклеиновой кислоты, могут включать дополнительно к специфическим последовательностям, кодирующие последовательности вплоть до десяти или даже вплоть до двадцати или более других белков или их частей или фрагментов, или могут включать функционально связанные регуляторные последовательности, которые контролируют экспрессию кодирующей области указанных последовательностей нуклеиновой кислоты, и/или могут включать векторные последовательности.
Используемый в описании настоящей патентной заявки термин «контрольные последовательности» относится к ДНК последовательностям, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Контрольные последовательности, подходящие для прокариотов, например, включают промотор, необязательно операторную последовательность и сайт связывания рибосом. Известно использование эукариотических клеток в качестве промоторов, сигналов полиаденирования полиаденилирования, и энхансеров.
Нуклеиновая кислота «функционально связана», когда она находится в функциональной связи с последовательностью другой нуклеиновой кислоты. Например, ДНК для предпоследовательности или секреторного лидера функционально связана с ДНК для полипептида, если он экспрессирован, как пребелок, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если это оказывает воздействие на транскрипцию последовательности; или сайт связывания рибосом функционально связан с кодирующей последовательностью, если он расположен таким образом, чтобы облегчить трансляцию. Как правило, «функционально связан» означает, что связанные ДНК последовательности являются перекрывающими sequences, и в случае секреторного линкера, перекрываются и в фазе считывания. Однако энхансеры не должны быть перекрывающимися. Связывание осуществляют лигированием в подходящих сайтах рестрикции. Если такие сайты отсутствуют, то используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с обычной практикой.
Используемые в описании настоящей патентной заявки термины «клетка», «клеточная линия» и «клеточная культура» взаимозаменяемы и все такие обозначения включают потомков. Следовательно, термины «трансформанты» и «трансформированные клетки» включают первичную клетку субъекта и культуры, полученные из нее, без учета количества переносов. Также понятно, что не все потомки будут иметь точное идентичное содержание ДНК в следствие преднамеренных или случайных мутаций. Включены мутировавшие потомки, которые имеют туже функцию или биологическую активность, как у прошедших отсев оригинально трансформированных клеток. Где предложены различные обозначения, будет ясно из контекста.
Используемый в описании настоящей патентной заявки термин «зародышевая последовательность» относится к не аранжированным последовательностям ДНК иммуноглобулина. Может быть использован любой подходящий источник не аранжированных последовательностей иммуноглобулина. Человеческие зародышевые последовательности могут быть получены, например, из баз данных зародышевых линий JOINSOLVER® на сайте the National Institute of Arthritis and Musculoskeletal and Skin Diseases of the United States National Institutes of Health. Мышиные зародышевые последовательности могут быть получены, например, как описано в Giudicelli et al. [Nucleic Acids Res. 33:D256-D261 (2005)].
Свойства канинизированного антитела
У относящихся к собачьим существует четыре тяжелые цепи IgG, указанные как A, B, C и D. Эти тяжелые цепи представляют четыре различных подкласса собачьего IgG, которые указаны, как IgGA, IgGB, IgGC и IgGD. ДНК и аминокислотные последовательности этих четырех тяжелых цепей были впервые идентифицированы Tang et al. [Vet. Immunol. Immunopathol. 80: 259-270 (2001)]. Аминокислотные последовательности и ДНК для этих четырех цепей также доступны из базы данных GenBank. Например, аминокислотная последовательность тяжелой цепи IgGТ имеет учетный номер AAL35301.1, IgGB имеет учетный номер AAL35302.1, IgGC имеет учетный номер AAL35303.1 и IgGD имеет учетный номер (AAL35304.1). Антитела собак также содержат два типа легких цепей, каппа и лямбда. Аминокислотная последовательность и ДНК этих легких цепей может быть получена из базы данных GenBank. Например, аминокислотная последовательность каппа легкой цепи имеет учетный номер ABY 57289.1, и лямбда легкой цепи имеет учетный номер ABY 55569.1. В настоящем изобретении аминокислотная последовательность для каждого из четырех Fc фрагментов собачьего IgG располагается на определенной границе доменов CH1 и CH2, как определено Tang et al., выше.
Разработка терапевтического моноклонального антитела представляет комплекс процессов, который влечет координацию совокупности действий для генерирования заданного антитела. Они включают оптимизацию специфичности антитела, аффиности, функциональной активности, уровня экспрессии в сконструированных клеточных линиях, долговременной стабильности, элиминации или усиления эффекторных функций и развитие коммерчески доступных способов получения и способов очистки. Принимая во внимание объекты настоящего изобретения независимо от способности активировать клетки иммунных систем человека, канинизированное или собачье моноклональное антитело против собачьего PD-1 оптимально имеет три дополнительных определяющих признака:
1. Отсутствие эффекторных функций, таких как антителозависимая цитотоксичность (ADCC) и комплиментзависимая цитотоксичность (CDC),
2. относительно длинный срок жизни in vivo; и
3. легкая очищаемость в большом масштабе при использовании промышленных стандартных технологий, таких как на основе хроматографии с белком A.
Ни один из встречающихся в природе подклассов собачьего IgG не удовлетворяет всем этим критериям. Например, IgGB может быть очищен при использовании белка A, но имеет высокий уровень ADCC активности. IgGC также имеет значительную ADCC активность. С другой стороны, IgGA слабо связывается с белком A, но имеет нежелательную ADCC активность. Дополнительно, ни IgGC, ни IgGD не могут быть очищены на колонках с белком A, хотя IgGD не имеет ADCC активности. Дополнительно IgGC имеет короткое время полужизни в кровяном русле, поскольку он не связывается с рецептором собачьего FcRn. Настоящее изобретение преодолевает эту трудность, обеспечивая модифицированное антитело собачьего IgG, специфическое к собачьим антигенам, например, собачьему PD-1; такое, как антитела с отсутствующими эффекторными функциями, такие как ADCC и CDC, и имеет относительно длинный срок полужизни, и может быть легко очищено при использовании промышленной стандартной хроматографии с белком A.
До настоящего времени не описывались генетически модифицированные собачьи IgGs, у которых отсутствуют обе, и DCC, и CDC эффекторные функции, и дополнительно они могут быть очищены при использовании хроматографии с белком A. Как приведено в описании настоящей патентной заявки, единственное замещение в позиции собачьего IgG, которое является аналогичным таковому в человеческом и мышином IgG, такое как N297A или D265A, не полностью элиминирует обе, и DCC, и CDC эффекторные функции в соответствующем собачьем антителе. Например, в то время, как каждое из замещений N297 и D265 в человеческом или мышином антителе в результате приводит к аннулированию связывания с Fcᵧ рецептором и C1q, ни одно замещение полностью не аннулирует связывание собачьего антитела с C1q. Вместо этого, как указано ниже, для элиминирования обоих, и DCC, и CDC в собачьем антителе подкаласса IgGB или IgGC доказана необходимость проведения двойного замещения в Fc собачьего антитела, комбинируя оба замещения, и аспарагин на аланин, и аспарагиновую кислоту на аланин. Дополнительно, совершенно неожиданно одно замещение, которое продемонстрировало снижение эффекторных функций в человеческом антителе, фактически привело к усилению связывания соответствующего собачьего IgG с FcᵧR и C1q.
Для получения вариантов собачьего IgGB и IgGC, не имеющих эффекторных функций, могут быть получены модифицированные тяжелые цепи собачьего IgGB или модифицированные тяжелые цепи собачьего IgGC. Всего для возможных замещений идентифицировано семь аминокислотных остатков, которые присутствуют в кристаллизуемых фрагментах обоих этих собачьих (cFcs). Эти семь аминокислотных остатков представляют: P4, D31, N63, G64, T65, A93 и P95 для обоих аминокислотных последовательностей SEQ ID NO: 130 для Fc собачьего IgGB; и аминокислотной последовательности SEQ ID NO: 132 для Fc собачьего IgGC. Соответственно, аминокислотная последовательность SEQ ID NO: 2 отличается от таковой SEQ ID NO: 130 наличием аминокислотных остатков в позициях: 4, 31, 63, 64, 65, 93 и 95, которые представляют пролин (P), аспарагиновую кислоту (D), аспарагин (N), глицин (G), треонин (T), аланин (A) и пролин (P), соответственно, в аминокислотной последовательности SEQ ID NO: 130 «X» (или «Xaa» в трехбуквенном коде) для всех семи позиций, означает, что эти семь аминокислотных позиций могут представлять любую из двадцати природных аминокислот (смотрите, в колонке 1 Таблицы 1 ниже). Аналогично, аминокислотная последовательность SEQ ID NO: 4 отличается от таковой SEQ ID NO: 132 наличием аминокислотных остатков в позициях 4, 31, 63, 64, 65, 93 и 95, приведенные как «X» (или «Xaa» в трехбуквенном коде) для всех семи позиций, обозначая эти семь аминокислотных позиций. Аминокислотная последовательность SEQ ID NO: 2 кодируется нуклеотидной последовательностью SEQ ID NO: 1, в то время аминокислотная последовательность SEQ ID NO: 4 кодируется нуклеотидной последовательностью SEQ ID NO: 3.
В одном варианте воплощения настоящего изобретения cFc содержит аминокислотную последовательность SEQ ID NO: 130 со следующими замещениями P4(A, G, или S), D31(A, G, или S) N63(A, G, или S), G64(A or P), T65(A, G, или S), A93(G или S) и P95(A, G, или S); в которой P4 (A G, или S) означает, что остаток пролина в позиции 4 замещен либо остатком аланина, глицина, либо остатком серина, и аналогично G64(P или A) означает, что остаток глицина в позиции 64 замещен либо остатком пролина, либо остатком аланина, и аналогичное им). В конкретном варианте воплощения настоящего изобретения cFc содержит аминокислотную последовательность SEQ ID NO: 130 со следующими замещениями: P4A, D31A, N63A, G64P, T65A, A93G и P95A.
В близком варианте воплощения настоящего изобретения cFc содержит аминокислотную последовательность SEQ ID NO: 4, которая содержит 7 аминокислот, обозначенных как Xaa следующими аминокислотными остатками: A4, A31, A63, G64, T65, G93, и 95, то есть аминокислотная последовательность SEQ ID NO: 132 со следующими пятью (5) изменениями аминокислотных остатков: P4A, D31A, N63A, A93G и P95A, сохранением двух аминокислотных остатков из семи, G64 и T65, неизменными из аминокислотной последовательности SEQ ID NO: 132.
Аминокислотные последовательности SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64и SEQ ID NO: 66 -все содержат «X» (или «Xaa» в трехбуквенном коде) в семи аминокислотных позициях, означая, что эти семь аминокислотных позиций могут представлять любую из двадцати природных аминокислот, приведеных в колонке 1 Таблицы 1 ниже. В частности, в соответствующих последовательностях SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64 и SEQ ID NO: 66 содержит либо аминокислотную последовательность SEQ ID NO: 2, либо таковую SEQ ID NO: 4. Конкретные примеры аминокислотных остатков в одной или более из этих семи позиций аминокислотных последовательностей приведены выше и ниже, следовательно, они включаются в семейство индивидуальных аминокислотных последовательностей SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64 и SEQ ID NO: 66, наряду с канинизированными антителами, которые содержат эти последовательности.
В приведенной ниже Таблице 10, в частности коррелируют семь аминокислотных позиций, как приведено в описании настоящей патентной заявки, Fc cIgGB (SEQ ID NO: 130 и SEQ ID NO: 2) и Fc cIgGC (SEQ ID NO: 132 и SEQ ID NO: 4) с таковыми полноразмерных тяжелых цепей собачьего, которые содержат эти аминокислотные последовательностей cFc, то есть, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, SEQ ID NO: 48, SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64 и SEQ ID NO: 66. Соответственно, фактическая позиция полноразмерной последовательности IgGB или IgGC может быть легко синхронизирована с таковой cFc, как приведено в Таблице 10, ниже.
В конкретных вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 40, 52, 56, или 64, содержащую (i) P, A, G или S в позиции 239, (ii) D, A, G или S в позиции 266, (iii) N, A, G или S в позиции 298, (iv) G, P или A в позиции 299, (v) T, A, G или S в позиции 300, (vi) A, G или S в позиции 328 и (vii) P, A, G или S в позиции 330. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 42, 54, 58 или 66, содержащую (i) P, A, G или S в позиции 237, (ii) D, A, G или S в позиции 264, (iii) N, A, G или S в позиции 296, (iv) G, P или A в позиции 297, (v) T, A, G или S в позиции 298, (vi) A, G или S в позиции 326и (vii) P, A, G или S в позиции 328. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 44, 50 или 60, содержащую (i) P, A, G или S в позиции 244, (ii) D, A, G или S в позиции 271, (iii) N, A, G или S в позиции 303, (iv) G, P или A в позиции 304, (v) T, A, G или S в позиции 305, (vi) A, G или S в позиции 333и (vii) P, A, G или S в позиции 335. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 46 или 62, содержащую (i) P, A, G или S в позиции 242, (ii) D, A, G или S в позиции 269, (iii) N, A, G или S в позиции 301, (iv) G, P или A в позиции 302, (v) T, A, G или S в позиции 303, (vi) A, G или S в позиции 331 и (vii) P, A, G или S в позиции 333. В других вариантах воплощения настоящего изобретения тяжелая цепь антитела содержит аминокислотную последовательность SEQ ID NO: 48, содержащую (i) P, A, G или S в позиции 246, (ii) D, A, G или S в позиции 273, (iii) N, A, G или S в позиции 305, (iv) G, P или A в позиции 306, (v) T, A, G или S в позиции 307, (vi) A, G или S в позиции 335 и (vii) P, A, G или S в позиции 337.
Настоящее изобретение также обеспечивает модифицированные собачьи IgGDs, которые содержат шарнирную область либо из IgGA, IgGB, либо из IgGC вместо природой шарнирной области IgGD. В качестве альтернативы, шарнирная область IgGD может быть генетически модифицирована замещением остатка серина на остаток пролина, как приведено в Таблице 5. Такие модификации приводят к тому, что у собачьго IgGD отсутствует изменение fab-области. Модифицированный собачий IgGDs может быть сконструирован при использовании стандартных способов технологии рекомбинантной ДНК [например, Maniatis et al., Molecular Cloning, A Laboratory Manual (1982)]. Для конструирования этих вариантов нуклеиновые кислоты, кодирующие аминокислотную последовательность собачьего IgGD, могут быть модифицированы, таким образом, что они будут кодировать модифицированные IgGDs. Модифицированные последовательности нуклеиновых кислот в таком случае клонируются в экспрессионных плазмидах для экспрессии белка. Нуклеиновые кислоты, кодирующие Fcs собачьего IgGD, с заменой шарнирной области в качестве примера представлены нуклеотидными последовательностями SEQ ID NO: 7, 9 и 11, которые кодируют аминокислотные последовательности SEQ ID NO: 8, 10 и 12. Нуклеиновая кислота, кодирующая Fc собачьего IgGD с модифицированной шарнирной областью IgGD, содержит нуклеотидную последовательность SEQ ID NO: 5, которая кодирует аминокислотную последовательность SEQ ID NO: 6.
Дополнительно настоящее изобретение обеспечивает полноразмерные собачьи тяжелые цепи, которые могут находиться в соответствии с соответствующими легкими цепями, с получением канинизированного антитела. Соответственно, дополнительно настоящее изобретение обеспечивает канинизированные мышиные антитела против собачьего антигена (включая выделенное канинизированное мышиное антитело против собачьего PD-1 антитела) и способы получения антитела или его антигенсвязывающих фрагментов для лечения заболеваний, например, лечения рака у относящихся к собачьим.
Дополнительно, настоящее изобретение обеспечивает канинизированное мышиное антитело против собачьего PD-1 или антигенсвязывающие фрагменты, которые связываются с собачьим PD-1 и блокируют связывание собачьего PD-1 с собачьим PD-L1. В конкретных вариантах воплощения настоящего изобретения канинизированное мышиное антитело против собачьего PD-1 содержит модифицированный Fc собачьего IgGB, модифицированный Fc собачьего IgGC, или у собачьего IgGD отсутствуют изменения fab-области, как приведено в описании настоящей патентной заявки.
Антитело или его антигенсвязывающий фрагмент, который связывается с собачьим антигеном, например, собачий PD-1, может содержать одну, две, три, четыре, пять или шесть областей, определяющих комплементарность (CDRs) мышиного антитела против собачьего, как приведено в описании настоящей патентной заявки. Одна, две, три, четыре, пять, или шесть CDRs могут быть независимо выбраны из CDR последовательностей, которые приведены ниже. В дополнительном варианте воплощения настоящего изобретения антитело или его антиген-связывающий фрагмент, который связывается с собачьим PD-1, содержит каппа легкую цепь собачьего антитела, содержащую CDR-1, CDR-2 и/или CDR-3 легкой цепи мышиного антитела, и тяжелую цепь собачьего IgG, содержащую CDR-1, CDR-2 и/или CDR 3 тяжелой цепи мышиного антитела. Соответственно, дополнительно настоящее изобретение обеспечивает полноразмерные собачьи тяжелые цепи, которые могут находиться в соответствии, например, с соответствующими легкими цепями, с получением канинизированного антитела [смотрите, Таблица 2 ниже, в которой приведены последовательности семи CDRs мышиного антитела против собачьего PD-1, например, 1B5, 2G9, 2H9, 3B6, 4D12, 5G5 и 7C9].
В других вариантах воплощения настоящее изобретение обеспечивает антитела или их антигенсвязывающие фрагменты, которые связываются специфически с PD-1 и имеют каппа легкие цепи собачьего антитела, содержащие от одной до шести различных CDRs, содержащих последовательность по меньшей мере на 80%, 85%, 90%, 95%, 98% или 99% идентичную аминокислотным последовательностям SEQ ID NOs: 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 и/или 26 и тяжелые цепи собачьего IgG, содержащие от одной до шести CDRs, содержащих последовательность по меньшей мере на 80%, 85%, 90%, 95%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NOs: 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38 и/или 146, продолжая при этом демонстрировать заданное связывание и функциональные свойства. В другом варианте воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент по настоящему изобретению содержит собачий каркас, содержащий комбинацию последовательности тяжелой цепи IgG с каппа легкой цепью, имеющей одну или более аминокислотную последовательность с 0, 1, 2, 3, 4 или 5 консервативными или не консервативными аминокислотными замещениями указанной выше CDR, продолжая при этом демонстрировать заданное связывание и функциональные свойства.
Идентичность последовательности относится к степени, до которой аминокислоты двух полипептидов идентичны в эквивалентных позициях, когда две последовательности оптимально выравнены. Используемый в описании настоящей патентной заявки термин одна аминокислотная последовательность на 100% «идентична» второй аминокислотной последовательности относится к случаю, когда аминокислотные остатки обеих последовательностей идентичны. Соответственно, аминокислотная последовательность на 50% «идентична» второй аминокислотной последовательности в случае, когда 50% аминокислотных остатков двух аминокислотных последовательностей идентичны. Сравнение последовательности проводят через перекрывающийся блок аминокислотных остатков, содержащихся в данном белке, например, сравниваемый белок или часть полипептида. В конкретном варианте воплощения настоящего изобретения во внимание принимаются выбранные делеции или вставки, которые могли бы в противном случае изменить сходство между двумя аминокислотными последовательностями.
Сходство последовательности включает идентичные остатки и не идентичные биохимически сходные аминокислоты. Биохимически сходные представляют таковые со сходными свойствами и взаимозаменяемые.
«Консервативно модифицированные варианты» или «консервативные замещения» относится к замещениям аминокислот в белке другими аминокислотами, имеющими сходные характеристики (например заряд, размер боковой цепи, гидрофобность/гидрофильность, конформация и жесткость скелета и, тому подобное), таким образом, что изменения могут быть часто сделаны без изменения биологической активности белка. Специалисту в области техники, к которой относится настоящее изобретение, известно, что как правило, одиночные аминокислотные замещения в несущественных областях полипептида по существу не изменяют биологическую активность [смотрите, например, Watson et al., Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th Ed.; 1987)]. Дополнительно, замещения структурно или функционально сходных аминокислот с меньшей вероятностью разрушить биологическую активность. Примеры консервативных замещений приведены в Таблице I непосредственно ниже.

Консервативный вариант функции антитела по настоящему изобретению также входит в объем притязаний настоящего изобретения. Используемый в описании настоящей патентной заявки термин «консервативный вариант функции» относится к антителу или фрагментам, в которых один или более аминокислотный остаток заменен без изменения заданного свойства, такого как аффиность и/или специфичность антигена, такие варианты включают без ограничения замену аминокислоты на таковую, имеющую сходные свойства, такую как консервативные аминокислотные замещения, приведенные в Таблице I выше.
Нуклеиновые кислоты
Дополнительно, настоящее изобретение включает нуклеиновые кислоты, кодирующие цепи иммуноглобулина канинизированного мышиного антитела против собачьего PD-1 и его антигенсвязывающие фрагменты, как приведено в описании настоящей патентной заявки (смотрите, Примеры ниже).
Также настоящее изобретение включает нуклеиновые кислоты, которые кодируют полипептиды иммуноглобулина, содержащие аминокислотные последовательности, которые по меньшей мере на около 70% идентичны, предпочтительно по меньшей мере на около 80% идентичны, более предпочтительно по меньшей мере на около 90% идентичны и наиболее предпочтительно по меньшей мере на около 95% идентичны (например, 95%, 96%, 97%, 98%, 99%, 100%) аминокислотным последовательностям CDRs и/или собачьих cFc's и/или антител по настоящему изобретению, когда сравнение проводят при использовании алгоритма BLAST, где параметры алгоритма, выбраны, таким образом, чтобы дать наибольшее совмещение между соответствующими последовательностями по всей длине соответствующих референсных последовательностей. Дополнительно, настоящее изобретение обеспечивает нуклеиновые кислоты, которые кодируют полипептиды иммуноглобулина, содержащие аминокислотные последовательности, которые по меньшей мере на около 70% сходны, предпочтительно по меньшей мере на около 80% сходны, более предпочтительно по меньшей мере на около 90% сходны и наиболее предпочтительно по меньшей мере на около 95% сходны (например, 95%, 96%, 97%, 98%, 99%, 100%) с любой референсной аминокислотной последовательностью, когда сравнение проводят при использовании алгоритма BLAST, где параметры алгоритма выбраны, таким образом, чтобы дать наибольшее совмещение между соответствующими последовательностями по всей длине соответствующих референсных последовательностей.
Используемый в описании настоящей патентной заявки термин процент идентичности нуклеотидной аминокислотной последовательности может быть определен при использовании C, MacVector (MacVector, Inc. Cary, NC 27519), Vector NTI (Informax, Inc. MD), Oxford Molecular Group PLC (1996) и алгоритма Clustal W с используемыми по умолчанию параметрами выравнивания для идентичности. Эти коммерчески доступные программы также могут быть использованы для определения сходства последовательности при использовании идентичных или аналогичных параметров по умолчанию. В качестве альтернативы, может быть использован поиск Advanced Blast на условиях фильтрации по умолчанию, например, при использовании GCG (Genetics Computer Group, Program Manual for the GCG Package, Version 7, Madison, Wisconsin) pileup program при использовании параметров по умолчанию.
Следующие ссылки относятся к часто используемым для анализа последовательностей алгоритмам BLAST: BLAST ALGORITHMS: Altschul, S.F., et al., J. Mol. Biol. 215:403-410 (1990); Gish, W., et al., Nature Genet. 3:266-272 (1993); Madden, T.L., et al., Meth. Enzymol. 266:131-141(1996); Altschul, S.F., et al., Nucleic Acids Res. 25:3389-3402 (1997); Zhang, J., et al., Genome Res. 7:649-656 (1997); Wootton, J.C., et al., Comput. Chem. 17:149-163 (1993); Hancock, J.M. et al., Comput. Appl. Biosci. 10:67-70 (1994); ALIGNMENT SCORING SYSTEMS: Dayhoff, M.O., et al., "A model of evolutionary change in proteins." in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3. M.O. Dayhoff (ed.), pp. 345-352, (1978); Natl. Biomed. Res. Found., Washington, DC; Schwartz, R.M., et al., "Matrices for detecting distant relationships." in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3." (1978), M.O. Dayhoff (ed.), pp. 353-358 (1978), Natl. Biomed. Res. Found., Washington, DC; Altschul, S.F., J. Mol. Biol. 219:555-565 (1991); States, D.J., et al., Methods 3:66-70(1991); Henikoff, S., et al., Proc. Natl. Acad. Sci. USA 89:10915-10919 (1992); Altschul, S.F., et al., J. Mol. Evol. 36:290-300 (1993); ALIGNMENT STATISTICS: Karlin, S., et al., Proc. Natl. Acad. Sci. USA 87:2264-2268 (1990); Karlin, S., et al., Proc. Natl. Acad. Sci. USA 90:5873-5877 (1993); Dembo, A., et al., Ann. Prob. 22:2022-2039 (1994); and Altschul, S.F. "Evaluating the statistical significance of multiple distinct local alignments." in Theoretical and Computational Methods in Genome Research (S. Suhai, ed.), pp. 1-14, Plenum, New York (1997).
Также настоящее изобретение обеспечивает векторы экспрессии, содержащие нуклеиновые кислоты (включая выделенные нуклеиновые кислоты) по настоящему изобретению, где нуклеиновая кислота функционально связана с контрольными последовательностями, которые распознаются клеткой-хозяином, когда трансфецируются вектором. Также обеспечены клетки-хозяева, содержащие вектор экспрессии по настоящему изобретению, и способы получения антитела или его антигенсвязывающего фрагмента, как приведено в описании настоящей патентной заявки, включающие культивирование клетки-хозяина, содержащей вектор экспрессии, кодирующий антитело или антигенсвязывающий фрагмент, в культуральной среде, и выделение антигена или его антигенсвязывающего фрагмента из клетки-хозяина или культуральной среды.
Канинизированное мышиное антитело против собачьего PD-1 может быть получено, например, рекомбинантно при использовании способов, известных из предшествующего уровня техники. Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии антитела или фрагментов по настоящему изобретению, хорошо известны из предшествующего уровня техники и включают множество иммортализованных клеточных линий, доступных от American Type Culture Collection (ATCC). Среди прочего они включают клетки яичника китайского хомячка (CHO), NSO, SP2 клетки, HeLa клетки, клетки почек новорожденного хомячка (BHK), клетки почек обезьяны (COS), клетки злокачественной гепатомы человека (например, Hep G2), A549 клетки, 3T3 клетки, HEK-293 клетки и множество других клеточных линий. Клетки-хозяева млекопитающих включают клетки человека, мышей, крыс, собак, обезьян, свиней, коз, жвачных животных, лошадей и хомяков. По существу предпочтительно клеточные линии выбирают, определяя, какие клеточные линии имеют высокие уровни экспрессии. Другие клеточные линии, которые могут быть использованы, представляют клеточные линии насекомых, такие как Sf9 клетки, клетки земноводных, бактериальные клетки, клетки растений и клетки грибов. В случае когда рекомбинантные векторы экспрессии, кодирующие тяжелую цепь или ее часть или антиген-связывающий фрагмент, легкую цепь и/или ее антиген-связывающий фрагмент вводят в клетки-хозяева млекопитающих, антитела получают культивированием клеток-хозяев в течение периода времени, достаточного для экспрессии антитела в клетках-хозяевах, или более предпочтительно секрецией антитела в культуральную среду, в которой выращивают клетки-хозяева.
Антитела могут быть выделены из культуральной среды при использовании стандартных методов очистки белка. Дополнительно, экспрессия антитела по настоящему изобретению (или других его фрагментов) из продуктивных клеточных линий может быть усилена при использовании множества известных технологий. Например, система генной экспрессии с глутаминсинтазой (GS система) представляет распространенный подход для усиления экспрессии при определенных условиях. GS система описана в целом или частично в европейских патентах № 0 216 846, 0 256 055и 0 323 997 и в европейской патентной заявкой № 89303964.4.
Как правило, гликобелки, полученные в определенных линиях клеток или от трансгенного животного, будут иметь профиль гликозилирования характерный для гликобелков, полученных в клеточной линии или от трансгенного животного. Следовательно, конкретный профиль гликозилирования будет зависеть от конкретной, используемой для получения клеточной линии или трансгенного животного. Однако, все антитела, кодируемые молекулами нуклеиновой кислоты по настоящему изобретению или содержащие аминокислотные последовательности по настоящему изобретению, входят в объем настоящего изобретения независимо от профиля гликозилирования, который имеет антитело. Аналогично, в конкретных вариантах воплощения настоящего изобретения антитела с профилем гликозилирования, содержащие только не фукозилированные N-гликаны, могут быть преимущественными, поскольку эти антитела показали, как правило, большую эффективность по сравнению с их фукозилированными аналогами, и in vitro, и in vivo [Смотрите, например, Shinkawa et al., J. Biol. Chem. 278: 3466-3473 (2003); патенты США №№ 6,946,292 и 7,214,775].
Дополнительно, настоящее изобретение включает фрагменты антител канинизированного мышиного антитела против собачьего PD-1 по настоящему изобретению. Фрагменты антитела включают F(ab)2 фрагменты, которые могут быть получены ферментативным расщеплением IgG, например, пепсином. Fab фрагменты могут быть получены, например, разделением F(ab)2 дитиотриэтолом или меркаптоэтиламином. Fab фрагмент представляет VL-CL цепь, присоединенную к VH-CH1 цепи дисульфидным мостиком. F(ab)2 фрагмент представляет два Fab фрагмента, которые в свою очередь прикреплены двумя дисульфидными мостиками. Fab часть F(ab)2 молекулы включает часть Fc области, между которыми расположены дисульфидные мостики. FV фрагмент представляет VL или VH область.
В одном варианте воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент содержит константную область тяжелой цепи, например, собачью константную область, такую как константная область тяжелой цепи собачьего IgGA, IgGB, IgGC и IgGD или ее вариант. В другом варианте воплощения настоящего изобретения антитело или антигенсвязывающий фрагмент содержит константную область легкой цепи, например, константную область собачей легкой цепи, такую как область лямбда или каппа собачей легкой цепи или ее вариант. Приведенная в качестве примера и не ограничивающая константная область собачей тяжелой цепи может представлять IgGB, а константная область собачей легкой цепи может представлять каппа.
Конструирование антитела
Канинизированное мышиное антитело против собачьего PD-1 по настоящему изобретению может быть сконструировано, таким образом, чтобы включать модификации в собачьем каркасе родительского (то есть относящийся к собачьим) моноклонального антитела, например, для улучшения свойств антитела, как детально описано ниже.
Приведенное в описании настоящей патентной заявки перекрестное блокирование канинизированного антитела и его антиген-связывающих фрагментов может быть определено на основе их способности перекрестно конкурировать с любым из IB5, 3B6, 4D12, 7C9, 2H9, 5G5, и/или 2G9 или при использовании стандартного анализа связывания (например, BIACore®, ELISA, как приведено в качестве примера ниже, или проточной цитометрии). Например, может быть использован стандартный анализ ELISA, в котором рекомбинантный собачий белок PD-1 иммобилизован на пластине, одно из антител флуоресцентно мечено и оценивается способность не меченного антитела конкурировать за связывание с меченным антителом. Дополнительно или в качестве альтернативы, может быть использован BIAcore® анализ для оценки способности антитела перекрестно конкурировать. Способность тестируемого антитела ингибировать связывание, например, IB5, 3B6, 4D12, 7C9, 2H9, 5G5 и/или 2G9 с собачьим PD-1 демонстрирует, что антитело может конкурировать с IB5, 3B6, 4D12, 7C9, 2H9, 5G5 и/или 2G9 за связывание с собачьим PD-1 и, следовательно, может в некоторых случаях связываться с тем же самым эпитопом на собачьем PD-1 как и IB5, 3B6, 4D12, 7C9, 2H9, 5G5 и/или 2G9. Как указано выше, антитела и фрагменты, которые связываются с одним и тем же эпитопом, как и любое из антител против собачьего PD-1 или фрагментов по настоящему изобретению, также является частью настоящего изобретения.
Фармацевтические композиции и введение
Для получения фармацевтических или стерильных композиций канинизированное мышиное антитело против собачьего PD-1 или его антигенсвязывающий фрагмент оно может быть смешано с фармацевтически приемлемым носителем или наполнителем. [Смотрите, например, Remington's Pharmaceutical Sciences и США Pharmacopeia: National Formulary, Mack Publishing Company, Easton, PA (1984)].
Составы терапевтических и диагностических агентов могут быть получены смешиванием с приемлемыми носителями, наполнителями или стабилизаторами в форме, например, лиофилизированных порошков, дисперсий, водных растворов или суспензий [смотрите, например, Hardman, et al. (2001) Goodman and Gilman's The Pharmacological Basis of Therapeutics, McGraw-Hill, New York, NY; Gennaro (2000) Remington: The Science and Practice of Pharmacy, Lippincott, Williams and Wilkins, New York, NY; Avis, et al. (eds.) (1993) Pharmaceutical Лекарственные формы: Parenteral Medications, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Лекарственные формы: Tablets, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Лекарственные формы: Disperse Systems, Marcel Dekker, NY; Weiner and Kotkoskie (2000) Excipient Toxicity and Safety, Marcel Dekker, Inc., New York, NY]. В одном варианте воплощения настоящего изобретения антитело против PD-1 по настоящему изобретению разводят до подходящей концентрации в растворе ацетата натрия pH 5-6 и для тоничности добавляют NaCl или сахарозу. Для усиления стабильности могут быть добавлены дополнительные агенты, такие как полисорбат 20 или полисорбат 80.
Токсичность и терапевтическая эффективность композиций антитела, вводимых как таковых или в комбинации с другим агентом, может быть определена при использовании стандартных фармацевтических процедур на клеточных культурах или экспериментальных животных, например, для определения LD50 (летальной дозы для 50% популяции) и ED50 (терапевтически эффективной дозы для 50% популяции). Соотношение дозы между токсическим и терапевтическим эффектом представляет терапевтический индекс (LD50/ ED50). В конкретных аспектах желательными являются антитела, демонстрирующие высокие терапевтические индексы. Данные, полученные из исследований клеточных культур и исследований на животных, могут быть использованы для составления диапазона доз для применения у животных, относящийся к собачьим. Предпочтительно доза таких соединений составляет в пределах циркулирующей концентрации, которая включает ED50 с малой токсичностью или без токсичности. Дозировка может варьировать в этих пределах в зависимости от используемой формы дозировки и способа введения.
Способ введения может варьировать. Подходящие способы введения включают оральный, ректальный, трансмукозальный, интестинальный, парентеральный, внутримышечный, подкожный, внутрикожный, интрамедуллярно, внутриоболочечно, напрямую внутрижелудочково, внутривенно, внутрибрюшинно, интраназально, интраокулярно, ингаляцией, инсуффляцией, местным нанесением, накожно, трансдермально или внутриартериально.
В конкретных вариантах воплощения настоящего изобретения канинизированное мышиное антитело против собачьего PD-1 или его антигенсвязывающий фрагмент может быть введен инвазивным способом, таким как инъекция. В других дополнительных вариантах воплощения настоящего изобретения мышиное антитело против собачьего PD-1 или его антигенсвязывающий фрагмент, или его фармацевтическую композицию вводят внутривенно, подкожно, внутримышечно, внутриартериально, внутриопухолево или ингаляцией, аэрозольным введением. Введение не инвазивными способами (например, орально; например, в пилюлях, капсулах или таблетках) также входит в объем притязаний настоящего изобретения.
Фармацевтические композиции по настоящему изобретению также могут быть введены капельно. Примеры хорошо известных имплантов и модульных форм введения фармацевтических композиций включают: Патент США № 4,487,603, в котором описывается имплантируемый микроинфузионный насос для дозирования лекарства с контролируемой скоростью; Патент США № 4,447,233, в котором описывается инфузионный насос для лекарственного средства для доставки лекарственного средства с точной скоростью инфузии; Патент США № 4,447,224, в котором описывается имплантируемое инфузионное устройство с регулятором расхода для непрерывной доставки лекарственного средства; Патент США № 4,439,196, в котором описывается осмотическая система доставки с камерой с множеством отделений. Многие другие такие импланты, системы доставки и модули хорошо известны специалисту в области техники, к которой относится настоящее изобретение.
В качестве альтернативы, введение канинизированного мышиного антитела против собачьего PD-1 может быть осуществлено локально а не системно, например, инъекцией антитела непосредственно в артритный сустав или патоген-индуцированное поражение, характеризующееся иммунопаталогией, часто в форме депо или состава с замедленным высвобождением. Дополнительно, можно вводить антитело в системе целевой доставки лекарственного средства, например, ткане-специфическое антитело с липосомальным покрытием, нацеленное, например, на артритный сустав или патоген-индуцированное поражение, характеризующееся иммунопатологией. Липосомы могут быть нацелены или селективно поглощены пораженными тканями.
Режим введения зависит от нескольких факторов, включая скорость метаболизма терапевтического антитела в сыворотке или тканях, уровень симптомов, иммуногенность терапевтического антитела и доступность целевых клеток в биологической матрице. Предпочтительно режим введения позволяет доставить достаточно терапевтических антител для достижения улучшения целевой стадии заболевания, при этом одновременно минимизируя нежелательные побочные эффекты. Соответственно, биологически доставленное количество зависит частично от конкретного терапевтического антитела и тяжести лечимого состояния. Доступно руководство по выбору подходящих доз терапевтического антитела [смотрите, например, Wawrzynczak Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK (1996); Kresina (ed.) Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, NY (1991); Bach (ed.) Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, NY (1993); Baert, et al. New Engl. J. Med. 348:601-608 (2003); Milgrom et al. New Engl. J. Med. 341:1966-1973 (1999); Slamon et al. New Engl. J. Med. 344:783-792 (2001); Beniaminovitz et al. New Engl. J. Med. 342:613-619 (2000); Ghosh et al. New Engl. J. Med. 348:24-32 (2003); Lipsky et al. New Engl. J. Med. 343:1594-1602 (2000)].
Определение подходящей дозы проводится ветеринаром, например, при использовании параметров или факторов, влияющих на лечение, известных или предполагаемых в области техники, к которой относится настоящее изобретение. Как правило, доза начинается с критерии включают таковые количества немного меньшего, чем оптимальная доза, и повышается при малом шаге приращения до достижения заданного или оптимального эффекта относительно любых негативных побочных эффектов. Важные диагностические симптомов, например, воспаление или уровень продуцированных воспалительных цитокинов.
Антитела или их антигенсвязывающие фрагменты по настоящему изобретению могут быть обеспечены непрерывной инфузией или введением доз, например, ежедневно, 1-7 раз в неделю, еженедельно, дважды в неделю, ежемесячно, дважды в месяц, ежеквартально, раз в полгода, ежегодно и тому подобное. Дозы могут быть обеспечены, например, внутривенно, подкожно, местным нанесением, орально, назально, ректально, внутримышечно, интрацеребрально, интраспинально или ингаляцией. Общая недельная доза, как правило, составляет по меньшей мере 0,05 μг/кг массы тела, более распространено по меньшей мере 0,2 μг/кг, 0,5 μг/кг, 1 μг/кг, 10 μг/кг, 100 μг/кг, 0,25 мг/кг, 1,0 мг/кг, 2,0 мг/кг, 5,0 мг/мл, 10 мг/кг, 25 мг/кг, 50 мг/кг или более [смотрите, например, Yang, et al. New Engl. J. Med. 349:427-434 (2003); Herold, et al. New Engl. J. Med. 346:1692-1698 (2002); Liu, et al. J. Neurol. Neurosurg. Psych. 67:451-456 (1999); Portielji, et al. Cancer Immunol. Immunother. 52:133-144 (2003)]. Также дозы могут быть обеспечены для достижения заранее определенной целевой концентрации канинизированного мышиного антитела против собачьего PD-1 в сыворотке субъекта, такой как 0,1, 0,3, 1, 3, 10, 30, 100, 300 μг/мл или более. В других вариантах воплощения настоящего изобретения канинизированное мышиное антитело против собачьего PD-1 по настоящему изобретению вводят подкожно или внутривенно, еженедельно, дважды в неделю, «каждые 4 недели», ежемесячно, дважды в месяц или ежеквартально из расчета 10, 20, 50, 80, 100, 200, 500, 1000 или 2500 мг/субъекту.
Используемый в описании настоящей патентной заявки термин «ингибирование» или «терапия», или «лечение» включает отсрочку развития симптомов, связанных с заболеванием, и/или снижение тяжести симптомов такого заболевания. Используемые в описании настоящей патентной заявки термины дополнительно включают облегчение существующих неконтролируемых или нежелательных симптомов, предотвращение дополнительных симптомов и облегчение или предотвращение первоначальных причин таких симптомов. Следовательно, используемые в описании настоящей патентной заявки термины обозначают, что положительный результат достигнут у позвоночного субъекта с заболеванием или болезнью, или симптомом, или с потенциальном для развития такого заболевания, болезни или симптома.
Используемые в описании настоящей патентной заявки термины «терапевтически эффективное количество», «терапевтически эффективная доза» и «эффективное количество» относятся к количеству канинизированного мышиного антитела против собачьего PD-1 или его антигенсвязывающих фрагментов по настоящему изобретению, которое при введении как такового или в комбинации с дополнительным терапевтическим агентом в клетку, ткань или субъекту, эффективно для достижения измеряемого улучшения одного или более симптома заболевания или состояния, или прогрессирования такого заболевания или состояния. Дополнительно, терапевтически эффективная доза относится к такому количеству связывающего соединения, которое достаточно для достижения по меньшей мере частичного ослабления симптомов, например, лечение, излечение, профилактика или улучшение релевантного медицинского состояния, или повышение скорости лечения, излечения, профилактики или ослабления таких состояний. При применении введением только индивидуального активного ингредиента, терапевтически эффективная доза относится к ингредиенту как таковому. При применении введением в комбинации, терапевтически эффективная доза относится к комбинированным количествам активных ингредиентов, которые приводят к терапевтическому эффекту, независимо от введения в комбинации периодически или одновременно. Терапевтически эффективное количество приводит к улучшению диагностичекого критерия или параметра по меньшей мере на 10%; как правило по меньшей мере на 20%; предпочтительно по меньшей мере на около 30%; более предпочтительно по меньшей мере 40% и наиболее предпочтительно по меньшей мере на 50%. Эффективное количество также может привести к улучшению субъективной оценки в случае, когда субъективные оценки используют для оценки тяжести заболевания.
Другая комбинированная терапия
Как указано выше, канинизированное мышиное антитело против собачьего PD-1 или его антигенсвязывающий фрагмент могут быть введены совместно с одним или более другим терапевтическим агентом (таким как химиотерапевтический агент). Антитело может быть связано с агентом (как иммунокомплекс) или может быть введено отдельно от агента. В последнем случае (раздельное введение) антитело может быть введено перед, после или одновременно с агентом или может быть совместно введено с другими известными лекарственным средством.
Наборы
Дополнительно обеспечены наборы, содержащие один или более компонент, которые включают без ограничения антитело или антигенсвязывающий фрагмент по настоящей заявке, который специфически связывается с PD-1 (например, канинизированное мышиное антитело против собачьего PD-1 или его антигенсвязывающий фрагмент) в сочетании с одним или более дополнительным компонентом, включая без ограничения фармацевтически приемлемый носитель и/или химиотерапевтический агент, как указано выше. Связывающая композиция и/или химиотерапевтическая композиция может быть составлена, как чистая композиция или в комбинации с фармацевтически приемлемым носителем в фармацевтической композиции.
В одном варианте воплощения настоящего изобретения набор включает связывающую композицию по настоящему изобретению (например, канинизированное мышиное антитело против собачьего PD-1 или его фармацевтическая композиция в одном контейнере (например, в стерильной стеклянной или пластиковой пробирке) и его фармацевтическую композицию и/или химиотерапевтический агент в другом контейнере (например, в стерильной стеклянной или пластиковой пробирке).
В случае, когда набор содержит фармацевтическую композицию для парентерального введения субъекту, набор также содержит устройство для проведения такого введения. Например, набор может содержать одну или более гиподермальную иглу или другие устройства для инъекции, как указано выше. Также набор содержит листок-вкладыш, содержащий информацию о фармацевтической композиции и лекарственных формах в наборе. Как правило, такая информация помогает владельцу животного и ветеринарам эффективно и безопасно использовать укупоренные фармацевтические композиции и лекарственные формы. Например, на вкладыш может быть помещена следующая информация, относящаяся к комбинации по настоящему изобретению: фармакокинетика, фармакодинамика, клинические исследования, показатели эффективности, показания к применению, противопоказания, предостережения и меры предосторожности, побочные реакции, передозировка, правильная дозировка и введение, форма выпуска, надлежащие условия хранения, референсные препараты, информация о производителе/дистрибьюторе и патентная информация.
Для удобства в наборе может быть обеспечено антитело или агент специфического связывания, то есть в виде комплекта реагентов в заранее определенных количествах с инструкциями для проведения диагностического анализа или анализа на обнаружение. В случае, когда антитело метят ферментом, набор будет содержать субстраты и кофакторы, требуемые для фермента (например, субстрат-предшественник, который обеспечивает детектируемый хромофор или флуорофор). Дополнительно, могут быть включены другие добавки, такие как стабилизаторы, буферы (например, блокирующий или лизисный буфер) и аналогичное им. Относительные количества различных реагентов могут широко варьировать для обеспечения концентраций в растворе реагентов, которые по существу оптимизируют чувствительность анализа. В частности, реагенты могут быть обеспечены в форме сухих порошков, как правило лиофилизованных, включая наполнители, которые при растворении будут обеспечивать раствор реагентов интрацеребрально в подходящей концентрации.
ПРИМЕРЫ
ПРИМЕР 1
СОБАЧИЙ PD-1 И PD-L1
Собачий PD-1 и PD-L1:
Предварительная заявка на патент США № 61/918,946, поданная 20 декабря 2013, введенная здесь ссылкой в полном объеме обеспечивает: полноразмерную нуклеотидную последовательность для собачьего PD-1 (cPD-1) SEQ ID NO: 113 [SEQ ID NO: 133 включает сигнальную последовательность]; соответствующую транслированную аминокислотную последовательность SEQ ID NO: 114 [SEQ ID NO: 134 включает сигнальную последовательность]; нуклеотидную последовательность, кодирующую внеклеточный домен (ECD) собачьего PD-1, SEQ ID NO: 115; аминокислотную последовательность ECD собачьего PD-1, SEQ ID NO: 116; нуклеотидную последовательность собачьего PD-1 ECD плюс GT линкер и Fc часть человеческого гена Fc IgG1, SEQ ID NO: 117; и аминокислотную последовательность собачьего PD-1 ECD плюс GT линкер и Fc часть человеческого гена Fc IgG1, SEQ ID NO: 118 [SEQ ID NO: 137 включает сигнальную последовательность].
Предварительная заявка на патент США № 61/918,946 дополнительно обеспечивает: полноразмерную нуклеотидную последовательность для собачьего PD-L1 (cPD-L1) SEQ ID NO: 119 [SEQ ID NO: 135 включает сигнальную последовательность]; соответствующую транслированную аминокислотную последовательность SEQ ID NO: 120 [SEQ ID NO: 136 включает сигнальную последовательность]; нуклеотидную последовательность, кодирующую внеклеточный домен (ECD) собачьего PD-L1, SEQ ID NO: 121; аминокислотную последовательность ECD собачьего PD-L1, SEQ ID NO: 122; нуклеотидную последовательность собачьего PD-L1 ECD плюс GT линкер и Fc часть человеческого гена Fc IgG1, SEQ ID NO: 123; и аминокислотную последовательность собачьего PD-L1 ECD плюс GT линкер и Fc часть человеческого гена Fc IgG1, SEQ ID NO: 124.
ПРИМЕР 2
МЫШИНЫЕ АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО PD-1
Получение моноклонального антитела против собачьего PD1:
В общей сложности трех Balb/c мышей иммунизировали несколько раз (каждый раз 10 μг) в течение 17 дневного периода. Иммунизирующий антиген представлял собачий слитый белок PD-1 ECD-Fc. После иммунизации собирали сыворотку каждой мыши, проводили тестирование на реактивность к целевому собачьему белку PD-1 ECD-HIS. Клетки печени мышей с самым высоким титром сыворотки против PD-1 ECD-HIS слили с клеточной линией миеломы P3X63Ag8.653. Через около 2 недели после слития супернатант клеток предполагаемой гибридомы протестировали при использовании ELISA на реактивность к целевому белкку PD-1 ECD-HIS. Гибридомы, продуцирующие резко-положительные сигналы, в ELISA субклонировали серийным разведением и снова протестировали на реактивность к целевому белку собачьего PD-1 ECD-HIS.
Подтверждение реактивности моноклональных антител против собачьего PD-1:
Реактивность антитела, секретированного гибридомами к ECD собачьего PD-1 подтверждали при использовании ELISA. Гибридомные клетки культивировали при использовании биореакторов CELLine (Integra-biosciences) в течение 10-30 дней. Сначала клетки выдерживали в DMEM с добавлением 4 мM на л глутамина и 10% Ultra Low IgG фетальной бычьей сыворотки (FBS) от Gibco. Гибридомные клетки высеяли в клеточные камеры биореактора CELLine с плотностью клеток около 2×106 клетки/мл в 15 мл той же самой среды с концентрацией FBS, повышенной до 20%. Внешнюю камеру заполнили 1 л питательной среды (DMEM с 4 мM на л глутамина и 2% стандартной FBS). Гибридомные клетки в клеточной камере размножились до около 2,5×107 клеток/мл в течение 3-7 дней. Затем из клеточной камеры собрали 10 мл клеточной суспензии и поместили на свежую среду для последующего размножения клеток и затем провели сбор клеток. Эту процедуру повторяли по необходимости для получения адекватных количеств mAb от каждого клона гибридомы. Суспензии собранных клеток центрифугировали и фильтровали супернатанты через 0,2 микронную фильтрационную мембрану. Для очистки антитела супернатант каждого клона очистили при использовании Protein G Sepharose 4 Fast flow 5 мл колонки (GE Healthcare) самотеком. После промывки буфером трис ЭДТА (TE) pH 8,0, связанные антитела элюировали при использовании 0,1 M глицинового буфера, pH 2,7, с последующей нейтрализацией pH при использовании 1 M трис, pH 8,0. Антитела концентрировали и заменили буфер на фосфатно-солевой буфер (PBS) при использовании установок - фильтрующих центрифуг Centriprep® YM-10, 10 кДа NMWL (Ultracel-10 представляет собой ультрафильтрационную мембрану с NMWL 10000 кДа из регенерированной целлюлозы,) (Millipore). Концентрации антител оценивали при использовании спектрофотометрии.
Очищенные mAbs против собачьего PD-1 протестировали на реактивность к HIS-целевым ECD домену собачьего PD-1 при использовании ELISA следующим образом: HIS-целевой ECD собачьего PD-1 белка развели до 10 μг/мл в буфере для сенсибилизации поверхностей (Carbonate/Bicarbonate pH 9,0) и распределили в 100 μл/лунку 96-луночных плоскодонных планшетов ELISA (NUNC). Планшеты инкубировали при температуре 4°C в течение ночи. Затем планшеты промыли три раза фосфатно-солевым буфером, содержащим 0,05% Tween® 20 (PBST). Затем добавили 200 μл блокирующего буфера (5% обезжиренного молока в PBST) в каждую лунку и инкубировали при температуре 37°C в течение 60 минут. Затем планшеты промыли три раза PBST. Далее 100 μл тестируемых mAbs, разведенных в блокирующем буфере, добавили в первые лунки соответствующих колонок. Затем тестируемые mAbs двухкратно развели в соответствующем положении планшета. После инкубирования планшетов при температуре 37°C в течение 60 минут, планшеты промыли три раза PBST. Далее в планшеты добавили 100 μl на лунку 1:2,000 разведения козий пероксидазный конъюгат хрена против IgG мыши (KPL), которые затем инкубировали при температуре 37°C в течение 60 минут. Затем планшеты промыли три раза PBST и добавили в планшеты 100 μл/лунку 3,3',5,5' тетраметил бензидина, (TMB) субстрат (из KPL). Позволили развиться цветной реакции в течение 5-20 минут при температуре 37°C перед измерением поглощения при 650 нм.
CHO клетки, экспрессирующие собачий PD-1 белок:
Полноразмерный собачий PD-1 ген клонировали в плазмиде p96793. В этой плазмиде провели экспрессию PD-1 белка при использовании промотора hCMV. CHO DXB11 клетки (dhfr-) выдерживали в MEM-альфа (Gibco) с добавлением 10% фетальной бычьей сыворотки. Трансфецировали CHO клетки плазмидой p96793 в 75 см2 колбах, содержащих около 6×106 клеток, липосомно-опосредованной доставкой генов при использовании липофектамина (Invitrogen). Через 48 часов клетки переместили в MEM-альфа среду без нуклеотидов с добавлением 10% FBS и 400 μг/мл гидромицина B (селективная среда). Провели клонирование методом серийных разведений в пуле dhfr+ гидромицин резистентных клеток. Провели оценку клонов на экспрессию собачьего PD-1 при использовании иммунофлуоресцентного анализа. Кратко, монослои клеток зафиксировали в 96 луночных планшетах 80% ацетоном. Зафиксированные и высушенные монослои клеток затем инкубировали в течение 1 часа с поликлональным козьим антителом против человеческого PD-1 (R&D Systems). Планшеты промыли PBS, затем инкубировали в течение 1 часа с флуоресцентно-меченным кроличьим антителом против козьего IgG (KPL). Планшеты промыли PBS. Клоны, демонстрировавшие флуоресценцию, размножили с получением запаса клеток.
Реактивность мышиных mAbs против собачьего PD-1 белка, экспрессированного CHO клетками:
Реактивность мышиных mAbs против собачьего PD-1 к собачим PD-1 на CHO клетках определили исследованием на клетках при использовании CHO клеток, экспрессирующих PD-1. Кратко, CHO клетки, экспрессирующие собачий PD-1, культивировали до 80-100% заселенности в 50 μл среды (DMEM/HAM's F12, 10% FBS). Далее 50 μл среды, содержащей различные концентрации очищенных mAbs, добавили на 1 час при температуре 37°C. Затем промыли три раза PBS-Tween, 100 μл козьего пероксидазного конъюгата хрена против мыши (HRP), разведенного 1:1000 в культуральной среде, добавили на один час при температуре 37°C. После трех дополнительных промывок PBS-Tween, связанные mAbs визуализировали при использовании пероксидазного субстрата (TMB). Увеличение поглощения света из-за активности пероксидазы 450 нм измерили с использованием считывающего устройства для микропланшетов.
Характеристика мышиного антитела против собачьего PD-1:
Как указано выше, наряду с Предварительной заявкой на патент США № 61/918,946, поданной 20 декабря 2013, введенной здесь ссылкой в полном объеме, мышиное антитело против собачьего PD-1 характеризуется множеством параметров, включая: реактивность к ECD собачьего PD-1 при использовании ELISA, реактивность к PD-1, экспрессированному на поверхности CHO клетки, способность блокировать связывание PD-1 с PD-L1 и способность связываться с PBMC клетками от здоровой собаки и от собаки с раком. Выбрали аминокислотные последовательности CDRs семи мышиных антител против собачьего PD-1 (обозначенные, как IB5, 2G9, 2H9, 3B6, 4D12, 5G5 и 7C9, соответственно), обладавшие значительной гомологией, как приведено в Таблице 2 ниже.
ТАБЛИЦА 2
АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ CDRs
VL CDR1 SEQ ID NO.
1B5 Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala 13
2G9 Arg Ser Ser Gln Asn Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu 14
2H9 His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser 15
3B6 Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala 13
4D12 Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala 13
5G5 His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser 15
7C9 Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu Ala 13
VL CDR2
1B5 Phe Ala Ser Thr Arg Val Ser 16
2G9 Lys Val Ser Asn Arg Phe Ser 17
2H9 Lys Ala Ser His Leu His Thr 18
3B6 Phe Ala Ser Ala Arg Val Ser 19
4D12 Phe Ala Ser Thr Arg Ile Ser 20
5G5 Lys Ala Ser Asn Leu His Thr 21
7C9 Phe Ala Ser Thr Arg Val Ser 16
VL CDR3
1B5 Gln Gln Tyr Phe Ser Thr Pro Leu Thr 22
2G9 Phe Gln Gly Ser His Val Pro Tyr Thr 23
2H9 Gln Gln Gly Gln Ser Trp Pro Leu Thr 24
3B6 Gln Gln Tyr Phe Ser Thr Pro Leu Thr 25
4D12 Gln Gln Tyr Phe Ser Thr Pro Leu Thr 25
5G5 Gln Gln Gly Gln Ser Tyr Pro Leu Thr 26
7C9 Gln Gln Tyr Phe Ser Thr Pro Leu Thr 22
VH CDR1
1B5 Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser 27
2G9 Gly Tyr Thr Phe Thr Arg Tyr Asn Met His 28
2H9 Gly Phe Asn Ile Lys Asn Thr Tyr Met His 29
3B6 Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser 27
4D12 Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser 27
5G5 Gly Phe Asn Ile Lys Asn Thr Tyr Met His 29
7C9 Gly Phe Ser Leu Thr Ser Tyr Gly Val His 30
VH CDR2
1B5 Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys Gly 31
2G9 Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe Lys Gly 32
2H9 Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe Gln Gly 33
3B6 Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys Gly 31
4D12 Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe Lys Gly 34
5G5 Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe Gln Gly 35
7C9 Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys Gly 31
VHCDR3
1B5 Phe Asp Gly Pro Asp Tyr 36
2G9 Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr 37
2H9 Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val 38
3B6 Phe Asp Gly Pro Asp Tyr 36
4D12 Phe Asp Gly Pro Asp Tyr 36
5G5 Ile Phe Tyr Asp Tyr Asp Gly Asp Ile Asp Val 146
7C9 Phe Asp Gly Pro Asp Tyr 36
Канонические структуры (классы) для VH цепи CDRs
mAbs: 4D12, 3B6, 7C9 и 1B5: CDR: H1-1; CDR2: H2-1; CDR3: H3-6
mAb: 5G5: CDR: H1-1; CDR2: H2-1; CDR3: H3-11
mAb: 2H9 CDR: H1-1; CDR2: H2-2A; CDR3: H3-11
mAb: 2G9 CDR: H1-1; CDR2: H2-2A; CDR3: H3-13
Канонические структуры (классы) для VL цепи CDRs
mAbs: 4D12, 3B6, 7C9, 1B5: CDRL: L1-3; CDR2: L2-1; CDR3: L3-1
mAb: 5G5: CDR: L1-2A; CDR2: L2-1; CDR3:L3-1
mAb: 2H9 CDR: L1-2A; CDR2: L2-1; CDR3:L3-1
mAb: 2G9 CDR: L1-4; CDR2: L2-1; CDR3:L3-1
ПРИМЕР 3
КАНИНИЗАЦИЯ И ХАРАКТЕРИСТИКА КАНИНИЗИРОВАННОГО АНТИТЕЛА
Для получения канинизированных антител необходимо идентифицировать ДНК последовательность, кодирующую тяжелую и легкую цепи собачьего IgG. Нуклеотидные и аминокислотные последовательности собачьей тяжелой цепи могут быть получены из NCBI гена и базы данных белка. Существует четыре известных подкласса собачьего IgG: IgGA, IgGB, IgGC и IgGD и два типа легких цепей: каппа и лямбда. В Таблице 7 приведены аминокислотная и нуклеотидная SEQ ID NO не модифицированных Fc фрагментов собачьего антитела.
Не ограничиваясь каким-либо конкретным подходом получения вариантов моноклональные антитела против PD-1 с различным содержанием собачьих и мышиных последовательностей, способ включен в следующую общую схему:
i) Определение нуклеотидной последовательности VH и VL цепей мышиных mAbs;
ii) Идентификация CDRs H и L цепи мышиных mAbs;
iii) Идентификация подходящей H и L цепи собачьего IgG;
iv) Определение нуклеотидной последовательности H и L цепей собачьего IgG;
v) Замена нуклеотидной последовательности, кодирующей эндогенные CDRs собачьих H и L цепей, нуклеотидными последовательностями, кодирующими соответствующие мышиные CDRs. Также необязательная замена некоторых остатков собачьего каркаса на выбранные остатки областей мышиного каркаса;
vi) Синтез нуклеотида со стадии (v) и вставка в подходящую экспрессионную плазмиду; Трансфекция плазмид в подходящие клетки, например, HEK 293 клетки;
vii) Очистка экспрессированного антитела от супернатанта HEK 293; и
viii)Тестирование очищенного антитела на связывание с собачьим PD-1.
Для получения ряда вараинтов канинизированных антител с различным содержанием собачьих и мышиных последовательностей провели ряд экспериментов, следуя указанным выше стадиям.
Реактивность канинизированных mAbs против собачьего PD-1 белка, экспрессированного на CHO клетках:
Реактивность мышиных mAbs канинизированных собачим PD-1 против собачьего PD-1 на CHO клетках определили исследованием на клетках при использовании CHO клеток, экспрессирующих PD-1. Кратко, CHO клетки, экспрессирующие собачий PD-1, культивировали до 80-100% заселенности в 50 μл среды (DMEM/HAM's F12, 10% FBS). Далее 50 μл среды, содержащей различные концентрации очищенных mAbs, добавили на 1 час при температуре 37°C. Затем промыли три раза PBS-Tween, 100 μл козьего пероксидазного конъюгата хрена против мыши (HRP), разведенного 1:1000 в культуральной среде добавили на один час при температуре 37°C. После трех дополнительных промывок PBS-Tween, связанные mAbs визуализировали с использованием пероксидазного субстрата (TMB). Увеличение поглощения света из-за активности пероксидазы 450 нм измерили с использованием считывающего устройства для микропланшетов.
Исследование связывания мышиных mAbs против собачьего PD-1 и канинизированных собачим PD-1 мышиных mAbs против собачьего PD-1
Около 70 резонансных единиц (RU) антигена собачьего PD-1 иммобилизовали прямым аминным спариванием. Измерение аффиности провели на основе технологии свободного от мечения поверхностного плазмонного резонанса (например, Biacore® T200) со временем ассоциации 300 секунд, временем диссоциации 1200 секунд, при наномолярных концентрациях 50, 100, 200 (x2) 400 и 800 (нM). Использовали подбор модели связываний 1:1. Антиген (собачий PD-1) иммобилизовали на сенсорном чипе через аминное спаривание и в качестве аналитов использовали четыре антитела, как указано в Таблице 14 ниже, которые пропускали по поверхности антигена. Результаты показали, что аффиность связывания антитела против собачьего PD-1 по настоящему изобретению для собачьего PD-1 антигена -сильная, с наномолярной и даже субмолярной константами диссоциации (Kd). Дополнительно моноклональное мышиное антитело против собачьего PD-1 и соответствующее канинизированное моноклональное мышиное антитело против собачье PD-1 из одного и того же клона дали поразительно схожие показатели Kd (смотрите, Таблица 14 ниже).
Определение констант связывания
M-1с-1
M
(RU2)
Блокировка лиганда канинизированными mAbs против собачьего PD1:
Для канинизированного антитела, реагирующего с собачьим PD-1 (cPD-1), использовали анализ на основе клеточного ELISA (CELISA), который основывается на клеточной линии CHO, экспрессирующей собачий PD-1. Кратко, cPD-1 CHO клетки поместили в 96-луночные планшеты при 4x104 клеток на лунку и инкубировали клетки при температуре 37°C в течение 18-24 часов до достижения 95-100% заселенности. Культуральную среду удалили отсасыванием, планшеты промыли 3x PBS плюс 0,05% Tween® 20 и 1x CHO средой. Получили 3-кратные серийные разведения канинизированных mAbs против cPD1 в CHO среде, и добавили в планшет, начиная с 30 μг/мл и 50 μл/лунку каждого разведения антитела. Затем планшеты инкубировали при температуре 37°C, 5% CO2 в течение 30 минут, при встряхивании. Добавили человеческий PD-L1-Fc в 4 μг/мл в CHO среду, 50 μл/лунку без удаления или промывки инкубированных mAbs против PD1, затем инкубировали при температуре 37°C, 5% CO2 в течение 45 минут, при встряхивании. Планшеты промыли 6x PBS плюс 0,05% Tween® 20, добавили 100 μл/лунку против человеческого Fc-HRP (Calbiochem) (1:2500) в CHO среде и инкубировали при температуре 37°C/5% CO2 в течение 30-60 минут (против человеческого Fc-HRP не связывается с собачьим Fc.). Планшеты промыли 5x PBS плюс 0,05% Tween® 20, добавили 100 μл/лунку TMB микролуночного субстрата и затем инкубировали при комнатной температуре в течение 10 минут. Реакцию остановили 100 μл/лунку 1,5 M фосфорной кислоты. Показатели A450-A620 на считывающем устройстве ELISA.
Выделение цитокина из собачьего PBMC:
PBMC получили из EDTA образцов крови, полученных от здоровых собак и от собак с раком при использовании разделения Фиколла. Клетки промыли 3 раза и ресуспендировали в полной среде для культуры тканей в концентрации 2,5×105 с трех кратным повторном лунок в 96-луночных планшетах. Клетки активировали конканавалином A в концентрации 1 μг/мл. Тестируемые антитела добавили в различных концентрациях и инкубировали культуры в течение 96 часов. Контроли включали клетки, инкубированные с conA и без антител или с conA и иррелевантными изотипически сходными антителами. Через 96 часов культивирования собрали супернатанты и провели анализ на выделение IFN-ᵧ при использовании коммерческого собачьего IFN-ᵧ набора ELISA (R & D Systems) [смотрите, Фигура 4].
ПРИМЕР 4
ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ СОБАЧЬИ IgGs
Для получения вариантов собачьего IgG с отсутствующими эффекторными функциями получили множество мутантных тяжелых цепей собачьего IgGB. Эти варианты могут включать одно из следующих: одиночного или комбинированного замещения в Fc части тяжелой цепи аминокислотной последовательности: P4A, D31A, N63A, G64P, T65A, A93G и P95A. Варианты тяжелых цепей (то есть, содержащих такие аминокислотные замещения) клонировали в экспрессионных плазмидах и трансфецировали в HEK 293 клетки вместе с плазмидой, содержащей ген, кодирующий легкую цепь. Экспрессированные интактные антитела очистили от HEK 293 клеток и провели оценку связывания с FcᵧRI и C1q для оценки потенциала опосредования иммунных эффекторных функций. В Таблице 3 приведены примеры плазмид, кодирующих тяжелые цепи генетически модифицированного канинизированного антитела, канинизированные тяжелые цепи; и генетические модификации в этих тяжелых цепях. Вариант тяжелой цепи использовали для оценки эффекторной функции в генетически модифицированных mAbs. Все тяжелые цепи содержали CDRs из 2H9 мышиного антитела против собачьего PD-1.
FcᵧRI связывание:
Связывание с FcRᵧI представляет критерий способности антитела опосредовать ADCC антитела. Для оценки этого свойства провели анализ канинизированного антитела для измерения связывания канинизированного антитела с FcᵧRI следующим образом: Покрыли пленкой 96-луночные планшеты 100 μл на лунку 2,5 μг/мл PD-1 HIS. Инкубировали при температуре 2-7°C в течение ночи. Планшеты доводили до комнатной температуры в течение 15 минут. Планшеты промыли 3X фосфатно-солевым буфером, содержащим 0,05% Tween® 20 (PBST), и затем блокировали лунки при использовании 200 μл/лунку 5% NFDM (нежирное сухое молоко). Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 3X PBST. Провели 2-кратное разведение антитела, начиная с 1 μг/мл в 5% NFDM. Добавили 100 μл/лунку разведенного антитела. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 6X PBST. Добавили 100 μл/лунку рекомбинантного человеческого CD64 белка (R&D systems), разведенного до 1 μг/мл. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 6X PBST. Добавили 100 μл/лунку биотинилированного антитела против CD64 (R&D systems), разведенного до 1:3000. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 6X PBST. Добавили 100 μл/лунку антитело+стрептавидин-HRP (R&D systems), разведенного до 1:7500. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 6X PBST. Добавили 100 μл/лунку субстрата TMB. Инкубировали в течение 10 минут при температуре 15-30°C. Для считывания планшетов использовали считывающее устройство планшетного типа ELISA при 450-540 нм.
Результаты: На Фигуре 5A показано, что канинизированное mAb, обозначенное, как can2H9 ADCC mut-1 VH4/VL3, которое имело генетическую модификацию D31A, или mAb, обозначенное, как can2H9 ADCC mut-2 VH4/VL3, которое имело генетическую модификацию N63A, продемонстрировали практически полное снижение связывания с FcᵧRI. С другой стороны, mab, обозначенное, как can2H9 ADCC (1062) VH4/VL3, которое содержит комбинированные генетические модификации D31A плюс N63A, не продемонстрировало детектируемое связывание с FcᵧRI. На Фигуре 5A can2H9 IgGD VH4/VL3, представляющий канинизированное антитело, которое содержит Fc из собачьего IgGD, и can3B6 VH4/VL4 IgGB, представляющее канинизированное антитело, которое не связывается с покрывающим антигеном (PD-1 HIS), и канинизированное mAb, которое обозначено, как can2H9 VH4/VL3, представляет антитело, которое содержит не модифицированный Fc IgGB. На Фигуре 5B показано, что канинизированное mAb, обозначенное, как can2H9 ADCC(1059) VH4/VL3, которое содержит генетическую модификацию P4A, и mAb, обозначенное, как can2H9 ADCC (1061) VH4/VL3, которое содержит генетическую модификацию P95A, продемонстрировало значительное снижение связывания FcᵧRI, при этом mAb, обозначенное, как can2H9 ADCC(1060) VH4/VL3, которое содержит генетическую модификацию A93G, продемонстрировало слабое снижение связывания с FcᵧRI. С другой стороны, mAb, обозначенное, как can2H9 IgGB ADCC (1068) VH4/VL3, которое содержит пять генетических модификаций (D31A, N63A, P4A, A93G, P95A), продемонстрировало полное отсутствие связывания с FcᵧRI.
C1q связывание:
Связывание с первым компонентом комплемента C1q представляет критерий способности антитела опосредовать CDC. Для оценки этого свойства провели анализ канинизированного антитела для измерения связывания канинизированного антитела с C1q следующим образом: Покрыли пленкой 96-луночные планшеты 100 μл на лунку 2,5 μг/мл PD-1 HIS. Инкубировали при температуре 2-7°C в течение ночи. Планшеты доводили до комнатной температуры в течение 15 минут. Планшеты промыли 3X PBST. Блокировали 200 μл/лунку 5% BSA. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 3X PBST. Провели 2-кратное разведение антитела, начиная с 1 μг/мл в 5% BSA. Добавили 100 μл/лунку разведенного антитела. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 6X PBST. Добавили 100 μл/лунку C1q белка до 4 μг/мл. Инкубировали в течение 60 минут при температуре 36-38°C. Добавили 100 μл/лунку покрывающего антитела против C1q, разведенного до 1:3000. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 6X PBST.
Добавили 100 μл/лунку ослиного против козьего антитела - HRP, разведенного до 1:10000. Инкубировали в течение 60 минут при температуре 36-38°C. Промыли 6X PBST. Добавили 100 μл/лунку субстрата TMB. Инкубировали в течение 10 минут при температуре 15-30°C. Для считывания планшетов использовали считывающее устройство планшетного типа ELISA при 450-540 нм.
Результаты: На Фигуре 6A показано, что канинизированное mAb, обозначенное, как can2H9VH4 IgGB ADCC (mut-1)/VL3, которое имеет генетическую модификацию D31A, или mAb, обозначенное, как can2H9 VH4 IgGB ADCC (mut-2)/VL3, которое имеет генетическую модификацию N63A, продемонстрировало значительное снижение связывания с C1q. С другой стороны, mAb, обозначенное, как can2H9 VH4 IgGB ADCC (1062)/VL3, которое содержит комбинированные генетические модификации D31A плюс N63A, не продемонстрировало детектируемое связывание с C1q.
На Фигуре 6A can2H9 VH4 IgGD/VL3, представляющее канинизированное антитело, которое содержит Fc из собачьего IgGD, и can3B6 VH4/VL4 IgGB, представляющее канинизированное антитело, которое не связывается с покрывающим антигеном (PD-1 HIS), и канинизированное mAb, обозначенное, как can2H9 VH4/VL3 IgGB, представляет антитело, которое содержит не мутированный IgGB Fc. На Фигуре 6B показано, что канинизированное mAb, обозначенное, как can2H9 VH4 IgGB ADCC(1059)/VL3, которое содержит замещение P4A, и mAb, обозначенное, как can2H9 VH4 IgGB ADCC (1061)/VL3, которая содержит замещение P95A, продемонстрировало значительное снижение связывание с C1q, при этом mAb, обозначенное, как can2H9 VH4 IgGB ADCC(1060)/VL3, которое содержит замещение A93G, продемонстрировало усиление связывания с C1q. С другой стороны, mAb, обозначенное, как can2H9 VH4 IgGB ADCC (1068) /VL3, которое содержит пять замещений (D31A, N63A, P4A, A93G, P95A), продемонстрировало полное отсутствие связывания с C1q. На Фигуре 6B mAb, обозначенное, как can3B6 VH4/VL4 IgGB, представляет канинизированное антитело, которое не связывается с покрывающим антигеном (PD-1 HIS), и канинизированное mAb, обозначенное, как can2H9 VH4/VL3 IgGB, представляет антитело, которое содержит не мутированный IgGB Fc.
МОДИФИЦИРОВАННЫЙ cFc ИЛИ НАТИВНЫЙ cFc С ШАРНИРНЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ
ПОСЛЕДОВАТЕЛЬНОСТИ ШАРНИРНЫХ ОБЛАСТЕЙ
ПОСЛЕДОВАТЕЛЬНОСТИ СОБАЧЬЕГО PD-1/PD-L1
ПОСЛЕДОВАТЕЛЬНОСТИ НАТИВНОГО cFc
АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ CDR
ОТДЕЛЬНЫЕ ЗАМЕЩЕННЫЕ КАНИНИЗИРОВАННЫЕ ТЯЖЕЛЫЕ ЦЕПИ
Потенциальные специфические замещения в P4, D31, N63, G64,T65, A93 и P95
КОРРЕЛЯЦИЯ ПОЗИЦИЙ АМИНОКИСЛОТНЫХ ОСТАТКОВ НАТИВНЫХ И ЗАМЕЩЕННЫХ cFc С ТАКОВЫМИ СООТВЕТСТВУЮЩИХ ЗАМЕЩЕННЫХ СОБАЧЬИХ ТЯЖЕЛЫХ ЦЕПЕЙ#
ОТДЕЛЬНЫЕ НЕЗАМЕЩЕННЫЕ КАНИНИЗИРОВАННЫЕ
ТЯЖЕЛЫЕ И ЛЕГКИЕ ЦЕПИ
ПРИМЕР 5
КАРТИРОВАНИЕ ЭПИТОПА АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО PD-1
Введение
Взаимодействие антитела с его распознанным белковым антигеном опосредованно через связывание специфических аминокислот (активных центров) антитела со специфическими аминокислотами (эпитопы) целевых антигенов. Эпитоп представляет антигенный детерминант, который вызывает специфическую реакцию иммуноглобулина. Он состоит из группы аминокислот на поверхности антигена.
Интересующий белок может содержать несколько эпитопов, которые распознаются различными антителами. Эпитопы, распознанные антителами, классифицируются как линейные или конформационные эпитопы. Линейные эпитопы формируются вытягиванием непрерывной последовательности аминокислот белка, в то время, как конформационные эпитопы состоят из аминокислот, которые являются дискретными (например, на большом расстоянии друг от друга) в первичной аминокислотной последовательности, но сходятся вместе в трехмерной укладке белка.
Картирование эпитопа относится к процессу идентификации аминокислотных последовательностей (то есть, эпитопов), которые распознаются антителом на их целевых антигенах. Идентификация эпитопов, распознанных моноклональными антителами (mAbs) на целевых антигенах, имеет важное значение. Например, это может помогать при разработке новых терапевтических средств, диагностических средств и вакцин. Картирование эпитопа также может помочь в выборе оптимальных терапевтических mAbs и помогает выяснить механизм их действия. Информация об эпитопе также может объяснить уникальные эпитопы рака и определить защитные или патогеннные эффекты вакцины.
Картирование эпитопа может быть проведено при использовании поликлональных или моноклональных антител, и для идентификации используют несколько способов в зависимости от предполагаемой природы эпитопа (то есть, линейный по сравнению с конформационным). Картирование линейных эпитопов более простое и относительно проще для выполнения. Для этой цели коммерческие организации для картирования линейных эпитопов часто используют сканирование пептидов. В таком случае химически синтезируют ряд перекрывающихся коротких пептидных последовательностей целевого белка и тестируют его на способность связываться с интересующим антителом. Такой подход является быстрым, имеет высокую пропускную способность и относительно недорог в осуществлении. С другой стороны, картирование прерывистого эпитопа технически более сложное и требует более специализированных методов, таких как рентгеновская кристаллография моноклонального антитела совместно с целевым белком, водород-дейтериевый (H/D) обмен, и/или масс-спектроскопия совместно с ферментативным расщеплением.
Картирование эпитопов PD-1 при использовании микроматриц ProImmune®:
Для идентификации аминокислот, которые образуют эпитопы для mAbs против PD1, химически синтезировали всего 28 пептидов, которые тмели 15 аминокислот в длину и перекрывались 10 аминокислотами. Эту библиотеку перекрывающихся пептидов разработали для охвата полноразмерного собачьего белка PD-1. Определение антитело-связывающего пептида провели связыванием образцов антитела с пептидной микроматрицей ProArray Ultra® с последующим инкубированием с вторичным флуоресцентно-меченным антителом. Все пептиды синтезировали отдельно и затем связывали с поверхностью слайда ProImmune® наряду с контролями мышиных IgG ProImmune®. Такой оптимизированный процесс гарантирует, что пептиды будут присутствовать на эррее, таким образом, что он близко иммитирует свойства соответствующей области белка, обходя природную физикохимическую вариацию свободных пептидов и делая совместимой комбинированную эррей платформу пептида и белка. Тестируемые аналиты (пептиды) расположили на слайде ProArray Ultra® в отдельных точках подходящих gal-файлов, что позволяет точно сгруппировать полученный массив признаков по отношению к внесенному аналиту. Слайды ProArray Ultra® блокировали при использовании валидированного блокирующего буфера для снижения неспецифического связывания mAbs. Затем их инкубировали с образцами mAb, с последующим инкубированием со специфическим вторичным флуоресцентно-меченным антителом. После нескольких стадий промывки, эррей ProArray Ultra® высушили и провели сканирование при использовании флуорисцентной сканирующей системы микроматриц с высоким разрешением. После сканирования флуорисцентно меченных слайдов ProArray Ultra® сканер записывает изображение, которое оценивают при использовании программного обеспечения для анализа изображений, позволяя интерпретировать и проводить количественную оценку уровней интенсивности флуоресценции, связанных с каждой флуоресцентной точкой на отсканированном слайде микроматрицы. Результаты этого эксперимента показали, что некоторые пептиды собачьего PD-1 были распознаны некоторыми из оцениваемых mAbs. Идентичность mAbs и аминокислотной последовательности, распознаваемой этими mAbs, приведена в Таблице 12. Это исследование показало, что mAb 2H9 распознает эпитоп, расположенный во внеклеточном домене собачьего PD-1, содержащем аминокислотную последовательность, представленную SEQ ID NO: 138, и что mAb 1A1 распознает эпитоп, содержащий аминокислотную последовательность, представленную SEQ ID NO: 138, и перекрывающая аминокислотная последовательность представлена аминокислотной последовательностью SEQ ID NO: 139.
Картирование эпитопов PD-1 при использовании масс-спектрометрии:
Для идентификации потенциально прерывистых эпитопов, распознанных против собачьего PD-1, использовали способ на основе химического перекрестного сшивания и детекцию масс-спектрометрией (CovalX® Instrument Incorporated). Применение этой технологии для картирования эпитопа собачьего PD-1 позволяет идентифицировать по меньшей мере части эпитопов, распознанные указанными mAbs, которые представляют приведенные в Таблице 13. Как видно из Таблицы 13, mAb 3B6 распознает по меньшей мере часть эпитопа, расположенного во внеклеточном домене собачьего PD-1 в аминокислотной последовательности, представленной SEQ ID NO: 140, и что mAb 2G9 распознает по меньшей мере часть эпитопа в аминокислотной последовательности, представленной SEQ ID NO: 141. С другой стороны, mAb 1E4 и mAb 1B5 распознают по меньшей мере часть эпитопа в аминокислотной последовательности, представленной SEQ ID NO: 142, в аминокислотной последовательности, представленной SEQ ID NO: 143, соответственно.
Как показано на Фигуре 7A, определение, проведенное при использовании химического перекрестного сшивания, детектора частиц с высокой массой в сочетании с масс-спектрометрией (High-Mass MALDI mass spectrometry) и не линейной хроматографии с орбитальной ионной ловушкой в сочетании с масс- спектрометрией (nLC-Orbitrap mass spectrometry) показало, что эпитоп на собачьем PD-1, распознанный канинизированным антителом 2G9, содержит R62, R69, R72 и R75 SEQ ID NO: 114. Аналогичное определение для эпитопа на собачьем PD-1, распознанного канинизированным антителом 3B6, содержит R75 и R90 SEQ ID NO: 114. Соответственно, R75 по-видимому оказывается очень важным важным аминокислотным остатком в одном или более эпитопе собачьего PD-1. Интересно, что после проведения этих анализов было установлено, что аминокислотная последовательность для CDRs 1A1 идентична таковой 2G9. Совместимость между областью на PD-1, с которой связывается 2G9, и установленная для 1A1, которая была получена этими двумя очень разными методами, указывает на то, что область содержит аминокислотные остатки, составляющие эпитоп PD-1, который распознается антителом против собачьего PD-1 (смотрите, Таблицы 12 и 13 ниже).
Дополнительно, область аминокислотной последовательности PD-1, которая распознается блокирующим протестированным антителом по настоящему изобретению, расположена во внеклеточном домене собачьего PD-1. Распознанная область состоит из следующего пептида (смотрите, Таблицы 12 и 13 ниже).
NQTDKLAAFQEDRIEPGRDRRFRVM*RLPNGRDFHMSIVAARLNDS (SEQ ID NO:144)
В этом пептиде представлен более короткий пептид, который выделен жирным шрифтом. Более короткий пептид был распознан при использовании микроматрицы ProImmune® (смотрите, Таблица 12).
DRIEPGRDRRFRVM*RLPNGR (SEQ ID NO:145).
Следует отметить, что R62, R69, R72 и R75 SEQ ID NO: 114 все входят в состав, как более длинного пептида (SEQ ID NO: 144), так и более короткого пептида (SEQ ID NO: 145), при этом R90 SEQ ID NO: 114 присутствует в более длинном пептиде. Эти пять аргининовых остатков по-видимому являются очень важными аминокислотными остатками в одном или более эпитопе собачьего PD-1. Как указано в Таблицах 6-8, помеченный звездочкой метиониновый остаток (*) также указывался как представляющий треониновый остаток been.
ЭПИТОПЫ PD-1, РАСПОЗНАННЫЕ MAABS ПРОТИВ СОБАЧЬЕГО PD-1
ПРИМЕНЕНИЕ МИКРОМАТРИЦЫ PROIMMUNE®
# CDRs 1A1 идентичны таковым 2G9.
ЭПИТОПЫ PD-1, РАСПОЗНАННЫЕ MAABS ПРОТИВ СОБАЧЬЕГО PD-1
ПРИМЕНЕНИЕ МАСС-СПЕКТРОМЕТРИИ
# CDRs 1E4 наиболее близки таковым 2G9.
Все ссылки, приведенные в описании настоящей патентной заявки, введены ссылками в том же объеме, как если бы это была отдельная публикация, значение в базе данных (например, Genbank sequences или GeneID entries), патентная заявка или патент был конкретно или отдельно указан вводимой ссылкой. Это утверждение о введение ссылкой, как предполагается заявителями, в соответствии с 37 C.F.R. §1.57(b)(1), относительно каждой и каждой отдельной публикации, значению в базе данных (например, Genbank sequences или GeneID entries), патентной заявке или патенту, каждое из которых четко определено в соответствии с 37 C.F.R. §1.57(b)(2), даже если с таким цитированием в непосредственной близости не приведено специального утверждение о введении ссылкой. Включение при использовании специальных утверждений о введении ссылкой, если такое имеет место быть, в описании никаким образом не ослабляет это общее утверждение о введении ссылкой. Цитирование ссылок в описании настоящей патентной заявки не следует рассматривать, как признание того, что ссылка относится к предшествующему уровню техники, а также не предполагает какого-либо признания в качестве содержимого или данных этих публикаций или документов.
Объем притязаний настоящего изобретения не ограничивается приведенными конкретными вариантами его воплощения. Естественным образом, специалисту в области техники, к которой относится настоящее изобретение, будут понятны различные модификации настоящего изобретения в дополнение к приведенному в указанном выше описании и приложенных фигурах. Такие модификации также входят в объем притязаний настоящего изобретения, изложенный в приложенной формуле изобретения.
Приведенное выше описание считается достаточным для возможности осуществления настоящего изобретения специалистом в области техники, к которой относится настоящее изобретение. Различные модификации настоящего изобретения дополнительно к приведенным и описанным выше будут ясны специалисту в области техники, к которой относится настоящее изобретение из приведенного выше описания, и входят в объем притязаний, изложенный в приложенной формуле изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Intervet Inc.
Morsey, Mohamad
Yuanzheng, Zhang
Tarpey, Ian
<120> КАНИНИЗИРОВАННЫЕ АНТИТЕЛА
<130> 23799
<150> 61/918946
<151> 2013-12-20
<150> 61/918847
<151> 2013-12-20
<150> 62/030812
<151> 2014-07-30
<160> 146
<170> PatentIn version 3.5
<210> 1
<211> 690
<212> ДНК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<220>
<221> misc_feature
<222> (10)..(12)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (91)..(93)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (187)..(195)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (277)..(279)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (283)..(285)
<223> n представляет a, c, g, или t
<400> 1
ctgggcggcn nnagcgtgtt tatttttccg ccgaaaccga aagataccct gctgattgcg
60
cgcaccccgg aagtgacctg cgtggtggtg nnnctggatc cggaagatcc ggaagtgcag
120
attagctggt ttgtggatgg caaacagatg cagaccgcga aaacccagcc gcgcgaagaa
180
cagtttnnnn nnnnntatcg cgtggtgagc gtgctgccga ttggccatca ggattggctg
240
aaaggcaaac agtttacctg caaagtgaac aacaaannnc tgnnnagccc gattgaacgc
300
accattagca aagcgcgcgg ccaggcgcat cagccgagcg tgtatgtgct gccgccgagc
360
cgcgaagaac tgagcaaaaa caccgtgagc ctgacctgcc tgattaaaga tttttttccg
420
ccggatattg atgtggaatg gcagagcaac ggccagcagg aaccggaaag caaatatcgc
480
accaccccgc cgcagctgga tgaagatggc agctattttc tgtatagcaa actgagcgtg
540
gataaaagcc gctggcagcg cggcgatacc tttatttgcg cggtgatgca tgaagcgctg
600
cataaccatt atacccagga aagcctgagc catagcccgg gcaaacataa ccattatacc
660
caggaaagcc tgagccatag cccgggcaaa
690
<210> 2
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (31)..(31)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (63)..(65)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (93)..(93)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (95)..(95)
<223> Xaa может представлять любую природную аминокислоту
<400> 2
Leu Gly Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
1 5 10 15
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Xaa Leu
20 25 30
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
35 40 45
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Xaa Xaa
50 55 60
Xaa Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
65 70 75 80
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser
85 90 95
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
100 105 110
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
115 120 125
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
130 135 140
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
145 150 155 160
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
165 170 175
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
180 185 190
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
195 200 205
Leu Ser His Ser Pro Gly Lys
210 215
<210> 3
<211> 690
<212> ДНК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<220>
<221> misc_feature
<222> (10)..(12)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (91)..(93)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (187)..(195)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (277)..(279)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (283)..(285)
<223> n представляет a, c, g, или t
<400> 3
ctgggcggcn nnagcgtgtt tatttttccg ccgaaaccga aagatattct ggtgaccgcg
60
cgcaccccga ccgtgacctg cgtggtggtg nnnctggatc cggaaaaccc ggaagtgcag
120
attagctggt ttgtggatag caaacaggtg cagaccgcga acacccagcc gcgcgaagaa
180
cagagcnnnn nnnnntatcg cgtggtgagc gtgctgccga ttggccatca ggattggctg
240
agcggcaaac agtttaaatg caaagtgaac aacaaannnc tgnnnagccc gattgaagaa
300
attattagca aaaccccggg ccaggcgcat cagccgaacg tgtatgtgct gccgccgagc
360
cgcgatgaaa tgagcaaaaa caccgtgacc ctgacctgcc tggtgaaaga tttttttccg
420
ccggaaattg atgtggaatg gcagagcaac ggccagcagg aaccggaaag caaatatcgc
480
atgaccccgc cgcagctgga tgaagatggc agctattttc tgtatagcaa actgagcgtg
540
gataaaagcc gctggcagcg cggcgatacc tttatttgcg cggtgatgca tgaagcgctg
600
cataaccatt atacccagat tagcctgagc catagcccgg gcaaacataa ccattatacc
660
cagattagcc tgagccatag cccgggcaaa
690
<210> 4
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (31)..(31)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (63)..(65)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (93)..(93)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (95)..(95)
<223> Xaa может представлять любую природную аминокислоту
<400> 4
Leu Gly Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile
1 5 10 15
Leu Val Thr Ala Arg Thr Pro Thr Val Thr Cys Val Val Val Xaa Leu
20 25 30
Asp Pro Glu Asn Pro Glu Val Gln Ile Ser Trp Phe Val Asp Ser Lys
35 40 45
Gln Val Gln Thr Ala Asn Thr Gln Pro Arg Glu Glu Gln Ser Xaa Xaa
50 55 60
Xaa Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
65 70 75 80
Ser Gly Lys Gln Phe Lys Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser
85 90 95
Pro Ile Glu Glu Ile Ile Ser Lys Thr Pro Gly Gln Ala His Gln Pro
100 105 110
Asn Val Tyr Val Leu Pro Pro Ser Arg Asp Glu Met Ser Lys Asn Thr
115 120 125
Val Thr Leu Thr Cys Leu Val Lys Asp Phe Phe Pro Pro Glu Ile Asp
130 135 140
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
145 150 155 160
Met Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
165 170 175
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
180 185 190
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Ile Ser
195 200 205
Leu Ser His Ser Pro Gly Lys
210 215
<210> 5
<211> 699
<212> ДНК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 5
ccgaaagaaa gcacctgcaa atgcattccg ccgtgcccgg tgccggaaag cctgggcggc
60
ccgagcgtgt ttatttttcc gccgaaaccg aaagatattc tgcgcattac ccgcaccccg
120
gaaattacct gcgtggtgct ggatctgggc cgcgaagatc cggaagtgca gattagctgg
180
tttgtggatg gcaaagaagt gcataccgcg aaaacccagc cgcgcgaaca gcagtttaac
240
agcacctatc gcgtggtgag cgtgctgccg attgaacatc aggattggct gaccggcaaa
300
gaatttaaat gccgcgtgaa ccatattggc ctgccgagcc cgattgaacg caccattagc
360
aaagcgcgcg gccaggcgca tcagccgagc gtgtatgtgc tgccgccgag cccgaaagaa
420
ctgagcagca gcgataccgt gaccctgacc tgcctgatta aagatttttt tccgccggaa
480
attgatgtgg aatggcagag caacggccag ccggaaccgg aaagcaaata tcataccacc
540
gcgccgcagc tggatgaaga tggcagctat tttctgtata gcaaactgag cgtggataaa
600
agccgctggc agcagggcga tacctttacc tgcgcggtga tgcatgaagc gctgcagaac
660
cattataccg atctgagcct gagccatagc ccgggcaaa
699
<210> 6
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 6
Pro Lys Glu Ser Thr Cys Lys Cys Ile Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Ser Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp
20 25 30
Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr Cys Val Val Leu Asp
35 40 45
Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly
50 55 60
Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg Glu Gln Gln Phe Asn
65 70 75 80
Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile Glu His Gln Asp Trp
85 90 95
Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn His Ile Gly Leu Pro
100 105 110
Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln
115 120 125
Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser
130 135 140
Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Glu
145 150 155 160
Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro Glu Pro Glu Ser Lys
165 170 175
Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu
180 185 190
Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Gln Gly Asp Thr
195 200 205
Phe Thr Cys Ala Val Met His Glu Ala Leu Gln Asn His Tyr Thr Asp
210 215 220
Leu Ser Leu Ser His Ser Pro Gly Lys
225 230
<210> 7
<211> 699
<212> ДНК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 7
tttaacgaat gccgctgcac cgataccccg ccgtgcccgg tgccggaacc gctgggcggc
60
ccgagcgtgt ttatttttcc gccgaaaccg aaagatattc tgcgcattac ccgcaccccg
120
gaaattacct gcgtggtgct ggatctgggc cgcgaagatc cggaagtgca gattagctgg
180
tttgtggatg gcaaagaagt gcataccgcg aaaacccagc cgcgcgaaca gcagtttaac
240
agcacctatc gcgtggtgag cgtgctgccg attgaacatc aggattggct gaccggcaaa
300
gaatttaaat gccgcgtgaa ccatattggc ctgccgagcc cgattgaacg caccattagc
360
aaagcgcgcg gccaggcgca tcagccgagc gtgtatgtgc tgccgccgag cccgaaagaa
420
ctgagcagca gcgataccgt gaccctgacc tgcctgatta aagatttttt tccgccggaa
480
attgatgtgg aatggcagag caacggccag ccggaaccgg aaagcaaata tcataccacc
540
gcgccgcagc tggatgaaga tggcagctat tttctgtata gcaaactgag cgtggataaa
600
agccgctggc agcagggcga tacctttacc tgcgcggtga tgcatgaagc gctgcagaac
660
cattataccg atctgagcct gagccatagc ccgggcaaa
699
<210> 8
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 8
Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Pro Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp
20 25 30
Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr Cys Val Val Leu Asp
35 40 45
Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly
50 55 60
Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg Glu Gln Gln Phe Asn
65 70 75 80
Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile Glu His Gln Asp Trp
85 90 95
Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn His Ile Gly Leu Pro
100 105 110
Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln
115 120 125
Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser
130 135 140
Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Glu
145 150 155 160
Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro Glu Pro Glu Ser Lys
165 170 175
Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu
180 185 190
Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Gln Gly Asp Thr
195 200 205
Phe Thr Cys Ala Val Met His Glu Ala Leu Gln Asn His Tyr Thr Asp
210 215 220
Leu Ser Leu Ser His Ser Pro Gly Lys
225 230
<210> 9
<211> 714
<212> ДНК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 9
ccgaaacgcg aaaacggccg cgtgccgcgc ccgccggatt gcccgaaatg cccggcgccg
60
gaaatgctgg gcggcccgag cgtgtttatt tttccgccga aaccgaaaga tattctgcgc
120
attacccgca ccccggaaat tacctgcgtg gtgctggatc tgggccgcga agatccggaa
180
gtgcagatta gctggtttgt ggatggcaaa gaagtgcata ccgcgaaaac ccagccgcgc
240
gaacagcagt ttaacagcac ctatcgcgtg gtgagcgtgc tgccgattga acatcaggat
300
tggctgaccg gcaaagaatt taaatgccgc gtgaaccata ttggcctgcc gagcccgatt
360
gaacgcacca ttagcaaagc gcgcggccag gcgcatcagc cgagcgtgta tgtgctgccg
420
ccgagcccga aagaactgag cagcagcgat accgtgaccc tgacctgcct gattaaagat
480
ttttttccgc cggaaattga tgtggaatgg cagagcaacg gccagccgga accggaaagc
540
aaatatcata ccaccgcgcc gcagctggat gaagatggca gctattttct gtatagcaaa
600
ctgagcgtgg ataaaagccg ctggcagcag ggcgatacct ttacctgcgc ggtgatgcat
660
gaagcgctgc agaaccatta taccgatctg agcctgagcc atagcccggg caaa
714
<210> 10
<211> 238
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 10
Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys Pro Lys
1 5 10 15
Cys Pro Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe Ile Phe Pro
20 25 30
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr
35 40 45
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
50 55 60
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg
65 70 75 80
Glu Gln Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile
85 90 95
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
100 105 110
His Ile Gly Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
115 120 125
Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
130 135 140
Glu Leu Ser Ser Ser Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp
145 150 155 160
Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro
165 170 175
Glu Pro Glu Ser Lys Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp
180 185 190
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
195 200 205
Gln Gln Gly Asp Thr Phe Thr Cys Ala Val Met His Glu Ala Leu Gln
210 215 220
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
225 230 235
<210> 11
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 11
gccaaggagt gcgagtgcaa gtgcaactgc aacaactgcc cctgccccgg ctgcggcctg
60
ctgggcggcc ccagcgtgtt catcttcccc cccaagccca aggacatcct gagaatcacc
120
agaacccccg agatcacctg cgtggtgctg gacctgggca gagaggaccc cgaggtgcag
180
atcagctggt tcgtggacgg caaggaggtg cacaccgcca agacccagcc cagagagcag
240
cagttcaaca gcacctacag agtggtgagc gtgctgccca tcgagcacca ggactggctg
300
accggcaagg agttcaagtg cagagtgaac cacatcggcc tgcccagccc catcgagaga
360
accatcagca aggccagagg ccaggcccac cagcccagcg tgtacgtgct gccccccagc
420
cccaaggagc tgagcagcag cgacaccgtg accctgacct gcctgatcaa ggacttcttc
480
ccccccgaga tcgacgtgga gtggcagagc aacggccagc ccgagcccga gagcaagtac
540
cacaccaccg ccccccagct ggacgaggac ggcagctact tcctgtacag caagctgagc
600
gtggacaaga gcagatggca gcagggcgac accttcacct gcgccgtgat gcacgaggcc
660
ctgcagaacc actacaccga cctgagcctg agccacagcc ccggcaag
708
<210> 12
<211> 236
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> модифицированная собачья последовательность
<400> 12
Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn Asn Cys Pro Cys Pro
1 5 10 15
Gly Cys Gly Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys
20 25 30
Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr Cys Val
35 40 45
Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser Trp Phe
50 55 60
Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg Glu Gln
65 70 75 80
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile Glu His
85 90 95
Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn His Ile
100 105 110
Gly Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln
115 120 125
Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys Glu Leu
130 135 140
Ser Ser Ser Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp Phe Phe
145 150 155 160
Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro Glu Pro
165 170 175
Glu Ser Lys Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp Gly Ser
180 185 190
Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Gln
195 200 205
Gly Asp Thr Phe Thr Cys Ala Val Met His Glu Ala Leu Gln Asn His
210 215 220
Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
225 230 235
<210> 13
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 13
Lys Ser Ser Gln Ser Leu Leu Asn Ser Val Asn Gln Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 14
<211> 16
<212> БЕЛОК
<213> Mus musculus
<400> 14
Arg Ser Ser Gln Asn Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 15
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 15
His Ala Ser Gln Asn Ile Asn Val Trp Leu Ser
1 5 10
<210> 16
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 16
Phe Ala Ser Thr Arg Val Ser
1 5
<210> 17
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 17
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 18
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 18
Lys Ala Ser His Leu His Thr
1 5
<210> 19
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 19
Phe Ala Ser Ala Arg Val Ser
1 5
<210> 20
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 20
Phe Ala Ser Thr Arg Ile Ser
1 5
<210> 21
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 21
Lys Ala Ser Asn Leu His Thr
1 5
<210> 22
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 22
Gln Gln Tyr Phe Ser Thr Pro Leu Thr
1 5
<210> 23
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 23
Phe Gln Gly Ser His Val Pro Tyr Thr
1 5
<210> 24
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 24
Gln Gln Gly Gln Ser Trp Pro Leu Thr
1 5
<210> 25
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 25
Gln Gln Tyr Phe Ser Thr Pro Leu Thr
1 5
<210> 26
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 26
Gln Gln Gly Gln Ser Tyr Pro Leu Thr
1 5
<210> 27
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 27
Gly Tyr Thr Phe Thr Thr Tyr Gly Met Ser
1 5 10
<210> 28
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 28
Gly Tyr Thr Phe Thr Arg Tyr Asn Met His
1 5 10
<210> 29
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 29
Gly Phe Asn Ile Lys Asn Thr Tyr Met His
1 5 10
<210> 30
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 30
Gly Phe Ser Leu Thr Ser Tyr Gly Val His
1 5 10
<210> 31
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 31
Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 32
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 32
Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 33
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 33
Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe Gln
1 5 10 15
Gly
<210> 34
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 34
Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 35
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 35
Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe Gln
1 5 10 15
Gly
<210> 36
<211> 6
<212> БЕЛОК
<213> Mus musculus
<400> 36
Phe Asp Gly Pro Asp Tyr
1 5
<210> 37
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 37
Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr
1 5 10
<210> 38
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 38
Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val
1 5 10
<210> 39
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (715)..(717)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (796)..(798)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (892)..(900)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (988)..(990)
<223> n представляет a, c, g, или t
<400> 39
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccagcaagac caaggtggac aagcccgtgc ccaagagaga gaacggcaga
660
gtgcccagac cccccgactg ccccaagtgc cccgcccccg agatgctggg cggcnnnagc
720
gtgttcatct tcccccccaa gcccaaggac accctgctga tcgccagaac ccccgaggtg
780
acctgcgtgg tggtgnnnct ggaccccgag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagc agatgcagac cgccaagacc cagcccagag aggagcagtt cnnnnnnnnn
900
tacagagtgg tgagcgtgct gcccatcggc caccaggact ggctgaaggg caagcagttc
960
acctgcaagg tgaacaacaa gnnnctgnnn agccccatcg agagaaccat cagcaaggcc
1020
agaggccagg cccaccagcc cagcgtgtac gtgctgcccc ccagcagaga ggagctgagc
1080
aagaacaccg tgagcctgac ctgcctgatc aaggacttct tcccccccga catcgacgtg
1140
gagtggcaga gcaacggcca gcaggagccc gagagcaagt acagaaccac ccccccccag
1200
ctggacgagg acggcagcta cttcctgtac agcaagctga gcgtggacaa gagcagatgg
1260
cagagaggcg acaccttcat ctgcgccgtg atgcacgagg ccctgcacaa ccactacacc
1320
caggagagcc tgagccacag ccccggcaag
1350
<210> 40
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (239)..(239)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (266)..(266)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (298)..(300)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (330)..(330)
<223> Xaa может представлять любую природную аминокислоту
<400> 40
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro
210 215 220
Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Xaa Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala
275 280 285
Lys Thr Gln Pro Arg Glu Glu Gln Phe Xaa Xaa Xaa Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe
305 310 315 320
Thr Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys
355 360 365
Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser
370 375 380
Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln
385 390 395 400
Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp
405 410 415
Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro
435 440 445
Gly Lys
450
<210> 41
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (709)..(711)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (790)..(792)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (886)..(894)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (976)..(978)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<400> 41
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagccaga gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta catccccgag cccgtgaccg tgagctggaa cagcgtgagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccaccaacac caaggtggac aagcccgtgg ccaaggagtg cgagtgcaag
660
tgcaactgca acaactgccc ctgccccggc tgcggcctgc tgggcggcnn nagcgtgttc
720
atcttccccc ccaagcccaa ggacatcctg gtgaccgcca gaacccccac cgtgacctgc
780
gtggtggtgn nnctggaccc cgagaacccc gaggtgcaga tcagctggtt cgtggacagc
840
aagcaggtgc agaccgccaa cacccagccc agagaggagc agagcnnnnn nnnntacaga
900
gtggtgagcg tgctgcccat cggccaccag gactggctga gcggcaagca gttcaagtgc
960
aaggtgaaca acaagnnnct gnnnagcccc atcgaggaga tcatcagcaa gacccccggc
1020
caggcccacc agcccaacgt gtacgtgctg ccccccagca gagacgagat gagcaagaac
1080
accgtgaccc tgacctgcct ggtgaaggac ttcttccccc ccgagatcga cgtggagtgg
1140
cagagcaacg gccagcagga gcccgagagc aagtacagaa tgaccccccc ccagctggac
1200
gaggacggca gctacttcct gtacagcaag ctgagcgtgg acaagagcag atggcagaga
1260
ggcgacacct tcatctgcgc cgtgatgcac gaggccctgc acaaccacta cacccagatc
1320
agcctgagcc acagccccgg caag
1344
<210> 42
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (237)..(237)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (264)..(264)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (296)..(298)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (326)..(326)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<400> 42
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Gln Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Ile Pro Glu Pro Val Thr Val Ser Trp Asn Ser Val Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Thr Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn
210 215 220
Asn Cys Pro Cys Pro Gly Cys Gly Leu Leu Gly Gly Xaa Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Val Thr Ala Arg Thr Pro
245 250 255
Thr Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asn Pro Glu Val
260 265 270
Gln Ile Ser Trp Phe Val Asp Ser Lys Gln Val Gln Thr Ala Asn Thr
275 280 285
Gln Pro Arg Glu Glu Gln Ser Xaa Xaa Xaa Tyr Arg Val Val Ser Val
290 295 300
Leu Pro Ile Gly His Gln Asp Trp Leu Ser Gly Lys Gln Phe Lys Cys
305 310 315 320
Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Glu Ile Ile Ser
325 330 335
Lys Thr Pro Gly Gln Ala His Gln Pro Asn Val Tyr Val Leu Pro Pro
340 345 350
Ser Arg Asp Glu Met Ser Lys Asn Thr Val Thr Leu Thr Cys Leu Val
355 360 365
Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly
370 375 380
Gln Gln Glu Pro Glu Ser Lys Tyr Arg Met Thr Pro Pro Gln Leu Asp
385 390 395 400
Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser
405 410 415
Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Ile Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 43
<211> 1365
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (730)..(732)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (811)..(813)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (907)..(915)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (997)..(999)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (1003)..(1005)
<223> n представляет a, c, g, или t
<400> 43
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgcccccg ccaacgtgga caccaagtac
180
gcccccaagt tccagggcaa ggccaccatc agcgccgaca ccgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt actactgcgt gctgatctac
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag caccagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacttcc ccgagcccgt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgtaca gcctgagcag catggtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggccca ccccgccagc aagaccaagg tggacaagcc cgtgcccaag
660
agagagaacg gcagagtgcc cagacccccc gactgcccca agtgccccgc ccccgagatg
720
ctgggcggcn nnagcgtgtt catcttcccc cccaagccca aggacaccct gctgatcgcc
780
agaacccccg aggtgacctg cgtggtggtg nnnctggacc ccgaggaccc cgaggtgcag
840
atcagctggt tcgtggacgg caagcagatg cagaccgcca agacccagcc cagagaggag
900
cagttcnnnn nnnnntacag agtggtgagc gtgctgccca tcggccacca ggactggctg
960
aagggcaagc agttcacctg caaggtgaac aacaagnnnc tgnnnagccc catcgagaga
1020
accatcagca aggccagagg ccaggcccac cagcccagcg tgtacgtgct gccccccagc
1080
agagaggagc tgagcaagaa caccgtgagc ctgacctgcc tgatcaagga cttcttcccc
1140
cccgacatcg acgtggagtg gcagagcaac ggccagcagg agcccgagag caagtacaga
1200
accacccccc cccagctgga cgaggacggc agctacttcc tgtacagcaa gctgagcgtg
1260
gacaagagca gatggcagag aggcgacacc ttcatctgcg ccgtgatgca cgaggccctg
1320
cacaaccact acacccagga gagcctgagc cacagccccg gcaag
1365
<210> 44
<211> 455
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (244)..(244)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (271)..(271)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (303)..(305)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (333)..(333)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (335)..(335)
<223> Xaa может представлять любую природную аминокислоту
<400> 44
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Ser Ala Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Leu Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Xaa Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Xaa Xaa
290 295 300
Xaa Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210> 45
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (724)..(726)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (805)..(807)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (901)..(909)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (991)..(993)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (997)..(999)
<223> n представляет a, c, g, или t
<400> 45
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgcccccg ccaacgtgga caccaagtac
180
gcccccaagt tccagggcaa ggccaccatc agcgccgaca ccgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt actactgcgt gctgatctac
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag ccagagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacatcc ccgagcccgt gaccgtgagc
480
tggaacagcg tgagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgtaca gcctgagcag catggtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggccca ccccgccacc aacaccaagg tggacaagcc cgtggccaag
660
gagtgcgagt gcaagtgcaa ctgcaacaac tgcccctgcc ccggctgcgg cctgctgggc
720
ggcnnnagcg tgttcatctt cccccccaag cccaaggaca tcctggtgac cgccagaacc
780
cccaccgtga cctgcgtggt ggtgnnnctg gaccccgaga accccgaggt gcagatcagc
840
tggttcgtgg acagcaagca ggtgcagacc gccaacaccc agcccagaga ggagcagagc
900
nnnnnnnnnt acagagtggt gagcgtgctg cccatcggcc accaggactg gctgagcggc
960
aagcagttca agtgcaaggt gaacaacaag nnnctgnnna gccccatcga ggagatcatc
1020
agcaagaccc ccggccaggc ccaccagccc aacgtgtacg tgctgccccc cagcagagac
1080
gagatgagca agaacaccgt gaccctgacc tgcctggtga aggacttctt cccccccgag
1140
atcgacgtgg agtggcagag caacggccag caggagcccg agagcaagta cagaatgacc
1200
cccccccagc tggacgagga cggcagctac ttcctgtaca gcaagctgag cgtggacaag
1260
agcagatggc agagaggcga caccttcatc tgcgccgtga tgcacgaggc cctgcacaac
1320
cactacaccc agatcagcct gagccacagc cccggcaag
1359
<210> 46
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (242)..(242)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (269)..(269)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (301)..(303)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (331)..(331)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (333)..(333)
<223> Xaa может представлять любую природную аминокислоту
<400> 46
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Ser Ala Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Leu Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Gln Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Ile Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Val Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Thr Asn Thr Lys Val Asp Lys Pro Val Ala Lys Glu Cys Glu Cys
210 215 220
Lys Cys Asn Cys Asn Asn Cys Pro Cys Pro Gly Cys Gly Leu Leu Gly
225 230 235 240
Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Val
245 250 255
Thr Ala Arg Thr Pro Thr Val Thr Cys Val Val Val Xaa Leu Asp Pro
260 265 270
Glu Asn Pro Glu Val Gln Ile Ser Trp Phe Val Asp Ser Lys Gln Val
275 280 285
Gln Thr Ala Asn Thr Gln Pro Arg Glu Glu Gln Ser Xaa Xaa Xaa Tyr
290 295 300
Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Ser Gly
305 310 315 320
Lys Gln Phe Lys Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile
325 330 335
Glu Glu Ile Ile Ser Lys Thr Pro Gly Gln Ala His Gln Pro Asn Val
340 345 350
Tyr Val Leu Pro Pro Ser Arg Asp Glu Met Ser Lys Asn Thr Val Thr
355 360 365
Leu Thr Cys Leu Val Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu
370 375 380
Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Met Thr
385 390 395 400
Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu
405 410 415
Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Ile Ser Leu Ser
435 440 445
His Ser Pro Gly Lys
450
<210> 47
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (736)..(738)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (817)..(819)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (913)..(921)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (1003)..(1005)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (1009)..(1011)
<223> n представляет a, c, g, или t
<400> 47
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc agatacaaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcacc atctaccccg gctacggcga caccagctac
180
aaccagaagt tcaagggcaa ggccaccctg agcgtggaca tcgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt acttctgcag cagagagttc
300
gccgacgact accccatccc ccccttcgac tactggggcc agggcaccct ggtgaccgtg
360
agcagcgcca gcaccaccgc ccccagcgtg ttccccctgg cccccagctg cggcagcacc
420
agcggcagca ccgtggccct ggcctgcctg gtgagcggct acttccccga gcccgtgacc
480
gtgagctgga acagcggcag cctgaccagc ggcgtgcaca ccttccccag cgtgctgcag
540
agcagcggcc tgtacagcct gagcagcatg gtgaccgtgc ccagcagcag atggcccagc
600
gagaccttca cctgcaacgt ggcccacccc gccagcaaga ccaaggtgga caagcccgtg
660
cccaagagag agaacggcag agtgcccaga ccccccgact gccccaagtg ccccgccccc
720
gagatgctgg gcggcnnnag cgtgttcatc ttccccccca agcccaagga caccctgctg
780
atcgccagaa cccccgaggt gacctgcgtg gtggtgnnnc tggaccccga ggaccccgag
840
gtgcagatca gctggttcgt ggacggcaag cagatgcaga ccgccaagac ccagcccaga
900
gaggagcagt tcnnnnnnnn ntacagagtg gtgagcgtgc tgcccatcgg ccaccaggac
960
tggctgaagg gcaagcagtt cacctgcaag gtgaacaaca agnnnctgnn nagccccatc
1020
gagagaacca tcagcaaggc cagaggccag gcccaccagc ccagcgtgta cgtgctgccc
1080
cccagcagag aggagctgag caagaacacc gtgagcctga cctgcctgat caaggacttc
1140
ttcccccccg acatcgacgt ggagtggcag agcaacggcc agcaggagcc cgagagcaag
1200
tacagaacca ccccccccca gctggacgag gacggcagct acttcctgta cagcaagctg
1260
agcgtggaca agagcagatg gcagagaggc gacaccttca tctgcgccgt gatgcacgag
1320
gccctgcaca accactacac ccaggagagc ctgagccaca gccccggcaa g
1371
<210> 48
<211> 457
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (246)..(246)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (273)..(273)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (305)..(307)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (335)..(335)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (337)..(337)
<223> Xaa может представлять любую природную аминокислоту
<400> 48
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Ser Val Asp Ile Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ser Arg Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr
130 135 140
Val Ala Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ser Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr
180 185 190
Val Pro Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala
195 200 205
His Pro Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu
210 215 220
Asn Gly Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro
225 230 235 240
Glu Met Leu Gly Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Xaa Leu Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp
275 280 285
Gly Lys Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe
290 295 300
Xaa Xaa Xaa Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp
305 310 315 320
Trp Leu Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Xaa Leu
325 330 335
Xaa Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His
340 345 350
Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys
355 360 365
Asn Thr Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp
370 375 380
Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys
385 390 395 400
Tyr Arg Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu
405 410 415
Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr
420 425 430
Phe Ile Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Glu Ser Leu Ser His Ser Pro Gly Lys
450 455
<210> 49
<211> 1365
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (730)..(732)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (811)..(813)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (907)..(915)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (997)..(999)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (1003)..(1005)
<223> n представляет a, c, g, или t
<400> 49
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc agatacaaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcacc atctaccccg gctacggcga caccagctac
180
aaccagaagt tcaagggcaa ggccaccctg agcgtggaca tcgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt acttctgcag cagagagttc
300
gccgacgact accccatccc ccccttcgac tactggggcc agggcaccct ggtgaccgtg
360
agcagcgcca gcaccaccgc ccccagcgtg ttccccctgg cccccagctg cggcagccag
420
agcggcagca ccgtggccct ggcctgcctg gtgagcggct acatccccga gcccgtgacc
480
gtgagctgga acagcgtgag cctgaccagc ggcgtgcaca ccttccccag cgtgctgcag
540
agcagcggcc tgtacagcct gagcagcatg gtgaccgtgc ccagcagcag atggcccagc
600
gagaccttca cctgcaacgt ggcccacccc gccaccaaca ccaaggtgga caagcccgtg
660
gccaaggagt gcgagtgcaa gtgcaactgc aacaactgcc cctgccccgg ctgcggcctg
720
ctgggcggcn nnagcgtgtt catcttcccc cccaagccca aggacatcct ggtgaccgcc
780
agaaccccca ccgtgacctg cgtggtggtg nnnctggacc ccgagaaccc cgaggtgcag
840
atcagctggt tcgtggacag caagcaggtg cagaccgcca acacccagcc cagagaggag
900
cagagcnnnn nnnnntacag agtggtgagc gtgctgccca tcggccacca ggactggctg
960
agcggcaagc agttcaagtg caaggtgaac aacaagnnnc tgnnnagccc catcgaggag
1020
atcatcagca agacccccgg ccaggcccac cagcccaacg tgtacgtgct gccccccagc
1080
agagacgaga tgagcaagaa caccgtgacc ctgacctgcc tggtgaagga cttcttcccc
1140
cccgagatcg acgtggagtg gcagagcaac ggccagcagg agcccgagag caagtacaga
1200
atgacccccc cccagctgga cgaggacggc agctacttcc tgtacagcaa gctgagcgtg
1260
gacaagagca gatggcagag aggcgacacc ttcatctgcg ccgtgatgca cgaggccctg
1320
cacaaccact acacccagat cagcctgagc cacagccccg gcaag
1365
<210> 50
<211> 455
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (244)..(244)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (271)..(271)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (303)..(305)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (333)..(333)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (335)..(335)
<223> Xaa может представлять любую природную аминокислоту
<400> 50
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Ser Val Asp Ile Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ser Arg Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Cys Gly Ser Gln Ser Gly Ser Thr
130 135 140
Val Ala Leu Ala Cys Leu Val Ser Gly Tyr Ile Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Val Ser Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ser Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr
180 185 190
Val Pro Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala
195 200 205
His Pro Ala Thr Asn Thr Lys Val Asp Lys Pro Val Ala Lys Glu Cys
210 215 220
Glu Cys Lys Cys Asn Cys Asn Asn Cys Pro Cys Pro Gly Cys Gly Leu
225 230 235 240
Leu Gly Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile
245 250 255
Leu Val Thr Ala Arg Thr Pro Thr Val Thr Cys Val Val Val Xaa Leu
260 265 270
Asp Pro Glu Asn Pro Glu Val Gln Ile Ser Trp Phe Val Asp Ser Lys
275 280 285
Gln Val Gln Thr Ala Asn Thr Gln Pro Arg Glu Glu Gln Ser Xaa Xaa
290 295 300
Xaa Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Ser Gly Lys Gln Phe Lys Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser
325 330 335
Pro Ile Glu Glu Ile Ile Ser Lys Thr Pro Gly Gln Ala His Gln Pro
340 345 350
Asn Val Tyr Val Leu Pro Pro Ser Arg Asp Glu Met Ser Lys Asn Thr
355 360 365
Val Thr Leu Thr Cys Leu Val Lys Asp Phe Phe Pro Pro Glu Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Met Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Ile Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210> 51
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (715)..(717)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (796)..(798)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (892)..(900)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (988)..(990)
<223> n представляет a, c, g, или t
<400> 51
gaggtgcagc tggtgcagag cggccccggc ctggtgaagc ccagccagag cctgagcctg
60
acctgcgtgg tgagcggctt cagcctgacc agctacggcg tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccagcaagac caaggtggac aagcccgtgc ccaagagaga gaacggcaga
660
gtgcccagac cccccgactg ccccaagtgc cccgcccccg agatgctggg cggcnnnagc
720
gtgttcatct tcccccccaa gcccaaggac accctgctga tcgccagaac ccccgaggtg
780
acctgcgtgg tggtgnnnct ggaccccgag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagc agatgcagac cgccaagacc cagcccagag aggagcagtt cnnnnnnnnn
900
tacagagtgg tgagcgtgct gcccatcggc caccaggact ggctgaaggg caagcagttc
960
acctgcaagg tgaacaacaa gnnnctgnnn agccccatcg agagaaccat cagcaaggcc
1020
agaggccagg cccaccagcc cagcgtgtac gtgctgcccc ccagcagaga ggagctgagc
1080
aagaacaccg tgagcctgac ctgcctgatc aaggacttct tcccccccga catcgacgtg
1140
gagtggcaga gcaacggcca gcaggagccc gagagcaagt acagaaccac ccccccccag
1200
ctggacgagg acggcagcta cttcctgtac agcaagctga gcgtggacaa gagcagatgg
1260
cagagaggcg acaccttcat ctgcgccgtg atgcacgagg ccctgcacaa ccactacacc
1320
caggagagcc tgagccacag ccccggcaag
1350
<210> 52
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (239)..(239)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (266)..(266)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (298)..(300)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (330)..(330)
<223> Xaa может представлять любую природную аминокислоту
<400> 52
Glu Val Gln Leu Val Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Val Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro
210 215 220
Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Xaa Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala
275 280 285
Lys Thr Gln Pro Arg Glu Glu Gln Phe Xaa Xaa Xaa Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe
305 310 315 320
Thr Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys
355 360 365
Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser
370 375 380
Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln
385 390 395 400
Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp
405 410 415
Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro
435 440 445
Gly Lys
450
<210> 53
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (709)..(711)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (790)..(792)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (886)..(894)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (976)..(978)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<400> 53
gaggtgcagc tggtgcagag cggccccggc ctggtgaagc ccagccagag cctgagcctg
60
acctgcgtgg tgagcggctt cagcctgacc agctacggcg tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagccaga gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta catccccgag cccgtgaccg tgagctggaa cagcgtgagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccaccaacac caaggtggac aagcccgtgg ccaaggagtg cgagtgcaag
660
tgcaactgca acaactgccc ctgccccggc tgcggcctgc tgggcggcnn nagcgtgttc
720
atcttccccc ccaagcccaa ggacatcctg gtgaccgcca gaacccccac cgtgacctgc
780
gtggtggtgn nnctggaccc cgagaacccc gaggtgcaga tcagctggtt cgtggacagc
840
aagcaggtgc agaccgccaa cacccagccc agagaggagc agagcnnnnn nnnntacaga
900
gtggtgagcg tgctgcccat cggccaccag gactggctga gcggcaagca gttcaagtgc
960
aaggtgaaca acaagnnnct gnnnagcccc atcgaggaga tcatcagcaa gacccccggc
1020
caggcccacc agcccaacgt gtacgtgctg ccccccagca gagacgagat gagcaagaac
1080
accgtgaccc tgacctgcct ggtgaaggac ttcttccccc ccgagatcga cgtggagtgg
1140
cagagcaacg gccagcagga gcccgagagc aagtacagaa tgaccccccc ccagctggac
1200
gaggacggca gctacttcct gtacagcaag ctgagcgtgg acaagagcag atggcagaga
1260
ggcgacacct tcatctgcgc cgtgatgcac gaggccctgc acaaccacta cacccagatc
1320
agcctgagcc acagccccgg caag
1344
<210> 54
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (237)..(237)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (264)..(264)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (296)..(298)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (326)..(326)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<400> 54
Glu Val Gln Leu Val Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Val Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Gln Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Ile Pro Glu Pro Val Thr Val Ser Trp Asn Ser Val Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Thr Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn
210 215 220
Asn Cys Pro Cys Pro Gly Cys Gly Leu Leu Gly Gly Xaa Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Val Thr Ala Arg Thr Pro
245 250 255
Thr Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asn Pro Glu Val
260 265 270
Gln Ile Ser Trp Phe Val Asp Ser Lys Gln Val Gln Thr Ala Asn Thr
275 280 285
Gln Pro Arg Glu Glu Gln Ser Xaa Xaa Xaa Tyr Arg Val Val Ser Val
290 295 300
Leu Pro Ile Gly His Gln Asp Trp Leu Ser Gly Lys Gln Phe Lys Cys
305 310 315 320
Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Glu Ile Ile Ser
325 330 335
Lys Thr Pro Gly Gln Ala His Gln Pro Asn Val Tyr Val Leu Pro Pro
340 345 350
Ser Arg Asp Glu Met Ser Lys Asn Thr Val Thr Leu Thr Cys Leu Val
355 360 365
Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly
370 375 380
Gln Gln Glu Pro Glu Ser Lys Tyr Arg Met Thr Pro Pro Gln Leu Asp
385 390 395 400
Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser
405 410 415
Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Ile Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 55
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (715)..(717)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (796)..(798)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (892)..(900)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (988)..(990)
<223> n представляет a, c, g, или t
<400> 55
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccagcaagac caaggtggac aagcccgtgc ccaagagaga gaacggcaga
660
gtgcccagac cccccgactg ccccaagtgc cccgcccccg agatgctggg cggcnnnagc
720
gtgttcatct tcccccccaa gcccaaggac accctgctga tcgccagaac ccccgaggtg
780
acctgcgtgg tggtgnnnct ggaccccgag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagc agatgcagac cgccaagacc cagcccagag aggagcagtt cnnnnnnnnn
900
tacagagtgg tgagcgtgct gcccatcggc caccaggact ggctgaaggg caagcagttc
960
acctgcaagg tgaacaacaa gnnnctgnnn agccccatcg agagaaccat cagcaaggcc
1020
agaggccagg cccaccagcc cagcgtgtac gtgctgcccc ccagcagaga ggagctgagc
1080
aagaacaccg tgagcctgac ctgcctgatc aaggacttct tcccccccga catcgacgtg
1140
gagtggcaga gcaacggcca gcaggagccc gagagcaagt acagaaccac ccccccccag
1200
ctggacgagg acggcagcta cttcctgtac agcaagctga gcgtggacaa gagcagatgg
1260
cagagaggcg acaccttcat ctgcgccgtg atgcacgagg ccctgcacaa ccactacacc
1320
caggagagcc tgagccacag ccccggcaag
1350
<210> 56
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (239)..(239)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (266)..(266)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (298)..(300)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (330)..(330)
<223> Xaa может представлять любую природную аминокислоту
<400> 56
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro
210 215 220
Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Xaa Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala
275 280 285
Lys Thr Gln Pro Arg Glu Glu Gln Phe Xaa Xaa Xaa Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe
305 310 315 320
Thr Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys
355 360 365
Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser
370 375 380
Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln
385 390 395 400
Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp
405 410 415
Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro
435 440 445
Gly Lys
450
<210> 57
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (709)..(711)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (790)..(792)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (886)..(894)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (976)..(978)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<400> 57
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagccaga gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta catccccgag cccgtgaccg tgagctggaa cagcgtgagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccaccaacac caaggtggac aagcccgtgg ccaaggagtg cgagtgcaag
660
tgcaactgca acaactgccc ctgccccggc tgcggcctgc tgggcggcnn nagcgtgttc
720
atcttccccc ccaagcccaa ggacatcctg gtgaccgcca gaacccccac cgtgacctgc
780
gtggtggtgn nnctggaccc cgagaacccc gaggtgcaga tcagctggtt cgtggacagc
840
aagcaggtgc agaccgccaa cacccagccc agagaggagc agagcnnnnn nnnntacaga
900
gtggtgagcg tgctgcccat cggccaccag gactggctga gcggcaagca gttcaagtgc
960
aaggtgaaca acaagnnnct gnnnagcccc atcgaggaga tcatcagcaa gacccccggc
1020
caggcccacc agcccaacgt gtacgtgctg ccccccagca gagacgagat gagcaagaac
1080
accgtgaccc tgacctgcct ggtgaaggac ttcttccccc ccgagatcga cgtggagtgg
1140
cagagcaacg gccagcagga gcccgagagc aagtacagaa tgaccccccc ccagctggac
1200
gaggacggca gctacttcct gtacagcaag ctgagcgtgg acaagagcag atggcagaga
1260
ggcgacacct tcatctgcgc cgtgatgcac gaggccctgc acaaccacta cacccagatc
1320
agcctgagcc acagccccgg caag
1344
<210> 58
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (237)..(237)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (264)..(264)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (296)..(298)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (326)..(326)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<400> 58
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Gln Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Ile Pro Glu Pro Val Thr Val Ser Trp Asn Ser Val Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Thr Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn
210 215 220
Asn Cys Pro Cys Pro Gly Cys Gly Leu Leu Gly Gly Xaa Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Val Thr Ala Arg Thr Pro
245 250 255
Thr Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asn Pro Glu Val
260 265 270
Gln Ile Ser Trp Phe Val Asp Ser Lys Gln Val Gln Thr Ala Asn Thr
275 280 285
Gln Pro Arg Glu Glu Gln Ser Xaa Xaa Xaa Tyr Arg Val Val Ser Val
290 295 300
Leu Pro Ile Gly His Gln Asp Trp Leu Ser Gly Lys Gln Phe Lys Cys
305 310 315 320
Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Glu Ile Ile Ser
325 330 335
Lys Thr Pro Gly Gln Ala His Gln Pro Asn Val Tyr Val Leu Pro Pro
340 345 350
Ser Arg Asp Glu Met Ser Lys Asn Thr Val Thr Leu Thr Cys Leu Val
355 360 365
Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly
370 375 380
Gln Gln Glu Pro Glu Ser Lys Tyr Arg Met Thr Pro Pro Gln Leu Asp
385 390 395 400
Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser
405 410 415
Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Ile Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 59
<211> 1365
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (730)..(732)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (811)..(813)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (907)..(915)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (997)..(999)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (1003)..(1005)
<223> n представляет a, c, g, или t
<400> 59
gaggtgcagc tggtgcagag cgtggccgag ctggtgaagc ccggcgccag cgtgaaggtg
60
agctgcaccg tgagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgaccccg ccaacgtgaa caccaagtac
180
gcccccaagt tccagggcag agccaccatc accgccgaca ccagcaccaa caccgcctac
240
atgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagaatctac
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag caccagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacttcc ccgagcccgt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgtaca gcctgagcag catggtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggccca ccccgccagc aagaccaagg tggacaagcc cgtgcccaag
660
agagagaacg gcagagtgcc cagacccccc gactgcccca agtgccccgc ccccgagatg
720
ctgggcggcn nnagcgtgtt catcttcccc cccaagccca aggacaccct gctgatcgcc
780
agaacccccg aggtgacctg cgtggtggtg nnnctggacc ccgaggaccc cgaggtgcag
840
atcagctggt tcgtggacgg caagcagatg cagaccgcca agacccagcc cagagaggag
900
cagttcnnnn nnnnntacag agtggtgagc gtgctgccca tcggccacca ggactggctg
960
aagggcaagc agttcacctg caaggtgaac aacaagnnnc tgnnnagccc catcgagaga
1020
accatcagca aggccagagg ccaggcccac cagcccagcg tgtacgtgct gccccccagc
1080
agagaggagc tgagcaagaa caccgtgagc ctgacctgcc tgatcaagga cttcttcccc
1140
cccgacatcg acgtggagtg gcagagcaac ggccagcagg agcccgagag caagtacaga
1200
accacccccc cccagctgga cgaggacggc agctacttcc tgtacagcaa gctgagcgtg
1260
gacaagagca gatggcagag aggcgacacc ttcatctgcg ccgtgatgca cgaggccctg
1320
cacaaccact acacccagga gagcctgagc cacagccccg gcaag
1365
<210> 60
<211> 455
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (244)..(244)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (271)..(271)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (303)..(305)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (333)..(333)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (335)..(335)
<223> Xaa может представлять любую природную аминокислоту
<400> 60
Glu Val Gln Leu Val Gln Ser Val Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Thr Val Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly
210 215 220
Arg Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met
225 230 235 240
Leu Gly Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Xaa Leu
260 265 270
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
275 280 285
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Xaa Xaa
290 295 300
Xaa Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
305 310 315 320
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser
325 330 335
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
340 345 350
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
355 360 365
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
370 375 380
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
385 390 395 400
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
405 410 415
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
420 425 430
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
435 440 445
Leu Ser His Ser Pro Gly Lys
450 455
<210> 61
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (724)..(726)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (805)..(807)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (901)..(909)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (991)..(993)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (997)..(999)
<223> n представляет a, c, g, или t
<400> 61
gaggtgcagc tggtgcagag cgtggccgag ctggtgaagc ccggcgccag cgtgaaggtg
60
agctgcaccg tgagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgaccccg ccaacgtgaa caccaagtac
180
gcccccaagt tccagggcag agccaccatc accgccgaca ccagcaccaa caccgcctac
240
atgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagaatctac
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag caccagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacttcc ccgagcccgt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgtaca gcctgagcag catggtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggccca ccccgccagc aagaccaagg tggacaagcc cgtggccaag
660
gagtgcgagt gcaagtgcaa ctgcaacaac tgcccctgcc ccggctgcgg cctgctgggc
720
ggcnnnagcg tgttcatctt cccccccaag cccaaggaca tcctggtgac cgccagaacc
780
cccaccgtga cctgcgtggt ggtgnnnctg gaccccgaga accccgaggt gcagatcagc
840
tggttcgtgg acagcaagca ggtgcagacc gccaacaccc agcccagaga ggagcagagc
900
nnnnnnnnnt acagagtggt gagcgtgctg cccatcggcc accaggactg gctgagcggc
960
aagcagttca agtgcaaggt gaacaacaag nnnctgnnna gccccatcga ggagatcatc
1020
agcaagaccc ccggccaggc ccaccagccc aacgtgtacg tgctgccccc cagcagagac
1080
gagatgagca agaacaccgt gaccctgacc tgcctggtga aggacttctt cccccccgag
1140
atcgacgtgg agtggcagag caacggccag caggagcccg agagcaagta cagaatgacc
1200
cccccccagc tggacgagga cggcagctac ttcctgtaca gcaagctgag cgtggacaag
1260
agcagatggc agagaggcga caccttcatc tgcgccgtga tgcacgaggc cctgcacaac
1320
cactacaccc agatcagcct gagccacagc cccggcaag
1359
<210> 62
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (242)..(242)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (269)..(269)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (301)..(303)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (331)..(331)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (333)..(333)
<223> Xaa может представлять любую природную аминокислоту
<400> 62
Glu Val Gln Leu Val Gln Ser Val Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Thr Val Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro
195 200 205
Ala Ser Lys Thr Lys Val Asp Lys Pro Val Ala Lys Glu Cys Glu Cys
210 215 220
Lys Cys Asn Cys Asn Asn Cys Pro Cys Pro Gly Cys Gly Leu Leu Gly
225 230 235 240
Gly Xaa Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Val
245 250 255
Thr Ala Arg Thr Pro Thr Val Thr Cys Val Val Val Xaa Leu Asp Pro
260 265 270
Glu Asn Pro Glu Val Gln Ile Ser Trp Phe Val Asp Ser Lys Gln Val
275 280 285
Gln Thr Ala Asn Thr Gln Pro Arg Glu Glu Gln Ser Xaa Xaa Xaa Tyr
290 295 300
Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Ser Gly
305 310 315 320
Lys Gln Phe Lys Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile
325 330 335
Glu Glu Ile Ile Ser Lys Thr Pro Gly Gln Ala His Gln Pro Asn Val
340 345 350
Tyr Val Leu Pro Pro Ser Arg Asp Glu Met Ser Lys Asn Thr Val Thr
355 360 365
Leu Thr Cys Leu Val Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu
370 375 380
Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Met Thr
385 390 395 400
Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu
405 410 415
Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Ile Ser Leu Ser
435 440 445
His Ser Pro Gly Lys
450
<210> 63
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (715)..(717)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (796)..(798)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (892)..(900)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (988)..(990)
<223> n представляет a, c, g, или t
<400> 63
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat gcccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccagcaagac caaggtggac aagcccgtgc ccaagagaga gaacggcaga
660
gtgcccagac cccccgactg ccccaagtgc cccgcccccg agatgctggg cggcnnnagc
720
gtgttcatct tcccccccaa gcccaaggac accctgctga tcgccagaac ccccgaggtg
780
acctgcgtgg tggtgnnnct ggaccccgag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagc agatgcagac cgccaagacc cagcccagag aggagcagtt cnnnnnnnnn
900
tacagagtgg tgagcgtgct gcccatcggc caccaggact ggctgaaggg caagcagttc
960
acctgcaagg tgaacaacaa gnnnctgnnn agccccatcg agagaaccat cagcaaggcc
1020
agaggccagg cccaccagcc cagcgtgtac gtgctgcccc ccagcagaga ggagctgagc
1080
aagaacaccg tgagcctgac ctgcctgatc aaggacttct tcccccccga catcgacgtg
1140
gagtggcaga gcaacggcca gcaggagccc gagagcaagt acagaaccac ccccccccag
1200
ctggacgagg acggcagcta cttcctgtac agcaagctga gcgtggacaa gagcagatgg
1260
cagagaggcg acaccttcat ctgcgccgtg atgcacgagg ccctgcacaa ccactacacc
1320
caggagagcc tgagccacag ccccggcaag
1350
<210> 64
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (239)..(239)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (266)..(266)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (298)..(300)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (330)..(330)
<223> Xaa может представлять любую природную аминокислоту
<400> 64
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro
210 215 220
Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Xaa Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala
275 280 285
Lys Thr Gln Pro Arg Glu Glu Gln Phe Xaa Xaa Xaa Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe
305 310 315 320
Thr Cys Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys
355 360 365
Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser
370 375 380
Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln
385 390 395 400
Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp
405 410 415
Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro
435 440 445
Gly Lys
450
<210> 65
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (709)..(711)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (790)..(792)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (886)..(894)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (976)..(978)
<223> n представляет a, c, g, или t
<220>
<221> misc_feature
<222> (982)..(984)
<223> n представляет a, c, g, или t
<400> 65
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat gcccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gcccaccccg ccagcaagac caaggtggac aagcccgtgg ccaaggagtg cgagtgcaag
660
tgcaactgca acaactgccc ctgccccggc tgcggcctgc tgggcggcnn nagcgtgttc
720
atcttccccc ccaagcccaa ggacatcctg gtgaccgcca gaacccccac cgtgacctgc
780
gtggtggtgn nnctggaccc cgagaacccc gaggtgcaga tcagctggtt cgtggacagc
840
aagcaggtgc agaccgccaa cacccagccc agagaggagc agagcnnnnn nnnntacaga
900
gtggtgagcg tgctgcccat cggccaccag gactggctga gcggcaagca gttcaagtgc
960
aaggtgaaca acaagnnnct gnnnagcccc atcgaggaga tcatcagcaa gacccccggc
1020
caggcccacc agcccaacgt gtacgtgctg ccccccagca gagacgagat gagcaagaac
1080
accgtgaccc tgacctgcct ggtgaaggac ttcttccccc ccgagatcga cgtggagtgg
1140
cagagcaacg gccagcagga gcccgagagc aagtacagaa tgaccccccc ccagctggac
1200
gaggacggca gctacttcct gtacagcaag ctgagcgtgg acaagagcag atggcagaga
1260
ggcgacacct tcatctgcgc cgtgatgcac gaggccctgc acaaccacta cacccagatc
1320
agcctgagcc acagccccgg caag
1344
<210> 66
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<220>
<221> misc_feature
<222> (237)..(237)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (264)..(264)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (296)..(298)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (326)..(326)
<223> Xaa может представлять любую природную аминокислоту
<220>
<221> misc_feature
<222> (328)..(328)
<223> Xaa может представлять любую природную аминокислоту
<400> 66
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys
195 200 205
Val Asp Lys Pro Val Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn
210 215 220
Asn Cys Pro Cys Pro Gly Cys Gly Leu Leu Gly Gly Xaa Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Val Thr Ala Arg Thr Pro
245 250 255
Thr Val Thr Cys Val Val Val Xaa Leu Asp Pro Glu Asn Pro Glu Val
260 265 270
Gln Ile Ser Trp Phe Val Asp Ser Lys Gln Val Gln Thr Ala Asn Thr
275 280 285
Gln Pro Arg Glu Glu Gln Ser Xaa Xaa Xaa Tyr Arg Val Val Ser Val
290 295 300
Leu Pro Ile Gly His Gln Asp Trp Leu Ser Gly Lys Gln Phe Lys Cys
305 310 315 320
Lys Val Asn Asn Lys Xaa Leu Xaa Ser Pro Ile Glu Glu Ile Ile Ser
325 330 335
Lys Thr Pro Gly Gln Ala His Gln Pro Asn Val Tyr Val Leu Pro Pro
340 345 350
Ser Arg Asp Glu Met Ser Lys Asn Thr Val Thr Leu Thr Cys Leu Val
355 360 365
Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly
370 375 380
Gln Gln Glu Pro Glu Ser Lys Tyr Arg Met Thr Pro Pro Gln Leu Asp
385 390 395 400
Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser
405 410 415
Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Ile Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 67
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 67
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gcacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgt tcaacgagtg cagatgcacc
660
gacacccccc cctgccccgt gcccgagccc ctgggcggcc ccagcgtgct gatcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg aggtgacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagag cagagagcag cagttcaacg gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcgacc tgcccagccc catcgagaga accatcagca aggccagagg cagagcccac
1020
aagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
agcatcacct gcctgatcaa ggacttctac ccccccgaca tcgacgtgga gtggcagagc
1140
aacggccagc aggagcccga gagaaagcac agaatgaccc ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
cccttcacct gcgccgtgat gcacgagacc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 68
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 68
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu His Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro
210 215 220
Cys Pro Val Pro Glu Pro Leu Gly Gly Pro Ser Val Leu Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Ser Arg
275 280 285
Glu Gln Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Asp Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Arg Ala His Lys Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Ser Ile Thr Cys Leu Ile Lys Asp
355 360 365
Phe Tyr Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Gln
370 375 380
Glu Pro Glu Arg Lys His Arg Met Thr Pro Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Pro Phe Thr Cys Ala Val Met His Glu Thr Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 69
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 69
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcaccg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgc ccaaggagag cacctgcaag
660
tgcatcagcc cctgccccgt gcccgagagc ctgggcggcc ccagcgtgtt catcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg agatcacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagcc cagagagcag cagttcaaca gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcggcc tgcccagccc catcgagaga accatcagca aggccagagg ccaggcccac
1020
cagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
accctgacct gcctgatcaa ggacttcttc ccccccgaga tcgacgtgga gtggcagagc
1140
aacggccagc ccgagcccga gagcaagtac cacaccaccg ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
accttcacct gcgccgtgat gcacgaggcc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 70
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 70
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Thr Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Glu Ser Thr Cys Lys Cys Ile Ser Pro
210 215 220
Cys Pro Val Pro Glu Ser Leu Gly Gly Pro Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg
275 280 285
Glu Gln Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Gly Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp
355 360 365
Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro
370 375 380
Glu Pro Glu Ser Lys Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Thr Phe Thr Cys Ala Val Met His Glu Ala Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 71
<211> 672
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 71
gacatcgtga tgacccagac ccccctgagc ctgagcgtga gccccggcga gcccgccagc
60
atgagctgca agagcagcca gagcctgctg aacagcgtga accagaagaa ctacctggcc
120
tggtacagac agaagcccgg ccagagcccc caggtgctgg tgtacttcgc cagcgccaga
180
gtgagcggcg tgcccgacag attcatcggc agcggcagcg gcaccgactt caccctgaga
240
atcagcagag tggaggccga cgacctgggc gtgtactact gccagcagta cttcagcacc
300
cccctgacct tcggccaggg caccaagctg gagctgaaga gaaacgacgc ccagcccgcc
360
gtgtacctgt tccagcccag ccccgaccag ctgcacaccg gcagcgccag cgtggtgtgc
420
ctgctgaaca gcttctaccc caaggacatc aacgtgaagt ggaaggtgga cggcgtgatc
480
caggacaccg gcatccagga gagcgtgacc gagcaggaca gcaaggacag cacctacagc
540
ctgagcagca ccctgaccat gagcagcacc gagtacctga gccacgagct gtacagctgc
600
gagatcaccc acaagagcct gcccagcacc ctgatcaaga gcttccagag aagcgagtgc
660
cagagagtgg ac
672
<210> 72
<211> 224
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 72
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Arg Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Val Leu Val Tyr Phe Ala Ser Ala Arg Val Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg
65 70 75 80
Ile Ser Arg Val Glu Ala Asp Asp Leu Gly Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Thr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Leu
100 105 110
Lys Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro
115 120 125
Asp Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser
130 135 140
Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile
145 150 155 160
Gln Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr
180 185 190
Leu Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro
195 200 205
Ser Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210> 73
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 73
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgcccccg ccaacgtgga caccaagtac
180
gcccccaagt tccagggcaa ggccaccatc agcgccgaca ccgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt actactgcgt gctgatctac
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag caccagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacttcc ccgagcccgt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgcaca gcctgagcag catggtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggtgca ccccgccagc aacaccaagg tggacaagcc cgtgttcaac
660
gagtgcagat gcaccgacac ccccccctgc cccgtgcccg agcccctggg cggccccagc
720
gtgctgatct tcccccccaa gcccaaggac atcctgagaa tcaccagaac ccccgaggtg
780
acctgcgtgg tgctggacct gggcagagag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagg aggtgcacac cgccaagacc cagagcagag agcagcagtt caacggcacc
900
tacagagtgg tgagcgtgct gcccatcgag caccaggact ggctgaccgg caaggagttc
960
aagtgcagag tgaaccacat cgacctgccc agccccatcg agagaaccat cagcaaggcc
1020
agaggcagag cccacaagcc cagcgtgtac gtgctgcccc ccagccccaa ggagctgagc
1080
agcagcgaca ccgtgagcat cacctgcctg atcaaggact tctacccccc cgacatcgac
1140
gtggagtggc agagcaacgg ccagcaggag cccgagagaa agcacagaat gacccccccc
1200
cagctggacg aggacggcag ctacttcctg tacagcaagc tgagcgtgga caagagcaga
1260
tggcagcagg gcgacccctt cacctgcgcc gtgatgcacg agaccctgca gaaccactac
1320
accgacctga gcctgagcca cagccccggc aag
1353
<210> 74
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 74
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Ser Ala Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Leu Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu His Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Val His Pro
195 200 205
Ala Ser Asn Thr Lys Val Asp Lys Pro Val Phe Asn Glu Cys Arg Cys
210 215 220
Thr Asp Thr Pro Pro Cys Pro Val Pro Glu Pro Leu Gly Gly Pro Ser
225 230 235 240
Val Leu Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Glu Val His Thr Ala
275 280 285
Lys Thr Gln Ser Arg Glu Gln Gln Phe Asn Gly Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn His Ile Asp Leu Pro Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Arg Ala His Lys Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp Thr Val Ser Ile Thr
355 360 365
Cys Leu Ile Lys Asp Phe Tyr Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Arg Lys His Arg Met Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asp Pro Phe Thr Cys Ala Val Met
420 425 430
His Glu Thr Leu Gln Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210> 75
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 75
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgcccccg ccaacgtgga caccaagtac
180
gcccccaagt tccagggcaa ggccaccatc agcgccgaca ccgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt actactgcgt gctgatctac
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag caccagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacttcc ccgagcccgt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgtaca gcctgagcag caccgtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggtgca ccccgccagc aacaccaagg tggacaagcc cgtgcccaag
660
gagagcacct gcaagtgcat cagcccctgc cccgtgcccg agagcctggg cggccccagc
720
gtgttcatct tcccccccaa gcccaaggac atcctgagaa tcaccagaac ccccgagatc
780
acctgcgtgg tgctggacct gggcagagag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagg aggtgcacac cgccaagacc cagcccagag agcagcagtt caacagcacc
900
tacagagtgg tgagcgtgct gcccatcgag caccaggact ggctgaccgg caaggagttc
960
aagtgcagag tgaaccacat cggcctgccc agccccatcg agagaaccat cagcaaggcc
1020
agaggccagg cccaccagcc cagcgtgtac gtgctgcccc ccagccccaa ggagctgagc
1080
agcagcgaca ccgtgaccct gacctgcctg atcaaggact tcttcccccc cgagatcgac
1140
gtggagtggc agagcaacgg ccagcccgag cccgagagca agtaccacac caccgccccc
1200
cagctggacg aggacggcag ctacttcctg tacagcaagc tgagcgtgga caagagcaga
1260
tggcagcagg gcgacacctt cacctgcgcc gtgatgcacg aggccctgca gaaccactac
1320
accgacctga gcctgagcca cagccccggc aag
1353
<210> 76
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 76
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Ala Pro Ala Asn Val Asp Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Ser Ala Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Leu Ile Tyr Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Thr Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Val His Pro
195 200 205
Ala Ser Asn Thr Lys Val Asp Lys Pro Val Pro Lys Glu Ser Thr Cys
210 215 220
Lys Cys Ile Ser Pro Cys Pro Val Pro Glu Ser Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg
245 250 255
Thr Pro Glu Ile Thr Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Glu Val His Thr Ala
275 280 285
Lys Thr Gln Pro Arg Glu Gln Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn His Ile Gly Leu Pro Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp Thr Val Thr Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Pro Glu Pro Glu Ser Lys Tyr His Thr Thr Ala Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asp Thr Phe Thr Cys Ala Val Met
420 425 430
His Glu Ala Leu Gln Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210> 77
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 77
gacatcgtga tgacccagac ccccctgagc ctgagcgtga gcctgggcga gcccgccagc
60
atcagctgcc acgccagcca gaacatcaac gtgtggctga gctggtacag acagaagccc
120
ggccagatcc cccagctgct gatctacaag gccagccacc tgcacaccgg cgtgcccgac
180
agattcagcg gcagcggcag cggcaccgac ttcaccctga gaatcagcag agtggaggcc
240
gacgacgccg gcgtgtacta ctgccagcag ggccagagct ggcccctgac cttcggccag
300
ggcaccaagg tggagatcaa gagaaacgac gcccagcccg ccgtgtacct gttccagccc
360
agccccgacc agctgcacac cggcagcgcc agcgtggtgt gcctgctgaa cagcttctac
420
cccaaggaca tcaacgtgaa gtggaaggtg gacggcgtga tccaggacac cggcatccag
480
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc
540
atgagcagca ccgagtacct gagccacgag ctgtacagct gcgagatcac ccacaagagc
600
ctgcccagca ccctgatcaa gagcttccag agaagcgagt gccagagagt ggac
654
<210> 78
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 78
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Leu Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Arg Gln Lys Pro Gly Gln Ile Pro Gln Leu Leu Ile
35 40 45
Tyr Lys Ala Ser His Leu His Thr Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val Glu Ala
65 70 75 80
Asp Asp Ala Gly Val Tyr Tyr Cys Gln Gln Gly Gln Ser Trp Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Asn Asp Ala Gln
100 105 110
Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr Gly
115 120 125
Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu Tyr
180 185 190
Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys Ser
195 200 205
Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210> 79
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 79
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc agatacaaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcacc atctaccccg gctacggcga caccagctac
180
aaccagaagt tcaagggcaa ggccaccctg agcgtggaca tcgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt acttctgcag cagagagttc
300
gccgacgact accccatccc ccccttcgac tactggggcc agggcaccct ggtgaccgtg
360
agcagcgcca gcaccaccgc ccccagcgtg ttccccctgg cccccagctg cggcagcacc
420
agcggcagca ccgtggccct ggcctgcctg gtgagcggct acttccccga gcccgtgacc
480
gtgagctgga acagcggcag cctgaccagc ggcgtgcaca ccttccccag cgtgctgcag
540
agcagcggcc tgcacagcct gagcagcatg gtgaccgtgc ccagcagcag atggcccagc
600
gagaccttca cctgcaacgt ggtgcacccc gccagcaaca ccaaggtgga caagcccgtg
660
ttcaacgagt gcagatgcac cgacaccccc ccctgccccg tgcccgagcc cctgggcggc
720
cccagcgtgc tgatcttccc ccccaagccc aaggacatcc tgagaatcac cagaaccccc
780
gaggtgacct gcgtggtgct ggacctgggc agagaggacc ccgaggtgca gatcagctgg
840
ttcgtggacg gcaaggaggt gcacaccgcc aagacccaga gcagagagca gcagttcaac
900
ggcacctaca gagtggtgag cgtgctgccc atcgagcacc aggactggct gaccggcaag
960
gagttcaagt gcagagtgaa ccacatcgac ctgcccagcc ccatcgagag aaccatcagc
1020
aaggccagag gcagagccca caagcccagc gtgtacgtgc tgccccccag ccccaaggag
1080
ctgagcagca gcgacaccgt gagcatcacc tgcctgatca aggacttcta cccccccgac
1140
atcgacgtgg agtggcagag caacggccag caggagcccg agagaaagca cagaatgacc
1200
cccccccagc tggacgagga cggcagctac ttcctgtaca gcaagctgag cgtggacaag
1260
agcagatggc agcagggcga ccccttcacc tgcgccgtga tgcacgagac cctgcagaac
1320
cactacaccg acctgagcct gagccacagc cccggcaag
1359
<210> 80
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 80
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Ser Val Asp Ile Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ser Arg Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr
130 135 140
Val Ala Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ser Val Leu Gln Ser Ser Gly Leu His Ser Leu Ser Ser Met Val Thr
180 185 190
Val Pro Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Val
195 200 205
His Pro Ala Ser Asn Thr Lys Val Asp Lys Pro Val Phe Asn Glu Cys
210 215 220
Arg Cys Thr Asp Thr Pro Pro Cys Pro Val Pro Glu Pro Leu Gly Gly
225 230 235 240
Pro Ser Val Leu Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Arg Ile
245 250 255
Thr Arg Thr Pro Glu Val Thr Cys Val Val Leu Asp Leu Gly Arg Glu
260 265 270
Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Glu Val His
275 280 285
Thr Ala Lys Thr Gln Ser Arg Glu Gln Gln Phe Asn Gly Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu Thr Gly Lys
305 310 315 320
Glu Phe Lys Cys Arg Val Asn His Ile Asp Leu Pro Ser Pro Ile Glu
325 330 335
Arg Thr Ile Ser Lys Ala Arg Gly Arg Ala His Lys Pro Ser Val Tyr
340 345 350
Val Leu Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp Thr Val Ser
355 360 365
Ile Thr Cys Leu Ile Lys Asp Phe Tyr Pro Pro Asp Ile Asp Val Glu
370 375 380
Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Arg Lys His Arg Met Thr
385 390 395 400
Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu
405 410 415
Ser Val Asp Lys Ser Arg Trp Gln Gln Gly Asp Pro Phe Thr Cys Ala
420 425 430
Val Met His Glu Thr Leu Gln Asn His Tyr Thr Asp Leu Ser Leu Ser
435 440 445
His Ser Pro Gly Lys
450
<210> 81
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 81
gaggtgcagc tggtgcagag cggcggcgac ctggtgaagc ccggcggcag cgtgagactg
60
agctgcgtgg ccagcggcta caccttcacc agatacaaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcacc atctaccccg gctacggcga caccagctac
180
aaccagaagt tcaagggcaa ggccaccctg agcgtggaca tcgccaagaa caccgcctac
240
atgcagctga acagcctgag agccgaggac accgccgtgt acttctgcag cagagagttc
300
gccgacgact accccatccc ccccttcgac tactggggcc agggcaccct ggtgaccgtg
360
agcagcgcca gcaccaccgc ccccagcgtg ttccccctgg cccccagctg cggcagcacc
420
agcggcagca ccgtggccct ggcctgcctg gtgagcggct acttccccga gcccgtgacc
480
gtgagctgga acagcggcag cctgaccagc ggcgtgcaca ccttccccag cgtgctgcag
540
agcagcggcc tgtacagcct gagcagcacc gtgaccgtgc ccagcagcag atggcccagc
600
gagaccttca cctgcaacgt ggtgcacccc gccagcaaca ccaaggtgga caagcccgtg
660
cccaaggaga gcacctgcaa gtgcatcagc ccctgccccg tgcccgagag cctgggcggc
720
cccagcgtgt tcatcttccc ccccaagccc aaggacatcc tgagaatcac cagaaccccc
780
gagatcacct gcgtggtgct ggacctgggc agagaggacc ccgaggtgca gatcagctgg
840
ttcgtggacg gcaaggaggt gcacaccgcc aagacccagc ccagagagca gcagttcaac
900
agcacctaca gagtggtgag cgtgctgccc atcgagcacc aggactggct gaccggcaag
960
gagttcaagt gcagagtgaa ccacatcggc ctgcccagcc ccatcgagag aaccatcagc
1020
aaggccagag gccaggccca ccagcccagc gtgtacgtgc tgccccccag ccccaaggag
1080
ctgagcagca gcgacaccgt gaccctgacc tgcctgatca aggacttctt cccccccgag
1140
atcgacgtgg agtggcagag caacggccag cccgagcccg agagcaagta ccacaccacc
1200
gccccccagc tggacgagga cggcagctac ttcctgtaca gcaagctgag cgtggacaag
1260
agcagatggc agcagggcga caccttcacc tgcgccgtga tgcacgaggc cctgcagaac
1320
cactacaccg acctgagcct gagccacagc cccggcaag
1359
<210> 82
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 82
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Asn Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Thr Ile Tyr Pro Gly Tyr Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Ser Val Asp Ile Ala Lys Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ser Arg Glu Phe Ala Asp Asp Tyr Pro Ile Pro Pro Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr
130 135 140
Val Ala Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ser Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Thr Val Thr
180 185 190
Val Pro Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Val
195 200 205
His Pro Ala Ser Asn Thr Lys Val Asp Lys Pro Val Pro Lys Glu Ser
210 215 220
Thr Cys Lys Cys Ile Ser Pro Cys Pro Val Pro Glu Ser Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Arg Ile
245 250 255
Thr Arg Thr Pro Glu Ile Thr Cys Val Val Leu Asp Leu Gly Arg Glu
260 265 270
Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Glu Val His
275 280 285
Thr Ala Lys Thr Gln Pro Arg Glu Gln Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu Thr Gly Lys
305 310 315 320
Glu Phe Lys Cys Arg Val Asn His Ile Gly Leu Pro Ser Pro Ile Glu
325 330 335
Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr
340 345 350
Val Leu Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp Thr Val Thr
355 360 365
Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu
370 375 380
Trp Gln Ser Asn Gly Gln Pro Glu Pro Glu Ser Lys Tyr His Thr Thr
385 390 395 400
Ala Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu
405 410 415
Ser Val Asp Lys Ser Arg Trp Gln Gln Gly Asp Thr Phe Thr Cys Ala
420 425 430
Val Met His Glu Ala Leu Gln Asn His Tyr Thr Asp Leu Ser Leu Ser
435 440 445
His Ser Pro Gly Lys
450
<210> 83
<211> 666
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 83
gacatcgtga tgacccagac ccccctgagc ctgcccgtga gcctgggcga gcccgccagc
60
atcagctgca gaagcagcca gaacatcgtg cacagcaacg gcaacaccta cctggagtgg
120
tacagacaga agcccggcca gagcccccag ctgctgatct acaaggtgag caacagattc
180
agcggcgtgc ccgacagatt cagcggcagc ggcagcggca ccgacttcac cctgagaatc
240
agcagagtgg aggccgacga cgccggcgtg tactactgct tccagggcag ccacgtgccc
300
tacaccttcg gccagggcac caaggtggag atcaagagag acgcccagcc cgccgtgtac
360
ctgttccagc ccagccccga ccagctgcac accggcagcg ccagcgtggt gtgcctgctg
420
aacagcttct accccaagga catcaacgtg aagtggaagg tggacggcgt gatccaggac
480
accggcatcc aggagagcgt gaccgagcag gacagcaagg acagcaccta cagcctgagc
540
agcaccctga ccatgagcag caccgagtac ctgagccacg agctgtacag ctgcgagatc
600
acccacaaga gcctgcccag caccctgatc aagagcttcc agagaagcga gtgccagaga
660
gtggac
666
<210> 84
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 84
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Asn Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Arg Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln
115 120 125
Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr
130 135 140
Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp
145 150 155 160
Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser
180 185 190
His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr
195 200 205
Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210> 85
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 85
gaggtgcagc tggtgcagag cggccccggc ctggtgaagc ccagccagag cctgagcctg
60
acctgcgtgg tgagcggctt cagcctgacc agctacggcg tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gcacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgt tcaacgagtg cagatgcacc
660
gacacccccc cctgccccgt gcccgagccc ctgggcggcc ccagcgtgct gatcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg aggtgacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagag cagagagcag cagttcaacg gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcgacc tgcccagccc catcgagaga accatcagca aggccagagg cagagcccac
1020
aagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
agcatcacct gcctgatcaa ggacttctac ccccccgaca tcgacgtgga gtggcagagc
1140
aacggccagc aggagcccga gagaaagcac agaatgaccc ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
cccttcacct gcgccgtgat gcacgagacc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 86
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 86
Glu Val Gln Leu Val Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Val Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu His Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro
210 215 220
Cys Pro Val Pro Glu Pro Leu Gly Gly Pro Ser Val Leu Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Ser Arg
275 280 285
Glu Gln Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Asp Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Arg Ala His Lys Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Ser Ile Thr Cys Leu Ile Lys Asp
355 360 365
Phe Tyr Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Gln
370 375 380
Glu Pro Glu Arg Lys His Arg Met Thr Pro Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Pro Phe Thr Cys Ala Val Met His Glu Thr Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 87
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 87
gaggtgcagc tggtgcagag cggccccggc ctggtgaagc ccagccagag cctgagcctg
60
acctgcgtgg tgagcggctt cagcctgacc agctacggcg tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcaccttc agcctggaca ccgccaagaa caccgcctac
240
ctgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcaccg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgc ccaaggagag cacctgcaag
660
tgcatcagcc cctgccccgt gcccgagagc ctgggcggcc ccagcgtgtt catcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg agatcacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagcc cagagagcag cagttcaaca gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcggcc tgcccagccc catcgagaga accatcagca aggccagagg ccaggcccac
1020
cagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
accctgacct gcctgatcaa ggacttcttc ccccccgaga tcgacgtgga gtggcagagc
1140
aacggccagc ccgagcccga gagcaagtac cacaccaccg ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
accttcacct gcgccgtgat gcacgaggcc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 88
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 88
Glu Val Gln Leu Val Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Val Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Thr Phe Ser Leu Asp Thr Ala Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Thr Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Glu Ser Thr Cys Lys Cys Ile Ser Pro
210 215 220
Cys Pro Val Pro Glu Ser Leu Gly Gly Pro Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg
275 280 285
Glu Gln Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Gly Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp
355 360 365
Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro
370 375 380
Glu Pro Glu Ser Lys Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Thr Phe Thr Cys Ala Val Met His Glu Ala Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 89
<211> 672
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 89
gacatcgtga tgacccagac ccccctgagc ctgagcgtga gccccggcga gcccgccagc
60
atgagctgca agagcagcca gagcctgctg aacagcgtga accagaagaa ctacctggcc
120
tggtacagac agaagcccgg ccagagcccc caggtgctgg tgtacttcgc cagcaccaga
180
gtgagcggcg tgcccgacag attcatcggc agcggcagcg gcaccgactt caccctgaga
240
atcagcagag tggaggccga cgacctgggc gtgtactact gccagcagta cttcagcacc
300
cccctgacct tcggccaggg caccaagctg gagctgaaga gaaacgacgc ccagcccgcc
360
gtgtacctgt tccagcccag ccccgaccag ctgcacaccg gcagcgccag cgtggtgtgc
420
ctgctgaaca gcttctaccc caaggacatc aacgtgaagt ggaaggtgga cggcgtgatc
480
caggacaccg gcatccagga gagcgtgacc gagcaggaca gcaaggacag cacctacagc
540
ctgagcagca ccctgaccat gagcagcacc gagtacctga gccacgagct gtacagctgc
600
gagatcaccc acaagagcct gcccagcacc ctgatcaaga gcttccagag aagcgagtgc
660
cagagagtgg ac
672
<210> 90
<211> 224
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 90
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Arg Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Val Leu Val Tyr Phe Ala Ser Thr Arg Val Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg
65 70 75 80
Ile Ser Arg Val Glu Ala Asp Asp Leu Gly Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Thr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Leu
100 105 110
Lys Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro
115 120 125
Asp Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser
130 135 140
Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile
145 150 155 160
Gln Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr
180 185 190
Leu Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro
195 200 205
Ser Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210> 91
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 91
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gcacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgt tcaacgagtg cagatgcacc
660
gacacccccc cctgccccgt gcccgagccc ctgggcggcc ccagcgtgct gatcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg aggtgacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagag cagagagcag cagttcaacg gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcgacc tgcccagccc catcgagaga accatcagca aggccagagg cagagcccac
1020
aagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
agcatcacct gcctgatcaa ggacttctac ccccccgaca tcgacgtgga gtggcagagc
1140
aacggccagc aggagcccga gagaaagcac agaatgaccc ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
cccttcacct gcgccgtgat gcacgagacc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 92
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 92
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu His Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro
210 215 220
Cys Pro Val Pro Glu Pro Leu Gly Gly Pro Ser Val Leu Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Ser Arg
275 280 285
Glu Gln Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Asp Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Arg Ala His Lys Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Ser Ile Thr Cys Leu Ile Lys Asp
355 360 365
Phe Tyr Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Gln
370 375 380
Glu Pro Glu Arg Lys His Arg Met Thr Pro Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Pro Phe Thr Cys Ala Val Met His Glu Thr Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 93
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 93
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat ccccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcgc cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcaccg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgc ccaaggagag cacctgcaag
660
tgcatcagcc cctgccccgt gcccgagagc ctgggcggcc ccagcgtgtt catcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg agatcacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagcc cagagagcag cagttcaaca gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcggcc tgcccagccc catcgagaga accatcagca aggccagagg ccaggcccac
1020
cagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
accctgacct gcctgatcaa ggacttcttc ccccccgaga tcgacgtgga gtggcagagc
1140
aacggccagc ccgagcccga gagcaagtac cacaccaccg ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
accttcacct gcgccgtgat gcacgaggcc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 94
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 94
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Ile Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Thr Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Glu Ser Thr Cys Lys Cys Ile Ser Pro
210 215 220
Cys Pro Val Pro Glu Ser Leu Gly Gly Pro Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg
275 280 285
Glu Gln Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Gly Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp
355 360 365
Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro
370 375 380
Glu Pro Glu Ser Lys Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Thr Phe Thr Cys Ala Val Met His Glu Ala Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 95
<211> 672
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 95
gacatcgtga tgacccagac ccccctgagc ctgagcgtga gccccggcga gcccgccagc
60
atgagctgca agagcagcca gagcctgctg aacagcgtga accagaagaa ctacctggcc
120
tggtacagac agaagcccgg ccagagcccc caggtgctgg tgtacttcgc cagcaccaga
180
gtgagcggcg tgcccgacag attcatcggc agcggcagcg gcaccgactt caccctgaga
240
atcagcagag tggaggccga cgacctgggc gtgtactact gccagcagta cttcagcacc
300
cccctgacct tcggccaggg caccaagctg gagctgaaga gaaacgacgc ccagcccgcc
360
gtgtacctgt tccagcccag ccccgaccag ctgcacaccg gcagcgccag cgtggtgtgc
420
ctgctgaaca gcttctaccc caaggacatc aacgtgaagt ggaaggtgga cggcgtgatc
480
caggacaccg gcatccagga gagcgtgacc gagcaggaca gcaaggacag cacctacagc
540
ctgagcagca ccctgaccat gagcagcacc gagtacctga gccacgagct gtacagctgc
600
gagatcaccc acaagagcct gcccagcacc ctgatcaaga gcttccagag aagcgagtgc
660
cagagagtgg ac
672
<210> 96
<211> 224
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 96
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Arg Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Val Leu Val Tyr Phe Ala Ser Thr Arg Val Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg
65 70 75 80
Ile Ser Arg Val Glu Ala Asp Asp Leu Gly Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Thr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Leu
100 105 110
Lys Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro
115 120 125
Asp Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser
130 135 140
Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile
145 150 155 160
Gln Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr
180 185 190
Leu Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro
195 200 205
Ser Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210> 97
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 97
gaggtgcagc tggtgcagag cgtggccgag ctggtgaagc ccggcgccag cgtgaaggtg
60
agctgcaccg tgagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgaccccg ccaacgtgaa caccaagtac
180
gcccccaagt tccagggcag agccaccatc accgccgaca ccagcaccaa caccgcctac
240
atgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagaatcttc
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag caccagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacttcc ccgagcccgt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgcaca gcctgagcag catggtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggtgca ccccgccagc aacaccaagg tggacaagcc cgtgttcaac
660
gagtgcagat gcaccgacac ccccccctgc cccgtgcccg agcccctggg cggccccagc
720
gtgctgatct tcccccccaa gcccaaggac atcctgagaa tcaccagaac ccccgaggtg
780
acctgcgtgg tgctggacct gggcagagag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagg aggtgcacac cgccaagacc cagagcagag agcagcagtt caacggcacc
900
tacagagtgg tgagcgtgct gcccatcgag caccaggact ggctgaccgg caaggagttc
960
aagtgcagag tgaaccacat cgacctgccc agccccatcg agagaaccat cagcaaggcc
1020
agaggcagag cccacaagcc cagcgtgtac gtgctgcccc ccagccccaa ggagctgagc
1080
agcagcgaca ccgtgagcat cacctgcctg atcaaggact tctacccccc cgacatcgac
1140
gtggagtggc agagcaacgg ccagcaggag cccgagagaa agcacagaat gacccccccc
1200
cagctggacg aggacggcag ctacttcctg tacagcaagc tgagcgtgga caagagcaga
1260
tggcagcagg gcgacccctt cacctgcgcc gtgatgcacg agaccctgca gaaccactac
1320
accgacctga gcctgagcca cagccccggc aag
1353
<210> 98
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 98
Glu Val Gln Leu Val Gln Ser Val Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Thr Val Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Phe Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu His Ser Leu Ser Ser Met Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Val His Pro
195 200 205
Ala Ser Asn Thr Lys Val Asp Lys Pro Val Phe Asn Glu Cys Arg Cys
210 215 220
Thr Asp Thr Pro Pro Cys Pro Val Pro Glu Pro Leu Gly Gly Pro Ser
225 230 235 240
Val Leu Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Glu Val His Thr Ala
275 280 285
Lys Thr Gln Ser Arg Glu Gln Gln Phe Asn Gly Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn His Ile Asp Leu Pro Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Arg Ala His Lys Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp Thr Val Ser Ile Thr
355 360 365
Cys Leu Ile Lys Asp Phe Tyr Pro Pro Asp Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Gln Glu Pro Glu Arg Lys His Arg Met Thr Pro Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asp Pro Phe Thr Cys Ala Val Met
420 425 430
His Glu Thr Leu Gln Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210> 99
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 99
gaggtgcagc tggtgcagag cgtggccgag ctggtgaagc ccggcgccag cgtgaaggtg
60
agctgcaccg tgagcggctt caacatcaag aacacctaca tgcactgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatcggcaga atcgaccccg ccaacgtgaa caccaagtac
180
gcccccaagt tccagggcag agccaccatc accgccgaca ccagcaccaa caccgcctac
240
atgcagctga gcagcctgag agccgaggac accgccgtgt actactgcgc cagaatcttc
300
tacgactacg acggcgacat cgacgtgtgg ggccagggca ccctggtgac cgtgagcagc
360
gccagcacca ccgcccccag cgtgttcccc ctggccccca gctgcggcag caccagcggc
420
agcaccgtgg ccctggcctg cctggtgagc ggctacttcc ccgagcccgt gaccgtgagc
480
tggaacagcg gcagcctgac cagcggcgtg cacaccttcc ccagcgtgct gcagagcagc
540
ggcctgtaca gcctgagcag caccgtgacc gtgcccagca gcagatggcc cagcgagacc
600
ttcacctgca acgtggtgca ccccgccagc aacaccaagg tggacaagcc cgtgcccaag
660
gagagcacct gcaagtgcat cagcccctgc cccgtgcccg agagcctggg cggccccagc
720
gtgttcatct tcccccccaa gcccaaggac atcctgagaa tcaccagaac ccccgagatc
780
acctgcgtgg tgctggacct gggcagagag gaccccgagg tgcagatcag ctggttcgtg
840
gacggcaagg aggtgcacac cgccaagacc cagcccagag agcagcagtt caacagcacc
900
tacagagtgg tgagcgtgct gcccatcgag caccaggact ggctgaccgg caaggagttc
960
aagtgcagag tgaaccacat cggcctgccc agccccatcg agagaaccat cagcaaggcc
1020
agaggccagg cccaccagcc cagcgtgtac gtgctgcccc ccagccccaa ggagctgagc
1080
agcagcgaca ccgtgaccct gacctgcctg atcaaggact tcttcccccc cgagatcgac
1140
gtggagtggc agagcaacgg ccagcccgag cccgagagca agtaccacac caccgccccc
1200
cagctggacg aggacggcag ctacttcctg tacagcaagc tgagcgtgga caagagcaga
1260
tggcagcagg gcgacacctt cacctgcgcc gtgatgcacg aggccctgca gaaccactac
1320
accgacctga gcctgagcca cagccccggc aag
1353
<210> 100
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 100
Glu Val Gln Leu Val Gln Ser Val Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Thr Val Ser Gly Phe Asn Ile Lys Asn Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Val Asn Thr Lys Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Phe Tyr Asp Tyr Asp Gly Asp Ile Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala
130 135 140
Leu Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Thr Val Thr Val Pro
180 185 190
Ser Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Val His Pro
195 200 205
Ala Ser Asn Thr Lys Val Asp Lys Pro Val Pro Lys Glu Ser Thr Cys
210 215 220
Lys Cys Ile Ser Pro Cys Pro Val Pro Glu Ser Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg
245 250 255
Thr Pro Glu Ile Thr Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro
260 265 270
Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Glu Val His Thr Ala
275 280 285
Lys Thr Gln Pro Arg Glu Gln Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe
305 310 315 320
Lys Cys Arg Val Asn His Ile Gly Leu Pro Ser Pro Ile Glu Arg Thr
325 330 335
Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu
340 345 350
Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp Thr Val Thr Leu Thr
355 360 365
Cys Leu Ile Lys Asp Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln
370 375 380
Ser Asn Gly Gln Pro Glu Pro Glu Ser Lys Tyr His Thr Thr Ala Pro
385 390 395 400
Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asp Thr Phe Thr Cys Ala Val Met
420 425 430
His Glu Ala Leu Gln Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser
435 440 445
Pro Gly Lys
450
<210> 101
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 101
gacatcgtga tgacccagac ccccctgagc ctgagcgtga gcctgggcga gcccgccagc
60
atcagctgcc acgccagcca gaacatcaac gtgtggctga gctggtacag acagaagccc
120
ggccagatcc cccagctgct gatctacaag gccagcaacc tgcacaccgg cgtgcccgac
180
agattcagcg gcagcggcag cggcaccgac ttcaccctga gaatcagcag agtggaggcc
240
gacgacgccg gcgtgtacta ctgccagcag ggccagagct accccctgac cttcggccag
300
ggcaccaagg tggagatcaa gagaaacgac gcccagcccg ccgtgtacct gttccagccc
360
agccccgacc agctgcacac cggcagcgcc agcgtggtgt gcctgctgaa cagcttctac
420
cccaaggaca tcaacgtgaa gtggaaggtg gacggcgtga tccaggacac cggcatccag
480
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc
540
atgagcagca ccgagtacct gagccacgag ctgtacagct gcgagatcac ccacaagagc
600
ctgcccagca ccctgatcaa gagcttccag agaagcgagt gccagagagt ggac
654
<210> 102
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 102
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Leu Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys His Ala Ser Gln Asn Ile Asn Val Trp
20 25 30
Leu Ser Trp Tyr Arg Gln Lys Pro Gly Gln Ile Pro Gln Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val Glu Ala
65 70 75 80
Asp Asp Ala Gly Val Tyr Tyr Cys Gln Gln Gly Gln Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Asn Asp Ala Gln
100 105 110
Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr Gly
115 120 125
Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu Tyr
180 185 190
Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys Ser
195 200 205
Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210> 103
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 103
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat gcccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gcacagcctg
540
agcagcatgg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgt tcaacgagtg cagatgcacc
660
gacacccccc cctgccccgt gcccgagccc ctgggcggcc ccagcgtgct gatcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg aggtgacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagag cagagagcag cagttcaacg gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcgacc tgcccagccc catcgagaga accatcagca aggccagagg cagagcccac
1020
aagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
agcatcacct gcctgatcaa ggacttctac ccccccgaca tcgacgtgga gtggcagagc
1140
aacggccagc aggagcccga gagaaagcac agaatgaccc ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
cccttcacct gcgccgtgat gcacgagacc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 104
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 104
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu His Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro
210 215 220
Cys Pro Val Pro Glu Pro Leu Gly Gly Pro Ser Val Leu Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Ser Arg
275 280 285
Glu Gln Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Asp Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Arg Ala His Lys Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Ser Ile Thr Cys Leu Ile Lys Asp
355 360 365
Phe Tyr Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Gln
370 375 380
Glu Pro Glu Arg Lys His Arg Met Thr Pro Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Pro Phe Thr Cys Ala Val Met His Glu Thr Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 105
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 105
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggcgccag cgtgaaggtg
60
agctgcaagg ccagcggcta caccttcacc acctacggca tgagctgggt gagacaggcc
120
cccggcaagg gcctgcagtg gatgggctgg atcaacatct acagcggcat gcccacctac
180
gccgacgact tcaagggcag attcgccctg agcctggaca ccagcaccag caccgcctac
240
atggagctga acagcctgag agccgaggac accgccgtgt actactgcac cagattcgac
300
ggccccgact actggggcca gggcaccctg gtgaccgtga gcagcgccag caccaccgcc
360
cccagcgtgt tccccctggc ccccagctgc ggcagcacca gcggcagcac cgtggccctg
420
gcctgcctgg tgagcggcta cttccccgag cccgtgaccg tgagctggaa cagcggcagc
480
ctgaccagcg gcgtgcacac cttccccagc gtgctgcaga gcagcggcct gtacagcctg
540
agcagcaccg tgaccgtgcc cagcagcaga tggcccagcg agaccttcac ctgcaacgtg
600
gtgcaccccg ccagcaacac caaggtggac aagcccgtgc ccaaggagag cacctgcaag
660
tgcatcagcc cctgccccgt gcccgagagc ctgggcggcc ccagcgtgtt catcttcccc
720
cccaagccca aggacatcct gagaatcacc agaacccccg agatcacctg cgtggtgctg
780
gacctgggca gagaggaccc cgaggtgcag atcagctggt tcgtggacgg caaggaggtg
840
cacaccgcca agacccagcc cagagagcag cagttcaaca gcacctacag agtggtgagc
900
gtgctgccca tcgagcacca ggactggctg accggcaagg agttcaagtg cagagtgaac
960
cacatcggcc tgcccagccc catcgagaga accatcagca aggccagagg ccaggcccac
1020
cagcccagcg tgtacgtgct gccccccagc cccaaggagc tgagcagcag cgacaccgtg
1080
accctgacct gcctgatcaa ggacttcttc ccccccgaga tcgacgtgga gtggcagagc
1140
aacggccagc ccgagcccga gagcaagtac cacaccaccg ccccccagct ggacgaggac
1200
ggcagctact tcctgtacag caagctgagc gtggacaaga gcagatggca gcagggcgac
1260
accttcacct gcgccgtgat gcacgaggcc ctgcagaacc actacaccga cctgagcctg
1320
agccacagcc ccggcaag
1338
<210> 106
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 106
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Met
35 40 45
Gly Trp Ile Asn Ile Tyr Ser Gly Met Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Leu Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Asp Gly Pro Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val
130 135 140
Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Thr Val Thr Val Pro Ser Ser Arg Trp Pro
180 185 190
Ser Glu Thr Phe Thr Cys Asn Val Val His Pro Ala Ser Asn Thr Lys
195 200 205
Val Asp Lys Pro Val Pro Lys Glu Ser Thr Cys Lys Cys Ile Ser Pro
210 215 220
Cys Pro Val Pro Glu Ser Leu Gly Gly Pro Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Ile Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr
245 250 255
Cys Val Val Leu Asp Leu Gly Arg Glu Asp Pro Glu Val Gln Ile Ser
260 265 270
Trp Phe Val Asp Gly Lys Glu Val His Thr Ala Lys Thr Gln Pro Arg
275 280 285
Glu Gln Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Ile
290 295 300
Glu His Gln Asp Trp Leu Thr Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
His Ile Gly Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg
325 330 335
Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser Pro Lys
340 345 350
Glu Leu Ser Ser Ser Asp Thr Val Thr Leu Thr Cys Leu Ile Lys Asp
355 360 365
Phe Phe Pro Pro Glu Ile Asp Val Glu Trp Gln Ser Asn Gly Gln Pro
370 375 380
Glu Pro Glu Ser Lys Tyr His Thr Thr Ala Pro Gln Leu Asp Glu Asp
385 390 395 400
Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asp Thr Phe Thr Cys Ala Val Met His Glu Ala Leu Gln
420 425 430
Asn His Tyr Thr Asp Leu Ser Leu Ser His Ser Pro Gly Lys
435 440 445
<210> 107
<211> 672
<212> ДНК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 107
gacatcgtga tgacccagac ccccctgagc ctgagcgtga gccccggcga gcccgccagc
60
atgagctgca agagcagcca gagcctgctg aacagcgtga accagaagaa ctacctggcc
120
tggtacagac agaagcccgg ccagagcccc caggtgctgg tgtacttcgc cagcaccaga
180
atcagcggcg tgcccgacag attcatcggc agcggcagcg gcaccgactt caccctgaga
240
atcagcagag tggaggccga cgacctgggc gtgtactact gccagcagta cttcagcacc
300
cccctgacct tcggccaggg caccaagctg gagctgaaga gaaacgacgc ccagcccgcc
360
gtgtacctgt tccagcccag ccccgaccag ctgcacaccg gcagcgccag cgtggtgtgc
420
ctgctgaaca gcttctaccc caaggacatc aacgtgaagt ggaaggtgga cggcgtgatc
480
caggacaccg gcatccagga gagcgtgacc gagcaggaca gcaaggacag cacctacagc
540
ctgagcagca ccctgaccat gagcagcacc gagtacctga gccacgagct gtacagctgc
600
gagatcaccc acaagagcct gcccagcacc ctgatcaaga gcttccagag aagcgagtgc
660
cagagagtgg ac
672
<210> 108
<211> 224
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> канинизированное мышиное антитело
<400> 108
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Val Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Arg Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Val Leu Val Tyr Phe Ala Ser Thr Arg Ile Ser Gly Val
50 55 60
Pro Asp Arg Phe Ile Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg
65 70 75 80
Ile Ser Arg Val Glu Ala Asp Asp Leu Gly Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Thr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Leu
100 105 110
Lys Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro
115 120 125
Asp Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser
130 135 140
Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile
145 150 155 160
Gln Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr
180 185 190
Leu Ser His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro
195 200 205
Ser Thr Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210> 109
<211> 17
<212> БЕЛОК
<213> Canis familiaris
<400> 109
Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Pro
<210> 110
<211> 22
<212> БЕЛОК
<213> Canis familiaris
<400> 110
Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys Pro Lys
1 5 10 15
Cys Pro Ala Pro Glu Met
20
<210> 111
<211> 20
<212> БЕЛОК
<213> Canis familiaris
<400> 111
Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn Asn Cys Pro Cys Pro
1 5 10 15
Gly Cys Gly Leu
20
<210> 112
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> modified canine
<400> 112
Pro Lys Glu Ser Thr Cys Lys Cys Ile Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Ser
<210> 113
<211> 795
<212> ДНК
<213> Canis familiaris
<400> 113
ctagactccc ctgacaggcc ctggagcccg ctcaccttct ccccggcgca gctcacggtg
60
caggagggag agaacgccac gttcacctgc agcctggccg acatccccga cagcttcgtg
120
ctcaactggt accgcctgag cccccgcaac cagacggaca agctggccgc cttccaggag
180
gaccgcatcg agccgggccg ggacaggcgc ttccgcgtca tgcggctgcc caacgggcgg
240
gacttccaca tgagcatcgt cgctgcgcgc ctcaacgaca gcggcatcta cctgtgcggg
300
gccatctacc tgccccccaa cacacagatc aacgagagtc cccgcgcaga gctctccgtg
360
acggagagaa ccctggagcc ccccacacag agccccagcc ccccacccag actcagcggc
420
cagttgcagg ggctggtcat cggcgtcacg agcgtgctgg tgggtgtcct gctactgctg
480
ctgctgacct gggtcctggc cgctgtcttc cccagggcca cccgaggtgc ctgtgtgtgc
540
gggagcgagg acgagcctct gaaggagggc cccgatgcag cgcccgtctt caccctggac
600
tacggggagc tggacttcca gtggcgagag aagacgccgg agcccccggc gccctgtgcc
660
ccggagcaga ccgagtatgc caccatcgtc ttcccgggca ggccggcgtc cccgggccgc
720
agggcctcgg ccagcagcct gcagggagcc cagcctccga gccccgagga cggacccggc
780
ctgtggcccc tctga
795
<210> 114
<211> 264
<212> БЕЛОК
<213> Canis familiaris
<400> 114
Leu Asp Ser Pro Asp Arg Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala
1 5 10 15
Gln Leu Thr Val Gln Glu Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu
20 25 30
Ala Asp Ile Pro Asp Ser Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro
35 40 45
Arg Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu
50 55 60
Pro Gly Arg Asp Arg Arg Phe Arg Val Met Arg Leu Pro Asn Gly Arg
65 70 75 80
Asp Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser Gly Ile
85 90 95
Tyr Leu Cys Gly Ala Ile Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu
100 105 110
Ser Pro Arg Ala Glu Leu Ser Val Thr Glu Arg Thr Leu Glu Pro Pro
115 120 125
Thr Gln Ser Pro Ser Pro Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly
130 135 140
Leu Val Ile Gly Val Thr Ser Val Leu Val Gly Val Leu Leu Leu Leu
145 150 155 160
Leu Leu Thr Trp Val Leu Ala Ala Val Phe Pro Arg Ala Thr Arg Gly
165 170 175
Ala Cys Val Cys Gly Ser Glu Asp Glu Pro Leu Lys Glu Gly Pro Asp
180 185 190
Ala Ala Pro Val Phe Thr Leu Asp Tyr Gly Glu Leu Asp Phe Gln Trp
195 200 205
Arg Glu Lys Thr Pro Glu Pro Pro Ala Pro Cys Ala Pro Glu Gln Thr
210 215 220
Glu Tyr Ala Thr Ile Val Phe Pro Gly Arg Pro Ala Ser Pro Gly Arg
225 230 235 240
Arg Ala Ser Ala Ser Ser Leu Gln Gly Ala Gln Pro Pro Ser Pro Glu
245 250 255
Asp Gly Pro Gly Leu Trp Pro Leu
260
<210> 115
<211> 438
<212> ДНК
<213> Canis familiaris
<400> 115
ctggattccc ccgacagacc ctggagccct ctcaccttct cccctgccca gctgaccgtc
60
caggaaggcg agaatgccac cttcacctgc agcctcgccg acatccccga cagcttcgtg
120
ctgaactggt acagactgag ccccaggaac cagaccgaca agctggccgc tttccaggag
180
gacaggatcg aacccggcag ggacaggagg tttagggtca tgaggctgcc caacggcagg
240
gacttccaca tgtccatcgt ggccgccaga ctgaacgact ccggcatcta cctgtgcggc
300
gctatctacc tgccccccaa cacccagatc aacgagagcc ccagggccga actgagcgtg
360
acagagagaa ccctggaacc tcccacccag agcccttccc ctcctcctag actgagcgga
420
cagctgcagg gcctggtg
438
<210> 116
<211> 146
<212> БЕЛОК
<213> Canis familiaris
<400> 116
Leu Asp Ser Pro Asp Arg Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala
1 5 10 15
Gln Leu Thr Val Gln Glu Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu
20 25 30
Ala Asp Ile Pro Asp Ser Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro
35 40 45
Arg Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu
50 55 60
Pro Gly Arg Asp Arg Arg Phe Arg Val Met Arg Leu Pro Asn Gly Arg
65 70 75 80
Asp Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser Gly Ile
85 90 95
Tyr Leu Cys Gly Ala Ile Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu
100 105 110
Ser Pro Arg Ala Glu Leu Ser Val Thr Glu Arg Thr Leu Glu Pro Pro
115 120 125
Thr Gln Ser Pro Ser Pro Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly
130 135 140
Leu Val
145
<210> 117
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> canine and human sequence
<400> 117
ctggattccc ccgacagacc ctggagccct ctcaccttct cccctgccca gctgaccgtc
60
caggaaggcg agaatgccac cttcacctgc agcctcgccg acatccccga cagcttcgtg
120
ctgaactggt acagactgag ccccaggaac cagaccgaca agctggccgc tttccaggag
180
gacaggatcg aacccggcag ggacaggagg tttagggtca tgaggctgcc caacggcagg
240
gacttccaca tgtccatcgt ggccgccaga ctgaacgact ccggcatcta cctgtgcggc
300
gctatctacc tgccccccaa cacccagatc aacgagagcc ccagggccga actgagcgtg
360
acagagagaa ccctggaacc tcccacccag agcccttccc ctcctcctag actgagcgga
420
cagctgcagg gcctggtggg taccgacaaa actcacacat gcccaccgtg cccagcacct
480
gaactcctgg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg
540
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag
600
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg
660
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac
720
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc
780
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc
840
ccatcccggg atgagctgac caagaaccag gtcagcctga cctgcctggt caaaggcttc
900
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag
960
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg
1020
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg
1080
cacaaccact acacgcagaa gagcctctcc ctgtctccgg gtaaatga
1128
<210> 118
<211> 375
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> canine and human sequence
<400> 118
Leu Asp Ser Pro Asp Arg Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala
1 5 10 15
Gln Leu Thr Val Gln Glu Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu
20 25 30
Ala Asp Ile Pro Asp Ser Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro
35 40 45
Arg Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu
50 55 60
Pro Gly Arg Asp Arg Arg Phe Arg Val Met Arg Leu Pro Asn Gly Arg
65 70 75 80
Asp Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser Gly Ile
85 90 95
Tyr Leu Cys Gly Ala Ile Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu
100 105 110
Ser Pro Arg Ala Glu Leu Ser Val Thr Glu Arg Thr Leu Glu Pro Pro
115 120 125
Thr Gln Ser Pro Ser Pro Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly
130 135 140
Leu Val Gly Thr Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
145 150 155 160
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
165 170 175
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
180 185 190
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
195 200 205
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
210 215 220
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
225 230 235 240
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
245 250 255
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
260 265 270
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
275 280 285
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
290 295 300
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
305 310 315 320
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
325 330 335
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
340 345 350
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
355 360 365
Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 119
<211> 816
<212> ДНК
<213> Canis familiaris
<400> 119
tttacgatca cagtttctaa ggacctgtat gtggtagagt atggtggcaa tgtgacaatg
60
gaatgcaaat tcccggtgga aaaacagtta aacttgtttg cactaatcgt ctactgggaa
120
atggaggata aaaaaattat acaatttgtg aatggaaagg aagacctgaa agttcagcac
180
agcagctaca gccagagggc tcagctattg aaggaccagc tcttcttggg gaaggctgcg
240
cttcagatca cagatgtgag attgcaggat gcaggggttt actgctgctt gatcggctat
300
ggcggtgctg actacaagcg gattactttg aaagttcatg ccccgtaccg caacatcagc
360
caaagaattt ctgtggatcc tgtcacctct gaacatgaac taatgtgtca ggctgagggt
420
taccctgagg ctgaagtcat ctggacaagc agtgaccacc gagtcctgag tggcaaaacc
480
accatcacta attccaatag ggaagagaag cttttcaatg tgaccagcac gctgaacatc
540
aatgcaacag ctaatgagat tttctactgc acttttcaaa gatcaggtcc tgaggaaaac
600
aatactgccg agttggtcat cccagaacga ctgcccgttc cagcaagtga gaggactcat
660
ttcatgattc tgggaccttt cctgttgctt cttggtgtag tcctggcagt cactttctgt
720
ctaaaaaaac atgggagaat gatggatgtg gaaaaatgtt gcacccgaga taggaactca
780
aagaaacgaa atgatataca atttgaagag acataa
816
<210> 120
<211> 271
<212> БЕЛОК
<213> Canis familiaris
<400> 120
Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr Gly Gly
1 5 10 15
Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asn Leu
20 25 30
Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile Ile Gln
35 40 45
Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser Tyr Ser
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp Pro Val
115 120 125
Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly Lys Thr
145 150 155 160
Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val Thr Ser
165 170 175
Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys Thr Phe
180 185 190
Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val Ile Pro
195 200 205
Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His Phe Met Ile Leu
210 215 220
Gly Pro Phe Leu Leu Leu Leu Gly Val Val Leu Ala Val Thr Phe Cys
225 230 235 240
Leu Lys Lys His Gly Arg Met Met Asp Val Glu Lys Cys Cys Thr Arg
245 250 255
Asp Arg Asn Ser Lys Lys Arg Asn Asp Ile Gln Phe Glu Glu Thr
260 265 270
<210> 121
<211> 660
<212> ДНК
<213> Canis familiaris
<400> 121
tttaccatca ccgtgtccaa ggacctgtac gtggtcgagt acggcggcaa tgtgaccatg
60
gagtgcaagt tccccgtgga gaagcagctg aacctgttcg ccctcatcgt gtactgggag
120
atggaggaca agaagatcat ccagttcgtg aacggcaagg aggacctgaa ggtgcagcac
180
tccagctact cccagagagc ccagctgctg aaggaccagc tgttcctggg caaggccgcc
240
ctgcagatca ccgacgtgag actgcaggac gccggcgtgt attgctgcct gatcggctac
300
ggaggcgccg actacaagag gatcaccctg aaggtgcatg caccctacag gaacatcagc
360
cagaggatca gcgtcgatcc cgtgaccagc gagcacgagc tgatgtgcca agccgagggc
420
tatcccgagg ccgaagtgat ctggaccagc agcgaccaca gggtcctgag cggcaagacc
480
accatcacca acagcaacag ggaggagaag ctgttcaacg tgaccagcac cctcaacatc
540
aacgccaccg ccaacgagat cttctactgc accttccaga ggagcggccc cgaagagaac
600
aacaccgccg agctggtgat ccccgagaga ctgcctgtgc ctgccagcga gaggacccac
660
<210> 122
<211> 220
<212> БЕЛОК
<213> Canis familiaris
<400> 122
Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr Gly Gly
1 5 10 15
Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asn Leu
20 25 30
Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile Ile Gln
35 40 45
Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser Tyr Ser
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp Pro Val
115 120 125
Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly Lys Thr
145 150 155 160
Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val Thr Ser
165 170 175
Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys Thr Phe
180 185 190
Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val Ile Pro
195 200 205
Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His
210 215 220
<210> 123
<211> 1350
<212> ДНК
<213> Canis familiaris
<400> 123
tttaccatca ccgtgtccaa ggacctgtac gtggtcgagt acggcggcaa tgtgaccatg
60
gagtgcaagt tccccgtgga gaagcagctg aacctgttcg ccctcatcgt gtactgggag
120
atggaggaca agaagatcat ccagttcgtg aacggcaagg aggacctgaa ggtgcagcac
180
tccagctact cccagagagc ccagctgctg aaggaccagc tgttcctggg caaggccgcc
240
ctgcagatca ccgacgtgag actgcaggac gccggcgtgt attgctgcct gatcggctac
300
ggaggcgccg actacaagag gatcaccctg aaggtgcatg caccctacag gaacatcagc
360
cagaggatca gcgtcgatcc cgtgaccagc gagcacgagc tgatgtgcca agccgagggc
420
tatcccgagg ccgaagtgat ctggaccagc agcgaccaca gggtcctgag cggcaagacc
480
accatcacca acagcaacag ggaggagaag ctgttcaacg tgaccagcac cctcaacatc
540
aacgccaccg ccaacgagat cttctactgc accttccaga ggagcggccc cgaagagaac
600
aacaccgccg agctggtgat ccccgagaga ctgcctgtgc ctgccagcga gaggacccac
660
ggtaccgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg
720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag
780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac
840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc
900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag
960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa
1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggatgagctg
1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc
1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg
1200
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag
1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag
1320
aagagcctct ccctgtctcc gggtaaatga
1350
<210> 124
<211> 449
<212> БЕЛОК
<213> Canis familiaris
<400> 124
Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr Gly Gly
1 5 10 15
Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asn Leu
20 25 30
Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile Ile Gln
35 40 45
Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser Tyr Ser
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp Pro Val
115 120 125
Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly Lys Thr
145 150 155 160
Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val Thr Ser
165 170 175
Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys Thr Phe
180 185 190
Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val Ile Pro
195 200 205
Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His Gly Thr Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 125
<211> 648
<212> ДНК
<213> Canis familiaris
<400> 125
ctgggcggcc ccagcgtgct gatcttcccc cccaagccca aggacatcct gagaatcacc
60
agaacccccg aggtgacctg cgtggtgctg gacctgggca gagaggaccc cgaggtgcag
120
atcagctggt tcgtggacgg caaggaggtg cacaccgcca agacccagag cagagagcag
180
cagttcaacg gcacctacag agtggtgagc gtgctgccca tcgagcacca ggactggctg
240
accggcaagg agttcaagtg cagagtgaac cacatcgacc tgcccagccc catcgagaga
300
accatcagca aggccagagg cagagcccac aagcccagcg tgtacgtgct gccccccagc
360
cccaaggagc tgagcagcag cgacaccgtg agcatcacct gcctgatcaa ggacttctac
420
ccccccgaca tcgacgtgga gtggcagagc aacggccagc aggagcccga gagaaagcac
480
agaatgaccc ccccccagct ggacgaggac ggcagctact tcctgtacag caagctgagc
540
gtggacaaga gcagatggca gcagggcgac cccttcacct gcgccgtgat gcacgagacc
600
ctgcagaacc actacaccga cctgagcctg agccacagcc ccggcaag
648
<210> 126
<211> 216
<212> БЕЛОК
<213> Canis familiaris
<400> 126
Leu Gly Gly Pro Ser Val Leu Ile Phe Pro Pro Lys Pro Lys Asp Ile
1 5 10 15
Leu Arg Ile Thr Arg Thr Pro Glu Val Thr Cys Val Val Leu Asp Leu
20 25 30
Gly Arg Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
35 40 45
Glu Val His Thr Ala Lys Thr Gln Ser Arg Glu Gln Gln Phe Asn Gly
50 55 60
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu
65 70 75 80
Thr Gly Lys Glu Phe Lys Cys Arg Val Asn His Ile Asp Leu Pro Ser
85 90 95
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Arg Ala His Lys Pro
100 105 110
Ser Val Tyr Val Leu Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp
115 120 125
Thr Val Ser Ile Thr Cys Leu Ile Lys Asp Phe Tyr Pro Pro Asp Ile
130 135 140
Asp Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Arg Lys His
145 150 155 160
Arg Met Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr
165 170 175
Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Gln Gly Asp Pro Phe
180 185 190
Thr Cys Ala Val Met His Glu Thr Leu Gln Asn His Tyr Thr Asp Leu
195 200 205
Ser Leu Ser His Ser Pro Gly Lys
210 215
<210> 127
<211> 648
<212> ДНК
<213> Canis familiaris
<400> 127
ctgggcggcc ccagcgtgtt catcttcccc cccaagccca aggacatcct gagaatcacc
60
agaacccccg agatcacctg cgtggtgctg gacctgggca gagaggaccc cgaggtgcag
120
atcagctggt tcgtggacgg caaggaggtg cacaccgcca agacccagcc cagagagcag
180
cagttcaaca gcacctacag agtggtgagc gtgctgccca tcgagcacca ggactggctg
240
accggcaagg agttcaagtg cagagtgaac cacatcggcc tgcccagccc catcgagaga
300
accatcagca aggccagagg ccaggcccac cagcccagcg tgtacgtgct gccccccagc
360
cccaaggagc tgagcagcag cgacaccgtg accctgacct gcctgatcaa ggacttcttc
420
ccccccgaga tcgacgtgga gtggcagagc aacggccagc ccgagcccga gagcaagtac
480
cacaccaccg ccccccagct ggacgaggac ggcagctact tcctgtacag caagctgagc
540
gtggacaaga gcagatggca gcagggcgac accttcacct gcgccgtgat gcacgaggcc
600
ctgcagaacc actacaccga cctgagcctg agccacagcc ccggcaag
648
<210> 128
<211> 216
<212> БЕЛОК
<213> Canis familiaris
<400> 128
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile
1 5 10 15
Leu Arg Ile Thr Arg Thr Pro Glu Ile Thr Cys Val Val Leu Asp Leu
20 25 30
Gly Arg Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
35 40 45
Glu Val His Thr Ala Lys Thr Gln Pro Arg Glu Gln Gln Phe Asn Ser
50 55 60
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Glu His Gln Asp Trp Leu
65 70 75 80
Thr Gly Lys Glu Phe Lys Cys Arg Val Asn His Ile Gly Leu Pro Ser
85 90 95
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
100 105 110
Ser Val Tyr Val Leu Pro Pro Ser Pro Lys Glu Leu Ser Ser Ser Asp
115 120 125
Thr Val Thr Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Glu Ile
130 135 140
Asp Val Glu Trp Gln Ser Asn Gly Gln Pro Glu Pro Glu Ser Lys Tyr
145 150 155 160
His Thr Thr Ala Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr
165 170 175
Ser Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Gln Gly Asp Thr Phe
180 185 190
Thr Cys Ala Val Met His Glu Ala Leu Gln Asn His Tyr Thr Asp Leu
195 200 205
Ser Leu Ser His Ser Pro Gly Lys
210 215
<210> 129
<211> 645
<212> ДНК
<213> Canis familiaris
<400> 129
ctgggcggcc ccagcgtgtt catcttcccc cccaagccca aggacaccct gctgatcgcc
60
agaacccccg aggtgacctg cgtggtggtg gacctggacc ccgaggaccc cgaggtgcag
120
atcagctggt tcgtggacgg caagcagatg cagaccgcca agacccagcc cagagaggag
180
cagttcaacg gcacctacag agtggtgagc gtgctgccca tcggccacca ggactggctg
240
aagggcaagc agttcacctg caaggtgaac aacaaggccc tgcccagccc catcgagaga
300
accatcagca aggccagagg ccaggcccac cagcccagcg tgtacgtgct gccccccagc
360
agagaggagc tgagcaagaa caccgtgagc ctgacctgcc tgatcaagga cttcttcccc
420
cccgacatcg acgtggagtg gcagagcaac ggccagcagg agcccgagag caagtacaga
480
accacccccc cccagctgga cgaggacggc agctacttcc tgtacagcaa gctgagcgtg
540
gacaagagca gatggcagag aggcgacacc ttcatctgcg ccgtgatgca cgaggccctg
600
cacaaccact acacccagga gagcctgagc cacagccccg gcaag
645
<210> 130
<211> 215
<212> БЕЛОК
<213> Canis familiaris
<400> 130
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
1 5 10 15
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu
20 25 30
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
35 40 45
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly
50 55 60
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
65 70 75 80
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
85 90 95
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
100 105 110
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
115 120 125
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
130 135 140
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
145 150 155 160
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
165 170 175
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
180 185 190
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
195 200 205
Leu Ser His Ser Pro Gly Lys
210 215
<210> 131
<211> 645
<212> ДНК
<213> Canis familiaris
<400> 131
ctgggcggcc ccagcgtgtt catcttcccc cccaagccca aggacatcct ggtgaccgcc
60
agaaccccca ccgtgacctg cgtggtggtg gacctggacc ccgagaaccc cgaggtgcag
120
atcagctggt tcgtggacag caagcaggtg cagaccgcca acacccagcc cagagaggag
180
cagagcaacg gcacctacag agtggtgagc gtgctgccca tcggccacca ggactggctg
240
agcggcaagc agttcaagtg caaggtgaac aacaaggccc tgcccagccc catcgaggag
300
atcatcagca agacccccgg ccaggcccac cagcccaacg tgtacgtgct gccccccagc
360
agagacgaga tgagcaagaa caccgtgacc ctgacctgcc tggtgaagga cttcttcccc
420
cccgagatcg acgtggagtg gcagagcaac ggccagcagg agcccgagag caagtacaga
480
atgacccccc cccagctgga cgaggacggc agctacttcc tgtacagcaa gctgagcgtg
540
gacaagagca gatggcagag aggcgacacc ttcatctgcg ccgtgatgca cgaggccctg
600
cacaaccact acacccagat cagcctgagc cacagccccg gcaag
645
<210> 132
<211> 215
<212> БЕЛОК
<213> Canis familiaris
<400> 132
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Ile
1 5 10 15
Leu Val Thr Ala Arg Thr Pro Thr Val Thr Cys Val Val Val Asp Leu
20 25 30
Asp Pro Glu Asn Pro Glu Val Gln Ile Ser Trp Phe Val Asp Ser Lys
35 40 45
Gln Val Gln Thr Ala Asn Thr Gln Pro Arg Glu Glu Gln Ser Asn Gly
50 55 60
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
65 70 75 80
Ser Gly Lys Gln Phe Lys Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
85 90 95
Pro Ile Glu Glu Ile Ile Ser Lys Thr Pro Gly Gln Ala His Gln Pro
100 105 110
Asn Val Tyr Val Leu Pro Pro Ser Arg Asp Glu Met Ser Lys Asn Thr
115 120 125
Val Thr Leu Thr Cys Leu Val Lys Asp Phe Phe Pro Pro Glu Ile Asp
130 135 140
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
145 150 155 160
Met Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
165 170 175
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
180 185 190
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Ile Ser
195 200 205
Leu Ser His Ser Pro Gly Lys
210 215
<210> 133
<211> 867
<212> ДНК
<213> Canis familiaris
<400> 133
atggggagcc ggcgggggcc ctggccgctc gtctgggccg tgctgcagct gggctggtgg
60
ccaggatggc tcctagactc ccctgacagg ccctggagcc cgctcacctt ctccccggcg
120
cagctcacgg tgcaggaggg agagaacgcc acgttcacct gcagcctggc cgacatcccc
180
gacagcttcg tgctcaactg gtaccgcctg agcccccgca accagacgga caagctggcc
240
gccttccagg aggaccgcat cgagccgggc cgggacaggc gcttccgcgt catgcggctg
300
cccaacgggc gggacttcca catgagcatc gtcgctgcgc gcctcaacga cagcggcatc
360
tacctgtgcg gggccatcta cctgcccccc aacacacaga tcaacgagag tccccgcgca
420
gagctctccg tgacggagag aaccctggag ccccccacac agagccccag ccccccaccc
480
agactcagcg gccagttgca ggggctggtc atcggcgtca cgagcgtgct ggtgggtgtc
540
ctgctactgc tgctgctgac ctgggtcctg gccgctgtct tccccagggc cacccgaggt
600
gcctgtgtgt gcgggagcga ggacgagcct ctgaaggagg gccccgatgc agcgcccgtc
660
ttcaccctgg actacgggga gctggacttc cagtggcgag agaagacgcc ggagcccccg
720
gcgccctgtg ccccggagca gaccgagtat gccaccatcg tcttcccggg caggccggcg
780
tccccgggcc gcagggcctc ggccagcagc ctgcagggag cccagcctcc gagccccgag
840
gacggacccg gcctgtggcc cctctga
867
<210> 134
<211> 288
<212> БЕЛОК
<213> Canis familiaris
<400> 134
Met Gly Ser Arg Arg Gly Pro Trp Pro Leu Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Trp Pro Gly Trp Leu Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Ser Pro Leu Thr Phe Ser Pro Ala Gln Leu Thr Val Gln Glu Gly Glu
35 40 45
Asn Ala Thr Phe Thr Cys Ser Leu Ala Asp Ile Pro Asp Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Leu Ser Pro Arg Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Gln Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg
85 90 95
Val Met Arg Leu Pro Asn Gly Arg Asp Phe His Met Ser Ile Val Ala
100 105 110
Ala Arg Leu Asn Asp Ser Gly Ile Tyr Leu Cys Gly Ala Ile Tyr Leu
115 120 125
Pro Pro Asn Thr Gln Ile Asn Glu Ser Pro Arg Ala Glu Leu Ser Val
130 135 140
Thr Glu Arg Thr Leu Glu Pro Pro Thr Gln Ser Pro Ser Pro Pro Pro
145 150 155 160
Arg Leu Ser Gly Gln Leu Gln Gly Leu Val Ile Gly Val Thr Ser Val
165 170 175
Leu Val Gly Val Leu Leu Leu Leu Leu Leu Thr Trp Val Leu Ala Ala
180 185 190
Val Phe Pro Arg Ala Thr Arg Gly Ala Cys Val Cys Gly Ser Glu Asp
195 200 205
Glu Pro Leu Lys Glu Gly Pro Asp Ala Ala Pro Val Phe Thr Leu Asp
210 215 220
Tyr Gly Glu Leu Asp Phe Gln Trp Arg Glu Lys Thr Pro Glu Pro Pro
225 230 235 240
Ala Pro Cys Ala Pro Glu Gln Thr Glu Tyr Ala Thr Ile Val Phe Pro
245 250 255
Gly Arg Pro Ala Ser Pro Gly Arg Arg Ala Ser Ala Ser Ser Leu Gln
260 265 270
Gly Ala Gln Pro Pro Ser Pro Glu Asp Gly Pro Gly Leu Trp Pro Leu
275 280 285
<210> 135
<211> 870
<212> ДНК
<213> Canis familiaris
<400> 135
atgagaatgt ttagtgtctt tacattcatg gcctactgcc atttgctaaa agcatttacg
60
atcacagttt ctaaggacct gtatgtggta gagtatggtg gcaatgtgac aatggaatgc
120
aaattcccgg tggaaaaaca gttaaacttg tttgcactaa tcgtctactg ggaaatggag
180
gataaaaaaa ttatacaatt tgtgaatgga aaggaagacc tgaaagttca gcacagcagc
240
tacagccaga gggctcagct attgaaggac cagctcttct tggggaaggc tgcgcttcag
300
atcacagatg tgagattgca ggatgcaggg gtttactgct gcttgatcgg ctatggcggt
360
gctgactaca agcggattac tttgaaagtt catgccccgt accgcaacat cagccaaaga
420
atttctgtgg atcctgtcac ctctgaacat gaactaatgt gtcaggctga gggttaccct
480
gaggctgaag tcatctggac aagcagtgac caccgagtcc tgagtggcaa aaccaccatc
540
actaattcca atagggaaga gaagcttttc aatgtgacca gcacgctgaa catcaatgca
600
acagctaatg agattttcta ctgcactttt caaagatcag gtcctgagga aaacaatact
660
gccgagttgg tcatcccaga acgactgccc gttccagcaa gtgagaggac tcatttcatg
720
attctgggac ctttcctgtt gcttcttggt gtagtcctgg cagtcacttt ctgtctaaaa
780
aaacatggga gaatgatgga tgtggaaaaa tgttgcaccc gagataggaa ctcaaagaaa
840
cgaaatgata tacaatttga agagacataa
870
<210> 136
<211> 289
<212> БЕЛОК
<213> Canis familiaris
<400> 136
Met Arg Met Phe Ser Val Phe Thr Phe Met Ala Tyr Cys His Leu Leu
1 5 10 15
Lys Ala Phe Thr Ile Thr Val Ser Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Gly Asn Val Thr Met Glu Cys Lys Phe Pro Val Glu Lys Gln Leu
35 40 45
Asn Leu Phe Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Lys Ile
50 55 60
Ile Gln Phe Val Asn Gly Lys Glu Asp Leu Lys Val Gln His Ser Ser
65 70 75 80
Tyr Ser Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Phe Leu Gly Lys
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Arg Leu Gln Asp Ala Gly Val Tyr
100 105 110
Cys Cys Leu Ile Gly Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu
115 120 125
Lys Val His Ala Pro Tyr Arg Asn Ile Ser Gln Arg Ile Ser Val Asp
130 135 140
Pro Val Thr Ser Glu His Glu Leu Met Cys Gln Ala Glu Gly Tyr Pro
145 150 155 160
Glu Ala Glu Val Ile Trp Thr Ser Ser Asp His Arg Val Leu Ser Gly
165 170 175
Lys Thr Thr Ile Thr Asn Ser Asn Arg Glu Glu Lys Leu Phe Asn Val
180 185 190
Thr Ser Thr Leu Asn Ile Asn Ala Thr Ala Asn Glu Ile Phe Tyr Cys
195 200 205
Thr Phe Gln Arg Ser Gly Pro Glu Glu Asn Asn Thr Ala Glu Leu Val
210 215 220
Ile Pro Glu Arg Leu Pro Val Pro Ala Ser Glu Arg Thr His Phe Met
225 230 235 240
Ile Leu Gly Pro Phe Leu Leu Leu Leu Gly Val Val Leu Ala Val Thr
245 250 255
Phe Cys Leu Lys Lys His Gly Arg Met Met Asp Val Glu Lys Cys Cys
260 265 270
Thr Arg Asp Arg Asn Ser Lys Lys Arg Asn Asp Ile Gln Phe Glu Glu
275 280 285
Thr
<210> 137
<211> 401
<212> БЕЛОК
<213> Canis familiaris
<400> 137
Met Asn Phe Leu Leu Ser Trp Val His Trp Ser Leu Ala Leu Leu Leu
1 5 10 15
Tyr Leu His His Ala Lys Trp Ser Gln Ala Leu Asp Ser Pro Asp Arg
20 25 30
Pro Trp Ser Pro Leu Thr Phe Ser Pro Ala Gln Leu Thr Val Gln Glu
35 40 45
Gly Glu Asn Ala Thr Phe Thr Cys Ser Leu Ala Asp Ile Pro Asp Ser
50 55 60
Phe Val Leu Asn Trp Tyr Arg Leu Ser Pro Arg Asn Gln Thr Asp Lys
65 70 75 80
Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg
85 90 95
Phe Arg Val Met Arg Leu Pro Asn Gly Arg Asp Phe His Met Ser Ile
100 105 110
Val Ala Ala Arg Leu Asn Asp Ser Gly Ile Tyr Leu Cys Gly Ala Ile
115 120 125
Tyr Leu Pro Pro Asn Thr Gln Ile Asn Glu Ser Pro Arg Ala Glu Leu
130 135 140
Ser Val Thr Glu Arg Thr Leu Glu Pro Pro Thr Gln Ser Pro Ser Pro
145 150 155 160
Pro Pro Arg Leu Ser Gly Gln Leu Gln Gly Leu Val Gly Thr Asp Lys
165 170 175
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
180 185 190
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
195 200 205
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
210 215 220
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
225 230 235 240
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
245 250 255
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
260 265 270
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
275 280 285
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
290 295 300
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
305 310 315 320
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
325 330 335
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
340 345 350
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
355 360 365
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
370 375 380
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
385 390 395 400
Lys
<210> 138
<211> 15
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa может быть Met или Thr
<400> 138
Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg
1 5 10 15
<210> 139
<211> 15
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa может быть Met или Thr
<400> 139
Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg
1 5 10 15
<210> 140
<211> 25
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa может быть Met или Thr
<400> 140
Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg Asp Phe His Met Ser
1 5 10 15
Ile Val Ala Ala Arg Leu Asn Asp Ser
20 25
<210> 141
<211> 26
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (20)..(20)
<223> Xaa может быть Met или Thr
<400> 141
Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg
1 5 10 15
Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg
20 25
<210> 142
<211> 31
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (15)..(15)
<223> Xaa может быть Met или Thr
<400> 142
Glu Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg
1 5 10 15
Leu Pro Asn Gly Arg Asp Phe His Met Ser Ile Val Ala Ala Arg
20 25 30
<210> 143
<211> 31
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (25)..(25)
<223> Xaa может быть Met или Thr
<400> 143
Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro
1 5 10 15
Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg
20 25 30
<210> 144
<211> 45
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (25)..(25)
<223> Xaa может быть Met или Thr
<400> 144
Asn Gln Thr Asp Lys Leu Ala Ala Phe Gln Glu Asp Arg Ile Glu Pro
1 5 10 15
Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu Pro Asn Gly Arg Asp
20 25 30
Phe His Met Ser Ile Val Ala Ala Arg Leu Asn Asp Ser
35 40 45
<210> 145
<211> 20
<212> БЕЛОК
<213> Canis familiaris
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa может быть Met или Thr
<400> 145
Asp Arg Ile Glu Pro Gly Arg Asp Arg Arg Phe Arg Val Xaa Arg Leu
1 5 10 15
Pro Asn Gly Arg
20
<210> 146
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 146
Ile Phe Tyr Asp Tyr Asp Gly Asp Ile Asp Val
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| КАНИНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CTLA-4 | 2020 |
|
RU2822460C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО CTLA-4 | 2020 |
|
RU2818586C2 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2840044C1 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2838943C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2795485C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2732604C2 |
| АНТИТЕЛА К ПАРВОВИРУСУ ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2830441C2 |
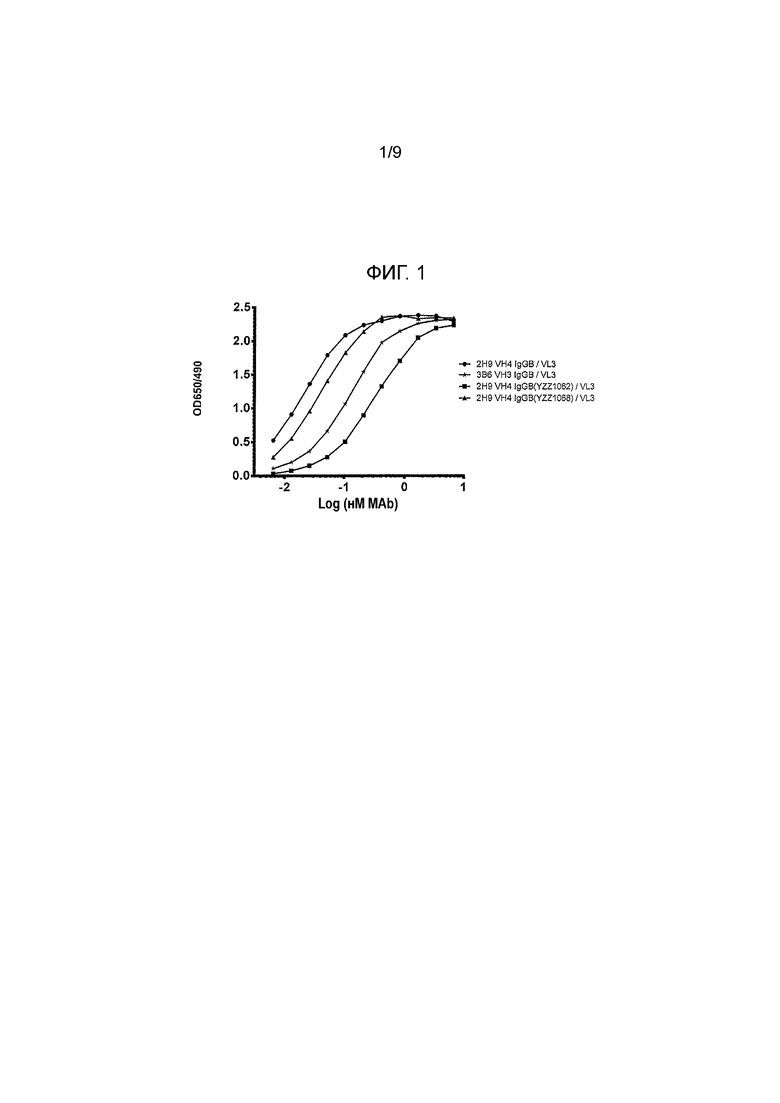
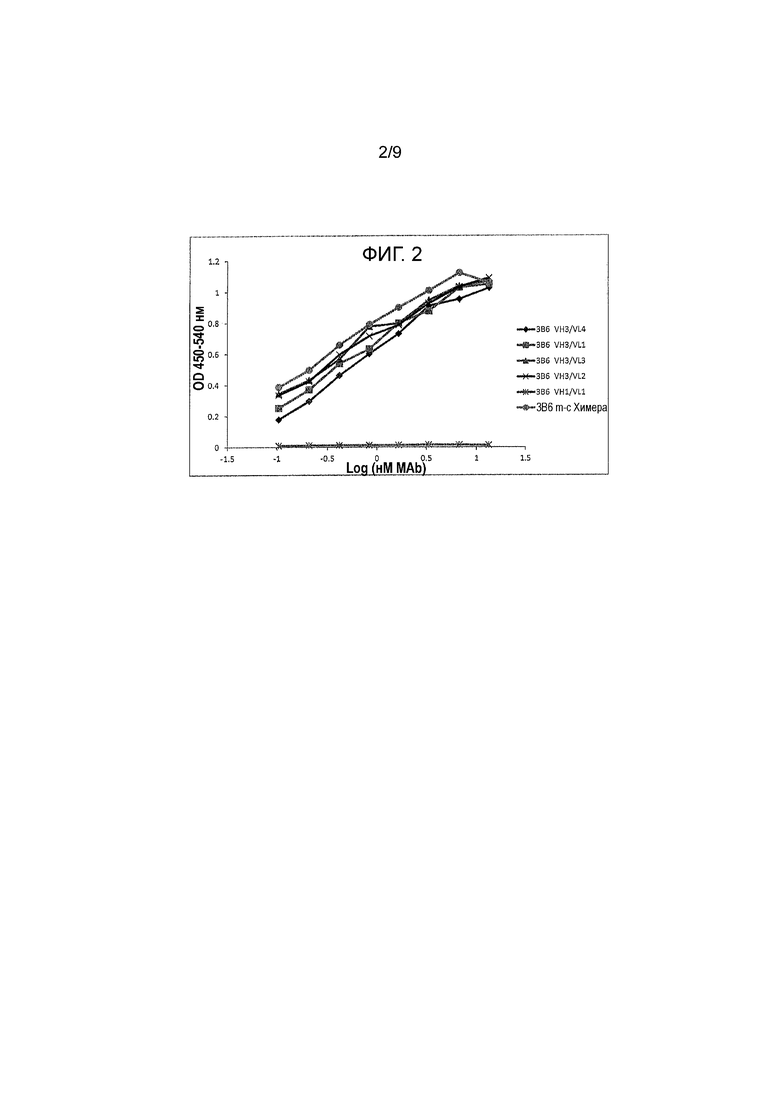


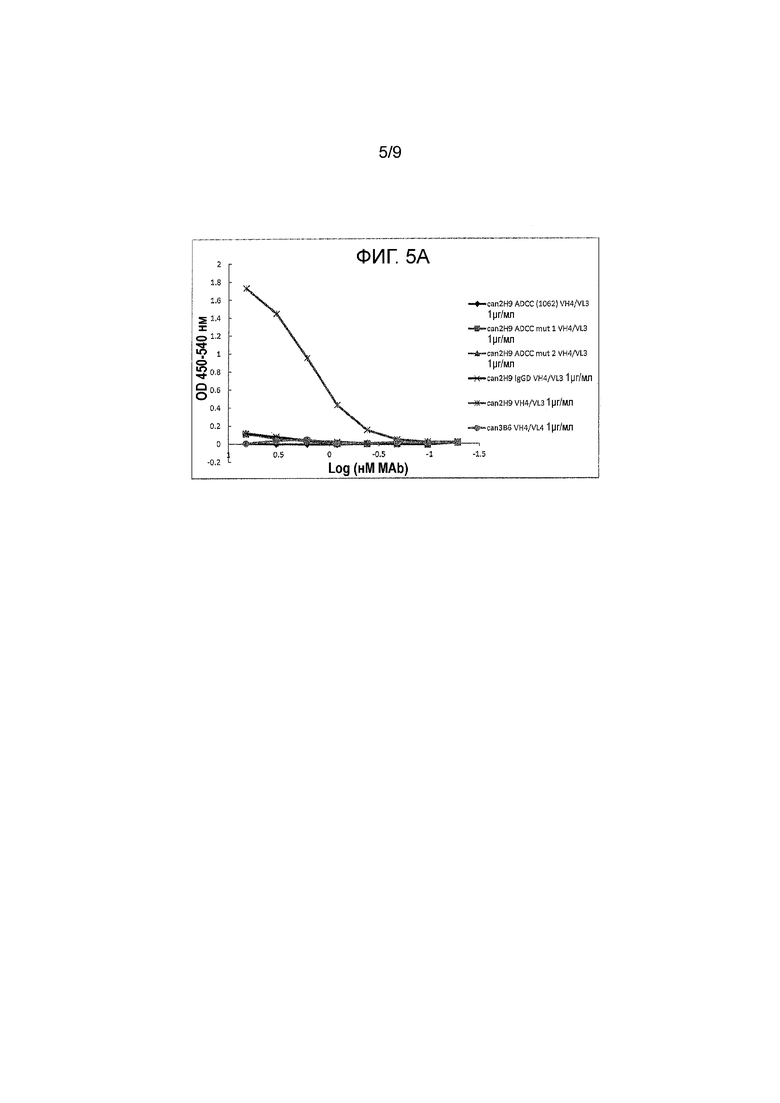



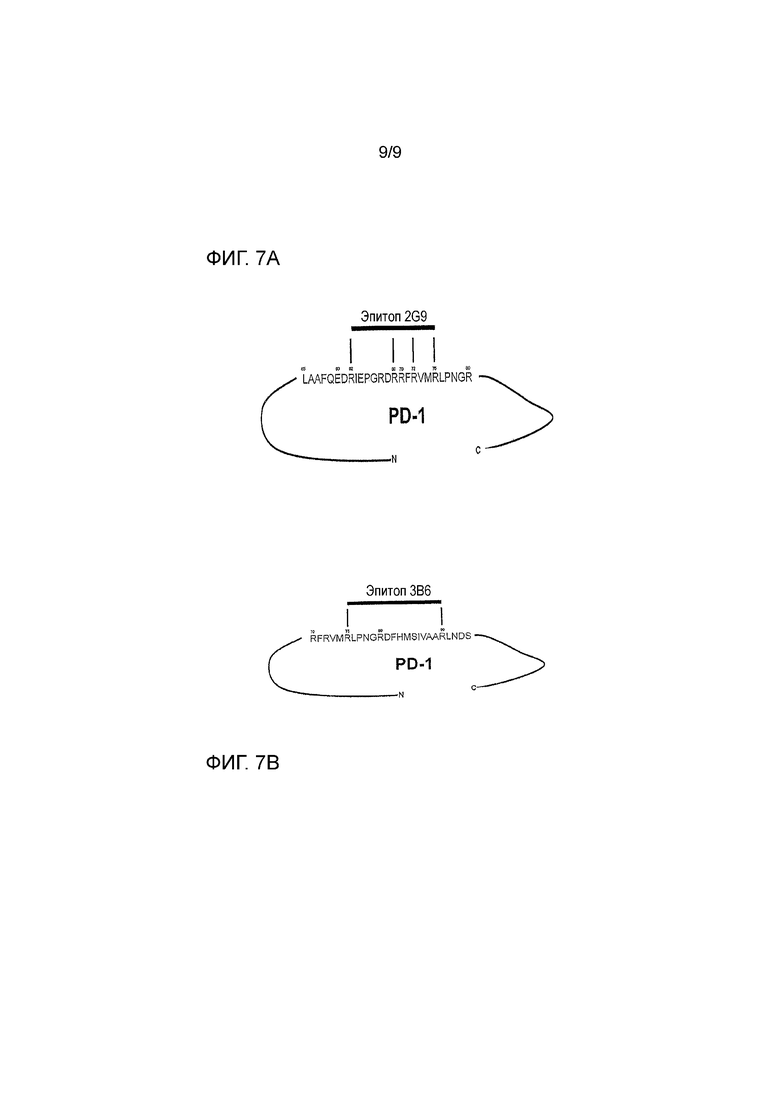
Изобретение относится к области биотехнологии, а именно к канинизированному антителу, включающему кристаллизуемую область фрагмента (cFc-область), шарнирную область и комбинацию комплементарно определяемых областей (CDR) тяжелой и легкой цепей, а также к содержащей его композиции. Также представлена нуклеиновая кислота, кодирующая тяжелую цепь и легкую вышеуказанного антитела, а также вектор, ее содержащий. Изобретение эффективно для лечения PD-1 опосредованных заболеваний. 6 н. и 4 з.п. ф-лы, 10 ил., 14 табл., 5 пр.
1. Канинизированное антитело для лечения PD-1 опосредованных заболеваний, включающее кристаллизуемую область фрагмента (cFc-область), шарнирную область и комбинации комплементарно определяемых областей (CDR) тяжелой и легкой цепей, выбранные из
аминокислотной последовательности SEQ ID NO: 15, 18, 24, 29, 33 и 38; и
аминокислотной последовательности SEQ ID NO: 13, 19, 25, 27, 31 и 36;
где тяжелая цепь содержит генетически модифицированную cFc-область, которая содержит аминокислотную последовательность, которая имеет 90% или более идентичность аминокислотной последовательности cFc-области, выбранной из группы, состоящей из SEQ ID NO: 130 и SEQ ID NO: 132;
где указанная генетически модифицированная cFc-область содержит аланиновый остаток в положении 31 последовательности SEQ ID NO: 130 вместо остатка аспарагиновой кислоты (D31A) и аланиновый остаток в положении 63 последовательности SEQ ID NO: 130 вместо остатка аспарагина (N63A); или
где указанная генетически модифицированная cFc область содержит аланиновый остаток в положении 31 последовательности SEQ ID NO: 132 вместо остатка аспарагиновой кислоты (D31A) и аланиновый остаток в положении 63 последовательности SEQ ID NO: 132 вместо остатка аспарагина (N63A).
2. Канинизированное антитело по п. 1, где шарнирная область содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 109, SEQ ID NO: 110, SEQ ID NO: 111 и SEQ ID NO: 112.
3. Канинизированное антитело по п. 1 или 2, где канинизированное антитело связывает рецептор запрограммированной гибели клеток 1 собаки со специфичностью.
4. Канинизированное антитело по любому из пп. 1-3, где легкая последовательность содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 72 и SEQ ID NO: 78.
5. Канинизированное антитело по п. 1, которое проявляет одно, два, три, четыре, пять или все из следующих свойств:
(i) связывание с PD-1 собаки с константой диссоциации (Kd), составляющей 1×10-5 - 1×10-12 M;
(ii) связывание с PD-1 собаки со скоростью ассоциации (kon), составляющей 1×102 - 1×107 M-1⋅c-1;
(iii) связывание с PD-1 собаки со скоростью диссоциации (koff), составляющей 1×10-3 - 1×10-8 c-1;
(iv) стимулирование памяти антигенспецифических ответов на опухоль или патоген;
(v) стимулирование гуморального иммунного ответа in vivo; и
(vi) стимулирование иммунного ответа у животного-субъекта.
6. Канинизированное антитело по любому из пп. 1-5, где при связывании с PD-1 собаки указанное антитело связывается с по меньшей мере одним аминокислотным остатком в рамках одной или более последовательностей, выбранных из группы, состоящей из SEQ ID NO: 138, SEQ ID NO: 139, SEQ ID NO: 140, SEQ ID NO: 141, SEQ ID NO: 142, SEQ ID NO: 143, SEQ ID NO: 144 и SEQ ID NO: 145, где антитело связывается с PD-1 собаки и блокирует связывание PD-1 собаки с рецептором запрограммированной гибели клеток 1 собаки.
7. Канинизированное антитело по п. 6, где указанное антитело связывается с одним или более аминокислотными остатками, выбранными из группы, состоящей из R62, R69, R72, R75 и R90 последовательности SEQ ID NO: 114.
8. Нуклеиновая кислота, которая кодирует тяжелую цепь и легкую цепь канинизированного антитела по п. 4.
9. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 8.
10. Фармацевтическая композиция для лечения PD-1 опосредованных заболеваний, содержащая эффективное количество канинизированного антитела по п. 4 и фармацевтически приемлемый носитель или разбавитель.
| WO 2012153126 A1, 15.11.2012 | |||
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К РЕЦЕПТОРУ ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2005 |
|
RU2402569C2 |
| WO 2010117760 A2, 14.10.2014 | |||
| WO 2008083174 A2, 10.07.2008. | |||
