ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет в соответствии с 35 U.S.C. § 119(e) предварительных заявок США с серийными номерами 63/015,209, поданной 24 апреля 2020 г., 63/015,220, поданной 24 апреля 2020 г., 62/951,778, поданной 20 декабря 2019, и 62/951,793, поданной 20 декабря 2019 г., содержание каждой из которых включено в настоящее описание во всей своей полноте посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к антителам к собачьему альфа-рецептору IL-4, демонстрирующим высокую аффинность связывания с собачьим альфа-рецептором IL-4, которые могут блокировать связывание собачьего IL-4 и/или IL-13 с собачьим альфа-рецептором IL-4. Настоящее изобретение также относится к применению антител по настоящему изобретению при лечении атопического дерматита у собак.
УРОВЕНЬ ТЕХНИКИ
Иммунная система состоит из сети резидентных и рециркулирующих специализированных клеток, которые функционируют совместно для защиты хозяина от инфекционных заболеваний и рака. Способность иммунной системы выполнять эту функцию в значительной степени зависит от биологической активности группы белков, секретируемых лейкоцитами, которые в совокупности называются интерлейкинами. Среди хорошо изученных интерлейкинов присутствуют четыре важные молекулы, идентифицированные как интерлейкин-4 (IL-4), интерлейкин-13 (IL-13), интерлейкин-31 (IL-31) и интерлейкин-22 (IL-22). IL-4 и IL-13 представляют собой близкородственные белки, которые могут секретироваться многими типами клеток, включая CD4+ Th2-клетки, естественные киллеры Т-клетки (NKT), макрофаги, тучные клетки и базофилы. IL-4 и IL-13 обладают многими перекрывающимися функциями и имеют решающее значение для развития Т-клеточно-зависимых гуморальных иммунных ответов. Известно, что IL-4 связывается с высокой аффинностью с двумя рецепторами, т.е. рецепторами IL-4 типа I и типа II. Рецептор IL-4 типа I состоит из α-цепи рецептора IL-4 и общей γ-С-цепи. Рецептор IL-4 типа II состоит из α-цепи рецептора IL-4 и α1-цепи рецептора IL-13. IL-13 связывается с рецептором IL-4 типа II и с уникальным рецептором, обозначенным как α2-рецептор IL-13. Связывание IL-13 с α2-рецептором IL-13 не передает сигнал, и этот рецептор также секретируется в растворимой форме. Соответственно, α2-рецептор IL-13 часто называют рецептором-ловушкой. Хотя IL-4, IL-13, IL-22 и IL-31 являются важными цитокинами для развития иммунных ответов, которые необходимы для защиты от внеклеточных патогенов (например, паразитов, обитающих в тканях или просвете), эти цитокины также участвуют в патогенезе аллергических заболеваний человека и животных, включая атопический дерматит.
Атопический дерматит (АД) представляет собой рецидивирующее зудящее и хроническое воспалительное заболевание кожи, характеризующееся нарушением регуляции иммунной системы и нарушениями эпидермального барьера у человека. Патологические и иммунологические признаки атопического дерматита были предметом обширных исследований [см. обзор Rahman et al. Inflammation & Allergy-drug target 10:486-496 (2011) и Harskamp et al., Seminar in Cutaneous Medicine and Surgery 32:132-139 (2013)]. Атопический дерматит также часто встречается у домашних животных, особенно у собак, причем его распространенность в собачьей популяции составляет примерно 10-15%. Патогенез атопического дерматита у собак и кошек [см. обзор Nuttall et al., Veterinary Records 172(8):201-207 (2013)] демонстрирует значительное сходство с патогенезом атопического дерматита у человека, включая инфильтрацию кожи различными иммунными клетками, и среду CD4+ Th2 поляризованных цитокинов, включая преобладание IL-4, IL-13 и IL-31. Кроме того, IL-22 вовлечен в усиленную пролиферацию эпителиальных клеток, приводящую к гиперплазии эпидермиса, что характерно для атопического дерматита.
Например, было показано, что антитела к собачьему IL-31 оказывают значительное влияние на зуд, ассоциированный с атопическим дерматитом у собак [US 8790651 B2; США 10,093,731 B2]. Кроме того, было протестировано антитело к человеческому альфа-рецептору IL-31 (IL-31RA), и обнаружено, что оно оказывает значимое влияние на зуд, ассоциированный с атопическим дерматитом у людей [Ruzicka, et al., New England Journal of Medicine, 376(9), 826-835 (2017)]. Соответственно, блокирование связывания IL-31 с рецептором IL-31RA приводит к облегчению зуда, ассоциированного с атопическим дерматитом.
Разработаны моноклональные антитела к человеческому альфа-рецептору IL-4 (IL-4 Rα), и некоторые из этих антител были тщательно протестированы на терапевтическое действие при лечении атопического дерматита у людей [см., например, US2015/0017176 A1]. Совсем недавно также были описаны канинизированные антитела к собачьему IL-4 Rα, которые блокируют связывание собачьего IL-4 с собачьим IL-4 Rα [US 2018/0346580A1, включенная в настоящее описание во всей полноте посредством ссылки]. Поскольку рецептор IL-4 типа II состоит из α-цепи рецептора IL-4 и α1-цепи рецептора IL-13, были получены антитела к собачьему IL-4 Rα, которые могут блокировать связывание как собачьего IL-4, так и собачьего IL-13 с собачьим рецептором IL-4 типа II, тем самым позволяя блокировать воспаление, ассоциированное с атопическим дерматитом [US2018/0346580A1].
Интерлейкин-22 (IL-22), также известный как IL-10 подобный T-клеточный индуцибельный фактор (IL-TIF), принадлежит к семейству цитокинов IL-10. IL-22 продуцируется нормальными Т-клетками при анти-CD3-стимуляции у людей. Экспрессия мышиного IL-22 также индуцируется в различных органах при инъекции липополисахаридов, что свидетельствует о том, что IL-22 может быть вовлечен в воспалительные реакции. IL-22 специфически связывается с рецепторным комплексом, состоящим из гетеродимерного комплекса IL-10R2 (также известного как IL-10R бета) и рецептора интерлейкина-22 (IL-22R), и передает через него сигналы [см. Lee et al., Pharmacology Research & Perspectives, pp. 1-13 (2018:e00434)]. Рецептор интерлейкина-22 также известен как интерлейкин-22R, альфа 1; IL-22RА1; IL-22R1; zcytor11; и CRF2-9 [Xu et al., Proc. Nat. Acad. Sci. 98(17):9511-9516 (2001); Gelebart and Lai, Atlas of Genetics and Cytogenetics 14(12):1106-1110 (2010)]. IL-22 индуцирует пролиферацию эпителиальных клеток во время заживления ран, и его дефицит может привести к неконтролируемой пролиферации и усилению развития опухоли [Huber et al., Nature 491:259-263 (2012)]. Было показано, что IL-22 активирует STAT-1 и STAT-3 в нескольких клеточных линиях гепатомы и усиливает продукцию белков острой фазы. Антитела к интерлейкину-22 и IL-22R действуют как антипролиферативные агенты, блокируя взаимодействие IL-22 с IL-22R и тем самым связанный с ним сигнальный путь, что приводит к пролиферации эпителиальных клеток.
Однако, несмотря на недавний успех в лечении атопического дерматита, ни один из используемых в настоящее время способов лечения не приводит к быстрому началу противозудного действия, сопровождающегося значительным влиянием на воспаление кожи с улучшением ее барьерной функции. Следовательно, существует необходимость в разработке более эффективных способов лечения, которые могли бы устранить один или более симптомов атопического дерматита.
Цитирование любой ссылки в настоящем описании не должно быть истолковано как признание того, что такая ссылка может использоваться в качестве «известного уровня техники» по отношению к настоящей заявке.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложены новые канинизированные антитела к собачьему IL-4R-альфа (IL-4Rα), которые выделены в конкретных вариантах осуществления и которые обладают превосходными свойствами по сравнению с антителами, известными из уровня техники, например, связываются более прочно, чем антитела к собачьему альфа-рецептору IL-4 предшествующего уровня техники. В конкретных вариантах осуществления настоящее изобретение относится к антителам млекопитающих или их антигенсвязывающим фрагментам, которые специфически связываются с собачьим альфа-рецептором интерлейкина-4, содержащим тяжелую цепь, которая содержит набор из трех определяющих комплементарность областей тяжелой цепи (CDR), CDR 1 тяжелой цепи (HCDR1), CDR 2 тяжелой цепи (HCDR2) и CDR 3 тяжелой цепи (HCDR3), где HCDR1 содержит аминокислотную последовательность SEQ ID NO: 12, HCDR2 содержит аминокислотную последовательность SEQ ID NO: 14, и HCDR3 содержит аминокислотную последовательность SEQ ID NO: 16. В родственных вариантах осуществления антитела млекопитающих или их антигенсвязывающие фрагменты, которые специфически связываются с собачьим α-рецептором интерлейкина-4 (IL-4Rα), дополнительно содержат легкую цепь, которая содержит набор из трех CDR легкой цепи: CDR 1 легкой цепи (LCDR1), CDR 2 легкой цепи (LCDR2) и CDR 3 легкой цепи (LCDR3), где LCDR1 содержит аминокислотную последовательность SEQ ID NO: 18, LCDR2 содержит аминокислотную последовательность SEQ ID NO: 20, и LCDR3 содержит аминокислотная последовательность SEQ ID NO: 22. В предпочтительных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент связывается с собачьим IL-4Rα и блокирует связывание собачьего IL-4Rα с собачьим интерлейкином-4 (cIL-4). В родственных вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент связывается с собачьим IL-4Rα и блокирует связывание собачьего IL-4Rα с собачьим интерлейкином-13 (cIL-13). В других вариантах осуществления антитело млекопитающего или его антигенсвязывающий фрагмент связывается с собачьим IL-4Rα и блокирует связывание собачьего IL-4Rα с cIL-4 и с cIL-13.
В конкретных вариантах осуществления антитело млекопитающего к собачьему IL 4R-альфа представляет собой мышиное антитело. В родственных вариантах осуществления антитело млекопитающего к собачьему IL 4R-альфа представляет собой канинизированное мышиное антитело. В конкретных вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит cFc IgG-D, но встречающаяся в природе шарнирная область IgG-D заменена шарнирной областью, содержащей аминокислотную последовательность SEQ ID NO: 6. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит cFc IgG-D, но встречающаяся в природе шарнирная область IgG-D заменена шарнирной областью, содержащей аминокислотную последовательность SEQ ID NO: 7. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит cFc IgG-D, но встречающаяся в природе шарнирная область IgG-D заменена шарнирной областью, содержащей аминокислотную последовательность SEQ ID NO: 8. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит cFc IgG D, но встречающаяся в природе шарнирная область IgG-D заменена шарнирной областью, содержащей аминокислотную последовательность SEQ ID NO: 9.
В конкретных вариантах осуществления канинизированное антитело содержит тяжелую цепь, содержащую модифицированный собачий IgG-B (IgG-Bm), содержащий аминокислотную последовательность SEQ ID NO: 10. В некоторых вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 40. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 41. В других вариантах осуществления канинизированное антитело содержит тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 42. В конкретных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 39. В альтернативных вариантах осуществления канинизированное антитело дополнительно содержит легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 44.
В конкретных вариантах осуществления канинизированные антитела или их антигенсвязывающие фрагменты связываются с SEQ ID NO: 46. В более конкретных вариантах осуществления канинизированные антитела или их антигенсвязывающие фрагменты связываются с одним, двумя или всеми тремя из следующих аминокислотных остатков собачьего IL-4Rα: К97, H112, T113 SEQ ID NO: 5. В родственных вариантах осуществления канинизированные антитела или их антигенсвязывающие фрагменты связываются с SEQ ID NO: 47. В более конкретных вариантах осуществления канинизированные антитела или их антигенсвязывающие фрагменты связываются с одним, двумя, тремя, четырьмя или всеми пятью из следующих аминокислотных остатков собачьего IL-4Rα: S164, T165, S171, Y172, S173 и R175 SEQ ID NO: 5. В еще более конкретных вариантах осуществления канинизированные антитела или их антигенсвязывающие фрагменты связываются с обеими SEQ ID NO: 46 и SEQ ID NO: 47. В еще более конкретных вариантах осуществления этого типа канинизированные антитела или их антигенсвязывающие фрагменты связываются с одним, двумя или всеми тремя из следующих аминокислотных остатков собачьего IL-4Rα: К97, H112, T113 SEQ ID NO: 5 и/или с одним, двумя, тремя, четырьмя или всеми пятью из следующих аминокислотных остатков собачьего IL-4Rα: S164, T165, S171, Y172, S173 и R175 SEQ ID NO: 5.
Настоящее изобретение также относится к нуклеиновым кислотам, включая выделенные нуклеиновые кислоты, которые кодируют CDR, тяжелые цепи канинизированных антител или их антигенсвязывающих фрагментов и/или легкие цепи канинизированных антител или их антигенсвязывающих фрагментов. Кроме того, настоящее изобретение относится к векторам экспрессии, которые содержат такие нуклеиновые кислоты, и клеткам-хозяевам, которые содержат такие векторы экспрессии.
Кроме того, настоящее изобретение относится к фармацевтическим композициям, которые содержат канинизированные антитела и их антигенсвязывающие фрагменты по настоящему изобретению вместе с фармацевтически приемлемым носителем и/или разбавителем. Настоящее изобретение дополнительно относится к способам лечения атопического дерматита, включающим введение одной из вышеупомянутых композиций представителю псовых, страдающему атопическим дерматитом. В конкретных вариантах осуществления настоящее изобретение относится к способам позволяющим блокировать воспаление, ассоциированное с атопическим дерматитом, включающим введение нуждающемуся в этом представителю псовых терапевтически эффективного количества фармацевтической композиции по настоящему изобретению.
Эти и другие аспекты настоящего изобретения будут лучше поняты со ссылкой на приведенное ниже краткое описание чертежей и подробное описание.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показаны два разных канинизированных моноклональных антитела к собачьему альфа-рецептору интерлейкина 4, обозначенные c4H3 [см. WO2016/156588] и c146E2-H3L3, которые оценивали на их способность ингибировать фосфорилирование STAT-6. Данные показывают, что оба антитела приводят к дозозависимому ингибированию фосфорилирования STAT-6 в присутствии собачьего интерлейкина-4. Контроль IL-4 в отсутствие антител к IL-4R-альфа (IL-4Rα) показан в верхней правой части графика.
На фиг.2 показаны два разных канинизированных моноклональных антитела к собачьему альфа-рецептору интерлейкина 4, обозначенных c4H3 [см. WO2016/156588] и c146E2-H3L3, которые оценивали на их способность ингибировать фосфорилирование STAT-6. Данные показывают, что оба антитела приводят к дозозависимому ингибированию фосфорилирования STAT-6 в присутствии собачьего интерлейкина-13. Контроль IL-13 в отсутствие антител к IL-4R-альфа (IL-4Rα) показан в верхней правой части графика.
На фиг.3 показано связывание канинизированных антител к собачьему IL-4Rα, содержащих легкие лямбда- или каппа-цепи, согласно результатам метода ELISA. Результаты показывают, что канинизированные антитела к собачьему IL-4Rα, содержащие легкие лямбда-цепи (c146ClL1-H1, c146ClL1-H2 и c146ClL1-H3), связываются с собачьим IL-4Rα так же, как и канинизированные антитела к собачьему IL-4Rα, содержащие такие же CDR, но с легкой каппа-цепью (c146E2-H3L3). 146mc представляет собой положительный контроль химерного мышино-собачьего антитела, а Iso-Ctr, служащий в качестве отрицательного контроля, является неродственным канинизированным антителом.
На фиг.4 показан эпитоп для антитела c146E2-H3L3, содержащий аминокислотные последовательности SEQ ID NO: 46 и SEQ ID NO: 47, на собачьем IL-4Rα.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В ответ на потребность в более эффективных способах лечения атопического дерматита в настоящем изобретении предлагаются канинизированные антитела, составы с канинизированными антителами и способы, которые могут оказывать значительное влияние на воспаление кожи, связанное с атопическим дерматитом.
СОКРАЩЕНИЯ
В подробном описании и примерах изобретения используются следующие сокращения:
ADCC - Антителозависимая клеточная цитотоксичность
CDC - Комплемент-зависимая цитотоксичность
CDR - Определяющая комплементарность область в вариабельных областях иммуноглобулина, определенная с помощью системы нумерации Кабат (Kabat)
EC50 - Концентрация, приводящая к 50% эффективности или связыванию
ELISA - Иммуноферментный анализ
FR - Каркасная область антитела: вариабельные области иммуноглобулина, за исключением областей CDR.
IC50 - Концентрация, приводящая к 50% ингибированию
IgG - Иммуноглобулин G
Kabat - Система выравнивания и нумерации иммуноглобулинов, впервые предложенная Элвином А. Кабатом (Elvin A. Kabat) [Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)]
mAb - Моноклональное антитело (также Mab или MAb)
V-область - Сегмент цепей IgG, последовательность которых различается у разных антител. Согласно нумерации Кабат, он простирается до остатка 109 в легкой цепи и 113 в тяжелой цепи.
VH - Вариабельная область тяжелой цепи иммуноглобулина
VL - Вариабельная область легкой цепи иммуноглобулина
VK - Вариабельная область легкой каппа-цепи иммуноглобулина
ОПРЕДЕЛЕНИЯ
Чтобы изобретение было более понятным, некоторые технические и научные термины определены ниже. Если в настоящем описании специально не определено иное, все другие используемые технические и научные термины имеют значение, обычно понимаемое специалистом в области техники, к которой относится настоящее изобретение.
Используемые в настоящем описании, включая прилагаемую формулу изобретения, указания на формы единственного числа, такие как «a», «an» и «the», включают соответствующие ссылки на множественное число, если из контекста в явном виде не следует иное.
«Введение» и «лечение» применительно к животному, например, представителю псовых, клетке, ткани, органу или биологической жидкости, относится к контакту экзогенного фармацевтического, терапевтического, диагностического агента или композиции с животным, например, представителем псовых, клеткой, тканью, органом или биологической жидкостью. Лечение клетки включает контакт реагента с клеткой, а также контакт реагента с жидкостью, причем жидкость находится в контакте с клеткой.
«Введение» и «лечение» также означают лечение in vitro и ex vivo, например, клетки реагентом, диагностическим средством, связывающим соединением или другой клеткой. Термин «субъект» включает любой организм, предпочтительно животное, более предпочтительно млекопитающее (например, представителя псовых, кошачьих или человека) и наиболее предпочтительно представителя псовых.
«Лечить» или «обработка» означает введение терапевтического агента, такого как композиция, содержащая любое из антител по настоящему изобретению, внутрь или наружно, например, представителю псовых или пациенту с одним или более симптомами или с подозрением на наличие состояния, в отношении которого указанный агент обладает терапевтической активностью. Как правило, агент вводят в количестве, эффективном для облегчения и/или ослабления одного или более симптомов заболевания/состояния у субъекта или популяции, получающей лечение, либо путем индуцирования регрессии, либо ингибирования прогрессирования такого(их) симптома(ов) на любую клинически измеримую величину. Количество терапевтического агента, которое является эффективным для ослабления любого конкретного симптома заболевания/состояния (также называемое «терапевтически эффективным количеством»), может меняться в зависимости от таких факторов, как тяжесть заболевания/состояния, возраст и вес пациента (например, представителя псовых), и способность фармацевтической композиции вызывать требуемый ответ у субъекта. Ослабление или облегчение симптома заболевания/состояния можно оценить с помощью любого клинического измерения, обычно используемого ветеринарами или другими квалифицированными медицинскими работниками для оценки тяжести или состояния прогрессирования этого симптома. Хотя вариант осуществления настоящего изобретения (например, способа лечения или промышленного изделия) может не быть эффективным для облегчения симптома(ов) целевого заболевания/состояния у каждого субъекта, он должен облегчать симптом(ы) целевого заболевания/состояния у статистически значимого количества субъектов, определяемого любым статистическим тестом, известным в данной области, таким как t-критерий Стьюдента, критерий хи-квадрат, U-критерий Манна-Уитни, критерий Краскела-Уоллиса (H-критерий), критерий Джонкхиера-Терпстра и критерий Уилкоксона.
«Лечение» применительно к человеку, в ветеринарии (например, представителю псовых) или субъекту исследования, относится к терапевтическому лечению, а также к исследовательским и диагностическим применениям. «Лечение» применительно к человеку, в ветеринарии (например, представителю псовых) или субъекту исследования, или клетке, ткани или органу включает контакт антител по настоящему изобретению, например, с организмом представителя псовых или другого животного, клеткой, тканью, физиологическим компартментом или физиологической жидкостью.
В контексте настоящего описания термин «собачий (псовый)» включает всех домашних собак, Canis lupus familiaris или Canis familiaris, если не указано иное.
В контексте настоящего описания термин «кошачий» относится к любому члену семейства кошачьих. Члены этого семейства включают диких кошек, кошек, содержащихся в зоопарке, и домашних кошек, включая домашних, чистопородных и/или беспородных кошек-компаньонов, выставочных кошек, лабораторных кошек, клонированных кошек, а также диких или одичавших кошек.
В контексте настоящего описания термин «собачий каркас» относится к аминокислотной последовательности тяжелой цепи и легкой цепи собачьего антитела, отличной от остатков гипервариабельной области, определенных в настоящем описании как остатки CDR. Что касается канинизированного антитела, то в большинстве вариантов осуществления аминокислотные последовательности нативных собачьих CDR заменены соответствующими чужеродными CDR (например, из мышиного антитела) в обеих цепях. Необязательно, тяжелые и/или легкие цепи собачьего антитела могут содержать некоторые чужеродные остатки, не относящиеся к остаткам CDR, например, с целью сохранения конформации чужеродных CDR внутри собачьего антитела и/или для модификации функции Fc, как показано ниже и/или раскрыто в US 10,106,607 B2, полностью включенном в настоящее описание посредством ссылки.
«Область кристаллизуемого фрагмента», сокращенно «Fc», соответствует части CH3-CH2 антитела, которая взаимодействует с рецепторами клеточной поверхности, называемыми Fc-рецепторами. Собачьи области кристаллизуемого фрагмента (cFc) каждого из четырех собачьих IgG были впервые описаны Tang et al. [Vet. Immunol. Immunopathol. 80:259-270 (2001); см. также Bergeron et al., Vet. Immunol. Immunopathol. 157:31-41 (2014) и U.S. 10,106,607 B2].
Собачья Fc (cFc) «IgG-Bm», используемая в настоящем описании, представляет собой собачью Fc IgG-B, содержащую две (2) замены аминокислотных остатков, D31A и N63A, в аминокислотной последовательности SEQ ID NO: 10 IgG-B (см. ниже), в которой отсутствует С-концевой лизин («К»). В IgG-Bm оба остатка, остаток аспарагиновой кислоты (D) в положении 31 SEQ ID NO: 10 и остаток аспарагина (N) в положении 63 SEQ ID NO: 10, замещены остатком аланина (A). Эти две замены аминокислотных остатков существенно снижают антителозависимую цитотоксичность (ADCC) и комплемент-зависимую цитотоксичность (CDC) встречающегося в природе собачьего IgG-B [см. U.S. 10,106,607 B2, содержание которого включено в настоящее описание во всей полноте посредством ссылки]. В IgG-Bm также предусмотрены дополнительные замены аминокислот, аналогичные заменам, которые могут быть параллельно введены в IgG-B и которые могут включать замены аминокислот, способствующие образованию гетеродимеров в биспецифических антителах. Аминокислотная последовательность IgG-B, SEQ ID NO: 45 является следующей:

Ниже представлена аминокислотная последовательность IgG-Bm, SEQ ID NO: 10.
lggpsvfifppkpkdtlliartpevtcvvvaldpedpevqiswfvdgkqmqtaktqpreeqfagtyrvvsvlpighqdwlkgkqftckvnnkalpspiertiskargqahqpsvyvlppsreelskntvsltclikdffppdidvewqsngqqepeskyrttppqldedgsyflysklsvdksrwqrgdtficavmhealhnhytqeslshspg
В контексте настоящего описания термин «замещение аминокислотного остатка» другим аминокислотным остатком в аминокислотной последовательности антитела, например, является эквивалентным термину «замена аминокислотного остатка» другим аминокислотным остатком и означает, что конкретный аминокислотный остаток в определенном положении аминокислотной последовательности заменен (или замещен) другим аминокислотным остатком. Такие замены могут быть специально предусмотренными, т.е. могут быть целенаправленной заменой аланина серином в определенном положении аминокислотной последовательности, например, выполненной методом рекомбинантной ДНК. Альтернативно, конкретный аминокислотный остаток или нить аминокислотных остатков антитела может быть заменен одним или более аминокислотными остатками посредством более естественных процессов отбора, например, основанных на способности антитела, продуцируемого клеткой, связываться с заданной областью на антигене, например, который содержит эпитоп или его часть, и/или путем отбора антитела, содержащего конкретную CDR с сохраненной канонической структурой, которая является такой же как у замененной ею CDR. Такие замещения/замены могут давать «вариантные» CDR и/или вариантные антитела.
В контексте настоящего описания термин «антитело» относится к любой форме антитела, которая проявляет требуемую биологическую активность. Антитело может быть мономером, димером или более крупным мультимером. Таким образом, он используется в самом широком смысле и, в частности, охватывает, помимо прочего, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, полиспецифические антитела (например, биспецифические антитела), канинизированные антитела, полностью собачьи антитела, химерные антитела и камелизованные (на основе антитела верблюдовых) однодоменные антитела. «Родительские антитела» представляют собой антитела, полученные путем воздействия иммунной системы на антиген до модификации антител для предполагаемого применения, такой как канинизация антитела для использования в качестве собачьего терапевтического антитела.
В контексте настоящего описания антитела по настоящему изобретению, которые «блокируют» или являются «блокирующими» или «блокируют связывание», например, собачьего рецептора с его партнером по связыванию (лигандом), представляют собой антитела, которые блокируют (частично или полностью) связывание собачьего рецептора с собачьим лигандом и наоборот, что определяется стандартными анализами связывания (например, BIACore®, ELISA или проточной цитометрией).
Как правило, антитело или его антигенсвязывающий фрагмент по изобретению сохраняет по меньшей мере 10% своей активности связывания собачьего антигена (по сравнению с исходным антителом), выраженной в молярных единицах. Предпочтительно антитело или антигенсвязывающий фрагмент по изобретению сохраняет по меньшей мере 20%, 50%, 70%, 80%, 90%, 95% или 100% или более аффинности исходного антитела в отношении связывания собачьего антигена. Также предполагается, что антитело или антигенсвязывающий фрагмент по изобретению может включать консервативные или неконсервативные аминокислотные замены (называемые «консервативными вариантами» или «вариантами с сохраненной функцией» антитела), которые существенно не изменяют его биологическую активность.
«Выделенное антитело» относится к статусу очистки и в этом контексте означает, что молекула практически свободна от других биологических молекул, таких как нуклеиновые кислоты, белки, липиды, углеводы или другой материал, такой как клеточный дебрис и среда для роста. Как правило, термин «выделенный» не относится к полному отсутствию такого материала или к отсутствию воды, буферов или солей, если только они не присутствуют в количествах, которые фактически мешают экспериментальному или терапевтическому применению раскрытого связывающего соединения.
В контексте настоящего описания термин «химерное антитело» представляет собой антитело, имеющее вариабельный домен первого антитела и константный домен второго антитела, при этом первое и второе антитела относятся к разным видам. [U.S. 4,816,567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)]. Обычно вариабельные домены получают из антитела экспериментального животного («родительское антитело»), такого как грызун, а последовательности константных доменов получают из антител животного-субъекта, например человека или представителя псовых, вследствие чего полученное химерное антитело будет с меньшей вероятностью вызывать неблагоприятный иммунный ответ у человека или представителя псовых, соответственно, по сравнению с родительским (например, грызуна) антителом.
В контексте настоящего описания термин «канинизированное антитело» относится к формам антител, которые содержат последовательности как собачьих, так и несобачьих (например, мышиных) антител. Как правило, канинизированное антитело будет содержать практически все из по меньшей мере одного или, как правило, двух вариабельных доменов, в которых все или практически все гипервариабельные петли соответствуют петлям несобачьего иммуноглобулина (например, содержит 6 CDR, как показано ниже), и все или по существу все каркасные (FR) области (и, как правило, все или по существу все оставшиеся каркасные участки) являются участками последовательности собачьего иммуноглобулина. Как показано в настоящем описании, канинизированное антитело содержит три CDR тяжелой цепи, и три CDR легкой цепи мышиного антитела к собачьему антигену вместе с собачьим каркасом или модифицированным собачьим каркасом. Модифицированный собачий каркас содержит одну или более аминокислотных замен, как показано в настоящем описании, которые позволяют дополнительно оптимизировать эффективность канинизированного антитела, например, путем увеличения его связывания с собачьим антигеном и/или способности блокировать связывание этого собачьего антигена с природным партнером по связыванию собачьего антигена.
Вариабельные области каждой пары легкая/тяжелая цепь образуют сайт связывания антитела. Таким образом, обычно интактное антитело имеет два сайта связывания. За исключением бифункциональных или биспецифических антител, два сайта связывания, как правило, являются одинаковыми. Как правило, вариабельные домены как тяжелой, так и легкой цепей содержат три гипервариабельные области, также называемые определяющими комплементарность областями (CDR), расположенные внутри относительно консервативных каркасных областей (FR). CDR обычно выровнены по каркасным областям, что обеспечивает связывание со специфическим эпитопом. Как правило, в направлении от N-конца к С-концу вариабельные домены как легкой, так и тяжелой цепи содержат FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Принадлежность аминокислот каждому домену, как правило, соответствует определениям, описанным в Sequences of Proteins of Immunological Interest, Kabat, et al.; National Institutes of Health, Bethesda, Md.; 5th ed.; NIH Publ. No. 91-3242 (1991); Kabat, Adv. Prot. Chem. 32:1-75 (1978); Kabat, et al., J. Biol. Chem. 252:6609-6616 (1977); Chothia, et al., J. Mol. Biol. 196:901-917 (1987) или Chothia, et al., Nature 342:878-883 (1989).
В контексте настоящего описания термин «гипервариабельная область» относится к аминокислотным остаткам антитела, ответственным за связывание антигена. Гипервариабельная область содержит аминокислотные остатки из «определяющей комплементарность области» или «CDR» (т.е. LCDR1, LCDR2 и LCDR3 в вариабельном домене легкой цепи и HCDR1, HCDR2 и HCDR3 в вариабельном домене тяжелой цепи). [См. Kabat et al. Sequences of Proteins of Immunological Interest, 5th ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), определение областей CDR антитела по последовательности; см. также Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987), определение областей CDR антитела по структуре]. В контексте настоящего описания термин «каркасные» или «FR» остатки относится к тем остаткам вариабельного домена, которые не относятся к остаткам гипервариабельной области, определенным в настоящем описании как остатки CDR.
Существует четыре известных подтипа тяжелых цепей IgG собачьего IgG, которые обозначаются как IgG-A, IgG-B, IgG-C и IgG-D. Два известных подтипа легкой цепи называются лямбда и каппа. В конкретных вариантах осуществления изобретения, помимо связывания и активации собачьих иммунных клеток, собачье или канинизированное антитело к антигену по настоящему изобретению в оптимальном варианте обладает двумя свойствами:
1) у него отсутствуют эффекторные функции, такие как антителозависимая цитотоксичность (ADCC) и комплемент-зависимая цитотоксичность (CDC), и
2) оно легко очищается в больших масштабах с помощью стандартных промышленных методов, таких как хроматография на основе белка А.
Ни один из встречающихся в природе собачьих изотипов IgG не удовлетворяет обоим критериям. Например, IgG-B может быть очищен с помощью белка А, но имеет высокий уровень активности ADCC. С другой стороны, IgG-A слабо связывается с белком А, а также проявляет активность ADCC. Более того, ни IgG-C, ни IgG-D не могут быть очищены на колонках с белком А, хотя IgG-D не проявляет ADCC-активность. (IgG-C обладает значительной ADCC-активностью). Одним из способов, которым настоящее изобретение решает эти проблемы, является предоставление модифицированных собачьих IgG-B антител по настоящему изобретению, специфичных к антигену по настоящему изобретению, которые лишены эффекторных функций, таких как ADCC, и могут быть легко очищены с помощью стандартной хроматографии с белком А.
В контексте настоящего описания «противовоспалительное антитело» представляет собой антитело, которое может действовать как противовоспалительное средство в организме животного, включая млекопитающих, таких как человек, представитель псовых и/или кошачьих, особенно в отношении атопического дерматита. В конкретных вариантах осуществления противовоспалительное антитело связывается со специфическими белками сигнального пути IL-4/IL-13, такими как IL-4 или рецептор IL-4Rα. Связывание противовоспалительного антитела с соответствующим ему антигеном (например, IL-4 или IL-4Rα) ингибирует связывание, например, IL-4 с IL-4Rα и препятствует передаче сигналов этого пути и/или предотвращает эту передачу, тем самым препятствуя или предотвращая появление хронического воспаления, связанного с атопическим дерматитом.
«Гомология», в контексте настоящего описания, относится к сходству двух полинуклеотидных последовательностей или двух полипептидных последовательностей в случае их оптимального выравнивания. Когда положение в обеих сравниваемых последовательностях занято одним и тем же основанием или аминокислотным остатком, например, если положение в каждой из двух молекул ДНК занято аденином, то молекулы гомологичны в этом положении. Процент гомологии представляет собой количество гомологичных положений, общих для двух последовательностей, деленное на общее количество сравниваемых положений × 100. Например, если 6 из 10 положений в двух последовательностях совпадают или гомологичны, когда последовательности оптимально выровнены, то эти две последовательности гомологичны на 60%. Как правило, сравнение проводят, когда две последовательности выровнены с целью получения максимального процента гомологии. Идентичность последовательностей относится к степени, в которой аминокислоты двух полипептидов одинаковы в эквивалентных положениях, когда две последовательности оптимально выровнены.
В контексте настоящего описания одна аминокислотная последовательность «идентична» второй аминокислотной последовательности на 100%, когда идентичны аминокислотные остатки обеих последовательностей. Соответственно, аминокислотная последовательность «идентична» второй аминокислотной последовательности на 50%, когда идентичны 50% аминокислотных остатков двух аминокислотных последовательностей. Сравнение последовательностей проводят на непрерывном блоке аминокислотных остатков, содержащихся в заданном белке, например белке или части полипептида, подлежащего сравнению. В конкретных вариантах осуществления учитываются выбранные делеции или вставки, которые в противном случае могли бы изменить соответствие между двумя аминокислотными последовательностями. Сходство последовательностей включает идентичные остатки и неидентичные биохимически родственные аминокислоты. Биохимически родственные аминокислоты являются аминокислотами, которые имеют сходные свойства и могут быть взаимозаменяемыми.
«Консервативно модифицированные варианты» или «консервативная замена» относятся к заменам аминокислот в белке другими аминокислотами, имеющими сходные характеристики (например, заряд, размер боковой цепи, гидрофобность/гидрофильность, конформацию и жесткость основной цепи и т.д.), настолько, что часто позволяют сделать такие изменения без изменения биологической активности белка. Специалистам в данной области известно, что, как правило, замены отдельных аминокислот в несущественных участках полипептида не приводят к существенному изменению биологической активности [см., например, Watson et al., Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th ed., 1987)]. Кроме того, замена структурно или функционально сходных аминокислот с меньшей вероятностью приведет к нарушению биологической активности. Примеры консервативных замен представлены в Таблице А ниже.
В настоящем изобретении также предполагаются функционально-консервативные варианты антител по изобретению. «Функционально-консервативные варианты», в контексте настоящего описания, относится к антителам или фрагментам, в которых заменен один или более аминокислотных остатков без изменения требуемого свойства, такого как аффинность и/или специфичность к антигену. Такие варианты включают, без ограничения, замену аминокислоты аминокислотой с аналогичными свойствами, например, консервативные замены аминокислот из Таблицы А выше.
«Молекула выделенной нуклеиновой кислоты» означает ДНК или РНК геномного, мРНК, кДНК или синтетического происхождения или некоторую их комбинацию, которая не связана со всем полинуклеотидом или частью полинуклеотида, в котором выделенный полинуклеотид встречается в естественных условиях, или которая связана с полинуклеотидом, с которым она не связана в естественных условиях. Для целей настоящего раскрытия следует понимать, что «молекула нуклеиновой кислоты, содержащая» конкретную нуклеотидную последовательность, не охватывает интактные хромосомы. Молекулы выделенных нуклеиновых кислот, «содержащие» определенные последовательности нуклеиновых кислот, могут включать, помимо указанных последовательностей, последовательности, кодирующие до десяти или даже до двадцати или более других белков или их частей или фрагментов, или могут включать функционально связанные регуляторные последовательности, которые контролируют экспрессию кодирующей области указанных последовательностей нуклеиновых кислот и/или могут включать векторные последовательности.
Настоящее изобретение относится к выделенным канинизированным антителам по настоящему изобретению, способам применения этих антител при лечении состояния, например, при лечении атопического дерматита у псовых. У псовых встречаются четыре тяжелые цепи IgG, обозначаемые как A, B, C и D. Эти тяжелые цепи представляют четыре разных подкласса IgG псовых, обозначаемые как IgG-A (или IgGA), IgG-B (или IgGB), IgG-C (или IgGC) и IgG-D (или IgGD). Каждая из двух тяжелых цепей состоит из одного вариабельного домена (VH) и трех константных доменов, обозначаемых как СН-1, СН-2 и СН-3. Домен СН-1 соединен с доменом СН-2 посредством аминокислотной последовательности, называемой «шарниром» или альтернативно «шарнирной областью».
Последовательности нуклеиновых кислот и аминокислот этих четырех тяжелых цепей впервые были идентифицированы Tang et al. [Vet. Immunol. Immunopathol. 80:259-270 (2001)]. Аминокислотные и нуклеиновые последовательности этих тяжелых цепей также доступны в базах данных GenBank. Например, аминокислотная последовательность тяжелой цепи IgGA доступна под номером AAL35301.1, IgGB доступна под номером AAL35302.1, IgGC доступна под номером AAL35303.1, и IgGD доступна под номером (AAL35304.1). Собачьи антитела также содержат два типа легких цепей, каппа и лямбда. Последовательность ДНК и аминокислот этих легких цепей можно получить из баз данных GenBank. Например, аминокислотная последовательность легкой каппа-цепи доступна под номером ABY 57289.1, и легкая лямбда-цепь доступна под номером ABY 55569.1.
В настоящем изобретении аминокислотная последовательность для каждого из четырех Fc-фрагментов собачьего IgG основана на идентифицированной границе доменов CH1 и CH2, как определено Tang et al., см. выше. Канинизированные мышиные анти-собачьи антитела, которые связываются с собачьим IL-4Rα, включают, без ограничения, антитела по настоящему изобретению, которые содержат собачьи тяжелые цепи и/или собачьи легкие каппа- или лямбда-цепи IgG-A, IgG-B, IgG-C и IgG-D вместе с мышиными CDR к собачьему IL-4Rα. Соответственно, настоящее изобретение относится к выделенным канинизированным мышиным анти-собачьим антителам по настоящему изобретению, которые связываются с собачьим IL-4Rα и блокируют связывание этого собачьего IL-4Rα с собачьими IL-4 и/или собачьих IL-13, естественными партнерами по связыванию.
Соответственно, настоящее изобретение дополнительно относится к канинизированным мышиным антителам и способам применения антител по настоящему изобретению при лечении состояния, например, при лечении атопического дерматита у собак.
В настоящем изобретении также предложены полноразмерные собачьи тяжелые цепи, которые могут быть спарены с соответствующими легкими цепями с получением канинизированного антитела. Соответственно, в настоящем изобретении дополнительно предлагаются канинизированные мышиные антитела к собачьему антигену (включая выделенные канинизированные мышиные антитела к собачьему антигену) по настоящему изобретению и способы применения антител по настоящему изобретению при лечении состояния, например, при лечении атопический дерматит у собак.
Настоящее изобретение также относится к антителам по настоящему изобретению, которые содержат собачью область кристаллизуемого фрагмента (cFc-область), причем cFc генетически модифицирована для увеличения, уменьшения или устранения одной или более эффекторных функций. В одном из аспектов настоящего изобретения генетически модифицированная cFc снижает или устраняет одну или более эффекторных функций. В другом аспекте изобретения генетически модифицированная cFc усиливает одну или более эффекторных функций. В некоторых вариантах осуществления генетически модифицированная cFc-область представляет собой генетически модифицированную Fc-область IgGB псовых. В другом таком варианте осуществления генетически модифицированная cFc-область представляет собой генетически модифицированную Fc-область IgGC псовых. В конкретном варианте осуществления эффекторная функция представляет собой антителозависимую цитотоксичность (ADCC), которая усилена, снижена или устранена. В другом варианте осуществления эффекторная функция представляет собой комплемент-зависимую цитотоксичность (CDC), которая усилена, снижена или устранена. В еще одном варианте осуществления cFc-область генетически модифицирована для увеличения, уменьшения или устранения как ADCC, так и CDC.
Для создания вариантов собачьего IgG, лишенного эффекторных функций, получали ряд мутантных собачьих тяжелых цепей IgGB. Эти варианты могут включать одну или более из следующих одиночных или комбинированных замен в Fc-части аминокислотной последовательности тяжелой цепи: P4A, D31A, N63A, G64P, T65A, A93G и P95A. Варианты тяжелых цепей (т.е. содержащие такие аминокислотные замены) клонировали в экспрессионные плазмиды и трансфицировали в клетки HEK 293 вместе с плазмидой, содержащей ген, кодирующий легкую цепь. Интактные антитела, экспрессированные и очищенные из клеток HEK 293, оценивали на связывание с FcγRI и C1q для оценки их эффективности в отношении опосредования иммунных эффекторных функций. [См. U.S. 10,106,607 B2, содержание которого включено в настоящее описание во всей полноте посредством ссылки.]
Настоящее изобретение также относится к модифицированным собачьим IgG-D, которые вместо природного шарнирного участка IgG-D содержат шарнирный участок из:
IgG A: fnecrctdtppcpvpep SEQ ID NO: 6
IgG B: pkrengrvprppdcpkcpapem SEQ ID NO: 7; или
IgG C: akececkcncnncpcpgcgl SEQ ID NO: 8.
Альтернативно, шарнирная область IgG-D может быть генетически модифицирована путем замены остатка серина остатком пролина, т.е. pkestckciPpcpvpes, SEQ ID NO: 9 (с остатком пролина (P), подчеркнутым и выделенным жирным шрифтом, на который заменен природный остатк серина). Такие модификации могут дать собачий IgG-D, у которого не будет происходить обмен fab-плечами. Модифицированные собачьи IgG-D могут быть сконструированы с помощью стандартных методов рекомбинантных ДНК [например, Maniatis et al., Molecular Cloning, A Laboratory Manual (1982)]. Для создания этих вариантов нуклеиновые кислоты, кодирующие аминокислотную последовательность собачьего IgG-D, можно модифицировать таким образом, чтобы она кодировала модифицированные IgG-D. Модифицированные последовательности нуклеиновых кислот затем клонируют в экспрессионные плазмиды для экспрессии белка.
Шесть областей, определяющих комплементарность (CDR) канинизированного мышиного анти-собачьего антитела, как описано в настоящем описании, могут содержать легкую каппа-цепь собачьего антитела, содержащую мышиные LCDR1, LCDR2 и LCDR3 легкой цепи, и тяжелую цепь собачьего антитела IgG, содержащую мышиные HCDR1, HCDR2 и HCDR3 тяжелой цепи.
Нуклеиновые кислоты
Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим антитела по настоящему изобретению (см., например, примеры ниже).
Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим полипептиды иммуноглобулина, включающие аминокислотные последовательности, идентичные на по меньшей мере примерно 70%, предпочтительно идентичные на по меньшей мере примерно 80%, более предпочтительно идентичные на по меньшей мере примерно 90% и наиболее предпочтительно идентичные на по меньшей мере примерно 95% (например, на 95%, 96%, 97%, 98%, 99%, 100%) аминокислотным последовательностям канинизированных антител, за исключением неизмененных CDR, представленным в настоящем описании, при сравнении с помощью алгоритма BLAST, где параметры алгоритма подобраны так, чтобы получить максимальное совпадение между соответствующими последовательностями по всей длине соответствующих референсных последовательностей. В настоящем изобретении также предложены нуклеиновые кислоты, кодирующие полипептиды иммуноглобулинов, которые содержат аминокислотные последовательности, идентичные на по меньшей мере примерно 70%, предпочтительно на по меньшей мере примерно 80%, более предпочтительно на по меньшей мере примерно 90% и наиболее предпочтительно на по меньшей мере примерно 95% (например, на 95%, 96%, 97%, 98%, 99%, 100%) любой из референсных аминокислотных последовательностей, при сравнении с помощью алгоритма BLAST, в котором параметры алгоритма подобраны так, чтобы получить максимальное совпадение между соответствующими последовательностями по всей длине соответствующих референсных последовательностей.
Используемый в настоящем описании процент идентичности нуклеотидной и аминокислотной последовательностей может быть определен с помощью алгоритма C. MacVector (MacVector, Inc. Cary, NC 27519), Vector NTI (Informax, Inc. MD), Oxford Molecular Group PLC (1996) и Clustal W. с параметрами выравнивания, установленными по умолчанию, и параметрами идентификации, установленными по умолчанию. Эти коммерчески доступные программы также можно использовать для определения сходства последовательностей с использованием таких же или аналогичных параметров по умолчанию. В качестве альтернативы можно использовать расширенный поиск Blast в условиях фильтрации по умолчанию, например, с помощью программы PILEUP GCG (Genetics Computer Group, Program Manual for the GCG Package, Version 7, Madison, Wisconsin) с параметрами по умолчанию.
Приведенные ниже ссылки относятся к алгоритмам BLAST, часто используемым для анализа последовательностей: BLAST ALGORITHMS: Altschul, S.F., et al., J. Mol. Biol. 215:403-410 (1990); Gish, W., et al., Nature Genet. 3:266-272 (1993); Madden, T.L., et al., Meth. Enzymol. 266:131-141(1996); Altschul, S.F., et al., Nucleic Acids Res. 25:3389-3402 (1997); Zhang, J., et al., Genome Res. 7:649-656 (1997); Wootton, J.C., et al., Comput. Chem. 17:149-163 (1993); Hancock, J.M. et al., Comput. Appl. Biosci. 10:67-70 (1994); ALIGNMENT SCORING SYSTEMS: Dayhoff, M.O., et al., "A model of evolutionary change in proteins." in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3. M.O. Dayhoff (ed.), pp. 345-352, (1978); Natl. Biomed. Res. Found., Washington, DC; Schwartz, R.M., et al., "Matrices for detecting distant relationships." in Atlas of Protein Sequence and Structure, vol. 5, suppl. 3." (1978), M.O. Dayhoff (ed.), pp. 353-358 (1978), Natl. Biomed. Res. Found., Washington, DC; Altschul, S.F., J. Mol. Biol. 219:555-565 (1991); States, D.J., et al., Methods 3:66-70(1991); Henikoff, S., et al., Proc. Natl. Acad. Sci. USA 89:10915-10919 (1992); Altschul, S.F., et al., J. Mol. Evol. 36:290-300 (1993); ALIGNMENT STATISTICS: Karlin, S., et al., Proc. Natl. Acad. Sci. USA 87:2264-2268 (1990); Karlin, S., et al., Proc. Natl. Acad. Sci. USA 90:5873-5877 (1993); Dembo, A., et al., Ann. Prob. 22:2022-2039 (1994); and Altschul, S.F. "Evaluating the statistical significance of multiple distinct local alignments." in Theoretical and Computational Methods in Genome Research (S. Suhai, ed.), pp. 1-14, Plenum, New York (1997).
Антитела по настоящему изобретению могут быть получены рекомбинантно способами, известными в данной области. Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии антител или фрагментов согласно настоящему описанию, хорошо известны в данной области и включают множество иммортализованных клеточных линий, доступных из Американской коллекции типовых культур (ATCC). К ним относятся, среди прочего, клетки яичника китайского хомяка (CHO), NSO, клетки SP2, клетки HeLa, клетки почек детенышей хомячка (BHK), клетки почки обезьяны (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2), клетки A549, клетки 3Т3, клетки НЕК-293 и ряд других клеточных линий. Клетки-хозяева млекопитающих включают клетки человека, мыши, крысы, собаки, обезьяны, свиньи, козы, быка, лошади и хомяка. Клеточные линии, которые являются особо предпочтительными, выбирают путем определения клеточных линий с высокими уровнями экспрессии. Другие клеточные линии, которые можно использовать, включают клеточные линии насекомых, такие как клетки Sf9, клетки амфибий, бактериальные клетки, клетки растений и клетки грибов. Когда рекомбинантные векторы экспрессии, кодирующие тяжелую цепь или ее антигенсвязывающую часть или фрагмент, легкую цепь и/или ее антигенсвязывающий фрагмент, вводят в клетки-хозяева млекопитающих, антитела продуцируют путем культивирования клеток-хозяев в течение периода времени, достаточного для обеспечения экспрессии антитела в клетках-хозяевах или, более предпочтительно, достаточного для секреции антитела в культуральную среду, в которой выращивают клетки-хозяева.
Антитела могут быть выделены из культуральной среды стандартными методами очистки белков. Кроме того, экспрессию антител по изобретению (или других их фрагментов) в продуцирующих клеточных линиях можно усилить несколькими известными способами. Например, система экспрессии гена глутаминсинтетазы (система GS) представляет собой распространенный подход к усилению экспрессии при определенных условиях. Полный или частичный обзор системы GS можно найти в европейских патентах №№ 0216846, 0256055 и 0323997 и заявке на европейский патент № 89303964.4.
Как правило, гликопротеины, продуцируемые в конкретной клеточной линии или в организме трансгенного животного, будут иметь тип гликозилирования, характерный для гликопротеинов, продуцируемых в этой клеточной линии или в организме трансгенного животного. Следовательно, конкретный характер гликозилирования антитела будет зависеть от конкретной клеточной линии или трансгенного животного, используемого для получения антитела. Однако в настоящее изобретение включены все антитела, кодируемые молекулами нуклеиновых кислот, представленными в настоящем описании, или содержащие аминокислотные последовательности, представленные в настоящем описании, независимо от паттерна гликозилирования, который могут иметь антитела. Аналогично, в конкретных вариантах осуществления предпочтительными могут быть антитела с паттерном гликозилирования, содержащим только нефукозилированные N-гликаны, поскольку было показано, что эти антитела обычно являются более эффективными, чем их фукозилированные аналоги, как in vitro, так и in vivo [см., например, Shinkawa et al., J. Biol. Chem. 278:3466-3473 (2003); патенты США №№ 6,946,292 и 7,214,775].
СОБАЧИЙ IL-4 РЕЦЕПТОР, АЛЬФА-РЕЦЕПТОР
кДНК, кодирующая предсказанную полноразмерную альфа-цепь собачьего рецептора IL-4 (SEQ ID NO: 1), идентифицировали путем поиска в базе данных Genbank (номер доступа XM_547077.4; см. также US 7208579 B2). Эта предсказанная кДНК кодирует 823 аминокислоты (SEQ ID NO: 2), включая лидерную последовательность из 25 аминокислот, и идентифицирована под номером доступа XP_547077.3. Предсказанный зрелый белок α-цепи собачьего рецептора IL-4 (SEQ ID NO: 4) идентичен на 65% α-цепи рецептора IL-4 человека (номер доступа NP_000409.1) и на 70% α-цепи рецептора IL-4 свиньи (номер доступа NP_999505.1). Предсказанный зрелый белок α-цепи собачьего рецептора IL-4 кодируется нуклеотидной последовательностью, идентифицированной как SEQ ID NO: 3. Сравнение предсказанной α-цепи зрелого рецептора IL-4 с известными последовательностями α-цепи рецептора IL-4 человека позволило идентифицировать внеклеточный домен (ECD) зрелого белка α-цепи собачьего рецептора IL-4, обозначенного как SEQ ID NO: 5. Все это ранее было описано в [US2018/0346580; включенной в настоящее описание во всей полноте].
Полноразмерная ДНК α-цепи собачьего рецептора IL-4 с сигнальной последовательностью [SEQ ID NO:1]
atgggcagactgtgcagcggcctgaccttccccgtgagctgcctggtgctggtgtgggtggccagcagcggcagcgtgaaggtgctgcacgagcccagctgcttcagcgactacatcagcaccagcgtgtgccagtggaagatggaccaccccaccaactgcagcgccgagctgagactgagctaccagctggacttcatgggcagcgagaaccacacctgcgtgcccgagaacagagaggacagcgtgtgcgtgtgcagcatgcccatcgacgacgccgtggaggccgacgtgtaccagctggacctgtgggccggccagcagctgctgtggagcggcagcttccagcccagcaagcacgtgaagcccagaacccccggcaacctgaccgtgcaccccaacatcagccacacctggctgctgatgtggaccaacccctaccccaccgagaaccacctgcacagcgagctgacctacatggtgaacgtgagcaacgacaacgaccccgaggacttcaaggtgtacaacgtgacctacatgggccccaccctgagactggccgccagcaccctgaagagcggcgccagctacagcgccagagtgagagcctgggcccagacctacaacagcacctggagcgactggagccccagcaccacctggctgaactactacgagccctgggagcagcacctgcccctgggcgtgagcatcagctgcctggtgatcctggccatctgcctgagctgctacttcagcatcatcaagatcaagaagggctggtgggaccagatccccaaccccgcccacagccccctggtggccatcgtgatccaggacagccaggtgagcctgtggggcaagagaagcagaggccaggagcccgccaagtgcccccactggaagacctgcctgaccaagctgctgccctgcctgctggagcacggcctgggcagagaggaggagagccccaagaccgccaagaacggccccctgcagggccccggcaagcccgcctggtgccccgtggaggtgagcaagaccatcctgtggcccgagagcatcagcgtggtgcagtgcgtggagctgagcgaggcccccgtggacaacgaggaggaggaggaggtggaggaggacaagagaagcctgtgccccagcctggagggcagcggcggcagcttccaggagggcagagagggcatcgtggccagactgaccgagagcctgttcctggacctgctgggcggcgagaacggcggcttctgcccccagggcctggaggagagctgcctgcccccccccagcggcagcgtgggcgcccagatgccctgggcccagttccccagagccggccccagagccgcccccgagggccccgagcagcccagaagacccgagagcgccctgcaggccagccccacccagagcgccggcagcagcgccttccccgagcccccccccgtggtgaccgacaaccccgcctacagaagcttcggcagcttcctgggccagagcagcgaccccggcgacggcgacagcgaccccgagctggccgacagacccggcgaggccgaccccggcatccccagcgccccccagccccccgagccccccgccgccctgcagcccgagcccgagagctgggagcagatcctgagacagagcgtgctgcagcacagagccgcccccgcccccggccccggccccggcagcggctacagagagttcacctgcgccgtgaagcagggcagcgcccccgacgccggcggccccggcttcggccccagcggcgaggccggctacaaggccttctgcagcctgctgcccggcggcgccacctgccccggcaccagcggcggcgaggccggcagcggcgagggcggctacaagcccttccagagcctgacccccggctgccccggcgcccccacccccgtgcccgtgcccctgttcaccttcggcctggacaccgagccccccggcagcccccaggacagcctgggcgccggcagcagccccgagcacctgggcgtggagcccgccggcaaggaggaggacagcagaaagaccctgctggcccccgagcaggccaccgaccccctgagagacgacctggccagcagcatcgtgtacagcgccctgacctgccacctgtgcggccacctgaagcagtggcacgaccaggaggagagaggcaaggcccacatcgtgcccagcccctgctgcggctgctgctgcggcgacagaagcagcctgctgctgagccccctgagagcccccaacgtgctgcccggcggcgtgctgctggaggccagcctgagccccgccagcctggtgcccagcggcgtgagcaaggagggcaagagcagccccttcagccagcccgccagcagcagcgcccagagcagcagccagacccccaagaagctggccgtgctgagcaccgagcccacctgcatgagcgccagc
Полноразмерный белок собачьего α-рецептора IL-4 с сигнальной последовательностью, выделенной жирным шрифтом [SEQ ID NO: 2]
MGRLCSGLTFPVSCLVLVWVASSGSVKVLHEPSCFSDYISTSVCQWKMDHPTNCSAELRLSYQLDFMGSENHTCVPENREDSVCVCSMPIDDAVEADVYQLDLWAGQQLLWSGSFQPSKHVKPRTPGNLTVHPNISHTWLLMWTNPYPTENHLHSELTYMVNVSNDNDPEDFKVYNVTYMGPTLRLAASTLKSGASYSARVRAWAQTYNSTWSDWSPSTTWLNYYEPWEQHLPLGVSISCLVILAICLSCYFSIIKIKKGWWDQIPNPAHSPLVAIVIQDSQVSLWGKRSRGQEPAKCPHWKTCLTKLLPCLLEHGLGREEESPKTAKNGPLQGPGKPAWCPVEVSKTILWPESISVVQCVELSEAPVDNEEEEEVEEDKRSLCPSLEGSGGSFQEGREGIVARLTESLFLDLLGGENGGFCPQGLEESCLPPPSGSVGAQMPWAQFPRAGPRAAPEGPEQPRRPESALQASPTQSAGSSAFPEPPPVVTDNPAYRSFGSFLGQSSDPGDGDSDPELADRPGEADPGIPSAPQPPEPPAALQPEPESWEQILRQSVLQHRAAPAPGPGPGSGYREFTCAVKQGSAPDAGGPGFGPSGEAGYKAFCSLLPGGATCPGTSGGEAGSGEGGYKPFQSLTPGCPGAPTPVPVPLFTFGLDTEPPGSPQDSLGAGSSPEHLGVEPAGKEEDSRKTLLAPEQATDPLRDDLASSIVYSALTCHLCGHLKQWHDQEERGKAHIVPSPCCGCCCGDRSSLLLSPLRAPNVLPGGVLLEASLSPASLVPSGVSKEGKSSPFSQPASSSAQSSSQTPKKLAVLSTEPTCMSAS
Зрелый полноразмерный белок собачьего α-рецептора IL-4 без сигнальной последовательности [SEQ ID NO: 4]
VKVLHEPSCFSDYISTSVCQWKMDHPTNCSAELRLSYQLDFMGSENHTCVPENREDSVCVCSMPIDDAVEADVYQLDLWAGQQLLWSGSFQPSKHVKPRTPGNLTVHPNISHTWLLMWTNPYPTENHLHSELTYMVNVSNDNDPEDFKVYNVTYMGPTLRLAASTLKSGASYSARVRAWAQTYNSTWSDWSPSTTWLNYYEPWEQHLPLGVSISCLVILAICLSCYFSIIKIKKGWWDQIPNPAHSPLVAIVIQDSQVSLWGKRSRGQEPAKCPHWKTCLTKLLPCLLEHGLGREEESPKTAKNGPLQGPGKPAWCPVEVSKTILWPESISVVQCVELSEAPVDNEEEEEVEEDKRSLCPSLEGSGGSFQEGREGIVARLTESLFLDLLGGENGGFCPQGLEESCLPPPSGSVGAQMPWAQFPRAGPRAAPEGPEQPRRPESALQASPTQSAGSSAFPEPPPVVTDNPAYRSFGSFLGQSSDPGDGDSDPELADRPGEADPGIPSAPQPPEPPAALQPEPESWEQILRQSVLQHRAAPAPGPGPGSGYREFTCAVKQGSAPDAGGPGFGPSGEAGYKAFCSLLPGGATCPGTSGGEAGSGEGGYKPFQSLTPGCPGAPTPVPVPLFTFGLDTEPPGSPQDSLGAGSSPEHLGVEPAGKEEDSRKTLLAPEQATDPLRDDLASSIVYSALTCHLCGHLKQWHDQEERGKAHIVPSPCCGCCCGDRSSLLLSPLRAPNVLPGGVLLEASLSPASLVPSGVSKEGKSSPFSQPASSSAQSSSQTPKKLAVLSTEPTCMSAS
Полноразмерная ДНК зрелого собачьего α-рецептора IL-4 без сигнальной последовательности [SEQ ID NO: 3]
gtgaaggtgctgcacgagcccagctgcttcagcgactacatcagcaccagcgtgtgccagtggaagatggaccaccccaccaactgcagcgccgagctgagactgagctaccagctggacttcatgggcagcgagaaccacacctgcgtgcccgagaacagagaggacagcgtgtgcgtgtgcagcatgcccatcgacgacgccgtggaggccgacgtgtaccagctggacctgtgggccggccagcagctgctgtggagcggcagcttccagcccagcaagcacgtgaagcccagaacccccggcaacctgaccgtgcaccccaacatcagccacacctggctgctgatgtggaccaacccctaccccaccgagaaccacctgcacagcgagctgacctacatggtgaacgtgagcaacgacaacgaccccgaggacttcaaggtgtacaacgtgacctacatgggccccaccctgagactggccgccagcaccctgaagagcggcgccagctacagcgccagagtgagagcctgggcccagacctacaacagcacctggagcgactggagccccagcaccacctggctgaactactacgagccctgggagcagcacctgcccctgggcgtgagcatcagctgcctggtgatcctggccatctgcctgagctgctacttcagcatcatcaagatcaagaagggctggtgggaccagatccccaaccccgcccacagccccctggtggccatcgtgatccaggacagccaggtgagcctgtggggcaagagaagcagaggccaggagcccgccaagtgcccccactggaagacctgcctgaccaagctgctgccctgcctgctggagcacggcctgggcagagaggaggagagccccaagaccgccaagaacggccccctgcagggccccggcaagcccgcctggtgccccgtggaggtgagcaagaccatcctgtggcccgagagcatcagcgtggtgcagtgcgtggagctgagcgaggcccccgtggacaacgaggaggaggaggaggtggaggaggacaagagaagcctgtgccccagcctggagggcagcggcggcagcttccaggagggcagagagggcatcgtggccagactgaccgagagcctgttcctggacctgctgggcggcgagaacggcggcttctgcccccagggcctggaggagagctgcctgcccccccccagcggcagcgtgggcgcccagatgccctgggcccagttccccagagccggccccagagccgcccccgagggccccgagcagcccagaagacccgagagcgccctgcaggccagccccacccagagcgccggcagcagcgccttccccgagcccccccccgtggtgaccgacaaccccgcctacagaagcttcggcagcttcctgggccagagcagcgaccccggcgacggcgacagcgaccccgagctggccgacagacccggcgaggccgaccccggcatccccagcgccccccagccccccgagccccccgccgccctgcagcccgagcccgagagctgggagcagatcctgagacagagcgtgctgcagcacagagccgcccccgcccccggccccggccccggcagcggctacagagagttcacctgcgccgtgaagcagggcagcgcccccgacgccggcggccccggcttcggccccagcggcgaggccggctacaaggccttctgcagcctgctgcccggcggcgccacctgccccggcaccagcggcggcgaggccggcagcggcgagggcggctacaagcccttccagagcctgacccccggctgccccggcgcccccacccccgtgcccgtgcccctgttcaccttcggcctggacaccgagccccccggcagcccccaggacagcctgggcgccggcagcagccccgagcacctgggcgtggagcccgccggcaaggaggaggacagcagaaagaccctgctggcccccgagcaggccaccgaccccctgagagacgacctggccagcagcatcgtgtacagcgccctgacctgccacctgtgcggccacctgaagcagtggcacgaccaggaggagagaggcaaggcccacatcgtgcccagcccctgctgcggctgctgctgcggcgacagaagcagcctgctgctgagccccctgagagcccccaacgtgctgcccggcggcgtgctgctggaggccagcctgagccccgccagcctggtgcccagcggcgtgagcaaggagggcaagagcagccccttcagccagcccgccagcagcagcgcccagagcagcagccagacccccaagaagctggccgtgctgagcaccgagcccacctgcatgagcgccagc
Внеклеточный домен α-цепи собачьего рецептора IL-4 [SEQ ID NO: 5]
vkvlhepscfsdyistsvcqwkmdhptncsaelrlsyqldfmgsenhtcvpenredsvcvcsmpiddaveadvyqldlwagqqllwsgsfqpskhvkprtpgnltvhpnishtwllmwtnpyptenhlhseltymvnvsndndpedfkvynvtymgptlrlaastlksgasysarvrawaqtynstwsdwspsttwlnyyepweqhlp
Белковая инженерия антител
В качестве примера, без ограничения, константная область тяжелой цепи псовых может происходить из IgG-B или модифицированного cFc, такого как IgG-Bm, используемого в настоящем описании [см. U.S. 10,106,607 B2, который включен в настоящее описание во всей полноте посредством ссылки], а константная область легкой цепи псовых может происходить из каппа-цепи.
Антитела могут быть сконструированы таким образом, чтобы они включали модификации собачьего каркаса и/или остатков собачьего каркаса в пределах вариабельных доменов родительского (т.е. мышиного) моноклонального антитела, например, для улучшения свойств антитела.
Фармацевтические композиции и введение
Для приготовления фармацевтических или стерильных композиций, содержащих антитела по настоящему изобретению, эти антитела можно смешать с фармацевтически приемлемым носителем или вспомогательным веществом. [См., например, Remington's Pharmaceutical Sciences and U.S. Pharmacopeia: National Formulary, Mack Publishing Company, Easton, PA (1984)].
Составы терапевтических и диагностических средств могут быть приготовлены путем смешивания с приемлемыми носителями, вспомогательными веществами или стабилизаторами в форме, например, лиофилизированных порошков, взвесей, водных растворов или суспензий [см., например, Hardman, et al. (2001) Goodman and Gilman’s The Pharmacological Basis of Therapeutics, McGraw-Hill, New York, NY; Gennaro (2000) Remington: The Science and Practice of Pharmacy, Lippincott, Williams, and Wilkins, New York, NY; Avis, et al. (eds.) (1993) Pharmaceutical Dosage Forms: Parenteral Medications, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Tablets, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Disperse Systems, Marcel Dekker, NY; Weiner and Kotkoskie (2000) Excipient Toxicity and Safety, Marcel Dekker, Inc., New York, NY]. В одном из вариантов осуществления антитела по настоящему изобретению разбавляют до соответствующей концентрации в растворе ацетата натрия, рН 5-6, и для тоничности добавляют NaCl или сахарозу. Для повышения стабильности могут быть добавлены дополнительные агенты, такие как полисорбат 20 или полисорбат 80.
Токсичность и терапевтическую эффективность композиций антител, вводимых отдельно или в комбинации с другим агентом, можно определить, используя стандартные фармацевтические процедуры, на культурах клеток или экспериментальных животных, например, для определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной у 50% населения). Соотношение токсических и терапевтических эффектов, вызываемых дозой, является терапевтическим индексом (LD50/ED50). В конкретных аспектах желательны антитела, демонстрирующие высокие терапевтические индексы. Данные, полученные в результате этих анализов клеточных культур и исследований на животных, могут быть использованы при определении диапазона доз для собак. Дозирование таких соединений предпочтительно находится в пределах диапазона циркулирующих концентраций, которые включают ED50 с небольшой токсичностью или без токсичности. Доза может варьировать в пределах этого диапазона в зависимости от используемой лекарственной формы и пути введения.
Способ введения может меняться. Подходящие пути введения включают пероральный, ректальный, чрезслизистый, кишечный, парентеральный; внутримышечный, подкожный, внутрикожный, интрамедуллярный, подоболочечный, непосредственно внутрь желудочка, внутривенный, внутрибрюшинный, интраназальный, интраокулярный, ингаляцию, инсуффляцию, топический, накожный, чрескожный или внутриартериальный. В конкретных вариантах осуществления антитела по настоящему изобретению можно вводить инвазивным путем, таким как инъекция. В дополнительных вариантах осуществления изобретения антитела по настоящему изобретению или их фармацевтические композиции вводят внутривенно, подкожно, внутримышечно, внутриартериально или путем ингаляции, или аэрозольной доставки. Введение неинвазивными путями (например, перорально; например, в виде пилюль, капсул или таблеток) также входит в объем настоящего изобретения.
Композиции можно вводить с помощью медицинских устройств, известных в данной области техники. Например, фармацевтическую композицию по изобретению можно вводить путем инъекции с помощью иглы для подкожных инъекций, включая, например, предварительно заполненный шприц или автоинжектор. Раскрытые в настоящем описании фармацевтические композиции также можно вводить с помощью безыгольного устройства для подкожных инъекций; например, с помощью устройств, раскрытых в патентах США № 6,620,135; 6,096,002; 5,399,163; 5,383,851; 5,312,335; 5,064,413; 4,941,880; 4,790,824 или 4,596,556.
Фармацевтические композиции, раскрытые в настоящем описании, также можно вводить путем инфузии. Примеры хорошо известных имплантатов и модулей для введения фармацевтических композиций включают, описанные в патенте США № 4,487,603, в котором описан имплантируемый микроинфузионный насос для дозирования лекарственных средств с контролируемой скоростью; патенте США № 4,447,233, в котором описан насос для инфузии лекарственных средств для доставки лекарственного средства с точно установленной скоростью инфузии; патенте США № 4,447,224, в котором раскрыт имплантируемый инфузионный аппарат с переменным потоком для непрерывной доставки лекарственного средства; патенте США № 4,439,196, в котором описана осмотическая система доставки лекарственных средств, имеющая многокамерные отсеки. Многие другие такие имплантаты, системы доставки и модули хорошо известны специалистам в данной области.
В качестве альтернативы антитела по настоящему изобретению можно вводить локально, а не системно, часто в виде депо или состава с замедленным высвобождением.
Режим введения зависит от нескольких факторов, включая скорость обмена терапевтических антител в сыворотке или ткани, тяжесть симптомов, иммуногенность терапевтических антител и доступность клеток-мишеней в биологической матрице. Предпочтительно режим введения позволяет доставлять терапевтические антитела в количестве, достаточном для облегчения целевого заболевания/состояния, одновременно сводя к минимуму нежелательные побочные эффекты. Соответственно, количество доставляемого биологического препарата частично зависит от конкретных терапевтических антител и тяжести состояния, подлежащего лечению. Доступны руководства по выбору соответствующих доз терапевтических антител [см., например, Wawrzynczak Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK (1996); Kresina (ed.) Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, NY (1991); Bach (ed.) Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, NY (1993); Baert, et al. New Engl. J. Med. 348:601-608 (2003); Milgrom et al. New Engl. J. Med. 341:1966-1973 (1999); Slamon et al. New Engl. J. Med. 344:783-792 (2001); Beniaminovitz et al. New Engl. J. Med. 342:613-619 (2000); Ghosh et al. New Engl. J. Med. 348:24-32 (2003); Lipsky et al. New Engl. J. Med. 343:1594-1602 (2000)].
Определение подходящей дозы выполняет ветеринарный врач, используя, например, параметры или факторы, о которых в данной области известно или предполагается, что они влияют на лечение. Как правило, дозирование начинают с количества, которое немного меньше оптимальной дозы, затем это количество увеличивают небольшими приращениями до тех пор, пока не будет достигнут требуемый или оптимальный эффект относительно любых негативных побочных эффектов. Важные диагностические измерения включают измерения симптомов.
Антитела, раскрытые в настоящем описании, можно вводить путем непрерывной инфузии или посредством доз, вводимых, например, ежедневно, 1-7 раз в неделю, еженедельно, раз в две недели, ежемесячно, раз в два месяца, ежеквартально, раз в полгода, ежегодно и т.д. Дозы можно вводить, например, внутривенно, подкожно, топически, перорально, назально, ректально, внутримышечно, интрацеребрально, интраспинально или путем ингаляции. Общая недельная доза обычно составляет по меньшей мере 0,05 мкг/кг массы тела, в более общем случае по меньшей мере 0,2 мкг/кг, 0,5 мкг/кг, 1 мкг/кг, 10 мкг/кг, 100 мкг/кг, 0,25 мг/кг, 1,0 мг/кг, 2,0 мг/кг, 5,0 мг/мл, 10 мг/кг, 25 мг/кг, 50 мг/кг или более [см., например, Yang, et al. New Engl. J. Med. 349:427-434 (2003); Herold, et al. New Engl. J. Med. 346:1692-1698 (2002); Liu, et al. J. Neurol. Neurosurg. Psych. 67:451-456 (1999); Portielji, et al. Cancer Immunol. Immunother. 52:133-144 (2003)]. Также могут быть предусмотрены дозы для достижения заранее определенной целевой концентрации антител по настоящему изобретению в собачьей сыворотке, например, 0,1, 0,3, 1, 3, 10, 30, 100, 300 мкг/мл или более. В других вариантах осуществления антитела по настоящему изобретению вводят подкожно или внутривенно еженедельно, раз в две недели, «каждые 4 недели», ежемесячно, раз в два месяца или ежеквартально по 10, 20, 50, 80, 100, 200, 500, 1000 или 2500 мг/субъекта.
В контексте настоящего описания термины «ингибировать», «лечить» или «лечение» включают отсрочку развития симптомов, связанных с расстройством, и/или снижение тяжести симптомов такого расстройства. Термины также включают облегчение существующих неконтролируемых или нежелательных симптомов, предотвращение появления дополнительных симптомов и улучшение или предотвращение основных причин появления таких симптомов. Таким образом, термины означают достижение положительного результата у позвоночного субъекта (например, представителя псовых), страдающего нарушением, состоянием и/или имеющего симптом или риск развития такого нарушения, заболевания или появления симптома.
В контексте настоящего описания термины «терапевтически эффективное количество», «терапевтически эффективная доза» и «эффективное количество» относятся к количеству антител по настоящему изобретению, которое при введении отдельно или в комбинации с дополнительным терапевтическим агентом в клетку, ткань или организм субъекта, например, представителю псовых, эффективно вызывает измеримое улучшение одного или более симптомов заболевания или состояния или подавляет прогрессирование такого заболевания или состояния. Терапевтически эффективная доза также относится к такому количеству антител, которое является достаточным для получения по меньшей мере частичного облегчения симптомов, например, лечения, излечения, профилактики или облегчения соответствующего медицинского состояния, или увеличения скорости лечения, заживления, профилактики или улучшения таких состояний. Применительно к комбинации терапевтически эффективная доза относится к комбинированным количествам активных ингредиентов, которые приводят к терапевтическому эффекту независимо от того, введены ли они в комбинации, последовательно или одновременно. Эффективное количество терапевтического средства приводит к улучшению диагностического измерения или параметра на по меньшей мере 10%; обычно на по меньшей мере 20%; предпочтительно на по меньшей мере примерно 30%; более предпочтительно на по меньшей мере 40% и наиболее предпочтительно на по меньшей мере 50%. Эффективное количество также может привести к улучшению субъективного показателя в случаях, когда для оценки тяжести состояния используются субъективные показатели.
ПРИМЕРЫ
ПРИМЕР 1
АНТИТЕЛА К АЛЬФА-РЕЦЕПТОРУ IL-4
Общие материалы и методы
Рекомбинантные белки получали путем предоставления аминокислотной последовательности выбранного белка коммерческому производителю (ATUM, Ньюарк, Калифорния), который, в свою очередь, подбирал подходящую нуклеотидную последовательность, кодирующую эту аминокислотную последовательность. Нуклеотидные последовательности также можно получить из общедоступных баз данных ДНК, таких как GenBank®. Затем коммерческий производитель химически синтезировал нуклеиновую кислоту, которую впоследствии клонировал с помощью ATUM в экспрессионную плазмиду (pD2610-v10; доступна от AUTM) с получением соответствующего рекомбинантного белка. Плазмиду помещали либо в клетки НЕК-293, либо в клетки СНО для экспрессии рекомбинантного белка, который затем выделяли обычными методами.
Мышей Balb/c иммунизировали несколько раз (по 10 мкг каждый раз) в течение 17 дней. Иммунизирующий антиген представлял собой слитый белок внеклеточного домена (ECD) собачьей альфа-цепи IL-4R и Fc человека. После иммунизации от каждой мыши собирали сыворотку и тестировали на реактивность с HIS-меченым белком ECD собачьей альфа-цепи рецептора IL-4. Клетки селезенки мыши с самым высоким титром ECD альфа-цепи рецептора IL-4 в сыворотке сливали с клеточной линией миеломы P3X63Ag8.653. Приблизительно через 2 недели после слияния надосадочную жидкость клеток предполагаемой гибридомы тестировали с помощью ELISA на реактивность по отношению к меченному белку ECD альфа-цепи рецептора IL-4. Гибридомы, генерирующие сильные положительные сигналы в ELISA, субклонировали методом предельного разведения и снова тестировали на реактивность к HIS-меченому белку ECD собачьей альфа-цепи рецептора IL-4.
Антитела к собачьему альфа-рецептору IL-4 включали антитело c152H11VL3-cCLk-s/c152H11VH3-cIgG-Bm и антитело c146E2VL3-cCLk-s/c146E2VH3-cIgG-Bm. Наборы из шести (6) CDR (трех отдельных последовательностей легких цепей (LC) и трех последовательностей тяжелых цепей (HC)) для этих двух антител представлены ниже в Таблицах 1A и 1B. В Таблице 1А представлены нуклеиновые кислоты, которые кодируют аминокислотные последовательности двенадцати CDR, перечисленных в Таблице 1В. Аминокислотные последовательности полноразмерных легких цепей и тяжелых цепей этих канинизированных антител представлены ниже непосредственно после Таблицы 1В.
c152H11VL3-cCLk-s (легкая каппа-цепь): [SEQ ID NO: 35]
EIVMTQSPASLSLSQEEKVTITCKASQNVGTNVAWYQQKPGQAPKLLIYSASYRYSGLPDRFSGSGSGTDFSFTISSLEPEDVAEFFCQQYNSYPYTFGQGTKLEIKRNDAQPAVYLFQPSPDQLHTGSASVVCLLNSFYPKDINVKWKVDGVIQDTGIQESVTEQDKDSTYSLSSTLTMSSTEYLSHELYSCEITHKSLPSTLIKSFQRSECQRVD
c152H11VH1-cIgGBm (тяжелая цепь): [SEQ ID NO: 36]
EVQLVESGGDLVKPGGSLRLSCAASGFTFSSYGMSWVRQAPGKGLQWVATISRGGDYTYYPDSVKGRFTISRDNAKNTLYLQMNSLRAEDTAMYYCAKGTLNNRGFACWGQGTLVTVSSASTTAPSVFPLAPSCGSTSGSTVALACLVSGYFPEPVTVSWNSGSLTSGVHTFPSVLQSSGLYSLSSMVTVPSSRWPSETFTCNVAHPASKTKVDKPVPKRENGRVPRPPDCPKCPAPEMLGGPSVFIFPPKPKDTLLIARTPEVTCVVVALDPEDPEVQISWFVDGKQMQTAKTQPREEQFAGTYRVVSVLPIGHQDWLKGKQFTCKVNNKALPSPIERTISKARGQAHQPSVYVLPPSREELSKNTVSLTCLIKDFFPPDIDVEWQSNGQQEPESKYRTTPPQLDEDGSYFLYSKLSVDKSRWQRGDTFICAVMHEALHNHYTQESLSHSPG
c152H11VH2-cIgGBm (тяжелая цепь): [SEQ ID NO: 37]
EVQLVESGGDLVKPGGSLRLSCAASGFTFSSYGMSWVRQAPDKRLQWVATISRGGDYTYYPDSVKGRFTISRDNAKNTLYLQMNSLRAEDTAMYYCARGTLNNRGFACWGQGTLVTVSSASTTAPSVFPLAPSCGSTSGSTVALACLVSGYFPEPVTVSWNSGSLTSGVHTFPSVLQSSGLYSLSSMVTVPSSRWPSETFTCNVAHPASKTKVDKPVPKRENGRVPRPPDCPKCPAPEMLGGPSVFIFPPKPKDTLLIARTPEVTCVVVALDPEDPEVQISWFVDGKQMQTAKTQPREEQFAGTYRVVSVLPIGHQDWLKGKQFTCKVNNKALPSPIERTISKARGQAHQPSVYVLPPSREELSKNTVSLTCLIKDFFPPDIDVEWQSNGQQEPESKYRTTPPQLDEDGSYFLYSKLSVDKSRWQRGDTFICAVMHEALHNHYTQESLSHSPG
c152H11VH3-cIgG-Bm (тяжелая цепь): [SEQ ID NO: 38]
EVQLVESGGDLVKPGGSLRLSCAASGFTFSSYGMSWVRQAPDKRLQWVATISRGGDYTYYPDSVKGRFTISRDNAKNTLYLQMNSLRAEDTAMYYCARGTLNNRGFASWGQGTLVTVSSASTTAPSVFPLAPSCGSTSGSTVALACLVSGYFPEPVTVSWNSGSLTSGVHTFPSVLQSSGLYSLSSMVTVPSSRWPSETFTCNVAHPASKTKVDKPVPKRENGRVPRPPDCPKCPAPEMLGGPSVFIFPPKPKDTLLIARTPEVTCVVVALDPEDPEVQISWFVDGKQMQTAKTQPREEQFAGTYRVVSVLPIGHQDWLKGKQFTCKVNNKALPSPIERTISKARGQAHQPSVYVLPPSREELSKNTVSLTCLIKDFFPPDIDVEWQSNGQQEPESKYRTTPPQLDEDGSYFLYSKLSVDKSRWQRGDTFICAVMHEALHNHYTQESLSHSPG
c146E2VL3-cCLk-s (легкая каппа-цепь): [SEQ ID NO: 39]
DIVLTQTPLSLSVSPGETASIYCRASESVDSYGNSFLNWYQQKPGQPPKLLIYRASNLASEIPDRFSGSGSRTEFTLKISRVEADDAGVYYCQQNYENPRTFGQGTKLEIKRNDAQPAVYLFQPSPDQLHTGSASVVCLLNSFYPKDINVKWKVDGVIQDTGIQESVTEQDKDSTYSLSSTLTMSSTEYLSHELYSCEITHKSLPSTLIKSFQRSECQRVD
c146E2VH1-cIgGBm (тяжелая цепь): [SEQ ID NO: 40]
EVQLVQSGAEVKKPGASVKVSCKASGYTFARYWMHWVRQAPGAGLDWMGMIHPDSGNINYNERFKTRVTLTADTSTSTAYMELSSLRAGDIAVYYCARQLRNAMDYWGQGTLVTVSSASTTAPSVFPLAPSCGSTSGSTVALACLVSGYFPEPVTVSWNSGSLTSGVHTFPSVLQSSGLYSLSSMVTVPSSRWPSETFTCNVAHPASKTKVDKPVPKRENGRVPRPPDCPKCPAPEMLGGPSVFIFPPKPKDTLLIARTPEVTCVVVALDPEDPEVQISWFVDGKQMQTAKTQPREEQFAGTYRVVSVLPIGHQDWLKGKQFTCKVNNKALPSPIERTISKARGQAHQPSVYVLPPSREELSKNTVSLTCLIKDFFPPDIDVEWQSNGQQEPESKYRTTPPQLDEDGSYFLYSKLSVDKSRWQRGDTFICAVMHEALHNHYTQESLSHSPG
c146E2VH2-cIgGBm (тяжелая цепь): [SEQ ID NO: 41]
EVQLVQSGAEVKKPGASVKVSCKASGYTFARYWMHWMKQAPGAGLDWIGMIHPDSGNINYNERFKTKATLTADTSTSTAYMELSSLRAGDIAVYYCARQLRNAMDYWGQGTLVTVSSASTTAPSVFPLAPSCGSTSGSTVALACLVSGYFPEPVTVSWNSGSLTSGVHTFPSVLQSSGLYSLSSMVTVPSSRWPSETFTCNVAHPASKTKVDKPVPKRENGRVPRPPDCPKCPAPEMLGGPSVFIFPPKPKDTLLIARTPEVTCVVVALDPEDPEVQISWFVDGKQMQTAKTQPREEQFAGTYRVVSVLPIGHQDWLKGKQFTCKVNNKALPSPIERTISKARGQAHQPSVYVLPPSREELSKNTVSLTCLIKDFFPPDIDVEWQSNGQQEPESKYRTTPPQLDEDGSYFLYSKLSVDKSRWQRGDTFICAVMHEALHNHYTQESLSHSPG
c146E2VH3-cIgG-Bm (тяжелая цепь): [SEQ ID NO: 42]
EVQLVQSGAEVKKPGASVKVSCKASGYTFARYWMHWMKQAPGAGLDWIGMIHPDSGNINYNERFKTKATLTVdkststaymelsslragdiavyycarqlrnamdywgqgtlvtvssasttapsvfplapscgstsgstvalaclvsgyfpepvtvswnsgsltsgvhtfpsvlqssglyslssmvtvpssrwpsetftcnvahpasktkvdkpvpkrengrvprppdcpkcpapemlggpsvfifppkpkdtlliartpevtcvvvaldpedpevqiswfvdgkqmqtaktqpreeqfagtyrvvsvlpighqdwlkgkqftckvnnkalpspiertiskargqahqpsvyvlppsreelskntvsltclikdffppdidvewqsngqqepeskyrttppqldedgsyflysklsvdksrwqrgdtficavmhealhnhytqeslshspg
Кроме того, также были сконструированы легкие цепи антител к собачьему альфа-рецептору IL-4 с легкой лямбда-цепью, как показано ниже.
c152H11LV1-cC1 (легкая лямбда-цепь) [SEQ ID NO: 43]
QSVLTQPASVSGSLGQRVTISCKASQNVGTNVAWYQQLPGTSPRTLIYSASYRYSGVPDRFSGSRSGSTATLTISGLQAEDEADYYCQQYNSYPYTFGGGTHLTVLGQPKASPSVTLFPPSSEELGANKATLVCLISDFYPSGVTVAWKADGSPVTQGVETTKPSKQSNNKYAASSYLSLTPDKWKSHSSFSCLVTHEGSTVEKKVAPAECS
c146E2LV1-cCl (легкая лямбда-цепь) [SEQ ID NO: 44]
QSVLTQPASVSGSLGQRVTISCRASESVDSYGNSFLNWYQQLPGKAPSLLIYRASNLASGVPERFSGSKSGSSATLTITGLQAEDEADYYCQQNYENPRTFGGGTHLTVLGQPKASPSVTLFPPSSEELGANKATLVCLISDFYPSGVTVAWKADGSPVTQGVETTKPSKQSNNKYAASSYLSLTPDKWKSHSSFSCLVTHEGSTVEKKVAPAECS
ПРИМЕР 2
ИНГИБИРОВАНИЕ SТАТ-6
Антитела к собачьему альфа-рецептору IL-4 тестировали на способность ингибировать фосфорилирование STAT-6 в клетках DH82 следующим образом:
Материалы
1. Активно растущие клетки DH82
2. Среда для выращивания клеток DH82 (ATCC® 302003™, минимальная основная среда Игла, содержащая инактивированную нагреванием фетальную бычью сыворотку с конечной концентрацией 15% мас./об.)
3. Набор для анализа AlphaLISA p-STAT6 (Tyr641): Perkin Elmer Каталог: ALSU-PST6-A-HV
4. Рекомбинантный собачий IL-4: R&D Systems, Каталог: 752-CL/CF
5. Рекомбинантный собачий IL-13: R&D Systems, Каталог: 5894-CL/CF
6. Perkin Elmer Envision
а. Канинизированные моноклональные антитела к собачьему IL-4Rα
b. c146Е2-H3L3
с. Канинизированные антитела 4H3 из US 2018/0346580
Антитела к собачьему альфа-рецептору IL-4 тестировали на способность ингибировать фосфорилирование STAT-6 в клетках DH82 следующим образом:
Методы
1. Два планшета для тканевых культур засевали клетками DH82 в количестве 8х104 на лунку (200 мкл с плотностью 4х105 клеток/мл) и инкубировали при 37°С в течение ночи.
2. Тестируемые антитела предварительно разводили до 500 мкг/мл, и затем осуществляли серийное разведение в 3 раза в среде для выращивания клеток DH82. Среду из планшетов для культивирования клеток удаляли, и в каждый планшет вносили по 50 мкл/лунку серийно разбавленного тестируемого образца.
3. Собачий IL-4 разводили до 5 нг/мл в среде для выращивания клеток DH82 и добавляли по 50 мкл в каждую лунку одного из планшетов. Собачий IL-13 разбавляли до 10 нг/мл в среде для выращивания клеток DH82 и добавляли по 50 мкл в каждую лунку второго планшета. Планшеты инкубировали в течение 15 мин при 37°С.
4. Среду из планшетов удаляли и в планшет добавляли по 100 мкл на лунку свежеприготовленного 1x буфера для лизиса из набора для анализа AlphaLISA p-STAT-6. Планшет встряхивали на шейкере для планшетов со скоростью 350 об/мин в течение 10 минут при комнатной температуре.
5. Акцепторную смесь готовили из набора для анализа AlphaLISA p-STAT6, и 15 мкл на лунку добавляли к 30 мкл клеточного лизата в 96-луночных планшетах 1/2 Area. Планшеты запечатывали, встряхивали в течение 2 минут при 350 об/мин и затем инкубировали в течение 2 часов при комнатной температуре.
6. Донорскую смесь готовили из набора для анализа AlphaLISA p-STAT6 при слабом лабораторном освещении, и в каждый планшет добавляли по 15 мкл на лунку. Планшеты запечатывали, накрывали фольгой, встряхивали в течение 2 минут при 350 об/мин и затем инкубировали в течение 2 часов при комнатной температуре.
7. Планшеты считывали, используя настройки AlphaScreen, на Perkin Elmer EnVison.
Два разных канинизированных моноклональных антитела к собачьему IL-4Rα, обозначенные c4H3 [WO2016/156588; US2018/0346580] и c146E2-H3L3 оценивали на их способность ингибировать фосфорилирование αSTAT-6 путем блокирования связывания собачьего IL-4 или собачьего IL-13 с собачьим IL-4Rα. Данные, представленные на фиг.1, демонстрируют, что оба антитела обеспечивают дозозависимое ингибирование фосфорилирования STAT-6 в присутствии IL-4. Однако связывание c146E2-H3L3 на удивление оказалось более прочным, чем связывание антитела предшествующего уровня техники c4H3 к собачьему альфа-рецептору IL-4 [WO2016/156588]. Контроль IL-4 в отсутствие антител к IL-4R-альфа (IL-4Rα) показан в верхней правой части графика. Данные, представленные на фиг.2, также демонстрируют, что оба антитела приводят к дозозависимому ингибированию фосфорилирования STAT-6 в присутствии IL-13, однако и в этом случае связывание c146E2-H3L3 было более прочным, чем связывание антитела предшествующего уровня техники c4H3 к собачьему альфа-рецептору IL-4. Контроль IL-13 в отсутствие антител к IL-4R-альфа (IL-4Rα) показан в верхней правой части графика. На фиг.3 показано, что замена легкой каппа-цепи на легкую лямбда-цепь не влияла на связывание c146E2-H3L3 с IL-4R-альфа.
ПРИМЕР 3
КАРТИРОВАНИЕ ЭПИТОПОВ
Взаимодействие антител с родственными им белками-антигенами опосредуется через связывание специфических аминокислот антител (паратопов) со специфическими аминокислотами (эпитопами) антигенов-мишеней. Эпитоп представляет собой антигенную детерминанту, вызывающую специфическую реакцию иммуноглобулина. Эпитоп состоит из группы аминокислот на поверхности антигена. Представляющий интерес белок может содержать несколько эпитопов, которые распознаются разными антителами. Эпитопы, распознаваемые антителами, классифицируют как линейные или конформационные эпитопы. Линейные эпитопы образованы участком непрерывной последовательности аминокислот в белке, в то время как конформационные эпитопы состоят из аминокислот, которые не следуют непрерывно друг за другом (например, расположены далеко друг от друга) в первичной аминокислотной последовательности, но сближаются друг с другом при формировании пространственной структуры белка.
Картирование эпитопов относится к процессу идентификации аминокислотных последовательностей (т.е. эпитопов), которые распознаются антителами на антигенах-мишенях. Идентификация эпитопов, распознаваемых моноклональными антителами (mAb) на антигенах-мишенях, имеет важные применения. Например, она может помочь в разработке новых терапевтических средств, диагностических средств и вакцин. Картирование эпитопов может также облегчить выбор оптимизированных терапевтических mAb и прояснить механизмы их действия. Информация об эпитопах альфа-рецептора IL-4 также может пролить свет на уникальные эпитопы и определить защитное или патогенное действие вакцин. Идентификация эпитопа также может привести к разработке субъединичных вакцин, основанных на химическом или генетическом связывании идентифицированного пептидного эпитопа с белком-носителем или другими иммуностимулирующими агентами.
Картирование эпитопов можно выполнять с использованием поликлональных или моноклональных антител, и для идентификации эпитопов применяют несколько методов в зависимости от предполагаемой природы эпитопа (т.е. линейного или конформационного). Картирование линейных эпитопов является более очевидным и относительно простым в осуществлении. С этой целью коммерческие службы, выполняющие картирование линейных эпитопов, часто используют сканирование пептидов. В этом случае химически синтезируют перекрывающийся набор коротких пептидных последовательностей целевого белка, которые тестируют на способность связывать представляющие интерес антитела. Стратегия является быстрой, высокопроизводительной и относительно недорогой в исполнении. С другой стороны, картирование прерывистого эпитопа является технически более сложным процессом и требует более специализированных методов, таких как рентгеновская сокристаллография моноклонального антитела с белком-мишенью, обмен водород-дейтерий (H/D), масс-спектрометрия в сочетании с ферментативным расщеплением, а также несколько других способов, известных специалистам в данной области.
Картирование эпитопов собачьего альфа-рецептора IL-4 с помощью масс-спектроскопии:
Для идентификации эпитопов, распознаваемых mAb к собачьему альфа-рецептору IL-4, использовали метод, основанный на химической сшивке, детектировании методом масс-спектрометрии и ковалентном мечении [CovalX Instruments Incorporated, расположенная по адресу: 999 Broadway, Suite 305, Saugus, MA 01906-4510 США).]
Применение этого метода для картирования эпитопа альфа-цепи собачьего рецептора IL-4 в ранее проведенном исследовании показало, что mAb распознают специфические пептидные эпитопы, которые присутствуют во внеклеточном домене собачьего альфа-рецептора IL-4 [US2018/0346580]. Аналогичный анализ, выполненный в отношении антитела c146E2-H3L3 к собачьему альфа-рецептору IL, представленный на фиг.4, привел к идентификации аминокислотных последовательностей SEQ ID NO: 46 и SEQ ID NO: 47 для эпитопа(ов), который(ые) имеет(ют) разумное сходство с ранее идентифицированными. Кроме того, как показано на фиг.4, аминокислотные остатки K97, H112, T113, S164, T165, S171, Y172, S173 и R175 были идентифицированы как конкретные точки контакта [см., например, SEQ ID NO: 5, для нумерации аминокислотных остатков].
--->
СПИСКИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ИНТЕРВЕТ ИНТЕРНЭШНЛ Б.В.
МОРСИ, Мохамад
ЧЖАН, Юаньчжэн
<120> АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4
<130> 24980
<150> US 63/015,209
<151> 2020-04-24
<150> US 63/015,220
<151> 2020-04-24
<150> US 62/951,778
<151> 2019-12-20
<150> US 62/951,793
<151> 2019-12-20
<160> 47
<170> PatentIn version 3.5
<210> 1
<211> 2469
<212> ДНК
<213> Canis familiaris
<400> 1
atgggcagac tgtgcagcgg cctgaccttc cccgtgagct gcctggtgct ggtgtgggtg 60
gccagcagcg gcagcgtgaa ggtgctgcac gagcccagct gcttcagcga ctacatcagc 120
accagcgtgt gccagtggaa gatggaccac cccaccaact gcagcgccga gctgagactg 180
agctaccagc tggacttcat gggcagcgag aaccacacct gcgtgcccga gaacagagag 240
gacagcgtgt gcgtgtgcag catgcccatc gacgacgccg tggaggccga cgtgtaccag 300
ctggacctgt gggccggcca gcagctgctg tggagcggca gcttccagcc cagcaagcac 360
gtgaagccca gaacccccgg caacctgacc gtgcacccca acatcagcca cacctggctg 420
ctgatgtgga ccaaccccta ccccaccgag aaccacctgc acagcgagct gacctacatg 480
gtgaacgtga gcaacgacaa cgaccccgag gacttcaagg tgtacaacgt gacctacatg 540
ggccccaccc tgagactggc cgccagcacc ctgaagagcg gcgccagcta cagcgccaga 600
gtgagagcct gggcccagac ctacaacagc acctggagcg actggagccc cagcaccacc 660
tggctgaact actacgagcc ctgggagcag cacctgcccc tgggcgtgag catcagctgc 720
ctggtgatcc tggccatctg cctgagctgc tacttcagca tcatcaagat caagaagggc 780
tggtgggacc agatccccaa ccccgcccac agccccctgg tggccatcgt gatccaggac 840
agccaggtga gcctgtgggg caagagaagc agaggccagg agcccgccaa gtgcccccac 900
tggaagacct gcctgaccaa gctgctgccc tgcctgctgg agcacggcct gggcagagag 960
gaggagagcc ccaagaccgc caagaacggc cccctgcagg gccccggcaa gcccgcctgg 1020
tgccccgtgg aggtgagcaa gaccatcctg tggcccgaga gcatcagcgt ggtgcagtgc 1080
gtggagctga gcgaggcccc cgtggacaac gaggaggagg aggaggtgga ggaggacaag 1140
agaagcctgt gccccagcct ggagggcagc ggcggcagct tccaggaggg cagagagggc 1200
atcgtggcca gactgaccga gagcctgttc ctggacctgc tgggcggcga gaacggcggc 1260
ttctgccccc agggcctgga ggagagctgc ctgccccccc ccagcggcag cgtgggcgcc 1320
cagatgccct gggcccagtt ccccagagcc ggccccagag ccgcccccga gggccccgag 1380
cagcccagaa gacccgagag cgccctgcag gccagcccca cccagagcgc cggcagcagc 1440
gccttccccg agcccccccc cgtggtgacc gacaaccccg cctacagaag cttcggcagc 1500
ttcctgggcc agagcagcga ccccggcgac ggcgacagcg accccgagct ggccgacaga 1560
cccggcgagg ccgaccccgg catccccagc gccccccagc cccccgagcc ccccgccgcc 1620
ctgcagcccg agcccgagag ctgggagcag atcctgagac agagcgtgct gcagcacaga 1680
gccgcccccg cccccggccc cggccccggc agcggctaca gagagttcac ctgcgccgtg 1740
aagcagggca gcgcccccga cgccggcggc cccggcttcg gccccagcgg cgaggccggc 1800
tacaaggcct tctgcagcct gctgcccggc ggcgccacct gccccggcac cagcggcggc 1860
gaggccggca gcggcgaggg cggctacaag cccttccaga gcctgacccc cggctgcccc 1920
ggcgccccca cccccgtgcc cgtgcccctg ttcaccttcg gcctggacac cgagcccccc 1980
ggcagccccc aggacagcct gggcgccggc agcagccccg agcacctggg cgtggagccc 2040
gccggcaagg aggaggacag cagaaagacc ctgctggccc ccgagcaggc caccgacccc 2100
ctgagagacg acctggccag cagcatcgtg tacagcgccc tgacctgcca cctgtgcggc 2160
cacctgaagc agtggcacga ccaggaggag agaggcaagg cccacatcgt gcccagcccc 2220
tgctgcggct gctgctgcgg cgacagaagc agcctgctgc tgagccccct gagagccccc 2280
aacgtgctgc ccggcggcgt gctgctggag gccagcctga gccccgccag cctggtgccc 2340
agcggcgtga gcaaggaggg caagagcagc cccttcagcc agcccgccag cagcagcgcc 2400
cagagcagca gccagacccc caagaagctg gccgtgctga gcaccgagcc cacctgcatg 2460
agcgccagc 2469
<210> 2
<211> 823
<212> Белок
<213> Canis familiaris
<400> 2
Met Gly Arg Leu Cys Ser Gly Leu Thr Phe Pro Val Ser Cys Leu Val
1 5 10 15
Leu Val Trp Val Ala Ser Ser Gly Ser Val Lys Val Leu His Glu Pro
20 25 30
Ser Cys Phe Ser Asp Tyr Ile Ser Thr Ser Val Cys Gln Trp Lys Met
35 40 45
Asp His Pro Thr Asn Cys Ser Ala Glu Leu Arg Leu Ser Tyr Gln Leu
50 55 60
Asp Phe Met Gly Ser Glu Asn His Thr Cys Val Pro Glu Asn Arg Glu
65 70 75 80
Asp Ser Val Cys Val Cys Ser Met Pro Ile Asp Asp Ala Val Glu Ala
85 90 95
Asp Val Tyr Gln Leu Asp Leu Trp Ala Gly Gln Gln Leu Leu Trp Ser
100 105 110
Gly Ser Phe Gln Pro Ser Lys His Val Lys Pro Arg Thr Pro Gly Asn
115 120 125
Leu Thr Val His Pro Asn Ile Ser His Thr Trp Leu Leu Met Trp Thr
130 135 140
Asn Pro Tyr Pro Thr Glu Asn His Leu His Ser Glu Leu Thr Tyr Met
145 150 155 160
Val Asn Val Ser Asn Asp Asn Asp Pro Glu Asp Phe Lys Val Tyr Asn
165 170 175
Val Thr Tyr Met Gly Pro Thr Leu Arg Leu Ala Ala Ser Thr Leu Lys
180 185 190
Ser Gly Ala Ser Tyr Ser Ala Arg Val Arg Ala Trp Ala Gln Thr Tyr
195 200 205
Asn Ser Thr Trp Ser Asp Trp Ser Pro Ser Thr Thr Trp Leu Asn Tyr
210 215 220
Tyr Glu Pro Trp Glu Gln His Leu Pro Leu Gly Val Ser Ile Ser Cys
225 230 235 240
Leu Val Ile Leu Ala Ile Cys Leu Ser Cys Tyr Phe Ser Ile Ile Lys
245 250 255
Ile Lys Lys Gly Trp Trp Asp Gln Ile Pro Asn Pro Ala His Ser Pro
260 265 270
Leu Val Ala Ile Val Ile Gln Asp Ser Gln Val Ser Leu Trp Gly Lys
275 280 285
Arg Ser Arg Gly Gln Glu Pro Ala Lys Cys Pro His Trp Lys Thr Cys
290 295 300
Leu Thr Lys Leu Leu Pro Cys Leu Leu Glu His Gly Leu Gly Arg Glu
305 310 315 320
Glu Glu Ser Pro Lys Thr Ala Lys Asn Gly Pro Leu Gln Gly Pro Gly
325 330 335
Lys Pro Ala Trp Cys Pro Val Glu Val Ser Lys Thr Ile Leu Trp Pro
340 345 350
Glu Ser Ile Ser Val Val Gln Cys Val Glu Leu Ser Glu Ala Pro Val
355 360 365
Asp Asn Glu Glu Glu Glu Glu Val Glu Glu Asp Lys Arg Ser Leu Cys
370 375 380
Pro Ser Leu Glu Gly Ser Gly Gly Ser Phe Gln Glu Gly Arg Glu Gly
385 390 395 400
Ile Val Ala Arg Leu Thr Glu Ser Leu Phe Leu Asp Leu Leu Gly Gly
405 410 415
Glu Asn Gly Gly Phe Cys Pro Gln Gly Leu Glu Glu Ser Cys Leu Pro
420 425 430
Pro Pro Ser Gly Ser Val Gly Ala Gln Met Pro Trp Ala Gln Phe Pro
435 440 445
Arg Ala Gly Pro Arg Ala Ala Pro Glu Gly Pro Glu Gln Pro Arg Arg
450 455 460
Pro Glu Ser Ala Leu Gln Ala Ser Pro Thr Gln Ser Ala Gly Ser Ser
465 470 475 480
Ala Phe Pro Glu Pro Pro Pro Val Val Thr Asp Asn Pro Ala Tyr Arg
485 490 495
Ser Phe Gly Ser Phe Leu Gly Gln Ser Ser Asp Pro Gly Asp Gly Asp
500 505 510
Ser Asp Pro Glu Leu Ala Asp Arg Pro Gly Glu Ala Asp Pro Gly Ile
515 520 525
Pro Ser Ala Pro Gln Pro Pro Glu Pro Pro Ala Ala Leu Gln Pro Glu
530 535 540
Pro Glu Ser Trp Glu Gln Ile Leu Arg Gln Ser Val Leu Gln His Arg
545 550 555 560
Ala Ala Pro Ala Pro Gly Pro Gly Pro Gly Ser Gly Tyr Arg Glu Phe
565 570 575
Thr Cys Ala Val Lys Gln Gly Ser Ala Pro Asp Ala Gly Gly Pro Gly
580 585 590
Phe Gly Pro Ser Gly Glu Ala Gly Tyr Lys Ala Phe Cys Ser Leu Leu
595 600 605
Pro Gly Gly Ala Thr Cys Pro Gly Thr Ser Gly Gly Glu Ala Gly Ser
610 615 620
Gly Glu Gly Gly Tyr Lys Pro Phe Gln Ser Leu Thr Pro Gly Cys Pro
625 630 635 640
Gly Ala Pro Thr Pro Val Pro Val Pro Leu Phe Thr Phe Gly Leu Asp
645 650 655
Thr Glu Pro Pro Gly Ser Pro Gln Asp Ser Leu Gly Ala Gly Ser Ser
660 665 670
Pro Glu His Leu Gly Val Glu Pro Ala Gly Lys Glu Glu Asp Ser Arg
675 680 685
Lys Thr Leu Leu Ala Pro Glu Gln Ala Thr Asp Pro Leu Arg Asp Asp
690 695 700
Leu Ala Ser Ser Ile Val Tyr Ser Ala Leu Thr Cys His Leu Cys Gly
705 710 715 720
His Leu Lys Gln Trp His Asp Gln Glu Glu Arg Gly Lys Ala His Ile
725 730 735
Val Pro Ser Pro Cys Cys Gly Cys Cys Cys Gly Asp Arg Ser Ser Leu
740 745 750
Leu Leu Ser Pro Leu Arg Ala Pro Asn Val Leu Pro Gly Gly Val Leu
755 760 765
Leu Glu Ala Ser Leu Ser Pro Ala Ser Leu Val Pro Ser Gly Val Ser
770 775 780
Lys Glu Gly Lys Ser Ser Pro Phe Ser Gln Pro Ala Ser Ser Ser Ala
785 790 795 800
Gln Ser Ser Ser Gln Thr Pro Lys Lys Leu Ala Val Leu Ser Thr Glu
805 810 815
Pro Thr Cys Met Ser Ala Ser
820
<210> 3
<211> 2394
<212> ДНК
<213> Canis familiaris
<400> 3
gtgaaggtgc tgcacgagcc cagctgcttc agcgactaca tcagcaccag cgtgtgccag 60
tggaagatgg accaccccac caactgcagc gccgagctga gactgagcta ccagctggac 120
ttcatgggca gcgagaacca cacctgcgtg cccgagaaca gagaggacag cgtgtgcgtg 180
tgcagcatgc ccatcgacga cgccgtggag gccgacgtgt accagctgga cctgtgggcc 240
ggccagcagc tgctgtggag cggcagcttc cagcccagca agcacgtgaa gcccagaacc 300
cccggcaacc tgaccgtgca ccccaacatc agccacacct ggctgctgat gtggaccaac 360
ccctacccca ccgagaacca cctgcacagc gagctgacct acatggtgaa cgtgagcaac 420
gacaacgacc ccgaggactt caaggtgtac aacgtgacct acatgggccc caccctgaga 480
ctggccgcca gcaccctgaa gagcggcgcc agctacagcg ccagagtgag agcctgggcc 540
cagacctaca acagcacctg gagcgactgg agccccagca ccacctggct gaactactac 600
gagccctggg agcagcacct gcccctgggc gtgagcatca gctgcctggt gatcctggcc 660
atctgcctga gctgctactt cagcatcatc aagatcaaga agggctggtg ggaccagatc 720
cccaaccccg cccacagccc cctggtggcc atcgtgatcc aggacagcca ggtgagcctg 780
tggggcaaga gaagcagagg ccaggagccc gccaagtgcc cccactggaa gacctgcctg 840
accaagctgc tgccctgcct gctggagcac ggcctgggca gagaggagga gagccccaag 900
accgccaaga acggccccct gcagggcccc ggcaagcccg cctggtgccc cgtggaggtg 960
agcaagacca tcctgtggcc cgagagcatc agcgtggtgc agtgcgtgga gctgagcgag 1020
gcccccgtgg acaacgagga ggaggaggag gtggaggagg acaagagaag cctgtgcccc 1080
agcctggagg gcagcggcgg cagcttccag gagggcagag agggcatcgt ggccagactg 1140
accgagagcc tgttcctgga cctgctgggc ggcgagaacg gcggcttctg cccccagggc 1200
ctggaggaga gctgcctgcc cccccccagc ggcagcgtgg gcgcccagat gccctgggcc 1260
cagttcccca gagccggccc cagagccgcc cccgagggcc ccgagcagcc cagaagaccc 1320
gagagcgccc tgcaggccag ccccacccag agcgccggca gcagcgcctt ccccgagccc 1380
ccccccgtgg tgaccgacaa ccccgcctac agaagcttcg gcagcttcct gggccagagc 1440
agcgaccccg gcgacggcga cagcgacccc gagctggccg acagacccgg cgaggccgac 1500
cccggcatcc ccagcgcccc ccagcccccc gagccccccg ccgccctgca gcccgagccc 1560
gagagctggg agcagatcct gagacagagc gtgctgcagc acagagccgc ccccgccccc 1620
ggccccggcc ccggcagcgg ctacagagag ttcacctgcg ccgtgaagca gggcagcgcc 1680
cccgacgccg gcggccccgg cttcggcccc agcggcgagg ccggctacaa ggccttctgc 1740
agcctgctgc ccggcggcgc cacctgcccc ggcaccagcg gcggcgaggc cggcagcggc 1800
gagggcggct acaagccctt ccagagcctg acccccggct gccccggcgc ccccaccccc 1860
gtgcccgtgc ccctgttcac cttcggcctg gacaccgagc cccccggcag cccccaggac 1920
agcctgggcg ccggcagcag ccccgagcac ctgggcgtgg agcccgccgg caaggaggag 1980
gacagcagaa agaccctgct ggcccccgag caggccaccg accccctgag agacgacctg 2040
gccagcagca tcgtgtacag cgccctgacc tgccacctgt gcggccacct gaagcagtgg 2100
cacgaccagg aggagagagg caaggcccac atcgtgccca gcccctgctg cggctgctgc 2160
tgcggcgaca gaagcagcct gctgctgagc cccctgagag cccccaacgt gctgcccggc 2220
ggcgtgctgc tggaggccag cctgagcccc gccagcctgg tgcccagcgg cgtgagcaag 2280
gagggcaaga gcagcccctt cagccagccc gccagcagca gcgcccagag cagcagccag 2340
acccccaaga agctggccgt gctgagcacc gagcccacct gcatgagcgc cagc 2394
<210> 4
<211> 798
<212> Белок
<213> Canis familiaris
<400> 4
Val Lys Val Leu His Glu Pro Ser Cys Phe Ser Asp Tyr Ile Ser Thr
1 5 10 15
Ser Val Cys Gln Trp Lys Met Asp His Pro Thr Asn Cys Ser Ala Glu
20 25 30
Leu Arg Leu Ser Tyr Gln Leu Asp Phe Met Gly Ser Glu Asn His Thr
35 40 45
Cys Val Pro Glu Asn Arg Glu Asp Ser Val Cys Val Cys Ser Met Pro
50 55 60
Ile Asp Asp Ala Val Glu Ala Asp Val Tyr Gln Leu Asp Leu Trp Ala
65 70 75 80
Gly Gln Gln Leu Leu Trp Ser Gly Ser Phe Gln Pro Ser Lys His Val
85 90 95
Lys Pro Arg Thr Pro Gly Asn Leu Thr Val His Pro Asn Ile Ser His
100 105 110
Thr Trp Leu Leu Met Trp Thr Asn Pro Tyr Pro Thr Glu Asn His Leu
115 120 125
His Ser Glu Leu Thr Tyr Met Val Asn Val Ser Asn Asp Asn Asp Pro
130 135 140
Glu Asp Phe Lys Val Tyr Asn Val Thr Tyr Met Gly Pro Thr Leu Arg
145 150 155 160
Leu Ala Ala Ser Thr Leu Lys Ser Gly Ala Ser Tyr Ser Ala Arg Val
165 170 175
Arg Ala Trp Ala Gln Thr Tyr Asn Ser Thr Trp Ser Asp Trp Ser Pro
180 185 190
Ser Thr Thr Trp Leu Asn Tyr Tyr Glu Pro Trp Glu Gln His Leu Pro
195 200 205
Leu Gly Val Ser Ile Ser Cys Leu Val Ile Leu Ala Ile Cys Leu Ser
210 215 220
Cys Tyr Phe Ser Ile Ile Lys Ile Lys Lys Gly Trp Trp Asp Gln Ile
225 230 235 240
Pro Asn Pro Ala His Ser Pro Leu Val Ala Ile Val Ile Gln Asp Ser
245 250 255
Gln Val Ser Leu Trp Gly Lys Arg Ser Arg Gly Gln Glu Pro Ala Lys
260 265 270
Cys Pro His Trp Lys Thr Cys Leu Thr Lys Leu Leu Pro Cys Leu Leu
275 280 285
Glu His Gly Leu Gly Arg Glu Glu Glu Ser Pro Lys Thr Ala Lys Asn
290 295 300
Gly Pro Leu Gln Gly Pro Gly Lys Pro Ala Trp Cys Pro Val Glu Val
305 310 315 320
Ser Lys Thr Ile Leu Trp Pro Glu Ser Ile Ser Val Val Gln Cys Val
325 330 335
Glu Leu Ser Glu Ala Pro Val Asp Asn Glu Glu Glu Glu Glu Val Glu
340 345 350
Glu Asp Lys Arg Ser Leu Cys Pro Ser Leu Glu Gly Ser Gly Gly Ser
355 360 365
Phe Gln Glu Gly Arg Glu Gly Ile Val Ala Arg Leu Thr Glu Ser Leu
370 375 380
Phe Leu Asp Leu Leu Gly Gly Glu Asn Gly Gly Phe Cys Pro Gln Gly
385 390 395 400
Leu Glu Glu Ser Cys Leu Pro Pro Pro Ser Gly Ser Val Gly Ala Gln
405 410 415
Met Pro Trp Ala Gln Phe Pro Arg Ala Gly Pro Arg Ala Ala Pro Glu
420 425 430
Gly Pro Glu Gln Pro Arg Arg Pro Glu Ser Ala Leu Gln Ala Ser Pro
435 440 445
Thr Gln Ser Ala Gly Ser Ser Ala Phe Pro Glu Pro Pro Pro Val Val
450 455 460
Thr Asp Asn Pro Ala Tyr Arg Ser Phe Gly Ser Phe Leu Gly Gln Ser
465 470 475 480
Ser Asp Pro Gly Asp Gly Asp Ser Asp Pro Glu Leu Ala Asp Arg Pro
485 490 495
Gly Glu Ala Asp Pro Gly Ile Pro Ser Ala Pro Gln Pro Pro Glu Pro
500 505 510
Pro Ala Ala Leu Gln Pro Glu Pro Glu Ser Trp Glu Gln Ile Leu Arg
515 520 525
Gln Ser Val Leu Gln His Arg Ala Ala Pro Ala Pro Gly Pro Gly Pro
530 535 540
Gly Ser Gly Tyr Arg Glu Phe Thr Cys Ala Val Lys Gln Gly Ser Ala
545 550 555 560
Pro Asp Ala Gly Gly Pro Gly Phe Gly Pro Ser Gly Glu Ala Gly Tyr
565 570 575
Lys Ala Phe Cys Ser Leu Leu Pro Gly Gly Ala Thr Cys Pro Gly Thr
580 585 590
Ser Gly Gly Glu Ala Gly Ser Gly Glu Gly Gly Tyr Lys Pro Phe Gln
595 600 605
Ser Leu Thr Pro Gly Cys Pro Gly Ala Pro Thr Pro Val Pro Val Pro
610 615 620
Leu Phe Thr Phe Gly Leu Asp Thr Glu Pro Pro Gly Ser Pro Gln Asp
625 630 635 640
Ser Leu Gly Ala Gly Ser Ser Pro Glu His Leu Gly Val Glu Pro Ala
645 650 655
Gly Lys Glu Glu Asp Ser Arg Lys Thr Leu Leu Ala Pro Glu Gln Ala
660 665 670
Thr Asp Pro Leu Arg Asp Asp Leu Ala Ser Ser Ile Val Tyr Ser Ala
675 680 685
Leu Thr Cys His Leu Cys Gly His Leu Lys Gln Trp His Asp Gln Glu
690 695 700
Glu Arg Gly Lys Ala His Ile Val Pro Ser Pro Cys Cys Gly Cys Cys
705 710 715 720
Cys Gly Asp Arg Ser Ser Leu Leu Leu Ser Pro Leu Arg Ala Pro Asn
725 730 735
Val Leu Pro Gly Gly Val Leu Leu Glu Ala Ser Leu Ser Pro Ala Ser
740 745 750
Leu Val Pro Ser Gly Val Ser Lys Glu Gly Lys Ser Ser Pro Phe Ser
755 760 765
Gln Pro Ala Ser Ser Ser Ala Gln Ser Ser Ser Gln Thr Pro Lys Lys
770 775 780
Leu Ala Val Leu Ser Thr Glu Pro Thr Cys Met Ser Ala Ser
785 790 795
<210> 5
<211> 208
<212> Белок
<213> Canis familiaris
<400> 5
Val Lys Val Leu His Glu Pro Ser Cys Phe Ser Asp Tyr Ile Ser Thr
1 5 10 15
Ser Val Cys Gln Trp Lys Met Asp His Pro Thr Asn Cys Ser Ala Glu
20 25 30
Leu Arg Leu Ser Tyr Gln Leu Asp Phe Met Gly Ser Glu Asn His Thr
35 40 45
Cys Val Pro Glu Asn Arg Glu Asp Ser Val Cys Val Cys Ser Met Pro
50 55 60
Ile Asp Asp Ala Val Glu Ala Asp Val Tyr Gln Leu Asp Leu Trp Ala
65 70 75 80
Gly Gln Gln Leu Leu Trp Ser Gly Ser Phe Gln Pro Ser Lys His Val
85 90 95
Lys Pro Arg Thr Pro Gly Asn Leu Thr Val His Pro Asn Ile Ser His
100 105 110
Thr Trp Leu Leu Met Trp Thr Asn Pro Tyr Pro Thr Glu Asn His Leu
115 120 125
His Ser Glu Leu Thr Tyr Met Val Asn Val Ser Asn Asp Asn Asp Pro
130 135 140
Glu Asp Phe Lys Val Tyr Asn Val Thr Tyr Met Gly Pro Thr Leu Arg
145 150 155 160
Leu Ala Ala Ser Thr Leu Lys Ser Gly Ala Ser Tyr Ser Ala Arg Val
165 170 175
Arg Ala Trp Ala Gln Thr Tyr Asn Ser Thr Trp Ser Asp Trp Ser Pro
180 185 190
Ser Thr Thr Trp Leu Asn Tyr Tyr Glu Pro Trp Glu Gln His Leu Pro
195 200 205
<210> 6
<211> 17
<212> Белок
<213> Canis familiaris
<400> 6
Phe Asn Glu Cys Arg Cys Thr Asp Thr Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Pro
<210> 7
<211> 22
<212> Белок
<213> Canis familiaris
<400> 7
Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys Pro Lys
1 5 10 15
Cys Pro Ala Pro Glu Met
20
<210> 8
<211> 20
<212> Белок
<213> Canis familiaris
<400> 8
Ala Lys Glu Cys Glu Cys Lys Cys Asn Cys Asn Asn Cys Pro Cys Pro
1 5 10 15
Gly Cys Gly Leu
20
<210> 9
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> модифицированная собачья
<400> 9
Pro Lys Glu Ser Thr Cys Lys Cys Ile Pro Pro Cys Pro Val Pro Glu
1 5 10 15
Ser
<210> 10
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> модифицированная собачья
<400> 10
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
1 5 10 15
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu
20 25 30
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
35 40 45
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly
50 55 60
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
65 70 75 80
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
85 90 95
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
100 105 110
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
115 120 125
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
130 135 140
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
145 150 155 160
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
165 170 175
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
180 185 190
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser
195 200 205
Leu Ser His Ser Pro Gly
210
<210> 11
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 11
agatactgga tgcac 15
<210> 12
<211> 5
<212> Белок
<213> Mus musculus
<400> 12
Arg Tyr Trp Met His
1 5
<210> 13
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 13
atgattcacc ccgacagcgg caacatcaac tacaacgagc ggttcaagac c 51
<210> 14
<211> 17
<212> Белок
<213> Mus musculus
<400> 14
Met Ile His Pro Asp Ser Gly Asn Ile Asn Tyr Asn Glu Arg Phe Lys
1 5 10 15
Thr
<210> 15
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 15
cagctgcgga acgccatgga ttat 24
<210> 16
<211> 8
<212> Белок
<213> Mus musculus
<400> 16
Gln Leu Arg Asn Ala Met Asp Tyr
1 5
<210> 17
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 17
agagccagcg agagcgtgga cagctacggc aacagcttcc tgaac 45
<210> 18
<211> 15
<212> Белок
<213> Mus musculus
<400> 18
Arg Ala Ser Glu Ser Val Asp Ser Tyr Gly Asn Ser Phe Leu Asn
1 5 10 15
<210> 19
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 19
agagccagca acctggcctc t 21
<210> 20
<211> 7
<212> Белок
<213> Mus musculus
<400> 20
Arg Ala Ser Asn Leu Ala Ser
1 5
<210> 21
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 21
cagcagaact acgagaaccc cagaacc 27
<210> 22
<211> 9
<212> Белок
<213> Mus musculus
<400> 22
Gln Gln Asn Tyr Glu Asn Pro Arg Thr
1 5
<210> 23
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 23
agctacggca tgagc 15
<210> 24
<211> 5
<212> Белок
<213> Mus musculus
<400> 24
Ser Tyr Gly Met Ser
1 5
<210> 25
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 25
acaatcagca gaggcggcga ctacacctac tatcccgaca gcgtgaaggg c 51
<210> 26
<211> 17
<212> Белок
<213> Mus musculus
<400> 26
Thr Ile Ser Arg Gly Gly Asp Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1 5 10 15
Gly
<210> 27
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 27
ggcaccctga acaaccgggg ctttgcttct 30
<210> 28
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> модифицированная мышиная
<400> 28
Gly Thr Leu Asn Asn Arg Gly Phe Ala Ser
1 5 10
<210> 29
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 29
aaggccagcc agaacgtggg caccaatgtg gcc 33
<210> 30
<211> 11
<212> Белок
<213> Mus musculus
<400> 30
Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala
1 5 10
<210> 31
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 31
agcgccagct accggtactc t 21
<210> 32
<211> 7
<212> Белок
<213> Mus musculus
<400> 32
Ser Ala Ser Tyr Arg Tyr Ser
1 5
<210> 33
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> сгенерированная компьютером на основе аминокислотной
последовательности CDR
<400> 33
cagcagtaca acagctaccc ctacacc 27
<210> 34
<211> 9
<212> Белок
<213> Mus musculus
<400> 34
Gln Gln Tyr Asn Ser Tyr Pro Tyr Thr
1 5
<210> 35
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 35
Glu Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ser Leu Ser Gln Glu
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Leu Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Ser Phe Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Val Ala Glu Phe Phe Cys Gln Gln Tyr Asn Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Asn Asp Ala Gln
100 105 110
Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln Leu His Thr Gly
115 120 125
Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr Pro Lys Asp Ile
130 135 140
Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp Thr Gly Ile Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His Glu Leu Tyr Ser
180 185 190
Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu Ile Lys Ser Phe
195 200 205
Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215
<210> 36
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 36
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Thr Ile Ser Arg Gly Gly Asp Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Lys Gly Thr Leu Asn Asn Arg Gly Phe Ala Cys Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu
130 135 140
Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser
180 185 190
Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala
195 200 205
Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg
210 215 220
Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp
260 265 270
Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln
275 280 285
Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys
305 310 315 320
Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro
325 330 335
Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser
340 345 350
Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val
355 360 365
Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val
370 375 380
Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr
385 390 395 400
Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys
405 410 415
Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys
420 425 430
Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu
435 440 445
Ser His Ser Pro Gly
450
<210> 37
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 37
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Asp Lys Arg Leu Gln Trp Val
35 40 45
Ala Thr Ile Ser Arg Gly Gly Asp Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Thr Leu Asn Asn Arg Gly Phe Ala Cys Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu
130 135 140
Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser
180 185 190
Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala
195 200 205
Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg
210 215 220
Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp
260 265 270
Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln
275 280 285
Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys
305 310 315 320
Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro
325 330 335
Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser
340 345 350
Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val
355 360 365
Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val
370 375 380
Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr
385 390 395 400
Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys
405 410 415
Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys
420 425 430
Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu
435 440 445
Ser His Ser Pro Gly
450
<210> 38
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 38
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Asp Lys Arg Leu Gln Trp Val
35 40 45
Ala Thr Ile Ser Arg Gly Gly Asp Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Thr Leu Asn Asn Arg Gly Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu
130 135 140
Ala Cys Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser
180 185 190
Ser Arg Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala
195 200 205
Ser Lys Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg
210 215 220
Val Pro Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp
260 265 270
Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln
275 280 285
Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys
305 310 315 320
Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro
325 330 335
Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser
340 345 350
Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val
355 360 365
Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val
370 375 380
Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr
385 390 395 400
Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys
405 410 415
Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys
420 425 430
Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu
435 440 445
Ser His Ser Pro Gly
450
<210> 39
<211> 221
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 39
Asp Ile Val Leu Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Thr Ala Ser Ile Tyr Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Glu Ile Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Glu Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Gln Gln Asn Tyr
85 90 95
Glu Asn Pro Arg Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln
115 120 125
Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr
130 135 140
Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp
145 150 155 160
Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His
180 185 190
Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu
195 200 205
Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
210 215 220
<210> 40
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 40
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ala Arg Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Ala Gly Leu Asp Trp Met
35 40 45
Gly Met Ile His Pro Asp Ser Gly Asn Ile Asn Tyr Asn Glu Arg Phe
50 55 60
Lys Thr Arg Val Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ala Gly Asp Ile Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Arg Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys
130 135 140
Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg
180 185 190
Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys
195 200 205
Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro
210 215 220
Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile
245 250 255
Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp Pro Glu
260 265 270
Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln
275 280 285
Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys
305 310 315 320
Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu
325 330 335
Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr
340 345 350
Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu
355 360 365
Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp
370 375 380
Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro
385 390 395 400
Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser
405 410 415
Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His
435 440 445
Ser Pro Gly
450
<210> 41
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 41
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ala Arg Tyr
20 25 30
Trp Met His Trp Met Lys Gln Ala Pro Gly Ala Gly Leu Asp Trp Ile
35 40 45
Gly Met Ile His Pro Asp Ser Gly Asn Ile Asn Tyr Asn Glu Arg Phe
50 55 60
Lys Thr Lys Ala Thr Leu Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ala Gly Asp Ile Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Arg Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys
130 135 140
Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg
180 185 190
Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys
195 200 205
Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro
210 215 220
Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile
245 250 255
Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp Pro Glu
260 265 270
Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln
275 280 285
Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys
305 310 315 320
Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu
325 330 335
Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr
340 345 350
Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu
355 360 365
Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp
370 375 380
Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro
385 390 395 400
Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser
405 410 415
Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His
435 440 445
Ser Pro Gly
450
<210> 42
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 42
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ala Arg Tyr
20 25 30
Trp Met His Trp Met Lys Gln Ala Pro Gly Ala Gly Leu Asp Trp Ile
35 40 45
Gly Met Ile His Pro Asp Ser Gly Asn Ile Asn Tyr Asn Glu Arg Phe
50 55 60
Lys Thr Lys Ala Thr Leu Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ala Gly Asp Ile Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Leu Arg Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Cys Gly Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys
130 135 140
Leu Val Ser Gly Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ser Leu Thr Ser Gly Val His Thr Phe Pro Ser Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg
180 185 190
Trp Pro Ser Glu Thr Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys
195 200 205
Thr Lys Val Asp Lys Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro
210 215 220
Arg Pro Pro Asp Cys Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile
245 250 255
Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Leu Asp Pro Glu
260 265 270
Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln
275 280 285
Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Ala Gly Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys
305 310 315 320
Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu
325 330 335
Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr
340 345 350
Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu
355 360 365
Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp
370 375 380
Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro
385 390 395 400
Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser
405 410 415
Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu Ser His
435 440 445
Ser Pro Gly
450
<210> 43
<211> 212
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 43
Gln Ser Val Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val
20 25 30
Ala Trp Tyr Gln Gln Leu Pro Gly Thr Ser Pro Arg Thr Leu Ile Tyr
35 40 45
Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Arg Ser Gly Ser Thr Ala Thr Leu Thr Ile Ser Gly Leu Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr His Leu Thr Val Leu Gly Gln Pro Lys Ala Ser
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gly Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Ser Gly Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Thr Gln Gly Val Glu
145 150 155 160
Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Asp Lys Trp Lys Ser His Ser Ser Phe Ser
180 185 190
Cys Leu Val Thr His Glu Gly Ser Thr Val Glu Lys Lys Val Ala Pro
195 200 205
Ala Glu Cys Ser
210
<210> 44
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> канинизированная мышиная
<400> 44
Gln Ser Val Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr Gly
20 25 30
Asn Ser Phe Leu Asn Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Ser
35 40 45
Leu Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Glu Arg
50 55 60
Phe Ser Gly Ser Lys Ser Gly Ser Ser Ala Thr Leu Thr Ile Thr Gly
65 70 75 80
Leu Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Gln Asn Tyr Glu
85 90 95
Asn Pro Arg Thr Phe Gly Gly Gly Thr His Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gly Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Ser Gly Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Thr
145 150 155 160
Gln Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Asp Lys Trp Lys Ser His
180 185 190
Ser Ser Phe Ser Cys Leu Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Lys Val Ala Pro Ala Glu Cys Ser
210 215
<210> 45
<211> 215
<212> Белок
<213> Canis familiaris
<400> 45
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
1 5 10 15
Leu Leu Ile Ala Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Leu
20 25 30
Asp Pro Glu Asp Pro Glu Val Gln Ile Ser Trp Phe Val Asp Gly Lys
35 40 45
Gln Met Gln Thr Ala Lys Thr Gln Pro Arg Glu Glu Gln Phe Asn Gly
50 55 60
Thr Tyr Arg Val Val Ser Val Leu Pro Ile Gly His Gln Asp Trp Leu
65 70 75 80
Lys Gly Lys Gln Phe Thr Cys Lys Val Asn Asn Lys Ala Leu Pro Ser
85 90 95
Pro Ile Glu Arg Thr Ile Ser Lys Ala Arg Gly Gln Ala His Gln Pro
100 105 110
Ser Val Tyr Val Leu Pro Pro Ser Arg Glu Glu Leu Ser Lys Asn Thr
115 120 125
Val Ser Leu Thr Cys Leu Ile Lys Asp Phe Phe Pro Pro Asp Ile Asp
130 135 140
Val Glu Trp Gln Ser Asn Gly Gln Gln Glu Pro Glu Ser Lys Tyr Arg
145 150 155 160
Thr Thr Pro Pro Gln Leu Asp Glu Asp Gly Ser Tyr Phe Leu Tyr Ser
165 170 175
Lys Leu Ser Val Asp Lys Ser Arg Trp Gln Arg Gly Asp Thr Phe Ile
180 185 190
Cys Ala Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
195 200 205
Leu Ser His Ser Pro Gly Lys
210 215
<210> 46
<211> 31
<212> Белок
<213> Canis familiaris
<400> 46
Phe Gln Pro Ser Lys His Val Lys Pro Arg Thr Pro Gly Asn Leu Thr
1 5 10 15
Val His Pro Asn Ile Ser His Thr Trp Leu Leu Met Trp Thr Asn
20 25 30
<210> 47
<211> 21
<212> Белок
<213> Canis familiaris
<400> 47
Arg Leu Ala Ala Ser Thr Leu Lys Ser Gly Ala Ser Tyr Ser Ala Arg
1 5 10 15
Val Arg Ala Trp Ala
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2840044C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| КАНИНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CTLA-4 | 2020 |
|
RU2822460C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2801209C2 |
| АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО CTLA-4 | 2020 |
|
RU2818586C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2795485C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИТЕЛА К ПАРВОВИРУСУ ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2830441C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2732604C2 |
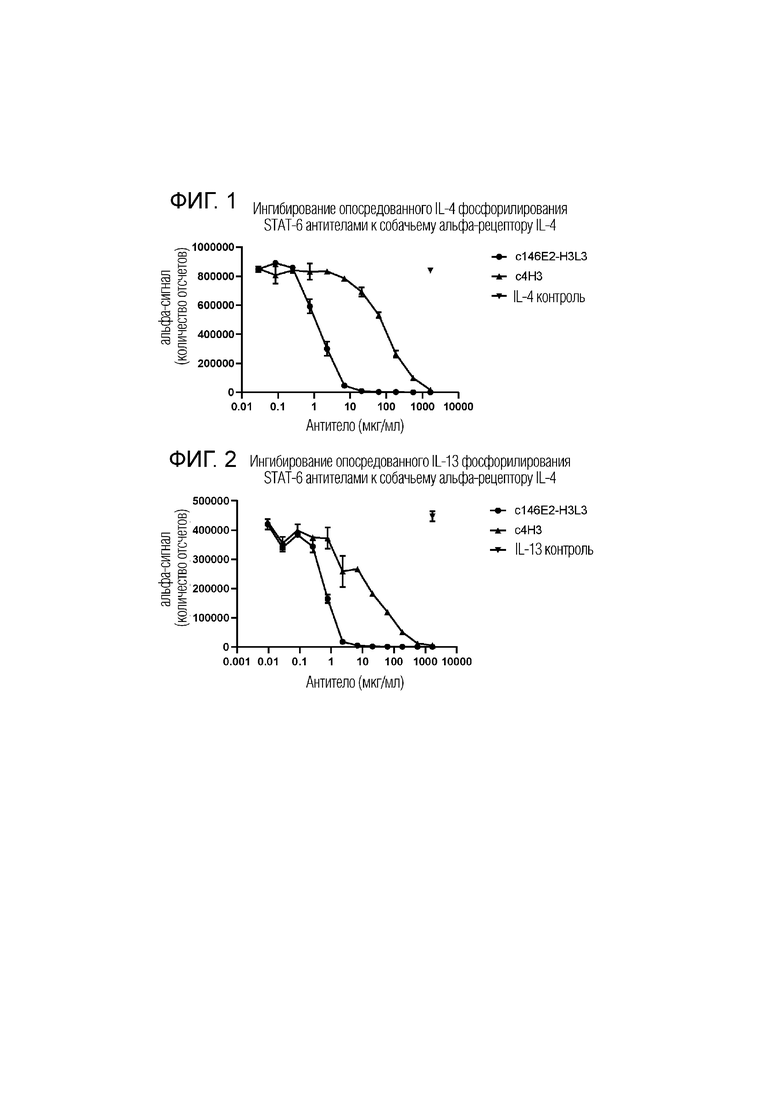
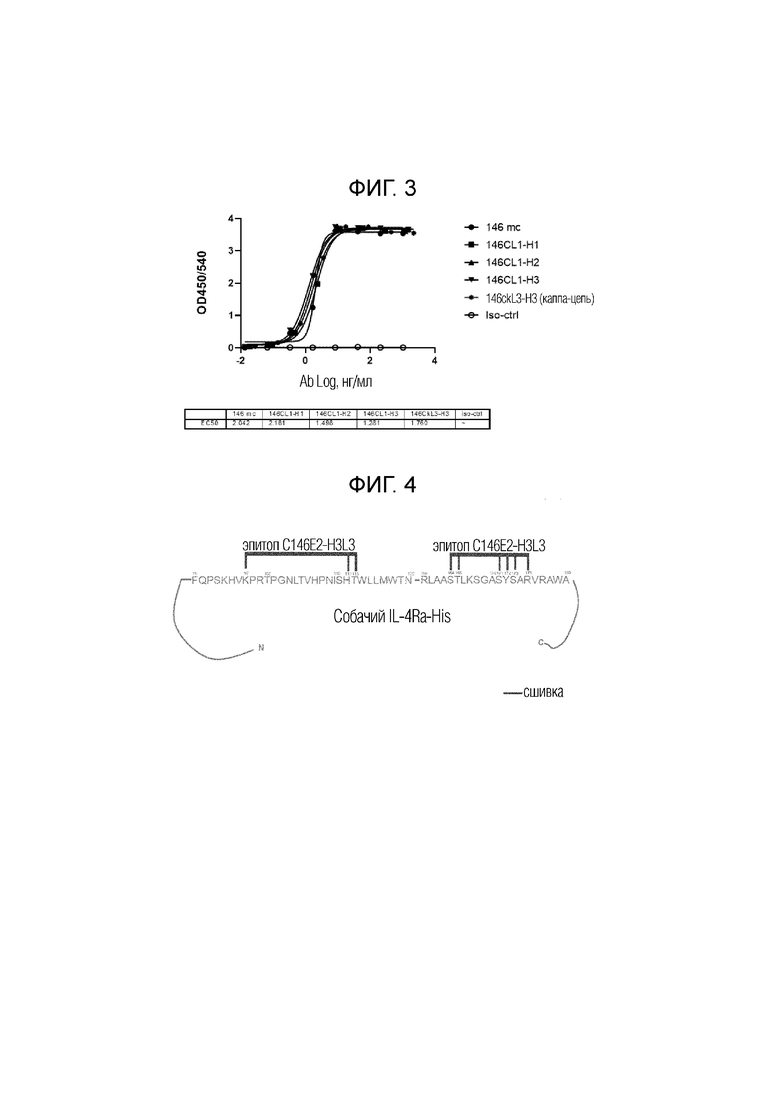
Изобретение относится к области биотехнологии. Предложены антитело млекопитающего или его антигенсвязывающий фрагмент, которые связываются с собачьим α-рецептором интерлейкина-4, а также кодирующие их нуклеиновые кислоты, векторы экспрессии и клетки-хозяева для получения антител. Кроме того, изобретение относится к применению антител для лечения атопического дерматита у собак. Изобретение обеспечивает высокую аффинность связывания с собачьим α-рецептором IL-4 и блокирование связывания собачьего IL-4 и/или IL-13 с собачьим α-рецептором IL-4. 11 н. и 7 з.п. ф-лы, 4 ил., 4 табл., 3 пр.
1. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, которое связывается с собачьим α-рецептором интерлейкина-4 (IL-4Rα), содержащее набор из шести определяющих комплементарность областей (CDR), три из которых представляют собой CDR тяжелой цепи: CDR 1 тяжелой цепи (HCDR1), CDR 2 тяжелой цепи (HCDR2) и CDR 3 тяжелой цепи (HCDR3) и три из которых представляют собой CDR легкой цепи: CDR 1 легкой цепи (LCDR1), CDR 2 легкой цепи (LCDR2) и CDR 3 легкой цепи (LCDR3); где
(i) HCDR1 содержит аминокислотную последовательность SEQ ID NO: 12;
(ii) HCDR2 содержит аминокислотную последовательность SEQ ID NO: 14;
(iii) HCDR3 содержит аминокислотную последовательность SEQ ID NO: 16;
(iv) LCDR1 содержит аминокислотную последовательность SEQ ID NO: 18;
(v) LCDR2 содержит аминокислотную последовательность SEQ ID NO: 20; и
(vi) LCDR3 содержит аминокислотную последовательность SEQ ID NO: 22.
2. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, по п. 1, где антитело, и его антигенсвязывающий фрагмент, связывается с собачьим IL-4Rα и блокирует связывание собачьего IL-4Rα с собачьим интерлейкином-4.
3. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, по п. 1 или 2, которое представляет собой канинизированное антитело, или его канинизированный антигенсвязывающий фрагмент.
4. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, по п. 3, которое содержит шарнирную область, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8 и SEQ ID NO: 9.
5. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, по п. 3, которое содержит тяжелую цепь, содержащую модифицированный собачий IgG-B (IgG-Bm), содержащий аминокислотную последовательность SEQ ID NO: 10.
6. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, по п. 5, которое содержит тяжелую цепь, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 40, SEQ ID NO: 41 и SEQ ID NO: 42.
7. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, по п. 6, которое содержит легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 39.
8. Выделенное антитело млекопитающего, или его антигенсвязывающий фрагмент, по п. 6, которое содержит легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 44.
9. Выделенная нуклеиновая кислота, кодирующая тяжелую цепь выделенного антитела млекопитающего или его антигенсвязывающего фрагмента по пп. 3, 4, 5, 6, 7 или 8.
10. Выделенная нуклеиновая кислота, кодирующая легкую цепь выделенного антитела млекопитающего или его антигенсвязывающего фрагмента по пп. 3, 4, 5, 6, 7 или 8.
11. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 9.
12. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 10.
13. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 9 и выделенную нуклеиновую кислоту по п. 10.
14. Клетка-хозяин, содержащая вектор экспрессии по п. 11, для получения тяжелой цепи выделенного антитела млекопитающего или его антигенсвязывающего фрагмента.
15. Клетка-хозяин, содержащая вектор экспрессии по п. 12, для получения легкой цепи выделенного антитела млекопитающего или его антигенсвязывающего фрагмента.
16. Клетка-хозяин, содержащая вектор экспрессии по п. 13, для получения антитела млекопитающего или его антигенсвязывающего фрагмента.
17. Фармацевтическая композиция для уменьшения воспаления, связанного с атопическим дерматитом, содержащая выделенное антитело млекопитающего или его антигенсвязывающий фрагмент по пп. 3, 4, 5, 6, 7 или 8 и фармацевтически приемлемый носитель или разбавитель.
18. Способ снижения воспаления, связанного с атопическим дерматитом, включающий введение пациенту терапевтически эффективного количества фармацевтической композиции по п. 17.
| WO 2016156588 A1, 06.10.2016 | |||
| US 7208579 B2, 24.04.2007 | |||
| WO 2017102920 A1, 22.06.2017 | |||
| СПОСОБЫ ЛЕЧЕНИЯ АЛЛЕРГИИ И ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АЛЛЕРГЕН-СПЕЦИФИЧЕСКОЙ ИММУНОТЕРАПИИ ПУТЕМ ВВЕДЕНИЯ ИНГИБИТОРА ИЛ-4Р | 2014 |
|
RU2679920C2 |
| AHMED N | |||
| et al., Identification and Characterization of a Novel IL-4 Receptor α Chain (IL-4Rα) Antagonist to Inhibit IL-4 Signalling, Cellular Physiology and Biochemistry, 2015, vol.36(3), pp.831-842. | |||

