Область техники
Настоящее изобретение имеет отношение к линиям клеток и маркерам отбора (селектируемым маркерам) для продукции белков.
Предпосылки создания изобретения
Для продуцирования рекомбинантных белков в промышленном масштабе требуется изоляция клонов, продуцирующих большие количества рекомбинантных белков. Введение гетерологичных генов в клетки-хозяева животных и скрининг на экспрессию добавленных генов является длительным и сложным процессом. Процесс включает трансфекцию и отбор клонов со стабильной длительной экспрессией, а также скрининг на клоны с высокими уровнями экспрессии соответствующего рекомбинантного белка.
При создании клонов, экспрессирующих рекомбинантный белок с экспрессионных векторов, клетки-хозяева обычно трансфицируют ДНК-вектором, кодирующим как представляющий интерес белок, так и селектируемый маркер в одном и том же векторе. Таким образом, такой экспрессионный вектор содержит селектируемый маркер, позволяющий отбирать клоны, в которых присутствует экспрессионный вектор. Такой селектируемый маркер может также приводить к коамплификации трансфицированной ДНК, что позволяет изолировать клоны-продуценты рекомбинантных белков на высоком уровне.
Большинство селектируемых маркеров представляют собой либо белок, придающий устойчивость к антибиотику или другому токсичному веществу, либо белок, необходимый для выживания клеток. В данной области техники известно несколько таких селектируемых маркеров, в том числе, например, G418, гигромицин, пуромицин, зеомицин, дигидрофолатредуктаза (DHFR), глутаминсинтетаза (GS) и гипоксантин-гуанинфосфорибозилтрансфераза (HPRT). В частности, GS широко используется в качестве селектируемого маркера в области промышленной продукции рекомбинантных белков в эукариотических клетках. Ген GS делает возможным синтез глутамина, необходимого для роста клеток, и ингибируется MSX (L-метионина сульфоксимином). В присутствии MSX выживают только клетки, экспрессирующие большее количество GS. После соответствующего скрининга можно отобрать клетки, продуцирующие экзогенные белки.
В предыдущей заявке WO2016/062837 авторы настоящего изобретения разработали экспрессионную систему, основанную на использовании дегидрооротатдегидрогеназы (DHODH) в качестве селектируемого маркера. DHODH представляет собой фермент, необходимый для синтеза пиримидина. Следовательно, соединения, которые ингибируют DHODH, подавляют синтез ДНК и, следовательно, пролиферацию клеток. Таким образом, этот маркер отбора включает экспрессионный вектор, кодирующий DHODH, используемый в сочетании с ингибитором DHODH, таким как лефлуномид и терифлуномид.
Однако большинство ингибиторов, используемых с указанными выше маркерами отбора, являются токсичными. В случае маркера отбора DHODH, терифлуномид является, например, мощным иммуносупрессором, и обработка им, особенно в больших масштабах, может быть проблематичной из соображений безопасности. В случае маркера отбора GS, MSX вызывает судороги в высоких дозах и, таким образом, может также вызвать проблемы при обработке им. В случае маркера отбора DHFR, метотрексат, как известно, проявляет гемопоэтическую и пищеварительную токсичность, что также вызывает проблемы при обработке им.
Соответственно, существует потребность в экспрессионных системах, в которых отбор клона, продуцирующего представляющий интерес белок, можно проводить без добавления сложного при обработке соединения.
Настоящее изобретение удовлетворяет эту потребность.
Краткое изложение сущности настоящего изобретения
Настоящее изобретение является результатом разработки авторами настоящего изобретения линии клеток, в случае которой клетки, продуцирующие представляющий интерес белок, могут быть отобраны в среде, лишенной уридина, благодаря частичной или полной инактивации гена DHODH в указанной линии клеток. Эту линию клеток, в которой ген DHODH частично или полностью инактивирован, выращивают, как правило, в среде, дополненной уридином, но при трансфекции экспрессионным вектором, содержащим нуклеотидную последовательность, кодирующую DHODH млекопитающего, в частности, кодирующую мутированную DHODH млекопитающего, и экспрессионную кассету для экспрессии представляющего интерес белка, культуральную среду, как правило, заменяют культуральной средой, лишенной уридина, тем самым отбирая клетки, продуцирующие представляющий интерес белок.
Такая экспрессионная система особенно выгодна, поскольку в результате избегания использования ингибиторов в качестве селекционного давления она увеличивает жизнеспособность продуцирующих клеток. Авторы настоящего изобретения, кроме того, продемонстрировали, что это снижение токсичности связано с высокой продуктивностью.
Таким образом, настоящее изобретение имеет отношение к линии клеток, содержащей эндогенный ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован.
В конкретном варианте осуществления указанная линия клеток представляет собой линию клеток яичника китайского хомячка (СНО).
В более конкретном варианте осуществления линия клеток получена путем
a) инактивации эндогенного гена DHODH в клетке, в частности, с помощью метода редактирования генов, такого как метод с использованием системы CRISPR-Cas9, и
b) культивирования клетки в культуральной среде, содержащей уридин, в условиях, подходящих для создания линии клеток, в которой эндогенный ген DHODH частично или полностью инактивирован.
В конкретном варианте осуществления все аллели эндогенного гена DHODH указанной линии клеток частично или полностью инактивированы.
В дальнейшем варианте осуществления указанная линия клеток содержит, кроме того, экспрессионный вектор, содержащий нуклеотидную последовательность, кодирующую экзогенную DHODH млекопитающего, и по крайней мере одну экспрессионную кассету для экспрессии рекомбинантного белка, причем указанная экзогенная DHODH включает последовательность, идентичную на по крайней мере 60% последовательности SEQ ID NO:2 или последовательности SEQ ID NO:4.
В его конкретном варианте осуществления указанная нуклеотидная последовательность включает последовательность SEQ ID NO: 1 или последовательность SEQ ID NO:3.
В его другом конкретном варианте осуществления указанный рекомбинантный белок представляет собой моноклональное антитело.
В его еще одном конкретном варианте осуществления указанный вектор содержит первую экспрессионную кассету, подходящую для клонирования легкой цепи антитела, и вторую экспрессионную кассету, подходящую для клонирования тяжелой цепи антитела.
Другим объектом настоящего изобретения является экспрессионная система, включающая:
(i) линию клеток, содержащую эндогенный ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован, как определено выше, и
(ii) экспрессионный вектор, содержащий нуклеотидную последовательность, кодирующую экзогенную DHODH млекопитающего, и по крайней мере одну экспрессионную кассету для экспрессии рекомбинантного белка, причем указанная экзогенная DHODH включает последовательность, идентичную на по крайней мере 60% последовательности SEQ ID NO:2 или последовательности SEQ ID NO:4.
В конкретном варианте осуществления указанная нуклеотидная последовательность включает последовательность SEQ ID NO:1 или последовательность SEQ ID NO:3.
В другом конкретном варианте осуществления указанный рекомбинантный белок представляет собой моноклональное антитело.
В еще одном конкретном варианте осуществления указанный вектор содержит первую экспрессионную кассету, подходящую для клонирования легкой цепи антитела, и вторую экспрессионную кассету, подходящую для клонирования тяжелой цепи антитела.
Настоящее изобретение, кроме того, имеет отношение к (i) линии клеток, определенной выше, или экспрессионной системе, определенной выше, и (ii) культуральной среды, лишенной уридина.
Другой объект настоящего изобретения относится к способу продуцирования рекомбинантного белка in vitro, включающему стадии:
А a1) предоставления линии клеток, определенной выше, содержащей, кроме того, экспрессионный вектор, содержащий нуклеотидную последовательность, кодирующую экзогенную DHODH млекопитающего, и по крайней мере одну экспрессионную кассету для экспрессии рекомбинантного белка, причем указанная экзогенная DHODH включает последовательность, идентичную на по крайней мере 60% последовательности SEQ ID NO:2 или последовательности SEQ ID NO:4;
или
а2) предоставления линии клеток, определенной выше, и
а2') введения экспрессионного вектора, определенного выше, в линию клеток, предоставленную на стадии а2);
или
a3) предоставления линии клеток, содержащей эндогенный ген DHODH,
a3') частичной или полной инактивации эндогенного гена DHODH в линии клеток, предоставленной на стадии a3), и
a3”) введения экспрессионного вектора, определенного выше, в линию клеток, содержащую частично или полностью инактивированный эндогенный ген DHODH, полученный на стадии а3');
B) культивирования указанной линии клеток в условиях, подходящих для продукции рекомбинантного белка; и
C) выделения и/или очистки указанного рекомбинантного белка.
В конкретном варианте осуществления стадию B) указанного способа проводят в культуральной среде, лишенной уридина.
В другом конкретном варианте осуществления указанный способ включает, кроме того, стадию D) составления указанного рекомбинантного белка в фармацевтическую композицию.
Настоящее изобретение, кроме того, имеет отношение к применению линии клеток, определенной выше, экспрессионной системы, определенной выше, или набора, определенного выше, для продуцирования рекомбинантного белка.
В конкретном варианте осуществления линию клеток, экспрессионную систему или набор используют в сочетании с культуральной средой, лишенной уридина.
Краткое описание чертежей
На фиг. 1 показана геномная структура гена DHODH человека, зарегистрированная под идентификатором гена: 100756632, доступным 21 декабря 2018 г. в Genbank NCBI.
На фиг. 2 показано совмещение последовательности №1 экзона 2 DHODH. PAM: последовательность примыкающего к протоспейсеру мотива (TGG).
На фиг. 3 показан скрининг различных клонов KO (с нокаутом по) DHODH на продукцию антител в присутствии различных концентраций терифлуномида в качестве селективного агента.
На фиг. 4 показано количество продуцируемого белка в мг/мл, используя различные варианты DHODH в качестве маркеров отбора.
На фиг. 5 показана продукция липазы в день 14, используя DHODH G202A человека или GS человека в качестве селектируемого маркера и DHODH KO или клетки CHO дикого типа.
На фиг. 6 показана продукция моноклональных антител, mAb-B, в день 14, используя DHODH G202A человека или GS человека в качестве селектируемого маркера и DHODH KO или клетки CHO дикого типа.
На фиг. 7 показана продукция биспецифических антител в день 14, используя DHODH G202A человека и/или GS человека в качестве селектируемого маркера и DHODH KO или клетки CHO дикого типа.
На фиг. 8 показана продукция триспецифических антител в день 14, используя DHODH G202A человека и GS человека в качестве селектируемых маркеров и DHODH KO или клетки CHO дикого типа.
Подробное описание настоящего изобретения
Дигидрооротатдегидрогеназа
Используемый здесь термин «дигидрооротатдегидрогеназа» или «DHODH» относится к полипептиду, способному катализировать превращение дигидрооротата (4,5-дигидрооротовой кислоты или 2,6-диоксо-1,3-диазинан-4-карбоновой кислоты) в оротат (оротовую кислоту или 1,2,3,6-тетрагидро-2,6-диоксо-4-пиримидинкарбоновую кислоту), как представлено следующей реакцией:
(S)-дигидрооротат+O2 ↔ оротат+H2O2
Такой полипептид классифицируется под номером 1.3.3.1 Комиссии по ферментам (ЕС). Полипептиды, способные катализировать вышеупомянутую выше реакцию, проявляют «активность DHODH».
Вышеупомянутая реакция является четвертой стадией синтеза de novo уридинмонофосфата (rUMP), необходимого для синтеза ДНК и РНК. Таким образом, ингибирование или инактивация DHODH имеет эффект ингибирования синтеза ДНК и РНК и, следовательно, ингибирует пролиферацию клеток.
Линия клеток
Настоящее изобретение имеет отношение к линии клеток, содержащей эндогенной ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован.
Линия клеток представляет собой линию эукариотических клеток, например линию клеток млекопитающих, такую как линия клеток яичника китайского хомячка (СНО), линия клеток обезьяны или линия клеток человека.
В конкретном варианте осуществления линия клеток представляет собой линию клеток CHO.
Линии клеток СНО обычно используются для промышленной продукции белков, и специалистам в данной области техники известно множество линий клеток СНО. Например, такие линии клеток CHO включают линии, которые общедоступны из Американской коллекции типовых культур, такие как линия клеток CHO-K1 (номер в АТСС: CCL-61), линия клеток CHO-S (продаваемая, например, Invitrogen и Gibco), линия клеток CHO DP-12 (с номерами в ATCC: CRL-12444 и 12445) и линия клеток CHO 1-15 (с номером в ATCC: CRL-9606). Другой линией клеток, подходящей для промышленной продукции белков, является линия клеток CHO 9E4. Линия клеток 9E4 была получена из клона линии клеток CHO-K1 посредством процесса клонирования потомства одной клетки. Получение линии клеток 9E4 более подробно представлено в примере 1. Линия клеток CHO-K1 была получена Puck в 1957 г. и депонирована в АТСС под номером CCL-61.
Клетки человека, такие как клетки HEK293 (с номером в ATCC: CRL-1573), HKB11 (с номером в ATCC: CRL-12568), PER-C6 (Crucell), HT1080 (с номером в ATCC: CRL-121), Jurkat, Daudi, Raji и CAP (с номером в ATCC: CRL-1098), также могут использоваться для продукции белков, чтобы получить встречающийся в природе профиль гликозилирования рекомбинантных белков человека.
В одном варианте осуществления линия клеток способна расти в бессывороточной среде (например, в среде определенного химического состава) и/или в суспензии. Такая линия клеток может быть легко получена специалистами в данной области техники путем адаптации родительской линии клеток к росту в бессывороточной среде и/или в суспензии (например, путем клонирования потомства одной клетки, путем постепенной адаптации и/или путем процесса «выдерживания в условиях голодания и спасения»).
Линия клеток по настоящему изобретению представляет собой линию клеток, содержащую эндогенной ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован.
Под «эндогенным геном DHODH» здесь подразумевается ген DHODH, обычно присутствующий в указанной конкретной клетке на конкретной стадии развития при определенных условиях окружающей среды.
«Эндогенный ген DHODH» отличается от «экзогенного гена DHODH», определенного ниже, тем, что указанный экзогенный ген DHODH предоставляется экспрессионным вектором, определенным ниже, который может присутствовать в линии клеток по настоящему изобретению, если указанный экспрессионный вектор был введен в указанную линию клеток.
Как будет понятно специалисту, эндогенный ген DHODH будет зависеть от линии клеток. Например, в линии клеток СНО эндогенный ген DHODH представляет собой ген DHODH китайского хомячка; в линии клеток человека эндогенный ген DHODH представляет собой ген DHODH человека.
Как правило, DHODH китайского хомячка дикого типа относится к последовательности, включающей или состоящей из SEQ ID NO:2, а также к ее вариантам, проявляющим активность DHODH. Такие варианты могут, например, соответствовать вариантам, которые встречаются в природе у видов хомяков (таким как аллельные варианты или варианты сплайсинга).
Как правило, DHODH человека дикого типа относится к последовательности, включающей или состоящей из SEQ ID NO:4, а также к ее вариантам, проявляющим активность DHODH. Такие варианты могут, например, соответствовать вариантам, которые встречаются в природе у вида человека (например, аллельным вариантам или вариантам сплайсинга).
Используемый здесь термин «ген» включает область ДНК, кодирующую продукт гена, а также все области ДНК, которые регулируют продукцию продукта гена, независимо от того, примыкают ли такие регуляторные последовательности к кодирующим и/или транскрибируемым последовательностям. Соответственно, ген включает промоторные последовательности, терминаторы, регулирующие трансляцию последовательности, такие как сайты связывания рибосом и внутренние участки посадки рибосом, энхансеры, сайленсеры, инсуляторы, граничные элементы, начала репликации, сайты прикрепления к матриксу и локус-контрольные области (регуляторные дальнодействующие области).
«Инактивация» гена относится к любому снижению экспрессии гена по сравнению с соответствующей клеткой дикого типа. Инактивация гена может быть полной (полной инактивацией или нокаутом) или частичной (например, гипоморфным геном (мутантным геном), уровень экспрессии с которого ниже нормального, или продуктом мутантного гена, который демонстрирует частичное снижение активности, на которую он влияет).
В конкретном варианте осуществления все аллели эндогенного гена DHODH частично или полностью инактивированы.
В конкретном варианте осуществления указанный эндогенный ген DHODH является полностью инактивированным.
В более конкретном варианте осуществления все аллели эндогенного гена DHODH являются полностью инактивированными.
В конкретном варианте осуществления эндогенный ген DHODH инактивирован методом с использованием системы CRISPR-Cas9, описанного в Aga et al. (2015) BMC Proceedings 9(suppl 9):P2.
Как хорошо известно специалисту, система CRISPR-Cas9 представляет собой прокариотическую систему адаптивного иммунного ответа, в которой используются некодирующие РНК, чтобы направлять нуклеазу Cas9 на вызов сайт-специфического расщепления ДНК. Репарация этого повреждения ДНК осуществляется с помощью клеточных механизмов репарации ДНК, либо через путь репарации ДНК посредством негомологичного соединения концов (NHEJ), либо через путь гомологически направленной репарации (HDR). Для создания нарушений генов одиночная направляющая РНК (gRNA), состоящая из последовательности crRNA, специфической для ДНК-мишени, и последовательности tracrRNA, которая взаимодействует с белком Cas9, связывается с рекомбинантной формой белка Cas9, обладающей ДНК-эндонуклеазной активностью. Полученный комплекс вызовет мишень-специфическое расщепление двухцепочечной ДНК. Репарация сайта расщепления будет осуществляться путем репарации ДНК посредством негомологичного соединения концов (NHEJ), подверженного ошибкам процесса, который может приводить к вставкам/делециям (INDEL), которые могут нарушать функцию гена.
В конкретном варианте осуществления по крайней мере один экзон гена DHODH намечен на инактивацию, в частности, с помощью метода редактирования генов, такого как метод с использованием системы CRIPR-Cas9. В более конкретном варианте осуществления часть гена DHODH, кодирующая N-концевую часть белка DHODH, намечена на инактивацию, в частности, с помощью метода редактирования гена, такого как метод с использованием системы CRISPR-Cas9. В еще одном варианте осуществления второй экзон гена DHODH намечен на инактивацию, в частности, с помощью метода редактирования генов, такого как метод с использованием системы CRISPR-Cas9.
В одном варианте осуществления 20-нуклеотидная последовательность с последовательностью CAAGGATGATGGCTGCATCC (SEQ ID NO:23) или с последовательностью GGATGCAGCCATCATCCTTG (SEQ ID NO:5) или любая последовательность, совместимая с нокаутом гена DHODH без уменьшения продолжительности существования CHO, используется в качестве соответствующего фрагмента ДНК для создания gRNA, которая нацелена на второй экзон гена DHODH. Эту gRNA, как правило, получают с использованием олигонуклеотидов с последовательностью CACCGCACCGGGATGCAGCCATCATCCTTG (SEQ ID NO:6) и AAAACCAAGGATGATGGCTGCATCC (SEQ ID NO:7) или с использованием олигонуклеотидов последовательности GGATGCAGCCATCATCCTTGGTTTT (SEQ ID NO:24) и CAAGGATGATGGCTGCATCCCGGTG (SEQ ID NO:25), обычно клонируют в уникальный рестрикционный сайт плазмиды, такой как сайт для BaeI плазмиды pCM3561 (превращенной в источник прибыли Invitrogen), так что клонированная последовательность ДНК находится под контролем промотора U6, и, как только указанная плазмида вводится в клетку, она транскрибируется в одну транскрипционную единицу, содержащую crRNA, слитую с tracrRNA, при этом часть crRNA является специфической для второго экзона гена DHODH, а часть tracrRNA распознается ферментом Cas9.
С целью идентификации линии клеток, инактивированной по гену DHODH, отдельные клетки, как правило, изолируют путем предельного разведения в планшетах с лунками, и после достижения соответствующего слияния, например 90% слияния, клетки разделяют на по крайней мере 2 условия, такие как одно в культуральной среде, дополненной уридином, а другое в культуральной среде, лишенной уридина. Представляющие интерес клоны, как правило, представляют собой клоны, чувствительные к недостатку уридина.
После изоляции эти представляющие интерес клетки можно культивировать в культуральной среде, содержащей пиримидиновое основание, в частности, в культуральной среде, содержащей уридин.
Под «пиримидиновым основанием» здесь подразумевается пиримидин как таковой и различные производные пиримидина, имеющие пиримидиновое ядро в качестве остова. Примеры таких пиримидиновых оснований включают вещества, имеющие отношение к урацилсодержащим компонентам нуклеиновых кислот, такие как урацил, уридин, уридинфосфаты, в частности уридинмонофосфат (UMP), уридиндифосфат (UDP) и уридинтрифосфат (UTP), дезоксиуридин, дезоксиуридинфосфаты, в частности, дезоксиуридинмонофосфат (dUMP), дезоксиуридиндифосфат (dUDP) и дезоксиуридинтрифосфат (dUTP); вещества, имеющие отношение к цитозинсодержащим компонентам нуклеиновых кислот, такие как цитозин, цитидин, цитидинфосфаты, в частности, цитидинмонофосфат (CMP), цитидиндифосфат (CDP), цитидинтрифосфат (CTP), дезоксицитидин, 2'-дезоксицитидин, дезоксицитидинфосфаты, в частности, дезоксицитидинмонофосфат (dCMP), дезоксицитидиндифосфат (dCDP) и дезоксицитидинтрифосфат (dCTP); тимин, тимидин, тимидинфосфаты, в частности тимидинмонофосфат (TMP), тимидиндифосфат (TDP) и тимидинтрифосфат (TTP), дезокситимидин, дезокситимидинфосфаты, в частности, дезокситимидинмонофосфат (dTMP), дезокситимидиндифосфат (dTTP) и дезокситимидинтрифосфат (dTTP) и оротат.
В конкретном варианте осуществления указанное пиримидиновое основание представляет собой уридин.
Под «уридином» здесь подразумевается нуклеозид следующей формулы:

Под «культуральной средой, лишенной уридина», подразумевается любая основная культуральная среда, подходящая для выращивания конкретной линии клеток, причем указанная среда содержит менее 1 мМ уридина, в частности указанная среда не содержит какой-либо уридин.
Под «культуральной средой, содержащей уридин» подразумевается любая основная культуральная среда, подходящая для выращивания конкретной линии клеток, причем указанная среда содержит, кроме того, от 1 до 25 мМ уридина, в частности от 5 до 10 мМ уридина.
Под «основной культуральной средой» здесь подразумевается среда без добавок, которая подходит для воздействия на клетки, например, клетки СНО. Как будет понятно специалисту, используемая основная культуральная среда будет зависеть от типа используемых клеток. Примеры основной культуральной среды включают среду CDCHO, среду OPTiCHOTM, среду Fecto CHOTM, среду FortiCHOTM, среду ExpiCHOTM, среду Ex-CellTM, среду ActiPROTM, среду MAM PF77TM и среду PowerCHOTM.
В конкретном варианте осуществления в основную культуральную среду дополнительно добавляют глутамин, обычно от 4 до 6 мМ глутамина.
Соответственно, в конкретном варианте осуществления линию клеток по настоящему изобретению создают путем
а) инактивации эндогенного гена DHODH в клетке, в частности, с помощью метода редактирования генов, такого как метод с использованием системы CRISPR-Cas9, и
b) культивирования клетки в культуральной среде, содержащей уридин, в условиях, подходящих для создания линии клеток, в которой эндогенный ген DHODH частично или полностью инактивирован.
Создание линии клеток СНО, содержащей эндогенный ген DHODH, который полностью или частично инактивирован с помощью подхода с использованием системы CRISPR-Cas9, подробнее проиллюстрировано в примерах 2 и 3.
Создание линии клеток, такой как линия клеток СНО, содержащей эндогенный ген DHODH, который полностью или частично инактивирован, можно осуществить с помощью ряда других методов молекулярной биологии, известных в данной области техники. Например, другие методы редактирования генов, применимые для создания линии клеток, содержащей эндогенный ген DHODH, который полностью или частично инактивирован, включают использование нуклеаз с цинковыми пальцами (ZFN) или эффекторных нуклеаз типа фактора транскрипции (TALEN). Метод с использованием системы ркомбинации Cre/Lox также может использоваться для нокаута одной или более или всех аллелей гена DHODH.
В конкретном варианте осуществления линия клеток по настоящему изобретению содержит, кроме того, экспрессионный вектор, определенный ниже в разделе «Экспрессионный вектор».
Указанный экспрессионный вектор может быть введен в линию клеток любым подходящим способом, хорошо известным специалисту, например путем трансфекции, в частности, путем электропорации или химической трансфекции, или трансдукции.
В конкретном варианте осуществления указанная линия клеток по настоящему изобретению может содержать, кроме того, дополнительный экспрессионный вектор, содержащий маркер отбора, отличный от экспрессионного вектора по настоящему изобретению, как правило, дополнительный экспрессионный вектор, содержащий последовательность, кодирующую глутаминсинтетазу.
Экзогенная DHODH
DHODH, кодируемая экспрессионным вектором, используемым в настоящем изобретении, (далее называемая «экзогенной DHODH»), может включать или состоять из последовательности, идентичной на по крайней мере 60%, 62%, 65%, 70%, 75%, 80%, 85%, 90%, 91%; 92%; 93%, 94%, 95%, 95,5%, 96%, 96,5%, 97%, 97,5%, 98%, 98,5%, 99%, 99,5% или 100% SEQ ID NO:2 или SEQ ID NO:4. Она также может включать или состоять из фрагмента из по крайней мере 100, 150, 200, 250, 300 или 350 следующих друг за другом аминокислот SEQ ID NO:2 или SEQ ID NO:4 при условии, что белок сохраняет активность DHODH.
В некоторых вариантах осуществления экзогенная DHODH в соответствии с настоящим изобретением включает или состоит из последовательности, идентичной на по крайней мере 60%, 62%, 65%, 70%, 75%, 80%, 85%, 90%, 91%; 92%; 93%, 94%, 95%, 95,5%, 96%, 96,5%, 97%, 97,5%, 98%, 98,5%, 99%, 99,5% или 100% как последовательности SEQ ID NO:2, так и последовательности SEQ ID NO:4.
В некоторых вариантах осуществления экзогенная DHODH в соответствии с настоящим изобретением представляет собой DHODH человека, т.е. DHODH человеческого происхождения.
Используемый здесь термин «DHODH человека» относится к белку с последовательностью, включающей или состоящей из SEQ ID NO:4, а также к его вариантам, проявляющим активность DHODH. Такие варианты могут, например, соответствовать вариантам, которые встречаются в природе у вида человека, (например, аллельным вариантам или вариантам сплайсинга). Альтернативно, такие варианты могут соответствовать вариантам, полученным с помощью генной инженерии. В одном варианте осуществления такие варианты отличаются от последовательности SEQ ID NO:4 только наличием самое большее 150, 140, 130, 120, 110, 100, 90, 80, 70, 60, 50, 40, 30, 25, 24, 23, 22, 21, 20, 19, 18,17,16,15,14,13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 изменения аминокислоты по сравнению с SEQ ID NO:4 (при этом указанные изменения включают замены, вставки и делеции).
В конкретном варианте осуществления указанная DHODH человека представляет собой вариант, содержащий мутацию G202A по сравнению с последовательностью дикого типа, обычно белок, включающий аминокислотную последовательность SEQ ID NO:26 или состоящий из нее.
В некоторых вариантах осуществления экзогенная DHODH представляет собой DHODH хомяка, т.е. DHODH хомячкового происхождения. DHODH хомяка может быть, например, DHODH китайского хомячка (Cetulus griseus).
Используемый здесь термин «DHODH китайского хомячка» относится к последовательности, включающей или состоящей из SEQ ID NO:2, а также к ее вариантам, проявляющим активность DHODH. Такие варианты могут, например, соответствовать вариантам, которые встречаются в природе у вида хомяков, (например, аллельным вариантам или вариантам сплайсинга). Альтернативно, такие варианты могут соответствовать вариантам, полученным с помощью генной инженерии. В одном варианте осуществления такие варианты отличаются от последовательности SEQ ID NO:2 только наличием самое большее 150, 140, 130, 120, 110, 100, 90, 80, 70, 60, 50, 40, 30, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 изменения аминокислоты по сравнению с SEQ ID NO:2 (при этом указанные изменения включают замены, вставки и делеции).
В другом варианте осуществления вариант DHODH будет обладать активностью DHODH, необязательно, таким же уровнем активности, что и белок дикого типа, или 50%, 60%, 70%, 80%, 90%, 100%, 110%, 120%. 130%, 140% или более от уровня активности белка дикого типа.
Под полипептидом, имеющим аминокислотную последовательность, идентичную на по крайней мере, например, на 95% запрашиваемой аминокислотной последовательности по настоящему изобретению, подразумевается, что аминокислотная последовательность рассматриваемого полипептида идентична запрашиваемой последовательности, за исключением того, что рассматриваемая полипептидная последовательность может включать до пяти изменений аминокислот на каждые 100 аминокислот запрашиваемой аминокислотной последовательности. Другими словами, для получения полипептида, имеющего аминокислотную последовательность, идентичную на по крайней мере 95% запрашиваемой аминокислотной последовательности, до 5% (5 из 100) аминокислотных остатков в рассматриваемой последовательности могут быть вставлены, делетированы или заменены другой аминокислотой.
Идентичность последовательности может быть определена по всей длине вариантной последовательности, по всей длине эталонной последовательности или по обоим параметрам. Например, процент идентичности может быть рассчитан с использованием глобального совмещения (т.е. две последовательности сравниваются по всей их длине). Способы сравнения идентичности и гомологии двух или более последовательностей хорошо известны в данной области техники. Программа «Needle», в которой используется алгоритм для глобального совмещения Needleman-Wunsch (Needleman and Wunsch (1970) J. Mol. Biol. 48:443-453), чтобы найти оптимальное совмещение (включая разрывы) двух последовательностей с учетом всей их длины, может, например, использоваться при выполнении глобального совмещения. Эта программа «Needle», например, доступна на всемирном веб-сайте ebi.ac.uk. Процент идентичности в соответствии с настоящим изобретением предпочтительно рассчитывается, используя программу EMBOSS::needle (global) с использованием параметра «Gap Open» (штрафа за открытие разрыва), равного 10,0, параметра «Gap Extend» (штрафа за удлинение разрыва), равного 0,5, и матрицы замен Blosum62.
Варианты эталонной последовательности могут содержать мутации, такие как делеции, вставки и/или замены, по сравнению с эталонной последовательностью. В случае замен замена предпочтительно соответствует консервативной замене, как указано в таблице ниже.
Экспрессионный вектор
Экспрессионный вектор используемый в контексте настоящего изобретения, подходит для продуцирования рекомбинантного белка и содержит последовательность, кодирующую дигидрооротатдегидрогеназу (DHODH).
Экспрессионный вектор предпочтительно представляет собой ДНК-вектор.
Экспрессионный вектор, используемый в контексте настоящего изобретения, содержит последовательность, кодирующую экзогенную DHODH, определенную выше в разделе «Экзогенная DHODH».
В конкретном варианте осуществления линия клеток, в которую должен быть введен экспрессионный вектор, представляет собой линию клеток CHO, и экзогенная DHODH имеет гетерологичное происхождение (т.е. экзогенная DHODH не является DHODH хомяка).
Последовательность, кодирующая такую экзогенную DHODH, может быть встречающейся в природе нуклеотидной последовательностью. Альтернативно, триплетные кодоны последовательности, кодирующей такую DHODH, могут быть смещены для экспрессии в клетках СНО. Программное обеспечение и алгоритмы для смещения кодонов в последовательности для получения оптимальной экспрессии известны в данной области техники и включают, например, алгоритм, описанный в Raab et al. (2010) Syst Synth Biol. 4:215-225. Этот алгоритм не только обеспечивает наилучшие доступные кодоны для экспрессии, но также учитывает содержание GC и отсутствие нежелательных мотивов ДНК.
Например, последовательность, кодирующая экзогенную DHODH, может включать или состоять из последовательности, идентичной на по крайней мере 60%, 62%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% последовательности SEQ ID NO:3 (т.е. последовательности, кодирующей DHODH человека с SEQ ID NO:4, которая была разработана для оптимальной экспрессии в клетках CHO) и/или последовательности SEQ ID NO:1 (т.е. последовательности, кодирующей DHODH хомяка с SEQ ID NO:2, которая была разработана для оптимальной экспрессии в клетках CHO).
В одном варианте осуществления последовательность, кодирующая экзогенную DHODH, включает или состоит из последовательности SEQ ID NO:1 или SEQ ID NO:3.
В экспрессионном векторе, используемом в контексте настоящего изобретения, последовательность, кодирующая экзогенную DHODH, определенную выше, может быть помещена под контроль любого промотора, известного специалистам в данной области техники.
Например, последовательность, кодирующая экзогенную DHODH, определенную выше, может, например, быть помещена под контроль промотора, подходящего для управления экспрессией DHODH, например промотора обезьяньего вакуолизирующего вируса 40 (SV40) (например, позднего или раннего промотора SV40), промотора CMV, промотора фактора 1 элонгации, промотора GAPDH, промотора RPL37, промотора актина. Ранний промотор SV40 описан, например, в Benoist and Chambon (1981) Nature 290:304-310 и в Moreau et al. (1981) Nucleic Acids Res. 9:6047-6068. В частности, указанный промотор SV40 является полноразмерным промотором. Указанный промотор SV40 может также иметь начало репликации, содержащее повтор из 72 пар оснований.
В некоторых вариантах осуществления указанный промотор SV40 не является промотором SV40, из которого были удалены положения с 128 по 270, т.е. указанный промотор SV40 не является промотором SV40, описанным в патенте Кореи с № 10-0267720, и трансформирующим трансформант E. coli, депонированный в Gene Bank, Institute of Bioengineering, KIST, 17 декабря 1997 г. под номером депонирования: KCTC 8860 P.
В других вариантах осуществления последовательность, кодирующая экзогенную DHODH, определенную выше, не находится под контролем промотора SV40.
Экспрессионные векторы, подходящие для продукции рекомбинантных белков, известны специалистам в данной области техники. Такие векторы, как правило, соответствуют экспрессионным векторам, которые содержат начало репликации и по крайней мере одну экспрессионную кассету, обеспечивающую клонирование и экспрессию рекомбинантного белка, продукция которого желательна. Экспрессионная кассета, как правило, содержит 5'-нетранслируемую область (включающую или состоящую из промотора и, необязательно, энхансерной последовательности), один или более сайтов рестрикции, позволяющих клонировать последовательность, кодирующую рекомбинантный белок, 3'-нетранслируемую область (например, сигнал (поли)А) и, необязательно, один или более интронов. Промоторная последовательность может соответствовать любому сильному промотору, хорошо известному в данной области техники, такому как, например, промотор CMV человека. Необязательно, экспрессионные векторы, используемые в контексте настоящего изобретения, содержат прокариотическое начало репликации (например, прокариотический репликон, такой как ColE1 в E. coli) и по крайней мере ген маркера отбора прокариот, также известный как прокариотический селектируемый маркер, так что эти векторы делают возможной репликацию в прокариотических клетках. Клетки, которые реплицируют векторы, также экспрессируют ген маркера отбора прокариот и, следовательно, могут быть идентифицированы и отобраны. Гены селективных в случае прокариот маркеров хорошо известны специалисту в данной области. Примерами генов маркеров отбора прокариот являются, например, последовательности нуклеиновой кислоты, кодирующие белок, придающий устойчивость к антибиотикам (например, последовательность, кодирующая белок, придающий устойчивость к ампициллину, хлорамфениколу, бластицидину или канамицину).
Рекомбинантный белок может соответствовать любому белку, который представляет интерес для специалистов в данной области техники.
Используемый здесь термин «белок», как подразумевается, охватывает пептиды (т.е. аминокислотные цепи из менее 50 аминокислот), полипептиды (т.е. аминокислотные цепи из не менее 50 аминокислот), мономерные белки (т.е. белки, состоящие из одной аминокислотной цепи) и мультимерные белки (т.е. белки, состоящие из двух или более аминокислотных цепей, такие как, например, моноклональные антитела).
Экспрессионный вектор, используемый в контексте настоящего изобретения, включает, как правило, ряд экспрессионных кассет, который идентичен ряду различных аминокислотных цепей, составляющих белок (например, одну экспрессионную кассету в случае мономерного белка или гомодимерного белка, две в случае гетеродимерного белка или моноклонального антитела и т.д.).
Альтернативно, экспрессионный вектор, используемый в контексте настоящего изобретения, может содержать только одну экспрессионную кассету, даже когда желательна продукция гетеродимерного белка или моноклонального антитела. В таком случае последовательность(и), кодирующая другую аминокислотную цепь(и) белка, присутствует(ют) в отдельном экспрессионном векторе, который котрансфицируется вместе с экспрессионным вектором в соответствии с настоящим изобретением в линию клеток-хозяев, в частности, в линию клеток CHO.
В этом случае дополнительные отдельные экспрессионные векторы могут содержать маркеры отбора, отличные от маркера отбора DHODH, описанного здесь, такие как DHFR, GS или HPRT.
В одном варианте осуществления экспрессионный вектор, используемый в контексте настоящего изобретения, может быть лишен экспрессионной кассеты. В таком случае экспрессионная кассета(ы), подходящая для экспрессии рекомбинантного белка, присутствует(ют) в отдельном векторе, который котрансфицируется вместе с экспрессионным вектором в соответствии с настоящим изобретением в линию клеток-хозяев, в частности, в линию клеток с инактивированным DHODH по настоящему изобретению, конкретнее, в линию клеток CHO с инактивированным DHODH по настоящему изобретению.
Таким образом, в некоторых вариантах осуществления экспрессионный вектор, используемый в контексте настоящего изобретения, содержит:
- последовательность, кодирующую экзогенную DHODH, определенную выше, помещенную под контроль раннего промотора SV40;
- первую экспрессионную кассету, в которой последовательность, кодирующая легкую цепь антитела, помещена под контролем промотора CMV;
- вторую экспрессионную кассету, в которой последовательность, кодирующая тяжелую цепь антитела, помещена под контролем промотора CMV;
- прокариотическое начало репликации; и
- селектируемый маркер для применения в прокариотических клетках, а именно последовательность, кодирующую белок, придающий устойчивость к ампициллину, помещенный под контроль своего природного промотора.
На протяжении настоящего описания термин «рекомбинантный белок» относится к любому рекомбинантному белку, продукция которого желательно. Он может, например, соответствовать терапевтическому и/или профилактическому белку, т.е. белку, предназначенному для применения в качестве лекарственного средства (в том числе вакцины). В конкретном варианте осуществления рекомбинантный белок, продукция которого желательна, не является DHODH. В другом конкретном варианте осуществления рекомбинантный белок, продукция которого желательно, представляет собой антитело, например моноклональное антитело. В еще одном конкретном варианте осуществления рекомбинантный белок, продукция которого желательна, представляет собой антигенный белок.
Термин «антитело» используется здесь в самом широком смысле и, в частности, охватывает моноклональные антитела (в том числе полноразмерные моноклональные антитела) любого изотипа, такого как IgG, IgM, IgA, IgD и IgE, поликлональные антитела, полиспецифические антитела (в том числе биспецифические и триспецифические антитела), фрагменты антител (такие как, например, Fv, scFv, dsFab, Fab' или F(ab)-фрагменты, однодоменные антитела и их фрагменты, а также слитые белки, содержащие фрагмент антитела. Антитело, реагирующее со специфическим антигеном, может создано с помощью рекомбинантных методов, таких как отбор из библиотек рекомбинантных антител в фаге или аналогичных векторах, или путем иммунизации животного антигеном или кодирующей антиген нуклеиновой кислотой.
«Моноклональное антитело», как здесь используется, представляет собой антитело, полученное из популяции по существу гомогенных антител, т.е. антитела, образующие эту популяцию, являются по существу идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Эти антитела направлены против одного эпитопа (или одной группы эпитопов в случае полиспецифических моноклональных антител) и поэтому обладают высокой специфичностью.
Типичное моноклональное антитело состоит из двух идентичных тяжелых цепей и двух идентичных легких цепей, соединенных дисульфидными связями. Каждая тяжелая и легкая цепь содержит константную область и вариабельную область. Каждая вариабельная область содержит три сегмента, называемых «определяющими комплементарность участками» («CDR») или «гипервариабельными участками», которые в основном отвечают за связывание с эпитопом антигена. Их обычно называют CDR1, CDR2 и CDR3, с нумерацией последовательно от N-конца (смотрите Kabat et al., Sequences of Proteins of Immunological Interest, 5th edition, National Institute of Health, Bethesda, MD, 1991). Более высококонсервативные части вариабельных областей называются «каркасными областями».
Моноклональное антитело может, например, представлять собой мышиное антитело, химерное антитело, гуманизированное антитело или полностью человеческое антитело.
Моноклональное антитело может быть моноспецифическим, биспецифическим или триспецифическим антителом.
Когда рекомбинантный белок, продукция которого желательна, представляет собой моноклональное антитело, экспрессионный вектор в соответствии с настоящим изобретением может содержать первую экспрессионную кассету, подходящую для клонирования легкой цепи антитела, и вторую экспрессионную кассету, подходящую для клонирования тяжелой цепи антитела.
В конкретном варианте осуществления каждая из указанных первой и второй экспрессионных кассет содержит промотор цитомегаловируса (CMV), например промотор CMV из CMV человека или мыши. Конкретнее, указанные первая и вторая экспрессионные кассеты могут содержать:
- немедленный ранний промотор-энхансера CMV (например, тот, который имеет последовательность, описанную в Teschendorf et al. (2002) Anticancer Res. 22:3325-3330); или
- область промотора/энхансера IE2 из CMV мыши (например, имеющую последовательность, описанную в Chatellard et al. (2007) Biotechnol Bioeng. 96:106-117); или
- регуляторный элемент hCMV-MIE (например, имеющий последовательность, описанную в WO 89/01036).
Термин «антигенный белок» используется здесь в самом широком смысле и охватывает любой белок, способный вызывать иммунный ответ, либо отдельно, либо в сочетании с адъювантом. Он может быть предназначен для применения либо в профилактической вакцине, либо в терапевтической вакцине. В конкретном варианте осуществления антигенный белок представляет собой вакцинный белок, т.е. белок, предназначенный для применения в профилактической вакцине.
Экспрессионный вектор либо может содержать по крайней мере одну последовательность, кодирующую представляющий интерес рекомбинантный белок (например, одну последовательность, кодирующую мономерный белок, одну последовательность, кодирующую цепь антитела, или две последовательности, кодирующие легкую цепь антитела и тяжелую цепь антитела, соответственно), либо он может быть пустым (т.е. лишенным такой последовательности, кодирующей представляющий интерес рекомбинантный белок).
Экспрессионная система, наборы, способы и применения
Настоящим изобретением предоставляется экспрессионная система, включающая:
(i) линию клеток, определенную в разделе «Линия клеток» выше, содержащую эндогенный ген DHODH, который частично или полностью инактивирован, как определено в разделе «Линия клеток» выше, и
(ii) экспрессионный вектор, определенный в разделе «Экспрессионный вектор» выше.
Экспрессионная система по настоящему изобретению может, кроме того, включать дополнительные отдельные экспрессионные векторы, каждый из которых содержит нуклеотидную последовательность, кодирующую маркер отбора, отличный от DHODH, такой как DHFR, GS или HPRT, и по крайней мере одну экспрессионную кассету для экспрессии рекомбинантного белка.
Альтернативно, экспрессионная система по настоящему изобретению может, кроме того, включать дополнительные экспрессионные векторы, определенные в разделе «Экспрессионный вектор» выше.
Настоящим изобретением предоставляется набор, включающий (i) линию клеток в соответствии с настоящим изобретением, содержащую экспрессионный вектор, определенный в разделе «Экспрессионный вектор» выше, или экспрессионную систему в соответствии с настоящим изобретением и (ii) культуральную среду, лишенную уридина, определенную выше.
Набор может включать экспрессионный вектор, кодирующий экзогенную DHODH, (в экспрессионной системе), как описано выше. В таком наборе вектор предпочтительно является пустым, поскольку это позволяет клонировать белок, представляющий интерес для специалистов в данной области техники. Кроме того, экспрессионный вектор предпочтительно выделяют из линии клеток в таком наборе.
Набор включает, кроме того, культуральную среду, лишенную уридина, как определено в разделе «Линия клеток» выше.
Набор может, кроме того, включать среду, подходящую для культивирования линии клеток, среду, подходящую для трансфекции вектора в линию клеток, упаковочный материал и/или инструкции по применению экспрессионной системы.
В конкретном варианте осуществления набор не содержит ингибитор DHODH.
Примеры ингибиторов DHODH включают бицинхониновую кислоту, бреквинар (6-фтор-2-(2'-фтор-1,1’-бифенил-4-ил)-3-метил-4-хинолинкарбоновую кислоту), производные нафтохинона, такие как дихлоралли-лаусон, производные изоксазола, такие как лефлуномид (5-метил-N-[4-(трифторметил)фенил]изоксазол-4-карбоксамид) и его активный метаболит терифлуномид ((2Z)-2-циано-3-гидрокси-N-[4- (трифторметил)фенил]бут-2-енамид), хинолонкарбоновые кислоты, нафтохиноны, изоксазолы, феноксихинолины, редоксаль и производные, лаусон, лапахол, атовахон и (8-хлор-4-(2-хлор-4-фторфенокси)хинолин). Ингибитор DHODH может подавлять активность DHODH на по крайней мере 20, 30, 40, 50, 60, 70, 80, 90, 95, 99 или 100%.
В конкретном варианте осуществления набор не содержит терифлуномид.
В настоящем изобретении, кроме того, предусматривается применение линии клеток в соответствии с настоящим изобретением, содержащей экспрессионный вектор, определенный в разделе «Экспрессионный вектор» выше, экспрессионной системы в соответствии с настоящим изобретением или набора в соответствии с настоящим изобретением для продукции рекомбинантного белка in vitro.
В конкретном варианте осуществления указанная линия клеток, экспрессионная система или набор применяются в сочетании с культуральной средой, лишенной уридина, как определено выше, конкретнее, в отсутствие ингибитора DHODH.
Кроме того, в настоящем изобретении также предусматривается применение экспрессионной системы в соответствии с настоящим изобретением, линии клеток в соответствии с настоящим изобретением, содержащей экспрессионный вектор, определенный в разделе «Экспрессионный вектор» выше, или набора в соответствии с настоящим изобретением для изоляции клеточного клона, который продуцирует рекомбинантный белок на высоком уровне, («продуцирующих на высоком уровне клонов») in vitro, в частности, в отсутствии ингибитора DHODH.
В контексте настоящего изобретения, термин «высокий уровень рекомбинантного белка», как подразумевается, означает, что в культуральной среде концентрация рекомбинантного белка составляет по крайней мере 0,05 г/л, предпочтительно по крайней мере 0,1 г/л, еще предпочтительнее по крайней мере 0,2 г/л, более предпочтительно от 0,3 до 1 г/л. Концентрация рекомбинантного белка может быть определена с помощью способов, которые хорошо известны специалисту в данной области техники, включающих, в частности, иммуноферментный анализ (ELISA), Вестерн-блоттинг, калипер-метод и диапазон концентраций очищенного белка, соответствующий рекомбинантному белку.
Настоящим изобретением, кроме того, предоставляется способ продукции рекомбинантного белка in vitro, включающий стадии:
А a1) предоставления линии клеток в соответствии с настоящим изобретением, содержащей экспрессионный вектор, определенный в разделе «Экспрессионный вектор» выше;
или
a2) предоставления линии клеток в соответствии с настоящим изобретением и
a2') введения экспрессионного вектора, определенного в разделе «Экспрессионный вектор» выше, в линию клеток, предоставленную на стадии a2);
или
a3) предоставления линии клеток, содержащей эндогенный ген DHODH,
a3') частичной или полной инактивации эндогенного гена DHODH в линии клеток, предоставленной на стадии a3), и
a3”) введения экспрессионного вектора, определенного в разделе «Экспрессионный вектор» выше, в линию клеток, содержащую частично или полностью инактивированный эндогенный ген DHODH, полученную на стадии а3');
В) культивирования указанной линии клеток в условиях, подходящих для продукции рекомбинантного белка; и
С) выделения и/или очистки указанного рекомбинантного белка.
В конкретном варианте осуществления стадия B) вышеуказанного способа проводится в культуральной среде, лишенной уридина, конкретнее также лишенной ингибитора DHODH, и, в частности, включает подстадию, заключающуюся в отборе трансфицированных клеток, которые растут, несмотря на отсутствие уридина, в частности, дополнительно в отсутствие ингибитора DHODH.
Настоящим изобретение, кроме того, предоставляется in vitro способ изоляции клеточного клона, который продуцирует рекомбинантный белок на высоком уровне, при этом указанный способ включает или состоит из следующих стадий:
А a1) предоставления линии клеток в соответствии с настоящим изобретением, содержащей экспрессионный вектор, определенный в разделе «Экспрессионный вектор» выше;
или
a2) предоставления линии клеток в соответствии с настоящим изобретением и
a2') введения экспрессионного вектора, определенного в разделе «Экспрессионный вектор» выше, в линию клеток, предоставленную на стадии a2);
или
a3) предоставления линии клеток, содержащей эндогенный ген DHODH,
a3') частичной или полной инактивация эндогенного гена DHODH в линии клеток, предоставленной на стадии a3), и
a3”) введения экспрессионного вектора, определенного в разделе «Экспрессионный вектор» выше, в линию клеток, содержащую частично или полностью инактивированный эндогенный ген DHODH, полученную на стадии а3');
В) культивирования указанной линии клеток в условиях, подходящих для продукции рекомбинантного белка; и
С) изоляции клона, который продуцирует рекомбинантный белок на высоком уровне.
В конкретном варианте осуществления стадия B) вышеуказанного способа проводится в культуральной среде, лишенной уридина, конкретнее также лишенной ингибитора DHODH, и, в частности, включает подстадию, заключающуюся в отборе трансфицированных клеток, которые растут, несмотря на отсутствие уридина, в частности, дополнительно в отсутствие ингибитора DHODH.
Указанный экспрессионный вектор может быть введен в указанную линию клеток на стадиях а2') или а3") любым способом, хорошо известным специалисту, например, путем трансфекции, в частности, путем электропорации, или химической трансфекции, или трансдукции.
Условия, подходящие для продуцирования рекомбинантных белков, хорошо известны специалистам в данной области техники. Например, можно использовать протоколы, описанные в разделе «Примеры».
В конкретном варианте осуществления культуральная среда, используемая на стадии B), содержит уменьшающиеся концентрации уридина. Это позволяет отбирать клоны, в которых произошла амплификация экзогенного гена DHODH векторного происхождения (и, таким образом, последовательности, кодирующей рекомбинантный белок).
Вышеупомянутые способы могут включать, кроме того, стадию составления рекомбинантного белка в фармацевтическую композицию.
По всему описанию такие термины, как «содержит (включает)», «содержащийся (включенный)» и «содержащий (включающий)», имеют значение, приписываемое им в большинстве патентных юрисдикций, предпочтительно в рассматриваемой юрисдикции; например, они могут означать «включает», «включенный», «включающий» и т.д. Такие термины, как «состоящий из», «по существу состоящий из» и «по существу состоит из», имеют значение, приписываемое им в большинстве патентных юрисдикций, предпочтительно в рассматриваемой юрисдикции; например, они подразумевают исключение всех, большинства или всех, за исключением незначительного количества других элементов, они допускают элементы, не перечисленные в явной форме, но исключают элементы, которые встречаются в предшествующем уровне техники, или которые влияют на основные или новые характеристики настоящего изобретения.
В тексте данного описания приводится несколько документов. Каждый из приведенных здесь документов (включая любую журнальную статью или реферат, опубликованную или неопубликованную заявку на патент, выданный патент, спецификации производителя, инструкции и т.д.) включен таким образом посредством ссылки. Однако нельзя допускать, что какой-либо документ, приведенный здесь, действительно является предшествующим уровнем техники относительно настоящего изобретения.
Далее настоящее изобретение будет описано со ссылкой на следующие чертежи и примеры, которые являются только иллюстративными и не предназначены для ограничения настоящего изобретения.
Настоящее изобретение определяется формулой изобретения, которую следует интерпретировать с помощью описания и чертежей.
Краткое описание последовательностей
Примеры
Пример 1: Получение линии клеток CHO 9E4
В этом примере описывается получение линии клеток CHO 9E4 из линии клеток CHO-K1, коммерчески доступной из ATCC под номером в ATCC: CCL-61.
1. Линия клеток CHO-K1
Флакон с клетками CHO-K1 (ATCC: CCL-61), замороженными в присутствии телячьей сыворотки в 1969 году, был получен из ATCC.
2. Оттаивание флакона в среде Ex-Cell™ 302 и приготовление банка CHO-LG-APF.
CHO-K1 во флаконе оттаивали непосредственно в среде Ex-Cell™ 302 (SAFC), дополненной 4 мМ глутамином, и размножали на неподвижной опоре, затем во вращающем устройстве. Полученный банк CHO-LG-APF замораживали в среде Ex-Cell™ 302 через 12 пассажей и 17,3 поколений.
3. Оттаивание банка CHO-LG-APF в среде Ex-Cell™ 302 и приготовление банка ABC-024 P22.
CHO-LG-APF во флаконе оттаивали в среде Ex-Cell™ 302 и размножали. Полученный банк ABC-024 P22 оттаивали через 18,5 поколений.
4. Адаптация банка CMV07-024 к среде CDCHO Fusion и приготовление банка ABC-003.
Банк CMV07-024 оттаивали и непосредственно адаптировали к среде Ex-cell™ CDCHO Fusion (SAFC), дополненной 4 мМ глутамином, и адаптировали на шейкере на протяжении более 12,5 поколений до замораживания банка ABC-003 в среде Ex-cell™ CDCHO Fusion.
5. Оттаивание банка ABC-003 в среде CDCHO Fusion и приготовление банка ABC-053 в среде CDCHO Fusion
ABC-003 во флаконе оттаивали в среде CDCHO Fusion, и после разведения банк ABC-53 замораживали через 4,2 поколения.
6. Оттаивание банка ABC-053 в среде CDCHO Fusion, отбор, клонирование и приготовление банка P15A11 в среде CDCHO Fusion
Банк ABC-053 оттаивали в среде Ex-cell™ CDCHO Fusion (SAFC), дополненной 4 мМ глутамином. После размножения культуры ее клонировали путем предельного разведения в планшетах, а затем размножали в среде CDCHO Fusion. Банк клона P15A11, полученный в результате этого клонирования, замораживали. Это клонирование и размножение соответствует приблизительно 94 поколениям.
7. Оттаивание банка P15A11 в среде CDCHO Fusion, адаптация банка путем прямого пассажа в CDCHO и приготовление банка CHOSP10-002 в среде CDCHO.
Банк P15A11 оттаивали в среде Ex-cell™ CDCHO Fusion (SAFC), дополненной 4 мМ глутамином, и после 2 пассажей в среде CDCHO Fusion клетки разводили в среде CDCHO. После 3 пассажей в среде CDCHO банк CHOSP10-002 замораживали через всего 15,9 поколений.
8. Оттаивание банка CHOSP10-002 в среде CDCHO, размножение, удаление масс центрифугированием и отбор путем субкультивирования без масс в 96-луночных планшетах, размножение в 6-луночных планшетах и на шейкере для приготовления банка CHOSP10-012 в среде CDCHO.
Банк CHOSP10-002 оттаивали в среде CDCHO (Invitrogen), дополненной 6 мМ глутамином, затем размножали. Культуру центрифугировали для удаления клеточных масс и продолжения культивирования только с использованием клеток, выделенных из супернатанта. На этой стадии, от оттаивания, прошло 11,3 поколения.
Эту культуру разделяли по 96-луночным планшетам по 10 клеток на лунку. Лунки с клетками, которые изолированно размножаются в суспензии, размножали в 6-луночных планшетах, затем на шейкере. До замораживания банка CHOSP10-012 прошло 23,2 дополнительных поколения.
9. Оттаивание банка CHOSP10-012, размножение и приготовление банка CHOSP11-008 (банка 9E4)
Банк CHOSP10-012 оттаивали в среде CDCHO (Invitrogen), дополненной 6 мМ глутамином, затем размножали, начиная со стадии в колбе Эрленмейера до 17-литрового биореактор.
Банк 9E4 замораживали через в общей сложности 10 поколений.
Пример 2: Получение линии клеток СНО, в которой ген DHODH является инактивированным.
A - Разработка и конструирование направляющей РНК (gRNA) для системы CRISPR-CAS9.
Для аннулирования гена DHDOH в клетках СНО авторы настоящего изобретения начали с получения последовательности гена DHODH хомячка и использования общедоступного программного обеспечения Tefor для разработки различных направляющих РНК (gRNA) для трансфекции вместе с CRISPR-Cas9 в геном CHO. Полноразмерная последовательность DHODH CHO, с интронами и экзонами, показана на фиг. 1.
Программа определила 8 последовательностей, которые могут быть нацелены на ген DHODH:
Последовательность 1
CACCGGGATGCAGCCATCATCCTTG (SEQ ID NO:6)
AAAACCAAGGATGATGGCTGCATCC (SEQ ID NO:7)
Последовательность 2
CACCGGATGCAGCCATCATCCTTGG (SEQ ID NO:8)
AAAACCCAAGGATGATGGCTGCATC (SEQ ID NO:9)
Последовательность 3
CACCGGCAGCCATCATCCTTGGGGG (SEQ ID NO:10)
AAAACCCCCCAAGGATGATGGCTGC (SEQ ID NO:11)
Последовательность 4
CACCGGCCATCATCCTTGGGGGAGG (SEQ ID NO:12)
AAAACCCTCCCCCAAGGATGATGGC (SEQ ID NO:13)
Последовательность 5
CACCGGCTATTCGCTTCACGTCCCT (SEQ ID NO:14)
AAAACAGGGACGTGAAGCGAATAGC (SEQ ID NO:15)
Последовательность 6
CACCGGCCTCTACAAACTGGGCTTT (SEQ ID NO:16)
AAAACAAAGCCCAGTTTGTAGAGGC (SEQ ID NO:17)
Последовательность 7
CACCGGGCTTTGGGTTTGTCGAGGT (SEQ ID NO:18)
AAAACACCTCGACAAACCCAAAGCC (SEQ ID NO:19)
Последовательность 8
CACCGGCTGGTCTGAGGAGCCTACA (SEQ ID NO:20)
AAAACTGTAGGCTCCTCAGACCAGC (SEQ ID NO:21)
Хотя 8 последовательностей были протестированы и клонированы, из 8 последовательностей были трансфицированы только четыре клонированных последовательности, и только одна оказалась успешной для вызова нокаута гена DHODH. Следующая последовательность из 20 нуклеотидов GGATGCAGCCATCATCCTTG (SEQ ID NO:5) была использована в качестве соответствующего фрагмента ДНК для создания gRNA, как показано на фиг. 2. Она нацелена на второй экзон гена DHODH. Для достижения транскрипции надлежащей gRNA, два олигонуклеотида CACCGGGATGCAGCCATCATCCTTG (олигонуклеотид 1, SEQ ID NO:6) и AAAACCAAGGATGATGGCTGCATCC (олигонуклеотид 2, SEQ ID NO:7) были синтезированы, подвергнуты отжигу и клонированы в уникальный сайт для BaeI pCM3561 (превращенной в источник прибыли Invitrogen).
Таким образом, клонированная последовательность ДНК находилась под контролем промотора U6, и после трансфекции ДНК в клетки СНО она транскрибировалась в единую транскрипционную единицу, содержащую crRNA, слитую с tracrRNA. Часть crRNA была специфической для второго экзона гена DHODH, в то время как tracrRNA распознавалась самим ферментом Cas9.
B-Подготовка материала для редактирования гена с помощью системы CRISPR-Cas9
Клетки CHO 9E4 были изолированы и отобраны из клеток CHO K-1, приобретенных в ATCC, как описано в примере 1, и выращены и сохранены в виде суспензионных культур в бессывороточной среде CDCHO с определенным химическим составом, оптимизированной для культивирования клеток яичника китайского хомячка (CHO), с добавлением 6 мМ L-глутамина при 37°C в термостате с 8% CO2 и 80% влажностью.
10 мкг вектора для экспрессии sgRNA (pCM3561) расщепляли 1 мкл фермента BaeI при добавлении 5 единиц/мкл с 20 мкМ S-аденозилметионином (SAM) при 25°C в течение 1 часа, затем расщепленную плазмиду разделяли с помощью электрофореза, используя 1% агарозный гель. Полученный вектор для клонирования sgRNA затем извлекали с помощью набора для экстракции из геля (Qiagen Kit).
Вектор для клонирования sgRNA и отожженные направляющие олигонуклеотиды лигировали с использованием фермента ДНК-лигазы Т4 (Biolabs) и инкубировали в течение 10 мин при комнатной температуре.
5 мкл продуктов лигирования добавляли к 50 мкл компетентных клеток E. coli DH5a (Invitrogen).
Клетки и ДНК инкубировали 30 мин на льду, а затем подвергали тепловому шоку при 42°C в течение 45 с. После добавления 500 мл среды SOC, 1-часовая инкубация при 37°C (при 800 об./мин) давала бактериям время для выработки белков устойчивости к антибиотикам, кодируемых в остове плазмиды. После инкубации каждую пробирку распределяли по одной чашке со слоем LB с добавлением 100 пг/мл ампициллина. Чашки инкубировали в течение ночи при 37°C. Отрицательные контроли (с водой вместо вставки ДНК) использовали для оценки успеха трансформации.
Для стадии амплификации были выбраны две колонии для каждой конструкции и засеяны в 2 мл среды LB, дополненной 100 мкг/мл ампициллина, в пробирке, помещенной в термостат на ночь (при 37°C, 700 об./мин). Культуру, подвергнутую инкубации в течение ночи, собирали путем центрифугирования. Набор QIAprep Miniprep Kit™ (QIAGEN) использовали для выделения амплифицированной ДНК (элюирования в буфер EB). Затем последовательность представляющих интерес направляющих олигонуклеотидов проверяли с помощью секвенирования по Сэнгеру (смыслового и антисмыслового секвенирования, GATC Company). После проверки путем совмещения в программном обеспечении Vector NTI (Thermofisher Scientific) соответствующие колонии использовали для засева 200 мл среды LB, дополненной 100 мкг/мл ампициллина. После инкубации в течение 24 часов бактерии собирали путем центрифугирования при 6000 х g в течение 15 минут при 4°C. Набор EndoFree Plasmid Maxi Kit™ (QIAGEN) использовали для приготовления MaxiPrep. ДНК осаждали путем добавления изопропанола при комнатной температуре. После центрифугирования в течение 1 часа (при 4°C, 8000 об./мин) осадок ДНК промывали 70% этанолом, не содержащим эндотоксинов, при комнатной температуре. После короткого нового центрифугирования осадок сушили на воздухе в течение 1 ч и повторно растворяли в подходящем объеме стерильной воды, не содержащей эндотоксинов, для получения концентрации ДНК, составляющей 5 мг/мл. Устройство NanoDrop использовали для измерения концентрации ДНК.
Были получены четыре различных плазмиды, а именно плазмида pBH6840 (KO DHODH SEQ1), плазмида pBH6841 (KO DHODH SEQ4), плазмида pBH6842 (KO DHODH SEQ5) и плазмида pBH6843 (KO DHODH SEQ7). Мишень этих плазмид в гене DHODH CHO показана в последовательности SEQ ID NO:22.
Секвенирование ДНК было выполнено субподрядчиком GATC - Eurofins Genomics Company.
С-Редактирование гена с помощью системы CRISPR-Cas9
Трансфекции были выполнены путем электропорации с использованием MaxCyte STX и его протокола, определенного для CHO. Они были выполнены в процесс-компоновках OC-100 (20 миллионов клеток на трансфекцию).
За день до трансфекции клетки засевали в количестве 1,5х106 клеток/мл в среду CDCHO, дополненную 6 мМ L-глутамином.
В день трансфекции клетки подсчитывали с помощью прибора ViCell (Beckman & Coulter). Необходимое количество клеток центрифугировали при 250 х g в течение 10 мин, и супернатант отбрасывали.
Для каждого условия трансфекции 20х106 клеток центрифугировали 10 мин при 250 х g. Осадок ресуспендировали в 70 мкл буфера Maxcyte. Добавляли 30 мкг ДНК, и смесь (клетки, буфер и ДНК) переносили в кассету для электропорации Maxcyte емкостью 100 мкл. Используемая процесс-компоновка представляла собой процесс-компоновку OC-100, приспособленную для 100-мкл кассеты, и была выбрана оптимизированная для CHO программа.
Были осуществлены следующие трансфекции.
После электропорации клетки были перенесены в колбы Эрленмейера с рабочим объемом=25 мл. Их помещали в термостат статического типа с температурой 37°C, 5% CO2 на 45 мин. Затем добавляли 25 мл среды CDCHO, дополненной 6 мМ L-глутамином, для ресуспендирования клеток, и колбы Эрленмейера помещали на шейкеры на 110 об./мин при 37°C, 5% CO2, 70% влажности.
На следующий день после электропорации по одной клетке в каждую лунку засевали путем предельного разведения из пулов, трансфицированных CH09E4, описанных выше. Приблизительно через 20 дней, когда клетки стали на приблизительно 90% сливающимися и стали выглядеть здоровыми при исследовании под микроскопом, клетки были разделены на 2 новых 96-луночных планшета с уридином или без него.
Несколько клонов было отобрано по их чувствительности к недостатку уридина. Эти клоны были адаптированы для культивирования в среде CDCHO, дополненной 6 мМ глутамином и 5 мМ уридином.
С целью подтверждения того, что редактирование гена было успешным, геномную ДНК экстрагировали из клеток клона CRISPR-Cas9 с помощью набора Qiagen DNeasy™ (Qiagen). Локус-мишень амплифицировали с помощью ПЦР, используя соответствующие праймеры для области локуса DHODH, на которую нацелена система CRISPR-Cas9, и ПЦР-продукты секвенировали с помощью NGS, используя ПЦР-фрагменты, покрывающие потенциально делетированные области.
Пример 3. Альтернативное получение линии клеток СНО, в которой ген DHODH является аннулированным.
A - Разработка и конструирование направляющей РНК (gRNA) для системы CRISPR-CAS9.
Для аннулирования гена DHDOH в клетках СНО авторы настоящего изобретения начали с получения последовательности гена DHODH хомячка и использования общедоступного программного обеспечения Tefor для разработки различных направляющих РНК (gRNA) для трансфекции вместе с CRISPR-Cas9 в геном CHO.
Программа определила 8 последовательностей, которые могут быть нацелены на ген DHODH:
Последовательность 1’
GGATGCAGCCATCATCCTTGGTTTT (SEQ ID NO:24)
CAAGGATGATGGCTGCATCCCGGTG (SEQ ID NO:25)
Последовательность 2’
GATGCAGCCATCATCCTTGGGTTTT (SEQ ID NO:27)
CCAAGGATGATGGCTGCATCCGGTG (SEQ ID NO:28)
Последовательность 3’
GCAGCCATCATCCTTGGGGGGTTTT (SEQ ID NO:29)
CCCCCAAGGATGATGGCTGCCGGTG (SEQ ID NO:30)
Последовательность 4’
GCCATCATCCTTGGGGGAGGGTTTT (SEQ ID NO:31)
CCTCCCCCAAGGATGATGGCCGGTG (SEQ ID NO:32)
Последовательность 5’
GCTATTCGCTTCACGTCCCTGTTTT (SEQ ID NO:33)
AGGGACGTGAAGCGAATAGCCGGTG (SEQ ID NO:34)
Последовательность 6’
GCCTCTACAAACTGGGCTTTGTTTT (SEQ ID NO:35)
AAAGCCCAGTTTGTAGAGGCCGGTG (SEQ ID NO:36)
Последовательность 7’
GGCTTTGGGTTTGTCGAGGTGTTTT (SEQ ID NO:37)
ACCTCGACAAACCCAAAGCCCGGTG (SEQ ID NO:38)
Последовательность 8’
GCTGGTCTGAGGAGCCTACAGTTTT (SEQ ID NO:39)
TGTAGGCTCCTCAGACCAGCCGGTG (SEQ ID NO:40)
Хотя 8 последовательностей были протестированы и клонированы, из 8 последовательностей были трансфицированы только четыре клонированных последовательности, и только одна оказалась успешной для вызова нокаута гена DHODH. Следующая последовательность из 20 нуклеотидов GGATGCAGCCATCATCCTTG (SEQ ID NO: 5) была использована в качестве соответствующего фрагмента ДНК для создания gRNA. Она нацелена на второй экзон гена DHODH. Для достижения транскрипции надлежащей gRNA, два олигонуклеотида GGATGCAGCCATCATCCTTGGTTTT (олигонуклеотид 1’, SEQ ID NO:24) и CAAGGATGATGGCTGCATCCCGGTG (олигонуклеотид 2’, SEQ ID NO:25) были синтезированы, подвергнуты отжигу и клонированы в уникальный сайт для BaeI плазмиды pCM3561 (превращенной в источник прибыли Invitrogen).
Таким образом, клонированная последовательность ДНК находилась под контролем промотора U6, и после трансфекции ДНК в клетки СНО она транскрибировалась в единую транскрипционную единицу, содержащую crRNA, слитую с tracrRNA. Часть crRNA была специфической для второго экзона гена DHODH, в то время как tracrRNA распознавалась самим ферментом Cas9.
B-Подготовка материала для редактирования гена с помощью системы CRISPR-Cas9
Клетки CHO 9E4 были изолированы и отобраны из клеток CHO K-1, приобретенных в ATCC, как описано в примере 1, и выращены и сохранены в виде суспензионных культур в бессывороточной среде CDCHO с определенным химическим составом, оптимизированной для культивирования клеток яичника китайского хомячка (CHO), с добавлением 6 мМ L-глутамина при 37°C в термостате с 8% CO2 и 80% влажностью.
10 мкг вектора для экспрессии sgRNA (pCM3561) расщепляли 1 мкл фермента BaeI при добавлении 5 единиц/мкл с 20 мкМ S-аденозилметионином (SAM) при 25°C в течение 1 часа, затем расщепленную плазмиду разделяли с помощью электрофореза, используя 1% агарозный гель. Полученный вектор для клонирования sgRNA затем извлекали с помощью набора для экстракции из геля (Qiagen Kit).
Вектор для клонирования sgRNA и отожженные направляющие олигонуклеотиды лигировали с использованием фермента ДНК-лигазы Т4 (Biolabs) и инкубировали в течение 10 мин при комнатной температуре.
5 мкл продуктов лигирования добавляли к 50 мкл компетентных клеток E. coli DH5a (Invitrogen).
Клетки и ДНК инкубировали 30 мин на льду, а затем подвергали тепловому шоку при 42°C в течение 45 с. После добавления 500 мл среды SOC, 1-часовая инкубация при 37°C (при 800 об./мин) давала бактериям время для выработки белков устойчивости к антибиотикам, кодируемых в остове плазмиды. После инкубации каждую пробирку распределяли по одной чашке со слоем LB с добавлением 100 пг/мл ампициллина. Чашки инкубировали в течение ночи при 37°C. Отрицательные контроли (с водой вместо вставки ДНК) использовали для оценки успеха трансформации.
Для стадии амплификации были выбраны две колонии для каждой конструкции и засеяны в 2 мл среды LB, дополненной 100 мкг/мл ампициллина, в пробирке, помещенной в термостат на ночь (при 37°C, 700 об./мин). Культуру, подвергнутую инкубации в течение ночи, собирали путем центрифугирования. Набор QIAprep Miniprep Kit™ (QIAGEN) использовали для выделения амплифицированной ДНК (элюирования в буфер EB). Затем последовательность представляющих интерес направляющих олигонуклеотидов проверяли с помощью секвенирования по Сэнгеру (смыслового и антисмыслового секвенирования, GATC Company). После проверки путем совмещения в программном обеспечении Vector NTI (Thermofisher Scientific) соответствующие колонии использовали для засева 200 мл среды LB, дополненной 100 мкг/мл ампициллина. После инкубации в течение 24 часов бактерии собирали путем центрифугирования при 6000 х g в течение 15 минут при 4°C. Набор EndoFree Plasmid Maxi Kit™ (QIAGEN) использовали для приготовления MaxiPrep. ДНК осаждали путем добавления изопропанола при комнатной температуре. После центрифугирования в течение 1 часа (при 4°C, 8000 об./мин) осадок ДНК промывали 70% этанолом, не содержащим эндотоксинов, при комнатной температуре. После короткого нового центрифугирования осадок сушили на воздухе в течение 1 ч и повторно растворяли в подходящем объеме стерильной воды, не содержащей эндотоксинов, для получения концентрации ДНК, составляющей 5 мг/мл. Устройство NanoDrop использовали для измерения концентрации ДНК.
Были получены четыре различных плазмиды, а именно плазмида pBH6840 (KO DHODH SEQ1), плазмида pBH6841 (KO DHODH SEQ4), плазмида pBH6842 (KO DHODH SEQ5) и плазмида pBH6843 (KO DHODH SEQ7).
Секвенирование ДНК было выполнено субподрядчиком GATC - Eurofins Genomics Company.
С-Редактирование гена с помощью системы CRISPR-Cas9
Трансфекции были выполнены путем электропорации с использованием MaxCyte STX и его протокола, определенного для CHO. Они были выполнены в процесс-компоновках OC-100 (20 миллионов клеток на трансфекцию).
За день до трансфекции клетки засевали в количестве 1,5х106 клеток/мл в среду CDCHO, дополненную 6 мМ L-глутамином.
В день трансфекции клетки подсчитывали с помощью прибора ViCell (Beckman & Coulter). Необходимое количество клеток центрифугировали при 250 х g в течение 10 мин, и супернатант отбрасывали.
Для каждого условия трансфекции 20х106 клеток центрифугировали 10 мин при 250 х g. Осадок ресуспендировали в 70 мкл буфера Maxcyte. Добавляли 30 мкг ДНК, и смесь (клетки, буфер и ДНК) переносили в кассету для электропорации Maxcyte емкостью 100 мкл. Используемая процесс-компоновка представляла собой процесс-компоновку OC-100, приспособленную для 100-мкл кассеты, и была выбрана оптимизированная для CHO программа.
Были осуществлены следующие трансфекции.
После электропорации клетки были перенесены в колбы Эрленмейера с рабочим объемом=25 мл. Их помещали в термостат статического типа с температурой 37°C, 5% CO2 на 45 мин. Затем добавляли 25 мл среды CDCHO, дополненной 6 мМ L-глутамином, для ресуспендирования клеток, и колбы Эрленмейера помещали на шейкеры на 110 об./мин при 37°C, 5% CO2, 70% влажности.
На следующий день после электропорации по одной клетке в каждую лунку засевали путем предельного разведения из пулов, трансфицированных CH09E4, описанных выше. Приблизительно через 20 дней, когда клетки стали на приблизительно 90% сливающимися и стали выглядеть здоровыми при исследовании под микроскопом, клетки были разделены на 2 новых 96-луночных планшета с уридином или без него.
Несколько клонов было отобрано по их чувствительности к недостатку уридина. Эти клоны были адаптированы для культивирования в среде CDCHO, дополненной 6 мМ глутамином и 5 мМ уридином.
С целью подтверждения того, что редактирование гена было успешным, геномную ДНК экстрагировали из клеток клона CRISPR-Cas9 с помощью набора Qiagen DNeasy™ (Qiagen). Локус-мишень амплифицировали с помощью ПЦР, используя соответствующие праймеры для области локуса DHODH, на которую нацелена система CRISPR-Cas9, и ПЦР-продукты секвенировали с помощью NGS, используя ПЦР-фрагменты, покрывающие потенциально делетированные области.
Пример 4: Использование дефицитной по DHODH линии клеток CHO для продуцирования рекомбинантных белков
Продукцию антител тестировали на подтвержденных, дефицитных по DHODH клонах CHO, полученных в примере 2 или 3, для проверки того, могут ли эти клоны экспрессировать антитела без терифлуномида.
Разработанные векторы были получены и приготовлены в концентрации, составляющей 5 мг/мл. Все они содержат ITR, позволяющие использовать систему транспозонов для интеграции плазмид в геном продуцирующих клеток, кроме pBH6209, которая является плазмидой, кодирующей транспозазу.
Используемыми линиями клеток были CHO 9E4_SP11 дикого типа и KО2 и KО19, нокаутные по DHODH.
CHO 9E4_SP11 культивировали в среде CDCHO с добавлением 6 мМ L-глутамина.
KО2 и KО19 культивировали в среде CDCHO с добавлением 6 мМ L-глутамина и 5 мМ уридина.
Вначале их культивировали в колбах Эрленмейера с рабочим объемом=25 мл и размножали до тех пор, пока не было достигнуто необходимое количество жизнеспособных клеток.
Различные белки были продуцированы с использованием высокоэффективного протокола электропорации, разработанного Maxcyte, на аппарате Maxcyte STX.
Клетки делили на порции в 1,5х106 за день до трансфекции.
В день трансфекции клетки котрансфицировали двумя векторами: плазмидным экспрессионным ДНК-вектором, содержащим кассеты для экспрессии тяжелой цепи (НС) и легкой цепи (LC) антитела против CD38 человека, и маркер отбора DHODH (как описано в WO2016/062837), которые были фланкированы сайтами распознавания PiggyBac (инвертированными концевыми повторами, ITR), и кодирующим транспозазу вектором от Transposagen, который катализирует мобилизацию транспозонов в геном СНО в сайты TTAA.
Для каждого условия трансфекции 80х106 клеток центрифугировали 10 мин при 250 х g. Осадок ресуспендировали в 250 мкл буфера Maxcyte. Добавляли 120 мкг ДНК, и смесь (клетки, буфер и ДНК) переносили в кассету для электропорации Maxcyte емкостью 400 мкл. Используемая процесс-компоновка представляла собой процесс-компоновку OC-400, приспособленную для 400-мкл кассеты, и была выбрана оптимизированная для CHO программа.
Для фазы восстановления трансфицированные клетки немедленно переносили в колбу на 125 мл при 37°C, 40 мин без перемешивания. Добавляли 25 мл предварительно нагретой среды CDCHO, дополненной 6 мМ глутамином (+5 мМ уридина для клеток KO), и трансфицированные культуры поддерживали при 37°C в термостате с 8% CO2 и 80% влажностью. Через 1 день после трансфекции клетки центрифугировали и ресуспендировали в селективной среде CD OPTiCHO™ (Gibco), дополненной 6 мМ глутамином, 30% FeedB (Gibco) и различными количествами терифлуномида (0, 5, 15 и 25 мкМ), до 1х106 клеток/мл.
В день 14 после трансфекции клетки центрифугировали при 200 х g в течение 10 мин при 25°C. Супернатант фильтровали через 0,22-мкм фильтр PES, и титр антител измеряли с помощью прибора Octet.
Как показано на фиг. 3, два клона (KО2 и KО19) продемонстрировали хорошую продукцию без терифлуномида, и геномное NGS показало, что эти два клона содержали вызывающую нокаут мутацию в двух аллелях локуса DHODH.
Примечательно, что все клоны КО продуцировали антитела даже в отсутствие терифлуномида в качестве селективного агента. Два клона KО2 и KО19 были выбраны для дальнейших исследований. Более того, эти клоны, на которые ссылаются, были единственными клонами, демонстрирующими гомозиготный нокаут гена DHODH.
Таким образом, этот пример показывает, что настоящим изобретением предоставляется ряд линий клеток и векторов, позволяющих продуцировать антитела без использования какого-либо селективного давления.
Пример 5: Оценка трех вариантов DHODH человека.
Для проверки того, увеличивает ли использование нарушенных форм DHODH человека число копий интеграции в геном СНО и тем самым обеспечивает большую продуктивность, были трансфицированы линии клеток DHODH KО2, KО19 и WT CHO 9E4 тремя вариантами кДНК для DHODH человека, описанными у пациентов с синдромом Миллера (R135C, G202A и R346W, смотрите, в частности, Fang et al. (2012) Biosci. 32:631-639). В качестве контрольного вектора трансфицировали плазмиду, содержащую кДНК, кодирующую DHODH человека дикого типа.
50 нг плазмидного вектора, кодирующего моноклональное антитело против CD38 человека, mAb-A (pBH6204), расщепленного рестрикционными ферментами SalI-BglII, смешивали с 37,5 нг очищенных SalI-BglII-фрагментов ДНК, соответствующих каждому варианту (R135C, G202A и R346W). После добавления 1 мкл Т4-ДНК-лигазы (BioLabs) и концентрированного буфера для лигирования реакцию лигирования (в конечном объеме 10 мкл) проводили в течение 10 минут при комнатной температуре.
Затем аликвоты этого пула ДНК использовали для трансформации компетентных клеток E. coli (Stellar™, Takara).
Приготовление плазмид в небольшом масштабе осуществляли с помощью коммерчески доступного набора для выполнения процедуры Min-Prep (Qiagen) в соответствии с рекомендациями производителя.
Секвенирование ДНК с использованием 603 смыслового (с последовательностью GTTGGCCTTCCAATGGCTT, SEQ ID NO:41) и 503 антисмыслового (с последовательностью GTTCCTTCACAAAGAT, SEQ ID NO:42) олигонуклеотидов было выполнено субподрядчиком GATC - Eurofins Genomics Company.
Трансфекцию вариантов DHODH проводили путем электропорации с использованием MaxCyte STX. Они были выполнены в процесс-компоновках OC-100, как описано выше.
Через один день после трансфекции клетки центрифугировали и ресуспендировали в селективной среде CD CHO, дополненной 6 мМ глутамином и 25 мкМ терифлуномидом для клеток 9E4 CHO, и без терифлуномида для клонов K02 и K019 в концентрации 1х106 клеток/мл.
После двух пассажей начинали продукцию mAb-A в среде CD OPTiCHO, дополненной 30% FeedB и 6 мМ глутамином и 25 мкМ терифлуномидом для клеток 9E4 CHO, и без терифлуномида для клонов K02 и K019 в концентрации 0,3х106 клеток/мл.
В день 14 после трансфекции клетки центрифугировали при 200 х g, 10 мин при 25°C. Супернатант фильтровали через 0,22-мкм фильтр PES, и титр антител измеряли с помощью прибора Octet.
Как показано на фиг. 4, не наблюдалось значительного эффекта мутаций R138C и R346W в DHODH на WT CHO 9E4 и клоны DHODH KO. С другой стороны, мутация G202A позволила получить продукцию антител в граммовом масштабе с использованием линии WT CHO 9E4 и повысить продуктивность клонов DHODH КО2 и KО19.
Пример 6: Продукция белка с использованием экспрессионной системы по настоящему изобретению.
Различные типы белков были продуцированы, используя экспрессионную систему по настоящему изобретению, в частности, используя кДНК для DHODH человека, в том числе описанную выше мутацию G202A, в подтвержденных клонах CHO 9E4 KО2 и KО19, описанных в примере 3 выше, чтобы подтвердить, что конечная продуктивность находится, по крайней мере, в том же диапазоне, что и в случае экспрессионной системы предшествующего уровня техники, в которой используется глутаминсинтетаза (GS) в качестве селектируемого маркера.
Были продуцированы следующие белки:
- Липаза:
используя моноцистронную кДНК PLBL2-His, и
используя плазмиду, кодирующую hDHODH G202A, или плазмиду, кодирующую hGS, в качестве селектируемых маркеров.
- Моноклональное антитело (mAb-B):
используя бицистронную кДНК, кодирующую VH- и VL-цепи антитела, и
используя плазмиду, кодирующую hDHODH G202A, или плазмиду, кодирующую hGS, в качестве селектируемого маркера.
- Биспецифическое антитело:
используя (i) бицистронную кДНК, кодирующую VH- и VL-цепи антитела, или (ii) две моноцистронные кДНК, кодирующие, соответственно, VH- и VL-цепи антитела, и
используя (i) плазмиду, кодирующую hDHODH G202A, (в случае бицистронной кДНК) или (ii) две плазмиды, кодирующих hDHODH G202A, или плазмиду, кодирующую hDHODH G202A, и плазмиду, кодирующую hGS, (в случае моноцистронных кДНК) в качестве селектируемых маркеров.
- Триспецифические антитела:
используя две бицистронные кДНК и
используя плазмиду, кодирующую hDHODH G202A, или плазмиду, кодирующую hGS, в качестве селектируемых маркеров.
За день до трансфекции FectoPRO® клетки разводили до 1,5х106 клеток/мл в среде CD CHO, дополненной 6 мМ глутамином и 5 мМ уридином.
В день трансфекции клеточную суспензию разводили до 1,1х106 клеток/мл в среде CD CHO, дополненной 6 мМ L-глутамином и 5 мМ уридином. Реагент FectoPRO® встряхивали в течение 5 с и прекращали вращение перед добавлением 25 мкл/пробирку в пустую 50-мл пробирку. Во второй 50-мл пробирке 12,5 мкг кДНК разводили в среде CD CHO, и разведенную ДНК сразу выливали в чистый реагент FectoPRO®. Раствор немедленно гомогенизировали и инкубировали в течение 10 мин. Смесь для трансфекции FectoPRO®/ДНК выливали на клетки, и культуру инкубировали при 37°C, 190 об./мин и уровнях CO2, составляющих 8%.
Через 24 часа после трансфекции клетки подсчитывали с помощью прибора Invitrogen™ Countess™. Цельноклеточную культуру центрифугировали при 200 х g, 10 мин. Осадок ресуспендировали в 2,5 мл предварительно нагретой среды для продуцирования в условиях, описанных ниже.
В случае сложных белков, содержащих 3 или более субъединиц, выполняют вторую трансфекцию, снова используя протокол трансфекции, описанный выше.
- Для липазы моноцистронный вектор (белок с His-меткой)
MSX: L-метионина сульфоксимин; Gln: глутамин; Uri: уридин; TNF: терифлуномид
- Для моноклонального антитела mAb-B бицистронный, кодирующий VH и VL вектор:
MSX: L-метионина сульфоксимин; Gln: глутамин; Uri: уридин; TNF: терифлуномид; FeedB: товарная подпитка.
- Для биспецифических антител бицистронные или два моноцистронные, кодирующие VH и VL вектора:
MSX: L-метионина сульфоксимин; Gln: глутамин; Uri: уридин; TNF: терифлуномид; FeedB: товарная подпитка.
- Для триспецифических антител, бицистронные, кодирующие VH и VL векторы
MSX: L-метионина сульфоксимин; Gln: глутамин; Uri: уридин; TNF: терифлуномид; FeedB: товарная подпитка.
Через 72 часа после трансфекции клетки подсчитывали с помощью прибора Invitrogen™ Countess™, и продукцию белка увеличивали с помощью еще одной замены среды. Кроме того, измеряли жизнеспособность трансфицированных клеток через 3 и 7 дней после трансфекции.
В день 14 после трансфекции клетки центрифугировали при 200 х g, 10 мин и 25°C. Супернатант фильтровали через 0,22-мкм фильтр PES, и титр белка измеряли с помощью прибора Octet.
Были получены следующие результаты.
a) Липаза
Продукция липазы в день 14 показана на фиг. 5.
Эти результаты показывают, что обе линии клеток KO DHODH обладают способностью продуцировать липазу в отсутствие селективного давления терифлуномида, что позволяет снизить токсичность по отношению к продуцирующим клеткам.
Кроме того, линии клеток KO DHODH были способны продуцировать липазу в том же диапазоне, что и в случае системы GS предшествующего уровня техники (в клетках СНО 9E4 дикого типа), т.е. 1 г/л в 25-мл масштабе.
b) Моноклональное антитело mAb-B
DHODH_G202A KO19
DHODH_G202A 9E4
Продукция моноклонального антитела mAb-B в день 14 показана на фиг. 6.
Эти результаты показывают, что линии клеток KO DHODH вели себя по-разному во время продукции этого конкретного антитела. Действительно, хотя оба клона и показали большую продуктивность, чем продуктивность в присутствии терифлуномида предшествующего уровня техники, клон KО19 показал значительно большую продуктивность, чем клон KО2.
В лучшей линии клеток KO (K019) продукция антител находилась в том же диапазоне, что и в случае системы GS предшествующего уровня техники (в клетках СНО 9E4 дикого типа), приблизительно 0,67 г/л в 25-мл масштабе.
Кроме того, увеличенная жизнеспособность наблюдалась при использовании линий клеток KO.
c) Биспецифическое антитело
Продукция биспецифического антитела в день 14 показана на фиг. 7.
Биспецифические антитела продуцируются не так эффективно, как моноспецифические антитела, во всех протестированных случаях. Однако, несмотря на трудности продуцирования биспецифических антител этого типа, продуктивность лучшей линии клеток KO была в том же диапазоне, что и в случае системы GS предшествующего уровня техники (в клетках 9E4 CHO дикого типа), приблизительно 0,145 г/л в 25-мл масштабе.
Кроме того, увеличенная жизнеспособность наблюдалась при использовании линий клеток KO.
d) Триспецифическое антитело
Продукция триспецифического антитела в день 14 показана на фиг. 8.
Во всех условиях две линии клеток КО показали результаты в том же диапазоне, что и система отбора предшествующего уровня техники, т.е. 0,5 г/л.
Таким образом, этот пример подтверждает, что клоны KO DHODH CHO подходят для экспрессии различных форм белков (белков, моноклональных антител, биспецифических антител, триспецифических антител). Эти клоны могут даже использоваться для двойной трансфекции с целью продукции сложных белков.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SANOFI
<120> Линия клеток, содержащая новый маркер отбора, и ее применение
для продуцирования белков
<130> BET 18P4548
<150> EP19305331.1
<151> 2019-03-19
<160> 45
<170> Патент в версии 3.5
<210> 1
<211> 1188
<212> ДНК
<213> Cricetus cricetus
<400> 1
atggcttggc ggcagatgcg gaagagagcc ctggacgccg ctatcatcct gggaggcgga 60
ggcctgctgt tcacctctta cctgacagcc accggcgacg accacttcta cgccgagtac 120
ctgatgcctg ccctgcagag actgctggac cctgagtctg cccaccggct ggccatcaga 180
ttcacctccc tgggcctgct gcccagagcc accttccagg actccgacat gctggaagtg 240
cgggtgctgg gccacaagtt cagaaacccc gtgggaatcg ccgctggctt cgacaagcac 300
ggcgaggctg tggacggcct gtacaagctg ggcttcggct tcgtggaagt gggctccgtg 360
acaccccagc cccaggaagg caaccccaga cctagagtgt tccggctgcc tgaggaccag 420
gccgtgatca acagatacgg cttcaactcc cacggcctgt ccgtggtgga acaccggctg 480
agagccagac agcagaagca gaacaagctg accgccgacg gcctgcccct gggcatcaat 540
ctgggcaaga acaagacctc cgaggacgct gccgccgact acgtggaagg cgtgcgagtg 600
ctgggacctc tggccgatta cctggtcgtg aacgtgtcct cccccaacac cgctggcctg 660
agaagcctgc agggaaaggc cgagctgaga aggctgctgg ccaaggtgct gcaggaacgg 720
gacgctctga agggcgccca gaaacctgcc gtgctcgtga agatcgcccc tgacctgacc 780
gcccaggaca aagaggatat cgcctccgtg gccagagagc tgggcatcga cggactgatc 840
gtgaccaaca ccaccgtgtc tcggcctacc ggactgcagg gcgctctgag atctgagatg 900
ggcggcctgt ctggcaagcc tctgagggac ctgtccaccc agaccatcag agagatgtac 960
accctgaccc agggccggat ccccatcatt ggagtgggcg gagtgtcctc tggccaggac
1020
gccctggaaa agatccaggc tggcgcctct ctggtgcagc tgtataccgc cctgaccttt
1080
ctgggccctc ccgtggtggt gcgagtgaag agagaactgg aagccctgct gaaagagcgg
1140
ggcttcaaca ccgtgaccga ggccatcggc gctgaccaca gaagatga 1188
<210> 2
<211> 395
<212> Белок
<213> Cricetus cricetus
<400> 2
Met Ala Trp Arg Gln Met Arg Lys Arg Ala Leu Asp Ala Ala Ile Ile
1 5 10 15
Leu Gly Gly Gly Gly Leu Leu Phe Thr Ser Tyr Leu Thr Ala Thr Gly
20 25 30
Asp Asp His Phe Tyr Ala Glu Tyr Leu Met Pro Ala Leu Gln Arg Leu
35 40 45
Leu Asp Pro Glu Ser Ala His Arg Leu Ala Ile Arg Phe Thr Ser Leu
50 55 60
Gly Leu Leu Pro Arg Ala Thr Phe Gln Asp Ser Asp Met Leu Glu Val
65 70 75 80
Arg Val Leu Gly His Lys Phe Arg Asn Pro Val Gly Ile Ala Ala Gly
85 90 95
Phe Asp Lys His Gly Glu Ala Val Asp Gly Leu Tyr Lys Leu Gly Phe
100 105 110
Gly Phe Val Glu Val Gly Ser Val Thr Pro Gln Pro Gln Glu Gly Asn
115 120 125
Pro Arg Pro Arg Val Phe Arg Leu Pro Glu Asp Gln Ala Val Ile Asn
130 135 140
Arg Tyr Gly Phe Asn Ser His Gly Leu Ser Val Val Glu His Arg Leu
145 150 155 160
Arg Ala Arg Gln Gln Lys Gln Asn Lys Leu Thr Ala Asp Gly Leu Pro
165 170 175
Leu Gly Ile Asn Leu Gly Lys Asn Lys Thr Ser Glu Asp Ala Ala Ala
180 185 190
Asp Tyr Val Glu Gly Val Arg Val Leu Gly Pro Leu Ala Asp Tyr Leu
195 200 205
Val Val Asn Val Ser Ser Pro Asn Thr Ala Gly Leu Arg Ser Leu Gln
210 215 220
Gly Lys Ala Glu Leu Arg Arg Leu Leu Ala Lys Val Leu Gln Glu Arg
225 230 235 240
Asp Ala Leu Lys Gly Ala Gln Lys Pro Ala Val Leu Val Lys Ile Ala
245 250 255
Pro Asp Leu Thr Ala Gln Asp Lys Glu Asp Ile Ala Ser Val Ala Arg
260 265 270
Glu Leu Gly Ile Asp Gly Leu Ile Val Thr Asn Thr Thr Val Ser Arg
275 280 285
Pro Thr Gly Leu Gln Gly Ala Leu Arg Ser Glu Met Gly Gly Leu Ser
290 295 300
Gly Lys Pro Leu Arg Asp Leu Ser Thr Gln Thr Ile Arg Glu Met Tyr
305 310 315 320
Thr Leu Thr Gln Gly Arg Ile Pro Ile Ile Gly Val Gly Gly Val Ser
325 330 335
Ser Gly Gln Asp Ala Leu Glu Lys Ile Gln Ala Gly Ala Ser Leu Val
340 345 350
Gln Leu Tyr Thr Ala Leu Thr Phe Leu Gly Pro Pro Val Val Val Arg
355 360 365
Val Lys Arg Glu Leu Glu Ala Leu Leu Lys Glu Arg Gly Phe Asn Thr
370 375 380
Val Thr Glu Ala Ile Gly Ala Asp His Arg Arg
385 390 395
<210> 3
<211> 1188
<212> ДНК
<213> Homo sapiens
<400> 3
atggcttggc ggcacctgaa gaagagggcc caggacgccg tgatcatcct gggaggcgga 60
ggcctgctgt tcgcctctta cctgatggct accggcgacg agcggttcta cgccgagcat 120
ctgatgccca cactgcaggg cctgctggac cctgagtctg cccatagact ggccgtgcgg 180
ttcacctccc tgggactgct gcctagagcc cggttccagg actccgacat gctggaagtg 240
cgggtgctgg gccacaagtt cagaaacccc gtgggaatcg ccgctggctt cgacaagcac 300
ggcgaggctg tggacggcct gtacaagatg ggcttcggct tcgtggaaat cggctccgtg 360
acccccaagc cccaggaagg caaccccaga cctcgggtgt tcagactgcc tgaggaccag 420
gctgtgatca acagatacgg cttcaactcc cacggcctgt ccgtggtgga acaccggctg 480
agagccagac agcagaagca ggccaagctg accgaggatg gcctgcctct gggagtgaac 540
ctgggcaaga acaagacctc cgtggacgcc gccgaggatt acgctgaagg cgtgcgagtg 600
ctgggacccc tggctgatta cctggtcgtg aacgtgtcct cccccaacac cgctggcctg 660
agatctctgc agggcaaggc cgagctgcgg agactgctga caaaggtgct gcaggaacgc 720
gacggcctgc ggagagtgca tagacctgcc gtgctcgtga agatcgcccc cgacctgacc 780
agccaggaca aagaggatat cgcctccgtc gtgaaagagc tgggcatcga cggactgatc 840
gtgaccaaca ccaccgtgtc taggcctgcc ggactgcagg gggctctgag atctgaaacc 900
ggcggactgt ccggcaagcc tctgagggat ctgtccaccc agaccatcag agagatgtac 960
gccctgaccc agggccgggt gccaatcatt ggagtgggcg gagtgtcctc cggccaggat
1020
gccctggaaa agatcagagc cggcgcttcc ctggtgcagc tgtacaccgc tctgaccttt
1080
tggggccctc ccgtcgtggg caaagtgaag agagagctgg aagccctgct gaaagagcag
1140
ggatttggcg gcgtgaccga tgccatcggc gctgaccaca gaagatga 1188
<210> 4
<211> 395
<212> Белок
<213> Homo sapiens
<400> 4
Met Ala Trp Arg His Leu Lys Lys Arg Ala Gln Asp Ala Val Ile Ile
1 5 10 15
Leu Gly Gly Gly Gly Leu Leu Phe Ala Ser Tyr Leu Met Ala Thr Gly
20 25 30
Asp Glu Arg Phe Tyr Ala Glu His Leu Met Pro Thr Leu Gln Gly Leu
35 40 45
Leu Asp Pro Glu Ser Ala His Arg Leu Ala Val Arg Phe Thr Ser Leu
50 55 60
Gly Leu Leu Pro Arg Ala Arg Phe Gln Asp Ser Asp Met Leu Glu Val
65 70 75 80
Arg Val Leu Gly His Lys Phe Arg Asn Pro Val Gly Ile Ala Ala Gly
85 90 95
Phe Asp Lys His Gly Glu Ala Val Asp Gly Leu Tyr Lys Met Gly Phe
100 105 110
Gly Phe Val Glu Ile Gly Ser Val Thr Pro Lys Pro Gln Glu Gly Asn
115 120 125
Pro Arg Pro Arg Val Phe Arg Leu Pro Glu Asp Gln Ala Val Ile Asn
130 135 140
Arg Tyr Gly Phe Asn Ser His Gly Leu Ser Val Val Glu His Arg Leu
145 150 155 160
Arg Ala Arg Gln Gln Lys Gln Ala Lys Leu Thr Glu Asp Gly Leu Pro
165 170 175
Leu Gly Val Asn Leu Gly Lys Asn Lys Thr Ser Val Asp Ala Ala Glu
180 185 190
Asp Tyr Ala Glu Gly Val Arg Val Leu Gly Pro Leu Ala Asp Tyr Leu
195 200 205
Val Val Asn Val Ser Ser Pro Asn Thr Ala Gly Leu Arg Ser Leu Gln
210 215 220
Gly Lys Ala Glu Leu Arg Arg Leu Leu Thr Lys Val Leu Gln Glu Arg
225 230 235 240
Asp Gly Leu Arg Arg Val His Arg Pro Ala Val Leu Val Lys Ile Ala
245 250 255
Pro Asp Leu Thr Ser Gln Asp Lys Glu Asp Ile Ala Ser Val Val Lys
260 265 270
Glu Leu Gly Ile Asp Gly Leu Ile Val Thr Asn Thr Thr Val Ser Arg
275 280 285
Pro Ala Gly Leu Gln Gly Ala Leu Arg Ser Glu Thr Gly Gly Leu Ser
290 295 300
Gly Lys Pro Leu Arg Asp Leu Ser Thr Gln Thr Ile Arg Glu Met Tyr
305 310 315 320
Ala Leu Thr Gln Gly Arg Val Pro Ile Ile Gly Val Gly Gly Val Ser
325 330 335
Ser Gly Gln Asp Ala Leu Glu Lys Ile Arg Ala Gly Ala Ser Leu Val
340 345 350
Gln Leu Tyr Thr Ala Leu Thr Phe Trp Gly Pro Pro Val Val Gly Lys
355 360 365
Val Lys Arg Glu Leu Glu Ala Leu Leu Lys Glu Gln Gly Phe Gly Gly
370 375 380
Val Thr Asp Ala Ile Gly Ala Asp His Arg Arg
385 390 395
<210> 5
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> соответствующий фрагмент ДНК для создания gRNA
<400> 5
ggatgcagcc atcatccttg 20
<210> 6
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность1)
<400> 6
caccgggatg cagccatcat ccttg 25
<210> 7
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность1)
<400> 7
aaaaccaagg atgatggctg catcc 25
<210> 8
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность2)
<400> 8
caccggatgc agccatcatc cttgg 25
<210> 9
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность2)
<400> 9
aaaacccaag gatgatggct gcatc 25
<210> 10
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность3)
<400> 10
caccggcagc catcatcctt ggggg 25
<210> 11
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность3)
<400> 11
aaaacccccc aaggatgatg gctgc 25
<210> 12
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность4)
<400> 12
caccggccat catccttggg ggagg 25
<210> 13
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность4)
<400> 13
aaaaccctcc cccaaggatg atggc 25
<210> 14
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность5)
<400> 14
caccggctat tcgcttcacg tccct 25
<210> 15
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность5)
<400> 15
aaaacaggga cgtgaagcga atagc 25
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность6)
<400> 16
caccggcctc tacaaactgg gcttt 25
<210> 17
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность6)
<400> 17
aaaacaaagc ccagtttgta gaggc 25
<210> 18
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность7)
<400> 18
caccgggctt tgggtttgtc gaggt 25
<210> 19
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность7)
<400> 19
aaaacacctc gacaaaccca aagcc 25
<210> 20
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность8)
<400> 20
caccggctgg tctgaggagc ctaca 25
<210> 21
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность8)
<400> 21
aaaactgtag gctcctcaga ccagc 25
<210> 22
<211> 14437
<212> ДНК
<213> Cricetulus griseus
<220>
<221> разный признак
<222> (1992)..(2015)
<223> мишень плазмид pBH6840 KO DHODH SEQ1 и pBH6841 KO DHODH
SEQ4
<220>
<221> разный признак
<222> (2129)..(2147)
<223> мишень плазмиды pBH6842 KO DHODH SEQ5
<220>
<221> разный признак
<222> (4015)..(4047)
<223> мишень плазмиды pBH6843 KO DHODH SEQ7
<400> 22
ccgagcccac attctccaat ggagtggagg cagaggcggg cccagtggca gaaggagcat 60
ggcgtggaga cagatgagag tgagtctgtt gcgggtgctt acgcttgttc ggaaaacagg 120
ttgggagtgg acgaggctgg gagattgaaa ggctggggcc cttgcagtgt gggctgcatg 180
tgtgctcagc taggggcatg caaaaagcaa gcgtgctggg cagaggctga gttgattgtg 240
agagatcgcg ggcttttttt tttttttttt tcttaaactg agctagtatc gggcccctgt 300
gagcacatgg tggctagcat tatggaaagg atccagggcg gaatgaaata agacacgcag 360
ccacccttcc agataaagtg tggttagaat gggagtcccg tgcgaagagt ccgtagattg 420
tgtccctggc ttgggaaaga gtgcgcagat attgtttctt gacacgtgag aatttctccc 480
tccagaggag ggagaatgaa tacttggttt gctgtacagg caaatctgat cacatcctct 540
acttccttat ttagtggaca aatcccatta aaaatataat cactgattgt ggtgaggggt 600
tgtgctgtga catgggagct ggaaccgaac tctgcactta accactgaac catctccagc 660
ccccaaatct gtattttggc tgacaccccc acatttctcc taccttactt taagaaatgc 720
tttcctttta gttccaaaca gacctgtctt cctctacccc attctctggc ccttctcatt 780
ggcttcagcc acctagagat tccttctcca gctgcttcct cctctttagc ttttgtgtct 840
ccttggaaag tcaaccttag cctggaagga gcctcaagat ccttccatta cttatctctt 900
cgtgtcagtt ttcttgggtt tgtgtttaca tggccattaa tttctctctt cattagatag 960
attacacact tctgaaggac agggacctgg tgtggtttgc ttgcctttgg tgaaggattg
1020
aatgaatgca ttagccactt ggcatctggc agtgataatt atcaaaaagc attgtgagcc
1080
tcctttgaac tagggtagtg attctctacc cagagtgtga agctatgttt gtattacaac
1140
cacttttata ttatacacat aggacatttc aggccagtaa cttttctagg tataatttta
1200
aaatatagac accatgtcct aactacagta taaaggagaa atgaaaacat gtatttcagt
1260
atgtagaagt cagaacacag ttcacctaca tagatggcca ggtgtttaag cctctattta
1320
gaaggtgtgc gttggtatac aaacagcact gtataccctt tctggtgggg ttttaggaac
1380
agtgaccaag tcttagtaat gttttgtgca aactgtacaa ttgtccttaa attacaaggt
1440
atctttattt ctgtaaaagt caccctgtac atgtaaacca tgtggttgtg tggtgaacta
1500
tagcttgctt cttgcctctc ctattatcct cagataatag gaagttttcc tcacataaat
1560
agcaatagca agttcaaaca tcatacagta catagggaaa ctagtgtgtt agctttaatt
1620
aaagaagaat attgacagga ctgatttctc cacaccaaat gtattagcat cattccctca
1680
ttaactagta aaaattttcc catgtgaccc ctagggggca gtactgcctg tcttgaaaat
1740
tccctgcctg gtccttagga aaatattcag ggctagaacc catgtaagca gtgtaggtac
1800
tgcatctacc tttgtagatg tgtaagtggg caggatatga agggggacag tgtgagcagg
1860
gaaggactgt cactttgtgt agttgttatg tcctctgggc aggtgtgaag ggtcaggaag
1920
ggtactgcag gctggtctga tggggttccc catggggcct tttctctccc ccactagaag
1980
cgggccctgg atgcagccat catccttggg ggaggaggac ttctcttcac ctcttacctg
2040
acagccacgg gcgatgacca tttctatgct gaatatctga tgccggccct gcagaggctg
2100
ctagacccag aatcagccca ccggctagct attcgcttca cgtccctggg gctccttcct
2160
cgagctacat ttcaggactc tgacatgctg gtaggtcccc cggtcaccct gtgttacatt
2220
tgtgtttgta ggggaggtca catccttcag cacttcggta ttcctctgct gccactgggc
2280
ttttgaaaac cagagctgca ggttcacaag cagctgggac ctggagccca gcccgaggaa
2340
gggacttttc ttctgtgtta caccctgtac ttacagaaca gcactcagca tatagccggt
2400
gcttacagct ttaatcctca cagataccct ttgagggagg tcctatctca tctccatcta
2460
caaatcagga aactgaggta cagactaagt aagtgacttg cccaaggtca cacagctggg
2520
aggtggcagg accaggattc aaacctatag agtttgactc cagtgtccaa ccttggtcac
2580
tggcctgcct tgctgagatc cctctgctga acaggaacat cttctggagt cattaagcag
2640
ctactggatg caggggacag ggtcagcagg gaagggctgt cacttgtaga tactgtgtcc
2700
tctgtctgga taggtgtgaa gtctcagagt gggtgctgca gcctggcctg atgggtccgt
2760
cctttccccc catcagaagc agaccctggc tgtagtcatc accctgaggg gtggaggact
2820
taagtcctag gcaacctcct gccccagcca cattcagcca caactcactc ctgtggccca
2880
cctggcacca tttataacag agccagtgtg gcccttctac tgtctctctg aatgctcccc
2940
ttttgagatg ggcctaggtt gtccaggttg gtctcaaact tggggtccca aatgatttca
3000
gaatctcagc ttcctgagta agtgggatca cacagtatgc accactgtgc gcagtggtcc
3060
ctgttccccg atagccaggc cggttgatac tcttagcccg attcaccttt ctcggttttc
3120
taagtcttca gttttctatc atgacgagac tctcctgggg gagggggatg gctgggtgct
3180
tctggaatgc ttggctgacc cggtctcctg attgaccggt tttaccataa tctcaaagaa
3240
gcctgacatc aacaccccag gctctgtcac acagccaggg atgagaactg gcttgagggt
3300
agcctcaagg aaggcctgtt ctagctgaac ttctcgcctc atggttatga agtgctgcgt
3360
gcacattttt accttccttg tgctcattct cgtctcctgt ggattttgag agcatgtagc
3420
tttccccttg gggactggcc tctaagagct cactggcagc accattgcct gccagtgtcc
3480
acccttcagc tctccatggt gcccatgggc tggcttcttg acagttatgg aggaagggcc
3540
agctagggct tgtatgctag gctggctaca ggctacctca aggctggtga caccaaggac
3600
tccttctcct gcccaggttg agtagatagg aagatacaca gtaattgtgc aagacagcac
3660
ctgtctcctc ttcacttgtg gtgggacact atctttctct ccccactgcc ttgagacccc
3720
ctggtgacag actccagcag acatcagaag cacctgcaga atgctgatgc agatagatac
3780
cccagggtgt ccttcctccc aaagatccct attttctggg cccagtgatg cttagcaagc
3840
tccaggggaa ttcttataca gtatgctggg atccctgctg tgagaaccac ttggtatagc
3900
tgcttatacc tctgacctgt ctccttccca ggaagtgaga gtcctgggcc ataaattccg
3960
aaatccggta gggattgctg cgggatttga caagcacggg gaagctgtag atggcctcta
4020
caaactgggc tttgggtttg tcgaggtagg aagtgtgact ccccagcctc aggaaggaaa
4080
ccccagaccc agagtgttcc gcctcccgga ggaccaagct gtcattaaca ggtaggtggt
4140
ggctcaatgt catagggaca tctcctcccc tgctgggatc tctgagatgg aacctgtttt
4200
gtgcttttct catatgatca gtggacagtc tgtcctttgg gatagtcagg agcacctttg
4260
aacacctgtt gtgtactagg cagctctcgc cccaaccttg ggaaacagct gttaggattt
4320
taccaggtat tcccactgag gctctgagag ctggtgctgg ttgcaaagaa cagcttggga
4380
aggcttgggc tgttgggctt cagtgtgagc ttttttacat tgttgctcta ttgcctttgt
4440
ccaatgttgg ctaacaggaa tataatgatc tcacatgtaa tttgaaatat tctaatgatg
4500
cattaaaaac tagtgaactt gataatacat ttataattat tatattttta attattatga
4560
tttaattata aacagtgtgt tctaaagtca ttttaacctg cagtcaatgt aaaaatcatt
4620
ggttaactat cgtcctcttt ttattggttt ccggtccccc aatgccttta cttctggttc
4680
ctctcacttg gccagggctg gtggccacaa gcttcagcag cgttggtact gtacaggtga
4740
cccttgcttt caatgaacag ggacatggta ttcaaatgta cagctgctct taggaggaga
4800
atccttcgac tgtatgaagc cttgcggttt gttgttctgt atagttagta taactgcaaa
4860
gggaagacct tcgtactttt tgctaggaga tttttggaga acctccttgt ggggattttg
4920
gcaggcctgt ggcagtgctt cctactgctt gtttcaggaa actgatttac aagtcataga
4980
tctttctgtt acattacagg cagggactga tgtctttagg actggccaga caaaatactt
5040
gtgtttctga tttggctgtt acaagtgact tcgggtctta ggatgatggt acttgggaca
5100
gggagtggtg agggacaggc acctgtgctc ttgggcatgt gcgaacttag ccttcttgct
5160
catatacaca gtgtgtgttc agtataggca tgctaacgga tgggtattta cacctgcaga
5220
cccacatgcc acaatccatg cagtgttgct ctggctgcaa gcaacaaagt gatgtaacca
5280
gatctcattt gttatcattc ttgtcactag cacaaaataa ctgacagcag gcagcttaag
5340
gaatggggtc aggtattttg gctcacagtt tgaggggaca cgccctcccg gtggcaggag
5400
cctgaggcag agggtcactc tgtacctaca gtgaggaagc agcaagggaa ggatgctggt
5460
gcttgggtct ctttctccgt cttgctcagc tggggtgtga atccatggtg ctgcccatgt
5520
ttgcggtggg tcttccctgc tccattcaac cttcccagaa acagtcttcc agacacacaa
5580
caggtgtgtt tccatggtga ctctaagtca catctagttg acaatgaagc ataaccagtg
5640
gtgagagcat ggaaatggtc tcaggtacgt tttccaaact gccacctttt aattccaccc
5700
cttacctgtg ggtttatgtg ctccaggtac ggattcaaca gccacgggct ctcagtggtg
5760
gaacacaggc tacgggccag acagcagaag cagaacaagc tcactgcagg taaactcagg
5820
tgttgtggga gggacttact aactctatta caaagaaaca tggggagggc atgggattca
5880
gcaataagaa acaaatgagc aaacaattaa aaccccacag aacaactcag gggattgtcc
5940
agcttctacc acagcctcaa aggtctcaac cagatggcat aattcagcaa aaccttgctg
6000
gtctctggca tccatggaac ctgaattgtg tcttaactta ggggcttgaa gatcagcaga
6060
gagcatgtgt ggagctaaga tgcgccccga gttcatcttc ccacatcccg ccctgctgct
6120
gtccttctgt tcactgtgta aatagtgtgt attagttctg gaagagctac agaagagaat
6180
agagttgtca cactcctgtc cttccactca ggggaccagc tgtcctcaaa tctcacttcg
6240
gaaacaacct ttgcttttgg tggattctca tggagacgct ccagtttgtc ctctgtggtc
6300
tgcatggctc tctgaagtgc attttggggt caaagccctg ggtggtggcc cccctcccca
6360
tgtctccctg tgtggcttct gcagtccatg gagatttttc tcctttgcct ctggaatgtc
6420
tttcaggagg tcttagtctc atttgatgtc cccatagtgt gacaattaag ccaaggactt
6480
ctgcatttat ttgaggagct aaggaagaga gggctgaagc atttgacgag tttagagggt
6540
gggaacagga aatgacaatg tttaaaaaaa cccctggttt tgacggaacc gggtctgcca
6600
gccctgtaga accttgaaca caatgtttgt ggtttgtggc gttctcctgc ctcctgcctt
6660
ccgtttcttc gttttaatgc cttctgtggg ctaggtttat tgccattgtc ctgtgtccac
6720
ccttcatcct cattctgtgc cctcacagtc actggagtga ggccaagttc tgtgtctttg
6780
ccctgcctac atgtgtctgc tgacagctag cagcctcata gaaacttttc atcagcacca
6840
tgcctaggaa ccctagtgga tagaagagac tgtgctttgt gtaaagtgta gttcagcctt
6900
cctgactcca gaactgtgac tgtgggcagg tgaccactaa tgaaaggctc agttgcctct
6960
tggaagccat actctcaatc cagtttactt gtgctaaagc ttacagtgat cctggcttgc
7020
tgggtgctag acttgaagct ttgagacctt gttctgtata tagtctaatc cctggctcca
7080
cccagcccac ttaggaaatt agttacacaa atcttacttt taagttgatg acccagaaga
7140
tgacgggttg tgagagatgg actgtgagaa gggggctagc aatacagtgg gggatgttgc
7200
ccgtgtcctc cctggcctgg tatgggtgag ctgacttggt gttctaggtc tggctggatt
7260
ggcagatgga cagcagcctg ggccactgac atactaggag ttgtaagaaa gttgagttgt
7320
aataaagatg aatccaggaa actcctttct gccaaagaga acatgagtct gaaagagcat
7380
ttttctagag gcttggcctt ttgagagtga cagggggcag gtgtgagtaa ttacacccag
7440
gccccaggag tgtcttgagg ctacagagat gtatcatcac cctaattata agttcttttc
7500
tcagagaaca agaaagattt attacacact tccatctact ttttaggcat gatagttgaa
7560
aacatttcct agtgttttcc tgtgtagctc aggctggcct cagacttaca gcaatcctcc
7620
tgcagactaa atttccatgt gtggcctcca gtgttttata cttttgtttt taatcatttc
7680
acatacagtc ccttgaccaa ggctcatgag attcaggttg ttcttgctga catgcccatc
7740
aatacctgcc tactctctct tttgttatct ctcaaaccac tgatctgtga atatagcaag
7800
tgtcattggg agtcatttta ttgctacata cttcaagaag agcagtagta tttgtttttt
7860
tccccatgtc cctgatttgt ctggactcag atccttggcc acccatgcag tcagtgttgg
7920
ggatgggttc catctcatgg attgggcctt aaatacaatc agattctggt tggttactcc
7980
cgcagctttg tgctactgtt tcactagtgc atccagggca ggtcatcatt gtagatcaaa
8040
agatgtgtag cggggttggt gtttaccttt ctcctttggt aacatgcaga atacctccca
8100
gtaccatgga caccaccaga gggatgaagg ctgtaggtag gcaccagctc gacttctccc
8160
tatttaataa cttgtgtagg tgttgtcttt agcaatagag ccttgtcagt tttcagagag
8220
caaccaatat ccttgacaat agcctgggtt gtttgggtat tctcatgggg cccctttggc
8280
caacaacttc tttgaaagtc tggtcaaatt cttcactaaa accatctggc actgggcatt
8340
ttttggttgg gaggctttta tgactgcttc tattttacta ggggttataa gtttgtttac
8400
attgcttatc cgatcttgat ttaactttag tagatggtat gtatcaagaa aattattcat
8460
ttattttcaa ttttccattt aggtagagta taggttttta aaacacatcc ctatgattct
8520
ttggatttcc ttggtgtctg ttgttatgtc actcttttca tctctaattt tgttaatttg
8580
gatctttcct ctttgccttt taattaattt ggctaagggt ttgtcaatct tactggtttt
8640
ctcaaagaac caactcttcg tttcattgat tctttgcatt atttttgtct gtttctattt
8700
cattgatttc agccctgagt ttgattattt cctgccatct actcctcctg ggtgtgactt
8760
cttgttgttc tagagctttc aggtgtgcta gtatgaaatc tctctaattt atgtaggcac
8820
ttagtgctac ataaagtttt gggtttttgt tttgttttga cctatttttc attcagtagt
8880
gagttgttca gtttcagttg ttcagtttcc atgagtttgt aagctttctg ttgtttttgt
8940
aggtgttgat attcagcttt aatctgtggt ggtatgatag ggtattattt cagttttctt
9000
gtatcttttg agacttgctt tatggcctag tatatggtca gttttggaga aagttccatg
9060
aggtgctgag aagaaacgtt ttgtgtttgg gtgaaatgtt ctataaatta ggtccatttg
9120
gtttaacaca tcatttagct ccatcatttc tttgttcagt ttttgtctgg atgacctgtc
9180
tattgtcgag agttggggag tatcaaagta tcccactatg tgtgtgtgtg tgcggagggg
9240
tcaatatgtg atttaagcta tagtagagtt tcttttatga acttggatgc ctttgtgctt
9300
ggtgcataga tgtttagaat ttcaatatcc tcttggtggg tttttccttt gatgagtatg
9360
tagtttcctt ccctatctat ctcttgatta gatttgattt gaaatctatt ctgtcatata
9420
ttaaaatggc ttcacccatt tgctttcttg tccatttgct tggaatatct ttttccatcc
9480
ttttactctg aggtgatatc tatccttgat gttaaagtat gtttcttgga ttcagttaaa
9540
gaatgaatcc tatttttgaa cccaattttt tagtctgtgt cttatttatt gggaaattga
9600
ggccactgat atcagtgttt gttgattcct gttatattat tgttatggcg taggcccctt
9660
cccttttgat ttgctggtct gagattattt attcaggctg aagttttcct tgtagtacct
9720
tctatcaagc tggatttgta gacagaaacc acttaagttt cattttattg tagattgtca
9780
ttctttcttc atctctgtga ttaaatgttt tgctggctat agtagtctgg gctggcatct
9840
gtggtagaat gtctgcccag gacctttgac ttttagagtc tcgattgaag tcaggggtta
9900
ttctaatagg tttgtctgcc gttatatgtt atctggtctt tctctattgt ggcttttggt
9960
attctttctt tgttctgtac attctgtttt ttgattatta tgtgttgtgg ggaatttatt
10020
tttggcccag tccgtttggt gttctatttg tttcttatac catgataggc acttccttct
10080
ttaggttagg taacttttca tctgtgattt tgttgaaaat attttctgtg cctttgacct
10140
gggtttcttc tccttttgct atccctgtta cttatagatt tttggtgttt tcatagtatc
10200
ctagatttcc tggatgtttt gtgcctggat tttgttttct ttcttttttt gtagatttaa
10260
cattttcttt gactgcagta tccttgtctc ctatcttgtc ctcagtgctt gagactctgt
10320
cttccatttc ttgtatgatt ttggtgaggc ttgcttctaa gatttctgtt ctagttccta
10380
aatttttcat ttccagcttt acctcaattt gggttttctt tagcaattcc atttcctctt
10440
tcatgtgttg aacaattttc ttcatttcat tgcactgttt gtgttttcat taatgggttt
10500
attcatatct tctttaggga ccttgaacat attcataata gctattctga gagccttgtc
10560
ttatgcttcc ctgtattgca tttctcggag cctgctgtgg tagggttgct gggctctagt
10620
ggagacatgt cgttctggct gttactgggt ttttacactg gtgtctaggc gactgggttt
10680
gagaagattg taattctagg tgctgatatc tggtcttgtt tttgttgggg tgatgttcag
10740
ttccttggtt tctgttgccc ccccccctca gggggggtgt ggtaactgga ttggttgcct
10800
ggtagggaat gcttctgaga tcctgccaga accctgccac tggcagtctt gggtagaaag
10860
tgtttctagg tgttgggagc tgacactaat gattgaggat gggttagaag cagtggtggc
10920
gggagagtcc acaggaggag gagagctggg tgtgccacca gggtctgcac agaggcctgg
10980
gaatgagaac agagatgaag gtgaggcctc agcaggtagt ctgctacaag gttgggataa
11040
ggctgggaga ttggaacttg gggacaggag ggagagtgaa gatcgcagac ccatcccctg
11100
gccaggtggg ggaagcctgg aggagaagat ctgtgtgatc tgctggaaat gggtccctta
11160
gtaacttctt gaagtttctt tgattttgag gctgctgttg ggggtcccct gtatgccaag
11220
catgtggtat atcactgaga tagaatctcc atagccctgc ttcttttttg agactgggtc
11280
tcatatcggc caggttggcc ttgaactagc tatgttgacc ttgaactttt ccttacctat
11340
acctgagaac tgggattaca agtgcccatc acacccagct tcctattgtt tttgcttttt
11400
tcctagtatt taagcttctg gcttccccat taaaatttaa aagatgaaag gcttagtaca
11460
cgggaagcat ccttgaaggt gcacacactt gccatataga gaggatgaag tggttctaag
11520
tcatcaggca gcagccccag gatcagacag ttcacattct ccatcatctg gttcagggga
11580
gccccacctg tactctgaat gttgcctggg aaactgtggg gacacactct tgatatttac
11640
agatgggctg cctctgggaa taaacctggg gaagaataag acttcggagg atgctgctgc
11700
agactatgta gagggtgttc gtgtcctggg ccccttggct gactacctgg tggtgaatgt
11760
gtccagtccc aacactgctg gtctgaggag cctacaggga aaggctgagc tgcgccgcct
11820
gctggccaag gtgtgtcatt acaccatcac atgcctgttg tccttactcc ttttcattct
11880
tcaggtgaag attcaggaca actgaggaga aactgttctt cacagctggc ctaggagccc
11940
tcatcacaca ttttccaagt accctctcat gtctcacagc cccggtcata cacaatagac
12000
agttcactgc tttaaaacca caggcaagca gagcagagca ggcggggctc ccatgtatca
12060
ctctgtcccc agaactgtga atgctcagca cttgtgacac acctccttgt ttgttttcag
12120
gaacaaaatc aaaacccttg agcagttatc tcggcggggg gtgcgggggt gggggaattg
12180
ctggtgcttt agttgacctt aggtgaaaat gctggcctgc actgcggggc tatgggctgg
12240
gtccccatct ctggcttgtc aacctaggtg ctgcaggaga gggacgcttt gaagggagcg
12300
cagaagccag cagtgctggt gaagatcgcc cccgacctca cggcccagga caaggaggac
12360
attgccagtg tggcgagaga ggtttgagtt ggggtggtcc agggcagggt gggggtagcc
12420
ttcatcgtcc actgctgctg ggataacaca gaagggcatg attggtgact tcctctgtgt
12480
gaagtggaca gctggttgat tgtctgtcat tgtatagttc atctggtagc aaacagtgag
12540
tttgaaaatg tgtttggtga gtctttttct tgcttctgat ctgtcttcat ccagaaagtc
12600
tgaggcctgt gctagtctct gccagtctca cctgggactt agaatggtgt ctgtcccttt
12660
ccagcttgtg taaacaccag gcttctctgg ctaaaaaggt agtaggaaca cagtctgctg
12720
ttctggactt cagtcttgtt ctactgtgta ctccctgaga tactcacaga cagcttgttc
12780
taagggtcaa actctgtgta ggcactgtga tgggttcagc aggagatgct gctgtgtctt
12840
cagcattgag aggctaattc tgatggcttc tccaatcaag atgtaggtga ggcctgtgag
12900
ggtcttgctc tgcagagctg gcccctggcc tggtggctgc ttcaatcctc aaaagacagt
12960
ttccttgagt acttcagatc catggcttaa ggctcttttt ctgtcttgtg ctgcagctgg
13020
gcattgatgg attaattgtc acaaacacca cagtgagtcg cccgactggc ctccaaggtg
13080
ctctgcgttc tgagatggga ggactgagcg ggaagccact ccgagatctg tcgacccaga
13140
ccatccggga gatgtacacc ctcactcaag gtaaggcttt tttgtttgtt tgtttgaggc
13200
aggatttctc tgtttaacac ctctggctgt cctggaactc acttggtaga ccaggctggc
13260
ctggaactca cagcttccct agtgctggga ttaaaggcat gtgccaccat tgcctggcta
13320
aggcaagact tctttggaca aatgtgaggt cctggatctt ccctcattca ctgtgtttgc
13380
tgagaactct ggttctgccc tcatcggctg tgtttgctgg gactgaggcc tgatggagcc
13440
ctgggtcttt agccctccct tcccggcctt gctatgtgct tctctccagg caggattccc
13500
attatcgggg ttggtggtgt gagcagtggg caagatgcgc tggagaagat ccaggcaggg
13560
gcctccctgg tgcagctgta cacggccctc accttcctgg ggccacccgt cgtggtcagg
13620
gtcaagcgtg agctggaggc acttctaaag tgagtagggt tcgatgcagc tgagacgtag
13680
aaagtgacac ttgtcatcag tctattgtgt actcccagag ggccggaggg aacactgagg
13740
gcatggtggg acgatttctc tgctgcagct ttggccaagg acaaacagtg ccagaggaat
13800
tcagaatgct tctgaggcag ggctcattac aaggcaggac ttgccacttg cccaggggct
13860
gggcatggag gatgaagtag taatttacat tgactcagtg tctggaagct gcaggttata
13920
aagtcacttc ccttcctgca cagaaggcct ggctgtacat atgtgcaggg gcctgggtga
13980
gggcagagta gcagtggttg taaggtgtgt tgggtgggac ggtgtggtaa gcggtgtgct
14040
catggtgagt gtgggcttcc ttaggacagg ctatttgttt tctgtccaga gagcggggtt
14100
ttaacacagt cacagaagcc attggagcag atcatcggag gtgacggttc ctgccagatg
14160
ccccatccag aacgtgccca ccaactcaag caagccttgt ggctgcatca taagaggaag
14220
atctgtctca agctatgtcc cttgactgtg tgacctggct ggactgcata agccagtcac
14280
ggttatcact agacagtaaa ggctttctct aatgagacca tgaactctac agtcactttc
14340
tggatctaag tcctgggatc cctcagtatt ataaggacat tggctctttg ggaggaaaaa
14400
tcatggagaa aataaagcca tttcaatctg ttttcaa 14437
<210> 23
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> соответствующий фрагмент ДНК для создания gRNA
<400> 23
caaggatgat ggctgcatcc 20
<210> 24
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность1')
<400> 24
ggatgcagcc atcatccttg gtttt 25
<210> 25
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность1')
<400> 25
caaggatgat ggctgcatcc cggtg 25
<210> 26
<211> 395
<212> Белок
<213> Искусственная последовательность
<220>
<223> аминокислотная последовательность DHODH G202A человека
<400> 26
Met Ala Trp Arg His Leu Lys Lys Arg Ala Gln Asp Ala Val Ile Ile
1 5 10 15
Leu Gly Gly Gly Gly Leu Leu Phe Ala Ser Tyr Leu Met Ala Thr Gly
20 25 30
Asp Glu Arg Phe Tyr Ala Glu His Leu Met Pro Thr Leu Gln Gly Leu
35 40 45
Leu Asp Pro Glu Ser Ala His Arg Leu Ala Val Arg Phe Thr Ser Leu
50 55 60
Gly Leu Leu Pro Arg Ala Arg Phe Gln Asp Ser Asp Met Leu Glu Val
65 70 75 80
Arg Val Leu Gly His Lys Phe Arg Asn Pro Val Gly Ile Ala Ala Gly
85 90 95
Phe Asp Lys His Gly Glu Ala Val Asp Gly Leu Tyr Lys Met Gly Phe
100 105 110
Gly Phe Val Glu Ile Gly Ser Val Thr Pro Lys Pro Gln Glu Gly Asn
115 120 125
Pro Arg Pro Arg Val Phe Arg Leu Pro Glu Asp Gln Ala Val Ile Asn
130 135 140
Arg Tyr Gly Phe Asn Ser His Gly Leu Ser Val Val Glu His Arg Leu
145 150 155 160
Arg Ala Arg Gln Gln Lys Gln Ala Lys Leu Thr Glu Asp Gly Leu Pro
165 170 175
Leu Gly Val Asn Leu Gly Lys Asn Lys Thr Ser Val Asp Ala Ala Glu
180 185 190
Asp Tyr Ala Glu Gly Val Arg Val Leu Ala Pro Leu Ala Asp Tyr Leu
195 200 205
Val Val Asn Val Ser Ser Pro Asn Thr Ala Gly Leu Arg Ser Leu Gln
210 215 220
Gly Lys Ala Glu Leu Arg Arg Leu Leu Thr Lys Val Leu Gln Glu Arg
225 230 235 240
Asp Gly Leu Arg Arg Val His Arg Pro Ala Val Leu Val Lys Ile Ala
245 250 255
Pro Asp Leu Thr Ser Gln Asp Lys Glu Asp Ile Ala Ser Val Val Lys
260 265 270
Glu Leu Gly Ile Asp Gly Leu Ile Val Thr Asn Thr Thr Val Ser Arg
275 280 285
Pro Ala Gly Leu Gln Gly Ala Leu Arg Ser Glu Thr Gly Gly Leu Ser
290 295 300
Gly Lys Pro Leu Arg Asp Leu Ser Thr Gln Thr Ile Arg Glu Met Tyr
305 310 315 320
Ala Leu Thr Gln Gly Arg Val Pro Ile Ile Gly Val Gly Gly Val Ser
325 330 335
Ser Gly Gln Asp Ala Leu Glu Lys Ile Arg Ala Gly Ala Ser Leu Val
340 345 350
Gln Leu Tyr Thr Ala Leu Thr Phe Trp Gly Pro Pro Val Val Gly Lys
355 360 365
Val Lys Arg Glu Leu Glu Ala Leu Leu Lys Glu Gln Gly Phe Gly Gly
370 375 380
Val Thr Asp Ala Ile Gly Ala Asp His Arg Arg
385 390 395
<210> 27
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность2')
<400> 27
gatgcagcca tcatccttgg gtttt 25
<210> 28
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность2')
<400> 28
ccaaggatga tggctgcatc cggtg 25
<210> 29
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность3')
<400> 29
gcagccatca tccttggggg gtttt 25
<210> 30
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность3')
<400> 30
cccccaagga tgatggctgc cggtg 25
<210> 31
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность4')
<400> 31
gccatcatcc ttgggggagg gtttt 25
<210> 32
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность4')
<400> 32
cctcccccaa ggatgatggc cggtg 25
<210> 33
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность5')
<400> 33
gctattcgct tcacgtccct gtttt 25
<210> 34
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность5')
<400> 34
agggacgtga agcgaatagc cggtg 25
<210> 35
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность6')
<400> 35
gcctctacaa actgggcttt gtttt 25
<210> 36
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность6')
<400> 36
aaagcccagt ttgtagaggc cggtg 25
<210> 37
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность7')
<400> 37
ggctttgggt ttgtcgaggt gtttt 25
<210> 38
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность7')
<400> 38
acctcgacaa acccaaagcc cggtg 25
<210> 39
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность8')
<400> 39
gctggtctga ggagcctaca gtttt 25
<210> 40
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид, используемый для получения gRNA,
(последовательность8')
<400> 40
tgtaggctcc tcagaccagc cggtg 25
<210> 41
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 603 смысловой олигонуклеотид
<400> 41
gttggccttc caatggctt 19
<210> 42
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 503 антисмысловой олигонуклеотид
<400> 42
gttccttcac aaagat 16
<210> 43
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность, включающая целевую последовательность и
PAM.
<400> 43
ggatgcagcc atcatccttg g 21
<210> 44
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Область смысловой последовательности экзона2 DHODH, включающая
последовательность n°1 для CrispR.
<400> 44
gacgaaacac cgggatgcag ccatcatcct tggttttaga 40
<210> 45
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Область антисмысловой последовательности экзона2 DHODH,
включающая последовательность n°1 для CrispR.
n°1
<400> 45
tctaaaacca aggatgatgg ctgcatcccg gtgtttcgtc 40
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ ДЛЯ РЕКОМБИНАНТНОЙ ЭКСПРЕССИИ ПРЕДСТАВЛЯЮЩЕГО ИНТЕРЕС ПРОДУКТА | 2014 |
|
RU2720525C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ПРЕДНАЗНАЧЕННЫЕ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2018 |
|
RU2834231C2 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2783313C1 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2021 |
|
RU2761879C1 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2760301C1 |
| МОДУЛЯЦИЯ АКТИВНОСТИ REP БЕЛКА ПРИ ПОЛУЧЕНИИ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (ЗКДНК) | 2020 |
|
RU2812850C2 |
| СВЯЗЫВАЮЩИЕСЯ С CD38 И PD-L1 МОЛЕКУЛЫ | 2017 |
|
RU2764201C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ ОПУХОЛИ | 2010 |
|
RU2563359C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок SMN1, и ее применение | 2020 |
|
RU2742837C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С TFR | 2019 |
|
RU2810756C2 |
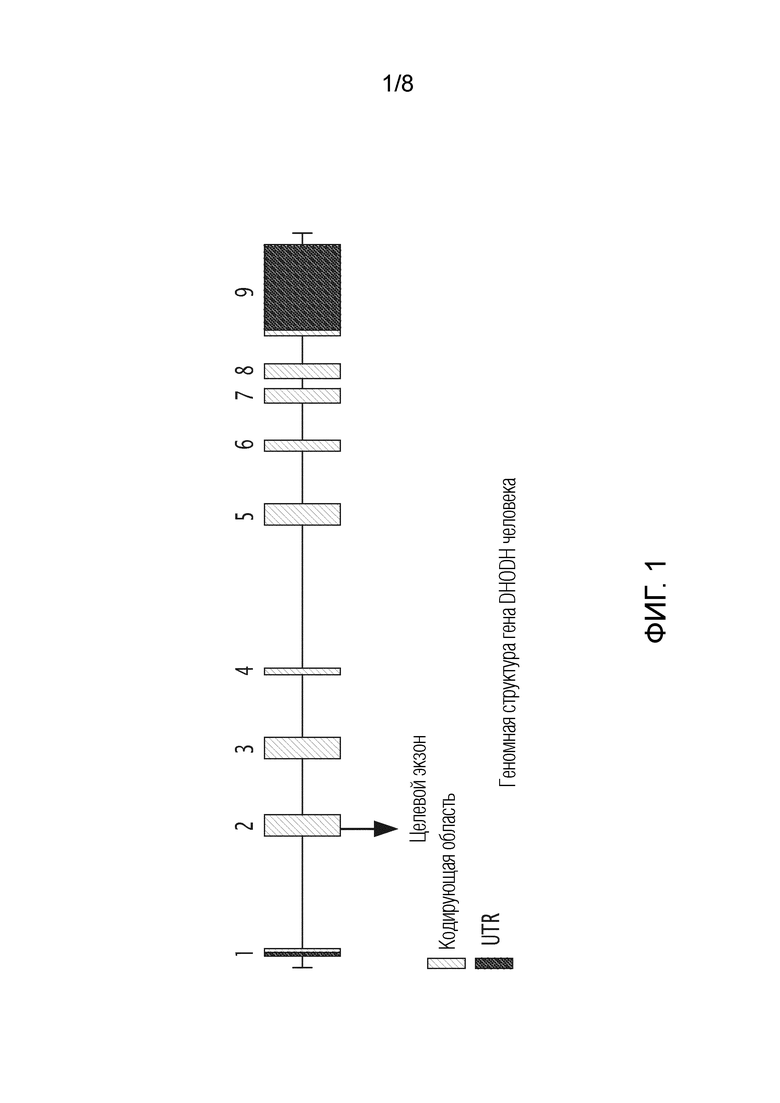
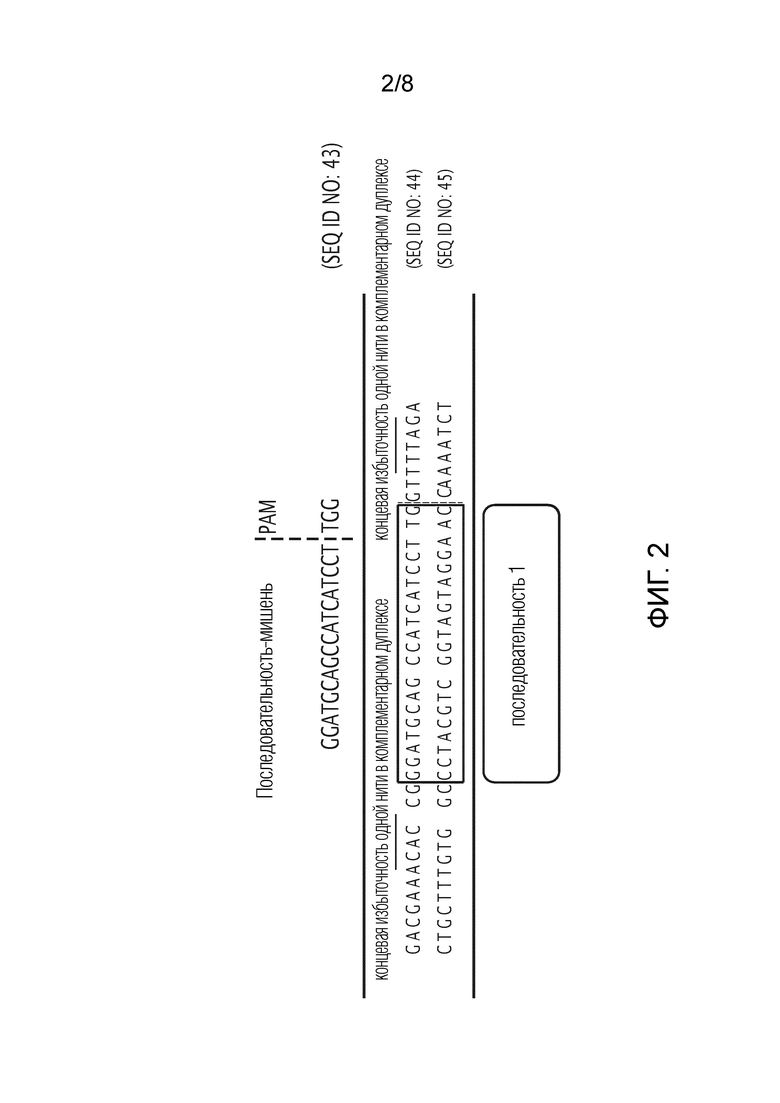
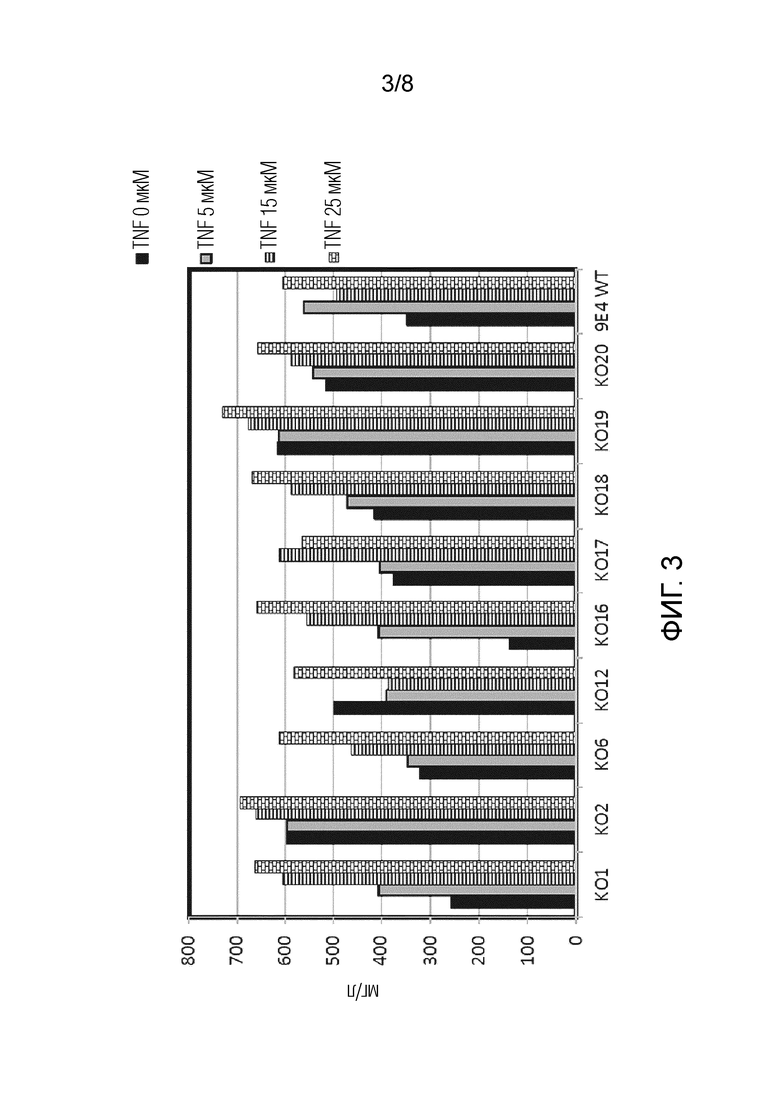
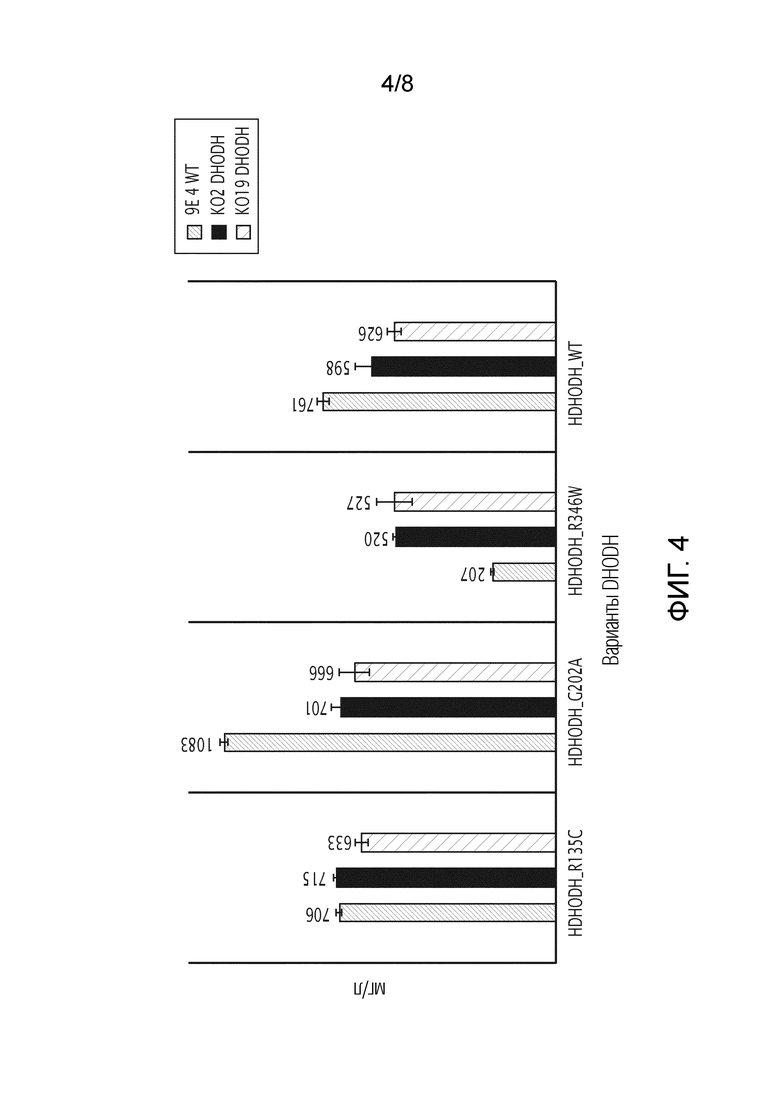
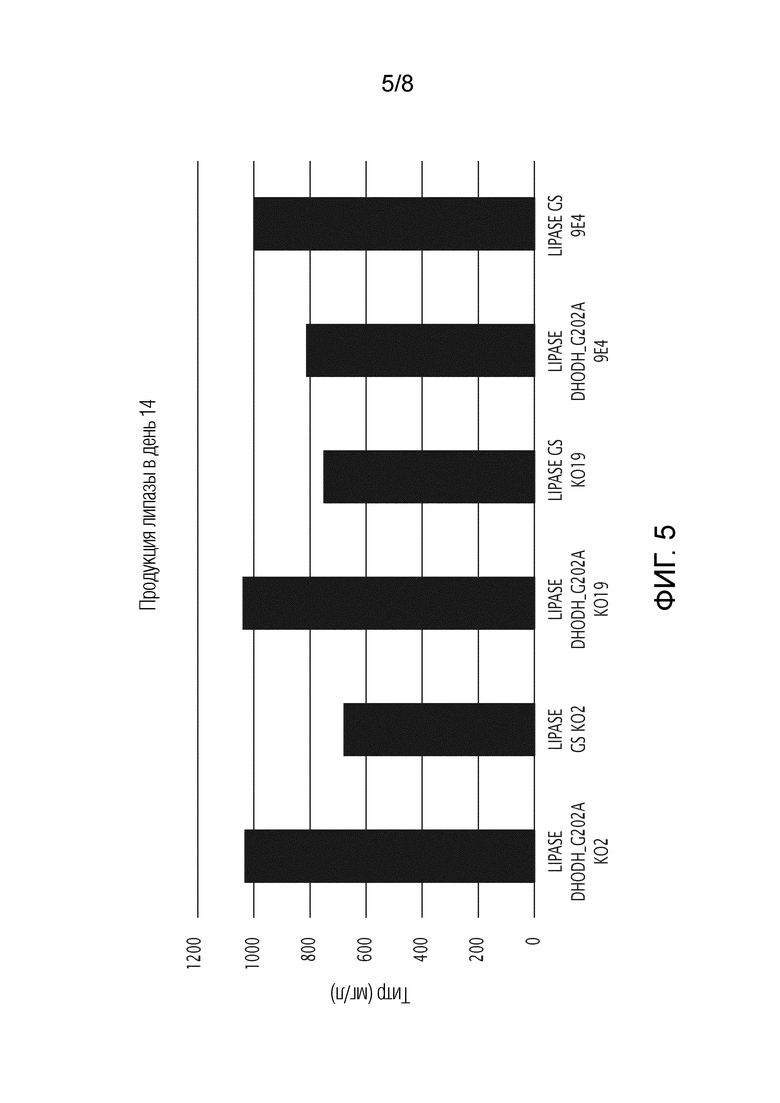
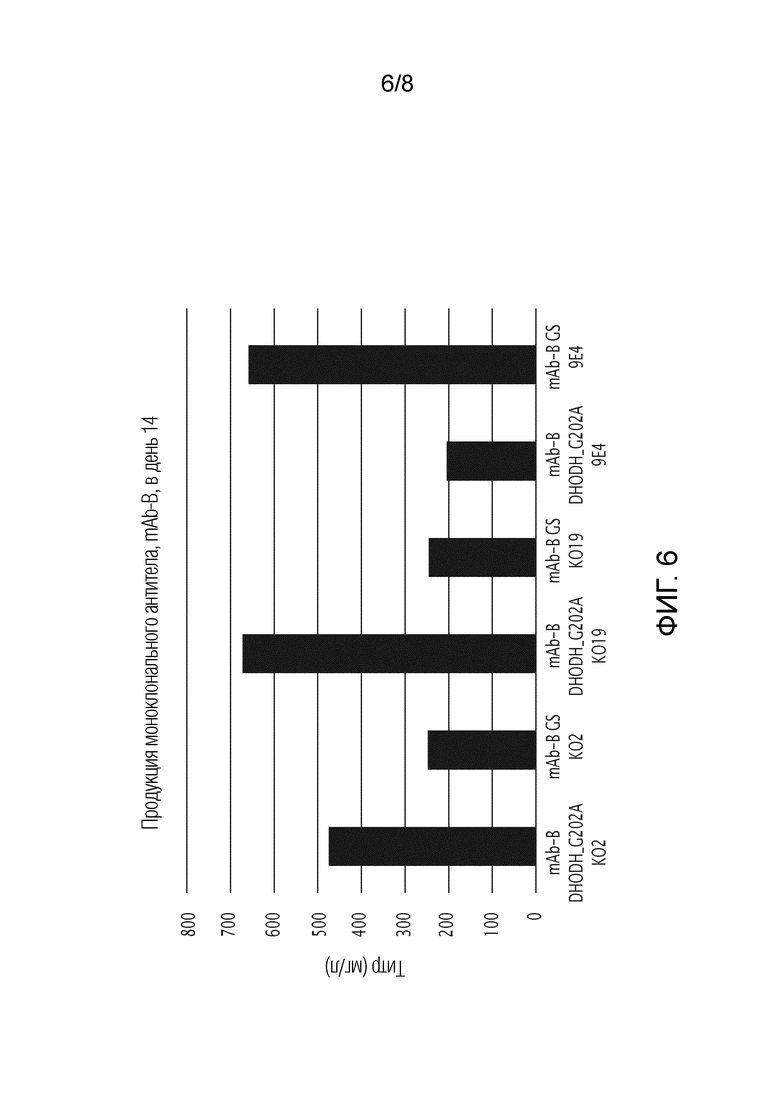
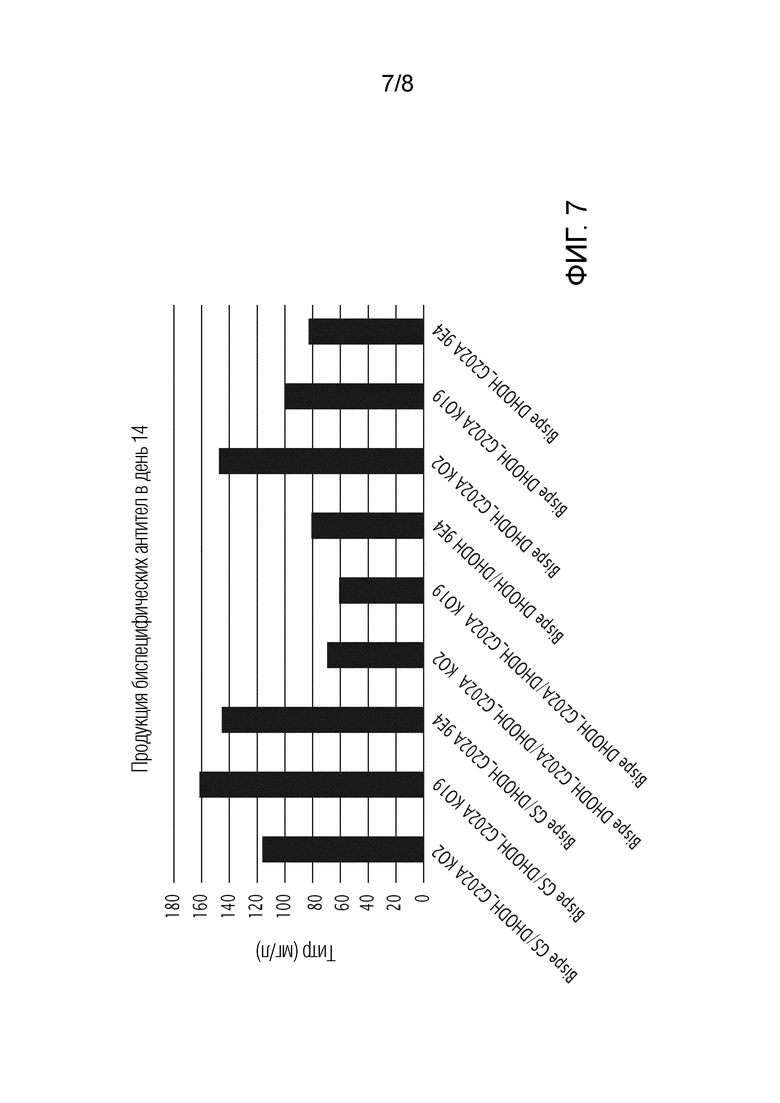
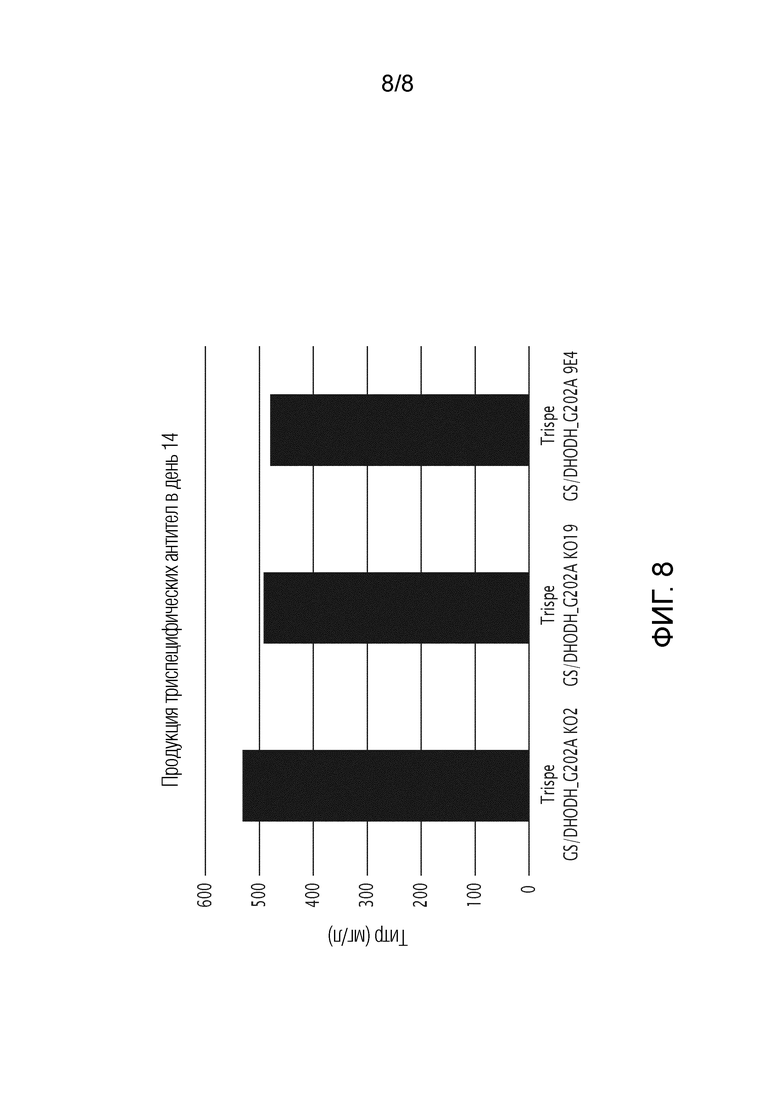
Изобретение относится к области биотехнологии, в частности к линии клеток млекопитающих, содержащей эндогенный ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован и экспрессионный вектор, подходящий для продуцирования рекомбинантного белка, где экспрессионный вектор включает последовательность, кодирующую DHODH, а также к экспрессионной системе, включающей линию клеток млекопитающих, включающую эндогенной ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован, и экспрессионный вектор, содержащий нуклеотидную последовательность, кодирующую DHODH млекопитающего, и по меньшей мере одну экспрессионную кассету. Также раскрыт набор, содержащий вышеуказанную линию клеток или экспрессионную систему, и культуральную среду, лишенную уридина. Изобретение эффективно для продуцирования рекомбинантного белка in vitro. 5 н. и 12 з.п. ф-лы, 8 ил., 6 пр.
1. Линия клеток млекопитающих для продуцирования рекомбинантного белка, содержащая эндогенный ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован и экспрессионный вектор, подходящий для продуцирования рекомбинантного белка, где экспрессионный вектор включает последовательность, кодирующую DHODH, и
где линия клеток выбрана из линий клеток яичника китайского хомячка (СНО), линий клеток HEK293, HKB11, PER-C6, HT1080, Jurkat, Daudi, Raji и CAP.
2. Линия клеток по п. 1, которая представляет собой линию клеток яичника китайского хомячка (СНО).
3. Линия клеток по п. 1 или 2, которая получена путем
a) инактивации эндогенного гена DHODH в клетке и
b) культивирования клетки в культуральной среде, содержащей уридин, в условиях, подходящих для создания линии клеток, в которой эндогенный ген DHODH частично или полностью инактивирован.
4. Линия клеток по п. 3, в которой эндогенный ген DHODH инактивирован с помощью метода редактирования генов.
5. Линия клеток по п. 4, в которой эндогенный ген DHODH инактивирован с помощью метода с использованием системы CRISPR-Cas9.
6. Линия клеток по любому из пп. 1-5, в которой одна или более или все аллели эндогенного гена DHODH частично или полностью инактивированы.
7. Линия клеток по любому из пп. 1-6, в которой экспрессионный вектор содержит нуклеотидную последовательность, кодирующую экзогенную DHODH млекопитающего, и по меньшей мере одну экспрессионную кассету для экспрессии рекомбинантного белка, причем указанная экзогенная DHODH включает последовательность, идентичную на по меньшей мере 60% последовательности SEQ ID NO: 2 или последовательности SEQ ID NO: 4.
8. Экспрессионная система, включающая:
(i) линию клеток млекопитающих, включающую эндогенной ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован, где линия клеток выбрана из линий клеток яичника китайского хомячка (СНО), линий клеток HEK293, HKB11, PER-C6, HT1080, Jurkat, Daudi, Raji и CAP, и
(ii) экспрессионный вектор, содержащий нуклеотидную последовательность, кодирующую DHODH млекопитающего, и по меньшей мере одну экспрессионную кассету для экспрессии рекомбинантного белка, причем указанная DHODH включает последовательность, идентичную на по меньшей мере 60% последовательности SEQ ID NO: 2 или последовательности SEQ ID NO: 4.
9. Линия клеток по любому из пп. 1-7 или экспрессионная система по п. 8, в которой указанная нуклеотидная последовательность, кодирующая DHODH, включает последовательность SEQ ID NO: 1 или последовательность SEQ ID NO: 3.
10. Линия клеток по любому из пп. 1-7 или 9 или экспрессионная система по п. 8 или 9, в которой указанный рекомбинантный белок представляет собой моноклональное антитело.
11. Линия клеток по любому из пп. 1-7, 9 и 10 или экспрессионная система по любому из пп. 8-10, в которой указанный вектор содержит первую экспрессионную кассету, подходящую для клонирования легкой цепи антитела, и вторую экспрессионную кассету, подходящую для клонирования тяжелой цепи антитела.
12. Набор для продуцирования рекомбинантного белка, включающий (i) линию клеток по любому из пп. 1-7 и 9-11, или экспрессионную систему по любому из пп. 8-11, и (ii) культуральную среду, лишенную уридина, в частности, лишенную, кроме того, ингибитора DHODH.
13. Способ продуцирования рекомбинантного белка in vitro, включающий стадии:
А a1) предоставления линии клеток по любому из пп. 1-7 и 9-11;
или
а2) предоставления линии клеток млекопитающих, включающей эндогенной ген дегидрооротатдегидрогеназы (DHODH), который частично или полностью инактивирован, где линия клеток выбрана из линий клеток яичника китайского хомячка (СНО), линий клеток HEK293, HKB11, PER-C6, HT1080, Jurkat, Daudi, Raji и CAP, и
а2') введения экспрессионного вектора, включающего нуклеотидную последовательность, кодирующую DHODH млекопитающих, и по меньшей мере одну экспрессионную кассету для экспрессии рекомбинантного белка, где указанная DHODH включает последовательность по меньшей мере на 60% идентичную последовательности SEQ ID NO: 2 или последовательности SEQ ID NO: 4, в линию клеток, предоставленную на стадии а2);
или
a3) предоставления линии клеток млекопитающих, содержащей эндогенный ген DHODH, где линия клеток выбрана из линий клеток яичника китайского хомячка (СНО), линий клеток HEK293, HKB11, PER-C6, HT1080, Jurkat, Daudi, Raji и CAP,
a3') частичной или полной инактивации эндогенного гена DHODH в линии клеток, предоставленной на стадии a3), и
a3'') введения экспрессионного вектора, включающего нуклеотидную последовательность, кодирующую DHODH млекопитающих, и по меньшей мере одну экспрессионную кассету для экспрессии рекомбинантного белка, где указанная DHODH включает последовательность по меньшей мере на 60% идентичная последовательности SEQ ID NO: 2 или последовательности SEQ ID NO: 4, в линию клеток, содержащую частично или полностью инактивированный эндогенный ген DHODH, полученный на стадии а3');
B) культивирования указанной линии клеток в условиях, подходящих для продукции рекомбинантного белка; и
C) выделения и/или очистки указанного рекомбинантного белка.
14. Способ по п. 13, в котором стадию B) проводят в культуральной среде, лишенной уридина, в частности, лишенной, кроме того, ингибитора DHODH.
15. Способ по п. 13 или 14, включающий, кроме того, стадию D) составления указанного рекомбинантного белка в фармацевтическую композицию.
16. Применение линии клеток по любому из пп. 1-7 и 9-11, экспрессионной системы по любому из пп. 9-11 или набора по п. 12 для продуцирования рекомбинантного белка.
17. Применение по п. 16, в случае которого линию клеток, экспрессионную систему или набор используют в сочетании с культуральной средой, лишенной уридина, в частности, в отсутствие ингибитора DHODH.
| DAVID B | |||
| SYKES, The emergence of dihydroorotate dehydrogenase (DHODH) as a therapeutic target in acute myeloid leukemia, Expert Opin Ther Targets | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| WO2016062837 A1, 28.04.2016 | |||
| YAMAZAKI H et al., Genetic transformation of a Rhizomucor pusillus mutant defective in asparagine-linked glycosylation: production of a | |||

