Изобретение относится к молекулам, связывающим CD38/PD-L1, в особенности к антителам, нацеленным на CD38 и PD-L1, способам получения данных молекул, композиций и их применению.
Уровень техники
Множественная миелома (MM) представляет собой третье наиболее распространенное гематологическое злокачественное новообразование с 114000 случаев в год по всему миру. Несмотря на успехи в лечении, MM остается одним из немногих гематологических злокачественных новообразований, где еще имеется неудовлетворённая медицинская потребность. Если после прогресса в ходе терапии первой линии у пациентов появилось рецидивирующее или рефрактерное (r/r) заболевание, варианты лечения очень ограничены. Однако в последние годы были разработаны анти-MM целевые опухолевые антигены (TAA). Одно анти-CD38 антитело, даратумумаб, было одобрено для лечения пациентов с рецидивирующей ММ, а другие анти-CD38 антитела разрабатываются в настоящее время (изатуксимаб и MOR-202, который описан в патенте США US 8,263,746). Однако, существует необходимость в улучшении ответов, которые в настоящее время находятся в диапазоне 30-35%. Было продемонстрировано, что активность анти-CD38 антител может быть повышена с помощью иммуномодулирующих терапевтических средств (например, леналидомида), которые стимулируют иммунную систему пациентов. Кроме того, было продемонстрировано, что один из механизмов резистентности опухолевых клеток ММ к лечению антителами связано с усилением сигнализации путей ингибиторов контрольных точек (например, PD-1/PD-L1). Таким образом, существует возможность усиления цитотоксичности анти-CD38 антител против опухолевых клеток MM и одновременной активации иммунной системы путем ингибирования путей ингибиторов контрольных точек (например, PD-1/PD-L1).
В физиологических условиях, PD-L1 играет главную роль в качестве защитника от аутоиммунной реакции путем понижающего регулирования иммунной системы. Он экспрессируется на иммунных «APC-подобных» клетках (T-клетках, NK-клетках, макрофагах, миелоидных DC, B-клетках, эпителиальных клетках, клетках эндотелия сосудов) и опухолевых клетках. PD-L1 связывается со своими рецепторами PD-1 и B7-1 и отрицательно регулирует иммунные клетки (T-клетки, NK-клетки и т.п.), ингибируя их пролиферацию и активацию.
В патологических условиях, PD-L1 сильно экспрессируется опухолевыми клетками (> 90% пациентов с MM) и связан с плохим прогнозом. Блокирующие антитела, нацеленные на пути точек контроля иммунного ответа (анти-PD-1, анти-CTLA-4, анти-PD-L1 и т.п.), продемонстрировали выдающуюся активность при различных типах рака (рака легкого, меланома и т.п.). Признаки эффективности наблюдали в MM, однако, активность этого класса перспективных терапевтических средств по-прежнему субоптимальна при MM. Одной из причин может быть то, что молекулы, которые обладают полезной активностью/профилем побочных эффектов (например, анти-PD-L1), требуют почти стехиометрического блокирования/насыщения их мишеней, чтобы вызвать максимальный иммуностимулирующий эффект на T-клетках.
Следовательно, специфическое нацеливание анти-PD-L1 антител в опухолевые локализации (например, нацеливание на раковые клетки CD38+) может помочь доставить анти-PD-L1 терапевтические средства к участку, в котором требуется иммунная стимуляция, и может привести к максимальной стимуляции иммунных клеток, что позволит осуществить полное блокирование PD-L1 на опухоли и клетках микроокружения. Такая направленная активация иммунных клеток в местоположениях опухоли также может уменьшить систематическую активацию иммунных клеток, предотвратить неблагоприятные побочные эффекты и позволить более высокую дозировку терапевтических антител.
Раскрытие изобретения
Для того чтобы использовать цитотоксический потенциал T-клеток, BK-клеток и других иммунных клеток для лечения множественной миеломы (MM) и других раков, предпочтительно CD38+ раков, были разработаны биспецифические молекулы с двумя сайтами связывания (специфические для CD38 и PD-L1, соответственно). Биспецифические молекулы изобретения снимают ингибирование иммунной системы, связанное с взаимодействием PD-1 с T-клетками и NK-клетками и PD-L1, экпрессированном на опухоли и клетках микроокружения опухоли. Такие молекулы полезны при лечении раков, в особенности, множественной миеломы или любых CD38+ раков, которые сверхэкспрессируют PD-L1 и растут в микроокружениях экспрессирующих PD-L1 иммунных клеток (плазмацитоидные дендритные клетки, миелоидные супрессорные клетки), что еще больше тормозит T-клетки и NK-клетки. Молекулы изобретения способствуют антитело-зависимой клеточно-опосредованной цитотоксичности (ADCC), фагоцитозу и комплемент-зависимой цитотоксичности (CDC) опухолевых клеток CD38+/PD-L1+, также как и клеток PD-L1+ из клеток микроокружения опухоли.
В одном из воплощений изобретения, были сконструированы несколько биспецифических молекул CD38/PD-L1, включавших анти-CD38 и анти-PD-L1 домены. Данные биспецифические молекулы CD38/PD-L1 были способны одновременно связываться с обоими антигенами.
Более конкретно, были сконструированы биспецифические антитела CD38/PD-L1, включающие анти-CD38 и анти-PD-L1 домены. Данные биспецифические CD38/PD-L1 антитела способны связываться одновременно с обоими антигенами.
В предпочтительном воплощении, биспецифические CD38/PD-L1 антитела экспрессируют в клетках CHO и очищают с помощью аффинной хроматографии, применяя смолы на основе белка А. Антитело-связывающие свойства характеризуют в in vitro анализах. Они одновременно связываются как с CD38, так и с PD-L1 в анализе ELISA.
Биспецифические тетравалентные антитела с четырьмя Fab, имеющие структуру, показанную на фиг. 1A или 1B, были названы BiXAb®, торговая марка Biomunex Therapeutics.
Антитело изобретения представляет собой биспецифическое и бивалентное для CD38 и PD-L1 антитело. Антиген-связывающие биспецифические антитела изобретения представляют собой полноразмерные биспецифические антитела, состоящие из непрерывной «составной тяжелой цепи» (сделанной из природной тяжелой цепи IgG из mAb1 с последующими линкерами и тяжелой цепь Fab из mAb2), которая построена из Fc (шарнир-CH2-CH3) с последующей тяжелой цепью Fab (CH1-VH) антитела 1 и последующей тяжелой цепью Fab (CH1-VH) антитела 2, последний присоединен с помощью полученной из шарнира полипептидной линкерной последовательности, и полученная в результате составная тяжелая цепь в ходе экспрессии белка соединяется с идентичной второй составной тяжелой цепью, в то время как совместно экспрессированные легкие цепи (LC) Fab антитела 2 и антитела 1 соединяются со своими соответствующими доменами тяжелых цепей, чтобы сформировать окончательный тандем молекулы F(ab')2-Fc; антитело 1 (Ab1) и антитело 2 (Ab2) представляют собой разные антитела, и их выбирают из группы, состоящей из анти-CD38 антител (даратумумаб, изатуксимаб, MOR-202 или любое другое анти-CD38 антитело), или их мутированных производных, и анти-PD-L1 антител (атезолизумаб, дурвалумаб, авелумаб, MDX-1105 или любое другое анти-PD-L1 антитело) или их мутированных производных.
Антитела BiXAb® способны связываться бивалентно как с CD38, так и с PD-L1.
Далее описан полипептид, который состоит из тяжелой цепи биспецифического антитела как определено выше, а также полинуклеотид, включающий последовательность, кодирующую указанный полипептид.
Клетка-хозяин, трансфицированная экспрессионным вектором, включающим указанный полинуклеотид, также описана.
Еще один объект изобретения представляет собой способ получения биспецифических антител изобретения.
Таким образом, обеспечивают способ получения биспецифического антитела изобретения, указанный способ, включающий следующие стадии: a) культивирование в подходящей среде и условиях культивирования клетки-хозяина, экспрессирующей тяжелую цепь антитела, как определено выше, и легкую цепь антитела, как определено выше; и b) извлечение указанного продуцированного антитела из культуральной среды или из указанных культивированных клеток.
В изобретении применяют рекомбинантные векторы, в особенности, экспрессионные векторы, включающие полинуклеотиды, кодирующие тяжелые и легкие цепи, определенные в настоящем документе, связанные с элементами, контролирующими транскрипцию и трансляцию, которые активны в выбранной клетке-хозяине. Векторы, которые могут быть применены для построения экспрессионных векторов в соответствии с изобретением, сами по себе известны и будут в частности выбраны в зависимости от клетки-хозяина, которую намереваются применять. Предпочтительно, указанную клетку-хозяина трансформируют полинуклеотидом, кодирующим тяжелую цепь, и двумя полинуклеотидами, кодирующими две разные легких цепи. Указанные полинуклеотиды могут быть вставлены в один и тот же экспрессионный вектор, или в разные экспрессионные векторы. Способ получения антитела изобретения включает культивирование такой клетки-хозяина и извлечение указанных антиген-связывающих фрагментов или антител из указанной культуры.
Каткое описание чертежей
Фиг. 1A и 1B представляют собой схематическими представления биспецифического антитела изобретения, которое включает две тяжелые цепи и четыре легкие цепи.
Фиг. 2 демонстрирует SDS-электрофорез в полиакриламидном геле биспецифических антител BiXAbs 4218, 4219 и 5104 в восстанавливающих условиях.
Фиг. 3 демонстрирует SDS-электрофорез в полиакриламидном геле BiXAbs 4218, 4219 и 5104 в невосстанавливающих условиях.
Фиг. 4 показывает анализ связывания ELISA для BiXAbs 4218 и 4219.
Фиг. 5 показывает SDS-электрофорез в полиакриламидном геле BiXAb-6567 в восстанавливающих и невосстанавливающих условиях. Дорожка 1: пробег BiXAb-6567 в восстанавливающих условиях; дорожка 2: маркеры молекулярной массы с указанной массой каждой полосы; дорожка 3: пробег BiXAb-6567 в невосстанавливающих условиях.
Фиг. 6 показывает анализ BiXAb-6567 способом гель-фильтрационной хроматографии.
Фиг. 7 показывает профили плавления двух родительских антител (анти-CD38 и анти-PD-L1) и BiXAb-6567, определенных с помощью цифровой сканирующей калориметрии.
Фиг. 8A показывает профили связывания двух родительских антител (анти-CD38 и анти-PD-L1) и BiXAb-6567 в анализе связывании антигена CD38 с помощью прямого ELISA. Фиг. 8B показывает профили связывания двух родительских антител (анти-CD38 и анти-PD-L1) и BiXAb-6567 в анализе связывании антигена PD-L1 с помощью прямого ELISA. Фиг. 8C показывает профиль связывания BiXAb-6567 в анализе связывании антигенов (PD-L1 и CD38) в двойном ELISA.
Фиг. 9A-9C показывают профили сортировки клеток с активированной флуоресценцией двух родительских mAbs (анти-CD38 и анти-PD-L1) и BiXAb-6567 на трех разных клеточных линиях, 9A: множественная миелома RPMI-8226, 9B: CHO клетки, стабильно трансфицированные полноразмерным CD38 и 9C: клеточная линия рака яичников SKOV-3.
Фиг. 10 показывает профили титрования связывания на клеточной линии CHO-CD38 двух родительских антител (анти-CD38 и анти-PD-L1), BiXAb-6567 и отрицательного контроля анти-CD20 антитела.
Фиг. 11 показывает профили цитотоксической активности двух родительских антител (анти-CD38 и анти-PD-L1), BiXAb-6567 и двух антител отрицательного контроля, анти-CD20 и анти-HER2, в анализе ADCC, применяющем клеточную линию множественной миеломы, RPMI-8226, в качестве клеток-мишеней и нефракционированные мононуклеарные клетки без предварительной активации в качестве эффекторных клеток.
Фиг. 12 показывает профили цитотоксической активности двух родительских антител (анти-CD38 и анти-PD-L1), BiXAb-6567 и двух антител отрицательного контроля, анти-CD20 и анти-HER2, в анализе ADCC, применяющем клеточную линию CHO-CD38 в качестве клеток-мишеней и нефракционированные мононуклеарные клетки без предварительной активации в качестве эффекторных клеток.
Фиг. 13 показывает профили цитотоксической активности двух родительских антител (анти-CD38 и анти-PD-L1), BiXAb-6567 и двух антител отрицательного контроля, анти-CD20 и анти-HER2, в анализе ADCC, применяющем клеточную линию SKOV-3 в качестве клеток-мишеней и обогащенные IL-12 предварительно активированные NK-клетки в качестве эффекторных клеток.
Фиг. 14 показывает профили цитотоксической активности двух родительских антител (анти-CD38 и анти-PD-L1), BiXAb-6567 и двух антител отрицательного контроля, анти-CD20 и анти-HER2, в анализе ADCC, применяющем клеточную линию SKOV-3 в качестве клеток-мишеней и обогащенные IL-15 предварительно активированные NK-клетки в качестве эффекторных клеток.
Осуществление изобретения
Определения
Базовая структура природной молекулы антитело представляет собой Y-образную тетрамерную четвертичную структуру, состоящую из двух идентичных тяжелых цепей и двух идентичных легких цепей, скрепленных совместно нековалентными взаимодействиями и межцепочечными дисульфидными связями.
У видов млекопитающих, существует пять типов тяжелой цепи: α, δ, ε, γ и μ, которые определяют класс (изотип) иммуноглобулина: IgA, IgD, IgE, IgG и IgM, соответственно. За N-концевым вариабельным доменом (VH) тяжелой цепи следует константная область, содержащая три домена (пронумерованные как CH1, CH2 и CH3 от N-конца к C-концу) в тяжелых цепях γ, α и δ, тогда как константная область тяжелых цепей µ и ε состоит из четырех доменов (пронумерованных как CH1, CH2, CH3 и CH4 от N-конца к C-концу). Домены CH1 и CH2 из IgA, IgG и IgD отделены друг от друга гибким шарниром, который варьируется по длине между разными классами и в случае IgA и lgG, между различными подтипами: lgG1, lgG2, IgG3 и IgG4 имеют, соответственно, шарниры размером в 15, 12, 62 (или 77) и 12 аминокислот, и IgA1 и IgA2 имеют, соответственно, шарниры размером в 20 и 7 аминокислот.
Существует два типа легких цепей: λ и κ, которые могут связываться с любым из изотипов тяжелых цепи, но обе представляют собой один и тот же тип в данной молекуле антитела. Обе легкие цепи, по-видимому, являются функционально идентичными. За их N-концевым вариабельным доменом (VL) следует константная область, состоящая из единственного домена, называемого CL.
Тяжелые и легкие цепи образуют пару путем белок/белок взаимодействий между доменами CH1 и CL и через взаимодействие VH/VL и две тяжелые цепи связаны белок/белок взаимодействиями между своими доменами CH3. Структура, показанная для молекулы иммуноглобулина, обычно стабилизируется межцепочечными дисульфидными связями между доменами CH1 и CL и между шарнирами.
Антиген-связывающие участки соответствуют плечам Y-образной структуры, которая состоят из каждой полной легкой цепи в паре с доменами VH и CH1 тяжелой цепи и называется Fab-фрагментами (означает фрагмент связывания антигена). Fab-фрагменты исходно создают из нативной молекулы иммуноглобулина путем расщепления папаином, который разрезает молекулу антитела в шарнирной области, на амино-концевой стороне межцепочечных дисульфидных связей, таким образом высвобождая два одинаковых антиген-связывающих плеча. Другие протеазы, такие как пепсин, также расщепляют молекулу антитела в шарнирной области, но на карбокси-концевой стороне межцепочечных дисульфидных связей, высвобождая фрагменты, состоящие из двух одинаковых Fab-фрагментов и оставаясь связанными дисульфидными связями; восстановление дисульфидных связей в F(ab')2 фрагментах приводит к образованию фрагментов Fab'.
Часть участка связывания антигена, соответствующая доменам VH и VL, называется Fv-фрагмент (означает вариабельный фрагмент); он содержит CDR (определяющие комплементарность области), которые формируют антиген-связывающий участок (также называемый паратопом).
Эффекторный участок антитела, который отвечает за его связывание с эффекторными молекулами или клетками, соответствует стеблю Y-образной структуры и содержит спаренные домены CH2 и CH3 тяжелой цепи (или домены CH2, CH3 и CH4, в зависимости от класса антитела) и называется участок Fc (означает кристаллизующийся фрагмент).
Благодаря идентичности двух тяжелых цепей и двух легких цепей, природные молекулы антитела имеют два идентичных антиген-связывающих участка и, таким образом, связываются одновременно с двумя идентичными эпитопами.
Антитело «специфически связывается» с антигеном-мишенью, если тот связывается с большей аффинностью, авидностью, быстрее и/или с большей продолжительностью, чем он связывается с другими веществами. «Специфическое связывание» или «преимущественное связывание» не обязательно требует (хотя может включать) эксклюзивное связывание. Как правило, но не обязательно, отсылка к связыванию означает преимущественное связывание.
Термины «субъект», «индивидуум» и «пациент» применяют в настоящем документе взаимозаменяемо, и они относятся к млекопитающему, оцениваемому для лечения и/или подвергаемого лечению. Субъекты могут представлять собой человека, но также включают других млекопитающих, в особенности, тех млекопитающих, которых применяют в качестве лабораторных моделей болезни человека, например, мышь, крысу, кролика, собаку и т.п.
Термин «лечение» относится к действию, применению или терапии, при которых субъекту, включая человека, оказывают медицинскую помощь с целью улучшения состояния субъекта, непосредственно или опосредованно. В особенности, термин относится к снижению или ослаблению симптомов, устранению рецидивов, предотвращению рецидивов, предотвращению случаев, улучшению симптомов, улучшению прогноза или их комбинации в некоторых воплощениях. Специалисту в данной области понятно, что лечение не обязательно приводит к полному отсутствию или устранению симптомов. Например, в отношении рака, «лечение» может относиться к замедлению роста неопластических или злокачественных клеток, пролиферации или метастазирования, предотвращению или задержке развития роста неопластических или злокачественных клеток, пролиферации, или метастазирования, или некоторых их комбинаций.
Конструкция предпочтительных биспецифических антител
Изобретение обеспечивает биспецифические тетравалентные антитела, включающие два сайта связывания с каждой из своих мишеней, и функциональный Fc-домен, позволяющий активировать эффекторные функции, такие как антитело-зависимую клеточную цитотоксичность (ADCC), фагоцитоз и комплемент-зависимую цитотоксичность (CDC).
Антитела изобретения представляют собой полноразмерные антитела. Они предпочтительно включают тяжелые цепи и легкие цепи из иммуноглобулинов человека, предпочтительно, IgG, еще более предпочтительно, IgG1.
Легкие цепи предпочтительно представляют собой легкие цепи kappa.
В предпочтительном воплощении линкер изобретения соединяет две пары Fab-доменов IgG в формате тетра-Fab биспецифического антитела, аминокислотная последовательность которого включает последовательности тяжелой цепи, по меньшей мере, двух Fab, соединенных с помощью линкера, за которыми следует нативная шарнирная последовательность, а затем последовательность Fc из IgG, совместно экспрессируемая с соответствующими последовательностями легкой цепи IgG.
Пример антител изобретения, который имеют IgG-подобную структуру, проиллюстрирован на фиг. 1A и 1B.
Биспецифические антитела изобретения, как правило, включают
- непрерывные тяжелые цепи, построенные из Fc (шарнир-CH2-CH3)
- за которыми следуют тяжелая цепь Fab (CH1-VH) антитела 1 и последующая тяжелая цепь Fab (CH1-VH) антитела 2, последняя присоединена с помощью полученной из шарнира полипептидной линкерной последовательности,
- и во время экспрессии белка полученные тяжелые цепи собираются в димеры, в то время как совместно экспрессированные легких цепи (VL-CL) антитела 1 и антитела 2 связываются со своими соответствующими тяжелыми цепями для того, чтобы сформировать конечный тандем молекулы F(ab)’2-Fc,
антитело 1 (Ab1) и антитело 2 (Ab2) представляют собой разные антитела.
Ab1 и Ab2, представляющие собой разные антитела, независимо выбирают из группы, состоящей из анти-CD38 антитела (такого как даратумумаб) и анти-PD-L1 антитела (такого как атезолизумаб).
Даратумумаб связывается уникальным эпитопом CD38 на С-концевом участке человеческого CD38, и особенно важны для связывания аминокислоты с 233 по 246 и с 267 по 280, с аминокислотами в положениях 272 и 274. Предпочтительно, Ab1 и/или Ab2 могут представлять собой антитела, которые связываются с тем же эпитопом, или перекрывающимся эпитопом (например, с перекрытием, по меньшей мере, в 4 аминокислоты) в отношении даратумумаба.
В другом воплощении, Ab1 и/или Ab2 могут представлять собой антитела, которые связываются с тем же эпитопом, или перекрывающимся эпитопом в отношении атезолизумаба.
В конкретном воплощении, биспецифическая молекула представляет собой биспецифическое антитело, которое включает, предпочтительно, состоит из, a) две тяжелые цепи, каждая включающая, предпочтительно состоящая из, SEQ ID NO: 1, и b) четыре легкие цепи, две включающие, предпочтительно, состоящие из, SEQ ID NO: 2, две другие включающие, предпочтительно, состоящие из, SEQ ID NO: 3. Такое биспецифическое антитело обозначают как BiXAb-4218.
В другом конкретном воплощении, биспецифическая молекула представляет собой биспецифическое антитело, которое включает, предпочтительно состоит из, a) две тяжелые цепи, каждая включающая, предпочтительно, состоящая из, SEQ ID NO: 4, и b) четыре легкие цепи, две включающие, предпочтительно, состоящие из, SEQ ID NO: 5, две другие включающие, предпочтительно состоящие из, SEQ ID NO: 6. Такое биспецифическое антитело обозначают как BiXAb-4219.
В другом конкретном воплощении, биспецифическая молекула представляет собой биспецифическое антитело, которое включает, предпочтительно состоит из, a) две тяжелые цепи, каждая включающая, предпочтительно, состоящая из, SEQ ID NO: 7, и b) четыре легкие цепи, две включающие, предпочтительно, состоящие из, SEQ ID NO: 8, две другие включающие, предпочтительно состоящие из, SEQ ID NO: 9. Такое биспецифическое антитело обозначают как BiXAb-5104.
В предпочтительном воплощении, биспецифическая молекула представляет собой биспецифическое антитело, которое включает, предпочтительно состоит из, a) две тяжелые цепи, каждая включающая, предпочтительно состоящая из, SEQ ID NO: 10, и b) четыре легкие цепи, две включающие, предпочтительно, состоящие из, SEQ ID NO: 11, две другие включающие, предпочтительно состоящие из, SEQ ID NO: 12. Такое биспецифическое антитело обозначают как BiXAb-6567.
Тяжелая цепь (SEQ ID NO: 10) включает
- VH даратумумаба (SEQ ID NO: 22)
- домен CH1 (человеческий IgG1 аллотипа G1m(3) с мутациями L124Q и S188V) Fab даратумумаба (SEQ ID NO: 23)
- линкер AP (SEQ ID NO: 15)
- VH атезолизумаба (SEQ ID NO: 24)
- домен CH1 (человеческий IgG1 аллотипа G1m(3) с мутацией T192D) Fab атезолизумаба (SEQ ID NO: 25)
- Шарнир человеческого IgG1 (SEQ ID NO: 26)
- домен CH2 человеческого IgG1 (SEQ ID NO: 27)
- домен CH3 человеческого IgG1 аллотипа G1m(3) (SEQ ID NO: 28)
Легкая цепь SEQ ID NO: 11 включает
- VL даратумумаба (SEQ ID NO: 29)
- домен Ckappa даратумумаба с мутациями V133T и S176V (SEQ ID NO: 30)
Легкая цепь SEQ ID NO: 12 включает
- VL атезолизумаба (SEQ ID NO: 31)
- домен Ckappa атезолизумаба с мутациями S114A и N137K (SEQ ID NO: 32)
Также были описаны биспецифические антитела с улучшенными свойствами, которые показали более высокую аффинность связывания к CD38 и/или к PD-L1. Например, такие биспецифические антитела могут проявлять Kd менее чем 1 x 10-7 M, 10-8 M, предпочтительно, менее чем 1 x 10-9 или 1 x 10-10 M, в отношении CD38 и/или PD-L1.
Конструкция линкеров
Полипептидный линкер, также обозначаемый как «полученная из шарнира полипептидная линкерная последовательность» или «псевдошарнирный линкер», включает всю или часть последовательности шарнирной области одного иммуноглобулина или нескольких иммуноглобулинов, выбираемых среди IgA, IgG и IgD, предпочтительно, человеческого происхождения. Указанный полипептидный линкер может включать всю или часть последовательности шарнирной области только одного иммуноглобулина. В этом случае, указанный иммуноглобулин может принадлежать к тому же изотипу и подклассу, что и иммуноглобулин, из которого получали примыкающий домен CH1, или к другому изотипу или подклассу. Альтернативно, указанный полипептидный линкер может включать всю или часть последовательности шарнирных областей, по меньшей мере, двух иммуноглобулинов разных изотипов или подклассов. В этом случае, N-концевая часть полипептидного линкера, который непосредственно следует за доменом CH1, предпочтительно, состоит из всей или части шарнирной области иммуноглобулина, принадлежащего к тому же изотипу и подклассу, что и иммуноглобулин, из которого получали указанный домен CH1.
Необязательно, указанный полипептидный линкер может дополнительно включать последовательность, состоящую из от 2 до 15, предпочтительно, от 5 до 10 N-концевых аминокислот домена CH2 иммуноглобулина.
Полипептидная линкерная последовательность, как правило, состоит менее чем из 80 аминокислот, предпочтительно, менее чем из 60 аминокислот, еще более предпочтительно, менее чем из 40 аминокислот.
В некоторых случаях, могут быть применены последовательности из нативных шарнирных областей; в других случаях, точечные мутации могут быть внесены в данные последовательности, в особенности, замена одного или нескольких цистеиновых остатков в нативных шарнирных последовательностях IgG1, IgG2 или IgG3 аланином или серином, для того, чтобы избежать образования нежелательных внутрицепочечных или межцепочечных дисульфидных связей.
В конкретном воплощении, полипептидная линкерная последовательность включает или состоит из аминокислотной последовательности EPKX1CDKX2HX3X4PPX5PAPELLGGPX6X7PPX8PX9PX10GG (SEQ ID NO: 13), в которой X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, идентичные или разные, представляют собой любую аминокислоту. В особенности, полипептидная линкерная последовательность может включать или состоять из последовательности, которую выбирают из группы, состоящей из
EPKSCDKTHTSPPAPAPELLGGPGGPPGPGPGGG (SEQ ID NO: 14);
EPKSCDKTHTSPPAPAPELLGGPAAPPAPAPAGG (SEQ ID NO: 15);
EPKSCDKTHTSPPAPAPELLGGPAAPPGPAPGGG (SEQ ID NO: 16);
EPKSCDKTHTCPPCPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 17) и EPKSCDKTHTSPPSPAPELLGGPSTPPTPSPSGG (SEQ ID NO: 18).
Не ограничивающий пример полученного из шарнира полипептидного линкера, который может быть применен в мультиспецифическом антигены-связывающем фрагменте изобретения, представляет собой полипептид, имеющий последовательность SEQ ID NO: 17. Указанный полипептид состоит из полноразмерной последовательности шарнира человеческого IgG1, за которым следуют 9 N-концевых аминокислот CH2 человеческого IgG1 (APELLGGPS, SEQ ID NO: 19), часть последовательности шарнира человеческого IgA1 (TPPTPSPS, SEQ ID NO: 20) и дипептид GG, добавленный для обеспечения дополнительной гибкости линкеру. В другом предпочтительном воплощении, полученная из шарнира полипептидная линкерная последовательность представляет собой SEQ ID NO: 15 или SEQ ID NO: 18.
В конкретном воплощении, X1, X2 и X3, идентичные или разные, представляют собой треонин (T) или серин (S).
В другом конкретном воплощении, X1, X2 и X3, идентичные или разные, выбирают из группы, состоящей из Ala (A), Gly (G), Val (V), Asn (N), Asp (D) и Ile (I), еще более предпочтительно X1, X2 и X3, идентичные или разные, могут представлять собой Ala (A) или Gly (G).
Альтернативно, X1, X2 и X3, идентичные или разные, могут представлять собой Leu (L), Glu (E), Gln (Q), Met (M), Lys (K), Arg (R), Phe (F), Tyr (T), His (H), Trp (W), предпочтительно, Leu (L), Glu (E), или Gln (Q).
В конкретном воплощении, X4 и X5, идентичные или разные, представляют собой любую аминокислоту, которую выбирают из группы, состоящей из серина (S), цистеин (C), аланин (A) и глицин (G).
В предпочтительном воплощении, X4 представляет собой серин (S) или цистеин (C).
В предпочтительном аспекте, X5 представляет собой аланин (A) или цистеин (C).
В конкретном воплощении, X6, X7, X8, X9, X10, идентичные или разные, представляют собой любую аминокислоту, отличную от треонина (T) или серина (S). Предпочтительно X6, X7, X8, X9, X10, идентичные или разные, выбирают из группы, состоящей из Ala (A), Gly (G), Val (V), Asn (N), Asp (D) и Ile (I).
Альтернативно, X6, X7, X8, X9, X10, идентичные или разные, могут представлять собой Leu (L), Glu (E), Gln (Q), Met (M), Lys (K), Arg (R), Phe (F), Tyr (T), His (H), Trp (W), предпочтительно, Leu (L), Glu (E) или Gln (Q).
В предпочтительном воплощении, X6, X7, X8, X9, X10, идентичные или разные, выбирают из группы, состоящей из Ala (A) и Gly (G).
В еще предпочтительном воплощении, X6 и X7 идентичны и их предпочтительно выбирают из группы, состоящей из Ala (A) и Gly (G).
В предпочтительном воплощении, полипептидная линкерная последовательность включает или состоит из последовательности SEQ ID NO: 13, в которой
X1, X2 и X3, идентичные или разные, представляют собой треонин (T), серин (S);
X4 представляет собой серин (S) или цистеин (C);
X5 представляет собой аланин (A) или цистеин (C);
X6, X7, X8, X9, X10, идентичные или разные, выбирают из группы, состоящей из Ala (A) и Gly (G).
В другом предпочтительном воплощении, полипептидная линкерная последовательность включает, или состоит из, последовательность SEQ ID NO: 13, в которой
X1, X2 и X3, идентичные или разные, представляют собой Ala (A) или Gly (G);
X4 представляет собой серин (S) или цистеин (C);
X5 представляет собой аланин (A) или цистеин (C);
X6, X7, X8, X9, X10, идентичные или разные, выбирают из группы, состоящей из Ala (A) и Gly (G).
Получение биспецифических антител
Специалист может обратиться к международной патентной заявке WO2013/005194, включенной в настоящем документе путем отсылки, для общих технологий экспрессии мультиспецифических антител.
Кроме того, в настоящем документе описан полинуклеотид, включающий последовательность, кодирующую белковую цепь молекулы или антитела изобретения. Указанный полинуклеотид также может включать дополнительные последовательности: в особенности, он может предпочтительно включать последовательность, кодирующую лидирующую последовательность или сигнальный пептид, делающие возможным секрецию указанной белковой цепи. Также раскрыты клетки-хозяева, трансформированные указанным полинуклеотидом.
Как правило, аминокислотные последовательности разных анти-CD38 и анти-PDL-1 моноклональных антител применяли для конструирования последовательности ДНК, необязательно, после оптимизации кодонов для экспрессии в млекопитающих. Для тяжелой цепи синтезировали ДНК, кодирующую сигнальные пептиды, вариабельную область и константный домен CH1 из Fab1, за которыми следует шарнирный линкер и вариабельная область, и константный домен CH1 из Fab2 с фланкирующими последовательностями для расщепления рестрикционными ферментами. Для легкой цепи синтезировали ДНК, кодирующую сигнальные пептиды и вариабельную область и константные области kappa-цепи.
Нуклеиновые кислоты, кодирующие тяжелые и легкие цепи антител изобретения, встраивали в экспрессионные векторы. Легкие и тяжелые цепи могут быть клонированы в одном и том же или в разных экспрессионных векторах. Сегменты ДНК, кодирующие цепи иммуноглобулина, функционально связаны с контрольными последовательностями в экспрессионном векторе (экспрессионных векторах), которые обеспечивают экспрессию полипептидов иммуноглобулина. Такие контрольные последовательности включают сигнальную последовательность, последовательность промотора, энхансера и терминации транскрипции. Экспрессионные векторы, как правило, способны к реплицикации в организмах-хозяевах либо как эписомы, или как интегральная часть хромосомной ДНК хозяина. Как правило, экспрессионные векторы содержат маркеры отбора, например, тетрациклин или неомицин, чтобы обеспечить обнаружение тех клеток, которые были трансформированы желаемыми последовательностями ДНК.
В одном из примеров, обе последовательности, кодирующие тяжелую и легкую цепи (например, последовательности, кодирующие VH и VL, VH-CH1 и VL-CL, или полноразмерную тяжелую цепь и полноразмерную легкую цепь) включают в один экспрессионный вектор. В другом примере, каждую из тяжелых и легких цепей антитела клонируют в индивидуальный вектор. В последнем случае, экспрессионные векторы, кодирующие тяжелые и легкие цепи, могут быть совместно трансфицированы в одну клетку-хозяина для экспрессии обеих цепей, которые могут быть собраны для формирования интактного антитела или in vivo, или in vitro. Альтернативно, экспрессионный вектор, кодирующий тяжелую цепь, и вектор или вектора, кодирующие легкие цепи, могут быть включены в разные клетки-хозяева для экспрессии каждой из тяжелой и легкой цепи, которые затем могут быть очищены и собраны для формирования интактного антитела in vitro.
В конкретном воплощении, клетку-хозяина совместно трансфицируют тремя независимыми экспрессионными векторами, такими как плазмиды, что приводит к совместной продукции всех трех цепей (а именно, тяжелая цепь HC и две легких цепи LC1 и LC2, соответственно) и к секреции биспецифического антитела.
Более конкретно, три вектора могут быть предпочтительно применены в следующем молекулярном отношении 2:1:1 (HC : LC1 : LC2).
Рекомбинантные векторы для экспрессии антитела, описанного в настоящем документе, как правило, содержат нуклеиновую кислоту, кодирующую аминокислотные последовательности антитела, функционально связанные с промотором, либо конститутивным, либо индуцируемым. Векторы может быть подходящим для репликации и для интеграции в прокариот, эукариот или в тех, и в других. Типичные векторы содержат терминаторы транскрипции и трансляции, последовательности инициации и промоторов, полезные для регуляции экспрессии нуклеиновой кислоты, кодирующей антитело. Векторы необязательно содержат универсальные экспрессионные кассеты, содержащие, по меньшей мере, одну независимую последовательность терминатора, последовательности, разрешающие репликацию кассеты, как у эукариот, так и у прокариот, т.е. челночные векторы, и маркеры отбора как для прокариотических, так и для эукариотических систем.
Биспецифические антитела, как описаны в настоящем документе, могут быть получены в прокариотических или эукариотических системах экспрессии, таких как клетки бактерий, дрожжей, мицелиальных грибов, растений, насекомых (например, с применением бакуловирусного вектора) и клетки млекопитающих. Не обязательно, чтобы рекомбинантный антитела изобретения были гликозилированы или экспрессированы в эукариотических клетках; однако, экспрессия в клетках млекопитающих обычно предпочтительна. Примеры применяемых линий клеток-хозяев млекопитающих представляют собой человеческую эмбриональную почечную линию (клетки 293), клетки почки новорождённого хомяка (клетки BHK), клетки яичника китайского хомячка /− или + DHFR (клетки CHO, CHO-S, CHO-DG44, Flp-in CHO), клетки почки африканской зеленой мартышки (клетки VERO) и клетки человеческой печени (клетки Hep G2).
Культура клеток тканей млекопитающих предпочтительна для экспрессии и продукции полипептидов, потому что в данной области уже разработан ряд подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины и включают клеточные линии CHO, различные клеточные линии Cos, клетки HeLa, предпочтительно, клеточные линии миеломы (такие как NS0), или трансформированные B-клетки или гибридомы.
В наиболее предпочтительном воплощении, биспецифические антитела изобретения получают, применяя клеточную линию CHO, наиболее предпочтительно, клеточные линии CHO-S или CHO-DG-44 или их производные.
Экспрессионные векторы для данных клеток могут включать последовательности контроля экспрессии, такие как точка начала репликации, промотор и энхансер, и необходимые сайты обработки информации, такие как сайты связывания рибосом, сайты сплайсинга РНК, сайты полиаденилирования и последовательности терминаторов транскрипции. Предпочтительные последовательности контроля экспрессии представляют собой промоторы, полученные из генов иммуноглобулина, SV40, аденовируса, вируса папилломы крупного рогатого скота, цитомегаловируса и т.п.
Векторы, содержащие представляющие интерес полинуклеотидные последовательности (например, последовательности, кодирующие тяжелую и легкую цепь и последовательности контроля экспрессии) могут быть трансфицированы в клетку-хозяина хорошо известными способами, которые варьируют в зависимости от типа клетки-хозяина. Например, обработка фосфатом кальция или электропорация могут быть применены для других клеток-хозяев (см. в целом, Sambrook et al., Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press, 2nd ed., 1989). Если тяжелые и легкие цепи клонируют в отдельные экспрессионные векторы, то векторы совместно трансфицируют для получения экспрессии и сборки интактного иммуноглобулины.
Клетки-хозяева трансформируют или трансфицируют векторами (например, путем химической трансфекции или электропорационными способами) и культивируют в обычных питательных средах (или модифицированных в зависимости от обстоятельств) для индукции промоторов, отбора трансформантов или амплификация генов, кодирующих желаемые последовательности.
Экспрессия антител может быть временной или стабильной.
Предпочтительно, биспецифические антитела получают способами стабильной экспрессии, в которых клеточные линии стабильно трансфицируют ДНК, кодирующей все полипептидные цепи биспецифического антитела, такого как BiXAb-6567, способные к устойчивой экспрессии, которая позволяет осуществлять производство терапевтических средств. Например, в особенности выгодна стабильная экспрессия в клеточной линии CHO.
После экспрессии, целые антитела, их димеры, индивидуальные легкие и тяжелые цепи, или другие формы иммуноглобулина настоящего изобретения могут быть дополнительно изолированы или очищены для получения препаратов, которые в значительной степени гомогенны для дальнейших анализов и применений. Могут быть применены стандартные способы очистки белка, известные в данной области техники. Например, подходящие процедуры очистки могут включать фракционирование на иммунноаффинных или ионообменных колонках, осаждение этанолом, высокоэффективную жидкостную хроматографию (HPLC), электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), осаждение сульфатом аммония и гель-фильтрацию (см. в целом, Scopes, Protein Purification (Springer-Verlag, N.Y., 1982). Для фармацевтического применения предпочтительны в значительной степени чистые иммуноглобулины с гомогенностью, равной, по меньшей мере, примерно от 90 до 95%, и наиболее предпочтительные с гомогенностью от 98 до 99% или более.
In vitro получение позволяет увеличивать масштаб производства, чтобы дать большое количество желаемых биспецифических антител изобретения. В таких способах могут применять гомогенную суспензионную культуру, например, в аэролифтном реакторе или в непрерывном перемешиваемом реакторе, или иммобилизованную культуру или культуру с захваченными клетками, например, в полых волокнах, микрокапсулах, на агарозном микрогранулах или керамических картриджах.
Мутированные производные и мутации
Полипептидные последовательности, которые связывают CD38, могут происходить из любого анти-CD38 антитела, например, их выбирают из группы, состоящей из даратумумаба, изатуксимаба, MOR-202 или их мутированных производных.
Полипептидные последовательности, которые связывают PD-L1, могут происходить из любого анти-PD-L1 антитела, например, их выбирают из группы, состоящей из атезолизумаба, дурвалумаба, авелумаба, MDX-1105 или их мутированных производных.
Термин «мутированное производное», «мутант» или «функциональный вариант» обозначает последовательность, которая отличается от родительской последовательности, к которой она относится, делецией, вставкой или заменой одной или нескольких аминокислот. Предпочтительно, мутированной производное предпочтительно демонстрирует, по меньшей мере, 80%, предпочтительно, по меньшей мере, 85%, еще более предпочтительно, по меньшей мере, 90% гомологии последовательности с нативной последовательностью. В конкретном воплощении, мутации в значительной степени не влияют на функцию антитела.
Мутированные производные, или функциональные варианты, могут включать цепь VH, которая включает аминокислотную последовательность, по меньшей мере, на 85% (например, на 90%, 92%, 94%, 95%, 96%, 97%, 98% или 99%) идентичную любой из базовых последовательностей, перечисленных в настоящем документе, цепь VL, которая имеет аминокислотную последовательность, по меньшей мере, на 85% (например, на 90%, 92%, 94%, 95%, 96%, 97%, 98% или 99%) идентичную любой из базовых последовательностей, перечисленных в настоящем документе, или обе. Данные варианты способны к связыванию с CD38 и PD-L1. В некоторых примерах, варианты обладают сходной антиген-связывающей аффинностью по отношению к базовым антителам, описанным выше (например, имеющим KD менее чем 1 x 10-7 M, 10-8 M, предпочтительно менее чем 1 x 10-9 или 1 x 10-10 M).
Аффинность связывания определяют в терминах ka (константа скорости ассоциации), kd (константа скорости диссоциации) или KD (равновесная диссоциация). Как правило, специфически связывающийся при применении по отношению к антителу относится к антителу, которое специфически связывается с («узнает») свою мишень(свои мишени) с величиной аффинности (KD) менее чем 10-7 M, предпочтительно, менее чем 10-8 M, например, менее чем 10-9 M или 10-10 M. Меньшее значение KD означает более высокую аффинность связывания (т.е. более сильное связывание), так что величина KD, равная 10-9, указывает на более высокую аффинность связывания, чем величина KD, равная 10-8.
«Процент идентичности» двух аминокислотных последовательностей определяют с помощью алгоритма, приведенного в работе Karlin and Altschul Proc. Natl. Acad. Sci. USA 87:2264-68, 1990, измененного в работе Karlin и Altschul Proc. Natl. Acad. Sci. USA 90:5873-77, 1993. Такой алгоритм включен в программы NBLAST и XBLAST (версия 2,0) в работе Altschul, et al. J. Mol. Biol. 215:403-10, 1990. Поиски BLAST белка могут быть выполнены с помощью программы XBLAST, балл = 50, длина слова = 3, для получения аминокислотной последовательности, гомологичной молекулам представляющего интерес белка. Там, где существуют пропуски между двумя последовательностями, может быть применена программа Gapped BLAST, как описано в работе Altschul et al., Nucleic Acids Res. 25(17):3389-3402, 1997. При применении программ BLAST и Gapped BLAST, могут быть применены параметры по умолчанию соответствующих программ (например, XBLAST и NBLAST).
В других воплощениях, функциональные варианты, описанные в настоящем документе, могут содержать одну или несколько мутации (например, консервативные замены), которые предпочтительно не происходят в остатках, для которых предсказано взаимодействие с одной или несколькими CDR.
В настоящем документе описаны мутированные производные, или функциональные варианты, который в значительной степени идентичны базовому антителу.
Термин «в значительной степени идентичны» или «несущественный» означает, что соответствующие аминокислотные последовательности (например, в каркасных областях (FRs), CDRs, домен VH или VL) варианта отличаются несущественно (например, включая консервативные аминокислотные замены) по сравнению с базовым антителом, так что вариант имеет в значительной степени сходные активности связывания (например, аффинность, специфичность или обе) и биоактивности по отношению к базовому антителу. Такие вариант может включать минорные аминокислотные изменения, например, 1 или 2 замены в 5-аминокислотной последовательности определенной области. В общем, больше замен может быть выполнено в FR-областях, в отличие от CDR-областей, до тех пор, пока они не будут оказывать неблагоприятного воздействия на функцию связывания антитела (например, снижение аффинности связывания на более чем 50% по сравнению с исходным антителом). В некоторых воплощениях, идентичность последовательности может составлять примерно 85%, 90%, 95%, 96%, 97%, 98%, 99% или более, между исходным и модифицированным антителом. В некоторых воплощениях, модифицированное антитело имеет ту же специфичность связывания и имеет, по меньшей мере, 50% аффинности исходного антитела.
Консервативные замены дают молекулы, имеющие функциональные и химические характеристики, сходные с теми молекулами, из которых такие модификации были сделаны. Например, «консервативная аминокислотная замена» может включать замену нативного аминокислотного остатка другим остатком, так что влияние на полярность или заряд аминокислотного остатка в этом положении будет незначительным или его не будет. Желаемые аминокислотные замены (консервативные или неконсервативные) могут быть определены специалистом в данной области техники. Например, аминокислотные замены могут быть применены для идентификации важных остатков последовательности молекулы, или для увеличения или снижения аффинности молекул, описанных в настоящем документе. Варианты, включающие одну или несколько консервативных аминокислотных замен, могут быть получены в соответствии со способами изменения полипептидной последовательности, известными специалисту обычной квалификации в данной области техники, такими как те, что могут быть найдены в ссылках, компилирующих такие способы, например, Molecular Cloning: A Laboratory Manual, J. Sambrook, et al., eds., Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, или Current Protocols in Molecular Biology, F.M. Ausubel, et al., eds., John Wiley & Sons, Inc., New York. Консервативные замены аминокислот включают замены, сделанный среди аминокислот в следующих группах: (a) M, I, L, V; (b) F, Y, W; (c) K, R, H; (d) A, G; (e) S, T; (f) Q, N; и (g) E, D.
Настоящее раскрытие также обеспечивает варианты антитела с улучшенными биологическими свойства антитела, такими как повышенная или сниженная аффинность связывания, или с измененным ADCC свойствами в отношении клеток, экспрессирующих CD38 и/или PD-L1.
Аминокислотная последовательность вариантов антитела может быть получена путем введения соответствующих нуклеотидных изменений в нуклеиновую кислоту антитела, или с помощью пептидного синтеза. Такие модификации включают, например, делеции из и/или вставки в и/или замены остатков в аминокислотной последовательности антитела. Любую комбинацию делеции, вставки и замены выполняют для достижения окончательной конструкции, при условии, что конечная конструкция обладает желаемыми характеристиками. Молекулы нуклеиновой кислоты, кодирующие аминокислотную последовательность вариантов антитела, могут быть получены множеством способов, известных в данной области техники. Данные способы включают, без ограничений, олигонуклеотид-опосредованный (или сайт-направленный) мутагенез, ПЦР-мутагенез и кассетный мутагенез из ранее полученного варианта или невариантной (природной) версии антитела. В одном из воплощений, величина равновесной константы диссоциации (KD) антител изобретения равна менее чем 10-7 M, в особенности, менее чем 10-8 M, 10-9 M или 10-10 M. Аффинность связывания может быть определена с применением технологий, известных в данной области техники, таких как ELISA или анализ биоспецифических взаимодействий (например, с помощью поверхностного плазмонного резонанса), или других технологий, известных в данной области техники.
Любая из молекул, описанная в настоящем документе, может быть исследована для определения ее свойств, таких как антиген-связывающая активность, антиген-связывающая специфичность и биологические функции, с помощью общепринятых способов.
Любая из молекул, описанная в настоящем документе, может быть изменена так, чтобы содержать дополнительные небелковые фрагменты, которые известны в данной области техники и легко доступны, например, путем присоединения ПЭГ, гипергликозилирования и т.п. Представляют интерес модификации, которые могут увеличить период полужизни в сыворотке.
На протяжении настоящего раскрытия, аминокислотные последовательности определяют в соответствии с руководством Kabat et al, Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991).
Мутации могут быть расположены в константных доменах. Биспецифические антитела действительно предпочтительно включают Fab-фрагменты, имеющие мутации на границе доменов CH1 и CL, указанные мутации способствуют родственному спариванию тяжелая цепь/легкая цепь и предотвращению их неправильного спаривания.
В предпочтительном воплощении, биспецифические антитела, описанные в настоящем документе, включают
• два Fab-фрагмента с разными доменами с мутированным CH1 и мутированным CL, состоящими из
a) Fab-фрагмента, имеющего мутированный CH1 и мутированный Ckappa домены, полученные из человеческого IgG1/Kappa, и доменов VH и VL из Ab1,
b) Fab-фрагмента, имеющего мутированный CH1 и мутированный Ckappa домены, полученные из человеческого IgG1/Kappa, и домены VH и VL из Ab2,
c) мутированную легкую цепь константного домена, которая получена из человеческого константного домена kappa,
Fab-фрагменты тандемно расположены в следующем порядке
- C-конец мутированного домена CH1 Fab-фрагмента из Ab1, соединенный с N-концом домена VH Fab-фрагмента из Ab2 через полипептидный линкер,
- шарнирная область человеческого IgG1, связывающая C-конец мутированного домена CH1 из фрагмента Ab2 с N-концом домена CH2,
- димеризованные домены CH2 и CH3 человеческого IgG1.
В конкретных примерах, описаны биспецифические антитела, в которых CH1-домен Fab одного из Ab1 или Ab2 представляет собой мутированный домен, который получают из домена CH1 иммуноглобулина путем замены остатка треонина в положении 192 указанного домена CH1 аспарагиновой кислотой, и соответствующий домен CL представляет собой мутированный домен, который получают из домена CL иммуноглобулина заменой остатка аспарагина в положении 137 указанного домена CL остатком лизина и заменой остатка серина в положении 114 указанного домена CL остатком аланина, и/или в котором домен CH1 Fab одного или другого из Ab1 или Ab2 представляет собой мутированный домен, который получают из домена CH1 иммуноглобулина путем замены остатка лейцина в положении 124 указанного домена CH1 глутамином и заменой остатка серина в положении 188 указанного домена CH1 остатком валина, и соответствующий домен CL представляет собой мутированный домен, который получают из домена CL иммуноглобулина путем замены остатка валина в положении 133 указанного домена CL остатком треонина и заменой остатка серина в положении 176 указанного домена CL остатком валина.
Антитела изобретения могут быть гликозилированными или негликозилированными, или могут демонстрировать разнообразные профили гликозилирования. В предпочтительном воплощении, антитела негликозилированы в вариабельной области тяжелой цепи, но гликозилированы в области Fc.
Определенные мутированные производные могут применять гуманизированные формы базового антитела. При гуманизации, определяющие комплементарность области (CDR) и определенные другие аминокислоты из донорских вариабельных областей мыши пересаживают на человеческие вариабельные акцепторные области и затем присоединяют к человеческим константным областям. Смотри, например, Riechmann et al., Nature 332:323-327 (1988); U.S. Pat. No. 5,225,539.
Терапевтическое применение
Биспецифическую молекулу, предпочтительно антитело, изобретения применяют в качестве лекарственного средства, в особенности, в лечении рака.
Термин «рак», как применен в настоящем документе включает любой рак, в особенности, злокачественное заболевание системы крови и любой другой рак, характеризуемый экспрессией или сверхэкспрессией CD38 или PD-L1, и в особенности, данные раки характеризуются совместной экспрессией обоих CD38 и PD-L1.
Примеры раков представляют собой лимфому или лейкемию, такие как неходжкинская лимфома (NHL), острая лимфоцитарная лейкемия (ALL), острая миелоидная лейкемия (AML), хроническая лимфоцитарная лейкемия (CLL), хроническая миелоидная лейкемия (CML) или множественная миелома (MM), рак молочной железы, рак яичников, рак головы и шей, рак мочевого пузыря, меланома, колоректальный рак, рак поджелудочной железы, рак легкого, лейомиома.
Таким образом, описан способ лечения пациента, страдающего от рака путем введения биспецифической молекулы в соответствии с изобретением указанному пациенту, нуждающемуся в таком лечении. Другой аспект изобретения, таким образом, заключается в применении биспецифической молекулы в соответствии с изобретением для производства лекарственного средства для лечения рака.
Один из аспектов изобретения представляет собой фармацевтическую композицию, включающую биспецифическую молекулу в соответствии с изобретением. Другой аспект изобретения представляет собой применение биспецифической молекулы в соответствии с изобретением для производства фармацевтической композиции. Дополнительный аспект изобретения представляет собой способ производства фармацевтической композиции, включающей биспецифическую молекулу в соответствии с изобретением.
В другом аспекте, настоящее изобретение обеспечивает композицию, например, фармацевтическую композицию, содержащую биспецифические молекулы, определенные в настоящем документе, составленные вместе с фармацевтическим носителем.
Как применен в настоящем документе, термин «фармацевтический носитель» включает любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические и задерживающие всасывание средства и т.п., которые физиологически совместимы. Предпочтительно, носитель подходит для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии).
Композиция настоящего изобретения может быть введена множеством способов, известных в данной области техники. Путь и/или способ введения будут изменяться в зависимости от желаемых результатов.
Для введения биспецифической молекулы или антитела изобретения с помощью определенных путей введения, может быть необходимо покрыть биспецифическую молекулу или антитело изобретения материалом, или совместно вводить биспецифическую молекулу или антитело изобретения с материалом для предупреждения их инактивации. Например, биспецифическая молекула или антитело изобретения могут быть введены субъекту в подходящем носителе, например, в липосомах или разбавителе. Фармацевтически приемлемые разбавители включают солевые и водные буферные растворы. Фармацевтические носители включают стерильные водные растворы или дисперсии и стерильные порошки для экстемпорального получения стерильных растворов для инъекций или дисперсии. Применение таких сред и средств для фармацевтически активных вещества известно в данной области техники.
Данные композиции также могут содержать вспомогательные вещества, такие как консерванты, смачивающие средства, эмульгаторы и диспергирующие средства. Предупреждение присутствия микроорганизмов могут обеспечивать как процедурами стерилизации, так и путем включения различных антибактериальных и противогрибковых средств, например, парабена, хлорбутанола, фенола, сорбиновой кислоты и т.п. Также может быть желательно включать в композиции изотонические средства, такие как хлорид натрия. Кроме того, пролонгированное всасывание инъекционных фармацевтических форм может быть вызвано путем включения средств, которые задерживают всасывание.
Фактические уровни дозировки активных ингредиентов в фармацевтических композициях настоящего изобретения можно варьировать, чтобы получить такое количество активного ингредиента, которое эффективно для достижения желаемого терапевтического ответа для конкретного пациента, композиции и способа введения которое не станет токсичным для пациента.
Выбранный уровень дозировки будет зависеть от разнообразных фармакокинетических факторов, включая активность конкретных примененных композиций настоящего изобретения, пути введения, время введения, продолжительность лечения, другие лекарственные препараты, соединения и/или материалы, примененные в комбинации с конкретными примененными композициями, возраст, пол, вес, состояние, общее состояние здоровья и предыдущая история болезни подвергаемого лечению пациента и тому подобные факторы, хорошо известные в области медицины. Например, биспецифическую молекулу или антитело изобретения можно вводить в дозировке, равной 0,2-20 мг/кг от 3 раз/неделю до 1 раза/месяц.
Настоящее изобретение, таким образом описанное выше в общем виде, будет легче понимать с отсылкой к следующим примерам, которые предусмотрены в качестве иллюстрации и не предназначены для ограничения настоящего изобретения.
Примеры
Пример 1. Получение биспецифических антител BiXAb-4218, BiXAb-4219 и BiXAb-5104
Синтез генов
Аминокислотные последовательности разных анти-CD38 и анти-PDL-1 моноклональных антител применяли для конструирования последовательности ДНК после оптимизации кодонов для экспрессии в млекопитающих с помощью программы GeneScript. Для тяжелой цепи, ДНК, кодирующую сигнальные пептиды, вариабельную область и константный домен CH1 из Fab1, за которыми следует шарнирный линкер и вариабельная область, и константный домен CH1 из Fab2 с фланкирующими последовательности для расщепления рестрикционными ферментами, синтезировали с помощью GeneScript. Для легкой цепи, ДНК, кодирующую сигнальные пептиды и вариабельную и константную kappa области, синтезировали с помощью GeneScript.
ПЦР-реакции осуществляли с помощью PfuTurbo Hot Start для амплификации вставок, которые затем расщепляли NotI + ApaI и NotI + HindIII для тяжелых и легких цепей, соответственно. Дважды расщепленные фрагменты тяжелых цепей лигировали с расщепленным NotI + ApaI экспрессионным вектором, принадлежавшим Evitria, в котором человеческий IgG1 CH1 + шарнир + CH2 + CH3 домены были уже вставлены. Дважды расщепленные фрагменты легкой цепи лигировали с вектором, принадлежавшим Evitria, обработанным NotI + HindIII. Плазмидные ДНК проверяли с помощью секвенирования последовательности двухцепочечной ДНК.
Экспрессия, очистка и характеристика
Для экспрессии в масштабе 50 мл, всего 50 мкг плазмидных ДНК в векторе, принадлежавшем Evitria (25 мкг тяжелых цепей + 12,5 мкг каждой из легкой цепи, LC1 и LC2), смешивали в 1,5 мл пробирке Eppendorf, добавляли 1 мл среды CHO SFM, содержавшей 25 мкл 3 мг/л PEI, pH 7,0, инкубировали при комнатной температуре в течение 20 мин. Смесь ДНК-PEI помещали в 49 мл клеток FreeStyle™ CHO-S при 1-2 x 106 клеток/мл в 125 мл встряхиваемых колбах. Клетки перебалтывали в течение дополнительных 6 дней. Супернатант собирали путем центрифугирования клеток при 3000 rpm в течение 15 мин. Собранный супернатант очищали на смоле с белком A. Электрофорез проводили в восстанавливающих условиях и невосстанавливающих условиях, применяя 4-15% гели Gel Biorad Stain-Free и соответствующий электродный буфер. Образцы получали путем объединения очищенных антител BiXAb® с 2X SDS-буфером для образцов и нагревания в течение 5 мин при 95°C. Получение восстановленных образцов включало добавление восстанавливающего агента NuPAGE перед нагреванием. Кажущиеся MW определяли с помощью смеси неокрашенных белков Ladder Precision Plus Protein Unstained Standards («Biorad»). Фиг. 2 демонстрирует образцы SDS-PAGE для антитела CD38/PD-L1 в восстанавливающих условиях. Наблюдали две полосы, соответствующие составным тяжелым цепям, и две совместно мигрирующим соответствующие легким цепям, которые имели ожидаемую молекулярную массу. Фиг. 3 демонстрирует образцы SDS-PAGE для антитела CD38/PD-L1 в невосстанавливающих условиях. Доминирующая полоса при 250 кДа, как и ожидалось, соответствует полной молекуле CD38/PD-L1 BiXAb®.
Для анализа с помощью ELISA связывания с планшетом двух антигенов применяли следующие реагенты: рекомбинантный человеческий CD38, Fc-меченый («Creative BioMart»); биотинилированный человеческий PD-L1, Avi Tag («AcroBiosystems»); стрептавидин-HRP, («Biotechne RD-Systems»). Планшеты «Maxisorp» покрывали человеческим слитым белком CD38-Fc, 2 мкг/мл в 1X PBS, pH7,4, в количестве 100 мкл/лунку при 4ºС в течение ночи. Планшеты промывали 5 раз с 1X PBS, содержавшем 0,05% Tween-20 (1X PBST), затем блокировали 3%-ным обезжиренным молоком/1X PBST, 200 мкл/лунку, при встряхивании при комнатной температуре в течение 1-го часа. Добавляли 100 мкл/лунку BiXAb® 4218 и BiXAb® 4219, концентрация исходного раствора 1 мг/мл, начиная с разведения 1/500 в 1X PBS в сериях разведений 1:3. Планшеты инкубировали при комнатной температуре в течение 1-го часа при встряхивании, за этим следовали 5 промывок с 1X PBST. Добавляли 100 мкл/лунку из 1 мкг/мл биотин-человеческий PD-L1 белок в 1X PBS, и планшеты встряхивали при комнатной температуре в течение 1-го часа. После 5 промывок с 1X PBST, добавляли 100 мкл/лунку 0,1 мкг/мл стрептавидин-конъюгированного HRP в 1XPBS. Планшеты встряхивали при комнатной температуре в течение 1-го часа, затем следовали 5 промывок с 1X PBST. Добавляли 100 мкл/лунку субстрата TMB в 1X PBS для развития окраски. Данные регистрировали при 405 нм, в течение 0,1 сек на лунку с помощью мультифункционального спектрофотометра для прочтения планшетов Victor II. Фиг. 4 демонстрирует профили связывания двойного антигена двух CD38/PD-L1 BiXAbs®. Данный профиль подтверждает, что оба типа доменов связывания данных молекул (анти-CD38 домены и анти-PD-L1 домены) связывают свои соответствующие антигены-мишени.
Пример 2. Получение биспецифического антитела изобретения BiXAb-6567
Синтез гена
Аминокислотную последовательность анти-CD38 (даратумумаб) и анти-PDL1 (атезолизумаб) применяли для конструирования последовательности ДНК, после оптимизации кодонов для экспрессии в млекопитающих, с помощью программы GeneScript. Данные антитела называют «родительскими» анти-CD38 и «родительскими» анти-PD-L1 mAbs.
ДНК-конструкцию тяжелой цепи конструировали следующим образом: сигнальный пептид (SEQ ID NO: 21), за которыми следует последовательность SEQ ID NO: 10 [состоящая из вариабельной области, за которой следует константный домен CH1 из Fab1 (анти-CD38), в который были введены мутации Leu на Gln и Ser на Val в положении по Кэбату 124 и 188, соответственно, за которым следует линкер, за которым следует вариабельная область, за которой следует константный домен CH1 из Fab2 (анти-PD-L1), в который была введена мутация Thr на Asp в положении 192 по Кэбату]; фланкирующие последовательности были введены для расщепления рестрикционными ферментами с обоих концов тяжелой цепи ДНК-конструкции. ДНК-конструкцию для легкой цепи конструировали следующим образом: сигнальный пептид (SEQ ID NO: 21), за которым следует вариабельная область, за которой следует константная kappa область. Для легкой цепи анти-CD38, мутации были введены в положениях 143 (Leu на Gln) и 188 (Ser на Val) по Кэбату в константный домен kappa. Для анти-PDL1 легкой цепи, мутации были введены в положении 133 (Val на Thr) и 176 (Ser на Val) по Кэбату в константны домен kappa. Все ДНК-конструкты синтезировали с помощью Gene Art.
ПЦР-реакции осуществляли с помощью PfuTurbo Hot Start для амплификации вставок, которые затем расщепляли с NotI и ApaI и NotI и HindIII для тяжелых и легких цепей, соответственно. Дважды расщепленные фрагменты тяжелой цепи лигировали с обработанным NotI и ApaI экспрессионным вектором pcDNA3.1 («Invitrogen»), в который уже был вставлен шарнир человеческого IgG1, за которыми следовали домены CH2-CH3. Дважды расщепленные фрагменты легкой цепи лигировали с обработанным NotI и HindIII экспрессионным вектором pcDNA3.1 («Invitrogen»). Плазмидные ДНК проверяли с помощью секвенирования последовательности двухцепочечной ДНК.
Экспрессия и очистка
Биспецифические антитело BiXAb-6567 получали, применяя временную экспрессию гена путем совместного трансфекции 3 генов, закодированных на отдельных векторах в молекулярном соотношении 2:1:1 = HC:LC1:LC2 (1 непрерывная тяжелая цепь (HC) и 2 легкие цепи (LC)) в клетках CHO-S, адаптированных к свободной от сыворотки среде в суспензии (среда CHO SFM-II, «Life Technologies»™). Как правило, для экспрессии в масштабе 50 мл, в общей сложности 50 мкг плазмидной ДНК (25 мкг тяжелой цепи, 12,5 мкг легкой цепи анти-CD38 и 12,5 мкг легкой цепи анти-PD-L1) смешивали в 1,5 мл пробирке Eppendorf, затем добавляли 1 мл среды CHO SFM, содержавшей 25 мкл 3 мг/л трансфекционного реагента PEI pH7,0 («Polyplus»), и реакцию инкубировали при комнатной температуре в течение 20 мин. Затем смесь ДНК-PEI добавляли к 49 мл клеток CHO-S «Life Technologies’ Invitrogen» FreeStyle™», в концентрации 1~2x106/мл в 125 мл встряхиваемой колбе. Клетки встряхивали в течение 6-ти дней. Супернатант собирали центрифугированием при 3000 rpm в течение 15 мин. Экспрессионный титр BiXAb-6567 в супернатанте определяли с помощью биосенсоров на белок А (ForteBio’s protein A («Octet® Systems»)). BiXAb-6567 затем очищали на аффинной с белком A (MabSelect SuRe, «GE Healthcare» Life Sciences). Антитело элюировали с белка A с помощью 0,1 M глицина pH 3,5 и элюат нейтрализовали 1 M TRIS. Очищенное антитело, в Дульбекко PBS («Lonza»), стерилизовали фильтрованием (0,2 мкМ фильтр для стерильной фильтрации, Techno Plastic Products AG) и конечную концентрацию определяли путем регистрации оптической плотности (OD) при 280 нм (Eppendorf BioSpectrometer®).
BiXAab-6567, как правило, демонстрирует хороший титр экспрессии (> 180 мг/литр) в транзиторной экспрессии в CHO. Данный уровень экспрессии сопоставим с уровнем экспрессии, наблюдаемым с обычными моноклональными антителами.
SDS-электрофорез в полиакриламидном геле
Для того чтобы оценить качество очищенных BiXAb-6567, мы проводили SDS-PAGE (автоматизированная система для проведения электрофореза Experion™, «BioRad»). В присутствии додецилсульфата натрия (SDS) в электродном буфере, скорость, с которой антитело мигрирует в геле, зависит, прежде всего, от его размера, что позволяет проводить определение молекулярной массы. Данный анализ был выполнен в невосстанавливающих условиях и в восстанавливающих условиях; последние приводят к разрушению дисульфидных связей и, следовательно, к визуализации индивидуальных полипептидных цепей (легких цепей и тяжелых цепей).
Результаты SDS-PAGE представлены на фиг. 5. В невосстанавливающих условиях, поддерживается четвертичная структура антитела, и наблюдаемая молекулярная масса должна представлять собой сумму молекулярных масс разных тяжелых и легких цепей. Биспецифическое антитело изобретения (BiXAb-6567) состоит из шести цепей: две тяжелые цепи и четыре легкие цепи. Теоретическая молекулярная масса BiXab-6567 равна 244,40 кДа, без учета пост-трансляционной модификации (PTM), например, N-гликозилирования в Fc на аспарагине 297. Гель был откалиброван с помощью смеси стандартов известной молекулярной массы. Данные в невосстанавливающих условиях демонстрировали основную полосу, движущуюся близко к стандарту молекулярной массы 250 кДа, что находится в соответствии с расчетной молекулярной массой и ожидаемым гликозилированием двух аспарагинов в положении 297 в домене Fc. В восстанавливающих условиях, дитиотреитол (DTT) дополнительно разрушает BiXAb-6567 в результате восстановления дисульфидных связей и нарушения четвертичной структуры, и, таким образом, шесть полипептидных цепей должны мигрировать в геле отдельно в соответствии с их молекулярной массой. Две идентичные тяжелые цепи BiXAb-6567 совместно мигрируют как единственная полоса, и две пары легких цепей, из-за их почти идентичной молекулярной массы, совместно мигрируют как вторая полоса. Следовательно, данные показывают две основные полосы, приблизительно при 75 кДа и 25 кДа, на основе подвижности стандартов молекулярной массы. Каждая тяжелая цепь обладала одним сайтом N-гликозилирования на аспарагине 297, что объясняет ширину полосы с более высокой молекулярной массой и то, что наблюдаемая молекулярная масса была немного выше расчетной (75,44 кДа); это уширение типично для гликозилированных белков. Рассчитанные молекулярные массы легких цепей анти-CD38 (23,40 кДа) и анти-PD-L1 (23.36 кДа) были очень похожи, что, таким образом, приводит к их совместной миграции.
В заключение, SDS-PAGE BiXAb-6567 продемонстрировал ожидаемые профили, как в невосстанавливающих и в восстанавливающих условиях и согласуется с рассчитанными теоретическими молекулярными массами, при учете существования сайта N-гликозилирования в тяжелой цепи.
Анализ с помощью гель-фильтрационной хроматографии
Агрегация белков часто наблюдается в молекулах белка, сконструированных генно-инженерными способами. Мы провели аналитическую гель-фильтрационную хроматографию (SEC) для анализа содержания форм с высокой молекулярной массой в одноступенчатом получении BiXAb-6567 с аффинной очисткой (смотри раздел Экспрессия и очистка вариантов). Мы применяли колонку SEC-s3000 (300x 7,8 мм) («BioSep») и систему Aktapurifier 10 («GE Healthcare»); анализ проводили при скорости элюции 1 мл/мин с помощью PBS-буфера, pH 7,4.
Хроматограмма гель-фильрации, представленная на фиг. 6, демонстрирует, что основной пик соответствует ожидаемому размеру мономерного BiXAb-6567; данный пик составил 98,2% от общего образца. Кроме того, наблюдали небольшой пик, соответствующий высокомолекулярным видам (возможно, димерам); этот пик составлял 1,8% от общего образца. Таким образом, мы пришли к выводу, что процентное содержание форм с высокой молекулярной массой минорно и сходно с ситуацией для обычных моноклональных антител, получаемых в системе экспрессии CHO. Узкая и симметричная форма мономерного пика предполагает, что BiXAb-6567 был правильно собран и представлен единственной формой.
Пример 3. Характеристика BiXAb-6567 с помощью дифференциальной сканирующей калориметрии
Дифференциальную сканирующую калориметрию (DSC) применяли для сравнения термической стабильности BiXAb-6567, родительского анти-CD38 mAb и родительского анти-PD-L1 mAb. Систему MicrocalTM VP-Capillary DSC («Malvern Instruments») применяли для выполнения экспериментов способом дифференциальной сканирующей калориметрии.
Все образцы центрифугировали (20000 x g, 5 мин, 4°C) и количественно определяли содержание в них белка перед анализом способом DSC с помощью спектрофотометра Nanodrop ND-1000 («Thermo Scientific»), применяя программу анализа Ig. Для анализа, все образцы разводили в PBS до конечной концентрации 1 мг/мл.
Время предварительного уравновешивания было равно 3 мин, и результирующие термограммы получали между 20 и 110°C со скоростью сканирования 60°C/час, с периодом фильтрации 25 сек и средним ответом. Перед анализом образца, 5 сканов буфер/буфер измеряли для стабилизации инструмента и сканирование буфер/буфер выполняли между каждым сканированием белок/буфер. Данные описывали моделью разворачивания не для двух устойчивых состояний, с пре- и постпереходом, скорректированным путем вычитания базовой линии.
Кривые DSC, представленные на фиг. 7 (охватывающие диапазон от 50 до 100°) демонстрируют способ, в котором индивидуальные Fv-области могут приводить к разным профилям разворачивания для Fab; данный эксперимент также демонстрирует, что Fv-области диктуют кажущиеся стабильности Fab. Профиль DSC для анти-CD38 mAb демонстрирует два перехода: большой пик, имеющий Cp max, равный 170 ккал/моль/°C, и Tm1, равную 70,9°C, соответствующие разворачиванию обоих доменов CH2 и Fab, и небольшой пик, имеющий Cp max, равный 20 ккал/моль/°C, и Tm2, равную 81,5oC, соответствующие разворачиванию домена CH3. Профиль DSC анти-PD-L1 mAb демонстрирует два перехода: небольшой пик, имеющий Cp max, равный 20 ккал/моль/°C, и Tm1, равную 69,9°C, соответствующие разворачиванию домена CH2, и большой пик, имеющий Cp max, равный 160 ккал/моль/°C, и Tm2, равную 83,4°C, соответствующие разворачиванию обоих доменов CH3 и Fab.
Профиль DSC BiXAb-6567 также демонстрирует два перехода с двумя большими пиками. Первый пик имел Cp max, равный 130 ккал/моль/°C, и Tm1, равную 71,5°C, и соответствовал разворачиванию доменов CH2 и Fab анти-CD38 mAb; второй пик имел Cp max, равный 170 ккал/моль/°C, и Tm2, равную 81,5°C, и соответствовал разворачиванию доменов CH3 и Fab анти-PD-L1 mAb. Таким образом, профиль DSC для BiXAb-6567 напоминал суперпозицию двух DSC-профилей двух родительских mAbs и проиллюстрировал отличную сборку и стабильность BiXAb-6567. Tonset для BiXAb-6567 (63,3°C) был сходным с таковым для родительского mAbs (анти-CD38 Tonset=63,5°C и анти-PD-L1 Tonset=63,2°C), указывая на то, что BiXAb-6567 обладал свойствами стабильности, сходными со свойствами родительского антитела. Рассчитанный ΔH для BiXAb-6567 был равен 1560 ккал/моль, что отражает больший размер биспецифической молекулы по сравнению с двумя родительскими антителами (для анти-CD38 ΔH=963 ккал/моль и для анти-PD-L1 ΔH =820 ккал/моль).
Определения
Tm или температура денатурации/плавления представляет собой точку, при которой концентрация развернутой и свернутой формы равна и представляет собой середину перехода разворачивания. В качестве параметра термин описывает восприимчивость белка к тепловой денатурации и, таким образом, он относится к стабильности белка. Чем выше Tm, тем более стабилен белок.
Tonset представляет собой температуру, при которой начинается переход разворачивания. Значения этого параметра обычно от 5 до 10°C ниже, чем Tm. Также это представляет собой параметр, описывающий стабильность белка, но с учетом сопротивления термической денатурации.
ΔH представляет собой калориметрическую энтальпию разворачивания и отражает нарушение внутримолекулярных взаимодействий в белке (т.е. разрушение внутри- и междудоменных взаимодействий). Процесс теплового разворачивания эндотермический и, таким образом, дает положительные значения энтальпии. Калориметрическая энтальпия (ΔH) представляет собой площадь под пиком перехода термического разворачивания.
Пример 4. Свойства бесклеточного связывания для BiXAb-6567
Прямой анализ ELISA связывания антигена CD38 на планшетах
100 мкл одного из родительских mAb, анти-CD38 или анти-PDL1, каждый в концентрации 3 мкг/мл, подготовленного разведением PBS pH 7,4, применяли для покрытия планшетов «Maxisorp» при 4°C в течение ночи. Кроме того, BiXAb-6567, в концентрации 5 мкг/мл, подготовленный разведением PBS, pH 7,4, применяли для покрытия планшетов «Maxisorp» при 4°C в течение ночи. Планшеты промывали 5 раз 1x PBS, содержавшем 0,05% Tween-20 (PBST), и затем блокировали 200 мкл/лунку 1% BSA в 1X PBS при комнатной температуре в течение 2 часов. Планшеты затем промывали 5 раз с 1X PBST. Готовили семиточечные серии 3-кратных разведений рекомбинантного His/Flag-меченого CD38 («Creative Biomart») в 1x PBS, начиная с 1 мкг/мл; 100 мкл каждой стадии разведения добавляли на лунку анализа. Планшеты инкубировали при комнатной температуре в течение 1-го часа и промывали 5 раз с 1X PBST. Добавляли 100 мкл/лунку анти-Flag-tag антитело-конъюгированного HRP (Abcam), разведенного в 10000 раз в 1x PBS, и планшеты инкубировали при комнатной температуре в течение 1-го часа. После 5 промывок с 1X PBST, добавляли 100 мкл/лунку субстрата TMB в 1X PBS для колориметрического считывания, и планшеты инкубировали в течение 15 мин при комнатной температуре для развития окрашивания. Данные анализа собрали, применяя считыватель для микропланшетов (Victor2 «Perkin Elmer») при 650 нм.
BiXAb-6567 демонстрировал кривую дозозависимого связывания очень сходную с кривой родительского анти-CD38 антитела (фиг. 8A). EC50 связывания CD38 для обоих антител были следующими: EC50[BiXAb-6567] = 171 нг/мл и EC50[анти-CD38] = 199 нг/мл. Данный результат предполагает, что BiXAb-6567 обладает правильно собранными анти-CD38 Fab-доменами, поскольку оно демонстрирует связывание, сходное со связыванием родительского анти-CD38 mAb. Родительское анти-PDL1 mAb, примененное в качестве отрицательного контроля, не продемонстрировало какого-либо связывания, как и ожидалось.
Прямой анализ ELISA связывания антигена PDL1 на планшетах
100 мкл биотинилированного человеческого белка PD-L1 («AcroBiosystems») в концентрации, равной 1 мкг/мл, подготовленного разведением в 1x PBS pH 7,4, применяли для покрытия планшетов «Maxisorp» при 4°C в течение ночи. Планшеты промывали 5 раз PBST и затем блокировали 200 мкл/лунку 1%-ным BSA в 1X PBS при комнатной температуре в течение 2 часов. Планшеты затем промывали 5 раз с 1X PBST. Подготавливали семиточечные серии с 3-кратным разведением или анти-CD38 mAb (начиная с 0,3 мг/мл), или анти-PD-L1 mAb (начиная с 0,3 мг/мл), или BiXAb-6567 (начиная с 0,5 мг/мл) в 1X PBS; 100 мкл каждой стадии разведения добавляли на лунку анализа. Планшеты инкубировали при комнатной температуре в течение 1-го часа и промывали 5 раз с 1X PBST. Добавляли 100 мкл/лунку антитела к человеческим антителам (IgG H&L), конъюгированные с HRP («Abliance»), разводили в 5000 раз в 1x PBS, и планшеты инкубировали при комнатной температуре в течение 1-го часа. После 5 промывок с 1X PBST, 100 мкл/лунку субстрата TMB в 1X PBS добавляли для колориметрического считывания, и планшеты инкубировали в течение 15 мин при комнатной температуре для развития окрашивания. Данные анализа собирали, применяя считыватель для микропланшетов Victor2 («Perkin Elmer») при 650 нм.
BiXAb-6567 демонстрировали кривую дозозависимого связывания, очень сходную с кривой родительского анти-PD-L1 антитела (фиг. 8B). EC50 связывания PD-L1 для обоих антител были следующими: EC50[BiXAb-6567] = 93 нг/мл и EC50[анти-PD-L1] = 72 нг/мл. Данный результат предполагает, что BiXAb-6567 обладал правильно собранными анти-PD-L1 Fab доменами, поскольку оно демонстрировало связывание, сходное со связыванием родительского анти-PD-L1 mAb. Родительское анти-CD38 mAb, примененное в качестве отрицательного контроля, не продемонстрировало какого-либо связывания, как и ожидалось.
Анализ ELISA связывания двойного антигена
100 мкл рекомбинантного человеческого Fc-меченого CD38 («Creative BioMart»), при 2 мкг/мл, подготовленного разведением в 1x PBS pH7,4, применяли для покрытия планшетов «Maxisorp» при 4°C в течение ночи. Планшеты промывали 5 раз с 1X PBST и затем блокировали 200 мкл/лунку 1% BSA в 1X PBS при комнатной температуре в течение 2-х часов. Планшеты промывали 5 раз с 1X PBST. Готовили семиточечные трехкратные серии разведений BiXAb-6567 в 1X PBS (начиная с 1 мкг/мл), и 100 мкл каждой стадии разведения добавляли на лунку анализа. Планшеты инкубировали при комнатной температуре в течение 1-го часа и затем промывали 5 раз с 1X PBST. Добавляли 100 мкл/лунку 1 мкг/мл биотинилированного человеческого PD-L1 («AcroBiosystems») в 1X PBS, и планшеты инкубировали при комнатной температуре в течение 1-го часа. После 5 промывок с 1X PBST, добавляли 100 мкл/лунку 0,1 мкг/мл стрептавидин-конъюгированного HRP («Biotechne»), подготовленного разведением в 1x PBS. Планшеты инкубировали при комнатной температуре в течение 1-го часа. После 5 промывок с 1X PBST, добавляли 100 мкл/лунку субстрата TMB в 1X PBS для колориметрического считывания, и планшеты инкубировали в течение 15 мин при комнатной температуре для развития окрашивания. Данные анализа собирали, применяя считыватель для микропланшетов Victor2 («Perkin Elmer») при 650 нм.
BiXAb-6567 демонстрировало кривую дозозависимого связывания в формате двойного ELISA, позволяя предположить, что он обладал правильно собранными доменами Fab анти-CD38 и анти-PD-L1 (фиг. 8C). Данные показывают, что BiXAb-6567 представляет собой биспецифическое антитело способное к связыванию CD38 и PD-L1 одновременно с EC50 = 144 нг/мл. Ни одно из двух родительских mAbs, анти-CD38 или анти-PDL1, не продемонстрировало какого-либо связывания в данном двойном формате ELISA, как и ожидалось.
Пример 5. Определение относительной активности связывания с помощью сортировки флуоресцентно-активированных клеток (FACS)
Клетки CHO-CD38 (клетки CHO, стабильно трансфицированные полноразмерным человеческим CD38) культивировали в среде DMEM-Glutamax-I, дополненной 100 мкг/мл пенициллина, 100 мкг/мл стрептомицина, 10%-ной эмбриональной сывороткой теленка и 500 мкг/мл генетицина. Клетки SKOV-3 и клетки RPMI-8226 культивировали в RPMI среде 1640-Glutamax-I, дополненной 100 мкг/мл пенициллина, 100 мкг/мл стрептомицина и 10%-ной эмбриональной сывороткой теленка.
Применяли 3x105 клеток (CHO-CD38, или SKOV-3, или RPMI-8226) на каждый образец. Клетки промывали 1x с раствором PBA (PBS, дополненной 1%BSA и 0,05% Na-азид). Для определения профилей FACS, клетки окрашивали соответствующими антителами в концентрации, равной 50 мкг/мл в объеме 30 мкл. Для титрования BiXAb-6567 и родительского анти-CD38 антитела и последующего определения параметров связывания, клетки CHO-CD38 окрашивали соответствующими антителами в указанных концентрациях в объеме 30 мкл. Клетки инкубировали в течение 30 мин на льду и затем промывали 2 раз с 1 мл раствора PBA. Клетки инкубировали с флуоресцентно мечеными специфическими вторичными антителами против kappa-цепи человека или против Fc гамма IgG человека на льду в темноте в течение 30 мин и затем промывали 2 раз с 1 мл раствора PBA; наконец, клетки ресуспендировали в конечном объеме 500 мкл раствора PBA. Образцы анализировали с помощью проточного цитометра или Epics-XL, или Navios («Beckman Coulter»). В каждом эксперименте получали 10000 событий.
Профили связывания BiXAb-6567 и родительского анти-CD38 и родительского анти-PD-L1 антител представлены на фиг. 9A-C. Мы решили протестировать клеточную линию множественной миеломы, RPMI-8226, которая экспрессирует высокие уровни CD38 и незначительные уровни PD-L1 (фиг. 9A); клеточная линия CHO-CD38, которая экспрессировала очень высокий уровень CD38 из-за стабильно трансфицируемого полноразмерного CD38 (фиг. 9B); и клеточная линия рака яичников SKOV-3, которая известна для экспрессии PD-L1 (фиг. 9C). Данные профили демонстрируют единственный пик для BiXAb-6567, который был очень сходен с профилями обеих родительских антител на 3 клеточные линии. Это позволяет предположить, что BiXAb-6567 был правильно сложен и обладал атрибутами связывания, сходными с атрибутами связывания родительского антитела. Как и ожидалось, CHO-CD38 экспрессировали только CD38 и не экспрессировали PD-L1, в то время как SKOV-3 экспрессировали только PD-L1 и не экспрессировали CD38.
Для того чтобы количественно подтвердить тот факт, что свойства связывания BiXAb-6567 сходны с таковыми родительского анти-CD38 антитела, проводили титрование BiXAb-6567 и родительского антитела анти-CD38, применяя клетки CHO-CD38, как представлено на фиг. 10. EC50 для BiXAb-6567 была определена равной 17,1 нм, а для родительского анти-CD38 – 8,5 нМ, что подтверждает сходные свойства связывания Fab-доменов анти-CD38 в BiXAb-6567 и в родительском анти-CD38 антителе. Отрицательные контроли в данном эксперименте, анти-PD-L1 и анти-CD20 антитела, не демонстрируют связывания с CHO-CD38 клетках, как и ожидалось.
Пример 6. Антитело-зависимая клеточная цитотоксичность (ADCC) с нефракционированными неактивированными моноядерными клетками (MNC)
Клетки CHO-CD38, SKOV-3 и RPMI-8226 культивировали, как описано выше в Примере 5.
Для получения MNC применяли следующую процедуру. Свежеприготовленную периферическую кровь обрабатывали цитратом для предотвращения коагуляции. Затем, на 5 мл раствора Ficoll-Paque PLUS наслаивали 6 мл цельной крови с антикоагулянтом. Образцы центрифугировали в течение 20 мин при 2500 rpm при комнатной температуре без последующего торможения центрифуги. MNC собирали с поверхности раздела плазма/Ficoll. Клеточную суспензию MNC разводили 1:10 в PBS и центрифугировали в течение 5 минут при 1800 rpm при комнатной температуре. Супернатант отбрасывали и эритроциты лизировали добавлением 45 мл ледяной дистиллированной воды к суспензии клеток в течение 30 секунд, после чего добавляли 5 мл 10x PBS. Клетки центрифугировали в течение 5 мин при 1800 rpm при комнатной температуре и промывали 1x PBS три раз для удаления тромбоцитов. Наконец, клетки ресуспендировали в 5 мл среды для клеточной культуры. Количество клеток доводили до достижения отношения 40:1= отношения эффекторная клетка : опухолевая клетка анализах ADCC.
Для определения ADCC в анализе выхода 51Хрома, 1x106 клеток-мишеней (RPMI 8226, SKOV-3 или CHO-CD38) инкубировали с 100 µCi 51Хром в 200 мкл PBS в течение 2-х часов при 37°C и 5% CO2. После 2-часовой инкубации, клетки промывали три раз 7 мл среды и наконец, ресуспендировали в концентрации, равной 0,1 x 106 клеток/мл. Клетки-мишени (5000 клеток/лунку) и MNC в присутствии антител инкубировали в 96-луночном планшете для микротитрования (анализируемый объем 200 мкл) в течение 3-х часов при 37°C и 5% CO2. Для определения максимального лизиса целевых клеток (=максимальный cpm) добавляли Triton X-100. Для определения базального выхода 51Хром (= базальный cpm) клетки-мишени не подвергали дополнительным манипуляциям. После 4-х часов инкубации, планшеты для микротитрования центрифугировали в течение 5 мин при 2000 rpm и 25 мкл супернатанта смешивали с 125 мкл Optiphase Supermix («Perkin Elmer») и инкубировали в вибрирующем инкубаторе в течение 1 мин. Образцы анализировали в бета-счетчике MicroBeta TriLux («Perkin Elmer»). Лизис клеток-мишеней рассчитывали с помощью следующего уравнения:
% лизиса = (экспериментальный cpm – базальный cpm)/(максимальный cpm – базальный cpm) x 100.
Все измерения выполняли в трех повторах.
Анализы ADCC клеток CD38+ (RPMI-8226 и CHO-CD38) проводили, применяя предварительно неактивированные MNC в качестве эффекторных клеток (фиг. 11 и 12). Анализы показали сильную цитотоксичность BiXAb-6567 и анти-CD38 антитела на клетки RPMI-8226 с EC50, равной 0,8 нм и 0,3 нМ, соответственно; на клетках CHO-CD38, цитотоксичность BiXAb-6567 и анти-CD38 антитело имела EC50, равные 0,2 нм и 0,07 нМ, соответственно. Анти-PD-L1 показал минимальную активность на обе клеточные линии; два отрицательных контроля mAbs, анти-CD20 и анти-HER2, не способствовали какому-либо лизису, как и ожидалось. Данные результаты демонстрируют сильную ADCC активность BMX-6567 против клеток CD38+, которая похожа на активность родительского анти-CD38 антитела.
Пример 7. ADCC с обогащенными предварительно активированными NK-клетками
Клетки SKOV3, RPMI 8226 и клетки CHO-CD38 культивировали, как описано в Примере 5. MNC получали как описано в Примере 6. NK-клетки были выделены из MNC путем негативной селекции, с помощью набора для выделения NK-клеток человека «NK cell isolation kit, human» («Miltenyi») в соответствии с инструкциями изготовителя. NK-клетки культивировали в течение ночи при плотности посева 2x106 клетки/мл в среде RPMI, дополненной 10%-ной фетальной сывороткой теленка. IL-12 или IL-15 добавляли до конечной концентрации 10 нг/мл. Анализы ADCC проводили, как указано в Примере 5, за исключением того, что отношение эффекторная клетка : опухолевая клетка поддерживали на уровне 10:1 и продолжительность реакции была уменьшена до 3 часов.
Свойства ADCC для группировки анти-PD-L1 в BiXAb-6567 анализировали на PD-L1+ клеточной линии SKOV-3, применяя обогащенные NK-клетки, предварительно активированные или IL-12, или IL-15. Результаты представлены на фиг. 13 и 14. Данный эксперимент сравнивает свойства ADCC для BiXAb-6567 с таковыми для родительского анти-PD-L1 антитела; в качестве положительного контроля применяли анти-HER2 антитело, и в качестве отрицательного контроля применяли анти-CD20 антитело и родительское анти-CD38, поскольку клетки SKOV-3 представляют собой PD-L1+/час ER2+/CD20-/CD38-. Фиг. 13 и 14 демонстрируют сильную активность ADCC для BiXAb-6567 и родительского анти-PD-L1 антитела, независимо от того применяли IL-12 или IL-15 при культивировании NK-клеток. EC50 для BiXAb-6567 и родительского анти-PD-L1 антитела были равны 0,007 нм и 0,03 нМ, соответственно, если применяли IL-12. Профили были еще более сходными, если применяли IL-15; однако аппроксимации кривых не сходились, таким образом, не давая возможности вычислить значения ЕС50. Данные результаты демонстрируют сильную активность ADCC для BMX-6567 против клеток PD-L1+, которая похожа на активность ADCC родительского анти-PD-L1 антитела.
--->
SEQUENCE LISTING
<110> Biomunex pharmaceuticals
<120> Binding molecules to CD38 and PD-L1
<130> B2210PC
<160> 32
<170> PatentIn version 3.5
<210> 1
<211> 709
<212> PRT
<213> Artificial Sequence
<220>
<223> BiXAb 4218 heavy chain
<400> 1
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Phe Asn Ser Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Lys Ile Leu Trp Phe Gly Glu Pro Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Lys
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser Gly Gly Glu Asn
245 250 255
Leu Tyr Phe Gln Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
260 265 270
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
275 280 285
Thr Phe Ser Asp Ser Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys
290 295 300
Gly Leu Glu Trp Val Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr
305 310 315 320
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser
325 330 335
Lys Asn Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
340 345 350
Ala Val Tyr Tyr Cys Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr
355 360 365
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
370 375 380
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
385 390 395 400
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
405 410 415
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
420 425 430
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
435 440 445
Glu Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
450 455 460
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
465 470 475 480
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
485 490 495
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
500 505 510
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
515 520 525
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
530 535 540
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
545 550 555 560
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
565 570 575
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
580 585 590
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
595 600 605
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
610 615 620
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
625 630 635 640
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
645 650 655
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
660 665 670
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
675 680 685
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
690 695 700
Leu Ser Pro Gly Lys
705
<210> 2
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> LC1
<400> 2
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Glu Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 3
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> LC2
<400> 3
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Lys Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 4
<211> 709
<212> PRT
<213> Artificial Sequence
<220>
<223> BiXAb 4219 Heavy chain
<400> 4
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Glu Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Thr Pro Pro Thr Pro Ser Pro Ser Gly Gly Glu Asn Leu Tyr Phe Gln
245 250 255
Gly Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
260 265 270
Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Phe Asn Ser
275 280 285
Phe Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
290 295 300
Val Ser Ala Ile Ser Gly Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser
305 310 315 320
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu
325 330 335
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe
340 345 350
Cys Ala Lys Asp Lys Ile Leu Trp Phe Gly Glu Pro Val Phe Asp Tyr
355 360 365
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
370 375 380
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
385 390 395 400
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
405 410 415
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
420 425 430
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
435 440 445
Lys Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
450 455 460
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
465 470 475 480
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
485 490 495
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
500 505 510
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
515 520 525
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
530 535 540
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
545 550 555 560
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
565 570 575
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
580 585 590
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
595 600 605
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
610 615 620
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
625 630 635 640
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
645 650 655
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
660 665 670
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
675 680 685
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
690 695 700
Leu Ser Pro Gly Lys
705
<210> 5
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> LC1
<400> 5
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Lys Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 6
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> LC2
<400> 6
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Glu Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 7
<211> 710
<212> PRT
<213> Artificial Sequence
<220>
<223> BiXAb 5104 Heavy chain
<400> 7
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Ala Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Trp Met Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Tyr Gly Pro Asp Gly Asp Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Lys Thr Val Tyr
65 70 75 80
Met His Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Tyr Tyr Gly Ser Asn Ser Leu Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Lys Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser Gly Gly Glu Asn Leu Tyr
245 250 255
Phe Gln Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
260 265 270
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
275 280 285
Ser Arg Tyr Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
290 295 300
Glu Trp Val Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val
305 310 315 320
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
325 330 335
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
340 345 350
Tyr Tyr Cys Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp
355 360 365
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
370 375 380
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
385 390 395 400
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
405 410 415
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
420 425 430
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
435 440 445
Val Glu Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
450 455 460
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
465 470 475 480
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
485 490 495
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
500 505 510
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
515 520 525
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
530 535 540
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
545 550 555 560
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
565 570 575
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
580 585 590
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
595 600 605
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
610 615 620
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
625 630 635 640
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
645 650 655
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
660 665 670
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
675 680 685
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
690 695 700
Ser Leu Ser Pro Gly Lys
705 710
<210> 8
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> LC1
<400> 8
Asp Ile Val Met Thr Gln Ser His Leu Ser Met Ser Thr Ser Leu Gly
1 5 10 15
Asp Pro Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Thr Val
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Arg Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Ile Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ala Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Glu Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 9
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> LC2
<400> 9
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Lys Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 10
<211> 702
<212> PRT
<213> Artificial Sequence
<220>
<223> BiXAB BMX101-AP-ML1-CC1
<400> 10
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Phe Asn Ser Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Lys Ile Leu Trp Phe Gly Glu Pro Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Gln Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Val Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Ser Pro Pro Ala Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ala Ala Pro Pro Ala Pro Ala Pro Ala Gly Gly Glu Val
245 250 255
Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
260 265 270
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser Trp Ile
275 280 285
His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp
290 295 300
Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
305 310 315 320
Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln
325 330 335
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
340 345 350
Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
355 360 365
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
370 375 380
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
385 390 395 400
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
405 410 415
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
420 425 430
Gly Leu Tyr Ser Leu Ser Ser Val Val Asp Val Pro Ser Ser Ser Leu
435 440 445
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
450 455 460
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
465 470 475 480
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
485 490 495
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
500 505 510
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
515 520 525
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
530 535 540
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
545 550 555 560
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
565 570 575
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
580 585 590
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
595 600 605
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
610 615 620
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
625 630 635 640
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
645 650 655
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
660 665 670
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
675 680 685
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
690 695 700
<210> 11
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Daratumumab LC
<400> 11
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Thr Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Val
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 12
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Atezolizumab-LC
<400> 12
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Lys Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 13
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 1
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (10)..(11)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (14)..(14)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (24)..(25)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (28)..(28)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (30)..(30)
<223> Xaa can be any naturally occurring amino acid
<220>
<221> misc_feature
<222> (32)..(32)
<223> Xaa can be any naturally occurring amino acid
<400> 13
Glu Pro Lys Xaa Cys Asp Lys Xaa His Xaa Xaa Pro Pro Xaa Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Xaa Xaa Pro Pro Xaa Pro Xaa Pro Xaa
20 25 30
Gly Gly
<210> 14
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 2
<400> 14
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ala Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Gly Gly Pro Pro Gly Pro Gly Pro Gly
20 25 30
Gly Gly
<210> 15
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 2
<400> 15
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ala Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ala Ala Pro Pro Ala Pro Ala Pro Ala
20 25 30
Gly Gly
<210> 16
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 3
<400> 16
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ala Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ala Ala Pro Pro Gly Pro Ala Pro Gly
20 25 30
Gly Gly
<210> 17
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 4
<400> 17
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
20 25 30
Gly Gly
<210> 18
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker 6
<400> 18
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Ser Pro Pro Ser Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
20 25 30
Gly Gly
<210> 19
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Nter IgG1 CH2
<400> 19
Ala Pro Glu Leu Leu Gly Gly Pro Ser
1 5
<210> 20
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Portion of human IgA1 hinge
<400> 20
Thr Pro Pro Thr Pro Ser Pro Ser
1 5
<210> 21
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> signal peptide
<400> 21
Met Asn Phe Gly Leu Arg Leu Ile Phe Leu Val Leu Thr Leu Lys Gly
1 5 10 15
Val Gln Cys
<210> 22
<211> 122
<212> PRT
<213> Homo sapiens
<400> 22
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Phe Asn Ser Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Lys Ile Leu Trp Phe Gly Glu Pro Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 23
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> CH1 domain of daratumumab
<400> 23
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Gln Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Val Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 24
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> VH of atezolizumab
<400> 24
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 25
<211> 98
<212> PRT
<213> Artificial Sequence
<220>
<223> CH1 domain of atezolizumab
<400> 25
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Asp Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 26
<211> 15
<212> PRT
<213> Homo sapiens
<400> 26
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 27
<211> 110
<212> PRT
<213> Homo sapiens
<400> 27
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 28
<211> 107
<212> PRT
<213> Homo sapiens
<400> 28
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
100 105
<210> 29
<211> 107
<212> PRT
<213> Homo sapiens
<400> 29
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 30
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> CKappa domain of daratumumab
<400> 30
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Thr Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Val Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 31
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL of atezolizumab
<400> 31
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 32
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> CKappa domain of atezolizumab
<400> 32
Arg Thr Val Ala Ala Pro Ala Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Lys Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УСТОЙЧИВЫЕ МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2018 |
|
RU2779602C2 |
| АНТИ-PD-L1 АНТИТЕЛО ДЛЯ ДЕТЕКЦИИ PD-L1 | 2018 |
|
RU2758723C2 |
| БИФУНКЦИОНАЛЬНЫЕ БЕЛКИ, КОМБИНИРУЮЩИЕ БЛОКАДУ КОНТРОЛЬНОЙ ТОЧКИ, ДЛЯ ТАРГЕТНОЙ ТЕРАПИИ | 2019 |
|
RU2756899C1 |
| МОНОСПЕЦИФИЧЕСКИЕ И БИСПЕЦИФИЧЕСКИЕ БЕЛКИ С РЕГУЛИРОВАНИЕМ ИММУННОЙ КОНТРОЛЬНОЙ ТОЧКИ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2793167C2 |
| АНТИ-PD-L1 АНТИТЕЛА | 2015 |
|
RU2722212C2 |
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| БЕЛКИ УПРАВЛЕНИЯ, НАВИГАЦИИ И КОНТРОЛЯ И СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2019 |
|
RU2811457C2 |
| АНТИТЕЛА К PD-1 И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2761640C2 |
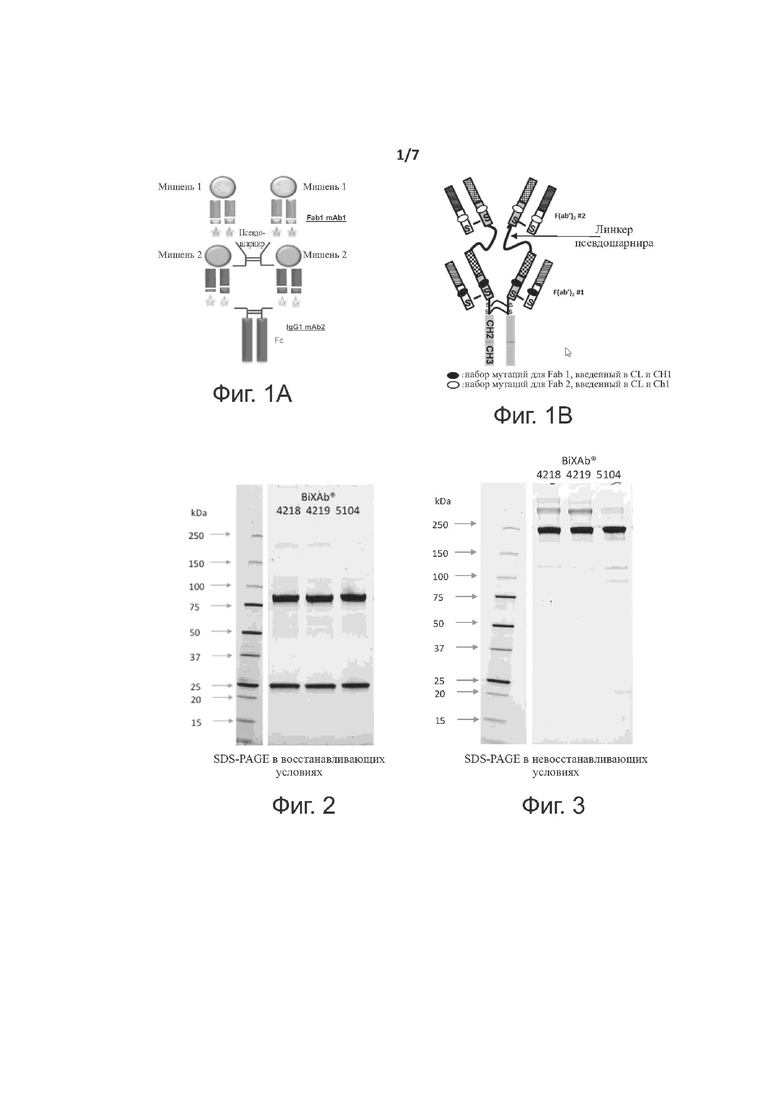
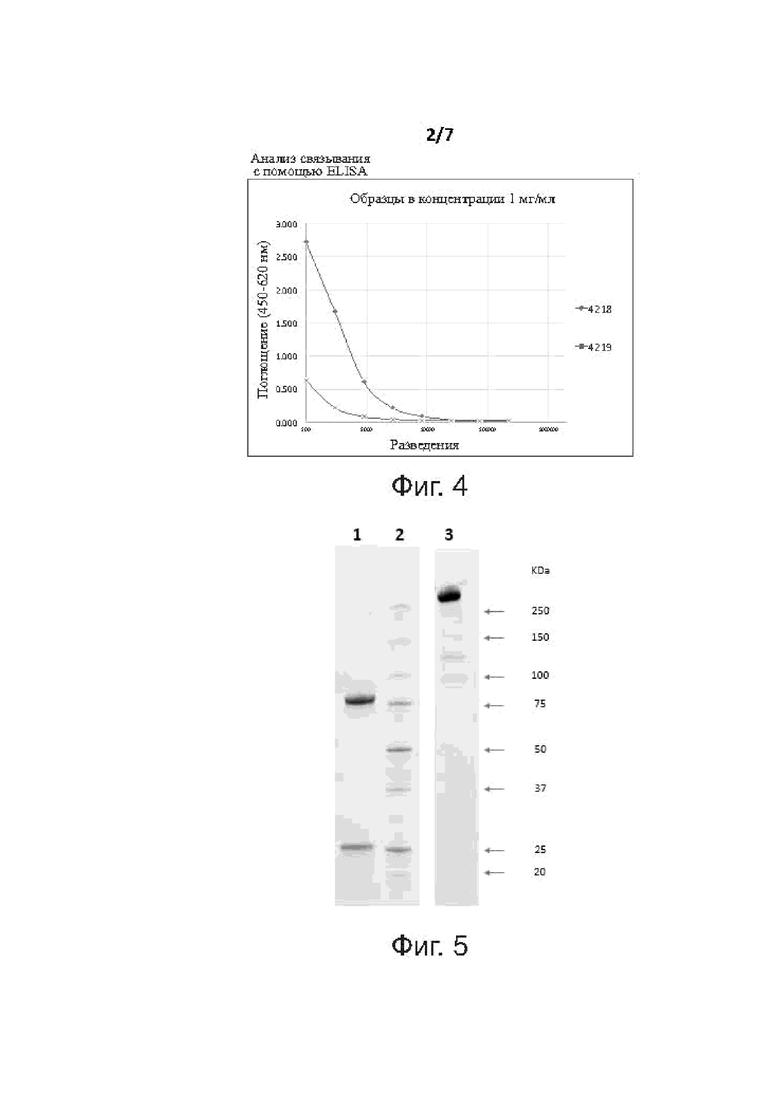
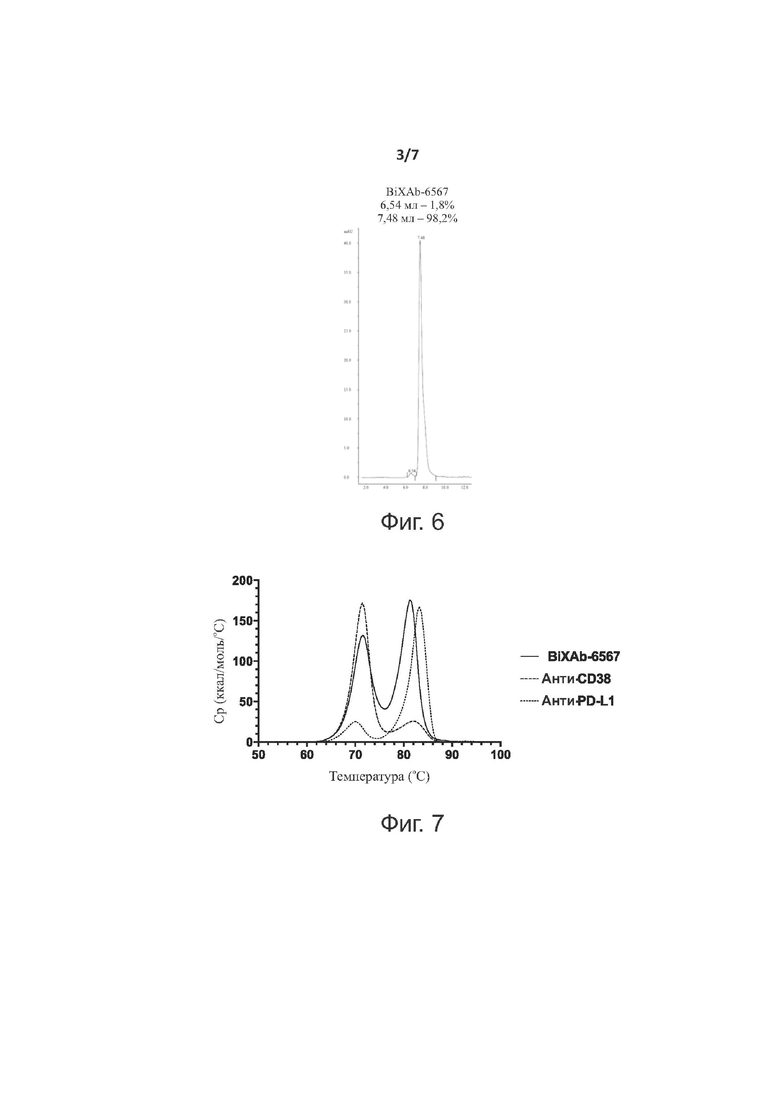
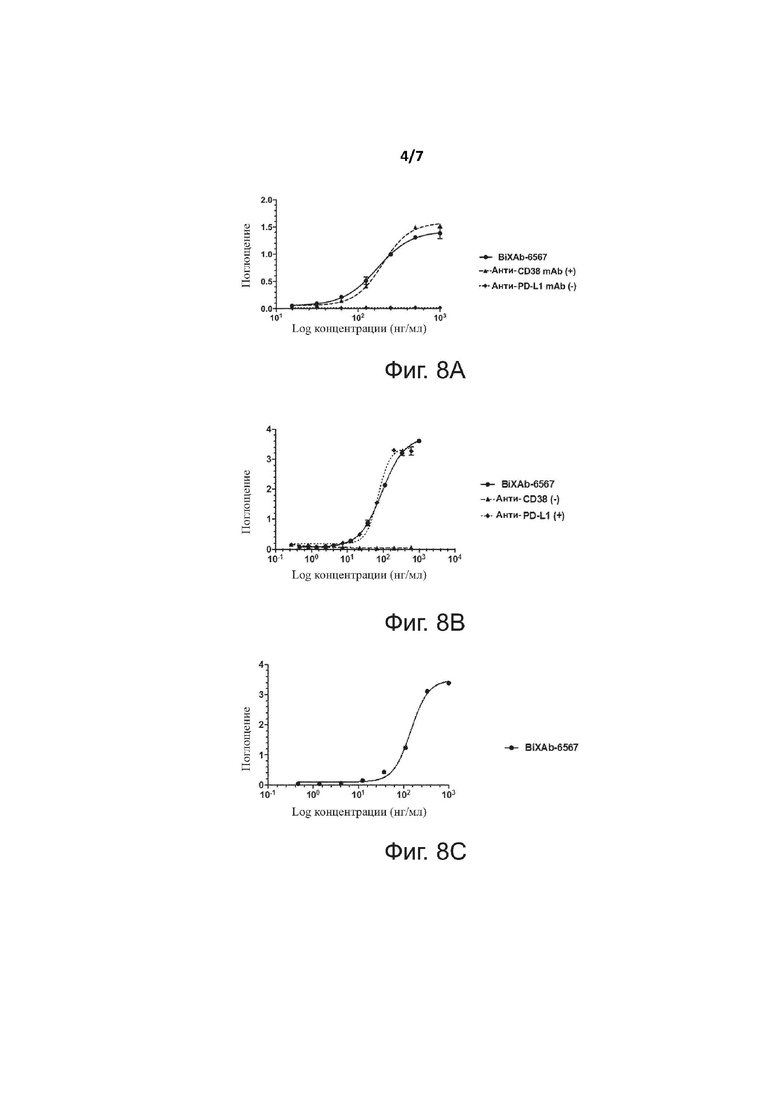
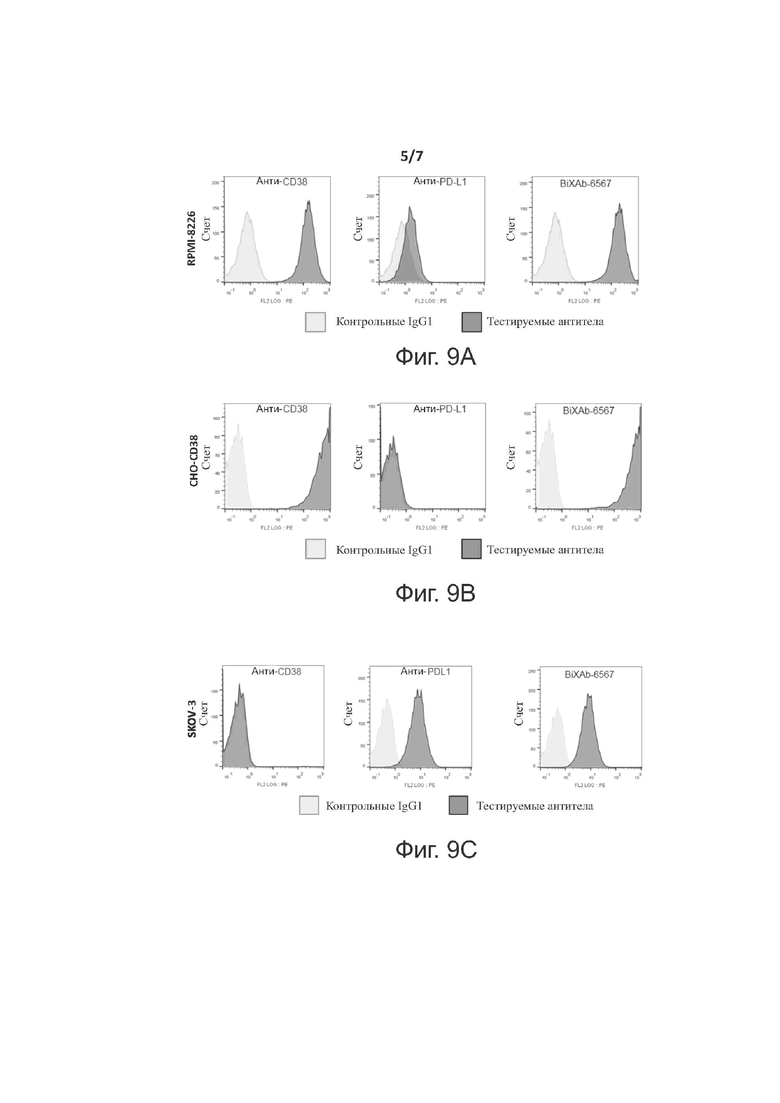
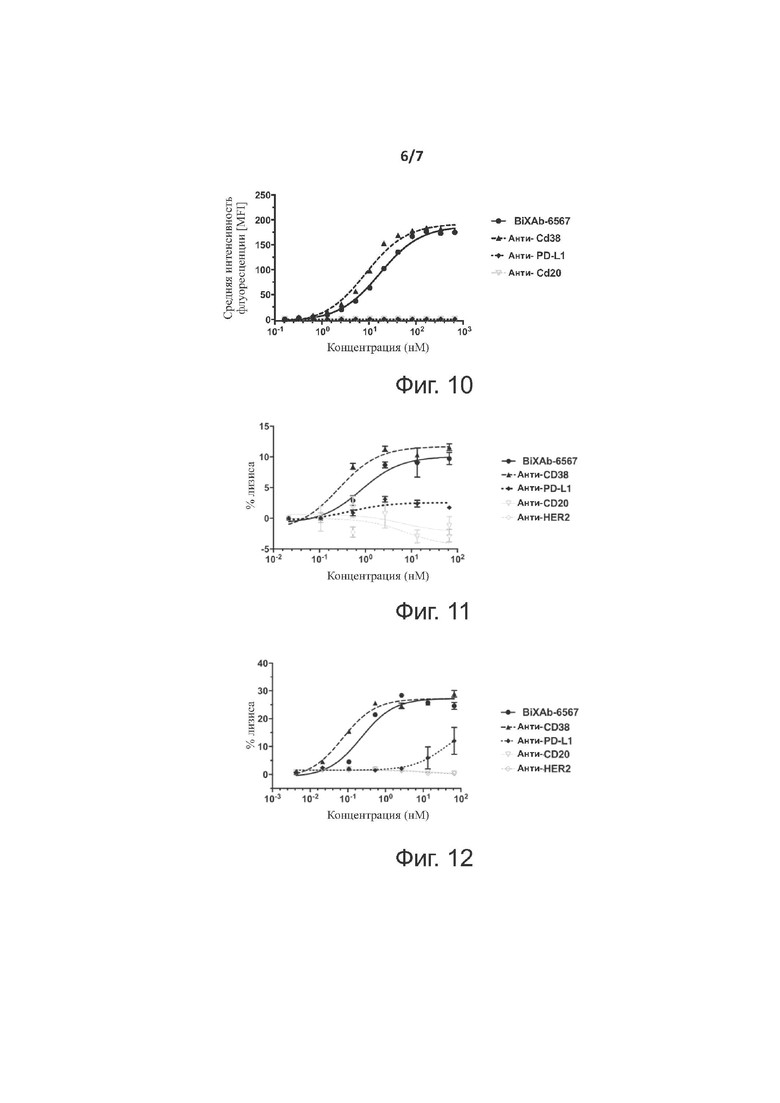
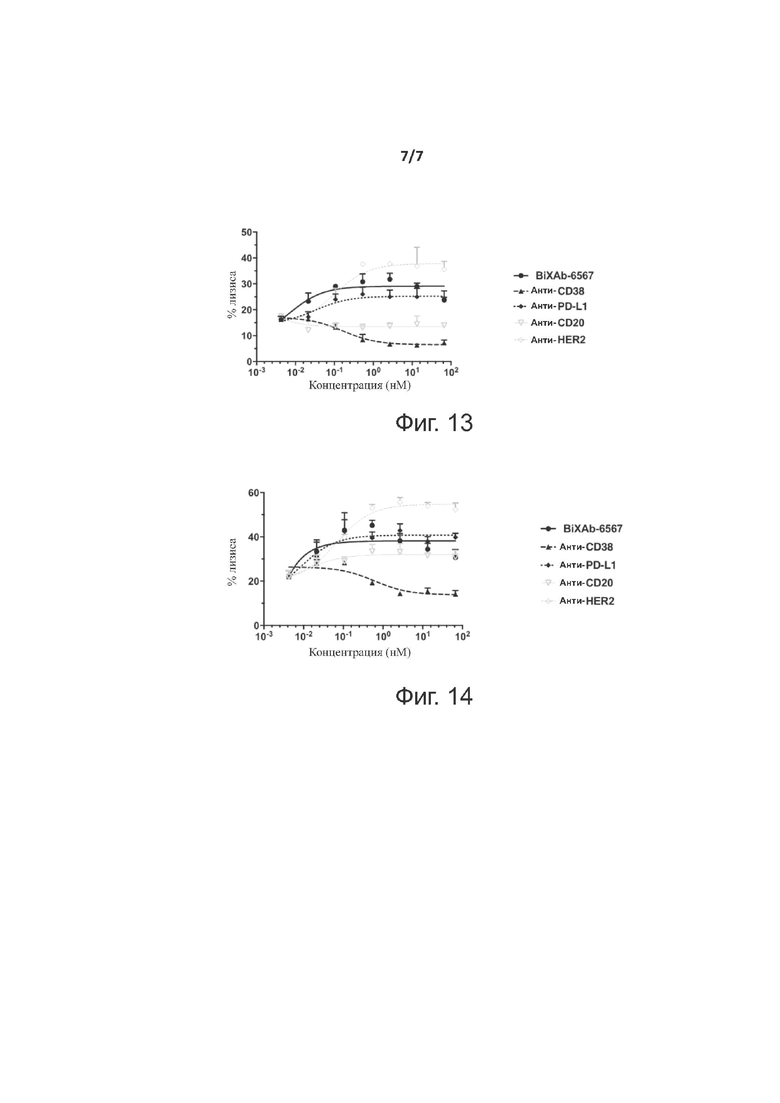
Изобретение относится к области биотехнологии, в частности к биспецифической молекуле, включающей, по меньшей мере, один анти-CD38 домен и, по меньшей мере, один анти-PD-L1 домен, которая способна одновременно связываться с антигеном CD38 и PD-L1. Изобретение эффективно для лечения рака, выбранного из группы, состоящей из множественной миеломы, неходжкинской лимфомы (NHL), рака молочной железы, меланомы и рака легкого. 9 з.п. ф-лы, 14 ил., 7 пр.
1. Биспецифическая молекула, включающая, по меньшей мере, один анти-CD38 домен и, по меньшей мере, один анти-PD-L1 домен, которая способна одновременно связываться с антигеном CD38 и PD-L1, соответственно,
которая представляет собой полноразмерное антитело, включающее две тяжелые цепи и четыре легкие цепи,
в котором каждая тяжелая цепь включает
a. область Fc, включающую шарнир-CH2-CH3 домены,
b. данная область Fc связана с тяжелой цепью Fab (CH1-VH) антитела 1 (Ab1),
c. которая, в свою очередь, связана с тяжелой цепью Fab (CH1-VH) антитела 2 (Ab2), с помощью полученной из шарнира полипептидной линкерной последовательности, в которой указанная полипептидная линкерная последовательность связывает N-конец указанной тяжелой цепи домена Fab VH из Ab1 с C-концом указанного домена CH1 из Ab2,
и четыре легкие цепи включают легкие цепи Fab (CL-VL) из Ab1 и легкие цепи Fab (CL-VL) из Ab2, связанные с тяжелыми цепями соответствующих доменов;
Ab1 и Ab2 представляют собой разные антитела, которые выбирают из группы, состоящей из анти-CD38 антител и анти-PD-L1 антител.
2. Биспецифическая молекула по п. 1, в которой Ab1 представляет собой анти-CD38 антитело и Ab2 представляет собой анти-PD-L1 антитело.
3. Биспецифическая молекула по п. 1, в которой Ab1 представляет собой анти-PD-L1 антитело и Ab2 представляет собой анти-CD38 антитело.
4. Биспецифическая молекула по любому из пп. 1-3, в которой анти-CD38 антитело выбирают из группы, состоящей из даратумумаба, изатуксимаба, MOR-202 или их мутированных производных.
5. Биспецифическая молекула по любому из пп. 1-4, в которой анти-PD-L1 антитело выбирают из группы, состоящей из атезолизумаба, дурвалумаба, авелумаба, MDX-1105 или их мутированных производных.
6. Биспецифическая молекула по любому из пп. 1-5, в которой домены CH1 и CL из Ab1 имеют последовательность, отличающуюся от доменов CH1 и CL из Ab2.
7. Биспецифическая молекула по любому из пп. 1-5, в которой домен CH1 Fab одного из Ab1 или Ab2 представляет собой мутированный домен, который получают из домена CH1 иммуноглобулина путем замены остатка треонина в положении 192 указанного домена CH1 аспарагиновой кислотой, и соответствующий домен CL представляет собой мутированный домен, который получают из домена CL иммуноглобулина заменой остатка аспарагина в положении 137 указанного домена CL остатком лизина и заменой остатка серина в положении 114 указанного домена CL остатком аланина, и/или в которой Fab домен CH1 одного или другого из Ab1 или Ab2 представляет собой мутированный домен, который получают из домена CH1 иммуноглобулина путем замены остатка лейцина в положении 124 указанного домена CH1 глутамином и заменой остатка серина в положении 188 указанного домена CH1 остатком валина, и соответствующий домен CL представляет собой мутированный домен, который получают из домена CL иммуноглобулина путем замены остатка валина в положении 133 указанного домена CL остатком треонина и заменой остатка серина в положении 176 указанного домена CL остатком валина.
8. Биспецифическая молекула по любому из пп. 1-7, которая включает или, предпочтительно, состоит из a) двух тяжелых цепей, каждая включающая или, предпочтительно, состоящая из SEQ ID NO: 10, и b) четырех легких цепей, две включающие или, предпочтительно, состоящие из SEQ ID NO: 11, две другие включающие или, предпочтительно, состоящие из SEQ ID NO: 12.
9. Биспецифическая молекула по любому из пп. 1-8, для применения в лечении рака, выбранного из группы, состоящей из множественной миеломы, неходжкинской лимфомы (NHL), рака молочной железы, меланомы и рака легкого.
10. Биспецифическая молекула по любому из пп. 1-8, для применения в лечении рака множественной миеломы.
| GREGORY L | |||
| MOORE, PHD3 et al., 1798 Tuning T Cell Affinity Improves Efficacy and Safety of Anti-CD38 ?Anti-CD3 Bispecific Antibodies in Monkeys - a Potential Therapy for Multiple Myeloma, 2015 | |||
| Genmab Announces Studies Of Daratumumab In Combination With Atezolizumab In A Solid Tumor And Multiple Myeloma, 21.03.2016 | |||
| US 8263746 B2, 11.09.2012 | |||
| WO |

