ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет по предварительной заявке на патент США № 62/463451, поданной 24 февраля 2017 г., и предварительной заявке на патент США № 62/583990, поданной 9 ноября 2017 г., содержание которых включено в данный документ посредством ссылки в полном объеме.
УРОВЕНЬ ТЕХНИКИ
Беспозвоночные организмы (например, насекомые, моллюски или нематоды) повсеместно распространены в окружающей человека среде. В некоторых случаях беспозвоночные выполняют полезные функции, такие как нематоды или членистоногие, используемые в сельском хозяйстве в целях опыления и борьбы с вредителями или в коммерческих целях для получения коммерческих продуктов (например, меда или шелка). В других случаях беспозвоночные могут быть вредными, в том числе некоторые виды моллюсков (например, улитки и слизни), нематоды или насекомые, которые могут представлять собой серьезных вредителей культурных растений или переносчиков заболевания. Таким образом, в данной области техники существует потребность в способах и композициях для модулирования приспособленности беспозвоночных, которые важны в сельском хозяйстве, коммерческой деятельности или здравоохранении.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В данном документе раскрыты композиции и способы модулирования приспособленности беспозвоночных, в том числе насекомых, нематод или моллюсков, с помощью изменения взаимодействий между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина.
В одном аспекте в данном документе предусмотрен способ снижения приспособленности насекомого, при этом способ включает доставку насекомому эффективного количества полинуклеотида, который содержит dsRNA, которая обеспечивает снижение экспрессии регуляторного гена бактериоцита насекомого или иммунорегуляторного гена насекомого в организме насекомого по сравнению с насекомым, которому dsRNA не была введена.
В некоторых вариантах осуществления ген кодирует белок из семейства BCR бактериоцит-специфических белков, богатых цистеином, белок из семейства SP секретируемых белков, BicD (белок bicaudal D), Cact (cactus), DIF (дорсальный иммунный фактор), Toll (Toll-взаимодействующий белок) или imd (белок иммунной недостаточности). В некоторых вариантах осуществления ген кодирует белок, характеризующийся по меньшей мере 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичностью аминокислотной последовательности по отношению к белку, приведенному в таблице 5, таблице 8 или таблице 9. В некоторых вариантах осуществления ген кодирует функциональный гомолог белка, приведенного в таблице 5, таблице 8 или таблице 9. Например, ген может кодировать cactus-подобный белок у тлей (например, любой из белков, описанный под №№ доступа в GenBank: XP_022175228.1, XP_016656687.1, NP_001156668.1, XP_008179071.1 или XP_016656686.1, соответствующие аминокислотные и нуклеотидные последовательности которых включены посредством ссылки).
В некоторых вариантах осуществления dsRNA является комплементарной 10-30 нуклеотидам гена у насекомого (например, 10-30 нуклеотидам, 12-28 нуклеотидам, 14-26 нуклеотидам, 16-24 нуклеотидам, 14-22 нуклеотидам или 18-20 нуклеотидам). В некоторых вариантах осуществления dsRNA является комплементарной 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидам гена у насекомого. В некоторых вариантах осуществления dsRNA по всей длине является комплементарной гену. В альтернативных вариантах осуществления лишь часть dsRNA является комплементарной гену.
В некоторых вариантах осуществления способ является эффективным для снижения экспрессии гена у насекомого, например, на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% или больше, по сравнению с насекомым, которому не был введен полинуклеотид. В некоторых вариантах осуществления способ является эффективным для снижения экспрессии гена у насекомого, например, на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% или больше по сравнению с эталонным уровнем (например, по сравнению с экспрессией одного или нескольких контрольных генов (например, гена домашнего хозяйства), экспрессией того же самого гена в другом образце (например, одном или нескольких контрольных образцах) или экспрессией одного и того же гена в одном и том же образце в одной или нескольких более ранних временных точках).
В некоторых вариантах осуществления способ является эффективным для снижения экспрессии гена у насекомого, например, в приблизительно 1,5x, 1,75x, 2x, 3x, 4x, 5x, 6x, 7x, 8x, 9x, 10x, 25x, 50x, 75x или 100x раз меньше, по сравнению с насекомым, которому не был введен полинуклеотид. В некоторых вариантах осуществления способ является эффективным для снижения экспрессии гена у насекомого, например, в приблизительно 1,5x, 1,75x, 2x, 3x, 4x, 5x, 6x, 7x, 8x, 9x, 10x, 25x, 50x, 75x или 100x раз меньше по сравнению с эталонным уровнем (например, по сравнению с экспрессией одного или нескольких контрольных генов (например, гена домашнего хозяйства), экспрессией того же самого гена в другом образце (например, одном или нескольких контрольных образцах) или экспрессией одного и того же гена в одном и том же образце в одной или нескольких более ранних временных точках).
В некоторых вариантах осуществления способ является эффективным для подавления экспрессии гена у насекомого или для снижения экспрессии гена до невыявляемого уровня.
В некоторых вариантах осуществления способ является эффективным для снижения уровня, разнообразия или метаболизма одного или нескольких микроорганизмов, обитающих в организме насекомого, по сравнению с насекомым, которому не был доставлен полинуклеотид. В некоторых вариантах осуществления способ является эффективным для снижения уровня, разнообразия или метаболизма одного или нескольких микроорганизмов, обитающих в организме насекомого, например, в приблизительно 1,5x, 1,75x, 2x, 3x, 4x, 5x, 6x, 7x, 8x, 9x, 10x, 25x, 50x, 75x или 100x раз меньше, по сравнению с насекомым, которому не был доставлен полинуклеотид. В некоторых вариантах осуществления способ является эффективным для снижения уровня, разнообразия или метаболизма одного или нескольких микроорганизмов, обитающих в организме насекомого, например, на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100%, по сравнению с насекомым, которому не был доставлен полинуклеотид. В определенных вариантах осуществления один или несколько микроорганизмов представляют собой Buchnera spp.
В некоторых вариантах осуществления способ является эффективным для снижения приспособленности насекомого по сравнению с насекомым, которому не был доставлен полинуклеотид. В некоторых вариантах осуществления способ является эффективным для снижения приспособленности насекомого, например, в приблизительно 1,5x, 1,75x, 2x, 3x, 4x, 5x, 6x, 7x, 8x, 9x, 10x, 25x, 50x, 75x или 100x раз меньше, по сравнению с насекомым, которому не был доставлен полинуклеотид. В некоторых вариантах осуществления способ является эффективным для снижения приспособленности насекомого, например, на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100%, по сравнению с насекомым, которому не был доставлен полинуклеотид.
В некоторых вариантах осуществления полинуклеотид доставляют в композиции, составленной для доставки насекомым. В некоторых вариантах осуществления доставка включает доставку полинуклеотида в по меньшей мере одну среду обитания, в которой насекомое-вредитель растет, живет, размножается, питается или осуществляет заражение. В некоторых вариантах осуществления доставка включает распыление противомикробного пептида на сельскохозяйственную культуру. В некоторых вариантах осуществления полинуклеотид доставляют в композиции, съедобной для насекомого, для заглатывания насекомым.
В некоторых вариантах осуществления полинуклеотид составляют с приемлемым с точки зрения сельского хозяйства носителем в виде жидкой, твердой, аэрозольной, пастообразной, гелеобразной или газообразной композиции.
В некоторых вариантах осуществления насекомое представляет собой тлю.
В другом аспекте в данном документе предусмотрена композиция, содержащая полинуклеотид, который содержит dsRNA, составленный для доставки насекомому, где dsRNA является комплементарной 15-30 нуклеотидам регуляторного гена бактериоцита насекомого или иммунорегуляторного гена насекомого. В некоторых вариантах осуществления ген кодирует белок, выбранный из группы, состоящей из белка из семейства BCR бактериоцит-специфических белков, богатых цистеином, белка из семейства SP секретируемых белков, BicD (белка bicaudal D), Cact (cactus), DIF (дорсального иммунного фактора), Toll (Toll-взаимодействующего белка) и imd (белка иммунной недостаточности). В некоторых вариантах осуществления ген кодирует белок, характеризующийся по меньшей мере 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичностью аминокислотной последовательности по отношению к белку, приведенному в таблице 5, таблице 8 или таблице 9. В некоторых вариантах осуществления ген кодирует функциональный гомолог белка, приведенного в таблице 5, таблице 8 или таблице 9. Например, ген может кодировать cactus-подобный белок у тлей (например, любой из белков, описанный под №№ доступа в GenBank: XP_022175228.1, XP_016656687.1, NP_001156668.1, XP_008179071.1 или XP_016656686.1, соответствующие аминокислотные и нуклеотидные последовательности которых включены посредством ссылки). В некоторых вариантах осуществления dsRNA является комплементарной 10-30 нуклеотидам гена у насекомого (например, 10-30 нуклеотидам, 12-28 нуклеотидам, 14-26 нуклеотидам, 16-24 нуклеотидам, 14-22 нуклеотидам или 18-20 нуклеотидам). В некоторых вариантах осуществления dsRNA является комплементарной 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидам гена у насекомого. В некоторых вариантах осуществления dsRNA по всей длине является комплементарной гену. В альтернативных вариантах осуществления лишь часть dsRNA является комплементарной гену.
В дополнительном аспекте в данном документе предусмотрено растение, предусматривающее местное нанесение композиций, описанных в данном документе.
В еще одном аспекте в данном документе предусмотрена клетка трансгенного растения, имеющая в своем геноме рекомбинантную конструкцию ДНК, при этом рекомбинантная конструкция ДНК содержит гетерологичный промотор, функционально связанный с ДНК, кодирующей РНК, содержащую по меньшей мере один двухнитевой участок РНК, по меньшей мере одна нить которого содержит нуклеотидную последовательность, которая является комплементарной 15-30 нуклеотидам, представляющим интерес, регуляторного гена бактериоцита насекомого или иммунорегуляторного гена насекомого. В некоторых вариантах осуществления ген кодирует белок, выбранный из группы, состоящей из белка из семейства BCR бактериоцит-специфических белков, богатых цистеином, белка из семейства SP секретируемых белков, BicD (белка bicaudal D), Cact (cactus), DIF (дорсального иммунного фактора), Toll (Toll-взаимодействующего белка) и imd (белка иммунной недостаточности). В некоторых вариантах осуществления ген кодирует белок, характеризующийся по меньшей мере 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичностью аминокислотной последовательности по отношению к белку, приведенному в таблице 5, таблице 8 или таблице 9. В некоторых вариантах осуществления ген кодирует функциональный гомолог белка, приведенного в таблице 5, таблице 8 или таблице 9. Например, ген может кодировать cactus-подобный белок у тлей (например, любой из белков, описанный под №№ доступа в GenBank: XP_022175228.1, XP_016656687.1, NP_001156668.1, XP_008179071.1 или XP_016656686.1, соответствующие аминокислотные и нуклеотидные последовательности которых включены посредством ссылки). В некоторых вариантах осуществления dsRNA является комплементарной 10-30 нуклеотидам гена у насекомого (например, 10-30 нуклеотидам, 12-28 нуклеотидам, 14-26 нуклеотидам, 16-24 нуклеотидам, 14-22 нуклеотидам или 18-20 нуклеотидам). В некоторых вариантах осуществления dsRNA является комплементарной 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидам гена у насекомого. В некоторых вариантах осуществления dsRNA по всей длине является комплементарной гену. В альтернативных вариантах осуществления лишь часть dsRNA является комплементарной гену.
В еще одном аспекте в данном документе предусмотрены композиции, которые содержат модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновую кислоту (например, ДНК, РНК (например, mRNA, gRNA или ингибирующую РНК (например, RNAi, shRNA, miRNA)), нуклеиновую кислоту CRISPR), малую молекулу (например, простагландин) или их комбинацию), которое модулирует (например, повышает или снижает) приспособленность беспозвоночного насекомого (например, насекомого, моллюска или нематоды), где модулирующее средство изменяет взаимодействия между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина.
В некоторых вариантах осуществления модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновая кислота (например, ДНК, РНК (например, mRNA, gRNA или ингибирующая РНК (например, RNAi, shRNA, miRNA)), нуклеиновая кислота CRISPR), малая молекула (например, простагландин) или их комбинация) целенаправленно воздействует на один или несколько путей хозяина, которые опосредуют взаимодействия между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина (например, взаимодействия хозяин-микробиота). В определенных вариантах осуществления целенаправленное воздействие (например, повышение регуляции, понижение регуляции или подавление) в отношении одного или нескольких путей хозяина изменяет уровень, разнообразие или функцию одного или нескольких микроорганизмов, обитающих в организме хозяина, по сравнению с организмом хозяина, которому не было введено модулирующее средство. В определенных вариантах осуществления целенаправленное воздействие (например, повышение регуляции, понижение регуляции или подавление) в отношении одного или нескольких путей хозяина повышает уровень, разнообразие или функцию одного или нескольких микроорганизмов, обитающих в организме хозяина, по сравнению с организмом хозяина, которому не было введено модулирующее средство. В альтернативных вариантах осуществления целенаправленное воздействие (например, повышение регуляции, снижение регуляции или ингибирование) в отношении одного или нескольких путей хозяина снижает уровень, разнообразие или функцию одного или нескольких микроорганизмов, обитающих в организме хозяина, по сравнению с организмом хозяина, которому не было введено модулирующее средство.
В некоторых вариантах осуществления путь хозяина представляет собой путь, который обеспечивает регуляцию функции или развития бактериоцитов. В некоторых вариантах осуществления целенаправленное воздействие на функцию или развитие бактериоцитов может повышать и/или понижать уровень, разнообразие и/или функцию одного или нескольких микроорганизмов, обитающих в бактериоците, по сравнению с организмом хозяина, которому не было введено модулирующее средство. В определенных вариантах осуществления целенаправленное воздействие на функцию или развитие бактериоцитов снижает уровень, разнообразие или функцию одного или нескольких микроорганизмов, обитающих в бактериоците (например, бактериоците тли), по сравнению с организмом хозяина, которому не было введено модулирующее средство. В определенных вариантах осуществления целенаправленное воздействие на функцию или развитие бактериоцитов повышает уровень, разнообразие или функцию одного или нескольких микроорганизмов, обитающих в бактериоците (например, бактериоците тли), по сравнению с организмом хозяина, которому не было введено модулирующее средство.
В некоторых вариантах осуществления путь хозяина представляет собой путь, который обеспечивает регуляцию иммунной системы хозяина. Например, в некоторых вариантах осуществления модулирующее средство активирует иммунный ответ против одного или нескольких микроорганизмов, обитающих в организме хозяина, со снижением тем самым уровня, разнообразия и/или функции одного или нескольких микроорганизмов по сравнению с организмом хозяина, которому не было введено модулирующее средство. Альтернативно, в некоторых вариантах осуществления модулирующее средство подавляет иммунный ответ против одного или нескольких микроорганизмов, обитающих в организме хозяина, с повышением тем самым уровня, разнообразия и/или функции одного или нескольких микроорганизмов по сравнению с организмом хозяина, которому не было введено модулирующее средство.
В некоторых вариантах осуществления модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновая кислота (например, ДНК, РНК (например, mRNA, gRNA или ингибирующая РНК (например, RNAi, shRNA, miRNA)), нуклеиновая кислота CRISPR), малая молекула (например, простагландин) или их комбинация) целенаправленно воздействует в отношении одного или нескольких путей хозяина с помощью изменения экспрессии генов в организме хозяина по сравнению с организмом хозяина, которому не было введено модулирующее средство. Например, модулирующее средство может обеспечивать повышение и/или снижение экспрессии генов у хозяина по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления модулирующее средство обеспечивает изменение экспрессии гена, который кодирует белок, приведенный в таблице 3, таблице 4, таблице 5, таблице 7, таблице 8 или таблице 9, по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления модулирующее средство обеспечивает снижение экспрессии гена, который кодирует белок, приведенный в таблице 3, таблице 4, таблице 5, таблице 7, таблице 8 или таблице 9, по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления модулирующее средство обеспечивает снижение экспрессии гена, который кодирует белок, приведенный в таблице 3, таблице 4, таблице 5, таблице 7, таблице 8 или таблице 9, по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления ген кодирует регуляторный пептид бактериоцита. К примеру, регуляторный пептид бактериоцита может представлять собой регуляторный пептид, приведенный в таблице 5 или таблице 8 (например, BCR1). В некоторых вариантах осуществления ген кодирует компонент иммунной системы. Например, компонент иммунной системы может представлять собой компонент иммунной системы, приведенный в таблице 9. В некоторых вариантах осуществления модулирующее средство целенаправленно воздействует на полипептид у хозяина. В некоторых вариантах осуществления полипептид представляет собой фермент или клеточный рецептор. В некоторых вариантах осуществления модулирующее средство повышает и/или снижает активность фермента по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления модулирующее средство повышает и/или снижает передачу сигнала с участием клеточного рецептора по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления белок хозяина представляет собой белок, приведенный в таблице 4, таблице 5, таблице 8 или таблице 9.
Модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновая кислота (например, ДНК, РНК (например, mRNA, gRNA или ингибирующая РНК (например, RNAi, shRNA, miRNA)), нуклеиновая кислота CRISPR), малая молекула (например, простагландин) или их комбинация) может дополнительно или альтернативно целенаправленно воздействовать в отношении одного или нескольких путей микроорганизма, которые опосредуют взаимодействия между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина. В некоторых вариантах осуществления модулирующее средство обеспечивает изменение экспрессии генов у одного или нескольких микроорганизмов, обитающих в организме хозяина, по сравнению с организмом хозяина, которому не было введено модулирующее средство. К примеру, модулирующее средство может обеспечивать повышение и/или снижение экспрессии генов у одного или нескольких микроорганизмов по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления модулирующее средство обеспечивает изменение (например, повышение или снижение) экспрессии гена, который кодирует белок, приведенный в таблице 3 или таблице 7, по сравнению с организмом хозяина, которому не было введено модулирующее средство. В некоторых вариантах осуществления модулирующее средство целенаправленно воздействует (например, связывается, антагонизирует и/или вызывает агонизацию) на полипептид у одного или нескольких микроорганизмов, обитающих у хозяина (например, белок, приведенный в таблице 3 или таблице 7).
В некоторых вариантах осуществления один или несколько микроорганизмов, обитающих у хозяина, представляют собой эндосимбиотический микроорганизм. В некоторых вариантах осуществления один или несколько микроорганизмов обитают в кишке хозяина. В некоторых вариантах осуществления один или несколько микроорганизмов обитают в бактериоците в организме хозяина. В некоторых вариантах осуществления один или несколько микроорганизмов, обитающих у хозяина, представляют собой гриб или бактерию. В некоторых вариантах осуществления бактерия, обитающая в организме хозяина, представляет собой по меньшей мере одну бактерию, выбранную из группы, состоящей из Candidatus spp, Buchenera spp, Blattabacterium spp, Baumania spp, Wigglesworthia spp, Wolbachia spp, Rickettsia spp, Orientia spp, Sodalis spp, Burkholderia spp, Cupriavidus spp, Frankia spp, Snirhizobium spp, Streptococcus spp, Wolinella spp, Xylella spp, Erwinia spp, Agrobacterium spp, Bacillus spp, Paenibacillus spp, Streptomyces spp, Micrococcus spp, Corynebacterium spp, Acetobacter spp, Cyanobacteria spp, Salmonella spp, Rhodococcus spp, Pseudomonas spp, Lactobacillus spp, Enterococcus spp, Alcaligenes spp, Klebsiella spp, Paenibacillus spp, Arthrobacter spp, Corynebacterium spp, Brevibacterium spp, Thermus spp, Pseudomonas spp, Clostridium spp и Escherichia spp. В некоторых вариантах осуществления гриб, обитающий в организме хозяина, представляет собой по меньшей мере один гриб, выбранный из группы, состоящей из Candida, Metschnikowia, Debaromyces, Starmerella, Pichia, Cryptococcus, Pseudozyma, Symbiotaphrina bucneri, Symbiotaphrina kochii, Scheffersomyces shehatae, Scheffersomyces stipites, Cryptococcus, Trichosporon, Amylostereum areolatum, Epichloe spp, Pichia pinus, Hansenula capsulate, Daldinia decipien, Ceratocytis spp, Ophiostoma spp и Attamyces bromatificus. В некоторых вариантах осуществления модулирующее средство обеспечивает изменение роста, деления, жизнеспособности, метаболизма и/или продолжительности жизни микроорганизма, обитающего в организме хозяина. В некоторых вариантах осуществления модулирующее средство обеспечивает снижение роста, деления, жизнеспособности, метаболизма и/или продолжительности жизни одного или нескольких микроорганизмов. В некоторых вариантах осуществления модулирующее средство обеспечивает повышение роста, деления, жизнеспособности, метаболизма и/или продолжительности жизни одного или нескольких микроорганизмов.
В некоторых вариантах осуществления модулирующее средство представляет собой полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновую кислоту (например, ДНК, РНК (например, mRNA, gRNA или ингибирующую РНК (например, RNAi, shRNA, miRNA)), нуклеиновую кислоту CRISPR), малую молекулу (например, простагландин) или любую их комбинацию.
В некоторых вариантах осуществления модулирующее средство представляет собой нуклеиновую кислоту. Нуклеиновая кислота может представлять собой молекулу ДНК, молекулу РНК (например, двухнитевую РНК (dsRNA) или однонитевую РНК (ssRNA)) или гибридную молекулу ДНК-РНК. В некоторых вариантах осуществления РНК представляет собой информационную РНК (mRNA), направляющую РНК (gRNA) или ингибирующую РНК. В некоторых вариантах осуществления ингибирующая РНК представляет собой RNAi, shRNA или miRNA. В некоторых вариантах осуществления нуклеиновая кислота кодирует полипептид. В некоторых вариантах осуществления нуклеиновая кислота представляет собой вектор экспрессии, кодирующий полипептид. В некоторых вариантах осуществления нуклеиновая кислота представляет собой нуклеиновую кислоту CRISPR.
В некоторых вариантах осуществления модулирующее средство представляет собой малую молекулу. В некоторых вариантах осуществления малая молекула представляет собой агонист, антагонист, ингибитор или активатор компонента пути иммунной системы хозяина или регуляторного пути бактериоцита. В некоторых вариантах осуществления малая молекула представляет собой простагландин.
В некоторых вариантах осуществления модулирующее средство представляет собой полипептид. В некоторых вариантах осуществления полипептид представляет собой антитело или фрагмент антитела. Например, антитело или фрагмент антитела может представлять собой агонист или антагонист фермента у хозяина (например, фермент иммунной системы или регуляторный фермент бактериоцита) или в микроорганизма, обитающего в организме хозяина, в том числе любой из белков, приведенных в таблице 5, таблице 7, таблице 8 или таблице 9.
В некоторых вариантах осуществления модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновая кислота (например, ДНК, РНК (например, mRNA, gRNA или ингибирующая РНК (например, RNAi, shRNA, miRNA)), нуклеиновая кислота CRISPR), малая молекула (например, простагландин) или их комбинация) модулирует приспособленность хозяина с помощью повышения или понижения восприимчивости хозяина к пестициду (например, пестициду, приведенному в таблице 11). В некоторых вариантах осуществления пестицид представляет собой бактерицид или фунгицид. В некоторых вариантах осуществления пестицид представляет инсектицид, моллюскоцид или нематоцид.
В некоторых вариантах осуществления композиция содержит несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более 10) различных модулирующих средств (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновую кислоту (например, ДНК, РНК (например, mRNA, gRNA или ингибирующую РНК (например, RNAi, shRNA, miRNA)), нуклеиновую кислоту CRISPR), малую молекулу (например, простагландин) или их комбинацию). В некоторых вариантах осуществления композиция содержит модулирующее средство и пестицид (например, пестицид, приведенный в таблице 11). В некоторых вариантах осуществления пестицид представляет собой бактерицид или фунгицид. В некоторых вариантах осуществления пестицид представляет инсектицид, моллюскоцид или нематоцид. В некоторых вариантах осуществления композиция содержит модулирующее средство и средство, повышающее рост сельскохозяйственной культуры.
В некоторых вариантах осуществления модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновая кислота (например, ДНК, РНК (например, mRNA, gRNA или ингибирующая РНК (например, RNAi, shRNA, miRNA)), нуклеиновая кислота CRISPR), малая молекула (например, простагландин) или любая их комбинация) связано со вторым фрагментом. В некоторых вариантах осуществления второй фрагмент выбран из группы, состоящей из модулирующего средства, нуклеиновой кислоты пептида, пептида, проникающего в клетку (CPP), и нацеливающегося домена. В некоторых вариантах осуществления модулирующее средство содержит линкер. В некоторых вариантах осуществления линкер представляет собой расщепляемый линкер. В некоторых вариантах осуществления CPP является любым, приведенным в таблице 10.
В некоторых вариантах осуществления композиция дополнительно содержит носитель. В некоторых вариантах осуществления носитель представляет собой приемлемый с точки зрения сельского хозяйства носитель. В некоторых вариантах осуществления композиция дополнительно содержит приманку для хозяина, липкое средство или их комбинацию. В некоторых вариантах осуществления приманка для хозяина представляет собой съедобное средство. В некоторых вариантах осуществления приманка для хозяина представляет собой хемоаттрактант.
В некоторых вариантах осуществления композиция составляет дозу, эффективную для модулирования приспособленности хозяина. В некоторых вариантах осуществления композиция составляет дозу, эффективную для повышения приспособленности хозяина. В некоторых вариантах осуществления композиция составляет дозу, эффективную для понижения приспособленности хозяина. В некоторых вариантах осуществления приспособленность хозяина измеряют по выживаемости, продолжительности жизни, размножению или метаболизму хозяина.
В некоторых вариантах осуществления композицию составляют для доставки в микроорганизм, обитающий в кишке хозяина. В некоторых вариантах осуществления композицию составляют для доставки в микроорганизм, обитающий в бактериоците хозяина. В некоторых вариантах осуществления композицию составляют для доставки в растение. В некоторых вариантах осуществления композицию составляют для применения в кормушке для хозяина. В некоторых вариантах осуществления композицию составляют в виде жидкости, порошка, гранул или наночастиц. В некоторых вариантах осуществления композицию составляют в виде композиции, выбранной из группы, состоящей из липосомы, полимера, бактерии, секретирующей пептид, и синтетической нанокапсулы. В некоторых вариантах осуществления синтетическая нанокапсула доставляет композицию в участок-мишень в организме хозяина. В некоторых вариантах осуществления участок-мишень представляет собой кишку хозяина. В некоторых вариантах осуществления участок-мишень представляет собой бактериоцит в организме хозяина.
В другом аспекте в данном документе предусмотрены растения, содержащие любую из предшествующих композиций (например, модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновую кислоту (например, ДНК, РНК (например, mRNA, gRNA или ингибирующую РНК (например, RNAi, shRNA, miRNA)), нуклеиновую кислоту CRISPR), малую молекулу (например, простагландин) или их комбинацию)). В некоторых вариантах осуществления растение содержит нуклеиновую кислоту, интегрированную в геном растения, при этом нуклеиновая кислота кодирует любое из предшествующих модулирующих средств (например, полипептид (например, антитело, бактериоцин (например, colA), противомикробный пептид, регуляторный пептид бактериоцита, нуклеиновую кислоту или малую молекулу). Модулирующее средство может быть неэндогенным в отношении растения. В некоторых вариантах осуществления растение дополнительно содержит съедобное средство для беспозвоночных (например, насекомого, моллюска или нематоды), при этом съедобное средство обеспечивает образование и/или перенос модулирующего средства. В некоторых вариантах осуществления съедобное средство содержит один или несколько компонентов растения. В некоторых вариантах осуществления один или несколько компонентов растения содержат корень, стебель, лист, цветок, сок, кору, древесину, шип, пыльцу, нектар, семя, плод или любую их комбинацию. К примеру, в некоторых вариантах осуществления растение продуцирует модулирующее средство, которое насекомое поглощает в результате поедания одного или нескольких компонентов растения.
В еще одном аспекте в данном документе предусмотрены хозяева, содержащие любую из предшествующих композиций (например, модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновую кислоту (например, ДНК, РНК (например, mRNA, gRNA или ингибирующую РНК (например, RNAi, shRNA, miRNA)), нуклеиновую кислоту CRISPR), малую молекулу (например, простагландин) или любую их комбинацию)). Хозяин может представлять собой беспозвоночное (например, насекомое, моллюска или нематоду). В некоторых вариантах осуществления беспозвоночное представляет собой насекомое. В некоторых вариантах осуществления насекомое представляет насекомое, содержащее бактериоцит. К примеру, в определенных вариантах осуществления насекомое, содержащее бактериоцит, может представлять собой тлю (например, тлю кукурузную листовую или тлю персиковую зеленую). В дополнительных вариантах осуществления насекомое представляет собой жука, долгоносика, муху, тлю, белокрылку, цикадку, червеца, моль, бабочку, кузнечика, сверчка, трипса или микроскопического клеща. В других вариантах осуществления беспозвоночное представляет собой моллюска. В некоторых вариантах осуществления моллюск представляет собой вид, принадлежащий к Veronicellidae, Ampullariidae, Achatinidae, Helicidae, Hydromiidae, Planobidae, Lymnaeidae, Urocyclidae, Bradybaenidae, Agriolimacidae, Arionidae или Milacidae. В другом варианте осуществления беспозвоночное может представлять собой нематоду. В некоторых вариантах осуществления нематода представляет собой вид, принадлежащий к Criconematidae, Belonolaimidae, Hoplolaimidae, Heteroderidae, Longidoridae, Pratylenchidae, Trichodoridae или Anguinidae.
В другом аспекте в данном документе предусмотрена система для модулирования (например, повышения или понижения) приспособленности хозяина. Указанная система содержит модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита), нуклеиновую кислоту (например, ДНК, РНК (например, mRNA, gRNA или ингибирующую РНК (например, RNAi, shRNA, miRNA)), нуклеиновую кислоту CRISPR), малую молекулу (например, простагландин) или любую их комбинацию), которое изменяет взаимодействия между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина, при этом система является эффективной для модулирования (например, повышения или понижения) приспособленности хозяина, и при этом хозяин представляет собой беспозвоночное (например, насекомое (например, тлю), моллюска или нематоду). В некоторых вариантах осуществления модулирующее средство системы представляет собой любую из предшествующих композиций. В некоторых вариантах осуществления модулирующее средство составляют в виде порошка. В некоторых вариантах осуществления модулирующее средство составляют в виде растворителя. В некоторых вариантах осуществления модулирующее средство составляют в виде концентрата. В некоторых вариантах осуществления модулирующее средство составляют в виде разбавителя. В некоторых вариантах осуществления модулирующее средство готовят для доставки посредством объединения любой из предыдущих композиций с носителем.
В другом аспекте в данном документе предусмотрены способы модулирования взаимодействия хозяин-микробиота, которые включают доставку любой из композиций, описанных в данном документе (например, модулирующего средства (например, полипептида (например, антитела, бактериоцина, противомикробного пептида или регуляторного пептида бактериоцита), нуклеиновой кислоты (например, ДНК, РНК (например, mRNA, gRNA или ингибирующей РНК (например, RNAi, shRNA, miRNA)), нуклеиновой кислоты CRISPR), малой молекулы (например, простагландина) или их комбинации), хозяину, при этом модулирующее средство модулирует одно или несколько взаимодействий между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина.
В другом аспекте в данном документе предусмотрены способы модулирования приспособленности хозяина-беспозвоночного (например, насекомого (например, тли), моллюска или нематоды), при этом способ включает доставку любой из композиций, описанных в данном документе (например, модулирующего средства (например, полипептида (например, антитела, бактериоцина, противомикробного пептида или регуляторного пептида бактериоцита), нуклеиновой кислоты (например, ДНК, РНК (например, mRNA, gRNA или ингибирующей РНК (например, RNAi, shRNA, miRNA)), нуклеиновой кислоты CRISPR), малой молекулы (например, простагландина) или их комбинации), хозяину, и при этом модулирующее средство изменяет взаимодействия между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина.
В некоторых вариантах осуществления любого из вышеуказанных способов один или несколько микроорганизмов, обитающих в организме хозяина, могут представлять собой гриб или бактерию. В некоторых вариантах осуществления один или несколько микроорганизмов представляют собой эндосимбиотический микроорганизм. В некоторых вариантах осуществления один или несколько микроорганизмов обитают в кишке хозяина. В некоторых вариантах осуществления один или несколько микроорганизмов обитают в бактериоците в организме хозяина. В некоторых вариантах осуществления один или несколько микроорганизмов требуются для приспособленности хозяина или выживаемости хозяина.
В некоторых вариантах осуществления любого из вышеуказанных способов модулирующее средство может целенаправленно воздействовать на один или несколько путей хозяина, которые опосредуют взаимодействия между хозяином и одним или несколькими микроорганизмами. В некоторых вариантах осуществления путь хозяина представляет собой путь, который обеспечивает регуляцию функции или развитие бактериоцитов насекомого (например, тли). В некоторых вариантах осуществления целенаправленное воздействие на функцию или развитие бактериоцитов хозяина снижает уровень, разнообразие и/или функцию одного или нескольких микроорганизмов, обитающих в бактериоците. В альтернативном случае целенаправленное воздействие на функцию или развитие бактериоцитов хозяина повышает уровень, разнообразие и/или функцию одного или нескольких микроорганизмов, обитающих в бактериоците. В некоторых вариантах осуществления путь хозяина представляет собой путь, который обеспечивает регуляцию иммунной системы хозяина.
В некоторых вариантах осуществления модулирующее средство активирует иммунный ответ против одного или нескольких микроорганизмов, обитающих в организме хозяина, со снижением тем самым уровня, разнообразия и/или функции одного или нескольких микроорганизмов. В некоторых вариантах осуществления модулирующее средство подавляет иммунный ответ против одного или нескольких микроорганизмов, обитающих в организме хозяина, с повышением тем самым уровня, разнообразия и/или функции одного или нескольких микроорганизмов.
В некоторых вариантах осуществления модулирующее средство целенаправленно воздействует на один или несколько путей микроорганизма, которые опосредуют взаимодействия между хозяином и одним или несколькими микроорганизмами.
В некоторых вариантах осуществления этап доставки включает предоставление модулирующего средства в дозе и в течение периода времени, достаточных для воздействия на один или несколько микроорганизмов с модулированием тем самым разнообразия микроорганизмов в организме хозяина. В некоторых вариантах осуществления этап доставки включает местное применение любой из предыдущих композиций в отношении растения. В некоторых вариантах осуществления этап доставки включает предоставление модулирующего средства с помощью генетически модифицированного, сконструированного или трансгенного растения (например, любого из растений, описанных в данном документе). В других вариантах осуществления этап доставки включает в себя предоставление модулирующего средства хозяину в виде съедобного средства для беспозвоночных (например. насекомого, моллюска или нематоды). В дополнительных вариантах осуществления этап доставки включает получение хозяина, несущего модулирующее средство. В некоторых вариантах осуществления хозяин, несущий модулирующее средство, может переносить модулирующее средство одному или нескольким дополнительным хозяевам.
Также предусмотрены скрининговые анализы для идентификации модулирующего средства, которое модулирует (например, повышает или снижает) приспособленность хозяина. Скрининговый анализ может включать этапы (a) воздействия на микроорганизм, который может обитать в организме хозяина, одного или нескольких кандидатных модулирующих средств и (b) идентификации модулирующего средства, которое повышает или снижает приспособленность хозяина. В некоторых вариантах осуществления модулирующее средство представляет собой микроорганизм, обитающий в организме хозяина. В некоторых вариантах осуществления микроорганизм представляет собой бактерию. В некоторых вариантах осуществления бактерия, обитая в организме хозяина, обеспечивает повышение приспособленности хозяина. В альтернативном случае бактерия, обитая в организме хозяина, обеспечивает снижение приспособленности хозяина. В некоторых вариантах осуществления модулирующее средство представляет собой любое из модулирующих средств, описанных в данном документе (например, модулирующее средство (например, полипептид (например, антитело, бактериоцин, противомикробный пептид или регуляторный пептид бактериоцита (например, колеоптерицин А)), нуклеиновую кислоту (например, ДНК, РНК (например, mRNA, gRNA или ингибирующую РНК (например, RNAi, shRNA, miRNA)), нуклеиновую кислоту CRISPR), малую молекулу (например, простагландин) или любую их комбинацию)). В некоторых вариантах осуществления модулирующее средство предоставляют посредством генетически модифицированного фага или бактерии. В некоторых вариантах осуществления приспособленность хозяина модулируют с помощью модулирования микробиоты хозяина.
Определения
Используемый в данном документе термин "бактериоцит" относится к специализированной клетке, встречающейся у беспозвоночных, например, насекомых, нематод или моллюсков, в которой обитают внутриклеточные бактерии со свойствами симбиотических бактерий. В некоторых случаях бактериоцит может кластеризоваться с другими бактериоцитами с образованием бактериома.
Используемый в данном документе термин "эффективное количество" относится к количеству модулирующего средства (например, полипептида, нуклеиновой кислоты, малой молекулы или их комбинаций) или композиции, содержащей указанное средство, достаточному для получения указанного результата, например, для повышения или понижения приспособленности организма-хозяина (например, насекомого, нематоды или моллюска); для достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации модулирующего средства в организме хозяина-мишени; для достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации модулирующего средства в кишке хозяина-мишени; для достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации модулирующего средства в бактериоците хозяина-мишени; для модулирования уровня или активности одного или нескольких микроорганизмов (например, эндосимбионтов) в организме хозяина-мишени.
Используемый в данном документе термин "приспособленность" относится к способности хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) выживать и/или давать жизнеспособное потомство. Приспособленность хозяина (например, насекомого, моллюска или нематоды) может быть измерена с помощью одного или нескольких параметров, в том числе без ограничения продолжительности жизни, скорости размножения, подвижности, массы тела или скорости метаболизма. В зависимости от хозяина, приспособленность может быть дополнительно измерена на основе показателей активности (например, укусов животных или людей, опыления растений), передачи заболеваний (например, переноса от переносчика переносчику или переноса от переносчика животному) или продуцирования (например, меда или шелка).
Используемый в данном документе термин "кишка" относится к любой части кишки хозяина, в том числе передней кишке, средне кишке или задней кишке хозяина.
Используемый в данном документе термин "хозяин" относится организму, такому как беспозвоночное (например, насекомое, моллюск или нематода), несущему обитающие в нем микроорганизмы (например, эндогенные микроорганизмы, эндосимбиотические микроорганизмы (например, первичные или вторичные эндосимбионты), организмы-комменсалы и/или патогенные микроорганизмы).
Используемый в данном документе термин "снижение приспособленности хозяина" относится к любому нарушению физиологических процессов хозяина или любой активности, осуществляемой указанным хозяином, вследствие введения модулирующего средства, в том числе без ограничения любому одному или нескольким из следующих желаемых эффектов: (1) уменьшению популяции хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (2) снижению скорости размножения (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (3) снижению подвижности хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (4) уменьшению массы тела хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (5) снижению скорости метаболизма или активности хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (6) снижению заражения растения хозяином (например, насекомым, моллюском или нематодой) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (7) снижению передачи заболеваний (например, растения, животного или человеческого патогена) хозяином (например, насекомым, моллюском или нематодой) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; или (8) снижению роста, повышению смертности нимф и/или повышению в зрелом возрасте стерильности хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше. Снижение приспособленности хозяина может быть определено по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
Используемый в данном документе термин "повышение приспособленности хозяина" или "способствование приспособленности хозяина" относится к любому благоприятному изменению физиологических процессов хозяина или любой активности, осуществляемой указанным хозяином, вследствие введения модулирующего средства, в том числе без ограничения любому одному или нескольким из следующих необходимых эффектов: (1) увеличению популяции хозяина на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (2) повышению скорости размножения хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (3) повышению подвижности хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (4) увеличению массы тела хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (5) повышению скорости метаболизма или активности хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (6) повышению опыления (например, числу растений, опыленных за определенное количество времени) хозяином (например, насекомым, моллюском или нематодой) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (7) повышению продуцирования побочных продуктов (например, меда или шелка) хозяина (например, насекомого, моллюска или нематоды) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; (8) повышению содержания питательного вещества хозяина (например, насекомого, моллюска или нематоды) (например, белок, жирные кислоты или аминокислоты) на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше; или (9) повышению устойчивости хозяина (например, насекомого, моллюска или нематоды) к пестициду на приблизительно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 99%, 100% или больше. Повышение приспособленности хозяина может быть определено по сравнению с организмом-хозяином, которому не было введено модулирующее средство
Как используется в данном документе, выражения "взаимодействия между хозяином и микроорганизмами, обитающими в хозяине" или "взаимодействия хозяин-микробиота" относятся к (i) любым путям (например, метаболическим путям, путям регуляции генов, передачи клеточных сигналов или иммуно-воспалительным путям) в организме хозяина, которые непосредственно или косвенно влияют на выживаемость, рост или метаболизм микроорганизмов, обитающих в организме хозяина (например, эндосимбиотических микроорганизмов), (ii) любым путям (например, метаболическим путям или путям передачи клеточных сигналов) в обитающем микроорганизме, которые непосредственно или косвенно влияют на приспособленность хозяина-беспозвоночного (например, насекомого, нематоды или моллюска), и/или (iii) любым путям (например, метаболическим путям, путям передачи клеточных сигналов или иммуно-воспалительных путям) в обитающем микроорганизме, которые непосредственно или косвенно влияют на выживаемость, рост или метаболизм хозяина, и/или (iv) любым путям (например, метаболическим путям, путям регуляции генов, передачи клеточных сигналов или иммуно-воспалительным путям) в организме хозяина, которые непосредственно или косвенно влияют на приспособленность обитающего микроорганизма.
Термин "насекомое" включает любой организм, принадлежащий к типу Arthropoda и классу Insecta или классу Arachnida, на любой стадии развития, т. е. неполовозрелых и взрослых насекомых.
Термин "моллюск" включает любой организм, принадлежащий к типу Mollusca, в том числе организмы класса Gastropoda (например, улитки и слизни), на любой стадии развития, т. е. неполовозрелых и взрослых моллюсков.
Термин "нематода" включает любой организм, принадлежащий к типу Nematoda (например, нематоды), на любой стадии развития, т. е. неполовозрелых и взрослых нематод.
Используемый в данном документе термин "микроорганизм" или "микробиота" относится к бактериям или грибам. Микроорганизмы могут относиться к микроорганизмам, обитающим в организме-хозяине (например, эндогенным микроорганизмам, эндосимбиотическим микроорганизмам (например, первичным или вторичным эндосимбионтам)), или микроорганизмам, экзогенным для хозяина, в том числе микроорганизмам, которые могут выступать в качестве модулирующих средств. Используемый в данном документе термин "микроорганизм-мишень" относится к микроорганизму, который обитает в организме хозяина и подвергается воздействию модулирующего средства, прямо или косвенно.
Используемый в данном документе термин "модулирующее средство" или "средство" относится к средству, которое способно изменять уровни и/или функционирование микроорганизмов, обитающих в организме-хозяине (например, беспозвоночном, например, насекомом, моллюске или нематоде), и тем самым модулировать (например, повышать или снижать) приспособленность организма-хозяина.
Определенный в данном документе термин "нуклеиновая кислота" и "полинуклеотид" являются взаимозаменяемыми и относятся к РНК или ДНК, которые являются линейными или разветвленными, одно- или двухнитевыми, или их гибридами, независимо от длины (например, по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 100, 150, 200, 250, 500, 1000 или более нуклеиновых кислот). Термин также охватывает гибриды РНК/ДНК. Нуклеотиды, как правило, в нуклеиновой кислоте связаны с помощью фосфодиэфирных связей, хотя термин "нуклеиновая кислота" также охватывает аналоги нуклеиновых кислот, имеющие другие типы связей или остовов (например, фосфорамидные, фосфотиоатные, фосфодитиоатные, O-метилфосфорамидатные, морфолино связи или остовы, связи или остовы по типу закрытых нуклеиновых кислот (LNA), по типу нуклеиновой кислоты с глицерином (GNA), по типу нуклеиновой кислоты с треозой (TNA) и по типу нуклеиновой кислоты с пептидом (PNA), среди прочего). Нуклеиновые кислоты могут быть однонитевыми, двухнитевыми или содержать однонитевую и двухнитевую последовательность. Нуклеиновая кислота может содержать любую комбинацию дезоксирибонуклеотидов и рибонуклеотидов, а также любую комбинацию оснований, в том числе, например, аденин, тимин, цитозин, урацил и модифицированные или неканонические основания (в том числе, например, гипоксантин, ксантин, 7-метилгуанин, 5,6-дигидроурацил, 5-метилцитозин и 5-гидроксиметилцитозин).
Используемый в данном документе термин "вредитель" относится к беспозвоночным (например, насекомым, нематодам или моллюскам), которые причиняют ущерб растениям или другим организмам или в иных отношениях являются вредоносными для людей, например, в отношении способов или продуктов сельского хозяйства людей.
Используемый в данном документе термин "пестицид" или "пестицидное средство" относится к веществу, которое можно применять для контроля вредителей, значимых для сельского хозяйства, окружающей среды и домашнего хозяйства/быта, таких как насекомые, моллюски, нематоды, грибы, бактерии и вирусы. Термин "пестицид" понимают как охватывающий встречающиеся в природе или синтетические инсектициды (ларвициды или адультициды), регуляторы роста насекомых, нематициды, моллюскоциды, акарициды (майтициды), нематоциды, эктопаразитициды, бактерициды, фунгициды или гербициды (вещества, которые можно применять в сельском хозяйстве для контроля или модификации роста растений). Дополнительные примеры пестицидов или пестицидных средств приведены в таблице 11. В некоторых случаях пестицид представляет собой аллелохимикат. Используемый в данном документе термин "аллелохимикат" или "аллелохимическое средство" означает вещество, продуцируемое организмом, которое может воздействовать на физиологическую функцию (например, зарождение, рост, выживаемость или размножение) другого организма (например, насекомого, моллюска или нематоды).
Используемый в данном документе термин "пептид", "белок" или "полипептид" охватывает любую цепь из встречающихся в природе или не встречающихся в природе аминокислот (как D-, так и L-аминокислот), независимо от длины (например, по меньшей мере 2, 3, 4, 5, 6, 7, 10, 12, 14, 16, 18, 20, 25, 30, 40, 50, 100 или больше аминокислот), наличия или отсутствия посттрансляционных модификаций (например, гликозилирования или фосфорилирования) или наличия, например, одной или нескольких не являющихся аминоацильными групп (например, углеводных, липидных и т. д.), ковалентно связанных с пептидом, и включает, например, встречающиеся в природе белки, синтетические или рекомбинантные полипептиды и пептиды, гибридные молекулы, пептоиды или пептидомиметики.
Как используется в данном документе, "процент идентичности" между двумя последовательностями определяют с помощью алгоритма BLAST 2.0, который описан в Altschul et al., J. Mol. Biol. 215:403-410, 1990. Программное обеспечение для проведения анализов BLAST общедоступно благодаря Национальному центру биотехнологической информации.
Используемый в данном документе термин "растение" относится к целым растениям, органам растений, растительным тканям, растительным клеткам, семенам или их потомству. Растительные клетки включают без ограничения клетки из семян, суспензионных культур, зародышей, участков меристемы, каллюсной ткани, листьев, корней, побегов, гаметофитов, спорофитов, пыльцы или микроспор. Части растений включают в себя дифференцированные и недифференцированные ткани, в том числе без ограничения следующее: корни, стебли, побеги, листья, пыльцу, семена, опухолевую ткань или различные формы клеток и культуры (например, отдельные клетки, протопласты, зародыши и каллюсную ткань). Растительная ткань может находиться в растении или в органе, ткани или культуре клеток растения.
Используемый в данном документе термин "трансгенное растение", "генетически сконструированное растение" или "генетически модифицированное растение" относится к растению, геном которого (например, хромосомная ДНК, хлоропластная ДНК или митохондриальная ДНК) был изменен с помощью стабильной интеграции рекомбинантной ДНК. Трансгенное растение включает растение, регенерированное из изначально трансформированной растительной клетки и потомков трансгенных растений из более поздних поколений или скрещиваний трансформированного растения. К примеру, трансгенное растение может быть генетически модифицированным таким образом, что в нем продуцируется гетерологично (например, неэндогенный) или негетерологично (например, эндогенный) кодируемый белок или РНК, например, любого из модулирующих средств в способах и композициях, описанных в данном документе. Любой вид растения может быть трансформирован с получением трансгенного растения. Трансгенное растение может быть двудольным растением или однодольным растением. Например, и без ограничения трансгенные растения композиций и способов, описанных в данном документе, могут быть получены из любого из следующих семейств двудольных растений: Leguminosae, в том числе растений, таких как горох, люцерна и соя; Umbelliferae, в том числе растений, таких как морковь и сельдерей; Solanaceae, в том числе растений, таких как томат, картофель, баклажан, табак и перец; Cruciferae, в частности, рода Brassica, который включает в себя растения, такие как масличный рапс, свеклу, кочанную капусту, цветную капусту и брокколи); и Arabidopsis thaliana; Compositae, которое включает в себя растения, такие как латук; Malvaceae, которое включает в себя хлопок; Fabaceae, которое включает в себя растения, такие как арахис, и т. п. Трансгенные растения по настоящему изобретению могут быть получены из однодольных растений, таких как, к примеру, пшеница, ячмень, сорго, просо, рожь, тритикале, маис, рис, овес, просо прутьевидное, мискантус и сахарный тростник. Также в трансгенные растения по настоящему изобретению включены деревья, такие как яблоня, груша, айва, слива, черешня, персик, нектарин, абрикос, папайя, манго и другие древесные виды, в том числе хвойные и лиственные деревья, такие как тополь, сосна, секвойя, кедр, дуб, ива и т. п.
Другие характеристики и преимущества настоящего изобретения будут очевидны из следующего подробного описания и формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигуры предусмотрены для иллюстрации одной или нескольких характеристик, аспектов или вариантов осуществления настоящего изобретения и не предполагаются как ограничивающие.
На фиг. 1 представлена панель графиков, на которых показано, что обработка с помощью P. pastoris приводила к задержке развития тлей. Тлей LSR-1 на стадии личинок первого и второго возраста помещали на листья, залитые водой (отрицательный контроль) или раствором P. pastoris в воде, и отслеживали стадию развития в каждой указанной временной точке во время эксперимента. Показаны средние значения в процентах тлей в каждой группе ± SD.
На фиг. 2 представлен график, на котором показано, что обработка с помощью P. pastoris приводила к гибели тлей. Тлей LSR-1 на стадии личинок первого и второго возраста обрабатывали водой (контроль) или P. pastoris посредством заливания листьев и отслеживали выживаемость ежедневно во время эксперимента. N=62-63 тлей/группа. Статистически значимые различия определяли с помощью логарифмического рангового критерия (Кокса-Мантеля). ****, p < 0,0001.
На фиг. 3 представлена панель графиков, на которых показано, что обработка с помощью P. pastoris посредством обрызгивания листьев не влияла в отношении развития тлей. Тлей LSR-1 на стадии личинок первого и второго возраста обрабатывали водой (контроль) или P. pastoris посредством распыления и отслеживали стадию развития с течением времени. Показаны средние значения в процентах ± SD мертвых или живых тлей на каждой стадии развития (личинка на стадии 1-го, 2-го, 3-го, 4-го или 5-го возраста) в каждой временной точке. N=две повторности из 30 тлей/группа.
На фиг. 4 представлен график, на котором показано, что обработка с помощью P. pastoris приводила к повышенной смертности тлей. Тлей LSR-1 на стадии личинок первого и второго возраста помещали на листья, на которые распыляли воду (контроль) или P. pastoris и отслеживали выживаемость с течением времени. N=60 тлей/группа обработки.
На фиг. 5A и 5B представлены графики, на которых показано, что распыление с помощью P. pastoris на листья конских бобов приводило к уменьшению количества эндосимбиотических Buchnera в тлях, кормящихся на листьях. Титр симбионтов определяли через 6 (A) и 9 (B) дней после обработки с помощью P. pastoris. Показаны средние соотношения копий Buchnera/тли ± SD. Число в прямоугольнике над указанной совокупностью данных обозначает медианное значение этой группы. Каждая точка представляет одну тлю.
На фиг. 6 представлен график, на котором показано, что микроинъекция PNA BCR-4 приводила к снижению экспрессии BCR-4. Тлям A. pisum на стадии личинок четвертого и пятого возраста вводили 20 нл воды или 321 нг/мкл PNA BCR-4, экстрагировали РНК из тлей через 7 дней и осуществляли RT-qPCR с целью измерения экспрессии BCR-4. Показаны средние соотношения копий BCR-4/актин ± SD. Каждая точка данных представляет одну тлю. Число в прямоугольнике над каждой совокупностью данных обозначает медиану этих данных.
На фиг. 7 представлен график, на котором показано снижение выживаемости насекомых после обработки с помощью PNA, целенаправленно воздействующей на BCR-4. Тлям A. pisum на четвертой и пятой возрастной стадиях вводили воду или PNA, целенаправленно воздействующую на BCR-4. Выживаемость отслеживали ежедневно в течение эксперимента. N=20 тлей/группа обработки.
На фиг. 8 представлен график, на котором показано, что введение тлям PNA, целенаправленно воздействующей на BCR-4, приводило к снижению плодовитости. Тлям A. pisum на четвертой и пятой возрастной стадиях вводили воду или PNA, целенаправленно воздействующую на BCR-4. Плодовитость измеряли с помощью подсчета количества потомков, полученных в каждой группе в каждой временной точке, и представляли в виде количества F1 (потомки первого поколения), полученных от F0 (взрослых) в день. N=20 тлей/группа обработки.
На фиг. 9 представлена панель графиков, на которых показано, что обработка PNA BCR-4 приводила к задержке развития тлей. Тлей LSR-1 на стадии личинок первого и второго возраста помещали на листья, залитые водой (отрицательный контроль) или раствором PNA BCR-4 в воде и отслеживали стадию развития в каждой указанной временной точке во время эксперимента. Показаны средние значения в процентах тлей в каждой группе ± SD.
На фиг. 10 представлен график, на котором показано, что обработка BCR-4 приводила к повышению смертности тлей. Тлей LSR-1 на стадии личинок первого и второго возраста обрабатывали водой (контроль) или PNA BCR-4 посредством заливания листьев и отслеживали выживаемость ежедневно во время эксперимента. N=60 тлей/группа. Статистически значимые различия определяли с помощью логарифмического рангового критерия (Кокса-Мантеля).
На фиг. 11 представлен график, на котором показано, что обработка с помощью PNA BCR-4, доставляемого посредством заливания листьев, приводила к увеличению титров Buchnera. Тлей LSR-1 на стадии личинок первого и второго возраста обрабатывали водой (контроль) или PNA BCR4 с помощью заливания листьев, а мертвых тлей собирали на дни 5 и 6 после обработки. ДНК экстрагировали и выполняли qPCR с целью определения количества копий ДНК Buchnera/тли. Показано среднее количество копий ДНК Buchnera/тлей ± SD для 6-7 тлей/группа.
На фиг. 12 представлен график, на котором показано, что обработка тлей PNA, направленной против BCR-4, с помощью заливания листьев приводила к снижению экспрессии BCR-4. Тлей LSR-1 на стадии личинок первого и второго возраста обрабатывали водой (контроль) или PNA BCR-4 с помощью заливания листьев и на 7-й день РНК экстрагировали из живых тлей и проводили RT-qPCR с целью количественного оценки экспрессии BCR-4 относительно экспрессии актина. Число в прямоугольнике обозначает медиану совокупности данных.
На фиг. 13 представлен график, на котором показано, что обработка dsRNA-ApGLNT1 приводила к нокдауну экспрессии ApGLNT1. Тлям A. pisum на стадии личинок первого возраста вводили воду или dsRNA-ApGLNT1 в воде. Через 2 дня после обработки экстрагировали общую РНК и выполняли RT-qPCR с целью определения относительной экспрессии гена ApGLNT1 (актина в качестве гена внутреннего стандарта). Показано среднее соотношение относительной экспрессии ApGLNT1/актин ± SD для 5-7 тлей/группа. Статистически значимые различия определяли с помощью t-критерия Стьюдента (**, p<0,01).
На фиг. 14 представлен график, на котором показано, что обработка с помощью dsRNA-ApGLNT1 приводила к повышению смертности тлей. Тлям A. pisum LSR-1 вводили воду или dsRNA-ApGLNT1 в воде и отслеживали выживаемость в течение эксперимента. N=40 тлей/группа. Статистически значимую разницу выявляли между двумя группами, определенную с помощью логарифмического рангового критерия (Кокса-Мантеля).
На фиг. 15 представлен график, на котором показано, что обработка с помощью dsRNA-ApGLNT1 приводила к снижению титров Buchnera. Тлям A. pisum LSR-1 вводили воду или dsRNA-ApGLNT1 в воде, ДНК экстрагировали из тлей через 5 дней после обработки и проводили qPCR для количественной оценки Buchnera. Показаны средние количества копий ДНК Buchnera/тлей ± SD. Каждая точка представляет отдельную тлю. Число в прямоугольнике над каждой совокупностью данных обозначает медиану этой группы.
На фиг. 16A и 16B представлена панель графиков, на которых показано, что потомство тлей, которым вводили микроинъекцию dsRNA-ApGLNT1, характеризовалось задержкой развития. Фиг. 16A: тлям A. pisum LSR-1 вводили воду или dsRNA-ApGLNT1 в воде и отслеживали стадии развития в указанной временной точке после введения. Показан среднее значение в процентах ± тлей, которые были мертвыми или живыми на каждой стадии личинок во время всего эксперимента. N=40 тлей/группа. Фиг. 16B: через 4 дня после сбора потомства получали фотографии каждой тли с целью определения площади каждой тли. Показана средняя площадь ± SD потомков, отобранных от тлей, которым вводили либо воду, либо dsRNA-ApGLNT1. Статистически значимые различия определяли с помощью t-критерия Стьюдента. Каждая точка данных представляет одну отдельную тлю.
На фиг. 17 представлена иллюстрация, на которой показана кассета экспрессии dsRNA. Промотор pCaMV 35S помещен выше последовательности, экспрессирующей dsRNA. Смысловая и антисмысловая нити участка гена-мишени тли помещены в тандеме с коротким спейсером, что будет выступать в качестве шпилечной петли. После экспрессии образованная РНК будет принимать двухнитевую конфигурацию в результате комплементарности последовательности.
На фиг. 18 представлена иллюстрация челночного вектора для конструкций для экспрессии dsRNA в N. tabacum. Плазмида содержит точки начала репликации, совместимые с E. coli и A. tumefaciens, маркеры устойчивости к канамицину и гентамицину, кассету экспрессии с зеленым флуоресцентным белком под контролем убиквитинового промотора петрушки и, наконец, кассету экспрессии dsRNA, управляемую pCaMV 35S.
На фиг. 19 представлена панель изображений, на которых показана экспрессия GFP в растении N. tabacum, инфильтрованном A. tumefaciens. Верхние панели представляют собой N. tabacum, инфильтрированный A. tumefaciens, содержащей плазмиду, которая конститутивно управляет экспрессией GFP в N. tabacum (верхняя левая панель представляет собой поле зрения высокой освещенности, а верхняя правая панель представляет собой зеленый канал). Нижние панели представляют собой листья отрицательного контроля, не инфильтрированные A. tumefaciens.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В данном документе предусмотрены способы и композиции для модулирования приспособленности хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) с помощью изменения взаимодействий между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина. В настоящем изобретении описана композиция, содержащая модулирующее средство (например, полипептид, нуклеиновую кислоту, малую молекулу или их комбинации), которое может опосредованно индуцировать изменения в микробиоте хозяина таким образом, что это модулирует (например, повышает или снижает) приспособленность хозяина. К примеру, модулирующее средство может целенаправленно воздействовать на пути хозяина (например, пути иммунной системы или бактериоцитов) или микробные пути, которые обеспечивают изменение (например, повышение или снижение) уровней микробов, активности микробов, метаболизма микробов и/или разнообразия микробов, и, в свою очередь, модулирует (например, повышает или снижает) приспособленность множества беспозвоночных (например, насекомого, моллюска или нематоды), которые важны для сельского хозяйства, коммерческой деятельности и/или здравоохранения.
Способы и композиции, описанные в данном документе, основаны отчасти на примерах, в которых проиллюстрировано то, как различные средства, к примеру, малые соединения (например, простагландин), ингибирующая РНК (например, dsRNA или PNA) и микроорганизмы (грибы или бактерии) можно применять для изменения ответа иммунной системы хозяина в отношении микроорганизмов, обитающих в организме хозяина. Способы и композиции, описанные в данном документе, также можно применять для изменения функции органов или клеток хозяина, в котором микроорганизмы обычно обитают. К примеру, РНК можно применять для нарушения функции бактериоцитов в организме тли с разрушением тем самым популяций эндосимбиотических микрорганизмов, обитающих в бактериоците тли. Нарушение эндосимбиотических популяций микроорганизмов (например, Buchnera spp.) в организме тли, в свою очередь, снижает приспособленность тли. Нуклеиновые кислоты, такие как РНК (например, dsRNA), или PNA, или малые молекулы (например, простагландин), являются представителями модулирующих средств, пригодных в настоящем изобретении, и другие модулирующие средства данного типа могут быть пригодны в настоящем изобретении. На этом основании в настоящем изобретении описано множество различных подходов для применения средств, которые модулируют (например, повышают или снижают) приспособленность хозяина-беспозвоночного (например, насекомого, моллюска или нематоды), при этом модулирующее средство изменяет взаимодействия между хозяином и одним или несколькими микроорганизмами, обитающими в организме хозяина.
I. Хозяева
i. Насекомые-хозяева
В некоторых случаях хозяин, описанный в данном документе, представляет собой организм, принадлежащий к типу Arthropoda. В некоторых случаях насекомое считается вредителем, например, сельскохозяйственным вредителем. В некоторых случаях насекомое несет бактерию или вирус, который считается вредителем растений, что вызывает заболевание у растения (например, Agrobacterium или вирус желтой курчавости листьев томата (TYLCV)). Хозяин может быть на любой стадии развития. Например, хозяин может представлять собой зародыш, личинку, куколку или взрослую особь.
В некоторых случаях насекомое может принадлежать к следующим отрядам: Acari, Araneae, Anoplura, Coleoptera, Collembola, Dermaptera, Dictyoptera, Diplura, Diptera (например, пестрокрылая Drosophila), Embioptera, Ephemeroptera, Grylloblatodea, Hemiptera (например, тля, белокрылка или щитник теплиц), Homoptera, Hymenoptera, Isoptera, Lepidoptera, Mallophaga, Mecoptera, Neuroptera, Odonata, Orthoptera, Phasmida, Plecoptera, Protura, Psocoptera, Siphonaptera, Siphunculata, Thysanura, Strepsiptera, Thysanoptera, Trichoptera или Zoraptera.
В некоторых случаях насекомое относится к классу Arachnida, к примеру, Acarus spp., Aceria sheldoni, Aculops spp., Aculus spp., Amblyomma spp., Amphitetranychus viennensis, Argas spp., Boophilus spp., Brevipalpus spp., Bryobia graminum, Bryobia praetiosa, Centruroides spp., Chorioptes spp., Dermanyssus gallinae, Dermatophagoides pteronyssinus, Dermatophagoides farinae, Dermacentor spp., Eotetranychus spp., Epitrimerus pyri, Eutetranychus spp., Eriophyes spp., Glycyphagus domesticus, Halotydeus destructor, Hemitarsonemus spp., Hyalomma spp., Ixodes spp., Latrodectus spp., Loxosceles spp., Metatetranychus spp., Neutrombicula autumnalis, Nuphersa spp., Oligonychus spp., Ornithodorus spp., Ornithonyssus spp., Panonychus spp., Phyllocoptruta oleivora, Polyphagotarsonemus latus, Psoroptes spp., Rhipicephalus spp., Rhizoglyphus spp., Sarcoptes spp., Scorpio maurus, Steneotarsonemus spp., Steneotarsonemus spinki, Tarsonemus spp., Tetranychus spp., Trombicula alfreddugesi, Vaejovis spp., Vasates lycopersici.
В некоторых случаях насекомое относится к классу Chilopoda, например, Geophilus spp., Scutigera spp.
В некоторых случаях насекомое относится к отряду Collembola, например, Onychiurus armatus.
В некоторых случаях насекомое относится к классу Diplopoda, например, Blaniulus guttulatus.
В некоторых случаях насекомое относится к классу Insecta, например, к отряду Blattodea, к примеру, Blattella asahinai, Blattella germanica, Blatta orientalis, Leucophaea maderae, Panchlora spp., Parcoblatta spp., Periplaneta spp., Supella longipalpa.
В некоторых случаях насекомое относится к отряду Coleoptera, к примеру, Acalymma vittatum, Acanthoscelides obtectus, Adoretus spp., Agelastica alni, Agriotes spp., Alphitobius diaperinus, Amphimallon solstitialis, Anobium punctatum, Anoplophora spp., Anthonomus spp., Anthrenus spp., Apion spp., Apogonia spp., Atomaria spp., Attagenus spp., Bruchidius obtectus, Bruchus spp., Cassida spp., Cerotoma trifurcata, Ceutorrhynchus spp., Chaetocnema spp., Cleonus mendicus, Conoderus spp., Cosmopolites spp., Costelytra zealandica, Ctenicera spp., Curculio spp., Cryptolestes ferrugineus, Cryptorhynchus lapathi, Cylindrocopturus spp., Dendroctonus spp. (например, Dendroctonus ponderosae), Dermestes spp., Diabrotica spp. (например, злаковый корневой червь), Dichocrocis spp., Dicladispa armigera, Diloboderus spp., Epilachna spp., Epitrix spp., Faustinus spp., Gibbium psylloides, Gnathocerus cornutus, Hellula undalis, Heteronychus arator, Heteronyx spp., Hylamorpha elegans, Hylotrupes bajulus, Hypera postica, Hypomeces squamosus, Hypothenemus spp. (Hypothenemus hampei), Lachnosterna consanguinea, Lasioderma serricorne, Latheticus oryzae, Lathridius spp., Lema spp., Leptinotarsa decemlineata, Leucoptera spp., Lissorhoptrus oryzophilus, Lixus spp., Luperodes spp., Lyctus spp., Megascelis spp., Melanotus spp., Meligethes aeneus, Melolontha spp., Migdolus spp., Monochamus spp., Naupactus xanthographus, Necrobia spp., Niptus hololeucus, Oryctes rhinoceros, Oryzaephilus surinamensis, Oryzaphagus oryzae, Otiorrhynchus spp., Oxycetonia jucunda, Phaedon cochleariae, Phyllophaga spp., Phyllophaga helleri, Phyllotreta spp., Popillia japonica, Premnotrypes spp., Prostephanus truncatus, Psylliodes spp., Ptinus spp., Rhizobius ventralis, Rhizopertha dominica, Sitophilus spp., Sitophilus oryzae, Sphenophorus spp., Stegobium paniceum, Sternechus spp., Symphyletes spp., Tanymecus spp., Tenebrio molitor, Tenebrioides mauretanicus, Tribolium spp., Trogoderma spp., Tychius spp., Xylotrechus spp., Zabrus spp.;
к отряду Diptera, к примеру, Aedes spp., Agromyza spp., Anastrepha spp., Anopheles spp., Asphondylia spp., Bactrocera spp., Bibio hortulanus, Calliphora erythrocephala, Calliphora vicina, Ceratitis capitata, Chironomus spp., Chrysomyia spp., Chrysops spp., Chrysozona pluvialis, Cochliomyia spp., Contarinia spp., Cordylobia anthropophaga, Cricotopus sylvestris, Culex spp., Culicoides spp., Culiseta spp., Cuterebra spp., Dacus oleae, Dasyneura spp., Delia spp., Dermatobia hominis, Drosophila spp., Echinocnemus spp., Fannia spp., Gasterophilus spp., Glossina spp., Haematopota spp., Hydrellia spp., Hydrellia griseola, Hylemya spp., Hippobosca spp., Hypoderma spp., Liriomyza spp., Lucilia spp., Lutzomyia spp., Mansonia spp., Musca spp. (например, Musca domestica), Oestrus spp., Oscinella frit, Paratanytarsus spp., Paralauterborniella subcincta, Pegomyia spp., Phlebotomus spp., Phorbia spp., Phormia spp., Piophila casei, Prodiplosis spp., Psila rosae, Rhagoletis spp., Sarcophaga spp., Simulium spp., Stomoxys spp., Tabanus spp., Tetanops spp., Tipula spp.
В некоторых случаях насекомое относится к отряду Heteroptera, к примеру, Anasa tristis, Antestiopsis spp., Boisea spp., Blissus spp., Calocoris spp., Campylomma livida, Cavelerius spp., Cimex spp., Collaria spp., Creontiades dilutus, Dasynus piperis, Dichelops furcatus, Diconocoris hewetti, Dysdercus spp., Euschistus spp., Eurygaster spp., Heliopeltis spp., Horcias nobilellus, Leptocorisa spp., Leptocorisa varicornis, Leptoglossus phyllopus, Lygus spp., Macropes excavatus, Miridae, Monalonion atratum, Nezara spp., Oebalus spp., Pentomidae, Piesma quadrata, Piezodorus spp., Psallus spp., Pseudacysta persea, Rhodnius spp., Sahlbergella singularis, Scaptocoris castanea, Scotinophora spp., Stephanitis nashi, Tibraca spp., Triatoma spp.
В некоторых случаях насекомое относится к отряду Hemiptera или подотряду Homoptera, к примеру, Acizzia acaciaebaileyanae, Acizzia dodonaeae, Acizzia uncatoides, Acrida turrita, Acyrthosipon spp., Acrogonia spp., Aeneolamia spp., Agonoscena spp., Aleyrodes proletella, Aleurolobus barodensis, Aleurothrixus floccosus, Allocaridara malayensis, Amrasca spp., Anuraphis cardui, Aonidiella spp., Aphanostigma pini, Aphis spp. (e.g., Apis gossypii), Arboridia apicalis, Arytainilla spp., Aspidiella spp., Aspidiotus spp., Atanus spp., Aulacorthum solani, Bemisia tabaci, Blastopsylla occidentalis, Boreioglycaspis melaleucae, Brachycaudus helichrysi, Brachycolus spp., Brevicoryne brassicae, Cacopsylla spp., Calligypona marginata, Carneocephala fulgida, Ceratovacuna lanigera, Cercopidae, Ceroplastes spp., Chaetosiphon fragaefolii, Chionaspis tegalensis, Chlorita onukii, Chondracris rosea, Chromaphis juglandicola, Chrysomphalus ficus, Cicadulina mbila, Coccomytilus halli, Coccus spp., Cryptomyzus ribis, Cryptoneossa spp., Ctenarytaina spp., Dalbulus spp., Dialeurodes citri, Diaphorina citri, Diaspis spp., Drosicha spp., Dysaphis spp., Dysmicoccus spp., Empoasca spp., Eriosoma spp., Erythroneura spp., Eucalyptolyma spp., Euphyllura spp., Euscelis bilobatus, Ferrisia spp., Geococcus coffeae, Glycaspis spp., Heteropsylla cubana, Heteropsylla spinulosa, Homalodisca coagulata, Homalodisca vitripennis, Hyalopterus arundinis, Icerya spp., Idiocerus spp., Idioscopus spp., Laodelphax striatellus, Lecanium spp., Lepidosaphes spp., Lipaphis erysimi, Macrosiphum spp., Macrosteles facifrons, Mahanarva spp., Melanaphis sacchari, Metcalfiella spp., Metopolophium dirhodum, Monellia costalis, Monelliopsis pecanis, Myzus spp., Nasonovia ribisnigri, Nephotettix spp., Nettigoniclla spectra, Nilaparvata lugens, Oncometopia spp., Orthezia praelonga, Oxya chinensis, Pachypsylla spp., Parabemisia myricae, Paratrioza spp., Parlatoria spp., Pemphigus spp., Peregrinus maidis, Phenacoccus spp., Phloeomyzus passerinii, Phorodon humuli, Phylloxera spp., Pinnaspis aspidistrae, Planococcus spp., Prosopidopsylla flava, Protopulvinaria pyriformis, Pseudaulacaspis pentagona, Pseudococcus spp., Psyllopsis spp., Psylla spp., Pteromalus spp., Pyrilla spp., Quadraspidiotus spp., Quesada gigas, Rastrococcus spp., Rhopalosiphum spp., Saissetia spp., Scaphoideus titanus, Schizaphis graminum, Selenaspidus articulatus, Sogata spp., Sogatella furcifera, Sogatodes spp., Stictocephala festina, Siphoninus phillyreae, Tenalaphara malayensis, Tetragonocephela spp., Tinocallis caryaefoliae, Tomaspis spp., Toxoptera spp., Trialeurodes vaporariorum, Trioza spp., Typhlocyba spp., Unaspis spp., Viteus vitifolii, Zygina spp.; к отряду Hymenoptera, к примеру, Acromyrmex spp., Athalia spp., Atta spp., Diprion spp., Hoplocampa spp., Lasius spp., Monomorium pharaonis, Sirex spp., Solenopsis invicta, Tapinoma spp., Urocerus spp., Vespa spp., Xeris spp. В определенных случаях насекомое представляет собой тлю (например, Rhopalosiphum maidis или Myzus persicae).
В некоторых случаях насекомое относится к отряду Isopoda, например, Armadillidium vulgare, Oniscus asellus, Porcellio scaber.
В некоторых случаях насекомое относится к отряду Isoptera, к примеру, Coptotermes spp., Cornitermes cumulans, Cryptotermes spp., Incisitermes spp., Microtermes obesi, Odontotermes spp., Reticulitermes spp.
В некоторых случаях насекомое относится к отряду Lepidoptera, к примеру, Achroia grisella, Acronicta major, Adoxophyes spp., Aedia leucomelas, Agrotis spp., Alabama spp., Amyelois transitella, Anarsia spp., Anticarsia spp., Argyroploce spp., Barathra brassicae, Borbo cinnara, Bucculatrix thurberiella, Bupalus piniarius, Busseola spp., Cacoecia spp., Caloptilia theivora, Capua reticulana, Carpocapsa pomonella, Carposina niponensis, Cheimatobia brumata, Chilo spp., Choristoneura spp., Clysia ambiguella, Cnaphalocerus spp., Cnaphalocrocis medinalis, Cnephasia spp., Conopomorpha spp., Conotrachelus spp., Copitarsia spp., Cydia spp., Dalaca noctuides, Diaphania spp., Diatraea saccharalis, Earias spp., Ecdytolopha aurantium, Elasmopalpus lignosellus, Eldana saccharina, Ephestia spp., Epinotia spp., Epiphyas postvittana, Etiella spp., Eulia spp., Eupoecilia ambiguella, Euproctis spp., Euxoa spp., Feltia spp., Galleria mellonella, Gracillaria spp., Grapholitha spp., Hedylepta spp., Helicoverpa spp., Heliothis spp., Hofmannophila pseudospretella, Homoeosoma spp., Homona spp., Hyponomeuta padella, Kakivoria flavofasciata, Laphygma spp., Laspeyresia molesta, Leucinodes orbonalis, Leucoptera spp., Lithocolletis spp., Lithophane antennata, Lobesia spp., Loxagrotis albicosta, Lymantria spp., Lyonetia spp., Malacosoma neustria, Maruca testulalis, Mamstra brassicae, Melanitis leda, Mocis spp., Monopis obviella, Mythimna separata, Nemapogon cloacellus, Nymphula spp., Oiketicus spp., Oria spp., Orthaga spp., Ostrinia spp., Oulema oryzae, Panolis flammea, Parnara spp., Pectinophora spp., Perileucoptera spp., Phthorimaea spp., Phyllocnistis citrella, Phyllonorycter spp., Pieris spp., Platynota stultana, Plodia interpunctella, Plusia spp., Plutella xylostella, Prays spp., Prodenia spp., Protoparce spp., Pseudaletia spp., Pseudaletia unipuncta, Pseudoplusia includens, Pyrausta nubilalis, Rachiplusia nu, Schoenobius spp., Scirpophaga spp., Scirpophaga innotata, Scotia segetum, Sesamia spp., Sesamia inferens, Sparganothis spp., Spodoptera spp., Spodoptera praefica, Stathmopoda spp., Stomopteryx subsecivella, Synanthedon spp., Tecia solanivora, Thermesia gemmatalis, Tinea cloacella, Tinea pellionella, Tineola bisselliella, Tortrix spp., Trichophaga tapetzella, Trichoplusia spp., Tryporyza incertulas, Tuta absoluta, Virachola spp.
В некоторых случаях насекомое относится к отряду Orthoptera или Saltatoria, к примеру, Acheta domesticus, Dichroplus spp., Gryllotalpa spp., Hieroglyphus spp., Locusta spp., Melanoplus spp., Schistocerca gregaria.
В некоторых случаях насекомое относится к отряду Phthiraptera, к примеру, Damalinia spp., Haematopinus spp., Linognathus spp., Pediculus spp., Ptirus pubis, Trichodectes spp.
В некоторых случаях насекомое относится к отряду Psocoptera, к примеру, Lepinatus spp., Liposcelis spp.
В некоторых случаях насекомое относится к отряду Siphonaptera, например, Ceratophyllus spp., Ctenocephalides spp., Pulex irritans, Tunga penetrans, Xenopsylla cheopis.
В некоторых случаях насекомое относится к отряду Thysanoptera, например, Anaphothrips obscurus, Baliothrips biformis, Drepanothrips reuteri, Enneothrips flavens, Frankliniella spp., Heliothrips spp., Hercinothrips femoralis, Rhipiphorothrips cruentatus, Scirtothrips spp., Taeniothrips cardamomi, Thrips spp.
В некоторых случаях насекомое относится к отряду Zygentoma (=Thysanura), например, Ctenolepisma spp., Lepisma saccharina, Lepismodes inquilinus, Thermobia domestica.
В некоторых случаях насекомое относится к классу Symphyla, например, Scutigerella spp.
В некоторых случаях насекомое представляет собой микроскопического клеща, включая без ограничения клещей из семейства Tarsonemidae, таких как Phytonemus pallidus, Polyphagotarsonemus latus, Tarsonemus bilobatus и т. п.; микроскопических клещей из семейства Eupodidae, таких как Penthaleus erythrocephalus, Penthaleus major и т. п.; паутинных клещиков, таких как Oligonychus shinkajii, Panonychus citri, Panonychus mori, Panonychus ulmi, Tetranychus kanzawai, Tetranychus urticae и т. п.; клещей из семейства Eriophyidae, таких как Acaphylla theavagrans, Aceria tulipae, Aculops lycopersici, Aculops pelekassi, Aculus schlechtendali, Eriophyes chibaensis, Phyllocoptruta oleivora и т. п.; клещей из семейства Acaridae, таких как Rhizoglyphus robini, Tyrophagus putrescentiae, Tyrophagus similis и т. п.; клещей, поражающих расплод пчел, таких как Varroa jacobsoni, Varroa destructor и т. п.; иксодовых клещей, таких как Boophilus microplus, Rhipicephalus sanguineus, Haemaphysalis longicornis, Haemophysalis flava, Haemophysalis campanulata, Ixodes ovatus, Ixodes persulcatus, Amblyomma spp., Dermacentor spp. и т. п.; Cheyletidae, таких как Cheyletiella yasguri, Cheyletiella blakei и т. п.; Demodicidae, таких как Demodex canis, Demodex cati и т. п.; Psoroptidae, таких как Psoroptes ovis и т. п.; Sarcoptidae, таких как Sarcoptes scabiei, Notoedres cati, Knemidocoptes spp. и т. п.
Способы и композиции, предусмотренные в данном документе, можно применять в отношении любого насекомого-хозяина, которое считается переносчиком патогена, способного вызывать заболевание у животных. Например, насекомое-хозяин может включать в себя без ограничения насекомых с колюще-сосущими ротовыми аппаратами, которые встречаются у Hemiptera и некоторых Hymenoptera и Diptera, таких как комары, пчелы, осы, галлицы, вши, муха цеце, блохи и муравьи, а также представителей Arachnidae, таких как иксодовые клещи и микроскопические клещи; в отряде, классе или семействе Acarina (иксодовых клещей и микроскопических клещей), например, у представителей семейств Argasidae, Dermanyssidae, Ixodidae, Psoroptidae или Sarcoptidae и представителей видов Amblyomma spp., Anocenton spp., Argas spp., Boophilus spp., Cheyletiella spp., Chorioptes spp., Demodex spp., Dermacentor spp., Dermanyssus spp., Haemophysalis spp., Hyalomma spp., Ixodes spp., Lynxacarus spp., Mesostigmata spp., Notoednes spp., Ornithodoros spp., Ornithonyssus spp., Otobius spp., Оtodectes spp., Pneumonyssus spp., Psoroptes spp., Rhipicephalus spp., Sancoptes spp. или Trombicula spp.; Anoplura (сосущих вшей и пухоедов), например, представителей видов Bovicola spp., Haematopinus spp., Linognathus spp., Menopon spp., Pediculus spp., Pemphigus spp., Phylloxera spp. или Solenopotes spp.; Diptera (мух), например, представителей видов Aedes spp., Anopheles spp., Calliphora spp., Chrysomyia spp., Chrysops spp., Cochliomyia spp., Culex spp., Culicoides spp., Cuterebra spp., Dermatobia spp., Gastrophilus spp., Glossina spp., Haematobia spp., Haematopota spp., Hippobosca spp., Hypoderma spp., Lucilia spp., Lyperosia spp., Melophagus spp., Oestrus spp., Phaenicia spp., Phlebotomus spp., Phormia spp., Sarcoptidae spp., Sarcophaga spp., Simulium spp., Stomoxys spp., Tabanus spp., Tannia spp. или Tipula spp.; Mallophaga (кусающих вшей), например, представителей видов Damalina spp., Felicola spp., Heterodoxus spp. или Trichodectes spp.; или Siphonaptera (бескрылых насекомых), например, представителей видов Ceratophyllus spp., Xenopsylla spp; Cimicidae (настоящих клопов), например, представителей видов Cimex spp., Tritominae spp., Rhodinius spp. или Triatoma spp. В некоторых случаях насекомое представляет собой кровососущее насекомое из отряда Diptera (например, подотряда Nematocera, например, семейства Colicidae). В некоторых случаях насекомое относится к подсемействам Culicinae, Corethrinae, Ceratopogonidae или Simuliidae. В некоторых случаях насекомое относится к Culex spp., Theobaldia spp., Aedes spp., Anopheles spp., Aedes spp., Forciponiyia spp., Culicoides spp. или Helea spp.
ii. Хозяева-моллюски
В некоторых случаях хозяин, описанный в данном документе, может представлять собой организм, принадлежащий к типу Mollusca. В некоторых случаях моллюск считается вредителем, например, сельскохозяйственным вредителем. К примеру, способы и композиции являются подходящими для контроля наземных брюхоногих моллюсков (например, слизней и улиток) в сельском хозяйстве и растениеводстве. Они включают в себя всех наземных слизней и улиток, которые главным образом встречаются в качестве многоядных вредителей на сельскохозяйственных и плодовых культурах.
В некоторых случаях моллюск принадлежит к семейству Achatinidae, Agriolimacidae, Ampullariidae, Arionidae, Bradybaenidae, Helicidae, Hydromiidae, Lymnaeidae, Milacidae, Urocyclidae или Veronicellidae.
К примеру, в некоторых случаях моллюск представляет собой Achatina spp., Agriolimax spp., Arion spp. (например, A. ater, A. circumscriptus, A. distinctus, A. fasciatus, A. hortensis, A. intermedius, A. rufus, A. subfuscus, A. silvaticus, A. lusitanicus), Biomphalaria spp., Bradybaena spp. (например, B. fruticum), Bulinus spp., Cantareus spp. (например, C. asperses), Cepaea spp. (например, C. hortensis, C. nemoralis), Cernuella spp., Cochlicella spp., Cochlodina spp. (например, C. laminata), Deroceras spp. (например, D. agrestis, D. empiricorum, D. laeve, D. panornimatum, D. reticulatum), Discus spp. (например, D. rotundatus), Euomphalia spp., Galba spp. (например, G. trunculata), Helicella spp. (например, H. itala, H. obvia), Helicigona spp. (например, H. arbustorum), Helicodiscus spp., Helix spp. (например, H. aperta, H. aspersa, H. pomatia), Limax spp. (например, L. cinereoniger, L. flavus, L. marginatus, L. maximus, L. tenellus), Lymnaea spp. (например, L. stagnalis), Milax spp. (например, M. gagates, M. marginatus, M. sowerbyi, M. budapestensis), Oncomelania spp., Opeas spp., Oxyloma spp. (например, O. pfeifferi), Pomacea spp. (например, P. canaliculata), Succinea spp., Tandonia spp. (например, T. budapestensis, T. sowerbyi), Theba spp., Vallonia spp. и Zonitoides spp. (например, Z. nitidus).
iii. Хозяева-нематоды
Хозяином в любой из композиций или способов, описанных в данном документе, также может быть любой организм, принадлежащий к типу Nematoda. В некоторых случаях нематода считается вредителем, например, сельскохозяйственным вредителем. К примеру, нематода может быть паразитической или вызывать проблемы со здоровьем у растения или у гриба (к примеру, виды из отрядов Aphelenchida, Meloidogyne, Tylenchida и др.) или у людей и животных (к примеру, видов из отрядов Trichinellida, Tylenchida, Rhabditina и Spirurida).
Растительные нематоды охватывают растительных паразитических нематод и нематод, живущих в почве. Растительные паразитические нематоды включают в себя без ограничения эктопаразитов, таких как Xiphinema spp., Longidorus spp. и Trichodorus spp.; полупаразитов, таких как Tylenchulus spp.; мигрирующих эндопаразитов, таких как Pratylenchus spp., Radopholus spp. и Scutellonema spp.; прикрепленных паразитов, таких как Heterodera spp., Globodera spp. и Meloidogyne spp., а также эндопаразитов стеблей и листьев, таких как Ditylenchus spp., Aphelenchoides spp. и Hirshmaniella spp. Особенно вредными корневыми паразитическими почвенными нематодами являются такие как цистообразующие нематоды родов Heterodera или Globodera и/или клубеньковые нематоды рода Meloidogyne. Вредоносными видами этих родов являются, например, Meloidogyne incognita, Heterodera glycines (соевая цистообразующая нематода), Globodera pallida и Globodera rostochiensis (картофельная цистообразующая нематода), эти виды эффективно контролируются с помощью модулирующих средств, описанных в данном документе. Однако применение модулирующих средств, описанных в данном документе, ни в коем случае не ограничено этими родами или видами, но также распространяется подобным образом на других нематод.
Растительные нематоды включают без ограничения, например, Aglenchus agricola, Anguina tritici, Aphelenchoides arachidis, Aphelenchoides fragaria и эндопаразитов стеблей и листьев Aphelenchoides spp. в целом, Belonolaimus gracilis, Belonolaimus longicaudatus, Belonolaimus nortoni, Bursaphelenchus cocophilus, Bursaphelenchus eremus, Bursaphelenchus xylophilus, Bursaphelenchus mucronatus и Bursaphelenchus spp. в целом, Cacopaurus pestis, Criconemella curvata, Criconemella onoensis, Criconemella ornata, Criconemella rusium, Criconemella xenoplax (=Mesocriconema xenoplax) и Criconemella spp. в целом, Criconemoides femiae, Criconemoides onoense, Criconemoides ornatum и Criconemoides spp. в целом, Ditylenchus destructor, Ditylenchus dipsaci, Ditylenchus myceliophagus и эндопаразитов стеблей и листьев Ditylenchus spp. в целом, Dolichodorus heterocephalus, Globodera pallida (=Heterodera pallida), Globodera rostochiensis (картофельная цистообразующая нематода), Globodera solanacearum, Globodera tabacum, Globodera virginia и прикрепленных цистообразующих паразитов Globodera spp. в целом, Helicotylenchus digonicus, Helicotylenchus dihystera, Helicotylenchus erythrine, Helicotylenchus multicinctus, Helicotylenchus nannus, Helicotylenchus pseudorobustus и Helicotylenchus spp. в целом, Hemicriconemoides, Hemicycliophora arenaria, Hemicycliophora nudata, Hemicycliophora parvana, Heterodera avenae, Heterodera cruciferae, Heterodera glycines (соевая цистообразующая нематода), Heterodera oryzae, Heterodera schachtii, Heterodera zeae и прикрепленных цистообразующих паразитов Heterodera spp. в целом, Hirschmaniella gracilis, Hirschmaniella oryzae Hirschmaniella spinicaudata и эндопаразитов стеблей и листьев Hirschmaniella spp. в целом, Hoplolaimus aegyptii, Hoplolaimus califomicus, Hoplolaimus columbus, Hoplolaimus galeatus, Hoplolaimus indicus, Hoplolaimus magnistylus, Hoplolaimus pararobustus, Longidorus africanus, Longidorus breviannulatus, Longidorus elongatus, Longidorus laevicapitatus, Longidorus vineacola и эктопаразитов Longidorus spp. в целом, Meloidogyne acronea, Meloidogyne africana, Meloidogyne arenaria, Meloidogyne arenaria thamesi, Meloidogyne artiella, Meloidogyne chitwoodi, Meloidogyne coffeicola, Meloidogyne ethiopica, Meloidogyne exigua, Meloidogyne fallax, Meloidogyne graminicola, Meloidogyne graminis, Meloidogyne hapla, Meloidogyne incognita, Meloidogyne incognita acrita, Meloidogyne javanica, Meloidogyne kikuyensis, Meloidogyne minor, Meloidogyne naasi, Meloidogyne paranaensis, Meloidogyne thamesi и прикрепленных паразитов Meloidogyne spp. в целом, Meloinema spp., Nacobbus aberrans, Neotylenchus vigissi, Paraphelenchus pseudoparietinus, Paratrichodorus allius, Paratrichodorus lobatus, Paratrichodorus minor, Paratrichodorus nanus, Paratrichodorus porosus, Paratrichodorus teres и Paratrichodorus spp. в целом, Paratylenchus hamatus, Paratylenchus minutus, Paratylenchus projectus и Paratylenchus spp. в целом, Pratylenchus agilis, Pratylenchus alleni, Pratylenchus andinus, Pratylenchus brachyurus, Pratylenchus cerealis, Pratylenchus coffeae, Pratylenchus crenatus, Pratylenchus delattrei, Pratylenchus giibbicaudatus, Pratylenchus goodeyi, Pratylenchus hamatus, Pratylenchus hexincisus, Pratylenchus loosi, Pratylenchus neglectus, Pratylenchus penetrans, Pratylenchus pratensis, Pratylenchus scribneri, Pratylenchus teres, Pratylenchus thornei, Pratylenchus vulnus, Pratylenchus zeae и мигрирующих эндопаразитов Pratylenchus spp. в целом, Pseudohalenchus minutus, Psilenchus magnidens, Psilenchus tumidus, Punctodera chalcoensis, Quinisulcius acutus, Radopholus citrophilus, Radopholus similis, мигрирующих эндопаразитов Radopholus spp. в целом, Rotylenchulus borealis, Rotylenchulus parvus, Rotylenchulus reniformis и Rotylenchulus spp. в целом, Rotylenchus laurentinus, Rotylenchus macrodoratus, Rotylenchus robustus, Rotylenchus uniformis и Rotylenchus spp. в целом, Scutellonema brachyurum, Scutellonema bradys, Scutellonema clathricaudatum и мигрирующих эндопаразитов Scutellonema spp. в целом, Subanguina radiciola, Tetylenchus nicotianae, Trichodorus cylindricus, Trichodorus minor, Trichodorus primitivus, Trichodorus proximus, Trichodorus similis, Trichodorus sparsus и эктопаразитов Trichodorus spp. в целом, Tylenchorhynchus agri, Tylenchorhynchus brassicae, Tylenchorhynchus clarus, Tylenchorhynchus claytoni, Tylenchorhynchus digitatus, Tylenchorhynchus ebriensis, Tylenchorhynchus maximus, Tylenchorhynchus nudus, Tylenchorhynchus vulgaris и Tylenchorhynchus spp. в целом, Tylenchulus semipenetrans и полупаразитов Tylenchulus spp. в целом, Xiphinema americanum, Xiphinema brevicolle, Xiphinema dimorphicaudatum, Xiphinema index и эктопаразитов Xiphinema spp. в целом.
Другие примеры хозяев-нематод включают в себя виды, принадлежащие к семейству Criconematidae, Belonolaimidae, Hoploaimidae, Heteroderidae, Longidoridae, Pratylenchidae, Trichodoridae или Anguinidae.
iv. Полезные хозяева
В некоторых случаях хозяин, описанный в данном документе, представляет собой полезного насекомого, моллюска или нематоду (например, опылителя, естественного конкурента вредителя или продуцента полезных веществ для людей). Термин "полезное насекомое", "полезный моллюск" или "полезная нематода", используемые в данном документе, относится к насекомому, моллюску или нематоде, которые приносят пользу (например, экономическую и/или экологическую) людям, животным, экосистеме и/или окружающей среде. К примеру, хозяин может представлять собой беспозвоночное (например, насекомого, моллюска или нематоду), которое участвует в продуцировании коммерческого продукта, в том числе без ограничения беспозвоночным, выращиваемым для продуцирования продукта питания (например, меда от медоносных пчел, например, Apis mellifera), материалов (таких как шелк от Bombyx mori) и/или веществ (например, лака от Laccifer lacca или пигментов от Dactylopius coccus и Cynipidae). Помимо этого, хозяин может включать беспозвоночных (например, насекомых, моллюсков или нематод), которых применяют в областях сельскохозяйственного применения, в том числе беспозвоночных (например, насекомых, моллюсков или нематод), которые способствуют опылению сельскохозяйственных культур, распространению семян или контролю вредителей. Кроме того, в некоторых случаях хозяин может представлять собой беспозвоночное (например, насекомого, моллюска или нематоду), которое является пригодным для удаления отходов и/или повторной переработки органики (например, земляные черви, термиты или личинки Diptera).
В некоторых случаях хозяева продуцируют пригодный для использования продукт (например, мед, шелк, пчелиный воск или шеллак). В некоторых случаях хозяин представляет собой пчелу. Иллюстративные роды пчел включают в себя без ограничения Apis, Bombus, Trigona и Osmia. В некоторых случаях пчела представляет собой медоносную пчелу (например, насекомое, принадлежащее к роду Apis). В некоторых случаях пчела представляет собой Apis mellifera (европейскую или западную медоносную пчелу), Apis cerana (азиатскую, восточную или гималайскую медоносную пчелу), Apis dorsata ("гигантскую" медоносную пчелу), Apis florea ("красную карликовую" медоносную пчелу), Apis andreniformis ("черную карликовую" медоносную пчелу) или Apis nigrocincta. В некоторых случаях хозяин представляет собой тутового шелкопряда. Тутовый шелкопряд может представлять собой вид в семействе Bombycidae или Saturniidae. В некоторых случаях тутовый шелкопряд представляет собой Bombyx mori. В некоторых случаях хозяин представляет собой лакового червеца. Лаковый червец может представлять собой вид в семействе Kerriidae. В некоторых случаях лаковый червец представляет собой Kerria lacca.
В некоторых случаях хозяин способствует опылению растения (например, пчелы, жуки, осы, мухи, бабочки и моли). В некоторых случаях хозяин, способствующий опылению растения, представляет собой жука. В некоторых случаях жук представляет собой вид в семействе Buprestidae, Cantharidae, Cerambycidae, Chrysomelidae, Cleridae, Coccinellidae, Elateridae, Melandryidae, Meloidae, Melyridae, Mordellidae, Nitidulidae, Oedemeridae, Scarabaeidae или Staphyllinidae. В некоторых случаях хозяин, способствующий опылению растения, представляет собой бабочку или моль (например, Lepidoptera). В некоторых случаях бабочка или моль представляет собой вид в семействе Geometridae, Hesperiidae, Lycaenidae, Noctuidae, Nymphalidae, Papilionidae, Pieridae или Sphingidae. В некоторых случаях хозяин, способствующий опылению растения, представляет собой муху (например, Diptera). В некоторых случаях муха находится в семействе Anthomyiidae, Bibionidae, Bombyliidae, Calliphoridae, Cecidomiidae, Certopogonidae, Chrionomidae, Conopidae, Culicidae, Dolichopodidae, Empididae, Ephydridae, Lonchopteridae, Muscidae, Mycetophilidae, Phoridae, Simuliidae, Stratiomyidae или Syrphidae. В некоторых случаях хозяин, способствующий опылению, представляет собой муравья (например, Formicidae), пилильщика (например, Tenthredinidae) или осу (например, Sphecidae или Vespidae). В некоторых случаях хозяин, способствующий опылению растения, представляет собой пчелу. В некоторых случаях пчела находится в семействе Andrenidae, Apidae, Colletidae, Halictidae или Megachilidae.
В некоторых случаях хозяин способствует контролю вредителей. В некоторых случаях хозяин, способствующий контролю вредителей, представляет собой хищную нематоду. В конкретных примерах нематода представляет собой вид Heterorhabditis или Steinernema. В некоторых случаях хозяин, способствующий контролю вредителей, представляет собой насекомое. К примеру, хозяин, способствующий контролю вредителей, может представлять собой вид, относящийся к семейству Braconidae (например, паразитоидные осы), Carabidae (например, жужелицы), Chrysopidae (например, златоглазки), Coccinellidae (например, божьи коровки), Hemerobiidae (например, гемеробы), Ichneumonidae (например, настоящие наездники), Lampyridae (например, светлячки), Mantidae (например, богомолы), Myrmeleontidae (например, муравьиные львы), Odonata (например, стрекозы и равнокрылые стрекозы) или Syrphidae (например, журчалки). В других случаях хозяин, способствующий контролю вредителей, представляет собой насекомое, которое конкурирует с насекомым, считающимся вредителем (например, сельскохозяйственным вредителем). К примеру, средиземноморская плодовая мушка, Ceratitis capitata, представляет собой распространенного вредителя плодов и фруктов во всем мире. Одним из способов контроля C. captitata является выпускание стерилизованных самцов насекомых в окружающую среду с целью конкуренции с дикими самцами за спаривание с самками. В других случаях хозяин может представлять собой стерилизованного самца, принадлежащего к виду, который обычно считается вредителем.
В некоторых случаях хозяин способствует разрушению отходов или органического материала. В некоторых случаях хозяин, способствующий разрушению отходов или органического материала, принадлежит к Coleoptera или Diptera. В некоторых случаях хозяин, принадлежащий к Diptera, находится в семействе Calliphoridae, Curtonotidae, Drosophilidae, Fanniidae, Heleomyzidae, Milichiidae, Muscidae, Phoridae, Psychodidae, Scatopsidae, Sepsidae, Sphaeroceridae, Stratiomyidae, Syrphidae, Tephritidae или Ulidiidae. В некоторых случаях хозяин, принадлежащий к Coleoptera, находится в семействе Carabidae, Hydrophilidae, Phalacaridae, Ptiliidae или Staphylinidae.
В некоторых случаях хозяином может быть насекомое или паукообразное, которое можно выращивать для потребляемого продукта (например, продукта питания или корма). К примеру, хозяином может быть моль, бабочка, муха, сверчок, паук или жук. В некоторых случаях хозяин принадлежит к отряду Anoplura, Araneae, Blattodea, Coleoptera, Dermaptera, Dictyoptera, Diplura, Diptera, Embioptera, Ephemeroptera, Grylloblatodea, Hemiptera, Homoptera, Hymenoptera, Isoptera, Lepidoptera, Mantodea, Mecoptera, Neuroptera, Odonata, Orthoptera, Phasmida, Plecoptera, Protura, Psocoptera, Siphonaptera, Siphunculata, Thysanura, Strepsiptera, Thysanoptera, Trichoptera или Zoraptera.
В некоторых примерах хозяин является черной львинкой (Hermetia illucens), комнатной мухой, смоляно-бурым хрущаком, муравьем-портным, шелкопрядом (Bombyx mori), кузнечиком, китайским кузнечиком (Acrida cinerea), мучным большым хрущаком (Clarias gariepinns), молью (Anaphe infracta или Bombyx mori), Spodoptera littoralis, домовым сверчком, термитом, пальмовым долгоносиком (Rhynchophorus ferruginens), белостоматидой (Lethocerus indicus), водяным жуком, термитом (Macrotermes subhyalinus), хлебным точильщиком (Stegobium paniceum), Imbrasia belina, Rhynchophorus phoenicis, Oryctes rhinoceros, Macrotermes bellicosus, Ruspolia differens, Oryctes Monoceros или Oecophylla smaragdina.
v. Снижение приспособленности хозяев
Способы и композиции, предусмотренные в данном документе, можно применять для снижения приспособленности любого из хозяев-беспозвоночных (например, насекомых, моллюсков или нематод), описанных в данном документе. Снижение приспособленности возникает в результате изменений в путях хозяина, которые опосредуют взаимодействия между хозяином и микроорганизмами, обитающими в организме хозяина, при этом изменения представляют собой следствие введения модулирующего средства и оказывают пагубные воздействия на хозяина.
В некоторых случаях снижение приспособленности хозяина может проявляться в виде нарушения или ухудшения физиологических процессов хозяина (например, ослабления здоровья или выживаемости) вследствие введения модулирующего средства. В некоторых случаях приспособленность организма может быть измерена с помощью одного или нескольких параметров, в том числе без ограничения скорости размножения, плодовитости, продолжительности жизни, подвижности, жизнеспособности, репродуктивной способности, развития хозяина, массы тела, скорости метаболизма или активности или выживаемости по сравнению с организмом-хозяином, которому модулирующее средство не было введено. Например, способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения общего состояния здоровья хозяина или для снижения общей выживаемости хозяина. В некоторых случаях снижение приспособленности хозяина составляет приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем 100% по сравнению с эталонным уровнем (например, уровня, обнаруженного у хозяина, который не получает модулирующее средство). В некоторых случаях способы и композиции являются эффективными для снижения интенсивности размножения хозяина (например, скорости размножения, плодовитости) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы и композиции являются эффективными для снижения других физиологических параметров, таких как подвижность, масса тела, продолжительность жизни, плодовитость или скорость метаболизма, на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях снижение приспособленности хозяина может проявляться в виде снижения продуцирования одного или нескольких питательных веществ в организме хозяина (например, витаминов, углеводов, аминокислот или полипептидов) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения продуцирования питательных веществ в организме хозяина (например, витаминов, углеводов, аминокислот или полипептидов) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство). В некоторых случаях способы или композиции, предусмотренные в данном документе, могут снижать содержание питательных веществ в организме хозяина путем снижения продуцирования питательных веществ одним или несколькими микроорганизмами (например, эндосимбионтами) в организме хозяина по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях снижение приспособленности хозяина может проявляться в виде повышения восприимчивости хозяина к пестицидному средству (например, пестициду, приведенному в таблице 11) и/или снижения устойчивости хозяина к пестицидному средству (например, пестициду, приведенному в таблице 11) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для повышения восприимчивости хозяина к пестицидному средству (например, пестициду, приведенному в таблице 11) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство). Пестицидное средство может представлять собой любое пестицидное средство, известное в данной области техники, в том числе инсектицидные средства. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут повышать восприимчивость хозяина к пестицидному средству (например, пестициду, упомянутому в таблице 11) посредством снижения способности хозяина метаболизировать или расщеплять пестицидное средство на пригодные к использованию субстраты по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях снижение приспособленности хозяина может проявляться в виде повышения восприимчивости хозяина к аллелохимическому средству и/или снижения устойчивости хозяина к аллелохимическому средству по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы и композиции, предусмотренные в данном документе, могут быть эффективными для снижения устойчивости хозяина к аллелохимическому средству на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство). В некоторых случаях аллелохимическое средство представляет собой кофеин, цистатин сои, фенитротион, монотерпены, дитерпеновые кислоты или фенольные соединения (например, таннины, флавоноиды). В некоторых случаях способы или композиции, предусмотренные в данном документе, могут повышать восприимчивость хозяина к аллелохимическому средству посредством снижения способности хозяина метаболизировать или расщеплять аллелохимическое средство на пригодные к использованию субстраты по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения устойчивости хозяина к паразитам или патогенам (например, грибным, бактериальным или вирусным патогенам или паразитам) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения устойчивости хозяина к патогену или паразиту (например, грибным, бактериальным или вирусным патогенам или паразитическим микроскопическим клещам) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения способности хозяина переносить или передавать патоген растений (например, вирус растений (например, TYLCV)) или бактерию растений (например, Agrobacterium spp)) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. К примеру, способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения способности хозяина переносить или передавать патоген растений (например, вирус растений (например, TYLCV)) или бактерию растений (например, Agrobacterium spp)) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях снижение приспособленности хозяина может проявляться в виде других недостатков приспособленности, таких как сниженная переносимость определенных факторов окружающей среды (например, переносимость высокой или низкой температуры), сниженная способность к выживанию в определенных средах обитания или сниженная способность к поддержанию определенного рациона по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения приспособленности хозяина множеством способов, описанных в данном документе. Кроме того, модулирующее средство может снижать приспособленность хозяина в любом количестве классов, отрядов, семейств, родов или видов хозяев (например, у 1 вида хозяев, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 200, 250, 500 или больше видов хозяев). В некоторых случаях модулирующее средство действует на один класс, отряд, семейство, род или вид хозяев.
Приспособленность хозяина может быть оценена с помощью любых стандартных способов в данной области техники. В некоторых случаях приспособленность хозяина может быть оценена с помощью оценки отдельного хозяина. В качестве альтернативы, приспособленность хозяина может быть оценена с помощью оценки популяции хозяев. Например, снижение приспособленности хозяина может проявляться в виде снижения успешного конкурирования по сравнению с другими насекомыми, что тем самым приводит к уменьшению размера популяции хозяев.
vi. Повышение приспособленности хозяев
Способы и композиции, предусмотренные в данном документе, можно применять для повышения приспособленности любого из хозяев, описанных в данном документе. Повышение приспособленности возникает в результате изменений в путях хозяина, которые опосредуют взаимодействия между хозяином и микроорганизмами, обитающими в организме хозяина, при этом изменения представляют собой следствие введения модулирующего средства и оказывают полезные воздействия на хозяина.
В некоторых случаях повышение приспособленности хозяина может проявляться в виде улучшения физиологических процессов хозяина (например, улучшения здоровья или выживаемости) вследствие введения модулирующего средства. В некоторых случаях приспособленность организма может быть измерена с помощью одного или нескольких параметров, в том числе без ограничения скорости размножения, продолжительности жизни, подвижности, плодовитости, массы тела, скорости метаболизма или активности или выживаемости по сравнению с организмом-хозяином, которому модулирующее средство не было введено. Например, способы или композиции, предусмотренные в данном документе, могут быть эффективными для улучшения общего состояния здоровья хозяина или для улучшения общей выживаемости хозяина по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях улучшение выживаемости хозяина составляет приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство). В некоторых случаях способы и композиции являются эффективными для повышения параметров размножения хозяина (например, скорости размножения) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы и композиции являются эффективными для повышения других физиологических параметров, таких как подвижность, масса тела, продолжительность жизни, плодовитость или скорость метаболизма, на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях повышение приспособленности хозяина может проявляться в виде повышения продуцирования продукта, образуемого указанным хозяином по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для повышения продуцирования продукта, образуемого хозяином, описанного в данном документе (например, меда, пчелиного воска, перги, прополиса, шелка или лака) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях повышение приспособленности хозяина может проявляться в виде повышения частоты или эффективности необходимой активности, осуществляемой хозяином (например, опыления, истребления вредителей, распространения семян или разрушения отходов или органического материала) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы и композиции, предусмотренные в данном документе, могут быть эффективными для повышения частоты или эффективности необходимой активности, осуществляемой хозяином (например, опыления, истребления вредителей, распространения семян или разрушения отходов или органического материала) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях повышение приспособленности хозяина может проявляться в виде повышения продуцирования одного или нескольких питательных веществ в организме хозяина (например, витаминов, углеводов, аминокислот или полипептидов) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для повышения продуцирования питательных веществ в организме хозяина (например, витаминов, углеводов, аминокислот или полипептидов) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство). В некоторых случаях способы или композиции, предусмотренные в данном документе, могут повышать содержание питательных веществ в организме хозяина путем повышения продуцирования питательных веществ одним или несколькими микроорганизмами (например, эндосимбионтами) в организме хозяина по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях повышение приспособленности хозяина может проявляться в виде снижения восприимчивости хозяина к пестицидному средству (например, пестициду, приведенному в таблице 11) и/или повышения устойчивости хозяина к пестицидному средству (например, пестициду, приведенному в таблице 11) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения восприимчивости хозяина к пестицидному средству (например, пестициду, приведенному в таблице 11) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство). Пестицидное средство может представлять собой любое пестицидное средство, известное в данной области техники, в том числе инсектицидные средства. В некоторых случаях пестицидное средство представляет собой неоникотиноид. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут снижать чувствительность хозяина к пестицидному средству (например, пестициду, приведенному в таблице 11) посредством повышения способности хозяина метаболизировать или расщеплять пестицидное средство на пригодные к использованию субстраты по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях повышение приспособленности хозяина может проявляться в виде снижения чувствительности хозяина к аллелохимическому средству и/или повышения устойчивости хозяина к аллелохимическому средству по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для повышения устойчивости хозяина к аллелохимическому средству на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство). В некоторых случаях аллелохимическое средство представляет собой кофеин, цистатин сои, фенитротион, монотерпены, дитерпеновые кислоты или фенольные соединения (например, таннины, флавоноиды). В некоторых случаях способы или композиции, предусмотренные в данном документе, могут снижать восприимчивость хозяина к аллелохимическому средству посредством повышения способности хозяина метаболизировать или расщеплять аллелохимическое средство на пригодные к использованию субстраты по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для повышения устойчивости хозяина к паразитам или патогенам (например, грибным, бактериальным или вирусным патогенам; или паразитическим микроскопическим клещам (например, микроскопический клещ Varroa destructor у пчел)) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для повышения устойчивости хозяина к патогену или паразиту (например, грибным, бактериальным или вирусным патогенам; или паразитическим микроскопическим клещам (например, микроскопический клещ Varroa destructor у пчел)) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях повышение приспособленности хозяина может проявляться в виде других преимуществ в отношении приспособленности, таких как улучшенная переносимость определенных факторов окружающей среды (например, переносимость высокой или низкой температуры), улучшенная способность к выживанию в определенных средах, или улучшенная способность к поддержанию определенного рациона (например, улучшенная способность метаболизировать сою по сравнению с кукурузой) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для повышения приспособленности хозяина множеством способов, описанных в данном документе. Кроме того, модулирующее средство может повышать приспособленность хозяина в любом количестве классов, отрядов, семейств, родов или видов хозяев (например, у 1 вида хозяев, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 200, 250, 500 или больше видов хозяев). В некоторых случаях модулирующее средство действует на один класс, отряд, семейство, род или вид хозяев.
Приспособленность хозяина может быть оценена с помощью любых стандартных способов в данной области техники. В некоторых случаях приспособленность хозяина может быть оценена с помощью оценки отдельного хозяина. В качестве альтернативы, приспособленность хозяина может быть оценена с помощью оценки популяции хозяев. Например, повышение приспособленности хозяина может проявляться в виде повышения успешного конкурирования по сравнению с другими насекомыми, что, тем самым, приводит к увеличению размера популяции хозяев.
vii. Хозяева в сельском хозяйстве
Модулирующие средства, описанные в данном документе, могут быть пригодны для способствования росту растений. Например, благодаря снижению приспособленности вредных беспозвоночных (например, насекомых, моллюсков или нематод) модулирующие средства, предусмотренные в данном документе, могут быть эффективными для способствования росту растений, которые, как правило, повреждаются хозяином. В альтернативном случае благодаря повышению приспособленности полезных беспозвоночных (например, насекомых, моллюсков или нематод) модулирующие средства, предусмотренные в данном документе, могут быть эффективными для способствования росту растений, которые получают пользу от указанных хозяев. Модулирующее средство может быть доставлено в растение с помощью любого из составов и способов доставки, описанных в данном документе, в количестве и в течение периода времени, эффективных для модулирования (например, повышения или снижения) приспособленности хозяина и тем самым оказания пользы растению, например, повышения роста сельскохозяйственных культур, повышения урожайности, снижения заражения вредителями и/или снижения вреда в отношении растений. Это может предусматривать или не предусматривать непосредственное применение модулирующего средства в отношении растения. Например, в случаях, когда первоначальная среда обитания хозяина отличается от региона, в котором растет растение, модулирующее средство может применяться в отношении одного из первоначальной среды обитания хозяина либо растений, представляющих интерес, или комбинации того и другого.
В некоторых случаях растения может представлять собой сельскохозяйственную продовольственную культуру, такую как злаковую, зерновую, бобовую, плодовую или овощную культуру, или непродовольственную культуру, например, травы, цветущие растения, хлопчатник, сено, коноплю. Композиции, описанные в данном документе, могут быть доставлены в сельскохозяйственную культуру в любое время до или после сбора злаковой, зерновой, бобовой, плодовой, овощной или другой сельскохозяйственной культуры. Урожайность представляет собой показатель, часто используемый для сельскохозяйственных культур и обычно измеряемый в метрических тоннах на гектар (или килограммах на гектар). Урожайность также относится к фактическому образованию семян из растения. В некоторых случаях модулирующее средство может быть эффективным для повышения урожайности (например, увеличения количества метрических тонн злаковых, зерновых, бобовых, плодовых или овощных культур на гектар и/или повышения образования семян) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или больше по сравнению с эталонным уровнем (например, сельскохозяйственной культурой, которой модулирующее средство не было введено).
В некоторых случаях у растения (например, сельскохозяйственной культуры) может иметься риск развития заражения вредителем (например, насекомым, моллюском или нематодой) или уже может развиться заражение вредителем. Способы и композиции, описанные в данном документе, можно применять для снижения или предупреждения заражения вредителем в отношении таких сельскохозяйственных культур с помощью снижения приспособленности беспозвоночных (например, насекомого, моллюска или нематоды), которые заражают растения. В некоторых случаях модулирующее средство может быть эффективным для снижения заражения сельскохозяйственных культур (например, снижения количества зараженных растений, уменьшения размера популяции вредителя, снижения повреждения растений) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или больше по сравнению с эталонным уровнем (например, сельскохозяйственной культурой, в которую модулирующее средство не было введено). В других случаях модулирующее средство может быть эффективным для предупреждения заражения или снижения вероятности заражения сельскохозяйственных культур на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или больше по сравнению с эталонным уровнем (например, сельскохозяйственной культурой, в которую модулирующее средство не было введено).
Композиции и способы, описанные в данном документе, могут оказывать пользу в любых подходящих растительных тканях, в том числе без ограничения в соматических зародышах, пыльце, листьях, стеблях, каллюсах, столонах, микроклубнях или побегах. Способы и композиции, описанные в данном документе, могут предусматривать обработку покрытосеменных или голосеменных растений, таких как акация, люцерна, яблоня, абрикос, артишок, ясень, аспарагус, авокадо, банан, овес, бобы, свекла, береза, бук, ежевика, голубика, брокколи, брюссельская капуста, кочанная капуста, канола, канталупа, морковь, кассава, цветная капуста, кедр, злаковое, сельдерей, каштан, черешня, пекинская капуста, цитрусовое растение, клементин, клевер, кофе, кукуруза, хлопчатник, хвойные деревья, вигна, огурец, кипарис, баклажан, вяз, цикорий, эвкалипт, конские бобы, фенхель, фуражные культуры, фиговые деревья, ель, плодовые и ореховые деревья, герань, виноград, грейпфрут, земляные орехи, фезалис, камедное дерево, тсуга, конопля, кария, кале, киви, кольраби, лиственница, латук, лук-порей, лимон, лайм, акация, сосна, адиантум, маис, манго, клен, дыня, просо, гриб, горчица, орехи, дуб, овес, окра, лук, апельсин, декоративное растение или цветок или дерево, папайя, пальма, петрушка, пастернак, горох, персик, арахис, груша, торф, перец, хурма, голубиный орех, сосна, ананас, плантан, слива, гранта, картофель, тыква, радиккио, редис, рапс, малина, рис, рожь, сорго, бредина, соя, шпинат, ель, кабачок, клубника, сахарная свекла, сахарный тростник, подсолнечник, батат, сахарная кукуруза, мандарин, чай, табак, томат, деревья, тритикале, мох, турнепс, виноградная лоза, грецкий орех, водяной кресс, арбуз, пшеница, ямс, тис или цуккини.
viii. Хозяин в получении кормов/продуктов питания
При достижении необходимой стадии развития хозяин может быть собран и, при необходимости, обработан для применения в изготовлении продукта, который можно потреблять. В некоторых случаях собранный хозяин-беспозвоночное (например, насекомое, моллюск или нематода) может быть распределен в цельной форме (например, в виде цельного, необработанного насекомого) в качестве продукта, который можно потреблять. В некоторых случаях цельный собранный хозяин является обработанным (например, измельченным) и распределенным в качестве продукта, который можно потреблять. В качестве альтернативы, одну или несколько частей хозяина (например, одну или несколько частей тела или одно или несколько веществ) можно извлекать из хозяина для применения в изготовлении продукта, который можно потреблять.
Продуктом, который можно потреблять, может быть любой продукт, безопасный для потребления человеком или животным (например, заглатывания). В некоторых случаях хозяин может применяться в изготовлении корма для животного. В некоторых случаях животное относится к домашнему скоту или представляет собой сельскохозяйственное животное (например, курицу, корову, лошадь или свинью). В некоторых случаях животное представляет собой птицу, рептилию, амфибию, млекопитающее или рыбу. В некоторых случаях хозяин может применяться в изготовлении продукта, который заменяет обычный корм для животного. В качестве альтернативы, хозяин может применяться в изготовлении продукта, который дополняет обычный корм для животного. Хозяин также может применяться в изготовлении продукта питания, пищевой добавки или пищевого ингредиента для людей. В некоторых случаях хозяина применяют в изготовлении биологически активной пищевой добавки (например, белковой добавки) для людей.
Хозяин может быть дикого типа или одомашненным хозяином. Кроме того, хозяин может быть на любой стадии развития во время доставки или применения композиций, описанных в данном документе. Дополнительно, хозяин может быть на любой стадии развития во время сбора хозяина для применения в изготовлении продукта, который можно потреблять. В некоторых случаях хозяин представляет собой личинку, куколку или взрослое насекомое во время сбора, применения, обработки или изготовления. Доставка модулирующего средства и стадии сбора могут происходить в одно и то же время или в различные моменты времени.
В некоторых случаях виды хозяев выбраны на основании их естественного питательного профиля. В некоторых случаях модулирующее средство применяют для улучшения питательного профиля насекомого, где модулирующее средство приводит к повышению продуцирования питательного вещества по сравнению с организмом-хозяином, которому модулирующее средство не было введено. Примеры питательных веществ включают витамины, углеводы, аминокислоты, полипептиды или жирные кислоты. В некоторых случаях повышение продуцирования может возникать в результате повышения продуцирования питательного вещества микроорганизмом, обитающим в организме хозяина. В качестве альтернативы, повышение продуцирования может возникать в результате повышения продуцирования питательного вещества самим насекомым-хозяином, где хозяин имеет повышенную приспособленность после доставки или введения модулирующего средства.
В некоторых случаях при конечной обработке первый вид насекомого объединяют со вторым видом насекомого, чей питательный профиль обеспечивает дополняющий благоприятный эффект к общей пищевой ценности пищевого или кормового продукта. Например, вид, имеющий профиль с высоким содержанием белка, может быть объединен с видом, имеющим профиль с высоким содержанием омега-3/6 жирной кислоты. Таким образом, белковая кормовая смесь хозяина может быть смешана на заказ для удовлетворения потребностей людей или различных видов животных.
ix. Насекомые-хозяева в переносе заболеваний
Благодаря снижению приспособленности насекомых-хозяев, которые несут патогены человека и/или животных, модулирующие средства, предусмотренные в данном документе, могут быть эффективными для снижения распространения заболеваний, передаваемых переносчиками. Модулирующее средство может быть доставлено хозяевами с помощью любого из составов и способов доставки, описанных в данном документе, в количестве и в течение периода времени, эффективных для снижения передачи заболевания, например, снижения вертикального или горизонтального переноса между переносчиками и/или снижения переноса людям и/или животным. Например, модулирующее средство, описанное в данном документе, может снижать вертикальную или горизонтальную передачу патогена, передаваемого переносчиком, на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или больше по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В качестве другого примера модулирующее средство, описанное в данном документе, может снижать способность к переносу у хозяина-переносчика на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или больше по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
Неограничивающие примеры заболеваний, которые можно контролировать с помощью композиций и способов, предусмотренных в данном документе, включают в себя заболевания, вызываемые вирусами Togaviridae (например, чикунгунья, лихорадку реки Росс, лихорадку Майаро, лихорадку Онионг-Нионг, лихорадку Синдбис, восточный лошадиный энцефаломиелит, западный лошадиный энцефаломиелит, венесуэльский лошадиный энцефаломиелит или лихорадку леса Барма); заболевания, вызываемые вирусами Flaviviridae (например, лихорадку Денге, желтую лихорадку, болезнь Кьясанурского леса, омскую геморрагическую лихорадку, японский энцефалит, энцефалит долины Муррея, энцефалит Росио, энцефалит Сент-Луис, энцефалит Западного Нила или клещевой энцефалит); заболевания, вызываемые вирусами Bunyaviridae (например, москитную лихорадку, лихорадку долины Рифт, энцефалит, вызываемый вирусом Ла Кросс, калифорнийский энцефалит. геморрагическую лихорадку Крым-Конго или лихорадку Оропуч); заболевания, вызываемые вирусами Rhabdoviridae (например, везикулярный стоматит); заболевания, вызываемые вирусами Orbiviridae (например, блутанг); заболевания, вызываемые бактериями (например, чуму, туляремию, Ку-лихорадку, пятнистую лихорадку Скалистых гор, мышиный тиф, марсельскую лихорадку, клещевой тиф Квинсленда, сибирский клещевой тиф, цуцугамуши, возвратную лихорадку или болезнь Лайма); или заболевания, вызываемые простейшими (например, малярию, африканский трипаносомоз, нагану, болезнь Шагаса, лейшманиоз, пироплазмоз, филяриоз Банкрофта или бругиоз).
II. Микроорганизмы-мишени
Микроорганизмы, на которые целенаправленно воздействует модулирующее средство, описанное в данном документе, могут включать в себя любой микроорганизм, обитающий в организме хозяина-беспозвоночного или на нем (например, насекомого, моллюска или нематоды), в том числе без ограничения любые бактерии и/или грибы, описанные в данном документе. Микроорганизмы, обитающие в организме хозяина, могут включать в себя, например, симбиотические микроорганизмы (например, эндосимбиотические микроорганизмы, которые предоставляют полезные питательные вещества или ферменты хозяину), микроорганизмы-комменсалы, патогенные или паразитические микроорганизмы. Симбиотический микроорганизм (например, бактерия или гриб) может являться облигатным симбионтом хозяина или факультативным симбионтом хозяина. Микроорганизмы, обитающие в организме хозяина, могут быть приобретены с помощью любого пути переноса, в том числе вертикального, горизонтального переноса или переноса множественного происхождения.
i. Бактерии
Иллюстративные бактерии, которые могут подвергаться целенаправленному воздействию в соответствии со способами и композициями, предусмотренными в данном документе, включают в себя без ограничения Xenorhabdus spp, Photorhabdus spp, Candidatus spp, Buchnera spp, Blattabacterium spp, Baumania spp, Wigglesworthia spp, Wolbachia spp, Rickettsia spp, Orientia spp, Sodalis spp, Burkholderia spp, Cupriavidus spp, Frankia spp, Snirhizobium spp, Streptococcus spp, Wolinella spp, Xylella spp (например, Xylella fastidiosa), Erwinia spp, Agrobacterium spp, Bacillus spp, Commensalibacter spp. (например, Commensalibacter intestini), Paenibacillus spp, Streptomyces spp, Micrococcus spp, Corynebacterium spp, Acetobacter spp (например, Acetobacter pomorum), Cyanobacteria spp, Salmonella spp, Rhodococcus spp, Pseudomonas spp (например, Psuedomonas fulva или Pseudomonas mandelii, Pseudomonas migulae), Pantoea spp. (например, Pantoea vagans), Lactobacillus spp (например, Lactobacillus plantarum), Lysobacter spp., Herbaspirillum spp., Enterococcus spp, Gluconobacter spp. (например, Gluconobacter morbifer), Alcaligenes spp, Hamiltonella spp., Klebsiella spp, Paenibacillus spp, Serratia spp. (например, Serratia marcescens), Rahnella spp. (например, Rahnella aquatilis), Arthrobacter spp, Azotobacter spp., Corynebacterium spp, Brevibacterium spp, Regiella spp. (например, Regiella insecticola), Thermus spp, Pseudomonas spp, Clostridium spp, Mortierella spp. (например, Mortierella elongata) и Escherichia spp. В некоторых случаях бактерии, подвергающиеся целенаправленному воздействию, представляют собой виды в родах Xenorhabdus spp., Photorhabdus spp. или Wolbachia spp. В некоторых случаях бактерии, подвергающиеся целенаправленному воздействию, находятся в отряде Streptomycetales, Rhizobiales, Pseudomonadales, Xanthomondadales, Sphingobacteriales, Chlorofelxales, Rhodospirllales, Enterobacteriales, Sphingomonadales, Gemmatimonadales, Micrococcales, Caulobacterales, Cytophagales, Firmicutes, Micromonosporales, Burkholderiales, Rickettsiales, Flavobacteriales, Acidimicroiales, Rhodocyclales или Bdellovibrionales. В некоторых случаях бактерии, подвергающиеся целенаправленному воздействию, представляют собой Armatimonadetes, Firmicutes, TM7, Bacteroidetes, Proteobacteria или Actinobacteria. В некоторых случаях бактерии, подвергающиеся целенаправленному воздействию, являются бактериями в родах Lactococcus spp., Aeromonas spp., Pseudomonas spp., Enterobacter spp., Citrobacter spp., Sulfurospillium spp., Phaeosphaeria spp. или Mycosphaerella spp. В некоторых случаях бактерии, подвергающиеся целенаправленному воздействию с помощью модулирующего средства, могут представлять собой бактерий, которые могут быть перенесены от хозяина (например, насекомого, моллюска или нематоды) растению, в том числе без ограничения бактериальные патогены растений (например, Agrobacterium spp.). Неограничивающие примеры бактерий, на которые можно целенаправленно воздействовать с помощью способов и композиций, предусмотренных в данном документе, приведены в таблице 1. В некоторых случаях последовательность 16S рРНК бактерий, на которые целенаправленно воздействуют с помощью модулирующего средства, на по меньшей мере 50%, 60%, 70%, 80%, 85%, 90%, 95%, 97%, 99%, 99,9% или 100% идентична последовательности, приведенной в таблице 1.
Таблица 1. Примеры бактерий-мишеней и насекомых-хозяев
(SEQ ID NO: 1)
(SEQ ID NO: 2)
(SEQ ID NO: 3)
(SEQ ID NO: 4)
(SEQ ID NO: 5)
(SEQ ID NO: 6)
(SEQ ID NO: 7)
(SEQ ID NO: 8)
(SEQ ID NO: 9)
(SEQ ID NO: 10)
(SEQ ID NO: 11)
(SEQ ID NO: 12)
(SEQ ID NO: 13)
(SEQ ID NO: 14)
(SEQ ID NO: 15)
(SEQ ID NO: 16)
(SEQ ID NO: 17)
(SEQ ID NO: 18)
(SEQ ID NO: 19)
(SEQ ID NO: 20)
(SEQ ID NO: 21)
Diceroprocta semicincta
(SEQ ID NO: 22)
Culex quinquefasciatus
(SEQ ID NO: 23)
(SEQ ID NO: 24)
(SEQ ID NO: 25)
(SEQ ID NO: 26)
(SEQ ID NO: 27)
(SEQ ID NO: 28)
(SEQ ID NO: 29)
(SEQ ID NO: 30)
(SEQ ID NO: 31)
(SEQ ID NO: 32)
(SEQ ID NO: 33)
(SEQ ID NO: 34)
(SEQ ID NO: 35)
(SEQ ID NO: 36)
(SEQ ID NO: 37)
(SEQ ID NO: 38)
(SEQ ID NO: 39)
(SEQ ID NO: 40)
(SEQ ID NO: 41)
На любое количество видов бактерий можно целенаправленно воздействовать с помощью композиций или способов, описанных в данном документе. Например, в некоторых случаях модулирующее средство может целенаправленно воздействовать на один вид бактерий. В некоторых случаях модулирующее средство может целенаправленно воздействовать на по меньшей мере приблизительно любой из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 500 или больше различных видов бактерий. В некоторых случаях модулирующее средство может целенаправленно воздействовать на любой из от приблизительно 1 до приблизительно 5, от приблизительно 5 до приблизительно 10, от приблизительно 10 до приблизительно 20, от приблизительно 20 до приблизительно 50, от приблизительно 50 до приблизительно 100, от приблизительно 100 до приблизительно 200, от приблизительно 200 до приблизительно 500, от приблизительно 10 до приблизительно 50, от приблизительно 5 до приблизительно 20 или от приблизительно 10 до приблизительно 100 различных видов бактерий. В некоторых случаях модулирующее средство может целенаправленно воздействовать на по меньшей мере приблизительно любые из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100 или больше типов, классов, порядков, семейств или родов бактерий.
В некоторых случаях модулирующее средство может обеспечивать увеличение популяции одной или нескольких бактерий на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше в организме хозяина. В некоторых случаях модулирующее средство может обеспечивать уменьшение популяции одной или нескольких бактерий на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше в организме хозяина. В некоторых случаях модулирующее средство может обеспечивать уничтожение популяции бактерии в организме хозяина.
В некоторых случаях модулирующее средство может изменять разнообразие бактерий и/или состав бактерий организма хозяина по сравнению с организмом хозяина, которому модулирующее средство не было введено. В некоторых случаях модулирующее средство может увеличивать разнообразие бактерий в организме хозяина относительно исходного разнообразия на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях модулирующее средство может уменьшать разнообразие бактерий в организме хозяина относительно исходного разнообразия на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях модулирующее средство может изменять функцию, активность, рост и/или деление одной или нескольких бактериальных клеток по сравнению с организмом хозяина, которому модулирующее средство не было введено. Например, модулирующее средство может изменять экспрессию одного или нескольких генов в бактерии. В некоторых случаях модулирующее средство может изменять функцию одного или нескольких белков в бактерии. В некоторых случаях модулирующее средство может изменять функцию одной или нескольких клеточных структур (например, клеточной стенки, внешней или внутренней мембраны) в бактерии. В некоторых случаях модулирующее средство может уничтожать (например, лизировать) бактерию.
Бактерия-мишень может обитать в одной или нескольких частях хозяина-беспозвоночного (например, насекомого, моллюска или нематоды). Кроме того, бактерия-мишень может быть внутриклеточной или внеклеточной. В некоторых случаях бактерии обитают в одной или нескольких частях кишки хозяина, в том числе, например, в передней кишке, средней кишке и/или задней кишке. В некоторых случаях бактерии обитают в качестве внутриклеточных бактерий в клетке хозяина. В некоторых случаях бактерии обитают в бактериоците хозяина-беспозвоночного (например, насекомого, моллюска или нематоды).
Изменения в отношении популяций бактерий в организме хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) могут быть определены с помощью любых способов, известных в данной области техники, таких как стандартные методики культивирования, подсчеты CFU, микроматричный анализ, полимеразная цепная реакция (ПЦР), ПЦР в реальном времени, проточная цитометрия, флуоресцентная микроскопия, трансмиссионная электронная микроскопия, флуоресцентная гибридизация in situ (например, FISH), спектрофотометрия, масс-спектрометрия с матрично-активированной лазерной десорбцией/ионизацией (MALDI-MS) или секвенирование ДНК. В некоторых случаях образец от хозяина, обработанного модулирующим средством, секвенируют (например, с помощью метагеномного секвенирования 16S рРНК или рДНК) для определения микробиоты хозяина после доставки или введения модулирующего средства. В некоторых случаях образец от хозяина, который не получал модулирующее средство, также секвенируют для получения эталона.
ii. Грибы и дрожжевые грибы
Иллюстративные грибы, на которые можно целенаправленно воздействовать в соответствии со способами и композициями, предусмотренными в данном документе, включают в себя без ограничения Amylostereum areolatum, Epichloe spp, Pichia pinus, Hansenula capsulate, Daldinia decipien, Ceratocytis spp, Ophiostoma spp и Attamyces bromatificus. Неограничивающие примеры дрожжевых и дрожжеподобных симбионтов, обнаруживаемых у беспозвоночных (например, насекомого, моллюска или нематоды), включают в себя Candida, Metschnikowia, Leucocoprinu (например, Leucocoprinus gongylophorus), Debaromyces, Scheffersomyces shehatae и Scheffersomyces stipites, Starmerella, Pichia, Trichosporon, Cryptococcus, Pseudozyma, а также дрожжеподобные симбионты из подтипа Pezizomycotina (например, Symbiotaphrina bucneri и Symbiotaphrina kochii). Неограничивающие примеры дрожжевых грибов, на которые можно целенаправленно воздействовать с помощью способов и композиций согласно данному документу, приведены в таблице 2.
Таблица 2. Примеры дрожжевых грибов у насекомых
H. sycophanta
На любое количество видов грибов можно целенаправленно воздействовать с помощью композиций или способов, описанных в данном документе. Например, в некоторых случаях, модулирующее средство может целенаправленно воздействовать на один вид грибов. В некоторых случаях модулирующее средство может целенаправленно воздействовать на по меньшей мере приблизительно любой из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 500 или больше различных видов грибов. В некоторых случаях модулирующее средство может целенаправленно воздействовать на любой из от приблизительно 1 до приблизительно 5, от приблизительно 5 до приблизительно 10, от приблизительно 10 до приблизительно 20, от приблизительно 20 до приблизительно 50, от приблизительно 50 до приблизительно 100, от приблизительно 100 до приблизительно 200, от приблизительно 200 до приблизительно 500, от приблизительно 10 до приблизительно 50, от приблизительно 5 до приблизительно 20 или от приблизительно 10 до приблизительно 100 различных видов грибов. В некоторых случаях модулирующее средство может целенаправленно воздействовать на по меньшей мере приблизительно любые из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100 или больше типов, классов, порядков, семейств или родов грибов.
В некоторых случаях модулирующее средство может обеспечивать увеличение популяции одного или нескольких грибов на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше в организме хозяина по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях модулирующее средство может обеспечивать уменьшение популяции одного или нескольких грибов на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше в организме хозяина по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях модулирующее средство может обеспечивать уничтожение популяции грибов в организме хозяина.
В некоторых случаях модулирующее средство может изменять разнообразие грибов и/или состав грибов хозяина. В некоторых случаях модулирующее средство может повышать разнообразие грибов в организме хозяина относительно исходного разнообразия на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях модулирующее средство может снижать разнообразие грибов в организме хозяина относительно исходного разнообразия на по меньшей мере приблизительно любое значение из 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
В некоторых случаях модулирующее средство может изменять функцию, активность, рост и/или деление одного или нескольких грибов. Например, модулирующее средство может изменять экспрессию одного или нескольких генов в грибе. В некоторых случаях модулирующее средство может изменять функцию одного или нескольких белков в грибе. В некоторых случаях модулирующее средство может изменять функцию одного или нескольких клеточных компонентов в грибе. В некоторых случаях модулирующее средство может уничтожать гриб.
Кроме того, гриб-мишень может обитать в одной или нескольких частях организма насекомого. В некоторых случаях гриб обитает в одной или нескольких частях кишки насекомого или на них, в том числе, например, в передней кишке, средней кишке и/или задней кишке. В некоторых случаях гриб живет внеклеточно в гемолимфе, жировых телах или в специализированных структурах в организме хозяина.
Изменения в отношении популяции грибов в организме хозяина могут быть определены с помощью любых способов, известных в данной области техники, таких как микроматричный анализ, полимеразная цепная реакция (ПЦР), ПЦР в реальном времени, проточная цитометрия, флуоресцентная микроскопия, трансмиссионная электронная микроскопия, флуоресцентная гибридизация in situ (например, FISH), спектрофотометрия, масс-спектрометрия с матрично-активированной лазерной десорбцией/ионизацией (MALDI-MS) и секвенирование ДНК. В некоторых случаях образец от хозяина, обработанного модулирующим средством, секвенируют (например, с помощью метагеномного секвенирования) для определения микробиоты хозяина после доставки или введения модулирующего средства. В некоторых случаях образец от хозяина, который не получал модулирующее средство, также секвенируют для получения эталона.
III. Модулирующие средства
Модулирующее средство из способов и композиций, предусмотренных в данном документе, может включать в себя полипептид, нуклеиновую кислоту, малую молекулу или любую их комбинацию. В некоторых случаях модулирующее средство представляет собой молекулу нуклеиновой кислоты (например, молекулу ДНК или молекулу РНК, например, mRNA, направляющую РНК (gRNA) или ингибирующую РНК (например, siRNA, shRNA или miRNA), или гибридную молекулу ДНК-РНК), малую молекулу, пептид или полипептид (например, молекулу антитела, например, антитело или его антигенсвязывающий фрагмент). Любое из этих средств можно применять для изменения микробиоты хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) с помощью целенаправленного воздействия на пути в организме хозяина и/или обитающих микроорганизмах (например, пути, которые опосредуют взаимодействия хозяин-микробиота, например, пути иммунной системы хозяина или пути бактериоцитов). К примеру, любые модулирующие средства, описанные в данном документе, можно применять для регуляции (например, индукции или подавления) гена или белка в организме хозяина или микроорганизме, обитающем в хозяине (например, белка или гена, кодирующего белок, приведенных в таблице 7, таблице 8 или таблице 9).
i. Полипептиды
Модулирующее средство, описанное в данном документе, может включать в себя полипептид (например, антитело). В некоторых случаях модулирующее средство, описанное в данном документе, включает в себя полипептид или функциональные фрагменты или их производные, которые целенаправленно воздействуют на пути хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, пути, которые опосредуют взаимодействия хозяин-микробиота, например, пути иммунной системы хозяина или пути бактериоцитов). В некоторых случаях средство представляет собой полипептид, приведенный в таблице 7, таблице 8 или таблице 9, при этом первичная последовательность полипептида, представляющего собой средство, представлена с помощью ссылки на его номер доступа.
Модулирующее средство, включающее в себя полипептид, описанный в данном документе, может быть приведено в контакт с хозяином-мишенью в количестве и в течение периода времени, достаточных для: (a) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации внутри хозяина-мишени; (b) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации внутри кишки хозяина-мишени; (c) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации внутри бактериоцита хозяина-мишени; (d) модулирования уровня или активности одного или нескольких микроорганизмов (например, эндосимбионтов) в организме хозяина-мишени или/и (e) модулирования приспособленности хозяина-мишени.
Полипептиды, включенные в данный документ, могут включать в себя встречающиеся в природе полипептиды или рекомбинантно получаемые варианты. В некоторых случаях полипептид может представлять собой функциональные фрагменты или их варианты (например, ферментативно активный фрагмент полипептида, приведенного в таблице 7, таблице 8 или таблице 9, или его вариант). Такие фрагменты или варианты могут быть получены и подвергнуты скринингу в отношении аналогичной активности, описанной в данном документе, и будут эквивалентными далее в данном документе в раскрытых способах и композициях). Например, полипептид может представлять собой функционально активный вариант любого из полипептидов, описанных в данном документе, на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичный, например, в определенной области или во всей последовательности, последовательности полипептида, описанного в данном документе, или встречающегося в природе полипептида. В некоторых случаях полипептид может характеризоваться по меньшей мере 50% (например, по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 97%, 99% или больше) идентичностью по отношению к аминокислотной последовательности белка, приведенного в таблице 7, таблице 8 или таблице 9 с представленной ссылкой на номер доступа.
Способы получения терапевтического полипетида являются стандартными в данной области техники. См., в целом, Smales & James (Eds.), Therapeutic Proteins: Methods and Protocols (Methods in Molecular Biology), Humana Press (2005); and Crommelin, Sindelar & Meibohm (Eds.), Pharmaceutical Biotechnology: Fundamentals and Applications, Springer (2013).
Способы получения полипептида предусматривают экспрессию в клетках млекопитающих, хотя рекомбинантные белки также могут продуцироваться с помощью клеток насекомых, дрожжей, бактерий или других клеток под контролем соответствующих промоторов. Векторы экспрессии млекопитающих могут содержать нетранскрибируемые элементы, такие как точка начала репликации, подходящий промотор и энхансер, и другие 5'- или 3'-фланкирующие нетранскрибируемые последовательности, и 5'- или 3'-нетранскрибируемые последовательности, такие как необходимые участки связывания рибосомы, сайт полиаденилирования, донорный сайт сплайсинга, акцепторный сайт сплайсинга и последовательности терминации. Последовательности ДНК, происходящие из генома вируса SV40, к примеру, точки начала репликации, раннего промотора, энхансера, сайтов сплайсинга и сайтов полиаденилирования mSV40 можно применять для обеспечения других генетических элементов, требуемых для экспрессии гетерологичной последовательности ДНК. Подходящие векторы клонирования и экспрессии для применения с клетками-хозяевами бактерий, грибов, дрожжей и млекопитающих описаны в Green & Sambrook, Molecular Cloning: A Laboratory Manual (Fourth Edition), Cold Spring Harbor Laboratory Press (2012).
Различные системы культивирования клеток млекопитающих можно применять для экспрессии и изготовления средства на основе рекомбинантного полипептида (например, приведенного в таблицах 4, 5 или 6). Примеры систем экспрессии млекопитающих включают клетки CHO, клетки COS, клеточные линии HeLA и BHK. Способы культивирования клеток-хозяев для получения терапевтических средств на основе белка описаны, например, в Zhou and Kantardjieff (Eds.), Mammalian Cell Cultures for Biologics Manufacturing (Advances in Biochemical Engineering/Biotechnology), Springer (2014). Очистка терапевтических средств на основе белка описана в Franks, Protein Biotechnology: Isolation, Characterization, and Stabilization, Humana Press (2013); и в Cutler, Protein Purification Protocols (Methods in Molecular Biology), Humana Press (2010). Составление терапевтических средств на основе белка описано в Meyer (Ed.), Therapeutic Protein Drug Products: Practical Approaches to formulation in the Laboratory, Manufacturing, and the Clinic, Woodhead Publishing Series (2012).
Модулирующие средства на основе полипептида, описанные далее в данном документе, а именно, антитела, бактериоцины, противомикробные пептиды и регуляторные пептиды бактериоцитов, можно применять для изменения путей у хозяина, которые опосредуют взаимодействия между хозяином и микроорганизмами, обитающими в организме хозяина, как указано в разделах в отношении повышения и снижения приспособленности хозяев.
(a) Антитела
В некоторых случаях модулирующее средство включает антитело или его антигенсвязывающий фрагмент. К примеру, средство, описанное в данном документе, может представлять собой антитело, которое блокирует или усиливает активность и/или функцию компонента пути иммунной системы хозяина или регуляторного пути бактериоцитов, приведенного в таблице 8 или таблице 9. Антитело может действовать в качестве антагониста или агониста полипептида (например, фермента или клеточного рецептора) в организме хозяина или микроорганизмах, обитающих в хозяине, в том числе любых белков, приведенных в таблице 7, таблице 8 или таблице 9.
Получение и применение антител против антигена-мишени (например, белков, которые опосредуют взаимодействия хозяин-микробиота, например, белков иммунной системы хозяев или белков бактериоцитов, например, белков, приведенных в таблице 7, таблице 8 или таблице 9) известно в данной области техники. См., например, Zhiqiang An (Ed.), Therapeutic Monoclonal Antibodies: From Bench to Clinic, 1st Edition, Wiley, 2009, а также Greenfield (Ed.), Antibodies: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory Press, 2013, в отношении способов получения рекомбинантных антител, в том числе конструирования антител, применения вырожденных олигонуклеотидов, 5'-RACE, фагового дисплея и мутагенеза; тестирования и характеристики антител; фармакокинетики и фармакодинамики антител; очистки и хранения антител; и методик скрининга и мечения.
(b) Бактериоцины
Модулирующее средство, описанное в данном документе, может включать в себя бактериоцин. В некоторых случаях бактериоцин продуцируется естественным образом грамположительными бактериями, такими как Pseudomonas, Streptomyces, Bacillus, Staphylococcus или молочнокислые бактерии (LAB, такие как Lactococcus lactis). В некоторых случаях бактериоцин продуцируется естественным образом грамотрицательными бактериями, такими как Hafnia alvei, Citrobacter freundii, Klebsiella oxytoca, Klebsiella pneumonia, Enterobacter cloacae, Serratia plymithicum, Xanthomonas campestris, Erwinia carotovora, Ralstonia solanacearum или Escherichia coli. Иллюстративные бактериоцины включают в себя без ограничения антибиотики LAB I-IV класса (такие как лантибиотики), колицины, микроцины и пиоцины. Неограничивающие примеры бактериоцинов приведены в таблице 3.
Таблица 3. Примеры бактериоцинов
Enterococcus,
Lactobacillus,
Lactococcus,
Leuconostoc,
Listeria,
Clostridium
(SEQ ID NO: 42)
(SEQ ID NO: 43)
Lactobacilli,
Leuconostoc,
Brochothrix thermosphacta,
Propionibacteria,
Bacilli,
Enterococci,
Staphylococci,
Listeria clostridia,
Listeria monocytogenes, Listeria innocua
(SEQ ID NO: 44)
(SEQ ID NO: 45)
(SEQ ID NO: 46)
Enterococcus faecalis
(SEQ ID NO: 47)
(SEQ ID NO: 48)
(SEQ ID NO: 49)
(SEQ ID NO: 50)
(SEQ ID NO: 51)
В некоторых случаях бактериоцин представляет собой колицин, пиоцин или микроцин, продуцируемые грамотрицательными бактериями. В некоторых случаях бактериоцин представляет собой колицин. Колицин может представлять собой колицин группы A (например, использует систему Tol для проникновения через внешнюю мембрану бактерии-мишени) или колицин группы B (например, использует систему Ton для проникновения через внешнюю мембрану бактерии-мишени). В некоторых случаях бактериоцин представляет собой микроцин. Микроцин может представлять собой микроцин класса I (например, < 5 кДа, имеет посттрансляционные модификации) или микроцин класса II (например, 5-10 кДа, с посттрансляционными модификациями или без них). В некоторых случаях микроцин класса II представляет собой микроцин класса IIa (например, требует более одного гена для синтеза и сборки функциональных пептидов) или микроцин класса IIb (например, линейные пептиды с посттрансляционными модификациями на С-конце или без них). В некоторых случаях бактериоцин представляет собой пиоцин. В некоторых случаях пиоцин представляет собой R-пиоцин, F-пиоцин или S-пиоцин.
В некоторых случаях бактериоцин представляет собой бактериоцин класса I, класса II, класса III или класса IV, продуцируемый грамположительными бактериями. В некоторых случаях модулирующее средство включает в себя бактериоцин класса I (например, антибиотики, содержащие лантионин (лантибиотики), продуцируемые грамположительными бактериями). Бактериоцины класса I или лантибиотики могут представлять собой пептид с низкой молекулярной массой (например, менее чем приблизительно 5 кДа) и могут иметь аминокислотные остатки с посттрансляционными модификациями (например, лантионин, β-метиллантионин или дегидрированные аминокислоты).
В некоторых случаях бактериоцин представляет собой бактериоцин класса II (например, соединения, не являющиеся лантибиотиками, продуцируемые грамположительными бактериями). Многие из них являются положительно заряженными не содержащими лантионина пептидами, которые, в отличие от лантибиотиков, не подвергаются обширной посттрансляционной модификации. Бактериоцин класса II может принадлежать к одному из следующих подклассов: ʺпедиоциноподобныеʺ бактериоцины (например, педиоцин PA-1 и карнобактериоцин X (класс IIa)); двухпептидные бактериоцины (например, лактацин F и ABP-118 (класс IIb)); круговые бактериоцины (например, карноциклин А и энтероцин AS-48 (класс IIc)) или немодифицированные линейные непедиоциноподобные бактериоцины (например, эпидермицин NI01 и лактококцин A (класс IId)).
В некоторых случаях бактериоцин представляет собой бактериоцин класса III (например, продуцируемый грамположительными бактериями). Бактериоцины класса III могут иметь молекулярную массу более 10 кДа и могут представлять собой термонестабильные белки. Бактериоцины класса III могут быть дополнительно подразделены на бактериоцины группы IIIA и группы IIIB. Бактериоцины группы IIIA включают в себя бактериолитические ферменты, которые уничтожают чувствительные штаммы посредством лизиса клеточной стенки, такие как энтеролизин А. Бактериоцины группы IIIB включают в себя нелитические белки, такие как казеицин 80, гельветицин J и лактацин B.
В некоторых случаях бактериоцин представляет собой бактериоцин класса IV (например, продуцируемый грамположительными бактериями). Бактериоцины класса IV представляют собой группу сложных белков, ассоциированных с другими липидными или углеводными фрагментами, которые, по-видимому, требуются для активности. Они являются относительно гидрофобными и термостабильными. Примерами бактериоцинов класса IV являются лейконоцин S, лактоцин 27 и лактоцин S.
В некоторых случаях бактериоцин представляет собой бактериоцин R-типа. Бактериоцины R-типа представляют собой сократительные бактериоцидные белковые комплексы. Некоторые бактериоцины R-типа имеют сократительную структуру, подобную отростку фага. C-концевая область белка нити отростка фага определяет специфичность связывания с мишенью. Они могут прикрепляться к клеткам-мишеням посредством белка, связывающегося с рецептором, например, нити отростка. За прикреплением следует сокращение наружного чехла и введение сердцевины через оболочку бактерии-мишени. Проникновение сердцевины приводит к быстрой деполяризации потенциала клеточной мембраны и мгновенной гибели клетки. Контакт с одной частицей бактериоцина R-типа может приводить к гибели клетки. Бактериоцин R-типа, например, может быть термолабильным, умеренно кислотостойким, устойчивым к трипсину, способным осаждаться в результате центрифугирования, быть различимым при использовании электронной микроскопии или обладать комбинацией этих свойств. Другие бактериоцины R-типа могут представлять собой сложные молекулы, включающие в себя многочисленные белки, полипептиды или субъединицы, и могут напоминать структуру отростка бактериофагов семейства Myoviridae. У встречающихся в природе бактериоцинов R-типа структуры субъединиц могут кодироваться бактериальным геномом, таким как геном C. difficile или P. aeruginosa, и образовывать бактериоцины R-типа, которые выступают в качестве природной защиты против других бактерий. В некоторых случаях бактериоцин R-типа представляет собой пиоцин. В некоторых случаях пиоцин представляет собой R-пиоцин, F-пиоцин или S-пиоцин.
В некоторых случаях бактериоцин представляет собой функционально активный вариант бактериоцинов, описанных в данном документе. В некоторых случаях вариант бактериоцина является на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичным, например, в определенной области или во всей последовательности, последовательности бактериоцина, описанного в данном документе, или встречающегося в природе бактериоцина.
В некоторых случаях бактериоцин можно создавать с помощью биоинженерии в соответствии со стандартными способами для модулирования его биологической активности, например, повышения, или снижения, или регулирования, или для определения его микроорганизмов-мишеней. В других случаях бактериоцин продуцируется трансляционным аппаратом (например, рибосомой и т. д.) микробной клетки. В некоторых случаях бактериоцин синтезируют химическим путем. Некоторые бактериоцины могут образовываться из полипептидного предшественника. Полипептидный предшественник может подвергаться расщеплению (например, процессингу под действием протеазы) с образованием самого полипептида бактериоцина. Таким образом, в некоторых случаях бактериоцин продуцируется из полипептида-предшественника. В некоторых других случаях бактериоцин включает в себя полипептид, который подвергся посттрансляционным модификациям, например, расщеплению или добавлению одной или нескольких функциональных групп.
Бактериоцины, описанные в данном документе, могут быть составлены в виде композиции для любого из путей применения, описанных в данном документе. Композиции, раскрытые в данном документе, могут содержать любые количество или тип (например, классы) бактериоцинов, как, например, по меньшей мере приблизительно любое количество из 1 бактериоцина, 2, 3, 4, 5, 10, 15, 20, 30, 40, 50, 100 или больше бактериоцинов. Подходящие концентрации каждого бактериоцина в композициях, описанных в данном документе, зависят от таких факторов, как эффективность, стабильность бактериоцина, количество различных типов бактериоцина в композициях, состав и способы применения композиции. В некоторых случаях концентрация каждого бактериоцина в жидкой композиции составляет от приблизительно 0,01 нг/мл до приблизительно 100 мг/мл. В некоторых случаях концентрация каждого бактериоцина в твердой композиции составляет от приблизительно 0,01 нг/г до приблизительно 100 мг/г. В некоторых случаях, если композиция содержит по меньшей мере два типа бактериоцинов, то концентрация каждого типа бактериоцинов может быть одной и той же или разной. В некоторых случаях бактериоцин предусмотрен в композиции, содержащей бактериальную клетку, которая секретирует бактериоцин. В некоторых случаях бактериоцин предусмотрен в композиции, содержащей полипептид (например, полипептид, выделенный из бактериальной клетки).
Бактериоцины могут нейтрализовать (например, уничтожать) по меньшей мере один микроорганизм, отличный от отдельной бактериальной клетки, в которой полипептид образуется, в том числе клетки, клонально родственные бактериальной клетке, и другие микробные клетки. Таким образом, бактериальная клетка может проявлять цитотоксические или ингибирующие рост эффекты в отношении множества микроорганизмов посредством секреции бактериоцинов. В некоторых случаях бактериоцин целенаправленно воздействует на один или несколько видов бактерий, обитающих в организме хозяина, и уничтожает их посредством образования пор в цитоплазматической мембране, разрушения клеточной стенки (например, активности пептидогликаназы) или активности нуклеазы (например, активности ДНКазы, активности 16S рРНКазы или активности тРНКазы).
В некоторых случаях бактериоцин характеризуется нейтрализующей активностью. Нейтрализующая активность бактериоцинов может включать в себя без ограничения остановку размножения микроорганизмов или цитотоксичность. Некоторые бактериоцины характеризуются цитотоксической активностью, и, таким образом, могут уничтожать микроорганизмы, например, бактерии, дрожжевые грибы, водоросли и т. п. Некоторые бактериоцины могут ингибировать размножение микроорганизмов, например, бактерий, дрожжевых грибов, водорослей и т. п., например, посредством остановки клеточного цикла.
В некоторых случаях бактериоцин характеризуется уничтожающей активностью. Механизм уничтожения у бактериоцинов является специфичным для каждой группы бактериоцинов. В некоторых случаях бактериоцин характеризуется биологической активностью узкого спектра. Бактериоцины известны своей очень высокой активностью в отношении своих штаммов-мишеней. Активность некоторых бактериоцинов ограничена штаммами, которые являются близкородственными по отношению к штамму-продуценту бактериоцина (биологическая активность узкого спектра). В некоторых случаях бактериоцин характеризуется биологической активностью широкого спектра в отношении широкого круга родов.
В некоторых случаях бактериоцины взаимодействуют с рецепторной молекулой или стыковочной молекулой на клеточной мембране бактерии-мишени. Например, низин является крайне активным в отношении своих штаммов бактерий-мишеней, демонстрируя противомикробную активность даже в одноразрядной наномолярной концентрации. Было показано, что молекула низина связывается с липидом II, который представляет собой основной переносчик субъединиц пептидогликанов из цитоплазмы в клеточную стенку.
В некоторых случаях бактериоцин характеризуется противогрибковой активностью. Был идентифицирован ряд бактериоцинов с противодрожжевой или противогрибковой активностью. Например, было показано, что бактериоцины из Bacillus характеризуются нейтрализующей активностью в отношении некоторых штаммов дрожжей (см., например, Adetunji and Olaoye, Malaysian Journal of Microbiology 9:130-13, 2013). В другом примере было показано, что пептид Enterococcus faecalis характеризуется нейтрализующей активностью в отношении видов Candida (см., например, Shekh and Roy, BMC Microbiology 12:132, 2012). В другом примере было показано, что бактериоцины из Pseudomonas характеризуются нейтрализующей активностью в отношении грибов, таких как Curvularia lunata, виды Fusarium, виды Helminthosporium и виды Biopolaris (см., например, Shalani and Srivastava, The Internet Journal of Microbiology Volume 5 Number 2, 2008). В другом примере было показано, что ботрицидин AJ1316 и алирин B1 из B. subtilis характеризуются противогрибковой активностью.
Модулирующее средство, включающее в себя бактериоцин, описанный в данном документе, может быть приведено в контакт с хозяином-мишенью в количестве и в течение времени, достаточных для: (a) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации бактериоцина внутри хозяина-мишени; (b) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации бактериоцина внутри кишки хозяина-мишени; (c) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации бактериоцина внутри бактериоцита хозяина-мишени; (d) модулирования уровня или активности одного или нескольких микроорганизмов (например, эндосимбионтов) в организме хозяина-мишени или/и (e) модулирования приспособленности хозяина-мишени.
Как показано с помощью примера 5, бактериоцины (например, colA, продуцируемый трансгенным растением) можно применять в качестве модулирующего средства, которое целенаправленно воздействует на путь хозяина (например, насекомого, например, тли), которое изменяет активность, уровни или метаболизм эндосимбиотических бактерий, обитающих в организме хозяина, таких как Buchnera spp., с целью модулирования (например, снижения) приспособленности хозяина.
(c) Противомикробные пептиды
Модулирующее средство, описанное в данном документе, может включать в себя противомикробный пептид (AMP). Можно применять любой AMP, подходящий для ингибирования микроорганизма, обитающего в организме хозяина. AMP представляют собой разнообразную группу молекул, которые разделяются на подгруппы на основании их аминокислотного состава и структуры. AMP может быть получен из любого организма или продуцироваться в любом организме, который естественным образом продуцирует AMP, в том числе AMP, получаемые из растений (например, копсин), насекомых (например, мастопаран, понератоксин, цекропин, морицин, меллитин), лягушек (например, магаинин, дермасептин, ауреин) и млекопитающих (например, кателицидины, дефензины и протегрины). Неограничивающие примеры AMP приведены в таблице 4.
Таблица 4. Примеры противомикробных пептидов
(SEQ ID NO: 52)
(SEQ ID NO: 53)
(SEQ ID NO: 54)
(SEQ ID NO: 55)
(SEQ ID NO: 56)
(SEQ ID NO: 57)
(SEQ ID NO: 58)
(SEQ ID NO: 59)
(SEQ ID NO: 60)
(SEQ ID NO: 61)
(SEQ ID NO: 62)
(SEQ ID NO: 63)
(SEQ ID NO: 64)
AMP может быть активным в отношении любого количества микроорганизмов-мишеней. В некоторых случаях AMP может характеризоваться формами антибактериальной и/или противогрибковой активности. В некоторых случаях AMP может характеризоваться биологической активностью узкого спектра или биологической активностью широкого спектра. Например, некоторые AMP целенаправленно воздействуют только на несколько видов бактерий или грибов и уничтожают их, тогда как другие являются активными в отношении как грамотрицательных, так и грамположительных бактерий, а также грибов.
Кроме того, AMP может функционировать посредством ряда известных механизмов действия. Например, цитоплазматическая мембрана представляет собой частую мишень для AMP, однако AMP могут также нарушать синтез ДНК и белка, сворачивание белка и синтез клеточной стенки. В некоторых случаях AMP с суммарным катионным зарядом и амфипатической природой разрушают мембраны бактерий, приводя к лизису клеток. В некоторых случаях AMP могут проникать в клетки и взаимодействовать с внутриклеточной мишенью с нарушением синтеза ДНК, РНК, белка или клеточной стенки. Помимо уничтожения микроорганизмов, AMP продемонстрировали ряд иммуномодулирующих функций, которые вовлечены в устранение инфекции, в том числе способность изменять экспрессию генов хозяев, выступать в роли хемокинов и/или индуцировать продуцирование хемокинов, ингибировать продуцирование провоспалительных цитокинов, индуцированное липополисахаридами, способствовать заживлению ран и модулировать ответы дендритных клеток и клеток, участвующих в адаптивном иммунном ответе.
В некоторых случаях AMP представляет собой функционально активный вариант AMP, описанных в данном документе. В некоторых случаях вариант AMP является на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичным, например, в определенной области или во всей последовательности, последовательности AMP, описанного в данном документе, или AMP природного происхождения.
В некоторых случаях AMP можно создавать с помощью биоинженерии для модулирования его биологической активности, например, повышения, или снижения, или регулирования, или для определения его микроорганизма-мишени. В некоторых случаях AMP продуцируется трансляционным аппаратом (например, рибосомой и т. д.) клетки. В некоторых случаях AMP синтезируют химическим путем. В некоторых случаях AMP образуется из полипептидного предшественника. Полипептидный предшественник может подвергаться расщеплению (например, процессингу под действием протеазы) с образованием самого полипептида AMP. Таким образом, в некоторых случаях AMP продуцируется из полипептида-предшественника. В некоторых случаях AMP включает в себя полипептид, который подвергся посттрансляционным модификациям, например, расщеплению или добавлению одной или нескольких функциональных групп.
AMP, описанные в данном документе, могут быть составлены в виде композиции для любого из путей применения, описанных в данном документе. Композиции, раскрытые в данном документе, могут содержать любые количество или тип (например, классы) AMP, как, например, по меньшей мере приблизительно любое количество из 1 AMP, 2, 3, 4, 5, 10, 15, 20 или больше AMP. Подходящая концентрация каждого AMP в композиции зависит от таких факторов, как эффективность, стабильность AMP, количество различных AMP в композиции, состав и способы применения композиции. В некоторых случаях концентрация каждого AMP в жидкой композиции составляет от приблизительно 0,1 нг/мл до приблизительно 100 мг/мл. В некоторых случаях концентрация каждого AMP в твердой композиции составляет от приблизительно 0,1 нг/г до приблизительно 100 мг/г. В некоторых случаях, если композиция содержит по меньшей мере два типа AMP, то концентрация каждого типа AMP может быть одной и той же или разной.
Модулирующее средство, включающее в себя AMP, описанный в данном документе, может быть приведено в контакт с хозяином-мишенью в количестве и в течение времени, достаточных для: (a) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации AMP внутри хозяина-мишени; (b) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации AMP внутри кишки хозяина-мишени; (c) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации AMP внутри бактериоцита хозяина-мишени; (d) модулирования уровня или активности одного или нескольких микроорганизмов (например, эндосимбионтов) в организме хозяина-мишени или/и (e) модулирования приспособленности хозяина-мишени.
(d) Регуляторные пептиды бактериоцитов
Модулирующее средство, описанное в данном документе, может включать в себя регуляторный пептид бактериоцитов (BRP). BRP представляют собой пептиды, экспрессируемые в бактериоцитах насекомых. Эти гены впервые экспрессируются в момент времени развития, совпадающий с включением симбионтов, а их специфичная для бактериоцитов экспрессия поддерживается в течение всей жизни насекомого. В некоторых случаях BRP имеет гидрофобный аминоконцевой домен, который, как предполагается, представляет собой сигнальный пептид. Помимо этого, некоторые BRP имеют домен, богатый цистеином. В некоторых случаях регуляторный пептид бактериоцитов представляет собой специфичный для бактериоцитов белок, богатый цистеином (BCR). Регуляторные пептиды бактериоцитов имеют длину от приблизительно 40 до 150 аминокислот. В некоторых случаях регуляторный пептид бактериоцитов имеет длину в диапазоне от приблизительно 45 до приблизительно 145, от приблизительно 50 до приблизительно 140, от приблизительно 55 до приблизительно 135, от приблизительно 60 до приблизительно 130, от приблизительно 65 до приблизительно 125, от приблизительно 70 до приблизительно 120, от приблизительно 75 до приблизительно 115, от приблизительно 80 до приблизительно 110, от приблизительно 85 до приблизительно 105 или в любом диапазоне в промежутке между этими значениями. Неограничивающие примеры BRP и форм их активности приведены в таблице 5.
Таблица 5. Примеры регуляторных пептидов бактериоцитов
(SEQ ID NO: 65)
(SEQ ID NO: 66)
(SEQ ID NO: 67)
(SEQ ID NO: 68)
(SEQ ID NO: 69)
(SEQ ID NO: 70)
(SEQ ID NO: 71)
(SEQ ID NO: 72)
(SEQ ID NO: 73)
(SEQ ID NO: 74)
(SEQ ID NO: 75)
(SEQ ID NO: 76)
(SEQ ID NO: 77)
В некоторых случаях BRP изменяет рост и/или активность одной или нескольких бактерий, обитающих в бактериоците хозяина. В некоторых случаях BRP можно создавать с помощью биоинженерии для модулирования его биологической активности (например, повышения, снижения или регулирования) или для определения его микроорганизма-мишени. В некоторых случаях BRP продуцируется трансляционным аппаратом (например, рибосомой и т. д.) клетки. В некоторых случаях BRP синтезируют химическим путем. В некоторых случаях BRP образуется из полипептидного предшественника. Полипептидный предшественник может подвергаться расщеплению (например, процессингу под действием протеазы) с образованием самого полипептида BRP. Таким образом, в некоторых случаях BRP продуцируется из полипептида-предшественника. В некоторых случаях BRP включает в себя полипептид, который подвергся посттрансляционным модификациям, например, расщеплению или добавлению одной или нескольких функциональных групп.
Функционально активные варианты BRP, описанные в данном документе, также пригодны в композициях и способах, описанных в данном документе. В некоторых случаях вариант BRP является на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичным, например, в определенной области или во всей последовательности, последовательности BRP, описанного в данном документе, или BRP природного происхождения.
BRP, описанный в данном документе, может быть составлен в виде композиции для любого из путей применения, описанных в данном документе. Композиции, раскрытые в данном документе, могут содержать любые количество или тип (например, классы) BRP, как, например, по меньшей мере приблизительно любое количество из 1 BRP, 2, 3, 4, 5, 10, 15, 20 или больше BRP. Подходящая концентрация каждого BRP в композиции зависит от таких факторов, как эффективность, стабильность BRP, количество различных BRP, состав и способы применения композиции. В некоторых случаях концентрация каждого BRP в жидкой композиции составляет от приблизительно 0,1 нг/мл до приблизительно 100 мг/мл. В некоторых случаях концентрация каждого BRP в твердой композиции составляет от приблизительно 0,1 нг/г до приблизительно 100 мг/г. В некоторых случаях, если композиция содержит по меньшей мере два типа BRP, то концентрация каждого типа BRP может быть одной и той же или разной.
Модулирующее средство, включающее в себя BRP, описанный в данном документе, может быть приведено в контакт с хозяином-мишенью в количестве и в течение времени, достаточных для: (a) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации BRP внутри хозяина-мишени; (b) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации BRP внутри кишки хозяина-мишени; (c) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации BRP внутри бактериоцита хозяина-мишени; (d) модулирования уровня или активности одного или нескольких микроорганизмов (например, эндосимбионтов) в организме хозяина-мишени или/и (e) модулирования приспособленности хозяина-мишени.
ii. Малые молекулы
В некоторых случаях модулирующее средство включает в себя малую молекулу. Многочисленные средства на основе малых молекул пригодны в способах и композициях, описанных в данном документе. Малые молекулы, обсуждаемые далее в данном документе, можно применять для изменения путей у хозяина, которые опосредуют взаимодействия между хозяином и микроорганизмами, обитающими в организме хозяина, как указано в разделах в отношении снижения приспособленности насекомых, таких как тли. Дополнительные средства на основе малых молекул также могут быть подвергнуты скринингу на основании их способности целенаправленно воздействовать на компоненты (например, полипептиды, например, ферменты или рецепторы поверхности клеток) путей у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, полипептиды, которые опосредуют взаимодействия хозяин-микробиота, например, пути иммунной системы хозяина или пути бактериоцитов). В некоторых случаях малая молекула включает в себя агонист, антагонист, ингибитор или активатор. К примеру, малая молекула, описанная в данном документе, может представлять собой агонист, антагонист, ингибитор или активатор, которые блокируют или усиливают активность и/или функцию компонента пути иммунной системы хозяина или пути бактериоцитов, приведенного в таблице 8 или таблице 9. Малая молекула может выступать в качестве антагониста или агониста полипептида (например, фермента или клеточного рецептора) в организме хозяина или микроорганизмах, обитающих в хозяине, в том числе любых белков, приведенные в таблице 7, таблице 8 или таблице 9.
Малые молекулы включают в себя без ограничения малые пептиды, пептидомиметики (например, пептоиды), аминокислоты, аналоги аминокислот, синтетические полинуклеотиды, аналоги полинуклеотидов, нуклеотиды, аналоги нуклеотидов, органические и неорганические соединения (в том числе гетероорганические и металлорганические соединения), как правило, характеризующиеся молекулярной массой менее чем приблизительно 5000 грамм на моль, например, органические или неорганические соединения, характеризующиеся молекулярной массой менее чем приблизительно 2000 грамм на моль, например, органические или неорганические соединения, характеризующиеся молекулярной массой менее чем приблизительно 1000 грамм на моль, например, органические или неорганические соединения, характеризующиеся молекулярной массой менее чем приблизительно 500 грамм на моль, а также соли, сложные эфиры и другие фармацевтически приемлемые формы таких соединений.
Малая молекула, описанная в данном документе, может быть составлена в композиции для любого из путей применения, описанных в данном документе. Композиции, раскрытые в данном документе, могут содержать любые количество или тип (например, классы) малых молекул, например, по меньшей мере приблизительно любое количество из 1 малой молекулы, 2, 3, 4, 5, 10, 15, 20 или более малых молекул. Подходящая концентрация каждой малой молекулы в композиции зависит от таких факторов, как эффективность, стабильность малой молекулы, количество различных малых молекул, состав и способы применения композиции. В некоторых случаях, если композиция содержит по меньшей мере два типа малых молекул, то концентрация каждого типа малой молекулы может быть одной и той же или разной.
Модулирующее средство, включающее в себя малую молекулу, описанную в данном документе, может быть приведено в контакт с хозяином-мишенью в количестве и в течение времени, достаточных для: (a) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации малой молекулы внутри хозяина-мишени; (b) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации малой молекулы внутри кишки хозяина-мишени; (c) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации малой молекулы внутри бактериоцита хозяина-мишени; (d) модулирования уровня или активности одного или нескольких микроорганизмов (например, эндосимбионтов) в организме хозяина-мишени или/и (e) модулирования приспособленности хозяина-мишени.
В некоторых случаях малая молекула активирует, стимулирует или повышает иммунный ответ хозяина по сравнению с организмом-хозяином, которому малая молекула не была введена. К примеру, малая молекула может представлять собой молекулу пептидогликана, которая активирует систему ROS у насекомых путем связывания с поверхностью эпителиальной клетки, что, в свою очередь, индуцирует ферментативную активность DUOX путем мобилизации внутриклеточного кальция. В другом примере молекула дигидроксифенилаланин (DOPA) представляет собой необходимый компонент для синтеза кутикулы. При получении кутикулы DOPA достигает высоких количеств ух насекомых, что вызывает апоптоз и активацию аутофагии. В другом случае иммунный ответ является эффективным для снижения уровня эндосимбионта или уничтожения эндосимбионта по сравнению с организмом-хозяином, которому малая молекула не была введена. В некоторых случаях малая молекула является эффективной для нарушения или снижения функции бактериоцитов по сравнению с организмом-хозяином, которому модулирующее средство не было введено. К примеру, молекулы, которые блокируют транспорт предшественников незаменимых аминокислот внутри бактериоцита, также нарушают продуцирование незаменимых аминокислот, например, аргинина. Это изменение в конечном итоге приводит к гибели эндосимбионта и, в конце концов, к гибели хозяина. Другие примеры модулирующих средств, которые можно применять для стимуляции иммунной системы хозяина и тем самым снижения уровней эндосимбионтов, обитающих в организме хозяина, включают в себя липополисахариды, рапамицин и β-глюкан.
В других случаях малая молекула снижает или повышает экспрессию генов обитающего микроорганизма путем связывания с некодирующей областью РНК. К примеру, малая молекула может представлять собой ингибитор рибопереключателя, такой как рибоцил, который связывается с регуляторным доменом 'рибопереключателя' в некодирующей области информационной РНК, которая кодирует фермент синтазу, участвующий в синтезе рибофлавина, с подавлением таким образом данного пути. В другом примере малая молекула является эффективной для повышения или снижения экспрессии генов, что приводит к уничтожению эндосимбионта. В некоторых случаях малая молекула является эффективной для нарушения функции бактериоцитов.
В некоторых случаях малая молекула изменяет гомеостаз хозяина. Например, малая молекула может представлять собой молекулу эйкозаиноида, такую как простагландин, которая активирует температурную реакцию в ответ на инфекцию, а также экзоцитоз белков в слюнных железах. Помимо постоянных видов активности в гомеостазе, определенные виды активности эйкозаноидов возникают в критических точках в жизненных циклах насекомых, таких как инфекционная атака и важные события при размножении. Эйкозаноиды опосредуют реакции клеточной защиты у насекомых. В одном из примеров подавление синтеза простагландинов в значительной степени нарушает способность насекомых к удалению бактерий из гемолимфы. В другом случае малая молекула является эффективной для повышения или снижения иммунного ответа у хозяина в отношении эндосимбионта или повышения или снижения температурной реакции у хозяина, что приводит к уничтожению эндосимбионта. В некоторых случаях малая молекула является эффективной для нарушения функции бактериоцитов.
Как показано с помощью примера 14, малые молекулы (например, простагландин) можно применять в качестве модулирующих средств, которые целенаправленно воздействуют на пути хозяина, что, в свою очередь, изменяет активность, уровни или метаболизм эндосимбиотических бактерий в организме хозяина и тем самым модулирует (например, снижает) приспособленность хозяина.
iii. Нуклеиновые кислоты
Многочисленные нуклеиновые кислоты являются пригодными в композициях и способах, описанных в данном документе. Композиции, раскрытые в данном документе, могут содержать любое число или тип (например, классы) нуклеиновых кислот (например, молекулу ДНК или молекулу РНК, например, молекулу mRNA, направляющей РНК (gRNA) или ингибирующей РНК (например, siRNA, shRNA или miRNA) или гибридную молекулу ДНК-РНК), например, по меньшей мере приблизительно 1 класс или вариант нуклеиновой кислоты, 2, 3, 4, 5, 10, 15, 20 или более классов или вариантов нуклеиновых кислот. Подходящая концентрация каждой нуклеиновой кислоты в композиции зависит от таких факторов, как эффективность, стабильность нуклеиновой кислоты, количество различных нуклеиновых кислот, состав и способы применения композиции. В некоторых случаях, если композиция содержит по меньшей мере два типа нуклеиновых кислот, то концентрация каждого типа нуклеиновой кислоты может быть одной и той же или разной.
Модулирующее средство, включающее в себя нуклеиновую кислоту, описанную в данном документе, может быть приведено в контакт с хозяином-мишенью в количестве и в течение времени, достаточных для: (a) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации нуклеиновой кислоты внутри хозяина-мишени; (b) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации нуклеиновой кислоты внутри кишки хозяина-мишени; (c) достижения целевого уровня (например, предварительно определенного или порогового уровня) концентрации нуклеиновой кислоты внутри бактериоцита хозяина-мишени; (d) модулирования уровня или активности одного или нескольких микроорганизмов (например, эндосимбионтов) в организме хозяина-мишени или/и (e) модулирования приспособленности хозяина-мишени.
Модулирующие средства на основе нуклеиновой кислоты, описанные далее в данном документе, в том числе нуклеиновые кислоты, кодирующие полипептиды, синтетическая РНК, ингибирующая РНК и системы редактирования генома, можно применять для изменения путей у хозяина, которые опосредуют взаимодействия между хозяином и микроорганизмами, обитающими в организме хозяина, как указано в разделах в отношении повышения и снижения приспособленности хозяев (например, тлей).
(a) Нуклеиновые кислоты, кодирующие полипептид
В некоторых случаях композиция содержит нуклеиновую кислоту, кодирующую любой из полипептидов, описанных в данном документе. Нуклеиновые кислоты, кодирующие полипептид, могут иметь длину от приблизительно 10 до приблизительно 50000 нуклеотидов (нукл.), от приблизительно 25 до приблизительно 100 нукл., от приблизительно 50 до приблизительно 150 нукл., от приблизительно 100 до приблизительно 200 нукл., от приблизительно 150 до приблизительно 250 нукл., от приблизительно 200 до приблизительно 300 нукл., от приблизительно 250 до приблизительно 350 нукл., от приблизительно 300 до приблизительно 500 нукл., от приблизительно 10 до приблизительно 1000 нукл., от приблизительно 50 до приблизительно 1000 нукл., от приблизительно 100 до приблизительно 1000 нукл., от приблизительно 1000 до приблизительно 2000 нукл., от приблизительно 2000 до приблизительно 3000 нукл., от приблизительно 3000 до приблизительно 4000 нукл., от приблизительно 4000 до приблизительно 5000 нукл., от приблизительно 5000 до приблизительно 6000 нукл., от приблизительно 6000 до приблизительно 7000 нукл., от приблизительно 7000 до приблизительно 8000 нукл., от приблизительно 8000 до приблизительно 9000 нукл., от приблизительно 9000 до приблизительно 10000 нукл., от приблизительно 10000 до приблизительно 15000 нукл., от приблизительно 10000 до приблизительно 20000 нукл., от приблизительно 10000 до приблизительно 25000 нукл., от приблизительно 10000 до приблизительно 30000 нукл., от приблизительно 10000 до приблизительно 40000 нукл., от приблизительно 10000 до приблизительно 45000 нукл., от приблизительно 10000 до приблизительно 50000 нукл. или любой диапазон между ними.
Модулирующее средство также может включать в себя функционально активные варианты нуклеиновых кислот, описанных в данном документе. В некоторых случаях вариант нуклеиновой кислоты является на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичным, например, в определенной области или во всей последовательности, последовательности нуклеиновых кислот, описанных в данном документе. В некоторых случаях настоящее изобретение предусматривает функционально активный полипептид, кодируемый вариантом нуклеиновой кислоты, описанным в данном документе. В некоторых случаях функционально активный полипептид, кодируемый вариантом нуклеиновой кислоты, характеризуется по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью, например, в определенной области или во всей аминокислотной последовательности, последовательности полипептида, описанного в данном документе, или последовательности полипептида природного происхождения.
Некоторые способы экспрессии нуклеиновой кислоты, кодирующей белок, могут предусматривать экспрессию в клетках, в том числе клетках хозяев (например, клетках насекомых, клетках моллюсков или клетках нематод), клетках дрожжевых грибов, клетках бактерий или других клетках под контролем соответствующих промоторов. Векторы экспрессии могут содержать нетранскрибируемые элементы, такие как точка начала репликации, подходящий промотор и энхансер, и другие 5'- или 3'-фланкирующие нетранскрибируемые последовательности, и 5'- или 3'-нетранскрибируемые последовательности, такие как необходимые участки связывания рибосомы, сайт полиаденилирования, донорный сайт сплайсинга, акцепторный сайт сплайсинга и последовательности терминации. Последовательности ДНК, происходящие из генома вируса SV40, к примеру, точки начала репликации, раннего промотора, энхансера, сайтов сплайсинга и сайтов полиаденилирования mSV40 можно применять для обеспечения других генетических элементов, требуемых для экспрессии гетерологичной последовательности ДНК. Подходящие векторы клонирования и экспрессии для применения с клетками-хозяевами бактерий, грибов, дрожжей и млекопитающих описаны в Green et al., Molecular Cloning: A Laboratory Manual, Fourth Edition, Cold Spring Harbor Laboratory Press, 2012.
Генетическая модификация с применением рекомбинантных способов является общеизвестной в данной области техники. Последовательность нуклеиновой кислоты, кодирующая необходимый ген, может быть получена с помощью рекомбинантных способов, известных в данной области техники, таких как, к примеру, скрининг библиотек на основе клеток, экспрессирующих ген, путем получения гена из вектора, который, как известно, содержит его, или путем выделения непосредственно из клеток и тканей, содержащих его, с помощью стандартных методик. В альтернативном случае ген, представляющий интерес, может быть получен синтетически, а не клонирован.
Экспрессия природных или синтетических нуклеиновых кислот, как правило, достигается с помощью функционального связывания нуклеиновой кислоты, кодирующей ген, представляющий интерес, с промотором, и включения данной конструкции в вектор экспрессии. Векторы экспрессии могут быть подходящими для репликации и экспрессии в бактериях. Векторы экспрессии также могут быть подходящими для репликации и интеграции в эукариотах. Типичные клонирующие векторы содержат терминаторы транскрипции и трансляции, последовательности инициации и промоторы, пригодные для экспрессии необходимой последовательности нуклеиновой кислоты.
Дополнительные промоторные элементы, например, энхансеры, регулируют частоту инициации транскрипции. Как правило, они расположены в области 30-110 пар оснований (п. о.) выше от сайта начала транскрипции, хотя, как недавно было показано, ряд промоторов также содержит функциональные элементы ниже сайта начала транскрипции. Область между промоторными элементами является гибкой, поэтому промоторная функция сохраняется, когда элементы инвертированы или перемещены друг по отношению к другу. В промоторе тимидинкиназы (tk) область между промоторными элементами может быть увеличена до 50 п. о. до того, как активность начинает снижаться. В зависимости от промотора, по-видимому, отдельные элементы могут функционировать совместно или независимо с целью активации транскрипции.
Одним примером подходящего промотора является последовательность промотора гена немедленно-раннего ответа цитомегаловируса (CMV). Данная промоторная последовательность представляет собой последовательность сильного конститутивного промотора, способного управлять высокими уровнями экспрессии любой полинуклеотидной последовательности, функционально связанной с ним. Другим примером подходящего промотора является фактор элонгации 1α (EF-1α). Однако также можно использовать другие последовательности конститутивных промоторов, в том числе без ограничения ранний промотор вируса обезьян 40 (SV40), промотор вируса рака молочной железы мыши (MMTV), промотор длинного концевого повтора (LTR) вируса иммунодефицита человека (HIV), промотор MoMuLV, промотор вируса лейкоза птиц, немедленно-ранний промотор вируса Эпштейна-Барр, промотор вируса саркомы Рауса, а также промоторы генов человека, такие как без ограничения промотор гена актина, промотор гена миозина, промотор гена гемоглобина и промотор гена креатинкиназы.
В альтернативном случае промотор может представлять собой индуцируемый промотор. Применение индуцируемого промотора предоставляет молекулярный переключатель, способный включать экспрессию полинуклеотидной последовательности, с которой он функционально связан, в случае когда такая экспрессия является желательной, или выключение экспрессии, в случае если экспрессия нежелательна. Примеры индуцируемых промоторов включают в себя без ограничения промотор гена металлотионина, промотор гена глюкокортикоидного рецептора, промотор гена прогестеронового рецептора и промотор гена устойчивости к тетрациклину.
Вектор экспрессии, подлежащий включению, также может содержать либо ген селективного маркера, либо репортерный ген, либо и то и другое с целью облегчения идентификации и отбора экспрессирующих клеток из популяции клеток, подлежащих трансфекции или инфицированию посредством вирусных векторов. В других аспектах селективный маркер может быть перенесен на отдельном участке ДНК и применен в процедуре котрансфекции. Как селектируемые маркеры, так и репортерные гены могут быть фланкированы подходящими регуляторными последовательностями с целью обеспечения экспрессии в клетках хозяев. Пригодные селектируемые маркеры включают в себя, к примеру, гены устойчивости к антибиотикам, такие как neo и т. п.
Репортерные гены можно применять для идентификации потенциально трансформированных клеток и для оценки функциональности регуляторных последовательностей. Как правило, репортерный ген представляет собой ген, который не присутствует в источнике реципиента или не кодируется им и который кодирует полипептид, экспрессия которого проявляется с помощью некоторого легко выявляемого свойства, например, ферментативной активности. Экспрессию репортерного гена анализируют в подходящее время после того, как ДНК была введена в клетки реципиента. Подходящие репортерные гены могут включать в себя гены, кодирующие люциферазу, бета-галактозидазу, хлорамфениколацетилтрансферазу, секретируемую щелочную фосфатазу, или ген зеленого флуоресцентного белка (например, Ui-Tei et al., FEBS Letters 479:79-82, 2000). Подходящие системы экспрессии хорошо известны и могут быть получены с помощью известных методик или приобретены коммерчески. Как правило, конструкция с минимальной 5'-фланкирующей областью, характеризующаяся наибольшим уровнем экспрессии репортерного гена, идентифицируется в качестве промотора. Такие промоторные области могут быть связаны с репортерным геном и применяться для оценки средств в отношении способности модулировать управляемую промотором транскрипцию.
В некоторых случаях организм может быть генетически модифицирован с целью изменения экспрессии одного или нескольких белков. Экспрессия одного или нескольких белков может быть модифицирована в течение определенного времени, например, состояния развития или дифференцировки организма. В одном случае в настоящем изобретении предусмотрена композиция для изменения экспрессии одного или нескольких белков, например, белков, которые влияют на активность, структуру или функцию. Экспрессия одного или нескольких белков может быть ограничена определенным(определенными) положением(положениями) или распространена по всему организму.
(b) Синтез mRNA
Модулирующее средство может включать в себя молекулу mRNA, например, синтетическую молекулу mRNA, кодирующую полипептид. В некоторых случаях молекула mRNA повышает уровень (например, уровень белка и/или mRNA) и/или активность средства, например, положительного регулятора функции, например, гена или продукта гена, приведенного в таблице 7, таблице 8 или таблице 9. В некоторых случаях молекула mRNA кодирует средство на основе полипептида или его фрагмент. К примеру, молекула mRNA может кодировать полипептид, характеризующийся по меньшей мере 50% (например, по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 97%, 99% или больше) идентичностью по отношению к аминокислотной последовательности средства, приведенного в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа. В других примерах молекула mRNA характеризуется по меньшей мере 50% (например, по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 97%, 99% или больше) идентичностью по отношению к последовательности нуклеиновой кислоты, кодирующей средство, приведенное в таблице 7, таблице 8 или таблице 9. В некоторых случаях молекула mRNA кодирует аминокислотную последовательность, отличающуюся на не более чем 30 (например, на не более чем 30, 20, 10, 5, 4, 3, 2 или 1) аминокислот по отношению к аминокислотной последовательности средства, приведенного в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа. В некоторых случаях молекула mRNA включает в себя последовательность, кодирующую фрагмент гена или продукт гена, приведенного в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа. К примеру, фрагмент содержит 10-20, 20-40, 40-60, 60-80, 80-100, 100-120, 120-140, 140-160, 160-180, 180-200, 200-250, 250-300, 300-400, 400-500, 500-600 или более аминокислот в длину. В некоторых случаях фрагмент представляет собой функциональный фрагмент, например, характеризующийся по меньше мере 20%, например, по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или больше активностью от полноразмерного гена или продукта гена, приведенного в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа. В некоторых случаях молекула mRNA повышает уровень и/или активность средства (или его фрагмента) или кодирует его.
Иллюстративная молекула mRNA включает в себя РНК, кодирующую любой полипептид, выбранный из таблицы 7, таблицы 8 или таблицы 9.
Синтетическая молекула mRNA может быть модифицированной, например, химически. Молекула mRNA может быть химически синтезирована или транскрибирована in vitro. Молекула mRNA может быть расположена в плазмиде, например, в вирусном векторе, бактериальном векторе или эукариотическом векторе экспрессии. В некоторых примерах молекула mRNA может быть доставлена в клетки с помощью трансфекции, электропорации или трансдукции (например, аденовирусной или лентивирусной трансдукции).
В некоторых случаях средство на основе модифицированной РНК, представляющее интерес, описанное в данном документе, имеет модифицированные нуклеозиды или нуклеотиды. Такие модификации известны и описаны, например, в WO 2012/019168. Дополнительные модификации описаны, например, в WO 2015/038892; WO 2015/038892; WO 2015/089511; WO 2015/196130; WO 2015/196118 и WO 2015/196128 A2.
В некоторых случаях модифицированная РНК, кодирующая полипептид, представляющий интерес, описанная в данном документе, имеет одну или несколько концевых модификаций, например, 5'-кэп-структуру и/или поли-A-хвост (например, от 100 до 200 нуклеотидов в длину). 5'-кэп-структура может быть выбрана из группы, состоящей из CapO, Capl, ARCA, инозина, Nl-метилгуанозина, 2'-фторгуанозина, 7-деазагуанозина, 8-оксогуанозина, 2-аминогуанозина, LNA-гуанозина и 2-азидогуанозина. В некоторых случаях модифицированные РНК также содержат 5'-UTR, содержащую по меньшей мере одну последовательность Козак, и 3'-UTR. Такие модификации известны и описаны, например, в WO 2012/135805 и WO 2013/052523. Дополнительные концевые модификации описаны, например, в WO 2014/164253 и WO 2016/011306, WO 2012/045075 и WO 2014/093924.
Химерные ферменты для синтеза кэппированных молекул РНК (например, модифицированной mRNA), которые могут содержать по меньшей мере одну химическую модификацию, описаны в WO 2014/028429.
В некоторых случаях модифицированная mRNA может быть циклизированной или конкатемеризованной с целью получения отвечающей требованиям трансляции молекулы для облегчения взаимодействий между поли-A-связывающими белками и 5'-конец-связывающими белками. Механизм циклизации или конкатемеризации может осуществляться посредством по меньшей мере 3 различных путей: 1) химического, 2) ферментативного и 3) катализируемого рибозимами. Вновь образованная 5'-/3'-связь может быть внутримолекулярной или межмолекулярной. Такие модификации описаны, например, в WO 2013/151736.
Способы получения и очистки модифицированных РНК известны и раскрыты в данной области техники. К примеру, модифицированные РНК получают с помощью только ферментативного синтеза с использованием транскрипции in vitro (IVT). Способы получения полинуклеотидов IVT известны в данной области техники и описаны в WO 2013/151666, WO 2013/151668, WO 2013/151663, WO 2013/151669, WO 2013/151670, WO 2013/151664, WO 2013/151665, WO 2013/151671, WO 2013/151672, WO 2013/151667 и WO 2013/151736. Способы очистки включают очистку РНК-транскрипта, содержащего поли-А-хвост, путем приведения образца в контакт с поверхностью, связанной со множеством тимидинов или их производных и/или множеством урацилов или их производных (polyT/U) в таких условиях, что РНК-транскрипт связывается с поверхностью, и элюирования очищенного РНК-транскрипта с поверхности (WO 2014/152031); применение ион(например, анион)обменной хроматографии, которая способствует разделению более длинных молекул РНК до 10000 нуклеотидов в длину включительно посредством масштабируемого способа (WO 2014/144767); и обработку образца модифицированной mRNA ДНКазой (WO 2014/152030).
Составы модифицированных РНК известны и описаны, например, в WO 2013/090648. К примеру, состав может представлять собой, но без ограничения, наночастицы, микросферы на основе сополимера молочной и гликолевой кислоты (PLGA), липидоиды, липоплекс, липосому, полимеры, углеводы (в том числе простые сахара, катионные липиды, фибриновый гель, фибриновый гидрогель, фибриновый клей, фибриновый герметик, фибриноген, тромбин, быстро элиминируемые липидные наночастицы (reLNP)) и их комбинации.
Модифицированные РНК, кодирующие полипептиды, в областях заболеваний человека, антител, вирусов и множества условий in vivo известны и раскрыты, к примеру, в таблице 6 международных публикаций №№ WO 2013/151666, WO 2013/151668, WO 2013/151663, WO 2013/151669, WO 2013/151670, WO 2013/151664, WO 2013/151665, WO 2013/151736; таблицах 6 и 7 международной публикации № WO 2013/151672; таблицах 6, 178 и 179 международной публикации № WO 2013/151671; таблицах 6, 185 и 186 международной публикации № WO 2013/151667. Любое из вышеизложенного может быть синтезировано в виде полинуклеотида IVT, химерного полинуклеотида или кольцевого нуклеотида, и каждое может включать в себя один или несколько модифицированных нуклеотидов или концевых модификаций.
(c) Ингибирующая РНК
В некоторых случаях модулирующее средство включает в себя молекулу ингибирующей РНК, например, которая действует с помощью пути РНК-интерференции (RNAi). К примеру, молекула ингибирующей РНК может включать в себя короткую интерферирующую РНК, короткую шпилечную РНК и/или microRNA, которые целенаправленно воздействуют на пути хозяина (например, пути, которые опосредуют взаимодействия хозяин-микробиота, например, пути иммунной системы хозяина или пути бактериоцитов, например, белки или гены, кодирующие белки, приведенные в таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа) у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды), и/или на пути обитающих микроорганизмов (например, белки или гены, кодирующие белки, приведенные в таблице 7, все из которых представлены со ссылкой на номер доступа). Определенные молекулы РНК могут подавлять экспрессию генов путем биологического процесса РНК-интерференции (RNAi). Молекулы для RNAi включают в себя РНК или РНК-подобные структуры, как правило, содержащие 15-50 пар оснований (например, приблизительно 18-25 пар оснований) и имеющие последовательность нуклеиновых оснований, идентичную (комплементарную) или почти идентичную (по сути комплементарную) кодирующей последовательности в экспрессируемом гене-мишени в клетке. Молекулы для RNAi включают в себя без ограничения короткие интерферирующие РНК (siRNA), двухнитевые РНК (dsRNA), короткие шпилечные РНК (shRNA), меродуплексы, субстраты dicer и мультивалентные интерферирующие РНК (патенты США №№ 8084599, 8349809, 8513207 и 9200276). shRNA представляет собой молекулу РНК, содержащую шпилечный оборот, которая обеспечивает снижение экспрессии генов-мишеней путем RNAi. shRNA могут быть доставлены в клетки в форме плазмид, например, вирусных или бактериальных векторов, например, с помощью трансфекции, электропорации или трансдукции). microRNA представляет собой некодирующую молекулу РНК, которая, как правило, имеет длину приблизительно 22 нуклеотида. MiRNA связываются с целевыми сайтами на молекулах mRNA и приводят к сайленсингу mRNA, например, вызывая расщепление mRNA, дестабилизацию mRNA или подавление трансляции mRNA. В некоторых случаях молекула ингибирующей РНК снижает уровень и/или активность отрицательного регулятора функции. В других вариантах осуществления молекула ингибирующей РНК снижает уровень и/или активность ингибитора положительного регулятора функции.
Молекулы для RNAi содержат последовательность, по сути комплементарную или полностью комплементарную всему гену-мишени или его фрагменту. Молекулы для RNAi могут комплементарно дополнять последовательности на границе между интронами и экзонами с целью предупреждения созревания вновь образованных ядерных РНК-транскриптов определенных генов в mRNA для транскрипции. Молекулы для RNAi, комплементарные определенным генам, могут гибридизироваться с mRNA для гена-мишени и предупреждать его трансляцию. Антисмысловая молекула может представлять собой ДНК, РНК или их производное или гибрид. Примеры таких производных молекул включают в себя без ограничения молекулы на основе нуклеиновой кислоты с пептидом (PNA) и фосфотиоата, такие как дезоксирибонуклеиновый гуанидин (DNG) или рибонуклеиновый гуанидин (RNG).
Молекулы для RNAi могут быть предусмотрены в виде "готовой к применению" РНК, синтезированной in vitro или в виде антисмыслового гена, трансфицированного в клетки, что приведет к образованию молекул для RNAi при транскрипции. Гибридизация с mRNA приводит к разрушению гибридизированной молекулы РНКазой H и/или подавлению образования трансляционных комплексов. И то и другое приводит к невозможности продуцировать продукт исходного гена.
Длина молекулы для RNAi, которая гибридизируется с транскриптом, представляющим интерес, может составлять около 10 нуклеотидов, от приблизительно 15 до 30 нуклеотидов или приблизительно 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более нуклеотидов. Степень идентичности антисмысловой последовательности с целевым транскриптом может составлять по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95%.
Молекулы для RNAi также могут содержать выступающие концы, т. е., как правило, неспаренные выступающие нуклеотиды, которые непосредственно не вовлечены в двойную спиральную структуру, обычно образующуюся коровыми последовательностями определенной в данном документе пары смысловой и антисмысловой нити. Молекулы для RNAi могут содержать 3'- и/или 5'-выступающие концы из приблизительно 1-5 оснований независимо на каждой из смысловых нитей и антисмысловых нитей. В некоторых случаях как смысловая нить, так и антисмысловая нить содержат 3'- и 5'-выступающие концы. В некоторых случаях один или несколько нуклеотидов 3'-выступающего конца одной нити спариваются основаниями с одним или несколькими нуклеотидами 5'-выступающего конца другой нити. В других случаях один или несколько нуклеотидов 3'-выступающего конца одной нити не спариваются основаниями с одним или несколькими нуклеотидами 5'-выступающего конца другой нити. Смысловая и антисмысловая нити молекулы для RNAi могут содержать или могут не содержать одинаковое количество нуклеотидных оснований. Антисмысловая и смысловая нити могут образовывать дуплекс, при этом только 5'-конец имеет тупой конец, только 3'-конец имеет тупой конец, как 5'-, так и 3'-концы заканчиваются тупым концом, или ни 5'-, ни 3'-конец не заканчивается тупым концом. В другом случае один или несколько нуклеотидов в выступающем конце содержат тиофосфат, фосфотиоат, дезоксинуклеотидный инвертированный (связанный в направлении от 3' до 3') нуклеотид или представляют собой модифицированный рибонуклеотид или дезоксинуклеотид.
Молекулы малой интерферирующей РНК (siRNA) содержат нуклеотидную последовательность, которая является идентичной от приблизительно 15 до приблизительно 25 смежных нуклеотидов целевой mRNA. В некоторых случаях последовательность siRNA начинается динуклеотидом AA, характеризуется содержанием GC приблизительно 30-70% (приблизительно 30-60%, приблизительно 40-60% или приблизительно 45%-55%) и не характеризуется высоким процентом идентичности по отношению к любой нуклеотидной последовательности, отличной от мишени в геноме, в который она подлежит введению, к примеру, определенной с помощью стандартного поиска BLAST.
siRNA и shRNA имеют сходство с промежуточными соединениями в пути процессинга генов эндогенной microRNA (miRNA) (Bartel, Cell 116:281-297, 2004). В некоторых случаях siRNA могут функционировать в качестве miRNA и наоборот (Zeng et al., Mol. Cell 9:1327-1333, 2002; Doench et al., Genes Dev. 17:438-442, 2003). Экзогенные siRNA снижают экспрессию mRNA с помощью затравочной области, комплементарной siRNA (Birmingham et al., Nat. Methods 3:199-204, 2006). Несколько целевых сайтов в 3'-UTR обеспечивают более сильное снижение экспрессии (Doench et al., Genes Dev. 17:438-442, 2003).
Известные эффективные последовательности siRNA и когнатные связывающие сайты также хорошо представлены в соответствующей литературе. Молекулы для RNAi легко конструируют и получают с помощью технологий, известных в данной области техники. Помимо этого, существуют компьютерные средства, которые повышают вероятность обнаружения эффективных и специфических мотивов последовательностей (Pei et al., Nat. Methods 3(9):670-676, 2006; Reynolds et al., Nat. Biotechnol. 22(3):326-330, 2004; Khvorova et al., Nat. Struct. Biol. 10(9):708-712, 2003; Schwarz et al., Cell 115(2):199-208, 2003; Ui-Tei et al., Nucleic Acids Res. 32(3):936-948, 2004; Heale et al., Nucleic Acids Res. 33(3):e30, 2005; Chalk et al., Biochem. Biophys. Res. Commun. 319(1):264-274, 2004; и Amarzguioui et al., Biochem. Biophys. Res. Commun. 316(4):1050-1058, 2004).
Молекула для RNAi модулирует экспрессию РНК, кодируемую геном. Поскольку многочисленные гены могут разделять некоторую степень гомологии последовательностей друг с другом, в некоторых случаях молекула RNAi может быть сконструирована таким образом, чтобы целенаправленно воздействовать на класс генов с достаточной гомологией последовательностей. В некоторых случаях молекула для RNAi может содержать последовательность, которая характеризуется комплементарностью по отношению к последовательностям, которые являются общими среди различных генных мишеней или являются уникальным в отношении определенной генной мишени. В некоторых случаях молекула для RNAi может быть сконструирована таким образом, чтобы целенаправленно воздействовать на консервативные области последовательности РНК, характеризующиеся гомологией между несколькими генами, тем самым целенаправленно воздействуя на несколько генов в семействе генов (например, различные изоформы генов, сплайс-варианты, мутантные гены и т. д.). В некоторых случаях молекула для RNAi может быть сконструирована с целью целенаправленного воздействия на последовательность, которая является уникальной в отношении определенной последовательности РНК одного гена.
В некоторых случаях молекула ингибирующей РНК обеспечивает снижение уровня и/или активности компонента хозяина (например, компонента в путях, которые опосредуют взаимодействия хозяин-микробиота, например, путях иммунной системы хозяина или путях бактериоцитов) и/или микробного компонента, в том числе генов, кодирующих белки, приведенные в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа. В некоторых случаях молекула ингибирующей РНК обеспечивает подавление экспрессии компонента хозяина (например, компонента в путях, которые опосредуют взаимодействия хозяин-микробиота, например, путях иммунной системы хозяина или путях бактериоцитов) или микробного компонента, например, генов, кодирующих белки, приведенные в таблице 7, таблице 8 или таблице 9 (например, подавляет трансляцию белка). В других случаях молекула ингибирующей РНК обеспечивает повышение разрушения компонента хозяина (например, компонента в путях, которые опосредуют взаимодействия хозяин-микробиота, например, путях иммунной системы хозяина или путях бактериоцитов) и/или микробного компонента, например, генов, кодирующих белки, приведенные в таблице 7, таблице 8 или таблице 9, и/или уменьшение стабильности (например, периода полужизни) компонента пути. Молекула ингибирующей РНК может быть химически синтезирована или транскрибирована in vitro.
В некоторых случаях композиции, описанные в данном документе, содержат молекулу для RNAi, например, siRNA, для регуляции (например, подавления) экспрессии гена, кодирующего любой из компонентов, описанных в данном документе, который регулирует иммунную систему хозяина. В некоторых случаях композиция содержит молекулу для RNAi, например, siRNA, для подавления экспрессии любого из генов, описанных в данном документе, которые регулируют (например, подавляют) развитие или функцию бактериоцита у хозяина. В некоторых случаях регуляция иммунной системы хозяина приводит к сокращению или уничтожению эндосимбиотического микроорганизма в организме хозяина и, в свою очередь, снижает приспособленность хозяина. В некоторых случаях регуляция развития и/или функции бактериоцитов приводит к сокращению или уничтожению эндосимбиотического микроорганизма в организме хозяина и, в свою очередь, снижает приспособленность хозяина. В некоторых случаях молекула для RNAi (например, siRNA) может обеспечивать подавление экспрессии Ubx с целью нарушения расположения бактериоцитов в симбионте. В некоторых случаях молекула для RNAi (например, siRNA) обеспечивает подавление экспрессии abd-A и Antp с целью нарушения целостности и расположения бактериома.
В некоторых случаях одна или несколько молекул для RNAi целенаправленно воздействуют на любой ген, кодирующий любой белок, описанный в данном документе, например, см., таблицу 7, таблицу 8 или таблицу 9. В некоторых случаях одна или несколько молекул для RNAi целенаправленно воздействуют на ген бактерии, описанный в данном документе. В некоторых случаях одна или несколько молекул для RNAi целенаправленно воздействуют на ген эндосимбионта, описанный в данном документе. В некоторых случаях одна или несколько молекул для RNAi целенаправленно воздействуют на ген бактериоцита, описанный в данном документе. В некоторых случаях одна или несколько молекул для RNAi целенаправленно воздействуют на ген хозяина, описанный в данном документе. В некоторых случаях одна или несколько молекул для RNAi целенаправленно воздействуют на ген иммунной системы у хозяина, описанный в данном документе.
Молекула ингибирующей РНК может быть модифицированной, например, с содержанием модифицированных нуклеотидов, например, 2'-фтор, 2'-o-метил, 2'-дезокси, замкнутой нуклеиновой кислоты, 2'-гидрокси, фосфотиоата, 2'-тиоуридина, 4'-тиоуридина, 2'-дезоксиуридина. Не ограничиваясь теорией, считается, что такие модификации могут повышать устойчивость к нуклеазе и/или стабильность в сыворотке крови или снижать иммуногенность.
В некоторых случаях молекула для RNAi связывается с полимером для доставки с помощью физиологически неустойчивой связи или линкера. Физиологически неустойчивый линкер выбирают таким образом, что он подвергается химической трансформации (например, расщеплению) при присутствии в определенных физиологических условиях (например, дисульфидная связь, расщепляемая в восстанавливающей среде цитоплазмы клетки). Высвобождение молекулы из полимера в результате расщепления физиологически неустойчивой связи облегчает взаимодействие молекулы с подходящими клеточными компонентами с целью активности.
Конъюгат молекула для RNAi-полимер может быть образован с помощью ковалентного связывания молекулы с полимером. Полимер полимеризуют или модифицируют таким образом, что он содержит реакционную группу A. Молекулу для RNAi также полимеризуют или модифицируют таким образом, что она содержит реакционную группу B. Реакционные группы A и B выбирают таким образом, что они могут связываться посредством обратимой ковалентной связи с помощью способов, известных в данной области техники.
Конъюгация молекулы для RNAi с полимером может быть выполнена в присутствии избытка полимера. Поскольку молекула для RNAi и полимер могут иметь разный заряд во время конъюгации, наличие избытка полимера может приводить к снижению или устранению агрегации конъюгата. В альтернативном случае можно применять избыток полимера-носителя, такого как поликатион. Избыток полимера может быть удален из конъюгированного полимера перед введением конъюгата. В альтернативном случае избыток полимера может быть введен совместно с конъюгатом.
Инъекция двухнитевой РНК (dsRNA) в организм материнских особей насекомых приводит к эффективному подавлению экспрессии генов их потомства во время эмбриогенеза, см., например, Khila et al., PLoS Genet. 5(7):e1000583, 2009; и Liu et al., Development 131(7):1515-1527, 2004. Matsuura et al. (PNAS 112(30):9376-9381, 2015) показали, что подавление Ubx приводит к устранению бактериоцитов и расположения бактериоцитов в симбионте.
Получение и применение ингибирующих средств на основе некодирующей РНК, такой как рибозимы, РНКаза P, молекулы для siRNA и miRNA, также известно в данной области техники, к примеру, как описано в Sioud, RNA Therapeutics: Function, Design, and Delivery (Methods in Molecular Biology). Humana Press (2010).
Другие примеры модулирующих средств на основе нуклеиновой кислоты, которые могут применяться в данном документе, включают в себя dsRNA, характеризующиеся по меньшей мере 50% (например, по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 97%, 99% или больше) идентичностью по отношению к последовательности под любым из SEQ ID NO: 148-150.
Как показано с помощью примеров 1-4 и 7-9, ингибирующая РНК (например, dsRNA или PNA) может быть использована в качестве модулирующего средства, которое целенаправленно воздействует на путь хозяина (например, насекомое, например, тлю), что, в свою очередь, изменяет активность, уровни или метаболизм эндосимбиотических бактерий, таких как Buchnera spp., обитающих в организме хозяина и, тем самым, модулирует (например, снижает) приспособленность хозяина.
(d) Редактирование генов
Модулирующие средства, описанные в данном документе, могут включать в себя компонент системы редактирования генов. К примеру, средство может обеспечивать введение изменения (например, вставки, делеции (например, нокаута), транслокации, инверсии, точечной мутации или другой мутации) в гене, связанном с иммунной системой или бактериоцитом хозяина-беспозвоночного (например, насекомого, моллюска или нематоды), или в гене у микроорганизма, обитающего в хозяине-беспозвоночном, например, гене фермента или рецептора, описанном в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа. Иллюстративные системы редактирования генов включают в себя нуклеазы с цинковыми пальцами (ZFN), нуклеазы на основе эффектора, подобного активаторам транскрипции (TALEN), и системы коротких палиндромных повторов, регулярно расположенных группами (CRISPR). Способы на основе ZFN, TALEN и CRISPR описаны, например, в Gaj et al., Trends Biotechnol. 31(7):397-405, 2013.
В типичной системе CRISPR/Cas эндонуклеаза направлена на целевую нуклеотидную последовательность (например, сайт в геноме, который подлежит редактированию последовательности) с помощью специфических в отношении последовательности некодирующих "направляющих РНК", которые целенаправленно воздействуют на одно- или двухнитевые последовательности ДНК. Были идентифицированы три класса (I-III) систем CRISPR. Системы CRISPR II класса используют одну эндонуклеазу Cas (вместо нескольких белков Cas). Система CRISPR II класса включает в себя эндонуклеазу Cas II типа, такую как Cas9, РНК CRISPR ("crRNA") и транс-активирующую crRNA ("tracrRNA"). crRNA содержит "направляющую РНК", т. е. как правило, последовательность РНК из приблизительно 20 нуклеотидов, которая соответствует целевой последовательности ДНК. crRNA также содержит область, которая связывается с tracrRNA с образованием частично двухнитевой структуры, которая расщепляется РНКазой III, приводя к образованию гибрида crRNA/tracrRNA. РНК выступают в качестве направляющих последовательностей для направления белков Cas к специфическим в отношении сайленсинга последовательностям ДНК/РНК, в зависимости от спейсерной последовательности. См., например, Horvath et al., Science 327:167-170, 2010; Makarova et al., Biology Direct 1:7, 2006; Pennisi, Science 341:833-836, 2013. Целевая последовательность ДНК должна, как правило, прилегать к "мотиву, примыкающему к протоспейсеру" ("PAM"), который является специфическим для определенной эндонуклеазы Cas; однако последовательности PAM встречаются по всему данному геному. Эндонуклеазы CRISPR, идентифицированные из различных прокариотических видов, обладают требованиями к уникальным последовательностям PAM; примеры последовательностей PAM включают в себя 5'-NGG (SEQ ID NO: 78) (Streptococcus pyogenes), 5'-NNAGAA (SEQ ID NO: 79) (Streptococcus thermophilus CRISPR1), 5'-NGGNG (SEQ ID NO: 80) (Streptococcus thermophilus CRISPR3) и 5'-NNNGATT (SEQ ID NO: 81) (Neisseria meningiditis). Некоторые эндонуклеазы, например, эндонуклеазы Cas9, ассоциированы с сайтами PAM с высоким содержанием G, например, 5'-NGG (SEQ ID NO: 78), и осуществляют расщепление на тупом конце целевой ДНК в положении на 3 нуклеотида выше от (от 5') сайта PAM. Другая система CRISPR II класса включает в себя эндонуклеазу Cpf1 V типа, которая является меньшего размера, чем Cas9; примеры включают в себя AsCpf1 (из Acidaminococcus sp.) и LbCpf1 (из Lachnospiraceae sp.). Cpf1-ассоциированные матрицы CRISPR подвергаются процессингу в зрелые crRNA без потребности в tracrRNA; другими словами, система Cpf1 требует только нуклеазы Cpf1 и crRNA для расщепления целевой последовательности ДНК. Эндонуклеазы Cpf1 ассоциированы с сайтами PAM с высоким содержанием T, например, 5'-TTN. Cpf1 также может распознавать мотив 5'-CTA PAM. Cpf1 расщепляет целевую ДНК в результате введения офсетного или ступенчатого двухнитевого разрыва с применением 4- или 5-нуклеотидного 5'-выступающего конца, к примеру, расщепление целевой ДНК с применением 5-нуклеотидного офсетного или ступенчатого разрыва, локализованного на 18 нуклеотидов ниже от (от 3') сайта PAM на кодирующей нити и на 23 нуклеотида ниже от сайта PAM на комлементарной нити; 5-нуклеотидный выступающий конец, который образуется в результате такого офсетного расщепления, обеспечивает более точное редактирование генома с помощью вставки ДНК путем гомологичной рекомбинации по сравнению со вставкой на тупом конце расщепляемой ДНК. См., например, Zetsche et al., Cell 163:759-771, 2015.
В целях редактирования генома матрицы CRISPR могут быть сконструированы таким образом, чтобы содержать одну или множество последовательностей направляющей РНК, соответствующих необходимой целевой последовательности ДНК; см., например, Cong et al., Science 339:819-823, 2013; Ran et al., Nature Protocols 8:2281-2308, 2013. По меньшей мере приблизительно 16 или 17 нуклеотидов из последовательности gRNA требуется Cas9 для того, чтобы произошло расщепление ДНК; в случае Cpf1 по меньшей мере приблизительно 16 нуклеотидов из последовательности gRNA необходимо для достижения детектируемого расщепления ДНК. Фактически, последовательности направляющей РНК, как правило, конструируют таким образом, чтобы они имели длину от 17 до 24 нуклеотидов (например, 19, 20 или 21 нуклеотидов) и комплементарность по отношению к последовательности гена-мишени или нуклеиновой кислоты. Генераторы и алгоритмы заданной пользователем gRNA доступны коммерчески для применения в конструировании эффективных направляющих РНК. Редактирование генома также было достигнуто с помощью химерных "одиночных направляющих РНК" ("sgRNA"), сконструированной (синтетической) одиночной молекулы РНК, которая имитирует встречающийся в природе комплекс crRNA-tracrRNA и содержит как tracrRNA (для связывания с нуклеазой), так и по меньшей мере одну crRNA (для направления нуклеазы к последовательности, в отношении которой происходит целенаправленное воздействие с целью редактирования). Также было продемонстрировано, что химически модифицированные sgRNA являются эффективными в редактировании геномов; см., к примеру, Hendel et al., Nature Biotechnol. 985-991, 2015.
В то время как Cas9 дикого типа приводит к образованию двухнитевых разрывов (DSB) в определенных последовательностях ДНК, в отношении которых происходит целенаправленное воздействие gRNA, доступен ряд эндонуклеаз CRISPR, имеющих модифицированные функциональные свойства, к примеру: версия "никаза" Cas9 приводит к образованию только однонитевого разрыва; каталитически неактивный Cas9 ("dCas9") не разрезает целевую ДНК, однако нарушает транскрипцию в результате стерического затруднения. dCas9 может быть дополнительно слит с эффектором для репрессии (CRISPRi) или активации (CRISPRa) экспрессии гена-мишени. К примеру, Cas9 может быть слит с транскрипционным репрессором (например, доменом KRAB) или транскрипционным активатором (например, слияние dCas9-VP64). Каталитически неактивный Cas9 (dCas9), слитый с нуклеазой FokI ("dCas9-FokI"), может быть слит с образованием DSB в целевых последовательностях, гомологичных двум gRNA. См., например, многочисленные плазмиды на основе CRISPR/Cas9, раскрытые в репозитории Addgene (Addgene, 75 Sidney St., Suite 550A, Cambridge, MA 02139; addgene.org/crispr/) и публично доступные из него. "Двойная никаза" Cas9, которая вводит два отдельных двухнитевых разрыва, каждый из которых направляется отдельной направляющей РНК, описана как достигающая более точного редактирования генома Ran et al., Cell 154:1380-1389, 2013.
Технология CRISPR для редактирования генов эукариот раскрыта в публикациях заявки на патент США US 2016/0138008 A1 и US 2015/0344912 A1, и в патентах США 8697359, 8771945, 8945839, 8999641, 8993233, 8895308, 8865406, 8889418, 8871445, 8889356, 8932814, 8795965 и 8906616. Эндонуклеаза Cpf1 и соответствующие направляющие РНК и сайты PAM раскрыты в публикации заявки на патент США 2016/0208243 A1.
В некоторых случаях необходимая модификация генома включает в себя гомологическую рекомбинацию, при этом один или несколько двухнитевых разрывов ДНК в целевой нуклеотидной последовательности образуются с помощью направляемой РНК нуклеазы и направляющих РНК с последующей репарацией разрыва(разрывов) путем механизма гомологичной рекомбинации ("репарации, направляемой гомологией"). В таких случаях донорная матрица, которая кодирует необходимую нуклеотидную последовательность, подлежащую вставке или нокированию в двухнитевом разрыве, предусмотрена для клетки или субъекта; примеры подходящих матриц включают в себя однонитевые матрицы ДНК и двухнитевые матрицы ДНК (например, связанные с полипептидом, описанным в данном документе). Как правило, донорная матрица, кодирующая нуклеотидное изменение на протяжении участка менее чем приблизительно 50 нуклеотидов, предусмотрена в форме однонитевой ДНК; более крупные донорные матрицы (например, более чем 100 нуклеотидов) часто предусмотрены в виде плазмид на основе двухнитевой ДНК. В некоторых случаях донорная матрица предусмотрена для клетки или субъекта в количестве, которое является достаточным для достижения необходимой репарации, направляемой гомологией, но она не сохраняется в клетке или у субъекта после определенного периода времени (например, после одного или нескольких циклов клеточного деления). В некоторых случаях донорная матрица имеет коровую нуклеотидную последовательность, которая отличается от целевой нуклеотидной последовательности (например, гомологичной эндогенной геномной области) на по меньшей мере 1, по меньшей мере 5, по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50 или более нуклеотидов. Указанная коровая последовательность фланкируется "гомологичными плечами" или областями высокой идентичности последовательности по отношению к целевой нуклеотидной последовательности; в некоторых случаях области высокой идентичности включают в себя по меньшей мере 10, по меньшей мере 50, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 300, по меньшей мере 400, по меньшей мере 500, по меньшей мере 600, по меньшей мере 750 или по меньшей мере 1000 нуклеотидов на каждой стороне коровой последовательности. В некоторых случаях, если донорная матрица находится в форме однонитевой ДНК, то коровая последовательность фланкируется гомологичными плечами, содержащими по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80 или по меньшей мере 100 нуклеотидов на каждой стороне коровой последовательности. В случаях, если донорная матрица находится в форме двухнитевой ДНК, то коровая последовательность фланкируется гомологичными плечами, содержащими по меньшей мере 500, по меньшей мере 600, по меньшей мере 700, по меньшей мере 800, по меньшей мере 900 или по меньшей мере 1000 нуклеотидов на каждой стороне коровой последовательности. В одном случае два отдельных двухнитевых разрыва вводят в клетку или целевую нуклеотидную последовательность субъекта с помощью "двойной никазы" Cas9 (см. Ran et al., Cell 154:1380-1389, 2013) с последующей доставкой донорной матрицы.
В некоторых случаях композиция содержит gRNA и целевую нуклеазу, например, Cas9, например, Cas9 дикого типа, никазу Cas9 (например, Cas9 D10A), неактивную форму Cas9 (dCas9), eSpCas9, Cpf1, C2C1 или C2C3 или нуклеиновую кислоту, кодирующую такую нуклеазу. Выбор нуклеазы и gRNA определяется тем, представляет ли собой целевая мутация делецию, замену или добавление нуклеотидов, например, делецию, замену или добавление нуклеотидов по отношению к целевой последовательности. Слияния каталитически неактивной эндонуклеазы, например, неактивной формы Cas9 (dCas9, например, D10A; H840A), связанной со всей или частью (например, биологически активной частью) (одного или нескольких) эффекторных доменов приводят к образованию химерных белков, которые могут быть слиты с полипептидом с целью направления композиции к определенным сайтам ДНК с помощью одной или нескольких последовательностей РНК (sgRNA) для модулирования активности и/или экспрессии одной или нескольких целевых последовательностей нуклеиновых кислот.
В некоторых случаях средство включает направляющую РНК (gRNA) для применения в системе CRISPR для редактирования генов. В некоторых случаях средство включает в себя нуклеазу с цинковыми пальцами (ZFN) или mRNA, кодирующую ZFN, которая целенаправленно воздействует (например, расщепляет) на последовательность нуклеиновой кислоты (например, последовательность ДНК) гена, связанного с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, гена, кодирующего белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа). В некоторых случаях средство включает в себя TALEN или mRNA, кодирующую TALEN, которая целенаправленно воздействует (например, расщепляет) на последовательность нуклеиновой кислоты (например, последовательность ДНК) в гене, связанном с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, в гене, кодирующем белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа).
К примеру, gRNA может быть использована в системе CRISPR для конструирования изменения в гене, связанном с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, в гене, кодирующем белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа). В других примерах ZFN и/или TALEN могут быть использованы в системе CRISPR для конструирования изменения в гене, связанном с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, в гене, кодирующем белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа). Иллюстративные изменения включают в себя вставки, делеции (например, нокауты), транслокации, инверсии, точечные мутации или другие мутации. Изменение может быть введено в ген в клетке, например, in vitro, ex vivo или in vivo. В некоторых примерах изменение обеспечивает повышение уровня и/или активности гена, связанного с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, гена, кодирующего белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа). В других примерах изменение обеспечивает снижение уровня и/или активности гена (например, нокдаунам или нокаутам), связанного с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, гена, кодирующего белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа). В еще одном примере изменение обеспечивает коррекцию дефекта (например, мутации, вызывающей дефект) в гене, связанном с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, в гене, кодирующем белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа).
В некоторых случаях система CRISPR применяется для редактирования (например, добавления или удаления пары оснований) гена-мишени, связанного с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, гена, кодирующего белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа). В других случаях система CRISPR применяется для введения преждевременного стоп-кодона, например, со снижением тем самым экспрессии гена-мишени. В еще одних случаях система CRISPR применяется для отключения гена-мишени обратимым образом, например, аналогично РНК-интерференции. В некоторых случаях система CRISPR применяется для направления Cas к промотору гена, со стерическим блокированием тем самым РНК-полимеразы.
В некоторых случаях система CRISPR может быть получена для редактирования гена, связанного с путями у хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) и/или обитающих микроорганизмов (например, путями, которые опосредуют взаимодействия хозяин-микробиота, например, путями иммунной системы хозяина или путями бактериоцитов, например, генов, кодирующих белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа) с помощью технологии, описанной, например, в публикации США № 20140068797, Cong, Science 339: 819-823, 2013; Tsai, Nature Biotechnol. 32:6 569-576, 2014; патентах США №№ 8871445; 8865406; 8795965; 8771945 и 8697359.
В некоторых случаях методику CRISPR-интерференции (CRISPRi) можно применять для репрессии транскрипции определенных генов, например, гена, кодирующего компонент иммунной системы хозяина или бактериоцитов (например, фермент или рецептор, описанные в данном документе). В случае CRISPRi сконструированный белок Cas9 (например, нулевая нуклеаза dCas9 или слитый белок dCas9, например, слияние dCas9-KRAB или dCas9-SID4X) может спариваться со специфической в отношении последовательности направляющей РНК (sgRNA). Комплекс Cas9-gRNA может блокировать РНК-полимеразу, тем самым препятствуя элонгации транскрипта. Данный комплекс также может блокировать инициацию транскрипции с помощью препятствования связыванию с транскрипционными факторами. Способ CRISPRi является специфическим с минимальными нецелевыми эффектами и мультиплексным, например, может одновременно приводить к репрессии более одного гена (например, с помощью нескольких gRNA). Кроме того, способ CRISPRi обеспечивает обратимую репрессию генов.
В некоторых случаях опосредованную CRISPR активацию генов (CRISPRa) можно применять для активации транскрипции, например, одного или нескольких генов, описанных в данном документе (например, гена, кодирующего любой из белков, приведенных в таблице 7, таблице 8 или таблице 9). В случае методики CRISPRa слитые белки dCas9 обеспечивают рекрутинг транскрипционных активаторов. К примеру, dCas9 может быть слит с полипептидами (например, доменами активации), такими как VP64 или доменом активации p65 (p65D) и применен совместно с sgRNA (например, одиночной sgRNA или множественными sgRNA) с целью активации гена или генов, например, эндогенного(эндогенных) гена(генов) у хозяина или микроорганизма, обитающего в организме хозяина. Многочисленные активаторы могут быть подвергнуты рекрутингу с помощью многочисленных sgRNA, это может обеспечивать повышение эффективности активации. Можно применять ряд доменов активации и одиночные или множественные домены активации. Помимо конструирования dCas9 с целью рекрутинга активаторов, также для рекрутинга активаторов могут быть сконструированы sgRNA. К примеру, РНК-аптамеры могут быть включены в sgRNA с целью рекрутинга белков (например, доменов активации), таких как VP64. В некоторых примерах системы посредников синергической активации (SAM) можно применять для активации транскрипции. В случае SAM MS2-аптамеры добавляют к sgRNA. MS2 способствует рекрутингу белка оболочки MS2 (MCP), слитого с p65AD и фактором теплового шока 1 (HSF1).
Методики CRISPRi и CRISPRa описаны более подробно, например, в Dominguez et al., Nat. Rev. Mol. Cell Biol. 17:5-15, 2016, включенном в данном документе посредством ссылки. Помимо этого, опосредованные dCas9 эпигенетические модификации и одновременная активация и репрессия с помощью систем CRISPR, описанных в Dominguez et al., могут быть использованы с целью модулирования компонента хозяина или микробного пути, описанных в данном документе (например, путей, которые опосредуют взаимодействия хозяин-микробиота, например, путей иммунной системы хозяина или путей бактериоцитов, например, гена, кодирующего белок, приведенный в таблице 7, таблице 8 или таблице 9, все из которых представлены со ссылкой на номер доступа).
iv. Гены- и белки-мишени
Любое из модулирующих средств, описанных в данном документе, можно применять для изменения (например, повышения или снижения) экспрессии генов, изменения (например, повышения или снижения) активности белка-мишени и/или изменения функции у хозяина или микроорганизма, обитающего в организме хозяина. Белки или гены, которые вовлечены во множество процессов, могут подвергаться целенаправленному воздействию, включая любой из функциональных классов, приведенных в таблице 6.
Таблица 6. Функциональные классы белков-мишеней
(a) Гены- и белки-мишени микроорганизмов
Любое из модулирующих средств, описанных в данном документе, можно применять с целью изменения экспрессии генов или целевых белков в микроорганизме, обитающем в организме хозяина. В некоторых случаях модулирующее средство (например, антитело) непосредственно целенаправленно воздействует на белок в микроорганизме, обитающем в организме хозяина, в том числе любой из белков, приведенных в таблице 7. В других случаях модулирующее средство (например, нуклеиновая кислота, например, RNAi) изменяет экспрессию генов (например, обеспечивает повышение или снижение экспрессии генов) в микроорганизме, обитающем в организме хозяина, в том числе генов, которые кодируют любой из белков, приведенных в таблице 7, по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
Таблица 7. Белки бактерий-мишеней
(b) Гены- и белки-мишени в организме хозяев
Любое из модулирующих средств, описанных в данном документе, можно применять с целью изменения экспрессии генов или белков-мишеней в организме хозяина. В некоторых случаях модулирующее средство (например, антитело) непосредственно целенаправленно воздействует на белок у хозяина, в том числе любой из белков, приведенных в таблице 8 или таблице 9. В других случаях модулирующее средство (например, нуклеиновая кислота, например, для RNAi) целенаправленно воздействует на ген в организме хозяина, в том числе гены, которые кодируют любой из белков, приведенных в таблице 8 или таблице 9. К примеру, нуклеиновые кислоты, описанные в данном документе, можно применять для изменения (например, повышения или снижения) экспрессии генов в организме хозяина (например, генов, которые регулируют функцию или развитие бактериоцитов (например, регуляторные пептиды бактериоцитов) или генов, которые регулируют иммунную систему), в том числе без ограничения любого из генов, приведенных в таблице 8 и таблице 9, по сравнению с организмом-хозяином, которому модулирующее средство не было введено.
Таблица 8. Белки, которые регулируют функцию бактериоцитов
В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения устойчивости хозяина к паразитам или патогенам (например, грибным, бактериальным или вирусным патогенам или паразитам) по сравнению с организмом-хозяином, которому модулирующее средство не было введено. В некоторых случаях способы или композиции, предусмотренные в данном документе, могут быть эффективными для снижения устойчивости хозяина к патогену или паразиту (например, грибным, бактериальным или вирусным патогенам или паразитическим микроскопическим клещам) на приблизительно 2%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более чем на 100% по сравнению с эталонным уровнем (например, уровнем, обнаруживаемым у хозяина, который не получает модулирующее средство).
В некоторых случаях модулирующее средство является эффективным для изменения врожденной иммунной системы хозяина с целью опосредованного изменения разнообразия микроорганизмов в организме хозяина относительно организма-хозяина, которому модулирующее средство не было введено. Беспозвоночные характеризуются многочисленными иммунными реакциями, некоторых из которых являются гомологичными по отношению к иммунным механизмам, встречающимся у млекопитающих. Общие принципы врожденного иммунитета у насекомых были представлены в других обзорах (Lemaitre et al., Annu. Rev. Immunol. 25:697-743, 2007; Charroux et al., Fly 4:40-47, 2010; Ganesan et al., Curr. Top. Microbiol. Immunol. 349:25-60, 2011; Chambers et al., Curr. Opin. Immunol. 24:10-14, 2012). К примеру, у D. melanogaster существуют два основных индуцируемых ответа, обеспечивающих локальный иммунитет в слое эпителиальных клеток кишечника: продуцирование AMP и синтез активных форм кислорода (ROS). Несмотря на то, что оба из этих индуцируемых ответов можно наблюдать в виде классических механизмов устойчивости, они оба включают в себя отрицательные обратные связи и модулирующие компоненты, которые могут придавать переносимость в отношении микробиоты кишки комменсала.
Колонизация кишки бактериями-комменсалами могла бы индуцировать явления иммунного примирования, приводящие к активации или изменению иммунного ответа не только в отношении повторной колонизации бактерий-комменсалов, но и против патогенов. Микробиота кишки является необходимой не только для примирования иммунной системы хозяина по отношению к бактериям, например, к Plasmodium, но также и для вызывания примирующего ответа при повторном заражении хозяев бактериями. Зависимый от бактерий примирующий ответ у комаров характеризуется дифференцировкой прогемоцитов в гранулоциты и присутствием повышенных количеств циркулирующих гранулоцитов с измененными морфологией и связывающими свойствами. В другом примере бактерии-симбионты мухи цеце, в том числе населяющая кишку гамма-протеобактерия S. glossinidius, являются необходимыми во время личиночного развития с целью того, чтобы взрослые мухи могли представлять устойчивый к трипаносомам фенотип. В этом случае бактерии, по-видимому, влияют на формирование и целостность перитрофической матрицы взрослой особи, тем самым опосредованно регулируя способность мухи выявлять присутствие трипаносом и отвечать на это.
В некоторых случаях иммунный ответ модулируется с помощью продуцирования модулирующего средства. К примеру, в случае системного иммунного ответа D. melanogaster Toll и IMD представляют собой два основных сигнальных пути, индуцирующих продуцирование противомикробного пептида (AMP). После воздействия патогенов лишь путь с участием IMD является активным и вызывает локальный ответ AMP. Активация происходит с помощью связывания различных вариантов бактериального пептидогликана (PGN) с вне- и внутриклеточными эпителиальными рецепторами, относящимися к семейству пептидогликан-распознающих белков (PGRP). Белок Pirk приводит к секвестрированию определенных PGN-связывающих рецепторов (PGRP-LC) в цитоплазме, тем самым снижая количество этих рецепторов, локализованных на клеточной поверхности и замедляющих передачу сигнала в пути с участием IMD. PGRP-LE обеспечивает иммунную переносимость по отношению к микробиоте комменсала посредством стимуляции амидаз и Pirk. Нисходящая передача сигнала посредством пути с участием IMD приводит к активации транскрипционного фактора Relish, который, в свою очередь, индуцирует экспрессию нескольких AMP и других связанных с иммунитетом генов. PGRP-LE индуцирует зависимый от Relish иммунный ответ в отношении патогенных бактерий. PGRP-LE также обеспечивает иммунную переносимость в отношении микробиоты комменсала посредством стимуляции амидаз и Pirk.
Транскрипционный фактор гомеобокса Caudal специфически репрессирует транскрипцию гена AMP в кишке путем связывания с промоторными областями. Недостаточность Caudal вызывает конститутивное продуцирование AMP и сдвиг в микробиоте кишечника. В одном варианте осуществления путь с участием IMD инактивируют с целью ограничения экспрессии одного или нескольких AMP и/или других генов, связанных с иммунитетом, например, недостаточностью caudal, тем самым активируя иммунный ответ в отношении микробиоты кишечника.
В некоторых случаях воздействие патогенов в кишке активирует образование ROS посредством системы мембраноассоциированной двойной оксидазы (DUOX). К примеру, у D. melanogaster PGN-зависимые и PGN-независимые сигнальные пути обеспечивают образование ROS, что вызывает окислительный стресс не только в бактериях, но также и в эпителиальных клетках хозяина. D. melanogaster устраняет избыточные ROS с помощью активации иммунореактивных каталаз. Данное продуцирование каталаз приводит к повышенной переносимости вследствие снижения самоповреждения, вызываемого индуцируемым бактериями иммунным ответом, возможно, в результате локального ограничения активности каталаз, например, близостью к эпителиальной поверхности. В одном варианте осуществления иммунореактивные каталазы инактивируют или репрессируют с целью поддержания продуцирования ROS с помощью системы DUOX и окислительного стресса с помощью PGN-зависимых и PGN-независимых сигнальных путей для активации иммунного ответа в отношении микробиоты кишечника.
В некоторых случаях модулирующее средство является эффективным для изменения (например, повышения или снижения) экспрессии генов у хозяина с целью повышения или снижения ответа иммунной системы хозяина или иммунорегуляторной передачи сигнала, например, ответа иммунной системы на микроорганизмы, обитающие в организме хозяина (например, микроорганизмы, обитающие в бактериоцитах хозяина), по сравнению с организмом, которому модулирующее средство не было введено. Неограничивающие примеры связанных с иммунной системой генов/белков в бактериоцитах показаны в таблице 9.
Таблица 9. Иммунорегуляторные белки, которые модулируют бактериоциты
v. Бактерии в качестве модулирующих средств
В некоторых случаях модулирующее средство, описанное в данном документе, включает в себя одну или несколько бактерий. Многочисленные бактерии являются пригодными в композициях и способах, описанных в данном документе. В некоторых случаях средство представляет собой вид бактерий, обнаруживаемый эндогенно в организме хозяина. В некоторых случаях бактериальное модулирующее средство представляет собой вид эндосимбиотических бактерий. В некоторых случаях бактериальное модулирующее средство представляет собой патоген в организме хозяина. Неограничивающие примеры бактерий, которые можно применять в качестве модулирующих средств, включают в себя все виды бактерий, описанные в данном документе в разделе II подробного описания, и виды бактерий, приведенные в таблице 1. Например, модулирующее средство может представлять собой вид бактерий из любого типа бактерий, присутствующего в кишках хозяев (например, насекомого, моллюска или нематоды), в том числе Gammaproteobacteria, Alphaproteobacteria, Betaproteobacteria, Bacteroidetes, Firmicutes (например, Lactobacillus и Bacillus spp.), Clostridia, Actinomycetes, Spirochetes, Verrucomicrobia и Actinobacteria.
В некоторых случаях бактерию можно применять в качестве модулирующего средства для стимуляции иммунного ответа у хозяина, что приводит к снижению уровня, разнообразия или метаболизма одного или нескольких микроорганизмов, обитающих в организме хозяина. В некоторых случаях бактерии доставляют в виде живых бактериальных клеток (например, клеток E. coli). В альтернативном случае бактерии могут быть доставлены в виде термоинактивированных бактериальных клеток (например, термоинактивированных клеток E. coli). В других случаях бактерии могут быть доставлены в виде лизатов (например, приготовленных из лизированных цельных клеток) или их фракций.
В некоторых случаях модулирующее средство включает в себя генетически модифицированные или трансформированные бактерии, описанные в данном документе. В некоторых случаях бактерии являются генетически модифицированными. К примеру, бактерия может быть модифицированной путем введения генетического материала в бактерию с помощью стандартных способов, известных в данной области техники (например, трансдукции, трансформации или конъюгации), с модификацией или изменением тем самым физиологии клетки и/или биохимии бактерии. В результате введения генетического материала модифицированная бактерия может приобретать новые функции или свойства. В некоторых случаях генетически модифицированные бактерии могут продуцировать и секретировать модулирующее средство, описанное в данном документе. К примеру, генетически модифицированные бактерии могут продуцировать полипептиды, малые молекулы или нуклеиновые кислоты, которые целенаправленно воздействуют на определенных эндосимбионтов хозяев или других микроорганизмов, обитающих в организме хозяина. В некоторых случаях модифицированные бактерии могут быть использованы для продуцирования продукта, который стимулирует иммунный ответ хозяина.
В некоторых случаях генетически сконструированные или трансформированные бактерии предусмотрены для придания новых функциональных свойств хозяину. Новые функциональные свойства могут включать в себя, к примеру, способность разрушать пестициды (например, инсектициды, моллюскоциды или нематициды; например, пестицид, приведенный в таблице 11), растительные аллелохимикаты или продуцировать питательные вещества. К примеру, генетически модифицированные бактерии могут быть получены из встречающихся в природе бактерий, выделенных из вредителей, устойчивых к пестицидам (например, инсектицидам, моллюскоцидам или нематицидам; например, пестициду, приведенному в таблице 11), например, штаммов Burkholderia из насекомого R. pedestris. Если бактерии из устойчивых к инсектицидам вредителей культивируют вместе с бактериями-комменсалами, выделенными из хозяина, представляющего интерес, например, пчел, то гены, придающие устойчивость к инсектицидам (например, способность применять инсектицид в качестве источника углерода), могут быть перенесены бактериям-комменсалам хозяина, представляющего интерес, например, пчел. Затем генетически модифицированные бактерии повторно вводят в организм хозяина, например, пчел, и устойчивых к инсектицидам насекомых отбирают с помощью их селекции в среде с высоким содержанием инсектицида. Любой из типов бактерий, который обычно присутствует в организме хозяина, может быть модифицирован таким образом, чтобы иметь новое функциональное свойство, в том числе без ограничения Gammaproteobacteria, Alphaproteobacteria, Betaproteobacteria, Bacteroidetes, Firmicutes including Lactobacillus and Bacillus species, Clostridia, Actinomycetes, Spirochetes, Verrucomicrobia, Actinobacteria и др.
Бактерия может быть генетически модифицирована с целью снижения приспособленности хозяина или уничтожения хозяина. В некоторых случаях бактерия может быть генетически модифицирована с целью повышения ее выживаемости в клетках хозяев, повышения ее токсичности в отношении клетки хозяина и/или обеспечения функции, которая снижает приспособленность хозяина (например, снижение устойчивости к пестициду, например, пестициду, приведенному в таблице 11). В некоторых случаях бактерию модифицируют таким образом, чтобы она больше не синтезировала необходимую молекулу, которая, как правило, предоставляется клетке хозяина. В некоторых случаях бактерию генетически модифицируют таким образом, что популяция бактерий увеличивается до уровня, который отрицательно влияет на хозяина, например, в результате избыточного использования необходимых веществ или системы функционирования или в результате конкурирования с полезными микроорганизмами, обитающими в организме хозяина. В некоторых случаях бактерия только уничтожает хозяина после последующего вмешательства человека.
Генетически модифицированные бактерии, описанные в данном документе, могут быть получены с помощью любых способов, известных в данной области техники. К примеру, способы доставки нуклеиновых кислот к бактериям включают в себя без ограничения различные химические, электрохимические и биологические подходы, к примеру, трансформацию тепловым шоком, электропорацию, трансфекцию, к примеру, опосредованную липосомами трансфекцию, опосредованную DEAE-декстраном трансфекцию или опосредованную фосфатом кальция трансфекцию. В некоторых случаях конструкцию на основе нуклеиновой кислоты, к примеру, экспрессионную конструкцию, содержащую по меньшей мере одну последовательность нуклеиновой кислоты, вводят в бактерии с помощью носителя или вектора с целью переноса генетического материала. Векторы для переноса генетического материала бактериям хорошо известны специалистам в данной области техники и включают в себя, например, плазмиды, хромосомы и вирусные векторы. Способы конструирования конструкций на основе нуклеиновых кислот, в том числе экспрессионных конструкций, содержащих конститутивные или индуцируемые гетерологичные промоторы, конструкции с нокаутом и нокдауном, а также способы и векторы для доставки нуклеиновой кислоты или конструкции на основе нуклеиновой кислоты бактериям хорошо известны специалистам в данной области техники и описаны, к примеру, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 2001; Amberg et al., Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual, Cold Spring Harbor Laboratory Press, 2005; Abelson et al., Guide to Yeast Genetics and Molecular Biology, Part A, Volume 194 (Methods in Enzymology Series, 194), Academic Press, 2004; Guthrie et al., Guide to Yeast Genetics and Molecular and Cell Biology, 1st edition, Part B, Volume 350 (Methods in Enzymology, Vol 350), Academic Press, 2002; Stephanopoulos et al., Metabolic Engineering: Principles and Methodologies, 1st edition, Academic Press, 1998; и Smolke, The Metabolic Pathway Engineering Handbook: Fundamentals, 1st edition, CRC Press, 2009, все из которых включены в данный документ посредством ссылки в полном объеме.
vi. Грибы в качестве модулирующих средств
В некоторых случаях модулирующее средство, описанное в данном документе, включает в себя один или несколько грибов. Многочисленные грибы являются пригодными в композициях и способах. К примеру, в некоторых случаях грибное модулирующее средство может представлять собой дрожжевой гриб, такой как Sacchromyces cerevisiae или Pichia pastoris.
В некоторых случаях грибы (например, S. cerevisiae или P. pastoris) можно применять в качестве модулирующих средств для стимуляции иммунного ответа в организме хозяина, что приводит к снижению уровней, разнообразия или метаболизма одного или нескольких микроорганизмов (например, бактерий или грибов), обитающих в организме хозяина. В некоторых случаях грибы доставляют в виде живых грибных клеток (например, клетки P. pastoris, см. пример 15). В альтернативном случае грибы могут быть доставлены в виде термоинактивированных грибных клеток (например, термоинактивированных клеток S. cerevisiae). В других случаях грибы могут быть доставлены в виде лизатов (например, приготовленных из лизированных цельных клеток) или их фракций.
В некоторых случаях модулирующее средство включает в себя генетически модифицированный или трансформированный гриб, описанный в данном документе. В некоторых случаях гриб является генетически модифицированным. К примеру, гриб может быть модифицированным путем введения генетического материала в гриб с помощью стандартных способов, известных в данной области техники, тем самым, с модификацией или изменением тем самым физиологии клетки и/или биохимии гриба. В результате введения генетического материала модифицированная гриб может приобретать новые функции или свойства. В некоторых случаях генетически модифицированный гриб может продуцировать и секретировать модулирующее средство, описанное в данном документе. В некоторых случаях генетически модифицированный гриб может быть использован для продуцирования продукта, который стимулирует иммунный ответ хозяина.
В некоторых случаях генетически сконструированный или трансформированный гриб предусмотрен для придания новых функциональных свойств хозяину. Новые функциональные свойства могут включать в себя, к примеру, способность разрушать пестициды (например, инсектициды), растительные аллелохимикаты или продуцировать питательные вещества. Любые типы грибов, которые обычно присутствуют в организме хозяина, могут быть генетически модифицированы таким образом, чтобы иметь новое функциональное свойство, в том числе без ограничения Candida, Metschnikowia, Debaromyces, Scheffersomyces shehatae и Scheffersomyces stipites, Starmerella, Pichia, Trichosporon, Cryptococcus, Pseudozyma дрожжеподобные симбионты из подтипаа Pezizomycotina (например, Symbiotaphrina bucneri и Symbiotaphrina kochii).
Как показано с помощью примера 15, дрожжевой гриб, такой как P. pastoris,можно применять в качестве модулирующего средства, которое стимулирует иммунный ответ хозяина (например, у насекомого, например, тли), что, в свою очередь, изменяет активность, уровни или метаболизм эндосимбиотических бактерий, таких как Buchnera spp., обитающих в организме хозяина и тем самым модулирует (например, снижает) приспособленность хозяина.
vii. Модификации в отношении модулирующих средств
В некоторых случаях молекула нуклеиновой кислоты, пептид, полипептид или молекула антитела могут быть модифицированы. К примеру, модификация может представлять собой химическую модификацию, например, конъюгацию с маркером, например, флуоресцентным маркером или радиоактивным маркером. В других случаях модификация может включать в себя конъюгацию или функциональную связь с фрагментом, что обеспечивает повышение в отношении стабильности, доставки, целенаправленного воздействия, доступности или периода полужизни средства, например, липида, гликана, полимера (например, PEG), катионного фрагмента.
(a) Продукты слияния
Любое из модулирующих средств, описанных в данном документе, может быть слито или связано с дополнительным фрагментом. В некоторых случаях модулирующее средство включает в себя продукт слияния одного или нескольких дополнительных фрагментов (например, 1 дополнительного фрагмента, 2, 3, 4, 5, 6, 7, 8, 9, 10 или больше дополнительных фрагментов). В некоторых случаях дополнительный фрагмент представляет собой любое из модулирующих средств, описанных в данном документе (например, пептид, полипептид, малую молекулу или антибиотик). В качестве альтернативы, дополнительный фрагмент может сам по себе не выступать в качестве модулирующего средства, однако вместо этого может выполнять вторичную функцию. Например, дополнительный фрагмент может облегчать доступ, связывание или активирование модулирующего средства в участке-мишени в организме хозяина (например, в кишке хозяина или бактериоците хозяина) или в микроорганизме-мишени, обитающем в организме хозяина.
В некоторых случаях дополнительный фрагмент может облегчать проникновение модулирующего средства в клетку хозяина-мишени или микроорганизм-мишень, обитающий в организме хозяина. Например, дополнительный фрагмент может включать в себя пептид, проникающий в клетку. Пептиды, проникающие в клетку (CPP), могут представлять собой природные последовательности, полученные из белков; химерные пептиды, которые образуются в результате слияния двух природных последовательностей; или синтетические CPP, которые представляют собой последовательности, сконструированные синтетическим путем на основе исследований взаимосвязи структуры и активности. В некоторых случаях CPP характеризуются способностью повсеместно проникать через клеточные мембраны (например, клеточные мембраны прокариот и эукариот) с ограниченной токсичностью. Кроме того, CPP могут характеризоваться способностью проникать через клеточные мембраны посредством энергозависимых и/или энергонезависимых механизмов без необходимости хирального распознавания специфическими рецепторами. Неограничивающие примеры CPP приведены в таблице 10.
Таблица 10. Примеры пептидов, проникающих в клетку (CPP)
(SEQ ID NO: 82)
(SEQ ID NO: 83)
(SEQ ID NO: 84)
(SEQ ID NO: 85)
(SEQ ID NO: 86)
(SEQ ID NO: 87)
(SEQ ID NO: 88)
(SEQ ID NO: 89)
В некоторых случаях дополнительный фрагмент представляет собой нуклеиновую кислоту с пептидом (PNA). Нуклеиновые кислоты с пептидами (PNA) включают в себя одну или несколько боковых цепей нуклеиновых кислот, связанных с амидным остовом. Одна или несколько аминокислотных единиц в PNA имеют амидсодержащий остов, например, аминоэтилглицин, аналогичный пептидному остову, с боковой цепью нуклеиновой кислоты вместо аминокислотной боковой цепи. Известно, что PNA комплементарно гибридизируются с ДНК и РНК с более высокой аффинностью, чем их олигонуклеотидные партнеры. Данный характер PNA не только делает полипептид по настоящему изобретению стабильным гибридом с боковым цепями нуклеиновых кислот, но в то же время нейтральный остов и гидрофобные боковые цепи приводят к образованию гидрофобной единицы в полипептиде. Примеры фрагментов PNA включают в себя молекулу, которая содержит пептид, такой как CPP (например, аминокислотную последовательность, характеризующуюся по меньшей мере 80% (например, 80%, 90%, 95%, 97%, 99% или больше) идентичностью по отношению к SEQ ID NO: 106) и нуклеотидную последовательность, характеризующуюся по меньшей мере 80% (например, 80%, 90%, 95%, 97%, 99% или больше) идентичностью по отношению к последовательности под одним из SEQ ID NO: 105 или 151-153.
Боковые цепи нуклеиновых кислот включают в себя без ограничения пуриновую или пиримидиновую боковую цепь, например, адениновую, цитозиновую, гуаниновую, тиминовую или урациловую. В одних случаях боковая цепь нуклеиновых кислот включает в себя нуклеозидный аналог, такой как 5-фторурацил; 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 4-метилбензимидазол, 5-(карбоксигидроксилметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, дигидроуридин, бета-D-галактозилквеуозин, инозин, N6-изопентениладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеуозин, 5'-метоксикарбоксиметилуарцил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, урацил-5-оксиуксусная кислота (v), вибутоксозин, псевдоурацил, квеуозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, сложный метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусную кислоту (v), 5-метил-2-тиоурацил, 3-(3-амино-3-N-2-карбоксипропил)урацил (acp3 U), 2,6-диаминопурин, 3-нитропиррол, инозин, тиоуридин, квеуозин, виозин, диаминопурин, изогуанин, изоцитозин, диаминопиримидин, 2,4-дифтортолуол, изохинолин, пирроло[2,3-β]пиридин и любые другие, которые могут спариваться основаниями с пуриновой или пиримидиновой боковой цепью.
В других случаях дополнительный фрагмент облегчает связывание модулирующего средства с микроорганизмом-мишенью (например, грибом или бактерий), обитающим в организме хозяина. Дополнительный фрагмент может включать в себя один или несколько нацеливающих доменов. В некоторых случаях нацеливающий домен может нацеливать модулирующее средство на один или несколько микроорганизмов (например, бактерию или гриб), обитающих в кишке хозяина. В некоторых случаях нацеливающий домен может нацеливать модулирующее средство на конкретную область в организме хозяина (например, кишку или бактериоцит хозяина) для доступа к микроорганизмам, которые обычно присутствуют в указанной области организма хозяина. Например, нацеливающий домен может нацеливать модулирующее средство на переднюю кишку, среднюю кишку или заднюю кишку хозяина. В других случаях нацеливающий домен может нацеливать модулирующее средство на бактериоцит в организме хозяина и/или одну или несколько бактерий, обитающих в бактериоците хозяина.
(b) Пре- или продомены
В некоторых случаях модулирующее средство может включать в себя аминокислотную пре- и пропоследовательность. Например, модулирующее средство может представлять собой неактивный белок или пептид, которые могут быть активированы посредством расщепления или посттрансляционной модификации пре- или пропоследовательности. В некоторых случаях модулирующее средство конструируют с инактивирующей пре- или пропоследовательностью. Например, пре- или пропоследовательность может закрывать сайт активации на модулирующем средстве, например, сайт связывания с рецептором, или может индуцировать конформационное изменение в модулирующем средстве. Таким образом, при отщеплении пре- или пропоследовательности модулирующее средство активируется.
В качестве альтернативы, модулирующее средство может включать в себя малую пре- или промолекулу, например, антибиотик. Модулирующее средство может представлять собой неактивную малую молекулу, описанную в данном документе, которая может быть активирована в среде-мишени внутри хозяина. Например, малая молекула может быть активирована при достижении определенного значения pH в кишке хозяина.
(с) Линкеры
В случаях, если модулирующее средство связано с дополнительным фрагментом, модулирующее средство может дополнительно содержать линкер. Например, линкер может представлять собой химическую связь, например, одну или несколько ковалентных связей или нековалентных связей. В некоторых случаях линкер может представлять собой пептидный линкер (например, длиной 2, 3, 4, 5, 6, 8, 10, 12, 14, 16, 20, 25, 30, 35, 40 или больше аминокислот). Линкер может включать в себя любые гибкие, жесткие или расщепляемые линкеры, описанные в данном документе.
Гибкий пептидный линкер может включать в себя любой из линкеров, широко применяемых в данной области техники, в том числе линкеры, имеющие последовательности, содержащие главным образом остатки Gly и Ser (линкер "GS"). Гибкие линкеры могут быть пригодны для соединения доменов, которые требуют определенной степени подвижности или взаимодействия, и могут содержать небольшие, неполярные (например, Gly) или полярные (например, Ser или Thr) аминокислоты.
В качестве альтернативы, пептидный линкер может представлять собой жесткий линкер. Жесткие линкеры пригодны для сохранения фиксированного расстояния между фрагментами и поддержания их независимых функций. Жесткие линкеры также могут быть пригодны, если пространственное разделение доменов является важным для сохранения стабильности или биологической активности одного или нескольких компонентов в продукте слияния. Жесткие линкеры, например, могут иметь альфа-спиральную структуру или Pro-богатую последовательность, (XP)n, при этом X обозначает любую аминокислоту, предпочтительно Ala, Lys или Glu.
В еще нескольких случаях пептидный линкер может представлять собой расщепляемый линкер. В некоторых случаях линкеры могут расщепляться в специфических условиях, таких как присутствие восстанавливающих реагентов или протеаз. Расщепляемые in vivo линкеры могут использовать обратимую природу дисульфидной связи. Один пример включает в себя чувствительную к тромбину последовательность (например, PRS) между двумя остатками Cys. Обработка тромбином CPRSC in vitro приводит к расщеплению чувствительной к тромбину последовательности, тогда как обратимая дисульфидная связь остается интактной. Такие линкеры известны и описаны, например, в Chen et al., Adv. Drug Deliv. Rev. 65(10):1357-1369, 2013. Расщепление линкеров в продуктах слияния также может осуществляться протеазами, которые экспрессируются в условиях in vivo в конкретных клетках или тканях хозяина или микроорганизмах, обитающих в организме хозяина. В некоторых случаях при достижении участка- или клетки-мишени в результате расщепления линкера может высвобождаться свободное функциональное модулирующее средство.
Продукты слияния, описанные в данном документе, в качестве альтернативы могут быть связаны со связывающей молекулой, в том числе гидрофобным линкером, таким как отрицательно заряженная сульфонатная группа; липидами, такими как поли(--CH2--)углеводородные цепи, такие как полиэтиленгликолевая (PEG) группа, их ненасыщенными вариантами, их гидроксилированными вариантами, амидированными или их в других отношениях N-содержащими вариантами, неуглеродными линкерами; углеводными линкерами; фосфодиэфирными линкерами, или другими молекулами, способными ковалентно связывать две или более молекул, например, модулирующих средств. Можно применять нековалентные линкеры, такие как гидрофобные липидные глобулы, с которыми связывается модулирующее средство, например, посредством гидрофобной области модулирующего средства или гидрофобного удлинения модулирующего средства, такого как ряд остатков, богатый лейцином, изолейцином, валином или, возможно, также аланином, фенилаланином или даже тирозином, метионином, глицином или другим гидрофобным остатком. Модулирующее средство может быть связано с помощью химического взаимодействия зарядов таким образом, что положительно заряженный фрагмент модулирующего средства связывается с отрицательно заряженным другим модулирующим средством или дополнительным фрагментом.
IV. Составы и композиции
Композиции, описанные в данном документе, могут быть составлены в чистой форме (например, композиция содержит модулирующее средство) либо совместно с одним или несколькими дополнительными средствами (такими как наполнитель, средство доставки, носитель, разбавитель, стабилизатор и т. д.) для облегчения применения или доставки композиций. Примеры подходящих наполнителей и разбавителей включают в себя без ограничения лактозу, декстрозу, сахарозу, сорбит, маннит, крахмалы, аравийскую камедь, фосфат кальция, альгинаты, трагакант, желатин, силикат кальция, микрокристаллическую целлюлозу, поливинилпирролидон, целлюлозу, воду, солевой раствор, сироп, метилцеллюлозу, метит- и пропилгидроксибензоаты, тальк, стеарат магния и минеральное масло.
В некоторых случаях композиция содержит средство доставки или носитель. В некоторых случаях средство доставки включает в себя наполнитель. Иллюстративные наполнители включают в себя без ограничения твердые или жидкие материалы-носители, растворители, стабилизаторы, наполнители для составов с замедленным высвобождением, красители и поверхностно-активные вещества (поверхностно-активные средства). В некоторых случаях средство доставки представляет собой стабилизирующее средство доставки. В некоторых случаях стабилизирующее средство доставки включает в себя стабилизирующий наполнитель. Иллюстративные стабилизирующие наполнители включают в себя без ограничения эпоксидированные растительные масла, противовспенивающие средства, например, силиконовое масло, консерванты, регуляторы вязкости, связующие средства и средства, придающие липкость. В некоторых случаях стабилизирующее средство доставки представляет собой буфер, подходящий для модулирующего средства. В некоторых случаях композиция является микроинкапсулированной в средство доставки на основе полимерных гранул. В некоторых случаях стабилизирующее средство доставки защищает модулирующее средство от УФ и/или кислых условий. В некоторых случаях средство доставки содержит pH-буфер. В некоторых случаях композиция составлена таким образом, что она имеет pH в диапазоне от приблизительно 4,5 до приблизительно 9,0, включая, например, любой из диапазонов pH от приблизительно 5,0 до приблизительно 8,0, от приблизительно 6,5 до приблизительно 7,5 или от приблизительно 6,5 до приблизительно 7,0.
В зависимости от предполагаемых целей и преобладающих обстоятельств композиция может быть составлена в виде концентратов эмульсий, концентратов суспензий, непосредственно распыляемых или разбавляемых растворов, паст, наносимых в виде покрытия, разбавленных эмульсий, распыляемых порошков, растворимых порошков, диспергируемых порошков, смачиваемых порошков, пылевидных препаратов, гранул, форм, инкапсулированных в полимерные вещества, микрокапсул, пен, аэрозолей, препаратов на основе диоксида углерода, таблеток, препаратов на основе смол, препаратов на основе бумаги, препаратов на основе нетканого полотна или препаратов на основе трикотажного или тканого полотна. В некоторых случаях композиция представляет собой жидкость. В некоторых случаях композиция представляет собой твердое вещество. В некоторых случаях композиция представляет собой аэрозоль, как, например, в находящемся под давлением аэрозольном баллончике. В некоторых случаях композиция присутствует в отходах (таких как экскременты) вредителя. В некоторых случаях композиция присутствует внутри или на поверхности тела живого вредителя.
В некоторых случаях средство доставки представляет собой пищу или воду для хозяина. В других случаях средство доставки представляет собой источник пищи для хозяина. В некоторых случаях средство доставки представляет собой пищевую приманку для хозяина. В некоторых случаях композиция представляет собой съедобное средство, поглощаемое хозяином. В некоторых случаях композиция доставляется хозяином второму хозяину и поглощается вторым хозяином. В некоторых случаях композиция поглощается хозяином или вторым хозяином, и композиция высвобождается в окружение хозяина или второго хозяина с отходами (такими как экскременты) хозяина или второго хозяина. В некоторых случаях модулирующее средство включено в пищевую приманку, предназначенную для поглощения хозяином или переноса назад в его колонию.
В некоторых случаях средство доставки представляет собой бактериальный вектор. Модулирующее средство может быть включено в бактериальный вектор с помощью любых подходящих способов и реагентов для клонирования, известных в данной области техники, таких как описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. "Бактериальный вектор", описанный в данном документе, относится к любому генетическому элементу, такому как плазмиды, векторы на основе бактериофагов, транспозоны, космиды и хромосомы, который способен к репликации внутри бактериальных клеток и который способен к переносу генов между клетками. Иллюстративные бактериальные векторы включают без ограничения векторную систему на основе фага лямбда gtl 1, gt WES.tB, Charon 4 и плазмидные векторы, такие как pBR322, pBR325, pACYC177, pACYC184, pUC8, pUC9, pUC18, pUC19, pLG339, pR290, pKC37, pKCIOI, SV 40, pBluescript II SK +/- или KS +/- (см. каталог "Stratagene Cloning Systems", Stratagene, La Jolla, California, 1993), серии pQE, pIH821, pGEX, pET (см. Studier et al., "Use of T7 RNA Polymerase to Direct Expression of Cloned Genes", Gene Expression Technology Vol. 185, 1990), а также любые их производные.
Каждый бактериальный вектор может кодировать одно или несколько модулирующих средств. В некоторых случаях бактериальный вектор содержит молекулу нуклеиновой кислоты, кодирующую полипептид, который должен экспрессироваться в симбиотической бактерии-мишени или бактерии-хозяине. В некоторых случаях бактериальный вектор содержит молекулу нуклеиновой кислоты, кодирующую бактериоцин, который должен экспрессироваться в бактерии-мишени. В некоторых случаях бактериальный вектор дополнительно содержит один или несколько регуляторных элементов, таких как промоторы, сигналы терминации и транскрипционные и трансляционные элементы. В некоторых случаях регуляторная последовательность функционально связана с нуклеиновой кислотой, кодирующей ген (например, любой из нуклеиновых кислот, описанных в данном документе), подлежащий экспрессии в симбиотической бактерии-мишени.
В некоторых случаях бактериальный вектор вводят в бактерию, которая должна быть поглощена хозяином или членом колонии хозяина. В некоторых случаях бактерия представляет собой симбиотическую бактерию-мишень. В некоторых случаях бактерия представляет собой встречающуюся в природе бактерию из кишки хозяина или ее генетически модифицированное производное, которое может быть легко введено хозяину посредством заглатывания. Иллюстративные бактерии для применения при переносе бактериального вектора включают в себя без ограничения Proteobacter, в том числе род Pseudomonas; Actinobacter, в том числе Priopionibacterium и Corynebacterium; Firmicutes, в том числе любой вид из родов Mycoplasma, Bacillus, Streptococcus, Staphylococcus; Fibrobacteres; Spirochaetes, в том числе Treponema и Borrelia; Bacteroides, в том числе роды Bacteroides и Flavobacterium. Также подходящими являются любые бактерии из Enterobacteriaceae, в том числе рода Serratia, в том числе без ограничения S. marcescens, S. entomophila, S. proteamaculans; любые виды Enterobacter, в том числе без ограничения E. cloacae, E. aerogenes, E. dissolvens, E. agglomerans, E. hafiiiae; и любые виды, принадлежащие к следующим родам: Citrobacter, Escherichia, Klebsiella, Kluyvera, Panotea, Proteus, Salmonella, Xenorhabdus и Yokenella.
В некоторых случаях модулирующее средство может составлять от приблизительно 0,1% до приблизительно 100% композиции, как, например, любое из количества от приблизительно 0,01% до приблизительно 100%, от приблизительно 1% до приблизительно 99,9%, от приблизительно 0,1% до приблизительно 10%, от приблизительно 1% до приблизительно 25%, от приблизительно 10% до приблизительно 50%, от приблизительно 50% до приблизительно 99% или от приблизительно 0,1% до приблизительно 90% активных ингредиентов (например, полипептида, нуклеиновой кислоты, малой молекулы или их комбинаций). В некоторых случаях композиция содержит по меньшей мере любое количество из 0,1%, 0,5%, 1%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или больше активных ингредиентов (например, полипептида, нуклеиновой кислоты, малой молекулы или их комбинаций). В некоторых случаях концентрированные средства являются предпочтительными в качестве коммерческих продуктов, а конечный потребитель, как правило, использует разведенные средства, которые имеют значительно более низкую концентрацию активного ингредиента.
Любой из составов, описанных в данном документе, можно применять в форме приманки, спирали, электрического коврика, дымящегося препарата, фумиганта или листа.
i. Жидкие составы
Композиции, предусмотренные в данном документе, могут находиться в форме жидкого состава. Жидкие составы, как правило, смешивают с водой, однако в некоторых случаях их можно применять с растительным маслом, дизельным топливом, керосином или другим легким маслом в качестве носителя. Количество активного ингредиента часто находится в диапазоне от приблизительно 0,5 до приблизительно 80 процентов по весу.
Состав в виде концентрата эмульсии может содержать жидкий активный ингредиент, один или несколько растворителей на нефтяной основе и средство, которое способствует смешиванию состава с водой с образованием эмульсии. Такие концентраты можно применять в составах для сельского хозяйства, декоративных и газонных растений, лесного хозяйства, обработки зданий, обработки пищевых продуктов, сельскохозяйственных животных и вредителей, значимых для общественного здравоохранения. Их можно адаптировать для эксплуатационного оборудования от небольших переносных опрыскивателей до гидравлических опрыскивателей, малообъемных наземных опрыскивателей, вентиляторных туманообразователей и малообъемных авиационных опрыскивателей. Некоторые активные ингредиенты легко растворяются в жидком носителе. При смешивании с носителем они образуют раствор, в котором не происходит осаждение и разделение, например, гомогенный раствор. Составы этого типа могут содержать активный ингредиент, носитель и один или несколько других ингредиентов. Растворы можно применять в любом типе распылителя, в закрытом помещении и на свежем воздухе.
В некоторых случаях композиция может быть составлена в виде обратной эмульсии. Обратная эмульсия представляет собой водорастворимый активный ингредиент, диспергированный в масляном носителе. Обратные эмульсии требуют эмульгатора, который способствуют смешиванию активного ингредиента с большим объемом носителя на нефтяной основе, обычно топливного масла. Обратные эмульсии способствуют снижению сноса. В случае других составов некоторый снос при распылении происходит, когда капли воды начинают испаряться до достижения целевых поверхностей; в результате этого капли могут стать очень маленькими и легковесными. Поскольку масло испаряется более медленно, чем вода, капли обратной эмульсии сжимаются меньше, и большее количество активного ингредиента достигает цели. Масло дополнительно способствует снижению поверхностного стока и повышает устойчивость к дождю. Оно дополнительно выступает в качестве адгезивного средства путем улучшения степени покрытия поверхности и всасывания. Поскольку капли являются сравнительно большими и тяжелыми, то им сложно пройти насквозь покрытие нижней стороны листьев. Обратные эмульсии наиболее широко применяются в полосах отчуждения, где снос на восприимчивые нецелевые площади может быть проблемой.
Текучий или жидкий состав объединяет многие из характеристик концентратов эмульсий и смачиваемых порошков. Изготовители используют эти составы, если активный ингредиент представляет собой твердое вещество, которое не растворяется ни в воде, ни в масле. Активный ингредиент, пропитанный веществом, таким как глина, измельчают до очень мелкого порошка. Затем порошок суспендируют в небольшом количестве жидкости. Образующийся в результате жидкий продукт является достаточно плотным. Текучие и жидкие составы имеют многие из характеристик концентратов эмульсий, а также они имеют аналогичные недостатки. Они требуют умеренного взбалтывания для поддержания их в виде суспензии и оставляют видимые остатки, аналогичные таковым у смачиваемых порошков.
Текучие/жидкие составы являются легкими в обращении и применении. Поскольку они представляют собой жидкости, то они подлежат разбрызгиванию и расплескиванию. Они содержат твердые частицы, поэтому они способствуют абразивному износу насадок и насосов. Текучие и жидкие суспензии осаждаются в своих контейнерах. Поскольку текучие и жидкие составы склонны к осаждению, упаковка в контейнеры по пять галлонов или меньше облегчает повторное смешивание.
Аэрозольные составы содержат один или несколько активных ингредиентов и растворитель. Большинство аэрозолей имеют низкое процентное содержание активных ингредиентов. Существует два типа аэрозольных составов - готовый к применению тип, широко доступный в находящихся под давлением запечатанных контейнерах, и продукты, используемые в электрических или работающих на бензине аэрозольных генераторах, которые высвобождают состав в виде дыма или тумана.
Готовые к применению аэрозольные составы обычно представляют собой небольшие автономные блоки, которые высвобождают состав, когда клапан насадки приводится в движение. Состав пропускается через тонкое отверстие с помощью инертного газа под давлением с образованием мелкодисперсных капель. Эти продукты используются в теплицах, на небольших площадях внутри зданий или в ограниченных площадях на открытом воздухе. Коммерческие модели, которые удерживают до 5 фунтов активного ингредиента, обычно являются многоразовыми.
Дымообразующие или туманообразующие аэрозольные составы не находятся под давлением. Они используются в машинах, которые разбивают жидкий состав на мелкодисперсный туман или густой туман (аэрозоль) с помощью быстро вращающегося диска или нагреваемой поверхности.
ii. Сухие или твердые составы
Сухие составы могут быть разделены на два типа: готовые к применению и концентраты, которые должны быть смешаны с водой для применения в качестве спрея. Большинство пылевидных составов являются готовыми к применению и имеют низкое процентное содержание активных ингредиентов (меньше чем приблизительно 10 процентов по весу) наряду с очень тонкодисперсным сухим инертным носителем, изготовленным из талька, мела, глины, ореховой скорлупы или вулканического пепла. Размер отдельных пылевидных частиц варьируется. Некоторые пылевидные составы представляют собой концентраты и имеют высокое процентное содержание активных ингредиентов. Перед применением их смешивают с сухими инертными носителями. Порошки всегда используются сухими и могут легко сноситься на нецелевые участки.
iii. Гранулированные или пеллетированные составы
В некоторых случаях композиция составлена в виде гранул. Гранулированные составы аналогичны пылевидным составам, за исключением того, что гранулированные частицы являются более крупными и тяжелыми. Крупнодисперсные частицы могут быть получены из таких материалов, как глина, кукурузные початки или скорлупа грецких орехов. Активный ингредиент покрывает наружную поверхность гранул или всасывается в них. Количество активного ингредиента может быть относительно малым, обычно находясь в диапазоне от приблизительно 0,5 до приблизительно 15 процентов по весу. Гранулированные составы наиболее часто используются для применения в отношении почвы, насекомых, моллюсков или нематод, живущих в почве, или всасывания растениями через корни. Гранулированные составы иногда применяют с использованием самолета или вертолета с целью сведения к минимуму сноса или проникновения в густые заросли. После применения гранулы могут медленно высвобождать активный ингредиент. Некоторые гранулы требуют почвенной влаги для высвобождения активного ингредиента. Гранулированные составы также используются для контроля личинок комаров и других водных вредителей. Гранулы используются в операциях контроля вредителей в сельском хозяйстве, при обработке зданий, на декоративных и газонных растениях, в водной среде, в полосах отчуждения, а также в общественном здравоохранении (в отношении жалящих насекомых).
В некоторых случаях композиция составлена в виде пеллет. Большинство пеллетированных составов являются весьма сходными с гранулированными составами; эти термины используются взаимозаменяемо. Однако в пеллетированном составе все частицы имеют одну и ту же массу и форму. Однородность частиц облегчает применение с использованием точного эксплуатационного оборудования.
iv. Порошки
В некоторых случаях композиция составлена в виде порошка. В некоторых случаях композиция составлена в виде смачиваемого порошка. Смачиваемые порошки представляют собой сухие тонкоизмельченные составы, которые выглядят как пылевидные препараты. Обычно они должны смешиваться с водой для применения в качестве спрея. Однако, немного продуктов можно применять в качестве пылевидного препарата или в качестве смачиваемого порошка - выбор остается за потребителем. Смачиваемые порошки имеют от приблизительно 1 до приблизительно 95 процентов по весу активного ингредиента; в некоторых случаях более чем приблизительно 50 процентов. Частицы не растворяются в воде. Они быстро осаждаются, если их постоянно не взбалтывать для поддержания в суспендированном состоянии. Их можно применять для решения большинства проблем, связанных с вредителями, и в большинстве типов распылительного оборудования, в которых возможно взбалтывание. Смачиваемые порошки обладают превосходной остаточной активностью. В связи с их физическими свойствами большинство состава остается на поверхности обработанных пористых материалов, таких как бетон, штукатурка и необработанная древесина. В таких случаях только вода проникает в материал.
В некоторых случаях композиция составлена в виде растворимого порошка. Растворимые порошковые составы выглядят как смачиваемые порошки. В то же время, при смешивании с водой растворимые порошки легко растворяются и образуют истинный раствор. После их тщательного смешивания дополнительное взбалтывание не требуется. Количество активного ингредиента в растворимых порошках находится в диапазоне от приблизительно 15 до приблизительно 95 процентов по весу; в некоторых случаях более чем приблизительно 50 процентов. Растворимые порошки имеют все преимущества смачиваемых порошков и ни одного из недостатков, за исключением опасности вдыхания во время смешивания.
В некоторых случаях композиция составлена в виде гранул, диспергируемых в воде. Гранулы, диспергируемые в воде, также известные как сухие текучие составы, подобны смачиваемым порошкам за исключением того, что являются пылевидными, и их составляют в виде небольших легко определяемых гранул. Гранулы, диспергируемые в воде, должны быть смешаны с водой перед применением. Оказавшись в воде, гранулы распадаются на мелкодисперсные частицы, аналогичные смачиваемым порошкам. Состав требует непрерывного взбалтывания для поддержания его в суспендированном состоянии в воде. Процентное содержание активного ингредиента является высоким, часто не менее 90 процентов по весу. Гранулы, диспергируемые в воде, имеют многие из тех же самых преимуществ и недостатков смачиваемых порошков, за исключением того, что они являются более легко определяемыми и смешиваемыми. В связи с низким содержанием пылевидных частиц они обуславливают меньшую опасность вдыхания для потребителя во время использования.
v. Приманка
В некоторых случаях композиция включает в себя приманку. Приманка может находиться в любой подходящей форме, такой как твердая, пастообразная, пеллетированная или порошкообразная форма. Приманка также может быть перенесена хозяином обратно в популяцию указанного хозяина (например, колонию или улей). Приманка может затем выступать в качестве источника пищи для других членов колонии, тем самым предоставляя эффективное модулирующее средство большому числу хозяев и потенциально всей колонии хозяина.
Приманки могут быть предоставлены в подходящем "домике" или "ловушке". Такие домики и ловушки являются коммерчески доступными, и существующие ловушки могут быть адаптированы для включения композиций, описанных в данном документе. Домик или ловушка могут быть предусмотрены, например, в форме коробки, и могут быть предусмотрены в готовом виде или могут быть изготовлены, например, из складывающегося картона. Подходящие материалы для домика или ловушки включают в себя пластмассу и картон, в частности, гофрированный картон. Внутренние поверхности ловушек могут быть выстланы липким веществом с целью ограничения движения хозяина при попадании в ловушку. Домик или ловушка могут содержать подходящую бороздку внутри, которая может удерживать приманку на месте. Ловушка отличается от домика, поскольку хозяин не может легко покинуть ловушку после попадания, тогда как домик выступает в качестве "кормушки", которая обеспечивает хозяина предпочтительной средой, в которой они могут кормиться и чувствовать себя в безопасности от хищников.
vi. Аттрактанты
В некоторых случаях композиция содержит аттрактант (например, хемоаттрактант). Аттрактант может привлекать взрослого хозяина или неполовозрелого хозяина (например, личинку) к месту поблизости композиции. Аттрактанты включают в себя феромоны, химические вещества, которые секретируются животным, особенно хозяином (например, насекомым, моллюском или нематодой), которые влияют на поведение или развитие других особей этого же вида. Другие аттрактанты включают в себя сахарные сиропы и сиропы на основе гидролизатов белков, дрожжевые грибы и гниющее мясо. Аттрактанты также можно объединять с активным ингредиентом и распылять на листья или другие предметы в обрабатываемой области.
Известны различные аттрактанты, которые влияют на поведение хозяина, например, поиск хозяином пищи, мест для откладывание яиц или спаривания или партнеров для спаривания. Аттрактанты, пригодные в способах и композициях, описанных в данном документе, включают в себя, например, эвгенол, фенэтилпропионат, этилдиметилизобутилциклопропанкарбоксилат, пропилбензодиоксанкарбоксилат, цис-7,8-эпокси-2-метилоктадекан, транс-8,транс-0-додекадиенол, цис-9-тетрадеценаль (и цис-11-гексадеценаль), транс-11-тетрадеценаль, цис-11-гексадеценаль, (Z)-11,12-гексадекадиеналь, цис-7-додеценилацетат, цис-8-додеценилацетат, цис-9-додеценилацетат, цис-9-тетрадеценилацетат, цис-11-тетрадеценилацетат, транс-11-тетрадеценилацетат (и цис-11), цис-9,транс-11-тетрадекадиенилацетат (и цис-9,транс-12), цис-9,транс-12-тетрадекадиенилацетат, цис-7,цис-11-гексадекадиенилацетат (и цис-7,транс-11), цис-3,цис-13-октадекадиенилацетат, транс-3,цис-13-октадекадиенилацетат, анетол и изоамилсалицилат.
Для привлечения хозяина (например, насекомого, моллюска или нематоды) также можно применять средства, отличные от хемоаттрактантов, в том числе осветительные устройства с различными длинами волн или цветами излучения.
vii. Нанокапсулы/микроинкапсулирование/липосомы
В некоторых случаях композиция предусмотрена в виде микроинкапсулированного состава. Микроинкапсулированные составы смешивают с водой и распыляют таким же образом, как и другие распыляемые составы. После распыления пластмассовые покрытия разрушаются и медленно высвобождают активный ингредиент.
viii. Носители
Любая из композиций, описанных в данном документе, может быть составлена с включением модулирующего средства, описанного в данном документе, и инертного носителя. Такой носитель может представлять собой твердый носитель, жидкий носитель, гелеобразный носитель и/или газообразный носитель. В определенных вариантах осуществления носитель может представлять собой покрытие для семян. Покрытие для семян представляет собой любой не встречающийся в природе состав, который прилипает, целиком или частично, к поверхности семени. Состав может дополнительно содержать вспомогательное вещество или поверхностно-активное вещество. Состав может также содержать одно или несколько модулирующих средств для расширения спектра действия.
Твердый носитель, используемый для состава, включает в себя мелкодисперсный порошок или гранулы глины (например, каолиновой глины, диатомовой земли, бентонитовой глины, глины Фубасами, кислой глины и т. д.), синтетический гидратированный диоксид кремния, тальк, керамические материалы, другие неорганические минералы (например, серицит, кварц, серу, активированный уголь, карбонат кальция, гидратированный диоксид кремния и т. д.), вещество, которое может быть сублимировано и находится в твердой форме при комнатной температуре (например, 2,4,6-триизопропил-1,3,5-триоксан, нафталин, п-дихлорбензол, камфору, адамантан и т. д.); шерсть; шелк; хлопок; пеньку; жом; синтетические смолы (например, полиэтиленовые смолы, такие как полиэтилен низкой плотности, немодифицированный полиэтилен низкой плотности и полиэтилен высокой плотности; сополимеры этилена и винилового сложного эфира, такие как сополимеры этилена и винилацетата; сополимеры этилена и сложного эфира метакриловой кислоты, такие как сополимеры этилена и метилметакрилата и сополимеры этилена и этилметакрилата; сополимеры сложного эфира этилена и акриловой кислоты, такие как сополимеры этилена и метилакрилата и сополимеры этилена и этилакрилата; сополимеры этилена и винилкарбоновой кислоты, такие как сополимеры этилена и акриловой кислоты; сополимеры этилена и тетрациклододецена; полипропиленовые смолы, такие как гомополимеры пропилена и сополимеры пропилена и этилена; поли-4-метилпентен-1, полибутен-1, полибутадиен, полистирол; смолы на основе сополимера акрилонитрила и стирола; стирольные эластомеры, такие как смолы на основе сополимера акрилонитрила, бутадиена и стирола, блок-сополимеры стирола и сопряженных диенов и гидриды юлок-сополимеров стирола и сопряженных диенов; фторкаучуки; акриловые смолы, такие как поли(метилметакрилат); полиамидные смолы, такие как нейлон 6 и нейлон 66; полиэфирные смолы, такие как полиэтилентерефталат, полиэтиленнафталат, полибутилентерефталат и полициклогексилендиметилентерефталат, поликарбонаты, полиацетали, полиакрилсульфоны, полиарилаты, полиэфиры гидроксибензойной кислоты, полиэфиримиды, полиэфиркарбонаты, полифениленэфирные смолы, поливинилхлорид, поливинилиденхлорид, полиуретан и пористые смолы, такие как пенополиуретан, пенополипропилен или пеноэтилен и т. д.), стекла, металлы, керамические материалы, волокнистые материалы, ткани, трикотажные полотна, листы, бумагу, пряжу, пену, пористые вещества и мультифиламенты.
Жидкий носитель может включать в себя, например, ароматические или алифатические углеводороды (например, ксилол, толуол, алкилнафталин, фенилксилилэтан, керосин, газойль, гексан, циклогексан и т. д.), галогензамещенные углеводороды (например, хлорбензол, дихлорметан, дихлорэтан, трихлорэтан и т. д.), спирты (например, метанол, этанол, изопропиловый спирт, бутанол, гексанол, бензиловый спирт, этиленгликоль и т. д.), эфиры (например, диэтиловый эфир, диметиловый эфир этиленгликоля, монометиловый эфир диэтиленгликоля, моноэтиловый эфир диэтиленгликоля, монометиловый эфир пропиленгликоля, тетрагидрофуран, диоксан и т. д.), сложные эфиры (например, этилацетат, бутилацетат и т. д.), кетоны (например, ацетон, метилэтилкетон, метилизобутилкетон, циклогексанон и т. д.), нитрилы (например, ацетонитрил, изобутиронитрил и т. д.), сульфоксиды (например, диметлисульфоксид и т. д.), амиды (например, N,N-диметилформамид, N,N-диметилацетамид, циклические имиды (например, N-метилпирролидон), алкилиденкарбонаты (например, пропиленкарбонат и т. д.), растительное масло (например, соевое масло, хлопковое масло и т. д.), растительные эфирные масла (например, апельсиновое масло, масло иссопа, лимонное масло и т. д.) или воду.
Газообразный носитель может включать в себя, например, газообразный бутан, газообразный политетрафторэтилен, сжиженный нефтяной газ (LPG), диметиловый эфир и газообразный диоксид углерода.
ix. Вспомогательные вещества
В некоторых случаях композиция, предусмотренная в данном документе, может содержать вспомогательное вещество. Вспомогательные вещества представляют собой химические вещества, которые не обладают активностью. Вспомогательные вещества предварительно смешивают в составе или добавляют к распылительному резервуару для улучшения смешивания или применения или для усиления функциональных характеристик. Они широко используются в продуктах, предназначенных для применения в отношении листьев. Вспомогательные вещества можно использовать для придания индивидуальных свойств составу в соответствии с конкретными нуждами и введения поправки на местные условия. Вспомогательные вещества могут быть предназначены для осуществления конкретных функций, в том числе смачивания, распределения, прилипания, снижения испарения, снижения улетучивания, буферизации, эмульгирования, диспергирования, снижения сноса при распылении и снижения пенообразования. Ни одно вспомогательное вещество не может осуществлять все эти функции само по себе, однако совместимые вспомогательные вещества часто можно объединять для осуществления нескольких функций одновременно.
Среди неограничивающих примеров вспомогательных веществ, включенных в состав, присутствуют связующие вещества, диспергирующие вещества и стабилизаторы, в частности, например, казеин, желатин, полисахариды (например, крахмал, аравийская камедь, производные целлюлозы, альгиновая кислота и т. д.), производные лигнина, бентонит, сахара, синтетические водорастворимые полимеры (например, поливиниловый спирт, поливинилпирролидон, полиакриловая кислота и т. д.), PAP (кислый изопропилфосфат), BHT (2,6-ди-трет-бутил-4-метилфенол), BHA (смесь 2-трет-бутил-4-метоксифенола и 3-трет-бутил-4-метоксифенола), растительные масла, минеральные масла, жирные кислоты и сложные эфиры жирных кислот.
x. Поверхностно-активные вещества
В некоторых случаях композиция, предусмотренная в данном документе, содержит поверхностно-активное вещество. Поверхностно-активные вещества, также называемые смачивающими средствами и распределителями, физически изменяют поверхностное натяжение капли спрея. Для того, чтобы состав надлежащим образом выполнял свою функцию, капля спрея должна быть способной смачивать листья и распределяться равномерно по листу. Поверхностно-активные вещества увеличивают площадь, покрываемую составом, тем самым усиливая воздействие химического вещества на вредителя. Поверхностно-активные вещества особенно важны при применении состава в отношении листьев, покрытых воском, или волосистых листьев. Без надлежащего смачивания и распределения капли спрея часто стекают или не могут покрыть поверхности листьев надлежащим образом. В то же время слишком большое количество поверхностно-активного вещества может вызвать избыточный поверхностный сток и снизить эффективность.
Поверхностно-активные вещества классифицируются на основе способа их ионизации или расщепления на электрически заряженные атомы или молекулы, называемые ионами. Поверхностно-активное вещество с отрицательным зарядом является анионным. Поверхностно-активное вещество с положительным зарядом является катионным, и поверхностно-активное вещество без электрического заряда является неионогенным. Активность состава в присутствии неионогенного поверхностно-активного вещества может значительно отличаться от активности в присутствии катионного или анионного поверхностно-активного вещества. Выбор неправильного поверхностно-активного вещества может снижать эффективность пестицидного продукта и наносить вред растению-мишени. Анионные поверхностно-активные вещества являются наиболее эффективными при использовании с контактными пестицидами (пестициды, которые контролируют вредителя путем непосредственного контакта, а не системного всасывания). Катионные поверхностно-активные вещества никогда не должны применяться в качестве самостоятельных поверхностно-активных веществ, поскольку они обычно являются фитотоксичными.
Неионогенные поверхностно-активные вещества, часто применяемые с системными пестицидами, облегчают проникновение пестицидных спреев через кутикулы растений. Неионогенные поверхностно-активные вещества совместимы с большинством пестицидов, и для большинства пестицидов, зарегистрированных EPA, которым необходимо поверхностно-активное вещество, рекомендуется неионогенный тип. Вспомогательные вещества включают в себя без ограничения клеящие вещества, сухие разбавители, вещества, обеспечивающие проникновение в растения, факторы совместимости, буферы или модификаторы pH, дополнительные компоненты для контроля сноса, противовспенивающие средства и загустители.
Среди неограничивающих примеров поверхностно-активных веществ, включенных в композиции, описанные в данном документе, присутствуют соли алкилсульфатных сложных эфиров, алкилсульфонаты, аклиларилсульфонаты, алкилариловые эфиры и их полиоксиэтиленированные продукты, простые эфиры полиэтиленгликоля, сложные эфиры многоатомных спиртов и производные сахарных спиртов.
xi. Комбинации
В составах и в применяемых формах, полученных из этих составов, модулирующее средство может находиться в смеси с другими активными соединениями, такими как пестицидные средства (например, инсектициды, стерилизаторы, акарициды, нематоциды, моллюскоциды или фунгициды; например, пестицид, приведенный в таблице 11), аттрактанты, вещества, регулирующие рост, или гербициды. Используемый в данном документе термин "пестицидное средство" относится к любому веществу или смеси веществ, предусмотренных для предупреждения воздействия, нарушения воздействия, отпугивания или ослабления воздействия любого вредителя. Пестицид может представлять собой химическое вещество или биологическое средство, применяемое против вредителей, в том числе насекомых, моллюсков, патогенов, сорняков, нематод и микробов, которые конкурируют с человеком за пищу, разрушают имущество, распространяют заболевания или представляют собой негативный раздражитель. Термин "пестицидное средство" может дополнительно охватывать другие биологически активные молекулы, такие как антибиотики, противовирусные пестициды, противогрибковые средства, антигельминтные средства, питательные вещества, пыльцу, сахарозу и/или средства, которые останавливают или замедляют движение насекомых.
В случаях, когда модулирующее средство применяют в отношении растений, смесь с другими известными соединениями, такими как гербициды, удобрения, регуляторы роста, антидоты, химические сигнальные вещества или другие средства для улучшения свойств растения, также является возможной.
V. Доставка
Хозяин, описанный в данном документе, может подвергаться воздействию любой из композиций, описанных в данном документе, любым подходящим образом, который обеспечивает доставку или введение композиции хозяину-беспозвоночному (например, насекомому, моллюску или нематоде). Модулирующее средство может быть доставлено в отдельности или в комбинации с другими активными или неактивными веществами и может применяться, например, посредством распыления, микроинъекции, через растения, посредством полива, погружения, в форме концентрированных жидкостей, гелей, растворов, суспензий, спреев, порошков, пеллет, брикетов, плиток и т. п., составленных для доставки эффективной концентрации модулирующего средства. Количества и места для применения композиций, описанных в данном документе, как правило, определяются условиями обитания хозяина, стадией жизненного цикла, на которой на микроорганизмы хозяина можно целенаправленно воздействовать с помощью модулирующего средства, участком, в котором применение должно выполняться, а также физическими и функциональными характеристиками модулирующего средства. Модулирующие средства, описанные в данном документе, можно вводить хозяину-беспозвоночному (например, насекомому, моллюску или нематоде) путем перорального заглатывания, однако также можно вводить с помощью средств, которые обеспечивают проникновение через кутикулу или проникновение в дыхательную систему хозяина (например, насекомого, моллюска или нематоды).
В некоторых случаях хозяин-беспозвоночное (например, насекомое, моллюск или нематода) может быть просто "намочен" или "опрыскан" раствором, содержащим модулирующее средство. В альтернативном случае модулирующее средство может быть связано с пищевым компонентом (например, съедобным) для хозяина-беспозвоночного (например, насекомого, моллюска или нематоды) для облегчения доставки и/или в целях повышения поглощения модулирующего средства хозяином. Способы перорального введения включают в себя, например, непосредственное смешивание модулирующего средства с пищей для хозяина, распыления модулирующего средства в среде обитания хозяина или поле, а также подходы на основе конструирования, в которых вид, который применяется в качестве пищи, конструируется с целью экспрессии модулирующего средства, после чего скармливается хозяину, подлежащему отрицательному воздействию. В некоторых случаях, например, композиция на основе модулирующего средства может быть включена в состав рациона хозяина или покрывать его сверху. К примеру, композицию на основе модулирующего средства можно распылять на поле с сельскохозяйственными культурами, на которых обитает хозяин.
В некоторых случаях композицию распыляют непосредственно на растение, например, сельскохозяйственные культуры, например, путем распыления из рюкзака, распыления с воздуха, опрыскивания/опыления растительных культур и т. д. В случаях, когда модулирующее средство доставляют в растение, растение, получающее модулирующее средство, может находиться на любой стадии роста растения. Например, составленные модулирующие средства можно применять в виде покрытия для семян или средства для обработки корней на ранних стадиях роста растения или в виде средства для полной обработки растения на более поздних стадиях цикла урожая. В некоторых случаях модулирующее средство можно применять в отношении растения в качестве местного средства, так чтобы хозяин-беспозвоночное (например, насекомое, моллюск или нематода) заглатывал или иным образом приходил в контакт с растением при взаимодействии с растением.
Кроме того, модулирующее средство можно применять (например, в отношении почвы, в которой растение растет, или в отношении воды, которую используют для полива растения) в качестве системного средства, которое поглощается и распределяется по тканям хозяина-растения или хозяина-животного таким образом, что хозяин-беспозвоночное (например, насекомое, моллюск или нематода), питающийся на нем, будет получать эффективную дозу модулирующего средства. В некоторых случаях растения или организмы, используемые в качестве пищи, могут быть генетически трансформированы для экспрессии модулирующего средства, так чтобы хозяин, питающийся на растении или организме, используемом в качестве пищи, заглатывал модулирующее средство.
Замедленное или непрерывное высвобождение также может достигаться посредством покрытия модулирующего средства или композиции с модулирующим(модулирующими) средством(средствами) растворимым или биоразрушаемым слоем покрытия, таким так желатин, при этом данное покрытие растворяется или разрушается в среде применения, что затем делает модулирующее средство доступным, или посредством диспергирования средства в растворимой или разрушаемой матрице. Такое непрерывное высвобождение и/или распределение означает, что устройства могут быть предпочтительно использованы для устойчивого поддержания эффективной концентрации одного или нескольких модулирующих средств, описанных в данном документе, в конкретной среде обитания хозяина.
В альтернативном случае модулирующее средство экспрессируется в бактериальной или грибной клетке, и бактериальная или грибная клетка захватывается или съедается видом хозяина-беспозвоночного (например, насекомого, моллюска или нематоды). Бактерии или грибы могут быть сконструированы с целью продуцирования любого из модулирующих средство, описанных в данном документе. В других случаях также можно применять вирус, такой как бакуловирус, который специфически инфицирует хозяев-беспозвоночных (например, насекомое, моллюска или нематоду). Это обеспечивает безопасность для млекопитающих, особенно людей и животных, поскольку указанный вирус не будет инфицировать млекопитающих.
Модулирующее средство также может быть включено в среду, в которой хозяин-беспозвоночное (например, насекомое, моллюск или нематода) растет, живет, размножается, питается или осуществляет заражение. Например, модулирующее средство может быть включено в контейнер для пищи, кормушку, защитную обертку или улей. В некоторых путях применения модулирующее средство может быть связано с твердой подложкой для применения в порошкообразной форме или в "ловушке" или "кормушке". В качестве примера, в областях применения, в которых композиция подлежит применению в ловушке или в виде приманки для определенного хозяина-беспозвоночного (например, насекомого, моллюска или нематоды), композиции также могут быть связаны с твердой подложкой или инкапсулированы в материал с медленным высвобождением. Например, композиции, описанные в данном документе, можно вводить посредством доставки композиции в по меньшей мере одну среду обитания, где сельскохозяйственный вредитель (например, тля) растет, живет, размножается или питается.
i. Сконструированные растения
Термины "генетически сконструированное растение" или "трансгенное растение" относятся к растительной клетке или растению, которые экспрессируют модулирующее средство. Трансгенные растения также включают в себя потомство (преемника, потомка и др.) из любого поколения такого трансгенного растения или семя из любого поколения всех таких трансгенных растений, при этом указанное потомство или семя содержит модулирующее средство.
Специалист в данной области техники признает, что в данной области техники существует большое разнообразие методик трансформации, и непрерывно становятся доступными новые методики. Можно применять любую методику, которая является подходящей для растения-хозяина, являющегося мишенью. К примеру, конструкции можно вводить в ряде форм, в том числе без ограничения в виде нити ДНК, в плазмиде или в искусственной хромосоме. Введение конструкций в клетки растений-мишеней можно осуществлять с помощью ряда методик, в том числе без ограничения опосредованной Agrobacterium трансформации, электропорации, микроинъекции, бомбардировки микрочастицами, копреципитации ДНК с фосфатом кальция или опосредованной липосомами трансформации гетерологичной нуклеиновой кислоты. Трансформация растения предпочтительно является постоянной, т.е. с помощью интеграции экспрессионных конструкций в геном хозяина-растения таким образом, что введенные конструкции передаются в последующие поколения растений.
Любой вид растения может быть трансформирован с получением трансгенного растения. Трансгенное растение может быть двудольным растением или однодольным растением. Например, и без ограничения трансгенные растения композиций и способов, описанных в данном документе, могут быть получены из любого из следующих семейств двудольных растений: Leguminosae, в том числе растений, таких как горох, люцерна и соя; Umbelliferae, в том числе растений, таких как морковь и сельдерей; Solanaceae, в том числе растений, таких как томат, картофель, баклажан, табак и перец; Cruciferae, в частности, рода Brassica, который включает в себя растения, такие как масличный рапс, свеклу, кочанную капусту, цветную капусту и брокколи); и Arabidopsis thaliana; Compositae, которое включает в себя растения, такие как латук; Malvaceae, которое включает в себя хлопок; Fabaceae, которое включает в себя растения, такие как арахис, и т. п. Трансгенные растения по настоящему изобретению могут быть получены из однодольных растений, таких как, к примеру, пшеница, ячмень, сорго, просо, рожь, тритикале, маис, рис, овес, просо прутьевидное, мискантус и сахарный тростник. Также в трансгенные растения по настоящему изобретению включены деревья, такие как яблоня, груша, айва, слива, черешня, персик, нектарин, абрикос, папайя, манго и другие древесные виды, в том числе хвойные и лиственные деревья, такие как тополь, сосна, секвойя, кедр, дуб, ива и т. п.
Можно применять любой промотор, способный направлять экспрессию в растении, представляющем интерес. Промотор может быть нативным, или аналогичным, или чужеродным, или гетерологичным по отношению к хозяину-растению. Выбор промоторов, подлежащих включению, зависит от нескольких факторов, в том числе без ограничения эффективности, избирательности, индуцируемости, необходимого уровня экспрессии или предпочтительной экспрессии в клетках или тканях. Это является стандартной задачей для специалиста в данной области техники с целью модулирования экспрессии последовательности с помощью подходящего отбора и расположения промоторов и других регуляторных областей относительно этой последовательности.
Особенно представляющимим интерес являются промоторы в фотосинтетической ткани с целью направления транскрипции в зеленых тканях, таких как листья и стебли. Наиболее подходящими являются промоторы, которые направляют экспрессию только или преимущественно в таких тканях. Промотор может обеспечивать экспрессию конститутивно во всем растении, или дифференциально по отношению к тканям зеленого цвета, или дифференциально по отношению к стадии развития зеленой ткани, в которой происходит экспрессия, или в ответ на внешний раздражитель.
Примеры таких промоторов включают в себя промоторы рибулозо-1,5-бисфосфаткарбоксилазы (RbcS), такие как промотор RbcS из лиственницы американской (Larix laricina), промотор сосны cab6 (Yamamoto et al., Plant Cell Physiol. 35:773-778, 1994), промотор гена Cab-1 из пшеницы (Fejes et al., Plant Mol. Biol. 15:921-932, 1990), промотор CAB-1 из шпината (Lubberstedt et al., Plant Physiol. 104:997-1006, 1994), промотор cab1 R из риса (Luan et al., Plant Cell 4:971-981, 1992), промотор пируватортофосфодикиназы (PPDK) из кукурузы (Matsuoka et al., Proc. Natl. Acad. Sci. USA 90:9586-9590, 1993), промотор Lhcb1*2 табака (Cerdan et al. (1997) Plant Mol. Biol. 33:245-255), промотор сахарозо-H+ симпортера SUC2 Arabidopsis thaliana (Truernit et al. Planta 196:564-570, 1995) и промотор мембранного белка тилакоидов из шпината (psaD, psaF, psaE, PC, FNR, atpC, atpD, cab, rbcS). Другие промоторы, которые управляют транскрипцией в стеблях, листьях и зеленой ткани, описаны в публикации патента США № 2007/0006346. Промотор TrpA представляет собой предпочтительный для сердцевины промотор и был описан в патенте США № 6018104.
Ген маиса, кодирующий фосфоенолкарбоксилазу (PEPC), был описан Hudspeth et al. (Plant Molec. Biol. 12:579-589, 1989). С помощью стандартных методик молекулярной биологии промотор для указанного гена может быть применен для управления экспрессией любого гена специфическим по отношению к зеленым тканям образом в трансгенных растениях.
В некоторых других случаях индуцируемые промоторы могут быть предпочтительными. Индуцируемые промоторы управляют транскрипцией в ответ на внешние раздражители, такие как химические средства или раздражители внешней среды. К примеру, индуцируемые промоторы могут обеспечивать транскрипцию в ответ на гормоны, такие как гиббереллиновая кислота или этилен, в ответ на свет или засуху.
VI. Скрининг
Любое из модулирующих средств, описанных в данном документе, может быть выделено в результате скринингового анализа, при этом библиотеку модулирующих средств (например, смесь вариантов исходного модулирующего средства) подвергают скринингу в отношении модулирующих средств (например, вариантов модулирующих средств), которые являются эффективными для изменения микробиоты хозяина (например, насекомого/моллюска/нематоды) и тем самым модулирования (например, повышения или снижения) приспособленности хозяев.
К примеру, скрининговые анализы, предусмотренные в данном документе, могут быть эффективными для идентификации одного или нескольких модулирующих средств (например, полипептида, нуклеиновой кислоты, малой молекулы или их комбинаций), которые целенаправленно воздействуют на симбиотические микроорганизмы, обитающие в организме хозяина, и тем самым снижают приспособленность хозяина. Например, идентифицированное модулирующее средство (например, полипептид, нуклеиновая кислота, малая молекула или их комбинации) может быть эффективным для снижения жизнеспособности микроорганизмов, которые расщепляют пестициды или аллелохимикаты (например, бактерий, например, бактерии, которая расщепляет пестицид, приведенный в таблице 11), с повышением тем самым восприимчивости хозяина к пестициду (например, восприимчивость к пестициду, приведенному в таблице 11) или аллелохимическому средству.
В альтернативном случае скрининговый анализ можно применять для идентификации модулирующего средства, эффективного для повышения приспособленности хозяина (например, приспособленности насекомого, моллюска или нематоды). Например, скрининговый анализ может быть применен для идентификации одного или нескольких модулирующих средств, которые целенаправленно воздействуют на определенные микроорганизмы и/или определенных хозяев. Дополнительно, скрининговые анализы можно применять для идентификации модулирующих средств, которые придают одному или нескольким микроорганизмам улучшенные функциональные свойства. К примеру, скрининговый анализ может быть эффективным для выделения модулирующих средств, которые придают одному или нескольким микроорганизмам улучшенную способность метаболизировать (например, расщеплять) пестицид (например, инсектицид, например, неоникотиноид) или аллелохимические вещества в растении (например, кофеин, цистатин сои, фенитротион, монотерпены, дитерпеновые кислоты или фенольные соединения (например, таннины, флавоноиды)). Доставка и колонизация выделенного микроорганизма в организме хозяина может повысить устойчивость хозяина к пестициду или растительному аллелохимикату, тем самым повышая приспособленность хозяина. Способы также могут быть пригодны для выделения модулирующих средств, которые придают микроорганизмам улучшенную способность колонизировать любого из хозяев, описанных в данном документе.
ПРИМЕРЫ
Далее представлен пример способов по настоящему изобретению. Следует понимать, что на практике могут осуществляться различные другие варианты осуществления c учетом общего описания, приведенного выше.
Пример 1. Создание библиотеки cDNA из личинок кукурузной тли (Rhopalosiphum maidis)
В данном примере демонстрируется получение библиотеки cDNA из личинок кукурузной тли (Rhopalosiphum maidis).
Схема эксперимента
С целью получения библиотеки РНК из 0,9 г целых личинок на стадии первого возраста (4-5 дней после вылупливания; содержали при 16°C) очищали с применением приведенного ниже способа на основе фенола/TRI REAGENT® (MOLECULAR RESEARCH CENTER, Цинциннати, Огайо). Личинок гомогенизировали при комнатной температуре в гомогенизаторе на 15 мл с использованием 10 мл TRI REAGENT® до получения однородной суспензии. После 5 мин. инкубации при комнатной температуре гомогенат распределяли в 1,5 мл микропробирки для центрифугирования (1 мл на пробирку), добавляли 200 мкл хлороформа и смесь энергично встряхивали в течение 15 секунд. После того, как экстракту давали возможность осесть при комнатной температуре в течение 10 мин., фазы разделяли путем центрифугирования при 12000×g при 4°C. Верхнюю фазу (составляющую приблизительно 0,6 мл) осторожно переносили в другую стерильную 1,5 мл пробирку и добавляли равный объем изопропанола комнатной температуры. После инкубации при комнатной температуре в течение 5-10 мин. смесь центрифугировали 8 мин. при 12000×g (4°C или 25°C).
Супернатант осторожно удаляли и отбрасывали, и осадок, содержащий РНК, дважды промывали путем встряхивания на вортексе с 75% этанолом с извлечением с помощью центрифугирования в течении 5 мин. при 7500×g (4°C или 25°C) после каждой промывки. Этанол осторожно удаляли, осадок оставляли для высыхания на воздухе 3-5 мин., а затем растворяли в стерильной воде, не содержащей нуклеаз. Концентрацию РНК определяли путем измерения поглощения (A) при 260 нм и 280 нм. Экстрагированную РНК хранили при -80°C до дальнейшей обработки, а качество РНК определяли путем прогонки аликвоты в 1% агарозном геле.
Общую РНК из личинок превращали в библиотеку cDNA с применением случайных праймеров. Библиотеку cDNA личинок секвенировали в масштабе планшета ½ с использованием системы GS FLX 454 Titanium™ в EUROFINS MWG Operon, в результате чего получали более 600000 ридов со средней длиной рида, составляющей 348 п.о. 350000 ридов собирали в более чем 50000 контигов. Как не подвергнутые сборке риды, так и контиги преобразовали в базы данных BLASTable с применением программы с открытым доступом FORMATDB (доступна от NCBI).
Пример 2. Получение и очистка dsRNABcr1
В данном примере демонстрируется получение и очистка синтетической dsRNA из библиотеки cDNA.
Схема эксперимента
Ген Bcr1 (ACYPI32128) представляет собой важнейший ген в регуляции и функции бактериоцитов у насекомых. Получали cDNA Bcr1 из общей РНК личинок, описанной в примере 1, и с целью синтеза dsRNA Bcr1 с помощью ПЦР использовали следующие пары праймеров: прямой 5'-aaactgctgcatggctttct-3' (SEQ ID NO: 90) и обратный праймеры 5'-acaggcctttcaggctttta-3' (SEQ ID NO: 91). С целью нацеливания на участок гена проводили две отдельные ПЦР-амплификации. С помощью первой ПЦР-амплификации вводили промоторную последовательность T7 на 5′-конце ((TTAATACGACTCACTATAGGGAGA; SEQ ID NO: 92) амплифицированных смысловых нитей. С помощью второй реакции встраивали промоторную последовательность T7 на 5′-концах антисмысловых нитей. Два ПЦР-амплифицированных фрагмента для каждого участка целевого гена Bcr1 затем смешивали в равных количествах, и данную смесь использовали в качестве транскрипционной матрицы для получения dsRNA. Двухнитевую РНК для биологического анализа насекомых синтезировали и очищали с применением набора для RNAi AMBION® MEGASCRIPT®, следуя инструкциям изготовителя (INVITROGEN), или набора для транскрипции In Vitro с T7 HiScribe®, следуя инструкциям изготовителя (New England Biolabs, Ипсуич, Масс.). Концентрацию dsRNA Bcr1 измеряли с применением спектрофотометра NANODROP™ 8000 (THERMO SCIENTIFIC, Уилмингтон, Дел.), и очищенную dsRNA Bcr1 подготавливали в TE-буфере.
Последовательность шпилечной dsRNA Bcr1 одной нити: AUGAAACUGCUGCAUGGCUUUCUGAUUAUUAUGCUGACCAUGCAUCUGAGCAUUCAGUAUGCGUAUGGCGGCCCGUUUCUGACCAAAUAUCUGUGCGAUCGCGUGUGCCAUAAACUGUGCGGCGAUGAAUUUGUGUGCAGCUGCAUUCAGUAUAAAAGCCUGAAAGGCCUGUGGUUUCCGCAUUGCCCGACCGGCAAAGCGAGCGUGGUGCUGCAUAACUUUCUGACCAGCCCGUUUUUUUUUUCGGGCUGGUCAGAAAGUUAUGCAGCACCACGCUCGCUUUGCCGGUCGGGCAAUGCGGAAACCACAGGCCUUUCAGGCUUUUAUACUGAAUGCAGCUGCACACAAAUUCAUCGCCGCACAGUUUAUGGCACACGCGAUCGCACAGAUAUUUGGUCAGAAACGGGCCGCCAUACGCAUACUGAAUGCUCAGAUGCAUGGUCAGCAUAAUAAUCAGAAAGCCAUGCAGCAGUUUCAU (SEQ ID NO: 93).
Пример 3. Обработка тлей (Rhopalosiphum maidis) с помощью dsRNA Bcr1
В данном примере демонстрируется способность уничтожать тлей Rhopalosiphum maidis или снижать их приспособленность посредством обработки раствором dsRNA за счет целенаправленного воздействия на экспрессию гена Bcr1 (ACYPI32128), который представляет собой важнейший ген в регуляции и функции бактериоцитов у насекомых.
Тли являются одними из наиболее важных насекомых-вредителей в сельском хозяйстве. Они обуславливают прямое повреждение сельскохозяйственных культур, вызванное поеданием, и служат в качестве переносчиков вирусов, поражающих растения. Помимо этого, медвяная падь тлей способствует росту плесневых налетов грибов и привлекает вредоносных муравьев. Применение средств химической обработки, к сожалению, по-прежнему остается широко распространенным, что приводит к отбору устойчивых особей, уничтожение которых становится все более сложным.
Структура терапевтического средства: растворы dsRNA составляли с концентрацией 0 (отрицательный контроль), 0,5, 1 или 5 мкг/мл dsRNA Bcr1 из примера 2 в 10 мл TE-буфера с 0,5% сахарозы и незаменимыми аминокислотами.
Схема эксперимента
Для подготовки к обработке тлей выращивали в лабораторных условиях и среде. В комнате с контролируемым климатом (фотопериод с 16 ч. света; 60±5% RH; 20±2°C) растения конских бобов выращивали в смеси вермикулита и перлита и заражали тлями. Для ограничения материнских эффектов или отличий в состоянии здоровья между растениями 5-10 взрослых особей с разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии личинок второго и третьего возраста собирали из здоровых растений и разделяли по видам обработки таким образом, что для каждого вида обработки получали примерно одно и то же число особей с каждого из растений коллекции.
Лунки 96-луночного планшета заполняли 200 мкл искусственного рациона для тлей (Febvay et al., Canadian Journal of Zoology 66(11):2449-2453, 1988), и планшет закрывали парафильмом для получения съедобного саше. Искусственный рацион смешивали или только с раствором TE-буфера (трис-HCl (1 мМ) с буфером на основе EDTA (0,1 мМ), pH 7,2) с 0,5% сахарозы и незаменимыми аминокислотами в качестве отрицательного контроля, или смешивали с растворами dsRNA, разведенными в TE-буфере, содержащем варьирующиеся концентрации dsRNA. Растворы dsRNA смешивали с искусственным рационом с получением конечных концентраций, составляющих от 0,5 до 5 мкг/мл.
Для каждой повторности обработки 30-50 тлей второй и третьей возрастной стадии помещали отдельно в лунки 96-луночного планшета, и планшет со съедобным саше переворачивали над ними, что давало возможность насекомым питаться через парафильм и ограничивать их отдельными лунками. Экспериментальных тлей содержали при тех же самых условиях среды, что и колонии тлей. После того, как тлей вскармливали в течение 24 часов, съедобное саше заменяли новым, содержащим стерильный искусственный рацион, и новое стерильное саше предоставляли каждые 24 часа в течение 4 дней. Когда саше заменяли, тлей также проверяли в отношении смертности. Тлю считали мертвой, если она становилась коричневой или располагалась на дне лунки и не двигалась во время наблюдения. Если тля находилась на парафильме съедобного саше, но не двигалась, она считалась кормящейся и живой.
Показатели выживаемости тлей, обработанных dsRNA Bcr1, сравнивали с тлями, обработанными отрицательным контролем. Показатели выживаемости тлей, обработанных dsRNA Bcr1, снижались по сравнению с тлями, обработанными контролем.
Пример 4. Получение трансгенных травянистых растений, экспрессирующих dsRNA Bcr1
В данном примере демонстрируются генетическая модификация и продуцирование dsRNA Bcr1 в трансгенном травянистом растении с целью доставки тлям.
Трансгенную кормовую траву бутелоуа изящную - Bouteloua gracilis, которая продуцирует молекулы dsRNA Bcr1 за счет экспрессии химерного гена, стабильно интегрированного в геном растения, получали путем бомбардировки микрочастицами с применением системы на основе эмбриогенной клеточной линии 'TIANSJ98' с высоким содержанием хлорофилла (Aguado-Santacruz et al., Theoretical and Applied Genetics 104(5):763-771, 2002).
Образование клеточной линии 'TIANSJ98' и ее поддержание. Эмбриогенную клеточную линию 'TIANSJ98' с высоким содержанием хлорофилла получали путем культивирования зеленых каллюсов апикальных меристем побега в жидкой среде MPC, как описано в (Aguado-Santacruz et al., Plant Cell Rep. 20:131-136, 2001). Данную клеточную линию пересевали каждые 20 дней путем переноса 1 мл клеточной суспензии в 24 мл свежей среды MPC. Обоснования для применения условий диффузного распределения эмбриогенных каллюсов были следующими: 1) синхронизация физиологической стадии с клетками-мишенями, 2) максимальное увеличение распределения тотипотентного материала на бумажных фильтрах (оптимизация покрытия побега плазмидами в ходе бомбардировки) и 3) облегчение идентификации независимых объектов, полученных в результате трансформации (зеленых точек), в пределах рассеянных клеточных кластеров при селекции.
Бомбардировка микрочастицами эмбриогенных клеток.
Бинарный трансформирующий вектор использовали с фрагментом матрицы для экспрессии dsRNA Bcr1. Данная плазмида содержала инвертированный повтор целевого гена Bcr1, находящийся под контролем двойного промотора 35S из вируса мозаики цветной капусты, и лидерную последовательность из вируса мозаики люцерны (Aguado-Santacruz et al., Theoretical and Applied Genetics 104(5):763-771, 2002).
Эмбриогенную клеточную линию 'TIANSJ98' с высоким содержанием хлорофилла использовали в качестве мишени в экспериментах с бомбардировкой микрочастицами. Клетки распределяли по поверхности дисков из бумажных фильтров диаметром 2,0 см (примерно 2 г клеток по весу сырой ткани). Смеси для бомбардировки готовили следующим образом: 50 мкл частиц вольфрама M10 (15 мг/мл), 10 мкл ДНК (1 мкг/мл), 50 мкл 2,5 M CaCl2 и 20 мкл 0,1 M спермидина последовательно смешивали, встряхивали на вортексе в течение 5 мин., а затем подвергали непродолжительной ультразвуковой обработке. Смесь центрифугировали при 10000 об./мин. в течение 10 с. Удаляли 60 мкл супернатанта, а остаток распределяли в 5-мкл аликвоты в случае каждого отдельного побега. Бомбардировки проводили с применением пушки для струйной бомбардировки Particle Inflow Gun (Finer et al., Plant Cell Rep. 11:323-328, 1992). Смесь частицы/ДНК размещали в центре шприцевого фильтра. Эмбриогенные клетки покрывали проницаемым барьером толщиной 500 мкм, размещали на расстоянии 10 см от сетки фильтра, содержащего частицы, и однократно бомбардировали в вакуумной камере при давлении 60 мм рт.ст. Тестировали две разные осмотические среды с целью обработки до и после бомбардировки (застывшая среда MPC, дополненная 0,4 и 1 M маннита) и три разных значения давления бомбардировки (60, 80 и 100 фунтов на кв. дюйм). Обработку до бомбардировки проводили за 24 ч. до обстрела. После завершения обстрела бумажные фильтры, несущие эмбриогенные клетки, выдерживали в течение дополнительных 3-х дней на той же осмотической среде, которую применяли при обработке до бомбардировки. Таким образом, в общей сложности оценивали девять вариантов обработки с десятью чашками клеток, бомбардированными на каждую обработку. В качестве контроля фильтры с суспензией материала бомбардировали с применением частиц без ДНК.
Отбор устойчивых трансформированных клонов и восстановление растений.
После бомбардировки и последующей 3-х дневной осмотической обработки на среде MPC, содержащей 0,4 или 1 M маннита, но не содержащей антибиотик, бумажные фильтровальные диски, несущие подвергнутые бомбардировке клетки, переносили на среду MPC, содержащую 140 мг/л канамицина, и инкубировали при 30 ± 1°C при освещении белым светом, обеспечиваемым флуоресцентными лампами холодного света. Той же процедуры придерживались и в случае клеток, подвергнутых бомбардировке, но не подвергнутых осмотической обработке. Через 2 месяца концентрацию канамицина повышали до 150 или 160 мг/л. Клетки пересевали каждые 3 недели и на фоне селекции поддерживали 8 месяцев. После завершения данного периода устойчивые к канамицину клоны переносили в среду для регенерации, содержащую полную среду MS, 3% сахарозы, 2,5% фитагеля (Sigma, Сент-Луис, Миссури), но не содержащую антибиотик. Регенерированные побеги переносили для укоренения в 1/2 MS, содержащую 3,0 мкМ (0,56 мг/л) α-нафталинуксусной кислоты, 2,5 мкМ (0,51 мг/л) индол-3-масляной кислоты и 2,5% фитагеля, и инкубировали при 30 ± 1°C под постоянным флуоресцентным освещением. Затем укоренившиеся проростки переносили в горшки, закаливали и выращивали до зрелости в теплице.
ПЦР-анализ для подтверждения трансформации.
Общую геномную ДНК выделяли из устойчивых к канамицину и нетрансформированных контрольных растений с применением следующего протокола: примерно 250 мг клеток собирали в 2-мл пробирки эппендорф и измельчали до тонкодисперсного порошка в жидком N2 с применением стеклянного пестика, присоединенного к гомогенизатору (Caframo, встряхиватель типа RZR). Порошкообразные клетки повторно суспендировали с 500 мкл буфера для экстрагирования (7,0 M мочевины, 0,35 M NaCl, 0,05 M трис-HCl, pH 8,0, 0,02 M EDTA, 1% саркозина) в течение по меньшей мере 45 мин. Гомогенат клеток экстрагировали с помощью 1 объема смеси фенол/хлороформ. Водную фазу отделяли путем центрифугирования, а затем осаждали с применением равного объема изопропилового спирта. Осажденную ДНК однократно промывали 70% этанолом и повторно суспендировали в TE-буфере (0,01 M трис-HCl, 0,01 M EDTA, pH 8,0).
В случае ПЦР-анализа использовали 100-150 нг для амплификаций геномной ДНК на один объем реакционной смеси, составляющий 25 мкл. Прямой 5'-aaactgctgcatggctttct-3′ (SEQ ID NO: 91) и обратный праймеры 5'-acaggcctttcaggctttta-3' (SEQ ID NO: 92) конструировали с целью амплификации внутреннего фрагмента из 169 п.о. Bcr1 смыслового и антисмыслового фрагмента-вставки. Реакции ПЦР проводили с применением амплификатора Perkin Elmer в течение 30 циклов. Температуры для реакций были следующими: денатурация при 95°C (2 мин.), отжиг при 56°C (30 с) и элонгация при 72°C (30 с). Каждый объем реакционной смеси, составляющий 25 мкл, содержал: 1 × буфер для ПЦР, 0,25 мМ dNTP, 2 мМ MgCl2, 0,2 мкМ праймеров и 2,5 ЕД Taq. Продукты амплификации анализировали с помощью электрофореза в гелях 1% агарoза/SYBR green.
Пример 5. Получение трансгенных травянистых растений, продуцирующих бактериоцин colA
В данном примере демонстрируется генетическая модификация и продуцирование регуляторного в отношении бактериоцитов пептида колеоптерицина А (colA) в трансгенном травянистом растении с целью доставки тлям.
Трансгенную кормовую траву бутелоуа изящную, Bouteloua gracilis, которая продуцирует колеоптерицин A за счет экспрессии химерного гена, стабильно интегрированного в геном растения, получали путем бомбардировки микрочастицами с применением системы, описанной в примере 4.
Эмбриогенную клеточную линию 'TIANSJ98' с высоким содержанием хлорофилла пересевали каждые 20 дней путем переноса 1 мл клеточной суспензии в 24 мл свежей среды MPC.
Колеоптерицин A (colA)
atgacccgcaccatgctgtttctggcgtgcgtggcggcgctgtatgtgtgcattagcgcgaccgcgggcaaaccggaagaatttgcgaaactgagcgatgaagcgccgagcaacgatcaggcgatgtatgaaagcattcagcgctatcgccgctttgtggatggcaaccgctataacggcggccagcagcagcagcagcagccgaaacagtgggaagtgcgcccggatctgagccgcgatcagcgcggcaacaccaaagcgcaggtggaaattaacaaaaaaggcgataaccatgatattaacgcgggctggggcaaaaacattaacggcccggatagccataaagatacctggcatgtgggcggcagcgtgcgctgg (SEQ ID NO: 94).
Трансформационную плазмиду конструировали с целью экспрессии колеоптерицина A (colA). Данная плазмида содержала нуклеиновую кислоту, кодирующую colA, находящуюся под контролем двойного промотора 35S из вируса мозаики цветной капусты, и лидерную последовательность из вируса мозаики люцерны (Aguado-Santacruz et al., Theoretical and Applied Genetics 104(5):763-771, 2002).
Эмбриогенную клеточную линию 'TIANSJ98' с высоким содержанием хлорофилла использовали в качестве мишени в экспериментах с бомбардировкой микрочастицами. Клетки распределяли по поверхности дисков из бумажных фильтров диаметром 2,0 см (примерно 2 г клеток по весу сырой ткани). Смеси для бомбардировки готовили следующим образом: 50 мкл частиц вольфрама M10 (15 мг/мл), 10 мкл ДНК (1 мкг/мл), 50 мкл 2,5 M CaCl2 и 20 мкл 0,1 M спермидина последовательно смешивали, встряхивали на вортексе в течение 5 мин., а затем подвергали непродолжительной ультразвуковой обработке. Смесь центрифугировали при 10000 об./мин. в течение 10 с. Удаляли 60 мкл супернатанта, а остаток распределяли в 5-мкл аликвоты в случае каждого отдельного побега. Бомбардировки проводили с применением пушки для струйной бомбардировки Particle Inflow Gun (Finer et al., Plant Cell Rep. 11:323-328, 1992). Смесь частицы/ДНК размещали в центре шприцевого фильтра. Эмбриогенные клетки покрывали проницаемым барьером толщиной 500 мкм, размещали на расстоянии 10 см от сетки фильтра, содержащего частицы, и однократно бомбардировали в вакуумной камере при давлении 60 мм рт.ст. Тестировали две разные осмотические среды с целью обработки до и после бомбардировки (застывшая среда MPC, дополненная 0,4 и 1 M маннита) и три разных значения давления бомбардировки (60, 80 и 100 фунтов на кв. дюйм). Обработку до бомбардировки проводили за 24 ч. до обстрела. После завершения обстрела бумажные фильтры, несущие эмбриогенные клетки, выдерживали в течение дополнительных 3-х дней на той же осмотической среде, которую применяли при обработке до бомбардировки. Таким образом, в общей сложности оценивали девять вариантов обработки с десятью чашками клеток, бомбардированными на каждую обработку. В качестве контроля фильтры с суспензией материала бомбардировали с применением частиц без ДНК.
Отбор устойчивых трансформированных клонов и восстановление растений.
После бомбардировки и последующей 3-х дневной осмотической обработки на среде MPC, содержащей 0,4 или 1 M маннита, но не содержащей антибиотик, бумажные фильтровальные диски, несущие подвергнутые бомбардировке клетки, переносили на среду MPC, содержащую 140 мг/л канамицина, и инкубировали при 30 ± 1°C при освещении белым светом, обеспечиваемым флуоресцентными лампами холодного света. Той же процедуры придерживались и в случае клеток, подвергнутых бомбардировке, но не подвергнутых осмотической обработке. Через 2 месяца концентрацию канамицина повышали до 150 или 160 мг/л. Клетки пересевали каждые 3 недели и на фоне селекции поддерживали 8 месяцев. После завершения данного периода устойчивые к канамицину клоны переносили в среду для регенерации, содержащую полную среду MS, 3% сахарозы, 2,5% фитагеля (Sigma, Сент-Луис, Миссури), но не содержащую антибиотик. Регенерированные побеги переносили для укоренения в 1/2 MS, содержащую 3,0 мкМ (0,56 мг/л) α-нафталинуксусной кислоты, 2,5 мкМ (0,51 мг/л) индол-3-масляной кислоты и 2,5% фитагеля, и инкубировали при 30 ± 1°C под постоянным флуоресцентным освещением. Затем укоренившиеся проростки переносили в горшки, закаливали и выращивали до зрелости в теплице.
ПЦР-анализ для подтверждения трансформации.
Общую геномную ДНК выделяли из устойчивых к канамицину и нетрансформированных контрольных растений с применением следующего протокола: примерно 250 мг клеток собирали в 2-мл пробирки эппендорф и измельчали до тонкодисперсного порошка в жидком N2 с применением стеклянного пестика, присоединенного к гомогенизатору (Caframo, встряхиватель типа RZR). Порошкообразные клетки повторно суспендировали с 500 мкл буфера для экстрагирования (7,0 M мочевины, 0,35 M NaCl, 0,05 M трис-HCl, pH 8,0, 0,02 M EDTA, 1% саркозина) в течение по меньшей мере 45 мин. Гомогенат клеток экстрагировали с помощью 1 объема смеси фенол/хлороформ. Водную фазу отделяли путем центрифугирования, а затем осаждали с применением равного объема изопропилового спирта. Осажденную ДНК однократно промывали 70% этанолом и повторно суспендировали в TE-буфере (0,01 M трис-HCl, 0,01 M EDTA, pH 8,0).
В случае ПЦР-анализа использовали 100-150 нг для амплификаций геномной ДНК на один объем реакционной смеси, составляющий 25 мкл. Прямой 5'-caacgatcaggcgatgtatg-3′ (SEQ ID NO: 95) и обратный праймеры 5'-ttaatttccacctgcgcttt-3' (SEQ ID NO: 96) конструировали с целью амплификации внутреннего фрагмента из 165 п.о. фрагмента-вставки гена colA. Реакции ПЦР проводили с применением амплификатора Perkin Elmer в течение 30 циклов. Температуры для реакций были следующими: денатурация при 95°C (2 мин.), отжиг при 56°C (30 с) и элонгация при 72°C (30 с). Каждый объем реакционной смеси, составляющий 25 мкл, содержал: 1 × буфер для ПЦР, 0,25 мМ dNTP, 2 мМ MgCl2, 0,2 мкМ праймеров и 2,5 ЕД Taq. Продукты амплификации анализировали с помощью электрофореза в гелях 1% агарoза/SYBR green.
Пример 6. Получение трансгенных травянистых растений, продуцирующих бактериоцин colA
В данном примере демонстрируются генетическая модификация и продуцирование бактериоцина colA в трансгенном травянистом растении с целью доставки тлям.
Трансгенную кормовую траву бутелоуа изящную - Bouteloua gracilis, которая продуцирует бактериоцин colA за счет экспрессии химерного гена, стабильно интегрированного в геном растения, получают путем бомбардировки микрочастицами с применением системы на основе эмбриогенной клеточной линии 'TIANSJ98' с высоким содержанием хлорофилла (Aguado- Santacruz et al., 2002. Theoretical and Applied Genetics, 104(5), 763-771).
Образование клеточной линии 'TIANSJ98' и ее поддержание. эмбриогенную клеточную линию 'TIANSJ98' с высоким содержанием хлорофилла получали путем культивирования зеленых каллюсов апикальных меристем побега в жидкой среде MPC, как описано в (Aguado- Santacruz et al., 2001. ex Steud. Plant Cell Rep 20:131-136). Данную клеточную линию пересевали каждые 20 дней путем переноса 1 мл клеточной суспензии в 24 мл свежей среды MPC. Обоснования для применения условий диффузного распределения эмбриогенных каллюсов были следующими: 1) синхронизация физиологической стадии с клетками-мишенями, 2) максимальное увеличение распределения тотипотентного материала на бумажных фильтрах (оптимизация покрытия побега плазмидами в ходе бомбардировки) и 3) облегчение идентификации независимых объектов, полученных в результате трансформации (зеленых точек), в пределах рассеянных клеточных кластеров при селекции.
Бомбардировка микрочастицами эмбриогенных клеток.
Трансформирующий вектор использовали с фрагментом матрицы, предназначенным для экспрессии бактериоцина colA.
Бактериоцин colA
atgacccgcaccatgctgtttctggcgtgcgtggcggcgctgtatgtgtgcattagcgcgaccgcgggcaaaccggaagaatttgcgaaactgagcgatgaagcgccgagcaacgatcaggcgatgtatgaaagcattcagcgctatcgccgctttgtggatggcaaccgctataacggcggccagcagcagcagcagcagccgaaacagtgggaagtgcgcccggatctgagccgcgatcagcgcggcaacaccaaagcgcaggtggaaattaacaaaaaaggcgataaccatgatattaacgcgggctggggcaaaaacattaacggcccggatagccataaagatacctggcatgtgggcggcagcgtgcgctgg (SEQ ID NO: 94).
Данная плазмида содержала нуклеиновую кислоту, кодирующую бактериоцин colA, находящуюся под контролем двойного промотора 35S из вируса мозаики цветной капусты, и лидерную последовательность из вируса мозаики люцерны (Aguado- Santacruz et al., 2002. Theoretical and Applied Genetics, 104(5), 763-771).
Эмбриогенную клеточную линию 'TIANSJ98' с высоким содержанием хлорофилла использовали в качестве мишени в экспериментах с бомбардировкой микрочастицами. Клетки распределяли по поверхности дисков из бумажных фильтров диаметром 2,0 см (примерно 2 г клеток по весу сырой ткани). Смеси для бомбардировки готовили следующим образом: 50 мкл частиц вольфрама M10 (15 мг/мл), 10 мкл ДНК (1 мкг/мл), 50 мкл 2,5 M CaCl2 и 20 мкл 0,1 M спермидина последовательно смешивали, встряхивали на вортексе в течение 5 мин., а затем подвергали непродолжительной ультразвуковой обработке. Смесь центрифугировали при 10000 об./мин. в течение 10 с. Удаляли 60 мкл супернатанта, а остаток распределяли в 5-мкл аликвоты в случае каждого отдельного побега.
Бомбардировки проводили с применением пушки для струйной бомбардировки Particle Inflow Gun (Finer et al.,1992. Plant Cell Rep 11:323-328). Смесь частицы/ДНК размещали в центре шприцевого фильтра. Эмбриогенные клетки покрывали проницаемым барьером толщиной 500 мкм, размещали на расстоянии 10 см от сетки фильтра, содержащего частицы, и однократно бомбардировали в вакуумной камере при давлении 60 мм рт.ст. Тестировали две разные осмотические среды с целью обработки до и после бомбардировки (застывшая среда MPC, дополненная 0,4 и 1 M маннита) и три разных значения давления бомбардировки (60, 80 и 100 фунтов на кв. дюйм). Обработку до бомбардировки проводили за 24 ч. до обстрела.
После завершения обстрела бумажные фильтры, несущие эмбриогенные клетки, выдерживали в течение дополнительных 3-х дней на той же осмотической среде, которую применяли при обработке до бомбардировки. Таким образом, в общей сложности оценивали девять вариантов обработки с десятью чашками клеток, бомбардированными на каждую обработку. В качестве контроля фильтры с суспензией материала бомбардировали с применением частиц без ДНК.
Отбор устойчивых трансформированных клонов и восстановление растений.
После бомбардировки и последующей 3-х дневной осмотической обработки на среде MPC с 0,4 или 1 M маннита, но не содержащей антибиотик, бумажные фильтровальные диски, несущие подвергнутые бомбардировке клетки, переносили на среду MPC, содержащую 140 мг/л канамицина, и инкубировали при 30 ± 1°C при освещении белым светом, обеспечиваемым флуоресцентными лампами холодного света. Той же процедуры придерживались и в случае клеток, подвергнутых бомбардировке, но не подвергнутых осмотической обработке. Через 2 месяца концентрацию канамицина повышали до 150 или 160 мг/л. Клетки пересевали каждые 3 недели и на фоне селекции поддерживали 8 месяцев. После завершению данного периода канамицин-устойчивые клоны переносили в среду для регенерации с полной средой MS, 3% сахарозы, 2,5% фитагеля (Sigma, Сент-Луис, Миссури), но не содержащей антибиотик. Регенерированные побеги переносили для укоренения в 1/2 MS с 3,0 мкМ (0,56 мг/л) α-нафталинуксусной кислоты, 2,5 мкМ (0,51 мг/л) индол-3-масляной кислоты и 2,5% фитагеля и инкубировали при 30 ± 1°C под постоянным флуоресцентным освещением. Затем укоренившиеся проростки переносили в горшки, закаливали и выращивали до зрелости в теплице.
ПЦР-анализ с целью подтверждения трансформации.
Общую геномную ДНК выделяли из устойчивых к канамицину и нетрансформированных контрольных растений с применением следующего протокола: примерно 250 мг клеток собирали в 2-мл пробирки эппендорф и измельчали до тонкодисперсного порошка в жидком N2 с применением стеклянного пестика, присоединенного к гомогенизатору (Caframo, встряхиватель типа RZR). Порошкообразные клетки повторно суспендировали с 500 мкл буфера для экстрагирования (7,0 M мочевины, 0,35 M NaCl, 0,05 M трис-HCl, pH 8,0, 0,02 M EDTA, 1% саркозина) в течение по меньшей мере 45 мин. Гомогенат клеток экстрагировали с помощью 1 объема смеси фенол/хлороформ. Водную фазу отделяли путем центрифугирования, а затем осаждали с применением равного объема изопропилового спирта. Осажденную ДНК однократно промывали 70% этанолом и повторно суспендировали в TE-буфере (0,01 M трис-HCl, 0,01 M EDTA, pH 8,0).
В случае ПЦР-анализа использовали 100-150 нг для амплификаций геномной ДНК на один объем реакционной смеси, составляющий 25 мкл. Прямой 5'-caacgatcaggcgatgtatg-3′ и обратный 5'-ttaatttccacctgcgcttt-3' праймеры конструировали с целью амплификации внутреннего фрагмента из 165 п.о. фрагмента-вставки гена ColA. Реакции ПЦР проводили с применением амплификатора Perkin Elmer в течение 30 циклов. Температуры для реакций были следующими: денатурация при 95°C (2 мин.), отжиг при 56°C (30 с) и элонгация при 72°C (30 с). Каждый объем реакционной смеси, составляющий 25 мкл, содержал: 1 × буфер для ПЦР, 0,25 мМ dNTP, 2 мМ MgCl2, 0,2 мкМ праймеров и 2,5 ЕД Taq. Продукты амплификации анализировали с помощью электрофореза в гелях 1% агарoза/SYBR green.
Пример 7. Получение библиотеки cDNA из тли персиковой зеленой (Myzus persicae)
В данном примере демонстрируется получение библиотеки cDNA из личинок тли персиковой зеленой (Myzus persicae).
Выделение общей РНК из личинок тли и получение библиотеки cDNA.
Общую РНК из 0,9 г целых личинок на стадии первого возраста (4-5 дней после вылупливания; содержали при 16°C) экстрагировали с применением приведенного ниже способа на основе фенола/TRI REAGENT® (MOLECULAR RESEARCH CENTER, Цинциннати, Огайо). Личинок гомогенизировали при комнатной температуре в гомогенизаторе на 15 мл с использованием 10 мл TRI REAGENT® до получения однородной суспензии. После 5 мин. инкубации при комнатной температуре гомогенат распределяли в 1,5 мл микропробирки для центрифугирования (1 мл на пробирку), добавляли 200 мкл хлороформа и смесь энергично встряхивали 15 секунд. После того, как экстракту давали возможность осесть при комнатной температуре в течение 10 мин., фазы разделяли путем центрифугирования при 12000×g при 4°C. Верхнюю фазу (составляющую приблизительно 0,6 мл) осторожно переносили в другую стерильную 1,5 мл пробирку и добавляли равный объем изопропанола комнатной температуры. После инкубации при комнатной температуре в течение 5-10 мин. смесь центрифугировали 8 мин. при 12000×g (4°C или 25°C).
Супернатант осторожно удаляли и отбрасывали, и осадок, содержащий РНК, дважды промывали путем встряхивания на вортексе с 75% этанолом с извлечением с помощью центрифугирования в течении 5 мин. при 7500×g (4°C или 25°C) после каждой промывки. Этанол осторожно удаляли, осадок оставляли для высыхания на воздухе 3-5 мин., а затем растворяли в стерильной воде, не содержащей нуклеаз. Концентрацию РНК определяли путем измерения поглощения (A) при 260 нм и 280 нм. РНК хранили при -80°C, а качество РНК определяли путем прогонки аликвоты в 1% агарозном геле.
Общую РНК из личинок превращали в библиотеку cDNA с применением случайных праймеров. Библиотеку cDNA личинок секвенировали в масштабе планшета ½ с использованием системы GS FLX 454 Titanium™ в EUROFINS MWG Operon, в результате чего получали более 600000 ридов со средней длиной рида, составляющей 348 п.о. 350000 ридов собирали в более чем 50000 контигов. Как не подвергнутые сборке риды, так и контиги преобразовали в базы данных BLASTable с применением программы с открытым доступом FORMATDB (доступна от NCBI).
Пример 8. Получение dsRNA Bcr1
В данном примере демонстрируется получение и очистка синтетической dsRNA из библиотеки cDNA.
Схема эксперимента
Ген Bcr1 (ACYPI32128) представляет собой важнейший ген в регуляции и функции бактериоцитов у насекомых. Описанную в примере 6 cDNA использовали для синтеза dsRNA Bcr1, полученной с помощью ПЦР с применением следующей пары праймеров: прямой 5'-aaactgctgcatggctttct-3' (SEQ ID NO: 90) и обратный 5'-acaggcctttcaggctttta-3' (SEQ ID NO: 91). С целью нацеливания на область гена проводили две отдельные ПЦР-амплификации. С помощью первой ПЦР-амплификации вводили промоторную последовательность T7 на 5′-конце ((TTAATACGACTCACTATAGGGAGA; SEQ ID NO: 92) амплифицированных смысловых нитей. С помощью второй реакции встраивали промоторную последовательность T7 на 5′-концах антисмысловых нитей. Два ПЦР-амплифицированных фрагмента для каждого участка целевого гена Bcr1 затем смешивали в равных количествах, и данную смесь использовали в качестве транскрипционной матрицы для получения dsRNA. Двухнитевую РНК для биологического анализа насекомых синтезировали и очищали с применением набора для RNAi AMBION® MEGASCRIPT®, следуя инструкциям изготовителя (INVITROGEN), или набора для транскрипции In Vitro с T7 HiScribe®, следуя инструкциям изготовителя (New England Biolabs, Ипсуич, Масс.). Концентрацию dsRNA к Bcr1 измеряли с применением спектрофотометра NANODROP™ 8000 (THERMO SCIENTIFIC, Уилмингтон, Дел.), и очищенную dsRNA подготавливали в TE-буфере.
Последовательность шпилечной dsRNA Bcr1 одной нити: AUGAAACUGCUGCAUGGCUUUCUGAUUAUUAUGCUGACCAUGCAUCUGAGCAUUCAGUAUGCGUAUGGCGGCCCGUUUCUGACCAAAUAUCUGUGCGAUCGCGUGUGCCAUAAACUGUGCGGCGAUGAAUUUGUGUGCAGCUGCAUUCAGUAUAAAAGCCUGAAAGGCCUGUGGUUUCCGCAUUGCCCGACCGGCAAAGCGAGCGUGGUGCUGCAUAACUUUCUGACCAGCCCGUUUUUUUUUUCGGGCUGGUCAGAAAGUUAUGCAGCACCACGCUCGCUUUGCCGGUCGGGCAAUGCGGAAACCACAGGCCUUUCAGGCUUUUAUACUGAAUGCAGCUGCACACAAAUUCAUCGCCGCACAGUUUAUGGCACACGCGAUCGCACAGAUAUUUGGUCAGAAACGGGCCGCCAUACGCAUACUGAAUGCUCAGAUGCAUGGUCAGCAUAAUAAUCAGAAAGCCAUGCAGCAGUUUCAU (SEQ ID NO: 93).
Пример 9. Обработка тлей (Myzus persicae) раствором dsRNA Bcr1
В данном примере демонстрируется способность уничтожать тлей Myzus persicae или снижать их приспособленность посредством обработки раствором dsRNA за счет целенаправленного воздействия на экспрессию гена Bcr1 (ACYPI32128), который представляет собой важнейший ген в регуляции и функции бактериоцитов у насекомых.
Тли являются одними из наиболее важных насекомых-вредителей в сельском хозяйстве. Они обуславливают прямое повреждение сельскохозяйственных культур, вызванное поеданием, и служат в качестве переносчиков вирусов, поражающих растения. Помимо этого, медвяная падь тлей способствует росту плесневых налетов грибов и привлекает вредоносных муравьев. Применение средств химической обработки, к сожалению, по-прежнему остается широко распространенным, что приводит к отбору устойчивых особей, уничтожение которых становится все более сложным.
Структура терапевтического средства: растворы dsRNA составляли с концентрацией 0 (отрицательный контроль), 0,5, 1 или 5 мкг/мл dsRNA Bcr1 из примера 7 в 10 мл TE-буфера с 0,5% сахарозы и незаменимыми аминокислотами.
Схема эксперимента
Для подготовки к обработке тлей выращивали в лабораторных условиях и среде. В комнате с контролируемым климатом (фотопериод с 16 ч. света; 60 ± 5% RH; 20±2°C) растения выращивали в смеси вермикулита и перлита и заражали тлями. Для ограничения материнских эффектов или отличий в состоянии здоровья между растениями 5-10 взрослых особей с разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии личинок второго и третьего возраста собирали из здоровых растений и разделяли по видам обработки таким образом, что для каждого вида обработки получали примерно одно и то же число особей с каждого из растений коллекции.
Лунки 96-луночного планшета заполняли 200 мкл искусственного рациона для тлей (Febvay et al., Canadian Journal of Zoology 66(11):2449-2453, 1988), и планшет закрывали парафильмом для получения съедобного саше. Искусственный рацион смешивали или только с раствором TE-буфера (трис-HCl (1 мМ) с буфером на основе EDTA (0,1 мМ), pH 7,2) с 0,5% сахарозы и незаменимыми аминокислотами в качестве отрицательного контроля, или смешивали с растворами dsRNA, разведенными в TE-буфере, содержащем варьирующиеся концентрации dsRNA. Растворы dsRNA смешивали с искусственным рационом с получением конечных концентраций, составляющих от 0,5 до 5 мкг/мл.
Для каждой повторности обработки 30-50 тлей второй и третьей возрастной стадии помещали отдельно в лунки 96-луночного планшета, и планшет со съедобным саше переворачивали над ними, что давало возможность насекомым питаться через парафильм и ограничивать их отдельными лунками. Экспериментальных тлей содержали при тех же самых условиях среды, что и колонии тлей. После того, как тлей вскармливали в течение 24 часов, съедобное саше заменяли новым, содержащим стерильный искусственный рацион, и новое стерильное саше предоставляли каждые 24 часа в течение 4 дней. Когда саше заменяли, тлей также проверяли в отношении смертности. Тлю считали мертвой, если она становилась коричневой или располагалась на дне лунки и не двигалась во время наблюдения. Если тля находилась на парафильме съедобного саше, но не двигалась, она считалась кормящейся и живой.
Показатели выживаемости тлей, обработанных dsRNA Bcr1, сравнивали с тлями, обработанными отрицательным контролем. Показатели выживаемости тлей, обработанных dsRNA Bcr1, снижались по сравнению с тлями, обработанными контролем.
Пример 10. Местная доставка в растения dsRNA, специфической в отношении бактериоцитов, с целью защиты культурных растений
В данном примере демонстрируется способность к доставке dsRNA, специфической в отношении бактериоцитов, путем местного применения на листьях табака. dsRNA целенаправленно воздействует на экспрессию гена Bcr1 (ACYPI32128), который является ключевым в регуляции и функции бактериоцитов у насекомых.
Структура терапевтического средства: распыляемые растворы dsRNA-LDH составляли с dsRNA Bcr1, полученной в примере 7, при соотношении 1:0 (отрицательный контроль), 1:1, 1:2 или 1:3 dsRNA:LDH при 125 мкл/см2 поверхности листа.
Получение и характеристика нанопластин на основе LDH.
Пластинчатые глинистые наночастицы, а именно положительно заряженный LDH, представляют собой отличные системы наноносителей для dsRNA для обеспечения RNAi в качестве стабильных распыляемых составов для защиты культурных растений (Mitter et al., Nature Plants 1-10, 2017). Конъюгаты на основе LDH прочно приклеиваются к поверхности листа даже после интенсивного споласкивания и повышают стабильность dsRNA в течение более длительного периода времени в условиях окружающей среды. Замедленное высвобождение dsRNA улучшается за счет образования угольной кислоты на поверхности листа из атмосферного CO2 и влажности, что способствует расщеплению нанопластинок на основе LDH.
Нанопластины на основе LDH получали в соответствии с (Mitter et al., Nature Plants 1-10, 2017) путем модифицированного безводного осаждения с последующими тепловой обработкой, очисткой и диспергированием в воде с получением частиц, средний размер которых составляет 45 нм. Характеристики частиц (образцы LDH в пяти повторах) получали с помощью прибора Nanosizer Nano ZS (Malvern Instruments) с получением Z-среднего размера и PDI, и их визуально анализировали с применением JEOL JSM-2010 TEM22. Химический состав и кристаллическую структуру определяли в пяти образцах LDH с помощью порошковой XRD и инфракрасной спектроскопии на основе преобразования Фурье в нарушенном режиме полного отражения, применяя соответственно дифрактометр Rigaku Miniflex X-Ray и Nicolet 6700 FT-IR (Thermo Electron Corporation).
Загрузка dsRNA на LDH.
Для определения оптимальности и полноты загрузки в нанопластины на основе LDH конструкции на основе шпилечной dsRNA, полученной в примере 7, несколько раз анализировали соотношение in vitro транскрибируемой dsRNA (500 нг) и LDH (dsRNA-LDH (вес/вес)) при 1:1, 1:2, 1:3, 1:4, 1:5 и 1:10. Инкубировали dsRNA и LDH в общем объеме 10 мкл при комнатной температуре в течение 30 мин. при осторожном перемешивании на орбитальном шейкере. Полноту загрузки dsRNA оценивали с помощью удержания комплексов dsRNA-LDH в лунке 1% агарозного геля. Соответствующие соотношения загрузки являются постоянными, независимо от требуемого масштаба увеличения.
Высвобождение dsRNA из комплексов dsRNA-LDH и стабильность высвобождаемой dsRNA тестировали так, как описано в (Mitter et al., Nature Plants 1-10, 2017).
Нозерн-блот-анализ с целью выявления dsRNA в листьях.
Для поглощения dsRNA растения N. tabacum (три повторности) опрыскивали с использованием либо LDH, Bcr1-dsRNA либо Bcr1-dsRNA-LDH в день 0. Соотношения dsRNA-LDH в тестируемом комплексе были следующими: 1:1, 1:2 или 1:3. Растения выращивали в грунте UQ23 в горшках диаметром 10 см в условиях теплицы (средняя температура 25°C с естественным освещением). Во время опрыскивания кончики таких растений накрывали изоляционной лентой. Собирали новые листья, появившиеся через 20 дней после опрыскивания. Общую РНК экстрагировали с помощью TRIzol и насыщали малыми РНК (Mitter et al., Am. Phytopathological Soc. 16:936-944, 2003). Малые РНК (20 мкг) для каждого варианта обработки разделяли в 15% (вес/об.) полиакриламидном геле, содержащем мочевину, в денатурирующих условиях (PAGE). ZR Small-RNA ladder (Zymo Research), меченую DIG на 3′-конце, использовали в качестве маркера молекулярной массы. Блоты переносили с помощью устройства для полусухого переноса Trans-Blot SD (Bio-Rad) на мембрану Hybond-N (Roche). Блоты обрабатывали согласно рекомендации изготовителя с использованием DIG-меченого 24-нуклеотидного зонда (5-atgctgaccatgcatctgagcatt) для Bcr1 в запатентованном буферном комплексе (Roche). После гибридизации фильтры выявляли с применением CSPD - хемилюминесцентного субстрата для щелочной фосфатазы. Количественный анализ проводили с применением программного обеспечения NIH Image 1.6.
Пример 11. Твердофазный синтез PNA
В данном примере демонстрируется твердофазный синтез PNA.
Структура терапевтического средства: конструкция на основе комплементарной антисмысловой PNA, целенаправленно воздействующей на целевой ген Bcr1:gaatgcagctgc бактериоцитов
Схема эксперимента
Антисмысловую PNA синтезировали автоматизированно (синтезатор пептидов MilliGen 9050) с помощью твердофазного способа с применением стандартной химии Fmoc (N-(9-флуоренил)метоксикарбонил) в режиме непрерывного потока.
Очистку антисмысловой PNA проводили с помощью обращено-фазовой высокоэффективной жидкостной хроматографии (RP-HPLC) с УФ-детекцией при 260 нм с применением полупрепаративной колонки C18 (10 мкМ, 300×7,7 мМ, Xterra Waters, 300 Å), элюирования водой, содержащей 0,1% TFA (элюент A), и ацетонитрилом, содержащим 0,1% TFA (элюент B); градиент элюирования от 100% A до 50% B в течение 30 мин., скорость потока: 4 мл/мин. Чистоту и идентичность очищенной антисмысловой PNA осуществляли с помощью сверхпроизводительной жидкостной хроматографии в сочетании с масс-спектрометрией (UPLC-MS; устройство Waters Acquity, оснащенное анализатором ESI-Q) с применением колонки Acquity UPLC BEH C18; 2,1×50 мМ, 1,7 мкМ. Ожидаемые массовые пики выявляли для правильной аминокислотной последовательности и последовательности нуклеиновых кислот.
Пример 12. Получение меченной Cy3 PNA
В данном примере демонстрируется связывание PNA, описанной в примере 10, с красителем Cy3 в качестве маркера, с целью мечения PNA посредством клик-химии.
Структура терапевтического средства: PNA с дибензилциклоoктиновой модификацией (DBCO) и красителем Cy3 с азидной модификацией: Cy3-gaatgcagctgc.
Схема эксперимента
С целью подготовки к клик-реакции PNA, синтезированную в примере 10, метили DBCO (Glen Research, Стерлинг, Вирджиния). Сложный эфир DBCO-сульфо-NHS растворяли при концентрации 5,2 мг в 60 мкл воды или безводного DMSO. Данный исходный раствор использовали для конъюгации аминомодифицированной PNA в конъюгационном буфере натрия карбонат/бикарбонат, pH=~9.
Для синтеза 0,2 мкмоль PNA растворяли PNA в 500 мкл конъюгационного буфера. Прим. 6-кратный избыток (6 мкл) раствора сложного эфира DBCO-сульфо-NHS добавляли в растворенную PNA. Смесь встряхивали на вортексе и инкубировали при комнатной температуре в течение периода от 2-4 часов до приблизительно всей ночи. Конъюгированную PNA обессоливали на колонке для обессоливания (Glen Research, Стерлинг, Вирджиния) с удалением солей и органических примесей.
Cy3-азид модифицированный краситель приобретали у Sigma Aldrich (777315). Для клик-реакции 1 мг Cy3-азида растворяли в 150 мкл DMSO. Раствор Cy3-азида добавляли к 10 OD PNA, конъюгированной с DBCO, в 100 мкл воды. Смесь инкубировали при комнатной температуре в течение ночи. Лигированные PNA обессоливали на колонке для обессоливания (Glen Research, Стерлинг, Вирджиния) с удалением солей и органических примесей.
Пример 13. Обработка тлей (Myzus persicae) с помощью раствора Cy3-PNA
В данном примере демонстрируется способность уничтожать тлей Myzus persicae или снижать их приспособленность посредством их обработки конструкциями Cy3-PNA, которые целенаправленно воздействуют на экспрессию гена Bcr1 (ACYPI32128), который представляет собой важнейший ген в регуляции функции бактериоцитов у насекомых.
Тли являются одними из наиболее важных насекомых-вредителей в сельском хозяйстве. Они обуславливают прямое повреждение сельскохозяйственных культур, вызванное поеданием, и служат в качестве переносчиков вирусов, поражающих растения. Помимо этого, медвяная падь тлей способствует росту плесневых налетов грибов и привлекает вредоносных муравьев. Применение средств химической обработки, к сожалению, по-прежнему остается широко распространенным, что приводит к отбору устойчивых особей, уничтожение которых становится все более сложным.
Структура терапевтического средства: растворы Cy3-PNA составляли с 0, 0,5, 1, 5 или 10 мг/л Cy3-PNA из примера 11 в 10 мл 0,5% сахарозы и незаменимых аминокислот.
Схема эксперимента
Для подготовки к обработке тлей выращивали в лабораторных условиях и среде. В комнате с контролируемым климатом (фотопериод с 16 ч. света; 60±5% RH; 20±2°C) растения выращивали в смеси вермикулита и перлита и заражали тлями. Для ограничения материнских эффектов или отличий в состоянии здоровья между растениями 5-10 взрослых особей с разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии личинок второго и третьего возраста собирали из здоровых растений и разделяли по видам обработки таким образом, что для каждого вида обработки получали примерно одно и то же число особей с каждого из растений коллекции.
Лунки 96-луночного планшета заполняли 200 мкл искусственного рациона для тлей (Febvay et al., Canadian Journal of Zoology 66(11):2449-2453, 1988), и планшет закрывали парафильмом для получения съедобного саше. Искусственный рацион смешивали или только с раствором стерильной воды с 0,5% сахарозой и незаменимыми аминокислотами в качестве отрицательного контроля, или смешивали с растворами Cy3-PNA. Растворы Cy3-PNA смешивали с искусственным рационом с получением конечных концентраций от 0,5 до 10 мг/мл.
Для каждой повторности обработки 30-50 тлей на стадии второго и третьего возраста помещали отдельно в лунки 96-луночного планшета, и планшет со съедобным саше переворачивали над ними, что давало возможность насекомым питаться через парафильм и ограничивать их отдельными лунками. Экспериментальных тлей содержали при тех же самых условиях среды, что и колонии тлей. После того, как тлей вскармливали в течение 24 часов, съедобное саше заменяли новым, содержащим стерильный искусственный рацион, и новое стерильное саше предоставляли каждые 24 часа в течение 4 дней. Когда саше заменяли, тлей также проверяли в отношении смертности. Тлю считали мертвой, если она становилась коричневой или располагалась на дне лунки и не двигалась во время наблюдения. Если тля находилась на парафильме съедобного саше, но не двигалась, она считалась кормящейся и живой.
Показатели выживаемости тлей, обработанных Cy3-PNA, специфичной в отношении Bcr1, сравнивали с тлями, обработанными отрицательным контролем. Показатели выживаемости тлей, обработанных Cy3-PNA, специфичными к Bcr1, снижались по сравнению с тлями, обработанными контролем.
Пример 14. Местная доставка в растения Cy3-PNA с целью защиты культурных растений
В данном примере демонстрируется способность к доставке Cy3-PNA за счет местного применения на листьях растений табака. Cy3-PNA целенаправленно воздействует на экспрессию гена Bcr1 (ACYPI32128), который представляет собой важнейший ген в регуляции и функции бактериоцитов у насекомых.
Структура терапевтического средства: распыляемые растворы Cy3-PNA-LDH составляли с Cy3-PNA, синтезированной в примере 11, при соотношении 1:0 (отрицательный контроль), 1:1, 1:2 или 1:3 Cy3-PNA:LDH при 125 мкл/см2 поверхности листа.
Получение и характеристика нанопластин на основе LDH.
Пластинчатые глинистые наночастицы, а именно положительно заряженный LDH, представляют собой отличные системы наноносителей для доставки нуклеиновых кислот в качестве стабильных распыляемых составов для защиты культурных растений (Mitter et al., Nature Plants 1-10, 2017). Конъюгаты на основе LDH прочно приклеиваются к поверхности листа даже после интенсивного споласкивания и повышают стабильность PNA в течение более длительного периода времени в условиях окружающей среды. Замедленное высвобождение PNA улучшается за счет образования угольной кислоты на поверхности листа из атмосферного CO2 и влажности, что способствует расщеплению нанопластин на основе LDH.
Нанопластины на основе LDH получали в соответствии с (Mitter et al., Nature Plants 1-10, 2017) путем модифицированного безводного осаждения с последующими тепловой обработкой, очисткой и диспергированием в воде с получением частиц, средний размер которых составляет 45 нм. Характеристики частиц (образцы LDH в пяти повторах) получали с помощью прибора Nanosizer Nano ZS (Malvern Instruments) с получением Z-среднего размера и PDI, и их визуально анализировали с применением JEOL JSM-2010 TEM22. Химический состав и кристаллическую структуру определяли в пяти образцах LDH с помощью порошковой XRD и инфракрасной спектроскопии на основе преобразования Фурье в нарушенном режиме полного отражения, применяя соответственно дифрактометр Rigaku Miniflex X-Ray и Nicolet 6700 FT-IR (Thermo Electron Corporation).
Загрузка Cy3-PNA на LDH.
Для определения оптимума и полноты загрузки Cy3-PNA, полученных в примере 11, в нанопластины на основе LDH несколько раз анализировали соотношение Cy3-PNA (0,5 мкг) и LDH (Cy3-PNA-LDH (вес/вес)) при 1:1, 1:2, 1:3, 1:4, 1:5 и 1:10. Cy3-PNA и LDH инкубировали в общем объеме 10 мкл при комнатной температуре в течение 30 мин. при осторожном перемешивании на орбитальном шейкере. Полноту загрузки Cy3-PNA оценивали с помощью удержания комплексов Cy3-PNA-LDH в лунке 1% агарозного геля. Соответствующие соотношения загрузки являются постоянными, независимо от требуемого масштаба увеличения.
Конфокальная визуализация с целью выявления Cy3-PNA в пределах листьев растений.
Семена Arabidopsis thaliana Col-0 со стерилизованной поверхностью подвергали яровизации, высевали и вертикально выращивали при 21°C (16 ч./8 ч. день/ночь) в течение 11 дней на твердых средах MS.
Для выявления поглощения Cy3-PNA проростки A. thaliana возрастом 11 дней в трех повторностях обрабатывали каплями по 3 мкл только Cy3, Cy3-PNA (1 мкг) и Cy3-PNA-LDH (1:3). Капли наносили на каждый отдельный лист. Через 48 ч. листья дважды споласкивали в течение 2 мин. в 3 мл воды с дальнейшим интенсивным пипетированием и визуальным осмотром эпидермиса, поскольку флуоресценция остается на поверхности в случае обработок с использованием LDH. Естественную флуоресценцию хлорофилла выявляли при помощи фильтра с полосой пропускания 650-800 нм. Серии срезов по оси Z использовали чтобы удостовериться, что флуоресценция, наблюдаемая в губчатом мезофиле и ксилеме, носит внутренний характер. Листья визуально осматривали под конфокальным лазерным сканирующим микроскопом Zeiss LSM510 META (Carl Zeiss). Флуоресценцию Cy3 визуально определяли по испусканию с помощью HeNe-лазера при 543 нм и выявляли с помощью фильтра с полосой пропускания 560-615 нм.
Пример 15. Обработка тлей с помощью раствора, повышающего температуру тела
В данном примере демонстрируется способность уничтожать тлей или снижать их приспособленность посредством их обработки простагландином. Простагландины представляют собой эйкозаноиды, связанные с иммунной функцией, которые быстро вызывают повышение температуры у позвоночных и беспозвоночных. В данном примере демонстрируется, что влияние простагландина на тлей опосредовано изменением популяций бактерий, эндогенных по отношению к тлям, которые являются чувствительными к повышению температуры тела, индуцированному простагландином. Одним целевым бактериальным штаммом являлся Buchnera.
Структура терапевтического средства: раствор простагландина E2 (PGE2) составляли с 0 (отрицательный контроль), 10, 20 или 50 мкг/мкл простагландина, составленного в растворе этанола и стерильной воды с 0,5% сахарозой и незаменимыми аминокислотами.
Схема эксперимента
Для подготовки к обработке тлей выращивали в лабораторных условиях и среде. В комнате с контролируемым климатом (фотопериод с 16 ч. света; 60 ± 5% RH; 20 ± 2°C) растения конских бобов выращивали в смеси вермикулита и перлита при 24°C при 16 ч. света и 8 ч. темноты. Для ограничения материнских эффектов или отличий в состоянии здоровья между растениями 5-10 взрослых особей с разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии личинок второго и третьего возраста собирали из здоровых растений и разделяли по видам обработки таким образом, что для каждого вида обработки получали примерно одно и то же число особей с каждого из растений коллекции.
Растворы простагландина получали путем растворения простагландина (SIGMA-ALDRICH, P5640) в воде и этаноле (1:1) с 0,5% сахарозой и незаменимыми аминокислотами. Лунки в 96-луночном планшете заполняли 200 мкл искусственного рациона для тлей (Febvay, et al., Canadian Journal of Zoology, 1988, 66(11): 2449-2453), а планшеты накрывали парафильмом с получением съедобного саше. Искусственный рацион смешивали либо со стерильной водой и 0,5% сахарозой и незаменимыми аминокислотами в качестве отрицательного контроля, либо раствором простагландина, содержащим простагландин в одной из определенных концентраций. Растворы простагландина смешивали с искусственным рационом с получением конечных концентраций от 10 до 50 мкг/мкл.
Для каждой повторности обработки 30-50 тлей второй и третьей возрастной стадий помещали отдельно в лунки 96-луночного планшета, и планшет со съедобным саше переворачивали над ними, что давало возможность насекомым питаться через парафильм и ограничивать их отдельными лунками. Экспериментальных тлей содержали при тех же самых условиях среды, что и колонии тлей. После того, как тлей вскармливали в течение 24 часов, съедобное саше заменяли новым, содержащим стерильный искусственный рацион, и новое стерильное саше предоставляли каждые 24 часа в течение четырех дней. Когда саше заменяли, тлей также проверяли в отношении смертности. Тлю считали мертвой, если она становилась коричневой или располагалась на дне лунки и не двигалась во время наблюдения. Если тля находилась на парафильме съедобного саше, но не двигалась, она считалась кормящейся и живой.
Статус Buchnera в образцах от тлей оценивали с помощью ПЦР. Общую ДНК выделяли из контрольных (не обработанных простагландином) и обработанных простагландином особей с помощью набора для выделения ДНК насекомых (OMEGA, Bio-tek) в соответствии с протоколом производителя. Праймеры для Buchnera: прямой праймер 5′-GTCGGCTCATCACATCC-3′ (SEQ ID NO: 97) и обратный праймер 5′-TTCCGTCTGTATTATCTCCT-3′ (SEQ ID NO: 98), конструировали на основе последовательностей 23S-5S рРНК, полученных из генома Buchnera (номер доступа: GCA_000009605.1) (Shigenobu, et al., Nature 200.407, 81-86) с применением программного обеспечения Primer 5.0 (Primer-E Ltd., Плимут, Великобритания). Циклы ПЦР-амплификации включали стадию начальной денатурации при 95°C в течение 5 мин., 35 циклов при 95°C в течение 30 с, при 55°C в течение 30 с и при 72°C в течение 60 с и конечную стадию элонгации в течение 10 мин. при 72°C. Продукты амплификации из обработанных простагландином и контрольных образцов анализировали в 1% агарозных гелях, окрашивали SYBR Safe и визуализировали с применением системы визуализации. У тлей, обработанных простагландином, продемонстрировано снижение экспрессии специфичных для Buchnera генов.
Показатели выживаемости тлей, обработанных раствором простагландина, сравнивали с тлями, обработанными отрицательным контролем. Показатели выживаемости тлей, обработанных раствором простагландина, снижались по сравнению с контролем.
Пример 16. Обработка тлей с помощью dsRNA для стимуляции иммунного ответа у насекомых с целью снижения приспособленности
В данном примере демонстрируется, что обработка тлей двухнитевой РНК (dsRNA) приводит к нокдауну иммунорегуляторных генов с индуцированием иммунного ответа и снижением приспособленности тлей. Индуцирование иммунного ответа путем подавления иммунорегуляторного гена, в частности, Cact, являющегося отрицательным регулятором сигнального пути Toll, который является первичным сигнальным путем иммунитета у тлей, с индуцированием активации сигнального пути Toll для экспрессии и секретирования антимикробных пептидов, лизоцимов профенолоксидазы, в конечном итоге негативно воздействует на популяции бактерий, являющиеся эндогенными по отношению к тлям. В данном примере демонстрируется, что эффект снижения уровней Cact у тлей с положительной регуляцией системных иммунных ответов приводит к нарушению функций у популяций бактерий, являющихся эндогенными в отношении тлей, которые являются чувствительными к повышению иммунных ответов у тлей, инициируемых за счет активации пути Toll. Одним целевым бактериальным штаммом являлся Buchnera.
Структура терапевтического средства: Тлей LSR-1 на стадии 5го возраста подвергали микроинъекции. Растворы для инъекций будут представлять собой или dd-воду (отрицательный контроль), или dsRNA, разбавленную в dd-воде в разных концентрациях (8 или 60 нг/тля; см. ниже).
Схема эксперимента
Тлей LSR-1 (которые переносят только Buchnera) A. pisum будут выращивать на растениях конских бобов (бобы Vroma vicia faba от Johnny's Selected Seeds) в инкубаторе с регулируемым климатом (фотопериод: 16 ч. света/8 ч. темноты; 60 ± 5% RH; 25 ± 2°C). Перед использованием для скармливания тлям растения конских бобов будут выращивать в почвенной смеси для горшечных культур при 24°C при 16 ч. света и 8 ч. темноты. Для ограничения материнских эффектов или отличий в состоянии здоровья между растениями 5-10 взрослых особей с разных растений будут распределять среди 10 растений в возрасте двух недель и оставлять размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии пятого возраста будут собирать на здоровых растениях и разделять на 2 разные группы обработки: 1) вода (отрицательный контроль) или 2) dsRNA, целенаправленно воздействующая на ApGLNT1 (в концентрациях, указанных в данном документе).
Доставка путем микроинъекции - схема эксперимента
Микроинъекцию будут осуществлять с помощью автоматического инъектора NanoJet III Auto-Nanoliter Injector с выдвижной иглой из боросиликатного стекла собственного изготовления (Drummond Scientific; № по кат. 3-000-203-G/XL). Тлей будут переносить с помощью малярной кисточки в систему трубок, соединенную с вакуумным насосом, что удерживает тлей на месте во время осуществления микроинъекции. Место инъекции будет располагаться в вентральной части груди тли. Объем инъекции будет составлять 20 нл для взрослой особи (при скорости 2 нл/сек). В каждой группе обработки будет примерно одинаковое количество получивших инъекцию особей с каждого из растений коллекции.
После инъекции тлей будут выпускать в чашки Петри на листья конских бобов, черешки которых находятся в пробирке эппендорф, заполненной 1 мл воды. Выживаемость тли будут контролировать ежедневно, а мертвых тлей удалять после их выявления. Подсчитывать количество потомства будут от каждой группы и оценивать плодовитость по количеству потомства (F1), полученного на одну тлю в каждой временной точке.
В выбранных экспериментах развитие будут оценивать в группах потомства, полученного от каждой группы обработки, путем ежедневной регистрации стадии развития потомства (стадии 1го, 2го, 3го, 4го и 5го возраста). Развитие также будут оценивать путем визуального осмотра тлей через 4 дня после сбора и определения их площади. Матрица для синтезирования dsRNA будет представлять собой cDNA, полученную путем обратной транскрипции с mRNA. РНК будут экстрагировать из одной особи A. pisum (линия LSR-1) на стадии 5го возраста и количественно определять с помощью Nanodrop (Thermo fisher scientific). ~100 мкг общей РНК будут добавлять в качестве матрицы в реакционную смесь для обратной транскрипции с применением набора Superscript IV (Thermo Fisher Scientific), следуя протоколу изготовителя.
С целью амлификации матриц для получения dsRNA cDNA будут разбавлять в 100 раз и в дальнейшем использовать в ПЦР. Реакционные смеси для ПЦР (конечный объем 25 мкл) содержали 12,5 мкл смеси 2X Go Taq Green (Promega), 0,2 мкл прямого праймера (таблица 12), 0,2 мкл обратного праймера (таблица 12) и 12,1 мкл cDNA, разбавленной в 100 раз. ПЦР-реакции проводят с применением следующий условий: 1) 95°C в течение 2 минут, 2) 95°C в течение 20 секунд, 3) 55°C в течение 15 секунд, 4) 72°C в течение 30 секунд, 5) повтор стадий 2-4 35x, 6) 72°C в течение 5 минут. Размеры ПЦР-ампликонов определяют с помощью электрофореза в 1,5% агарозном геле, и полосы ожидаемого размера вырезают и очищают с помощью набора для очистки ДНК QIAquick (Qiagen). Будут синтезировать dsRNA in vitro с применением набора T7 MEGAscript (Ambion, Thermo Fisher Scientific; № по кат. AM1334), следуя протоколу изготовителя. Концентрацию dsRNA измеряют с помощью Nanodrop (Thermo Fisher Scientific).
Таблица 12. Последовательности dsRNA Cact
Статус Buchnera в образцах тлей будут оценивать с помощью ПЦР. Взрослых тлей из групп отрицательного контроля и обработки фагом вначале будут подвергать поверхностной стерилизации 70% этанолом в течение 1 мин., 10% отбеливателем в течение 1 мин. и тремя промывками сверхчистой водой в течение 1 мин. Общую ДНК будут экстрагировать из каждой особи (всего тела) с помощью набора для выделения ДНК насекомых (OMEGA, Bio-tek) в соответствии с протоколом производителя. Праймеры для Buchnera: прямой праймер 5′-GTCGGCTCATCACATCC-3′ (SEQ ID NO: 97) и обратный праймер 5′-TTCCGTCTGTATTATCTCCT-3′ (SEQ ID NO: 98), конструировали на основе последовательностей 23S-5S рРНК, полученных из генома Buchnera (номер доступа: GCA_000009605.1) (Shigenobu, et al., Nature 200.407, 81-86) с применением программного обеспечения Primer 5.0 (Primer-E Ltd., Плимут, Великобритания). Циклы ПЦР-амплификации включают стадию начальной денатурации при 95°C в течение 5 мин., 35 циклов при 95°C в течение 30 с, при 55°C в течение 30 с и при 72°C в течение 60 с и конечную стадию элонгации в течение 10 мин. при 72°C. Продукты амплификации из обработанных рифампицином и контрольных образцов будут анализировать в 1% агарозных гелях, окрашивать с помощью SYBR Safe и визуально анализировать с применением системы визуализации. Предполагается, что у тлей, обработанных dsRNA, будет продемонстрировано снижение экспрессии генов, специфичных для Buchnera.
Экспрессию Cact и показатели выживаемости тлей, обработанных Cactus, сравнивают с тлями, обработанными отрицательным контролем. Ожидается, что экспрессия Cact и показатели выживаемости тлей, обработанных dsRNA Cact, будут снижаться по сравнению с тлями, обработанными контролем.
Пример 17. Обработка тлей раствором грибов, стимулирующим иммунный ответ у насекомых, с целью снижения приспособленности
В данном примере демонстрируется обработка тлей раствором из Pichia pastoris, штамма дрожжей, что в результате снижает приспособленность тлей. Хотя ряд сигнальных путей системы врожденного иммунитета, присутствующих у насекомых, у тлей отсутствует, ключевые компоненты, которые распознают грибы и индуцируют иммунные ответы к грибам, остаются интактными. В частности, присутствие грибов индуцирует расщепление сериновой протеазой у тли, Persephone, которая затем активирует сигнальный путь Toll. Затем Cactus фосфорилируется и Dorsal высвобождается и перемещается в ядро, что активирует экспрессию и секретирование антимикробных пептидов, лизоцимов и профенолоксидазы, которые в конечном итоге оказывают воздействие на популяцию бактерий, являющихся эндогенными по отношению к тлям. В данном примере демонстрируется, что влияние раствора из Pichia pastoris на тлей опосредовано модуляцией популяций бактерий, эндогенных по отношению к тлям, которые являются чувствительными к повышению иммунного ответа тлей, индуцируемого присутствием дрожжей. Одним целевым бактериальным штаммом являлся Buchnera.
Тли представляют собой сельскохозяйственных насекомых-вредителей, причиняющих непосредственный вред растению в результате кормления и выступающих в качестве переносчиков вирусов растений. Помимо этого, медвяная падь тлей способствует росту плесневых налетов грибов и привлекает вредоносных муравьев. Применение средств химической обработки, к сожалению, по-прежнему остается широко распространенным, что приводит к отбору устойчивых особей, уничтожение которых становится все более сложным.
Структура терапевтического средства
P. pastoris доставляли с применением двух разных способов: перфузии в листья конских бобов и опрыскивание листьев конских бобов с помощью пульверизатора. Для каждого эксперимента получали две экспериментальные группы: 1) группу обработки водой в качестве отрицательного контроля и 2) группу обработки P. pastoris в воде. Способы и дозы обработки описаны в данном документе в разделе "Схема эксперимента".
Схема эксперимента с перфузией в листья
Тлей (линии LSR-1, Acyrthosiphon pisum) выращивали на растениях конских бобов (бобы Vroma vicia от Johnny's Selected Seeds) в инкубаторе с контролируемым климатом (фотопериод: 16 ч. света/8 ч. темноты; 60 ± 5% RH; 25 ± 2°C). Перед использованием для скармливания тлям растения конских бобов выращивали в почвенной смеси для горшечных культур при 24°C при 16 ч. света и 8 ч. темноты. Для ограничения материнских эффектов или различий в состоянии здоровья между растениями 5-10 взрослых особей из разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии первого и второго возраста собирали на здоровых растениях и разделяли на две разные группы обработки: 1) которым обеспечивали кормление на листьях, подвергнутых перфузии водой, и 2) которым обеспечивали кормление на листьях, подвергнутых перфузии раствором P. pastoris в воде.
В экспериментах использовали штамм с нокаутом по ade2 "протеазы дикого типа" (штамм 1) из системы Pichia Pink (Thermo Fisher Scientific). P. pastoris выращивали в YPD в течение ночи при 30°C, а на следующий день культуру промывали однократно водой и повторно суспендировали в воде до достижения конечного показателя 0,918 OD600 и использовали для перфузии в лист в день 0, или 5,58 OD600 и использовали для перфузии в лист в день 3 эксперимента. В случае каждой перфузии в лист примерно 1 мл раствора из P. pastoris или воды (отрицательного контроля) инъецировали в лист конских бобов. Черешок листа затем помещали в 1,5 мл пробирку эппендорф, которую герметично закрывали парафильмом. Стебель листа затем помещали в глубокую чашку петри (Fisher Scientific, № по кат. FB0875711), а на листья растения помещали тлей для обеспечения кормления. Листья меняли в день 3 эксперимента, а старые листья заменяли новыми листьями, которых подвергли перфузии раствором из P. pastoris (при указанной OD600) или водой.
Для каждой обработки на каждый лист помещали 62-63 тли. Ежедневно осуществляли контроль тлей в отношении их выживаемости, и мертвых тлей удаляли в случае их обнаружения. Помимо этого, стадию развития (1-ю, 2-ю, 3-ю, 4-ю и 5-ю возрастные стадии) определяли ежедневно на протяжении всего эксперимента.
После 6 дней обработки ДНК экстрагировали из нескольких тлей из каждой группы обработки. Вкратце, поверхность тела тлей стерилизовали путем погружения тли в 6% раствор отбеливателя примерно на 5 секунд. Затем тлей промывали в стерильной воде, и ДНК экстрагировали из каждой отдельной тли с помощью набора для экстракции ДНК (Qiagen, набор DNeasy) в соответствии с инструкциями производителя. Концентрацию ДНК измеряли посредством количественной оценки нуклеиновой кислоты с использованием NanoDrop, и число копий ДНК Buchnera и тлей измеряли с помощью qPCR. Праймерами, используемыми для Buchnera, были Buch_groES_18F (CATGATCGTGTGCTTGTTAAG; SEQ ID NO: 101) и Buch_groES_98R (CTGTTCCTCGAGTCGATTTCC; SEQ ID NO: 102) (Chong and Moran, 2016 PNAS). Праймерами, используемыми для тлей, были ApEF1a 107F (CTGATTGTGCCGTGCTTATTG; SEQ ID NO: 103) и ApEF1a 246R (TATGGTGGTTCAGTAGAGTCC; SEQ ID NO: 104) (Chong and Moran, 2016 PNAS). Проводили qPCR с применением скорости линейного изменения для qPCR-амплификации 1,6 градуса C/с и следующих условий: 1) 95°C в течение 10 минут, 2) 95°C в течение 15 секунд, 3) 55°C в течение 30 секунд, 4) повторение стадий 2-3 40x, 5) 95°C в течение 15 секунд, 6) 55°C в течение 1 минуты, 7) изменение скорости линейного изменения до 0,15 градуса C/с, 8) 95°C в течение 1 секунды. Данные qPCR анализировали с помощью аналитического программного обеспечения (Thermo Fisher Scientific, QuantStudio Design and Analysis).
Обработка раствором дрожжей обеспечивала замедление развития тли
Тлей LSR-1 первой и второй возрастных стадий разделяли на две группы обработки, как определено в разделе Схема эксперимента с перфузией в листья. Тлей ежедневно контролировали и определяли количество тлей на каждой стадии развития. Ко дню 6 эксперимента примерно 30% тлей, питающихся на листьях, в отношении которых осуществляли перфузию водой, достигали стадии личинки пятого возраста (фиг. 1). Для сравнения примерно 3% тлей, питающихся на листьях, в отношении которых осуществляли перфузию с использованием P. рastoris,достигали стадии личинки пятого возраста ко дню 6 эксперимента (фиг. 1). Большая часть тлей, питающихся листьями, в отношении которых осуществляли перфузию с использованием P. pastoris, которые выжили ко дню 6, находились на 4ой возрастной стадии (~5%) (фиг. 1). Эти данные указывают на то, что обработка с использованием P. pastoris обеспечивала замедление развития тли.
Обработка с применением раствора дрожжей обеспечивала повышение смертности тли
Показатель выживаемости тлей также измеряли в ходе обработок. Примерно 55,5% тлей, питающихся листьями, в отношении которых осуществляли перфузию с использованием воды, выживали ко дню 6 эксперимента (фиг. 2). Для сравнения тли, питающиеся на листьях, в отношении которых осуществляли перфузию с использованием P. рastoris,, начинали быстро погибать со дня 2 после обработки, и ко дню 6 лишь 11% тлей оставались живыми (фиг. 2). Эти данные указывают на то, что обработка с использованием P. рastoris,, доставляемых посредством перфузии в листья, обеспечивала в результате значительное (p<0,0001) повышение показателя смертности тли.
В частности, для исследования того, приводила ли обработка P. pastoris к потере Buchnera у тлей и влияла ли эта потеря на жизнеспособность тлей, ДНК экстрагировали из тлей в каждой группе обработки после 6 дней обработки, и проводили qPCR для определения количества копий Buchnera/тлей. Результаты были неубедительными по причине низкого количества тлей, оставшихся в группе обработки P. pastoris ко дню 6, и отсутствия экстрагируемой ДНК Buchnera.
Схема эксперимента с опрыскиванием с применением пульверизатора.
Тлей (линии LSR-1, Acyrthosiphon pisum) выращивали на растениях конских бобов, как описано в данном документе. Для экспериментов тлей на стадии первого и второго возраста собирали на здоровых растениях и разделяли на 2 разные группы обработки: 1) тли, помещенные на листья, опрысканные водой (отрицательным контролем), и 2) тли, помещенные на листья, опрысканные P. pastoris.
В экспериментах использовали штамм с нокаутом по ade2 "протеазы дикого типа" (штамм 1) из системы Pichia Pink (Thermo Fisher Scientific). P. pastoris выращивали в YPD в течение ночи при 30°C, а на следующий день культуру промывали однократно водой и повторно суспендировали в воде до достижения конечного показателя 0,918 OD600 и использовали для опрыскивания первого и второго листьев (в день 0 и 3), и 5,58 OD600 и использовали для завершающего опрыскивания листьев в день 6 эксперимента. С целью обработок листья конских бобов отрезали и черешки помещали в 1,5 мл пробирку эппендорф со стерильной водой и опрыскивали с обеих сторон водой или P. pastoris с помощью пульверизатора в концентрации, указанной выше. Листья затем помещали в глубокую чашку Петри (Fisher Scientific, № по кат. FB0875711), а на листья растения помещали тлей для обеспечения кормления. В дни 3 и 6 эксперимента старые листья заменяли новыми, которые недавно опрыскивали.
Для каждой группы обработки получали примерно одно и то же число особей с каждого из растений коллекции. Для каждой обработки на каждый лист помещали 30 тлей. Каждую обработку проводили в двухкратной повторности с общим количеством 60 тлей/группа обработки. Ежедневно осуществляли контроль тлей в отношении их выживаемости, и мертвых тлей удаляли в случае их обнаружения. Помимо этого, стадию развития (1-ю, 2-ю, 3-ю, 4-ю и 5-ю возрастные стадии) определяли ежедневно на протяжении всего эксперимента.
Через 6 и 9 дней после обработки экстрагировали ДНК из нескольких тлей из каждой группы обработки, и проводили qPCR для количественной оценки уровней Buchnera так, как описано в данном документе.
Обработка с помощью раствора дрожжей не оказывала значительного влияния в отношении развития тли
Личинок тлей LSR-1 первой и второй возрастной стадии распределяли в две группы обработки, как определено в разделе Схема эксперимента с опрыскиванием с применением пульверизатора, описанном в данном документе. Тлей ежедневно контролировали и определяли количество тлей на каждой стадии развития. К дню 6 после обработки тли из обеих групп обработки достигали 5ой возрастной стадии (фиг. 3). К дню 9 после обработки практически все оставшиеся тли в каждой группе достигали зрелого возраста (фиг. 3). Эти данные свидетельствуют о том, что имело место незначительное отличие в развитии тлей, обработанных водой или P. pastoris.
Обработка с помощью раствора дрожжей посредством опрыскивания приводила к повышенной смертности тли
Показатель выживаемости тлей также измеряли в ходе экспериментов по опрыскиванию листьев. В ходе проведения эксперимента лишь несколько тлей, которые кормились на листьях, опрысканных водой, погибли, при этом большее количество тлей погибло в каждой временной точке в группе обработки P. рastoris (фиг. 4). В частности, в день 2 после обработки 91% из обработанных водой тлей оставались живыми, тогда как лишь 80% из тлей, обработанных P. рastoris, оставались живыми. Данная тенденция сохранялась в течение 3, 6 и 7 дней эксперимента, где 82%, 66% и 60% из обработанных водой тлей оставались живыми в каждой временной точке соответственно, в отличие от всего лишь 73%, 55% и 49% тлей, обработанных P. рastoris, которые оставались живыми в каждой временной точке соответственно (фиг. 4). Эти данные свидетельствовали о том, что обработка с помощью P. pastoris путем доставки с применением опрыскивания пульверизатором приводила к более высокой смертности по сравнению с обработкой только водой.
Обработка с помощью раствора дрожжей обеспечивала снижение количества титров Buchnera у тлей
В частности, для исследования того, приводили ли P. рastoris,, доставляемые посредством опрыскивания пульверизатором, к потере Buchnera у тлей и влияла ли эта потеря на жизнеспособность тлей, экстрагировали ДНК из тлей в каждой группе обработки через 6 и 9 дней обработки и проводили qPCR для определения количества копий Buchnera/тлей. В день 6 после обработки средние значения соотношения Buchnera/тлей составляли 47,5 у тлей, которые кормились на листьях, подвергнутых опрыскиванию водой (фиг. 5). Для сравнения, средние значения соотношения Buchnera/тлей были в примерно 1,2 раза ниже (~39) у тлей, которые кормились на листьях, подвергнутых опрыскиванию P. pastoris (фиг. 5). Однако через 9 дней после обработки соотношения копий ДНК Buchnera/тли были сходными в обоих группах обработки (фиг. 5).
Такие данные указывают на то, что нанесение P. pastoris с помощью опрыскивания на листья конских бобов обеспечивает снижение количества эндосимбиотических Buchnera у тлей через 6 дней после обработки. Существовала возможность того, что обработанные P. pastoris тли, которые выжили через 9 дней после обработки, смогли преодолеть неблагоприятные эффекты повышенного иммунного ответа, и это могло дать объяснение, почему у этих тлей имели место титры Buchnera, аналогичные титрам у контрольных особей, обработанных водой. Тем не менее, оценка медианного значения копий Buchnera/тлей в день 9 выявила тенденцию к снижению Buchnera в группе обработки P. pastoris.
Подтверждение того, что P. pastoris обеспечивают положительную регуляцию иммунного ответа у тлей
Принимая во внимание то, что обработка с использованием P. pastoris в результате приводила к снижению приспособленности тли и к понижению титров эндосимбионта тли, Buchnera, целью следующего эксперимента было подтверждение того, что P. pastoris обеспечивает положительную регуляцию иммунного ответа. Чтобы это проверить, из тлей выделяли РНК в каждом из описанных в данном документе экспериментов (схема эксперимента с перфузией в листья и схема эксперимента с опрыскиванием с применением пульверизатора) и измеряли экспрессию генов, вовлеченных в иммунный ответ на грибы (Persephone, Cactus, and Dorsal) как описано в Gerardo et al., 2010, Genome Biology, 11:R21, https://doi.org/10.1186/gb-2010-11-2-r21. Предполагалось выявить положительную регуляцию генов, вовлеченных в данный путь.
В своей совокупности эти данные, описанные в этих примерах, демонстрируют способность уничтожать эндогенные бактериальные популяции и снижать их развитие, продолжительность жизни и количество, например, приспособленность у тли, за счет их обработки с использованием P. рastoris,, что, очевидно, приводит в результате к активации иммунного ответа у тли.
Пример 18. Обработка тлей с помощью раствора нуклеиновой кислоты с пептидом
В данном примере демонстрируется, что обработка тлей нуклеиновой кислотой с пептидом (PNA), целенаправленно воздействующей на BCR-4, cлитой с проникающим в клетку пептидом (CPP) (Cermenati et al., 2011; Zhou et al., 2015), в данном документе называемой PNA, целенаправленно воздействующей на BCR-4, или PNA BCR-4, приводит в результате к нокдауну экспрессии гена BCR-4 и снижению приспособленности тли.
BCR-4 является одним из нескольких богатых цистеином секретируемых пептидов, которые экспрессируются в бактериоцитах тли гороховой Acyrthosiphon pisum. Облигатный бактериальный симбионт Buchnera у тли обитает внутри бактериоцитов. BCR-4 характеризуется последовательностью, гомологичной по отношению к множеству специфичных для клубеньков богатых цистеином пептидов (NCR), вовлеченных в поддержание количества бактериальных симбионтов у проверяемых растений (Pan and Wang, 2017 Nature Plants and Durgo et al., 2015 Proteomics). С учетом сходства последовательностей BCR-4 и NCR, в данном примере демонстрируется, что BCR-4 выполняет аналогичную функцию в поддержании гомеостаза эндосимбионтов внутри бактериоцита за счет блокирования BCR-4 посредством нокдауна гена с нарушением регуляции Buchnera, за счет чего осуществляется отрицательное воздействие на приспособленность тли.
Структура терапевтического средства
PNA BCR-4 доставляли посредством либо микроинъекции, либо перфузии в листья конских бобов. Для экспериментов с микроинъекцией инъекционные растворы представляли собой либо воду (отрицательный контроль), либо PNA BCR-4 в воде. В исследованиях с перфузией в листья конских бобов обрабатывали либо с помощью перфузии водой (отрицательным контролем), либо PNA BCR-4 в воде. Каждая схема доставки в эксперименте подробно пояснена ниже.
Доставка путем микроинъекции - схема эксперимента
Микроинъекцию осуществляли с помощью автоматического инъектора NanoJet III Auto-Nanoliter Injector с выдвижной иглой из боросиликатного стекла собственного изготовления (Drummond Scientific; № по кат. 3-000-203-G/XL). Тлей переносили с использованием малярной кисточки в систему трубок, соединенную с вакуумным насосом, что удерживает их на месте во время осуществления инъекции. Место инъекции находилось в вентральной части груди тли. Растворами для инъекций были вода (отрицательный контроль) или 321 нг/мкл PNA BCR-4 в воде. Объем инъекции составлял 20 нл для взрослой особи (стадии 4 и 5 возраста) при скорости 2 нл/сек. В каждой группе обработки было примерно одинаковое количество получивших инъекцию особей с каждого из растений коллекции. После инъекции тлей выпускали в чашки Петри на листья конских бобов, черешки которых находились в пробирке эппендорф, заполненной 1 мл воды. Выживаемость и плодовитость контролировали ежедневно в течение эксперимента.
Вскармливание и содержание тли.
Тлей LSR-1 (которые переносят только Buchnera) A. pisum выращивали на растениях конских бобов (бобы Vroma vicia от Johnny's Selected Seeds) в инкубаторе с контролируемым климатом (фотопериод: 16 ч. света/8 ч. темноты; 60 ± 5% RH; 25 ± 2°C). Перед использованием для скармливания тлям растения конских бобов выращивали в почвенной смеси для горшечных культур при 24°C при 16 ч. света и 8 ч. темноты. Для ограничения материнских эффектов или различий в состоянии здоровья между растениями 5-10 взрослых особей из разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии четвертого и пятого возраста собирали на здоровых растениях и разделяли на две разные группы обработки: 1) вода (отрицательный контроль) или 2) PNA BCR-4 в воде.
Синтез PNA BCR-4
BCR-4-CPP синтезировали с помощью PNA bio, и последовательность представляла собой YGRKKRRQRRR-CGTACAATAATCTCATGG; SEQ ID NO: 105 и 106. Последовательность CPP (Tat) представляла собой YGRKKRRQRRR; SEQ ID NO: 106. PNA растворяли в смеси 80% ацетонитрила и 20% воды, обогащенной 0,1% трифторуксусной кислотой (TFA). После растворения PNA разделяли на аликвоты [32,1 мкг/5 нмоль на аликвоту], высушивали на воздухе и хранили при -20°C. Растворы с рабочими концентрациями PNA получали в воде при 50 мкM.
Через 7 дней обработки РНК экстрагировали из оставшихся тлей в каждой группе обработки. Вкратце, поверхность тела тлей стерилизовали путем погружения тли в 6% раствор отбеливателя примерно на 5 секунд. Затем тлей промывали в стерильной воде, и общую РНК экстрагировали из каждой отдельной тли с помощью набора для экстрагирования РНК (Qiagen, набор miRNeasy) в соответствии с инструкциями производителя. Концентрации РНК измеряли с использованием количественного определения нуклеиновой кислоты с помощью NanoDrop. Относительную экспрессию BCR-4 измеряли с помощью RT-qPCR. Использовали следующие праймеры: ApBCR-4F (CTCTGTCAACCACCATGAGATTA; SEQ ID NO: 107) и ApBCR-4R (TGCAGACTACAGCACAATACTT; SEQ ID NO: 108). Праймерами для внутреннего референтного гена были праймеры для актина (гена домашнего хозяйства). Последовательность прямого праймера представляла собой GATCAGCAGCCACACACAAG; SEQ ID NO: 109, а последовательность обратного праймера представляла собой TTTGAACCGGTTTACGACGA; SEQ ID NO: 110. RT-qPCR проводили с применением скорости линейного изменения для qPCR-амплификации 1,6 градуса C/с и следующих условий: 1) 48°C в течение 30 мин, 2) 95°C в течение 10 минут, 3) 95°C в течение 15 секунд, 4) 60°C в течение 1 минуты, 5) повторение стадий 3-4 40x, 6) 95°C в течение 15 секунд, 7) 60°C в течение 1 минуты, 8) изменение скорости линейного изменения до 0,15 градуса C/с, 9) 95°C в течение 1 секунды. Данные RT-qPCR анализировали с помощью аналитического программного обеспечения (Thermo Fisher Scientific, QuantStudio Design and Analysis).
Микроинъекция PNA, направленной против гена BCR-4, в результате приводит к снижению экспрессии BCR-4
Чтобы проверить, будет ли PNA, направленная против BCR-4, доставляемая с помощью микроинъекции, приводить в результате к снижению экспрессии гена BCR-4 у тлей, тлям вводили воду (контроль) или PNA BCR-4. Через 7 дней обработки РНК экстрагировали из тлей в каждой группе обработки и проводили RT-qPCR. Тли, которым с помощью микроинъекции вводили воду, демонстрировали примерно 2-кратное повышение экспрессии BCR-4 по сравнению с тлями, которым вводили PNA BCR-4 (фиг. 6), что указывает на то, что инъекция PNA BCR-4 приводила в результате к нокдауну BCR-4.
Обработка с помощью PNA, целенаправленно воздействующей на BCR-4, обеспечивает повышение смертности тли
Показатель выживаемости тлей также измеряли в ходе обработок. В большей части временных точек в ходе эксперимента живых тлей наблюдалось больше среди контрольных (инъекция воды) тлей по сравнению с тлями, которым вводили PNA, целенаправленно воздействующую на BCR-4 (фиг. 7). Эти данные указывают на то, что имело место незначительное снижение выживаемости при инъекции PNA, целенаправленно воздействующей на BCR-4.
Обработка с помощью PNA, целенаправленно воздействующей на BCR-4, обеспечивает снижение плодовитости тли
Количество потомства, полученного от тлей в каждой группе обработки, также оценивали в ходе проведения эксперимента, и плодовитость выражали в виде числа потомков, полученных от одной взрослой особи в каждую оцениваемой временной точке. В общем наблюдали тенденцию, заключающуюся в том, что контрольные (которым инъекцировали воду) тли давали большее количество потомства по сравнению с теми, которым инъецировали PNA, целенаправленно воздействующую на BCR-4. В частности, в дни 3 и 7 после обработки тли в группе обработки водой производили в среднем 5 потомков/взрослая особь, при этом тли в группе обработки PNA BCR-4 производили лишь 4 потомков/взрослая особь (фиг. 8). Эти данные указывают на то, что обработка с использованием PNA BCR-4 в результате приводила к незначительному снижению плодовитости у тлей.
Схема эксперимента с перфузией в листья
Тлей (линии LSR-1, Acyrthosiphon pisum) выращивали на растениях конских бобов (бобы Vroma vicia от Johnny's Selected Seeds) в инкубаторе с контролируемым климатом (фотопериод: 16 ч. света/8 ч. темноты; 60 ± 5% RH; 25 ± 2°C). Перед использованием для скармливания тлям растения конских бобов выращивали в почвенной смеси для горшечных культур при 24°C при 16 ч. света и 8 ч. темноты. Для ограничения материнских эффектов или отличий в состоянии здоровья между растениями 5-10 взрослых особей с разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии первого и второго возраста собирали на здоровых растениях и разделяли на две разные группы обработки: 1) которым обеспечивали кормление на листьях, подвергнутых перфузии водой, и 2) которым обеспечивали кормление на листьях, подвергнутых перфузии раствором PNA BCR-4 в воде.
PNA BCR-4, слитую с CPP, синтезировали, как описано в данном документе (см. "Синтез PNA BCR-4 для схемы эксперимента с доставкой посредством микроинъекции").
Для каждой перфузии в лист примерно 1 мл воды (отрицательного контроля) или 1 мкM раствора PNA BCR-4 инъецировали в лист конских бобов. Черешок листа затем помещали в 1,5 мл пробирку эппендорф, которую герметично закрывали парафильмом. Стебель листа затем помещали в глубокую чашку Петри (Fisher Scientific, № по кат. FB0875711), а на листья растения помещали тлей для обеспечения кормления. В ходе проведения эксперимента каждые 2-3 дня старые листья заменяли новыми, которые недавно подвергли инъекции. Для каждой обработки на каждый лист помещали 60 тлей. Ежедневно осуществляли контроль тлей в отношении их выживаемости, и мертвых тлей удаляли в случае их обнаружения. Помимо этого, стадию развития (1-ю, 2-ю, 3-ю, 4-ю и 5-ю возрастные стадии) определяли ежедневно на протяжении всего эксперимента.
Через 5 и 6 дней обработки экстрагировали ДНК из мертвых тлей из каждой группы обработки. Вкратце, поверхность тела тлей стерилизовали путем погружения тли в 6% раствор отбеливателя примерно на 5 секунд. Затем тлей промывали в стерильной воде, и ДНК экстрагировали из каждой отдельной тли с помощью набора для экстракции ДНК (Qiagen, набор DNeasy) в соответствии с инструкциями производителя. Концентрацию ДНК измеряли посредством количественной оценки нуклеиновой кислоты с использованием NanoDrop, и число копий ДНК Buchnera и тлей измеряли с помощью qPCR. Праймерами, используемыми для Buchnera, были Buch_groES_18F (CATGATCGTGTGCTTGTTAAG; SEQ ID NO: 101) и Buch_groES_98R (CTGTTCCTCGAGTCGATTTCC; SEQ ID NO: 102) (Chong and Moran, 2016 PNAS). Праймерами, используемыми для тлей, были ApEF1a 107F (CTGATTGTGCCGTGCTTATTG; SEQ ID NO: 103) и ApEF1a 246R (TATGGTGGTTCAGTAGAGTCC; SEQ ID NO: 104) (Chong and Moran, 2016 PNAS). Проводили qPCR с применением скорости линейного изменения для qPCR-амплификации 1,6 градуса C/с и следующих условий: 1) 95°C в течение 10 минут, 2) 95°C в течение 15 секунд, 3) 55°C в течение 30 секунд, 4) повторение стадий 2-3 40x, 5) 95°C в течение 15 секунд, 6) 55°C в течение 1 минуты, 7) изменение скорости линейного изменения до 0,15 градуса C/с, 8) 95°C в течение 1 секунды. Данные qPCR анализировали с помощью аналитического программного обеспечения (Thermo Fisher Scientific, QuantStudio Design and Analysis).
В день 7 после обработки РНК экстрагировали из живых тлей и проводили RT-pPCR, как описано выше (см. "Схема эксперимента с доставкой с помощью микроинъекции"), для количественной оценки экспрессии BCR-4.
Обработка посредством перфузии в листья PNA BCR-4 обеспечивала замедление развития тлей
Тлей LSR-1 первой и второй возрастных стадий разделяли на две группы, как определено в разделе "Схема эксперимента с перфузией в листья". Тлей ежедневно контролировали и определяли количество тлей на каждой стадии развития. В некоторых временных точках в течение эксперимента у тлей, обработанных PNA BCR-4, развитие замедлялось. Например, в день 2 после обработки примерно 19% тлей, которые кормились на листьях, обработанных посредством перфузии воды, находились на 3-й возрастной стадии (фиг. 9). Для сравнения, лишь 8% тлей, которые кормились на листьях, обработанных посредством перфузии PNA, целенаправленно воздействующей на BCR-4, находились на 3-й возрастной стадии. Эти данные указывают на то, что обработка с использованием PNA целенаправленно воздействующей на BCR-4, замедляла развитие тлей.
Обработка посредством перфузии в листья PNA BCR-4 обеспечивала повышение смертности тлей
Выживаемость также отслеживали на в ходе всей обработки. Через 7 дней после обработки 53% тлей в группе обработки водой (контролем) оставались живыми (фиг. 10). Для сравнения, лишь 30% тлей в группе обработки PNA BCR-4 оставались живыми в день 7 (фиг. 10). Эти данные показали, что обработка PNA, целенаправленно воздействующей на BCR-4, приводила в результате к повышенной смертности тлей.
Обработка посредством перфузии в листья PNA BCR-4 обеспечивала повышение титров Buchnera у тлей
Для исследования того, оказывала ли, в частности, обработка с помощью PNA BCR-4 путем перфузии в лист влияние на Buchnera у тлей, и оказывало ли это воздействие на приспособленность тлей, экстрагировали ДНК из тлей в каждой группе обработки через 5 и 6 дней после обработки и проводили qPCR для определения титров Buchnera в каждой группе обработки. Хотя у тлей, которые кормились на листьях, подвергнутых перфузии водой, среднее соотношение копий Buchnera/тлей составляло примерно 20, тли, которые кормились на листьях, подвергнутых перфузии PNA, целенаправленно воздействующей на BCR-4, характеризовались значительно большим соотношением копий Buchnera/тлей (примерно 57), см. фиг. 11. Эти данные указывают на то, что обработка с использованием PNA, целенаправленно воздействующей на BCR-4, приводит к нарушению равновесия титров Buchnera.
Обработка с помощью PNA, целенаправленно воздействующей на BCR-4, посредством перфузии в лист обеспечивает нокдаун BCR-4
Чтобы оценить, будет ли экспрессия BCR-4 снижаться у тлей, которые кормились на листьях, подвергнутых перфузии PNA, целенаправленно воздействующей на BCR-4, РНК выделяли из живых тлей через 7 дней обработки и проводили RT-qPCR. Медианное значение уровней транскрипта BCR-4 у тлей, обработанных PNA, целенаправленно воздействующей на BCR-4, было в примерно 3 раза ниже по сравнению с тлями, обработанными только водой (фиг. 12), что подтверждает нокдаун экспрессии BCR-4 за счет PNA, целенаправленно воздействующей на BCR-4.
В своей совокупности эти данные, описанные в этих примерах, демонстрируют способность уничтожать тлей и снижать их развитие, плодовитость и продолжительность жизни (например, приспособленность) с помощью их обработки с использованием PNA, целенаправленно воздействующей на ген, экспрессируемый в бактериоцитах (BCR-4), с целью контроля популяции эндосимбионта Buchnera
Пример 19. Обработка тлей с помощью раствора dsRNA, направленной против транспортеров бактериоцитов
В данном примере демонстрируется, что обработка тлей двухнитевой РНК (dsRNA) приводит к нокдауну ряда важнейших генов, включая ген, кодирующий транспортeр 1 глутамина (ApGLNT1), который был идентифицированный в бактериоцитах у тли, Acyrthosiphon pisum. Транспортер глутамина является ответственным за захват глутамина из гемолимфы тли с доставкой в бактериоциты, где располагается облигатный эндосимбионт Buchnera aphidicola. Объединенная биосинтетическая способность голофита (A. pisum и Buchnera) является достаточной для биосинтеза всех двадцати кодирующих белки аминокислот, включая аминокислоты, которые у тлей не могут синтезироваться сами. Блокирование захвата глутамина путем инактивации или сайленсинга (например, РНК-интерференции с помощью dsRNA) транспортeра глутамина отрицательно воздействует на синтез аминокислот и белков в бактериоцитах и во всем организме тли, что тем самым оказывает отрицательное воздействие на их приспособленность.
Структура терапевтического средства
Тлей LSR-1 на стадии 5го возраста подвергали микроинъекции. Растворы для инъекций представляли собой или dd-воду (отрицательный контроль), или dsRNA, разбавленную в dd-воде в разных концентрациях (8 или 60 нг/тля; см. ниже).
Вскармливание и содержание тли.
Тлей LSR-1 (которые переносят только Buchnera) A. pisum выращивали на растениях конских бобов (бобы Vroma vicia faba от Johnny's Selected Seeds) в инкубаторе с контролируемым климатом (фотопериод: 16 ч. света/8 ч. темноты; 60 ± 5% RH; 25 ± 2°C). Перед использованием для скармливания тлям растения конских бобов выращивали в почвенной смеси для горшечных культур при 24°C при 16 ч. света и 8 ч. темноты. Для ограничения материнских эффектов или различий в состоянии здоровья между растениями 5-10 взрослых особей из разных растений распределяли среди 10 растений в возрасте двух недель и оставляли размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей пятой возрастной стадии собирали на здоровых растениях и разделяли на 2 разные группы обработки: 1) вода (отрицательный контроль) или 2) dsRNA, целенаправленно воздействующая на ApGLNT1 (в концентрациях, указанных в данном документе).
Доставка путем микроинъекции - схема эксперимента
Микроинъекцию осуществляли с помощью автоматического инъектора NanoJet III Auto-Nanoliter Injector с выдвижной иглой из боросиликатного стекла собственного изготовления (Drummond Scientific; № по кат. 3-000-203-G/XL). Тлей переносили с использованием малярной кисточки в систему трубок, соединенную с вакуумным насосом, что удерживает тлей на месте во время осуществления микроинъекции. Место инъекции находилось в вентральной части груди тли. Объем инъекции составлял 20 нл для взрослой особи (при скорости 2 нл/сек). В каждой группе обработки было примерно одинаковое количество получивших инъекцию особей с каждого из растений коллекции..
После инъекции тлей выпускали в чашки Петри на листья конских бобов, черешки которых находились в пробирке эппендорф, заполненной 1 мл воды. Выживаемость тли контролировали ежедневно и мертвых тлей удаляли после их выявления. Подсчитывали количество потомства от каждой группы, а плодовитость оценивали по количеству потомства (F1), полученного на одну тлю в каждой временной точке.
В выбранных экспериментах развитие оценивали в группах потомства, полученного от каждой группы обработки, путем ежедневной регистрации стадии развития потомства (стадии 1го, 2го, 3го, 4го и 5го возраста). Развитие также оценивали путем визуального осмотра тлей через 4 дня после сбора и определения их площади. Матрица для синтезирования dsRNA представляла собой cDNA, полученную путем обратной транскрипции с mRNA. РНК экстрагировали из одной особи на стадии 5го возраста A. pisum (линия LSR-1) и количественно определяли с помощью Nanodrop (Thermo fisher scientific). ~100 мкг общей РНК добавляли в качестве матрицы в реакционную смесь для обратной транскрипции с применением набора Superscript IV (Thermo Fisher Scientific), следуя протоколу изготовителя.
Для амлификации матриц для получения dsRNA cDNA разбавляли в 100 раз и в дальнейшем использовали в ПЦР. Реакционные смеси для ПЦР (конечный объем 25 мкл) содержали 12,5 мкл смеси 2X Go Taq Green (Promega), 0,2 мкл прямого праймера (таблица 13), 0,2 мкл обратного праймера (таблица 13) и 12,1 мкл cDNA, разбавленной в 100 раз. ПЦР-реакции проводили с использованием следующий условий: 1) 95°C в течение 2 минут, 2) 95°C в течение 20 секунд, 3) 55°C в течение 15 секунд, 4) 72°C в течение 30 секунд, 5) повтор стадий 2-4 35x, 6) 72°C в течение 5 минут. Размеры амплифицированных продуктов ПЦР определяли с помощью электрофореза в 1,5% агарозном геле и полосы ожидаемого размера вырезали и очищали с помощью набора для очистки ДНК QIAquick (Qiagen). Синтезировали dsRNA in vitro с применением набора T7 MEGAscript (Ambion, Thermo Fisher Scientific; № по кат. AM1334), следуя протоколу изготовителя. Концентрацию dsRNA измеряли с помощью Nanodrop (Thermo Fisher Scientific).
Таблица 13. Номера доступа и праймеры, предназначенные для синтеза dsRNA
В указанные временные точки после обработки ДНК и/или РНК экстрагировали из тлей в каждой группе обработки. Вкратце, поверхность тела тлей стерилизовали путем погружения тли в 6% раствор отбеливателя примерно на 5 секунд. Затем тлей промывали в стерильной воде, и ДНК или общую РНК экстрагировали из каждой отдельной тли с помощью набора для экстрагирования ДНК или РНК (соответственно набор DNeasy или miRNeasy) в соответствии с инструкциями производителя. Концентрации ДНК и РНК измеряли с применением количественного определения нуклеиновой кислоты с помощью NanoDrop. Количество копий ДНК Buchnera и тли измеряли с помощью qPCR. Праймерами, используемыми для Buchnera, были Buch_groES_18F (CATGATCGTGTGCTTGTTAAG; SEQ ID NO: 101) и Buch_groES_98R (CTGTTCCTCGAGTCGATTTCC; SEQ ID NO: 102) (Chong and Moran, 2016 PNAS). Праймерами, используемыми для тлей, были ApEF1a 107F (CTGATTGTGCCGTGCTTATTG; SEQ ID NO: 103) и ApEF1a 246R (TATGGTGGTTCAGTAGAGTCC; SEQ ID NO: 104) (Chong and Moran, 2016 PNAS). Проводили qPCR с применением скорости линейного изменения для qPCR-амплификации 1,6 градуса C/с и следующих условий: 1) 95°C в течение 10 минут, 2) 95°C в течение 15 секунд, 3) 55°C в течение 30 секунд, 4) повторение стадий 2-3 40x, 5) 95°C в течение 15 секунд, 6) 55°C в течение 1 минуты, 7) изменение скорости линейного изменения до 0,15 градуса C/с, 8) 95°C в течение 1 секунды. Данные qPCR анализировали с помощью аналитического программного обеспечения (Thermo Fisher Scientific, QuantStudio Design and Analysis). Относительную экспрессию ApGLNT1 измеряли с помощью RT-qPCR. Праймерами, используемыми для ApGLNT1, были ACYPI001018-fwd (CCTGAAATCGACGGGGTCC; SEQ ID NO: 113) и ACYPI001018-rev (AGATCGGCAACATCTGTTCGT; SEQ ID NO: 114) (оба праймера были сконструированы с помощью NCBI Pick Primers). Внутренним референтным геном выступал ген актина (ген домашнего хозяйства). Праймерами, используемыми для актина, были Actin-F (GATCAGCAGCCACACACAAG; SEQ ID NO: 109) и Actin-R (TTTGAACCGGTTTACGACGA; SEQ ID NO: 110). RT-qPCR проводили с использованием скорости линейного изменения для qPCR-амплификации 1,6 градуса C/с и следующих условий: 1) 48°C в течение 30 мин, 2) 95°C в течение 10 минут, 3) 95°C в течение 15 секунд, 4) 60°C в течение 1 минуты, 5) повторение стадий 3-4 40x, 6) 95°C в течение 15 секунд, 7) 60°C в течение 1 минуты, 8) изменение скорости линейного изменения до 0,15 градуса C/с, 9) 95°C в течение 1 секунды. Данные RT-qPCR анализировали с помощью аналитического программного обеспечения (Thermo Fisher Scientific, QuantStudio Design and Analysis).
Микроинъекция с использованием dsRNA обеспечивала нокдаун экспрессии гена, кодирующего транспортер, в бактериоците у тлей
В предварительном эксперименте оценивали, будет ли инъецирование dsRNA тлям в результате приводить к понижению экспрессии гена. Взрослым особям тли инъецировали 8 нг dsRNA или воды (в качестве отрицательного контроля). Через два дня после инъекции РНК экстрагировали из остальных тлей в каждой группе обработки и RT-qPCR проводили для количественного определения экспрессии ApGLNT1. У тлей, которым посредством микроинъекции вводили раствор отрицательного контроля (воду), экспрессия гена ApGLNT1 была относительно высокой. Для сравнения, у взрослых особей тлей, которым посредством микроинъекции вводили dsRNA, направленную против ApGLNT1, наблюдалось резкое и значительное снижение экспрессии гена ApGLNT1 (фиг. 13), указывающее на то, что обработка посредством микроинъекции dsRNA снижала экспрессию гена ApGLNT1.
Микроинъекция с использованием dsRNA-ApGLNT1 приводила к повышенной смертности тлей
Для оценки влияния микроинъекции с помощью dsRNA-ApGLNT1 на приспособленность насекомых тлям LSR-1 пятой возрастной стадии инъецировали 60 нг dsRNA и контролировали их выживаемость в течение 5 дней. В день 3 после инъекции примерно 72% тлей из группы, которым инъецировали воду, оставались живыми (фиг. 14). Для сравнения, только 52% тлей, которым инъецировали dsRNA, оставались живыми (фиг. 14). В дни 4 и 5 после инъекции оставалось значительно большее количество живых тлей в группе, которым инъецировали воду, по сравнению с группой, которым вводили dsRNA. Примерно 62% и 51% тлей, которым инъецировали воду, оставались живыми соответственно в дни 4 и 5, при этом только 30% и 12,5% тлей, которым вводили dsRNA, оставались живыми соответственно в день 4 и 5 (фиг. 14). Эти данные указывают на то, что dsRNA, направленные против ApGLNT1, в значительной степени (p=0,0004) повышали смертность тли по сравнению с контролями, которым вводили воду.
Микроинъекция с помощью dsRNA-ApGLNT1 в результате приводит к снижению титров Buchnera
Чтобы проверить, было ли снижение выживаемости и приспособленности обусловлено пониженным количеством эндосимбионтов, ДНК экстрагировали из живых тлей через 5 дней после инъекции и проводили qPCR с целью подсчета количества Buchnera, присутствующих у тлей. У тлей, которым вводили воду путем микроинъекции, среднее соотношение копий Buchnera/тлей составляло примерно 11 (фиг. 15). Для сравнения, количество копий Buchnera/тлей было в примерно 1,28 раза ниже у тлей, которым вводили dsRNA, направленную против ApGLNT1 (фиг. 15). Эти данные показали, что микроинъекция с помощью dsRNA-ApGLNT1 обеспечивала в результате снижение титров Buchnera, что приводит к снижению приспособленности тли.
Развитие замедлялось у потомства тлей, которым вводили с помощью микроинъекции dsRNA-ApGLNT1
В день 3 после инъекции 40 потомков (первой возрастной стадии) из каждой группы обработки переносили на их собственную искусственную систему кормления система (чашку петри, содержащую листья конских бобов, черешок которых погружен в 1,5 мл пробирку эппендорф, которую герметично закрывали парафильмом) и развитие контролировали в динамике. В целом развитие замедлялось у потомства, полученного от взрослых особей, которым вводили dsRNA-ApGLNT1. К дню 4 после переноса потомства примерно 22,5% потомства тлей, которым инъецировали воду, начали достигать 5ой возрастной стадии (фиг. 4A). Для сравнения, в день 4 только 7,5% потомков взрослых особей, которым вводили dsRNA-ApGLNT1, достигли 5ой возрастной стадии (фиг. 16A). Кроме того, измеряли площади тела тлей в день 4 путем визуального осмотра каждой тли в каждой группе. Хотя средний размер потомков от взрослых особей, которым инъецировали воду, составлял 0,55 мм2, размер потомков взрослых особей, которым инъецировали dsRNA-ApGLNT1, являлся в значительной степени меньшим (p=0,009) и в среднем составлял 0,4 мм2 (фиг. 16B). Эти данные указывают на то, что обработка взрослых особей с помощью dsRNA-ApGLNT1 в результате обеспечивает получение потомства, которое значительно отстает в развитии.
Пример 20. Получение трансгенных растений, экспрессирующих dsRNA, которые целенаправленно воздействуют на несколько путей, с целью нарушения гомеостаза насекомое-симбионт посредством обработки тлей трансгенными растениями
В данном примере демонстрируется возможность генетически модифицировать Nicotiana tabacum, чтобы продуцировать dsRNA для доставки тлям с целью отрицательного воздействия на гомеостаз насекомое-симбионт. Генетические конструкции, которые стабильно экспрессируют dsRNA в Nicotiana tabacum, доставляют в растение с применением трансгенной Agrobacterium tumefaciens, которая будет переносить плазмиду в растение.
Схема эксперимента
Несколько генов будут подвергаться целенаправленному воздействию с целью нокдауна во многих сигнальных путях, которые являются ключевыми для симбиотических взаимоотношений между тлями и их облигатными эндосимбиотическими бактериями Buchnera. В частности, целенаправленное воздействие будет осуществляться в отношении транспортера глутамина бактериоцитов (GlnT1), ультрабиторакса (Ubx), бета-аланин-синтазы (bAS) и cactus (Cact). GlnT1 представляет собой транспортер глутамина, который применяется для доставки глутамина в бактериоциты, и следующие в нисходящем направлении продукты глутамина являются ключевыми в синтезе незаменимых аминокислот с участием Buchnera. Ubx представляет собой ген, вовлеченный как в общее развитие тли, так и в образование бактериоцитов, которые будут содержать Buchnera. bAS представляет собой ген тли, необходимый для синтеза бета-аланина, который является предшественником в синтезе витамина B5 с участием Buchnera. Cact представляет собой отрицательный регулятор сигнального пути Toll, который является путем первичного иммунитета у тлей. Снижение уровней Cact может положительно регулировать системные иммунные ответы, что приводит к нарушению регуляции уровней Buchnera.
Получение плазмиды, содержащей кассету для экспрессии dsRNA.
Челночный вектор между E. coli и A. tumefaciens будет применяться для переноса кассеты для экспрессии dsRNA, которая включает промотор 35S вируса мозаики цветной капусты (pCaMV 35S) выше последовательности, экспрессирующей dsRNA. Последовательность, экспрессирующая dsRNA, включает смысловую последовательность, за которой следует антисмысловая последовательность участка гена-мишени тли, связанного с малой последовательностью шпилечной петли (фиг. 17).
Кассету для экспрессии dsRNA затем помещали в челночный вектор для E. coli и A. tumefaciens (фиг. 18). Плазмида также содержала кассеты устойчивости как к канамицину, так и к гентамицину, которые можно применять в качестве селективных маркеров. Последовательность терминатора транскрипции размещали после транскрипта, полученного в результате экспрессии dsRNA, для устранения безудержной транскрипции.
Экспрессирущие dsRNA последовательности, целенаправленно воздействующие на разные гены (GlnT1, Ubx, bAS и Cact), можно было вводить в вектор посредством сборки по Гибсону. Сперва с применением подходящих праймеров в ПЦР будут получены ампликоны смысловой и антисмысловой нитей с липкими концами, которые соответствуют прилегающим участкам в плазмиде. В частности, левый липкий конец смысловой нити будет содержать участок гомологии (~30 п.о.) с pCaMV 35S, а правый липкий конец антисмысловой нити будет содержать участок гомологии (~30 п.о.) с 35S терминаторным участком. Правый липкий конец смысловой нити и левый липкий конец антисмысловых нитей будут иметь перекрывающиеся друг с другом части с включением небольшой шпилечной области (acacgt, SEQ ID NO: 115). Последовательности праймеров, предназначенные для получения ампликонов, показаны в таблице 14.
Таблица 14. Перечень праймеров для получения ампликонов, предназначенных для сборки по Гибсону для получения последовательностей шпилечной dsRNA, направленной против генов A. pisum. Участок каждого гена-мишени будет амплифицирован или в виде смысловой, или антисмысловой нити с фланкирующими участками, которые гомологичны плазмидному остову, для сборки по Гибсону.
Таблица 15. Перечень праймеров для получения ампликонов, предназначенных для сборки по Гибсону для получения последовательностей шпилечной dsRNA, направленной против генов Myzus persicae. Участок каждого гена-мишени будет амплифицирован или в виде смысловой, или антисмысловой нити с фланкирующими участками, которые гомологичны плазмидному остову, для сборки по Гибсону.
Как только получают ампликоны смысловой и антисмысловой нитей, плазмида будет расщеплена в двух местах с получением липких концов на конце pCaMV 35S и в начале терминатора 35S. Расщепленные плазмида, ампликоны смысловой и антисмысловой нитей будут собранв вместе с помощью сборки по Гибсону с применением наборов для сборки по Гибсону (такого, как набор для сборки по Гибсону SGI). E. coli (DH5α, электрокомпетентные клетки) затем будут трансформировать плазмидами и выращивать в чашках с LB, содержащей 50 мг/мл канамицина. Устойчивые колонии будут использовать для получения плазмиды, содержащей кассету для экспрессии dsRNA. Плазмиды будут называться pGlnT1dsRNA, pUbxdsRNA, pbASdsRNA и pCactdsRNA в случае каждой из четырех разных вставок Эти плазмиды будут использовать для трансформации A. tumefaciens посредством электропорации. При отбора на среде LB, содержащей канамицин (50мг/мл) и гентамицин (50мг/мл), устойчивые колонии будут выделять и поддерживать на чашках для отбора.
Трансформированными A. tumefaciens инфильтрировали N. tabacum.
Трансформированные A. tumefaciens выращивали на протяжении ночи в среде LB с антибиотиками для отбора до достижения 0,6 при OD 600. Клетки осаждали, повторно суспендировали в инфильтрационной среде (10 мМ MES, 150 мкМ ацетосирингона и 10 мМ MgCl2, pH 5.5), и доводили до 0,6 OD600. Клетки инкубировали при комнатной температуре в течение 2-4 часов. Клеточной суспензией инфильтрировали здоровые листья N. tabacum. Способ инфильтрации осуществляли путем размещения обрезанного открытого конца 1 мл шприца с нижней стороны листа и принудительного нагнетания клеточной суспензии в лист. Площади инфильтратов можно было легко отличить от необработанных областей, и для наглядности их разграничивали с использованием маркера. Затем растения накрывали на 24 ч. с целью создания условий повышенной увлажненности.
В листьях экспрессировался зеленый флуоресцентный белок, и его визуально анализировали с помощью эпифлуоресцентного микроскопа через 1-2 дня, см. фиг. 19. Генетический материал, который переносили в растение посредством A. tumefaciens, содержал кассету для экспрессии зеленого флуоресцентного белка (GFP), которая управлялась убиквитиновым промотором, который являлся активным в N. tabacum. Экспрессию GFP будут применять в качестве индикатора экспрессии dsRNA.
Получение устойчивого клона N. tabacum, экспрессирующего dsRNA
Инфильтрированные листья, которые демонстрируют экспрессию GFP, будут выделять и вырезать участки листа, расположенные рядом с центральной жилкой, в которых очень сильно экспрессируется GFP. Затем листовые диски будут тщательно стерилизовать с применением стерилизующего раствора (2% гипохлорита, 0,01% твин 20) в течение 10 мин. при взбалтывании. Стерильные листовые диски будут помещать на чашки Петри, содержащие среду для роста побегов (2,15 г/л солей по Мурасиге и Скугу (без IAA, кинетина или сахарозы), 0,8% (вес/об.) агара, 3,0% (вес/об.) сахарозы, 0,1 мг/л индолмасляной кислоты, 0,8 мг/л 6-бензиламинопурина, 0,1 мг/л карбенициллина, 0,2 мг/л тикарциллина/клавулановой кислоты и подходящий селективный маркер для бинарного вектора, несущего слитую конструкцию на основе FP) при 25°C при цикле 16 ч:8 ч света:темноты до появления всходов. Новые побеги затем будут переносить на планшеты, содержащие среду для укоренения (2,15 г/л солей по Мурасиге и Скугу, 0,8% (вес/об.) агар, 3,0% (вес/об.) сахарозы, 0,5 мг/л индолмасляной кислоты, 0,1 мг/л карбенициллина, 0,2 мг/л тикарциллина/клавулановой кислоты) при 25°C при цикле 16 ч.:8 ч. света:темноты до появления корней. Эти новые растения будут переносить в ящики для выращивания растений с целью получения более крупных корней, в результате чего их можно будет пересадить в почву. Экспрессию GFP у всех новых клонов будут проверять для того, чтобы удостовериться в стабильной экспрессии.
Обработка тлей с помощью dsRNA путем их разведения на N. tabacum, экспрессирующем dsRNA
Тлей будут растить на растениях N. tabacum 10-недельного возраста в инкубаторе с регулируемом климатом (цикл свет:темнота 16 ч.:8 ч., 60% влажность, 25°C). Для ограничения материнских эффектов или отличий в состоянии здоровья между растениями 5-10 взрослых особей с разных растений будут распределять среди 10 растений в возрасте двух недель и оставлять размножаться до высокой плотности в течение 5-7 дней. Для экспериментов тлей на стадии личинок первого и второго возраста будут собирать со здоровых растений и разделять на две разные группы обработки: 1) которым обеспечивают кормление на листьях, которые экспрессируют dsRNA, и 2) которым обеспечивают кормление на контрольных листьях, которые не экспрессируют dsRNA.
В каждом эксперименте с кормлением с растения будут брать листья и помещать в 1,5 мл пробирки эппендорф, которые запечатывают парафильмом. Стебель листа затем будут помещать в глубокую чашку Петри (Fisher Scientific, № по кат. FB0875711), а на листья растения будут помещать тлей для обеспечения кормления. В ходе проведения эксперимента каждые 2-3 дня старые листья будут заменять новыми, которые недавно подвергли инъекции. Для каждой обработки на каждый лист будут помещать 60 тлей. Выживаемость тлей будут контролировать ежедневно и мертвых тлей удалять после их выявления. Помимо этого, стадию развития (1-ю, 2-ю, 3-ю, 4-ю и 5-ю возрастные стадии) будут определять ежедневно на протяжении всего эксперимента.
Через 5 и 6 дней обработки будут экстрагировать ДНК из мертвых тлей из каждой группы обработки. Вкратце, поверхность тела тлей будут стерилизовать путем погружения тлей в 6% раствор отбеливателя примерно на 5 секунд. Затем тлей будут промывать в стерильной воде и экстрагировать ДНК из каждой отдельной тли с помощью набора для экстрагирования ДНК (Qiagen, набор DNeasy) в соответствии с инструкциями производителя. Концентрацию ДНК будут измерять посредством количественной оценки нуклеиновой кислоты с применением NanoDrop, а число копий ДНК Buchnera и тлей измерять с помощью qPCR. Праймерами, которые будут использованы для Buchnera, являются Buch_groES_18F (CATGATCGTGTGCTTGTTAAG; SEQ ID NO: 101) и Buch_groES_98R (CTGTTCCTCGAGTCGATTTCC; SEQ ID NO: 102) (Chong and Moran, 2016 PNAS). Праймерами, которые будут использованы для тлей, являются ApEF1a 107F (CTGATTGTGCCGTGCTTATTG; SEQ ID NO: 103) и ApEF1a 246R (TATGGTGGTTCAGTAGAGTCC; SEQ ID NO: 104) (Chong and Moran, 2016 PNAS). qPCR будут проводить с применением скорости линейного изменения для qPCR-амплификации 1,6 градуса C/с и следующих условий: 1) 95°C в течение 10 минут, 2) 95°C в течение 15 секунд, 3) 55°C в течение 30 секунд, 4) повторение стадий 2-3 40x, 5) 95°C в течение 15 секунд, 6) 55°C в течение 1 минуты, 7) изменение скорости линейного изменения до 0,15 градуса C/с, 8) 95°C в течение 1 секунды. Данные qPCR будут анализировать с помощью аналитического программного обеспечения (Thermo Fisher Scientific, QuantStudio Design and Analysis).
В день 7 после обработки будут экстрагировать РНК из живых тлей и проводить RT-pPCR для количественной оценки экспрессии BCR-4. Вкратце, поверхность тела тлей будут стерилизовать путем погружения тлей в 6% раствор отбеливателя примерно на 5 секунд. Затем тлей будут промывать в стерильной воде и будут экстрагировать общую РНК из каждой отдельной тли с помощью набора для экстрагирования РНК (Qiagen, набор miRNeasy) в соответствии с инструкциями производителя. Концентрации РНК будут измерять с применением количественного определения нуклеиновой кислоты с помощью NanoDrop. Относительную экспрессию BCR-4 будут измерять с помощью RT-qPCR. Будут использовать следующие праймеры: ApBCR-4F (CTCTGTCAACCACCATGAGATTA; SEQ ID NO: 107) и ApBCR-4R (TGCAGACTACAGCACAATACTT; SEQ ID NO: 108). Праймерами для внутреннего референтного гена были праймеры для актина (гена домашнего хозяйства). Последовательность прямого праймера представляет собой GATCAGCAGCCACACACAAG; SEQ ID NO: 109, а последовательность обратного праймера представляет собой TTTGAACCGGTTTACGACGA; SEQ ID NO: 110. RT-qPCR будут проводить с применением скорости линейного изменения для qPCR-амплификации 1,6 градуса C/с и следующих условий: 1) 48°C в течение 30 мин, 2) 95°C в течение 10 минут, 3) 95°C в течение 15 секунд, 4) 60°C в течение 1 минуты, 5) повторение стадий 3-4 40x, 6) 95°C в течение 15 секунд, 7) 60°C в течение 1 минуты, 8) изменение скорости линейного изменения до 0,15 градуса C/с, 9) 95°C в течение 1 секунды. Данные RT-qPCR анализировали с помощью аналитического программного обеспечения (Thermo Fisher Scientific, QuantStudio Design and Analysis).
Как показано в предыдущих примерах, ожидается, что тли, питающиеся на листьях, экспрессирующих dsRNA, целенаправленно воздействующую на гены-мишени тлей, будут характеризоваться более низкой выживаемостью, более медленным развитием, содержать меньшее количество Buchnera и обладать пониженной экспрессией генов-мишеней по сравнению с тлями, которых растили на контрольных листьях.
Вместе взятые, эти данные, описанные в данном документе, демонстрируют возможность уничтожения тлей и снижения показателей их развития и продолжительности жизни (например, приспособленности) за счет их обработки с помощью растений, экспрессирующих dsRNA, целенаправленно воздействующую на ключевой(ключевые) ген(гены) (например, транспортeр глутамина ApGLNT1) в бактериоцитах в организме тлей.
ДРУГИЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ
Несмотря на то, что вышеизложенное изобретение было достаточно подробно описано с помощью иллюстраций и примеров в целях ясности понимания, описания и примеры не следует истолковывать как ограничивающие объем настоящего изобретения. Раскрытия всех источников патентной и научной литературы, цитируемых в данном документе, явным образом включено в их полном объеме посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> FLAGSHIP PIONEERING, INC.
<120> КОМПОЗИЦИИ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ МОДУЛИРОВАНИЯ
ЭНДОСИМБИОНТОВ
<130> 51215-008WO3
<150> US 62/583,990
<151> 2017-11-09
<150> US 62/463,451
<151> 2017-02-24
<160> 153
<170> PatentIn версия 3.5
<210> 1
<211> 1526
<212> ДНК
<213> Carsonella ruddii
<400> 1
tatccagcca caggttcccc tacagctacc ttgttacgac ttcaccccag ttacaaatca
60
taccgttgta atagtaaaat tacttatgat acaatttact tccatggtgt gacgggcggt
120
gtgtacaagg ctcgagaacg tattcaccgt aacattctga tttacgatta ctagcgattc
180
caacttcatg aaatcgagtt acagatttca atccgaacta agaatatttt ttaagattag
240
cattatgttg ccatatagca tataactttt tgtaatactc attgtagcac gtgtgtagcc
300
ctacttataa gggccatgat gacttgacgt cgtcctcacc ttcctccaat ttatcattgg
360
cagtttctta ttagttctaa tatattttta gtaaaataag ataagggttg cgctcgttat
420
aggacttaac ccaacatttc acaacacgag ctgacgacag ccatgcagca cctgtctcaa
480
agctaaaaaa gctttattat ttctaataaa ttctttggat gtcaaaagta ggtaagattt
540
ttcgtgttgt atcgaattaa accacatgct ccaccgcttg tgcgagcccc cgtcaattca
600
tttgagtttt aaccttgcgg tcgtaatccc caggcggtca acttaacgcg ttagcttttt
660
cactaaaaat atataacttt ttttcataaa acaaaattac aattataata tttaataaat
720
agttgacatc gtttactgca tggactacca gggtatctaa tcctgtttgc tccccatgct
780
ttcgtgtatt agtgtcagta ttaaaataga aatacgcctt cgccactagt attctttcag
840
atatctaagc atttcactgc tactcctgaa attctaattt cttcttttat actcaagttt
900
ataagtatta atttcaatat taaattactt taataaattt aaaaattaat ttttaaaaac
960
aacctgcaca ccctttacgc ccaataattc cgattaacgc ttgcacccct cgtattaccg
1020
cggctgctgg cacgaagtta gccggtgctt cttttacaaa taacgtcaaa gataatattt
1080
ttttattata aaatctcttc ttactttgtt gaaagtgttt tacaacccta aggccttctt
1140
cacacacgcg atatagctgg atcaagcttt cgctcattgt ccaatatccc ccactgctgc
1200
cttccgtaaa agtttgggcc gtgtctcagt cccaatgtgg ttgttcatcc tctaagatca
1260
actacgaatc atagtcttgt taagctttta ctttaacaac taactaattc gatataagct
1320
cttctattag cgaacgacat tctcgttctt tatccattag gatacatatt gaattactat
1380
acatttctat atacttttct aatactaata ggtagattct tatatattac tcacccgttc
1440
gctgctaatt atttttttaa taattcgcac aacttgcatg tgttaagctt atcgctagcg
1500
ttcaatctga gctatgatca aactca
1526
<210> 2
<211> 1536
<212> ДНК
<213> Portiera aleyrodidarum BT-B
<400> 2
aagagtttga tcatggctca gattgaacgc tagcggcaga cataacacat gcaagtcgag
60
cggcatcata caggttggca agcggcgcac gggtgagtaa tacatgtaaa tatacctaaa
120
agtggggaat aacgtacgga aacgtacgct aataccgcat aattattacg agataaagca
180
ggggcttgat aaaaaaaatc aaccttgcgc ttttagaaaa ttacatgccg gattagctag
240
ttggtagagt aaaagcctac caaggtaacg atccgtagct ggtctgagag gatgatcagc
300
cacactggga ctgagaaaag gcccagactc ctacgggagg cagcagtggg gaatattgga
360
caatgggggg aaccctgatc cagtcatgcc gcgtgtgtga agaaggcctt tgggttgtaa
420
agcactttca gcgaagaaga aaagttagaa aataaaaagt tataactatg acggtactcg
480
cagaagaagc accggctaac tccgtgccag cagccgcggt aagacggagg gtgcaagcgt
540
taatcagaat tactgggcgt aaagggcatg taggtggttt gttaagcttt atgtgaaagc
600
cctatgctta acataggaac ggaataaaga actgacaaac tagagtgcag aagaggaagg
660
tagaattccc ggtgtagcgg tgaaatgcgt agatatctgg aggaatacca gttgcgaagg
720
cgaccttctg ggctgacact gacactgaga tgcgaaagcg tggggagcaa acaggattag
780
ataccctggt agtccacgct gtaaacgata tcaactagcc gttggattct taaagaattt
840
tgtggcgtag ctaacgcgat aagttgatcg cctggggagt acggtcgcaa ggctaaaact
900
caaatgaatt gacgggggcc cgcacaagcg gtggagcatg tggtttaatt cgatgcaacg
960
cgcaaaacct tacctactct tgacatccaa agtactttcc agagatggaa gggtgcctta
1020
gggaactttg agacaggtgc tgcatggctg tcgtcagctc gtgttgtgaa atgttgggtt
1080
aagtcccgta acgagcgcaa cccttgtcct tagttgccaa cgcataaggc gggaacttta
1140
aggagactgc tggtgataaa ccggaggaag gtggggacga cgtcaagtca tcatggccct
1200
taagagtagg gcaacacacg tgctacaatg gcaaaaacaa agggtcgcaa aatggtaaca
1260
tgaagctaat cccaaaaaaa ttgtcttagt tcggattgga gtctgaaact cgactccata
1320
aagtcggaat cgctagtaat cgtgaatcag aatgtcacgg tgaatacgtt ctcgggcctt
1380
gtacacaccg cccgtcacac catggaagtg aaatgcacca gaagtggcaa gtttaaccaa
1440
aaaacaggag aacagtcact acggtgtggt tcatgactgg ggtgaagtcg taacaaggta
1500
gctgtagggg aacctgtggc tggatcacct ccttaa
1536
<210> 3
<211> 1540
<212> ДНК
<213> Buchnera aphidicola, штамм APS (Acyrthosiphon pisum)
<400> 3
agagtttgat catggctcag attgaacgct ggcggcaagc ctaacacatg caagtcgagc
60
ggcagcgaga agagagcttg ctctctttgt cggcaagcgg caaacgggtg agtaatatct
120
ggggatctac ccaaaagagg gggataacta ctagaaatgg tagctaatac cgcataatgt
180
tgaaaaacca aagtggggga ccttttggcc tcatgctttt ggatgaaccc agacgagatt
240
agcttgttgg tagagtaata gcctaccaag gcaacgatct ctagctggtc tgagaggata
300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat
360
attgcacaat gggcgaaagc ctgatgcagc tatgccgcgt gtatgaagaa ggccttaggg
420
ttgtaaagta ctttcagcgg ggaggaaaaa aataaaacta ataattttat ttcgtgacgt
480
tacccgcaga agaagcaccg gctaactccg tgccagcagc cgcggtaata cggagggtgc
540
aagcgttaat cagaattact gggcgtaaag agcgcgtagg tggtttttta agtcaggtgt
600
gaaatcccta ggctcaacct aggaactgca tttgaaactg gaaaactaga gtttcgtaga
660
gggaggtaga attctaggtg tagcggtgaa atgcgtagat atctggagga atacccgtgg
720
cgaaagcggc ctcctaaacg aaaactgaca ctgaggcgcg aaagcgtggg gagcaaacag
780
gattagatac cctggtagtc catgccgtaa acgatgtcga cttggaggtt gtttccaaga
840
gaagtgactt ccgaagctaa cgcattaagt cgaccgcctg gggagtacgg ccgcaaggct
900
aaaactcaaa tgaattgacg ggggcccgca caagcggtgg agcatgtggt ttaattcgat
960
gcaacgcgaa aaaccttacc tggtcttgac atccacagaa ttctttagaa ataaagaagt
1020
gccttcggga gctgtgagac aggtgctgca tggctgtcgt cagctcgtgt tgtgaaatgt
1080
tgggttaagt cccgcaacga gcgcaaccct tatcccctgt tgccagcggt tcggccggga
1140
actcagagga gactgccggt tataaaccgg aggaaggtgg ggacgacgtc aagtcatcat
1200
ggcccttacg accagggcta cacacgtgct acaatggttt atacaaagag aagcaaatct
1260
gcaaagacaa gcaaacctca taaagtaaat cgtagtccgg actggagtct gcaactcgac
1320
tccacgaagt cggaatcgct agtaatcgtg gatcagaatg ccacggtgaa tacgttcccg
1380
ggccttgtac acaccgcccg tcacaccatg ggagtgggtt gcaaaagaag caggtatcct
1440
aaccctttaa aaggaaggcg cttaccactt tgtgattcat gactggggtg aagtcgtaac
1500
aaggtaaccg taggggaacc tgcggttgga tcacctcctt
1540
<210> 4
<211> 1552
<212> ДНК
<213> Buchnera aphidicola, штамм Sg (Schizaphis graminum)
<400> 4
aaactgaaga gtttgatcat ggctcagatt gaacgctggc ggcaagccta acacatgcaa
60
gtcgagcggc agcgaaaaga aagcttgctt tcttgtcggc gagcggcaaa cgggtgagta
120
atatctgggg atctgcccaa aagaggggga taactactag aaatggtagc taataccgca
180
taaagttgaa aaaccaaagt gggggacctt ttttaaaggc ctcatgcttt tggatgaacc
240
cagacgagat tagcttgttg gtaaggtaaa agcttaccaa ggcaacgatc tctagctggt
300
ctgagaggat aaccagccac actggaactg agacacggtc cagactccta cgggaggcag
360
cagtggggaa tattgcacaa tgggcgaaag cctgatgcag ctatgccgcg tgtatgaaga
420
aggccttagg gttgtaaagt actttcagcg gggaggaaaa aattaaaact aataatttta
480
ttttgtgacg ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat
540
acggagggtg cgagcgttaa tcagaattac tgggcgtaaa gagcacgtag gtggtttttt
600
aagtcagatg tgaaatccct aggcttaacc taggaactgc atttgaaact gaaatgctag
660
agtatcgtag agggaggtag aattctaggt gtagcggtga aatgcgtaga tatctggagg
720
aatacccgtg gcgaaagcgg cctcctaaac gaatactgac actgaggtgc gaaagcgtgg
780
ggagcaaaca ggattagata ccctggtagt ccatgccgta aacgatgtcg acttggaggt
840
tgtttccaag agaagtgact tccgaagcta acgcgttaag tcgaccgcct ggggagtacg
900
gccgcaaggc taaaactcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg
960
tttaattcga tgcaacgcga aaaaccttac ctggtcttga catccacaga attttttaga
1020
aataaaaaag tgccttcggg aactgtgaga caggtgctgc atggctgtcg tcagctcgtg
1080
ttgtgaaatg ttgggttaag tcccgcaacg agcgcaaccc ttatcccctg ttgccagcgg
1140
ttcggccggg aactcagagg agactgccgg ttataaaccg gaggaaggtg gggacgacgt
1200
caagtcatca tggcccttac gaccagggct acacacgtgc tacaatggtt tatacaaaga
1260
gaagcaaatc tgtaaagaca agcaaacctc ataaagtaaa tcgtagtccg gactggagtc
1320
tgcaactcga ctccacgaag tcggaatcgc tagtaatcgt ggatcagaat gccacggtga
1380
atacgttccc gggccttgta cacaccgccc gtcacaccat gggagtgggt tgcaaaagaa
1440
gcagatttcc taaccacgaa agtggaaggc gtctaccact ttgtgattca tgactggggt
1500
gaagtcgtaa caaggtaacc gtaggggaac ctgcggttgg atcacctcct ta
1552
<210> 5
<211> 1566
<212> ДНК
<213> Buchnera aphidicola, штамм Bp (Baizongia pistaciae)
<400> 5
acttaaaatt gaagagtttg atcatggctc agattgaacg ctggcggcaa gcttaacaca
60
tgcaagtcga gcggcatcga agaaaagttt acttttctgg cggcgagcgg caaacgggtg
120
agtaacatct ggggatctac ctaaaagagg gggacaacca ttggaaacga tggctaatac
180
cgcataatgt ttttaaataa accaaagtag gggactaaaa tttttagcct tatgctttta
240
gatgaaccca gacgagatta gcttgatggt aaggtaatgg cttaccaagg cgacgatctc
300
tagctggtct gagaggataa ccagccacac tggaactgag atacggtcca gactcctacg
360
ggaggcagca gtggggaata ttgcacaatg ggctaaagcc tgatgcagct atgccgcgtg
420
tatgaagaag gccttagggt tgtaaagtac tttcagcggg gaggaaagaa ttatgtctaa
480
tatacatatt ttgtgacgtt acccgaagaa gaagcaccgg ctaactccgt gccagcagcc
540
gcggtaatac ggagggtgcg agcgttaatc agaattactg ggcgtaaaga gcacgtaggc
600
ggtttattaa gtcagatgtg aaatccctag gcttaactta ggaactgcat ttgaaactaa
660
tagactagag tctcatagag ggaggtagaa ttctaggtgt agcggtgaaa tgcgtagata
720
tctagaggaa tacccgtggc gaaagcgacc tcctaaatga aaactgacgc tgaggtgcga
780
aagcgtgggg agcaaacagg attagatacc ctggtagtcc atgctgtaaa cgatgtcgac
840
ttggaggttg tttcctagag aagtggcttc cgaagctaac gcattaagtc gaccgcctgg
900
ggagtacggt cgcaaggcta aaactcaaat gaattgacgg gggcccgcac aagcggtgga
960
gcatgtggtt taattcgatg caacgcgaag aaccttacct ggtcttgaca tccatagaat
1020
tttttagaga taaaagagtg ccttagggaa ctatgagaca ggtgctgcat ggctgtcgtc
1080
agctcgtgtt gtgaaatgtt gggttaagtc ccgcaacgag cgcaacccct atcctttgtt
1140
gccatcaggt tatgctggga actcagagga gactgccggt tataaaccgg aggaaggtgg
1200
ggatgacgtc aagtcatcat ggcccttacg accagggcta cacacgtgct acaatggcat
1260
atacaaagag atgcaactct gcgaagataa gcaaacctca taaagtatgt cgtagtccgg
1320
actggagtct gcaactcgac tccacgaagt aggaatcgct agtaatcgtg gatcagaatg
1380
ccacggtgaa tacgttcccg ggccttgtac acaccgcccg tcacaccatg ggagtgggtt
1440
gcaaaagaag caggtagctt aaccagatta ttttattgga gggcgcttac cactttgtga
1500
ttcatgactg gggtgaagtc gtaacaaggt aaccgtaggg gaacctgcgg ttggatcacc
1560
tcctta
1566
<210> 6
<211> 828
<212> ДНК
<213> Buchnera aphidicola BCc
<400> 6
atgagatcat taatatataa aaatcatgtt ccaattaaaa aattaggaca aaatttttta
60
cagaataaag aaattattaa tcagataatt aatttaataa atattaataa aaatgataat
120
attattgaaa taggatcagg attaggagcg ttaacttttc ctatttgtag aatcattaaa
180
aaaatgatag tattagaaat tgatgaagat cttgtgtttt ttttaactca aagtttattt
240
attaaaaaat tacaaattat aattgctgat attataaaat ttgatttttg ttgttttttt
300
tctttacaga aatataaaaa atataggttt attggtaatt taccatataa tattgctact
360
atattttttt taaaaacaat taaatttctt tataatataa ttgatatgca ttttatgttt
420
caaaaagaag tagcaaagag attattagct actcctggta ctaaagaata tggtagatta
480
agtattattg cacaatattt ttataagata gaaactgtta ttaatgttaa taaatttaat
540
ttttttccta ctcctaaagt agattctact tttttacgat ttactcctaa atattttaat
600
agtaaatata aaatagataa acatttttct gttttagaat taattactag attttctttt
660
caacatagaa gaaaattttt aaataataat ttaatatctt tattttctac aaaagaatta
720
atttctttag atattgatcc atattcaaga gcagaaaatg tttctttaat tcaatattgt
780
aaattaatga aatattattt gaaaagaaaa attttatgtt tagattaa
828
<210> 7
<211> 921
<212> ДНК
<213> Buchnera aphidicola (Cinara tujafilina)
<400> 7
ttatcttatt tcacatatac gtaatattgc gctgcgtgca cgaggatttt tttgaatttc
60
agatatattt ggtttaatac gtttaataaa acgtattttt ttttttattt ttcttatttg
120
caattcagta ataggaagtt ttttaggtat atttggataa ttactgtaat tcttaataaa
180
gttttttaca atcctatctt caatagaatg aaaactaata atagcaattt ttgatccgga
240
atgtaatatg ttaataataa tttttaatat tttatgtaat tcatttattt cttggttaat
300
atatattcga aaagcttgaa atgttctcgt agctggatgt ttaaatttgt catattttgg
360
gattgatttt tttatgattt gaactaactc taacgtgctt gttatggttt ttttttttat
420
ttgtaatatg atggctcggg atattttttt tgcgtatttt tcttcgccaa aattttttat
480
tacctgttct attgtttttt ggtttgtttt ttttaaccat tgactaactg atattccaga
540
tttagggttc atacgcatat ctaaaggtcc atcattcata aatgaaaatc ctcggatact
600
agaatttaac tgtattgaag aaatacctaa atctaataat attccatcta ttttatctct
660
atttttttct ttttttaata ttttttcaat attagaaaat ttacctaaaa atattttaaa
720
tcgcgaatct tttatttttt ttccgatttt tatagattgt gggtcttgat caatactata
780
taactttcca ttaaccccta attcttgaag aattgctttt gaatgaccac cacctccaaa
840
tgtacaatca acatatgtac cgtctttttt tatttttaag tattgtatga tttcttttgt
900
taaaacaggt ttatgaatca t
921
<210> 8
<211> 822
<212> ДНК
<213> Buchnera aphidicola, штамм G002 (Myzus persicae)
<400> 8
atgaaaagta taaaaacttt taaaaaacac tttcctgtga aaaaatatgg acaaaatttt
60
cttattaata aagagatcat aaaaaatatt gttaaaaaaa ttaatccaaa tatagaacaa
120
acattagtag aaatcggacc aggattagct gcattaactg agcccatatc tcagttatta
180
aaagagttaa tagttattga aatagactgt aatctattat attttttaaa aaaacaacca
240
ttttattcaa aattaatagt tttttgtcaa gatgctttaa actttaatta tacaaattta
300
ttttataaaa aaaataaatt aattcgtatt tttggtaatt taccatataa tatctctaca
360
tctttaatta tttttttatt tcaacacatt agagtaattc aagatatgaa ttttatgctt
420
caaaaagaag ttgctgcaag attaattgca ttacctggaa ataaatatta cggtcgtttg
480
agcattatat ctcaatatta ttgtgatatc aaaattttat taaatgttgc tcctgaagat
540
ttttggccta ttccgagagt tcattctata tttgtaaatt taacacctca tcataattct
600
ccttattttg tttatgatat taatatttta agccttatta caaataaggc tttccaaaat
660
agaagaaaaa tattacgtca tagtttaaaa aatttatttt ctgaaacaac tttattaaat
720
ttagatatta atcccagatt aagagctgaa aatatttctg tttttcagta ttgtcaatta
780
gctaattatt tgtataaaaa aaattatact aaaaaaaatt aa
822
<210> 9
<211> 822
<212> ДНК
<213> Buchnera aphidicola, штамм Ak (Acyrthosiphon kondoi)
<400> 9
attataaaaa attttaaaaa acattttcct ttaaaaaggt atggacaaaa ttttcttgtc
60
aatacaaaaa ctattcaaaa gataattaat ataattaatc caaacaccaa acaaacatta
120
gtggaaattg gacctggatt agctgcatta acaaaaccaa tttgtcaatt attagaagaa
180
ttaattgtta ttgaaataga tcctaattta ttgtttttat taaaaaaacg ttcattttat
240
tcaaaattaa cagtttttta tcaagacgct ttaaatttca attatacaga tttgttttat
300
aagaaaaatc aattaattcg tgtttttgga aacttgccat ataatatttc tacatcttta
360
attatttctt tattcaatca tattaaagtt attcaagata tgaattttat gttacagaaa
420
gaggttgctg aaagattaat ttctattcct ggaaataaat cttatggccg tttaagcatt
480
atttctcagt attattgtaa aattaaaata ttattaaatg ttgtacctga agattttcga
540
cctataccga aagtgcattc tgtttttatc aatttaactc ctcataccaa ttctccatat
600
tttgtttatg atacaaatat cctcagttct atcacaagaa atgcttttca aaatagaagg
660
aaaattttgc gtcatagttt aaaaaattta ttttctgaaa aagaactaat tcaattagaa
720
attaatccaa atttacgagc tgaaaatatt tctatctttc agtattgtca attagctgat
780
tatttatata aaaaattaaa taatcttgta aaaatcaatt aa
822
<210> 10
<211> 822
<212> ДНК
<213> Buchnera aphidicola, штамм Ua (Uroleucon ambrosiae)
<400> 10
atgatactaa ataaatataa aaaatttatt cctttaaaaa gatacggaca aaattttctt
60
gtaaatagag aaataatcaa aaatattatc aaaataatta atcctaaaaa aacgcaaaca
120
ttattagaaa ttggaccggg tttaggtgcg ttaacaaaac ctatttgtga atttttaaat
180
gaacttatcg tcattgaaat agatcctaat atattatctt ttttaaagaa atgtatattt
240
tttgataaat taaaaatata ttgtcataat gctttagatt ttaattataa aaatatattc
300
tataaaaaaa gtcaattaat tcgtattttt ggaaatttac catataatat ttctacatct
360
ttaataatat atttatttcg gaatattgat attattcaag atatgaattt tatgttacaa
420
caagaagtgg ctaaaagatt agttgctatt cctggtgaaa aactttatgg tcgtttaagt
480
attatatctc aatattattg taatattaaa atattattac atattcgacc tgaaaatttt
540
caacctattc ctaaagttaa ttcaatgttt gtaaatttaa ctccgcatat tcattctcct
600
tattttgttt atgatattaa tttattaact agtattacaa aacatgcttt tcaacataga
660
agaaaaatat tgcgtcatag tttaagaaat tttttttctg agcaagattt aattcattta
720
gaaattaatc caaatttaag agctgaaaat gtttctatta ttcaatattg tcaattggct
780
aataatttat ataaaaaaca taaacagttt attaataatt aa
822
<210> 11
<211> 816
<212> ДНК
<213> Buchnera aphidicola (Aphis glycines)
<400> 11
atgaaaaagc atattcctat aaaaaaattt agtcaaaatt ttcttgtaga tttgagtgtg
60
attaaaaaaa taattaaatt tattaatccg cagttaaatg aaatattggt tgaaattgga
120
ccgggattag ctgctatcac tcgacctatt tgtgatttga tagatcattt aattgtgatt
180
gaaattgata aaattttatt agatagatta aaacagttct cattttattc aaaattaaca
240
gtatatcatc aagatgcttt agcatttgat tacataaagt tatttaataa aaaaaataaa
300
ttagttcgaa tttttggtaa tttaccatat catgtttcta cgtctttaat attgcattta
360
tttaaaagaa ttaatattat taaagatatg aattttatgc tacaaaaaga agttgctgaa
420
cgtttaattg caactccagg tagtaaatta tatggtcgtt taagtattat ttctcaatat
480
tattgtaata taaaagtttt attgcatgtg tcttcaaaat gttttaaacc agttcctaaa
540
gtagaatcaa tttttcttaa tttgacacct tatactgatt atttccctta ttttacttat
600
aatgtaaacg ttcttagtta tattacaaat ttagcttttc aaaaaagaag aaaaatatta
660
cgtcatagtt taggtaaaat attttctgaa aaagttttta taaaattaaa tattaatccc
720
aaattaagac ctgagaatat ttctatatta caatattgtc agttatctaa ttatatgata
780
gaaaataata ttcatcagga acatgtttgt atttaa
816
<210> 12
<211> 1463
<212> ДНК
<213> Annandia pinicola
<400> 12
agattgaacg ctggcggcat gccttacaca tgcaagtcga acggtaacag gtcttcggac
60
gctgacgagt ggcgaacggg tgagtaatac atcggaacgt gcccagtcgt gggggataac
120
tactcgaaag agtagctaat accgcatacg atctgaggat gaaagcgggg gaccttcggg
180
cctcgcgcga ttggagcggc cgatggcaga ttaggtagtt ggtgggataa aagcttacca
240
agccgacgat ctgtagctgg tctgagagga cgaccagcca cactggaact gagatacggt
300
ccagactctt acgggaggca gcagtgggga atattgcaca atgggcgcaa gcctgatgca
360
gctatgtcgc gtgtatgaag aagaccttag ggttgtaaag tactttcgat agcataagaa
420
gataatgaga ctaataattt tattgtctga cgttagctat agaagaagca ccggctaact
480
ccgtgccagc agccgcggta atacgggggg tgctagcgtt aatcggaatt actgggcgta
540
aagagcatgt aggtggttta ttaagtcaga tgtgaaatcc ctggacttaa tctaggaact
600
gcatttgaaa ctaataggct agagtttcgt agagggaggt agaattctag gtgtagcggt
660
gaaatgcata gatatctaga ggaatatcag tggcgaaggc gaccttctgg acgataactg
720
acgctaaaat gcgaaagcat gggtagcaaa caggattaga taccctggta gtccatgctg
780
taaacgatgt cgactaagag gttggaggta taacttttaa tctctgtagc taacgcgtta
840
agtcgaccgc ctggggagta cggtcgcaag gctaaaactc aaatgaattg acgggggcct
900
gcacaagcgg tggagcatgt ggtttaattc gatgcaacgc gtaaaacctt acctggtctt
960
gacatccaca gaattttaca gaaatgtaga agtgcaattt gaactgtgag acaggtgctg
1020
catggctgtc gtcagctcgt gttgtgaaat gttgggttaa gtcccgcaac gagcgcaacc
1080
cttgtccttt gttaccataa gatttaagga actcaaagga gactgccggt gataaactgg
1140
aggaaggcgg ggacgacgtc aagtcatcat ggcccttatg accagggcta cacacgtgct
1200
acaatggcat atacaaagag atgcaatatt gcgaaataaa gccaatctta taaaatatgt
1260
cctagttcgg actggagtct gcaactcgac tccacgaagt cggaatcgct agtaatcgtg
1320
gatcagcatg ccacggtgaa tatgtttcca ggccttgtac acaccgcccg tcacaccatg
1380
gaagtggatt gcaaaagaag taagaaaatt aaccttctta acaaggaaat aacttaccac
1440
tttgtgactc ataactgggg tga
1463
<210> 13
<211> 1554
<212> ДНК
<213> Moranella endobia
<400> 13
tctttttggt aaggaggtga tccaaccgca ggttccccta cggttacctt gttacgactt
60
caccccagtc atgaatcaca aagtggtaag cgccctccta aaaggttagg ctacctactt
120
cttttgcaac ccacttccat ggtgtgacgg gcggtgtgta caaggcccgg gaacgtattc
180
accgtggcat tctgatccac gattactagc gattcctact tcatggagtc gagttgcaga
240
ctccaatccg gactacgacg cactttatga ggtccgctaa ctctcgcgag cttgcttctc
300
tttgtatgcg ccattgtagc acgtgtgtag ccctactcgt aagggccatg atgacttgac
360
gtcatcccca ccttcctccg gtttatcacc ggcagtctcc tttgagttcc cgaccgaatc
420
gctggcaaaa aaggataagg gttgcgctcg ttgcgggact taacccaaca tttcacaaca
480
cgagctgacg acagccatgc agcacctgtc tcagagttcc cgaaggtacc aaaacatctc
540
tgctaagttc tctggatgtc aagagtaggt aaggttcttc gcgttgcatc gaattaaacc
600
acatgctcca ccgcttgtgc gggcccccgt caattcattt gagttttaac cttgcggccg
660
tactccccag gcggtcgatt taacgcgtta actacgaaag ccacagttca agaccacagc
720
tttcaaatcg acatagttta cggcgtggac taccagggta tctaatcctg tttgctcccc
780
acgctttcgt acctgagcgt cagtattcgt ccagggggcc gccttcgcca ctggtattcc
840
tccagatatc tacacatttc accgctacac ctggaattct acccccctct acgagactct
900
agcctatcag tttcaaatgc agttcctagg ttaagcccag ggatttcaca tctgacttaa
960
taaaccgcct acgtactctt tacgcccagt aattccgatt aacgcttgca ccctccgtat
1020
taccgcggct gctggcacgg agttagccgg tgcttcttct gtaggtaacg tcaatcaata
1080
accgtattaa ggatattgcc ttcctcccta ctgaaagtgc tttacaaccc gaaggccttc
1140
ttcacacacg cggcatggct gcatcagggt ttcccccatt gtgcaatatt ccccactgct
1200
gcctcccgta ggagtctgga ccgtgtctca gttccagtgt ggctggtcat cctctcagac
1260
cagctaggga tcgtcgccta ggtaagctat tacctcacct actagctaat cccatctggg
1320
ttcatctgaa ggtgtgaggc caaaaggtcc cccactttgg tcttacgaca ttatgcggta
1380
ttagctaccg tttccagcag ttatccccct ccatcaggca gatccccaga ctttactcac
1440
ccgttcgctg ctcgccggca aaaaagtaaa cttttttccg ttgccgctca acttgcatgt
1500
gttaggcctg ccgccagcgt tcaatctgag ccatgatcaa actcttcaat taaa
1554
<210> 14
<211> 1539
<212> ДНК
<213> Ishikawaella capsulata Mpkobe
<400> 14
aaattgaaga gtttgatcat ggctcagatt gaacgctagc ggcaagctta acacatgcaa
60
gtcgaacggt aacagaaaaa agcttgcttt tttgctgacg agtggcggac gggtgagtaa
120
tgtctgggga tctacctaat ggcgggggat aactactgga aacggtagct aataccgcat
180
aatgttgtaa aaccaaagtg ggggacctta tggcctcaca ccattagatg aacctagatg
240
ggattagctt gtaggtgggg taaaggctca cctaggcaac gatccctagc tggtctgaga
300
ggatgaccag ccacactgga actgagatac ggtccagact cctacgggag gcagcagtgg
360
ggaatcttgc acaatgggcg caagcctgat gcagctatgt cgcgtgtatg aagaaggcct
420
tagggttgta aagtactttc atcggggaag aaggatatga gcctaatatt ctcatatatt
480
gacgttacct gcagaagaag caccggctaa ctccgtgcca gcagccgcgg taacacggag
540
ggtgcgagcg ttaatcggaa ttactgggcg taaagagcac gtaggtggtt tattaagtca
600
tatgtgaaat ccctgggctt aacctaggaa ctgcatgtga aactgataaa ctagagtttc
660
gtagagggag gtggaattcc aggtgtagcg gtgaaatgcg tagatatctg gaggaatatc
720
agaggcgaag gcgaccttct ggacgaaaac tgacactcag gtgcgaaagc gtggggagca
780
aacaggatta gataccctgg tagtccacgc tgtaaacaat gtcgactaaa aaactgtgag
840
cttgacttgt ggtttttgta gctaacgcat taagtcgacc gcctggggag tacggccgca
900
aggttaaaac tcaaatgaat tgacgggggt ccgcacaagc ggtggagcat gtggtttaat
960
tcgatgcaac gcgaaaaacc ttacctggtc ttgacatcca gcgaattata tagaaatata
1020
taagtgcctt tcggggaact ctgagacgct gcatggctgt cgtcagctcg tgttgtgaaa
1080
tgttgggtta agtcccgcaa cgagcgccct tatcctctgt tgccagcggc atggccggga
1140
actcagagga gactgccagt attaaactgg aggaaggtgg ggatgacgtc aagtcatcat
1200
ggcccttatg accagggcta cacacgtgct acaatggtgt atacaaagag aagcaatctc
1260
gcaagagtaa gcaaaactca aaaagtacat cgtagttcgg attagagtct gcaactcgac
1320
tctatgaagt aggaatcgct agtaatcgtg gatcagaatg ccacggtgaa tacgttctct
1380
ggccttgtac acaccgcccg tcacaccatg ggagtaagtt gcaaaagaag taggtagctt
1440
aacctttata ggagggcgct taccactttg tgatttatga ctggggtgaa gtcgtaacaa
1500
ggtaactgta ggggaacctg tggttggatt acctcctta
1539
<210> 15
<211> 1561
<212> ДНК
<213> Baumannia cicadellinicola
<400> 15
ttcaattgaa gagtttgatc atggctcaga ttgaacgctg gcggtaagct taacacatgc
60
aagtcgagcg gcatcggaaa gtaaattaat tactttgccg gcaagcggcg aacgggtgag
120
taatatctgg ggatctacct tatggagagg gataactatt ggaaacgata gctaacaccg
180
cataatgtcg tcagaccaaa atgggggacc taatttaggc ctcatgccat aagatgaacc
240
cagatgagat tagctagtag gtgagataat agctcaccta ggcaacgatc tctagttggt
300
ctgagaggat gaccagccac actggaactg agacacggtc cagactccta cgggaggcag
360
cagtggggaa tcttgcacaa tgggggaaac cctgatgcag ctataccgcg tgtgtgaaga
420
aggccttcgg gttgtaaagc actttcagcg gggaagaaaa tgaagttact aataataatt
480
gtcaattgac gttacccgca aaagaagcac cggctaactc cgtgccagca gccgcggtaa
540
gacggagggt gcaagcgtta atcggaatta ctgggcgtaa agcgtatgta ggcggtttat
600
ttagtcaggt gtgaaagccc taggcttaac ctaggaattg catttgaaac tggtaagcta
660
gagtctcgta gaggggggga gaattccagg tgtagcggtg aaatgcgtag agatctggaa
720
gaataccagt ggcgaaggcg cccccctgga cgaaaactga cgctcaagta cgaaagcgtg
780
gggagcaaac aggattagat accctggtag tccacgctgt aaacgatgtc gatttgaagg
840
ttgtagcctt gagctatagc tttcgaagct aacgcattaa atcgaccgcc tggggagtac
900
gaccgcaagg ttaaaactca aatgaattga cgggggcccg cacaagcggt ggagcatgtg
960
gtttaattcg atacaacgcg aaaaacctta cctactcttg acatccagag tataaagcag
1020
aaaagcttta gtgccttcgg gaactctgag acaggtgctg catggctgtc gtcagctcgt
1080
gttgtgaaat gttgggttaa gtcccgcaac gagcgcaacc cttatccttt gttgccaacg
1140
attaagtcgg gaactcaaag gagactgccg gtgataaacc ggaggaaggt gaggataacg
1200
tcaagtcatc atggccctta cgagtagggc tacacacgtg ctacaatggt gcatacaaag
1260
agaagcaatc tcgtaagagt tagcaaacct cataaagtgc atcgtagtcc ggattagagt
1320
ctgcaactcg actctatgaa gtcggaatcg ctagtaatcg tggatcagaa tgccacggtg
1380
aatacgttcc cgggccttgt acacaccgcc cgtcacacca tgggagtgta ttgcaaaaga
1440
agttagtagc ttaactcata atacgagagg gcgcttacca ctttgtgatt cataactggg
1500
gtgaagtcgt aacaaggtaa ccgtagggga acctgcggtt ggatcacctc cttacactaa
1560
a
1561
<210> 16
<211> 1464
<212> ДНК
<213> Sodalis-подобные
<400> 16
attgaacgct ggcggcaggc ctaacacatg caagtcgagc ggcagcggga agaagcttgc
60
ttctttgccg gcgagcggcg gacgggtgag taatgtctgg ggatctgccc gatggagggg
120
gataactact ggaaacggta gctaataccg cataacgtcg caagaccaaa gtgggggacc
180
ttcgggcctc acaccatcgg atgaacccag gtgggattag ctagtaggtg gggtaatggc
240
tcacctaggc gacgatccct agctggtctg agaggatgac cagtcacact ggaactgaga
300
cacggtccag actcctacgg gaggcagcag tggggaatat tgcacaatgg gggaaaccct
360
gatgcagcca tgccgcgtgt gtgaagaagg ccttcgggtt gtaaagcact ttcagcgggg
420
aggaaggcga tggcgttaat agcgctatcg attgacgtta cccgcagaag aagcaccggc
480
taactccgtg ccagcagccg cggtaatacg gagggtgcga gcgttaatcg gaattactgg
540
gcgtaaagcg tacgcaggcg gtctgttaag tcagatgtga aatccccggg ctcaacctgg
600
gaactgcatt tgaaactggc aggctagagt ctcgtagagg ggggtagaat tccaggtgta
660
gcggtgaaat gcgtagagat ctggaggaat accggtggcg aaggcggccc cctggacgaa
720
gactgacgct caggtacgaa agcgtgggga gcaaacagga ttagataccc tggtagtcca
780
cgctgtaaac gatgtcgatt tgaaggttgt ggccttgagc cgtggctttc ggagctaacg
840
tgttaaatcg accgcctggg gagtacggcc gcaaggttaa aactcaaatg aattgacggg
900
ggcccgcaca agcggtggag catgtggttt aattcgatgc aacgcgaaga accttaccta
960
ctcttgacat ccagagaact tggcagagat gctttggtgc cttcgggaac tctgagacag
1020
gtgctgcatg gctgtcgtca gctcgtgttg tgaaatgttg ggttaagtcc cgcaacgagc
1080
gcaaccctta tcctttattg ccagcgattc ggtcgggaac tcaaaggaga ctgccggtga
1140
taaaccggag gaaggtgggg atgacgtcaa gtcatcatgg cccttacgag tagggctaca
1200
cacgtgctac aatggcgcat acaaagagaa gcgatctcgc gagagtcagc ggacctcata
1260
aagtgcgtcg tagtccggat tggagtctgc aactcgactc catgaagtcg gaatcgctag
1320
taatcgtgga tcagaatgcc acggtgaata cgttcccggg ccttgtacac accgcccgtc
1380
acaccatggg agtgggttgc aaaagaagta ggtagcttaa ccttcgggag ggcgcttacc
1440
actttgtgat tcatgactgg ggtg
1464
<210> 17
<211> 1465
<212> ДНК
<213> Hartigia pinicola
<400> 17
agatttaacg ctggcggcag gcctaacaca tgcaagtcga gcggtaccag aagaagcttg
60
cttcttgctg acgagcggcg gacgggtgag taatgtatgg ggatctgccc gacagagggg
120
gataactatt ggaaacggta gctaataccg cataatctct gaggagcaaa gcaggggaac
180
ttcggtcctt gcgctatcgg atgaacccat atgggattag ctagtaggtg aggtaatggc
240
tcccctaggc aacgatccct agctggtctg agaggatgat cagccacact gggactgaga
300
cacggcccag actcctacgg gaggcagcag tggggaatat tgcacaatgg gcgaaagcct
360
gatgcagcca tgccgcgtgt atgaagaagg ctttagggtt gtaaagtact ttcagtcgag
420
aggaaaacat tgatgctaat atcatcaatt attgacgttt ccgacagaag aagcaccggc
480
taactccgtg ccagcagccg cggtaatacg gagggtgcaa gcgttaatcg gaattactgg
540
gcgtaaagcg cacgcaggcg gttaattaag ttagatgtga aagccccggg cttaacccag
600
gaatagcata taaaactggt caactagagt attgtagagg ggggtagaat tccatgtgta
660
gcggtgaaat gcgtagagat gtggaggaat accagtggcg aaggcggccc cctggacaaa
720
aactgacgct caaatgcgaa agcgtgggga gcaaacagga ttagataccc tggtagtcca
780
tgctgtaaac gatgtcgatt tggaggttgt tcccttgagg agtagcttcc gtagctaacg
840
cgttaaatcg accgcctggg ggagtacgac tgcaaggtta aaactcaaat gaattgacgg
900
gggcccgcac aagcggtgga gcatgtggtt taattcgatg caacgcgaaa aaccttacct
960
actcttgaca tccagataat ttagcagaaa tgctttagta ccttcgggaa atctgagaca
1020
ggtgctgcat ggctgtcgtc agctcgtgtt gtgaaatgtt gggttaagtc ccgcaacgag
1080
cgcaaccctt atcctttgtt gccagcgatt aggtcgggaa ctcaaaggag actgccggtg
1140
ataaaccgga ggaaggtggg gatgacgtca agtcatcatg gcccttacga gtagggctac
1200
acacgtgcta caatggcata tacaaaggga agcaacctcg cgagagcaag cgaaactcat
1260
aaattatgtc gtagttcaga ttggagtctg caactcgact ccatgaagtc ggaatcgcta
1320
gtaatcgtag atcagaatgc tacggtgaat acgttcccgg gccttgtaca caccgcccgt
1380
cacaccatgg gagtgggttg caaaagaagt aggtaactta accttatgga aagcgcttac
1440
cactttgtga ttcataactg gggtg
1465
<210> 18
<211> 1571
<212> ДНК
<213> Tremblaya phenacola
<400> 18
aggtaatcca gccacacctt ccagtacggc taccttgtta cgacttcacc ccagtcacaa
60
cccttacctt cggaactgcc ctcctcacaa ctcaaaccac caaacacttt taaatcaggt
120
tgagagaggt taggcctgtt acttctggca agaattattt ccatggtgtg acgggcggtg
180
tgtacaagac ccgagaacat attcaccgtg gcatgctgat ccacgattac tagcaattcc
240
aacttcatgc actcgagttt cagagtacaa tccgaactga ggccggcttt gtgagattag
300
ctcccttttg caagttggca actctttggt ccggccattg tatgatgtgt gaagccccac
360
ccataaaggc catgaggact tgacgtcatc cccaccttcc tccaacttat cgctggcagt
420
ctctttaagg taactgacta atccagtagc aattaaagac aggggttgcg ctcgttacag
480
gacttaaccc aacatctcac gacacgagct gacgacagcc atgcagcacc tgtgcactaa
540
ttctctttca agcactcccg cttctcaaca ggatcttagc catatcaaag gtaggtaagg
600
tttttcgcgt tgcatcgaat taatccacat catccactgc ttgtgcgggt ccccgtcaat
660
tcctttgagt tttaaccttg cggccgtact ccccaggcgg tcgacttgtg cgttagctgc
720
accactgaaa aggaaaactg cccaatggtt agtcaacatc gtttagggca tggactacca
780
gggtatctaa tcctgtttgc tccccatgct ttagtgtctg agcgtcagta acgaaccagg
840
aggctgccta cgctttcggt attcctccac atctctacac atttcactgc tacatgcgga
900
attctacctc cccctctcgt actccagcct gccagtaact gccgcattct gaggttaagc
960
ctcagccttt cacagcaatc ttaacaggca gcctgcacac cctttacgcc caataaatct
1020
gattaacgct cgcaccctac gtattaccgc ggctgctggc acgtagtttg ccggtgctta
1080
ttctttcggt acagtcacac caccaaattg ttagttgggt ggctttcttt ccgaacaaaa
1140
gtgctttaca acccaaaggc cttcttcaca cacgcggcat tgctggatca ggcttccgcc
1200
cattgtccaa gattcctcac tgctgccttc ctcagaagtc tgggccgtgt ctcagtccca
1260
gtgtggctgg ccgtcctctc agaccagcta ccgatcattg ccttgggaag ccattacctt
1320
tccaacaagc taatcagaca tcagccaatc tcagagcgca aggcaattgg tcccctgctt
1380
tcattctgct tggtagagaa ctttatgcgg tattaattag gctttcacct agctgtcccc
1440
cactctgagg catgttctga tgcattactc acccgtttgc cacttgccac caagcctaag
1500
cccgtgttgc cgttcgactt gcatgtgtaa ggcatgccgc tagcgttcaa tctgagccag
1560
gatcaaactc t
1571
<210> 19
<211> 1535
<212> ДНК
<213> Tremblaya princeps
<400> 19
agagtttgat cctggctcag attgaacgct agcggcatgc attacacatg caagtcgtac
60
ggcagcacgg gcttaggcct ggtggcgagt ggcgaacggg tgagtaacgc ctcggaacgt
120
gccttgtagt gggggatagc ctggcgaaag ccagattaat accgcatgaa gccgcacagc
180
atgcgcggtg aaagtggggg attctagcct cacgctactg gatcggccgg ggtctgatta
240
gctagttggc ggggtaatgg cccaccaagg cttagatcag tagctggtct gagaggacga
300
tcagccacac tgggactgag acacggccca gactcctacg ggaggcagca gtggggaatc
360
ttggacaatg ggcgcaagcc tgatccagca atgccgcgtg tgtgaagaag gccttcgggt
420
cgtaaagcac ttttgttcgg gatgaagggg ggcgtgcaaa caccatgccc tcttgacgat
480
accgaaagaa taagcaccgg ctaactacgt gccagcagcc gcggtaatac gtagggtgcg
540
agcgttaatc ggaatcactg ggcgtaaagg gtgcgcgggt ggtttgccaa gacccctgta
600
aaatcctacg gcccaaccgt agtgctgcgg aggttactgg taagcttgag tatggcagag
660
gggggtagaa ttccaggtgt agcggtgaaa tgcgtagata tctggaggaa taccgaaggc
720
gaaggcaacc ccctgggcca tcactgacac tgaggcacga aagcgtgggg agcaaacagg
780
attagatacc ctggtagtcc acgccctaaa ccatgtcgac tagttgtcgg ggggagccct
840
ttttcctcgg tgacgaagct aacgcatgaa gtcgaccgcc tggggagtac gaccgcaagg
900
ttaaaactca aaggaattga cggggacccg cacaagcggt ggatgatgtg gattaattcg
960
atgcaacgcg aaaaacctta cctacccttg acatggcgga gattctgccg agaggcggaa
1020
gtgctcgaaa gagaatccgt gcacaggtgc tgcatggctg tcgtcagctc gtgtcgtgag
1080
atgttgggtt aagtcccata acgagcgcaa cccccgtctt tagttgctac cactggggca
1140
ctctatagag actgccggtg ataaaccgga ggaaggtggg gacgacgtca agtcatcatg
1200
gcctttatgg gtagggcttc acacgtcata caatggctgg agcaaagggt cgccaactcg
1260
agagagggag ctaatcccac aaacccagcc ccagttcgga ttgcactctg caactcgagt
1320
gcatgaagtc ggaatcgcta gtaatcgtgg atcagcatgc cacggtgaat acgttctcgg
1380
gtcttgtaca caccgcccgt cacaccatgg gagtaagccg catcagaagc agcctcccta
1440
accctatgct gggaaggagg ctgcgaaggt ggggtctatg actggggtga agtcgtaaca
1500
aggtagccgt accggaaggt gcggctggat tacct
1535
<210> 20
<211> 1450
<212> ДНК
<213> Nasuia deltocephalinicola
<400> 20
agtttaatcc tggctcagat ttaacgcttg cgacatgcct aacacatgca agttgaacgt
60
tgaaaatatt tcaaagtagc gtataggtga gtataacatt taaacatacc ttaaagttcg
120
gaataccccg atgaaaatcg gtataatacc gtataaaagt atttaagaat taaagcgggg
180
aaaacctcgt gctataagat tgttaaatgc ctgattagtt tgttggtttt taaggtaaaa
240
gcttaccaag actttgatca gtagctattc tgtgaggatg tatagccaca ttgggattga
300
aataatgccc aaacctctac ggagggcagc agtggggaat attggacaat gagcgaaagc
360
ttgatccagc aatgtcgcgt gtgcgattaa gggaaactgt aaagcacttt tttttaagaa
420
taagaaattt taattaataa ttaaaatttt tgaatgtatt aaaagaataa gtaccgacta
480
atcacgtgcc agcagtcgcg gtaatacgtg gggtgcgagc gttaatcgga tttattgggc
540
gtaaagtgta ttcaggctgc ttaaaaagat ttatattaaa tatttaaatt aaatttaaaa
600
aatgtataaa ttactattaa gctagagttt agtataagaa aaaagaattt tatgtgtagc
660
agtgaaatgc gttgatatat aaaggaacgc cgaaagcgaa agcatttttc tgtaatagaa
720
ctgacgctta tatacgaaag cgtgggtagc aaacaggatt agataccctg gtagtccacg
780
ccctaaacta tgtcaattaa ctattagaat tttttttagt ggtgtagcta acgcgttaaa
840
ttgaccgcct gggtattacg atcgcaagat taaaactcaa aggaattgac ggggaccagc
900
acaagcggtg gatgatgtgg attaattcga tgatacgcga aaaaccttac ctgcccttga
960
catggttaga attttattga aaaataaaag tgcttggaaa agagctaaca cacaggtgct
1020
gcatggctgt cgtcagctcg tgtcgtgaga tgttgggtta agtcccgcaa cgagcgcaac
1080
ccctactctt agttgctaat taaagaactt taagagaaca gctaacaata agtttagagg
1140
aaggagggga tgacttcaag tcctcatggc ccttatgggc agggcttcac acgtcataca
1200
atggttaata caaaaagttg caatatcgta agattgagct aatctttaaa attaatctta
1260
gttcggattg tactctgcaa ctcgagtaca tgaagttgga atcgctagta atcgcggatc
1320
agcatgccgc ggtgaatagt ttaactggtc ttgtacacac cgcccgtcac accatggaaa
1380
taaatcttgt tttaaatgaa gtaatatatt ttatcaaaac aggttttgta accggggtga
1440
agtcgtaaca
1450
<210> 21
<211> 1536
<212> ДНК
<213> Zinderia insecticola CARI
<400> 21
atataaataa gagtttgatc ctggctcaga ttgaacgcta gcggtatgct ttacacatgc
60
aagtcgaacg acaatattaa agcttgcttt aatataaagt ggcgaacggg tgagtaatat
120
atcaaaacgt accttaaagt gggggataac taattgaaaa attagataat accgcatatt
180
aatcttagga tgaaaatagg aataatatct tatgctttta gatcggttga tatctgatta
240
gctagttggt agggtaaatg cttaccaagg caatgatcag tagctggttt tagcgaatga
300
tcagccacac tggaactgag acacggtcca gacttctacg gaaggcagca gtggggaata
360
ttggacaatg ggagaaatcc tgatccagca ataccgcgtg agtgatgaag gccttagggt
420
cgtaaaactc ttttgttagg aaagaaataa ttttaaataa tatttaaaat tgatgacggt
480
acctaaagaa taagcaccgg ctaactacgt gccagcagcc gcggtaatac gtagggtgca
540
agcgttaatc ggaattattg ggcgtaaaga gtgcgtaggc tgttatataa gatagatgtg
600
aaatacttaa gcttaactta agaactgcat ttattactgt ttaactagag tttattagag
660
agaagtggaa ttttatgtgt agcagtgaaa tgcgtagata tataaaggaa tatcgatggc
720
gaaggcagct tcttggaata atactgacgc tgaggcacga aagcgtgggg agcaaacagg
780
attagatacc ctggtagtcc acgccctaaa ctatgtctac tagttattaa attaaaaata
840
aaatttagta acgtagctaa cgcattaagt agaccgcctg gggagtacga tcgcaagatt
900
aaaactcaaa ggaattgacg gggacccgca caagcggtgg atgatgtgga ttaattcgat
960
gcaacacgaa aaaccttacc tactcttgac atgtttggaa ttttaaagaa atttaaaagt
1020
gcttgaaaaa gaaccaaaac acaggtgctg catggctgtc gtcagctcgt gtcgtgagat
1080
gttgggttaa gtcccgcaac gagcgcaacc cttgttatta tttgctaata aaaagaactt
1140
taataagact gccaatgaca aattggagga aggtggggat gacgtcaagt cctcatggcc
1200
cttatgagta gggcttcaca cgtcatacaa tgatatatac aatgggtagc aaatttgtga
1260
aaatgagcca atccttaaag tatatcttag ttcggattgt agtctgcaac tcgactacat
1320
gaagttggaa tcgctagtaa tcgcggatca gcatgccgcg gtgaatacgt tctcgggtct
1380
tgtacacacc gcccgtcaca ccatggaagt gatttttacc agaaattatt tgtttaacct
1440
ttattggaaa aaaataatta aggtagaatt catgactggg gtgaagtcgt aacaaggtag
1500
cagtatcgga aggtgcggct ggattacatt ttaaat
1536
<210> 22
<211> 1423
<212> ДНК
<213> Hodgkinia
<400> 22
aatgctggcg gcaggcctaa cacatgcaag tcgagcggac aacgttcaaa cgttgttagc
60
ggcgaacggg tgagtaatac gtgagaatct acccatccca acgtgataac atagtcaaca
120
ccatgtcaat aacgtatgat tcctgcaaca ggtaaagatt ttatcgggga tggatgagct
180
cacgctagat tagctagttg gtgagataaa agcccaccaa ggccaagatc tatagctggt
240
ctggaaggat ggacagccac attgggactg agacaaggcc caaccctcta aggagggcag
300
cagtgaggaa tattggacaa tgggcgtaag cctgatccag ccatgccgca tgagtgattg
360
aaggtccaac ggactgtaaa actcttttct ccagagatca taaatgatag tatctggtga
420
tataagctcc ggccaacttc gtgccagcag ccgcggtaat acgaggggag cgagtattgt
480
tcggttttat tgggcgtaaa gggtgtccag gttgctaagt aagttaacaa caaaatcttg
540
agattcaacc tcataacgtt cggttaatac tactaagctc gagcttggat agagacaaac
600
ggaattccga gtgtagaggt gaaattcgtt gatacttgga ggaacaccag aggcgaaggc
660
ggtttgtcat accaagctga cactgaagac acgaaagcat ggggagcaaa caggattaga
720
taccctggta gtccatgccc taaacgttga gtgctaacag ttcgatcaag ccacatgcta
780
tgatccagga ttgtacagct aacgcgttaa gcactccgcc tgggtattac gaccgcaagg
840
ttaaaactca aaggaattga cggagacccg cacaagcggt ggagcatgtg gtttaattcg
900
aagctacacg aagaacctta ccagcccttg acataccatg gccaaccatc ctggaaacag
960
gatgttgttc aagttaaacc cttgaaatgc caggaacagg tgctgcatgg ctgttgtcag
1020
ttcgtgtcgt gagatgtatg gttaagtccc aaaacgaaca caaccctcac ccatagttgc
1080
cataaacaca attgggttct ctatgggtac tgctaacgta agttagagga aggtgaggac
1140
cacaacaagt catcatggcc cttatgggct gggccacaca catgctacaa tggtggttac
1200
aaagagccgc aacgttgtga gaccgagcaa atctccaaag accatctcag tccggattgt
1260
actctgcaac ccgagtacat gaagtaggaa tcgctagtaa tcgtggatca gcatgccacg
1320
gtgaatacgt tctcgggtct tgtacacgcc gcccgtcaca ccatgggagc ttcgctccga
1380
tcgaagtcaa gttacccttg accacatctt ggcaagtgac cga
1423
<210> 23
<211> 1504
<212> ДНК
<213> Wolbachia sp. wPip
<400> 23
aaatttgaga gtttgatcct ggctcagaat gaacgctggc ggcaggccta acacatgcaa
60
gtcgaacgga gttatattgt agcttgctat ggtataactt agtggcagac gggtgagtaa
120
tgtataggaa tctacctagt agtacggaat aattgttgga aacgacaact aataccgtat
180
acgccctacg ggggaaaaat ttattgctat tagatgagcc tatattagat tagctagttg
240
gtggggtaat agcctaccaa ggtaatgatc tatagctgat ctgagaggat gatcagccac
300
actggaactg agatacggtc cagactccta cgggaggcag cagtggggaa tattggacaa
360
tgggcgaaag cctgatccag ccatgccgca tgagtgaaga aggcctttgg gttgtaaagc
420
tcttttagtg aggaagataa tgacggtact cacagaagaa gtcctggcta actccgtgcc
480
agcagccgcg gtaatacgga gagggctagc gttattcgga attattgggc gtaaagggcg
540
cgtaggctgg ttaataagtt aaaagtgaaa tcccgaggct taaccttgga attgctttta
600
aaactattaa tctagagatt gaaagaggat agaggaattc ctgatgtaga ggtaaaattc
660
gtaaatatta ggaggaacac cagtggcgaa ggcgtctatc tggttcaaat ctgacgctga
720
agcgcgaagg cgtggggagc aaacaggatt agataccctg gtagtccacg ctgtaaacga
780
tgaatgttaa atatggggag tttactttct gtattacagc taacgcgtta aacattccgc
840
ctggggacta cggtcgcaag attaaaactc aaaggaattg acggggaccc gcacaagcgg
900
tggagcatgt ggtttaattc gatgcaacgc gaaaaacctt accacttctt gacatgaaaa
960
tcatacctat tcgaagggat agggtcggtt cggccggatt ttacacaagt gttgcatggc
1020
tgtcgtcagc tcgtgtcgtg agatgttggg ttaagtcccg caacgagcgc aaccctcatc
1080
cttagttgcc atcaggtaat gctgagtact ttaaggaaac tgccagtgat aagctggagg
1140
aaggtgggga tgatgtcaag tcatcatggc ctttatggag tgggctacac acgtgctaca
1200
atggtgtcta caatgggctg caaggtgcgc aagcctaagc taatccctaa aagacatctc
1260
agttcggatt gtactctgca actcgagtac atgaagttgg aatcgctagt aatcgtggat
1320
cagcatgcca cggtgaatac gttctcgggt cttgtacaca ctgcccgtca cgccatggga
1380
attggtttca ctcgaagcta atggcctaac cgcaaggaag gagttattta aagtgggatc
1440
agtgactggg gtgaagtcgt aacaaggtag cagtagggga atctgcagct ggattacctc
1500
ctta
1504
<210> 24
<211> 1532
<212> ДНК
<213> Uzinura diaspidicola
<400> 24
aaaggagata ttccaaccac accttccggt acggttacct tgttacgact tagccctagt
60
catcaagttt accttaggca gaccactgaa ggattactga cttcaggtac ccccgactcc
120
catggcttga cgggcggtgt gtacaaggtt cgagaacata ttcaccgcgc cattgctgat
180
gcgcgattac tagcgattcc tgcttcatag agtcgaattg cagactccaa tccgaactga
240
gactggtttt agagattagc tcctgatcac ccagtggctg ccctttgtaa ccagccattg
300
tagcacgtgt gtagcccaag gcatagaggc catgatgatt tgacatcatc cccaccttcc
360
tcacagttta caccggcagt tttgttagag tccccggctt tacccgatgg caactaacaa
420
taggggttgc gctcgttata ggacttaacc aaacacttca cagcacgaac tgaagacaac
480
catgcagcac cttgtaatac gtcgtataga ctaagctgtt tccagcttat tcgtaataca
540
tttaagcctt ggtaaggttc ctcgcgtatc atcgaattaa accacatgct ccaccgcttg
600
tgcgaacccc cgtcaattcc tttgagtttc aatcttgcga ctgtacttcc caggtggatc
660
acttatcgct ttcgctaagc cactgaatat cgtttttcca atagctagtg atcatcgttt
720
agggcgtgga ctaccagggt atctaatcct gtttgctccc cacgctttcg tgcactgagc
780
gtcagtaaag atttagcaac ctgccttcgc tatcggtgtt ctgtatgata tctatgcatt
840
tcaccgctac accatacatt ccagatgctc caatcttact caagtttacc agtatcaata
900
gcaattttac agttaagctg taagctttca ctactgactt aataaacagc ctacacaccc
960
tttaaaccca ataaatccga ataacgcttg tgtcatccgt attgccgcgg ctgctggcac
1020
ggaattagcc gacacttatt cgtatagtac cttcaatctc ctatcacgta agatatttta
1080
tttctataca aaagcagttt acaacctaaa agaccttcat cctgcacgcg acgtagctgg
1140
ttcagagttt cctccattga ccaatattcc tcactgctgc ctcccgtagg agtctggtcc
1200
gtgtctcagt accagtgtgg aggtacaccc tcttaggccc cctactgatc atagtcttgg
1260
tagagccatt acctcaccaa ctaactaatc aaacgcaggc tcatcttttg ccacctaagt
1320
tttaataaag gctccatgca gaaactttat attatggggg attaatcaga atttcttctg
1380
gctatacccc agcaaaaggt agattgcata cgtgttactc acccattcgc cggtcgccga
1440
caaattaaaa atttttcgat gcccctcgac ttgcatgtgt taagctcgcc gctagcgtta
1500
attctgagcc aggatcaaac tcttcgttgt ag
1532
<210> 25
<211> 1470
<212> ДНК
<213> Sulcia muelleri
<400> 25
ctcaggataa acgctagcgg agggcttaac acatgcaagt cgaggggcag caaaaataat
60
tatttttggc gaccggcaaa cgggtgagta atacatacgt aactttcctt atgctgagga
120
atagcctgag gaaacttgga ttaatacctc ataatacaat tttttagaaa gaaaaattgt
180
taaagtttta ttatggcata agataggcgt atgtccaatt agttagttgg taaggtaatg
240
gcttaccaag acgatgattg gtagggggcc tgagaggggc gttcccccac attggtactg
300
agacacggac caaacttcta cggaaggctg cagtgaggaa tattggtcaa tggaggaaac
360
tctgaaccag ccactccgcg tgcaggatga aagaaagcct tattggttgt aaactgcttt
420
tgtatatgaa taaaaaattc taattataga aataattgaa ggtaatatac gaataagtat
480
cgactaactc tgtgccagca gtcgcggtaa gacagaggat acaagcgtta tccggattta
540
ttgggtttaa agggtgcgta ggcggttttt aaagtcagta gtgaaatctt aaagcttaac
600
tttaaaagtg ctattgatac tgaaaaacta gagtaaggtt ggagtaactg gaatgtgtgg
660
tgtagcggtg aaatgcatag atatcacaca gaacaccgat agcgaaagca agttactaac
720
cctatactga cgctgagtca cgaaagcatg gggagcaaac aggattagat accctggtag
780
tccatgccgt aaacgatgat cactaactat tgggttttat acgttgtaat tcagtggtga
840
agcgaaagtg ttaagtgatc cacctgagga gtacgaccgc aaggttgaaa ctcaaaggaa
900
ttgacggggg cccgcacaat cggtggagca tgtggtttaa ttcgatgata cacgaggaac
960
cttaccaaga cttaaatgta ctacgaataa attggaaaca atttagtcaa gcgacggagt
1020
acaaggtgct gcatggttgt cgtcagctcg tgccgtgagg tgtaaggtta agtcctttaa
1080
acgagcgcaa cccttattat tagttgccat cgagtaatgt caggggactc taataagact
1140
gccggcgcaa gccgagagga aggtggggat gacgtcaaat catcacggcc cttacgtctt
1200
gggccacaca cgtgctacaa tgatcggtac aaaagggagc gactgggtga ccaggagcaa
1260
atccagaaag ccgatctaag ttcggattgg agtctgaaac tcgactccat gaagctggaa
1320
tcgctagtaa tcgtgcatca gccatggcac ggtgaatatg ttcccgggcc ttgtacacac
1380
cgcccgtcaa gccatggaag ttggaagtac ctaaagttgg ttcgctacct aaggtaagtc
1440
taataactgg ggctaagtcg taacaaggta
1470
<210> 26
<211> 1761
<212> ДНК
<213> Symbiotaphrina buchneri
<220>
<221> другой_признак
<222> (30)..(30)
<223> n представляет собой a, g, c или t
<220>
<221> другой_признак
<222> (40)..(40)
<223> n представляет собой a, g, c или t
<400> 26
agattaagcc atgcaagtct aagtataagn aatctatacn gtgaaactgc gaatggctca
60
ttaaatcagt tatcgtttat ttgatagtac cttactacat ggataaccgt ggtaattcta
120
gagctaatac atgctaaaaa ccccgacttc ggaaggggtg tatttattag ataaaaaacc
180
aatgcccttc ggggctcctt ggtgattcat gataacttaa cgaatcgcat ggccttgcgc
240
cggcgatggt tcattcaaat ttctgcccta tcaactttcg atggtaggat agtggcctac
300
catggtttta acgggtaacg gggaattagg gttcgattcc ggagagggag cctgagaaac
360
ggctaccaca tccaaggaag gcagcaggcg cgcaaattac ccaatcccga cacggggagg
420
tagtgacaat aaatactgat acagggctct tttgggtctt gtaattggaa tgagtacaat
480
ttaaatccct taacgaggaa caattggagg gcaagtctgg tgccagcagc cgcggtaatt
540
ccagctccaa tagcgtatat taaagttgtt gcagttaaaa agctcgtagt tgaaccttgg
600
gcctggctgg ccggtccgcc taaccgcgtg tactggtccg gccgggcctt tccttctggg
660
gagccgcatg cccttcactg ggtgtgtcgg ggaaccagga cttttacttt gaaaaaatta
720
gagtgttcaa agcaggccta tgctcgaata cattagcatg gaataataga ataggacgtg
780
cggttctatt ttgttggttt ctaggaccgc cgtaatgatt aatagggata gtcgggggca
840
tcagtattca attgtcagag gtgaaattct tggatttatt gaagactaac tactgcgaaa
900
gcatttgcca aggatgtttt cattaatcag tgaacgaaag ttaggggatc gaagacgatc
960
agataccgtc gtagtcttaa ccataaacta tgccgactag ggatcgggcg atgttattat
1020
tttgactcgc tcggcacctt acgagaaatc aaagtctttg ggttctgggg ggagtatggt
1080
cgcaaggctg aaacttaaag aaattgacgg aagggcacca ccaggagtgg agcctgcggc
1140
ttaatttgac tcaacacggg gaaactcacc aggtccagac acattaagga ttgacagatt
1200
gagagctctt tcttgattat gtgggtggtg gtgcatggcc gttcttagtt ggtggagtga
1260
tttgtctgct taattgcgat aacgaacgag accttaacct gctaaatagc ccggtccgct
1320
ttggcgggcc gctggcttct tagagggact atcggctcaa gccgatggaa gtttgaggca
1380
ataacaggtc tgtgatgccc ttagatgttc tgggccgcac gcgcgctaca ctgacagagc
1440
caacgagtaa atcaccttgg ccggaaggtc tgggtaatct tgttaaactc tgtcgtgctg
1500
gggatagagc attgcaatta ttgctcttca acgaggaatt cctagtaagc gcaagtcatc
1560
agcttgcgct gattacgtcc ctgccctttg tacacaccgc ccgtcgctac taccgattga
1620
atggctcagt gaggccttcg gactggcaca gggacgttgg caacgacgac ccagtgccgg
1680
aaagttggtc aaacttggtc atttagagga agtaaaagtc gtaacaaggt ttccgtaggt
1740
gaacctgcgg aaggatcatt a
1761
<210> 27
<211> 1801
<212> ДНК
<213> Symbiotaphrina kochii
<220>
<221> другой_признак
<222> (1753)..(1755)
<223> n представляет собой a, g, c или t
<400> 27
tacctggttg attctgccag tagtcatatg cttgtctcaa agattaagcc atgcaagtct
60
aagtataagc aatctatacg gtgaaactgc gaatggctca ttaaatcagt tatcgtttat
120
ttgatagtac cttactacat ggataaccgt ggtaattcta gagctaatac atgctaaaaa
180
cctcgacttc ggaaggggtg tatttattag ataaaaaacc aatgcccttc ggggctcctt
240
ggtgattcat gataacttaa cgaatcgcat ggccttgcgc cggcgatggt tcattcaaat
300
ttctgcccta tcaactttcg atggtaggat agtggcctac catggtttca acgggtaacg
360
gggaattagg gttcgattcc ggagagggag cctgagaaac ggctaccaca tccaaggaag
420
gcagcaggcg cgcaaattac ccaatcccga cacggggagg tagtgacaat aaatactgat
480
acagggctct tttgggtctt gtaattggaa tgagtacaat ttaaatccct taacgaggaa
540
caattggagg gcaagtctgg tgccagcagc cgcggtaatt ccagctccaa tagcgtatat
600
taaagttgtt gcagttaaaa agctcgtagt tgaaccttgg gcctggctgg ccggtccgcc
660
taaccgcgtg tactggtccg gccgggcctt tccttctggg gagccgcatg cccttcactg
720
ggtgtgtcgg ggaaccagga cttttacttt gaaaaaatta gagtgttcaa agcaggccta
780
tgctcgaata cattagcatg gaataataga ataggacgtg tggttctatt ttgttggttt
840
ctaggaccgc cgtaatgatt aatagggata gtcgggggca tcagtattca attgtcagag
900
gtgaaattct tggatttatt gaagactaac tactgcgaaa gcatttgcca aggatgtttt
960
cattaatcag tgaacgaaag ttaggggatc gaagacgatc agataccgtc gtagtcttaa
1020
ccataaacta tgccgactag ggatcgggcg atgttattat tttgactcgc tcggcacctt
1080
acgagaaatc aaagtctttg ggttctgggg ggagtatggt cgcaaggctg aaacttaaag
1140
aaattgacgg aagggcacca ccaggagtgg agcctgcggc ttaatttgac tcaacacggg
1200
gaaactcacc aggtccagac acattaagga ttgacagatt gagagctctt tcttgattat
1260
gtgggtggtg gtgcatggcc gttcttagtt ggtggagtga tttgtctgct taattgcgat
1320
aacgaacgag accttaacct gctaaatagc ccggtccgct ttggcgggcc gctggcttct
1380
tagagggact atcggctcaa gccgatggaa gtttgaggca ataacaggtc tgtgatgccc
1440
ttagatgttc tgggccgcac gcgcgctaca ctgacagagc caacgagtac atcaccttgg
1500
ccggaaggtc tgggtaatct tgttaaactc tgtcgtgctg gggatagagc attgcaatta
1560
ttgctcttca acgaggaatt cctagtaagc gcaagtcatc agcttgcgct gattacgtcc
1620
ctgccctttg tacacaccgc ccgtcgctac taccgattga atggctcagt gaggccttcg
1680
gactggcaca gggacgttgg caacgacgac ccagtgccgg aaagttcgtc aaacttggtc
1740
atttagagga agnnnaagtc gtaacaaggt ttccgtaggt gaacctgcgg aaggatcatt
1800
a
1801
<210> 28
<211> 1490
<212> ДНК
<213> Burkholderia sp. SFA1
<400> 28
agtttgatcc tggctcagat tgaacgctgg cggcatgcct tacacatgca agtcgaacgg
60
cagcacgggg gcaaccctgg tggcgagtgg cgaacgggtg agtaatacat cggaacgtgt
120
cctgtagtgg gggatagccc ggcgaaagcc ggattaatac cgcatacgac ctaagggaga
180
aagcggggga tcttcggacc tcgcgctata ggggcggccg atggcagatt agctagttgg
240
tggggtaaag gcctaccaag gcgacgatct gtagctggtc tgagaggacg accagccaca
300
ctgggactga gacacggccc agactcctac gggaggcagc agtggggaat tttggacaat
360
gggggcaacc ctgatccagc aatgccgcgt gtgtgaagaa ggcttcgggt tgtaaagcac
420
ttttgtccgg aaagaaaact tcgtccctaa tatggatgga ggatgacggt accggaagaa
480
taagcaccgg ctaactacgt gccagcagcc gcggtaatac gtagggtgcg agcgttaatc
540
ggaattactg ggcgtaaagc gtgcgcaggc ggtctgttaa gaccgatgtg aaatccccgg
600
gcttaacctg ggaactgcat tggtgactgg caggctttga gtgtggcaga ggggggtaga
660
attccacgtg tagcagtgaa atgcgtagag atgtggagga ataccgatgg cgaaggcagc
720
cccctgggcc aactactgac gctcatgcac gaaagcgtgg ggagcaaaca ggattagata
780
ccctggtagt ccacgcccta aacgatgtca actagttgtt ggggattcat ttccttagta
840
acgtagctaa cgcgtgaagt tgaccgcctg gggagtacgg tcgcaagatt aaaactcaaa
900
ggaattgacg gggacccgca caagcggtgg atgatgtgga ttaattcgat gcaacgcgaa
960
aaaccttacc tacccttgac atggtcggaa ccctgctgaa aggtgggggt gctcgaaaga
1020
gaaccggcgc acaggtgctg catggctgtc gtcagctcgt gtcgtgagat gttgggttaa
1080
gtcccgcaac gagcgcaacc cttgtcctta gttgctacgc aagagcactc taaggagact
1140
gccggtgaca aaccggagga aggtggggat gacgtcaagt cctcatggcc cttatgggta
1200
gggcttcaca cgtcatacaa tggtcggaac agagggttgc caagccgcga ggtggagcca
1260
atcccagaaa accgatcgta gtccggatcg cagtctgcaa ctcgactgcg tgaagctgga
1320
atcgctagta atcgcggatc agcatgccgc ggtgaatacg ttcccgggtc ttgtacacac
1380
cgcccgtcac accatgggag tgggtttcac cagaagtagg tagcctaacc gcaaggaggg
1440
cgcttaccac ggtgggattc atgactgggg tgaagtcgta acaaggtagc
1490
<210> 29
<211> 1408
<212> ДНК
<213> Burkholderia sp. KM-A
<400> 29
gcaaccctgg tggcgagtgg cgaacgggtg agtaatacat cggaacgtgt cctgtagtgg
60
gggatagccc ggcgaaagcc ggattaatac cgcatacgat ctacggaaga aagcggggga
120
tccttcggga cctcgcgcta taggggcggc cgatggcaga ttagctagtt ggtggggtaa
180
aggcctacca aggcgacgat ctgtagctgg tctgagagga cgaccagcca cactgggact
240
gagacacggc ccagactcct acgggaggca gcagtgggga attttggaca atgggggcaa
300
ccctgatcca gcaatgccgc gtgtgtgaag aaggccttcg ggttgtaaag cacttttgtc
360
cggaaagaaa acgtcttggt taatacctga ggcggatgac ggtaccggaa gaataagcac
420
cggctaacta cgtgccagca gccgcggtaa tacgtagggt gcgagcgtta atcggaatta
480
ctgggcgtaa agcgtgcgca ggcggtctgt taagaccgat gtgaaatccc cgggcttaac
540
ctgggaactg cattggtgac tggcaggctt tgagtgtggc agaggggggt agaattccac
600
gtgtagcagt gaaatgcgta gagatgtgga ggaataccga tggcgaaggc agccccctgg
660
gccaacactg acgctcatgc acgaaagcgt ggggagcaaa caggattaga taccctggta
720
gtccacgccc taaacgatgt caactagttg ttggggattc atttccttag taacgtagct
780
aacgcgtgaa gttgaccgcc tggggagtac ggtcgcaaga ttaaaactca aaggaattga
840
cggggacccg cacaagcggt ggatgatgtg gattaattcg atgcaacgcg aaaaacctta
900
cctacccttg acatggtcgg aagtctgctg agaggtggac gtgctcgaaa gagaaccggc
960
gcacaggtgc tgcatggctg tcgtcagctc gtgtcgtgag atgttgggtt aagtcccgca
1020
acgagcgcaa cccttgtcct tagttgctac gcaagagcac tctaaggaga ctgccggtga
1080
caaaccggag gaaggtgggg atgacgtcaa gtcctcatgg cccttatggg tagggcttca
1140
cacgtcatac aatggtcgga acagagggtt gccaagccgc gaggtggagc caatcccaga
1200
aaaccgatcg tagtccggat cgcagtctgc aactcgactg cgtgaagctg gaatcgctag
1260
taatcgcgga tcagcatgcc gcggtgaata cgttcccggg tcttgtacac accgcccgtc
1320
acaccatggg agtgggtttc accagaagta ggtagcctaa ccgcaaggag ggcgcttacc
1380
acggtgggat tcatgactgg ggtgaagt
1408
<210> 30
<211> 1383
<212> ДНК
<213> Burkholderia sp. KM-G
<400> 30
gcaaccctgg tggcgagtgg cgaacgggtg agtaatacat cggaacgtgt cctgtagtgg
60
gggatagccc ggcgaaagcc ggattaatac cgcatacgac ctaagggaga aagcggggga
120
tcttcggacc tcgcgctata ggggcggccg atggcagatt agctagttgg tggggtaaag
180
gcctaccaag gcgacgatct gtagctggtc tgagaggacg accagccaca ctgggactga
240
gacacggccc agactcctac gggaggcagc agtggggaat tttggacaat gggggcaacc
300
ctgatccagc aatgccgcgt gtgtgaagaa ggccttcggg ttgtaaagca cttttgtccg
360
gaaagaaaac ttcgaggtta atacccttgg aggatgacgg taccggaaga ataagcaccg
420
gctaactacg tgccagcagc cgcggtaata cgtagggtgc gagcgttaat cggaattact
480
gggcgtaaag cgtgcgcagg cggtctgtta agaccgatgt gaaatccccg ggcttaacct
540
gggaactgca ttggtgactg gcaggctttg agtgtggcag aggggggtag aattccacgt
600
gtagcagtga aatgcgtaga gatgtggagg aataccgatg gcgaaggcag ccccctgggc
660
caacactgac gctcatgcac gaaagcgtgg ggagcaaaca ggattagata ccctggtagt
720
ccacgcccta aacgatgtca actagttgtt ggggattcat ttccttagta acgtagctaa
780
cgcgtgaagt tgaccgcctg gggagtacgg tcgcaagatt aaaactcaaa ggaattgacg
840
gggacccgca caagcggtgg atgatgtgga ttaattcgat gcaacgcgaa aaaccttacc
900
tacccttgac atggtcggaa gtctgctgag aggtggacgt gctcgaaaga gaaccggcgc
960
acaggtgctg catggctgtc gtcagctcgt gtcgtgagat gttgggttaa gtcccgcaac
1020
gagcgcaacc cttgtcctta gttgctacgc aagagcactc taaggagact gccggtgaca
1080
aaccggagga aggtggggat gacgtcaagt cctcatggcc cttatgggta gggcttcaca
1140
cgtcatacaa tggtcggaac agagggttgc caagccgcga ggtggagcca atcccagaaa
1200
accgatcgta gtccggatcg cagtctgcaa ctcgactgcg tgaagctgga atcgctagta
1260
atcgcggatc agcatgccgc ggtgaatacg ttcccgggtc ttgtacacac cgcccgtcac
1320
accatgggag tgggtttcac cagaagtagg tagcctaacc tgcaaaggag ggcgcttacc
1380
acg
1383
<210> 31
<211> 538
<212> ДНК
<213> Xiphinematobacter sp.
<400> 31
gcaagtcgaa cggagtggaa cctgcagtaa tgcagattcg attcagtggc gtacgggtgc
60
gtaacacgtg agtgatctac cggtaagtgg gggataaccc gccgaaaggc gaattaatac
120
cgcatgtggc tagggatgcc ttcatcctgt agctaaagtc gattttgacg ctttctgatg
180
agctcgcggc ctatcagctt gttggtggag gtaatggccc accaaggcaa tgacgggtag
240
ctggtctgag aggacgatca gccacactgg aactgagaca cggtccagac acctacgggt
300
ggcagcagtc gagaattttt cacaatgggg gaaaccctga tgaagcaacg ccgcgtggag
360
gatgaagggc ttcgcgctcg taaactcctg tcaagcggga acaagaaagt gatagtaccg
420
ctagaggaag agacggctaa ctctgtgcca gcagccgcgg taatacagag gtctcgagcg
480
ttgttcggat ttattgggcg taaagggtgc gtaggcggtg tggcaagtca agtgtgaa
538
<210> 32
<211> 975
<212> ДНК
<213> Wolbachia sp. wOo
<400> 32
atgacacaca taccggtttt actaaaagaa atgctatcgc aactttcacc acaaaatggt
60
agtgtatatg tggatgccac atttggagct ggaggatata gtaaagcaat attggagtca
120
gctgattgca gagtgtatgc aatcgacaga gatgaaacgg ttattaaatt ttataatagt
180
ttgaatacca agtaccacgg taaaataaaa ctatttattg aaaagtttag caatattcaa
240
actatactaa acagtagtaa tctcaaacac tttacagaac cttccgtcat tgtttcagct
300
ggaattcaga aaaaaaatgc aaggtcaagc accgagatga tacaaagtaa taccgtagat
360
ggagttgtgt tcgatatagg agtatcgtct atgcagcttg atgaagaaaa tagaggattt
420
tcatttttac ataacagtcc gcttgatatg cgcatggata cctcttctca cattaacgct
480
tcaatatttg ttaatgcctt acgcgaagaa gaaattgcaa acactatata tagctatgga
540
ggtgaacgtt attctcgcaa aattgcaaga gcaatagtga acgtacgtaa gaaaaaaact
600
atcgacacta catttgagct tgcagacatt gtacgttccg tggtatctcg cggaaaaagc
660
aagattgatc ctgcaactag gacatttcaa gcaatcagaa tatgggtaaa cgatgagctt
720
agagagcttg aaaagggtat taaagctgca tccaaaatct taaataggaa tggcaagctg
780
attgtcatta cttttcattc cttggaagat cgtatagtca agaccttttt taaaggctta
840
tgtgagccaa aattcaccaa ctgtagaacg ttttctcttc tgaataaaaa agtaatcaag
900
gcaagcgcag aagaaataag tgcaaatcca cgtgcgcgtt cagcaaaact aagagctata
960
caaaggttat tatga
975
<210> 33
<211> 1505
<212> ДНК
<213> Snodgrassella alvi
<400> 33
gagagtttga tcctggctca gattgaacgc tggcggcatg ccttacacat gcaagtcgaa
60
cggcagcacg gagagcttgc tctctggtgg cgagtggcga acgggtgagt aatgcatcgg
120
aacgtaccga gtaatggggg ataactgtcc gaaaggatgg ctaataccgc atacgccctg
180
agggggaaag cgggggatcg aaagacctcg cgttatttga gcggccgatg ttggattagc
240
tagttggtgg ggtaaaggcc taccaaggcg acgatccata gcgggtctga gaggatgatc
300
cgccacattg ggactgagac acggcccaaa ctcctacggg aggcagcagt ggggaatttt
360
ggacaatggg gggaaccctg atccagccat gccgcgtgtc tgaagaaggc cttcgggttg
420
taaaggactt ttgttaggga agaaaagccg ggtgttaata ccatctggtg ctgacggtac
480
ctaaagaata agcaccggct aactacgtgc cagcagccgc ggtaatacgt agggtgcgag
540
cgttaatcgg aattactggg cgtaaagcga gcgcagacgg ttaattaagt cagatgtgaa
600
atccccgagc tcaacttggg acgtgcattt gaaactggtt aactagagtg tgtcagaggg
660
aggtagaatt ccacgtgtag cagtgaaatg cgtagagatg tggaggaata ccgatggcga
720
aggcagcctc ctgggataac actgacgttc atgctcgaaa gcgtgggtag caaacaggat
780
tagataccct ggtagtccac gccctaaacg atgacaatta gctgttggga cactagatgt
840
cttagtagcg aagctaacgc gtgaaattgt ccgcctgggg agtacggtcg caagattaaa
900
actcaaagga attgacgggg acccgcacaa gcggtggatg atgtggatta attcgatgca
960
acgcgaagaa ccttacctgg tcttgacatg tacggaatct cttagagata ggagagtgcc
1020
ttcgggaacc gtaacacagg tgctgcatgg ctgtcgtcag ctcgtgtcgt gagatgttgg
1080
gttaagtccc gcaacgagcg caacccttgt cattagttgc catcattaag ttgggcactc
1140
taatgagact gccggtgaca aaccggagga aggtggggat gacgtcaagt cctcatggcc
1200
cttatgacca gggcttcaca cgtcatacaa tggtcggtac agagggtagc gaagccgcga
1260
ggtgaagcca atctcagaaa gccgatcgta gtccggattg cactctgcaa ctcgagtgca
1320
tgaagtcgga atcgctagta atcgcaggtc agcatactgc ggtgaatacg ttcccgggtc
1380
ttgtacacac cgcccgtcac accatgggag tgggggatac cagaattggg tagactaacc
1440
gcaaggaggt cgcttaacac ggtatgcttc atgactgggg tgaagtcgta acaaggtagc
1500
cgtag
1505
<210> 34
<211> 1541
<212> ДНК
<213> Gilliamella apicola
<400> 34
ttaaattgaa gagtttgatc atggctcaga ttgaacgctg gcggcaggct taacacatgc
60
aagtcgaacg gtaacatgag tgcttgcact tgatgacgag tggcggacgg gtgagtaaag
120
tatggggatc tgccgaatgg agggggacaa cagttggaaa cgactgctaa taccgcataa
180
agttgagaga ccaaagcatg ggaccttcgg gccatgcgcc atttgatgaa cccatatggg
240
attagctagt tggtagggta atggcttacc aaggcgacga tctctagctg gtctgagagg
300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg
360
aatattgcac aatgggggaa accctgatgc agccatgccg cgtgtatgaa gaaggccttc
420
gggttgtaaa gtactttcgg tgatgaggaa ggtggtgtat ctaataggtg catcaattga
480
cgttaattac agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg
540
tgcgagcgtt aatcggaatg actgggcgta aagggcatgt aggcggataa ttaagttagg
600
tgtgaaagcc ctgggctcaa cctaggaatt gcacttaaaa ctggttaact agagtattgt
660
agaggaaggt agaattccac gtgtagcggt gaaatgcgta gagatgtgga ggaataccgg
720
tggcgaaggc ggccttctgg acagatactg acgctgagat gcgaaagcgt ggggagcaaa
780
caggattaga taccctggta gtccacgctg taaacgatgt cgatttggag tttgttgcct
840
agagtgatgg gctccgaagc taacgcgata aatcgaccgc ctggggagta cggccgcaag
900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc
960
gatgcaacgc gaagaacctt acctggtctt gacatccaca gaatcttgca gagatgcggg
1020
agtgccttcg ggaactgtga gacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa
1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccatc ggttaggccg
1140
ggaactcaaa ggagactgcc gttgataaag cggaggaagg tggggacgac gtcaagtcat
1200
catggccctt acgaccaggg ctacacacgt gctacaatgg cgtatacaaa gggaggcgac
1260
ctcgcgagag caagcggacc tcataaagta cgtctaagtc cggattggag tctgcaactc
1320
gactccatga agtcggaatc gctagtaatc gtgaatcaga atgtcacggt gaatacgttc
1380
ccgggccttg tacacaccgc ccgtcacacc atgggagtgg gttgcaccag aagtagatag
1440
cttaaccttc gggagggcgt ttaccacggt gtggtccatg actggggtga agtcgtaaca
1500
aggtaaccgt aggggaacct gcggttggat cacctcctta c
1541
<210> 35
<211> 1528
<212> ДНК
<213> Bartonella apis
<400> 35
aagccaaaat caaattttca acttgagagt ttgatcctgg ctcagaacga acgctggcgg
60
caggcttaac acatgcaagt cgaacgcact tttcggagtg agtggcagac gggtgagtaa
120
cgcgtgggaa tctacctatt tctacggaat aacgcagaga aatttgtgct aataccgtat
180
acgtccttcg ggagaaagat ttatcggaga tagatgagcc cgcgttggat tagctagttg
240
gtgaggtaat ggcccaccaa ggcgacgatc catagctggt ctgagaggat gaccagccac
300
attgggactg agacacggcc cagactccta cgggaggcag cagtggggaa tattggacaa
360
tgggcgcaag cctgatccag ccatgccgcg tgagtgatga aggccctagg gttgtaaagc
420
tctttcaccg gtgaagataa tgacggtaac cggagaagaa gccccggcta acttcgtgcc
480
agcagccgcg gtaatacgaa gggggctagc gttgttcgga tttactgggc gtaaagcgca
540
cgtaggcgga tatttaagtc aggggtgaaa tcccggggct caaccccgga actgcctttg
600
atactggata tcttgagtat ggaagaggta agtggaattc cgagtgtaga ggtgaaattc
660
gtagatattc ggaggaacac cagtggcgaa ggcggcttac tggtccatta ctgacgctga
720
ggtgcgaaag cgtggggagc aaacaggatt agataccctg gtagtccacg ctgtaaacga
780
tgaatgttag ccgttggaca gtttactgtt cggtggcgca gctaacgcat taaacattcc
840
gcctggggag tacggtcgca agattaaaac tcaaaggaat tgacgggggc ccgcacaagc
900
ggtggagcat gtggtttaat tcgaagcaac gcgcagaacc ttaccagccc ttgacatccc
960
gatcgcggat ggtggagaca ccgtctttca gttcggctgg atcggtgaca ggtgctgcat
1020
ggctgtcgtc agctcgtgtc gtgagatgtt gggttaagtc ccgcaacgag cgcaaccctc
1080
gcccttagtt gccatcattt agttgggcac tctaagggga ctgccggtga taagccgaga
1140
ggaaggtggg gatgacgtca agtcctcatg gcccttacgg gctgggctac acacgtgcta
1200
caatggtggt gacagtgggc agcgagaccg cgaggtcgag ctaatctcca aaagccatct
1260
cagttcggat tgcactctgc aactcgagtg catgaagttg gaatcgctag taatcgtgga
1320
tcagcatgcc acggtgaata cgttcccggg ccttgtacac accgcccgtc acaccatggg
1380
agttggtttt acccgaaggt gctgtgctaa ccgcaaggag gcaggcaacc acggtagggt
1440
cagcgactgg ggtgaagtcg taacaaggta gccgtagggg aacctgcggc tggatcacct
1500
cctttctaag gaagatgaag aattggaa
1528
<210> 36
<211> 1390
<212> ДНК
<213> Parasaccharibacter apium
<220>
<221> другой_признак
<222> (643)..(756)
<223> n представляет собой a, g, c или t
<400> 36
ctaccatgca agtcgcacga aacctttcgg ggttagtggc ggacgggtga gtaacgcgtt
60
aggaacctat ctggaggtgg gggataacat cgggaaactg gtgctaatac cgcatgatgc
120
ctgagggcca aaggagagat ccgccattgg aggggcctgc gttcgattag ctagttggtt
180
gggtaaaggc tgaccaaggc gatgatcgat agctggtttg agaggatgat cagccacact
240
gggactgaga cacggcccag actcctacgg gaggcagcag tggggaatat tggacaatgg
300
gggcaaccct gatccagcaa tgccgcgtgt gtgaagaagg tcttcggatt gtaaagcact
360
ttcactaggg aagatgatga cggtacctag agaagaagcc ccggctaact tcgtgccagc
420
agccgcggta atacgaaggg ggctagcgtt gctcggaatg actgggcgta aagggcgcgt
480
aggctgtttg tacagtcaga tgtgaaatcc ccgggcttaa cctgggaact gcatttgata
540
cgtgcagact agagtccgag agagggttgt ggaattccca gtgtagaggt gaaattcgta
600
gatattggga agaacaccgg ttgcgaaggc ggcaacctgg ctnnnnnnnn nnnnnnnnnn
660
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn
720
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnngagc taacgcgtta agcacaccgc
780
ctggggagta cggccgcaag gttgaaactc aaaggaattg acgggggccc gcacaagcgg
840
tggagcatgt ggtttaattc gaagcaacgc gcagaacctt accagggctt gcatggggag
900
gctgtattca gagatggata tttcttcgga cctcccgcac aggtgctgca tggctgtcgt
960
cagctcgtgt cgtgagatgt tgggttaagt cccgcaacga gcgcaaccct tgtctttagt
1020
tgccatcacg tctgggtggg cactctagag agactgccgg tgacaagccg gaggaaggtg
1080
gggatgacgt caagtcctca tggcccttat gtcctgggct acacacgtgc tacaatggcg
1140
gtgacagagg gatgctacat ggtgacatgg tgctgatctc aaaaaaccgt ctcagttcgg
1200
attgtactct gcaactcgag tgcatgaagg tggaatcgct agtaatcgcg gatcagcatg
1260
ccgcggtgaa tacgttcccg ggccttgtac acaccgcccg tcacaccatg ggagttggtt
1320
tgaccttaag ccggtgagcg aaccgcaagg aacgcagccg accaccggtt cgggttcagc
1380
gactggggga
1390
<210> 37
<211> 1583
<212> ДНК
<213> Lactobacillus kunkeei
<400> 37
ttccttagaa aggaggtgat ccagccgcag gttctcctac ggctaccttg ttacgacttc
60
accctaatca tctgtcccac cttagacgac tagctcctaa aaggttaccc catcgtcttt
120
gggtgttaca aactctcatg gtgtgacggg cggtgtgtac aaggcccggg aacgtattca
180
ccgtggcatg ctgatccacg attactagtg attccaactt catgcaggcg agttgcagcc
240
tgcaatccga actgagaatg gctttaagag attagcttga cctcgcggtt tcgcgactcg
300
ttgtaccatc cattgtagca cgtgtgtagc ccagctcata aggggcatga tgatttgacg
360
tcgtccccac cttcctccgg tttatcaccg gcagtctcac tagagtgccc aactaaatgc
420
tggcaactaa taataagggt tgcgctcgtt gcgggactta acccaacatc tcacgacacg
480
agctgacgac aaccatgcac cacctgtcat tctgtccccg aagggaacgc ccaatctctt
540
gggttggcag aagatgtcaa gagctggtaa ggttcttcgc gtagcatcga attaaaccac
600
atgctccacc acttgtgcgg gcccccgtca attcctttga gtttcaacct tgcggtcgta
660
ctccccaggc ggaatactta atgcgttagc tgcggcactg aagggcggaa accctccaac
720
acctagtatt catcgtttac ggcatggact accagggtat ctaatcctgt tcgctaccca
780
tgctttcgag cctcagcgtc agtaacagac cagaaagccg ccttcgccac tggtgttctt
840
ccatatatct acgcatttca ccgctacaca tggagttcca ctttcctctt ctgtactcaa
900
gttttgtagt ttccactgca cttcctcagt tgagctgagg gctttcacag cagacttaca
960
aaaccgcctg cgctcgcttt acgcccaata aatccggaca acgcttgcca cctacgtatt
1020
accgcggctg ctggcacgta gttagccgtg gctttctggt taaataccgt caaagtgtta
1080
acagttactc taacacttgt tcttctttaa caacagagtt ttacgatccg aaaaccttca
1140
tcactcacgc ggcgttgctc catcagactt tcgtccattg tggaagattc cctactgctg
1200
cctcccgtag gagtctgggc cgtgtctcag tcccaatgtg gccgattacc ctctcaggtc
1260
ggctacgtat catcgtcttg gtgggctttt atctcaccaa ctaactaata cggcgcgggt
1320
ccatcccaaa gtgatagcaa agccatcttt caagttggaa ccatgcggtt ccaactaatt
1380
atgcggtatt agcacttgtt tccaaatgtt atcccccgct tcggggcagg ttacccacgt
1440
gttactcacc agttcgccac tcgctccgaa tccaaaaatc atttatgcaa gcataaaatc
1500
aatttgggag aactcgttcg acttgcatgt attaggcacg ccgccagcgt tcgtcctgag
1560
ccaggatcaa actctcatct taa
1583
<210> 38
<211> 1395
<212> ДНК
<213> Lactobacillus Firm-4
<400> 38
acgaacgctg gcggcgtgcc taatacatgc aagtcgagcg cgggaagtca gggaagcctt
60
cgggtggaac tggtggaacg agcggcggat gggtgagtaa cacgtaggta acctgcccta
120
aagcggggga taccatctgg aaacaggtgc taataccgca taaacccagc agtcacatga
180
gtgctggttg aaagacggct tcggctgtca ctttaggatg gacctgcggc gtattagcta
240
gttggtggag taacggttca ccaaggcaat gatacgtagc cgacctgaga gggtaatcgg
300
ccacattggg actgagacac ggcccaaact cctacgggag gcagcagtag ggaatcttcc
360
acaatggacg caagtctgat ggagcaacgc cgcgtggatg aagaaggtct tcggatcgta
420
aaatcctgtt gttgaagaag aacggttgtg agagtaactg ctcataacgt gacggtaatc
480
aaccagaaag tcacggctaa ctacgtgcca gcagccgcgg taatacgtag gtggcaagcg
540
ttgtccggat ttattgggcg taaagggagc gcaggcggtc ttttaagtct gaatgtgaaa
600
gccctcagct taactgagga agagcatcgg aaactgagag acttgagtgc agaagaggag
660
agtggaactc catgtgtagc ggtgaaatgc gtagatatat ggaagaacac cagtggcgaa
720
ggcggctctc tggtctgtta ctgacgctga ggctcgaaag catgggtagc gaacaggatt
780
agataccctg gtagtccatg ccgtaaacga tgagtgctaa gtgttgggag gtttccgcct
840
ctcagtgctg cagctaacgc attaagcact ccgcctgggg agtacgaccg caaggttgaa
900
actcaaagga attgacgggg gcccgcacaa gcggtggagc atgtggttta attcgaagca
960
acgcgaagaa ccttaccagg tcttgacatc tcctgcaagc ctaagagatt aggggttccc
1020
ttcggggaca ggaagacagg tggtgcatgg ttgtcgtcag ctcgtgtcgt gagatgttgg
1080
gttaagtccc gcaacgagcg caacccttgt tactagttgc cagcattaag ttgggcactc
1140
tagtgagact gccggtgaca aaccggagga aggtggggac gacgtcaaat catcatgccc
1200
cttatgacct gggctacaca cgtgctacaa tggatggtac aatgagaagc gaactcgcga
1260
ggggaagctg atctctgaaa accattctca gttcggattg caggctgcaa ctcgcctgca
1320
tgaagctgga atcgctagta atcgcggatc agcatgccgc ggtgaatacg ttcccgggcc
1380
ttgtacacac cgccc
1395
<210> 39
<211> 1549
<212> ДНК
<213> Enterococcus
<400> 39
aggtgatcca gccgcacctt ccgatacggc taccttgtta cgacttcacc ccaatcatct
60
atcccacctt aggcggctgg ctccaaaaag gttacctcac cgacttcggg tgttacaaac
120
tctcgtggtg tgacgggcgg tgtgtacaag gcccgggaac gtattcaccg cggcgtgctg
180
atccgcgatt actagcgatt ccggcttcat gcaggcgagt tgcagcctgc aatccgaact
240
gagagaagct ttaagagatt tgcatgacct cgcggtctag cgactcgttg tacttcccat
300
tgtagcacgt gtgtagccca ggtcataagg ggcatgatga tttgacgtca tccccacctt
360
cctccggttt gtcaccggca gtctcgctag agtgcccaac taaatgatgg caactaacaa
420
taagggttgc gctcgttgcg ggacttaacc caacatctca cgacacgagc tgacgacaac
480
catgcaccac ctgtcacttt gtccccgaag ggaaagctct atctctagag tggtcaaagg
540
atgtcaagac ctggtaaggt tcttcgcgtt gcttcgaatt aaaccacatg ctccaccgct
600
tgtgcgggcc cccgtcaatt cctttgagtt tcaaccttgc ggtcgtactc cccaggcgga
660
gtgcttaatg cgtttgctgc agcactgaag ggcggaaacc ctccaacact tagcactcat
720
cgtttacggc gtggactacc agggtatcta atcctgtttg ctccccacgc tttcgagcct
780
cagcgtcagt tacagaccag agagccgcct tcgccactgg tgttcctcca tatatctacg
840
catttcaccg ctacacatgg aattccactc tcctcttctg cactcaagtc tcccagtttc
900
caatgaccct ccccggttga gccgggggct ttcacatcag acttaagaaa ccgcctgcgc
960
tcgctttacg cccaataaat ccggacaacg cttgccacct acgtattacc gcggctgctg
1020
gcacgtagtt agccgtggct ttctggttag ataccgtcag gggacgttca gttactaacg
1080
tccttgttct tctctaacaa cagagtttta cgatccgaaa accttcttca ctcacgcggc
1140
gttgctcggt cagactttcg tccattgccg aagattccct actgctgcct cccgtaggag
1200
tctgggccgt gtctcagtcc cagtgtggcc gatcaccctc tcaggtcggc tatgcatcgt
1260
ggccttggtg agccgttacc tcaccaacta gctaatgcac cgcgggtcca tccatcagcg
1320
acacccgaaa gcgcctttca ctcttatgcc atgcggcata aactgttatg cggtattagc
1380
acctgtttcc aagtgttatc cccctctgat gggtaggtta cccacgtgtt actcacccgt
1440
ccgccactcc tctttccaat tgagtgcaag cactcgggag gaaagaagcg ttcgacttgc
1500
atgtattagg cacgccgcca gcgttcgtcc tgagccagga tcaaactct
1549
<210> 40
<211> 1541
<212> ДНК
<213> Delftia
<400> 40
cagaaaggag gtgatccagc cgcaccttcc gatacggcta ccttgttacg acttcacccc
60
agtcacgaac cccgccgtgg taagcgccct ccttgcggtt aggctaccta cttctggcga
120
gacccgctcc catggtgtga cgggcggtgt gtacaagacc cgggaacgta ttcaccgcgg
180
catgctgatc cgcgattact agcgattccg acttcacgca gtcgagttgc agactgcgat
240
ccggactacg actggtttta tgggattagc tccccctcgc gggttggcaa ccctctgtac
300
cagccattgt atgacgtgtg tagccccacc tataagggcc atgaggactt gacgtcatcc
360
ccaccttcct ccggtttgtc accggcagtc tcattagagt gctcaactga atgtagcaac
420
taatgacaag ggttgcgctc gttgcgggac ttaacccaac atctcacgac acgagctgac
480
gacagccatg cagcacctgt gtgcaggttc tctttcgagc acgaatccat ctctggaaac
540
ttcctgccat gtcaaaggtg ggtaaggttt ttcgcgttgc atcgaattaa accacatcat
600
ccaccgcttg tgcgggtccc cgtcaattcc tttgagtttc aaccttgcgg ccgtactccc
660
caggcggtca acttcacgcg ttagcttcgt tactgagaaa actaattccc aacaaccagt
720
tgacatcgtt tagggcgtgg actaccaggg tatctaatcc tgtttgctcc ccacgctttc
780
gtgcatgagc gtcagtacag gtccagggga ttgccttcgc catcggtgtt cctccgcata
840
tctacgcatt tcactgctac acgcggaatt ccatccccct ctaccgtact ctagccatgc
900
agtcacaaat gcagttccca ggttgagccc ggggatttca catctgtctt acataaccgc
960
ctgcgcacgc tttacgccca gtaattccga ttaacgctcg caccctacgt attaccgcgg
1020
ctgctggcac gtagttagcc ggtgcttatt cttacggtac cgtcatgggc cccctgtatt
1080
agaaggagct ttttcgttcc gtacaaaagc agtttacaac ccgaaggcct tcatcctgca
1140
cgcggcattg ctggatcagg ctttcgccca ttgtccaaaa ttccccactg ctgcctcccg
1200
taggagtctg ggccgtgtct cagtcccagt gtggctggtc gtcctctcag accagctaca
1260
gatcgtcggc ttggtaagct tttatcccac caactaccta atctgccatc ggccgctcca
1320
atcgcgcgag gcccgaaggg cccccgcttt catcctcaga tcgtatgcgg tattagctac
1380
tctttcgagt agttatcccc cacgactggg cacgttccga tgtattactc acccgttcgc
1440
cactcgtcag cgtccgaaga cctgttaccg ttcgacttgc atgtgtaagg catgccgcca
1500
gcgttcaatc tgagccagga tcaaactcta cagttcgatc t
1541
<210> 41
<211> 1502
<212> ДНК
<213> Pelomonas
<220>
<221> другой_признак
<222> (192)..(193)
<223> n представляет собой a, g, c или t
<220>
<221> другой_признак
<222> (832)..(833)
<223> n представляет собой a, g, c или t
<400> 41
atcctggctc agattgaacg ctggcggcat gccttacaca tgcaagtcga acggtaacag
60
gttaagctga cgagtggcga acgggtgagt aatatatcgg aacgtgccca gtcgtggggg
120
ataactgctc gaaagagcag ctaataccgc atacgacctg agggtgaaag cgggggatcg
180
caagacctcg cnngattgga gcggccgata tcagattagg tagttggtgg ggtaaaggcc
240
caccaagcca acgatctgta gctggtctga gaggacgacc agccacactg ggactgagac
300
acggcccaga ctcctacggg aggcagcagt ggggaatttt ggacaatggg cgcaagcctg
360
atccagccat gccgcgtgcg ggaagaaggc cttcgggttg taaaccgctt ttgtcaggga
420
agaaaaggtt ctggttaata cctgggactc atgacggtac ctgaagaata agcaccggct
480
aactacgtgc cagcagccgc ggtaatacgt agggtgcaag cgttaatcgg aattactggg
540
cgtaaagcgt gcgcaggcgg ttatgcaaga cagaggtgaa atccccgggc tcaacctggg
600
aactgccttt gtgactgcat agctagagta cggtagaggg ggatggaatt ccgcgtgtag
660
cagtgaaatg cgtagatatg cggaggaaca ccgatggcga aggcaatccc ctggacctgt
720
actgacgctc atgcacgaaa gcgtggggag caaacaggat tagataccct ggtagtccac
780
gccctaaacg atgtcaactg gttgttggga gggtttcttc tcagtaacgt anntaacgcg
840
tgaagttgac cgcctgggga gtacggccgc aaggttgaaa ctcaaaggaa ttgacgggga
900
cccgcacaag cggtggatga tgtggtttaa ttcgatgcaa cgcgaaaaac cttacctacc
960
cttgacatgc caggaatcct gaagagattt gggagtgctc gaaagagaac ctggacacag
1020
gtgctgcatg gccgtcgtca gctcgtgtcg tgagatgttg ggttaagtcc cgcaacgagc
1080
gcaacccttg tcattagttg ctacgaaagg gcactctaat gagactgccg gtgacaaacc
1140
ggaggaaggt ggggatgacg tcaggtcatc atggccctta tgggtagggc tacacacgtc
1200
atacaatggc cgggacagag ggctgccaac ccgcgagggg gagctaatcc cagaaacccg
1260
gtcgtagtcc ggatcgtagt ctgcaactcg actgcgtgaa gtcggaatcg ctagtaatcg
1320
cggatcagct tgccgcggtg aatacgttcc cgggtcttgt acacaccgcc cgtcacacca
1380
tgggagcggg ttctgccaga agtagttagc ctaaccgcaa ggagggcgat taccacggca
1440
gggttcgtga ctggggtgaa gtcgtaacaa ggtagccgta tcggaaggtg cggctggatc
1500
ac
1502
<210> 42
<211> 34
<212> БЕЛОК
<213> Lactococcus lactis
<400> 42
Ile Thr Ser Ile Ser Leu Cys Thr Pro Gly Cys Lys Thr Gly Ala Leu
1 5 10 15
Met Gly Cys Asn Met Lys Thr Ala Thr Cys His Cys Ser Ile His Val
20 25 30
Ser Lys
<210> 43
<211> 22
<212> БЕЛОК
<213> Staphylococcus epidermis
<400> 43
Ile Ala Ser Lys Phe Ile Cys Thr Pro Gly Cys Ala Lys Thr Gly Ser
1 5 10 15
Phe Asn Ser Tyr Cys Cys
20
<210> 44
<211> 44
<212> БЕЛОК
<213> Pediococcus acidilactici
<400> 44
Lys Tyr Tyr Gly Asn Gly Val Thr Cys Gly Lys His Ser Cys Ser Val
1 5 10 15
Asp Trp Gly Lys Ala Thr Thr Cys Ile Ile Asn Asn Gly Ala Met Ala
20 25 30
Trp Ala Thr Gly Gly His Gln Gly Asn His Lys Cys
35 40
<210> 45
<211> 44
<212> БЕЛОК
<213> Enterococcus faecium
<400> 45
Ala Thr Arg Ser Tyr Gly Asn Gly Val Tyr Cys Asn Asn Ser Lys Cys
1 5 10 15
Trp Val Asn Trp Gly Glu Ala Lys Glu Asn Ile Ala Gly Ile Val Ile
20 25 30
Ser Gly Trp Ala Ser Gly Leu Ala Gly Met Gly His
35 40
<210> 46
<211> 39
<212> БЕЛОК
<213> Streptococcus lactis
<400> 46
Gly Thr Trp Asp Asp Ile Gly Gln Gly Ile Gly Arg Val Ala Tyr Trp
1 5 10 15
Val Gly Lys Ala Met Gly Asn Met Ser Asp Val Asn Gln Ala Ser Arg
20 25 30
Ile Asn Arg Lys Lys Lys His
35
<210> 47
<211> 48
<212> БЕЛОК
<213> Lactobacillus johnsonii
<400> 47
Asn Arg Trp Gly Asp Thr Val Leu Ser Ala Ala Ser Gly Ala Gly Thr
1 5 10 15
Gly Ile Lys Ala Cys Lys Ser Phe Gly Pro Trp Gly Met Ala Ile Cys
20 25 30
Gly Val Gly Gly Ala Ala Ile Gly Gly Tyr Phe Gly Tyr Thr His Asn
35 40 45
<210> 48
<211> 70
<212> БЕЛОК
<213> Enterococcus faecalis
<400> 48
Met Ala Lys Glu Phe Gly Ile Pro Ala Ala Val Ala Gly Thr Val Leu
1 5 10 15
Asn Val Val Glu Ala Gly Gly Trp Val Thr Thr Ile Val Ser Ile Leu
20 25 30
Thr Ala Val Gly Ser Gly Gly Leu Ser Leu Leu Ala Ala Ala Gly Arg
35 40 45
Glu Ser Ile Lys Ala Tyr Leu Lys Lys Glu Ile Lys Lys Lys Gly Lys
50 55 60
Arg Ala Val Ile Ala Trp
65 70
<210> 49
<211> 51
<212> БЕЛОК
<213> Staphylococcus aureus
<400> 49
Met Ser Trp Leu Asn Phe Leu Lys Tyr Ile Ala Lys Tyr Gly Lys Lys
1 5 10 15
Ala Val Ser Ala Ala Trp Lys Tyr Lys Gly Lys Val Leu Glu Trp Leu
20 25 30
Asn Val Gly Pro Thr Leu Glu Trp Val Trp Gln Lys Leu Lys Lys Ile
35 40 45
Ala Gly Leu
50
<210> 50
<211> 43
<212> БЕЛОК
<213> Lactococcus garvieae
<400> 50
Ile Gly Gly Ala Leu Gly Asn Ala Leu Asn Gly Leu Gly Thr Trp Ala
1 5 10 15
Asn Met Met Asn Gly Gly Gly Phe Val Asn Gln Trp Gln Val Tyr Ala
20 25 30
Asn Lys Gly Lys Ile Asn Gln Tyr Arg Pro Tyr
35 40
<210> 51
<211> 103
<212> БЕЛОК
<213> Escherichia coli
<400> 51
Met Arg Thr Leu Thr Leu Asn Glu Leu Asp Ser Val Ser Gly Gly Ala
1 5 10 15
Ser Gly Arg Asp Ile Ala Met Ala Ile Gly Thr Leu Ser Gly Gln Phe
20 25 30
Val Ala Gly Gly Ile Gly Ala Ala Ala Gly Gly Val Ala Gly Gly Ala
35 40 45
Ile Tyr Asp Tyr Ala Ser Thr His Lys Pro Asn Pro Ala Met Ser Pro
50 55 60
Ser Gly Leu Gly Gly Thr Ile Lys Gln Lys Pro Glu Gly Ile Pro Ser
65 70 75 80
Glu Ala Trp Asn Tyr Ala Ala Gly Arg Leu Cys Asn Trp Ser Pro Asn
85 90 95
Asn Leu Ser Asp Val Cys Leu
100
<210> 52
<211> 48
<212> БЕЛОК
<213> Homo sapiens
<400> 52
Ser Ser Leu Leu Glu Lys Gly Leu Asp Gly Ala Lys Lys Ala Val Gly
1 5 10 15
Gly Leu Gly Lys Leu Gly Lys Asp Ala Val Glu Asp Leu Glu Ser Val
20 25 30
Gly Lys Gly Ala Val His Asp Val Lys Asp Val Leu Asp Ser Val Leu
35 40 45
<210> 53
<211> 37
<212> БЕЛОК
<213> Hyalophora cecropia
<400> 53
Lys Trp Lys Leu Phe Lys Lys Ile Glu Lys Val Gly Gln Asn Ile Arg
1 5 10 15
Asp Gly Ile Ile Lys Ala Gly Pro Ala Val Ala Val Val Gly Gln Ala
20 25 30
Thr Gln Ile Ala Lys
35
<210> 54
<211> 62
<212> БЕЛОК
<213> Drosophila teissieri
<400> 54
Met Lys Tyr Phe Ser Val Leu Val Val Leu Thr Leu Ile Leu Ala Ile
1 5 10 15
Val Asp Gln Ser Asp Ala Phe Ile Asn Leu Leu Asp Lys Val Glu Asp
20 25 30
Ala Leu His Thr Gly Ala Gln Ala Gly Phe Lys Leu Ile Arg Pro Val
35 40 45
Glu Arg Gly Ala Thr Pro Lys Lys Ser Glu Lys Pro Glu Lys
50 55 60
<210> 55
<211> 66
<212> БЕЛОК
<213> Bombyx mori
<400> 55
Met Asn Ile Leu Lys Phe Phe Phe Val Phe Ile Val Ala Met Ser Leu
1 5 10 15
Val Ser Cys Ser Thr Ala Ala Pro Ala Lys Ile Pro Ile Lys Ala Ile
20 25 30
Lys Thr Val Gly Lys Ala Val Gly Lys Gly Leu Arg Ala Ile Asn Ile
35 40 45
Ala Ser Thr Ala Asn Asp Val Phe Asn Phe Leu Lys Pro Lys Lys Arg
50 55 60
Lys His
65
<210> 56
<211> 71
<212> БЕЛОК
<213> Ceratitis capitata
<400> 56
Met Ala Asn Leu Lys Ala Val Phe Leu Ile Cys Ile Val Ala Phe Ile
1 5 10 15
Ala Leu Gln Cys Val Val Ala Glu Pro Ala Ala Glu Asp Ser Val Val
20 25 30
Val Lys Arg Ser Ile Gly Ser Ala Leu Lys Lys Ala Leu Pro Val Ala
35 40 45
Lys Lys Ile Gly Lys Ile Ala Leu Pro Ile Ala Lys Ala Ala Leu Pro
50 55 60
Val Ala Ala Gly Leu Val Gly
65 70
<210> 57
<211> 53
<212> БЕЛОК
<213> Apis mellifera
<400> 57
Met Lys Val Val Ile Phe Ile Phe Ala Leu Leu Ala Thr Ile Cys Ala
1 5 10 15
Ala Phe Ala Tyr Val Pro Leu Pro Asn Val Pro Gln Pro Gly Arg Arg
20 25 30
Pro Phe Pro Thr Phe Pro Gly Gln Gly Pro Phe Asn Pro Lys Ile Lys
35 40 45
Trp Pro Gln Gly Tyr
50
<210> 58
<211> 283
<212> БЕЛОК
<213> Apis mellifera
<400> 58
Lys Asn Phe Ala Leu Ala Ile Leu Val Val Thr Phe Val Val Ala Val
1 5 10 15
Phe Gly Asn Thr Asn Leu Asp Pro Pro Thr Arg Pro Thr Arg Leu Arg
20 25 30
Arg Glu Ala Lys Pro Glu Ala Glu Pro Gly Asn Asn Arg Pro Val Tyr
35 40 45
Ile Pro Gln Pro Arg Pro Pro His Pro Arg Leu Arg Arg Glu Ala Glu
50 55 60
Pro Glu Ala Glu Pro Gly Asn Asn Arg Pro Val Tyr Ile Pro Gln Pro
65 70 75 80
Arg Pro Pro His Pro Arg Leu Arg Arg Glu Ala Glu Leu Glu Ala Glu
85 90 95
Pro Gly Asn Asn Arg Pro Val Tyr Ile Ser Gln Pro Arg Pro Pro His
100 105 110
Pro Arg Leu Arg Arg Glu Ala Glu Pro Glu Ala Glu Pro Gly Asn Asn
115 120 125
Arg Pro Val Tyr Ile Pro Gln Pro Arg Pro Pro His Pro Arg Leu Arg
130 135 140
Arg Glu Ala Glu Leu Glu Ala Glu Pro Gly Asn Asn Arg Pro Val Tyr
145 150 155 160
Ile Ser Gln Pro Arg Pro Pro His Pro Arg Leu Arg Arg Glu Ala Glu
165 170 175
Pro Glu Ala Glu Pro Gly Asn Asn Arg Pro Val Tyr Ile Pro Gln Pro
180 185 190
Arg Pro Pro His Pro Arg Leu Arg Arg Glu Ala Glu Pro Glu Ala Glu
195 200 205
Pro Gly Asn Asn Arg Pro Val Tyr Ile Pro Gln Pro Arg Pro Pro His
210 215 220
Pro Arg Leu Arg Arg Glu Ala Glu Pro Glu Ala Glu Pro Gly Asn Asn
225 230 235 240
Arg Pro Val Tyr Ile Pro Gln Pro Arg Pro Pro His Pro Arg Leu Arg
245 250 255
Arg Glu Ala Lys Pro Glu Ala Lys Pro Gly Asn Asn Arg Pro Val Tyr
260 265 270
Ile Pro Gln Pro Arg Pro Pro His Pro Arg Ile
275 280
<210> 59
<211> 228
<212> БЕЛОК
<213> Sus scrofa
<400> 59
Met Glu Thr Gln Arg Ala Ser Leu Cys Leu Gly Arg Trp Ser Leu Trp
1 5 10 15
Leu Leu Leu Leu Ala Leu Val Val Pro Ser Ala Ser Ala Gln Ala Leu
20 25 30
Ser Tyr Arg Glu Ala Val Leu Arg Ala Val Asp Arg Leu Asn Glu Gln
35 40 45
Ser Ser Glu Ala Asn Leu Tyr Arg Leu Leu Glu Leu Asp Gln Pro Pro
50 55 60
Lys Ala Asp Glu Asp Pro Gly Thr Pro Lys Pro Val Ser Phe Thr Val
65 70 75 80
Lys Glu Thr Val Cys Pro Arg Pro Thr Arg Arg Pro Pro Glu Leu Cys
85 90 95
Asp Phe Lys Glu Asn Gly Arg Val Lys Gln Cys Val Gly Thr Val Thr
100 105 110
Leu Asp Gln Ile Lys Asp Pro Leu Asp Ile Thr Cys Asn Glu Gly Val
115 120 125
Arg Arg Phe Pro Trp Trp Trp Pro Phe Leu Arg Arg Pro Arg Leu Arg
130 135 140
Arg Gln Ala Phe Pro Pro Pro Asn Val Pro Gly Pro Arg Phe Pro Pro
145 150 155 160
Pro Asn Val Pro Gly Pro Arg Phe Pro Pro Pro Asn Phe Pro Gly Pro
165 170 175
Arg Phe Pro Pro Pro Asn Phe Pro Gly Pro Arg Phe Pro Pro Pro Asn
180 185 190
Phe Pro Gly Pro Pro Phe Pro Pro Pro Ile Phe Pro Gly Pro Trp Phe
195 200 205
Pro Pro Pro Pro Pro Phe Arg Pro Pro Pro Phe Gly Pro Pro Arg Phe
210 215 220
Pro Gly Arg Arg
225
<210> 60
<211> 144
<212> БЕЛОК
<213> Bos taurus
<400> 60
Met Gln Thr Gln Arg Ala Ser Leu Ser Leu Gly Arg Trp Ser Leu Trp
1 5 10 15
Leu Leu Leu Leu Gly Leu Val Val Pro Ser Ala Ser Ala Gln Ala Leu
20 25 30
Ser Tyr Arg Glu Ala Val Leu Arg Ala Val Asp Gln Leu Asn Glu Leu
35 40 45
Ser Ser Glu Ala Asn Leu Tyr Arg Leu Leu Glu Leu Asp Pro Pro Pro
50 55 60
Lys Asp Asn Glu Asp Leu Gly Thr Arg Lys Pro Val Ser Phe Thr Val
65 70 75 80
Lys Glu Thr Val Cys Pro Arg Thr Ile Gln Gln Pro Ala Glu Gln Cys
85 90 95
Asp Phe Lys Glu Lys Gly Arg Val Lys Gln Cys Val Gly Thr Val Thr
100 105 110
Leu Asp Pro Ser Asn Asp Gln Phe Asp Leu Asn Cys Asn Glu Leu Gln
115 120 125
Ser Val Ile Leu Pro Trp Lys Trp Pro Trp Trp Pro Trp Arg Arg Gly
130 135 140
<210> 61
<211> 149
<212> БЕЛОК
<213> Sus scrofa
<400> 61
Met Glu Thr Gln Arg Ala Ser Leu Cys Leu Gly Arg Trp Ser Leu Trp
1 5 10 15
Leu Leu Leu Leu Ala Leu Val Val Pro Ser Ala Ser Ala Gln Ala Leu
20 25 30
Ser Tyr Arg Glu Ala Val Leu Arg Ala Val Asp Arg Leu Asn Glu Gln
35 40 45
Ser Ser Glu Ala Asn Leu Tyr Arg Leu Leu Glu Leu Asp Gln Pro Pro
50 55 60
Lys Ala Asp Glu Asp Pro Gly Thr Pro Lys Pro Val Ser Phe Thr Val
65 70 75 80
Lys Glu Thr Val Cys Pro Arg Pro Thr Arg Gln Pro Pro Glu Leu Cys
85 90 95
Asp Phe Lys Glu Asn Gly Arg Val Lys Gln Cys Val Gly Thr Val Thr
100 105 110
Leu Asp Gln Ile Lys Asp Pro Leu Asp Ile Thr Cys Asn Glu Val Gln
115 120 125
Gly Val Arg Gly Gly Arg Leu Cys Tyr Cys Arg Arg Arg Phe Cys Val
130 135 140
Cys Val Gly Arg Gly
145
<210> 62
<211> 17
<212> БЕЛОК
<213> Tachypleus gigas
<400> 62
Lys Trp Cys Phe Arg Val Cys Tyr Arg Gly Ile Cys Tyr Arg Arg Cys
1 5 10 15
Arg
<210> 63
<211> 102
<212> БЕЛОК
<213> Anopheles gambiae
<400> 63
Met Lys Cys Ala Thr Ile Val Cys Thr Ile Ala Val Val Leu Ala Ala
1 5 10 15
Thr Leu Leu Asn Gly Ser Val Gln Ala Ala Pro Gln Glu Glu Ala Ala
20 25 30
Leu Ser Gly Gly Ala Asn Leu Asn Thr Leu Leu Asp Glu Leu Pro Glu
35 40 45
Glu Thr His His Ala Ala Leu Glu Asn Tyr Arg Ala Lys Arg Ala Thr
50 55 60
Cys Asp Leu Ala Ser Gly Phe Gly Val Gly Ser Ser Leu Cys Ala Ala
65 70 75 80
His Cys Ile Ala Arg Arg Tyr Arg Gly Gly Tyr Cys Asn Ser Lys Ala
85 90 95
Val Cys Val Cys Arg Asn
100
<210> 64
<211> 70
<212> БЕЛОК
<213> Drosophila melanogaster
<400> 64
Met Met Gln Ile Lys Tyr Leu Phe Ala Leu Phe Ala Val Leu Met Leu
1 5 10 15
Val Val Leu Gly Ala Asn Glu Ala Asp Ala Asp Cys Leu Ser Gly Arg
20 25 30
Tyr Lys Gly Pro Cys Ala Val Trp Asp Asn Glu Thr Cys Arg Arg Val
35 40 45
Cys Lys Glu Glu Gly Arg Ser Ser Gly His Cys Ser Pro Ser Leu Lys
50 55 60
Cys Trp Cys Glu Gly Cys
65 70
<210> 65
<211> 78
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 65
Met Lys Leu Leu His Gly Phe Leu Ile Ile Met Leu Thr Met His Leu
1 5 10 15
Ser Ile Gln Tyr Ala Tyr Gly Gly Pro Phe Leu Thr Lys Tyr Leu Cys
20 25 30
Asp Arg Val Cys His Lys Leu Cys Gly Asp Glu Phe Val Cys Ser Cys
35 40 45
Ile Gln Tyr Lys Ser Leu Lys Gly Leu Trp Phe Pro His Cys Pro Thr
50 55 60
Gly Lys Ala Ser Val Val Leu His Asn Phe Leu Thr Ser Pro
65 70 75
<210> 66
<211> 77
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 66
Met Lys Leu Leu Tyr Gly Phe Leu Ile Ile Met Leu Thr Ile His Leu
1 5 10 15
Ser Val Gln Tyr Phe Glu Ser Pro Phe Glu Thr Lys Tyr Asn Cys Asp
20 25 30
Thr His Cys Asn Lys Leu Cys Gly Lys Ile Asp His Cys Ser Cys Ile
35 40 45
Gln Tyr His Ser Met Glu Gly Leu Trp Phe Pro His Cys Arg Thr Gly
50 55 60
Ser Ala Ala Gln Met Leu His Asp Phe Leu Ser Asn Pro
65 70 75
<210> 67
<211> 86
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 67
Met Ser Val Arg Lys Asn Val Leu Pro Thr Met Phe Val Val Leu Leu
1 5 10 15
Ile Met Ser Pro Val Thr Pro Thr Ser Val Phe Ile Ser Ala Val Cys
20 25 30
Tyr Ser Gly Cys Gly Ser Leu Ala Leu Val Cys Phe Val Ser Asn Gly
35 40 45
Ile Thr Asn Gly Leu Asp Tyr Phe Lys Ser Ser Ala Pro Leu Ser Thr
50 55 60
Ser Glu Thr Ser Cys Gly Glu Ala Phe Asp Thr Cys Thr Asp His Cys
65 70 75 80
Leu Ala Asn Phe Lys Phe
85
<210> 68
<211> 69
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 68
Met Arg Leu Leu Tyr Gly Phe Leu Ile Ile Met Leu Thr Ile Tyr Leu
1 5 10 15
Ser Val Gln Asp Phe Asp Pro Thr Glu Phe Lys Gly Pro Phe Pro Thr
20 25 30
Ile Glu Ile Cys Ser Lys Tyr Cys Ala Val Val Cys Asn Tyr Thr Ser
35 40 45
Arg Pro Cys Tyr Cys Val Glu Ala Ala Lys Glu Arg Asp Gln Trp Phe
50 55 60
Pro Tyr Cys Tyr Asp
65
<210> 69
<211> 77
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 69
Met Arg Leu Leu Tyr Gly Phe Leu Ile Ile Met Leu Thr Ile His Leu
1 5 10 15
Ser Val Gln Asp Ile Asp Pro Asn Thr Leu Arg Gly Pro Tyr Pro Thr
20 25 30
Lys Glu Ile Cys Ser Lys Tyr Cys Glu Tyr Asn Val Val Cys Gly Ala
35 40 45
Ser Leu Pro Cys Ile Cys Val Gln Asp Ala Arg Gln Leu Asp His Trp
50 55 60
Phe Ala Cys Cys Tyr Asp Gly Gly Pro Glu Met Leu Met
65 70 75
<210> 70
<211> 108
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 70
Met Lys Leu Phe Val Val Val Val Leu Val Ala Val Gly Ile Met Phe
1 5 10 15
Val Phe Ala Ser Asp Thr Ala Ala Ala Pro Thr Asp Tyr Glu Asp Thr
20 25 30
Asn Asp Met Ile Ser Leu Ser Ser Leu Val Gly Asp Asn Ser Pro Tyr
35 40 45
Val Arg Val Ser Ser Ala Asp Ser Gly Gly Ser Ser Lys Thr Ser Ser
50 55 60
Lys Asn Pro Ile Leu Gly Leu Leu Lys Ser Val Ile Lys Leu Leu Thr
65 70 75 80
Lys Ile Phe Gly Thr Tyr Ser Asp Ala Ala Pro Ala Met Pro Pro Ile
85 90 95
Pro Pro Ala Leu Arg Lys Asn Arg Gly Met Leu Ala
100 105
<210> 71
<211> 178
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 71
Met Val Ala Cys Lys Val Ile Leu Ala Val Ala Val Val Phe Val Ala
1 5 10 15
Ala Val Gln Gly Arg Pro Gly Gly Glu Pro Glu Trp Ala Ala Pro Ile
20 25 30
Phe Ala Glu Leu Lys Ser Val Ser Asp Asn Ile Thr Asn Leu Val Gly
35 40 45
Leu Asp Asn Ala Gly Glu Tyr Ala Thr Ala Ala Lys Asn Asn Leu Asn
50 55 60
Ala Phe Ala Glu Ser Leu Lys Thr Glu Ala Ala Val Phe Ser Lys Ser
65 70 75 80
Phe Glu Gly Lys Ala Ser Ala Ser Asp Val Phe Lys Glu Ser Thr Lys
85 90 95
Asn Phe Gln Ala Val Val Asp Thr Tyr Ile Lys Asn Leu Pro Lys Asp
100 105 110
Leu Thr Leu Lys Asp Phe Thr Glu Lys Ser Glu Gln Ala Leu Lys Tyr
115 120 125
Met Val Glu His Gly Thr Glu Ile Thr Lys Lys Ala Gln Gly Asn Thr
130 135 140
Glu Thr Glu Lys Glu Ile Lys Glu Phe Phe Lys Lys Gln Ile Glu Asn
145 150 155 160
Leu Ile Gly Gln Gly Lys Ala Leu Gln Ala Lys Ile Ala Glu Ala Lys
165 170 175
Lys Ala
<210> 72
<211> 311
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 72
Met Lys Thr Ser Ser Ser Lys Val Phe Ala Ser Cys Val Ala Ile Val
1 5 10 15
Cys Leu Ala Ser Val Ala Asn Ala Leu Pro Val Gln Lys Ser Val Ala
20 25 30
Ala Thr Thr Glu Asn Pro Ile Val Glu Lys His Gly Cys Arg Ala His
35 40 45
Lys Asn Leu Val Arg Gln Asn Val Val Asp Leu Lys Thr Tyr Asp Ser
50 55 60
Met Leu Ile Thr Asn Glu Val Val Gln Lys Gln Ser Asn Glu Val Gln
65 70 75 80
Ser Ser Glu Gln Ser Asn Glu Gly Gln Asn Ser Glu Gln Ser Asn Glu
85 90 95
Gly Gln Asn Ser Glu Gln Ser Asn Glu Val Gln Ser Ser Glu His Ser
100 105 110
Asn Glu Gly Gln Asn Ser Lys Gln Ser Asn Glu Gly Gln Asn Ser Glu
115 120 125
Gln Ser Asn Glu Val Gln Ser Ser Glu His Ser Asn Glu Gly Gln Asn
130 135 140
Ser Glu Gln Ser Asn Glu Val Gln Ser Ser Glu His Ser Asn Glu Gly
145 150 155 160
Gln Asn Ser Lys Gln Ser Asn Glu Gly Gln Asn Ser Lys Gln Ser Asn
165 170 175
Glu Val Gln Ser Ser Glu His Trp Asn Glu Gly Gln Asn Ser Lys Gln
180 185 190
Ser Asn Glu Asp Gln Asn Ser Glu Gln Ser Asn Glu Gly Gln Asn Ser
195 200 205
Lys Gln Ser Asn Glu Gly Gln Asn Ser Lys Gln Ser Asn Glu Asp Gln
210 215 220
Asn Ser Glu Gln Ser Asn Glu Gly Gln Asn Ser Lys Gln Ser Asn Glu
225 230 235 240
Val Gln Ser Ser Glu Gln Ser Asn Glu Gly Gln Asn Ser Lys Gln Ser
245 250 255
Asn Glu Gly Gln Ser Ser Glu Gln Ser Asn Glu Gly Gln Asn Ser Lys
260 265 270
Gln Ser Asn Glu Val Gln Ser Pro Glu Glu His Tyr Asp Leu Pro Asp
275 280 285
Pro Glu Ser Ser Tyr Glu Ser Glu Glu Thr Lys Gly Ser His Glu Ser
290 295 300
Gly Asp Asp Ser Glu His Arg
305 310
<210> 73
<211> 431
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 73
Met Lys Thr Ile Ile Leu Gly Leu Cys Leu Phe Gly Ala Leu Phe Trp
1 5 10 15
Ser Thr Gln Ser Met Pro Val Gly Glu Val Ala Pro Ala Val Pro Ala
20 25 30
Val Pro Ser Glu Ala Val Pro Gln Lys Gln Val Glu Ala Lys Pro Glu
35 40 45
Thr Asn Ala Ala Ser Pro Val Ser Asp Ala Lys Pro Glu Ser Asp Ser
50 55 60
Lys Pro Val Asp Ala Glu Val Lys Pro Thr Val Ser Glu Val Lys Ala
65 70 75 80
Glu Ser Glu Gln Lys Pro Ser Gly Glu Pro Lys Pro Glu Ser Asp Ala
85 90 95
Lys Pro Val Val Ala Ser Glu Ser Lys Pro Glu Ser Asp Pro Lys Pro
100 105 110
Ala Ala Val Val Glu Ser Lys Pro Glu Asn Asp Ala Val Ala Pro Glu
115 120 125
Thr Asn Asn Asp Ala Lys Pro Glu Asn Ala Ala Ala Pro Val Ser Glu
130 135 140
Asn Lys Pro Ala Thr Asp Ala Lys Ala Glu Thr Glu Leu Ile Ala Gln
145 150 155 160
Ala Lys Pro Glu Ser Lys Pro Ala Ser Asp Leu Lys Ala Glu Pro Glu
165 170 175
Ala Ala Lys Pro Asn Ser Glu Val Pro Val Ala Leu Pro Leu Asn Pro
180 185 190
Thr Glu Thr Lys Ala Thr Gln Gln Ser Val Glu Thr Asn Gln Val Glu
195 200 205
Gln Ala Ala Pro Ala Ala Ala Gln Ala Asp Pro Ala Ala Ala Pro Ala
210 215 220
Ala Asp Pro Ala Pro Ala Pro Ala Ala Ala Pro Val Ala Ala Glu Glu
225 230 235 240
Ala Lys Leu Ser Glu Ser Ala Pro Ser Thr Glu Asn Lys Ala Ala Glu
245 250 255
Glu Pro Ser Lys Pro Ala Glu Gln Gln Ser Ala Lys Pro Val Glu Asp
260 265 270
Ala Val Pro Ala Ala Ser Glu Ile Ser Glu Thr Lys Val Ser Pro Ala
275 280 285
Val Pro Ala Val Pro Glu Val Pro Ala Ser Pro Ser Ala Pro Ala Val
290 295 300
Ala Asp Pro Val Ser Ala Pro Glu Ala Glu Lys Asn Ala Glu Pro Ala
305 310 315 320
Lys Ala Ala Asn Ser Ala Glu Pro Ala Val Gln Ser Glu Ala Lys Pro
325 330 335
Ala Glu Asp Ile Gln Lys Ser Gly Ala Val Val Ser Ala Glu Asn Pro
340 345 350
Lys Pro Val Glu Glu Gln Lys Pro Ala Glu Val Ala Lys Pro Ala Glu
355 360 365
Gln Ser Lys Ser Glu Ala Pro Ala Glu Ala Pro Lys Pro Thr Glu Gln
370 375 380
Ser Ala Ala Glu Glu Pro Lys Lys Pro Glu Ser Ala Asn Asp Glu Lys
385 390 395 400
Lys Glu Gln His Ser Val Asn Lys Arg Asp Ala Thr Lys Glu Lys Lys
405 410 415
Pro Thr Asp Ser Ile Met Lys Lys Gln Lys Gln Lys Lys Ala Asn
420 425 430
<210> 74
<211> 160
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 74
Met Asn Gly Lys Ile Val Leu Cys Phe Ala Val Val Phe Ile Gly Gln
1 5 10 15
Ala Met Ser Ala Ala Thr Gly Thr Thr Pro Glu Val Glu Asp Ile Lys
20 25 30
Lys Val Ala Glu Gln Met Ser Gln Thr Phe Met Ser Val Ala Asn His
35 40 45
Leu Val Gly Ile Thr Pro Asn Ser Ala Asp Ala Gln Lys Ser Ile Glu
50 55 60
Lys Ile Arg Thr Ile Met Asn Lys Gly Phe Thr Asp Met Glu Thr Glu
65 70 75 80
Ala Asn Lys Met Lys Asp Ile Val Arg Lys Asn Ala Asp Pro Lys Leu
85 90 95
Val Glu Lys Tyr Asp Glu Leu Glu Lys Glu Leu Lys Lys His Leu Ser
100 105 110
Thr Ala Lys Asp Met Phe Glu Asp Lys Val Val Lys Pro Ile Gly Glu
115 120 125
Lys Val Glu Leu Lys Lys Ile Thr Glu Asn Val Ile Lys Thr Thr Lys
130 135 140
Asp Met Glu Ala Thr Met Asn Lys Ala Ile Asp Gly Phe Lys Lys Gln
145 150 155 160
<210> 75
<211> 415
<212> БЕЛОК
<213> Buchnera aphidicola
<400> 75
Met His Leu Phe Leu Ala Leu Gly Leu Phe Ile Val Cys Gly Met Val
1 5 10 15
Asp Ala Thr Phe Tyr Asn Pro Arg Ser Gln Thr Phe Asn Gln Leu Met
20 25 30
Glu Arg Arg Gln Arg Ser Ile Pro Ile Pro Tyr Ser Tyr Gly Tyr His
35 40 45
Tyr Asn Pro Ile Glu Pro Ser Ile Asn Val Leu Asp Ser Leu Ser Glu
50 55 60
Gly Leu Asp Ser Arg Ile Asn Thr Phe Lys Pro Ile Tyr Gln Asn Val
65 70 75 80
Lys Met Ser Thr Gln Asp Val Asn Ser Val Pro Arg Thr Gln Tyr Gln
85 90 95
Pro Lys Asn Ser Leu Tyr Asp Ser Glu Tyr Ile Ser Ala Lys Asp Ile
100 105 110
Pro Ser Leu Phe Pro Glu Glu Asp Ser Tyr Asp Tyr Lys Tyr Leu Gly
115 120 125
Ser Pro Leu Asn Lys Tyr Leu Thr Arg Pro Ser Thr Gln Glu Ser Gly
130 135 140
Ile Ala Ile Asn Leu Val Ala Ile Lys Glu Thr Ser Val Phe Asp Tyr
145 150 155 160
Gly Phe Pro Thr Tyr Lys Ser Pro Tyr Ser Ser Asp Ser Val Trp Asn
165 170 175
Phe Gly Ser Lys Ile Pro Asn Thr Val Phe Glu Asp Pro Gln Ser Val
180 185 190
Glu Ser Asp Pro Asn Thr Phe Lys Val Ser Ser Pro Thr Ile Lys Ile
195 200 205
Val Lys Leu Leu Pro Glu Thr Pro Glu Gln Glu Ser Ile Ile Thr Thr
210 215 220
Thr Lys Asn Tyr Glu Leu Asn Tyr Lys Thr Thr Gln Glu Thr Pro Thr
225 230 235 240
Glu Ala Glu Leu Tyr Pro Ile Thr Ser Glu Glu Phe Gln Thr Glu Asp
245 250 255
Glu Trp His Pro Met Val Pro Lys Glu Asn Thr Thr Lys Asp Glu Ser
260 265 270
Ser Phe Ile Thr Thr Glu Glu Pro Leu Thr Glu Asp Lys Ser Asn Ser
275 280 285
Ile Thr Ile Glu Lys Thr Gln Thr Glu Asp Glu Ser Asn Ser Ile Glu
290 295 300
Phe Asn Ser Ile Arg Thr Glu Glu Lys Ser Asn Ser Ile Thr Thr Glu
305 310 315 320
Glu Asn Gln Lys Glu Asp Asp Glu Ser Met Ser Thr Thr Ser Gln Glu
325 330 335
Thr Thr Thr Ala Phe Asn Leu Asn Asp Thr Phe Asp Thr Asn Arg Tyr
340 345 350
Ser Ser Ser His Glu Ser Leu Met Leu Arg Ile Arg Glu Leu Met Lys
355 360 365
Asn Ile Ala Asp Gln Gln Asn Lys Ser Gln Phe Arg Thr Val Asp Asn
370 375 380
Ile Pro Ala Lys Ser Gln Ser Asn Leu Ser Ser Asp Glu Ser Thr Asn
385 390 395 400
Gln Gln Phe Glu Pro Gln Leu Val Asn Gly Ala Asp Thr Tyr Lys
405 410 415
<210> 76
<211> 126
<212> БЕЛОК
<213> Sitophilus zeamais
<400> 76
Met Thr Arg Thr Met Leu Phe Leu Ala Cys Val Ala Ala Leu Tyr Val
1 5 10 15
Cys Ile Ser Ala Thr Ala Gly Lys Pro Glu Glu Phe Ala Lys Leu Ser
20 25 30
Asp Glu Ala Pro Ser Asn Asp Gln Ala Met Tyr Glu Ser Ile Gln Arg
35 40 45
Tyr Arg Arg Phe Val Asp Gly Asn Arg Tyr Asn Gly Gly Gln Gln Gln
50 55 60
Gln Gln Gln Pro Lys Gln Trp Glu Val Arg Pro Asp Leu Ser Arg Asp
65 70 75 80
Gln Arg Gly Asn Thr Lys Ala Gln Val Glu Ile Asn Lys Lys Gly Asp
85 90 95
Asn His Asp Ile Asn Ala Gly Trp Gly Lys Asn Ile Asn Gly Pro Asp
100 105 110
Ser His Lys Asp Thr Trp His Val Gly Gly Ser Val Arg Trp
115 120 125
<210> 77
<211> 220
<212> БЕЛОК
<213> Acyrthosiphon pisum
<400> 77
Met Lys Glu Thr Thr Val Val Trp Ala Lys Leu Phe Leu Ile Leu Ile
1 5 10 15
Ile Leu Ala Lys Pro Leu Gly Leu Lys Ala Val Asn Glu Cys Lys Arg
20 25 30
Leu Gly Asn Asn Ser Cys Arg Ser His Gly Glu Cys Cys Ser Gly Phe
35 40 45
Cys Phe Ile Glu Pro Gly Trp Ala Leu Gly Val Cys Lys Arg Leu Gly
50 55 60
Thr Pro Lys Lys Ser Asp Asp Ser Asn Asn Gly Lys Asn Ile Glu Lys
65 70 75 80
Asn Asn Gly Val His Glu Arg Ile Asp Asp Val Phe Glu Arg Gly Val
85 90 95
Cys Ser Tyr Tyr Lys Gly Pro Ser Ile Thr Ala Asn Gly Asp Val Phe
100 105 110
Asp Glu Asn Glu Met Thr Ala Ala His Arg Thr Leu Pro Phe Asn Thr
115 120 125
Met Val Lys Val Glu Gly Met Gly Thr Ser Val Val Val Lys Ile Asn
130 135 140
Asp Arg Lys Thr Ala Ala Asp Gly Lys Val Met Leu Leu Ser Arg Ala
145 150 155 160
Ala Ala Glu Ser Leu Asn Ile Asp Glu Asn Thr Gly Pro Val Gln Cys
165 170 175
Gln Leu Lys Phe Val Leu Asp Gly Ser Gly Cys Thr Pro Asp Tyr Gly
180 185 190
Asp Thr Cys Val Leu His His Glu Cys Cys Ser Gln Asn Cys Phe Arg
195 200 205
Glu Met Phe Ser Asp Lys Gly Phe Cys Leu Pro Lys
210 215 220
<210> 78
<211> 3
<212> ДНК
<213> Streptococcus pyogenes
<220>
<221> другой_признак
<222> (1)..(1)
<223> n представляет собой a, g, c или t
<400> 78
ngg
3
<210> 79
<211> 6
<212> ДНК
<213> Streptococcus thermophilus
<220>
<221> другой_признак
<222> (1)..(2)
<223> n представляет собой a, g, c или t
<400> 79
nnagaa
6
<210> 80
<211> 5
<212> ДНК
<213> Streptococcus thermophilus
<220>
<221> другой_признак
<222> (1)..(1)
<223> n представляет собой a, g, c или t
<220>
<221> другой_признак
<222> (4)..(4)
<223> n представляет собой a, g, c или t
<400> 80
nggng
5
<210> 81
<211> 7
<212> ДНК
<213> Neisseria meningitidis
<220>
<221> другой_признак
<222> (1)..(3)
<223> n представляет собой a, g, c или t
<400> 81
nnngatt
7
<210> 82
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Пенетратин
<400> 82
Arg Gln Ile Lys Ile Trp Phe Gln Asn Arg Arg Met Lys Trp Lys Lys
1 5 10 15
<210> 83
<211> 13
<212> БЕЛОК
<213> Вирус иммунодефицита человека типа 1
<400> 83
Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Pro Pro Gln
1 5 10
<210> 84
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pVEC
<400> 84
Leu Leu Ile Ile Leu Arg Arg Arg Ile Arg Lys Gln Ala His Ala His
1 5 10 15
Ser Lys
<210> 85
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Транспортан
<400> 85
Gly Trp Thr Leu Asn Ser Ala Gly Tyr Leu Leu Gly Lys Ile Asn Leu
1 5 10 15
Lys Ala Leu Ala Ala Leu Ala Lys Lys Ile Leu
20 25
<210> 86
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> MPG
<400> 86
Gly Ala Leu Phe Leu Gly Phe Leu Gly Ala Ala Gly Ser Thr Met Gly
1 5 10 15
Ala Trp Ser Gln Pro Lys Lys Lys Arg Lys Val
20 25
<210> 87
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Pep-1
<400> 87
Lys Glu Thr Trp Trp Glu Thr Trp Trp Thr Glu Trp Ser Gln Pro Lys
1 5 10 15
Lys Lys Arg Lys Val
20
<210> 88
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> MAP
<400> 88
Lys Leu Ala Leu Lys Leu Ala Leu Lys Ala Leu Lys Ala Ala Leu Lys
1 5 10 15
Leu Ala
<210> 89
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> R6W3
<400> 89
Arg Arg Trp Trp Arg Arg Trp Arg Arg
1 5
<210> 90
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для Bcr1
<400> 90
aaactgctgc atggctttct
20
<210> 91
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для Bcr1
<400> 91
acaggccttt caggctttta
20
<210> 92
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность промотора T7
<400> 92
ttaatacgac tcactatagg gaga
24
<210> 93
<211> 478
<212> РНК
<213> Acyrthosiphon pisum
<400> 93
augaaacugc ugcauggcuu ucugauuauu augcugacca ugcaucugag cauucaguau
60
gcguauggcg gcccguuucu gaccaaauau cugugcgauc gcgugugcca uaaacugugc
120
ggcgaugaau uugugugcag cugcauucag uauaaaagcc ugaaaggccu gugguuuccg
180
cauugcccga ccggcaaagc gagcguggug cugcauaacu uucugaccag cccguuuuuu
240
uuuucgggcu ggucagaaag uuaugcagca ccacgcucgc uuugccgguc gggcaaugcg
300
gaaaccacag gccuuucagg cuuuuauacu gaaugcagcu gcacacaaau ucaucgccgc
360
acaguuuaug gcacacgcga ucgcacagau auuuggucag aaacgggccg ccauacgcau
420
acugaaugcu cagaugcaug gucagcauaa uaaucagaaa gccaugcagc aguuucau
478
<210> 94
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ген колеоптерицина A (colA)
<400> 94
atgacccgca ccatgctgtt tctggcgtgc gtggcggcgc tgtatgtgtg cattagcgcg
60
accgcgggca aaccggaaga atttgcgaaa ctgagcgatg aagcgccgag caacgatcag
120
gcgatgtatg aaagcattca gcgctatcgc cgctttgtgg atggcaaccg ctataacggc
180
ggccagcagc agcagcagca gccgaaacag tgggaagtgc gcccggatct gagccgcgat
240
cagcgcggca acaccaaagc gcaggtggaa attaacaaaa aaggcgataa ccatgatatt
300
aacgcgggct ggggcaaaaa cattaacggc ccggatagcc ataaagatac ctggcatgtg
360
ggcggcagcg tgcgctgg
378
<210> 95
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для ColA
<400> 95
caacgatcag gcgatgtatg
20
<210> 96
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для ColA
<400> 96
ttaatttcca cctgcgcttt
20
<210> 97
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для Buchnera
<400> 97
gtcggctcat cacatcc
17
<210> 98
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для Buchnera
<400> 98
ttccgtctgt attatctcct
20
<210> 99
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для Mpcactus
<400> 99
taatacgact cactataggg tacacccatt gtgtgcacct
40
<210> 100
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для Mpcactus
<400> 100
taatacgact cactataggg ccactgtcca aggcaatttt
40
<210> 101
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Buch_groES_18F
<400> 101
catgatcgtg tgcttgttaa g
21
<210> 102
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Buch_groES_98R
<400> 102
ctgttcctcg agtcgatttc c
21
<210> 103
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ApEF1a 107F
<400> 103
ctgattgtgc cgtgcttatt g
21
<210> 104
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ApEF1a 246R
<400> 104
tatggtggtt cagtagagtc c
21
<210> 105
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> BCR4 (из BCR4-CPP)
<400> 105
cgtacaataa tctcatgg
18
<210> 106
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Tat
<400> 106
Tyr Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg
1 5 10
<210> 107
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ApBCR-4F
<400> 107
ctctgtcaac caccatgaga tta
23
<210> 108
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ApBCR-4R
<400> 108
tgcagactac agcacaatac tt
22
<210> 109
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для актина
<400> 109
gatcagcagc cacacacaag
20
<210> 110
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для актина
<400> 110
tttgaaccgg tttacgacga
20
<210> 111
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для ApGLNT1
<400> 111
taatacgact cactataggg caattacaaa aggacggcag
40
<210> 112
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для ApGLNT1
<400> 112
taatacgact cactataggg ccgctctagg aacaccgtat
40
<210> 113
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для ACYPI001018
<400> 113
cctgaaatcg acggggtcc
19
<210> 114
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для ACYPI001018
<400> 114
agatcggcaa catctgttcg t
21
<210> 115
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Малая шпилька
<400> 115
acacgt
6
<210> 116
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити ApGlnT1
<400> 116
ctacaaatct atctctccta ggcaattaca aaaggacggc ag
42
<210> 117
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити ApGlnT1
<400> 117
ttacaaactg ggaagaacct ggagacgtgc cgctctagga acaccgtat
49
<210> 118
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити ApGlnT1
<400> 118
ttacaaactg ggaagaacct ggaacacgtc ccgctctagg aacaccgtat
50
<210> 119
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити ApGlnT1
<400> 119
agaaactaga gcttgtcgat cgttaattaa caattacaaa aggacggcag
50
<210> 120
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити ApUbx
<400> 120
ctacaaatct atctctccta ggttttaccg tcacaggcat ca
42
<210> 121
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити ApUbx
<400> 121
acgagtgctg aagtccctag ccagacgtgt tcgtgctcgt taccaaatgt
50
<210> 122
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити ApUbx
<400> 122
acgagtgctg aagtccctag ccaacacgtc tcgtgctcgt taccaaatgt
50
<210> 123
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити ApUbx
<400> 123
agaaactaga gcttgtcgat cgttaattaa ttttaccgtc acaggcatca
50
<210> 124
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити ApbAs
<400> 124
ctacaaatct atctctccta ggggtgtcac catcgagacg tt
42
<210> 125
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити ApbAs
<400> 125
aaaaaccaca tacctcgagt ggggacgtgt catgactctg gcagttgaag tt
52
<210> 126
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити ApbAs
<400> 126
aaaaaccaca tacctcgagt gggacacgtc catgactctg gcagttgaag tt
52
<210> 127
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити ApbAs
<400> 127
agaaactaga gcttgtcgat cgttaattaa ggtgtcacca tcgagacgtt
50
<210> 128
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити Apcactus
<400> 128
ctacaaatct atctctccta gggtcgtcgt cgtcgtcgta gt
42
<210> 129
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити Apcactus
<400> 129
accaaaattg ccttggacag tgggacgtgt gcacgcacgg aaaacattta
50
<210> 130
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити Apcactus
<400> 130
accaaaattg ccttggacag tggacacgtc gcacgcacgg aaaacattta
50
<210> 131
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити Apcactus
<400> 131
agaaactaga gcttgtcgat cgttaattaa gtcgtcgtcg tcgtcgtagt
50
<210> 132
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити MpGlnT1
<400> 132
ctacaaatct atctctccta ggttggaagg gattggtgtt gtaatgcc
48
<210> 133
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити MpGlnT1
<400> 133
ttacaaactg ggaagaacct ggagacgtgt tccaggttct tcccagtttg taactagatc
60
g
61
<210> 134
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити MpGlnT1
<400> 134
ttacaaactg ggaagaacct ggaacacgtc tccaggttct tcccagtttg taactagatc
60
g
61
<210> 135
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити MpGlnT1
<400> 135
agaaactaga gcttgtcgat cgttaattaa ttggaaggga ttggtgttgt aatgcc
56
<210> 136
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити MpUbx
<400> 136
ctacaaatct atctctccta ggtcgtgtgg agcaagtaca gcg
43
<210> 137
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити MpUbx
<400> 137
acgagtgctg aagtccctag ccagacgtgt tggctaggga cttcagcact cg
52
<210> 138
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити MpUbx
<400> 138
acgagtgctg aagtccctag ccaacacgtc tggctaggga cttcagcact cg
52
<210> 139
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити MpUbx
<400> 139
agaaactaga gcttgtcgat cgttaattaa tcgtgtggag caagtacagc gg
52
<210> 140
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити MpbAs
<400> 140
ctacaaatct atctctccta gggaggaact ccaactgcca gagtcatg
48
<210> 141
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити MpbAs
<400> 141
aaaaaccaca tacctcgagt ggggacgtgt cccactcgag gtatgtggtt tttcctatg
59
<210> 142
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити MpbAs
<400> 142
aaaaaccaca tacctcgagt gggacacgtc cccactcgag gtatgtggtt tttcc
55
<210> 143
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити MpbAs
<400> 143
agaaactaga gcttgtcgat cgttaattaa gaggaactcc aactgccaga gtcatg
56
<210> 144
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для смысловой нити Mpcactus
<400> 144
ctacaaatct atctctccta ggtacaccca ttgtgtgcac ctgagtac
48
<210> 145
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для смысловой нити Mpcactus
<400> 145
accaaaattg ccttggacag tgggacgtgt ccactgtcca aggcaatttt ggttg
55
<210> 146
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для антисмысловой нити Mpcactus
<400> 146
accaaaattg ccttggacag tggacacgtc ccactgtcca aggcaatttt ggttg
55
<210> 147
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для антисмысловой нити Mpcactus
<400> 147
agaaactaga gcttgtcgat cgttaattaa tacacccatt gtgtgcacct gagtac
56
<210> 148
<211> 172
<212> ДНК
<213> Искусственная последовательность
<220>
<223> C002
<400> 148
ctgaaccgta cgatgagcag gaagaagcgt ctgtcgaatt accgatggag caccgtcagt
60
gcgatgaata caaatcgaag atctgggaca aagcatttag caaccaggag gctatgcagc
120
tgatggaact aacgtttaat acaggtaagg aattaggctc ccacgaagtg tg
172
<210> 149
<211> 266
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ApGLNT1
<400> 149
caattacaaa aggacggcag cgatacaact ctcatacaga cagaacagca atattagcgt
60
tgtcaattac agctattagt gtccagtaaa attgttttta aacaaaaaaa cgacaataaa
120
tattttgtac tgattaacta gggtgcaaca gtgaattttg aaaaaaatat aaaaacaaaa
180
tcatggattt tagatattag tcttttatct ttggtcatca gtttttgagt attatattat
240
tatattatac ggtgttccta gagcgg
266
<210> 150
<211> 191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> РНКаза
<400> 150
attgcaccga gcagaagact acctataccc ataataatat tacaaggaag aaacctgagg
60
actgtactta gacgtgttat atcataagac ctaataacgt acatgctcga aaatgcattc
120
aactgcatta attactgcag tatgtttagt catttgttcg tgtcttcttc cgaccatcaa
180
tgccaagaaa a
191
<210> 151
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ApGLNT1 (из ApGLNT1-CPP)
<400> 151
tgatgcgaca tagc
14
<210> 152
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ApC002 (из ApC002-CPP)
<400> 152
taacttccca tgttg
15
<210> 153
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> BCR-4 (из BCR-4-CPP)
<400> 153
tacaataatc tcatgg
16
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ ДЛЯ СЕЛЬСКОГО ХОЗЯЙСТВА | 2018 |
|
RU2783258C2 |
| КОМПОЗИЦИИ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ КОНТРОЛЯ ЗАБОЛЕВАНИЙ, ПЕРЕДАВАЕМЫХ ПЕРЕНОСЧИКАМИ | 2018 |
|
RU2777518C2 |
| КОМПОЗИЦИИ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ КОНТРОЛЯ ЗАБОЛЕВАНИЙ, ПЕРЕДАВАЕМЫХ ПЕРЕНОСЧИКАМИ | 2018 |
|
RU2804136C2 |
| СПОСОБЫ И СООТВЕТСТВУЮЩИЕ КОМПОЗИЦИИ ДЛЯ ИЗГОТОВЛЕНИЯ ПРОДУКТОВ ПИТАНИЯ И КОРМОВ | 2018 |
|
RU2780586C2 |
| КОМПОЗИЦИИ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ ДЛЯ СЕЛЬСКОГО ХОЗЯЙСТВА | 2018 |
|
RU2805081C2 |
| ПОДАВЛЕНИЕ ЭКСПРЕССИИ ГЕНОВ У НАСЕКОМЫХ-ВРЕДИТЕЛЕЙ | 2018 |
|
RU2725953C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2019 |
|
RU2810220C2 |
| КОНТРОЛЬ ЖЕСТКОКРЫЛЫХ ВРЕДИТЕЛЕЙ С ПРИМЕНЕНИЕМ МОЛЕКУЛ РНК | 2017 |
|
RU2783144C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765304C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765722C2 |
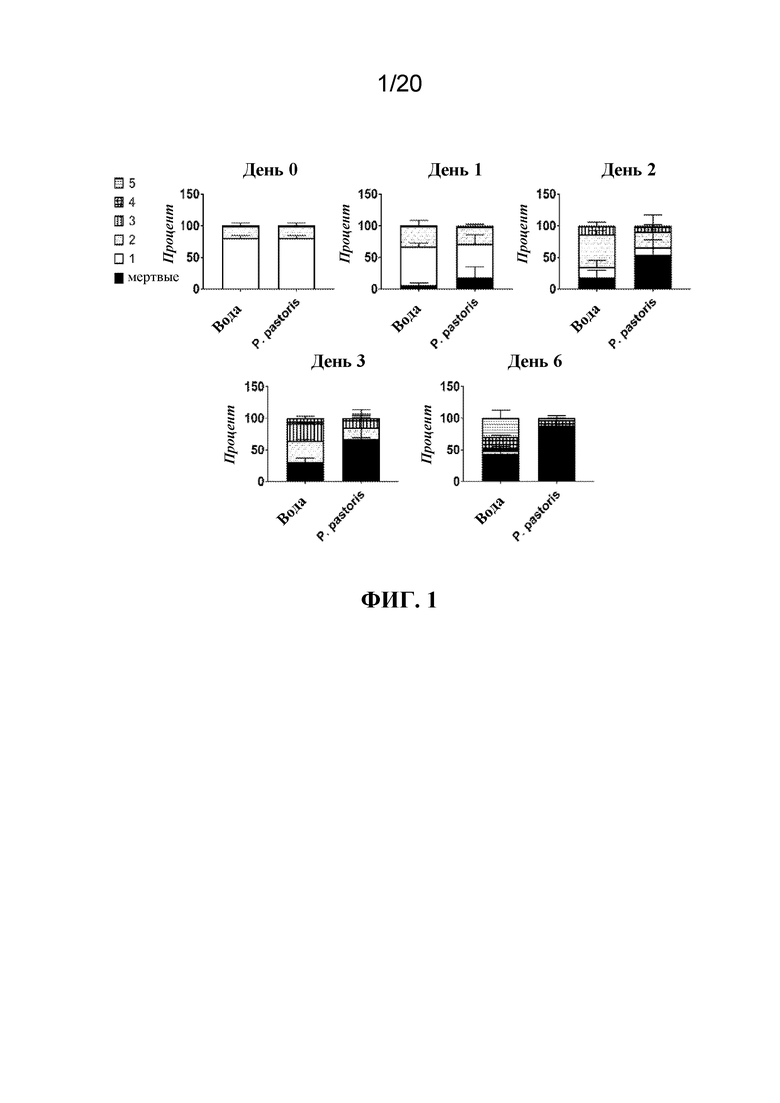
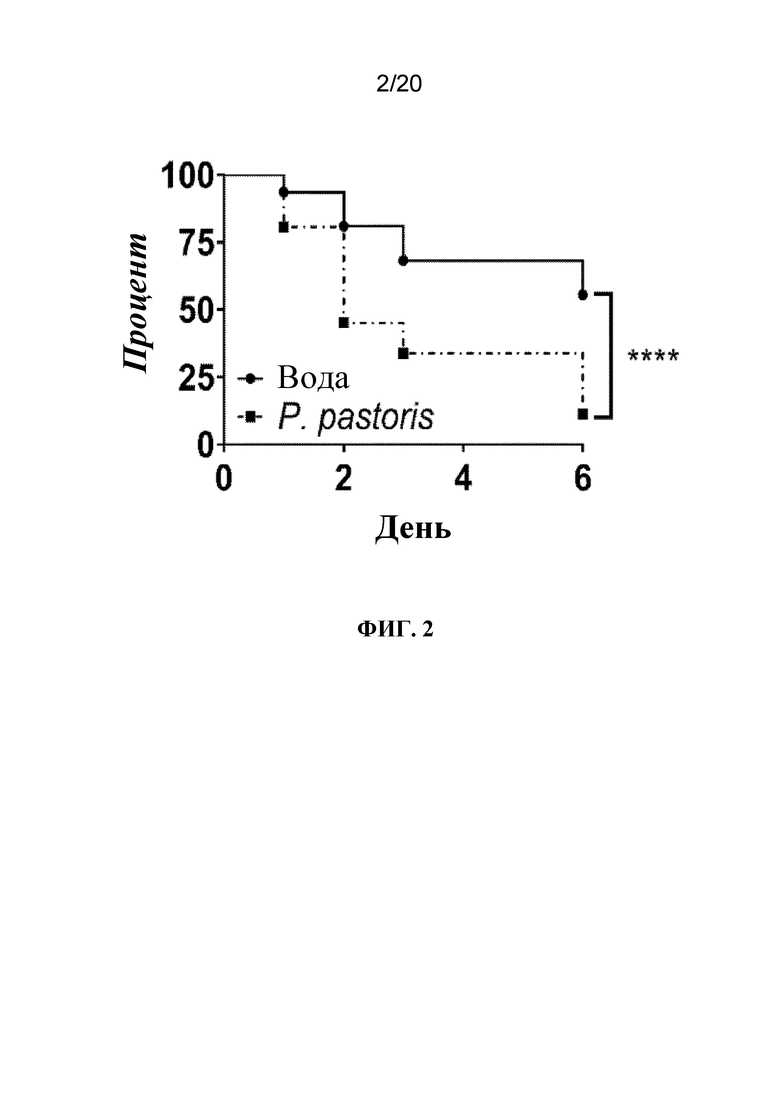
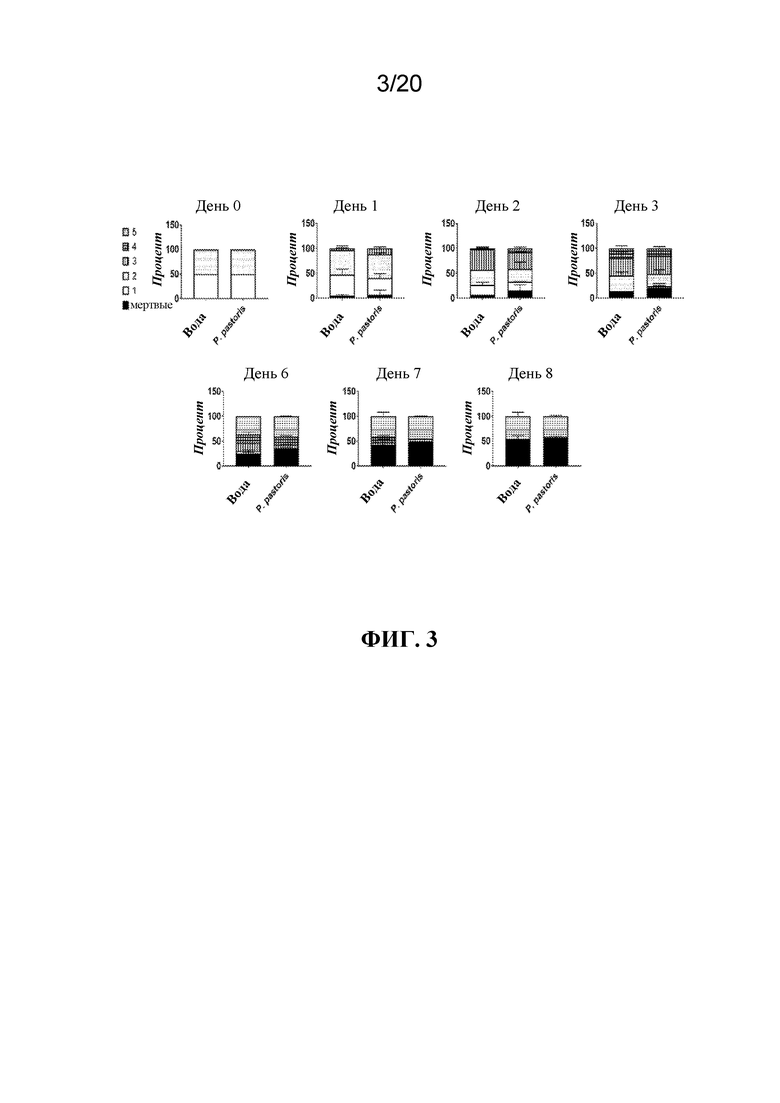
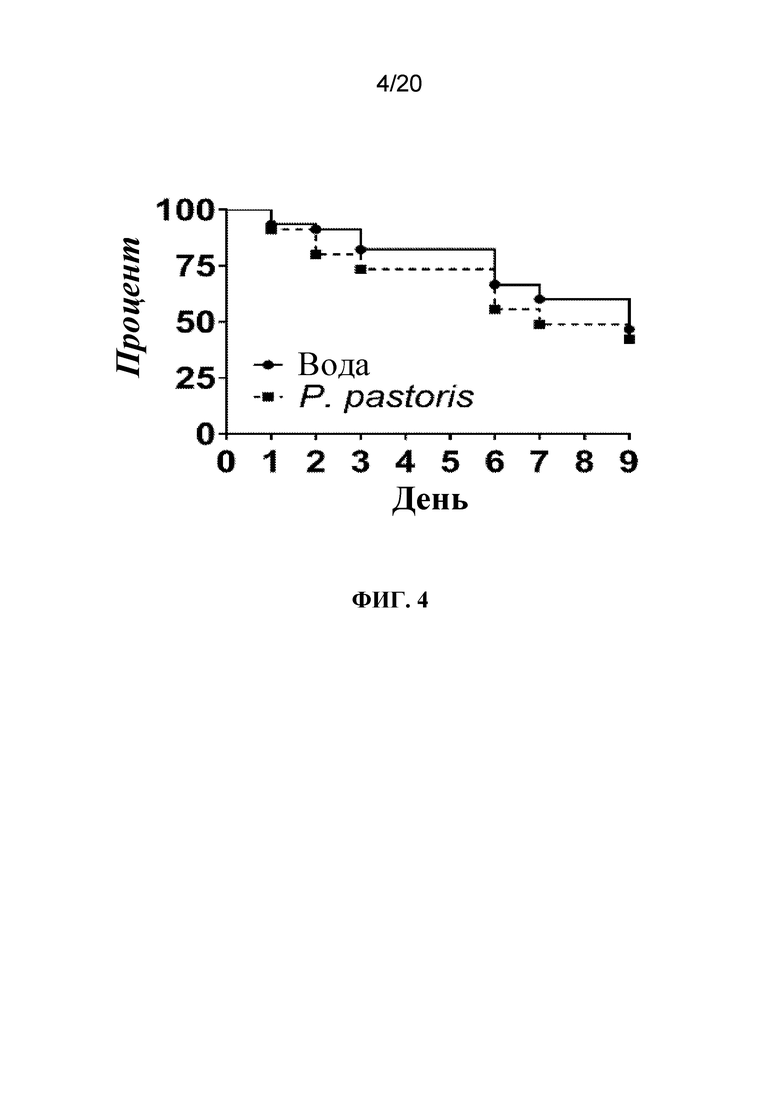
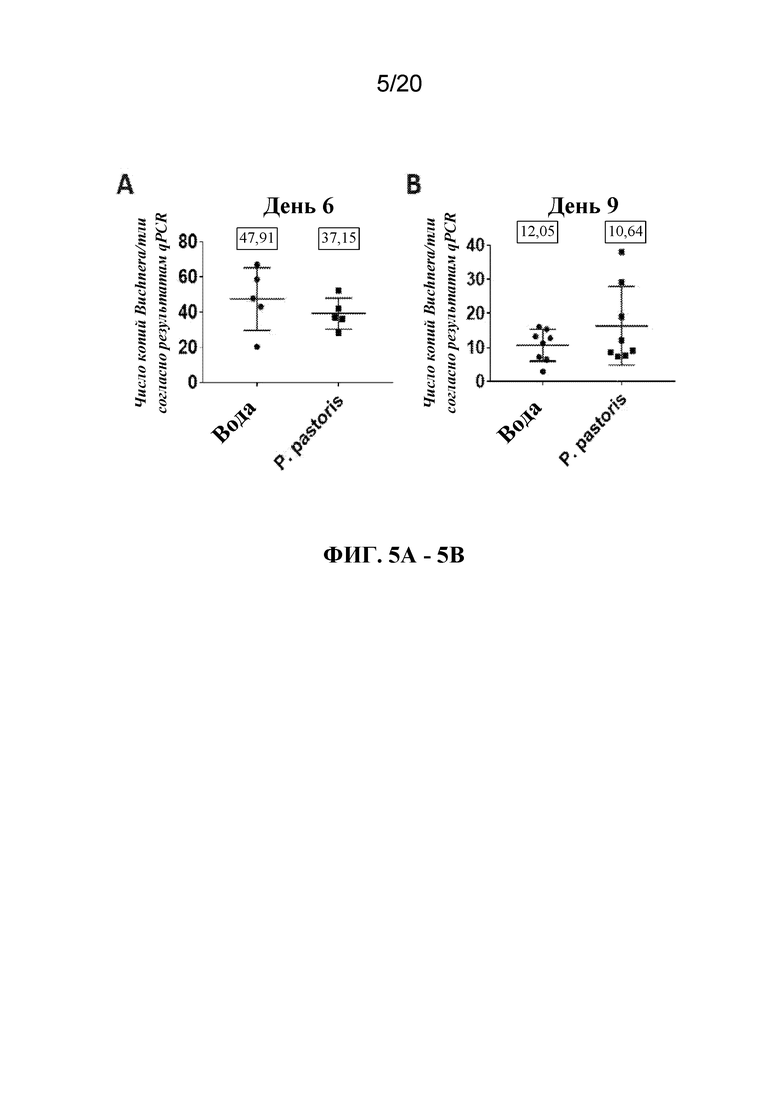
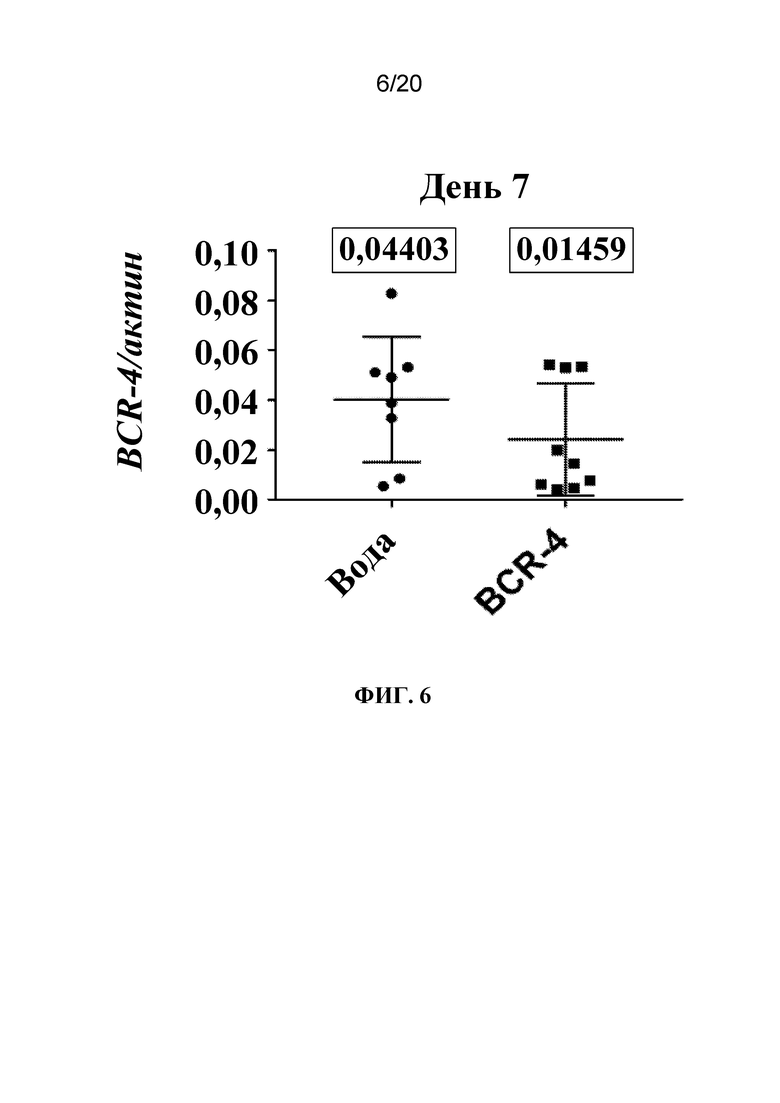
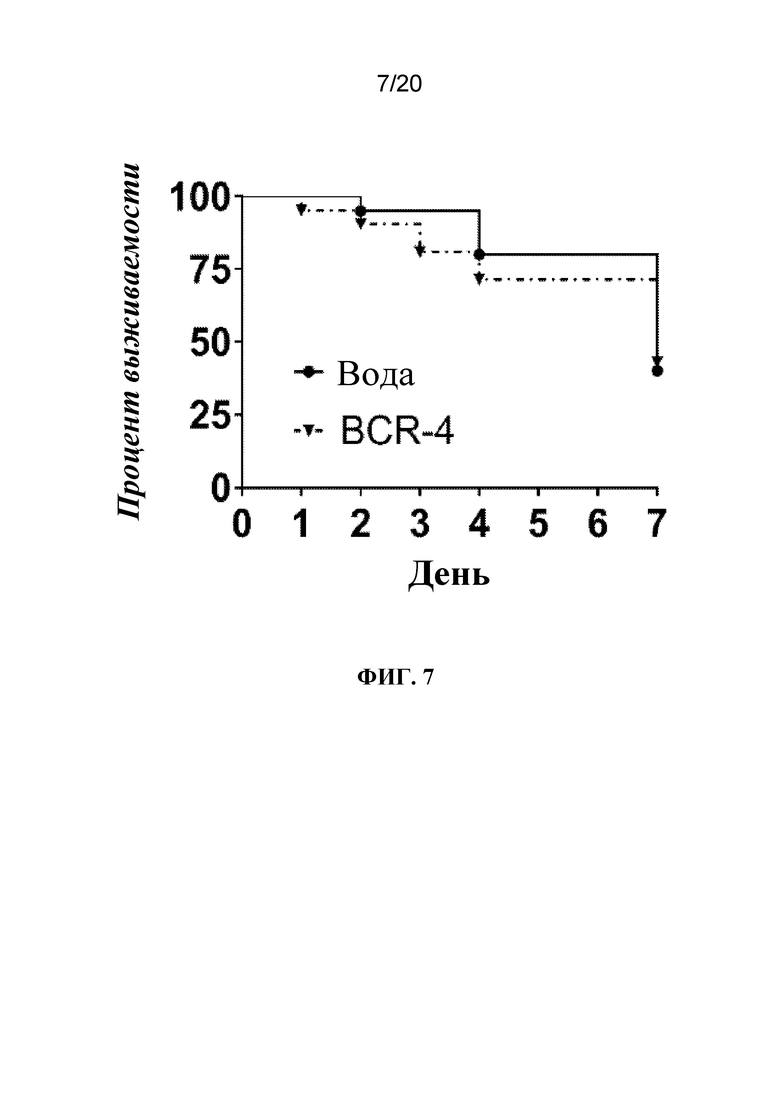
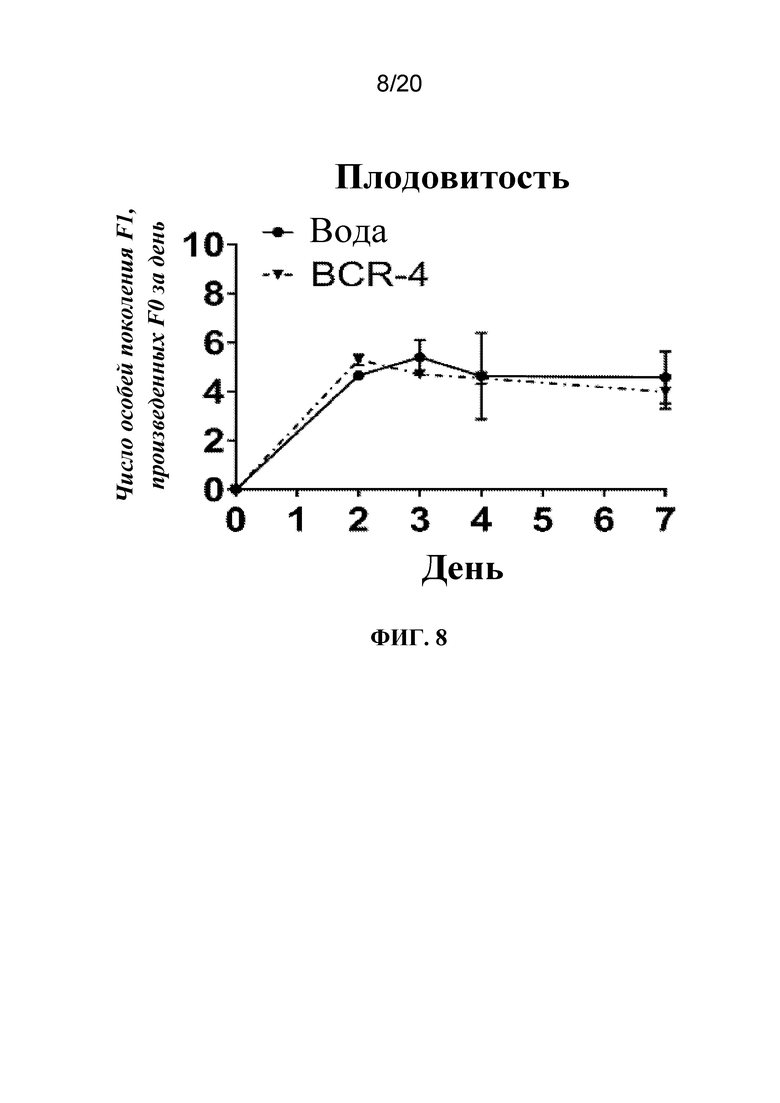
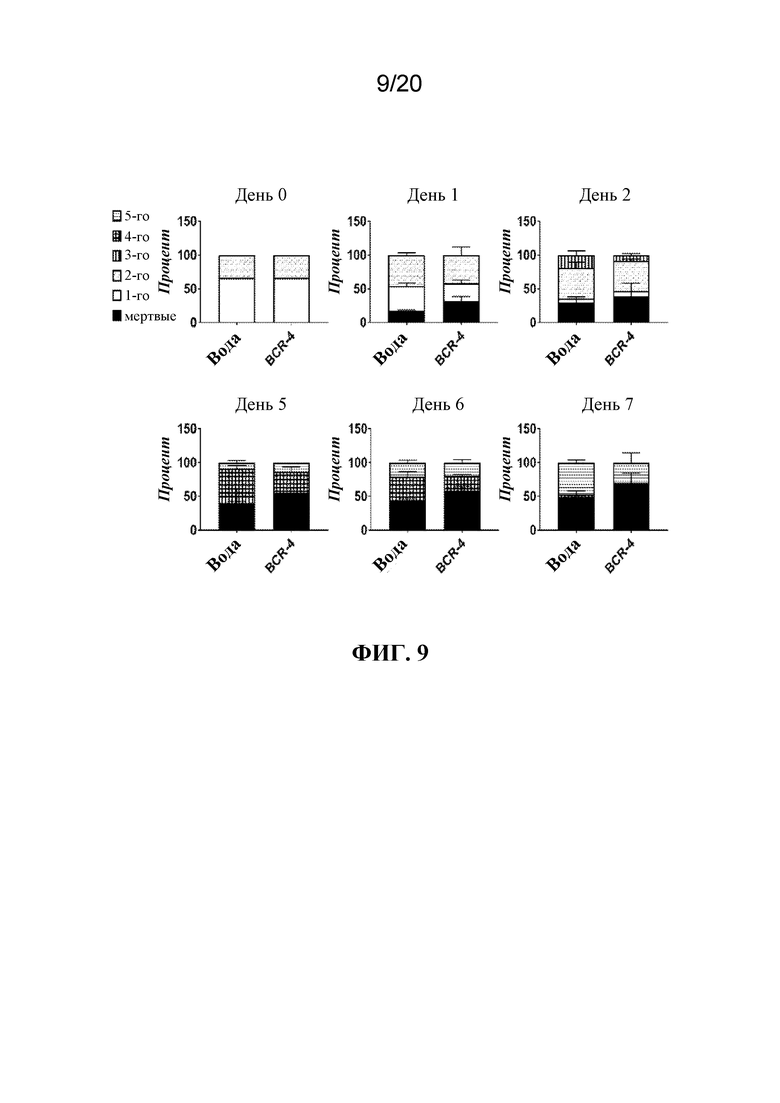
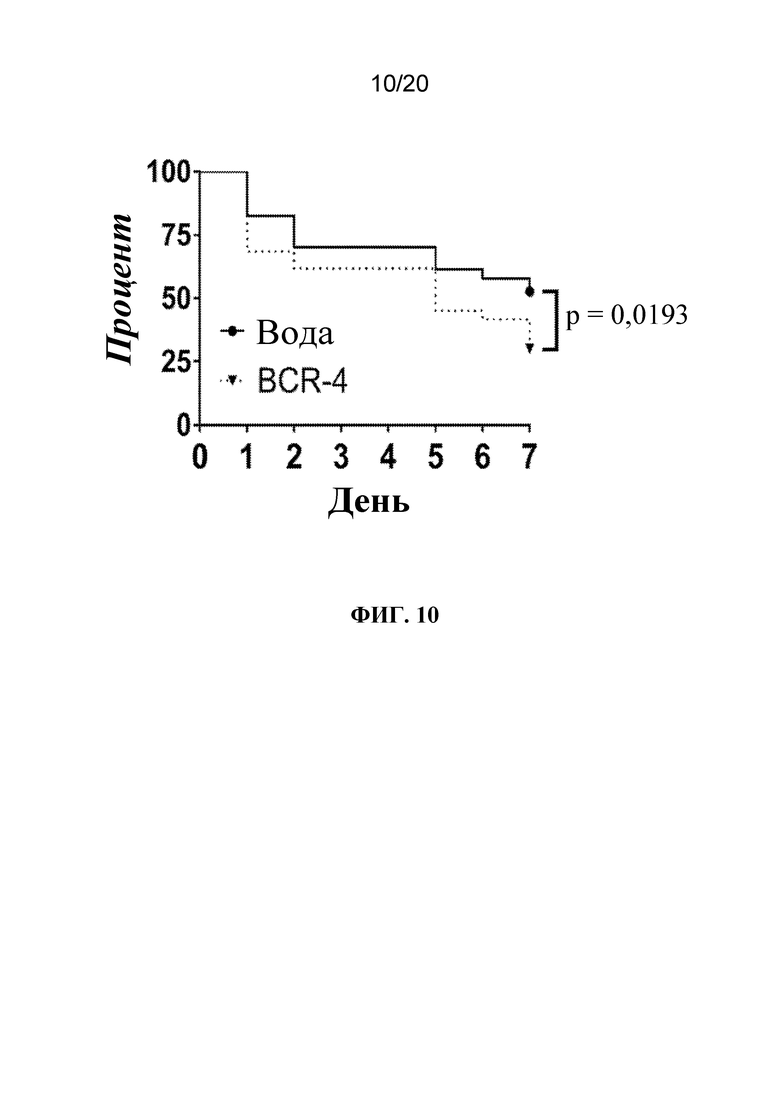
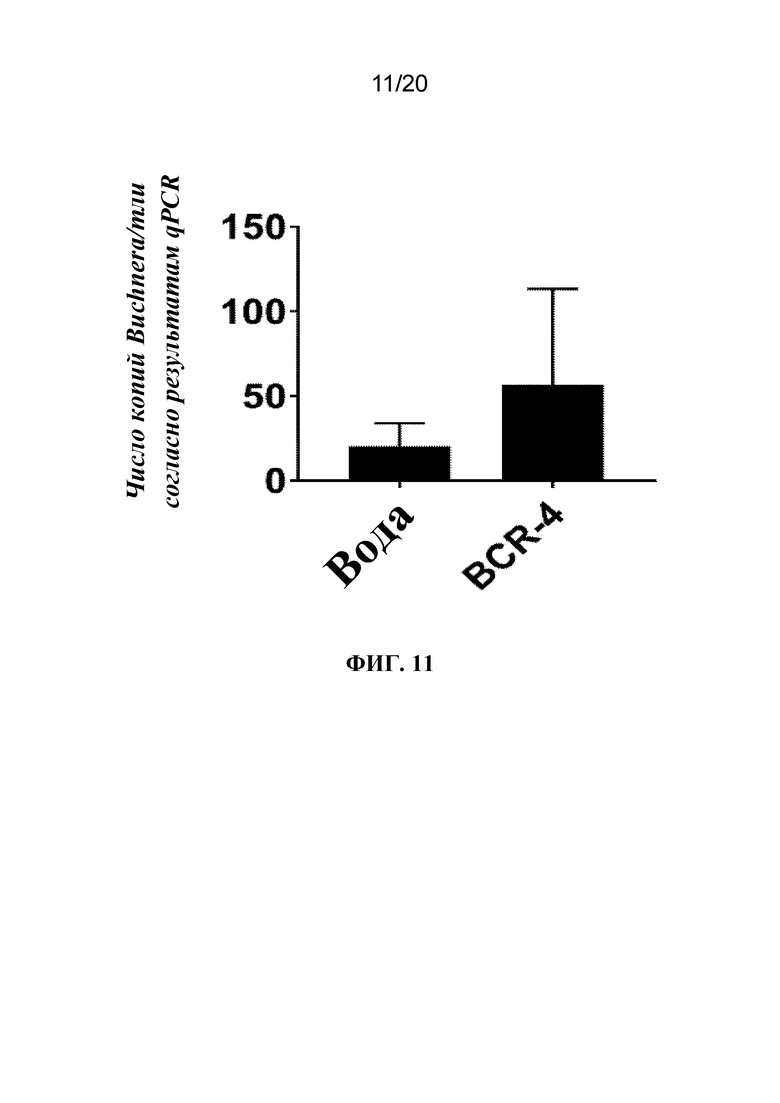
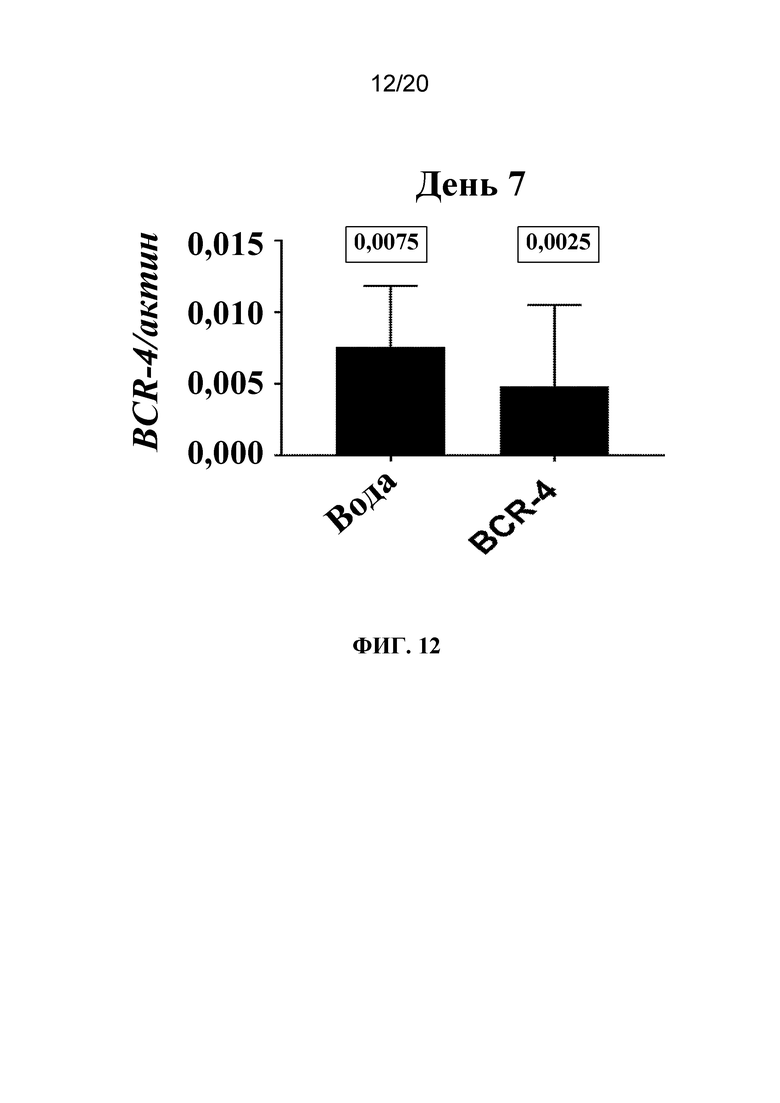
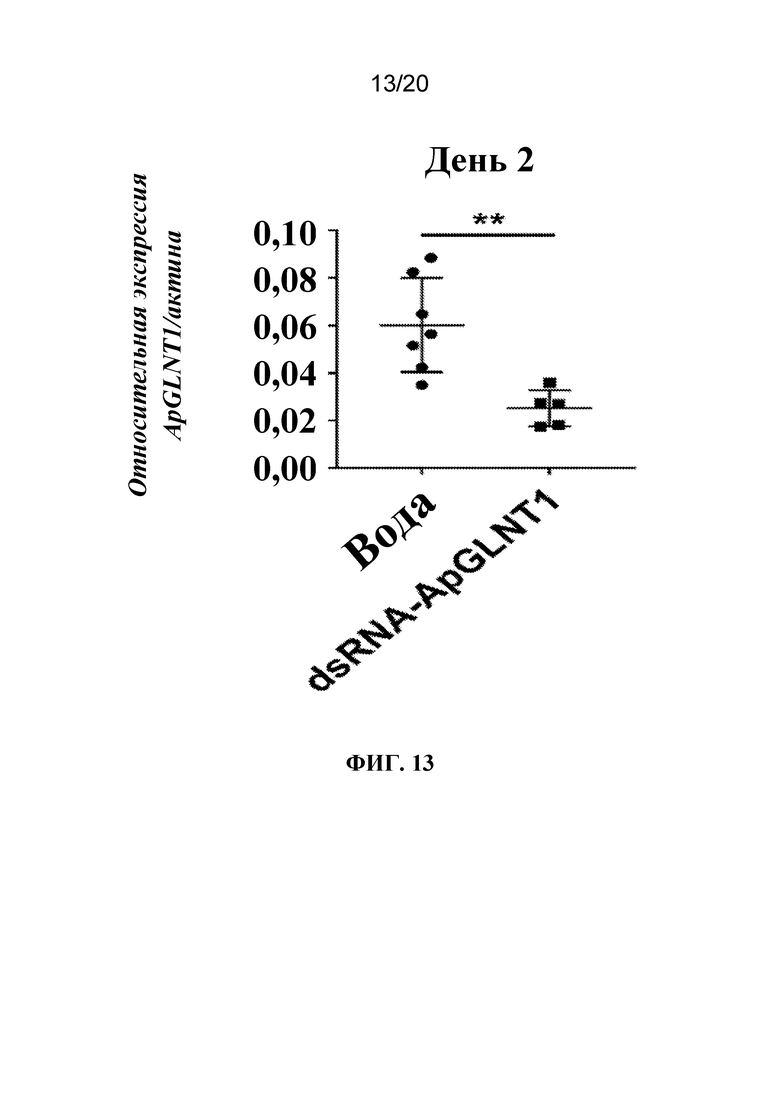
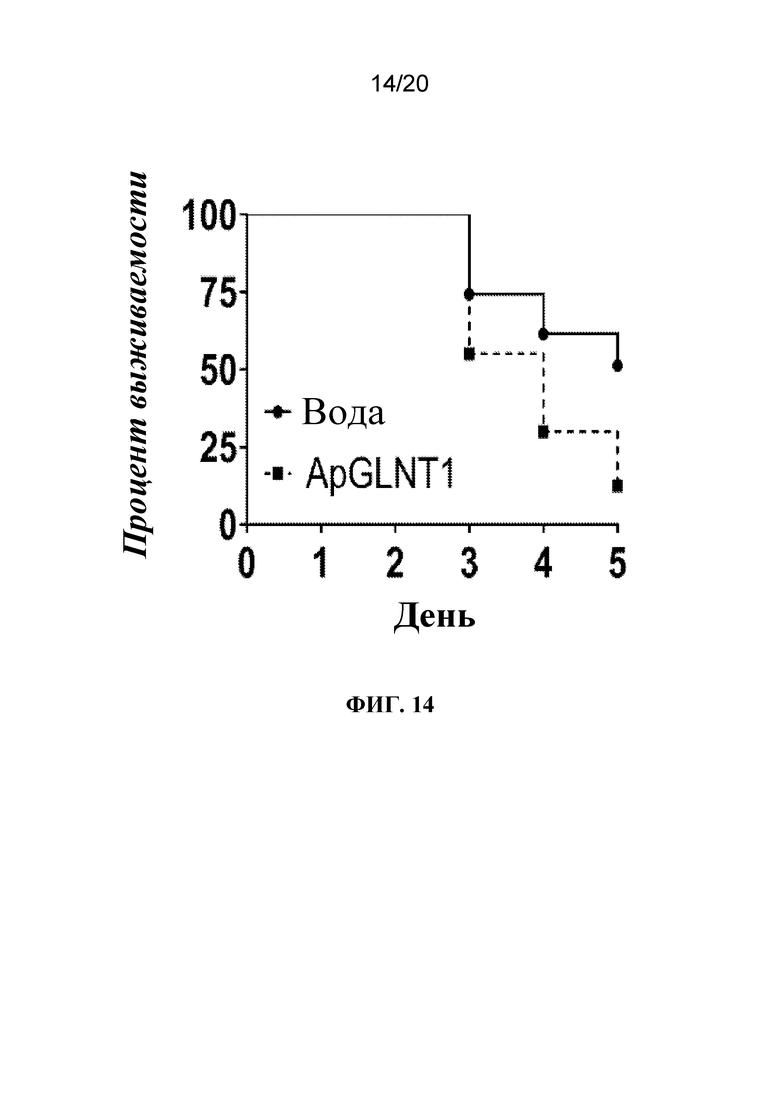
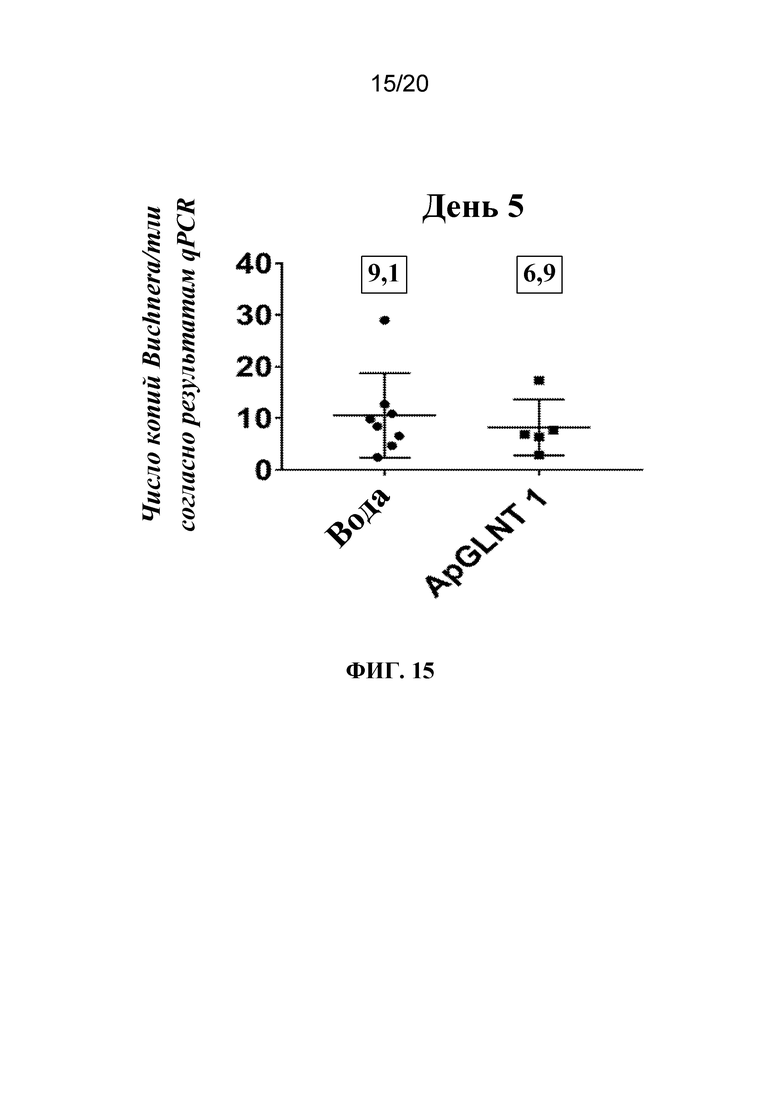
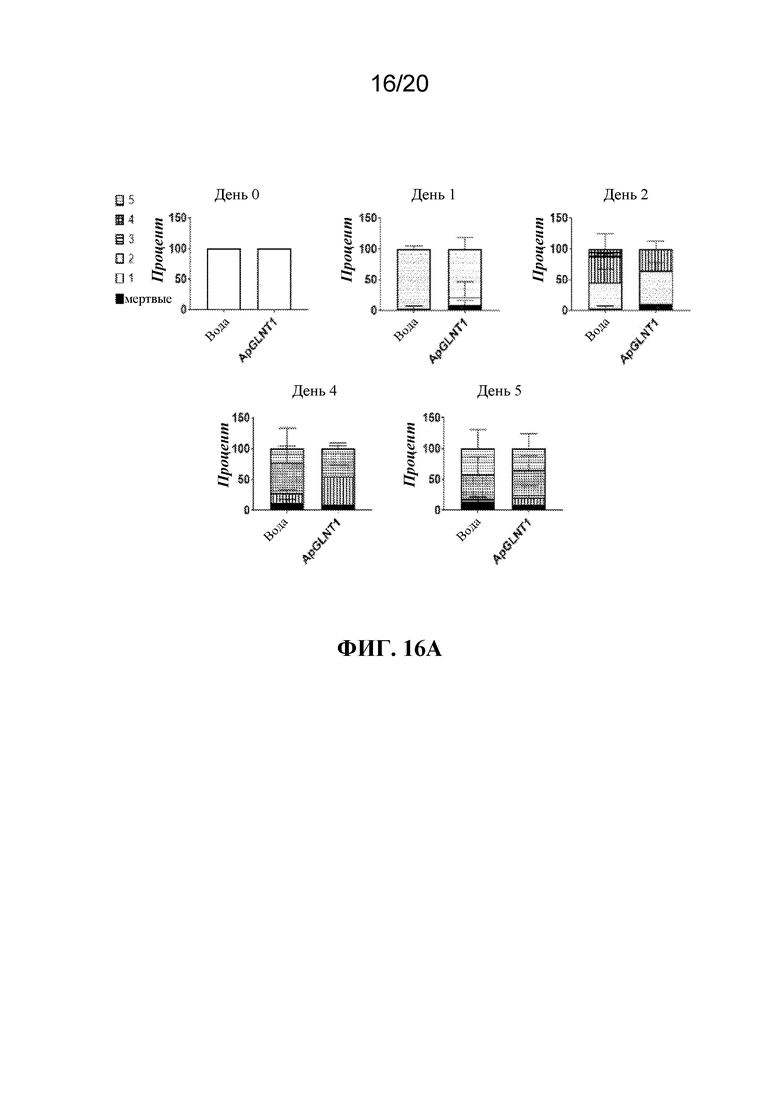
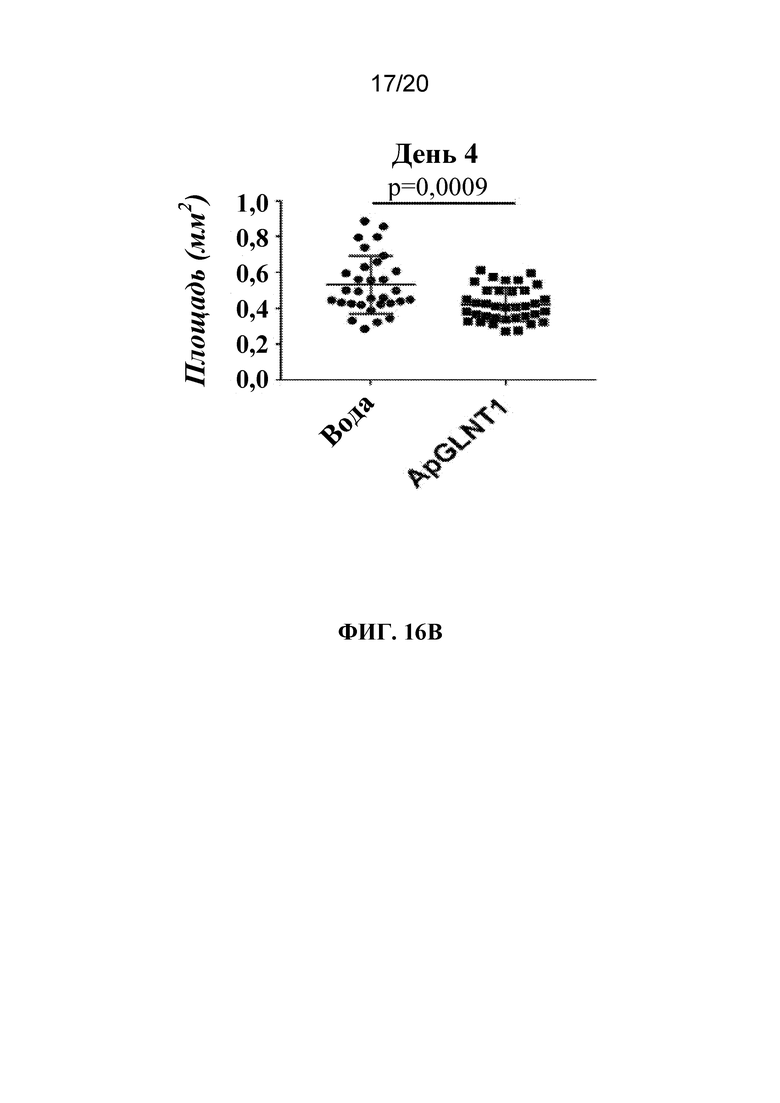
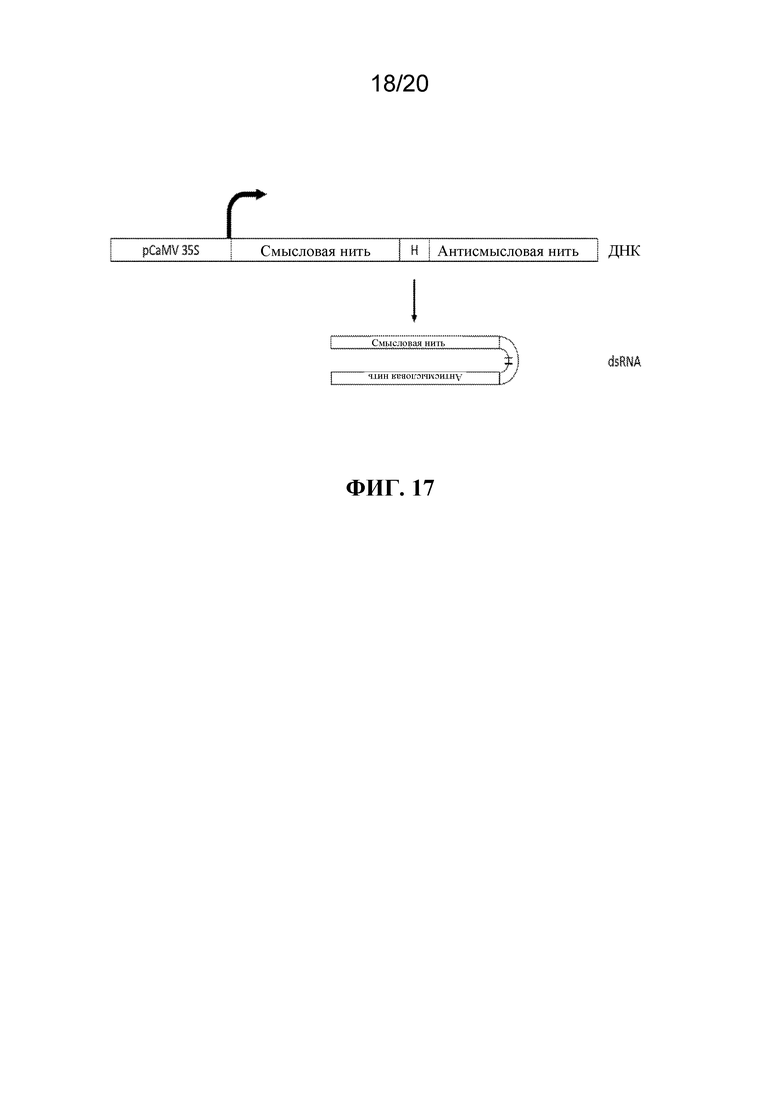
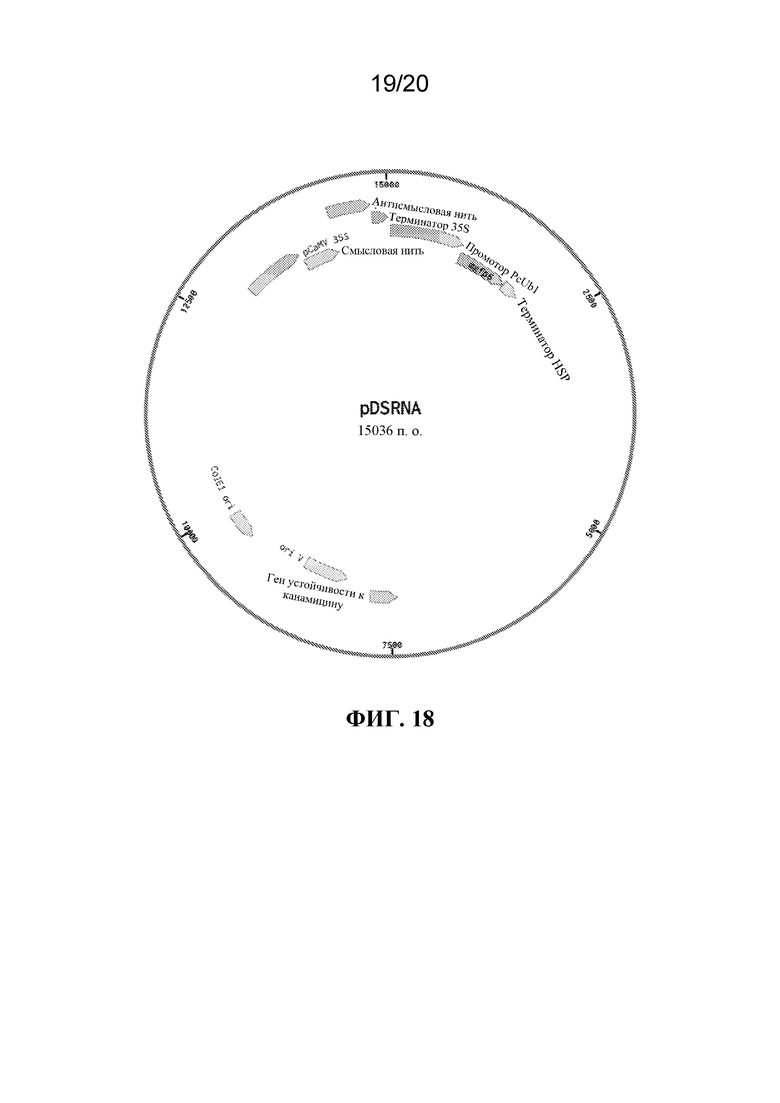

Изобретение относится к области биохимии, в частности к способу снижения приспособленности насекомого. Также раскрыты композиция, содержащая указанный полинуклеотид; конструкция рекомбинантной DNA, содержащая указанный нуклеотид. Раскрыто применение указанной композиции. Изобретение позволяет эффективно бороться с насекомыми. 4 н. и 11 з.п. ф-лы, 19 ил., 15 табл., 20 пр.
1. Способ снижения приспособленности насекомого отряда Hemiptera, при этом способ включает доставку насекомому отряда Hemiptera эффективного количества полинуклеотида, содержащего dsRNA, которая нацеливается и обеспечивает снижение экспрессии гена ApGLNT1 с SEQ ID NO: 149 в организме насекомого по сравнению с насекомым, которому dsRNA не была введена, таким образом снижая приспособленность насекомого отряда Hemiptera.
2. Способ по п. 1, где dsRNA является комплементарной 15-30 нуклеотидам указанного гена ApGLNT1 в организме насекомого.
3. Способ по п. 1, где способ является эффективным для подавления экспрессии гена в организме насекомого отряда Hemiptera.
4. Способ по п. 1, где способ является эффективным для снижения уровня, разнообразия или метаболизма микробиоты, обитающей в организме насекомого отряда Hemiptera, по сравнению с насекомым отряда Hemiptera, которому полинуклеотид не был доставлен.
5. Способ по п. 4, где микробиота содержит Buchnera spp.
6. Способ по любому из пп. 1-5, где способ является эффективным для снижения приспособленности насекомого отряда Hemiptera по сравнению с насекомым отряда Hemiptera, которому полинуклеотид не был доставлен.
7. Способ по любому из пп. 1-6, где полинуклеотид доставляют в композиции, составленной для доставки насекомым отряда Hemiptera.
8. Способ по любому из пп. 1-7, где доставка включает осуществление доставки полинуклеотида в по меньшей мере одну среду обитания, в которой насекомое отряда Hemiptera растет, живет, размножается, питается или осуществляет заражение.
9. Способ по любому из пп. 1-8, где доставка включает распыление полинуклеотида на сельскохозяйственную культуру.
10. Способ по любому из пп. 1-9, где полинуклеотид доставляют в виде композиции, съедобной для насекомого, для заглатывания насекомым отряда Hemiptera.
11. Способ по любому из пп. 1-10, где полинуклеотид составляют с приемлемым с точки зрения сельского хозяйства носителем в виде жидкой, твердой, аэрозольной, пастообразной, гелеобразной или газообразной композиции.
12. Способ по любому из пп. 1-11, где насекомое отряда Hemiptera представляет собой тлю.
13. Композиция для снижения приспособленности насекомого отряда Hemiptera, содержащая полинуклеотид, который содержит dsRNA, составленная для доставки насекомому отряда Hemiptera, где dsRNA является комплементарной 15-30 нуклеотидам гена ApGLNT1 с SEQ ID NO: 149 и где dsRNA нацеливается на ген ApGLNT1 и снижает его экспрессию.
14. Применение композиции по п. 13 для местного нанесения на растение.
15. Конструкция рекомбинантной DNA для экспрессии dsRNA в растении, содержащая гетерологичный промотор, функционально связанный с DNA, кодирующей RNA, содержащую по меньшей мере один участок dsRNA, по меньшей мере одна цепь которого содержит нуклеотидную последовательность, комплементарную 15-30 нуклеотидам гена ApGLNT1 с SEQ ID NO: 149, и где dsRNA нацеливается на ген ApGLNT1 и снижает его экспрессию.
| ПЕСТИЦИДНЫЙ БЕЛОК (ВАРИАНТЫ) | 2000 |
|
RU2267535C2 |
| РАСТЕНИЯ, ИМЕЮЩИЕ УСИЛЕННЫЕ ПРИЗНАКИ, СВЯЗАННЫЕ С УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2008 |
|
RU2503721C2 |
| РАСТИТЕЛЬНЫЕ КЛЕТКИ И ОРГАНИЗМЫ РАСТЕНИЙ С МОДИФИЦИРОВАННЫМ КЛЕТОЧНЫМ РОСТОМ, РАЗВИТИЕМ И ДИФФЕРЕНЦИРОВКОЙ | 2004 |
|
RU2349642C2 |

