Область техники
[0001] Настоящая заявка относится к связыванию слитого белка с белком CD47 и к его применению. Указанный слитый белок может специфически блокировать взаимодействие между белком CD47 и SIRPα, не вызывая реакцию свертывания крови, и может ингибировать рост и/или пролиферации опухолей или опухолевых клеток.
Уровень техники
[0002] Белок CD47 представляет собой трансмембранный гликопротеин, который является членом иммуноглобулинового суперсемейства и экспрессируется на поверхности различных клеток, включая эритроциты. Лиганды CD47 включают интегрины, тромбоспондин-1 и сигнальные белки (SIRP). CD47 влияет на ряд биологических функций, включая клеточную миграцию, Т-клетки, активацию дендритных клеток, развитие аксонов и т.д. Кроме того, взаимодействуя с SIRPα, CD47 может ингибировать макрофагальный фагоцитоз и защищать нормальные клетки, такие как клетки крови и т.п., от фагоцитоза макрофагами. Исследования показали, что в дополнение к экспрессии CD47 клетками нормальных тканей многие опухолевые клетки сверхэкспрессируют CD47 и предотвращают фагоцитоз опухолевых клеток макрофагами путем комбинации с SIRPα на поверхности макрофагов. Это рассматривается как механизм, с помощью которого опухоли ускользают от иммунного надзора организма. Блокирование взаимодействия между белком CD47 и SIRPα может подавлять рост опухолей. (Theocharides APA, et al., 2012).
[0003] Однако, существующие реагенты для блокирования взаимодействия между белком CD47 и SIRPα обладают ограниченной активностью распознавания. Они имеют тенденцию к недостаточной аффинности с белком CD47 и ограниченную способность ингибировать опухоли. В другом аспекте, существующие лекарственные средства с антителами, нацеленными на CD47, обладают побочными эффектами, вызывающими анемию или тромбоцитопению (Yinpeng Bai et al., Chin J Clin Oncol., 2017 Vol 44. No. 7). Существует острая необходимость в разработке новой терапии, которая может эффективно блокировать взаимодействие между белком CD47 и SIRPα с меньшим количеством побочных эффектов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0004] Настоящая заявка относится к связыванию слитого белка с белком CD47 и к его применению. Слитый белок может специфически связываться с белком CD47. Слитый белок по настоящей заявке имеет, по меньшей мере, одну из следующих характеристик: 1) специфическое связывание с белком CD47 с относительно высокой аффинностью; 2) специфическое блокирование взаимодействия белка CD47 с SIRPα; 3) не вызывает реакцию свертывания крови; 4) ингибирование роста и/или пролиферации опухолей или опухолевых клеток; 5) блокирование апоптотического сигнала, индуцированного взаимодействием CD47/SIRPα; и/или 6) безопасен для человека и не имеет побочных эффектов, наносящих вред организму. Настоящая заявка дополнительно относится к способу получения и применению слитого белка.
[0005] В одном из аспектов, настоящая заявка относится к слитому белку, способному специфически связываться с белком CD47, и имеющему, по меньшей мере, одну из следующих характеристик: 1) связывание с белком CD47 с величиной KD 1×10-8 M или меньше; 2) специфическое блокирование взаимодействия белка CD47 с SIRPα; 3) не вызывает реакцию свертывания крови; и 4) ингибирование роста и/или пролиферации опухолей или опухолевых клеток.
[0006] В некоторых вариантах осуществления указанный белок CD47 представляет собой человеческий белок CD47.
[0007] В некоторых вариантах осуществления указанный белок CD47 представляет собой белок CD47, экспрессирующийся на поверхности клеток.
[0008] В некоторых вариантах осуществления указанные опухоли или опухолевые клетки являются CD47-положительными.
[0009] В некоторых вариантах осуществления указанные опухоли выбраны из группы, состоящей из CD47-положительных гематологических опухолей и/или CD47-положительных солидных опухолей.
[0010] В некоторых вариантах осуществления указанный слитый белок содержит домен человеческого SIRPα, который может специфически связываться с указанным белком CD47 и Fc-областью иммуноглобулина, где указанный домен человеческого SIRPα напрямую или опосредованно связан с Fc-областью иммуноглобулина.
[0011] В некоторых вариантах осуществления указанный домен человеческого SIRPα содержит внеклеточный домен человеческого SIRPα, его фрагмент или его вариант, который содержит одну или более замен аминокислот. В некоторых вариантах осуществления указанный домен человеческого SIRPα содержит домен IgV человеческого SIRPα, его фрагмент или его вариант, который содержит одну или более замен аминокислот. В некоторых вариантах осуществления указанный домен человеческого SIRPα содержит домен человеческого SIRPα варианта 1, его фрагмент или его вариант, который содержит одну или более замен аминокислот. В некоторых вариантах осуществления указанный домен человеческого SIRPα содержит домен IgV человеческого SIRPα варианта 1, его фрагмент или его вариант, который содержит одну или более замен аминокислот. В некоторых вариантах осуществления указанный домен человеческого SIRPα содержит аминокислотные остатки в положениях 33-149 человеческого SIRPα варианта 1, его фрагмента или его варианта, который содержит одну или более замен аминокислот.
[0012] В некоторых вариантах осуществления домен человеческого SIRPα по настоящей заявке содержит аминокислотную последовательность, указанную в одной из SEQ ID NO: 1-20, 62-65, и аминокислотную последовательность, имеющую с ней, по меньшей мере, 80% (например, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98%, по меньшей мере, 99%, или, по меньшей мере, 100%) гомологии последовательности.
[0013] В некоторых вариантах осуществления слитый белок по настоящей заявке содержит аминокислотную последовательность, указанную в одной из SEQ ID NO: 21-61, и аминокислотную последовательность, имеющую с ней, по меньшей мере, 80% (например, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98%, по меньшей мере, 99%, или, по меньшей мере, 100%) гомологии последовательности.
[0014] В некоторых вариантах осуществления указанный домен человеческого SIRPα, его фрагмент или его вариант, который содержит одну или более замен аминокислот, включает замены, делеции или добавления одного или более аминокислотных остатков.
[0015] В некоторых вариантах осуществления указанный мутант содержит замены аминокислот по одному или более остаткам, выбранным из группы, состоящей из I61, V63, E77, Q82, K83, E84, V93, D95, D96, K98, N100, R107, G109 и V132.
[0016] В некоторых вариантах осуществления указанный мутант содержит одну или более замен аминокислот, выбранных из группы, состоящей из R22C, I29L, I61L/V/F, V63I, E77I/N/Q/K/H/M/R/N/V/L, Q82S/R/G/N, K83R, E84K/H/D/R/G, V93L/A, D95H/R/E, D96S/T, K98R, N100G/K/D/E, R107N/S, G109R/H и V132L/R/I/S.
[0017] В некоторых вариантах осуществления указанный домен человеческого SIRPα содержит аминокислотную последовательность, указанную в одной из SEQ ID NO: 1-20, 62-65.
[0018] В некоторых вариантах осуществления указанная Fc-область иммуноглобулина содержит Fc-область IgG.
[0019] В некоторых вариантах осуществления указанный IgG представляет собой человеческий IgG. В некоторых вариантах осуществления указанный IgG выбран из группы, состоящей из IgG1 и/или IgG4.
[0020] В некоторых вариантах осуществления указанный домен человеческого SIRPα расположен у N-конца указанной Fc-области иммуноглобулина.
[0021] В некоторых вариантах осуществления указанный домен человеческого SIRPα связан с Fc-областью иммуноглобулина через линкер.
[0022] В некоторых вариантах осуществления указанная Fc-область иммуноглобулина содержит аминокислотную последовательность, указанную в одной из SEQ ID NO: 67-68.
[0023] В некоторых вариантах осуществления указанный слитый белок содержит аминокислотную последовательность, указанную в одной из SEQ ID NO: 21-61.
[0024] В другом аспекте настоящая заявка относится к молекуле нуклеиновой кислоты, кодирующей слитый белок по настоящей заявке.
[0025] В другом аспекте настоящая заявка относится к вектору, содержащему молекулу нуклеиновой кислоты по настоящей заявке.
[0026] В другом аспекте настоящая заявка относится к клетке-хозяину, содержащей молекулу нуклеиновой кислоты по настоящей заявке или вектор по настоящей заявке.
[0027] В другом аспекте настоящая заявка относится к способу получения слитого белка по настоящей заявке, способу, включающему культивирование клетки-хозяина по настоящей заявке в условиях, которые позволяют экспрессировать слитый белок.
[0028] В другом аспекте настоящая заявка относится к композиции, содержащей слитый белок, молекулу нуклеиновой кислоты, вектор и/или клетку-хозяина по настоящей заявке, и необязательно фармацевтически приемлемые адъюванты.
[0029] В другом аспекте настоящая заявка относится к применению слитого белка, молекулы нуклеиновой кислоты, вектора, клетки-хозяина и/или композиции по настоящей заявке для получения лекарственного средства и/или набора, где указанное лекарственное средство и/или набор предназначены для профилактики или лечения опухолей или аутоиммунных заболеваний. В некоторых вариантах осуществления опухоли выбраны из группы, состоящей из CD47-положительных гематологических опухолей и/или CD47-положительных солидных опухолей. В некоторых вариантах осуществления аутоиммунные заболевания выбраны из группы, состоящей из болезни Крона, аллергической астмы и ревматоидного артрита.
[0030] В другом аспекте настоящая заявка относится к способу блокирования взаимодействия между белком CD47 и SIRPα, способу, включающему введение слитого белка или композиции по настоящей заявке.
[0031] В другом аспекте настоящая заявка относится к способу ингибирования роста и/или пролиферации опухолей или опухолевых клеток, способу, включающему контакт слитого белка или композиции по настоящей заявке с опухолями или опухолевыми клетками. В некоторых вариантах осуществления контакт происходит in vitro.
[0032] В другом аспекте настоящая заявка относится к способу профилактики или лечения опухолей или аутоиммунных заболеваний у индивидуума, способу, включающему введение индивидууму эффективного количества слитого белка или композиции по настоящей заявке. В некоторых вариантах осуществления опухоли выбраны из группы, состоящей из CD47-положительных гематологических опухолей и/или CD47-положительных солидных опухолей. В некоторых вариантах осуществления аутоиммунные заболевания выбраны из группы, состоящей из болезни Крона, аллергической астмы и ревматоидного артрита.
[0033] Специалисты в данной области могут легко узнать о других аспектах и преимуществах настоящего изобретения из подробного последующего описания. Подробное описание ниже показывает и описывает только иллюстративные варианты осуществления настоящего изобретения. Как будет очевидно специалистам в данной области, содержание настоящего изобретения позволяет специалистам в данной области изменять конкретные описанные варианты осуществления без отклонения от сущности и объема изобретения, которые изложены в настоящей заявке. Соответственно, чертежи и описание по настоящей заявке являются лишь иллюстративными, а не ограничительными.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0034] Конкретные признаки изобретения, относящиеся к настоящей заявке, показаны в прилагаемой формуле изобретения. Ссылаясь на иллюстративные варианты осуществления, подробно описанные ниже, и прилагаемые чертежи, можно лучше понять особенности и преимущества настоящего изобретения. Краткое описание прилагаемых чертежей выглядит следующим образом:
[0035] ФИГ. 1 показывает физическое структурное схематическое изображение вектора pTM.
[0036] ФИГ. 2 показывает схематическое изображение способа детекции взаимодействий между укороченными доменами SIRPα и их мутантами и CD47.
[0037] ФИГ. 3 показывает результаты скрининга с обогащением для мутантов укороченного домена SIRPα путем проточной цитометрии.
[0038] ФИГ. 4 показывает выравнивание последовательностей укороченных доменов SIRPα и их вариантов.
[0039] ФИГ. 5 показывает результаты распознавания слитым белком по настоящей заявке белка CD47.
[0040] ФИГ. 6A-6B показывают специфичность слитого белка по настоящей заявке относительно распознавания человеческого белка CD47.
[0041] ФИГ. 7 показывает распознавание слитым белком по настоящей заявке человеческого белка CD47 и других белков.
[0042] ФИГ. 8 показывает, что слитый белок по настоящей заявке и TTI-621 конкурентно блокируют связывание белка CD47 с его лигандом SIRPα.
[0043] ФИГ. 9A-9C показывают результаты распознавания слитым белком по настоящей заявке клеток Raji, клеток Jurkat и белка CD47 на поверхности клеток A549.
[0044] ФИГ. 10A-10C показывают результаты распознавания слитым белком по настоящей заявке и TTI-621 клеток Raji, клеток Jurkat и белка CD47 на поверхности клеток A549.
[0045] ФИГ. 11 показывает результаты слитого белка по настоящей заявке и Hu5F9-G4 по устойчивости к реакции свертывания крови.
[0046] ФИГ. 12A-12B показывают опухольингибирующую активность слитого белка по настоящей заявке.
[0047] ФИГ. 13A-13B показывают воздействия слитого белка по настоящей заявке на эритроциты и тромбоциты по сравнению с TTI-621.
[0048] ФИГ. 14 показывает результаты распознавания слитым белком по настоящей заявке белка CD47.
[0049] ФИГ. 15 показывает, что слитый белок по настоящей заявке конкурентно блокирует связывание белка CD47 с его лигандом SIRPα.
[0050] ФИГ. 16 показывает результаты слитого белка по настоящей заявке и Hu5F9-G4 по устойчивости к реакции свертывания крови.
[0051] ФИГ. 17 показывает, что слитый белок по настоящей заявке эффективно блокирует CD47-Fc-индуцированный апоптоз клеток Jurkat-CSR.
[0052] ФИГ. 18A показывает воздействия слитого белка по настоящей заявке на уровень эритроцитов в периферической крови мышей.
[0053] ФИГ. 18B показывает воздействия слитого белка по настоящей заявке на уровень тромбоцитов в периферической крови мышей.
[0054] ФИГ. 19 показывает ингибирующее действие слитого белка по настоящей заявке на рост опухоли у мышей.
[0055] ФИГ. 20 показывает воздействия слитого белка по настоящей заявке на массу тела мышей.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[0056] Далее в настоящем документе варианты осуществления изобретения, использованные в настоящей заявке, описаны посредством конкретных примеров. Специалисты в данной области могут легко понять другие преимущества и эффекты изобретения, описанного в настоящей заявке, из содержания, раскрытого в настоящем описании.
СЛИТЫЙ БЕЛОК
[0057] В одном из аспектов, настоящая заявка относится к слитому белку, который может специфически связываться с белком CD47 со значением KD 1×10-8 M или меньше, например, со значением KD не более чем 9×10-9 M, не более чем 8×10-9 M, не более чем 7×10-9 M, не более чем 6,2×10-9 M, не более чем 6×10-9 M, не более чем 5×10-9 M, не более чем 4,8×10-9 M, не более чем 4,5×10-9 M, не более чем 2×10-9 M, не более чем 1,5×10-9 M, или не более чем 1×10-10 M или меньше.
[0058] В некоторых случаях слитый белок по настоящей заявке может также специфически блокировать взаимодействие между белком CD47 и SIRPα, тем самым активируя макрофаги для фагоцитоза опухолевых клеток или ингибируя апоптотический сигнал некоторых специфических клеток. Кроме того, слитый белок по настоящей заявке может не вызывать реакцию свертывания крови. Например, указанный слитый белок и раствор эритроцитов добавляли в тестовый планшет для гемагглютинации, а затем эритроциты погружались на дно лунки вместо того, чтобы уплощаться в форме сетки. Слитый белок может дополнительно ингибировать рост и/или пролиферацию опухолей или опухолевых клеток, например, он может уменьшать площадь или размер опухоли или может увеличивать коэффициент выживаемости индивидуума с опухолью. Слитый белок также безопасен для введения индивидууму, потому что он не повлияет отрицательно на массу тела и/или уровень смертности индивидуума. Кроме того, слитый белок по настоящей заявке легко приготовить и получить, не ограничиваясь только источником Fc-области специфического иммуноглобулина.
[0059] В настоящей заявке, термин «слитый белок», в основном, относится к сложному полипептиду, то есть, к одной непрерывной последовательности аминокислот, состоящей из двух или более полипептидов. Слитый белок, как правило, можно получать искусственно с помощью рекомбинантной нуклеиновой кислоты или химического синтеза.
[0060] В настоящей заявке термин «белок CD47» также известен как интегринсвязанный белок (IAP), который относится к суперсемейству иммуноглобулинов. Белок CD47 может связываться с мембранными интегринами, тромбоспондином-1 (TSP-1) или сигнальным регуляторным белком альфа (SIRPα). Белок CD47 можно экспрессировать на поверхности клеточной мембраны. Указанный белок CD47 может представлять собой супрамолекулярный комплекс, состоящий из определенных IAP, G-белков и холестерина. В настоящей заявке указанный белок CD47 может быть человеческим белком CD47 с регистрационным номером CEJ95640.1 в базе данных GenBank. В настоящей заявке белок CD47 может включать аминокислотную последовательность, показанную в SEQ ID NO: 66.
[0061] В настоящей заявке термин "CD47-положительный" в основном относится к выражению в организме или на поверхности клеток характеристики белка CD47, его фрагмента или его варианта, который содержит одну или более замен. CD47-положительные клетки могут быть клетками, сверхэкспрессирующими CD47. CD47-положительная клетка в основном может служить признаком заболеваний. Например, в случае заболевания, плотность указанного белка CD47 на поверхности CD47-положительной клетки будет превышать плотность белка CD47 того же типа в клетках в нормальных условиях. В некоторых вариантах осуществления, указанные опухоли или опухолевые клетки могут быть CD47-положительными. Например, указанные опухоли могут быть выбраны из группы, состоящей из CD47-положительных гематологических опухолей и/или CD47-положительных солидных опухолей.
[0062] В настоящей заявке термин «KD» может использоваться взаимозаменяемо с «KD», и, в основном, относится к константе диссоциации специфического взаимодействия антиген-антитело на единицу M (моль/л). KD можно рассчитать по концентрации материала AB и диссоциированных материалов A и B: KD=c(A)×c(B)/c(AB). Из формулы видно, что чем больше значение KD, тем больше диссоциация и тем слабее аффинность материалов A и B; напротив, чем ниже значение KD, тем меньше диссоциация и тем сильнее аффинность материалов A и B.
[0063] В настоящей заявке термин «SIRPα» в основном относится к гликопротеину регуляторной мембраны из семейства SIRP. Указанный SIRPα может распознавать белок CD47 и может служить лигандом белка CD47. SIRPα представляет собой трансмембранный белок, который может иметь в своей внеклеточной области три области, подобные областям иммуноглобулинового семейства, где область на N-конце опосредует связывание с CD47. Указанный SIRPα исходно экспрессируется на поверхности макрофагов, дендритных клеток и нервных клеток. Цитоплазматический участок указанного SIRPα высоко консервативен у крыс. Указанный SIRPα проявляет полиморфизм, который, однако, не влияет на его распознавание и связывание с белком CD47.
[0064] В настоящей заявке, термин «домен человеческого SIRPα», в основном, включает человеческий SIRPα, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. В настоящей заявке, указанный домен человеческого SIRPα может содержать внеклеточный домен человеческого SIRPα, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. Указанный домен человеческого SIRPα может содержать домен IgV человеческого SIRPα, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. Указанный домен человеческого SIRPα может содержать домен человеческого SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. Указанный домен человеческого SIRPα может включать аминокислотные остатки в положениях 33-149 человеческого SIRPα варианта 1, его фрагмента, или его варианта, который содержит одну или более замен аминокислот. У человека, белок SIRPα исходно имеет два типа, один тип (человеческий SIRPα, вариант 1 или Тип V1) имеет аминокислотную последовательность с номером доступа GeneBank NP_542970.1 (его аминокислотная последовательность изложена в SEQ ID NO: 62, где аминокислотные остатки в положениях 31-504 могут составлять домен зрелого SIRPα). Другой тип (Вариант 2 или Тип V2) на 13 аминокислот отличается от варианта 1 или Типа V1, и его аминокислотная последовательность имеет номер доступа GeneBank CAA71403.1.
[0065] В настоящей заявке, указанный домен человеческого SIRPα может содержать внеклеточный домен человеческого SIRPα, его фрагмент, или его вариант, который содержит одну или более замен аминокислот.
[0066] В настоящей заявке термин «внеклеточный домен», в основном, относится к функциональной структурной области белка, расположенной снаружи от клеточной мембраны. В некоторых вариантах осуществления указанный внеклеточный домен может относиться к домену человеческого SIRPα, его фрагменту, или его варианту, который содержит одну или более замен аминокислот. Например, указанный внеклеточный домен из доменов человеческого SIRPα может содержать 3 домена иммуноглобулинового суперсемейства (IgSF) и множество участков гликозилирования. Указанный внеклеточный домен из доменов человеческого SIRPα может связываться с конкретным лигандом (например, белком CD47), включая функцию трансдукции сигнала. Указанный внеклеточный домен из доменов человеческого SIRPα также может быть активирован рядом митогенов, таких как, сыворотка, инсулин, факторы роста, EGF, PDGF и нейротрофические факторы, и фосфорилирован.
[0067] В настоящей заявке, указанный домен человеческого SIRPα может содержать домен IgV человеческого SIRPα, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. В настоящей заявке, указанный домен человеческого SIRPα может включать человеческий SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. Например, указанный домен человеческого SIRPα может включать аминокислотные остатки в положениях 33-149 человеческого SIRPα варианта 1, его фрагмента, или его варианта, который содержит одну или более замен аминокислот.
[0068] В настоящей заявке, термин «домен IgV», в основном, относится к IgV-подобному домену, который аналогичен вариабельному домену антитела. Иммуноглобулиновый домен может быть подразделен на четыре класса: IgV, IgC1, IgC2 и IgI. Домен IgV может присутствовать у различных семейств белков, и содержит легкую цепь и тяжелую цепь, T-клеточный рецептор иммуноглобулина. Человеческий SIRPα может быть высокополиморфным в домене IgV. Например, указанный домен IgV человеческого SIRPα варианта 1 может опосредовать связывание домена человеческого SIRPα с человеческим белком CD47 (Seiffert, M. et al. (2001) Blood 97,2741-9; Vernon-ffilson, E.F. et al. (2000) Eur J Immunol 30, 2130-7).
[0069] Например, указанный человеческий SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот, может содержать аминокислотную последовательность, изложенную в SEQ ID NO: 62. Например, указанный домен человеческого SIRPα может содержать домен IgV из человеческого SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. Например, указанный домен IgV из человеческого SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот, может содержать аминокислотную последовательность, изложенную в SEQ ID NO: 65 (т.е. остатки в положениях 38-145 аминокислотной последовательности, изложенной в SEQ ID NO: 62). Альтернативно, например, указанный домен человеческого SIRPα может содержать укороченный домен человеческого SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот. Указанный укороченный домен человеческого SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот, может содержать аминокислотную последовательность, изложенную в SEQ ID NO: 63 (т.е. остатки в положениях 33-149 аминокислотной последовательности, изложенной в SEQ ID NO: 62).
[0070] Домен человеческого SIRPα по настоящей заявке может содержать аминокислотную последовательность, изложенную в любой одной последовательности, выбранной из группы, состоящей из SEQ ID NO: 1-20 и 62-65.
[0071] В настоящей заявке термин «мутант», в основном, относится к белку, полипептиду или аминокислотной последовательности, в которой происходит мутация. Указанная мутация может относиться к различию по сравнению с диким типом. Например, указанная мутация может быть структурным изменением аминокислотной последовательности, которое происходит на основе дикого типа. Например, указанный дикий тип может быть типичным фенотипом без указанного структурного изменения. Например, указанный мутант можно получить при мутации домена человеческого SIRPα, его фрагмента (содержит домен IgV человеческого SIRPα, его фрагмент, домен человеческого SIRPα варианта 1, его фрагмент или аминокислотные остатки в положениях 33-149 человеческого SIRPα вариант 1, его фрагмента), который рассматривается как дикий тип.
[0072] В настоящей заявке, мутант может содержать аминокислотные замены по одному или более остаткам, выбранным из группы, состоящей из I61, V63, E77, Q82, K83, E84, V93, D95, D96, K98, N100, R107, G109, и V132. Указанные положения аминокислотных остатков для замен аминокислот могут представлять собой точный номер остатка при использовании в качестве эталона аминокислотной последовательности, изложенной в SEQ ID NO: 62, где «остаток Xn» относится к остатку X, который соответствует положению n в аминокислотной последовательности, изложенной в SEQ ID NO: 62, где n является положительным целым числом, X является аббревиатурой любого аминокислотного остатка. Например, «остаток I61» относится к аминокислотному остатку I, который соответствует положению 61 в аминокислотной последовательности, изложенной в SEQ ID NO:62.
[0073] В настоящей заявке, «замена аминокислоты Xn» относится к замене аминокислоты, находящейся в аминокислотном остатке X в положении n аминокислотной последовательности, изложенной в SEQ ID NO: 62, где n является положительным целым числом, X является аббревиатурой любого аминокислотного остатка. Например, «замены аминокислот I61» относится к замене аминокислоты, находящейся в аминокислотном остатке I, который соответствует положению 61 в аминокислотной последовательности, изложенной в SEQ ID NO:62.
[0074] В настоящей заявке, определенный аминокислотный остаток в определенной аминокислотной последовательности "соответствующий" определенному аминокислотному остатку в другой аминокислотной последовательности, в основном относится к соответствующему родству аминокислотного остатка, полученному выравниванием аминокислотной последовательности в оптимальных условиях. Указанное выравнивание последовательностей можно проводить способами, которые понятны специалистам в данной области, например, с использованием программного обеспечения BLAST, BLAST-2, ALIGN, NEEDLE или Megalign (DNASTAR) и т.д. Специалисты в данной области могут определить подходящие параметры для применения для выравнивания, включающие любой алгоритм, необходимый для достижения оптимального выравнивания в сравниваемой полноразмерной последовательности.
[0075] Замены аминокислот по настоящей заявке могут быть неконсервативными заменами. Указанные неконсервативные замены могут включать замену аминокислотных остатков в целевом белке или полипептиде неконсервативным образом, например, замену аминокислотного остатка, имеющего определенный размер боковой цепи или определенную характеристику (например, гидрофильный), на аминокислотный остаток, имеющего другой размер боковой цепи или другую характеристику (например, гидрофобный).
[0076] Указанные замены аминокислот также могут быть консервативными заменами. Указанные консервативные замены могут включать замену аминокислотных остатков в целевом белке или полипептиде консервативным образом, например, замену аминокислотного остатка, имеющего определенный размер боковой цепи или определенную характеристику (например, гидрофильный) на аминокислотный остаток, имеющий такой же или подобный размер боковой цепи или такую же или похожую характеристику (например, все еще гидрофильный). Такие консервативные замены в основном не оказывают достоверного воздействия на структуру или функцию продуцируемого белка. В настоящей заявке, вариант аминокислотной последовательности, который является мутантом слитого белка, его фрагментом, или его вариантом, который содержит одну или более замен аминокислот, может содержать консервативные замены аминокислот, которые не могут заметно изменить структуру или функцию белка (например, способность блокировать связывание CD47 с его лигандом).
[0077] Например, консервативные замены в настоящей заявке можно рассматривать как консервативные замены между аминокислотами в каждой из следующих групп:
Группа аминокислот с неполярной боковой цепью/цепями: аланин, валин, лейцин, изолейцин, пролин, фенилаланин, триптофан и метионин.
Группа незаряженных аминокислот с полярными боковыми цепями: глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин.
Группа отрицательно заряженных аминокислот с полярными боковыми цепями: аспарагиновая кислота и глутаминовая кислота.
Группа положительно заряженных основных аминокислот: лизин, аргинин и гистидин.
Группа аминокислот с фенилом: фенилаланин, триптофан и тирозин.
[0078] В некоторых вариантах осуществления указанный мутант может содержать одну или более аминокислотных замен, выбранных из группы, состоящей из I61L/V/F, V63I, E77I/N/Q/K/H/M/R/N/V/L, Q82S/R/G/N, K83R, E84K/H/D/R/G, V93L/A, D95H/R/E, D96S/T, K98R, N100G/K/D/E, R107N/S, G109R/H и V132L/R/I/S.
[0079] В настоящей заявке, замена аминокислот "XnY/Z» означает, что остаток X, соответствующий положению n в аминокислотной последовательности, изложенной в SEQ ID NO: 62, замещен аминокислотным остатком Y или аминокислотным остатком Z, где n является положительным целым числом, X, Y и Z независимо являются аббревиатурой любого аминокислотного остатка, соответственно, и X отличается от Y или Z. Например, аминокислотная замена «I61L/V/F» означает, что остаток I, соответствующий положению 61 в аминокислотной последовательности, изложенной в SEQ ID NO: 62, замещен аминокислотным остатком L, V или F.
[0080] Например, слитый белок по настоящей заявке может содержать замену аминокислотной группы, выбранной из группы, включающей:
(1) I61L, V63I, E77I, E84K, V93L, L96S, K98R, N100G и V132 L;
(2) I61V, E77N, Q82S, K83R и E84H;
(3) I61F, V63I, K83R, E84K и V132I;
(4) I61L, E77Q, E84D, R107N и V132I;
(5) I61L, V63I, E77K, K83R, E84D и N100G;
(6) I61V, E77H, Q82R, K83R, E84H и R107S;
(7) I61L, E77I, Q82G, E84R, V93L, L96T, N100G, R107S, G109R и V132R;
(8) I61L, E77M, Q82G, K83R, E84D и V132L;
(9) I61L;
(10) I61F, D95H, L96S, G109H и V132S;
(11) I61F, D95H, L96S, K98R, G109H и V132S;
(12) I61L, E77Q, E84D, V93A, R107N и V132I;
(13) E77K, L96S, N100K, G109H и V132L;
(14) I61L, V63I, Q82G, E84G, D95R, L96S, N100D и V132I;
(15) I61L, E77R, Q82N, K83R, E84G, V93L, D95E, L96T, K98R, N100D и V132L;
(16) I61V, E77N, Q82S, K83R, E84H и V93A;
(17) I61V, V63I, E77V, K83R, E84D, D95E, L96T, K98R и N100E;
(18) I61L, V63I, E77V, K83R, D95E, L96S, K98R, N100D и G109R;
(19) I61V, E77L, Q82G, E84G, V93L, D95E, L96T, K98R и N100G; и
(20) I61L, V63I, E77N, Q82G и E84G.
[0081] В настоящей заявке, варианты домена SIRPα, соответственно содержащего одну группу аминокислотных замен из вышеуказанных от (1) до (20) на основе укороченного домена человеческого SIRPα варианта 1 (аминокислотная последовательность, изложенная в SEQ ID NO: 63, то есть остатки в положениях 33-149 аминокислотной последовательности, изложенной в SEQ ID NO: 62), могут быть последовательно названы M1, M5, M12, M35, M37, M41, M57, M67, M81, M82, M84, M91, M99, M102, M111, M122, M126, M130, M135 и M145. Эти мутанты могут последовательно содержать аминокислотные последовательности, показанные в одной из SEQ ID NO: с 1 до 20.
[0082] В настоящей заявке, слитый белок, содержащий укороченный домен человеческого SIRPα варианта 1 (аминокислотная последовательность, изложенная в SEQ ID NO: 63, а именно, остатки в положениях 33-149 аминокислотной последовательности, изложенной в SEQ ID NO: 62) и Fc человеческого IgG1 (аминокислотная последовательность, изложенная в SEQ ID NO: 67), может быть последовательно назван SS002, который содержит аминокислотную последовательность, изложенную в SEQ ID NO: 61.
[0083] В настоящей заявке термин «Fc-область иммуноглобулина» в основном относится к основной области Y-образной структуры из структуры антитела, которую также называют областью кристаллизующегося фрагмента (Fc-областью). В изотипах антитела IgG, IgA и IgD Fc-область может состоять из двух идентичных белковых фрагментов, полученных из второго и третьего константных доменов двух тяжелых цепей антитела. Fc-области IgM и IgE могут содержать три константных домена тяжелой цепи в каждой полипептидной цепи. Fc-область IgG имеет высококонсервативный сайт N-гликозилирования. В некоторых вариантах осуществления указанный иммуноглобулин Fc-область может содержать Fc-область IgG. В некоторых вариантах осуществления указанная Fc-область иммуноглобулина может включать области CH2 и CH3 константной области тяжелой цепи. В некоторых вариантах осуществления указанная Fc-область иммуноглобулина может содержать шарнирную область. Например, указанная Fc-область иммуноглобулина может содержать аминокислотную последовательность, выбранную из любой из следующих последовательностей: SEQ ID NO: 67-68.
[0084] В настоящей заявке термин «IgG» в основном относится к иммуноглобулину G (Иммуноглобулин G). IgG является одним из иммуноглобулинов человека. По разнице в антигенности гамма цепи в молекулах IgG, IgG человека имеет четыре подтипа: IgG1, IgG2, IgG3 и IgG4. В настоящей заявке термин «IgG1» в основном относится к одному подтипу с наибольшей долей IgG, который имеет относительно высокую аффинность с Fc-рецептором. Например, указанный IgG может быть человеческим IgG. Альтернативно, например, указанный IgG может быть выбранн из группы, состоящей из IgG1 и/или IgG4.
[0085] В настоящей заявке, указанный слитый белок содержит домен человеческого SIRPα, который может специфически связываться с белком CD47 и Fc-областью иммуноглобулина, где человеческий домен SIRPα может быть прямо или опосредованно связан с Fc-областью иммуноглобулина. Например, домен человека SIRPα может располагаться у N-конца Fc-области иммуноглобулина. Например, указанный C-конец домена человеческого SIRPα может быть прямо или опосредованно связан с N-концом Fc иммуноглобулина. Например, указанный домен человеческого SIRPα может быть связан с указанной Fc иммуноглобулина через линкер. В настоящей заявке, указанный линкер может быть пептидным линкером.
[0086] В настоящей заявке, слитый белок по настоящей заявке, соответственно содержащий одну группу из аминокислотных замен из вышеизложенных (1)-(20) на основе указанного SS002, соответственно может быть последовательно назван SS002M1, SS002M5, SS002M12, SS002M35, SS002M37, SS002M41, SS002M57, SS002M67, SS002M81, SS002M82, SS002M84, SS002M91, SS002M99, SS002M102, SS002M111, SS002M122, SS002M126, SS002M130, SS002M135 и SS002M145. Эти слитые белки могут последовательно включать аминокислотную последовательность, изложенную в SEQ ID NO: 40.
[0087] В настоящей заявке, слитый белок по настоящей заявке, соответственно содержащий одну группу из аминокислотных замен из вышеизложенных (1)-(20) и Fc человеческого IgG4 (аминокислотная последовательность, изложенная в SEQ ID NO: 68) на основе усеченного домена человеческого SIRPα вариант 1 (содержащего аминокислотную последовательность из SEQ ID NO: 63, то есть, остатки в положениях 33-149 в аминокислотной последовательности, изложенной в SEQ ID NO: 62), может быть последовательно назван SS002M1G4, SS002M5G4, SS002M12G4, SS002M35G4, SS002M37G4, SS002M41G4, SS002M57G4, SS002M67G4, SS002M81G4, SS002M82G4, SS002M84G4, SS002M91G4, SS002M99G4, SS002M102G4, SS002M111G4, SS002M122G4, SS002M126G4, SS002M130G4, SS002M135G4 и SS002M145G4. Эти слитые белки могут последовательно включать аминокислотную последовательность, изложенную в одной из SEQ ID NO: с 41 по SEQ ID NO: 60.
[0088] В некоторых вариантах осуществления слитый белок по настоящей заявке может содержать аминокислотную последовательность, указанную в одной из SEQ ID NO: 21-SEQ ID NO: 61.
[0089] Белок, полипептид и/или полипептид, включенный в настоящую заявку, также можно понимать как охватывающий, по меньшей мере, следующий объем: варианты или гомологи с такой же или сходной функцией, как и указанный белок или полипептид.
[0090] В настоящей заявке, указанный варианты могут быть белком или полипептидами, образованными в результате замены, делеции или добавления одной или более аминокислот по сравнению с аминокислотной последовательностью указанного белка и указанного его полипептида (например, домена человеческого SIRPα, его фрагмента, или его варианта, который содержит одну или более замен аминокислот; или указанного слитого белка). Например, указанный функциональный вариант может включать белки или полипептиды с аминокислотными изменениями путем замены, или вставки, по меньшей мере, одной аминокислоты, например, 1-30, 1-20 или 1-10, альтернативно, например, 1, 2, 3, 4 или 5 замен, делеций и/или вставок. Указанный функциональный вариант может по существу сохранять биологические характеристики указанного белка или указанного полипептида до изменения (например, замены, делеции или вставки). Например, указанный функциональный вариант может сохранять, по меньшей мере, 60%, 70%, 80%, 90% или 100% биологической активности (например, способности специфически связываться с белком CD47) от активности указанного белка или указанного полипептида до изменения.
[0091] В настоящей заявке, указанный гомолог может быть белком или полипептидом, который имеет, по меньшей мере, приблизительно 80% (например, по меньшей мере, приблизительно 85%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или больше) гомологии последовательности с аминокислотной последовательностью указанного белка и/или указанного полипептида (например, домена человеческого SIRPα, его фрагмента, или его варианта, который содержит одну или более замен аминокислот; или указанного слитого белка).
[0092] В настоящей заявке, указанная гомология, в основном, относится к подобию, аналогии или ассоциации между двумя или более последовательностями. «Процент гомологии последовательностей» можно рассчитать путем сравнения двух последовательностей, которые должны быть выровнены в окне сравнения, чтобы определить количество положений, в которых присутствуют одно и то же основание нуклеиновой кислоты (например, A, T, C, G, I) или одинаковый аминокислотный остаток (например, Ala, Pro, Ser, Thr, Gly, Val, Leu, Ile, Phe, Tyr, Trp, Lys, Arg, His, Asp, Glu, Asn, Gln, Cys и Met), и, таким образом, получить число совпадающих положений, а затем разделить количество совпадающих позиций на общее количество положений в окне сравнения (т.е. размер окна) и умножить результат на 100, чтобы получить процент гомологии последовательностей. Выравнивание для определения процента гомологии последовательностей можно проводить рядом способов, известных в данной области, например, с использованием общедоступных компьютерных программ, таких как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить подходящие параметры для выравнивания последовательностей, включая любой алгоритм, необходимый для достижения максимального выравнивания в пределах сравниваемой полноразмерной последовательности или в пределах области целевой последовательности. Указанную гомологию можно также определить следующими способами: FASTA и BLAST. Алгоритм FASTA описан, например, в W. R. Pearson and D. J. Lipman's "Improved Tool for Biological Sequence Comparison", Proc. Natl. Acad. Sci., 85: 2444-2448, 1988; и D, J. Lipman and W. R. Pearson’s "Fast and Sensitive Protein Similarity Search", Science, 227:1435-1441, 1989. Для описания алгоритма BLAST, пожалуйста, обращайтесь к S. Altschul, W. Gish, W. Miller, E. W. Myers and D. Lipman, "A Basic Local Alignment Search Tool", Journal of Molecular Biology, 215: 403-410, 1990.
МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, ВЕКТОР, КЛЕТКА-ХОЗЯИН
[0093] В другом аспекте настоящая заявка относится к одной или более молекулам нуклеиновой кислоты, способным кодировать слитый белок по настоящей заявке.
[0094] В некоторых вариантах осуществления указанная молекула нуклеиновой кислоты может полностью кодировать слитый белок по настоящей заявке. Например, указанный слитый белок можно получать при использовании только одного типа молекулы нуклеиновой кислоты. В некоторых вариантах осуществления указанная молекула нуклеиновой кислоты может кодировать часть слитого белка по настоящей заявке. Например, указанный слитый белок можно получать при использовании более чем двух типов различных указанных молекул нуклеиновой кислоты. Например, указанная молекула нуклеиновой кислоты может кодировать указанные домены человеческого SIRPα слитого белка по настоящей заявке (например, указанный внеклеточный домен человеческого SIRPα, его фрагмент, или его вариант, который содержит одну или более замен аминокислот, указанный домен IgV человеческого SIRPα, его фрагмент, или его вариант, который содержит одну или более замен аминокислот, указанный домен человеческого SIRPα варианта 1, его фрагмент, или его вариант, который содержит одну или более замен аминокислот). Альтернативно, например, указанная молекула нуклеиновой кислоты может кодировать Fc-область иммуноглобулина из слитого белка.
[0095] В другом аспекте настоящая заявка относится к одному или более векторам, которые могут содержать одну или более молекул нуклеиновой кислоты по настоящей заявке. В другом аспекте, настоящая заявка относится к клетке (например, клетке-хозяину), которая может содержать молекулу нуклеиновой кислоты по настоящей заявке или вектор по настоящей заявке.
[0096] В настоящей заявке, термин «молекула нуклеиновой кислоты» в основном относится к выделенной форме нуклеотида, дезоксирибонуклеотида или рибонуклеотида или их аналогам любой длины, выделенным из их природного окружения или искусственно синтезированным. Молекулы нуклеиновой кислоты по настоящей заявке можно выделять. Например, их можно получать или синтезировать следующими способами: (i) амплификация in vitro, такая как амплификация путем полимеразной цепной реакции (ПЦР), (ii) клональная рекомбинация, (iii) очистка, например, фракционирование путем расщепления рестрикционными ферментами и электрофореза в геле, или (iv) синтез, например, химический синтез. В некоторых вариантах осуществления указанная выделенная нуклеиновая кислота представляет собой молекулу нуклеиновой кислоты, полученную при помощи технологии рекомбинантных ДНК. В настоящей заявке, нуклеиновую кислоту, кодирующую указанное антитело или его антигенсвязывающий фрагмент, можно получать рядом способов, известных в данной области. Эти способы в качестве неограничивающих примеров включают ПЦР с достройкой перекрывающихся участков с использованием манипуляций с рестрикционными фрагментами или синтетическими олигонуклеотидами. Конкретные манипуляции можно найти в Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989; и Ausube et al. Current Protocols in Molecular Biology, Greene Publishing and Wiley-Interscience, New York NY, 1993.
[0097] В настоящей заявке термин «вектор» в основном относится к нуклеиновой молекуле, способной к саморепликации в подходящем хозяине, которая переносит вставленную молекулу нуклеиновой кислоты в клетку-хозяина и/или между клетками-хозяевами. Указанный вектор может включать вектор, который в основном используется для вставки ДНК или РНК в клетки, вектор, который в основном используется для репликации ДНК или РНК, и вектор, который в основном используется для транскрипции ДНК или РНК и/или экспрессии трансляции. Указанный вектор также включает вектор с несколькими функциями, описанными выше. Указанный вектор может быть полинуклеотидом, который может быть расшифрован и переведен в полипептид при введении в подходящую клетку-хозяина. В основном, при культивировании подходящей клетки-хозяина, содержащей указанный вектор, указанный вектор может производить желаемый продукт экспрессии. В настоящей заявке, указанный вектор может включать одну или более указанных молекул нуклеиновой кислоты. Например, указанный вектор может содержать все молекулы нуклеиновой кислоты, необходимые для кодирования указанного слитого белка. В этом случае для получения слитого белка по настоящей заявке требуется только один вектор. В некоторых вариантах осуществления указанный вектор может содержать молекулу нуклеиновой кислоты, кодирующую часть указанного слитого белка, например, молекулу нуклеиновой кислоты, кодирующую домен человеческого SIRPα в слитом белке по настоящей заявке. Альтернативно, указанный вектор может содержать, например, молекулу нуклеиновой кислоты, кодирующую указанную Fc-область иммуноглобулина из указанного слитого белка. В настоящее время для получения слитого белка по настоящей заявке необходимы два или более различных векторов.
[0098] Кроме того, указанный вектор может также содержать другие гены, такие как маркерный ген, что позволяет проводить селекцию вектора в подходящей клетке-хозяине и в подходящем состоянии. Кроме того, указанный вектор может также включать элемент контроля экспрессии, который позволяет кодирующей области правильно экспрессироваться в подходящем хозяине. Такой элемент контроля хорошо известен специалистам в данной области. Например, они могут содержать промоторы, рибосомные участки связывания, энхансеры и другие элементы контроля, регулирующие транскрипцию гена или трансляцию мРНК. В некоторых вариантах осуществления указанная последовательность контроля экспрессии представляет собой регуляторный элемент. Конкретная структура указанной последовательности контроля экспрессии может варьироваться в зависимости от функции видов или типов клеток, но, как правило, содержит 5'-нетранскрибируемые последовательности и 5'- и 3'-нетранслируемые последовательности, участвующих в инициации транскрипции и трансляции, такие как TATA-боксы, последовательности для кэпирования, последовательности CAAT и т.д. Например, 5'-нетранскрибируемая последовательность для контроля экспрессии может содержать промоторную область, а промоторная область может содержать промоторную последовательность для транскрипционного контроля функционально связанной нуклеиновой кислоты. В настоящей заявке, указанный вектор может быть вектором pTM.
[0099] В настоящей заявке термины «клетка-хозяин», «клетка» и «хозяин» используют взаимозаменяемо, и в основном они относятся к плазмиде или векторам, которые могут включать или включают молекулу нуклеиновой кислоты по настоящей заявке, или могут относиться к отдельным клеткам, клеточным линиям или клеточным культурам со слитым белком по настоящей заявке, его фрагментами или его вариантами Указанная клетка-хозяин может включать потомство одной клетки-хозяина. Из-за естественных, случайных или преднамеренных мутаций клетки-потомки и исходные родительские клетки не обязательно могут быть полностью идентичными по морфологии или геному при условии, что они могут экспрессировать антитела по настоящей заявке или их антигенсвязывающие фрагменты. Указанную клетку-хозяина можно получить путем трансфекции клетки in vitro вектором по настоящей заявке. Указанная клетка-хозяин может быть прокариотической клеткой (например, Escherichia coli) или эукариотической клеткой (например, дрожжевые клетки, например, клетки COS, клетки китайского хомяка (CHO), клетки HeLa, клетки HEK293, клетки COS-1, клетки NS0 или миеломные клетки). В настоящей заявке, указанная клетка-хозяин может быть клеткой CHO.
КОМПОЗИЦИЯ, СПОСОБ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ
[00100] В другом аспекте настоящая заявка относится к способу получения указанного слитого белка, включающему культивирование клетки-хозяина в условиях, позволяющих экспрессировать указанный слитый белок.
[00101] В другом аспекте настоящая заявка относится к композиции, содержащей указанный слитый белок, указанную молекулу нуклеиновой кислоты, указанный вектор и/или указанную клетку-хозяина, и необязательно фармацевтически приемлемый адъювант.
[00102] В настоящей заявке, термин «фармацевтически приемлемый адъювант» может включать буферы, антиоксиданты, консерванты, низкомолекулярные полипептиды, белки, гидрофильные полимеры, аминокислоты, сахара, хелатирующие средства, противоионы, комплексные соединения с металлами и/или неионные поверхностно-активные вещества, и т.д.
[00103] Указанный фармацевтически приемлемый может включать буферы, антиоксиданты, консерванты, низкомолекулярные полипептиды, белки, гидрофильные полимеры, аминокислоты, сахара, хелатирующие средства, противоионы, комплексные соединения с металлами и/или неионные поверхностно-активные вещества, и т.д.
[00104] В настоящей заявке, указанную фармацевтическую композицию можно формулировать с фармацевтически приемлемым носителем или разбавителем и любыми другими известными адъювантами и эксципиентами в соответствии с общепринятыми в данной области техническими способами, например, следуя способам в Remington: The Science и Practice of Pharmacy, девятнадцатое издание, издано Gennaro, Mack Publishing Co., Easton, PA, 1995.
[00105] В настоящей заявке, указанную композицию можно формулировать для перорального введения, внутривенного введения, внутримышечного введения, введения in situ в опухолевый участок, ингаляции, ректального введения, вагинального введения, трансдермального введения или лекарственное средство вводят через подкожное депо.
[00106] В настоящей заявке, указанную композицию можно использовать для ингибирования роста опухоли. Например, композиция по настоящей заявке может ингибировать или задерживать развитие или прогрессирование заболеваний (например, опухолей или аутоиммунных заболеваний) (например, уменьшать размер опухоли или даже по существу устранять опухоли) и/или может уменьшать и/или стабилизировать состояние заболевания.
[00107] Фармацевтическая композиция по настоящей заявке может содержать терапевтически эффективное количество указанного слитого белка. Указанное терапевтически эффективное количество представляет собой дозу, необходимую для предотвращения и/или лечения (по меньшей мере частично) заболеваний (например, опухолей или аутоиммунных заболеваний) и/или любых их осложнений у индивидуума с заболеванием или с риском возникновения заболевания.
[00108] В другом аспекте настоящая заявка относится к применению слитого белка, молекулы нуклеиновой кислоты, вектора, клетки-хозяина и/или композиции по настоящей заявке для получения лекарственного средства и/или набора для применения для профилактики и/или лечения опухолей или аутоиммунных заболеваний.
[00109] В настоящей заявке термин «опухоль», как правило, относится к новообразованиям, образующимся в результате пролиферации клеток местных тканей организма. Поскольку такие новообразования в основном представлены в виде объемных выпуклостей, их также называют неоплазиями. В соответствии с клеточными характеристиками новообразований и степенью повреждения организма, опухоли делятся на два класса, то есть доброкачественные опухоли и злокачественные опухоли. Рак - это общий термин для злокачественных опухолей. Опухоли по настоящей заявке можно выбирать из группы CD47-положительных гематологических опухолей и/или CD47-положительных солидных опухолей.
[00110] В настоящей заявке термин «CD47-положительная гематологическая опухоль» в основном относится к гематологическим опухолям, сверхэкспрессирующим CD47, которые могут включать ряд лейкозов, лимфом и миелом. Указанный «лейкоз» в основном относится к злокачественной опухоли крови, при которой вырабатывается слишком много лейкоцитов, неэффективных в борьбе с инфекцией и вытесняющих другие части крови, такие как тромбоциты и эритроциты. Лейкозы можно разделить на острые и хронические лейкозы. Некоторые формы лейкоза могут представлять собой, например, острый лимфоцитарный лейкоз (ОЛЛ), острый миелолейкоз (ОМЛ), хронический лимфоцитарный лейкоз (ХЛЛ), хронический миелогенный лейкоз (ХМЛ), миелопролиферативное нарушение/опухоль (MPDS) и миелодиспластический синдром. Указанная «лимфома» может относиться к лимфоме Ходжкина, вялотекущим и агрессивным неходжкинским лимфомам, лимфоме Беркитта, фолликулярной лимфоме (мелкоклеточной и крупноклеточной) и т.д. Указанная миелома может относиться к множественной миеломе (ММ), гигантоклеточной миеломе, миеломе тяжелых цепей, миеломе легких цепей или миеломе Бенс-Джонса.
[00111] В настоящей заявке термин «CD47-положительная солидная опухоль» в основном относится к солидной опухоли или видимой опухоли, которая сверхэкспрессирует CD47, что может быть обнаружено клиническим обследованием, таким как рентгеновский снимок, сканирование при помощи компьютерной томографии, B-ультразвук или пальпация. Основные категории могут включать карциномы и саркомы. Например, указанные CD47-положительные солидные опухоли могут включать саркому Юинга, остеосаркому, рабдомиосаркому, рак мочевого пузыря, рак яичников, рак предстательной железы, рак легких, рак толстой кишки, рак молочной железы, рак поджелудочной железы, астроцитарную карциному, глиобластому, карциному почечных клеток и т.д.
[00112] В настоящей заявке, указанные аутоиммунные заболевания могут включать болезнь Крона, аллергическую астму и ревматоидный артрит.
[00113] В настоящей заявке, термин "болезнь Крона" в основном относится к воспалительному заболеванию тонкого кишечника неизвестного генеза, которое может возникать в любой части желудочно-кишечного тракта. Указанная болезнь Крона и хронический неспецифический язвенный колит вместе обозначают как воспалительное заболевание кишечника (IBD).
[00114] В настоящей заявке, термин «аллергическая астма» в основном относится к хроническому воспалению дыхательных путей, в которое вовлечен ряд клеток, в частности, тучные клетки, эозинофилы и T-лимфоциты.
[00115] В настоящей заявке, термин «ревматоидный артрит» в основном относится к хроническому системному аутоиммунному заболеванию, в котором преобладает поражение суставов.
[00116] В другом аспекте слитый белок, молекулу нуклеиновой кислоты, вектор, клетку-хозяина и/или композицию по настоящей заявке можно использовать для профилактики или лечения указанных опухолей или указанных аутоиммунных заболеваний.
[00117] В другом аспекте настоящая заявка относится к способу для профилактики или лечения опухолей или аутоиммунных заболеваний, включающему введение слитого белка, молекулы нуклеиновой кислоты, вектора, клетки-хозяина и/или композиции по настоящей заявке индивидууму.
[00118] В другом аспекте настоящая заявка относится к способу блокирования взаимодействия белка CD47 и SIRPα, включающему введение слитого белка или композиции по настоящей заявке (например, введение индивидууму или клетке или биологическому образцу, которым это необходимо).
[00119] В другом аспекте настоящая заявка относится к способу ингибирования роста и/или пролиферации опухолей или опухолевых клеток, включающему контакт слитого белка или композиции по настоящей заявке с опухолями или опухолевыми клетками. Например, указанный контакт может происходить in vitro.
[00120] В настоящей заявке, термин «индивидуум» в основном относится к любому человеку или не являющемуся человеком животному. Термин «не являющееся человеком животное» может включать всех позвоночных, таких как млекопитающие и не-млекопитающие, например, не являющиеся человеком приматы, козы, овцы, собаки, коровы, курицы, амфибии, пресмыкающиеся, и т.д.
[00121] В настоящей заявке термин «приблизительно» в основном относится к отклонениям в пределах 0,5-10% от указанного значения, например, в пределах 0,5%, 1%, 1,5%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5%, 5%, 5,5%, 6%, 6,5%, 7%, 7,5%, 8%, 8,5%, 9%, 9,5% или 10% от указанного значения.
[00122] В настоящей заявке, термин «содержащий», как правило, означает «включающий», «состоящий», «имеющий» или «охватывающий». В некоторых случаях, он также относится к значению «являющийся» или «состоящий из».
[00123] Без ограничения какой-либо теорией, следующие примеры приведены только с целью иллюстрации режимов работы устройства, способа и системы по настоящей заявке, а не для ограничения объема изобретения, заявленного в настоящей заявке.
Примеры
Пример 1: Скрининг вариантов
[00124] Брали укороченный домен человеческого SIRPα, вариант 1 (NP_542970.1) с аминокислотной последовательностью, изложенной в SEQ ID NO: 63 (т.е. остатки в положениях 33-149 в SEQ ID NO: 62) использовали программное обеспечение Discovery Studio (Neotrident) для конструирования структуры в укороченном домене, который взаимодействует с человеческим CD47 (CEJ95640.1). Участки взаимодействия в двух белках и способы их взаимодействия были теоретически проанализированы, чтобы определить, что сайты аминокислот в укороченных доменах, которые прямо или опосредованно участвовали во взаимодействии с CD47, были I61, V63, E77, Q82, K83, E84, V93, D95, D96 , K98, N100, R107, G109 и V132 (в которых положения аминокислотных остатков в в аминокислотных заменах были пронумерованы с использованием аминокислотной последовательности, изложенной в SEQ ID NO: 62 в качестве эталонной). Эти сайты действия были случайным образом мутированы, и конструировали библиотеку мутантов. Затем библиотеку мутантов клонировали в вектор pTM. Указанный вектор pTM содержал последовательности сигнального пептида и трансмембранной области (как показано на ФИГ. 1), которая могла отображать ген, клонированный в векторе, на клеточной поверхности.
[00125] Экспрессирующим вектором со сконструированной библиотеой мутантов трансфицировали клетки СНО (АТСС), чтобы отображать и экспрессировать библиотеку мутантов на клеточной поверхности. Затем белок CD47 (Yiqiao Shenzhou) флуоресцентно метили FITC для получения CD47-FITC. В соответствии с различием активности связывания между CD47-FITC и мутантами укороченного домена на поверхности клетки CHO, мутанты, связывающиеся с CD47-FITC, были обогащены и исследованы с помощью проточной цитометрии. Конкретный принцип скрининга можно видеть на ФИГ. 2, где укороченный домен и мутант связаны с белком CD47 с флуоресцентной молекулой, таким образом, результаты связывания могут быть отражены уровнем флуоресцентной молекулы.
[00126] После четырех раундов скрининга и обогащения собирали клетки, которые прочно связывались с CD47-FITC (как показано на ФИГ. 3). Затем выделяли мРНК и проводили обратную транскрипцию с получением кДНК, и секвенировали мутантный усеченный домен (как показано на ФИГ. 4). Результаты секвенирования показали, что указанные положения I61, V63, E77, Q82, K83, E84, V93, D95, D96, K98, N100, R107, G109, V132 имеют разные комбинации сайтов мутации.
[00127] Из результатов видно, что мутант усеченного домена человеческого SIRPα, вариант 1, который может специфически распознавать CD47, можно получить путем введения различных комбинаций сайтов мутации по остаткам I61, V63, E77, Q82, K83, E84, V93, D95, D96. , K98, N100, R107, G109 и/или V132.
[00128] Дальнейший анализ дал возможные типы мутированных аминокислот в указанных сайтах мутации: I61L/V/F, V63I, E77I/N/Q/K/H/M/R/N/V/L, Q82S/R/G/N, K83R, E84K/H/D/R/G, V93L/A, D95H/R/E, D96S/T, K98R, N100G/K/D/E, R107N/S, G109R/H, V132 L/R/I/S.
[00129] Мутанты, содержащие эти мутации, были названы M1, M5, M12, M35, M37, M41, M57, M67, M81, M82, M84, M91, M99, M102, M111, M122, M126, M130, M135 и M145, и содержали следующие аминокислотные последовательности, последовательно указанные в одной из SEQ ID NO: 1-20, соответственно. В то же время эти мутанты включали последовательно следующие комбинации аминокислотных мутаций на основе аминокислотной последовательности, изложенной в SEQ ID NO: 63:
(1) I61L, V63I, E77I, E84K, V93L, L96S, K98R, N100G и V132 L;
(2) I61V, E77N, Q82S, K83R и E84H;
(3) I61F, V63I, K83R, E84K и V132I;
(4) I61L, E77Q, E84D, R107N и V132I;
(5) I61L, V63I, E77K, K83R, E84D и N100G;
(6) I61V, E77H, Q82R, K83R, E84H и R107S;
(7) I61L, E77I, Q82G, E84R, V93L, L96T, N100G, R107S, G109R и V132R;
(8) I61L, E77M, Q82G, K83R, E84D и V132L;
(9) I61L;
(10) I61F, D95H, L96S, G109H и V132S;
(11) I61F, D95H, L96S, K98R, G109H и V132S;
(12) I61L, E77Q, E84D, V93A, R107N и V132I;
(13) E77K, L96S, N100K, G109H и V132L;
(14) I61L, V63I, Q82G, E84G, D95R, L96S, N100D и V132I;
(15) I61L, E77R, Q82N, K83R, E84G, V93L, D95E, L96T, K98R, N100D и V132L;
(16) I61V, E77N, Q82S, K83R, E84H и V93A;
(17) I61V, V63I, E77V, K83R, E84D, D95E, L96T, K98R и N100E;
(18) I61L, V63I, E77V, K83R, D95E, L96S, K98R, N100D и G109R;
(19) I61V, E77L, Q82G, E84G, V93L, D95E, L96T, K98R и N100G; и (20) I61L, V63I, E77N, Q82G и E84G.
Пример 2: Измерение активности связывания слитого белка
[00130] Укороченный домен человеческого SIRPα, вариант 1 (или укороченный домен так называемого дикого типа SIRPα) в Примере 1 и мутант домена человеческого SIRPα, полученный в Примере 1 (т.е. M1, M5, M12, M35, M37, M41, M57 , M67, M81, M82, M84, M91, M99, M102, M111, M122, M126, M130, M135 и M145) были слиты и экспрессированы с человеческой IgG1-Fc (аминокислотная последовательность которой изложена в SEQ ID NO: 67) соответственно, чтобы получить соответствующий слитый белок из мутанта 1 SIRPα, состоящий из укороченного домена-человеческой Fc (в кратком изложении, слитый белок). Эти слитые белки были названы SS002, SS002M1, SS002M5, SS002M12, SS002M35, SS002M37, SS002M41, SS002M57, SS002M67, SS002M81, SS002M82, SS002M84, SS002M91, SS002M99, SS002M102, SS002M111, SS002M122, SS002M126, SS002M130, SS002M135 и SS002M145, соответственно. Эти слитые белки состояли из аминокислотной последовательности, указанной в одной из SEQ ID NO: 61 и 21-40, соответственно.
[00131] Для анализа биологической активности в качестве примера выбирали слитые белки SS002, SS002M12, SS002M5, SS002M82, SS002M84, SS002M91, SS002M102 и SS002M130.
[00132] Аффинность различных слитых белков, включая SS002, SS002M5, SS002M12, SS002M82, SS002M84, SS002M91, SS002M102, SS002M130 и подобных к CD47 определяли при помощи способа ELISA.
[00133] Планшет для ELISA покрывали 1 г/мл целевого антигена CD47-His при 4°C в течение ночи. После промывания PBST добавляли 10% эмбриональную телячью сыворотку, и смесь блокировали при 37°C в течение 1 часа. Затем добавляли SS002, SS002M5, SS002M82, SS002M84, SS002M91, SS002M102, и SS002M130 соответственно и проводили реакцию при 37°C в течение 1 часа. Затем реакционную смесь отмывали с PBST, и добавляли меченное пероксидазой хрена козлиное антитело к человеческому IgG (Thermo Fisher Scientific) и проводили реакцию при комнатной температуре в течение 30 минут. Затем планшет повторно промывали 5 раз PBST и сушили для удаления остатков жидкости, по возможности, промокательной бумагой. Затем в каждую лунку добавляли 100 мл TMB (eBioscience) и выдерживали при температуре (20±5°C) в темноте в течение 1-5 минут. В каждую лунку добавляли 100 мл 2N H2SO4 для гашения реакции субстрата. Значение OD считывали на 450 нм с помощью микроспектрофотометра для чтения планшетов, и анализировали аффинность между каждым слитым белком и молекулой CD47 (как показано на ФИГ. 5).
[00134] Результаты на ФИГ. 5 показали, что слитые белки, включая SS002, SS002M5, SS002M12, SS002M82, SS002M84, SS002M91, SS002M102, SS002M130 и т.п. могут эффективно распознавать молекулу CD47.
Пример 3: Анализ аффинности
[00135] В качестве примера использовали способ с Biacore для измерения аффинности каждого слитого белка, включая SS002, SS002M5, SS002M12, SS002M82, SS002M84, SS002M91, SS002M102, SS002M130 и т.п. к молекуле CD47, и результаты показаны в таблице 2 ниже.
[00136] Результаты в таблице 2 показывают, что SS002, SS002M5, SS002M12, SS002M82, SS002M84, SS002M91, SS002M102, SS002M130 и другие слитые белки могут распознавать молекулы CD47 с высокой аффинностью.
Пример 4: Специфичность видового распознавания для слитого белка
[00137] Для проведения анализа специфической активности раcпознавания в качестве примера брали слитые белки SS002 и SS002M91.
[00138] Для проведения видового анализа слитых белков, покрывали планшеты для ELISA 1 мг/мл человеческого CD47 и мышиного CD47 (Beijing Yiqiao Shenzhou Biotechnology Co., Ltd.), соответственно, и выдерживали при 4°C в течение ночи. После промывания PBST добавляли 10% эмбриональную телячью сыворотку и блокировали смесь при 37°C в течение 1 часа. Затем добавляли SS002 и SS002M91, соответственно, и проводили реакцию при 37°C в течение 1 часа. После промывания PBST добавляли меченное пероксидазой хрена козлиное антитело к человеческому IgG (Thermo Fisher Scientific) и проводили реакцию при комнатной температуре в течение 30 минут. Затем планшет повторно промывали 5 раз PBST и сушили для удаления остаточных капель жидкости, по возможности, промокательной бумагой. Затем в каждую лунку добавляли 100 мл TMB (eBioscience) и выдерживали при комнатной температуре (20± 5°C) в темноте в течение 1-5 минут. В каждую лунку добавляли 100 мл 2N H2SO4 для гашения реакции субстрата. Значение OD считывали при 450 нм с помощью микроспектрофотометра для чтения планшетов, и анализировали способность связывания слитого белка с различными видами CD47 (экспериментальные результаты для слитого белка SS002 и SS002M91 показаны на фиг. 6A и 6B, соответственно).
[00139] Результаты на фиг. 6A-6B показали, что как SS002, так и SS002M91 могут специфически распознавать молекулы CD47 человека, но не могут распознавать молекулы CD47 мыши.
Пример 5: Слитые белки специфически распознают целевой антиген
[00140] Слитые белки SS002 и SS002M91 были взяты в качестве примера. Покрывали планшеты для ELISA 1 мг/мл SS002, SS002M91, а также молока (Beijing Bomed Biotechnology Co., Ltd.), BSA (BOVOGEN), CD19 (Beijing Yiqiao Shenzhou Biotechnology Co., Ltd.), TROP2 (Beijing Yiqiao Shenzhou Biotechnology Co., Ltd.), CD47 (Beijing Magppel Biotech Co., Ltd.), CD38 (Beijing Yiqiao Shenzhou Biotechnology Co., Ltd.), Gas6 (R&D) и других белков, и AXL (ACRO Biosystems), соответственно, и оставляли при 4°C в течение ночи. После промывания PBST добавляли 10% эмбриональную телячью сыворотку, и блокировали смесь при 37°C в течение 1 часа. Затем добавляли SS002 и SS002M91, соответственно, и проводили реакцию при 37°C в течение 1 часа. После промывания PBST добавляли меченное пероксидазой хрена козлиное антитело к человеческому IgG (Thermo Fisher Scientific) и проводили реакцию при комнатной температуре в течение 30 минут. Затем пластину повторно промывали 5 раз PBST и сушили для удаления остаточных капель жидкости, по возможности, промокательной бумагой. Затем в каждую лунку добавляли 100 мл TMB (eBioscience) и выдерживали при комнатной температуре (20±5°C) в темноте в течение 1-5 минут. В каждую лунку добавляли 100 мл 2N H2SO4 для гашения реакции субстрата. Величину OD считывали на 450 нм с помощью микроспектрофотометра для чтения планшетов и анализировали способность слитого белка связываться с различными вышеуказанными белками (как показано на ФИГ. 7).
[00141] Результаты на ФИГ. 7 показали, что слитые белки SS002 и SS002M91 легко распознают молекулу CD47 и не участвуют в перекрестной реакции с другими различными белками.
Пример 6: Слитые белки специфически блокируют взаимодействие CD47/SIRPα
[00142] В качестве примера для проведения анализа специфического блокирования активности взаимодействия CD47/SIRPα был использован SS002M91, а экспрессию американского аналога TTI-621 (см. CN105073780A) использовали в качестве положительного контроля.
[00143] Покрывали планшеты для ELISA 1 мг/мл SIRPα-His и оставляли при 4°C в течение ночи. После промывания PBST добавляли 10% эмбриональную телячью сыворотку, и блокировали смесь при 37°C в течение 1 часа. SS002M91 и TTI-621 серийно разбавляли 10% фетальной телячьей кровью, соответственно, и Биотин-Fc-CD47 добавляли в образцы до конечной концентрации 2 мкг/мл. Смесь предварительно инкубировали при 37°C в течение 30 минут для применения в качестве первичного антитела. После того, как планшет ELISA отмывали PBST, добавляли первичное антитело и инкубировали при 37°C в течение 1 часа. Затем после 5-кратной промывки PBST добавляли меченный пероксидазой хрена авидин (Стрептавидин-HRP, Jiaxuan Bio) и инкубировали при 37°C в течение 30 минут. После 5-кратной промывки PBST в каждую лунку добавляли 100 мкл TMB (eBioscience) и оставляли при комнатной температуре (20±5°C) в темноте в течение 1-5 минут. В каждую лунку добавляли 100 мл 2N H2SO4 для гашения реакции субстрата. Значение OD считывали при 450 нм с помощью микроспектрофотометра для чтения планшетов, и анализировали блокирующий эффект слитого белка SIRPα на CD47/SIRPα (как показано на ФИГ. 8).
[00144] Результаты на ФИГ. 8 показали, что как SS002M91, так и TTI-621 могут конкурентно блокировать связывание CD47 с его лигандом SIRPα. Однако слитый белок SS002M91 обладает значительно более высокой блокирующей активностью, чем TTI-621. Значение IC50 для SS002M91 составляет 5,47 мкг/мл, а значение IC50 для TTI-621 составляет 493,5 мкг/мл.
Пример 7: Слитый белок специфически распознает молекулу CD47 на поверхности опухолевых клеток
[00145] В качестве примеров для анализа активности распознавания молекулы CD47 на поверхности опухолевых клеток брали слитые белки SS002 и SS002M91.
[00146] Использовали проточную цитометрию (BD Calibur) для обнаружения соответствующим образом активности SS002 и SS002M91, которые специфически распознают молекулы CD47 на поверхности клеток Raji, клеток Jurkat и клеток A549. Собирали вышеуказанные клетки в логарифмической фазе роста, соответственно, с корректировкой клеточной плотности до 5×106 клеток/мл, и предварительно охлаждали на льду в течение 10 минут. Слитые белки SIRPα SS002 и SS002M91 разбавляли до различной концентрации предварительно охлажденным физиологическим раствором, содержащим 2% FBS. Добавляли 100 мкл клеток в равный объем вышеуказанного разбавленного слитого белка SIRPα, и проводили реакцию при 4°C в темноте в течение 30 минут. После завершения клетки отмывали дважды предварительно охлажденным физиологическим раствором, содержащим 2% FBS. Клетки ресуспендировали в 100 мкл разбавленного вторичного антитела козы, меченого PE, к человеческой IgG-Fc (eBioscience), и проводили реакцию при 4°C в темноте в течение 30 минут. После завершения реакции клетки отмывали дважды предварительно охлажденным физиологическим раствором, содержащим 2% FBS, и ресуспендировали в 400 мкл 1% параформальдегида. Использовали проточную цитометрию (BD Calibur) для анализа связывающей способности слитого белка с CD47 на поверхности клетки (как показано на фиг. 9A-9C, где слитые белки SS002 и SS002M91 специфически распознают поверхностный CD47 на клетках Raji, клетках Jurkat и клетках A549, соответственно).
[00147] Результаты показали, что слитый белок SS002M91 может специфически распознавать CD47 на поверхности клеток Raji, клеток Jurkat и клеток A549 с активностью распознавания, которая была значительно выше, чем у SS002, и зависела от дозы. Среди них значение связывания ЕС50 с клетками Raji составляло 197,0 нг/мл для SS002M91 и 1140,0 нг/мл для SS002 (как показано на ФИГ. 9A). Значение связывания ЕС50 с клетками Jurkat для SS002M91 составляло 796,0 нг/мл, а для SS002 составляло 4529,0 нг/мл (как показано на ФИГ. 9B). Значение связывания ЕС50 с клетками A549 для SS002M91 составляло 321,9 нг/мл, а для SS002 составляло 1655,0 нг/мл (как показано на ФИГ. 9C).
[00148] Аналогичным образом, способ из этого примера применяли для сравнения активности SS002M91 и TTI-621 по распознаванию молекул CD47 поверхностях клеток Raji (как показано на ФИГ. 10A), клеток Jurkat (как показано на ФИГ. 10B) и клеток A549 (как показано на ФИГ. 10С). Результаты показали, что в одном из аспектов, максимальная интенсивность флуоресценции для SS002M91, который специфически распознает молекулы CD47 на поверхности опухолевых клеток, была значительно выше, чем у TTI-621: максимальная интенсивность флуоресценции связывания SS002M91 с клетками Raji была приблизительно 1200, в то время как максимальная интенсивность флуоресценции связывания TTI-621 с клетками Raji была приблизительно 600; максимальная интенсивность флуоресценции связывания SS002M91 с клетками Jurkat была приблизительно 5000, в то время как максимальная интенсивность флуоресценции связывания TTI-621 с клетками Jurkat была приблизительно 3000; максимальная интенсивность флуоресценции связывания SS002M91 с клетками A549 была приблизительно 180, в то время как максимальная интенсивность флуоресценции связывания TTI-621 с клетками A549 была приблизительно 60. В другом аспекте полуоптимальная доза SS002M91, которая специфически распознает молекулы CD47 на поверхности опухолевых клеток, была значительно лучше, чем у TTI-621: значение ЕС50 для связывания SS002M91 с клетками Raji составляло 13,06 нг/мл, тогда как значение EC50 для связывания TTI-621 с клетками Raji составляло 40,37 нг/мл; значение ЕС50 для связывания SS002M91 с клетками Jurkat составляло 28,09 нг/мл, тогда как значение ЕС50 для связывания TTI-621 с клетками Jurkat было 53,92 нг/мл; значение ЕС50 для связывания SS002M91 с клетками A549 составляло 26,95 нг/мл, а значение ЕС50 для связывания TTI-621 с клетками A549 составляло 1003 нг/мл. Можно видеть, что SS002M91, который специфически распознает молекулы CD47 на поверхности опухолевых клеток, значительно лучше, чем TT1-621.
Пример 8: Детекция реакции свертывания крови
[00149] SS002 и SS002M91 были взяты в качестве примеров для проведения анализа свертывающей активности с использованием антитела Hu5F9-G4 к CD47 в качестве контроля (см. Guerriero JL, Sotayo A, Ponichtera HE, et al. Class IIa HDAC inhibition reduces breast tumors and metastases through anti-tumour macrophages. [J] Nature, 2017, 543(7645): 428.432 и Gholamin S, Mitra SS, Feroze AH et al. Disrupting the CD47-SIRPα antiphagocytic axis by a humanized anti-CD47 antibody is an efficacious treatment for malignant pediatric brain tumors. Sci. Transl. Med 2017).
[00150] Цельную кровь от здоровых доноров использовали для получения эритроцитов человека (собрана из периферической крови добровольцев). Цельную кровь разбавляли в 5 раз при помощи PBS, 3 раза промывали и готовили свежий 1% раствор эритроцитов. В каждую лунку планшета для гемагглютинации добавляли 50 мкл различной концентрации слитых белков SIRPα SS002, SS002M91 и антитела к CD47 Hu5F9-G4, в каждую лунку добавляли 50 мкл 1% раствора эритроцитов. Смесь осторожно перемешивали до однородности и инкубировали при 37°C в атмосфере 5% CO2 в течение ночи. Затем планшет фотографировали для интерпретации с использованием стандартов, согласно которым все эритроциты коагулировали, опускались на дно лунки и сплющивались в форме сетки при 100%-ной коагуляции (++++), и явление, когда эритроциты опускались на дно и выглядели как точки, принимали как отсутствие коагуляции (-) (как показано на ФИГ. 11).
[00151] Результаты на ФИГ. 11 показали, что слитые белки SS002M91 и SS002 не вызывают агглютинации эритроцитов, тогда как антитело CD47 Hu5F9-G4 может в значительной степени вызывать агглютинацию эритроцитов в определенном диапазоне доз.
Пример 9: Выявление опухольингибирующей активности in vivo
[00152] В качестве примера для анализа опухольингибирующей активности in vivo использовали слитый белок SS002M91.
[00153] Мышам B-NSG инокулировали клетки Raji-Luc для создания модели опухоли для оценки опухольингибирующей активности антитела SS002M91. Самок мышей B-NSG в возрасте 8 недель (Beijing Biocytogene Biotechnology Co., Ltd.) брали в качестве экспериментальных животных, а для тестирования выбирали клетки Raji-Luc (Beijing Biocytogene Biotechnology Co., Ltd.). Клетки Raji-Luc переносили в стабильную клеточную линию, полученную с репортерным геном флуоресцеина. После реанимации и культивирования до необходимого количества, клетки собирали в лог-фазе роста и суспендировали до 5×106 клеток/0,2 мл. Затем мышам B-NSG инокулировали клетки через хвостовую вену в концентрации 0,2 мл/мышь. После вакцинации рост опухоли и массу тела наблюдали на Сутки 0 и Сутки 3 с помощью визуализатора для мелких животных. И на Сутки 3 выбирали 12 мышей с умеренным сигналом визуализации опухоли (приблизительно 1,00×106 P/S) и случайным образом делили на 2 группы (по 6 мышей на группу), то есть контрольную группу с растворителем (G1, физиологический раствор) и экспериментальную группу (G2, SS002M91). Экспериментальной группе вводили дозу 10 мг/кг на Сутки 0 и D3 после распределения по группам, всего дважды. Наблюдали рост опухоли и коэффициент выживаемости мышей (как показано на РИС. 12A и 12B соответственно).
[00154] Результаты показали, что на Сутки 10 после распределения по группам средняя интенсивность флуоресценции опухолей в контрольной группе составляла 6,75×108 P/S, в то время как в группе дозирования средняя интенсивность флуоресценции опухолей составляла 1,76×106 P/S, а степень ингибирования приблизительно 95%.
Пример 10: Выявления воздействия на эритроциты и тромбоциты
[00155] Для проведения предварительной оценки безопасности in vivo в качестве примера использовали слитый белок SS002M91, а в качестве модели - мышей B-NSG.
[00156] Были выбраны восемнадцать самок 8-недельных мышей B-NSG (Beijing Biocytogene Biotechnology Co., Ltd.) и случайным образом разделены на 3 группы, то есть, контрольную группу с растворителем (вводили физиологический раствор), экспериментальную группу (вводили слитый белок SS002M91) и группу положительного контроля (вводили TTI-621) (6 мышей на группу). Мышам вводили дозу 10 мг/кг на Сутки 0, Сутки 3 и Сутки 7 после распределения по группам, всего 3 раза. На следующие сутки после третьего введения (т.е. Сутки 8) анализировали в периферической крови мышей уровни эритроцитов и тромбоцитов (уровень эритроцитов и уровень тромбоцитов показаны на ФИГ. 13A и ФИГ. 13B, соответственно).
[00157] Результаты показали, что по сравнению с контрольной группой SS002M91 не вызывает значимого снижения эритроцитов (P=0,4483) и тромбоцитов (P=0,9199); в то время как TTI-621 оказывал небольшое влияние на тромбоциты (P=0,9447), но вызывал снижение уровня эритроцитов (P=0,0246).
Пример 11: Эффект слияния с различными подтипами Fc IgG на активность слитого белка
[00158] В соответствии со способом конструирования слитого белка в Примере 2, мутанты M1, M5, M12, M35, M37, M41, M57, M67, M81, M82, M84, M91, M99, M102, M111, M122, M126, M130, M135 и M145 домена SIRPα, полученного в Примере 1, были слиты и экспрессированы с IgG4-Fc человека (аминокислотные последовательности которой были указаны в SEQ ID NO: 68), соответственно, для получения соответствующего слитого белка: укороченные домены SIRPα мутанта 1-человеческая Fc (именуемого слитым белком). Эти слитые белки были названы SS002M1G4, SS002M5G4, SS002M12G4, SS002M35G4, SS002M37G4, SS002M41G4, SS002M57G4, SS002M67G4, SS002M81G4, SS002M82G4, SS002M84G4, SS002M91G4, SS002M99G4, SS002M102G4, SS002M111G4, SS002M122G4, SS002M126G4, SS002M130G4, SS002M135G4 и SS002M145G4 (аминокислотные последовательности которых были изложены в SEQ ID NO: 41-60, соответственно).
[00159] Для анализа биологической активности в качестве примера выбирали слитый белок SS002M91G4.
[00160] Анализировали активность связывания SS002M91G4 с антигеном CD47 в соответствии со способом измерения активности связывания в Примере 2, и результаты показаны на ФИГ. 14. Результаты на ФИГ. 14 показали, что SS002M91G4 обладает хорошей активностью связывания с антигеном CD47 со значением EC50 0,0157 мкг/мл, что по существу согласуется с EC50 (0,0195 мкг/мл) для SS002M91.
[00161] В соответствии со способом анализа специфического блокирования взаимодействия CD47/SIRPα в Примере 6 анализировали активность SS002M91G4 по блокированию взаимодействия CD47 SIRPα, и результаты показаны на ФИГ. 15. Результаты на ФИГ. 15 показали, что SS002M91G4 обладает хорошей активностью по блокированию взаимодействия CD47/SIRPα со значением IC50 3,46 мкг/мл, что по существу согласуется со значением IC50 (5,47 мкг/мл) для SS002M91.
[00162] Из приведенных выше результатов видно, что слияние с различными подтипами Fc IgG не оказывает достоверного воздействия на активность слитого белка, сконструированного по настоящей заявке.
Пример 12: Выявление реакции свертывания крови для слитого белка
[00163] Оценивали реакцию свертывания крови для слитого белка на основе IgG4 Fc, используя в качестве примера SS002M91G4 и в соответствии со способом обнаружения реакции свертывания крови в Примере 8. Цельную кровь от здоровых доноров использовали для получения эритроцитов человека. Цельную кровь разбавляли в 5 раз при помощи PBS, 3 раза промывали и готовили свежий 1% раствор эритроцитов. В каждую лунку планшета для гемагглютинации добавляли 50 мкл различной концентрации слитых белков SS002M91G4, положительного контроля TTI-621 и антитела к CD47 Hu5F9-G4, в каждую лунку добавляли 50 мкл 1% раствора эритроцитов. Смесь осторожно перемешивали до однородности и инкубировали при 37°C в атмосфере 5% CO2 в течение ночи. Затем планшет фотографировали для интерпретации с использованием стандартов, согласно которым все эритроциты коагулировали, опускались на дно лунки и сплющивались в форме сетки при 100%-ной коагуляции (++++), и явление, когда эритроциты опускались на дно и выглядели как точки, принимали как отсутствие коагуляции (-).
[00164] Результаты показаны на ФИГ. 16. Результаты показали, что слитые белки SS002M91G4 и TTI-621 на основе Fc IgG4 не вызывают агглютинации эритроцитов, тогда как антитело CD47 Hu5F9-G4 достоверно вызывает агглютинацию эритроцитов в определенном диапазоне доз.
Пример 13: Анализ биологической активности слитого белка
[00165] Для оценки биологической активности указанного слитого белка на основе Fc IgG4 брали в качестве примера SS002M91G4, использовали клеточную систему Jurkat-CSR (Immune OncoBiomedical Technology (Shanghai) Co., Ltd.), и TTI-621 в качестве положительного контроля.
[00166] Белок CD47-Fc (Каталожный номер 12283-H02H, Sino Biological) разбавляли до 0,2 мкг/мл, и добавляли 50 мкл разбавителя в каждую лунку. Затем TTI-621 и SS002M91G4 разбавляли до 0,4 мг/мл, соответственно, а затем последовательно разбавляли до различной концентрации. Добавляли 50 мкл в каждую лунку и инкубировали совместно с CD47-Fc в инкубаторе при 37°C, 5% CO2 в течение 45 минут. Брали клетки Jurkat-CSR и корректировали плотность до 5×105/мл, в каждую лунку добавляли 100 мкл клеточной суспензии, и устанавливали контрольную «пустую» группу. Суспензию культивировали совместно со смесью белков в инкубаторе при 37°C, 5% CO2 в течение 20 часов. После завершения культивирования 20 мкл CCK-8 (Dojindo, Японский институт химии) добавляли в каждую лунку и культивировали в инкубаторе с температурой 37°C, 5% CO2 в течение 4 часов. Значение OD измеряли при длине волны 450 нм с микроспектрофотометром для чтения планшетов. Скорость ингибирования роста клеток рассчитывали в соответствии со следующей формулой: Процент ингибирования = (OD450 (образца)-OD450 (пустой)/(OD450 (Jurkat-CSR)-OD450 (пустой))×100.
[00167] Согласно принципу клеточной системы Jurkat-CSR, взаимодействие CD47/SIRPα может индуцировать апоптоз клетки-мишени, а добавление ингибитора, который ингибирует взаимодействие CD47/SIRPα, может блокировать сигнал апоптоза. Чем сильнее действие ингибитора, тем полнее блокируется апоптотический сигнал. Результат показан на ФИГ. 17. Результаты показали, что SS002M91G4 может значительно блокировать апоптоз клеток Jurkat-CSR, индуцированный CD47-Fc, блокирующий эффект сильнее, чем у TTI-621, и IC50 составляет примерно 1/100 от TTI-621. Можно видеть, что биологическая активность SS002M91G4 при блокировании взаимодействия CD47/SIRPα значительно лучше, чем у TTI-621.
Пример 14: Опухольингибирующая активность in vivo Fc слитого белка из различных подтипов IgG и воздействие на эритроциты и тромбоциты
[00168] В качестве примера брали слитые белки SS002M91 и SS002M91G4 и анализировали опухолевую активность и влияние на эритроциты и тромбоциты in vivo.
[00169] Мышам NOD/SCID подкожно инокулировали клетки Raji и создавали модель трансплантированной подкожной опухоли из человеческой лимфомы для оценки активности SS002M91 и SS002M91G4 по ингибированию опухоли in vivo.
[00170] Выбирали самок мышей NOD/SCID возрастом 6-7 недель (Shanghai Lingchang Biotechnology Co., Ltd.), и культивировали клетки Raji в среде RPMI1640, содержащей 10% эмбриональной телячьей сыворотки. Собирали 1×107 клетки Raji в фазе экспоненциального роста и ресуспендировали до подходящей концентрации в PBS, а затем смешивали с матригелем (BD Matrigel™) в соотношении 1:1 для подкожной инокуляции опухоли мышам. После инокуляции, когда средний объем опухолей составляет примерно 98,6 мм3, мыши были случайным образом разделены на 4 группы (по 6 мышей на группу) в соответствии с размером опухоли (то есть контрольная группа с растворителем, группа SS002M91, группа SS002M91G4 и группа TTI-621). Мышам делали интраперитонеальную инъекцию в дозе 10 мг/кг за раз (где вышеупомянутым четырем группам вводили раствор PBS, SS002M91, SS002M91G4 и TTI-621, соответственно) с частотой дважды в неделю в течение двух недель.
[00171] Эксперимент закончили на сутки 6 после последнего введения и взяли кровь на плановое тестирование. Анализировали уровни эритроцитов и тромбоцитов в периферической крови мышей (уровень эритроцитов и уровень тромбоцитов показаны на ФИГ. 18A и ФИГ. 18B, соответственно). Во время введения наблюдали за ростом опухоли. Терапевтический эффект оценивали в соответствии с относительным ингибированием роста опухоли (TGI) (результаты показаны на ФИГ. 19), и оценивали безопасность на основе изменения массы животных и смертности (результаты показаны на ФИГ. 20). Формула для расчета TGI (%), то есть относительного ингибирования роста опухоли, была следующей: TGI% = (1-T/C)×100%. Из них T и C представляют собой относительный объем опухоли (RTV) или массу опухоли (TW) групп лечения (например, группы SS002M91, группы SS002M91G4 и группы TTI-621) и контрольной группы (т.е. контрольной группы с растворителем) в определенный момент времени.
[00172] Результаты показали, что группа SS002M91, группа SS002M91G4 и группа TTI-621 (10 мг/кг) продемонстрировали значительный эффект ингибирования опухоли на Сутки 6 после отмены лекарственного средства и продемонстрировали относительное ингибирование роста опухоли TGI (%), которое составило 74,09%, 66,65% и 54,75%, соответственно. По сравнению с контрольной группой с растворителем есть статистически значимые отличия (все значения p менее 0,01). Группа SS002M91 и группа SS002M91G4 имеют сходное опухольингибирующее действие, лучшее, чем у группы положительного контроля TTI-621.
[00173] Во время лечения ни одно животное не погибло или не проявило явной лекарственной токсичности, и все они хорошо переносили лекарство. Результаты планового анализа крови показали, что по сравнению с группой положительного контроля TTI-621, эритроциты и тромбоциты группы SS002M91 были снижены, тромбоциты группы TTI-621 также немного уменьшились, но эритроциты по существу не пострадали от SS002M91G4.
[00174] Вышеуказанное подробное описание было предоставлено в качестве иллюстрации и примеров, и не предназначено для ограничения объема прилагаемой формулы изобретения. Сейчас различные варианты осуществления, перечисленные в настоящем документе, очевидны специалистам в данной области и остаются в объеме прилагаемой формулы изобретения и ее эквивалентных решений.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> HANGZHOU SUMGEN BIOTECH CO., LTD.; SUMGEN MAB (BEIJING)
BIOTECH CO., LTD.
<120> СВЯЗЫВАНИЕ СЛИТОГО БЕЛКА С БЕЛКОМ CD47 И ЕГО ПРИМЕНЕНИЕ
<130> 0070-PA-009
<160> 68
<170> PatentIn version 3.5
<210> 1
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M1
<400> 1
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gln Lys Lys Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Ser
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 2
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M5
<400> 2
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 3
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M12
<400> 3
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Arg Lys Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 4
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M35
<400> 4
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 5
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M37
<400> 5
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 6
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M41
<400> 6
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg His Leu Ile Tyr
35 40 45
Asn Arg Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Ser Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 7
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M57
<400> 7
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gly Lys Arg Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Thr
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Ser Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Arg Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 8
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M67
<400> 8
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Met Leu Ile Tyr
35 40 45
Asn Gly Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 9
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M81
<400> 9
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 10
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M82
<400> 10
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 11
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M84
<400> 11
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Arg Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 12
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M91
<400> 12
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 13
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99
<400> 13
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Ser
50 55 60
Thr Lys Arg Lys Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 14
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M102
<400> 14
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Arg Ser
50 55 60
Thr Lys Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 15
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M111
<400> 15
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Arg Leu Ile Tyr
35 40 45
Asn Asn Arg Gly Gly His Phe Pro Arg Val Thr Thr Leu Ser Glu Thr
50 55 60
Thr Arg Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 16
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M122
<400> 16
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 17
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M126
<400> 17
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Thr
50 55 60
Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 18
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M130
<400> 18
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Ser
50 55 60
Thr Arg Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 19
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M135
<400> 19
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Leu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Leu Ser Glu Thr
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 20
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> M145
<400> 20
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 21
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M1
<400> 21
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gln Lys Lys Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Ser
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 22
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M5
<400> 22
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 23
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M12
<400> 23
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Arg Lys Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 24
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M35
<400> 24
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 25
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M37
<400> 25
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 26
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M41
<400> 26
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg His Leu Ile Tyr
35 40 45
Asn Arg Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Ser Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 27
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M57
<400> 27
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gly Lys Arg Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Thr
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Ser Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Arg Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 28
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M67
<400> 28
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Met Leu Ile Tyr
35 40 45
Asn Gly Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 29
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M81
<400> 29
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 30
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M82
<400> 30
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 31
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M84
<400> 31
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Arg Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 32
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M91
<400> 32
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 33
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M99
<400> 33
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Ser
50 55 60
Thr Lys Arg Lys Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 34
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M102
<400> 34
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Arg Ser
50 55 60
Thr Lys Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 35
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M111
<400> 35
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Arg Leu Ile Tyr
35 40 45
Asn Asn Arg Gly Gly His Phe Pro Arg Val Thr Thr Leu Ser Glu Thr
50 55 60
Thr Arg Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 36
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M122
<400> 36
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 37
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M126
<400> 37
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Thr
50 55 60
Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 38
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M130
<400> 38
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Ser
50 55 60
Thr Arg Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 39
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M135
<400> 39
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Leu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Leu Ser Glu Thr
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 40
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M145
<400> 40
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 41
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M1G4
<400> 41
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gln Lys Lys Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Ser
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 42
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M5G4
<400> 42
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 43
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M12G4
<400> 43
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Arg Lys Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 44
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M35G4
<400> 44
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 45
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M37G4
<400> 45
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 46
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M41G4
<400> 46
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg His Leu Ile Tyr
35 40 45
Asn Arg Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Ser Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 47
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M57G4
<400> 47
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gly Lys Arg Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Thr
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Ser Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Arg Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 48
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M67G4
<400> 48
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Met Leu Ile Tyr
35 40 45
Asn Gly Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 49
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M81G4
<400> 49
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 50
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M82G4
<400> 50
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 51
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M84G4
<400> 51
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Arg Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 52
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M91G4
<400> 52
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 53
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M99G4
<400> 53
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Ser
50 55 60
Thr Lys Arg Lys Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 54
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M102G4
<400> 54
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Arg Ser
50 55 60
Thr Lys Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 55
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M111G4
<400> 55
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 56
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M122G4
<400> 56
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 57
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M126G4
<400> 57
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Thr
50 55 60
Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 58
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M130G4
<400> 58
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Ser
50 55 60
Thr Arg Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 59
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M135G4
<400> 59
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Leu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Leu Ser Glu Thr
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 60
<211> 346
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002M145G4
<400> 60
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
115 120 125
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
130 135 140
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
145 150 155 160
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
165 170 175
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
180 185 190
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
195 200 205
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
210 215 220
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
225 230 235 240
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
245 250 255
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
260 265 270
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
275 280 285
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
290 295 300
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn
305 310 315 320
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
325 330 335
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
340 345
<210> 61
<211> 350
<212> Белок
<213> Искусственная последовательность
<220>
<223> SS002
<400> 61
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<210> 62
<211> 504
<212> Белок
<213> Искусственная последовательность
<220>
<223> Человеческий SIRP ¦Б вариант 1
<400> 62
Met Glu Pro Ala Gly Pro Ala Pro Gly Arg Leu Gly Pro Leu Leu Cys
1 5 10 15
Leu Leu Leu Ala Ala Ser Cys Ala Trp Ser Gly Val Ala Gly Glu Glu
20 25 30
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
35 40 45
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
50 55 60
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
65 70 75 80
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
85 90 95
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
100 105 110
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
115 120 125
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
130 135 140
Arg Ala Lys Pro Ser Ala Pro Val Val Ser Gly Pro Ala Ala Arg Ala
145 150 155 160
Thr Pro Gln His Thr Val Ser Phe Thr Cys Glu Ser His Gly Phe Ser
165 170 175
Pro Arg Asp Ile Thr Leu Lys Trp Phe Lys Asn Gly Asn Glu Leu Ser
180 185 190
Asp Phe Gln Thr Asn Val Asp Pro Val Gly Glu Ser Val Ser Tyr Ser
195 200 205
Ile His Ser Thr Ala Lys Val Val Leu Thr Arg Glu Asp Val His Ser
210 215 220
Gln Val Ile Cys Glu Val Ala His Val Thr Leu Gln Gly Asp Pro Leu
225 230 235 240
Arg Gly Thr Ala Asn Leu Ser Glu Thr Ile Arg Val Pro Pro Thr Leu
245 250 255
Glu Val Thr Gln Gln Pro Val Arg Ala Glu Asn Gln Val Asn Val Thr
260 265 270
Cys Gln Val Arg Lys Phe Tyr Pro Gln Arg Leu Gln Leu Thr Trp Leu
275 280 285
Glu Asn Gly Asn Val Ser Arg Thr Glu Thr Ala Ser Thr Val Thr Glu
290 295 300
Asn Lys Asp Gly Thr Tyr Asn Trp Met Ser Trp Leu Leu Val Asn Val
305 310 315 320
Ser Ala His Arg Asp Asp Val Lys Leu Thr Cys Gln Val Glu His Asp
325 330 335
Gly Gln Pro Ala Val Ser Lys Ser His Asp Leu Lys Val Ser Ala His
340 345 350
Pro Lys Glu Gln Gly Ser Asn Thr Ala Ala Glu Asn Thr Gly Ser Asn
355 360 365
Glu Arg Asn Ile Tyr Ile Val Val Gly Val Val Cys Thr Leu Leu Val
370 375 380
Ala Leu Leu Met Ala Ala Leu Tyr Leu Val Arg Ile Arg Gln Lys Lys
385 390 395 400
Ala Gln Gly Ser Thr Ser Ser Thr Arg Leu His Glu Pro Glu Lys Asn
405 410 415
Ala Arg Glu Ile Thr Gln Asp Thr Asn Asp Ile Thr Tyr Ala Asp Leu
420 425 430
Asn Leu Pro Lys Gly Lys Lys Pro Ala Pro Gln Ala Ala Glu Pro Asn
435 440 445
Asn His Thr Glu Tyr Ala Ser Ile Gln Thr Ser Pro Gln Pro Ala Ser
450 455 460
Glu Asp Thr Leu Thr Tyr Ala Asp Leu Asp Met Val His Leu Asn Arg
465 470 475 480
Thr Pro Lys Gln Pro Ala Pro Lys Pro Glu Pro Ser Phe Ser Glu Tyr
485 490 495
Ala Ser Val Gln Val Pro Arg Lys
500
<210> 63
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Укороченный домен человеческого SIRP ¦Б вариант 1
<400> 63
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 64
<211> 373
<212> Белок
<213> Искусственная последовательность
<220>
<223> Внеклеточный домен человеческого SIRP ¦Б домен
<400> 64
Met Glu Pro Ala Gly Pro Ala Pro Gly Arg Leu Gly Pro Leu Leu Cys
1 5 10 15
Leu Leu Leu Ala Ala Ser Cys Ala Trp Ser Gly Val Ala Gly Glu Glu
20 25 30
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
35 40 45
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
50 55 60
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
65 70 75 80
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
85 90 95
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
100 105 110
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
115 120 125
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
130 135 140
Arg Ala Lys Pro Ser Ala Pro Val Val Ser Gly Pro Ala Ala Arg Ala
145 150 155 160
Thr Pro Gln His Thr Val Ser Phe Thr Cys Glu Ser His Gly Phe Ser
165 170 175
Pro Arg Asp Ile Thr Leu Lys Trp Phe Lys Asn Gly Asn Glu Leu Ser
180 185 190
Asp Phe Gln Thr Asn Val Asp Pro Val Gly Glu Ser Val Ser Tyr Ser
195 200 205
Ile His Ser Thr Ala Lys Val Val Leu Thr Arg Glu Asp Val His Ser
210 215 220
Gln Val Ile Cys Glu Val Ala His Val Thr Leu Gln Gly Asp Pro Leu
225 230 235 240
Arg Gly Thr Ala Asn Leu Ser Glu Thr Ile Arg Val Pro Pro Thr Leu
245 250 255
Glu Val Thr Gln Gln Pro Val Arg Ala Glu Asn Gln Val Asn Val Thr
260 265 270
Cys Gln Val Arg Lys Phe Tyr Pro Gln Arg Leu Gln Leu Thr Trp Leu
275 280 285
Glu Asn Gly Asn Val Ser Arg Thr Glu Thr Ala Ser Thr Val Thr Glu
290 295 300
Asn Lys Asp Gly Thr Tyr Asn Trp Met Ser Trp Leu Leu Val Asn Val
305 310 315 320
Ser Ala His Arg Asp Asp Val Lys Leu Thr Cys Gln Val Glu His Asp
325 330 335
Gly Gln Pro Ala Val Ser Lys Ser His Asp Leu Lys Val Ser Ala His
340 345 350
Pro Lys Glu Gln Gly Ser Asn Thr Ala Ala Glu Asn Thr Gly Ser Asn
355 360 365
Glu Arg Asn Ile Tyr
370
<210> 65
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен IgV
<400> 65
Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly Glu Thr Ala Thr Leu
1 5 10 15
Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly Pro Ile Gln Trp Phe
20 25 30
Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr Asn Gln Lys Glu Gly
35 40 45
His Phe Pro Arg Val Thr Thr Val Ser Asp Leu Thr Lys Arg Asn Asn
50 55 60
Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr Pro Ala Asp Ala Gly
65 70 75 80
Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser Pro Asp Asp Val Glu
85 90 95
Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val Arg
100 105
<210> 66
<211> 293
<212> Белок
<213> Искусственная последовательность
<220>
<223> человеческий CD47
<400> 66
Met Trp Pro Leu Val Ala Ala Leu Leu Leu Gly Ser Ala Cys Cys Gly
1 5 10 15
Ser Ala Gln Leu Leu Phe Asn Lys Thr Lys Ser Val Glu Phe Thr Phe
20 25 30
Cys Asn Asp Thr Val Val Ile Pro Cys Phe Val Thr Asn Met Glu Ala
35 40 45
Gln Asn Thr Thr Glu Val Tyr Val Lys Trp Lys Phe Lys Gly Arg Asp
50 55 60
Ile Tyr Thr Phe Asp Gly Ala Leu Asn Lys Ser Thr Val Pro Thr Asp
65 70 75 80
Phe Ser Ser Ala Lys Ile Glu Val Ser Gln Leu Leu Lys Gly Asp Ala
85 90 95
Ser Leu Lys Met Asp Lys Ser Asp Ala Val Ser His Thr Gly Asn Tyr
100 105 110
Thr Cys Glu Val Thr Glu Leu Thr Arg Glu Gly Glu Thr Ile Ile Glu
115 120 125
Leu Lys Tyr Arg Val Val Ser Trp Phe Ser Pro Asn Glu Asn Ile Leu
130 135 140
Ile Val Ile Phe Pro Ile Phe Ala Ile Leu Leu Phe Trp Gly Gln Phe
145 150 155 160
Gly Ile Lys Thr Leu Lys Tyr Arg Ser Gly Gly Met Asp Glu Lys Thr
165 170 175
Ile Ala Leu Leu Val Ala Gly Leu Val Ile Thr Val Ile Val Ile Val
180 185 190
Gly Ala Ile Leu Phe Val Pro Gly Glu Tyr Ser Leu Lys Asn Ala Thr
195 200 205
Gly Leu Gly Leu Ile Val Thr Ser Thr Gly Ile Leu Ile Leu Leu His
210 215 220
Tyr Tyr Val Phe Ser Thr Ala Ile Gly Leu Thr Ser Phe Val Ile Ala
225 230 235 240
Ile Leu Val Ile Gln Val Ile Ala Tyr Ile Leu Ala Val Val Gly Leu
245 250 255
Ser Leu Cys Ile Ala Ala Cys Ile Pro Met His Gly Pro Leu Leu Ile
260 265 270
Ser Gly Leu Ser Ile Leu Ala Leu Ala Gln Leu Leu Gly Leu Val Tyr
275 280 285
Met Lys Phe Val Glu
290
<210> 67
<211> 233
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG1-FC
<400> 67
Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 68
<211> 229
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG4-FC
<400> 68
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800923C2 |
| СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2811120C2 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| АНТИТЕЛА К БЕЛКУ CD38 И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2776795C2 |
| ВАРИАНТЫ СЛИТОГО БЕЛКА SIRP-альфа-4-1BBL И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2815515C2 |
| СЛИТЫЙ БЕЛОК ИЗ БЕЛКА DCTN1 С БЕЛКОМ RET | 2018 |
|
RU2813996C2 |
| ИММУНОМОДУЛИРУЮЩИЕ СЛИТЫЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2755227C2 |
| ХИМЕРНЫЕ БЕЛКИ НА ОСНОВЕ TIGIT И LIGHT | 2018 |
|
RU2775490C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ СЛИТОГО БЕЛКА РЕЦЕПТОРА ТРАНСФОРМИРУЮЩЕГО ФАКТОРА РОСТА БЕТА И ЕЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2791683C2 |
| АНТИТЕЛО К SIRPα И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2822496C1 |
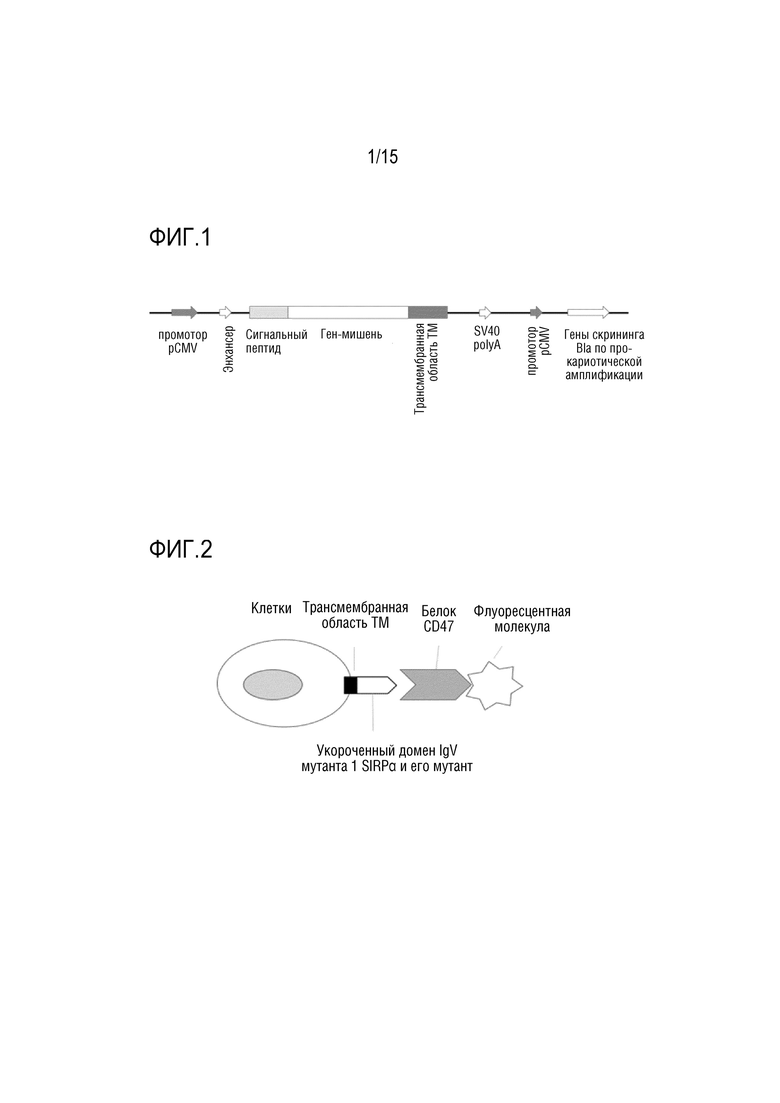
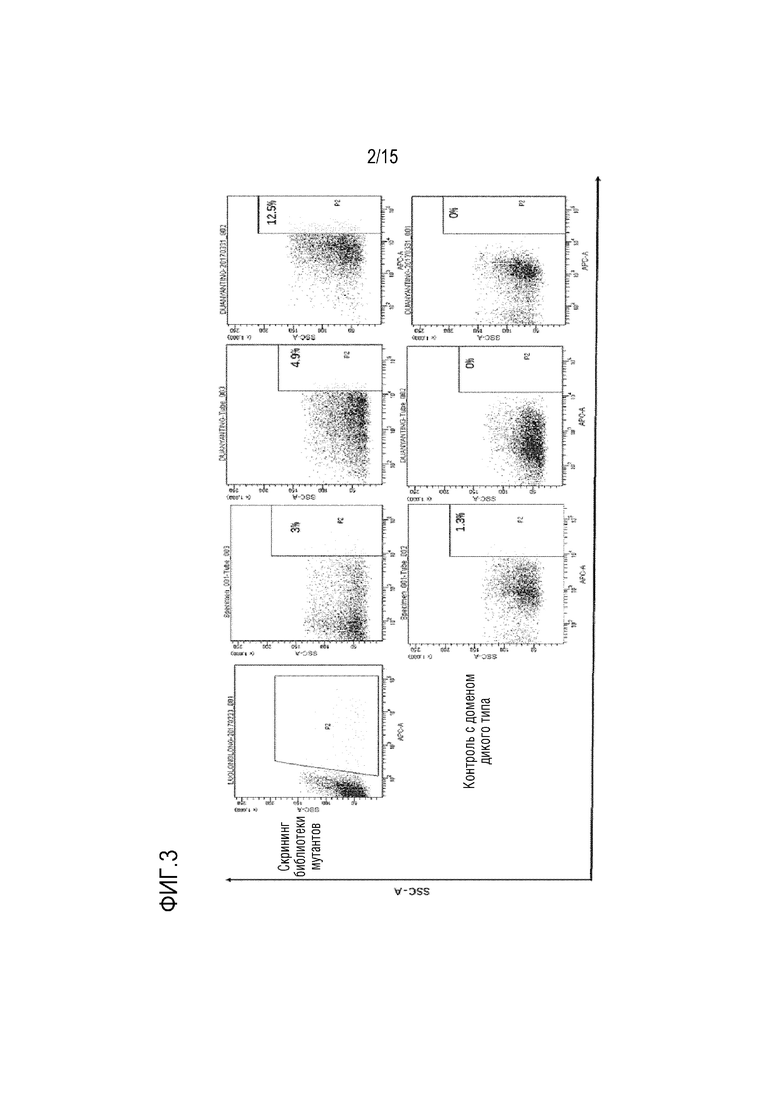

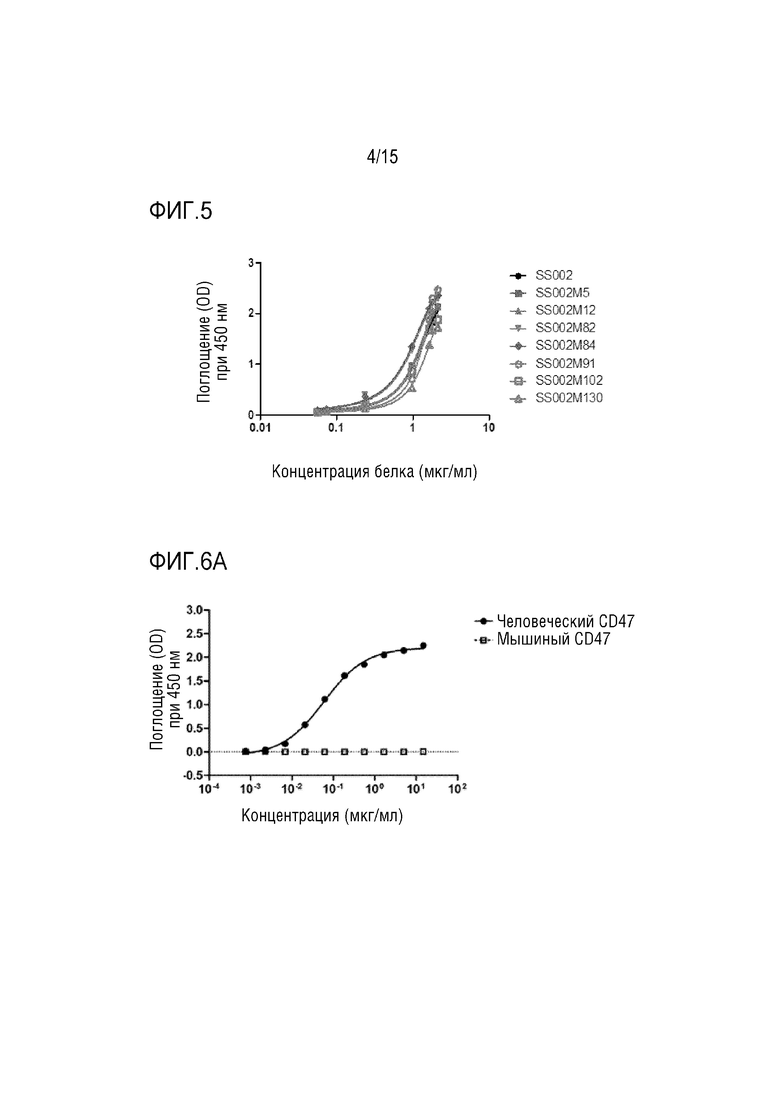
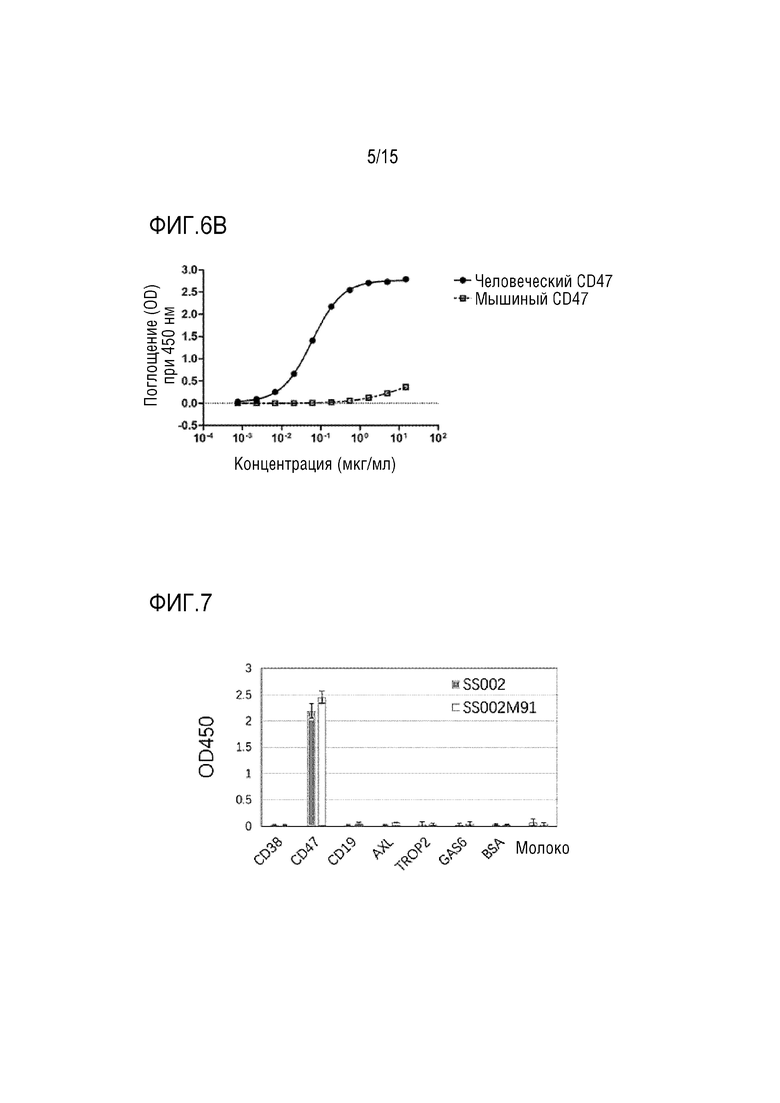
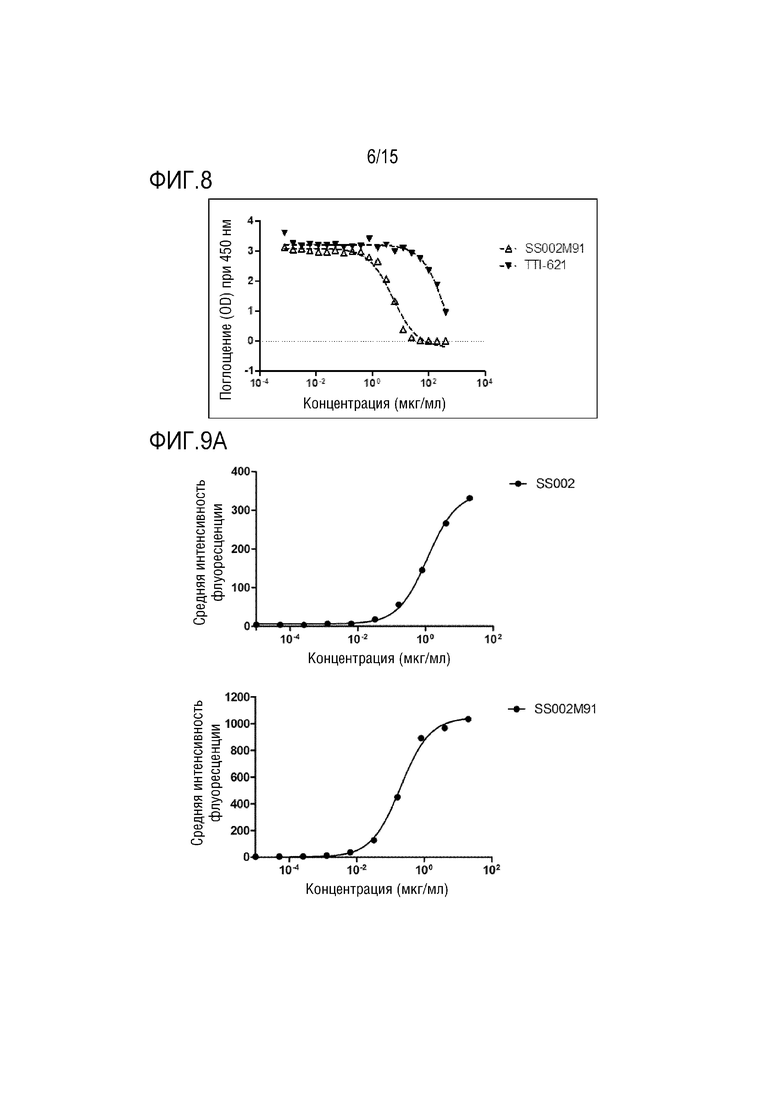
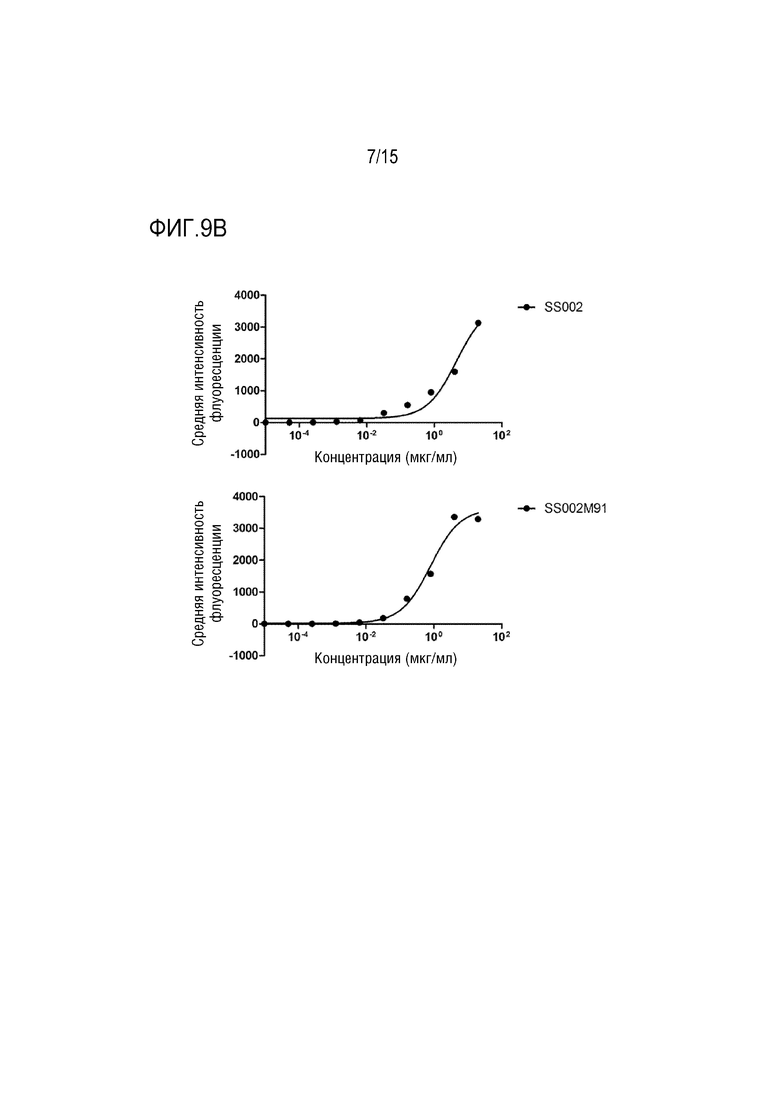
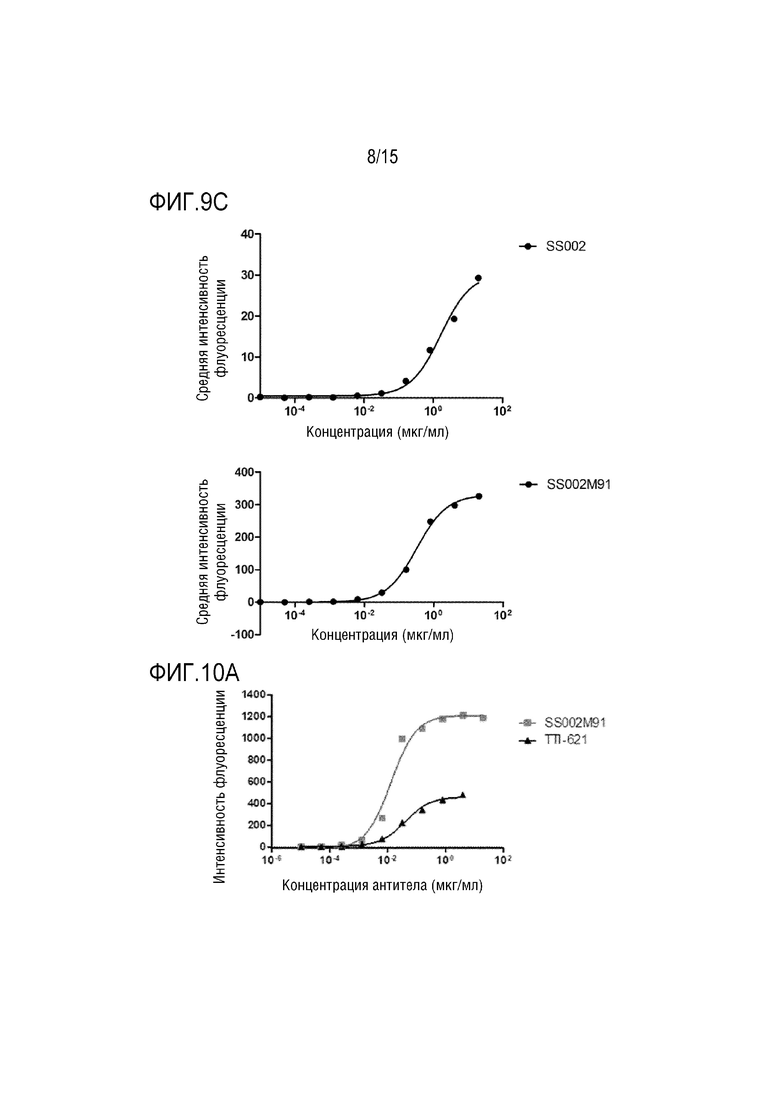
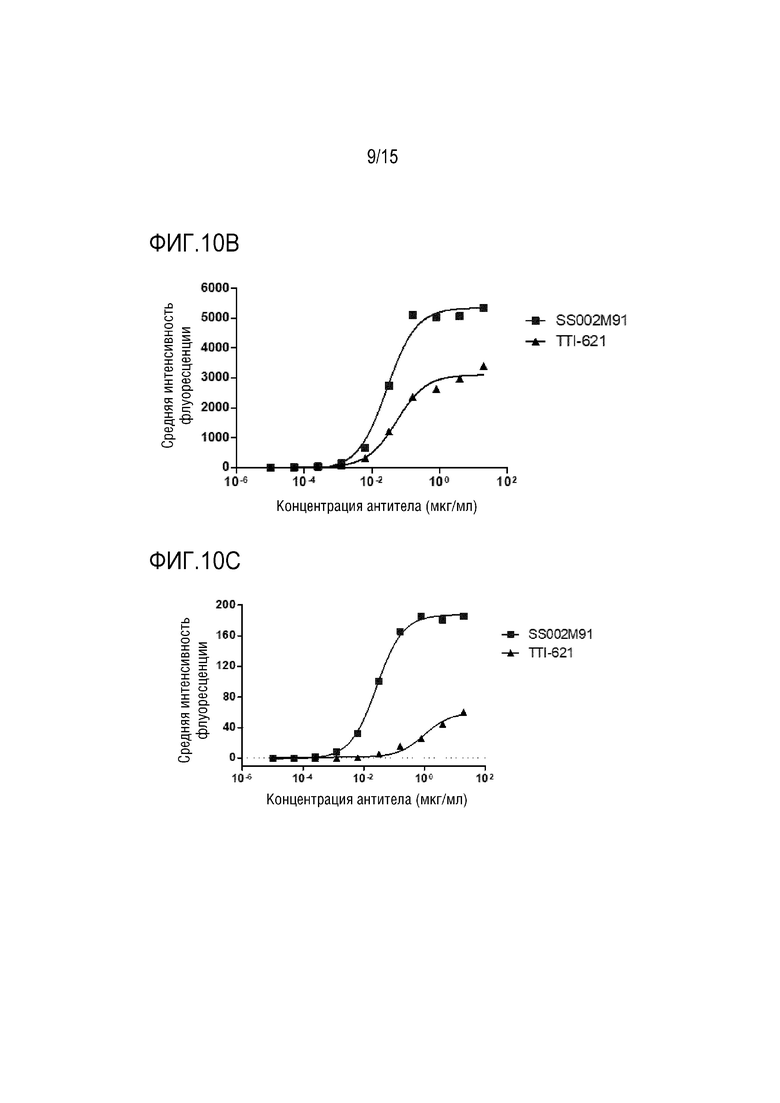
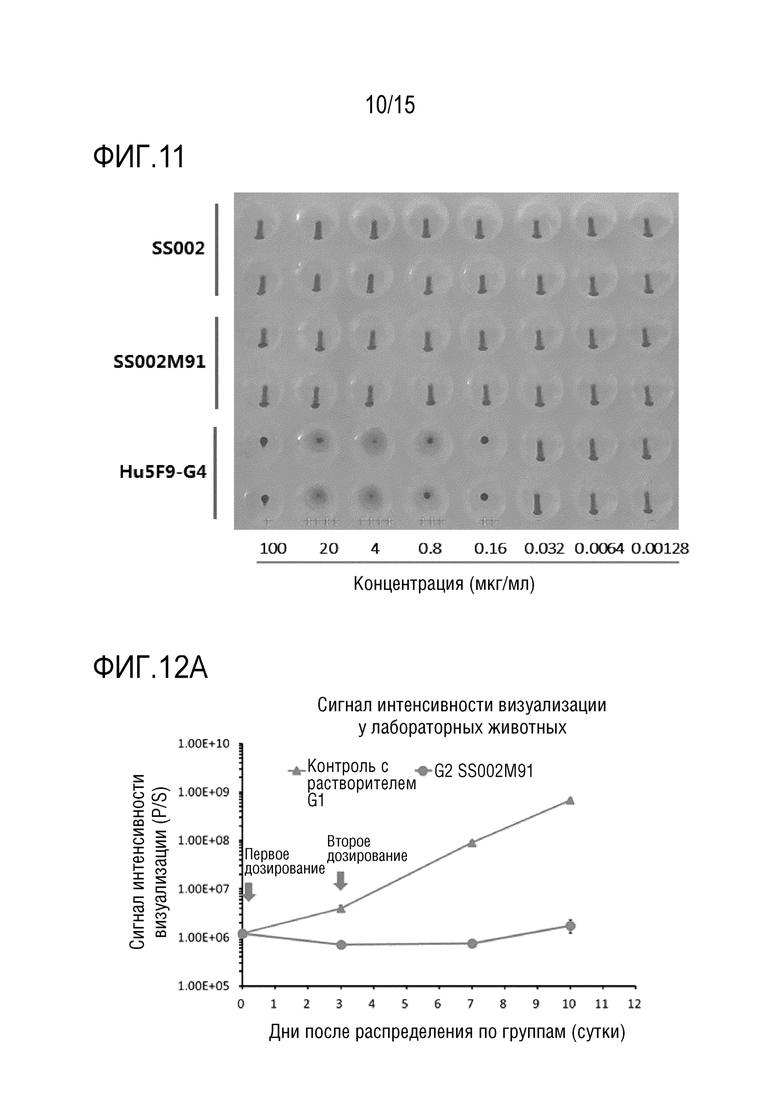
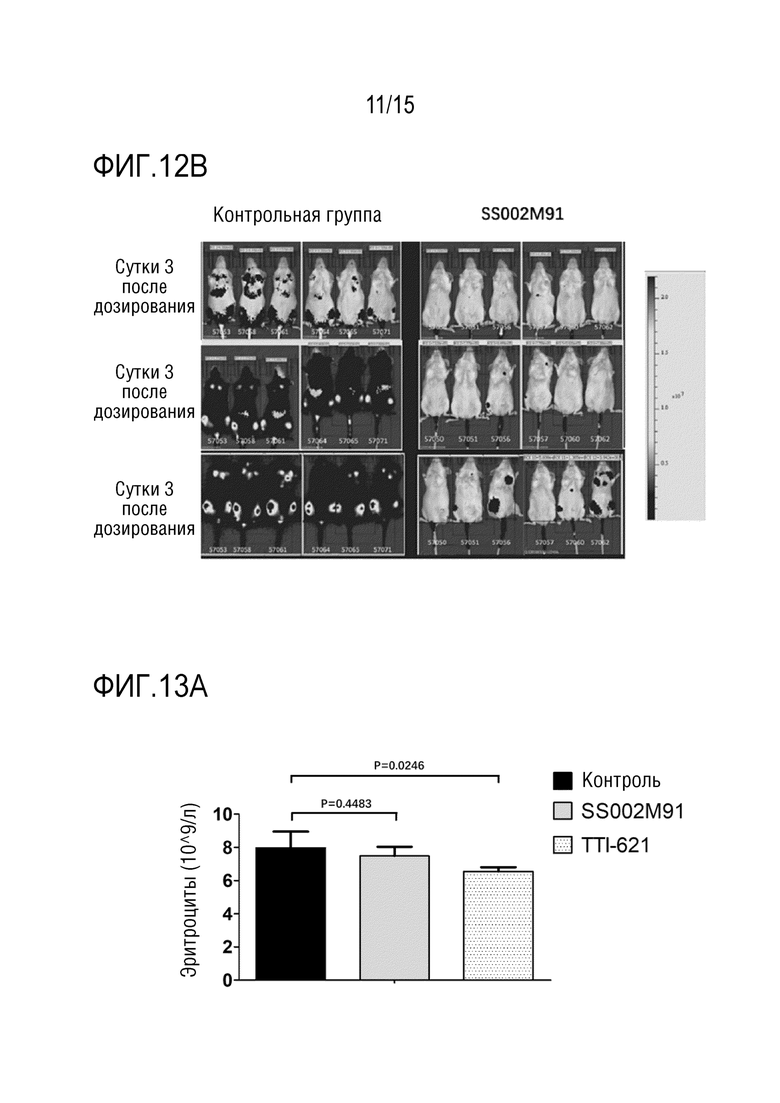
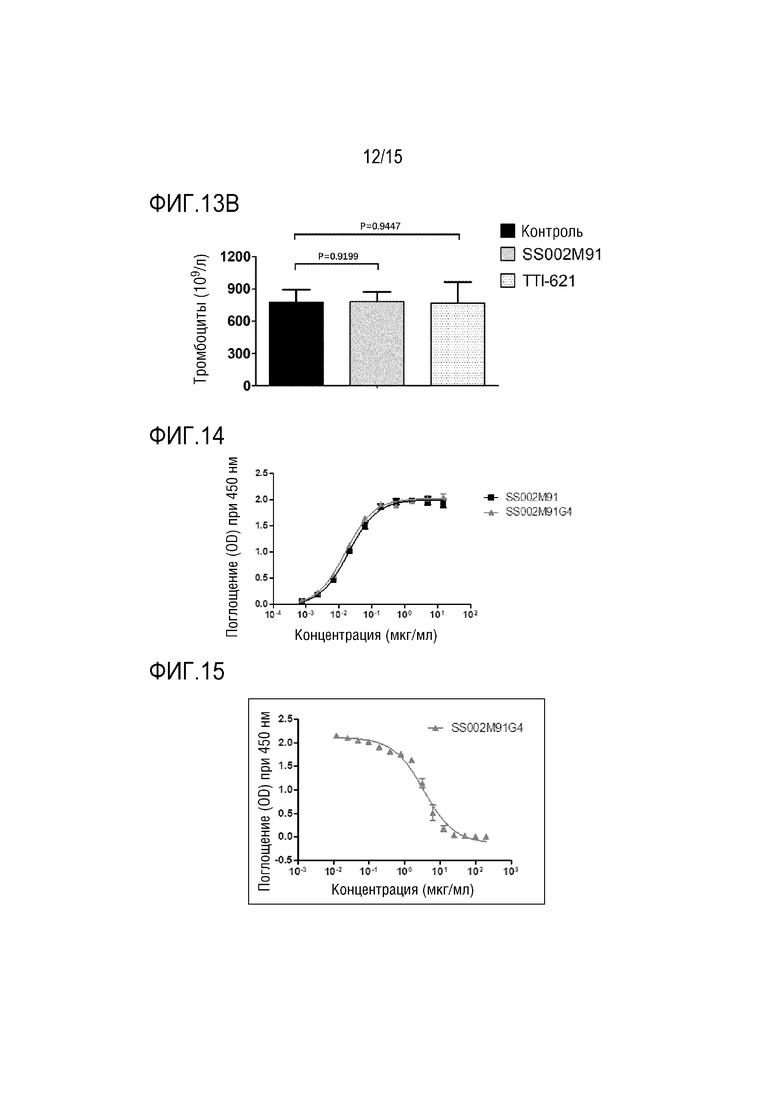
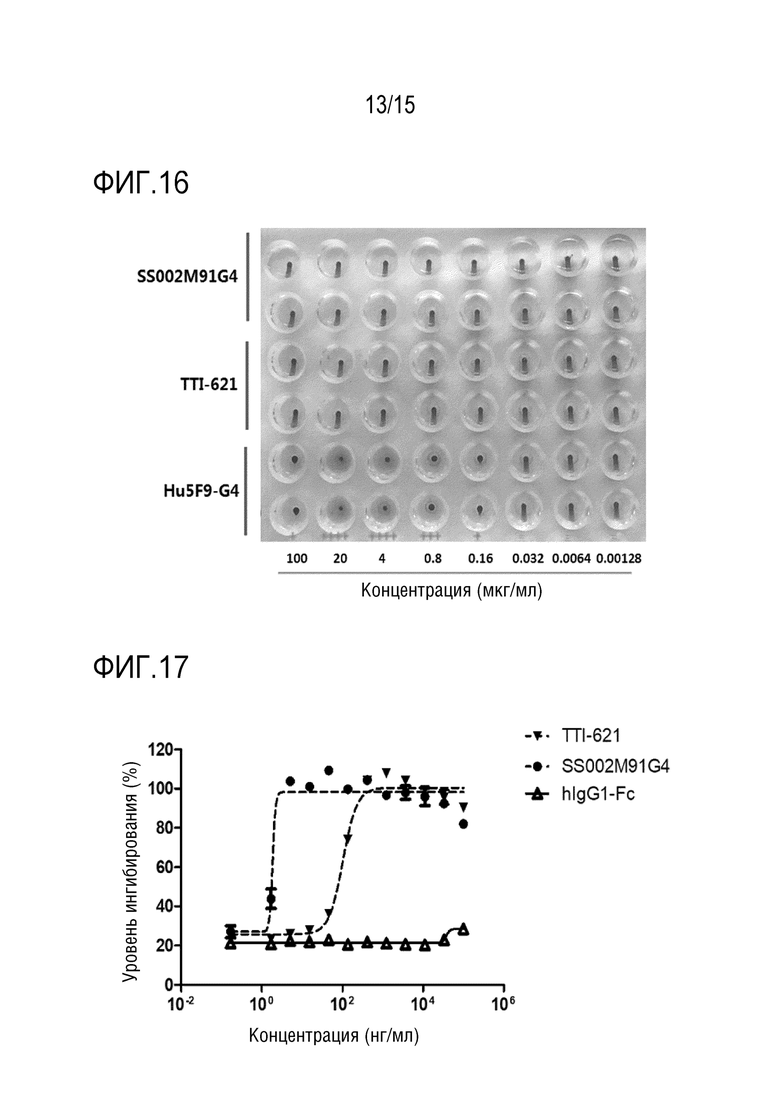
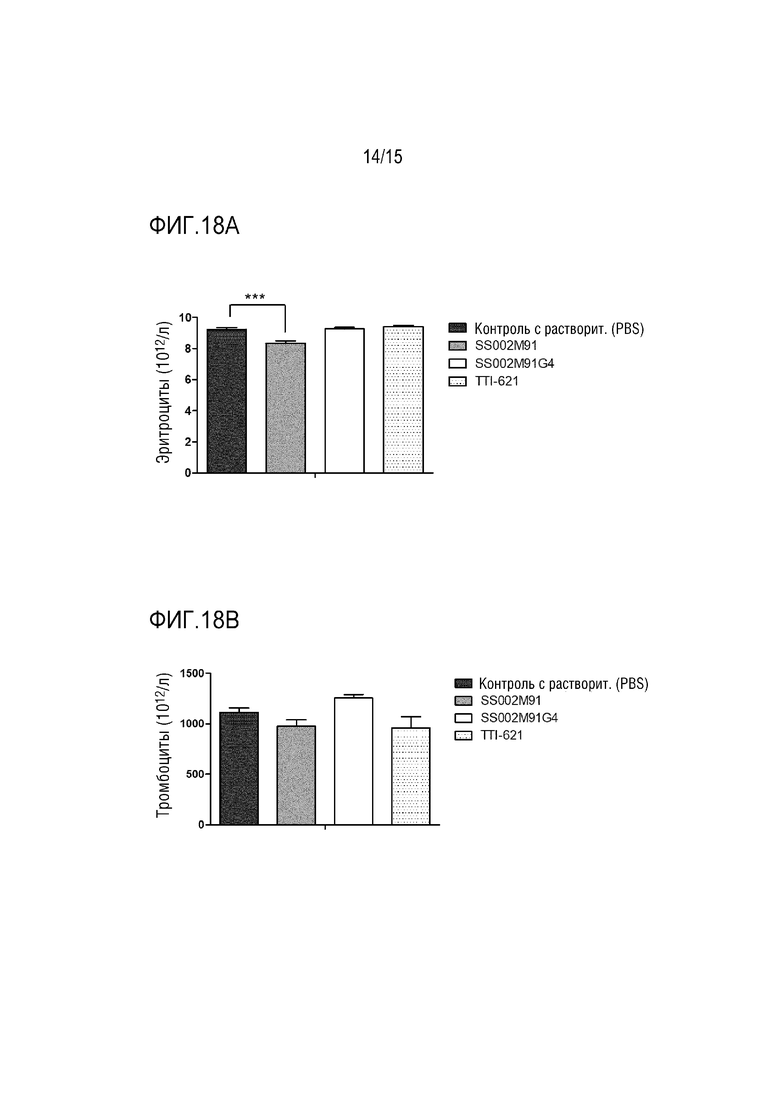
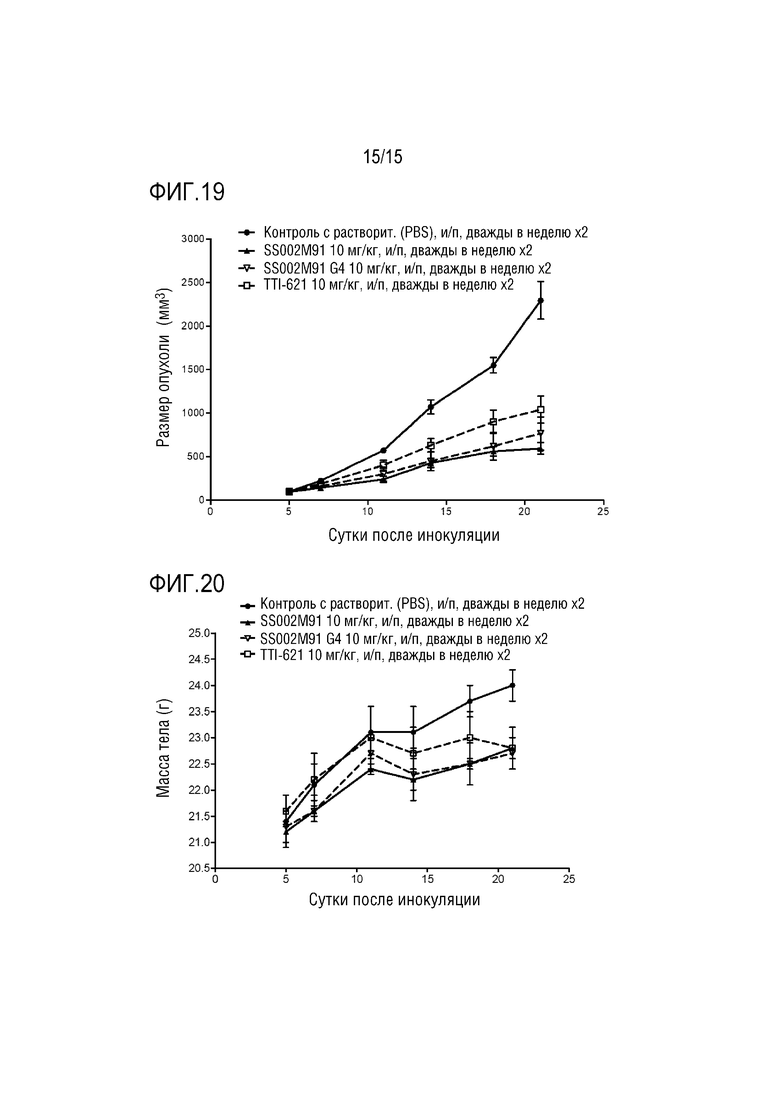
Настоящее изобретение относится к области биотехнологии, конкретно к связывающим CD47 белкам, и может быть использовано в медицине для терапии CD47-положительных гематологических опухолей и CD47-положительных солидных опухолей, а также аутоиммунных заболеваний, таких как болезнь Крона, аллергическая астма и ревматоидный артрит. Предложенное изобретение обеспечивает получение белка, который обладает следующими характеристиками: 1) специфическое связывание с белком CD47 с относительно высокой аффинностью; 2) специфическое блокирование взаимодействия белка CD47 с SIRPальфа; 3) не вызывает реакцию свертывания крови; 4) ингибирование роста и/или пролиферации опухолей или опухолевых клеток; 5) блокирование апоптотического сигнала, индуцированного взаимодействием CD47/SIRPальфа; и/или 6) безопасен для человека и не имеет побочных эффектов, наносящих вред организму. 8 н. и 7 з.п. ф-лы, 20 ил., 2 табл., 14 пр.
1. Мутант домена человеческого SIRPα, который специфически связывается с белком CD47, где указанный домен человеческого SIRPα содержит аминокислотные остатки в положениях 33-149 варианта 1 человеческого SIRPα, указанные аминокислотные остатки в положениях 33-149 варианта 1 человеческого SIRPα представлены в виде SEQ ID NO: 63, и указанный мутант домена человеческого SIRPα содержит группу аминокислотных замен, выбранную из группы, состоящей из:
(1) I61L, V63I, E77I, E84K, V93L, L96S, K98R, N100G и V132L;
(2) I61V, E77N, Q82S, K83R и E84H;
(3) I61F, V63I, K83R, E84K и V132I;
(4) I61L, E77Q, E84D, R107N и V132I;
(5) I61L, V63I, E77K, K83R, E84D и N100G;
(6) I61V, E77H, Q82R, K83R, E84H и R107S;
(7) I61L, E77I, Q82G, E84R, V93L, L96T, N100G, R107S, G109R и V132R;
(8) I61L, E77M, Q82G, K83R, E84D и V132L;
(9) I61L;
(10) I61F, D95H, L96S, G109H и V132S;
(11) I61F, D95H, L96S, K98R, G109H и V132S;
(12) I61L, E77Q, E84D, V93A, R107N и V132I;
(13) E77K, L96S, N100K, G109H и V132L;
(14) I61L, V63I, Q82G, E84G, D95R, L96S, N100D и V132I;
(15) I61L, E77R, Q82N, K83R, E84G, V93L, D95E, L96T, K98R, N100D и V132L;
(16) I61V, E77N, Q82S, K83R, E84H и V93A;
(17) I61V, V63I, E77V, K83R, E84D, D95E, L96T, K98R и N100E;
(18) I61L, V63I, E77V, K83R, D95E, L96S, K98R, N100D и G109R;
(19) I61V, E77L, Q82G, E84G, V93L, D95E, L96T, K98R и N100G; и
(20) I61L, V63I, E77N, Q82G и E84G.
2. Мутант домена человеческого SIRPα по п.1, где указанный мутант домена человеческого SIRPα содержит аминокислотную последовательность, представленную в любой из SEQ ID NO: 1-20.
3. Мутант домена человеческого SIRPα по любому из пп.1-2, имеющий, по меньшей мере, одну из следующих характеристик:
(a) связывается с белком CD47 с величиной KD 1×10-8 M или меньше;
(b) специфически блокирует взаимодействие между белком CD47 и SIRPα;
(c) не вызывает реакцию свертывания крови; и
(d) ингибирует рост и/или пролиферацию опухолей или опухолевых клеток.
4. Слитый белок, который специфически связывается с белком CD47, содержащий указанный мутант домена человеческого SIRPα по любому из пп.1-3 и Fc-область иммуноглобулина, где указанный домен человеческого SIRPα прямо или опосредованно связан с Fc-областью иммуноглобулина.
5. Слитый белок по п.4, где указанный мутант домена человеческого SIRPα расположен у N-конца Fc-области иммуноглобулина.
6. Слитый белок по п.4, где указанная Fc-область иммуноглобулина содержит Fc-область IgG1 человека.
7. Слитый белок по п.4, где указанная Fc-область иммуноглобулина содержит Fc-область IgG4 человека.
8. Слитый белок по п.4, где указанная Fc-область иммуноглобулина содержит аминокислотную последовательность, указанную в одной из SEQ ID NO: 67-68.
9. Слитый белок по любому из пп.4-8, содержащий аминокислотную последовательность, указанную в одной из SEQ ID NO: 21-60.
10. Композиция, которая специфически связывается с белком CD47, содержащая эффективное количество мутанта домена человеческого SIRPα по любому из пп.1-3 или эффективное количество слитого белка по любому из пп.4-9 и необязательно фармацевтически приемлемый адъювант.
11. Применение мутанта домена человеческого SIRPα по любому из пп.1-3 или слитого белка по любому из пп.4-9 в получении лекарственного средства, где указанное лекарственное средство предназначено для профилактики или лечения опухолей, где указанные опухоли выбраны из группы, состоящей из CD47-положительных гематологических опухолей и/или CD47-положительных солидных опухолей.
12. Применение мутанта домена человеческого SIRPα по любому из пп.1-3 или слитого белка по любому из пп.4-9 в получении набора, где указанный набор предназначен для профилактики или лечения опухолей, где указанные опухоли выбраны из группы, состоящей из CD47-положительных гематологических опухолей и/или CD47-положительных солидных опухолей.
13. Применение мутанта домена человеческого SIRPα по любому из пп.1-3 или слитого белка по любому из пп.4-9 в получении лекарственного средства, где указанное лекарственное средство предназначено для профилактики или лечения аутоиммунных заболеваний, где указанные аутоиммунные заболевания выбраны из группы, состоящей из болезни Крона, аллергической астмы и/или ревматоидного артрита.
14. Применение мутанта домена человеческого SIRPα по любому из пп.1-3 или слитого белка по любому из пп.4-9 в получении набора, где указанный набор предназначен для профилактики или лечения аутоиммунных заболеваний, где указанные аутоиммунные заболевания выбраны из группы, состоящей из болезни Крона, аллергической астмы и/или ревматоидного артрита.
15. Способ блокирования взаимодействия между белком CD47 и SIRPα, указанный способ включает введение слитого белка по любому из пп.4-9 или композиции по п.10 индивидууму или клетке или биологическому образцу, которым это необходимо.
| WO 2016024021 A1, 18.02.2016, WO 2013109752 A), 25.07.2013, WO 2014094122 A1, 26.06.2014, EA 201100947 A1, 28.02.2012, WO 2017027422 A1, 16.02.2017, KONTERMANN R | |||
| E | |||
| et al., Bispecific antibodies, Drug Discovery Today, 2015, V | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| et al., Single variable domain-IgG fusion: a novel recombinant approach to Fc | |||

