В рамках настоящего изобретения предложены растворимые мультидоменные связывающие молекулы, содержащие Т-клеточные рецепторы (TCR), обладающие специфичностью в отношении антигена, Fc-домен иммуноглобулина или фрагмента связывающий альбумин; и иммунный эффекторный домен. Такие мультидоменные связывающие молекулы имеют преимущество, поскольку они демонстрируют увеличенный период полувыведения при сохранении функции.
Уровень техники
Многие терапевтические средства на основе белков, включая фрагменты антител и слитые белки, быстро выводятся из организма после введения. Их короткий период полувыведения из системы кровообращения обычно объясняется небольшим размером, который обеспечивает эффективное выведение при помощи почечной фильтрации, а также отсутствием защиты от внутриклеточного расщепления. В таких случаях для поддержания эффективной концентрации лекарственного средства в течение длительных периодов требуется частое введение или продолжительные инфузии. Для улучшения режима дозирования было использовано несколько стратегий с целью увеличить период полувыведения из кровотока. К ним относятся увеличение гидродинамического радиуса белка за счет присоединения гибких гидрофильных молекул, таких как углеводы или ПЭГ (полиэтиленгликоль), и применение рециркуляции, опосредуемой неонатального рецептора Fc (FcRn), а также присоединение Fc-доменов антитела или сывороточного альбумина (Konnteman, Curr Opin Biotechnol. 2011 Dec; 22 (6): 868-76).
Стратегии, в которых применяют рециркуляцию, опосредуемую FcRn, особенно привлекательны благодаря возможности достижения более низкого риска индукции иммуногенности in vivo и достижения длительного увеличения периода полувыведения. Например, сообщают, что период полувыведения биспецифичного антитела формата BiTE®, привлекающего Т-клетки, после присоединения домена Fc превышает 200 часов (Lorenczewski и др., Blood 2017. 130 (Suppl 1), 2815). Аналогичным образом, биспецифичные антитела формата TriTac®, которые включают связывающий альбумин домен, имеют период полувыведения более четырех дней (Wesche et al., Cancer Res 2018; 78 (13 Suppl): Abstract nr 3814).
Слитые белки, содержащие растворимый Т-клеточный рецептор, слитый с фрагментом анти-CD3-антитела, представляют собой новую категорию биспецифичных слитых белков, привлекающих Т-клетки, с периодом полувыведения in vivo приблизительно 6-8 часов (Sato et al., 2018 J Clin One 2018 36, №15_suppl 9521-9521; Миддлтон и др., J Clin One 2016 34, №15_suppl 3016-3016). В отличие от традиционных антител, рецепторы Т-клеток предназначены для распознавания коротких пептидов, происходящих из внутриклеточных антигенов и презентированных на поверхности клетки лейкоцитарным антигеном человека (пептид-HLA). Эффективное формирование иммунного синапса между комплексом пептид-HLA на антигенпрезентирующей клетке и Т-клеткой зависит от фиксированной геометрии взаимодействия, которая нарушается увеличением внутримембранного расстояния (Choudhuri et al., 2005 Nature Jul 28; 436 (7050): 578-82).
Описание изобретения
Существует потребность в биспецифичных белках, привлекающих Т-клетки (взаимодействующих с Т-клетками), с увеличенным периодом полувыведения, которые способны опосредовать эффективное формирование иммунного синапса. Вопреки ожиданиям в данной области техники, авторы настоящего изобретения обнаружили, что слияние слитого белка TCR-анти-CD3 с Fc-областью антитела или фрагментом, связывающим альбумин, неожиданно привело к эффективному образованию иммунного синапса.
В первом аспекте предложена мультидоменная связывающая молекула, содержащая:
i) фрагмент, связывающий пептид-главный комплекс гистосовместимости (рМНС), связанный с иммунным эффектором, привлекающим Т-клетки; и
ii) домен, увеличивающий период полувыведения, содержащий Fc иммуноглобулина или домен, связывающий альбумин.
Предпочтительно, фрагмент, связывающий рМНС, представляет собой рецептор Т-клеток (TCR) или TCR-подобное антитело, содержащее вариабельные домены TCR и/или антитела и по меньшей мере один константный домен. Предпочтительно фрагмент, связывающий рМНС, содержит по меньшей мере один константный домен иммуноглобулина. Предпочтительно константный домен может соответствовать константному домену из альфа-цепи TCR или бета-цепи TCR (TRAC или TRBC соответственно). В качестве альтернативы, константный домен TCR фрагмента, связывающего рМНС, может быть заменен константным доменом легкой или тяжелой цепи антитела (CL, CH1, СН2, СН3 или СН4). Константный домен может быть полноразмерным или усеченным. Константный домен TCR может быть усечен для удаления трансмембранного домена. Если константный домен усечен, предпочтительно удаляют только связанные с мембраной части. Дополнительные мутации могут быть введены в аминокислотную последовательность константного домена относительно природного константного домена. Константная область может также включать остатки, встречающиеся в природе или введенные искусственно, которые допускают димеризацию, например, за счет дисульфидной связи между двумя остатками цистеина.
Авторы настоящего изобретения неожиданно обнаружили, что мультидоменная связывающая молекула, содержащая фрагмент, связывающий рМНС, иммунный эффекторный домен и либо Fc-домен иммуноглобина, либо связывающий альбумин фрагмент, сохраняет свои функции. Это особенно удивительно с учетом знаний в данной области техники о том, что взаимодействие TCR-pMHC основывается на фиксированной геометрии связывания и что активация TCR чувствительна к увеличению межмембранного расстояния (Garboczi et al., Nature. 1996 Nov 14; 384 (6605): 134-41; Choudhuri et al., 2005 Nature Jul 28; 436 (7050): 578-82; Rudolph et al., Annu Rev Immunol. 2006; 24: 419-66). Действительно, модель кинетической сегрегации предполагает, что активация TCR является результатом тетеринга комплекса TCR-CD3 в зонах тесного контакта, в которых предпочтительно происходит фосфорилирование тирозина из-за зависящего от размера исключения тирозинфосфатаз, таких как CD45 (Choudhuri et al., 2005 Nature Jul 28; 436 (7050): 578-82; Davis et al., Nat Immunol. 2006 августа; 7 (8): 803-9). Таким образом, считают, что небольшие размеры комплекса TCR-pMHC (приблизительно 100 А) и фиксированная геометрия связывания важны для образования иммунного синапса и активации TCR. На основании этих сведений специалист в данной области техники понял бы, что применение антигенсвязывающих полипептидов согласно настоящему изобретению согласно ожиданию приведет к снижению образования иммунного синапса, нарушению геометрии связывания TCR-pMHC и, в конечном итоге, к неэффективной активации TCR.
Фрагмент, связывающий рМНС, может представлять собой TCR-подобное антитело. Предпочтительно фрагмент, связывающий рМНС, включает вариабельные домены TCR-подобного антитела. Антитела естественным образом не распознают рМНС; однако известно, что могут быть сконструированы антитела со специфичностью к рМНС. Такие антитела называют TCR-подобными или TCR-мимиками (Chang et al., Expert Opin Biol Ther. 2016 Aug; 16 (8): 979-87 и Dahan et al., Expert Rev Mol Med. 2012 Feb 24; 14: e6).
TCR может представлять собой пару гетеродимерных полипептидов альфа/бета или гамма/дельта TCR. В других случаях TCR может представлять собой одноцепочечный полипептид альфа/бета или гамма/дельта TCR. Аминокислотная последовательность вариабельных доменов TCR может соответствовать тем, которые встречаются в природе, или они могут содержать одну или более мутаций относительно природного TCR. Такие мутации могут быть введены для увеличения аффинности TCR к выбранному антигену. Дополнительно или в качестве альтернативы могут быть включены мутации, улучшающие стабильность и производственную технологичность.
TCR может связываться с МНС в комплексе с пептидным антигеном. Предпочтительно, пептидный антиген представляет собой любой антиген, связанный с заболеванием. Предпочтительно пептидный антиген представляет собой любой антиген, связанный с опухолью. Пептидный антиген может представлять собой пептид, происходящий из GP100, NYESO, MAGEA4 или PRAME, как описано в WO 2011001152, WO 2017109496, WO 2017175006 и WO 2018234319.
TCR может иметь аминокислотную последовательность, как определено в WO 2011001152, WO 2017109496, WO 2017175006 и WO 2018234319.
Иммунный эффекторный домен, привлекающий Т-клетки, может представлять собой эффекторный домен CD3. Привлекающий Т-клетки иммунный эффектор может представлять собой антитело scFv (или антитело-подобный каркас аналогичного размера), который активирует Т-клетку посредством взаимодействия с CD3 и/или комплексом TCR/CD3. Эффекторы CD3 включают, но не ограничиваются ими, анти-CD3-антитела или фрагменты антител, в частности анти-CD3 scFv или антитело-подобные каркасы. Дополнительные иммунные эффекторы включают, но не ограничиваются ими, цитокины, такие как ИЛ-2 и ФНО-β; суперантигены и их мутанты; хемокины, такие как ИЛ-8, фактор тромбоцитов 4, белок, стимулирующий рост меланомы; антитела, включая их фрагменты, производные и варианты, которые связываются с антигенами иммунных клеток, таких как Т-клетки или NK-клетки (например, анти-CD28 или анти-CD16 или любыми молекулами, которые располагаются в иммунном синапсе), и активаторы комплемента.
Домен, увеличивающий период полувыведения, может быть связан с С- или N-концом фрагмента, связывающего рМНС, или с С- или N-концом иммунного эффектора, привлекающего Т-клетки.
Домен, увеличивающий период полувыведения, может содержать Fc иммуноглобулина. Fc-домен иммуноглобулина может представлять собой Fc-область любого антитела. Область Fc представляет собой хвостовую область антитела, которая взаимодействует с рецепторами Fc на клеточной поверхности и некоторыми белками системы комплемента. Область Fc обычно содержит две полипептидные цепи, обе из которых имеют два или три константных домена тяжелой цепи (называемые СН2, СН3 и СН4), и шарнирную область. Две цепи связаны дисульфидными связями с шарнирной областью. Fc-домены подклассов иммуноглобулинов IgG1, IgG2 и IgG4 связываются с FcRn и подвергаются опосредованной им рециркуляции, что обеспечивает длительный период полувыведения из кровотока (3-4 недели). Взаимодействие IgG с FcRn локализовано в области Fc, охватывающей части домена СН2 и СН3. Предпочтительный Fc-домен иммуноглобулина для использования в настоящем изобретении включает, но не ограничивается, Fc-домены из IgG1 или IgG4. Предпочтительно домен Fc является производным последовательностей человека. Область Fc может также предпочтительно включать мутации KiH, которые облегчают димеризацию, а также мутации для предотвращения взаимодействия с активирующими рецепторами, то есть функционально молчащие молекулы. Fc-домен иммуноглобулина может быть соединен с С- или N-концом других доменов (т.е. TCR или иммунного эффектора). Fc иммуноглобулина может быть слит с другими доменами (TCR или иммунным эффектором) через линкер, в качестве альтернативы линкер не используют. Линкерные последовательности обычно гибкие, поскольку они состоят в основном из аминокислот, таких как глицин, аланин и серии, которые не имеют объемных боковых цепей, ограничивающих гибкость. В качестве альтернативы могут быть предпочтительны линкеры с большей жесткостью. Применимые или оптимальные длины линкерных последовательностей можно легко определить. Часто такая последовательность будет иметь длину менее 12, например, менее 10 или от 2 до 10 аминокислот. Линкер может иметь 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 аминокислот в длину. Примеры подходящих линкеров, которые можно использовать в мультидоменных связывающих молекулах согласно настоящему изобретению, включают, но не ограничиваются: GGGSGGGG, GGGGS, GGGSG, GGSGG, GSGGG, GSGGGP, GGEPS, GGEGGGP и GGEGGGSEGGGS (как описано в WO 282010/1338). Когда Fc иммуноглобулина слит с TCR, он может быть слит либо с альфа-, либо с бета-цепями, либо с альфа- и бета-цепями, с линкером или без него. Кроме того, отдельные цепи Fc могут быть слиты с отдельными цепями TCR.
Предпочтительно область Fc может происходить из подкласса IgG1 или IgG4. Две цепи могут содержать константные домены СН2 и СН3 и всю или часть шарнирной области. Шарнирная область может в значительной степени или частично соответствовать шарнирной области IgG1, IgG2, IgG3 или IgG4. Шарнир может включать полностью или частично основной шарнирный домен и полностью или частично нижнюю шарнирную область. Предпочтительно шарнирная область содержит по меньшей мере одну дисульфидную связь, связывающую две цепи.
Область Fc может содержать мутации относительно последовательности Fc дикого типа. Мутации включают замены, вставки и делеции. Такие мутации могут быть сделаны с целью придания желаемых терапевтических свойств. Например, для облегчения гетеродимерации мутации «выступы во впадины» (KiH) могут быть встроены в домен СН3. В этом случае одну цепь проектируют так, чтобы она содержала громоздкий выступающий остаток (то есть выступ), такой как Y, а другую так, чтобы она содержала дополнительный карман (то есть впадину). В данной области техники известны подходящие положения для мутаций KiH. Дополнительно или в качестве альтернативы могут быть введены мутации, которые предотвращают или уменьшают связывание с рецепторами Fey и/или увеличивают связывание с FcRn, и/или предотвращают обмен Fab-плеча, или удаляют сайты протеазы. Дополнительно или в качестве альтернативы мутации могут быть введены по технологическим причинам, например, для удаления или замены аминокислот, которые могут подвергаться посттрансляционным модификациям, таким как гликозилирование.
Примеры включают:
IgG4 (подчеркнутые остатки указывают на мутации по сравнению с последовательностью дикого типа)
«Выступ»

«Впадина»

IgG1 (подчеркнутые остатки указывают на мутации по сравнению с последовательностью дикого типа)
«Выступ»

«Впадина»

Домен, увеличивающий период полувыведения, может содержать домен, связывающий альбумин. Как известно в данной области, альбумин имеет длительный период полувыведения из кровотока, составляющий 19 дней, отчасти из-за своего размера, превышающего почечный порог, а также из-за специфичного взаимодействия и рециркуляции, опосредуемой FcRn. Присоединение к альбумину представляет собой хорошо известную стратегию увеличения периода полувыведения терапевтической молекулы в кровотоке in vivo. Альбумин может быть присоединен нековалентно с помощью специфичного домена, связывающего альбумин или ковалентно с помощью конъюгации или прямого генетического слияния. Примеры терапевтических молекул, в которых задействуют присоединение к альбумину для увеличения периода полувыведения, приведены в Sleep et al., Biochim Biophys Acta. 2013 декабрь; 1830 (12): 5526-34.
Домен, связывающий альбумин, может представлять собой любой фрагмент, способный связываться с альбумином, включая любой известный альбумин-связывающий фрагмент. Домены, связывающие альбумин, могут быть выбраны из эндогенных или экзогенных лигандов, небольших органических молекул, жирных кислот, пептидов и белков, которые специфично связывают альбумин. Примеры предпочтительных доменов, связывающих альбумин, включают короткие пептиды, например, описанные в Dennis et al., J Biol Chem. 2002 Sep 20; 277 (38): 35035-43 (например, пептид QRLMEDICLPRWGCLWEDDF); белки, сконструированные для связывания альбумина, например, антитела, их фрагменты и антитело-подобные каркасы, например, Albudab® (O'Connor-Semmes et al., Clin Pharmacol Ther. 2014 Dec; 96 (6): 704-12), коммерчески поставляемые GSK и Nanobody® (Van Roy et al., Arthritis Res Ther. 2015 May 20; 17: 135), коммерчески поставляемые Ablynx; и белки на основе связывающих альбумин доменов, встречающиеся в природе, такие как белок стрептококка Белок G (Stork et al., Eng Des Sel. 2007 Nov; 20 (11): 569-76), например, Albumod®, коммерчески поставляемый Affibody. Предпочтительно альбумин представляет собой сывороточный альбумин человека (HSA). Аффинность связывающего альбумин домена к альбумину человека может находиться в диапазоне от пикомолярного до микромолярного. Учитывая чрезвычайно высокую концентрацию альбумина в сыворотке крови человека (35-50 мг/мл, приблизительно 0,6 мМ), подсчитано, что по существу все домены, связывающие альбумин, будут связаны с альбумином in vivo.
Фрагмент, связывающий альбумин, может быть связан с С- или N-концом других доменов (то есть с TCR или иммунным эффектором). Фрагмент, связывающий альбумин, может быть связан с другими доменами (т.е. с TCR или иммунным эффектором) через линкер. Линкерные последовательности обычно гибкие, поскольку они состоят в основном из аминокислот, таких как глицин, аланин и серии, которые не имеют объемных боковых цепей, ограничивающих гибкость. В качестве альтернативы могут быть предпочтительны линкеры с большей жесткостью. Применимые или оптимальные длины линкерных последовательностей можно легко определить. Часто линкерная последовательность будет иметь длину менее 12, например, менее 10 или от 2 до 10 аминокислот. Она может иметь 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 аминокислот в длину. Примеры подходящих линкеров, которые можно использовать в мультидоменных связывающих молекулах согласно настоящему изобретению, включают, но не ограничиваются: GGGSGGGG, GGGGS, GGGSG, GGSGG, GSGGG, GSGGGP, GGEPS, GGEGGGP и GGEGGGSEGGGS (как описано в WO 282010/1338). Если фрагмент, связывающий альбумин, связан с TCR, он может быть связан либо с альфа- или бета-цепями, либо с альфа- и бета-цепями, с линкером или без него.
Мультидоменная связывающая молекула согласно первому аспекту может применяться в качестве лекарственного средства.
В дополнительном аспекте предложена фармацевтическая композиция, содержащая мультидоменную связывающую молекулу в соответствии с первым аспектом.
В еще одном дополнительном аспекте предложена нуклеиновая кислота, кодирующая мультидоменную связывающую молекулу в соответствии с первым аспектом. Также предложен вектор экспрессии, содержащий нуклеиновую кислоту этого аспекта. Кроме того, предложена клетка-хозяин, содержащая нуклеиновую кислоту или вектор этого аспекта, где нуклеиновая кислота, кодирующая мультидоменную связывающую молекулу, присутствует в виде одной открытой рамки считывания или двух отдельных открытых рамок считывания, кодирующих альфа-цепь и бета-цепь соответственно.
Также в дополнительном аспекте предложен способ получения мультидоменной связывающей молекулы в соответствии с первым аспектом, включающий поддержание клетки-хозяина, описанной выше, в оптимальных условиях для экспрессии нуклеиновой кислоты и выделения мультидоменного антигенсвязывающего полипептида.
В еще одном дополнительном аспекте предложен способ лечения, включающий введение мультидоменной связывающей молекулы в соответствии с первым аспектом пациенту, нуждающемуся в этом.
В рамки изобретения входят фенотипически «молчащие» варианты любой раскрытой в настоящей заявке молекулы. Используемый в настоящей заявке термин «фенотипически молчащие варианты» относится к варианту, который включает в себя одну или более дополнительных аминокислотных замен, включая замены, вставки и делеции, в дополнение к тем, что изложены выше, причем этот вариант имеет фенотип, аналогичный фенотипу соответствующей молекулы без указанных изменений. Для целей данной заявки фенотип включает аффинность связывания (KD и/или период полувыведения из связи) и специфичность. Предпочтительно, фенотип растворимой мультидоменной связывающей молекулы включает в себя способность активации иммунной системы и выход при очистке, в дополнение к аффинности связывания и специфичности.
Фенотипически молчащие варианты могут содержать одну или более консервативных замен и/или одну или несколько нейтральных замен. Под нейтральными заменами подразумевают те замены, которые не подпадают под определение консервативных, как указано ниже, но, тем не менее, фенотипически молчащие. Специалисту известно, что различные аминокислоты имеют схожие свойства и, следовательно, являются «консервативными». Одна или несколько таких аминокислот белка, полипептида или пептида часто могут быть заменены одной или более другими такими аминокислотами без устранения желаемой активности этого белка, полипептида или пептида.
Таким образом, аминокислоты глицин, аланин, валин, лейцин и изолейцин часто могут быть заменены друг на друга (аминокислоты с алифатическими боковыми цепями). Из этих возможных замен предпочтительно, чтобы глицин и аланин заменяли друг друга (поскольку они имеют относительно короткие боковые цепи), а валин, лейцин и изолейцин заменяли друг друга (поскольку они имеют более крупные гидрофобные алифатические боковые цепи). Другие аминокислоты, которые часто могут быть заменены друг на друга, включают: фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи); лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи); аспартат и глутамат (аминокислоты с кислотными боковыми цепями); аспарагин и глутамин (аминокислоты, имеющие боковые амидные цепи); и цистеин и метионин (аминокислоты с серосодержащими боковыми цепями). Следует понимать, что замены аминокислот в рамках настоящего изобретения могут быть выполнены с использованием встречающихся в природе или не встречающихся в природе аминокислот. Например, в настоящей заявке предполагается, что метальная группа в аланине может быть заменена этильной группой и/или что в остов пептида могут быть внесены незначительные изменения. Независимо от того, используют ли природные или синтетические аминокислоты, предпочтительно, чтобы присутствовали только L-аминокислоты.
Замены такого рода часто называют «консервативными» или «полуконсервативными» аминокислотными заменами. Таким образом, настоящее изобретение распространяется на применение молекулы, содержащей любую аминокислотную последовательность, описанную выше, но с одной или несколькими консервативными заменами и/или одной или несколькими нейтральными заменами в последовательности, так что аминокислотная последовательность TCR имеет не менее 90% идентичности, а именно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности, с последовательностями TCR, раскрытыми в настоящей заявке.
«Идентичность», как известно в данной области техники, представляет собой взаимоотношение между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, определенное путем их сравнения. В данной области техники идентичность также означает степень родства последовательностей между полипептидными или полинуклеотидными последовательностями, в зависимости от обстоятельств, что определяется соответствием между нитями таких последовательностей. Хотя существует ряд методов для измерения идентичности между двумя полипептидными или двумя полинуклеотидными последовательностями, методы, обычно используемые для определения идентичности, кодифицированы в компьютерных программах. Предпочтительные компьютерные программы для определения идентичности двух последовательностей включают, помимо прочего, программный пакет GCG (Devereux, et al., Nucleic Acids Research, 12, 387 (1984), BLASTP, BLASTN и FASTA (Atschul et al., J. Molec. Biol., 215, 403 (1990)).
Для сравнения аминокислотных последовательностей можно использовать такую программу, как CLUSTAL. Эта программа сравнивает аминокислотные последовательности и находит оптимальное выравнивание, вставляя пробелы в любую последовательность, если это необходимо. Можно рассчитать идентичность или сходство аминокислот (идентичность плюс сохранение типа аминокислоты) для оптимального выравнивания. Программа, подобная BLASTx, выровняет самый длинный отрезок похожих последовательностей и присвоит значение совпадению. Таким образом, можно получить сравнение, в котором обнаружено несколько областей сходства, каждая из которых имеет свою оценку. Оба типа анализа идентичности рассматриваются в настоящем изобретении.
Процент идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот определяют путем выравнивания последовательностей для оптимального сравнения (например, в первую последовательность могут быть введены пробелы для наилучшего выравнивания со второй) и сравнения аминокислотных остатков или нуклеотидов на соответствующих положениях. «Лучшее выравнивание» - это выравнивание двух последовательностей, которое дает наивысший процент идентичности. Процент идентичности определяется количеством идентичных аминокислотных остатков или нуклеотидов в сравниваемых последовательностях (т.е. % идентичности = количество идентичных положений/общее количество положений х 100).
Определение процента идентичности между двумя последовательностями может быть выполнено с использованием математического алгоритма, известного специалистам в данной области техники. Примером математического алгоритма для сравнения двух последовательностей является алгоритм Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87: 2264-2268, изменено как у Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-5877. Программы BLASTn и BLASTp Altschul et al. (1990) J. Mol. Biol. 215: 403-410 включают такой алгоритм. Определение процента идентичности между двумя нуклеотидными последовательностями можно выполнить с помощью программы BLASTn. Определение процента идентичности между двумя последовательностями белка может быть выполнено с помощью программы BLASTp. Чтобы получить выравнивания с пробелами в целях сравнения, можно применять Gapped BLAST, как описано в Altschul et al. (1997) Nucleic Acids Res. 25: 3389-3402. В качестве альтернативы можно применять для выполнения повторяющегося поиска PSI-Blast, который обнаруживает отдаленные отношения между молекулами (Id.). При использовании программ BLAST, Gapped BLAST и PSI-Blast можно применять параметры по умолчанию соответствующих программ (например, BLASTp и BLASTp). См. Http://www.ncbi.nlm.nih.gov. Общие параметры по умолчанию могут включать, например, Word Size=3, Expect Threshold=10. Параметры могут быть выбраны для автоматической настройки для коротких входных последовательностей. Другим примером математического алгоритма, применяемого для сравнения последовательностей, является алгоритм Майерса и Миллера, CABIOS (1989). Программа ALIGN (версия 2.0), которая является частью пакета программного обеспечения для выравнивания последовательностей CGC, включает такой алгоритм. Другие алгоритмы анализа последовательности, известные в данной области, включают ADVANCE и ADAM, как описано в Torellis and Robotti (1994) Comput. Appl. Biosci., 10: 3-5; и FASTA, описанные в Pearson and Lipman (1988) Proc. Natl. Acad. Sci. 85: 2444-8. В FASTA ktup - это опция управления, которая устанавливает чувствительность и скорость поиска. В целях оценки процента идентичности в рамках настоящего изобретения в качестве методологии сравнения применяют BLASTp с параметрами по умолчанию. Кроме того, когда указанный процент идентичности дает значение не целого числа для аминокислот (т.е. последовательность из 25 аминокислот, имеющая 90% идентичности последовательности, дает значение «22,5», полученное значение округляется в меньшую сторону до следующего целого числа, то есть, «22»). Соответственно, в представленном примере последовательность, имеющая 22 совпадения из 25 аминокислот, находится в пределах 90% идентичности.
Как будет понятно специалистам в данной области техники, можно усечь или удлинить последовательности, представленные на их С-конце и/или N-конце, на 1, 2, 3, 4, 5 или более остатков без существенного влияния на функциональные характеристики молекулы, например, на часть TCR. Последовательности, представленные на их С-конце и/или N-конце, могут быть усечены или удлинены на 1, 2, 3, 4 или 5 остатков. Все данные варианты охватываются настоящим изобретением.
Мутации, включая консервативные и нейтральные замены, вставки и делеции, могут быть введены в последовательности, полученные с использованием любого подходящего метода, включая, но не ограничиваясь ими, методы, основанные на полимеразной цепной реакции (ПЦР), клонирование на основе рестрикционных ферментов или процедуры независимого от лигирования клонирования (LIC). Эти методы подробно описаны во многих стандартных работах по молекулярной биологии. Для получения дополнительных сведений о полимеразной цепной реакции (ПЦР) и клонировании на основе рестрикционных ферментов см. Sambrook& Russell, (2001) Molecular Cloning - A Laboratory Manual (3rd Ed.) CSHL Press. Дополнительную информацию о процедурах независимого от лигирования клонирования (LIC), можно найти в Rashtchian, (1995) Curr Opin Biotechnol 6 (1): 30-6. Последовательности TCR, предложенные в настоящем изобретении, могут быть получены твердофазным синтезом или любым другим подходящим способом, известным в данной области техники.
Молекулы согласно настоящему изобретению могут иметь идеальный профиль безопасности для применения в качестве терапевтических реагентов. Идеальный профиль безопасности означает, что помимо демонстрации хорошей специфичности молекулы согласно настоящему изобретению могли пройти дополнительные доклинические испытания на безопасность. Примеры таких тестов включают анализы цельной крови для подтверждения минимального высвобождения цитокинов в присутствии цельной крови и, следовательно, низкого риска возникновения потенциального синдрома высвобождения цитокинов in vivo, а также тесты аллореактивности для подтверждения низкого потенциала распознавания альтернативных типов HLA.
Молекулы согласно настоящему изобретению можно изменять для очистки с высоким выходом. Выход может быть определен на основе количества материала, удерживаемого в процессе очистки (т.е. количества правильно фолдированного материала, полученного в конце процесса очистки, по отношению к количеству солюбилизированного материала, полученного до рефолдинга), также расчет выхода может быть основан на соотношении количества правильно фолдированного материала, полученного в конце процесса очистки, к исходному объему культуры. Высокий выход означает более 1% или, что более предпочтительно, более 5% или более высокий выход. Высокий выход означает более 1 мг/мл, или, что более предпочтительно, более 3 мг/мл, или более 5 мг/мл, или более высокий выход.
Способы определения аффинности связывания (обратно пропорциональной константе равновесия KD) и периода полувыведения (выраженного как Т½) известны специалистам в данной области. В предпочтительном варианте аффинность связывания и период полувыведения определяют с использованием поверхностного плазменного резонанса (SPR) или биослойной интерферометрии (BLI), например, с использованием прибора BIAcore или прибора Octet, соответственно. Следует принять во внимание, что удвоение аффинности приводит к уменьшению KD вдвое. Т½ рассчитывают как ln2, деленный на коэффициент отклонения (koff). Следовательно, удвоение Т½ приводит к уменьшению koff вдвое. Значения KD и koff для TCR обычно измеряют для растворимых форм TCR, то есть форм, которые усечены для удаления остатков цитоплазматических и трансмембранных доменов. Чтобы учесть различия между независимыми измерениями и, в частности, влияние времени диссоциации, превышающего 20 часов, аффинность связывания и/или период полувыведения данного TCR можно измерить несколько раз, например, 3 или более раз, с использованием одного и того же протокол анализа и взять среднее значение полученных результатов. Для сравнения данных связывания между двумя образцами (т.е. двумя разными TCR и/или двумя препаратами одного и того же TCR) предпочтительно, чтобы измерения проводили в одних и тех же условиях анализа (например, температуры). Методы измерения, описанные в отношении TCR, также могут быть применены к мультидоменным антигенсвязывающим полипептидам, описанным в настоящей заявке.
Некоторые предпочтительные мультидоменные связывающие молекулы согласно настоящему изобретению способны генерировать высокоэффективный Т-клеточный ответ in vitro в отношении антигенположительных клеток, в частности тех клеток, которые презентируют низкий уровень антигена, типичного для раковых клеток (т.е. порядка 5-100, например, 50 антигенов на клетку (Bossi et al., (2013) Oncoimmunol. 1; 2 (11): e26840; Purbhoo et al., (2006). J. Immunol. 176 (12): 7308-7316.)). Такие TCR могут подходить для включения в описанные в настоящей заявке мультидоменные антигенсвязывающие полипептиды. Измеряемый ответ Т-клеток может представлять собой высвобождение маркеров активации Т-клеток, таких как интерферон-γ, гранзим В, уничтожения клеток-мишеней, или другого показателя активации Т-клеток, такого как пролиферация Т-клеток. Предпочтительно высокоэффективным ответом является ответ со значением ЕС50 в диапазоне нМ - пМ, предпочтительно 500 нМ или ниже, наиболее предпочтительно, 1 нМ или ниже или 500 пМ или ниже.
Части молекул TCR согласно настоящему изобретению могут представлять собой ар гетеродимеры. Части альфа-бета-гетеродимерных TCR молекул данного изобретения обычно содержат последовательность константного домена TRAC альфа-цепи и/или последовательность константного домена TRBC1 или TRBC2 бета-цепи. Константные домены могут быть в растворимом виде (т.е. не иметь трансмембранных или цитоплазматических доменов). Один или оба константных домена могут содержать мутации, замены или делеции относительно нативных последовательностей TRAC и/или TRBC1/2. Термин TRAC и TRBC1/2 также охватывает природные полиморфные варианты, например, N-K в положении 4 TRAC (Bragado et al. International Immunology. 1994 Feb; 6 (2): 223-30).
Последовательности константных доменов альфа- и бета-цепей можно модифицировать путем усечения или замены для удаления нативной дисульфидной связи между Cys4 экзона 2 TRAC и Cys2 экзона 2 TRBC1 или TRBC2. Последовательности константных доменов альфа- и/или бета-цепи могут иметь введенную дисульфидную связь между остатками соответствующих константных доменов, как описано, например, в WO 03/020763 и WO 06000830. Константные домены альфа и бета могут быть модифицированы заменой остатков цистеина в положении Thr 48 TRAC и положении Ser 57 TRBC1 или TRBC2, причем указанные цистеины образуют дисульфидную связь между константными доменами альфа и бета TCR. TRBC1 или TRBC2 могут дополнительно включать мутацию цистеина в аланин в положении 75 константного домена и мутацию аспарагина в аспарагиновую кислоту в положении 89 константного домена. Один или оба внеклеточных константных домена, присутствующие в гетеродимере αβ, могут быть усечены на С-конце или С-концах, например, до 15, 10, или до 8 или меньше аминокислот. С-конец внеклеточного константного домена альфа-цепи может быть усечен на 8 аминокислот.
Части молекул TCR согласно настоящему изобретению могут быть представлены в одноцепочечном формате. Такие одноцепочечные форматы включают полипептиды αβ TCR типов Vα-L-Vβ, Vβ-L-Vα, Vα-Cα-L-Vβ, Vα-L-Vβ-Cβ или Vα-Cα-L-Vβ-Cβ, но не ограничиваются ими, при этом Vα и Vβ представляют собой вариабельные области TCR α и β соответственно, Сα и Сβ представляют собой константные области TCR α и β соответственно, a L представляет собой линкерную последовательность (Weidanz et al., (1998) J Immunol Methods. Dec. 1; 221 (1-2): 59-76; Epel et al., (2002), Cancer Immunol Immunother. Nov; 51 (10): 565-73; WO 2004/033685; WO 9918129). Одноцепочечные TCR могут иметь введенную дисульфидную связь между остатками соответствующих константных доменов, как описано в WO 2004/033685. Одноцепочечные TCR дополнительно описаны в WO 2004/033685; WO 98/39482; WO 01/62908; Weidanz et al. (1998) J Immunol Methods 221 (1-2): 59-76; Hoo et al. (1992) Proc Natl Acad Sci U S. A 89 (10): 4759-4763; Schodin (1996) Mol Immunol 33 (9): 819-829).
Терапевтические агенты, которые могут быть связаны с молекулами согласно настоящему изобретению, включают иммуномодуляторы и эффекторы, радиоактивные соединения, ферменты (например, перфорин) или химиотерапевтические средства (например, цисплатин). Чтобы гарантировать, что токсические эффекты проявляются в желаемом месте, токсин может находиться внутри липосомы, связанной с мультидоменным антигенсвязывающим полипептидом, описанным в настоящей заявке, так, чтобы соединение высвобождалось медленно. Это предотвратит повреждающее действие во время транспортировки в организме и обеспечит максимальный эффект токсина после связывания описанного в настоящей заявке мультидоменного антигенсвязывающего полипептида с соответствующими антигенпрезентирующими клетками.
Примеры подходящих терапевтических агентов включают, но не ограничиваются ими:
• низкомолекулярные цитотоксические агенты, т.е. соединения, способные убивать клетки млекопитающих, с молекулярной массой менее 700 Дальтон. Такие соединения также могут содержать токсичные металлы, способные оказывать цитотоксическое действие. Кроме того, следует понимать, что эти низкомолекулярные цитотоксические агенты также включают пролекарства, то есть соединения, которые распадаются или превращаются в физиологических условиях с высвобождением цитотоксических агентов. Примеры таких агентов включают цисплатин, производные майтанзина, рахелмицин, калихеамицин, доцетаксел, этопозид, гемцитабин, ифосфамид, иринотекан, мелфалан, митоксантрон, порфимер натрия, фотофрин II, темозоломид, топотекан, триметрексата арбурат, ауристатин Е, винкристин и доксорубицин;
• пептидные цитотоксины, т.е. белки или их фрагменты, обладающие способностью убивать клетки млекопитающих. Например, рицин, дифтерийный токсин, бактериальный экзотоксин А синегнойной палочки, ДНКаза и РНКаза;
• радионуклиды, т.е. нестабильные изотопы элементов, распадающиеся с одновременным испусканием одной или нескольких а или р частиц или у лучей. Например, йод 131, рений 186, индий 111, иттрий 90, висмут 210 и 213, актиний 225 и астатин 213; хелатирующие агенты могут быть использованы для облегчения связывания этих радионуклидов с высокоаффинными TCR или их мультимерами;
• Иммуностимуляторы, т.е. иммунные эффекторные молекулы, которые стимулируют иммунный ответ. Например, цитокины, такие как ИЛ-2 и ФНО-γ,
• Супер антигены и их мутанты;
• Слитые молекулы TCR-HLA, например слитая молекула с комплексом пептид-HLA, где указанный пептид получен из общего патогена человека, такого как вирус Эпштейна-Барра (EBV);
• хемокины, такие как ИЛ-8, фактор тромбоцитов 4, белок, стимулирующий рост меланомы, и т.д.;
• антитела или их фрагменты, включая антитела против детерминант Т-клеток или NK-клеток (например, анти-CD3, анти-CD28 или анти-CD16);
• антитела или их фрагменты, которые связываются с молекулами, находящимися в иммунном синапсе
• альтернативные белковые каркасы с антителоподобными характеристиками связывания
• активаторы комплемента;
• ксеногенные белковые домены, аллогенные белковые домены, вирусные/бактериальные белковые домены, вирусные/бактериальные пептиды.
Особенно предпочтительным иммунным эффектором является анти-CD3-антитело, или функциональный фрагмент или вариант указанного анти-CD3-антитела. Используемый в настоящей заявке термин «антитело» охватывает такие фрагменты и варианты. Примеры антител к CD3 включают, но не ограничиваются OKT3, UCHT-1, ВМА-031 и 12F6. Фрагменты и варианты/аналоги антител, которые подходят для использования в описанных в настоящей заявке композициях и способах, включают мини-тела, Fab-фрагменты, F(ab')2-фрагменты, dsFv и scFv-фрагменты, Nanobodies™ (эти конструкции, продаваемые Ablynx (Бельгия), содержат синтетический единичный вариабельный домен тяжелой цепи иммуноглобулина, полученный из антитела верблюдовых (например, верблюда или ламы)) и доменные антитела (Domantis (Бельгия), содержащие единичный вариабельный домен тяжелой цепи или вариабельный домен легкой цепи иммуноглобулина с созревшей аффинностью) или альтернативные белковые каркасы, которые проявляют характеристики связывания, подобные антителам, такие как Affibodies (Affibody (Швеция), включающие каркас из сконструированного белка А) или Anticalins (Pieris (Германия)), включающие спроектированные антикалины), и многие другие, иммунный эффектор связан с TCR-частью мультидоменного антигенсвязывающего полипептида, где иммунный эффектор предпочтительно представляет собой анти-CD3-антитело.
Связывание отдельных компонентов мультидоменной связывающей молекулы может происходить посредством ковалентного или нековалентного присоединения. Ковалентное присоединение может быть прямым или непрямым через линкерную последовательность. Линкерные последовательности обычно гибкие, поскольку они состоят в основном из аминокислот, таких как глицин, аланин и серии, которые не имеют объемных боковых цепей, ограничивающих гибкость. В качестве альтернативы могут быть предпочтительны линкеры с большей жесткостью. Применимые или оптимальные длины линкерных последовательностей можно легко определить. Часто линкерная последовательность будет иметь длину менее 12, например, менее 10 или от 2 до 10 аминокислот. Примеры подходящих линкеров, которые можно использовать в молекулах данного изобретения, включают, но не ограничиваются: GGGSGGGG, GGGGS, GGGSG, GGSGG, GSGGG, GSGGGP, GGEPS, GGEGGGP и GGEGGGSEGGGS (как описано в WO 2010/133828).
В дополнительном аспекте настоящего изобретения предложена нуклеиновая кислота, кодирующую мультидоменную связывающую молекулу согласно настоящему изобретению. В некоторых вариантах реализации нуклеиновая кислота представляет собой кДНК. В некоторых вариантах реализации нуклеиновая кислота может представлять собой мРНК. Нуклеиновая кислота может не встречаться в природе, и/или может быть очищенной, и/или сконструированной. Последовательность нуклеиновой кислоты может быть кодон-оптимизирована в соответствии с используемой системой экспрессии. Как известно специалистам в данной области, системы экспрессии могут включать бактериальные клетки, такие как E. coli, или дрожжевые клетки, или клетки млекопитающих, или клетки насекомых, или они могут быть бесклеточными системами экспрессии.
В рамках настоящего изобретения также предложены конструкции в форме плазмид, векторов, кассет транскрипции или экспрессии, которые содержат по меньшей мере одну нуклеиновую кислоту, как описано выше. В настоящем изобретении также предложена рекомбинантная клетка-хозяин, которая содержит одну или несколько конструкций, указанных выше. Как указано, нуклеиновая кислота, кодирующая специфичную связывающую молекулу согласно настоящему изобретению, составляет аспект настоящего изобретения, как и способ получения специфичной связывающей молекулы, который включает экспрессию из кодирующей ее нуклеиновой кислоты. Экспрессия может быть легко проведена путем культивирования в подходящих условиях рекомбинантных клеток-хозяев, содержащих нуклеиновую кислоту. После продуцирования путем экспрессии специфичная связывающая молекула может быть выделена и/или очищена с применением любого подходящего метода, а затем использована при необходимости.
Системы для клонирования и экспрессии полипептида в различных клетках-хозяевах хорошо известны. Подходящие клетки-хозяева включают бактерии, клетки млекопитающих, дрожжевые и бакуловирусные системы. Клеточные линии млекопитающих, доступные в данной области для экспрессии гетерологичного полипептида, включают клетки яичников китайского хомячка, клетки HeLa, клетки почек детенышей хомячка, клетки меланомы мыши NSO и многие другие. Обычным предпочтительным бактериальным хозяином является Е. coli. Экспрессия антител и фрагментов антител в прокариотических клетках, таких как Е. coli, хорошо известна в данной области. Для обзора см., например, Pltickthun, Bio/Technology 9: 545-551 (1991). Экспрессия в эукариотических клетках в культуре также доступна специалистам в данной области в качестве варианта продукции специфичной связывающей молекулы, см. недавний обзор, например, Reff, Curr. Opinion Biotech. 4: 573-576 (1993); Trill et al., Curr. Opinion Biotech. 6: 553-560 (1995).
Можно выбрать или сконструировать подходящие векторы, содержащие соответствующие регуляторные последовательности, включая промоторные последовательности, терминаторные последовательности, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие соответствующие последовательности. Векторы могут быть плазмидами, вирусными, например, фаг или фагмида, в зависимости от случая. Для получения дополнительной информации см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual: 2nd Edition, Cold Spring Harbor Laboratory Press (1989). Многие известные методы и протоколы для манипуляции с нуклеиновой кислотой, например, при получении конструкций нуклеиновых кислот, мутагенезе, секвенировании, введении ДНК в клетки и экспрессии генов, а также анализе белков, подробно описаны в Ausubel et al. ред., Short Protocols in Molecular Biology, 2-nd edition, John Wiley & Sons (1992).
Таким образом, в дополнительном аспекте настоящего изобретения предложена клетка-хозяин, содержащая нуклеиновую кислоту, как описано в настоящей заявке. Еще один аспект относится к способу, включающему введение такой нуклеиновой кислоты в клетку-хозяина. Для введения можно использовать любую доступную технику. Для эукариотических клеток подходящие методы могут включать трансфекцию фосфатом кальция, DEAE-декстран, электропорацию, опосредованную липосомами трансфекцию и трансдукцию с использованием ретровируса или другого вируса, например, оспы, или, в случае клеток насекомых, бакуловируса. Для бактериальных клеток подходящие методы могут включать трансформацию хлоридом кальция, электропорацию и трансфекцию с использованием бактериофага. После введения может последовать активация экспрессии нуклеиновой кислоты, например, путем культивирования клеток-хозяев в условиях экспрессии гена.
Нуклеиновая кислота согласно настоящему изобретению может быть интегрирована в геном (например, хромосому) клетки-хозяина. Интеграции может способствовать включение последовательностей, которые способствуют рекомбинации с геномом, в соответствии со стандартными методами.
Как хорошо известно в данной области техники, молекулы могут подвергаться посттрансляционным модификациям. Гликозилирование является одной из таких модификаций, которая включает ковалентное присоединение олигосахаридных фрагментов к определенным аминокислотам в цепи TCR. Например, остатки аспарагина или остатки серина/треонина являются хорошо известными местами присоединения олигосахаридов. Статус гликозилирования конкретного белка зависит от ряда факторов, включая последовательность белка, конформацию белка и доступность определенных ферментов. Кроме того, статус гликозилирования (то есть тип олигосахарида, ковалентная связь и общее количество связей) может влиять на функцию белка. Следовательно, при производстве рекомбинантных белков часто желателен контроль гликозилирования. Контролируемое гликозилирование использовали для улучшения терапевтических средств на основе антител. (Джефферис и др., (2009) Nat Rev Drug Discov Mar; 8 (3): 226-34). Для частей молекул TCR согласно настоящему изобретению гликозилирование можно контролировать, используя, например, определенные клеточные линии (включая, линии клеток млекопитающих, такие как клетки яичника китайского хомячка (СНО) или клетки эмбриональной почки человека (НЕК), но не ограничиваясь ими), или путем химической модификации. Такие модификации могут быть желательными, поскольку гликозилирование может улучшить фармакокинетику, снизить иммуногенность и помочь более точно имитировать нативный белок человека (Sinclair and Elliott, (2005) Pharm Sci. Aug; 94 (8): 1626-35).
Для введения пациентам молекулы согласно настоящему изобретению (предпочтительно связанные с детектируемой меткой или терапевтическим агентом), нуклеиновые кислоты, векторы экспрессии или клетки согласно настоящему изобретению могут быть предложены как часть стерильной фармацевтической композиции вместе с одним или несколькими фармацевтически приемлемыми носителями или вспомогательными веществами. Эта фармацевтическая композиция может быть в любой подходящей форме (в зависимости от желаемого способа введения пациенту). Она может быть предложена в виде однократной формы дозирования, обычно предлагаемой в запечатанном контейнере и может быть предложена в виде набора. Такой набор обычно (хотя и не обязательно) включает инструкции по применению. Он может включать множество указанных однократных форм дозирования.
Фармацевтическая композиция может быть адаптирована для введения любым подходящим путем, таким как парентеральный (включая подкожный, внутримышечный, интратекальный или внутривенный), энтеральный (включая пероральный или ректальный), ингаляционный или интраназальный пути. Такие композиции могут быть приготовлены любым способом, известным в области фармации, например, путем смешивания активного ингредиента с носителем(-ями) или наполнителем(-ями) в стерильных условиях.
Дозировки молекул согласно настоящему изобретению могут варьировать в широких пределах, в зависимости от заболевания или расстройства, подлежащего лечению, возраста и состояния индивидуума, подлежащего лечению, и т.д. Подходящий диапазон доз для молекулы данного изобретения может находиться в диапазоне диапазон от 25 нг/кг до 50 мкг/кг или от 1 мкг до 1 г. Применяемые подходящие дозировки в конечном итоге определит врач.
Мультидоменные антигенсвязывающие полипептиды, фармацевтические композиции, векторы, нуклеиновые кислоты и клетки согласно настоящему изобретению могут быть предложены по существу в чистом виде, например, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% чистые.
Также в настоящем изобретении предложены:
• Мультидоменный антигенсвязывающий полипептид, нуклеиновая кислота, фармацевтическая композиция или клетка согласно настоящему изобретению для примеения в медицине, предпочтительно для применения с целью лечения рака или опухоли;
• применение мультидоменного антигенсвязывающего полипептида, нуклеиновой кислоты, фармацевтической композиции или клетки согласно настоящему изобретению в производстве лекарственного средства для лечения рака или опухоли;
• способ лечения рака или опухоли у пациента, включающий введение пациенту мультидоменного антигенсвязывающего полипептида, нуклеиновой кислоты, фармацевтической композиции или клетки согласно настоящему изобретению;
• лекарственная форма для инъекций для введения пациенту, содержащая мультидоменный антигенсвязывающий полипептид, нуклеиновую кислоту, фармацевтическую композицию или клетку согласно настоящему изобретению.
Способ лечения может дополнительно включать отдельное, последовательное или комбинированное введение дополнительного противоопухолевого агента. Примеры таких агентов известны в данной области техники и могут включать иммуноактивирующие агенты и/или агенты, модулирующие Т-клетки.
Предпочтительные признаки каждого аспекта изобретения такие же, как и для каждого из других аспектов, с соответствующими изменениями. Упомянутые в настоящей заявке документы предшествующего уровня техники включены посредством ссылки в максимально возможной степени, разрешенной законом.
Описание чертежей
Фигура 1 А) Дизайн слитого белка TCR-антиCD3-Fc; В) Пример последовательности слитого белка TCR-антиCD3-Fc по настоящему изобретению.
Фигура 2 показывает, что слитая молекула TCR-анти-CD3, содержащая домен Fc, способна опосредовать активацию Т-клеток в присутствии антиген-положительных клеток-мишеней. Показаны данные для трех слитых молекул IgG1-Fc.
На Фигуре 3 А) показано, что слитая молекула TCR-анти-CD3, содержащая пептид, связывающий альбумин, способна опосредовать активацию Т-клеток в присутствии антиген-положительных клеток-мишеней. Показаны три формата, в которых пептид, связывающий альбумин, присоединен к С-альфа (F1), N-альфа (F2) или С-бета (F3). В) показывает, что слитая молекула TCR-анти-CD3, содержащая нанотело, связывающее альбумин, способна опосредовать активацию Т-клеток в присутствии антиген-положительных клеток-мишеней. Показаны два формата, в которых пептид, связывающий альбумин, присоединен либо к С-альфа (R), либо к С-бета (Y). С) показывает, что слитая молекула TCR-анти-CD3, содержащая доменное антитело, связывающее альбумин (Albudab®), способна опосредовать активацию Т-клеток в присутствии антиген-положительных клеток-мишеней. Показан один формат, в котором Albudab® присоединен к С-альфа слитой молекулы TCR-анти-CD3.
Фигура 4 А) показывает PK-характеристики слитой молекулы TCR-анти-CD3-Albudab® в сыворотке мышей и В) показывает теоретическую PK слитой молекулы TCR-анти-CD3-Albudab® у человека на основе оценки у мышей.
Примеры
В следующих примерах описаны мультидоменные связывающие молекулы согласно настоящему изобретению, которые можно обозначать как слитые белки TCR-анти-CD3-Fc или слитые белки TRC-анти-CD3-альбумин-связывающий фрагмент.
Пример 1 (Слитая молекула с Fc)
а) Дизайн слитых белков TCR-анти-CD3-Fc
В этом примере использовали слитой белок TCR-анти-CD3, содержащий TCR с высокой аффинностью, который связывается с пептидом, рестриктированным по HLA-A*02 из PRAME. Примеры таких молекул представлены в WO 2018234319.
Fc-домен IgG1 человека был слит через линкер с С-концом TCR-anti-CD3 (см. схему на Фигуре 1А). Были получены еще две конструкции, содержащие функциональные варианты IgG1 Fc человека, известные в данной области техники. Вариант 1 не связывается с рецепторами Fcγ (FcγRs) или белком комплемента C1q и поэтому функционально является «молчащим». Вариант 2 демонстрирует повышенное связывание с FcRn, что может привести к увеличению периода полувыведения in vivo. В каждом случае домен Fc содержит известные мутации «выступ-во-впадину» Y86T и T22Y для облегчения гетеродимеризации.
На Фигуре 1В показана последовательность слитой молекулы TCR-анти-CD3-Fc, содержащей функционально молчащий Fc IgG1 человека (вариант 1).
б) Экспрессия и очистка спитых белков TCR-антиCD3-Fc.
Экспрессию слитых молекул с Fc проводили с применением системы временной экспрессии на основе адаптированных к суспензии клеток яичника китайского хомячка (СНО) (система экспрессии ExpiCHO, Thermo Fisher). Клетки трансфицировали в соответствии с инструкциями производителя с использованием плазмид экспрессии млекопитающих, содержащих соответствующие цепи TCR, соединенные с различными доменами Fc Ig. После сбора супернатанты клеточных культур осветляли центрифугированием клеток при 4000-5000 g в течение 30 минут в охлаждаемой центрифуге. Супернатанты фильтровали через фильтр 0,22 мкм и собирали для дальнейшей очистки.
Для очистки слитые молекулы Fc предварительно корректировали буфером с применением предварительно упакованных колонок mAbselect Sure (GE Healthcare или эквивалентные смолы) в соответствии с инструкциями производителя. Конкретные фракции, содержащие белок, объединяли и дополнительно очищали эксклюзионной хроматографией с использованием соответствующих колонок (GE Healthcare) в физиологически релевантных буферах. Конкретные фракции, содержащие белок, объединяли и концентрировали для последующего тестирования и хранения.
с) Сильная активация Т-клеток слитыми белками TCR-анти-CD3-Fc.
Слитые белки TCR-анти-CD3-Fc оценивали по их способности опосредовать сильное перенаправление CD3+ Т-клеток на антигенпрезентирующие Т2-клетки. Высвобождение интерферона-γ (IFN-γ) использовали для учета активации Т-клеток.
Анализы выполняли с применением набора ELISPOT IFN-γ человека (BD Biosciences) в соответствии с инструкциями производителя. Вкратце, клетки Т2 использовали в качестве клеток-мишеней и нагружали 5 нМ пептида PRAME. Клетки-мишени получали при плотности 1×106/мл в среде для анализа (RPMI 1640, содержащей 10% инактивированного нагреванием FBS и 1% пенициллин-стрептомицин-L-глутамина) и высевали по 50000 клеток на лунку в объеме 50 мкл. Мононуклеарные клетки периферической крови (РВМС), выделенные из свежей донорской крови, использовали в качестве эффекторных клеток и высевали по 35000 клеток на лунку в объеме 50 мкл. Слитые белки TCR-анти-CD3-Fc титровали до конечных концентраций от 10 нМ до 0,0001 нМ и добавляли в лунку в объеме 50 мкл.
Планшеты были приготовлены в соответствии с инструкциями производителя. Лунки, содержащие клетки-мишени, эффекторные клетки и слитые белки, доводили до конечного объема 200 мкл с помощью среды для анализа. Все реакции проводили в трех повторах. Также были приготовлены контрольные лунки без слитого белка, эффекторных клеток или клеток-мишеней. Планшеты инкубировали в течение ночи (37°С/5% CO2). На следующий день планшеты трижды промывали промывочным буфером (пакет 1xPBS, содержащий 0,05% Твин-20, приготовленный в деионизированной воде). Затем в каждую лунку добавляли антитела для первичного обнаружения в объеме 50 мкл. Планшеты инкубировали при комнатной температуре в течение 2 часов перед повторной трехкратной промывкой. Вторичное обнаружение выполняли путем добавления 50 мкл разбавленного стрептавидина-HRP в каждую лунку и инкубации при комнатной температуре в течение 1 часа, после чего этап промывки повторяли. Не более чем за 15 минут до использования к каждому 1 мл субстрата АЕС добавляли одну каплю (20 мкл) хромогена АЕС, перемешивали и добавляли по 50 мкл в каждую лунку. Появление пятен регулярно отслеживали, и планшеты промывали водопроводной водой для прекращения реакции проявления. Затем планшетам давали высохнуть при комнатной температуре в течение не менее 2 часов перед подсчетом пятен с использованием анализатора CTL с программным обеспечением Immunospot (Cellular Technology Limited). Данные были подготовлены и проанализированы с использованием программного обеспечения PRISM.
Результаты, представленные на Фиг. 2, демонстрируют, что слитые белки TCR-анти-CD3-Fc опосредуют эффективную активацию Т-клеток в присутствии антиген-положительных клеток-мишеней. Значения Ес50 находились в диапазоне пМ (238 пМ, 257 пМ и 25 пМ соответственно для слитых молекул с IgG1-Fc, IgG1-Fc-вариант 1 и IgG1-Fc-вариант 2 соответственно). Контрольные эксперименты с антиген-отрицательными клетками-мишенями продемонстрировали, что функционально молчащий вариант 2 IgG1 дает незначительную фоновую активность, поэтому функционально молчащие домены Fc сочли наиболее предпочтительными для терапевтического применения.
Пример 2 (связывание с альбумином)
а) Дизайн слитых белков TCR-антиСD3-альбумин-связывающий фрагмент
В первой конструкции слитой белок TCR-анти-CD3 содержал TCR с высокой аффинностью, который связывается с пептидом, рестриктированным по HLA-A*02, из gp100. Аминокислотная последовательность таких молекул раскрыта в WO 2011001152. В частности, слитая молекула TCR-антиСD3 включает альфа-цепь SEQ ID NO: 45 из WO 2011001152, в которой аминокислоты 1-109 заменены на SEQ ID NO: 8 из WO 2011001152, а аминокислота в положении 1 представляет собой А, исходя из нумерации SEQ ID NO: 45; и бета-цепь с SEQ ID NO: 36 WO 2011001152, в которой остатки 259-370 соответствуют SEQ ID NO: 27 WO 2011001152, аминокислоты в положениях 1 и 2 представляют собой А и I соответственно. Пептид, связывающий альбумин, имеющий аминокислотную последовательность QRLMEDICLPRWGCLWEDDF (как описано у Dennis et al., J Biol Chem. 2002 Sep 20; 277 (38): 35035-43), был присоединен к слитой молекуле TCR-анти-CD3 через линкер. Подходящим линкером является GGGGS. Были приготовлены три варианта, в которых пептид, связывающий альбумин, был соединен в трех разных сайтах связывания: С-альфа (F1), N-альфа (F2) или С-бета (F3).
Во второй конструкции нанотело, связывающее альбумин, было присоединено к той же слитой молекуле TCR-анти-CD3, которую применяли в первой конструкции. Нанотело, связывающее альбумин, имеющее последовательность SEQ ID NO: 52 в WO 2006122787, было присоединено к слитой молекуле TCR-анти-CD3 через линкер. Подходящим линкером является GGGGS. Было приготовлено два варианта, в которых нанотело, связывающее альбумин, было соединено в двух разных сайтах связывания: С-альфа (R) или С-бета (Y).
В третьей конструкции альбумин-связывающее доменное антитело было присоединено к С-концу альфа-цепи слитой молекулы TCR-анти-CD3. Антитело принадлежит платформе Albudab®. Применяли два варианта доменного антитела; DOM 7h-10-14 dAb и DOM 7h-11-15 dAb, представленные SEQ ID NO: 26 и 27 соответственно в WO 201010893. Антитело было прикреплено через линкер и напрямую (т.е. без линкера). Подходящим линкером является GGGGS. Слитой белок TCR-анти-CD3 содержал TCR с высокой аффинностью, который связывается с пептидом, рестриктированным по HLA-A*01, из MAGEA3. Аминокислотные последовательности таких молекул представлены в WO 2013041865.
б) Экспрессия и очистка слитых белков TCR-анти-CD3- альбумин-связывающий фрагмент
Слитые белки TCR-анти-CD3-альбумин-связывающий фрагмент, были экспрессированы в E. coli в виде телец включения и впоследствии подвергнуты рефолдингу и очистке с использованием той же методологии, которая известна в данной области для слитых белков TCR-анти-CD3 (например, см. WO 2011001152, пример 2)
с) Сильная активация Т-клеток слитыми белками TCR-анти-CD3-альбумин-связывающий фрагмент
Слитые белки TCR-анти-CD3-альбумин-связывающий фрагмент, оценивали по их способности опосредовать сильное перенаправление CD3+ Т-клеток на антиген-положительные раковые клетки. Высвобождение интерферона-γ (ИФН-γ) использовали для учета активации Т-клеток.
Анализы выполняли с использованием набора ELISPOT IFN-γ человека (BD Biosciences) в соответствии с инструкциями производителя. Вкратце, для слитой молекулы, содержащей пептид, связывающий альбумин, в качестве клеток-мишеней применяли клетки меланомы Ме152. Клетки-мишени получали при плотности 1×106/мл в среде для анализа (RPMI 1640, плюс 150 мкМ сывороточного альбумина человека (HSA) и 1% пенициллин-стрептомицин-L-глутамина) и высевали по 50000 клеток на лунку в объеме 50 мкл. Мононуклеарные клетки периферической крови (РВМС), выделенные из свежей донорской крови, использовали в качестве эффекторных клеток и высевали по 30 000 клеток на лунку в объеме 50 мкл. Слитые белки TCR-анти-CD3-Fc титровали до конечных концентраций от 10 нМ до 0,0001 нМ и добавляли в лунку в объеме 50 мкл.
Для слитых молекул, содержащих Albudab® в качестве клеток-мишеней применяли клетки миеломы EJM. Клетки-мишени получали при плотности 1×106/мл в среде для анализа (RPMI 1640, плюс 45 мкМ HSA и 1% пенициллин-стрептомицин-L-глутамина) и высевали по 50000 клеток на лунку в объеме 50 мкл. Мононуклеарные клетки периферической крови (РВМС), выделенные из свежей донорской крови, использовали в качестве эффекторных клеток и высевали по 30 000 клеток на лунку в объеме 50 мкл. Слитые белки TCR-анти-CD3-Fc титровали до конечных концентраций от 10 нМ до 0,0001 нМ и добавляли в лунку в объеме 50 мкл.
Планшеты были подготовлены и проявлены, как описано в Примере 1 с.
На Фиг. 3А-С показано, что слияние фрагмента, связывающего альбумин, с слитым белком TCR-анти-CD3 опосредует эффективную активацию Т-клеток в присутствии антиген-положительных клеток-мишеней. Значения Ес50 находились в диапазоне пМ (137,4/178,0/137,1).
Пример 3 (увеличенный период полувыведения)
а) PK-оценка слитых белков TCR-анти-CD3- альбумин-связывающий фрагмент
Фармакокинетические (РК) характеристики слитых молекул TCR-анти-CD3-Albudab исследовали в сыворотке мышей.
Мышам вводили 0,1 мг/кг слитого белка путем внутривенной болюсной инъекции, и образцы сыворотки отбирали через равные промежутки времени в течение 120 часов. Оценка PK проводилась с использованием анализа на основе ИФА. Вкратце, биотинилиро ванный комплекс pHLA прикрепляли к планшетам, покрытым стрептавидином, и затем добавляли образцы сыворотки. Обнаружение осуществляли с использованием первичного антитела козы к CD3 scFv и конъюгированного с HRP IgG против антитела козы, активированного для колориметрического обнаружения с помощью ТМВ при 450 нм. Полученные результаты использовали для подтверждения присутствия и связывающей активности слитой молекулы TCR-анти-CD3-Albudab с использованием серии разведений и анализа по стандартной кривой. Результаты представлены в виде % активности и их используют для построения графика Cmax в зависимости от времени.
Полученные данные PK показаны на Фигуре 4А. Обратите внимание, что образцы, обозначенные «TCR-Alb (10-14)» и «TCR-Alb (11-15)», относятся к слитым молекулам TCR-анти-CD3, описанным в Примере 2а. «TCR-Alb (D)» представляет собой контрольный образец, соединенный с не связывающим альбумин Albudab, a «TCR» относится к TCR-anti-CD3 без Albudab.
PK-данные из Фиг. 4А использовали для расчета теоретического PK-профиля для слитых молекул TCR-анти-CD3-Albudab у людей (Фиг. 4В). Предполагается, что соединение с Albudab продлит период полувыведения in vivo с 7 до 264 часов.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ИММУНОКОР ЛИМИТЕД
<120> Мультидоменные связывающие молекулы
<130> P121630WO
<150> GB1901306.9
<151> 2019-01-30
<160> 16
<170> PatentIn версии 3.5
<210> 1
<211> 8
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 1
Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 2
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 2
Gly Gly Gly Gly Ser
1 5
<210> 3
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 3
Gly Gly Gly Ser Gly
1 5
<210> 4
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 4
Gly Gly Ser Gly Gly
1 5
<210> 5
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 5
Gly Ser Gly Gly Gly
1 5
<210> 6
<211> 6
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 6
Gly Ser Gly Gly Gly Pro
1 5
<210> 7
<211> 5
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 7
Gly Gly Glu Pro Ser
1 5
<210> 8
<211> 7
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 8
Gly Gly Glu Gly Gly Gly Pro
1 5
<210> 9
<211> 12
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Последовательность линкера
<400> 9
Gly Gly Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser
1 5 10
<210> 10
<211> 226
<212> БЕЛОК
<213> homo sapiens
<400> 10
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly
1 5 10 15
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
20 25 30
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
35 40 45
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
50 55 60
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
65 70 75 80
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
85 90 95
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
100 105 110
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
115 120 125
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
130 135 140
Tyr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
145 150 155 160
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
165 170 175
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
180 185 190
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
195 200 205
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
210 215 220
Gly Lys
225
<210> 11
<211> 226
<212> БЕЛОК
<213> homo sapiens
<400> 11
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly
1 5 10 15
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
20 25 30
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
35 40 45
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
50 55 60
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
65 70 75 80
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
85 90 95
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
100 105 110
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
115 120 125
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
130 135 140
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
145 150 155 160
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
165 170 175
Leu Asp Ser Asp Gly Ser Phe Phe Leu Thr Ser Arg Leu Thr Val Asp
180 185 190
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
195 200 205
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
210 215 220
Gly Lys
225
<210> 12
<211> 223
<212> БЕЛОК
<213> homo sapiens
<400> 12
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
100 105 110
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Tyr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 13
<211> 223
<212> БЕЛОК
<213> homo sapiens
<400> 13
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
100 105 110
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Thr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 14
<211> 20
<212> БЕЛОК
<213> homo sapiens
<400> 14
Gln Arg Leu Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10 15
Glu Asp Asp Phe
20
<210> 15
<211> 450
<212> БЕЛОК
<213> homo sapiens
<400> 15
Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu Glu
1 5 10 15
Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr Asp
20 25 30
Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr Val
35 40 45
Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu Ala
50 55 60
Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala Thr
65 70 75 80
Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His Ser
85 90 95
Arg Leu Gly Asn Tyr Ile Ala Thr Phe Gly Lys Gly Thr Lys Leu Ser
100 105 110
Val Ile Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg
115 120 125
Asp Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp
130 135 140
Ser Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr
145 150 155 160
Asp Lys Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser
165 170 175
Ala Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe
180 185 190
Asn Asn Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser
195 200 205
Ser Thr Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Arg
210 215 220
Ser Gly Thr Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Tyr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 16
<211> 743
<212> БЕЛОК
<213> homo sapiens
<400> 16
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala Asp Thr Gly Gly Gly Gly Ser Gly Gly Gly Gly
500 505 510
Ser Gly Gly Gly Arg Ser Gly Thr Val Glu Cys Pro Pro Cys Pro Ala
515 520 525
Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
530 535 540
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
545 550 555 560
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
565 570 575
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
580 585 590
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
595 600 605
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
610 615 620
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
625 630 635 640
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
645 650 655
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
660 665 670
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
675 680 685
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Thr Ser
690 695 700
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
705 710 715 720
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
725 730 735
Leu Ser Leu Ser Pro Gly Lys
740
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| СПЕЦИФИЧНЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2825837C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| ВЫСОКОАФФИННЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР ДЛЯ РАСПОЗНАВАНИЯ АНТИГЕНА AFP | 2019 |
|
RU2836365C2 |
| ИММУНОМОДУЛИРУЮЩИЕ СЛИТЫЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2755227C2 |
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2017 |
|
RU2775623C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| НОВЫЕ БИСПЕЦИФИЧНЫЕ ПОЛИПЕПТИДНЫЕ КОМПЛЕКСЫ ПРОТИВ CD3/CD19 | 2018 |
|
RU2788127C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
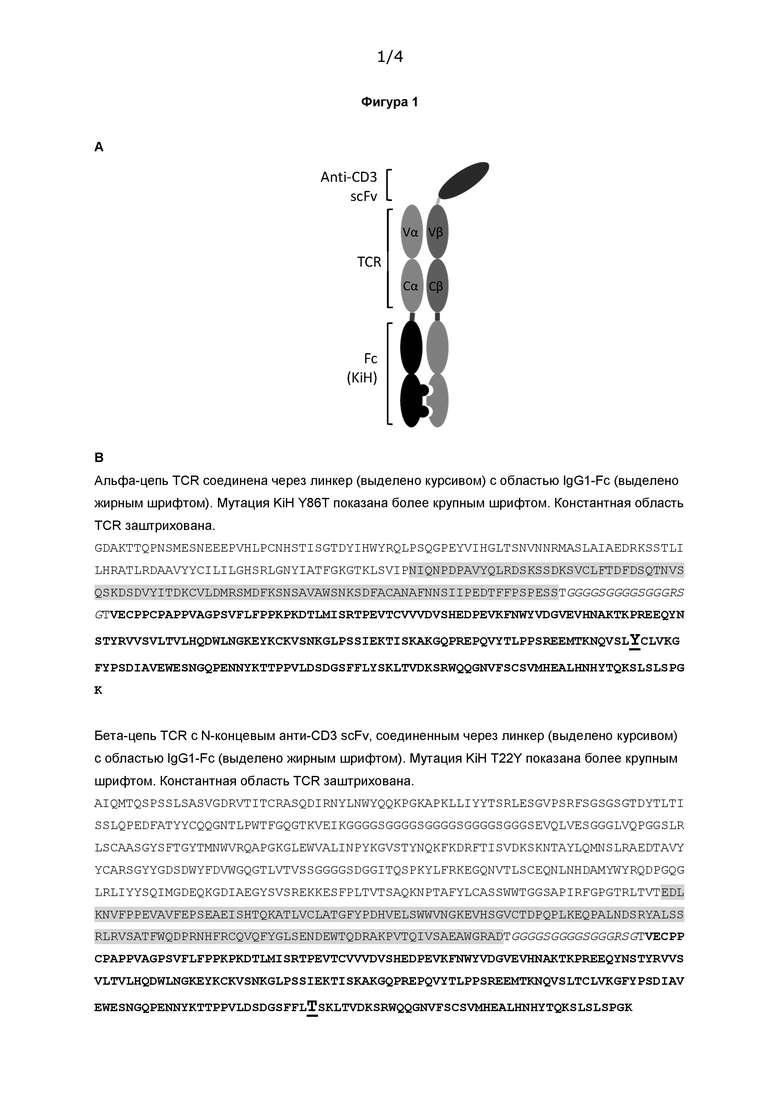
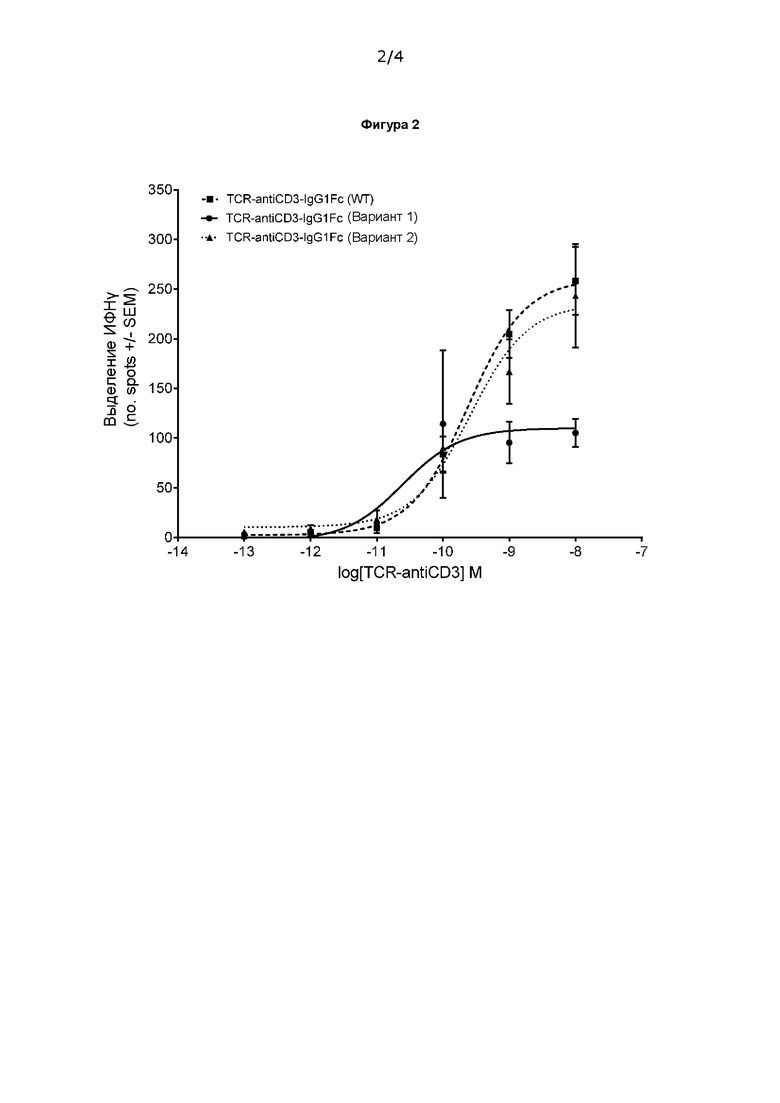
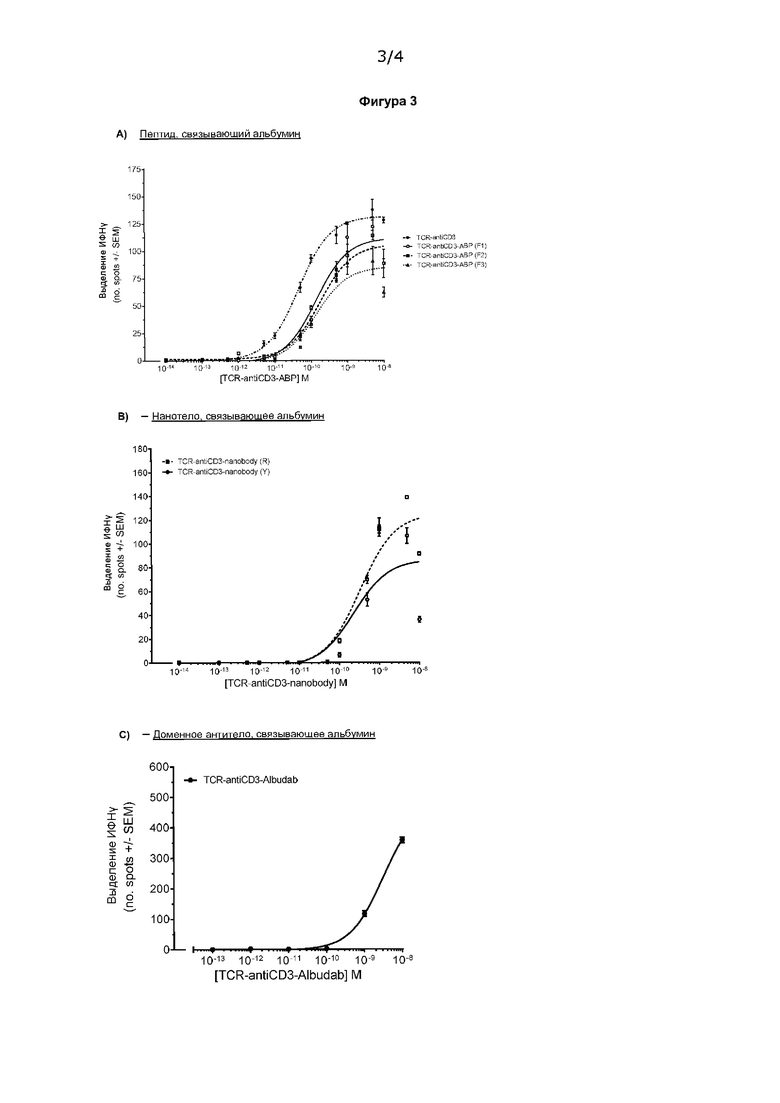
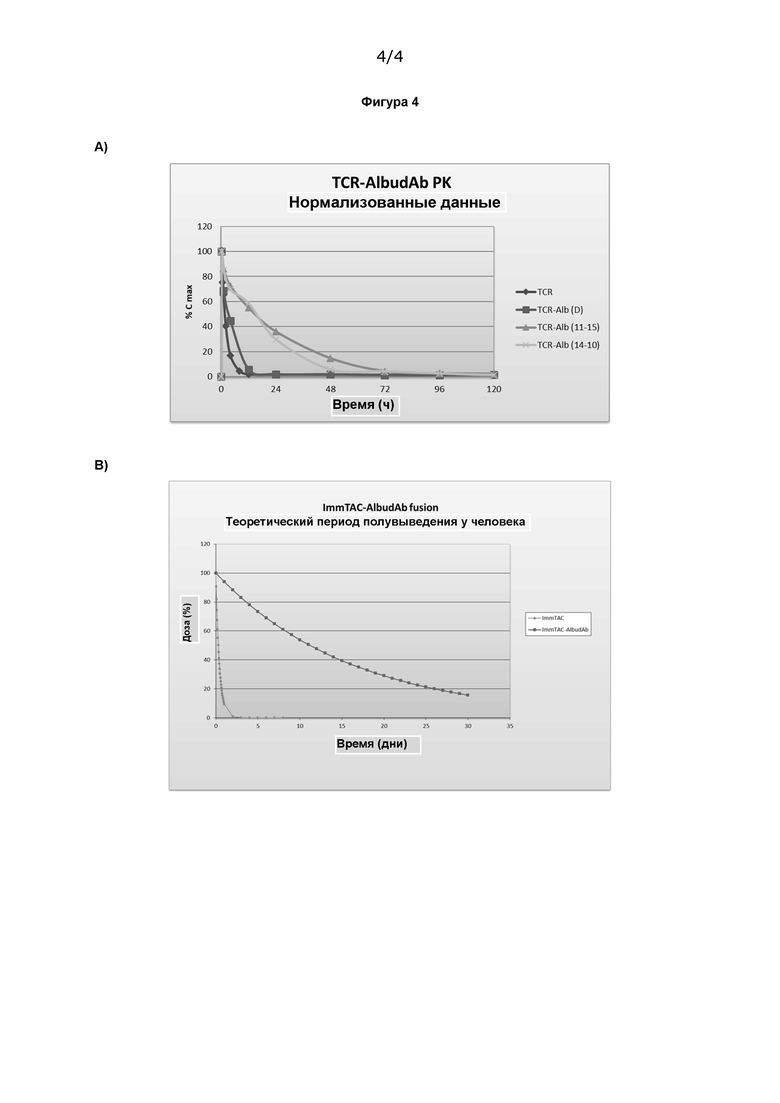
Изобретение относится к области биотехнологии, а именно к мультидоменной связывающей молекуле, способной связывать пептид-главный комплекс гистосовместимости (pMHC), которая содержит фрагмент, связывающий pMHC, который связан с иммунным эффектором, привлекающим Т-клетки, и домен, увеличивающий период полувыведения, содержащий Fc иммуноглобулина или домен, связывающий альбумин, содержащей ее композиции, а также к способу ее получения. Также раскрыты экспрессионный вектор, содержащий нуклеиновую кислоту, кодирующую вышеуказанную мультидоменную связывающую молекулу, а также клетка, его содержащая. Изобретение эффективно для лечения рака или опухоли. 5 н. и 9 з.п. ф-лы, 4 ил., 3 пр.
1. Мультидоменная связывающая молекула, способная связывать пептид-главный комплекс гистосовместимости (pMHC), которая содержит:
i) фрагмент, связывающий pMHC, который связан с иммунным эффектором, привлекающим Т-клетки; и
ii) домен, увеличивающий период полувыведения, содержащий Fc иммуноглобулина или домен, связывающий альбумин;
причем указанный домен, увеличивающий период полувыведения, связан с С- или N-концом указанного фрагмента, связывающего pMHC, или с С- или N-концом указанного иммунного эффектора, привлекающего Т-клетки,
причем указанный фрагмент, связывающий pMHC, представляет собой Т-клеточный рецептор (TCR), содержащий вариабельные домены TCR;
при этом указанный иммунный эффектор, привлекающий Т-клетки, представляет собой эффекторный домен CD3, который активирует Т-клетку посредством взаимодействия с CD3 и/или комплексом TCR/CD3.
2. Мультидоменная связывающая молекула по п. 1, отличающаяся тем, что указанный фрагмент, связывающий pMHC, содержит по меньшей мере один константный домен.
3. Мультидоменная связывающая молекула по п.1, отличающаяся тем, что указанный TCR представляет собой пару гетеродимерных полипептидов альфа/бета TCR.
4. Мультидоменная связывающая молекула по п.1, отличающаяся тем, что указанный TCR представляет собой одноцепочечный полипептид альфа/бета TCR.
5. Мультидоменная связывающая молекула по любому из пп. 1-4, отличающаяся тем, что указанный эффекторный домен CD3 содержит scFv антитела или антитело-подобный каркас.
6. Мультидоменная связывающая молекула по любому из пп. 1-5, отличающаяся тем, что указанный домен, увеличивающий период полувыведения, представляет собой домен Fc иммуноглобулина.
7. Мультидоменная связывающая молекула по любому из пп. 1-5, отличающаяся тем, что указанный домен, увеличивающий период полувыведения, содержит домен, связывающий альбумин.
8. Мультидоменная связывающая молекула по любому из пп. 1-6, отличающаяся тем, что указанный домен, увеличивающий период полувыведения, соединен с указанным фрагментом, связывающим pMHC, или с указанным иммунным эффектором, привлекающим Т-клетки, через линкер.
9. Мультидоменная связывающая молекула по любому из предшествующих пунктов для применения в качестве лекарственного средства при лечении рака или опухоли.
10. Фармацевтическая композиция для применения при лечении рака или опухоли, содержащая мультидоменную связывающую молекулу по любому из пп. 1-8 и по меньшей мере один фармацевтически приемлемый носитель и/или фармацевтически приемлемое вспомогательное вещество.
11. Экспрессионный вектор, содержащий нуклеиновую кислоту, кодирующую мультидоменную связывающую молекулу по любому из пп. 1-8.
12. Клетка-хозяин для экспрессии мультидоменной связывающей молекулы, содержащая вектор по п. 11, при этом нуклеиновая кислота, кодирующая мультидоменную связывающую молекулу, представлена в указанном векторе в виде одной открытой рамки считывания или двух отдельных открытых рамок считывания, кодирующих альфа-цепь и бета-цепь, соответственно.
13. Способ получения мультидоменной связывающей молекулы по п. 1, включающий поддержание клетки-хозяина по п. 12 в оптимальных условиях для экспрессии нуклеиновой кислоты и выделение мультидоменного антигенсвязывающего полипептида.
14. Способ лечения рака или опухоли, включающий введение мультидоменной связывающей молекулы по любому из пп. 1-8 или фармацевтической композиции по п.10 пациенту, который в этом нуждается.
| WO 2018234319 A1, 27.12.2018 | |||
| WO 2018132597 A1, 19.07.2018 | |||
| WO 2017208018 A1, 07.12.2017 | |||
| GAOWEI FAN et al | |||
| Bispecific antibodies and their applications, J Hematol Oncol, 2015, Vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Реверсивный дисковый культиватор для тросовой тяги | 1923 |
|
SU130A1 |
| ВДОВИН А.С.и др | |||
| Т-лимфоциты с модифицированной специфичностью в терапии злокачественных заболеваний, МОЛЕКУЛЯРНАЯ БИОЛОГИЯ, 2017, том 51, N 6, с | |||
| ПЛУГ ДЛЯ РАЗГРУЗКИ БАЛЛАСТНЫХ ПЛАТФОРМ | 1923 |
|
SU1008A1 |

