Изобретение относится к области молекулярной биологии, биотехнологии и регенеративной медицины и может быть использовано при экспериментальном моделировании репаративной регенерации in vivo, в частности, регенерации кожных ран.
Известно, что регенерация кожных ран млекопитающих является сложным многокомпонентным процессом. У млекопитающих, в том числе и у человека, сильно снижена способность к репаративной регенерации по сравнению с другими классами позвоночных животных. Низкие темпы заживления ран у человека составляют предпосылки для развития современных технологий регенеративной медицины. В здравоохранении 21 века проблемы ранозаживления постепенно начинают занимать все большее место, их называют "молчащей эпидемией" [Lindholm С, Searle R. Wound management for the 21st century: combining effectiveness and efficiency. Int Wound J. 2016 Jul;13 Suppl 2(Suppl 2):5-15. doi: 10.1111/iwj.12623]. Было показано, что на 1 миллион жителей приходится около 3500 человек с ранами, из них у 525 человек раны сохраняются дольше 1 года (длительно не заживающие раны). Помимо этого, существуют проблемы возникновения келоидных и гипертрофических рубцов как частое осложнение при заживлении ран, возникающих после травм, хирургических вмешательств, ожогов и любых процессов, вызывающих воспаление кожи.
Для детального изучения особенностей регенерации используют экспериментальные модели лабораторных животных. Процесс ранозаживления у млекопитающих после рождения сильно отличается от того, что происходит с раневой поверхностью на эмбриональной стадии развития, когда заживление происходит без образования рубцов [Satish L, Kathju S. Cellular and molecular characteristics of scarless versus fibrotic wound healing. Dermatol Res Pract. 2010;2010:790234. doi: 10.1155/2010/790234]. Изучение опыта применение хирургического метода внутриутробного удаления амниотических перетяжек, например, конечностей плода крупных млекопитающих, показывает что заживление раны происходит без образования рубцов вплоть до 60 дня гестации [Soldado F, Aguirre М,  Fontecha CG, Esteves M, Velez R,
Fontecha CG, Esteves M, Velez R, Fetal surgery of extremity amniotic bands: an experimental model of in utero limb salvage in fetal lamb. J Pediatr Orthop.2009 Jan-Feb;29(l):98-102. doi: 10.1097/ВРО.0b013e318192196e]. После рождения у млекопитающих происходит переключение молекулярных механизмов управляющих процессом регенерации таким образом, что регенеративный потенциал снижается.
Fetal surgery of extremity amniotic bands: an experimental model of in utero limb salvage in fetal lamb. J Pediatr Orthop.2009 Jan-Feb;29(l):98-102. doi: 10.1097/ВРО.0b013e318192196e]. После рождения у млекопитающих происходит переключение молекулярных механизмов управляющих процессом регенерации таким образом, что регенеративный потенциал снижается.
Механизмы регенерации и ее переключения являются предметом активного изучения, в том числе на моделях эмбрионов мыши [Wilgus ТА. А murine incisional fetal wound-healing model to study scarless and fibrotic skin repair. Methods Mol Biol. 2021;2193:13-21. doi: 10.1007/978-1-0716-0845-6 2]. После рождения у млекопитающих остается способность к регенерации конечностей лишь на уровне дистальных частей фаланги пальцев [Fernando WA, Leininger Е, Simkin J, Li N, Malcom CA, Sathyamoorthi S, Han M, Muneoka K. Wound healing and blastema formation in regenerating digit tips of adult mice. Dev Biol. 2011 Feb 15;350(2):301-10. doi: 10.1016/j.ydbio. 2010.11.035]. Показано, что наличие грануляционной ткани и привлечение миофибробластов, осуществляющих усиленное депонирование коллагеновых волокон и альфа-актина играет ключевую роль в фиброзном перерождении ткани после заживления раны. При этом существует большое разнообразие гипотез в поддержку того или иного терапевтического подхода, направленного на снижение процессов формирования патологических рубцов при ранозаживлении [Haverstock BD. Hypertrophic scars and keloids. Clin Podiatr Med Surg. 2001 Jan;18(l): 147-59. PMID: 11344975].
Регуляторный каскад AGR играет важную роль в контроле регенерации у низших позвоночных [Ivanova AS, Shandarin IN, Ermakova GV, Minin AA, Tereshina MB, Zaraisky AG. The secreted factor Agl missing in higher vertebrates regulates fins regeneration in Danio rerio. Sci Rep. 2015 Jan 29;5:8123. doi: 10.1038/srep08123; Kumar A, Godwin JW, Gates PB, Garza-Garcia AA, Brockes JP. Molecular basis for the nerve dependence of limb regeneration in an adult vertebrate. Science. 2007 Nov 2;318(5851):772-7. doi: 10.1126/science.1147710]. У млекопитающих, характеризующихся снижением или потерей способности к регенерации, часть генов этого каскада утеряна в процессе эволюции. Показано, что белок xAG является ключевым фактором, ускоряющим регенерацию головастиков шпорцевой лягушки, однако для оценки влияния xAg на регенеративный потенциал млекопитающих необходимо создать адекватную модель изучения [Ivanova AS, Tereshina MB, Araslanova KR, Martynova NY, Zaraisky AG. The secreted protein disulfide isomerase Agl lost by ancestors of poorly regenerating vertebrates is required for Xenopus laevis tail regeneration. Front Cell Dev Biol. 2021 Oct 5;9:738940. doi: 10.3389/fcell.2021.738940]. Поскольку экспрессия чужеродного белка может оказывать непредсказуемое воздействие на организм, в том числе и в процессе эмбриогенеза, важным подходом при проектировании животной модели, является создание системы индуцибельной экспрессии целевого гена.
Из уровня техники известны технологии получения линий трансгенных мышей, используемых для тестирования вакцин против COVID-19, а также для детального изучения механизмов проникновения вируса SARS-CoV-2 (RU 2 757 114 С1, ФГБУН Институт биологии гена Российской академии наук, 11.10.2021) (1). Известный способ получения трансгенных мышей включает операции: получение генетической конструкции с CAG-промотором и STOP-кассетой, терминирующей транскрипцию с данного промотора и фланкированной LoxP-сайтам, получение линии трансгенных мышей, путем инъекции экспрессионного вектора в пронуклеус и последующей подсадке трансгенных эмбрионов самкам-реципиентам. В результате была получена мышь, в геном которой включен ген hACE2 Homo sapiens, экспрессия которого индуцируется тканеспецифично путем введения Cre-рекомбиназы. Однако в данной системе не предусмотрена система индуцибельной активации транскрипции, позволяющей регулировать экспрессию целевого трансгена. Кроме того, известный способ не касается исследование репаративной регенерации кожных ран.
Самым распространенным подходом для ускорения регенерации человека является применение широкого спектра лекарственных средств различного механизма действия. Так, известен способ стимуляции регенерации тканей при моделировании кожной раны у мыши с помощью наружного применения двух факторов: SDF-1 и G-CSF (RU 2 634 576, С1). В другом патенте хлоргидрат N-изопропиламид-2-(1-фенилэтил)аминоэтансульфокислоты указан как агент, ускоряющий процессы клеточной регенерации (RU 2 270 007, С2).
В обоих случаях стимуляция регенерации осуществляется за счет повторяющего введения заявленных веществ, а не за счет увеличения регенеративного потенциала организма.
Настоящее изобретение направлено на решение технической проблемы создания трансгенных мышей с индуцированной экспрессией генов, активных в регенерационных процессах низших позвоночных.
Нами поставлена задача получить и использовать генетически модифицированного грызуна как экспериментальной модели для анализа репаративной регенерации in vivo.
Технический результат, достигаемый при осуществлении изобретения заключается в достижении стимуляции регенеративных способностей созданного трансгенного животного (модели) под воздействием эндогенной экспрессии факторов регенерации, активных в организмах низших позвоночных.
Сущность изобретения заключается в следующем.
Предложено применение трансгенной мыши с индуцируемой экспрессией гена xAG шпорцевой лягушки Xenopus laevis в качестве модельного животного для исследования репаративной регенерации кожной раны.
Созданная модель может быть использована для изучения процессов репаративной регенерации после индукции экспрессии полипептида xAG шпорцевой лягушки, активных в процессе регенерации у личиночных форм. Изобретение может быть использовано для проверки регенеративного потенциала тканей in vivo, в частности, регенерации кожных ран.
Существо изобретения поясняется на следующих фигурах.
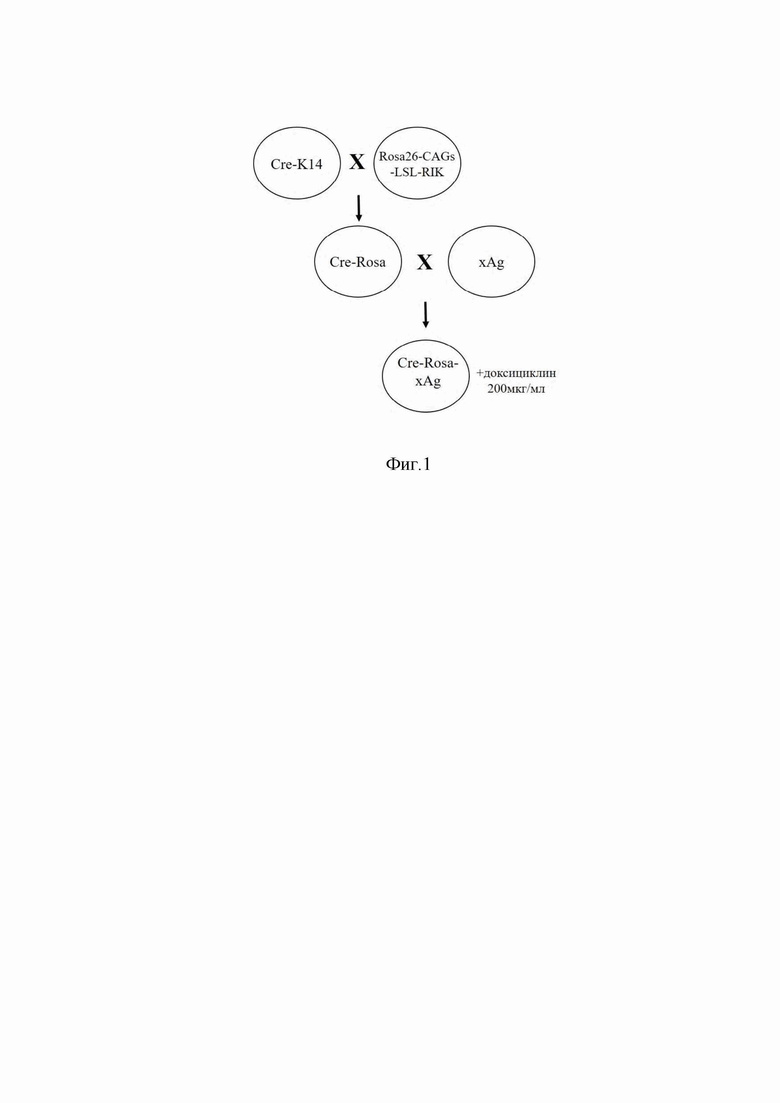
Фиг. 1 показывает схему получения трансгенных мышей, способных к индуцированной активации экспрессии xAG.
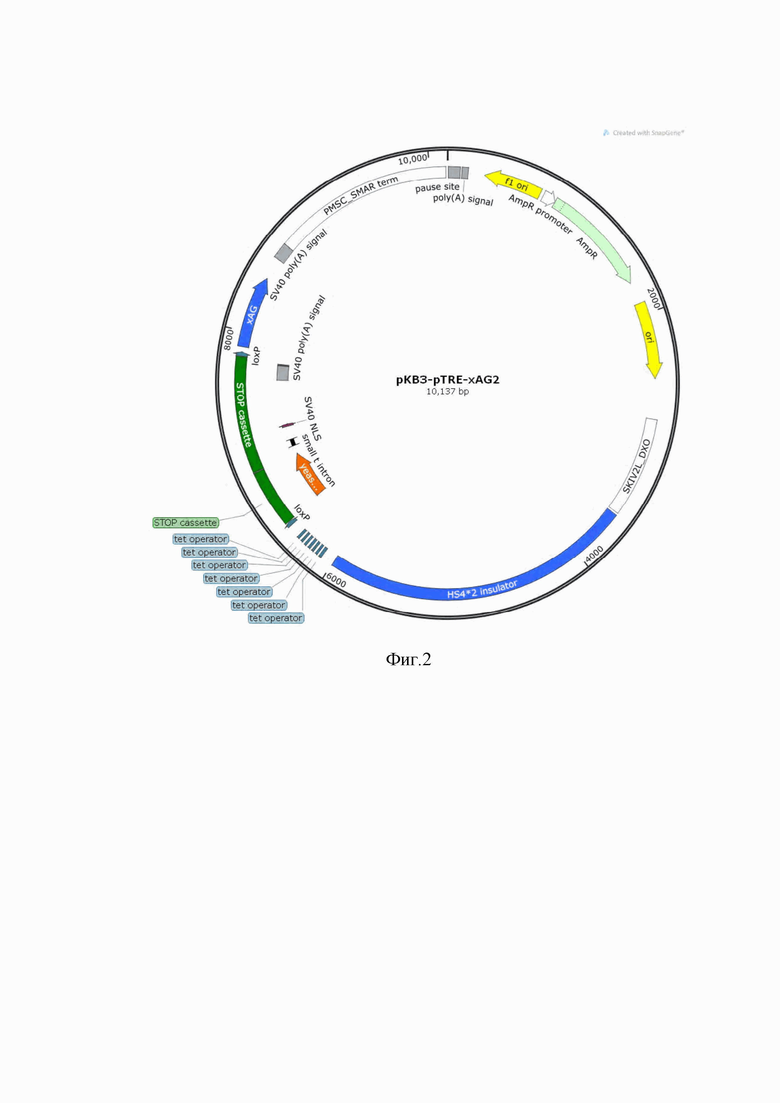
Фиг. 2. показывает схему плазмиды pKB2-xAG.
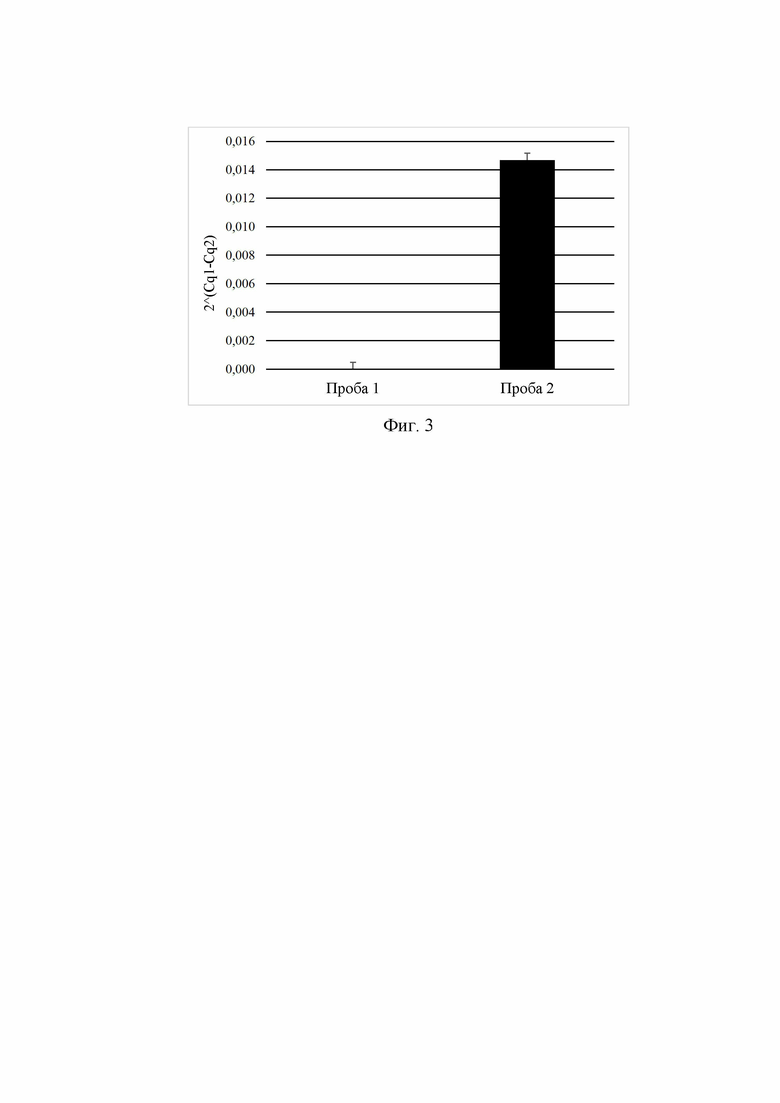
Фиг. 3. иллюстрирует оценку уровня экспрессии мРНК трансгена xAG в тканях мышей до и после активации доксициклином.
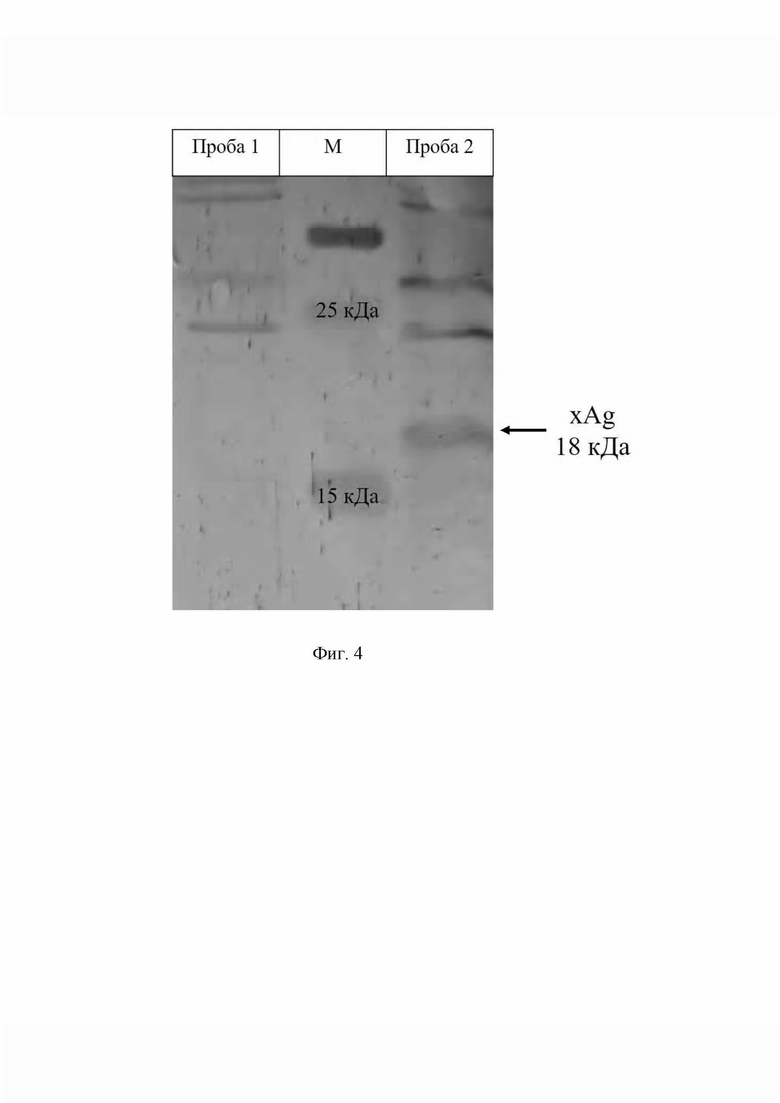
Фиг. 4. иллюстрирует оценку уровня экспрессии xAG в тканях трансгенных мышей.
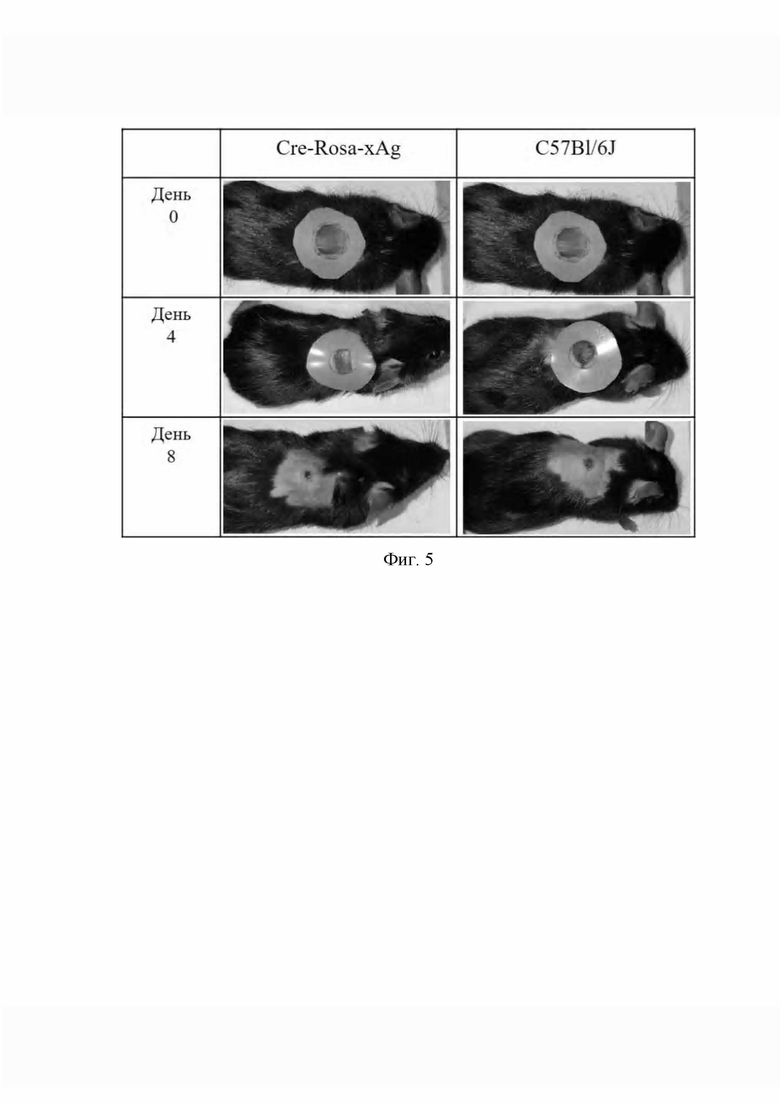
Фиг. 5. демонстрирует модель полнослойной раны у мышей.
Фиг. 6. демонстрирует модель эксцизионной ушной раны мыши.
Патентуемое изобретение предусматривает получение трансгенных мышей с последовательным проведением следующих этапов.
1. Получение линеаризованного вектора pKB2-xAG с последовательностью SEC ID NO: 1 на основе плазмиды NIF (1), с геном устойчивости к ампициллину, с инсуляторами и терминатором для защиты трансгена от эффекта положения, STOP-кассетой, фланкированной LoxP сайтами, с внесенным множественным сайтом клонирования кДНК xAG, под контролем Tet-On промотора. Рестрикция вектора pKB2-xAG эндонуклеазой PvuI с удалением части бактериальных последовательностей.
2. Создание мышей, несущих ген xAg, по следующей схеме:
а) проведение микроинъекции плазмидной ДНК из п. 1 в мужской пронуклеус зиготы мыши гибрида F1(CBA X C57BL/6) на стадии двух пронуклеусов;
б) выявление и отбор жизнеспособных эмбрионов;
в) подсадка выживших эмбрионов из п. 26 псевдобеременным самкам-реципиентам линии ICR;
г) получение новорожденных мышей на 19 день после пересадки зигот из п. 2в;
д) ДНК-анализ мышей из п. 2 г на наличие последовательности xAg с использованием прямого праймера FW-xAG, имеющего последовательность SEC ID NO: 2 и обратного праймера RV-xAG, имеющего последовательность SEC ID NO: 3;
е) отбор мышей, содержащих xAg.
3. Получение линии мышей - двойных трансгенов Cre-Rosa:
а) скрещивание мышей линии B6N.Cg-Tg(KRT14-cre)lAmc/J Stock No: 018964 с мышами линии B6.Cg-Gt(ROSA)26Sortm2 (CAG-rtTA3,-mKate2)Slowe/J Stock No: 029633;
б) получение новорожденных мышей на 21-23 сутки после скрещивания;
в) ДНК-анализ мышей из п. 36 на наличие последовательности Cre и Rtta с использованием с использованием прямого праймера FW-Cre, имеющего последовательность SEC ID NO: 4, обратного праймера RV-Cre, имеющего последовательность SEC ID NO: 5, прямого праймера FW-Rtta, имеющего последовательность SEC ID NO: 6, обратного праймера RV-Rtta, имеющего последовательность SEC ID NO: 7;
г) отбор трансгенных мышей Cre-Rosa.
4. Получение итоговой линии мышей - тройных трансгенов Cre-Rosa- xAg:
а) скрещивание мышей, несущих Cre-Rosa с трансгенными мышами xAg;
б) получение новорожденных мышей на 21-23 сутки после ссадки;
в) ДНК-анализ мышей из п. 4б на наличие последовательности Cre, Rtta и xAg с использованием праймеров, указанных в п. 2д и п. 3в;
г) отбор трансгенных мышей Cre-Rosa-xAg.
5. Активация экспрессии xAg in vivo путем администрации доксициклина мышам Cre-Rosa-xAg из п. 4 г. Схема проведения скрещивания для получения целевого трансгена Cre-Rosa-xAg и индукции экспрессии xAg представлена на фиг.1.
6. Подтверждение экспрессии xAg с помощью ОТ-ПНР.
Способ осуществляется следующим образом.
Для создания генетических конструкций получали кДНК гена xAg, на основе суммарной мРНК, выделенной из ткани бластемы регенерирующего хвоста головастика шпорцевой лягушки Xenopus laevis, диссоциированной в тризоле. Осаждение проводили по стандартной процедуре на основе этанола в присутствии гликогена (Fermentas). Обратную транскрипцию проводили с использованием обратной транскриптазы M-MLV (Promega) в присутствии oligo-dT праймера (Евроген). Для амплификации гена xAg использовали прямой праймер FW-xAG1 Agel, имеющий последовательность SEC ID NO: 8, обратный праймер RV-xAG1Mlul STOP, имеющий последовательность SEC ID NO: 9, и Q5 HF ДНК-полимеразу (СкайДжин). Полученный фрагмент затем разрезали рестриктазами MluI-FD и BshTI-FD (СибЭнзим) и лигировали в вектор. Схема плазмиды pKB2-xAG представлена на фиг. 2. Tet-On промотор содержит 7 операторов (tet operator), активация работы этого промотора происходит под действием тетрациклина/доксициклина. STOP-кассета содержит интрон и сигнал ядерной локализации. Фрагмент кодирующей части гена xAG отделен от Tet-On промотора STOP-кассетой, в присутствии STOP-кассеты экспрессии трансгена не происходит.LoxP последовательности фланкируют STOP-кассету, что позволяет избирательно удалить STOP-кассету с помощью Cre рекомбиназы. SV40polyA и PMSC SMARterm - сигнал полиаденилирования и терминатор транскрипции трансгена. Ген устойчивости к ампициллину AmpR выделен зеленым цветом. Ori репликации плазмиды выделен желтым цветом. HS4 insulator - последовательность инсулятора, выделенная синим, стабилизирующего экспрессию трансгена от воздействия генного окружения. Последовательность SKIV 2L DXO, выделенная белым, также является инсулятором.
Лигазной смесью трансформировали компетентные клетки DH5a. Полученные колонии анализировали, отбирали и наращивали культуру для выделения плазмидной ДНК. Перед использованием плазмиду проверяли на соответствие с использованием прямого праймера FW-xAG, имеющего последовательность SEC ID NO: 2 и обратного праймера RV-xAG, имеющего последовательность SEC ID NO: 3, после чего линеаризовали с использованием эндонуклеазы рестрикции PvuI, очищали в агарозном геле и выделяли с помощью набора QIAquick Gel Extraction Kit (Qiagen), и разводили до рабочей концентрации - 1нг/мкл.
Эмбрионы мыши для инъекции плазмиды выделяли из неполовозрелых самок-гибридов F1(CBA×C57BL/6) в возрасте 3-3,5 недель после стимуляции суперовуляции по схеме: гормон PMS инъецировали животным в 15:00, через 48-50 часов вводили HCG; через час после последней инъекции гормонов самок подсаживали к самцам той же линии; на следующее утро после ссадки отбирали самок с влагалищными пробками, образующимися после прошедшего коитуса. Животные содержались в помещении с автоматизированным освещением, обеспечивающим световой цикл 12/12 с 8.00 до 20.00. Зиготы выделяли на следующее утро после последней инъекции гормонов. Самок умертвляли путем дислокации шейных позвонков, вырезали комплекс яичника с яйцеводом и рогом матки. Из ампулы яйцевода выделяли зиготы с клетками кумулюса в среду М2 (Sigma-Aldrich, США) с добавлением гиалуронидазы (Sigma-Aldrich, США). Зиготы последовательно отмывали в 3 сменах среды М2.
Микроинъекции зигот проводили в среде М2 (Sigma-Aldrich, США) под микроскопом Zeiss Axiovert 200М при помощи микроманипуляторов Narishige. После проведения микроинъекций зиготы культивировали в течение 1 часа, после чего проводили отбор жизнеспособных эмбрионов.
Для получения вазектомированных самцов использовали животных линии ICR возрастом 12-18 недель. Самцов наркотизировали внутрибрюшинным введением раствора авертина (2,2,2-бромэтанола) (Sigma-Aldrich, США) в концентрации 250 мг на кг веса. На брюшной стороне делали продольный разрез кожи и брюшины и извлекали семенник с придатками из полости тела. Семявыносящий проток отсепаровывали от соединительной ткани, обрезали участок протока размером около 5-8 мм и прижигали образовавшиеся кончики. Семенник с придатками возвращали обратно в полость тела и повторяли процедуру для второго семявыносящего протока. После операции края брюшины и кожи послойно зашивали резорбируемым шовным материалом (Ethicon, США).
Для получения псевдобеременных самок-реципиентов самок линии ICR возрастом 12-18 недель, находящихся в фазе позднего проэструса, либо начала эструса, ссаживали с вазектомированными самцами той же линии. На следующий день, до 12 часов, проводили осмотр и самок с влагалищными пробками, образующимися после прошедшего коитуса, отбирали для последующих подсадок.
Для подсадки эмбрионов самок-реципиентов наркотизировали внутрибрюшинным введением раствора авертина (2,2,2-бромэтанола) (Sigma-Aldrich, США) в концентрации 250 мг на кг веса. Через последовательные разрезы кожи и брюшины со спинной стороны в воронку яйцевода вводили выжившие после инъекции эмбрионы по 6-10 штук на животное.
Новорожденных мышей получали при помощи кесарева сечения. На 19 сутки после подсадки микроинъецированных эмбрионов самку-реципиента умерщвляли путем дислокации шейных позвонков и проводили кесарево сечение, после чего выживших детенышей помещали к заранее подготовленной самке-кормилице. Через 14-21 сутки после рождения у мышат брали образец ткани, который инкубировали в растворе протеиназы К в течение 12-15 часов при 56°С, после чего выделяли ДНК и анализировали наличие трансгена методом ПНР.
Анализ наличия xAg в геноме мышей проводили при помощи метода ПНР. ДНК выделяли из тканей по стандартному протоколу. Амплификацию фрагментов ДНК проводили с помощью набора GenPak™ PCR Core (Isogene) в присутствии 10 пМ прямого праймера FW-xAG, имеющего последовательность SEC ID NO: 2, и обратного праймера RV-xAG, имеющего последовательность SEC ID NO: 3, в следующих режимах: денатурация 95°С -3 мин; далее следовало 35 циклов: 95°С - 40 сек, 58°С - 40 сек; 72°С - 40 сек; и последний синтез 72°С - 2 мин.
Для получения двойных трансгенов Cre-Rosa мышей линии B6N.Cg-Tg(KRT14-cre)lAmc/J Stock No: 018964 скрещивали с мышами линии B6.Cg-Gt(ROSA)26Sortm2 (CAG-rtTA3,-mKate2)Slowe/J Stock No: 029633 и через 14-21 сутки после наступления естественных родов мышат анализировали на наличие последовательности Cre и Rtta методом ПЦР, описанном выше, с использованием прямого праймера FW-Cre, имеющего последовательность SEC ID NO: 4, обратного праймера RV-Cre, имеющего последовательность SEC ID NO: 5, прямого праймера FW-Rtta, имеющего последовательность SEC ID NO: 6, обратного праймера RV-Rtta, имеющего последовательность SEC ID NO: 7.
Для получения целевых трансгенов Cre-Rosa-xAg отобранных ранее трансгенных мышей xAg скрещивали с мышами, несущими Cre-Rosa, и через 14-21 сутки после наступления естественных родов мышат также анализировали на наличие последовательности Cre, Rtta и xAG методом ПНР, описанном выше, с использованием прямого праймера FW-xAG, имеющего последовательность SEC ID NO: 2, обратного праймера RV-xAG, имеющего последовательность SEC ID NO: 3, прямого праймера FW-Cre, имеющего последовательность SEC ID NO: 4, обратного праймера RV-Cre, имеющего последовательность SEC ID NO: 5, прямого праймера FW-Rtta, имеющего последовательность SEC ID NO: 6, обратного праймера RV-Rtta, имеющего последовательность SEC ID NO: 7.
Процесс активации экспрессии xAg проходил в два этапа. Под воздействием Cre-рекомбиназы, экспрессирующейся под промотором К14, полученной от мышей линии B6N.Cg-Tg(KRT14-cre)lAmc/J Stock No: 018964, произошло вырезание СТОП-кассеты в кератиноцитах целевых трансгенов Cre-Rosa-xAg. Оверэкспрессия xAg была индуцирована посредством доксициклина, добавленного в питьевую воду животных в концентрации 200 мкг/мл.
Проверка экспрессии xAG проводилась с помощью метода ОТ-ПЦР и вестерн-блота. Для проведения ОТ-ПЦР суммарную РНК из тканей отобранных мышей Cre-Rosa-xAg выделяли с помощью стандартного протокола с использованием тризола. Для снижения количества геномной ДНК в образце проводили обработку ДНКазой с последующим ее ингибированием ЭДТА. Синтез первой цепи кДНК проводили с помощью обратной транскриптазы MMLV (Евроген) с использованием случайной затравки. В качестве контроля для ОТ-ПЦР использовали образец РНК до обратной транскрипции (NRT образец), образец без добавления матрицы (NTC образец).
В ПЦР в реальном времени оценивали уровень экспрессии гена xAG у мышей до и после активации экспрессии - через 7 суток после администрации доксициклина. Для оценки экспрессии гена xAG использовали прямой праймер FW-xAG-RT, имеющий последовательность SEC ID NO: 16 и обратный праймер RV-xAG-RT, имеющий последовательность SEC ID NO: 17, и вычисляли значение Cq1. В качестве нормировки уровня экспрессии использовали гены «домашнего хозяйства» GAPDH, ODC и EF1 (для них подсчитали усредненное значение Cq2), для амплификации которых были использованы следующие праймеры: прямой праймер FW-GAPDH, имеющий последовательность SEC ID NO: 10, обратный праймер RV-GAPDH, имеющий последовательность SEC ID NO: 11, прямой праймер FW-ODC, имеющий последовательность SEC ID NO: 12, обратный праймер RV-ODC, имеющий последовательность SEC ID NO: 13, прямой праймер FW-EF1, имеющий последовательность SEC ID NO: 14, обратный праймер RV-EF1, имеющий последовательность SEC ID NO: 15. Уровень экспрессии оценивали по формуле: 2Cq1-Cq2. На 3 показано возникновение экспрессии xAg в тканях мыши Cre-Rosa-xAg после получения животными доксициклина (проба 2) при полном остутсвии мРНК xAg у животных, в отсутствии доксициклина (проба 1).
Для проведения вестерн-блота были получены биоптаты тканей мыши Cre-Rosa-xAg до и после активации экспрессии - через 7 суток после администрации доксициклина. Образцы диссоциировали в лизирующем буфере, белки разделяли электрофорезом в полиакриламидном геле в присутствии додецилсульфата натрия, переносили на мембрану и инкубировали с антителами к xAg. На фиг. 4 представлены результаты анализа растворимой фракции суммарных лизатов. Показано, что только в пробе под номером 2, соответствующей лизату тканей мыши Cre-Rosa-xAg после активации экспрессии доксициклином, присутствует белок размером 18кДа (размер определен в соответствии с нанесенным в средней колонке маркером, обозначенным М), что соответствует белку xAg. В то время как в контрольной пробе под номером 1, полученной от неактивированной мыши, отсутствует полоса, соответствующая белку xAg.
Изучение регенеративного потенциала трансгенных мышей Cre-Rosa-xAg, полученных по способу, описанному на страницах 7-11, может быть осуществлено следующим образом.
Пример 1. Модель заживления полнослойной кожной раны мыши.
Эксперимент проводили на мышах в возрасте 10-12 недель на момент начала эксперимента. В качестве контроля выступали мыши линии C57B1/6J, являющиеся генетическим бэкграундом для используемых трансгенных линий мышей. На протяжении всего эксперимента все животные получали доксициклин.
Животных взвешивали, и наркотизировали внутрибрюшинным введением 2,2,2,-трибромэтанола (250 мг/кг), обезболивали капрофеном (5 мг/кг), операционное поле обрабатывали антисептическим 2% раствором йода и этиловым спиртом. На дорсальной стороне между лопаток вырезали округлый участок кожи диаметром 7-10 мм. На рану накладывали шину из силикона в виде кольца с отверстием, соответствующим диаметру раны, предварительно смазанного клеем. Размеры раны замеряли сразу после нанесения и далее через день до полного закрытия ран в контрольной группе. Скорость закрытия раны была оценена в процентах от первоначального размера нанесенной раны, принятого за 100%. На фиг. 5 показано, что к 8 суткам в опытной группе, состоящей из активированных мышей Cre-Rosa-xAg, средние размеры раны были меньше в сравнении с контрольной группой мышей C57B1/6J (27,91% против 43,75% по результатам измерений). Полное закрытие раны происходило к 13 суткам эксперимента. Далее животных умерщвляли путем передозировки трехкратной дозой 2,2,2,-трибромэтанола и забирали ткани для выделения РНК и гистологического исследования.
Пример 2. Модель эксцизионной ушной раны.
Экспериментальные группы животных были сформированы аналогично тому, как описано в Примере 1. В центре каждого уха пробивали сквозное отверстие диаметром 2 мм с помощью пробойника для клинической биопсии. Размеры раны замеряли сразу после нанесения и раз в неделю до полной остановки процессов зарастания раны. Скорость закрытия раны была оценена в процентах от первоначального размера нанесенной раны, принятого за 100%. Было показано ускоренное закрытие пробоя в экспериментальной группе в первую неделю после нанесения повреждения в сравнении с контрольной группой, однако полного закрытия повреждения не происходило. Через 7 суток после операции средняя площадь раны ушной раковины у активированных мышей Cre-Rosa-xAg, составила 58,62%, в противоположность контрольным мышам C57B1/6J с показателем в 90,48%.
Изменения размеров ушной раны с момента нанесения и на 7 сутки после операции представлены на фиг. 6.
Таким образом, созданный нами генетически модифицированный грызун может быть использован как экспериментальная модель для анализа репаративной регенерации in vivo.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения линии гуманизированных мышей, трансгенных по hACE2 | 2020 |
|
RU2757114C1 |
| Способ получения генно-модифицированных лабораторных животных с индуцируемой системной и тканеспецифической экспрессией циклофилин А человека | 2022 |
|
RU2800264C1 |
| Способ получения генно-модифицированных мышей, экспрессирующих миниген антитромбина III человека, с помощью микроинъекций TelN-линеаризованного фрагмента ДНК | 2022 |
|
RU2806568C1 |
| Набор для определения копийности гена АРР в геноме человека | 2021 |
|
RU2789799C1 |
| Набор для определения копийности вставки интересующей конструкции в AAVS1 локус генома человека | 2021 |
|
RU2786396C1 |
| ГЕННО-ИНЖЕНЕРНАЯ КОНСТРУКЦИЯ pGoatcasGCSF, ОБЕСПЕЧИВАЮЩАЯ ПРОДУКЦИЮ ГРАНУЛОЦИТ-КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА ЧЕЛОВЕКА В МОЛОКО ТРАНСГЕННЫХ ЖИВОТНЫХ | 2010 |
|
RU2422529C1 |
| Линия мышей, трансгенных по альфа-цепи Т-клеточного рецептора клеток памяти, для изучения их функциональной активности | 2017 |
|
RU2691484C2 |
| ПОЛУЧЕНИЕ АНТИТЕЛ ИЗ ОДНИХ ТЯЖЕЛЫХ ЦЕПЕЙ В ТРАНСГЕННЫХ ЖИВОТНЫХ | 2007 |
|
RU2435784C2 |
| ТРАНСГЕННОЕ ЖИВОТНОЕ, ОТЛИЧНОЕ ОТ ЧЕЛОВЕКА, И ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2579701C2 |
| Способ получения мышиной модели для изучения синдрома Леша-Нихена путем внесения делеции p.Val8del в ген hprt1 | 2021 |
|
RU2768048C1 |

Изобретение относится к области биотехнологии, в частности к применению трансгенной мыши с индуцируемой экспрессией гена xAG шпорцевой лягушки Xenopus laevis со STOP-кассетой и под контролем Tet-On промотора в качестве модельного животного для исследования репаративной регенерации раны кожи. Изобретение обеспечивает достижение стимуляции регенеративных способностей созданного трансгенного животного (модели) под воздействием эндогенной экспрессии факторов регенерации, активных в организмах низших позвоночных. 6 ил., 2 пр.
Применение трансгенной мыши с индуцируемой экспрессией гена xAG шпорцевой лягушки Xenopus laevis со STOP-кассетой и под контролем Tet-On промотора в качестве модельного животного для исследования репаративной регенерации раны кожи.
| Бункер уборочной машины | 1987 |
|
SU1498423A1 |
| MASAHITO IKAWA et al., Generation of transgenic mice using lentiviral vectors: a novel preclinical assessment of lentiviral vectors for gene therapy, Molecular Therapy, 2003,Vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Комнатная печь | 1923 |
|
SU666A1 |
| IVANOVA A | |||
| S | |||
| et al, The Secreted Protein Disulfide Isomerase Ag1 Lost by Ancestors of Poorly Regenerating Vertebrates Is | |||

