1. Перекрестная ссылка на родственные заявки
В настоящей заявке испрашивается приоритет предварительной заявки США №62/699667, поданной 17 июля 2018 г, которая тем самым является включенной посредством ссылки во всей ее полноте.
2. Перечень последовательностей
Настоящая заявка содержит перечень последовательностей, который был подан посредством файловой системы электронного архивирования (EFS - от англ. electronic filing system) и включен в данный документ посредством ссылки во всей его полноте. Указанная копия ASCII, созданная 1 июля 2019 г, называется 37536US_CRF_sequencelisting.txt и имеет размер 130163 байт.
3. Предшествующий уровень техники
Нейропатия представляет собой хроническое патологическое состояние, возникающее из-за повреждения нервов. Нейропатия является обычным последствием диабета, причем нейропатия у диабетического пациента конкретно называется диабетической нейропатией. Нейропатия также может быть вызвана повреждением нервов, вызванным инфекциями (например, герпесом с ассоциированной нейропатией, возникающей после инфекции, известной как постгерпетическая невралгия; ВИЧ (вирус иммунодефицита человека)/СПИД (синдром приобретенного иммунодефицита человека); болезнью Лайма; лепрой; сифилисом и опоясывающим герпесом); аутоиммунными заболеваниями (например, ревматоидный артрит, системная волчанка и синдром Гийена-Барре); генетическими или наследственными расстройствами (например, наследственная атаксия Фридрейха и болезнь Шарко-Мари-Тута); амилоидозом; уремией; воздействием токсинов, ядов или лекарственных средств; травмами или повреждением. В некоторых случаях данная причина неизвестна, причем в данном случае нейропатия называется идиопатической нейропатией.
Независимо от причины нейропатия ассоциирована с характерными симптомами, которые отчасти зависят от анатомического места повреждения нерва (например, периферическая нейропатия, краниальная нейропатия, автономная нейропатия, очаговая нейропатия), такими как боль (невропатическая боль), другие сенсорные нарушения (например, анестезии, включающие частичную или полную потерю ощущения; и парестезии, включающие онемение, покалывание и т.д.), моторные дефекты (например, слабость, потеря рефлексов, потеря мышечной массы, спазмы, потеря точной координации и т.д.) и автономная дисфункция (например, тошнота, рвота, импотенция, головокружение, запор, диарея и т.д.).
Нейропатию традиционно лечат с использованием мер, которые управляют ассоциированными симптомами, и, когда этиология известна, посредством лечения лежащей в основе причины нейропатии. Например, использовали обезболивающие лекарственные средства или медицинские способы лечения диабета, аутоиммунных заболеваний, инфекций или недостаточностей витаминов. Однако данные способы не лечат само повреждение нервов.
Следовательно, существует потребность в эффективном способе лечения, который может предупреждать или осуществлять репарацию повреждения нервов, ассоциированного с нейропатией.
Разные факторы роста предлагали в качестве возможных средств для лечения нейропатии, и Kessler с коллегами недавно описали успешное двойное слепое, плацебо-контролируемое клиническое испытание на человеке, фаза 2, невирусной генотерапии фактором роста гепатоцитов (HGF - от англ. hepatocyte growth factor) при диабетической периферической нейропатии. Kessler et a/., Annals Clin. Transl. Neurology 2(5):465-478 (2015). См. также патент США №9963493, включенный в данный документ посредством ссылки во всей его полноте.
Принимая во внимание широкий спектр этиологий, которые вызывают нейропатию, и широкий спектр клинических проявлений нейропатии, несмотря на клинические успехи в лечении диабетической периферической нейропатии с использованием ДНК-конструкций, экспрессирующих HGF, остается потребность в дополнительных лечениях, включая лечения с использованием HGF с другими терапевтическими средствами.
4. Краткое изложение сущности изобретения
Настоящее изобретение основывается на новых данных о том, что введение ДНК-конструкции, кодирующей инсулиноподобный фактор роста 1 (IGF-1 - от англ. insulin-like growth factor 1), способной экспрессировать человеческую изоформу IGF-1, в комбинации с ДНК-конструкцией, кодирующей HGF, способной экспрессировать человеческую изоформу HGF, является эффективным в лечении симптома, ассоциированного с нейропатией. Было продемонстрировано, что эффекты лечения двумя данными ДНК-конструкциями в комбинации больше, чем эффекты лечения самой ДНК-конструкцией, кодирующей HGF (например, VM202 или pCK-HGF728). Согласно настоящему изобретению дополнительно предложены разные ДНК-конструкции, кодирующие изоформу IGF-1 или изоформу HGF, которые можно использовать для комбинированной терапии. Кроме того, в данном документе предложены способы введения ДНК-конструкций, продемонстрированных как эффективные в лечении симптомов, ассоциированных с нейропатией in vivo.
Таким образом, согласно настоящему изобретению предложена новая комбинированная терапия с использованием изоформ IGF-1 и HGF для лечения нейропатии.
В частности, в одном аспекте согласно настоящему изобретению предложен способ лечения нейропатии, включающий следующие стадии: (1) введение субъекту, имеющему нейропатию, терапевтически эффективного количества первой ДНК-конструкции, кодирующей IGF-1, способной экспрессировать человеческую изоформу IGF-1; и (2) введение данному субъекту терапевтически эффективного количества первой ДНК-конструкции, кодирующей HGF, способной экспрессировать человеческую изоформу HGF.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, способна экспрессировать белок IGF-1Ea класса I, содержащий полипептид SEQ ID NO 14 или белок IGF-1Ec класса I, содержащий полипептид SEQ ID NO 16. В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, не способна экспрессировать как белок IGF-1Ea Класса II, содержащий полипептид SEQ ID NO 18, так и белок IGF-1Еb Класса I, содержащий полипептид SEQ ID NO 20.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 15. В некоторых воплощениях данный способ дополнительно включает стадию введения субъекту второй ДНК-конструкции, кодирующей IGF-1, где данная вторая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 17.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 17. В некоторых воплощениях данный способ дополнительно включает стадию введения субъекту второй ДНК-конструкции, кодирующей IGF-1, где данная вторая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 15.
В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения второй ДНК-конструкции, кодирующей IGF-1, проводятся одновременно. В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения второй ДНК-конструкции, кодирующей IGF-1, проводятся последовательно.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, кодирует более чем одну изоформу человеческого IGF-1. В некоторых воплощениях более чем одна изоформа человеческого IGF-1 содержит полипептид SEQ ID NO 14 и полипептид SEQ ID NO 16.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит: первый полинуклеотид IGF SEQ ID NO 1 (экзоны 1, 3, 4) или его вырожденную последовательность; второй полинуклеотид IGF SEQ ID NO 2 (интрон 4) или его фрагмент; третий полинуклеотид IGF SEQ ID NO 3 (экзоны 5 и 6-1) или его вырожденную последовательность; четвертый полинуклеотид IGF SEQ ID NO 4 (интрон 5) или его фрагмент; и пятый полинуклеотид IGF SEQ ID NO 5 (экзон 6-2) или его вырожденную последовательность, где первый полинуклеотид, второй полинуклеотид, третий полинуклеотид, четвертый полинуклеотид и пятый полинуклеотид связываются в последовательном порядке от 5' к 3'.
В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 6. В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 7. В некоторых воплощениях четвертый полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 8.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит плазмидный вектор. В некоторых воплощениях данный плазмидный вектор представляет собой рСK. В некоторых воплощениях данный плазмидный вектор представляет собой рТх.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 10. В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 9.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, и первая ДНК-конструкция, кодирующая HGF, вводятся в достаточном количестве для уменьшения у субъекта боли. В некоторых воплощениях данный субъект имеет диабетическую нейропатию.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, и первая ДНК-конструкция, кодирующая HGF, вводятся посредством множества внутримышечных инъекций.
В некоторых воплощениях человеческая изоформа HGF представляет собой fIHGF SEQ ID NO 11 или dHGF SEQ ID NO 12.
В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, кодирует более чем одну человеческую изоформу HGF. В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, кодирует две человеческие изоформы HGF, где две данные человеческие изоформы HGF представляют собой fIHGF SEQ ID NO 11 и dHGF SEQ ID NO 12.
В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, содержит плазмидный вектор, возможно где данный плазмидный вектор представляет собой вектор рСK или вектор рТх.
В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, содержит первый полинуклеотид HGF экзонов 1-4 SEQ ID NO 22 или его вырожденную последовательность; второй полинуклеотид HGF интрона 4 SEQ ID NO 25 или его функциональный фрагмент; и третий полинуклеотид HGF экзонов 5-18 SEQ ID NO 23 или его вырожденную последовательность, где данный второй полинуклеотид HGF расположен между первым полинуклеотидом HGF и третьим полинуклеотидом HGF, и первая ДНК-конструкция, кодирующая HGF, кодирует две изоформы человеческого HGF.
В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, содержит полинуклеотид SEQ ID NO 13.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, и первая ДНК-конструкция, кодирующая HGF, вводятся совместно. В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, и первая ДНК-конструкция, кодирующая HGF, вводятся совместно посредством внутримышечной инъекции.
В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения первой ДНК-конструкции, кодирующей HGF, проводятся раздельно. В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения первой ДНК-конструкции, кодирующей HGF, проводятся с разделением во времени по меньшей мере три недели.
В некоторых воплощениях данный способ дополнительно включает стадию введения субъекту второй ДНК-конструкции, кодирующей HGF, способной экспрессировать человеческую изоформу HGF, выбранную из fIHGF SEQ ID NO 11 и dHGF SEQ ID NO 12.
В некоторых воплощениях данный способ включает следующие стадии: введение субъекту, имеющему нейропатию, ДНК-конструкции, кодирующей HGF, содержащей полинуклеотид SEQ ID NO 13, и введение субъекту ДНК-конструкции, кодирующей IGF-1, содержащей полинуклеотид SEQ ID NO 10 или полинуклеотид SEQ ID NO 9, где стадия введения ДНК-конструкции, кодирующей HGF, и стадия введения ДНК-конструкции, кодирующей IGF-1, проводятся с разделением во времени по меньшей мере три недели.
В некоторых воплощениях данный способ включает следующие стадии: введение субъекту, имеющему нейропатию, ДНК-конструкции, кодирующей HGF, содержащей полинуклеотид SEQ ID NO 33, и введение данному субъекту ДНК-конструкции, кодирующей IGF-1, содержащей полинуклеотид SEQ ID NO 10 или полинуклеотид SEQ ID NO 9, где стадия введения ДНК-конструкции, кодирующей HGF, и стадия введения ДНК-конструкции, кодирующей IGF-1, проводятся с разделением во времени по меньшей мере три недели.
В некоторых воплощениях следующие стадии: введение субъекту, имеющему нейропатию, ДНК-конструкции, кодирующей HGF, содержащей полинуклеотид SEQ ID NO 13, и введение данному субъекту первой ДНК-конструкции, кодирующей IGF-1, содержащей полинуклеотид SEQ ID NO 15, и второй ДНК-конструкции, кодирующей IGF-1, содержащей полинуклеотид SEQ ID NO 17, где стадия введения ДНК-конструкции, кодирующей HGF, и стадия введения первой ДНК-конструкции, кодирующей IGF-1, и второй ДНК-конструкции, кодирующей IGF-1, проводятся с разделением во времени по меньшей мере три недели.
В другом аспекте согласно настоящему изобретению предложена фармацевтическая композиция, содержащая: ДНК-конструкцию, кодирующую IGF-1, способную экспрессировать по меньшей мере одну изоформу человеческого IGF-1; ДНК-конструкцию, кодирующую HGF, способную экспрессировать по меньшей мере одну изоформу человеческого HGF, и фармацевтически приемлемый эксципиент.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, кодирует белок IGF-1Ea класса I, содержащий полипептид SEQ ID NO 14 или белок IGF-1Ec класса I, содержащий полипептид SEQ ID NO 16.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, кодирует более чем одну изоформу человеческого IGF-1. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, кодирует две изоформы человеческого IGF-1, где данные две изоформы человеческого IGF-1 представляют собой белок IGF-1Ea класса I, содержащий полипептид SEQ ID NO 14, и белок IGF-1Ec Класса I, содержащий полипептид SEQ ID NO 16.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит: первый полинуклеотид IGF SEQ ID NO 1 (экзоны 1, 3, 4) или его вырожденную последовательность; второй полинуклеотид IGF SEQ ID NO 2 (интрон 4) или его фрагмент; третий полинуклеотид IGF SEQ ID NO 3 (экзоны 5 и 6-1) или его вырожденную последовательность; четвертый полинуклеотид IGF SEQ ID NO 4 (интрон 5) или его фрагмент; и пятый полинуклеотид IGF SEQ ID NO 5 (экзон 6-2) или его вырожденную последовательность, где первый полинуклеотид, второй полинуклеотид, третий полинуклеотид, четвертый полинуклеотид и пятый полинуклеотид связываются в последовательном порядке от 5' к 3'.
В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 6. В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 7. В некоторых воплощениях четвертый полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 8.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, дополнительно содержит плазмидный вектор. В некоторых воплощениях данный плазмидный вектор представляет собой рСK. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, выбрана из группы, состоящей из pCK-IGF-1X6 и pCK-IGF-1X10. В некоторых воплощениях данный плазмидный вектор представляет собой рТх. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, выбрана из группы, состоящей из pTx-IGF-1X6 и pTx-IGF-1X10.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 9. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 10.
В некоторых воплощениях по меньшей мере одна человеческая изоформа HGF представляет собой fIHGF SEQ ID NO 11 или dHGF SEQ ID NO 12. В некоторых воплощениях ДНК-конструкция, кодирующая HGF, способна экспрессировать и fIHGF SEQ ID NO 11, и dHGF SEQ ID NO 12.
В некоторых воплощениях ДНК-конструкция, кодирующая HGF, содержит: первый полинуклеотид HGF SEQ ID NO 22 (экзоны 1-4) или его вырожденную последовательность; второй полинуклеотид HGF SEQ ID NO 25 (интрон 4) или его функциональный фрагмент; и третий полинуклеотид HGF SEQ ID NO 23 (экзоны 5-18) или его вырожденную последовательность, где второй полинуклеотид HGF расположен между первым полинуклеотидом HGF и третьим полинуклеотидом HGF, и первая ДНК-конструкция, кодирующая HGF, кодирует две изоформы человеческого HGF.
В некоторых воплощениях ДНК-конструкция, кодирующая HGF, содержит полинуклеотид любой из SEQ ID NO: 26-32 и 13. В некоторых воплощениях ДНК-конструкция, кодирующая HGF, содержит полинуклеотид SEQ ID NO 13.
В некоторых воплощениях данная фармацевтическая композиция содержит полинуклеотид SEQ ID NO 13 и полинуклеотид SEQ ID NO 9. В некоторых воплощениях данная фармацевтическая композиция содержит полинуклеотид SEQ ID NO 13 и полинуклеотид SEQ ID NO 10. В некоторых воплощениях данная фармацевтическая композиция содержит полинуклеотид SEQ ID NO 13, полинуклеотид SEQ ID NO 15 или полинуклеотид SEQ ID NO 17. В некоторых воплощениях данная фармацевтическая композиция содержит полинуклеотид SEQ ID NO 13, полинуклеотид SEQ ID NO 15 и полинуклеотид SEQ ID NO 17.
В некоторых воплощениях данная фармацевтическая композиция содержит полинуклеотид SEQ ID NO 33 и полинуклеотид SEQ ID NO 9, SEQ ID NO 10, SEQ ID NO 15 или SEQ ID NO 17.
В еще одном другом аспекте согласно настоящему изобретению предложен набор для лечения нейропатии, содержащий: первую фармацевтическую композицию, содержащую ДНК-конструкцию, кодирующую IGF-1, способную экспрессировать по меньшей мере одну изоформу человеческго IGF-1, и первый фармацевтически приемлемый эксципиент; и вторую фармацевтическую композицию, содержащую ДНК-конструкцию, кодирующую HGF, способную экспрессировать по меньшей мере одну изоформу человеческго HGF, и второй фармацевтически приемлемый эксципиент.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, кодирует белок IGF-1Ea класса I, содержащий полипептид SEQ ID NO 14, или белок IGF-1Ec класса I, содержащий полипептид SEQ ID NO 16. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, кодирует более чем одну человеческую изоформу IGF-1. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, кодирует две человеческие изоформы IGF-1, где данные две человеческие изоформы IGF-1 представляют собой белок IGF-1Ea класса I, содержащий полипептид SEQ ID NO 14, и белок IGF-1Ec класса I, содержащий полипептид SEQ ID NO 16.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит: первый полинуклеотид IGF SEQ ID NO 1 (экзоны 1, 3, 4) или его вырожденную последовательность; второй полинуклеотид IGF SEQ ID NO 2 (интрон 4) или его фрагмент; третий полинуклеотид IGF SEQ ID NO 3 (экзоны 5 и 6-1) или его вырожденную последовательность; четвертый полинуклеотид IGF SEQ ID NO 4 (интрон 5) или его фрагмент; и пятый полинуклеотид IGF SEQ ID NO 5 (экзон 6-2) или его вырожденную последовательность, где первый полинуклеотид, второй полинуклеотид, третий полинуклеотид, четвертый полинуклеотид и пятый полинуклеотид связываются в последовательном порядке от 5' к 3'.
В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 6. В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 7. В некоторых воплощениях четвертый полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 8.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, дополнительно содержит плазмидный вектор. В некоторых воплощениях данный плазмидный вектор представляет собой рСK. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит pCK-IGF-1X6 или pCK-IGF-1X10. В некоторых воплощениях данный плазмидный вектор представляет собой рТх. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит pTx-IGF-1X6 или pTx-IGF-1X10.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 9. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 10.
В некоторых воплощениях по меньшей мере одна человеческая изоформа HGF представляет собой fIHGF SEQ ID NO 11 или dHGF SEQ ID NO 12. В некоторых воплощениях ДНК-конструкция, кодирующая HGF, способна экспрессировать и fIHGF SEQ ID NO 11, и dHGF SEQ ID NO 12.
В некоторых воплощениях ДНК-конструкция, кодирующая HGF, содержит: первый полинуклеотид HGF SEQ ID NO 22 (экзоны 1-4) или его вырожденную последовательность; второй полинуклеотид HGF SEQ ID NO 25 (интрон 4) или его функциональный фрагмент; и третий полинуклеотид HGF SEQ ID NO 23 (экзоны 5-18) или его вырожденную последовательность, где второй полинуклеотид HGF расположен между первым полинуклеотидом HGF и третьим полинуклеотидом HGF, и первая ДНК-конструкция, кодирующая HGF, кодирует две изоформы человеческого HGF.
В некоторых воплощениях ДНК-конструкция, кодирующая HGF, содержит полинуклеотид любой из SEQ ID NO: 26-32 и 13. В некоторых воплощениях ДНК-конструкция, кодирующая HGF, содержит полинуклеотид SEQ ID NO 13.
В некоторых воплощениях первая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 9; и вторая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 13.
В некоторых воплощениях первая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 10; и вторая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 13.
В некоторых воплощениях первая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 15 и полинуклеотид SEQ ID NO 17; и вторая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 13.
В некоторых воплощениях первая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 9, SEQ ID NO 10, SEQ ID NO 15 или SEQ ID NO 17, и вторая фармацевтическая композиция содержит полинуклеотид SEQ ID NO 33.
В другом аспекте согласно настоящему раскрытию предложена первая ДНК-конструкция, кодирующая IGF-1, способная экспрессировать человеческую изоформу IGF-1 для применения в медицинском способе лечения нейропатии, причем данный медицинский способ включает следующие стадии: введение субъекту, имеющему нейропатию, терапевтически эффективного количества первой ДНК-конструкции, кодирующей IGF-1, и введение данному субъекту терапевтически эффективного количества первой ДНК-конструкции, кодирующей HGF, способной экспрессировать человеческую изоформу HGF.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, способна экспрессировать белок IGF-1Ea Класса I, содержащий полипептид SEQ ID NO 14, или белок IGF-1Ec Класса I, содержащий полипептид SEQ ID NO 16. В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, не способна экспрессировать как белок IGF-1Ea Класса II, содержащий полипептид SEQ ID NO 18, так и белок IGF-1Еb Класса I, содержащий полипептид SEQ ID NO 20.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 15. В некоторых воплощениях данный медицинский способ дополнительно включает стадию введения субъекту второй ДНК-конструкции, кодирующей IGF-1, где данная вторая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 17.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 17. В некоторых воплощениях данный медицинский способ дополнительно включает стадию введения субъекту второй ДНК-конструкции, кодирующей IGF-1, где данная вторая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 15.
В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения второй ДНК-конструкции, кодирующей IGF-1, проводятся одновременно. В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения второй ДНК-конструкции, кодирующей IGF-1, проводятся последовательно.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, кодирует более чем одну человеческую изоформу IGF-1. В некоторых воплощениях более чем одна человеческая изоформа IGF-1 содержит полипептид SEQ ID NO 14 и полипептид SEQ ID NO 16.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, содержит: первый полинуклеотид IGF SEQ ID NO 1 (экзоны 1, 3, 4) или его вырожденную последовательность; второй полинуклеотид IGF SEQ ID NO 2 (интрон 4) или его фрагмент; третий полинуклеотид IGF SEQ ID NO 3 (экзоны 5 и 6-1) или его вырожденную последовательность; четвертый полинуклеотид IGF SEQ ID NO 4 (интрон 5) или его фрагмент; и пятый полинуклеотид IGF SEQ ID NO 5 (экзон 6-2) или его вырожденную последовательность, где первый полинуклеотид, второй полинуклеотид, третий полинуклеотид, четвертый полинуклеотид и пятый полинуклеотид связываются в последовательном порядке от 5' к 3'.
В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 6. В некоторых воплощениях второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 7. В некоторых воплощениях четвертый полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 8.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит плазмидный вектор. В некоторых воплощениях данный плазмидный вектор представляет собой рСK. В некоторых воплощениях данный плазмидный вектор представляет собой рТх.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 10. В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, содержит полинуклеотид SEQ ID NO 9.
В некоторых воплощениях первую ДНК-конструкцию, кодирующую IGF-1, и первую ДНК-конструкцию, кодирующую HGF, вводят в достаточном количестве для уменьшения у субъекта боли. В некоторых воплощениях данный субъект имеет диабетическую нейропатию. В некоторых воплощениях первую ДНК-конструкцию, кодирующую IGF-1, и первую ДНК-конструкцию, кодирующую HGF, вводят посредством целого ряда внутримышечных инъекций.
В некоторых воплощениях человеческая изоформа HGF представляет собой fIHGF SEQ ID NO 11 или dHGF SEQ ID NO 12. В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, кодирует более чем одну человеческую изоформу HGF. В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, кодирует две человеческие изоформы HGF, где данные две человеческие изоформы HGF представляют собой fIHGF SEQ ID NO 11 и dHGF SEQ ID NO 12.
В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, содержит плазмидный вектор, возможно, где данный плазмидный вектор представляет собой вектор рСK или вектор рТх. В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, содержит первый полинуклеотид HGF SEQ ID NO 22 (экзоны 1-4) или его вырожденную последовательность; второй полинуклеотид HGF SEQ ID NO 25 (интрон 4) или его функциональный фрагмент; и третий полинуклеотид HGF SEQ ID NO 23 (экзоны 5-18) или его вырожденную последовательность, где второй полинуклеотид HGF расположен между первым полинуклеотидом HGF и третьим полинуклеотидом HGF, и первая ДНК-конструкция, кодирующая HGF, кодирует две изоформы человеческого HGF.
В некоторых воплощениях первая ДНК-конструкция, кодирующая HGF, содержит полинуклеотид SEQ ID NO 13.
В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, и первая ДНК-конструкция, кодирующая HGF, вводятся совместно. В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, и первая ДНК-конструкция, кодирующая HGF, вводятся совместно посредством внутримышечной инъекции. В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения первой ДНК-конструкции, кодирующей HGF, осуществляются раздельно. В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения первой ДНК-конструкции, кодирующей HGF, осуществляются с разделением во времени по меньшей мере три недели.
В некоторых воплощениях данный медицинский способ дополнительно включает стадию введения субъекту второй ДНК-конструкции, кодирующей HGF, способной экспрессировать человеческую изоформу HGF, выбранную из fIHGF SEQ ID NO 11 и dHGF SEQ ID NO 12.
В другом аспекте согласно настоящему раскрытию дополнительно предложена первая ДНК-конструкция, кодирующая HGF, способная экспрессировать человеческую изоформу HGF для применения в медицинском способе лечения нейропатии, причем данный методологический способ включает следующие стадии: введение субъекту, имеющему нейропатию, терапевтически эффективного количества первой ДНК-конструкции, кодирующей HGF, и введение данному субъекту терапевтически эффективного количества ДНК-конструкции, кодирующей IGF-1, способной экспрессировать человеческую изоформу IGF-1.
5. Краткое описание графических материалов
Фиг. 1 представляет собой схематическое представление человеческого гена IGF-1, включающего сайты альтернативной инициации транскрипции и сайты альтернативного сплайсинга. Изоформы IGF-1, которые образуются в природе от гена IGF-1, включают Еc класса I (изоформа №1); Еа класса II (изоформа №2); Еb класса I (изоформа №3) и Еа класса I (изоформа №4).
На Фиг. 2а описан экспериментальный протокол для анализа терапевтической эффективности одновременного введения ДНК-конструкции, кодирующей HGF (VM202), и ДНК-конструкции, кодирующей одну изоформу IGF-1, в модели хронической компрессии (CCI - от англ. chronic constriction injury).
Фиг. 2в представляет собой график, показывающий частоту (%) отдергивания лапы, измеренную у мышей CCI или у плацебо мышей в эксперименте, описанном на ФИГ. 2А. Мышам CCI инъецировали ДНК-конструкцию - (i) вектор рСK («рСK»). (ii) VM202 («VM202») или (iii) VM202 плюс (+) ДНК-конструкцию, кодирующую IGF-1 - VM202 и pCK-IGF-1#1 («1»), VM202 и pCK-IGF-1#2 («2»), VM202 и pCK-IGF-1#3 («3») или VM202 и pCK-IGF-1#4 («4»).
На Фиг. 3а описан экспериментальный протокол для анализа терапевтической эффективности одновременного введения ДНК-конструкции, кодирующей HGF (VM202), и одной или двух ДНК-конструкций, кодирующих одну изоформу IGF-1, в модели хронической компрессии (CCI).
Фиг. 3в представляет собой график, показывающий частоту (%) отдергивания лапы, измеренную у мышей CCI или у плацебо мышей в эксперименте, описанном на ФИГ. 3А. Мышам CCI инъецировали ДНК-конструкцию - (i) вектор рСK («рСK»), (ii) VM202 («VM202») или (iii) VM202 и ДНК-конструкцию, кодирующую IGF-1 - VM202 и pCK-IGF-1#1 («1»), VM202 и pCK-IGF-1#4 («4») или VM202, pCK-IGF-1#1 и pCK-IGF-1#4 («1+4»).
На Фиг. 4а описан экспериментальный протокол для анализа терапевтической эффективности серийного введения ДНК-конструкции, кодирующей HGF (VM202), и двух ДНК-конструкций, кодирующих IGF-1 - pCK-IGF-1#1 и pCK-IGF-1#4, в модели хронической компрессии (CCI).
Фиг. 4в представляет собой график, показывающий частоту (%) отдергивания лапы, измеренную у мышей CCI, в эксперименте, описанном на ФИГ. 4А. Мышам CCI дважды инъецировали одну или более чем одну ДНК-конструкцию - (i) вектор рСK в 1-ой инъекции и вектор рСK во 2-ой инъекции ("рСK"), (ii) рСK-IGF-1#1 и pCK-IGF-1#4 в 1-ой инъекции, и pCK-IGF-1#1 и pCK-IGF-1#4 во 2-ой инъекции ("IGF-1 -> IGF-1"), (iii) VM202 в 1-ой инъекции и вектор рСK во 2-ой инъекции ("VM202 ->р СK"), (iv) pCK-IGF-1#1 и pCK-IGF-1#4 в 1-ой инъекции, и вектор рСK во 2-ой инъекции ("IGF-1 -> рСK"), (v) pCK-IGF-1#1 и pCK-IGF-1#4 в 1-ой инъекции и VM202 во 2-ой инъекции ("IGF-1 -> VM202"), (vi) VM202 в 1-ой инъекции и VM202 во 2-ой инъекции ("VM202 -> VM202"), или (vii) VM202 в 1-ой инъекции и pCK-IGF-1#1, и pCK-IGF-1#4 во 2-ой инъекции ("VM202 -> изоформы IGF-1").
На Фиг. 5а описан экспериментальный протокол, используемый в Примере 3 для оценки экспрессии in vivo изоформ IGF-1 от разных ДНК-конструкций.
На Фиг. 5в показаны результаты иммуноферментного твердофазного анализа (ИФА), измеряющего общее количество человеческих изоформ IGF-1, экспрессируемых после инъекции ДНК-конструкции, не кодирующей IGF (только вектор, «рСK»); pCK-IGF-1#1 ("1"); pCK-IGF-1#4 ("4"); pCK-IGF-1 #1 и pCK-IGF-1#4 ("1+4"); и двойных экспрессионных конструкций pCK-IGF-1X6 ("Х6") и pCK-IGF-1X10 ("Х10").
На Фиг. 6а показано положение прямого («F») и обратного («R») праймеров, используемых в ПЦР-ОТ (полимеразная цепная реакция, сопряженная с обратной транскрипцией), для различения экспрессии изоформ №1 (изоформа Еc класса I) и №4 (изоформа Еа класса I) IGF-1.
На Фиг. 6в показан электрофорез в агарозном геле продуктов ПЦР-ОТ, демонстрирующий экспрессию изоформ №1 и №4 от конструкций двойной экспрессии pCK-IGF-1X6 и pCK-IGF-1X10. И pCK-IGF-1X6, и pCK-IGF-1X10 индуцировали высокий уровень экспрессии обеих изоформ.
На Фиг. 7а описан протокол, используемый в Примере 3 для оценки экспрессии белка от ДНК-конструкций, кодирующих IGF-1, в клетках 293Т in vitro.
На Фиг. 7в показаны результаты вестерн-блоттинга, демонстрирующие экспрессию изоформ №1 и/или №4 IGF-1 после трансфекции in vitro (i) pCK-IGF-1#1 ("1"), (ii) pCK-IGF-1#4 ("4"), (iii) двух одиночных экспрессионных конструкций: pCK-IGF-1#1 и pCK-IGF-1#4 ("1+4"), (iv) двойной экспрессионной конструкции рСK-IGF-1X6 ("Х6") или (v) двойной экспрессионной конструкции pCK-IGF-1X10 ("Х10").
На Фиг. 8а описан экспериментальный протокол, используемый в Примере 4 для анализа эффективности одновременного введения конструкции, кодирующей HGF - VM202, и разных ДНК-конструкций, кодирующих IGF-1, в уменьшении механической аллодинии в животной CCI модели.
Фиг. 8в представляет собой график, показывающий частоту отдергивания лапы, измеренную у плацебо мышей или мышей CCI, в эксперименте, описанном на ФИГ. 8А. Мышам CCI инъецировали одну или более чем одну ДНК-конструкцию - (i) вектор рСK ("рСK"), (ii) VM202 ("VM202"), (iii) VM202, pCK-IGF-1#1, и pCK-IGF-1#4 ("IGF-1#1+#4"), (iv) VM202 и pCK-IGF-1X6 ("IGF-1 X6"), и (v) VM202 и pCK-IGF-1X10 ("IGF-1 X10").
На Фиг. 9a описан экспериментальный протокол, используемый в Примере 5 для анализа эффективности одновременного введения конструкции, экспрессирующей HGF728, и разных ДНК-конструкций, кодирующих IGF-1, в уменьшении механической аллодинии в животной модели CCI.
Фиг. 9в представляет собой график, показывающий частоту отдергивания лапы, измеренную у плацебо мышей или мышей CCI, в эксперименте, описанном на ФИГ. 9А.
Фиг. 9с представляет собой график, показывающий порог отдергивания лапы, измеренный у плацебо мышей или мышей CCI, в эксперименте, описанном на ФИГ. 9А. Мышам CCI инъецировали одну или более чем одну ДНК-конструкцию - один вектор ("CCI-pCK") или (i) pCK-HGF728 ("CCI-HGF728"), (ii) pCK-HGF728 и pCK-IGF-1#1 ("CCI-HGF728+IGF-1#1"), (iii) pCK-HGF728 и pCK-IGF-1#4 ("CCI-HGF728+IGF-1#4"), или (iv) pCK-HGF728 и pCK-IGF-1X10 ("CCI-HGF728+IGF-1X10").
Данные графические материалы показывают разные воплощения настоящего изобретения лишь в целях иллюстрации. Специалист в данной области легко узнает из следующего обсуждения то, что можно использовать альтернативные воплощения структур и способов, проиллюстрированных в данном документе, без отступления от принципов изобретения, описанного в данном документе.
6. Подробное описание изобретения
6.1 Определения
Если не определено иначе, все технические и научные термины, используемые в данном документе, имеют значение, обычно понятное специалисту в области, к которой принадлежит данное изобретение. Следующие термины в том виде, в котором они используются в данном документе, имеют значения, приписываемые им ниже.
Термины «изоформа IGF-1» или «человеческая изоформа IGF-1» в том виде, в котором они используются в данном документе, используются взаимозаменяемо для названия полипептида, имеющего аминокислотную последовательность, которая является по меньшей мере на 80% идентичной аминокислотной последовательности одного из встречающихся в природе пре-прополипептидов человеческого IGF-1 или их аллельного варианта, образующегося в результате сплайсинга варианта или образующегося в результате делеции вариантов. Встречающиеся в природе полипептиды пре-про-IGF-1 включают изоформы Еc класса I (SEQ ID NO 16); Еа класса II (SEQ ID NO 18); Eb класса I (SEQ ID NO 20) и Еа класса I (SEQ ID NO 14).
Термины «изоформа №1», «изоформа Еc класса I», «изоформа Еc IGF-1 класса I» или «Еc IGF-1 класса I» используются в данном документе взаимозаменяемо для названия полипептида SEQ ID NO 16.
Термины «изоформа №2», «изоформа Еа класса II», «изоформа Еа IGF-1 класса II» или «Еа IGF-1 класса II» используются в данном документе взаимозаменяемо для названия полипептида SEQ ID NO 18.
Термины «изоформа №3», «изоформа Еb класса I», «изоформа Eb IGF-1 класса I» или «Еb IGF-1 класса I» используются в данном документе взаимозаменяемо для названия полипептида SEQ ID NO 20.
Термины «изоформа №4», «изоформа Еа класса I», «изоформа Еа IGF-1 класса I» или «Еа IGF-1 класса I» используются в данном документе взаимозаменяемо для названия полипептида SEQ ID NO 14.
Термин «лечение» в том виде, в котором он используется в данном документе, относится ко всем актам (а) подавления симптома нейропатии; (b) облегчения симптома нейропатии; и (с) устранения симптома нейропатии. В некоторых воплощениях композиция по настоящему изобретению может лечить нейропатию посредством роста нейронов или подавления гибели нейронов.
Термин «VM202» в том виде, в котором он используется в данном документе, относится к плазмидной ДНК, также именуемой pCK-HGF-X7, содержащей вектор рСK (SEQ ID NO 24) и HGF-X7 (SEQ ID NO 13), клонированный в вектор рСK. VM202 депонировали согласно условиям Будапештского соглашения в Корейский центр культуры микроорганизмов (KССМ) под номером доступа KССМ-10361 12 марта 2002 г.
Термин «изоформы HGF» в том виде, в котором он используется в данном документе, относится к полипептиду, имеющему аминокислотную последовательность, которая по меньшей мере на 80% идентична аминокислотной последовательности встречающегося в природе у животного, включая человека, полипептида HGF. Данный термин включает полипептиды, имеющие аминокислотную последовательность, которая по меньшей мере на 80% идентична любому полноразмерному полипептиду HGF дикого типа, и включает полипептиды, имеющие аминокислотную последовательность, которая по меньшей мере на 80% идентична встречающемуся в природе аллельному варианту HGF, варианту, образующемуся в результате сплайсинга, или варианту, образующемуся в результате делеции. Предпочтительные для применения в настоящем изобретении изоформы HGF включают две или более чем две изоформы, выбранные из группы, состоящей из полноразмерного HGF (fIHGF) (синонимично, fHGF), делетированного варианта HGF (dHGF), NK1, NK2 и NK4. Согласно более предпочтительному воплощению настоящего изобретения изоформы HGF, используемые в способах, описанных в данном документе, включают fIHGF (SEQ ID NO 11) и dHGF (SEQ ID NO 12).
Термины «человеческий fIHGF», «fIHGF» и «fHGF» используются в данном документе взаимозаменяемо для названия белка, состоящего из аминокислот 1-728 белка человеческого HGF. Последовательность fIHGF приводится в SEQ ID NO 11.
Термины «человеческий dHGF» или «dHGF» используются в данном документе взаимозаменяемо для названия делетированного варианта белка HGF, продуцируемого альтернативным сплайсингом человеческого гена HGF. В частности, «человеческий dHGF» или «dHGF» относится к человеческому белку HGF с делецией пяти аминокислот (F, L, Р, S и S) в первом домене «двойная петля» альфа цепи из последовательности полноразмерного HGF. Человеческий dHGF имеет 723 аминокислоты в длину. Аминокислотная последовательность человеческого dHGF приводится в SEQ ID NO 12.
Термин «терапевтически эффективная доза» или «эффективное количество» в том виде, в котором он используется в данном документе, относится к дозе или количеству, которое дает желательный эффект, для которого оно вводится. В контексте настоящих способов терапевтически эффективное количество представляет собой эффективное количество для лечения симптома нейропатии. Данное количество может представлять собой эффективное количество для лечения симптома нейропатии само по себе или в комбинации с другим терапевтическим средством.
Термин «достаточное количество» в том виде, в котором он используется в данном документе, относится к достаточному количеству, для того, чтобы давать желательный эффект. Данное количество может быть достаточным количеством для получения желательного эффекта самим по себе или в комбинации с другим терапевтическим средством.
Термин «вырожденная последовательность» в том виде, в котором он используется в данном документе, относится к последовательности нуклеиновой кислоты, которая может транслироваться с получением идентичной аминокислотной последовательности относительно аминокислотной последовательности, транслируемой от эталонной последовательности нуклеиновой кислоты.
6.2. Другие договоренности по интерпретации
Понятно, что перечисленные в данном документе интервалы представляют собой сокращенную запись для всех значений в пределах данного интервала, включая перечисленные конечные точки. Например, понятно то, что интервал от 1 до 50 включает любое число, комбинацию чисел или подинтервал из группы, состоящей из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, и 50.
6.3. Способы лечения нейропатии
В первом аспекте предложены способы лечения нейропатии. Данные способы включают введение субъекту, имеющему нейропатию, терапевтически эффективного количества первой ДНК-конструкции, кодирующей IGF-1, способной экспрессировать человеческую изоформу IGF-1; и терапевтически эффективного количества первой ДНК-конструкции, кодирующей HGF, способной экспрессировать человеческую изоформу HGF.
6.3.1. ДНК-конструкции, кодирующие IGF-1
В способах, предложенных в данном документе, используются ДНК-конструкции, способные экспрессировать по меньшей мере одну человеческую изоформу IGF-1.
Как проиллюстрировано на Фиг. 1, человеческий ген IGF-1 содержит шесть экзонов (экзоны 1, 2, 3, 4, 5 и 6 (6-1 и 6-2)), охватывающих почти 90 т.п.н. геномной ДНК. Экзоны 1 и 2 представляют собой взаимоисключающие лидерные экзоны, причем каждый имеет многочисленные промоторные сайты, которые используются по-разному. Кроме того, ген IGF-1 может подвергаться дифференциальному сплайсингу с созданием множества вариантов транскриптов. Каждый вариант транскрипта кодирует отличный белок пре-про-IGF-1 («изоформа IGF-1»), обладающий вариабельными последовательностями лидерного сигнального пептида. Тем не менее, все изоформы транскрипта приводят к одинаковому зрелому 70-аминокислотному пептиду IGF-1, который после процессинга использует одинаковый рецептор.
Пептиды пре-про-IGF-1 отличаются по их лидерным или сигнальным последовательностям и их карбокси(С)-концу. Включение экзона 1 или экзона 2 является взаимоисключающим, и один из них служит в качестве лидерной последовательности пептида пре-про-IGF-1; причем разные лидерные экзоны создают разные 5'-UTR (от англ. untranslated region - нетранслируемая область). Данные полипептиды пре-про-IGF-1 подвергаются посттрансляционному протеолитическому расщеплению с удалением лидера и карбоксиконца Е-пептида, приводя к зрелому 70-аминокислотному IGF-1.
Транскрипты, содержащие экзон 1, называются транскриптами класса 1 (например, Еc класса I; Еb класса I и Еа класса I на Фиг. 1), тогда как транскрипты, содержащие экзон 2, называются транскриптами класса 2 (например, Еа класса II на Фиг. 1). Почти все пре-пропептиды включают 27 аминокислот в сигнальном пептиде, происходящем от экзона 3, причем остальные сигнальные последовательности происходят от включения экзона 1 или 2. У меньшинства транскриптов используется отличный сайт инициации транскрипции в пределах экзона 3, генерируя более короткий сигнальный пептид из 22 аминокислот. Экзоны 3 и 4 являются инвариантными и кодируют домены В, С, А и D зрелого пептида IGF-1; экзон 4 кодирует две третьих зрелого пептида IGF-1. Человеческий пептид Еb состоит только из экзонов 4 и 5, тогда как Еc содержит экзоны 4, 5 и 6 (Фиг. 1).
Альтернативный сплайсинг и взаимоисключающая инициация транскрипции, которые приводят к образованию разных полипептидов пре-про-IGF-1 (т.е. изоформ IGF-1), проиллюстрированы на Фиг. 1. В частности, изоформа IGF-1 Еc класса I (SEQ ID NO 16), содержащая по меньшей мере фрагмент экзонов 1, 3/4, 5 и 6, генерируется из транскрипта, содержащего последовательность SEQ ID NO 17. Изоформа IGF-1 Еа класса II (SEQ ID NO 18), содержащая по меньшей мере фрагмент экзонов 2, 3/4 и 6, генерируется из транскрипта, содержащего последовательность SEQ ID NO 19. Изоформа IGF-1 Еb класса I (SEQ ID NO 20), содержащая по меньшей мере фрагмент экзонов 1, 3/4 и 5, генерируется из транскрипта, содержащего последовательность SEQ ID NO 21. Изоформа IGF-1 Еа класса I (SEQ ID NO 14), содержащая по меньшей мере фрагмент экзонов 1, 3/4 и 6, генерируется из транскрипта, содержащего последовательность SEQ ID NO 15.
Несмотря на то, что зрелый белок IGF-1, образующийся от разных транскриптов, не отличается, предполагали то, что разные изоформы транскриптов имеют разные регуляторные функции. Формы вариантов обладают разными стабильностями, партнерами связывания и активностью, указывая на ключевую регуляторную роль для данных изоформ. Биологическая значимость данных изоформ остается неясной, хотя и предполагали то, что изоформы класса I с экзоном 1 являются аутокринными/паракринными формами, тогда как изоформы класса II с экзоном 2 являются секретируемыми эндокринными формами. Это основывается на данных о том, что транскрипты класса II включают типичный мотив сигнального пептида, ассоциированный с эффективной секрецией, тогда как транскрипты класса I имеют более длинный сигнальный пептид, который возможно может препятствовать секреции.
Считается, что большинство тканей использует транскрипты класса I, хотя в печени используются обе формы, и печеночные транскрипты класса II предпочтительно усиливаются во время развития. Имеется множество изменений в количестве транскриптов IGF-1 на протяжении развития. Обнаружили то, что Еа класса 1 является самой преобладающей формой во время фазы активного роста, а Еb класса 1 также однородно экспрессируется, хотя и на более низких уровнях, по всей ростовой пластинке на протяжении ранних фаз роста.
В данном документе предложены ДНК-конструкции, способные экспрессировать по меньшей мере одну изоформу человеческого IGF-1. Такая одиночная экспрессионная конструкция включает pCK-IGF-1#1, который представляет собой вектор рСK, содержащий кодирующую последовательность для изоформы №1 IGF-1; pCK-IGF-1#2, который представляет собой вектор рСK, содержащий кодирующую последовательность для изоформы №2 IGF-1; pCK-IGF-1#3, который представляет собой вектор рСK, содержащий кодирующую последовательность для изоформы №3 IGF-1, и pCK-IGF-1#4, который представляет собой вектор рСK, содержащий кодирующую последовательность для изоформы №4 IGF-1, но не ограничивается ими. В некоторых воплощениях используется более чем одна ДНК-конструкция, причем каждая кодирует другую изоформу IGF-1. Например, совместно используются первая конструкция, кодирующая изоформу Еc класса I (изоформа №1), и вторая конструкция, кодирующая изоформу Еа класса I (изоформа №4). Например, можно использовать совместно pCK-IGF-1#1 и pCK-IGF-1#4.
Такая конструкция одиночной экспрессии дополнительно включает pTx-IGF-1#1, который представляет собой вектор рТх, содержащий кодирующую последовательность изоформы №1 IGF-1; pTx-IGF-1#2, который представляет собой вектор рТх, содержащий кодирующую последовательность изоформы №2 IGF-1; pTx-IGF-1#3, который представляет собой вектор рТх, содержащий кодирующую последовательность изоформы №3 IGF-1, и pTx-IGF-1#4, который представляет собой вектор рТх, содержащий кодирующую последовательность изоформы №4 IGF-1, но не ограничивается ими. В некоторых воплощениях используется более чем одна ДНК-конструкция, причем каждая кодирует другую изоформу IGF-1. Например, совместно используются первая конструкция, кодирующая изоформу Еc класса I (изоформа №1), и вторая конструкция, кодирующая изоформу Еа класса I (изоформа №4). Например, можно совместно использовать pTx-IGF-1#1 и pTx-IGF-1#4.
В некоторых воплощениях ДНК-конструкция содержит кодирующую последовательность одной из изоформ IGF-1. Например, данная ДНК-конструкция может содержать последовательность, кодирующую Еа класса I (изоформа №4) (SEQ ID NO 15); Eb класса I (изоформа №3) (SEQ ID NO 21); Еc класса I (изоформа №1) (SEQ ID NO 17); или Еа класса II (изоформа №2) (SEQ ID NO 19).
В некоторых воплощениях ДНК-конструкция представляет собой конструкцию двойной экспрессии - ДНК-конструкцию, которая может экспрессировать более чем одну изоформу IGF-1 посредством содержания регуляторной последовательности экспрессии для каждой последовательности, кодирующей изоформу (CDS - от англ. coding sequence). В некоторых воплощениях данная конструкция содержит участок внутренней посадки рибосомы (IRES - от англ. internal ribosomal entry site) между двумя кодирующими последовательностями, например, в порядке: (1) регуляторная последовательность экспрессии - (2) кодирующая последовательность первой изоформы - (3) IRES - (4) кодирующая последовательность второй изоформы - (5) последовательность терминации транскрипции. IRES обеспечивает начало трансляции в последовательности IRES, обеспечивая, посредством этого, экспрессию двух белковых продуктов от одного транскрипта. В других воплощениях совместно используется множество конструкций, причем каждая кодирует одну изоформу IGF-1, для индукции экспрессии более чем одной изоформы IGF-1 у субъекта, которому они вводятся.
В предпочтительных воплощениях данная ДНК-конструкция способна одновременно экспрессировать две или более чем две изоформы IGF-1 - например, (i) изоформу Еc класса I (изоформа №1) и изоформу Еа класса II (изоформа №2); (ii) изоформу Еc класса I (изоформа №1) и изоформу Еb класса I (изоформа №3); (iii) изоформу Еc класса I (изоформа №1) и изоформу Еа класса I (изоформа №4); (iv) изоформу Еа Класса II (изоформа №2) и изоформу Еb Класса I (изоформа №3); (v) изоформу Еа класса II (изоформа №2) и изоформу Еа класса I (изоформа №4); (vi) изоформу Еb класса I (изоформа №3) и изоформу Еа класса I (изоформа №4) - посредством содержания сайта альтернативного сплайсинга.
Например, данная ДНК-конструкция может содержать (i) первую последовательность, содержащую экзоны 1, 3 и 4 человеческого гена IGF-1 (SEQ ID NO 1), или вырожденную последовательность данной первой последовательности; (ii) вторую последовательность, содержащую интрон 4 человеческого гена IGF-1 (SEQ ID NO 2), или фрагмент данной второй последовательности; (iii) третью последовательность, содержащую экзоны 5 и 6-1 человеческого гена IGF-1 (SEQ ID NO 3), или вырожденную последовательность данной третьей последовательности; (iv) четвертую последовательность, содержащую интрон 5 человеческого гена IGF-1 (SEQ ID NO 4) или фрагмент данной второй последовательности; и (v) пятую последовательность, содержащую экзон 6-2 человеческого гена IGF-1 (SEQ ID NO 5), или вырожденную последовательость данной пятой последовательности. Интроны 4 и 5 могут подвергаться альтернативному сплайсингу, приводя к продукции двух изоформ IGF-1 (например, Еc класса I и Еа класса I).
В некоторых воплощениях данную ДНК-конструкцию анализируют in vitro и/или in vivo в отношении ее способности экспрессировать одну или более чем одну изоформу IGF-1. В предпочтительных воплощениях выбирают ДНК-конструкции, способные экспрессировать и изоформы Еc класса I, и изоформы Еа класса I IGF-1.
В некоторых воплощениях данная конструкция содержит полную последовательность интрона 4 (SEQ ID NO 2) или его фрагмент. В предпочтительных воплощениях данная конструкция содержит фрагмент интрона 4, имеющий последовательность SEQ ID NO 6 или SEQ ID NO 7.
В некоторых воплощениях данная конструкция содержит полную последовательность интрона 5 (SEQ ID NO 4) или его фрагмент. В предпочтительных воплощениях данная конструкция содержит фрагмент интрона 5, имеющий последовательность SEQ ID NO 8.
Разные ДНК-конструкции, содержащие последовательности, соответствующие (i) экзонам 1-6 человеческого гена IGF-1 и (ii) интронам 4 и 5 человеческого гена IGF-1 или разные фрагменты интронов 4 и 5, называются «IGF-1Х» с последующим уникальным номером. Проанализированные заявителем конструкции IGF-1X включают IGF-1X1, IGF-1X2, IGF-1X3, IGF-1X4, IGF-1X5, IGF-1X6, IGF-1X7, IGF-1X8, IGF-1X9 и IGF-1X10, но не ограничиваются ими. Конструкции IGF-1X1, клонированные в вектор рСK, называются pCK-IGF-1X1, pCK-IGF-1X2, pCK-IGF-1X3, pCK-IGF-1X4, pCK-IGF-1X5, pCK-IGF-1X6, pCK-IGF-1X7, pCK-IGF-1X8, pCK-IGF-1X9 и pCK-IGF-1X10 соответственно. Среди проанализированных конструкций идентифицировали то, что pCK-IGF-1X6 и рСK-IGF-1X10 экспрессируют и изоформу Еc класса I, и Еа класса I IGF-1. Конструкции IGF-1X, клонированные в вектор рТх, называются pTx-IGF-1X1, pTx-IGF-1X2, рТх-IGF-1X3, pTx-IGF-1X4, pTx-IGF-1X5, pTx-IGF-1X6, pTx-IGF-1X7, pTx-IGF-1X8, pTx-IGF-1X9 и pTx-IGF-1X10 соответственно. pTx-IGF-1X6 и pTx-IGF-1X10 экспрессируют и изоформу Еc класса I, и Еа класса I IGF-1.
В предпочтительных воплощениях используются IGF-1X6 (SEQ ID NO 9) или IGF-1X10 (SEQ ID NO 10). IGF-1X6 (SEQ ID NO 9) и IGF-1X10 (SEQ ID NO 10), клонированные в вектор рСK, называются pCK-IGF-1X6 и pCK-IGF-1X10 соответственно. Клетки Е. coli, трансформированные pCK-IGF-1X6 ("DH5α_pCK-IGF1 Х6"), депонировали согласно условиям Будапештского соглашения в Корейскую коллекцию типовых культур (KСТС (от англ. Korea Collection for Type Cultures), Корейский исследовательский институт биологических наук и биотехнологии (KRIBB - от англ. Korea Research Institute of Bioscience and Biotechnology) 181, Ipsin-gil, Jeongeup-si, Jeollabuk-do, 56212, Республика Корея) с номером доступа KСТС 13539 ВР 30 мая 2018 г. Клетки E. coli, трансформированные pCK-IGF-1X10 ("DH5α_pCK-IGF1 Х10"), депонировали согласно условиям Будапештского соглашения в Корейскую коллекцию типовых культур (KСТС, Korea Research Institute of Bioscience and Biotechnology (KRIBB) 181, Ipsin-gil, Jeongeup-si, Jeollabuk-do, 56212, Республика Корея) с номером доступа KСТС 13540 ВР 30 мая 2018 г.
В других предпочтительных воплощениях используют IGF-1X6 (SEQ ID NO 9) и IGF-1X10 (SEQ ID NO 10), клонированные в вектор рТх (SEQ ID NO 38). Данные конструкции IGF называются pTx-IGF-1X6 и pTx-IGF-1X10 (SEQ ID NO 39) соответственно.
Изоформы IGF-1 или ДНК-конструкции, кодирующие изоформы IGF-1, описанные в данном документе, могут включать модификации от человеческих изоформ IGF-1 дикого типа. Данные модифицированные последовательности могут включать последовательности с по меньшей мере 80%-ной идентичностью, более предпочтительно по меньшей мере 90%-ной идентичностью и наиболее предпочтительно по меньшей мере 95%-ной идентичностью при максимальном выравнивании данных модифицированных последовательностей с последовательностями человеческой изоформы IGF-1 дикого типа. Способы выравнивания последовательностей для сравнения хорошо известны в данной области. Способы выравнивания последовательностей для сравнения являются хорошо известными в данной области. В частности, алгоритм выравнивания, раскрытый на сайте Средства поиска основного локального выравнивания (BLAST - от англ. Basic Local Alignment Search Tool) NCBI Национального центра биотехнологической информации (NBCI - от англ. National Center for Biotechnological Information, Bethesda, Md.) и используемый в связи с программами анализа последовательностей blastp, blasm, blastx, tblastn и tblastx, можно использовать для определения процента идентичности.
6.3.2. ДНК-конструкции, кодирующие HGF
В способах, предложенных в данном документе, используются ДНК-конструкции, способные экспрессировать по меньшей мере одну изоформу человеческого HGF.
Фактор роста гепатоцитов (HGF) представляет собой гепаринсвязывающий гликопротеин, также известный как рассеивающий фактор или гепатопоэтин-А. HGF имеет многочисленные биологические эффекты, такие как митогенез, мотогенез и морфогенез разных типов клеток. HGF кодируется геном, содержащим 18 экзонов и 17 интронов, расположенным на хромосоме 7q21.1.
Ген HGF кодирует две изоформы HGF посредством альтернативного сплайсинга между экзоном 4 и экзоном 5 - две данные изоформы включают: (1) предшественник полноразмерного полипептида HGF («fIHGF»), содержащий 728 аминокислот (SEQ ID NO 11), со следующими доменами: сериновая протеаза, инактивированная N-концевой петлей шпильки-двойной петлей 1-двойной петлей 2-двойной петлей 3-двойной петлей 4, и (2) делетированный вариант HGF («dHGF»), содержащий 723 аминокислоты (SEQ ID NO 12) с делецией пяти аминокислот в первом домене «двойная петля» альфа-цепи (т.е. F, L, Р, S и S). fIHGF и dHGF имеют несколько общих биологических функций, но отличаются в показателях иммунологических характеристик и нескольких биологических свойств. Было продемонстрировано, что данные две изоформы HGF являются эффективными в лечении диабетической нейропатии, как раскрыто в публикации США №20140296142, включенной в данный документ посредством ссылки во всей ее полноте.
Согласно некоторым воплощениям настоящего изобретения предложен способ введения конструкции, кодирующей одну или более чем одну изоформу HGF. В некоторых воплощениях используется конструкция, кодирующая и fIHGF, и dHGF. В некоторых воплощениях используется конструкция, кодирующая либо fIHGF, либо dHGF. В частности, может использоваться конструкция, содержащая полинуклеотид SEQ ID NO 33. Данные конструкции могут содержать вектор с одной или более чем одной регуляторной последовательностью (например, промотором или энхансером), связанной функциональным образом с кодирующей последовательностью, кодирующей fIHGF, dHGF или и ту, и другую. Данная регуляторная последовательность может регулировать экспрессию изоформы HGF.
В некоторых воплощениях конструкция может кодировать две или более чем две изоформы HGF посредством содержания регуляторной последовательности экспрессии для кодирующей последовательности (CDS) каждой изоформы. В качестве альтернативы, данная конструкция может содержать участок внутренней посадки рибосомы (IRES) между двумя кодирующими последовательностями, например, в порядке: (1) регуляторная последовательность экспрессии - (2) кодирующая последовательность первой изоформы - (3) IRES - (4) кодирующая последовательность второй изоформы - (5) последовательность терминации транскрипции. IRES обеспечивает начало трансляции в последовательности IRES, обеспечивая, посредством этого, экспрессию двух интересующих генов от одной конструкции. В качестве альтернативы, для индукции экспрессии более чем одной изоформы HGF в мишени можно использовать более чем одну конструкцию, причем каждая кодирует одну изоформу HGF.
В предпочтительных воплощениях используется конструкция, которая одновременно экспрессирует две или более чем две разные изоформы HGF - т.е. fIHGF и dHGF - посредством включения сайта альтернативного сплайсинга. В патенте США №7812146, включенном в данный документ посредством ссылки во всей его полноте, ранее было продемонстрировано то, что конструкция, кодирующая две изоформы HGF (fIHGF и dHGF), имеет значительно большую (почти в 250 раз большую) эффективность экспрессии, чем конструкция, кодирующая одну изоформу HGF (либо fIHGF, либо dHGF).
Данная конструкция может включать кДНК, соответствующую экзону 1-18 человеческого HGF и интрону 4 гена человеческого HGF или его фрагмента, который вставлен между экзоном 4 и экзоном 5 кДНК. От данной конструкции могут быть генерированы две изоформы HGF (fIHGF и dHGF) посредством альтернативного сплайсинга между экзоном 4 и экзоном 5. В некоторых воплощениях данная конструкция содержит полную последовательность интрона 4 (SEQ ID NO 25). В некоторых воплощениях данная конструкция содержит фрагмент интрона 4.
Конструкция, содержащая кДНК, соответствующую экзону 1-18 человеческого HGF и интрону 4 человеческого гена HGF или его фрагмента, может кодировать две изоформы HGF посредством альтернативного сплайсинга в интроне 4 или его фрагменте. В частности, данная конструкция может содержать нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO 13 и SEQ ID NO 26 - SEQ ID NO 32. Нуклеотидная последовательность SEQ ID NO 26 имеет 7113 п. н. и соответствует конструкции, содержащей полную последовательность интрона 4. Нуклеотидная последовательность SEQ ID NO 13 и 27-32 соответствует конструкциям, содержащим разные фрагменты интрона 4.
Разные ДНК-конструкции, содержащие кДНК, соответствующую экзону 1-18 человеческого HGF и интрону 4 человеческого гена HGF или его фрагмента, называются «HGF-Х» с последующим уникальным номером. HGF-X, которая может использоваться для разных воплощений настоящего изобретения, включает HGF-Х1 (SEQ ID NO 26), HGF-X2 (SEQ ID NO 27), HGF-X3 (SEQ ID NO 28), HGF-X4 (SEQ ID NO 29), HGF-X5 (SEQ ID NO 30), HGF-X6 (SEQ ID NO 31), HGF-X7 (SEQ ID NO 13; кодирующая последовательность HGF в VM202) и HGF-X8 (SEQ ID NO 32), но не ограничивается ими.
Продемонстрировали то, что pCK-HGF-X7 (т.е. VM202) имеет наивысшую эффективность экспрессии, как раскрыто в патенте США №7812146. Соответственно, в предпочтительных воплощениях настоящего изобретения может использоваться ДНК-конструкция, содержащая HGF-X7.
Конструкции, используемые в данном изобретении, могут включать по существу идентичные нуклеотидные последовательности относительно последовательностей человеческих изоформ HGF дикого типа. Существенная идентичность включает последовательности с по меньшей мере 80%-ной идентичностью, более предпочтительно по меньшей мере 90%-ной идентичностью и наиболее предпочтительно по меньшей мере 95%-ной идентичностью при измерении с использованием одного из алгоритмов сравнения последовательностей, где аминокислотная последовательность или нуклеотидная последовательность человеческой изоформы HGF дикого типа максимально выравнивается с последовательностью. Способы выравнивания последовательностей для сравнения хорошо известны в данной области. Разные программы и алгоритмы выравнивания описываются в Smith and Waterman, Adv. Appl. Math. 2: 482 (1981); Needleman and Wunsch, J. Mol. Bio. 48: 443 (1970); Pearson and Lipman, Methods in Mol. Biol. 24: 307-31 (1988); Higgins and Sharp, Gene 73: 15 237-44 (1988); Higgins and Sharp, CABIOS 5: 151-3 (1989) Corpet et al., Nuc. Acids Res. 16: 10881-90 (1988); Huang et al., Соmр. Appl. BioSci. 8: 155-65 (1992) и Pearson et al., Meth. Mol. Biol. 24: 307-31 (1994). Basic Local Alignment Search Tool (BLAST) от NCBI [Altschul 20 et al., J. Mol. Biol. 215: 403-10 (1990) доступен из нескольких источников, включающих Национальный центр биологической информации (NBCI, Bethesda, Md.) и интернет, для применения в связи с программами анализа последовательностей blastp, blasm, blastx, tblastn и tblastx.
6.3.3. Вектор
ДНК-конструкции, экспрессирующие изоформу IGF-1 или изоформу HGF, используемые в способах, описанных в данном документе, типично содержат вектор с одной или более чем одной регуляторной последовательностью (например, промотором или энхансером), связанной функциональным образом с экспрессируемыми последовательностями. Данные регуляторные последовательности регулируют экспрессию изоформ IGF-1 или изоформ HGF.
Предпочтительным является то, что в экспрессионной конструкции полинуклеотид, кодирующий одну или более чем одну изоформу IGF-1 или изоформу HGF, связан функциональным образом с промотором. Термин «связанный функциональным образом» относится к функциональной связи между последовательностью контроля экспрессии нуклеиновой кислоты (такой как промотор, сигнальная последовательность или группа сайтов связывания транскрипционных факторов) и второй последовательностью нуклеиновой кислоты, где последовательность контроля экспрессии влияет на транскрипцию и/или трансляцию нуклеиновой кислоты, соответствующей второй последовательности.
В типичных воплощениях промотор, связанный с данным полинуклеотидом, является функциональным предпочтительно у животных, более предпочтительно в клетках млекопитающих, для контроля транскрипции полинуклеотида, включая промоторы, происходящие из генома клеток млекопитающих или из вирусов млекопитающих, например, промотор цитомеголовируса (CMV - от англ. cytomegalovirus), поздний промотор аденовируса, промотор вируса осповакцины 7.5K, промотор SV40, промотор tk HSV (от англ. Herpes Simplex Virus - вирус простого герпеса), промотор RSV (от англ. respiratory syncytial virus - вирус саркомы Рауса), промотор EF1 альфа, промотор металлотионеина, промотор бета-актина, промотор гена IL-2 (от англ. interleukin - интерлейкин) человека, промотор гена IFN (от англ. interferon - интерферон) человека, промотор гена IL-4 человека, промотор гена лимфотоксина человека и промотор гена GM-CSF (от англ. Granulocyte-macrophage colony-stimulating factor - гранулоцитарно-макрофагальный колониестимулирующий фактор) человека, но не ограничиваясь ими. Более предпочтительно, полезный в данном изобретении промотор представляет собой промотор, происходящий из IE (немедленный ранний) гена человеческого CMV (hCMV), или промотор EF1 альфа, наиболее предпочтительно промотор/энхансер, происходящий из IE гена hCMV, и 5'-UTR (нетранслируемая область), содержащая всю последовательность экзона 1 и экзона 2, охватывающую последовательность непосредственно перед инициирующим кодоном ATG.
Экспрессионная кассета, используемая в данном изобретении, может содержать последовательность полиаденилирования, например, включающую терминатор коровьего гормона роста (Gimmi, Е.R., et al., Nucleic Acids Res. 17:6983-6998 (1989)), происходящую из SV40 последовательность полиаденилирования (Schek, N, et al., Mol. Cell Biol. 12:5386-5393 (1992)), полиА ВИЧ-1 (вирус иммунодефицита человека-1) (Klasens, В. I. F., et al., Nucleic Acids Res. 26:1870-1876 (1998)), полиА β-глобина (Gil, A., et al, Cell 49:399-406 (1987)), полиА ТK HSV (Cole, C.N. and T.P. Stacy, Mol. Cell. 5 Biol. 5: 2104-2113 (1985)) или полиА вируса полиомы (Batt, D. Band G.G. Carmichael, Mol. Cell. Biol. 15:4783-4790 (1995)), но не ограничивающуюся ими.
6.3.3.1. Невирусный вектор
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, способная экспрессировать человеческую изоформу IGF-1, и/или ДНК-конструкция, кодирующая HGF, способная экспрессировать человеческую изоформу HGF, представляет собой невирусный вектор, способный экспрессировать одну или более чем одну изоформу IGF-1 или одну или более чем одну изоформу HGF.
В типичных воплощениях невирусный вектор представляет собой плазмиду. В предпочтительных в настоящее время воплощениях данная плазмида представляет собой рСK, рСР, pVAXI, рТх или pCY. В особенно предпочтительных воплощениях, данная плазмида представляет собой рСK, подробности о которой можно найти в WO 2000/040737 и Lee et al., Biochem. Biophys. Res. Comm. 272:230-235 (2000), которые обе включаются в данный документ посредством ссылки во всей их полноте. Е. coli, трансформированную рСK (Тор10-рСK), депонировали в Корейский центр культуры микроорганизмов (KССМ) согласно условиям Будапештского соглашения 21 марта 2003 г. (№ доступа: KССМ-10476). Е. coli, трансформированную pCK-VEGF165 (т.е. вектор рСK с кодирующей последовательностью VEGF - Top10-pCK/VEGF165'), депонировали в Корейский центр культуры микроорганизмов (KССМ) согласно условиям Будапештского соглашения 27 декабря 1999 г. (№ доступа: KССМ-10179).
Вектор рСK конструируется таким образом, что экспрессия гена, например, гена IGF-1 или гена HGF, регулируется под энхансером/промотором человеческого цитомегаловируса (HCMV), как подробно раскрыто в Lee et al., Biochem, Biophys, Res. Commun. 272: 230 (2000); WO 2000/040737, которые обе включаются посредством ссылки во всей их полноте. Вектор рСK использовали для клинических испытаний в человеческом организме, и его безопасность и эффективность были подтверждены (Henry et al., Gene Ther. 18:788 (2011)).
В предпочтительных воплощениях плазмида рСK содержит кодирующую последовательность для изоформы IGF-1 Еc класса I и/или изоформы IGF-1 Еа класса I. В особенно предпочтительных воплощениях плазмида рСK содержит IGF-1X6 (т.е. pCK-IGF-1X6) или IGF-1 Х10 (т.е. pCK-IGF-1X10).
В предпочтительных воплощениях плазмида рСK содержит кодирующую последовательность для изоформ fIHGF и/или dHGF. В особенно предпочтительных воплощениях плазмида рСK содержит HGF-X7 (т.е. pCK-HGF-Х7 или VM202).
В других предпочтительных воплощениях данная плазмида представляет собой рТх (SEQ ID NO 38) - плазмидный вектор, происходящий из рСK. рТх получали посредством двух последовательных раундов мутагенеза рСK. Первый делеционный мутагенез проводили для удаления ненеобходимой последовательности между геном устойчивости к канамицину и СоIЕ1 рСK. В частности, делеционный ПЦР-мутагенез проводили с использованием первой пары праймеров (SEQ ID NO: 34 и 35). Делецию 228 пар оснований между геном устойчивости к канамицину и СоIЕ1 подтверждали секвенированием плазмиды. Затем проводили второй делеционный ПЦР-мутагенез с использованием второй пары праймеров (SEQ ID NO: 36 и 37) для оптимизации размера последовательности интрона HCMV. Последовательность интрона HCMV (421 пара оснований) между экзоном 1 и экзоном 2 IE1 подвергали делеции, и данную делецию подтверждали секвенированием.
В конкретных воплощениях плазмида рТх содержит IGF-1X6 (т.е. pTx-IGF-1X6) или IGF-1 Х10 (т.е. pTx-IGF-1X10). Например, рТх-1Х10 (SEQ ID NO 39) получали посредством лигирования IGF-1X10 в рТх, расщепленную ферментом Clal на 5' и ферментом Sal1 на 3'.
6.3.3.2. Вирусный вектор
В других воплощениях для доставки и экспрессии одной или более чем одной изоформы IGF-1 и/или одной или более чем одной изоформы HGF по настоящему изобретению можно использовать разные вирусные векторы, известные в данной области. Например, для некоторых воплощений настоящего изобретения можно использовать векторы, разработанные с использованием ретровирусов, лентивирусов, аденовирусов или аденоассоциированных вирусов.
(а) Ретровирус
Ретровирусы, способные нести относительно большие экзогенные гены, использовали в качестве вирусных векторов доставки генов в том смысле, что они интегрируют их геном в геном хозяина и имеют широкий спектр хозяев.
Для того чтобы сконструировать ретровирусный вектор, полинуклеотид по изобретению (например, кодирующую последовательность одной или более чем одной изоформы IGF-1) вставляют в вирусный геном вместо определенных вирусных последовательностей для получения дефектного по репликации вируса. Для получения вирионов конструируют упаковочную линию клеток, содержащую гены gag, pol и env, но без компонентов LTR (от англ. long terminal repeat - длинный концевой повтор) и W (Mann et al., Cell, 33:153-159 (1983)). Когда рекомбинантную плазмиду, содержащую полинуклеотид по изобретению, в данную линию клеток вводят LTR и W, последовательность W обеспечивает упаковку РНК-транскрипта рекомбинантной плазмиды в вирусные частицы, которые затем секретируются в культуральную среду (Nicolas and Rubinstein "Retroviral vectors," In: Vectors: A survey of molecular cloning vectors and their uses, Rodriguez and Denhardt (eds.), Stoneham: Butterworth, 494-513(1988)). Среду, содержащую рекомбинантные ретровирусы, затем отбирают, возможно концентрируют и используют для доставки генов.
Был описан успешный перенос генов с использованием ретровирусного вектора второго поколения. Kasahara et al. (Science, 266:1373-1376 (1994)) получили варианты вируса мышиного лейкоза Молони, в котором вместо области оболочки вставлена последовательность ЕРО (от англ. erythropoietin - эритропоэтин), получая впоследствии химерные белки, имеющие новые свойства связывания. Подобным образом, настоящую систему доставки генов можно конструировать согласно стратегиям конструирования для ретровирусного вектора второго поколения.
(b) Лентивирусы
В некоторых воплощениях настоящего изобретения также можно использовать лентивирусы. Лентивирусы представляют собой подкласс ретровирусов. Однако лентивирус может интегрироваться в геном неделящихся клеток, тогда как ретровирусы могут инфицировать только делящиеся клетки.
Лентивирусные векторы обычно продуцируются из упаковочной линии клеток, обычно НЕK293, трансформированной несколькими плазмидами. Данные плазмиды включают (1) упаковочные плазмиды, кодирующие вирионные белки, такие капсид и обратную транскриптазу, (2) плазмиду, содержащую экзогенный ген (например, кодирующую последовательность одной или более чем одной изоформы IGF-1, или одной или более чем одной изоформы HGF), подлежащий доставке в мишень.
При поступлении данного вируса в клетку вирусный геном в виде РНК подвергается обратной транскрипции с получением ДНК, которая затем вставляется в геном посредством фермента вирусной интегразы. Таким образом, экзогенно доставленное с использованием лентивирусного вектора может оставаться в геноме и передаваться потомству клетки при ее делении.
(c) Аденовирус
Аденовирус обычно использовали в качестве системы доставки генов из-за его генома среднего размера, легкости манипуляции, высокого титра, широкого интервала клеток-мишеней и высокой инфекционности. Оба конца данного вирусного генома содержат ITR (от англ. inverted terminal repeats инвертированные концевые повторы) из 100-200 п.н., которые представляют собой цис-элементы, необходимые для репликации и упаковки вирусной ДНК. Область EI (EIA и EIB) кодирует белки, ответственные за регуляцию транскрипции вирусного генома и нескольких клеточных генов. Экспрессия области Е2 (Е2А и Е2В) приводит к синтезу белков для репликации вирусной ДНК.
Из аденовирусных векторов, разработанных до настоящего времени, обычно используют некомпетентный в отношении репликации аденовирус, имеющий делетированную область EI. Делетированная область Е3 в аденовирусных векторах может обеспечивать сайт вставки для трансгенов (Thimmappaya, В. еt al., Cell, 31:543-551(1982) и Riordan, J.R. et al., Science, 245:1066- 1073 (1989)). Следовательно, предпочтительным является то, что нуклеотидную последовательность, кодирующую декорин, вставляют либо в делетированную область EI (область EIA и/или область EIB 5, предпочтительно область EIB), либо в делетированную область Е3. Полинуклеотид по изобретению может быть вставлен в делетированную область Е4. Термин «делеция» по отношению к вирусным геномным последовательностям охватывает полную делецию, а также частичную делецию. В природе аденовирус может упаковывать приблизительно 105% от генома дикого типа, обеспечивая вместительность для примерно 2 дополнительных т.п.н. ДНК (Ghosh-Choudhury et al., EMBO J. 6:1733-1 739 (1987)). В данном отношении чужеродные последовательности, описанные выше, вставленные в аденовирус, могут быть дополнительно вставлены в геном аденовируса дикого типа.
Данный аденовирус может быть любого известного серотипа или подгруппы A-F. Аденовирус типа 5 подгруппы С является самым предпочтительным исходным материалом для конструирования аденовирусной системы доставки генов по данному изобретению. Известно значительное количество биохимической и генетической информации об аденовирусе типа 5. Чужеродные гены, доставляемые аденовирусной системой доставки генов, являются эписомными и генотоксичные для клеток-хозяев. Следовательно, генотерапия с использованием аденовирусной системы доставки генов может быть в значительной степени безопасной.
(d) Аденоассоциированный вирус (AAV, от англ - adeno-associated virus)
Аденоассоциированные вирусы способны инфицировать неделящиеся клетки и разные типы клеток, делая их полезными в конструировании системы доставки генов по данному изобретению. Подробные описания для применения и получения вектора на основе AAV находятся в патентах США №10308958; 10301650; 10301648; 10266846; 10265417; 10208107; 10167454; 10155931; 10149873; 10144770; 10138295; 10137176; 10113182; 10041090; 9890365; 9790472; 9770011; 9738688; 9737618; 9719106; 9677089; 9617561; 9597363; 9593346; 9587250; 9567607; 9493788; 9382551; 9359618; 9217159; 9206238; 9163260; 9133483; 8962332, раскрытия которых включаются в данный документ посредством ссылки во всей их полноте, и патентах США №5139941 и 4797368, раскрытия которых включаются в данный документ посредством ссылки во всей их полноте.
Результаты исследования для AAV в качестве систем доставки генов раскрываются в LaFace et al., Viology, 162: 483486 (1988), Zhou et al., Exp. Hematol. (NY), 21:928-933(1993), Walsh et al., J. Clin. Invest., 94:1440-1448(1994) и Flotte et al., Gene Therapy, 2:29-37(1995). Типично рекомбинантный вирус AAV получают сотрансфицированием плазмиды, содержащей интересующий ген (т.е. интересующую нуклеотидную последовательность, подлежащую доставке, например, кодирующую последовательность изоформы IGF-1), фланкированный двумя концевыми повторами AAV (McLaughlin et al., 1988; Samulski et al., 1989), и экспрессией плазмиды, содержащей кодирующие последовательности AAV дикого типа без концевых повторов (McCarty et al., J. Viral., 65:2936-2945 (1991)).
(e) Другие вирусные векторы
В настоящем изобретении можно использовать другие вирусные векторы в качестве системы доставки генов. В настоящих системах доставки для трансфицирования в клетки полинуклеотида по изобретению можно использовать векторы, полученные из таких вирусов, как вирус осповакцины (Puhlmann М. et al., Human Gene Therapy 10:649-657(1999); Ridgeway, "Mammalian expression vectors," In: Vectors: A survey of molecular cloning vectors and their uses. Rodriguez and Denhardt, eds. Stoneham: Butterworth, 467-492 (1988); Baichwal and Sugden, "Vectors for gene transfer derived from animal DNA viruses: Transient and stable expression of transferred genes," In: Kucherlapati R, ed. Gene transfer. New York: Plenum Press, 117-148 (1986) и Coupar et al., Gene, 68:1-10(1988)), лентивирус (Wang G. et al., J. Clin. Invest. 104 (11): RS 5-62 (1999)) и вирус простого герпеса (Chamber R., et al., Proc. Natl. 10 15 Acad. Sci USA 92:1411-1415 (1995)).
6.3.4. Способы введения
Для введения ДНК-конструкции, кодирующей IGF-1, и ДНК-конструкции, кодирующей HGF, можно использовать разные способы.
6.3.4.1.1. Инъекция
В некоторых воплощениях ДНК-конструкция вводится посредством инъекции жидкой фармацевтической композиции. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, и ДНК-конструкция, кодирующая HGF, вводятся совместно посредством одной инъекции. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, и ДНК-конструкция, кодирующая HGF, вводятся совместно посредством многих инъекций. В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, и ДНК-конструкция, кодирующая HGF, вводятся индивидуально посредством многих инъекций.
В предпочтительных в настоящее время воплощениях ДНК-конструкция вводится посредством внутримышечной инъекции. Типично ДНК-конструкция вводится посредством внутримышечной инъекции близко к месту повреждения нерва, месту боли или ощущаемому пациентом месту боли, или месту другого симптома, ассоциированного с нейропатическим заболеванием. В некоторых воплощениях ДНК-конструкции вводятся в мышцы кистей, ступней или рук субъекта.
В некоторых воплощениях данную конструкцию инъецируют подкожно или внутрикожно. В некоторых воплощениях ДНК-конструкция вводится посредством внутрисосудистой доставки. В некоторых воплощениях данную конструкцию инъецируют посредством ретроградной внутривенной инъекции.
6.3.4.1.2. Электропорация
Эффективность трансформации плазмидной ДНК в клетки in vivo в некоторых случаях может улучшаться посредством осуществления инъекции с последующей электропорацией. Таким образом, в некоторых воплощениях ДНК-конструкция вводится посредством инъекции, с последующей электропорацией. В конкретных воплощениях введение элекропорацией осуществляется с использованием системы доставки TriGrid™ (Ichor Medical Systems, Inc., Сан-Диего, США).
6.3.4.1.3. Сонопорация
В некоторых воплощениях для усиления эффективности трансформации ДНК-конструкции по настоящему изобретению используют сонопорацию. В сонопорации используется ультразвуковая волна для временной пермеабилизации клеточной мембраны для обеспечения клеточного поглощения ДНК. ДНК-конструкции могут включаться в микропузырьки и вводиться в большой круг кровообращения, с последующим внешним приложением ультразвука. Ультразвук индуцирует кавитацию микропузырька в пределах ткани-мишени, что приводит к высвобождению и трансфекции конструкций.
6.3.4.1.4. Магнитофекция
В некоторых воплощениях для усиления эффективности трансформации ДНК-конструкции по настоящему изобретению используют магнитофекцию. Данная конструкция вводится после связывания с магнитной наночастицей. Приложение высокоградиентных внешних магнитов вызывает захват и удержание комплекса в мишени. Данная ДНК-конструкция может высвобождаться ферментативным расщеплением сшивающей молекулы, взаимодействием зарядов или деградацией матрикса.
6.3.4.1.5. Липосома
В некоторых воплощениях ДНК-конструкции по настоящему изобретению могут доставляться посредством липосом. Липосомы спонтанно образуются при суспендировании фосфолипидов в избытке водной среды. Опосредованная липосомами доставка ДНК была успешной, как описано в Dos Santos Rodrigues et al., Int. J. Pharm. 566:717-730 (2019); Rasoulianboroujeni et al., Mater Sci Eng С Mater Biol Appl. 75:191-197 (2017); Xiong et al., Pharmazie 66(3):158-164 (2011); Nicolau and Sene, Biochim. Biophys. Acta, 721:185-190 (1982) и Nicolau et al., Methods Enzymol., 149:157-176 (1987). Пример имеющихся в продаже реактивов для трансфицирования животных клеток с использованием липосом включает липофектамин (Gibco BRL). Липосомы, захватывающие ДНК-конструкции по изобретению, взаимодействуют с клетками посредством таких механизмов, как эндоцитоз, адсорбция и слияние, и затем переносят данные последовательности в клетки.
6.3.4.1.6. Трансфекция
При использовании вирусного вектора для доставки ДНК-конструкции, кодирующей IGF-1, или ДНК-конструкции, кодирующей HGF, данная конструкция может доставляться в клетки разными способами вирусной инфекции, известными в данной области. Инфекция клеток-хозяев с использованием вирусных векторов известна в данной области.
Фармацевтическая композиция по данному изобретению может вводиться парентерально. Для неперорального введения можно использовать внутривенную инъекцию, внутрибрюшинную инъекцию, внутримышечную инъекцию, подкожную инъекцию или местную инъекцию. Например, данная фармацевтическая композиция может инъецироваться ретроградной внутривенной инъекцией.
Предпочтительно фармацевтическая композиция по настоящему изобретению может вводиться в мышцу. В некоторых воплощениях данное введение нацелено в мышцу, пораженную нейропатией (например, невропатической болью или другими симптомами).
6.3.5. Доза
ДНК-конструкция, кодирующая IGF-1, и ДНК-конструкция, кодирующая HGF, вводятся в терапевтически эффективных количествах. В описанных в данном документе способах терапевтически эффективное количество или доза ДНК-конструкции представляет собой эффективную дозу для лечения нейропатии у субъекта самой по себе или в комбинации с другой ДНК-конструкцией, или в комбинации с другим терапевтическим средством.
В некоторых воплощениях способов, описанных в данном документе, каждая из ДНК-конструкций (ДНК-конструкция, кодирующая IGF-1, и ДНК-конструкция, кодирующая HGF) вводится в общей дозе от 1 мкг до 200 мг, от 1 мг до 200 мг, от 1 мг до 100 мг, от 1 мг до 50 мг, от 1 мг до 20 мг, от 2 мг до 10 мг, 16 мг, 8 мг, 4 мг или 2 мг.
В некоторых воплощениях общая доза ДНК-конструкции, кодирующей IGF-1, и общая доза ДНК-конструкции, кодирующей HGF, которые вводятся субъекту, являются одинаковыми. В некоторых воплощениях общая доза ДНК-конструкции, кодирующей IGF-1, и общая доза ДНК-конструкции, кодирующей HGF, являются разными. В некоторых воплощениях общая доза ДНК-конструкции, кодирующей IGF-1, корректируется в зависимости от общей дозы ДНК-конструкции, кодирующей HGF. В некоторых воплощениях общая доза ДНК-конструкции, кодирующей HGF, корректируется в зависимости от общей дозы ДНК-конструкции, кодирующей IGF-1.
В типичных воплощениях общая доза каждой ДНК-конструкции делится на целый ряд индивидуальных инъекционных доз. В некоторых воплощениях общая доза делится на целый ряд равных инъекционных доз. В некоторых воплощениях общая доза делится на неравные инъекционные дозы.
В разных воплощениях с раздельными дозами общая доза каждой ДНК-конструкции вводится в 4, 8, 16, 24, 32 или 64 разных инъекционных сайтов.
В некоторых воплощениях доза каждой ДНК-конструкции на инъекцию составляет от 0,1 до 20 мг, от 1 до 10 мг, от 2 до 8 мг или от 3 до 8 мг. В определенных воплощениях доза каждой ДНК-конструкции на инъекцию составляет 0,1 мг, 0,15 мг, 0,2 мг, 0,25 мг, 0,3 мг, 0,35 мг, 0,4 мг, 0,45 мг, 0,5 мг, 1 мг, 2 мг, 4 мг, 8 мг, 16 мг или 32 мг.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, и ДНК-конструкция, кодирующая HGF, вводятся совместно. В данных случаях доза двух ДНК-конструкций в комбинации составляет от 0,1 до 20 мг, от 1 до 10 мг, от 2 до 8 мг или от 3 до 8 мг на инъекцию. В некоторых воплощениях доза двух ДНК-конструкций в комбинации составляет 0,1 мг, 0,15 мг, 0,2 мг, 0,25 мг, 0,3 мг, 0,35 мг, 0,4 мг, 0,45 мг, 0,5 мг, 1 мг, 2 мг, 4 мг, 8 мг, 16 мг или 32 мг на инъекцию.
Общая доза каждой ДНК-конструкции или обеих ДНК-конструкций в комбинации может вводиться во время одного посещения или на протяжении двух или более чем двух посещений.
В типичных воплощениях с раздельными дозами все из целого ряда инъекционных доз вводятся в пределах 1 часа друг от друга. В некоторых воплощениях все из целого ряда инъекционных доз вводятся в пределах 1,5; 2; 2,5 или 3 часов друг от друга.
В разных воплощениях данных способов общая доза каждой ДНК-конструкции или общая доза двух ДНК-конструкций в комбинации, независимо от того, вводится ли она в виде единичной однократной дозы или разделенной на множество инъекционных доз, вводится субъекту только один раз.
В некоторых воплощениях один цикл может включать введение общей дозы каждой ДНК-конструкции или двух ДНК-конструкций в комбинации в целый ряд инъекционных сайтов на протяжении одного, двух, трех или четырех посещений. Например, один цикл может включать введение 64 мг, 32 мг, 16 мг, 8 мг, 4 мг или 2 мг каждой ДНК-конструкции в целый ряд инъекционных сайтов на протяжении двух посещений. Данные два посещения могут быть разделены 3, 5, 7, 14, 21 или 28 сутками.
В некоторых воплощениях один цикл может включать введение ДНК-конструкции, кодирующей IGF-1, и введение ДНК-конструкции, кодирующей HGF, в целый ряд инъекционных сайтов на протяжении одного, двух, трех или четырех посещений. Например, один цикл может включать введение 64 мг, 32 мг, 16 мг, 8 мг, 4 мг или 2 мг ДНК-конструкции, кодирующей IGF-1, в целый ряд инъекционных сайтов и введение 64 мг, 32 мг, 16 мг, 8 мг, 4 мг или 2 мг ДНК-конструкции, кодирующей HGF, в целый ряд инъекционных сайтов на протяжении двух посещений. Данные два посещения могут быть разделены 3, 5, 7, 14, 21 или 28 сутками.
В некоторых воплощениях данный цикл может быть повторен. Данный цикл можно повторять дважды, три раза, четыре раза, пять раз, шесть раз или более.
В некоторых воплощениях данный цикл может быть повторен через 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более, чем 12 месяцев после предыдущего цикла.
В некоторых воплощениях общая доза, введенная в последующем цикле, является такой же, как и общая доза, введенная в предыдущем цикле. В некоторых воплощениях общая доза, введенная в последующем цикле, отличается от общей дозы, введенной в предыдущем цикле.
В предпочтительных воплощениях ДНК-конструкция (ДНК-конструкция, кодирующая IGF-1, или ДНК-конструкция, кодирующая HGF) вводится в дозе 8 мг на пораженную конечность, равномерно разделенной на целый ряд внутримышечных инъекций и целый ряд посещений, где каждая из целого ряда инъекций в любое одиночное посещение осуществляется в отдельное место инъекции. В некоторых воплощениях ДНК-конструкция (ДНК-конструкция, кодирующая IGF-1, или ДНК-конструкция, кодирующая HGF) вводится в дозе 8 мг на пораженную конечность, равномерно разделенной на первую дозу в 4 мг на конечность в сутки 0 и вторую дозу в 4 мг на конечность в сутки 14, где каждая из первой и второй дозы равномерно разделяется на целый ряд инъекционных доз.
В некоторых воплощениях ДНК-конструкция, кодирующая IGF-1, и ДНК-конструкция, кодирующая HGF, вводится одновременно или раздельно в общей дозе 16 мг на пораженную конечность, равномерно разделенной на целый ряд внутримышечных инъекий и целый ряд посещений, где каждая из целого ряда инъекий в любое одиночное посещение осуществляется в отдельное место инъекции. В некоторых воплощениях один цикл составляет введение ДНК-конструкции, кодирующей IGF-1, в дозе 8 мг на пораженную конечность и введение ДНК-конструкции, кодирующей HGF, в дозе 8 мг на пораженную конечность. Данный цикл можно повторять один раз, дважды, три раза или более.
Фактическое введенное количество, скорость и динамика введения будут зависеть от природы и тяжести нейропатии, которую лечат. В типичных воплощениях одна или более чем одна ДНК-конструкция вводится в эффективном количестве для уменьшения симптомов нейропатии, например, нейропатической боли. В некоторых воплощениях данное количество является эффективным для уменьшения симптома нейропатии в пределах 1 недели введения. В некоторых воплощениях данное количество является эффективным для уменьшения симптома нейропатии в пределах 2 недель, 3 недель или 4 недель введения.
В некоторых воплощениях вместе вводят два разных типа ДНК-конструкций, кодирующих IGF-1, или два разных типа ДНК-конструкций, кодирующих HGF. В некоторых воплощениях для индукции экспрессии двух изоформ IGF-1 или HGF доставляется конструкция для двойной экспрессии.
Согласно традиционным методикам, известным специалистам в данной области, данная фармацевтическая композиция может быть приготовлена с фармацевтически приемлемым носителем и/или наполнителем, как описано выше, обеспечивая, наконец, несколько видов стандартной леарственной формы и многодозовой формы. Неограничивающие примеры данных препаратов включают раствор, суспензию или эмульсию в масле или водной среде, экстракт, эликсир, поршок, гранулу, таблетку и капсулу, но не ограничиваются ими, и могут дополнительно содержать диспергирующее средство или стабилизатор.
Для помощи в идентификации оптимальных интервалов дозировки возможно можно использовать анализы in vivo и/или in vitro. Точная доза, подлежащая применению в препарате, также будет зависеть от пути введения, серьезности состояния и должна быть определена согласно решению практика и обстоятельствам каждого субъекта. Эффективные дозы могут быть экстраполированы из кривых доза-ответ, полученных от опытных систем животных моделей in vitro.
Данные ДНК-конструкции могут вводиться сами по себе или в комбинации с другими лечениями, либо одновременно, либо последовательно.
6.3.6. Пациенты с нейропатией
В описанных в данном документе способах выбранные для лечения пациенты имеют нейропатию. Данные пациенты могут иметь периферическую нейропатию, черепную нейропатию, автономную нейропатию или очаговую нейропатию. Нейропатия может быть вызвана заболеваниями, повреждениями, инфекциями или состояниями недостаточности витаминов. Например, нейропатия может быть вызвана диабетом, недостаточностями витаминов, аутоиммунными заболеваниями, генетическими или наследственными расстройствами, амилоидозом, уремией, токсинами или ядами, травмой или повреждением, опухолями или может быть идиопатической. В некоторых воплощениях данные пациенты имеют диабетическую периферическую нейропатию.
Данные пациенты могут иметь один или более чем один симптом, ассоциированный с нейропатией, такой как боль (невропатическая боль), другие сенсорные дефекты (например, потеря ощущения, онемение, покалывание и т.д.), моторные дефекты (например, слабость, потеря рефлексов, потеря мышечной массы, хромание, потеря точных координированных движений и т.д.) и автономная дисфункция (например, тошнота, рвота, импотенция, головокружение, запор, диарея и т.д.).
Данных пациентов можно лечить одним или более чем одним способом лечения, известным в данной области, помимо лечения способом, предложенным в данном документе.
Способы лечения по настоящему изобретению можно использовать для лечения человеческого пацента или животного с нейропатией.
6.3.7. Порядок введения
Описанные в данном документе способы включают стадии введения терапевтически эффективного количества первой ДНК-конструкции, кодирующей IGF-1, способной экспрессировать человеческую изоформу IGF-1, и введения терапевтически эффективного количества первой ДНК-конструкции, кодирующей HGF, способной экспрессировать человеческую изоформу HGF. Терапевтически эффективное количество представляет собой эффективное количество в лечении данного заболевания в комбинации или индивидуально.
Стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения первой ДНК-конструкции, кодирующей HGF, могут проводиться одновременно или последовательно. В некоторых воплощениях введение первой ДНК-конструкции, кодирующей IGF-1, и введение первой ДНК-конструкции, кодирующей HGF, проводится раздельно, по меньшей мере с интервалом в несколько минут друг от друга, несколько часов друг от друга, один месяц друг от друга, два месяца друг от друга, три месяца друг от друга или шесть месяцев друг от друга. В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей HGF, проводится до стадии введения первой ДНК-конструкции, кодирующей IGF-1. В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, проводится до стадии введения первой ДНК-конструкции, кодирующей HGF.
В некоторых воплощениях стадия введения первой ДНК-конструкции, кодирующей IGF-1, стадия введения первой ДНК-конструкции, кодирующей HGF, или и та, и другая повторяются. В некоторых воплощениях данная стадия повторяется дважды, три раза или более.
Данная первая ДНК-конструкция, кодирующая IGF-1, может представлять собой любую из ДНК-конструкций, кодирующих IGF-1, предложенных в данном документе, или ее модификацию. Она может экспрессировать одну или более чем одну изоформу IGF-1. Она может представлять собой ДНК-конструкцию, кодирующую одну изоформу IGF-1: изоформы Еc класса I, (SEQ ID NO 16); Еа класса II (SEQ ID NO 18); Eb класса I (SEQ ID NO 20) или Еа класса I (SEQ ID NO 14). Она может представлять собой ДНК-конструкцию двойной экспрессии, кодирующую две изоформы IGF-1. В некоторых воплощениях данная ДНК-конструкция может кодировать изоформы Еc класса I (SEQ ID NO 16) и Еа класса I (SEQ ID NO 14).
Данная первая ДНК-конструкция, кодирующая HGF, может представлять собой любую из ДНК-конструкций, кодирующих HGF, предложенных в данном документе, или ее модификацию. Она может экспрессировать одну или более чем одну изоформу HGF. Она может представлять собой ДНК-конструкцию, кодирующую одну изоформу HGF: fIHGF (SEQ ID NO 11) или dHGF (SEQ ID NO 12). Она может представлять собой ДНК-конструкцию двойной экспрессии, кодирующую две изоформы HGF. В предпочтительных воплощениях данная ДНК-конструкция содержит полинуклеотид SEQ ID NO 13. Она может представлять собой VM202.
Данный способ может дополнительно включать стадию введения второй ДНК-конструкции, кодирующей IGF-1. Данная вторая ДНК-конструкция, кодирующая IGF-1, может быть такой же или отличной от первой ДНК-конструкции, кодирующей IGF-1. Данная вторая ДНК-конструкция, кодирующая IGF-1, может представлять собой любую из ДНК-конструкций, кодирующих IGF-1, предложенных в данном документе, или ее модификацию. Стадия введения первой ДНК-конструкции, кодирующей IGF-1, и стадия введения второй ДНК-конструкции, кодирующей IGF-1, могут осуществляться одновременно или последовательно. В некоторых воплощениях первая ДНК-конструкция, кодирующая IGF-1, способная экспрессировать Еc класса I (SEQ ID NO 16), и вторая ДНК-конструкция, кодирующая IGF-1, способная экспрессировать Еа класса I (SEQ ID NO 14), вводятся одновременно. В некоторых воплощениях введение первой ДНК-конструкции, кодирующей IGF-1, и введение второй ДНК-конструкции, кодирующей IGF-1, проводится раздельно, по меньшей мере с интервалом в несколько минут друг от друга, несколько часов друг от друга, одни сутки друг от друга, двое суток друг от друга, трое суток друг от друга, одна неделя друг от друга, две недели друг от друга, три недели друг от друга, один месяц друг от друга, два месяца друг от друга, три месяца друг от друга или шесть месяцев друг от друга.
Данный способ может дополнительно включать стадию введения второй ДНК-конструкции, кодирующей HGF. Данная вторая ДНК-конструкция, кодирующая HGF, может быть такой же или отличной от первой ДНК-конструкции, кодирующей HGF. Данная вторая ДНК-конструкция, кодирующая HGF, может представлять собой любую из ДНК-конструкций, кодирующих HGF, предложенных в данном документе, или ее модификацию. Стадия введения первой ДНК-конструкции, кодирующей HGF, и стадия введения второй ДНК-конструкции, кодирующей HGF, могут осуществляться одновременно или последовательно. Например, первая ДНК-конструкция, кодирующая HGF, способная экспрессировать fIHGF (SEQ ID NO 11), и вторая ДНК-конструкция, кодирующая HGF, способная экспрессировать dHGF (SEQ ID NO 12), могут вводиться одновременно. В некоторых воплощениях введение первой ДНК-конструкции, кодирующей HGF, и введение второй ДНК-конструкции, кодирующей HGF, проводится раздельно, по меньшей мере с интервалом в несколько минут друг от друга, несколько часов друг от друга, одни сутки друг от друга, двое суток друг от друга, трое суток друг от друга, одна неделя друг от друга, две недели друг от друга, три недели друг от друга, один месяц друг от друга, два месяца друг от друга, три месяца друг от друга или шесть месяцев друг от друга.
В некоторых воплощениях данный способ включает введение VM202 наряду с pCK-IGF-1X6 или pCK-IGF-1X10. В некоторых воплощениях данный способ включает введение другой ДНК-конструкции, кодирующей HGF (например, конструкции, содержащей полинуклеотид SEQ ID NO 33), наряду с pCK-IGF-1X6 или pCK-IGF-1X10.
В некоторых воплощениях данный способ включает введение VM202, наряду с pTx-IGF-1X6 или pTx-IGF-1X10. В некоторых воплощениях данный способ включает введение другой ДНК-конструкции, кодирующей HGF (например, конструкции, содержащей полинуклеотид SEQ ID NO 33), наряду с pTx-IGF-1X6 или pTx-IGF-1X10.
В некоторых воплощениях данный способ включает введение VM202 с последующим введением pCK-IGF-1X6 или pCK-IGF-1X10. В некоторых воплощениях данный способ включает введение другой ДНК-конструкции, кодирующей HGF (например, pCK-HGF728, которая представляет собой конструкцию, содержащую полинуклеотид SEQ ID NO 33), с последующим введением pCK-IGF-1X6 или pCK-IGF-1X10. В некоторых воплощениях данный способ включает введение pCK-IGF-1X6 или pCK-IGF-1X10 с последующим введением VM202 или другой ДНК-конструкции, кодирующей HGF (например, рСK-HGF728).
В некоторых воплощениях данный способ включает введение VM202 с последующим введением pTx-IGF-1X6 или pTx-IGF-1X10. В некоторых воплощениях данный способ включает введение другой ДНК-конструкции, кодирующей HGF (например, pCK-HGF728, которая представляет собой конструкцию, содержащую полинуклеотид SEQ ID NO 33), с последующим введением pTx-IGF-1X6 или pTx-IGF-1X10. В некоторых воплощениях данный способ включает введение pTx-IGF-1X6 или pTx-IGF-1X10 с последующим введением VM202 или другой ДНК-конструкции, кодирующей HGF (например, рСK-HGF728).
6.4. Фармацевтическая композиция, содержащая ДНК-конструкцию, кодирующую IGF-1, и ДНК-конструкцию, кодирующую HGF-1
В другом аспекте предложена фармацевтическая композиция, содержащая ДНК-конструкцию, кодирующую IGF-1, и ДНК-конструкцию, кодирующую HGF.
6.4.1. Фармацевтические композиции и стандартные лекарственные формы, адаптированные для инъекции
Для внутривенной, внутримышечной, внутрикожной или подкожной инъекции ДНК-конструкция может находиться в форме парентерально приемлемого водного раствора, который является непирогенным и имеет подходящий рН, изотоничность и стабильность. Специалисты релевантной квалификации в данной области легко способны готовить подходящие растворы с использованием, например, изотоничных носителей, таких как инъекционный раствор хлорида натрия, инъекционный раствор Рингера, инъекционный раствор Рингера с лактатом. Могут быть включены консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки по потребности.
В некоторых воплощениях данная фармацевтическая композиция содержит ДНК-конструкцию, кодирующую одну изоформу IGF-1. Например, данная ДНК-конструкция может экспрессировать изоформу Еc класса I (изоформа №1); изоформу Еа класса II (изоформа №2); изоформу Еb класса I (изоформа №3) или изоформу Еа класса I (изоформа №4). Данные ДНК-конструкции могут представлять собой pCK-IGF-1#1, pCK-IGF-1#2, pCK-IGF-1#3 или pCK-IGF-1#4. В некоторых воплощениях данные ДНК-конструкции могут представлять собой рТх-IGF-1#1, pTx-IGF-1#2, pTx-IGF-1#3 или pTx-IGF-1#4.
В некоторых воплощениях данная фармацевтическая композиция содержит более чем одну ДНК-конструкцию, причем каждая кодирует одну изоформу IGF-1. Например, данная фармацевтическая композиция может содержать (i) первую ДНК-конструкцию, кодирующую изоформу Еc класса I (изоформа №1) и вторую ДНК-конструкцию, кодирующую изоформу Еа класса II (изоформа №2); (ii) первую ДНК-конструкцию, кодирующую изоформу Еc класса I (изоформа №1) и вторую ДНК-конструкцию, кодирующую изоформу Еb класса I (изоформа №3); (iii) первую ДНК-конструкцию, кодирующую изоформу Еc класса I (изоформа №1) и вторую ДНК-конструкцию, кодирующую изоформу Еа класса I (изоформа №4); (iv) первую ДНК-конструкцию, кодирующую изоформу Еа класса II (изоформа №2) и вторую ДНК-конструкцию, кодирующую изоформу Еb класса I (изоформа №3); (v) первую ДНК-конструкцию, кодирующую изоформу Еа класса II (изоформа №2) и вторую ДНК-конструкцию, кодирующую изоформу Еа класса I (изоформа №4); (vi) первую ДНК-конструкцию, кодирующую изоформу Еb класса I (изоформа №3) и вторую ДНК-конструкцию, кодирующую изоформу Еа класса I (изоформа №4).
В некоторых воплощениях данная фармацевтическая композиция содержит конструкцию двойной экспрессии, причем ДНК-конструкция может экспрессировать более чем одну изоформу IGF-1. Например, данная фармацевтическая композиция может содержать конструкцию двойной экспрессии, которая может экспрессировать (i) изоформу Еc класса I (изоформа №1) и изоформу Еа класса II (изоформа №2); (ii) изоформу Еc класса I (изоформа №1) и изоформу Еb класса I (изоформа №3); (iii) изоформу Еc класса I (изоформа №1) и изоформу Еа класса I (изоформа №4); (iv) изоформу Еа класса II (изоформа №2) и изоформу Еb класса I (изоформа №3); (v) изоформу Еа класса II (изоформа №2) и изоформу Еа класса I (изоформа №4); (vi) изоформу Еb класса I (изоформа №3) и изоформу Еа класса I (изоформа №4).
В некоторых воплощениях данная фармацевтическая композиция содержит конструкцию двойной экспрессии: pCK-IGF-1X6 или pCK-IGF-1X10. В некоторых воплощениях данная фармацевтическая композиция содержит конструкцию двойной экспрессии: pTx-IGF-1X6 или pTx-IGF-1X10. В некоторых воплощениях данная фармацевтическая композиция содержит две конструкции двойной экспрессии, например, включающие и pCK-IGF-1X6, и pCK-IGF-1X10. В некоторых воплощениях данная фармацевтическая композиция содержит две конструкции двойной экспрессии, например, включающие и pTx-IGF-1X6, и pTx-IGF-1X10.
В некоторых воплощениях данная фармацевтическая композиция дополнительно содержит ДНК-конструкцию, кодирующую одну изоформу HGF. Например, данная ДНК-конструкция может экспрессировать fIHGF или dHGF. В некоторых воплощениях данная фармацевтическая композиция содержит более чем одну ДНК-конструкцию, причем каждая кодирует одну изоформу HGF. Например, данная фармацевтическая композиция может содержать первую ДНК-конструкцию, кодирующую fIHGF, и вторую ДНК-конструкцию, кодирующую dHGF.
В некоторых воплощениях данная фармацевтическая композиция содержит конструкцию двойной экспрессии - ДНК-конструкцию, которая может экспрессировать более чем одну изоформу HGF. Например, данная фармацевтическая композиция может содержать конструкцию двойной экспрессии, которая может экспрессировать и fIHGF, и dHGF.
В предпочтительных воплощениях данная фармацевтическая композиция содержит конструкцию двойной экспрессии - pCK-HGF-X7 (VM202). В некоторых воплощениях данная фармацевтическая композиция содержит две ДНК-конструкции, кодирующих HGF, причем каждая кодирует fIHGF или dHGF. В некоторых воплощениях данная фармацевтическая композиция содержит одну ДНК-конструкцию, кодирующую HGF, способную экспрессировать fIHGF (рСK-HGF728).
В некоторых воплощениях данная фармацевтическая композиция дополнительно содержит другое терапевтическое средство. Например, данная фармацевтическая композиция может дополнительно содержать другое эффективное в лечение нейропатии терапевтическое средство.
В разных воплощениях одна или более чем одна ДНК-конструкция присутствует в жидкой композиции в концентрации 0,01 мг/мл, 0,05 мг/мл, 0,1 мг/мл, 0,25 мг/мл, 0,45 мг/мл, 0,5 мг/мл или 1 мг/мл, индивидуально или в комбинации. В некоторых воплощениях стандартная лекарственная форма представляет собой флакон, содержащий 2 мл фармацевтической композиции с одной или более чем одной ДНК-конструкцией в концентрации 0,01 мг/мл, 0,1 мг/мл, 0,5 мг/мл или 1 мг/мл, индивидуально или в комбинации. В некоторых воплощениях стандартная лекарственная форма представляет собой флакон, содержащий 1 мл фармацевтической композиции с одной или более чем одной ДНК-конструкцией в концентрации 0,01 мг/мл, 0,1 мг/мл, 0,5 мг/мл или 1 мг/мл, индивидуально или в комбинации. В некоторых воплощениях стандартная лекарственная форма представляет собой флакон, содержащий меньше, чем 1 мл фармацевтической композиции с одной или более чем одной ДНК-конструкцией в концентрации 0,01 мг/мл, 0,1 мг/мл, 0,5 мг/мл или 1 мг/мл, индивидуально или в комбинации.
В некоторых воплощениях стандартная лекарственная форма представляет собой флакон, ампулу, бутыль или предварительно заполненный шприц. В некоторых воплощениях стандартная лекарственная форма содержит 0,01 мг, 0,1 мг, 0,2 мг, 0,25 мг, 0,5 мг, 1 мг, 2,5 мг, 5 мг, 8 мг, 10 мг, 12,5 мг, 16 мг, 24 мг, 25 мг, 50 мг, 75 мг, 100 мг, 150 мг или 200 мг одной или более чем одной ДНК-конструкции по настоящему изобретению.
В некоторых воплощениях фармацевтическая композиция в стандартной лекарственной форме находится в жидкой форме. В разных воплощениях стандартная лекарственная форма содержит от 0,1 мл до 50 мл фармацевтической композиции. В некоторых воплощениях стандартная лекарственная форма содержит 0,25 мл, 0,5 мл, 1 мл, 2,5 мл, 5 мл, 7,5 мл, 10 мл, 25 мл или 50 мл фармацевтической композиции.
В конкретных воплощениях стандартная лекарственная форма представляет собой флакон, содержащий 0,5 мл, 1 мл, 1,5 мл или 2 мл фармацевтической композиции, в подходящих воплощениях стандартной лекарственной формы для подкожного, внутрикожного или внутримышечного введения, включая предварительно загруженные шприцы, аутоинъекторы и аутоинъекционные шприцы-ручки, причем каждый содержит заданное количество фармацевтической композиции, описанной в данном документе выше.
В разных воплощениях стандартная лекарственная форма представляет собой предварительно заполенный шприц, включающий шприц и заданное количество фармацевтической композиции. В некоторых воплощениях предварительно заполенного шприца данный шприц приспособлен для подкожного введения. В некоторых воплощениях данный шприц подходит для самостоятельного введения. В конкретных воплощениях предварительно заполненный шприц представляет собой одноразовый шприц.
В разных воплощениях предварительно заполненный шприц содержит от примерно 0,1 мл до примерно 0,5 мл фармацевтической композиции. В некоторых воплощениях данный шприц содержит примерно 0,5 мл фармацевтической композиции. В конкретных воплощениях данный шприц содержит примерно 1,0 мл фармацевтической композиции. В конкретных воплощениях данный шприц содержит примерно 2,0 мл фармацевтической композиции.
В некоторых воплощениях стандартная лекарственная форма представляет собой шприц-ручку для автоинъекции. Данная шприц-ручка для автоинъекции включает шприц-ручку для автоинъекции, содержащую фармацевтическую композицию, как описано в данном документе. В некоторых воплощениях данная шприц-ручка для автоинъекции доставляет заданный объем фармацевтической композиции. В других воплощениях данная шприц-ручка для автоинъекции сконфигурирована для доставки установленного пользователем объема фармацевтической композиции.
В разных воплощениях данная шприц-ручка для автоинъекции содержит от примерно 0,1 мл до примерно 5,0 мл фармацевтической композиции. В конкретных воплощениях данная шприц-ручка для автоинъекции содержит примерно 0,5 мл фармацевтической композиции. В конкретных воплощениях данная шприц-ручка для автоинъекции содержит примерно 1,0 мл фармацевтической композиции. В других воплощениях данная шприц-ручка для автоинъекции содержит примерно 5,0 мл фармацевтической композиции.
6.4.2. Лиофилизированные препараты ДНК
В некоторых воплощениях ДНК-конструкции по настоящему изобретению приготовлены в виде лиофилизированной композиции. В конкретных воплощениях ДНК-конструкции лиофилизируют, как описано в патенте США №8389492, включенном в данный документ посредством ссылки во всей его полноте.
В некоторых воплощениях ДНК-конструкции готовят с некоторыми эксципиентами, например, углеводом и солью, перед лиофилизацией. Стабильность ДНК-конструкции, подлежащей применению в качестве диагностического или терапевтического средства, может быть увеличена посредством приготовления данной ДНК-конструкции перед лиофилизацией с водным раствором, содержащим стабилизирующее количество углевода.
Данный углевод может представлять собой моно-, олиго- или полисахарид, такой как сахароза, глюкоза, лактоза, трегалоза, арабиноза, пентоза, рибоза, ксилоза, галактоза, гексоза, идоза, манноза, талоза, гептоза, фруктоза, глюконовая кислота, сорбит, маннит, метил-α-глюкопиранозид, мальтоза, изоаскорбиновая кислота, аскорбиновая кислота, лактон, сорбоза, глюкаровая кислота, эритроза, треоза, аллоза, альтроза, гулоза, эритрулоза, рибулоза, ксилулоза, псикоза, тагатоза, глюкуроновая кислота, галактуроновая кислота, маннуроновая кислота, глюкозамин, галактозамин, нейраминовая кислота, арабинаны, фруктаны, фуканы, галактаны, галактуронаны, глюканы, маннаны, ксиланы, леван, фукоидан, каррагенан, галактокаролоза, пектины, пектиновые кислоты, амилоза, пуллулан, гликоген, амилопектин, целлюлоза, декстран, циклодекстрин, пустулан, хитин, агароза, кератин, хондроитин, дерматан, гиалуроновая кислота, альгиновая кислота, ксантановая камедь или крахмал.
В одной серии воплощений данный углевод представляет собой маннит или сахарозу.
Раствор углевода перед лиофилизацией может соответствовать углеводу в одной воде, или может быть включен буфер. Примеры таких буферов включают PBS (от англ. phosphate buffered saline - фосфатно-солевой буферный раствор), HEPES (от англ. (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) - 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота), TRIS или TRIS/EDTA (от англ. Ethylenediaminetetraacetic acid - этилендиаминтетрауксусная кислота). Типично раствор углевода объединяют с ДНК-конструкцией до конечной концентрации от примерно 0,05% до примерно 30% сахарозы, типично от 0,1% до примерно 15% сахарозы, как, например, от 0,2% до примерно 5%, 10% или 15% сахарозы, предпочтительно примерно от 0,5% до 10% сахарозы, от 1% до 5% сахарозы, от 1% до 3% сахарозы и наиболее предпочтително примерно 1,1% сахарозы.
Препарат ДНК по данному изобретению также может включать соль, например, NaCl или KCl. В некоторых воплощениях данная соль представляет собой NaCl. В некоторых воплощениях соль препарата ДНК находится в количестве, выбранном из группы, состоящей из от примерно 0,001% до примерно 10%, примерно от 0,1% до 5%, примерно от 0,1% до 4%, примерно от 0,5% до 2%, примерно от 0,8% до 1,5%, примерно от 0,8% до 1,2 масс/об. %. В некоторых воплощениях соль препарата ДНК находится в количестве примерно 0,9 масс/об. %.
Конечная концентрация одной или более чем одной ДНК-конструкции в жидких композициях, растворенных из лиофилизированных препаратов, может составлять от примерно 1 нг/мл до примерно 30 мг/мл. Например, конечная концентрация может составлять примерно 1 нг/мл, примерно 5 нг/мл, примерно 10 нг/мл, примерно 50 нг/мл, примерно 100 нг/мл, примерно 200 нг/мл, примерно 500 нг/мл, примерно 1 мкг/мл, примерно 5 мкг/мл, примерно 10 мкг/мл, примерно 50 мкг/мл, примерно 100 мкг/мл, примерно 200 мкг/мл, примерно 400 мкг/мл, примерно 500 мкг/мл, примерно 600 мкг/мл, примерно 800 мкг/мл, примерно 1 мг/мл, примерно 2 мг/мл, примерно 2,5 мг/мл, примерно 3 мг/мл, примерно 3,5 мг/мл, примерно 4 мг/мл, примерно 4,5 мг/мл, примерно 5 мг/мл, примерно 5,5 мг/мл, примерно 6 мг/мл, примерно 7 мг/мл, примерно 8 мг/мл, примерно 9 мг/мл, примерно 10 мг/мл, примерно 20 мг/мл или примерно 30 мг/мл, индивидуально или в комбинации. В некоторых воплощениях данного изобретения конечная концентрация одной или более чем одной ДНК-конструкции составляет от примерно 100 мкг/мл до примерно 2,5 мг/мл индивидуально или в комбинации. В конкретных воплощениях изобретения конечная концентрация одной или более чем одной ДНК-конструкции составляет от примерно 0,5 мг/мл до 1 мг/мл индивидуально или в комбинации.
Препарат ДНК по изобретению лиофилизируют при стандартных условиях, известных в данной области. Способ лиофилизации препарата ДНК по изобретению может включать (а) загрузку контейнера (например, флакона) с препаратом ДНК (например, препаратом ДНК, содержащим одну или более чем одну ДНК-конструкцию по настоящему изобретению), солью и углеводом в лиофилизатор, где данный лиофилизатор имеет исходную температуру от примерно 5°С до примерно -50°С; (b) охлаждение препарата ДНК до температур ниже нуля (например, от -10°С до -50°С); и (с) существенную сушку ДНК-препарата. Условия для лиофилизации, например, температуру и продолжительность, препарата ДНК по изобретению может корректировать обычный специалист в данной области, принимая во внимание факторы, которые влияют на параметры лиофилизации, например, используемый тип лиофилизатора, используемое количество ДНК и используемый размер контейнера.
Контейнер, содержащий лиофилизированный препарат ДНК, можно затем запечатывать и хранить в течение длительного периода времени при разных температурах (например, от комнатной температуры до примерно -180°С, предпочтительно от примерно 2-8°С до примерно -80°С, более предпочтительно от примерно -20°С до примерно -80°С и наиболее предпочтительно - примерно -20°С). В некоторых аспектах данные лиофилизированные препараты ДНК предпочтительно являются стабильными в пределах интервала от примерно 2-8°С до примерно -80°С в течение периода по меньшей мере 6 месяцев без потери значительной активности. Препарат плазмидной ДНК стабильного хранения также может соответствовать хранению плазмидной ДНК в стабильной форме в течение длительных периодов времени перед применением в качестве таковых для исследования или терапии на основе плазмиды. Срок хранения может составлять до нескольких месяцев, 1 года, 5 лет, 10 лет, 15 лет или вплоть до 20 лет. Предпочтительно данный препарат является стабильным в течение периода по меньшей мере примерно 3 лет.
6.5. Наборы для комбинированной терапии
В другом аспекте согласно настоящему изобретению предложен набор для комбинированной терапии с ДНК-конструкцией, кодирующей IGF-1, и ДНК-конструкцией, кодирующей HGF.
Данный набор может содержать первую фармацевтическую композицию, содержащую ДНК-конструкцию, кодирующую IGF-1, и вторую фармацевтическую композицию, содержащую ДНК-конструкцию, кодирующую HGF. В некоторых воплощениях первая фармацевтическая композиция и вторая фармацевтическая композиция представляют собой ту же самую фармацевтическую композицию в одном контейнере. В некоторых воплощениях первая фармацевтическая композиция и вторая фармацевтическая композиция представляют собой отдельные фармацевтические композиции в двух или более чем двух отдельных контейнерах.
Данная первая фармацевтическая композиция может содержать любую предложенную в данном документе ДНК-конструкцию, кодирующую IGF-1. Например, данная ДНК-конструкция, кодирующая IGF-1, может представлять собой ДНК-конструкцию одиночной экспрессии, способную экспрессировать одну изоформу IGF-1, или ДНК-конструкцию двойной экспрессии, экспресирующую две изоформы IGF-1. Вторая фармацевтическая композиция может содержать любую предложенную в данном документе ДНК-конструкцию, кодирующую HGF. Например, данная ДНК-конструкция, кодирующая HGF, может представлять собой ДНК-конструкцию одиночной экспрессии, способную экспрессировать одну изоформу HGF, или ДНК-конструкцию двойной экспрессии, экспресирующую две изоформы HGF.
Данный набор может содержать одну или более чем одну разовую дозу ДНК-конструкции, кодирущей IGF-1, ДНК-конструкции, кодирущей HGF, или и той, и другой.
Данный набор может дополнительно содержать инструкцию, объясняющую способ введения ДНК-конструкции, кодирущей IGF-1, ДНК-конструкции, кодирущей HGF, или и той, и другой. Данный способ может представлять собой любой из способов введения, предложенный в данном документе.
6.6. Примеры
Следующие примеры используются таким образом, чтобы предоставлять обычным специалистам в данной области полное раскрытие и описание того, как сделать и применять настоящее изобретение, и не предназначены ни для того, чтобы ограничивать объем того, что авторы данного изобретения рассматривают как их изобретение, ни для того, чтобы представлять, что описанные ниже эксперименты представляют собой все или единственные проведенные эксперименты. Были предприняты усилия для обеспечения точности в отношении используемых чисел (например, количеств, температуры и т.д.), но следует учитывать некоторые экспериментальные ошибки и отклонения. Если не указано иное, части представляют собой части по массе, молекулярные массы представляют собой среднемассовую молекулярную массу, температура приводится в градусах Цельсия, и давление является атмосферным или околоатмосферным. Можно использовать стандартные сокращения, например, п. н. - пара(ры) нукпеотидов; т.п.н. - тысяча(чи) пар нукпеотидов; пл - пиколитр; с - секунда(ды); мин - минута(ты); ч - час(сы); ак - аминокислота(ты); нт - нукпеотид(ды) и тому подобные.
В воплощении на практике настоящего изобретения будут применяться, если не указано иное, традиционные способы белковой химии, биохимии, методики генной инженерии и фармакологии, находящиеся в пределах квалификации в данной области.
6.6.1. Пример 1: терапевтическая эффективность совместного введения ДНК-конструкций, кодирущих IGF-1, и ДНК-конструкции, кодирущей HGF, в мышиной модели нейропатии CCI
Терапевтические эффекты совместного введения конструкции, кодирующей IGF-1, и конструкции, кодирующей HGF, анализировали в модели хронического компрессионного повреждения (CCI) у мышей, которая представляет собой широко принятую модель для исследования нейропатии. Мышиную модель CCI получали посредством хронического компрессионного повреждения (CCI) седалищного нерва, которое, как известно, инициирует несколько процессов, которые приводят к хроническому повреждению нервов на периферии. Также известно, что мыши CCI имеют невропатическую боль.
В частности, покупали 4-недельных самцов мышей ICR (с массой тела от 24 до 26 г) и использовали для получения модели CCI и анализа эффектов. Все хирургические и экспериментальные процедуры были одобрены Институциональным комитетом по уходу и использованию животных в Сеульском национальном университете. Для осуществления CCI у мышей, у них делали тупые диссекции примерно 1 см длины для экспонирования правого седалищного нерва, который обычно находится между ягодичной мышцей и двуглавой мышцей бедра. Как только экспонировался седалищный нерв рядом с местом разделения на три ветви, на него три раза накладывали слабые лигатуры на расстоянии 0,5 мм друг от друга с использованием шелковых нитей 6-0 (Ethicon). Данные лигатуры слегка подтягивали, пока не было заметной судороги правой задней конечности. Ложнооперированным мышам делали такие же диссекции в правом бедре, но они не получали лигатур на седалищном нерве.
В сутки CCI внутримышечно инъецировали суммарно 200 мкг ДНК-конструкций, и измеряли болевую чувствительность в отношении механических стимулов посредством нити фон Фрея. Типы и количества ДНК-конструкций, инъецированных каждой группе, обобщаются ниже в ТАБЛИЦЕ 1.

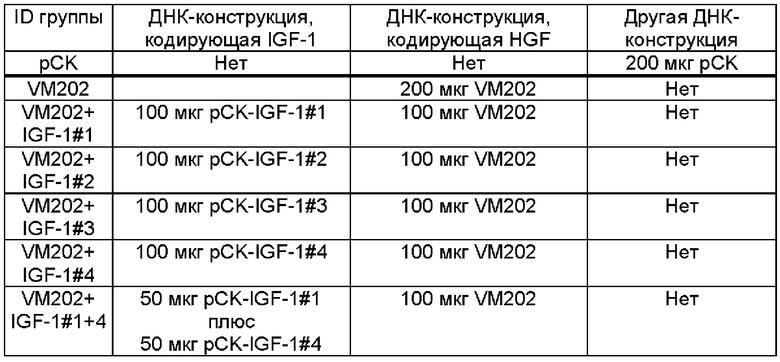
Каждая группа состояла из 6 мышей, и проводили больше, чем 2 независимых эксперимента (среднее плюс/минус стандартная ошибка среднего (SEM - от англ. standard error mean); * - р меньше 0,05, ** - р меньше 0,01, *** - р меньше 0,001).
В ТАБЛИЦЕ 1 pCK-IGF-1#1, pCK-IGF-1#2, pCK-IGF-1#3 и pCK-IGF-1#4 представляют собой ДНК-конструкции, кодирующие индивидуальную человеческую изоформу IGF-1, клонированную в вектор рСK. Данные ДНК-конструкции конструировали в векторе рСK с использованием стандартных методик молекулярного клонирования. В частности, получали четыре полинуклеотида (SEQ ID NO: 15, 17, 19 и 21) посредством кастомизированного способа синтеза ДНК, обеспечиваемого Bioneer (Корея). Данные полинукпетиды синтезировали с 5'-линкером, Cla I и 3'-линкером, Sal I. Вектор рСK и полинуклеотиды подвергали рестрикции Cla I и Sal I. IGF-1 №1, кодирующую Еc класса I (изоформа №1) получали вставкой полинуклеотида SEQ ID NO 17, который представляет собой кодирующую последовательность изоформы Еc класса I и содержит по меньшей мере часть экзонов 1, 3/4, 5 и 6 гена IGF-1, в сайт клонирования вектора рСK. IGF-1 №2, кодирующую Еа класса II (изоформа №2) получали вставкой полинуклеотида SEQ ID NO 19, который представляет собой кодирующую последовательность изоформы Еа класса II и содержит по меньшей мере часть экзонов 2, 3/4 и 6 гена IGF-1, в сайт клонирования вектора рСK. IGF-1 №3, кодирующую Еb класса I (изоформа №3) получали вставкой полинуклеотида SEQ ID NO 21, который представляет собой кодирующую последовательность изоформы Еb класса I и содержит по меньшей мере часть экзонов 1, 3/4 и 5 гена IGF-1, в сайт клонирования вектора рСK. IGF-1 №4, кодирующую Еа класса I (изоформа №4) получали вставкой полинуклеотида SEQ ID NO 15, и содержит по меньшей мере часть экзонов 1, 3/4 и 6, в сайт клонирования вектора рСK. Экспрессию каждой изоформы IGF-1 от каждой плазмиды анализировали и подтверждали как in vitro, так и in vivo.
Через одну неделю после хирургии CCI и введения ДНК-конструкций оценивали развитие механической аллодинии с использованием нити фон Фрея, и симптомы боли оценивали еженедельно, как приводится на Фиг. 2А. Анализ с нитью фон Фрея проводили для измерения механической чувствительности мышей. Вкратце, животных индивидуально помещали в цилиндр над полом из металлической сетки для адаптации. Частоту механической чувствительности мышей оценивали посредством стимулирования задней лапы с использованием постоянной толщины нити (калибр 0,16).
Фиг. 2В представляет собой график, обобщающий частоту (%) отдергивания лапы, измеренную в эксперименте CCI, описанном на Фиг. 2А. Частота (%)Фиг. 2В представляет собой среднее измерений, сделанных в 1 неделю - 4 недели после хирургии CCI. Данный результат демонстрирует то, что инъекция одной VM202 или VM202 в комбинации с разными ДНК-конструкциями, кодирующими IGF-1, обеспечивает значимое снижение частоты отдергивания лапы по сравнению с одним вектором (рСK). Кроме того, инъекция VM202 в комбинации с pCK-IGF-1#1 или pCK-IGF-1#4 (т.е. ДНК-конструкция, способная экспрессировать изоформу №1 IGF-1 или изоформу №4 IGF-1) обеспечивала более значительное снижение, чем инъекция одного VM202 или VM202 в комбинации с IGF-1#2 или IGF-1#3. Данные свидетельствуют о том, что изоформа №1 IGF (Еc класса I) и изоформа №4 IGF (Еа класса I) являются особенно эффективными в лечении нейропатии при введении совместно с VM202.
IGF-1#1 и IGF-1#4, которые оказались самыми эффективными в данных, приведенных на Фиг. 2 В, анализировали далее для того, чтобы понять, могут ли быть усилены их эффекты при совместном введении. В частности, 50 мкг IGF-1 #1 и 50 мкг IGF-1#4 вводили мышам CCI совместно с VM202, и частоту отдергивания их лапы измеряли, как обобщено на Фиг. ЗА. Результат (среднее от 1 недели до 4 недель), приведенный на Фиг. ЗВ, демонстрирует то, что инъекция VM202 в комбинации как с pCK-IGF-1#1, так и с pCK-IGF-1#4 давала даже более значительное уменьшение частоты отдергивания лапы по сравнению с VM202 в комбинации с одним pCK-IGF-1#1 или с VM202 в комбинации с одним pCK-IGF-1#4. Данные свидетельстуют о том, что изоформа №1 IGF (Еc класса I) или изоформа №4 IGF (Еа класса I) в комбинации имеют даже большую терапевтическую эффективность при введении совместно с VM202.
6.6.2. Пример 2: терапевтическая эффективность серийного введения ДНК-конструкций, кодирующих IGF-1, и ДНК-конструкции, кодирующей HGF, в мышиной модели нейропатии CCI
CCI нейропатию мышей получали, как приводится в Примере 1, и их делили на семь групп, как приводится в ТАБЛИЦЕ 2. Суммарно 200 мкг ДНК-конструкций внутримышечно инъецировали мышам CCI в сутки хирургии CCI (1-ая инъекция), а другую инъекцию осуществляли в неделю 3 (2-ая инъекция). ДНК-конструкции, введенные в 1-ую инъекцию и 2-ую инъекцию для каждой группы, обобщаются ниже в ТАБЛИЦЕ 2. Каждая группа состояла из 6 мышей, и проводили больше, чем 2 независимых эксперимента (среднее плюс/минус SEM (стандартная ошибка среднего); * - р меньше 0,05; ** - р меньше 0,01; *** - р меньше 0,001).

Терапевтические эффекты данных инъекций анализировали посредством измерения механической аллодинии на еженедельной основе посредством анализа с нитями фон Фрея на протяжении всего эксперимента. Экспериментальная методика обобщается на Фиг. 4А. Вкратце, животных индивидуально помещали в цилиндр над полом из металлической сетки для адаптации. Частоту механической чувствительности мышей оценивали посредством стимулирования задней лапы с использованием постоянной толщины нити (калибр 0,16). Данный анализ затем повторяли еженедельно.
Результаты обобщаются на Фиг. 4В, предоставляя частоту (%) отдергивания лапы, измеренную в каждой группе на еженедельной основе. Данный результат подтверждает то, что инъекция ДНК-конструкции, кодирующей IGF-1 (т.е. IGF-1 #1 и IGF-1#4), или инъекция ДНК-конструкции, кодирующей HGF (т.е. VM202), обеспечивает значительное уменьшение частоты отдергивания лапы по сравнению с одним вектором (рСK). Кроме того, продемонстрировали то, что инъекция ДНК-конструкций, кодирующих IGF-1 (т.е. IGF-1#1 и IGF-1 #4), после инъекции ДНК-конструкции, кодирующей HGF (т.е. VM202) дополнительно уменьшает частоту отдергивания лапы (VM202 -> изоформы IGF). Это дополнительное снижение посредством 2-ой инъекции не наблюдали в группе, где IGF-1#1 и IGF-1#4 инъецировали до 2-ой инъекции VM202 (изоформы IGF -> VM202) или VM202 инъецировали перед 2-ой инъекцией VM202 (VM202 -> VM202). Следовательно, инъекция VM202 с последующей инъекцией IGF-1#1 и IGF-1#4 (VM202 -> изоформы IGF) давала самое значительное уменьшение частоты отдергивания лапы.
6.6.3. Пример 3: ДНК-конструкции, способные экспрессировать и изоформу №1 IGF-1 (Еc класса I), и изоформу №4 (Еа класса I)
Поскольку первая ДНК-конструкция, кодирующая IGF-1, экспрессирующая IGF-1 Еc класса I (IGF-1#1) и вторая ДНК-конструкция, кодирующая IGF-1, экспрессирующая Еа класса I (IGF-1#4) совместно имели статистически значимо большую способность уменьшать механическую аллодинию в поведенческих экспериментах, описанных выше, авторы данного изобретения сконструировали несколько плазмид, разработанных для одновременной экспрессии обеих изоформ IGF-1 - Еc класса I (изоформа №1) и Еа класса I (изоформа №4), через альтернативный сплайсинг РНК-транскрипта. В частности, ДНК-конструкции получали так, чтобы они содержали последовательности экзонов 1, 3/4, 5 и 6, и интронов гена IGF-1 или их фрагменты. Несколько ДНК-конструкций, включая разные вариации, получали и анализировали на их способность экспрессировать и изоформу IGF-1 Еc класса I (изоформа №1), и изоформу Еа класса I (изоформа №4).
Каждую плазмиду конструировали с использованием рСK в качестве остова плазмиды так, чтобы она содержала вставку, связанную функциональным образом с контрольными последовательностями экспрессии рСK. Данную вставку создавали посредством объединения (1) первого полинуклеотида, кодирующего экзоны 1, 3 и 4 человеческого IGF-1 (SEQ ID NO 1); (2) второго полинуклеотида, кодирующего либо интрон 4 IGF-1 (SEQ ID NO 2), либо его фрагмент; (3) третьего полинуклеотида, кодирующего экзоны 5 и 6-1 (SEQ ID NO 3); (4) четвертого полинуклеотида, кодирующего либо интрон 5 (SEQ ID NO 4), либо его фрагмент; и (5) пятого полинуклеотид, кодирующего экзон 6-2 (SEQ ID NO 5), в которой первый полинуклеотид, второй полинуклеотид, третий полинуклеотид, четвертый полинуклеотид и пятый полинуклеотид были связаны с последовательном порядке от 5' к 3'. Данные плазмиды отличались по размеру фрагмента интрона 4 и/или интрона 5. В частности, SEQ ID NO 6 дает нукпеотидную последовательность фрагмента интрона 4, используемого в векторе pCK-IGF-1X6, и SEQ ID NO 7 дает нукпеотидную последовательность фрагмента интрона 4, используемого в векторе pCK-IGF-1-X10. SEQ ID NO 8 дает нукпеотидную последовательность фрагмента интрона 5, используемого в векторе pCK-IGF-1X6 и pCK-IGF1X10.
Для анализа экспрессии изоформы №1 (Еc класса I) и изоформы №4 (Еа класса I) от разных конструкций in vivo, 9-месячным самцам мышей C57BL/6 инъецировали 50 мкг плазмиды в их мышцу Т.А. (передняя большеберцовая мышца). Их скелетные мышцы ТА получали через 5 суток после инъекции. Данные скелетные мышцы затем гомогенизировали в лизисном буфере, содержащем ингибитор протеазы, смесь ингибиторов фосфатаз (Roche Diagnostic Ltd.) и PMSF (фенилметилсульфонилфторид) (Sigma), с использованием полипропиленовых пестиков (Bel-Art Scienceware). Данные образцы центрифугировали при 12000 об./мин в течение 15 минут при 4°С, и супернатанты, содержащие общий белок, подвергали ИФА на человеческий IGF-1 (R&D Systems), как изложено в протоколе изготовителя. Выявленный уровень IGF-1 нормировали к общему количеству белковых экстрактов из ткани при измерении посредством набора для анализа белка ВСА (Thermo, IL, США). Данная экспериментальная процедура обобщается на Фиг. 5А.
Как показано на Фиг. 5Б, посредством ИФА был определен общий уровень экспрессии человеческих белков IGF-1 в мышиной мышце Т.А. Независимо от того, получала ли мышь 50 мкг конструкции, экспрессирующей одну изоформу («1» (Еc класса I) или «4» (Еа класса I)), 25 мкг первой конструкции, экспрессирующей изоформу №1 (Еc класса I), плюс 25 мкг второй конструкции, экспрессирующей изоформу №4 (Еа класса I) («1+4»), или 50 мкг одной из конструкций, экспрессирующих обе изоформы - pCK-IGF-1X6 («Х6») или pCK-IGF-1X10 («Х10»), общие уровни экспрессии белка - человеческого IGF-1 - были аналогичными.
Авторы данного изобретения использовали ПЦР-ОТ (полимеразная цепная реакция, сопряженная с обратной транскрипцией) для определения того, экспрессировали ли данные конструкции зрелые транскрипты и для изоформы №1, и для изоформы №4 одновременно. Реакции ПЦР-ОТ проводили с прямым праймером (F), который связывается с экзоном 3/4, и обратным праймером (R), который связывается с экзоном 6. Как дополнительно объясняется на Фиг. 6А, ПЦР-ОТ транскрипта изоформы №1 (Еc класса I) генерировала бы два ампликона - ампликон из 178 п. н. и ампликон из 259 п. н., тогда как ПЦР-ОТ транскрипта изоформы №4 (Еа класса I) генерировала бы один ампликон из 129 п.н.
Для ПЦР-ОТ отбирали скелетные мышцы, механически гомогенизировали с использованием полипропиленовых пестиков (Bel-Art Scienceware) и экстрагировали в RNAiso plus (Takara). Количественное измерение РНК осуществляли с использованием прибора nanodrop. Для синтеза кДНК использовали равные количества РНК с использованием обратной транскриптазы XL (AMV) (Takara), и ПЦР проводили с использованием прямого (TGA ТСТ AAG GAG GCT GGA) (SEQ ID NO 40) и обратного (СТА СТТ GCG ТТС ТТС AAA TG) (SEQ ID. NO: 41) праймеров, показанных на Фиг. 6А.
Как проиллюстрировано на Фиг. 6В, pCK-IGF-1X6 и pCK-IGF-1X10 экспрессировали зрелые транскрипты как для изоформы №1 (полосы 178 п. н. и 259 п. н.), так и для изоформы №4 (полоса 129 п.н.). Экспрессию зрелых транскриптов как для изоформы №1, так и для изоформы №4 не выявляли от конструкций, отличных от pCK-IGF-1X6 и pCK-IGF-1X10, причем эти данные не приводятся в данном документе.
Для того чтобы подтвердить то, что оба транскрипта двух данных изоформ эффективно транслировались в белок, авторы данного изобретения трансфицировали клетки 293Т pCK-IGF-1X6 и pCK-IGF-1X10, как проиллюстрировано на Фиг. 7А. Для иммуноблоттинга клетки получали через 2 суток (48 часов) после трансфекции плазмидной ДНК, с последующим лизисом с использованием буфера RIPA со смесью ингибиторов протеазы и фосфатазы (Roche Diagnostic Ltd.). Равные количества белка разделяли на 10%-ном SDS полиакриламидном геле (от англ. sodium dodecyl sulphate-polyacrylamide gel -полиакриламидный гель в присутствии додецилсульфата натрия) и переносили на мембрану для вестерн-блоттинга (PVDF - от англ. polyvinylidenfluoride - поливинилиденфторид). Данную мембрану блокировали 1% BSA (от англ. bovine serum albumin - бычий сывороточный альбумин) (Invitrogen-Gibco) в смеси трис-забуференного физиологического раствора и полисорбата 20 (TBST - от англ. tris-buffered saline (TBS) and Polysorbate 20 (Tween 20)) (20 мМ Tris-HCl, pH 7,4, 0,9% NaCl и 0,1% Tween 20) в течение 1 часа и зондировали с использованием первичных антител, разведенных в блокирующем растворе, при 4°С в течение ночи. Первичные антитела, используемые для проверки уровня изоформы 1 и изоформы 4 IGF-1, были предоставлены Abclon (Корея), а антитела для IGF-1 и β-актина были приобретены у Abeam (Великобритания) и Sigma-Aldrich (США). После промывки TBST мембраны инкубировали с конъюгированным с HRP (от англ. horseradish peroxidase - пероксидаза хрена) вторичным антителом козы против мышиного или кроличьего IgG (Sigma) при комнатной температуре в течение 1 часа. Блоты затем три раза промывали TBST, и полосы белка визуализировали с использованием системы усиленной хемилюминисценции (Millipore). β-актин использовали в качестве контроля загрузки.
Данные вестерн-блоттинга, показанные на Фиг. 7 В, подтверждают то, что плазмиды pCK-IGF-1X6 и pCK-IGF-1X10 экспрессируют обе изоформы IGF-1 на уровне белка.
6.6.4. Пример 4: терапевтическая эффективность совместного введения ДНК-конструкций, кодирующих HGF (VM202), и ДНК-конструкции, кодирующей IGF-1, экспрессирующей как изоформу №1 (Еc класса I), так и изоформу №4 (Еа класса I) IGF-1 в мышиной модели CCI нейропатии
Как обсуждалось выше в Примерах 1 и 2, одновременное или серийное введение VM202 и ДНК-конструкций, кодирующих IGF-1 - pCK-IGF-1#1 и pCK-IGF-1#4, (т.е. ДНК-конструкции, кодирующей IGF-1, способной экспрессировать изоформу №1 IGF-1 или изоформу №4 IGF-1), обеспечивало значимое уменьшение механической аллодинии в мышиной модели CCI нейропатии.
Авторы данного изобретения анализировали, могут ли обеспечиваться одинаковые эффекты с использованием ДНК-конструкции двойной экспрессии, способной экспрессировать и изоформу №1 IGF-1, и изоформу №4 IGF-1, полученной в Примере 3. В частности, pCK-IGF-1X6 и pCK-IGF-1X10 анализировали в комбинации с VM202 в мышиной модели CCI нейропатии.
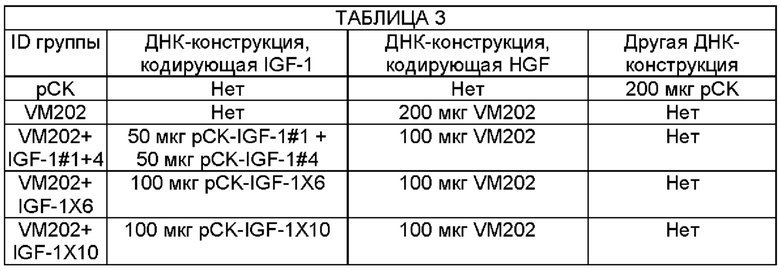
Мышей CCI делили на пять групп и вводили им суммарно 200 мкг ДНК-конструкции (как приводится в ТАБЛИЦЕ 3) посредством внутримышечных инъекций в сутки CCI. Болевую чувствительность в отношении механических стимулов измеряли в подходящие моменты времени посредством нити фон Фрея. Каждая группа состояла из 6 мышей, и осуществляли больше, чем 2 независимых эксперимента (среднее плюс/минус SEM; * - р меньше 0,05; ** - р меньше 0,01; *** - р меньше 0,001). Через одну неделю после хирургии CCI оценивали развитие механической аллодинии с использованием нити фон Фрея, и болевые симптомы оценивали еженедельно. Экспериментальная методика обобщается на Фиг. 8А.
Частоты отдергивания лапы, измеренные через одну неделю после хирургии CCI, приводятся на Фиг. 8В. Данные демонстрируют статистически значимые уменьшения механической аллодинии после одновременной внутримышечной инъекции VM202 и конструкций, кодирующих изоформу №1 и №4 IGF-1 (т.е. IGF-1 №1 и IGF-1 №4; IGF-1X6 и IGF-1X10). В частности, влияние на механическую аллодинию было лучше при введении мышам VM202 одновременно с двумя ДНК-конструкциями, кодирующими IGF-1, причем каждая кодирует изоформу №1 или №4 IGF-1, или с конструкцией двойной экспрессии pCK-IGF-1X10. Данные эффекты согласованно наблюдали до последнего измерения в четыре недели после хирургии CCI.
6.6.5. Пример 5: терапевтическая эффективность совместного введения ДНК-конструкций, кодирующих HGF (HGF728), и ДНК-конструкции, кодирующей IGF-1, экспрессирующей и изоформу №1 (Еc класса I), и изоформу №4 IGF-1 (Еа класса I), в мышиной модели CCI нейропатии Как обсуждалось выше в Примерах 1 -4, одновременное или серийное введение VM202 и ДНК-конструкций, кодирующих IGF-1, давало значительное снижение механической аллодинии в мышиной модели CCI нейропатии. Терапевтические эффекты другой ДНК-конструкции, кодирующей HGF - HGF728 - дополнительно тестировали в комбинации с ДНК-конструкциями, кодирующими IGF-1.
В частности, мышей с CCI нейропатией получали, как приводится в Примере 1, и делили на пять групп. Как схематически показано на Фиг. 9А, внутримышечно инъецировали суммарно 200 мкг плазмидной ДНК в сутки CCI хирургии. ДНК-конструкции, введенные в каждой группе, обобщаются в ТАБЛИЦЕ 4.
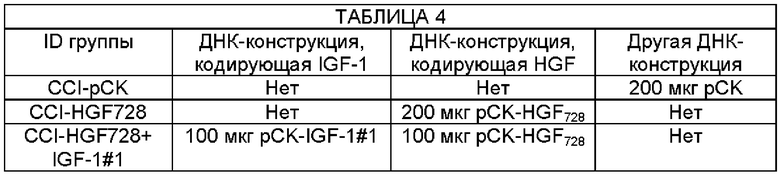

Через одну неделю после хирургии CCI оценивали развитие механической аллодинии с использованием анализа с нитью фон Фрея. Вкратце, животных индивидуально помещали в цилиндр над полом из металлической сетки для адаптации. Для проверки частоты механической чувствительности мышей оценивали посредством стимулирования задней лапы с использованием постоянной толщины нити (калибр 0,16). Для определения механического порога, мышей стимулировали разной толщиной нити (калибр 0,04 ~ 2,0). Данный анализ затем повторяли еженедельно.
Частоты отдергивания лапы и механический порог, измеренные через одну неделю после CCI хирургии, приводятся на Фиг. 9В-9С, причем на Фиг. 9В приводится частота (%), и на Фиг. 9С приводится порог отдергивания лапы. Все значения приводятся в виде среднего плюс/минус стандартная ошибка среднего (SEM) из трех независимых экспериментов. Различия между значениями определеляли посредством однофакторного ANOVA (дисперсионный анализ), с последующим апостериорным критерием Тьюки или критерием множественных сравнений Бонферрони.
Данные показывают то, что инъекция pCK-HGF728 значимо уменьшает частоту отдергивания лапы и увеличивает порог отдергивания лапы. При обработке животных pCK-HGF728 и pCK-IGF-1X10 наблюдали большее уменьшение частоты отдергивания лапы и наблюдали большее увеличение порога механической чувствительности, которое является статистически значимым. Данные результаты демонстрируют то, что терапевтические эффекты pCK-HGF728 также могут быть усилены совместным введением pCK-IGF-1X10. Данные эффекты согласованно наблюдали до последнего измерения в две недели после CCI хирургии.
7. Включение посредством ссылки
Все публикации, патенты, патентные заявки и другие документы, процитированные в данной заявке, являются тем самым включенными посредством ссылки во всей их полноте для всех целей в той же самой степени, как если бы каждая индивидуальная публикация, патент, патентная заявка или другой документ были индивидуально указанными как включенные посредством ссылки для всех целей.
8. Эквиваленты
В то время как разные конкретные воплощения были проиллюстрированы и описаны, приведенное выше описание изобретения не является ограничивающим. Будет понятно, что могут быть сделаны разные изменения без отступления от сущности и объема данного(ных) изобретения(ний). Многие вариации станут очевидными для специалистов в данной области при обзоре данного описания изобретения.













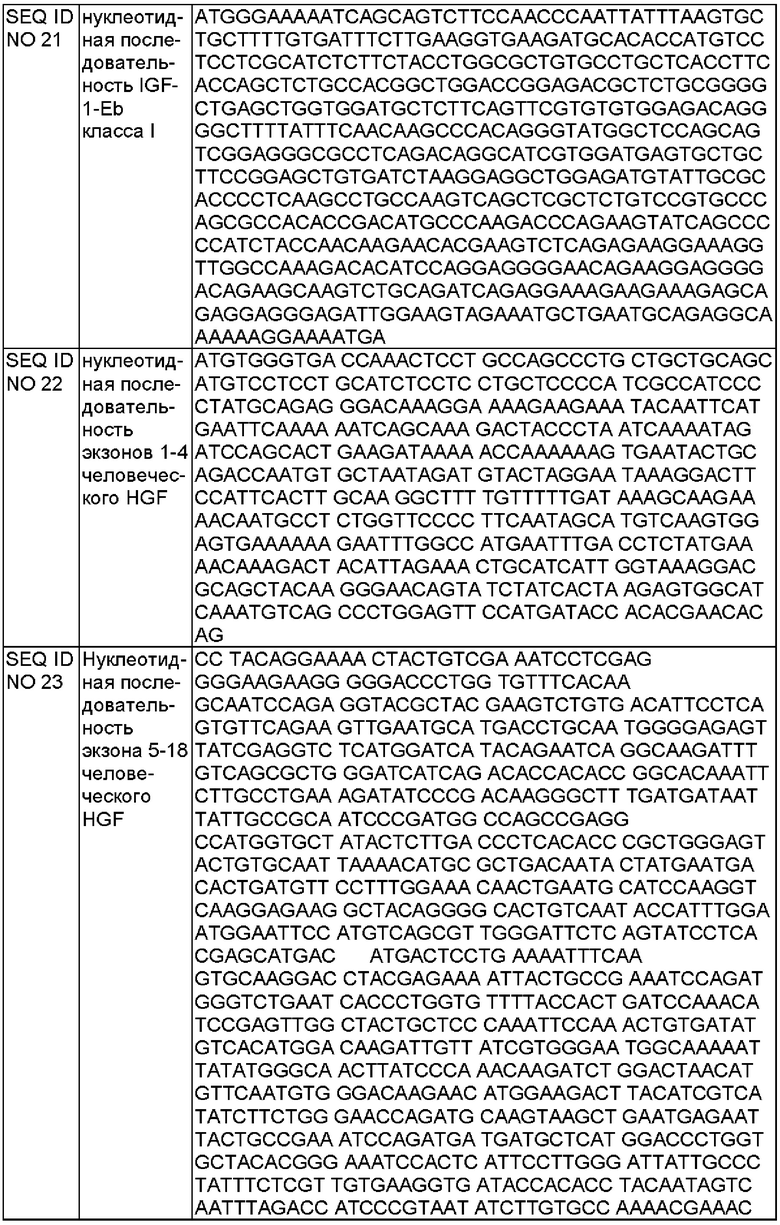


























--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> HELIXMITH CO., LTD.
<120> ЛЕЧЕНИЕ НЕЙРОПАТИИ ДНК-КОНСТРУКЦИЯМИ, КОДИРУЮЩИМИ IGF-1, И
ДНК-КОНСТРУКЦИЯМИ, КОДИРУЮЩИМИ HGF
<130> 33730-37536/WO (012WO)
<140>
<141>
<150> 62/699,667
<151> 2018-07-17
<160> 41
<170> PatentIn версия 3.5
<210> 1
<211> 402
<212> ДНК
<213> Homo sapiens
<400> 1
atgggaaaaa tcagcagtct tccaacccaa ttatttaagt gctgcttttg tgatttcttg 60
aaggtgaaga tgcacaccat gtcctcctcg catctcttct acctggcgct gtgcctgctc 120
accttcacca gctctgccac ggctggaccg gagacgctct gcggggctga gctggtggat 180
gctcttcagt tcgtgtgtgg agacaggggc ttttatttca acaagcccac agggtatggc 240
tccagcagtc ggagggcgcc tcagacaggc atcgtggatg agtgctgctt ccggagctgt 300
gatctaagga ggctggagat gtattgcgca cccctcaagc ctgccaagtc agctcgctct 360
gtccgtgccc agcgccacac cgacatgccc aagacccaga ag 402
<210> 2
<211> 1505
<212> ДНК
<213> Homo sapiens
<400> 2
gtaagcccac ctgggtggga tccagccatc ctcaagtggt ctctctcttg tgcatgtggg 60
tgggccaagc agaaatcctg ccccatagtc tcctggctta caagtcagaa aagctccttt 120
gcaccaaagg gatggattac atccccatct ctttggtcac tctgcattgc aaatttcccc 180
tcccaccgct atggacgatg tgatgattgg aagatgttac aaaacagtgg ctaaacaaac 240
atgggctttg gtgtcagaca aaagtgaagt cctggctttc tcacacacca gcttagagcc 300
cttggcaaat aatgtgatgt acccaagcct cagtttcatc agtaacattg ggataataat 360
aatatctacc acatcagttt gttgtcaaaa ttaagtagct catgcatata ctttgagatg 420
cttttcacat gcctgcataa agtaattgtt ggaccatcgt taatgtctgc cataattgca 480
cttaataaca aagcttgtaa cctttcaagt tctgagattc tacaatcttc caaagaaaat 540
aaaaggctaa tgggaactat tcaaaattca tattcagtag caagcataat taaacatgaa 600
acattaaaaa tagaaatttc tgtttggcta taagaatgcc tagacatttg taatgatcaa 660
aatctgcagg catcattttc taagagctag actgtaaaca aacctcagag gtaccaacta 720
tgccatcagt agtacataaa acatctgatg cacatttagt cacttgatcg atttctcttg 780
aatgagtgaa cgaatgaaca aatgaatata agagattaaa attttagcca ttaagtagaa 840
agaataagaa ctaaagagaa ggtaaaggag gaaaaagaga aggcaaggaa gttgagtaag 900
ggaagaaata gctctcgttt aagtattttg gggactctgt tgaaaaaaga aatgccaaca 960
tgtggtttta atctttggag ctagaactaa taatattgtg caaaagcaca agatgagaga 1020
tcaagaagtt caccatgaca ccttcgctgc ttcctggtct taaacctcag ctgaggctgg 1080
aagaggacca tggtggctta ttggagatgt gaccccaggg agcccctctg aaggatggaa 1140
ggggactggg caagacccaa cacacacaga acacagtagc cactggccag gcaggaagca 1200
aggatctcag aaaagacttt taggtgaatg tggcaggaaa gcgtgcttgc tggggcaaag 1260
gcagattcat tctttctctt cccaggtgac ccagcgcctc ttggtttcta actggggagg 1320
gggtaggtgt caagagatga gtcccaaagt tctggaatgg tgggtcttgt gactgaggtc 1380
tagacccctc tccagcatga gtgctgtctc ctgcatcata tggagcctgg gcattctgag 1440
ctcattcaaa gggacaccat gggaaccact tgttctcaat gcaattattt ttgtgatgtt 1500
tacag 1505
<210> 3
<211> 75
<212> ДНК
<213> Homo sapiens
<400> 3
tatcagcccc catctaccaa caagaacacg aagtctcaga gaaggaaagg aagtacattt 60
gaagaacgca agtag 75
<210> 4
<211> 15250
<212> ДНК
<213> Homo sapiens
<400> 4
aggacaggag gattaaacag acagaggcaa ggatgatgag agaggagcag acagcaagaa 60
tgaaaagcag aaaatacaat agaggaaatg aagaaaagta ggcctgctgg agctagatga 120
tgatgtgatg gaaatagaag taacctttta gagaatctcg ctaagaaaca tggagaaaac 180
ggaaaagaaa aatgtaatgc cctagaaagc gcaaagaaag acagtggcaa aaatgaaaaa 240
aaaaaataaa aattataaaa gaggcaaaaa aagacacact attctctgcc tctaaaacac 300
aattaaataa aagaatttaa ataaaaatta aggcttctat atgcattttt aaattttgta 360
tgaatctgtt atggaagaat tgcctatgtc aatatatgtt cagagttaaa tattagcccc 420
aaatgctcag caagactgaa ttgtgtcata gaagttccca gattcccttt tcccgcaatg 480
tcattggagg ctgcatttct tagtcaagtc cagggtttag gccaaagggc atccggtatt 540
gcctaaaacc ctgtgaggtc tgtgaggtaa cttttgagaa gaggtcactg cactcttcat 600
cttttttgca ctttggaatc agatataaaa gatgtataag tttgctaggg ctgccataac 660
aaagtatcat aggctaggta gtttaaacca cagaaattga ttttttcata gttctgggag 720
ttgaaagtcc aaaatcaaag tatcagccct tgcaagggcc ttagagaagg ctctgtcatg 780
ggctcctccc ctcggcttgt aggtggcctc cttcttctcc ccctgtgtct tcacttcatc 840
ttccctccat acatatctct gtgtctaaac atcctctgtg tgaaacaaca ccagccaggt 900
tggatttggg cccaccccac tgacctcatt ttaacttaat tatctctgta aagactctgt 960
ctccaaatac agtcatattt tgacgtactg ggagttaggg cttcaacaca tgaatttgga 1020
cacaattcag ccagtgacag aagacttctg atctctgatg ataaccactg cattttgatt 1080
acagctccta gaaaacactc ccctccacca ccccaccaca gatctatttt tatatctgaa 1140
accctgagtt tctgctccat gagaacccca ggaacatact atgttagatc tggaagaagc 1200
ctcagaaatc cccttatttt gaagactagg acactgagat ccagaagtgg gtaaagatgt 1260
gcttgggttc taagctgctc ttcttttggc caggagacaa cagcacataa tcaaagtggg 1320
tcaactaaga aagaattcca gaaggaaaag agagggcaga aatgaaggga gagaatgaga 1380
gcaaaagtgc tggatttccc tgagggtgaa gaaaagttaa atagaatcac agaattcaga 1440
ttttagagat cttctccttc agatcccttg gtttaatcag taggattggg gtcttcatag 1500
ataataaagc aaaaactctc gccatcctcc aagttgtgaa ttagaagagc tgagaaaggg 1560
tacaagacgg aagttctcta ccaaacaaat ggtgacattt tggggtaaga atatgactaa 1620
cccagaagtg aagcatttca tccaagtagt ctattttgaa gatgtcatgg tataaaggaa 1680
cctcctttct gcctggtcct ccatgcctct gccatgcttt ttactccagg atcacccttt 1740
ctagtggttc actgaaaacc caggattact taaatatgat ggacatgttc acggctcaat 1800
ccaggaggaa aaggtcgaac tgaaagcatg ccaaagcccc acatgggagc caagccactg 1860
ctgctgtggt tgcaaagtgg atcctggctt atcagagcag agagaagcca ggctcgtgcc 1920
ttagcccaag tggccagtca ccttattcag gagatactaa gttctccagc taagacatcc 1980
atgctttggg accagctgca gacagaagcc aattcctact acaaccatca ccttagagta 2040
gcatatagac acagatggct cttcaaagga ccacagttcc atggaataac taagaattca 2100
tgtcctgtgg aaaggtttga ataaactata attataccca atcataaatt tcattcaaga 2160
agaactaaag caaaggcaaa gacagagaga agaaggaagg aaggagggag ggagggaggg 2220
aaggaaggaa ggaaggaagg aaggaaggaa ggaaggaagg aaggaaggaa ggaaagggaa 2280
ggaagaacaa aaagactttc tagttaaaga atgcttaact agcaaactat gtactataag 2340
acagttcttt tcggaatgag ttttatcaac tctaaagcaa ttatcttgaa tgcctacatg 2400
tgattactga ataatatgaa ccaagaaaac agaaagaatc tatattatct ttccatttcc 2460
ttctttccag tatcaatacc caagcctcta gtgatacatg gcatataatg ttggatggat 2520
ggatggatgg atggatggat ggatggatgg atggatggat gaatggatgg ttggatggac 2580
aaatgagtaa cataggctga tgaatagtgg tagaaagaca caccataaaa acaagtggca 2640
cttctgagat gaaatgattc ctattctcct acacaagaca gtgaggcaag tacagagtaa 2700
aaaaggaaag gcataggagc tatgcttata caagtattgt atgtttggaa tttccttcgc 2760
tggccaaatt gaaattgttc aaggacctat tgctacaggt ggcaactggc taagaatttc 2820
atagtgaata ttatacacct attactcccc ttaatgtttc tttgaagtaa gcagaatatt 2880
aataatcatt taaaattcca gtgtttcaac ttcaattgtt tcctagggca aattgataat 2940
tgtgtgtaaa actaattgga atatgtatgg aataatcatc ctgaaataaa attggtgaaa 3000
agtatttgtt attgggcatc tacaatgtgc aaacctctgt actaggcatg aacaagagtt 3060
ataagcattg gagaggctaa aatatagtcc ttaaggctgg gcacagtggc tcatgcctgt 3120
aatcctagca ctttgggagg ccaaggcggg cagattgcct gagctcagga gttcaagacc 3180
agcctgggca acatagcgaa accccatctc tactaaaaat acaaaaaaat tacctgggca 3240
tggtggcacg cacctgtaat cccagctact caggaggctg aggcatgaga attccttgaa 3300
cctgggaggc agaggttgca gcgagccgag atcctgccgc tgcatcccag cttgggtgac 3360
agagtgagac tctgtctcaa aaaaaaatta aataataaat aaatagtaaa atacagtcat 3420
taagagtaca aaatgtagat tcagactacc tgggttcaaa tcttggctct tacttgcatt 3480
gtggctttgg gcagatcatg taacttatgt gtgcctcagt ttcctcatct gttaaatagg 3540
ggcaacaact gaatctacct tattcagttg ttgtgagggt ttattgagat tgtgtgtgtg 3600
tatgtgtgtg agtgtagtgt gtgcatgtgt gtgtctgtgc aaggagtggg aggtgtatat 3660
tcagagacac atattacagc acttaaaatg gtatctagca cttagtaagc attattcaag 3720
ttttagttaa cattatttta cttacctctg aaaattggag ctatgtgaaa aagaagttgg 3780
tctcctgaag tagaagccag tcttgtgtca ccaaaaactt caagcccaag cttgccaacg 3840
cttttccatg atgtggtagt agagtttcaa gcatgtggta ggataagaga actcaatgac 3900
ctaagaacca ttccaaccca gagaacccct ggttctatga ataattccaa cttaaatagg 3960
tagcttggct ctcccaagtg agagccattg cttctgtttc cgggtcatat aatgaacttt 4020
cagaaaacca ccatttttct caaccagtta aaattaagtg taatacgtgc tttcatttca 4080
tggtgcctgg ggaaaattta attgtagtat gaactccagt tattggtagt cttaagtaaa 4140
attgccaaaa taaatagaaa tgcaggatat ttctgggctc acacagcttc cgggacactt 4200
tagtttcttg ggctgccaat ccagtgcctt tcacaagcat ttgatctttt ttcaaacatc 4260
tcttgaaaac aaacaaaacc tcacacagct tctaatgtgt gcactgttcg aatgtaaggg 4320
tggaaaagga ggcaaagaaa tgagctccca aagagcaatt ccccttctct cgcctccatc 4380
ccttgacgac ctccctccca ctaaagggaa acattgtttt cttaggtaat aaattctgca 4440
atttctcaag tccattaaca tccactgggc aagatgagat ctattctttt tatttgccca 4500
taggaaaaga atagtgcttt tttgcaatat tcactagata acacagagtt gacttttaat 4560
ccaagggcaa cattgatagt ctctagttaa aggggaagcc ttcaggagca atgaaaagat 4620
taatagtttt agatgaagca gaatccaaat ccctttttat gagttttgaa atatccagtt 4680
tgtatgctca cctcaatact taaagcccag ttactgattc ctttggccta agcaagacag 4740
gtcaattttt aaagagggag tagctgaggt tagcaaaaat tctccaggtc cacaaaactt 4800
ccagacctgc aaggtgaaaa tcagcttttc tgtcatccct aaaggcctaa ctggaatcag 4860
aacttttccc tgatgcccac atatttggag gtcctttttt aatgggactc cttaatgcct 4920
ttagtgccat cccattttca tccagtgtcc aaaagaaatg atttaaaaat ataaacgtat 4980
gtttaaattc cagaagagag aaatggagat tgagaacaat agggaaatga tgagagctat 5040
gggaaaagag gtttatgagt ccatgtctga ttcttccaga gagcccctaa gaaagttctt 5100
atcataccag gaactcaatt ataactttca ttgcctattg ttagatgagt aacaggagct 5160
agaaaacatt ttggaaattc ccatctttat ttttttaact aatatgatta tagttttaag 5220
aaccattggt caagaagcta actttttaaa aagtggaagt atgatggtta gaaataagaa 5280
tgctaaaggt gcatcaagct gattttaatt ctaaatgtcc ttggcagcaa tttagaatct 5340
gtaataaact acaccaaaca gttttgaggg gaaggggatt agtttctccc cttccttcgt 5400
gtgtgtgtgt gcgcgtgtgt gtgtgtgcac ctttgtgttc tagcattgtt gcacccatta 5460
cagagctggg gggaactatt ttccaaaatt ataggtgaga acagtttctt ggattgtctt 5520
tcagtgaagg taaattcctc tgtaaaaact aaccatcatt cagtaaaaac tgcaggattc 5580
ctttgtcttc tcaaaagcct gtttctcatc ctaaattaaa aattattcag gaaatagaga 5640
ggacattatt ggaggggtgg aaataagttg gttttctttt tattgtatct tttgaggatc 5700
cagggacttc taccatttcc catctaacat acagagaagg attctctagg tccctgtcta 5760
tagactgcag taactttcct atagaaccaa tttgcaattt tagaaatttc taggtctaat 5820
tattgaccca ttacaaccaa aggtcaatgc atccagccaa tcttccttct atcatcccct 5880
gcccttactt ctattaggga ctgggattac aggcaaaacc catcaaatgc ctcttctacc 5940
actttcccat ttcttaacca ttagcctcta acttcctcta ttcagtttct catatgcttt 6000
catgcccatt gggtcagata aaggaacatt catttatttg agtaggcatc tgttatgatc 6060
actccggaaa aaagatgaca atgggttacc ttgtcctcct gggcttctct aactgacatg 6120
gtcaaaatgc ccatatgaag ataagatgtt aagagcaaga tttatgaaaa gctgagtatg 6180
atggcagctc ttgtctcata aaataactcg aaagttccca gtgaaagacc aagaaatttt 6240
acatcaaacc caaaccggcc aaatggtcca agcttccaag ctgggatcca tggctaaagt 6300
ttctacaaaa ttctgggtac aatgtataaa cattcacttg gggctttctg tctagccagc 6360
accaagaggt caagtaatca aggaccaact agccctgcca tctgtgaaaa tatgtgctat 6420
tttcacggct ttagttcaca attatggcaa gacaaaagtt ccaaataatt aggagcaaga 6480
ccatggcagg ttgacggttg agtaaggttc tcaatcagcc gacaattgta gagttgggga 6540
tgtgcaatgt ttatgtcatg gtgtaagtat gtggcatgct tgactagctt gtgaggcact 6600
ggaagactag aaggaatgaa aaatatgaat gaatcaataa atgcatagta taattactgt 6660
tattttgtca gtattgtttt acctaggtca ctattgaatg ctctgatttg tctctttata 6720
aataataata tgttttcttc ttcaaaagaa cactaggatg aaggtagagg tgcttttggc 6780
acaatgccac aattctgatt tttttaaaac tgtatgcatg cataaaatgt tcttgagcca 6840
ttctctgcct tggaatagca ctggctggca ttctgcatgt ttacttttat atgctgaagg 6900
cccccatcaa cctcaaacag aggcaaatca atttaacttc tcatagtgtt attttgttca 6960
tcctaaaagt tcaagagagc cttccaaact tccaaaattt ctctcaattc agtgaggagg 7020
aaaattcaga acacagcatt tgaatgttct gcccagattt gtcacacaca caaggaatga 7080
gtgaaagagg gcaacaccct ttcctcctaa ccctgtgaac tcatcactat tgcattgaaa 7140
tgacaccaaa aggtaaaaac cctaggcctc acatctccca agaacactgc aataggagtt 7200
actgcataca ccagtttaag taactctagc ataaattgta tgtcagatga aacaatggca 7260
ttttggaggc ttaagagaaa aagaataatc aaatccagtt tttaggtact aatgtgctga 7320
atctttagca catagcagca aaattgctag aatctggtgt ttcacttttt aaaataccac 7380
atttgaacct ttcagcaatt ccaaaatcaa ctccctctgc gaaagataat aagcttaaac 7440
attttttaaa tttaaaaatg taacacaaac aaacagctaa gcaaacaagc tgcccataaa 7500
atcaacagtc tggggagccc tgatcctgaa gtattttaca acatccttca tgactattaa 7560
aggcaacata aacacctctt gtcagcaagg gaaactaccc ttggcatttt tttttctttg 7620
ttccccaggc ttttaaacca ttttgataga gattttttac atcacaggca gaaatatttg 7680
aaatagagtc aggtggtagt ctttaaaaga gtaagaaagt tgctaagtca agataatctt 7740
ggaataaagt cctctgattc ctggggattc ctagggatgc cccagtcact agaaaacaga 7800
gctgtaagtc cactctccca gcactcaacg gagctccgga aaccaaggag ctagctactg 7860
tttccccaca ttcagccaga gaaagggcag cactctagca tgcaaactgc tttgacaata 7920
gtaacaatta aaaagtaaat taaaaagaat cataatagct gatattgatt aggtacttgc 7980
cctgtggcaa gagctatagg gaatcacctc atttaatctt cacatgaagc ttgcagagtg 8040
agtaccacaa ttatcactat tgtatagaca ggaaaactca ggctgagtat ggctaagtgt 8100
cttgccaacg tcttgggcta acaagcggtc aagcagaatc caaacccgag atagatagac 8160
cacagtgtgc taatcaagca ctgcactctc tcctgcattt cttagttgat atttaccata 8220
tacaatctgt cacttgtatg agatggcagg gggttctgtg ctatttgtcc ttgtagagaa 8280
taccacagga agaaagtaag cagccatgca atatttgctg ttgacctgaa ctccattcca 8340
tcattcctgc aggaaattcg catccattaa atgagcattt cctggtttgc cactttgctc 8400
aaacactttg cttggatctg gagaggatat agaagtgaag gaaatatgct acctgctctc 8460
aaggaactta tgttttagtg gagagacaaa catgcagaat ttactctaca gaacatcaat 8520
gcttgagcaa atgtagaccc agagagggct cttacagcac acaagccaga acagactgat 8580
ggtgctaaca attaggttca aggtttttct aaacagtaga ctctcctgca tacaactata 8640
ccgcatgcca ggtaaatgac tgagggttat tacatccaat tataacacca ctgtgatgta 8700
ggtgctctta ccccacactt tcattttaca gaagaggaaa ttgaggacag cacaatgtag 8760
tgattatcaa aggtcacacg actactgtgt gggagagcta ggatttaaac cagatgcata 8820
agatgaggtc ctccaagaaa cagaagatga gaaggtgtta aatgagcagg ggttttatta 8880
gggggaatta atgtgtgaac agaaataggg gaggataggc aaagccatca gattgcaagg 8940
caagcctaac cccaagggaa ggagagagag agagtagatt ggttggaaac atttttggtg 9000
ggtctatggt ctaaggaaag ttcagcaaag tcatcatgga gtttttgagc caaagttggg 9060
caatacagtt gcccaacaaa tttctgtgtt tctcagaaat aggtctgcct caatgtcccc 9120
accatacttg gtcactggct cttgggaggg gcctgccctg ttccaatcca ctagagccaa 9180
agaagagccg ttgtactggc agggggtggg ggaattccta caaccacata aaaagtgggg 9240
tgaggtttcc agaaaaaaac gtgatgctgg gctaaccaaa actgtgtcca gtaagtacat 9300
atccctcact ctgttaaaga agcagccaca taaacaagga gtacacgttt ctcaaaatgt 9360
gcaccttgtt ctttggtttt gaagtcacat cccaaagtgc tgagtagatc gcatgaccct 9420
cgctttgcct ggctgccaga gaggaaaggc tgatccaact ctcctggaat ttgaacttgt 9480
gattccctga agtaaagaga tatcaaagtt gatactgaga catctaaatc atcctccacc 9540
atttcacatg tccccaggcc aagccagcaa aattgctata gcacatccct ttcaacaggt 9600
aaagggctga tatctgagcc ctctttccaa tcatccactg ctcttttctt ctcattttgc 9660
cctttttggg agcaggtcaa tgctgagtta gtactttatg ctgtacaata agctgctgat 9720
attccatgct ggacagaatt ttcccagtat tttttataga gtgccaggct tttcctagac 9780
ttcatgtcat acaatactta acttgtttgg agtgggtgga gatggaaaca tagtctattg 9840
aaaacatcac tgcttcctcc ctgaagttta aagagcctat ttttatcctt ttagattcta 9900
tctctcaggc aaaatctcat aaagataagt ggggaggaaa aaaagggggt tataatacct 9960
agggagtttg cttttgctaa ttgaatactg tgctcctaga cttctataaa taccattaca 10020
aatgggtccc agcttgtggt aatactcacc ctcctcattg agtcttctgt cccatggcac 10080
agcctttccc tccaaactag catctacccc catctggaag catgggcagc tcatgatatt 10140
atcaactatt gctattggaa agtgatttgg acttgaaagc actagatatt ttttacctct 10200
tggggaggca gtttagcaga gtggttaact ggtgagctcc agaatcagaa ggaataggtc 10260
caaattccaa ccactattac atctccatca taagaaatta ggcaagttgt ttatcctaag 10320
tttcagattc cttaaagata aaacagtcaa gacagtagta cttatccctg agagaagtat 10380
aggaaacaag aaaatatatg caatttacat acatactaca atccccagca catgacaaat 10440
gttcaagtaa tgggaactgt tattatttta gccctttgtc tatcagtttg ttcctctgtg 10500
acctcaagca cattactaaa tgttagcgag cttcagcttg tacgtgggac tgacaggaat 10560
aacaccgcat cacctcatgt ggtgattgta aggattcagt gatattattt tgtaaactgt 10620
aaagcctttg caaatgttaa gcaagattat tattattgcc gttgttatta gtcctcagtg 10680
atcttttttt tttttttttt tttttttttt tttttttttt ttggagacag agttttactc 10740
tgtcgccaag gctggaatgc agtggcacaa tctcagctca ctgcaacctc cgcctcctgg 10800
gttcaagcaa ttttcctgcc tcagcctcct gagtagctga aactacaggc acacgccacc 10860
acaccaggct aattttttgt atttttagta gagacggggt ttcaccatgt tggccaggct 10920
ggtctccagc tcctgacctc aagtgatctg cccacctcgg cctcccaaag tgctgggatt 10980
acaggtgtga gccaccacac ctggcacagt aatcttaatt gaaaagtctg tggatagctt 11040
tccaaaggaa agcttggagc ttggataaga accaagagat aatgggagaa ggtgaatggc 11100
ctcttcaggg ccttttctag caccctaaat atgcgtgtct gtccataatg ggtaatcata 11160
tatatcacaa atcaaaccct ccacaaactt atttcctaat gtgtttgtta acctttcctt 11220
ctaaagggta aacttcttta accaacccca gtgagctgga ggatcaatgt tttcttaata 11280
gtcttacctt cgttggtgtc aataggaaac agtatttact cactactgtt ttccttttaa 11340
aaatctgtct agttgcatac tagaaacagt ttcagctggt ttgtttgtat tggacaagct 11400
gctgaagtga aaagtttttg cttgactgaa tgtgagacag tttcataact cttcaagaag 11460
tgcaccaaag gtgggtgcca gctctgatga cggctgcttc taacatgcct ccacttgccg 11520
cccattgtca agggtggctg gcgtaattaa gttaagacaa tgagcaaagc aacagatgca 11580
actgagacct agtccctgag tgcttttgtt ttgtcactgt cattgtctgc aacaaagaag 11640
tcacatgtga cagcctggga agagagccaa atgcaaacca gacgatatcc cagctggttt 11700
gaatggcctc caccgtgcac gtgtgtgcat gggaatcatg ctacttggta cagcatctgc 11760
ttcactcaag tgagtttcag cccatggctt tgctgtgatg ctgagacaga cccagaagaa 11820
acagaccagg gaatccctcc gctcagactt tacactttat accttgtgct ttgagagaaa 11880
agaaaaagaa tctctctatt ggagacaaaa aataggatgt atgtggttgg tcaatctaac 11940
ctcaattctt tttgctatag ccccccgcta atttaaagag tgaagcatag atggtatctt 12000
aatgttttct tgtagaaatt tgggattaat ttggcttgag aggaagaatg gagattaaac 12060
gctttatgag gctttctttt aatttgttcc catttcattc ctgaatattt tcttagtttg 12120
ggcattgcag atgtttaaag aacttcttat tttgagctgg tatgcctctt aaacagaaaa 12180
acaaaaggta aaattcaaat tagtgtgttt ctccgcctgt taattaattt ggttagtagt 12240
taggcagaga gatggcatcc ttaataatat ctattttgcg ggtttgatca gctacagacc 12300
atcaacagtg ttgattgaga attgaacaaa aacatttcaa ggagtttggg aacattaggg 12360
atgctattct gtggccccat gtgtccttct ctcatttttc tagagaactc ctataagaaa 12420
gcagaacacg gccaggcatg atggctcatg cctgtaatcc cagcacttca ggaggctgag 12480
gcaggcagat cacctgaggt caggagttca agaccagcct ggccaacatg gtgaaaccct 12540
atctctatta aaaatacaaa aaattagctg ggcatgatgg cgcgtgcctg taatcccagc 12600
tacttgggag gctgaggcag gagaatcact tgaactggga ggcaaaggtt gcagtgagcc 12660
tagatcacac cactgcactc cagcctgggt gacagagtga gactccaact caaaaaaaag 12720
aaagaaagaa agaaagaaag cagaacccaa tggaagatta agaacacaca tttagcttac 12780
gcctgtaata ccagcacttt gggaggccaa ggcgggtgga tcacaaggtc agaagttcga 12840
gaccaacctg gccaatatgg tgaaacccca tctctactaa aaagtacaaa aattagccat 12900
gcatggtggc aggcgtctgt aatcccagct actacagagg ctgaggcagg agaatcactt 12960
gaacccggga ggcagaggtt gcagtgagct gagaacgcgc cactgcactc cagcctgggt 13020
gacagagcga gactccatct caaaaaaaaa aaacaacaaa aaaaaacaaa acacaagttt 13080
actgggaact tagcagtaga tgctttgcac cacaacaaat gtatcttaag tggtcttttg 13140
tgatatttga gggaaagtgc cagaatttaa aacaaatggc atttcaagtt attctataca 13200
aatgcccagt ttctttctac catctttttt tcctttttgc agtggtcact gagctatttt 13260
agtgaatgtt tttacacaat gatgccatct tccttctact cagtcagtac aagatgttga 13320
ccatcgactc ataaaacact agctaccttt catgaaggac ttggtgataa ctctcatgtt 13380
ccaagtagaa ccggaaaaca tgtgtaagaa aacctgccga tccctatggg ccttggccaa 13440
taggtattat tcccaagggg tggcagttta tctttttccc cagccttcat attaaaacct 13500
ctcaccttct ccaggtctca ggtctgtgta atctcaaatg tgctttagct cctcacaata 13560
ttgtaactgt gtgggtgttc attaccttag ccagaagaca gtttacagat tccaggtctc 13620
atggagagaa cttttgtttt tggttatgaa cctcactgta taccaataat tatccattac 13680
atccttctgt agagggctct ctggctagag ataaaaccaa aaaaagaagt acctcaggtt 13740
tatgcatata aatgccagtt cctccttgat tttatttcaa aactcctgtc tacatacttt 13800
gcaatttaaa tacattcaag gataaagtaa taactgtagg aaaagtatta taatataatg 13860
acttagttct gcacatcaca agggggtccc tcatactcat tcattcattt cactcatttt 13920
acagatattt attgagcacc tgcaataacc tgcacactgc tctagacact gggactataa 13980
cagtaaacag acagatacat ctctggtctc acagggcttc tattctaagc aaaactcaat 14040
atccaggccg ggtgcagtgg ctcatgcctg gaatgccagc actttgggag accaaggcca 14100
ggcagatcac ctgagcccac tagttgaaga ccagcctggg caatatagca aaaccccgtc 14160
tctacaaaaa aaaaaaaaaa aaaaaaaaaa aaattgtcaa ggcatggtgg catgcgcctg 14220
tggtcccagc tacttaggag gctgaggcag gaggattgtg taagcctggg aggcagaggt 14280
tgcagtgacc tgagatggca ccaccacact ccagcctggg caacagagtg agaccctgtc 14340
caaaaaaaaa aaaccctcac tatccttaag ataacatcat tgcttgttga tgagtgaatg 14400
ttaacaccaa attaggaacc caggactttt agtcttggca tggttacttt ccaataaaga 14460
tgacaatact aagaagagaa aaatgattta ataatgataa tagtggctaa tacttatgta 14520
gtgcttacca tgtgccaggt ctattgtaag tacttttata tatattaatt atttaatctt 14580
tgatcctata aggtagatat tattgttacc ctagtttata gatgaagaaa cggaaacaca 14640
agagattgcc actcatacaa gtttacacag ccagaaaata gaaaagctac gagttgagct 14700
cagcccagta tgtctatgat tttacagact caaaattaat tataagattt cctaatcttc 14760
gatttctgaa actctgcctt gctctagagg aaaacaagaa aaacaatgaa aaataaatgt 14820
ctctttttta caaaaattaa aacagaacaa actgcaataa aacaacagag gatgaatcca 14880
gaatgtgatt gatttttttt cttactagga aaggatctag aggccagaag gctggatttt 14940
tcaggatctc ctttcaatca atgaatctgt gatagaagca gatgaatcaa atctcatctt 15000
tgtgtgatta taaagctgtc tgtggtattc acgccaccag gggtacatag aagatgcctg 15060
agtgaggttt ggcaaaagta ctaagggcct gtccacctat acatgccctt ctcaggaaaa 15120
ccaaggttca agctctctat tagctcaact ggtaaggcgt aagacatgga aggttgaggc 15180
ccaatgttag aaatagatgg atacataaaa cttcatcaag ttaatgtcac tttttctcct 15240
ttatttatag 15250
<210> 5
<211> 60
<212> ДНК
<213> Homo sapiens
<400> 5
gaagtacatt tgaagaacgc aagtagaggg agtgcaggaa acaagaacta caggatgtag 60
<210> 6
<211> 375
<212> ДНК
<213> Homo sapiens
<400> 6
gtaagcccac ctgggtggga tccagccatc ctcaagtggt ctctctcttg tgcatgtggg 60
tgggccaagc agaaatcctg ccccatagtc tcctggctta caagtcagaa aagctccttt 120
gcaccaaagg gatggattac atccccatct ctttggtcac tctgcattgc aaatttcccc 180
tcccaccgct atggacgatg tgatgattgg aagatgttac aaaacagtgg ctaaacaaac 240
atgggctttg gtgtcagaca aaagtgaagt cctggctttc tcacacacca gcttagagag 300
aaaagacttt taggtgaatg tggcaggaaa gcgtgcttgc tggggcaaag gcagattcat 360
tctttctctt cccag 375
<210> 7
<211> 300
<212> ДНК
<213> Homo sapiens
<400> 7
gtaagcccac ctgggtggga tccagccatc ctcaagtggt ctctctcttg tgcatgtggg 60
tgggccaagc agaaatcctg ccccatagtc tcctggctta caagtcagaa aagctccttt 120
gcaccaaagg gatggattac atccccatct ctttgctaaa caaacatggg ctttggtgtc 180
agacaaaagt gaagtcctgg ctttctcaca caccagctta gagagaaaag acttttaggt 240
gaatgtggca ggaaagcgtg cttgctgggg caaaggcaga ttcattcttt ctcttcccag 300
<210> 8
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 8
ctttttctcc tttatttata g 21
<210> 9
<211> 933
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 9
atgggaaaaa tcagcagtct tccaacccaa ttatttaagt gctgcttttg tgatttcttg 60
aaggtgaaga tgcacaccat gtcctcctcg catctcttct acctggcgct gtgcctgctc 120
accttcacca gctctgccac ggctggaccg gagacgctct gcggggctga gctggtggat 180
gctcttcagt tcgtgtgtgg agacaggggc ttttatttca acaagcccac agggtatggc 240
tccagcagtc ggagggcgcc tcagacaggc atcgtggatg agtgctgctt ccggagctgt 300
gatctaagga ggctggagat gtattgcgca cccctcaagc ctgccaagtc agctcgctct 360
gtccgtgccc agcgccacac cgacatgccc aagacccaga aggtaagccc acctgggtgg 420
gatccagcca tcctcaagtg gtctctctct tgtgcatgtg ggtgggccaa gcagaaatcc 480
tgccccatag tctcctggct tacaagtcag aaaagctcct ttgcaccaaa gggatggatt 540
acatccccat ctctttggtc actctgcatt gcaaatttcc cctcccaccg ctatggacga 600
tgtgatgatt ggaagatgtt acaaaacagt ggctaaacaa acatgggctt tggtgtcaga 660
caaaagtgaa gtcctggctt tctcacacac cagcttagag agaaaagact tttaggtgaa 720
tgtggcagga aagcgtgctt gctggggcaa aggcagattc attctttctc ttcccagtat 780
cagcccccat ctaccaacaa gaacacgaag tctcagagaa ggaaaggaag tacatttgaa 840
gaacgcaagt agctttttct cctttattta taggaagtac atttgaagaa cgcaagtaga 900
gggagtgcag gaaacaagaa ctacaggatg tag 933
<210> 10
<211> 858
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 10
atgggaaaaa tcagcagtct tccaacccaa ttatttaagt gctgcttttg tgatttcttg 60
aaggtgaaga tgcacaccat gtcctcctcg catctcttct acctggcgct gtgcctgctc 120
accttcacca gctctgccac ggctggaccg gagacgctct gcggggctga gctggtggat 180
gctcttcagt tcgtgtgtgg agacaggggc ttttatttca acaagcccac agggtatggc 240
tccagcagtc ggagggcgcc tcagacaggc atcgtggatg agtgctgctt ccggagctgt 300
gatctaagga ggctggagat gtattgcgca cccctcaagc ctgccaagtc agctcgctct 360
gtccgtgccc agcgccacac cgacatgccc aagacccaga aggtaagccc acctgggtgg 420
gatccagcca tcctcaagtg gtctctctct tgtgcatgtg ggtgggccaa gcagaaatcc 480
tgccccatag tctcctggct tacaagtcag aaaagctcct ttgcaccaaa gggatggatt 540
acatccccat ctctttgcta aacaaacatg ggctttggtg tcagacaaaa gtgaagtcct 600
ggctttctca cacaccagct tagagagaaa agacttttag gtgaatgtgg caggaaagcg 660
tgcttgctgg ggcaaaggca gattcattct ttctcttccc agtatcagcc cccatctacc 720
aacaagaaca cgaagtctca gagaaggaaa ggaagtacat ttgaagaacg caagtagctt 780
tttctccttt atttatagga agtacatttg aagaacgcaa gtagagggag tgcaggaaac 840
aagaactaca ggatgtag 858
<210> 11
<211> 728
<212> Белок
<213> Homo sapiens
<400> 11
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Phe Leu Pro Ser Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr
165 170 175
Cys Arg Asn Pro Arg Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser
180 185 190
Asn Pro Glu Val Arg Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu
195 200 205
Val Glu Cys Met Thr Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp
210 215 220
His Thr Glu Ser Gly Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro
225 230 235 240
His Arg His Lys Phe Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp
245 250 255
Asp Asn Tyr Cys Arg Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr
260 265 270
Thr Leu Asp Pro His Thr Arg Trp Glu Tyr Cys Ala Ile Lys Thr Cys
275 280 285
Ala Asp Asn Thr Met Asn Asp Thr Asp Val Pro Leu Glu Thr Thr Glu
290 295 300
Cys Ile Gln Gly Gln Gly Glu Gly Tyr Arg Gly Thr Val Asn Thr Ile
305 310 315 320
Trp Asn Gly Ile Pro Cys Gln Arg Trp Asp Ser Gln Tyr Pro His Glu
325 330 335
His Asp Met Thr Pro Glu Asn Phe Lys Cys Lys Asp Leu Arg Glu Asn
340 345 350
Tyr Cys Arg Asn Pro Asp Gly Ser Glu Ser Pro Trp Cys Phe Thr Thr
355 360 365
Asp Pro Asn Ile Arg Val Gly Tyr Cys Ser Gln Ile Pro Asn Cys Asp
370 375 380
Met Ser His Gly Gln Asp Cys Tyr Arg Gly Asn Gly Lys Asn Tyr Met
385 390 395 400
Gly Asn Leu Ser Gln Thr Arg Ser Gly Leu Thr Cys Ser Met Trp Asp
405 410 415
Lys Asn Met Glu Asp Leu His Arg His Ile Phe Trp Glu Pro Asp Ala
420 425 430
Ser Lys Leu Asn Glu Asn Tyr Cys Arg Asn Pro Asp Asp Asp Ala His
435 440 445
Gly Pro Trp Cys Tyr Thr Gly Asn Pro Leu Ile Pro Trp Asp Tyr Cys
450 455 460
Pro Ile Ser Arg Cys Glu Gly Asp Thr Thr Pro Thr Ile Val Asn Leu
465 470 475 480
Asp His Pro Val Ile Ser Cys Ala Lys Thr Lys Gln Leu Arg Val Val
485 490 495
Asn Gly Ile Pro Thr Arg Thr Asn Ile Gly Trp Met Val Ser Leu Arg
500 505 510
Tyr Arg Asn Lys His Ile Cys Gly Gly Ser Leu Ile Lys Glu Ser Trp
515 520 525
Val Leu Thr Ala Arg Gln Cys Phe Pro Ser Arg Asp Leu Lys Asp Tyr
530 535 540
Glu Ala Trp Leu Gly Ile His Asp Val His Gly Arg Gly Asp Glu Lys
545 550 555 560
Cys Lys Gln Val Leu Asn Val Ser Gln Leu Val Tyr Gly Pro Glu Gly
565 570 575
Ser Asp Leu Val Leu Met Lys Leu Ala Arg Pro Ala Val Leu Asp Asp
580 585 590
Phe Val Ser Thr Ile Asp Leu Pro Asn Tyr Gly Cys Thr Ile Pro Glu
595 600 605
Lys Thr Ser Cys Ser Val Tyr Gly Trp Gly Tyr Thr Gly Leu Ile Asn
610 615 620
Tyr Asp Gly Leu Leu Arg Val Ala His Leu Tyr Ile Met Gly Asn Glu
625 630 635 640
Lys Cys Ser Gln His His Arg Gly Lys Val Thr Leu Asn Glu Ser Glu
645 650 655
Ile Cys Ala Gly Ala Glu Lys Ile Gly Ser Gly Pro Cys Glu Gly Asp
660 665 670
Tyr Gly Gly Pro Leu Val Cys Glu Gln His Lys Met Arg Met Val Leu
675 680 685
Gly Val Ile Val Pro Gly Arg Gly Cys Ala Ile Pro Asn Arg Pro Gly
690 695 700
Ile Phe Val Arg Val Ala Tyr Tyr Ala Lys Trp Ile His Lys Ile Ile
705 710 715 720
Leu Thr Tyr Lys Val Pro Gln Ser
725
<210> 12
<211> 723
<212> Белок
<213> Homo sapiens
<400> 12
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr Cys Arg Asn Pro Arg
165 170 175
Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser Asn Pro Glu Val Arg
180 185 190
Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu Val Glu Cys Met Thr
195 200 205
Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp His Thr Glu Ser Gly
210 215 220
Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro His Arg His Lys Phe
225 230 235 240
Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp Asp Asn Tyr Cys Arg
245 250 255
Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr Thr Leu Asp Pro His
260 265 270
Thr Arg Trp Glu Tyr Cys Ala Ile Lys Thr Cys Ala Asp Asn Thr Met
275 280 285
Asn Asp Thr Asp Val Pro Leu Glu Thr Thr Glu Cys Ile Gln Gly Gln
290 295 300
Gly Glu Gly Tyr Arg Gly Thr Val Asn Thr Ile Trp Asn Gly Ile Pro
305 310 315 320
Cys Gln Arg Trp Asp Ser Gln Tyr Pro His Glu His Asp Met Thr Pro
325 330 335
Glu Asn Phe Lys Cys Lys Asp Leu Arg Glu Asn Tyr Cys Arg Asn Pro
340 345 350
Asp Gly Ser Glu Ser Pro Trp Cys Phe Thr Thr Asp Pro Asn Ile Arg
355 360 365
Val Gly Tyr Cys Ser Gln Ile Pro Asn Cys Asp Met Ser His Gly Gln
370 375 380
Asp Cys Tyr Arg Gly Asn Gly Lys Asn Tyr Met Gly Asn Leu Ser Gln
385 390 395 400
Thr Arg Ser Gly Leu Thr Cys Ser Met Trp Asp Lys Asn Met Glu Asp
405 410 415
Leu His Arg His Ile Phe Trp Glu Pro Asp Ala Ser Lys Leu Asn Glu
420 425 430
Asn Tyr Cys Arg Asn Pro Asp Asp Asp Ala His Gly Pro Trp Cys Tyr
435 440 445
Thr Gly Asn Pro Leu Ile Pro Trp Asp Tyr Cys Pro Ile Ser Arg Cys
450 455 460
Glu Gly Asp Thr Thr Pro Thr Ile Val Asn Leu Asp His Pro Val Ile
465 470 475 480
Ser Cys Ala Lys Thr Lys Gln Leu Arg Val Val Asn Gly Ile Pro Thr
485 490 495
Arg Thr Asn Ile Gly Trp Met Val Ser Leu Arg Tyr Arg Asn Lys His
500 505 510
Ile Cys Gly Gly Ser Leu Ile Lys Glu Ser Trp Val Leu Thr Ala Arg
515 520 525
Gln Cys Phe Pro Ser Arg Asp Leu Lys Asp Tyr Glu Ala Trp Leu Gly
530 535 540
Ile His Asp Val His Gly Arg Gly Asp Glu Lys Cys Lys Gln Val Leu
545 550 555 560
Asn Val Ser Gln Leu Val Tyr Gly Pro Glu Gly Ser Asp Leu Val Leu
565 570 575
Met Lys Leu Ala Arg Pro Ala Val Leu Asp Asp Phe Val Ser Thr Ile
580 585 590
Asp Leu Pro Asn Tyr Gly Cys Thr Ile Pro Glu Lys Thr Ser Cys Ser
595 600 605
Val Tyr Gly Trp Gly Tyr Thr Gly Leu Ile Asn Tyr Asp Gly Leu Leu
610 615 620
Arg Val Ala His Leu Tyr Ile Met Gly Asn Glu Lys Cys Ser Gln His
625 630 635 640
His Arg Gly Lys Val Thr Leu Asn Glu Ser Glu Ile Cys Ala Gly Ala
645 650 655
Glu Lys Ile Gly Ser Gly Pro Cys Glu Gly Asp Tyr Gly Gly Pro Leu
660 665 670
Val Cys Glu Gln His Lys Met Arg Met Val Leu Gly Val Ile Val Pro
675 680 685
Gly Arg Gly Cys Ala Ile Pro Asn Arg Pro Gly Ile Phe Val Arg Val
690 695 700
Ala Tyr Tyr Ala Lys Trp Ile His Lys Ile Ile Leu Thr Tyr Lys Val
705 710 715 720
Pro Gln Ser
<210> 13
<211> 3679
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 13
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgga tcctgggtag gaaacacatt tgaatggtat ttactaagat actaaaatcc 780
ttggacttca ctctaatttt agtgccattt agaactcaag gtctcagtaa aagtagaaat 840
aaagcctgtt aacaaaacac aaactgaata ttaaaaatgt aactggattt tcaaagaaat 900
gtttactggt attacctgta gatgtatatt ctttattatg atcttttgtg taaagtctgg 960
cagacaaatg caatatctaa ttgttgagtc caatatcaca agcagtacaa aagtataaaa 1020
aagacttggc cttttctaat gtgttaaaat actttatgct ggtaataaca ctaagagtag 1080
ggcactagaa attttaagtg aagataatgt gttgcagtta ctgcactcaa tggcttacta 1140
ttataaacca aaactgggat cactaagctc cagtcagtca aaatgatcaa aattattgaa 1200
gagaataagc aattctgttc tttattagga cacagtagat acagactaca aagtggagtg 1260
tgcttaataa gaggtagcat ttgttaagtg tcaattactc tattatccct tggagcttct 1320
caaaataacc atataaggtg taagatgtta aaggttatgg ttacactcag tgcacaggta 1380
agctaatagg ctgagagaag ctaaattact tactggggtc tcacagtaag aaagtgagct 1440
gaagtttcag cccagattta actggattct gggctcttta ttcatgttac ttcatgaatc 1500
tgtttctcaa ttgtgcagaa aaaagggggc tatttataag aaaagcaata aacaaacaag 1560
taatgatctc aaataagtaa tgcaagaaat agtgagattt caaaatcagt ggcagcgatt 1620
tctcagttct gtcctaagtg gccttgctca atcacctgct atcttttagt ggagctttga 1680
aattatgttt cagacaactt cgattcagtt ctagaatgtt tgactcagca aattcacagg 1740
ctcatctttc taacttgatg gtgaatatgg aaattcagct aaatggatgt taataaaatt 1800
caaacgtttt aaggacagat ggaaatgaca gaattttaag gtaaaatata tgaaggaata 1860
taagataaag gatttttcta ccttcagcaa aaacataccc actaattagt aaaattaata 1920
ggcgaaaaaa agttgcatgc tcttatactg taatgattat cattttaaaa ctagcttttt 1980
gccttcgagc tatcggggta aagacctaca ggaaaactac tgtcgaaatc ctcgagggga 2040
agaaggggga ccctggtgtt tcacaagcaa tccagaggta cgctacgaag tctgtgacat 2100
tcctcagtgt tcagaagttg aatgcatgac ctgcaatggg gagagttatc gaggtctcat 2160
ggatcataca gaatcaggca agatttgtca gcgctgggat catcagacac cacaccggca 2220
caaattcttg cctgaaagat atcccgacaa gggctttgat gataattatt gccgcaatcc 2280
cgatggccag ccgaggccat ggtgctatac tcttgaccct cacacccgct gggagtactg 2340
tgcaattaaa acatgcgctg acaatactat gaatgacact gatgttcctt tggaaacaac 2400
tgaatgcatc caaggtcaag gagaaggcta caggggcact gtcaatacca tttggaatgg 2460
aattccatgt cagcgttggg attctcagta tcctcacgag catgacatga ctcctgaaaa 2520
tttcaagtgc aaggacctac gagaaaatta ctgccgaaat ccagatgggt ctgaatcacc 2580
ctggtgtttt accactgatc caaacatccg agttggctac tgctcccaaa ttccaaactg 2640
tgatatgtca catggacaag attgttatcg tgggaatggc aaaaattata tgggcaactt 2700
atcccaaaca agatctggac taacatgttc aatgtgggac aagaacatgg aagacttaca 2760
tcgtcatatc ttctgggaac cagatgcaag taagctgaat gagaattact gccgaaatcc 2820
agatgatgat gctcatggac cctggtgcta cacgggaaat ccactcattc cttgggatta 2880
ttgccctatt tctcgttgtg aaggtgatac cacacctaca atagtcaatt tagaccatcc 2940
cgtaatatct tgtgccaaaa cgaaacaatt gcgagttgta aatgggattc caacacgaac 3000
aaacatagga tggatggtta gtttgagata cagaaataaa catatctgcg gaggatcatt 3060
gataaaggag agttgggttc ttactgcacg acagtgtttc ccttctcgag acttgaaaga 3120
ttatgaagct tggcttggaa ttcatgatgt ccacggaaga ggagatgaga aatgcaaaca 3180
ggttctcaat gtttcccagc tggtatatgg ccctgaagga tcagatctgg ttttaatgaa 3240
gcttgccagg cctgctgtcc tggatgattt tgttagtacg attgatttac ctaattatgg 3300
atgcacaatt cctgaaaaga ccagttgcag tgtttatggc tggggctaca ctggattgat 3360
caactatgat ggcctattac gagtggcaca tctctatata atgggaaatg agaaatgcag 3420
ccagcatcat cgagggaagg tgactctgaa tgagtctgaa atatgtgctg gggctgaaaa 3480
gattggatca ggaccatgtg agggggatta tggtggccca cttgtttgtg agcaacataa 3540
aatgagaatg gttcttggtg tcattgttcc tggtcgtgga tgtgccattc caaatcgtcc 3600
tggtattttt gtccgagtag catattatgc aaaatggata cacaaaatta ttttaacata 3660
taaggtacca cagtcatag 3679
<210> 14
<211> 153
<212> Белок
<213> Homo sapiens
<400> 14
Met Gly Lys Ile Ser Ser Leu Pro Thr Gln Leu Phe Lys Cys Cys Phe
1 5 10 15
Cys Asp Phe Leu Lys Val Lys Met His Thr Met Ser Ser Ser His Leu
20 25 30
Phe Tyr Leu Ala Leu Cys Leu Leu Thr Phe Thr Ser Ser Ala Thr Ala
35 40 45
Gly Pro Glu Thr Leu Cys Gly Ala Glu Leu Val Asp Ala Leu Gln Phe
50 55 60
Val Cys Gly Asp Arg Gly Phe Tyr Phe Asn Lys Pro Thr Gly Tyr Gly
65 70 75 80
Ser Ser Ser Arg Arg Ala Pro Gln Thr Gly Ile Val Asp Glu Cys Cys
85 90 95
Phe Arg Ser Cys Asp Leu Arg Arg Leu Glu Met Tyr Cys Ala Pro Leu
100 105 110
Lys Pro Ala Lys Ser Ala Arg Ser Val Arg Ala Gln Arg His Thr Asp
115 120 125
Met Pro Lys Thr Gln Lys Glu Val His Leu Lys Asn Ala Ser Arg Gly
130 135 140
Ser Ala Gly Asn Lys Asn Tyr Arg Met
145 150
<210> 15
<211> 462
<212> ДНК
<213> Homo sapiens
<400> 15
atgggaaaaa tcagcagtct tccaacccaa ttatttaagt gctgcttttg tgatttcttg 60
aaggtgaaga tgcacaccat gtcctcctcg catctcttct acctggcgct gtgcctgctc 120
accttcacca gctctgccac ggctggaccg gagacgctct gcggggctga gctggtggat 180
gctcttcagt tcgtgtgtgg agacaggggc ttttatttca acaagcccac agggtatggc 240
tccagcagtc ggagggcgcc tcagacaggc atcgtggatg agtgctgctt ccggagctgt 300
gatctaagga ggctggagat gtattgcgca cccctcaagc ctgccaagtc agctcgctct 360
gtccgtgccc agcgccacac cgacatgccc aagacccaga aggaagtaca tttgaagaac 420
gcaagtagag ggagtgcagg aaacaagaac tacaggatgt ag 462
<210> 16
<211> 158
<212> Белок
<213> Homo sapiens
<400> 16
Met Gly Lys Ile Ser Ser Leu Pro Thr Gln Leu Phe Lys Cys Cys Phe
1 5 10 15
Cys Asp Phe Leu Lys Val Lys Met His Thr Met Ser Ser Ser His Leu
20 25 30
Phe Tyr Leu Ala Leu Cys Leu Leu Thr Phe Thr Ser Ser Ala Thr Ala
35 40 45
Gly Pro Glu Thr Leu Cys Gly Ala Glu Leu Val Asp Ala Leu Gln Phe
50 55 60
Val Cys Gly Asp Arg Gly Phe Tyr Phe Asn Lys Pro Thr Gly Tyr Gly
65 70 75 80
Ser Ser Ser Arg Arg Ala Pro Gln Thr Gly Ile Val Asp Glu Cys Cys
85 90 95
Phe Arg Ser Cys Asp Leu Arg Arg Leu Glu Met Tyr Cys Ala Pro Leu
100 105 110
Lys Pro Ala Lys Ser Ala Arg Ser Val Arg Ala Gln Arg His Thr Asp
115 120 125
Met Pro Lys Thr Gln Lys Tyr Gln Pro Pro Ser Thr Asn Lys Asn Thr
130 135 140
Lys Ser Gln Arg Arg Lys Gly Ser Thr Phe Glu Glu Arg Lys
145 150 155
<210> 17
<211> 477
<212> ДНК
<213> Homo sapiens
<400> 17
atgggaaaaa tcagcagtct tccaacccaa ttatttaagt gctgcttttg tgatttcttg 60
aaggtgaaga tgcacaccat gtcctcctcg catctcttct acctggcgct gtgcctgctc 120
accttcacca gctctgccac ggctggaccg gagacgctct gcggggctga gctggtggat 180
gctcttcagt tcgtgtgtgg agacaggggc ttttatttca acaagcccac agggtatggc 240
tccagcagtc ggagggcgcc tcagacaggc atcgtggatg agtgctgctt ccggagctgt 300
gatctaagga ggctggagat gtattgcgca cccctcaagc ctgccaagtc agctcgctct 360
gtccgtgccc agcgccacac cgacatgccc aagacccaga agtatcagcc cccatctacc 420
aacaagaaca cgaagtctca gagaaggaaa ggaagtacat ttgaagaacg caagtag 477
<210> 18
<211> 137
<212> Белок
<213> Homo sapiens
<400> 18
Met Ile Thr Pro Thr Val Lys Met His Thr Met Ser Ser Ser His Leu
1 5 10 15
Phe Tyr Leu Ala Leu Cys Leu Leu Thr Phe Thr Ser Ser Ala Thr Ala
20 25 30
Gly Pro Glu Thr Leu Cys Gly Ala Glu Leu Val Asp Ala Leu Gln Phe
35 40 45
Val Cys Gly Asp Arg Gly Phe Tyr Phe Asn Lys Pro Thr Gly Tyr Gly
50 55 60
Ser Ser Ser Arg Arg Ala Pro Gln Thr Gly Ile Val Asp Glu Cys Cys
65 70 75 80
Phe Arg Ser Cys Asp Leu Arg Arg Leu Glu Met Tyr Cys Ala Pro Leu
85 90 95
Lys Pro Ala Lys Ser Ala Arg Ser Val Arg Ala Gln Arg His Thr Asp
100 105 110
Met Pro Lys Thr Gln Lys Glu Val His Leu Lys Asn Ala Ser Arg Gly
115 120 125
Ser Ala Gly Asn Lys Asn Tyr Arg Met
130 135
<210> 19
<211> 414
<212> ДНК
<213> Homo sapiens
<400> 19
atgattacac ctacagtgaa gatgcacacc atgtcctcct cgcatctctt ctacctggcg 60
ctgtgcctgc tcaccttcac cagctctgcc acggctggac cggagacgct ctgcggggct 120
gagctggtgg atgctcttca gttcgtgtgt ggagacaggg gcttttattt caacaagccc 180
acagggtatg gctccagcag tcggagggcg cctcagacag gcatcgtgga tgagtgctgc 240
ttccggagct gtgatctaag gaggctggag atgtattgcg cacccctcaa gcctgccaag 300
tcagctcgct ctgtccgtgc ccagcgccac accgacatgc ccaagaccca gaaggaagta 360
catttgaaga acgcaagtag agggagtgca ggaaacaaga actacaggat gtag 414
<210> 20
<211> 195
<212> Белок
<213> Homo sapiens
<400> 20
Met Gly Lys Ile Ser Ser Leu Pro Thr Gln Leu Phe Lys Cys Cys Phe
1 5 10 15
Cys Asp Phe Leu Lys Val Lys Met His Thr Met Ser Ser Ser His Leu
20 25 30
Phe Tyr Leu Ala Leu Cys Leu Leu Thr Phe Thr Ser Ser Ala Thr Ala
35 40 45
Gly Pro Glu Thr Leu Cys Gly Ala Glu Leu Val Asp Ala Leu Gln Phe
50 55 60
Val Cys Gly Asp Arg Gly Phe Tyr Phe Asn Lys Pro Thr Gly Tyr Gly
65 70 75 80
Ser Ser Ser Arg Arg Ala Pro Gln Thr Gly Ile Val Asp Glu Cys Cys
85 90 95
Phe Arg Ser Cys Asp Leu Arg Arg Leu Glu Met Tyr Cys Ala Pro Leu
100 105 110
Lys Pro Ala Lys Ser Ala Arg Ser Val Arg Ala Gln Arg His Thr Asp
115 120 125
Met Pro Lys Thr Gln Lys Tyr Gln Pro Pro Ser Thr Asn Lys Asn Thr
130 135 140
Lys Ser Gln Arg Arg Lys Gly Trp Pro Lys Thr His Pro Gly Gly Glu
145 150 155 160
Gln Lys Glu Gly Thr Glu Ala Ser Leu Gln Ile Arg Gly Lys Lys Lys
165 170 175
Glu Gln Arg Arg Glu Ile Gly Ser Arg Asn Ala Glu Cys Arg Gly Lys
180 185 190
Lys Gly Lys
195
<210> 21
<211> 588
<212> ДНК
<213> Homo sapiens
<400> 21
atgggaaaaa tcagcagtct tccaacccaa ttatttaagt gctgcttttg tgatttcttg 60
aaggtgaaga tgcacaccat gtcctcctcg catctcttct acctggcgct gtgcctgctc 120
accttcacca gctctgccac ggctggaccg gagacgctct gcggggctga gctggtggat 180
gctcttcagt tcgtgtgtgg agacaggggc ttttatttca acaagcccac agggtatggc 240
tccagcagtc ggagggcgcc tcagacaggc atcgtggatg agtgctgctt ccggagctgt 300
gatctaagga ggctggagat gtattgcgca cccctcaagc ctgccaagtc agctcgctct 360
gtccgtgccc agcgccacac cgacatgccc aagacccaga agtatcagcc cccatctacc 420
aacaagaaca cgaagtctca gagaaggaaa ggttggccaa agacacatcc aggaggggaa 480
cagaaggagg ggacagaagc aagtctgcag atcagaggaa agaagaaaga gcagaggagg 540
gagattggaa gtagaaatgc tgaatgcaga ggcaaaaaag gaaaatga 588
<210> 22
<211> 482
<212> ДНК
<213> Homo sapiens
<400> 22
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
ag 482
<210> 23
<211> 1675
<212> ДНК
<213> Homo sapiens
<400> 23
cctacaggaa aactactgtc gaaatcctcg aggggaagaa gggggaccct ggtgtttcac 60
aagcaatcca gaggtacgct acgaagtctg tgacattcct cagtgttcag aagttgaatg 120
catgacctgc aatggggaga gttatcgagg tctcatggat catacagaat caggcaagat 180
ttgtcagcgc tgggatcatc agacaccaca ccggcacaaa ttcttgcctg aaagatatcc 240
cgacaagggc tttgatgata attattgccg caatcccgat ggccagccga ggccatggtg 300
ctatactctt gaccctcaca cccgctggga gtactgtgca attaaaacat gcgctgacaa 360
tactatgaat gacactgatg ttcctttgga aacaactgaa tgcatccaag gtcaaggaga 420
aggctacagg ggcactgtca ataccatttg gaatggaatt ccatgtcagc gttgggattc 480
tcagtatcct cacgagcatg acatgactcc tgaaaatttc aagtgcaagg acctacgaga 540
aaattactgc cgaaatccag atgggtctga atcaccctgg tgttttacca ctgatccaaa 600
catccgagtt ggctactgct cccaaattcc aaactgtgat atgtcacatg gacaagattg 660
ttatcgtggg aatggcaaaa attatatggg caacttatcc caaacaagat ctggactaac 720
atgttcaatg tgggacaaga acatggaaga cttacatcgt catatcttct gggaaccaga 780
tgcaagtaag ctgaatgaga attactgccg aaatccagat gatgatgctc atggaccctg 840
gtgctacacg ggaaatccac tcattccttg ggattattgc cctatttctc gttgtgaagg 900
tgataccaca cctacaatag tcaatttaga ccatcccgta atatcttgtg ccaaaacgaa 960
acaattgcga gttgtaaatg ggattccaac acgaacaaac ataggatgga tggttagttt 1020
gagatacaga aataaacata tctgcggagg atcattgata aaggagagtt gggttcttac 1080
tgcacgacag tgtttccctt ctcgagactt gaaagattat gaagcttggc ttggaattca 1140
tgatgtccac ggaagaggag atgagaaatg caaacaggtt ctcaatgttt cccagctggt 1200
atatggccct gaaggatcag atctggtttt aatgaagctt gccaggcctg ctgtcctgga 1260
tgattttgtt agtacgattg atttacctaa ttatggatgc acaattcctg aaaagaccag 1320
ttgcagtgtt tatggctggg gctacactgg attgatcaac tatgatggcc tattacgagt 1380
ggcacatctc tatataatgg gaaatgagaa atgcagccag catcatcgag ggaaggtgac 1440
tctgaatgag tctgaaatat gtgctggggc tgaaaagatt ggatcaggac catgtgaggg 1500
ggattatggt ggcccacttg tttgtgagca acataaaatg agaatggttc ttggtgtcat 1560
tgttcctggt cgtggatgtg ccattccaaa tcgtcctggt atttttgtcc gagtagcata 1620
ttatgcaaaa tggatacaca aaattatttt aacatataag gtaccacagt catag 1675
<210> 24
<211> 3756
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 24
cgcgttgaca ttgattattg actagttatt aatagtaatc aattacgggg tcattagttc 60
atagcccata tatggagttc cgcgttacat aacttacggt aaatggcccg cctggctgac 120
cgcccaacga cccccgccca ttgacgtcaa taatgacgta tgttcccata gtaacgccaa 180
tagggacttt ccattgacgt caatgggtgg agtatttacg gtaaactgcc cacttggcag 240
tacatcaagt gtatcatatg ccaagtccgc cccctattga cgtcaatgac ggtaaatggc 300
ccgcctggca ttatgcccag tacatgacct tacgggactt tcctacttgg cagtacatct 360
acgtattagt catcgctatt accatggtga tgcggttttg gcagtacacc aatgggcgtg 420
gatagcggtt tgactcacgg ggatttccaa gtctccaccc cattgacgtc aatgggagtt 480
tgttttggca ccaaaatcaa cgggactttc caaaatgtcg taataacccc gccccgttga 540
cgcaaatggg cggtaggcgt gtacggtggg aggtctatat aagcagagct cgtttagtga 600
accgtcagat cgcctggaga cgccatccac gctgttttga cctccataga agacaccggg 660
accgatccag cctccgcggc cgggaacggt gcattggaac gcggattccc cgtgccaaga 720
gtgacgtaag taccgcctat agactctata ggcacacccc tttggctctt atgcatgcta 780
tactgttttt ggcttggggc ctatacaccc ccgcttcctt atgctatagg tgatggtata 840
gcttagccta taggtgtggg ttattgacca ttattgacca ctcccctatt ggtgacgata 900
ctttccatta ctaatccata acatggctct ttgccacaac tatctctatt ggctatatgc 960
caatactctg tccttcagag actgacacgg actctgtatt tttacaggat ggggtcccat 1020
ttattattta caaattcaca tatacaacaa cgccgtcccc cgtgcccgca gtttttatta 1080
aacatagcgt gggatctcca cgcgaatctc gggtacgtgt tccggacatg ggctcttctc 1140
cggtagcggc ggagcttcca catccgagcc ctggtcccat gcctccagcg gctcatggtc 1200
gctcggcagc tccttgctcc taacagtgga ggccagactt aggcacagca caatgcccac 1260
caccaccagt gtgccgcaca aggccgtggc ggtagggtat gtgtctgaaa atgagctcgg 1320
agattgggct cgcaccgctg acgcagatgg aagacttaag gcagcggcag aagaagatgc 1380
aggcagctga gttgttgtat tctgataaga gtcagaggta actcccgttg cggtgctgtt 1440
aacggtggag ggcagtgtag tctgagcagt actcgttgct gccgcgcgcg ccaccagaca 1500
taatagctga cagactaaca gactgttcct ttccatgggt cttttctgca gtcaccgtcc 1560
ttgacacgaa gcttggtacc gagctcggat ccactagtcc agtgtggtgg aattctgcag 1620
atatccagca cagtggcggc cgctcgagtc tagagggccc gtttaaaccc gctgatcagc 1680
ctcgactgtg ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt 1740
gaccctggaa ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca 1800
ttgtctgagt aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga 1860
ggattgggaa gacaatagca ggcatgctgg ggagtcgaaa ttcagaagaa ctcgtcaaga 1920
aggcgataga aggcgatgcg ctgcgaatcg ggagcggcga taccgtaaag cacgaggaag 1980
cggtcagccc attcgccgcc aagctcttca gcaatatcac gggtagccaa cgctatgtcc 2040
tgatagcggt ccgccacacc cagccggcca cagtcgatga atccagaaaa gcggccattt 2100
tccaccatga tattcggcaa gcaggcatcg ccatgggtca cgacgagatc ctcgccgtcg 2160
ggcatgctcg ccttgagcct ggcgaacagt tcggctggcg cgagcccctg atgctcttcg 2220
tccagatcat cctgatcgac aagaccggct tccatccgag tacgtgctcg ctcgatgcga 2280
tgtttcgctt ggtggtcgaa tgggcaggta gccggatcaa gcgtatgcag ccgccgcatt 2340
gcatcagcca tgatggatac tttctcggca ggagcaaggt gagatgacag gagatcctgc 2400
cccggcactt cgcccaatag cagccagtcc cttcccgctt cagtgacaac gtcgagcaca 2460
gctgcgcaag gaacgcccgt cgtggccagc cacgatagcc gcgctgcctc gtcttgcagt 2520
tcattcaggg caccggacag gtcggtcttg acaaaaagaa ccgggcgccc ctgcgctgac 2580
agccggaaca cggcggcatc agagcagccg attgtctgtt gtgcccagtc atagccgaat 2640
agcctctcca cccaagcggc cggagaacct gcgtgcaatc catcttgttc aatcatgcga 2700
aacgatcctc atcctgtctc ttgatcagat cttgatcccc tgcgccatca gatccttggc 2760
ggcaagaaag ccatccagtt tactttgcag ggcttcccaa ccttaccaga gggcgcccca 2820
gctggcaatt ccggttcgct tgctgtccat aaaaccgccc agtctagcta tcgccatgta 2880
agcccactgc aagctacctg ctttctcttt gcgcttgcgt tttcccttgt ccagatagcc 2940
cagtagctga cattcatccg gggtcagcac cgtttctgcg gactggcttt ctacgtgaaa 3000
aggatctagg tgaagatcct ttttgataat ctcatgacca aaatccctta acgtgagttt 3060
tcgttccact gagcgtcaga ccccgtagaa aagatcaaag gatcttcttg agatcctttt 3120
tttctgcgcg taatctgctg cttgcaaaca aaaaaaccac cgctaccagc ggtggtttgt 3180
ttgccggatc aagagctacc aactcttttt ccgaaggtaa ctggcttcag cagagcgcag 3240
ataccaaata ctgttcttct agtgtagccg tagttaggcc accacttcaa gaactctgta 3300
gcaccgccta catacctcgc tctgctaatc ctgttaccag tggctgctgc cagtggcgat 3360
aagtcgtgtc ttaccgggtt ggactcaaga cgatagttac cggataaggc gcagcggtcg 3420
ggctgaacgg ggggttcgtg cacacagccc agcttggagc gaacgaccta caccgaactg 3480
agatacctac agcgtgagct atgagaaagc gccacgcttc ccgaagggag aaaggcggac 3540
aggtatccgg taagcggcag ggtcggaaca ggagagcgca cgagggagct tccaggggga 3600
aacgcctggt atctttatag tcctgtcggg tttcgccacc tctgacttga gcgtcgattt 3660
ttgtgatgct cgtcaggggg gcggagccta tggaaaaacg ccagcaacgc ggccttttta 3720
cggttcctgg ccttttgctg gccttttgct cacatg 3756
<210> 25
<211> 4956
<212> ДНК
<213> Homo sapiens
<400> 25
gtaagaacag tatgaagaaa agagatgaag cctctgtctt ttttacatgt taacagtctc 60
atattagtcc ttcagaataa ttctacaatc ctaaaataac ttagccaact tgctgaattg 120
tattacggca aggtttatat gaattcatga ctgatattta gcaaatgatt aattaatatg 180
ttaataaaat gtagccaaaa caatatctta ccttaatgcc tcaatttgta gatctcggta 240
tttgtgaaat aataacgtaa acttcgttta aaaggattct tcttcctgtc tttgagaaag 300
tacggcactg tgcaggggga gaggttgatt gtgaaaaatc agaggtagat gagaatctta 360
ctgagggctg agggttcttt aaccttggtg gatctcaaca ttggttgcac attaaaatca 420
cctgctgcaa gcccttgacg aatcttactt agaagatgac aacacagaac aattaaatca 480
gaatctctgg ggagaatagg gcaccagtat tttttgagct cccaccatga ttccaaagtg 540
cagccaaatt tgagaaccac tgctaaaagc tcaagcttca gattgaccag cttttccatc 600
tcacctatcg cctaaagacc aaattggata aatgtgttca ttacgacaga tgggtactat 660
ttaaagatga gtaaacacaa tatacttagg ctcgtcagac tgagagtttt aatcatcact 720
gaggaaaaac atagatatct aatactgact ggagtattag tcaaggctta tttcacacac 780
aattttatca gaaaccaaag tagtttaaaa cagctctccc cttattagta atgcattgga 840
gggtttactt taccatgtac cttgctgagc actgtacctt gttaatctca tttacttgta 900
atgagaacca cacagcgggt agttttattg gttctatttt acctacatga caaaactgaa 960
gcataaaaac acttagtaag ttttcagtgt catgcacaac taggaagtga catggccaga 1020
atataagccc agtcaccatc actctataac ctgcgctttt aacaacttca gggcatgaca 1080
catttggccg gtcagtagaa cccatgctgt gatttgtttt tgcagtggtg gtgatgactg 1140
ccttgttgaa tccacttttt attctattcc attttgggga cacaattctg caagatgatt 1200
cttcattagg aaacagagat gagttattga ccaacacaga aagaaaaaga gtttgttgct 1260
ccacactggg attaaaccta tgatcttggc ctaattaaca ctagctagta agtgtccaag 1320
ctgatcatct ctacaacatt tcaataacag aaaacaacaa ttttcaaaat tagttactta 1380
caattatgta gaaatgcctc taaaacacag tattttcctt atattacaaa aacaaaaatt 1440
ataattggtt ttgtcctctt ttgagagttt gcatggtgtt actccctgca tagtgaagaa 1500
aacattttat ttaagtagat ggatctaagt ttttcatgaa caaaggaatg acatttgaaa 1560
tcaatcctac cctagtccag gagaatgcat tagattaacc tagtagaggt cttatttcac 1620
cctgagtttt ctatgatcgt gattctctgc tggaggagta attgtgaaat agatctctct 1680
gggaactggc ttcctagtcc aatcagctct tttaccaatg aacacttcct tgtgatatag 1740
atgtttatgg ccgagaggat ccagtatatt aataaaatcc ctttttgtat tcaatgaggg 1800
aaacacataa ttttcatcaa ttagcagctt attggaatat ctgcatgatg gtttaacact 1860
tttaagtgtt gactaaagat taattttaca gaaaatagaa aaagaaatat gtttctgtct 1920
ggaggaatga tttattgttg acccctaaat tgaaatattt tactagtggc ttaatggaaa 1980
gatgatgaaa gatgatgaaa ttaatgtaga agcttaacta gaaaatcagg tgacctgata 2040
tctacatctg tatccttcat tggccaccca gcattcatta atgaatcaga tgatggaata 2100
gatcaagttt cctaggaaca cagtgaatat taaaagaaaa caaagggagc ctagcaccta 2160
gaagacctag tttatatttc aaagtatatt tggatgtaac ccaattttaa acatttcctc 2220
acttgtctct cttaaagcct tgccaacagc aaggacagag aaccaaaaat agtgtatata 2280
tgaataaatg cttattacag aatctgctga ctggcacatg ctttgtgtgt aatgggttct 2340
cataaacact tgttgaatga acacacataa gtgaaagagc atggctaggc ttcatccctt 2400
ggtcaaatat ggggtgctaa agaaaagcag gggaaataca ttgggacact aacaaaaaaa 2460
aacagttaat ttaggtaaaa gataaaatac accacagaat gaagaaaaga gatgacccag 2520
actgctcttt aaccttcatg tcctagagag gtttttgata tgaattgcat tcagaattgt 2580
ggaaaggagc ccatcttttc tcttcatttt gattttatta actccaatgg gggaatttta 2640
ttcgtgtttt ggccatatct acttttgatt tctacattat tctctcttcc tttctacctg 2700
tatttgtcct aataaattgt tgacttatta attcactact tcctcacagc ttttttttgg 2760
ctttacaaat ccactggaaa ggtatatggg tgtatcactt tgtgtatttc ggtgtgcatg 2820
tgtagagggg acaaaaatcc tctctcaaac tataaatatt gagtatttgt gtattgaaca 2880
tttgctataa ctactaggtt tcttaaataa tcttaatata taaaatgata tagaaaaagg 2940
gaaattatag ttcgtattat tcatctaagt gaagagatta aaacccaggg agtaaataaa 3000
ttgtctaagg actaaggttg tatactattt aggtgataga tatggggcaa ccgtatgggt 3060
tttatgatta acaaataaac ttctcaccac tctaccatat caacttttcc ataaaagaga 3120
gctatagtat tctttgctta aataaatttg attagtgcat gacttcttga aaacatataa 3180
agcaaaagtc acatttgatt ctatcagaaa agtgagtaag ccatggccca aacaaaagat 3240
gcattaaaat attctggaat gatggagcta aaagtaagaa aaatgacttt ttaaaaaagt 3300
ttactgttag gaattgtgaa attatgctga attttagttg cattataatt tttgtcagtc 3360
atacggtctg acaacctgtc ttatttctat ttccccatat gaggaatgct agttaagtat 3420
ggatattaac tattactact tagatgcatt gaagttgcat aatatggata atacttcact 3480
ggttccctga aaatgtttag ttagtaataa gtctcttaca ctatttgttt tgtccaataa 3540
tttatatttt ctgaagactt aactctagaa tacactcatg tcaaaatgaa agaatttcat 3600
tgcaaaatat tgcttggtac atgacgcata cctgtatttg ttttgtgtca caacatgaaa 3660
aatgatggtt tattagaagt ttcattgggt aggaaacaca tttgaatggt atttactaag 3720
atactaaaat ccttggactt cactctaatt ttagtgccat ttagaactca aggtctcagt 3780
aaaagtagaa ataaagcctg ttaacaaaac acaaactgaa tattaaaaat gtaactggat 3840
tttcaaagaa atgtttactg gtattacctg tagatgtata ttctttatta tgatcttttg 3900
tgtaaagtct ggcagacaaa tgcaatatct aattgttgag tccaatatca caagcagtac 3960
aaaagtataa aaaagacttg gccttttcta atgtgttaaa atactttatg ctggtaataa 4020
cactaagagt agggcactag aaattttaag tgaagataat gtgttgcagt tactgcactc 4080
aatggcttac tattataaac caaaactggg atcactaagc tccagtcagt caaaatgatc 4140
aaaattattg aagagaataa gcaattctgt tctttattag gacacagtag atacagacta 4200
caaagtggag tgtgcttaat aagaggtagc atttgttaag tgtcaattac tctattatcc 4260
cttggagctt ctcaaaataa ccatataagg tgtaagatgt taaaggttat ggttacactc 4320
agtgcacagg taagctaata ggctgagaga agctaaatta cttactgggg tctcacagta 4380
agaaagtgag ctgaagtttc agcccagatt taactggatt ctgggctctt tattcatgtt 4440
acttcatgaa tctgtttctc aattgtgcag aaaaaagggg gctatttata agaaaagcaa 4500
taaacaaaca agtaatgatc tcaaataagt aatgcaagaa atagtgagat ttcaaaatca 4560
gtggcagcga tttctcagtt ctgtcctaag tggccttgct caatcacctg ctatctttta 4620
gtggagcttt gaaattatgt ttcagacaac ttcgattcag ttctagaatg tttgactcag 4680
caaattcaca ggctcatctt tctaacttga tggtgaatat ggaaattcag ctaaatggat 4740
gttaataaaa ttcaaacgtt ttaaggacag atggaaatga cagaatttta aggtaaaata 4800
tatgaaggaa tataagataa aggatttttc taccttcagc aaaaacatac ccactaatta 4860
gtaaaattaa taggcgaaaa aaagttgcat gctcttatac tgtaatgatt atcattttaa 4920
aactagcttt ttgccttcga gctatcgggg taaaga 4956
<210> 26
<211> 7113
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 26
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgaa ataataacgt aaacttcgtt taaaaggatt cttcttcctg tctttgagaa 780
agtacggcac tgtgcagggg gagaggttga ttgtgaaaaa tcagaggtag atgagaatct 840
tactgagggc tgagggttct ttaaccttgg tggatctcaa cattggttgc acattaaaat 900
cacctgctgc aagcccttga cgaatcttac ttagaagatg acaacacaga acaattaaat 960
cagaatctct ggggagaata gggcaccagt attttttgag ctcccaccat gattccaaag 1020
tgcagccaaa tttgagaacc actgctaaaa gctcaagctt cagattgacc agcttttcca 1080
tctcacctat cgcctaaaga ccaaattgga taaatgtgtt cattacgaca gatgggtact 1140
atttaaagat gagtaaacac aatatactta ggctcgtcag actgagagtt ttaatcatca 1200
ctgaggaaaa acatagatat ctaatactga ctggagtatt agtcaaggct tatttcacac 1260
acaattttat cagaaaccaa agtagtttaa aacagctctc cccttattag taatgcattg 1320
gagggtttac tttaccatgt accttgctga gcactgtacc ttgttaatct catttacttg 1380
taatgagaac cacacagcgg gtagttttat tggttctatt ttacctacat gacaaaactg 1440
aagcataaaa acacttagta agttttcagt gtcatgcaca actaggaagt gacatggcca 1500
gaatataagc ccagtcacca tcactctata acctgcgctt ttaacaactt cagggcatga 1560
cacatttggc cggtcagtag aacccatgct gtgatttgtt tttgcagtgg tggtgatgac 1620
tgccttgttg aatccacttt ttattctatt ccattttggg gacacaattc tgcaagatga 1680
ttcttcatta ggaaacagag atgagttatt gaccaacaca gaaagaaaaa gagtttgttg 1740
ctccacactg ggattaaacc tatgatcttg gcctaattaa cactagctag taagtgtcca 1800
agctgatcat ctctacaaca tttcaataac agaaaacaac aattttcaaa attagttact 1860
tacaattatg tagaaatgcc tctaaaacac agtattttcc ttatattaca aaaacaaaaa 1920
ttataattgg ttttgtcctc ttttgagagt ttgcatggtg ttactccctg catagtgaag 1980
aaaacatttt atttaagtag atggatctaa gtttttcatg aacaaaggaa tgacatttga 2040
aatcaatcct accctagtcc aggagaatgc attagattaa cctagtagag gtcttatttc 2100
accctgagtt ttctatgatc gtgattctct gctggaggag taattgtgaa atagatctct 2160
ctgggaactg gcttcctagt ccaatcagct cttttaccaa tgaacacttc cttgtgatat 2220
agatgtttat ggccgagagg atccagtata ttaataaaat ccctttttgt attcaatgag 2280
ggaaacacat aattttcatc aattagcagc ttattggaat atctgcatga tggtttaaca 2340
cttttaagtg ttgactaaag attaatttta cagaaaatag aaaaagaaat atgtttctgt 2400
ctggaggaat gatttattgt tgacccctaa attgaaatat tttactagtg gcttaatgga 2460
aagatgatga aagatgatga aattaatgta gaagcttaac tagaaaatca ggtgacctga 2520
tatctacatc tgtatccttc attggccacc cagcattcat taatgaatca gatgatggaa 2580
tagatcaagt ttcctaggaa cacagtgaat attaaaagaa aacaaaggga gcctagcacc 2640
tagaagacct agtttatatt tcaaagtata tttggatgta acccaatttt aaacatttcc 2700
tcacttgtct ctcttaaagc cttgccaaca gcaaggacag agaaccaaaa atagtgtata 2760
tatgaataaa tgcttattac agaatctgct gactggcaca tgctttgtgt gtaatgggtt 2820
ctcataaaca cttgttgaat gaacacacat aagtgaaaga gcatggctag gcttcatccc 2880
ttggtcaaat atggggtgct aaagaaaagc aggggaaata cattgggaca ctaacaaaaa 2940
aaaacagtta atttaggtaa aagataaaat acaccacaga atgaagaaaa gagatgaccc 3000
agactgctct ttaaccttca tgtcctagag aggtttttga tatgaattgc attcagaatt 3060
gtggaaagga gcccatcttt tctcttcatt ttgattttat taactccaat gggggaattt 3120
tattcgtgtt ttggccatat ctacttttga tttctacatt attctctctt cctttctacc 3180
tgtatttgtc ctaataaatt gttgacttat taattcacta cttcctcaca gctttttttt 3240
ggctttacaa atccactgga aaggtatatg ggtgtatcac tttgtgtatt tcggtgtgca 3300
tgtgtagagg ggacaaaaat cctctctcaa actataaata ttgagtattt gtgtattgaa 3360
catttgctat aactactagg tttcttaaat aatcttaata tataaaatga tatagaaaaa 3420
gggaaattat agttcgtatt attcatctaa gtgaagagat taaaacccag ggagtaaata 3480
aattgtctaa ggactaaggt tgtatactat ttaggtgata gatatggggc aaccgtatgg 3540
gttttatgat taacaaataa acttctcacc actctaccat atcaactttt ccataaaaga 3600
gagctatagt attctttgct taaataaatt tgattagtgc atgacttctt gaaaacatat 3660
aaagcaaaag tcacatttga ttctatcaga aaagtgagta agccatggcc caaacaaaag 3720
atgcattaaa atattctgga atgatggagc taaaagtaag aaaaatgact ttttaaaaaa 3780
gtttactgtt aggaattgtg aaattatgct gaattttagt tgcattataa tttttgtcag 3840
tcatacggtc tgacaacctg tcttatttct atttccccat atgaggaatg ctagttaagt 3900
atggatatta actattacta cttagatgca ttgaagttgc ataatatgga taatacttca 3960
ctggttccct gaaaatgttt agttagtaat aagtctctta cactatttgt tttgtccaat 4020
aatttatatt ttctgaagac ttaactctag aatacactca tgtcaaaatg aaagaatttc 4080
attgcaaaat attgcttggt acatgacgca tacctgtatt tgttttgtgt cacaacatga 4140
aaaatgatgg tttattagaa gtttcattgg gtaggaaaca catttgaatg gtatttacta 4200
agatactaaa atccttggac ttcactctaa ttttagtgcc atttagaact caaggtctca 4260
gtaaaagtag aaataaagcc tgttaacaaa acacaaactg aatattaaaa atgtaactgg 4320
attttcaaag aaatgtttac tggtattacc tgtagatgta tattctttat tatgatcttt 4380
tgtgtaaagt ctggcagaca aatgcaatat ctaattgttg agtccaatat cacaagcagt 4440
acaaaagtat aaaaaagact tggccttttc taatgtgtta aaatacttta tgctggtaat 4500
aacactaaga gtagggcact agaaatttta agtgaagata atgtgttgca gttactgcac 4560
tcaatggctt actattataa accaaaactg ggatcactaa gctccagtca gtcaaaatga 4620
tcaaaattat tgaagagaat aagcaattct gttctttatt aggacacagt agatacagac 4680
tacaaagtgg agtgtgctta ataagaggta gcatttgtta agtgtcaatt actctattat 4740
cccttggagc ttctcaaaat aaccatataa ggtgtaagat gttaaaggtt atggttacac 4800
tcagtgcaca ggtaagctaa taggctgaga gaagctaaat tacttactgg ggtctcacag 4860
taagaaagtg agctgaagtt tcagcccaga tttaactgga ttctgggctc tttattcatg 4920
ttacttcatg aatctgtttc tcaattgtgc agaaaaaagg gggctattta taagaaaagc 4980
aataaacaaa caagtaatga tctcaaataa gtaatgcaag aaatagtgag atttcaaaat 5040
cagtggcagc gatttctcag ttctgtccta agtggccttg ctcaatcacc tgctatcttt 5100
tagtggagct ttgaaattat gtttcagaca acttcgattc agttctagaa tgtttgactc 5160
agcaaattca caggctcatc tttctaactt gatggtgaat atggaaattc agctaaatgg 5220
atgttaataa aattcaaacg ttttaaggac agatggaaat gacagaattt taaggtaaaa 5280
tatatgaagg aatataagat aaaggatttt tctaccttca gcaaaaacat acccactaat 5340
tagtaaaatt aataggcgaa aaaaagttgc atgctcttat actgtaatga ttatcatttt 5400
aaaactagct ttttgccttc gagctatcgg ggtaaagacc tacaggaaaa ctactgtcga 5460
aatcctcgag gggaagaagg gggaccctgg tgtttcacaa gcaatccaga ggtacgctac 5520
gaagtctgtg acattcctca gtgttcagaa gttgaatgca tgacctgcaa tggggagagt 5580
tatcgaggtc tcatggatca tacagaatca ggcaagattt gtcagcgctg ggatcatcag 5640
acaccacacc ggcacaaatt cttgcctgaa agatatcccg acaagggctt tgatgataat 5700
tattgccgca atcccgatgg ccagccgagg ccatggtgct atactcttga ccctcacacc 5760
cgctgggagt actgtgcaat taaaacatgc gctgacaata ctatgaatga cactgatgtt 5820
cctttggaaa caactgaatg catccaaggt caaggagaag gctacagggg cactgtcaat 5880
accatttgga atggaattcc atgtcagcgt tgggattctc agtatcctca cgagcatgac 5940
atgactcctg aaaatttcaa gtgcaaggac ctacgagaaa attactgccg aaatccagat 6000
gggtctgaat caccctggtg ttttaccact gatccaaaca tccgagttgg ctactgctcc 6060
caaattccaa actgtgatat gtcacatgga caagattgtt atcgtgggaa tggcaaaaat 6120
tatatgggca acttatccca aacaagatct ggactaacat gttcaatgtg ggacaagaac 6180
atggaagact tacatcgtca tatcttctgg gaaccagatg caagtaagct gaatgagaat 6240
tactgccgaa atccagatga tgatgctcat ggaccctggt gctacacggg aaatccactc 6300
attccttggg attattgccc tatttctcgt tgtgaaggtg ataccacacc tacaatagtc 6360
aatttagacc atcccgtaat atcttgtgcc aaaacgaaac aattgcgagt tgtaaatggg 6420
attccaacac gaacaaacat aggatggatg gttagtttga gatacagaaa taaacatatc 6480
tgcggaggat cattgataaa ggagagttgg gttcttactg cacgacagtg tttcccttct 6540
cgagacttga aagattatga agcttggctt ggaattcatg atgtccacgg aagaggagat 6600
gagaaatgca aacaggttct caatgtttcc cagctggtat atggccctga aggatcagat 6660
ctggttttaa tgaagcttgc caggcctgct gtcctggatg attttgttag tacgattgat 6720
ttacctaatt atggatgcac aattcctgaa aagaccagtt gcagtgttta tggctggggc 6780
tacactggat tgatcaacta tgatggccta ttacgagtgg cacatctcta tataatggga 6840
aatgagaaat gcagccagca tcatcgaggg aaggtgactc tgaatgagtc tgaaatatgt 6900
gctggggctg aaaagattgg atcaggacca tgtgaggggg attatggtgg cccacttgtt 6960
tgtgagcaac ataaaatgag aatggttctt ggtgtcattg ttcctggtcg tggatgtgcc 7020
attccaaatc gtcctggtat ttttgtccga gtagcatatt atgcaaaatg gatacacaaa 7080
attattttaa catataaggt accacagtca tag 7113
<210> 27
<211> 6190
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 27
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgaa ataataacgt aaacttcgtt taaaaggatt cttcttcctg tctttgagaa 780
agtacggcac tgtgcagggg gagaggttga ttgtgaaaaa tcagaggtag atgagaatct 840
tactgagggc tgagggttct ttaaccttgg tggatctcaa cattggttgc acattaaaat 900
cacctgctgc aagcccttga cgaatcttac ttagaagatg acaacacaga acaattaaat 960
cagaatctct ggggagaata gggcaccagt attttttgag ctcccaccat gattccaaag 1020
tgcagccaaa tttgagaacc actgctaaaa gctcaagctt cagattgacc agcttttcca 1080
tctcacctat cgcctaaaga ccaaattgga taaatgtgtt cattacgaca gatgggtact 1140
atttaaagat gagtaaacac aatatactta ggctcgtcag actgagagtt ttaatcatca 1200
ctgaggaaaa acatagatat ctaatactga ctggagtatt agtcaaggct tatttcacac 1260
acaattttat cagaaaccaa agtagtttaa aacagctctc cccttattag taatgcattg 1320
gagggtttac tttaccatgt accttgctga gcactgtacc ttgttaatct catttacttg 1380
taatgagaac cacacagcgg gtagttttat tggttctatt ttacctacat gacaaaactg 1440
aagcataaaa acacttagta agttttcagt gtcatgcaca actaggaagt gacatggcca 1500
gaatataagc ccagtcacca tcactctata acctgcgctt ttaacaactt cagggcatga 1560
cacatttggc cggtcagtag aacccatgct gtgatttgtt tttgcagtgg tggtgatgac 1620
tgccttgttg aatccacttt ttattctatt ccattttggg gacacaattc tgcaagatga 1680
ttcttcatta ggaaacagag atgagttatt gaccaacaca gaaagaaaaa gagtttgttg 1740
ctccacactg ggattaaacc tatgatcttg gcctaattaa cactagctag taagtgtcca 1800
agctgatcat ctctacaaca tttcaataac agaaaacaac aattttcaaa attagttact 1860
tacaattatg tagaaatgcc tctaaaacac agtattttcc ttatattaca aaaacaaaaa 1920
ttataattgg ttttgtcctc ttttgagagt ttgcatggtg ttactccctg catagtgaag 1980
aaaacatttt atttaagtag atggatctaa gtttttcatg aacaaaggaa tgacatttga 2040
aatcaatcct accctagtcc aggagaatgc attagattaa cctagtagag gtcttatttc 2100
accctgagtt ttctatgatc gtgattctct gctggaggag taattgtgaa atagatctct 2160
ctgggaactg gcttcctagt ccaatcagct cttttaccaa tgaacacttc cttgtgatat 2220
agatgtttat ggccgagagg atctcttcct ttctacctgt atttgtccta ataaattgtt 2280
gacttattaa ttcactactt cctcacagct tttttttggc tttacaaatc cactggaaag 2340
gtatatgggt gtatcacttt gtgtatttcg gtgtgcatgt gtagagggga caaaaatcct 2400
ctctcaaact ataaatattg agtatttgtg tattgaacat ttgctataac tactaggttt 2460
cttaaataat cttaatatat aaaatgatat agaaaaaggg aaattatagt tcgtattatt 2520
catctaagtg aagagattaa aacccaggga gtaaataaat tgtctaagga ctaaggttgt 2580
atactattta ggtgatagat atggggcaac cgtatgggtt ttatgattaa caaataaact 2640
tctcaccact ctaccatatc aacttttcca taaaagagag ctatagtatt ctttgcttaa 2700
ataaatttga ttagtgcatg acttcttgaa aacatataaa gcaaaagtca catttgattc 2760
tatcagaaaa gtgagtaagc catggcccaa acaaaagatg cattaaaata ttctggaatg 2820
atggagctaa aagtaagaaa aatgactttt taaaaaagtt tactgttagg aattgtgaaa 2880
ttatgctgaa ttttagttgc attataattt ttgtcagtca tacggtctga caacctgtct 2940
tatttctatt tccccatatg aggaatgcta gttaagtatg gatattaact attactactt 3000
agatgcattg aagttgcata atatggataa tacttcactg gttccctgaa aatgtttagt 3060
tagtaataag tctcttacac tatttgtttt gtccaataat ttatattttc tgaagactta 3120
actctagaat acactcatgt caaaatgaaa gaatttcatt gcaaaatatt gcttggtaca 3180
tgacgcatac ctgtatttgt tttgtgtcac aacatgaaaa atgatggttt attagaagtt 3240
tcattgggta ggaaacacat ttgaatggta tttactaaga tactaaaatc cttggacttc 3300
actctaattt tagtgccatt tagaactcaa ggtctcagta aaagtagaaa taaagcctgt 3360
taacaaaaca caaactgaat attaaaaatg taactggatt ttcaaagaaa tgtttactgg 3420
tattacctgt agatgtatat tctttattat gatcttttgt gtaaagtctg gcagacaaat 3480
gcaatatcta attgttgagt ccaatatcac aagcagtaca aaagtataaa aaagacttgg 3540
ccttttctaa tgtgttaaaa tactttatgc tggtaataac actaagagta gggcactaga 3600
aattttaagt gaagataatg tgttgcagtt actgcactca atggcttact attataaacc 3660
aaaactggga tcactaagct ccagtcagtc aaaatgatca aaattattga agagaataag 3720
caattctgtt ctttattagg acacagtaga tacagactac aaagtggagt gtgcttaata 3780
agaggtagca tttgttaagt gtcaattact ctattatccc ttggagcttc tcaaaataac 3840
catataaggt gtaagatgtt aaaggttatg gttacactca gtgcacaggt aagctaatag 3900
gctgagagaa gctaaattac ttactggggt ctcacagtaa gaaagtgagc tgaagtttca 3960
gcccagattt aactggattc tgggctcttt attcatgtta cttcatgaat ctgtttctca 4020
attgtgcaga aaaaaggggg ctatttataa gaaaagcaat aaacaaacaa gtaatgatct 4080
caaataagta atgcaagaaa tagtgagatt tcaaaatcag tggcagcgat ttctcagttc 4140
tgtcctaagt ggccttgctc aatcacctgc tatcttttag tggagctttg aaattatgtt 4200
tcagacaact tcgattcagt tctagaatgt ttgactcagc aaattcacag gctcatcttt 4260
ctaacttgat ggtgaatatg gaaattcagc taaatggatg ttaataaaat tcaaacgttt 4320
taaggacaga tggaaatgac agaattttaa ggtaaaatat atgaaggaat ataagataaa 4380
ggatttttct accttcagca aaaacatacc cactaattag taaaattaat aggcgaaaaa 4440
aagttgcatg ctcttatact gtaatgatta tcattttaaa actagctttt tgccttcgag 4500
ctatcggggt aaagacctac aggaaaacta ctgtcgaaat cctcgagggg aagaaggggg 4560
accctggtgt ttcacaagca atccagaggt acgctacgaa gtctgtgaca ttcctcagtg 4620
ttcagaagtt gaatgcatga cctgcaatgg ggagagttat cgaggtctca tggatcatac 4680
agaatcaggc aagatttgtc agcgctggga tcatcagaca ccacaccggc acaaattctt 4740
gcctgaaaga tatcccgaca agggctttga tgataattat tgccgcaatc ccgatggcca 4800
gccgaggcca tggtgctata ctcttgaccc tcacacccgc tgggagtact gtgcaattaa 4860
aacatgcgct gacaatacta tgaatgacac tgatgttcct ttggaaacaa ctgaatgcat 4920
ccaaggtcaa ggagaaggct acaggggcac tgtcaatacc atttggaatg gaattccatg 4980
tcagcgttgg gattctcagt atcctcacga gcatgacatg actcctgaaa atttcaagtg 5040
caaggaccta cgagaaaatt actgccgaaa tccagatggg tctgaatcac cctggtgttt 5100
taccactgat ccaaacatcc gagttggcta ctgctcccaa attccaaact gtgatatgtc 5160
acatggacaa gattgttatc gtgggaatgg caaaaattat atgggcaact tatcccaaac 5220
aagatctgga ctaacatgtt caatgtggga caagaacatg gaagacttac atcgtcatat 5280
cttctgggaa ccagatgcaa gtaagctgaa tgagaattac tgccgaaatc cagatgatga 5340
tgctcatgga ccctggtgct acacgggaaa tccactcatt ccttgggatt attgccctat 5400
ttctcgttgt gaaggtgata ccacacctac aatagtcaat ttagaccatc ccgtaatatc 5460
ttgtgccaaa acgaaacaat tgcgagttgt aaatgggatt ccaacacgaa caaacatagg 5520
atggatggtt agtttgagat acagaaataa acatatctgc ggaggatcat tgataaagga 5580
gagttgggtt cttactgcac gacagtgttt cccttctcga gacttgaaag attatgaagc 5640
ttggcttgga attcatgatg tccacggaag aggagatgag aaatgcaaac aggttctcaa 5700
tgtttcccag ctggtatatg gccctgaagg atcagatctg gttttaatga agcttgccag 5760
gcctgctgtc ctggatgatt ttgttagtac gattgattta cctaattatg gatgcacaat 5820
tcctgaaaag accagttgca gtgtttatgg ctggggctac actggattga tcaactatga 5880
tggcctatta cgagtggcac atctctatat aatgggaaat gagaaatgca gccagcatca 5940
tcgagggaag gtgactctga atgagtctga aatatgtgct ggggctgaaa agattggatc 6000
aggaccatgt gagggggatt atggtggccc acttgtttgt gagcaacata aaatgagaat 6060
ggttcttggt gtcattgttc ctggtcgtgg atgtgccatt ccaaatcgtc ctggtatttt 6120
tgtccgagta gcatattatg caaaatggat acacaaaatt attttaacat ataaggtacc 6180
acagtcatag 6190
<210> 28
<211> 5190
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 28
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgaa ataataacgt aaacttcgtt taaaaggatt cttcttcctg tctttgagaa 780
agtacggcac tgtgcagggg gagaggttga ttgtgaaaaa tcagaggtag atgagaatct 840
tactgagggc tgagggttct ttaaccttgg tggatctcaa cattggttgc acattaaaat 900
cacctgctgc aagcccttga cgaatcttac ttagaagatg acaacacaga acaattaaat 960
cagaatctct ggggagaata gggcaccagt attttttgag ctcccaccat gattccaaag 1020
tgcagccaaa tttgagaacc actgctaaaa gctcaagctt cagattgacc agcttttcca 1080
tctcacctat cgcctaaaga ccaaattgga taaatgtgtt cattacgaca gatgggtact 1140
atttaaagat gagtaaacac aatatactta ggctcgtcag actgagagtt ttaatcatca 1200
ctgaggaaaa acatagatat ctaatactga ctggagtatt agtcaaggct tatttcacac 1260
acaattttat cagaaaccaa agtagtttaa aacagctctc cccttattag taatgcattg 1320
gagggtttac tttaccatgt accttgctga gcactgtacc ttgttaatct catttacttg 1380
taatgagaac cacacagcgg gtagttttat tggttctatt ttacctacat gacaaaactg 1440
aagcataaaa acacttagta agttttcagt gtcatgcaca actaggaagt gacatggcca 1500
gaatataagc ccagtcacca tcactctata acctgcgctt ttaacaactt cagggcatga 1560
cacatttggc cggtcagtag aacccatgct gtgatttgtt tttgcagtgg tggtgatgac 1620
tgccttgttg aatccacttt ttattctatt ccattttggg gacacaattc tgcaagatga 1680
ttcttcatta ggaaacagag atgagttatt gaccaacaca gaaagaaaaa gagtttgttg 1740
ctccacactg ggattaaacc tatgatcttg gcctaattaa cactagctag taagtgtcca 1800
agctgatcat ctctacaaca tttcaataac agaaaacaac aattttcaaa attagttact 1860
tacaattatg tagaaatgcc tctaaaacac agtattttcc ttatattaca aaaacaaaaa 1920
ttataattgg ttttgtcctc ttttgagagt ttgcatggtg ttactccctg catagtgaag 1980
aaaacatttt atttaagtag atggatctaa gtttttcatg aacaaaggaa tgacatttga 2040
aatcaatcct accctagtcc aggagaatgc attagattaa cctagtagag gtcttatttc 2100
accctgagtt ttctatgatc gtgattctct gctggaggag taattgtgaa atagatctct 2160
ctgggaactg gcttcctagt ccaatcagct cttttaccaa tgaacacttc cttgtgatat 2220
agatgtttat ggccgagagg atcctgggta ggaaacacat ttgaatggta tttactaaga 2280
tactaaaatc cttggacttc actctaattt tagtgccatt tagaactcaa ggtctcagta 2340
aaagtagaaa taaagcctgt taacaaaaca caaactgaat attaaaaatg taactggatt 2400
ttcaaagaaa tgtttactgg tattacctgt agatgtatat tctttattat gatcttttgt 2460
gtaaagtctg gcagacaaat gcaatatcta attgttgagt ccaatatcac aagcagtaca 2520
aaagtataaa aaagacttgg ccttttctaa tgtgttaaaa tactttatgc tggtaataac 2580
actaagagta gggcactaga aattttaagt gaagataatg tgttgcagtt actgcactca 2640
atggcttact attataaacc aaaactggga tcactaagct ccagtcagtc aaaatgatca 2700
aaattattga agagaataag caattctgtt ctttattagg acacagtaga tacagactac 2760
aaagtggagt gtgcttaata agaggtagca tttgttaagt gtcaattact ctattatccc 2820
ttggagcttc tcaaaataac catataaggt gtaagatgtt aaaggttatg gttacactca 2880
gtgcacaggt aagctaatag gctgagagaa gctaaattac ttactggggt ctcacagtaa 2940
gaaagtgagc tgaagtttca gcccagattt aactggattc tgggctcttt attcatgtta 3000
cttcatgaat ctgtttctca attgtgcaga aaaaaggggg ctatttataa gaaaagcaat 3060
aaacaaacaa gtaatgatct caaataagta atgcaagaaa tagtgagatt tcaaaatcag 3120
tggcagcgat ttctcagttc tgtcctaagt ggccttgctc aatcacctgc tatcttttag 3180
tggagctttg aaattatgtt tcagacaact tcgattcagt tctagaatgt ttgactcagc 3240
aaattcacag gctcatcttt ctaacttgat ggtgaatatg gaaattcagc taaatggatg 3300
ttaataaaat tcaaacgttt taaggacaga tggaaatgac agaattttaa ggtaaaatat 3360
atgaaggaat ataagataaa ggatttttct accttcagca aaaacatacc cactaattag 3420
taaaattaat aggcgaaaaa aagttgcatg ctcttatact gtaatgatta tcattttaaa 3480
actagctttt tgccttcgag ctatcggggt aaagacctac aggaaaacta ctgtcgaaat 3540
cctcgagggg aagaaggggg accctggtgt ttcacaagca atccagaggt acgctacgaa 3600
gtctgtgaca ttcctcagtg ttcagaagtt gaatgcatga cctgcaatgg ggagagttat 3660
cgaggtctca tggatcatac agaatcaggc aagatttgtc agcgctggga tcatcagaca 3720
ccacaccggc acaaattctt gcctgaaaga tatcccgaca agggctttga tgataattat 3780
tgccgcaatc ccgatggcca gccgaggcca tggtgctata ctcttgaccc tcacacccgc 3840
tgggagtact gtgcaattaa aacatgcgct gacaatacta tgaatgacac tgatgttcct 3900
ttggaaacaa ctgaatgcat ccaaggtcaa ggagaaggct acaggggcac tgtcaatacc 3960
atttggaatg gaattccatg tcagcgttgg gattctcagt atcctcacga gcatgacatg 4020
actcctgaaa atttcaagtg caaggaccta cgagaaaatt actgccgaaa tccagatggg 4080
tctgaatcac cctggtgttt taccactgat ccaaacatcc gagttggcta ctgctcccaa 4140
attccaaact gtgatatgtc acatggacaa gattgttatc gtgggaatgg caaaaattat 4200
atgggcaact tatcccaaac aagatctgga ctaacatgtt caatgtggga caagaacatg 4260
gaagacttac atcgtcatat cttctgggaa ccagatgcaa gtaagctgaa tgagaattac 4320
tgccgaaatc cagatgatga tgctcatgga ccctggtgct acacgggaaa tccactcatt 4380
ccttgggatt attgccctat ttctcgttgt gaaggtgata ccacacctac aatagtcaat 4440
ttagaccatc ccgtaatatc ttgtgccaaa acgaaacaat tgcgagttgt aaatgggatt 4500
ccaacacgaa caaacatagg atggatggtt agtttgagat acagaaataa acatatctgc 4560
ggaggatcat tgataaagga gagttgggtt cttactgcac gacagtgttt cccttctcga 4620
gacttgaaag attatgaagc ttggcttgga attcatgatg tccacggaag aggagatgag 4680
aaatgcaaac aggttctcaa tgtttcccag ctggtatatg gccctgaagg atcagatctg 4740
gttttaatga agcttgccag gcctgctgtc ctggatgatt ttgttagtac gattgattta 4800
cctaattatg gatgcacaat tcctgaaaag accagttgca gtgtttatgg ctggggctac 4860
actggattga tcaactatga tggcctatta cgagtggcac atctctatat aatgggaaat 4920
gagaaatgca gccagcatca tcgagggaag gtgactctga atgagtctga aatatgtgct 4980
ggggctgaaa agattggatc aggaccatgt gagggggatt atggtggccc acttgtttgt 5040
gagcaacata aaatgagaat ggttcttggt gtcattgttc ctggtcgtgg atgtgccatt 5100
ccaaatcgtc ctggtatttt tgtccgagta gcatattatg caaaatggat acacaaaatt 5160
attttaacat ataaggtacc acagtcatag 5190
<210> 29
<211> 4241
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 29
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgaa ataataacgt aaacttcgtt taaaaggatt cttcttcctg tctttgagaa 780
agtacggcac tgtgcagggg gagaggttga ttgtgaaaaa tcagaggtag atgagaatct 840
tactgagggc tgagggttct ttaaccttgg tggatctcaa cattggttgc acattaaaat 900
cacctgctgc aagcccttga cgaatcttac ttagaagatg acaacacaga acaattaaat 960
cagaatctct ggggagaata gggcaccagt attttttgag ctcccaccat gattccaaag 1020
tgcagccaaa tttgagaacc actgctaaaa gctcaagctt cagattgacc agcttttcca 1080
tctcacctat cgcctaaaga ccaaattgga taaatgtgtt cattacgaca gatgggtact 1140
atttaaagat gagtaaacac aatatactta ggctcgtcag actgagagtt ttaatcatca 1200
ctgaggaaaa acatagatat ctaatactga ctggagtatt agtcaaggct tatttcacac 1260
acaattttat cagaaaccaa agtagtttaa aacagctctc cccttattag taatgcattg 1320
gagggtttac tttaccatgt accttgctga gcactgtacc ttgttaatct catttacttg 1380
taatgagaac cacacagcgg gtagttttat tggttctatt ttacctacat gacaaaactg 1440
aagcataaaa acacttagta agttttcagt gtcatgcaca actaggaagt gacatggcca 1500
gaatataagc ccagtcacca tcactctata acctgcgctt ttaacaactt cagggcatga 1560
cacatttggc cggtcagtag aacccatgct gtgatttgtt tttgcagtgg tggtgatgac 1620
tgccttgttg aatccacttt ttattctatt ccattttggg gacacaattc tgcaagatga 1680
ttcttcatta ggaaacagag atgagttatt gaccaacaca gaaagaaaaa gagtttgttg 1740
ctccacactg ggattaaacc tatgatcttg gcctaattaa cactagctag taagtgtcca 1800
agctgatcat ctctacaaca tttcaataac agaaaacaac aattttcaaa attagttact 1860
tacaattatg tagaaatgcc tctaaaacac agtattttcc ttatattaca aaaacaaaaa 1920
ttataattgg ttttgtcctc ttttgagagt ttgcatggtg ttactccctg catagtgaag 1980
aaaacatttt atttaagtag atggatctaa gtttttcatg aacaaaggaa tgacatttga 2040
aatcaatcct accctagtcc aggagaatgc attagattaa cctagtagag gtcttatttc 2100
accctgagtt ttctatgatc gtgattctct gctggaggag taattgtgaa atagatctct 2160
ctgggaactg gcttcctagt ccaatcagct cttttaccaa tgaacacttc cttgtgatat 2220
agatgtttat ggccgagagg atccttatgt ttcagacaac ttcgattcag ttctagaatg 2280
tttgactcag caaattcaca ggctcatctt tctaacttga tggtgaatat ggaaattcag 2340
ctaaatggat gttaataaaa ttcaaacgtt ttaaggacag atggaaatga cagaatttta 2400
aggtaaaata tatgaaggaa tataagataa aggatttttc taccttcagc aaaaacatac 2460
ccactaatta gtaaaattaa taggcgaaaa aaagttgcat gctcttatac tgtaatgatt 2520
atcattttaa aactagcttt ttgccttcga gctatcgggg taaagaccta caggaaaact 2580
actgtcgaaa tcctcgaggg gaagaagggg gaccctggtg tttcacaagc aatccagagg 2640
tacgctacga agtctgtgac attcctcagt gttcagaagt tgaatgcatg acctgcaatg 2700
gggagagtta tcgaggtctc atggatcata cagaatcagg caagatttgt cagcgctggg 2760
atcatcagac accacaccgg cacaaattct tgcctgaaag atatcccgac aagggctttg 2820
atgataatta ttgccgcaat cccgatggcc agccgaggcc atggtgctat actcttgacc 2880
ctcacacccg ctgggagtac tgtgcaatta aaacatgcgc tgacaatact atgaatgaca 2940
ctgatgttcc tttggaaaca actgaatgca tccaaggtca aggagaaggc tacaggggca 3000
ctgtcaatac catttggaat ggaattccat gtcagcgttg ggattctcag tatcctcacg 3060
agcatgacat gactcctgaa aatttcaagt gcaaggacct acgagaaaat tactgccgaa 3120
atccagatgg gtctgaatca ccctggtgtt ttaccactga tccaaacatc cgagttggct 3180
actgctccca aattccaaac tgtgatatgt cacatggaca agattgttat cgtgggaatg 3240
gcaaaaatta tatgggcaac ttatcccaaa caagatctgg actaacatgt tcaatgtggg 3300
acaagaacat ggaagactta catcgtcata tcttctggga accagatgca agtaagctga 3360
atgagaatta ctgccgaaat ccagatgatg atgctcatgg accctggtgc tacacgggaa 3420
atccactcat tccttgggat tattgcccta tttctcgttg tgaaggtgat accacaccta 3480
caatagtcaa tttagaccat cccgtaatat cttgtgccaa aacgaaacaa ttgcgagttg 3540
taaatgggat tccaacacga acaaacatag gatggatggt tagtttgaga tacagaaata 3600
aacatatctg cggaggatca ttgataaagg agagttgggt tcttactgca cgacagtgtt 3660
tcccttctcg agacttgaaa gattatgaag cttggcttgg aattcatgat gtccacggaa 3720
gaggagatga gaaatgcaaa caggttctca atgtttccca gctggtatat ggccctgaag 3780
gatcagatct ggttttaatg aagcttgcca ggcctgctgt cctggatgat tttgttagta 3840
cgattgattt acctaattat ggatgcacaa ttcctgaaaa gaccagttgc agtgtttatg 3900
gctggggcta cactggattg atcaactatg atggcctatt acgagtggca catctctata 3960
taatgggaaa tgagaaatgc agccagcatc atcgagggaa ggtgactctg aatgagtctg 4020
aaatatgtgc tggggctgaa aagattggat caggaccatg tgagggggat tatggtggcc 4080
cacttgtttg tgagcaacat aaaatgagaa tggttcttgg tgtcattgtt cctggtcgtg 4140
gatgtgccat tccaaatcgt cctggtattt ttgtccgagt agcatattat gcaaaatgga 4200
tacacaaaat tattttaaca tataaggtac cacagtcata g 4241
<210> 30
<211> 5602
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 30
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgga tccagtatat taataaaatc cctttttgta ttcaatgagg gaaacacata 780
attttcatca attagcagct tattggaata tctgcatgat ggtttaacac ttttaagtgt 840
tgactaaaga ttaattttac agaaaataga aaaagaaata tgtttctgtc tggaggaatg 900
atttattgtt gacccctaaa ttgaaatatt ttactagtgg cttaatggaa agatgatgaa 960
agatgatgaa attaatgtag aagcttaact agaaaatcag gtgacctgat atctacatct 1020
gtatccttca ttggccaccc agcattcatt aatgaatcag atgatggaat agatcaagtt 1080
tcctaggaac acagtgaata ttaaaagaaa acaaagggag cctagcacct agaagaccta 1140
gtttatattt caaagtatat ttggatgtaa cccaatttta aacatttcct cacttgtctc 1200
tcttaaagcc ttgccaacag caaggacaga gaaccaaaaa tagtgtatat atgaataaat 1260
gcttattaca gaatctgctg actggcacat gctttgtgtg taatgggttc tcataaacac 1320
ttgttgaatg aacacacata agtgaaagag catggctagg cttcatccct tggtcaaata 1380
tggggtgcta aagaaaagca ggggaaatac attgggacac taacaaaaaa aaacagttaa 1440
tttaggtaaa agataaaata caccacagaa tgaagaaaag agatgaccca gactgctctt 1500
taaccttcat gtcctagaga ggtttttgat atgaattgca ttcagaattg tggaaaggag 1560
cccatctttt ctcttcattt tgattttatt aactccaatg ggggaatttt attcgtgttt 1620
tggccatatc tacttttgat ttctacatta ttctctcttc ctttctacct gtatttgtcc 1680
taataaattg ttgacttatt aattcactac ttcctcacag cttttttttg gctttacaaa 1740
tccactggaa aggtatatgg gtgtatcact ttgtgtattt cggtgtgcat gtgtagaggg 1800
gacaaaaatc ctctctcaaa ctataaatat tgagtatttg tgtattgaac atttgctata 1860
actactaggt ttcttaaata atcttaatat ataaaatgat atagaaaaag ggaaattata 1920
gttcgtatta ttcatctaag tgaagagatt aaaacccagg gagtaaataa attgtctaag 1980
gactaaggtt gtatactatt taggtgatag atatggggca accgtatggg ttttatgatt 2040
aacaaataaa cttctcacca ctctaccata tcaacttttc cataaaagag agctatagta 2100
ttctttgctt aaataaattt gattagtgca tgacttcttg aaaacatata aagcaaaagt 2160
cacatttgat tctatcagaa aagtgagtaa gccatggccc aaacaaaaga tgcattaaaa 2220
tattctggaa tgatggagct aaaagtaaga aaaatgactt tttaaaaaag tttactgtta 2280
ggaattgtga aattatgctg aattttagtt gcattataat ttttgtcagt catacggtct 2340
gacaacctgt cttatttcta tttccccata tgaggaatgc tagttaagta tggatattaa 2400
ctattactac ttagatgcat tgaagttgca taatatggat aatacttcac tggttccctg 2460
aaaatgttta gttagtaata agtctcttac actatttgtt ttgtccaata atttatattt 2520
tctgaagact taactctaga atacactcat gtcaaaatga aagaatttca ttgcaaaata 2580
ttgcttggta catgacgcat acctgtattt gttttgtgtc acaacatgaa aaatgatggt 2640
ttattagaag tttcattggg taggaaacac atttgaatgg tatttactaa gatactaaaa 2700
tccttggact tcactctaat tttagtgcca tttagaactc aaggtctcag taaaagtaga 2760
aataaagcct gttaacaaaa cacaaactga atattaaaaa tgtaactgga ttttcaaaga 2820
aatgtttact ggtattacct gtagatgtat attctttatt atgatctttt gtgtaaagtc 2880
tggcagacaa atgcaatatc taattgttga gtccaatatc acaagcagta caaaagtata 2940
aaaaagactt ggccttttct aatgtgttaa aatactttat gctggtaata acactaagag 3000
tagggcacta gaaattttaa gtgaagataa tgtgttgcag ttactgcact caatggctta 3060
ctattataaa ccaaaactgg gatcactaag ctccagtcag tcaaaatgat caaaattatt 3120
gaagagaata agcaattctg ttctttatta ggacacagta gatacagact acaaagtgga 3180
gtgtgcttaa taagaggtag catttgttaa gtgtcaatta ctctattatc ccttggagct 3240
tctcaaaata accatataag gtgtaagatg ttaaaggtta tggttacact cagtgcacag 3300
gtaagctaat aggctgagag aagctaaatt acttactggg gtctcacagt aagaaagtga 3360
gctgaagttt cagcccagat ttaactggat tctgggctct ttattcatgt tacttcatga 3420
atctgtttct caattgtgca gaaaaaaggg ggctatttat aagaaaagca ataaacaaac 3480
aagtaatgat ctcaaataag taatgcaaga aatagtgaga tttcaaaatc agtggcagcg 3540
atttctcagt tctgtcctaa gtggccttgc tcaatcacct gctatctttt agtggagctt 3600
tgaaattatg tttcagacaa cttcgattca gttctagaat gtttgactca gcaaattcac 3660
aggctcatct ttctaacttg atggtgaata tggaaattca gctaaatgga tgttaataaa 3720
attcaaacgt tttaaggaca gatggaaatg acagaatttt aaggtaaaat atatgaagga 3780
atataagata aaggattttt ctaccttcag caaaaacata cccactaatt agtaaaatta 3840
ataggcgaaa aaaagttgca tgctcttata ctgtaatgat tatcatttta aaactagctt 3900
tttgccttcg agctatcggg gtaaagacct acaggaaaac tactgtcgaa atcctcgagg 3960
ggaagaaggg ggaccctggt gtttcacaag caatccagag gtacgctacg aagtctgtga 4020
cattcctcag tgttcagaag ttgaatgcat gacctgcaat ggggagagtt atcgaggtct 4080
catggatcat acagaatcag gcaagatttg tcagcgctgg gatcatcaga caccacaccg 4140
gcacaaattc ttgcctgaaa gatatcccga caagggcttt gatgataatt attgccgcaa 4200
tcccgatggc cagccgaggc catggtgcta tactcttgac cctcacaccc gctgggagta 4260
ctgtgcaatt aaaacatgcg ctgacaatac tatgaatgac actgatgttc ctttggaaac 4320
aactgaatgc atccaaggtc aaggagaagg ctacaggggc actgtcaata ccatttggaa 4380
tggaattcca tgtcagcgtt gggattctca gtatcctcac gagcatgaca tgactcctga 4440
aaatttcaag tgcaaggacc tacgagaaaa ttactgccga aatccagatg ggtctgaatc 4500
accctggtgt tttaccactg atccaaacat ccgagttggc tactgctccc aaattccaaa 4560
ctgtgatatg tcacatggac aagattgtta tcgtgggaat ggcaaaaatt atatgggcaa 4620
cttatcccaa acaagatctg gactaacatg ttcaatgtgg gacaagaaca tggaagactt 4680
acatcgtcat atcttctggg aaccagatgc aagtaagctg aatgagaatt actgccgaaa 4740
tccagatgat gatgctcatg gaccctggtg ctacacggga aatccactca ttccttggga 4800
ttattgccct atttctcgtt gtgaaggtga taccacacct acaatagtca atttagacca 4860
tcccgtaata tcttgtgcca aaacgaaaca attgcgagtt gtaaatggga ttccaacacg 4920
aacaaacata ggatggatgg ttagtttgag atacagaaat aaacatatct gcggaggatc 4980
attgataaag gagagttggg ttcttactgc acgacagtgt ttcccttctc gagacttgaa 5040
agattatgaa gcttggcttg gaattcatga tgtccacgga agaggagatg agaaatgcaa 5100
acaggttctc aatgtttccc agctggtata tggccctgaa ggatcagatc tggttttaat 5160
gaagcttgcc aggcctgctg tcctggatga ttttgttagt acgattgatt tacctaatta 5220
tggatgcaca attcctgaaa agaccagttg cagtgtttat ggctggggct acactggatt 5280
gatcaactat gatggcctat tacgagtggc acatctctat ataatgggaa atgagaaatg 5340
cagccagcat catcgaggga aggtgactct gaatgagtct gaaatatgtg ctggggctga 5400
aaagattgga tcaggaccat gtgaggggga ttatggtggc ccacttgttt gtgagcaaca 5460
taaaatgaga atggttcttg gtgtcattgt tcctggtcgt ggatgtgcca ttccaaatcg 5520
tcctggtatt tttgtccgag tagcatatta tgcaaaatgg atacacaaaa ttattttaac 5580
atataaggta ccacagtcat ag 5602
<210> 31
<211> 4680
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 31
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgga tctcttcctt tctacctgta tttgtcctaa taaattgttg acttattaat 780
ytcactactt cctcacagct tttttttggc tttacaaatc cactggaaag gtatatgggt 840
gtatcacttt gtgtatttcg gtgtgcatgt gtagagggga caaaaatcct ctctcaaact 900
ataaatattg agtatttgtg tattgaacat ttgctataac tactaggttt cttaaataat 960
cttaatatat aaaatgatat agaaaaaggg aaattatagt tcgtattatt catctaagtg 1020
aagagattaa aacccaggga gtaaataaat tgtctaagga ctaaggttgt atactattta 1080
ggtgatagat atggggcaac cgtatgggtt ttatgattaa caaataaact tctcaccact 1140
ctaccatatc aacttttcca taaaagagag ctatagtatt ctttgcttaa ataaatttga 1200
ttagtgcatg acttcttgaa aacatataaa gcaaaagtca catttgattc tatcagaaaa 1260
gtgagtaagc catggcccaa acaaaagatg cattaaaata ttctggaatg atggagctaa 1320
aagtaagaaa aatgactttt taaaaaagtt tactgttagg aattgtgaaa ttatgctgaa 1380
ttttagttgc attataattt ttgtcagtca tacggtctga caacctgtct tatttctatt 1440
tccccatatg aggaatgcta gttaagtatg gatattaact attactactt agatgcattg 1500
aagttgcata atatggataa tacttcactg gttccctgaa aatgtttagt tagtaataag 1560
tctcttacac tatttgtttt gtccaataat ttatattttc tgaagactta actctagaat 1620
acactcatgt caaaatgaaa gaatttcatt gcaaaatatt gcttggtaca tgacgcatac 1680
ctgtatttgt tttgtgtcac aacatgaaaa atgatggttt attagaagtt tcattgggta 1740
ggaaacacat ttgaatggta tttactaaga tactaaaatc cttggacttc actctaattt 1800
tagtgccatt tagaactcaa ggtctcagta aaagtagaaa taaagcctgt taacaaaaca 1860
caaactgaat attaaaaatg taactggatt ttcaaagaaa tgtttactgg tattacctgt 1920
agatgtatat tctttattat gatcttttgt gtaaagtctg gcagacaaat gcaatatcta 1980
attgttgagt ccaatatcac aagcagtaca aaagtataaa aaagacttgg ccttttctaa 2040
tgtgttaaaa tactttatgc tggtaataac actaagagta gggcactaga aattttaagt 2100
gaagataatg tgttgcagtt actgcactca atggcttact attataaacc aaaactggga 2160
tcactaagct ccagtcagtc aaaatgatca aaattattga agagaataag caattctgtt 2220
ctttattagg acacagtaga tacagactac aaagtggagt gtgcttaata agaggtagca 2280
tttgttaagt gtcaattact ctattatccc ttggagcttc tcaaaataac catataaggt 2340
gtaagatgtt aaaggttatg gttacactca gtgcacaggt aagctaatag gctgagagaa 2400
gctaaattac ttactggggt ctcacagtaa gaaagtgagc tgaagtttca gcccagattt 2460
aactggattc tgggctcttt attcatgtta cttcatgaat ctgtttctca attgtgcaga 2520
aaaaaggggg ctatttataa gaaaagcaat aaacaaacaa gtaatgatct caaataagta 2580
atgcaagaaa tagtgagatt tcaaaatcag tggcagcgat ttctcagttc tgtcctaagt 2640
ggccttgctc aatcacctgc tatcttttag tggagctttg aaattatgtt tcagacaact 2700
tcgattcagt tctagaatgt ttgactcagc aaattcacag gctcatcttt ctaacttgat 2760
ggtgaatatg gaaattcagc taaatggatg ttaataaaat tcaaacgttt taaggacaga 2820
tggaaatgac agaattttaa ggtaaaatat atgaaggaat ataagataaa ggatttttct 2880
accttcagca aaaacatacc cactaattag taaaattaat aggcgaaaaa aagttgcatg 2940
ctcttatact gtaatgatta tcattttaaa actagctttt tgccttcgag ctatcggggt 3000
aaagacctac aggaaaacta ctgtcgaaat cctcgagggg aagaaggggg accctggtgt 3060
ttcacaagca atccagaggt acgctacgaa gtctgtgaca ttcctcagtg ttcagaagtt 3120
gaatgcatga cctgcaatgg ggagagttat cgaggtctca tggatcatac agaatcaggc 3180
aagatttgtc agcgctggga tcatcagaca ccacaccggc acaaattctt gcctgaaaga 3240
tatcccgaca agggctttga tgataattat tgccgcaatc ccgatggcca gccgaggcca 3300
tggtgctata ctcttgaccc tcacacccgc tgggagtact gtgcaattaa aacatgcgct 3360
gacaatacta tgaatgacac tgatgttcct ttggaaacaa ctgaatgcat ccaaggtcaa 3420
ggagaaggct acaggggcac tgtcaatacc atttggaatg gaattccatg tcagcgttgg 3480
gattctcagt atcctcacga gcatgacatg actcctgaaa atttcaagtg caaggaccta 3540
cgagaaaatt actgccgaaa tccagatggg tctgaatcac cctggtgttt taccactgat 3600
ccaaacatcc gagttggcta ctgctcccaa attccaaact gtgatatgtc acatggacaa 3660
gattgttatc gtgggaatgg caaaaattat atgggcaact tatcccaaac aagatctgga 3720
ctaacatgtt caatgtggga caagaacatg gaagacttac atcgtcatat cttctgggaa 3780
ccagatgcaa gtaagctgaa tgagaattac tgccgaaatc cagatgatga tgctcatgga 3840
ccctggtgct acacgggaaa tccactcatt ccttgggatt attgccctat ttctcgttgt 3900
gaaggtgata ccacacctac aatagtcaat ttagaccatc ccgtaatatc ttgtgccaaa 3960
acgaaacaat tgcgagttgt aaatgggatt ccaacacgaa caaacatagg atggatggtt 4020
agtttgagat acagaaataa acatatctgc ggaggatcat tgataaagga gagttgggtt 4080
cttactgcac gacagtgttt cccttctcga gacttgaaag attatgaagc ttggcttgga 4140
attcatgatg tccacggaag aggagatgag aaatgcaaac aggttctcaa tgtttcccag 4200
ctggtatatg gccctgaagg atcagatctg gttttaatga agcttgccag gcctgctgtc 4260
ctggatgatt ttgttagtac gattgattta cctaattatg gatgcacaat tcctgaaaag 4320
accagttgca gtgtttatgg ctggggctac actggattga tcaactatga tggcctatta 4380
cgagtggcac atctctatat aatgggaaat gagaaatgca gccagcatca tcgagggaag 4440
gtgactctga atgagtctga aatatgtgct ggggctgaaa agattggatc aggaccatgt 4500
gagggggatt atggtggccc acttgtttgt gagcaacata aaatgagaat ggttcttggt 4560
gtcattgttc ctggtcgtgg atgtgccatt ccaaatcgtc ctggtatttt tgtccgagta 4620
gcatattatg caaaatggat acacaaaatt attttaacat ataaggtacc acagtcatag 4680
<210> 32
<211> 2730
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 32
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgga tccttatgtt tcagacaact tcgattcagt tctagaatgt ttgactcagc 780
aaattcacag gctcatcttt ctaacttgat ggtgaatatg gaaattcagc taaatggatg 840
ttaataaaat tcaaacgttt taaggacaga tggaaatgac agaattttaa ggtaaaatat 900
atgaaggaat ataagataaa ggatttttct accttcagca aaaacatacc cactaattag 960
taaaattaat aggcgaaaaa aagttgcatg ctcttatact gtaatgatta tcattttaaa 1020
actagctttt tgccttcgag ctatcggggt aaagacctac aggaaaacta ctgtcgaaat 1080
cctcgagggg aagaaggggg accctggtgt ttcacaagca atccagaggt acgctacgaa 1140
gtctgtgaca ttcctcagtg ttcagaagtt gaatgcatga cctgcaatgg ggagagttat 1200
cgaggtctca tggatcatac agaatcaggc aagatttgtc agcgctggga tcatcagaca 1260
ccacaccggc acaaattctt gcctgaaaga tatcccgaca agggctttga tgataattat 1320
tgccgcaatc ccgatggcca gccgaggcca tggtgctata ctcttgaccc tcacacccgc 1380
tgggagtact gtgcaattaa aacatgcgct gacaatacta tgaatgacac tgatgttcct 1440
ttggaaacaa ctgaatgcat ccaaggtcaa ggagaaggct acaggggcac tgtcaatacc 1500
atttggaatg gaattccatg tcagcgttgg gattctcagt atcctcacga gcatgacatg 1560
actcctgaaa atttcaagtg caaggaccta cgagaaaatt actgccgaaa tccagatggg 1620
tctgaatcac cctggtgttt taccactgat ccaaacatcc gagttggcta ctgctcccaa 1680
attccaaact gtgatatgtc acatggacaa gattgttatc gtgggaatgg caaaaattat 1740
atgggcaact tatcccaaac aagatctgga ctaacatgtt caatgtggga caagaacatg 1800
gaagacttac atcgtcatat cttctgggaa ccagatgcaa gtaagctgaa tgagaattac 1860
tgccgaaatc cagatgatga tgctcatgga ccctggtgct acacgggaaa tccactcatt 1920
ccttgggatt attgccctat ttctcgttgt gaaggtgata ccacacctac aatagtcaat 1980
ttagaccatc ccgtaatatc ttgtgccaaa acgaaacaat tgcgagttgt aaatgggatt 2040
ccaacacgaa caaacatagg atggatggtt agtttgagat acagaaataa acatatctgc 2100
ggaggatcat tgataaagga gagttgggtt cttactgcac gacagtgttt cccttctcga 2160
gacttgaaag attatgaagc ttggcttgga attcatgatg tccacggaag aggagatgag 2220
aaatgcaaac aggttctcaa tgtttcccag ctggtatatg gccctgaagg atcagatctg 2280
gttttaatga agcttgccag gcctgctgtc ctggatgatt ttgttagtac gattgattta 2340
cctaattatg gatgcacaat tcctgaaaag accagttgca gtgtttatgg ctggggctac 2400
actggattga tcaactatga tggcctatta cgagtggcac atctctatat aatgggaaat 2460
gagaaatgca gccagcatca tcgagggaag gtgactctga atgagtctga aatatgtgct 2520
ggggctgaaa agattggatc aggaccatgt gagggggatt atggtggccc acttgtttgt 2580
gagcaacata aaatgagaat ggttcttggt gtcattgttc ctggtcgtgg atgtgccatt 2640
ccaaatcgtc ctggtatttt tgtccgagta gcatattatg caaaatggat acacaaaatt 2700
attttaacat ataaggtacc acagtcatag 2730
<210> 33
<211> 2187
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 33
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
agctttttgc cttcgagcta tcggggtaaa gacctacagg aaaactactg tcgaaatcct 540
cgaggggaag aagggggacc ctggtgtttc acaagcaatc cagaggtacg ctacgaagtc 600
tgtgacattc ctcagtgttc agaagttgaa tgcatgacct gcaatgggga gagttatcga 660
ggtctcatgg atcatacaga atcaggcaag atttgtcagc gctgggatca tcagacacca 720
caccggcaca aattcttgcc tgaaagatat cccgacaagg gctttgatga taattattgc 780
cgcaatcccg atggccagcc gaggccatgg tgctatactc ttgaccctca cacccgctgg 840
gagtactgtg caattaaaac atgcgctgac aatactatga atgacactga tgttcctttg 900
gaaacaactg aatgcatcca aggtcaagga gaaggctaca ggggcactgt caataccatt 960
tggaatggaa ttccatgtca gcgttgggat tctcagtatc ctcacgagca tgacatgact 1020
cctgaaaatt tcaagtgcaa ggacctacga gaaaattact gccgaaatcc agatgggtct 1080
gaatcaccct ggtgttttac cactgatcca aacatccgag ttggctactg ctcccaaatt 1140
ccaaactgtg atatgtcaca tggacaagat tgttatcgtg ggaatggcaa aaattatatg 1200
ggcaacttat cccaaacaag atctggacta acatgttcaa tgtgggacaa gaacatggaa 1260
gacttacatc gtcatatctt ctgggaacca gatgcaagta agctgaatga gaattactgc 1320
cgaaatccag atgatgatgc tcatggaccc tggtgctaca cgggaaatcc actcattcct 1380
tgggattatt gccctatttc tcgttgtgaa ggtgatacca cacctacaat agtcaattta 1440
gaccatcccg taatatcttg tgccaaaacg aaacaattgc gagttgtaaa tgggattcca 1500
acacgaacaa acataggatg gatggttagt ttgagataca gaaataaaca tatctgcgga 1560
ggatcattga taaaggagag ttgggttctt actgcacgac agtgtttccc ttctcgagac 1620
ttgaaagatt atgaagcttg gcttggaatt catgatgtcc acggaagagg agatgagaaa 1680
tgcaaacagg ttctcaatgt ttcccagctg gtatatggcc ctgaaggatc agatctggtt 1740
ttaatgaagc ttgccaggcc tgctgtcctg gatgattttg ttagtacgat tgatttacct 1800
aattatggat gcacaattcc tgaaaagacc agttgcagtg tttatggctg gggctacact 1860
ggattgatca actatgatgg cctattacga gtggcacatc tctatataat gggaaatgag 1920
aaatgcagcc agcatcatcg agggaaggtg actctgaatg agtctgaaat atgtgctggg 1980
gctgaaaaga ttggatcagg accatgtgag ggggattatg gtggcccact tgtttgtgag 2040
caacataaaa tgagaatggt tcttggtgtc attgttcctg gtcgtggatg tgccattcca 2100
aatcgtcctg gtatttttgt ccgagtagca tattatgcaa aatggataca caaaattatt 2160
ttaacatata aggtaccaca gtcatag 2187
<210> 34
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
праймер
<400> 34
agctggcaat tccggttcgc ttgctgcgtc agaccccgta 40
<210> 35
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
праймер
<400> 35
tacggggtct gacgcagcaa gcgaaccgga attgccagct 40
<210> 36
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
праймер
<400> 36
ctaatccata acatggctct agacttaagg cagcggcaga 40
<210> 37
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
праймер
<400> 37
tctgccgctg ccttaagtct agagccatgt tatggattag 40
<210> 38
<211> 3250
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 38
cagggtcgga acaggagagc gcacgaggga gcttccaggg ggaaacgcct ggtatcttta 60
tagtcctgtc gggtttcgcc acctctgact tgagcgtcga tttttgtgat gctcgtcagg 120
ggggcggagc ctatggaaaa acgccagcaa cgcggccttt ttacggttcc tggccttttg 180
ctggcctttt gctcacatgc gcgttgacat tgattattga ctagttatta atagtaatca 240
attacggggt cattagttca tagcccatat atggagttcc gcgttacata acttacggta 300
aatggcccgc ctggctgacc gcccaacgac ccccgcccat tgacgtcaat aatgacgtat 360
gttcccatag taacgccaat agggactttc cattgacgtc aatgggtgga gtatttacgg 420
taaactgccc acttggcagt acatcaagtg tatcatatgc caagtccgcc ccctattgac 480
gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt acatgacctt acgggacttt 540
cctacttggc agtacatcta cgtattagtc atcgctatta ccatggtgat gcggttttgg 600
cagtacacca atgggcgtgg atagcggttt gactcacggg gatttccaag tctccacccc 660
attgacgtca atgggagttt gttttggcac caaaatcaac gggactttcc aaaatgtcgt 720
aataaccccg ccccgttgac gcaaatgggc ggtaggcgtg tacggtggga ggtctatata 780
agcagagctc gtttagtgaa ccgtcagatc gcctggagac gccatccacg ctgttttgac 840
ctccatagaa gacaccggga ccgatccagc ctccgcggcc gggaacggtg cattggaacg 900
cggattcccc gtgccaagag tgacgtaagt accgcctata gactctatag gcacacccct 960
ttggctctta tgcatgctat actgtttttg gcttggggcc tatacacccc cgcttcctta 1020
tgctataggt gatggtatag cttagcctat aggtgtgggt tattgaccat tattgaccac 1080
tcccctattg gtgacgatac tttccattac taatccataa catggctcta gacttaaggc 1140
agcggcagaa gaagatgtag gcagctgagt tgttgtattc tgataagagt cagaggtaac 1200
tcccgttgcg gtgctgttaa cggtggaggg cagtgtagtc tgagcagtac tcgttgctgc 1260
cgcgcgcgcc accagacata atagctgaca gactaacaga ctgttccttt ccatgggtct 1320
tttctgcagt caccgtcctt gacacgaagc ttatcgatgt cgacctcgag tctagagggc 1380
ccgtttaaac ccgctgatca gcctcgactg tgccttctag ttgccagcca tctgttgttt 1440
gcccctcccc cgtgccttcc ttgaccctgg aaggtgccac tcccactgtc ctttcctaat 1500
aaaatgagga aattgcatcg cattgtctga gtaggtgtca ttctattctg gggggtgggg 1560
tggggcagga cagcaagggg gaggattggg aagacaatag caggcatgct ggggagtcga 1620
aattcagaag aactcgtcaa gaaggcgata gaaggcgatg cgctgcgaat cgggagcggc 1680
gataccgtaa agcacgagga agcggtcagc ccattcgccg ccaagctctt cagcaatatc 1740
acgggtagcc aacgctatgt cctgatagcg gtccgccaca cccagccggc cacagtcgat 1800
gaatccagaa aagcggccat tttccaccat gatattcggc aagcaggcat cgccatgggt 1860
cacgacgaga tcctcgccgt cgggcatgct cgccttgagc ctggcgaaca gttcggctgg 1920
cgcgagcccc tgatgctctt cgtccagatc atcctgatcg acaagaccgg cttccatccg 1980
agtacgtgct cgctcgatgc gatgtttcgc ttggtggtcg aatgggcagg tagccggatc 2040
aagcgtatgc agccgccgca ttgcatcagc catgatggat actttctcgg caggagcaag 2100
gtgagatgac aggagatcct gccccggcac ttcgcccaat agcagccagt cccttcccgc 2160
ttcagtgaca acgtcgagca cagctgcgca aggaacgccc gtcgtggcca gccacgatag 2220
ccgcgctgcc tcgtcttgca gttcattcag ggcaccggac aggtcggtct tgacaaaaag 2280
aaccgggcgc ccctgcgctg acagccggaa cacggcggca tcagagcagc cgattgtctg 2340
ttgtgcccag tcatagccga atagcctctc cacccaagcg gccggagaac ctgcgtgcaa 2400
tccatcttgt tcaatcatgc gaaacgatcc tcatcctgtc tcttgatcag atcttgatcc 2460
cctgcgccat cagatccttg gcggcaagaa agccatccag tttactttgc agggcttccc 2520
aaccttacca gagggcgccc cagctggcaa ttccggttcg cttgctgcgt cagaccccgt 2580
agaaaagatc aaaggatctt cttgagatcc tttttttctg cgcgtaatct gctgcttgca 2640
aacaaaaaaa ccaccgctac cagcggtggt ttgtttgccg gatcaagagc taccaactct 2700
ttttccgaag gtaactggct tcagcagagc gcagatacca aatactgttc ttctagtgta 2760
gccgtagtta ggccaccact tcaagaactc tgtagcaccg cctacatacc tcgctctgct 2820
aatcctgtta ccagtggctg ctgccagtgg cgataagtcg tgtcttaccg ggttggactc 2880
aagacgatag ttaccggata aggcgcagcg gtcgggctga acggggggtt cgtgcacaca 2940
gcccagcttg gagcgaacga cctacaccga actgagatac ctacagcgtg agctatgaga 3000
aagcgccacg cttcccgaag ggagaaaggc ggacaggtat ccggtaagcg gcagggtcgg 3060
aacaggagag cgcacgaggg agcttccagg gggaaacgcc tggtatcttt atagtcctgt 3120
cgggtttcgc cacctctgac ttgagcgtcg atttttgtga tgctcgtcag gggggcggag 3180
cctatggaaa aacgccagca acgcggcctt tttacggttc ctggcctttt gctggccttt 3240
tgctcacatg 3250
<210> 39
<211> 4108
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 39
cagggtcgga acaggagagc gcacgaggga gcttccaggg ggaaacgcct ggtatcttta 60
tagtcctgtc gggtttcgcc acctctgact tgagcgtcga tttttgtgat gctcgtcagg 120
ggggcggagc ctatggaaaa acgccagcaa cgcggccttt ttacggttcc tggccttttg 180
ctggcctttt gctcacatgc gcgttgacat tgattattga ctagttatta atagtaatca 240
attacggggt cattagttca tagcccatat atggagttcc gcgttacata acttacggta 300
aatggcccgc ctggctgacc gcccaacgac ccccgcccat tgacgtcaat aatgacgtat 360
gttcccatag taacgccaat agggactttc cattgacgtc aatgggtgga gtatttacgg 420
taaactgccc acttggcagt acatcaagtg tatcatatgc caagtccgcc ccctattgac 480
gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt acatgacctt acgggacttt 540
cctacttggc agtacatcta cgtattagtc atcgctatta ccatggtgat gcggttttgg 600
cagtacacca atgggcgtgg atagcggttt gactcacggg gatttccaag tctccacccc 660
attgacgtca atgggagttt gttttggcac caaaatcaac gggactttcc aaaatgtcgt 720
aataaccccg ccccgttgac gcaaatgggc ggtaggcgtg tacggtggga ggtctatata 780
agcagagctc gtttagtgaa ccgtcagatc gcctggagac gccatccacg ctgttttgac 840
ctccatagaa gacaccggga ccgatccagc ctccgcggcc gggaacggtg cattggaacg 900
cggattcccc gtgccaagag tgacgtaagt accgcctata gactctatag gcacacccct 960
ttggctctta tgcatgctat actgtttttg gcttggggcc tatacacccc cgcttcctta 1020
tgctataggt gatggtatag cttagcctat aggtgtgggt tattgaccat tattgaccac 1080
tcccctattg gtgacgatac tttccattac taatccataa catggctcta gacttaaggc 1140
agcggcagaa gaagatgtag gcagctgagt tgttgtattc tgataagagt cagaggtaac 1200
tcccgttgcg gtgctgttaa cggtggaggg cagtgtagtc tgagcagtac tcgttgctgc 1260
cgcgcgcgcc accagacata atagctgaca gactaacaga ctgttccttt ccatgggtct 1320
tttctgcagt caccgtcctt gacacgaagc ttatcgatat gggaaaaatc agcagtcttc 1380
caacccaatt atttaagtgc tgcttttgtg atttcttgaa ggtgaagatg cacaccatgt 1440
cctcctcgca tctcttctac ctggcgctgt gcctgctcac cttcaccagc tctgccacgg 1500
ctggaccgga gacgctctgc ggggctgagc tggtggatgc tcttcagttc gtgtgtggag 1560
acaggggctt ttatttcaac aagcccacag ggtatggctc cagcagtcgg agggcgcctc 1620
agacaggcat cgtggatgag tgctgcttcc ggagctgtga tctaaggagg ctggagatgt 1680
attgcgcacc cctcaagcct gccaagtcag ctcgctctgt ccgtgcccag cgccacaccg 1740
acatgcccaa gacccagaag gtaagcccac ctgggtggga tccagccatc ctcaagtggt 1800
ctctctcttg tgcatgtggg tgggccaagc agaaatcctg ccccatagtc tcctggctta 1860
caagtcagaa aagctccttt gcaccaaagg gatggattac atccccatct ctttgctaaa 1920
caaacatggg ctttggtgtc agacaaaagt gaagtcctgg ctttctcaca caccagctta 1980
gagagaaaag acttttaggt gaatgtggca ggaaagcgtg cttgctgggg caaaggcaga 2040
ttcattcttt ctcttcccag tatcagcccc catctaccaa caagaacacg aagtctcaga 2100
gaaggaaagg aagtacattt gaagaacgca agtagctttt tctcctttat ttataggaag 2160
tacatttgaa gaacgcaagt agagggagtg caggaaacaa gaactacagg atgtaggtcg 2220
acctcgagtc tagagggccc gtttaaaccc gctgatcagc ctcgactgtg ccttctagtt 2280
gccagccatc tgttgtttgc ccctcccccg tgccttcctt gaccctggaa ggtgccactc 2340
ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca ttgtctgagt aggtgtcatt 2400
ctattctggg gggtggggtg gggcaggaca gcaaggggga ggattgggaa gacaatagca 2460
ggcatgctgg ggagtcgaaa ttcagaagaa ctcgtcaaga aggcgataga aggcgatgcg 2520
ctgcgaatcg ggagcggcga taccgtaaag cacgaggaag cggtcagccc attcgccgcc 2580
aagctcttca gcaatatcac gggtagccaa cgctatgtcc tgatagcggt ccgccacacc 2640
cagccggcca cagtcgatga atccagaaaa gcggccattt tccaccatga tattcggcaa 2700
gcaggcatcg ccatgggtca cgacgagatc ctcgccgtcg ggcatgctcg ccttgagcct 2760
ggcgaacagt tcggctggcg cgagcccctg atgctcttcg tccagatcat cctgatcgac 2820
aagaccggct tccatccgag tacgtgctcg ctcgatgcga tgtttcgctt ggtggtcgaa 2880
tgggcaggta gccggatcaa gcgtatgcag ccgccgcatt gcatcagcca tgatggatac 2940
tttctcggca ggagcaaggt gagatgacag gagatcctgc cccggcactt cgcccaatag 3000
cagccagtcc cttcccgctt cagtgacaac gtcgagcaca gctgcgcaag gaacgcccgt 3060
cgtggccagc cacgatagcc gcgctgcctc gtcttgcagt tcattcaggg caccggacag 3120
gtcggtcttg acaaaaagaa ccgggcgccc ctgcgctgac agccggaaca cggcggcatc 3180
agagcagccg attgtctgtt gtgcccagtc atagccgaat agcctctcca cccaagcggc 3240
cggagaacct gcgtgcaatc catcttgttc aatcatgcga aacgatcctc atcctgtctc 3300
ttgatcagat cttgatcccc tgcgccatca gatccttggc ggcaagaaag ccatccagtt 3360
tactttgcag ggcttcccaa ccttaccaga gggcgcccca gctggcaatt ccggttcgct 3420
tgctgcgtca gaccccgtag aaaagatcaa aggatcttct tgagatcctt tttttctgcg 3480
cgtaatctgc tgcttgcaaa caaaaaaacc accgctacca gcggtggttt gtttgccgga 3540
tcaagagcta ccaactcttt ttccgaaggt aactggcttc agcagagcgc agataccaaa 3600
tactgttctt ctagtgtagc cgtagttagg ccaccacttc aagaactctg tagcaccgcc 3660
tacatacctc gctctgctaa tcctgttacc agtggctgct gccagtggcg ataagtcgtg 3720
tcttaccggg ttggactcaa gacgatagtt accggataag gcgcagcggt cgggctgaac 3780
ggggggttcg tgcacacagc ccagcttgga gcgaacgacc tacaccgaac tgagatacct 3840
acagcgtgag ctatgagaaa gcgccacgct tcccgaaggg agaaaggcgg acaggtatcc 3900
ggtaagcggc agggtcggaa caggagagcg cacgagggag cttccagggg gaaacgcctg 3960
gtatctttat agtcctgtcg ggtttcgcca cctctgactt gagcgtcgat ttttgtgatg 4020
ctcgtcaggg gggcggagcc tatggaaaaa cgccagcaac gcggcctttt tacggttcct 4080
ggccttttgc tggccttttg ctcacatg 4108
<210> 40
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
праймер
<400> 40
tgatctaagg aggctgga 18
<210> 41
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
праймер
<400> 41
ctacttgcgt tcttcaaatg 20
<---
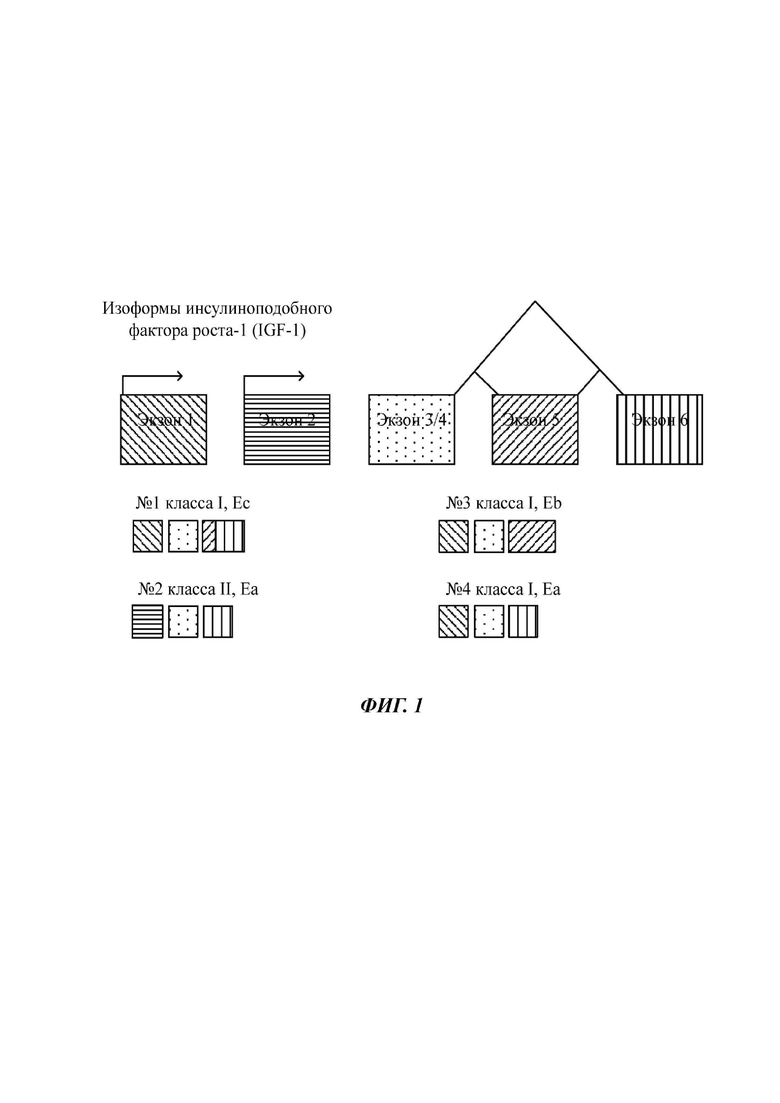
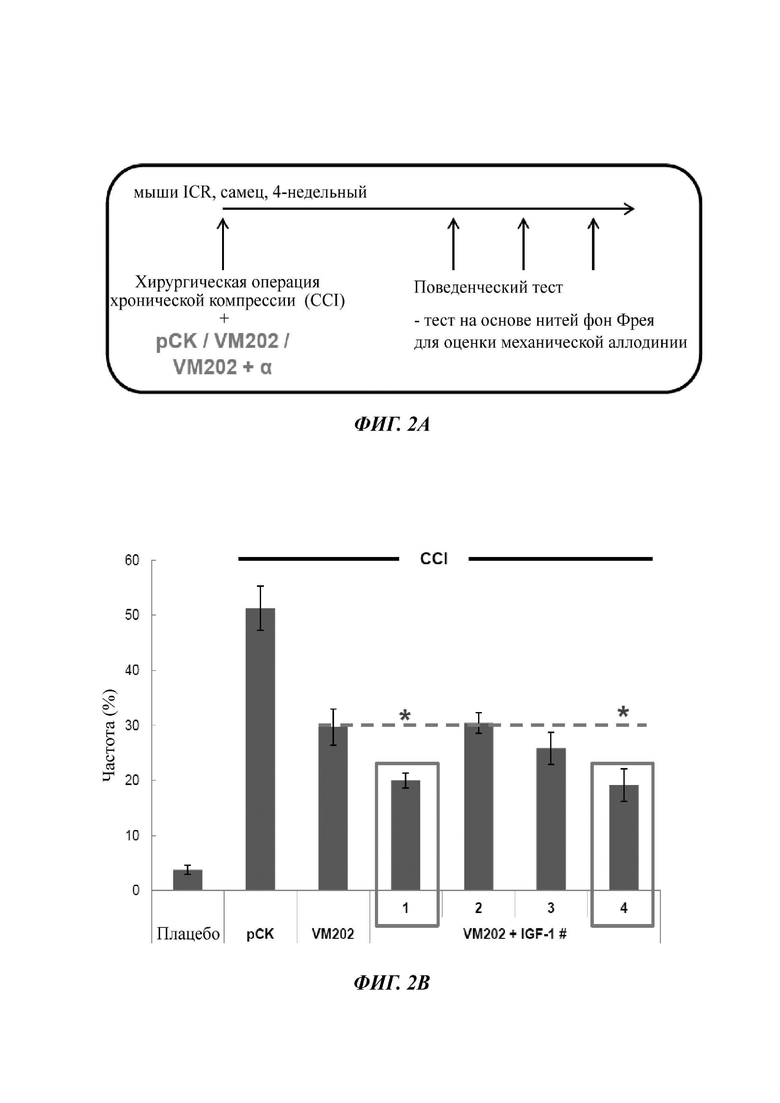
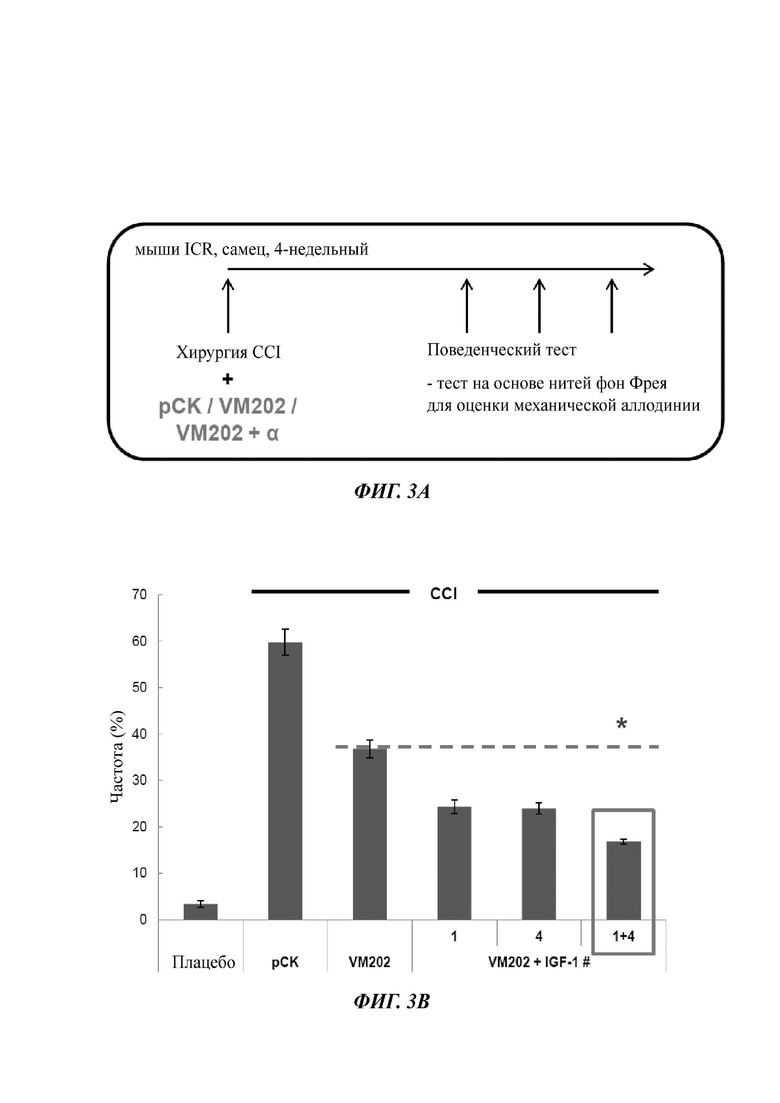





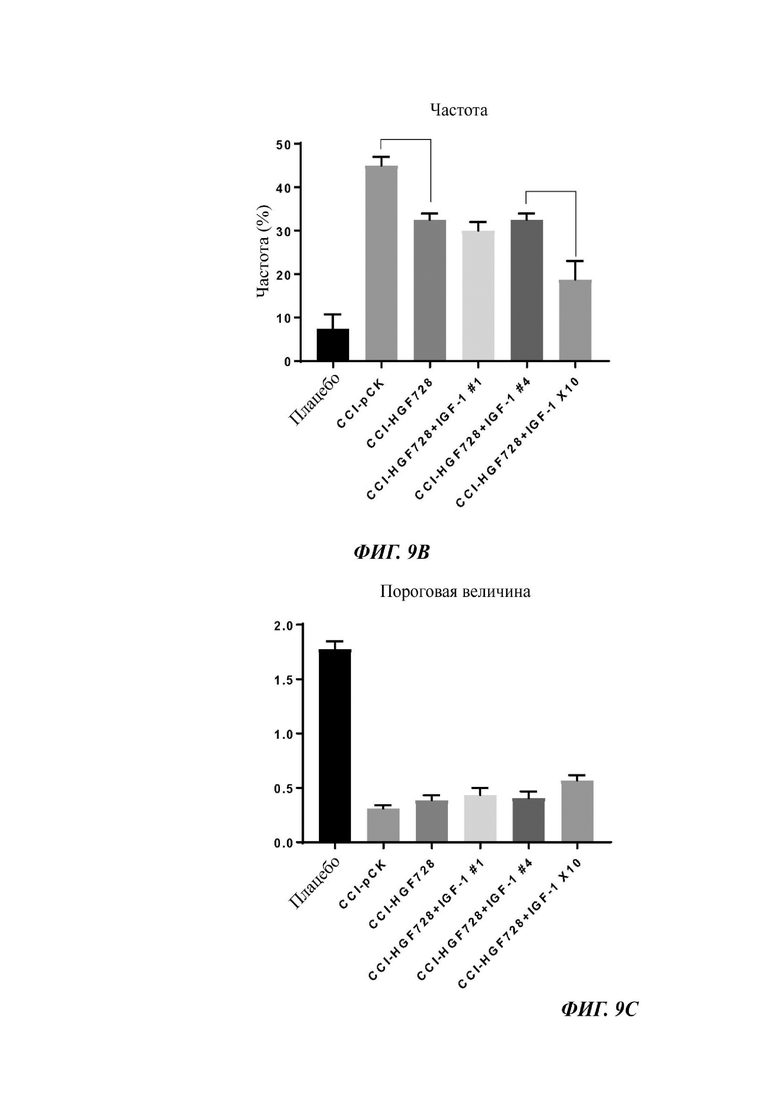
Настоящее изобретение относится к области биотехнологии, конкретно к терапевтическим ДНК-конструкциям, и может быть использовано в медицине для лечения нейропатии. Предложены фармацевтические композиции, включающие экспрессионные ДНК-конструкции, кодирующие человеческую изоформу инсулиноподобного фактора роста-1 (IGF-1) и человеческую изоформу фактора роста гепатоцитов (HGF). Изобретение обеспечивает эффективное лечение нейропатических пациентов. 2 н. и 8 з. п. ф-лы, 18 ил., 4 табл., 5 пр.
1. Фармацевтическая композиция для лечения нейропатии, содержащая эффективное количество:
ДНК-конструкцию, кодирующую инсулиноподобный фактор роста-1 (IGF-1), способную экспрессировать по меньшей мере одну изоформу человеческого IGF-1, выбранную из группы, состоящей из белка IGF-1Ea класса I, содержащего аминокислотную последовательность SEQ ID NO 14, и белка IGF-1Ec класса I, содержащего аминокислотную последовательность SEQ ID NO 16; и
ДНК-конструкцию, кодирующую фактор роста гепатоцитов (HGF), способную экспрессировать по меньшей мере одну человеческую изоформу HGF, выбранную из группы, состоящей из полноразмерного фактора роста гепатоцитов (flHGF), содержащего аминокислотную последовательность SEQ ID NO 11, и человеческого фактора роста гепатоцитов с делециями (dHGF), содержащего аминокислотную последовательность SEQ ID NO 12.
2. Фармацевтическая композиция по п. 1, в которой ДНК-конструкция, кодирующая IGF-1, содержит:
первый полинуклеотид IGF SEQ ID NO 1 (экзоны 1, 3, 4) или его вырожденную последовательность;
второй полинуклеотид IGF SEQ ID NO 2 (интрон 4) или его фрагмент;
третий полинуклеотид IGF SEQ ID NO 3 (экзоны 5 и 6-1) или его вырожденную последовательность;
четвертый полинуклеотид IGF SEQ ID NO 4 (интрон 5) или его фрагмент; и
пятый полинуклеотид IGF SEQ ID NO 5 (экзон 6-2) или его вырожденную последовательность,
где первый полинуклеотид, второй полинуклеотид, третий полинуклеотид, четвертый полинуклеотид и пятый полинуклеотид связываются в последовательном порядке от 5' к 3'.
3. Фармацевтическая композиция по п. 2, в которой второй полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 6 или полинуклеотид SEQ ID NO 7.
4. Фармацевтическая композиция по п. 2, в которой четвертый полинуклеотид IGF представляет собой полинуклеотид SEQ ID NO 8.
5. Фармацевтическая композиция по п. 1, в которой ДНК-конструкция, кодирующая IGF-1, дополнительно содержит плазмидный вектор.
6. Фармацевтическая композиция по п. 5, в которой плазмидный вектор представляет собой рCK, содержащий нуклеотидную последовательность SEQ ID NO 24, или pTx, содержащий нуклеотидную последовательность SEQ ID NO 38.
7. Фармацевтическая композиция по п. 6, в которой ДНК-конструкция, кодирующая IGF-1, представляет собой рCK или pTx, в который клонирован IGF-1X6 или IGF-1X10, где IGF-1X6 представлен SEQ ID NO 9, а IGF-1X10 представлен SEQ ID NO: 10.
8. Фармацевтическая композиция по любому из пп. 1-6, в которой ДНК- конструкция, кодирующая HGF, содержит:
первый полинуклеотид HGF SEQ ID NO 22 (экзоны 1-4) или его вырожденную последовательность;
второй полинуклеотид HGF SEQ ID NO 25 (интрон 4) или его функциональный фрагмент; и
третий полинуклеотид HGF SEQ ID NO 23 (экзоны 5-18) или его вырожденную последовательность,
где второй полинуклеотид HGF расположен между первым полинуклеотидом HGF и третьим полинуклеотидом HGF, и первая ДНК-конструкция, кодирующая HGF, кодирует две человеческие изоформы HGF.
9. Фармацевтическая композиция по любому из пп. 1-7, в которой ДНК- конструкция, кодирующая HGF, содержит полинуклеотид любой из SEQ ID NO: 26-32 и 13.
10. Фармацевтическая композиция для лечения нейропатии, содержащая эффективное количество
a) полинуклеотида SEQ ID NO 13 и i) полинуклеотида SEQ ID NO 10, ii) полинуклеотида SEQ ID NO 15, iii) полинуклеотида SEQ ID NO 17, или iv) полинуклеотида SEQ ID NO 15 и полинуклеотида SEQ ID NO 17; или
b) полинуклеотида SEQ ID NO 33 и i) SEQ ID NO 10 или ii) SEQ ID NO: 39.
| US 2014296142 А1, 02.10.2014 | |||
| US 2006058239 А1, 16.03.2006 | |||
| US 9587240 В2, 07.03.2017 | |||
| US 2016120942 A1, 05.05.2016 | |||
| US 2017333527 A1, 23.11.2017 | |||
| US 5473054 A, 05.12.1995 | |||
| US 8715660 В2, 06.05.2014 | |||
| 0 |
|
SU260957A1 | |
| Сдвоенная папиросно-набивная машина | 1925 |
|
SU3176A1 |

