Область техники
Группа изобретений относится к области к биотехнологии, иммунологии и вирусологии и касается интраназальной вакцины на основе вирусоподобных частиц, состоящая из смеси вирусоподобных частиц и фармацевтически приемлемого буфера, где вирусоподобные частицы состоят из четырёх структурных белков S, M, N, E SARS-CoV-2, где S белок выбран из вариантов 19А, Альфа, Дельта, Омикрон SARS-CoV-2. Также разработан способ профилактики COVID-19, включающий введение интраназально трижды с интервалом между введениями 21 день.
Уровень техники
Отмена Всемирной организацией здравоохранения режима чрезвычайной ситуации в области здравоохранения, введенного из-за пандемии COVID-19 более чем на 3 года, не означает, что коронавирусная инфекция перестала представлять угрозу. Одна из основных проблем - многообразие генетических вариантов SARS-CoV-2. Накопление мутаций, возникающих в результате последующих репликации вируса, является естественным процессом. Известно, что вирус SARS-CoV-2 эволюционирует со скоростью примерно 1,1 × 10 - 3 замены на участок в год. ВОЗ в конце 2020 года для растущего числа вариантов SARS-CoV-2 предложила классифицировать новые штаммы SARS-CoV-2 следующим образом:
• варианты, представляющие интерес (VOIs) - варианты с мутациями, которые приводят к изменениям в связывании с рецепторами, снижению эффективности лечения, снижению нейтрализации антителами и потенциальному увеличению тяжести заболевания и/или трансмиссивности
• варианты, вызывающие озабоченность (VOCs) - варианты, в отношении которых могут быть убедительные доказательства увеличения трансмиссивности, большей тяжести заболевания, заметного снижения нейтрализации вырабатываемыми антителами и, следовательно, снижения реакции на лечение и вакцины.
Кроме того, для универсальной номенклатуры, облегчающей упрощенное отслеживание каждого из новых вариантов SARS-CoV-2, ВОЗ рекомендовала использовать греческий алфавит для однозначной идентификации каждого нового варианта.
Несмотря на многочисленные исследования, эффективного специфического лечения нет, а основа лечения - противовирусные препараты, нацеленные на широкий спектр вирусов, иммуномодуляторы. Поэтому основа профилактики и защиты от COVID-19 является вакцинация.
Различные платформы вакцинации против COVID-19 представлены векторными, мРНКовыми, цельновирусными инактивированными и белковыми вакцинами, каждая из которых имеет свои преимущества и недостатки. В их основе лежит формирование иммунитета на спайковый гликопротеин SARS-CoV-2. В результате постоянных и быстрых мутаций у новых штаммов может происходить изменение эпитопов в S-белке и соответственно их прогнозируемый выход из-под действия вакциноиндуцированных антител.
Прогнозирование влияния новых VOCs на эффективность вакцин против COVID-19 должно лежать в основе подходов к разработке и оптимизации вакцин, которые не позволят эволюционирующему вирусу избегать или подавлять иммунные реакции человека.
Одним из перспективных подходов создания эффективных, а главное безопасных вакцин, является технология создания нереплицирующихся вакцин на основе вирусоподобных частиц (VLP). VLP - это ансамбли вирусных структурных белков, способных к самосборке, имитирующих структуру нативных вирионов, но не несущие в себе вирусного генома. В отличие от субъединичных вакцин VLP-вакцины презентуют эпитоп в конформации, более похожей на ту, которую имеет нативный вирус, и это значительно усиливает иммунный ответ. VLP-вакцины одобрены и находятся на разных стадиях клинических исследований - NVX-CoV2373 (Novavax), Covifenz (Medicago), ABNCoV2 (Radboud University) и LYB001 (Yantai Patronus Biotech Co Ltd). Большинство используемых вакцин против COVID-19, в том числе и на основе VLP, вводятся путем внутримышечной инъекции, и большинство из них содержат в своем составе адъюванты различной природы для повышения иммуногенности, и перечисленные выше вакцины не исключение.
В качестве ближайшего аналога можно рассмотреть документ US10953089B1, опуб. 23.03.2021, в котором раскрывается иммуногенная композиция основанная на наночастицах, образованных пептидами S белка коронавируса (например, гликопротеин BV2373 Spike), фармацевтически приемлемый буфер и сапониновый адъювант (например, MATRIX-M™). Показана иммуногенность полученной вакцины на модели яванского макака с инфекцией SARS-CoV-2. Недостатком решения является то, что данные наночастицы не имитируют вирион SARS-CoV-2, а также то, что в отличии от приведенного аналога, в заявленном техническом решении присутствуют 4 белка коронавируса - М, N, Е и S, поэтому спектр антител, после иммунизации данной вакциной значительно шире.
Таким образом, в уровне техники существует потребность в создании новой удобной в применении вакцины, способной обеспечить защиту от инфекций, вызываемых вирусом SARS-CoV-2.
Раскрытие сущности изобретения
Техническая задача заявленной группы изобретений заключается в разработке вакцины против COVID-19, которая обладает иммуногенностью, достаточной для формирования иммунного ответа против COVID-19.
Технический результат заключается в создании интраназальной вакцины, не содержащей адъюванта, и при этом обладающей иммуногенностью, достаточной для формирования иммунного ответа против COVID-19, способностью индуцировать гуморальный и клеточный иммунный ответ, стимулировать защитный иммунитет слизистых оболочек, а также имеющей дополнительное преимущество простоты введения по сравнению с инъекционными формами. Также технический результат заключается в том, что создана вакцина, которая обладает иммуногенностью против различных штаммов SARS-CoV-2.
Технический результат достигается тем, что создана интраназальная вакцина на частиц для профилактики COVID-19, состоящая из рекомбинантных вирусоподобных частиц, содержащих на поверхности S белок, вариантов19А, Альфа, Дельта, Омикрон SARS-CoV-2, синтезированных в бакуловирусной системе экспрессии - 80-120 мкг; калия дигидрофосфат - 0,63 мг; динатрия гидрофосфат - 0,65 мг; натрия хлорид - 3,84 мг; калия хлорид - 0,09 мг; кальция хлорид - 0,02 мг; трис(гидроксиметил)аминометан-HCl - 0,03 мг; тиомерсал - 4,00 мкг; вода для инъекций - до 0,5 мл.
В частном случае выполнения вакцина не содержит адъюванта.
В частном случае выполнения вакцины количество антигена в вакцине на дозу выбрано из 80, 90, 100, 110, 120 мкг.
Также технический результат достигается тем, что разработан способ профилактики COVID-19, включающий введение разработанной вакцины интраназально трижды с интервалом между введениями 21 день.
Краткое описание чертежей
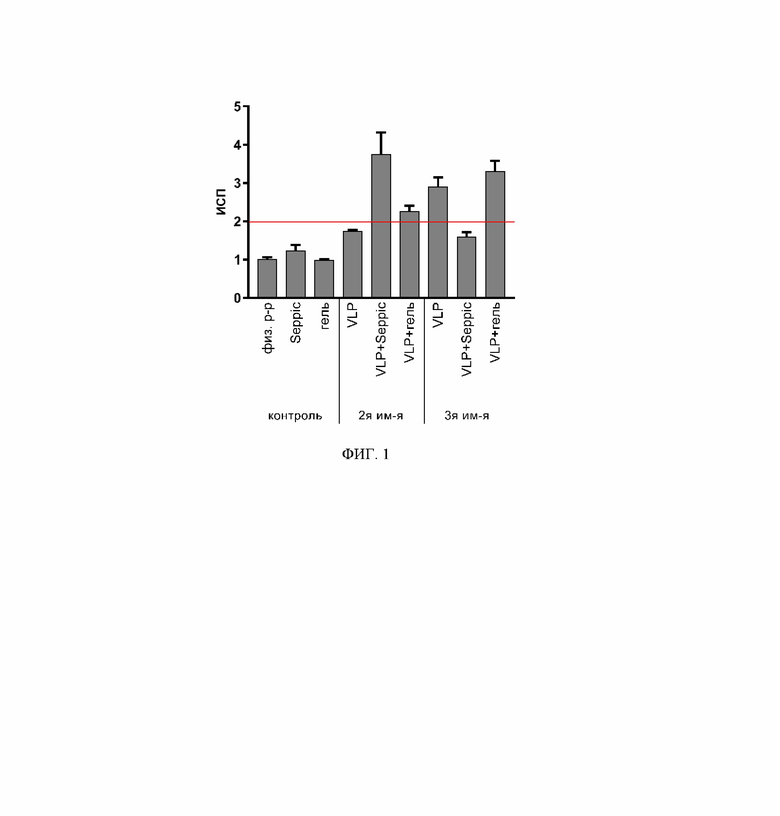
На фиг. 1 представлены результаты реакции бласттрансформации лимфоцитов мышей линии hACE2 Ac70 на 7 сутки после 2-й и 3-й иммунизации. По оси абсцисс указаны группы мышей, которым вводили интраназально: Плацебо (физ. р-р); адъювант SEPPIC с буферным раствором (Seppic); адъювант ГЕЛЬ с буферным раствором (гель); VLP вакцину 80 мкг/доза без адъюванта (VLP); VLP вакцину 80 мкг/доза с адъювантом SEPPIC (VLP+Seppic); VLP вакцину 80 мкг/доза с адъювантом Гель (VLP+гель). По оси ординат приведены средние значения ИСП.
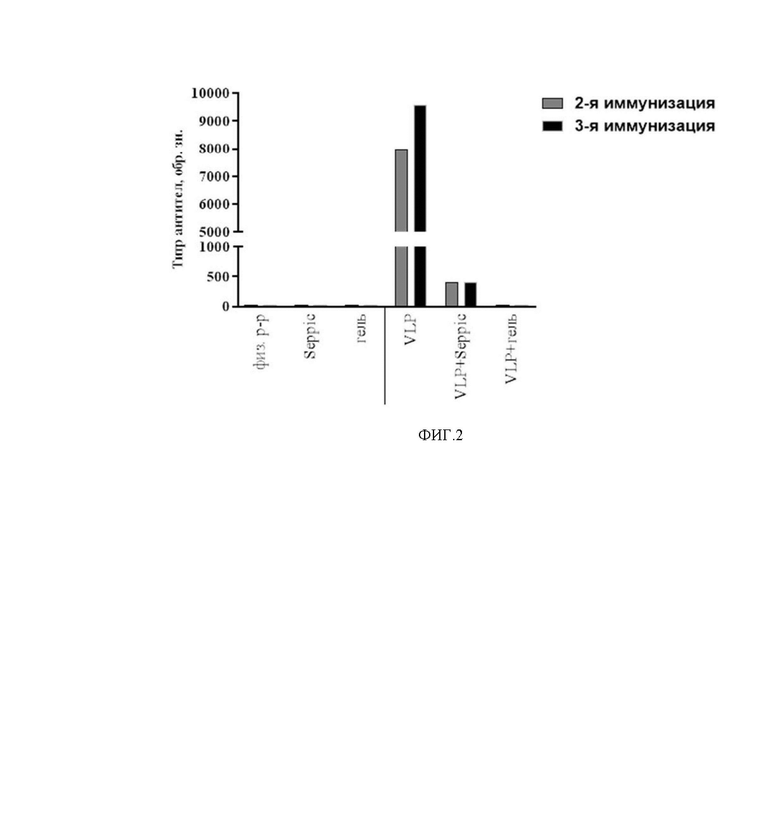
На фиг. 2 представлено определение титра антител к S-белку вируса SARS-CoV-2 у мышей линии hACE2 Ac70 после 2-й и 3-й иммунизации. По оси абсцисс указаны группы мышей, которым вводили интраназально: Плацебо (физ. р-р); адъювант SEPPIC с буферным раствором (Seppic); адъювант ГЕЛЬ с буферным раствором (гель); VLP вакцину 80 мкг/доза без адъюванта (VLP); VLP вакцину 80 мкг/доза с адъювантом SEPPIC (VLP+Seppic); VLP вакцину 80 мкг/доза с адъювантом Гель (VLP+гель). По оси ординат приведены обратные значения титра антител.
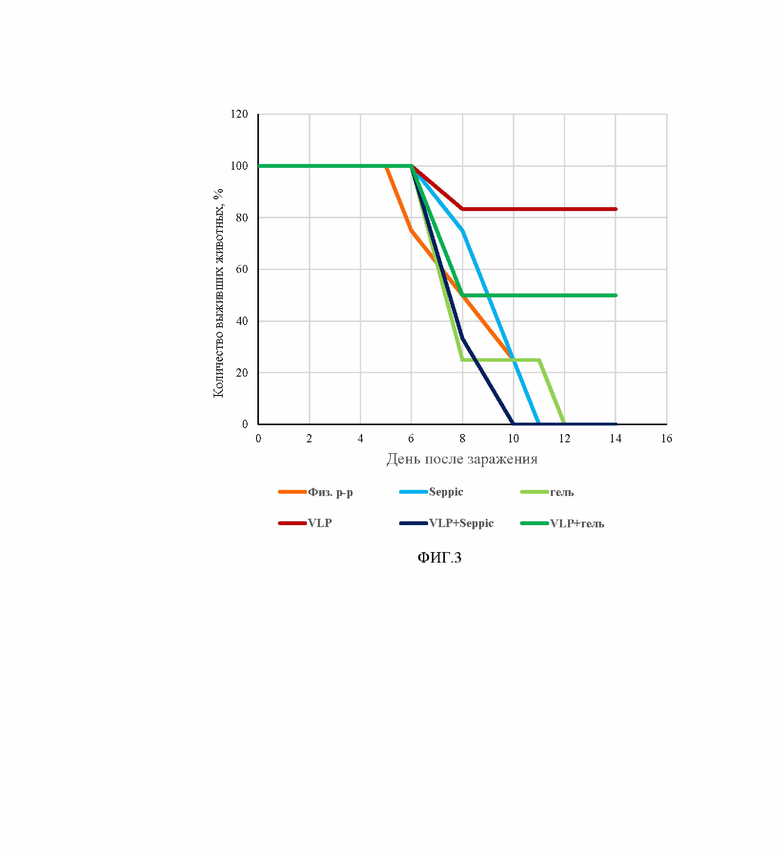
На фиг. 3 представлена выживаемость мышей линии hACE2 AC70 опытных и контрольных групп после инфицирования вирусом SARS-CoV-2 (100 LD50). Группы мышей, которым вводили интраназально: Плацебо (физ. р-р); адъювант SEPPIC с буферным раствором (Seppic); адъювант ГЕЛЬ с буферным раствором (гель); VLP вакцину 80 мкг/доза без адъюванта (VLP); VLP вакцину 80 мкг/доза с адъювантом SEPPIC (VLP+Seppic); VLP вакцину 80 мкг/доза с адъювантом Гель (VLP+гель). По оси абсцисс указаны сутки после заражения. По оси ординат количество выживших животных, выраженное в %.
Осуществление изобретения
Пример 1. Получение VLP с использованием рекомбинантных бакуловирусов и получение состава вакцины.
Активным компонентом разработанной вакцины является рекомбинантные вирусоподобные частицы (VLP), которые состоят из четырёх структурных белков S, M, N, E SARS-CoV-2, где S белок выбран из вариантов 19А, Альфа, Дельта, Омикрон SARS-CoV-2, содержащие нуклеотидные последовательности SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7. Указанные частицы синтезированы в бакуловирусной системе экспрессии, как это указано в примерах в публикации RU 2769223 С1.
pFastBac донорная плазмида содержит экспрессионную кассету, в которой помимо клонированных генов находится фланкирующие последовательности транспозона Tn7. Рекомбинантная трансферная плазмида используется для трансформации клеток DH10Bac E.coli, которые содержат модифицированный бакуловирусный геном в виде большой плазмиды (бакмиды) и вектор-помощник, кодирующий фермент транспозазу. В трансформированных клетках DH10Bac транспозаза осуществляет сайт-специфический перенос экспрессионной кассеты из трансферного вектора в модифицированный бакуловирусный геном. Отбор рекомбинатных клонов DH10Bac осуществляется методом цветного теста, в выбранных колониях белого цвета перенос экспрессионной кассеты в бакуловирусный геном подтверждается методом ПЦР с праймерами, один из которых специфичен к клонированной последовательности, а другой - к геному бакуловируса.
Для дальнейшей работы отбирали клон, из которых выделили бакмиды. Выделенные рекомбинантные бакмиды смешивают с липосомным агентом Cellfectin для трансфекции клеток насекомых Sf-9 или Sf-21. Проникшая в клетку кольцевая молекула бакмидной ДНК инфекционна и запускает жизненный цикл бакуловируса в клетке. Таким образом, получаются рекомбинантные бакуловирусы. Трансфекцию перевиваемой линии клеток Spodoptera frugiperda Sf-21 проводили очищенными препаратами бакмидной ДНК, содержащей оптимизированные гены коронавируса,с использованием катионного липосомного агента Cellfectin (Invitrogen, США), для каждой конструкции использовали по два клона (посевная концентрация клеток 5х105/мл, 10 мкл бакмиды).После трансфекции проводили еще два пассажа на клетках Sf-9 или Sf-21. Таким образом, получили рекомбинантные бакуловирусы.
Концентрацию белка в растворах определяли с использованием коммерческого набора “Micro BCA Protein Assay Kit” (Thermo, США). Анализ структурных белков Sars-CoV-2 проводили методом электрофореза в 12% полиакриламидном геле с додецилсульфатом натрия (ПААГ-ДСН) по методу Laemmli (1970) Электрофорез проводили в пластинах полиакриламидного геля размером 70 × 100 × 0,75 на приборе Mini-PROTEAN II (Bio-Rad, США) в восстанавливающих условиях при по-стоянном напряжении 200 V. Разделяющий гель содержал 12% акриламида, 0,5% N,N-метилен-бис-акриламида, 0,375 М трис-HCl pH 8,8 и 0,1% ДСН. Фокусирующий гель содержал 4% акриламида, 10% N,N-метилен- бисакриламида в 0,125 М трис-HCl буфере pH 6,8. Для полимеризации в оба геля вносили по 0,025% персульфата аммония и 0,075% TEMED. Электродный буфер содержал 0,025 М трис-HCl, 0,192 М глицина, pH 8,3 и 0,1% ДСН. Все испытуемые пробы содержали лизирующий буфер с восстановителем (0,125 М трис-HCl, pH 6,8, 5% ДСН, 0,5% β-меркаптоэтанола, 10,8% глицерина, 0,01% бромфенолового синего) и были прогреты в течение 5 минут при 100 °C. Заливку геля и подготовку аппарата для электрофореза к работе проводили согласно рекомендациямизготовителя. Белки в гелях окрашивали в течение 1 часа 0,1% раствором Кумасси ярко-голубого (CBB R-350) в водном растворе, содержащем 10% уксусной кислоты и 30% метанола. Избыток красителя отмывали 10% раствором уксусной кислотой такое же время с несколькими его сменами. В качестве белков-маркеров молекулярной массы использовали β-галактозидазу- 116 кД; фосфорилазу В - 94 кД, БСА- 66 кД, овальбумин- 45 кД, карбонат ангидразу-30 кД, ингибитор трипсина- 20,1 кД. Для определения антигенной активности структурных белков SARS-CoV-2 использовали сэндвич-ИФАвестерн-блот.
Таким образом, было определено, что были получены структурные белки коронавируса M, E, N и S.
VLP получали методом коинфекции, то есть одновременного заражения перевиваемой линии клеток насекомых T.ni различными сочетаниями рекомбинантных бакуловирусов. Перевиваемую культуру клеток насекомых Trichoplusia ni, культивировали в течение 4 суток после заражения. Был использован 2 пассаж рекомбинантных бакуловирусов.
Культуральную жидкость, содержащую вирус или вирусоподобные частицы, подвергали низкоскоростному центрифугированию, освобождаясь от клеток и клеточного дебриса при 1000 об/мин в течение 5 минут и при 6000 об/мин в течение 20 минут соответственно (+4°С, ротор Sorval® SS34). Распределение SARS-VLP в процессе предварительной очистки представлено на фиг. 5. Фракция 5 использовалась для выделения и очистки SARS-VLP методом ультрацентрифугирования. Полученные осветлённые суспензии наслаивали на 6 мл 25% или 35% (w/v) сахарозы, приготовленной на буфере TNC (10 mM Tris-HCl, 140 mM NaCl, 10 mM CaCl2 рН 7.4). Центрифугировали в течение 2 часов при 28 000 об/мин (центрифуга Optima XE-100, ротор SW 32Ti, Beckman Coulter, +4°С). Полученные осадки ресуспендировали в буфере TNC, и хранили при температуре +4°С.
Таким образом, был получен препарат, содержащий очищенные VLP.
Была получена вакцина следующего состава:
- рекомбинантные вирусоподобные частиц, содержащие на поверхности S белок, вариантов19А, Альфа, Дельта, Омикрон SARS-CoV-2, синтезированных в бакуловирусной системе экспрессии - 80-120 мкг;
калия дигидрофосфат - 0,63 мг;
динатрия гидрофосфат - 0,65 мг;
натрия хлорид - 3,84 мг;
калия хлорид - 0,09 мг;
кальция хлорид - 0,02 мг;
трис(гидроксиметил)аминометан-HCl - 0,03 мг;
тиомерсал - 4,00 мкг;
вода для инъекций - до 0,5 мл.
Активный компонент (вирусоподобные частицы) в буфере TNC (10 mM Tris-HCl, 140 mM NaCl, 10 mM CaCl2 рН 7.4) ресуспендируют в фосфатно-солевом буферном растворе (0,01М КН2РО4, 0,01М Na2HPO4; 0,137М NaCl; 0,0027М KCl, рН 7,4) до необходимой концентрации, и затем перемешивают в течение 30 минут при скорости вращения 250-300 оборотов в минуту.
Вакцина разливается в стерильные флаконы. Хранение производят в защищенном от света месте, при температуре 2-8°С.
Пример 2. Исследование Т-клеточного иммунного ответа
1. Реакция бласттрансформации лимфоцитов
У мышей в стерильных условиях извлекали селезенки и гомогенизировали в 3 мл чистой среды RPMI-1640. Суспензию клеток центрифугировали на одноступенчатом градиенте плотности фиколла, выделяли фракцию мононуклеарных клеток, отмывали дважды в чистой среде RPMI-1640 и помещали в 96-луночные культуральные планшеты с концентрацией 105 клеток в 100 мкл в лунку. Антигены-стимуляторы добавлялись по 100 мкл в лунку к клеткам до конечных концентраций. В качестве положительного контроля служили спленоциты, активированные конковалином А. В качестве отрицательных контролей использовали: клеточные культуры из селезенок неиммунизированных мышей; нестимулированные клеточные культуры из селезенок мышей; культуры, стимулированные неспецифическим антигеном - инактивированного вируса Конго-крымской геморрагической лихорадки. Клетки культивировали в полноростовой среде RPMI-1640, с 20% эмбриональной телячьей сыворотки, 2 мМ глутамина, 4.5 г/л глюкозы, 50 мкг/мл гентамицина, 0.2 ед/мл инсулина при 37°С в атмосфере 5% СО2.
Пролиферацию спленоцитов оценивали в РБТЛ через 4-5 суток с помощью инвертированного микроскопа (увеличение x400). Результаты РБТЛ (фиг.1) выражали в виде индекса стимуляции пролиферации (ИСП) - отношения среднего количества бластов в присутствии стимуляторов к среднему количеству бластов в отсутствие стимуляторов. Положительным считали результат, если ИСП превышает 2.
2. Определение количества клеток, синтезирующих IFN-γ, методом ELISpot
Количественное определение клеток, секретирующих IFN-γ, проводили с помощью набора мышиных IFN-γ ELISpot («BD Biosciences», США). Окрашивание клеток проводили в соответствии с инструкцией производителя. Визуализацию окрашенных пятен «spots» проводили на стереомикроскопе МБС-10 (ЛОМЗ, Россия). Результаты были выражены как разница в количестве пятен (пятнообразующих единиц, SFU) на 106 клеток между лунками, стимулированными специфическим антигеном и средними контрольными лунками без специфических стимуляторов (только питательная среда).
Таблица 1. Формирование IFN-γ-секретирующих лимфоцитов в культуре клеток, выделенных из селезенок мышей, иммунизированных интраназально VLP вакциной на 7 сутки после второй и третьей иммунизации.
Результаты исследования Т-клеточного иммунного ответа показали, что мыши, получившие три дозы вакцины для профилактики COVID-19 на основе VLP, потенциально способны к количественному эффективному клеточно-опосредованному иммунному ответу. Клеточный ответ является защитным, что подтверждается высоким ИСП в реакции РБТЛ при двукратном интраназальном введении вакцины в сочетании с адъювантом Seppic (ИСП 3,75±0,57) и после третьей дозы вакцины без адъюванта (2,9±0,24) и в сочетании с адъювантом гель (ИСП 3,3±0,28) (фиг. 1), коррелирующим с формированием IFN-γ секретирующих клеток (табл. 1).
Пример 3. Исследование гуморального иммунного ответа.
1. Определение антител к S-белку вируса SARS-CoV-2
Уровень антител к S-белку вируса SARS-CoV-2 после иммунизации определяли методом непрямого ИФА. Для этого рекомбинантный белок RBD (иммуногенный фрагмент S-белка вируса SARS-CoV-2) сорбировали в лунках микропланшета в 0,1 М карбонатном буфере, рН 9,5 в концентрации 5 мкг/мл по 100 мкл в лунку в течение ночи при 4°С. После 4-кратной отмывки планшета в лунки вносили по 100 мкл исследуемой сыворотки, разведенной в фосфатно-солевом буфере рН 7.5, содержащим 0.01% Твин 20 и 0.5% желатина (Gerbu). Инкубировали планшет 1 час при +37°С. После промывания планшета вносили по 0.1 мл пероксидазного конъюгата антител к IgG мыши или хомяка. Инкубировали планшет 1 час при +37°С. После промывания планшета вносили по 0.1 мл субстратного раствора с тетраметилбензидином. Инкубировали 15 мин при комнатной температуре в темноте и добавляли 0.1 мл 1 М H2SO4 для остановки реакции. Оптическую плотность при 450 нм (А450) измеряли на спектрофотометре с вертикальным лучом Multiscan FX (Thermo, США).
После 2-ой и 3-ей иммунизации у животных групп, получавших вакцину на основе VLP c адъювантом Seppic и без адъюванта определяли специфические антитела к RBD белка S SARS-CoV-2 в сыворотках крови методом непрямого ИФА. Результаты представлены на фиг. 2. Указаны средние уровни обратной величины титра специфических антител по группам.
2. Реакция нейтрализации (РН)
Для проведения реакции нейтрализации штамм SARS-CoV-2 PMVL-12 (клайд 20В) накопили, пулировали и заморозили при -70°С. Определение lg ТЦИД50 пула вируса проводили методом конечных разведений, для чего вирус титровали на клетках Vero E6 от 10-1 до 10-8. Подсчет lg ТЦИД50 делали по методике Рида и Менча.
Реакцию нейтрализации проводили микрометодом в 96-луночных планшетах (Costar, США) на перевиваемой культуре клеток Vero E6, выращенной на среде Игла с добавлением 5% эмбриональной телячьей сыворотки. Поддерживающая среда содержала 1% сыворотки. При постановке реакции нейтрализации использовали разведение вируссодержащей культуральной жидкости, содержащее 100 ТЦИД50 в 100 мкл.
Из сывороток, готовили разведения от 1:5 до 1:1280 и вносили по 100 мкл в лунки 96-ти луночного планшета. К разведениям сывороток добавляли по 100 мкл вируссодержащей суспензии и инкубировали смесь в течение часа при 37°С. Затем смесь переносили в 96-ти луночный планшет с монослоем клеток Vero E6. Через 72 часа реакцию учитывали, просматривая лунки планшета в микроскоп. Если в сыворотке крови есть нейтрализующие вирус антитела, вирус не будет вызывать ЦПД клеток. Титром сыворотки (последним нейтрализующим разведением) считали разведение, при котором обеспечивается 100% защита клеток (нет ЦПД).
Таблица 2. Результаты РН с сыворотками мышей линии hACE2 Ac70 при интраназальном введении вакцины после после 2-й и 3-й иммунизации.
При исследовании гуморального иммунитета выявлено, что после третьей иммунизации интраназально у мышей линии hACE2 Ac70, наблюдалась выработка специфических антител к RBD домену белка S SARS-CoV-2. При этом, в группе, иммунизированной вакциной 80 мкг/доза с адъювантом Seppic, средний титр специфических антител составлял 1/400, а в группе, иммунизированной вакциной 80 мкг/доза без адъюванта - средний титр специфических антител был существенно выше (в 24 раза) и составлял выше 1/9600. В группе животных, вакциной 80 мкг/доза с адъювантом гель, специфических антител не выявлено.
После заражения мышей hACE2 Ac70 SARS-CoV-2 не зарегистрировано низкоавидных антител, что также может быть косвенной проверкой на проявление потенциального ADE-эффекта.
Показано также, что после второй и третьей иммунизации интраназально, у животных вырабатываются нейтрализующие антитела против вируса SARS-CoV-2, количество вируснейтрализующих антител было максимальным в группе, в которой животных иммунизировали VLP вакциной 80 мкг/доза без адъюванта.
Пример 4. Исследование протективности вакцины Гам-VLP-Мультивак
Протективную эффективность вакцины оценивали путем учета потребления корма и воды животными, количества павших животных опытных и контрольных групп. После заражения потребление корма в контрольных группах, получавших буфер с адъювантами и плацебо было меньше практически в 2 раза, по сравнению с группами, получавшими вакцину с адъювантом и без адъюванта. Снижение потребление корма связано с гибелью животных после инфицирования. Также показано, что в группе животных, получивших вакцину 80 мкг/доза без адъюванта, потребление корма было максимальное, в том числе и за счет минимальной гибели животных.
Потребление воды определяли количественно для клетки содержания еженедельно замеряя начальный объем и спустя 24 часа. Потребление воды животными контрольных групп, получавших буфер с адъювантами и плацебо, сократилось, начиная с 6 дня практически в 2, а затем в 3 раза, тогда как в группах, получавшими вакцину с адъювантом и без адъюванта животными потребление воды сократилось, но гораздо меньше. Потребление воды в группе животных, получивших вакцину 80 мкг/доза без адъюванта было максимальное, в том числе и за счет того, что гибель животных была минимальная.
Учет выживаемости инфицированных животных проводили в течение 14 дней после инфицирования вирусом SARS-CoV-2 дозой 100LD50. Результаты учета представлены на фиг. 3.
Гибель животных отмечали, начиная с 6-го дня после инфицирования. В контрольных группах гибель мышей составила 100%. В группах, получавших вакцину VLP с адъювантом Гель и без адъюванта выживаемость животных составила 50% и 83,4% соответственно.
Средняя продолжительность жизни мышей в группе, получавшей VLP вакцину 80 мкг/доза с адъювантом Гель составила 11 дней. Максимальная продолжительность жизни животных 13 дней наблюдалась в группе, получавшей VLP вакцину 80 мкг/доза без адъюванта. В контрольных группах средняя продолжительность жизни животных была существенно меньше и составила от 8,5 до 9,75 дней.
При интраназальной вакцинации наибольшую протективность показала VLP вакцина 80 мкг/доза без адъюванта. Выживаемость животных составила 83,4%.
Пример 5. Исследование острой и субхронической токсичности вакцины при однократном и многократном введении.
Доклинические исследования острой токсичности проводили на 60 половозрелых аутбредных мышах (30 самцах и 30 самках). Животным вводилась интраназально одна прививочная доза, превышающая дозу для человека более чем в 2000 раз. Введение проводилось дробно с интервалом 1,5 часа в максимально возможном объёме для однократного интраназального введения - 25 мкл. В течение 14 суток за животными велось ежедневное наблюдение, на 15 сутки - эвтаназия с последующей некропсией и морфометрической оценкой внутренних органов. На протяжении всего исследования у животных не было установлено клинической картины интоксикации, гибели не зарегистрировано. Масса животных на протяжении эксперимента значимо не различалась между группами (табл. 3).
Таблица 3. Динамика массы тела мышей, гр. (M±m).
У некоторых животных появилось покраснение и припухлость носового зеркальца (исчезало в течение суток) при многократных инстилляциях, как вакцины, так и носителя. При вскрытии не выявлено отрицательного воздействия вакцины на структуру и массу органов (табл. 4).
Таблица 4. Средние групповые показатели относительной массы внутренних органов мышей, % (M±m).
Доклинические исследования субхронической токсичности проводили на 90 аутбредных половозрелых крысах (45 самцах и 45 самках). Вакцину вводили интраназально трижды с интервалом между введениями 21 день. Такой режим введения приближен к планируемому клиническому применению. Использовали одну прививочную дозу для человека (80 мкг) и удвоенную (160 мкг). Из-за невозможности разового введения таких доз крысам, ведение производилось дробно с интервалом 3 часа. Через сутки после последнего введения вакцины (43-44 день эксперимента) проводили эвтаназию части животных из каждой группы, оставшихся животных подвергали эвтаназии на 58 день эксперимента. При исследовании использовали общепринятые методы: оценка интегральных и физиологических показателей, исследования клинических и биохимических показателей (кровь, моча), морфометрическая и гистологическая оценка внутренних органов и тканей.
При наблюдении за животными в течение всего исследования не было каких-либо отклонений в поведении (тест «Открытое поле») и общем состоянии крыс. Гибели животных на протяжении исследования не зарегистрировано.
Оценка динамики массы тела крыс показало, что интраназальное введение вакцины в исследуемых дозах не оказывает негативного влияния на данный показатель (табл. 5).
Таблица 5. Динамика массы тела крыс, гр. (M±m).
Средние групповые значения клинического анализа крови представлены на табл. 6. Анализ данных гематологических показателей выявил обратимое увеличение количества тромбоцитов (менее чем в 2 раза) у самцов крыс на 43 день эксперимента (через сутки, после последней иммунизации), получавших удвоенную прививочную дозу. Через 14 суток данный показатель в опытных группах не отличался от контрольных групп. Остальные показатели клинического анализа крови находились в пределах физиологической нормы для крыс и не отличались между группами.
Таблица 6. Средние значения показателей клинического анализа крови крыс (M±m).
** статистически значимое различие в сравнении с контролем по t-критерию Стьюдента (р<0,05)
Показатели гемостаза (время рекальцификации, АЧТВ, фибриноген), биохимического анализа крови (АЛТ, АСТ, ЩФ, билирубин общий, холестерин, триглицериды, общий белок, мочевина) и анализа мочи (удельный вес, рН, белок, кетоны) у животных, получавших интраназально вакцину в обеих исследуемых дозах, значимо не отличался от аналогичных показателей у крыс контрольных групп.
У крыс не обнаружены проявления интоксикации и патологических отклонений в структуре внутренних органов. Морфометрический анализ внутренних органов крыс представлен в табл 7.
Таблица 7. Средние групповые показатели относительной массы внутренних органов крыс на 1 и 14 сутки после последней вакцинации (43-44 и 58 сутки эксперимента, соответственно), % (M±m).
яичники
У крыс, получавших двойную прививочную дозу, через 14 дней после последней вакцинации выявлено статистически значимое увеличение (~ в 1,5 раза) по сравнению с контрольными крысами легких у самцов, а селезенок у самок, при этом значения не превышают физиологические нормы для лабораторных крыс.
Визуальная оценка и цитогистоархитектонический анализ не выявил местно-раздражающего действия места введения (носовые ходы) в рамках данного исследования.
Пример 6. Изучение иммунотоксичности
Исследование иммунотоксичности вакцины проводили путем изучения иммунного ответа на стандартный антиген (KLH) при введении крысам SD (Sprague-Dawley) 30 самцах и 30 самках.
Все животные осматривались дважды в день на наличие смертности и умирающих животных. Подробный клинический осмотр с измерением массы тела и регистрацией потребления корма выполняли непосредственно после введения препарата и далее еженедельно в течение исследования. В конце прижизненной фазы животные были подвергнуты эвтаназии путем анестезии (Телазол®/Ксила®) с последующим терминальным взятием крови для определения наличия иммунотоксических эффектов (43-й день исследования).
Анализ сыворотки крови на наличие иммуноглобулинов класса М (IgM) и G (IgG) к препарату KLH проводился методом конкурентного иммуноферментного анализа с помощью планшетного спектрофотометра” Multiscan Go” (Thermo Fisher Scientific Oy, Финляндия) с использованием соответствующих тест-систем («Rat keyhole limpet hemocyanin antibody (KLH-IgM) Elisa kit» и «Rat keyhole limpet hemocyanin antibody (IgG) (KLH-IgG) Elisa kit»).
Принцип метода основан на конкуренции KLH-Ig из образцов и конъюгата KLH-Ig-HRP за сайт связывания антител против KLH-Ig. Поскольку количество сайтов-связывания ограничено, чем больше сайтов занято KLH-Ig из образца, тем меньше сайтов остается для связывания конъюгата KLH-Ig-HRP. При этом происходит изменении интенсивности окраски в зависимости от концентрации антител в образцах или стандартных растворах. Строится стандартная калибровочная кривая, отражающая зависимость интенсивности окраски от концентрации стандартных растворов. Концентрация антител в каждом образце интерполируется по данному калибровочному графику.
На протяжении всего исследования ни в одной экспериментальной группе гибели животных не было. Ни у самцов, ни у самок крыс SD в острой фазе после первой инъекции вакцины не наблюдалось никаких клинических отклонений, связанных с тестируемым объектом. На 1-2 день исследования у самцов и самок животных в группах, получавших тестируемый препарат в двух дозах, было выявлено статистически значимое изменение потребления корма относительно контрольной группы, получавшей носитель. Во всех случаях зарегистрированные отклонения проявлялись в виде снижения потребления корма. Уменьшенное потребление корма не привело к изменениям в показателе прироста массы тела, следовательно, отклонение в потребление корма было краткосрочным и не является критичным. В дальнейший период наблюдения на 6-7 день исследования и далее - отличий в потреблении корма не выявлено. Статистически значимых межгрупповых различий показателей массы тела и прироста массы тела ни у самцов, ни у самок, на протяжении текущего периода наблюдения выявлено не было. Введение тестируемого объекта не изменило показатели иммунного ответа (KLH-IgM) у самок крыс во все дни проведения анализа. В то же время уровень KLH-IgG был достоверно увеличен на 42й день исследования, однако это было временное и преходящее изменение, и на 35й и 49й дни исследования данный показатель не отличался от показателей животных контрольной группы. Введение тестируемого объекта не изменило показатели иммунного ответа (KLH-IgM и KLH-IgG) на введение KLH самцам крыс. Следовательно, иммунотоксического действия не выявлено в исследуемых дозах: предполагаемой клинической (40 мкг) и в 4 раза большей (160 мкг).
Исследования иммунотоксичности также проводили на здоровых половозрелых мышах самцах и самках линий СВА, C57BL/6 и мышах-гибридах первого поколения (СВА × C57BL/6)F1 с помощью определения массы и количества ядросодержащих клеток в органах иммунной системы, оценки клеточного и гуморального иммунного ответа и активности фагоцитов. Было показано, что вакцина стимулирует повышенную выработку антителообразующих клеток (АОК) и содержание антител в сыворотке крови мышей, иммунизированных эритроцитами барана. Введение адъюванта приводит к усилению иммунного ответа на эритроциты барана, происходит значимое увеличение количества АОК и титров антител в реакции гемагглютинации. Вакцина и адъювант также приводят к стимуляции клеточного звена иммунной системы и не индуцируют реакции гиперчувствительности замедленного типа к корпускулярному антигену (ЭБ). Вакцина и адъювант не оказывают значимого влияния на фагоцитарную активность перитонеальных макрофагов. В результате проведенных гистологических исследований установлено, что вакцина и адъювант вызывают функциональные изменения в органах иммунной системы: изменение соотношения зон селезенки: увеличение площади белой пульпы и сокращение площади красной пульпы; увеличение митозов в тимусе и лимфатическом узле по сравнению с контрольной группой.
Таким образом, результаты проведенного исследования показали, что вакцина против COVID-19, вызываемого вирусом SARS-CoV-2, на основе вирусоподобных частиц и адъювант обладают иммунологической активностью, что непосредственно связано с механизмом их действия.
Пример 7. Изучение генотоксичности
Исследование генотоксичности вакцины проводилось в тесте in vivo на хромосомы и/или митотический аппарат в тесте in vivo на образование микроядер в эритробластах крыс SD (30 самцов и 30 самок). Через 24 часа после последнего введения животные подвергались эвтаназии в CO2-камере. Сразу после эвтаназии у крыс забирается костный мозг из бедренной кости аспирацией, и готовится 2 мазка с последующей их фиксацией и окраской по Романовскому и Май-Грюнвальду. В мазках подсчитывается частота встречаемости микроядерных полихромных (незрелых) эритроцитов (анализ 4000 полихромных эритроцитов) и соотношение между полихромными и зрелыми эритроцитами (подсчет 500 эритроцитов). Максимальная вводимая доза не должна обладать выраженной токсичностью на систему гемопоэза: соотношение незрелых эритроцитов к общему количеству эритроцитов не должно отличаться более чем на 20% от соотношения в контрольной группе с растворителем. Статистически значимое увеличение частоты встречаемости микроядер в полихромных эритроцитах, по крайней мере, в одной из получавших препарат групп по сравнению с контрольной считается основанием предполагать у тестируемого препарата наличие генотоксического действия. Кроме того, у животных в прижизненной фазе исследования контролируется проявление токсических эффектов вводимого препарата: масса тела, потребление корма, внешние клинические признаки. В конце исследования после эвтаназии и взятия костного мозга выполняляется некропсия тел с осмотром и взвешиванием органов.
Таблица 8. Суммарные данные встречаемости микроядер в эритроцитах костного мозга
Исследуемые дозы не оказывали общетоксического действия и токсического действия на гемопоэз. Результаты исследования свидетельствуют об отсутствии способности тестируемого объекта оказывать повреждающее действие на хромосомы или митотический аппарат эритробластов у используемой животной модели, при введении в предполагаемой клинической дозе (40 мкг) и при введении в дозах, превышающих предполагаемую клиническую в 4 и 8 раз (160 мкг и 320 мкг, соответственно).
Исследование генотоксичности вакцины также проводилось в тесте Эймса на аутбредных крысах SD (30 самцах и 30 самках). Тестируемый образец рассматривают как обладающим мутагенной активностью, если он вызывает воспроизводимое, зависящее от дозы и статистически достоверное увеличение частоты мутаций по сравнению с соответствующими отрицательными контролями у одного или более штаммов в условиях метаболической активации или без нее, по меньшей мере, в двух независимых экспериментах. При этом число ревертантных колоний на чашку в опыте должно в 2 или более раз превышать число спонтанных ревертантных колоний в отрицательном контроле в случае штаммов ТА97, ТА98, ТА100, ТА102 и в 3 или более раз в случае ТА1535. Положительные результаты в тесте оценки обратных мутации на бактериях свидетельствуют о том, что вещество индуцирует точковые мутации типа замены пар оснований или сдвига рамки считывания в геноме Salmonella typhimurium.
Неопределенный результат отмечается тогда, когда при одной или более дозах, по меньшей мере, на одном штамме наблюдается повышенное количество ревертантных колоний на чашку в опыте, по сравнению с отрицательным контролем, но отсутствует отчетливая зависимость доза-эффект. Испытуемое вещество, результаты исследования которого не соответствуют вышеупомянутым критериям, рассматривается как не мутагенное в данном тесте. Статистический анализ проводили, используя критерий Даннетта t и ранговый метод Спирмена (при α=0,05).
Тест Даннетта t показал отсутствие значимого повышения частоты обратных генных мутаций при действии вакцины и адъюванта по сравнению с отрицательным контролем при α = 0,05 в условиях эксперимента. Методом Спирмена не выявлено значимой монотонной зависимости повышения количества ревертантов от концентрации ни для одного из штаммов при α = 0,05.
Полученные результаты свидетельствуют о том, что число ревертантных колоний на чашках при действии вакцины и адъюванта, сопоставимо с числом спонтанных ревертантных колоний на чашках в отрицательных контрольных вариантах, как в условиях метаболической активации, так и в ее отсутствии. Таким образом, вакцина против COVID-19, вызываемого вирусом SARS-CoV-2, на основе вирусоподобных частиц и адъювант не проявляли способности к индукции точковых мутаций у исследуемых штаммов Salmonella typhimurium в концентрациях 0,05-5,0 мкл/чашка, что свидетельствуют об отсутствии способности тестируемых образцов вызывать генные мутации в условиях проведенных экспериментов.
Таким образом, разработанная вакцина обладает высоким уровнем безопасности, вызывает гуморальный и Т-клеточный иммунитет и может быть использована для вакцинации и ревакцинации.
| название | год | авторы | номер документа |
|---|---|---|---|
| Вакцина на основе вирусоподобных частиц (VLP) для профилактики COVID-19 для парентерального применения | 2024 |
|
RU2834060C1 |
| Вирусоподобные химерные частицы для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2, содержащие белки коронавируса и ротавируса | 2022 |
|
RU2779810C1 |
| Рекомбинантные вирусоподобные частицы для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2 | 2021 |
|
RU2769224C1 |
| Иммунобиологическое средство на основе вирусоподобных частиц для индукции специфического иммунитета против инфекции, вызываемой ротавирусом А человека | 2022 |
|
RU2795055C1 |
| Вакцинная композиция против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743595C1 |
| Пептидные иммуногены и вакцинная композиция против коронавирусной инфекции COVID-19 с использованием пептидных иммуногенов | 2020 |
|
RU2743593C1 |
| Пептидные иммуногены, используемые в качестве компонентов вакцинной композиции против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743594C1 |
| Пептидные иммуногены и вакцинная композиция против коронавирусной инфекции COVID-19 с использованием пептидных иммуногенов | 2020 |
|
RU2738081C1 |
| Вакцина против ротавирусной инфекции человека на основе вирусоподобных частиц ротавируса | 2024 |
|
RU2829862C1 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ КОРОНАВИРУСА БЛИЖНЕВОСТОЧНОГО РЕСПИРАТОРНОГО СИНДРОМА (MERS-COV) И СПОСОБЫ | 2014 |
|
RU2685185C2 |
Группа изобретений относится к области к биотехнологии, иммунологии и вирусологии. Описана интраназальная вакцина на основе вирусоподобных частиц, состоящая из смеси вирусоподобных частиц и фармацевтически приемлемого буфера. Вирусоподобные частицы состоят из четырёх структурных белков S, M, N, E SARS-CoV-2, где S белок выбран из вариантов 19А, Альфа, Дельта, Омикрон SARS-CoV-2. Также разработан способ профилактики COVID-19, включающий введение вакцины интраназально трижды с интервалом между введениями 21 день. Группа изобретений обеспечивает создание интраназальной вакцины, обладающей иммуногенностью, достаточной для формирования иммунного ответа против COVID-19, способностью индуцировать гуморальный и клеточный иммунный ответ, стимулировать защитный иммунитет слизистых оболочек, а также имеющей дополнительное преимущество простоты введения по сравнению с инъекционными формами. 2 н. и 2 з.п. ф-лы, 3 ил., 8 табл., 8 пр.
1. Интраназальная вакцина на основе вирусоподобных частиц для профилактики COVID-19, состоящая из:
- рекомбинантных вирусоподобных частиц, содержащих на поверхности S белок, вариантов 19А, Альфа, Дельта, Омикрон SARS-CoV-2, синтезированных в бакуловирусной системе экспрессии – 80-120 мкг;
- калия дигидрофосфата – 0,63 мг;
- динатрия гидрофосфата – 0,65 мг;
- натрия хлорида – 3,84 мг;
- калия хлорида – 0,09 мг;
- кальция хлорида – 0,02 мг;
- трис(гидроксиметил)аминометана-HCl – 0,03 мг;
- тиомерсала – 4,00 мкг;
- воды для инъекций – до 0,5 мл.
2. Интраназальная вакцина по п. 1, где вакцина не содержит адъюванта.
3. Интраназальная вакцина по любому из пп. 1-2, где количество антигена в вакцине на дозу выбрано из 80, 90, 100, 110, 120 мкг.
4. Способ профилактики COVID-19, включающий введение вакцины по любому из пп. 1-3 интраназально трижды с интервалом между введениями 21 день.
| WO 2022046633 A1, 03.03.2022 | |||
| Вакцина против гриппа типа А, гриппа типа B и COVID-19 | 2021 |
|
RU2751485C1 |
| WO 2021248145 A2, 09.12.2021 | |||
| WO 2021229311 A1, 18.11.2021 | |||
| Ю.А.БЕЛИКОВА, Ю.В.САМСОНОВ, Е.В.АБАКУШИНА, СОВРЕМЕННЫЕ ВАКЦИНЫ И КОРОНАВИРУСНЫЕ ИНФЕКЦИИ, Исследования и практика в медицине, 2020, т.7, N4, с | |||
| Способ обделки поверхностей приборов отопления с целью увеличения теплоотдачи | 1919 |
|
SU135A1 |
| Е.В | |||
| ЗУЕВ, О.Л | |||
| ЕВДОКИМОВА и др., Сравнительная оценка безопасности | |||

