Настоящее изобретение относится к способам лечения комплемент-опосредуемых расстройств, а в частности, расстройств, ассоциированных с гиперактивностью цикла обратной связи C3b комплемента (например, возрастной дегенерации желтого пятна (ВДЖП)), методом генотерапии для повышения уровней фактора I комплемента (или его фрагментов или производных); к рекомбинантным вирусным векторам, кодирующим фактор I (или его фрагменты или производные); к рекомбинантным вирусным частицам, икапсидирующим эти векторы, и к их применению в способах лечения.
Система комплемента образует защиту первого ряда против инфекций посредством запуска воспалительных ответов, подающих сигнал иммунной системе об опасности. Эта система играет важную роль в мечении микробов и инфицированных или поврежденных клеток для стимуляции их уничтожения посредством лизиса или фагоцитарного клиренса. Система комплемента может активироваться по трем различным механизмам: по классическому пути, по лектиновому пути и по альтернативному пути (см. фигуру 1). Все три пути сводятся к расщеплению белка С3, присутствующего в плазме в избыточном количестве, что приводит к активации конечных стадий комплемента, а также к запуску других эффекторных механизмов комплемента.
В настоящее время хорошо известно, что альтернативный путь активируется либо одним из двух путей комплемента, способствующих амплификации положительной обратной связи, либо iC3 (или C3(H2O)), который приводит к спонтанному гидролизу внутренней тиоэфирной связи в C3, либо C3b, который постоянно продуцируется в сыворотке в очень низких концентрациях. Этот «механизм холостого хода» обеспечивает нужное минимальное количество активированного C3, который может связываться с фактором B, который, в свою очередь, будет расщепляться фактором D с образованием исходного C3-конвертазного комплекса iC3Bb, который может превращать нативный C3 в C3b. C3b подвергается структурной реаранжировке, что обеспечивает связывание фактора B, то есть, предшественника сериновой протеазы, с C3b. Фактор D может затем связываться с комплексом и расщеплять фактор B с образованием Bb и Ba. C3bBb представляет собой C3-конвертазу альтернативного пути. Фактор D не расщепляет фактор B в сыворотке, если только фактор B не связывается с C3b или iC3. Только что образовавшийся C3b может ковалентно связываться с поверхностью патогенов (или с любой другой близко расположенной поверхностью), где он может связываться с другой молекулой фактора B. Такое связывание стабилизируется пропердином, который способствует увеличению времени полужизни C3-конвертазы альтернативного пути посредством ингибирования функциональной активности фактора H.
Альтернативный путь регулируется балансом между двумя конкурирующими циклами, которые действуют на C3b. Такими циклами являются циклы обратной связи C3b и циклы ингибирования C3b, которые лежат в основе активации комплемента (см. фигуру 2). После образования iC3 или C3b и их отложения на поверхности, две различные реакции могут происходить, главным образом, независимо от механизма продуцирования C3b: он может быть связан либо с фактором B (посредством обратной связи), либо с фактором H (или с кофакторами CD46 или CD35 FI, связанными с мембраной) (посредством расщепления).
Связывание C3b (или C3(H2O)) приводит либо к амплификации C3-конвертазы (в присутствии фактора B, фактора D и пропердина) и инициации сборки комплекса, атакующего мембрану (MAC), либо к инактивации C3b (в присутствии фактора H и фактора I). Наличие амплификации или инактивации зависит только от природы поверхности, с которой связывается C3b. На так называемых «защищенных поверхностях» наблюдается снижение уровня связывания с фактором H (и следовательно, также расщепления C3b фактором I). Так, например, уровень связывания фактора H с C3b на липополисахариде (ЛПС) Escherichia coli 04 гораздо ниже, чем уровень связывания фактора B.
Ингибирование C3 во время активации комплемента показано на фигуре 3. C3 (185 кДа) состоит из двух цепей, а именно, α- и β-цепей, а в C3b, N-концевой пептид C3a (9 кДа) расщепляется C3-конвертазой. Расщепление C3b достигается посредством разрезания, опосредуемого двумя факторами I в α'-цепи C3b. Небольшой фрагмент, обозначаемый С3f (3 кДа), высвобождается, что приводит к расщеплению α'-цепи на фрагменты 68 и 43 кДа. Такой подвергнутый протеолизу C3 обозначается здесь iC3b, и он больше не участвует в пути расщепления C3- или C5-конвертазой. Однако, он является важдым медиатором воспаления, индуцируемого комплементом, что обусловлено его способностью реагировать с рецептором CR3 комплемента (CD11b,CD18) на нейтрофилах. Такое первое расщепление с образованием iC3b происходит очень быстро, и в нем участвует фактор H в качестве кофактора, а второе расщепление iC3b происходит гораздо медленнее. iC3b может снова расщепляться фактором I только в присутствии рецептора кофактора 1 комплемента (CR1). Фактор I снова расщепляет iC3b во фрагменте 68 кДа, который высвобождает большую часть молекулы, называемой C3c. C3dg остается связанным с поверхностью посредством ковалентной связи (либо сложноэфирной связи с сахаром, либо амидной связи с белком). C3dg не реагирует с CR3 и не является медиатором воспаления.
Генетические изменения в белках комплемента, которые влияют на их функцию, ассоциируются с резистентностью к инфекционным заболеваниям и восприимчивостью к различными воспалительным заболеваниям. Хорошо охарактеризованный полиморфизм ассоциируется с возрастной дегенерацией желтого пятна (ВДЖП), с болезнью плотных отложений (БПО) и с атипическим гемолитическим уремическим синдромом (АГУС).
ВДЖП представляет собой наиболее распространенное заболевание, вызываемое полиморфизмом в альтернативном пути. Такое заболевание ассоциируется с несколькими общеизвестными модификациями в факторе H и FH-родственных белках: в FH гаплотипа 1, который включает fHY402H SNP (Haines, et al., Science, 308(5720):419-421, April 2005; Edwards, et al., Science, 308(5720):421-424, April 2005; Klein, et al., Science, 308(5720):385-389, April 2005); FH гаплотипа 2, который включает fHV62I SNP (Hageman et al., Proc. Natl. Acad. Sci. U.S.A., 102(20):7227-7232, May 2005); и FH гаплотипа 3, который включает делецию CFHR1/CFHR3 и является протективным (Hughes et al., Nat. Genet., 38(10):1173-1177, October 2006). Проведенные исследования показали, что наследование двух копий FH гаплотипа 1 в семь раз повышает риск развития ВДЖП. Другие ВДЖП-ассоциированные генетичесике модификации присутствуют в факторе B и в C3 (C3 R102G (Heurich, et al., Proc. Natl. Acad. Sci. U.S.A., 108(21):8761-8766, May 2011)). Совсем недавно было обнаружено, что редко встречающиеся мутации фактора I, которые могут снижать уровни фактора I в сыворотке, ассоциируются со значительным увеличением риска развития прогрессирующей ВДЖП (Kavanagh, et al., Hum. Mol. Genet., 24(13):3861-3870, July 2015). Вышеупомянутый полиморфизм представлен ниже в таблице 1.

Болезнь плотных отложений, БПО, клинически ассоциирована с ВДЖП, но у пациентов с БПО также наблюдается изменение сетчатки, как и в случае ВДЖП, но в гораздо более раннем возрасте (если они доживают до этого возраста), чем у пациентов, страдающих «только ВДЖП». Фактически, полиморфизм в гаплотипах H1 и H2 и полиморфизм в C3 также способствуют повышению или снижению риска развития БПО, соответственно, как это наблюдалось в случае ВДЖП (Pickering, et al., J. Exp. Med., 204(6):1249-1256, June 2007). Комбинации нескольких гаплотипов риска значительно повышают риск развития БПО (Abrera-Abeleda, et al., J. Am. Soc. Nephrol., 22(8):1551-1559, August 2011). Исследования на мышах показали, что развитие БПО полностью зависит от способности генерировать iC3b (Rose, et al., J. Clin. Invest., 118(2):608-618, February 2008). Для развития заболевания абсолютно необходимо присутствие фактора I. Полное отсутствие фактора I устраняет риск развития БПО и ВДЖП благодаря отсутствию медиатора воспаления iC3b.
ВДЖП и БПО ассоциируются с изменениями регуляции альтернативного пути жидкой фазы, а поэтому они рассматриваются как системные заболевания, а не только глазные или почечные заболевания. Причиной конкретных поражений этих двух органов, вероятно, является равномерное распределение белков-регуляторов комплемента и строение этих органов. В обоих органах наблюдается разделение эритроцитов и плазмы и базальной мембраны, которая непосредственно контактирует с плазмой, что приводит к защите от атаки комплемента, где такая атака в гораздо большей степени зависит от активности кофактора фактора H и фактора I. Большинство полиморфизмов, которые могут вызывать предрасположенность к ВДЖП и БПО, наблюдается в областях фактора H, ответственных за связывание с C3.
Атипический гемолитический уремический синдром (АГУС) также ассоциируется с мутациями в генах альтернативного пути, а в частности, в гене фактора H (в половине всех случаев АГУС), но, в отличие от ВДЖП и БПО, эти мутации обычно присутствуют у С-конца фактора H, то есть, области, ответственной за секвестрирование фактора H на поверхностях (Manuelian, et al., J. Clin. Invest., 111(8):1181-1190, April 2003). Другой полиморфизм, вызывающий предрасположенность к заболеванию, был обнаружен в гене C3 или CFB (Goicoechea de Jorge, et al., Proc. Natl. Acad. Sci. U.S.A., 104(1):240-245, January 2007;  et al., Blood, 112(13):4948-4952, December 2008). В итоге, риск АГУС значительно увеличивается в присутствии ассоциированного с риском полиморфизма, который стимулирует несоответствующую активацию альтернативного пути на клеточных поверхностях (
et al., Blood, 112(13):4948-4952, December 2008). В итоге, риск АГУС значительно увеличивается в присутствии ассоциированного с риском полиморфизма, который стимулирует несоответствующую активацию альтернативного пути на клеточных поверхностях ( et al., Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease, 1812(1):12-22, January 2011).
et al., Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease, 1812(1):12-22, January 2011).
Исследования по ассоциации геномов широкого ряда выявили дополнительный полиморфизм, который, например, присутствует в гене CR1, кластерине (оба эти полиморфизма вызывают предрасположенность к болезни Альцгеймера (БА) у пациентов, у которых уже присутствует одна или две копии аллеля APOE-ε4, вызывающего предрасположенность к БА) (Harold, et al., Nat. Genet., 41(10):1088-1093, October 2009; Lambert, et al., Nat. Genet., 41(10):1094-1099, October 2009), и в C5 (ассоциированном с ревматоидным артритом) (Kurreeman, et al., PLoS Med., 4(9):e278, September 2007). Также следует отметить, что амплификация комплемента и ее эффекты в цикле обратной связи наблюдаются во всех путях комплемента, а поэтому их модификация может также приводить к ослаблению других патофизиологических процессов.
Большинство вышеупомянутых полиморфизмов имеют общие свойства, заключающиеся в том, что ассоциированный с заболеванием аллель стимулирует повышение активности обратной связи в альтернативном пути, что приводит к амплификации всех путей комплемента независимо от инициирующих раздражителей (Lachmann, Adv. Immunol., 104:115-149, 2009). Таким образом, гипервоспалительный комплотип (определенный как представляющий паттерн генетических вариантов в генах комплемента, наследуемых у пациента, у которого наблюдается изменение степени риска развития воспалительных расстройств и инфекционных заболеваний, вызываемых комплементом) защищает от инфекций, а в частности, у детей в раннем возрасте, но истощается в более позднем возрасте, когда у пациента уже вырабатываются антитела IgG против наиболее распространенных инвазивных патогенов, что вызывает предрасположенность к развитию воспалительных заболеваний. Поскольку список заболеваний, вызываемых комплотипом, постоянно растет по мере проведения генетических исследований известных или новых полиморфизмов комплемента (Harris, et al., Trends Immunol., 33(10):513-521, October 2012), то в настоящее время особый интерес представляет разарботка новых способов устранения этой гиперактивности цикла обратной связи C3b.
Баланс петли обратной связи, который определяет количество продуцированного C3b, может быть достигнут с обоих концов. Большее количество C3b может быть продуцировано либо путем амплификации петли обратной связи, либо путем ингибирования механизма расщепления C3b. Петля обратной связи может быть амплифицирована путем увеличения количества исходного C3b (например, под действием большего числа C3-конвертаз классических/лектиновых путей), путем ускорения реакций обратной связи (например, увеличения количества функциональных мутантов фактора B или увеличения количества фактора D или ионов магния), путем добавления фактора яда кобры (который обладает C3b-подобными свойствами и может связываться с фактором B и образовывать C3-конвертазу после расщепления фактора B под действием фактора D) или путем стабилизации C3-конвертазы альтернативного пути (например, под действием пропердина или нефритных факторов). Расщепление C3b может быть предотвращено в отсутствии или при низких концентрациях фактора H и фактора I или мембранного белка-кофактора (MCP) или при восприимчивом C3 определенного генотипа (например, в C3F, SNP: R102G имеет низкую аффинность по отношению к фактору H, чем C3S, а поэтому он более медленно расщепляется фактором I). Расщепление C3 также влияет на защищенные поверхности. Негативная регуляция цикла обратной связи C3b достигается посредством стимуляции расщепления C3b. Это может быть осуществлено либо путем повышения концентрации фактора H или фактора I, либо путем ингибирования фактора D, фактора B или пропердина. Конкретными мишенями для негативной регуляции цикла обратной связи C3b являются фактор D, фактор H и фактор I.
Способ лечения региональной атрофии, то есть, ВДЖП, на более поздней стадии, уже был разработан и в настоящее время применяется в клиническом испытании фазы III (в исследовании MAHALO Genentech/Roche). В настоящее время, для прекращения прогрессирования региональной атрофии используется гуманизованное моноклональное ингибирующее антитело против фактора D (лампализумаб), вводимое путем инъекции в стекловидное тело. Фактор D присутствует в сыворотке в очень низких концентрациях и является важным фактором для альтернативного пути. Тем не менее фактор D из-за его небольшого размера (27 кДа), быстро выводится из почек и быстро подвергается повторному синтезу. Таким образом, лечение антителом против фактора D применяется для локального ингибирования фактора D в небольшом компартменте, таком как сетчатка, но не является системным. Способ введения, а именно, инъекция в стекловидное тело все же ассоциируется с некоторым риском, что также является недостатком такого способа лечения.
Что касается других двух белков-мишеней, то уже в 70-е годы было известно, что цикл обратной связи C3b может ингибироваться путем повышения концентрации фактора H и I в плазме (Nydegger, et al., J Immunol, 120(4):1404-1408, January 1978; Lachmann and Halbwachs, Clin. Exp. Immunol., 21(1):109-114, July 1975). В настоящее время появляется все больше данных о геномах, указывающих на связь комплемента и/или нарушения альтернативного пути с рядом заболеваний (например, Harris et al., 2012 (supra); Haines, et al., 2005 (supra); Hageman, et al., 2005 (supra); Kavanagh, et al., Hum. Mol. Genet., 24(13):3861-3870, July 2015), что подтверждает возможность применения фактора I или фактора H для лечения расстройства, ассоциированного с нерегулируемыми путями. Тем не менее, фактор I является лучшей мишенью по нескольким причинам. Прежде всего, он присутствует в гораздо меньших концентрациях, чем фактор H (≈35 мкг/мл (FI) и ≈200-500 мкг/мл (FH), соответственно). Меньшие количества легче поддаются обработке, что также снижает стоимость лечения. Кроме того, имеются варианты фактора H, называемые белками, родственными фактору H 1-5 (FHR1-5), которые конкурируют с фактором H за связывание с поверхностями. FHR1, 2 и 5 содержат общий мотив димеризации, что способствует образованию трех гомодимеров и трех гетеродимеров, которые значительно повышают авидность и предотвращают конкуренцию с фактором Н в физиологически релевантных концентрациях. Увеличение концентрации фактора H, вероятно, не предотвращает доминантные патогенные эффекты этих конкурирующих антагонистов. Последней и самой главной причиной является то, что фактор I представляет собой лишь регулятор, который не только стимулирует расщепление C3b с образованием iC3b, но также и ускоряет расщепление iC3b. В физиологических условиях, аффинность фактора H по отношению к iC3b является слишком низкой для образования комплекса. Поскольку реакция iC3b с рецептором CR3 комплемента является основным механизмом, посредством которого активация комплемента способствует развитию воспаления, то расщепление iC3b с образованием C3dg играет важную роль в снижении степени воспаления, индуцированного комплементом (Lachmann, Adv. Immunol., 104:115-149, 2009).
Исследования ВДЖП затруднены из-за отсутствия оптимальных животных, которые могли бы стать моделью человеческого заболевания, а в частности, мышей из-за отсутствия у них желтого пятна. Однако, авторам настоящего изобретения известно, что ВДЖП представляет собой системное, а не только глазное заболевание, на что указывает все увеличивающийся список ВДЖП-ассоциированных полиморфизмов в белках и регуляторах комплемента. Авторы настоящего изобретения считают, что ключ к лечению ВДЖП может быть найден в регуляции системного альтернативного пути. Негативная регуляция цикла обратной связи C3b альтернативного пути может быть достигнута путем повышения уровней фактора I в плазме. В WO 2010/103291 описаны методы лечения ВДЖП путем введения очищенного от плазмы или рекомбинантного фактора I.
Влияние введения фактора I (в присутствии зимозана как активатора альтернативного пути) на образование iC3b и его последующего расщепления до C3dg было оценено для трех различных комплотипов (Lay, et al., Clin. Exp. Immunol., 181(2):314-322, August 2015; Lachmann, et al., Clin. Exp. Immunol., September 2015). Выбранные комплотипы были подразделены на восприимчивые или протективные гомозиготные и гетерозиготные комплотипы по трем общим полиморфизмам, ассоциированным с ВДЖП, C3 S/FR102G, fHY402H и fHV62I. Полученные результаты указывали не только на то, что восприимчивые комплотипы замедляли и гораздо меньше способствовали превращению iC3b в C3dg, чем протективные и гетерозиготные группы, но также и на то, что при увеличении концентрации общего фактора I, восприимчивый генотип может быть «превращен» в протективный генотип, и может устранять нежелательный полиморфизм в CFH и C3. Добавление 22 мкг/мл фактора I (в концентрации, меньшей, чем обычная концентрация FI) превращает генотип «с риском» в защищенный генотип (Lachmann et al., 2015 (см. выше)). Следовательно, присутствие лишь трех локусов оказывает огромное влияние на регуляцию активации комплемента альтернативного пути под действием фактора I, и такое влияние заключается в том, что гипервоспалительный комплотип становится более резистентным к негативной регуляции при увеличении концентрации фактора I. При увеличении концентрации фактора I в плазме, эффекты нежелательного комплотипа могут быть устранены и прогрессирование заболевания замедляется.
Генотерапия представляет собой способ доставки генетического материала пациенту в терапевтических целях. В случае генотерапии in vivo, пациенту вводят вектор, проникающий в клетки-мишени, в которых затем экспрессируется трансген, передаваемый этим вектором. Вначале, генотерапия была направлена на лечение редко встречающихся заболеваний с моногенетическими дефектами, которые требуют прямого, но сравнительно простого подхода, например, тяжелого комбинированного иммунодефицита (СПИД'а). Успех лечения на ранней стадии наблюдается и в случае приобретенных заболеваний, таких как рак, сердечно-сосудистые заболевания, нейродегенеративные расстройства и инфекционные заболевания. Значительные успехи были достигнуты при лечении гемофилии В и заболевания, ассоциированного с дефицитом липопротеин-липазы. Однако, несмотря на множество проведенных клинических испытаний, этот продукт для генотерапии пока еще не был апробирован специалистами Управления по контролю за качеством пищевых продуктов и лекарственных средств (FDA). В Европе, первый продукт для генотерапии, Glybera, был апробирован специалистами Европейского Фармацевтического Агенства в 2012 г. Glybera представляет собой средство для лечения редкого моногенетического заболевания, а именно, дефицита липопротеин-липазы (LPLD), и состоит из дефицитного гена, упакованного в рекомбинантный вирусный вектор, который вводят путем внутримышечной инъекции.
Современная генотерапия основана на замене дефектного гена, а не на достижении сверхэкспрессии имеющегося функционального гена. Авторами настоящего изобретения было неожиданно обнаружено, что уровни фактора I в плазме могут быть значительно повышены у мышей посредством генотерапии с использованием рекомбинантного вирусного вектора.
Настоящее изобретение основано на обнаружении того факта, что негативная регуляция цикла обратной связи C3b комплемента может быть достигнута посредством in vivo сверхэкспрессии фактора I в плазме с помощью генотерапии. Достигнутое восстановление равновесия петли обратной связи альтернативного пути будет стимулировать расщепление C3b и iC3b и тем самым удаление основных факторов заболевания при лечении комплемент-опосредованных расстройств, а в частности, расстройств, в основе которых лежит дефект регуляции альтернативного пути.
В соответствии с этим, настоящее изобретение относится к способу профилактики, лечения или ослабления комплемент-опосредуемого расстройства у пациента, где указанный способ включает введение пациенту рекомбинантного вирусного вектора, содержащего нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, которые сохраняют C3b-инактивирующую и iC3b-расщепляющую активность, так, чтобы терапевтически эффективное количество кодируемого фактора I или его фрагмента или производного экспрессировалось нуклеиновой кислотой у пациента и приводило к повышению уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента.
Способы согласно изобретению могут быть применены для достижения системного повышения уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности увеличивается до уровня, превышающего нормальный уровень.
Таким образом, в одном из своих вариантов, настоящее изобретение относится к способу профилактики, лечения или ослабления комплемент-опосредуемого расстройства у пациента, где указанный способ включает введение пациенту рекомбинантного вирусного вектора, содержащего нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, которые сохраняют C3b-инактивирующую и iC3b-расщепляющую активность, так, чтобы терапевтически эффективное количество кодируемого фактора I или его фрагмента или производного экспрессировалось нуклеиновой кислотой у пациента и приводило к повышению уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента до уровня, превышающего нормальный уровень.
В конкретных вариантах осуществления изобретения, пациенту вводят рекомбинантную вирусную частицу, которая инкапсидирует рекомбинантный вирусный вектор.
Настоящее изобретение также относится к рекомбинантному вирусному вектору, который содержит нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, которые сохраняют C3b-инактивирующую и iC3b-расщепляющую активность.
Настоящее изобретение также относится к рекомбинантному вирусному вектору, который содержит вирусный капсид, содержащий рекомбинантный вирусный вектор согласно изобретению.
Вирусный геном состоит из генов и цис-действующих регуляторных последовательностей, которые, в большинстве вирусов имеют пространственное распределение. Такое распределение используют для конструирования вирусных векторов. Вирусные гены, которые ответственны за репликацию, или капсидные/оболочечные белки заменяют терапевтическими трансгенами, которые затем фланкируются по обоим концам регуляторными цис-действующими последовательностями (Thomas, et al., Nat. Rev. Genet., 4(5):346-358, May 2003). Делетированные гены действуют in trans и могут доставлены либо посредством упаковывающей клеточной линии, которая имеет вирусные гены, включенные в геном, или посредством гетерологичных плазмид, котрансфецированных вирусным вектором (Kay et al., Nat. Med., 7(1):33-40, January 2001). Таким образом могут быть получены дефицитные по репликации вирусные частицы, содержащие трансген и способные передавать его клетке-мишени.
«Рекомбинантный вирусный вектор» означает рекомбинантный полинуклеотидный вектор, содержащий одну или более гетерологичных последовательностей, то есть, последовательностей нуклеиновой кислоты без вирусного ориджина. Рекомбинантный вирусный вектор может быть представлен в любой форме, включая плазмиды, линейные искусственные хромосомы, образующие комплекс с липидами, инкапсулированные в липосомы и инкапсидированные в вирусную частицу. Рекомбинантный вирусный вектор может быть упакован в вирусный капсид для получения рекомбинантной вирусной частицы. Термин «рекомбинантная вирусная частица» означает вирусную частицу, состоящую по меньшей мере из одного вирусного капсидного белка и инкапсидированного генома рекомбинантного вирусного вектора.
Рекомбинантными вирусными векторами, подходящими для их применения в соответствии с настоящим изобретением, являются рекомбинантные вирусные векторы, происходящие от ретровирусов, лентивирусов, вирусов простого герпеса-1 (HSV-1), аденовирусов и аденоассоциированных вирусов (AAV) (Thomas et al., supra - см. Таблицу 1 этого документа для сравнения относительных преимуществ и недостатков применения этих векторов в генотерапии).
Вирусные векторы могут быть подразделены на интергирующиеся и неинтегрирующиеся векторы. Интергирующиеся векторы, такие как ретровирусы и лентивирусы, перманентно встраивают трансген в хромосому клетки-хозяина. Неинтегрирующиеся векторы, такие как аденовирусные векторы и векторы на основе HSV, опосредуют экспрессию трансгена из эписом. Векторы на основе AAV преимущественно являются неинтегрирующимися векторами (интергирующиеся на <10%). Неинтегрирующиеся векторы, по сравнению с интергирующимися векторами, которые присутствуют в каждой дочерней клетке, будут быстро истощаться в быстро делящихся клетках, но они также не будут приводить к риску инсерционного мутагенеза. Поэтому, ретровирусы обычно используют для трансфекции клеток, которые подвергаются быстрому делению и дифференцировке (например, гемопоэтических стволовых клеток), а неинтегрирующиеся вирусы используют для введения в ткань после митоза (например, в печень, мышцы или глаза).
В конкретных вариантах осуществления изобретения, рекомбинантным вирусным вектором является неинтегрирующийся (или преимущественно неинтегрирующийся вектор, то есть, имеющий менее чем 50%, например, менее, чем 40%, 30%, 20% или 10% персистентных векторных геномов, интегрирующихся in vivo) эписомный вирусный вектор.
В конкретных вариантах осуществления изобретения, рекомбинантная вирусная частица инфицирует неделящиеся клетки, например, клетки печени (гепатоциты).
В других вариантах осуществления изобретения, рекомбинантная вирусная частица инфицирует делящиеся клетки, например, B-лимфоциты.
В конкретных вариантах осуществления изобретения, рекомбинантная вирусная частица инфицирует клетки, которые экспрессируют и секретируют кодируемый фактор I или его фрагмент или производное в кровотоке пациента. Подходящими клетками являются клетки печени (гепатоциты) и B-лимфоциты.
Рекомбинантная вирусная частица должна обладать способностью к переносу клеток, которые являются мишенью для экспрессии кодируемого фактора I или его фрагмента или производного. В конкретных вариантах осуществления изобретения, рекомбинантная вирусная частица способна переноситься в клетки печени (в частности, в гепатоциты).
В некоторых вариантах осуществления изобретения, рекомбинантный вирусный вектор содержит нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, фланкированные вирусными цис-действующими регуляторными последовательностями, такими как инвертированные концевые повторы (ITR).
В некоторых вариантах осуществления изобретения, нуклеиновая кислота, кодирующая фактор I или его фрагмент или производное, функционально присоединена к промотору. Репрезентативными промоторами являются, но не ограничиваются ими, предранний промотор цитомегаловируса (CMV), LTR RSV, LTR MoMLV, промотор фосфоглицерат-киназы-1 (PGK), промотор обезьяньего вируса 40 (SV40) и промотор CK6, промотор транстиретина (TTR), промотор ΤΚ, промотор чувствительности к тетрациклину (TRE), промотор HBV, промотор человеческого альфа 1-антитрипсина (hAAT), промотор альбумина, печень-специфический промотор (LSP), химерные печень-специфические промоторы (LSP), промотор E2F, промотор теломеразы (hTERT); промотор энхансера цитомегаловируса/куриного бета-актина/кроличьего β-глобина (промотор CAG; Niwa et al., Gene, 1991, 108(2): 193-9) и промотор фактора элонгации 1-альфа (EFI-альфа) (Kim et al., Gene, 1990, 91(2):217-23 и Guo et al., Gene Ther., 1996, 3(9):802-10). В некоторых вариантах осуществления изобретения, промотором является человеческой промотор β-глюкуронидазы, промотор β-актина, промотор куриного β-актина (CBA), промотор энхансера цитомегаловируса, присоединенного к CBA, промотор LTR ретровируса саркомы Рауса (RSV) или промотор дигидрофолатредуктазы.
В некоторых вариантах осуществления изобретения, промотор стимулирует экспрессию нуклеиновой кислоты, кодирующей фактор I или его фрагмент или производное, в клетках печени, например, в гепатоцитах.
Примерами печень-специфических промоторов, подходящих для их использования в настоящем изобретении, являются HLP, LP1, HLP2, hAAT, HCR-hAAT, ApoE-hAAT и LSP.
Эти печень-специфические промоторы более подробно описаны в следующих работах:
HLP: Mcintosh J. et al., Blood 2013 Apr 25, 121(17):3335-44, and WO 2011/005968;
LP1: Nathwani et al., Blood 2006 April 1, 107(7): 2653-2661, and WO 06/036502;
HCR-hAAT: Miao et al., Mol Ther. 2000; 1: 522-532;
ApoE-hAAT: Okuyama et al., Human Gene Therapy, 7, 637-645 (1996); LSP: Wang et al., Proc Natl Acad Sci USA 1999 March 30, 96(7): 3906-3910; и
HLP2: WO 2016/075473.
Промотор может быть конститутивным, индуцибельным или репрессируемым. Репрезентативные промоторы и их описание можно найти, например, в заявке на патент США 2014/0335054.
Примерами конститутивных промоторов являются, но не ограничиваются ими, промотор LTR ретровируса саркомы Рауса (RSV) (необязательно с энхансером RSV), промотор цитомегаловируса (CMV) (необязательно с энхансером CMV) (см., например, Boshart et al., Cell, 41:521-530 (1985)), промотор SV40, промотор дигидрофолатредуктазы, промотор 13-актина, промотор фосфоглицерин-киназы (PGK) и промотор EF1a (Invitrogen).
Индуцибельные промоторы регулируют экспрессию гена и могут регулироваться экзогенно доставляемыми соединениями, факторами окружающей среды, такими как температура, или конкретным физиологическим состоянием, например, острой фазой, конкретной стадией дифференцировки клетки или только реплицирующимися клетками. Индуцибельные промоторы и индуцибельные системы поставляются различными коммерческими источниками, включая, но не ограничиваясь ими, Invitrogen, Clontech и Ariad. Были описаны и многие другие системы, которые могут быть легко выбраны специалистом. Примерами индуцибельных промоторов, регулируемых экзогенно доставляемыми промоторами, являются овечий промотор металлотионеина, индуцируемый цинком (ΜΤ); промотор вируса опухоли молочной железы мышей, индуцируемый дексаметазоном (Dex) (MMTV); промоторная система полимеразы Τ7 (WO 98/10088); промотор экдизона насекомых (No et al., Proc. Natl. Acad. Sci. USA, 93:3346-3351 (1996)); система, репрессируемая тетрациклином (Gossen et al., Proc. Natl. Acad. Sci. USA, 89:5547-5551 (1992)); система, индуцируемая тетрациклином (Gossen et al., Science, 268:1766-1769 (1995), см. также Harvey et al., Curr. Opin. Chem. Biol., 2:512-518 (1998)); RU486-индуцируемая система (Wang et al., Nat. Biotech., 15:239-243 (1997) и Wang et al., Gene Ther., 4:432-441 (1997)); и система, индуцируемая рапамицином (Magari et al., J. Clin. Invest., 100:2865-2872 (1997)). Индуцибельными промоторами других типов, которые могут быть использованы в этом контексте, являются промоторы, которые регулируются конкретным физиологическим состоянием, например, острой фазой, конкретной стадией дифференцировки клетки или только реплицирующимися клетками.
В другом варианте осуществления изобретения может быть использован нативный промотор или его фрагмент для фактора I или его фрагмента или производного. Нативный промотор может быть использован в том случае, когда желательно, чтобы экспрессия фактора I или его фрагмента или производного имитировала нативную экспрессию (при условии, что после такой экспрессии, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента будет увеличиваться или превышать нормальный уровень). Нативный промотор может быть использован в том случае, когда экспрессия трансгена должна быть времяспецифической, или фазоспецифической, или тканеспецифической, или должна регулироваться в ответ на специфический стимулятор транскрипции. В другом варианте осуществления изобретения, для имитации нативной экспрессии могут быть также использованы и другие элементы регуляции нативной экспрессии, такие как энхансерные элементы, сайты полиаденилирования или консенсусные последовательности Козака.
Рекомбинантный вирусный вектор может включать любую регуляторную последовательность, необходимую для экспрессии кодируемого фактора I или его фрагмента или производного, в ткани или в клетке-мишени. Так, например, рекомбинантный вирусный вектор может содержать экспрессионный кластер, включающий нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, функционально присоединенные к промотору и сайту распознавания полиаденилирования. Рекомбинантный вирусный вектор может также включать и другие регуляторные последовательности для экспрессии фактора I или его фрагмента или производного, такие как элемент, связывающийся с рибосомой, терминатор, энхансер, селективный маркер, интрон, сигнал polyA и/или ориджин репликации или последовательность посттранскрипционного регуляторного элемента (WPRE) вируса гепатита сурка (WHP) или мутированная последовательность WPRE.
В некоторых вариантах осуществления изобретения, регуляторные последовательности влияют на тканеспецифическую экспрессию гена. В некоторых случаях, тканеспецифические регуляторные последовательности связываются с тканеспецифическими факторами транскрипции, которые индуцируют тканеспецифическую транскрипцию. Такие тканеспецифические регуляторные последовательности (например, промоторы, энхансеры и т.п.) хорошо известны специалистам.
В некоторых вариантах осуществления изобретения, промоторная последовательность может повсеместно экспрессироваться у пациента, и таким образом, она может экспрессироваться в одной или более клетках ткани-мишени (например, в печени) благодаря ее доставке в эту ткань. В других вариантах осуществления изобретения может быть использована промоторная последовательность, которая специфически экспрессируется в ткани-мишени (например, в печени) или в субсерии одной или более клеток ткани-мишени (например, в гепатоцитах).
В некоторых вариантах осуществления изобретения, фактор I или его фрагмент или производное экспрессируются исключительно в конкретных клетках ткани-мишени (например, в гепатоцитах), но не в других клетках этой ткани.
В конкретных вариантах осуществления изобретения, кодируемый фактор I или его фрагмент или производное экспрессируются из промотора, который является специфичным для клеток или тканей конкретных типов, представляющих собой мишень для экспрессии колируемого фактора I или его фрагмента или производного. Так, например, промотором может быть печень-специфический промотор, такой как промотор hAAT.
В одном из вариантов осуществления изобретения, промотором является печень-специфический промотор, выбранный из HLP, LP1, HLP2, hAAT, HCR-hAAT, ApoE-hAAT и LSP.
В одном из вариантов осуществления изобретения, промотор содержит, или состоит из нее, нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности SEQ ID NO: 8. В одном из вариантов осуществления изобретения, промотор содержит, или состоит из нее, нуклеотидную последовательность SEQ ID NO: 8.
В одном из вариантов осуществления изобретения, промотор содержит, или состоит из нее, нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности SEQ ID NO: 9. В одном из вариантов осуществления изобретения, промотор содержит, или состоит из нее, нуклеотидную последовательность SEQ ID NO: 9.
В одном из вариантов осуществления изобретения, промотор содержит, или состоит из нее, нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности SEQ ID NO: 10. В одном из вариантов осуществления изобретения, промотор содержит, или состоит из нее, нуклеотидную последовательность SEQ ID NO: 10.
В некоторых вариантах осуществления изобретения, рекомбинантный вирусный вектор или экспрессионный кластер содержит печень-специфический энхансер, такой как печень-специфический энхансер аполипопротеина E (ApoE). В конкретных вариантах осуществления изобретения, рекомбинантный вирусный вектор или экспрессионный кластер содержит печень-специфический промотор, такой как промотор hAAT и печень-специфический энхансер, такой как печень-специфический энхансер аполипопротеина E (ApoE).
В некоторых вариантах осуществления изобретения, экспрессионный кластер может быть фланкирован вирусными цис-действующими регуляторными последовательностями, такими как инвертированные концевые повторы (ITR).
В конкретных вариантах осуществления изобретения, рекомбинантная вирусная частица инфицирует печень (например, гепатоциты) пациента после введения, что приводит к экспрессии фактора I или его фрагмента или производного в печени (например, в гепатоцитах) и к секреции фактора I или его фрагмента или производного в кровоток пациента. В таких вариантах осуществления изобретения, кодируемый фактор I или его фрагмент или производное могут экспрессироваться из печень-специфического промотора, такого как промотор человеческого альфа-1-антитрипсина (hAAT) или промотор альбумина.
В конкретном аспекте изобретения, рекомбинантным вирусным вектором является рекомбинантный вектор AAV (rAAV). AAV представляет собой небольшой безоболочечный вирус, который состоит из линейного одноцепочечного генома ДНК с упаковывающей емкостью приблизительно 4,7 т.п.о. Его репликативный цикл зависит от совместного инфицирования вирусом-помощником, способным комплементировать делетированные гены для репликации AAV (Buller, et al., J. Virol., 40(1):241-247, October 1981), например, аденовирусом (возможно, природным вирусом-помощником) (Urabe, et al., J. Virol., 80(4):1874-1885, February 2006), герпесвирусом (Afione, et al., J. Virol., 70(5):3235-3241, May 1996) или бакуловирусом (Samulski, et al., EMBO J., 10(12):3941-3950, December 1991). Тот факт, что AAV представляет собой «природный дефектный» вирус, является дополнительным баррьером, обеспечивающим безопасность, то есть, предупреждающим нежелательное распространение вирусного вектора при клиническом применении (Nakai, et al., J. Virol., 75(15):6969-6976, August 2001). Рекомбинантные векторы AAV (rAAV) могут быть получены путем инсерции трансгена (то есть, нуклеиновой кислоты, кодирующей фактор I или его фрагмент или производное) между двумя инвертированными концевыми повторами (ITR). Рекомбинантные частицы AAV могут быть получены, например, путем котрансфекции таких векторов одной или более плазмидами, кодирующими все другие основные гены. Они могут инфицировать делящиеся и покоящиеся клетки и присутствуют, главным образом, в эписомной форме, в отличие от AAV дикого типа, который интегрируется, предпочтительно, в специфический сайт человеческой хромосомы 19. Если они присутствуют в делящейся клетке, то эписомный геном AAV быстро истощается в процессе деления клетки.
Термин «рекомбинантный вектор AAV (вектор rAAV)» означает полинуклеотидный вектор, содержащий нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное и фланкированную по меньшей мере одной последовательностью инвертированного концевого повтора AAV (ITR). В конкретных вариантах осуществления изобретения, нуклеиновая кислота, кодирующая фактор I или его фрагмент или производное, фланкирована двумя ITR AAV. Такие векторы rAAV могут реплицироваться и упаковываться в инфекционные вирусные частицы, если они присутствуют в клетке-хозяине, инфицированной, например, подходящим вирусом-помощником (или вирусом, который экспрессирует подходящие хелперные функции), и другой плазмидой, которая представляет собой продукты экспрессии генов rep и cap AAV (то есть белки Rep и Cap AAV), как более подробно объясняется ниже.
В некоторых вариантах осуществления изобретения, вектор rAAV содержит экспрессионный кластер, фланкированный по меньшей мере одной последовательностью ITR AAV, где экспрессионный кластер содержит нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное и функционально присоединенную к промотору и к сайту распознавания полиаденилирования, и необязательно, к последовательности WPRE или к мутантной последовательности WPRE. В конкретных вариантах осуществления изобретения, экспрессионный кластер фланкируется двумя последовательностями ITR AAV. В некоторых вариантах осуществления изобретения, экспрессионный кластер может также содержать регуляторные последовательности, включающие последовательности инициации и терминации транскрипции.
Термин «вирус-помощник» для AAV означает вирус, который позволяет AAV (то есть, дефектному парвовирусу) реплицироваться в клетке-хозяине и упаковываться в эту клетку. Был идентифицирован ряд таких вирусов-помощников, включая аденовирусы, гепресвирусы и поксвирусы, такие как вирус коровьей оспы. Аденовирусы включает ряд различных подгрупп, хотя чаще всего используется аденовирус типа 5 подгруппы C (Ad5). Множество аденовирусов, происходящих от человека, млекопитающего, не являющегося человеком, и от птиц, являются известными и хранятся в депозитариях, таких как ATCC. Вирусами, принадлежащими к семейству герпесвирусов, которые также хранятся в депозитариях, таких как ATCC, являются, например, вирусы простого герпеса (HSV), вирусы Эпштейна-Барра (EBV), цитомегаловирусы (CMV) и вирусы псевдобешенства (PRV).
Термины «инвертированный концевой повтор» или «последовательность ITR» хорошо известны специалистам и означатт относительно короткие последовательности, присутствующие на концах вирусных геномов в противоположной ориентации. Термин «последовательность инвертированного концевого повтора AAV (ITR)» также хорошо известен специалистам и означает последовательность, состоящую приблизительно из 145 нуклеотидов, которые присутствуют по обоим концам нативного одноцепочечного генома AAV. Самые крайние 125 нуклеотидов ITR могут присутствовать в любых двух альтернативных ориентациях, сообщающих гетерогенность между различными геномами AAV и между двумя концами одного генома AAV. Самые крайние 125 нуклеотидов также содержат несколько более коротких областей с аутокомплементарностью (обозначенных как области A, A', Β, B', C, C' и D), что позволяет образовывать пары оснований внутри цепи в этой части ITR.
Последовательность(и) ITR AAV должна(ы) выполнять функцию «спасения», репликации и упаковки вириона AAV (см., Davidson et al., PNAS, 2000, 97(7)3428-32; Passini et al., J. Virol., 2003, 77(12):7034-40; и Pechan et al., Gene Ther., 2009, 16:10-16). ITR AAV, если они используются в векторах согласно изобретению, необязательно должны иметь нуклеотидную последовательность дикого типа (например, как описано Kotin, Hum. Gene Ther., 1994, 5:793-801), и могут быть модифицированы путем инсерции, делеции или замены нуклеотидов, либо ITR AAV могут происходить от AAV любых нескольких серотипов (см., Gao et al., PNAS, 2002, 99(18): 11854-6; Gao et al., PNAS, 2003, 100(10):6081-6; and Bossis et al., J. Virol., 2003, 77(12):6799-810.
ITR для использования в векторах согласно изобретению могут содержать нуклеотидную последовательность природного ITR или состоять из нее, либо они могут содержать, или состоять из нее, нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR. В частности, ITR AAV для использования в векторах согласно изобретению могут содержать, или состоять из нее, нуклеотидную последовательность природного ITR AAV, или они могут содержать или состоять из нее, нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR AAV, такого как ITR AAV, имеющего следующие серотипы AAV: AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12.
Векторы AAV могут интегрироваться в человеческий геном, хотя скорость их интергации in vivo является низкой. Первичным источником rAAV-опосредуемой экспрессии гена является внехромосомные неинтегрированные геномы. В модели мышиной гепатектомии, менее чем 10% персистентных векторных геномов интегрируются в печени (Miller, et al., Nat. Genet., 30(2):147-148, February 2002).
AAV продемонстрировал впечатляющий рекорд по безопасности и не был ассоциирован ни с одним из известных заболеваний человека или животных, хотя большинство людей (>70%) являются серопозитивными по одному или более серотипам. Векторы AAV не ассоциируются с токсичностью или с воспалительным ответом, хотя в данном случае, наблюдается продуцирование нейтрализующих антител, которые могут давать эффект после введения. Однако такие иммунные ответы могут быть снижены путем модификации капсидной последовательности (Mingozzi and High, Blood, 122(1):23-36, July 2013).
В конкретных вариантах осуществления изобретения, рекомбинантной вирусной частицей является рекомбинантная частица аденоассоциированного вируса (rAAV). Термин «частица rAAV» означает вирусную частицу, состоящую по меньшей мере из одного капсидного белка AAV и из инкапсидированного генома рекомбинантного вирусного вектора, а в частности, инкапсидированного генома вектора rAAV.
Основные стадии метода генотерапии на основе AAV согласно изобретению проиллюстрированы на фигуре 4. Рекомбинантный вектор AAV, кодирующий фактор I или его фрагмент или производное, используют для продуцирования частиц rAAV, инкапсидирующих вектор. После введения частиц rAAV пациенту, например, путем инъекции, частицы rAAV инфицируют клетки-мишени, а фактор I или его фрагмент или производное, кодируемые вектором, экспрессируются в клетках-мишенях, что приводит к секреции фактора I или его фрагмента или производного в кровоток пациента. Такие методы более подробно описаны ниже.
Известно, что протеогликан на основе сульфата гепарана (HSPG) действует как клеточный рецептор для частиц AAV серотипа 2 (AAV2) (Summerford, C. and Samulski, R.J. (1998) J. Virol. 72(2):1438-45). Связывание частицы AAV2 с HSPG в клеточной мембране служит для присоединения частицы к клетке. Другие белки клеточной поверхности, такие как рецептор фактора роста фибробластов и интегрин ανβ5 могут также облегчать инфицирование клеток. После связывания, частица AAV2 может проникать в клетку по определенным механизмам, включая опосредуемый рецептором эндоцитоз в полости, покрытой клатрином. Частица AAV2 может высвобождаться из эндоцитарной везикулы после подкисления эндосомы. Это позволяет частице AAV2 проникать в околоядерную область, а затем поступать в клеточное ядро.
Известно, что капсид AAV включает три капсидных белка: VP1, VP2 и VP3. Эти белки содержат значительное количество перекрывающихся аминокислотных последовательностей и уникальные N-концевые последовательности. Капсид AAV2 включает 60 субъединиц, имеющих икосаэдрическую симметрию (Xie, Q., et al. (2002) Proc. Natl. Acad. Sci. 99(16): 10405-10). Было обнаружено, что VP1, VP2 и VP3 присутствуют в отношении 1:1:10. Связывание капсидных белков AAV2 с HSPG происходит посредством электростатических взаимодействий между основными остатками капсидного белка AAV2 и отрицательно заряженными гликозаминогликановыми остатками (Opie, SR et al., (2003) J. Virol. 77:6995-7006; Kern, A et al., (2003) J. Virol. 11:11072-11081). Специфическими капсидными остатками, участвующими в таких взаимодействиях, являются R484, R487, Κ532, R585 и R588. Вирусы инфицируют природную клетку-хозяина наиболее эффективно. Псевдотипирование включает замену поверхностных белков, которые опосредуют проникновение в клетку, белками другого вируса для модификации вирусного тропизма. Примером псевдотипирования является псевдотипирование лентивируса G-белком вируса везикулярного стоматита (G-псевдотипированного лентивируса VSV), который может проникать почти во все клетки (Akkina, et al., J. Virol., 70(4):2581-2585, April 1996; Willett and Bennett, Front Immunol, 4:261, 2013; Sharland, et al., Discov Med, 9(49):519-527, June 2010). В других случаях, тропизм вируса ограничивается только клеткой-мишенью. Это позволяет снижать дозу вводимого вектора и предотвращать экспрессию трансгена за пределами клетки соответствующего типа, например, когда экспрессия должна осуществляться только в глазах (Grimm, et al., Blood, 102(7):2412-2419, October 2003) или в печени (Thomas, et al., J. Virol., 78(6):3110-3122, March 2004; Lisowski, et al., Nature, 506(7488):382-386, February 2014; Niidome and Huang. Gene Ther., 9(24):1647-1652, December 2002). Псевдотипы могут быть также получены искусственно, например, путем создания библиотек, состоящих из капсидов AAV, в которых была сделана перестановка ДНК для повышения тропизма по отношению к человеческим гепатоцитам (Li and Huang. Gene Ther., 13(18):1313-1319, September 2006). Это позволяет предотвращать слабое поглощение вектора и недостаточную экспрессию трансгена.
В соответствии с настоящим изобретением, рекомбинантная вирусная частица может быть псевдотипирована для сообщения тропизма по отношению к клетке(ам) определенного типа, которая(ые) должна(ы) быть инфицирована(ы). Так, например, в вариантах настоящего изобретения, в которых рекомбинантная вирусная частица инфицирует клетки печени (например, гепатоциты), рекомбинантная вирусная частица может быть псевдотипирована для сообщения тропизма по отношению к печени, а в частности, к гепатоцитам. В других вариантах осуществления изобретения, в которых рекомбинантная вирусная частица инфицирует B-лимфоциты, тропизм по отношению к B-лимфоцитам может быть достигнут с использованием gp220/350 или его фрагмента, происходящих от вируса Эпштейна-Барра.
В некоторых вариантах осуществления изобретения, рекомбинантной вирусной частицей является частица rAAV, которая была псевдотипирована для сообщения тропизма по отношению к печени, а в частности, к гепатоцитам.
Было идентифицировано свыше 100 различных серотипов AAV (Atchison, et al., Science, 149(3685):754-756, August 1965; Warrington, et al., J. Virol., 78(12):6595-6609, June 2004). Псевдотипирование может быть легко достигнуто для вирусных частиц rAAV, например, путем упаковки капсидной последовательности другого серотипа в хелперную плазмиду. Для повышения тропизма и повышения уровня экспрессии трансгенов может быть проведено несколько стадий: i) конструирования капсидных библиотек, состоящих из рандомизированных капсидных последовательностей (Li and Huang, Gene Ther., 13(18):1313-1319, September 2006); ii) инсерции аминокислотных последовательностей в капсид AAV2 (размером до 30 кДа, возможно для инсерции последовательности ScFv для нацеливания AAV-мишени на клетку конкретного типа) (Calcedo, et al., J. Infect. Dis., 199(3):381-390, February 2009); и iii) рационального конструирования путем объединения данных о механизмах доставки с данными структурных анализов AAV (Calcedo, et al., Clin. Vaccine Immunol., 18(9):1586-1588, September 2011).
В объем настоящего изобретения входит применение AAV любого серотипа. В некоторых вариантах осуществления изобретения, вектором rAAV является вектор, происходящий от серотипа AAV, включая ITR AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11 или AAV12. Частица rAAV может содержать капсидный белок, происходящий от AAV любого серотипа, включая капсид AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAVrh10, AAV11 или AAV12. Для оптимизации трансдукции конкретных клеток-мишеней или для нацеливания на клетки конкретных типов в конкретной ткани-мишени (например, в ткани печени) используют AAV различных серотипов. Частица rAAV может содержать вирусные белки и вирусные нуклеиновые кислоты одного и того же серотипа или смешанных серотипов.
Капсидный белок рекомбинантной вирусной частицы согласно изобретению может включать или состоять из нее, аминокислотную последовательность природного капсидного белка (например, природного капсидного белка AAV, такого как капсидный белок серотипа AAV: AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAVrh10, AAV11 или AAV12), либо он может представлять собой производное природного капсидного белка, которое включает одну или более аминокислотных замен, делеций или добавлений по сравнению с аминокислотной последовательностью природного капсидного белка, например, для сообщения повышенного тропизма по отношению к тканям или клеткам нужных типов (таким как печень, или тропизма по отношению к гепатоцитам) или для снижения иммуногенности рекомбинантной вирусной частицы.
В некоторых вариантах осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению имеет аминокислотную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности природного капсидного белка. В некоторых вариантах осуществления изобретения, капсидный белок AAV частицы rAAV согласно изобретению имеет аминокислотную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности природного капсидного белка AAV, например, природного капсидного белка AAV серотипа AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAVrh10, AAV11 или AAV12.
В некоторых вариантах осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению представляет собой неприродный капсидный белок, например, неприродный капсидный белок AAV, такой как сконструированный или химерный капсидный белок.
Химерными капсидными белками являются белки, полученные путем рекомбинации между двумя или более капсид-кодирующими последовательностями природных серотипов AAV. Это может быть осуществлено, например, методом «спасения» маркера, где неинфекционные капсидные последовательности одного серотипа котрансфецируют капсидными последовательностями другого серотипа и проводят прямой отбор на капсидные последовательности, имеющие нужные свойства. Капсидные последовательности различных серотипов могут быть модифицированы путем гомологичной рекомбинации в клетке для продуцирования новых химерных капсидных белков.
Химерными капсидными белками также являются белки, полученные путем конструирования последовательностей капсидного белка для переноса специфических доменов капсидного белка, поверхностных петель или специфических аминокислотных остатков, расположенных между двумя или более капсидными белками, например, между двумя или более капсидными белками различных серотипов.
Капсидные белки, полученные путем перестановки, или химерные капсидные белки, могут быть также получены путем перестановки ДНК или с помощью ПЦР с вероятностью ошибки. Гибридные капсидные гены AAV могут быть получены путем рандомизированной фрагментации последовательностей родственных генов AAV, например, генов, кодирующих капсидные белки множества различных серотипов, с последующей повторой сборкой фрагментов посредством полимеразной реакции с аутопраймированием, которая может также вызывать кроссинговер в областях гомологии последовательностей. Библиотека гибридных генов AAV, созданная таким путем перестановки капсидных генов некоторых серотипов, может быть скринирована для идентификации вирусных клонов, имеющих нужные функциональные свойства. Аналогичным образом, ПЦР с вероятностью ошибки может быть использована для рандомизированной мутации капсидных генов AAV в целях создания библиотек с разнообразными вариантами, которые могут быть затем отобраны на нужные свойства.
Последовательности капсидных генов могут быть также генетически модифицированы для введения специфических делеций, замен или инсерций в нативную последовательность дикого типа. В частности, капсидные гены могут быть модифицированы путем инсерции последовательности неродственного белка или пептида в открытую рамку считывания капсид-кодирующей последовательности или у N- и/или C-конца капсид-кодирующей последовательности.
Примеры неприродных капсидных белков, подходящих для их использования в настоящем изобретении, описаны в WO 2016/181123 и WO 2013/029030.
В одном из вариантов осуществления изобретения, капсидным белком рекомбинантной вирусной частицы согласно изобретению является капсидный белок MutC (например, капсидный белок Mut C, описанный в WO 2016/181123). В одном из вариантов осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению имеет аминокислотную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 11. В одном из вариантов осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению имеет аминокислотную последовательность, которая включает, или состоит из нее, аминокислотную последовательность SEQ ID NO: 11.
В одном из вариантов осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению представляет собой капсидный белок LK03 (например, капсидный белок LK03, описанный в WO 2013/029030). В одном из вариантов осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению имеет аминокислотную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 12. В одном из вариантов осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению имеет аминокислотную последовательность, которая включает, или состоит из нее, аминокислотную последовательность SEQ ID NO: 12.
В одном из вариантов осуществления изобретения, капсидный белок рекомбинантной вирусной частицы согласно изобретению представляет собой белок AAVrh10, описанный Wang et al. Mol. Ther. 2015 Dec; 23(12): 1877-1887 или его производное, имеющие аминокислотную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности описанного белка AAVrh10.
В конкретных вариантах осуществления изобретения, рекомбинантным вирусным вектором является вирусный вектор, происходящий от AAV2, то есть, рекомбинантный вирусный вектор, в котором нуклеиновая кислота, кодирующая фактор I или его фрагмент или производное, фланкирована по меньшей мере одной последовательностью ITR AAV2 (Kotterman and Schaffer, Nat. Rev. Genet., 15(7):445-451, July 2014), или ее производным, имеющими нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR AAV2. В некоторых вариантах осуществления изобретения, нуклеиновая кислота, кодирующая фактор I или его фрагмент или производное, фланкирована двумя ITR AAV2 или производными ITR AAV2 (где каждое производное ITR AAV2 содержит нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR AAV2).
AAV2 имеет широкий тропизм и относительно эффективно инфицирует клетки многих типов, включая гепатоциты in vivo. Псевдотипирование рекомбинантного вектора AAV с различными капсидными белками может значительно влиять на тропизм. AAV8 и AAV9 обладают гораздо большей аффинностью по отношению к гепатоцитам по сравнению с AAV2. В частности, AAV8 может инфицировать в 3-4 раза большее число гепатоцитов и доставлять в 3-4 раза большее число геномов на трансдуцировнную клетку по сравнению с AAV2.
В одном из вариантов осуществления изобретения, рекомбинантный вирусный вектор не является вирусным вектором, происходящим от AAV2.
В некоторых вариантах осуществления изобретения, частица rAAV согласно изобретению содержит капсидный белок AAV8 или AAV9 (или его производное, аминокислотная последовательность которого по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности природного капсидного белка AAV8 или AAV9). В конкретных вариантах осуществления изобретения, частица rAAV инкапсидирует вирусный вектор, происходящий от AAV2, и была псевдотипирована капсидом AAV8 (rAAV2/8) или капсидом AAV9 (rAAV2/9) (или его производным, аминокислотная последовательность которого по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности природного капсидного белка AAV8 или AAV9). В других вариантах осуществления изобретения, частица rAAV инкапсидирует вирусный вектор, происходящий от AAV2, и была псевдотипирована капсидом Mut C (rAAV2/Mut C) или капсидом LK03 (rAAV2/LK03) или капсидом AAVrh10 (rAAV2/rh10).
Специалистам известно множество методов продуцирования частиц rAAV, включая системы трансфекции, системы стабильного продуцирования клеточных линий и системы продуцирования инфекционных гибридных вирусов, которые включают гибриды «аденовирус-AAV», гибриды «герпесвирус-AAV» (Conway, et al., (1997) J. Virology 71(11):8780-8789) и гибриды «бакуловирус-AAV».
Для всех rAAV-продуцирующих культур, используемых для продуцирования вирусных частиц rAAV, необходимы: i) подходящие клетки-хозяева, включая, например, человеческие клеточные линии, такие как клетки HeLa, Α549, 293 или 293T, или клеточные линии насекомых, такие как SF-9, в случае систем продуцирования бакуловирусов; ii) подходящая хелперная функция, сообщаемая аденовирусом дикого типа или мутантным аденовирусом (таким как термочувствительный аденовирус), герпесвирусом, бакуловирусом или плазмидной конструкцией с хелперными функциями; iii) гены rep и cap AAV; iv) нуклеиновая кислота, кодирующая фактор I или его фрагмент или производное и фланкированная по меньшей мере одной последовательностью ITR AAV; и v) среда и компоненты среды, подходящие для поддержания продуцирования rAAV.
Для продуцирования частиц rAAV могут быть использованы подходящие среды, известные специалистам. Такими средами являются, но не ограничиваются ими, среды, получаемые в лабораториях Hyclone Laboratories и JRH, включая модифицированную среду Игла (MEM), модифицированную по способу Дульбекко среду Игла (DMEM) с высоким содержанием глюкозы, специально приготовленные композиции, такие как композиции, описанные в патенте США 6566118, и среды Sf-900 II SFM, описанные в патенте США 6723551.
Частицы rAAV могут быть получены известными методами. См., например, патенты США 6566118, 6989264 и 6995006. При осуществлении настоящего изобретения, клетками-хозяевами для продуцирования частиц rAAV являются клетки млекопитающих, клетки насекомых, клетки растений, микроорганизмы и дрожжи. Клетками-хозяевами могут быть также упаковывающие клетки, в которых гены rep и cap AAV стабильно поддерживались в клетке-хозяине, или клетки-продуценты, в которых стабильно поддерживается геном вектора AAV. Репрезентативные упаковывающие клетки и клетки-продуценты происходят от клеток 293, 293T, Α549 или HeLa. Векторы AAV очищают и приготавливают стандартными методами, известными специалистам.
В некоторых вариантах осуществления изобретения, частицы rAAV могут быть получены методом тройной трансфекции, таким как репрезентативный метод тройной трансфекции, описанный ниже. Вкратце, плазмида, содержащая ген rep и капсидный ген, хелперная аденовирусная плазмида и плазмида, содержащая трансген (то есть, ген, кодирующий фактор I или его фрагмент или производное) могут быть перенесены (например, методом с использованием фосфата кальция или с использованием полиэтиленимина) в клеточную линию (например, в клетки HEK-293), а затем вирус может быть собран и необязательно очищен.
В некоторых вариантах осуществления изобретения, частицы rAAV могут быть получены методом с использованием клеточной линии-продуцента, таким как репрезентативный метод с использованием клеточной линии-продуцента, описанный ниже (см. также публикацию Martin et al., (2013) Human Gene Therapy Methods 24:253-269). Вкратце, клеточная линия (например, клеточная линия HeLa) может быть стабильно трансфицирована плазмидой, содержащей ген rep, капсидный ген и последовательность промотора-трансгена. Клеточные линии могут быть скринированы для отбора главного клона на продуцирование rAAV, а затем эти клеточные линии могут быть размножены в биореакторе и инфицированы аденовирусом (например, аденовирусом дикого типа) в качестве вируса-помощника для инициации продуцирования rAAV. Затем вирус может быть собран, после чего аденовирус может быть инактивирован (например, путем нагревания) и/или удален, и частицы rAAV могут быть очищены.
В некоторых своих аспектах, настоящее изобретение относится к способу продуцирования частицы rAAV согласно изобретению, включающему: (a) культивирование клетки-хозяина в условиях, способствующих продуцированию частиц rAAV, где клетка-хозяин содержит (i) один или более упаковывающих генов AAV, где каждый упаковывающий ген AAV кодирует белок репликации и/или инкапсидирования AAV; (ii) вектор rAAV, содержащий нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, которые сохраняют C3b-инактивирующую и iC3b-расщепляющую активность, где указанная нуклеиновая кислота фланкирована по меньшей мере одним ITR AAV (предпочтительно, двумя ITR AAV), и (iii) AAV с хелперной функцией (то есть, гены, необходимые для поддержания продуктивного клеточного цикла AAV, например, аденовируса, герпесвируса или бакуловируса); и (b) выделение частиц rAAV, продуциованных клеткой-хозяином. В некоторых вариантах осуществления изобретения, гены (i), (ii) и (iii) могут быть получены на трех отдельных плазмидах, например, как описано ниже в примере 1.
В некоторых вариантах осуществления изобретения, один или каждый ITR AAV содержит, или состоит из нее, нуклеотидную последовательность природного ITR AAV, например, любого из нижеследующих серотипов ITR AAV: AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV 10, AAV11, AAV12, например, ITR AAV2, или содержит, или состоит из нее, нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR AAV, такого как ITR AAV, имеющий любой из нижеследующих серотипов: AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12.
В некоторых вариантах осуществления изобретения, инкапсидирующим белком является инкапсидирующий белок AAV8 или AAV9 или Mut C или LK03 или AAVrh10.
В подходящую культуральную среду для продуцирования rAAV могут быть добавлены сыворотка или сывороточные рекомбинантные белки на уровне 0,5%-20% (об/об или масс/об). Альтернативно, как известно специалистам, частицы rAAV могут быть получены в бессывороточных условиях, что может также означать, что среда не содержит продуктов животного происхождения. Для среднего специалиста в данной области очевидно, что в коммерчески доступные или специально приготовленные среды для поддержания продуцирования частиц rAAV могут быть также добавлены один или более компонентов клеточной культуры, известных специалистам, включая, но не ограничиваясь ими, глюкозу, витамины, аминокислоты и/или факторы роста, для повышения титра rAAV в культурах-продуцентах.
Культуры для продуцирования rAAV могут быть культивированы в различных условиях (при температурах в широких пределах, в течение различных периодов времени и т.п.), подходящих для использования конкретной клетки-хозяина. Как известно специалистам, культуры для продуцирования rAAV включают зависящие от связывания культуры, которые могут быть культивированы в подходящих зависящих от связывания сосудах, таких как, например, гиперколбы, роллер-флаконы, фильтры из полого волокна, микроносители и биореакторы пакетного типа или биореакторы с псевдоожиженным слоем. Культуры для продуцирования вектора rAAV могут также включать адаптированные для суспензий клетки-хозяева, такие как клетки HeLa, 293, 293T и SF-9, которые могут быть культивированы различными способами, например, в центрифужных колбах, в биореакторах с резервуаром для перемешивания и в одноразовых системах, таких как пакетная система волнового типа.
Частицы rAAV согласно изобретению могут быть собраны из культур для продуцирования rAAV путем лизиса клеток-хозяев в культуре-продуценте или путем удаления истощенной среды из культуры-продуцента при условии, что клетки будут культивированы в условиях, известных специалистам и подходящих для высвобождения частиц rAAV в среду из интактных клеток, как более подробно описано в патенте США 6566118. Подходящие методы лизиса клеток также известны специалистам и включают, например, множество циклов замораживания/оттаивания, обработку ультразвуком, микрофлюидизацию и обработку химическими веществами, такими как детергенты и/или протеазы.
В другом варианте осуществления изобретения, частицы rAAV являются очищенными. Используемый здесь термин «очищенный» относится к препарату частиц rAAV, не содержащих по меньшей мере некоторых других компонентов, которые могут присутствовать в природных частицах rAAV или в исходных частицах. Так, например, выделенные частицы rAAV могут быть получены методом очистки для обогащения из смеси источника, такой как лизат культуры или супернатант культуры-продуцента. Обогащение может быть оценено различными способами, например, по количеству резистентных к ДНКазе частиц (DRP) или геномных копий (гк), присутствующих в растворе, или по инфекционности, либо оно может быть оценено по наличию другого потенциально опасного вещества, присутствующего в смеси источника, такого как примеси, включая примеси культуры-продуцента, или примеси, образовавшиеся в процессе культивирования, включая вирус-помощник или компоненты среды.
В некоторых вариантах осуществления изобретения, культуру для продуцирования rAAV осветляют для удаления дебриса клеток-хозяев. В некоторых вариантах осуществления изобретения, полученную культуру-продуцент осветляют путем фильтрации через серию фильтров для глубокой фильтрации, включая, например, корпусный фильтр DOHC Millipore Millistak+HC, корпусный фильтр A1HC Millipore Millistak+HC и 0,2 мкм-фильтр с гидрофильной мембраной Opticap XL1O Millipore Express SHC. Осветление может быть также достигнуто различными другими стандартными методами, известными специалистам, такими как центрифугирование или фильтрация через любой фильтр из ацетата целлюлозы размером 0,2 мкм или еще большим размером пор, как известно специалистам.
В некоторых вариантах осуществления изобретения, собранную культуру для продуцирования rAAV дополнительно обрабатывают бензоназой® для расщепления любой высокомолекулярной ДНК, присутствующей в культуре для продуцирования. В некоторых вариантах осуществления изобретения, расщепление бензоназой® осуществляют в стандартных условиях, известных специалистам, включая, например, конечную концентрацию 1-2,5 единиц/мл бензоназы® при температуре в пределах от температуры окружающей среды до 37°C в течение периода времени от 30 минут до нескольких часов.
Частицы rAAV могут быть выделены или очищены путем проведения одной или более из нижеследующих стадий очистки: центрифугирования в градиенте плотности хлорида цезия (например, как описано Cooper et al., Molecular Therapy (2005) 11, Supplement 1, S53-S54), или иодоксанола; равновесного центрифугирования; проточной анионообменной фильтрации; фильтрации в тангенциальном потоке (TFF) для концентрирования частиц rAAV; захвата rAAV с помощью хроматографии на апатитах; термоинактивации вируса-помощника; захвата rAAV с помощью гидрофобной хроматографии; буферного обмена с помощью эксклюзионной хроматографии (ЭХ); нанофильтрации; и захвата rAAV с помощью анионообменной хроматографии, катионообменной хроматографии или аффинной хроматографии. Эти стадии могут быть проведены отдельно, в различных комбинациях или в различном порядке. Методы очистки частиц rAAV описаны, например, Xiao et al., (1998) Journal of Virology 72:2224-2232; в патентах США 6989264; 8137948; и в заявке WO 2010/148143.
Рекомбинантная вирусная частица может быть введена пациенту любым подходящим способом. В конкретных вариантах осуществления изобретения, рекомбинантную вирусную частицу вводят пациенту внутривенно.
В конкретных вариантах осуществления изобретения, рекомбинантную вирусную частицу не вводят пациенту путем локального (или местного) введения.
В конкретных вариантах осуществления изобретения, рекомбинантную вирусную частицу не вводят в глаза. Таким образом, в конкретных вариантах осуществления изобретения, введение рекомбинантной вирусной частицы не включает внутриглазное введение, такое как внутриглазная, например, субретинальная инъекция.
Рекомбинантная вирусная частица согласно изобретению может быть введена пациенту путем системного введения.
Специалисту в данной области известно множество способов системного введения и множество подходящих схем такого введения.
Так, например, системное введение может быть осуществлено парентерально. Примерами парентерального введения являются внутривенное введение, внутриартериальное введение, внутримышечное введение, интратекальное введение и подкожное введение.
Таким образом, в конкретных вариантах осуществления изобретения, рекомбинантную вирусную частицу согласно изобретению вводят пациенту системно.
Предпочтительным способом введения рекомбинантной вирусной частицы согласно изобретению является внутривенное введение.
В одном из вариантов осуществления изобретения, рекомбинантную вирусную частицу согласно изобретению вводят в печеночную воротную вену пациента. Предпочтительно, чтобы введение рекомбинантной вирусной частицы в печеночную воротную вену осуществлялось путем непосредственного введения рекомбинантной вирусной частицы в печень.
Как описано выше, в конкретных вариантах осуществления изобретения, рекомбинантная вирусная частица инфицирует печень (например, гепатоциты) пациента после введения, что приводит к экспрессии фактора I или его фрагмента или производного из печени (например, из гепатоцитов) и к секреции фактора I или его фрагмента или производного в кровоток пациента. В таких вариантах осуществления изобретения, кодируемый фактор I или его фрагмент или производное могут экспрессироваться из печень-специфического промотора.
Фактор I или его фрагмент или производное, секретируемые в кровоток пациента из печени, могут оказывать терапевтический эффект в ткани-мишени или в клетке-мишени, находящихся в любом участке организма. Фактор I или его фрагмент или производное, секретируемые в кровоток пациента из печени, могут приводить к системному увеличению уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента.
AAV-опосредуемая генотерапия путем непосредственной доставки в печень обсуждается в публикациях Sands Methods Mol Biol. 2011; 807: 141-157, и Sharland et al., Discovery Medicine, 9(49):519-527, June 2010. Так, например, в зависимости от дозы, AAV8 может переносить до 90-95% гепатоцитов в печень мышей после инъекции в воротную вену. Интересно отметить, что сравнимые уровни трансдукции могут быть достигнуты после внутривенной инъекции. Прямая инъекция вектора AAV в паренхиму также опосредует относительно длительную экспрессию на высоком уровне. Дополнительная специфичность может сообщаться с использованием печень-специфических промоторов в комбинации с капсидными белками AAV8.
Эффективное количество рекомбинантного вирусного вектора (в некоторых вариантах осуществления изобретения, инкапсидированного рекомбинантными вирусными частицами) вводят в зависимости от целей лечения. Так, например, если нужный терапевтический эффект может достигаться при небольшом проценте трансдукции, то тогда цель лечения обычно достигается при таком уровне или более высоком трансдукции. В некоторых случаях, такой уровень трансдукции может достигаться путем переноса лишь приблизительно от 1 до 5% клеток-мишеней, в некоторых вариантах осуществления изобретения, по меньшей мере приблизительно 20% клеток ткани нужного типа, в некоторых вариантах осуществления изобретения, по меньшей мере приблизительно 50%, в некоторых вариантах осуществления изобретения, по меньшей мере приблизительно 80%, в некоторых вариантах осуществления изобретения, по меньшей мере приблизительно 95%, а в некоторых вариантах осуществления изобретения, по меньшей мере приблизительно 99% клеток ткани нужного типа.
В некоторых вариантах осуществления изобретения, доза вирусных частиц, вводимых пациенту, составляет от 1 × 108 до 1 × 1013 геномных копий/кг массы тела.
В некоторых вариантах осуществления изобретения, общее количество вирусных частиц, вводимых пациенту, составляет от 1 × 109 до 1 × 1015 геномных копий.
Для вариантов осуществления изобретения, в которых вирусные частицы вводят пациенту, в качестве примера можно сказать, что число вирусных частиц, вводимых за одну инъекцию, обычно составляет от 1 × 106 до 1 × 1014 частиц, от 1 × 107 до 1 × 1013 частиц, от 1 × 109 до 1 × 1012 частиц или 1 × 1011 частиц.
Рекомбинантные вирусные частицы могут быть введены за одну или более инъекций, либо путем проведения той же самой процедуры, либо с перерывом в несколько дней, недель, месяцев или лет. В некоторых вариантах осуществления изобретения, для лечения пациента может быть использовано множество векторов.
В некоторых вариантах осуществления изобретения, по меньшей мере 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70% или 75%-100% клеток-мишеней ткани (например, гепатоцитов печени) являются трансдуцированными. В некоторых вариантах осуществления изобретения, по меньшей мере 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70% или 75%-100% клеток ткани-мишени (например, гепатоцитов печени) являются трансдуцированными.
Методы идентификации клеток, трансдуцированных рекомбинантными вирусными частицами, содержащими капсид рекомбинантных вирусных частиц, известны специалистам. Так, например, для детектирования трансдукции рекомбинантных вирусных частиц может быть применен иммуногистохимический метод или маркер, такой как активированный белок, флуоресцирующий в зеленом диапазоне спектра.
В некоторых вариантах осуществления изобретения, рекомбинантные вирусные частицы вводят (например, путем инъекции) в один или более участков нужной ткани (например, печени). В некоторых вариантах осуществления изобретения, рекомбинантные вирусные частицы вводят (например, путем инъекции) в любой один или любой из двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти или более участков ткани.
В некоторых вариантах осуществления изобретения, рекомбинантные вирусные частицы вводят одновременно или последовательно в более чем один участок. В некоторых вариантах осуществления изобретения, множество инъекций рекомбинантных вирусных частиц вводят с интервалом не более, чем в один час, два часа, три часа, четыре часа, пять часов, шесть часов, девять часов, двенадцать часов или 24 часа.
Пациенту могут быть введены один или более дополнительных терапевтических средств для лечения комплемент-опосредуемых расстройств. Такое(ие) средство(а) может (могут) быть введено(ы) вместе с вирусными частицами или последовательно. Интервал между последовательными введениями может составлять по меньшей мере (или, альтернативно, менее, чем) в течение нескольких минут, часов или дней.
Следует отметить, что C3b-инактивирующая и iC3b-расщепляющая активность у пациента после экспрессии фактора I или его фрагмента или производного из нуклеиновой кислоты рекомбинантного вирусного вектора может включать C3b-инактивирующую и iC3b-расщепляющую активность из эндогенного фактора I пациента (то есть, где фактор I пациента не продуцируется посредством экспрессии из рекомбинантного вирусного вектора) и C3b-инактивирующую и iC3b-расщепляющую активность, продуцируемую в результате экспрессии из рекомбинантного вирусного вектора, так, чтобы общий уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента был еще выше (по сравнению с уровнем до введения вектора) или превышал нормальный уровень.
Уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента может представлять собой уровень такой активности в сыворотке пациента. Для человека, нормальный уровень C3b-инактивирующей и iC3b-расщепляющей активности эквивалентен уровню, достигаемому при введении 30-40 мкг/мл фактора I в сыворотке пациента.
До введения рекомбинантной вирусной частицы согласно изобретению, у пациента может наблюдаться нормальный уровень эндогенного фактора I. В других вариантах осуществления изобретения, у пациента может наблюдаться уровень фактора I ниже нормального уровня, например, количество фактора I в сыворотке пациента может составлять менее чем 30 мкг/мл и более чем 0 мкг/мл.
В другом примере, количество фактора I в сыворотке пациента может составлять менее чем 25 мкг/мл и более чем 0 мкг/мл, или менее чем 20 мкг/мл и более чем 0 мкг/мл, или менее чем 15 мкг/мл и более чем 0 мкг/мл, или менее чем 10 мкг/мл и более чем 0 мкг/мл или менее чем 5 мкг/мл и боле, чем 0 мкг/мл.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, превышающего нормальный уровень.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который по меньшей мере на 5%, 10%, 15%, 20% или 25% превышает нормальный уровень.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который в два раза превышает нормальный уровень или на 80%, 60%, 40% или 20% превышает нормальный уровень.
Так, например, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента может повышаться до уровня, который на 5-100%, 5-80%, 5-60%, 5-40%, 5-20%, 10-100%, 10-80%, 10-60%, 10-40%, 10-20%, 15-100%, 15-80%, 15-60%, 15-40%, 15-20%, 20-100%, 20-80%, 20-60%, 20-40%, 25-100%, 25-80%, 25-60% или 25-40% превышает нормальный уровень.
Как описано выше, уровень эндогенного фактора I в сыворотке пациента может быть ниже нормального уровня. Таким образом, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента до введения ему рекомбинантной вирусной частицы согласно изобретению может быть ниже нормального уровня. Терапевтические эффекты у такого пациента могут быть достигнуты путем увеличения уровня C3b-инактивирующей и iC3b-расщепляющей активности до уровня, который превышает базовый уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента до введения ему рекомбинантной вирусной частицы согласно изобретению. Указанный повышенный уровень C3b-инактивирующей и iC3b-расщепляющей активности может, но необязательно, сохраняться на уровне ниже нормального уровня.
Таким образом, в конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, превышающего базовый уровень у пациента до введения ему рекомбинантной вирусной частицы согласно изобретению.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который по меньшей мере на 5%, 10%, 15%, 20% или 25% превышает базовый уровень у пациента до введения ему рекомбинантной вирусной частицы согласно изобретению.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который в два раза превышает базовый уровень у пациента до введения ему рекомбинантной вирусной частицы согласно изобретению или на 80%, 60%, 40% или 20% превышает базовый уровень у пациента до введения ему рекомбинантной вирусной частицы согласно изобретению.
Так, например, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента может повышаться до уровня, который на 5-100%, 5-80%, 5-60%, 5-40%, 5-20%, 10-100%, 10-80%, 10-60%, 10-40%, 10-20%, 15-100%, 15-80%, 15-60%, 15-40%, 15-20%, 20-100%, 20-80%, 20-60%, 20-40%, 25-100%, 25-80%, 25-60% или 25-40% превышает базовый уровень у пациента до введения ему рекомбинантной вирусной частицы согласно изобретению.
Комплемент-опосредуемым расстройством может быть расстройство, ассоциированное с дефектом регуляции альтернативного пути, а в частности, с гиперактивностью цикла обратной связи C3b комплемента.
Комплемент-опосредуемым расстройством может быть расстройство, характеризующееся наличием симптомов, которые могут быть ослаблены посредством повышения уровней C3b-инактивирующей и iC3b-расщепляющей активности у пациента.
Примерами комплемент-опосредуемых расстройств, которые могут быть подвергнуты профилактике или лечению в соответствии с настоящим изобретением, являются возрастная дегенерация желтого пятна (ВДЖП) (а в частности, ВДЖП на ранней стадии (сухой глаз), или региональная атрофия), болезнь плотных отложений (БПО), атипический гемолитический уремический синдром (АГУС), C3-гломерулопатии, мембранопролиферативный гломерулонефрит типа 2 (МПГН2 или МПГН типа II), атеросклероз, хроническое сердечно-сосудистое заболевание, болезнь Альцгеймера, системный васкулит, пароксизмальная ночная гемоглобинурия (ПНГ), и воспалительные или аутовоспалительные заболевания у пациентов пожилого возраста.
В одном из вариантов осуществления изобретения осуществляют профилактику или ослабление тяжести региональной атрофии. В другом варианте осуществления изобретения снижается тяжесть региональной атрофии.
C3-гломерулопатиями, которые могут быть подвергнуты профилактике или лечению в соответствии с настоящим изобретением, являются C3-гломерулонефрит и болезнь плотных отложений (также известная как мембранопролиферативный гломерулонефрит (МПГН) типа II).
Другими примерами комплемент-опосредуемых расстройств, которые могут быть подвергнуты профилактике или лечению в соответствии с настоящим изобретением, являются мембранопролиферативный гломерулонефрит типа I (МПГН типа I), мембранопролиферативный гломерулонефрит типа III (МПГН типа III), синдром Гийена-Барре, пурпура Геноха-Шенлейна, IgA-нефропатия и мембранозный гломерулонефрит. Мембранопролиферативный гломерулонефрит (МПГН) также известен как мезангиокапиллярный гломерулонефрит.
В конкретных вариантах осуществления изобретения, комплемент-опосредуемое расстройство, которое подвергают профилактике, лечению или ослаблению в соответствии с настоящим изобретением, выбрано из БПО, АГУС, C3-гломерулопатии, атеросклероза, хронического сердечно-сосудистого заболевания, болезни Альцгеймера, системного васкулита, ПНГ, воспалительных или аутовоспалительных заболеваний у пациентов пожилого возраста, МПГН типа I, МПГН типа III, синдрома Гийена-Барре, пурпуры Геноха-Шенлейна, IgA-нефропатии и мембранозного гломерулонефрита.
В одном из вариантов осуществления изобретения, указанные профилактика, лечение или ослабление комплемент-опосредуемого расстройства у пациента включают снижение указанной требуемой дозы, вводимой пациенту, и/или частоты введения дозы в соответствии с уже существующей схемой лечения, например снижение указанной требуемой дозы, вводимой пациенту, и/или частоты введения дозы ингибитора компонента пути комплемента (такого как ингибитор белка C5 комплемента, например, анти-C5 антитело) или противовоспалительного средства (такого как стероид).
В конкретных вариантах осуществления изобретения, комплемент-опосредуемым расстройством, которое подвергают профилактике или лечению в соответствии с настоящим изобретением, не является глазное заболевание.
В конкретных вариантах осуществления изобретения, комплемент-опосредуемым расстройством, которое подвергают профилактике или лечению в соответствии с настоящим изобретением, не является глазное комплемент-опосредуемое расстройство.
В конкретных вариантах осуществления изобретения, комплемент-опосредуемым расстройством, которое подвергают профилактике или лечению в соответствии с настоящим изобретением, не является возрастная дегенерация желтого пятна (ВДЖП) или региональная атрофия.
В конкретных вариантах осуществления изобретения, комплемент-опосредуемым расстройством, которое подвергают профилактике или лечению в соответствии с настоящим изобретением, не является диабетическая ретинопатия.
У пациента может наблюдаться риск развития комплемент-опосредуемого расстройства. Так, например, пациент может быть гомозиготным или гетерозиготным по одному или более SNP, ассоциированным с комплемент-опосредуемым расстройством.
В конкретных вариантах осуществления изобретения, у пациента наблюдается риск развития ВДЖП. Так, например, пациент может быть гомозиготным или гетерозиготным по одному или более SNP, ассоциированным с ВДЖП. Примерами таких SNP являются rs1061170 (кодирующий Y402H) фактора H, rs800292 (кодирующий V62I) фактора H, rs641153 (кодирующий R32Q) фактора B или rs2230199 (кодирующий R102G) C3, а в частности, rs1061170 фактора H, rs800292 фактора H или rs2230199 C3. Другие примеры редких мутаций в CFI, ассоциированных с прогрессирующей ВДЖП, которые обычно приводят к снижению уровней фактора I в сыворотке, описаны Kavanagh et al., Hum Mol Genet. 2015 Jul 1;24(13):3861-70. Эти мутации включают следующие SNP: rs144082872 (кодирующий P50A); 4:110687847 (кодирующий P64L); rs141853578 (кодирующий G119R); 4:110685721 (кодирующий V152M); 4:110682846 (кодирующий G162D); 4:110682801 (кодирующий N177I); rs146444258 (кодирующий A240G); rs182078921 (кодирующий G287R); rs41278047 (кодирующий K441R); rs121964913 (кодирующий R474).
Способы согласно изобретению могут также включать определение наличия у пациента риска развития комплемент-опосредуемого расстройства (например ВДЖП), например, путем обнаружения наличия у пациента гомозиготности или гетерозиготности по одному или более SNP, ассоциированным с комплемент-опосредуемым расстройством (например, путем обнаружения наличия у пациента гомозиготности или гетерозиготности по одному или более SNP, ассоциированным с перечисленными выше ВДЖП).
В исследовании, описанном Lay et al. (Clin Exp Immunol 181(2):314-322 Aug 2015), сыворотки, взятые у большого числа здоровых пациентов, были типированы по трем общим полиморфизмам, одному в С3 (R102G) и двум в факторе H (V62I и Y402H), которые влияют на предрасположенность к развитию ВДЖП и некоторых форм заболевания почек. Были протестированы сыворотки трех групп, то есть, группы, гомозиготной по трем аллелям с риском заболевания; группы, гетерозиготной по всем трем аллелям; и группы, гомозиготной по аллелям с низким риском заболевания. Эти группы варьировались по их ответу на добавление экзогенного фактора I, если альтернативный путь комплемента был активирован зимозаном. При этом наблюдалось снижение максимального количества образовавшегося iC3b и скорости превращения iC3b в C3dg. В случае использования комплотипа с риском развития заболевания, для достижения аналогичной негативной регуляции в обеих реакциях требуются более высокие дозы фактора I. Поскольку реакция взаимодействия iC3b с рецептором CR3 комплемента представляет собой основной механизм, посредством которого активация комплемента вызывает воспаление, то расщепление iC3b с образованием C3dg может рассматриваться как главный признак снижения воспаления, индуцируемого комплементом. Эти данные впервые продемонстрировали, что сыворотки, взятые у пациентов с различными аллелями комплемента, имеют такие же свойства, как это было предсказано в in vitro анализе на негативную регуляцию альтернативного пути комплемента после увеличения концентрации фактора I. Эти результаты подтвердили возможность использования экзогенного фактора I в качестве терапевтического средства для ингибирования гиперактивности цикла обратной связи C3b и для разработки способа лечения ВДЖП и некоторых форм заболевания почек.
В исследовании, описанном Lachmann et al. (Clinical & Experimental Immunology, Volume 183, Issue 1, pages 150-156, January 2016), количество фактора I, необходимое для превращения группы с наивысшим риском (гомозиготной по трем аллелям с риском развития ВДЖП, C3 (R102G), фактора H (V621 и Y402H)) в группу с наименьшим риском (гомозиготную по аллелям с низким риском), составляет приблизительно 22 мкг/мл фактора I; количество, необходимое для превращения группы с наивысшим риском в гетерозиготную группу (гетерозиготную по всем трем аллелям с риском развития заболевания) составляет 12,5 мкг/мл, а количество, необходимое для превращения гетерозиготной группы в группу с наименьшим риском, составляет 5 мкг/мл.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который по меньшей мере на 10% превышает нормальный уровень, если у пациента наблюдается гомозиготность по одному или более SNP, ассоциированным с ВДЖП. В конкретных вариантах осуществления изобретения, SNP, ассоциированными с ВДЖП, являются rs1061170 фактора H, rs800292 фактора H или rs2230199 C3.
В конкретных вариантах осуществления изобретения, уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который по меньшей мере на 50% превышает нормальный уровень, если у пациента наблюдается гомозиготность по одному или более SNP, ассоциированным с ВДЖП. В конкретных вариантах осуществления изобретения, SNP, ассоциированными с ВДЖП, являются rs1061170 фактора H, rs800292 фактора H или rs2230199 C3.
Способы согласно изобретению могут также включать определение уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента по меньшей мере через один, два, три, четыре, пять или шесть дней или по меньшей мере через одну, две или три недели или по меньшей мере через один, два, три, четыре, пять или шесть месяцев или по меньшей мере через один год после введения, и повторное введение, если уровень активности равен нормальному уровню или ниже этого уровня. Если было обнаружено, что уровень активности превышает нормальный уровень через один год после первого введения, то могут быть проведены последующие ежегодные анализы для подтверждения того, что уровень активности превышает нормальный уровень.
In vivo, фактор I комплемента (FI) экспрессируется, главным образом, в печени гепатоцитами, хотя было также обнаружено, что он экспрессируется in vitro в моноцитах, фибробластах, кератиноцитах и в эндотелиальных клетках пупочной вены человека. FI представляет собой гетеродимер, состоящий из тяжелой и легкой цепи, ковалентно связанных дисульфидной связью. FI экспрессируется как одна пропептидная цепь. После трансляции, белок подвергается нескольким модификациям, включая N-гликозилирование в эндоплазматическом ретикулуме и в аппарате Гольджи. Каждая цепь может быть гликозилирована в трех аспарагиновых остатках; эти молекулы сахара гликана с высокой молекулярной массой составляют 20-25% от кажущейся молекулярной массы белка. Кроме того, FI протеолитически расщепляется в сети всего аппарата Гольджи под действием фурина (или фермента, расщепляющего спаренные основные аминокислоты, PACE), который расщепляет четыре положительно заряженных аминокислоты, а именно RRKR. Эти четыре аминокислоты присутствуют на границе раздела тяжелой и легкой цепей и образуют линкерный пептид. После их удаления, профермент процессируется с образованием ферментативно активной расщепленной гетеродимерной формы. Хотя про-FI содержит 28 спаренных основных аминокислотных остатков по всей последовательности, однако, фурин распознает только сайт Arg-Arg-Lys-Arg и расщепляет только в этом положении. FI имеет N-концевую лидерную последовательность из 18-остатков, которая отщепляется до секреции с последующим высвобождением зрелого белка в кровоток (Nilsson, et al., Mol. Immunol., 44(8):1835-1844, March 2007).
FI состоит из нескольких доменов (Nilsson et al., 2011, Mol. Immunol. 48(14):1611-1620). Тяжелая цепь состоит из N-концевого домена комплекса, атакующего мембрану FI (FIMAC), акцепторного богатого цистеином домена рецептора (SRCR, также известного как домен CD5), домена рецептора липопротеина низкой плотности 1 и 2 и небольшой области неизвестной гомологии, иногда называемой D-областью. Легкая цепь FI представляет собой ферментативно активный домен, который состоит из домена сериновой протеазы, подобной химотрипсину (SP), где указанный домен содержит остатки, образующие каталитическую триаду His-Asp-Ser.
Было высказано предположение, что тяжелая цепь фактора I связывается с субстратом и ориентирует домен SP интактного FI на взаимодействие с двумя сайтами расщепления в C3b, что приводит к образованию iC3b. В 2011 году была опубликована кристаллическая структура FI, что позволило получить еще больше информации о расположении тяжелой и легкой цепей в комплексе C3b-FH-FI (Tsiftsoglou and Sim, J. Immunol., 173(1):367-375, July 2004). Было высказано предположение, что субстрат (то есть, комплекс C3b-FH) индуцирует структурное ремоделирование в активном сайте фактора I. Это было также подтверждено данными, указывающими на то, что ингибитор протеазы, диизопропилфторфосфат, способен взаимодействовать только с серином активного сайта, если FI был предварительно инкубирован в C3b. Наложение FI на кристаллическую структуру комплекса C3b-FH(1-4) показало, что в отличие от домена SRCR, тяжелая цепь плотно упакована вместе с C3b и кофактором. Это связывание нейтрализует аллостерические ингибирующие эффекты тяжелой цепи и индуцирует ремоделирование домена SP, который становится более активным и расщепляет C3b. Важное значение доступности домена FIMAC и FI в функции, присущей FI, было наглядно продемонстрировано в серии экспериментов по мутагенезу (Schlott, et al., J. Mol. Biol., 318(2):533-546, April 2002).
Фактор I или его фрагмент или производное, кодируемые рекомбинантным вирусным вектором согласно изобретению, могут включать сигнальную последовательность из нативных 18 аминокислотных остатков, что будет гарантировать правильный процессинг и секрецию зрелого фактора I. Альтернативно, может быть использована гетерологичная сигнальная последовательность (то есть, сигнальная последовательность, которая не кодируется нативным геном), либо эта сигнальная последовательность может быть удалена.
В конкретных вариантах осуществления изобретения, фактором I является человеческий фактор I, например, человеческий фактор I с аминокислотной последовательностью SEQ ID NO: 2 (вариант 2, с N-концевой сигнальной последовательностью из 18 остатков, эталонная последовательность NCBI: NM_000204.4), или с последовательностью SEQ ID NO: 4 (вариант 2, без N-концевой сигнальной последовательности из 18 остатков).
В других вариантах осуществления изобретения могут быть использованы и другие варианты фактора I, такие как человеческий фактор I, вариант 1 (этот вариант представляет собой более длинную изоформу и включает альтернативный экзон с сохранением рамки считывания в центральной кодирующей области по сравнению с вариантом 2). Последовательность этого варианта представляет собой эталонную последовательность: NCBI NM_001318057.1 и может быть использована вместе с N-концевой сигнальной последовательностью из 18 остатков или без этой последовательности.
Фрагмент или производное фактора I может представлять собой полипептид, который сохраняет C3b-инактивирующую и iC3b-расщепляющую активность и имеет аминокислотную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична человеческому фактору I с аминокислотной последовательностью SEQ ID NO: 2 или 4. Этот фрагмент или это производное могут содержать одну или более аминокислотных добавлений, делеций или замен, например, одну или более консервативных аминокислотных замен.
Консервативные аминокислотные замены, вероятно, оказывают минимальное влияние на активность полученного белка. Дополнительную информацию о консервативных заменах можно найти, например, у Ben Bassat et al. (J. Bacterial., 169:751-757, 1987), O'Regan et al. (Gene, 77:237-251, 1989), Sahin-Toth et al. (Protein Sci., 3:240-247, 1994), Hochuli et al. (Bio/Technology, 6:1321-1325, 1988) и в широко известных руководствах по генетике и молекулярной биологии. Матрицы Blosum обычно применяются для определения родства полипептидных последовательностей. Матрицы Blosum были созданы с использованием крупной базы данных по доверительному выравниванию (базы данных BLOCKS), где было оценено попарное выравнивание родственных последовательностей с меньшим процентом пороговой идентичности (Henikoff et al., Proc. Natl. Acad. Sci. USA, 89:10915-10919, 1992). Порог 90%-ной идентичности может быть использован для оценки частоты встречаемости для высококонсервативных мишеней в матрице BLOSUM90. Порог 65%-идентичности был использован для матрицы BLOSUM65. Оценка «ноль» и выше в матрицах Blosum рассматривается как «консервативные замены» при выбранном проценте идентичности. В нижеследующей таблице проиллюстрировано репрезентативные консервативные аминокислотные замены.
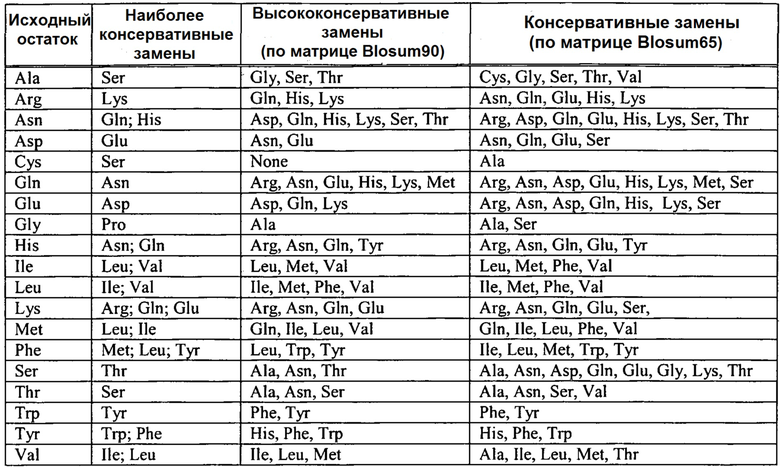
Фрагмент или производное фактора I может сохранять по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% или 100% C3b-инактивирующей и iC3b-расщепляющей активности нативного фактора I. C3b-инактивирующая и iC3b-расщепляющая активность фрагмента или производного фактора I и нативного фактора I может быть определена любым подходящим методом, известным специалистам. Так, например, измерение протеолитической активности фактора I описано Hsiung et al. (Biochem. J. (1982) 203, 293-298) на странице 295 в левом столбце. Анализы на гемолитическую и конглютинирующую активность FI описаны Lachmann PJ & Hobart MJ (1978) «Complement Technology» в руководстве по экспериментальной иммунологии в 3-м издании научной публикации Ed DM Weir Blackwells в главе 5A, стр. 17. Более подробное описание, также включающее анализ на протеолитическую активность, можно найти в публикации Harrison RA(1996) «Weir's Handbook of Experimental Immunology» 5th Edition Eds; Herzenberg Leonore A'Weir DM, Herzenberg Leonard A & Blackwell C Blackwells Scientific Publications Chapter 75 36-37. Анализ на конглютинацию является в высокой степени чувствительным и может быть использован для детектирования первого (двойного) захвата, превращающего фиксированный C3b в iC3b, и приобретения способности реагировать с конглютинином; и для детектирования конечного связывания с C3dg путем инициации реакции с фиксированным iC3b с последующим скринингом на потерю способности реагировать с конглютинином. Анализ на гемолитическую активность проводят для детектирования превращения C3b в iC3b, а анализ на протеолитическую активность позволяет детектировать все эти связывания.
В некоторых вариантах осуществления изобретения, нуклеиновая кислота, кодирующая фактор I, кодирует человеческий фактор I с аминокислотной последовательностью SEQ ID NO: 2 (вариант 2, с N-концевой сигнальной последовательностью из 18 остатков, эталонная последовательность NCBI: NM_000204.4) или с последовательностью SEQ ID NO: 4 (вариант 2, без N-концевой сигнальной последовательности из 18 остатков).
В некоторых вариантах осуществления изобретения, нуклеиновая кислота, кодирующая фактор I, содержит нуклеотидную последовательность SEQ ID NO: 1 (кодирующую область CDS фактора I, вариант транскрипта 2, мРНК, из эталонной последовательности NCBI: NM_000204.4, с N-концевой сигнальной последовательностью из 18 остатков) или SEQ ID NO: 3 (кодирующую область CDS фактора I, вариант транскрипта 2, мРНК, без N-концевой сигнальной последовательности из 18 остатков).
В некоторых вариантах осуществления изобретения может быть использована оптимизированная по кодонам нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное. Подходящие оптимизированные по кодонам последовательности могут быть получены, например, с использованием компьютерной программы по оптимизации кодонов, такой как SnapGene.
Примером оптимизированной по кодонам нуклеотидной последовательности, кодирующей фактор I, является SEQ ID NO: 7. Таким образом, в некоторых вариантах осуществления изобретения, нуклеиновая кислота, кодирующая фактор I, содержит, или состоит из нее, нуклеотидную последовательность SEQ ID NO: 7.
В некоторых вариантах осуществления изобретения, нуклеиновая кислота, кодирующая фактор I или его фрагмент или производное, может содержать последовательность нуклеиновой кислоты, которая по всей своей длине по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеиновой кислоте, кодирующей человеческий фактор I с нуклеотидной последовательностью SEQ ID NO: 1 или 3.
Термин «процент идентичности (%) последовательностей» по сравнению с эталонной полипептидной последовательностью или последовательностью нуклеиновой кислоты определен здесь как процент аминокислотных остатков или нуклеотидов в последовательности, которые идентичны аминокислотным остаткам или нуклеотидам в эталонной полипептидной последовательности или последовательности нуклеиновой кислоты после выравнивания и введения «пробелов», если это необходимо для достижения максимального процента идентичности последовательностей. Выравнивание в целях определения процента идентичности аминокислотных последовательностей или последовательностей нуклеиновой кислоты может быть осуществлено различными способами, известными специалистам, например, с использованием общедоступных компьютерных программ, таких как программы, описанные в руководствах Current Protocols in Molecular Biology (Ausubel et al., eds., 1987), Приложение 30, раздел 7.7.18, таблица 7.7.1, и такие программы включают программы BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Примером программы выравнивания является ALIGN Plus (Scientific and Educational Software, Pennsylvania). Специалист в данной области может самостоятельно определить соответствующие параметры для оценки выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Для этих целей, процент идентичности данной аминокислотной последовательности А с данной аминокислотной последовательностью В (которая, альтернативно, может быть описана как данная аминокислотная последовательность А, которая имеет или включает некоторый % идентичности с данной аминокислотной последовательностью Β) вычисляется следующим образом: 100 × дробь Χ/Υ, где X представляет собой число аминокислотных остатков, оцененных как идентичные в программе выравнивания последовательностей А и В, и где Υ означает общее число аминокислотных остатков в последовательности Β. Следует отметить, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то процент идентичности аминокислотной последовательности А и В не равен проценту идентичности аминокислотных последовательностей В и А. В этих целях, процент идентичности данной последовательности нуклеиновой кислоты С и данной последовательности нуклеиновой кислоты D (которая, альтернативно, может быть описана как данная последовательность нуклеиновой кислоты С, которая имеет или включает некоторый % идентичности с данной последовательностью нуклеиновой кислоты D) вычисляется следующим образом: 100 × дробь W/Z, где W означает число нуклеотидов, оцененных как идентичные в программе выравнивания последовательностей С и D, и где Ζ означат общее число нуклеотидов в D. Следует отметить, что если длина последовательности нуклеиновой кислоты С не равна длине последовательности нуклеиновой кислоты D, то процент идентичности последовательности нуклеиновой кислоты С и D не равен проценту идентичности последовательностей нуклеиновой кислоты D и C.
Термин «пациент» означает млекопитающее. Подходящими примерами являются домашние животные (такие как коровы, овцы, кошки, собаки, лошади), приматы (например, человек и приматы, не являющиеся человеком, такие как обезьяны), кролики и грызуны (например, мыши и крысы). В некоторых вариантах осуществления изобретения, пациентом является человек.
Используемые здесь термины «лечить» или «лечение» означают способ получения благоприятных или желаемых клинических результатов. В соответствии с настоящим изобретением, благоприятные или желаемые клинические результаты включают ослабление симптомов, уменьшение степени заболевания, стабилизацию (например, отсутствие ухудшения) заболевания, предотвращение распространения заболевания, отсрочку или замедление прогрессирования заболевания, ослабление или снижение тяжести патологического состояния, снижение требуемой дозы и/или частоты введения при уже проводимом курсе терапии и ремиссию (частичную или полную) независимо от того, является ли она детектируемой или недетектируемой. Термин «лечение» может также означать увеличение продолжительности жизни по сравнению с ожидаемой продолжительностью жизни без лечения. Термин «лечение» может быть применим и к периоду времени и/или к частоте введения, достаточным для достижения благоприятных или желаемых клинических результатов.
Используемый здесь термин «предупреждение» или «профилактика» относится к лечению, если известно, что у пациента имеется или, возможно, имеется риск развития расстройства, но при этом, у него не наблюдается никаких симптомов или минимальных симптомов расстройства. Пациентом, проходящим профилактическое лечение, может быть пациент, проходящий лечение до начала проявления симптомов.
«Терапевтически эффективное количество» означает количество, достаточное для достижения благоприятных или желаемых результатов, включая клинические результаты (например, ослабление симптомов и достижение клинических конечных точек). Терапевтически эффективное количество может быть введено за одно или более введений. В случае патологического состояния, терапевтически эффективным количеством является количество, достаточное для ослабления симптомов, стабилизации или замедления развития заболевания.
Следует отметить, что рекомбинантный вирусный вектор согласно изобретению обычно представляет собой выделенный рекомбинантный вирусный вектор, то есть, вектор, отделенный от компонентов его природного окружения и/или не содержащий таких компонентов. Аналогичным образом, рекомбинантная вирусная частица согласно изобретению обычно представляет собой выделенную рекомбинантную вирусную частицу, то есть, частицу, отделенную от компонентов ее природного окружения и/или не содержащую таких компонентов.
Настоящее изобретение также относится к фармацевтической композиции, которая включает: рекомбинантный вирусный вектор согласно изобретению или рекомбинантную вирусную частицу согласно изобретению и фармацевтически приемлемый носитель, наполнитель или разбавитель.
Фармацевтическая композиция согласно изобретению может быть подходящей для любого соответствующего способа ее введения, например, внутривенного введения или для любого подходящего способа вышеописанного введения.
Фармацевтические композиции согласно изобретению могут быть подходящими для их введения человеку.
Настоящее изобретение также относится к набору, который включает: рекомбинантный вирусный вектор согласно изобретению или рекомбинантную вирусную частицу согласно изобретению и фармацевтически приемлемый носитель, наполнитель или разбавитель.
В некоторых вариантах осуществления изобретения, набор согласно изобретению может также включать инструкции по лечению описанного здесь комплемент-опосредуемого расстройства с применением любых способов, векторов или вирусных частиц согласно изобретению. Набор согласно изобретению может включать фармацевтически приемлемый носитель, наполнитель или разбавитель, подходящий для инъекции пациенту, нуждающемуся в лечении. Набор согласно изобретению может также включать любые из следующих компонентов, таких как буфер, фильтр, игла, шприц и вкладыш в упаковку с инструкциями по введению пациенту. Могут быть также включены подходящие упаковочные материалы, и такими материалами могут быть любые материалы, известные специалистам, включая, например, флаконы (такие как герметичные флаконы), сосуды, ампулы, бутыли, кувшины, жесткие упаковки (например, герметично закрытые пакеты Милара или пластиковые пакеты). Эти упаковочные материалы могут быть также стерилизованы и/или герметично запаяны.
Фармацевтическая композиция или набор согласно изобретению могут быть упакованы в виде одной унифицированной дозы или в форме с множеством доз. Фармацевтическая композиция согласно изобретению и содержимое набора согласно изобретению обычно приготавливают в виде стерильного и, в основном, изотонического раствора.
Фармацевтически приемлемые носители, наполнители и разбавители представляют собой относительно инертные вещества, которые облегчают введение фармакологически эффективного вещества и могут быть получены в виде жидких растворов или суспензий, в виде эмульсий или в виде твердых веществ, подходящих для растворения или суспендирования в жидкости перед их использованием. Так, например, наполнитель может придавать форму или консистенцию или действовать как разбавитель. Подходящими наполнителями являются, но не ограничиваются ими, стабилизирующие агенты, смачивающие и эмульгирующие агенты, соли для изменения осмомолярности, инкапсулирующие агенты, рН-забуферивающие вещества и буферы. Такие наполнители включает любое фармацевтическое средство, подходящее для прямой доставки пациенту (например, внутривенно или в печень), которая может быть осуществлена без излишней токсичности. Фармацевтически приемлемыми наполнителями являются, но не ограничиваются ими, сорбит, любые различные соединения TWEEN и жидкости, такие как вода, физиологический раствор, глицерин и этанол. Могут быть также включены фармацевтически приемлемые соли, такие как, например, соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты, сульфаты и т.п.; и соли органических кислот, такие как ацетаты, пропионаты, малонаты или бензоаты.
В некоторых вариантах осуществления изобретения, фармацевтически приемлемыми наполнителями могут быть фармацевтически приемлемые носители. Такими фармацевтически приемлемыми носителями могут быть стерильные жидкости, такие как вода и масло, включая жидкости, полученные из нефти и продуктов животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло и минеральное масло. Физиологические растворы и водные растворы декстрозы, полиэтиленгликоля (ПЭГ) и глицерина могут быть также использованы в виде жидких носителей, а в частности, носителей для приготовления растворов для инъекций. Могут быть также использованы дополнительные ингредиенты, например, консерванты, буферы, агенты, придающие тоничность, антиоксиданты и стабилизаторы, неионные смачивающие или осветляющие агенты или агенты, повышающие вязкость.
Подробное обсуждение фармацевтически приемлемых наполнителей и носителей приводится в руководстве Remington's Pharmaceutical Sciences (Mack Pub. Co., N.J. 1991).
В некоторых вариантах осуществления изобретения, титр вируса в фармацевтической композиции согласно изобретению составляет по меньшей мере 5 × 1012, 6 × 1012, 7 × 1012, 8 × 1012, 9 × 1012, 10 × 1012, 11 × 1012, 15 × 1012, 20 × 1012, 25 × 1012, 30 × 1012 или 50 × 1012 геномных копий/мл. В некоторых вариантах осуществления изобретения, титр вируса в фармацевтической композиции согласно изобретению составляет от 5 × 1012 до 10 × 1012, от 10 × 1012 до 25 × 1012 или от 25 × 1012 до 50 × 1012 геномных копий/мл. В некоторых вариантах осуществления изобретения, титр вируса в фармацевтической композиции согласно изобретению составляет от 5 × 1012 до 6 × 1012, от 6 × 1012 до 7 × 1012, от 7 × 1012 до 8 × 1012, от 8 × 1012 до 9 × 1012, от 9 × 1012 до 10 × 1012, от 10 × 1012 до 11 × 1012, от 11 × 1012 до 15 × 1012, от 15 × 1012 до 20 × 1012, от 20 × 1012 до 25 × 1012, от 25 × 1012 до 30 × 1012, от 30 × 1012 до 50 × 1012 или от 50 × 1012 до 100 × 1012 геномных копий/мл.
В некоторых вариантах осуществления изобретения, титр вируса в фармацевтической композиции согласно изобретению составляет по меньшей мере 5 × 109, 6 × 109, 7 × 109, 8 × 109, 9 × 109, 10 × 109, 11 × 109, 15 × 109, 20 × 109, 25 × 109, 30 × 109 или 50 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления изобретения, титр вируса в композиции составляет от 5 × 109 до 10 × 109, от 10 × 109 до 15 × 109, от 15 × 109 до 25 × 109 или от 25 × 109 до 50 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления изобретения, титр вируса в композиции составляет от 5 × 109 до 6 × 109, от 6 × 109 до 7 × 109, от 7 × 109 до 8 × 109, от 8 × 109 до 9 × 109, от 9 × 109 до 10 × 109, от 10 × 109 до 11 × 109, от 11 × 109 до 15 × 109, от 15 × 109 до 20 × 109, от 20 × 109 до 25 × 109, от 25 × 109 до 30 × 109, от 30 × 109 до 50 × 109 или от 50 × 109 до 100 × 109 трансдуцирующих единиц/мл.
Используемый здесь термин «трансдуцирующая единица», если он относится к титру вируса, означает число инфекционных частиц рекомбинантного вектора, необходимое для получения функционального трансгенного продукта, оцениваемого в функциональных анализах, таких как анализы, описанные в примерах в WO 2015/168666 или например, в публикациях Xiao et al. (1997) Exp. Neurobiol., 144:113-124; или Fisher et al. (1996) J. Virol., 70:520-532 (анализ LFU).
В некоторых вариантах осуществления изобретения, титр вируса в фармацевтической композиции согласно изобретению составляет по меньшей мере 5 × 1010, 6 × 1010, 7 × 1010, 8 × 1010, 9 × 1010, 10 × 1010, 11 × 1010, 15 × 1010, 20 × 1010, 25 × 1010, 30 × 1010, 40 × 1010 или 50 × 1010 инфекционных единиц/мл. В некоторых вариантах осуществления изобретения, титр вируса в композиции составляет по меньшей мере от 5 × 1010 до 10 × 1010, от 10 × 1010 до 15 × 1010, от 15 × 1010 до 25 × 1010 или от 25 × 1010 до 50 × 1010 инфекционных единиц/мл. В некоторых вариантах осуществления изобретения, титр вируса в композиции составляет по меньшей мере от 5 × 1010 до 6 × 1010, от 6 × 1010 до 7 × 1010, от 7 × 1010 до 8 × 1010, от 8 × 1010 до 9 × 1010, от 9 ×× 1010 до 10 × 1010, от 10 × 1010 до 11 × 1010, от 11 × 1010 до 15 × 1010, от 15 × 1010 до 20 × 1010, от 20 × 1010 до 25 × 1010, от 25 × 1010 до 30 × 1010, от 30 × 1010 до 40 × 1010, от 40 × 1010 до 50 × 1010 или от 50 × 1010 до 100 × 1010 инфекционных единиц/мл.
Используемый здесь термин «инфекционная единица», если он относится к титру вируса, означает число инфекционных и компетентных по репликации частиц рекомбинантного вектора, оцениваемое с помощью анализа, проводимого в Инфекционном центре и также известного как анализ на репликацию в Биологическом центре, описанный, например, McLaughlin et al. (1988) J. Virol., 62:1963-1973.
Настоящее изобретение также относится к набору для продуцирования частиц rAAV, включающему вектор rAAV согласно изобретению; одну или более хелперных плазмид, содержащих нуклеиновую кислоту, кодирующую белки репликации AAV и капсидные белки; и гены, необходимые для осуществления продуктивного жизненного цикла AAV.
В некоторых вариантах осуществления изобретения, набор согласно изобретению для продуцирования частиц rAAV включает первую хелперную плазмиду, содержащую нуклеиновую кислоту, кодирующую белки репликации AAV и капсидные белки; и вторую хелперную плазмиду, содержащую нуклеиновую кислоту, включающую гены, необходимые для осуществления продуктивного жизненного цикла AAV.
Гены, необходимые для осуществления продуктивного жизненного цикла AAV, могут представлять собой гены аденовируса, герпесвируса или бакуловируса. В конкретных вариантах осуществления изобретения, генами являются гены аденовируса, а в частности, РНК-гены аденовируса E1a, E1b, E2a, E4 и VA.
Настоящее изобретение также относится к рекомбинантному вирусному вектору согласно изобретению, к рекомбинантной вирусной частице согласно изобретению или к фармацевтической композиции согласно изобретению, используемой в качестве лекарственного средства.
Настоящее изобретение также относится к рекомбинантному вирусному вектору согласно изобретению, к рекомбинантной вирусной частице согласно изобретению или к фармацевтической композиции согласно изобретению, используемой для лечения комплемент-опосредуемого расстройства.
Настоящее изобретение также относится к применению рекомбинантного вирусного вектора согласно изобретению, рекомбинантной вирусной частицы согласно изобретению или фармацевтической композиции согласно изобретению в целях приготовления лекарственного средства для лечения комплемент-опосредуемого расстройства.
Комплемент-опосредуемым расстройством может быть расстройство, ассоциированное с дефектом регуляции альтернативного пути, а в частности, с гиперактивностью цикла обратной связи C3b комплемента.
Примерами комплемент-опосредуемых расстройств, которые могут быть подвергнуты профилактике или лечению в соответствии с настоящим изобретением, являются возрастная дегенерация желтого пятна (ВДЖП) (а в частности, ВДЖП на ранней стадии (сухой глаз), или региональная атрофия), болезнь плотных отложений (БПО), атипический гемолитический уремический синдром (АГУС), C3-гломерулопатии, мембранопролиферативный гломерулонефрит типа 2 (МПГН2), атеросклероз, хроническое сердечно-сосудистое заболевание, болезнь Альцгеймера, пароксизмальная ночная гемоглобинурия (ПНГ) и аутовоспалительные заболевания у пациентов пожилого возраста.
Другими примерами комплемент-опосредуемых расстройств, которые могут быть подвергнуты профилактике или лечению в соответствии с настоящим изобретением, являются МПГН типа I, МПГН типа III, синдром Гийена-Барре, пурпура Геноха-Шенлейна, IgA-нефропатия и мембранозный гломерулонефрит.
Представленные здесь варианты осуществления изобретения описаны лишь в качестве примеров со ссылками на прилагаемые чертежи, где:
На фигуре 1 проиллюстрирована схема стадий активации комплемента.
На фигуре 2 проиллюстрирована петля обратной связи альтернативного пути комплемента у позвоночных.
На фигуре 3 проиллюстрирован протеолиз C3 в процессе активации комплемента. C3 состоит из α- и β-цепи. β-цепь не является модифицированной, а α-цепь подвергалась расщеплению несколько раз: i) C3a отщеплялась под действием C3-конвертазы, и оставшийся белок был обозначен C3b; ii) FI расщеплялся в двух сайтах, что приводило к высвобождению пептида C3f (FH как кофактора), обозначаемого iC3b или C3bi; iii) FI снова расщеплялся под действием CR1 в качестве кофактора с образованием фрагмента iC3b размером 68 кДа. При этом C3c подвергался диффузии. C3dg оставался связанным с клеточной поверхностью и мог впоследствии расщепляться трипсин-подобными протеазами.
На фигуре 4 схематически представлена генотерапия на основе AAV.
На фигуре 5 представлена конструкция AAV2/8, используемая для сверхэкспрессии фактора I в сыворотке. Трансген, называемый здесь фактором I, был встроен между промотором и сайтом распознавания полиаденилирования. На обеих сторонах были фланкированы два инвертированных концевых повтора (ITR). Капсидная последовательность и другие аденовирусные гены были упакованы в другие плазмиды.
На фигуре 6 показаны повышенные уровни фактора I, измеренные с помощью вестерн-блот-анализа. Мышиную сыворотку разводили до 5%, и 10 мкл разделяли с помощью электрофореза в ДСН-ПААГ. FI детектировали с использованием антитела против α-FI (1:500), которое взаимодействует с тяжелой цепью mFI. Этот эксперимент проводили два раза.
На фигуре 7 представлены репрезентативные результаты ELISA на ингибирование. (a) Данные анализа калибровали по известным концентрациям очищенного рекомбинантного mFI. Концентрации ниже 0,25 мкг/мл давали позитивный сигнал. (b) Нормальная мышиная сыворотка (NMS) была позитивной при концентрациях ниже 0,625%. (c-f) ELISA на ингибирование проводили с использованием сыворотки мышей, которым вводили AAV-конструкции. На этой фигуре показана только одна проба сыворотки на группу. Этот эксперимент проводили два раза с тремя повторностями.
На фигуре 8 представлены результаты анализа на расщепление C3b и iC3b in vitro для оценки функциональной активности сверхэкспрессируемого фактора I. Субстрат, то есть, C3α'1-цепь, продуцировался быстро и имел сравнимую скорость расщепления, то есть, расщепления с образованием C3dg. Расщепление C3 детектировали с использованием α-hC3dg (1 мкг/мл). Этот эксперимент проводили два раза; и
На фигуре 9 представлены результаты анализа на отложение iC3b для тестирования функциональной активности сверхэкспрессируемого фактора I. Сыворотку мыши, которой вводили инъекцию, разводили до 25% в буфере для фиксации комплемента альтернативного пути и загружали на планшет, покрытый ЛПС. После инкубирования в течение 1 часа при 37°C, связанный iC3b детектировали с использованием антитела против человеческого α-C3c. Поглощение измеряли на 415 нм. Этот эксперимент проводили два раза с двумя повторностями.
Пример 1
AAV-экспрессионная система
В этом примере описано получение рекомбинантного вирусного вектора, кодирующего мышиный фактор I, сверхэкспрессирующийся в мышиных гепатоцитах, и получение вирусных частиц (вирионов), инкапсидирующих этот вектор.
Была использована конструкция аденоассоциированного вируса (AAV). Эта конструкция состоит из остова вируса AAV2, который был псевдотипирован капсидным белком AAV8 для сообщения тропизма к печени. Поэтому, этот вирус инифицрует, главным образом, печень, а трансген FI сверхэкспрессируется в природном сайте. Для дополнительного подавления экспрессии трансгена фактора I вне печени использовали промотор α-1-антитрипсина (AAT) с двумя дополнительными регуляторными областями ApoE в печени (см. фигуру 5). Все гены, необходимые для продуцирования вируса, распределяли между тремя плазмидами, которые были котрансфецированы в клеточную линию, которая вводила остальные недостающие гены для упаковки AAV как подробно описано ниже.
AAV (дикого типа): 4,7 т.п.о.-геном AAV дикого типа характеризуется наличием двух инвертированных концевых повторов (ITR) и двух серий открытых рамок считывания (ОРС), которые кодируют белки репликации (Rep) и капсидные белки (Cap). ОРС Rep кодирует четыре белка (78, 68, 52 и 40 кДа), функция которых заключается, главным образом, в регуляции репликации и интеграции AAV. ОРС Cap кодирует три структурных белка (85 кДа (VP1), 72 кДа (VP2) и 61 кДа (VP3)). Соотношение VP1:VP2:VP3 в капсиде составляет приблизительно 1:1:8 или 1:1:10.
Рекомбинантный AAV (rAAV): rAAV, используемый в этом примере, представляет собой AAV2, псевдотипированный капсидом AAV8 (rAAV2/8) для сообщения тропизма к печени. Вирионы rAAV2/8 упаковывали в клетки HEK293 путем тройной трансфекции с использованием трех плазмид, описанных ниже:
1. pAM2AA (ITR и кластер для экспрессии трансгена): В этой плазмиде, 145 п.о. инвертированных концевых повторов (ITR) от AAV2 фланкируют трансгенный кластер. Два ITR представляют собой лишь цис-элементы, необходимые для осуществления всех стадий жизненного цикла AAV. Они функционируют как ориджин репликации ДНК, обеспечивают передачу сигналов упаковки и интеграции и служат в качестве регуляторных элементов для экспрессии гена AAV дикого типа. Кластер pAM2AA включает промотор человеческого α-1-антитрипсина (hAAT) с двумя энхансерами ApoE, за которыми следует кДНК, кодирующая трансген. 3'-нетранслируемая область (UTR) содержит посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE) и сигнал полиаденилирования бычьего гормона роста (BGH polyA). Упаковывающая способность AAV дикого типа составляет приблизительно 4,7 т.п.о. В pAM2AA, регуляторные области включают ≈2390 п.о., а остальные ≈2310 п.о. ответственны за инсерцию трансгенов.
2. p5E18VD/8 (хелперные последовательности AAV): Делетированные вирусные кодирующие последовательности AAV2, включая гены Rep и Cap, присутствуют в плазмиде, запускаемой промотором p5 AAV. В данном случае, последовательности, кодирующие капсид AAV2, заменяют последовательностями, кодирующими капсид AAV8. При этом, какой-либо гомологии между векторными и хелперными последовательностями не наблюдалось, что снижает вероятность продуцирования рекомбинантных AAV дикого типа.
3. pXX6 (аденовирусные хелперные функции): Аденовирусные гены, играющие важную роль в продуктивном жизненном цикле AAV, представляют собой гены РНК E1a, E1b, E2a, E4 и VA. E1a служит в качестве трансактиватора, который активирует транскрипцию аденовирусных генов, а также генов Rep и Cap AAV. E1b взаимодействует с E4, что облегчает перенос вирусных мРНК. E4 также участвует в репликации ДНК AAV. РНК E2a и VA способствуют повышению стабильности вирусной мРНК и повышают эффективность трансляции, в частности, транскриптов Cap. pXX6 содержит главне хелперные гены аденовируса, но не содержит структурных генов и генов репликации аденовируса. Главными хелперными генами, включенными в pXX6, являются гены E2a, E4 и VA. E1a и E1b были делетированы, и эти отсутствующие гены E1a и E1b были комплементированы клетками HEK293.
Получение рекомбинантного вектора AAV, кодирующего мышиный фактор I (AAV_mFI)
Для клонирования фактора I в экспрессионный вектор pAM2AA, рестрикционные сайты были заменены совместимыми фермент-распознающими сайтами (с использованием праймерных последовательностей: FI-AAV-F: 5'-TCT AGA GGA TCC GCC ACC ATG AAG CTC-3' (SEQ ID NO: 5); и FI-AAV-R: 5'-AAG CTT GGC GGC CGC TCA GAC ATT GTG TTG AGA AAC AAG AGA CCT TC-3'(SEQ ID NO:6). Новые последовательности были присоединены к мышиной кДНК фактора I посредством ПЦР-амплификации. После очистки амплифицированной конструкции, она была лигирована в pAM2AA. кДНК мышиного фактора I комплемента клонировали в pAM2AA с использованием рестрикционных сайтов XbaI и HindIII. После получения вектора, вектор AAV_GFP обрабатывали тем же самым способом, а затем проводили стадии упаковки и очистки вируса. Это гарантировало отсутствие ошибок после экспрессии GFP в печени, который можно легко наблюдать визуально под флуоресцентным микроскопом.
Получение вирионов rAAV2/8
Вектор pAM2AA с кДНК фактора I мышиного комплемента переносили в клетки SURE2 для предотвращения нежелательных событий рекомбинации. Клетки HEK293 трансфецировали всеми тремя плазмидами с использованием CaPO4. Через два дня после трансфекции, клетки собирали и подвергали лизису путем проведения повторяющихся циклов замораживания/оттаивания в бане со смесью сухого льда/100% этанола, а затем хранили при -80°C. Вирионы AAV очищали в градиенте хлорида цезия, и вирусные частицы количественно оценивали с помощью ПЦР в реальном времени.
Пример 2
In vivo сверхэкспрессия фактора I комплемента посредством системы доставки аденоассоциированного вируса
В этом примере описаны: инъекция мышам вирионов, инкапсидирующих вектор AAV_mFI, полученный как описано в Примере 1; определение уровней фактора I в плазме после инъекции вирионов; и оценка негативной регуляции альтернативного пути комплемента у мышей после инъекции.
Инъекция мышам
12 мышей распределяли по 4 группам, каждой из которых вводили различные концентрации AAV_mFI или препарат контрольного вируса:
- низкая доза mFI=5 × 109 вирионов
- средняя доза mFI=5 × 1010 вирионов (=100% трансдукция в печень в предыдущих экспериментах)
- высокая доза mFI=5 × 1011 вирионов
- GFP=5 × 1011 вирионов.
Мышам вводили внутривенную инъекцию в хвостовую вену. Кровь брали через 4 недели, а еще через 8 недель мышей умерщвляли и их сыворотки анализировали.
Результаты
Количественная оценка повышенных уровней фактора I
Для оценки уровней FI заказывали специально приготовленную поликлональную антисыворотку α-mFI. При этом осуществляли вестерн-блот-анализ. В данном случае, антитело против α-FI (Santa Cruz, sc-69465) использовали в качестве детектирующего антитела для приблизительной оценки уровней FI. Сыворотку разводили в несколько последовательных стадий для снижения погрешностей при пипетировании. Относительную концентрацию FI определяли следующим образом: AAV_GFP < AAV_FI в низкой дозе < AAV_FI в средней дозе < AAV_FI в высокой дозе (см. фигуру 6).
После получения антисыворотки проводили гораздо более чувствительный анализ, то есть, ELISA на ингибирование. Сыворотку предварительно инкубировали с антисывороткой α-mFI с заранее определенной концентрацией, а затем инкубировали на микротитрационном планшете, покрытом рекомбинантным mFI. Получение и специфичность связывания α-mFI-антисыворотки описаны в разделе «Материалы и методы», приведенном ниже. На этом планшете, только несвязанные антитела были способны взаимодействовать с иммобилизованным антигеном. Поэтому, анализ давал положительные результаты только в том случае, когда количество α-mFI-антител в антисыворотке превышало количество mFI, присутствующего в сыворотке. Данные этого анализа калибровали по известным концентрациям фактора I.
Было обнаружено, что концентрации антисыворотки, превышающие 75 мкг/мл, давали сильный положительный сигнал, если микротитрационный планшет был покрыт 1 мкг mFI на лунку (не показано). На фигуре 7 представлены репрезентативные результаты ELISA на ингибирование. Этот анализ давал положительный сигнал при концентрациях менее, чем 0,25 мкг/мл (фигура 7a). С использованием такой калибровки можно определить концентрацию FI в пробе сыворотки. Сначала был протестирован нормальный образец, взятый у мыши дикого типа, и было обнаружено, что положительный сигнал наблюдался при концентрациях менее, чем 0,625% (фигура 7b). После определения порога положительного сигнала в этом анализе при 0,25 мкг/мл mFI могла быть вычислена концентрация mFI в 100% сыворотке, и эта концентрация составляла 40 мкг/мл. Такие же вычисления были сделаны для каждой сыворотки мыши, которой вводили AAV-конструкции (фигура 7c-f), и определяли интервалы концентраций на группу (см. таблицу 2).
Таблица 2. Количественная оценка фактора I после сверхэкспрессии посредством системы экспрессии аденоассоциированного вируса
Концентрации даны в следующем порядке: 20-40 мкг/мл FI у контрольной группы, которой вводили GFP; 40-80 мкг/мл у группы, которой вводили низкую дозу FI; 80 мкг/мл у группы, которой вводили среднюю дозу FI; и 80-160 мкг/мл FI у группы, которой вводили высокую дозу FI. Был сделан вывод, что при проведении генотерапии с использованием AAV, концентрация в сыворотке могла увеличиваться в четыре раза по сравнению с нормальными уровнями.
Функциональный анализ в сыворотке с повышенными уровнями фактора I
Сыворотку трансгенных мышей анализировали таким же способом, как и рекомбинантный фактор I. Поскольку теоретически сверхэкспрессируемый белок процессировался и секретировался как эндогенный FI, то сыворотки мышей можно было непосредственно сравнивать друг с другом. Затем повышение уровня FI было подтверждено с помощью иммуноблотинга и ELISA на ингибирование, после чего был проведен анализ на функциональную активность сверхэкспрессируемого фермента. Следовательно, сыворотки анализировали на функциональную активность FI с помощью анализа на расщепление C3b in vitro, анализа на гемолиз и анализа на отложение iC3b.
Расщепление C3b и iC3b оценивали в анализе in vitro. Человеческий C3 гидролизовали до C3b под действием трипсина и инкубировали с hFH и сывороткой от трансгенных мышей. Через 30 минут, 1 мкг человеческого C3 загружали на ДСН-гель и подвергали блоттингу с использованием клона 9. C3b сначала расщеплялся с образованием iC3b в быстрой реакции (антитело реагировало с цепью C3α'1 iC3b), а затем с образованием C3dg в более медленной реакции. Эта вторая реакция могла ускоряться за счет повышения концентрации FI. Было показано, что человеческий C3b, инкубированный с человеческим фактором H и с сывороткой, взятой у мышей различных групп, более эффективно расщеплялся с образованием C3dg за период инкубирования у группы со средней дозой AAV_FI и у группы с высокой дозой AAV_FI (см. фигуру 8, дорожки 10-14). У мышей, которым вводили только контрольную плазмиду, почти не наблюдалось полосы C3dg за этот период инкубирования.
Следующая стадия была проведена для того, чтобы определить, является ли сверхэкспрессируемый фактор I также функционально активным в цельной сыворотке. Для этого, микротитрационный планшет покрывали ЛПС и оценивали отложение C3b/iC3b после инкубирования с сывороткой трансгенных мышей при 37°C. Результаты представлены на фигуре 9. Как и в случае рекомбинантного белка, у мышей, которым вводили AAV_mFI, наблюдался меньший уровень отложения C3b/iC3b на ЛПС, поскольку петля положительной обратной связи в альтернативном пути прерывалась фактором I. Отложение C3b/iC3b снижалось на 11% у группы, которой вводили низкую дозу AAV_FI; на 38% у группы, которой вводили среднюю дозу AAV_FI; и на 50% у группы, которой вводили высокую дозу AAV_FI.
Был сделан вывод, что фактор I комплемента мог сверхэкспрессироваться при проведении генотерапии с использованием вирусного вектора до уровней, которые способствуют значительному ингибированию альтернативного пути комплемента.
Обсуждение
Генотерапия представляет собой новый и перспективный терапевтический способ лечения различных заболеваний. Этот способ позволяет осуществлять in vivo экспрессию белков после трансфекции соответствующих клеток конструкцией, несущей последовательность ДНК, кодирующую представляющий интерес белок. Теоретически, генотерапия может быть применена для экспрессии любого белка, однако, на практике, обычно существуют ограничения, такие как, например, длина последовательности. Применение генотерапии позволяет заменять отсутствующие белки. Однако это всегда связано с риском иммуногенности. Авторы считают, что такой риск можно устранить при повышении концентраций уже существующих (эндогенных) белков. В этом исследовании, авторами была поставлена задача выяснить, может ли фактор I комплемента сверхэкспрессироваться посредством генотерапии до уровней, которые будут приводить к значительному ингибированию альтернативного пути комплемента.
Авторами был успешно продуцирован вектор для трансгенной экспрессии мышиного фактора I комплемента. Используемая вирусная экспрессионная система была получена на основе AAV2, а его капсид происходит от AAV8. Вместе с этим, эта конструкция AAV2/8, главным образом, способствует трансдукции в печень мышей, хотя вирусные частицы могут также присутствовать в ткани за пределами печени. Для повышения уровня печень-специфической экспрессии, трансген должен находиться под контролем промотора α-1-антитрипсина с двумя контрольными областями ApoE печени, что будет обеспечивать высокий уровень и специфичность экспрессии в гепатоцитах. Следовательно, экспрессия трансгена должна осуществляться только в гепатоцитах (хотя, иногда очень незначительный уровень экспрессии трансгена также может наблюдаться в ткани поджелудочной железы, которая представляет собой очень близкородственную ткань).
Авторами было продемонстрировано, что сверхэкспрессия FI под действием используемой экспрессионной системы AAV оказывает заметное влияние на негативную регуляцию комплемента, и во-вторых, этот эффект может быть проанализирован путем титрования и становится более заметным по мере повышения дозы вектора. Для измерения повышения уровня общего FI у кроликов увеличивают уровень поликлонального антитела против мышиного фактора I, поскольку коммерчески доступные антитела против α-FI являются неподходящими для измерения концентраций мышиного FI в ELISA (они не реагируют с нативным FI, а реагируют только с денатурированным белком в вестерн-блот-анализе). С использованием этого специально приготовленного поликлонального антитела стало возможным увеличение концентраций в сыворотке в четыре раза по сравнению с нормальными уровнями, то есть с 20-40 мкг/мл у контрольных мышей до 80-160 мкг/мл FI у трансгенных мышей.
Другой представляющий интерес вопрос заключается в том, может ли экспрессия трансгена дополнительно увеличиваться в реакции острой фазы. Поскольку в используемой конструкции FI находится под контролем промотора α-1-антитрипсина (α-1-антитрипсин и FI оба являются реагентами острой позитивной фазы), то можно ожидать, что экспрессия трансгена будет также повышаться в реакции острой фазы. Это может быть проанализировано в прямом эксперименте на животных, который может быть легко проведен путем инъекции ЛПС или IL6 мышам дикого типа и трансгенным мышам с последующим сравнением скорости увеличения уровня FI. Принимая во внимание то, что экспрессия эндогенного и трансгенного FI будет увеличиваться, то можно снизить исходную вирусную нагрузку при введении AAV, но при этом увеличить уровень экспрессии, если это необходимо.
Такие факторы, как иммунитет хозяина, позволяют предотвратить широкое распространение AAV у человека. После AAV-инфицирования, у человека вырабатываются нейтрализующие антитела, которые блокируют доставку гена. Однако путем конструирования капсидных белков могут быть получены новые варианты, которые имеют более эффективную трансдукцию и не распознаются нейтрализующими антителами. Поэтому, в настоящее время ведется разработка новых способов ограничения этих иммунных ответов (Mingozzi, et al., Science Translational Medicine, 5(194):194ra92-194ra92, July 2013). Недавно проведенные клинические исследования продемонстрировали, что одно успешное вливание AAV-вектора в течение двух лет давало терапевтические уровни фактора IX у пациента с тяжелой гемофилией B (Mingozzi and High, Blood, 122(1):23-36, July 2013), хотя следует также обратить внимание на то, что увеличение лишь на 1% от нормальных уровней FIX будет приводить к значительному снижению фенотипа, ассоциированного с сильным кровотечением у пациентов с гемофилией B (Nathwani, et al., N. Engl. J. Med., 371(21):1994-2004, November 2014). Ранее было показано, что экспрессия трансгена AAV FIX в скелетной мышце продолжалась в течение 10 лет, хотя в этом случае, FIX в кровотоке оставался на субтерапевтическом уровне (<1%) (Buchlis, et al., Blood, 119(13):3038-3041, March 2012).
Для лечения человека необходимо повышение уровня FI максимум на 50%. При использовании AAV_mFI-конструкции у мышей, этот уровень был значительно выше, и анализ на отложение iC3b указывал на снижение уровня отложения C3b менее чем на 50% при высокой дозе AAV_mFI. Независимо от этого, авторами было показано, что увеличение уровня FI более чем на 50% превращает активность комплотипа высокого риска в активность комплотипа низкого риска (Lay et al., 2015 (см. выше)). Поскольку оба эти комплотипа встречаются крайне редко, и у большинства людей они присутствуют в очень незначительных количествах, то для снижения риска у большинства людей требуется увеличение уровня FI только лишь на 50%.
Материалы и методы
Мышиный α-фактор I
Поликлональное антитело против этого фактора является коммерчески доступным и поставляется от Santa Cruz Biotechnology, Inc (# sc-69465). Это антитело продуцируется у коз против пептида в тяжелой цепи мышиного фактора I и распознает тяжелую цепь в вестерн-блот-анализах или в ELISA, проводимых в восстанавливающих и невосстанавливающих условиях. Это антитело, если оно является интактным, также распознает профермент. Тем не менее это антитело не является преципитирующим, поскольку пептид, используемый для иммунизации, является слишком коротким.
Нормальная рабочая концентрация 1:500.
Антисыворотка, содержащая мышиный α-фактор I
Это антитело не является коммерчески доступным и было специально заказано у Absea Biotechnology Ltd. для осаждения антитела против mFI, что позволяет легко определить уровни фактора I в анализе на двойную иммунодиффузию Охтерлони. Кроликов иммунизировали пептидным фрагментом (203-510 а.к.) рекомбинантного мышиного фактора I, который продуцировался в бактериях. После тестирования, антитело превращалось в не-преципитирующее, а поэтому был разработан ELISA на ингибирование для измерения уровней фактора I в сыворотке. Для получения поливалентного антитела против мышиного фактора I, которое преципитирует FI, фактор I, выделенный из клеточной культуры млекопитающего, отправляли в лабораторию Absea Biotechnology Ltd., но период иммунизации (8 недель) этой антисывороткой, содержащей «второе» антитело против мышиного α-фактора I, не был завершен, но мог быть продолжен в последующих экспериментах.
Клон 9 антитела
Клон 9 представляет собой крысиное антитело против человеческого α-C3g, распознающее неоэпитоп в C3g, который становится доступным только при расщеплении C3 фактором I с образованием iC3b или C3dg (Lachmann, et al., Immunology, 41(3):503-515, November 1980). В природных условиях, клон 9 реагирует только с iC3b или C3dg человека, тогда как в денатурирующих условиях, он распознает α,α'-цепь и 68 кДа-фрагмент человеческого C3, поскольку все эти три фрагмента включают эпитоп в C3g. Нормальная рабочая концентрация для детектирования в вестерн-блот-анализе составляет 0,5 мкг/мл, а концентрация антитела для захвата, используемого для нанесения покрытий, составляет 1,35 мкг/мл. Клон 9 и антитело против альфа-hC3g являются синонимами.
Клон 4 антитела
Клон 4 представляет собой крысиное моноклональное анти-C3c антитело, которое распознает конформационный эпитоп в C3c и реагирует с C3, C3b, iC3b и C3c (Lachmann, et al., Immunology, 41(3):503-515, November 1980). Поэтому, оно не связывается с Cdg или C3g из-за отсутствия эпитопа в C3c. Нормальная рабочая концентрация составляет 5 мкг/мл.
Человеческий α-C3c
Это антитело против α-C3c закупали у Dako и использовали для детектирования отложения C3b и iC3b. C3b и iC3b использовали в концентрации 1:5000 и детектировали с использованием «второго» кроличьего антитела против α-цепи.
Твердофазный иммуноферментный анализ (ELISA)
Анализ на отложение iC3b - «анализ с использованием клона 9». Планшеты Maxisorb Nunc покрывали в течение ночи при 4°C клоном 9 в концентрации 1,25 мкг/лунку в буфере для нанесения покрытий. На следующий день, планшеты промывали 5 раз промывочным буфером (PBS-0,05% Твином 20) и блокировали 4% сухим молоком Марвеля в PBS-T в течение 2 часов при комнатной температуре. Планшеты 5 раз промывали, и разведенные пробы сыворотки (1:2000 в PBS/желатине) добавляли в каждую лунку (получение проб сыворотки описано ниже). После инкубирования в течение 1 часа, планшеты промывали и инкубировали в течение 1 часа с биотинилированным клоном 4 в концентрации 5 мкг/мл в PBS/желатине. Планшеты 5 раз промывали и инкубировали с экстравидином-ПХ, 1:1000, в PBS/желатине. И наконец, после интенсивной промывки добавляли 100 мкл субстрата TMB (Invitrogen), и планшеты инкубировали на тарельчатом шейкере в течение 30 минут. Реакцию прекращали добавлением 50 мкл H2SO4 и планшеты считывали на микропланшет-ридере (на 450 нм).
Анализ на отложение C3b. Планшеты Maxisorb Nunc покрывали в течение ночи маннаном (Sigma, маннаном Saccharomyces cerevisiae) или липополисахаридом (Sigma) при плотности 1 мкг/лунку и при 4°C в буфере для нанесения покрытий. На следующий день, планшеты промывали в TBS-0,05% Твине-20 и блокировали 1% BSA в TBS-T при комнатной температуре. Через 2 часа, планшеты промывали и инкубировали с серийными разведениями рекомбинантных белков и/или сыворотки. После инкубирования в течение 1 часа при 37°C, планшеты промывали TBS-T и инкубировали в течение 1 часа с поликлональным кроличьим антителом против человеческого α-C3c комплемента (Dako), используемого в соотношении 1:5000 в TBS-T. После интенсивной промывки в течение 1 часа добавляли кроличий α-IgG (полноразмерную молекулу), конъюгированный со щелочной фосфатазой (продуцированный у коз, Sigma), 1:5000, в TBS-T. Планшеты 4 раза промывали TBS-T, и анализируемые планшеты проявляли с использованием таблеток, содержащих быстрый п-нитрофинилфосфатный краситель (Sigma), и проводили измерения (на 405 нм). Альтернативно, реакцию проявления в данном анализе завершали путем добавления 3M NaOH, а затем считывали.
ELISA на ингибирование. В ELISA на ингибирование, микротитрационные планшеты покрывали антигеном и после блокирования определяли концентрацию антисыворотки, которая давала сильный сигнал в присутствии «второго» антитела для детектирования. Затем эиу концентрацию использовали в анализе на ингибирование и смешивали с серийными разведениями образца с неизвестной концентрацией антигена. В это же время приготавливали серийные разведения антигена в известной концентрации. Во время этой стадии предварительного инкубирования образовывались иммунные комплексы, и после загрузки смеси на микротитрационный планшет с покрытием, положительный сигнал давали только те образцы, в которых концентрация антигена не превышала количество специфического антитела. В случае избытка антигена не образовывалось каких-либо свободных антител, которые могли бы связываться с иммобилизованным антигеном на микротитрационном планшете. Поэтому положительный результат в анализе на ингибирование указывал на разведение образца, в котором количество свободного антигена является ограничивающим, и путем сравнения со стандартными разведениями антигена в известных концентрациях может быть определена неизвестная концентрация.
Для определения концентрации мышиного фактора I в пробе сыворотки, планшеты Maxisorb Nunc покрывали в течение ночи при 4°C очищенным рекомбинантным мышиным фактором I (1 мкг/лунку) в буфере для нанесения покрытий. На следующий день, планшеты блокировали 1% BSA в TBS-T в течение 2 часов при комнатной температуре. Во время этого инкубирования приготавливали разведения мышиной сыворотки (снижение на 5%) или очищенного мышиного фактора I (снижение на 2 мкг/мл) в 1% BSA/150 мM NaCl. Затем, разведения смешивали 1:1 с определенными концентрациями очищенных и биотинилированных α-mFI IgG, то есть, 150 мкг/мл. Образцы инкубировали при комнатной температуре в течение 1 часа на тарельчатом шейкере, а затем загружали на планшет с покрытием. После инкубирования в течение одного часа, планшет интенсивно промывали, и связанное биотинилированное антитело против α-фактора I детектировали с использованием экстравидин-щелочной фосфатазы в концентрации 1:5000. Планшеты 4 раза промывали TBS-T, и анализируемые планшеты проявляли с использованием таблеток, содержащих быстрый п-нитрофинилфосфатный краситель (Sigma), и проводили измерения (на 405 нм).
Вестерн-блот-анализ
Вестерн-блот-анализ проводили для детектирования специфических белков или для подтверждения их присутствия в образце. Сначала проводили гель-электрофорез для разделения белков по размерам. Образцы белка кипятили в 4× буфере для загрузки в целях их денатурации, а если для анализа требовался восстановленный белок, то добавляли β-меркаптоэтанол. Электрофорез в ДСН-геле осуществляли в геле с 4-12% бис-трис-белком (Invitrogen) в течение 50 минут при 200 В. После разделения белков осуществляли влажный перенос для доставки отдельных белков на ПВДФ-мембрану, где белки мишени могут быть детектированы с использованием специфических антител.
Блот-анализ проводили путем комплексного переноса бумаги для блоттинга, уравновешенной буфером для переноса; геля и мембраны (сначала активированной MeOH) в кластер для переноса. Этот перенос осуществляли в течение 1 часа при 350 мA, 60 Вт.
Затем, мембрану блокировали в течение 30 минут на роторе в блокирующем буфере. «Первое» и «второе» антитело инкубировали путем встряхивания в течение 1 часа, а в перерывах, мембрану 3 раза промывали в течение 5 минут в промывочном буфере. Последние промывки проводили в TBS-T, pH 8, поскольку оба используемых субстрата давали более сильные сигналы, если эти промывки проводились при слабощелочном рН. В зависимости от конъюгата «второго» антитела были использованы различные субстраты: белки детектировали в хромогенных условиях путем добавления 3 мл субстрата TMB или AP (Life Technologies), или, в случае конъюгированных с пероксидазой антител, белки также детектировали хемилюминесцентным методом после добавления ЭХЛ-реагента.
Приготовление сыворотки
Приготовление сыворотки осуществляли, как описано в литературе (Lachmann, J. Immunol. Methods, 352(1-2):195-197, January 2010). Кровь брали в стерильных условиях и оставляли для свертывания при комнатной температуре. Первое центрифугирование обычно проводили в настольной центрифуге приблизительно при 3000×g в течение приблизительно 5-10 минут при комнатной температуре. Затем брали сыворотку и проводили второе высокоскоростное центрифугирование при 20000×g в течение 2-5 минут. Эта стадия необходима для удаления всех фрагментов эритроцитов, которые затем могут искажать результаты. Затем сыворотку, полученную после второго центрифугирования, охлаждали, разделяли на аликвоты и замораживали для проведения последующего эксперимента. Следует отметить, что сыворотка, используемая для функциональных анализов, никогда не должна храниться при температуре выше -80°C. При приготовлении аликвот нельзя проводить повторные циклы замораживания/оттаивания.
In vitro анализы
Расщепление профактора I фурином. Клетки Hek и CHO секретировали только частично процессированный mFI, а поэтому большинство mFI представляют собой неактивный про-фермент фактора I. Для получения активного фактора I, очищенный про-фермент гидролизовали протеазой фурином. Вкратце, очищенный mFI диализовали против TBS, а затем доводили до 10 мM CaCl2. Была добавлена 1 единица фурина на 25 мкг mFI. Реакцию осуществляли в течение ночи при 30°C. На следующий день, фактор I либо использовали сразу, либо разделяли на аликвоты и хранили при температуре -80°C.
Анализ на расщепление C3b. В анализе на расщепление C3b, C3b сначала получали путем лимитированного гидролиза трипсином, как описано в литературе (Bokisch, et al., J. Exp. Med., 129(5):1109-1130, May 1969). Вкратце, 2 мкл человеческого C3 инкубировали с 0,8 мкл трипсина (10 мкг/мл) ровно 60 секунд при 20°C. Реакцию прекращали путем добавления 2,4 мкл ингибитора соевого трипсина (10 мкг/мл). Такой ограниченный гидролиз приводил к образованию C3b из C3 посредством расщепления C3a. Затем, C3b смешивали с различными количествами мышиного фактора I или человеческого фактора I и с 2 мкл человеческого фактора H, а затем реакционный объем доводили до 10 мкл путем добавления TBS. Альтернативно, мышиную сыворотку добавляли в качестве источника фактора I. Реакцию осуществляли в течение 30-60 минут при 37°C, а затем завершали путем добавления восстанавливающего 4× буфера для загрузки и кипячения. Образцы либо анализировали с помощью вестерн-блот-анализа, либо хранили при 4°C.
Анализ в каждый период времени. Для анализа способности рекомбинантного мышиного фактора I расщеплять C3b и iC3b был разработан анализ для оценки в каждый период времени. Для этого, сыворотку быстро оттаивали при 37°C, подвергали быстрому вихревому перемешиванию, а затем помещали на лед. После этого, сыворотку смешивали с рекомбинантным человеческим или мышиным фактором I, а затем добавляли зимозан до конечной концентрации 5%. В этом анализе, в качестве буфера использовали буфер для альтернативного пути. Смесь, после ее получения, встряхивали в инкубаторе при 37°C, и через определенные промежутки времени брали 40 мкл каждого образца в 10 мкл 50 мМ EDTA. Образцы брали через 0, 30, 60, 120, 180, 240, 480, 600 и 1440 минут. Через каждый промежуток времени, образцы подвергали микроцентрифугированию для удаления зимозана и хранили при -80°C до проведения ELISA. Альтернативно, это анализ также проводили с использованием ЛПС в качестве комплемент-активирующего реагента, преимущество которого заключается в его растворимости.
Все цитируемые здесь документы, включая публикации, патенты и патентные заявки вводится в настоящее описание посредством ссылки так, как если бы каждый из этих документов во всей своей полноте был отдельно и конкретно введен в настоящее описание посредством ссылки. Обсуждаемые здесь документы должны быть поданы для раскрытия только до даты подачи настоящей заявки. Ничто здесь не должно быть истолковано как признание того, что любой документ порочит новизну или неочевидность описанных здесь вариантов осуществления изобретения.
--->
Список последовательностей
SEQ ID NO: 1
Кодирующая область CDS для фактора I комплемента
Homo sapiens (CFI), вариант 2 транскрипта, мРНК, эталонная
последовательность из NCBI: NM_000204.4:
atgaagcttc ttcatgtttt cctgttattt ctgtgcttcc acttaaggtt ttgcaaggtc 60
acttatacat ctcaagagga tctggtggag aaaaagtgct tagcaaaaaa atatactcac 120
ctctcctgcg ataaagtctt ctgccagcca tggcagagat gcattgaggg cacctgtgtt 180
tgtaaactac cgtatcagtg cccaaagaat ggcactgcag tgtgtgcaac taacaggaga 240
agcttcccaa catactgtca acaaaagagt ttggaatgtc ttcatccagg gacaaagttt 300
ttaaataacg gaacatgcac agccgaagga aagtttagtg tttccttgaa gcatggaaat 360
acagattcag agggaatagt tgaagtaaaa cttgtggacc aagataagac aatgttcata 420
tgcaaaagca gctggagcat gagggaagcc aacgtggcct gccttgacct tgggtttcaa 480
caaggtgctg atactcaaag aaggtttaag ttgtctgatc tctctataaa ttccactgaa 540
tgtctacatg tgcattgccg aggattagag accagtttgg ctgaatgtac ttttactaag 600
agaagaacta tgggttacca ggatttcgct gatgtggttt gttatacaca gaaagcagat 660
tctccaatgg atgacttctt tcagtgtgtg aatgggaaat acatttctca gatgaaagcc 720
tgtgatggta tcaatgattg tggagaccaa agtgatgaac tgtgttgtaa agcatgccaa 780
ggcaaaggct tccattgcaa atcgggtgtt tgcattccaa gccagtatca atgcaatggt 840
gaggtggact gcattacagg ggaagatgaa gttggctgtg caggctttgc atctgtggct 900
caagaagaaa cagaaatttt gactgctgac atggatgcag aaagaagacg gataaaatca 960
ttattaccta aactatcttg tggagttaaa aacagaatgc acattcgaag gaaacgaatt 1020
gtgggaggaa agcgagcaca actgggagac ctcccatggc aggtggcaat taaggatgcc 1080
agtggaatca cctgtggggg aatttatatt ggtggctgtt ggattctgac tgctgcacat 1140
tgtctcagag ccagtaaaac tcatcgttac caaatatgga caacagtagt agactggata 1200
caccccgacc ttaaacgtat agtaattgaa tacgtggata gaattatttt ccatgaaaac 1260
tacaatgcag gcacttacca aaatgacatc gctttgattg aaatgaaaaa agacggaaac 1320
aaaaaagatt gtgagctgcc tcgttccatc cctgcctgtg tcccctggtc tccttaccta 1380
ttccaaccta atgatacatg catcgtttct ggctggggac gagaaaaaga taacgaaaga 1440
gtcttttcac ttcagtgggg tgaagttaaa ctaataagca actgctctaa gttttacgga 1500
aatcgtttct atgaaaaaga aatggaatgt gcaggtacat atgatggttc catcgatgcc 1560
tgtaaagggg actctggagg ccccttagtc tgtatggatg ccaacaatgt gacttatgtc 1620
tggggtgttg tgagttgggg ggaaaactgt ggaaaaccag agttcccagg tgtttacacc 1680
aaagtggcca attattttga ctggattagc taccatgtag gaaggccttt tatttctcag 1740
tacaatgtat aa 1752
SEQ ID NO: 2
Трансляция кодирующей области CDS для фактора I комплемента
Homo sapiens (CFI), вариант 2 транскрипта, мРНК, эталонная
последовательность из NCBI: NM_000204.4:
MKLLHVFLLF LCFHLRFCKV TYTSQEDLVE KKCLAKKYTH LSCDKVFCQP WQRCIEGTCV 60
CKLPYQCPKN GTAVCATNRR SFPTYCQQKS LECLHPGTKF LNNGTCTAEG KFSVSLKHGN 120
TDSEGIVEVK LVDQDKTMFI CKSSWSMREA NVACLDLGFQ QGADTQRRFK LSDLSINSTE 180
CLHVHCRGLE TSLAECTFTK RRTMGYQDFA DVVCYTQKAD SPMDDFFQCV NGKYISQMKA 240
CDGINDCGDQ SDELCCKACQ GKGFHCKSGV CIPSQYQCNG EVDCITGEDE VGCAGFASVA 300
QEETEILTAD MDAERRRIKS LLPKLSCGVK NRMHIRRKRI VGGKRAQLGD LPWQVAIKDA 360
SGITCGGIYI GGCWILTAAH CLRASKTHRY QIWTTVVDWI HPDLKRIVIE YVDRIIFHEN 420
YNAGTYQNDI ALIEMKKDGN KKDCELPRSI PACVPWSPYL FQPNDTCIVS GWGREKDNER 480
VFSLQWGEVK LISNCSKFYG NRFYEKEMEC AGTYDGSIDA CKGDSGGPLV CMDANNVTYV 540
WGVVSWGENC GKPEFPGVYT KVANYFDWIS YHVGRPFISQ YNV 583
SEQ ID NO: 3
Кодирующая область CDS для фактора I комплемента Homo
sapiens (CFI), вариант 2 транскрипта, мРНК, эталонная
последовательность из NCBI: NM_000204.4, без
последовательности, кодирующей N-концевую сигнальную
последовательность из 18 остатков:
aaggtcactt atacatctca agaggatctg gtggagaaaa agtgcttagc aaaaaaatat 60
actcacctct cctgcgataa agtcttctgc cagccatggc agagatgcat tgagggcacc 120
tgtgtttgta aactaccgta tcagtgccca aagaatggca ctgcagtgtg tgcaactaac 180
aggagaagct tcccaacata ctgtcaacaa aagagtttgg aatgtcttca tccagggaca 240
aagtttttaa ataacggaac atgcacagcc gaaggaaagt ttagtgtttc cttgaagcat 300
ggaaatacag attcagaggg aatagttgaa gtaaaacttg tggaccaaga taagacaatg 360
ttcatatgca aaagcagctg gagcatgagg gaagccaacg tggcctgcct tgaccttggg 420
tttcaacaag gtgctgatac tcaaagaagg tttaagttgt ctgatctctc tataaattcc 480
actgaatgtc tacatgtgca ttgccgagga ttagagacca gtttggctga atgtactttt 540
actaagagaa gaactatggg ttaccaggat ttcgctgatg tggtttgtta tacacagaaa 600
gcagattctc caatggatga cttctttcag tgtgtgaatg ggaaatacat ttctcagatg 660
aaagcctgtg atggtatcaa tgattgtgga gaccaaagtg atgaactgtg ttgtaaagca 720
tgccaaggca aaggcttcca ttgcaaatcg ggtgtttgca ttccaagcca gtatcaatgc 780
aatggtgagg tggactgcat tacaggggaa gatgaagttg gctgtgcagg ctttgcatct 840
gtggctcaag aagaaacaga aattttgact gctgacatgg atgcagaaag aagacggata 900
aaatcattat tacctaaact atcttgtgga gttaaaaaca gaatgcacat tcgaaggaaa 960
cgaattgtgg gaggaaagcg agcacaactg ggagacctcc catggcaggt ggcaattaag 1020
gatgccagtg gaatcacctg tgggggaatt tatattggtg gctgttggat tctgactgct 1080
gcacattgtc tcagagccag taaaactcat cgttaccaaa tatggacaac agtagtagac 1140
tggatacacc ccgaccttaa acgtatagta attgaatacg tggatagaat tattttccat 1200
gaaaactaca atgcaggcac ttaccaaaat gacatcgctt tgattgaaat gaaaaaagac 1260
ggaaacaaaa aagattgtga gctgcctcgt tccatccctg cctgtgtccc ctggtctcct 1320
tacctattcc aacctaatga tacatgcatc gtttctggct ggggacgaga aaaagataac 1380
gaaagagtct tttcacttca gtggggtgaa gttaaactaa taagcaactg ctctaagttt 1440
tacggaaatc gtttctatga aaaagaaatg gaatgtgcag gtacatatga tggttccatc 1500
gatgcctgta aaggggactc tggaggcccc ttagtctgta tggatgccaa caatgtgact 1560
tatgtctggg gtgttgtgag ttggggggaa aactgtggaa aaccagagtt cccaggtgtt 1620
tacaccaaag tggccaatta ttttgactgg attagctacc atgtaggaag gccttttatt 1680
tctcagtaca atgtataa 1698
SEQ ID NO: 4
Трансляция кодирующей области CDS для фактора I комплемента
Homo sapiens (CFI), вариант 2 транскрипта, мРНК, эталонная
последовательность из NCBI: NM_000204.4, без N-концевой
сигнальной последовательности из 18 остатков:
KVTYTSQEDL VEKKCLAKKY THLSCDKVFC QPWQRCIEGT CVCKLPYQCP KNGTAVCATN 60
RRSFPTYCQQ KSLECLHPGT KFLNNGTCTA EGKFSVSLKH GNTDSEGIVE VKLVDQDKTM 120
FICKSSWSMR EANVACLDLG FQQGADTQRR FKLSDLSINS TECLHVHCRG LETSLAECTF 180
TKRRTMGYQD FADVVCYTQK ADSPMDDFFQ CVNGKYISQM KACDGINDCG DQSDELCCKA 240
CQGKGFHCKS GVCIPSQYQC NGEVDCITGE DEVGCAGFAS VAQEETEILT ADMDAERRRI 300
KSLLPKLSCG VKNRMHIRRK RIVGGKRAQL GDLPWQVAIK DASGITCGGI YIGGCWILTA 360
AHCLRASKTH RYQIWTTVVD WIHPDLKRIV IEYVDRIIFH ENYNAGTYQN DIALIEMKKD 420
GNKKDCELPR SIPACVPWSP YLFQPNDTCI VSGWGREKDN ERVFSLQWGE VKLISNCSKF 480
YGNRFYEKEM ECAGTYDGSI DACKGDSGGP LVCMDANNVT YVWGVVSWGE NCGKPEFPGV 540
YTKVANYFDW ISYHVGRPFI SQYNV 565
SEQ ID NO: 5
Праймерная последовательность: FI-AAV-F:
tctagaggat ccgccaccat gaagctc 27
SEQ ID NO: 6
Праймерная последовательность: FI-AAV-R:
aagcttggcg gccgctcaga cattgtgttg agaaacaaga gaccttc 47
SEQ ID NO: 7
Нуклеотидная последовательность, кодирующая фактор I
комплемента Homo sapiens (CFI) и оптимизированная по
кодонам для экспрессии в человеческих клетках:
ATGAAGCTGCTGCATGTCTTTCTGCTGTTTCTGTGCTTCCATCTGCGGTTCTGTAAAGT GACCTATACTAGCCAGGAGGATCTGGTGGAGAAGAAGTGTCTGGCCAAGAAGTACACACACCTG AGCTGCGACAAGGTGTTCTGTCAGCCTTGGCAGCGGTGCATCGAGGGCACCTGCGTGTGCAAGC TGCCTTACCAGTGCCCAAAGAACGGCACCGCCGTGTGCGCCACAAATCGGAGATCTTTTCCAAC ATATTGCCAGCAGAAGAGCCTGGAGTGTCTGCACCCCGGCACCAAGTTCCTGAACAATGGCACC TGCACAGCCGAGGGCAAGTTTTCTGTGAGCCTGAAGCACGGCAACACAGATAGCGAGGGCATCG TGGAGGTGAAGCTGGTGGACCAGGATAAGACCATGTTCATCTGTAAGAGCTCCTGGTCCATGAG GGAGGCAAACGTGGCATGCCTGGATCTGGGATTCCAGCAGGGAGCAGACACACAGAGGCGCTTT AAGCTGTCCGACCTGTCTATCAATAGCACCGAGTGCCTGCACGTGCACTGTAGGGGCCTGGAGA CATCCCTGGCAGAGTGCACCTTCACAAAGCGGAGAACCATGGGCTACCAGGACTTTGCCGACGT GGTGTGCTATACCCAGAAGGCCGATAGCCCCATGGACGATTTCTTTCAGTGCGTGAACGGCAAG TATATCTCCCAGATGAAGGCCTGCGACGGCATCAATGACTGTGGCGATCAGTCTGACGAGCTGT GCTGTAAGGCCTGTCAGGGCAAGGGCTTCCACTGCAAGAGCGGCGTGTGCATCCCTTCCCAGTA CCAGTGCAACGGCGAGGTGGATTGTATCACAGGAGAGGACGAAGTGGGATGCGCAGGATTTGCA TCTGTGGCACAGGAGGAGACAGAGATCCTGACAGCCGACATGGATGCCGAGAGGCGCCGGATCA AGTCTCTGCTGCCTAAGCTGAGCTGTGGCGTGAAGAATCGGATGCACATCAGAAGGAAGCGCAT CGTGGGAGGCAAGAGGGCACAGCTGGGCGATCTGCCATGGCAGGTGGCCATCAAGGACGCCTCT GGCATCACCTGCGGCGGCATCTACATCGGAGGATGTTGGATCCTGACCGCAGCACACTGCCTGA GAGCAAGCAAGACACACAGGTATCAGATCTGGACCACAGTGGTGGATTGGATCCACCCAGACCT GAAGAGAATCGTGATCGAGTACGTGGATAGGATCATCTTTCACGAGAACTACAATGCCGGCACA TATCAGAACGACATCGCCCTGATCGAGATGAAGAAGGATGGCAATAAGAAGGACTGTGAGCTGC CCAGATCCATCCCTGCATGCGTGCCATGGAGCCCCTATCTGTTCCAGCCCAACGATACCTGCAT CGTGTCCGGATGGGGAAGGGAGAAGGACAATGAGCGGGTGTTTTCTCTGCAGTGGGGCGAGGTG AAGCTGATCTCCAACTGTTCTAAGTTCTACGGCAATAGGTTTTATGAGAAGGAGATGGAGTGCG CCGGCACCTACGATGGCAGCATCGACGCCTGTAAGGGCGATTCCGGAGGACCACTGGTGTGCAT GGACGCAAACAATGTGACATACGTGTGGGGAGTGGTGTCCTGGGGAGAGAACTGCGGCAAGCCA GAGTTCCCCGGCGTATATACCAAGGTGGCCAATTATTTTGATTGGATTTCCTACCACGTCGGCA GGCCCTTTATTTCCCAGTATAATGTCTAA
SEQ ID NO: 8
Нуклеотидная последовательность печень-специфического
промотора HLP:
TGTTTGCTGC TTGCAATGTT TGCCCATTTT AGGGTGGACA CAGGACGCTG TGGTTTCTGA 60
GCCAGGGGGC GACTCAGATC CCAGCCAGTG GACTTAGCCC CTGTTTGCTC CTCCGATAAC 120
TGGGGTGACC TTGGTTAATA TTCACCAGCA GCCTCCCCCG TTGCCCCTCT GGATCCACTG 180
CTTAAATACG GACGAGGACA GGGCCCTGTC TCCTCAGCTT CAGGCACCAC CACTGACCTG 240
GGACAGTGAA T 251
SEQ ID NO: 9
Нуклеотидная последовательность печень-специфического
промотора LP1:
CCCTAAAATG GGCAAACATT GCAAGCAGCA AACAGCAAAC ACACAGCCCT CCCTGCCTGC
TGACCTTGGA GCTGGGGCAG AGGTCAGAGA CCTCTCTGGG CCCATGCCAC CTCCAACATC
CACTCGACCC CTTGGAATTT CGGTGGAGAG GAGCAGAGGT TGTCCTGGCG TGGTTTAGGT
AGTGTGAGAG GGGAATGACT CCTTTCGGTA AGTGCAGTGG AAGCTGTACA CTGCCCAGGC
AAAGCGTCCG GGCAGCGTAG GCGGGCGACT CAGATCCCAG CCAGTGGACT TAGCCCCTGT
TTGCTCCTCC GATAACTGGG GTGACCTTGG TTAATATTCA CCAGCAGCCT CCCCCGTTGC
CCCTCTGGAT CCACTGCTTA AATACGGACG AGGACAGGGC CCTGTCTCCT CAGCTTCAGG
CACCACCACT GACCTGGGAC AGTGAAT
SEQ ID NO: 10
Нуклеотидная последовательность печень-специфического
промотора HLP2:
CCCTAAAATG GGCAAACATT GCAAGCAGCA AACAGCAAAC ACACAGCCCT CCCTGCCTGC 60
TGACCTTGGA GCTGGGGCAG AGGTCAGACA CCTCTCTGGG CCCATGCCAC CTCCAACTGG 120
ACACAGGACG CTGTGGTTTC TGAGCCAGGG GGCGACTCAG ATCCCAGCCA GTGGACTTAG 180
CCCCTGTTTG CTCCTCCGAT AACTGGGGTG ACCTTGGTTA ATATTCACCA GCAGCCTCCC 240
CCGTTGCCCC TCTGGATCCA CTGCTTAAAT ACGGACGAGG ACAGGGCCCT GTCTCCTCAG 300
CTTCAGGCAC CACCACTGAC CTGGGACAGT GAATGATCCC CCTGATCTGC GGCC 354
SEQ ID NO: 11
Аминокислотная последовательность капсидного белка MutC:
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Gln Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Asp Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Val Gly
145 150 155 160
Lys Ser Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Thr Ser Leu Gly Ser Asn Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Lys Leu Ser Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg Thr
435 440 445
Gln Gly Thr Thr Ser Gly Thr Thr Asn Gln Ser Arg Leu Leu Phe Ser
450 455 460
Gln Ala Gly Pro Gln Ser Met Ser Leu Gln Ala Arg Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Leu Ser Lys Thr Ala Asn Asp Asn
485 490 495
Asn Asn Ser Asn Phe Pro Trp Thr Ala Ala Ser Lys Tyr His Leu Asn
500 505 510
Gly Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Glu Lys Phe Phe Pro Met His Gly Asn Leu Ile Phe Gly
530 535 540
Lys Glu Gly Thr Thr Ala Ser Asn Ala Glu Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln
565 570 575
Tyr Gly Thr Val Ala Asn Asn Leu Gln Ser Ser Asn Thr Ala Pro Thr
580 585 590
Thr Arg Thr Val Asn Asp Gln Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Met Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Thr Thr Phe Ser Pro Ala Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
SEQ ID NO: 12
Аминокислотная последовательность капсидного белка LK03:
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Gln Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Asp Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Val Gly
145 150 155 160
Lys Ser Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Thr Ser Leu Gly Ser Asn Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Lys Leu Ser Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg Thr
435 440 445
Gln Gly Thr Thr Ser Gly Thr Thr Asn Gln Ser Arg Leu Leu Phe Ser
450 455 460
Gln Ala Gly Pro Gln Ser Met Ser Leu Gln Ala Arg Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Leu Ser Lys Thr Ala Asn Asp Asn
485 490 495
Asn Asn Ser Asn Phe Pro Trp Thr Ala Ala Ser Lys Tyr His Leu Asn
500 505 510
Gly Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Glu Lys Phe Phe Pro Met His Gly Asn Leu Ile Phe Gly
530 535 540
Lys Glu Gly Thr Thr Ala Ser Asn Ala Glu Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln
565 570 575
Tyr Gly Thr Val Ala Asn Asn Leu Gln Ser Ser Asn Thr Ala Pro Thr
580 585 590
Thr Arg Thr Val Asn Asp Gln Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Met Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Thr Thr Phe Ser Pro Ala Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CAMBRIDGE ENTERPRISE LIMITED
THE SYDNEY CHILDREN'S HOSPITALS NETWORK (RANDWICK AND
WESTMEAD) (INCORPORATING THE ROYAL ALEXANDRA HOSPITAL FOR
CHILDREN)
<120> Лечение комплемент-опосредованных заболеваний
<130> P/74985.WO01
<150> GB1608046.7
<151> 2016-05-09
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 1752
<212> ДНК
<213> Homo sapiens
<400> 1
atgaagcttc ttcatgtttt cctgttattt ctgtgcttcc acttaaggtt ttgcaaggtc 60
acttatacat ctcaagagga tctggtggag aaaaagtgct tagcaaaaaa atatactcac 120
ctctcctgcg ataaagtctt ctgccagcca tggcagagat gcattgaggg cacctgtgtt 180
tgtaaactac cgtatcagtg cccaaagaat ggcactgcag tgtgtgcaac taacaggaga 240
agcttcccaa catactgtca acaaaagagt ttggaatgtc ttcatccagg gacaaagttt 300
ttaaataacg gaacatgcac agccgaagga aagtttagtg tttccttgaa gcatggaaat 360
acagattcag agggaatagt tgaagtaaaa cttgtggacc aagataagac aatgttcata 420
tgcaaaagca gctggagcat gagggaagcc aacgtggcct gccttgacct tgggtttcaa 480
caaggtgctg atactcaaag aaggtttaag ttgtctgatc tctctataaa ttccactgaa 540
tgtctacatg tgcattgccg aggattagag accagtttgg ctgaatgtac ttttactaag 600
agaagaacta tgggttacca ggatttcgct gatgtggttt gttatacaca gaaagcagat 660
tctccaatgg atgacttctt tcagtgtgtg aatgggaaat acatttctca gatgaaagcc 720
tgtgatggta tcaatgattg tggagaccaa agtgatgaac tgtgttgtaa agcatgccaa 780
ggcaaaggct tccattgcaa atcgggtgtt tgcattccaa gccagtatca atgcaatggt 840
gaggtggact gcattacagg ggaagatgaa gttggctgtg caggctttgc atctgtggct 900
caagaagaaa cagaaatttt gactgctgac atggatgcag aaagaagacg gataaaatca 960
ttattaccta aactatcttg tggagttaaa aacagaatgc acattcgaag gaaacgaatt 1020
gtgggaggaa agcgagcaca actgggagac ctcccatggc aggtggcaat taaggatgcc 1080
agtggaatca cctgtggggg aatttatatt ggtggctgtt ggattctgac tgctgcacat 1140
tgtctcagag ccagtaaaac tcatcgttac caaatatgga caacagtagt agactggata 1200
caccccgacc ttaaacgtat agtaattgaa tacgtggata gaattatttt ccatgaaaac 1260
tacaatgcag gcacttacca aaatgacatc gctttgattg aaatgaaaaa agacggaaac 1320
aaaaaagatt gtgagctgcc tcgttccatc cctgcctgtg tcccctggtc tccttaccta 1380
ttccaaccta atgatacatg catcgtttct ggctggggac gagaaaaaga taacgaaaga 1440
gtcttttcac ttcagtgggg tgaagttaaa ctaataagca actgctctaa gttttacgga 1500
aatcgtttct atgaaaaaga aatggaatgt gcaggtacat atgatggttc catcgatgcc 1560
tgtaaagggg actctggagg ccccttagtc tgtatggatg ccaacaatgt gacttatgtc 1620
tggggtgttg tgagttgggg ggaaaactgt ggaaaaccag agttcccagg tgtttacacc 1680
aaagtggcca attattttga ctggattagc taccatgtag gaaggccttt tatttctcag 1740
tacaatgtat aa 1752
<210> 2
<211> 583
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Met Lys Leu Leu His Val Phe Leu Leu Phe Leu Cys Phe His Leu Arg
1 5 10 15
Phe Cys Lys Val Thr Tyr Thr Ser Gln Glu Asp Leu Val Glu Lys Lys
20 25 30
Cys Leu Ala Lys Lys Tyr Thr His Leu Ser Cys Asp Lys Val Phe Cys
35 40 45
Gln Pro Trp Gln Arg Cys Ile Glu Gly Thr Cys Val Cys Lys Leu Pro
50 55 60
Tyr Gln Cys Pro Lys Asn Gly Thr Ala Val Cys Ala Thr Asn Arg Arg
65 70 75 80
Ser Phe Pro Thr Tyr Cys Gln Gln Lys Ser Leu Glu Cys Leu His Pro
85 90 95
Gly Thr Lys Phe Leu Asn Asn Gly Thr Cys Thr Ala Glu Gly Lys Phe
100 105 110
Ser Val Ser Leu Lys His Gly Asn Thr Asp Ser Glu Gly Ile Val Glu
115 120 125
Val Lys Leu Val Asp Gln Asp Lys Thr Met Phe Ile Cys Lys Ser Ser
130 135 140
Trp Ser Met Arg Glu Ala Asn Val Ala Cys Leu Asp Leu Gly Phe Gln
145 150 155 160
Gln Gly Ala Asp Thr Gln Arg Arg Phe Lys Leu Ser Asp Leu Ser Ile
165 170 175
Asn Ser Thr Glu Cys Leu His Val His Cys Arg Gly Leu Glu Thr Ser
180 185 190
Leu Ala Glu Cys Thr Phe Thr Lys Arg Arg Thr Met Gly Tyr Gln Asp
195 200 205
Phe Ala Asp Val Val Cys Tyr Thr Gln Lys Ala Asp Ser Pro Met Asp
210 215 220
Asp Phe Phe Gln Cys Val Asn Gly Lys Tyr Ile Ser Gln Met Lys Ala
225 230 235 240
Cys Asp Gly Ile Asn Asp Cys Gly Asp Gln Ser Asp Glu Leu Cys Cys
245 250 255
Lys Ala Cys Gln Gly Lys Gly Phe His Cys Lys Ser Gly Val Cys Ile
260 265 270
Pro Ser Gln Tyr Gln Cys Asn Gly Glu Val Asp Cys Ile Thr Gly Glu
275 280 285
Asp Glu Val Gly Cys Ala Gly Phe Ala Ser Val Ala Gln Glu Glu Thr
290 295 300
Glu Ile Leu Thr Ala Asp Met Asp Ala Glu Arg Arg Arg Ile Lys Ser
305 310 315 320
Leu Leu Pro Lys Leu Ser Cys Gly Val Lys Asn Arg Met His Ile Arg
325 330 335
Arg Lys Arg Ile Val Gly Gly Lys Arg Ala Gln Leu Gly Asp Leu Pro
340 345 350
Trp Gln Val Ala Ile Lys Asp Ala Ser Gly Ile Thr Cys Gly Gly Ile
355 360 365
Tyr Ile Gly Gly Cys Trp Ile Leu Thr Ala Ala His Cys Leu Arg Ala
370 375 380
Ser Lys Thr His Arg Tyr Gln Ile Trp Thr Thr Val Val Asp Trp Ile
385 390 395 400
His Pro Asp Leu Lys Arg Ile Val Ile Glu Tyr Val Asp Arg Ile Ile
405 410 415
Phe His Glu Asn Tyr Asn Ala Gly Thr Tyr Gln Asn Asp Ile Ala Leu
420 425 430
Ile Glu Met Lys Lys Asp Gly Asn Lys Lys Asp Cys Glu Leu Pro Arg
435 440 445
Ser Ile Pro Ala Cys Val Pro Trp Ser Pro Tyr Leu Phe Gln Pro Asn
450 455 460
Asp Thr Cys Ile Val Ser Gly Trp Gly Arg Glu Lys Asp Asn Glu Arg
465 470 475 480
Val Phe Ser Leu Gln Trp Gly Glu Val Lys Leu Ile Ser Asn Cys Ser
485 490 495
Lys Phe Tyr Gly Asn Arg Phe Tyr Glu Lys Glu Met Glu Cys Ala Gly
500 505 510
Thr Tyr Asp Gly Ser Ile Asp Ala Cys Lys Gly Asp Ser Gly Gly Pro
515 520 525
Leu Val Cys Met Asp Ala Asn Asn Val Thr Tyr Val Trp Gly Val Val
530 535 540
Ser Trp Gly Glu Asn Cys Gly Lys Pro Glu Phe Pro Gly Val Tyr Thr
545 550 555 560
Lys Val Ala Asn Tyr Phe Asp Trp Ile Ser Tyr His Val Gly Arg Pro
565 570 575
Phe Ile Ser Gln Tyr Asn Val
580
<210> 3
<211> 1698
<212> ДНК
<213> Homo sapiens
<400> 3
aaggtcactt atacatctca agaggatctg gtggagaaaa agtgcttagc aaaaaaatat 60
actcacctct cctgcgataa agtcttctgc cagccatggc agagatgcat tgagggcacc 120
tgtgtttgta aactaccgta tcagtgccca aagaatggca ctgcagtgtg tgcaactaac 180
aggagaagct tcccaacata ctgtcaacaa aagagtttgg aatgtcttca tccagggaca 240
aagtttttaa ataacggaac atgcacagcc gaaggaaagt ttagtgtttc cttgaagcat 300
ggaaatacag attcagaggg aatagttgaa gtaaaacttg tggaccaaga taagacaatg 360
ttcatatgca aaagcagctg gagcatgagg gaagccaacg tggcctgcct tgaccttggg 420
tttcaacaag gtgctgatac tcaaagaagg tttaagttgt ctgatctctc tataaattcc 480
actgaatgtc tacatgtgca ttgccgagga ttagagacca gtttggctga atgtactttt 540
actaagagaa gaactatggg ttaccaggat ttcgctgatg tggtttgtta tacacagaaa 600
gcagattctc caatggatga cttctttcag tgtgtgaatg ggaaatacat ttctcagatg 660
aaagcctgtg atggtatcaa tgattgtgga gaccaaagtg atgaactgtg ttgtaaagca 720
tgccaaggca aaggcttcca ttgcaaatcg ggtgtttgca ttccaagcca gtatcaatgc 780
aatggtgagg tggactgcat tacaggggaa gatgaagttg gctgtgcagg ctttgcatct 840
gtggctcaag aagaaacaga aattttgact gctgacatgg atgcagaaag aagacggata 900
aaatcattat tacctaaact atcttgtgga gttaaaaaca gaatgcacat tcgaaggaaa 960
cgaattgtgg gaggaaagcg agcacaactg ggagacctcc catggcaggt ggcaattaag 1020
gatgccagtg gaatcacctg tgggggaatt tatattggtg gctgttggat tctgactgct 1080
gcacattgtc tcagagccag taaaactcat cgttaccaaa tatggacaac agtagtagac 1140
tggatacacc ccgaccttaa acgtatagta attgaatacg tggatagaat tattttccat 1200
gaaaactaca atgcaggcac ttaccaaaat gacatcgctt tgattgaaat gaaaaaagac 1260
ggaaacaaaa aagattgtga gctgcctcgt tccatccctg cctgtgtccc ctggtctcct 1320
tacctattcc aacctaatga tacatgcatc gtttctggct ggggacgaga aaaagataac 1380
gaaagagtct tttcacttca gtggggtgaa gttaaactaa taagcaactg ctctaagttt 1440
tacggaaatc gtttctatga aaaagaaatg gaatgtgcag gtacatatga tggttccatc 1500
gatgcctgta aaggggactc tggaggcccc ttagtctgta tggatgccaa caatgtgact 1560
tatgtctggg gtgttgtgag ttggggggaa aactgtggaa aaccagagtt cccaggtgtt 1620
tacaccaaag tggccaatta ttttgactgg attagctacc atgtaggaag gccttttatt 1680
tctcagtaca atgtataa 1698
<210> 4
<211> 565
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Lys Val Thr Tyr Thr Ser Gln Glu Asp Leu Val Glu Lys Lys Cys Leu
1 5 10 15
Ala Lys Lys Tyr Thr His Leu Ser Cys Asp Lys Val Phe Cys Gln Pro
20 25 30
Trp Gln Arg Cys Ile Glu Gly Thr Cys Val Cys Lys Leu Pro Tyr Gln
35 40 45
Cys Pro Lys Asn Gly Thr Ala Val Cys Ala Thr Asn Arg Arg Ser Phe
50 55 60
Pro Thr Tyr Cys Gln Gln Lys Ser Leu Glu Cys Leu His Pro Gly Thr
65 70 75 80
Lys Phe Leu Asn Asn Gly Thr Cys Thr Ala Glu Gly Lys Phe Ser Val
85 90 95
Ser Leu Lys His Gly Asn Thr Asp Ser Glu Gly Ile Val Glu Val Lys
100 105 110
Leu Val Asp Gln Asp Lys Thr Met Phe Ile Cys Lys Ser Ser Trp Ser
115 120 125
Met Arg Glu Ala Asn Val Ala Cys Leu Asp Leu Gly Phe Gln Gln Gly
130 135 140
Ala Asp Thr Gln Arg Arg Phe Lys Leu Ser Asp Leu Ser Ile Asn Ser
145 150 155 160
Thr Glu Cys Leu His Val His Cys Arg Gly Leu Glu Thr Ser Leu Ala
165 170 175
Glu Cys Thr Phe Thr Lys Arg Arg Thr Met Gly Tyr Gln Asp Phe Ala
180 185 190
Asp Val Val Cys Tyr Thr Gln Lys Ala Asp Ser Pro Met Asp Asp Phe
195 200 205
Phe Gln Cys Val Asn Gly Lys Tyr Ile Ser Gln Met Lys Ala Cys Asp
210 215 220
Gly Ile Asn Asp Cys Gly Asp Gln Ser Asp Glu Leu Cys Cys Lys Ala
225 230 235 240
Cys Gln Gly Lys Gly Phe His Cys Lys Ser Gly Val Cys Ile Pro Ser
245 250 255
Gln Tyr Gln Cys Asn Gly Glu Val Asp Cys Ile Thr Gly Glu Asp Glu
260 265 270
Val Gly Cys Ala Gly Phe Ala Ser Val Ala Gln Glu Glu Thr Glu Ile
275 280 285
Leu Thr Ala Asp Met Asp Ala Glu Arg Arg Arg Ile Lys Ser Leu Leu
290 295 300
Pro Lys Leu Ser Cys Gly Val Lys Asn Arg Met His Ile Arg Arg Lys
305 310 315 320
Arg Ile Val Gly Gly Lys Arg Ala Gln Leu Gly Asp Leu Pro Trp Gln
325 330 335
Val Ala Ile Lys Asp Ala Ser Gly Ile Thr Cys Gly Gly Ile Tyr Ile
340 345 350
Gly Gly Cys Trp Ile Leu Thr Ala Ala His Cys Leu Arg Ala Ser Lys
355 360 365
Thr His Arg Tyr Gln Ile Trp Thr Thr Val Val Asp Trp Ile His Pro
370 375 380
Asp Leu Lys Arg Ile Val Ile Glu Tyr Val Asp Arg Ile Ile Phe His
385 390 395 400
Glu Asn Tyr Asn Ala Gly Thr Tyr Gln Asn Asp Ile Ala Leu Ile Glu
405 410 415
Met Lys Lys Asp Gly Asn Lys Lys Asp Cys Glu Leu Pro Arg Ser Ile
420 425 430
Pro Ala Cys Val Pro Trp Ser Pro Tyr Leu Phe Gln Pro Asn Asp Thr
435 440 445
Cys Ile Val Ser Gly Trp Gly Arg Glu Lys Asp Asn Glu Arg Val Phe
450 455 460
Ser Leu Gln Trp Gly Glu Val Lys Leu Ile Ser Asn Cys Ser Lys Phe
465 470 475 480
Tyr Gly Asn Arg Phe Tyr Glu Lys Glu Met Glu Cys Ala Gly Thr Tyr
485 490 495
Asp Gly Ser Ile Asp Ala Cys Lys Gly Asp Ser Gly Gly Pro Leu Val
500 505 510
Cys Met Asp Ala Asn Asn Val Thr Tyr Val Trp Gly Val Val Ser Trp
515 520 525
Gly Glu Asn Cys Gly Lys Pro Glu Phe Pro Gly Val Tyr Thr Lys Val
530 535 540
Ala Asn Tyr Phe Asp Trp Ile Ser Tyr His Val Gly Arg Pro Phe Ile
545 550 555 560
Ser Gln Tyr Asn Val
565
<210> 5
<211> 27
<212> ДНК
<213> Искусственная
<220>
<223> Праймерная последовательность: FI-AAV-F
<400> 5
tctagaggat ccgccaccat gaagctc 27
<210> 6
<211> 47
<212> ДНК
<213> Искусственная
<220>
<223> Праймерная последовательность: FI-AAV-R
<400> 6
aagcttggcg gccgctcaga cattgtgttg agaaacaaga gaccttc 47
<210> 7
<211> 1752
<212> ДНК
<213> Homo sapiens
<400> 7
atgaagctgc tgcatgtctt tctgctgttt ctgtgcttcc atctgcggtt ctgtaaagtg 60
acctatacta gccaggagga tctggtggag aagaagtgtc tggccaagaa gtacacacac 120
ctgagctgcg acaaggtgtt ctgtcagcct tggcagcggt gcatcgaggg cacctgcgtg 180
tgcaagctgc cttaccagtg cccaaagaac ggcaccgccg tgtgcgccac aaatcggaga 240
tcttttccaa catattgcca gcagaagagc ctggagtgtc tgcaccccgg caccaagttc 300
ctgaacaatg gcacctgcac agccgagggc aagttttctg tgagcctgaa gcacggcaac 360
acagatagcg agggcatcgt ggaggtgaag ctggtggacc aggataagac catgttcatc 420
tgtaagagct cctggtccat gagggaggca aacgtggcat gcctggatct gggattccag 480
cagggagcag acacacagag gcgctttaag ctgtccgacc tgtctatcaa tagcaccgag 540
tgcctgcacg tgcactgtag gggcctggag acatccctgg cagagtgcac cttcacaaag 600
cggagaacca tgggctacca ggactttgcc gacgtggtgt gctataccca gaaggccgat 660
agccccatgg acgatttctt tcagtgcgtg aacggcaagt atatctccca gatgaaggcc 720
tgcgacggca tcaatgactg tggcgatcag tctgacgagc tgtgctgtaa ggcctgtcag 780
ggcaagggct tccactgcaa gagcggcgtg tgcatccctt cccagtacca gtgcaacggc 840
gaggtggatt gtatcacagg agaggacgaa gtgggatgcg caggatttgc atctgtggca 900
caggaggaga cagagatcct gacagccgac atggatgccg agaggcgccg gatcaagtct 960
ctgctgccta agctgagctg tggcgtgaag aatcggatgc acatcagaag gaagcgcatc 1020
gtgggaggca agagggcaca gctgggcgat ctgccatggc aggtggccat caaggacgcc 1080
tctggcatca cctgcggcgg catctacatc ggaggatgtt ggatcctgac cgcagcacac 1140
tgcctgagag caagcaagac acacaggtat cagatctgga ccacagtggt ggattggatc 1200
cacccagacc tgaagagaat cgtgatcgag tacgtggata ggatcatctt tcacgagaac 1260
tacaatgccg gcacatatca gaacgacatc gccctgatcg agatgaagaa ggatggcaat 1320
aagaaggact gtgagctgcc cagatccatc cctgcatgcg tgccatggag cccctatctg 1380
ttccagccca acgatacctg catcgtgtcc ggatggggaa gggagaagga caatgagcgg 1440
gtgttttctc tgcagtgggg cgaggtgaag ctgatctcca actgttctaa gttctacggc 1500
aataggtttt atgagaagga gatggagtgc gccggcacct acgatggcag catcgacgcc 1560
tgtaagggcg attccggagg accactggtg tgcatggacg caaacaatgt gacatacgtg 1620
tggggagtgg tgtcctgggg agagaactgc ggcaagccag agttccccgg cgtatatacc 1680
aaggtggcca attattttga ttggatttcc taccacgtcg gcaggccctt tatttcccag 1740
tataatgtct aa 1752
<210> 8
<211> 251
<212> ДНК
<213> Искусственная
<220>
<223> Печень-специфическая промоторная последовательность HLP
<400> 8
tgtttgctgc ttgcaatgtt tgcccatttt agggtggaca caggacgctg tggtttctga 60
gccagggggc gactcagatc ccagccagtg gacttagccc ctgtttgctc ctccgataac 120
tggggtgacc ttggttaata ttcaccagca gcctcccccg ttgcccctct ggatccactg 180
cttaaatacg gacgaggaca gggccctgtc tcctcagctt caggcaccac cactgacctg 240
ggacagtgaa t 251
<210> 9
<211> 447
<212> ДНК
<213> Искусственная
<220>
<223> Печень-специфическая промоторная последовательность LP1
<400> 9
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt cggtggagag gagcagaggt tgtcctggcg tggtttaggt 180
agtgtgagag gggaatgact cctttcggta agtgcagtgg aagctgtaca ctgcccaggc 240
aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact tagcccctgt 300
ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct cccccgttgc 360
ccctctggat ccactgctta aatacggacg aggacagggc cctgtctcct cagcttcagg 420
caccaccact gacctgggac agtgaat 447
<210> 10
<211> 354
<212> ДНК
<213> Искусственная
<220>
<223> Печень-специфическая промоторная последовательность HLP2
<400> 10
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaca cctctctggg cccatgccac ctccaactgg 120
acacaggacg ctgtggtttc tgagccaggg ggcgactcag atcccagcca gtggacttag 180
cccctgtttg ctcctccgat aactggggtg accttggtta atattcacca gcagcctccc 240
ccgttgcccc tctggatcca ctgcttaaat acggacgagg acagggccct gtctcctcag 300
cttcaggcac caccactgac ctgggacagt gaatgatccc cctgatctgc ggcc 354
<210> 11
<211> 736
<212> БЕЛОК
<213> Искусственная
<220>
<223> Последовательность капсидного белка MutC
<400> 11
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln Lys Gln Asp Asp Gly Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Gln Ala Gly Asp Asn Pro Tyr Leu Arg Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Gln Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Gly Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Asp Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Val Gly
145 150 155 160
Lys Ser Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Thr Ser Leu Gly Ser Asn Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Lys Leu Ser Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg Thr
435 440 445
Gln Gly Thr Thr Ser Gly Thr Thr Asn Gln Ser Arg Leu Leu Phe Ser
450 455 460
Gln Ala Gly Pro Gln Ser Met Ser Leu Gln Ala Arg Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Leu Ser Lys Thr Ala Asn Asp Asn
485 490 495
Asn Asn Ser Asn Phe Pro Trp Thr Ala Ala Ser Lys Tyr His Leu Asn
500 505 510
Gly Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Glu Lys Phe Phe Pro Met His Gly Asn Leu Ile Phe Gly
530 535 540
Lys Glu Gly Thr Thr Ala Ser Asn Ala Glu Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln
565 570 575
Tyr Gly Thr Val Ala Asn Asn Leu Gln Ser Ser Asn Thr Ala Pro Thr
580 585 590
Thr Arg Thr Val Asn Asp Gln Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Met Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Thr Thr Phe Ser Pro Ala Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 12
<211> 736
<212> БЕЛОК
<213> Искусственная
<220>
<223> Последовательность капсидного белка LK03
<400> 12
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Gln Pro Gly Ala Pro Lys Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Asp Gln Ser Pro Gln Glu Pro Asp Ser Ser Ser Gly Val Gly
145 150 155 160
Lys Ser Gly Lys Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ser Glu Ser Val Pro Asp Pro Gln Pro Leu Gly Glu Pro Pro
180 185 190
Ala Ala Pro Thr Ser Leu Gly Ser Asn Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Lys Leu Ser Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Asn Arg Thr
435 440 445
Gln Gly Thr Thr Ser Gly Thr Thr Asn Gln Ser Arg Leu Leu Phe Ser
450 455 460
Gln Ala Gly Pro Gln Ser Met Ser Leu Gln Ala Arg Asn Trp Leu Pro
465 470 475 480
Gly Pro Cys Tyr Arg Gln Gln Arg Leu Ser Lys Thr Ala Asn Asp Asn
485 490 495
Asn Asn Ser Asn Phe Pro Trp Thr Ala Ala Ser Lys Tyr His Leu Asn
500 505 510
Gly Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Asp Asp Glu Glu Lys Phe Phe Pro Met His Gly Asn Leu Ile Phe Gly
530 535 540
Lys Glu Gly Thr Thr Ala Ser Asn Ala Glu Leu Asp Asn Val Met Ile
545 550 555 560
Thr Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln
565 570 575
Tyr Gly Thr Val Ala Asn Asn Leu Gln Ser Ser Asn Thr Ala Pro Thr
580 585 590
Thr Arg Thr Val Asn Asp Gln Gly Ala Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu
625 630 635 640
Lys His Pro Pro Pro Gln Ile Met Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asn Pro Pro Thr Thr Phe Ser Pro Ala Lys Phe Ala Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Pro Leu
725 730 735
<---

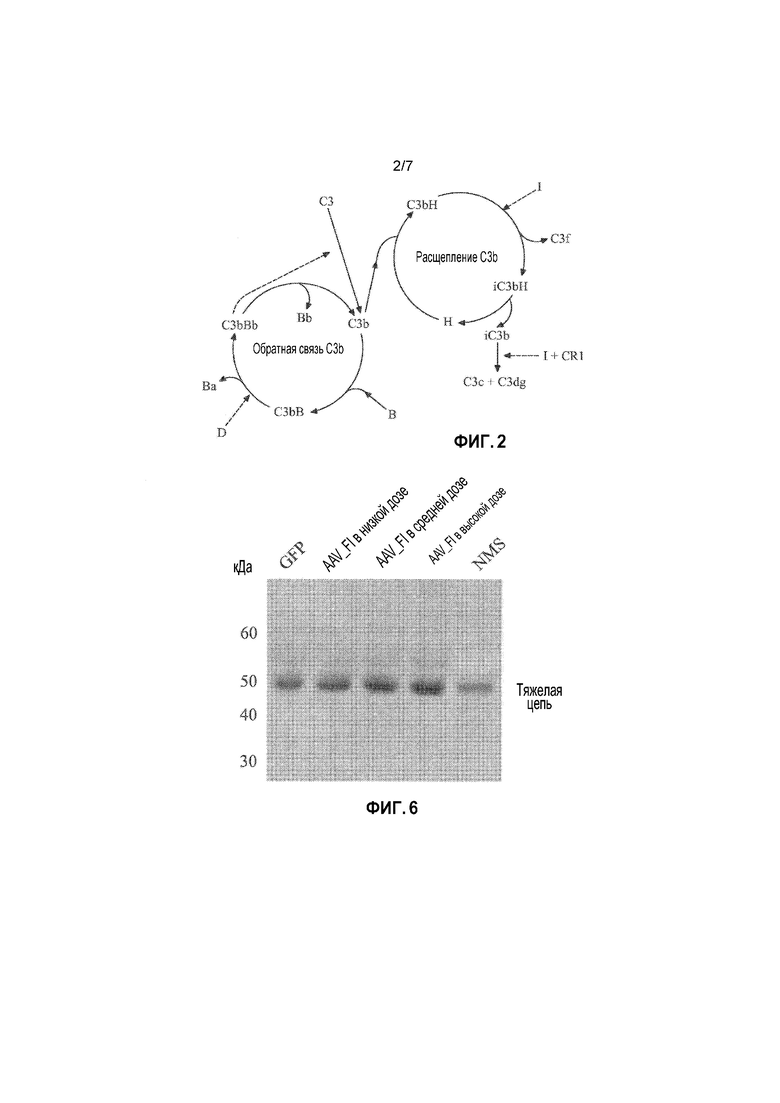
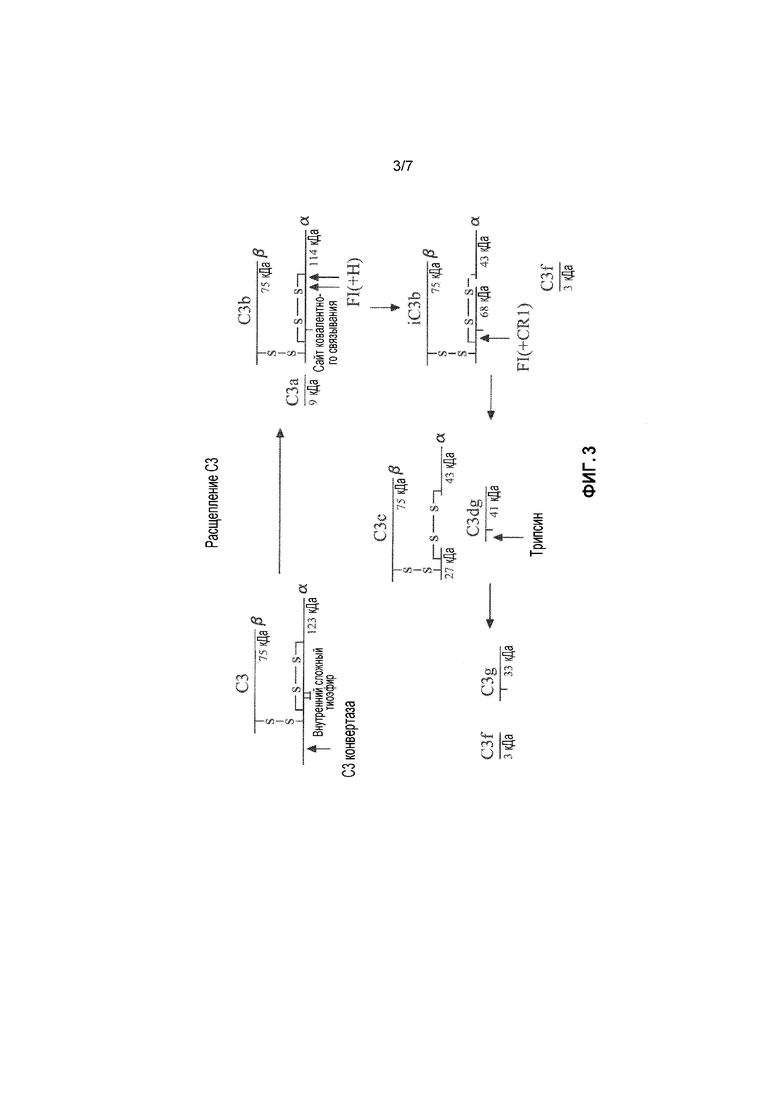
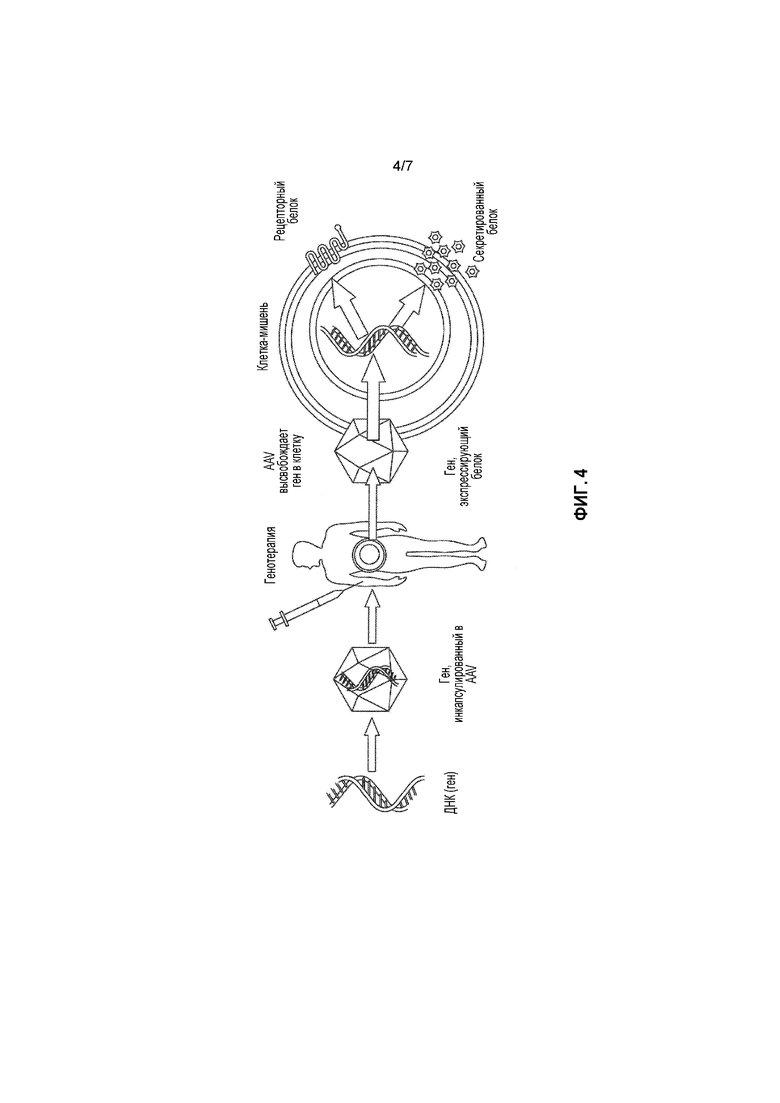
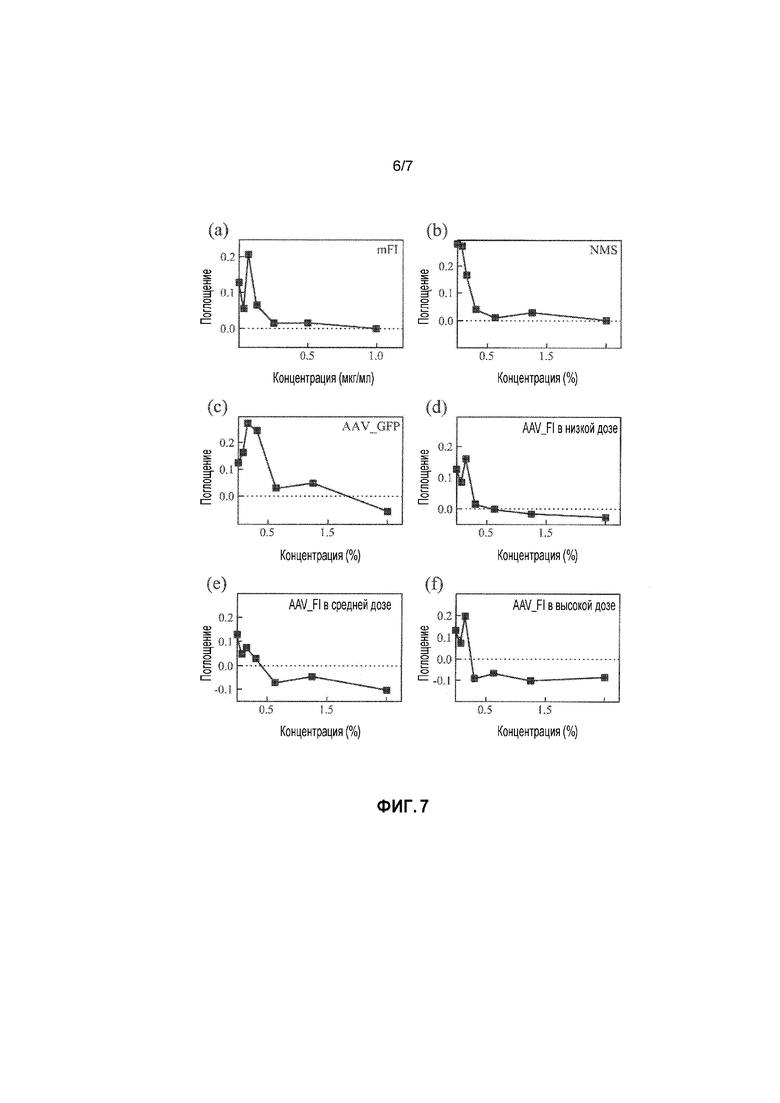
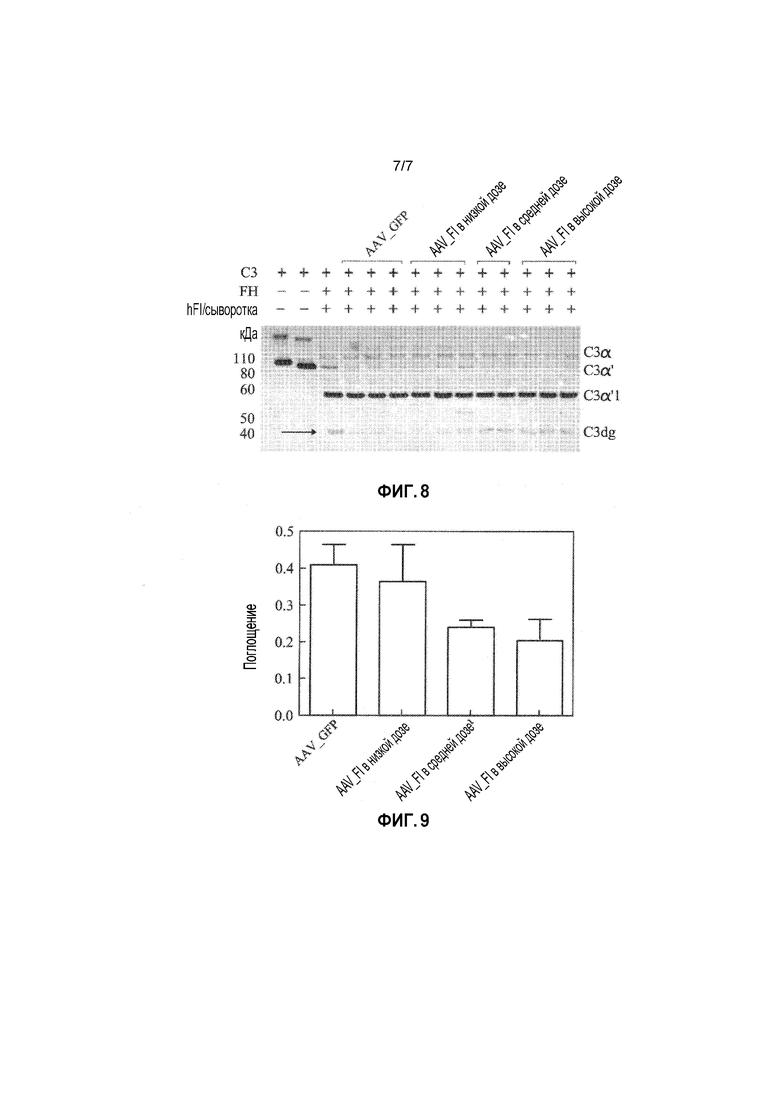
Изобретение относится к области биотехнологии. Предложен способ профилактики, лечения или ослабления комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента, у пациента, а также предложены фармацевтическая композиция и набор для осуществления указанного способа. Также описаны рекомбинантные вирусные векторы, кодирующие фактор I, рекомбинантные вирусные частицы, икапсидирующие эти векторы, и их применение в способах лечения. Изобретение обеспечивает значительное повышение уровня фактора I в плазме, что приводит к достижению системного повышения уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента. 9 н. и 35 з.п. ф-лы, 9 ил., 2 табл., 2 пр.
1. Способ профилактики, лечения или ослабления комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента, у пациента, где указанный способ включает введение пациенту рекомбинантной частицы аденоассоциированного вируса (rAAV)вектора, где частица rAAV является псевдотипированной для сообщения тропизма к печени, где частица rAAV инкапсидирует вектор rAAV, где вектор rAAV содержит нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, которые сохраняют C3b-инактивирующую и iC3b-расщепляющую активность, так, чтобы терапевтически эффективное количество кодируемого фактора I или его фрагмента или производного экспрессировалось нуклеиновой кислотой у пациента и приводило к повышению уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента, где кодируемый фактор I или его фрагмент или производное экспрессируется из печень-специфического промотора, и где частицу rAAV вводят пациенту внутривенно, и где частица rAAV инфицирует печень пациента после введения, что приводит к экспрессии фактора I или его фрагмента или производного из печени.
2. Способ по п. 1, где уровень C3b-инактивирующей и iC3b-расщепляющей активности увеличивается до уровня, превышающего нормальный уровень.
3. Способ по любому из предшествующих пунктов, где частица rAAV содержит один или два ITR AAV2 или их производные, где каждое производное ITR AAV2 содержит нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR AAV2, и где частица rAAV была псевдотипирована капсидным белком AAV8 (rAAV2/8), или частица AAV2 была псевдотипирована капсидным белком AAV9 (rAAV2/9) или его производным, аминокислотная последовательность которого по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности природного капсидного белка AAV8 или AAV9.
4. Способ по любому из предшествующих пунктов, отличающийся тем, что частица rAAV содержит капсидный белок, аминокислотная последовательность которого имеет по меньшей мере 98% или по меньшей мере 99% аминокислотную идентичность по всей своей длине с аминокислотной последовательностью SEQ ID NO: 11.
5. Способ по любому из предшествующих пунктов, где печень-специфический промотор представляет собой человеческий промотор альфа-1-антитрипсина (hAAT).
6. Способ по любому из пп.1-4, отличающийся тем, что печень-специфический промотор представляет собой HLP, LP1, HLP2, hAAT, HCR-hAAT, ApoE-hAAT или LSP.
7. Способ по любому из пп.1-4 или 6, в котором печень-специфический промотор содержит нуклеотидную последовательность, которая по меньшей мере на 90% или по меньшей мере 95% идентична по всей своей длине нуклеотидной последовательности SEQ ID NO: 10.
8. Способ по любому из предшествующих пунктов, где уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, в два раза превышающего нормальный уровень.
9. Способ по любому из предшествующих пунктов, где уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который на 80% или на 60% превышает нормальный уровень.
10. Способ по любому из предшествующих пунктов, где уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который на 40% или на 20% превышает нормальный уровень.
11. Способ по любому из предшествующих пунктов, где уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который по меньшей мере на 5%, 10%, 15%, 20% или 25% превышает нормальный уровень.
12. Способ по любому из предшествующих пунктов, где уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента представляет собой уровень в сыворотке пациента.
13. Способ по любому из предшествующих пунктов, где нормальный уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента эквивалентен уровню, обеспечиваемому 30-40 мкг/мл фактора I в сыворотке пациента.
14. Способ по любому из предшествующих пунктов, где комплемент-опосредуемым расстройством является возрастная дегенерация желтого пятна (ВДЖП) (а в частности, ВДЖП на ранней стадии (сухой глаз) или региональная атрофия), болезнь плотных отложений (БПО), атипический гемолитический уремический синдром (АГУС), C3-гломерулопатии, мембранопролиферативный гломерулонефрит типа 2 (МПГН2), атеросклероз, хроническое сердечно-сосудистое заболевание, болезнь Альцгеймера, системный васкулит, пароксизмальная ночная гемоглобинурия (ПНГ), воспалительные или аутовоспалительные заболевания у пациентов пожилого возраста, мембранопролиферативный гломерулонефрит типа I (МПГН типа I), мембранопролиферативный гломерулонефрит типа III (МПГН типа III), синдром Гийена-Барре, пурпура Геноха-Шенлейна, IgA-нефропатия или мембранозный гломерулонефрит.
15. Способ по п. 14, где у пациента имеется риск развития ВДЖП.
16. Способ по п. 15, где пациент является гомозиготным или гетерозиготным по одному или более SNP, ассоциированным с ВДЖП.
17. Способ по п. 15 или 16, который также включает определение риска развития ВДЖП у пациента.
18. Способ по п. 17, где наличие риска развития ВДЖП у пациента определяют по наличию у пациента гомозиготности или гетерозиготности по одному или более SNP, ассоциированным с ВДЖП.
19. Способ по п. 16 или 18, где уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который по меньшей мере на 10% превышает нормальный уровень, если пациент является гетерозиготным по одному или более SNP, ассоциированным с ВДЖП.
20. Способ по п. 16 или 18, где уровень C3b-инактивирующей и iC3b-расщепляющей активности у пациента повышается до уровня, который по меньшей мере на 50% превышает нормальный уровень, если пациент является гомозиготным по одному или более SNP, ассоциированным с ВДЖП.
21. Способ по любому из предшествующих пунктов, который также включает определение уровня C3b-инактивирующей и iC3b-расщепляющей активности у пациента по меньшей мере через неделю после введения, и повторение введения, если уровень активности равен нормальному уровню или ниже.
22. Способ по любому из предшествующих пунктов, где фактором I является человеческий фактор I с аминокислотной последовательностью SEQ ID NO: 2 или 4.
23. Способ по любому из предшествующих пунктов, где фрагмент или производное фактора I представляет собой полипептид, аминокислотная последовательность которого по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности человеческого фактора I с аминокислотной последовательностью SEQ ID NO: 2 или 4.
24. Способ по любому из предшествующих пунктов, где пациентом является человек.
25. Рекомбинантный вектор аденоассоциированного вируса (rAAV) для профилактики, лечения или ослабления комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента, который включает нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное, которые сохраняют C3b-инактивирующую и iC3b-расщепляющую активность, где нуклеиновая кислота, кодирующая фактор I или его фрагмент или производное, функционально присоединена к промотору, и где промотором является печень-специфический промотор.
26. Вектор rAAV по п. 25, где печень-специфическм промотором является человеческий промотор альфа-1-антитрипсина (hAAT).
27. Вектор rAAV по п. 25, где печень-специфический промотор представляет собой HLP, LP1, HLP2, hAAT, HCR-hAAT, ApoE-hAAT или LSP.
28. Вектор rAAV по п. 25 или 27, в котором печень-специфический промотор содержит нуклеотидную последовательность, которая по меньшей мере на 90% или по меньшей мере 95% идентична по всей своей длине нуклеотидной последовательности SEQ ID NO: 10.
29. Вектор rAAV по любому из пп. 25-28, который включает экспрессионный кластер, фланкированный инвертированными концевыми повторами AAV (ITR), где экспрессионный кластер содержит нуклеиновую кислоту, кодирующую фактор I или его фрагмент или производное и функционально присоединенную к печень-специфическому промотору и к сайту распознавания полиаденилирования.
30. Вектор rAAV по п. 29, где ITR представляют собой ITR AAV2 или их производные, где каждое производное ITR AAV2 содержит нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR AAV2.
31. Частица rAAV для профилактики, лечения или ослабления комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента, которая включает вирусный капсид, инкапсидирующий вектор rAAV по любому из пп. 25-30, и где частица rAAV является псевдотипированной для сообщения тропизма к печени.
32. Частица rAAV по п. 31, которая способна проникать в гепатоциты.
33. Частица rAAV по п. 31 или 32, где частица rAAV содержит капсидный белок AAV8 или AAV9 или его производное, содержащие аминокислотную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности природного капсидного белка AAV8 или AAV9.
34. Частица rAAV по п. 33, где частица rAAV содержит один или два ITR AAV2 или их производные, где каждое производное ITR AAV2 содержит нуклеотидную последовательность, которая по всей своей длине по меньшей мере на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности природного ITR AAV2.
35. Частица rAAV по любому из пп. 31 или32, отличающаяся тем, что частица rAAV содержит капсидный белок, аминокислотная последовательность которого имеет по меньшей мере 98% или по меньшей мере 99% аминокислотную идентичность по всей своей длине с аминокислотной последовательностью SEQ ID NO: 11.
36. Фармацевтическая композиция для профилактики, лечения или ослабления комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента, которая включает: терапевтически эффективное количество вектора rAAV по любому из пп. 25-30 или терапевтически эффективное количество частицы rAAV по любому из пп. 31-35, и фармацевтически приемлемый носитель, наполнитель или разбавитель.
37. Фармацевтическая композиция по п.36, подходящая для внутривенного введения.
38. Набор для профилактики, лечения или ослабления комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента, который включает: вектор rAAV по любому из пп. 25-30 или частицу rAAV по любому из пп. 31-35 и фармацевтически приемлемый носитель, наполнитель или разбавитель.
39. Набор для продуцирования частиц rAAV по любому из пп. 31-35, который включает: вектор rAAV по любому из пп. 25-30; первую хелперную плазмиду, содержащую нуклеиновую кислоту, кодирующую белки репликации AAV и капсидные белки, которые сообщают тропизм к печени, и вторую хелперную плазмиду, содержащую нуклеиновую кислоту, кодирующую гены, необходимые для продуктивного жизненного цикла AAV.
40. Применение вектора rAAV по любому из пп. 25-30, частицы rAAV по любому из пп. 31-35 или фармацевтической композиции по п. 36 или 37 в качестве лекарственного средства для профилактики, лечения или ослабления комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента.
41. Применение вектора rAAV по любому из пп. 25-30, частицы rAAV по любому из пп. 31-35 или фармацевтическая композиция по п. 36 или 37, для лечения комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента.
42. Применение вектора rAAV по любому из пп. 25-30, частицы rAAV по любому из пп. 31-35 или фармацевтической композиции по п. 36 или 37 в получении лекарственного средства для лечения комплемент-опосредуемого расстройства, ассоциированного с гиперактивностью цикла обратной связи C3b комплемента.
43. Применение вектора, частицы или композиции по п. 41 или 42, где комплемент-опосредуемым расстройством является возрастная дегенерация желтого пятна (ВДЖП) (а в частности, ВДЖП на ранней стадии (сухой глаз) или региональная атрофия), болезнь плотных отложений (БПО), атипический гемолитический уремический синдром (АГУС), C3-гломерулопатии, мембранопролиферативный гломерулонефрит типа 2 (МПГН2), атеросклероз, хроническое сердечно-сосудистое заболевание, болезнь Альцгеймера, системный васкулит, пароксизмальная ночная гемоглобинурия (ПНГ), воспалительные или аутовоспалительные заболевания у пациентов пожилого возраста, мембранопролиферативный гломерулонефрит типа I (МПГН типа I), мембранопролиферативный гломерулонефрит типа III (МПГН типа III), синдром Гийена-Барре, пурпура Геноха-Шенлейна, IgA-нефропатия или мембранозный гломерулонефрит.
44. Применение вектора, частицы или композиции по п. 41 или 42, где комплемент-опосредуемым расстройством является возрастная дегенерация желтого пятна (ВДЖП).
| WO 2010103291 A2, 16.09.2010 | |||
| WO 2015162302 A2, 29.10.2015 | |||
| US 6632670 B1, 14.10.2003 | |||
| THOMAS C.E | |||
| et al., Rapid uncoating of vector genomes is the key to efficient liver transduction with pseudotyped adeno-associated virus vectors, JOURNAL OF VIROLOGY, 2004, vol | |||
| Парный автоматический сцепной прибор для железнодорожных вагонов | 0 |
|
SU78A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| АНТИТЕЛА ПРОТИВ C3b И СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С КОМПЛЕМЕНТОМ НАРУШЕНИЙ | 2008 |
|
RU2473563C2 |

