ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к рекомбинантным вирусам герпеса, в которые вставлены по меньшей мере две рекомбинантные нуклеотидные последовательности, и к их применению. Изобретение особенно подходит для получения поливалентных вирусов или композиций, таких как вакцины, которые могут индуцировать защитный иммунитет против птичьих патогенов или болезней.
УРОВЕНЬ ТЕХНИКИ
Мясо птицы и яйца являются важными источниками пищи, потребление которых постоянно увеличивается из-за роста численности населения и отличного соотношения цены и качества. Чтобы обеспечить здоровье птицы, а также технику безопасности и охрану труда на предприятиях и безопасность пищевых продуктов, технология вакцины для птицы стала предметом озабоченности во всем мире.
В качестве вакцин для домашней птицы против патогенов-мишеней обычно используются вирусные векторы, экспрессирующие белки патогенов. Вакцины, включающие такие вирусные векторы, индуцируют экспрессию чужеродных белков патогенов в инфицированных хозяевах, что может приводить к получению защитного иммунитета.
В качестве векторов-кандидатов для вакцинации птиц были исследованы многие различные классы вирусов, такие как аденовирусы, адено-ассоциированные вирусы, вирусы оспы домашней птицы, вирусы герпеса и т.п.
Были определены три типа вирусов герпеса: MDV1, MDV2 и MDV3 (также известный как вирус герпеса индейки (HVT)). Между указанными вирусами существует большое сходство (см. Kingham et al., Journal of General Virology (2001) 82, 1123-1135), и все они были использованы для получения рекомбинантных вирусов, в которые интегрирован чужеродный ген, полученный из патогена, для применения в качестве вакцины для птиц, особенно домашней птицы, такой как курица.
Хотя такие вакцинные препараты обеспечивают эффективные результаты для вакцинации видов птиц от многих заболеваний, в случаях, когда птицам вводят два или более рекомбинантных вирусов герпеса, между патогенами, каждый из которых кодирует свой антиген, может возникать конкуренция и иммуносупрессия.
Чтобы преодолеть такое взаимодействие, а также облегчить вакцинацию против множества заболеваний, были предприняты различные попытки получить поливалентные вирусы герпеса, кодирующие несколько антигенов.
В первых исследованиях было вставлено несколько генов в один сайт клонирования в геном вируса герпеса (см., например, EP1026246). Однако такие конструкции либо не обеспечивали требуемый уровень защитного иммунитета, либо оказывались нестабильными, все или часть чужеродных генов удалялись во время повторяющихся пассажей в культуре клеток.
В WO2013/144355 сообщается о стабильных вирусах герпеса, кодирующих несколько чужеродных антигенов, полученных с использованием комбинаций сайтов клонирования, расположенных в некодирующих областях вирусного генома.
В WO2013/057236, WO2013/082327 и WO2013/082317 сообщается о другом подходе к созданию поливалентного HVT путем клонирования по меньшей мере одного гена в кодирующей последовательности US2 вирусов герпеса. Согласно этим применениям, US2 не является необходимым для репликации вируса, хотя это свойство и не было доказано.
Учитывая количество патогенов и видов, в данной области существует потребность в дополнительных рекомбинантных поливалентных вирусах герпеса, которые могут стабильно экспрессировать несколько генов in vivo и подходят для вакцинации птиц, особенно домашней птицы.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к рекомбинантным вирусам птичьего герпеса, содержащим несколько рекомбинантных нуклеотидных последовательностей по меньшей мере в двух отдельных участках вирусного генома.
Более конкретно, изобретение относится к рекомбинантным вирусам герпеса индейки (HVT), содержащим по меньшей мере первую и вторую рекомбинантные нуклеотидные последовательности, причем каждая рекомбинантная нуклеотидная последовательность кодирует полипептид, где первая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположеную между HVT053 (UL45) и HVT054 (UL46), и где вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT064 (LORF3) и HVT070, в частности между HVT064 (LORF3) и HVT065 (UL55), между HVT065 (UL55) и HVT066 (LORF4), между HVT066 (LORF4) и HVT067 (LORF5), между HVT067 (LORF5) и HVT069 (LORF6) или между HVT069 (LORF6) и HVT070, даже более конкретно, между HVT065 (UL55) и HVT066 (LORF4).
Каждый рекомбинантный нуклеотид может кодировать один и тот же или другой полипептид, в частности антиген, цитокин, адъювант, гормон и т.п. В конкретном варианте осуществления каждый рекомбинантный нуклеотид кодирует антиген, который может быть одинаковым или различным (или идентичными или различными частями одного и того же антигена), и может происходить от одного и того же или от различных патогенов. Антиген(ы) может быть выбран или получен, например, из поверхностных белков, секретируемых белков или структурных белков указанного патогена(ов) или их иммуногенных фрагментов.
Изобретение также относится к нуклеиновой кислоте, содержащей геном рекомбинантного HVT, как определено выше, и к вектору (например, плазмиде), содержащему такую нуклеиновую кислоту.
Изобретение также относится к клетке, содержащей рекомбинантный HVT, или нуклеиновую кислоту, или вектор, как определено выше.
Еще одной целью изобретения является композиция, содержащая рекомбинантный HVT, как определено выше, и подходящий наполнитель или разбавитель.
Еще одной целью изобретения является композиция, содержащая нуклеиновую кислоту или клетку, как определено выше, и подходящий наполнитель или разбавитель.
Другая цель изобретения заключается в вакцине, которая содержит эффективное иммунизирующее количество рекомбинантного HVT, нуклеиновой кислоты и/или клетки, как определено выше.
Дополнительная цель изобретения заключается в рекомбинантном HVT, нуклеиновой кислоте или клетке, как определено выше, для использования для иммунизации птицы, такой как домашняя птица, против патогена.
Следующая цель изобретения заключается в рекомбинантном HVT, нуклеиновой кислоте или клетке, как определено выше, для использования для защиты птицы, такой как домашняя птица, от заболевания, вызываемого патогеном.
Еще одна цель изобретения заключается в вакцине, как определено выше, для использования для вакцинации птицы, такой как домашняя птица, против одного или нескольких патогенов.
Еще одна цель изобретения заключается в способе вакцинации птиц, включающем введение птице композиции, вакцины или вируса, как определено выше.
Еще одна цель изобретения заключается в способе индукции иммунного ответа на антиген у птицы, включающем введение птице композиции, вакцины или вируса, как определено выше.
В изобретении также предлагается набор, содержащий вакцину, для иммунизации птиц, который включает следующие компоненты:
а. эффективное количество композиции или вакцины, как определено выше, и
б. средство для введения указанной композиции или вакцины указанной птице.
Изобретение может быть использовано для вакцинации любой птицы против любого птичьего патогена. Он особенно подходит для вакцинации домашней птицы, например, кур.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
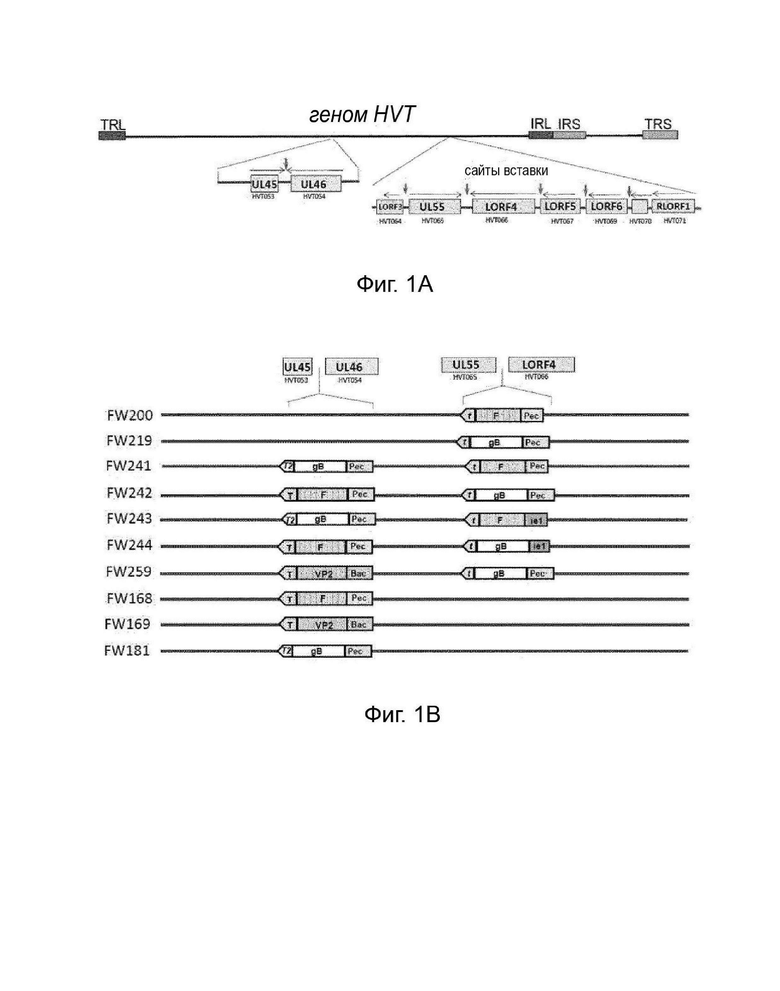
На Фигуре 1 представлена схематическая диаграмма генома HVT (1A) . Отмечено расположение Unique Long (UL) 45 (HVT053) и UL 46 (HVT054), а также расположение Unique Long HVT064-070. Рекомбинантные нуклеотидные последовательности могут быть вставлены в созданные ПЦР сайты SfiI между HVT053 (UL45) и HVT054 (UL46), и между HVT064 и 065, и/или HVT065 и 066, и/или HVT066 и 067, и/или HVT 067 и 069 и/или HVT 069 и 070. Также представлены конструкции HVT, объединяющие различные кластеры нуклеотидных последовательностей и промоторов, в соответствии с конкретными вариантами осуществления изобретения (1B).
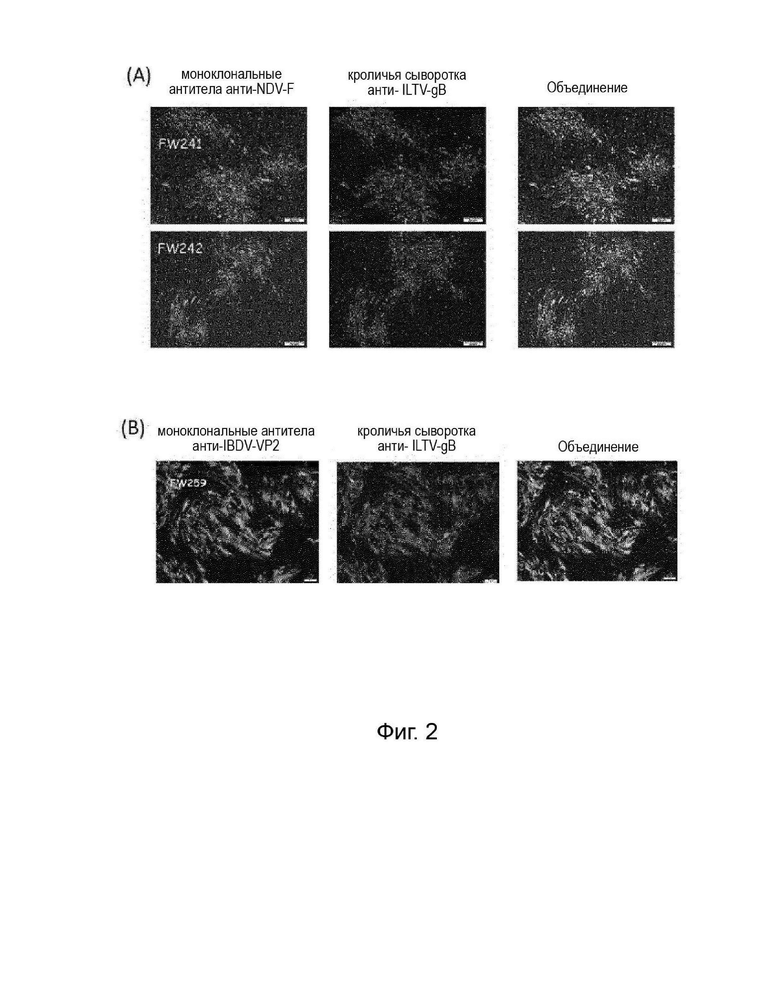
На Фигуре 2А показано иммунофлуоресцентное окрашивание клеток CEF, инфицированных двойными рекомбинантными HVT согласно вариантам осуществления изобретения (FW241 и FW242), коэкспрессирующих NDV-F и ILTV-gB (клетки, инфицированные rHVT/ND/ILT). Экспрессию белка F выявляли с помощью моноклонильных антител анти-F (77-2) и Alexa Flour 488. Экспрессию белка gB выявляли с помощью кроличьей сыворотки против gBN4 и Alexa Flour 546. Результаты показывают, что обе клетки, инфицированные FW241 или FW242, экспрессируют как встроенный белок NDV-F, так и встроенный белок ILTV-gB. Их наблюдали с помощью флуоресцентного микроскопа при 40-кратном увеличении.
На Фигуре 2B показано иммунофлуоресцентное окрашивание клеток CEF, инфицированных двойными рекомбинантными HVT в соответствии с вариантами осуществления изобретения (FW259), коэкспрессирующих VP2 IBDV и ILTV-gB (клетки, инфицированные rHVT/IBD/ILT). Экспрессию белка VP2 выявляли с помощью моноклональных антител анти-VP2 (R63) и Alexa Flour 488. Экспрессию белка gB выявляли с помощью кроличьей сыворотки против gBN4 и Alexa Flour 546. Результаты показывают, что клетки, инфицированные FW259, экспрессируют как встроенный белок IBDV-VP2, так и встроенный белок ILTV-gB. Их наблюдали с помощью флуоресцентного микроскопа при 100-кратном увеличении.
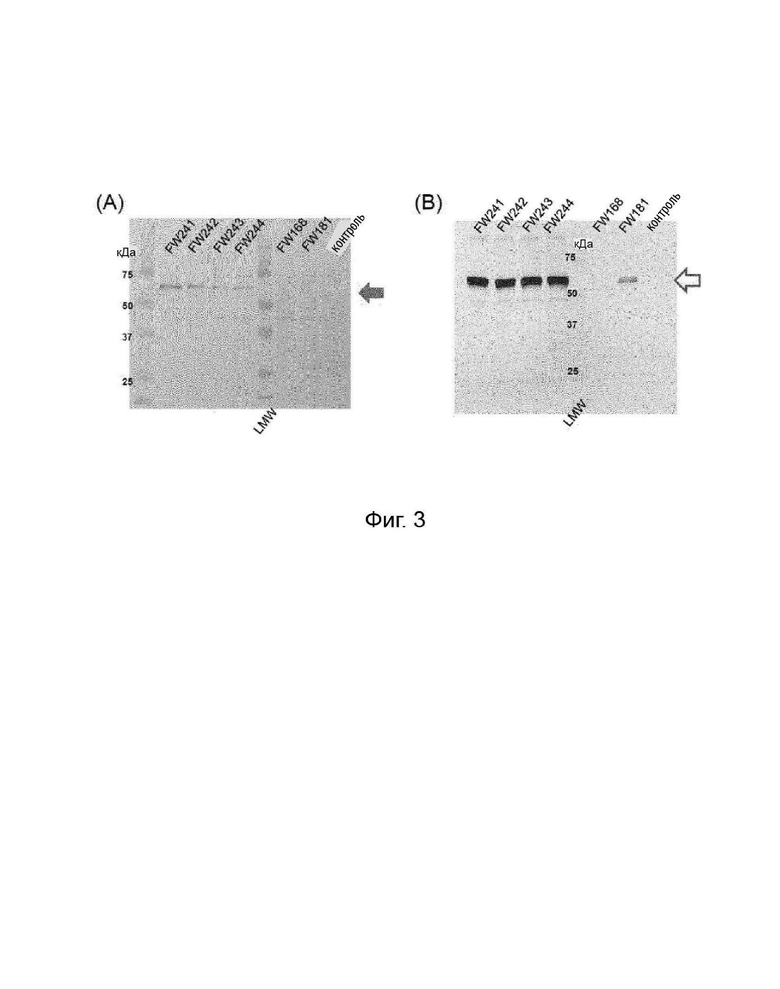
Фигуры 3A и 3B представляют собой анализ вестерн-блоттинг, показывающий экспрессию белка F и/или белка gB в клетках CEF, инфицированных rHVT по настоящему изобретению. Как показано на Фигуре 3А, полоса белка в 60 килодальтон (кДа) наблюдалась только в полосе с инфицированными rHVT/ND/ILT клетками, что соответствовало ожидаемому размеру белка F  Полосы на дорожке rHVT/45-46BacVP2 (FW181) отсутствовали. Как показано на Фигуре 3B, белок gB наблюдался при 54 килодальтонах (kd) в дорожках каждого rHVT/ND/ILT
Полосы на дорожке rHVT/45-46BacVP2 (FW181) отсутствовали. Как показано на Фигуре 3B, белок gB наблюдался при 54 килодальтонах (kd) в дорожках каждого rHVT/ND/ILT  Напротив, полоса на дорожке rHVT/45-46 PecF (FW168) отсутствовала. Двойные rHVT по изобретению экспрессировали как NDV-F, так и ILTV-gB.
Напротив, полоса на дорожке rHVT/45-46 PecF (FW168) отсутствовала. Двойные rHVT по изобретению экспрессировали как NDV-F, так и ILTV-gB.
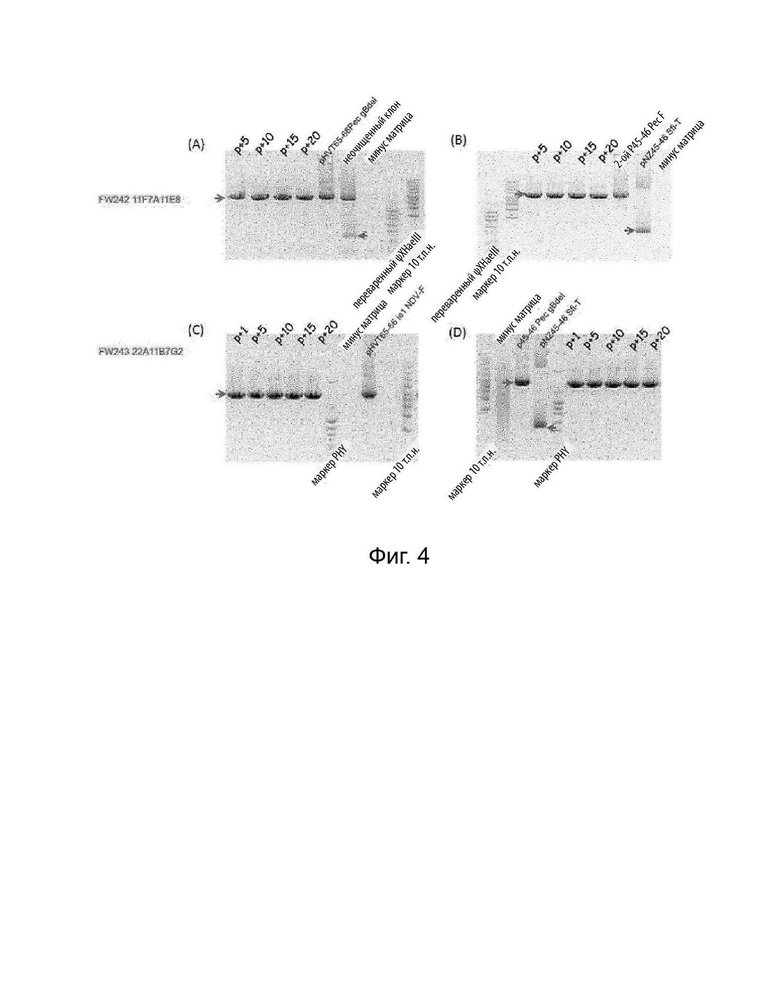
На Фигурах 4A и 4B показаны результаты ПЦР-анализа для проверки стабильности рекомбинантного HVT FW242 в последовательных пассажах, что указывает на то, что после 20 пассажей ген F и ген gB стабильно экспрессировались в rHVT FW242 по настоящему изобретению.
На Фигурах 4C и 4D показаны результаты ПЦР-анализа для проверки структуры генома очищенного FW243 и проверки стабильности FW243 в последовательных пассажах. Они указали, что FW243, двойной рекомбинантный HVT/ND/ILT по изобретению, имел ожидаемую геномную структуру и после 20 пассажей ген F и ген gB стабильно экспрессировались в rHVT FW243 по изобретению.
На фигурах 5A и 5B показаны результаты анализа вестерн-блоттинга для проверки стабильности рекомбинантного HVT FW244 в последовательных пассажах, указывающие на то, что после 20 пассажей белок F и белок gB стабильно экспрессировались в CEF, инфицированном rHVT FW244 по настоящему изобретению.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение в целом относится к рекомбинантным вирусам птичьего герпеса, содержащим несколько рекомбинантных нуклеотидных последовательностей, их изготовлению, композициям, содержащим их, и их применениям.
Определения
Настоящее раскрытие будет лучше всего понято со ссылкой на следующие определения:
Термин «рекомбинантный» по отношению к вирусу герпеса относится к вирусу герпеса, геном которого был модифицирован путем вставки по меньшей мере одной нуклеотидной последовательности (например, ДНК, такой как ген), которая в природе не встречается в геноме вируса герпеса, или которая в природе встречается в указанном геноме, но в другой форме или в другом положении. Будет понятно, что рекомбинантный вирус герпеса может быть получен различными способами, такими как технология рекомбинантной ДНК, как описано здесь, и, однажды полученный, может быть воспроизведен без дальнейшего использования технологии рекомбинантной ДНК. Таким образом, структура «рекомбинантного вируса герпеса» описывается с точки зрения вставки ДНК.
В настоящем описании термины «нуклеиновая кислота», «нуклеиновая последовательность» и «нуклеотидная последовательность» используются взаимозаменяемо и относятся к молекуле нуклеиновой кислоты, имеющей определенную последовательность, которая может представлять собой дезоксирибонуклеотиды и/или рибонуклеотиды. Нуклеотидная последовательность может быть сначала получена, например, рекомбинантными, ферментативными и/или химическими способами, а затем реплицирована в клетке-хозяине или системе in vitro. Нуклеотидная последовательность предпочтительно включает открытую рамку считывания, кодирующую молекулу (например, пептид или белок). Нуклеотидная последовательность может содержать дополнительные последовательности, такие как промотор, терминатор транскрипции, сигнальный пептид, IRES и т.д. Предпочтительно открытая рамка считывания в рекомбинантной нуклеиновой кислоте не содержит интрона.
«Рекомбинантная нуклеотидная последовательность» обозначает последовательность, которая не встречается в природе в геноме вируса герпеса или которая в природе встречается в указанном геноме, но в другой форме или в другом положении. Типичные «рекомбинантные нуклеотидные последовательности» представляют собой гены, предпочтительно кодирующие молекулы, которые не продуцируются в природе вирусом птичьего герпеса, например молекулы из другого вируса или из клетки. «Ген» в контексте такой «рекомбинантной нуклеотидной последовательности» обозначает любую молекулу нуклеиновой кислоты, содержащую открытую рамку считывания, такую как нуклеиновая кислота, состоящая или состоящая по существу из открытой рамки считывания. Ген может дополнительно содержать регуляторные элементы, такие как, например, промотор или терминатор.
В настоящем описании термины «полипептид», «пептид» и «белок» используются взаимозаменяемо и относятся к любой молекуле, содержащей полимер из по меньшей мере 2 последовательных аминокислот.
Термин «межгенная область» хорошо известен в данной области и относится к любой области вирусного генома, которая расположена между двумя указанными открытыми рамками считывания (ОРС) вируса. Таким образом, межгенная область между UL45 (HVT053) и UL46 (HVT054) обозначает область, начинающуюся сразу с 3' стоп-кодона UL45 и заканчивающуюся сразу на 5' стоп-кодоне UL46 (поскольку обе ОРС имеют противоположную ориентацию). Межгенная область между HVT064 и HVT065 обозначает область, начинающуюся сразу с 3' старт-кодона HVT064 и заканчивающуюся непосредственно на 5' старт-кодоне HVT065 (поскольку обе ОРС имеют обратную ориентацию). Точно так же межгенная область между HVT065 и HVT066 обозначает область, начинающуюся сразу с 3' стоп-кодона HVT065 и заканчивающуюся сразу на 5' стоп-кодоне HVT066 (поскольку обе ОРС имеют противоположную ориентацию). Межгенная область может включать регуляторные области, такие как терминаторы или промоторы.
Термин «виды птиц» предназначен для обозначения всех видов птиц, таких как птицы класса Aves, т.е. пернатых, крылатых, двуногих, эндотермических и откладывающих яйца позвоночных животных. В контексте изобретения птицы или виды птиц относятся, в частности, к птицам с экономическим и/или агрономическим значением, таким как домашняя птица (например, куры и индейки), водоплавающая птица (например, утки и гуси) и декоративные птицы (такие как как лебеди и попугаи).
Используемый здесь термин «вакцина» обозначает агент, который можно использовать для того, чтобы вызывать, стимулировать или усиливать иммунный ответ в организме.
Термин «поливалентный», используемый здесь в отношении рекомбинантного вируса герпеса, вектора или вакцины по изобретению, относится к рекомбинантному вирусу герпеса или вакцине, которая содержит по меньшей мере две рекомбинантные нуклеотидные последовательности, как определено выше, причем указанные последовательности являются последовательностями того же или другого патогена, и от того же или другого патогена.
Поливалентный рекомбинантный HVT
Изобретение относится к рекомбинантному HVT, содержащему несколько чужеродных генов в определенных местах. Более конкретно, изобретение показывает, что стабильный и эффективный поливалентный рекомбинантный HVT может быть получен, когда:
. по меньшей мере одна рекомбинантная нуклеотидная последовательность клонирована в межгенной области генома, расположенной между HVT053 и HVT054, и
. по меньшей мере одна рекомбинантная нуклеотидная последовательность клонирована в межгенной области генома, расположенной между HVT064 и HVT070.
Как показано в примерах, при использовании такой комбинации сайтов клонирования рекомбинантный HVT стабилен в течение по меньшей мере 10, предпочтительно по меньшей мере 15, более предпочтительно по меньшей мере 20 пассажей в клетках CEF. Кроме того, с использованием такой комбинации сайтов клонирования достигается сильная и продолжительная экспрессия генов in vivo, достаточная для обеспечения высокого защитного иммунитета.
В рекомбинантном вирусе герпеса существует множество потенциальных сайтов клонирования. В частности, в геноме вируса герпеса имеется около 397 ОРС, 99 из которых, по оценкам, кодируют функциональные белковые продукты (см. Afonso et al., J. Virology 75 (2), 2001, 971). Кроме того, некоторые сайты клонирования, подходящие для использования в контексте моновалентных вирусов, показали нестабильность при использовании в контексте поливалентных вирусов, особенно когда два разных сайта клонирования используются в одном и том же вирусе. Изобретение показывает, что комбинация межгенной области между HVT053 и HVT054 с межгенной областью между HVT064 и HVT070 обеспечивает стабильность и подходящий уровень экспрессии обеих клонированных рекомбинантных нуклеотидных последовательностей in vitro. Таким образом, такие конкретные рекомбинантные вирусы представляют собой новые эффективные агенты для индукции сильного защитного иммунитета in vivo.
В конкретном варианте осуществления изобретение относится к рекомбинантному HVT, содержащему несколько рекомбинантных нуклеиновых кислот (например, чужеродные гены), где:
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT053 и HVT054, и
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT064 и HVT065.
В другом конкретном варианте осуществления изобретение относится к рекомбинантному HVT, содержащему несколько рекомбинантных нуклеиновых кислот (например, чужеродные гены), где:
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT053 и HVT054, и
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT065 и HVT066.
В другом конкретном варианте осуществления изобретение относится к рекомбинантному HVT, содержащему несколько рекомбинантных нуклеиновых кислот (например, чужеродные гены), где:
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT053 и HVT054, и
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT066 и HVT067.
В другом конкретном варианте осуществления изобретение относится к рекомбинантному HVT, содержащему несколько рекомбинантных нуклеиновых кислот (например, чужеродные гены), где:
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT053 и HVT054, и
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT067 и HVT069. Такая межгенная область может в ограниченной степени перекрываться с ОРС HVT068, которая, однако, расположена на комплементарной цепи. Таким образом, клонирование в такой межгенной области может изменять кодирующую последовательность HVT068.
В другом конкретном варианте осуществления изобретение относится к рекомбинантному птичьему HVT, содержащему несколько рекомбинантных нуклеиновых кислот (например, чужеродные гены), где:
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT053 и HVT054, и
. по меньшей мере одна рекомбинантная нуклеиновая кислота клонируется в межгенной области генома, расположенной между HVT069 и HVT070.
Рекомбинантный HVT по изобретению может быть получен из любого HVT, предпочтительно непатогенного HVT. Примером непатогенного штамма HVT (MDV3), подходящего для использования в изобретении, является штамм FC126.
Геномная последовательность штамма FC126 доступна в данной области (Afonso et al., см. выше; Kingham et al., см. выше). Расположение указанных межгенных областей может быть легко идентифицировано квалифицированным специалистом с использованием положений настоящей заявки, общеизвестных знаний и информации о последовательностях, доступных в литературе. Например, в Kingham et al., выше, представлена нуклеотидная последовательность эталонного штамма FC 126, а также расположение большинства ОРС в указанном геноме.
Что касается полного генома FC126 (GenBank: AF291866.1), межгенная область между HVT053 и HVT054 предпочтительно соответствует нуклеотидам 95323-95443 генома HVT, межгенная область между HVT064 и HVT065 соответствует нуклеотидам 111304-111499 HVT генома HVT, межгенная область между HVT065 и HVT066 соответствует нуклеотидам 112010-112207 генома HVT, межгенная область между HVT066 и HVT067 соответствует нуклеотидам 112766-113107 генома HVT, межгенная область между HVT067 и HVT069 соответствует нуклеотидам 113906-114078 генома HVT, а межгенная область между HVT069 и HVT070 соответствует нуклеотидам 116731-116831 генома HVT.
В настоящей заявке ссылка на полный геном штамма FC126 является ссылкой на указанный штамм, как описано в Genbank под номером доступа AF291866.1 на дату подачи настоящей заявки.
Рекомбинантные нуклеотидные последовательности, встроенные в геном, могут иметь любую ориентацию.
В предпочтительном варианте рекомбинантный HVT по изобретению содержит:
. по меньшей мере одну рекомбинантную нуклеотидную последовательность, клонированную в межгенной области генома между HVT053 и HVT054, и
. по меньшей мере одну рекомбинантную нуклеотидную последовательность, клонированную в межгенной области генома между HVT065 и HVT066.
В конкретном варианте осуществления нуклеиновая кислота, вставленная в межгенную область между HVT053 и HVT054, имеет ту же ориентацию, что и HVT054.
В другом конкретном варианте осуществления рекомбинантная нуклеотидная последовательность, вставленная в межгенную область между HVT065 и HVT066, имеет ту же ориентацию, что и HVT066.
Рекомбинантная нуклеотидная последовательность или встроенная последовательность нуклеиновой кислоты может содержать (или быть функционально связанной с) регуляторные(ми) последовательности(ями), такими как промотор и/или терминатор. Используемый промотор может быть синтетическим или природным, эндогенным или гетерологичным промотором. В принципе можно использовать любой промотор, если он может эффективно функционировать в клетках-мишенях или в хозяине. В этом отношении промотор может быть эукариотическим, прокариотическим, вирусным или синтетическим промотором, способным управлять транскрипцией гена в птичьих клетках в контексте поливалентного вектора. Также каждая рекомбинантная нуклеотидная последовательность может содержать промотор, который может быть одинаковым или отличаться один от другого.
Предпочтительно промотор, используемый для каждой рекомбинантной нуклеотидной последовательности, выбран из промотора куриного бета-актина (Bac), промотора Pec, промотора цитомегаловируса мыши (Mcmv) ie1, промотора цитомегаловируса человека (Hcmv), промотора обезьяньего вируса 40 (SV40) и промотора вируса саркомы Рауса (RSV) или любых их фрагментов, которые сохраняют промоторную активность.
Последовательность нуклеиновой кислоты куриного промотора Bac показана в SEQ ID NO: 1, последовательность промотора Pec показана в SEQ ID NO: 2, а последовательность промотора Mcmv ie1 показана в SEQ ID NO: 3. Следует отметить, что варианты таких последовательностей, кодирующих функциональные промоторы, известны и/или могут быть разработаны/протестированы квалифицированным специалистом для использования в настоящем изобретении.
В предпочтительном варианте рекомбинантная нуклеотидная последовательность, вставленная в межгенную область, расположенную между HVT053 и HVT054, содержит промотор Pec или промотор Bac. Результаты, полученные авторами изобретения, показывают, что такие промоторы эффективны при размещении в указанном сайте клонирования в контексте поливалентного вектора по настоящему изобретению.
В другом предпочтительном варианте осуществления чужеродный ген, вставленный в межгенную область, расположенную между HVT065 и HVT066, содержит промотор Pec или Mcmv ie1. Результаты, полученные авторами изобретения, показывают, что такие промоторы особенно эффективны при размещении в указанном сайте клонирования в контексте поливалентного вектора по настоящему изобретению.
Особенно предпочтительный рекомбинантный HVT по изобретению содержит (i) рекомбинантную нуклеиновую кислоту, вставленную в межгенную область между HVT065 и HVT066 под контролем промотора Pec или Mcmv ie1, и (ii) рекомбинантную нуклеиновую кислоту, вставленную в межгенную область между HVT053 и HVT054 под контроль промотора Pec.
Рекомбинантная нуклеотидная последовательность
Рекомбинантные нуклеотидные последовательности могут кодировать любой интересующий полипептид. Рекомбинантные нуклеотидные последовательности могут кодировать полипептиды, такие как, например, антигены, цитокины, гормоны или адъюванты.
В конкретном варианте осуществления одна или каждая из указанных по меньшей мере 2 рекомбинантных нуклеотидных последовательностей кодирует антиген патогена или его иммуногенный фрагмент.
Они могут происходить или быть получены из любого патогенного организма, способного вызывать инфекцию у птиц. Примеры патогенов, вызывающих инфекцию у птиц, включают вирусы, бактерии, грибки и простейшие.
Антигенами могут быть любые иммуногенные пептиды или белки патогена, такие как пептид или белок, выбранный из поверхностных белков, секретируемых белков или структурных белков указанного патогена или их фрагментов или полученные из них.
Предпочтительные рекомбинантные нуклеотидные последовательности для использования в настоящем изобретении кодируют антиген вируса птичьего гриппа, птичьего парамиксовируса типа 1 (также называемого вирусом болезни Ньюкасла (NDV)), птичьего метапневмовируса, вируса болезни Марека, вируса болезни Гамборо (также называемого вирусом инфекционной бурсальной болезни (IBDV)), вируса инфекционного ларинготрахеита (ILVT), вируса инфекционного бронхита (IBV), Escherichia coli, Salmonella, Pasteurella multocida, Riemerella anatipestifer, Ornithobacterium rhinotracheale, Mycoplasma gallascobacterium, Mycoplasma synoviae, микроорганизмов Mycoplasma, инфицирующих птичьи виды и/или кокцидий.
Предпочтительно, по меньшей мере одна из рекомбинантных нуклеотидных последовательностей, встроенных в вирусный геном, кодирует антиген, выбранный из белка F NDV, белка VP2 IBDV, белка gB ILTV, белка 40K Mycoplasma gallisepticum и поверхностного белка гемагглютинина (НА) вируса птичьего гриппа или их иммуногенные фрагменты. Предпочтительно обе рекомбинантные нуклеотидные последовательности кодируют такой антиген, который может быть одинаковым или нет.
Иммуногенные фрагменты антигена обозначают любой фрагмент, который может вызывать иммунный ответ против указанного антигена in vivo, предпочтительно любой фрагмент, содержащий эпитоп. Иммуногенные фрагменты обычно содержат от 5 до 50 последовательных аминокислотных остатков антигена, например от 5 до 40 или от 10 до 40.
Можно рассмотреть различные комбинации антигенных пептидов.
В одном варианте осуществления рекомбинантный HVT по изобретению содержит рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, и нуклеотидную последовательность, кодирующую белок VP2 IBDV или его иммуногенный фрагмент.
В другом варианте осуществления рекомбинантный HVT по изобретению содержит нуклеотидную последовательность, кодирующую белок VP2 IBDV или его иммуногенный фрагмент, и нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент.
В предпочтительном варианте осуществления рекомбинантный HVT по изобретению содержит нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, и нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент.
В другом варианте рекомбинантный HVT по изобретению экспрессирует два или более антигенов от одного и того же патогена. Указанные антигены могут быть одинаковыми или разными.
В дополнительном варианте по меньшей мере одна из рекомбинантных нуклеотидных последовательностей кодирует активную молекулу, такую как цитокин или иммуномодулятор, адъювант, гормон, противопаразитарный агент, антибактериальный агент и т.п., а другая кодирует, например, антиген.
Согласно дополнительному варианту осуществления в вирусный геном вставляют три или более рекомбинантные нуклеотидные последовательности.
Способ получения конструкций
Рекомбинантный HVT по изобретению может быть получен с использованием методик, известных в данной области техники, таких как рекомбинантная технология, гомологичная рекомбинация, сайт-специфическая вставка, мутагенез и тому подобное.
Клонирование генов и конструирование плазмид хорошо известны обычному специалисту в данной области техники и могут быть в основном выполнены стандартными методиками молекулярной биологии (Molecular Cloning: A Laboratory Manual. 4th Edition, Cold Spring Harbor Laboratory Press, Woodbury, N.Y. 2012).
Вирусы герпеса можно размножать в любой подходящей клетке-хозяине и среде. Хозяин и условия для размножения вирусов герпеса включают, например, клетки, полученные из курицы, такие как CEF (фибробласты куриного эмбриона), почечные клетки курицы и т.п. Такие клетки можно культивировать в культуральной среде, такой как Eagle's MEM, культуральная среда Leibowitz-L-15/McCoy 5A (смесь 1: 1) при температуре около 37°C в течение 3-4 дней.
Геномная ДНК может быть извлечена из инфицированных вирусом клеток любой обычной методикой. В частности, после денатурирования белков в буфере для лизиса и удаления ДНК можно экстрагировать фенолом и этанолом.
Обычно рекомбинантные вирусы могут быть получены путем гомологичной рекомбинации между вирусным геномом и конструкцией (например, плазмидой), содержащей рекомбинантную нуклеотидную последовательность или нуклеиновую кислоту, которая должна быть вставлена, фланкирована нуклеотидами от сайта вставки, позволяя рекомбинацию. Вкратце, последовательность, содержащую межгенную область-мишень, сначала клонируют в плазмиду или другой подходящий вектор. Примеры плазмид включают pBR322, pBR325, pBR327, pBR328, pUC18, pUC19, pUC7, pUC8 и pUC9, примеры фагов включают фаг лямбда и фаг M13, а пример космид включает pHC79. Клонированная область предпочтительно должна иметь достаточную длину, чтобы при вставке чужеродного гена последовательности, фланкирующие чужеродный ген, имели подходящую длину, чтобы обеспечить гомологичную рекомбинацию in vitro с вирусным геномом. Предпочтительно каждая фланкирующая последовательность должна иметь длину по меньшей мере приблизительно 50 нуклеотидов.
Чтобы вставить одну или несколько рекомбинантных нуклеотидных последовательностей в межгенную область, можно провести мутацию в конкретном сайте межгенной области, чтобы создать сайт расщепления для рестрикционного фермента. Способ осуществления мутации может быть обычной методикой, и можно использовать методику, обычно используемую специалистом в данной области, такой как мутагенез in vitro и ПЦР. Таким образом, в методе ПЦР выполняется мутация, такая как делеция, замена или добавление 1-2 нуклеотидов в праймере ПЦР, и затем праймер используется для создания мутации. В качестве альтернативы можно использовать существующие в природе сайты рестрикции. Затем чужеродный ген (и промотор) вставляют в сайт встраивания вирусного генома в плазмиду.
Полученная плазмида может быть введена в клетки, инфицированные HV, или клетки, трансфицированные геномом HV, с использованием любой подходящей методики (например, электропорации, фосфата кальция, методики на основе липофектина и т.п.). Когда количество вводимой плазмиды находится в диапазоне от 0,1 до 1000 мкг, эффективность генерации рекомбинантных вирусов путем рекомбинации между гомологичными областями генома HV и плазмидой в клетках становится высокой. Это приводит к событию рекомбинации между плазмидой и вирусным геномом, что приводит к встраиванию рекомбинантной нуклеотидной последовательности в вирус.
Полученный рекомбинантный вирус может быть выбран генотипически или фенотипически с использованием известных методик отбора, например, путем гибридизации, обнаружения активности фермента, кодируемого геном, совместно интегрированным с последовательностями рекомбинантной нуклеиновой кислоты, или обнаружения антигенного пептида, иммунологически экспрессируемого рекомбинантным вирусом герпеса.
Выбранный рекомбинантный вирус герпеса можно культивировать в крупном масштабе в культуре клеток. После создания вирус может размножаться в подходящих клетках.
Предпочтительный вариант осуществления
Нижеследующие рекомбинантные HVT являются предпочтительными конкретными вариантами осуществления изобретения. Как показано в примерах, они обеспечивают сильный иммунный ответ in vivo против антигенов, кодируемых каждой рекомбинантной нуклеотидной последовательностью.
Особенно предпочтительный рекомбинантный HVT по изобретению включает: (i) вставку в межгенную область между HVT065 и HVT066, рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Mcmv ie1 и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec (FW243).
Другой особенно предпочтительный рекомбинантный HVT по изобретению представляет собой рекомбинантный вирус птичьего герпеса, который включает: (i) вставленную в межгенную область между HVT065 и HVT066 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec (FW242).
Другой особенно предпочтительный рекомбинантный HVT по изобретению представляет собой рекомбинантный вирус птичьего герпеса, который включает: (i) вставленную в межгенную область между HVT065 и HVT066 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно находящийся под контролем. промотора Mcmv ie1 и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec (FW244).
Другой особенно предпочтительный рекомбинантный HVT по изобретению включает: (i) вставку в межгенную область между HVT065 и HVT066, рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec (FW241).
Другой рекомбинантный HVT по изобретению включает: (i) вставку в межгенную область между HVT065 и HVT066, рекомбинантную нуклеотидную последовательность, кодирующую белок VP2 IBDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Mcmv ie1 или промотора Pec и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec.
Другой рекомбинантный HVT по изобретению включает: (i) вставку в межгенную область между HVT065 и HVT066, рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем промотора Mcmv ie1 или промотора Pec и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок VP2 IBDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Pec (FW259).
Рекомбинантный HVT по настоящему изобретению можно размножать в культурах клеток. В предпочтительных вариантах осуществления CEF, яйца с зародышем, клетки куриных почек и т.п. используются в качестве клеток-хозяев для размножения рекомбинантного HVT. Мультивалентный рекомбинантный HVT по настоящему изобретению можно культивировать в такой культуральной среде, как Eagle's MEM, культуральная среда Leibowitz-L-15/McCoy 5A (смесь 1: 1) при температуре около 37°C в течение 3-4 дней. Полученные таким образом инфицированные клетки суспендируют в культуральной среде, содержащей 10% диметилсульфоксид (ДМСО), и хранят замороженными в атмосфере жидкого азота.
Рекомбинантный поливалентный HVT по изобретению обеспечивает преимущество, состоящее в высоком уровне стабильности при пассажах, что соответствует коэкспрессии рекомбинантных нуклеотидных последовательностей в клетках видов птиц, предпочтительно клетках CEF, даже после 10 или более пассажей, предпочтительно после 15 пассажей, еще предпочтительнее после 20 пассажей. В контексте изобретения «пассаж» или «пассаж клеток» означает культуру клеток в подходящих условиях для обеспечения их роста и поддержания их жизни до тех пор, пока они не станут конфлюэнтными от 90% до 100%. Стадия пассирования заключается в переносе небольшого количества клеток предыдущей конфлюэнтной культуры в новую культуральную среду. Аликвоту предыдущей конфлюэнтной культуры, содержащей несколько клеток, можно разбавить большим объемом свежей среды.
Вирусы можно собирать или очищать с использованием обычных методик. Они могут храниться в любой подходящей среде в замороженном и/или лиофилизированном виде.
Нуклеиновые кислоты и клетки
Другая цель изобретения относится к любой нуклеиновой кислоте, содержащейся в вирусе, как определено выше. Нуклеиновые кислоты могут быть одно- или двухцепочечными, ДНК или РНК или их вариантами.
Изобретение также относится к вектору (например, плазмиде, космиде, искусственной хромосоме и т.д.), содержащему нуклеиновую кислоту по изобретению.
Изобретение также относится к клетке, содержащей рекомбинантный HVT, нуклеиновую кислоту или вектор по изобретению. Клетки обычно представляют собой эукариотические клетки, такие как клетки птиц или прокариотические клетки (если вектор подходит для репликации или поддержания в таком типе клеток).
Вакцинные композиции
Изобретение также относится к композициям, таким как вакцины, которые содержат поливалентный рекомбинантный HVT по изобретению, нуклеиновую кислоту по изобретению или клетку по изобретению.
Вакцины по изобретению обычно содержат иммунологически эффективное количество рекомбинантного HVT, как описано выше, в фармацевтически приемлемом носителе.
Композиции и вакцины согласно настоящему изобретению обычно содержат подходящий растворитель, разбавитель или наполнитель, такой как, например, водный буфер или фосфатный буфер. Композиции также могут содержать добавки, такие как белки или пептиды, полученные от животных (например, гормоны, цитокины, костимуляторные факторы), нуклеиновые кислоты, полученные из вирусов и других источников (например, двухцепочечная РНК, CpG) и тому подобное, которые вводятся с вакциной в количестве, достаточном для усиления иммунного ответа. Кроме того, любое количество комбинаций вышеупомянутых веществ может обеспечивать эффект иммунопотенцирования и, следовательно, может образовывать иммунопотенциатор по настоящему изобретению.
В состав вакцин по настоящему изобретению могут быть дополнительно включены одна или несколько дополнительных добавок для поддержания изотоничности, физиологического pH и стабильности, например, буфера, такого как физиологический раствор (0,85%), фосфатно-солевой буфер (PBS), цитратные буферы, Трис(гидроксиметиламинометан (TRIS), забуференный трис-физиологический раствор и т.п., либо антибиотик, например неомицин или стрептомицин и т.п.
Путь введения может быть любым, включая пероральный, глазной (например, с помощью глазных капель), окуло-назальное введение с использованием аэрозоля, интраназальный, клоакальный с кормом, в воде или в виде спрея, in ovo, местный или путем инъекции (например, внутривенная, подкожная, внутримышечная, внутриглазничная, внутриглазная, внутрикожная и/или внутрибрюшинная) вакцинация. Специалист легко адаптирует состав вакцинной композиции для каждого типа введения.
Каждая доза вакцины может содержать подходящую дозу, достаточную для того, чтобы вызвать защитный иммунный ответ у видов птиц. Оптимизация такой дозы хорошо известна в данной области. Количество антигена на дозу может быть определено известными методами с использованием реакций антиген/антитело, например методом ELISA.
Вакцины по изобретению можно вводить в виде однократных или многократных доз, в зависимости от протокола вакцинации.
Вакцины по настоящему изобретению обладают дополнительным преимуществом, состоящим в том, что они обеспечивают видам птиц до 80% защиты от целевых птичьих патогенов после 3 недель вакцинации.
Настоящее изобретение также относится к применению описанной выше вакцины для иммунизации видов птиц, таких как домашняя птица, против патогена.
Настоящее изобретение также относится к способу иммунизации видов птиц путем введения иммунологически эффективного количества вакцины согласно изобретению. Вакцину можно преимущественно вводить внутрикожно, подкожно, внутримышечно, перорально, in ovo, через слизистую оболочку или через окуло-назальное введение.
Настоящее изобретение также относится к наборам для вакцинации для иммунизации видов птиц, которые содержат эффективное количество поливалентной вакцины, как описано выше, и средства для введения указанных компонентов указанным видам. Например, такой набор включает инъекционное устройство, заполненное поливалентной вакциной согласно изобретению, и инструкции для внутрикожной, подкожной, внутримышечной инъекции или инъекции in ovo. Альтернативно, набор включает спрей/аэрозоль или устройство для глазных капель, заполненное поливалентной вакциной согласно изобретению, и инструкции по окуло-назальному введению, пероральному введению или введению через слизистые оболочки.
Дополнительные аспекты и преимущества настоящей заявки теперь будут раскрыты в следующих примерах, которые иллюстрируют изобретение.
ПРИМЕРЫ
В экспериментальной части было создано несколько рекомбинантных HVT (моновалентных или поливалентных согласно изобретению), обозначенных следующим образом (HVT/первый сайт вставки - первый чужеродный ген/второй сайт вставки - второй чужеродный ген):
Моновалентный:
FW200: HVT/65-66 Pec F
FW219: HVT/65-66 Pec gB
FW168: HVT/45-46 Pec F
FW169: HVT/45-46 Bac VP2
FW181: HVT/45-46 Pec gB
Мультивалентный:
FW241: HVT/65-66 Pec F/45-46 Pec gB
FW242: HVT/65-66 Pec gB/45-46 Pec F
FW243: HVT/65-66 Mcmv ie1 F/45-46 Pec gB
FW244: HVT/65-66 Mcmv ie1 gB/45-46 Pec F
FW259: HVT/65-66 Pec gB/45-46 Bac VP2
Пример 1: Конструирование векторов гомологии
Конструирование плазмиды в основном выполнялось стандартными методиками молекулярной биологии (Molecular Cloning: A Laboratory Manual. 4th Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 2012). Фрагменты рестрикции ДНК подвергали электрофорезу на агарозных гелях и очищали с помощью набора Plasmid plus Midi (QIAGEN, № по каталогу 12945).
ДНК HVT получали из клеток CEF, инфицированных штаммом HVT FC126, в соответствии с методом Lee et al. (J. Gen. Virol., 51: 245-253, 1980). HVT064-071 амплифицировали с помощью ПЦР с использованием полученной ДНК HVT в качестве матрицы. Использовали два праймера: один был SEQ ID NO: 4 (5'-GGATGTCCAATTCGCACATG -3'), а другой был SEQ ID NO: 5 (5-GCTACAGTTACGGGATTCATAGG-3'). Каждый праймер был разработан на основе информации GenBank AF291866.1.
Конструкция pHVT64-65
Используя HVT064-071 в качестве матрицы, ПЦР выполняли с двумя парами праймеров. Фрагмент ДНК, имеющий сайт SfiI между двумя ОРС, HVT064 (LORF3) и HVT065 (UL55), получали с помощью ПЦР и клонировали в pUC18.
Первой парой была SEQ ID NO: 6 (5'-GGGGGAATTCACTACTTTTAATTCTCTTTA-3') и SEQ ID NO: 7 (5'-GGGGGCCAATAAGGCCGCTAGCGGCCGCCTAACACCCCCGAATATTAGTC-3').
Вторая пара была SEQ ID NO: 8 (5'-GGGGCGGCCGCTAGCGGCCTTATTGGCCTCAC GTGTAGCCCATTGTGTGCATATAAC -3') и SEQ ID NO: 9 (5'-GGGAAGCTTAG ATCTGAAATAACGCAGTTG-3').
Первый полученный фрагмент переваривали EcoRI и NheI. Второй полученный фрагмент переваривали NheI и HindIII. Эти расщепленные фрагменты были интегрированы в pUC18, расщепленную EcoRI и HindIII, в результате чего была получена плазмида pHVT 64-65.
Конструкция pHVT65-66
Используя HVT064-071 в качестве матрицы, ПЦР выполняли с двумя парами праймеров. Фрагмент ДНК, имеющий сайт SfiI между двумя ORF, HVT065 (UL55) и HVT066 (LORF4), получали с помощью ПЦР и клонировали в pUC18.
Первой парой была SEQ ID NO: 10 (5'-GGGGAATTCGCCAGATATCCAAAGTACAGC-3') и SEQ ID NO: 11 (5'-GGGGGCCAATAAGGCCGCTAGCGGCCGCCAATTATTTTATTTAATAACATAT-3').
Вторая пара - это SEQ ID NO: 12 (5'-GGGGCGGCCGCTAGCGGCCTTATTGGCCACCAGTGAACAATTT GTTTAATGTTA-3') и SEQ ID NO: 13 (5'-GGGAAGCTTGGGTCTGTCCTAGCGATATAA-3').
Первый полученный фрагмент переваривали EcoRI и NheI. Второй полученный фрагмент переваривали NheI и HindIII. Эти расщепленные фрагменты были интегрированы в pUC18, расщепленную EcoRI и HindIII, в результате чего была получена плазмида pHVT 65-66.
Конструкция pHVT66-67
ПЦР выполняли с двумя парами праймеров, используя HVT064-071 в качестве матрицы,. Фрагмент ДНК, имеющий сайт SfiI между двумя ОРС, HVT066 (LORF4) и HVT067 (LORF5), получали с помощью ПЦР и клонировали в pUC18.
Первой парой была SEQ ID NO: 14 (5'-GGGGGAATTCTCCAGATTGTTGGATATCTG-3') и SEQ ID NO: 15 (5'-GGGGGCCAATAAGGCCGCTAGCGGCCGCCTTATTGATTTATAAAAACATACATGCAGTG-3').
Вторая пара - это SEQ ID NO: 16 (5'-GGGGCGGCCGCTAGCGGCCTTATTGGCCAGTACATAATTTATTACGTATCATTTCCG-3') и SEQ ID NO: 17 (5'-GGGAAG CTTCCTGCAAGACCTCATACGGAA-3').
Первый полученный фрагмент переваривали EcoRI и NheI. Второй полученный фрагмент переваривали NheI и HindIII. Эти расщепленные фрагменты были интегрированы в pUC18, расщепленную EcoRI и HindIII, в результате чего была получена плазмида pHVT 66-67.
Конструкция pHVT67-69
ПЦР выполняли с двумя парами праймеров, используя HVT064-071 в качестве матрицы. Фрагмент ДНК, имеющий сайт SfiI между двумя ОРС, HVT067 (LORF5) и HVT069 (LORF6), получали с помощью ПЦР и клонировали в pUC18.
Первой парой была SEQ ID NO: 18 (5'-GGGGGAATTCATTTCTTCATTGCAACGACG-3') и SEQ ID NO: 19 (5'-GGGGGCCAATAAGGCCGCTAGCGGCCGCATGATC GTGCTCATTACTGCATCG-3').
Вторая пара была SEQ ID NO: 20 (5'-GGGGCGGCCGCTAGCGGCCTTATTG GCCGGG CGGGGCGATGACGTTCTATTTGC-3') и SEQ ID NO: 21 (5'-GGGAAGCTTAA TACGCAGATTCTTTTCGG-3').
Первый полученный фрагмент переваривали EcoRI и NheI. Второй полученный фрагмент переваривали NheI и HindIII. Эти расщепленные фрагменты были интегрированы в pUC18, расщепленную EcoRI и HindIII, в результате чего была получена плазмида pHVT 67-69.
Конструкция pHVT69-70
ПЦР выполняли с двумя парами праймеров, используя HVT064-071 в качестве матрицы. Фрагмент ДНК, имеющий сайт SfiI между двумя ОРС, HVT069 (LORF6) и HVT070, получали с помощью ПЦР и клонировали в pUC18.
Первой парой была SEQ ID NO: 22 (5'-GGGGGAATTCTAAAGAATCGTACATGAGCG-3') и SEQ ID NO: 23 (5'-GGGGGCCAATAAGGCCGCTAGCGGCCGCCTGAT GTATAAGATTGCCGAAAAG-3').
Вторая пара была SEQ ID NO: 24 (5'-GGGGCGGCCGCTAGCGGCCTTATTGGCCCGGGTTGCGTGAATACTGGTCACAG-3') и SEQ ID NO: 25 (5'-GGGAAGCTTACGATCTGGCAAAAGGGTCC-3').
Первый полученный фрагмент переваривали EcoRI и NheI. Второй полученный фрагмент переваривали NheI и HindIII. Эти расщепленные фрагменты были интегрированы в pUC18, расщепленную EcoRI и HindIII, в результате чего была получена плазмида pHVT 69-70.
Конструирование вектора гомологии pHVT65-66 Pec F
Расщепленный SfiI pHVT65-66 дефосфорилировали с использованием щелочной фосфатазы Shewanella sp., рекомбинантный штамм S1B1 (PAP) (Funakoshi # DE110). Фрагмент был лигирован с расщепленным BglI p45/46Pec F (WO2003 001066), в результате была получена плазмида pHVT65-66 Pec F. Синтезированная короткая сигнальная последовательность polyA (SPA: SEQ ID NO: 26 CTGCAGGCGGCCGCTCTAGAGTCGACAATAAAAGATCTTTATTTTCATTAGATCTGTGTGTTGGTTTTTTGTGTGGCCAATAAGGCC) была интегрирована в pHVT65-66 PecF, расщепленный с помощью SalI и SfiI, с получением гомологичной плазмиды pHVT65-66 Pec F SPА.
Конструирование вектора гомологии pHVT65-66 Pec gB
Вектор гомологии pHVT65-66 Pec F SPA расщепляли XbaI и NotI. Фрагмент длиной 5210 п.н. лигировали с расщепленным Xbal/Notl pHVT87-88 Pec ILgB del (WO2013 144355) (1506 п.н.), получая гомологичную плазмиду pHVT65-66 Pec gB SPA.
Химически синтезированный промотор Mcmv ie1
Промотор Mcmv ie1 (SEQ ID NO: 3) был синтезирован на основе информации о 4191-4731 п.н. в Gene Bank L06816.1, представленной Koszinowski U.H. Синтезированный промотор Mcmv ie1 был сконструирован таким образом, что спереди были добавлены сайты BglI-PstI и в конце были добавлены сайты XbaI-NotI.
Пример 2: Очистка рекомбинантного HVT в клетках CEF, трансфицированных каждым вектором для переноса
Вирусную ДНК HVT дикого типа, штамм FC126 (wt-HVT) получали, как описано Morgan et al. (Avian Diseases, 34:345-351, 1990). Вирусные ДНК FW168 (rHVT/45-46PecF), FW169 (rHVT/45-46BacVP2) и FW181 (rHVT/45-46 PecgBdel) (WO2005 093070) получали аналогичным способом. Первый характерный участок двойного rHVT состоял в том, что клетки CEF трансфицировали приготовленной ДНК wt-HVT и pHVT65-66 Pec F (например, FW200). Второй характерный участок состоял в том, что клетки CEF трансфицировали приготовленной ДНК FW168 и pHVT65-66 Pec gB (например, FW242). Третий характерный участок состоял в том, что клетки CEF трансфицировали приготовленной ДНК FW181 и pHVT65-66 Pec F (например, FW241). Четвертый характерный участок состоял в том, что CEF трансфицировали приготовленной ДНК FW169 и pHVT65-66 Pec gB (например, FW259). Полученные рекомбинантные вирусы очищали из бляшек путем окрашивания бляшек антителом против gB ILTV, антителом против NDV-F или антителом против IBDV-VP2.
Вкратце, 107 первичных клеток CEF суспендировали в 100 мкл MEF-1 (Lonza LNJVD-1004) и котрансфицировали путем электропорации 1 мкг вектора гомологии, например, pHVT65-66 PecF и pHVT65-66 Mcmv ie1 gB, и 2 мкг ДНК HVT, например FC126, FW181 и FW168. Электропорацию проводили с помощью Nucleofector II. Трансфицированные клетки разводили в 20 мл среды L-15 Лейбовица (GIBCO BRL, кат. № 41300-39), McCoy's 5A Medium (GIBCO BRL, кат. № 21500-061) (1: 1) и 4% телячьей сыворотки (указанный раствор среды LM (+)) и распределяли по 100 мкл на лунку 96-луночного планшета.
Инкубируют при 37°C в 5% CO2 до тех пор, пока бляшки не станут видимыми, клетки отделяют от планшетов трипсинизацией, разводят в свежеприготовленных вторичных клетках CEF, переносят поровну в два 96-луночных планшета и инкубируют в течение 3 дней для визуализации бляшек. Затем один из двух планшетов окрашивают антителом, с помощью которого мы выявляли встроенный ген, содержащийся в каждом векторе для переноса в качестве первичного антитела. NDV-F выявляли кроличьей сывороткой анти-Fc или моноклональными антителами Mab77-2. ILTV-gB выявляли кроличьей сывороткой анти-ILTV gBN4 или моноклональными антителами Mab №1_B4_7. IBDV-VP2 выявляли моноклональными антителами анти-VP2 R63 (ATCC №: HB-9490). После выявления лунок, содержащих окрашенные рекомбинантные бляшки, были извлечены клетки из соответствующей лунки другого планшета, разбавлены свежими вторичными клетками CEF и перенесены в равных количествах в два 96-луночных планшета для завершения первого цикла очистки. Процедуру очистки повторяли до тех пор, пока каждая полученная бляшка не окрашивалась положительно каждым поликлональным антителом или моноклональным антителом. После этого кандидат двойного rHVT был окрашен другим антителом. Наконец, экспрессия белков всеми бляшками кандидата rHVT была подтверждена двойным окрашиванием IFA. CEF, инфицированные каждым rHVT, фиксировали холодным ацетон-метанолом (2: 1), промывали PBS, проводили реакцию со смесью антител (разведенная 1: 1000 кроличья сыворотка против gBN4 (или Mab#1_B4_1) и кроличья сыворотка анти-Fc (Mab77-2) или мышиные моноклональные антитела R63 против VP2) при 37°C в течение 60 минут. После 3-кратной промывки PBS проводили реакцию клеток со смесью флуоресцентных антител (разведенные 1: 1000 Alexa Fluor488 против кролика и Alexa Fluor546 против мыши, предоставленные Invitrogen) при 37°C в течение 60 минут. После 3-кратной промывки PBS их наблюдали с помощью флуоресцентного микроскопа при увеличении в 40 или 100 раз.
Экспрессию белка gB выявляли с помощью кроличьей сыворотки анти-gBN4 и Alexa Flour 546. Экспрессию белка F выявляли с помощью моноклональных антител анти-F Mab 77-2 и Alexa Flour 488. Экспрессию белка VP2 выявляли с помощью моноклональных антител анти-VP2 Mab (R63) и Alexa Flour 546. Когда все бляшки экспрессировали как F, так и gB (как gB, так и VP2), мы пришли к выводу, что очистка завершена. На Фигурах 2A и B показаны некоторые примеры двойного иммуноферментного анализа.
Очищенный рекомбинантный HVT был обозначен как rHVT/ND/ILT или rHVT/ILT/IBDV.
В таблице 1 ниже показана экспрессия gB и белка F, полученных от различных rHVT/ND/ILT.
Штамм FW168 (HVT/45-46 PecF) и FW200 (HVT/65-66 PecF) соответствуют моновалентному рекомбинантному вирусу герпеса, используемому в качестве контроля для экспрессии белка F, и FW181 (HVT/45-46 Pec gBdel) и FW219 (HVT/65-66 Pec gBdel) соответствуют моновалентному рекомбинантному вирусу герпеса, используемому в качестве контроля для экспрессии белка gB, а FW169 (HVT/45-46 Bac VP2) соответствует моновалентному рекомбинантному вирусу герпеса, используемому в качестве контроля для экспрессии VP2.
Результаты показывают, что двухвалентные конструкции по изобретению экспрессируют оба антигена.
Таблица 1. Экспрессия встроенного гена gB NDV-F и ILTV с помощью rHVT/ND/ILT (выявление флуоресценции)
Пример 3: Ко-экспрессия двух белков в клетках CEF, инфицированных двойными рекомбинантными HVT.
2 мл, содержащих 2X105 клеток CEF, инфицировали рекомбинантными HVT и инкубировали при 37°C в 5% CO2 в течение 3 дней.
Затем культуру центрифугировали при 300 g в течение 3 минут и осажденные клетки ресуспендировали в 100 мкл. К суспензии клеток добавляли буфер Лэммли (100 мкл). Полученную смесь затем кипятили в течение 5 мин и 5 мкл подвергали электрофорезу в 10% SDS-полиакриламидном геле. Разделенные электрофорезом белки переносили из SDS-геля на PVDF-мембрану (Immobilon-P, Millipore), которую блокировали в 1% в/о обезжиренном сухом молоке в PBS при комнатной температуре в течение одного часа.
Для обнаружения F (Фигура 3A) обработанную мембрану затем подвергали реакции с кроличьей антисывороткой против Fc в 500-кратном разведении при комнатной температуре в течение одного часа, трижды промывали PBS и инкубировали в течение одного часа с биотинилированной анти-кроличьей козьей антисывороткой.
Для обнаружения gB (Фигура 3B) обработанную мембрану затем подвергали реакции с кроличьей сывороткой против gBN4 в 500-кратном разведении при комнатной температуре в течение одного часа, трижды промывали PBS и инкубировали в течение одного часа с биотинилированной анти-кроличьей козьей антисывороткой.
После трехкратного промывания при помощи PBS мембрану инкубировали в течение одного часа с комплексом авидин-щелочная фосфатаза, трижды промывали PBS и один раз TBS (трис-буферный солевой раствор) и проводили реакцию с BCIP-NBT (субстрат щелочной фосфатазы). Как показано на Фигуре 3A, полоса белка 60 килодальтон (кДа) наблюдалась только в дорожке с rHVT/ND/ILT инфицированными клетками, что было ожидаемым для размера белка F (). Полосы на дорожке rHVT/45-46 Pec gB (FW181) отсутствовали.
На Фигуре 3B показано, что белок gB наблюдался при 54 килодальтон (кД) на дорожках каждого rHVT/ND/ILT  . Напротив, полосы на дорожке rHVT/45-46 PecF (FW168) отсутствовали. 54 кДа представляет собой усеченный белок gB (469aa; WO2005/093070), который вставлен в rHVT/ND/ILT или rHVT/ILT/IBD.
. Напротив, полосы на дорожке rHVT/45-46 PecF (FW168) отсутствовали. 54 кДа представляет собой усеченный белок gB (469aa; WO2005/093070), который вставлен в rHVT/ND/ILT или rHVT/ILT/IBD.
Двойные рекомбинантные HVT согласно изобретению экспрессировали как NDV-F, так и ILTV-gB.
Пример 4: Проверка геномной структуры
ПЦР анализ
Очищенный rHVT/ND/ILT размножали на клетках CEF одного 6-луночного планшета для получения конфлюэнтных бляшек. Клетки извлекали из чашек соскабливанием, переносили в пробирки Falcon и подвергали центрифугированию при 300 xg в течение 5 мин. Одну десятую собранных клеток (из одной лунки) суспендировали в 0,1 мл буфера для лизиса (20 мМ трис-HCl, 100 мМ NaCl, 5 мМ EDTA, 0,1% SDS) и лизировали путем встряхивания в течение 1 мин. Лизаты инкубировали при 60°C в течение 10 мин с 2 мкл протеазы K (10 мг/мл). Полученную смесь дважды обрабатывали фенол-хлороформом. Водную фазу (вирусную ДНК) использовали в качестве матрицы.
Для каждого сайта вставки были разработаны наборы праймеров.
Для анализа около HVT65/66
65R (SEQ ID NO: 27): 5'-CCACTGCCACTGTGATGATAAG-3'
66F (SEQ ID NO: 28): 5'- GCCTACTATGCACATTGTTACTCCT-3'
Для анализа около с UL45/46
45-46F-K (SEQ ID NO: 29): 5'-GGGGAAGTCTTCCGGTTAAGGGAC-3'
45-46R (SEQ ID NO: 30): 5'-GGTGCAATTCGTAAGACCGATGGG-3'
ПЦР проводили с использованием ДНК-полимеразы Tks Gflex (TAKARA BIO Inc. № R060) в соответствии с инструкцией производителя.
Результат ПЦР на Фигуре 4C показывает, что фрагмент длиной 2550 п.н. был амплифицирован с набором праймеров 66F/65R в ДНК из очищенного FW243 (p+1). Фрагмент, амплифицированный в плазмиде переноса pHVT65-66 ie1 NDV-F, имел такой же размер. Кроме того, фрагмент длиной 2281 п.н. амплифицировали с другим набором праймеров 45-46-K/45-46R в ДНК из очищенного FW243 (p+1) (Фигура 4D). Такая же полоса была обнаружена в p45-46 Pec gBdel. Нижний фрагмент (546 п.н.) амплифицировали в pNZ45-46 Sfi-T.
Фигура 4C-D (p + 1) показывает, что полученные двойные рекомбинанты HVT/ND/ILT имеют ожидаемую геномную структуру. Было подтверждено, что рекомбинантные вирусы являются чистыми, поскольку полоса, соответствующая родительским штаммам, не амплифицировалась.
Пример 5: Стабильность рекомбинантных HVT при пассаже
ПЦР анализ
Каждый rHVT/ND/ILT 20 раз пассировали в клетках CEF и подвергали ПЦР-анализу, как описано в Эксперименте 4. Результаты были такими же, как и в Эксперименте 4, что указывает на то, что рекомбинантный вирус был стабильным даже после 20 пассажей.
Результат ПЦР на Фигуре 4А показал, что фрагмент длиной 2118 п.н. амплифицировали с набором праймеров 66F/65R в ДНК из FW242 (p+5, +10, +15, +20) и плазмидой переноса pHVT65-66 Pec gBdel. Напротив, нижний фрагмент (252 п.н.) амплифицировали в неочищенном клоне.
На Фигуре 4B показано, что фрагмент длиной 3083 п.н. был амплифицирован с другим набором праймеров 45-46-K/45-46R в FW242 (p+5, +10, +15, +20) и 2-ой p45-46 Pec F. Нижняя полоса (547 пар оснований) была амплифицирована в pNZ45-46 Sfi-T (US 7569365).
На Фигуре 4С показано, что фрагмент длиной 2550 п.н. был амплифицирован при пассировании FW243 (p+5, +10, +15, +20). На Фигуре 4D показано, что фрагмент длиной 2281 п.н. был амплифицирован при пассировании FW243 (p+5, +10, +15, +20).
ПЦР-анализ с 65R/66F и 45-46-K/45-46R показал, что ген F или ген gB стабильно сохраняется в сайте вставки HVT65/66 или UL45/46 соответственно в FW242 и FW243.
Вестерн-блоттинг
Двойные рекомбинантные HVT пассировали серийно (до 20 раз) на фибробластах куриных эмбрионов (CEF). Затем клеточные лизаты подвергали вестерн-блоттингу. На первой панели (Фигура. 5A) блот подвергали реакции с кроличьей сывороткой против Fc (1: 500). На второй панели (Фигура 5B) блот подвергался реакции с кроличьей сывороткой против gBN4 (1: 500).
Контроль: незараженный CEF
M: Стандарты протеина Precision Plus Bio Rad # 161-0374
После 20 пассажей белок NDV-F  и ILTV-gB были стабильно экспрессированы в CEF, инфицированном двойным рекомбинантным HVT.
и ILTV-gB были стабильно экспрессированы в CEF, инфицированном двойным рекомбинантным HVT.
Пример 6: Индукция антител против NDV у цыплят, инокулированных двойными рекомбинантными HVT
Двойные рекомбинантные HVT инокулировали подкожно в спину десяти однодневных цыплят SPF (LineM, NISSEIKEN) с использованием шприца калибра 20. Дозы инокуляции показаны в таблице 2. Сыворотку собирали у вакцинированных птиц, и титры антител против NDV измеряли с помощью коммерческого набора для ELISA (IDVET, ID screen Newcastle Disease Indirect kit для диагностики болезни Ньюкасла). Цыплятам группы отрицательного контроля (не иммунизированные) вакцина не вводилась.
Таблица 2 показывает титр анти-NDV по значению S/P, и результаты ясно показывают, что двойной рекомбинантный HVT согласно изобретению индуцировал антитела против NDV уже через три недели после инокуляции и увеличивал количество антител против NDV после этого.
Таблица 2 Индукция антител против NDV у кур SPF, инокулированных двойными рекомбинантами.
Пример 7: Эффективность rHVT/ND/ILT у кур SPF против ILTV
Эффективность rHVT/ND/ ILT (FW241, FW242, FW243, FW244) в качестве вакцины против ILT оценивали путем заражения штаммом ILTV USDA.
Двойные рекомбинантные HVT подкожно инокулировали в спину однодневных цыплят SPF. Через четыре недели после инокуляции вакцинированных цыплят интратрахеально (ИТ) заражали 103 EID50/птицу штамма ILTV USDA. За зараженными цыплятами наблюдали ежедневно для проверки смертности и выявления любых симптомов инфекционного ларинготрахеита, таких как подавленность, затрудненное дыхание, хрипы, кровянистая мокрота, чихание, удушье, взмахи головой, растянутая шея или взъерошенные перья. Результаты на 10-й день после заражения суммированы в таблице 3.
Таблица 3. Эксперименты по заражению цыплят SPF, вакцинированных rHVT/ND/ILT, вирулентным штаммом ILTV US
Как показано в таблице 3, все двойные рекомбинантные HVT согласно изобретению индуцировали у цыплят защитный иммунитет против заражения вирулентным ILTV.
Пример 8: Эффективность rHVT/ND/ILT у кур SPF против NDV
Оценивали эффективность rHVT/ND/ILT (FW241, FW242, FW243, FW244) в качестве вакцины против ND.
Подкожно вводили в спину 2000 БОЕ/200 мкл rHVT/ND ILT на птицу пятнадцати однодневным цыплятам SPF (LineM, Japan Biological Laboratories) с использованием шприца калибра 20. Через три недели после вакцинации у вакцинированных птиц собирали сыворотку и измеряли титр антител против NDV с помощью коммерческого набора для ELISA (IDVET, ID screen Newcastle Disease Indirect kit для диагностики болезни Ньюкасла).
Цыплятам группы отрицательного контроля не вводили никакой вакцины (НИ: не иммунизированы).
В возрасте 49 дней (48 дней после вакцинации) цыплят всех пяти групп заражали 103 EID50 штамма NDV-TexasGB, который часто используется в качестве контрольного штамма в исследованиях эффективности, путем внутримышечной инъекции в бедренную область. За зараженными цыплятами наблюдали ежедневно для проверки смертности и выявления любых симптомов болезни Ньюкасла, таких как депрессия, затрудненное дыхание, неврологические симптомы и умирание. Результаты на 10-й день после заражения представлены в таблице 4.
Таблица 4. Эксперименты по заражению цыплят SPF, вакцинированных rHVT/ND/ILT, вирулентным NDV
Как показано в таблице 4, все двойные рекомбинантные HVT согласно изобретению индуцировали у цыплят защитный иммунитет против заражения NDV.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CEVA SANTE ANIMALE
<120> РЕКОМБИНАНТНЫЕ ВИРУСЫ ПТИЧЬЕГО ГЕРПЕСА, СОДЕРЖАЩИЕ НЕСКОЛЬКО
ЧУЖЕРОДНЫХ ГЕНОВ
<130> B2916
<160> 30
<170> PatentIn version 3.5
<210> 1 <211> 1506 <212> ДНК
<213> Искусственная последовательность
<220>
<223> Bac промотор <400> 1
tgcagctcag tgcatgcacg ctcattgccc atcgctatcc ctgcctctcc tgctggcgct 60
ccccgggagg tgacttcaag gggaccgcag gaccacctcg ggggtggggg gagggctgca 120
cacgcggacc ccgctccccc tccccaacaa agcactgtgg aatcaaaaag gggggagggg 180
ggatggaggg gcgcgtcaca cccccgcccc acaccctcac ctcgaggtga gccccacgtt 240
ctgcttcact ctccccatct cccccccctc cccaccccca attttgtatt tatttatttt 300
ttaattattt tgtgcagcga tgggggcggg gggggggggg gcgcgcgcca ggcggggcgg 360
ggcggggcca ggggcggggc ggggcgaggc ggagaggtgc ggcggcagcc aatcagagcg 420
gcgcgctccg aaagtttcct tttatggcga ggcggcggcg gcggcggccc tataaaaagc 480
gaagcgcgcg gcgggcggga gtcgctgcgc gctgccttcg ccccgtgccc cgctccgccg 540
ccgcctcgcg ccgcccgccc cggctctgac tgaccgcgtt actcccacag gtgagcgggc 600
gggacggccc ttctcctccg ggctgtaatt agcgcttggt ttaatgacgg ctcgtttctt 660
ttctgtggct gcgtgaaagc cttaaagggc tccgggaggg ccctttgtgc gggggggagc 720
ggctcggggg gtgcgtgcgt gtgtgtgtgc gtggggagcg ccgcgtgcgg ctccgcgctg 780
cccggcggct gtgagcgctg cgggcgcggc gcggggcttt gtgcgctccg cagtgtgcgc 840
gaggggagcg cggccggggg cggtgccccg cggtgcgggg ggggctgcga ggggaacaaa 900
ggctgcgtgc ggggtgtgtg cgtggggggg tgagcagggg gtgtgggcgc ggcggtcggg 960
ctgtaacccc cccctgcacc cccctccccg aagttgctga gcacggcccg gcttcgggtg
1020
cggggctccg tgcggggcgt ggcgcggggc tcgccgtgcc gggcgggggg tggcggcagg
1080
tgggggtgcc gggcggggcg gggccgcctc gggccgggga gggctcgggg gaggggcgcg
1140
gcggcccccg gagcgccggc ggctgtcgag gcgcggcgag ccgcagccat tgccttttat
1200
ggtaatcgtg cgagagggcg cagggacttc ctttgtccca aatctgtgcg gagccgaaat
1260
ctgggaggcg ccgccgcacc ccctctagcg ggcgcggggc gaagcggtgc ggcgccggca
1320
ggaaggaaat gggcggggag ggccttcgtg cgtcgccgcg ccgccgtccc cttctccatc
1380
tccagcctcg gggctgtccg cagggggacg gctgccttcg ggggggacgg ggcagggcgg
1440
ggttcggctt ctggcgtgtg accggcgggg tttatatctt cccttctctg ttcctccgca
1500
gccccc 1506
<210> 2
<211> 557
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Peo промотор
<400> 2
tgcagagtta ttaatagtaa tcaattacgg ggtcattagt tcatagccca tatatggagy 60
tccgcgttac ataacttacg gtaaatggcc cgccggctga ccgcccaacg acccccgccc 120
attgacgtca ataatgacgt atgytcccat agtaacgcca atagggactt tccattgacg 180
tcaatgggtg gagtayttac ggtaaactgc ccattggcag tacatcaagt gtatcatatg 240
ccaagtacgc cccctattga cgtcaatgac ggtaaatgga tgcagtattt tgtgcagcga 300
tgggggcggg gggggggggc gcgcgccagg cggggcgggg cggggcgagg ggcggggcgg 360
ggcgaggcgg agaggtgcgg cggcagccaa tcagagcggc gcgctccgaa agtttccttt 420
tatggcgagg cggcggcggc ggcggcccta taaaaagcga agcgcgcggc gggcgggagt 480
cgctgcgcgc tgccttcgcc ccgtgccccg ctccgccgcc gcctcgcgcc gcccgccccg 540
gctctgactg accgcgt 557
<210> 3 <211> 572 <212> ДНК
<213> Искусственная последовательность
<220>
<223> Промотор mcmv ie1 <400> 3
ggccaataag gctgcagtac tgagtcatta gggactttcc aatgggtttt gcccagtaca 60
taaggtcaat aggggtgaat caacaggaaa gtcccattgg agccaagtac actgagtcaa 120
tagggacttt ccattgggtt ttgcccagta caaaaggtca atagggggtg agtcaatggg 180
tttttcccat tattggcacg tacataaggt caataggggt gagtcattgg gtttttccag 240
ccaatttaat taaaacgcca tgtactttcc caccattgac gtcaatgggc tattgaaact 300
aatgcaacgt gacctttaaa cggtactttc ccatagctga ttaatgggaa agtaccgttc 360
tcgagccaat acacgtcaat gggaagtgaa agggcagcca aaacgtaaca ccgccccggt 420
tttcccctgg aaattccata ttggcacgca ttctattggc tgagctgcgt tctacgtggg 480
tataagaggc gcgaccagcg tcggtaccgt cgcagtcttc ggtctgacca ccgtagaacg 540
cagagctcct cgctgcaggc ggccgctcta ga 572
<210> 4
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 4
ggatgtccaa ttcgcacatg 20
<210> 5
<211> 19
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 5
gctacagtta cgggattca 19
<210> 6
<211> 30
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 6
gggggaattc actactttta attctcttta 30
<210> 7
<211> 50
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 7
gggggccaat aaggccgcta gcggccgcct aacacccccg aatattagtc 50
<210> 8
<211> 57
<212> ДНК
<213> Искусственная последовательность <220>
8
<223> олигонуклеотид
<400>
ggggcggccg ctagcggcct tattggcctc acgtgtagcc cattgtgtgc atataac 57
<210> 9
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 9
gggaagctta gatctgaaat aacgcagttg tagg 34
<210> 10
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 10
ggggaattcg ccagatatcc aaagtacagc 30
<210> 11
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 11
gggggccaat aaggccgcta gcggccgcca attattttat ttaataacat at 52
<210> 12
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 12
ggggcggccg ctagcggcct tattggccac cagtgaacaa tttgtttaat gtta 54
<210> 13
<211> 30
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид <400> 13
gggaagcttg ggtctgtcct agcgatataa 30
<210> 14
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 14
gggggaattc tccagattgt tggatatctg 30
<210> 15
<211> 59
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 15
gggggccaat aaggccgcta gcggccgcct tattgattta taaaaacata catgcagtg 59
<210> 16
<211> 57
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 16
ggggcggccg ctagcggcct tattggccag tacataattt attacgtatc atttccg 57
<210> 17
<211> 30
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 17
gggaagcttc ctgcaagacc tcatacggaa 30
<210> 18
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 18
gggggaattc atttcttcat tgcaacgacg 30
<210> 19
<211> 52
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 19
gggggccaat aaggccgcta gcggccgcat gatcgtgctc attactgcat cg 52
<210> 20
<211> 54
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 20
ggggcggccg ctagcggcct tattggccgg gcggggcgat gacgttctat ttgc 54
<210> 21
<211> 29
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 21
gggaagctta atacgcagat tcttttcgg 29
<210> 22
<211> 30
<212> ДНК
<213> Искусственная последовательность <220>
22
<223> олигонуклеотид
<400>
gggggaattc taaagaatcg tacatgagcg 30
<210> 23
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 23
gggggccaat aaggccgcta gcggccgcct gatgtataag attgccgaaa ag 52
<210> 24
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 24
ggggcggccg ctagcggcct tattggcccg ggttgcgtga atactggtca cag 53
<210> 25
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид
<400> 25
gggaagctta cgatctggca aaagggtcc 29
<210> 26 <211> 87 <212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид <400> 26
ctgcaggcgg ccgctctaga gtcgacaata aaagatcttt attttcatta gatctgtgtg 60
ttggtttttt gtgtggccaa taaggcc 87
<210> 27 <211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид <400> 27
ccactgccac tgtgatgata ag 22
<210> 28
<211> 25
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 28
gcctactatg cacattgtta ctcct 25
<210> 29
<211> 24
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 29
ggggaagtct tccggttaag ggac 24
<210> 30
<211> 24
<212> ДНК
<213> Искусственная последовательность <220>
<223> олигонуклеотид
<400> 30
ggtgcaattc gtaagaccga tggg 24
<---
Изобретение относится к биотехнологии. Предложен рекомбинантный вирус герпеса индейки (HVT), подходящий для вакцинации птицы против патогена. Рекомбинантный вирус содержит по меньшей мере первую и вторую рекомбинантные нуклеотидные последовательности, каждая рекомбинантная нуклеотидная последовательность кодирует полипептид, причем первая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT053 и HVT054, а вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT064 и HVT070. Предложены: нуклеиновая кислота, подходящая для вакцинации птицы против патогена, содержащая геном рекомбинантного HVT; клетка, содержащая рекомбинантный HVT, подходящая для вакцинации птицы против патогена; композиция, подходящая для вакцинации птицы против патогена, содержащая рекомбинантный вирус птичьего герпеса или нуклеиновую кислоту или клетку и подходящий наполнитель или разбавитель; мультивалентная вакцина; применение рекомбинантного HVT для изготовления мультивалентной вакцины для иммунизации птицы, такой как домашняя птица, против патогенна; применение мультивалентной вакцины, которая содержит эффективное иммунизирующее количество рекомбинантного вируса птичьего герпеса, нуклеиновую кислоту по п. 16 и/или клетку, подходящая для вакцинации птицы против патогена; набор для вакцинации для иммунизации птиц, состоящий из следующих компонентов: эффективное количество мультивалентной вакцины и средство для введения указанной вакцины указанной птице. Изобретение обеспечивает стабильность экспрессии белка в клетках CEF, инфицированных двойным рекомбинантным HVT. 9 н. и 14 з.п. ф-лы, 6 ил., 4 табл., 8 пр.
1. Рекомбинантный вирус герпеса индейки (HVT), подходящий для вакцинации птицы против патогена, который содержит по меньшей мере первую и вторую рекомбинантные нуклеотидные последовательности, каждая рекомбинантная нуклеотидная последовательность кодирует полипептид, причем первая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT053 и HVT054, а вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT064 и HVT070.
2. Рекомбинантный HVT по п. 1, отличающийся тем, что вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT064 и HVT065.
3. Рекомбинантный HVT по п. 1, отличающийся тем, что вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT065 и HVT066.
4. Рекомбинантный HVT по п. 1, отличающийся тем, что вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT066 и HVT067.
5. Рекомбинантный HVT по п. 1, отличающийся тем, что вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT067 и HVT069.
6. Рекомбинантный HVT по п. 1, отличающийся тем, что вторая рекомбинантная нуклеотидная последовательность вставлена в межгенную область вирусного генома, расположенную между HVT069 и HVT070.
7. Рекомбинантный HVT по любому из пп. 1-6, отличающийся тем, что каждая рекомбинантная нуклеотидная последовательность кодирует антиген птичьего патогена или его иммуногенный фрагмент.
8. Рекомбинантный HVT по п. 7, отличающийся тем, что каждый антиген выбран из поверхностных белков, секретируемых белков и структурных белков указанного птичьего патогена или его иммуногенного фрагмента.
9. Рекомбинантный HVT по п. 7 или 8, отличающийся тем, что каждая рекомбинантная нуклеотидная последовательность кодирует антиген, выбранный из антигена птичьего парамиксовируса типа 1, предпочтительно белка F вируса болезни Ньюкасла (NDV) или его иммуногенного фрагмента, антигена вируса болезни Гамборо, предпочтительно белок VP2 вируса инфекционной бурсальной болезни (IBDV) или его иммуногенный фрагмент, антиген вируса инфекционного ларинготрахеита (ILTV), предпочтительно белок gB или его иммуногенный фрагмент, антиген Mycoplasma gallisepticum, предпочтительно 40К белок или его иммуногенный фрагмент и антиген вируса птичьего гриппа, предпочтительно поверхностный белок гемагглютинин (НА) или его иммуногенный фрагмент.
10. Рекомбинантный HVT по любому из предшествующих пунктов, отличающийся тем, что первая и вторая рекомбинантные нуклеотидные последовательности кодируют разные антигены одного и того же или разных патогенов или их иммуногенные фрагменты.
11. Рекомбинантный HVT по любому из пп. 1-10, отличающийся тем, что каждая рекомбинантная нуклеотидная последовательность находится под контролем промотора.
12. Рекомбинантный HVT по п. 11, отличающийся тем, что каждый промотор, контролирующий экспрессию рекомбинантной нуклеотидной последовательности, выбран из промотора куриного бета-актина (Вас), промотора Рес, немедленного-раннего (ie) промотора 1 цитомегаловируса мыши (Mcmv), промотора цитомегаловируса человека (Hcmv), промотора обезьяньего вируса (SV) 40 и промотора вируса саркомы Рауса (RSV), или любых их фрагментов, которые сохраняют промоторную активность.
13. Рекомбинантный HVT по п. 1, который содержит: (i) вставленную в межгенную область между HVT065 и HVT066 рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, предпочтительно под контролем ie промотора 1 Mcmv и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем промотора Рес.
14. Рекомбинантный HVT по п. 1, который содержит: (i) вставленную в межгенную область между HVT065 и HVT066 рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Рес и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем промотора Рес.
15. Рекомбинантный HVT по п. 1, который содержит: (i) вставленную в межгенную область между HVT065 и HVT066 рекомбинантную нуклеотидную последовательность, кодирующую белок gB ILTV или его иммуногенный фрагмент, предпочтительно под контролем ie промотра 1 Mcmv и (ii) вставленную в межгенную область между HVT053 и HVT054 рекомбинантную нуклеотидную последовательность, кодирующую белок F NDV или его иммуногенный фрагмент, предпочтительно под контролем промотора Рес.
16. Нуклеиновая кислота, подходящая для вакцинации птицы против патогена, содержащая геном рекомбинантного HVT по любому из пп. 1-15.
17. Клетка, содержащая рекомбинантный HVT по любому из пп. 1-15 или нуклеиновую кислоту по п. 16, подходящая для вакцинации птицы против патогена.
18. Композиция, подходящая для вакцинации птицы против патогена, содержащая рекомбинантный вирус птичьего герпеса по любому из пп. 1-15 и подходящий наполнитель или разбавитель.
19. Композиция, подходящая для вакцинации птицы против патогена, содержащая нуклеиновую кислоту по п. 16 или клетку по п. 17 и подходящий наполнитель или разбавитель.
20. Мультивалентная вакцина, которая содержит эффективное иммунизирующее количество рекомбинантного вируса птичьего герпеса по любому из пп. 1-15, нуклеиновую кислоту по п. 16 и/или клетку по п. 17, подходящая для вакцинации птицы против патогена.
21. Применение рекомбинантного HVT по любому из пп. 1-15 для изготовления мультивалентной вакцины для иммунизации птицы, такой как домашняя птица, против патогена.
22. Применение мультивалентной вакцины по п. 20 для одновременной вакцинации птицы против по меньшей мере двух патогенов.
23. Набор для вакцинации для иммунизации птиц, состоящий из следующих компонентов:
c. эффективное количество мультивалентной вакцины по п. 20, и
d. средство для введения указанной вакцины указанной птице.
| WO 2013144355 A1, 03.10.2013 | |||
| CN 106701693 A, 24.05.2017 | |||
| WO 2013082327 A1, 06.06 | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| ИСАЕНКО Е.Ю | |||
| и др | |||
| Вакцины нового поколения, Вестник Смоленской государственной медицинской академии 2015, Т | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |
