Настоящее изобретение относится к генетически модифицированной бактерии и ее промышленному применению, в частности, для 1,2-дегидрирования стероидов.
1,2-дегидрирование стероидов (также называемое Δ1-дегидрированием) может осуществляться путем биотрансформации с помощью бактерии, имеющей соответствующую ферментную систему. Например, превращение может катализироваться микроорганизмом семейства Nocardioidaceae, который готовят и используют в виде бактериальной суспензии или в качестве супернатанта после лизиса бактериальной суспензии. К сожалению, микроорганизм также экспрессирует фермент, который катализирует вторую (нежелательную) реакцию молекулы стероида, которая представляет собой восстановление кетогруппы в положении 20 и приводит к образованию побочных продуктов, называемых здесь «примесью 20-ОН». Эти две реакции схематически описаны ниже на примере гидрокортизона.
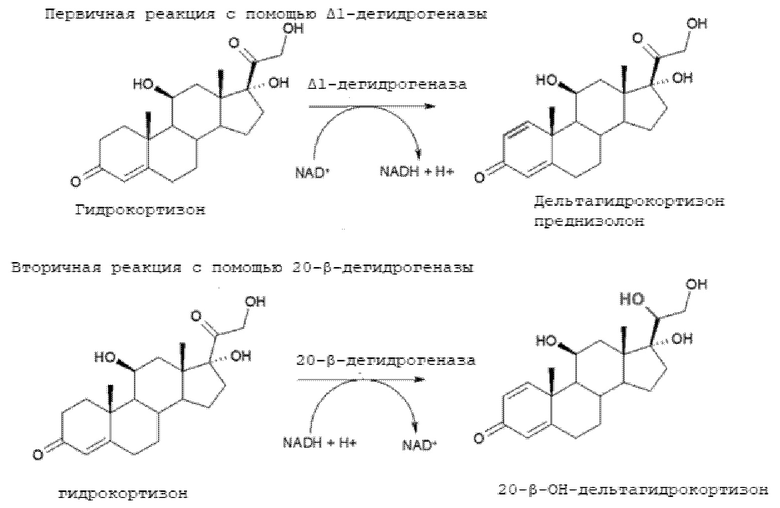
Было установлено, что за эту побочную реакцию отвечает фермент 20β-HSDH (20-бета-гидроксистероид дегидрогеназа), который приводит к кето-восстановлению стероидов в положении 20.
Теперь авторы изобретения идентифицировали ген, ответственный за активность побочных реакций, и генетически сконструировали микроорганизм, лишенный этой активности. При использовании в превращении стероидов он больше не производит нежелательный побочный продукт, и указанный генетически сконструированный микроорганизм согласно изобретению может быть непосредственно использован в превращении стероидов без какой-либо потенциальной стадии (стадий) созревания. Дополнительные стадии очистки продукта от нежелательного побочного продукта также могут быть опущены. Продукт биотрансформации можно использовать сразу после производства. В заключение, это приводит к более высокому выходу продукта, более высокому качеству продукта и более простому производственному процессу.
Описание изобретения
Таким образом, первый аспект изобретения относится к генетически модифицированной бактерии, где экспрессия гена, кодирующего 20β-HSDH, снижена или подавлена по сравнению с соответствующей немодифицированной бактерией, где соответствующая немодифицированная бактерия обладает активностью Δ1-дегидрогеназы и активностью 20-β-дегидрогеназы.
Используемый в настоящем описании термин «соответствующая немодифицированная бактерия» обозначает тот же штамм бактерии, что и немодифицированная бактерия, которая затем подвергается генетической модификации.
Термин «бактерия» означает прокариотический микроорганизм. Немодифицированная бактерия обладает активностью 20β-HSDH и активностью Δ1-дегидрогеназы. В одном варианте осуществления бактерия принадлежит к отряду, выбранному из отряда Actinomycetales, отряда Clostridiales, отряда Aeromonadales, отряда Pasteurellales, отряда Bacillales, отряда Lactobacillales, отряда Bifidobacteriales и отряда Coriobacteriales. В одном предпочтительном варианте осуществления бактерия принадлежит к отряду Actinomycetales. Например, бактерия может быть Nocardiodiaceae, как например Nocardioides simplex, Streptomycetaceae, как например Streptomyces albus, или Mycobacteriaceae, как например Mycobacterium avium и, в частности, Mycobacterium avium subsp. paratuberculosis MAP4. В конкретном варианте осуществления бактерия принадлежит к семейству Nocardioidaceae, предпочтительно Nocardioides и наиболее предпочтительно бактерия представляет собой Nocardioides simplex.
Термин «бактерия из семейства Nocardioidaceae» означает любую бактерию из этого семейства, обладающую активностью 20β-HSDH и активностью Δ1-дегидрогеназы, такую как, например, бактерия рода Actinopolymorpha, Aeromicrobium, Flindersiella, Friedmanniella, Kribbella, Marmoricola, Micropruina, Mumia, Nocardioides, Pimelobacter, Propionicella, Propionicimonas, Tenggerimyces или Thermasporomyces. Предпочтительно бактерия из семейства Nocardioidaceae представляет собой Nocardioides simplex (ранее называвшуюся Arthrobacter simplex или Corynebacterium simplex или Pimelobacter simplex). Предпочтительно бактерия представляет собой Nocardioides simplex ATCC6946. Другие предпочтительные бактерии включают Friedmanniella flava (ID таксономии: 1036181), Friedmanniella sagamiharensis (ID таксономии: 546874), Nocardioides dokdonensis FR1436 (ID таксономии: 1300347), Nocardioides lianchengensis (ID таксономии: 1045774), Nocardioides lianchengensis (ID таксономии: 1045774), Nocardioides, Nocardioides szechwanensis (ID таксономии: 1005944), Corynebacterium coyleae (ID таксономии: 53374), Corynebacterium glyciniphilum AJ 3170 (ID таксономии: 1404245), Corynebacterium kroppenstedtii DSM 44385 (ID таксономии), Corynebacterium kroppenstedtii DSM 44385 (ID таксономии 13), Corynebacterium kroppenstedtii DSM 44385 (ID таксономии), Corynebacterium spheniscorum (ID таксономии: 185761), Corynebacterium vitaeruminis DSM 20294 (ID таксономии: 1224164), Aeromicrobium marinum DSM 15272 (ID таксономии: 585531), Nocardioides luteus (ID таксономии: 1844), Corynebacterium YIM halotolera 1121362), Corynebacterium maris DSM 45190 (ID таксономии: 1224163), Corynebacterium riegelii (ID таксономии: 156976).
Термин «ген» означает последовательность ДНК, которая кодирует или соответствует определенной последовательности аминокислот, которая включает весь или часть одного или более белков или ферментов, и может включать или не включать регуляторные последовательности ДНК, такие как промоторные последовательности, которые определяют, например, условия, при которых экспрессируется ген. В частности, термин «ген» может означать геномную последовательность, кодирующую белок, то есть последовательность, содержащую регуляторные, промоторные, интронные и экзонные последовательности.
Используемый в настоящем описании термин «ген, кодирующий 20β-HSDH», соответствует гену, кодирующему 20β-HSDH любой бактерии. Предпочтительно ген, кодирующий 20β-HSDH, имеет по меньшей мере 60% идентичности с последовательностью SEQ ID NO: 1, более предпочтительно он имеет более чем 70%, 75%, 80%, 85%, 90%, 95% идентичности и наиболее предпочтительно более чем 99% идентичности с последовательностью SEQ ID NO: 1. Согласно варианту осуществления, ген, кодирующий 20β-HSDH, имеет по меньшей мере 60% идентичности с последовательностью SEQ ID NO: 3, более предпочтительно он имеет более чем 70%, 75%, 80%, 85%, 90%, 95% идентичности и наиболее предпочтительно более чем 99% идентичности с последовательностью SEQ ID NO: 3.
Под «геном, имеющим, по меньшей мере, x% идентичности с эталонной последовательностью» подразумевается, что последовательность нуклеиновой кислоты отличается от эталонной последовательности до 100-х нуклеотидных изменений на каждые 100 нуклеотидов эталонной последовательности по всей ее длине. Другими словами, для получения нуклеиновой кислоты, имеющей последовательность, по меньшей мере, на х% идентичности с эталонной последовательностью, до 100-х% нуклеотидов в рассматриваемой последовательности могут быть вставлены, удалены или заменены другой аминокислотой или нуклеотидом.
Последовательность нуклеиновой кислоты, «идентичная по меньшей мере на 60%, 70%, 75%, 80%, 85%, 90%, 95% или 99%» эталонной последовательности, может содержать мутацию(и), такую как делеция(и), вставки и/или замены по сравнению с эталонной последовательностью. В случае замены замена предпочтительно соответствует «молчащей» замене или замене, ведущей к консервативной замене в транслируемой аминокислотной последовательности.
Методы сравнения идентичности двух или более последовательностей хорошо известны в научном сообществе. Например, программы, доступные в пакете анализа последовательностей «Wisconsin Sequence Analysis Package», версия 9.1, например программы BESTFIT и GAP, могут использоваться для определения процента идентичности между двумя последовательностями. BESTFIT использует алгоритм «локальной гомологии» Смита и Уотермана и находит лучшую единственную область сходства между двумя последовательностями. Другие программы для определения идентичности между последовательностями также известны в данной области, например программа Needle, которая основана на алгоритме Нидлмана и Вунша, описанном Needleman and Wunsch (1970) J. Mol Biol. 48: 443-453, например, со следующими параметрами для сравнения полинуклеотидных последовательностей: матрица сравнения: DNAFULL; штраф за открытие пробела=10, штраф за удлинение пробела=0,5, штраф за концевой пробел: штраф за концевой, ложный пробел=10, штраф за удлинение пробела=0,5.
Термин «генетически модифицированный» означает любое детектируемое изменение генетического материала, например ДНК, РНК, кДНК или любой процесс, механизм или результат такого изменения. Сюда входят генные мутации, при которых изменяется структура (например, последовательность ДНК) гена, любой ген или ДНК, возникающие в результате любого процесса мутации, и любой продукт экспрессии (например, белок или фермент), экспрессируемый модифицированным геном или последовательностью ДНК. Обычно мутацию идентифицируют у пациента путем сравнения последовательности нуклеиновой кислоты или полипептида, экспрессируемых указанным пациентом, с соответствующей нуклеиновой кислотой или полипептидом, экспрессируемыми в контрольной популяции.
Термин «экспрессия гена, кодирующего 20β-HSDH, снижена или подавлена» означает снижение или подавление уровня экспрессии матричной РНК (мРНК) и/или белка, кодируемого геном, кодирующим 20β-HSDH. Экспрессия гена, кодирующего 20β-HSDH, может быть снижена по меньшей мере на 75%, 80%, 85%, 90% или 95% по сравнению с немодифицированной бактерией. Предпочтительно экспрессия гена, кодирующего 20β-HSDH, снижена на 100%, что соответствует подавлению экспрессии.
Уровень экспрессии гена или нуклеиновой кислоты можно измерить способами, которые хорошо известны специалистам в данной области, включая, в частности, анализы на основе прямой гибридизации и анализы на основе амплификации.
Экспрессия гена, кодирующего 20β-HSDH, может быть снижена или подавлена по сравнению с немодифицированной бактерией путем инактивации или делеции этого гена. В конкретном варианте осуществления ген, кодирующий 20β-HSDH, мутирован и предпочтительно укорочен.
Предпочтительно генетически модифицированная бактерия не содержит экзогенной ДНК.
Лабораторные методы инактивации, делеции или укорачивания гена хорошо известны в данной области.
Как предполагается в настоящем описании, когда ген, кодирующий 20β-HSDH, мутирован и/или укорочен, он может не кодировать полностью функциональный белок 20β-HSDH, но кодирует белок 20β-HSDH, имеющий активность ниже, чем у соответствующего природного белка 20β-HSDH, измеренную в тех же условиях, в частности, кодируемый белок 20β-HSDH может не иметь активности.
Генетически модифицированная бактерия согласно изобретению предпочтительно обладает активностью Δ1-дегидрогеназы, но имеет пониженную активность 20-β-дегидрогеназы или лишена ее по сравнению с немодифицированной бактерией.
Таким образом, как совершенно очевидно из данной заявки, изобретение относится к генетически модифицированной бактерии, где:
- соответствующая немодифицированная бактерия обладает активностью Δ1-дегидрогеназы и активностью 20-β-дегидрогеназы,
- по меньшей мере, одна генетическая модификация индуцирует сниженную или подавленную экспрессию гена, кодирующего 20-бета-гидроксистероид дегидрогеназу (20β-HSDH), и
- предпочтительно указанная генетически модифицированная бактерия сохраняет свою активность Δ1-дегидрогеназы.
Предпочтительно, генетически модифицированная бактерия согласно изобретению предпочтительно обладает активностью Δ1-дегидрогеназы, но имеет пониженную активность 20-β-дегидрогеназы или лишена ее по сравнению с немодифицированной бактерией.
Второй аспект изобретения относится к способу получения генетически модифицированной бактерии согласно изобретению, включающему:
а) предоставление бактерии, которая обладает активностью Δ1-дегидрогеназы и активностью 20-β-дегидрогеназы; и
b) инактивацию или делецию гена, кодирующего 20β-HSDH.
Стадия b способа получения генетически модифицированной бактерии согласно изобретению может состоять из мутации, укорачивания или делеции гена, кодирующего 20β-HSDH. Мутация или укорачивание могут быть реализованы любыми способами, известными специалистам в данной области, и предпочтительно двойным кроссинговером гена, кодирующего 20β-HSDH.
Предпочтительно полученная генетически модифицированная бактерия не содержит экзогенной ДНК.
В другом аспекте изобретение относится к способу получения 1,2-дегидрированного стероида, включающему:
а) предоставление стероида, который не является 1,2-дегидрированным;
b) контактирование указанного стероида, который не является 1,2-дегидрированным, с бактерией согласно изобретению или ее экстрактом в условиях, достаточных для получения 1,2-дегидрированного стероида.
Предпочтительно стероид, который не является 1,2-дегидрированным, используемый в способе получения 1,2-дегидрированного стероида согласно изобретению, представляет собой кортикостероид.
В конкретном варианте осуществления стероид, который не является 1,2-дегидрированным, используемый в способе, представляет собой прогестерон, тестостерон, андростендион или производное этих молекул, которые содержат стероидный каркас, содержащий кольцо А из шести атомов углерода, не дегидрированное между положениями 1,2.
В конкретном варианте осуществления стероид, который не является 1,2-дегидрированным, представляет собой:
- 11бета 17,21-тригидроксипрегн-4-ен-3,20-дион (гидрокортизон), и указанный способ предназначен для получения 11бета, 17альфа, 21-тригидрокси-1,4-прегнадиен-3,20-диона (преднизолона); или
- 17альфа, 21-дигидрокси-4-прегнен-3,11,20-трион (кортизон), и указанный способ предназначен для получения 17альфа, 21-дигидрокси-1,4-прегнадиен-3,11,20-триона (преднизона); или
- прегн-4-ен-3,20-дион, и указанный способ предназначен для получения прегна-1,4-диен-3,20-диона; или
- 11бета,17,21-тригидрокси-6-альфа-1,4-прегнен-3,20-дион (6-метил-гидрокортизон) для получения 11бета,17альфа,21-тригидрокси-6-альфа-метил-1,4-прегнадиен-3,20-диона(6-метилпреднизолона); или
- 9,11-дегидрокортексолон и указанный способ предназначен для получения 1,2-9,11-дегидрокортексолона; или
- 9,11-дегидрокортексолон-17,21-диацетат, и указанный способ предназначен для получения 1,2-9,11-дегидрокортексолона; или
- 9,11-16,17-дегидрокортексолон-21-ацетат, и указанный способ предназначен для получения 1,2-9,11-16,17-дегидрокортексолон-21-ацетата; или
- 16-альфа-метилгидрокортизон и указанный способ предназначен для получения 16-альфа-метилпреднизолона.
В одном варианте осуществления стадия b) способа получения 1,2-дегидрированного стероида выполняется с экстрактом бактерии согласно изобретению. Указанный экстракт предпочтительно обладает активностью Δ1-дегидрогеназы и предпочтительно имеет пониженную активность 20-β-дегидрогеназы или лишен ее. В частности, такой экстракт можно получить путем лизиса клеток бактерии согласно изобретению и, необязательно, путем фракционирования продукта лизиса клеток, чтобы сконцентрировать активность Δ1-дегидрогеназы или облегчить очистку активности Δ1-дегидрогеназы.
Способ получения 1,2-дегидрированного стероида может дополнительно включать выделение и очистку указанного 1,2-дегидрированного стероида.
Предпочтительно способ получения 1,2-дегидрированного стероида не включает стадию удаления 20-β-дегидрированного стероида. Действительно, эта стадия может быть ненужной, если экспрессия гена, кодирующего 20β-HSDH, значительно снижена.
Кроме того, способ получения 1,2-дегидрированного стероида может содержать включение 1,2-дегидрированного стероида в состав фармацевтической композиции.
В другом аспекте изобретение относится к применению бактерии или ее экстракта согласно изобретению для получения 1,2-дегидрированного стероида из стероида, который не является 1,2-дегидрированным.
Указанный экстракт предпочтительно обладает активностью Δ1-дегидрогеназы и предпочтительно имеет пониженную активность 20-β-дегидрогеназы или лишен ее. В частности, такой экстракт можно получить путем клеточного лизиса бактерии согласно изобретению и, необязательно, после фракционирования продукта клеточного лизиса, чтобы сконцентрировать активность Δ1-дегидрогеназы или облегчить очистку активности Δ1-дегидрогеназы.
Предпочтительно стероид, который не является 1,2-дегидрированным, представляет собой кортикостероид.
В конкретном варианте осуществления стероид, который не является 1,2-дегидрированным, представляет собой прогестерон, тестостерон, андростендион или производное этих молекул, которые содержат стероидный каркас, содержащий кольцо A из шести атомов углерода, не дегидрированное между положениями 1,2.
В конкретном варианте осуществления стероид, который не является 1,2-дегидрированным, представляет собой:
- 11бета,17,21-тригидроксипрегн-4-ен-3,20-дион (гидрокортизон) и получают 11бета,17альфа,21-тригидрокси-1,4-прегнадиен-3,20-дион (преднизолон); или
- 17альфа,21-дигидрокси-4-прегнен-3,11,20-трион (кортизон) и получают 17альфа,21-дигидрокси-1,4-прегнадиен-3,11,20-трион (преднизон); или
- прегн-4-ен-3,20-дион и получают прегна-1,4-диен-3,20-дион; или
- 11бета,17,21-тригидрокси-6альфа-1,4-прегнен-3,20-дион (6-метил-гидрокортизон) и получают 11бета,17альфа,21-тригидрокси-6альфа-метил-1,4-прегнадиен-3,20-дион (6-метилпреднизолон); или
- 9,11-дегидрокортексолон и получают 1,2-9,11-дегидрокортексолон; или
- 9,11-дегидрокортексолон-17,21-диацетат и получают 1,2-9,11-дегидрокортексолон; или
- 9,11-16,17-дегидрокортексолон-21-ацетат и получают 1,2-9,11-16,17-дегидрокортексолон-21-ацетат; или
- 16-альфа-метилгидрокортизон и получают 16-альфа-метилпреднизолон.
Используемый в настоящем описании термин «фармацевтическая композиция» относится к препарату 1,2-дегидрированного стероида с другими химическими компонентами, такими как физиологически подходящие носители и вспомогательные вещества. Назначение фармацевтической композиции состоит в том, чтобы облегчить введение указанного 1,2-дегидрированного стероида в организм.
Фармацевтические композиции по настоящему изобретению могут быть получены способами, хорошо известными в данной области, например, с помощью обычных процессов смешивания, растворения, гранулирования, измельчения, пульверизации, изготовления драже, измельчения, эмульгирования, инкапсулирования, улавливания или лиофилизации.
Далее изобретение будет проиллюстрировано следующими чертежами и примерами.
ЧЕРТЕЖИ
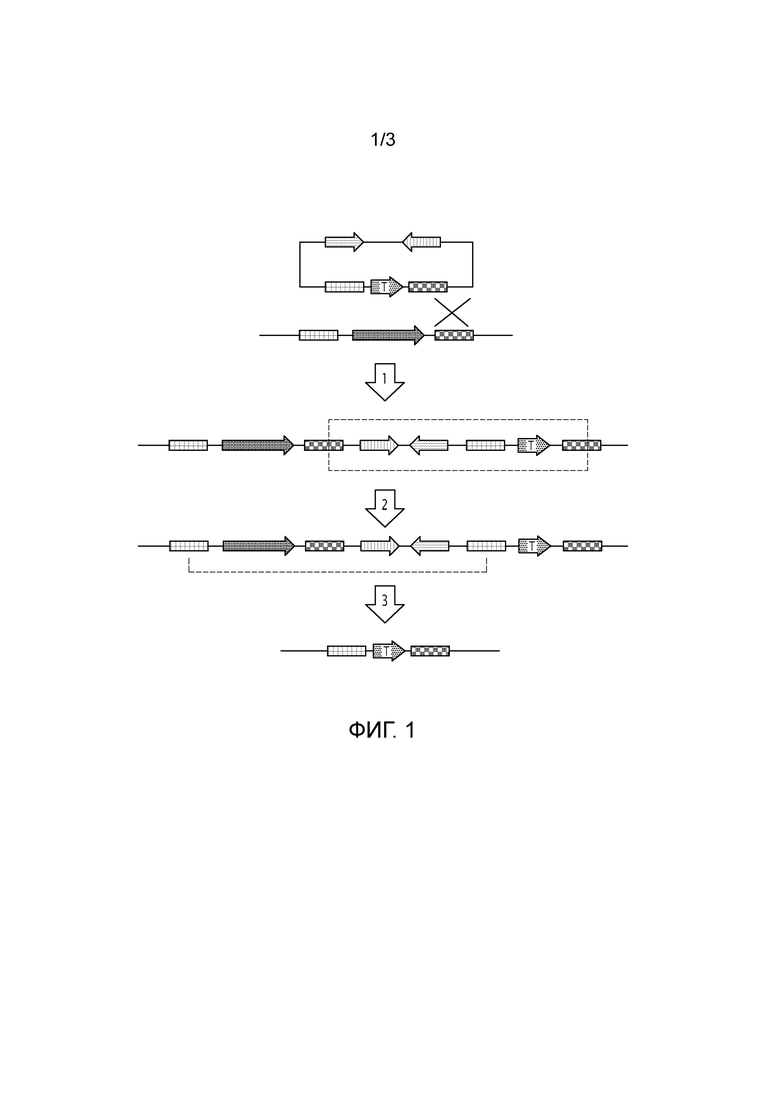
Фигура 1: Схематическое изображение инактивации гена двойным кроссинговером. Плазмида с укороченной формой интересующего гена (стрелка, содержащая букву «Т»), фланкированная двумя гомологичными плечами (прямоугольники с квадратами) и генами устойчивости к апрамицину и β-глюкуронидазы (стрелки, состоящие из вертикальных и волнистых линий), вступает в контакт с хромосомой бактерии и одно из гомологичных плеч рекомбинирует с соответствующей последовательностью в хромосоме (стадия 1). Эта стадия приводит к формированию промежуточной конструкции одиночного кроссинговера, вся плазмида интегрируется в геном (обведена пунктирной линией). Тогда штамм содержит два варианта интересующего гена и проявляет устойчивость к апрамицину и активность β-глюкуронидазы. После нескольких циклов культивирования (стадия 2) два оставшихся гомологичных плеча взаимодействуют друг с другом (пунктирная линия), и происходит двойной кроссинговер (стадия 3). Тогда полученный штамм содержит только укороченную форму гена (стрелка с буквой «Т»).
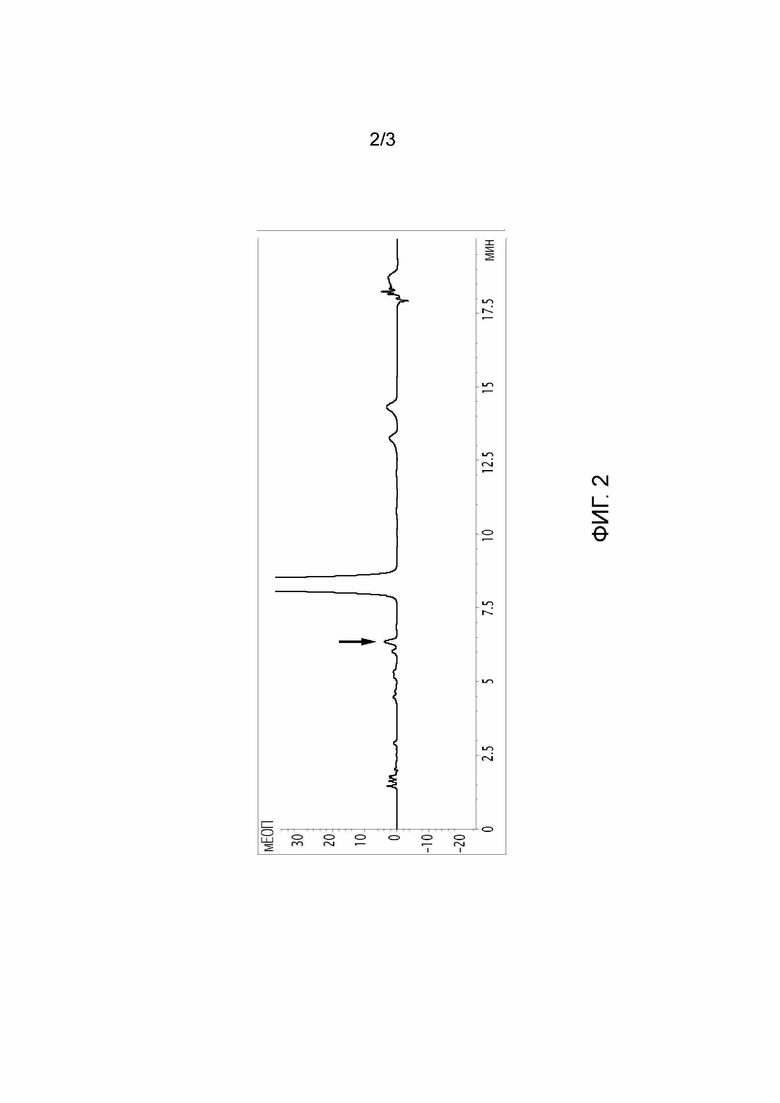
Фигура 2: ВЭЖХ-хроматограмма полученного продукта после классического процесса получения преднизолона из гидрокортизона с помощью N. simplex. Стрелка указывает на примеси 20-ОН преднизолона.
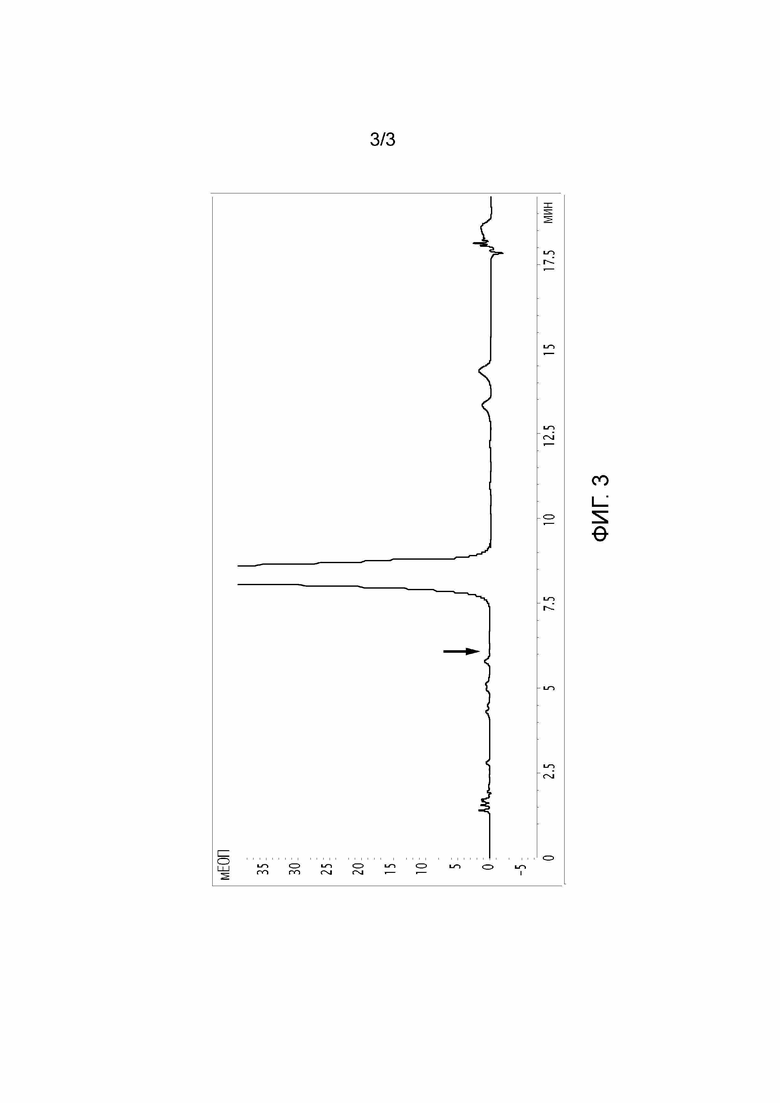
Фигура 3: ВЭЖХ-хроматограмма полученного продукта после классического процесса получения преднизолона из гидрокортизона с мутантами с двойным кроссинговером NOSIM5169 N. simplex. Стрелка указывает на место, где обычно наблюдаются примеси 20-ОН преднизолона.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ:
SEQ ID NO: 1 соответствует последовательности NOSIM3735 N. simplex.
SEQ ID NO: 2 соответствует последовательности NOSIM3735 N. simplex.
SEQ ID NO: 3 соответствует последовательности гена 20β-HSDH Mycobacterium avium subsp. Paratuberculosis MAP4.
ПРИМЕР
Анализируя геном N. simplex, были обнаружены два возможных гена для 20β-HSDH, названные NOSIM5169 (соответствует последовательности SEQ ID NO: 1) и NOSIM3735 (соответствует последовательности SEQ ID NO: 2). Оба они имеют сходство с последовательностью гена, ответственного за 20β-HSDH Mycobacterium avium subsp. Paratuberculosis MAP4 (соответствует последовательности SEQ ID NO: 3).
1. Конструкция плазмиды (NOSIM5169)
Плазмиды для инактивации NOSIM5169 были сконструированы следующим образом. Гомологичные плечи размером 2 и 1,7 т.п.н. амплифицировали с помощью ПЦР. Праймеры, используемые для амплификации одного плеча, содержали сайты рестрикции Xbal и EcoRV, а праймеры для второго гомологичного плеча были сконструированы так, чтобы они содержали сайты рестрикции EcoRV и EcoRI.
Впоследствии плечи были клонированы в первую плазмиду.
Для получения новой второй плазмиды ген устойчивости к гигромицину был клонирован по тупому концу в сайт рестрикции EcoRV.
Плазмида без гена устойчивости к гигромицину была названа «вторая плазмида-hyg».
2. Протокол электропорации для N. simplex
Протокол основан на процедуре, описанной для Arthrobacter (Zhang et al., 2011). Оптимизированная процедура получения электрокомпетентных клеток была следующей: клетки N. simplex инокулировали в 20 мл LB+Ph (LB означает бульон для лизогенизации, а Ph означает фосфомицин) и культивировали до достижения стационарной фазы. Затем 1% посевной материал из прекультуры вносили в свежую среду LB и культивировали до тех пор, пока оптическая плотность не приближалась к 0,4-0,5 при 600 нм. Затем добавляли ампициллин (Ap) и глицин до конечной концентрации 30 мкг/л и 5 г/л соответственно, и культивирование продолжали. Через 3 часа культуры помещали на лед и собирали центрифугированием (4500 об/мин, 6 минут, 4 °C). После трех промывок ледяным буфером для электропорации (0,5 М сорбит, 10% глицерин) клетки концентрировали центрифугированием в 100 раз. Наконец, суспензию распределяли на аликвоты по 60 мкл и хранили на льду (этот протокол использовали для создания N. simplex NOSIM5169-hyg).
В качестве альтернативы может применяться протокол для S. albus ATCC 21838 (Izumikawa et al., 2003). В этом случае процедура получения электрокомпетентных клеток была изменена, чтобы соответствовать физиологии N. simplex: 1% инокулята из прекультуры N. simplex в стационарной фазе инокулировали в свежую среду LB+Ph и культивировали. Когда оптическая плотность приближалась к 0,4-0,5 при 600 нм, культуры собирали центрифугированием (4500 об/мин, 6 мин, 4 °C) и трижды промывали стерильной ледяной водой. Затем мицелий суспендировали в 25 мл холодного буфера для электропорации (10% сахарозы, 15% глицерина) и добавляли лизоцим (0,5 мг/мл). После 20-минутной инкубации при 37°C клетки собирали центрифугированием и концентрировали в 100 раз в буфере для электропорации. Наконец, суспензию распределяли на аликвоты по 60 мкл и хранили на льду.
Оптимизированная процедура электропорации в обоих случаях была следующей. Электрокомпетентные клетки смешивали с 500 нг ДНК (полученной из E. coli GB2005) и смесь переносили в предварительно охлажденную кювету для электропорации (электрод 0,1 см). На смесь подавали одиночный импульс с использованием импульсного генератора Eppendorf Pulser с напряжением 2,1 кВ/см. Сразу после электрического импульса клеточные суспензии переносили в пробирку Eppendorf, содержащую 800 мкл восстановительной среды при комнатной температуре (SOC), и инкубировали при встряхивании при 850 об/мин в течение 12 часов при 30 °C. Для отбора трансформированных клеток последовательные разведения бактерий наносили на чашки с агаром LB, содержащим апрамицин (Am). В каждой серии экспериментов проводили отрицательные контроли, не добавляя плазмидную ДНК.
3. Мутанты с одиночным кроссинговером (NOSIM5169)
После электропорации штамма N. simplex второй плазмидой был получен одиночный трансформант кроссинговера и нанесен на чашки с агаром LB+Am+X-Gluc (схематически см. Фигуру 1) (X-Gluc обозначает моногидрат, X-GlcA). Способность расти на апрамицине и выполнять реакцию β-глюкуронидазы (GUS) (например, Myronovskyi et al., 2011) подтвердила, что трансформант был мутантом с одиночным кроссинговером. После этого мутант инокулировали в 100 мл среды LB+Ph и инкубировали в течение 48 часов при 28 °C, 200 об/мин.
4. Мутанты с двойным кроссинговером (NOSIM5169)
Затем 1 мл культуры переносили в свежую среду LB+Ph и инкубировали еще 48 ч при тех же условиях. Эту стадию повторяли несколько раз. После последней инкубации были произведены последовательные разведения мутантов на чашках с агаром LB+X-Gluc.
Мутанты, которые не проявляли GUS-активности, переносили на новый LB+X-Gluc и проверяли еще раз. Отсутствие GUS-активности означало, что у этих мутантов произошел двойной кроссинговер, и они потеряли остов плазмиды.
Мутанты, прошедшие второй тест, выращивали в LB, их гДНК выделяли и затем анализировали с помощью ПЦР с использованием праймеров, гомологичных хромосомной ДНК N. simplex.
5. Мутанты с двойным кроссинговером без чужеродной ДНК (NOSIM5169)
Вместо второй плазмиды для проведения трансформации использовали «вторую плазмиду-hyg». Присутствие гена устойчивости к антибиотикам (устойчивость к гигромицину) может облегчить дальнейшую идентификацию мутантов с двойным кроссинговером. Однако этот ген устойчивости оказался неактивным в геноме N. simplex.
Таким образом, предполагается, что после двойного кроссинговера полученный мутант не будет содержать экзогенной ДНК и будет доступен для дальнейшего промышленного применения (Фигура 1).
После введения «второй плазмиды -hyg» в N. simplex была получена колония, устойчивая к апрамицину.
Этот мутант был пропущен через шесть культивирований в среде LB+Ph, а затем были сделаны последовательные разведения культуры на LB+X-Gluc.
Полученные колонии, которые не могли расти на апрамицине и не имели GUS-активности, подвергали дальнейшему анализу с помощью ПЦР. Амплификацию выполняли с использованием двух праймеров, гомологичных участкам гДНК N. simplex, соседним с NOSIM5169.
6. Инактивация NOSIM3735
Для конструирования штаммов с инактивированным NOSIM3735 ранее описанные методы нельзя было применить, поскольку область ДНК перед NOSIM3735 была недоступна для ее амплификации с помощью ПЦР. Чтобы преодолеть эту проблему, была упорядочена и секвенирована библиотека космид, содержащая фрагменты генома N. simplex.
Чтобы инактивировать NOSIM3735, была идентифицирована соответствующая космида из космидной библиотеки, содержащая ген (секвенирование по Сэнгеру приблизительно 1500 космид GATC Biotech, Констанц), и ген был заменен геном устойчивости к хлорамфениколу (Cm), cat, с использованием λ-опосредованной рекомбинации. в E. coli (например, Murphy, 1998). Затем модифицированную космиду гидролизовали эндонуклеазой BamHI. Таким способом была получена смесь фрагментов и среди них фрагмент, который содержал ген cat вместо NOSIM3735, фланкированный двумя плечами размером 3 т.п.н. для гомологичной рекомбинации с геномом N. simplex. Фрагменты разделяли электрофорезом в агарозном геле, фракцию 7-8 т.п.н. очищали и лигировали в линеаризованную плазмиду BamHI. Для отбора мутантов, содержащих конструкцию с кошачьим геном, LB+Am+Cm использовали в качестве ростовой среды. Полученные мутанты анализировали с помощью ПЦР и секвенирования. Одна из полученных плазмид в дальнейшем была использована для инактивации NOSIM3735.
Инактивацию проводили согласно протоколу, ранее адаптированному для инактивации NOSIM5169 (см. пункт 2 и последующие).
Этот мутант несколько раз культивировали в среде LB+Ph, а затем проводили последовательные разведения культуры на LB+X-Gluc.
Полученные колонии, которые не могли расти на апрамицине и не имели GUS-активности, подвергали дальнейшему анализу с помощью ПЦР. Амплификацию проводили с использованием двух праймеров, гомологичных участкам гДНК N. simplex, прилегающим к NOSIM3735.
7. Использование NOSIM3735 и NOSIM5169 на гидрокортизоне
Клоны используются в классическом процессе производства преднизолона из гидрокортизона.
Вкратце, гидрокортизон смешивали с раствором менадиона в тетрагидрофуране. Затем эту суспензию смешивали с фосфатным буфером, доведенным до pH 8. Затем добавляли бактериальную суспензию. После инкубации при 30°C при перемешивании реакцию останавливали добавлением фосфорной кислоты. Увеличение преднизолона измеряли и анализировали с помощью ВЭЖХ.
ВЭЖХ-хроматограммы получали на колонке Waters-Xbrigde RP18 с использованием градиента вода/ацетонитрил в качестве подвижной фазы (хроматограммы показаны на фигурах 2-3).
В качестве альтернативы был проведен анализ активности продукции 20-ОН-побочного продукта. Преднизолон суспендировали в фосфатном буфере при pH 8,0 и инкубировали с бактериальной суспензией. Через 60 мин реакцию останавливали добавлением фосфорной кислоты. Потенциальное увеличение количества продукта, восстановленного 20-ОН-кето, затем измеряли с помощью ВЭЖХ.
Укорачивание NOSIM5169 позволяет полностью подавить активность 20β-HSDH и, таким образом, получить преднизолон без примесей 20-ОН (фигура 3).
Ген NOSIM3735 был нокаутирован, но штаммы показывают те же реакции, что и штаммы дикого типа, нежелательную побочную реакцию нельзя было отключить путем нокаута этого гена.
Список литературы:
Izumikawa, M.; Murata, M.; Tachibana, K.; Ebizuka, Y. and Fujii, I. (2003): Cloning of Modular Type I Polyketide Synthase Genes from Salinomycin Producing Strain of Streptomyces albus. Bioorganic & Medicinal Chemistry 11: 3401-3405.
Murphy, K.C. (1998): Use of bacteriophage λ recombination functions to promote gene replacement in Escherichia coli. J. Bacteriol. 180: 2063-2071.
Myronovskyi, M.; Welle, E.; Fedorenko, V. and Luzhetskyy, A. (2011): Glucuronidase as a Sensitive and Versatile Reporter in Actinomycetes. Applied and Environmental Microbiology, Aug. 2011, 5370-5383.
Zhang, H.; Li, Y.; Chen, X.; Sheng, H. and An, L. (2011): Optimization of electroporation conditions for Arthrobacter with plasmid PART2, Journal of Microbiological Methods, 84: 114-120.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SANOFI
<120> БИОТЕХНОЛОГИЧЕСКАЯ ОПТИМИЗАЦИЯ МИКРООРГАНИЗМОВ
<130> BET17P1750
<160> 3
<170> PatentIn version 3.5
<210> 1
<211> 748
<212> ДНК
<213> Nocardioides simplex
<400> 1
ggtcgtcctc gtcaccggag gcgcccgcgg gctgggcgcc gccttcgccc gcggcatcgt 60
cgccgccggc ggccgggtcg tcatcggcga cctgctggac gacgagggcg ccgccgtcgc 120
cgacgagctc ggcgccgcgg cccgctacgt ccacctcgac gtcaccagcg aggagtcctg 180
ggaggccgcc gtcgccgcga gcgtcgacgc cttcggccgg ctcgacgggc tggtcaacaa 240
cgccggcatc tcggccaccg gccagctgac cgccgacgag ccgaccgacg tcttccgccg 300
gatcatcgag atcaacctga tcgccgtcca caccgggctg cgcgccgtcg tccccgcgat 360
gcggacggcg ggcggcggct cgatcgtcaa catctcgtcg gccgcgggcc tgatgggcat 420
ggcgatgacc agcggctacg gcgccgccaa gtggggcgtg cgcgggctca gcaagatcgc 480
ggcggtcgag ctcggtcgcg accggatccg ggtcaactcg gtgcaccccg gcatggtcct 540
gacgccgatg accgccccga ccgggatcgt cgccgacgag ggcgccttcc cgaacaaccc 600
gtaccagcgg gtcggccggc ccgaggagct cgtgggtgcg gtcgtccacc tgctgagcga 660
cgccgcgtcg tacacgaccg gcgccgagct cgccgtcgac ggcggctgga ccgccggtcc 720
gtcggtggag tacatcgcag gtcggtga 748
<210> 2
<211> 825
<212> ДНК
<213> Nocardioides simplex
<400> 2
gtgccgctgg ccggcaccct gcccgcgcgc ccgcgcggac ggcgcttcat tggggccatg 60
acccagatcc aggccccgac gaacccgtcc accactcccg gccgcgtcgc cgaccgcgtc 120
gtcatcgtga cgggaggagc ccgcggcatc ggcgccgcct gcgtgcgcgc actcgtcgcc 180
gagggcgccc gggtggtggt cgccgacgtc ctcgaggccg aggcgagcgc cctcgtcgcc 240
gcgctggggg agcggacggc gtacgtcccg ctcgacgtga cgagcgagga ggcgtggcag 300
cacgcggtcg cggcggccga ggagcgcttc gggccggtct cgggactcgt caacaacgcc 360
ggcatcgtgc acatcgaccc gatcgagacc ctgagcgagg ccgactaccg gcgggtgatc 420
gacgtcaacc aggtcggcgt cttcctcggc atgaaggcgg tcatcgggtc gatgcggcgc 480
gcggggggcg gctcgatcgt caacatctcc tcgaccggcg ggctggtcgc ctactcccgg 540
atcctcgggt acgtcgcctc gaagtgggcc gtgcgcggca tgaccaagac cgccgcgcag 600
gagctcggcc ccgacggcat ccgggtcaac tcggtgcacc ccggcatcgt cgcctccgcg 660
atgaccgcga gctccgaccg ctcccacgag caggtcagga cccagccgct ggcgcgggcg 720
gcggacccgt ccgagatcgg cgcgctcgtg ctcttcctga tctcggagga gtccagctac 780
agcaccggct cggagttcgt cgccgacggg ggcttcacct cgctc 825
<210> 3
<211> 741
<212> ДНК
<213> Mycobacterium avium
<400> 3
gtgggacggg tagacggcaa ggttgcgctg atcagcgggg gcgcccgcgg catgggcgcc 60
gagcacgcgc ggctgctggc ggccgagggc gccaaggtgg tgatcggcga catcctcgac 120
gacgagggca aggccgtggc cgacgagatc ggcgactcgg tgcgctacgt ccacctggac 180
gtcacccagc ccgaccagtg ggacgccgcc gtcgaaaccg ccgtcggcga attcggcaag 240
ctcaacgtgt tggtcaacaa cgccggcacc gtcgcgctcg ggccgctgaa gagcttcgat 300
ctggccaagt ggcagaaggt gattgacgtc aacctgaccg gcaccttcct gggcatgcgg 360
gtggccgtcg agccgatgat cgcggccggc ggcggctcga tcatcaacat ctcctccatc 420
gaggggctgc gcggtgcgcc catggtgcac ccctacgtcg cctccaagtg gggcgtgcgc 480
ggcctggcga agtccgcggc gctggagctg gcgccgcaca acatccgggt caactccgtg 540
caccccggct tcatccgcac cccgatgacc aaacacctgc ccgacgacat ggtgaccgtc 600
ccgctcggcc gtccggccga gtcccgcgag gtgtcgacgt tcgtcttgtt cctggccagc 660
gacgagtcgt cgtacgcgac cggcagcgaa ttcgtgatgg acggcggact ggtcaccgac 720
gtgccgcaca agcagttcta g 741
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ КОНСТРУИРОВАНИЯ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННОГО ШТАММА МИКРООРГАНИЗМА | 2000 |
|
RU2268935C2 |
| МИКРОБИОЛОГИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ 1,2-ДЕГИДРИРОВАННЫХ ПРОИЗВОДНЫХ Δ-3-КЕТОСТЕРОИДОВ РЯДА АНДРОСТАНА В ВОДНО-ОРГАНИЧЕСКИХ СРЕДАХ | 2010 |
|
RU2447154C1 |
| СПОСОБ ПОЛУЧЕНИЯ 6-МЕТИЛЕНАНДРОСТ-4-ЕН-3,17-ДИОНА ИЗ АНДРОСТ-4-ЕН-3,17-ДИОНА, СПОСОБ ПОЛУЧЕНИЯ 6-МЕТИЛЕНАНДРОСТА-1,4-ДИЕН-3,17-ДИОНА (ЭКСЕМЕСТАНА) С ИСПОЛЬЗОВАНИЕМ ПОЛУЧЕННОГО 6-МЕТИЛЕНАНДРОСТ-4-ЕН-3,17-ДИОНА | 2010 |
|
RU2425052C1 |
| СПОСОБ ПОЛУЧЕНИЯ 1,2-ДЕГИДРОПРОИЗВОДНЫХ 4-ДЕЛЬТА-3-КЕТОСТЕРОИДОВ | 1998 |
|
RU2156302C1 |
| Способ получения 2,4-дигидроксимасляной кислоты | 2011 |
|
RU2626531C2 |
| Микроорганизм рода Corynebacterium, обладающий L-изолейцин-продуцирующей способностью, и способ получения L-изолейцина с использованием этого микроорганизма | 2016 |
|
RU2698394C1 |
| НОВАЯ АЛЬДОЛАЗА, ДНК, КОДИРУЮЩАЯ АЛЬДОЛАЗУ, КЛЕТКИ, ТРАНСФОРМИРОВАННЫЕ ДНК, СПОСОБ ПОЛУЧЕНИЯ АЛЬДОЛАЗЫ И СПОСОБ ПОЛУЧЕНИЯ 4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА (ВАРИАНТЫ) | 2006 |
|
RU2338784C2 |
| ПОЛИНУКЛЕОТИД, ОБЛАДАЮЩИЙ ПРОМОТОРНОЙ АКТИВНОСТЬЮ, И ПРИМЕНЕНИЕ ПОЛИНУКЛЕОТИДА ДЛЯ ПОЛУЧЕНИЯ АМИНОКИСЛОТЫ | 2021 |
|
RU2815942C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ОРНИТИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, СВЕРХЭКСПРЕССИРУЮЩИХ LYSE | 2011 |
|
RU2571932C2 |
| ШТАММ PIMELOBACTER SIMPLEX, ПРОЯВЛЯЮЩИЙ СТЕРОИД-1,2-ДЕГИДРОГЕНАЗНУЮ АКТИВНОСТЬ | 2001 |
|
RU2215038C2 |
Изобретение относится к биотехнологии. Предложена генетически модифицированная бактерия для применения для 1,2-дегидрирования стероидов, способ ее получения и способ получения 1,2-дегидрированного стероида. Изобретение обеспечивает получение стероидов без примеси 20-ОН-побочного продукта. 3 н. и 9 з.п. ф-лы, 3 ил., 1 пр.
1. Генетически модифицированная бактерия для применения для 1,2-дегидрирования стероидов, где генетически модифицированная бактерия лишена активности 20-бета-гидроксистероид-дегидрогеназы (20β-HSDH) или имеет сниженную активность 20β-HSDH вследствие генетической модификации по сравнению с соответствующей немодифицированной бактерией, где генетически модифицированная бактерия обладает активностью Δ1-дегидрогеназы,
где ген, кодирующий указанный 20β-HSDH, инактивирован или удален,
где соответствующая немодифицированная бактерия обладает активностью Δ1-дегидрогеназы и 20-β-дегидрогеназы.
2. Генетически модифицированная бактерия по п. 1, где ген, кодирующий 20β-HSDH, мутирован.
3. Генетически модифицированная бактерия по любому из пп. 1-2, где ген, кодирующий 20β-HSDH, укорочен.
4. Генетически модифицированная бактерия по любому из пп. 1-3, где бактерия принадлежит к семейству Nocardioidaceae.
5. Способ получения генетически модифицированной бактерии по любому из пп. 1-4, включающий:
а) предоставление бактерии, которая обладает активностью Δ1-дегидрогеназы и активностью 20-β-дегидрогеназы; и
b) инактивацию или делецию гена, кодирующего 20β-HSDH.
6. Способ получения 1,2-дегидрированного стероида, включающий:
а) предоставление стероида, который не является 1,2-дегидрированным;
b) контактирование указанного стероида, который не является 1,2-дегидрированным,
с бактерией по любому из пп. 1-4 или
ее экстрактом, где указанный экстракт обладает активностью Δ1-дегидрогеназы и имеет сниженную активность 20-β-дегидрогеназы или лишен такой активности в условиях, достаточных для получения 1,2-дегидрированного стероида.
7. Способ по п. 6, где стероид, который не является 1,2-дегидрированным, представляет собой кортикостероид.
8. Способ по п. 6 или 7 где указанный стероид, который не является 1,2-дегидрированным, представляет собой гидрокортизон, и указанный способ предназначен для получения преднизолона.
9. Способ по любому из пп. 6-8, где указанный способ дополнительно включает выделение и очистку указанного 1,2-дегидрированного стероида.
10. Способ по п. 9, который не включает стадию удаления 20 β-дегидрированного стероида.
11. Способ по любому из пп. 6-10, который дополнительно содержит включение 1,2-дегидрированного стероида в состав фармацевтической композиции.
12. Способ по любому из пп. 6-11, где указанный экстракт получают с помощью клеточного лизиса бактерии по одному из пп. 1-4 и необязательно фракционирования продукта клеточного лизиса для концентрирования Δ1-дегидрогеназной активности или для облегчения очистки Δ1-дегидрогеназной активности.
| COVEY D.F | |||
| et al | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Пневматический водоподъемный аппарат-двигатель | 1917 |
|
SU1986A1 |
| WO 2006124141 A2, 23.11.2006 | |||
| СПОСОБ ПОЛУЧЕНИЯ 1,2-ДЕГИДРОПРОИЗВОДНЫХ 4-ДЕЛЬТА-3-КЕТОСТЕРОИДОВ | 1998 |
|
RU2156302C1 |
| US 3047469 A, 31.07.1962. | |||

