ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0001] Изобретение относится к области биотехнологии, а именно к рекомбинантным генетическим векторам, созданным на базе аденоассоциированного вируса, кодирующим широко нейтрализующие антитела против вируса иммунодефицита человека (ВИЧ-1), а также к иммунобиологическим препаратам на их основе для лечения и профилактики ВИЧ-инфекции за счет длительной персистенции широко нейтрализующих антител против ВИЧ-1 в организме человека.
УРОВЕНЬ ТЕХНИКИ
[0002] Инфекция, вызванная ВИЧ-1, продолжает оставаться неизлечимым заболеванием и является одной из острых проблем здравоохранения. По данным Всемирной организации здравоохранения (ВОЗ) число людей, живущих с ВИЧ-1, в 2022 г. составило 39000000 человек [UN AIDS [Электронный ресурс], https://www.unaids.org/ru/resources/fact-sheet].
[0003] Терапия антиретровирусными препаратами не приводит к излечению, поэтому препараты должны приниматься ежедневно и пожизненно. Длительный прием препаратов вызывает токсические побочные эффекты, снижает приверженность пациентов к терапии, что в свою очередь приводит к развитию лекарственной устойчивости к принимаемым препаратам [WHO [Электронный ресурс], https://www.who.int/news-room/fact-sheets/detail/hiv-drugresistance. (дата обращения 23.11.2021)]. Все это обуславливает необходимость поиска новых подходов к лечению ВИЧ-инфекции.
[0004] Одним из таких подходов может быть использование моноклональных широко нейтрализующих антител против ВИЧ-1 (broadly neutralizing antibodies, bNAbs). Впервые такие антитела, способные нейтрализовать вирусы, принадлежащие к разным субтипам ВИЧ-1, были описаны в 2008 году [Scheid J.F. [и др.]. Broad diversity of neutralizing antibodies isolated from memory В cells in HIV-infected individuals // Nature. 2009. №7238 (458). C. 636-640]. С тех пор список широко нейтрализующих антител существенно увеличился и на сегодняшний день известно уже более 90 различных bNAbs против ВИЧ-1, причем широко нейтрализующие антитела нового поколения являются более активными и способны обеспечивать защиту в отношении широкого спектра вирусов [Glazkova, D.V.; Bogoslovskaya, E.V.; Shipulin, G.A.; Yudin, S.M. «Broadly neutralizing antibodies for the treatment of HIV infection». HIV Infect. Immunosuppr. Disord. 2021, 13, 81-95].
[0005] На настоящий момент проведены исследования, показывающие возможность использования широко нейтрализующих антител для лечения ВИЧ-инфекции. Не менее 10 bNAbs находятся на различных стадиях клинических испытаний, которые показали их высокий потенциал и способность снижать вирусную нагрузку у пациентов [Caskey М. et al. «Antibody 10-1074 suppresses viremia in HIV-1-infected individuals», Nature Medicine. 2017. №2 (23). C. 185-191], [Caskey M., Klein F., Nussenzweig M.C. «Broadly neutralizing anti-HIV-1 monoclonal antibodies in the clinic», Nature Medicine. 2019. №4 (25). C. 547-553].
[0006] Однако были выявлены недостатки такой терапии, а именно формирование устойчивых штаммов вируса при использовании одного антитела и быстрое падение концентрации антител после их внутривенного введения. Появление устойчивых штаммов - ожидаемая проблема, поскольку вирус обладает высокой изменчивостью, и по аналогии с высокоактивной антиретровирусной терапией решением проблемы может стать использование комбинаций антител. В нескольких работах было продемонстрировано in vitro, что для максимальной защиты клеток целесообразно использование комбинаций антител, распознающих разные эпитопы вирусного белка Env. В этом случае увеличивается эффективность и широта нейтрализации [Kochina, Е.А.; Urusov, F.A.; Kruglov, А.А.; Glazkova, D.V.; Shipulin, G.A.; Bogoslovskaya, E.V. «Double and Triple Combinations of Broadly Neutralizing Antibodies Provide Efficient Neutralization of All HIV-1 Strains from the Global Panel». Viruses 2022,14, 1910], [Mendoza, P.; Gruell, H.; Nogueira, L.; Pai, J.A.; Butler, A.L.; Millard, K.; Lehmann, C; Suarez, I.; Oliveira, T.Y.; Lorenzi, C.C.; et al. «Combination therapy with anti-HIV-1 antibodies maintains viral suppression», Nature 2018, 561, 479-484], [Wagh K., Seaman M.S. «Divide and conquer: broadly neutralizing antibody combinations for improved HIV-1 viral coverage», Curr Opin HIV AIDS. 2023 Jul 1; 18(4): 164-170].
[0007] В то же время быстрое снижение концентрации антител в крови потребует частых повторных введений препарата, что усложняет лечение, а с учетом необходимости пожизненной терапии делает его очень дорогостоящим. Кроме того, постепенное падение концентрации вводимых антител, увеличивает вероятность образования лекарственно устойчивых вариантов вируса. Перспективным подходом, позволяющим увеличить продолжительность персистирования антител в организме, является доставка ДНК-последовательностей, кодирующих желаемые антитела с помощью рекомбинантного генетического вектора, полученного на базе аденоассоциированного вируса (Adeno-Associated Virus, AAV) (далее по тексту AAV-вектор), который может обеспечить экспрессию антител in vivo в течение года или больше. Такой подход поможет повысить приверженность к терапии, удешевить лечение и позволит снизить дозу антиретровирусных препаратов или вообще отказаться от их применения, что может решить проблему токсичности антиретровирусной терапии.
РАСКРЫТИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0008] Техническая задача, на решение которой направлено заявляемое изобретение, заключается в разработке иммунобиологического препарата для лечения ВИЧ-инфекции путем пассивной иммунизации с использованием различных комбинаций рекомбинантных генетических векторов, кодирующих разные широко нейтрализующие антитела против ВИЧ-1.
[0009] Технический результат, достигаемый при реализации заявляемого изобретения, заключается в создании иммунобиологического препарата для лечения ВИЧ-инфекции на основе различных комбинаций рекомбинантных генетических векторов на основе аденоассоциированного вируса серотипа 9, кодирующих широко нейтрализующие антитела против ВИЧ-1.
[0010] Заявляемый технический результат достигается за счет использования набора рекомбинантных генетических векторов, кодирующих широко нейтрализующие антитела против ВИЧ, направленные на разные эпитопы вирусного белка Env, для получения иммунобиологического препарата для лечения ВИЧ-инфекции, где в качестве рекомбинантного генетического вектора используется аденоассоциированный вектор серотипа 9.
[0011] Кроме того, в частном случае реализации изобретения, иммунобиологический препарат включает в себя рекомбинантные генетические векторы, кодирующие широко нейтрализующие антитела против ВИЧ-1, выбраны из группы:
i. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074;
ii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10Е8;
iii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело N6;
iv. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело VRC07-523;
v. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело PGDM1400;
[0012] В частном случае реализации изобретения аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело VRC07-523, имеет нуклеотидную последовательность со сниженным содержанием CG динуклеотидов, аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеет нуклеотидную последовательность со сниженным содержанием CG динуклеотидов, аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело PGDM1400, имеющий нуклеотидную последовательность со сниженным содержанием CG динуклеотидов.
[0013] Кроме того, в частном случае реализации изобретения рекомбинантные генетические векторы, кодирующие широко нейтрализующие антитела против ВИЧ-1, имеют следующие нуклеотидные последовательности:
i. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеющий нуклеотидную последовательность SEQ ID NO: l;
ii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10Е8, имеющий нуклеотидную последовательность SEQ ID NO: 2;
iii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело N6, имеющий нуклеотидную последовательность SEQ ID NO: 3;
iv. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело VRC07-523, имеющий нуклеотидную последовательность SEQ ID NO: 4;
v. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеющий нуклеотидную последовательность SEQ ID NO: 5;
vi. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело PGDM1400, имеющий нуклеотидную последовательность SEQ ID NO: 6;
[0014] Также технический результат достигается за счет того, что иммунобиологический препарат для лечения и профилактики ВИЧ-инфекции, содержит комбинацию нескольких аденоассоциированных векторов серотипа 9, выбранных из приведенного выше набора векторов.
[0015] Кроме того, в частном случае реализации изобретения иммунобиологический препарат содержит комбинацию из аденоассоциированных векторов серотипа 9 (i), (ii), (iii) или комбинацию из аденоассоциированных векторов серотипа 9 (iv), (v), (vi).
[0016] Кроме того, в частном случае реализации изобретения иммунобиологический препарат представляет собой смесь аденоассоциированных векторов серотипа 9, входящих в комбинацию, при эквимолярном соотношении компонентов или набор отдельных аденоассоциированных векторов серотипа 9, входящих в комбинацию.
[0017] Также технический результат достигается за счет того, что способ применения иммунобиологического препарата характеризуется тем, что препарат вводят внутримышечно в виде смеси аденоассоциированных векторов серотипа 9 или каждый аденоассоциированный вектор серотипа 9 вектор, входящий в состав комбинации, вводят по отдельности.
[0018] Использование комбинации из нескольких AAV-векторов, кодирующих широко нейтрализующие антитела, направленные на разные эпитопы поверхностного белка Env ВИЧ-1, позволяет с одной стороны нейтрализовать все многообразие штаммов вируса, с другой стороны может предотвратить появление устойчивых вариантов ВИЧ-1.
[0019] Механизм действия препарата основан на том, что после введения AAV-векторов внутримышечно, AAV-векторы проникают в клетки, где происходит экспрессия трансгенов, кодирующих широко нейтрализующие антитела, и секреция данных антител в кровоток. В результате в организме начинают циркулировать антитела против ВИЧ-1 в концентрациях, достаточных для нейтрализации вируса. Применяемый AAV-вектор серотипа 9 способен обеспечить активную длительную экспрессию антител. Безопасность и эффективность AAV-векторов была неоднократно продемонстрирована в клинических испытаниях [Машуа, S., Sarangi, Р. & Jayandharan, G.R. «Safety of Adeno-associated virus-based vector-mediated gene therapy-impact of vector dose». Cancer Gene Ther 29, 1305-1306 (2022)].
[0020] Используемые в рамках настоящего изобретения широко нейтрализующие антитела против ВИЧ, а именно антитела 10-1074, VRC07-253, N6, 10Е8 и PGDM1400 описаны ранее, их аминокислотные последовательности известны. Например, антитело 10-1074 в виде белкового препарата моноклонального антитела было изучено в клиническом исследовании [Cohen Y.Z., Butler A.L., Millard K., et al. «Safety, pharmacokinetics, and immunogenicity of the combination of the broadly neutralizing anti-HIV-1 antibodies 3BNC117 and 10-1074 in healthy adults: A randomized, phase 1 study», PLoS One. 2019; 14(8): e0219142. Published 2019 Aug 8].
[0021] Антитело VRC07-253 было также исследовано в клинических испытаниях. Показана безопасность и хорошая переносимость у пациентов, получавших внутривенные или подкожные инъекции препарата вышеуказанного антитела [Gaudinski M.R. et al, «Safety and pharmacokinetics of broadly neutralising human monoclonal antibody VRC07-523LS in healthy adults: a phase 1 dose-escalation clinical trial». Lancet HIV. 2019 Oct; 6(10): e667-e679].
[0022] Антитело N6 характеризуется максимальной широтой действия, что выгодно отличает его от других широко нейтрализующих антител против ВИЧ-1 [Sok D., Burton D.R. «HIV Broadly Neutralizing Antibodies: Taking Good Care Of The 98», Immunity. 2016 Nov 15; 45(5): 958-960].
[0023] Использование AAV-векторов для экспрессии широко нейтрализующих антител против ВИЧ-1 также известно, однако для этой цели использовали AAV-векторы серотипа 1 и 8. При введении широко нейтрализующего антитела против ВИЧ PG9 с помощью AAV-вектора серотипа 1 антитела в крови не обнаруживали [Priddy F.H., Lewis D.J.M., Gelderblom Н.С., et al. «Adeno-associated virus vectored immunoprophylaxis to prevent HIV in healthy adults: a phase 1 randomised controlled trial», Lancet HIV. 2019 Apr 3].
[0024] При введении антитела VRC07-253 с помощью AAV серотипа 8 концентрация в крови была низкой [Casazza J.P., Cale Е.М., Narpala S., et al. «Safety and tolerability of AAV8 delivery of a broadly neutralizing antibody in adults living with HIV: a phase 1, dose-escalation trial», Nat Med. 2022; 28(5): 1022-1030].
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
[0025] Реализация заявляемого изобретения, подтверждается приведенными ниже примерами, но не ограничивается ими.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0026] Для пояснений сущности изобретения используются графические материалы, на которых изображены:
фиг. 1 - схема конструкции плазмиды, предназначенной для сборки AAV-вектора, кодирующего IgG;
фиг. 2 - графики концентрации антител (мкг/мл) в сыворотке мышей через 4 недели после введения препарата (Д - доза, М - номер мыши);
фиг. 3 - концентрации IgG в сыворотках мышей спустя 4, 8 и 12 недель после введения препарата «КомбиМаб1», мкг/мл (точки соответствуют индивидуальным значениям; пороговое значение чувствительности использованного ИФА набора - 0,01 мкг/мл);
фиг. 4 - графики нейтрализующей активности пула сывороток мышей, получивших препарат «КомбиМаб1», против 11 псевдовирусов глобальной панели ВИЧ-1;
фиг. 5 - концентрации IgG в сыворотках мышей через различные промежутки времени (до 22 месяцев) после введения препарата «КомбиМаб1», мкг/мл (точки соответствуют индивидуальным значениям);
фиг. 6 - графики концентрации IgG в сыворотках мышей (мкг/мл) через 2, 4, 6, 8 недель (2W, 4W, 6W, 8W) после введения препарата «КомбиМаб1» мышам C57BL/6. **р0,01 (0,0079); 1 site - введение в одну лапу, 3 site - введение в разные лапы;
фиг. 7 - графики нейтрализации псевдовирусов глобальной панели ВИЧ-1 пулами сывороток, полученных от мышей, которым проводилось введение препарата «КомбиМаб1» в одну лапу;
фиг. 8 - графики нейтрализации псевдовирусов глобальной панели ВИЧ-1 пулами сывороток, полученных от мышей, которым проводилось введение препарата «КомбиМаб1» в три лапы;
фиг. 9 - концентрации человеческого антитела 10-1074 в крови лабораторных животных через 2, 4, 6 и 8 недель после введения вектора AAV-10-1074 или вектора AAV-10-1074CpG-low. - Р<0,05 по критерию Манна-Уитни;
фиг. 10 -концентрация IgG человека в сыворотках мышей (мкг/мл) через 4 недели после введения мышам векторов AAV-VRC-07CpG-low, AAV-PGDM1400CpG-low и AAV-10-1074CpG-low или их смеси (препарат «КомбиМаб2») в однократной и в тройной дозе (линии указывают средние значения для n=8);
фиг. 11 - графики нейтрализующей активности пула сывороток мышей С57 Black, получивших смесь AAV-векторов, AAV-9-VRC07-523CpG-low, AAV-9-10-1074CpG-low и AAV-9-PGDM1400CpG-low, против 11 вирусов глобальной панели ВИЧ-1.
[0027] На поверхности белка Env ВИЧ-1 известно пять основных эпитопов, антитела к которым могут обладать широким спектром действия [McCoy L.E., Burton D.R. «Identification and specificity of broadly neutralizing antibodies against HIV», Immunol Rev. 2017 Jan; 275(1): 11-20]:
1) Сайт связывания CD4 (CD4bs, CD4-binding site);
2) Вариабельная область V3 (V3 loop) и сопутствующий комплекс гликанов;
3) Прилегающая к мембране внешняя область gp41 (MPER, membrane proximal external region);
4) Присутствующий только в составе тримера gp120 эпитоп, образованный областями V1 и V2, расположенный на верхушке Env (V1/V2 loop);
5) Эпитоп, образующийся в области контакта (интерфейса) между gp120 и gp41.
[0028] Наибольшее число обнаруженных и хорошо изученных антител против ВИЧ-1 - это антитела к первым 4-м эпитопам.
[0029] Вариабельность нуклеотидных последовательностей белка Env между штаммами вируса различных субтипов достигает 30%. Однако, не все области одинаково вариабельны. В структуре белка gp120 выделяют пять консервативных (С1-С5) и пять вариабельных областей (V1-V5) Консервативные области располагаются преимущественно внутри белка Env, в то время как вариабельные области образуют петли на поверхности белка Env [Merk A., Subramaniam S. «HIV-1 envelope glycoprotein structure», Curr Opin Struct Biol. 2013 Apr; 23(2): 268-76], [Klasse P.J., Ozorowski G., Sanders R.W., Moore J.P., «Env Exceptionalism: Why Are HIV-1 Env Glycoproteins Atypical Immunogens?», Cell Host Microbe. 2020 Apr 8; 27(4): 507-518].
[0030] Функционально важные и высоко консервативные структуры располагаются в труднодоступных местах четвертичной структуры Env. Сайт связывания CD4 (CD4bs), являющийся единственным полностью пептидным эпитопом Env, окружен по периметру гликанами (в частности, связанными с петлей V5), формирующими своего рода воронку. Это окружение уменьшает доступную для связывания поверхность эпитопа, таким образом, препятствуя одновременному связыванию этого сайта двумя вариабельными доменами антитела - легкой и тяжелой цепей. Однако, этой площади вполне достаточно для контакта с рецептором CD4.
[0031] Антитела, направленные против сайта связывания CD4 (CD4bs) на gp120 и эпитопа MPER в проксимальной области мембраны gp41, демонстрируют наибольшую широту и активность. Некоторые антитела против CD4bs нейтрализуют все вирусы из стандартных панелей вирусов ВИЧ-1 [deCamp A. et al. «Global panel of HIV-1 Env reference strains for standardized assessments of vaccine-elicited neutralizing antibodies», Journal of virology. - 2014. - T. 88. - №. 5. - C. 2489-2507]. Несколько меньшая широта нейтрализации наблюдается у антител против эпитопов, содержащих гликаны в петле gp120 V1/V2, петле gp120 V3 и интерфейсе gp120-gp41, при этом эти bNAb имеют большую активность по сравнению с антителами, обладающими большей широтой нейтрализации.
[0032] В рамках заявляемого изобретения для формирования комбинации AAV-векторов были выбраны следующие широко нейтрализующие антитела против ВИЧ: N6, 10-1074, 10Е8, VRC07-523, PGDM1400. В табл. 1 указаны эпитопы белка Env, с которыми связываются данные антитела.

I. ДИЗАЙН ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ПЛАЗМИДНЫХ ВЕКТОРОВ, КОДИРУЮЩИХ АНТИТЕЛА
[0033] В качестве основы для создания плазмидных векторов, кодирующих выбранные широко нейтрализующие антитела против ВИЧ-1 (N6, VRC07-523, 10-1074, PGDM1400, 10Е8), была использована плазмида pAAV-GFP (CellBioLabs, Inc., каталожный номер AAV-400), в которую были добавлены нуклеотидные последовательности, кодирующие легкую и тяжелую цепь указанных антител, и регулирующие экспрессию элементы. На фиг. 1 представлена универсальная схема плазмиды, кодирующей IgG антитело и предназначенной для получения AAV-вектора, согласно которой конструировались все плазмидные ДНК для выбранных антител.
[0034] В структуре экспрессионной кассеты последовательно расположены следующие элементы:
1) Гибридный промотор CMV-EF1α, обеспечивающий длительную и стабильную экспрессию трансгена в клетках эукариот;
2) Транслируемая часть, объединяющая в одной рамке считывания стартовый кодон ATG (в контексте последовательности Козак GCCGCCACC), последовательность сигнального пептида человеческого гормона роста (HGH сигнал) для обеспечения секреции тяжелой цепи антител, тяжелая цепь иммуноглобулина (вариабельная -HEAVY var) и константная часть (HEAVY const), сайт разрезания фурина, пептид GT2A, последовательность сигнального пептида человеческого гормона роста легкой цепи (HGH сигнал), легкая цепь иммуноглобулина (вариабельная - LIGHT var) и константная часть (LIGHT const), дублированный стоп-кодон TGA. Согласно предложенной конфигурации кассеты, тяжелая и легкая цепь антител находятся в одной рамке считывания и между ними находятся пептид GT2A, обеспечивающий разделение белковой цепочки в процессе трансляции, и сайт, разрезаемый протеазой фурин. Таким образом, в результате процессинга единой полипептидной цепи получаются отдельные тяжелая и легкая цепь антител, которые затем объединяются в функционально активное антитело. Для обеспечения секреции антитела, к каждой из цепей непосредственно к N-концу вариабельного домена добавлена последовательность сигнального пептида человеческого гормона роста (HGH-сигнал). Аминокислотные последовательности HGH-сигнала у тяжелой и легкой цепи антител одинаковы, однако кодирующие их нуклеотидные последовательности сигнального пептида отличаются друг от друга для уменьшения вероятности рекомбинационных событий в пределах вектора.
3) Последовательность посттранскрипционного регуляторного элемента вируса гепатита В сурков - WPRE (Woodchuck Posttranscriptional Regulatory Element), необходимая для повышения стабильности мРНК и усиления экспрессии трансгена.
4) Сигнал полиаденилирования человеческого гормона роста (PolyA сигнал).
5) Энхансер SV40 для усиления предшествующего сигнала полиаденилирования (SV40 энхнасер).
Все элементы фланкированы инвертированными терминальными повторами (ITR), необходимыми для сборки AAV-вектора.
[0035] На первом этапе на основе плазмиды pAAV-GFP (Cell Biolabs, США) был собран плазмидный вектор pAAVcore. Для этого фрагмент вектора pAAV-GFP/NotI, содержащий последовательности ITR AAV2 и бактериальную часть, соединили со следующими фрагментами:
1) фрагмент, кодирующий промотор CMV_EF1A, амплифицированный с плазмиды pD2610-v3 (Atum.bio),
2) интрон Ubc, амплифицированный с геномной ДНК человека с праймерами cagaacacaggtaagtccggcctccgcgccgggttttgg и ATTATAGATCTGGTGGCGGCATCGATGCTGTTCGTCACCCAGGACCTG,
3) последовательность WPRE, амплифицированная с плазмиды AAV Ef1a-EGFP-WPRE (Addgene)
4) сигнал полиаденилирования вируса SV40, амплифицированная с плазмиды pD2610-v3 (Atum.bio).
[0036] Между интроном Ubc и последовательностью WPRE были встроены сайты рестрикции ClaI, XbaI, BglII для удобства последующих стадий клонирования.
[0037] Фрагменты, кодирующие тяжелую и легкую цепь антител N6, 10Е8, 10-1074, амплифицировали из плазмид, описанных в работе [Kochina, Е.А.; Urusov, F.A.; Kruglov, А.А.; Glazkova, D.V.; Shipulin, G.A.; Bogoslovskaya, E.V. «Double and Triple Combinations of Broadly Neutralizing Antibodies Provide Efficient Neutralization of All HIV-1 Strains from the Global Panel», Viruses 2022, 14, 1910], добавляя необходимые сайты рестрикции. Также с помощью праймеров в начало последовательности легкой цепи была добавлена последовательность, кодирующая 2А-пептид и сайт узнавания протеазы фурин. В плазмидный вектор pAAVcore по сайтам ClaI-XbaI одновременно вставляли два фрагмента: последовательность, кодирующую тяжелую цепь, фланкированную сайтами ClaI-BamHI, и последовательность, кодирующую легкую цепь, фланкированную сайтами BamHI-XbaI. Корректность полученных плазмид подтверждали секвенированием.
[0038] На основании вышеприведенной универсальной схемы были сконструированы следующие плазмиды, кодирующие широко нейтрализующее антитела против ВИЧ-1 и предназначенные для получения AAV-векторов:
Плазмидный вектор, кодирующий широко нейтрализующее антитело 10-1074;
Плазмидный вектор, кодирующий широко нейтрализующее антитело 10Е8;
Плазмидный вектор, кодирующий широко нейтрализующее антитело N6.
II. НАРАБОТКА AAV-ВЕКТОРОВ
1. Получение AAV-частиц в клетках-продуцентах
[0039] Для наработки AAV-векторов (вирусных частиц) использовали клеточную линию HEK293FT (Invitrogen, США).
[0040] Линию HEK293T культивировали в среде DMEM (Thermo Scientific, США) + 10% эмбриональной телячьей сыворотки (FBS) (Gibco, США) + 1% пенициллин/стрептомицин (Thermo Scientific, США) с пассированием один 1 раз в 2 суток при достижении 70-80% конфлюэнтности. За сутки до трансфекции клетки высаживали в количестве 70000-75000 клеток на см2 площади культурального флакона. За 2-4 часа до трансфекции проводилась замены среды на свежую среду DMEM + 2,5% FBS + 1% пенициллин/стрептомицин. Трансфекция культуры проводилась 3 (тремя) плазмидами в эквимолярном соотношении: 1) плазмидным вектором, кодирующим антитела 2) плазмидой pAAV-Helper (Cell Biolabs, США), 3) плазмидой pAAV-RC-9 (Cell Biolabs, США), кодирующей капсидные белки, из расчета 0,35 мкг ДНК на см2 площади флакона. В качестве трансфицирующего агента использовался полиэтиленимин PEI MAX 40К (Polyscience, Франция) в соотношении 1 мкг дезоксирибонуклеиновой кислоты (ДНК) на 3 мкг полиэтиленимина (ПЭИ). Приготовление трансфекционной смеси проводилось из расчета 5% от финального объема наработки в среде DMEM. После смешивания ДНК и ПЭИ, полученный раствор инкубировали в течение 10 минут. Трансфекционную смесь добавляли в культуральные флаконы с HEK-293FT. Через 24 ч после трансфекции ростовую среду меняли на бессывороточную среду OptiMEM (Gibco) и инкубировали при 37°С и 5% СО2 в течение 48 ч.
[0041] Через 48 часов клетки снимались с поверхности флакона, центрифугировались 5 мин при 2000g. Полученный клеточный осадок, растворенный в однократном фосфатно-солевом буфере (PBS) (Thermo Scientific, США) с 2 мМ MgCl2, подвергался 3 (трем) циклам заморозки и разморозки при температурах -80°С и 37°С. Клеточный лизат обрабатывали бензоназой в концентрации 50 ед/мл (Merck, Германия) при 37°С в течение 30 мин и осветляли центрифугированием при 2300g в течение 10 мин. Полученный супернатант фильтровали через мембраны с размером пор 0.45 мкм (ТРР).
2. Очистка AAV-векторов в градиенте йодиксанола
[0042] Протокол для очистки AAV-векторов с помощью ультрацентрифугирования в градиенте йодиксанола основан на протоколе, описанном в книге Fredric Manfredsson «Gene therapy for neurological disorders. Methods and protocols», Springer Science + Business Media New York 2016.
[0043] Растворы с разными концентрациями йодиксанола готовили в стерильных условиях разведением медицинского контрастирующего препарата «ВИЗИПАК» (GE Healthcare, Норвегия) в пропорциях, указанных в табл. 2. Буферный раствор 5Х TD состоял из 5-кратного раствора PBS (Gibco, США), 5 мМ MgCl2 (Хеликон, Россия) и 12,5 мМ KCl (Хеликон, Россия).
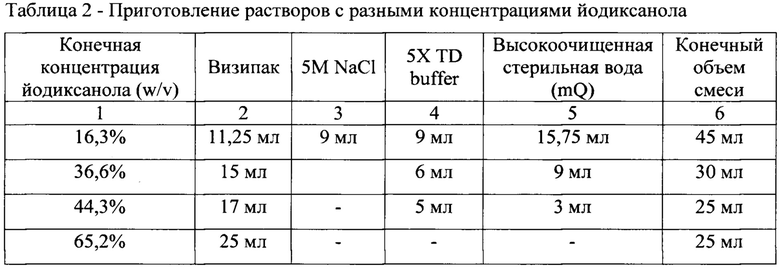
[0044] С использованием шприца с иглой Зельдингера (М. Schilling Gmbh, Германия), опущенной до дна пробирки, последовательно наслаивали в центрифужные пробирки Beckman (Beckman Coulter Life Sciences, США) 25.4×82.3 мм следующие растворы: 9 мл осветленного лизата клеток-продуцентов, обработанного бензоназой (Merck Millipore, Германия), затем 9 мл 16,3% раствора йодиксанола, далее 6 мл 36,6% раствора йодиксанола, далее 5 мл 44,3% раствора йодиксанола, далее 5 мл 65,2% раствора йодиксанола. Таким образом, каждый следующий, более плотный слой, вытеснял предыдущий слой вверх. Пробирки уравновешивали с помощью аналитических весов, и запаивали. Далее пробирки центрифугировали в ультрацентрифуге при 18°С с ускорением не менее 174000 g в течение 2 часов. После центрифугирования со дна ультрацентрифужной пробирки сначала отбирали 5 мл «пустой» фракции, а затем в отдельную пробирку 11 мл фракции, содержащей AAV-частицы.
3. Определение физического титра AAV-векторов
[0045] Для определения физического титра векторов AAV использовали метод капельной цифровой ПЦР (кцПЦР, droplet digital PCR (ddPCR). Последовательности праймеров и зонда были комплементарны последовательностям ITR AAV (табл. 3).


[0046] ДНК выделяли из образцов AAV-векторов объемом 100 мкл (предварительно разведенного в 106 раз препарата) с использованием набора реагентов для выделения РНК/ДНК «АмплиТест РИБО-преп» (ФГБУ «ЦСП» ФМБА России).
[0047] Для проведения ddPCR использовали систему с автоматической загрузкой образцов в генератор капель QX200™ AutoDG™ (BioRad) и амплификатор С1000 Touch Thermal Cycler (Bio-Rad). Для проведения анализа использовали набор реагентов ddPCR™ для подготовки образцов с зондами TaqMan QX100/QX200™ (Bio-Rad).
III. ВЫБОР ОПТИМАЛЬНОГО КАПСИДА AAV-ВЕКТОРА ДЛЯ IN VIVO ДОСТАВКИ ДНК ПОСЛЕДОВАТЕЛЬНОСТЕЙ АНТИТЕЛ
[0048] Для эффективной и безопасной экспрессии антител in vivo необходимо обеспечить доставку последовательностей, кодирующих антитела, с помощью AAV-вектора. После внутримышечного введения данного вектора должны наблюдаться:
1) Продолжительная и высокая экспрессия антител in vivo;
2) Максимальная концентрация вектора в месте введения;
3) Ограниченное распределение вектора в другие ткани.
[0049] Биораспределение и эффективность трансдукции AAV-векторов при введении в организм в большой степени определяется структурой капсида (серотипа AAV). Поэтому мы исследовали доставку антитела с помощью разных капсидов - 8, 9 и DJ. Для этого были наработаны AAV-частицы, несущие ДНК-последовательности, кодирующие антитело 10-1074 с использованием разных упаковочных плазмид (pAAV-RC-9, pAAV-RC-8, pAAV-RC-DJ).
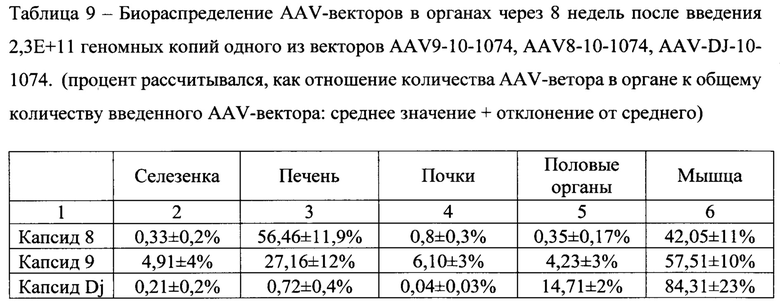
[0050] Полученные AAV-векторы (AAV9-10-1074, AAV8-10-1074, AAV-DJ-10-1074) вводили мышам Balb/c внутримышечно в двух дозах Д1 (2,3Е + 11 вирусных геномов на мышь) и Д2 (5,0Е + 10 вирусных геномов на мышь) в объеме 50 мкл/мышь. Животные контрольной группы получали контрольное вещество - 0,9% NaCl.
[0051] Далее была проведена оценка способности трех вариантов AAV-частиц (AAV9-10-1074, AAV8-10-1074, AAV-DJ-10-1074) обеспечивать экспрессию антитела in vivo. Оценивая концентрацию антител в сыворотке мышей, было установлено, что AAV-векторы на основе 8 и 9 серотипа и DJ-капсида способны доставлять в организм трансгены, кодирующие антитела, но эффективность их экспрессии в каждом случае разная (фиг. 2). Наибольшая концентрация антител наблюдалась при введении AAV-вектора на основе капсидов 9 и DJ. Для этих векторов прослеживается прямая зависимость от дозы: при увеличении дозы повышается концентрация антител.
[0052] Помимо эффективности экспрессии трансгена изучаемые векторы продемонстрировали различия в биораспределении, которое оценивалось методом ПЦР по количеству ДНК AAV-вектора в образцах ДНК, выделенных из органов животных.
[0053] Для изучения биораспределения через 8 недель после введения препарата животных умерщвляли методом кардиальной пункции и производили отбор крови и органов (печень, селезенка, почки, матка и мышца - место инъекции). Образцы тканей взвешивали, гомогенизировали с использованием жидкого азота в фарфоровой ступке.
[0054] ДНК из гомогенизированных органов выделяли с помощью набора «ДНК-Сорб-С-М» (ФБУН ЦНИИ Эпидемиологии Роспотребнадзора) согласно инструкции производителя.
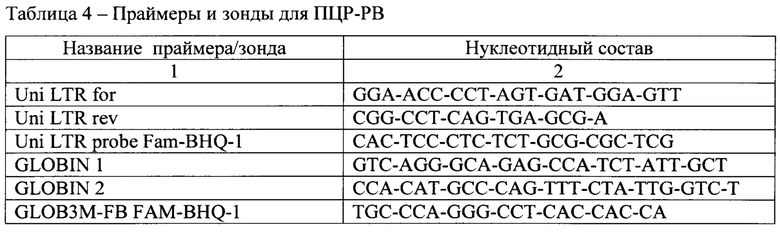
[0055] Полученную ДНК разводили в 3 раза. Содержание вектора в тканях и органах определяли методом ПЦР в реальном времени (ПЦР-РВ). В качестве внутреннего контроля ПЦР и для нормализации числа копий вектора на число клеток использовали ген глобина. Праймеры и зонды приведены в табл. 4.
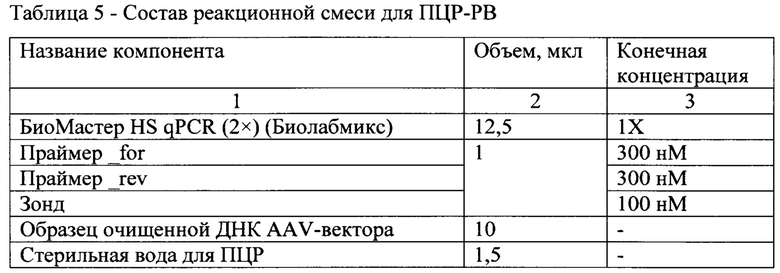
[0056] Смесь для ПЦР-РВ готовили согласно табл. 5.
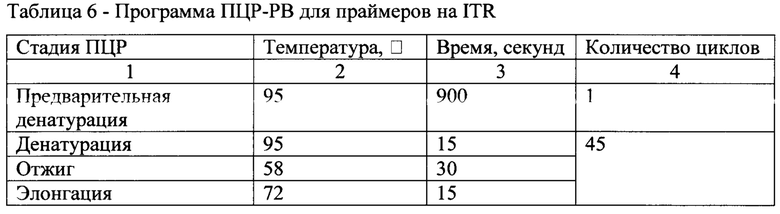
[0057] ПЦР-РВ осуществляли с использованием прибора RotorGene Q согласно следующей программам, описанным в табл. 6, 7.
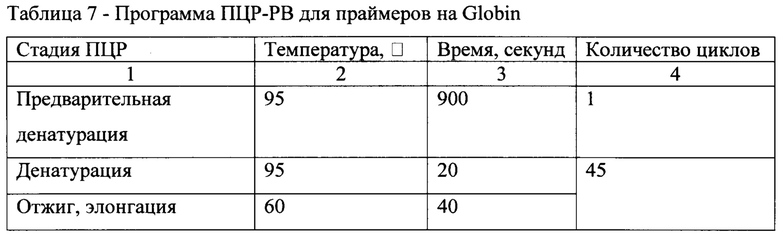
[0058] Измерение флуоресценции проводили на стадии отжига праймеров. Для построения калибровочной кривой и последующего расчета концентрации ДНК-фрагментов AAV-вектора и ампликонов гена глобина использовали плазмиды, содержащие последовательности ITR AAV и Globin, в которых концентрация ДНК была измерена методом цифровой ПЦР (Digital PCR) с использованием системы с автоматической загрузкой образцов в генератор капель QX200™ AutoDG™ (BioRad) и амплификатора С1000 Touch Thermal Cycler (Bio-Rad) с использованием праймеров и зондов к гену устойчивости к ампицилину (табл. 8).

[0059] По данным проведенного ПЦР-анализа было установлено, что AAV-векторы на основе капсида DJ в большей степени остаются в месте введения, т.е. в мышце. AAV-векторы с капсидом 9 имеет иное распределение: примерно 2/3 приходится на мышцу и 1/3 попадает в печень. AAV-вектор с капсидом 8 почти равномерно распределяется между печенью и мышцей (табл. 9).
[0060] На основании данных проведенных экспериментов был выбран капсид 9, который оказался способен обеспечить максимальный уровень экспрессии антител in vivo.
IV. ПОЛУЧЕНИЕ КОМБИНАЦИИ AAV-ВЕКТОРОВ СЕРОТИПА 9, КОДИРУЮЩИХ ШИРОКО НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА ПРОТИВ ВИЧ
[0061] Предварительно был наработан каждый AAV-вектор 9 серотипа (далее по тексту - AAV-9), как описано в разделе II: AAV-вектор 9 серотипа, кодирующий широко нейтрализующее антитело против ВИЧ-1 10-1074 (SEQ ID NO.T), AAV-вектор 9 серотипа, кодирующий широко нейтрализующее антитело против ВИЧ-1 10Е8 (SEQ ID NO: 2) и AAV-вектор 9 серотипа, кодирующий широко нейтрализующее антитело N6 против ВИЧ-1 (SEQ ID NO: 3).
[0062] Была подготовлена смесь, включающая следующие AAV-векторы 9 серотипа: AAV-вектор 9 серотипа, кодирующий широко нейтрализующее антитело против ВИЧ-1 10-1074 (SEQ ID NO: l), AAV-вектор 9 серотипа, кодирующий широко нейтрализующее антитело против ВИЧ 10Е8 (SEQ ID NO: 2) и AAV-вектор 9 серотипа, кодирующий широко нейтрализующее антитело N6 против ВИЧ-1 (SEQ ID NO: 3) в соотношении 1:1:1 каждого вектора (далее по тексту - препарат «КомбиМаб1»).
[0063] Смесь вышеприведенных векторов AAV-9 была подготовлена простым смешением суспензий векторов, входящих в их состав.
[0064] Также в одном из вариантов реализации изобретения может быть подготовлен набор AAV-векторов 9 серотипа (SEQ ID NO: 1), (SEQ ID NO: 2), (SEQ ID NO: 3), где каждый вектор может вводиться по отдельности.
V. ОЦЕНКА ЭФФЕКТИВНОСТИ КОМБИНАЦИЙ ИЗ AAV - ВЕКТОРОВ 9 СЕРОТИПА, КОДИРУЮЩИЕ РАЗНЫЕ АНТИТЕЛА НА МЫШАХ IN VIVO
1. Оценка эффективности комбинации из AAV-векторов 9 серотипа, кодирующих антитела N6, 10-1074 и 10Е8, на мышах линии CBAxC57Black
[0065] Было проведено исследование по оценке специфической активности комбинации AAV-9 векторов, кодирующих широко нейтрализующие антитела против ВИЧ - N6, 10-1074 и 10Е8 (далее по тексту - препарат «КомбиМаб1»). Для этого мышам линии СВАхС57 В1аск вводили однократно дозу 2×1011 геномных копий каждого AAV-вектора в объеме 100 мкл. Через 4, 8 и 12 недель после введения препарата «КомбиМаб1» была измерена концентрация IgG в индивидуальных образцах сыворотки крови мышей в группе «КомбиМаб1» и в группе контроля (фиг. 3).
[0066] Результаты измерения показали, что средняя концентрация человеческих антител в сыворотке крови мышей составляла от 2,4 до 3,7 мкг/мл в разное время после введения смеси антител, при этом экспрессия оказалась весьма неоднородной, наблюдались большие различия в концентрации антител от 0,1 до 16 мкг/мл, в сыворотках некоторых животных концентрация антител была ниже уровня детекции.
[0067] Для оценки нейтрализующей активности сывороток, полученных от животных после введения «КомбиМаб1», были использованы 11 (одиннадцать) псевдовирусов из глобальной панели ВИЧ-1 [De Camp, А.; Hraber, P.; Bailer, R.T.; Seaman, M.S.; Ochsenbauer, C; Kappes, J.; Gottardo, R.; Edlefsen, P.; Self, S.; Tang, H.; et al. «Global Panel of HIV-1 Env Reference Strains for Standardized Assessments of Vaccine-Elicited Neutralizing Antibodies», J. Virol. 2014,88, 2489-2507]. Считается, что варианты гликопротеинов Env глобальной панели представляют основные генетические варианты вируса. Способность антитела (или комбинации антител) нейтрализовать псевдовирусы глобальной панели отражает его потенциальную широту действия в отношении большинства циркулирующих в мире штаммов ВИЧ-1. Таким образом, глобальная панель позволяет сократить число вирусов в исследовании широты антител.
[0068] Для получения псевдовирусных частиц ВИЧ-1 клетки HEK293FT засевали в культуральный флакон площадью 75 см2 и трансфицировали смесью плазмид, содержащей вектор pSG3ΔENV (6,9 мкг) и плазмиду, кодирующую соответствующий белок Env из глобальной панели вирусов ВИЧ-1, в количестве 3,5 мкг. Глобальная панель (cat# 12670) была получена в рамках NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH, from Dr. David Montefiori. Для трансфекции использовали линейный полиэтиленимин (PEI МАХ 40000; Polysciences Inc., США). Соотношение ДНК:РЕ1 составляло 1:3. Процедура трансфекции и сбора клеточного супернатанта, содержащего псевдовирусные частицы, описана в статье [Omelchenko, D.O.; Glazkova, D.V.; Bogoslovskaya, E.V.; Urusov, F.A.; Zhogina, Y.A.; Tsyganova, G.M.; Shipulin, G.A. «Protection of Lymphocytes Against HIV using Lentivirus Vector Carrying a Combination of TRIM5α-HRH Genes and microRNA Against CCR5», Mol. Biol. 2018, 52, 251-261]. После сбора клеточный супернатант объемом 15 мл концентрировали до 1,5 мл с помощью Amicon Ultra-15 (100K) (Merck Millipore, Германия). Концентрировнный раствор псевдовирусных частиц хранили в виде малых аликвот (50 мкл) при температуре минус 70°С.
[0069] Титровку псевдовирусов проводили на клетках TZM-bl в 96-луночных планшетах по протоколу, описанному в статье [Sarzotti-Kelsoe, М.; Bailer, R.T.; Turk, Е.; Lin, С.; Bilska, М.; Greene, K.М.; Gao, Н.; Todd, С.А.; Ozaki, D.A.; Seaman, M.S.; et al. «Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1», J. Immunol. Methods 2014, 409, 131-146]. В лунки добавляли по 100 мкл среды DMEM (Gibco, США). Далее вносили по 25 мкл титруемого вируса и делали пятикратные разведения. Сверху наносили суспензию клеток TZM-bl в количестве 10000 клеток на лунку в полной среде DMEM (Gibco, США) + 10% FBS (Gibco, США) + 25 мМ Hepes (Gibco, США) с полибреном (Sigma, Германия) в концентрации 5 мкг/мл. Через 24 часа после трансдукции среду меняли на полную среду DMEM (Gibco, США) + 10% FBS (Gibco, США) + 25 мМ Hepes (Gibco, США). Через 72 часа после трансдукции из лунок удаляли 100 мкл среды и сверху добавляли 100 мкл реагента Bright-Glow (Promega, США). Результат оценивали с помощью люминометра Тесап Infinite 200 Pro.
[0070] Реакцию нейтрализации проводили в 96-луночных планшетах. В лунки добавляли по 100 мкл среды DMEM (Gibco, США). Далее в лунки вносили исследуемые антитела в объеме 20 мкл в трех повторах каждое и делали пятикратные разведения, сверху добавляли псевдовирус в количестве 200 TCID50 на лунку. Инкубировали полтора часа при температуре 37°С. Сверху наносили суспензию клеток TZM-bl в количестве 10000 клеток на лунку в полной среде DMEM (Gibco, США) + 10% FBS (Gibco, США) + 25 мМ Hepes (Gibco, США) с полибреном (Sigma, Германия) в концентрации 5 мкг/мл. Через 24 часа после трансдукции среду меняли на полную среду DMEM (Gibco, США) + 10% FBS (Gibco, США) + 25 мМ Hepes (Gibco, США). Через 72 часа после трансдукции из лунок удаляли 100 мкл среды и сверху добавляли 100 мкл реагента Bright-Glow (Promega, США). Результат оценивали с помощью люминометра Tecan Infinite 200 Pro.
[0071] Расчет TCID50 проводился в программе Excel по руководству, представленному лабораторией Montefiori DC [Montefiori, D.C. «Measuring HIV Neutralization in a Luciferase Reporter Gene Assay», Methods Mol. Biol. 2009, 485, 395-405]. Значения ID50 (50% inhibitory dilution) и ID80 были рассчитаны с помощью нелинейной регрессионной модели, основанной на зависимости процента нейтрализации от разведения сывороток в логарифмической шкале. Значения IC50 (50% inhibitory concentration) и IC80 определяли с использованием количественного определения общего человеческого IgG в образцах сывороток и ID50 и ID80. Статистически значимыми считали различия между показателями, если параметр р не превышал 0,05.
[0072] Для всех вирусов глобальной панели ВИЧ-1 проводили реакцию нейтрализации, используя пятикратные разведения пула сывороток, в которых обнаруживались IgG по данным ИФА. Кривые зависимости уровня нейтрализации от разведения пула сывороток для разных псевдовирусов ВИЧ-1 приведены на фиг. 4.
[0073] На фиг. 4 можно видеть, что сыворотки в разведении от 1:5 до 1:125 успешно нейтрализовали все изученные псевдовирусы ВИЧ-1. Наиболее «сложным» для нейтрализации оказался вирус BJXO2000 из глобальной панели, тем не менее, сыворотки в разведениях 1:5 и 1:25 были способны нейтрализовать и данный вирус.
[0074] Продемонстрированная высокая нейтрализующая активность сыворотки мышей против псевдовирусов из глобальной панели свидетельствует об эффективности препарата «КомбиМаб1» в отношении широкого спектра вирусов, который отражает глобальная панель.
[0075] В рамках доклинического исследования был проведен анализ длительности персистирования антител в крови животных после введения препарата. Концентрацию антител человека в крови животных определяли через 4, 8, 12 недель, и 9, 10, 12, 18 и 22 месяца после введения смеси векторов в суммарной дозе 6×1011 геномных копий на мышь, (фиг. 5). Средние концентрации антител в сыворотках мышей в разное время после введения AAV-векторов 9 серотипа, кодирующих антитела N6, 10-1074 и 10Е8 представлены в табл. 10. Антитела в крови обнаруживались по крайней мере в течение 22 месяцев.
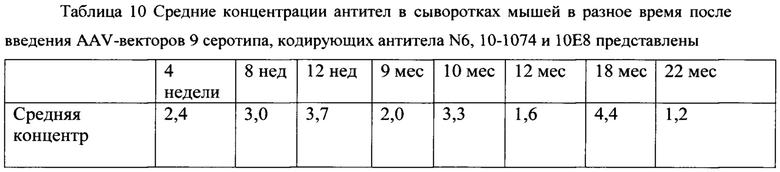
[0076] Таким образом, применение AAV-векторов 9 серотипа, кодирующих антитела N6, 10-1074 и 10Е8 для длительной экспрессии антител является эффективной стратегией.
2. Оценка эффективности комбинации из AAV-векторов 9 серотипа, кодирующих антитела N6, 10-1074 и 10Е8, у мышей линии C57BL/6
2.1 Экспрессия антител у мышей C57BL/6 после введения смеси AAV-векторов 9 серотипа, кодирующих антитела N6, 10Е8 и 10-1074.
[0077] Животным вводили комбинацию AAV-векторов, кодирующих антитела N6, 10Е8 и 10-1074, по 2*1011 гк каждого вектора на мышь. Было изучено два способа введения: в одной группе мышам вводили по 100 мкл смеси всех векторов в бедренную мышцу. Животным второй группы каждый AAV-вектор вводили отдельно: 100 мкл AAV9-N6 вектора в левое бедро, 100 мкл AAV9-10-1074 вектора в правое бедро и по 50 мкл AAV9-10E8 вектора в правую и левую передние лапы. Через 2, 4, 6, 8 недель анализировали концентрацию антител в крови (фиг. 6).
[0078] В течение 8 недель титр антител в крови увеличивался в обоих группах. Во всех точках средняя концентрация антител у животных, которым вводили AAV-вектора в разные лапы, была больше, достоверное отличие между группами наблюдалось на 6-й неделе (р<0,05 Манн Уитни тест).
2.2 Оценка нейтрализующей активности сывороток мышей линии C57BL/6 после введения смеси AAV-векторов 9 серотипа, кодирующих антитела N6, 10Е8 и 10-1074
[0079] Была измерена нейтрализующая активность сывороток мышей линии C57BL/6, получивших смесь AAV-9 векторов, кодирующих антитела N6, 10Е8 и 10-1074, с использованием разных способов введения. Для этого пулировали сыворотки животных из группы 1, которым вводили препарат в одну лапу (концентрация антител в пуле 4,0 мкг/мл), и отдельно пулировали сыворотки, полученные от мышей, введение векторов которым проводили в разные лапы (группа 2, концентрация антител в пуле 9,5 мкг/мл). Для определения нейтрализующей активности пулов использовали глобальную панель ВИЧ-1. Результаты нейтрализующей активности приведены на фиг. 7 (введение в одну лапу) и фиг. 8 (введение в три лапы). Хуже всего нейтрализовались вирусы BJOX20000 и CNE8. Эффективность нейтрализации этих вирусов сыворотками, разведенными в 5 раз составляла более 50%, но не достигала 80%. Для остальных вирусов сыворотка в разведении 1:5 обеспечивала нейтрализацию выше 80%.
[0080] Нейтрализация псевдовирусов сыворотками мышей, которым вводили препарат «КомбиМаб1» в три разные лапы, была более эффективной, в этом случае сыворотка в разведении 1:5 нейтрализовала все вирусы с эффективностью более 80%. Но, как и в случае введения в одну лапу, эффективность нейтрализации псевдовирусов BJOX20000 и CNE8 была ниже, чем эффективность нейтрализации других псевдовирусов.
VI. ОПТИМИЗАЦИЯ AAV-ВЕКТОРА С ЦЕЛЬЮ УМЕНЬШЕНИЯ ИММУНОГЕННОСТИ
[0081] Повышенная иммуногенность AAV-векторов может приводить к снижению эффективности экспрессии трансгена вследствие индукции CD8+Т-клеточного ответа и элиминации клеток, которые экспрессируют антитела [Manno C.S., Pierce G.F., Arruda V.R., Glader В., Ragni M., et al. «Successful transduction of liver in hemophilia by AAV-Factor IX and limitations imposed by the host immune response», Nat Med 12: 342-347], [Federico Mingozzi et.al., «AAV-1-mediated gene transfer to skeletal muscle in humans results in dose-dependent activation of capsid-specific T cells» (2009), 10.1182/blood-2008-07-167510]. Ключевую роль в запуске такого ответа играет эндосомальный внутриклеточный рецептор TRL9 (toll-подобный рецептор 9), выполняющий роль ДНК-сенсора в цитоплазме клетки [Eicke Latz et. al., «TLR9 signals after translocating from the ER to CpG DNA in the lysosome», Nature Immunology volume 5, pages 190-198 (2004)].
[0082] Внутриклеточный рецептор TLR9 способен обнаруживать ДНК патогенов, узнавая неметилированные CpG-мотивы. Таким образом, CpG-мотивы являются триггером активации TLR9-пути и последующего развития ответа CD8+ Т-клеток [Jiangao Zhu 1, Xiaopei Huang, Yiping Yang, «The TLR9-MyD88 pathway is critical for adaptive immune responses to adeno-associated virus gene therapy vectors in mice» J Clin Invest 2009 Aug], [Geoffrey L Rogers et.al. «Plasmacytoid and conventional dendritic cells cooperate in crosspriming AAV capsid-specific CD8+ T cells», Blood 2017 Jun 15; 129(24): 3184-3195].
[0083] Уменьшение количества CpG-мотивов в AAV-геноме позволяет снизить иммуногенность вектора [Susan М Faust et. al., «CpG-depleted adeno-associated virus vectors evade immune detection», J Clin Invest 2013 Jul; 123(7): 2994-3001].
[0084] Для исследования данного подхода были сконструированы векторные плазмиды 10-1074CpG-low, VRC07-523CpG-low, PGDM 1400CpG-low. Последовательности, кодирующие тяжелые и легкие цепи антител 10-1074CpG-low, VRC07-523CpG-low, PGDM 1400CpG-low, в которых число CpG динуклеотидов было уменьшено за счет внесения синонимичных замен в кодоны, образующие CpG, были синтезированы de novo.
[0085] Затем фрагменты, кодирующие тяжелую и легкую цепь этих антител, амплифицировали, добавляя необходимые сайты рестрикции. Также с помощью праймеров в начало последовательности легкой цепи была добавлена последовательность, кодирующая 2А-пептид и сайт узнавания протеазы фурин. В плазмидный вектор pAAVcore (получение этого плазмидного вектора описано в разделе I настоящей заявки) по сайтам ClaI-XbaI одновременно вставляли два фрагмента: последовательность, кодирующую тяжелую цепь, фланкированную сайтами ClaI-BamHI и последовательность, кодирующую легкую цепь, фланкированную сайтами BamHI-XbaI. Корректность полученных плазмид подтверждали секвенированием.
[0086] На основании вышеприведенной схемы были сконструированы следующие плазмидные векторы, кодирующие широко нейтрализующее антитела против ВИЧ-1:
Плазмидный вектор VRC07-523CpG-low, кодирующий широко нейтрализующее антитело VRC07-523;
Плазмидный вектор 10-1074CpG-low, кодирующий широко нейтрализующее антитело 10-1074;
Плазмидный вектор PGDM 1400CpG-low, кодирующий широко нейтрализующее антитело PGDM 1400;
[0087] Для изучения влияния CpG динуклеотидов на экспрессию антител in vivo были использованы две конструкции плазмидного вектора, кодирующие антитело 10-1074. Одна из них - плазмидный вектор 10-1074. Другая конструкция - плазмидный вектор 10-1074CpG-low, который является производным от плазмидного вектора 10-1074, в которой содержание CpG динуклеотидов уменьшено за счет внесения синонимичных замен в кодоны, образующие CpG.
[0088] Полученные конструкции плазмидных векторов были исследованы in vivo. Для этого были наработаны AAV-частицы 9-го серотипа и очищены в градиенте йодиксанола. После чего полученные AAV-вектора были внутримышечно введены мышам линии C57BL/6 в количестве 2*1011 vg/мышь. Наблюдение за концентрацией человеческих IgG в сыворотках экспериментальных животных проводилось в течение 8 недель. В результате было показано (фиг. 9), что конструкция AAV-вектора 10-1074CpG-low обеспечивает достоверно более высокие концентрации антитела 10-1074 в крови за счет проведенной оптимизации трансгена на 4 и 6 неделю эксперимента по сравнению с исходным вектором.
VII. ОЦЕНКА ЭФФЕКТИВНОСТИ КОМБИНАЦИИ ИЗ AAV - ВЕКТОРОВ VRC07-523CPG-LOW, PGDM 1400CPG-LOW И 10-1074CPG-LOW
[0089] Для оценки эффективности была составлена новая комбинация AAV - векторов, кодирующих широко нейтрализующие антитела человека против ВИЧ-1. В состав новой комбинации были включены AAV-вектора 9 серотипа, кодирующие антитела VRC07-523, PGDM 1400 и 10-1074, распознающие соответственно CD4-связывающий сайт оболочечного белка ВИЧ-1, эпитоп V1/V2 и эпитоп V3.
[0090] Были получены AAV-вектора, как описано в разделе II: AAV-9-VRC07-523CpG-low, AAV-9-10-1074CpG-low и AAV-9-PGDM1400CpG-low, несущие последовательности, кодирующие антитела VRC07-523, 10-1074 и PGDM1400, с нуклеотидными последовательностями SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6 соответственно.
[0091] Была получена смесь, включающая вектор AAV-9-VRC07-523CpG-low, кодирующий широко нейтрализующее антитело против ВИЧ-1 VRC07-523 (SEQ ID NO: 4), вектор AAV-9-10-1074CpG-low, кодирующий широко нейтрализующее антитело против ВИЧ 10-1074 (SEQ ID NO: 5) и вектор AAV-9-PGDM1400CpG-low, кодирующий широко нейтрализующее антитело против ВИЧ-1 PGDM 1400 (SEQ ID NO: 6) в соотношении 1:1:1 каждого вектора (далее по тексту - препарат «КомбиМаб2»).
[0092] Смесь AAV-векторов была подготовлена простым смешением суспензий векторов, входящих в их состав.
[0093] Также в одном из вариантов реализации изобретения может быть подготовлен набор AAV - 9 векторов (SEQ ID NO: 4), (SEQ ID NO: 5), (SEQ ID NO: 6), где каждый из AAV-9 векторов в наборе может вводиться по отдельности.
[0094] AAV-вектора, кодирующие отдельные антитела, а также их смесь вводили мышам линии С57 В1аск внутримышечно. Отдельные вектора вводили по 2*1011 гк на мышь в объеме 100 мкл, смесь векторов вводили в объеме 100 мкл в двух вариантах: в одном варианте смесь содержала по 2*1011 каждого вектора (доза 1, суммарно 6*1011), в другом 7*1010 каждого вектора (доза 2, суммарно 2*1011). Через 4 недели измеряли концентрацию антител в сыворотках крови мышей (фиг. 10).
[0095] Концентрация антител после введения векторов AAV-9-VRC07-523CpG-low и AAV-9-10-1074CpG-low находилась на одном уровне (средние 17,5 и 20,8 мкг/мл соответственно), экспрессия антитела после введения AAV-9-PGDM1400CpG-low была ниже, хотя статистически не отличалась (среднее 9,6 мкг/мл). При введении смеси векторов в дозе 1 (6*1011) средняя концентрация составила 28 мкг/мл, при введении в три раза меньшей дозы - 8,5 мкг/мл.
[0096] Далее была измерена нейтрализующая активность сывороток мышей линии C57BL/6, получивших «КомбиМаб2». Для этого сыворотки животных, получивших «КомбиМаб2» в дозе 1 пулировали (концентрация IgG в пуле составила 24,2 мкг/мл) и измеряли нейтрализующую активность пулов по отношению к псевдовирусам глобальной панели ВИЧ-1. Результаты исследования представлены на фиг. 11.
[0097] Пул сывороток показал высокую нейтрализующую активность: в разведении 1:125 наблюдалась нейтрализация всех вирусов глобальной панели вирусов ВИЧ с эффективностью более 80%. В разведении 1:625 сыворотки нейтрализовали 5 вирусов с эффективностью 80%, 4 вируса с эффективностью 50% и один вирус с эффективностью чуть ниже 50%.
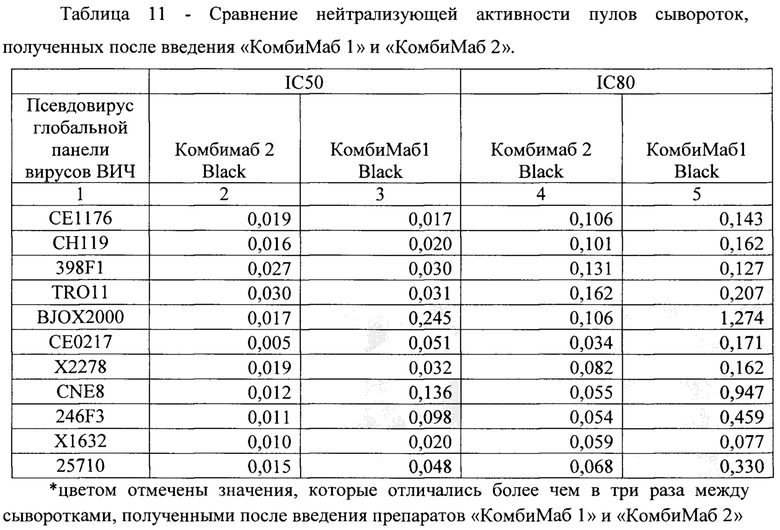
[0098] Чтобы сравнить нейтрализующую активность антител в пулах сывороток от мышей, которым вводили «КомбиМаб1» и «КомбиМаб2», исключив влияние разности концентрации антител, провели сравнение вычисленных для этих пулов значений IC50 и IC80 (табл. 11). Для «КомбиМаб2» IC50 не превышало 0,03 мкг/мл, IC80 - 0,16 мкг/мл. Для 5-ти вирусов глобальной панели ВИЧ-1 (BJOX2000, СЕ0217, CNE8, 246F3,25710) эффективность нейтрализации сыворотками мышей, получивших «КомбиМаб2», существенно превышала эффективность нейтрализации сыворотками мышей, получивших «КомбиМаб1».
[0099] Широко нейтрализующие антитела против ВИЧ-1 являются перспективным направлением для разработки терапевтического препарата для лечения ВИЧ-инфекции, и уже показали свою эффективность в клинических исследованиях. Однако для использования антител как терапевтического средства необходимо решить проблемы короткого времени жизни антител в крови после внутривенного вливания и быстрой выработки резистентности при использовании одного антитела. В заявленном изобретении предложена возможность использования комбинации антител против ВИЧ-1, доставляемых с помощью AAV-векторов серотипа 9, которая в дальнейшем может лечь в основу создания препарата от ВИЧ.
VIII. ПЕРЕЧЕНЬ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ









--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3"
fileName="antiHIV_AB_sequences.xml" softwareName="WIPO Sequence"
softwareVersion="2.3.0" productionDate="2024-05-24">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate>2024-05-24</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>123456789</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение "Центр стратегического планирования и
управления медико-биологическими рисками здоровью" Федерального
медико-биологического агентства, RU</ApplicantName>
<ApplicantNameLatin>Federal'noye gosudarstvennoye byudzhetnoye
uchrezhdeniye "Tsentr strategicheskogo planirovaniya i
upravleniya mediko-biologicheskimi riskami zdorov'yu"
Federal'nogo mediko-biologicheskogo agentstva,
RU</ApplicantNameLatin>
<InventionTitle languageCode="ru">НАБОР РЕКОМБИНАНТНЫХ ГЕНЕТИЧЕСКИХ
ВЕКТОРОВ, КОДИРУЮЩИХ ШИРОКО НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА ПРОТИВ ВИЧ-1,
ИММУНОБИОЛОГИЧЕСКИЙ ПРЕПАРАТ ДЛЯ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ВИЧ-ИНФЕКЦИИ
И СПОСОБ ЕГО ПРИМЕНЕНИЯ</InventionTitle>
<SequenceTotalQuantity>6</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>4557</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4557</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcgtc
gggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcac
taggggttcctgcggccgcgctctggagacgcgttacataacttacggtaaatggcccgcctggctgacc
gcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttc
cattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgc
caagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgacctt
atgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatgggtgaggctccggtg
cccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaac
cggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttccc
gagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccg
ccagaacacaggtaagtccggcctccgcgccgggttttggcgcctcccgcgggcgcccccctcctcacgg
cgagcgctgccacgtcagacgaagggcgcaggagcgttcctgatccttccgcccggacgctcaggacagc
ggcccgctgctcataagactcggccttagaaccccagtatcagcagaaggacattttaggacgggacttg
ggtgactctagggcactggttttctttccagagagcggaacaggcgaggaaaagtagtcccttctcggcg
attctgcggagggatctccgtggggcggtgaacgccgatgatgcctctactaaccatgttcatgttttct
ttttttttctacaggtcctgggtgacgaacagcatcgatgccgccaccatggctactggctctcgaacca
gcctcctgctggcatttgggttgctgtgtctgccatggctgcaggagggctctgcccaggtgaggctgag
ccagtctggaggtcagatgaagaagcctggagacagcatgagaatttcttgtagagctagtggatatgaa
tttattaattgtccaataaattggattagactggccccaggaaaaaggcctgagtggatgggatggatga
agcctaggcatggggcagtcagttatgcaagacaacttcagggcagagtgaccatgactagggacatgta
ttctgagacagcctttttggagctcagatccttgacaagtgatgacacagctgtctatttttgtactaga
ggaaaatattgcactgccagggactattataattgggactttgaacactggggccagggcaccccagtca
cagtcagctcagctagcaccaagggcccaagtgtcttccccctggcaccctcctccaagagcacctctgg
gggcacagctgccctgggctgcctggtcaaggactacttcccagaaccagtgacagtgagctggaactca
ggagccctgaccagcggagtgcacaccttcccagctgtccttcagtcctcaggactctactccctcagct
ctgtggtgacagtgccctccagcagcttgggcacccagacctacatctgcaatgtgaatcacaagcccag
caacaccaaggtggacaagaaagcagagcccaaatcttgtgacaaaactcacacatgcccaccctgccca
gcacctgaactcctggggggaccctcagtcttcctcttccccccaaaacccaaggacaccctcatgatct
ccaggacccctgaggtcacatgtgtggtggtggatgtgagccatgaagaccctgaggtcaagttcaactg
gtatgtggatggagtggaggtgcataatgccaagacaaagccaagagaggagcagtacaacagcacatat
agagtggtctctgtcctcacagtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtct
ccaacaaagccctcccagcccccatagagaaaaccatctccaaagccaaagggcagcccagggaaccaca
ggtgtacaccctgcccccatccagggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaa
ggcttctatccctctgacattgcagtggagtgggagagcaatgggcagccagagaacaactacaagacca
cccctccagtgctggactctgatggctccttcttcctctacagcaagctcacagtggacaagagcaggtg
gcagcaggggaatgtcttctcatgctctgtgctccatgaggctctgcacagccactacacccagaagagc
ctgtccctgtctccagggaaaaggaagagaaggggatctggagagggcagaggaagtctgctgacatgtg
gtgatgtcgaggagaatcctggcccaatggctaccggcagcagaacaagcctgctgctggcttttggact
gctctgtctcccctggttgcaagaaggctctgcctccttgacacagtctccaggcaccctgtctttgtct
ccaggggaaacagccatcatctcttgtaggaccagtcagtatggttccctggcctggtatcaacagaggc
caggccaggcccccaggctggtcatctattctggctctactagagctgctggcatcccagacaggttctc
tggcagtaggtgggggccagactacaatctcaccatcagcaacctggagtctggagattttggtgtttat
tattgccagcagtatgaattttttggccaggggaccaaggtccaggttgacattaagaggagaactgtgg
ctgcaccatctgtcttcatcttcccaccatctgatgagcagttgaaatctggaactgcctctgttgtgtg
cctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataatgccctccaatctggt
aactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgaccc
tgagcaaagcagactatgagaaacacaaagtctatgcctgtgaagtcacccatcagggcctgagcagccc
agtcacaaagagcttcaacaggggagagtgttgatgatctagatgagatctataatcaacctctggatta
caaaatttgtgaaagattgactggtattcttaactatgttgctccttttacgctatgtggatacgctgct
ttaatgcctttgtatcatgctattgcttcccgtatggctttcattttctcctccttgtataaatcctggt
tgctgtctctttatgaggagttgtggcccgttgtcaggcaacgtggcgtggtgtgcactgtgtttgctga
cgcaacccccactggttggggcattgccaccacctgtcagctcctttccgggactttcgctttccccctc
cctattgccacggcggaactcatcgccgcctgccttgcccgctgctggacaggggctcggctgttgggca
ctgacaattccgtggtgttgtcggggaaatcatcgtcctttccttggctgctcgcctgtgttgccacctg
gattctgcgcgggacgtccttctgctacgtcccttcggccctcaatccagcggaccttccttcccgcggc
ctgctgccggctctgcggcctcttccgcgtcttcgccttcgccctcagacgagtcggatctccctttggg
ccgcctccccgcatcctcgaggctcgctgatcagcctcgactgtgccttctagttgccagccatctgttg
tttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatga
ggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaag
ggggaggattgggaagacaatagcaggcatgctggggatctgagtgtcgataaggatctgaacgatggag
cggagaatgggcggaactgggcggagttaggggcgggatgggcggagttaggggcgggactatggttgct
gactaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacacctggttg
ctgactaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacaccctaa
ctgacacacattccacagcgcggccgcaggaacccctagtgatggagttggccactccctctctgcgcgc
tcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtg
agcgagcgagcgcgcagctgcctgcagg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>4593</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4593</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcgtc
gggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcac
taggggttcctgcggccgcgctctggagacgcgttacataacttacggtaaatggcccgcctggctgacc
gcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttc
cattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgc
caagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgacctt
atgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatgggtgaggctccggtg
cccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaac
cggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttccc
gagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccg
ccagaacacaggtaagtccggcctccgcgccgggttttggcgcctcccgcgggcgcccccctcctcacgg
cgagcgctgccacgtcagacgaagggcgcaggagcgttcctgatccttccgcccggacgctcaggacagc
ggcccgctgctcataagactcggccttagaaccccagtatcagcagaaggacattttaggacgggacttg
ggtgactctagggcactggttttctttccagagagcggaacaggcgaggaaaagtagtcccttctcggcg
attctgcggagggatctccgtggggcggtgaacgccgatgatgcctctactaaccatgttcatgttttct
ttttttttctacaggtcctgggtgacgaacagcatcgatgccgccaccatggctactggctctcgaacca
gcctcctgctggcatttgggttgctgtgtctgccatggctgcaggagggctctgcccaggtgcaactaca
agagtcaggccctggcttagtgaagccctcagagacactgagtgtaacatgctcagtgagtggggattcc
atgaataattattattggacctggattaggcagtctcctggaaaaggtttagaatggataggttatatta
gtgacagagagagtgctacatacaacccttctctgaattccagagtggtcatttccagagatacttcaaa
aaatcagctgagcctgaaattaaacagtgttacccctgctgatactgcagtgtattattgtgccacagcc
aggagagggcagaggatttatggagttgtgtcttttggagaattcttctactattactccatggatgtgt
ggggtaaaggcactacagtgactgtgtcctcagctagcaccaagggcccaagtgtcttccccctggcacc
ctcctccaagagcacctctgggggcacagctgccctgggctgcctggtcaaggactacttcccagaacca
gtgacagtgagctggaactcaggagccctgaccagcggagtgcacaccttcccagctgtccttcagtcct
caggactctactccctcagctctgtggtgacagtgccctccagcagcttgggcacccagacctacatctg
caatgtgaatcacaagcccagcaacaccaaggtggacaagaaagcagagcccaaatcttgtgacaaaact
cacacatgcccaccctgcccagcacctgaactcctggggggaccctcagtcttcctcttccccccaaaac
ccaaggacaccctcatgatctccaggacccctgaggtcacatgtgtggtggtggatgtgagccatgaaga
ccctgaggtcaagttcaactggtatgtggatggagtggaggtgcataatgccaagacaaagccaagagag
gagcagtacaacagcacatatagagtggtctctgtcctcacagtcctgcaccaggactggctgaatggca
aggagtacaagtgcaaggtctccaacaaagccctcccagcccccatagagaaaaccatctccaaagccaa
agggcagcccagggaaccacaggtgtacaccctgcccccatccagggatgagctgaccaagaaccaggtc
agcctgacctgcctggtcaaaggcttctatccctctgacattgcagtggagtgggagagcaatgggcagc
cagagaacaactacaagaccacccctccagtgctggactctgatggctccttcttcctctacagcaagct
cacagtggacaagagcaggtggcagcaggggaatgtcttctcatgctctgtgctccatgaggctctgcac
agccactacacccagaagagcctgtccctgtctccagggaaaaggaagagaaggggatctggagagggca
gaggaagtctgctgacatgtggtgatgtcgaggagaatcctggcccaatggctaccggcagcagaacaag
cctgctgctggcttttggactgctctgtctcccctggttgcaagaaggctctgcctcatatgtgagacct
ttgtctgtggcattgggagaaactgcaagaatcagctgtgggaggcaagccctggggagtagggctgtgc
agtggtatcagcacagaccaggacaggctcctatcttgctgatctataacaaccaggacagacccagtgg
tatacctgagagattttctggcactccagatatcaactttggcaccagggctacccttaccatcagtggt
gttgaggctggggatgaggcagactactactgccatatgtgggacagcagatcaggcttttcctggtcct
ttggaggggccactagacttactgtgctgggacagcccaaggctgccccctcagtcactctgttcccacc
ctctagtgaggagcttcaagccaacaaggccacactggtgtgtctcataagtgacttctacccaggagca
gtgacagtggcctggaaggcagatagcagccctgtcaaggcaggagtggagaccaccacaccctccaaac
aaagcaacaacaagtatgctgccagcagctacctgagcctgacacctgagcagtggaagtcccacagaag
ctacagctgccaggtcacccatgaagggagcacagtggagaagacagtggcccctacagaatgttcatga
tgatctagatgagatctataatcaacctctggattacaaaatttgtgaaagattgactggtattcttaac
tatgttgctccttttacgctatgtggatacgctgctttaatgcctttgtatcatgctattgcttcccgta
tggctttcattttctcctccttgtataaatcctggttgctgtctctttatgaggagttgtggcccgttgt
caggcaacgtggcgtggtgtgcactgtgtttgctgacgcaacccccactggttggggcattgccaccacc
tgtcagctcctttccgggactttcgctttccccctccctattgccacggcggaactcatcgccgcctgcc
ttgcccgctgctggacaggggctcggctgttgggcactgacaattccgtggtgttgtcggggaaatcatc
gtcctttccttggctgctcgcctgtgttgccacctggattctgcgcgggacgtccttctgctacgtccct
tcggccctcaatccagcggaccttccttcccgcggcctgctgccggctctgcggcctcttccgcgtcttc
gccttcgccctcagacgagtcggatctccctttgggccgcctccccgcatcctcgaggctcgctgatcag
cctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctgga
aggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcat
tctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctg
gggatctgagtgtcgataaggatctgaacgatggagcggagaatgggcggaactgggcggagttaggggc
gggatgggcggagttaggggcgggactatggttgctgactaattgagatgcatgctttgcatacttctgc
ctgctggggagcctggggactttccacacctggttgctgactaattgagatgcatgctttgcatacttct
gcctgctggggagcctggggactttccacaccctaactgacacacattccacagcgcggccgcaggaacc
cctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtc
gcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg</INSD
Seq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>4635</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4635</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcgtc
gggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcac
taggggttcctgcggccgcgctctggagacgcgttacataacttacggtaaatggcccgcctggctgacc
gcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttc
cattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgc
caagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgacctt
atgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatgggtgaggctccggtg
cccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaac
cggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttccc
gagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccg
ccagaacacaggtaagtccggcctccgcgccgggttttggcgcctcccgcgggcgcccccctcctcacgg
cgagcgctgccacgtcagacgaagggcgcaggagcgttcctgatccttccgcccggacgctcaggacagc
ggcccgctgctcataagactcggccttagaaccccagtatcagcagaaggacattttaggacgggacttg
ggtgactctagggcactggttttctttccagagagcggaacaggcgaggaaaagtagtcccttctcggcg
attctgcggagggatctccgtggggcggtgaacgccgatgatgcctctactaaccatgttcatgttttct
ttttttttctacaggtcctgggtgacgaacagcatcgatgccgccaccatggctactggctctcgaacca
gcctcctgctggcatttgggttgctgtgtctgccatggctgcaggagggctctgcccaggtgcatctgac
tcagtctgggcctgaggtgaggaagcctgggacctcagtaaaggtctcctgcaaggcccctggaaacaca
ttgaagacttatgatctacactgggtgagatctgtccctggacaaggccttcagtggatgggatggataa
gccatgagggagacaagaaggtcattgtggaaagattcaaggccaaagtcaccattgattgggacaggtc
caccaatactgcctatctccaactgagtggcctcacatctggagacactgctgtctattattgtgcaaaa
ggctcaaaacacaggctgagagattatgctctctatgatgatgatggagcattgaattgggctgtagatg
ttgactacctttcaaacttggaattctggggccaagggacagctgtcacagtctcttcagctagcaccaa
gggcccaagtgtcttccccctggcaccctcctccaagagcacctctgggggcacagctgccctgggctgc
ctggtcaaggactacttcccagaaccagtgacagtgagctggaactcaggagccctgaccagcggagtgc
acaccttcccagctgtccttcagtcctcaggactctactccctcagctctgtggtgacagtgccctccag
cagcttgggcacccagacctacatctgcaatgtgaatcacaagcccagcaacaccaaggtggacaagaaa
gcagagcccaaatcttgtgacaaaactcacacatgcccaccctgcccagcacctgaactcctggggggac
cctcagtcttcctcttccccccaaaacccaaggacaccctcatgatctccaggacccctgaggtcacatg
tgtggtggtggatgtgagccatgaagaccctgaggtcaagttcaactggtatgtggatggagtggaggtg
cataatgccaagacaaagccaagagaggagcagtacaacagcacatatagagtggtctctgtcctcacag
tcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccc
catagagaaaaccatctccaaagccaaagggcagcccagggaaccacaggtgtacaccctgcccccatcc
agggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatccctctgacattg
cagtggagtgggagagcaatgggcagccagagaacaactacaagaccacccctccagtgctggactctga
tggctccttcttcctctacagcaagctcacagtggacaagagcaggtggcagcaggggaatgtcttctca
tgctctgtgctccatgaggctctgcacagccactacacccagaagagcctgtccctgtctccagggaaaa
ggaagagaaggggatctggagagggcagaggaagtctgctgacatgtggtgatgtcgaggagaatcctgg
cccaatggctaccggcagcagaacaagcctgctgctggcttttggactgctctgtctcccctggttgcaa
gaaggctctgccgattttgtcctgactcagtctccacactctctgtctgtcacccctggagagtcagcct
ccatctcctgcaagtctagtcacagcctcattcatggtgataggaacaattatttggcttggtatgtaca
gaagccagggagatctccacaactcctgatctatttggcttccagtagagcctcaggggtccctgacagg
ttcagtggcagtggatcagacaaagattttacactgaagatcagcagagtggagactgaggatgttggga
catattactgcatgcaaggtagagaaagtccctggacttttggccaagggaccaaggtggacatcaaaag
aactgtggctgcaccatctgtcttcatcttcccaccatctgatgagcagttgaaatctggaactgcctct
gttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataatgccctcc
aatctggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcac
cctgaccctgagcaaagcagactatgagaaacacaaagtctatgcctgtgaagtcacccatcagggcctg
agcagcccagtcacaaagagcttcaacaggggagagtgttgatgatctagatgagatctataatcaacct
ctggattacaaaatttgtgaaagattgactggtattcttaactatgttgctccttttacgctatgtggat
acgctgctttaatgcctttgtatcatgctattgcttcccgtatggctttcattttctcctccttgtataa
atcctggttgctgtctctttatgaggagttgtggcccgttgtcaggcaacgtggcgtggtgtgcactgtg
tttgctgacgcaacccccactggttggggcattgccaccacctgtcagctcctttccgggactttcgctt
tccccctccctattgccacggcggaactcatcgccgcctgccttgcccgctgctggacaggggctcggct
gttgggcactgacaattccgtggtgttgtcggggaaatcatcgtcctttccttggctgctcgcctgtgtt
gccacctggattctgcgcgggacgtccttctgctacgtcccttcggccctcaatccagcggaccttcctt
cccgcggcctgctgccggctctgcggcctcttccgcgtcttcgccttcgccctcagacgagtcggatctc
cctttgggccgcctccccgcatcctcgaggctcgctgatcagcctcgactgtgccttctagttgccagcc
atctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaa
taaaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcagg
acagcaagggggaggattgggaagacaatagcaggcatgctggggatctgagtgtcgataaggatctgaa
cgatggagcggagaatgggcggaactgggcggagttaggggcgggatgggcggagttaggggcgggacta
tggttgctgactaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccaca
cctggttgctgactaattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttcca
caccctaactgacacacattccacagcgcggccgcaggaacccctagtgatggagttggccactccctct
ctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcgg
cctcagtgagcgagcgagcgcgcagctgcctgcagg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>4593</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4593</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcgtc
gggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcac
taggggttcctgcggccgcgctctggagacgcgttacataacttacggtaaatggcccgcctggctgacc
gcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttc
cattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgc
caagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgacctt
atgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatgggtgaggctccggtg
cccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaac
cggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttccc
gagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccg
ccagaacacaggtaagtccggcctccgcgccgggttttggcgcctcccgcgggcgcccccctcctcacgg
cgagcgctgccacgtcagacgaagggcgcaggagcgttcctgatccttccgcccggacgctcaggacagc
ggcccgctgctcataagactcggccttagaaccccagtatcagcagaaggacattttaggacgggacttg
ggtgactctagggcactggttttctttccagagagcggaacaggcgaggaaaagtagtcccttctcggcg
attctgcggagggatctccgtggggcggtgaacgccgatgatgcctctactaaccatgttcatgttttct
ttttttttctacaggtcctgggtgacgaacagcatcgatgccgccaccatggctactggctctcgaacca
gcctcctgctggcatttgggttgctgtgtctgccatggctgcaggagggctctgcccaggtgcaactaca
agagtccggccctggcttagtgaagccctccgagacactgagtgtaacatgcagcgtgagcggggattcc
atgaataattattattggacctggattaggcagtctcctggaaaaggtttagaatggatcggttatatta
gcgaccgtgagagtgcgacgtacaacccttctctgaattcccgcgtggtcatttcccgggatacttcaaa
aaatcagctgagcctgaaattaaacagcgttacccccgctgataccgcagtgtattattgcgccaccgcc
aggagagggcagaggatttacggcgttgtgtctttcggagaattcttctactattactccatggatgtgt
ggggtaaaggcacgacagtgaccgtgtcctcagctagcaccaagggcccatcggtcttccccctggcacc
ctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccc
gtgaccgtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccagctgtccttcagtcct
caggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctg
caacgtgaatcacaagcccagcaacaccaaggtggacaagaaagcagagcccaaatcttgtgacaaaact
cacacatgcccaccgtgcccagcacctgaactcctggggggaccctcagtcttcctcttccccccaaaac
ccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaaga
ccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccacgggag
gagcagtacaacagcacatatcgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggca
aggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaa
agggcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtc
agcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagc
cggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagct
caccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgctccatgaggctctgcac
agccactacacgcagaagagcctctccctgtctcccgggaaatggaagagaaggggatctggagagggca
gaggaagtctgctgacatgtggtgatgtcgaggagaatcctggcccaatggctaccggcagcagaacaag
cctgctgctcgcttttggactgctctgtctcccctggttgcaagaaggcagcgcctcatatgtgcggccg
ttgtctgtggcattgggagaaactgcacgaatcagctgtgggaggcaagccctggggagtagggccgtgc
agtggtatcagcacagaccaggacaggcgcctatcttgctgatctataacaaccaggaccgtcccagtgg
tatacctgagagattttccggcactccagatatcaactttggcaccagggctacccttaccatcagcggc
gtcgaggctggggacgaggccgactactactgccatatgtgggacagcagatccggcttttcctggtcct
ttggaggggccacgcggcttaccgtgctgggacagcccaaggctgccccctcggtcactctgttcccgcc
ctcgagtgaggagcttcaagccaacaaggccacactggtgtgtctcataagtgacttctacccgggagcc
gtgacagtggcctggaaggcagatagcagccccgtcaaggcgggagtggagaccaccacaccctccaaac
aaagcaacaacaagtacgcggccagcagctacctgagcctgacgcctgagcagtggaagtcccacagaag
ctacagctgccaggtcacgcatgaagggagcaccgtggagaagacagtggcccctacagaatgttcatga
tgatctagatgagatctataatcaacctctggattacaaaatttgtgaaagattgactggtattcttaac
tatgttgctccttttacgctatgtggatacgctgctttaatgcctttgtatcatgctattgcttcccgta
tggctttcattttctcctccttgtataaatcctggttgctgtctctttatgaggagttgtggcccgttgt
caggcaacgtggcgtggtgtgcactgtgtttgctgacgcaacccccactggttggggcattgccaccacc
tgtcagctcctttccgggactttcgctttccccctccctattgccacggcggaactcatcgccgcctgcc
ttgcccgctgctggacaggggctcggctgttgggcactgacaattccgtggtgttgtcggggaaatcatc
gtcctttccttggctgctcgcctgtgttgccacctggattctgcgcgggacgtccttctgctacgtccct
tcggccctcaatccagcggaccttccttcccgcggcctgctgccggctctgcggcctcttccgcgtcttc
gccttcgccctcagacgagtcggatctccctttgggccgcctccccgcatcctcgaggctcgctgatcag
cctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctgga
aggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcat
tctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctg
gggatctgagtgtcgataaggatctgaacgatggagcggagaatgggcggaactgggcggagttaggggc
gggatgggcggagttaggggcgggactatggttgctgactaattgagatgcatgctttgcatacttctgc
ctgctggggagcctggggactttccacacctggttgctgactaattgagatgcatgctttgcatacttct
gcctgctggggagcctggggactttccacaccctaactgacacacattccacagcgcggccgcaggaacc
cctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtc
gcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg</INSD
Seq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>4593</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4593</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q10">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcgtc
gggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcac
taggggttcctgcggccgcgctctggagacgcgttacataacttacggtaaatggcccgcctggctgacc
gcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttc
cattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgc
caagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgacctt
atgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatgggtgaggctccggtg
cccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaac
cggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttccc
gagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccg
ccagaacacaggtaagtccggcctccgcgccgggttttggcgcctcccgcgggcgcccccctcctcacgg
cgagcgctgccacgtcagacgaagggcgcaggagcgttcctgatccttccgcccggacgctcaggacagc
ggcccgctgctcataagactcggccttagaaccccagtatcagcagaaggacattttaggacgggacttg
ggtgactctagggcactggttttctttccagagagcggaacaggcgaggaaaagtagtcccttctcggcg
attctgcggagggatctccgtggggcggtgaacgccgatgatgcctctactaaccatgttcatgttttct
ttttttttctacaggtcctgggtgacgaacagcatcgatgccgccaccatggctactggctctcgaacca
gcctcctgctggcatttgggttgctgtgtctgccatggctgcaggagggctctgccgaggtgcagctggt
ggagtctgggggaggcttggtgaagcctggaggatcccttagactctcatgttcagcctctggtttcgac
ttcgataacgcctggatgacttgggtccgccagcctccagggaagggcctcgaatgggttggtcgtatta
cgggtccaggtgaaggttggtcagtggactatgctgcacccgtggaaggcagatttaccatctcgagact
caattcaataaatttcttatatttggagatgaacaatttaagaatggaagactcaggcctttacttctgt
gcccgcacgggaaaatattatgatttttggagtggctatccgccgggagaagaatacttccaagactggg
gccggggcaccctggtcaccgtctcctcagctagcaccaagggcccatcggtcttccccctggcaccctc
ctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaacccgtg
accgtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccagctgtccttcagtcctcag
gactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaa
cgtgaatcacaagcccagcaacaccaaggtggacaagaaagcagagcccaaatcttgtgacaaaactcac
acatgcccaccgtgcccagcacctgaactcctggggggaccctcagtcttcctcttccccccaaaaccca
aggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccc
tgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccacgggaggag
cagtacaacagcacatatcgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaagg
agtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagg
gcagccccgagaaccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagc
ctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccgg
agaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcac
cgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgctccatgaggctctgcacagc
cactacacgcagaagagcctctccctgtctcccgggaaatggaagagaaggggatctggagagggcagag
gaagtctgctgacatgtggtgatgtcgaggagaatcctggcccaatggctaccggcagcagaacaagtct
gctgctcgcttttggactgctctgtctcccctggttgcaagaaggcagcgcctcatacgaactgactcag
gagactggcgtctctgtggcactggggaggactgtgactattacttgccgaggcgactcactgcggagcc
actacgcttcctggtatcagaagaaacccggccaggcacctattctgctgttctacggaaagaacaatag
gccatctggcgtccccgaccgcttttctggcagtgcatcagggaaccgagccagtctgaccatttccggc
gcccaggctgaggacgacgccgaatactattgcagctcccgggataagagcggctccagactgagcgtgt
tcggaggaggaactaaactgaccgtcctcagtcagcccaaggctgccccctcggtcactctgttcccgcc
ctcgagtgaggagcttcaagccaacaaggccacactggtgtgtctcataagtgacttctacccgggagcc
gtgacagtggcctggaaggcagatagcagccccgtcaaggcgggagtggagaccaccacaccctccaaac
aaagcaacaacaagtacgcggccagcagctacctgagcctgacgcctgagcagtggaagtcccacagaag
ctacagctgccaggtcacgcatgaagggagcaccgtggagaagacagtggcccctacagaatgttcatga
tgatctagatgagatctataatcaacctctggattacaaaatttgtgaaagattgactggtattcttaac
tatgttgctccttttacgctatgtggatacgctgctttaatgcctttgtatcatgctattgcttcccgta
tggctttcattttctcctccttgtataaatcctggttgctgtctctttatgaggagttgtggcccgttgt
caggcaacgtggcgtggtgtgcactgtgtttgctgacgcaacccccactggttggggcattgccaccacc
tgtcagctcctttccgggactttcgctttccccctccctattgccacggcggaactcatcgccgcctgcc
ttgcccgctgctggacaggggctcggctgttgggcactgacaattccgtggtgttgtcggggaaatcatc
gtcctttccttggctgctcgcctgtgttgccacctggattctgcgcgggacgtccttctgctacgtccct
tcggccctcaatccagcggaccttccttcccgcggcctgctgccggctctgcggcctcttccgcgtcttc
gccttcgccctcagacgagtcggatctccctttgggccgcctccccgcatcctcgaggctcgctgatcag
cctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctgga
aggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcat
tctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctg
gggatctgagtgtcgataaggatctgaacgatggagcggagaatgggcggaactgggcggagttaggggc
gggatgggcggagttaggggcgggactatggttgctgactaattgagatgcatgctttgcatacttctgc
ctgctggggagcctggggactttccacacctggttgctgactaattgagatgcatgctttgcatacttct
gcctgctggggagcctggggactttccacaccctaactgacacacattccacagcgcggccgcaggaacc
cctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtc
gcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg</INSD
Seq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>4551</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4551</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q12">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcgtc
gggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcac
taggggttcctgcggccgcgctctggagacgcgttacataacttacggtaaatggcccgcctggctgacc
gcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttc
cattgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgc
caagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatgacctt
atgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatgggtgaggctccggtg
cccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaac
cggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttccc
gagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccg
ccagaacacaggtaagtccggcctccgcgccgggttttggcgcctcccgcgggcgcccccctcctcacgg
cgagcgctgccacgtcagacgaagggcgcaggagcgttcctgatccttccgcccggacgctcaggacagc
ggcccgctgctcataagactcggccttagaaccccagtatcagcagaaggacattttaggacgggacttg
ggtgactctagggcactggttttctttccagagagcggaacaggcgaggaaaagtagtcccttctcggcg
attctgcggagggatctccgtggggcggtgaacgccgatgatgcctctactaaccatgttcatgttttct
ttttttttctacaggtcctgggtgacgaacagcatcgatgccgccaccatggctactggctctcgaacca
gcctcctgctggcatttgggttgctgtgtctgccatggctgcaggagggctctgcccgagcgcacctggt
acaatcagggactgcgatgaagaaaccgggggcctcagtaagagtctcctgccagacctctggatacacc
tttaccgcccacatattattttggttccgacaggcccccgggcgaggacttgagtgggtggggtggatca
agccacaatatggggccgtgaattttggtggtggttttcgggacagggtcacattgactcgagacgtata
tagagagattgcgtacatggacatcagaggccttaaacctgacgacacggccgtctattactgtgcgaga
gaccgttcctatggcgactcctcttgggccttagatgcctggggacagggaacgacggtcgtcgtctccg
cggctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagc
ggccctgggctgcctggtcaaggactacttccccgaacccgtgaccgtgtcgtggaactcaggcgccctg
accagcggcgtgcacaccttcccagctgtccttcagtcctcaggactctactccctcagcagcgtggtga
ccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaa
ggtggacaagaaagcagagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaa
ctcctggggggaccctcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggaccc
ctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtgga
cggcgtggaggtgcataatgccaagacaaagccacgggaggagcagtacaacagcacatatcgtgtggtc
agcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaag
ccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacac
cctgcccccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctat
cccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccg
tgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggg
gaacgtcttctcatgctccgtgctccatgaggctctgcacagccactacacgcagaagagcctctccctg
tctcccgggaaatggaagagaaggggatctggagagggcagaggaagtctgctgacatgtggtgatgtcg
aggagaatcctggcccaatggctaccggcagcagaacaagcctgctgctcgcttttggactgctctgtct
cccctggttgcaagaaggcagcgcctacatccacgtgacccagtctccgtcctccctgtctgtgtctatt
ggagacagagtcaccatcaattgccagacgagtcagggtgttggcagtgacctacattggtatcaacaca
aaccggggagagcccctaaactcttgatccaccatacctcttctgtggaagacggtgtcccctcaagatt
cagcggctctggatttcacacatcttttaatctgaccatcagcgacctacaggctgacgacattgccaca
tattactgtcaagttttacaatttttcggccgagggagtcgactccatattaaacgaactgtggctgcac
catctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgct
gaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcc
caggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagca
aagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcac
aaagagcttcaacaggggagagtgttgatgatctagatgagatctataatcaacctctggattacaaaat
ttgtgaaagattgactggtattcttaactatgttgctccttttacgctatgtggatacgctgctttaatg
cctttgtatcatgctattgcttcccgtatggctttcattttctcctccttgtataaatcctggttgctgt
ctctttatgaggagttgtggcccgttgtcaggcaacgtggcgtggtgtgcactgtgtttgctgacgcaac
ccccactggttggggcattgccaccacctgtcagctcctttccgggactttcgctttccccctccctatt
gccacggcggaactcatcgccgcctgccttgcccgctgctggacaggggctcggctgttgggcactgaca
attccgtggtgttgtcggggaaatcatcgtcctttccttggctgctcgcctgtgttgccacctggattct
gcgcgggacgtccttctgctacgtcccttcggccctcaatccagcggaccttccttcccgcggcctgctg
ccggctctgcggcctcttccgcgtcttcgccttcgccctcagacgagtcggatctccctttgggccgcct
ccccgcatcctcgaggctcgctgatcagcctcgactgtgccttctagttgccagccatctgttgtttgcc
cctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaat
tgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggag
gattgggaagacaatagcaggcatgctggggatctgagtgtcgataaggatctgaacgatggagcggaga
atgggcggaactgggcggagttaggggcgggatgggcggagttaggggcgggactatggttgctgactaa
ttgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacacctggttgctgact
aattgagatgcatgctttgcatacttctgcctgctggggagcctggggactttccacaccctaactgaca
cacattccacagcgcggccgcaggaacccctagtgatggagttggccactccctctctgcgcgctcgctc
gctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgag
cgagcgcgcagctgcctgcagg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИНА ПРОТИВ ВИЧ/СПИД | 2010 |
|
RU2475264C2 |
| Искусственный ген YkuJ-MPER, кодирующий химерный белок-иммуноген YkuJ-MPER, рекомбинантная плазмидная ДНК pET21a-YkuJ-MPER, обеспечивающая экспрессию искусственного гена YkuJ-MPER, и химерный белок-иммуноген YkuJ-MPER, являющийся носителем мембранно-проксимальной области ВИЧ-1 и направленный на индукцию в организме широконейтрализующих антител | 2017 |
|
RU2679076C1 |
| Экспрессионный вектор на основе аденоассоциированного вируса, несущий гены рекомбинантных антител, и его применение для профилактики заболеваний, вызываемых вирусом гриппа А и вирусом SARS-CoV-2 | 2023 |
|
RU2817792C1 |
| СПОСОБ БЫСТРОГО ОТБОРА ВАРИАНТОВ GP-120 ВИЧ | 2012 |
|
RU2603732C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2648791C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ХИМЕРНЫЙ ПОЛИПЕПТИД ВИЧ, ПОЛИПЕПТИДЫ И ТЕСТ-СИСТЕМА ДЛЯ ОБНАРУЖЕНИЯ АНТИТЕЛ К ВИЧ | 2001 |
|
RU2214274C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2721274C2 |
| ВЕКТОРЫ И ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2011 |
|
RU2588667C2 |
| Повышение тканеспецифической доставки генов путем модификации капсида | 2019 |
|
RU2801217C2 |
| ПРОФИЛАКТИЧЕСКАЯ ВАКЦИНА ПРОТИВ ВИЧ, ОСНОВАННАЯ НА ВИЧ-СПЕЦИФИЧЕСКИХ АНТИТЕЛАХ | 2008 |
|
RU2505604C2 |
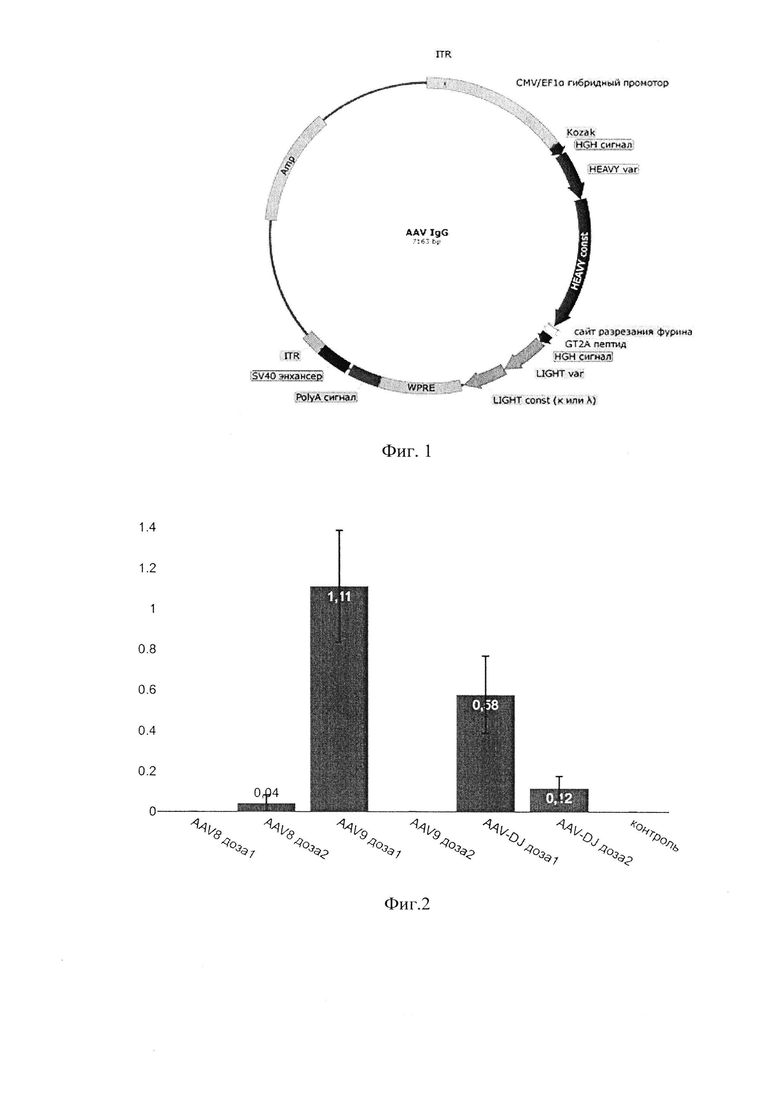
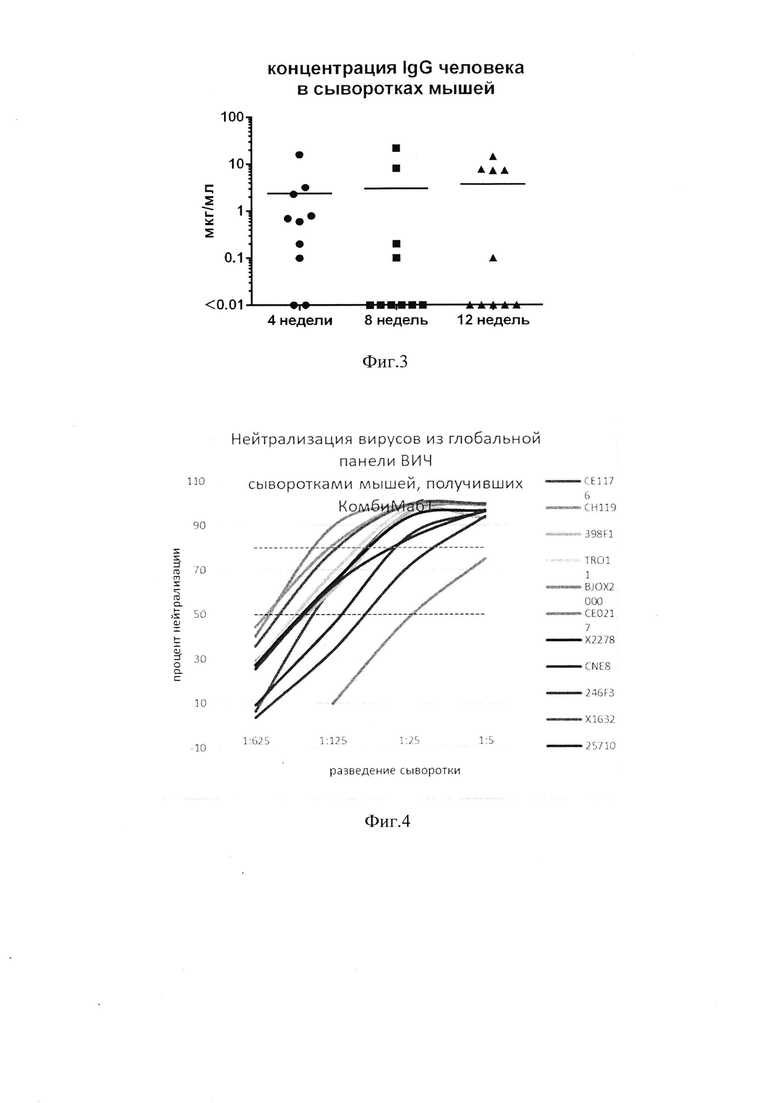
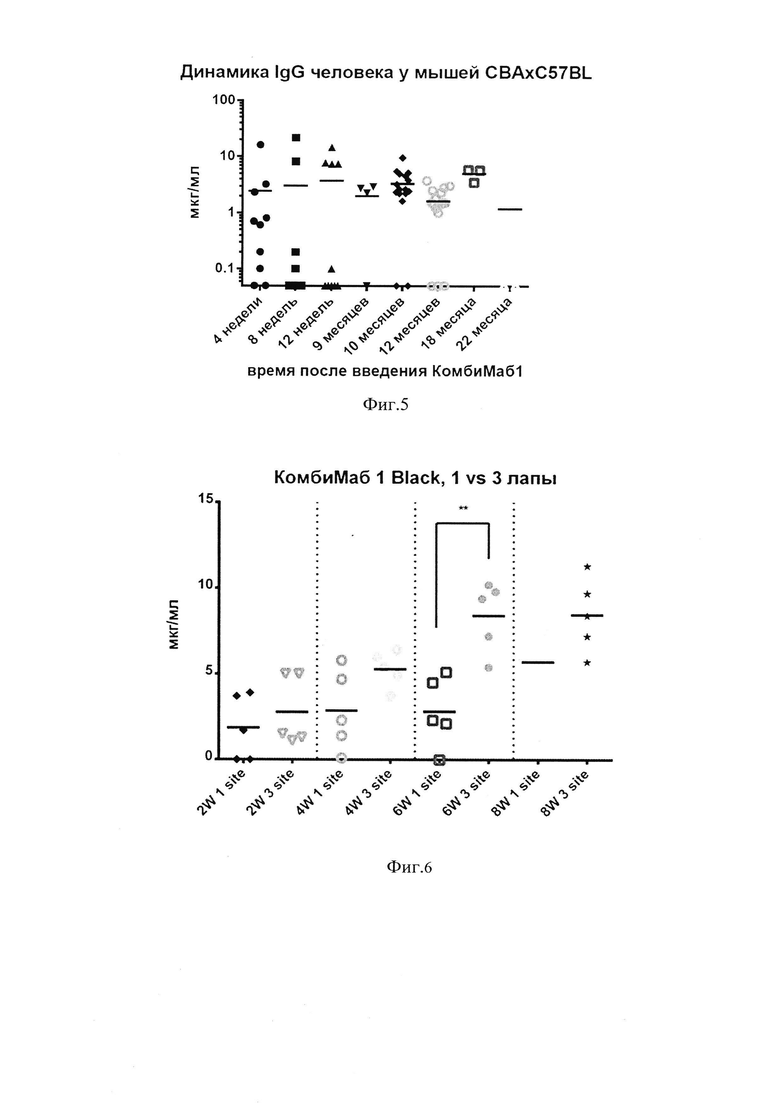
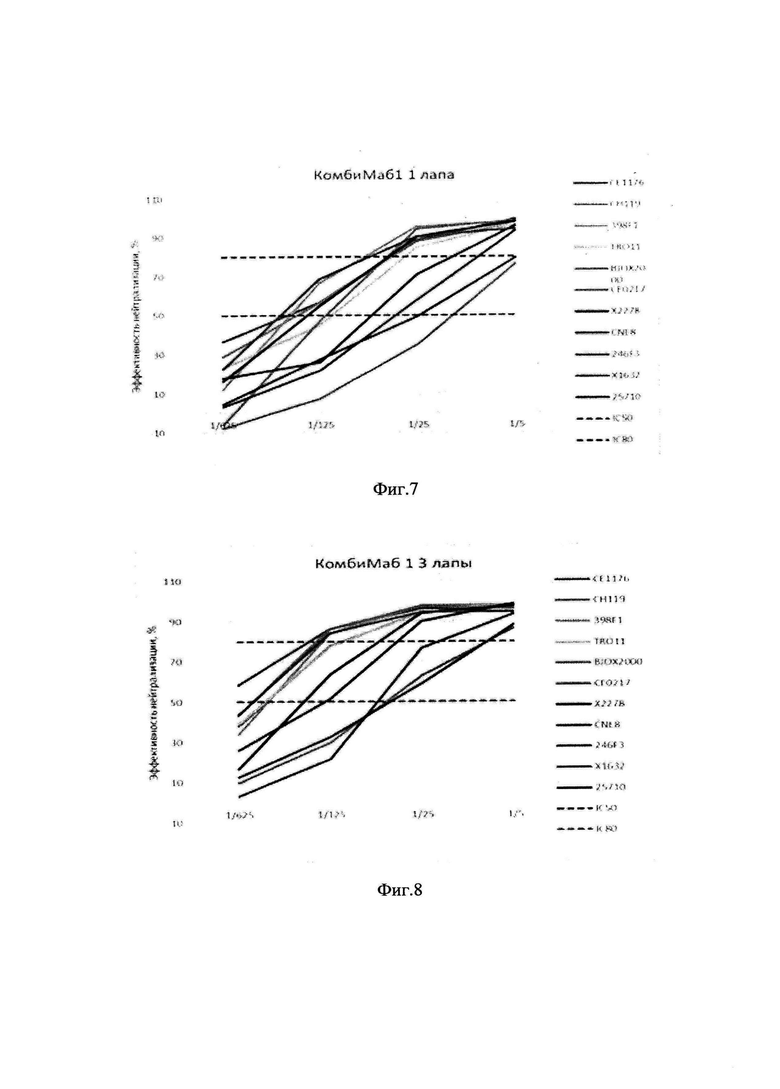
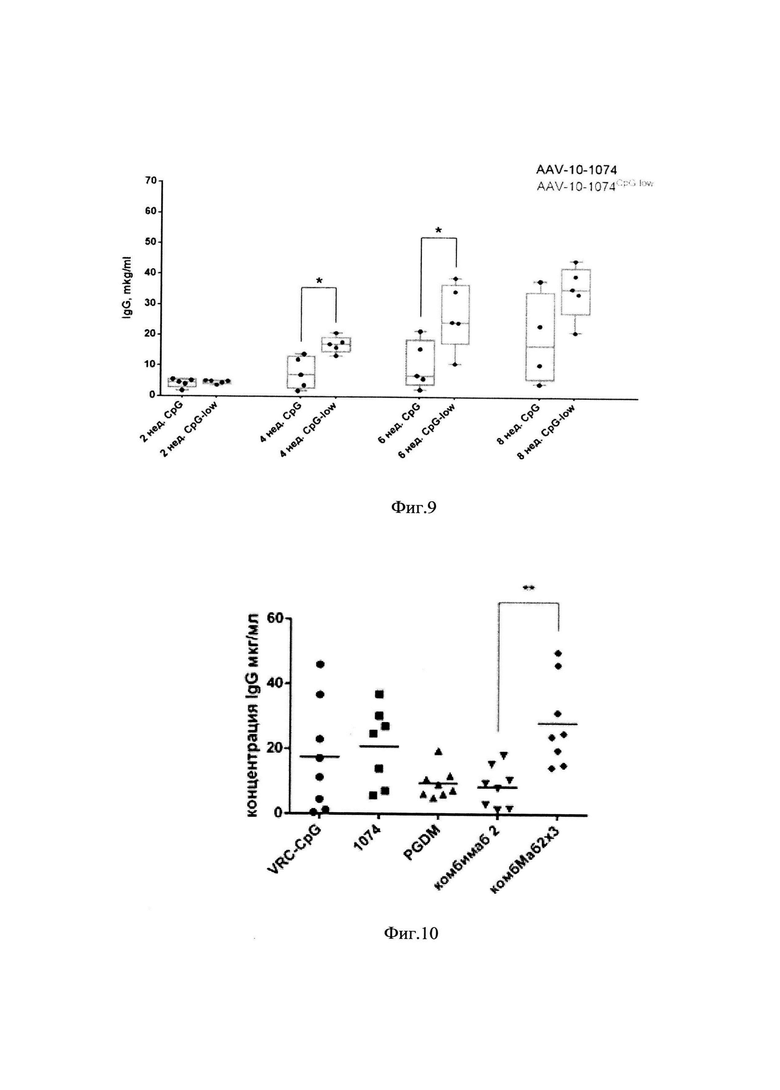
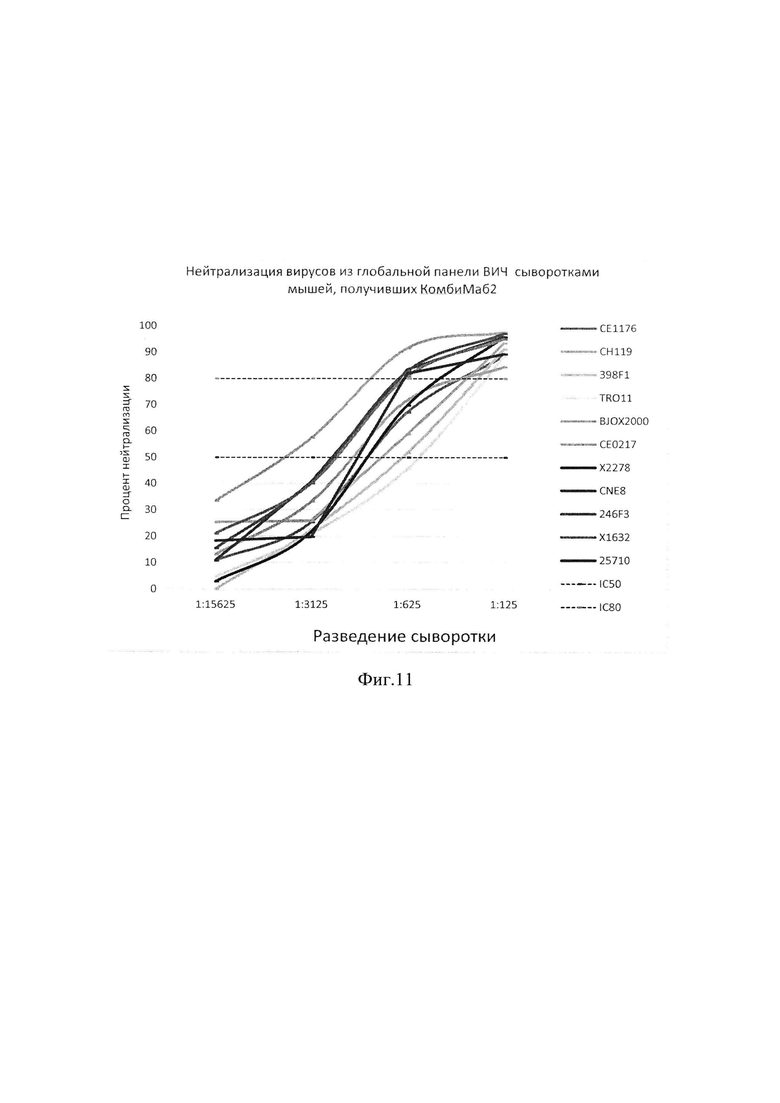
Изобретение относится к области биотехнологии, а именно к рекомбинантным генетическим векторам на базе аденоассоциированного вируса, кодирующим широко нейтрализующие антитела против вируса иммунодефицита человека (ВИЧ-1), а также к иммунобиологическим препаратам на их основе для лечения и профилактики ВИЧ-инфекции за счет обеспечения длительной персистенции широко нейтрализующих антител против ВИЧ-1. Описан набор рекомбинантных генетических векторов, кодирующих широко нейтрализующие антитела против ВИЧ-1, направленные на разные эпитопы вирусного белка Env, где в качестве рекомбинантного генетического вектора используется аденоассоциированный вектор серотипа 9. Представлен иммунобиологический препарат для лечения или профилактики ВИЧ-1-инфекции, содержащий комбинацию нескольких аденоассоциированных векторов серотипа 9, выбранных из набора. Также раскрыт способ применения иммунобиологического препарата, характеризующийся тем, что препарат вводят внутримышечно в виде смеси аденоассоциированных векторов серотипа 9 или каждый аденоассоциированный вектор серотипа 9, входящий в состав комбинации, вводят по отдельности. Технический результат - создание иммунобиологического препарата для лечения и профилактики ВИЧ-инфекции на основе различных комбинаций рекомбинантных генетических векторов на основе аденоассоциированного вируса серотипа 9, экспрессирующих широко нейтрализующие антитела против ВИЧ-1. 3 н. и 5 з.п. ф-лы, 11 ил., 11 табл.
1. Набор рекомбинантных генетических аденоассоциированных векторов серотипа 9, кодирующих широко нейтрализующие антитела против ВИЧ-1, направленные на разные эпитопы вирусного белка Env, для получения иммунобиологического препарата для лечения ВИЧ-инфекции.
2. Набор рекомбинантных генетических аденоассоциированных векторов серотипа 9 по п. 1, где рекомбинантные генетические вектора, кодирующие широко нейтрализующие антитела против ВИЧ-1, выбранные из группы:
i. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074;
ii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10Е8;
iii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело N6;
iv. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело VRC07-523;
v. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело PGDM1400.
3. Набор рекомбинантных генетических аденоассоциированных векторов серотипа 9 по п. 2, где аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело VRC07-523, имеет нуклеотидную последовательность со сниженным содержанием CG последовательностей, аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеет нуклеотидную последовательность со сниженным содержанием CG последовательностей, аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело PGDM1400, имеет нуклеотидную последовательность со сниженным содержанием CG последовательностей.
4. Набор рекомбинантных генетических аденоассоциированных векторов серотипа 9 по п. 2 или 3, где рекомбинантные генетические вектора, кодирующие широко нейтрализующие антитела против ВИЧ, имеют следующие нуклеотидные последовательности:
i. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеет нуклеотидную последовательность SEQ ID NO: 1;
ii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10Е8, имеет нуклеотидную последовательность SEQ ID NO: 2;
iii. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело N6, имеет нуклеотидную последовательность SEQ ID NO: 3;
iv. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело VRC07-523, имеет нуклеотидную последовательность SEQ ID NO: 4;
v. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеет нуклеотидную последовательность SEQ ID NO: 5;
vi. аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело PGDM1400, имеет нуклеотидную последовательность SEQ ID NO: 6.
5. Иммунобиологический препарат для лечения или профилактики ВИЧ-1-инфекции, содержащий комбинацию нескольких аденоассоциированных векторов серотипа 9, выбранных из набора по п. 4.
6. Иммунобиологический препарат по п. 5, отличающийся тем, что содержит комбинацию из аденоассоциированных векторов серотипа 9: аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеет нуклеотидную последовательность SEQ ID NO: 1; аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10Е8, имеет нуклеотидную последовательность SEQ ID NO: 2; аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело N6, имеет нуклеотидную последовательность SEQ ID NO: 3; или комбинацию из аденоассоциированных векторов серотипа 9: аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело VRC07-523, имеет нуклеотидную последовательность SEQ ID NO: 4; аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело 10-1074, имеет нуклеотидную последовательность SEQ ID NO: 5; аденоассоциированный вектор серотипа 9, кодирующий широко нейтрализующее антитело PGDM1400, имеет нуклеотидную последовательность SEQ ID NO: 6.
7. Иммунобиологический препарат по п. 6, отличающийся тем, представляет собой смесь аденоассоциированных векторов серотипа 9, входящих в комбинацию векторов, при эквимолярном соотношении компонентов или набор отдельных аденоассоциированных векторов серотипа 9, входящих в комбинацию.
8. Способ применения иммунобиологического препарата по пп. 5-7, характеризующий тем, что препарат по любому из пп. 5-7 вводят внутримышечно в виде смеси аденоассоциированных векторов серотипа 9 или каждый аденоассоциированный вектор серотипа 9, входящий в состав комбинации, вводят по отдельности.
| RU 2016104614 A, 18.08.2017 | |||
| WO 2020223240 A1, 05.11.2020 | |||
| С.В | |||
| КОРОБОВА, Г.В | |||
| КОРНИЛАЕВА и др., ИММУНОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА СИНТЕТИЧЕСКИХ ПЕПТИДОВ, КОПИРУЮЩИХ АКТУАЛЬНЫЕ АНТИГЕННЫЕ ДЕТЕРМИНАНТЫ ВИЧ, Медицинская иммунология, 2016, т | |||
| Способ использования делительного аппарата ровничных (чесальных) машин, предназначенных для мериносовой шерсти, с целью переработки на них грубых шерстей | 1921 |
|
SU18A1 |
| Способ запрессовки не выдержавших гидравлической пробы отливок | 1923 |
|
SU51A1 |

