Изобретение относится к искусственному гену YkuJ-MPER, кодирующему химерный белок-иммуноген YkuJ-MPER, рекомбинантной плазмидной ДНК рЕТ21a-YkuJ-MPER, обеспечивающей экспрессию искусственного гена YkuJ-MPER, и химерному белку-иммуногену YkuJ-MPER, являющемуся носителем мембранно-проксимальной области ВИЧ-1 и направленному на индукцию в организме широконейтрализующих антител и может быть использовано в качестве компонента вакцины против ВИЧ-1 в области биотехнологии, генной инженерии и медицины.
Общее число носителей ВИЧ-инфекции в мире на данный момент составляет более 36 миллионов [1]. В России ситуация с ВИЧ-инфекцией остается по-прежнему одной из самых неблагополучных в Восточной Европе и Азии. Российская Федерация - одна из стран в мире, где число новых заражений ВИЧ-инфекцией и число смертей, связанных с ВИЧ/СПИД, продолжают возрастать. На 31 декабря 2016 года в России официально зарегистрировано 1114815 ВИЧ-инфицированных людей [2]. Вакцины считаются одним из самых эффективных средств борьбы с инфекционными заболеваниями. В случае вируса иммунодефицита человека классические подходы по созданию вакцины, такие как использование аттенуированного или инактивированного вируса, оказались не эффективны. В связи с этим разрабатываются альтернативные технологии создания вакцин.
С момента открытия вируса иммунного дефицита человека было проведено огромное количество исследований и экспериментов, касающихся происхождения, патогенеза, разработки антиретровирусных препаратов и вакцин против ВИЧ-1. Для создания вакцин против ВИЧ-1 было испробовано множество подходов, включая использование инактивированных, аттенуированных форм вируса, субъединичных вакцин, состоящих из отдельных вирусных гликопротеинов, создание генно-инженерных конструкций, ДНК-вакцин, а также препаратов на основе рекомбинантных вирусных и бактериальных векторов [3]. Однако ни одна из кандидатных вакцин не продемонстрировала достаточный для широкого применения в клинике уровень эффективности.
Относительным успехом в области создания анти-ВИЧ вакцин можно считать испытание вакцины RV144 в 2009 году (совместная разработка компаний Мерк и Санофи Пастер) в Таиланде [4, 5]. Вакцинный препарат ALVAC-HIV vCP1521/AIDSVAX В/Е, используемый в испытаниях RV144, включал в свой состав иммуногены, предназначенные для одновременной стимуляции В- и Т-клеточного иммунного ответа. В качестве праймирующего антигена использовался рекомбинантный вирус оспы канареек, экспрессирующий гены env, gag и протеазы ВИЧ-1 (ALVAC-HIV vCP1521), а бустерную вакцинацию осуществляли с помощью препарата AIDSVAX В/Е (Vax003) на основе гликопротеина gp120 ВИЧ-1. Данное сочетание обеспечило снижение риска инфицирования на 31,2% [4]. Более детальный анализ результатов показал, что в первый год испытаний вакцина обеспечивала защиту на уровне 60%, после чего в крови вакцинированных начинала падать концентрация ВИЧ-специфических антител. Дальнейшие исследования позволили установить причину невысоких показателей защиты от ВИЧ, связанных, по-видимому, с бустирующим компонентом AIDSVAX, ответственным за индукцию гуморального ответа. Выяснилось, что антитела, образующиеся в ответ на введение вакцины, либо не способны нейтрализовать вирус, либо обладают слабой нейтрализующей активностью [6, 7].
Несмотря на то, что ВИЧ-1 без приема антиретровирусной терапии является смертельным, скорость развития СПИДа и сила иммунного ответа у отдельных людей могут сильно различаться. За прошедшие с момента открытия ВИЧ-1 годы был описан феномен ВИЧ-инфицированных долгожителей (non-progressors) - людей, у которых, несмотря на ВИЧ-инфекцию, не развиваются симптомы СПИДа даже без соответствующего лечения. В этой группе ВИЧ-инфицированных описана когорта людей, антитела сывороток которых способны нейтрализовать широкий спектр первичных изолятов ВИЧ-1. Моноклональные антитела, полученные на основе разнообразия иммуноглобулинов иммунной системы от таких ВИЧ-инфицрованных доноров, получили название широконейтрализующих (broadly neutralizing antibodies, bnAbs). По всей видимости, длительное аффинное созревание (которое может продолжаться более года) приводит к тому, что у большинства людей bnAbs появляются в организме с сильным опозданием, и к этому моменту уже неспособны существенно снизить вирусную нагрузку. Считается, что появление подобных антител в результате вакцинации способно предотвратить заражение ВИЧ-1. В связи с этим в настоящее время значительная часть исследований в области вакцинации против ВИЧ-1 направлена на разработку стратегий иммунизации и иммуногенов, которые будут вызывать защитные антитела, в том числе широконейтрализующие (bnAbs). Эта цель подтверждается данными о способности широконейтрализующих mAb (monoclonal antibodies) при воздействии на слизистую оболочку, предотвращать у макак заражение химерным вирусом SHIV, несущим env ВИЧ [8, 9, 10, 11]. Нейтрализующие поликлональные антитела в высоких концентрациях могут блокировать проникновение вируса в клетку и, как было показано, уменьшают тяжесть заболевания на модели приматов [12, 13, 14].
Однако использование моноклональных широконейтрализующих антител возможно только в качестве терапевтического, а не профилактического средства. Кроме того, из-за высокой стоимости моноклональных антител данная терапия может быть доступна лишь очень небольшому проценту населения. Тем не менее, учитывая уникальные свойства таких антител, очень перспективной выглядит идея создания иммуногена, способного индуцировать их наработку. Предложен ряд подходов для достижения этой цели.
Один из распространенных подходов, использующихся при создании подобных иммуногенов, основан на концепции рационального дизайна поверхностного гликопротеина gp120 ВИЧ-1, в составе которого находится сайт связывания вируса с клеточным рецептором CD4 (CD4-binding site, CD4bs), модифицированного таким образом, чтобы он содержал только те эпитопы, которые узнаются нейтрализующими антителами широкого спектра действия. Для этого вносят изменения, позволяющие стабилизировать структуру молекулы (точечные мутации, введение модификаций, приводящих к образованию дисульфидных связей), а также удаляют вариабельные фрагменты, которые маскируют эпитопы bnAbs [15].
Другой подход заключается в создании не отдельных фрагментов gp120, а цельных тримеров со стабилизированной структурой, способных имитировать нативную конформацию поверхностного белка Env ВИЧ-1 [16, 17, 18]. В качестве альтернативного варианта было предложено использовать мономерную стабилизированную форму оболочечного гликопротеина вируса для индукции антител к CD4bs - ключевому участку gp120, ответственному за связывание с CD4. Для получения иммуногена в соответствии с таким подходом сердцевина белка Env модифицируется с помощью направленного мутагенеза таким образом, чтобы полученная конструкция имитировала CD4bs сайт [19, 20, 21].
Однако в ходе многочисленных исследований было установлено, что иммунизация целым комплексом Env ВИЧ-1 или его субъединицами, такими как мономерные gp120, не обеспечивают воспроизводимого образования bnAbs. Более того, вследствие дороговизны и сложности данных методов пока не было создано ни одного вакцинного препарата, основанного на данной концепции.
Помимо описанных подходов и экспериментальных вакцин известен вакцинный препарат против ВИЧ-1 «ВИЧРЕПОЛ» (ГНЦ Институт иммунологии) [22]. Основной компонент вакцины - химерный белок rec (24-41), состоящий из фрагментов иммуногенных белков р24, gp41 и р17 ВИЧ-1, конъюгированный с иммуностимулятором - носителем полиоксидонием (фармакологически активное соединение с молекулярной массой 100 кД с выраженной иммуномодулирующей активностью) [23]. Первая фаза клинических испытаний показала безопасность, иммуногенность и антигенную активность представленной вакцины. Кроме того, при иммунизации лабораторных животных данная вакцина вызывает наработку специфических антител к белкам р24 и gp41 ВИЧ-1, входящих в состав вакцины. Однако эти антитела обладают низкой вирус-нейтрализующей активностью: индекс нейтрализации составляет 54,9% при разведении иммунной сыворотки 1:50.
Одним из перспективных и технологически более простым подходом является использование каркасных молекул, которые экспонируют эпитопы ВИЧ-1, узнаваемые нейтрализующими антителами широкого спектра действия. Подобные стратегии сосредоточены на консервативных нейтрализующих эпитопах Env ВИЧ-1, таких как мембрано-проксимальная наружная область gp41 (membrane-proximal external region, MPER). MPER участвует в слиянии мембран вируса и клетки-мишени и содержит сайты связывания для bnAbs ВИЧ-1 2F5, 4Е10, 10Е8 и др [24, 25]. Некоторые из этих MPER-специфических bnAbs, такие как 10Е8, также обладают сильной антителозависимой клеточно-опосредованной цитотоксичностью (ADCC) [26]. При этом даже когда варианты ВИЧ-1 развивают более высокую устойчивость к bnAbs, нацеленным на gp120, их чувствительность к bnAbs, нацеленная на MPER, остается неизменной [27]. Соответственно, MPER рассматривается как одна из наиболее перспективных мишеней разработки вакцин для индукции bnAbs ВИЧ-1. Так как прямое использование аминокислотной последовательности MPER в качестве пептидного иммуногена не принесло успеха, необходимым является корректная презентация эпитопов bnAbs MPER иммунной системе. Способом презентации этого региона иммунной системе могут быть искусственные иммуногены, полученные при помощи рационального дизайна.
Так как индуцировать широконейтрализующие антитела с помощью одного иммуногена с использованием типичных подходов к вакцинации на данный момент практически невозможно, Banerjee с сотрудниками [28] разработали стратегию, которая называется инкрементальной (поэтапной) фазированной антигенной стимуляцией быстрого созревания антител (Incremental, Phased Antigenic Stimulation for Rapid Antibody Maturation, or IPAS-RAM). Основная идея данной концепции состоит в том, чтобы направить эволюцию иммунной системы с использованием небольшого пептида, полученного из MPER, для стимуляции индукции широконейтрализующих антител против MPER, а затем выборочно амплифицировать те, которые связывают нативную структуру. Уникальность стратегии IPAS-RAM заключается в том, что иммунная система подвергается воздействию разных, но взаимосвязанных иммуногенов поэтапно, так что В-клетки, стимулированные более мелким иммуногеном, могут одновременно взаимодействовать с общими эпитопами на более крупном иммуногене. Предполагается, что, используя цепочку иммуногенов, можно "нацелить" иммунную систему на нейтрализующие эпитопы MPER, при этом созревание антител будет проходить более эффективно, чем обычно при инфекции. Была оценена стратегия вакцинации IPAS-RAM, где использовалось три иммуногена у кроликов: полипептид на основе MPER, мини-белок gp41, связанный с мембраной, и полноразмерный gp160. Предполагалось, что прайминг-антиген на основе пептида будет высокоэффективным в индукции антител против MPER, и что последующие иммунизации более крупными и «нативными» антигенами позволит селективным антителам созревать в bnAbs, способным связывать gp41, как это видно на нативном тримере. На основании проведенного исследования, данной группе не удалось выявить нейтрализующие антитела, но их исследования демонстрируют потенциальные преимущества последовательной иммунизации несколькими иммуногенами поэтапно.
Наиболее близким аналогом (прототипом) является исследования автора Sun с сотрудниками [29]. На основе участка связывания антитела 10Е8 они спроектировали иммуноген, содержащий четыре копии этого эпитопа (10Е8-4Р) и пептид, содержащий одну копию данного эпитопа (10Е8-1Р). Затем, была проведена оценка их способности индуцировать антитела с нейтрализующей активностью. Для этого самок новозеландских белых кроликов иммунизировали 50 мкг пептида 10Е8-1Р и иммуногеном 10Е8-4Р в комплексе с полным адъювантом Фрейнда. Повторные иммунизации проводили с интервалом в две недели с увеличивающейся концентрацией 10Е8-4Р в комплексе с неполным адъювантом Фрейнда. Методом иммуноферментного анализа было показано, что в организме кроликов нарабатываются антитела, связывающиеся как с пептидом 10Е8-1Р, так и с иммуногеном 10Е8-4Р. При этом антитела из сывороток кроликов, иммунизированных 10Е8-4Р, связываются как с пептидом, так и с иммуногеном, а антитела из сывороток кроликов, иммунизированных 10Е8-1Р, связываются только с пептидом 10Е8-1Р. В тесте вируснейтрализации было показано, что антитела в сыворотках кроликов иммунизированных 10Е8-4Р обладают слабой нейтрализующей активностью, в то же время антитела из сывороток после иммунизации только пептидом 10Е8-1Р не проявляли никакой нейтрализующей активности [29]. Недостаток данного подхода заключался в том, что при создании описываемой конструкции не учитывался тот факт, что аминокислотное окружение эпитопа, при нахождении его в составе гликопротеина gp41, оказывает влияние на его конформацию. Данное обстоятельство влияет на образование характерных (10Е8-подобных) антител. Также, недостатком данной разработки является использование одного эпитопа широконейтрализующего антитела, входящего в MPER ВИЧ-1.
Техническим результатом заявляемого изобретения является разработка рекомбинантного химерного белка-иммуногена YkuJ-MPER на основе белка В. subtilis YkuJ, содержащего два участка MPER ВИЧ-1, использование которого при подборе оптимальной стратегии иммунизации может обеспечить эффективную индукцию широконейтрализующих анти-ВИЧ-1 антител.
Указанный технический результат достигается тем, что искусственный ген YkuJ-MPER, кодирующий бело к-иммуноген YkuJ-MPER против ВИЧ-1, имеет следующую нуклеотидную последовательность SEQ ID NO: 1 длиной 360 п. н.:
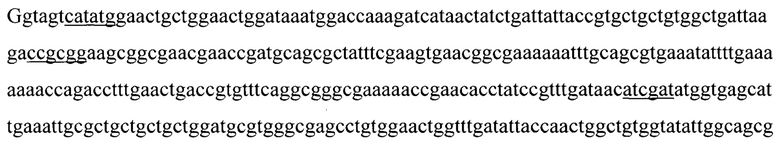
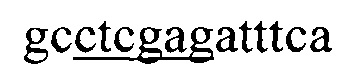 (360 п. н.).
(360 п. н.).
Указанный технический результат достигается также тем, что рекомбинантная плазмидная ДНК рЕТ21a-YkuJ-MPER, имеющая размер 5700 п. н., молекулярную массу 3,8×103 кДа, содержит целевой ген по п. 1, кодирующий химерный белок YkuJ-MPER, находящийся под контролем промотора РНК-полимеразы фага Т7, обеспечивающего его экспрессию в клетках Е. coli и состоящая из следующих фрагментов:
- NdeI-XhoI - векторного фрагмента ДНК плазмиды рЕТ21а(-) размером 5340 п.н., содержащего промотор и терминатор РНК-полимеразы фага Т7 и регуляторную область лактозного оперона, обеспечивающие индуцибельную экспрессию гена YkuJ-MPER в клетках Е. coli; промотор AmpR, обеспечивающий эффективную транскрипцию мРНК гена bla, ген устойчивости к ампициллину (bla), кодирующий белок бета-лактамазу, и (ori) точка начала репликации ColE1, обеспечивающие селекцию и размножение целевой плазмиды в клетках бактерий Escherichia coli; С-терминальный His-tag (His×6) в рамке считывания с геном YkuJ-MPER.
- NdeI-XhoI - фрагмента размером 360 п.н., содержащего ген YkuJ-MPER с инициирующим кодоном ATG, полученного путем обработки рестриктазами NdeI-XhoI плазмиды рЕТ21а;
- уникальных сайтов рестрикции, существенных для генно-инженерных манипуляций: NdeI - 5203, SacII - 5287, ClaI - 5434, XhoI - 5538, и имеющая следующее положение генов:
Указанный технический результат достигается также тем, что искусственный белок-иммуноген YkuJ-MPER, состоящий из участка белка В. subtilis YkuJ и двух участков MPER ВИЧ-1, имеет молекулярную массу 14,2 кДа и следующую аминокислотную последовательность SEQ ID NO: 2:

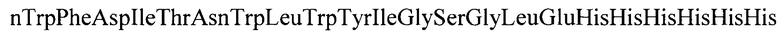 (119 а.о)
(119 а.о)
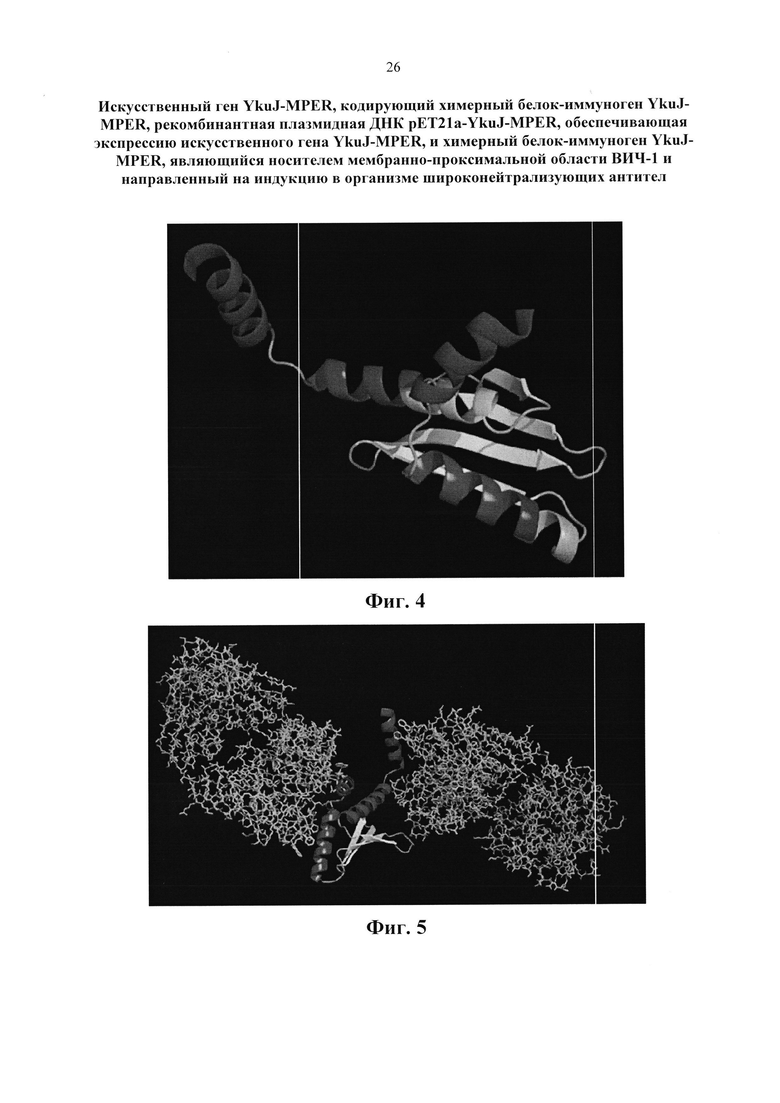
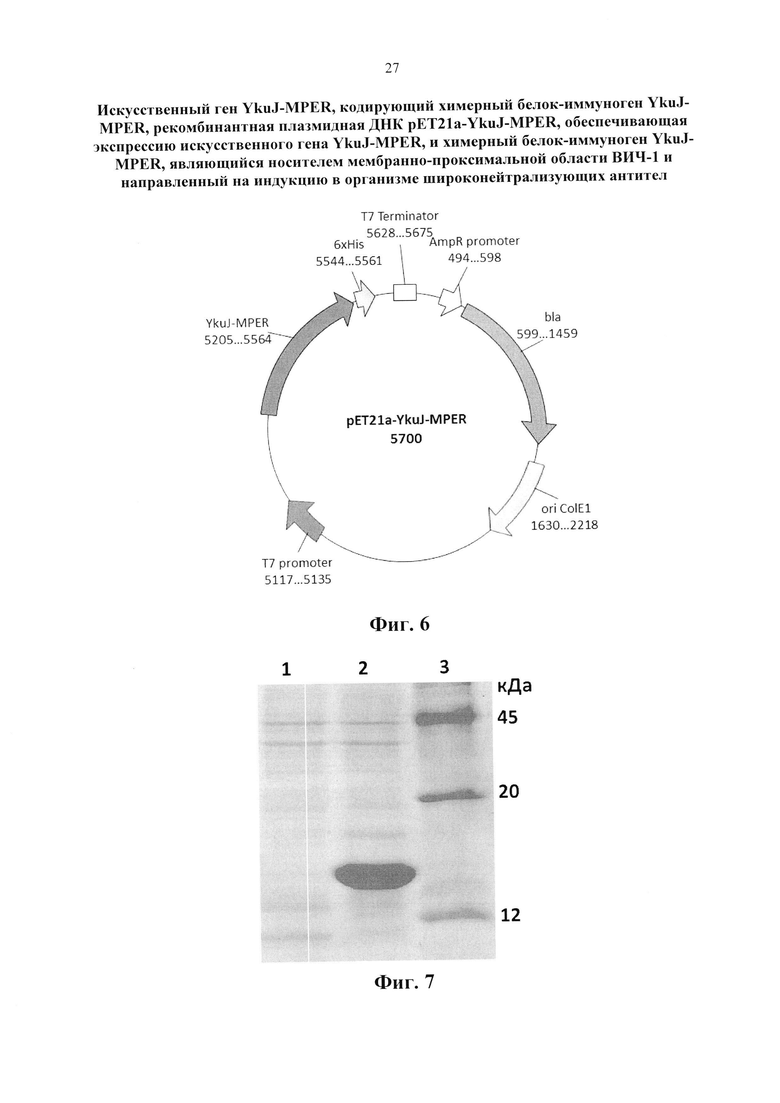
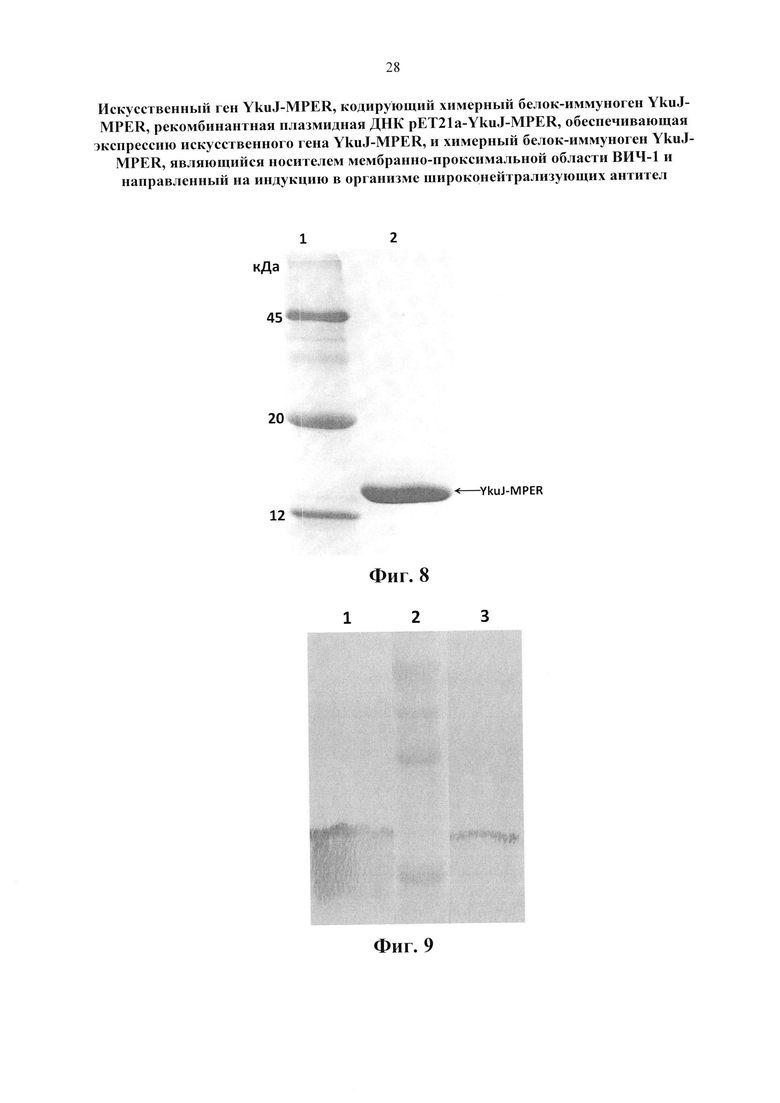
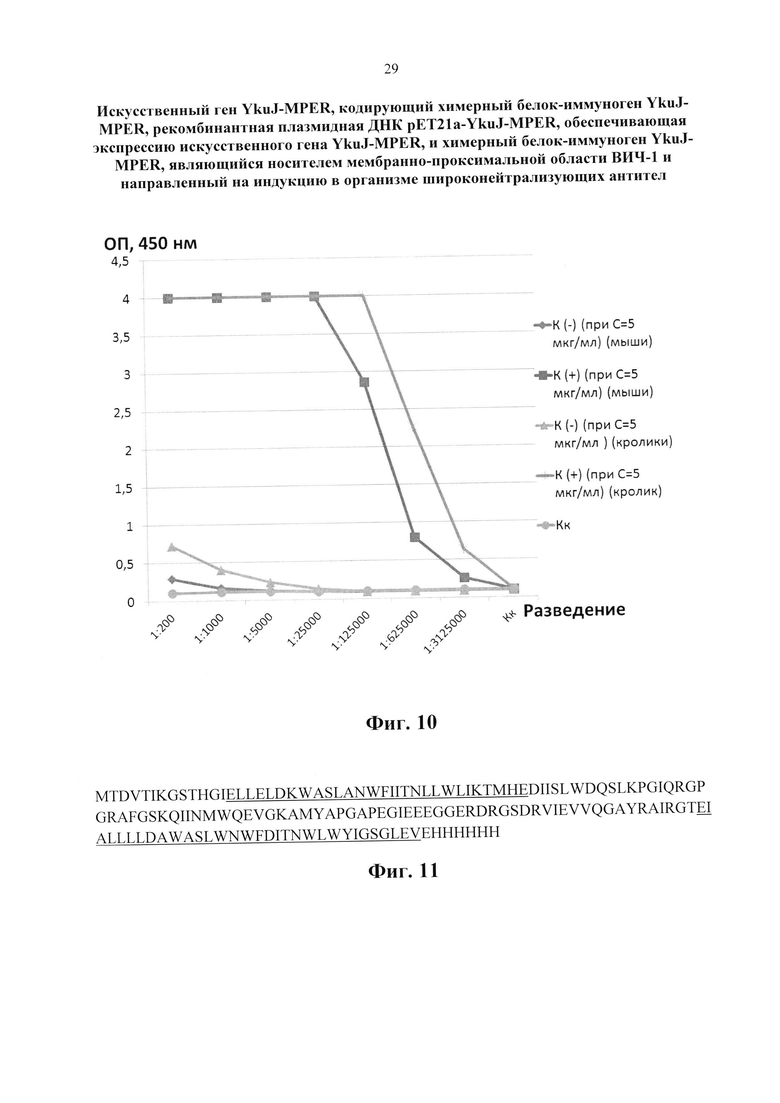
Изобретение поясняется следующими графическими материалами. На фиг. 1 представлена модель каркасного белка В. subtilis YkuJ. На фиг. 2 приведена нуклеотидная последовательность гена YkuJ-MPER, где подчеркиванием выделены сайты рестрикции NdeI, SacII, ClaI, XhoI соответственно. На фиг. 3 представлена аминокислотная последовательность белка, кодируемого геном YkuJ-MPER, а на фиг. 4 - модель химерного белка YkuJ-MPER. Участки MPER в составе каркасного белка В. subtilis YkuJ имеют альфа-спиральную структуру. На фиг. 5 представлена модель взаимодействия конструкции YkuJ-MPER с Fab-фрагментами антитела 10Е8. Альфа-спиралями обозначены MPER ВИЧ-1 в составе белка В. subtilis YkuJ. По левую и правую сторону от альфа-спирали обозначены Fab-фрагменты антитела 10Е8. На фиг. 6 изображена карта рекомбинантной плазмиды pET21a-YkuJ-MPE. На фиг. 7 представлена электрофореграмма лизатов клеток Е. coli ВL21/pYkuJ-MPER в 15% ПААГ, где: 1 - отрицательный контроль (культура Е. coli BL21 без плазмиды); 2 - лизат клеток Е. coli BL21/pYkuJ-MPER; 3 - маркер молекулярной массы (12, 20, 45 кДА). На фиг. 8 представлена электрофореграмма очищенного препарата белка YkuJ-MPER, где: 1 - маркер молекулярной массы (12, 20, 45 кДА), 2 - Препарат белка YkuJ-MPER. На фиг. 9 представлен вестерн-блот белка YkuJ-MPER после разделения в 15% ПААГ с моноклональным антителом 2F5 (дорожка 1) и 10Е8 (дорожка 3), дорожка 2 - маркер молекулярной массы (12, 20, 45 и 66 кДА). На фиг. 10 приведены результаты иммуноферментного анализа сывороток крови иммунизированных YkuJ-MPER лабораторных животных (мыши и кролики), где в качестве антигена был сорбирован белок YkuJ-MPER. На фиг. 11 приведена аминокислотная последовательность белка, используемого для определения титра антител сывороток животных, иммунизированных YkuJ-MPER, специфически взаимодействующих с MPER ВИЧ-1 (MPER-TBI), где подчеркиванием выделены участки MPER ВИЧ-1 (MPER-TBI). На фиг. 12 приведены результаты иммуноферментного анализа сывороток крови иммунизированных YkuJ-MPER лабораторных животных (мыши и кролики), где в качестве антигена был сорбирован белок MPER-TBI.
Заявляемый химерный полипептид-иммуноген, направленный на индукцию нейтрализующих ВИЧ-1 антител широкого спектра действия, получают следующим образом.
При разработке химерного иммуногена YkuJ-MPER за основу были взяты белок В. subtilis YkuJ и участок белка gp41 ВИЧ-1 MPER. Данный белок В. subtilis был выбран потому, что, имея небольшой размер, позволяет встроить два района MPER таким образом, что антитела могут связаться одновременно с двумя районами, не мешая друг другу. При этом районы, находясь на концах белка, могут принимать конфирмацию, характерную для эпитопов различных известных моноклональных антител: 2F5 и Z13 (конформация без регулярной вторичной структуры), 4Е10 и 10Е8 (альфа-спиральная конформация). Предполагается, что данное условие может обеспечить образование специфических в отношении данного региона антител, которые будут способны нейтрализовать ВИЧ-1.
Спроектированный ген клонировали в составе вектора рЕТ21а по сайтам, узнаваемым эндонуклеазами рестрикции XhoI и NdeI. Полученной конструкцией трансформировали клетки Е. coli BL21 (DE3), биомассу трансформированных клеток наращивали в течение ночи на среде YT×2. Полученную биомассу обрабатывали с помощью ультразвукового дезинтегратора, центрифугировали и ресуспендировали в лизирующем буфере, содержащем 8 М мочевину. Очистку целевого белка проводили с помощью металл-хелатной хроматографии на колонке с Ni-NTA-агарозой. Для перевода белка в нативное состояние использовали ступенчатый диализ против буферов со снижающейся концентрацией мочевины. Наличие фракции целевого белка и степень очистки оценивали с помощью электрофореза в 15% полиакриламидном геле (ПААГ) по Лэммли [30].
Для подтверждения того, что эпитопы широконейтрализующих антител, входящие в MPER ВИЧ-1 в составе белка YkuJ-MPER, связываются с известными моноклональными антителами против этого региона, такими как 2F5 и 10Е8, проводили вестерн-блот анализ. Он подтвердил, что эпитопы в составе химерного белка YkuJ-MPER распознаются МКА 2F5 и 10Е8, что подтверждает теоретическую структуру иммуногена.
Для анализа иммуногенности химерного белка очищенным препаратом YkuJ-MPER были иммунизированы мыши линии BALB/c. Было установлено, что полученный препарат не токсичен для лабораторных животных.
Полученные сыворотки лабораторных животных после иммунизации были протестированы в иммуноферментном анализе для оценки иммуногенности химерного белка YkuJ-MPER. Было установлено, что антитела из сывороток иммунизированных животных специфически взаимодействуют с рекомбинантным белком YkuJ-MPER. Титр сывороток иммунизированных животных для мышей составил 1:625 тысяч, для кроликов 1:3 миллиона.
Для анализа наличия антител из сывороток животных иммунизированных YkuJ-MPER, специфически взаимодействующих с MPER ВИЧ-1, был использован другой белок, также содержащий два участка MPER ВИЧ-1 (TBI-MPER). Анализ показал, что титр сывороток животных, иммунизированных YkuJ-MPER, в ИФА к TBI-MPER составляет для мышей 1:125 тысяч, для кроликов 1:625 тысяч. Исходя, из полученных результатов можно сделать вывод о том, что в организме лабораторных животных образуются антитела, специфичные к MPER ВИЧ-1 (Фиг. 12).
Пример 1. Проектирование гена YkuJ-MPER
Для конструирования химерного белка иммуногена YkuJ-MPER с помощью классификатора белковых структур SCOP был выбран глобулярный белок В. subtilis YkuJ (номер PDB 2FFG) с известной структурой (Фиг. 1). Данный белок был выбран потому, что при небольшом размере его пространственная структура позволяет встроить два района MPER таким образом, что антитела могут связаться одновременно с двумя районами, не мешая друг другу. Его концевые участки являются спиральными, что соответствует конформации эпитопов bnAbs 4Е10 и 10Е8. Способность связываться одновременно с двумя антителами без стерических проблем была подтверждена методами молекулярного моделирования. (Фиг.4, Фиг. 5).
При проектировании гена YkuJ-MPER, N- и С-концы белка YkuJ были частично заменены на две последовательности MPER ВИЧ-1 консенсусной последовательности субтипа В. В состав нуклеотидной последовательности гена YkuJ-MPER были заложены уникальные сайты рестрикции NdeI, SacII, ClaI, XhoI, фланкирующие нуклеотидную последовательность MPER, для того, чтобы в последующем каждый из этих участков было нетрудно заменить на MPER других генетических вариантов ВИЧ-1, а так же для клонирования гена в составе плазмидных векторов.
Ниже приведена нуклеотидная последовательность гена YkuJ-MPER, где подчеркиванием выделены сайты рестрикции Ndel, SacII, Clal, Xhol соответственно (SEQ ID NO: 1):
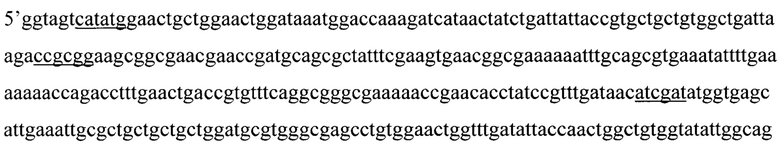
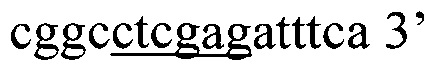 (360 п.н.), (Фиг. 2).
(360 п.н.), (Фиг. 2).
Аминокислотная последовательность белка, кодируемого геном YkuJ-MPER (SEQ ID N0:2) имеет следующий вид:

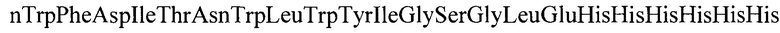 (119 а.о)
(119 а.о)
Пример 2. Получение плазмиды рЕТ21а- YkuJ-MPER
В качестве экспрессионного вектора была выбрана плазмида рЕТ21а (Novagen). Конструирование плазмиды проводили с помощью стандартных методов молекулярного клонирования [31]. Теоретически рассчитанный ген, кодирующий белок YkuJ-MPER, был химически синтезирован и клонирован в составе плазмиды pGH компанией «Евроген».
После этого ген химерного белка YkuJ-MPER амплифицировали с плазмиды pGH с использованием пары праймеров, содержащих сайты гидролиза рестриктаз NdeI и XhoI (таблица 1).

Реакцию проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия), при следующих параметрах реакции: начальная денатурация 2 мин при 94°C, последующие 25 циклов 15 с - 94°C, 30 с - 55°C, 90 с - 72°C, один цикл 1 мин - 72°C.
Затем ПЦР-продукт и плазмиду рЕТ21а обрабатывали изошизомерами эндонуклеаз рестрикции NdeI и XhoI «СибЭнзим», полученные продукты ДНК выделяли с помощью набора Cleanup Standard «Евроген». Очищенный вектор и ген смешивали в эквимолярных концентрациях и лигировали ДНК-лигазой бактериофага Т4 («СибЭнзим»). Реакция проводилась при +37°C в течение одного часа. Полученной лигазной смесью трансформировали клетки Е. coli JM103 методом электропорации согласно методике, описанной в [31]. Трансформированные клетки высевали на чашки с LB агаром, содержащим 100 мкг/мл ампициллина. Чашки помещали в термостат 37°C и инкубировали в течении ночи. На следующий день были идентифицированы отдельные колонии JM103/рЕТ21a-YkuJ-MPER.
После трансформации отдельные клоны засевали в жидкой среде LB с антибиотиком ампициллином 100 мкг/мл для наработки плазмидной ДНК. Плазмидную ДНК из бактериальной массы выделяли с помощью набора Plasmid Miniprep «Евроген». Наличие вставки гена YkuJ-MPER в рекомбинантной плазмиде подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). Секвенирование плазмидной ДНК клонов в районе встройки гена YkuJ-MPER позволило отобрать клоны с отсутствием дефектов встраиваемого гена (вставки, делении, замены), после чего из отобранных клонов была наработана и выделена плазмидная ДНК для дальнейших работ. Карта рекомбинантной плазмиды приведена на фигуре 6.
Таким образом, ген химерного белка YkuJ-MPER был клонирован в составе плазмидного вектора рЕТ21а по сайтам рестрикции NdeI и XhoI в общей рамке считывания полигистидиновой метки (6xHis). Это позволило включить в состав рекомбинантного белка YkuJ-MPER фрагмент 6xHis для очистки при помощи металл-хелатной хроматографии. Как показало моделирование пространственной структуры белка, этот фрагмент не препятствует связыванию антител.
Пример 3. Получение продуцента химерного белка YkuJ-MPER Для получения продуцента рекомбинантного белка YkuJ-MPER плазмидой рЕТ21-YkuJ-MPER трансформировали штамм Е. coli BL-21 (DE3) методом, описанном в примере 2. Полученные клоны Е. coli штамма BL21 (DE3) после трансформации рекомбинантной плазмидой рЕТ21a-YkuJ-MPER наращивали в 5 мл среды YT×2 в течение ночи. Полученную ночную культуру добавляли по 1,5 мл в колбы объемом 750 мл, содержащих 150 мл среды YT×2 и растили до плотности ODλ600=0,8. После этого в культуры вносили ИПТГ (Изопропил-(3-β-тиогалактопиранозид) до конечной концентрации 1 мкМ для индукции промотора фага Т7 и продолжали культивирование в течение 5 часов при температуре 37°C, 170 об/мин. Биомассу бактериальных клеток осаждали центрифугированием (7000 об/мин). Наличие целевого белка контролировали в 15% ПААГ (Фиг. 7). Затем биомассу гомогенизировали в лизирующем буфере (20 мМ Трис, 300 мМ NaCl, 0,1% Twin 20, рН=8,0) с использованием ультразвукового гомогенизатора Soniprep 150 Plus. Дебрис (осадок, формируемый в результате центрифугирования суспензии гомогенизированной ткани и содержащий полуразрушенные клетки) отделяли центрифугированием при 16000 rpm (31000 g) в течение 15 мин, при 4°C.
После разделения обе фракции были проанализированы в 15% ПААГ на наличие целевого белка. Результаты показали, что весь целевой белок находится в нерастворимой фракции.
Пример 4. Очистка химерного белка YkuJ-MPER
Так как целевой белок находится в тельцах включения (пример 3), для последующей очистки дебрис растворяли в буфере (20 мМ Трис, 300 мМ NaCl, рН=8), содержащем 8 М мочевину, и очищали с помощью металл-хелатной хроматографии на Ni-NTA агарозе.
Колонку, содержащую Ni-NTA агарозу, уравновешивали базовым буфером: 20 мМ Трис, 300 мМ NaCl, 8М мочевина, рН=8,0. Растворенный дебрис наносили на колонку. Промывали колонку 10 объемами промывочного буфера (базовый буфер с 8 М мочевиной + 20 мМ имидазола). Десорбцию связанных с сорбентом белков, проводили базовым буфером со ступенчато (шаг 50 мМ) повышающейся (20-300 мМ) концентрацией имидазола. Было установлено, что белок элюируется при концентрации имидазола 150 мМ. Рефолдинг белка, связанного с металл-хелатным сорбентом, и его очистку проводили диализом против физ. раствора. Степень очистки рекомбинантного белка YkuJ-MPER контролировали с помощью электрофореза в 15% ПААГ. После электрофореза гель фиксировали в 40%-ном спиртовом растворе, содержащем 10% уксусной кислоты, отмывали в дистиллированной воде и окрашивали Кумасси G-250. В конечном препарате чистота белка составляла более 98% (Фиг. 8).
Пример 5. Анализ иммунохимических свойств YkuJ-MPER
Для подтверждения способности эпитопов широконейтрализующих антител, входящих в состав YkuJ-MPER, связываться с широконейтрализующими ВИЧ-1 моноклональными антителами, был проведен вестерн-блот анализ. Для этого электрофоретически разделенные препараты белков YkuJ-MPER переносили на нитроцеллюлозную мембрану методом полусухого переноса [31]. Мембрану блокировали 1% раствором бычьего сывороточного альбумина в PBS (0,002 М KH2PO4, 0,01 М Na2HPO4×7H2O, 0,137 М NaCl, 0,0027 М KCl, pH 7,4) для предотвращения неспецифического связывания. Иммунодетекцию проводили согласно протоколу для системы «SNAP i.d.» (Merck Millipore, США) с первичными моноклональными человеческими антителами 2F5 и 10Е8. Связывание специфического антитела визуализировали с использованием вторичных антител rabbit anti-human IgG alkaline phosphatase (АР) в разведениях, рекомендованных производителем, и добавлением хромогенного субстрата BCIP/NBT. Результаты вестерн-блот анализа, представленные на (Фиг. 9), демонстрируют способность эпитопов, включенных в состав YkuJ-MPER, специфически взаимодействовать с соответствующими МКА.
Пример 6. Анализ иммуногенных свойств белка YkuJ-MPER
Для проверки иммуногенных свойств YkuJ-MPER очищенным препаратом белка иммунизировали лабораторных животных: мышей линии BALB/c и беспородных кроликов, затем проводили иммунологические тесты с использованием иммунных сывороток. Иммунизацию проводили по следующей схеме. Для иммунизации были отобраны 50 мышей линии BALB/c, 10 из которых были трехкратно иммунизированы по схеме: 1 иммунизация - 50 мкг препарата с полным адъювантом Фрейнда, 2 иммунизация - 50 мкг препарата с неполным адъювантом Фрейнда, 3 иммунизация - 100 мкг чистым белком. Иммунизация проводилась с интервалом 2 недели. Вторая группа мышей в количестве 10 особей служила в качестве отрицательного контроля - вместо YkuJ-MPER вводился физиологический раствор (0,9%-й), по той же схеме. По схеме, описанной выше, также были иммунизированы беспородные кролики, с разницей в концентрации вводимого препарата, на одного кролика - 500 мкг иммуногена, в качестве отрицательного контроля служила сыворотка этих же кроликов, взятая перед иммунизацией. Все животные содержались в питомнике согласно правилам лабораторной практики и получали должный уход.
Полученные сыворотки лабораторных животных после иммунизации были протестированы в иммуноферментном анализе для оценки антигенности химерного белка YkuJ-MPER. Было показано, что антитела из сывороток иммунизированных животных специфически взаимодействуют с рекомбинантным белком YkuJ-MPER. Титр сывороток иммунизированных животных в случаи мышей составил более 1:600 тысяч, в случаи кроликов составил более 1:3 миллионам (Фиг. 10).
Таким образом, было показано, что химерный белок-иммуноген YkuJ-MPER безопасен при введении лабораторным животным и обладает высокой иммуногенностью.
Для анализа наличия антител в сыворотках животных иммунизированных YkuJ-MPER, специфически взаимодействующих с MPER ВИЧ-1, был использован другой белок, также содержащий два участка MPER ВИЧ-1 (TBI-MPER, Фигура 11). Для получения белка TBI-MPER в полиэпитопном иммуногене TBI (компонент вакцины «КомбиВИЧвак», разработанной ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора) N-и С-концевые участки были заменены на MPER ВИЧ-1. Анализ вторичной структуры TBI-MPER показал, что встроенные участки имеют альфа-спиральную структуру, характерную для MPER ВИЧ-1.
Анализ показал, что титр сывороток животных, иммунизированных YkuJ-MPER, в ИФА к TBI-MPER составляет для мышей 1:125 тысяч, для кроликов 1:625 тысяч. Исходя из полученных результатов можно сделать вывод о том, что в организме лабораторных животных образуются антитела, специфичные к MPER ВИЧ-1 (Фигура 12).
Источники патентной и научно-технической информации:
1. Объединенная программа ООН по ВИЧ/СПИД [Электронный ресурс]. - URL: http://www.unaids.org/ru (дата обращения 20.09.2017).
2. Федеральный научно-методический Центр по профилактике и борьбе со СПИДом [Электронный ресурс]. - URL: http://www.hivrussia.ru/ (дата обращения 20.09.2017).
3. Gorry P.R., McPhee D.A., Verity Е., Dyer W.B., Wesselingh S.L., Learmont J., Sullivan J.S., Roche M., Zaunders J.J., Gabuzda D., Crowe S.M., Mills J., Lewin S.R., Brew B.J., Cunningham A.L., Churchill M.J. Pathogenicity and immunogenicity of attenuated, nef-deleted HIV-1 strains in vivo // Retrovirology. - 2007. - V. 4. - N3. - P.1
4. Rerks-Ngarm S., Pitisuttithum P., Nitayaphan S., Kaewkungwal J., Chiu J., Paris R., Premsri N., Namwat C., de Souza M., Adams E., Benenson M., Gurunathan S., Tartaglia J., McNeil J.G., Francis D.P., Stablein D., Birx D.L., Chunsuttiwat S., Khamboonruang C., Thongcharoen P., Robb M.L., Michael N.L., Kunasol P., Kim J.H. Vaccination with ALVAC and AIDSVAX to Prevent HIV-1 Infection in Thailand // New England Journal of Medicine. - 2009. - V. 361. - N. 23. - P. 2209-2220.
5. Chung A.W., Ghebremichael M., Robinson H., Brown E., Choi I., Lane S., Dugast A.S., Schoen M.K., Rolland M., Suscovich T.J., Mahan AE, Liao L, Streeck H., Andrews C., Rerks-Ngarm S., Nitayaphan S., de Souza M.S., Kaewkungwal J., Pitisuttithum P., Francis D., Michael N.L., Kim J.H., Bailey-Kellogg C., Ackerman M.E., Alter G. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines // Science Translational Medicine. - 2014. - V. 6. - N. (228). - P. 1-11.
6. Kim J.H., Excler J.L., Michael N.L. Lessons from the RV144 Thai Phase III HIV-1 Vaccine Trial and the Search for Correlates of Protection // Annual Review of Medicine. - 2015. - V. 66. - P. 423-427.
7. Montefiori D.C., Karnasuta C., Huang Y., Ahmed H., Gilbert P., De Souza M.S., McLinden R., Tovanabutra S., Laurence-Chenine A., Sanders-Buell E., Moody M.A., Bonsignori M., Ochsenbauer C., Kappes J., Tang H., Greene K., Gao H., Labranche C.C., Andrews C., Polonis V.R., Rerks-Ngarm S., Pitisuttithum P., Nitayaphan S., Kaewkungwal J., Self S.G., Berman P.W., Francis D., Sinangil F., Lee C, Tartaglia J., Robb M.L., Haynes B.F., Michael N.L., Kim J.H. Magnitude and breadth of the neutralizing antibody response in the RV144 and Vax003 HIV-1 vaccine efficacy trials // Journal of Infectious Diseases. - 2012. - V. 206. - N 3. P. 431-441.
8. Hessell A.J., Poiqnard P., Hunter M., Hanqartner L., Tehrani D.M., Bleeker W.K., Parren P.W., Marx P.A., Burton D.R. Effective, low-titer antibody protection against low-dose repeated mucosal SHIV challenge in macaques // Nature medicine. - 2009. - V. 15. - №. 8.-P. 951-954.
9. Mascola J.R., Lewis M.G., Stiegler G., Harris D., VanCott T.C., Hayes D., Louder M.K., Brown C.R., Sapan C.V., Frankel S.S., Lu Y., Robb ML., Katinger H., Birx D.L. Protection of macaques against pathogenic simian/human immunodeficiency virus 89.6 PD by passive transfer of neutralizing antibodies // Journal of virology. - 1999. - V. 73. - №. 5. - P. 4009-4018.
10. Moldt В., Rakasz E., Schultz N., Chan-Hui P., Swiderek K., Weisgrau K., Piaskowski Sh., Bergman Z., Watkins D., Poignard P., Burton D. Highly potent HIV-specific antibody neutralization in vitro translates into effective protection against mucosal SHIV challenge in vivo // Proceedings of the National Academy of Sciences. - 2012. - V. 109. - №. 46. - P. 18921-18925.
11. Parren P., Marx P., Hessell A., Luckay A., Harouse J., Cheng-Mayer C., Moore J., Burton D. Antibody protects macaques against vaginal challenge with a pathogenic R5 simian/human immunodeficiency virus at serum levels giving complete neutralization in vitro // Journal of virology. - 2001. - V. 75. - №. 17. - P. 8340-8347.
12. Ng С.Т., Jaworski J. P., Jayaraman P., Sutton W. F., Delio P., Kuller L. et al. Passive neutralizing antibody controls SHIV viremia and enhances В cell responses in infant macaques // Nature Medicine. - 2010. - V. 16. - P. 1117-1119.
13. Nishimura, Y. et al. Generation of the pathogenic R5-tropic simian/human immunodeficiency virus SHIVAD8 by serial passaging in rhesus macaques // Journal of Virology. - 2010. - V. 84. - P. 4769-1781.
14. Hessell A. J. et al. Early short-term treatment with neutralizing human monoclonal antibodies halts SHIV infection in newborn macaques // Nature medicine. - 2016. - V. 22. - №. 4. - P. 362.
15. Gilbert PB, Peterson ML, Follmann D, Hudgens MG, Francis DP, Gurwith M, et al. Correlation between immunologic responses to a recombinant glycoprotein 120 vaccine and incidence of HIV-1 infection in a phase 3 HIV-1 preventive vaccine trial // Journal of Infectious Diseases. - 2005. - V. 191. - P. 666-677.
16. Kovacs J.M., Nkolola J.P., Peng H., Cheung A., Perry J., Miller C.A., Seaman M.S., Barouch D.H., Chen B. HIV-1 envelope trimer elicits more potent neutralizing antibody responses than monomeric gp120 // Proceedings of the National Academy of Sciences. - 2012. - V. 109. - N. 30. - P. 12111-12116
17. Yang X., Lee J., Mahony E.M., Kwong P.D., Wyatt R., Sodroski J. Highly stable trimers formed by human immunodeficiency virus type 1 envelope glycoproteins fused with the trimeric motif of T4 bacteriophage fibritin // Journal of Virology. - 2002. - V. 76. - N 9. - P. 4634-4642.
18. Yasmeen A., Ringe R., Derking R., Cupo A., Julien J.P., Burton D.R., Ward A.B., Wilson I.A., Sanders R.W., Moore J.P., Klasse P.J. Differential binding of neutralizing and non-neutralizing antibodies to native-like soluble HIV-1 Env trimers, uncleaved Env proteins, and monomeric subunits // Retro virology. - 2014. - V. 11. - N 1.DOI: 10.1186/1742-4690-11-41.
19. Wu X.L., Yang Z.Y., Li Y.X., Hogerkorp С.М., Schief W.R., Seaman M.S., Zhou T.Q., Schmidt S.D., Wu L., Xu L., Longo N.S., McK.ee K.,  , Louder M.K., Wycuff D.L., Feng Y., Nason M., Doria-Rose N., Connors M., Kwong P.D., Roederer M., Wyatt R.T., Nabel G.J., Mascola J.R. Rational Design of Envelope Identifies Broadly Neutralizing Human Monoclonal Antibodies to HIV-1 // Science. - 2010. - V. 329. - N 5993. - P. 856-861.
, Louder M.K., Wycuff D.L., Feng Y., Nason M., Doria-Rose N., Connors M., Kwong P.D., Roederer M., Wyatt R.T., Nabel G.J., Mascola J.R. Rational Design of Envelope Identifies Broadly Neutralizing Human Monoclonal Antibodies to HIV-1 // Science. - 2010. - V. 329. - N 5993. - P. 856-861.
20. Zhou T.Q., Xu L., Dey В., Hessell A.J., Van Ryk D., Xiang S.H., Yang X.Z., Zhang M.Y., Zwick M.B., Arthos J., Burton D.R., Dimitrov D.S., Sodroski J., Wyatt R., Nabel G.J., Kwong P.D. Structural definition of a conserved neutralization epitope on HIV-1 gp120 // Nature. - 2007. - V. 445. - N 7129. - P. 732-737.
21. Zhou T.Q., Georgiev I., Wu X.L., Yang Z.Y., Dai K.F., Finzi A., Kwon Y.D., Scheid J.F., Shi W., Xu L., Yang Y.P., Zhu J.A., Nussenzweig M.C., Sodroski J., Shapiro L., Nabel G.J., Mascola J.R., Kwong P.D. Structural Basis for Broad and Potent Neutralization of HIV-1 by Antibody VRC01 // Science. - 2010. - V. 329. - N5993. - P. 811-817.
22. Патент РФ №2475264, кл. A61K 39/395, C07K 14/155, опубл.: 20.02.2013
23. Патент РФ №2214274, кл. А61K 39/21, опубл. 20.10.2003 г.
24. Huang J., Ofek G., Laub L. Broad and potent neutralization of HIV-1 by a gp41-specific human antibody // Nature. - 2012. - V. 491. - P. 406-412
25. Montero M., van Houten N.E., Wang X. The membrane-proximal external region of the human immunodeficiency virus type 1 envelope: dominant site of antibody neutralization and target for vaccine design // Microbiology and Molecular Biology Reviews. - 2008. - V. 72. - P. 54-84.
26. Bruel Т., Guivel-Benhassine. F, Amraoui S. Elimination of HIV-1-infected cells by broadly neutralizing antibodies // Nature Communications. - 2016. - V. 7. - P. 10844.
27. Bouvin-Pley M., Morgand M., Meyer L. Drift of the HIV-1 envelope glycoprotein gp120 toward increased neutralization resistance over the course of the epidemic: a comprehensive study using the most potent and broadly neutralizing monoclonal antibodies // Journal Virology. - 2014. - V. 88. - P. 13910-13917.
28. Banerjee S., Shi H., Banasik M., Moon H., Lees W., Qin Y., Harley A., Shepherd A., Cho M.W. Evaluation of a novel multi-immunogen vaccine strategy for targeting 4E10/10E8 neutralizing epitopes on HIV-1 gp41 membrane proximal external region // Virology. 2017. - V. 505. - P. 113-126.
29. Sun Zh., Zhu Yu., Wang Q., Ye L., Dai Ya., Su Sh., Yu F., YingT., Yang Ch., Jiang Sh., Lu L. An immunogen containing four tandem 10E8 epitope repeats with exposed key residues induces antibodies that neutralize HIV-1 and activates an ADCC reporter gene // Emerging Microbes and Infections. - 2016. - V. 5. - P. 1-11 (прототип).
30. Laemmli U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4 // Nature. - 1970. - V. 227. - P. 680-685.
31. Sambrook J., Fritsch E. F., Maniatis T. Molecular cloning // New York: Cold spring harbor laboratory press. - 1989. - V. 2. - P. 14-23.
Приложение
<110> Федеральное бюджетное учреждение науки «Государственный научный центр вирусологии и биотехнологии «Вектор» Федеральной службы по надзору в сфере защиты прав потребителей и благополучия человека (ФБУН ГНЦ В Б «Вектор» Роспотребнадзора) Роспотребнадзора» (ФБУН ГНЦ ВБ "Вектор" Роспотребнадзора)
<120> Искусственный ген YkuJ-MPER, кодирующий химерный белок-иммуноген YkuJ-MPER, рекомбинантная плазмидная ДНК рЕТ21a-YkuJ-MPER, обеспечивающая экспрессию искусственного гена YkuJ-MPER, и химерный белок-иммуноген YkuJ-MPER, являющийся носителем мембранно-проксимальной области ВИЧ-1 и направленный на индукцию в организме широконейтрализующих антител
<160> SEQ ID NO 2
<210> SEQ ID NO: 1
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность искусственного гена YkuJ-MPER, кодирующего белок-иммуноген YkuJ-MPER против ВИЧ-1
<400>1

<210> SEQ ID NO: 2
<211> 119<212>DNA
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность искусственного белка-иммуногена YkuJ-MPER.
<400> 2

(119 аминокислотных остатка).
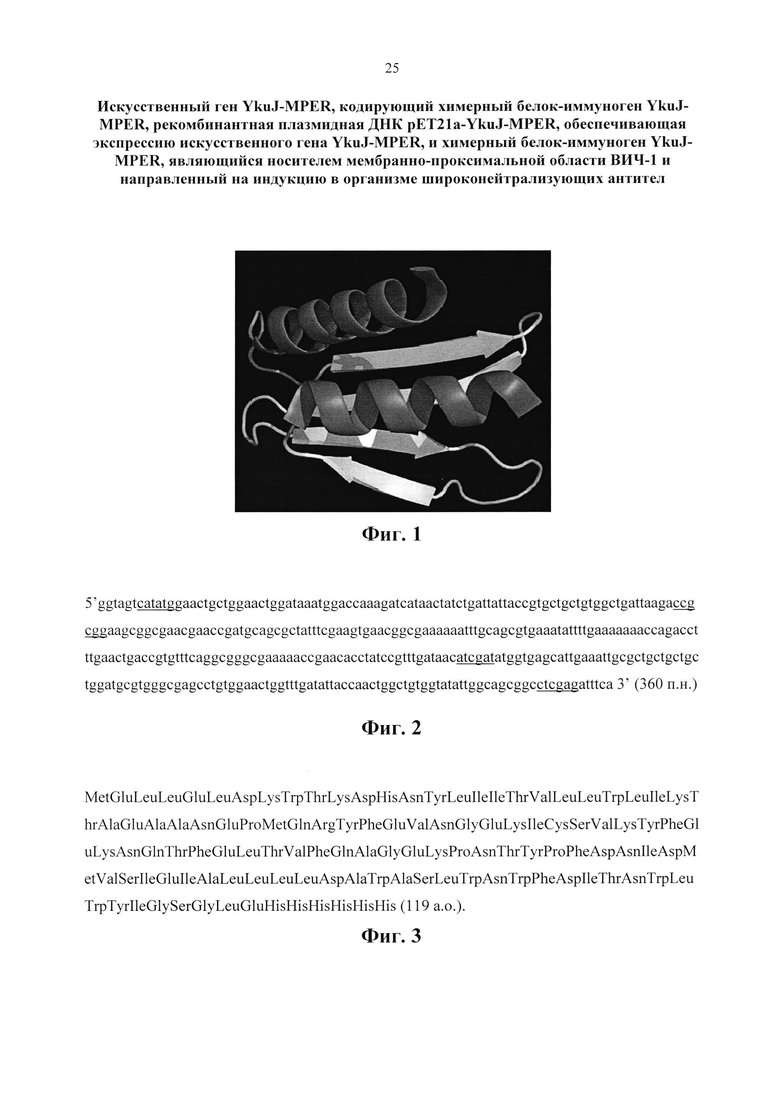

Группа изобретений относится к области биотехнологии, генной инженерии и медицины. Предложен искусственный ген YkuJ-MPER, кодирующий белок-иммуноген YkuJ-MPER против ВИЧ-1, имеющий нуклеотидную последовательность SEQ ID NO:1 длиной 360 п.н. Предложена также рекомбинантная плазмидная ДНК pET21a-YkuJ-MPER, предназначенная для получения белка-иммуногена, обеспечивающего индукцию ВИЧ-специфического В-клеточного ответа, имеющая размер 5700 п.н., молекулярную массу 3,8·103 кДа, содержащая ген YkuJ-MPER, кодирующий белок-иммуноген YkuJ-MPER против ВИЧ-1, находящийся под контролем промотора РНК-полимеразы фага Т7, обеспечивающего его экспрессию в клетках Е. coli. Предложен также искусственный белок-иммуноген YkuJ-MPER, состоящий из участка белка В. subtilis YkuJ и двух участков MPER ВИЧ-1, имеющий молекулярную массу 14,2 кДа и аминокислотную последовательность SEQ ID NO:2. Группа изобретений обеспечивает эффективную индукцию широконейтрализующих анти-ВИЧ-1 антител при подборе оптимальной стратегии иммунизации. 3 н.п. ф-лы, 12 ил., 1 табл., 6 пр.
1. Искусственный ген YkuJ-MPER, кодирующий белок-иммуноген YkuJ-MPER против ВИЧ-1, имеющий следующую нуклеотидную последовательность SEQ ID NO:1 длиной 360 п.н.:
5'ggtagtcatatggaactgctggaactggataaatggaccaaagatcataactatctgattattaccgtgctgctgtggctgattaagaccgcggaagcggcgaacgaaccgatgcagcgctatttcgaagtgaacggcgaaaaaatttgcagcgtgaaatattttgaaaaaaaccagacctttgaactgaccgtgtttcaggcgggcgaaaaaccgaacacctatccgtttgataacatcgatatggtgagcattgaaattgcgctgctgctgctggatgcgtgggcgagcctgtggaactggtttgatattaccaactggctgtggtatattggcagcggcctcgagatttca 3'.
2. Рекомбинантная плазмидная ДНК pET21a-YkuJ-MPER, предназначенная для получения белка-иммуногена, обеспечивающего индукцию ВИЧ-специфического В-клеточного ответа, имеющая размер 5700 п.н., молекулярную массу 3,8×103 кДа, содержащая в соответствии с физической и генетической картой, представленной на фиг. 3, целевой ген по п. 1, кодирующий химерный белок YkuJ-MPER, находящийся под контролем промотора РНК-полимеразы фага Т7, обеспечивающего его экспрессию в клетках Е. coli, и состоящая из следующих фрагментов:
- NdeI-XhoI - векторного фрагмента ДНК плазмиды рЕТ21а(-) размером 5340 п.н., содержащего промотор и терминатор РНК-полимеразы фага Т7, обеспечивающие индуцибельную экспрессию гена YkuJ-MPER в клетках Е. coli; промотор AmpR, обеспечивающий эффективную транскрипцию мРНК гена bla, ген устойчивости к ампициллину (bla), кодирующий белок бета-лактамазу, и (ori) точка начала репликации ColEl, обеспечивающие селекцию и размножение целевой плазмиды в клетках бактерий Escherichia coli; С-терминальный His-tag (His×6) в рамке считывания с геном YkuJ-MPER;
- NdeI-XhoI - фрагмента размером 360 п.н., содержащего ген YkuJ-MPER с инициирующим кодоном ATG, полученного путем обработки рестриктазами NdeI-XhoI плазмиды рЕТ21а;
- уникальных сайтов рестрикции, существенных для генно-инженерных манипуляций: NdeI - 5203, SacII - 5287, ClaI - 5434, XhoI - 5538, и имеющая следующее положение генов:
3. Искусственный белок-иммуноген YkuJ-MPER, состоящий из участка белка В. subtilis YkuJ и двух участков MPER ВИЧ-1, имеющий молекулярную массу 14,2 кДа и аминокислотную последовательность SEQ ID NO:2:
MetGluLeuGluLeuAspLysTrpThrLysAspHisAsnTyrLeuIleIleThrValLeuLeuTrpLeuIleLysThrAlaGluAlaAlaAsnGluProMetGlnArgTyrPheGluValAsnGlyGluLysIleCysSerValLysTyrPheGluLysAsnGlnThrPheGluLeuThrValPheGlnAlaGlyGluLysProAsnThrTyrProPheAspAsnlleAspMetValSerlleGluIleAlaLeuLeuLeuLeuAspAlaTrpAlaSerLeuTrpAsnTrpPheAsplleThrAsnTrpLeuTrpTyrlleGlySerGlyLeuGluHisHisHisHisHisHis.
| SUN Z | |||
| ET AL | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| РУДОМЕТОВ А.П | |||
| и др | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| III Международная конференция молодых ученых: биотехнологов, молекулярных биологов и вирусологов | |||
| Токарный резец | 1924 |
|
SU2016A1 |
| тез | |||
| / Новосиб | |||
| Гос | |||
| Ун-т | |||
| - Новосибирск: ИПЦ НГУ, 2016 | |||
| Способ переработки сплавов меди и цинка (латуни) | 1922 |
|
SU328A1 |
| ФОРМА ДЛЯ БРИКЕТОВ | 1919 |
|
SU286A1 |
| РУДОМЕТОВ А.П | |||
| и др | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Тезисы Пятой конференции по ВИЧ/СПИДу в Восточной Европе и Центральной Азии | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| Способ обработки грубых шерстей на различных аппаратах для мериносовой шерсти | 1920 |
|
SU113A1 |

