ОБЛАСТЬ
[0001] Настоящее изобретение относится к генам C-глюкозилтрансферазы (CGT), полученным из гречихи, или их гомологам, а также к способу создания трансгенных растений с голубым цветом цветка, включающему стадию применения генов для получения совместного присутствия антоцианов дельфинидинового типа и флавоновых моно-C-гликозидов в растительных клетках.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[0002] Роза, хризантема и гвоздика являются промышленно важными декоративными цветковыми растениями во всем мире. В частности, роза, будучи самым популярным цветковым растением, выращивается с древних времен и подвергается искусственному скрещиванию на протяжении сотен лет. Одна из проблем, однако, заключается в том, что ни один из родственных видов, поддающихся гибридизации, не имеет диких сортов с голубым цветом цветков, и поэтому было трудно создать сорта роз с голубым цветом цветков путем общепринятого скрещивания и мутационной селекции. Создание совершенно новых цветов синего цвета должно привести к новому спросу на еще более широкое использование декоративных цветковых растений, а также должно способствовать увеличению их производства и потребления. Таким образом, путем генетической инженерии была предпринята попытка создать розы с голубыми цветами.
[0003] Известно, например, что цветки от фиолетового до синего цвета в изобилии содержат антоцианы дельфинидинового типа, имеющие дельфинидиновый, петунидиновый и мальвидиновый остовы, но поскольку декоративные цветковые растения, такие как роза, не могут продуцировать такие антоцианы дельфинидинового типа, продолжаются исследования с целью искусственного получения дельфинидинов путем экспрессии гена флавоноидной 3',5'-гидроксилазы, необходимого для их синтеза (NPL 1). Однако даже когда фермент подвергается искусственной модификации для экспрессии гена, который продуцирует интересующее вещество в рекомбинантном растении, интересующее вещество часто присутствует в малом количестве или совсем не накапливается.
[0004] Кроме того, окраска цветка изменяется не только за счет структуры самих антоцианов как основных пигментов, но и за счет совместно присутствующих флавоноидов (также известных как копигменты), ионов металлов и pH вакуолей. Флавоны или флавонолы являются типичными копигментами, которые образуют сэндвич-подобные слои с антоцианами и окрашивают антоцианы в синий цвет, производя эффект интенсивности (NPL 2). Он известен как «эффект копигмента». Известно, в частности, что флавоны обладают мощным копигментным эффектом, а анализ генетически рекомбинантных гвоздик, например, показал, что флавоны проявляют значительный копигментирующий эффект (NPL 3). Было опубликовано, что для голландского ириса более высокое отношение общего содержания флавонов к общему содержанию дельфинидина приводит к более сильному эффекту копигментации и более синему цвету (NPL 4).
[0005] Однако, не все растения могут производить флавоны, и известно, что розы и петунии не запасают флавоны. Таким образом, были предприняты попытки модифицировать окраску цветков путем экспрессии в растениях различных генов, кодирующих белки, обладающие активностью синтеза флавонов из флаванонов (PTL 1).
[0006] В растениях флавоны распределяются не только в свободной форме, но и в виде гликозидов, причем в первую очередь образуются флавоновые О-гликозиды и флавоновые С-гликозиды, причем известно, что флавоновые С-гликозиды проявляют особенно мощный копигментный эффект. Например, сообщалось, что изовитексин, как один из типов флавонового C-гликозида, проявляет копигментный эффект с антоцианами японского садового ириса (Iris ensata Thunb.) и придает голубой цвет цветку за счет стабилизации антоцианов (NPL 5). Одним из известных путей биосинтеза флавоновых С-гликозидов является синтез из флаванонов посредством реакции, катализируемой флаванон-2-гидроксилазой (F2H), С-глюкозилтрансферазой (CGT) и дегидратазой (FDH) (NPL 6).
[0007] Ранее было опубликовано, что розы с голубым цветом цветков были созданы путем введения гена F3',5'H, полученного из колокольчика, и гена MT, полученного из торении, гена F2H, полученного из солодки, гена CGT, полученного из риса, и гена FDH, полученного из Lotus japonicus, для того чтобы вызвать совместное присутствие антоцианов дельфинидинового типа и флавоновых C-гликозидов в растительных клетках (PTL 2). Однако было обнаружено, что окраска цветков сортов роз, созданных таким образом, имеет сильный красноватый оттенок, и, таким образом, по-прежнему желательно разработать способы контроля экспрессии синего цвета, чтобы обеспечить однородное и стабильное создание роз с более синей окраской цветков.
[СПИСОК ЦИТИРОВАНИЯ]
[ПАТЕНТНАЯ ЛИТЕРАТУРА]
[0008] [PTL 1] Японская нерассмотренная патентная публикация No. 2000-279182
[PTL 2] Международная патентная публикация No. 2019/069946
[PTL 3] Международная патентная публикация No. 2008/156206
[НЕПАТЕНТНАЯ ЛИТЕРАТУРА]
[0009] [NPL 1] Phytochemistry Reviews 5, 283-291
[NPL 2] Prog. Chem. Org. Natl. Prod. 52
[NPL 3] Phytochemistry, 63, 15-23(2003)
[NPL 4] Plant Physiol. Bioch. 72, 116-124(2013)
[NPL 5] Euphytica 115, 1-5(2000)
[NPL 6] FEBS Lett. 589, 182-187(2015)
СУЩНОСТЬ
[ТЕХНИЧЕСКАЯ ЗАДАЧА]
[0010] Задача, решаемая изобретением, заключается в создании трансгенного растения, имеющего равномерную и стабильную синюю окраску цветков (RHS Color Chart 5th Edition: Violet-Blue group/Blue group и/или угол оттенка: 339,7°-270,0°), на основании исследования причин покраснения окраски цветков.
[РЕШЕНИЕ ЗАДАЧИ]
[0011] В результате интенсивных исследований и большого количества экспериментов по этой проблеме авторы настоящего изобретения обнаружили, что причиной покраснения являются флавоновые ди-С-гликозиды, и что копигментирующий эффект флавоновых моно-С-гликозидов с дельфинидиновым типом антоцианов значительно выше, чем у флавоновых ди-С-гликозидов. Путем переноса гена CGT, полученного из гречихи, который был выбран из различных генов CGT, авторам настоящего изобретения удалось добиться значительного накопления только флавоновых моно-C-гликозидов в лепестках растений. После этих результатов изобретение было завершено.
[0012] В частности, настоящее изобретение относится к следующему.
[1] Ген CGT, полученный из гречихи, или его гомолог, где:
ген CGT, полученный из гречихи, или его гомолог выбран из группы, состоящей из:
(1-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 11;
(1-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 11, при жестких условиях, и имеет ту же активность, что и полинуклеотид из (1-a); и
(1-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 12.
[2] Ген CGT, полученный из гречихи, или его гомолог согласно [1], к которому добавлена нетранслируемая область (5'-UTR), полученная из гена ADH Arabidopsis thaliana (SEQ ID NO: 15), или нетранслируемая область (5'-UTR), полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13).
[3] Вектор, содержащий ген CGT, полученный из гречихи, или его гомолог, где ген CGT, полученный из гречихи, или его гомолог выбран из группы, состоящей из:
(1-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 11;
(1-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 11 при жестких условиях, и имеет ту же активность, что и полинуклеотид из (1-a);
(1-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 12;
(1-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 12, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (1-с); и
(1-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичностью в отношении аминокислотной последовательности, указанной как SEQ ID NO: 12, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (1-c).
[4] Вектор согласно [3], где нетранслируемая область (5'-UTR), полученная из гена ADH Arabidopsis thaliana (SEQ ID NO: 15), или нетранслируемая область, полученная из гена HSPRO Arabidopsis thaliana (5'-UTR) (SEQ ID NO: 13) была добавлена к гену CGT, полученному из гречихи, или его гомологу.
[5] Вектор согласно [3] или [4], который дополнительно содержит ген флаванон-2-гидроксилазы (F2H) или его гомолог и ген дегидратазы (FDH) или его гомолог.
[6] Вектор согласно [5], где
ген F2H или его гомолог выбран из группы, состоящей из:
(2-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 5;
(2-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 5 при жестких условиях, и имеет ту же активность, что и полинуклеотид из (2-a);
(2-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 6;
(2-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 6, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом из (2-с); и
(2-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичности в отношении аминокислотной последовательности, указанной как SEQ ID NO: 6, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (2-c), и
ген FDH или его гомолог выбран из группы, состоящей из:
(3-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 9;
(3-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 9 при жестких условиях, и имеет ту же активность, что и полинуклеотид из (3-a);
(3-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 10;
(3-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 10, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (3-с); и
(3-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичности в отношении аминокислотной последовательности, указанной как SEQ ID NO: 10, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (3-c).
[7] Вектор по любому из [3]-[6], который дополнительно включает ген флавоноидной F3',5' гидроксилазы (F3',5'H) или его гомолог, и ген метилтрансферазы (MT) или его гомолог.
[8] Вектор согласно [7], где
ген F3',5'H или его гомолог выбран из группы, состоящей из:
(4-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 1;
(4-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 1, при жестких условиях и имеет ту же активность, что и полинуклеотид из (4-а);
(4-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 2;
(4-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 2, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом из (4-с); и
(4-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичности в отношении аминокислотной последовательности, указанной как SEQ ID NO: 2, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (4-c), и
ген MT или его гомолог выбран из группы, состоящей из:
(5-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 3;
(5-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 3, при жестких условиях и имеет ту же активность, что и полинуклеотид из (5-a);
(5-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 4;
(5-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 4, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (5-с); и
(5-e) полинуклеотид, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичности в отношении аминокислотной последовательности, указанной как SEQ ID NO: 4, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (5-c).
[9] Трансгенное растение, содержащее ген CGT, полученный из гречихи, или его гомолог согласно [1] или [2], или вектор по любому из [3]-[8], или его инбредное или аутбредное потомство.
[10] Трансгенное растение согласно [9], или его инбредное или аутбредное потомство, где растение выбрано из розы, хризантемы, гвоздики или лилии.
[11] Трансгенное растение согласно [10], или его инбредное или аутбредное потомство, где растение является розой.
[12] Выводковые почки, части растительного организма, ткань или клетки трансгенного растения по любому из [9]-[11] или его инбредное или аутбредное потомство.
[13] Срезанные цветы трансгенного растения по любому из [9]-[11], или его инбредное или аутбредное потомство, или переработанная форма, созданная из срезанных цветов.
[14] Способ создания трансгенных растений с голубым цветом цветка, включающий стадию переноса гена C-глюкозилтрансферазы (CGT) из гречихи или его гомолога в растение-хозяин для получения совместного присутствия антоцианов дельфинидинового типа и флавоновых моно-C-гликозидов в растительных клетках.
[15] Способ согласно [14], где флавоновый моно-С-гликозид представляет собой апигенин 6-С-глюкозид, лютеолин 6-С-глюкозид, трицетин 6-С-глюкозид, апигенин 8-С-глюкозид, лютеолин 8-C-глюкозид или трицетин 8-C-глюкозид или его производное.
[16] Способ согласно [14] или [15], где антоцианы дельфинидинового типа выбраны из группы, состоящей из мальвидинов, дельфинидинов, петунидинов и их комбинации.
[17] Способ по любому из [14]-[16], который дополнительно включает перенос гена CGT, полученного из гречихи, или его гомолога по [1] или [2] или вектора по любому из [3]-[8] в растительные клетки хозяина.
[18] Способ по любому из [14]-[17], где растение представляет собой розу, хризантему, гвоздику или лилию.
[19] Способ согласно [18], где растение является розой.
[ПОЛЕЗНЫЕ ЭФФЕКТЫ ПО ИЗОБРЕТЕНИЮ]
[0013] По изобретению можно однородно и стабильно создавать сорта растений, имеющие синюю окраску цветков (RHS Color Chart 5th Edition: Violet-Blue group/Blue group и/или угол оттенка: 339,7°-270,0°).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
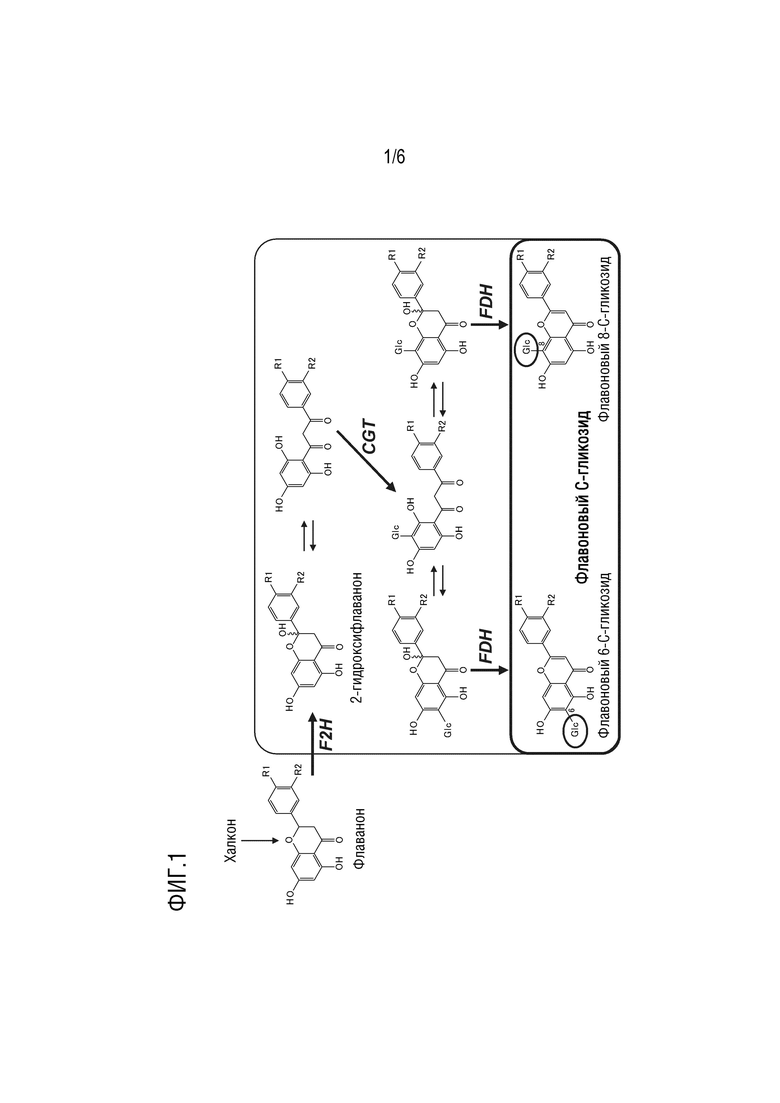
[0014] На фиг. 1 представлен путь биосинтеза флавонового моно-С-гликозида в растении.
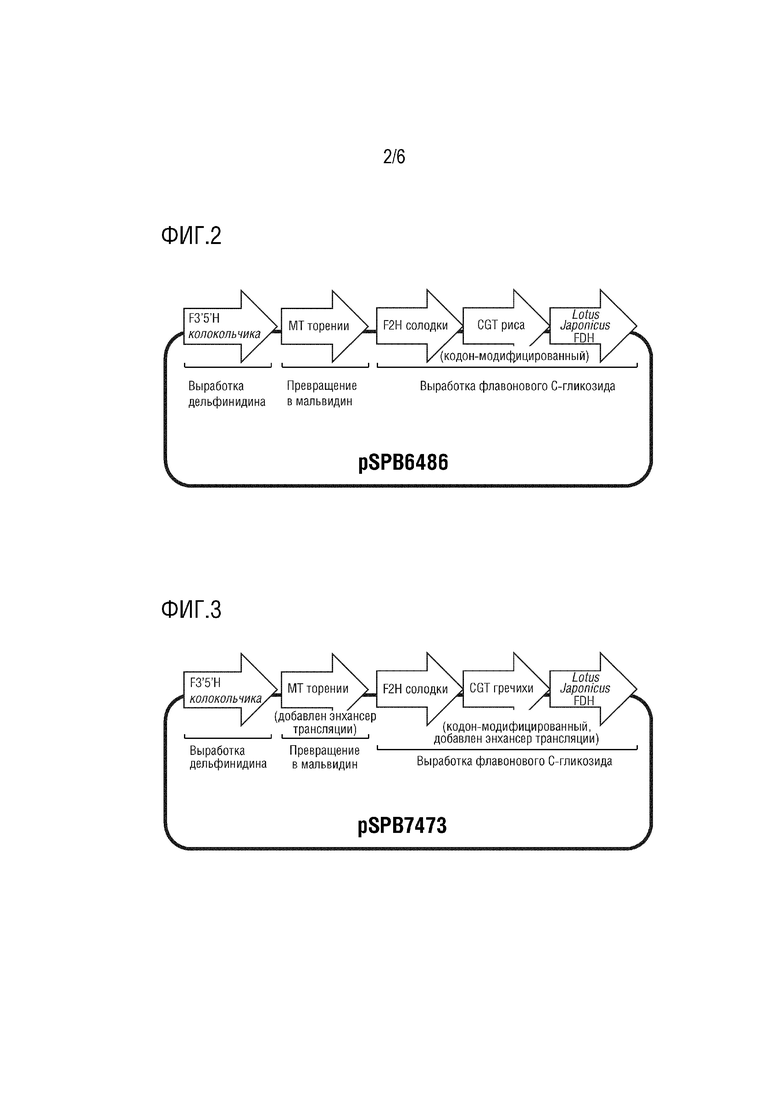
На фиг. 2 представлена структура pSPB6486.
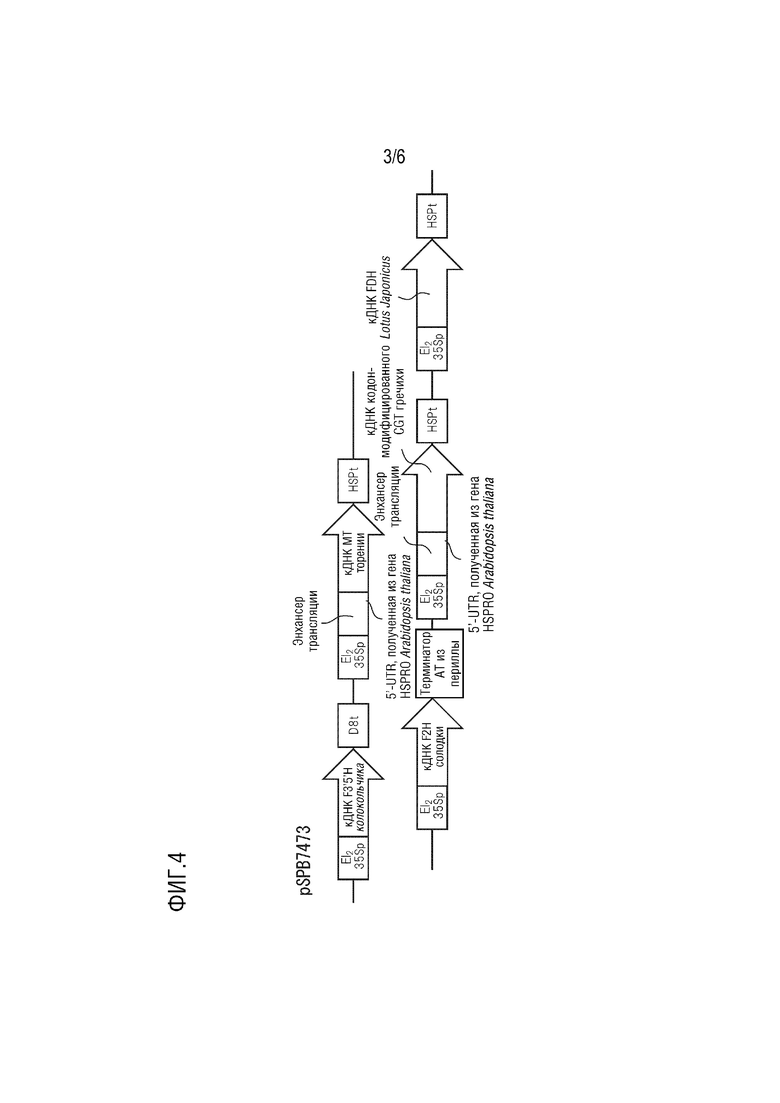
На фиг. 3 представлена структура pSPB7473.
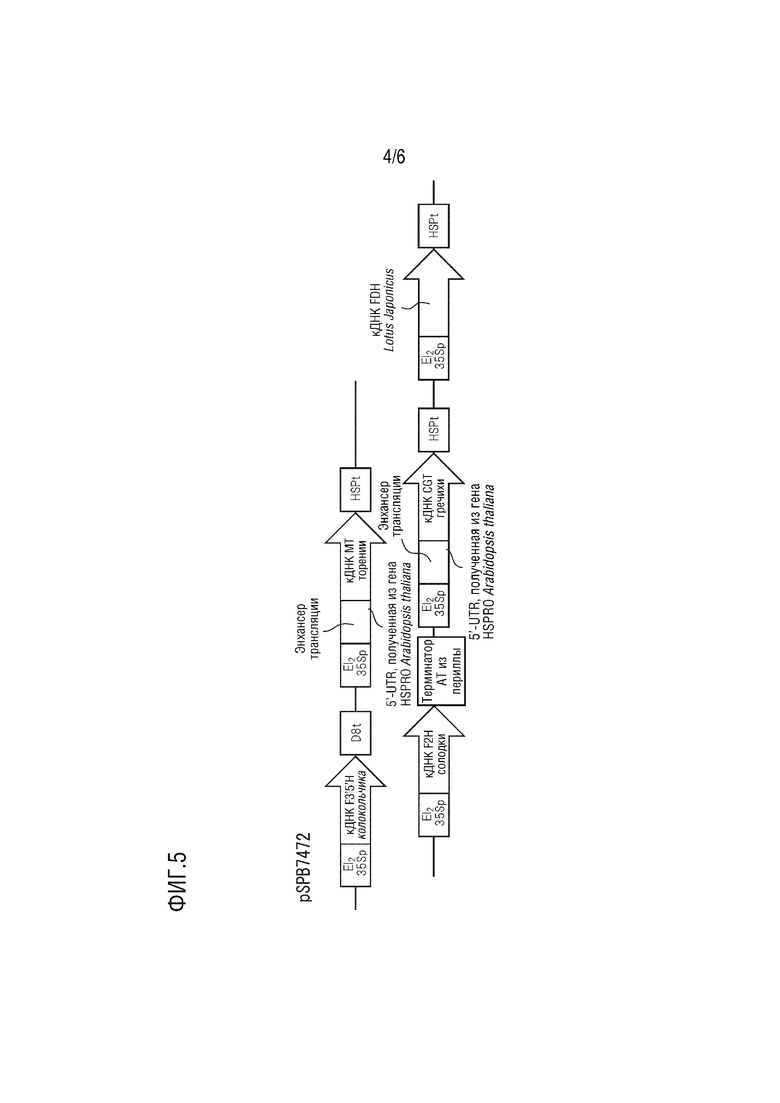
На фиг. 4 представлена подробная структура pSPB7473.
На фиг. 5 представлена подробная структура pSPB7472.
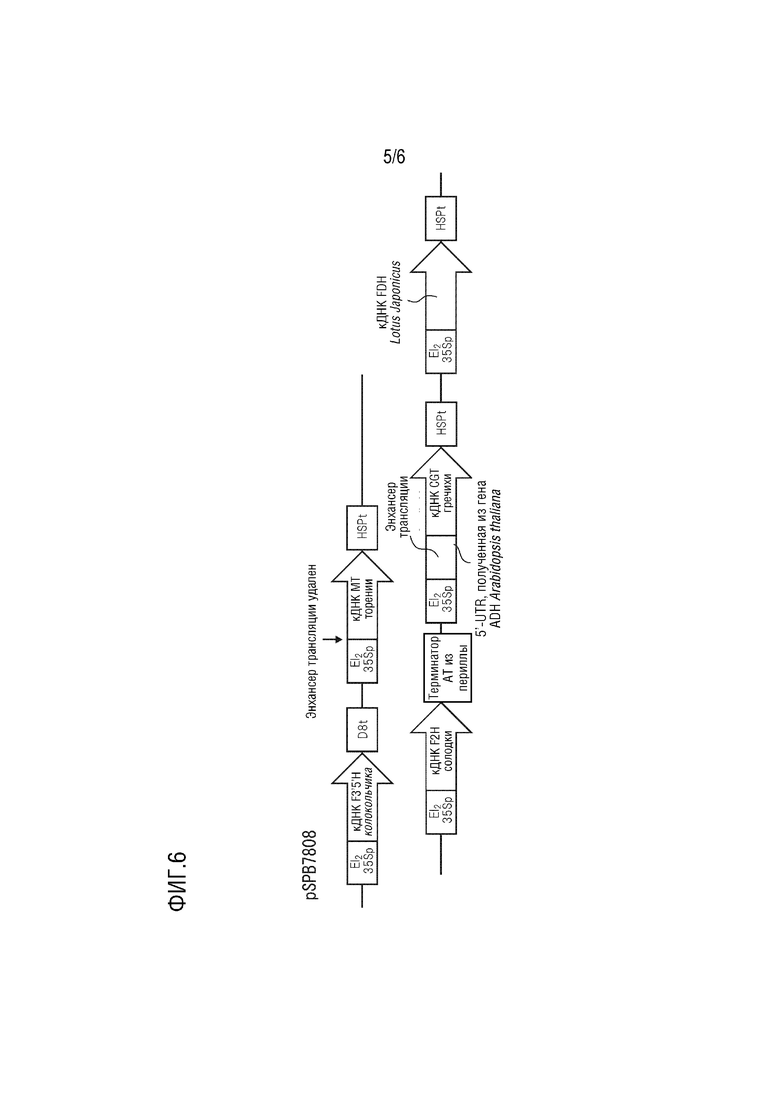
На фиг. 6 представлена подробная структура pSPB7808.
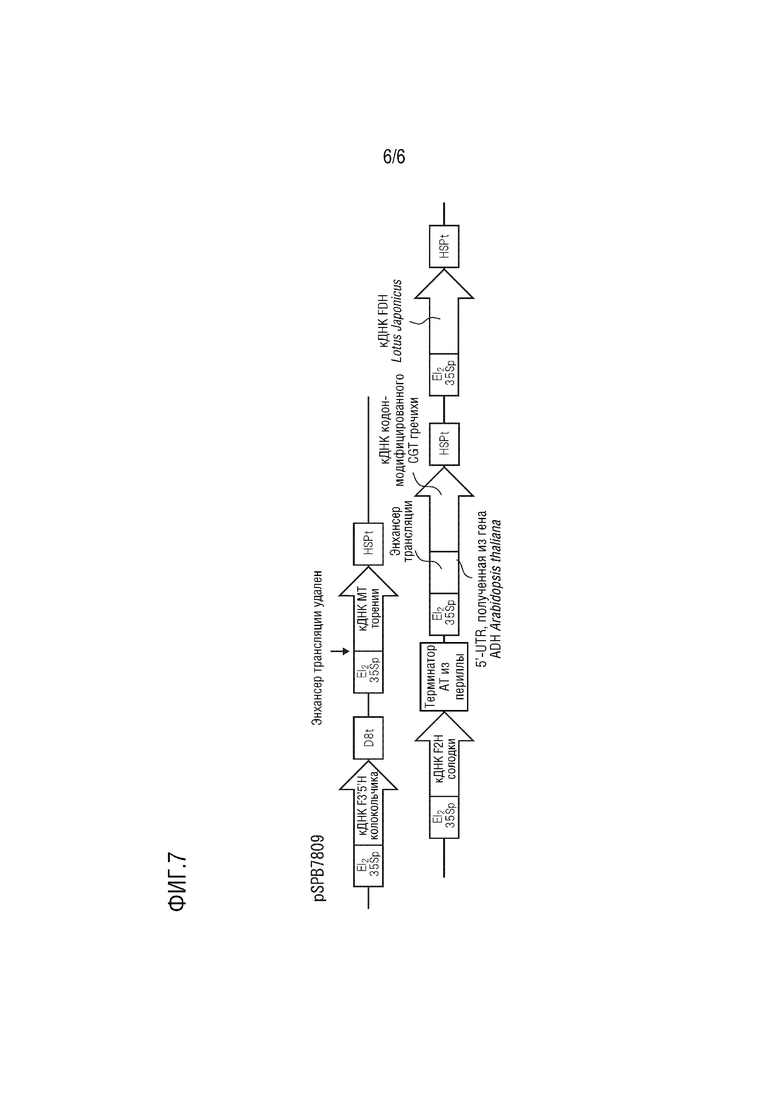
На фиг. 7 представлена подробная структура pSPB7809.
[0015] Антоцианы представляют собой группу пигментов, которые широко распространены в растениях, и известно, что они проявляют красный, синий и пурпурный цвета цветков. Они подразделяются на 3 типа: пеларгонидин, цианидин и дельфинидин, в зависимости от количества гидроксильных групп в B-кольце антоцианидина, в виде агликоновой формы. Хромофорная группа представляет собой агликоновую часть с антоцианами пеларгонидинового типа, имеющими оранжевый цвет, антоцианами цианидинового типа, имеющими красный цвет, и антоцианами дельфинидинового типа, имеющими цвет от фиолетового до синего. На всем протяжении настоящего описания «антоцианы дельфинидинового типа» также включают их производные, имеющие дельфинидиновую, мальвидиновую или петунидиновую основу, причем мальвидин является предпочтительным.
[0016] Когда антоцианы дельфинидинового типа присутствуют вместе с такими веществами, как флавоны, флавонолы, органические кислоты, сложные эфиры и танины, их молекулярное взаимодействие часто дает голубоватые цвета. Это явление известно как «копигментация», а вещества, вызывающие это явление, известны как копигменты. Копигментация включает в себя не только эффект углубления цвета, вызывающий образование синего цвета, но также эффект насыщенности цвета или эффект повышения устойчивости цвета. Авторы изобретения подтвердили, что копигментация между антоцианами дельфинидинового типа и флавоновыми С-гликозидами вызывает проявление синей окраски лепестков розы (PTL 2).
[0017] Флавоны являются органическими соединениями, которые представляют собой полученные из флавана циклические кетоны, и в растениях они в основном существуют в виде гликозидов. Флавон, в строгом определении, относится к 2,3-дидегидрофлаван-4-ону, представляющему собой соединение с химической формулой C15H10O2 и молекулярной массой 222,24, но в более широком смысле флавоны представляют собой категорию флавоноидов, причем флавоноид классифицируется как «флавон», если он имеет флавоновую структуру в качестве основного остова, а также не имеет гидроксильной группы в 3-м положении. Как применяют в настоящем документе, «флавоновый С-гликозид» означает гликозид флавона в широком смысле, т.е. производное, подпадающее под определение флавонов, где агликон непосредственно связан с аномерным углеродом альдозы. Флавоновые C-гликозиды в качестве неограничивающих примеров включают лютеолин C-гликозид, трицетин C-гликозид, апигенин C-гликозид и акацетин C-гликозид. Флавоновые С-гликозиды также включают производные гликозидов апигенина, лютеолина, трицетина и акацетина. Одним из известных путей биосинтеза флавоновых С-гликозидов в растениях является путь, показанный на фиг. 1. В этом пути флавоновый С-гликозид образуется при помощи F2H, CGT и FDH.
[0018] В этом пути синтеза флавоновые ди-C-гликозиды, такие как виценин-2 (апигенин 6,8-ди-C-глюкозид), также синтезируются в дополнение к флавоновым моно-C-гликозидам, но авторы настоящего изобретения неожиданно обнаружили, что флавоновые ди-C-гликозиды являются причиной покраснения и что флавоновые моно-C-гликозиды обладают более высоким эффектом копигментации с антоцианами дельфинидинового типа, чем флавоновые ди-C-гликозиды. Для однородного и стабильного получения трансгенного растения с синей окраской цветков необходимо свести к минимуму накопление флавоновых ди-С-гликозидов, а также аккумулировать в лепестках значительные количества только флавоновых моно-С-гликозидов. Флавоновые моно-C-гликозиды, как правило, представляют собой флавоновые 6-C-глюкозиды или флавоновые 8-C-глюкозиды, но чаще всего являются флавоновыми 6-C-глюкозидами. Примеры флавоновых моно-C-гликозидов включают апигенин 6-C-глюкозид (изовитексин), лютеолин 6-C-глюкозид (изоориентин), трицетин 6-C-глюкозид, апигенин 8-C-глюкозид (витексин), лютеолин 8-C-глюкозид (ориентин) и трицетин 8-С-глюкозид, или их производные.
[0019] Накопление флавоновых С-гликозидов в растительных клетках может быть достигнуто путем трансформации растений-хозяев генами, необходимыми для указанных выше путей синтеза (т.е. геном F2H, геном CGT и геном FDH), или их гомологами. Если используемый ген CGT представляет собой ген CGT, полученный из гречихи, или его гомолог, и, в частности, ген CGT, полученный из гречихи, или его гомолог с нетранслируемой областью (5'-UTR), полученной из гена ADH Arabidopsis thaliana (SEQ ID NO: 15), или с нетранслируемой областью (5'-UTR), полученной из гена HSPRO из Arabidopsis thaliana (SEQ ID NO: 13), добавленной в качестве последовательности энхансера трансляции, возможно накопление значительно большего количества флавоновых моно-C-гликозидов в лепестках, чем флавоновых ди-C-гликозидов.
[0020] Ген CGT, полученный из гречихи, или его гомолог выбран из группы, состоящей из следующих полинуклеотидов:
(1-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 11;
(1-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 11 при жестких условиях, и имеет ту же активность, что и полинуклеотид из (1-a);
(1-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 12;
(1-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 12, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (1-с); и
(1-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичностью в отношении аминокислотной последовательности, указанной как SEQ ID NO: 12, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (1-c).
[0021] Источник гена F2H или его гомолога конкретно не ограничен при условии, что он обладает желаемой функцией, но предпочтительно он является полученным из солодки геном F2H или его гомологом, выбранным из группы, состоящей из следующих полинуклеотидов:
(2-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 5;
(2-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 5 при жестких условиях, и имеет ту же активность, что и полинуклеотид из (2-a);
(2-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 6;
(2-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 6, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом из (2-с); и
(2-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичностью в отношении аминокислотной последовательности, указанной как SEQ ID NO: 6, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (2-c).
[0022] Источник гена FDH или его гомолога конкретно не ограничен при условии, что он обладает желаемой функцией, но предпочтительно он является полученным из Lotus japonicus геном FDH или его гомологом, и выбран из группы, состоящей из следующих полинуклеотидов:
(3-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 9;
(3-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 9, при жестких условиях, и имеет ту же активность, что и полинуклеотид из (3-a);
(3-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 10;
(3-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 10, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (3-с); и
(3-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичностью в отношении аминокислотной последовательности, указанной как SEQ ID NO: 10, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (3-c).
[0023] Накопление антоцианов дельфинидинового типа в растительных клетках может быть достигнуто путем включения гена флавоноидной F3',5' гидроксилазы (F3',5'H) или его гомолога и гена метилтрансферазы (МТ) или его гомолога в растение-хозяин (PTL 3). Путем трансформации растения-хозяина вектором, дополнительно содержащим ген F3',5'H или его гомолог и ген MT или его гомолог в дополнение к гену, необходимому для пути синтеза указанных флавоновых моно-C-гликозидов или их гомологов, возможно заставить антоцианин дельфинидинового типа и флавоновый моно-C-гликозид сосуществовать в клетках растения-хозяина.
[0024] Источник гена F3',5'H или его гомолога конкретно не ограничен при условии, что он обладает желаемой функцией, но предпочтительно он является полученным из колокольчика геном F3',5'H или его гомологом, и выбран из группы, состоящей из:
(4-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 1;
(4-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 1, при жестких условиях и имеет ту же активность, что и полинуклеотид из (4-a);
(4-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 2;
(4-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 2, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом из (4-с); и
(4-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичностью в отношении аминокислотной последовательности, указанной как SEQ ID NO: 2, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (4-c).
[0025] Источник гена МТ или его гомолога конкретно не ограничен при условии, что он обладает желаемой функцией, но предпочтительно он является полученным из торении геном МТ или его гомологом, выбранным из группы, состоящей из:
(5-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 3;
(5-b) полинуклеотида, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, указанной как SEQ ID NO: 3, при жестких условиях и имеет ту же активность, что и полинуклеотид из (5-a);
(5-c) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 4;
(5-d) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, которая представляет собой аминокислотную последовательность, указанную как SEQ ID NO: 4, имеющую делецию, замену, вставку и/или добавление одной или нескольких аминокислот, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (5-с); и
(5-e) полинуклеотида, который кодирует белок, имеющий аминокислотную последовательность, по меньшей мере, с 90% идентичностью в отношении аминокислотной последовательности, указанной как SEQ ID NO: 4, и имеющий ту же активность, что и белок, кодируемый полинуклеотидом (5-c).
[0026] К гену МТ или его гомологу также может быть добавлена нетранслируемая область (5'-UTR), полученная из гена ADH Arabidopsis thaliana (SEQ ID NO: 15), или нетранслируемая область (5'-UTR), полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13), в качестве последовательности энхансера трансляции.
[0027] На всем протяжении настоящего описания термин «полинуклеотид» относится к ДНК или РНК.
Как применяют в настоящем документе, термин «жесткие условия» относится к условиям, которые позволяют проводить специфическое связывание между полинуклеотидом или олигонуклеотидом и геномной ДНК избирательным и поддающимся обнаружению образом. Жесткие условия определяются соответствующей комбинацией концентрации соли, органического растворителя (например, формамида), температуры и других известных условий. В частности, жесткость повышается за счет снижения концентрации соли, повышения концентрации органического растворителя или повышения температуры гибридизации. На жесткость также влияют условия отмывания после гибридизации. Условия отмывания определяются концентрацией и температурой, и жесткость отмывания повышается за счет снижения концентрации соли и повышения температуры. Таким образом, термин «жесткие условия» означает такие условия, при которых специфическая гибридизация происходит только между нуклеотидными последовательностями с высокой идентичностью, такими как степень «идентичности» между нуклеотидными последовательностями в среднем приблизительно 80% или больше, предпочтительно приблизительно 90% или больше, более предпочтительно приблизительно 95% или больше, даже более предпочтительно приблизительно 97% или больше и наиболее предпочтительно приблизительно 98% или больше. «Жесткими условиями» могут быть, например, температура от 60°C до 68°C, концентрация натрия от 150 до 900 мМ и предпочтительно от 600 до 900 мМ, pH от 6 до 8, с конкретными примерами, включающими гибридизацию в условиях 5× SSC (750 мМ NaCl, 75 мМ трицитрат натрия), 1% SDS, 5× раствора Денхардта, 50% формальдегида, 42°C, отмывание в условиях 0,1× SSC (15 мМ NaCl, 1,5 мМ трицитрат натрия), 0,1% ДСН, 55°С.
[0028] Гибридизацию можно проводить с помощью общеизвестного в данной области способа или аналогичного способа, описанного в Current Protocols in Molecular Biology (под редакцией Frederick M. Ausubel et al., 1987). При использовании коммерческой доступной библиотеки гибридизацию можно проводить в соответствии со способом, описанным в прилагаемых инструкциях по использованию. Ген, отобранный с помощью гибридизации, может быть получен природным путем, как из растений, так и не из растений. Геном, выбранным путем гибридизации, может быть кДНК, геномная ДНК или химически синтезированная ДНК.
[0029] Фраза «аминокислотная последовательность, имеющая делецию, замену, вставку и/или добавление одной или нескольких аминокислот», как применяют в настоящем документе, означает аминокислотную последовательность, имеющую делецию, замену, вставку и/или добавление от 1 до 20, предпочтительно от 1 до 5 и более предпочтительно от 1 до 3 произвольных аминокислот. Сайт-специфический мутагенез является полезным методом генетической инженерии, поскольку он позволяет вводить специфические мутации в указанные сайты, и его можно проводить способом, описанным в Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989. Экспрессируя мутантную ДНК с помощью подходящей экспрессирующей системы, можно получить белок, состоящий из аминокислот, последовательность которых включает делецию, замену, вставку и/или добавление одной или нескольких аминокислот.
Полинуклеотид можно получить способом, который широко известен специалистам в данной области, таким как способ химического синтеза с использованием фосфорамидитного способа, или способ амплификации нуклеиновой кислоты с использованием образца растительной нуклеиновой кислоты в качестве матрицы, и праймеров, разработанных на основе нуклеотидной последовательности целевого гена.
[0030] На всем протяжении настоящего описания термин «идентичность» означает для полипептидных последовательностей (или аминокислотных последовательностей) или полинуклеотидных последовательностей (или нуклеотидных последовательностей) количество (число) составляющих их аминокислотных остатков или нуклеотидов, которые можно определить как идентичные между двумя цепями, в смысле взаимного согласования между ними, то есть степень корреляции последовательности между двумя полипептидными последовательностями или двумя полинуклеотидными последовательностями, и эта «идентичность» может быть легко вычислена. Известно множество способов измерения идентичности между двумя полинуклеотидными последовательностями или полипептидными последовательностями, а термин «идентичность» хорошо известен специалистам в данной области (например, см. Lesk, A.M. (Ed.), Computational Molecular Biology, Oxford University Press, New York, (1988); Smith, D.W. (Ed.), Biocomputing: Informatics and Genome Projects, Academic Press, New York, (1993); Grifin, A.M. & Grifin, H.G. (Ed.), Computer Analysis of Sequence Data: Part I, Human Press, New Jersey, (1994); von Heinje, G., Sequence Analysis in Molecular Biology, Academic Press, New York, (1987); Gribskov, M. & Devereux, J. (Ed.), Sequence Analysis Primer, M-Stockton Press, New York, (1991) и в других местах).
[0031] Кроме того, числовые значения для «идентичности», используемые в настоящем описании, если не указано иное, могут быть числовыми значениями, рассчитанными с использованием программы поиска идентичности, известной специалистам в данной области, но они предпочтительно являются числовыми значениями, рассчитанными с использованием программы ClustalW из приложения MacVector (версия 9.5, Oxford Molecular Ltd., Oxford, England). По изобретению, степень «идентичности» между полинуклеотидными последовательностями или аминокислотными последовательностями составляет, например, приблизительно 90% или больше, предпочтительно приблизительно 95% или больше, более предпочтительно приблизительно 97% или больше, и наиболее предпочтительно приблизительно 98% или больше.
[0032] Полинуклеотид (нуклеиновая кислота, ген) по изобретению «кодирует» интересующий белок. Здесь «кодирует» означает, что он позволяет экспрессировать интересующий белок в состоянии, в котором он проявляет свою активность. Кроме того, термин «кодирует» включает как кодирование структурной последовательности (экзон), которая представляет собой непрерывный участок интересующего белка, так и кодирование промежуточной последовательности (интрон).
[0033] Ген с природной нуклеотидной последовательностью можно получить, например, путем анализа с использованием секвенатора ДНК. Кроме того, ДНК, кодирующая фермент с модифицированной аминокислотной последовательностью, может быть синтезирована с использованием обычного сайт-специфического мутагенеза или ПЦР на основе ДНК, имеющей природную нуклеотидную последовательность. Например, фрагмент ДНК, подлежащий модификации, можно получить путем рестрикционного ферментативного преобразования природной кДНК или геномной ДНК и использовать в качестве матрицы для сайт-специфического мутагенеза или ПЦР с использованием праймеров с желаемой мутацией для получения фрагмента ДНК, имеющего желаемую модификацию. Фрагмент ДНК, имеющий мутацию, затем может быть связан с фрагментом ДНК, кодирующим другую часть целевого фермента.
Альтернативно, для того, чтобы получить ДНК, кодирующую фермент, состоящий из укороченной аминокислотной последовательности, ДНК, кодирующая аминокислотную последовательность, более длинную, чем целевая аминокислотная последовательность, такую как полноразмерная аминокислотная последовательность, может быть обрезана желаемым ферментом рестрикции, и если полученный фрагмент ДНК не имеет код полной целевой аминокислотной последовательности, то фрагмент ДНК, состоящий из последовательности недостающей части, может быть синтезирован и связан с ним.
[0034] Путем экспрессии полученного полинуклеотида с использованием системы экспрессии гена в Escherichia coli или дрожжах и измерения активности фермента можно подтвердить, что полученный полинуклеотид кодирует белок с желаемой активностью.
[0035] Настоящее изобретение относится к (рекомбинантному) вектору, и особенно к экспрессирующему вектору, включая указанный выше полинуклеотид, и к растениям хризантемы, трансформированным вектором.
[0036] Вектор по изобретению также содержит область контроля экспрессии, такую как промотор, терминатор и точку начала репликации, которые зависят от типа растения-хозяина, в которое вводят вектор. Примеры промоторов, которые конститутивно экспрессируют полинуклеотиды в растительных клетках, включают промотор 35S вируса мозаики цветной капусты, промотор El235S, имеющий два энхансерных участка промотора 35S, связанных вместе, и промотор гена rd29A, промотор rbcS и промотор mac-1. Для тканеспецифической экспрессии гена, можно использовать промотор для гена, экспрессируемого конкретно в этой ткани.
[0037] Вектор может быть создан обычным способом с использованием фермента рестрикции и лигазы. Трансформация растения-хозяина с помощью экспрессирующего вектора также может осуществляться по общепринятому способу.
[0038] На современном уровне технологий можно использовать способы введения полинуклеотида в растение и конститутивно или тканеспецифически экспрессировать полинуклеотид. Перенос ДНК в растение можно проводить способом, известным специалистам в данной области, таким как способ с Agrobacterium, способ бинарного вектора, способ электропорации, способ с ПЭГ или способом генной пушки.
[0039] Растения, используемые в качестве растений-хозяев по настоящему изобретению, конкретно не ограничены и могут представлять собой растения, принадлежащие к роду Rosaceae Rosa, Compositae Chrysanthemum, Caryophyllaceae Dianthus (например, гвоздика) или Liliaceae Lilium, среди которых сорт розы Rosaceae Rosa (научное название: Rosa hybrida) является наиболее предпочтительным. Как применяют в настоящем документе термин «растение розы» представляет собой сорт розы Rosaceae Rosa (научное название: Rosa hybrida), что является его таксономической классификацией. Розы в основном классифицируются как чайно-гибридные розы, розы Флорибунда и Полианта на основе их древовидной формы и размера цветка, при этом основной пигмент (антоциан) в лепестках всех линий относится к двум типам, цианидиновому типу и пеларгонидиновому типу. Тип растения розы, используемого в качестве растения-хозяина по настоящему изобретению, конкретно не ограничен, и подходят любые из этих разновидностей или линий. Примеры сортов роз, которые можно использовать в качестве хозяев, включают Ocean Song, Noblesse, Rita Perfumera, Cool Water, Fame, Topless и Peach Avalanche.
[0040] С помощью способов по настоящему изобретению можно однородно и стабильно создавать трансгенные растения, предпочтительно Rosaceae Rosa, Compositae Chrysanthemum и Caryophyllaceae Dianthus (гвоздика), и наиболее предпочтительно растения роз, имеющие голубую окраску цветков. Когда полученное трансгенное растение представляет собой растение розы, оно проявляет окраску цветка в синей группе или фиолетово-синей группе в соответствии с таблицей цветов RHS, и/или с углом оттенка от 339,7° до 270,0°, и более предпочтительно 315° или меньше в цветовой системе CIEL*a*b*.
[0041] Изобретение дополнительно также относится к срезанным цветкам полученного трансгенного растения или его инбредного или аутбредного потомства, или выводковым почкам, частям растительного организма, тканям или клеткам, или обработанной форме, созданной из срезанных цветов (особенно обработанных срезанных цветов). Упомянутые здесь обработанные срезанные цветы включают прессованные цветы, сформированные из срезанных цветов, или консервированные цветы, сухие цветы или продукты, запечатанные смолой, не ограничиваясь вышеуказанными.
[0042] Настоящее изобретение теперь будет объяснено более подробно с помощью примеров.
ПРИМЕРЫ
[0043] [Пример 1: Имитация копигментного эффекта флавонового С-гликозида с антоцианом (мальвином)]
Получали антоциан (мальвин) и флавоновый С-гликозид для имитации копигментного эффекта флавонового С-гликозида на мальвин. Мальвин (мальвидин 3,5-диглюкозид) и флавоновые С-гликозиды (витексин (апигенин 8-С-глюкозид), изовитексин (апигенин 6-С-глюкозид), ориентин (лютеолин 8-С-глюкозид), изоориентин (лютеолин 6-C-глюкозид) и виценин-2 (апигенин 6,8-ди-C-глюкозид)), используемые в экспериментах, были приобретены у Nacalai Tesque, Inc.
Каждый флавоновый С-гликозид (витексин, изовитексин, ориентин, изоориентин или виценин-2) добавляли к полученному мальвину в 5-молярном эквиваленте концентрации в буферном растворе при рН 5,0, и измеряли спектры поглощения. Концентрация мальвина составила 0,5 мМ.
[0044] [Таблица 1]
Максимум поглощения (λmax) и угол оттенка (°) раствора мальвина после добавления флавонового C-гликозида
(λmax)
[0045] Добавление флавонового С-гликозида увеличивало оптическую абсорбцию раствора мальвина и сдвигало максимум поглощения (λmax) в сторону более длинной длины волны по сравнению с одним мальвином, независимо от того, какой флавоновый С-гликозид добавляли. Этот эффект проявляется в виде большего смещения максимума поглощения в сторону длинной длины волны в следующем порядке: изовитексин > изоориентин > ориентин > витексин > виценин-2. Также было подтверждено, что угол оттенка меньше в следующем порядке: изовитексин > изоориентин > ориентин > витексин > виценин-2. Таким образом, было показано, что копигментный эффект флавоновых моно-С-гликозидов выше, чем у флавоновых ди-С-гликозидов. Кроме того, показано, что копигментный эффект флавоновых моно-С-гликозидов особенно высок с флавоновыми 6-С-гликозидами.
[0046] [Пример 2: Перенос гена F3',5'H, полученного из колокольчика, гена MT, полученного из торении, гена F2H, полученного из солодки, гена CGT с модифицированной частотой использования кодонов, полученного из риса, и гена FDH, полученного из Lotus japonicus, в сорт розы «Ocean Song»]
Плазмида pSPB6486 имеет в качестве основы pBINPLUS и содержит следующие четыре экспрессирующие кассеты.
(1) промотор El235S, полноразмерная кДНК F3',5'H, полученного из колокольчика (SEQ ID NO: 1), и терминатор D8;
(2) промотор El235S, полноразмерная кДНК MT, полученного из торении (SEQ ID NO: 3), и терминатор NOS;
(3) промотор 35S, полноразмерная кДНК F2H, полученного из солодки (SEQ ID NO: 5), и терминатор AT, полученный из периллы;
(4) промотор 35S, полноразмерная кДНК CGT с модифицированной частотой использования кодонов, полученного из риса (SEQ ID NO: 7), и терминатор HSP, полученный из Arabidopsis thaliana;
(5) промотор 35S, полноразмерная кДНК FDH, полученного из Lotus japonicus (SEQ ID NO: 9), и терминатор HSP, полученный из Arabidopsis thaliana.
Эта плазмида конститутивно экспрессирует в растениях ген F3',5'H колокольчика, ген MT торении, ген F2H солодки, ген CGT с модифицированной частотой использования кодонов из риса и ген FDH Lotus japonicus.
[0047] Сконструированная плазмида pSPB6486 была введена в сорт голубой розы «Ocean Song», всего было получено 27 трансформантов. При анализе пигментов запасы мальвидина восстановились в 26 трансформантах с максимальным содержанием мальвидина 74,5% (среднее: 57,0%). Флавоновые С-гликозиды изовитексин (апигенин 6-С-глюкозид), витексин (апигенин 8-С-глюкозид), изоориентин (лютеолин 6-С-глюкозид), ориентин (лютеолин 8-С-глюкозид) и виценин-2 (апигенин 6,8-ди-С-глюкозид) также были идентифицированы и определены количественно. Флавоновые С-гликозиды были обнаружены во всех трансформантах, в которых были обнаружены мальвидины, при этом флавоновый ди-С-гликозид виценин-2 был основным обнаруженным компонентом. Количество продукции возрастало в следующем порядке: виценин-2 > изовитексин > витексин > изоориентин > ориентин, с максимальным общим количеством 1,563 мг на 1 г свежей лепестковой массы. Общее количество флавоновых С-гликозидов более чем в 10 раз превышало количество мальвидинов.
[0048] Измеренные значения для типичных трансформантов показаны в таблице 2 ниже.
[Таблица 2]
(%)
Хозяин: Ocean Song
Del: дельфинидин, Cya: цианидин, Pet: петунидин, Pel: пеларгонидин, Mal: малвидин
M: мирицетин, Q: кверцетин, K: камферол
Tri: трицетин, Lut: лютеолин, Api: апигенин, Vic2: виценин-2, VX: витексин, IVX: изовитексин, Ori: ориентин, Iori: изоориентин
Mal(%): доля мальвидина в общем количестве антоцианидинов
[0049] [Пример 3: Перенос гена F3',5'H, полученного из колокольчика, гена MT, полученного из торении, гена F2H, полученного из солодки, гена CGT с модифицированной частотой использования кодонов, полученного из гречихи, и гена FDH, полученного из Lotus japonicus, в сорт розы «Ocean Song»]
Плазмида pSPB7473 имеет в качестве основы pBINPLUS и содержит следующие четыре экспрессирующие кассеты.
(1) промотор El235S, полноразмерная кДНК F3',5'H, полученного из колокольчика (SEQ ID NO: 1), и терминатор D8;
(2) промотор El235S, полноразмерная кДНК MT, полученного из торении (SEQ ID NO: 3), (область 5'-UTR, полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13) и добавленная к 5'-концу) и полученный из Arabidopsis thaliana терминатор HSP;
(3) промотор El235S, полноразмерная кДНК F2H, полученного из солодки (SEQ ID NO: 5), и терминатор AT, полученный из периллы;
(4) промотор El235S, полноразмерная кДНК CGT с модифицированной частотой использования кодонов, полученного из гречихи (SEQ ID NO: 11), (область 5'-UTR, полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13) и добавленная к 5'-концу) и полученный из Arabidopsis thaliana терминатор HSP;
(5) промотор El235S, полноразмерная кДНК FDH, полученного из Lotus japonicus (SEQ ID NO: 9), и терминатор HSP, полученный из Arabidopsis thaliana.
Эта плазмида конститутивно экспрессирует в растениях ген F3',5'H колокольчика, ген MT торении, ген F2H солодки, ген CGT с модифицированной частотой использования кодонов из гречихи и ген FDH Lotus japonicus.
[0050] Сконструированная плазмида pSPB7473 была введена в сорт голубой розы «Ocean Song», всего было получено 35 трансформантов. При анализе пигментов запасы мальвидина восстановились в 21 трансформанте с максимальным содержанием мальвидина 20,8% (в среднем: 9,1%). Флавоновые С-гликозиды изовитексин (апигенин 6-С-глюкозид), витексин (апигенин 8-С-глюкозид), изоориентин (лютеолин 6-С-глюкозид), ориентин (лютеолин 8-С-глюкозид) и виценин-2 (апигенин 6,8-ди-С-глюкозид) также были идентифицированы и определены количественно. Флавоновые С-гликозиды были обнаружены во всех трансформантах, в которых были обнаружены мальвидины, при этом флавоновый 6-С-гликозид изовитексин был основным обнаруженным компонентом. Количество продукции возрастало в следующем порядке: изовитексин > витексин > виценин-2 > изоориентин > ориентин, и максимальное общее количество составило 7,418 мг на 1 г свежей лепестковой массы, что указывает на очень высокое содержание. Кроме того, по сравнению с линией OS/6486, описанной в примере 2, общее количество флавоновых С-гликозидов было высоким - 3 мг или более на 1 г массы свежих лепестков у большинства трансформантов, что было, по меньшей мере, примерно в 100 раз больше по сравнению с дельфинидинами.
[0051] Измеренные значения для типичных трансформантов показаны в таблице 3 ниже.
[Таблица 3]
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(%)
(мг/г свежей массы)
Хозяин: Ocean Song
Del: дельфинидин, Cya: цианидин, Pet: петунидин, Pel: пеларгонидин, Mal: малвидин
M: мирицетин, Q: кверцетин, K: камферол
Tri: трицетин, Lut: лютеолин, Api: апигенин, Vic2: виценин-2, VX: витексин, IVX: изовитексин, Ori: ориентин, Iori: изоориентин
Mal(%): доля мальвидина в общем количестве антоцианидинов
[0052] [Пример 4: Перенос гена F3',5'H, полученного из колокольчика, гена MT, полученного из торении, гена F2H, полученного из солодки, гена CGT, полученного из гречихи, и гена FDH, полученного из Lotus japonicus, в сорт розы «Ocean Song»]
Плазмида pSPB7472 имеет в качестве базовой основы pBINPLUS и содержит следующие четыре экспрессирующие кассеты.
(1) промотор El235S, полноразмерная кДНК F3',5'H, полученного из колокольчика (SEQ ID NO: 1), и терминатор D8;
(2) промотор El235S, полноразмерная кДНК MT, полученного из торении (SEQ ID NO: 3), (область 5'-UTR, полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13) и добавленная к 5'-концу) и полученный из Arabidopsis thaliana терминатор HSP;
(3) промотор El235S, полноразмерная кДНК F2H, полученного из солодки (SEQ ID NO: 5), и терминатор AT, полученный из периллы;
(4) промотор El235S, полноразмерная кДНК CGT, полученного из гречихи (SEQ ID NO: 14), (область 5'-UTR, полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13) и добавленная к 5'-концу) и полученный из Arabidopsis thaliana терминатор HSP;
(5) промотор El235S, полноразмерная кДНК FDH, полученного из Lotus japonicus (SEQ ID NO: 9) и терминатор HSP, полученный из Arabidopsis thaliana.
Эта плазмида конститутивно экспрессирует в растениях ген F3',5'H колокольчика, ген MT торении, ген F2H солодки, ген CGT гречихи и ген FDH Lotus japonicus.
[0053] Сконструированная плазмида pSPB7472 была введена в сорт голубой розы «Ocean Song», всего было получено 33 трансформанта. Затем идентифицировали флавоновые С-гликозиды изовитексин (апигенин 6-С-глюкозид), витексин (апигенин 8-С-глюкозид), изоориентин (лютеолин 6-С-глюкозид), ориентин (лютеолин 8-С-глюкозид) и виценин-2 (апигенин 6,8-ди-С-глюкозид) и антоцианидины дельфинидин, цианидин, петунидин, пеларгонидин и мальвидин, и определяли их количество. В результате накопление флавоновых С-гликозидов и мальвидина подтверждено в 11 трансформантах. Среднее содержание флавоновых С-гликозидов составило 3,75 мг на 1 г свежей лепестковой массы, при этом в качестве основного компонента выявлен флавоновый 6-С-гликозид изовитексин. Содержание мальвидина составляло максимум 15,6% (среднее: 8,8%).
[0054] Таким образом, среднее содержание флавоновых С-гликозидов на 1 г свежей лепестковой массы было выше у линии OS/7473, описанной в примере 3, и составило 4,19 мг. Другими словами, можно более эффективно производить флавоновые С-гликозиды с кодон-модифицированным геном CGT, чем с исходным геном CGT, полученным из гречихи.
[0055] Измеренные значения для типичных трансформантов показаны в таблице 4 ниже.
[Таблица 4]
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(%)
(мг/г свежей массы)
Хозяин: Ocean Song
Del: дельфинидин, Cya: цианидин, Pet: петунидин, Pel: пеларгонидин, Mal: малвидин
M: мирицетин, Q: кверцетин, K: камферол
Tri: трицетин, Lut: лютеолин, Api: апигенин, Vic2: виценин-2, VX: витексин, IVX: изовитексин, Ori: ориентин, Iori: изоориентин
Mal(%): доля мальвидина в общем количестве антоцианидинов
[0056] [Пример 5: Перенос гена F3',5'H, полученного из колокольчика, гена MT, полученного из торении, гена F2H, полученного из солодки, гена CGT, полученного из гречихи, и гена FDH, полученного из Lotus japonicus, в сорт розы «Ocean Song»]
Плазмида pSPB7808 имеет в качестве базовой основы pBINPLUS и содержит следующие четыре экспрессирующие кассеты.
(1) промотор El235S, полноразмерная кДНК F3',5'H, полученного из колокольчика (SEQ ID NO: 1), и терминатор D8;
(2) промотор El235S, полноразмерная кДНК MT, полученного из торении (SEQ ID NO: 3), и терминатор HSP, полученный из Arabidopsis thaliana;
(3) промотор El235S, полноразмерная кДНК F2H, полученного из солодки (SEQ ID NO: 5) и терминатор AT, полученный из периллы;
(4) промотор El235S, полноразмерная кДНК CGT, полученного из гречихи (SEQ ID NO: 14), (область 5'-UTR, полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13) и добавленная к 5'-концу) и полученный из Arabidopsis thaliana терминатор HSP;
(5) промотор El235S, полноразмерная кДНК FDH, полученного из Lotus japonicus (SEQ ID NO: 9), и терминатор HSP, полученный из Arabidopsis thaliana.
Эта плазмида конститутивно экспрессирует в растениях ген F3',5'H колокольчика, ген MT торении, ген F2H солодки, ген CGT гречихи и ген FDH Lotus japonicus.
[0057] Сконструированная плазмида pSPB7808 была введена в сорт голубой розы «Ocean Song», всего было получено 65 трансформантов. Затем идентифицировали флавоновые С-гликозиды изовитексин (апигенин 6-С-глюкозид), витексин (апигенин 8-С-глюкозид), изоориентин (лютеолин 6-С-глюкозид), ориентин (лютеолин 8-С-глюкозид) и виценин-2 (апигенин 6,8-ди-С-глюкозид) и антоцианидины дельфинидин, цианидин, петунидин, пеларгонидин и мальвидин, и определяли их количество. В результате накопление флавоновых С-гликозидов и мальвидина подтверждено у 32 трансформантов. Среднее содержание флавоновых С-гликозидов составило 3,00 мг на 1 г свежей лепестковой массы, при этом в качестве основного компонента выявлен флавоновый 6-С-гликозид изовитексин. Содержание мальвидина составляло максимум 69,2% (среднее: 43,9%).
[0058] Измеренные значения для типичных трансформантов показаны в таблице 5 ниже.
[Таблица 5]
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(%)
(мг/г свежей массы)
Хозяин: Ocean Song
Del: дельфинидин, Cya: цианидин, Pet: петунидин, Pel: пеларгонидин, Mal: малвидин
M: мирицетин, Q: кверцетин, K: камферол
Tri: трицетин, Lut: лютеолин, Api: апигенин, Vic2: виценин-2, VX: витексин, IVX: изовитексин, Ori: ориентин, Iori: изоориентин
Mal(%): доля мальвидина в общем количестве антоцианидинов
[0059] [Пример 6: Перенос гена F3',5'H, полученного из колокольчика, гена MT, полученного из торении, гена F2H, полученного из солодки, гена CGT с модифицированной частотой использования кодонов, полученного из гречихи, и гена FDH, полученного из Lotus japonicus, в сорт розы «Ocean Song»]
Плазмида pSPB7809 имеет в качестве базовой основы pBINPLUS и содержит следующие четыре экспрессирующие кассеты.
(1) промотор El235S, полноразмерная кДНК F3',5'H, полученного из колокольчика (SEQ ID NO: 1), и терминатор D8;
(2) промотор El235S, полноразмерная кДНК MT, полученного из торении (SEQ ID NO: 3), и терминатор HSP, полученный из Arabidopsis thaliana;
(3) промотор El235S, полноразмерная кДНК F2H, полученного из солодки (SEQ ID NO: 5), и терминатор AT, полученный из периллы;
(4) промотор El235S, полноразмерная кДНК CGT с модифицированной частотой использования кодонов, полученного из гречихи (SEQ ID NO: 11), (область 5'-UTR, полученная из гена HSPRO Arabidopsis thaliana (SEQ ID NO: 13) и добавленная к 5'-концу) и полученный из Arabidopsis thaliana терминатор HSP;
(5) промотор El235S, полноразмерная кДНК FDH, полученного из Lotus japonicus (SEQ ID NO: 9), и терминатор HSP, полученный из Arabidopsis thaliana.
Эта плазмида конститутивно экспрессирует в растениях ген F3',5'H колокольчика, ген MT торении, ген F2H солодки, ген CGT с модифицированной частотой использования кодонов из гречихи и ген FDH Lotus japonicus.
[0060] Сконструированная плазмида pSPB7809 была введена в сорт голубой розы «Ocean Song», всего было получено 143 трансформанта. Затем идентифицировали флавоновые С-гликозиды изовитексин (апигенин 6-С-глюкозид), витексин (апигенин 8-С-глюкозид), изоориентин (лютеолин 6-С-глюкозид), ориентин (лютеолин 8-С-глюкозид) и виценин-2 (апигенин 6,8-ди-С-глюкозид) и антоцианидины дельфинидин, цианидин, петунидин, пеларгонидин и мальвидин, и определяли их количество. В результате накопление флавоновых С-гликозидов произошло у 58 трансформантов. Среднее содержание флавоновых С-гликозидов составило 3,24 мг на 1 г свежей лепестковой массы, при этом в качестве основного компонента выявлен флавоновый 6-С-гликозид изовитексин. Содержание мальвидина составляло максимум 80,3% (среднее: 46,6%).
[0061] Таким образом, среднее содержание флавоновых С-гликозидов на 1 г свежей лепестковой массы было выше у линии OS/7809, описанной в примерах. Аналогично результатам, полученным в примерах 3 и 4, показано, что можно более эффективно продуцировать флавоновые С-гликозиды с помощью кодон-модифицированного гена CGT, чем с исходным геном CGT, полученным из гречихи.
[0062] Измеренные значения для типичных трансформантов показаны в таблице 6 ниже.
[Таблица 6-1]
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(мг/г свежей массы)
(%)
(мг/г свежей массы)
[Таблица 6-2]
Хозяин: Ocean Song
Del: дельфинидин, Cya: цианидин, Pet: петунидин, Pel: пеларгонидин, Mal: малвидин
M: мирицетин, Q: кверцетин, K: камферол
Tri: трицетин, Lut: лютеолин, Api: апигенин, Vic2: виценин-2, VX: витексин, IVX: изовитексин, Ori: ориентин, Iori: изоориентин
Mal(%): доля мальвидина в общем количестве антоцианидинов
[0063] [Пример 7: Оценка окраски цветков роз, содержащих флавоновые С-гликозиды]
Трансформанты, созданные в Примерах 2 и 3 (с сортом розы «Ocean Song» в качестве хозяина), измеряли для определения цветовых оттенков соответствующих лепестков с помощью спектроколориметра CM-2022 (Minolta) с полем зрения 10° и источником света D65, и анализировали с помощью программного обеспечения для управления цветом SpectraMagicTM (Minolta).
При сравнении средних значений углов оттенка не было обнаружено различий в угле оттенка лепестков между розами с генами CGT, полученными из гречихи (Пример 3), и розами с генами CGT, полученными из риса (Пример 2). Однако на индивидуальном уровне большее количество трансформантов демонстрировало угол оттенка 315° или меньше среди роз с перенесенным геном CGT, полученным из гречихи, причем одна из них был изменена до самого синего цвета, который был получен, со значением 294,5°. Эти результаты подтвердили, что использование гена CGT, полученного из гречихи, значительно увеличило количество продукции флавоновых С-гликозидов, и особенно количество моно-С-гликозидов, изменяя оттенок лепестков до синего оттенка за счет их совместного присутствия с антоцианами.
[0064] Результаты показаны в таблице 7.
[Таблица 7]
Накапливает цианидин в качестве основного пигмента, совершенно не содержит флавонового C-гликозида
Имеет высокое накопление дельфинидина в качестве основного пигмента, совершенно не содержит флавонового C-гликозида
Самая синяя точка: 333,15°
Имеет высокое накопление мальвидина в качестве основного пигмента, содержит флавоновый C-гликозид
Самая синяя точка: 318,89°
Имеет высокое накопление дельфинидина в качестве основного пигмента, содержит флавоновый C-гликозид
Самая синяя точка: 294,53°
| название | год | авторы | номер документа |
|---|---|---|---|
| РОЗА, СОДЕРЖАЩАЯ ФЛАВОН И МАЛЬДИВИН, И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2008 |
|
RU2463348C2 |
| НОВЫЙ ГЕН ГЛИКОЗИЛТРАНСФЕРАЗЫ И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2636463C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2706789C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2772947C2 |
| НОВЫЙ ГЕН ФЛАВОНОИД 3,5-ГИДРОКСИЛАЗЫ КОЛОКОЛЬЧИКА И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2640248C2 |
| КОДОН-ОПТИМИЗИРОВАННЫЕ СИНТЕТИЧЕСКИЕ НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩИЕ БЕЛОК Cry2Ai, И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2799821C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2016 |
|
RU2745306C2 |
| РЕГУЛИРУЮЩИЙ ВЫСОТУ РАСТЕНИЙ ГЕН И ЕГО ПРИМЕНЕНИЯ | 2008 |
|
RU2458132C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2016 |
|
RU2759224C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2741103C2 |
Группа изобретений относится к биотехнологии. Предложены ген C-глюкозилтрансферазы (CGT) с модифицированной частотой использования кодонов, полученный из гречихи, для создания трансгенного растения с синей окраской цветка; вектор, содержащий ген CGT с модифицированной частотой использования кодонов, полученный из гречихи; трансгенное растение с синей окраской цветка, содержащее заявленный ген; выводковые почки, части растений трансгенного растения или его инбредное или аутбредное потомство для создания трансгенного растения с синей окраской цветка; ткань трансгенного растения, клетка трансгенного растения, срезанный цветок трансгенного растения или его инбредное или аутбредное потомство для создания трансгенного растения с синей окраской цветка; способ создания трансгенных растений с синей окраской цветка, включающий стадию переноса гена или вектора в растение-хозяин. Изобретения обеспечивают создание трансгенных растений с голубым цветом цветка. 9 н. и 11 з.п. ф-лы, 7 ил., 7 табл., 3 пр.
1. Ген C-глюкозилтрансферазы (CGT) с модифицированной частотой использования кодонов, полученный из гречихи, для создания трансгенного растения с синей окраской цветка, где:
ген CGT, полученный из гречихи, представляет собой:
(1-a) полинуклеотид, состоящий из нуклеотидной последовательности, указанной как SEQ ID NO: 11.
2. Вектор, содержащий ген CGT с модифицированной частотой использования кодонов, полученный из гречихи, для создания трансгенного растения с синей окраской цветка, где ген CGT, полученный из гречихи, выбран из группы, состоящей из:
(1-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 11; и
(1-b) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 12.
3. Вектор по п. 2, который дополнительно содержит ген флаванон-2-гидроксилазы (F2H) и ген дегидратазы (FDH).
4. Вектор по п. 3, где
ген F2H выбран из группы, состоящей из:
(2-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 5; и
(2-b) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 6, и
ген FDH выбран из группы, состоящей из:
(3-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 9; и
(3-b) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 10.
5. Вектор по любому из пп. 2-4, который дополнительно включает ген флавоноидной F3',5' гидроксилазы (F3',5'H) и ген метилтрансферазы (МТ).
6. Вектор по п. 5,
ген F3',5'H выбран из группы, состоящей из:
(4-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 1; и
(4-b) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 2, и
ген MT выбран из группы, состоящей из:
(5-a) полинуклеотида, состоящего из нуклеотидной последовательности, указанной как SEQ ID NO: 3; и
(5-b) полинуклеотида, который кодирует белок, состоящий из аминокислотной последовательности, указанной как SEQ ID NO: 4.
7. Трансгенное растение с синей окраской цветка, содержащее ген по п. 1, или вектор по любому из пп. 2-6, или его инбредное или аутбредное потомство.
8. Трансгенное растение по п. 7 или его инбредное или аутбредное потомство, где растение выбрано из розы, хризантемы, гвоздики или лилии.
9. Трансгенное растение по п. 8 или его инбредное или аутбредное потомство, где растение представляет собой розу.
10. Выводковые почки трансгенного растения по любому из пп. 7-9 или его инбредное или аутбредное потомство, для создания трансгенного растения с синей окраской цветка, содержащие ген по п. 1 или вектор по п. 2.
11. Части растений трансгенного растения по любому из пп. 7-9 или его инбредное или аутбредное потомство, для создания трансгенного растения с синей окраской цветка, содержащие ген по п. 1 или вектор по п. 2.
12. Ткань трансгенного растения по любому из пп. 7-9 или его инбредное или аутбредное потомство, для создания трансгенного растения с синей окраской цветка, содержащие ген по п. 1 или вектор по п. 2.
13. Клетка трансгенного растения по любому из пп. 7-9 или его инбредное или аутбредное потомство, для создания трансгенного растения с синей окраской цветка, содержащая ген по п. 1 или вектор по п. 2.
14. Срезанный цветок трансгенного растения по любому из пп. 7-9, или его инбредное или аутбредное потомство, или обработанная форма, созданная из срезанных цветов, для получения трансгенного растения с синей окраской цветка, содержащие ген по п. 1 или вектор по п. 2.
15. Способ создания трансгенных растений с синей окраской цветка, включающий стадию переноса гена по п. 1 или вектора по п. 2 в растение-хозяин, чтобы вызвать совместное присутствие антоцианов дельфинидинового типа и флавоновых моно-С-гликозидов в растительных клетках.
16. Способ по п. 15, где флавоновый моно-С-гликозид представляет собой апигенин 6-С-глюкозид, лютеолин 6-С-глюкозид, трицетин 6-С-глюкозид, апигенин 8-С-глюкозид, лютеолин 8-С-глюкозид или трицетин 8-С-глюкозид или его производное.
17. Способ по п. 15 или 16, где антоцианы дельфинидинового типа выбраны из группы, состоящей из мальвидинов, дельфинидинов, петунидинов и их комбинации.
18. Способ по любому из пп. 15-17, который дополнительно включает перенос гена CGT, полученного из гречихи, по п. 1 или вектора по любому из пп. 2-6 в растительные клетки хозяина.
19. Способ по любому из пп. 15-18, где растение представляет собой розу, хризантему, гвоздику или лилию.
20. Способ по п. 19, где растение представляет собой розу.
| NAGATOMO, Y | |||
| et al., Purification, molecular cloning and functional characterization of flavonoid C-glucosyltransferases from Fagopyram esculentum M | |||
| (buckwheat) cotyledon, The Plant Journal, 2014, v | |||
| Капельная масленка с постоянным уровнем масла | 0 |
|
SU80A1 |
| WO 2019069946 Al, 11.04.2019 | |||
| ИДЕНТИФИКАЦИЯ И ПРИМЕНЕНИЕ МУТАНТНЫХ KRP У РАСТЕНИЙ | 2012 |
|
RU2631790C2 |
| ПРОСЛЕЖИВАЕМОСТЬ СЕМЯН ТРАНСГЕННЫХ РАСТЕНИЙ ПРИ ПРЕДВАРИТЕЛЬНОЙ И ПОСЛЕДУЮЩЕЙ ОБРАБОТКЕ | 2005 |
|
RU2397250C2 |

