Область техники, к которой относится изобретение
Настоящее изобретение имеет отношение к полинуклеотиду, который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
Уровень техники
Цветы, имеющие новые признаки, всегда ценятся в цветочной индустрии. В частности, в промышленном отношении считается важным развитие растений, у которых был изменен "цвет", который считается наиболее важным признаком цветка, и были разработаны различные окраски цветков путем селекции с использованием классического метода кроссбридинга. Хотя кроссбридинг является эффективным методом селекции, поскольку существуют уникальные для растений генетические ограничения, этот метод имеет недостаток, будучи в состоянии использовать только генетические ресурсы, которыми обладают родственные виды, обладающие способностью к кроссбридингу. Например, несмотря на многие годы селекции, до сих пор не получены сорта роз, гвоздик, хризантем и лилий, имеющие цвет от фиолетового до синего, яркие красные сорта горечавок, ирисов и желтые сорта ипомей (вьюнков пурпурных утренняя слава).
Окраска цветка образуется из четырех типов пигментов, представленных флавоноидами, каротиноидами, хлорофиллом и беталаином. Среди них флавоноиды обладают широким спектром цветов, например, желтого, красного, фиолетового и синего. Группа, которая проявляет красный, фиолетовый и синий цвет, в общем называется антоцианами, и разнообразная структура антоцианов является одной из причин широкого диапазона окраски цветка. В целом антоцианы могут быть разделены на три группы, согласно структуре их агликона, вследствие их (антоцианов) пути биосинтеза. Цветки, имеющие яркий красный цвет, наподобие гвоздик и гераней, содержат большое количество антоцианов на основе пеларгонидина, тогда как цветки, имеющие синий или фиолетовый цвет, содержат большое количество антоцианов на основе дельфинидина. Причина отсутствия синего или фиолетового цвета у сортов роз, гвоздик, хризантем и лилий состоит в том, что эти растения не обладают способностью синтезировать антоцианы на основе дельфинидина.
Считается, что в дополнение к накоплению дельфинидина, для возникновения синей окраски цветка требуется любое из: (i) модификации антоциана одной или несколькими ароматическими ацильными группами, (ii) присутствия копигмента, такого как флавон или флавонол вместе с антоцианом, (ш) присутствия ионов железа или ионов алюминия вместе с антоцианом, (iv) повышения рН вакуолей, где локализован антоциан, от нейтрального до слабощелочного, или (v) образования комплекса из антоциана, копигмента и ионов металлов (и этот тип антоциана называется металлоантоцианинами) (см. непатентный документ 1).
Было проведено значительное исследование по биосинтезу флавоноидов и антоцианов и были идентифицированы связанные с биосинтезом ферменты и гены, кодирующие эти ферменты (см. непатентный документ 2 и фиг. 1). Например, из множества растений получен ген флавоноид-3ʹ,5ʹ-гидроксилазы (F3ʹ5ʹH), которая гидроксилирует В-кольцо флавоноидов, необходимых для биосинтеза дельфинидина. Кроме того, были созданы трансгенные растения, которые накапливают дельфинидин в своих лепестках, вызывая изменение окраски цветка на синюю (см. непатентный документ 4) введением этих генов F3ʹ5ʹH в гвоздики (см. патентный документ 1), розы (см. непатентный документ 3 и патентные документы 2 и 3) и хризантемы (см. патентный документ 4). Такие гвоздики и розы доступны в продаже.
Флавон является типом органического соединения, которое является циклическим кетоном производных флавана, и в узком смысле, относится к соединению 2,3-дигидрофлаван-4-он, представленного химической формулой С15Н10О2, и имеющего молекулярную массу 222.24. В широком смысле, производные, классифицированные как флавоны, в общем называют "флавонами". Флавон в широком смысле (флавоны) относится к категории флавоноидов, которые классифицируются как, имеющие флавоновую структуру основного скелета, но не имеющие гидроксильную группу в 3-положении. Примеры типичных флавонов включают апигенин (4ʹ,5,7-тригидроксифлавон) и лютеолин (3ʹ,4ʹ,5,7-тетрагидроксифлавон). В описании настоящей заявки термин "флавон" относится к флавонам в широком смысле, а именно производным, классифицированным как флавоны.
Гены флавонсинтаз (FNS), требуемые для биосинтеза флавона, получены из множества растений. Известно, что флавоны, если присутствуют с антоцианами, обладают эффектом изменения цвета антоцианов в темно-синий цвет, и эти гены FNS привлекли внимание для модификации окраски цветка. Одновременно с накоплением дельфинидина, в цветочных лепестках также накапливались флавоны, и окраска цветка изменялась даже на еще более темный синий цвет в результате введения гена F3ʹ5ʹH и FNS в розы, не обладающие способностью синтезировать флавоны (см. патентный документ 5). Кроме того, поскольку флавоны также поглощают ультрафиолетовые лучи, в дополнение к тому, что являются причиной синей окраски цветка, флавоны защищают растения от ультрафиолетовых лучей или действуют в качестве визуального сигнала для насекомых в случае цветков, опыляемых насекомыми. Кроме того, флавоны также участвуют во взаимодействии между растениями и почвенными микроорганизмами. Кроме того, флавоны также используют в качестве ингредиента в пищевых продуктах и косметических средствах в качестве полезных для здоровья компонентов. Например, считается, что флавоны обладают противораковой активностью и употребление продуктов, содержащих большие количества флавонов, было показано для лечения и предотвращения рака.
Кроме того, из множества растений также были получены гены, которые модифицируют антоцианы и флавоны. Хотя они включают гены гликозилтрансфераз, ацилтрансфераз и метилтрансфераз, последующее обеспечивает описание гликозилтрансфераз (GT), которые катализируют реакции гликозилирования. Например, ген, кодирующий белок, обладающий активностью переноса глюкозы на гидроксильную группу в 3-положении антоциана, был выделен из растений, таких как, горечавка, перилла, петуния, роза и львиный зев (см. непатентные документы 4-6 и патентный документ 6). Ген, кодирующий белок, обладающий активностью переноса глюкозы на гидроксильную группу в 5-положении антоциана, был выделен из таких растений, как перилла, петуния, горечавка, вербена или торения (см. непатентные документы 5-7 и патентный документ 7). Ген, кодирующий белок, обладающий активностью переноса глюкозы на гидроксильную группу в 7-положении флавона, был выделен из Arabidopsis thaliana (thale cress) (см. непатентный документ 8). Ген, кодирующий белок, обладающий активностью переноса глюкозы на гидроксильную группу в 7-положении байкалеина, был выделен из шлемника байкальского, и сообщалось, что белок, экспрессируемый этим геном в Escherichia coli, катализирует реакцию, демонстрируя активность переноса глюкозы на гидроксильную группу в 7-положении флавоноидов (см. непатентный документ 9). Ген, кодирующий белок, обладающий активностью переноса глюкозы на гидроксильную группу в 3-положении антоциана, был выделен из горечавки, мотылькового горошка и цинерии (cineria) (см. патентный документ 8). Кроме того, ген, кодирующий белок, обладающий активностью последовательного переноса глюкозы на гидроксильные группы в двух различных положениях А-кольца и С-кольца антоциана, был выделен из розы (см. патентный документ 9). Ген, кодирующий белок, обладающий активностью последовательного переноса глюкозы на гидроксильные группы в двух различных положениях В-кольца антоциана, был выделен из мотылькового горошка (см. патентный документ 10).
Хотя вышеупомянутые гликозилтрансферазы используют УДФ-глюкозу в качестве донора сахара, недавно были идентифицированы гликозилтрансферазы, которые используют ацилглюкозу в качестве донора сахара. Ген, кодирующий белок, обладающий активностью переноса глюкозы на гидроксильную группу в 5-положении антоциан-3-глюкозида, был выделен из гвоздики, тогда как ген, кодирующий белок, обладающий активностью переноса глюкозы на гидроксильную группу в 7-положении, был выделен из дельфиниума (см. непатентный документ 10).
Таким образом, большое количество белков существует в качестве гликозилтрансфераз, обладающих активностью переноса глюкозы на различные гидроксильные группы.
Однако считается, что остается большое число гликозилтрансфераз, для которых функция еще должна быть установлена. Например, ген, кодирующий белок, обладающий активностью переноса сахара в 4-положение флавоноида, и ген, кодирующий белок, обладающий активностью последовательного переноса сахара на гидроксильные группы в двух различных положениях А-кольца и В-кольца флавоноида, еще должны быть идентифицированы. Хотя сообщалось, что ген гликозилтрансферазы, полученный из мезембриантемум Ливингстона, демонстрирует активность, в отношении переноса глюкозы на одну из гидроксильных групп в 4ʹ-положении или в 7-положении флавоноида in vitro, активность, присущая этой гликозилтрансферазе растений, обеспечивает перенос глюкозы на гидроксильную группу в 5-положении бетанидина (см. непатентный документ 11).
Однако металлоантоцианины, представленные пигментами коммелины, василька, шалфея и немофилы, состоят из шести молекул антоцианов, шести молекул флавонов и двух атомов ионов металлов, и каждый компонент собран с образованием стабильного синего пигмента (см. фиг. 2 и непатентный документ 1). Например, металлоантоцианин немофилы образуется из немофилина (см. фиг. 3), малонил-апигенин 4ʹ,7ʹ-диглюкозида (см. фиг. 4), Mg2+ и Fe3+. Металлоантоцианин шалфея образуется из цианосальвианина (cyanosalvianin) (см. фиг. 5), апигенин-4,7ʹ-диглюкозида (см. фиг. 6) и Mg2+. Согласно предшествующему исследованию все голубые цветки, в которых образуются металлоантоцианины, биосинтезируют флавон, в котором сахар присоединяется к гидроксильным группам как в 4ʹ-положении, так и в 7-положении, и было установлено, что сахар, присоединенный к этому флавону играет важную роль в молекулярном узнавании на протяжении образования металлоантоцианина. Было показано, что сахар, скоординированный в 4ʹ-положении флавона, является важным в молекулярном распознавании в ходе образования, тогда как сахар в 7-положении участвует в стабильности флавона (см. непатентный документ 1). Только после добавления этих двух Сахаров к флавону образуется металлоантоцианин, который приводит к экспрессии привлекательного голубого цвета. Кроме того, лепестки голубого голландского ириса содержат флавон, в котором сахар был добавлен в 4-положение. Кроме того, поскольку в результате присоединения двух Сахаров к флавонам повышается их растворимость и изменяются физические свойства, то ожидается, что расширится применение флавонов в пищевых продуктах для здорового питания, лекарственных препаратах и в виде косметических ингредиентов.
Документы предыдущего уровня техники
Патентные документы
Патентный документ 1: Международная публикация WO 2006/105598
Патентный документ 2: Международная публикация WO 2010/122849
Патентный документ 3: Международная публикация WO 2005/017147
Патентный документ 4: Международная публикация WO 2009/062253
Патентный документ 5: Международная публикация WO 2008/156211
Патентный документ 6: Международная публикация WO 2007/094521
Патентный документ 7: Международная публикация WO 99/05287
Патентный документ 8: Международная публикация WO 001/092509
Патентный документ 9: Японская нерассмотренная патентная публикация №2006-149293
Патентный документ 10: Японская нерассмотренная патентная публикация №2005-95005
Непатентные документы
Непатентный документ 1: Natural Product Reports (2009), 26, 884-915
Непатентный документ 2: Biosci. Biotechnol. Biochem. (2010), 74(9), 1760-1769
Непатентный документ 3: Plant Cell Physiol. (2007), 48(11), 1589-1600
Непатентный документ 4: Plant Cell Physiol. (1996), 37(5), 711-716
Непатентный документ 5: J. Biol. Chem. (1999), 274(11), 7405-7411
Непатентный документ 6: Plant Molecular Biology (2002), 48,401-411
Непатентный документ 7: Journal of Experimental Botany (2008), 59(6), 1241-1252
Непатентный документ 8: Biosci. Biotechnol. Biochem. (2006), 70(6), 1471-1477
Непатентный документ 9: Planta (2010), 210,1006-1013
Непатентный документ 10: Plant Cell (2010), 22(10), 3374-89
Непатентный документ 11: The Plant Journal (1999), 19(5), 509-519
Раскрытие сущности изобретения
Проблемы, подлежащие разрешению данным изобретением
Изменение физических свойств флавонов необходимо для изменения окраски цветка и разработки материалов для пищевых продуктов, лекарственных и косметических препаратов. Например, хотя гвоздики, розы и хризантемы, которые накапливают дельфинидин, имеют синевато-фиолетовый цвет, проводятся исследования, чтобы сделать этот цвет еще более синим.
В соответствии с вышеизложенной целью, задачей настоящего изобретения является обеспечение полинуклеотида, который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, и применение полинуклеотида.
Способы решения поставленных задач
В результате проведения обширных исследований и экспериментов для решения вышеупомянутых задач, авторы настоящего изобретения выделили полинуклеотид, который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, и подтвердили применимость полипептида, что приводит к созданию настоящего изобретения.
А именно, настоящее изобретение представляет собой как описано ниже.
[1] Полинуклеотид, выбранный из группы, состоящей из следующих от (а) до (е):
(a) полинуклеотида, состоящего из последовательности оснований SEQ ID NO:1;
(b) полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, состоящим из последовательности оснований, комплементарной последовательности оснований SEQ ID NO:1, и который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона;
(c) полинуклеотида, кодирующего белок, состоящий из аминокислотной последовательности SEQ ID NO:2;
(d) полинуклеотида, кодирующего белок, состоящий из аминокислотной последовательности, в которой одна или несколько аминокислот были делетированы, замещены, встроены и/или добавлены в аминокислотную последовательность SEQ ID NO:2, и обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона; и,
(e) полинуклеотида, кодирующего белок, имеющий аминокислотную последовательность, имеющую 90% или большую идентичность, с аминокислотной последовательностью SEQ ID NO:2, и обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
[2] Полинуклеотид, описанный в [1] выше, который является (а) полинуклеотидом, состоящим из последовательности оснований SEQ ID NO:1.
[3] Полинуклеотид, описанный в [1] выше, который является (с) полинуклеотидом, кодирующим белок, состоящий из аминокислотной последовательности SEQ ID NO:2.
[4] Полинуклеотид, описанный в [1] выше, который является (f) полинуклеотидом, кодирующим белок, имеющий аминокислотную последовательность, имеющую 95% или большую идентичность, с аминокислотной последовательностью SEQ ID NO:2, и обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
[5] Полинуклеотид, описанный в [4] выше, который является (g) полинуклеотидом, кодирующим белок, имеющий аминокислотную последовательность, имеющую 97% или большую идентичность, с аминокислотной последовательностью SEQ ID NO:2, и обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
[6] Полинуклеотид, описанный в [5] выше, который является (h) полинуклеотидом, кодирующим белок, имеющий аминокислотную последовательность, имеющую 98% или большую идентичность, с аминокислотной последовательностью SEQ ID NO:2, и обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
[7] Полинуклеотид, описанный в любом из с [1] по [6] выше, который является ДНК.
[8] Белок, кодируемый полинуклеотидом, описанным в любом из с [1] по [7] выше.
[9] Вектор, содержащий полинуклеотид, описанный в любом из с [1] по [7] выше.
[10] Не относящийся к человеку хозяин, со вставленным вектором, описанным в [9] выше.
[11] Способ добавления сахара к гидроксильной группе в 4ʹ-положении флавона с использованием полинуклеотида, описанного в любом из с [1] по [7] выше.
[12] Растение, потомство растения, или часть, или ткань растения, с введенным полинуклеотидом, описанным в любом из с [1] по [7] выше, и содержащие этот полинуклеотид.
[13] Часть растения, описанного в [12] выше, которая является срезанным цветком.
[14] Процессированный срезанный цветок, где использован срезанный цветок, описанный в [13] выше.
[15] Способ получения белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, содержащий следующие стадии:
культивирования или выращивания не относящегося к человеку хозяина, описанного в [10] выше, и
получения белка из не относящегося к человеку хозяина, белка, который обладает активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
[16] Способ получения флавона, у которого сахар был добавлен к гидроксильной группе в его 4ʹ-положении, содержащий следующие стадии:
культивирования или выращивания не относящегося к человеку хозяина, описанного в [10] выше, и
получение флавона, у которого сахар был добавлен к гидроксильной группе в его 4ʹ-положении, из не относящегося к человеку хозяина.
[17] Продукт питания, содержащий флавон, полученный согласно способу, описанному в [16] выше, у которого сахар был добавлен к гидроксильной группе в его 4ʹ-положении.
[18] Фармацевтический препарат, содержащий флавон, полученный согласно способу, описанному в [16] выше, у которого сахар был добавлен к гидроксильной группе в его 4ʹ-положении.
[19] Косметическое средство, содержащие флавон, полученный согласно способу, описанному в [16] выше, у которого сахар был добавлен к гидроксильной группе в его 4ʹ-положении.
Эффекты изобретения
Полинуклеотид настоящего изобретения обеспечивает возможность получения белка, обладающего активностью специфического переноса сахара на гидроксильную группу в 4ʹ-положении флавона, экспрессируемого в подходящих клетках хозяина. По изобретению, настоящее изобретение может быть использовано для изменения окраски цветка путем конститутивной или тканеспецифической экспрессии в растении белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
Флавон, у которого сахар был добавлен к гидроксильным группам как в 4ʹ-положении, так и в 7-положении, образуется введением белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, в растение, заведомо обладающее активностью переноса сахара на гидроксильную группу в 7-положении флавона, например, наподобие розы. Альтернативно, флавон, у которого сахар был добавлен к гидроксильным группам как в 4ʹ-положении, так и в 7-положении, образуется экспрессией в растении белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, вместе с белком, обладающим активностью переноса сахара на гидроксильную группу в 7-положении флавона.
Кроме того, по изобретению также обеспечивают способ получения флавона, у которого сахар был добавлен к гидроксильной группе в 4ʹ-положении, и продукт питания, фармацевтический препарат или косметическое средство и т.п., полученное согласно этому способу получения.
Краткое описание чертежей
Фиг. 1 является чертежом для объяснения пути биосинтеза антоцианов.
Фиг. 2 является схематическим чертежом структуры металлоантоцианина.
Фиг. 3 демонстрирует структурную формулу антоциана, полученного из немофилы (немофилина).
Фиг. 4 демонстрирует структурную формулу флавона, полученного из немофилы (малонилапигенин-4ʹ,7ʹ-диглюкозида).
Фиг. 5 демонстрирует структурную формулу флавона, полученного из шалфея (цианосальвианина) (cyanosalvianin).
Фиг. 6 демонстрирует структурную формулу флавона, полученного из шалфея (апигенин-4ʹ,7-диглюкозида).
Фиг. 7 является хроматограммой, полученной после ферментативной реакции между экстрактом из лепестка цветка и апигенином с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ).
Фиг. 8 является чертежом для объяснения пути биосинтеза апигенин-4ʹ,7-диглюкозида.
Фиг. 9 является хроматограммой, полученной после ферментативной реакции между раствором белка NmGT8 и апигенином с помощью высокоэффективной жидкостной хроматографии.
Фиг. 10 является хроматограммой, полученной после ферментативной реакции между раствором белка NmGT8 и апигенин-7-глюкозидом с помощью высокоэффективной жидкостной хроматографии.
Фиг. 11 является чертежом, суммирующим реакционные способности раствора белка NmGT8 и фермента, в который был добавлен сахар в 5-положении байкалеина, при различных субстратах флавоноидов.
Фиг. 12-1 является диаграммой выравнивания для сравнения аминокислотных последовательностей NmGT8 и NmG13.
Фиг. 12-2 является диаграммой выравнивания для сравнения аминокислотных последовательностей NmGT8 и NmGT4.
Фиг. 12-3 является диаграммой выравнивания для сравнения аминокислотных последовательностей NmGT8, NmGT3 и NmGT4.
Фиг. 13 является диаграммой выравнивания для сравнения аминокислотных последовательностей NmGT8 и фермента, у которого сахар был добавлен в 4ʹ-положение, халкона львиного зева.
Фиг. 14 является диаграммой выравнивания для сравнения аминокислотных последовательностей NmGT8 и фермента, у которого сахара были добавлены в 3-положение и 5-положение, антоцианидина розы.
Фиг. 15 является филогенетическим древом, указывающим на родство между NmGT8 настоящего изобретения и указанных выше ферментов.
Фиг. 16 демонстрирует конструкцию, содержащую NmGT8, введенную в торению (pSPB-4583).
Фиг. 17 демонстрирует конструкции, содержащие NmGT8, введенную в петунию (pSPB5424 и pSPB5428).
Фиг. 18 является высокоэффективной жидкостной хроматограммой экстракта пестиков цветка рекомбинантной петунии со вставленным NmGT8.
Фиг. 19 является хроматограммой экстрагированных положительных ионов (с отношением масса/заряд (m/z) от 250 m/z до 1250 m/z) из экстракта лепестков цветков рекомбинантной петунии с введенным NmGT8.
Фиг. 20 демонстрирует конструкцию, содержащую NmGT8, введенную в гвоздику (pSPB5433).
Фиг. 21 демонстрирует конструкции, содержащие NmGT8, введенные в розу (pSPB4577, pSPB4578, pSPB5437 и pSPB5440).
Фиг. 22 является высокоэффективной жидкостной хроматограммой экстракта пестиков цветка рекомбинантной розы со вставленным NmGT8.
Фиг. 23 является масс-спектром экстрагированных положительных ионов (с отношением масса/заряд (m/z) от 250 m/z до 1250 m/z) из экстракта лепестков цветков рекомбинантной розы с введенным NmGT8.
Лучший способ осуществления изобретения
Настоящее изобретение имеет отношение к полинуклеотиду, выбранному из группы, состоящей из следующих от (а) до (е):
(a) полинуклеотида, состоящего из последовательности оснований SEQ ID NO:1;
(b) полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, состоящим из последовательности оснований, комплементарной последовательности оснований SEQ ID NO:1, и который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона;
(c) полинуклеотида, кодирующего белок, состоящий из аминокислотной последовательности SEQ ID NO:2;
(d) полинуклеотида, кодирующего белок, состоящий из аминокислотной последовательности, в которой одна или несколько аминокислот были делегированы, замещены, встроены и/или добавлены в аминокислотную последовательность SEQ ID NO:2, и обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона; и,
(e) полинуклеотида, кодирующего белок, имеющий аминокислотную последовательность, имеющую 90% или большую идентичность, с аминокислотной последовательностью SEQ ID NO:2, и обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
В настоящем описании термин "полинуклеотид" относится к ДНК или РНК.
В настоящем описании термин "жесткие условия" относится к условиям, при которых полинуклеотид или олигонуклеотид и геномная ДНК связываются избирательно и специфически, чтобы иметь возможность быть обнаруженными. Жесткие условия определяют подходящей комбинацией концентрации соли, концентрации органического растворителя (такого как формальдегид), температуры и других известных условий. А именно, жесткость повышают снижением концентрации соли, увеличением концентрации органического растворителя или повышением температуры гибридизации. Кроме того, условия отмывки после гибридизации также влияют на жесткость. Эти условия отмывки также определяются концентрацией соли и температурой, и жесткость отмывки повышают снижением концентрации соли или повышением температуры. Таким образом, термин "жесткие условия" относится к условиям, при которых специфическая гибридизация происходит только между последовательностями оснований, имеющими высокую гомологию, так что степень идентичности или гомология между каждой последовательностью оснований составляет, например, в среднем около 80% или более, предпочтительно около 90% или более, более предпочтительно около 95% или более, даже более предпочтительно около 97% или более и наиболее предпочтительно 98%) или более. Пример "жестких условий" состоит из концентрации натрия от 150 мМ до 900 мМ, и предпочтительно от 600 мМ до 900 мМ и рН от 6 до 8 при температуре от 60°С до 68°С, и в частности, состоит из выполнения гибридизации в условиях, состоящих из 5 × SSC (750 мМ NaCl, 75 мМ натрия лимоннокислого трехзамещенного), 1% SDS (додецилсульфата натрия (ДСН)), 5 × раствора Денхардта, 50% формальдегида и 42°С, с последующей отмывкой в условиях, состоящих из 0,1 × SSC (15 mM NaCl, 1,5 тМ натрия лимоннокислого трехзамещенного), 0,1 × SDS и 55°С.
Гибридизация может быть выполнена в соответствии с методом известным в данной области, или методом, удовлетворяющим требованиям, таким как метод, описанный в Current Protocols in Molecular Biology (edited by Frederick M. Ausubel, et al, 1987). Кроме того, в случае использования коммерчески доступной библиотеки, гибридизация может быть выполнена в соответствии с методом, описанным в руководстве, предоставленном вместе с ней (библиотекой). Гены, выбранные с использованием такой гибридизации, могут быть природными генами, такими как гены, произведенные из растений, или гены, не произведенные из растений. Кроме того, гены, отобранные гибридизацией, могут быть кДНК, геномной ДНК или химически синтезированной ДНК.
Вышеупомянутая "аминокислотная последовательность, у которой одна или множество аминокислот были делегированы, заменены, встроены и/или добавлены" относится к аминокислотной последовательности, у которой было делегировано, заменено, вставлено и/или добавлено произвольное число аминокислот, такое как от 1 до 20, предпочтительно от 1 до 5 и даже более предпочтительно от 1 до 3. Может оказаться полезной такая генно-инженерная технология, как сайт-специфический мутагенез, поскольку он позволяет вводить специфическую мутацию в специфический сайт, и может быть выполнен в соответствии с методом, таким как описанный в Molecular Cloning: А Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989. Экспрессией этой мутантной ДНК с использованием подходящей экспрессирующей системы, может быть получен белок, который состоит из аминокислотной последовательности, у которой одна или множество аминокислот были делегированы, заменены, встроены и/или добавлены.
Кроме того, ДНК по изобретению может быть получена методом, известным среднему специалисту в данной области техники, таким как метод, состоящий из химического синтеза согласно фосфоамидному методу, или методом амплификации нуклеиновых кислот с использованием пробы нуклеиновой кислоты из растения в качестве матрицы, и с использованием праймеров, сконструированных на основе нуклеотидной последовательности гена-мишени.
В настоящем описании термины "идентичность" и "гомология" относятся к количеству (числу), которое дает возможность определить, что соответствующие аминокислотные остатки или основания, образующие две цепи в полипептидной последовательности (или аминокислотной последовательности) или полинуклеотидной последовательности (или последовательности оснований), идентичны исходя из отношения взаимной совместимости между двумя цепями, и "идентичность" и "гомология" могут быть легко вычислены. Известны многочисленные методы для измерения гомологии между двумя полинуклеотидными последовательностями или полипептидными последовательностями, и термины "идентичность" и "гомология" широко известны обычным специалистам в данной области (см., например, Lesk, A.M. (ed.), Computational Molecular Biology, Oxford University Press, New York (1988); Smith, D.W. (ed.), Biocomputing: Informatics and Genome Projects, Academic Press, New York (1993); Grifin, A.M. & Grifin, H.G. (ed.), Computer Analysis of Sequence Data: Part I, Human Press, New Jersey (1994); von Heinje, G., Sequence Analysis in Molecular Biology, Academic Press, New York (1987); и Gribskov, M. & Devereux, J. (ed.), Sequence Analysis Primer, M-Stockton Press, New York (1991)).
Кроме того, хотя описанные в настоящем описании значения "идентичности" и "гомологии" могут быть значениями, вычисленными с использованием программы поиска гомологий, известной специалистам с обычной квалификацией в данной области техники, если специально не указано иное, они предпочтительно являются значениями, рассчитанными с использованием программы Clustal W Program с приложением MacVector (версия 9.5, Oxford Molecular Ltd., Оксфорд, Англия)
Полинуклеотид (нуклеиновая кислота или ген) настоящего изобретения это тот, который "кодирует" представляющий интерес белок. Здесь, "кодирует" относится к экспрессии представляющего интерес белка в состоянии сохранения его (белка) активности. Кроме того, "кодирует" включает как значение кодирующий в виде непосредственных составляющих последовательности (экзонов), где представляющий интерес белок является сплошным, так и кодирующий с использованием промежуточных последовательностей (интронов).
Как будет описано в примерах, которые впоследствии будут описаны, ген, имеющий природную последовательность оснований получают, например, анализом с использованием секвенатора ДНК. Кроме того, ДНК, кодирующая фермент, имеющий модифицированную аминокислотную последовательность, может быть синтезирована с использованием обычного сайт-специфического мутагенеза или ПЦР, с использованием ДНК, имеющей природную последовательность в качестве основы. Например, после получения необходимого фрагмента ДНК, подлежащего модификации обработкой кДНК рестриктазой или рестриктазой для геномной ДНК, сайт-специфический мутагенез или ПЦР выполняют с использованием фрагмента ДНК в качестве матрицы и с использованием праймеров, введенных необходимой мутацией для получения необходимого модифицированного фрагмента ДНК. Затем фрагмент ДНК, введенный этой мутацией, связывают с фрагментом ДНК, кодирующим другой сегмент фермента-мишени.
Альтернативно, для получения ДНК, кодирующей фермент, состоящий из укороченной аминокислотной последовательности, ДНК, кодирующую аминокислотную последовательность более длинную, чем аминокислотная последовательность-мишень, такую как ДНК, кодирующую полноразмерную аминокислотную последовательность, расщепляют рестриктазой, и в случае, если полученный фрагмент ДНК не кодирует всю аминокислотную последовательность-мишень, то синтезируют фрагмент ДНК, состоящий из неполного участка последовательности, и связывают с ним.
Кроме того, измерением ферментативной активности после экспрессии полученного полинуклеотида в Escherichia coli или дрожжах с использованием системы экспрессии генов, может быть подтверждено, что полученный полинуклеотид пригоден для кодирования белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона. Кроме того, полинуклеотидный продукт в форме белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, может быть получен экспрессией этого полинуклеотида. Альтернативно, белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, также может быть получен с использованием антитела к полипептиду, состоящего из аминокислотной последовательности, описанной в SEQ ID NO:2, и происходящий из другого организма полинуклеотид, кодирующий белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, также может быть клонирован с использованием этого антитела.
Настоящее изобретение также имеет отношение к (рекомбинантному) вектору, содержащему указанный выше полинуклеотид, и в частности к экспрессирующему вектору, а также хозяину, трансформированному этим вектором.
Прокариоты или эукариоты могут быть использованы в качестве хозяина. Примеры прокариот включают обычно используемые прокариоты-хозяева, такие как бактерии, в том числе бактерии, принадлежащие к роду Escherichia, такие как Escherichia coli, и бактерии, принадлежащие к роду Bacillus, такие как Bacillus subtilis. Примеры эукариот, которые могут быть использованы, включают низшие эукариоты, такие как эукариотические организмы наподобие грибов, такие как дрожжи или мицелиальные грибы.
Примеры дрожжей включают виды Saccharomyces, такие как Saccharomyces cerevisiae, и примеры мицелиальных грибов включают виды Aspergillus, такие как Aspergillus oryzae или Aspergillus niger, и виды Penicillium. Животные клетки или растительные клетки также могут быть использованы в качестве хозяина, примеры используемых систем животных клеток включают клетки мыши, хомяка, обезьяны или человека и клетки насекомых, такие как клетки тутового шелкопряда или взрослых особей тутового шелкопряда в чистом виде, также используют в качестве хозяев.
Экспрессирующий вектор по изобретению содержит регуляторную область экспрессии, такую как промотор, терминатор или репликативную точку, зависящую от типа хозяина, в который вводят экспрессирующий вектор. Обычно используемый промотор, такой как trc-промотор, tac-промотор или lac-промотор, используют в качестве промотора бактериального экспрессирующего вектора, промотор глицеральдегид-3-фосфат дегидрогеназы или промотор РН05, например, используют в качестве дрожжевого промотора, и амилазный промотор или trpC-промотор, например, используют в качестве промотора мицелиальных грибов. Кроме того, примеры промоторов клеток-хозяев животного включают вирусные промоторы, такие как ранний промотор SV40 поздний промотор SV40.
Примеры промоторов, которые конститутивно экспрессируют полинуклеотид в растительных клетках, включают промотор 35S РНК вируса мозаики цветной капусты, промотор гена rd29A, промотор rbcS и промотор тас-1. Кроме того, промотор гена, который специфически экспрессируется в этой ткани, может быть использован для тканеспецифической экспрессии гена.
Экспрессирующий вектор может быть получен в соответствии с обычными методами с использованием рестриктаз, лигаз и т.п. Кроме того, трансформация хозяина экспрессирующим вектором также может быть выполнена в соответствии с обычными методами.
Белок-мишень может быть получен культивированием, культивированием или выращиванием хозяина, трансформированного вышеуказанным экспрессирующим вектором, и извлечением и очисткой из культуры или среды в соответствии с обычными методами, такими как фильтрация, разделение центрифугированием, разрушение клеток, гель-фильтрационная хроматография или ионообменная хроматография.
Хотя настоящее описание описывает полученный из немофилы ген, кодирующий белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, полинуклеотид по изобретению не ограничивается геном, полученным из немофилы, но скорее может быть применен для изменения окраски цветка растения независимо от того, источником гена, кодирующего белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, является ли растение, животное или микроорганизм, при условии, что источник гена обладает активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
Настоящее изобретение также относится к растению, его потомству или его части или ткани, полученными введением экзогенного полинуклеотида, кодирующего белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, приводящее к тому, что экзогенный полинуклеотид должен содержаться в растении. Примером формы части ткани является срезанный цветок. 4ʹ-положение флавона может быть гликозилированным, или гликозилирование 4ʹ-положения флавона может быть ингибировано при помощи полинуклеотида по изобретению, который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, и в результате этого может быть изменена окраска цветка растения.
На данном уровне развития технологии, может быть применена технология, которая обеспечивает полинуклеотид, подлежащий введению в растение, которая и обеспечивает возможность полинуклеотиду экспрессироваться конститутивно или тканеспецифически. Введение ДНК в растение может быть выполнено способом, известным специалистам с обычной квалификацией в данной области техники, таким как метод с использованием Agrobacterium, (с использованием) бинарных векторов, метод электропорации, метод (на основе) ПЭГ или метод с использованием генной пушки.
Примеры растений, которые могут быть трансформированы, включают, без ограничения перечисленным, розу, гвоздику, хризантему, львиный зев, цикламен, орхидею, колокольчик, фрезию, герберу, гладиолус, перекати-поле, каланхоэ, лилию, пеларгонию, герань, петунию, торению, тюльпан, антуриум, орхидею-мотылек, рис, ячмень, пшеницу, рапс, картофель, томат, тополь, банан, эвкалипт, сладкий картофель, сою, люцерну, базилик рубиновый, кукурузу, цветную капусту и георгин.
Настоящее изобретение также имеет отношение к процессированному продукту, где использован вышеуказанный срезанный цветок (процессированный срезанный цветок). Здесь, примеры процессированных срезанных цветков включают, без ограничения перечисленным, прессованные цветы, законсервированные цветы, высушенные цветы и залитые смолой цветы с использованием этих же срезанных цветков.
Кроме того, полученный согласно способу получения по изобретению флавон, в котором сахар был добавлен к гидроксильной группе в 4ʹ-положение флавона, может найти различные применения, например в способах получения пищевых продуктов, в лекарственных препаратах или косметических средствах.
В настоящем изобретении, экспрессия гена-мишени в растении может быть подавлена антисмысловым методом, методом косупрессии или методом РНК-интерференции. Подавление экспрессии (супрессия) гена-мишени может быть выполнена в соответствии с методом, известным специалистам с обычной квалификацией в данной области техники, и их примеры включают технологию антисмысловых РНК/ДНК (Bioscience and Industry, 50, 322 (1992); Chemistry, 46, 681 (1991); Biotechnology, 9, 358 (1992); Trends in Biotechnology, 10, 152 (1992); Cellular Engineering, 16, 1463 (1997)), и технологию тройной спирали (триплексов) (Trends in Biotechnology, 10, 132 (1992)). Например, подавление экспрессии гена выполняют с использованием одноцепочечной молекулы нуклеиновой кислоты, содержащей всю или часть нуклеотидной последовательности, идентичной последовательности антисмысловой цепи гена по изобретению. Этот тип метода известен как антисмысловой метод. В антисмысловом методе экспрессию гена-мишени подавляют экспрессией высокого уровня РНК, имеющей последовательность, комплементарную гену, для которого необходимо подавление экспрессии. В этом методе может быть использована одноцепочечная РНК, которая содержит всю нуклеотидную последовательность, идентичную антисмысловой цепи полинуклеотида (гена) по изобретению. Кроме того, в указанном выше методе также может быть использована одноцепочечная РНК, которая содержит часть нуклеотидной последовательности, идентичную антисмысловой цепи гена по изобретению. Хотя этот тип неполной одноцепочечной РНК может быть подходящим образом сконструирован специалистом с обычной квалификацией в данной области техники при условии, что она способна подавлять экспрессию гена по изобретению, она предпочтительно является специфической для гена по изобретению, и ее длина составляет предпочтительно от 5 нуклеотидов до 100 нуклеотидов, более предпочтительно от 5 нуклеотидов до 50 нуклеотидов и даже более предпочтительно от 10 нуклеотидов до 20 нуклеотидов.
Подавление экспрессии гена выполняют с использованием одноцепочечной молекулы нуклеиновой кислоты, содержащей всю или часть нуклеотидной последовательности, идентичной смысловой цепи гена по изобретению. То есть, эта смысловая одноцепочечная нуклеиновая кислота может быть использована для подавления экспрессии гена по изобретению таким же образом, как и указанная выше антисмысловая одноцепочечная нуклеиновая кислота. В этом методе может быть использована одноцепочечная РНК, которая содержит полную нуклеотидную последовательность, идентичную смысловой цепи гена по изобретению. Кроме того, в указанном выше методе может быть использована одноцепочечная РНК, которая содержит часть нуклеотидной последовательности, идентичной смысловой цепи гена. Хотя этот тип частичной одноцепочечной РНК может быть подходящим образом сконструирован специалистом с обычной квалификацией в данной области техники при условии, что она способна подавлять экспрессию гена по изобретению, она предпочтительно является специфической для гена по изобретению, и ее длина составляет предпочтительно от 5 нуклеотидов до 100 нуклеотидов, более предпочтительно от 5 нуклеотидов до 50 нуклеотидов и даже более предпочтительно от 10 нуклеотидов до 20 нуклеотидов.
Кроме того, подавление экспрессии гена выполняют с использованием двухцепочечной молекулы нуклеиновой кислоты, содержащей всю или часть нуклеотидной последовательности, идентичной гену по изобретению. Например, антисмысловая или смысловая одноцепочечная нуклеиновая кислота гена по изобретению может быть супрессирована (подавлена) у покрытосемянных с использованием этой двухцепочечной молекулы нуклеиновой кислоты. Двухцепочечной молекулой нуклеиновой кислоты по изобретению предпочтительно является ДНК, и ее длина цепи и специфическая нуклеотидная последовательность соответствуют длине цепи и нуклеотидной последовательности одноцепочечной молекулы-мишени нуклеиновой кислоты. Например, в случае экспрессии указанной выше антисмысловой одноцепочечной нуклеиновой кислоты, двухцепочечная молекула нуклеиновой кислоты по изобретению содержит антисмысловую цепь гена по изобретению в качестве кодирующей цепи. Кроме того в случае экспрессии указанной выше смысловой одноцепочечной нуклеиновой кислоты, двухцепочечная молекула нуклеиновой кислоты по изобретению содержит смысловую цепь гена по изобретению в качестве кодирующей цепи.
Двухцепочечная молекула нуклеиновой кислоты может быть экспрессирована в растении с использованием метода, известного специалистам с обычной квалификацией в данной области техники. Например, двухцепочечная молекула нуклеиновой кислоты может быть экспрессирована введением экспрессирующего вектора, содержащего промотор, двухцепочечную молекулу нуклеиновой кислоты по изобретению и терминатор транскрипции, в растение-мишень с последующим культивированием полученного растительного организма. Введение экспрессирующего вектора в растение может быть выполнено в соответствии с методом, известным специалистам с обычной квалификацией в данной области техники, таким как метод с использованием Agrobacterium, (с использованием) бинарных векторов, метод электропорации, метод (на основе) ПЭГ или метод с использованием генной пушки.
Другим примером метода подавления экспрессии гена (супрессии) с использованием молекулы нуклеиновой кислоты по изобретению является метод косупрессии. В этом методе смысловую двухцепочечную ДНК, имеющую полную нуклеотидную последовательность гена по изобретению, вводят в растение-хозяин. В результате, экспрессируется смысловая одноцепочечная РНК по изобретению и экспрессия гена существенно подавляется этой РНК (Plant Cell, 9: 1357-1368, 1997).
Примеры
Нижеследующее обеспечивает подробное объяснение изобретения в соответствии с его примерами.
Пример 1: Обнаружение активности переноса сахара на гидроксильную группу в 4ʹ-положении флавона в лепестках цветков немофилы (Nemophila)
Лепестки цветков немофилы (Nemophila menziesii) собирали и делили на стадиях развития, определенных в порядке, указанном ниже, с последующим замораживанием жидким азотом и хранением в морозильной камере при -80°С.
Стадия 1: Неокрашенный, плотно закрытой бутон (приблизительно от 2 до 5 мм)
Стадия 2: Окрашенный, плотно закрытой бутон (приблизительно от 2 до 5 мм)
Стадия 3: Окрашенный, закрытый бутон с почти открытой чашечкой (приблизительно от 5 до 10 мм)
Стадия 4: Бутон с почти открытыми цветочными лепестками (приблизительно от 10 до 15 мм)
Стадия 5: Полностью открытый цветок Получение экстракта из лепестка цветка Nemophila
Ожидается, что активность гликозилтрансферазы флавона должна быть обнаружена в лепестках цветков на стадии 1 и стадии 2 до биосинтеза антоциана. Таким образом, экстракты из лепестков цветков готовили с использованием лепестков цветков на стадии 1 и стадии 2. 500 мг пробы лепестков цветков (250 мг каждого из лепестков цветков на стадиях 1 и 2 хранили при -80°С) измельчали в ступке с пестиком при охлаждении жидким азотом с последующим растворением в 1,5 мл буфера для экстракции (состав: фосфатно-калийный буфер (рН 7,5): 100 мМ, дитиотреитола (DTT): 1 мМ, поливинилпирролидона 40: 50 мг/мл, сахарозы: 100 мг/мл). Полученный раствор белка центрифугировали (10000 об/мин, 4°С, 10 минут), и к полученному супернатанту добавляли сульфат аммония до концентрации 30% от насыщения. После перемешивания в течение 1 часа при 4°С раствор центрифугировали (10000 об/мин, 4°С, 10 минут) с последующим получением супернатанта. К полученному супернатанту добавляли сульфат аммония до концентрации 70% от насыщения, и после перемешивания в течение 1 часа при 4°С раствор центрифугировали (10000 об/мин, 4°С, 10 минут) для получения осадка. Этот осадок растворяли в 500 мкл буфера для элюции (состав: Tris НС1 (рН 7,5): 2,5 мМ, DTT: 1 мМ, амидинофенилметансульфонилфторида гидрохлорид (APMSF): 10 мкМ) с последующей очисткой на колонке с использованием колонок NAP-5 с Sephadex G-25 DNA Grade (со степенью чистоты для ДНК) (GE Healthcare Inc.) и для удаления сульфата аммония. Затем этот (полученный после колонки) раствор использовали в качестве "экстракта из лепестков цветка". Для центрифугирования использовали (центрифугу) Avanti НР-26ХР (ротор: JA-2) (Beckman Coulter Inc.).
Определение ферментативной активности
Реакционную жидкость, приготовленную смешиванием 40 мкл экстракта из лепестков цветка, 20 мкл 5 мМ УДФ-глюкозы, 20 мкл 1 М Tris HCl (рН 7,5) и 1 мкл апигенина 500 нг/мкл на бане со льдом и доведением объема до 200 мкл водой, выдерживали в течение 1 часа при 30°С. Затем, добавляли 200 мкл буфера для остановки (90%) водный раствор ацетонитрила, содержащий 0,1% трифторуксусной кислоты (ТФУ)) для остановки реакции с последующим анализом реакционной жидкости высокоэффективной жидкостной хроматографией (Prominence (Shimadzu Corp.)). Для обнаружения использовали (спектрофотометр) Shimadzu PDA SPD-M10AVP и флавон обнаруживали при 330 нм. В качестве колонки использовали колонку Shim-Pack ODS 150 мм × 4,6 мм (Shimadzu Corp.). Раствор А (0,1% водный раствор ТФУ) и раствор В (90% водный раствор ацетонитрила, содержащий 0,1% ТФУ) использовали для элюции. Элюцию выполняли с использованием линейного градиента концентраций из смеси двух растворов 8:2 до смеси двух растворов 3:7 в течение 10 минут, за которым следует смесь 3:7 в течение 5 минут. Скорость потока составляла 0,6 мл/мин. Реакционную жидкость, полученную при предоставлении возможности ферменту взаимодействовать в одних и тех же условиях с использованием экстракта из лепестков цветка, полученного тепловой обработкой экстракта из лепестков цветка в течение 20 минут при 100°С, использовали в качестве контроля.
Как результат, флавон, демонстрирующий такое же время удерживания и такой же максимум поглощения, как и очищенный апигенин-4ʹ,7-диглюкозид и стандарт апигенин-7-глюкозид, биосинтезировался в дополнение к флавону, демонстрирующему время удерживания близкое к времени удерживания апигенин-7-глюкозида (см. фиг. 7). Ни один из этих продуктов не образовывался, когда ферменту давали возможность взаимодействовать без добавления УДФ-глюкозы.
Пример 2: Определение времени удерживания и максимума поглощения апигенин-4ʹ-глюкозида
Принимая во внимание путь биосинтеза апигенин-4ʹ,7-диглюкозида в лепестках цветков немофилы, апигенин-4ʹ-глюкозид и апигенин-7-глюкозид должны быть образованы в виде промежуточных продуктов в процессе биосинтеза апигенин-4ʹ,7-диглюкозида (см. фиг. 8). Исходя из этого, флавон, демонстрирующий время удерживания близкое к времени удерживания апигенин-7-глюкозида, обнаруженное в примере 1, был признан апигенин-4ʹ-глюкозидом (см. фиг. 7). Смогли определить время удерживания и максимум поглощения апигенин-4ʹ-глюкозида.
На основании этих результатов, было четко определено, что белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении и 7-положении флавона, которая зависит от УДФ-глюкозы, присутствует в лепестках цветков немофилы. Были рассмотрены две возможности гликозилирования гидроксильных групп в 4ʹ-положении и 7-положении флавона, одна возможность состоит в том, что один фермент выполняет гликозилирование обеих гидроксильных групп, и другая возможность состоит в том, что гликозилирование гидроксильных групп 4ʹ-положении и 7-положении соответственно, выполняется различными ферментами.
Пример 3: Обнаружение генов-кандидатов для гена, кодирующего белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона
Выделение тотальной РНК
Тотальную РНК выделяли из лепестков цветков немофилы стадии 1 и стадии 2 с использованием набора Plant RNAeasy Kit (Qiagen Corp.) в соответствии с протоколом, рекомендованным производителем.
Анализ экспрессии кДНК, полученной из лепестков цветков немофилы
Реакцию обратной транскрипции выполняли на 30 мкг тотальной РНК, полученной из лепестков цветков немофилы с последующим получением нормализованной библиотеки кДНК. После амплификации полученной библиотеки для каждого клона эмульсионной ПЦР, последовательности оснований определяли с использованием геномного секвенатора FLX Genome Sequencer (Roche Diagnostics K.K.). Кроме того, данные о полученной последовательности транслировали в аминокислотные последовательности с последующей экстракций тех последовательностей, которые демонстрировали гомологию с аминокислотной последовательностью антоцианин-3ʹ-гликозилтрансферазы (anthocyanin 3ʹ-glycosyltransferase) львиного зева. Затем эти последовательности были скомпонованы для получения генов-кандидатов, кодирующих гликозилтрансферазу.
Пример 4: Получение полноразмерной последовательности кДНК генов-кандидатов, кодирующих белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона
25 типов последовательностей гена гликозилтрансферазы было получено в примере 3. Эксперименты проводили для получения полноразмерных последовательностей кДНК для 10 из этих последовательностей (с NmGTO по NmGT9).
Получение полноразмерной кДНК выполняли с использованием (набора) GeneRacer (Invitrogen Corp.) в соответствии с протоколом, рекомендованным производителем. Области, специфические к клонам, выбирали из неполных последовательностей кДНК, полученных в примере 3 и праймеры для (проведения) RACE (пер., быстрой амплификации концов кДНК) конструировали на основе последовательностей этих областей для получения 5ʹ- и 3ʹ-концевых последовательностей RACE ПЦР (пер., ПЦР с быстрой амплификацией концов кДНК). Праймеры для амплификации полноразмерной последовательности кДНК конструировали на основе этих последовательностей, и ПЦР выполняли с использованием 50 мкл кДНК немофилы в качестве матрицы и с использованием полимеразы KOD-Plus Polymerase (Toyobo Co., Ltd.) в соответствии с протоколом, рекомендованным производителем (состоящим из повторения 30 циклов выдерживания при 94°С в течение 2 минут, при 94°С в течение 15 секунд, при 55°С в течение 30 секунд и при 68°С в течение 2 минут с последующим выдерживанием при 4°С). кДНК немофилы синтезировали с использованием тотальной РНК, выделенной в примере 2 с использованием обратной транскриптазы Superscript II (Invitrogen Corp.) в соответствии с протоколом, рекомендованным производителем. Праймеры конструировали таким образом, чтобы они содержали рестриктазные сайты на обоих концах полноразмерной кДНК, для обеспечения введения генов с NmGTO по NmGT9 в экспрессирующий вектор pET15b Е. coli (Novagen Inc.). Плазмиды, содержащие полноразмерные гены NmGT (с рТОРО-NmGTO по pTOPO-NmGT9) были получены с использованием набора для секвенирования Zero Blunt ТОРО PCR Cloning Kit for Sequencing (Invitrogen Corp.) в соответствии с протоколом, рекомендованным производителем. Последовательности оснований, вставленные с помощью плазмид, анализировали для получения последовательности полноразмерной кДНК (NmGT8: SEQ ID NO:1) из числа генов-кандидатов (с NmGTO по NmGT9) для кодирования гена, кодирующего белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона.
Пример 5: Эксперимент по измерению ферментативной активности белков-андидатов, обладающих активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона (случай использования неочищенного фермента)
Получение экспрессирующих конструкций Е. coli
3 мкг аликвот каждого из (генов) с pTOPO-NmGTO по pTOPO-NmGT9 обрабатывали соответствующими им рестриктазами с последующим извлечением полученных фрагментов ДНК (длиной) приблизительно 1,5 т.п.н.. 2 мкг вектора pET15b также обрабатывали рестриктазой и лигировали с полученными фрагментами ДНК для получения экспрессирующих конструкций Е. coli (с pET-NmGTO по pET-NmGT9).
Экспрессия гликозилтрансферазы в Е. coli
(Конструкции) с pET-NmGTO по pET-NmGT9 вводили в штамм BL2 Е. coli с использованием One Shot BL21 (DE3) (Invitrogen Corp.) в соответствии с протоколом, рекомендованным производителем для получения трансформированной Е. coli. Затем эту Е. coli культивировали с использованием системы Overnight Express Autoinduction System 1 (Novagen Inc.) в соответствии с протоколом, рекомендованным производителем. Трансформированную Е. coli культивировали при 37°С до значения OD600 0,5 в 2 мл подготовленной культуральной жидкости (приблизительно 4 часа). Затем эту культуральную жидкость Е. coli использовали в качестве предкультуральной жидкости и добавляли в 50 мл культуральной жидкости с последующим окончательным культивированием в течение ночи при 27°С.
После окончательного культивирования в течение ночи культуру Е. coli центрифугировали (3000 об/мин, 4°С, 15 минут), собранные клетки суспендировали в 5 мл буфера для ультразвуковой обработки (состав: Tris HCl (рН 7,0): 2,5 мМ, дитиотреитол (DTT): 1 мМ, амидинофенилметансульфонилфторида гидрохлорид (APMSF): 10 мкМ), и после разрушения Е. coli ультразвуковой обработкой, разрушенные клетки центрифугировали (15000 об/мин, 4°С, 10 минут) с последующим восстановлением супернатанта. Этот супернатант использовали в качестве жидкости неочищенного фермента. Для центрифугирования использовали Avanti НР-26ХР (ротор: JA-2) (Beckman Coulter Inc.).
Определение ферментативной активности
Реакционную жидкость, приготовленную смешиванием 80 мкл жидкости неочищенного фермента, 20 мкл 5 мМ УДФ-глюкозы, 20 мкл 1 М Tris HCl (рН 7,5) и 1 мкл апигенина 500 нг/мкл на бане со льдом и доведением объема до 200 мкл водой, выдерживали в течение 1 часа при 30°С. Затем, добавляли 200 мкл буфера для остановки (90% водный раствор ацетонитрила, содержащий 0,1% ТФУ) для остановки реакции с последующим анализом реакционной жидкости высокоэффективной жидкостной хроматографией (Prominence (Shimadzu Corp.)). Для обнаружения использовали (спектрофотометр) Shimadzu PDA SPD-M10AVP и флавон обнаруживали при 330 нм. В качестве колонки использовали колонку Shim-Pack ODS 150 мм × 4,6 мм (Shimadzu Corp.). Раствор А (0,1% водный раствор ТФУ) и раствор В (90% водный раствор ацетонитрила, содержащий 0,1% ТФУ) использовали для элюции. Элюцию выполняли с использованием линейного градиента концентраций из смеси двух растворов 8:2 до смеси двух растворов 3:7 в течение 10 минут, за которым следует смесь 3:7 в течение 5 минут. Скорость потока составляла 0,6 мл/мин. Реакционную жидкость, полученную при предоставлении возможности ферменту взаимодействовать в одних и тех же условиях с использованием жидкости неочищенного фермента Е. coli со вставленным вектором рЕТ без встроенной вставки (инсерта), использовали в качестве контроля.
В результате, для NmGT8 наблюдали пик, отличающийся от пика для субстрата.
Описания, начиная с описания примера 6, описывают применительно к NmGT8 (SEQ ID NO:1).
Пример 6: Эксперимент по измерению ферментативной активности белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона (случай использования белка с добавленными His-Tag
Экспрессия гликозилтрансферазы в Е. coli и очистка белка
Штамм BL2 Е. coli со вставленным pET-NmGT8, описанный в примере 5, культивировали с использованием системы Overnight Express Autoinduction System 1 (Novagen Inc.) очистки His-Tag-белка. Трансформированную E.coli культивировали при 37°C до значения OD600 0,5 в 8 мл приготовленной культуральной жидкости (приблизительно 4 часа). Затем эту культуральную жидкость Е. coli использовали в качестве предкультуральной жидкости и добавляли в 200 мл культуральной жидкости с последующим окончательным культивированием в течение ночи при 25°С.
После окончательного культивирования в течение ночи культуральную жидкость Е. coli центрифугировали (1000 × g, 4°С, 10 минут), собранные клетки суспендировали в 20 мл буфера для экстракции (состав: буфер (KCl: 300 мМ, KH2PO4: 50 мМ, имидазол: 5 мМ) (рН 8,0), амидинофенилметансульфонилфторида гидрохлорид (APMSF): 10 мкМ), и после разрушения Е. coli ультразвуковой обработкой, разрушенные клетки центрифугировали (1400 × g, 4°С, 20 минут) с последующим восстановлением супернатанта. Этот супернатант пропускали через фильтр 0,45 мкм и очищали с использованием (системы) Profinia (Bio-Rad Laboratories, Inc.) для очистки His-Tag-белка в соответствии с протоколом, рекомендованным производителем. Полученный раствор очищенного белка центрифугировали (7500 × g, 4°С, 15 минут) с использованием центрифужных фильтров (Ultracel-10K, Amicon Corp.) и концентрированный раствор белка называли "раствор белка NmGT8". Для центрифугирования использовали Avanti НР-26ХР (rotor: JA-2) (Beckman Coulter Inc.).
Измерение ферментативной активности
Реакционную жидкость, приготовленную смешиванием 10 мкл раствора белка, 2 мкл 50 мМ УДФ-глюкозы, 10 мкл 1 М Tris HCl (рН 7,5) и 5 мкл 1 мМ апигенина на бане со льдом и доведением объема до 100 мкл водой, выдерживали в течение 20 минут при 30°С. Затем, добавляли 100 мкл буфера для остановки (90% водный раствор ацетонитрила, содержащий 0,1% ТФУ) для остановки реакции с последующим анализом реакционной жидкости высокоэффективной жидкостной хроматографией (Prominence (Shimadzu Corp.)). Для обнаружения использовали (спектрофотометр) Shimadzu PDA SPD-M10AVP и флавон обнаруживали при 330 нм. В качестве колонки использовали колонку Shim-Pack ODS 150 мм × 4,6 мм (Shimadzu Corp.). Раствор А (0,1% водный раствор ТФУ) и раствор В (90% водный раствор метанола, содержащий 0,1% ТФУ) использовали для элюции. Элюцию выполняли с использованием линейного градиента концентраций из смеси двух растворов 8:2 до смеси двух растворов 3:7 в течение 10 минут, за которым следует смесь 3:7 в течение 6 минут. Скорость потока составляла 0,6 мл/мин.
В результате был биосинтезирован флавон, демонстрирующий то же время удерживания и тот же максимум поглощения, что и очищенный апигенин-4ʹ-глюкозид (см. фиг. 9). Лютеолин-4ʹ-глюкозид был биосинтезирован в случае выполнения ферментативной реакции в тех же самых условиях замещением субстрата на другой флавон в форме лютеолина (см. фиг. 11). С другой стороны, апигенин-4ʹ,7-дигюкозид не был биосинтезирован в случае выполнения ферментативной реакции в тех же самых условиях замещением субстрата на 1 мМ апигенин-7-глюкозид (см. фиг. 10). Подобным образом, лютеолин-4ʹ,7-дигюкозид также не был биосинтезирован в случае выполнения ферментативной реакции в тех же самых условиях замещением субстрата на 1 мМ лютеолин-7-глюкозид (см. фиг. 11). На этом основании, было четко определено, что белок NmGT8 проводит гликозилирование 4ʹ-положения до момента гликозилирования 7-положения апигенина и лютеолина (см. фиг. 8). Кроме того, при исследовании реакционной способности различных типов соединений флавонов, описанных на фиг. 11, и батанидина, было четко определено, что белок NmGT8 демонстрирует высокую субстратную специфичность и избирательно гликозилирует 4ʹ-положение флавонов, наподобие апигенина и лютеолина (см. фиг. 11).
Кроме того, хотя ген гликозилтрансферазы, полученный из мезембриантемума Ливингстона (Dbs5GT) переносит глюкозу на гидроксильную группу в 5-положении бетанидина, сообщалось также, что он демонстрирует активность in vitro переноса глюкозы на гидроксильную группу в одном из положений, в 4ʹ-положении или в 7-положении флавоноида. Было четко определено, что эта гликозилтрансфераза, полученная из мезембриантемума Ливингстона, существенно отличается от белка NmGT8 настоящего изобретения в отношении реакционной способности к соединениям флавоноидов и бетанидина (см. фиг. 11).
Идентичность аминокислотных последовательностей между NmGT8 и NmGT3 и между NmGT8 и NmGT4 составляла 32% и 32% соответственно (см. фиг. с 12-1 до 12-3). Программу Clustal W Program of the MacVector Application (версия 11.02, Oxford Molecular Ltd., Оксфорд, Англия) использовали для этого анализа. Среди ранее идентифицированных гликозилтрансфераз, аминокислотной последовательностью, имеющей самую высокую идентичность с NmGT8, была аминокислотная последовательность фермента, который добавляет сахар в 3-положение и 5-положение антоцианидинов розы ((номер в банке генов) GenBank Accession No. Q4R119), и демонстрирует 52% идентичность аминокислотной последовательности (см. фиг. 14). Затем, аминокислотной последовательностью, имеющей высокую идентичность с NmGT8, была аминокислотная последовательность фермента, который добавляет сахар в 4ʹ-положение халконов львиного зева (описанных в PCT/JP2004/019461), и демонстрирует 51% идентичность (см. фиг. 13).
Кроме того, фиг. 15 демонстрирует филогенетическое древо, указывающее на родство между NmGT8 настоящего изобретения и указанных выше ферментов.
Пример 7: Экспрессия гена, кодирующего пептид, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона у торении
Бинарный вектор pSPB4583 для экспрессии NmGT8 конструировали и вводили в торению (сорта Summer Wave) для подтверждения транслирует ли (или не транслирует) ген NmGT8 настоящего изобретения белок, обладающий активностью переноса сахара на гидроксильную группу в 4-положении флавона у растений. Детали введенной конструкции описываются ниже (см. фиг. 16).
Получение конструкции
Бинарный вектор pBINPLUS для введения в растения (van Engel, et al., Transgenic Research, 4, p.288) использовали для основного скелета pSPB4583, и в нем (бинарном векторе) содержались: промотор EI235S, имеющий две расположенные впереди (по ходу) повторности энхансерной последовательности из 35S промотора вируса мозаики цветной капусты (Mitsuhara, et al., (1996) Plant Cell Physiol., 37, p.49), полноразмерная кДНК NmGT8 и терминатор mas.
Анализ экспрессии гена
Анализ экспрессии гена выполняли путем формирования желобков в селективных средах, содержащих канамицин, акклиматизации индивидуумов, у которых наблюдалось укоренение, и использования лепестков бутонов каждой рекомбинантной торении, у которой чашечка еще не раскололась. Выделение тотальной РНК выполняли тем же самым способом, который описан в примере 3, и синтез кДНК выполняли тем же самым способом, который описан в примере 4. ПЦР с обратной транскрипцией выполняли в 30 мкл с использованием кДНК в качестве матрицы и с использованием полимеразы ExTaq (Takara Co., Ltd.) в соответствии с протоколом, рекомендованным производителем (состоящую из повторения 25 циклов выдерживания при 94°С в течение 2 минут, 94°С в течение 1 минуты, 55°С в течение 1 минуты и 72°С в течение 2 минут с последующим выдерживанием при 4°С). Буферы создавали с тем, чтобы специфически амплифицировалась каждая полноразмерная кДНК. В результате было подтверждено, что NmGT8 транскрибируется в торении.
Пример 8: Экспрессия гена, кодирующего белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона у петунии
Бинарные векторы pSPB5424 и pSBP5428 для экспрессии NmGT8 конструировали и вводили в петунию (Surfinia Bouquet Red). Поскольку петуния не биосинтезирует флавоны естественным путем, то векторы вводили вместе с флавонсинтазой торении и оценивали обладает ли (или не обладает) NmGT8 активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона у петунии. Детали введенных конструкций описывают ниже (см. фиг. 17).
Получение конструкций
pBINPLUS использовали для основного скелета pSPB5424, и в нем (векторе) содержалось три экспрессирующих кассеты (состоящих из: 1) промотора EI235S, полноразмерной кДНК F3ʹ5ʹH анютиных глазок (описанной в PCT/JP2004/011958, SEQ ID NO:3) и HSP терминатора (Plant Cell Physiology (2010), 51, 328-332); 2) промотора EI235S, полноразмерной кДНК флавонсинтазы торении и HSP терминатора; и 3) промотора EI235S, полноразмерной кДНК NmGT8 HSP-терминатора).
pBINPLUS также использовали для основного скелета pSPB5428, и в нем (векторе) содержалось три экспрессирующих кассеты (состоящих из: 1) промотора EI235S, полноразмерной кДНК флавонсинтазы торении и HSP терминатора; и 2) промотора EI235S, полноразмерной кДНК NmGT8 и HSP терминатора).
Анализ экспрессии гена
Анализ экспрессии гена выполняли тем же самым способом, что и описанный в примере 7 путем формирования желобков в селективных средах, содержащих канамицин, акклиматизации индивидуумов, у которых наблюдалось укоренение, и использования лепестков цветков каждой рекомбинантной петунии, которые полностью открылись. В результате было подтверждено, что NmGT8 транскрибируется в петунии.
Анализ пигментов лепестков цветка
Пигменты в лепестках цветка анализировали для полноразмерной кДНК флавонсинтазы торении и полноразмерной кДНК NmGT8 в тех штаммах, для которых были подтверждены продукты транскрипции. 0,2 г полностью раскрытых лепестков цветков были лиофильно высушены в течение 24 часов или более и тонко измельчены с помощью шпателя, с последующим добавлением 4 мл буфера для экстракции (состав: 50% раствор водного ацетонитрила, содержащий 0,1% ТФУ) и подверганием обработке ультразвуком в течение 20 минут. Экстракт лепестков цветка анализировали высокоэффективной жидкостной хроматографией (Prominence (Shimadzu Corp.)). Анализ выполняли в тех же самых условиях и тем же самым способом, что описаны в примере 5. Нерекомбинантную петунию, петунию без вставленных генов, и рекомбинантную петунию со вставленной только флавонсинтазой торении, которая биосинтезирует флавон, анализировали тем же самым способом, что и контроли (фиг. 18). Кроме того, разбавленный в 50 раз экстракт лепестков цветков, также анализировали высокоэффективной жидкостной хроматографией (Shimadzu Corp.). Shimadzu LCMS-IF-TOF (гибридный жидкостной хроматомасс-спектрометр) использовали для обнаружения и флавоны детектировали при 433,1057 нм ([Api-Glc+H]) и 449,1084 нм ([Lut-Glc+H]). В качестве колонки использовали Inertsil ODS-4 (250×4,6 мм, 5 мкм, Shimadzu Corp.). Раствор А (0,1% водный раствор муравьиной кислоты) и раствор В (90% водный раствор ацетонитрила, содержащего 0,1% муравьиную кислоту) использовали для элюции. Элюцию выполняли с использованием линейного градиента концентраций из смеси 9:1 двух растворов до смеси 11:9 двух растворов в течение 35 минут и линейного градиента концентраций из смеси 11:9 двух растворов до смеси 0:10 в течение 10 минут, за которым следует смесь 0:10 в течение 5 минут. Скорость потока составляла 0,6 мл/мин (фиг. 19).
В результате, в рекомбинантной петунии, со вставленными флавонсинтазой и NmGT8, обнаруживали апигенин-4ʹ-глюкозид и лютеолин-4ʹ-глюкозид (фиг. 18), и было определено, что флавон-4ʹ-глюкозид составляет 95,6% биосинтезированных флавонов. Было определено, что оставшиеся 4,4% состоят из флавон-7-глюкозида, биосинтезированного присущей петунии активностью, которая переносит сахар на гидроксильную группу в 7-положении флавона (фиг. 19). С другой стороны, флавоны не были обнаружены в нерекомбинантной петунии, и было определено, что флавон-7-глюкозид составляет 82,8% биосинтезированных флавонов в рекомбинантной петунии, только с введенной флавонсинтазой торении. На основании этого было четко продемонстрировано, что NmGT8 является белком, обладающим активностью переноса сахара на гидроксильную группу в 4-положении флавона, который функционирует предпочтительно более сильно, чем белок, обладающий активностью переноса сахара на гидроксильную группу в 7-положении флавона, который является присущим петунии. Применение NmGT8 обеспечивает возможность эффективного биосинтеза флавон-4ʹ-глюкозида в петунии.
Пример 9: Экспрессия гена, кодирующего белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона у гвоздики
Бинарный вектор pSPB5433 для экспрессии NmGT8 конструировали и вводили в гвоздику (Cream Cinderella). Поскольку гвоздика не биосинтезирует флавоны естественным путем, то вектор вводили вместе с флавонсинтазой торении и оценивали обладает ли (или не обладает) NmGT8 активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона у гвоздики. Детали введенных конструкций описываются ниже (см. фиг. 20).
Бинарный вектор pWTT2132 для введения в растения (DNA Plant Technologies, USA=DNAP) использовали для основного скелета pSPB5433, и в нем (бинарном векторе) содержалось четыре экспрессирующих кассеты (состоящие из: 1) синтазного промотора халкона львиного зева (описанного в PCT/AU94/00265), полноразмерной кДНК F3ʹ5ʹH анютиных глазок и HSP-терминатора; 2) синтазного промотора халкона львиного зева, полноразмерной кДНК флавонсинтазы торении и HSP-терминатора; 3) синтазного промотора антоциана гвоздики (описанного в PCT/AU/2009/001659), полноразмерной кДНК NmGT8 и HSP-терминатора; и 4) синтазного промотора антоциана гвоздики, полноразмерной кДНК NmGT3 (Патентная заявка Японии №2011-006317) и HSP-терминатора).
Пример 10: Экспрессия гена, кодирующего белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона у розы
Бинарные векторы pSPB4577, pSBP4578, pSBP5437 и pSBP5440 для экспрессии NmGT8 конструировали и вводили в розу (Noblesse, Ritapa Humera). Поскольку роза не биосинтезирует флавоны естественным путем, то векторы вводили вместе с флавонсинтазой торении и оценивали, обладает ли (или не обладает) NmGT8 активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона у розы. Детали введенных конструкций описывают ниже (см. фиг. 21).
pBINPLUS использовали для основного скелета pSPB5477, и в нем содержалось три экспрессирующих кассеты (состоящие из: 1) промотора EI235S, полноразмерной кДНК F3ʹ5ʹH анютиных глазок и терминатора mas; 2) промотора EI235S, полноразмерной кДНК флавонсинтазы торении и терминатора mas; и 3) промотора EI235S, полноразмерной кДНК NmGT8 и терминатора mas).
pBINPLUS использовали для основного скелета pSPB4578, и в нем содержалось три экспрессирующих кассеты (состоящие из: 1) промотора антоцианин-3-ацилтрансферазы (anthocyanin 3-acyltransferase) периллы (описанного в PCT/JP2010/053909), полноразмерной кДНК F3ʹ5ʹH анютиных глазок и терминатора mas; 2) промотора EI235S, полноразмерной кДНК флавонсинтазы торении и терминатора mas; и 3) промотора EI235S, полноразмерной кДНК NmGT8 и терминатора mas).
pBINPLUS использовали для основного скелета pSPB5437, и в нем содержалось пять экспрессирующих кассет (состоящих из: 1) промотора EI235S, полноразмерной кДНК F3ʹ5ʹH анютиных глазок и HSP-терминатора; 2) хромоген-антоцианин-3-ацилтрансферазы периллы (описанной в PCT/JP2010/053886, см. SEQ ID NO:7); 3) промотора EI235S, полноразмерной кДНК флавонсинтазы торении и HSP-терминатора; 4) промотора EI235S, полноразмерной кДНК NmGT8 и HSP-терминатора); и 5) промотора EI235S, полноразмерной кДНК NmGT3 и HSP-терминатора).
pBINPLUS использовали для основного скелета pSPB5440, и в нем содержалось пять экспрессирующих кассет (состоящих из: 1) промотора EI235S, полноразмерной кДНК F3ʹ5ʹH анютиных глазок и HSP-терминатора; 2) промотора EI235S, кДНК антоцианин-3-ацилтрансферазы лаванды (описанной в PCT/JP1996/000348, см., SEQ ID NO:8); 3) промотора EI235S, полноразмерной кДНК флавонсинтазы торении и HSP-терминатора; 4) промотора EI235S, полноразмерной кДНК NmGT8 и HSP-терминатора); и 5) промотора EI235S, полноразмерной кДНК NmGT3 и HSP-терминатора).
Анализ экспрессии гена
Анализ экспрессии гена выполняли тем же самым способом, что и описанный в примере 7 путем формирования желобков в селективных средах, содержащих канамицин, акклиматизации индивидуумов, у которых наблюдалось укоренение, и использования лепестков цветков каждой рекомбинантной розы, которые полностью открылись. В результате было подтверждено, что NmGT8 транскрибируется в розе.
Анализ пигментов лепестков цветка
Пигменты в лепестках цветка анализировали для полноразмерной кДНК флавонсинтазы торении и полноразмерной кДНК NmGT8 в тех штаммах, для которых были подтверждены продукты транскрипции. 0,2 г полностью раскрытых лепестков цветков были лиофильно высушены в течение 24 часов или более и тонко измельчены с помощью шпателя, с последующим добавлением 4 мл буфера для экстракции (состав: 50% раствор водного ацетонитрила, содержащий 0,1% ТФУ) и подверганием обработке ультразвуком в течение 20 минут. Экстракт лепестков цветка анализировали высокоэффективной жидкостной хроматографией (Prominence (Shimadzu Corp.)). Анализ выполняли в тех же самых условиях и тем же самым способом, что описаны в примере 5. Нерекомбинантную розу, без вставленных генов, анализировали тем же самым способом, что и контроль (фиг. 22). Кроме того, разбавленный в 50 раз экстракт лепестков цветков, также анализировали высокоэффективной жидкостной хроматографией (Shimadzu Corp.). Shimadzu LCMS-IF-TOF (гибридный жидкостной хроматомасс-спектрометр) использовали для обнаружения и флавоны детектировали при 433,1057 нм ([Api-Glc+H]) и 449,1084 нм ([Lut-Glc+H]). В качестве колонки использовали Inertsil ODS-4 (250×4,6 мм, 5 мкм, Shimadzu Corp.). Раствор A (0,1% водный раствор муравьиной кислоты) и раствор В (90% водный раствор ацетонитрила, содержащего 0,1% муравьиной кислоты) использовали для элюции. Элюцию выполняли с использованием линейного градиента концентраций из смеси 9:1 двух растворов до смеси 11:9 двух растворов в течение 35 минут, и линейного градиента концентраций из смеси 11:9 двух растворов до смеси 0:10 двух растворов в течение 10 минут, за которым следует смесь 0:10 в течение 5 минут. Скорость потока составляла 0,6 мл/мин. (фиг. 23).
В результате, в рекомбинантной розе со вставленными флавонсинтазой и NmGT8 обнаруживали апигенин-4ʹ-глюкозид и лютеолин-4ʹ-глюкозид (фиг. 22), и было определено, что флавон-4ʹ-глюкозид составляет 97,0% от биосинтезированных флавонов. Было определено, что оставшиеся 3,0% состоят из флавон-7-глюкозида, биосинтезированного присущей розе активностью, которая переносит сахар на гидроксильную группу в 7-положении флавона (фиг. 23). С другой стороны, флавоны не были обнаружены в нерекомбинантной розе. На основании этого было четко продемонстрировано, что NmGT8 является белком, обладающим активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона, который функционирует предпочтительно более сильно, чем белок, обладающий активностью переноса сахара на гидроксильную группу в 7-положении флавона, который является присущим розе. Применение NmGT8 обеспечивает возможность эффективного биосинтеза флавон-4ʹ-глюкозида в розе.
Промышленная применимость
В настоящем изобретении впервые идентифицировали полинуклеотид, кодирующий белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона. Экспрессия полинуклеотида настоящего изобретения в подходящих клетках-хозяевах обеспечивает возможность получения белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона. По изобретению, конститутивная или тканеспецифическая экспрессия белка, обладающего активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона в растении, может быть применена для изменения окраски цветка. Кроме того, по изобретению, обеспечивают способ получения флавона, в котором сахар был присоединен к гидроксильной группе в 4ʹ-положении флавона, и пищевые продукты, лекарственные и косметические препараты, полученные по способу получения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕН C-ГЛИКОЗИЛТРАНСФЕРАЗЫ ИЗ ГРЕЧИХИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2833446C1 |
| НОВЫЙ ГЕН ФЛАВОНОИД 3,5-ГИДРОКСИЛАЗЫ КОЛОКОЛЬЧИКА И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2640248C2 |
| РОЗА, СОДЕРЖАЩАЯ ФЛАВОН И МАЛЬДИВИН, И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2008 |
|
RU2463348C2 |
| КОМПОЗИЦИЯ ДЛЯ ПОЛУЧЕНИЯ РАСТИТЕЛЬНОГО ОРГАНИЗМА С УЛУЧШЕННЫМ СОДЕРЖАНИЕМ САХАРА И ЕЕ ПРИМЕНЕНИЕ | 2008 |
|
RU2446688C2 |
| РЕГУЛИРУЮЩИЙ ВЫСОТУ РАСТЕНИЙ ГЕН И ЕГО ПРИМЕНЕНИЯ | 2008 |
|
RU2458132C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИЛИЙ, СОДЕРЖАЩИХ В ЛЕПЕСТКАХ ДЕЛФИНИДИН | 2011 |
|
RU2590722C2 |
| МОДУЛЯЦИЯ УРОВНЯ ПРЕДШЕСТВЕННИКОВ АРОМАТА КОФЕ В СЫРЫХ (НЕОБЖАРЕННЫХ) КОФЕЙНЫХ ЗЕРНАХ | 2004 |
|
RU2348693C2 |
| СПОСОБЫ УВЕЛИЧЕНИЯ УРОЖАЯ И СТРЕССОУСТОЙЧИВОСТИ У РАСТЕНИЯ | 2012 |
|
RU2632569C2 |
| ТРАНСГЕННЫЕ РАСТЕНИЯ С УЛУЧШЕННЫМИ ХАРАКТЕРИСТИКАМИ РОСТА | 2009 |
|
RU2582260C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ И СПОСОБЫ ПОЛУЧЕНИЯ СЕМЯН, ИМЕЮЩИХ В ЗАРОДЫШЕ ПОЛНЫЙ ДИПЛОИДНЫЙ ГЕНОМ, КОМПЛЕМЕНТАРНЫЙ МАТЕРИНСКОМУ | 2006 |
|
RU2438297C2 |


























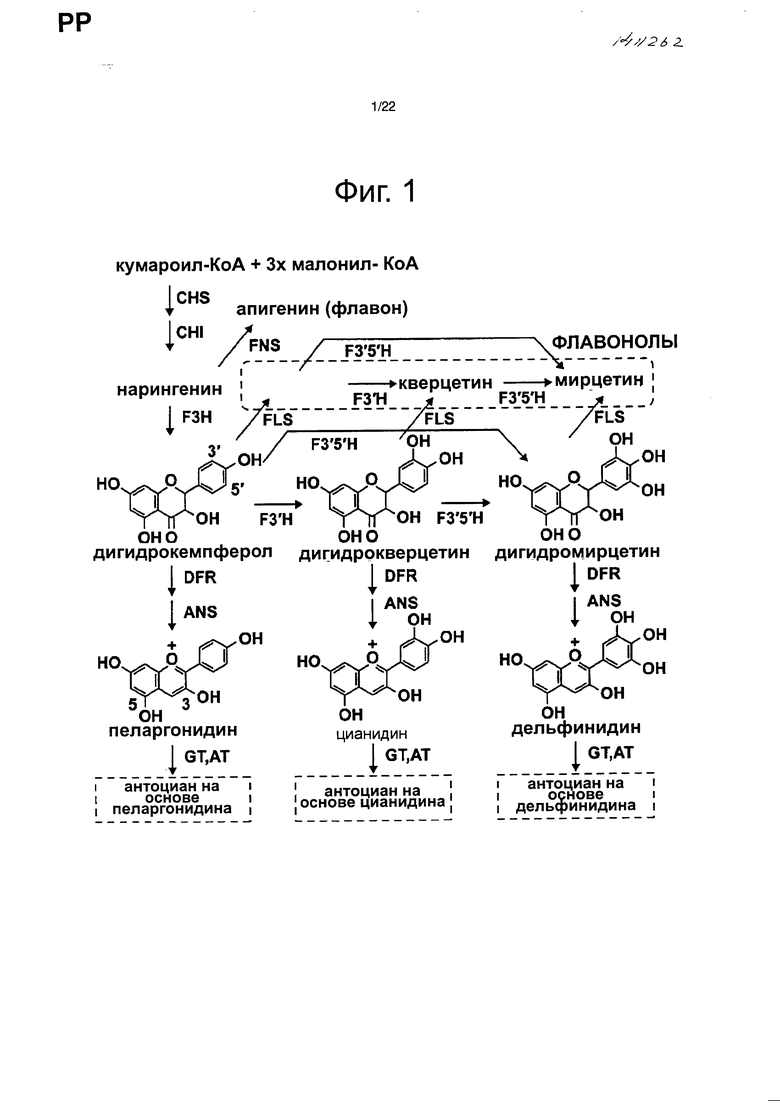
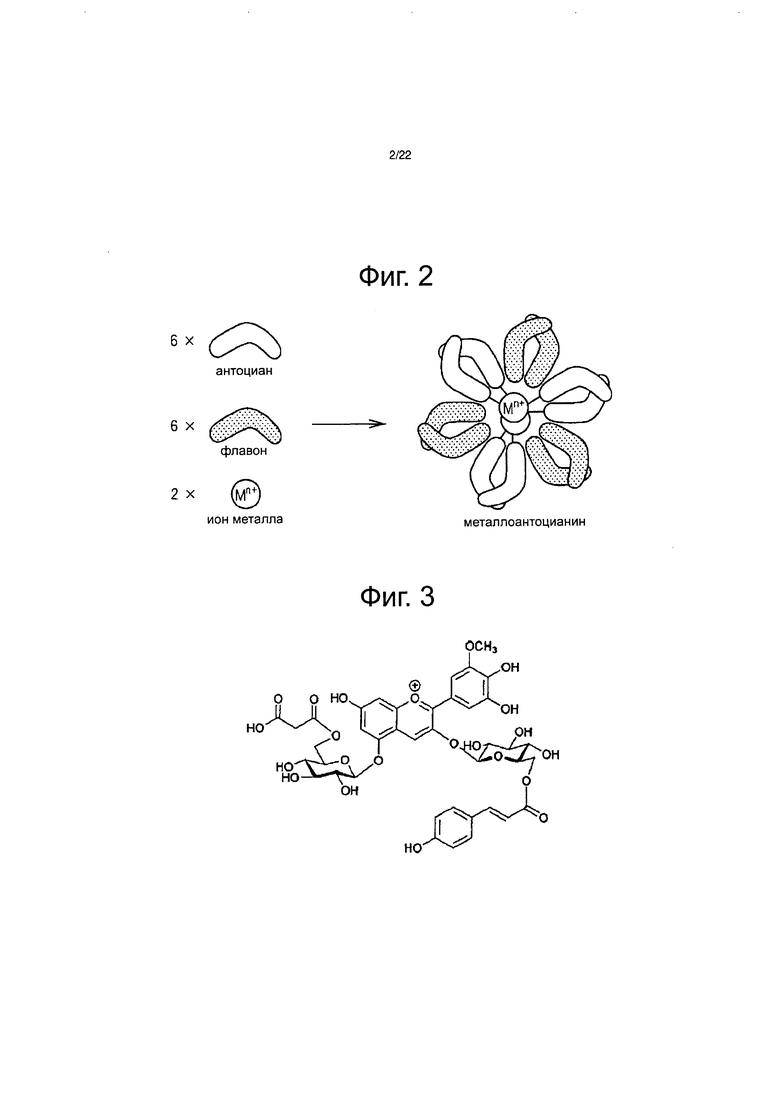
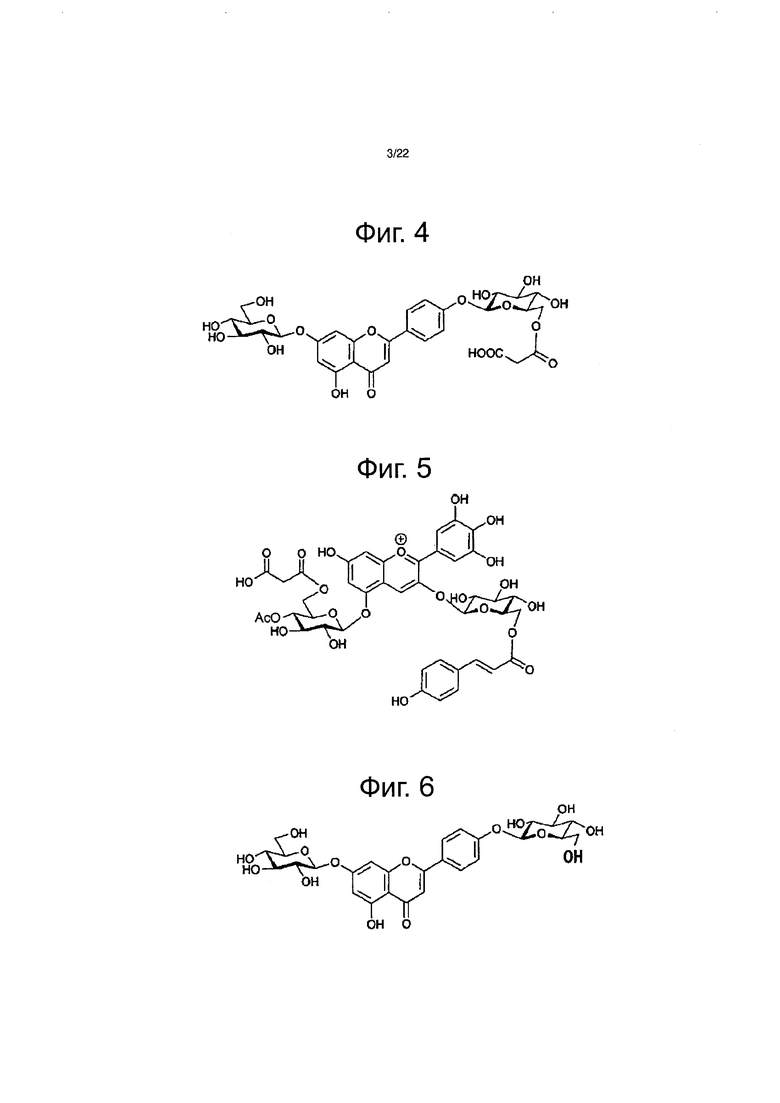
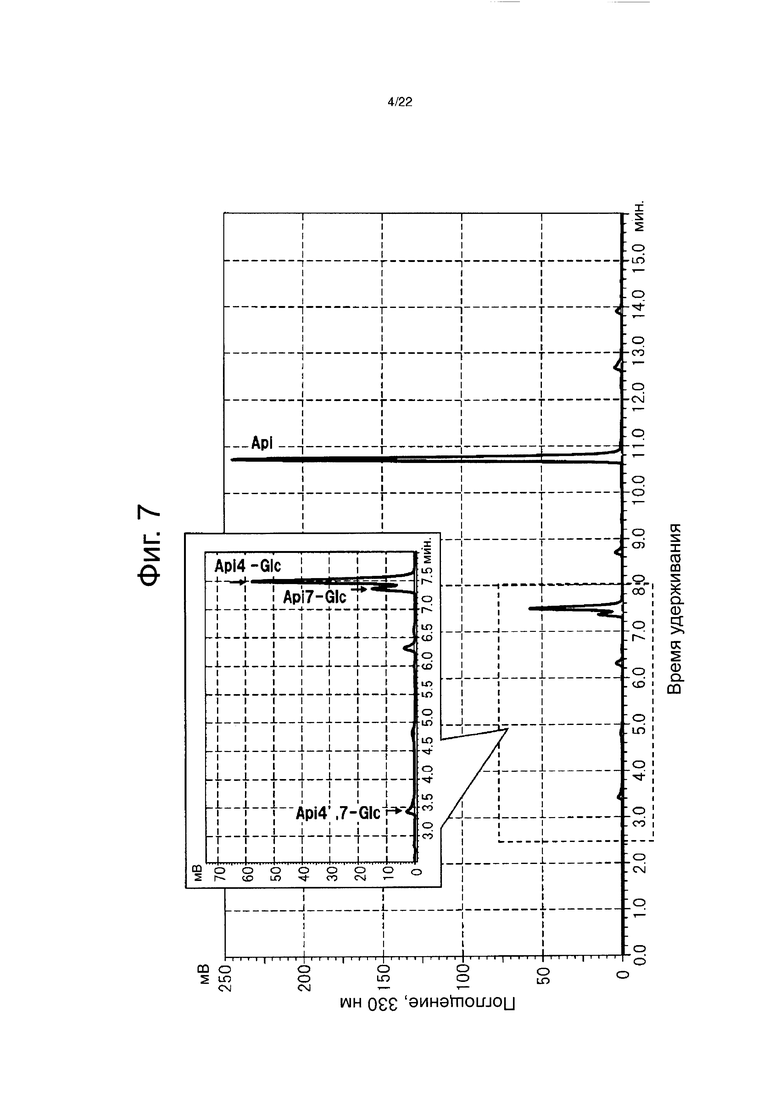
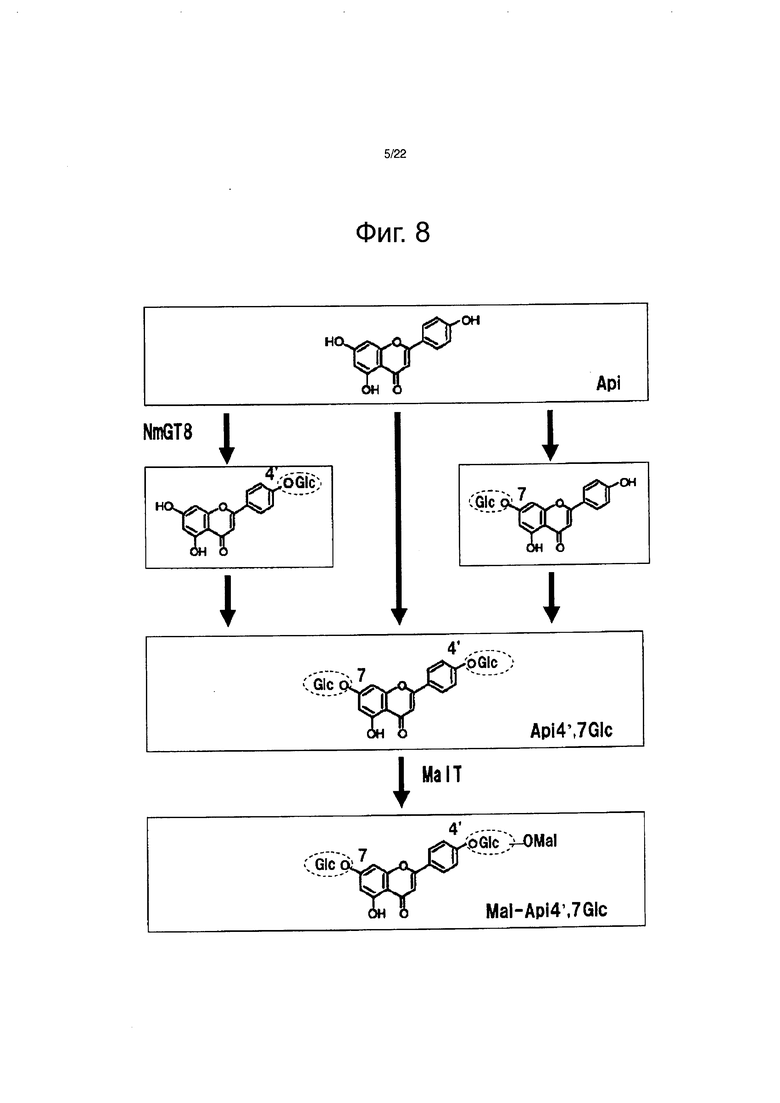
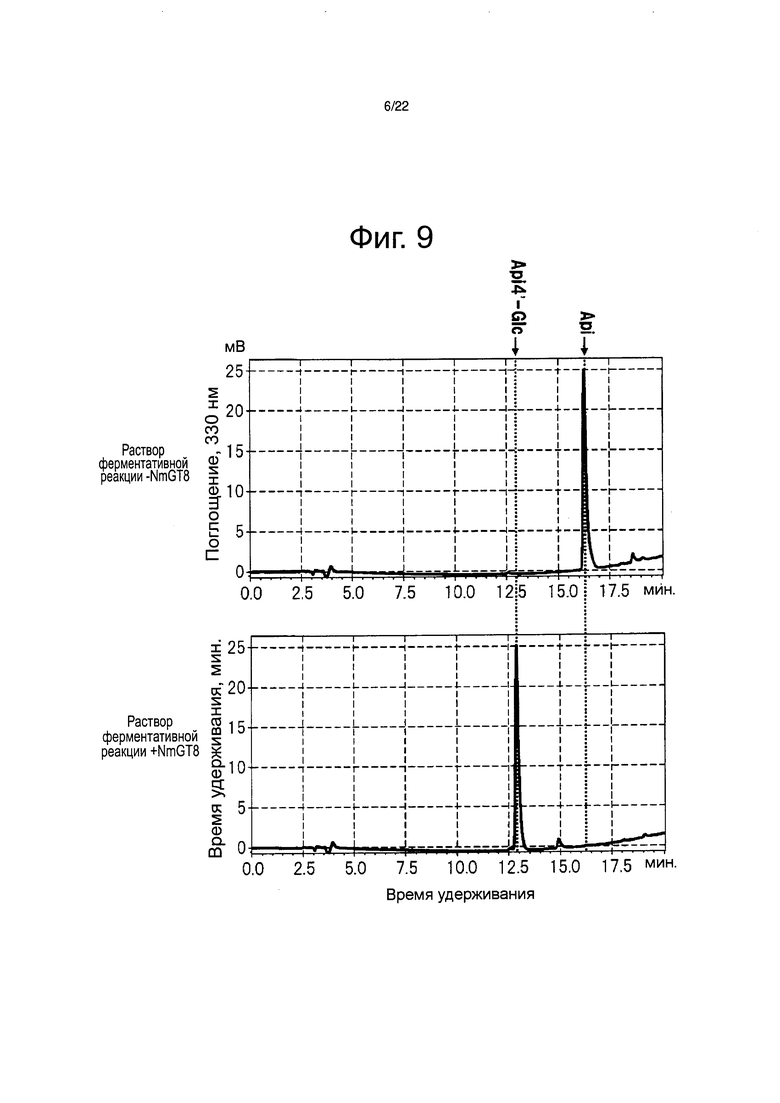
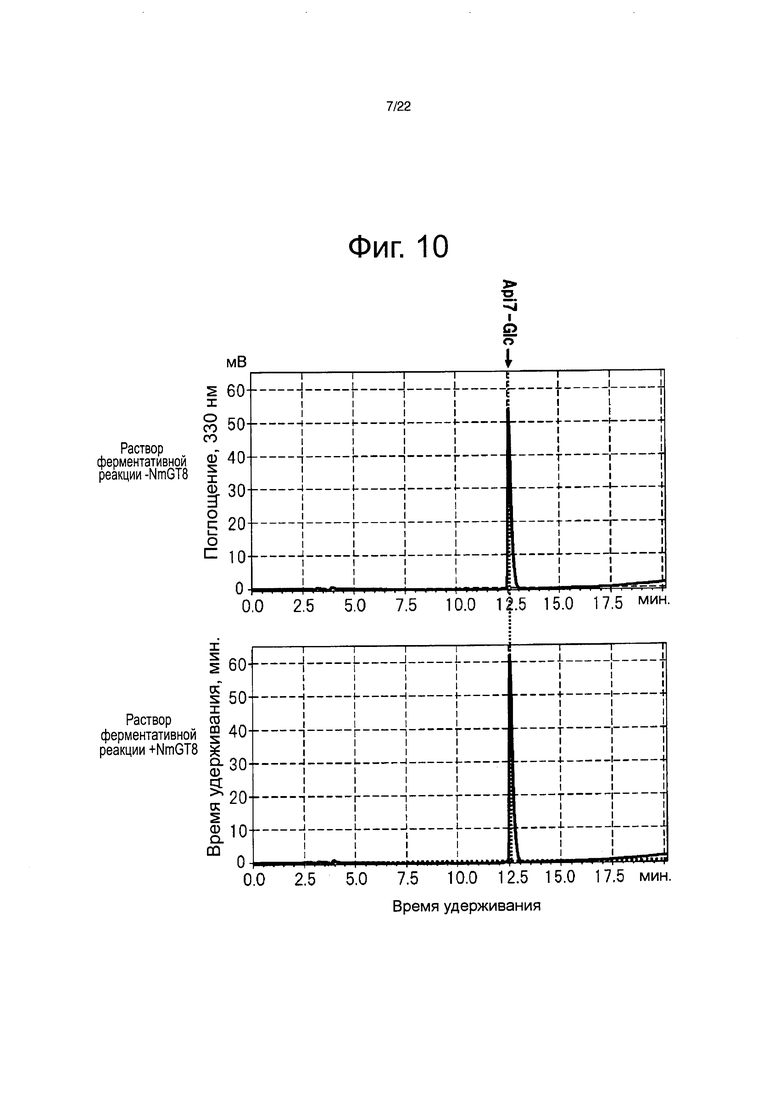




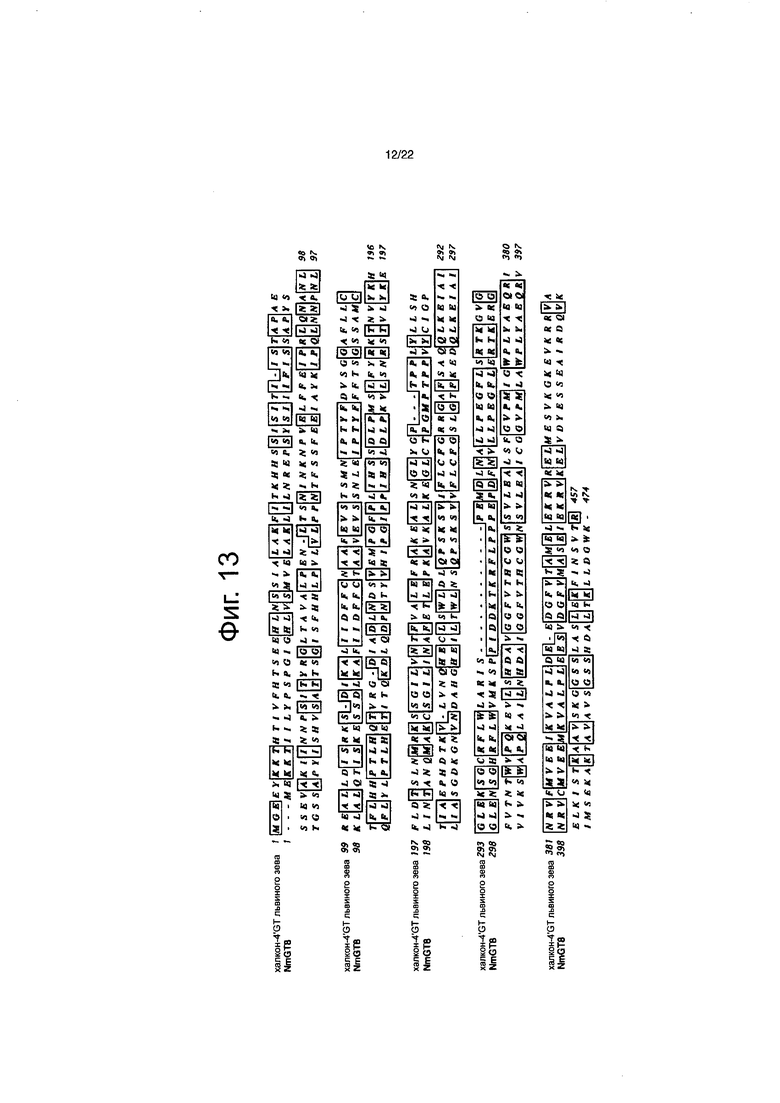
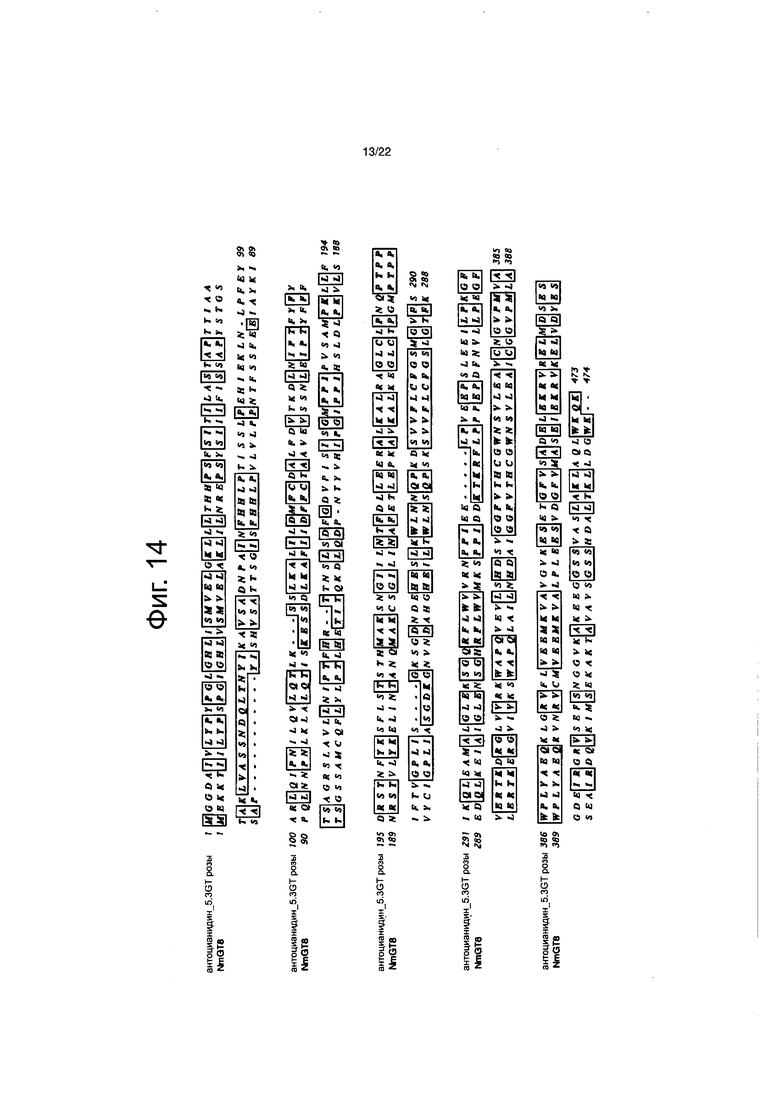

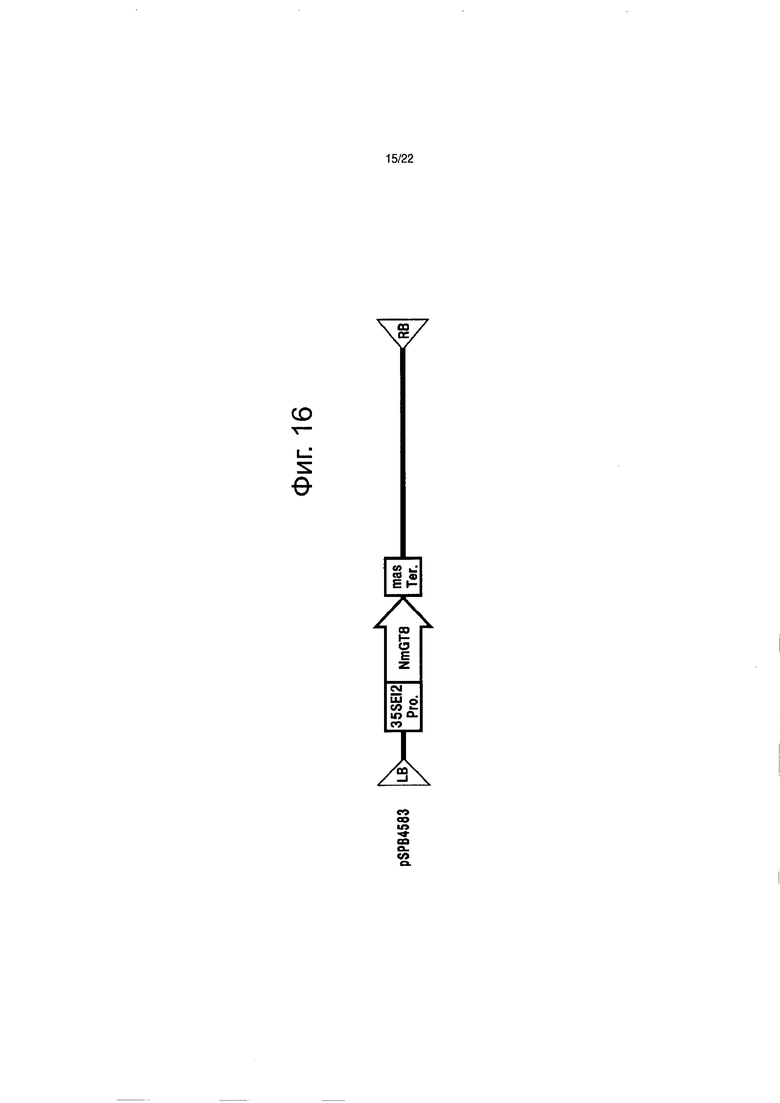

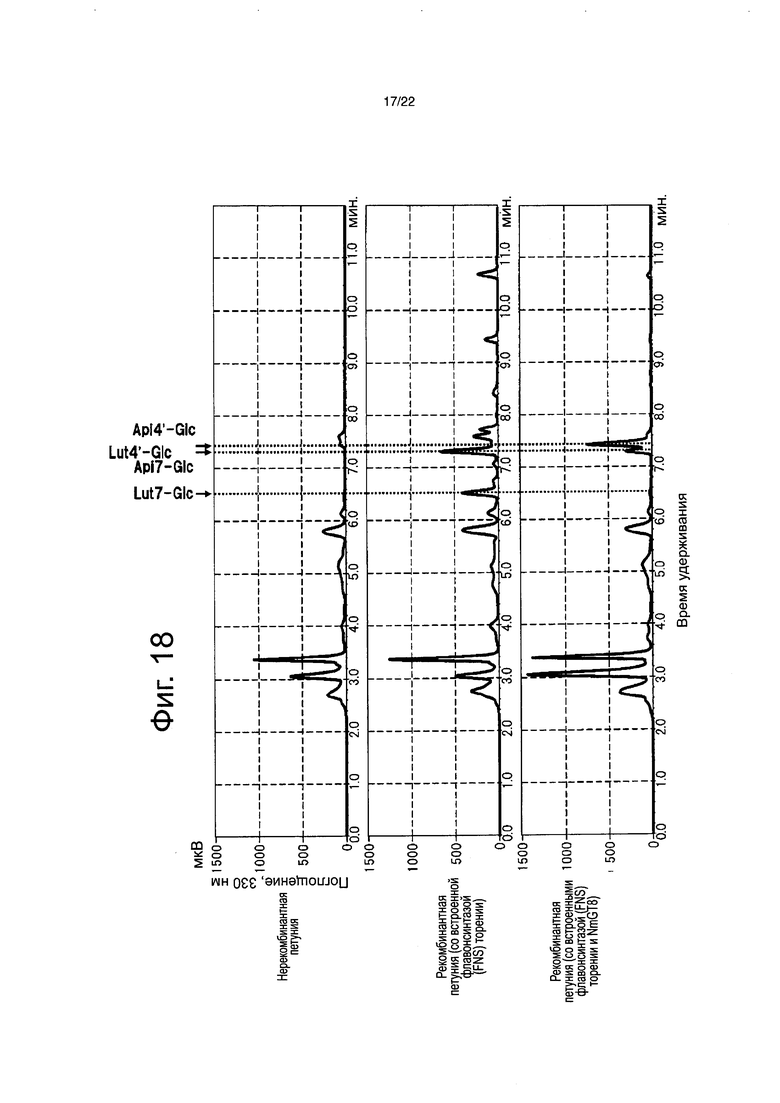
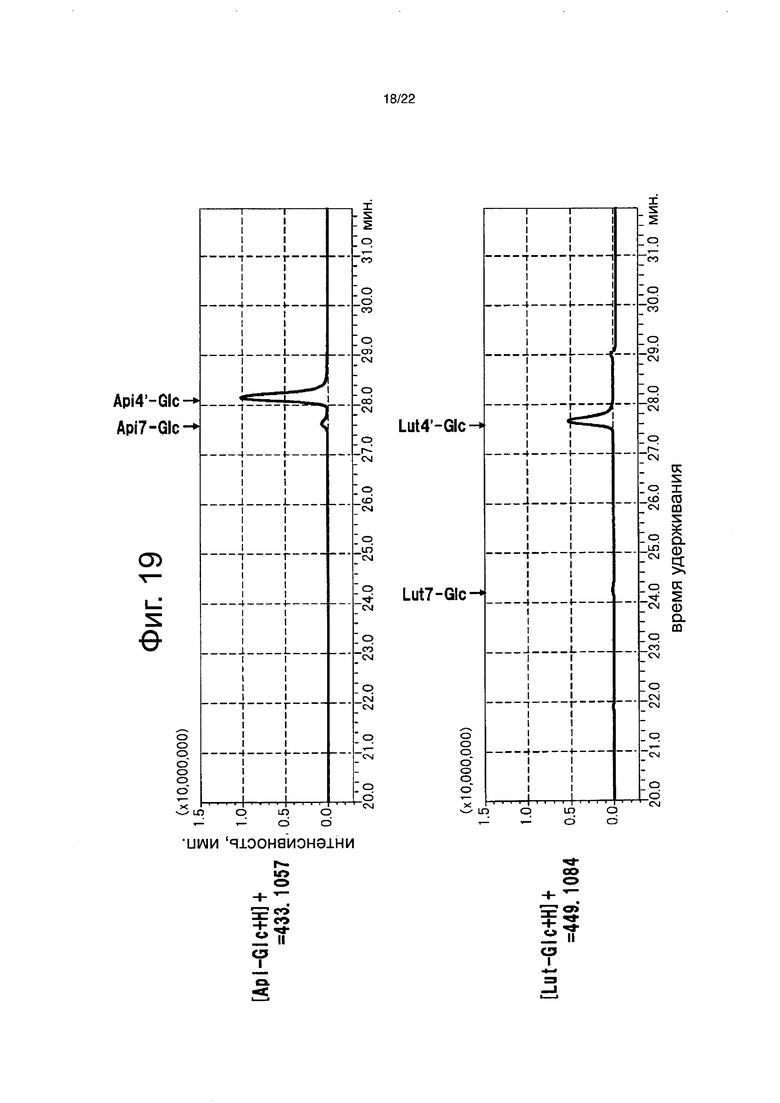
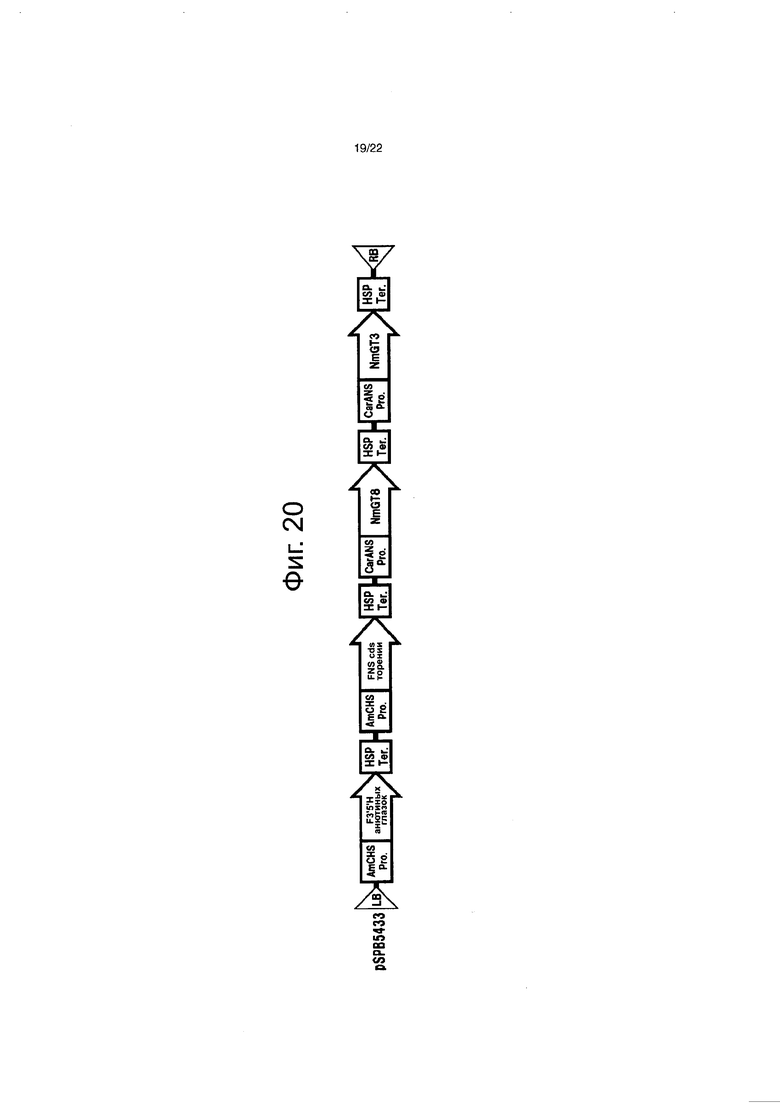

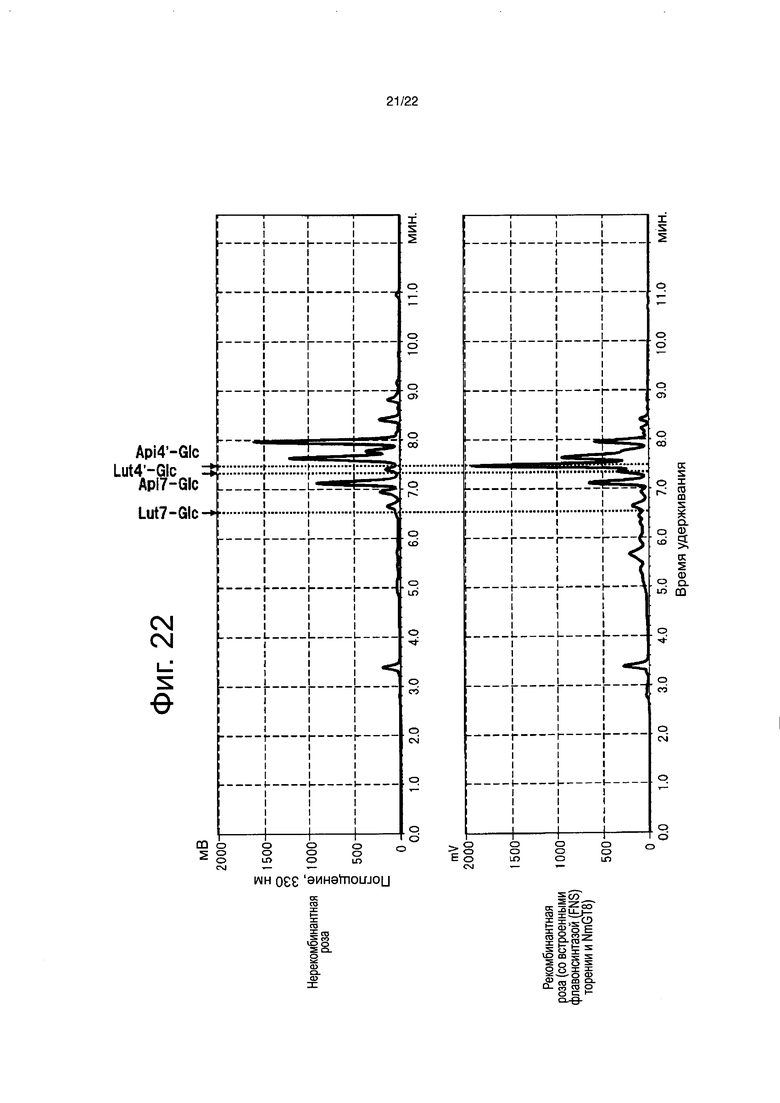
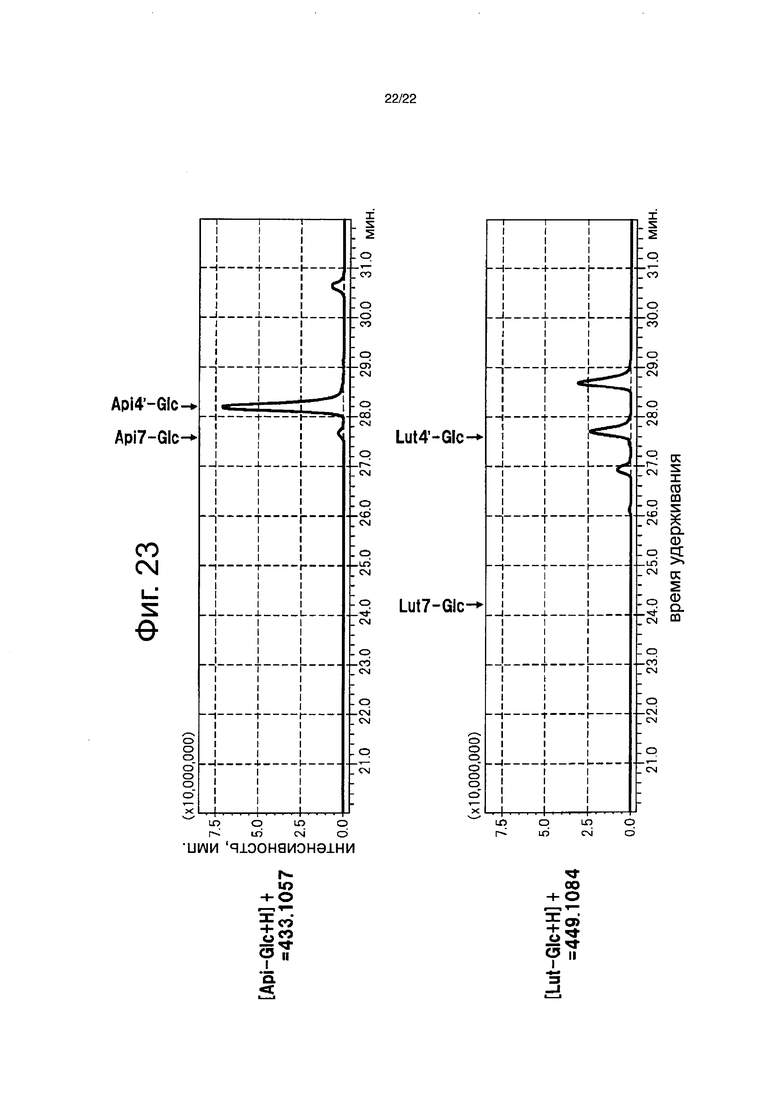
Изобретение относится к области биохимии, а именно представлен полинуклеотид, который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона. Полинуклеотид выбран из группы, состоящей из: (а) полинуклеотида, состоящего из последовательности оснований SEQ ID NO:1; (b) полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, состоящим из последовательности оснований, коплементарной последовательности оснований SEQ ID NO:1, и кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона; (с) полинуклеотида, кодирующего белок, состоящий из аминокислотной последовательности SEQ ID NO:2. Изобретение позволяет эффективно получать полинуклеотид, который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4ʹ-положении флавона. 8 н. и 4 з.п. ф-лы, 23 ил., 10 пр.
1. Полинуклеотид, который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4’-положении флавона, где полинуклеотид выбран из группы, состоящей из следующих от (а) до (c):
(a) полинуклеотида, состоящего из последовательности оснований SEQ ID NO:1;
(b) полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, состоящим из последовательности оснований, комплементарной последовательности оснований SEQ ID NO:1, и который кодирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4’-положении флавона;
(c) полинуклеотида, кодирующего белок, состоящий из аминокислотной последовательности SEQ ID NO:2.
2. Полинуклеотид по п. 1, который является (а) полинуклеотидом, состоящим из последовательности оснований SEQ ID NO:1.
3. Полинуклеотид по п. 1, который является (с) полинуклеотидом, кодирующим белок, состоящий из аминокислотной последовательности SEQ ID NO:2.
4. Полинуклеотид по любому из пп. 1-3, который является ДНК.
5. Белок, кодируемый полинуклеотидом по любому из пп. 1-4, где белок обладает активностью переноса сахара на гидроксильную группу в 4’-положении флавона.
6. Вектор, содержащий полинуклеотид по любому из пп. 1-4, где вектор является вектором экспрессии.
7. Не относящийся к человеку хозяин с введенным вектором по п. 6, для получения белка, обладающего активностью переноса сахара на гидроксильную группу в 4’-положении флавона, где не относящийся к человеку хозяин является клеткой бактерии, клеткой гриба, клеткой животного, клеткой насекомого или растительной клеткой.
8. Растение, содержащее полинуклеотид по любому из пп. 1-4, где растение продуцирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4’-положении флавона.
9. Часть растения, содержащая полинуклеотид по любому из пп. 1-4, где часть растения продуцирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4’-положении флавона.
10. Часть растения по п. 9, которая является срезанным цветком.
11. Ткань растения, содержащая полинуклеотид по любому из пп. 1-4, где ткань растения продуцирует белок, обладающий активностью переноса сахара на гидроксильную группу в 4’-положении флавона.
12. Способ получения белка, обладающего активностью переноса сахара на гидроксильную группу в 4’-положении флавона, содержащий следующие стадии:
культивирования или выращивания не относящегося к человеку хозяина по п. 7, и
сбора белка из не относящегося к человеку хозяина, который обладает активностью переноса сахара на гидроксильную группу в 4’-положении флавона.
| CN 104473178 A, 20.09.2005 | |||
| БИОЛОГИЧЕСКИ АКТИВНАЯ ДОБАВКА | 2007 |
|
RU2350122C1 |
| US 2007148284 A1, 28.06.2007 | |||
| DE 19923838 A1, 30.11.2000 | |||
| US 2008227829 A1, 18.09.2008 | |||
| US 2009111761 A1, 30.04.2009 | |||
| КОСМЕТИЧЕСКОЕ СРЕДСТВО НА ОСНОВЕ ФИТОЭСТРОГЕННОГО БИОКОМПЛЕКСА | 2009 |
|
RU2414885C1 |
| WO 2008156211 A1, 24.12.2008 | |||
| WO 2008156214 A1, 24.12.2008 | |||
| WO 2010026666 A1, 11.03.2010. | |||

