Область техники, к которой относится изобретение
Данное изобретение относится к искусственно полученной розе, содержащей флавон и мальвидин. Это изобретение дополнительно относится к способу модификации окраски лепестков при помощи эффекта копигментации, который получают введением флавона и мальвидина методами генной инженерии, и, в частности, к способу изменения окраски лепестков в направлении синего цвета.
Уровень техники
Цветки являются репродуктивными органами растений, которые необходимы для образования семян для последующих генераций. Образование семян требует адгезии пыльцы на пестиках и оплодотворения. Пыльца часто переносится насекомыми, такими как пчелы и бабочки, колибри и изредка летучими мышами. Роль лепестков цветка заключается в приманивании этих организмов, которые несут пыльцу, и для этой цели растения развили модификации окраски цветков, формы и характера окраски цветков.
Поскольку окраска цветков является также наиболее важным признаком цветков для декоративных целей, цветки различных окрасок традиционно выводили скрещиванием. Однако один сорт растений редко имеет разные окраски цветков, и, например, скрещивание не давало каких-либо пурпурных - синих сортов для розы (Rosa hybrida), гвоздики (Dianthus caryophyllus), хризантемы (Chrysanthemum morifolium) или лилии (Lilium spp.) или ярко-красных сортов для Японского садового ириса (Iris ensata Thumb.) или горечавки (Gentiana triflora).
Светло-желтая - красная или синяя окраски цветков обычно обусловлены присутствием флавоноидов и антоцианинов (окрашенных глюкозидов, принадлежащих к классу флавоноидов). Флавоноиды являются вторичными метаболитами растений, и они имеют основной скелет макромолекулы C6C3C6 и синтезируются из фенилаланина и малонил-СоА, как показано ниже. Они классифицируются как флавоны, флавонолы и т.д. в соответствии с окислительными состояниями С-колец.
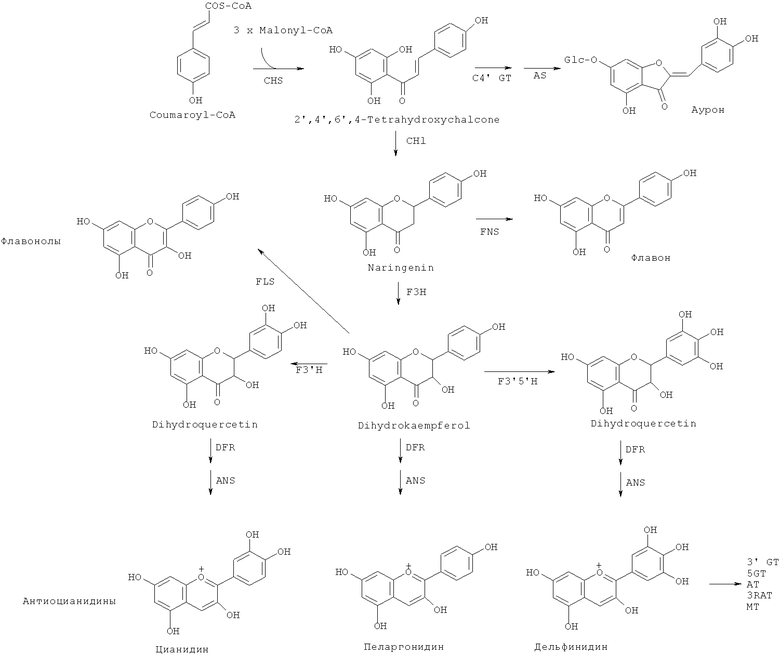
3×Malonyl-CoA-3 × Малонил-СоА; Coumaroyl-CoA-Кумароил-СоА;
2',4',6',4-Tetrahydroxychalcone-2',4',6',4-тетрагидроксихалькон; Naringenin-Нарингенин;
Dihydroquercetin-Дигидрокверцетин, Dihydrokaempferol-Дигидрокампферол;
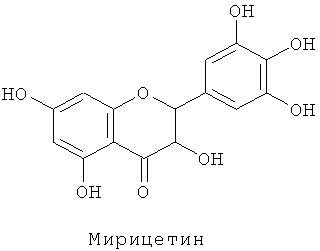
Dihydromyricetin-Дигидромирицетин

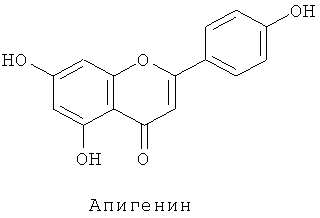
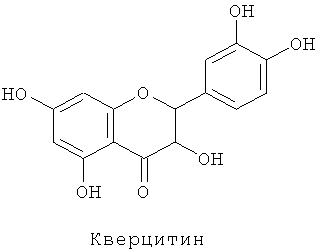
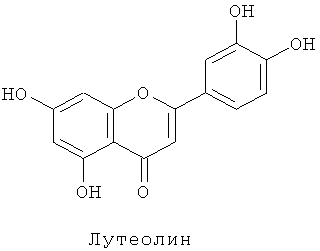

Флавоноиды поглощают ультрафиолетовые лучи и удаляют радикалы, и, следовательно, считается, что они являются защитой растительных организмов от различных форм стресса. В последние годы они также привлекли внимание как компоненты, важные для здоровья (см. Harborne and Williams 2000 Phytochemistry 55, 481-504).
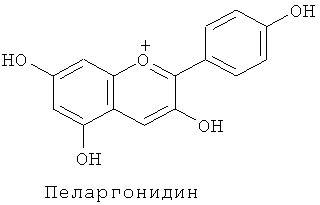
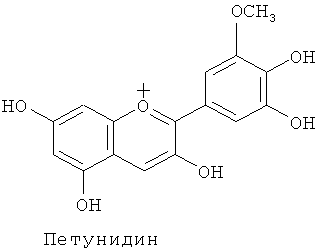
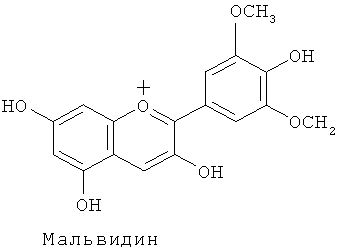
Известны несколько сотен молекулярных разновидностей окрашенных антоцианинов, и среди хромофорных антоцианидинов наиболее обычными являются следующие 6 типов: (1) пеларгонидин, изобилующий в оранжевых - красных цветках, (2) цианидин и пеонидин, изобилующие в красных - темно-красных (малиновых) цветках, и (3) дельфинидин, петунидин и мальвидин, изобилующие в фиолетовых - синих цветках.



Структура антоцианина оказывает большое влияние на окраску. Увеличенное количество гидроксильных групп на В-кольце антоцианина приводит к большей степени синего цвета. Антоцианины типа дельфинидина являются более синими, чем антоцианины типа пеларгонидина и антоцианины типа цианидина. Биосинтез флавоноидов, в том числе антоцианинов, является высококонсервативным среди видов растений. Флавоноиды биосинтезируются в цитозоле и после добавления сахаров и ацильных групп они транспортируются в вакуоли и накапливаются (см. Tanaka et al. 2005 Plant Cell, Tissue and Organ Culture 80, 1-24 и Tanaka and Brugliera 2006 Ainsworth, ed. Flowering and its manipulation, pp.201-239, Blackwell Publishing Ltd.).
Все структурные гены ферментов, участвующих в этом биосинтезе, были клонированы. Таким образом, создание рекомбинантных растений делает возможной модификацию структур и количеств флавоноидов, которые накапливаются в цветках, посредством искусственной экспрессии их генов, изменяя посредством этого окраску цветков (Tanaka et al., Plant Cell, Tissue and Organ Culture 80, 1-24, Tanaka and Brugliera 2006 Ainsworth, ed. Flowering and its manipulation, pp.201-239, Blackwell Publishing Ltd.). Например, для гвоздик или роз, которые не могут продуцировать дельфинидин в лепестках, ген флавоноид-3',5'-гидроксилазы (далее сокращаемой здесь как "F3'5'H"), необходимый для синтеза дельфинидина, экспрессировали с получением дельфинидина для получения искусственного синего цветка (см. Tanaka 2006 Phytochemistry Reviews 5, 283-291).
Такие способы искусственной модификации метаболизма растений называют иногда «метаболической инженерией». Для модификации метаболизма для накапливания представляющего интерес вещества возможна экспрессия гена для фермента, который продуцирует это представляющее интерес вещество в рекомбинантном растении, но во многих случаях конкуренция с эндогенными ферментами того же самого растения приводит к малому накапливанию или к абсолютному отсутствию накапливания представляющего интерес вещества, и это часто приводит к редкому применению в промышленности.
Например, петунии (Petunia hybrida) не накапливают пеларгонидин вследствие специфичности дигидрофлавонолредуктазы (далее сокращаемой здесь как "DFR") и, следовательно, не существуют природные сорта с окрашенными в оранжевый цвет цветками.
Хотя сообщались оранжевые петунии, которые накапливают пеларгонидин посредством переноса гена DFR из роз или т.п., накапливание пеларгонидина требует применения сортов петунии, не имеющих генов для флавоноид-3'-гидроксилазы (далее сокращаемой здесь как "F3'H"), F3'5'H и флавонолсинтазы (далее сокращаемой здесь как "FLS"), которые конкурирует с DFR, так как не наблюдается изменение в фенотипе, когда ген DFR розы переносят в петунии, которые не лишены этих генов (см. Tanaka and Brugliera 2006 Ainsworth, ed. Flowering and its manipulation, pp.201-239, Blackwell Publishing Ltd.). Таким образом, нельзя предсказать, будет ли представляющее интерес соединение накапливаться с проявлением желаемого фенотипа просто вследствие переноса представляющего интерес гена.
Кроме того, метаболическая инженерия часто производит непредсказуемые результаты. Например, при ингибировании экспрессии гена флавонсинтазы в торении (Torenia hybrida) содержание флавонов уменьшалось и наблюдалось накапливание флаванонов. Можно было бы ожидать, что накапливание флаванонов приведет к увеличенному содержанию антоцианина, но в действительности содержание антоцианина уменьшалось (Ueyma et al. Plant Science, 163, 253-263, 2002). Таким образом, трудно предсказать изменения в метаболитах и необходимы стойкие модификации для получения желаемых фенотипов.
Антоцианины, связанные с большими количествами ароматических ацильных групп, являются, по-видимому, более синими вследствие эффекта внутримолекулярного копигмента. Антоцианины с двумя или более ароматическими ацильными группами известны как полиацилированные антоцианины, и они проявляют стабильную синюю окраску (см. Harborne and Williams 2000 Phytochemistry 55, 481-504).
Окраска цветка изменяется не только посредством структуры самих антоцианиновых пигментов в качестве незаменимых пигментов, но обусловлена также одновременно присутствующими флавоноидами (также известными как копигменты), ионами металлов и pH вакуолей. Например, флавоны или флавонолы являются типичными копигментами, которые образуют сэндвич-подобную упаковку с антоцианинами и делают антоцианины более синими и усиливают окрашивающие эффекты (см. Goto (1987 Prog. Chem. Org. Natl. Prod. 52). Таким образом, флавоны могут рассматриваться как бесцветные копигментные компоненты. Например, изовитексин, тип флавона, обнаруживает копигментный эффект в отношении антицианинов в Японском садовом ирисе (Iris ensata Thunb.). Изовитексин также стабилизирует антоцианины, производя таким образом стабилизирующее действие, эффект на окраску цветков Японского садового ириса (см. Yabuya et al. Euphytica 2000 115, 1-5).
Флавоны обычно проявляют более сильные копигментные эффекты, чем флавонолы. Например, анализ генетически модифицированных гвоздик показал более сильный копигментный эффект для флавонов, чем для флавонолов (см. Fukui et al. Phytochemistry, 63, 15-23, 2003). Таким образом, накапливание флавонов является важным для создания синей окраски цветков. Однако не все растения могут продуцировать флавоны, и известно, что розы и петунии не накапливают флавоны. Известно, что кроме окраски цветков флавоны играют роль в поглощении ультрафиолетовых лучей, препятствуя различным типам стресса, и взаимодействии с микроорганизмами, и что растения с новыми признаками могут быть получены посредством синтеза флавонов (в качестве патентного документа, касающегося гена, кодирующего флавонсинтазу, см. публикацию нерассмотренного Японского патента №2000-279182). Однако примеры флавон-экспрессирующих роз до сих пор еще неизвестны.
Флавоны синтезируются из флаванонов реакцией, катализируемой флавонсинтазой. Конкретно, апигенин синтезируется из нарингенина, лутеолин синтезируется из эриодиктиола и трицетин синтезируется из пентагидроксифлаванона. Флавонсинтаза существует в двух формах, флавонсинтаза I и флавонсинтаза II. Обе катализируют одну и ту же реакцию, но являются ферментами разного типа. Флавонсинтаза I является зависимой от 2-оксоглутаровой кислоты диоксигеназой (см. British et al. (1981) Z.Natutforsch 36 с pp.742-750 и British (1990) Arch. Biochem. Biphys. 282 pp.152-160), тогда как флавонсинтаза II является монооксигеназой типа цитохрома Р450. Структурный ген флавонсинтазы II может быть получен из торении, львиного зева, периллы (Perilla frutescens), герберы (Gerbera hybrida) и горечавки (см. Tanaka and Brugliera 2006 Ainsworth, ed. Flowering and its manipulation, pp.201-239, Blackwell Publishing Ltd.).
Синтез флавонов является предсказуемым, когда ген флавонсинтазы экспрессируется в генетически модифицированных растениях, которые не продуцируют флавоны. Однако при экспрессии гена флавонсинтазы торении в петуниях сообщалось, что глубокая фиолетовая окраска цветка становится слабой (Tsuda et al. Plant Biotechnology, 21, 377-386, 2004). Сообщалось также, что экспрессия полученного из горечавки гена флавонсинтазы в табаке приводит к синтезу флавонов, но также приводит к более слабой окраске цветков (Nakatsuka et al. 2006, Molecular Breeding 17:91-99). Таким образом, синюю окраску цветков не всегда получают даже при синтезе флавонов. Причиной отсутствия копигментного эффекта могло бы быть неподходящее соотношение содержаний антоцианина и флавона или неподходящая модификация этих антоцианинов и флавонов сахарами и ацильными группами. Эти результаты предполагают, что невозможно увеличивать синюю окраску цветков просто экспрессией гена флавонсинтазы и накапливанием флавонов.
Розы являются наиболее популярными среди цветущих растений и их культивировали с древних времен. Искусственно модифицированные сорта также получили в последние несколько сотен лет. Таким образом, были получены розы, содержащие флавоноиды, такие как пеларгонидин, цианидин и флавонолы. В недавние годы также создавали розы способами генетической модификации для получения дельфинидина, который природно не обнаруживается в розах. Однако накапливающие флавон розы еще не были получены ни скрещиванием, ни генетической модификацией. Кроме того, еще не были получены розы, которые накапливают как флавон, так и мальвидин.
Сущность изобретения
Главным преимуществом использования генетической модификации для селекции растений является то, что в отличие от скрещивания она делает возможными модификации растений, которые не могут быть достигнуты скрещиванием, и модификации с использованием генов из других организмов. То есть генетическая модификация позволяет переносить любой ген организма другого вида в растение, такое как роза, для придания новой способности этому растению. Однако в отличие от модельных растений, таких как Arabidopsis (Arabidopsis thaliana) и табак (Nicotiana tabacum L.), функционирование перенесенных генов в розах в значительной степени зависит от используемых источника этого гена и промотора.
Согласно WO 2005/017147 перенос гена флавоноид-3',5'-гидроксилазы (F3'5'H) в розы не приводил к экспрессии в генетически модифицированной розе и к детектированию дельфинидина, когда этот ген получали из петунии или горечавки, но, неожиданно, при получении этого гена из фиалки трехцветной (анютиных глазок) он экспрессировался и придавал розам новую способность продуцировать дельфинидин. Таким образом, в розах невозможно легко сделать вывод о том, какие гены и полученные из каких сортов растений гены будут функционировать при переносе.
При перенесении гена в хризантемы также трудно предсказать, какой ген будет функционировать в хризантеме, и известно, что перенесенные гены теряют их функцию при старении рекомбинантной хризантемы. Сообщалось, что промотор 35S вируса мозаичной болезни цветной капусты, который часто используют для переноса чужеродных генов в рекомбинантных растениях, функционирует в горечавке (см. Mishiba et al. Plant Journal 2005, 44:541-556).
Хотя можно предположить, что синтез флавонов в розах может быть легко достигнут экспрессией гена флавонсинтазы, нелегко предсказать, будет ли экспрессироваться флавонсинтаза диоксигеназного типа или типа цитохрома Р450, или какое растение-источник должно быть использовано для гена флавонсинтазы, и, следовательно, обязательным является способ проб и ошибок (способ подбора). Этот копигментный эффект является феноменом, получаемым при одновременном присутствии антоцианинов и флавонов или флавонолов в определенной концентрации в вакуолях, и было продемонстрировано, что это требует подвергания флавоновых или флавоноловых копигментов гликозилированию или другой модификации, более приспособленной к условиям гликозилирования, или другой модификации источников антоцианиновой окраски (см. Nature, 2005 Aug 11; 436 (7052):791 и Nature, 358, 515-518 (1992)).
Для экспрессии необходимого тона окраски необходимо, чтобы антоцианины и флавоны/флавонолы находились в оптимальной структурной комбинации, и это требует использования способа проб и ошибок в отношении того, какой сорт модификаций должен существовать в одновременно присутствующих антоцианинах и флавонах/флавонолах. Кроме того, поскольку флаваноны, такие как нарингенин, быстро гидроксилируются флаванон-3-гидроксилазой (далее сокращаемой здесь как "F3H") в природных розах, флавоны необязательно синтезируются из флаванонов, даже если флавонсинтаза функционирует в этой розе.
Таким образом, целью данного изобретения является обеспечение роз, содержащих подходящие пигменты, для экспрессии желаемых оттенков окраски в этих розах.
В результате обширного исследования, направленного на решение вышеупомянутых проблем, авторы данного изобретения выполнили это изобретение после обнаружения, что экспрессия желаемого оттенка окраски может выполняться искусственным добавлением флавонов и мальвидина розам.
Конкретно, данное изобретение обеспечивает следующее:
1. Роза, содержащая флавон и мальвидин, включенные с помощью генетической модификации.
2. Роза, которая содержит флавон и мальвидин вследствие экспрессии флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы фиалки трехцветной (Viola × wittrockiana).
3. Роза, которая содержит мальвидин, флавон и дельфинидин вследствие экспрессии гена антоцианин-метилтрансферазы, гена флавонсинтазы и гена флавоноид-3',5'-гидроксилазы фиалки трехцветной (Viola wittrockiana).
4. Роза, в которой ген флавонсинтазы является геном флавонсинтазы, полученным из семейства Scrophulariaceae.
5. Роза, в которой ген флавонсинтазы, полученный из семейства Scrophulariaceae, является геном флавонсинтазы, полученным из львиного зева семейства Scrophulariaceae (Scrophulariaceae, Antirrihnum majus).
6. Роза, в которой ген флавонсинтазы, полученный из семейства Scrophulariaceae, является геном флавонсинтазы, полученным из торении семейства Scrophulariaceae (Scrophulariaceae, Torenia hybrida).
7. Роза, в которой ген, полученный из львиного зева семейства Scrophulariaceae, является геном, кодирующим:
(1) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 2,
(2) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 2, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами,
(3) флавонсинтазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 2, или
(4) флавонсинтазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 1, при условиях высокой строгости.
8. Роза, в которой ген, полученный из торении семейства Scrophulariaceae, является геном, кодирующим:
(1) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 4,
(2) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 4, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами,
(3) флавонсинтазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 4, или
(4) флавонсинтазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 3, при условиях высокой строгости.
9. Роза, в которой ген флавоноид-3',5'-гидроксилазы фиалки трехцветной является геном, кодирующим:
(1) флавоноид-3',5'-гидроксилазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 8,
(2) флавоноид-3',5'-гидроксилазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 8, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой другими аминокислотами,
(3) флавоноид-3',5'-гидроксилазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 8, или
(4) флавоноид-3',5'-гидроксидазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 7, при условиях высокой строгости.
10. Роза, в которой ген антоцианин-метилтрансферазы является геном, кодирующим:
(1) метилтрансферазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 10,
(2) метилтрансферазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 10, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой другими аминокислотами,
(3) метилтрансферазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 10, или
(4) метилтрансферазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 9, при условиях высокой строгости.
11. Роза, в которой окраска цветков изменена относительно хозяина перед переносом гена антоцианин-метилтрансферазы, гена флавонсинтазы и гена флавоноид-3',5'-гидроксилазы фиалки трехцветной.
12. Роза, в которой изменение окраски цветков является изменением в направлении синей окраски.
13. Роза, в которой изменение окраски цветков является таким изменением, что угол θ цветового тона (оттенка) в соответствии с диаграммой цветности системы цвета L*a*b* приближается к 270°, что является синей осью цветового тона.
14. Роза, в которой изменение окраски цветков является таким изменением, что минимальная величина спектра отражения лепестка смещается в направлении стороны более длинных длин волн.
15. Часть розы, потомок, ткань, вегетативное тело или клетка, имеющие те же самые свойства, что и роза, охарактеризованная выше.
16. Способ модификации окраски цветков розы посредством эффекта копигментации, продуцируемого введением флавона и мальвидина с помощью генетической модификации.
17. Способ, в котором эффект копигментации является эффектом изменения окраски цветков в направлении синего цвета.
Раскрытие изобретения
Термин "роза", в применении ко всему данному описанию, является общим названием декоративного растения, которое является лиственным кустарником отряда Rosales, семейства Rosaceae, рода Rose, и он не ограничивается каким-либо конкретным сортом и включает в себя цельное растение или его части, обычно содержащие цветок.
Ссылка на часть, потомка, ткань, вегетативное тело или клетку "розы", в применении ко всему данному описанию, обозначает любую часть, происходящую из "розы", пока она сохраняет желаемый генетический признак "розы" в соответствии с данным изобретением, и она не ограничивается какой-либо конкретной частицей.
Фраза "условия высокой строгости", в применении ко всему данному описанию, обозначает, например, условия нагревания антисмысловой цепи и нуклеиновой кислоты-мишени на протяжении ночи при 55°С в растворе, содержащем 6×SSC (состав 1×SSC: 0,15 М NaCl, 0,015 М цитрат натрия, pH 7,0), 0,5% ДСН, 5 × среда Денхардта, 100 мкг/мл денатурированной фрагментированной ДНК спермы лосося и 50% формамид, и промывание при условиях 0,1 × SSC при 60°С или более, и, конкретно, она относится к любым условиям, при которых неспецифический сигнал фона по существу отсутствует.
Фраза "угол цветового тона (оттенка) θ в соответствии с диаграммой цветности системы цвета L*a*b*", в контексте всего данного описания, относится к углу цветового тона (θ), стандартизованного в 1976 году Commission Internationale de l'eclairage (CIE) и заимствованного в Японии в виде JIS8729, где 0° является красным направлением, 90°С является желтым направлением, 180°С является зеленым направлением и 270°С является синим направлением. Окраска цветков может быть представлена комбинацией этих углов цветового тона и данными цветовой диаграммы (таблицы цветов) RNS (Royal Horticultural Society).
Перенос гена флавонсинтазы, гена флавоноид-3',5'-гидроксилазы и гена антоцианин-метилтрансферазы
Ген флавонсинтазы II, происходящий из периллы, переносили в розу посредством известной процедуры, вместе с геном F3'5'H фиалки трехцветной (анютиных глазок). В результате, флавоны не были детектированы в розах, в которые был перенесен ген флавонсинтазы II периллы, что указывает на то, что этот ген не функционирует в розе. С другой стороны, флавон детектировали в розах, в которые были перенесены гены флавонсинтазы II торении или львиного зева, что указывает на то, что эти гены действительно функционируют в розе.
Таким образом, были созданы накапливающие флавоны розы, не обнаруживаемые в известном уровне техники. Содержание флавона (%) от общих флавоноидов может быть равно 1% или более, предпочтительно 5% или более, более предпочтительно 10% или более и наиболее предпочтительно 30% или более. Розы, накапливающие как антоцианины, так и флавоны, имеют относительно более синие окраски в сравнении с розами, содержащими только те же самые антоцианины, что предполагает, следовательно, что накапливание флавонов вносит вклад в этот новый признак синего цвета (голубизны).
Кроме того, было обнаружено, что, когда присутствуют одновременно антоцианин с метилированным кольцом В (антоцианины, включающие в себя мальвидин) и флавон, обнаруживается более высокий эффект копигментации, чем в том случае, когда присутствуют одновременно дельфинидинсодержащий антоцианин и флавон, и что перенесением гена метилтрансферазы для кольца B, наряду с геном флавонсинтазы и геном F3'5'H фиалки трехцветной можно накапливать метилированные антоцианины типа дельфинидина и флавоны в лепестках розы, что приводит к более синей окраске лепестков розы.
Посредством гибридизационного теста было обнаружено, что признак накапливания как антоцианинов типа дельфинидина, так и флавонов передается потомству.
Эти открытия указывают на то, что в растениях, которые не накапливают флавоны или не накапливают метилированные антоцианины, такие как мальвидины, в лепестках, более синий оттенок цвета лепестков может быть получен вызыванием одновременного накапливания этих соединений.
Ферменты, ассоциированные с этим изобретением, являются обычно ферментами, имеющими конкретные аминокислотные последовательности, перечисленные в Списке последовательностей. Однако, хорошо известно, что желаемая активность фермента может сохраняться не только природной аминокислотной последовательностью фермента, но также той же самой аминокислотной последовательностью, имеющей модификации в районах, других, чем районы, ассоциированные с активностью фермента. Таким образом, ферменты этого изобретения включают в себя белки, имеющие аминокислотные последовательности, представленные этими номерами SEQ ID NO:, которые модифицированы добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами, с сохранением все еще активностей исходного фермента, а также белки, имеющие аминокислотные последовательности по меньшей мере с 90% идентичностью последовательности относительно конкретных аминокислотных последовательностей, представленных этими номерами SEQ ID NO:, с сохранением все еще активностей исходного фермента.
Известно, что для любого гена, кодирующего определенный фермент, имеется высокая вероятность того, что нуклеиновая кислота, которая гибридизуется с этим геном при условиях высокой строгости, будет кодировать фермент, имеющий ту же самую активность, что и этот фермент. Таким образом, ферменты, кодируемые нуклеиновыми кислотами, которые гибридизуются с нуклеиновыми кислотами, имеющими нуклеотидные последовательности, указанные этими номерами SEQ ID NO:, при условиях высокой строгости, имеющие желаемые активности фермента, также включены в качестве ферментов по данному изобретению.
Таким образом, следующие гены могут быть упомянуты в качестве генов ферментов, находящихся в объеме этого изобретения.
(A) Ген флавонсинтазы львиного зева (Antirrihnum majus)
Ген, кодирующий:
(1) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 2,
(2) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 2, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами,
(3) флавонсинтазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 2, или
(4) флавонсинтазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 1, при условиях высокой строгости.
(B) Ген флавонсинтазы торении (Torenia hybrida)
Ген, кодирующий:
(1) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 4,
(2) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 4, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами,
(3) флавонсинтазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 4, или
(4) флавонсинтазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 3, при условиях высокой строгости.
(C) Ген флавонсинтазы периллы (Perilla frutescens)
Ген, кодирующий:
(1) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 6,
(2) флавонсинтазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 6, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами,
(3) флавонсинтазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 6, или
(4) флавонсинтазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 5, при условиях высокой строгости.
(D) Ген 3',5'-гидроксилазы фиалки трехцветной (анютиных глазок) (Viola × wittrockiana)
Ген, кодирующий:
(1) 3',5'-гидроксилазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 8,
(2) 3',5'-гидроксилазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 8, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами,
(3) 3',5'-гидроксилазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 8, или
(4) 3',5'-гидроксилазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 7, при условиях высокой строгости.
(E) Ген метилтрансферазы торении (Torenia hybrida)
Ген, кодирующий:
(1) метилтрансферазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 10,
(2) метилтрансферазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 10, модифицированную добавлением или делецией одной или нескольких аминокислот и/или заменой одной или нескольких аминокислот другими аминокислотами,
(3) метилтрансферазу, имеющую аминокислотную последовательность по меньшей мере с 90% идентичностью относительно аминокислотной последовательности, представленной в списке как SEQ ID NO: 10, или
(4) метилтрансферазу, кодируемую нуклеиновой кислотой, которая гибридизуется с нуклеиновой кислотой, имеющей нуклеотидную последовательность SEQ ID NO: 9, при условиях высокой строгости.
Сведения, подтверждающие возможность осуществления изобретения
Данное изобретение будет теперь объяснено более подробно следующими примерами. Однако эти примеры представлены только для цели иллюстрации этого изобретения и не предназначены для ограничения каким бы то ни было образом этого изобретения.
Пример 1: Имитация копигментного эффекта флавона с антоцианинами
Антоцианины готовили сначала для имитации копигментного эффекта флавона с антоцианинами. Цианин экстрагировали и очищали из лепестков сорта розы "Красная роза" (культивара (cv.) розы "Rote Rose"). Дельфин получали щелочным гидролизом пигмента, экстрагированного из лепестков вербены "Tapien Violet" (культивара (cv.) вербены "Tapien Violet" или сорта вербены Sunmaref TP-V ("Tapien Violet") ("Tapien" является товарной маркой, зарегистрированной в Японии)), с последующей очисткой. Мальвин и лутеолин-7-О-глюкозид покупали из Funakoshi Corp.
Флавон (лутеолин-7-О-глюкозид) добавляли к каждому антоцианину, полученному таким образом, в концентрациях 0, 1, 2 и 4 молярных эквивалента в буферящем растворе при pH 5,0 и измеряли абсорбционные спектры (спектры поглощения). Использованными антоцианинами были цианин (цианидин-3,5-диглюкозид), дельфин (дельфинидин-3,5-диглюкозид) и мальвин (мальвидин-3,5-диглюкозид). Концентрации антоцианина для цианина, дельфина и мальвина были 1 мМ.
Как показано в таблицах 1 и 2, добавление флавона увеличивало оптическую плотность водных растворов антоцианина, и степень изменения (коэффициент оптической плотности) был наибольшим с мальвином. Максимумы поглощения (Амакс) также смещались в сторону длинных длин волн добавлением флавона. Степень изменения была наиболее высокой с мальвином и затем с дельфином. После оценивания величины оттенка цвета на основе системы цвета L*a*b* было обнаружено, что добавление флавона производит более синий оттенок цвета и увеличенную интенсивность окраски. Этот эффект был наиболее заметным с мальвином. То есть было продемонстрировано, что эффект копигментации лутеолин-7-О-глюкозида проявлялся в наивысшей степени с мальвином.


Пример 2 (ссылочный пример): Перенос гена F3'5'H#40 фиалки трехцветной (анютиных глазок) и гена флавонсинтазы периллы в сорт розы "Лаванда"
Содержащую ген флавонсинтазы периллы плазмиду pYFS3, описанную в публикации нерассмотренного Японского патента №2000-279182, расщепляли XbaI и затем концы затупляли и дополнительно расщепляли BamHI с получением фрагмента гена флавонсинтазы приблизительно 1 т.п.н. Отдельно, плазмиду pSPB906, описанную в WO 2005/017147, расщепляли XhoI и затем концы затупляли и дополнительно расщепляли BamHI. Этот фрагмент гена флавонсинтазы периллы инсертировали между тупыми концами и сайтом расщепления BamHI с получением плазмиды 906-pYES3. Плазмида 906-pYES3 содержит ген флавонсинтазы периллы между промотором Eb35S и терминатором D8 (оба описаны в WO 2005/017147).
Плазмиду, полученную инсертированием фрагмента гена F3'5'H#40 фиалки трехцветной (анютиных глазок), вырезанную из pCGP1961, описанной в WO 2005/017147, частичным расщеплением BamHI и XhoI, в сайтах BamHI и SaiI pSPB 176, сообщенной Ueyama et al. (Ueyama et al., Plant Science, 163, 253-263, 2002), обозначали как pSPB575. В сайте AscI этой плазмиды была инсертирована кассета экспрессии флавонсинтазы периллы приблизительно 3,4 т.п.н., полученная расщеплением вышеупомянутой плазмиды 906-pYES3 с использованием AscI. Из полученных плазмид, вектор, имеющий экспрессионную кассету гена F3'5'H#40 и экспрессионную кассету флавонсинтазы периллы, связанные в одном и том же направлении, обозначали как pSPB1310. Эта плазмида конститутивно экспрессирует ген F3'5'H#40 фиалки трехцветной и ген флавонсинтазы периллы в растениях.
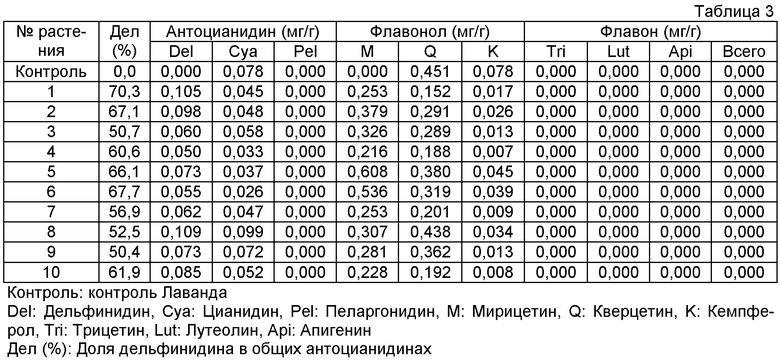
Плазмиду pSPB1310, сконструированную таким образом, переносили в розовато-лиловый сорт розы "Лаванда" и получали 55 трансформантов. Накапливание дельфинидина подтверждали в 49 из 50 анализированных на пигмент трансформантов, с максимальным содержанием дельфинидина 70% (среднее: 26%). Однако флавоны абсолютно не детектировались, и, следовательно, был сделан вывод, что ген флавонсинтазы периллы не функционирует в клетках розы.
Величины этого анализа для репрезентативных трансформантов показаны в таблице 3 ниже.
Пример 3: Перенос гена F3'5'H#40 фиалки трехцветной (анютиных глазок) и гена флавонсинтазы торении в сорт розы "Лаванда"
Плазмиду, полученную инсертированием гена флавонсинтазы торении, сообщенного Akashi et al. (Plant Cell Physiol 40, 1182-1186, 1999), в сайтах EcoPI и XhoI плазмиды pBlueScript II SK(-), обозначали как pSPB426. После расшепления этой плазмиды KpnI ее концы затупляли и затем ее дополнительно расщепляли BamHI с получением фрагмента гена флавонсинтазы торении приблизительно 1,7 т.п.н. Отдельно, pSPB906, описанную в WO 2005/017147, расщепляли XhoI и затем концы затупляли и плазмиду дополнительно расщепляли BamHI. Этот фрагмент гена флавонсинтазы торении инсертировали между тупыми концами и сайтом расщепления BamHI с получением плазмиды 906-426.
Плазмиду, полученную инсертированием фрагмента гена F3'5'H#40 фиалки трехцветной, вырезанного из pCGP1961, описанной в WO 2005/017147, частичным расщеплением BamHI и XhoI, в сайтах BamHI и SalI pSPB176, сообщенной Ueyama et al. (Ueyama et al., Plant Science, 163, 253-263, 2002), обозначали как pSPB575. В сайте AscI этой плазмиды была инсертирована кассета экспрессии флавонсинтазы торении приблизительно 3,3 т.п.н., полученная расщеплением вышеупомянутой плазмиды 906-426 с использованием AscI. Из полученных плазмид вектор, имеющий экспрессионную кассету гена F3'5'H#40 и экспрессионную кассету флавонсинтазы торении, связанные в одном и том же направлении, обозначали как pSPB1309. Эта плазмида конститутивно экспрессирует ген F3'5'H#40 фиалки трехцветной (анютиных глазок) и ген флавонсинтазы торении в растениях.
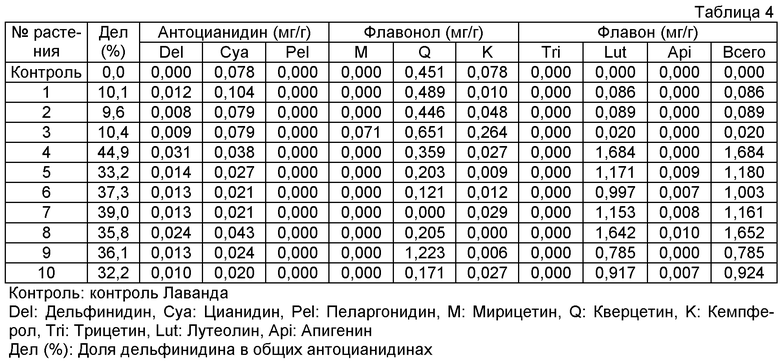
Плазмиду pSPB1309, сконструированную таким образом, переносили в розовато-лиловый сорт розы "Лаванда" и получали 50 трансформантов. Накапливание дельфинидина подтверждали в 36 из 38 анализированных на пигмент трансформантов, с максимальным содержанием дельфинидина 45% (среднее: 12%). Подтверждали также новое накапливание флавонов (лутеолина и апигенина) в 35 трансформантах вследствие действия гена флавонсинтазы торении. Максимально, общее количество флавонов было высоким содержанием 1,68 мг на 1 г сырой массы лепестков.
Величины этого анализа для репрезентативных трансформантов показано в таблице 4 ниже.
Пример 4: Перенос гена F3'5'H#40 фиалки трехцветной (анютиных глазок) и гена флавонсинтазы торении в сорт розы "WKS124"
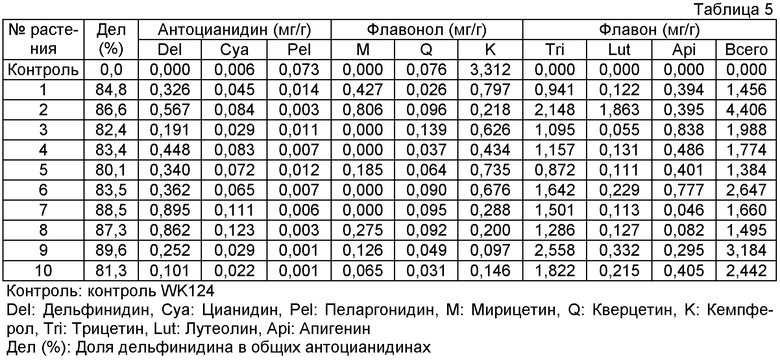
Плазмиду pSPB1309, описанную в примере 3, переносили в оранжево-розовый сорт розы "WK124" и получали 40 трансформантов. Накапливание дельфинидина подтверждали в 26 из 28 анализированных на пигмент трансформантов, с максимальным содержанием дельфинидина 96% (среднее: 81%). Подтверждали также новое накапливание флавонов (трицетина, лутеолина и апигенина) в 26 трансформантах вследствие действия гена флавонсинтазы торении. Максимально, общее количество флавонов было высоким содержанием 4,41 мг на 1 г сырой массы лепестков.
Величины этого анализа для репрезентативных трансформантов показано в таблице 5 ниже.
Пример 5: Перенос гена F3'5'H#40 фиалки трехцветной (анютиных глазок), гена флавонсинтазы торении и гена антоцианин-метилтрансферазы в сорт розы "WKS124"
Плазмиду pSPB1309, описанную в примере 3, обрабатывали PacI для расщепления в сайте PacI, присутствующем вблизи точки связывания между экспрессионной кассетой флавонсинтазы торении и экспрессионной кассетой гена F3'5'H#40 фиалки трехцветной (более конкретно, расположенной вблизи 3'-конца терминатора D8 экспрессионной кассеты флавонсинтазы), и в сайте PacI в векторе множественного клонирования, для вырезания экспрессионной кассеты F3'5'H#40 фиалки трехцветной.
Отдельно, бинарный вектор pSPB1530, имеющий экспрессионную кассету гена метилтрансферазы торении, описанную в WO 2003-062428, разрезали PacI и вышеупомянутую экспрессионную кассету F3'5'H#40 фиалки трехцветной инрсертировали в нее в том же самом направлении, что и направление в экспрессионной кассете гена метилтрансферазы. Эту плазмиду обозначали как ТМТ-ВР40.
Отдельно, плазмиду pSPB1309 расщепляли AscI для вырезания экспрессионной кассеты флавонсинтазы торении. Ее инсертировали в сайт AscI ТМТ-ВР40 в том же самом направлении, что и направления предыдущих экспрессионных кассет, и полученную плазмиду обозначали как pSEL535. Эта плазмида конститутивно экспрессирует ген F3'5'H#40 фиалки трехцветной, ген метилтрансферазы торении и ген флавонсинтазы торении в растениях.
Плазмиду pSEL535, полученную таким образом, переносили в оранжево-розовый сорт розы "WK124" и получали 173 трансформанта. Накапливание мальвидина (антоцианидина, который был метилирован в 3'- и 5'-положениях дельфинидина) подтверждали в 88 из 98 анализированных на антоцианидин трансформантов, и присутствие продукта указывало на то, что ген F3'5'H#40 фиалки трехцветной и ген антоцианин-метилтрансферазы торении функционировали в лепестках розы. Содержание мальвидина было равно максимально 84% (среднее: 50%).
Подтверждали также новое накапливание флавонов (трицетина, лутеолина и апигенина) в 77 трансформантах вследствие действия гена флавонсинтазы торении. Максимально, общее количество флавонов было высоким содержанием 4,58 мг на 1 г сырой массы лепестков. Метилированный трицетин детектировали в 51 трансформанте.
Величины этого анализа для репрезентативных трансформантов показано в таблице 6 ниже.

Пример 6: Перенос гена F3'5'H#40 фиалки трехцветной, гена флавонсинтазы торении и гена антоцианин-метилтрансферазы торении в сорт розы "Лаванда"
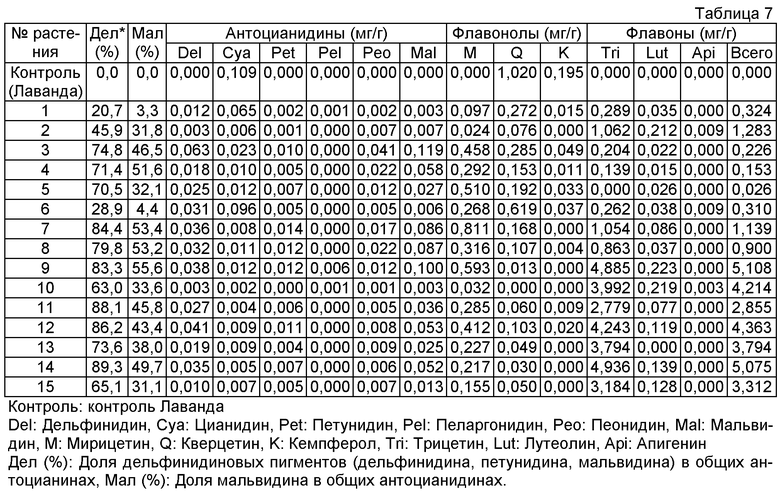
Плазмиду pSEL535, описанную в примере 5, переносили в розовато-лиловый сорт розы "Лаванда" и получали 130 трансформантов. Накапливание мальвидина (антоцианидина, который был метилирован в 3'- и 5'-положениях дельфинидина) подтверждали в 37 из 118 анализированных на антоцианидин трансформантов, и присутствие продукта указывало на то, что ген F3'5'H#40 фиалки трехцветной и ген антоцианин-метилтрансферазы торении функционировали в лепестках розы. Содержание мальвидина было равно максимально 55,6% (среднее: 20,5%).
Подтверждали также новое накапливание флавонов (трицетина, лутеолина и апигенина) в 78 трансформантах вследствие действия гена флавонсинтазы торении. Максимально, общее количество флавонов было высоким содержанием 5,11 мг на 1 г сырой массы лепестков. Кроме того, метилированный трицетин или лутеолин детектировали в 20 из продуцирующих флавон трансформантов.
Величины этого анализа для репрезентативных трансформантов показано в таблице 7 ниже.
Пример 7: Перенос гена F3'5'H#40 фиалки трехцветной (анютиных глазок), гена флавонсинтазы торении и гена антоцианин-метилтрансферазы торении в сорт розы "WKS82"
Плазмиду pSEL535, описанную в примере 5, переносили в розовато-лиловый сорт розы "WKS82" и получали 250 трансформантов. Накапливание мальвидина (антоцианидина, который был метилирован в 3'- и 5'-положениях дельфинидина) подтверждали в 110 из 232 анализированных на антоцианидин трансформантов и присутствие продукта указывало на то, что ген F3'5'H#40 фиалки трехцветной и ген антоцианин-метилтрансферазы торении функционировали в лепестках розы. Содержание мальвидина было равно максимально 65,2% (среднее: 19,7%).
Подтверждали также новое накапливание флавонов (трицетина, лутеолина и апигенина) в 125 трансформантах вследствие действия гена флавонсинтазы торении. Максимально, общее количество флавонов было высоким содержанием 4,71 мг на 1 г сырой массы лепестков. Кроме того, метилированный трицетин или лутеолин детектировали в 80 из продуцирующих флавон трансформантов.
Величины этого анализа для репрезентативных трансформантов показано в таблице 8 ниже.
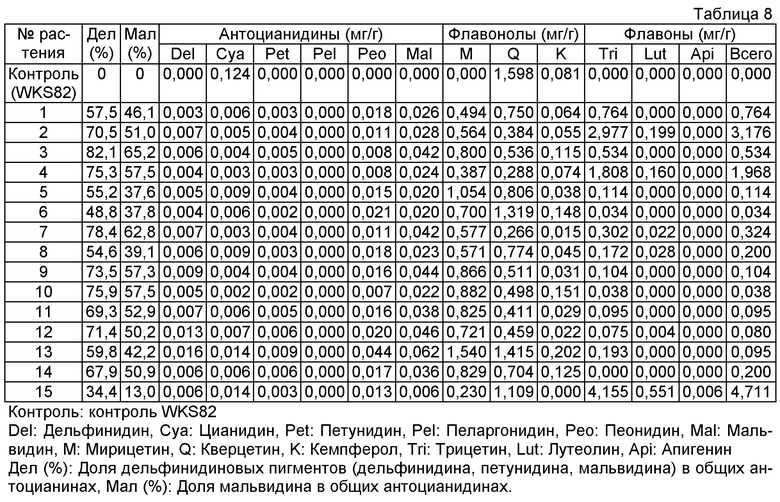
Пример 8: Перенос гена F3'5'H#40 фиалки трехцветной, гена флавонсинтазы торении и гена антоцианин-метилтрансферазы торении в сорт розы "WKS82"
Плазмиду pSEL535, описанную в примере 5, переносили в розовато-лиловый сорт розы "WKS82" и получали 74 трансформанта. Накапливание мальвидина (антоцианидина, который был метилирован в 3'- и 5'-положениях дельфинидина) подтверждали в 20 из 74 анализированных на антоцианидин трансформантов и присутствие продукта указывало на то, что ген F3'5'H#40 фиалки трехцветной и ген антоцианин-метилтрансферазы торении функционировали в лепестках розы. Содержание мальвидина было равно максимально 51,3% (среднее: 33,5%).
Подтверждали также новое накапливание флавонов (трицетина, лутеолина и апигенина) в 29 трансформантах вследствие действия гена флавонсинтазы торении. Максимально, общее количество флавонов было высоким содержанием 3,04 мг на 1 г сырой массы лепестков. Кроме того, метилированный трицетин или лутеолин детектировали в 20 из продуцирующих флавон трансформантов.
Величины этого анализа для репрезентативных трансформантов показано в таблице 9 ниже.
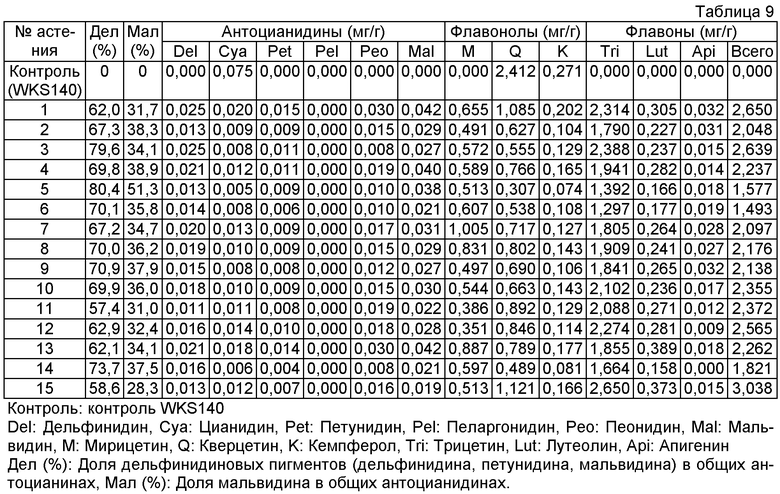
Пример 9: Распространение способности синтеза флавона и мальвидина на потомство - Гибридизация между культивируемыми розами и полученными генной инженерией розами, содержащими перенесенные ген F3'5'H#40 фиалки трехцветной, ген флавонсинтазы торении и ген антоцианин-метилтрансферазы
Для исследования способа наследования потомством способности синтеза флавонов в розах проводили скрещивание с использованием мальвидин- и флавон-продуцирующей розы, созданной в примере 5 (растение №6 в таблице 6), в качестве опылителя. В качестве родителя, образующего семена, использовали культивируемую розу «Медео» среднего размера (сорт Rose floribunda «Medeo»).
Накапливание мальвидина подтверждали в 7 из 10 анализированных на пигмент гибридных потомков F1 трансформантов, которые были получены, и присутствие продукта показало, что ген F3'5'H#40 фиалки трехцветной и ген антоцианин-метилтрансферазы торении функционируют в лепестках розы. Содержание мальвидина было равно максимально 68,2% (среднее: 46,6%).
С другой стороны, новое накапливание флавонов (трицетина, лутеолина и апигенина) подтверждали в 8 потомках трансформантов, вследствие действия гена флавонсинтазы торении. Максимально, общее количество флавонов было крайне высоким содержанием 7,35 мг на 1 г сырой массы лепестков. Кроме того, метилированный трицетин или лутеолин детектировали в 6 продуцирующих флавон потомках трансформантов.
Величины этого анализа для репрезентативных трансформантов показано в таблице 10 ниже.
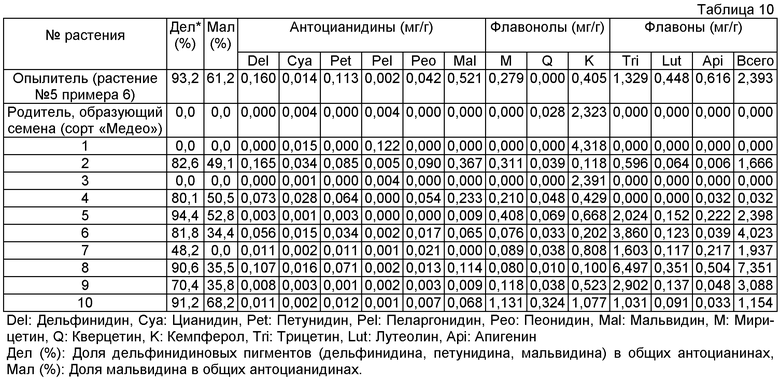
Пример 10: Распространение способности синтеза флавона и мальвидина на потомство
Скрещивание сорта розы "WKS123", содержащего перенесенный ген F3'5'H#40 фиалки трехцветной и ген антоцианин-метилтрансферазы торении, с сортом розы "Лаванда", содержащим перенесенный ген F3'5'H#40 фиалки трехцветной и ген флавонсинтазы торении.
Для исследования способа наследования потомством способности синтеза флавонов в розах проводили скрещивание с использованием флавон-продуцирующей линии, созданной в примере 3 (растение №4 в таблице 4), в качестве опылителя. В качестве родителя, образующего семена, использовали трансформант WKS124/1532-1-2-1 (описанный в WO 2003/062428), с высоким накапливанием мальвидина в лепестках вследствие переноса pSPB1532 в сорт розы WKS124 и получения действий гена F3'5'H#40 фиалки трехцветной и гена антицианин-метилтрансферазы торении.
После анализа на пигменты 149 из полученных потомков-трансформантов накапливание флавонов (трицетина, лутеолина, апигенина) подтверждали в 88 индивидуальных растений. Максимально, общее количество флавонов было высоким содержанием 4,09 мг на 1 г сырой массы лепестков. Кроме того, метилированный трицетин детектировали в 42 потомках, тогда как метилированный лутеолин (хризоэриол (3-Met-Lut)) детектировали в 11 потомках. Накапливание мальвидина подтверждали в 129 из 149 анализированных на пигменты потомков. Максимально содержание мальвидина было равно 79% (среднее: 36%).
Величины этого анализа для репрезентативных трансформантов показано в таблице 11 ниже.
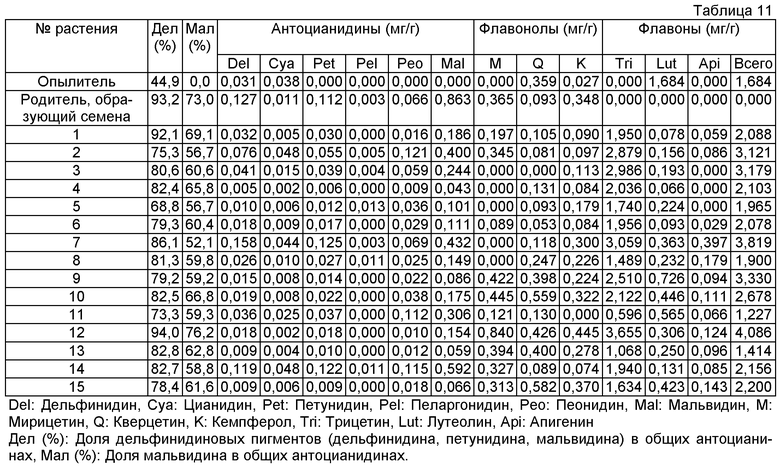
Пример 13: Оценивание окраски цветков флавонсодержащих роз
Трансформанты, созданные в примерах 4 и 5 (хозяин: сорт розы "WKS124"), делили на 5 группы: (1) трансформанты, накапливающие дельфинидин в качестве основного пигмента и не содержащие флавонов, (2) трансформанты, накапливающие дельфинидин в качестве основного пигмента и содержащие флавоны, (3) трансформанты, накапливающие мальфидин в высокой степени в качестве основного пигмента и не содержащие флавонов, (4) трансформанты, накапливающией в высокой степени мальвидин и содержащие флавоны, и (5) хозяин (накапливающий пеларгонидин в качестве основного пигмента), и оттенок окраски лепестков оценивали с использованием спектроколориметра (n=10).
Как в розах с дельфинидином в качестве основного пигмента, так и розах с мальвидином в качестве основного пигмента происходило смещение угла цветового тона (оттенка) лепестков в направлении синего цвета, когда одновременно присутствовали флавоны. Эта тенденция была более выраженной в розах с мальвидином в качестве основного пигмента, и минимум спектра отражения (Амин) также смещался значимо в направлении стороны длинных длин волн. Эти результаты подтвердили, что оттенок лепестков смещался к синему цвету одновременным присутствием флавонов. Эти результаты показаны в таблице 12 ниже.
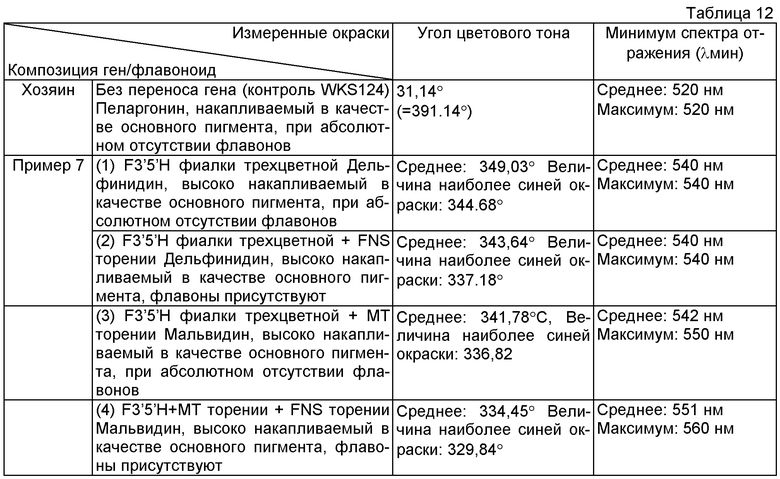
Угол цветового тона (цветовой тон): Смещение угла для цветового тона в направлении против часовой стрелки (влево) от оси цветового тона а* (красное направление) в качестве 0° в системе цвета L*a*b*, для указания положения цвета. Угол 90° является желтым направлением, угол 180° является зеленым направлением, угол 270° является синим направлением и угол 0° (=360°) является красным направлением. Другими словами, численная величина, приближающаяся к 270°, представляет более синий цветовой тон (оттенок).
Промышленная применимость
Согласно этому изобретению можно добавлять флавоны и дельфинидин генетической модификацией в розы в качестве популярных цветущих растений, используемых для декоративных целей, для изменения окраски цветков розы в направлении синего цвета посредством эффекта копигментаций. Ожидается, что розы с синей окраской цветков будут иметь высокий коммерческий спрос в качестве декоративных растений.
Все патентные документы и непатентные технические документы, цитируемые в данном описании, включены здесь в качестве ссылки либо по отдельности, либо в виде целого.
Это завершает объяснение данного изобретения, но это изобретение должно интерпретироваться как включающее в себя любые изменения или модификации, такие, которые не отклоняются от сущности этого изобретения, и объем этого изобретения не должен рассматриваться как объем, основанный на описании в примерах, а скорее определяется объемом прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ ГЕН ФЛАВОНОИД 3,5-ГИДРОКСИЛАЗЫ КОЛОКОЛЬЧИКА И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2640248C2 |
| ГЕН C-ГЛИКОЗИЛТРАНСФЕРАЗЫ ИЗ ГРЕЧИХИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2833446C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИЛИЙ, СОДЕРЖАЩИХ В ЛЕПЕСТКАХ ДЕЛФИНИДИН | 2011 |
|
RU2590722C2 |
| НОВЫЙ ГЕН ГЛИКОЗИЛТРАНСФЕРАЗЫ И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2636463C2 |
| НОВОЕ СОЕДИНЕНИЕ, СОДЕРЖАЩЕЕСЯ В ГОЛУБОЙ РОЗЕ | 2010 |
|
RU2507206C2 |
| ВЫДЕЛЕННЫЙ ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ПОЛИПЕПТИД, ВОВЛЕЧЕННЫЙ В БИОСИНТЕЗ ПИРИПИРОПЕНА А, ВЕКТОР И КЛЕТКА-ХОЗЯИН СОДЕРЖАЩИЕ ТАКОЙ ПОЛИНУКЛЕОТИД И СПОСОБ ПОЛУЧЕНИЯ ПРЕДШЕСТВЕННИКА ПИРИПИРОПЕНА А (ВАРИАНТЫ) | 2009 |
|
RU2540017C2 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ОСНОВАННОГО НА РЕКОМБИНАЦИИ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ ШАФРАНА | 2012 |
|
RU2676730C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ, СОДЕРЖАЩИЕ КЛАСТЕР ГЕНОВ БИОСИНТЕЗА ПИРИПИРОПЕНА И МАРКЕРНЫЙ ГЕН | 2011 |
|
RU2576001C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГИАЛУРОНАНА В РЕКОМБИНАНТНОЙ КЛЕТКЕ-ХОЗЯИНЕ | 2002 |
|
RU2346049C2 |
| ТРАНСГЕННЫЕ РАСТЕНИЯ С УЛУЧШЕННЫМИ ХАРАКТЕРИСТИКАМИ РОСТА | 2009 |
|
RU2582260C2 |













Изобретение относится к области биохимии. Представлена роза, цвет которой изменяется к синему, содержащая ген флавонсинтазы, полученный из семейства Scrophulariaceae, ген флавоноид-3',5'-гидроксилазы фиалки трехцветной (Viola × wittrockiana) и ген антоцианин-метилтрансферазы, аминокислотные последовательности которых представлены в описании, и где роза содержит: (a) флавон, вырабатываемый вследствие экспрессии гена флавонсинтазы, (b) делфинидин, вырабатываемый геном флавоноид-3',5'-гидроксилазы, и (c) мальвидин, вырабатываемый геном антоцианин-метилтрансферазы. Описаны часть, ткань, вегетативное тело и клетка розы, имеющие указанные гены. Предложен способ изменения окраски цветков розы к синему цвету, включающий введение в розу гена флавонсинтазы, полученного из семейства Scrophulariaceae, гена флавоноид-3',5'-гидроксилазы, полученного из фиалки трехцветной (Viola × wittrockiana), и гена антоцианин-метилтрансферазы, и включающий экспрессию генов флавонсинтазы, флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы. Изобретение позволяет изменить окраску лепестков розы в направлении синего цвета. 6 н. и 1 з.п. ф-лы, 12 табл., 13 пр.
1. Роза, чей цвет изменяется к синему, содержащая ген флавонсинтазы, полученный из семейства Scrophulariaceae, ген флавоноид-3',5'-гидроксилазы фиалки трехцветной (Viola × wittrockiana) и ген антоцианин-метилтрансферазы,
где ген флавонсинтазы, полученный из торении, кодирует:
(a) флавонсинтазу, имеющую аминокислотную последовательность SEQ ID NO: 4,
(b) флавонсинтазу, имеющую аминокислотную последовательность, по меньшей мере, с 50% идентичностью относительно аминокислотной последовательности SEQ ID NO: 4 и имеющую биологическую активность как у флавонсинтазы, определенной в (a), или
(c) флавонсинтазу, кодируемую нуклеиновой кислотой, которая в очень строгих условиях комплементарно гибридизуется с нуклеиновой кислотой с нуклеотидной последовательностью SEQ ID NO: 3, и имеющую такую же биологическую активность, что и флавонсинтаза, определенная в (a),
где ген флавоноид-3',5'-гидроксилазы кодирует:
(a) флавоноид-3',5'-гидроксилазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 8,
(b) флавоноид-3',5'-гидроксилазу, имеющую аминокислотную последовательность, по меньшей мере, с 50% идентичностью относительно аминокислотной последовательности SEQ ID NO: 8 и имеющую биологическую активность как у флавоноид-3',5'-гидроксилазы, определенной в (a), или
(c) флавоноид-3',5'-гидроксилазу, кодируемую нуклеиновой кислотой, которая в очень строгих условиях комплементарно гибридизуется с нуклеиновой кислотой с нуклеотидной последовательностью SEQ ID NO: 7, и имеющую такую же биологическую активность, что и флавоноид-3',5'-гидроксилаза, определенная в (a),
где ген антоцианин-метилтрансферазы является геном, кодирующим:
(a) метилтрансферазу, имеющую аминокислотную последовательность SEQ ID NO: 10,
(b) метилтрансферазу, имеющую аминокислотную последовательность, по меньшей мере, с 90% идентичностью аминокислотной последовательности SEQ ID NO: 10 и имеющую такую же биологическую активность, что и метилтрансфераза, определенная в (a), или
(c) метилтрансферазу, кодируемую нуклеиновой кислотой, которая в очень строгих условиях комплементарно гибридизуется с нуклеиновой кислотой с нуклеотидной последовательностью SEQ ID NO: 9, и имеющую такую же биологическую активность, что и метилтрансфераза, определенная в (a), и где роза содержит:
(a) флавон, вырабатываемый вследствие экспрессии гена флавонсинтазы,
(b) делфинидин, вырабатываемый геном флавоноид-3',5'-гидроксилазы, и
(c) мальвидин, вырабатываемый геном антоцианин-метилтрансферазы.
2. Часть розы, имеющая те же самые гены флавонсинтазы, флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы, что и роза по п.1.
3. Ткань розы, имеющая те же самые гены флавонсинтазы, флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы, что и роза по п.1.
4. Вегетативное тело розы, имеющее те же самые гены флавонсинтазы, флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы, что и роза по п.1.
5. Клетка розы, имеющая те же самые гены флавонсинтазы, флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы, что и роза по п.1.
6. Способ изменения окраски цветков розы к синему цвету, включающий введение в розу гена флавонсинтазы, полученного из семейства Scrophulariaceae, гена флавоноид-3',5'-гидроксилазы, полученного из фиалки трехцветной (Viola × wittrockiana), и гена антоцианин-метилтрансферазы, и включающий экспрессию генов флавонсинтазы, флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы,
где ген флавонсинтазы получен из торении, которая кодирует:
(a) флавонсинтазу, имеющую аминокислотную последовательность SEQ ID NO: 4,
(b) флавонсинтазу, имеющую аминокислотную последовательность, по меньшей мере, с 90% идентичностью аминокислотной последовательности SEQ ID NO: 4 и имеющую такую же биологическую активность, что и флавонсинтаза, определенная в (a), или
(c) флавонсинтазу, кодируемую нуклеиновой кислотой, которая в очень строгих условиях комплементарно гибридизуется с нуклеиновой кислотой с нуклеотидной последовательностью SEQ ID NO: 3, и имеющую такую же биологическую активность, что и флавонсинтаза, определенная в (a), где ген флавоноид-3',5'-гидроксилазы кодирует:
(a) флавоноид-3',5'-гидроксилазу, имеющую аминокислотную последовательность, представленную в списке как SEQ ID NO: 8,
(b) флавоноид-3',5'-гидроксилазу, имеющую аминокислотную последовательность, по меньшей мере, с 90% идентичностью с аминокислотной последовательностью SEQ ID NO: 8 и имеющую такую же биологическую активность, что и флавоноид-3',5'-гидроксилаза, определенная в (a), или
(c) флавоноид-3',5'-гидроксилазу, кодируемую нуклеиновой кислотой, которая в очень строгих условиях комплементарно гибридизуется с нуклеиновой кислотой с нуклеотидной последовательностью SEQ ID NO: 7 и имеющую такую же биологическую активность, что и флавоноид-3',5'-гидроксилаза, определенная в (a),
где ген антоцианин-метилтрансферазы является геном, кодирующим:
(a) метилтрансферазу, имеющую аминокислотную последовательность SEQ ID NO: 10,
(b) метилтрансферазу, имеющую аминокислотную последовательность, по меньшей мере, с 90% идентичностью аминокислотной последовательности SEQ ID NO: 10 и имеющую такую же биологическую активность, что и метилтрансфераза, определенная в (a), или
(c) метилтрансферазу, кодируемую нуклеиновой кислотой, которая в очень строгих условиях комплементарно гибридизуется с нуклеиновой кислотой с нуклеотидной последовательностью SEQ ID NO: 9, и имеющую такую же биологическую активность, что и метилтрансфераза, определенная в (a), и где роза содержит:
(a) флавон, вырабатываемый вследствие экспрессии гена флавонсинтазы,
(b) делфинидин, вырабатываемый геном флавоноид-3',5'-гидроксилазы, и
(c) мальвидин, вырабатываемый геном антоцианин-метилтрансферазы.
7. Способ по п.6, где роза в соответствии с диаграммой цветности системы цвета L·a·b демонстрирует угол цветового тона (Θ) более близко к 270°, чем роза, которая не содержит гены флавонсинтазы, флавоноид-3',5'-гидроксилазы и антоцианин-метилтрансферазы.
| FUKUI, et al., "A rationale for the shift in colour towards blue in transgenic carnation flowers expressing the flavonoid 3',5'-hydroxylase gene", Phytochemistry, 63, 2003, pp.15-23 | |||
| SHIONO, et al., "Phytochemistry: structure of the blue cornflower pigment", Nature, 2005, Aug., 11; 436 (7052): 791 | |||
| СПОСОБЫ И СОСТАВЫ ДЛЯ ИЗМЕРЕНИЯ УРОВНЯ СОДЕРЖАНИЯ ВТОРИЧНЫХ ПРОДУКТОВ ОБМЕНА ВЕЩЕСТВ В РАСТЕНИЯХ | 1999 |
|
RU2260050C2 |

