Область техники, к которой относится изобретение
Изобретение относится к применению фитоэкдизонов и полусинтетических производных фитоэкдизонов для лечения нервно-мышечных заболеваний, в частности спинальной мышечной атрофии детского возраста и бокового амиотрофического склероза.
Уровень техники
Нервно-мышечные заболевания характеризуются нарушением функции двигательных единиц, состоящих из моторных нейронов, нервно-мышечных синапсов и скелетных мышц. Независимо от происхождения заболевания, будь то нервные заболевания, такие как спинальная мышечная атрофия детского возраста или боковой амиотрофический склероз, или мышечные заболевания, все они вызывают изменение двигательной функции пациентов, которое может варьироваться от инвалидности до преждевременной смерти при поражении жизненно важных мышц. Лечение этих нервно-мышечных заболеваний на сегодняшний день все еще является симптоматическим и влечет существенные экономические затраты с учетом уровня инвалидности пациентов, прогрессирования этих патологий, а также связанных с этих потребностей в персонале и материалах. Исследования и разработка способов лечения, позволяющих облегчить моторные симптомы и зависимость пациентов, представляют определенный социальный и экономический интерес.
Среди этих нервно-мышечных заболеваний два описаны как специфически поражающие моторные нейроны: спинальная мышечная атрофия детского возраста (или СМА), симптомы которой проявляются в детстве, и боковой амиотрофический склероз (или БАС), симптомы которого возникают во взрослом возрасте. Эти два нейродегенеративных заболевания с разными причинами и клиническими проявлениями характеризуются общей прогрессирующей мышечной денервацией, ответственной за амиотрофию (Al-Chalabi и Hardiman, 2013; Crawford и Pardo, 1996).
Спинальная мышечная атрофия детского возраста представляет собой наиболее частую причину детской смертности, имеющую генетическое происхождение, с распространенностью от 1/6000 до 1/10 000 рождений (Crawford и Pardo, 1996). Описаны три основных типа по тяжести заболевания, в зависимости от возраста появления симптомов и прогрессирования клинических проявлений, от 1-го, наиболее тяжелого, до 3-го типа, продолжительность жизни при котором может составлять более 40 лет. Пациенты, страдающие СМА, имеют симметричное поражение скелетных мышц из-за атрофии мышечных волокон, изолированных или сгруппированных в пучки. Практически все виды СМА имеют проксимальный характер, т.е. преимущественно поражают мышцы туловища и мышцы, прилегающие к туловищу. Постепенно моторный дефицит распространяется на первой стадии на мышцы нижних конечностей, затем на второй стадии на мышцы верхних конечностей, преимущественно поражая мышцы-разгибатели. Несмотря на большую клиническую неоднородность, генетический анализ показал, что все формы СМА обусловлены мутацией гена выживания моторных нейронов SMN1, расположенного в хромосомной области 5q13.
В геноме человека имеется инвертированная центромерная копия этого гена, ген SMN2, который может быть обнаружен в нескольких копиях (Lorson et al., 1998), но который позволяет лишь частично компенсировать потерю функции гена SMN1. Действительно, SMN2 имеет 5 нуклеотидных отличий от SMN1, одно из которых находится в экзоне 7 и способствует его удалению путем сплайсинга 90% мРНК, продуцируемой геном SMN2. Этот альтернативный сплайсинг приводит к продукции усеченного и нестабильного белка SMNΔ7. Поэтому только 10% белков, продуцируемых геном SMN2, являются полными и функциональными (Vitte et al., 1997; Lefebvre et al., 1997). Таким образом, была продемонстрирована связь между количеством копий гена SMN2, их уровнем экспрессии и тяжестью заболевания. Белок SMN - это небольшой повсеместно распространенный белок, расположенный в дискретных доменах ядер, называемых «gems» от «Gemini of Coiled Bodies» («близнецы свернутых телец»), и в цитоплазме клеток. Хотя белок SMN выполняет специфические нейрональные функции, такие как рост аксонов (McWhorter et al., 2003) и аксональный транспорт (Akten et al., 2011; Peter et al., 2011), он также участвует в общих функциях, таких как биогенез ядерных рибонуклеопротеинов (Buhler et al., 1999; Liu et al., 1997; Zhang et al., 2008) или регуляция трансляции мРНК (Sanchez et al., 2013). Таким образом, ни экспрессия белка SMN, ни его клеточные функции не позволяют напрямую объяснить специфическую дегенерацию моторных нейронов.
Однако разработка различных животных моделей и успехи в фундаментальных исследованиях позволили продемонстрировать, что условная делеция гена Smn только в нейронах не воспроизводит полные симптомы СМА или дегенерацию моторных нейронов (Frugier et al., 2000). В других исследованиях описана роль белка SMN в правильном функционировании астроцитов (Rindt et al., 2015), клеток Шванна (Hunter et al., 2014), клеток сердца (Bevan et al., 2010; Heier et al., 2010; Shabadi et al., 2010; Biondi et al., 2010), клеток печени (Vitte et al., 2004) и сосудов (Somers et al., 2016), что позволяет сделать предположение о роли энергетического метаболизма в прогрессировании и тяжести заболевания. Другие дополнительные исследования были сосредоточены на двигательных единицах при СМА и выявили специфические изменения нервно-мышечных синапсов (Kariya et al., 2008; Kong et al., 2009; Muray et al., 2008; Biondi et al., 2008), важного фактора для выживания моторных нейронов. Действительно, было продемонстрировано, что белок SMN участвует в правильном функционировании нервно-мышечных синапсов, воздействуя на пул везикул нейротрансмиттеров, синаптическую активность (Torres-Benito et al., 2011), а также в созревании синапсов, что является важным шагом, позволяющим достичь функциональности и поддерживать ее (Kariya et al., 2008; Biondi et al., 2008). С другой стороны, отключение гена Smn только в мышечных клетках приводит к серьезным мышечным изменениям (Cifuentes-Diaz et al., 2001; Rajendra et al., 2007; Lee et al., 2011), таким как тяжелая дистрофия (Cifuentes-Diaz et al., 2001), плохая организация саркомерной структуры, обеспечивающей сокращение (Walker et al., 2008), снижение правильного слияния мышечных стволовых клеток (Nicole et al., 2003) и, следовательно, их дифференцировки (Shafey et al., 2005). Кроме того, миогенные клетки пациентов, страдающих СМА, бедны определенными факторами, такими как CANP (нейтральная протеаза, активируемая кальцием), необходимая для установления нервно-мышечных контактов (Fidzianska et al., 1984; Vrbova et al., 1989). Наконец, in vitro было обнаружено, что моторные нейроны при совместном культивировании с мышечными клетками SMA дегенерируют быстрее (Vrbova et al., 1989).
На сегодняшний день были рассмотрены очень разные подходы к разработке лечения СМА. Текущие стратегии исследований сосредоточены, по большей части, на модификации экспрессии SMN напрямую или опосредованно, либо путем повышения экспрессии гена SMN2, либо путем модификации сплайсинга транскриптов SMN2, или на генных терапиях, направленных на восстановление правильного гена. Некоторые исследования сосредоточены на подходах, независимых от SMN, путем разработки клеточной терапии, или cфокусированы на активирующей нейропротекции (олезоксим) или на улучшении мышечной функции (активатор тропонина).
Несмотря на продолжающиеся десятки клинических испытаний СМА, на сегодняшний день одобрены только два препарата.
Первый препарат, активатор тропонина (CK-2127107), направлен на повышение чувствительности саркомеров к кальцию, чтобы улучшить выносливость и производительность мышечной функции (Hwee et al., 2015). Это вещество недавно было признано контролирующими органами США орфанным лекарством от СМА (май 2017).
Второй препарат, антисмысловой олигонуклеотид (ASO), направлен на стимуляцию включения экзона 7 транскриптов, продуцируемых генами SMN2, что позволяет осуществлять сверхэкспрессию полного белка SMN клетками спинного мозга после интратекальных инъекций (Hache et al., 2016; Chiriboga et al., 2016). Этот препарат, разработанный компанией Biogen и получивший название Нусинерсен (Nusinersen), был одобрен американскими и европейскими регуляторными органами и представляет собой первый препарат для терапевтического применения, выпущенный на рынок для лечения тяжелой спинальной мышечной атрофии детского возраста 1-го и 2-го типа. Первые результаты, полученные с помощью ASO, нацеленного на интрон 7, на животных моделях (Hua et al., 2010; Passini et al., 2011) или во время клинических испытаний II и III фазы у пациентов с тяжелой СМА (Finkel et al., 2016), являются очень впечатляющими и многообещающими для этой смертельной болезни. Однако остается несколько недоисследованных областей в отношении использования олигонуклеотида in vivo, вводимого только в центральную нервную систему и/или в течение длительного времени. Прежде всего, научному сообществу не хватает ретроспективного взгляда на долгосрочную толерантность пациентов к этому типу экзогенных молекул, нацеленных на экспрессию генов. Кроме того, дополнительные вопросы относятся к рискам, присущим значительной и неконтролируемой сверхэкспрессии белка SMN, функция которого связана с пролиферацией клеток (Grice et al., 2011). Кроме того, эффективность нусинерсена кажется еще более важной, поскольку начало лечения связано с преждевременным вмешательством в прогрессирование заболевания, т.е. с клинической ситуацией, которой трудно добиться с учетом времени постановки диагноза и семейного анамнеза. Наконец, при СМА необходимо отметить важность роли, которую играют мульти-тканевые изменения в прогрессировании заболевания. Тем не менее, нусинерсен - это вещество, которое не проходит гематоэнцефалический барьер, что требует введения интратекальным путем, позволяя нацеливаться только на нейроны и глиальные клетки, не затрагивая другие органы, такие как мышцы, печень, поджелудочная железа и сосудистая система. Следовательно, их изменения могут ограничивать терапевтические эффекты нусинерсена. Наконец, для пациентов, страдающие СМА 3 типа, у которых уровень экспрессии белка SMN намного выше, и прогноз выживания редко зависит от него, эта терапия еще не получила разрешения, поэтому лечение заболевания этого типа не осуществляется.
Таким образом, представляется важным разработать физиологические и/или фармакологические подходы, дополняющие нусинерсен, чтобы усилить его защитные эффекты и максимально ограничить клинический курс у пациентов, их зависимость и клиническую помощь им.
Фитоэкдизоны представляют собой большое семейство полигидроксилированных стеринов. Эти вещества вырабатываются различными видами растений (папоротники, голосеменные, покрытосеменные) и участвуют в защите этих растений от вредителей. Главным фитоэкдизоном в растительном царстве является 20-гидроксиэкдизон.
В патенте FR3021318 раскрывается, что фитоэкдизоны и, в частности, 20-гидроксиэкдизон (20E) были предметом множества фармакологических исследований. Эти исследования выявили антидиабетические и анаболические свойства этого вещества. Его стимулирующее действие на синтез белка в мышцах наблюдается у крыс in vivo (Syrov et al., 2000; Toth et al., 2008; Lawrence et al., 2012) и на мышечные трубочки клеток C2C12 у мышей in vitro (Gorelick-Feldman et al., 2008). Некоторые из описанных выше эффектов на животных моделях были обнаружены в клинических исследованиях, но это все еще является редкостью. Так, 20-гидроксиэкдизон способствует увеличению мышечной массы у молодых спортсменов (Simakin et al., 1988). Наконец, во французском патенте FR3021318 дополнительно описано использование 20-гидроксиэкдизона и производных 20-гидроксиэкдизона для лечения и профилактики саркопении и саркопенического ожирения (Lafont et al., 2017).
Изложение сущности изобретения
Целью настоящего изобретения является ограничение потери моторных нейронов, связанной с нервно-мышечными заболеваниями, а также последствий их дегенерации.
Фитоэкдизоны представляют собой большое семейство фитополигидроксилированных стеринов, структурно схожих с гормонами линьки насекомых. Эти молекулы вырабатываются многими видами растений и участвуют в их защите от насекомых-вредителей. Главным фитоэкдизоном является 20-гидроксиэкдизон.
Авторы изобретения неожиданно обнаружили, что фитоэкдизоны и полусинтетические производные фитоэкдизонов значительно улучшают выживаемость, а также изменение массы тела у млекопитающих, страдающих спинальной мышечной атрофией. Кроме того, фитоэкдизоны и их полусинтетические производные ограничивают мышечную атрофию и аплазию, присутствующие при этой патологии, и значительно ограничивают потерю моторных нейронов млекопитающих, страдающих спинальной мышечной атрофией.
Для этой цели изобретение относится к композиции, содержащей 20-гидроксиэкдизон и/или по меньшей мере одно полусинтетическое производное 20-гидроксиэкдизона, для применения в лечении специфического расстройства моторных нейронов у млекопитающих, страдающих нервно-мышечным заболеванием, включая изменение мышечной функции из-за специфического расстройства моторных нейронов.
В конкретных вариантах осуществления изобретение дополнительно соответствует следующим характеристикам, реализованным по отдельности или в любой из их технически возможных комбинаций.
20-гидроксиэкдизон и его производные предпочтительно очищают до фармацевтической степени чистоты.
Используемый 20-гидроксиэкдизон предпочтительно находится в форме растительного экстракта, богатого 20-гидроксиэкдизоном, или композиции, содержащей 20-гидроксиэкдизон в качестве активного ингредиента. Экстракты растений, богатые 20-гидроксиэкдизоном, представляют собой, например, экстракты Stemmacantha carthamoides (также называемой Leuzea carthamoides), Cyanotis arachnoidea и Cyanotis vaga.
Полученные экстракты предпочтительно очищают до фармацевтической степени чистоты.
В одном варианте осуществления 20-гидроксиэкдизон находится в форме экстракта растения или части растения, где указанное растение выбрано из растений, содержащих по меньшей мере 0,5% 20-гидроксиэкдизона по сухой массе указанного растения, при этом указанный экстракт включает по меньшей мере 95%, и предпочтительно по меньшей мере 97% 20-гидроксиэкдизона. Указанный экстракт предпочтительно очищают до фармацевтической степени чистоты.
В дальнейшем указанный экстракт упоминается как BIO101. Он отличается тем, что включает от 0 до 0,05% примесей по сухой массе экстракта в качестве второстепенных соединений, способных повлиять на безопасность, доступность или эффективность фармацевтического применения указанного экстракта.
Согласно одному варианту осуществления изобретения примеси представляют собой соединения с 19 или 21 атомом углерода, такие как рубростерон, дигидрорубростерон или постстерон.
Растение, из которого получают BIO101, предпочтительно выбирают из Stemmacantha carthamoides (также называемой Leuzea carthamoides), Cyanotis arachnoidea и Cyanotis vaga.
Производные 20-гидроксиэкдизона получают полусинтетическим способом, и они могут быть получены, в частности, способом, описанным в заявке на европейский патент ЕР 15732785.9.
Согласно предпочтительному варианту осуществления изменение мышечной функции обусловлено изменением функции моторных нейронов или их дегенерацией.
В одном варианте осуществления измененная мышечная функция представляет собой функцию поперечно-полосатых скелетных мышц или миокарда.
В одном варианте осуществления изменение мышечной функции связано с аплазией и/или атрофией.
В одном конкретном варианте осуществления расстройство моторных нейронов является результатом генетического изменения у млекопитающих, страдающих нервно-мышечным заболеванием.
Термин «генетическое изменение» означает мутацию, такую как замена или вставка нуклеотида (нуклеотидов) или делеция нуклеотида (нуклеотидов).
В одном конкретном варианте осуществления изобретение направлено на композицию для применения у млекопитающих при лечении спинальной мышечной атрофии детского возраста (СМА) или бокового амиотрофического склероза (БАС).
В одном конкретном варианте осуществления изобретение направлено на композицию для применения у млекопитающих при лечении спорадического нервно-мышечного заболевания (связанного со случайной мутацией причинного гена или одного или нескольких генов восприимчивости) или семейной формы, при которой обнаруживается мутация по меньшей мере одного гена, выбранного из SMN1, который вовлечен в структуру СМА, SOD1, TARDBP, кодирующего TAR ДНК-связывающий белок 43, VCP (валозин-содержащего белка), FUS/TLS (слитый в саркоме/транслоцированный в липосаркоме) и C9ORF72 (открытая рамка считывания 72 хромосомы 9), вовлеченного в структуру БАС.
В одном конкретном варианте осуществления лечение специфического расстройства моторных нейронов включает улучшение выживаемости моторных нейронов и/или ускорение созревания нервно-мышечных синапсов.
В одном конкретном варианте осуществления фитоэкдизоны применяют у людей в дозе от 3 до 15 миллиграммов на килограмм в сутки. Термин «фитоэкдизон» в настоящей заявке означает как фитоэкдизоны в целом, так и их производные, 20-гидроксиэкдизон (в частности, в форме экстракта) и его производные.
Фитоэкдизоны предпочтительно вводят в дозе от 200 до 1000 мг/сутки в один или несколько приемов у взрослого человека и в дозе от 5 до 350 мг/сутки в один или несколько приемов у ребенка или младенца. Под фитоэкдизоном здесь понимаются как фитоэкдизоны в целом, так и их производные, 20-гидроксиэкдизон (в частности, в форме экстракта) и его производные.
В некоторых вариантах осуществления композиция включает по меньшей мере одно соединение, которое считается производным фитоэкдизона, имеющее общую формулу (I):
Хим. Формула 1
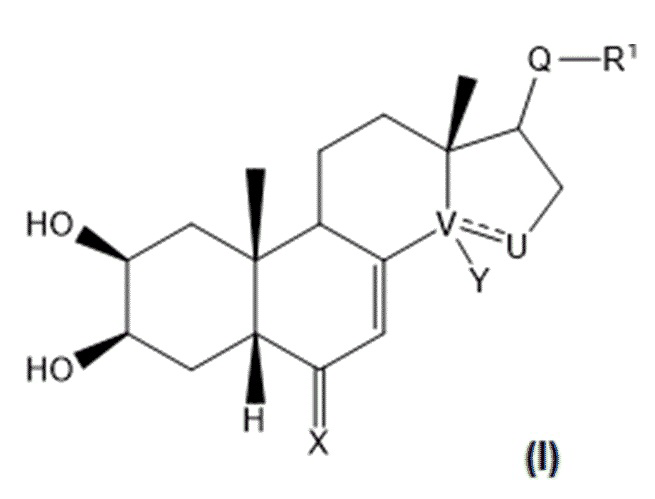
где:
V-U представляет собой одинарную углерод-углеродную связь, а Y представляет собой гидроксильную группу или атом водорода, или V-U представляет собой этиленовую связь C=C;
X представляет собой атом кислорода;
Q представляет собой карбонильную группу;
R1 выбран из: группы (C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)CO2(C1-C6); группы (C1-C6)А, где A представляет собой гетероцикл, при необходимости замещенный группой типа OH, OMe, (C1-C6), N(C1-C6), CO2(C1-C6); группы CH2Br;
W представляет собой гетероатом, выбранный из N, O и S, предпочтительно O и еще более предпочтительно S.
В контексте настоящего изобретения «(C1-C6)» означает любую линейную или разветвленную алкильную группу с 1-6 атомами углерода, в частности метильную, этильную, н-пропильную, изопропильную, н-бутильную, изобутильную, втор-бутильную, трет-бутильную, н-пентильную и н-гексильную группы. Предпочтительно она включает метильную, этильную, изопропильную или трет-бутильную группу, в частности метильную или этильную группу, более предпочтительно метильную группу.
В предпочтительном варианте осуществления в формуле (I):
Y представляет собой гидроксильную группу;
R1 выбран из: группы (C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)CO2(C1-C6); группы (C1-C6)А, где A представляет собой гетероцикл, при необходимости замещенный группой типа OH, OMe, (C1-C6), N(C1-C6), CO2(C1-C6);
W представляет собой гетероатом, выбранный из N, O и S, предпочтительно O и более предпочтительно S.
В некоторых вариантах осуществления композиция включает по меньшей мере одно соединение, выбранное из следующих соединений:
№1: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-10,13-диметил-17-(2-морфолиноацетил)-2,3,4,5, 9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-он;
№2: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-(3-гидроксипирролидин-1-ил)ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a] фенантрен-6-он;
№3: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-(4-гидрокси-1-пиперидил)ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a] фенантрен-6-он;
№4: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-[4-(2-гидроксиэтил)-1-пиперидил]ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a] фенантрен-6-он;
№5: (2S,3R,5R,10R,13R,14S,17S)-17-[2-(3-диметиламинопропил(метил)амино) ацетил]-2,3,14-тригидрокси-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-он;
№6: 2-[2-оксо-2-[(2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-10,13-диметил-6-оксо-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-17-ил]этил]этил сульфанилацетат;
№7: (2S,3R,5R,10R,13R,14S,17S)-17-(2-этилсульфанилацетил)-2,3,14-тригидрокси-10,13-диметил-2,3,4,5, 9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-он;
№8: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-(2-гидроксиэтилсульфанил)ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-он.
В некоторых вариантах осуществления композиция включает по меньшей мере одно соединение, которое считается производным фитоэкдизона, имеющее общую формулу (II):
Хим. Формула 2
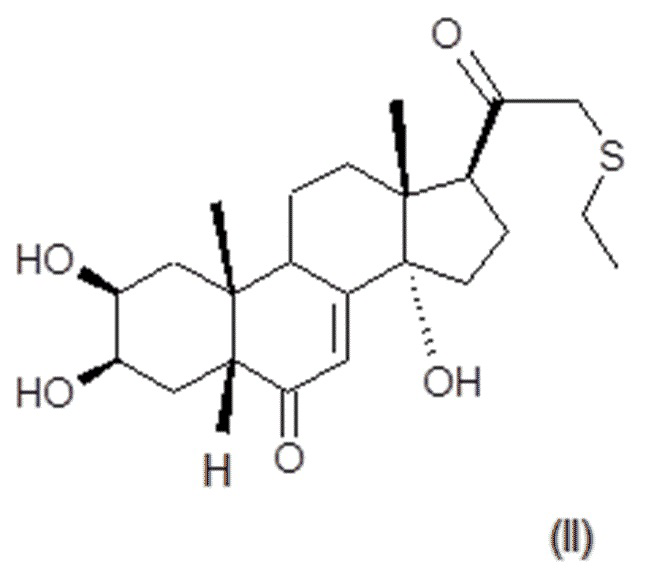
Соединение формулы (II) в дальнейшем упоминается как BIO103.
В некоторых вариантах осуществления композиция включена в фармацевтически приемлемый состав, который можно применять перорально.
В контексте настоящего изобретения «фармацевтически приемлемый» означает то, что можно использовать при приготовлении фармацевтической композиции и что обычно безопасно, нетоксично и приемлемо для ветеринарии, а также для фармацевтического применения у человека.
Краткое описание чертежей
Изобретение будет более понятно при чтении следующего описания, приведенного в качестве неограничивающего примера и со ссылкой на фигуры, на которых:
На фиг. 1A показана кривая изменения массы тела мышей со СМА, получавших носитель или BIO101, от рождения (P0) до 11 дней после рождения (P11). Здесь и в остальной части описания P соответствует количеству дней после рождения (постнатальных), n соответствует размеру выборки, а p соответствует «значению p», используемому для количественной оценки статистической значимости результата.
Фиг. 1В представляет собой кривые выживаемости Каплана-Мейера до P11 мышей со СМА, получавших носитель или BIO101, от P0 до P11.
Фиг. 2 представляет собой гистограмму, показывающую общее количество мышечных волокон передней большеберцовой мышцы (Tibialis) здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель), или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 3А представляет собой изображение, показывающее поперечный срез передней большеберцовой мышцы с окрашиванием гематоксилином и эозином, от здоровых контрольных мышей (контроль).
Фиг. 3В представляет собой изображение, показывающее поперечный срез передней большеберцовой мышцы с окрашиванием гематоксилином и эозином, от мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11.
Фиг. 3C представляет собой изображение, показывающее поперечный срез передней большеберцовой мышцы с окрашиванием гематоксилином и эозином, от мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 3D представляет собой гистограмму, показывающую поверхность поперечного среза мышечных волокон передней большеберцовой мышцы (Tibialis) здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 3E представляет собой гистограмму, показывающую поверхность поперечного среза мышечных волокон подошвенной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 3F представляет собой гистограмму, показывающую поверхность поперечного среза мышечных волокон камбаловидной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 4A представляет собой гистограмму, показывающую распределение мышечных волокон передней большеберцовой мышцы (Tibialis) в соответствии с их поверхностью поперечного среза, у мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 4В представляет собой гистограмму, показывающую распределение мышечных волокон подошвенной мышцы в соответствии с их поверхностью поперечного среза у мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 4C представляет собой гистограмму, показывающую распределение мышечных волокон камбаловидной мышцы в соответствии с их поверхностью поперечного среза у мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 5A представлен вестерн-блоттинг, показывающий фосфорилирование AKT подошвенной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей с СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 5B представлена гистограмма, показывающая количественное определение с помощью денситометрии фосфорилирования AKT подошвенной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 5C представлен вестерн-блоттинг, показывающий фосфорилирование ERK подошвенной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 5D представлена гистограмма, показывающая количественное определение с помощью денситометрии фосфорилирования ERK подошвенной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 5C представлен вестерн-блоттинг, показывающий уровень экспрессии белка SMN подошвенной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 5F представлена гистограмма, показывающая количественную оценку с помощью денситометрии белка SMN в подошвенной мышце здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 6А представлен вестерн-блоттинг, показывающий фосфорилирование AKT в спинном мозге здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 6B представлена гистограмма, показывающая количественную оценку с помощью денситометрии фосфорилирования AKT в спинном мозге здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 6C представлен вестерн-блоттинг, показывающий фосфорилирование ERK в спинном мозге здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 6D представлена гистограмма, показывающая количественную оценку с помощью денситометрии фосфорилирования ERK в спинном мозге здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 6E представлен вестерн-блоттинг, показывающий уровень экспрессии белка SMN в спинном мозге здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 6F представлена гистограмма, показывающая количественную оценку с помощью денситометрии белка SMN на поясничном уровне спинного мозга здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 7А представляет изображение, показывающее иммунофлуоресцентную метку моторных нейронов (анти-холинацетилтрансферазу) на поперечном срезе спинного мозга в поясничной области здоровых контрольных мышей (контроль, носитель).
Фиг. 7В представляет изображение, показывающее иммунофлуоресцентную метку моторных нейронов (анти-холинацетилтрансферазу) на поперечном срезе спинного мозга в поясничной области мышей со СМА, получавших носитель от P0 до P11 (SMA, носитель).
Фиг. 7C представляет изображение, показывающее иммунофлуоресцентную метку моторных нейронов (анти-холинацетилтрансферазу) на поперечном срезе спинного мозга в поясничной области мышей со СМА, получавших BIO101 от P0 до P11 (SMA BIO101).
На фиг. 7D показана гистограмма общего количества моторных нейронов на срез в брюшном отделе спинного мозга здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, и мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
На фиг. 7E показана гистограмма количества латеральных и дистальных моторных нейронов на срез в брюшном отделе спинного мозга здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 7F представляет гистограмму, показывающую распределение моторных нейронов в соответствии с поверхностью их тел, на срез в брюшном отделе спинного мозга у здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель), от P0 до P11, мышей со СМА, получавших BIO101 (SMA BIO101) от P0 до P11.
Фиг. 8 представляет собой гистограмму, показывающую распределение количества различных типов моторных нейронов (медленных, промежуточных и быстрых) в группах здоровых контрольных мышей (контроль, носитель, n ≥ 5), мышей со СМА, получавших носитель (SMA, носитель, n = 5) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101, n≥6) от P0 до P11. Медленные моторные нейроны характеризуются экспрессией маркеров ChAT+ERRβ+, быстрые моторные нейроны - экспрессией маркеров ChAT+MMP9+, а промежуточные моторные нейроны - экспрессией маркеров ChAT+ERRβ-MMP9-. Достоверность считается достигнутой при p <0,05. Символ «*» указывает на достоверную разницу по сравнению с группой здоровых мышей, которые получали носитель. Символ «#» указывает на достоверную разницу по сравнению с группой мышей со СМА, получавших носитель.
На фиг. 9А показаны фотографии иммунофлуоресцентной метки нервно-мышечного синапса различными маркерами пресинаптической стороны (нейрофиламент и SNAP25) и постсинаптической стороны (α-бунгаротоксин). Окрашивание бис-бензимидом позволяет идентифицировать ядра. Масштабный отрезок представляет 10 мкм.
Фиг. 9В представляет собой гистограмму, показывающую распределение процентного содержания различных типов нервно-мышечных синапсов в соответствии с их состоянием созревания (так называемой формы «кренделя» - зрелые; перфорированные - в процессе созревания; или однородные - незрелые). в подошвенной мышце в группах здоровых контрольных мышей (контроль, носитель, n = 4), мышей со СМА, получавших носитель (SMA, носитель, n = 4) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101, n = 4 ) от P0 до P11. Символ «*» указывает на достоверную разницу (p <0,05) по сравнению с группой контрольных мышей, получавших носитель.
Фиг. 9C представляет собой гистограмму, показывающую распределение процентного содержания различных типов нервно-мышечных синапсов в соответствии с их состоянием созревания (так называемой формы «кренделя» - зрелые; перфорированные - в процессе созревания; или однородные - незрелые) в камбаловидной мышце в группах здоровых контрольных мышей (контроль, носитель, n = 4), мышей со СМА, получавших носитель (SMA, носитель, n = 4) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101, n = 4) от P0 до P11. Символ «*» указывает на достоверную разницу (p <0,05) по сравнению с контрольной группой мышей, получавших носитель. Символ «#» указывает на достоверную разницу по сравнению с группой мышей со СМА, получавших носитель.
Фиг. 9D представляет собой гистограмму, показывающую распределение процентного содержания различных типов нервно-мышечных соединений в соответствии с их состоянием созревания (так называемой формы «кренделя» - зрелые; перфорированные - в процессе созревания; или однородные - незрелые) в передней большеберцовой мышце в группах здоровых контрольных мышей (контроль, носитель, n = 4), мышей со СМА, получавших носитель (SMA, носитель, n = 4) от P0 до P11, или мышей со СМА, получавших BIO101 (SMA BIO101, n = 4) от P0 до P11. Символ «*» указывает на достоверную разницу (p <0,05) по сравнению с группой контрольных мышей, получавших носитель. Символ «#» указывает на достоверную разницу по сравнению с группой мышей со СМА, получавших носитель.
На фиг. 10A показаны двигательные характеристики, оцененные с помощью теста открытого поля в дни P5, P7 и P9 у здоровых контрольных мышей (контроль, носитель, n = 13), мышей с СМА, получавших носитель (SMA, носитель, n = 11), от P0 до P9, или мышей со СМА, получавших BIO101 (SMA BIO101, n = 12) от P0 до P9. Символ «*» указывает на достоверную разницу по сравнению с группой контрольных мышей, получавших носитель (* = p <0,05, ** = p <0,01, *** = p <0,001 и **** = p <0,0001).
На фиг. 10B показана мышечная утомляемость, оцененная с помощью теста на захват в дни P5, P7 и P9 у здоровых контрольных мышей (контроль, носитель, n = 17), мышей со СМА, получавших носитель (SMA, носитель, n = 11) от P0 до P9, или мышей со СМА, получавших BIO101 (SMA BIO101, n = 10) от P0 до P9. Символ «*» указывает на достоверную разницу по сравнению с группой контрольных мышей, получавших носитель (* = p <0,05, ** = p <0,01 и *** = p <0,001).
Фиг. 11A показывает кривую массы тела мышей со СМА, получавших носитель или BIO103, от P0 до P11.
Фиг. 11В представляет собой кривую Каплана-Мейера выживаемости до P11 мышей со СМА, получавших носитель или BIO101, от P0 до P11.
Фиг. 12A представляет собой гистограмму, показывающую поверхность поперечного среза мышечных волокон передней большеберцовой мышцы (Tibialis) здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO103 (SMA BIO101) от P0 до P11.
Фиг. 12В представляет собой гистограмму, показывающую поверхность поперечного среза мышечных волокон подошвенной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO103 (SMA BIO101) от P0 до P11.
Фиг. 12C представляет собой гистограмму, показывающую поверхность поперечного среза мышечных волокон камбаловидной мышцы здоровых контрольных мышей (контроль, носитель), мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO103 (SMA BIO101) от P0 до P11.
Фиг. 13A представляет собой гистограмму, показывающую распределение мышечных волокон передней большеберцовой мышцы (Tibialis) в соответствии с поверхностью поперечного среза у мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO103 (SMA BIO103) от P0 до P11.
Фиг. 13B представляет собой гистограмму, показывающую распределение мышечных волокон подошвенной мышцы в соответствии с поверхностью поперечного среза у мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO103 (SMA BIO103) от P0 до P11.
Фиг. 13C представляет собой гистограмму, показывающую распределение мышечных волокон камбаловидной мышцы в соответствии с поверхностью поперечного среза у мышей со СМА, получавших носитель (SMA, носитель) от P0 до P11, или мышей со СМА, получавших BIO103 (SMA BIO103) от P0 до P11.
Описание вариантов осуществления изобретения
Далее изобретение будет описано в конкретном контексте некоторых из его предпочтительных и неограничивающих областей применения.
1. Способ очистки BIO101.
BIO101 получают полусинтетическим способом из 20-гидроксиэкдизона чистотой 90% в соответствии со следующими этапами:
i) горячее растворение 20-гидроксиэкдизона с чистотой 90% в метаноле, фильтрация и частичное концентрирование;
ii) добавление 3 объемов ацетона;
iii) охлаждение до температуры от 0 до 5°C при перемешивании;
iv) фильтрация полученного осадка;
v) последовательное промывание ацетоном и водой; и
vi) сушка.
Эта очистка использует процесс перекристаллизации, подходящий для этого вещества, который можно проводить в промышленном масштабе.
Фильтрацию на этапе (i) выполняют с использованием фильтра, отфильтровывающего частицы с размером 0,2 мкм.
Частичное концентрирование на этапе (i) предпочтительно проводят вакуумной перегонкой при температуре порядка 50°C в присутствии MeOH.
Этап сушки (vi) проводят в вакууме при температуре порядка 50°C.
2. Способ синтеза BIO103.
BIO103 получают полусинтетическим путем из 20-гидроксиэкдизона с последующей очисткой до фармацевтического качества в соответствии со следующим способом получения:
Хим. Формула 3
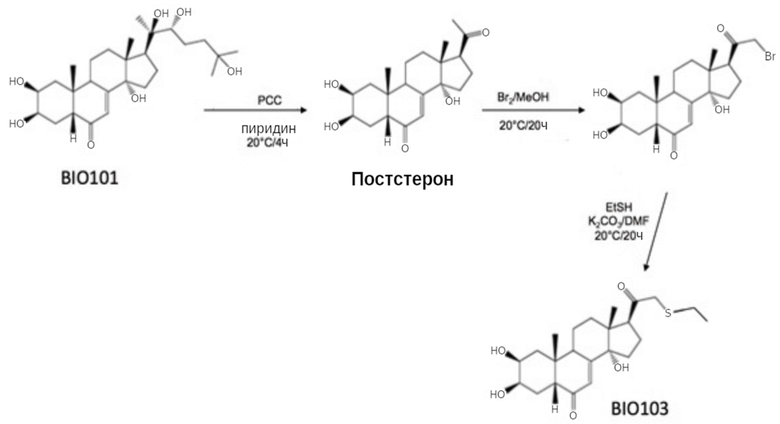
Схема синтеза BIO103 в 3 этапа:
I) окислительное расщепление боковой цепи 20-гидроксиэкдизона между атомами углерода C20 и C22 с целью получения постстерона (протокол, известный специалистам в данной области техники);
II) введение атома брома в положение C21;
III) реакция полученного таким образом соединения брома с этантиолом.
3. Биологическая активность BIO101 и BIO103.
а. Фенотипический анализ эффектов BIO101
Использовали модель тяжелой СМА на мышах на генном пуле FVB/NRj, характеризующемся инактивацией экзона 7 мышиного гена Smn и экспрессией 2 копий человеческого трансгена SMN2 (SmnΔ7/Δ7; huSMN2+/+) (Hsieh et al., 2000). Мыши, полученные в результате этих скрещиваний, имеющие генотип «FVB/NRj-SmnΔ7/Δ7 huSMN2+/+ 2 копии», описаны как «SMA». Эти мыши характеризуются прогрессирующей задержкой роста, которая наблюдается, начиная с 4 дней после рождения, и имеют дегенерацию около 50% моторных нейронов вентрального рога спинного мозга в конце жизни, прогрессирующую мышечную атрофию и среднюю продолжительность жизни около 12 дней (Hsieh et al., 2000). Мыши, имеющие генотип «FVB/NRj-Smn+/е huSMN2+/+ 2 копии», не имеют определенного фенотипа, и были использованы в качестве так называемых «контрольных» мышей. Мышам ежедневно вводили через зонд в дозе 50 мг/кг либо BIO101 в комплексе с носителем (в данном случае циклодекстрином), либо один носитель (VH). Массу тела и выживаемость анализировали ежедневно до P11. В остальной части описания n соответствует размеру выборки, а p соответствует «значению p», используемому для количественной оценки статистической значимости результата.
Результаты демонстрируют, что один лишь BIO101 (n = 18, где n - размер выборки) при ежедневном пероральном применении с момента рождения мышей позволяет значительно ограничить (p <0,05) потерю массы тела животных, заметную спустя 9 дней после рождения, по сравнению с животными, получавшими носитель (n = 22) (фиг. 1A), и значительно снижает (p <0,05) смертность животных, получавших лечение до P11 (фиг. 1B).
б. Анализ мышечно-трофических эффектов BIO101
На P11, через час после последнего введения желудочного зонда, получавших лечение мышей анестезировали пентобарбиталом 1% в дозе 6 мкл/г массы тела мыши, затем брали образцы камбаловидных мышц-разгибателей смешанной типологии, подошвенных мышц-разгибателей быстрой типологии и большеберцовых мышц-сгибателей быстрой типологии для проведения гистологических или молекулярных исследований.
После отбора образцы камбаловидной, подошвенной и большеберцовой мышцы по отдельности помещали в консервирующую среду, а затем замораживали в охлажденном изопентане. Для каждой мышцы были выполнены медиальные срезы толщиной 10 мкм. Эти срезы окрашивали гематоксилином-эозином, дегидратировали и помещали в среду для включения. Изображения этих срезов были получены с помощью микроскопа (с увеличением x200). Для получения изображений с оттиском рельефа использовали методику дифференциально-интерференционного контраста. На основе этих изображений количество мышечных волокон каждой мышцы и площадь поперечного сечения 20% этих волокон подсчитывали с использованием программного обеспечения для обработки изображений (фиг. 2).
Гистологический анализ поперечных срезов отобранных мышц, окрашенных гематоксилином-эозином, показывает положительное влияние BIO101 на аплазию мышц (количество мышечных волокон). Действительно, в большеберцовой мышце количество мышечных волокон у мышей со СМА, получавших BIO101, значительно увеличивается по сравнению с количеством волокон у животных, получавших носитель (соответственно, 2358 и 2069 волокон (+ 14%), p <0,05).
Интересно, что мышечная атрофия, присутствующая у мышей со СМА, ограничивается при лечении, независимо от природы и типологии мышц. Действительно, лечение BIO101 позволяет значительно ограничить атрофию трех исследованных мышц: сгибателей и разгибателей.
Атрофия мышечных волокон у мышей со СМА (фиг. 3А) по сравнению со здоровыми контрольными мышами (фиг. 3В) действительно видна на гистологических срезах большеберцовой мышцы, и количественная оценка площади поперечного сечения мышечных волокон показывает, что эта атрофия составляет 56,1% в большеберцовой мышце мышей со СМА (n = 4) по сравнению с контрольными мышами (n = 4) (фиг. 3D). Эта атрофия значительно снижается (p <0,05) до 32,7% на P11 благодаря лечению BIO101 (n = 4) (фиг. 3C и фиг. 3D). То же самое относится и к подошвенной мышце, где у мышей со СМА наблюдается значительная атрофия мышечных волокон на 46% по сравнению со здоровыми контрольными мышами (p <0,05) и где лечение BIO101 значительно ограничивает эту атрофию до 17,9% (p <0,05) (фиг. 3E), и в камбаловидной мышце (50,2% для мышей со СМА по сравнению с контролем (p <0,01), против 37,8% при лечении BIO101, p <0,05) (фиг. 3F).
Для проведения более точного анализа влияния BIO101 на атрофию было оценено распределение мышечных волокон по категориям поверхности поперечного сечения. По своей природе мышечные волокна имеют различную поверхность на поперечном срезе. Волокна типа I, характеризующиеся медленным сокращением, имеют размер меньше, чем волокна типа II, характеризующиеся быстрым сокращением. Значительное снижение атрофии препаратом BIO101, наблюдаемое при лечении мышей со СМА от P0 до P11, приводит к значительному влиянию этого лечения на распределение волокон в соответствии с их площадью поперечного сечения в трех мышцах, изученных у мышей со СМА, получавших BIO101, по сравнению с мышами со СМА, получавшими носитель (фиг. 4A, 4B и 4C). В большеберцовой мышце мышей со СМА, получавших BIO101 от P0 до P11, наблюдается значительное увеличение доли волокон с площадью поперечного сечения от 400 до 800 мкм2, в то время как доля волокон с поверхностью поперечного сечения в пределах от 100 до 200 мкм2 значительно снижается (p <0,05), по сравнению с группой мышей со СМА, получавших носитель (фиг. 4A). В подошвенной мышце и камбаловидной мышце мышей со СМА, получавших BIO101 от P0 до P11, наблюдается увеличение доли волокон с площадью поперечного сечения от 200 до 400 мкм2, сопровождающееся уменьшением доли волокон, которые имеют площадь поперечного сечения от 0 до 100 мкм2 для подошвенной мышцы (фиг. 4B) и волокон, которые имеют площадь поперечного сечения от 100 до 200 мкм2 для камбаловидной мышцы (фиг. 4C).
c. Молекулярный анализ
Предыдущие исследования позволили продемонстрировать недостаточную активацию пути AKT/CREB и чрезмерную активацию пути ERK/Elk-1, связанную с низкой экспрессией белка SMN в вентральном роге спинного мозга трансгенных мышей «FVB/NRj-SmnΔ7/Δ7 huSMN2+/+ 2 копии», используемых в качестве мышиной модели тяжелой спинальной мышечной атрофии типа II, что предполагает роль этих путей в заболевании (Branchu et al., 2013).
Замороженную подошвенную мышцу гомогенизировали в буфере для экстракции путем механического измельчения. Отбирали пробы надосадочной жидкости, содержащей белковый экстракт, затем дозировали белковые экстракты в соответствии с анализом белка Лоури. Проводили электрофорез на геле ДСН-ПААГ, затем разделенные белки переносили на мембраны. Использовали следующие первичные антитела: мышиные моноклональные анти-SMN (1:5000), кроличьи поликлональные анти-Ser 473 фосфо-AKT (1:1000), кроличьи поликлональные анти-AKT (1:100), кроличьи моноклональные анти-фосфо-ERK 1/2 (1:500), анти-MAP киназа 1/2 (ERK 1/2) (1:1000). После промывания мембраны инкубировали с вторичными антителами против белков мыши (1:5,000) или против белков кролика (1:5,000), конъюгированными с пероксидазой. После использования первичных антител комплексы антитело-антиген разрушали путем инкубации в растворе для диссоциации, затем мембраны снова инкубировали с кроличьими антителами против AKT и против MAP-киназы 1/2 (ERK 1/2) (1:1000). Комплексы антител выявляли с помощью хемилюминесценции и визуализировали с помощью устройства получения цифрового изображения образца геля, мембраны или пленки. Оптическую плотность каждой конкретной полосы количественно оценивали с помощью программного обеспечения для обработки изображений путем вычитания фона и нормализации с оптической плотностью полос β-актина. Для контролей, получавших носитель, полученные значения были определены как 1, а значения других групп были нормализованы по отношению к этим контролям и выражены как относительная величина. Результаты для животных из каждой группы получали путем проведения независимых экспериментов и с использованием различных мембран, на которых каждую группу сравнивали с контролем. Количественные значения pAKT для подошвенной мышцы представляют 3 мыши на группу, получавшую носитель (контрольную или со СМА), и 4 мыши, получавших BIO101. Количественные значения pERK для подошвенной мышцы представляют не менее 4 мышей на группу. Количественные значения SMN составляют n = 2. Что касается количественных значений pAKT в спинном мозге, они составляют n = 2 мыши на группу для pAKT и pERK, тогда как для уровня SMN они составляют n = 4 на группу.
Молекулярный анализ подошвенной мышцы подтверждает, что путь AKT действительно недостаточно активирован (фиг. 5A и 5B) в пользу пути ERK (фиг. 5C и 5D) у мышей со СМА по сравнению с контрольными мышами, получавшими носитель (контроль, носитель) (Branchu et al., 2013). 11-дневное применение BIO101 позволяет обратить эту взаимосвязь в мышцах мышей, получавших лечение от P0 до P11, за счет существенного увеличения фосфорилирования AKT (pAKT) (фиг. 5B), в то время как уровень фосфорилирования ERK (pERK) сильно уменьшается (фиг. 5D). Эта полезная и уже продемонстрированная благоприятная ситуация связана со сверхэкспрессией белка SMN (Branchu et al., 2013). Как и ожидалось, белок SMN практически не экспрессируется у животных со СМА по сравнению со здоровыми контрольными животными. Однако, и полностью неожиданно, после применения BIO101 не наблюдалось никаких изменений в уровне экспрессии SMN, что свидетельствует об оригинальной молекулярной регуляции (фиг. 5E и 5F).
Этот баланс сигнальных путей между ERK и AKT также обнаруживается в спинном мозге мышей со СМА. Действительно, наблюдается явное снижение фосфорилирования AKT (pAKT) у мышей со СМА (фиг. 6A и 6B) по сравнению со здоровыми контрольными мышами (контроль, носитель), в то время как фосфорилирование ERK (pERK) увеличивается (фиг. 6C и 6D). Ежедневное лечение мышей со СМА в течение 11 дней с помощью BIO101 частично восстанавливает этот баланс в спинном мозге животных за счет увеличения уровня pAKT и снижения уровня pERK (фиг. 6B и 6D). Ка и ожидалось, в этой модели белок SMN не экспрессируется у мышей со СМА. Как наблюдалось на подошвенной мышце, применение BIO101 не позволяет восстановить уровень SMN у мышей со СМА, получавших лечение в течение 11 дней (фиг. 6E и 6F).
d. Анализ моторных нейронов
Количественное и качественное исследование популяций моторных нейронов на толстых поперечных срезах поясничной области спинного мозга (L1-L5) здоровых контрольных мышей или мышей со СМА, получавших носитель или BIO101 в течение 11 дней, проводили с помощью иммунофлуоресцентного мечения холинацетилтрансферазой (ChAT), как описано выше (Biondi et al., 2008; Boyer et al., 2013). Затем анализ был уточнен путем изучения различных субпопуляций моторных нейронов по характеристике их расположения в спинном мозге (латеральное или медиальное положение), а также распределения размеров их тел.
Анестезированным мышам проводили внутрисердечную инфузию ФБР. У мышей брали спинной мозг, фиксировали, а затем промывали. Поясничный отдел спинного мозга (L1-L5) покрывали 4% раствором агарозы. С помощью вибратома были сделаны поперечные срезы 50 мкм по всей длине образца. Затем для иммуногистохимических анализов использовали один срез на каждые 5 срезов спинного мозга. После насыщения 0,1 М глицином ткани затем пермеабилизировали, блокировали, а затем метили козьим поликлональным первичным антителом против холинацетилтрансферазы (ChAT) (1/400e). Затем срезы промывали, затем инкубировали с поликлональным вторичным анти-козьим антителом, соединенным с Цианином 3 (1:400e). Ядра маркировали с использованием бис-бензимида (1/1000e), затем срезы снова промывали перед установкой с фотообесцвечивающим ингибитором флуоресцентных красителей. Специфичность маркировки подтверждали по контрольной маркировке, проводимой в отсутствие первичного антитела.
Изображения были получены с использованием камеры, установленной на микроскопе с увеличением x200 и соединенной с центральным процессором типа микрокомпьютера, содержащим, в частности, подходящее программное обеспечение для получения изображений. Все подсчеты проводили с использованием программного обеспечения для обработки изображений.
При иммунофлуоресценции моторные нейроны, меченные анти-ChAT антителом, действительно могут быть идентифицированы светло-серым цветом на изображениях вентрального рога спинного мозга (фиг. 7A, 7B и 7C). Количество моторных нейронов на срез вентрального отдела спинного мозга определяли для каждой из трех групп мышей. Как ожидалось, количество моторных нейронов (n = 5) мышей со СМА значительно уменьшается (фиг. 7B) по сравнению с количеством моторных нейронов (n = 5), обнаруженных в группе здоровых контрольных мышей (фиг. 7A). Действительно, разница дегенерации моторных нейронов между этими двумя группами составляет 25% (p <0,05) (фиг. 7D). Очень интересно то, что через 11 дней ежедневного лечения BIO101 отмечено, что количество моторных нейронов значительно выше (p <0,05) у этих мышей со СМА (фиг. 7C), чем у мышей со СМА, получавших носитель (n = 5). Таким образом, отмечается, что лечение BIO101 значительно ограничивает дегенерацию моторных нейронов, вызванную патологией, и что лечение оказывает значительный нейропротекторный эффект, ограничивающий потерю моторных нейронов на 13% в группе, получавшей BIO101 (фиг. 7D).
Затем количественный анализ был уточнен, с одной стороны, путем изучения различных субпопуляций моторных нейронов по анализу их расположения в спинном мозге (латеральное или медиальное положение), а с другой стороны, путем изучения распределения поверхности их тел.
Наблюдается, что количество латеральных моторных нейронов (которые иннервируют дистальные мышцы) повышено у мышей со СМА, получавших BIO101, по сравнению с мышами со СМА, которые получали носитель, в то время как значительного воздействия на медиальные моторные нейроны не наблюдалось (фиг. 7E).
Анализ поверхности тел моторных нейронов показывает, как и ожидалось, атрофию моторных нейронов у мышей со СМА, получавших носитель, по сравнению со здоровыми контрольными мышами, со значительным увеличением количества моторных нейронов, имеющих поверхность тел менее 600 мкм2 (p <0,05). Параллельно у мышей со СМА наблюдается потеря моторных нейронов, имеющих тела с большей поверхностью, чем 900 мкм2 (p <0,05) (фиг. 7F). Применение BIO101 позволяет ограничить эту атрофию моторных нейронов, значительно снижая количество малых моторных нейронов с поверхностью тела менее 300 мкм2 (p <0,05) (фиг. 7F). Важно отметить, что в рамках БАС было описано, что двигательные единицы не подвергаются однородному воздействию патологического процесса. В самом деле, пресимптоматическая модель заболевания на мышах выявила предпочтительную дегенерацию моторных единиц типа FF (быстро утомляемых), заставляя действовать моторные нейроны с телом с большой поверхностью (Pun et al., 2006). О подобной дифференциальной дегенерации также сообщалось и у пациентов (Dengler et al., 1990; Theys, Peeters и Robberecht, 1999). Таким образом, уменьшение доли моторных нейронов с малой поверхностью в пользу тех, у которых поверхность тела больше, может быть интересным подходом в рамках такой патологии, как БАС.
В дополнение к анализу количества моторных нейронов и размера их тел, а также качественному исследованию их местоположения (медиального или латерального в вентральном спинном мозге) мы изучили влияние лечения BIO101 на защиту субпопуляций моторных нейронов (медленные моторные нейроны, промежуточные моторные нейроны и быстрые моторные нейроны). Этот качественный анализ субпопуляций моторных нейронов был проведен на толстых поперечных срезах поясничной области спинного мозга (L1-L5) контрольных мышей или мышей со СМА, получавших или не получавших BIO101, путем иммунофлуоресцентного мечения холинацетилтрансферазой, как было описано выше (Biondi et al., 2008; Branchu et al., 2013), а также анализа типа моторных нейронов с помощью иммунофлуоресцентного мечения с эстроген-связанным рецептором-β (ERRβ), специфическим маркером медленных моторных нейронов, или с помощью матричной металлопептидазы 9 (MMP9), специфического маркера быстрых моторных нейронов.
Анестезированным мышам проводили внутрисердечную инфузию ФБР. У мышей брали спинной мозг, фиксировали, а затем промывали. Поясничный отдел спинного мозга (L1-L5) покрывали 4% раствором агарозы. На вибратоме были сделаны поперечные срезы 50 мкм по всей длине образца. Затем для иммуногистохимических анализов использовали один срез на каждые 5 срезов спинного мозга. После насыщения 0,1 М глицином ткани пермеабилизировали, блокировали, а затем метили следующими первичными антителами: козье антитело против ChAT (1/400e), мышиное антитело против ERRβ (1/400e), кроличье антитело против MMP9 (1/600e). После трех промываний со срезами инкубировали следующие антитела: ослиное анти-козье антитело с Cy5 (1/400e), ослиное анти-мышиное антитело с Alexa 488 (1/400e), ослиное анти-кроличье антитело с Cy3 (1/400e). Ядра маркировали с использованием бис-бензимида (1/1000e), затем срезы снова промывали перед установкой с фотообесцвечивающим ингибитором флуоресцентных красителей. Специфичность маркировки подтверждали по контрольной маркировке, проводимой в отсутствие первичного антитела.
Изображения были получены с использованием камеры, установленной на микроскопе с увеличением x200 и соединенной с центральным процессором типа микрокомпьютера, содержащим, в частности, подходящее программное обеспечение для получения изображений. Все подсчеты проводили с использованием программного обеспечения для обработки изображений.
Количество медленных моторных нейронов (ChAT+ERRβ+) существенно не меняется в группах, будь то здоровые мыши, получавшие носитель, мыши со СМА, получавшие носитель, или мыши со СМА, получавшие BIO101 (фиг. 8). В группе животных со СМА, получавших носитель, наблюдалась значительное снижение среднего числа быстрых моторных нейронов (ChAT+MMP9+) по сравнению с группой здоровых контрольных мышей (11 против 16, соответственно; p <0,05). Эта потеря происходит в пользу моторных нейронов промежуточного типа (ChAT+ERRb-MMP9-), количество которых значительно выше в группе мышей со СМА, получавших носитель, по сравнению с количеством, присутствующим у здоровых контрольных мышей (7 против 4; р <0,05).
Интересно, что лечение мышей со СМА BIO101 предпочтительно способствует выживанию моторных нейронов быстрого типа (ChAT+MMP9+). Действительно, в группе мышей со СМА, получавших BIO101, количество быстрых моторных нейронов значительно выше, чем в группе мышей со СМА, получавших носитель (14 против 11; p <0,05).
Следовательно, лечение BIO101 ограничивает потерю моторных нейронов, наблюдаемую в этой модели тяжелой СМА, в частности, путем защиты мышей от потери быстрых моторных нейронов.
е. Анализ нервно-мышечных синапсов
СМА характеризуется специфическим изменением нервно-мышечных синапсов, обусловленным отсутствием белка SMN и денервацией, вызванной специфической гибелью моторных нейронов (Kariya et al., 2008; Biondi et al., 2008; Chali et al., 2016). Мы провели морфологическое исследование нервно-мышечных синапсов с целью определения степени созревания и фрагментации зрелой структуры так называемого «кренделя». Для этого мы провели специальную маркировку пресинаптических (синаптофизин и нейрофиламент) и постсинаптических (α-бунгаротоксин) сторон с помощью иммунофлуоресценции (Leroy et al., 2014) на срезах мышечных волокон камбаловидной, подошвенной и подошвенной мышц мышей со СМА, получавших или не получавших лечение BIO101.
Продольные срезы толщиной 75 мкм получали с помощью вибратома. Затем срезы насыщали 0,1 М глицином при слабом перемешивании, затем промывали ФБР. Затем их обрабатывали блокирующим раствором и повышали проницаемость с помощью раствора ФБР с 4% БСА, 5% козьей сыворотки, 0,5% тритона. Первичные антитела против нейрофиламентов (1/800e) и против синаптофизина (SNAP25; 1/200e) (для идентификации пресинаптической поверхности нервно-мышечного синапса) инкубировали в течение 48 часов и проявляли вторичным антителом (анти-кроличьим антителом с AlexaFluor® 647; 1/400e), затем промывали. Наконец, срезы инкубировали с анти-α-бунгаротоксином, непосредственно связанным с AlexaFluor® 555 (1/500e). Срезы промывали, ядра маркировали бис-бензимидом (1/1000e), затем помещали между покровным стеклом и предметными стеклами с фотообесцвечивающим ингибитором флуоресцентных красителей для наблюдения в микроскопической эпифлуоресцентной визуализации (фиг. 9A).
Нервно-мышечные синапсы определяли и количественно оценивали по трем категориям, от самых незрелых до наиболее зрелых: однородные пластинки, перфорированные или имеющие форму «кренделя».
Во всех исследованных мышцах (подошвенной, камбаловидной и большеберцовой) процент однородных нервно-мышечных синапсов у животных со СМА был значительно увеличен по сравнению со здоровыми контрольными мышами. Эта задержка созревания нервно-мышечных соединений ожидаема и уже описана в литературе (Biondi et al., 2008). Действительно, в быстром разгибателе подошвенной мышце процент незрелых нервно-мышечных синапсов на Р10 составляет 89,7% по сравнению с 49,7% у здоровых контрольных мышей (p <0,05) (фиг. 9В), в медленном разгибателе камбаловидной мышце процент незрелых нервно-мышечных синапсов составляет 84,3% по сравнению с 45,7% у здоровых контрольных мышей (p <0,05) (фиг. 9C), в быстром сгибателе большеберцовой мышце процент незрелых нервно-мышечных синапсов составляет 69,3% по сравнению с 28,7% у здоровых контрольных мышей (p <0,05) (фиг. 9D). Когда мышей со СМА ежедневно лечили BIO101, от их рождения до P10, наблюдается более существенное созревание нервно-мышечных синапсов во всех протестированных мышцах с уменьшением процента незрелых пластин в пользу перфорированных пластин, что является свидетельством ускорения созревания. Действительно, в подошвенной мышце 16,3% синапсов имеют перфорированный тип по сравнению с 10,4% у животных со СМА, которым вводили носитель (p = n.s) (фиг. 9B). Это различие является значительным в камбаловидной мышце (26,5% перфорированных соединений в группе мышей со СМА по сравнению с 15,7% в группе мышей со СМА, получавших носитель, p<0,05, фиг. 9C) и большеберцовой мышце (45,7% перфорированных синапсов в группе получавших лечение мышей со СМА, по сравнению с 30,7% в группе мышей со СМА, получавших носитель, p <0,05, фиг. 9D).
Таким образом, эти результаты показывают, что применение BIO101 ускоряет созревание нервно-мышечных синапсов.
f. Анализ двигательных способностей на модели мышей с тяжелой СМА
Были проведены фенотипические анализы мышей с тяжелой формой СМА 2 типа, получавших или не получавших BIO101, начиная с P0. Параллельно каждые два дня тестировали двигательные способности мышей от P5 до P9. Авторы оценивали способность к самостоятельному передвижению с помощью теста открытого поля, а также мышечную утомляемость с помощью теста захвата, как описано выше (Biondi et al., 2008; Branchu et al., 2013; Chali et al., 2016).
Устройство, используемое для теста в открытом поле, различалось в зависимости от возраста мышей. Для животных от P0 до P6 оно представляло собой пластиковую коробку размером 15×15×5 см с сетчатой структурой, разделенной на 25 квадратов размером 3 см × 3 см. Для животных от P7 до P21 оно представляло собой пластиковую коробку размером 28 × 28 × 5 см с сетчатой структурой, разделенной на 16 квадратов 7×7 см. Мышей тестировали индивидуально, и устройство для оценки промывали после каждого сеанса. Каждая мышь, изначально помещенная в центр поля, могла свободно перемещаться в течение 5 минут. В течение этих 5 минут экспериментатор регистрировал поведенческие измерения и записывал общее количество пересеченных квадратов.
Как и ожидалось, во все время тестирования (P5, P7 и P9) мыши со СМА, получавшие носитель, демонстрировали значительно сниженные двигательные характеристики по сравнению со здоровыми контрольными мышами (фиг. 10A). Действительно, количество квадратов, которые они могут пересечь, составляет 10 на P5, 6 на P7 и 8 на P9, по сравнению с 17 на P5, 12 на P7 и 24 на P9 в группе здоровых контрольных мышей (с p <0,01, p <0,0001 и p <0,001, соответственно). На P5 лечение BIO101 мышей со СМА не улучшало их двигательные характеристики (11 квадратов) по сравнению с мышами со СМА, получавшими носитель (10 квадратов). На P7 подвижность мышей, получавших BIO101, значительно возрастала (пересечено 10 квадратов) по сравнению с мышами со СМА, получившими носитель (пересечено 6 квадратов, с p <0,05). На P9 это различие незначительно, но лечение BIO101 имеет тенденцию к проявлению положительного эффекта у мышей, которые способны пересекать 11 квадратов в группе мышей BIO101 со СМА, по сравнению с 8 квадратами в группе мышей со СМА, получавших носитель, p = n.s).
Чтобы оценить мышечную утомляемость, силу захвата задних лап мышей испытывали от P5 до P9 (тест захвата). Мышей подвешивали за задние лапы на тонком металлическом стержне, подвешенном в воздухе горизонтально. Регистрировали время, проведенное в подвешенном состоянии. Каждую мышь подвергали пяти последовательным попыткам с периодом отдыха в одну минуту между двумя испытаниями. Для оценки мышечных функций оставляли только наилучшие результаты.
Мышечная утомляемость, проверенная с помощью теста на захват, ожидаемо показала, что мыши со СМА, получавшие носитель, имеют значительно сниженную мышечную функцию по сравнению со здоровыми контрольными мышами (фиг. 10B). Действительно, время, в течение которого они могут оставаться подвешенными на металлическом стержне, составляет 0,2 секунды на P5, 6,2 секунды на P7 и 5,6 секунды на P9 против 3,3 секунды на P5, 11,4 секунды на P7 и 16,1 секунды на P9 в группе здоровых контрольных мышей (с p <0,01, p <0,01 и p <0,001, соответственно). На P5 лечение BIO101 мышей со СМА значительно улучшает их мышечные характеристики (2,3 секунды) по сравнению с мышами со СМА, получавшими носитель (0,2 секунды; p <0,05). На P7 лечение BIO101 имеет тенденцию к увеличению, но незначительному, времени пребывания в подвешенном состоянии мышей со СМА, по сравнению с мышами со СМА, получившими носитель (12,2 секунды против 6,2 секунды, соответственно, при p = n.s). Наконец, на P9 BIO101 существенно улучшает этот параметр у мышей, получавших BIO101 (20,9 секунды), по сравнению с мышами со СМА, получавшими носитель (5,6 секунды, с p <0,01).
В заключение, BIO101 оказывает благотворное влияние на произвольную моторику, а также на мышечную утомляемость животных со СМА.
g. Фенотипический анализ эффектов BIO103
Ту же модель мышей типа II использовали для характеристики фенотипических эффектов препарата BIO103 на модели тяжелой СМА. Мышам вводили ежедневно через зонд дозу 50 мг/кг либо BIO103 в комплексе с носителем (циклодекстрином в данном случае), либо один носитель (VH). Массу тела и выживаемость проверяли ежедневно до P11.
Результаты показывают, что BIO103 (n = 12) при ежедневном пероральном применении с момента рождения мышей имеет тенденцию ограничивать потерю массы тела животных, хотя разница не достигает порога значимости по сравнению с животными, получавшими носитель (n = 22) (фиг. 11A). С другой стороны, BIO103 значительно (p <0,05) увеличивает выживаемость животных, получавших лечение до P11 (выживаемость 66,6% на P11), по сравнению с мышами, получавшими носитель (выживаемость 36,3% на P11) (фиг. 11B).
f. Анализ мышечно-трофических эффектов BIO103
Гистологический анализ окрашенных гематоксилином/эозином поперечных срезов отобранных мышц показывает положительное влияние BIO103 на мышечную атрофию (поперечное сечение мышечных волокон).
Так же, как и в случае с молекулой BIO101, мышечная атрофия, присутствующая у мышей со СМА, ограничивается при лечении, независимо от природы и типологии мышц. Действительно, лечение BIO103 позволяет значительно ограничить атрофию трех исследованных мышц.
Количественная оценка поверхности мышечных волокон ожидаемо показывает, что эта атрофия сильно выражена и составляет 56,1% в большеберцовой мышце мышей со СМА (n = 4) по сравнению с контрольными мышами (n = 4). Эта атрофия значительно снижается (p <0,05) до 22,1% на P11 благодаря лечению BIO103 (n = 4) (фиг. 12A). То же самое относится и к подошвенной мышце, для которой наблюдается значительная атрофия мышечных волокон на 46% у мышей со СМА по сравнению со здоровыми контрольными мышами (p <0,05), а лечение BIO103 значительно ограничивает эту атрофию до 15,3% (p <0,05) (фиг. 12B), и к камбаловидной мышце (50,2% для мышей со СМА по сравнению с контролем (p <0,01) по сравнению с 37,1% при лечении BIO103, p <0,05) (фиг. 12C).
Чтобы провести более точный анализ воздействия BIO103 на атрофию, было оценено распределение мышечных волокон по категориям поверхности поперечного среза. Значительное уменьшение атрофии при воздействии BIO103, наблюдаемое при лечении мышей со СМА от P0 до P11, приводит к значительному влиянию этого лечения на распределение волокон в соответствии с их поперечной поверхностью в трех мышцах мышей со СМА, получавших BIO103, по сравнению с мышами со СМА, получавшими носитель (фиг. 13A, 13B и 13C). В большеберцовой мышце мышей со СМА, получавших BIO103 от P0 до P11, наблюдается значительное увеличение доли волокон, имеющих площадь поперечного сечения более 400 мкм2, в то время как доля мелких волокон, имеющих площадь поперечного сечения менее 200 мкм2, значительно снижается (p <0,05) по сравнению с группой мышей со СМА, получавших носитель (фиг. 13A). В подошвенной мышце (фиг. 13B) и камбаловидной мышце (фиг. 13C) мышей со СМА, получавших BIO101 от P0 до P11, наблюдается увеличение доли мышечных волокон с площадью поперечного сечения более 300 мкм2, сопровождающееся уменьшением доли волокон, имеющих площадь поперечного сечения менее 200 мкм2.
4. Вывод
Таким образом, в свете свойств BIO101 и BIO103 в отношении аплазии, мышечной атрофии и дегенерации моторных нейронов млекопитающих, страдающих детской спинальной мышечной атрофией, можно предложить использование фитоэкдизонов, в частности BIO101 и BIO103, по отдельности или в качестве дополнения к лечению, направленному на коррекцию последствий генетического изменения, чтобы сохранить мышечную ткань, а также моторные нейроны, и в результате замедлить развитие нервно-мышечных заболеваний, которые, как следствие, приводят к ухудшению мышечной функции и/или потере моторных нейронов. Нервно-мышечные заболевания включают, в частности, боковой амиотрофический склероз, а также спинальную мышечную атрофию.
В более общем плане следует отметить, что способы реализации и осуществления изобретения, рассмотренного выше, были описаны посредством неограничивающих примеров, и что, следовательно, могут быть рассмотрены другие альтернативные варианты.
Библиографические ссылки
Akten B, Kye MJ, Hao le T, Wertz MH, Singh S; et al. Interaction of survival of motor neuron (SMN) and HuD proteins with mRNA cpg15 rescues motor neuron axonal deficits. Proc. Natl. Acad. Sci. USA (2011); 108:10337–42.
Al-Chalabi A & Hardiman O. The epidemiology of ALS: a conspiracy of genes, environment and time. Nat. Rev. Neurol. (2013); 9:617–28.
Bevan AK, Hutchinson KR, Foust KD, Braun L, McGovern VL, et al. Early heart failure in the SMNDelta7 model of spinal muscular atrophy and correction by postnatal scAAV9-SMN delivery. Hum. Mol. Genet. (2010); 19:3895–905.
Biondi O, Grondard C, Lécolle S, Deforges S, Pariset C, et al. Exercise-induced activation of NMDA receptor promotes motor unit development and survival in a type 2 spinal muscular atrophy model mouse. J. Neurosci. (2008); 28:953–62.
Biondi O, Lopes P, Desseille C, Branchu J, Chali F, et al. Physical exercise reduces cardiac defects in type 2 spinal muscular atrophy-like mice. J. Physiol. (2012); 590:5907–25.
Boyer JG, Murray LM, Scott K, De Repentigny Y, Renaud JM, Kothary R. Early onset muscle weakness and disruption of muscle proteins in mouse models of spinal muscular atrophy. Skelet. Muscle (2013); 3:24.
Branchu J, Biondi O, Chali F, Collin T, Leroy F, et al. Shift from extracellular signal-regulated kinase to AKT/cAMP response element-binding protein pathway increases survival-motor-neuron expression in spinal-muscular-atrophy-like mice and patient cells. J. Neurosci. (2013); 33:4280–94.
Buhler D, Raker V, Luhrmann R & Fischer U. Essential role for the tudor domain of SMN in spliceosomal U snRNP assembly: implications for spinal muscular atrophy. Hum. Mol. Genet. (1999); 8:2351–7.
Chali, F. et al. Long-term exercise-specific neuroprotection in spinal muscular atrophy-like mice. J Physiol 594, 1931-1952 (2016).
Chiriboga CA, Swoboda KJ, Darras BT, Iannaccone ST, Montes J, et al. Results from a phase 1 study of nusinersen (ISIS-SMN(Rx)) in children with spinal muscular atrophy. Neurology (2016); 86:890-7.
Cifuentes-Diaz C, Frugier T, Tiziano FD, Lacène E, Robiot N, et al. Deletion of murine SMN exon 7 directed to skeletal muscle leads to severe muscular dystrophy. J. Cell Biol. (2001); 152:1107–14.
Crawford TO & Pardo CA. The neurobiology of childhood spinal muscular atrophy. Neurobiol. Dis. (1996); 3:97–110.
Dengler R, Konstanzer A, Küther G, Hesse S, Wolf W, Struppler A. Amyotrophic lateral sclerosis: macro-EMG and twitch forces of single motor units. Muscle Nerve (1990); 13 (6):545–50.
Fidzianska A, Rafalowska J, Glinka Z. Ultrastructural study of motoneurons in Werdnig-Hoffmann disease. Clin. Neuropathol. (1984); 3:260–5.
Finkel RS, Chiriboga CA, Vaisar J, Day JW, Montes J, et al. Treatment of infantile-onset spinal muscular atrophy with nusinersen: a phase 2, open-label, dose-escalation study. Lancet (2016); 388:3017–26.
Frugier T, Tiziano FD, Cifuentes-Diaz C, Miniou P, Roblot N, et al. Nuclear targeting defect of SMN lacking the C-terminus in a mouse model of spinal muscular atrophy. Hum. Mol. Genet. (2000); 9:849–58.
Gorelick-Feldman J, MacLean D, Ilic N, Poulev A, Lila MA, Cheng D, Raskin I. Phytoecdysteroids increase protein synthesis in skeletal muscle cells. J. Agric. Food Chem. (2008); 56:3532–37.
Greensmith L & Vrbova G. Alterations of nerve-muscle interaction during postnatal development influence motoneurone survival in rats. Brain Res. Dev. Brain Res. (1992); 69(1):125–31 (1992).
Grice SJ & Liu JL. Survival motor neuron protein regulates stem cell division, proliferation, and differentiation in Drosophila. PLoS Genetics (2011) 7:e1002030.
Hache M, Swoboda KJ, Sethna N, Farrow-Gillespie A, Khandji A, et al. Intrathecal Injections in Children With Spinal Muscular Atrophy: Nusinersen Clinical Trial Experience. J. Child Neurol. (2016); 31:899–906.
Heier CR, Satta R, Lutz C, DiDonato CJ. Arrhythmia and cardiac defects are a feature of spinal muscular atrophy model mice. Hum. Mol. Genet. (2010); 19:3906–18.
Hsieh-Li HM, Chang JG, Jong YJ, Wu MH, Wang NM, et al. A mouse model for spinal muscular atrophy. Nat. Genet. (2000); 24:66-70.
Hua Y, Sahashi K, Hung G, Rigo F, Passini MA, et al. Antisense correction of SMN2 splicing in the CNS rescues necrosis in a type III SMA mouse model. Genes Dev. (2010); 24:1634–44.
Hunter G, Aghamaleky Sarvestany A, Roche SL, Symes RC, Gillingwater TH. SMN-dependent intrinsic defects in Schwann cells in mouse models of spinal muscular atrophy. Hum. Mol. Genet. (2014); 23:2235–50.
Hwee DT, Kennedy AR, Hartman JJ, Ryans J, Durham N, et al. The small-molecule fast skeletal troponin activator, CK-2127107, improves exercise tolerance in a rat model of heart failure. J. Pharmacol. Exp. Ther. (2015); 353:159–68.
Kariya S, Park GH, Maeno-Hikichi Y, Leykekhman O, Lutz C, Arkovitz MS et al. Reduced SMN protein impairs maturation of the neuromuscular junctions in mouse models of spinal muscular atrophy. Hum. Mol. Genet. (2008); 17(16):2552–69.
Kong L, Wang X, Choe DW, Polley M, Burnett BG, et al. Impaired synaptic vesicle release and immaturity of neuromuscular junctions in spinal muscular atrophy mice. J. Neurosci. (2009); 29:842–51.
Lawrence MM. Ajuga turkestanica as a countermeasure against sarcopenia and dynapenia. MS thesis, Appalachian State University (2012).
Lee YI, Mikesh M, Smith I, Rimer M, Thompson W. Muscles in a mouse model of spinal muscular atrophy show profound defects in neuromuscular development even in the absence of failure in neuromuscular transmission or loss of motor neurons. Dev. Biol. (2011); 356:432–44.
Lefebvre S, Burlet P, Liu Q, Beltrandy S, Clermont O, et al. Correlation between severity and SMN protein level in spinal muscular atrophy. Nat. Genet. (1997); 16:265–9.
Leroy, F. et al. Early intrinsic hyperexcitability does not contribute to motoneuron degeneration in amyotrophic lateral sclerosis eLife. 3, e04046 (2014).
Liu Q, Fischer U, Wang F, Dreyfuss G. The spinal muscular atrophy disease gene product, SMN, and its associated protein SIP1 are in a complex with spliceosomal snRNP proteins. Cell (1997); 90:1013–21.
Lorson CL, Strasswimmer J, Yao JM, Baleja JD, Hahnen E,
et al. SMN oligomerization defect correlates with spinal muscular atrophy severity. Nat. Genet. (1998); 19:63–6.
McWhorter ML, Monani UR, Burghes AH, Beattie CE. Knockdown of the survival motor neuron (Smn) protein in zebrafish causes defects in motor axon outgrowth and pathfinding. J. Cell Biol. (2003); 162:919–31.
Murray LM, Comley LH, Thomson D, Parkinson N, Talbot K, Gillingwater TH.. Selective vulnerability of motor neurons and dissociation of pre- and post-synaptic pathology at the neuromuscular junction in mouse models of spinal muscular atrophy. Hum. Mol. Genet. (2008); 17:949–62.
Nicole S, Desforges B, Millet G, Lesbordes J, Cifuentes-Diaz C, et al. Intact satellite cells lead to remarkable protection against Smn gene defect in differentiated skeletal muscle. J. Cell Biol. ‘2003); 161:571–82.
Passini MA, Bu J, Richards AM, Kinnecom C, Sardi SP, et al. Antisense oligonucleotides delivered to the mouse CNS ameliorate symptoms of severe spinal muscular atrophy. Sci. Transl. Med. (2011); 3:72ra18.
Peter CJ, Evans M, Thayanithy V, Taniguchi-Ishigaki N, Bach I, et al. The COPI vesicle complex binds and moves with survival motor neuron within axons. Hum. Mol. Genet. (2011); 20:1701–11.
Pun S, Santos AF, Saxena S, Xu L, Caroni P. Selective Vulnerability and Pruning of Phasic Motoneuron Axons in Motoneuron Disease Alleviated by CNTF. Nature Neuroscience (2006); 9 (3):408–19.
Rajendra TK, Gonsalvez GB, Walker MP, Shpargel KB, Salz HK, Matera AG. A Drosophila melanogaster model of spinal muscular atrophy reveals a function for SMN in striated muscle. J. Cell Biol. (2007); 176:831–41.
Rindt H, Feng Z, Mazzasette C, Glascock JJ, Valdivia D, et al. Astrocytes influence the severity of spinal muscular atrophy. Hum. Mol. Genet. (2015); 24:4094–102.
Sanchez G, Dury AY, Murray LM, Biondi O, Tadesse H, et al. A novel function for the survival motoneuron protein as a translational regulator. Hum. Mol. Genet. (2013); 22:668–84.
Shababi M, Habibi J, Yang HT, Vale SM, Sewell WA, Lorson CL. Cardiac defects contribute to the pathology of spinal muscular atrophy models. Hum. Mol. Genet. (2010); 19:4059–71.
Shafey D, Cote PD, Kothary R. Hypomorphic Smn knockdown C2C12 myoblasts reveal intrinsic defects in myoblast fusion and myotube morphology. Exp. Cell Res. (2005); 311:49–61.
Simakin SYu, Panyushkin VV, Portugalov SN, Kostina LV, Martisorov EG. Combined application of preparation Ecdysten. Science Bulletin (1998); N°2, 29–31.
Somers E, Lees RD, Hoban K, Steigh JN, Zhou H, et al. Vascular Defects and Spinal Cord Hypoxia in Spinal Muscular Atrophy. Ann. Neurol. (2016); 79:217–30.
Syrov VN. Comparative experimental investigations of the anabolic activity of ecdysteroids and steranabols. Pharm. Chem. J. (2000); 34(4):193-197.
Theys PA, Peeters E, Robberecht W. Evolution of Motor and Sensory Deficits in Amyotrophic Lateral Sclerosis Estimated by Neurophysiological Techniques. J. Neurol. (1999); 246 (6):438–42.
Torres-Benito L, Neher MF, Cano R, Ruiz R, Tabares L. SMN requirement for synaptic vesicle, active zone and microtubule postnatal organization in motor nerve terminals. PLoS One (2011); 6:e26164.
Tóth N, Szabó A, Kacsala P, Héger J, Zádor E. 20-Hydroxyecdysone increases fiber size in a muscle-specific fashion in rat. Phytomedicine (2008); 15: 691-8.
Vitte J, Fassier C, Tiziano FD, Dalard C, Soave S, et al. Refined characterization of the expression and stability of the SMN gene products. Am J Pathol (2007); 171:1269-80.
Vitte JM, Davoult B, Roblot N, Mayer M, Joshi V, et al. Deletion of murine Smn exon 7 directed to liver leads to severe defect of liver development associated with iron overload. Am. J. Pathol. (2004); 165:1731-41.
Vrbova G, Fisher TJ. The Effect of Inhibiting the Calcium Activated Neutral Protease, on Motor Unit Size after Partial Denervation of the Rat Soleus Muscle. Eur. J. Neurosci. (1989); 1:616–25.
Walker MP, Rajendra TK, Saieva L, Fuentes JL, Pellizzoni L, Matera AG. SMN complex localizes to the sarcomeric Z-disc and is a proteolytic target of calpain. Hum. Mol. Genet. (2008); 17:3399–410.
Zhang Z, Lotti F, Dittmar K, Younis I, Wan L, et al. SMN deficiency causes tissue-specific perturbations in the repertoire of snRNAs and widespread defects in splicing. Cell (2008); 133:585-600.
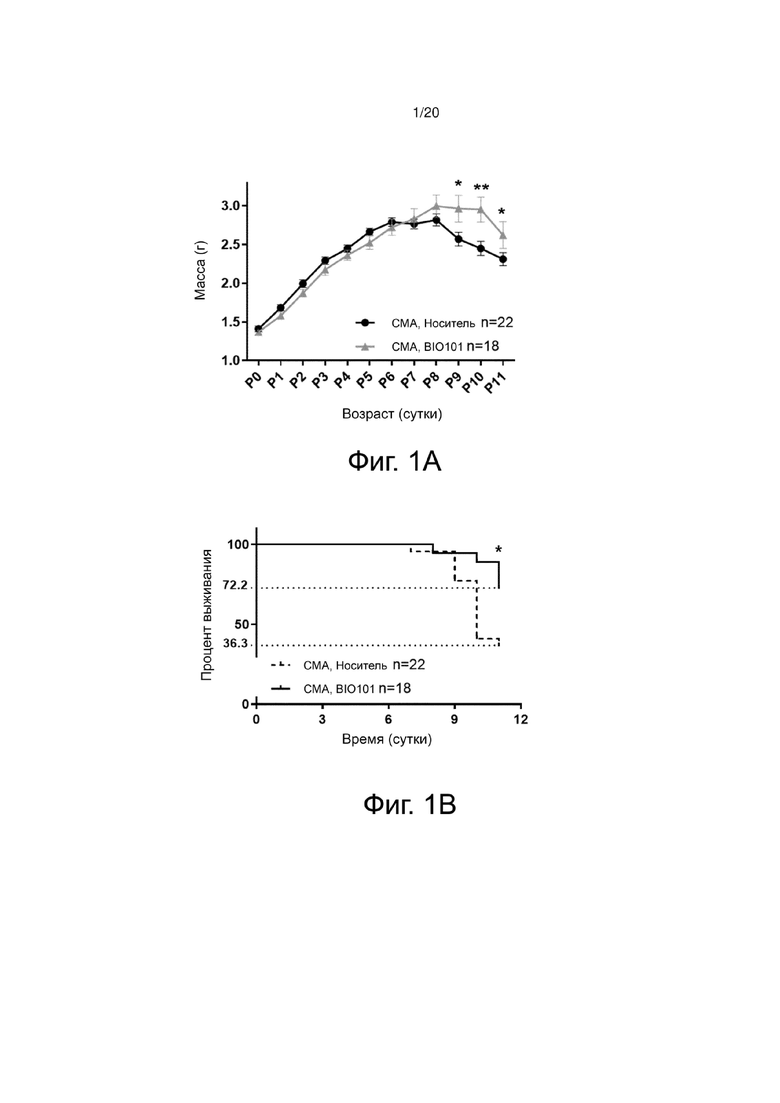
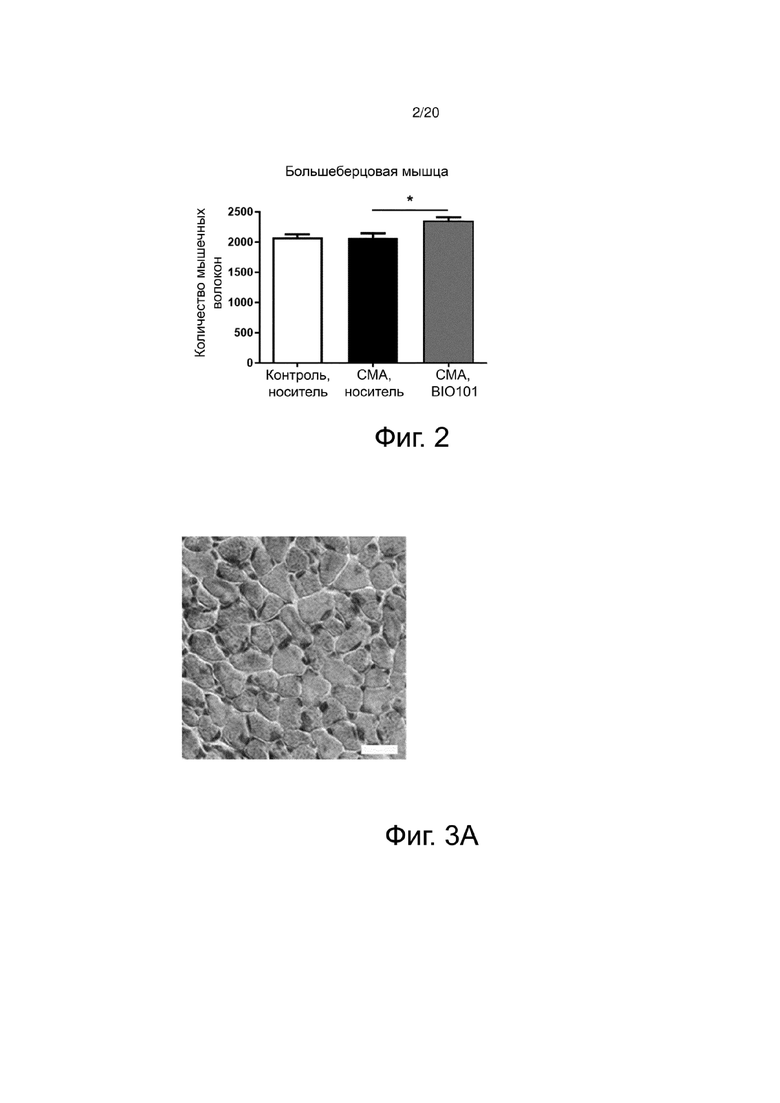

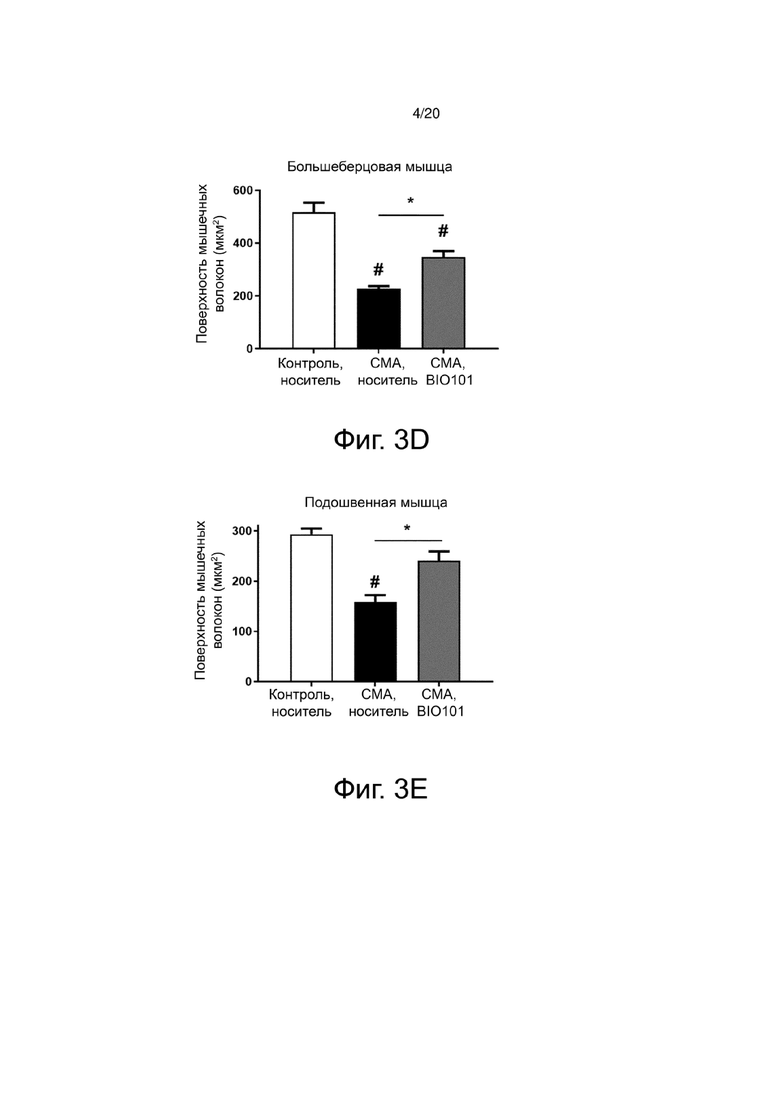
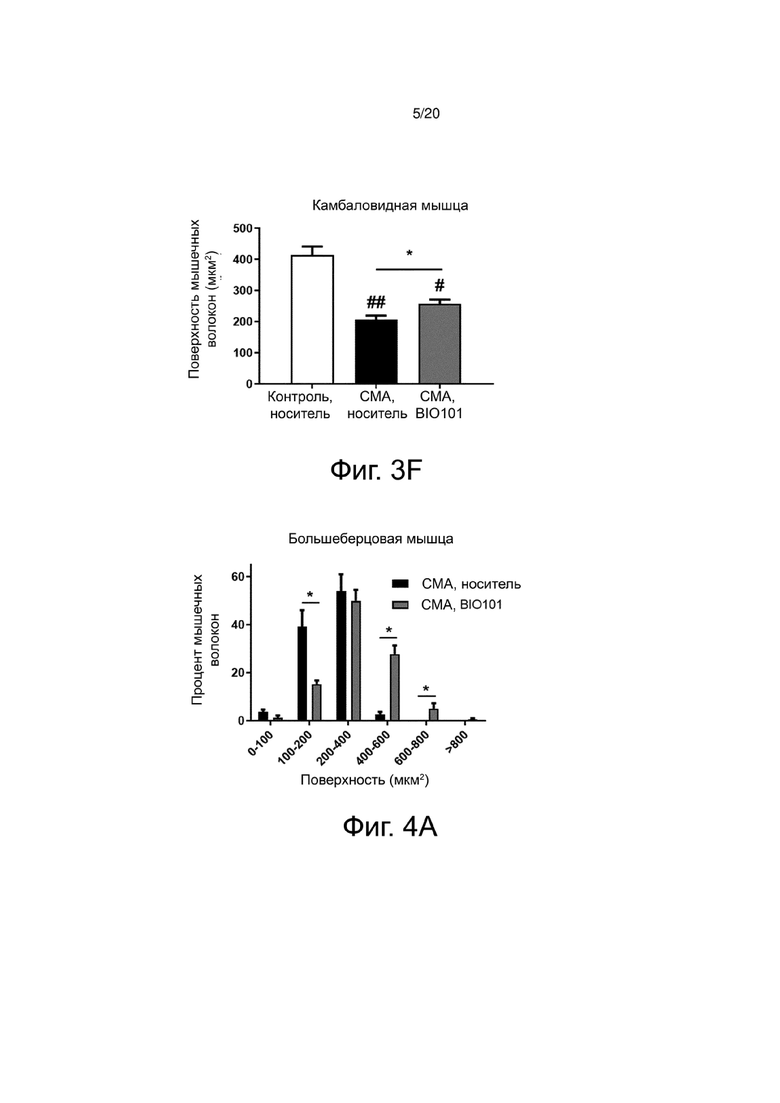
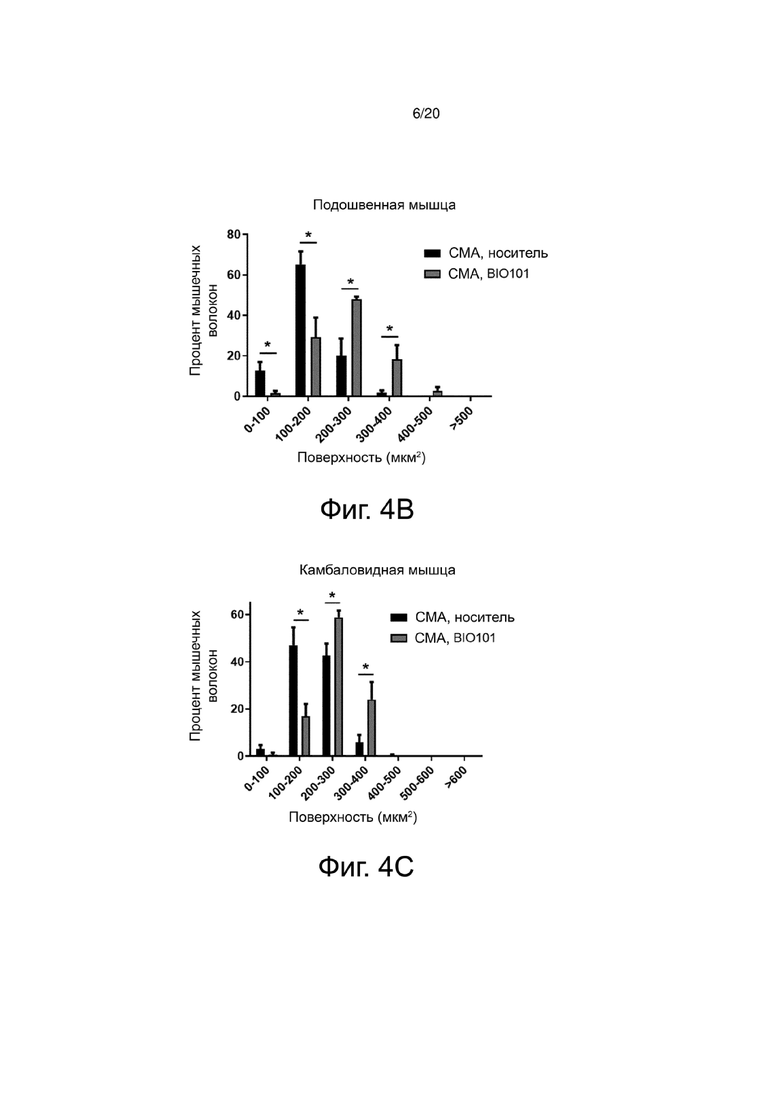
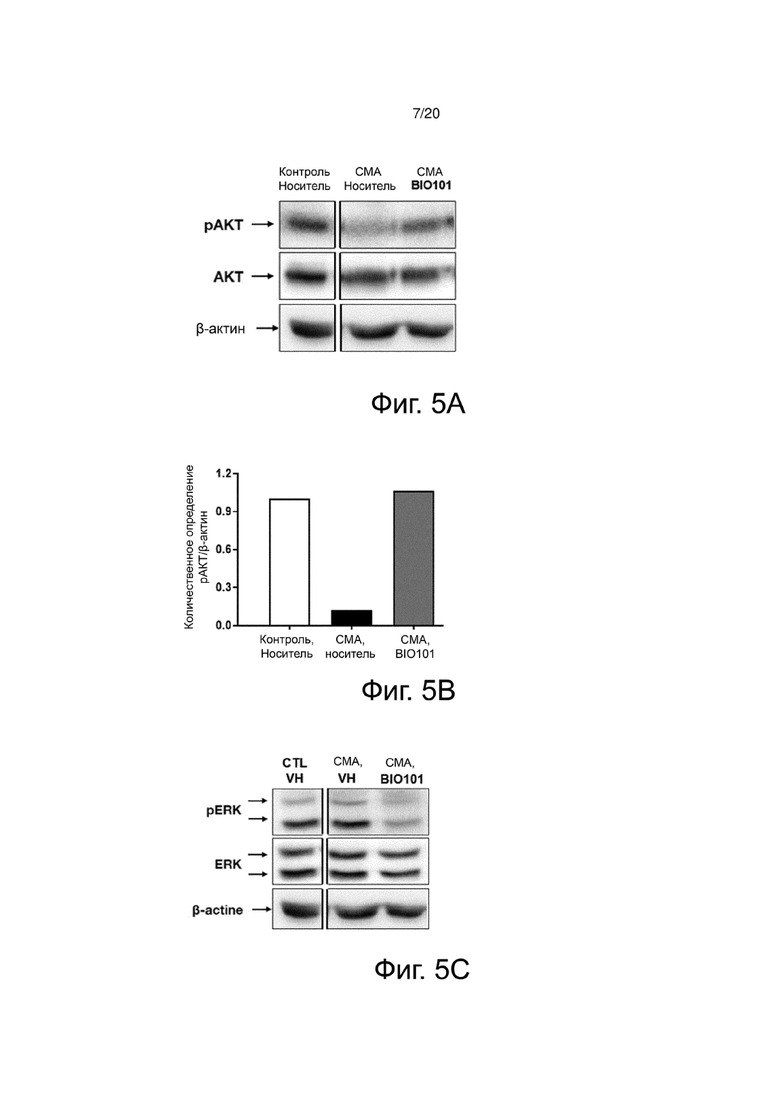
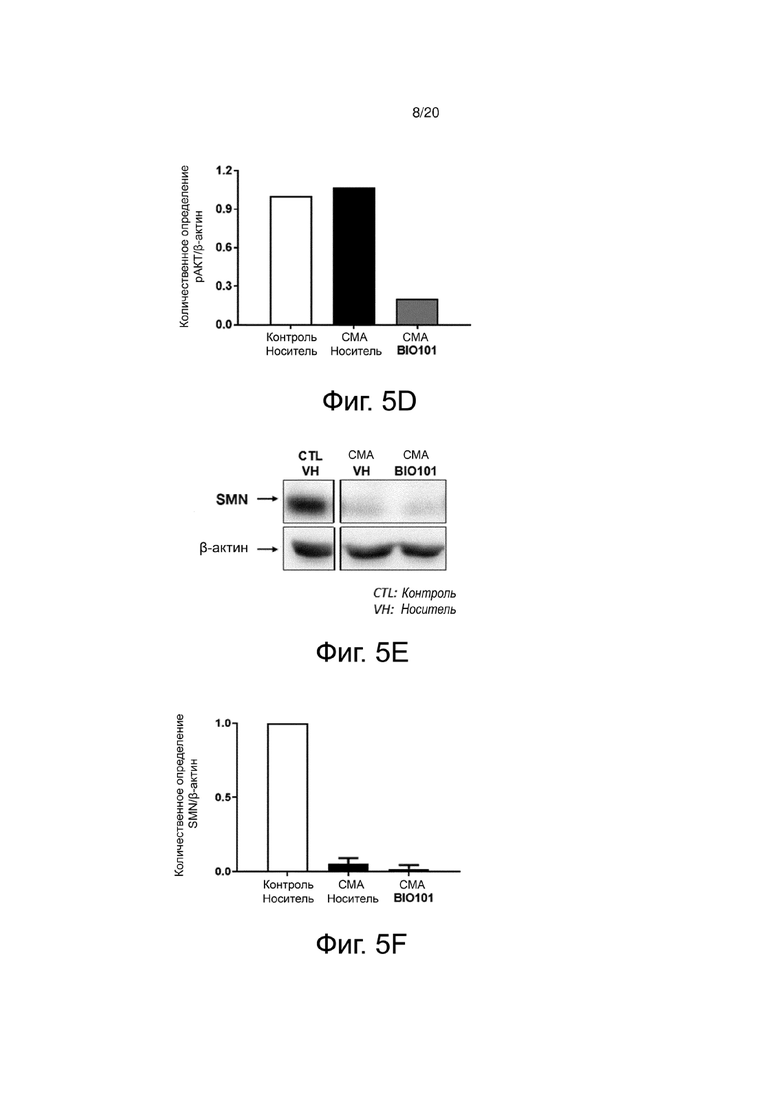
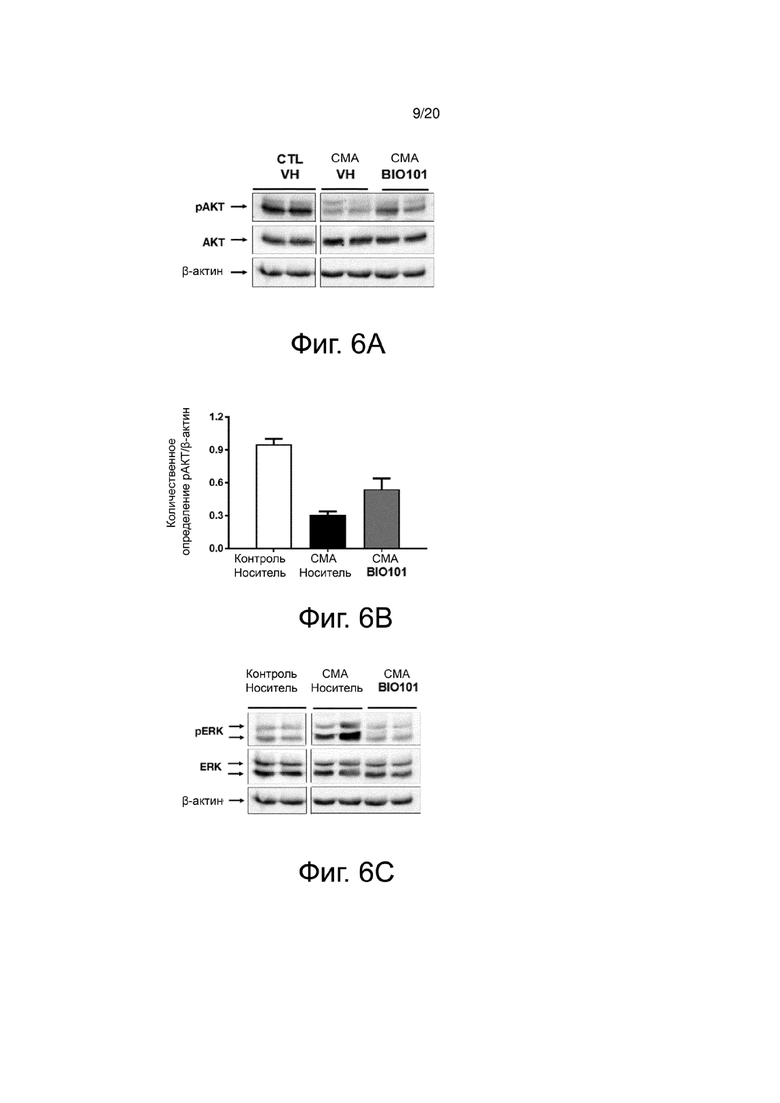
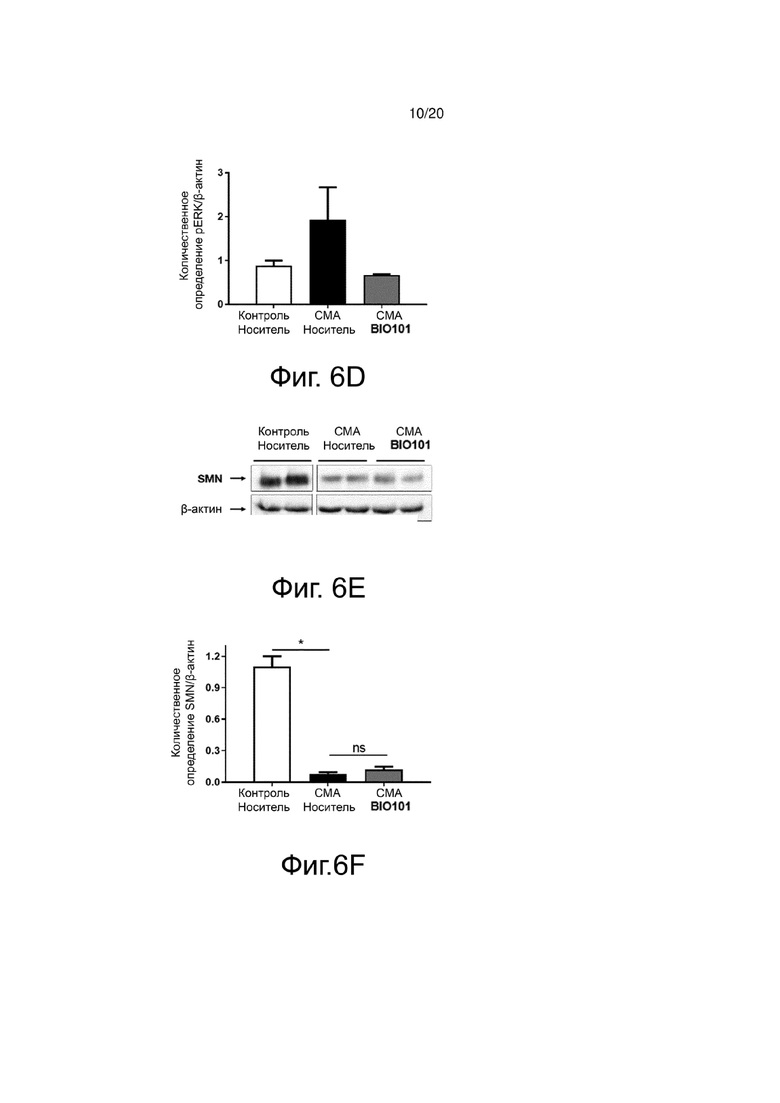
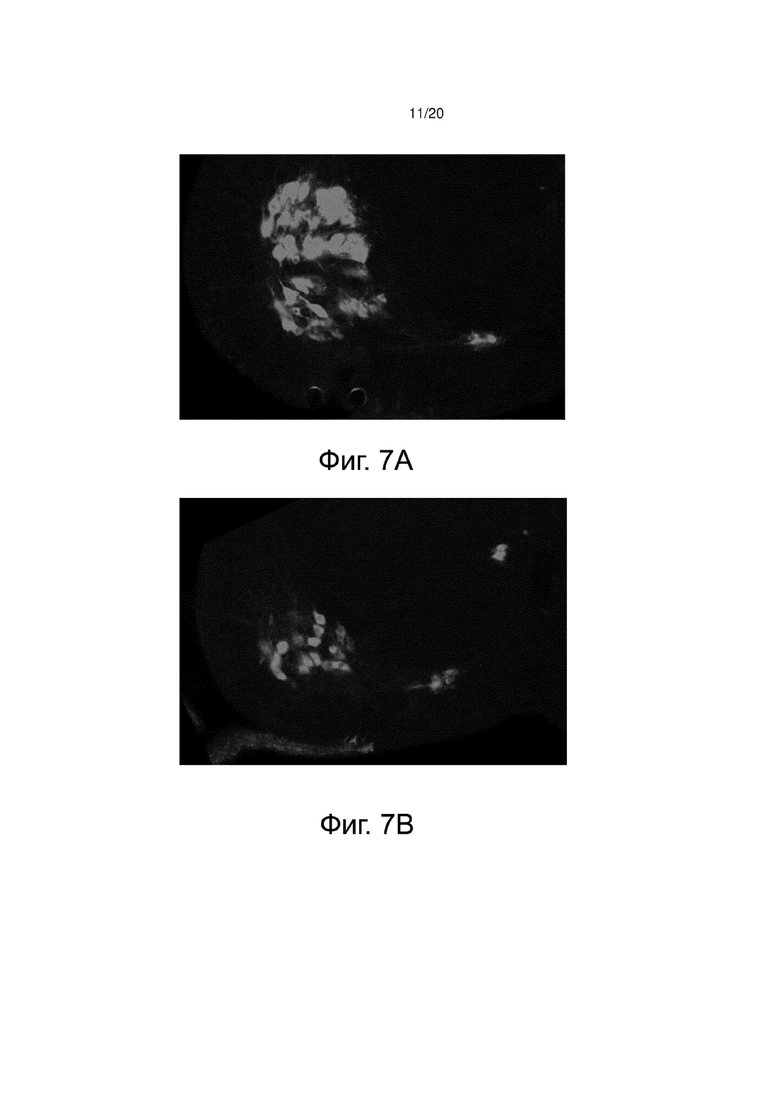
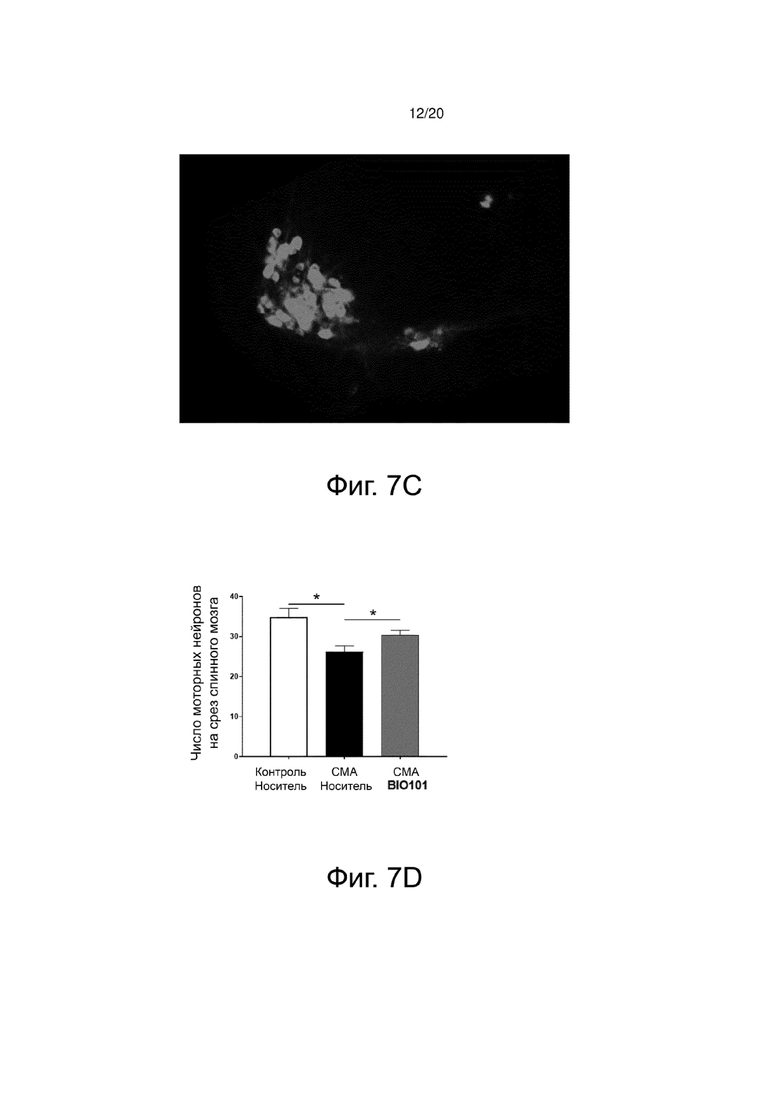
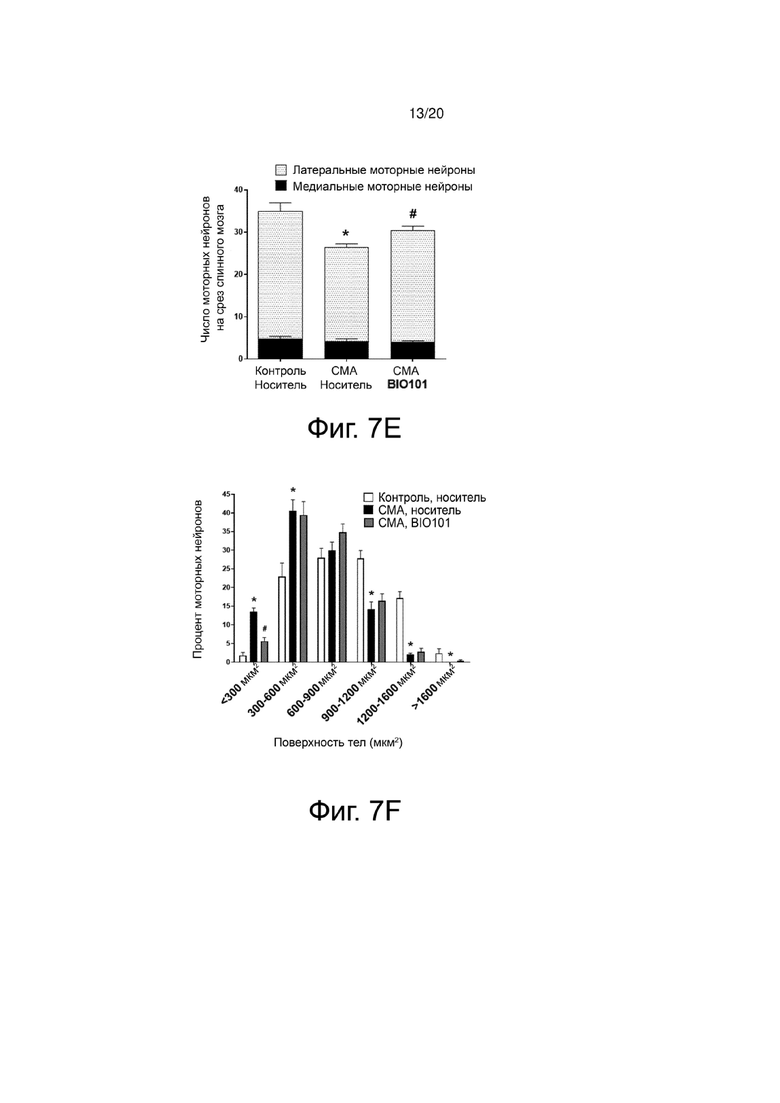
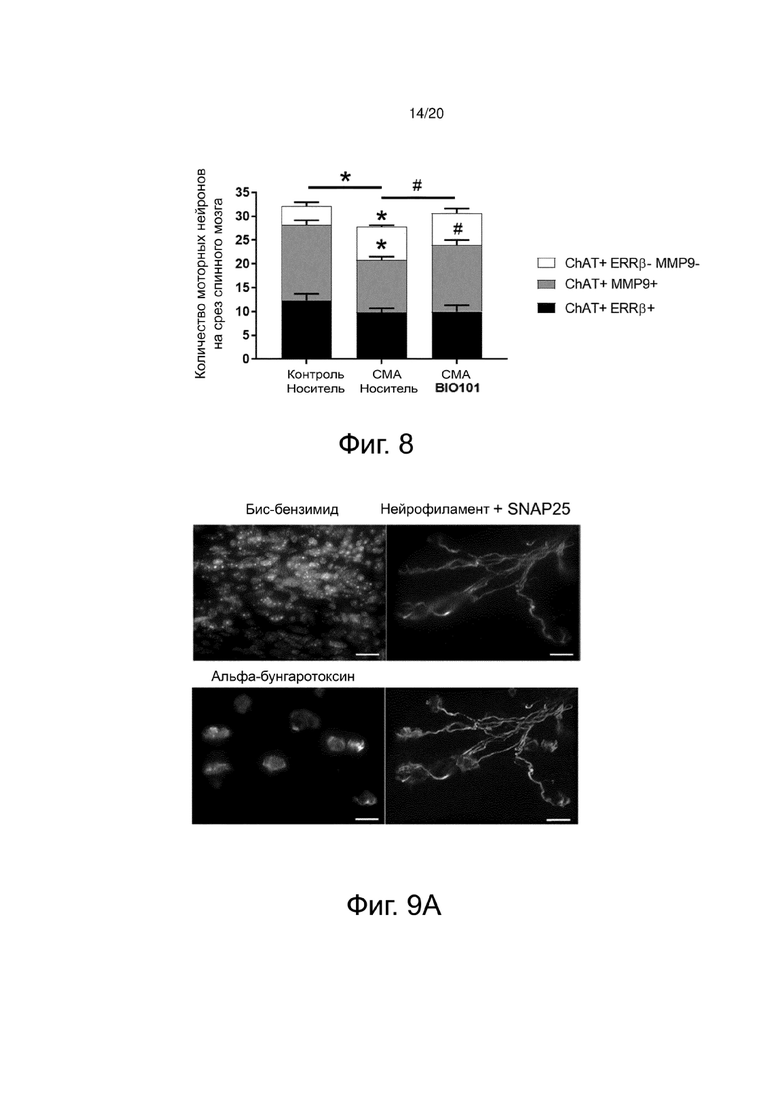
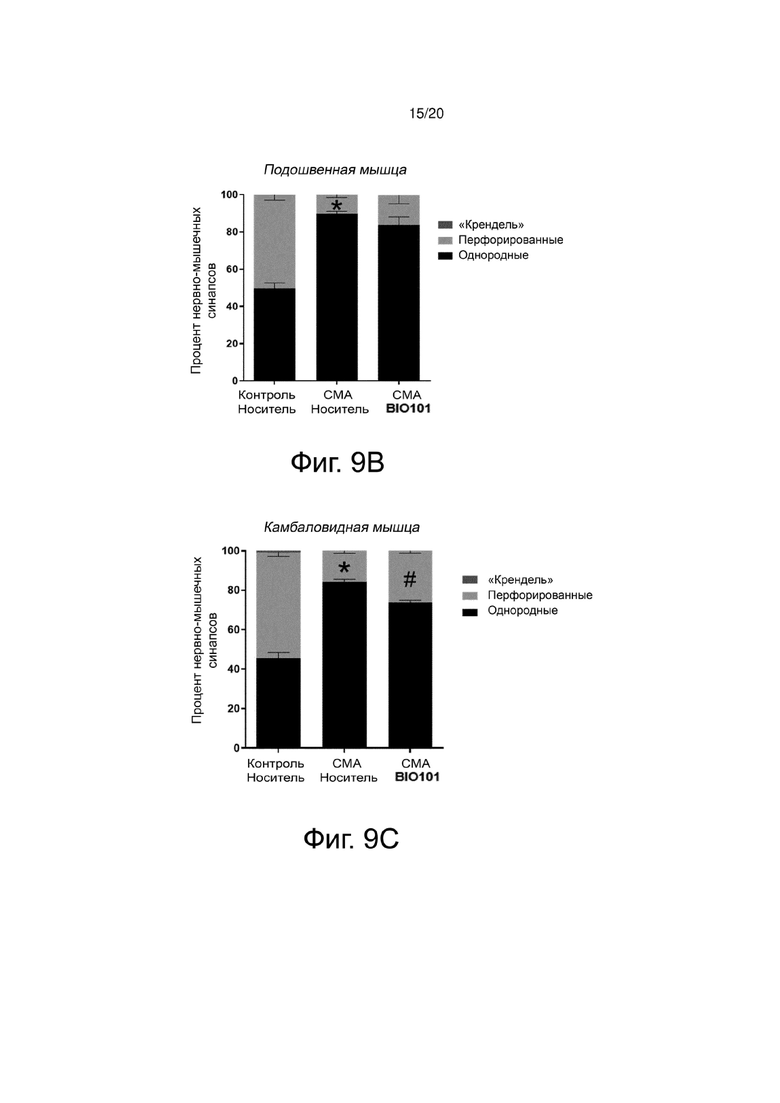
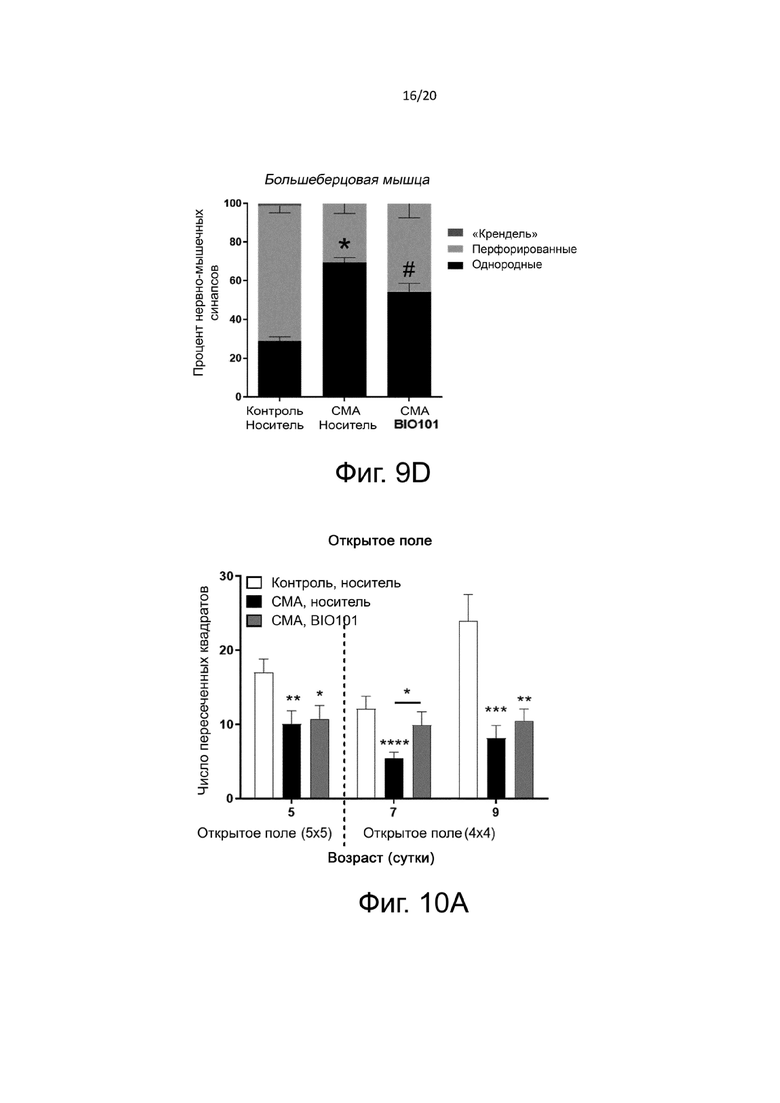
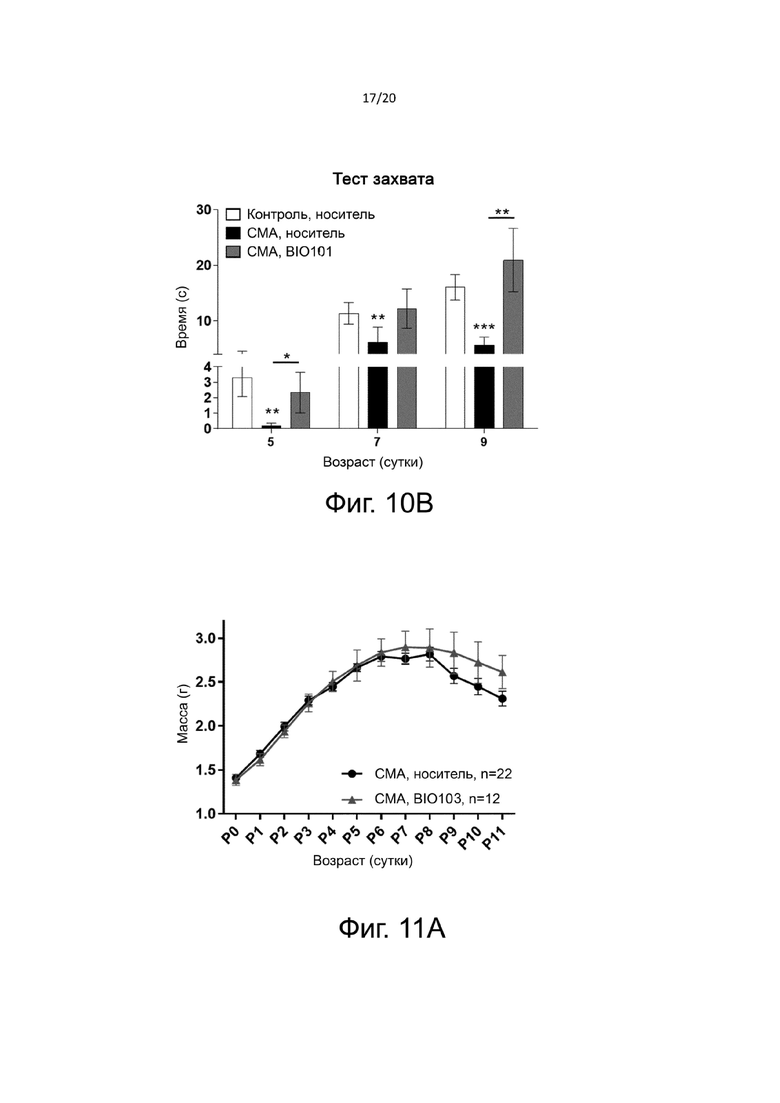
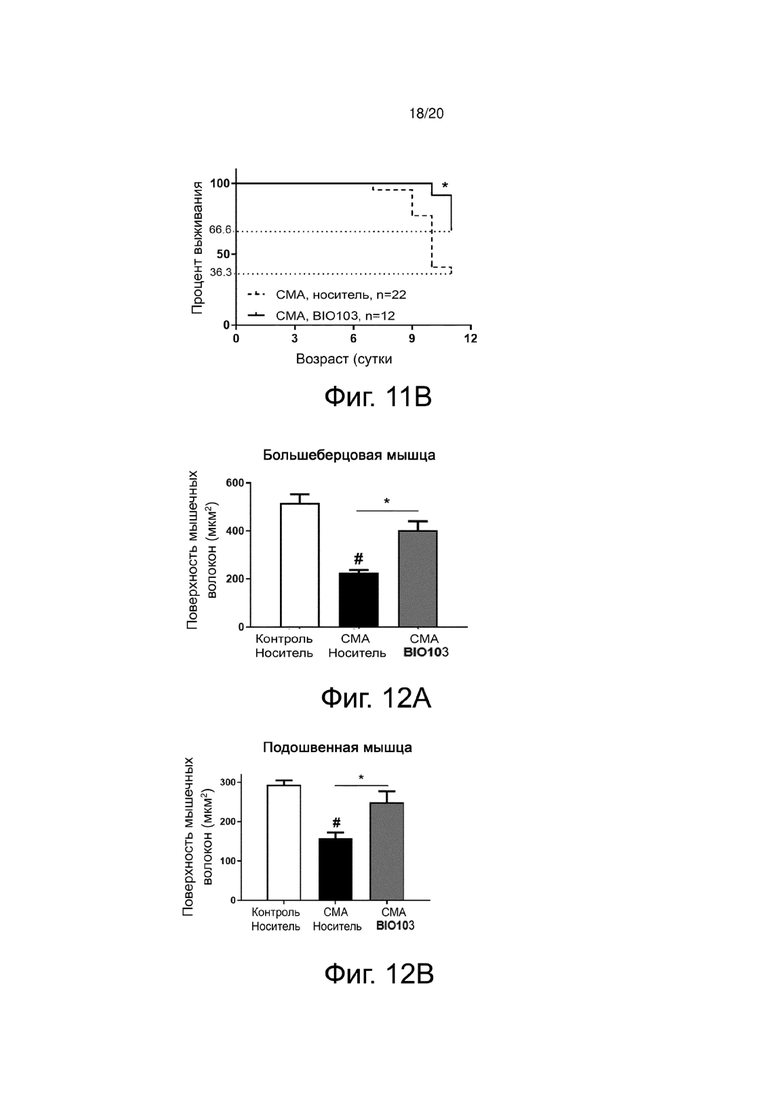
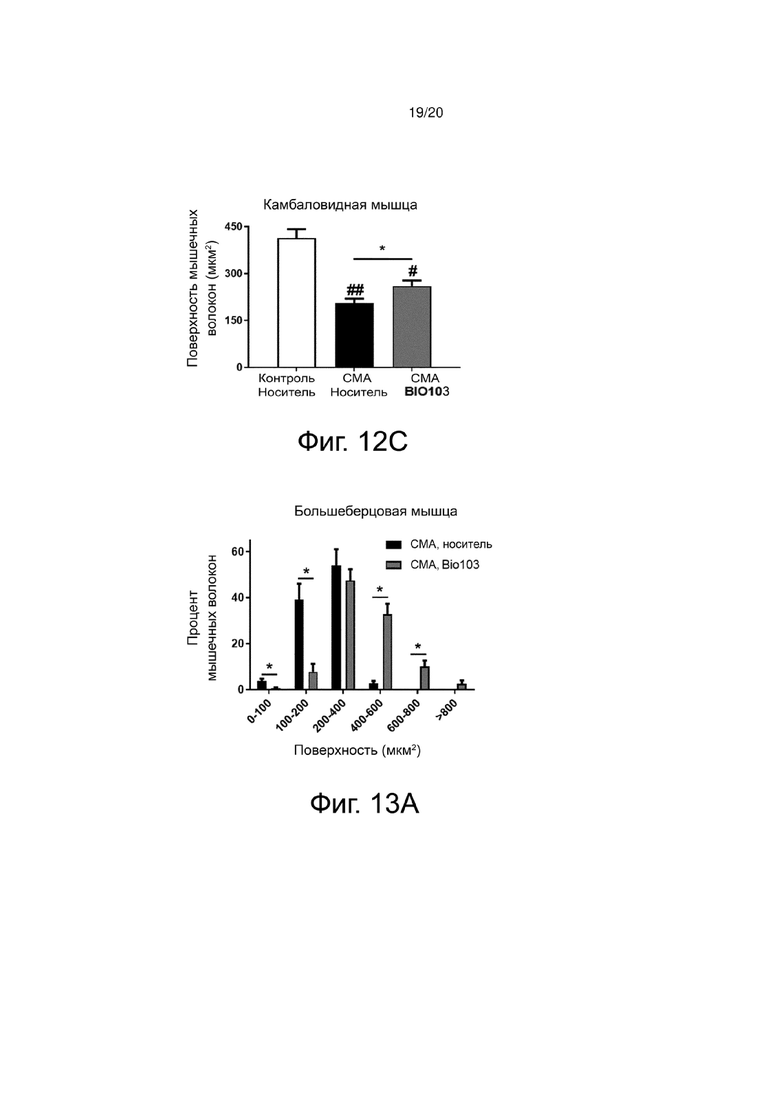
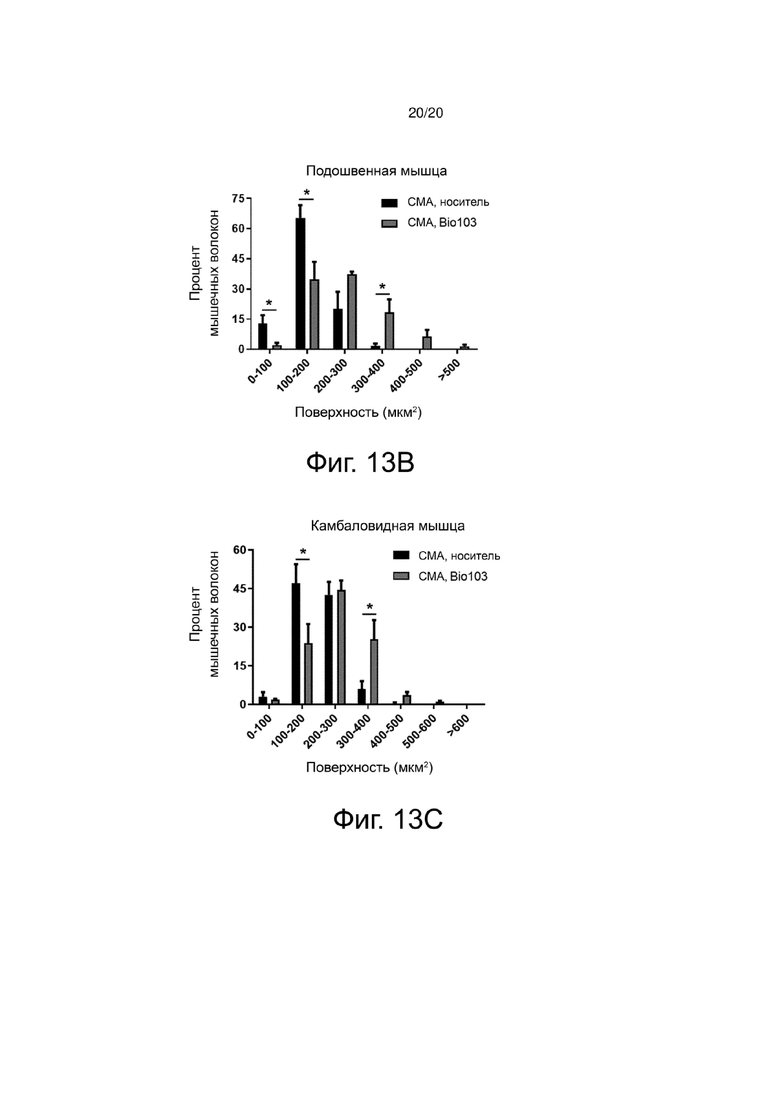
Изобретение относится к применению композиции, содержащей по меньшей мере 20-гидроксиэкдизон и/или по меньшей мере одно полусинтетическое производное 20-гидроксиэкдизона, для лечения специфического расстройства моторных нейронов у млекопитающих, страдающих нервно-мышечным заболеванием, включающим изменение мышечной функции из-за специфического нарушения моторных нейронов, причем указанное заболевание представляет собой спинальную мышечную атрофию (СМА) и указанное полусинтетическое производное 20-гидроксиэкдизона выбрано из соединения формулы (I) или соединения общей формулы (II). В общей формуле (I) R1 выбран из: группы (C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)CO2(C1-C6); группы (C1-C6)A, причем A представляет собой гетероцикл, необязательно замещенный группой, выбранной из типа OH, OMe, (C1-C6), N(C1-C6), CO2(C1-C6), группой CH2Br; W является гетероатомом, выбранным из N, O и S, предпочтительно O, но более предпочтительно S. Использование изобретения позволяет эффективно лечить спинальную мышечную атрофию (СМА). 13 з.п. ф-лы, 13 ил.
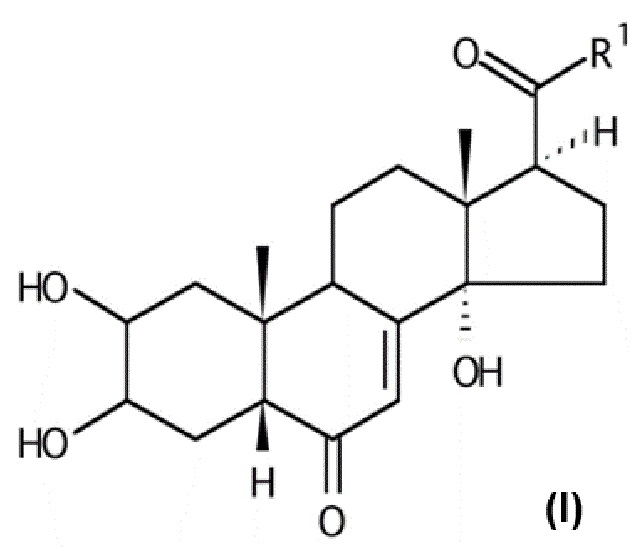
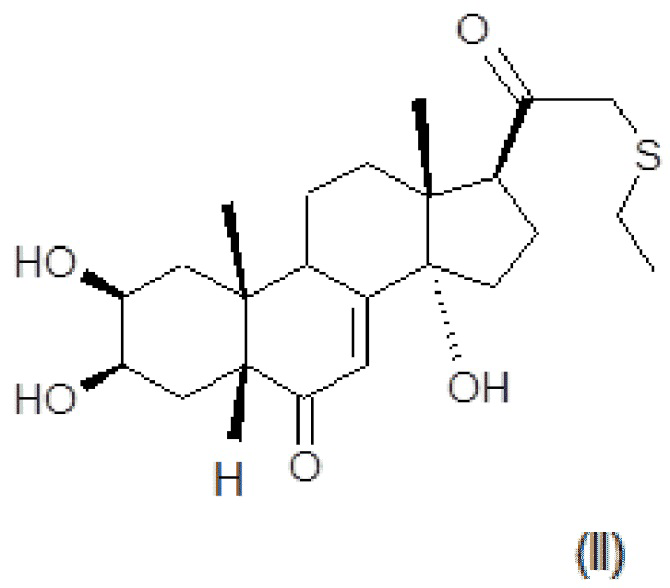
1. Применение композиции, содержащей по меньшей мере 20-гидроксиэкдизон и/или по меньшей мере одно полусинтетическое производное 20-гидроксиэкдизона, для лечения специфического расстройства моторных нейронов у млекопитающих, страдающих нервно-мышечным заболеванием, включающим изменение мышечной функции из-за специфического нарушения моторных нейронов, причем указанное заболевание представляет собой спинальную мышечную атрофию (СМА) и указанное полусинтетическое производное 20-гидроксиэкдизона выбрано из:
- соединения формулы (I)
[соединение 1]
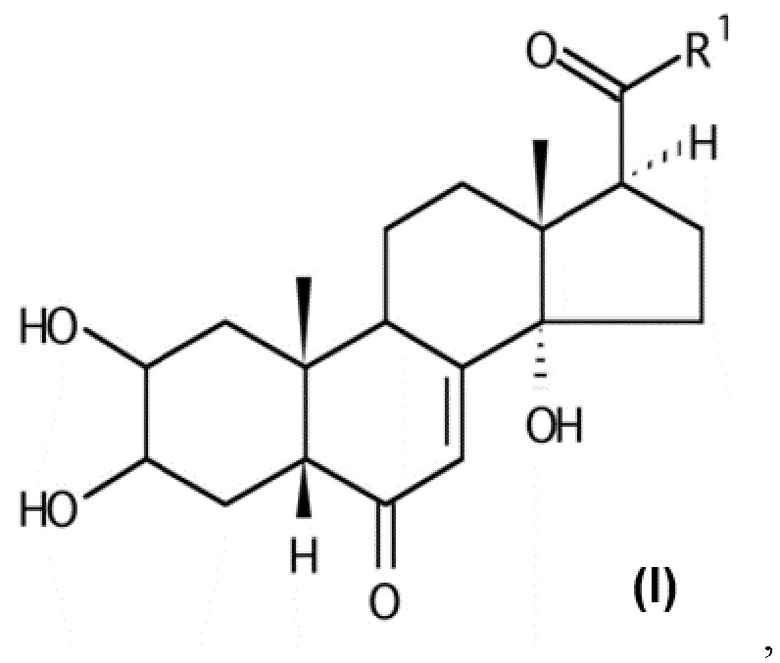
где R1 выбран из: группы (C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)W(C1-C6); группы (C1-C6)W(C1-C6)CO2(C1-C6); группы (C1-C6)A, причем A представляет собой гетероцикл, необязательно замещенный группой, выбранной из типа OH, OMe, (C1-C6), N(C1-C6), CO2(C1-C6), группой CH2Br;
W является гетероатомом, выбранным из N, O и S, предпочтительно O, но более предпочтительно S;
- соединения общей формулы (II)
[соединение 2]
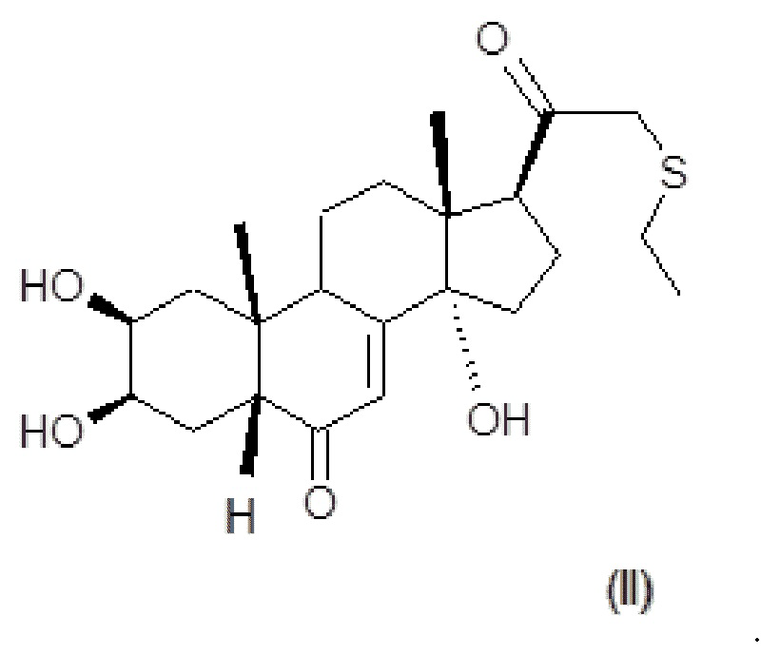
2. Применение по п. 1, где 20-гидроксиэкдизон находится в форме экстракта растения или экстракта части растения, где указанное растение выбрано из растений, содержащих по меньшей мере 0,5% 20-гидроксиэкдизона по сухой массе указанного растения, причем указанный экстракт включает по меньшей мере 95%, а предпочтительно по меньшей мере 97% 20-гидроксиэкдизона.
3. Применение по п. 2, где экстракт включает от 0 до 0,05% по сухой массе экстракта примесей, способных повлиять на безопасность, доступность или эффективность фармацевтического применения указанного экстракта.
4. Применение по любому из пп. 2, 3, где растение выбрано из Stemmacantha carthamoides, Cyanotis arachnoidea и Cyanotis vaga.
5. Применение по любому из пп. 1-4, где изменение мышечной функции вызвано изменением функции моторных нейронов или их дегенерацией.
6. Применение по любому из пп. 1-5, где расстройство моторных нейронов является результатом генетического изменения у млекопитающих, страдающих нервно-мышечным заболеванием.
7. Применение по любому из пп. 1-6, где измененной мышечной функцией является функция поперечно-полосатых скелетных мышц или миокарда.
8. Применение по любому из пп. 1-7, где изменение мышечной функции связано с аплазией и/или атрофией.
9. Применение по любому из пп. 1-8, где 20-гидроксиэкдизон и/или указанное по меньшей мере одно полусинтетическое производное 20-гидроксиэкдизона используют для лечения по меньшей мере одного генетического изменения, ответственного за СМА.
10. Применение по любому из пп. 1-9, где нервно-мышечное заболевание возникает в результате мутации гена SMN1.
11. Применение по любому из пп. 1-10, где лечение специфического расстройства моторных нейронов включает улучшение выживаемости моторных нейронов и/или ускорение созревания нервно-мышечных синапсов.
12. Применение по любому из пп. 1-11, где 20-гидроксиэкдизон и/или указанное по меньшей мере одно полусинтетическое производное 20-гидроксиэкдизона применяют в дозе от 3 до 15 мг на 1 кг в сутки у людей.
13. Применение по любому из пп. 1-12, где 20-гидроксиэкдизон и/или указанное по меньшей мере одно полусинтетическое производное 20-гидроксиэкдизона применяют в дозе от 200 до 1000 мг/сутки, разделенной на одну или несколько доз, для взрослых людей и в дозе от 5 до 350 мг/сутки, разделенной на одну или несколько доз, для детей или младенцев.
14. Применение по любому из пп. 1-13, где указанное по меньшей мере одно соединение общей формулы (I) выбрано из:
№1: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-10,13-диметил-17-(2-морфолиноацетил)-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-она;
№2: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-(3-гидроксипирролидин-1-ил)ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-она;
№3: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-(4-гидрокси-1-пиперидил)ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-она;
№4: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-[4-(2-гидроксиэтил)-1-пиперидил]ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-она;
№5: (2S,3R,5R,10R,13R,14S,17S)-17-[2-(3-диметиламинопропил(метил)амино) ацетил]-2,3,14-тригидрокси-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-она;
№6: 2-[2-оксо-2-[(2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-10,13-диметил-6-оксо-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-17-ил]этил]этил сульфанилацетата;
№7: (2S,3R,5R,10R,13R,14S,17S)-17-(2-этилсульфанилацетил)-2,3,14-тригидрокси-10,13-диметил-2,3,4,5, 9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-она;
№8: (2S,3R,5R,10R,13R,14S,17S)-2,3,14-тригидрокси-17-[2-(2-гидроксиэтилсульфанил)ацетил]-10,13-диметил-2,3,4,5,9,11,12,15,16,17-декагидро-1H-циклопента[a]фенантрен-6-она.
| WO 2018197708 A1, 01.11.2018 | |||
| US 20160220624 A1, 04.08.2016 | |||
| RASHIN MOHSENI et al., Overexpression of SMN2 Gene in Motoneuron-Like Cells Differentiated from Adipose-Derived Mesenchymal Stem Cells by Ponasterone A, Journal of Molecular Neuroscience | |||
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| US 20170226151 A1, 10.08.2017 | |||
| KOKONTIS L, | |||

