Область техники
[0001] Настоящее изобретение относится к области медицины и касается метабиотической композиции, обладающей антагонистической активностью по отношению к Helicobacter pylori. В частности данная композиция направлена на лечение и предупреждение расстройств желудочно-кишечного тракта, вызванных Helicobacter pylori, а также на улучшение пищеварения и поддержку здоровой микрофлоры пищеварительной системы пациента.
Уровень техники
[0002] Грамотрицательная спиралевидная Helicobacter pylori (H. pylori) вызывает широкий ряд заболеваний желудочно-кишечного тракта (ЖКТ), таких как гастрит, язва желудка и двенадцатиперстной кишки, синдром раздраженного кишечника, рак желудка и другие. Примерно около половины населения земного шара инфицировано данной бактерией [1]. Известно, что при попадании бактерии H. pylori в желудочно-кишечный тракт происходит колонизация слизистой оболочки желудка и последующая выработка аммиака с повышением значений pH. Это приводит к повышенной секреции соляной кислоты и пепсина, которые начинают разъедать поврежденную слизистую желудка с развитием острого гастрита [2]. Профилактическое лечение пациентов позволяет снизить риски возникновения большинства желудочно-кишечных патологий, в том числе и предотвратить возникновение рака желудка [3]. В ряде случаев прямая связь между присутствием H. pylori и заболеванием не до конца выяснена, как, например, в случае синдрома раздраженного кишечника.
[0003] При лечении организма от инвазии H. pylori применяется стандартная тройная эрадикационная терапия, которая использует сочетание трех направленных препаратов - ингибитора протонной помпы для подавления выработки соляной кислоты, и антибиотиков, например, кларитромицина и амоксициллина, направленных на эрадикацию (уничтожение) бактерии в организме [4]. Поскольку для лечения используются антибиотики широкого спектра действия, лечение может сопровождаться дисбактериозом кишечника. В дополнение к этому, эрадикационная терапия способствует повышению антибиотикорезистентности патогенов, что приводит к появлению бактериальных заболеваний, тяжело поддающихся стандартному лечению [5]. Для H. pylori известны случаи возникновения устойчивости к кларитромицину, метронидазолу и левофлоксацину, из-за чего эффективность эрадикационной терапии по всему миру снижается [6].
[0004] В ЖКТ обитает множество разнообразных микроорганизмов, таких как бактерии, грибы и простейшие, которые формируют так называемую микрофлору (микробиоту) кишечника [7]. Микрофлора кишечника обладает разнообразными функциями и играет важную роль в поддержании гомеостаза. При лечении антибиотиками в организме наблюдается дисбактериоз - нарушение качественного и количественного состава микрофлоры ЖКТ, что приводит к нарушению нормальной работы органов пищеварительного тракта. Помимо этого, микрофлора кишечника играет важную роль в функционировании иммунитета посредством формирования колонизационной резистентности, то есть защиты организма от патогенной микрофлоры. Таким образом, в результате нарушения естественного индивидуального количественного и качественного состава микрофлоры кишечника может происходить ослабление иммунитета всего организма [8].
[0005] Для лечения и профилактики дисбактериозов применяют пробиотики и пребиотики. Наиболее часто для лечения и профилактики дисбактериоза кишечника человека в качестве пробиотиков используют живые бактерии, принадлежащие к родам Bifidobacterium, Lactobacillus и Bacillus [9]. В качестве пребиотиков часто используют различные олиго- и полисахариды - например, инулин, клетчатка, лактулоза и другие. За счет своих свойств пребиотики усиливают эффект пробиотиков, поэтому их часто назначают в комплексе [10].
[0006] Пробиотики, являясь компонентами нормальной микрофлоры кишечника, могут проявлять противомикробные свойства относительно патогенных микроорганизмов. Противомикробные свойства могут быть обусловлены выделением веществ, непосредственно подавляющих рост патогенных микроорганизмов. Кроме того, пробиотики могут влиять на кислотность среды, создавая тем самым неблагоприятные условия для размножения патогенных микроорганизмов [11, 12].
[0007] Пробиотические композиции активно используют для лечения ряда заболеваний ЖКТ, вызванных или ассоциированных с H. pylori. Так, известна композиция, включающая бактерии семейства Lactobacillus, ингибирующие рост патогенных бактерий, в том числе, H. pylori (US2 0200069749 A1; опубл. 05.03.2020; МПК A61K 35/747, A61K 35/66, A61K 35/74, A61K 35/744, C12N 1/20, A61K 2035/115, Y02A 50/30).
[0008] Указанная композиция, направлена на уменьшение кислотности желудка, вызванной приемом ингибитора протонной помпы при эрадикации H. pylori. Композиция включает несколько пробиотических штаммов бактерий из семейства Lactobacillus. В частности, в одном из вариантов исполнения, указанная композиция включает, как минимум, один штамм Lactobacillus reuteri (L. reuteri) (EP 2753343 B1, опубл. 16.07.2014; МПК A23L 33/135, A61K 35/747, A23L 33/40, A61K 31/198, A61K 35/74, A61K 35/741, A61K 35/744, A61K 35/745, A61K 38/47, A61K 45/06, A61P 1/00, Y02A 50/30).
[0009] Пробиотические штаммы L. reuteri известны своими антагонистическими свойствами в отношении H. pylori. Было показано, что L. reuteri специфически распознает и связывается с белками на поверхности H. pylori, что способствует выведению H. pylori из ЖКТ за счет перистальтики. Данная особенность позволяет снизить нагрузку патогенной флоры на желудок [13]. В сочетании с тройной терапией использование L. reuteri улучшает эффективность эрадикации H. pylori до 20%, и общая картина симптомов у пациентов смягчается [14]. Также отмечается, что стандартное лечение антибиотиками в сочетании с данными пробиотическими бактериями показывает меньше побочных эффектов.
[0010] Однако обе указанные выше композиции включают живые бактерии семейства Lactobacillus. Данные композиции могут значительно терять эффективность при сочетании их с антибиотиками за счет гибели пробиотиков. Кроме того, использование лишь одной пробиотической культуры в составе вышеуказанных композиций увеличивает шанс развития устойчивости организма и при длительном применении пробиотических препаратов возможно развитие конкуренции вносимых микроорганизмов с собственной микрофлорой, которая является уникальной для каждого пациента. Синбиотическое лечение с использованием пребиотиков и метабиотиков, (продуктов жизнедеятельности бактерий и неживых клеток) выступает в качестве альтернативы известным пробиотическим композициям, что особенно важно в терапии H.pylori.
[0011] Для лечения синдрома раздраженного кишечника описан способ с использованием пробиотического штамма Lactobacillus paracasei (L. paracasei) (RU 2490325 C2; опубл. 20.08.2013; МПК A61K 35/74, A61K 35/747, A23L 33/135, A61K 35/745, A61P 1/00, A61P 1/06, A61P 1/12, A61P 29/00, A61P 31/00). В указанном способе описана возможность использования не живых бактерий, а и «полученного из пробиотика материала». Однако указанный «материал» описан лишь общими терминами без конкретизации состава или способа получения. Кроме того, хотя H. pylori упоминается как возможный патоген, вызывающий синдром раздраженного кишечника, нет никаких данных об эффективности указанной композиции в отношении H. pylori.
[0012] Известна композиция на основе инактивированных (т.е. неспособных к размножению) пробиотиков, обладающая антагонистическим действием в отношении H. pylori (CN 113577218 A; опубл. 02.11.2021; МПК: A61K 35/747, A61K 36/9064, A61K 8/9789, A61K 8/9794, A61K 8/99, A61P 1/02, A61P 11/04, A61P 31/04, A61Q 11/00). Однако данная композиция помимо нескольких пробиотических штаммов включает еще девять компонентов, относящихся к традиционной китайской медицине. Столь сложный и специфический состав усложняет производство композиции.
[0013] Известна композиция на основе инактивированных клеток L. reuteri, обладающая антагонистическим действием в отношении H. pylori. Данная композиция представляет собой высушенный штамм L. reuteri с известной коаггрегационной активностью относительно H. pylori, в одном из вариантов исполнения это штамм DSM 17648 (EP 2717890 B1; опубл. 08.06.2012; A61K 35/747, A61P 1/00, A61P 1/04, A61P 1/06, A61P 1/08, A61P 1/12, A61P 1/14, A61P 31/04, A61P 35/00). Однако данная композиция включает в себя лишь один пробиотический штамм, что делает композицию более узкоспецифичной относительно патогена. Таким образом, в настоящее время существует необходимость в создании метабиотической композиции с показанной эффективностью ингибирования H. pylori, способствующей профилактике дисбактериоза кишечника, доступной в производстве.
Термины и сокращения
[0014] Дисбактериоз - это качественное и/или количественное изменение бактериальной микрофлоры в организме, перемещение различных ее представителей в несвойственные им места обитания, сопровождающееся метаболическими и иммунными нарушениями. Причем указанные нарушения не исчезают после устранения неблагоприятного фактора, вызвавшего дисбактериоз. Чаще всего под дисбактериозом подразумевают нарушение микрофлоры кишечника [15,16].
[0015] Культуральная жидкость - жидкая среда, получаемая при культивировании различных про- и эукариотических клеток in vitro и содержащая остаточные питательные вещества и продукты метаболизма этих клеток [17].
[0016] Культуральная суспензия - жидкая среда, получаемая при культивировании различных про- и эукариотических клеток in vitro и содержащая культивируемые клетки и/или споры.
[0017] Метабиотики - вещества, являющиеся структурными компонентами пробиотических микроорганизмов и/или их метаболиты, которые способны оптимизировать физиологические функции, метаболические, эпигенетические, информационные, регуляторные, транспортные, иммунные, нейрогормональные, и/или поведенческие реакции, связанные с деятельностью симбиотической (индигенной) микрофлоры организма-хозяина [18].
[0018] Микрофлора (микробиота) - совокупность различных видов микроорганизмов, населяющих определенную среду обитания. Постоянно живущие в симбиозе с хозяином микроорганизмы называются нормальной, или симбиотической, микрофлорой [19].
[0019] Пребиотики - химические соединения, способные стимулировать рост и развитие групп бактерий, входящих в состав естественной микрофлоры организма [10].
[0020] Пробиотики - жизнеспособные микроорганизмы, способствующие улучшению функций пищеварения за счет восстановления и поддержания нормального количественно-качественного состава микрофлоры [11].
[0021] ЖК - жирные кислоты.
[0022] ЖКТ - желудочно-кишечный тракт.
[0023] кДа - килодальтон.
[0024] КОЕ - колониеобразующая единица.
[0025] B. subtilis - Bacillus subtilis.
[0026] E. coli - Escherichia coli.
[0027] L. reuteri - Lactobacillus reuteri.
[0028] CD64 - мембранный белок Fc-гамма-рецептор 1, моноцитарный маркер, также провоспалительный маркер макрофагов.
[0029] CDX-2 - белок гомеобокса каудального типа 2, маркер кишечной дифференцировки.
[0030] COX-2 - белок циклооксигеназы-2, белок-маркер воспаления.
[0031] IL1b - интерлейкин 1 бета, провоспалительный цитокин.
[0032] IL6 - интерлейкин 6, про- и противовоспалительный цитокин, фактор регенерации, фактор активации макрофагов, фактор стимуляции лимфоцитов.
[0033] IL10 - интерлейкин 10, противовоспалительный цитокин.
[0034] IL-23 - интерлейкин 23, провоспалительный цитокин.
[0035] MRSA - метициллин-резистентные штаммы S. Aureus.
[0036] RI - индекс удерживания.
[0037] RT - время удерживания.
[0038] TNFa - фактор некроза опухолей альфа, фактор активации макрофагов.
[0039] TGFb - трансформирующий фактор роста бета, блокирует активацию макрофагов, подавляет воспалительный иммунный ответ.
Сущность изобретения
[0040] Задачей настоящего изобретения является создание метабиотической композиции для лечения и профилактики болезней желудочно-кишечного тракта, ассоциированных с наличием, а также эрадикацией H. pylori.
[0041] Данная задача решается заявляемым изобретением за счет достижения такого технического результата, как создание метабиотической композиции, обладающей антагонистическими свойствами по отношению к патогенным и условно-патогенным микроорганизмам, в том числе к H. pylori, стимулирующим действием в отношении нормальной микрофлоры кишечника, а также обладающей иммуномодулирующим потенциалом.
[0042] Заявленный технический результат достигается за счет того, что заявленная композиция включает инактивированные клетки L. reuteri, комплекс метаболитов B. subtilis и наполнитель. При этом соотношение компонентов, мас. %:
[0043] В рамках заявленного изобретения инактивированные клетки L. reuteri представляют собой высушенные клетки, не способные к размножению. Инактивированные клетки пробиотического штамма L. reuteri не составляют конкуренцию нормальной микрофлоре пациента, при этом выполняя антагонистическую роль в отношении ряда патогенных бактерий. В предпочтительном варианте реализации инактивированные клетки пробиотического штамма L. reuteri являются неживыми, например, умерщвленными посредством термической обработки. В качестве пробиотического штамма L. reuteri могут быть использованы штаммы L. reuteri, обладающие коаггрегационным эффектом по отношению к H. pylori, например, DSM-17648, SALR01 или ATCC-55730. В предпочтительном варианте реализации пробиотический штамм L. reuteri представляет собой L. reuteri GMNL-0902. В предпочтительном варианте реализации инактивированные клетки пробиотического штамма L. reuteri являются высушенными.
[0044] Комплекс метаболитов B. subtilis представляет собой вещества, содержащиеся в смеси культуральной жидкости, вегетативных клеток и спор пробиотичсекого штамма B. subtilis. В качестве пробиотического штамма B. subtilis могут быть использованы штаммы B. subtilis ВКПМ №В-2335 (3), B. subtilis 534, B. subtilis ТПИ 13 - 07.06.21 - ДЕП/ВГНКИ, B. subtilis ВКПМ №В4759 (3), B. subtilis р ВМВ105, B. subtilis ТПИ 9; B. subtilis рВМВ 105. В предпочтительном варианте реализации пробиотический штамм B. subtilis представляет собой B. subtilis ВКМ В-3536D и/или ВКПМ №В-2335 (3).
[0045] Метаболиты B. subtilis способны селективно подавлять рост широкого ряда патогенных и условно патогенных микроорганизмов. Кроме того, комплекс метаболитов B. subtilis обладает также противовоспалительными свойствами, что может позволить дополнительно облегчить состояние пациентов с заболеваниями желудочно-кишечного тракта. Сочетание вегетативных клеток и метаболитов пробиотического штамма B. subtilis позволяет комплексно воздействовать на патогенные микроорганизмы без потери эффективности при терапии антибиотиками, что позволяет использовать ее для лечения и профилактики дисбактериоза для широкого круга пациентов.
[0046] Наполнитель представляет собой одно или несколько веществ, которые могут способствовать сохранению целостности композиции, сохранению биологической активности указанных метаболитов и/или могут оказывать полезный физиологический эффект, усиливая полезное действие композиции.
[0047] В предпочтительном варианте реализации заявленная композиция в качестве наполнителя включает один или несколько пребиотиков, которые могут представлять собой ди-, олиго- и полисахариды. В еще более предпочтительном варианте реализации один или несколько пребиотиков могут иметь растительное происхождение. Например, пребиотики могут быть представлены такими полифруктозанами, как, инулин, олигофруктоза или их смесь. Пребиотики стимулируют рост симбиотической микрофлоры кишечника, оказывают иммуномодулирующее и гепатопротекторное действие.
[0048] В предпочтительном варианте реализации заявленная композиция включает в состав наполнителя источник цинка. В качестве источника цинка может выступать соль цинка, предпочтительно, органическая соль цинка, например, например, цитрат цинка.
[0049] Технический результат также достигается за счет разработки доступного способа получения заявленной композиции. Способ включает следующие этапы:
а) выращивание пробиотического штамма B. subtilis в жидкой питательной среде.
В предпочтительном варианте среда имеет исходный рН 7,0±0,2. Выращивание могут проводить путем глубинного выращивания в ферментере с коэффициентом заполнения 60-70% при температуре 36-38°С при постоянном перемешивании и аэрации. При этом выращивание могут заканчивать, когда рН культуральной суспензии достигает 8,5-8,9 и/или до достижения созревания культуры не менее 90%. Этот этап выращивания соответствует максимальной концентрации метаболитов B. subtilis в культуральной суспензии. Указанный способ выращивания позволяет максимально эффективно использовать оборудование для получения метаболитов B. subtilis;
б) путем центрифугирования культуральной суспензии на проточной центрифуге отделяют вегетативные клетки и споры пробиотического штамма B. subtilis от культуральной жидкости;
в) высушивают полученную культуральную жидкость. В предпочтительном варианте реализации культуральную жидкость высушивают методом лиофилизации;
г) инактивируют вегетативные клетки и споры B. subtilis, отделенные от культуральной жидкости;
д) смешивают лиофилизированную культуральную жидкость, инактивированные вегетативные клетки и споры B. subtilis в соотношении;
е) к полученному комплексу метаболитов B. subtilis добавляют инактивированные клетки L. reuteri, а также наполнитель.
[0050] Заявленный способ обеспечивает получение композиции, включающей как метаболиты L. reuteri, так и практически весь спектр метаболитов B. subtilis. Благодаря этому заявленная композиция способствует эрадикации H. pylori, усиливает иммунитет и позволяет избежать или снизить эффект дисбактериоза, который может быть вызван антибиотиками, используемыми при лечении заболеваний ЖКТ, ассоциированных с H. pylori.
[0051] Способ может дополнительно включать высушивание инактивированных вегетативных клеток и спор B. subtilis. В предпочтительном варианте реализации инактивированные клетки и споры B. subtilis высушивают методом лиофилизации.
[0052] В предпочтительном варианте реализации для получения композиции все компоненты используют в сухом виде. Это увеличивает срок хранения композиции без потери ее эффективности. Это связано с сохранением активности метаболитов и с минимизацией риска загрязнения композиции какими-либо активными микроорганизмами.
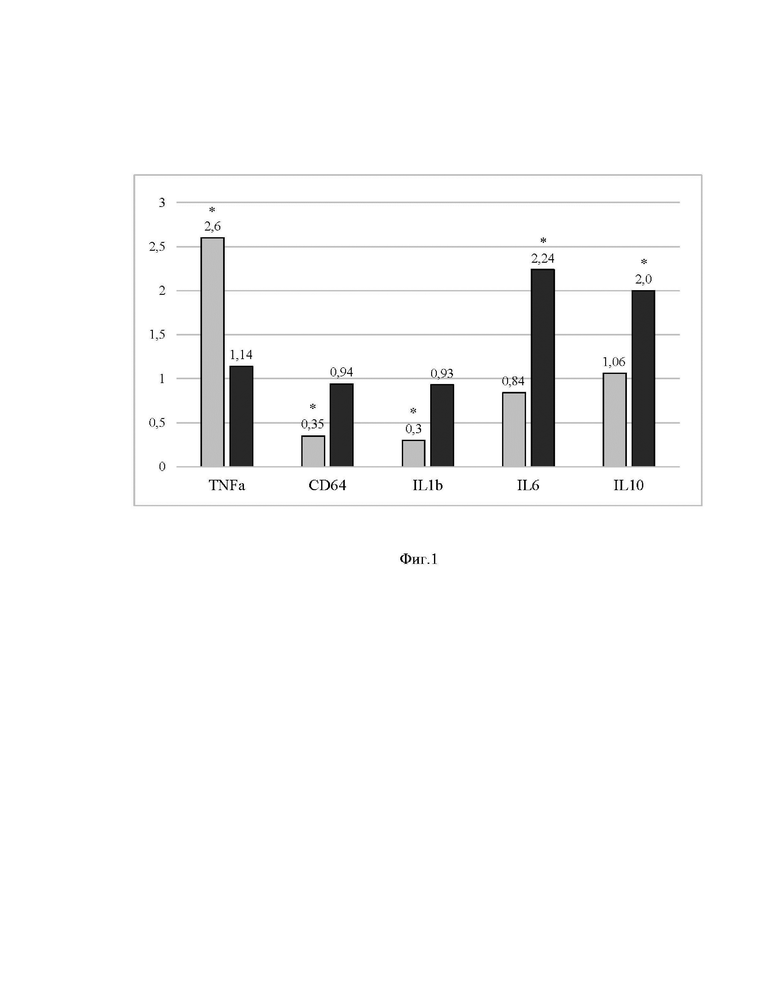
Описание чертежей
[0053] На Фиг. 1 представлены результаты исследования иммуномодулирующих свойств комплекса метаболитов B. subtilis.
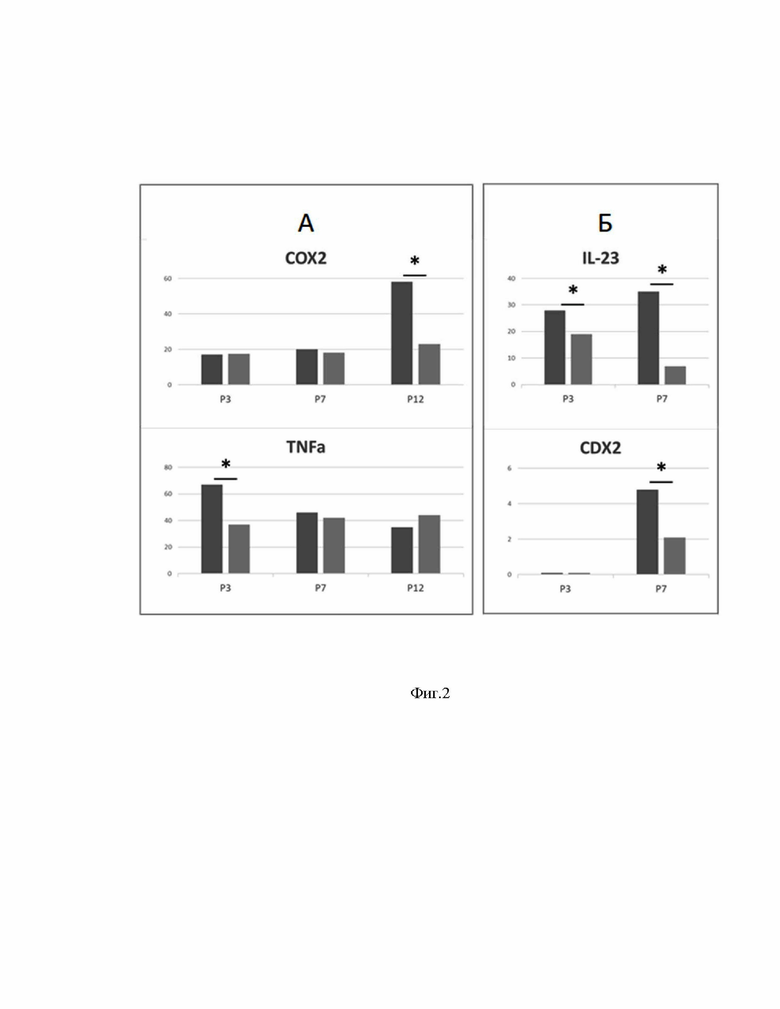
[0054] На Фиг.2А представлены результаты исследования экспрессии маркеров воспаления в культуре клеток желудка человека.
[0055] На Фиг.2Б представлены результаты исследования экспрессии маркеров воспаления в культуре клеток кишечника человека.
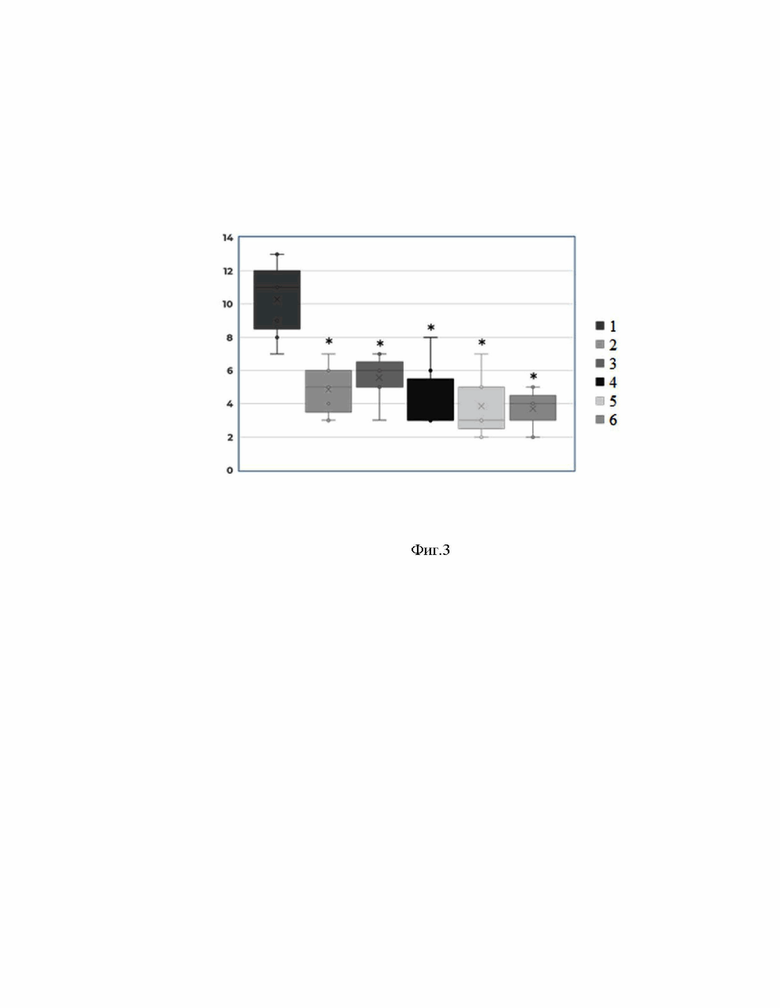
[0056] На Фиг. 3 представлены результаты исследования по оценке эффективности применения заявленной композиции при лечении хеликобактериоза на модели монгольских песчанок.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0057] В приведенном ниже подробном описании реализации изобретения приведены многочисленные детали реализации, призванные обеспечить отчетливое понимание настоящего изобретения. Однако, квалифицированному в предметной области специалисту, очевидно, каким образом можно использовать настоящее изобретение, как с данными деталями реализации, так и без них. В других случаях хорошо известные методы, процедуры и компоненты не описаны подробно, чтобы не затруднять излишне понимание особенностей настоящего изобретения.
[0058] Кроме того, из приведенного изложения ясно, что изобретение не ограничивается приведенной реализацией. Многочисленные возможные модификации, изменения, вариации и замены, сохраняющие суть и форму настоящего изобретения, очевидны для квалифицированных в предметной области специалистов.
[0059] Заявленная композиция включает метаболиты клетки L. reuteri, комплекс метаболитов B. subtilis и наполнитель. При этом соотношение компонентов, мас. %:
[0060] В заявленной композиции инактивированные клетки L. reuteri представляют собой высушенные клетки, не способные к размножению. Под инактивированными клетками подразумеваются клетки, не способные к размножению. В предпочтительном варианте реализации клетки пробиотического штамма L. reuteri являются неживыми, например, умерщвленными посредством термической обработки. В качестве пробиотического штамма L. reuteri могут быть использованы штаммы L. reuteri, обладающие коаггрегационным эффектом по отношению к H. pylori - SALR01, DSM-17648 или ATCC - 55730. В предпочтительном варианте реализации пробиотический штамм L. reuteri представляет собой L. reuteri GMNL-0902.
[0061] Использование инактивированных клеток L. reuteri в концентрации менее 16 мас. % приводит к существенному снижению эффективности композиции. Увеличение содержания метаболитов более 70 мас. % нерационально, так как увеличивает стоимость производства и при этом не оказывает значительного влияния на эффективность композиции.
[0062] Комплекс метаболитов B. subtilis представляет собой вещества, содержащиеся в культуральной суспензии пробиотического штамма B. subtilis. В предпочтительном варианте реализации комплекс метаболитов представляет собой вещества, содержащиеся в смеси культуральной жидкости, вегетативных клеток и спор пробиотического штамма B. subtilis. Указанная смесь включает практически весь спектр метаболитов пробиотического штамма B. subtilis. Экспериментальные данные показывают, что метаболиты B. subtilis способны селективно подавлять рост широкого ряда патогенных и условно-патогенных микроорганизмов, например: Staphylococcus aureus, Listeria monocytogenes, Pseudomonas aeruginosa, Salmonella typhimurium, Staphylococcus epidermidis, Streptococcus agalactiae, Enterococcus faecium, Enterococcus faecalis, Streptococcuss salivarius, Streptococcus mitis, Escherichia coli (Пр. 2 и 4).
[0063] Метаболиты B. subtilis включают разнообразные биологически активные вещества, которые обладают противомикробным, пробиотическим, иммунномодулирующим действием. Например, экспериментальные данные (Пр. 3) показывают, что метаболиты B. subtilis включают амикумацины А и В, которые относятся к классу продуктов дигидроизокумарина природного происхождения и проявляют антибактериальную, противогрибковую, противоопухолевую и противовоспалительную активность. Механизм их действия заключается в том, чтобы остановить транслокацию рибосом, тем самым блокируя синтез белка в клетках. Амикумацин A обладает антагонистической активностью в отношении H. pylori [20]. Кроме того, амикумацины активны в отношении устойчивых к антибиотикам штаммов возбудителя S. aureus (MRSA - метициллин-резистентные штаммы S. aureus), поэтому представляет особый интерес для борьбы с растущей устойчивостью штаммов возбудителя к антибиотикам. Амикумацин A активен против грамположительных бактерий Salmonella sp. и Shigella sp. [20, 21, 22, 23, 24, 25].
[0064] Также метаболиты B. subtilis включают циклические дипептиды (2,5-дикетопиперазины) проявляют противоопухолевую, и нейропротекторную и антигипергликемическую активность. Кроме того, циклические дипептиды проявляют противирусную, противогрибковую и антибактериальную активность. В частности, цикло(лейцилпролил) (L-Phe-D-Pro лактам) и цикло(D-фенилаланин-L-пролил) показывают антагонистическую активность в отношении S. aureus, E. coli, P. aeruginosa и C. albicans. При этом P. aeruginosa - один из самых распространенных возбудителей внутрибольничных инфекций, приобретающий устойчивость к обычно используемым антибиотикам., а C. albicans - частая причина возникновения грибковых инфекций [26].
[0065] Молочная кислота также относится к метаболитам B. subtilis и обладает противовоспалительным эффектом за счет стимуляции противовоспалительного цитокина IL-10 и подавления активности провоспалительного цитокина IL-12. Также молочная кислота и ее производные (например, 3-фенилмолочная кислота, 4-гидроксифенилмолочная кислота) обладают антибактериальным и противогрибковым эффектом. Например, 3-фенилмолочная кислота (самостоятельно или в сочетании с молочной кислотой) проявляет - подавляет рост таких патогенов, как L. monocyotgenes, E. coli, S. typhimurium, B. cereus, E. faecalis [27, 28, 29, 30,31, 32, 33].
[0066] а-Гидроксиизокапроновая кислота обладает широким антибактериальным действием, включая такие бактерии, как MRSA (метициллин-резистентные штаммы S. aureus), устойчивые к множеству системно применяемых противомикробных препаратов [34].
[0067] Индол представляет собой гетероциклическое конденсированное ароматическое соединение, образующееся в результате разложения аминокислоты триптофана. Индол обладает пробиотическими свойствами и поддерживает нормальный количественный и качественный состав микробиоты кишечника. Также индол поддерживает функцию эпителиального барьера и способствует иммунной толерантности для поддержания микробного комменсализма при защите от патогенных инфекций. Кроме того, индол обладает антикарциногенным, антиоксидантным и противовоспалительным действием. Последнее, вероятно, за счет стимуляции выработки цитокина IL-22 иммунными клетками кишечника [35,36].
[0068] Длинноцепочечные мононенасыщенные жирные кислоты (т.н. Омега-9 жирные кислоты), например, миристолеиновая (C14:1), пальмитолеиновая (C16:1), олеиновая (C18:1) и элаидиновая (C18:1) кислоты увеличивают разнообразие кишечной микробиоты [37]. Кроме того, миристолеиновая, пальмитолеиновая и олеиновая кислоты обладают антагонистической активностью в отношении H. pylori.
[0069] Некоторые насыщенные жирные кислоты с длинной цепью (ДЦЖК, LCFAs) обладают противомикробным и противовоспалительным действием. Например, пальмитиновая кислота обладает антогонистической активностью в отношении S.mutans, S.gordonii, S.sanquis, P.ginbivalis, F.nucleatum. Пентадекановая и маргариновая кислоты оказывают противовоспалительное действие [38,39].
[0070] Насыщенные жирные кислоты со средней длиной цепи, например, каприловая (C8), каприновая (C10), ундециловая (C11) и лауриновая (C12) кислоты, улучшают функцию кишечного эпителия, повышение активности ферментов микроворсинок слизистой оболочки в тонкой кишке. Указанные кислоты обладают антибактериальным, противогрибковым, противовоспалительным действием. Например, лауриновая кислота является наиболее эффективной в отношении Campylobacter jejuni, каприновая и лауриновая кислоты обладают антибактериальным действием в отношении H. pylori [37,40, 41, 42].
[0071] Витамин B5 (пантотеновая кислота) замедляет рост бактерий посредством регуляции врожденного иммунитета и адаптивного иммунитета у мышей, инфицированных Mycobacterium tuberculosis [43].
[0072] Аминокислоты поддерживают функцию кишечного барьера и эндокринную функцию кишечника. Аминокислоты, попадающие в кишечник извне, метаболизируются кишечной микробиотой, которая использует их для синтеза собственных белков и катаболических путей. Показано, что пищевые аминокислоты могут регулировать состав микробиоты. Например, лизин поддерживает комменсальную (пробиотическую) микробиоту в кишечнике. Кроме того, лизин способствует поддержанию иммунной системы [44, 45].
[0073] Культуральная жидкость пробиотического штамма B. subtilis в составе заявленной композиции может быть высушенной до такого состояния, что влажность сухой культуральной жидкости составляет 4-6 мас. % При этом вегетативные клетки и споры пробиотического штамма B. subtilis могут быть инактивированными, то есть неспособными к размножению, предпочтительно неживыми. В одном из вариантов реализации вегетативные клетки и споры пробиотического штамма B. subtilis могут быть инактивированы путем нагрева.
[0074] В качестве пробиотического штамма B. subtilis могут быть использованы штаммы B. subtilis ВКПМ № В-2335 (3), B. subtilis 534, B. subtilis ТПИ 13 - 07.06.21 - ДЕП/ВГНКИ, B. subtilis ВКПМ № В4759 (3), B. subtilis р ВМВ105, B. subtilis ТПИ 9; B. subtilis рВМВ 105. В предпочтительном варианте реализации пробиотический штамм B. subtilis представляет собой B. subtilis ВКМ В-3536D и/или ВКПМ № В-2335 (3).
[0075] Использование комплекса метаболитов B. subtilis в концентрации менее 1 мас. % приводит к существенному снижению эффективности композиции. Увеличение содержания метаболитов более 7 мас. % нерационально, так как увеличивает стоимость производства и при этом не оказывает значительного влияния на эффективность композиции.
[0076] Благодаря отсутствию живых микроорганизмов, заявленная композиция не теряет эффективности при сочетанном приеме с антибиотиками, что позволяет использовать ее для лечения и профилактики заболеваний желудочно-кишечного тракта у широкого круга пациентов. Кроме того, инактивированные вегетативные клетки и споры B. subtilis, а также инактивированные клетки L. reuteri, не создают конкуренции микрофлоре кишечника и не нарушают ее баланс, уникальный для каждого пациента.
[0077] Заявленная композиция включает наполнитель, представляющий собой одно или несколько веществ, которые могут способствовать сохранению целостности композиции, сохранению биологической активности указанных метаболитов и/или могут оказывать полезный физиологический эффект, усиливая полезное действие композиции. В качестве наполнителя могут быть использованы, например, но не ограничиваясь этим: мальтодекстрин, пищевые волокна, производные крахмала, пектины, целлюлоза, микрокристаллическая целлюлоза и другие производные целлюлозы, а также другие технологически приемлемые наполнители (например, выбранные из списка, представленного на сайте https://lexparency.org/eu/32008R1333/ANX_III/). Кроме того, в качестве наполнителя может использоваться один или несколько пребиотиков, которые могут представлять собой ди-, олиго- и полисахариды, устойчивые к гидролизу пищеварительными ферментами человека и не абсорбирующиеся в верхних отделах пищеварительного тракта. Например, это могут быть пищевые волокна злаковых, овощей, фруктов, трав, например, инулин и/или олигофруктоза, или синтетические полисахариды, такие как лактулоза, или их сочетания. В еще более предпочтительном варианте реализации один или несколько пребиотиков могут иметь растительное происхождение. Например, в качестве пребиотиков могут быть использованы полифруктозаны (фруктаны), такие как инулин и/или олигофруктоза. При этом полифруктозаны могут быть получены, например, из цикория, топинамбура, георгина, артишока, свеклы и др. растений.
[0078] Наполнитель также может включать источник цинка. Цинк играет важную роль в развитии и поддержании иммунной системы. Применение цинка подавляет рост патогенных микроорганизмов, например, Mycobacterium, а также препятствует развитию вирусной инфекции [46]. Источник цинка в композиции может составлять не менее 3 мас. % В предпочтительном варианте реализации источником цинка является органическая соль цинка. Это обусловлено тем, что органические соли обладают большей биодоступностью, чем неорганические соли цинка. В одном варианте реализации цинк может быть включен в композицию в хелатной форме, например, в виде соединения с аминокислотой. Однако такие соединения на рынке сравнительно дороги, что значительно увеличивает себестоимость композиции. В предпочтительном варианте реализации источником цинка является цитрата цинка. Цитрат цинка широко доступен на рынке и обеспечивает оптимальную стоимость производства композиции без потери ее эффективности.
[0079] Заявленная композиция может быть использована для изготовления пищевой добавки или фармацевтического препарата в виде порошка, таблетки или капсулы.
[0080] Сочетание бактериальных метабиотиков L. reuteri и B. subtilis способствует лечению заболеваний ЖКТ, ассоциированных с H. pylori. Кроме того, указанное сочетание позволяет проводить профилактику дисбактериоза кишечника, вызванного применением антибиотиков при эрадикации H. pylori. Профилактика дисбактериоза, в свою очередь, способствует предупреждению повторной инфекции.
[0081] Важной частью заявленного изобретения является способ получения заявленной композиции, включающий следующие этапы. На этапе а) проводят глубинное выращивание пробиотического штамма B. subtilis в жидкой питательной среде, имеющей исходный рН 7,0±0,2. В предпочтительном варианте реализации глубинное выращивание проводят в ферментере. При этом коэффициент заполнения ферментера может составлять от приблизительно 60% до приблизительно 70%. Указанный коэффициент заполнения, с одной стороны, позволяет избежать нежелательного пенообразования, и с другой стороны, позволяет максимально эффективно использовать оборудование для получения культуральной суспензии, насыщенной метаболитами пробиотического штамма B. subtilis. Глубинное выращивание проводят при температуре 36-38°С при постоянном перемешивании и аэрации до тех пор, когда рН культуральной суспензии достигнет 8,5-8,9 и/или до достижения созревания культуры не менее 90%. На данном этапе выращивания происходит переход культуры в стационарную фазу, когда клетки перестают делиться, те количество клеточной массы не увеличивается. Этот этап выращивания соответствует максимальной концентрации метаболитов B. subtilis в культуральной жидкости.
[0082] На этапе б) проводят отделение культуральной жидкости от клеток и спор пробиотического штамма B. subtilis. В одном варианте реализации культуральную жидкость отделяют от вегетативных клеток и спор путем фильтрации. В предпочтительном варианте реализации культуральную жидкость отделяют от вегетативных клеток и спор путем центрифугирования культуральной суспензии на проточной центрифуге до получения прозрачного супернатанта, представляющего собой культуральную жидкость. При этом осадок в основном содержит вегетативные клетки и споры пробиотического штамма B. subtilis.
[0083] На этапе в) высушивают культуральную жидкость, полученную на этапе б). Культуральная жидкость может быть высушена любым подходящим способом, известным из уровня техники. В предпочтительном варианте реализации культуральную жидкость высушивают методом лиофилизации. При этом культуральную жидкость высушивают до такого состояния, что влажность сухой культуральной жидкости составляет 4-6 мас. %
[0084] На этапе г) инактивируют вегетативные клетки и споры пробиотического штамма B. subtilis путем нагревания осадка, полученного на этапе б), до температуры 60-140°С. Осадок инкубируют при указанной температуре 5-120 минут. Инактивация путем нагревания обусловливает необходимость отделения вегетативных клеток и спор от культуральной жидкости. Дело в том, что указанная температура инактивации может критически снизить биологическую активность, по меньшей мере, части метаболитов B. subtilis, содержащихся в культуральной жидкости.
[0085] После этого смешивают высушенную культуральную жидкость B. subtilis с инактивированными вегетативными клетками и спорами.
[0086] Далее к полученному комплексу метаболитов B. subtilis добавляют инактивированные клетки пробиотического штамма L. reuteri, а также наполнитель.
[0087] Заявленный способ обеспечивает получение композиции, включающей широкий спектр метаболитов пробиотических штаммов L. reuteri и B. subtilis. Отделение вегетативных клеток и спор B. subtilis от культуральной жидкости перед нагревом вегетативных клеток и спор B. subtilis позволяет получить композицию с максимально широким спектром биологически активных метаболитов B. subtilis - как секретируемых вегетативными клетками в культуральную жидкость, так и содержащихся в самих вегетативных клетках и спорах. При этом нагревание вегетативных клеток и спор B. subtilis в указанном выше режиме позволяет эффективно инактивировать вегетативные клетки и споры.
[0088] В одном из вариантов реализации способ дополнительно включает высушивание инактивированных вегетативных клеток и спор пробиотического штамма B. subtilis. При этом инактивированные вегетативные клетки и споры B. subtilis могут быть высушены любым подходящим способом, известным из уровня техники. В предпочтительном варианте реализации инактивированные вегетативные клетки и споры B. subtilis высушивают путем лиофилизации. В одном варианте реализации инактивированные вегетативные клетки и споры B. subtilis сначала смешивают с культуральной жидкостью, а затем полученную смесь высушивают, например, лиофилизируют с использованием подходящего носителя, например, мальтодекстрина. В предпочтительном варианте реализации инактивированные вегетативные клетки и споры B. subtilis высушивают отдельно от культуральной жидкости. В любом случае инактивированные клетки и споры B. subtilis высушивают до такого состояния, что их влажность составляет 6-8 мас. % Использование высушенных вегетативных клеток и спор B. subtilis позволяет получать композицию в сухом виде.
[0089] В предпочтительном варианте реализации получают композицию в сухом виде, например, в виде порошка. Это увеличивает срок хранения композиции без потери ее эффективности. Это связано и с сохранением активности метаболитов, и с минимизацией риска загрязнения композиции какими-либо активными микроорганизмами. Кроме того, сухая композиция менее требовательна к условиям хранения и проще в транспортировке, что увеличивает ее доступность для широкого круга пользователей.
ОПИСАНИЕ РЕАЛИЗАЦИИ ИЗОБРЕТЕНИЯ
Пример 1. Получение комплекса метаболитов B. subtilis
[0090] В данном примере получали метаболиты штамма B. subtilis ВКМ В-3536D. Процесс глубинного культивирования вели в ферментере Р1000 фирмы Bioengineering (Швейцария) объемом 1000 дм3 с коэффициентом заполнения 0,6. В процессе культивирования поддерживали температуру 37±1°С. Скорость перемешивания составляля 250 об/мин. Начальная скорость аэрации составляла 0,2 об/мин, через 16 часов роста скорость изменяли до значения 0,5 об/мин. Процесс глубинного культивирования вели в течение 32 часов.
[0091] Приблизительно 30 л культуральной суспензии B. subtilis ВКМ В-3536D подвергали центрифугированию на проточной суперцентрифуге Sharpless AS-26 до получения приблизительно 20 л прозрачного супернатанта. При этом супернатант представлял собой культуральную жидкость, а осадок преимущественно включал в себя вегетативные клетки и споры B. subtilis.
[0092] Полученную культуральную жидкость лиофилизировали с добавлением 2,5 мас. % мальтодекстрина в качестве носителя. В результате получали 560 г порошка, представляющего собой лиофилизированную культуральную жидкость.
[0093] Осадок, включающий вегетативные клетки и споры, инкубировали при температуре 118°С в течение 40 минут. Таким образом инактивировали вегетативные клетки и споры B. subtilis. Затем инактивированные вегетативные клетки и споры подвергали лиофилизации с добавлением 2,5 мас. % мальтодекстрина в качестве носителя.
[0094] Смешивали лиофилизированную культуральную жидкость с лиофилизированными инактивированными вегетативными клетками и спорами B. subtilis. В результате получали весь комплекс метаболитов, присутствовавший в культуральной суспензии B. subtilis.
Пример 2. Исследование состава культуральной жидкости B. subtilis
[0095] В данном примере количественно исследовали состав культуральной жидкости пробиотического штамма B. subtilis ВКМ В-3536D. Исследование было выполнено в Федеральном государственном унитарном предприятии «Научно-исследовательский институт гигиены, профпатологии и экологии человека» федерального медико-биологического агентства (ФГУП «НИИ ГПЭЧ» ФМБА РОССИИ).
[0096] Образец культуральной жидкости после перемешивания был разделен на пробы, которые хранились до анализа в замороженном виде при -18°С. В дальнейшем пробы были разделены на аликвоты, в которых было проведено измерение массовых концентраций аминокислот, витаминов, свободных и этерифицированных жирных кислот. Определены содержания макро/микро элементов, а также тяжелых металлов. Результаты исследования представлены в Табл. 2-5.
[0097]
[0098]
[0099]
[00100]
[00101] Также был проведен общий скрининг с целью идентификации априори не предсказанных соединений в культуральной жидкости пробиотического штамма B. subtilis. Общий скрининг выполнялся методом газовой хроматомасс-спектрометрии с использованием баз данных масс-спектров и индексов удерживания в режиме автоматической интерпретации хроматомасс-спектрометрических данных. Группы органических соединений, идентифицированных при проведении хроматомасс-спектрометрическом скрининге образцов, представлены в табл. 6-10.
[00102]
[00103]
RT - Время удерживания, RI - индекс удерживания, эксп. - в эксперименте, лит. - по литературным данным;
х - количественная оценка содержания идентифицированного соединения невозможна.
[00104]
RT - Время удерживания, RI - индекс удерживания, эксп. - в эксперименте, лит. - по литературным данным;
х - количественная оценка содержания идентифицированного соединения невозможна.
[00105]
(2-пирролидон)
(Амид эруковой кислоты)
RT - Время удерживания, RI - индекс удерживания, эксп. - в эксперименте, лит. - по литературным данным;
х - количественная оценка содержания идентифицированного соединения невозможна.
[00106]
Цикло(лейцилпролил) (L-Phe-D-Pro лактам) **
Цикло(лейцилпролил)**
Цикло(лейцилпролил) (L-Phe-D-Pro лактам) **
3-Метил-6-(фенилметил)-2,5-пиперазиндион (Цикло(Ala-Phe)) **
3-Бензиогексагидропирроло[1,2-a]пиразин-1,4-дион (Цикло(D-фенилаланин-L-пролил))**
3-Бензиогексагидропирроло[1,2-a]пиразин-1,4-дион (Цикло(D-фенилаланин-L-пролил))**
RT - Время удерживания, RI - индекс удерживания, эксп. - в эксперименте, лит. - по литературным данным;
х - количественная оценка содержания идентифицированного соединения невозможна.
[00107] Также проводили скрининг с использованием высокоэффективной жидкостной хроматомасс-спектрометрии высокого разрешения (ВЭЖХ-МС ВР). В результате скрининга методом ВЭЖХ-МС ВР были идентифицированы соединения, представленные в Табл. 11. Отличие теоретической массой от практической составляет 0,7 ppm, таким образом можно утверждать о достоверной идентификации.
[00108]
RT - Время удерживания, ЖК - жирная кислота.
Пример 3. Изучение воздействия культуральной суспензии B. subtilis на ингибирование роста патогенной и условно-патогенной микрофлоры
[00109] В данном примере методом перпендикулярных штрихов изучали антагонистическую активность культуральной суспензии пробиотического штамма B. subtilis. В качестве пробиотического штамма использовали штамм B. subtilis ВКМ В-3536D. На поверхность агаризованной среды Луриа-Бертани в чашке Петри наносили штрихи культуральной суспензии штамма B. subtilis ВКМ В-3536D. Затем указанную чашку Петри на сутки помещали в термостат при температуре 37°С. После диффузии метаболитов B. subtilis в агаризованную среду, перпендикулярно к выросшему штриху подсевали штрихами тест-культуры, начиная от краев чашки Петри. Затем указанную чашку Петри на 48 часов помещали в термостат при температуре 37°С. Наличие антагонистической активности фиксировали, измеряя зоны задержки роста тест-культур.
[00110]
В результате исследования было установлена выраженная способность культуральной суспензии пробиотического штамма B. subtilis к подавлению роста патогенных и условно-патогенных микроорганизмов.
Пример 4. Изучение воздействия метаболитов B. subtilis на ингибирование роста патогенной и условно-патогенной микрофлоры
[00111] В данном примере изучали способность метаболитов пробиотического штамма B. subtilis ВКМ В-3536D с молекулярным весом более 5 кДа подавлять рост индикаторных культур. Исследование проводили с использованием адаптированного метода стандартных дисков для определения чувствительности микроорганизмов к антибиотикам. Использованные в данном примере индикаторные культуры представлены патогенными и условно-патогенными для человека микроорганизмами. Для реализации метода индикаторные культуры помещали на поверхность агаризованной питательной среды Луриа-Бертани в чашке Петри в количестве 107 КОЕ. После застывания чашки Петри подсушивали в течение 15 минут при 37°С. Затем на поверхность питательной среды с индикаторными культурами в виде капель диаметром 5±1 мм наносили по 10 мкл растворов, включающих метаболиты B. subtilis с молекулярным весом более 5 кДа. Указанные растворы были получены методом тангенциальной ультрафильтрации. При этом раствор №1 включал метаболиты с молекулярным весом более 100 кДа, а раствор №2 включал метаболиты с молекулярным весом 5-100 кДа.
[00112] Чашки Петри с нанесенными растворами оставляли при комнатной температуре до полного впитывания капель и затем инкубировали при 37°С в течение 24-48 часов. Детекцию результатов проводили путем измерения зоны задержки роста индикаторных культур вокруг области нанесения растворов, включающих метаболиты B. subtilis. Появление зон, свободных от индикаторных культур, вокруг области нанесения указанных растворов свидетельствуют об антагонистической активности метаболитов B. subtilis в отношении индикаторных культур, указанных в Табл. 2. Величина зон, свободных от индикаторных культур, указывает на эффективность антагонистического действия метаболитов B. subtilis.
[00113]
(> 100 кДа)
(5-100 кДа)
* - бактериостатическое действие;
«-» - задержка роста не наблюдалась.
[00114] Из Табл. 12 видно, что способностью к подавлению роста обладают как высокомолекулярные метаболиты весом более 100кДа (раствор №1), так и метаболиты весом 5-100кДа (раствор №2). При этом в воздействии этих двух вариантов метаболитов есть существенная разница: высокомолекулярные метаболиты весом >100 кДа вызывают задержку роста E. coli, а метаболиты весом 5-100 кДа не вызывают. При этом культуральная суспензия штамма B. subtilis ВКМ В-3536D, включающая все метаболиты, также вызывала значительную задержку роста E. coli (Пр. 3). Стоит отметить, что E. coli является одним из условно-патогенных микроорганизмов, часто обнаруживаемых в ЖКТ при развитии воспалительных заболеваний. Таким образом, использование всего спектра метаболитов B. subtilis обеспечивает эффективность заявленной композиции в отношении максимально широкого спектра патогенных и условно-патогенных микроорганизмов.
Пример 5. Исследование цитотоксичности метаболитов B. subtilis в отношении клеток млекопитающих
[00115] В данном примере исследовали потенциальную цитотоксичность метаболитов пробиотического штамма B. subtilis. Цитотоксичность оценивали согласно рекомендациям Европейского Агентства по безопасности пищевых продуктов (EFSA) для характеризации микроорганизмов используемых в качестве кормовых добавок или штаммов-продуцентов [47]. Штамм B. subtilis ВКМ В-3536D выращивали в сердечно-мозговом бульоне при 30°С в течение 6 ч. Клетки отделяли от культуральной жидкости центрифугированием.
[00116] Анализ цитотоксичности на клетках Vero (клеточная линия почечного эпителия зеленой мартышки) проводили по методу [48] с модификациями. Клетки Vero инкубировали с культуральной жидкостью. Степень поврежденности клеток Vero оценивали по выделению клетками фермента лактат-дегидрогеназы. Цитотоксичность выражали в процентах от токсичности детергента Triton-X100, вызывающего выход из клеток лактат-дегидрогеназы за счет перфорации клеточных мембран. В качестве контрольных штаммов использовали цитотоксичный и нецитотоксичный штаммы рода Bacillus. В Табл. 3 представлены средние значения для штамма ВКМ В-3536D и двух контрольных штаммов. Каждый эксперимент проводился в двух повторах.
[00117]
** Интерпретация результатов: 0-20% - не токсичный, 20-100% - токсичный.
[00118]
Из представленных результатов видно, что цитотоксичность культуральной жидкости штамма B. subtilis ВКМ В-3536D не превышает границы в 20%. Таким образом, метаболиты штамма B. subtilis ВКМ В-3536D, содержащиеся в исследованной культуральной жидкости, не являются токсичными в отношении клеток Vero, и могут считаться безопасными для клеток млекопитающих.
Пример 6. Оценка иммуномодулирующих свойств метаболитов B. subtilis
[00119] В данном примере исследовали иммуномодулирующие свойства комплекса метаболитов, включающего инактивированные вегетативные клетки и споры пробиотического штамма B. subtilis ВКМ В-3536D, а также вещества из культуральной жидкости с молекулярным весом более 5 кДа. Указанные вещества с молекулярным весом более 5 кДа были получены методом тангенциальной ультрафильтрации культуральной жидкости пробиотического штамма B. subtilis ВКМ В-3536D, смешаны с инактивированными клетками и спорами, а затем лиофилизированы на мальтодекстрине в качестве носителя. Указанный комплекс метаболитов добавляли в культуру клеток моноцитов человека ТНР-1. Сравнивали эффект добавления метаболитов по отношению к контролю без метаболитов 1) на стадии дифференцировки моноцитов в макрофаги с помощью форбол-миристат-ацетата и 2) на стадии поляризации ранее дифференцированных моноцитов. Эффект оценивали методом обратной транскрипции (RT-PCR), по экспрессии мРНК маркерных генов, кодирующих интерлейкины и другие факторы: IL1b, IL6, IL10, TGFb, TNFa и CD64.
[00120] На Фиг. 1 показаны значения экспрессии указанных маркерных генов при добавлении метаболитов, нормализованные к контролю без добавления метаболитов. Указаны средние значения двух экспериментов, по три повтора в каждом эксперименте. Статистически значимые (р≤0,05) количественные изменения (не менее чем в два раза по сравнению с контролем) в обоих экспериментах отмечены звездочкой (*). Светло-серые столбцы отражают уровень экспрессии соответствующего гена на стадии дифференцировки, темно-серые столбцы отражают уровень экспрессии на стадии поляризации.
[00121] Видно, что на стадии дифференцировки исследованные метаболиты значительно усиливали экспрессию гена TNFa, и в то же время значительно снижали экспрессию гена CD64. Это указывает на усиление процесса дифференцировки моноцитов в макрофаги. Также значительно снижалась экспрессия цитокина IL1b, что указывает на противовоспалительный эффект. На стадии поляризации исследуемые метаболиты значительно повышали экспрессию цитокинов IL6 и IL10. IL10 - классический противовоспалительный цитокин, и повышение его экспрессии свидетельствует об активации противовоспалительной поляризации макрофагов. IL6 - важный регуляторный цитокин, высокие концентрации которого также ассоциированы с противовоспалительным эффектом. При этом метаболиты не оказывали достоверно значимого эффекта на экспрессию противовоспалительного цитокина TGFb, что указывает на избирательность их действия.
[00122] Полученные данные указывают на то, что исследованные метаболиты обладают иммуномодулирующими свойствами. Они могут способствовать дифференцировке моноцитов в макрофаги, а также способствовать противовоспалительной поляризации макрофагов, которая необходима для окончания воспаления и регенерации тканей.
Пример 7. Оценка противовоспалительных и анти-неопластических свойств метаболитов B. subtilis
[00123] В данном примере исследовали влияние очищенной метаболитной фракции штамма B.subtilis ВКПМ В-2335 (5-100 кДа) на экспрессию белков - маркеров воспаления: циклооксигеназы 2 (COX-2) и фактора некроза опухоли α (TNF-α) в клетках эпителия желудка, и интерлейкина 23 (IL-23) в первичных клеточных культурах эпителия кишечника и желудка человека. Кроме того, в клетках кишечного эпителия изучали влияние метаболитов B. subtilis на экспрессию маркера кишечной дифференцировки - белка гомеобокса каудального типа 2 (CDX-2).
[00124] Первичные клеточные культуры получали из тканей биопсий тонкого кишечника и желудка. Пассажи (пересевание) культур производили каждые 2 дня. Всего культуры проходили 7-12 пассажей. К опытным культурам добавляли метаболиты B.subtilis в концентрации 0.1 мкг/мл. Экспрессию маркеров определяли методом иммуноцитохимии (ИЦХ) с использованием первичных специфических антител к изучаемым белкам. Детекцию проводили с помощью вторичных флюоресцентно меченых антител (метка Alexa Fluor 647), связывающихся с первичными антителами, с последующей визуализацией на инвертированном конфокальном микроскопе Olympus FluoView FV1000-IX70 с апохроматическим объективом 606 UPlan (Япония) при увеличении 400×. Регистрировали по четыре поля зрения на каждый препарат. Изображения оценивали по параметру относительной площади экспрессии в %, которую высчитывали по формуле: Sотн=Sэкспр/Sобщ*100%, где S отн - относительная площадь экспрессии, S экспр - площадь изображения с экспрессией маркера, S общ - общая площадь препарата на изображении. Значимость различий оценивали по t-критерию Стьюдента (р≤0,05).
[00125] Результаты оценки экспрессии исследованных маркеров на клетках эпителия желудка представлены на Фиг.2А. Результаты оценки экспрессии исследованных маркеров на клетках кишечного эпителия представлены на Фиг.2Б Темно-серым цветом обозначена контрольная культура, серым - культура с добавлением метаболитов B. subtilis. По оси абсцисс обозначен номер пассажа культуры, по оси ординат - относительная площадь экспрессии маркера в %. Статистически значимые (р≤0,05) количественные изменения (не менее чем в два раза по сравнению с контролем) в экспериментах отмечены звездочкой (*).
[00126] Метаболиты B. subtilis достоверно снижали экспрессию маркера воспаления COX-2 после 12 пассажа культуры и экспрессию TNF-α на 3 пассаже в клетках желудка. Экспрессия IL-23 в клетках кишечника понижалась под действием метаболитов B. subtilis как на 3, так и на 7 пассаже. Маркер метаплазии CDX-2 заметно экспрессировался на 7 пассаже культуры клеток кишечника, и также снижался в культуре с добавлением метаболитов B. subtilis.
[00127] Полученные данные демонстрируют противовоспалительное и антиметапластическое действие метаболитов B. subtilis. В случае маркеров COX-2 и TNF-α эффект метаболитов B. subtilis зависел от количества пассажей, пройденных культурами, т.е. от «зрелости» культуры. В условиях in vivo клетки эпителия ЖКТ находятся на разных стадиях дифференцировки («созревания»), но метаболиты B. subtilis могут потенциально одновременно воздействовать на эти клетки, благодаря своим различным молекулярным мишеням.
Пример 8. Оценка анти-хеликобактерного эффекта заявленной композиции на модели монгольских песчанок
[00128] В данном примере исследовали влияние заявленной композиции метабиотиков на эффективность эрадикации H. pylori на модели хеликобактериоза у монгольских песчанок. Монгольские песчанки представляют собой эффективную модель изучения патогенеза H. pylori, отражающую многие черты воспаления тканей желудка, вызываемых H. pylori у человека [49]. Животных заражали двукратным (через день) внутрижелудочным введением бактериальной суспензии H. pylori. После двукратного заражения за животными наблюдали в течение 11-ти недель. К 12-й неделе после заражения у животных наблюдалось развитие гастрита и язвенных поражений тканей желудка. С 12-й недели начинали введение исследуемых субстанций в соответствии со схемой (Табл. 14). Исследуемые субстанции представляли собой:
1) смесь инактивированных клеток штамма L. reuteri DSM17648 (метаболиты L. reuteri) и наполнителя;
2) смесь инактивированных клеток штамма L. reuteri DSM17648 (метаболиты L. reuteri), метаболитов 5 -100 кДа штамма B. subtilis ВКПМ В-2335 и наполнителя.
При этом наполнитель представлял собой крахмал в количестве 1 мас. % В качестве позитивного контроля использовали препараты стандартной эрадикационной терапии H. pylori у человека, которая включает в себя ингибитор протонной помпы омепразол и антибиотики кларитромицин и амоксициллин [50]. В качестве негативного контроля использовали просто наполнитель. Также исследовали воздействие указанных субстанций 1 и 2 в сочетании с препаратами позитивного контроля.
[00129]
+ позитивный контроль
+ позитивный контроль
[00130] Наполнитель, препараты позитивного контроля и исследуемые субстанции, вводили песчанкам внутрижелудочно, поскольку этот способ является аналогом перорального способа применения у человека. По окончании эксперимента животных эвтаназировали и проводили гистологическое исследование тканей желудка и двенадцатиперстной кишки. Оценка состояния желудка проводилась по следующим параметрам: площадь воспаления; макроскопические изменения слизистой; морфологические изменения (степень воспаления; активность воспаления; атрофия желудочных желез; метаплазия; обсеменение слизистой оболочки H. pylori). Для морфологической оценки степени выраженности патологии использовали визуально-аналоговые шкалы, предложенные в Сиднейской системе [49]. В данном случае применяли балльную систему оценки:
0 - норма;
1 - слабо выраженная;
2 - умеренная;
3 - выраженная.
[00131] При макроскопической оценке тканей желудка и двенадцатиперстной кишки во всех экспериментальных группах видимых патологических изменений не обнаружили. Результаты микроскопических гистологических исследований приведены в Табл. 15.
[00132]
(метаболиты L.reuteri + наполнитель)
(метаболиты L.reuteri, метаболиты+ B.subtilis
+ наполнитель)
+
позитивный контроль
+
позитивный контроль
(0,0; 0,0)
(1,0; 3,0) а
(1,0; 1,0)b
(1,0; 1,0) b
(1,0; 1,0) b
(1,0; 1,0)
(1,0; 1,0) b
(0,0; 0,0)
(2,0; 2,0)а
(1,0; 2,0)b
(1,0; 2,0)
(1,0; 2,0) b
(1,0; 2,0) b
(1,0; 1,0) b
(0,0; 0,0)
(2,0; 3,0)а
(1,0; 2,0)b
(1,0; 2,0) b
(1,0; 2,0) b
(0,0; 1,0) b
(0,0; 1,0) b
(0,0; 0,0)
(2,0; 3,0)а
(0,0; 1,0)b
(1,0; 1,0) b
(0,0; 1,0) b
(0,0; 1,0) b
(0,0; 1,0) b
(0,0; 0,0)
(1,0; 2,0)а
(0,0; 1,0)b
(0,0; 1,0) b
(0,0; 1,0) b
(0,0; 1,0) b
(0,0; 1,0) b
[00133] Медиана, верхний и нижний квартиль (Me (Q1; Q3)) указаны в белых ячейках, сумма баллов в группе указана в серых ячейках; а - достоверное отличие от интактной группы мышей, (p<0,05); b - достоверное отличие от группы негативного контроля (p<0,05). В интактной группе мышей, не подвергавшихся заражению и лечению, строение стенки желудка соответствовало норме. В группе негативного контроля у всех животных были выявлены морфологические изменения слизистой оболочки желудка, характерные для катарального гастрита различной степени выраженности: от слабого до умеренного, с тенденцией к хронизации процесса. Развитие инфекции у животных группы негативного контроля привело к формированию преимущественно пангастрита (площадь воспаления 3, средний балл 15, Табл. 15). В данной группе отмечали достоверное увеличение степени выраженности патологических изменений, степени и активности воспаления, атрофии желудочных желез и обсеменения слизистой оболочки желудка H. pylori по сравнению с интактной группой (p<0,05). В группе позитивного контроля происходило значимое снижение показателей по всем исследованным критериям по сравнению с группой негативного контроля (Табл. 15).
[00134] На Фиг.3. представлены диаграммы размаха суммарных показателей выраженности патологических симптомов для исследуемых субстанций Негативному контролю соответствует бокс-плот 1. Позитивному контролю соответствует бокс-плот 2. Субстанции 1 (метаболиты L. reuteri + наполнитель) соответствует бокс-плот 3. Субстанции 2 (метаболиты L. reuteri+ метаболиты B. subtilis + наполнитель) соответствует бокс-плот 4. Сочетание субстанции 1 и позитивного контроля отвечает бокс-плоту 5. Сочетание субстанции 2 и позитивного контроля соответствует бокс-плоту 6. По оси ординат высота диаграмм соответствует суммарной выраженности патологических симптомов в баллах визуально-аналоговой шкалы. Указаны средние значения и медиана экспериментов, по семь повторов в каждом эксперименте. Статистически значимые (р≤0,05) количественные изменения (не менее чем в два раза по сравнению с негативным контролем) в экспериментах отмечены звездочкой (*).
[00135] При монотерапии субстанцией 1 (метаболиты L. reuteri + наполнитель) наблюдалось достоверное снижение показателей площади и активности воспаления, морфологических изменений желудка и степени обсеменения слизистой оболочки желудка H. pylori по сравнению с негативным контролем. Однако по большей части критериев монотерапия субстанцией 1 уступала в эффективности позитивному контролю (Табл. 15).
[00136] Монотерапия субстанцией 2 (метаболиты L. reuteri + метаболиты B. subtilis + наполнитель) не уступала в эффективности позитивному контролю, а по некоторым параметрам даже превосходила его. Так, при монотерапии субстанцией 2 атрофия желудочных желез, а также степень обсеменения слизистой оболочки желудка H. pylori были меньше, чем в группе позитивного контроля (Табл. 15).
[00137] Комбинированная терапия субстанцией 1 вместе с препаратами позитивного контроля или субстанцией 2 с препаратами позитивного контроля превосходила позитивный контроль по некоторым показателям. Так, добавление исследуемых субстанций к препаратам позитивного контроля снижало активность воспаления, выраженность атрофии желудочных желез, а также степень обсеменения слизистой оболочки желудка H. pylori (Табл. 15). При этом комбинированная терапия субстанцией 2 с препаратами позитивного контроля показывала наименьший разброс данных и ошибку среднего значения (Фиг.3). Таким образом, комбинированная терапия субстанцией 2 с препаратами позитивного контроля наиболее сильно и статистически достоверно снижала общую выраженность патологических изменений по сравнению с другими исследованными вариантами терапии.
[00138] Полученные данные показывают, что заявленная композиция, включающая метаболиты L. reuteri и B. subtilis, более эффективна при лечении хеликобактериоза, чем композиция, включающая только метаболиты L. reuteri. Причем эта закономерность наблюдалась как при монотерапии исследуемыми субстанциями 1 и 2, так и при комбинировании этих субстанций с препаратами стандартной эрадикационной терапии H. pylori. Это говорит о том, что добавление метаболитов B. subtilis к метаболитам L. reuteri обусловливает эффективность заявленной композиции в отношении H.pylori и сопутствующего воспаления. Последнее может быть связано с противовоспалительным и иммуномодулирующим действием метаболитов B. subtilis, показанном в Пр. 6 и 7.
Пример 9. Оценка эффективности применения заявленной композиции в терапии у пациентов с хроническим гастритом или функциональной диспепсией, ассоциированных с инфекцией H. pylori
[00139] В данном примере проводили открытое рандомизированное проспективное контролируемое исследование эффективности и безопасности заявленной композиции с участием пациентов с функциональной диспепсией и/или хроническим гастритом, ассоциированных с инфекцией H. pylori. Использованная в примере композиция включала инактивированные клетки пробиотического штамма L. reuteri GMNL-0902, комплекс метаболитов пробиотического штамма B. subtilis ВКМ B-3536D, цитрат тригидрата цинка, а также фруктоолигосахариды. В исследование было включено 40 пациентов обоих полов в возрасте от 18 до 65 лет включительно с подтвержденным диагнозом функциональная диспепсия или хронический гастрит, и с положительным анализом на наличие ДНК H. pylori в кале методом ПЦР в реальном времени (Real-Time PCR). Отдельным требованием для пациентов было отсутствие в анамнезе ранее проведенной эрадикационной терапии менее чем за год до скрининга.
[00140] Все пациенты были поровну разбиты на экспериментальную группу 1 и контрольную группу 2. Пациенты из экспериментальной группы 1 в течение 14 дней получали заявленную композицию 2 раза в сутки утром и вечером после еды дополнительно к стандартной трехкомпонентной эрадикационной терапии первой линии (эзомепразол 20 мг перорально 2 раза в сутки утром и вечером после еды, амоксициллин 1000 мг 2 раза в сутки утром и вечером после еды, кларитромицин 500 мг 2 раза в сутки утром и вечером после еды). Следующие 14 дней после окончания терапии пациенты данной группы получали заявленную композицию 2 раза в сутки утром и вечером после еды в качестве монопрепарата. Пациенты из контрольной группы 2 получали в течение 14 дней эрадикационную терапию, включающую эзомепразол 20 мг перорально 2 раза в сутки утром и вечером после еды, амоксициллин 1000 мг перорально 2 раза в сутки утром и вечером после еды, кларитромицин 500 мг перорально 2 раза в сутки утром и вечером после еды. Выраженность симптомов, характерных для хронического гастрита или функциональной диспепсии оценивали с помощью опросников, которые все пациенты заполняли перед началом эксперимента и через 28 дней от начала эксперимента (Табл. 16).
[00141] Статистическое сравнение эффектов терапии в экспериментальной и контрольной группах показало, что применение заявленной композиции достоверно облегчало такие симптомы, как избыточное отхождение газов, урчание в животе и склонность к диарее. При этом в экспериментальной группе частота данных симптомов снизилась после курса терапии по сравнению с моментом начала терапии. В то же время в контрольной группе указанные симптомы стали встречаться чаще после прохождения терапии, чем вначале. Это свидетельствует о том, что стандартная эрадикационная терапия нарушает работу ЖКТ. Вероятнее всего, это связано с дисбактериозом, вызванным антибиотиками широкого спектра. При этом применение заявленной композиции вместе с препаратами стандартной эрадикационной терапии снижало частоту практически всех симптомов после прохождения терапии. Это может быть связано с облегчением или предотвращением дисбактериоза, благодаря благотворному влиянию заявленной композиции на представителей нормальной микрофлоры кишечника.
[00142] В настоящих материалах заявки представлено предпочтительное раскрытие осуществления заявленного технического решения, которое не должно использоваться как ограничивающее иные, частные воплощения его реализации, которые не выходят за рамки испрашиваемого объема правовой охраны и являются очевидными для специалистов в соответствующей области техники.
Список цитируемой литературы:
1. Brown L.M. Helicobacter pylori: epidemiology and routes of transmission. Epidemiol Rev. 2000; 22 (2): 283-97.
2. Warren J.R, Marshall B. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet. 1983 Jun 4; 1 (8336): 1273-5.
3. Watari J., Chen N., Amenta P.S., Fukui H., Oshima T., Tomita T., Miwa H., Lim K.J., Das K.M. Helicobacter pylori associated chronic gastritis, clinical syndromes, precancerous lesions, and pathogenesis of gastric cancer development. World J Gastroenterol. 2014 May 14; 20 (18): 5461-73.
4. Molina-Infante J., Gisbert J.P. Optimizing clarithromycin-containing therapy for Helicobacter pylori in the era of antibiotic resistance. World J Gastroenterol. 2014 Aug 14; 20 (30): 10338-47.
5. Намазова-Баранова Лейла Сеймуровна, Баранов А.А. «Антибиотикорезистентность в современном мире». Педиатрическая фармакология, vol. 14, no. 5, 2017, pp. 341-354.
6. Ahmetova D.G., Baltabekova A.Zh., Shustov A.V. Antibiotic resistance of Helicobacter pylori: review of epidemiological trends and problems of treatment // RMJ. Medical Review. 2018. № 7(I). P. 13-18.
7. Shah T., Baloch Z., Shah Z., Cui X., Xia X. «The Intestinal Microbiota: Impacts of Antibiotics Therapy, Colonization Resistance and Diseases» // Int. J. Mol. Sci. 2021, 22, 6597.
8. Бондаренко В.М., Грачева Н.М., Мацулевич Т.В. «Дисбактериозы кишечника у взрослых» // М.: КМК. - 2003. - 224 с.
9. Sullivan A., Nord C.E. Probiotics and gastrointestinal diseases. J Intern Med. 2005; 257: 78-92.
10. Shenderov B.A. Probiotics and Functional Foods / B.A. Shenderov // Food Engineering, Encyclopedia of Life Support Systems (EOLSS). Developed under the Auspices of the UNESCO. - Eolss Publishers: Oxford, UK, 2011.
11. Воробейчиков Е.В., Василенко А.Ж., Синица А.В и др. «Методологические аспекты применения пробиотиков и антибиотиков для экстренной профилактики инфекционных заболеваний» // Сб. научн. тр. III съезда общества биотехнологов России им. Ю.А. Овчинникова. - М: МаксПресс, 2005. - С. 35-36.
12. Homan M., Orel R. Are probiotics useful in Helicobacter pylori eradication? World J Gastroenterol. 2015 Oct 7; 21 (37): 10644-53.
13. Parth K., Prudhivi R., Palatheeya S., Abbas S.K., Varsha K., Niharika B. et al. (2021). Efficacy of lactobacillus reuteri supplementation in eradication of h. pylori: A comparison study with triple drug therapy. J. Pharm. Res. Int.
14. Lin R., Liu W., Piao M., Zhu H. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids. 2017 Dec; 49 (12): 2083-2090. doi: 10.1007/s00726-017-2493-3.
15. Бондаренко В.М., Боев Б.В., Лыкова Е.А. и др. Дисбактериозы желудочно-кишечного тракта // Российский журнал гастроэнтерологии, гепатологии, колопроктологии. 1999. №1. С. 66-70 [Bondarenko V.M., Boev B.V., Lykova E.A. i dr. Disbakteriozy zheludochno-kishechnogo trakta // Rossijskij zhurnal gastroehnterologii, gepatologii, koloproktologii. 1999. № 1. S. 66-70 (in Russian)].
16. Хурса Р.В., Месникова И.Л., Микша Я.С. «Кишечная микрофлора: роль в поддержании здоровья и развитии патологии, возможности коррекции» // Минск БГМУ, 2017.
17. Жучкова Т.В., «Современные представления о микрофлоре человека: что такое дисбактериоз?» // Медицинская энциклопедия Видаль: Гастроэнтерология, 2015.
18. Тарантула В.З. «Словарь биотехнологических терминов» // Москва: Роспатент, 2005.
19. Шендеров Б.А., Ткаченко Е.И., Лазебник Л.Б., Ардатская М.Д., Синица А.В., Захарченко М.М. «Метабиотики - новая технология профилактики и лечения заболеваний, связанных с микроэкологическими нарушениями в организме человека» // Экспериментальная и клиническая гастроэнтерология. 2018; 151 (3): 83-92.
20. Фирсов Н.Н., «Микробиология: словарь терминов» // М: Дрофа, 2006.
21. Maksimova E.M., Vinogradova D.S., Osterman I.A., Kasatsky P.S., Nikonov O.S., Milón P., Dontsova O.A., Sergiev P.V., Paleskava A., Konevega A.L. Multifaceted Mechanism of Amicoumacin A Inhibition of Bacterial Translation. Front Microbiol. 2021 Feb 12; 12: 618857.
22. Kaspar F., Neubauer P., Gimpel M. Bioactive Secondary Metabolites from Bacillus subtilis: A Comprehensive Review. J Nat Prod. 2019, Jul 26; 82 (7): 2038-2053. doi: 10.1021/acs.jnatprod.9b00110.
23. Park H.B., Perez C.E., Perry E.K. and Crawford J.M. Activating and Attenuating the Amicoumacin Antibiotics.// Molecules.2016. 21 (7).
24. Prokhorova I.V., Akulich K.A., Makeeva D.S., Osterman I.A., Skvortsov D.A., Sergiev P.V., Dontsova O.A., Yusupova G., Yusupov M.M., Dmitriev S.E. Amicoumacin A induces cancer cell death by targeting the eukaryotic ribosome. Sci Rep. 2016 Jun 14; 6: 27720. doi: 10.1038/srep27720.
25. Tosco A., Chobelet A., Bathany K., Schmitter J.-M, Urdaci M. C., Buré C. Characterization by Tandem Mass Spectrometry of Biologically Active Compounds Produced by Bacillus Strains // journal of Applied bioanalysis. 2015. Jan., P. 19-25
26. Hamdache A., Lamarti A.; Aleu J.; Collado I.G. Non-peptide metabolites from the genus Bacillus // J. Nat. Prod. 2011. V. 74, P. 893-899.
27. Zhao K., Xing R., Yan X. Cyclic dipeptides: Biological activities and self-assembled materials. Peptide Science, 2020, https://doi.org/10.1002/pep2.24202.
28. Ning Y., Fu Y., Hou L., Ma M., Wang Z., Li X., Jia Y. iTRAQ-based quantitative proteomic analysis of synergistic antibacterial mechanism of phenyllactic acid and lactic acid against Bacillus cereus. Food Res Int. 2021 Jan; 139:109562. doi: 10.1016/j.foodres.2020.109562. Epub 2020 Nov 1. PMID: 33509445.
29. Liu F., Sun Z., Wang F., Liu Y., Zhu Y., Du L., Wang D., Xu W. Inhibition of biofilm formation and exopolysaccharide synthesis of Enterococcus faecalis by phenyllactic acid. Food Microbiol. 2020 Apr; 86: 103344. doi: 10.1016/j.fm.2019.103344.
30. Zheng R., Zhao T., Hung Y.C., Adhikari K. Evaluation of Bactericidal Effects of Phenyllactic Acid on Escherichia coli O157:H7 and Salmonella Typhimurium on Beef Meat. J Food Prot. 2019 Dec; 82 (12): 2016-2022. doi: 10.4315/0362-028X.JFP-19-217.
31. Wang F., Wu H., Jin P., Sun Z., Liu F., Du L., Wang D., Xu W. Antimicrobial Activity of Phenyllactic Acid Against Enterococcus faecalis and Its Effect on Cell Membrane. Foodborne Pathog Dis. 2018 Oct; 15 (10): 645-652. doi: 10.1089/fpd.2018.2470.
32. Ning Y., Yan A., Yang K., Wang Z., Li X., Jia Y. Antibacterial activity of phenyllactic acid against Listeria monocytogenes and Escherichia coli by dual mechanisms. Food Chem. 2017 Aug. 1; 228: 533-540. doi: 10.1016/j.foodchem.2017.01.112. Epub 2017 Jan 25. PMID: 28317760.
33. Sun S., Li H., Chen J., Qian Q. Lactic Acid: No Longer an Inert and End-Product of Glycolysis. Physiology (Bethesda). 2017 Nov; 32 (6): 453-463. doi: 10.1152/physiol.00016.2017.
34. Narayana K.J., Prabhakar P., Vijayalakshmi M., Venkateswarlu Y., Krishna P.S. Biological activity of phenylpropionic acid isolated from a terrestrial Streptomycetes. Pol J Microbiol. 2007; 56 (3): 191-7.
35. Sakko M., Tjäderhane L., Sorsa T., Hietala P., Järvinen A., Bowyer P., Rautemaa R. 2-Hydroxyisocaproic acid (HICA): a new potential topical antibacterial agent. Int J Antimicrob Agents. 2012 Jun; 39 (6): 539-40. doi: 10.1016/j.ijantimicag.2012.02.006.
36. Guimarães A., Venancio A., Abrunhosa L. Antifungal effect of organic acids from lactic acid bacteria on Penicillium nordicum. Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2018 Sep; 35.(9):.1803-1818. doi: 10.1080/19440049.2018.1500718.
37. Zhang, L.S., Davies, S.S. Microbial metabolism of dietary components to bioactive metabolites: opportunities for new therapeutic interventions. Genome Med 8, 46, 2016. https://doi.org/10.1186/s13073-016-0296-x.
38. Machate D.J., Figueiredo P.S., Marcelino G., Guimarães R.C.A., Hiane P.A, Bogo D., Pinheiro V.A.Z., Oliveira L.C.S., Pott A. Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis. Int J Mol Sci. 2020 Jun 8; 21 (11): 4093.
39. Venn-Watson, S., Lumpkin, R. & Dennis, E.A. Efficacy of dietary odd-chain saturated fatty acid pentadecanoic acid parallels broad associated health benefits in humans: could it be essential?. Sci Rep 10, 8161, 2020. https://doi.org/10.1038/s41598-020-64960-y.
40. Huang C.B., Alimova Y., Myers T.M., Ebersole J.L. Short- and medium-chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch Oral Biol. 2011 Jul; 56 (7): 650-4. doi: 10.1016/j.archoralbio.2011.01.011.
41. Bae Y.S., Rhee M.S. Short-Term Antifungal Treatments of Caprylic Acid with Carvacrol or Thymol Induce Synergistic 6-Log Reduction of Pathogenic Candida albicans by Cell Membrane Disruption and Efflux Pump Inhibition. Cell Physiol Biochem. 2019; 53 (2): 285-300. doi: 10.33594/000000139.
42. Jung S.W., Lee S.W. The antibacterial effect of fatty acids on Helicobacter pylori infection. Korean J Intern Med. 2016 Jan; 31 (1): 30-5. doi: 10.3904/kjim.2016.31.1.30. Epub 2015 Dec 28. PMID: 26767854; PMCID: PMC4712431.
43. Omura Y., O'Young B., Jones M., Pallos A., Duvvi H., Shimotsuura Y. Caprylic acid in the effective treatment of intractable medical problems of frequent urination, incontinence, chronic upper respiratory infection, root canalled tooth infection, ALS, etc., caused by asbestos & mixed infections of Candida albicans, Helicobacter pylori & cytomegalovirus with or without other microorganisms & mercury. Acupunct Electrother Res. 2011; 36 (1-2): 19-64. doi: 10.3727/036012911803860886.
44. He W., Hu S., Du X., Wen Q., Zhong X.P., Zhou X., Zhou C., Xiong W., Gao Y., Zhang S., Wang R., Yang J., Ma L. Vitamin B5 Reduces Bacterial Growth via Regulating Innate Immunity and Adaptive Immunity in Mice Infected with Mycobacterium tuberculosis. Front Immunol. 2018 Feb 26; 9: 365. doi: 10.3389/fimmu.2018.00365.
45. Levine M, Lohinai Z.M. Resolving the Contradictory Functions of Lysine Decarboxylase and Butyrate in Periodontal and Intestinal Diseases. Journal of Clinical Medicine. 2021; 10 (11): 2360. https://doi.org/10.3390/jcm10112360.
46. Read S.A., Obeid S., Ahlenstiel C., Ahlenstiel G. The Role of Zinc in Antiviral Immunity. Adv Nutr. 2019 Jul 1; 10 (4): 696-710. doi: 10.1093/advances/nmz013.
47. Rychen G., Aquilina G., Azimonti G., Bampidis V., Bastos M.L., Bories G., Chesson A., Cocconcelli P.S., Flachowsky G., Gropp J., Kolar B., Kouba M., Lopez-Alonso M., Lopez Puente S., Mantovani A., Mayo B., Ramos F., Saarela M., Villa R.E., Wallace R.J., Wester P., Glandorf B., Herman L., Kärenlampi S., Aguilera J., Anguita M., Brozzi R., Galobart J. «EFSA FEEDAP Panel (EFSA Panel on Additives and Products or Substances used in Animal Feed): Guidance on the characterisation of microorganisms used as feed additives or as production organisms» // EFSA Journal, 2018, 16 (3), 5206, 24 pp. https://doi.org/10.2903/j.efsa.2018.5206.
48. Roberts P.H., Davis K.C., Garstka W., Bhunia A.K. «Lactate dehydrogenase release assay from Vero cells to distinguish verotoxin producing Escherichia coli from non-verotoxin producing strains» // Journal of Microbiological, 2001, Methods, 43: 171-181.
49. Noto, Jennifer M., Judith Romero-Gallo, M. Blanca Piazuelo, and Richard M. Peek. 2016. “The Mongolian Gerbil: A Robust Model of Helicobacter Pylori-Induced Gastric Inflammation and Cancer.”, Methods in molecular biology (Clifton, N.J.), 1422, pp. 263-280.
50. Ивашкин В.Т., Маев И.В., Лапина Т.Л. Клинические рекомендации Российской гастроэнтерологической ассоциации по диагностике и лечению инфекции Helicobacter pylori у взрослых // Российский журнал Гастроэнтерологии, Гепатологии, Колопроктологии. - 2018. - №28 (1). - С. 55-70.
51. Dixon M.F., Genta R.M., Yardley J.H., Correa P. Classification and grading of gastritis: the updated Sydney system // The American journal of surgical pathology. - 1996. - Vol. 20. - №10. - P. 1161-1181.
Группа изобретений относится к биотехнологии и медицине, а именно к композициям на основе метаболитов пробиотических организмов, направленных на лечение и профилактику заболеваний желудочно-кишечного тракта, вызванных Helicobacter pylori. Предложена метабиотическая композиция, включающая инактивированные клетки Lactobacillus reuteri, метаболиты Bacillus subtilis и наполнитель, при этом соотношение компонентов, мас. %: инактивированные клетки Lactobacillus reuteri - 16-70; комплекс метаболитов Bacillus subtilis - 1-7; наполнитель - 23-83. Комплекс метаболитов Bacillus subtilis представляет собой вещества, содержащиеся в смеси культуральной жидкости, вегетативных клеток и спор пробиотического штамма Bacillus subtilis. Предложены также применение указанной композиции для лечения и профилактики болезней желудочно-кишечного тракта, ассоциированных с Helicobacter pylori, и способ получения указанной композиции. Предложенная композиция обладает антагонистическим действием в отношении патогенных микроорганизмов Helicobacter pylori, а также иммуномодулирующим действием. Прием предложенной композиции положительно влияет на состояние желудочно-кишечного тракта и в сочетании с терапией антибиотиками позволяет повысить эффективность эрадикации H.pylori. 3 н. и 17 з.п. ф-лы, 3 ил., 16 табл., 9 пр.
1. Метабиотическая композиция, отличающаяся тем, что включает в себя инактивированные клетки Lactobacillus reuteri, метаболиты Bacillus subtilis и наполнитель, при этом соотношение компонентов, мас. %:
где комплекс метаболитов Bacillus subtilis представляет собой вещества, содержащиеся в смеси культуральной жидкости, вегетативных клеток и спор пробиотического штамма Bacillus subtilis.
2. Метабиотическая композиция по п. 1, отличающаяся тем, что пробиотический штамм В. subtilis представляет собой ВКМ B-3536D и/или ВКПМ В-2335.
3. Метабиотическая композиция по п. 1, отличающаяся тем, что вегетативные клетки и споры пробиотического штамма В. subtilis являются инактивированными.
4. Метабиотическая композиция по п. 1, отличающаяся тем, что наполнитель включает пребиотик.
5. Метабиотическая композиция по п. 4, отличающаяся тем, что пребиотик представляет собой полифруктозан.
6. Метабиотическая композиция по п. 1, отличающаяся тем, что наполнитель включает в себя источник цинка в содержании не менее 3 мас. % от массы композиции.
7. Метабиотическая композиция по п. 6, отличающаяся тем, что источник цинка представляет собой цитрат цинка.
8. Применение композиции по п. 1 для лечения и профилактики болезней желудочно-кишечного тракта, ассоциированных с Helicobacter pylori.
9. Применение композиции по п. 8 для лечения и профилактики хронического гастрита, ассоциированного с Helicobacter pylori.
10. Применение композиции по п. 8 для лечения и профилактики функциональной диспепсии, ассоциированной с Helicobacter pylori.
11. Способ получения композиции по п. 1, включающий следующие этапы:
а) выращивают Bacillus subtilis в жидкой питательной среде;
б) центрифугируют культуральную суспензию Bacillus subtilis на проточной центрифуге с получением культуральной жидкости Bacillus subtilis и осадка, включающего вегетативные клетки и споры Bacillus subtilis;
в) высушивают полученную культуральную жидкость Bacillus subtilis;
г) инактивируют вегетативные клетки и споры Bacillus subtilis;
д) смешивают высушенную культуральную жидкость Bacillus subtilis и инактивированные вегетативные клетки и споры Bacillus subtilis с получением комплекса метаболитов Bacillus subtilis;
е) добавляют инактивированные клетки Lactobacillus reuteri и наполнитель к полученному комплексу метаболитов Bacillus subtilis.
12. Способ по п. 11, в котором культуральную жидкость Bacillus subtilis высушивают до влажности 4-6% мас.
13. Способ по п. 11, в котором культуральную жидкость Bacillus subtilis высушивают путем лиофилизации.
14. Способ по п. 11, в котором вегетативные клетки и споры Bacillus subtilis инактивируют путем инкубации осадка, полученного на этапе центрифугирования, при температуре 60-140°С.
15. Способ по п. 14, в котором осадок инкубируют в течение 5-120 минут.
16. Способ по п. 11, по которому после инактивации вегетативные клетки и споры Bacillus subtilis дополнительно высушивают так, что их влажность составляет не более 6-8% мас.
17. Способ по п. 16, где инактивированные клетки и споры Bacillus subtilis высушивают путем лиофилизации.
18. Способ по п. 11, в котором используют наполнитель, включающий пребиотик.
19. Способ по п. 11, в котором используют наполнитель, включающий источник цинка.
20. Способ по п. 19, где в качестве источника цинка используют цитрат цинка.
| ПРЕПАРАТ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА "БАКТИСТАТИН" | 2005 |
|
RU2287335C1 |
| ПРЕПАРАТ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА "ТоксиБиоВит" | 2011 |
|
RU2475254C1 |
| СИНБИОТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ КОРРЕКЦИИ ДИСБИОТИЧЕСКИХ НАРУШЕНИЙ МИКРОБИОЦЕНОЗА ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА | 2015 |
|
RU2592988C1 |
| МЕТАБИОТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ КОЛОНИЗАЦИОННОЙ РЕЗИСТЕНТНОСТИ МИКРОБИОЦЕНОЗА КИШЕЧНИКА ЧЕЛОВЕКА | 2015 |
|
RU2589818C1 |
| Способ подземного аккумулирования тепла или холода | 2019 |
|
RU2717890C1 |
| CN 105995972 A, 12.10.2016 | |||
| M.H | |||
| EMARA et al | |||
| Lactobacillus reuteri in management of Helicobacter pylori infection in dyspeptic patients: a double-blind placebo-controlled randomized clinical trial | |||
| Therapeutic | |||

