Изобретение относится к области биотехнологии, в частности к новым противогрибковым средствам на основе липосом.
Уровень техники
Грибковые патогены представляют собой серьезную угрозу для здравоохранения, поскольку они становятся все более распространенными и устойчивыми к лечению (Rodrigues, Nosanchuk, 2020). Инвазивные формы грибковых инфекций часто поражают тяжелобольных пациентов и пациентов со значительными сопутствующими заболеваниями, связанными с иммунной системой, что приводит к высокому уровню смертности (Kainz et al., 2020). В 2022 году ВОЗ опубликовала отчет со списком высокоприоритетных грибковых патогенов, авторы которого выразили обеспокоенность тем, что грибковым инфекциям уделяется очень мало внимания, особенно при изучении моделей устойчивости к противогрибковым препаратам (https://www.who.int/publications/i/item/9789240060241, дата обращения 02.06.2024).
Во время недавней пандемии COVID-19 наблюдалась оппортунистическая грибковая инфекция у COVID-положительных пациентов. При этом наибольшая тяжесть и смертность, связанные с этими инфекциями, наблюдаются у COVID-19-позитивных пациентов с предрасполагающими факторами, такими как острый респираторный дистресс синдром, сахарный диабет, искусственная вентиляция легких, пациентов с трансплантацией органов, получавших антибиотики и иммуносупрессивную терапию (Ventoulis et al., 2020). Случаи аспергиллеза у пациентов с COVID-19 были зарегистрированы в нескольких ретроспективных исследованиях, проведенных в Китае в первой половине 2020 г. (Wang et al., 2020). В Индии во время второй волны COVID-19 было зарегистрировано более 14000 случаев мукормикоза - так называемого «черного гриба» (Raut, Huy, 2021). Более того, заболеваемость и смертность, связанные с аспергиллезом, кандидозом и пневмоцистной пневмонией у пациентов с COVID-19, принимала более серьезный характер в европейских странах, а именно в Великобритании, Франции, Нидерландах, Испании и Германии (Silva et al., 2020). Таким образом, заболеваемость оппортунистическими грибковыми инфекциями привлекла внимание исследователей по всему миру.
По данным на 2011 год, общая заболеваемость микозами в России составила 2,7 млн человек, при этом на поверхностные микозы приходилось 2,2 млн, на аллергический аспергиллез и бронхиальную астму 406 тыс., а на системные микозы - 76 тыс. человек. В исследованиях на COVID-положительных пациентах с ИВЛ-ассоциированной пневмонией было установлено, что системные микозы в 41% случаев являлись причиной пневмонии и сопровождались летальным исходом в 100% случаев (Martynova et al., 2023).
Глубокую озабоченность вызывает не только широкое распространение грибковых инфекций как на глобальном, так и на национальном уровне, но и существенный рост устойчивости. Несмотря на отсутствие централизованного подхода к оценке устойчивости, существует большое количество исследований по отдельным видам грибов. Показано, что устойчивость к азолам возрастает у Candida albicans и Aspergillus fumigatus, а устойчивость к эхинокандину у С. glabrata (Wiederhold, 2017). В 2012 году в крови пациентов были обнаружены штаммы С. glabrata перекрестно устойчивые к флуконазолу и эхинокандинам (Pfaller et al., 2012). Другим видом, который недавно привлек большое внимание, является Candida auris, причем до 90% изолятов демонстрируют пониженную чувствительность к флуконазолу, 35% - к амфотерицину Б и 7% - к эхинокандинам (Lockhart et al., 2017). Некоторые виды грибов по своей природе устойчивы ко всем основным группам противогрибковых препаратов, например, Lomentospora prolificans (Biswas et al., 2018).
Таким образом, существует потребность в новых противогрибковых препаратах, предпочтительно с новым механизмом действия, чтобы избежать перекрестной резистентности и/или перекрестной токсичности. Кроме того, наличие пероральных лекарственных форм позволило бы проводить амбулаторное лечение, что привело бы к повышению комфорта пациента и приверженности терапии. Разработка противогрибковых препаратов долгое время находилась в стагнации. Так в период с 2007 по 2017 год появилось только одно новое противогрибковое средство - завуконазол (Gonzalez-Lara et al., 2017). В дальнейшем ситуация немного улучшилась с появлением таких препаратов как резафунгин, ибрексафунгерп, олорофим и фосманогепикс (The Lancet Infectious Diseases, 2023). Тем не менее число противогрибковых препаратов, выходящих на рынок, остается низким, что обусловлено препятствиями, которые необходимо преодолеть, прежде чем противогрибковое средство может быть коммерциализировано: необходимо определить спектр противогрибковой активности и показания, хорошо охарактеризовать токсичность, провести длительные испытания на выборке достаточного размера, и так далее, при этом стоимость разработки должна быть приемлемой (Perfect, 2017). Следовательно, не каждая молекула, обладающая противогрибковой активностью, приведет к созданию одобренного лекарственного средства: приблизительно 80% потенциальных противогрибковых препаратов, опубликованных в литературе, не получили дальнейшего развития из-за обнаружения нежелательных свойств (Pouliot, Jeanmart, 2016).
Эхинокандины, один из пяти основных классов противогрибковых препаратов, являются относительно новыми и рекомендованы Американским обществом инфекционных заболеваний в качестве препаратов первой линии (Pappas et al., 2016). Такие препараты высокоэффективны при лечении инвазивных грибковых инфекций (Htet et al., 2023) и против грибковых биопленок (Tóth et al., 2023). В 1974 году в Швейцарии был открыт первый противогрибковый препарат этой группы - эхинокандин В, который продемонстрировал хорошие противогрибковые свойства, но в то же время показал сильный гемолитический эффект. Чтобы противодействовать этому, был синтезирован цилофунгин, полусинтетический аналог эхинокандина В с боковой цепью из 4-октилоксибензоата. Это соединение значительно снижает гемолитическую активность, сохраняя при этом противогрибковые свойства (Hüttel, 2021). Однако цилофунгин был исключен из клинических испытаний II фазы из-за плохой растворимости в воде и токсичности (Patil, Majumdar, 2017). Важным шагом в исследованиях эхинокандинов стало открытие пневмокандина А0 и пневмокандина В0. В 1992 году из пневмокандина В0 был впервые синтезирован ацетат каспофунгина и одобрен для клинических испытаний (Balkovec et al., 2014). Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) одобрило каспофунгин в январе 2001 года в качестве лекарственного средства для профилактики грибковых инфекций у взрослых пациентов. Соединение под номером FR901379 было обнаружено в Японии компанией Fujisawa Pharmaceutical. Это вещество вызывало лизис ретикулоцитов, поэтому компания исследовала различные химические модификации FR901379. В результате было получено соединение FK463, которое получило название микафунгин, оно обладало пониженной гемолитической активностью и было эффективным в отношении видов Candida и Aspergillus (Hashimoto, 2009). Микафунгин содержит сульфатную группу в боковой цепи остатка дигидроксигомотирозина в структуре соединения, что увеличивает его растворимость в воде (Hashimoto, 2009). Препарат был одобрен в 2005 году для лечения инвазивного кандидоза у пациентов (Taormina et al., 2021).
Эхинокандины неконкурентно ингибируют (1,3) бета-глюкансинтазу, что приводит к нарушению синтеза клеточной стенки грибов (Szymanski et al., 2024). Структура этого транс мембранного белка была недавно определена с помощью криоэлектронной микроскопии в двух независимых исследованиях (Hu et al., 2023; Zhao et al., 2023). Авторы этих работ установили, что очаги мутаций, устойчивых к эхинокандину, тесно связаны с мембранным окружением. Ху и соавторы (Hu et al., 2023) заметили, что горячие точки были окружены несколькими связанными липидными молекулами, которые меняют конформацию при мутации в (1,3) бета-глюкансинтазе, и пришли к выводу, что либо сайт связывания эхинокандина находится внутри мембранной части (1,3) бета-глюкансинтазы, либо лекарства действуют через мембранные изменения. Чжао и соавторы (Zhao et al., 2023) указали, что эхинокандины, вероятно, ингибируют транслокацию глюканов, воздействуя на транспортный канал в мембране. Более того, эта группа авторов обнаружила, что связанные липиды, очищенные совместно с (1,3) бета-глюкансинтазой, содержат эндогенный эргостерин, который помогает стабилизировать структуру (1,3) бета-глюкансинтазы.
Хотя нет никаких сомнений в том, что противогрибковый эффект эхинокандинов связан с ингибированием (1,3) бета-глюкансинтазы, существуют некоторые мутации, не связанные с (1,3) бета-глюкансинтазой, которые приводят к устойчивости к эхинокандину. Хили и др. наблюдали, что мутации, нарушающие биосинтез сфинголипидов, приводят к устойчивости к каспофунгину у Candida glabrata (Healey et al., 2012). Мутации в четырех разных генах привели к одинаковым изменениям в составе мембран: накоплению длинноцепочечных оснований дигидросфингозина и фитосфингозина. Кроме того, внешнее добавление этих веществ вызывало аналогичную устойчивость. Более того, Сатиш и соавторы продемонстрировали, что стресс-индуцированная устойчивость к эхинокандину у Aspergillus fumigatus вызывает аналогичные мембранные изменения, а экзогенный дигидросфингозин и фитосфингозин, добавленные к очищенной (1,3) бета-глюкансинтазе, ингибируют ее каталитическую активность (Satish et al., 2019). Другой косвенный признак поражения мембран был продемонстрирован в исследовании зависимости чувствительности к каспофунгину от температуры у Candida albicans без мутаций в канонических генах (Zheng et al., 2023). Эти данные вместе с упомянутыми выше структурными данными указывают на то, что свойства мембран чрезвычайно важны для правильного функционирования (1,3) бета-глюкансинтазы. Прямое влияние эхинокандинов на мембраны клеток-мишеней пока не изучено.
Недавнее поисковое исследование чувствительных к каспофунгину штаммов Cryptococcus neoformans выявило некоторые заметные мутации, вызывающие такую чувствительность (Moreira-Walsh et al., 2022). Две из четырех выявленных делеций находились в генах, связанных с синтезом эргостерина, и приводили к повышенной чувствительности к мембранному стрессу и повышенной чувствительности к каспофунгину при более высоких температурах без явных дефектов клеточной стенки. Более того, микроскопия показала, что основной причиной этой чувствительности была потеря целостности мембраны, что способствует либо увеличению проницаемости для каспофунгина, либо изменению местоположения или ориентации (1,3) бета-глюкансинтазы, что делает его более восприимчивым к ингибированию (Moreira-Walsh et al., 2022). Эти данные вместе с упомянутыми выше структурными данными указывают на то, что свойства мембран чрезвычайно важны для правильного функционирования (1,3) бета-глюкансинтазы. Прямое влияние эхинокандинов на мембраны клеток-мишеней пока не изучено.
Несмотря на то, что эхинокандины хорошо переносятся, у них есть два существенных недостатка, ограничивающих их применение. Первый - наличие у некоторых грибов естественной устойчивости к этим молекулам. Например, виды Basidiomycetes, Mucormycetes и Fusarium проявляют устойчивость к эхинокандинам, поскольку β-1,3-глюкан составляет лишь второстепенный компонент их клеточных стенок (Lockhart et al., 2023). Второй - быстрое появление грибных изолятов с приобретенной устойчивостью. Один из способов преодолеть устойчивость к эхинокандинам - объединить их с малыми растительными молекулами. Трипати с соавторами показали, что комбинация каспофунгина с сесквитерпенхиноном морского происхождения, пуупехеноном, была высокоэффективной против устойчивых к каспофунгину С. albicans, С. glabrata и С. neoformans, возможно, за счет ингибирования активности белка теплового шока 90 (Tripathi et al., 2020). В другом исследовании синтетическая небольшая молекула L-269289 смогла потенцировать активность эхинокандина у резистентных С. albicans, C. tropicis и С. parapsilosis, воздействуя на геранилгеранил-трансферазу типа I (GGTase I) и блокируя мембранную локализацию Rho1, регулятора субъединицы (1,3) бета-глюкансинтазы (Sun et al., 2020). Представляется рациональным комбинировать эхинокандины с небольшими молекулами, уже обладающими противогрибковой активностью.
Флоретин - это флавоноид из подкласса дигидрохалконов, содержащийся в яблоках и клубнике и известный своими мощными антиоксидантными, противоопухолевыми и противовоспалительными свойствами (Cheon et al., 2019). Было показано, что флоретин проявляет противогрибковую активность in vitro в отношении некоторых фитопатогенных грибов, включая Phytophthora capsici, Alternaria panax и Sclerotinia sclerotiorum (Liu et al., 2021). В исследовании с С. albicans флоретин ингибировал рост грибных клеток и образование биопленок, а также эффективно боролся с кандидозом полости рта в моделях in vivo (Liu et al., 2021). Флоретин снижал температуру фазового перехода, что говорит о повышении текучести мембран в его присутствии (Valenta et al., 2004). Более того, было выявлено увеличение порообразующей активности противогрибковых макролидных антибиотиков при его добавлении (Ostroumova et al., 2012а; Ostroumova et al., 2012b; Ostroumova et al., 2014).
Образование биопленок многими патогенными грибами существенно затрудняет противогрибковое лечение, особенно у пациентов с ослабленным иммунитетом (Pierce et al., 2013). Например, Candida spp. в настоящее время являются третьими наиболее распространенными микроорганизмами, связанными с катетер-ассоциированными инфекциями кровотока (Cramp, Collignon, 2000). Инкапсулирование противогрибковых препаратов в состав липосом может улучшить их противогрибковую активность, растворимость лекарственного препарата, снизить токсичность и преодолеть резистентность. Например, более эффективное проникновение липосомальной формы флуконазола приводило к ингибированию роста резистентных изолятов С. albicans, образующих биопленки (Hassanpour et al., 2021). Липосомальная форма противогрибкового амфотерицина Б была разработана в основном для устранения его высокой токсичности, однако также наблюдалось быстрое накопление этой формы препарата в месте инфекции, включая биопленки (Maertens et al., 2022). На сегодняшний день из эхинокандинов только анидулафунгин изучен в липосомных наночастицах (Vera-González et al., 2020). Хотя минимальная ингибирующая концентрация (МИК) против С. albicans была эквивалентна свободному лекарственному средству, липосомальная композиция продемонстрировала превосходное разрушение биопленки, снижая грибковую нагрузку на 99%. Таким образом, липосомальная форма эхинокандинов может преодолеть грибковую резистентность, связанную с биопленками.
Технической проблемой, на решение которой направлено настоящее изобретение, являлось расширение арсенала противогрибковых средств, в том числе создание препарата на основе микафунгина, обладающего повышенной противогрибковой активностью.
Сущность изобретения
Техническая проблема решена тем, что созданы однослойные липосомы, содержащие микафунгин в эквимолярном соотношении с липидом.
В одном из вариантов осуществления изобретения липосомы дополнительно содержат флоретин в эквимолярном соотношении с липидом.
В одном из вариантов осуществления изобретения в качестве липида используют смесь 1,2-диолеоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 80:20 мол.%, либо смесь 1,2-дипальмитоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 85:15 мол.%, либо смесь 1,2-дипальмитоил-sn-глицеро-3-фосфохолина и эргостерина в соотношении 85:15 мол.%, либо смесь 1-пальмитоил-2-олеоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 67:33 моль.%, либо 1,2-дипальмитоил-sn-глицеро-3-фосфохолин.
В другом варианте осуществления изобретения в качестве липида используют смесь 1-пальмитоил-2-олеоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 67:33 моль.
Изобретение иллюстрируется следующими графическими материалами:
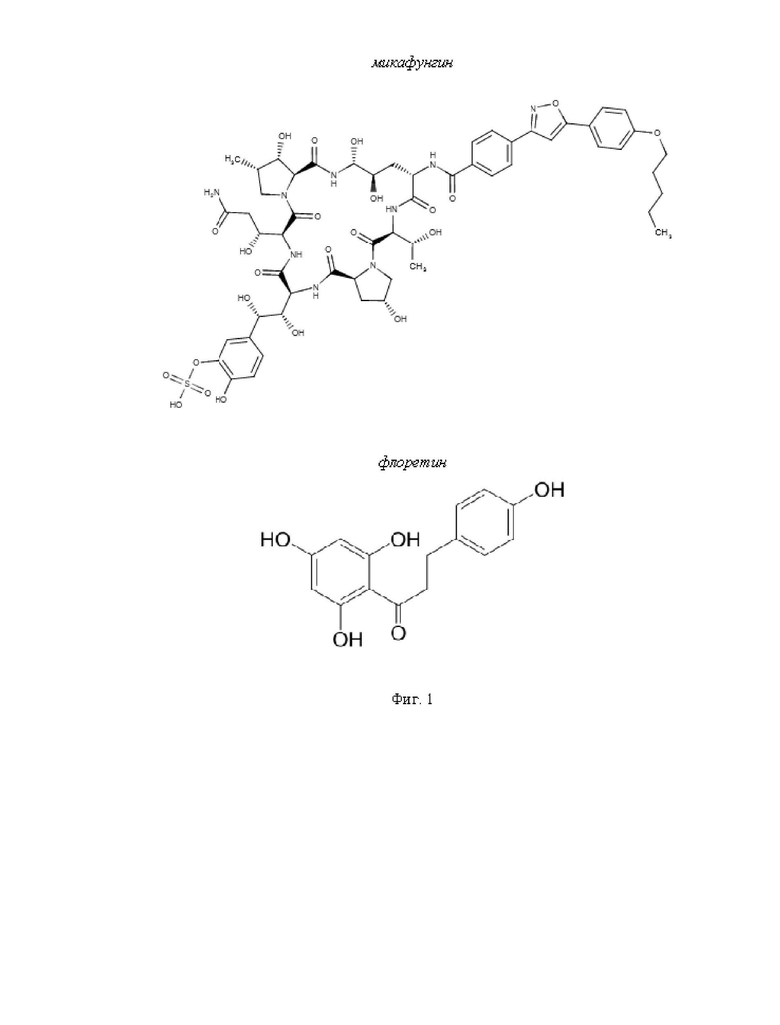
На Фиг. 1 представлены химические структуры молекул микафунгина и флоретина.
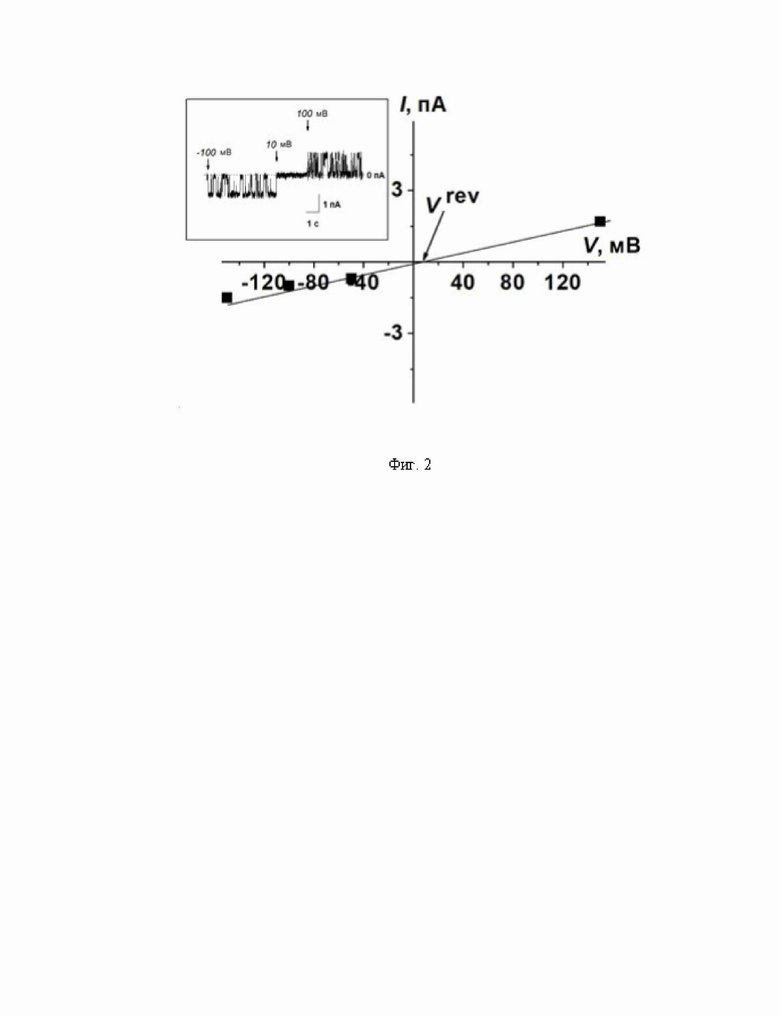
На Фиг. 2 представлена вольт-амперная характеристика модифицированной микафунгином мембраны, сформированной из смеси ПОФХ:ЭРГ (67:33 мол.%) в асимметричных по концентрации электролита омывающих растворах: 0,025 М NaCl (цис-отсек) и 0,15 М NaCl (транс-отсек) (10 мМ HEPES, рН 7,4). Стрелка указывает на потенциал реверсии (трансмембранное напряжение, отвечающее нулевому току) (Vrev). Вставка: пример записи со ступенеобразными флуктуациями трансмембранного тока, соответствующими открыванию и закрыванию одиночных ион-проницаемых транс мембранных пор, индуцированных микафунгином при различных транс мембранных напряжениях.
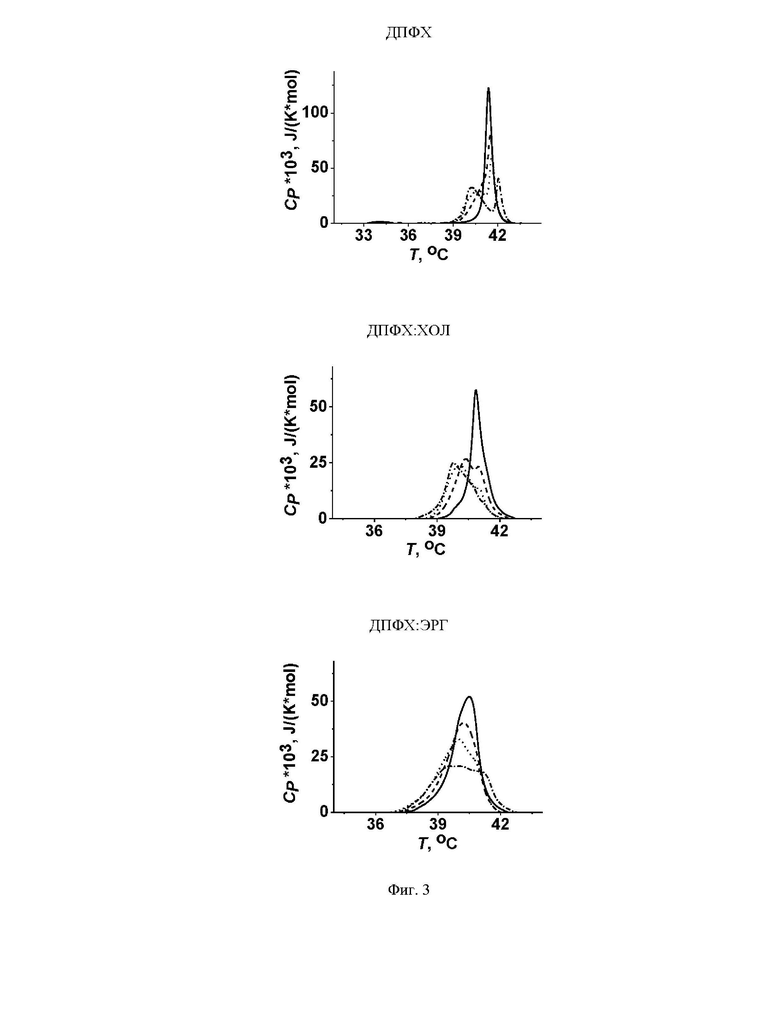
На Фиг. 3 представлены термограммы плавления ДПФХ (верхняя панель), ДПФХ:ХОЛ (85:15 мол.%) (средняя панель) и ДПФХ:ЭРГ (85:15 мол.%) (нижняя панель) в отсутствие (контроль, черные кривые) и в присутствии микафунгина при соотношении липид: антибиотик 50:1 (пунктирные кривые), 25:1 (точечные кривые) и 10:1 (пунктирно-точечные кривые).
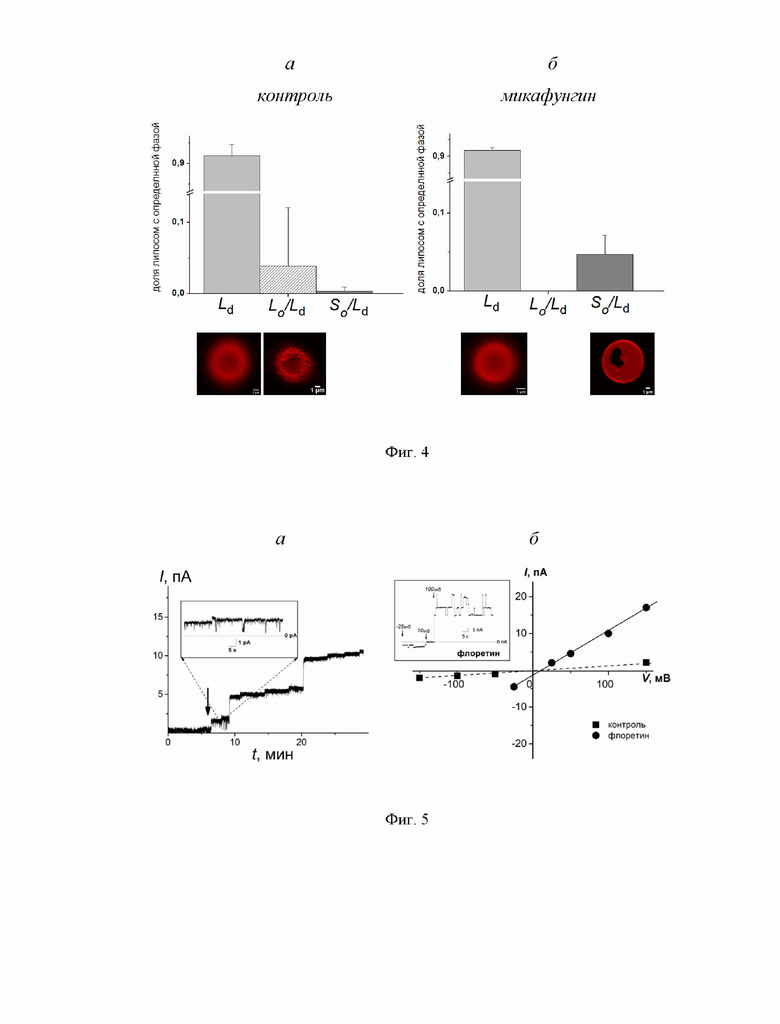
На Фиг. 4 представлены гистограммы, иллюстрирующие процентное содержание в суспензии липосом с определенным паттерном фазового разделения в мембране: Ld - жидкая неупорядоченная фаза, Lo/Ld - сосуществование жидких упорядоченной и неупорядоченной фаз, So/Ld - сосуществование квази-гель и жидкой неупорядоченной фаз до (а) и после введения 50 мкМ микафунгина (б). Нижняя панель представляет примеры микрофотографий гигантских однослойных липосом с разными паттернами фазового разделения в мембранах (масштабная линейка равна 1 мкм). Тип фазового разделения определяли на основании морфологии неокрашенных флуоресцентной меткой липидных доменов (неокрашенные домены круглой и нерегулярной формы были отнесены к Lo и So фазе соответственно). Для подтверждения статистической значимости использовали ANOVA (p-значение <0,05).
На Фиг. 5 представлено потенцирующее действие флоретина на порообразующую способность микафунгина. Мембраны сформированы из смеси ПОФХ:ЭРГ (67:33 мол.%) и омываются растворами 0,025 М NaCl с цис-стороны и 0,15 М NaCl с транс-стороны (10 мм HEPES, рН 7,4). (а) Влияние флоретина на стационарный трансмембранный микафунгин-индуцированный ток. Микафунгин добавлен в омывающий мембрану раствор до концентрации 5 мкМ в начальный момент времени. Момент введения флоретина в омывающий мембрану раствор до концентрации 20 мкМ. Трансмембранное напряжение составляет 100 мВ. Вставка: Ступенеобразные флуктуации тока, протекающего через поры, сформированные микафунгином, после введения флоретина. (б) Вольт-амперные характеристики модифицированной микафунгином мембраны до (пунктирная линия) и после введения флоретина (сплошная линия). Вставка: Ступенеобразные флуктуации тока, протекающего через поры, сформированные микафунгином, после введения флоретина при различных трансмембранных напряжениях.
Детальное описание изобретения
Если не указано иначе, предполагается, что все термины, обозначения и другие научные термины, используемые в данной заявке, имеют значения, которые обычно понимают специалисты в области, к которой относится настоящее изобретение. В некоторых случаях определения терминов с общепринятыми значениями приведены в данной заявке для ясности и/или для быстрой справки и понимания, и включение таких определений в настоящее описание не должно истолковываться как наличие существенного отличия значения термина от обычно подразумеваемого в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белка и нуклеиновых кислот, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Ферментативные реакции и способы очистки осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
В настоящем изобретении предлагаются однослойные липосомы, содержащие микафунгин в эквимолярном соотношении с липидом.
Однослойные липосомы представляют собой липосомы размером от примерно сотен нанометров до нескольких микрометров, состоящие из одного билипидного слоя. Однослойные липосомы известны из уровня техники и могут быть получены любым известным из уровня техники способом (см., например, Dua et al., 2012; Akbarzadeh et al., 2013). Конкретный липидный состав липосом не влияет на достижение технического результата, липосомы могут быть получены из традиционно используемых для этого липидов и их композиций липидов (см., например, Liu et al., 2020; D'Angelo et al., 2022; Pande, 2023). Включение в состав липосом дополнительных веществ, как гидрофильных, так и гидорофобных или амфифильных, достигается известными из уровня техники способами (Lu et al., 2009; Babazadeh et al., 2017; Sharma et al., 2019; Barani et al., 2021; Dwivedi et al., 2023).
Микафунгин (CAS Number: 235114-32-6, Фиг. 1) - противогрибковое соединение, получаемое из грибов Coleophoma empetri, которые синтезируют и выделяют во внешнюю среду это вещество с целью подавления окружающей микофлоры. Микофунгин может быть получен известными из уровня техники способами (см., например, Hashimoto, 2009), в том числе можно приобрести коммерчески доступный микафунгин (см, например, https://www.sigmaaldrich.com/RU/en/product/aablocksinc/aabh97d2d3b3?context=bbe, дата обращения 06.06.2024).
В одном из вариантов осуществления изобретения липосома дополнительно содержит флоретин в эквимолярном соотношении с липидом.
Флоретин (CAS Number: 60-82-2, Фиг. 1) - полифенольное соединение, производное дигидрохалкона. Флоретин может быть получен известными из уровня техники способами, в том числе можно приобрести коммерчески доступный флоретин (см, например, https://www.sigmaaldrich.com/RU/en/product/supelco/phl82624, дата обращения 06.07.2024).
В одном из вариантов осуществления изобретения в качестве липида используют смесь 1,2-диолеоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 80:20 мол.%, либо из смеси 1,2-дипальмитоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 85:15 мол.%, либо из смеси 1,2-дипальмитоил-sn-глицеро-3-фосфохолина и эргостерина в соотношении 85:15 мол.%, либо из смеси 1-пальмитоил-2-олеоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 67:33 моль.%, либо из чистого 1,2-дипальмитоил-sn-глицеро-3-фосфохолина.
В другом варианте осуществления изобретения в качестве липида используют смесь 1-пальмитоил-2-олеоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 67:33 моль.
В нашей работе мы реализовали комплексный подход к исследованию действия микафунгина (Фиг. 1) на модельные липидные мембраны, включая регистрацию ионных токов, протекающих через плоские липидные бислои, дифференциальную сканирующую микрокалориметрию фазовых переходов липидов, конфокальную флуоресцентную микроскопию липидных доменов в липосомах и молекулярно-динамическое моделирование. Основные результаты образования трансмембранных пор с катион/анионой селективностью и усиления сегрегации липидной фазы. Более того, было обнаружено, что растительный полифенол флоретин (Фиг. 1) усиливает порообразующую способность микафунгина в обогащенных эргостерином бислоях. Потенциальный эффект сочетания микафунгина с флоретином оценивали против Candida spp. Таким образом, наши результаты выявили возможный альтернативный противогрибковый механизм действия микафунгина, который может быть связан с изменением липидного микроокружения (1,3) бета-глюкансинтазы и увеличением проницаемости мембран. Комбинация микафунгина с флоретином представляется подходом к повышению эффективности противогрибкового агента за счет потенциации альтернативного механизма действия.
Техническим результатом настоящего изобретения является реализация назначения липосом по настоящему изобретению: показана их противогрибковая активность, что делает возможным реализацию назначения настоящего изобретения, иными словами, липосомы по настоящему изобретению могут быть использованы в качестве противогрибкового средства. Противогрибковая активность липосом по настоящему изобретению значительно выше активности известного из уровня техники амфотерицина Б, что подтверждается значительно более низкой минимальной ингибирующей концентрацией (МИК) липосом по настоящему изобретению по сравнению с амфотерицином Б. Так, МИК однослойных липосом с микафунгином в отношении устойчивых к флуконазолу клинических изолятов видов Candida в 8-60 раз ниже, чем МИК амфотерицина Б. Добавление флоретина в липосомы с микафунгином дополнительно уменьшает МИК в 1-4 раз.
Сущность и практическая применимость настоящего изобретения поясняется следующими примерами:
Пример 1. Материалы и методы
Пример 1.1. Материалы
1-Пальмитоил-2-олеоил-sn-глицеро-3-фосфохолин (ПОФХ), холестерин (ХОЛ), 1,2-диолеоил-sn-глицеро-3-фосфохолин (ДОФХ), 1,2-дипальмитоил-sn-глицеро-3-фосфохолин (ДПФХ), 1,2-дипальмитоил-sn-глицеро-3-фосфоэтаноламин-N-(лиссамино-родамин В-сульфонил) (Род-ДПФЭ) были получены от Avanti Polar Lipids Inc (Алабастер, Алабама, США). Эргостерин (ЭРГ), микафунгин, флоретин, амфотерицин Б, пентан, этанол, диметилсульфоксид (ДМСО), метанол, хлороформ, сорбитол, ЭДТА, NaCl, HEPES, КОН, КС1 и NaOH были приобретены у компании Sigma-Aldrich Company Ltd (Гиллингем, Великобритания). Химические структуры микафунгина и флоретина представлены на Фиг. 1.
Все реагенты были высокого качества. В экспериментах использовалась дистиллированная вода. Эксперименты проводились при комнатной температуре (25°С).
Пример 1.2. Формирование плоских липидных бислоев и их модификация микафунгином и флоретином
Для регистрации токов, протекающих через ион-проницаемые поры, индуцированные микафунгином в плоских липидных бислоях, использовали метод фиксации потенциала. Модельные липидные мембраны были сформированы по методу Монтала и Мюллера путем сведения конденсированных монослоев на отверстии в тефлоновой пленке, разделяющей камеру на два отсека (Montal, Muller, 1972). Объем каждого отсека составлял 1,5 мл, толщина пленки - 10 мкм, диаметр отверстия - 50-100 мкм. Перед началом формирования мембраны отверстие в тефлоновой пленке обрабатывали раствором гексадекана. Микафунгин из исходного 4 мМ раствора в ДМСО добавляли к водной фазе цис-отделения камеры. Для подачи трансмембранного напряжения и отведения сигнала использовали хлорсеребряные электроды, соединенные с растворами камеры через мостики, содержащие 1,5% агарозы. Положительным считали потенциал, вызывающий поток катионов из цис- в транс- отсек камеры. Регистрацию и оцифровку трансмембранных токов проводили в режиме фиксации потенциала по падению напряжения на сопротивлении обратной связи с помощью имеющегося Лабораторного аппаратно-измерительного комплекса для регистрации и анализа токов Axopatch 200В и Digidata 1440А (Axon Instruments, США). Обработку записей трансмембранных токов осуществляли с использованием программного пакета Clampfit 9.0 (Axon Instruments, США). Статистический анализ полученных данных производили при помощи программы Origin 8.0 (OriginLab, США).
Пример 1.3. Катион-анионная селективность ион-проницаемых пор, индуцированных микафунгином
Использовали плоские липидные бислои, состоящие из смеси 67 моль.% ПОФХ и 33 мол.% стерина (ЭРГ или ХОЛ), и буферные растворы с шестикратной разницей в концентрации электролита: 0,025 М NaCl (цис-отсек) и 0,15 М NaCl (транс-отсек). После того, как мембрана была полностью сформирована и стабилизирована, в цис-отсек добавляли микафунгин до конечной концентрации 0,5-2 мкМ.
Число переноса катионов (t+) было определено согласно (Morf, 1977):
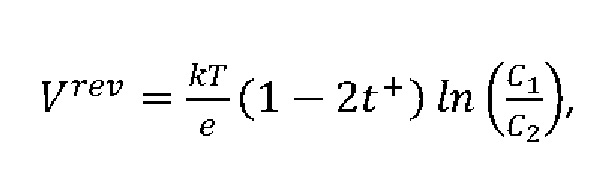
где Vrev - потенциал реверсии (напряжение, при котором трансмембранный ток равен 0); е - заряд электрона; k - постоянная Больцмана; Т - температура в Кельвинах; C1 и C2 - концентрация NaCl в буферном растворе в цис- и транс-отсеке соответственно.
Значение числа переноса катионов было усреднено по результатам 3-5 независимых экспериментов и представлено в виде среднего значения ± стандартная ошибка (р≤0,05).
Пример 1.4. Дифференциальная сканирующая микрокалориметрия фазового перехода липидов
Гигантские однослойные липосомы были получены из чистого ДПФХ и смеси ДПФХ:ХОЛ (85:15 мол.%) и ДПФХ:ЭРГ (85:15 мол.%) методом электроформации с помощью аппарата Vesicle Pre Pro® (Nanion Technologies, Мюнхен, Германия) (стандартный протокол, 3 В, 10 Гц, 58 мин, 55°С). Полученная суспензия липосом содержала 5 мМ липида в буфере (5 мМ HEPES-KOH при рН 7,4). Микафунгин вводили в суспензию до достижения молярного соотношения липид:агент 50:1, 25:1 или 10:1. Затем суспензию нагревали и охлаждали с постоянными скоростями 0,2 и 0,3°С/мин соответственно на микрокалориметре μDSC 7EVO (Setaram, Caluire-et-Cuire, Франция) в температурном диапазоне 30-50°С. Обратимость термических переходов оценивали путем повторного нагрева образца сразу после этапа охлаждения, проведенного при предыдущем сканировании.
Полученные термограммы характеризовали температурой пред-перехода (только в случае ДПФХ), средней температурой плавления (температура, при которой избыточная теплоемкость достигает максимума, Tm) и энтальпией основного фазового перехода (равной площади под основным пиком, ΔHcal). С целью характеристики кооперативности фазового перехода из геля в жидкокристаллическую фазу определяли разницу температур между левой (начало) и правой границей (завершение) основного фазового перехода, ΔTb. Разложение основного пика на составляющие в присутствии исследуемых веществ проводили с использованием программного обеспечения Calisto. Были определены температура максимума для каждого (i-го) компонента, Tm_i, и его процентный вклад в общую площадь, 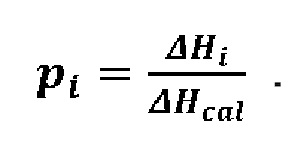
Для каждой липидной смеси было проведено по меньшей мере два независимых эксперимента, чтобы убедиться в воспроизводимости полученных результатов.
Пример 1.5. Конфокальная микроскопия гигантских однослойных липосом
Гигантские однослойные липосомы формировали из смеси ДОФХ:ХОЛ (80:20 мол.%) методом электроформации, как описано в Примере 1.4. (стандартный протокол, 3 В, 10 Гц, 58 мин, 45°С). Для визуализации липосом использовали флуоресцентный липидный зонд Род-ДПФЭ (1 мол.%). Полученную суспензию липосом, содержащую 0,8 мМ липида и 1,5 М сорбитола, разделяли на аликвоты по 30 мкл. Контрольные образцы липосом не содержали микафунгина. Микафунгин вводили в суспензию липосом до концентрации 50 мкМ. Суспензию липосом с микафунгином инкубировали в течение 30 мин при комнатной температуре (25±2°С). 10 мкл полученной суспензии липосом помещали на стандартное предметное стекло микроскопа и закрывали защитным стеклом. Изображения липосом получали с помощью масляного иммерсионного объектива (65×/1,4НСХ PL) Olympus (Гамбург, Германия). Температуру во время наблюдения контролировали с помощью системы нагрева/охлаждения воздуха в теплоизолированной камере. Для возбуждения Род-ДПФЭ использовали гелий-неоновый лазер (543 нм).
Известно, что Род-ДПФЭ преимущественно распределяется в жидкую неупорядоченную липидную фазу (Ld), в то время как жидкая упорядоченная (Lo) и гель (So) фазы остаются неокрашенными (Juhasz et al., 2010). Наличие сил поверхностного натяжения, стремящихся сократить длину границы между Lo и Ld фазами, определяет круглую форму Lo-доменов. Неокрашенные домены сложной формы принято относить к гелеобразной So фазе. Преимущественно, контрольные липосомы из смеси ДОФХ:ХОЛ (80:20 мол.%) были однородно окрашены Род-ДПФЭ, т.е. не демонстрировали фазового разделения (Ld). Небольшое количество липосом содержало небольшие неокрашенные домены круглой формы, которые могут быть отнесены к обогащенным стерином Lo-доменам (Suga, Umakoshi, 2013). В каждом образце в пределах одного поля зрения определяли число липосом без видимого фазового разделения (Ld) и с видимым фазовым разделением различного типа (Lo/Ld - сосуществование жидких упорядоченной и неупорядоченной фаз, So/Ld - сосуществование квази-гель и жидкой неупорядоченной фаз). Было проанализировано несколько соседних полей зрения.
Количество липосом с различным фазовым разделением в каждой тестируемой системе рассчитывали, как отношение количества липосом, демонстрирующих определенный паттерн фазового разделения, к общему количеству липосом. Было проведено восемь независимых экспериментов со средним количеством липосом для каждого образца 125. Различия между группами оценивали с помощью одностороннего дисперсионного анализа (ANOVA) с уровнем значимости р≤0,05.
Пример 1.6. Компьютерное моделирование взаимодействия микафунгина с мембранами
Параметры топологии микафунгина генерировали на основании 3D структуры с помощью CGenFF (Vanommeslaeghe, MacKerell, 2012).
Модельные мембраны были собраны в CHARMM-GUI Membrane Builder (Jo et al., 2009). GROMACS 2023.2 (Abraham et al., 2015) был использован для выполнения компьютерных симуляций с использованием обще-атомного силового поля CHARMM36M (Lee et al., 2016). Минимизация энергии проводилась с помощью алгоритма наискорейшего спуска. Уравновешивание было проведено в шесть ступеней путем постепенного снятия ограничений положения липидных молекул. Все расчеты проводили при постоянной температуре 25°С (298,15 К) и давлении 1 бар с использованием метода Nosé-Hoover и полуизотропного сопряжения давления с использованием баростата C-rescale (Martyna et al., 1992; Bernetti, Bussi, 2020). Постоянная времени сопряжения для температуры и давления составляла 1 и 5 пс соответственно. Метод Particle Mesh Ewald использовали для расчета электростатического взаимодействия на больших расстояниях с ограничением на коротких расстояниях 1,2 нм (Darden et al., 1993); алгоритм смещенного потенциала Леннарда-Джонса использовали для расчета ван-дер-ваальсовых взаимодействий с общим ограничением 1,2 нм и смещенным ограничением 1,0 нм. Траектория в моделях во время компьютерного симулирования регистрировалась каждые 10 пс. Время симулирования составляло 100 не. Площадь на липидную молекулу (APL) была вычислена на MEMBPLUGIN (Guixà-González et al., 2014) и в мембране ПОФХ составляла 63,1 Å2, что хорошо согласовывалось с экспериментальными данными (62,7 Å2 при 20°С, 64,3 Å2 при 30°С (Kučerka et al., 2011)) и данными молекулярной динамики (63,5 Å2 при 303,15 К (Saito et al., 2018)).
Водный раствор ионизировали NaCl до концентрации 0,15 М. Для оценки влияния микафунгина на свойства мембраны использовали модели размером 6×6×8 нм, содержащие (1) 120 ПОФХ; (2) 100 ПОФХ, 50 ХОЛ; (3) 100 ПОФХ, 50 ЭРГ и тот же буферный раствор. Одна молекула микафунгина была помещена в модельную мембрану таким образом, что ее головная часть находилась в области головных групп липидов, а гидрофобная часть молекулы - в области хвостов липидов.
Визуализацию проводили с помощью VMD (Humphrey et al., 1996). Связи между молекулами оценивали с помощью Hydrogen Bonds VMD, причем определяли все связи, которые имеют донорно-акцепторное расстояние менее 6 А: как водородные, так и ван-дер-ваальсовы связи. Взаимодействие на малых расстояниях Леннарда Джонса рассчитывали с помощью GROMACS energy tool. APL вычисляли на MEMBPLUGIN (Guixà-González et al., 2014).
Пример 1.8. Исследование противогрибковой активности in vitro
Приготовление липосомалъной формы микафунгина
Для приготовления липосомальной формы микафунгина была использована методика (Yamskov et al., 2008) с модификациями. Липидную смесь (67 моль.% ПОФХ и 33 моль.% ХОЛ) растворяли в смеси хлороформа (67 об.%) и метанола (33 об.%). Загрузку микафунгина и флоретина проводили таким образом, чтобы достичь эквимолярного соотношения с липидом. Полученный раствор выпаривали в вакуумно-роторном испарителе при комнатной температуре в течение 120 мин. Далее липидную пленку диспергировали в буфере (0,15 М NaCl, 10 мМ HEPES, 1 мМ ЭДТА, рН 7,4) и подвергали воздействию ультразвука в течение 10 минут. Концентрацию микафунгина в образцах, используемых для противогрибковых тестов, оценивали путем получения УФ-спектра согласно (Martens-Lobenhoffer et al., 2011). Спектроскопию поглощения проводили с использованием спектрофлуориметра Флюорат-02-Панорама (Люмэкс, Санкт-Петербург). Все образцы сканировали в диапазоне от 240 до 340 нм с шагом 1 нм.
Тестируемые микроорганизмы
Клинические изоляты устойчивых к флуконазолу штаммов Candida albicans 604М, Candida tropicalis 56-05, Candida krusei 72-05, Candida glabrata 61L и чувствительные к флуконазолу штаммы Candida neoformans для скрининга противогрибковых препаратов были любезно предоставлены А.Б. Кулько (Научно-клинический противотуберкулезный центр Департамента здравоохранения правительства Москвы, Москва).
Подготовка образцов
Образцы разводили в среде RPMI1640 до концентрации 8 мкг/мл в зависимости от содержания активного компонента. Амфотерицин Б, который использовали в качестве контрольного соединения, растворяли в ДМСО до концентрации 10 000 мкг/мл в соответствии с процентным содержанием основного вещества (95%) и дополнительно разбавляли в питательной среде. Диапазон рабочей концентрации протестированных образцов составлял 4-0,03 мкг/мл. Диапазон рабочей концентрации амфотерицина Б составлял 32-0,25 мкг/мл.
Параметры эксперимента
Противогрибковую активность изучали методом последовательных микроразведений в питательной среде в соответствии с руководством EUCAST E.DEF 7.3.2 (EUCAST Definitive document E.DEF 7.3.2. Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts, апрель 2020 г.). Питательную среду RPMI 1640 (с L-глютамином и индикатором рН, но без бикарбоната) готовили в соответствии с нормативной документацией из сухого порошка (партия SIGMA SLBZ6264, St. Лоиус, США) с добавлением глюкозы в конечной концентрации 2% и MOPS (3-(N-морфолино)пропансульфоновой кислоты, ПанЭко, Россия) до конечной концентрации 0,165 моль/л. рН доводили до 7,0 с помощью 1 М/л гидроксида натрия.
Каждую культуру Candida из отдельных колоний, выращенных на 48-часовом агаре Sabouraud, суспендировали в растворе PBS и определяли титр клеточного посева в соответствии со стандартом McFarland, равный 5×106 КОЕ/мл, микробной суспензии с использованием денситометра Den-1B (детектора мутности суспензии, Латвия). После серии разведений тестируемых образцов противогрибковых препаратов в 96-луночных планшетах (Медполимер, Россия) добавляли инокулят культуры Candida до конечного титра около 2×105 КОЕ/мл. Планшеты инкубировали без встряхивания при температуре 35±2°C в аэробных условиях.
Противогрибковую активность оценивали визуально через 24 и 48 часов. Значения МИК соответствовали минимальной концентрации, при которой не наблюдалось видимого роста тестируемого микроорганизма.
Пример 2. Результаты и обсуждения
Пример 2.1. Оценка влияния микафунгина на ионную проницаемость модельных липидных мембран
Исследована способность микафунгина увеличивать ионную проницаемость липидных бислоев. Введение микафунгина в омывающий раствор до концентрации 1-2 мкМ с цис-стороны ПОФХ:ЭРГ (67:33 мол,%) бислоя приводило к появлению скачкообразных флуктуаций трансмембранного тока различной амплитуды в пикоамперном диапазоне. На вставке к Фиг. 2 показаны примеры ступенеобразных флуктуаций, соответствующих открыванию и закрыванию одиночных ион-проницаемых пор, индуцированных микафунгином при различных трансмембранных напряжениях. Была определена катион-анионная специфичность индуцированных микафунгином трансмембранных дефектов. Потенциал реверсии (Vrev), при котором сила тока, протекающего через модифицированную микафунгином мембрану, была равна нулю, составлял около 10 мВ (Фиг. 2). Среднее значение потенциала реверсии соответствовало числу переноса для катионов Na+, равному 0,6±0,1. Наиболее вероятное объяснение преимущественной катионной селективности микафугиновых пор может быть связано с частичными отрицательными зарядами на карбонильных группах в его пептидном кольце.
Пример 2.2. Модификация свойств липидного матрикса микафунгином
Для исследования фазовых переходов липидов использовали дифференциальную сканирующую микрокалориметрию. В отсутствие микафунгина температура плавления ДПФХ, ДПФХ:ХОЛ (85:15 мол.%) и ДПФХ:ЭРГ (85:15 мол.%), Tm, была равна 41,5±0,1°C, 40,6±0,2°C и 40,5±0,1°C соответственно. Ширина пика, связанного с основным переходом ДПФХ, ДПФХ:ХОЛ (85:15 мол.%) и ДПФХ:ЭРГ (85:15 мол.%), ΔTb, была равна 2,6±0,3°C, 3,8±0,2°C и 4,9±0,2°C соответственно. Было показано, что микафунгин влияет на термотропные свойства чистого ДПФХ и его смесей с различными стеринами. На Фиг. 3 показаны репрезентативные термограммы нагрева липосом из ДПФХ, ДПФХ:ХОЛ (85:15 мол.%) и ДПФХ:ЭРГ (85:15 мол.%) до и после добавления микафунгина в суспензию до различных молярных соотношений липид:антибиотик. Микафунгин полностью подавляет пред-переход, наблюдаемый для чистого ДПФХ (Фиг. 3) при температуре 35,1±0,5°C, и увеличивает ширину основного перехода для всех тестируемых липидных систем,
ΔTb. Эти факты указывают на погружение микафунгина в область липидных головок. Основные пики на термограммах в присутствии микафунгина приобретают сложный профиль, характеризующийся наличием двух перекрывающихся компонентов (Фиг. 3). Это свидетельствует о существовании нескольких липидных фаз с различной концентрацией микафунгина.
В Таблице 1 представлены результаты деконволюционного анализа термограмм ДПФХ, ДПФХ:ХОЛ (85:15 мол.%) и ДПФХ:ЭРГ (85:15 мол.%) в присутствии микафунгина.
Таблица 1 - Деконволюционный анализ основного пика, соответствующего плавлению мембранных липидов в присутствии микафунгина
Введение микафунгина вызывало дозозависимое снижение температуры перехода низкотемпературной компоненты во всех тестируемых липидных системах. Более того, с ростом концентрации микафунгина происходило перераспределение энтальпии между двумя компонентами в пользу низкотемпературной. Снижение температуры плавления липидов и уменьшение кооперативности фазового перехода (выраженное в увеличении ΔTb) в присутствии микафунгина указывало на его разупорядывающее действие на мембранообразующие липиды путем погружения в мембрану и увеличения APL.
Нет возможности напрямую сравнить результаты дифференциальной сканирующей калориметрии с данными молекулярной динамики, представленными в Таблице 2, где APL оценивался в ходе молекулярно-динамической симуляции в течение 100 нс с одной молекулой лекарственного средства, вставленной в модельную мембрану. Вызванное микафунгином увеличение значения APL в бислоях из ПОФХ и ПОФХ:ХОЛ и ПОФХ:ЭРГ (Таблица 2) соответствовало результатам дифференциальной сканирующей микрокалориметрии, демонстрирующим разупорядочивающее действие микафунгина (Фиг. 3). Можно заметить, что меньшее изменение в APL в мембране из ПОФХ:ЭРГ (Таблица 2) хорошо согласовывалось с меньшим относительным увеличением ширины основного пика на термограмме ПОФХ:ЭРГ по сравнению с другими тестируемыми системами (Фиг. 3).
Таблица 2 - Площадь, приходящаяся на одну липидную молекулу в мембране в отсутствие и в присутствии микафунгина
На основании данных дифференциальной сканирующей микрокалориметрии было выдвинуто предположение о влиянии микафунгина на фазовое разделение в мембране. Для проверки предположения была выполнена конфокальная флуоресцентная микроскопия гигантских моноламеллярных везикул из ДОФХ:ХОЛ (80:20 мол.%) до и после добавки микафунгина (Фиг. 4). Подавляющее большинство липосом, как в контрольных образцах с необработанными липосомами, так и после введения микафунгина, были гомогенно окрашены, т.е. липиды находились в жидкой неупорядоченной фазе (Ld). На нижней панели к Фиг. 4 показаны типичные микрофотографии однородно окрашенных гигантских липосом до (а) и после (б) введения микафунгина. В контрольных образцах обнаруживалось небольшое количество липосом с маленькими неокрашенными доменами округлой формы, которые могли быть отнесены к стерин-обогащенной упорядоченной Lo фазе (Фиг. 4а, нижняя панель). В присутствии микафунгина наблюдалось изменение морфологии неокрашенных доменов: появлялись липосомы с более крупными неокрашенными доменами нерегулярной формы, которые были отнесены к квази-гель (So) фазе (Фиг. 4б). Было высказано предположение, что адсорбция молекул микафунгина, имеющего крупную полярную пептидную «головку» и только один гидрофобный «хвост», приводит к уменьшению поверхностного натяжения на границе раздела липидных фаз. Известно, что глюкансинтаза встроена в липидные рафты (Satish et al., 2019), и реорганизация липидных доменов при взаимодействии микафунгина с мембраной может нарушать функционирование фермента.
Опираясь на результаты, представленные выше, можно утверждать, что микафунгин характеризуется выраженным действием на липидный бислой мембран. Влияние микафунгина на фазовое разделение в мембране может приводить к ингибированию глюкансинтазы. Помимо этого, микафунгин обладает способностью формировать ион- проницаемые трансмембранные дефекты. Этот вид активности особенно интересен в рамках задачи преодоления антибиотикорезистентности. Усиление порообразующей способности микафунгина в таргетных мембранах может позволить преодолеть резистентность, вызванную мутациями в ферменте.
Пример 2.3. Усиление порообразующей способности микафунгина флоретином
Для усиления порообразующей активности микафунгина был выбран флавоноид флоретин, который, как известно, существенно влияет на физические свойства мембран (Cseh, Benz, 1999; Ostroumova et al., 2013). Литературные данные свидетельствуют, что стратегия успешно работает с полиеновыми противогрибковыми препаратами, амфотерицином Б и нистатином, противогрибковым липопептидом сирингомицином Е и антибактериальным липопептидом-антибиотиком полимиксином Б (Ostroumova et al., 2008; Chulkov et al., 2015; Chulkov, Ostroumova, 2016; Zakharova et al., 2022).
Фиг. 5а демонстрирует влияние 20 мкМ флоретина на трансмембранный ток, индуцируемый микафунгином в мембранах из ПОФХ:ЭРГ (67:33 мол.%). Флоретин приводил к десятикратному росту стационарного трансмембранного тока, индуцированного микафунгином. При этом, флоретин не изменял величину Vrev бислоев, модифицированных микафунгином (Фиг. 5б). В присутствии флоретина также наблюдались ступенеобразные флуктуации трансмембранного тока (Фиг. 5, вставки), которые следует отнести к функционированию ион-проницаемых пор, образующихся под действием микафунгина. Эти данные наглядно демонстрируют, что флоретин способен усиливать порообразующую активность микафунгина.
В работе было показано, что липосомальная форма анидулафунгина более эффективна по сравнению со свободным антибиотиком (Vera-González et al., 2020). Поэтому для оценки потенциальной клинической значимости усиления порообразующей активности микафунгина флоретином, был изучен противогрибковый эффект липосомальных форм микафунгина без и с флоретином. Были использованы клинические изоляты видов Candida, устойчивых к флуконазолу, за исключением штамма C. neoformans (чувствительного к флуконазолу). В качестве положительного контрольного образца использовали амфотерицин Б, растворенный в ДМСО. Липосомы без микафунгина и флоретина и липосомы только с флоретином не проявляли какой-либо противогрибковой активности в отношении тестируемых микроорганизмов в использованных концентрациях. Из Таблицы 3 видно, что комбинированная липосомальная форма микафунгина с флоретином характеризовалась меньшими минимальными ингибирующими концентрациями антибиотика в отношении Candida albicans 604M, Candida tropicalis 56-05 и Candida krusei 72-05 по сравнению с липосомальной формой микафунгина без флоретина.
Таблица 3 - Противогрибковая активность, оцененная визуально после 24/48 часов инкубации при 35°C. Значения минимальной ингибирующей концентрации соответствовали минимальной концентрации антибиотика, при которой не наблюдалось видимого роста тестируемого микроорганизма
Таким образом, можно полагать, что комбинация микафунгина с флоретином, повышающим порообразующую активность антибиотика, может служить основой для разработки инновационных форм микафунгина, которые обладают двумя различными механизмами действия, направленными как на ингибирование глюкансинтазы, так и на образование пор в мембранах патогенных грибов.
В результате проведенных экспериментов получено представление о возможной критической роли мембранной активности микафунгина. Впервые продемонстрирована способность микафунгина образовывать ион-проницаемые дефекты в модельных липидных мембранах. Данные находки могут свидетельствовать о дополнительном, ранее не описанном, механизме противогрибкового действия микафунгина. Впервые показано, что микафунгин влияет на организацию мембранных доменов и термотропные характеристики стерин-содержащих мембран, что приводит к образованию липидной фазы, сходной с гель- состоянием. Эти эффекты могут играть определенную роль в функционировании (1,3) бета- глюкансинтазы. Обнаружен способ усиления порообразующей активности микафунгина, путем его комбинирования в составе липосомальной формы с растительным полифенолом флоретином, что привело к значительному снижению минимальной ингибирующей концентрации в отношении устойчивых к флуканозолу Candida albicans, Candida tropicalis и Candida krusei. Обнаруженная мембранная активность микафунгина может способствовать разработке новых терапевтических средств для борьбы с устойчивостью к антимикотикам.
Список литературы
Abraham, M.J., Murtola, Т., Schulz, R., Páll, S., Smith, J.C., Hess, В., & Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX, (1-2), 19-25 (2015). DOI: 10.1016/j.softx.2015.06.001.
Akbarzadeh, A., Rezaei-Sadabady, R., Davaran, S., Joo, S.W., Zarghami, N., Hanifehpour, Y., Samiei, M., Kouhi, M., Nejati-Koshki, K. Liposome: classification, preparation, and applications. Nanoscale Res. Lett. 8(1), 102 (2013). DOI: 10.1186/1556-276X-8-102.
Babazadeh, A., Ghanbarzadeh, В., Hamishehkar, H. Phosphatidylcholine-rutin complex as a potential nanocarrier for food applications. J. Func. Foods, 33, 134-141 (2017). DOI: 10.1016/j.jff.2017.03.038.
Balkovec, J.M., Hughes, D.L., Masurekar, P.S., Sable, C.A., Schwartz, R.E., & Singh, S.B. Discovery and development of first in class antifungal caspofungin (CANCIDAS®)-a case study. Natural Product Reports. 31(1), 15-34. (2014). DOI: 10.1039/C3NP70070D.
Barani, M., Sangiovanni, E., Angarano, M., Rajizadeh, M.A., Mehrabani, M., Piazza, S., Gangadharappa, H.V., Pardakhty, A., Mehrbani, M., Dell'Agli, M., Nematollahi, M.H. Phytosomes as innovative delivery systems for phytochemicals: a comprehensive review of literature. Int. J. Nanomed. 16, 6983-7022 (2021). DOI: 10.2147/IJN.S318416.
Bernetti, M., & Bussi, G. Pressure control using stochastic cell rescaling. J Chem Phys, 153(11), 114107. (2020). DOI: 10.1063/5.0020514.
Biswas, C., Law, D., Birch, M., Halliday, C., Sorrell, Т.C., Rex, J., Slavin, M., & Chen, S.C.A. In vitro activity of the novel antifungal compound F901318 against Australian Scedosporium and Lomentospora fungi. Medical Mycology. 56(8), 1050-1054. (2018). DOI: 10.1093/MMY/MYX161.
Cheon, D., Kim, J., Jeon, D., Shin, H.C., & Kim, Y. Target Proteins of Phloretin for Its Anti-Inflammatory and Antibacterial Activities Against Propionibacterium acnes-Induced Skin Infection. Molecules. 24(7), 1319 (2019). DOI: 10.3390/molecules24071319.
Chulkov, E.G., & Ostroumova, O.S. Phloretin modulates the rate of channel formation by polyenes. Biochim Biophys Acta. 1858(2), 289-294(2016). DOI: 10.1016/j.bbamem.2015.
Chulkov, E.G., Schagina, L.V., & Ostroumova, O.S. Membrane dipole modifiers modulate single-length nystatin channels via reducing elastic stress in the vicinity of the lipid mouth of a pore. Biochim Biophys Acta. 1848(lPtA), 192-199 (2015). DOI: 10.1016/j.bbamem.2014.09.004.
Crump, J.A., & Collignon, P.J. Intravascular catheter-associated infections. Eur J Clin Microbiol Infect Dis. 19(1), 1-8. (2000). DOI: 10.1007/sl00960050001.
Cseh, R., & Benz, R. Interaction of phloretin with lipid monolayers: relationship between structural changes and dipole potential change. Biophys J. 77(3), 1477-1488 (1999). DOI: 10.1016/S0006-3495(99)76995-X.
D'Angelo, N.A., Noronha, M.A., Câmara, M.C.C, Kurnik, I.S., Feng, C, Araujo, V.H.S., Santos, J.H.P.M., Feitosa, V., Molino, J.V.D., Rangel-Yagui, CO., Chorilli, M., Но, E.A., Lopes, A.M. Doxorubicin nanoformulations on therapy against cancer: an overview from the last 10 years. Biomater. Adv. 133, 112623 (2022). DOI: 10.1016/j.msec.2021.112623.
Darden, Т., York, D., & Pedersen, L. Particle mesh Ewald: An N-log(N) method for Ewald sums in large systems. J. Chem. Phys. 98(12), 10089-10092 (1993). DOI: 10.1063/1.464397.
Dua, J.S., Rana A.C., Bhandari, A.K. Liposome: methods of preparation and applications. Int. J. Pharm. Stud. Res. 3, 14-20 (2012).
Dwivedi, J., Sachan, P., Wal, P., Kosey, S., Khan, M.M.U. Progressive journey of phytosomes: preparation, characterization, patents, clinical trials and commercial products. J. Res. Pharm. 27, 1687-1733 (2023). DOI: 10.29228/jrp.457.
Gonzalez-Lara, M.F., Sifuentes-Osornio, J., & Ostrosky-Zeichner, L. Drugs in Clinical Development for Fungal Infections. Drugs, 77(14), 1505-1518. (2017). DOI: 10.1007/S40265-017-0805-2.
Guixà-González, R., Rodriguez-Espigares, I., Ramfrez-Anguita, J.M., Carrió-Gaspar, P., Martinez-Seara, H., Giorgino, Т., & Selent, J. MEMBPLUGIN: studying membrane complexity in VMD. Bioinformatics, 30(10), 1478-1480 (2014). DOI: 10.1093/bioinformatics/btu037.
Hashimoto, S. Micafungin: a sulfated echinocandin. The Journal of Antibiotics. 62(1), 27-35. (2009). DOI: 10.1038/JA.2008.3.
Hassanpour, P., Hamishehkar, H., Bahari Baroughi, В., Baradaran, В., Sandoghchian Shotorbani, S., Mohammadi, M., Shomali, N., Aghebati-Maleki, L., & Nami, S. Antifungal Effects of Voriconazole-Loaded Nano-Liposome on Fluconazole-Resistant Clinical Isolates of Candida albicans, Biological Activity and ERG11, CDR1, and CDR2 Gene Expression. Assay Drug Dev Technol. 19(7), 453-462. (2021). DOI: 10.1089/adt.2020.1057.
Healey, K.R., Katiyar, S.K., Raj, S., & Edlind, T.D. CRS-MIS in Candida glabrata: sphingolipids modulate echinocandin-Fks interaction. Mol Microbiol. 86(2), 303-313 (2012). DOI: 10.1111/j.l365-2958.2012.08194.x.
Htet, L.L., Wang, L.N., & Liew, Y.X. Efficacy and safety of echinocandins versus triazoles or amphotericin В in the treatment of invasive fungal infections in paediatric patients: a systematic review. Singapore Med J. 2023. DOI: 10.4103/singaporemedj.SMJ-2021-173.
Hu, X., Yang, P., Chai, C, Liu, J., Sun, H., Wu, Y., Zhang, M., Zhang, M.,Liu, X., & Yu H. Structural and mechanistic insights into fungal β-l,3-glucan synthase FKS1. Nature. 616(7955), 190-198 (2023). DOI: 10.1038/s41586-023-05856-5.
Humphrey, W., Dalke, A., & Schulten, K. VMD: Visual molecular dynamics. Journal of Molecular Graphics. 14(1), 33-38. (1996). DOI: 10.1016/0263-7855(96)00018-5.
Hüttel, W. Echinocandins: structural diversity, biosynthesis, and development of antimycotics. Applied Microbiology and Biotechnology. 105(1), 55-66. (2021). DOI: 10.1007/S00253-020-11022-Y
Jo, S., Lim, J.B., Klauda, J.B., & Im, W. CHARMM-GUI Membrane Builder for mixed bilayers and its application to yeast membranes. Biophys J, 97(1), 50-58 (2009). DOI: 10.1016/j.bpj.2009.04.013.
Juhasz, J., Davis, J.H., & Sharom, F.J. Fluorescent probe partitioning in giant unilamellar vesicles of 'lipid raft' mixtures. BiochemJ, 430(3), 415-423 (2010). DOI: 10.1042/BJ20100516.
Kainz, K., Bauer, M.A., Madeo, F., & Carmona-Gutierrez, D. Fungal infections in humans: the silent crisis. Microb Cell. 7(6), 143-145 (2020). DOI: 10.15698/mic 2020.06.718.
Kučerka, N., Nieh, M.P., & Katsaras, J. Fluid phase lipid areas and bilayer thicknesses of commonly used phosphatidylcholines as a function of temperature. Biochim Biophys Acta, 1808(11), 2761-2771 (2011). DOI: 10.1016/j.bbamem.2011.07.022.
Lee, J., Cheng, X., Swails, J.M., Yeom, M.S., Eastman, P.K., Lemkul, J.A., Wei, S., Buckner, J., Jeong, J.C., Qi, Y., Jo, S., Pande, V.S., Case, D.A., & Brooks, C.L. 3rd; MacKerell, AD.Jr.; Klauda, J.B.; Im, W. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J С hem Theory Comput, 12(1), 405-413 (2016). DOI: 10.1021/acs.jctc.5b00935.
Liu, N., Zhang, N., Zhang, S., Zhang, L., & Liu, Q. Phloretin inhibited the pathogenicity and virulence factors against Candida albicans. Bioengineered. 12(1), 2420. (2021). DOI: 10.1080/21655979.2021.1933824.
Liu, Y., Mei, Z., Mei, L., Tang, J., Yuan, W., Srinivasan, S., Ackermann, R., Schwendeman, A.S. Analytical method development and comparability study for AmBisome® and generic Amphotericin В liposomal products. Eur. J. Pharm. Biopharm. 157, 241-249 (2020). DOI: 10.1016/j.ejpb.2020.09.008.
Lockhart, S.R., Chowdhary, A., & Gold, J.A. W. The rapid emergence of antifungal-resistant human-pathogenic fungi. Nat Rev Microbiol. 21(12), 818-832 (2023). DOI: 10.103 8/s41579-023-00960-9.
Lockhart, S.R., Etienne, K.A., Vallabhaneni, S., Farooqi, J., Chowdhary, A., Govender, N.P., Colombo, A.L., Calvo, В., Cuomo, C.A., Desjardins, C.A., Berkow, E.L., Castanheira, M., Magobo, R.E., Jabeen, K., Asghar, R.J., Meis, J.F., Jackson, В., Chiller, Т., & Litvintseva, A.P. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America. 64(2), 134-140. (2017). DOI: 10.1093/CID/CIW691.
Lu, M.; Qiu, Q.; Luo, X.; Liu, X.; Sun, J.; Wang, C; Lin, X.; Deng, Y.; Song, Y. Phyto-phospholipid complexes (phytosomes): a novel strategy to improve the bioavailability of active constituents. Asian J. Pharm. Sci. 14, 265-274(2019). DOI: 10.1016/j.ajps.2018.05.011.
Maertens, J., Pagano, L., Azoulay, E., & Warris, A. Liposomal amphotericin B-the present. J Antimicrob Chemother. 77(Suppl_2), ii 11-ii20 (2022). DOI: 10.1093/jac/dkac352.
Martens-Lobenhoffer, J., Rupprecht, V., & Bode-Boger, S.M. Determination of micafungin and anidulafungin in human plasma: UV- or mass spectrometric quantification? J Chromatogr В Analyt Technol Biomed Life Sci. 879(22), 2051-2056. (2011). DOI: 10.1016/j.jchromb.2011.05.033.
Martyna, G.J., Klein, M.L., & Tuckerman, M. Nose-Hoover chains: The canonical ensemble via continuous dynamics. J. Chem. Phys. 97(4), 2635-2643. (1992). DOI: 10.1063/1.463940.
Martynova, A.V., Pavlova, O.S., & Yusupova, E.P. Эпидемиологический анализ системных микозов при COVID-19. Медицинский Совет.10(13), 326-331. (2023). DOI: 10.21518/MS2023-178.
Montal, М., & Mueller, P. Formation of bimolecular membranes from lipid monolayers and a study of their electrical properties. Proc Natl Acad Sci USA, 69, 3561-3566 (1972). DOI: 10.1073/pnas.69.12.3561.
Moreira-Walsh, В., Ragsdale, A., Lam, W., Upadhya, R., Xu, E., Lodge, J.K., & Donlin, M.J. Membrane Integrity Contributes to Resistance of Cryptococcus neoformans to the Cell Wall Inhibitor Caspofungin. mSphere. 7(4), e0013422 (2022). DOI: 10.1128/msphere.00 134-22.
Morf, W. Calculation of liquid-junction potentials and membrane potentials on the basis of the Planck theory. Analytical Chemistry, 49(6), 810-813 (1977). DOI: 10.1021/ac50014a035.
Ostroumova, O.S., Efimova, S.S., & Schagina, L.V. Phloretin-induced reduction in dipole potential of sterol-containing bilayers. J Membr Biol. 246(12), 985-991 (2013). DOI: 10.1007/s00232-013-9603-2.
Ostroumova, O.S., Efimova, S.S., & Schagina, L.V. Probing amphotericin В single channel activity by membrane dipole modifiers. PloS One, 7(1), e30261. (2012). DOI: 10.1371/journal.pone.0030261.
Ostroumova, O.S., Efimova, S.S., Chulkov, E.G., & Schagina, L.V. The interaction of dipole modifiers with polyene-sterol complexes. PloS One, 7(9), e45135. (2012). DOI: 10.1371/journal.pone.0045135.
Ostroumova, O.S., Efimova, S.S., Mikhailova, E.V., & Schagina, L.V. The interaction of dipole modifiers with amphotericin-ergosterol complexes. Effects of phospholipid and sphingolipid membrane composition. European Biophysics Journal. 43(4-5), 207-215. (2014). DOI: 10.1007/S00249-014-0946-0.
Ostroumova, O.S., Shchagina, L.V, & Malev, V.V. The effect of dipole potential of lipid bilayers on the properties of ion channels formed by cyclic lipodepsipeptide syringomycin E. Biochem. (Moscow) Suppl. Ser. A: Membr. Cell Biol. 2, 259-270 (2008). DOI: 10.1134/s 1990747808030100.
Pande S. Liposomes for drug delivery: review of vesicular composition, factors affecting drug release and drug loading in liposomes. Artif. Cells Nanomed. Biotechnol. 51(1), 428-440 (2023). DOI: 10.1080/21691401.2023.2247036.
Pappas, P.G, Kauffman, C.A., Andes, D.R., Clancy, C.J., Marr, K.A., Ostrosky-Zeichner, L., Reboli, A.C., Schuster, M.G, Vazquez, J.A., Walsh, T.J., Zaoutis, Т.Е., & Sobel, J.D. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin Infect Dis. 62(4), el-50 (2016). DOI: 10.1093/cid/civ933.
Patil, A., & Majumdar, S. Echinocandins in antifungal pharmacotherapy. The Journal of Pharmacy and Pharmacology. 69(12), 1635-1660. (2017). DOI: 10.1111/JPHP.12780.
Perfect, J.R. The antifungal pipeline: a reality check. Nature Reviews. Drug Discovery. 16(9), 603-616. (2017). DOI: 10.1038/NRD.2017.46.
Pfaller, M.A., Castanheira, M., Lockhart, S.R., Ahlquist, A.M., Messer, S.A., & Jones, R. N. Frequency of decreased susceptibility and resistance to echinocandins among fluconazole-resistant bloodstream isolates of Candida glabrata. Journal of Clinical Microbiology. 50(4), 1199-1203. (2012). DOI: 10.1128/JCM.06112-11.
Pierce, C.G., Srinivasan, A., Uppuluri, P., Ramasubramanian, A.K., & López-Ribot, J.L. Antifungal therapy with an emphasis on biofilms. Curr Opin Pharmacol. 13(5), 726-730 (2013). DOI: 10.1016/j.coph.2013.08.008.
Pouliot, M., & Jeanmart, S. Pan Assay Interference Compounds (PAINS) and Other Promiscuous Compounds in Antifungal Research. Journal of Medicinal Chemistry. 59(2), 497-503. (2016). DOI: 10.1021/ACS.JMEDCHEM.5B00361.
Raut, A., & Huy, N.T. Rising incidence of mucormycosis in patients with COVID-19: another challenge for India amidst the second wave? The Lancet. Respiratory Medicine. 9(8), e77. (2021). DOI: 10.1016/S2213-2600(21)00265-4.
Rodrigues, M.L., & Nosanchuk, J.D. Fungal diseases as neglected pathogens: A wake-up call to public health officials. PLoS Negl Trop Dis. 14(2), e0007964 (2020). DOI: 10.1371/journal.pntd.0007964.
Saito, H., Morishita, Т., Mizukami, Т., Nishiyama, K., Kawaguchi, K., & Nagao, H. Molecular dynamics study of binary POPC bilayers: molecular condensing effects on membrane structure and dynamics. Phys. Conf. Ser, 1136, 012022 (2018). DOI: 10.1088/1742-6596/1136/1/012022.
Satish, S., Jiménez-Ortigosa, C, Zhao, Y., Lee, M.H., Dolgov, E., Krüger, T.;, Park, S., Denning, D.W., Kniemeyer, O., Brakhage, A.A., & Perlin, D.S. Stress-Induced Changes in the Lipid Micro environment of β-(l,3)-d-Glucan Synthase Cause Clinically Important Echinocandin Resistance in Aspergillus fumigatus. mBio, 10(3), e00779-19 (2019). DOI: 10.1128/mBio.00779-19.
Sharma, P.; Dogra, L.; Thakur, D.; Smily, O.; Chauhan, D. Phytosomes, a novel strategy to improve the bioavailability of flavonoids: a review. Eur. J. Biomed. Pharm. Sci. 6, 209-218 (2019).
Silva, L.N., de Mello, T.P., de Souza Ramos, L., Branquinha, M.H., Roudbary, M., & dos Santos, A.L.S. Fungal Infections in COVID-19-Positive Patients: A Lack of Optimal Treatment Options. Current Topics in Medicinal Chemistry. 20(22), 1951-1957. (2020). DOI: 10.2174/156802662022200917110102.
Singh, A., Yadav, V, & Prasad, R. Comparative lipidomics in clinical isolates of Candida albicans reveal crosstalk between mitochondria, cell wall integrity and azole resistance. PLoS One. 7(6), e39812 (2012). DOI: 10.1371/journal.pone.0039812.
Suga, K., & Umakoshi, H. Detection of nanosized ordered domains in DOPC/DPPC and DOPC/Ch binary lipid mixture systems of large unilamellar vesicles using a TEMPO quenching method. Langmuir, 29(15), 4830-4838 (2013). DOI: 10.1021/1a304768f.
Sun, Q., Xiong, K., Yuan, Y., Yu, J., Yang, L., Shen, C, Su, C, Lu, Y. Inhibiting Fungal Echinocandin Resistance by Small-Molecule Disruption of Geranylgeranyltransferase Type I Activity. Antimicrob Agents Chemother. 64(2), e02046-19 (2020). DOI: 10.1128/AAC.02046-19.
Szymański, M., Chmielewska, S., Czyżewska, U., Malinowska, M., & Tylicki, A. Echinocandins - structure, mechanism of action and use in antifungal therapy. J Enzyme Inhib Med Chem. 37(1), 876-894 (2022). DOI: 10.1080/14756366.2022.2050224.
Taormina, G., Gopinath, R., Moore, J., Yasinskaya, Y., Colangelo, P., Reynolds, K., & Nambiar, S.A Regulatory Review Approach for Evaluation of Micafungin for Treatment of Neonatal Candidiasis. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America. 73(12), 2335-2340. (2021). DOI: 10.1093/CID/CIAB025.
The Lancet Infectious Diseases. An exciting time for antifungal therapy. The Lancet. Infectious Diseases, 23(7), 763. (2023). DOI: 10.1016/S1473-3099(23)00380-8.
Tóth, Z., Bozó, A., Kovacs, R., Balogh, В., Balázs, В., Forgács, L., Kelentey, В., & Majoros, L. The In Vitro Activity of Fluconazole, Amphotericin В and Echinocandins Against Cyberlindnerafabianii Planktonic Cells and Biofilms. Mycopathologia, 188(1-2), 111-118 (2023). DOI: 10.1007/s11046-022-00688-9.
Tripathi, S.K., Feng, Q., Liu, L., Levin, D.E., Roy, K.K., Doerksen, R.J., Baerson, S.R., Shi, X., Pan, X., Xu, W.H., Li, X.C., Clark, A.M., & Agarwal, A.K. Puupehenone, a Marine-Sponge-Derived Sesquiterpene Quinone, Potentiates the Antifungal Drug Caspofungin by Disrupting Hsp90 Activity and the Cell Wall Integrity Pathway. mSphere. 5(1), e00818-19 (2020). DOI: 10.1128/mSphere.00818-19.
van Meer, G., & de Kroon, A.I. Lipid map of the mammalian cell. J Cell Sci. 124(Pt 1), 5-8 (2011). DOI: 10.1242/jcs.071233.
Vanommeslaeghe, K., & MacKerell, A.D. Automation of the CHARMM General Force Field (CGenFF) I: bond perception and atom typing. J Chem InfModel, 52(12), 3144-3154 (2012). DOI: 10.1021/ci300363c.
Ventoulis, L, Sarmourli, Т., Amoiridou, P., Mantzana, P., Exindari, M., Gioula, G., & Vyzantiadis, T.A. Bloodstream Infection by Saccharomyces cerevisiae in Two COVID-19 Patients after Receiving Supplementation of Saccharomyces in the ICU. Journal of Fungi. 6(3), 1-7. (2020). DOI: 10.3390/JOF6030098.
Vera-González, N., Bailey-Hytholt, С.M., Langlois, L., de Camargo Ribeiro, F., de Souza Santos, E.L., Junqueira, J.C, & Shukla, A. Anidulafungin liposome nanoparticles exhibit antifungal activity against planktonic and biofilm Candida albicans. J Biomed Mater Res A. 108(11), 2263-2276. (2020). DOI: 10.1002/jbm.a.36984.
Wang, J., Yang, Q., Zhang, P., Sheng, J., Zhou, J., Qu, Т., & Qu, T. Clinical characteristics of invasive pulmonary aspergillosis in patients with COVID-19 in Zhejiang, China: a retrospective case series. Critical Care, 24(1), 299. (2020). DOI: 10.1186/S13054-020-03046-7.
WHO fungal priority pathogens list to guide research, development and public health action. Geneva: World Health Organization. 2022. Licence: CC BY-NC-SA 3.0 IGO.
Wiederhold, N.P. Antifungal resistance: current trends and future strategies to combat. Infection and Drug Resistance. 10, 249-259. (2017). DOI: 10.2147/IDR.S 124918.
Yamskov, I.A., Kuskov, A.N., Babievskiĭ, K.K., Berezin, B.B., Kraiukhina, M.A., Samoĭlova, N.A., Tikhonov, V.E., & Shtil'man, M.I. New liposomal forms of antifungal antibiotics, modified by amphiphilic polymers. Appl Biochem Microbiol, 44(6), 688-693 (2008).
Zakharova, A.A., Efimova, S.S., & Ostroumova, O.S. Lipid Microenvironment Modulates the Pore-Forming Ability of Polymyxin B. Antibiotics. 11(10), 1445 (2022). DOI: 10.3390/antibiotics 11101445.
Zhao, C.R., You, Z.L., Chen, D.D., Hang, J., Wang, Z.B., Ji, M., Wang, L.X., Zhao, P., Qiao, J., Yun, C.H., & Bai, L. Structure of a fungal 1,3-β-glucan synthase. Sci Adv. 9(37), eadh7820 (2023). DOI: 10.1126/sciadv.adh7820.
Zheng, L., Xu, Y., Wang, C, Yang, F., Dong, Y., & Guo, L. Susceptibility to caspofungin is regulated by temperature and is dependent on calcineurin in Candida albicans. Microbiol Spectr. 11(6), e0179023 (2023). DOI: 10.1128/spectrum.01790-23.
| название | год | авторы | номер документа |
|---|---|---|---|
| Применение липопептидов в качестве ингибиторов слияния мембран | 2022 |
|
RU2802823C1 |
| Химерный рекомбинантный интерферон альфа-2б, обладающий противогрибковой активностью | 2024 |
|
RU2823607C1 |
| КОМПОЗИЦИЯ НА ОСНОВЕ ГИБРИДНОГО РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА ДЛЯ ЗАЩИТЫ РАСТЕНИЙ ОТ ЗАБОЛЕВАНИЙ, ВЫЗЫВАЕМЫХ ООМИЦЕТАМИ | 2021 |
|
RU2786706C2 |
| Противоопухолевый липосомальный препарат и способ его получения | 2017 |
|
RU2663291C1 |
| КОМПОЗИЦИИ ЛИПИДНЫХ ВЕЗИКУЛ И СПОСОБЫ ПРИМЕНЕНИЯ | 2011 |
|
RU2595872C2 |
| Композиция на основе миРНК для ингибирования Gal-9 (LGALS9), препарат на ее основе и способ ее применения в качестве таргетного иммуностимулятора при анти-PD-L1 терапии | 2023 |
|
RU2832798C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭФИРНОГО МАСЛА ПЛОДОВ БОРЩЕВИКА СОСНОВСКОГО, ОБЛАДАЮЩЕГО ПРОТИВОГРИБКОВОЙ АКТИВНОСТЬЮ | 2024 |
|
RU2835487C1 |
| ИНЪЕКЦИОННАЯ ЛЕКАРСТВЕННАЯ ФОРМА ОЛИГОПЕПТИДНОГО ПРЕПАРАТА ДЛЯ ЛЕЧЕНИЯ РАССЕЯННОГО СКЛЕРОЗА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2010 |
|
RU2561582C2 |
| Способ лечения пролиферативной витреоретинопатии с помощью мелфалана, ассоциированного с липосомами, в эксперименте | 2021 |
|
RU2772520C1 |
| Способ получения липосомальной лекарственной формы ацетилцистеина | 2024 |
|
RU2836760C1 |
Изобретение относится к области биотехнологии, в частности к новым противогрибковым средствам на основе липосом. Предлагаются однослойные липосомы, содержащие микафунгин. Дополнительно липосомы могут содержать флоретин в эквимолярном соотношении с липидом. Противогрибковая активность липосом по настоящему изобретению значительно выше активности известного из уровня техники амфотерицина Б, что подтверждается их значительно более низкой минимальной ингибирующей концентрацией (МИК) по сравнению с амфотерицином Б. Так, МИК однослойных липосом с микафунгином в отношении устойчивых к флуконазолу клинических изолятов видов Candida в 8-60 раз ниже, чем МИК амфотерицина Б. Добавление флоретина в липосомы с микафунгином уменьшает МИК в 14 раз. 3 з.п. ф-лы, 5 ил., 3 табл., 2 пр.
1. Противогрибковое средство в виде сформированных с участием фосфатидилхолина однослойных липосом, содержащих микафунгин.
2. Противогрибковое средство по п. 1, в котором в качестве фосфатидилхолина используют 1,2-диолеоил-sn-глицеро-3-фосфохолин, либо 1,2-дипальмитоил-sn-глицеро-3-фосфохолин, либо 1-пальмитоил-2-олеоил-sn-глицеро-3-фосфохолин.
3. Противогрибковое средство по п. 1, дополнительно содержащее флоретин.
4. Противогрибковое средство по пп. 1-3, в котором в качестве липида используют смесь 1-пальмитоил-2-олеоил-sn-глицеро-3-фосфохолина и холестерина в соотношении 67:33 моль.
| US 5874104 A, 1999.02.23 | |||
| WO 2017028811 A1, 2017.02.23 | |||
| WO 1988006442 A1, 1988.09.07 | |||
| Maria Chountoulesi et al | |||
| The significance of drug-to-lipid ratio to the development of optimized liposomal formulation, Journal of Liposome Research, 2017, 28(3): 249-258 | |||
| KV Clemons et al | |||
| Comparative efficacies of conventional amphotericin B, liposomal |

