Изобретение относится к области биотехнологии и молекулярной биологии, а именно к применению антимикробных циклических липопептидов (ЦЛП) по новому назначению в качестве ингибиторов слияния мембран.
Оболочечные вирусы представляют собой патогены, капсид которых окружен двойным липидным бислоем с включенными в него белками (Yuan et al., 2018). К представителям этого класса относятся множество вирусов: ВИЧ, вирус гриппа, кори, Эболы, MERS-CoV-2, SARS-CoV, SARS-CoV-2, вирус Денге, Западного Нила, желтой лихорадки, везикулярного стоматита и др. (Rey and Lok, 2018). Критической стадией жизненного цикла оболочечного вируса является слияние его липидного бислоя с клеточной мембраной, после чего вирусный геном попадает внутрь клетки (Teissier et al., 2010). Слияние двух контактирующих липидных бислоев требует не только сильного сближения их апикальных частей, но и ряда топологических перестроек. Промежуточные состояния, характеризующиеся локальным минимумом энергии, называются интермедиатами слияния. Для перехода между ними необходимо преодолеть соответствующие энергетические барьеры (Risselada et al., 2014). Энергию для подобных преобразований обеспечивает трансформация вирусных белков слияния (Kielian, 2014). В последнее время частота возникновения заболеваний, вызванных оболочечными вирусами, растет (Buchmann and Holmes, 2015). Причиной такой тенденции, без сомнений, является высокая мутационная активность вирусов (Pizzorno et al., 2011). Это снижает эффективность применения существующих противовирусных препаратов, механизм действия которых направлен на различные белковые вирусные мишени (Klebe and Schlitzer, 2011). Кроме того, данные фармакологические средства имеют узкий спектр противовирусной активности (Wei et al., 2002). Требуется развитие новых терапевтических стратегий, в основе которых будет лежать воздействие на консервативные вирусные элементы. Подобной альтернативой может стать применение ингибиторов вирусного слияния с мембрано- или липидоопосредованным механизмом действия. Известно, что молекулы, объем гидрофильной части/частей которых значительно преобладает над объемом гидрофобных частей, способствуют образованию выпуклых липидных поверхностей (с положительной кривизной) и могут успешно подавлять слияние вирусов с клетками за счет увеличения энергии формирования промежуточных состояний, характеризующихся вогнутыми липидными поверхностями (с отрицательной кривизной) (St Vincent et al., 2010).
ЦЛП характеризуются наличием объемной гидрофильной пептидной «головки» и гидрофобного углеводородного «хвоста», что может указывать на возможность индукции ими положительной спонтанной кривизны (Xia et al., 2021; Schneider et al., 2014; Fewer et al., 2021). Циклическая структура олигопептида увеличивает стабильность ЦЛП in vivo в связи с более низким протеолизом относительно линейных аналогов, однако, это ограничивает конформационную подвижность пептида (Hamley et al., 2015). Указанные соединения обладают быстрой биоразлагаемостью (Makovitzki et al., 2007) и относительной безопасностью применения (Kanlayavattanakul and Lourith, 2010). По происхождению выделяют природные, полусинтетические и синтетические ЦЛП. Основными продуцентами природных ЦЛП являются грибы, например, Aspergillus, и бактерии, в том числе, Streptomyces, Pseudomonas и Bacillus (Raaijmakers et al., 2010). В состав пептидной головки могут входить неканонические аминокислоты, в частности, D -аминокислоты (Xia et al., 2021). Это, прежде всего, связано с тем, что ЦЛП синтезируются вне рибосом с помощью специальных пептид с интетаз (Biniarz et al., 2017). К данному классу можно отнести такие липопептиды как: даптомицин (Miao et al., 2005), сурфактин (Arima et al., 1968), итурин A (Maget-Dana and Peypoux, 1994), фенгицин (Deleu et al., 2008), полимиксин Б (Velkov et al., 2013) и другие. На основании проведенных структурно-функциональных исследований были созданы полусинтетические липопептиды. Примерами полусинтетических ЦЛП являются эхинокандины, полученные путем модификации вторичных метаболитов грибов. В частности, каспофунгин и анидулафунгин были синтезированы на основе пневмокандина ВО (Glarea lozoyensis) (Rahway, 2005) и эхинокандина ВО (Aspergillus nidulans) (Murdoch and Plosker, 2004). Следующим шагом стало создание полностью синтетических липопептидов, в основе которых не лежат природные молекулярные матрицы. Представителями класса синтетических ЦЛП могут являться вещества, полученные в работе Вила с соавторами: ВРС194, ВРС590, ВРС504, ВРС528, ВРС588, ВРС676, ВРС684, ВРС702, ВРС710, ВРС728 ( et al., 2016). Авторы, взяв за основу синтетический циклический пептид ВРС194, путем присоединения различных жирнокислотных остатков получили панель соединений, которые демонстрировали антибактериальную и противогрибковую активность, в том числе против: Xanthomonas axonopodis pv. vesicatoria, Pseudomonas syringae pv. syringae, Fusarium oxysporum ( et al., 2016).
et al., 2016). Авторы, взяв за основу синтетический циклический пептид ВРС194, путем присоединения различных жирнокислотных остатков получили панель соединений, которые демонстрировали антибактериальную и противогрибковую активность, в том числе против: Xanthomonas axonopodis pv. vesicatoria, Pseudomonas syringae pv. syringae, Fusarium oxysporum ( et al., 2016).
Акулеацин противогрибковый антибиотик, продуцентом которого является Aspergillus aculeatus. Состоит из циклического гексапептида и связанного с ним остатка пальмитиновой кислоты (Yamaguchi et al., 1985). Акулеацин проявляет высокую активность против Candida и демонстрирует малую эффективность в отношении диморфных, дематиевых и дерматофитных грибов (Iwata et al., 1982). Не обладает антибактериальной активностью (Iwata et al., 1982). Механизм действия акулеацина основан на блокаде β-(1,3)глюкансинтазы и ингибировании синтеза р-глюкана (Yamaguchi et al., 1985).
Эхинокандины, каспофунгин и анидулафунгин, представляют собой амфифильные циклические гексапептиды с N-связанными ацил-липидными боковыми цепями (Mikamo et al., 2000). Эхинокандины демонстрируют высокую активность в отношении Candida, включая С. parapsilosis, С. lusitaniae и С. guilliermondii (Pfaller et al., 2005; Pfaller et al., 2006), эффективно подавляют развитие Pneumocystis carinii (Ito et al., 2000), Alternaria sp., Curvularia sp., Acremonium sp., Bipolaris sp. и Trichoderma sp. H др. (Kahn et al., 2006). Механизм действия эхинокандинов основан на ингибировании 1,3-β- (Deresinski and Stevens, 2003) и 1,6-β-D-глюкансинтазы (Stevens et al., 2006).
Даптомицин - липопептид, содержащий деканоильную липидную цепь, присоединенную к пептидному участку, состоящему из 13 аминокислот, циклизованных с образованием 10-членного макролактонового кольца с тремя экзоциклическими остатками (Miao et al., 2005). Пептидная часть даптомицина содержит семь протеиногенных и шесть непротеиногенных аминокислот (D-Asn2, Orn6, D-Ala8, D-Serll, MeGlu12n Kyn13) (Liao et al., 2012). Даптомицин содержит кальций-связывающий домен с 7 по 10 аминокислотный остаток (Liao et al., 2012), который играет важную роль при активности ЦЛП (Jung et al., 2008). Предполагается, что образование комплекса липопептида с кальцием способствует компенсации общего отрицательного заряда молекулы (-3), позволяя даптомицину взаимодействовать с анионной бактериальной мембраной (Schneider et al., 2014). Продуцентом даптомицина является грамположительный актиномицет Streptomyces roseoporous (Miao et al., 2005). Даптомицин демонстрирует выраженную активность в отношении широкого спектра грамположительных бактерий, включая Staphylococcus aureus, Enterococcus faecalis, Streptococcus pyogenes, S. agalactiae, S. dysgalactiae и др (Streit et al., 2004). Предполагается несколько механизмов его действия (Silverman et al., 2003). По одной из версий активность ЦЛП связана со способностью ингибировать синтез липотейхоевой кислоты (Silverman et al., 2003). Другое предложение базируется на способности даптомицина индуцировать деполяризацию мембраны за счет формирования пор (Zhang et al., 2014). Считается, что липопептид взаимодействует с фосфатидилглицерином, что приводит к мембранным перестройкам и последующему нарушению архитектуры бислоя (Jung et al., 2008).
Фенгицин - липопептид, синтезируемый несколькими штаммами Bacillus subtilis (Zhao et al., 2017). Он состоит из жирнокислотного остатка, длина которого может варьировать от 14 до 17 атомов углерода (Pathak et al., 2012), и пептидной части, включающей 10 аминокислотных остатков (Акра et al., 2001) Восемь из них участвуют в образовании кольца посредством лактоновой связи между Tyr-3 и Ile-10 (Акра et al., 2001). В шестом положении фенгицина А и Б содержится Ala и Val соответственно (Meena and Kanwar, 2015; Tang et al., 2014). Фенгицин активен в отношении нитевидных грибов, в том числе Rhizoctonia solani (Guo et al., 2014). Механизм действия этого ЦЛП основан на его способности изменять физико-химические свойства таргетных мембран. В высоких концентрациях фенгицин действует как детергент (Deleu et al., 2005). В низких концентрациях ЦЛП образует олигомеры, которые способствуют локальному возмущению бислоя, индукции положительной кривизны, формированию пор и повреждению мембраны (Deleu et al., 2005; Horn et al., 2013). Присутствие ионов кальция, как и в случае даптомицина, усиливает активность липопептида за счет конформационных изменений декапептидной области (Nasir et al., 2013).
Итурин А и микосубтилин являются членами одного семейства липопептидов - итуринов. Продуцентом обоих является В. subtilis (Raaijmakers et al., 2010). Пептидное кольцо итурина состоит из семи аминокислотных остатков и связано с жирнокислотным «хвостом» в области Ser-7 с образованием макролактамного кольца (Raaijmakers et al., 2010; Zhao et al., 2017). Длина хвостовой части для гомологов итурина А, С, D, Е отличается и варьирует от 14 до 17 углеродов (Zhao et al., 2017). Микосубитилин и итурин являются близкими гомологами и отличаются последовательностью и стероизомеризацией серина и аспарагина в шестом и седьмом положениях (Nasir et al., 2012). Итурины характеризуются широким спектром биологической активности. Антибактериальное действие включает патогены растений Xanthomonas campestris pv. cucurbitae, Pectobacterium carotovorum subsp. carotovorum, R. solani, Fusarium graminearum и др. (Zeriouh et al., 2011, Gong et al., 2015). Противогрибковая активность отмечена в отношении Colletotrichum demiatium (Hiradate et al., 2002), Penicillium roqueforti (Chitarra et al., 2003), A. flavus (Moyne et al., 2001), R. solani (Yu et al., 2002). Механизм действия итурина А основан на способности липопептида формировать трансмембранные поры (Maget-Dana and Peypoux, 1994). Считается, что во взаимодействии с клеточной мембраной критическую роль играет Tyr (Harnois et al., 1989). Стоит отметить, что присутствие холестерина в липидном бислое значительно увеличивает вероятность открывания итурин-индуцированных пор (Regine et al., 1985). В свою очередь, эргостерин, присутствующий в мембранах дрожжей или грибов, также может служить мишенью для действия итуринов. Ассоциация ЦЛП с эргостерином приводит к изменению в организации и морфологии таргетных мембран (Nasir and Besson, 2012).
Полимиксины - класс циклических липопептидов, продуцентами которых являются различные виды рода Paenibacillus, такие как P. amylolyticus, P. polymyxa (De Crescenzo Henriksen et al., 2007; Yoshino et al., 2013). Полимиксины состоят из пептидного участка, в котором семь остатков образуют цикл, а три экзоциклические аминокислоты соединяются с остатком жирной кислоты (Schneider et al., 2014). Значительный положительный заряд молекулы обеспечивает наличие 2,4-диаминомасляной кислоты (Storm et al., 1977). В качестве липидного «хвоста» в структуре ЦЛП могут присутствовать 6-метилоктановая кислота или 6-метилгептановая кислота (Schneider et al., 2014). Липопептиды, относящиеся к классу полимиксинов, эффективны против грамотрицательных бактерий, например P. aeruginosa, Acinetobacter baumannii, Klebsiella pneumonia (Arnold et al., 2011; Kanj and Kanafani, 2011). Высокая избирательность связана с механизмом действия, в основе которого лежит ассоциация пентакатионного пептидного фрагмента с бактериальным липополисахаридом. Это приводит к истончению мембраны, нарушению ее целостности, нарушению водно-электролитного баланса и, как следствие, гибели бактерии (Clausell et al., 2007). Кроме того, гидрофобные боковые цепи в структуре полимиксина могут взаимодействовать с липидными цепями липополисахаридов (Mares et al., 2009), способствуя внедрению ЦЛП во внешнюю мембрану бактерии (Hermsen et al., 2003, Evans et al., 1999). Делеция липидного фрагмента приводит к потере антибактериальных свойств полимиксина (Deris et al., 2014).
Сирингостатин А и сиринготоксин Б относятся к группе липопептидов продуцируемых Pseudomonas syringae-pv. syringae (Dalla Serra et al., 1999). Сиринготоксин Б состоит из циклической пептидной части и сшитого с ней посредством амидной связи гидрофобного хвоста в виде 3-гидрокситетрадекановой жирной кислоты. Пептидная часть молекулы имеет вид Ser-Dab-Gly-Hse-Orn-aThr-Dhb-(3-OH)Asp-(4-Cl)Thr. Четыре аминокислоты являются нестандартными: Dab2, Dhb7, (3-OH)Asp8, (4-Cl)Thr9. Лактоновое кольцо образуется между гидроксильной группой боковой цепи Serl и карбоксилатной группой (4-Cl)Thr9 (Matyus et al., 2008). Пептидная часть сирингостатина А отличается от сиринготоксина Б одной заменой Gly3 на Dab3, что придает молекуле более положительный заряд (Ballio et al., 1990). Данные соединения демонстрируют широкую противогрибковую активность, в том числе против С. albicans, С. kefyr, С. tropicalis, Saccharomyces cerevisiae, A. fumigatus и др. Эффективность ЦЛП в отношении дрожжевых грибов была выше относительно мицелиальных грибов (Sorensen et al., 1996). Механизм действия веществ, по всей видимости, связан с их способностью повышать проницаемость мембраны посредством образования олигомерных каналов (Serra et al., 1999; Gur'nev et al., 2002). Включение стеринов в состав таргетного бислоя повышает активность сиринготоксина Б (Serra et al., 1999).
Сурфактин имеет в структуре пептидного участка семь аминокислот. Хвостовая часть - β-гидроксижирная кислота с длиной цепи от 12 до 16 атомов углерода, - образует вместе с пептидной лактоновый цикл (Seydlova et al., 2011). Гомологичные варианты сурфактина могут отличаться как по аминокислотному составу, так и по длине гидрофобной части (Korenblum et al., 2012).
Как уже отмечено выше, биологическая активность липопептидов многообразна, а, следовательно, и области их применения многочисленны. Они используются в медицине в качестве противомикробных агентов, и в частности, против резистентных форм бактерий (Fowler et al., 2006). Некоторые липопептиды обладают выраженной противогрибковой активностью (Romero et al., 2007). Имеются сообщения и о противовирусной активности липопептидов (Dey et al., 2021; Jeon et al., 2020; Barrows et al., 2016; Elmorsy et al., 2022). В ходе in vitro скрининга Барроус с соавторами показали, что добавление даптомицина снижает уровень вирусного потомства в клетках HuH-7, HeLa и НАЕС (Barrows et al., 2016). Чен с соавторами показали успешное применение анидулафунгина и каспофунгина в отношении вируса Денге (Chen et al., 2021). В работе Лу с соавторами (Lu et al., 2021) было показано, что анидулафунгин эффективно подавляет развитие вируса Зика. Поскольку применение ЛП было эффективным на ранней стадии инфекции, авторы предположили, что основной механизм действия связан с прямым нарушением целостности вириона (Lu et al., 2021). Противовирусная активность анидулафунгина в отношении SARS-CoV-2 была показана в работе (Jeon et al., 2020). При этом механизмы противовирусной активности ЦЛП остаются спорными. В работе (Ahamad et al., 2022) было показано, что анидулафунгин значительно подавляет формирование синцитиев из клеток Vero, экспрессирующих S-белок SARS-CoV-2. С использованием методов молекулярной динамики авторы предположили, что анидулафунгин препятствует взаимодействию S-белка с АСЕ2-рецепторами. На основании результатов молекулярного докинга Дей с коллегами склонны связывать анти-S ARS-CoV-2-действие анидулафунгина с взаимодействием липопептида с белком nsp12, который управляет РНК-зависимой РНК-полимеразной активностью SARS-CoV-2 (Dey et al., 2021). Методами молекулярного докинга Анваар с коллегами (Anwaar et al., 2022) продемонстрировали возможность связывания анидулафунгина с рядом вирусных белков SARS-CoV-2: РНК-зависимой РНК полимераза (RdRp), геликазой, экзонуклеазой, поверхностными гликопротеинами S и N, при этом флуоресцентный анализ не выявил значимого взаимодействия липопептида с папаин-подобной протеиназой вируса (PLpro). Верготен и Байи также с использованием молекулярного докинга показали взаимодействие другого эхинокандина, каспофунгина, с главной протеазой SARS-CoV-2 (Mpro) (Vergoten and Bailly, 2021). Молекулярный докинг, проведенный в работах (Anwaar et al., 2022; Elmorsy et al., 2022), выявил возможность связывания полимиксина Б с Mpro, RdRp, S-, N-белками и комплексом nsp16-nsp10. Анализ имеющихся литературных данных обнажает необходимость изучения молекулярных механизмов противовирусного действия ЦЛП, так как большинство сведений получены исключительно in silico.
В своей работе Kang с соавторами зарегистрировали противовирусный эффект фенгицина и сурфактина в отношении вируса огуречной мозаики (Kang et al., 2021). Сурфактин также демонстрирует эффективность в отношении вируса леса Семлики, простого герпеса, везикулярного стоматита, иммунодефицита обезьян, калицивируса кошек, вируса энцефаломиокардита мышей (Vollenbroich et al., 1997). Следует отметить, что наибольшую активность ЦЛП проявлял в отношении оболочечных вирусов (Vollenbroich et al., 1997). Установлено, что количество атомов углерода в жирнокислотной цепи сурфактина определяет его противовирусную активность (Kracht et al., 1999). В работах Юань с соавторами (Yuan et al., 2018; Yuan et al., 2019) продемонстрирована высокая активность сурфактина и его линейных аналогов в отношении вирусов эпидемической диареи и трансмиссивного гастроэнтерита свиней. Авторам удалось связать противовирусную активность сурфактина с его способностью индуцировать положительную кривизну липидных поверхностей, акцентировав внимание на связи формы молекулы липопептида (соотношении полярной/заряженной и гидрофобной частей) и способности подавлять слияние липидной оболочки вириона с клеточной мембраной. Эти данные позволяют предположить, что механизм противовирусного действия ЦЛП может быть связан с липидной оболочкой патогена.
Одной из задач настоящего изобретения являлось получение средства, ингибирующего слияние клеточных мембран. Еще одной возможной задачей настоящего изобретения являлось получение средства, ингибирующего слияние вирусных частиц SARS-CoV-2 с клетками.
Задача решена тем, что было обнаружено, что липопептид, выбранный из группы, содержащей акулеацин А, анидулафунгин, каспофунгин, даптомицин, фенгицин, итурин А, микосубтилин полимиксин Б, сиринготоксин Б, оказывает ингибирующее влияние на слияние мембран.
В том числе было обнаружено, что липопептид, выбранный из группы, содержащей акулеацин А, анидулафунгин, итурин А, микосубтилин, оказывает ингибирующее влияние на слияние вирусных частиц SARS-CoV-2 с клетками.
Для выявления ингибиторов слияния была проведена оценка способности ЦЛП подавлять слияние липидных везикул, запущенное введением кальция, полиэтиленглиголя с молекулярной массой 8000 и фрагмента пептида слияния SARS-CoV-2. Были протестированы одиннадцать ЦЛП, имеющих разное происхождение: акулеацин А, анидулафунгин, каспофунгин, даптомицин, фенгицин, итурин А, микосубтилин, полимиксин Б, сирингостатин А, сиринготоксин Б, сурфактин. Для in vitro тестирования противовирусной активности липопептидов была использована клеточная линия эпителия почки африканской зеленой обезьяны - Vero, с успехом применяемая при оценке цитопатогенного эффекта SARS-CoV-2 (Park et al., 2020; Riva et al., 2020).
Изобретение иллюстрируется следующими графическими материалами:
На Фиг. 1 представлены графики временной зависимости относительной флуоресценции кальцеина (% слияния), вытекшего при слиянии ДОФХ/ДОФГ/ХОЛ (40/40/20 мол. %), ДОФХ/ХОЛ (80/20 мол. %) и ПОФХ/СМ/ХОЛ (60/20/20 мол. %) липосом, запущенном 40 мМ CaCl2 (а), 20 вес .% ПЭГ-8000 (б) или 150 мкМ FP-SARS-CoV-2 (в) соответственно. Черные линии соответствуют процессу слияния в отсутствие циклических липопептидов, соответствие цвета линии конкретному липопептиду указано на рисунке. Концентрации липопептидов приведены в Таблице 1.
На Фиг. 2 представлены индексы ингибирования (ИИ) тестируемыми липопептидами слияния везикул из ДОФХ/ДОФГ/ХОЛ (40/40/20 мол. %) (а), ДОФХ/ХОЛ (80/20 мол. %) (б) и ПОФХ/СМ/ХОЛ (60/20/20 мол. %) (в) под действием 40 мМ CaCl2, 20 вес. % ПЭГ-8000- или 150 мкМ FP-SARS-CoV-2 соответственно. Соответствие между цифрами и липопептидами указано на рисунке.
На Фиг. 3 представлено графическое изображение химической структуры молекул липопептидов.
На Фиг. 4 представлены термограммы нагревания липосом ДПФХ/ХОЛ (90/10 мол. %) в отсутствие (контроль, черные линии) и в присутствии акулеацина А (а), анидулафунгина (б), каспофунгина (в), фенгицина (г), итурина А (д), сурфактина (е), даптомицина (ж) и полимиксина Б (з). Молярное соотношение липид : липопептид равно 50:1 (красные линии), 25:1 (зеленые линии) и 10:1 (синие линии).
На Фиг. 5 представлены термограммы перехода ПОФЭ из ламеллярной в инвертированную гексагональную фазу в отсутствие (черная линия) и в присутствии акулеацина А, анидулафунгина, каспофунгина, фенгицина, даптомицина и полимиксина Б. Молярное соотношение липид : липопептид равно 25:1.
Сущность и практическая применимость настоящего изобретения поясняется следующими примерами:
Пример 1. Материалы и методы
Пример 1.1. Материалы
Биологические мембраны представляют собой сложные структуры, многокомпонентность которых существенно затрудняет интерпретацию полученных при их изучении данных (Khan et al., 2013; Marrink et al., 2019). Это определяет целесообразность использования модельных липидных мембран, плоских липидных бислоев и липидных везикул, для идентификации молекулярных механизмов действия соединений с фармакологической активностью (Munusamy et al., 2020; Andersson and Koper, 2016).
Были использованы следующие реактивы: 1-пальмитоил-2-олеил-sn-глицеро-3-фосфохолин (ПОФХ), 1,2-диолеил-sn-глицеро-3-фосфохолин (ДОФХ), 1,2-диолеоил-sn-глицеро-3-фосфо-(1'-rac-глицерин) (ДОФГ), 1,2-дипальмитоил-sn-глицеро-3-фосфохолин (ДПФХ), 1-пальмитоил-2-олеил-sn-глицеро-3-фосфоэтаноламин (ПОФЭ), холестерин (ХОЛ) и сфингомиелин (из свиного мозга) (СМ) производства Avanti Polar Lipids (Avanti Polar Lipids, Inc., USA). Хлорид натрия (NaCl), HEPES, EDTA, NaOH, этанол, диметилсульфоксид (ДМСО), тритон Х-100, сефадекс G-50, кальцеин, нонактин, хлорид кальция (CaCl2) и полиэтиленгликоль с молекулярной массой 8 кДа (ПЭГ-8000), липопептиды: акулеацин А, анидулафунгин, каспофунгин, даптомицин, фенгицин, итурин А, микосубтилин, полимиксин Б, сурфактин производства Sigma-Aldrich Company Ltd. (Gillingham, United Kingdom). Сирингостатин А, сиринготоксин Б были любезно предоставлены доктором Д. Такемото (Utah State University, США).
Выбранный фрагмент пептида слияния FP-SARS-CoV-2 (SFIEDLLFNKVT) был синтезирован IQ Chemical (Россия). Чистота пептида составляла >98%.
Для анализа противовирусной активности соединений использовали эпителиальные клетки почки обезьяны Vero (АТСС CCL81).
Культуральные среды DMEM и DMEM/F12, эмбриональную бычью сыворотку, раствор трипсина, фосфатный буферный раствор Дульбекко были произведены Gibco (Шотландия).
В работе использовали материалы и оборудование ЦКП «Коллекция культур клеток позвоночных».
Пример 1.2. Флуориметрия утечки кальцеина из липосом
Определение процента слияния больших моноламеллярных липосом проводили с помощью измерения относительной интенсивности флуоресценции кальцеина. Флуоресцентный зонд - кальцеин, в концентрации 35 мМ внутри липидных везикул обладает эффектом самотушения. При смешении внутреннего пространства сливающихся липосом происходит утечка красителя в окружающий везикулы раствор. Таким образом, флуоресценция кальцеина в омывающем растворе зависит от интенсивности слияния липосом (Pattnaik and Chakraborty, 2018). Выбор липидов для формирования везикул обусловлен высоким содержанием в вирусной оболочке некоторых фосфолипидов, холестерина и сфингомиелина (Gerl at al., 2012; Kalvodova et al., 2009). Тестируемые липидные везикулы были изготовлены из смеси ДОФГ/ХОЛ (80/20 мол. %), ДОФХ/ДОФГ/ХОЛ (40/40/20 мол. %) и ПОФХ/СМ/ХОЛ (60/20/20 мол. %). Липосомы заданного диаметра (до 100 нм) получали методом экструзии, используя миниэкструдер Avanti Polar Lipids® (Avanti Polar Lipids, Inc., USA). Хлороформ из липидной суспензии удаляли потоком азота. Полученную липидную пленку гидратировали буфером (35 мМ кальцеина, 10 мМ HEPES, рН 7.4) и подвергали пятикратному замораживанию-размораживанию. Образовавшуюся суспензию мультиламеллярных липосом 13 раз пропускали через поликарбонатную мембрану с диаметром пор 100 нм (США). Свободный кальцеин удаляли из раствора путем гель-фильтрации с помощью колонки, заполненной сефадексом G-50. Элюентом выступал буферный раствор без красителя (10 мМ HEPES, рН 7.4). Исходная концентрация липида в суспензии составила 3 мМ.
В качестве индукторов слияния использовали CaCl2 (в концентрации 40 мМ), ПЭГ-8000 (в концентрации 20 вес. %) и фрагмент пептида слияния SARS-CoV/SARS-CoV-2 (SFIEDLLFNKVT, FP-SARS-CoV-2) (в концентрации 150 мкМ) (Tang et al., 2020). Молекулы кальция и полиэтиленгликоля являются широко признанными индукторами слияния, механизм действия которых декларирован во множестве работ (Leventis et al., 1986; Allolio et al., 2021; Lentz, 2007).
Используемый фрагмент пептида слияния (SFIEDLLFNKVT) характеризуется высокой степенью консервативности и α-спирализации (Lai and Freed, 2021). В работе (Shekunov et al., 2021) мы продемонстрировали высокую фузогенную активность его удлиненного аналога RSFIEDLLFNKVT. Ранее для фрагмента пептида слияния MERS-CoV (RSARSAIEDLLFDKV) была продемонстрирована высокая фузогенная активность в отношении везикул, сформированных из смеси дипальмитоилфосфатидилхолин / СМ / ХОЛ (Alsaadi et al., 2019). Было установлено, что LLF-последовательность играет ключевую роль в мембранных перестройках при пептид-ассоциированном слиянии (Lai et al., 2017).
Для оценки антифузогенной активности липопептиды добавляли к липосомам в концентрациях, которые в отсутствие индуктора слияния вызывали утечку кальцеина не более 20%, и инкубировали 30±10 минут при комнатной температуре. Каспофунгин, даптомицин, микосубтилин, сирингостатин А, сиринготоксин Б и сурфактин вносили в концентрации 33, 100, 25, 20, 20 и 0,01 мкМ, независимо от липидного состава везикул. Акулеацин А (2, 2,5 и 1 мкМ), анидулафунгин (20, 5 и 10 мкМ), фенгицин (10, 10 и 2 мкМ), итурин А (20, 20 и 40 мкМ) и полимиксин Б (1, 25, 10 и 10 мкМ) добавляли к ДОФХ/ДОФГ/ХОЛ, ДОФХ/ХОЛ, и ПОФХ/СМ/ХОЛ липосомам соответственно (Таблица 1).
Измерение интенсивности флуоресценции вытекающего кальцеина осуществляли с помощью спектрофлюориметра «Флюорат-02-Панорама» (Россия) при длине волны возбуждения 490 нм, эмиссии 520 нм. В конце эксперимента для полного высвобождения флуоресцентного маркера в исследуемые образцы добавляли водный раствор ТХ-100 до концентрации 1%, после чего измеряли интенсивность общего количества кальцеина.
Расчет относительной величины утечки кальцеина, отражающей процент слившихся липосом (% слияния), определяли по формуле:
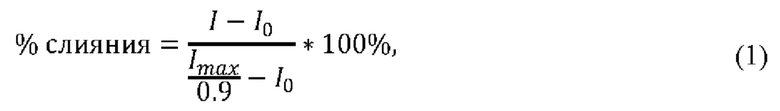
где I и I0 - интенсивность флуоресценции в присутствии (за исключением утечки маркера, вызванной липопептидом в отсутствии индуктора слияния) и в отсутствие ЦЛП,
Imax - интенсивность после добавления тритона Х-100. Коэффициент 0,9 введен для учета разбавления пробы детергентом. Процент слияния представляли как среднее арифметическое значение ± стандартная ошибка среднего в двух-пяти биологических повторностях.
Для сравнительной оценки эффективности липопептидов был введен индекс ингибирования (ИИ), который рассчитывали по формуле:
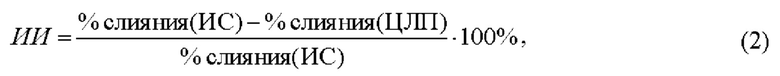
где % слияния (ИС) и % слияния (ЦЛП) - максимальная относительная флуоресценция кальцеина, возникшая в результате индуцированного слияния до и после инкубирования везикул с липопептидом соответственно. Величину ИИ определяли в каждом эксперименте по отношению к контролю с этой же липосомальной суспензией, а затем вычисляли среднее арифметическое значение ± стандартная ошибка среднего в двух-пяти биологических повторностях.
Пример 1.3. Оценка противовирусной активности in vitro
Анализ противовирусной активности исследуемых веществ проводили с использованием эпителиальных клеток почки обезьяны Vero (АТСС CCL81) и вируса SARS-CoV-2 (изолят 17612) с инфекционным титром 10 TCID 50/мл (инфекционная доза для тканевой культуры). Клетки выращивали в культуральной среде DMEM (Шотландия) с добавлением 10% эмбриональной бычьей сыворотки (Шотландия). Итоговую концентрацию клеток доводили до 5×105 клеток/мл и высевали в 96-луночные культуральные планшеты из расчета 0,1 мл на лунку. Планшеты инкубировали в течение 24 ч при 37°С в присутствии 5% CO2. Липопептиды растворяли в ДМСО, после чего готовили двукратные серийные разведения от 200 до 1,56 мкг/мл в бессывороточной среде DMEM/F12. Далее в каждую лунку с клетками вносили по 100 мкл каждого разведения и инкубировали в течение 1 часа при 37°С в присутствии 5% СО2. В контрольные лунки вместо липопептидов добавляли среду DMEM/F12 без сыворотки. После в лунки вносили по 100 мкл вируссодержащей жидкости SARS-CoV-2 и инкубировали в течение 1 часа при 37°С в присутствии 5% СО2. В лунки вирусного контроля добавляли вирусе о держащую среду. После инкубации вирус и тестируемые вещества удаляли, а клетки трехкратно промывали фосфатным буферным раствором Дульбекко (DPBS). Далее в отмытые лунки вносили по 100 мкл каждого разведения липопептидов (200-1,56 мкг/мл) и инкубировали в течение 24 часов при 37°С в присутствии 5% СО2. В контрольные лунки добавляли бессывороточную среду DMEM/F12. Через 24 часа супернатант из опытных и контрольных лунок собирали и титровали TCID50 с использованием клеток Vero. Результаты титрования оценивали через 72 ч с дальнейшим определением цитопатогеиного действия (ЦПД) вируса. Оценку ЦПД проводили с помощью светового инвертированного микроскопа Olympus СКХ41 (Япония).
Пример 1.4. Измерение изменения граничного потенциала липидных бислоев Плоские липидные бислои были сформированы по методу Монтала и Мюллера (Montal, Muller, 1972): путем сведения конденсированных липидных монослоев в двух отсеках тефлоновой камеры (цис- и транс-), сообщающихся между собой с помощью отверстия диаметром 50 мкМ в тефлоновой пленке толщиной 10 мкМ. Отверстие было предварительно обработано гексадеканом. Мембраны формировали из смеси ПОФХ/СМ/ХОЛ (60/20/20 моль %). Эксперименты проводили при одинаковом ионном составе разделяемых мембраной водных растворов электролита (0,1 М KCl). Кислотность растворов (рН 7,4) поддерживали буферной смесью 5 мМ HEPES-KOH. После испарения пентана на поверхности водного раствора оставался конденсированный липидный монослой. Подъем уровней жидкости в обоих отсеках камеры выше отверстия в тефлоновой пленке приводил к образованию бислойной мембраны. Измерения тока, протекающего через бислойную липидную мембрану, осуществляли в режиме фиксации потенциала. Подача трансмембранного напряжения (V) и отведение сигнала с мембраны осуществлялась с помощью хлорсеребряных электродов (Ag/AgCl), соединенных с растворами в отсеках камеры через агарозные мостики (1,5% агарозы в 2 М KCl).
Акулеацин А, анидулафунгин, каспофунгин, фенгицин, итурин А, сурфактин, сирингостатин А, сиринготоксин Б, даптомицин и полимиксин Б добавляли в липосомальную суспензию до концентрации 1, 10, 33, 2, 50, 0,01, 20, 20, 100 и 10 мкМ, соответственно, после модификации мембраны ионофором нонактивном.
Измеряли K+-нонактин-индуцированную проводимость мембраны (G) при постоянном трансмембранном напряжении (V=50 мВ). Изменения электрического потенциала на границе мембрана/водный раствор при введении тестируемых липопептидов (Δϕb) определяли согласно распределению Больцмана (Andersen et al., 1976):
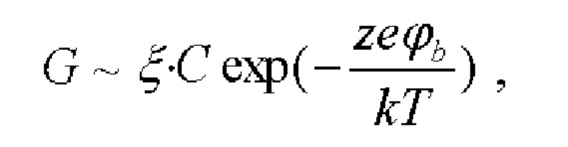
где ξ - подвижность ионов, ze - заряд иона, k - постоянная Больцмана, а Т - абсолютная температура.
Величину Δϕk представляли как среднее арифметическое значение ± стандартная ошибка среднего как минимум для трех независимых экспериментов с каждым липопептидом.
Ток измеряли с помощью усилителя Axopatch 200 В (Molecular Devices, LLC, США). Данные были оцифрованы с использованием Digidata 1440А и проанализированы с использованием pClamp 10.0 (Molecular Devices, LLC, США) и Origin 8.0 (OriginLab Corporation, США).
Пример 1.5. Дифференциальная сканирующая микрокалориметрия липидных везикул, модифицированных липопептидами.
Гигантские одноламеллярные липосомы формировали из смеси ДПФХ/ХОЛ (90/10 моль %) методом электроформации с помощью прибора «Nanion vesicle prep pro» (Германия). На стекла подавали переменное напряжение с амплитудой 3 В и частотой 10 Гц в течение 1 ч при температуре 55°С. Концентрация липида составляла 3 мМ. В экспериментальные образцы вводили липопептиды (акулеацин А, анидулафунгин, каспофунгин, фенгицин, итурин А, сурфактин, даптомицин и полимиксин Б) до достижения молярного соотношения липид : липопептид, равного 50:1, 25:1 и 10:1. Термограммы липосомальных суспензий получали при помощи дифференциального сканирующего микрокалориметра μDSC7 («Setaram», Франция). Суспензию липосом нагревали и охлаждали с постоянной скоростью 0,2 и 0,3°С/мин, соответственно.
Воспроизводимости температурной зависимости теплоемкости достигали путем повторного нагревания образца сразу после охлаждения. Термограммы липосомальных суспензий в отсутствие и в присутствии липопептидов анализировали с помощью Calisto Processing (Setaram, Caluire-et-Cuire, France).
Основными анализируемыми характеристиками термограммы ДПФХ/ХОЛ (90/10 моль %) служили максимальная температура основного фазового перехода (Tm) и полуширина основного пика (ширина на полувысоте) (Т1/2). Изменение указанных параметров позволяет судить о влиянии ЦЛП на плотность упаковки мембранных липидов. Величины ΔTm и ΔТ1/2 представляли как среднее арифметическое значение ± стандартное отклонение для двух независимых экспериментов с каждым липопептидом.
Суспензию ПОФЭ липосом изготавливливали путем растворения 8-9 мг сухого липида без или с 0,35-0,45 мг липопептида в теплом буфере (5 мМ HEPES при рН 7,4). После этого суспензию встряхивали на водяной бане при 40°С в течение 60 мин и обрабатывали ультразвуком с частотой 40 кГц в течение 1-3 мин. Полученную суспензию ПОФЭ выдерживали в холодильнике при 8°С не менее 8 часов перед проведением калориметрических измерений. Термограммы ПОФЭ характеризовались температурой ламеллярной инвертированной гексагональной фазы (ТНП). В экспериментальные образцы вводили липопептиды (акулеацин А, анидулафунгин, каспофунгин, фенгицин, даптомицин и полимиксин Б) до достижения молярного соотношения липид : липопептид, равного 25:1.
Пример 2. Результаты и обсуждения
Пример 2.1. Антифузогенный эффект липопептидов в отношении СаС2-, ПЭГ-8000-, FP-S ARS-CoV-2-индуцированного слияния
Исследована способность липопептидов ингибировать слияние липидных везикул вследствие добавления различных индукторов с применением метода регистрации утечки флуоресцентного маркера кальцеина из везикул. На Фиг. 1 представлены временные зависимости относительной интенсивности флуоресценции красителя, вытекшего из липосом различного состава при их слиянии в отсутствие и в присутствии тестируемых ЦЛП (% слияния). Липосомы формировали из незаряженных и отрицательно заряженных фосфолипидов, сфинголипидов и стеринов: ДОФХ/ДОФГ/ХОЛ (40/40/20 мол. %), ДОФХ/ХОЛ (80/20 мол. %) и ПОФХ/СМ/ХОЛ (60/20/20 мол. %). Контрольные образцы не модифицировали ЦЛП (черная линия, Фиг. 1). В экспериментальные образцы добавляли липопептиды (акулеацин А, анидулафунгин, каспофунгин, фенгицин, итурин А, микосубтилин, сурфактин, сирингостатин А, сиринготоксин Б, даптомицин и полимиксин Б) в концентрациях, указанных в Таблице 1, и инкубировали в течении 30±10 минут при комнатной температуре. Из представленных данных видно, что некоторые ЦЛП способны подавлять слияние липосом различного состава. Их антифузогенная активность зависит от используемого индуктора слияния. В Таблице 1 представлены средние значения максимальной утечки кальцеина при слиянии липосом в отсутствие и в присутствии липопептидов (% слияния).
Видно, что слияние не модифицированных липопептидами везикул под действием CaCl2, ПЭГ-8000 и FP-SARS-CoV-2 составляет около 80%, 75% и 80%, соответственно (Таблица 1). Снижение или рост этой величины в присутствии тестируемых липопептидов указывает на ингибирование (антифузогенную активность) или активацию слияния липосом соответственно.
Из Фиг. 1а и Таблицы 1 видно, что при CaCl2-ассоциированном слиянии ДОФХ/ДОФГ/ХОЛ-везикул % слияния значительно снижается в присутствии акулеацина А, анидулафунгина, каспофунгина, фенгицина и микосубтилина (не более 15%), умеренно в присутствии итурина А, сиринготоксина Б и полимиксина Б (от 35% до 51%) и практически не изменяется по сравнению с контрольным значением при добавлении в липосомальную суспензию сирингостатина А, сурфактина и даптомицина (от 76% до 92%).
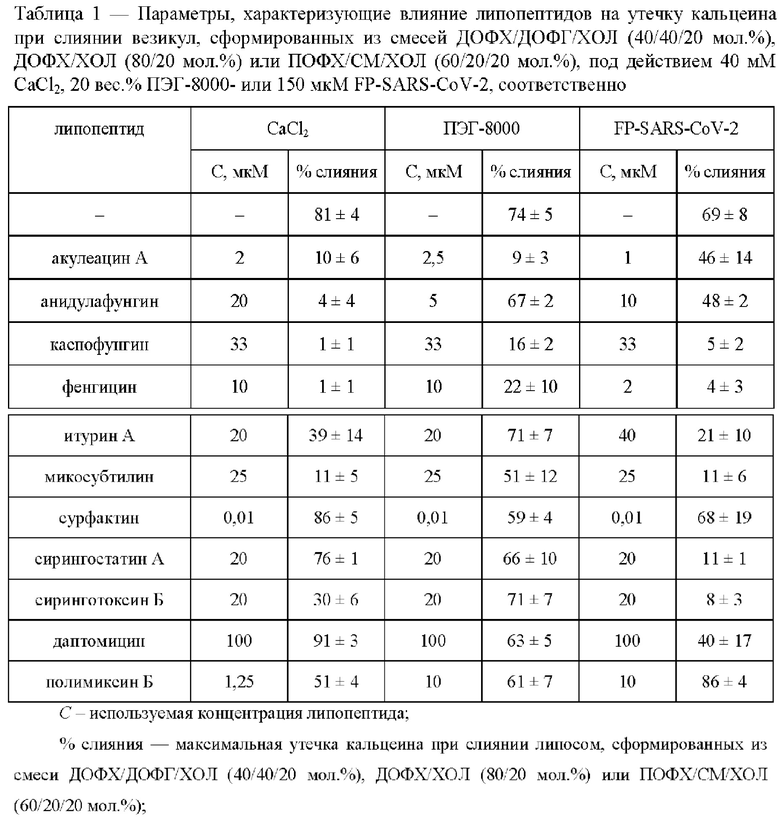
Замена липидной смеси для изготовления везикул, ДОФХ/ДОФГ/ХОЛ на ДОФХ/ХОЛ, и индуктора слияния, CaCl2 на ПЭГ-800, приводит к изменению эффективности действия ЦЛП. В случае ПЭГ-ассоциированного слияния ДОФХ/ХОЛ-липосом % слияния значительно снижается при введении акулеацина А и каспофунгина (не более 16%). Менее эффективно подавляют слияние везикул фенгицин и микосубтилин (от 35 до 51%) (Фиг. 1б, Таблица 1). При этом сурфактин, полимиксин Б, даптомицин, сирингостатин А, анидулафунгин, итурин А и сиринготоксин Б проявляют малую или практически не имеют антифузогенной активности в этой системе (от 59 до 71%, Фиг. 1б, Таблица 1).
Каспофунгин, фенгицин, микосубтилин, сирингостатин А, сиринготоксин Б и итурин А значительно ингибируют слияние ПОФХ/СМ/ХОЛ-везикул под действием FP-SARS-CoV-2 (% слияния не превышает 21%) (Фиг. 1в, Таблица 1). FP-SARS-CoV-2-ассоциированное слияние липосом значительно снижается в присутствии даптомицина, акулеацина А и анидулафунгина (от 40 до 48%) (Фиг. 1в, Таблица 1). Сурфактин и полимиксин Б не эффективны в отношении ингибирования FP-SARS-CoV-2-индуцированного слияния ПОФХ/СМ/ХОЛ-везикул (% слияния превышает 60%) (Фиг. 1в, Таблица 1).
Для сравнения эффективности антифузогенного действия липопептидов был рассчитан ИИ (Фиг. 2). Из Фиг. 2а видно, что способность подавлять слияние ДОФХ/ДОФГ/ХОЛ-везикул под действием СаС12 возрастает в ряду полимиксин Б (ИИ около 40%) < итурин А ≈ сиринготоксин Б (ИИ около 60%) < акулеацин А ≈ микосубтилин (ИИ около 85%) < анидулафунгин ≈ каспофунгин ≈ фенгицин (ИИ превышает 95%). При этом только акулеацин А, каспофунгин и фенгицин эффективно подавляют ПЭГ-опосредованное слияние незаряженных липосом (ИИ превышает 70%) (Фиг. 2б), а активность микосубтилина существенно снижается по сравнению с ДОФХ/ДОФГ/ХОЛ-везикулами (ИИ составляет около 30%). Каспофунгин, фенгицин, микосубтилин, сирингостатин А и сиринготоксин Б существенно ингибируют слияние сфинголипид-обогащенных мембран под действием FP-SARS-CoV-2 (ИИ более 70%), анидулафунгин, итурин А и даптомицин характеризуются умеренной эффективностью (ИИ около 30-50%) (Фиг. 2в). Интересно, что сирингостатин А и даптомицин способны подавлять слияние везикул, индуцированное FP-SARS-CoV-2, в отличие от слияния везикул, запущенного кальцием и ПЭГ-8000 (Фиг. 2а, б). Сурфактин и полимиксин В практически не характеризуются способностью подавлять слияние липосом во всех используемых липосомальных суспензиях (Фиг. 2).
Длина углеводородного «хвоста» возрастает в ряду: полимиксин Б (8 углеродных атомов) < даптомицин (10 углеродных атомов) < итурин А ≈ сурфактин (11 углеродных атомов) < каспофунгин ≈ сирингостатин А ≈ сиринготоксин Б (14 углеродных атомов) < микосубтилин (15 последовательно соединенных углеродных атомов) < акулеацин А ≈ фенгицин (16 последовательно соединенных углеродных атомов) < анидулафунгин (18 углеродных атомов). Коэффициент корреляции Пирсона между длиной «хвоста» ЦЛП и его ИИ составляет 0,7, 0,5 и 0,4 для СаС12-, ПЭГ-8000- и FP-SARS-CoV-2-индуцированного слияния ДОФХ/ДОФГ/ХОЛ-, ДОФХ/ХОЛ- и ПОФХ/СМ/ХОЛ-везикул, соответственно. Заметная корреляция в двух из трех липосомальных суспензий указывает на значимость длины углеводородного фрагмента в антифузогенной активности ЦЛП. По всей вероятности, это связано с зависимостью способности липопептидов погружаться в липидный бислой, разупорядочивать мембранные липиды и/или индуцировать положительную кривизну от длины их углеводородного хвоста. Следует отметить, что влияние липопептидов на эластические свойства мембраны также должно зависеть от размера пептидной «головки» и наличия в ней гидрофобных аминокислотных остатков. Отсутствие выраженной корреляции между длиной хвоста и ИИ при пептид-индуцированном слиянии можно связать с доменной организацией ПОФХ/СМ/ХОЛ мембран одновременном сосуществовании в мембране липидов в трех различных агрегатных состояниях (гель, жидкой упорядоченной и жидкой неупорядоченной фазах). Вероятно, распределение ЦЛП между липидными доменами в мембране зависит как от структуры агента, так и плотности упаковки липидов в фазе.
Пример 2.2. Оценка противовирусной активности липопептидов в условиях in vitro.
Таблица 2 демонстрирует цитотоксичность и противовирусную активность тестируемых агентов в отношении SARS-CoV-2. При оценке токсичности липопептидов было установлено, что даптомицин, полимиксин Б и сурфактин вызывали гибель клеток Vero при всех используемых концентрациях и поэтому были исключены из дальнейшего исследования. Акулеацин и микосубтилин были токсичны в концентрации выше 12,5 мкг/мл, анидулафунгин - 6,25 мкг/мл, каспофунгин - 100 мкг/мл, и итурин А - 50 мкг/мл.
Для определения инфекционной активности вирусного потомства готовили десятикратные разведения супернатантов, которые собирали из лунок с присутствием и отсутствием (вирусный контроль) липопептидов в бессывороточной среде DMEM/F12. Полученные суспензии вносили в 96-луночные культуральные планшеты с 80-90% монослоем клеток Vero и инкубировали в течение 72 ч при 37°С в присутствии 5% CO2. После этого проводили визуальную оценку вирусспецифического цитопатогенного действия (ЦПД) с помощью инвертированного микроскопа CKX41 (Olympus, Япония).
Титр вируса в контрольных образцах составил 106 ТС ID 50/мл. Инкубация клеток с акулеацином (в концентрации 12,5 мкг/мл), микосубтилином (12,5 мкг/мл) и итурином А (50 мкг/мл), позволила снизить титр вируса до 103 TCID 50/мл. В свою очередь, анидулафунгин (в концентрации 6,25 мкг/мл) демонстрирует более выраженное противовирусное действие и снижает вирусный титр до 10 TCID 50/мл. Каспофунгин оказался неактивным против SARS-CoV-2 даже при самых высоких используемых концентрациях.
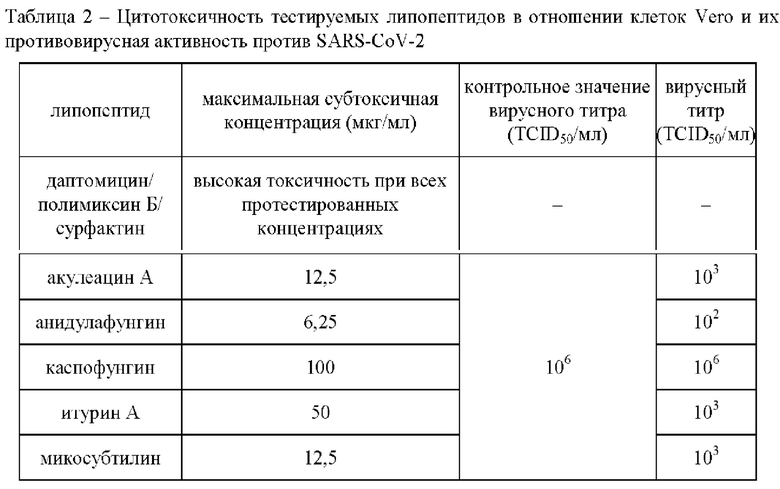
По итогам экспериментов можно отметить, что тестируемые липопептиды относительно токсичны для клеток Vero и демонстрируют умеренную активность против SARS-CoV-2.
Пример 2.3. Оценка изменения физических свойств мембран при введении липопептидов
Для понимания взаимосвязи между способностью липопептидов ингибировать слияние липидных везикул и их способностью модифицировать физико-химические свойства мембран было изучено действие липопептидов на электрические и эластические свойства модельных липидных мембран.
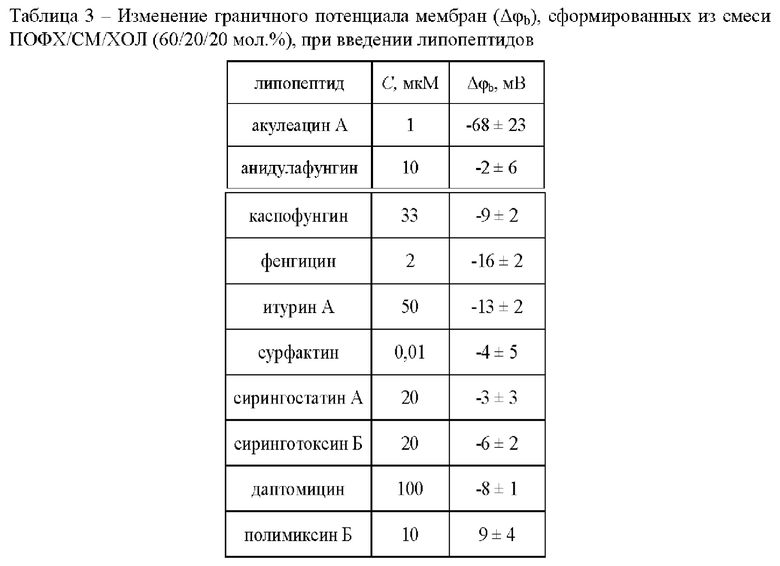
Для установления влияния тестируемых липопептидов на трансмембранное распределение электрического потенциала проведена оценка изменений граничного потенциала липидных бислоев при введении тестируемых липопептидов в концентрациях, используемых в экспериментах по слиянию (1 мкМ акулеацина А, 10 мкМ анидулафунгина, 33 мкМ каспофунгина, 100 мкМ даптомицина, 2 мкМ фенгицина, 40 мкМ итурина А, 25 мкМ микосубтилина, 10 мкМ полимиксина Б, 20 мкМ сирингостатина А, 20 мкМ сиринготоксина Б и 0,01 мкМ сурфактина). Граничный потенциал мембраны - это межфазный скачок электрического потенциала на границе мембрана/водный раствор. Он включает в себя поверхностный потенциал (падение потенциала в диффузной части электрического двойного слоя), связанный с поверхностной плотностью заряда мембраны, и так называемую дипольную компоненту граничного потенциала, определяемую взаимной специфической ориентацией диполей мембранных липидов и молекул воды на границе раздела фаз (Ермаков, 2000). Учитывая, что некоторые тестируемые липопептиды имеют электрический заряд и включают высокополярные аминокислотные остатки, то при их адсорбции на мембране могут изменяться как поверхностная, так и дипольная составляющая граничного потенциала мембраны. В таблице 3 представлены средние величины изменения граничного потенциала бислоев из ПОФХ/СМ/ХОЛ (60/20/20 мол. %) в присутствии липопептидов. Можно отметить, что изменение граничного потенциала мембран после добавления анидулафунгина, каспофунгина, даптомицина, фенгицина, полимиксина Б, сирингостатина А, сиринготоксина Б и сурфактина не превышает 20 мВ по абсолютному значению. Учитывая высокое значение граничного потенциала незаряженных липидных бислоев, связанное с существованием дипольной компоненты граничного потенциала (280 мВ (Brockman, 1994)), полученные величины указывают на неспособность этих липопептидов в указанных концентрациях значительно влиять на граничный потенциал мембраны. Единственным модулятором граничного потенциала среди тестируемых липопептидов является акулеацин А, который приводит к уменьшению дипольного потенциала на 100 мВ. Отсутствие корреляции между способностью липопептидов ингибировать пептид-индуцированное слияние ПОФХ/СМ/ХОЛ (60/20/20 мол. %) везикул (Таблица 2) и их действием на трансмембранное распределение электрического потенциала ПОФХ/СМ/ХОЛ (60/20/20 мол. %) (Таблица 3) указывает на то, что изменение граничного потенциала при введении липопептидов не является ключевым фактором, обеспечивающим их способность подавлять слияние мембран.
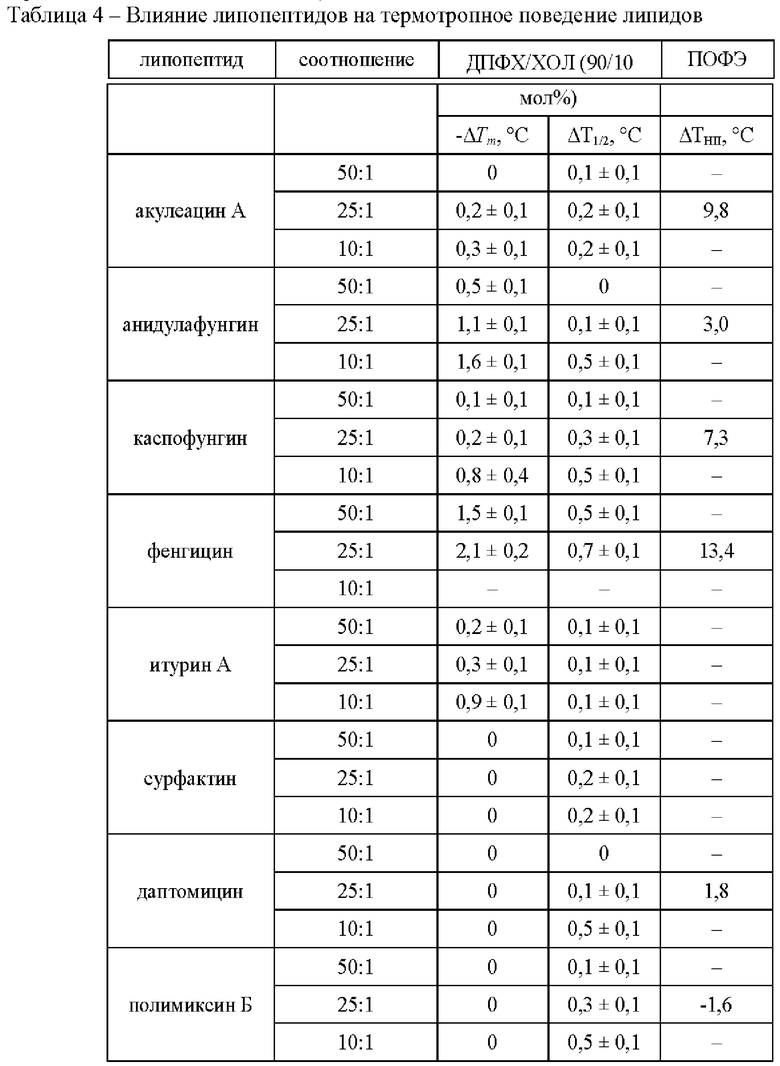
Для понимания взаимосвязи между влиянием ЦЛП на слияние липосом и их способностью модифицировать эластические свойства мембран было изучено термотропное поведение мембранообразующих липидов с использованием дифференциальной сканирующей микрокалориметрии везикул, сформированных из смеси дипальмитоилфосфатидилхолина и холестерина - ДПФХ/ХОЛ (90/10 мол. %), до и после введения в липосомальную суспензию акулеацина А, анидулафунгина, каспофунгина, фенгицина, итурина А, сурфактина, даптомицина и полимиксина Б.
В отсутствие липопептидов температура главного фазового перехода (плавления) смеси ДПФХ/ХОЛ (90/10 мол %), Tm, составляет около 40,6°С, а полуширина соответствующего пика, характеризующая величину, обратную кооперативности фазового перехода, Т1/'2, равна 0,6°С (Фиг. 4). В Таблице 3 представлены эти величины в присутствии липопептидов акулеацина А, анидулафунгина, каспофунгина, фенгицина, итурина А, сурфактина, даптомицина и полимиксина Б. Акулеацин А, анидулафунгин, каспофунгин, фенгицин и итурин А проявляют дозозависимый эффект (Таблица 4). При молярном соотношении липид : липопептид 50 к 1 акулеацин А, каспофунгин и итурин А мало влияют на основные характеристики термограммы плавления ДПФХ/ХОЛ: температура плавления и полуширина пика практически не меняются: изменения составляют 0,1-0,2°С (Фиг. 4а, в, г, Таблица 4). Повышение доли этих липопептидов в смеси с липидами до молярного соотношения липид : липопептид 25:1 приводит к снижению Tm на 0,2-0,4°С, при этом величина T1/2 увеличивается на 0,1-0,3°С (Фиг. 4а, в, г, Таблица 4). При молярном соотношении липид: липопептид 10:1 акулеацин А, каспофунгин и итурин А характеризуются более выраженным эффектом в отношении плавления ДПФХ/ХОЛ (90/10 мол %): Tm уменьшается на 0,3-1,0°С, Т1/2 увеличивается менее значительно (на 0,1-0,5°С) (Фиг. 4а, в, г, таблица 3). Анидулафунгин и фенгицин демонстрируют более выраженное влияние на фазовый переход ДПФХ/ХОЛ (Фиг. 4б, г, Таблица 4). С ростом концентрации анидулафунгина абсолютная величина ΔTm увеличивается от 0,5 до 1,6°С, а значение ΔТ1/2 возрастает от 0 до 0,5°С (Фиг. 4б, Таблица 4). При самой большой концентрации липопептида в смеси с липидом: в молярном соотношении липид : липопептид 50:1, фенгицин приводит к уменьшению Tm на 1,5°С и увеличению Т1/2 на 0,5°С (Фиг. 4г, Таблица 4). С увеличением концентрации фенгицина в 2 раза его модифицирующий эффект увеличивается на 40% (Фиг. 4г, Таблица 4). Сурфактин (Фиг. 4е), даптомицин (Фиг. 4ж) и полимиксин Б (Фиг. 4з) практически не влияют на основные характеристики термограммы плавления ДПФХ/ХОЛ при всех тестируемых молярных соотношениях липид : липопептид (50:1, 25:1 и 10:1).
Значимая корреляция между ИИ липопептидов и ΔTm в их присутствии обнаруживается для кальций-индуцированного слияния ДОФХ/ДОФГ/ХОЛ-липосом при всех исследованных молярных соотношениях липид : липопептид и для пептид-индуцированного слияния ПОФХ/СМ/ХОЛ-везикул только при большой концентрации липопептида в смеси с липидом: в молярном соотношении липид : липопептид 50:1 и 25:1). Коэффициент корреляции Пирсона между ИИ липопептидами кальций-, ПЭГ- и пептид-индуцированного слияния везикул (Таблица 1) и величинами ΔТ1/2 (Таблица 4) при различных молярных соотношениях липид : липопептид составляет 0,5-0,9. Наблюдаемая корреляция между величиной ΔТ1/2, связанной с кооперативностью плавления мембранообразующих липидов в присутствии липопептидов, и их индексом ингибирования может указывать на взаимосвязь между способностью тестируемых липопептидов разупорядочивать мембранные липиды и ингибировать слияние липосом.
Другим возможным механизмом антифузогенного действия липопептидов может быть способность липопептидов индуцировать образование положительной кривизны липидных монослоев. Эта гипотеза была проверена с помощью термограммы плавления гексагональной фазы ПОФЭ. Переход ПОФЭ из ламеллярной в инвертированную гексагональную фазу оценивали с помощью дифференциальной сканирующей микрокалориметрии (Фиг. 5, контроль). Акулеацин А, анидулафунгин, каспофунгин, фенгицин и даптомицин при молярном соотношении липид : липопептид 25:1 повышают температуру перехода ПОФЭ (ТНП) на 9,8, 3,0, 7,3, 13,4 и 1,8°С соответственно (Фиг. 5, Таблица 4). В то же время полимиксин Б снижает Тип ПОФЭ на 1,6°С. Это указывает на то, что липопептиды подавляют образование структур с высокой отрицательной кривизной, и эта способность коррелирует с их ингибирующим действием на слияние мембран, запускаемое CaCl2 и ПЭГ-8000 (коэффициенты корреляции Пирсона равны 0,7 и 0,9 соответственно).
Работа выполнена в рамках гранта РНФ (№22-15-00417).
Заявитель просит рассмотреть представленные материалы заявки «Применение липопептидов в качестве ингибиторов слияния мембран» на предмет выдачи патента на изобретение.
Список литературы
1. Ермаков Ю.А., Распределение электрического потенциала на границах липидных мембран. Автореферат диссертации на соискание ученой степени доктора физико-математических наук, 2000 г.
2. Ahamad, S., АН, Н., Secco, I., Giacca, М., Gupta, D. (2022) Anti-fungal drug anidulafungin inhibits SARS-CoV-2 spike-induced syncytia formation by targeting ACE2-spike protein interaction. Frontiers in Genetics, 13, 866474. https://doi.org/10.3389/fgene.2022.866474.
3. Акра, E., Jacques, P., Wathelet, В., Paquot, M., Fuchs, R., Budzikiewicz, H., & Thonart, P. (2001). Influence of culture conditions on lipopeptide production by Bacillus subtilis. Applied biochemistry and biotechnology, 91-93, 551-561. https://doi.Org/10.1385/abab:91-93:l-9:551.
4. Allolio, C, & Harries, D. (2021). Calcium Ions Promote Membrane Fusion by Forming Negative-Curvature Inducing Clusters on Specific Anionic Lipids. ACS nano, 10.1021/acsnano.0c08614. Advance online publication, https://doi.org/10.1021/acsnano.0c08614.
5. Alsaadi, E., Neuman, B. W., & Jones, I. M. (2019). A Fusion Peptide in the Spike Protein of MERS Coronavirus. Viruses, 11(9), 825. https://doi.org/10.3390/v11090825.
6. Andersen, O.S., Finkelstein A., Katz I., Cass A. Effect of phloretin on the permeability of thin lipid membranes. Journal of General Physiology. 1976, 67(6), 749-71. https://doi: 10.1085/jgp.67.6.749.
7. Andersson, J., &  (2016). Tethered and Polymer Supported Bilayer Lipid Membranes: Structure and Function. Membranes, 6(2), 30. https://doi.org/10.3390/membranes6020030.
(2016). Tethered and Polymer Supported Bilayer Lipid Membranes: Structure and Function. Membranes, 6(2), 30. https://doi.org/10.3390/membranes6020030.
8. Anwaar, M. U., Adnan, F., Abro, A., Khan, R. A., Rehman, A. U., Osama, M., Rainville, C., Kumar, S., Sterner, D. E., Javed, S., Jamal, S. В., Baig, A., Shabbir, M. R., Ahsan, W., Butt, T. R., & Assir, M. Z. (2022). Combined deep learning and molecular docking simulations approach identifies potentially effective FDA approved drugs for repurposing against SARS-CoV-2. Computers in biology and medicine, 141, 105049. https://doi.org/10.1016/j.compbiomed.2021.105049.
9. Arima, K., Kakinuma, A., & Tamura, G. (1968). Surfactin, a crystalline peptidelipid surfactant produced by Bacillus subtilis: isolation, characterization and its inhibition of fibrin clot formation. Biochemical and biophysical research communications, 31(3), 488 494. https://doi.org/10.1016/0006-291x(68)90503-2.
10. Arnold, R. S., Thorn, K. A., Sharma, S., Phillips, M., Kristie Johnson, J., & Morgan, D. J. (2011). Emergence of Klebsiella pneumoniae carbapenemase-producing bacteria. Southern medical journal, 104(1), 40-45. https://doi.org/10.1097/SMJ.0b013e3181fd7d5a.
11. Ballio, A., Bossa, F., Collina, A., Gallo, M., Iacobellis, N. S., Paci, M., … & Simmaco, M. (1990). Structure of syringotoxin, a bioactive metabolite of Pseudomonas syringae pv. syringae. FEBS letters, 269(2), 377-380.
12. Barrows, N. J., Campos, R. K., Powell, S. Т., Prasanth, K. R., Schott-Lerner, G., Soto-Acosta, R.,  G., McGrath, E. L., Urrabaz-Garza, R., Gao, J., Wu, P., Menon, R., Saade, G., Fernandez-Salas, I., Rossi, S. L., Vasilakis, N., Routh, A., Bradrick, S. S., & Garcia-Blanco, M. A. (2016). A Screen of FDA-Approved Drugs for Inhibitors of Zika Virus Infection. Cell host & microbe, 20(2), 259-270. https://doi.Org/10.1016/j.chom.2016.07.004.
G., McGrath, E. L., Urrabaz-Garza, R., Gao, J., Wu, P., Menon, R., Saade, G., Fernandez-Salas, I., Rossi, S. L., Vasilakis, N., Routh, A., Bradrick, S. S., & Garcia-Blanco, M. A. (2016). A Screen of FDA-Approved Drugs for Inhibitors of Zika Virus Infection. Cell host & microbe, 20(2), 259-270. https://doi.Org/10.1016/j.chom.2016.07.004.
13. Biniarz, P., Lukaszewicz, M., & Janek, T. (2017). Screening concepts, characterization and structural analysis of microbial-derived bioactive lipopeptides: a review. Critical reviews in biotechnology, 37(3), 393-410. https://doi.org/10.3109/07388551.2016.1163324.
14. Brockman H. Dipole potential of lipid membranes. Chemistry and Physics of Lipids, 1994, vol. 73, no. 1-2. pp. 57-79.
15. Buchmann, J. P., & Holmes, E. C. (2015). Cell Walls and the Convergent Evolution of the Viral Envelope. Microbiology and molecular biology reviews: MMBR, 79(4), 403-418. https://doi.org/10.1128/MMBR.00017-15.
16. Cancidas PI. Cancidas package insert [online] Rahway, NJ: Merck; 2005. Accessed on 3 January2006. URL: http://www.cancidas.com/cancidas/shared/documents/english/pi.pdf.
17. Chen, Y. C, Lu, J. W., Yeh, С. Т., Lin, T. Y., Liu, F. С, & Ho, Y. J. (2021). Micafungin Inhibits Dengue Virus Infection through the Disruption of Virus Binding, Entry, and Stability. Pharmaceuticals (Basel, Switzerland), 14(4), 338. https://doi.org/10.3390/phl4040338.
18. Chitarra, G. S., Breeuwer, P., Nout, M. J., van Aelst, A. C., Rombouts, F. M., & Abee, T. (2003). An antifungal compound produced by Bacillus subtilis YM 10-20 inhibits germination of Penicillium roqueforti conidiospores. Journal of applied microbiology, 94(2), 159-166. https://doi.Org/10.1046/j.1365-2672.2003.01819.x.
19. Clausell, A., Garcia-Subirats, M., Pujol, M., Busquets, M. A., Rabanal, F., & Cajal, Y. (2007). Gram-negative outer and inner membrane models: insertion of cyclic cationic lipopeptides. The journal of physical chemistry. B, 111(3), 551 563. https://doi.org/10.1021/jp064757+.
20. Dalla Serra, M., Fagiuoli, G., Nordera, P., Bernhart, L, Delia Volpe, C., Di Giorgio, D., Ballio, A., & Menestrina, G. (1999). The interaction of lipodepsipeptide toxins from Pseudomonas syringae pv. syringae with biological and model membranes: a comparison of syringotoxin, syringomycin, and two syringopeptins. Molecular plant-microbe interactions: MPMI, 12(5), 391-400. https://doi.Org/10.1094/MPMI.1999.12.5.391.
21. De Crescenzo Henriksen, E., Phillips, D. R., & Peterson, J. D. (2007). Polymyxin E production by P. amylolyticus. Letters in applied microbiology, 45(5), 491-496.
22. Deleu, M., Paquot, M., & Nylander, T. (2005). Fengycin interaction with lipid monolayers at the air-aqueous interface-implications for the effect of fengycin on biological membranes. Journal of colloid and interface science, 283(2), 358-365. https://doi.Org/10.1016/j.jcis.2004.09.036
23. Deleu, M., Paquot, M., & Nylander, T. (2008). Effect of fengycin, a lipopeptide produced by Bacillus subtilis, on model biomembranes. Biophysical journal, 94(7), 2667 2679. https://doi.org/10.1529/biophysj.107.114090.
24. Deresinski, S. C., & Stevens, D. A. (2003). Caspofungin. Clinical infectious diseases: an official publication of the Infectious Diseases Society of America, 36(11), 1445 1457. https://doi.org/10.1086/375080.
25. Deris, Z. Z., Swarbrick, J. D., Roberts, K. D., Azad, M. A., Akter, J., Home, A. S., Nation, R. L., Rogers, K. L., Thompson, P. E., Velkov, Т., & Li, J. (2014). Probing the penetration of antimicrobial polymyxin lipopeptides into gram-negative bacteria. Bioconjugate chemistry, 25(4), 750-760. https://doi.org/10.1021/bc500094d.
26. Dey, S. K., Saini, M., Dhembla, C., Bhatt, S., Rajesh, A. S., Anand, V., Das, H. K., & Kundu, S. (2021). Suramin, penciclovir, and anidulafungin exhibit potential in the treatment of COVID-19 via binding to nspl2 of SARS-CoV-2. Journal of biomolecular structure & dynamics, 1-17. Advance online publication, https://doi.org/10.1080/07391102.2021.2000498.
27. Elmorsy, M. A., El-Baz, A. M., Mohamed, N. H., Almeer, R., Abdel-Daim, M. M., & Yahya, G. (2022). In silico screening of potent inhibitors against COVID-19 key targets from a library of FDA-approved drugs. Environmental science and pollution research international, 29(8), 12336-12346. https://doi.org/10.1007/sll356-021-16427-4.
28. Evans, M. E., Feola, D. J., & Rapp, R. P. (1999). Polymyxin В sulfate and colistin: old antibiotics for emerging multiresistant gram-negative bacteria. The Annals of pharmacotherapy, 33(9), 960 967. https://doi.org/10.1345/aph.18426.
29. Fewer, D. P., Jokela, J., Heinila, L., Aesoy, R., Sivonen, K., Galica, Т., Hrouzek, P., & Herfindal, L. (2021). Chemical diversity and cellular effects of antifungal cyclic lipopeptides from cyanobacteria. Physiologiaplantarum, 173(2), 639-650. https://doi.org/10.1111/ppl.13484.
30. Fowler, V. G., Jr, Boucher, H. W., Corey, G. R., Abrutyn, E., Karchmer, A. W., Rupp, M. E., Levine, D. P., Chambers, H. F., Tally, F. P., Vigliani, G. A., Cabell, С. FL, Link, A. S., DeMeyer, I., Filler, S. G., Zervos, M., Cook, P., Parsormet, J., Bernstein, J. M., Price, C. S., Forrest, G. N., … S. aureus Endocarditis and Bacteremia Study Group (2006). Daptomycin versus standard therapy for bacteremia and endocarditis caused by Staphylococcus aureus. The New England journal of medicine, 355(7), 653-665. https://doi.org/10.1056/NEJMoa053783.
31. Gerl, M. J., Sampaio, J. L., Urban, S., Kalvodova, L., Verbavatz, J. M., Binnington, В., Lindemann, D., Lingwood, C. A., Shevchenko, A., Schroeder, C., & Simons, K. (2012). Quantitative analysis of the lipidomes of the influenza virus envelope and MDCK cell apical membrane. The Journal of cell biology, 196(2), 213-221. https://doi.org/10.1083/jcb.201108175.
32. Gong, A. D., Li, H. P., Yuan, Q. S., Song, X. S., Yao, W., He, W. J., Zhang, J. В., & Liao, Y. C. (2015). Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PloS one, 10(2), e0116871. https://doi.org/10.1371/journal.pone.0116871.
33. Guo, Q., Dong, W., Li, S., Lu, X., Wang, P., Zhang, X., Wang, Y., & Ma, P. (2014). Fengycin produced by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling damping-off disease. Microbiological research, 169(7-8), 533-540. https://doi.Org/10.1016/j.micres.2013.12.001.
34. Gur'nev, F. A., Kaulin, I., Tikhomirova, A. V., Wangspa, R., Takemoto, D., Malev, V. V., & Shchagina, L. V. (2002). Aktivnost' toksinov, produtsiruemykh bacteriiami Pseudomonas syringae pv. syringae, v model'nykh i kletochnykh membranakh [Activity of toxins produced by Pseudomonas syringae pv. syringae in model and cell membranes]. Tsitologiia, 44(3), 296-304.
35. Hamley I. W. (2015). Lipopeptides: from self-assembly to bioactivity. Chemical communications (Cambridge, England), 51(41), 8574-8583. https://doi.org/10.1039/c5cc01535a.
36. Harnois, I., Maget-Dana, R., & Ptak, M. (1989). Methylation of the antifungal lipopeptide iturin A modifies its interaction with lipids. Biochimie, 71(1), 111-116.
37. Hermsen, E. D., Sullivan, C. J., & Rotschafer, J. C. (2003). Polymyxins:: Pharmacology, pharmacokinetics, pharmacodynamics, and clinical applications. Infectious Disease Clinics, 17(3), 545-562.
38. Hiradate, S., Yoshida, S., Sugie, H., Yada, H., & Fujii, Y. (2002). Mulberry anthracnose antagonists (iturins) produced by Bacillus amyloliquefaciens RC-2. Phytochemistry, 61(6), 693-698. https://doi.org/10.1016/s0031-9422(02)00365-5
39. Horn, J. N., Cravens, A., & Grossfield, A. (2013). Interactions between fengycin and model bilayers quantified by coarse-grained molecular dynamics. Biophysical journal, 105(7), 1612-1623. https://doi.Org/10.1016/j.bpj.2013.08.034.
40. Ito, M., Nozu, R., Kuramochi, Т., Eguchi, N., Suzuki, S., Hioki, K., Itoh, Т., & Ikeda, F. (2000). Prophylactic effect of FK463, a novel antifungal lipopeptide, against Pneumocystis carinii infection in mice. Antimicrobial agents and chemotherapy, 44(9), 2259-2262. https://doi.Org/10.1128/AAC.44.9.2259-2262.2000.
41. Iwata, K., Yamamoto, Y., Yamaguchi, H., & Hiratani, T. (1982). In vitro studies of aculeacin A, a new antifungal antibiotic. The Journal of antibiotics, 35(2), 203-209. https://doi.org/10.7164/antibiotics.35.203.
42. Jeon, S., Ко, M., Lee, J., Choi, I., Byun, S. Y., Park, S., Shum, D., & Kim, S. (2020). Identification of Antiviral Drug Candidates against SARS-CoV-2 from FDA-Approved Drugs. Antimicrobial agents and chemotherapy, 64(7), e00819-20. https://doi.org/10.1128/AAC.00819-20.
43. Jung, D., Powers, J. P., Straus, S. K., & Hancock, R. E. (2008). Lipid-specific binding of the calcium-dependent antibiotic daptomycin leads to changes in lipid polymorphism of model membranes. Chemistry and physics of lipids, 154(2), 120-128. https://doi.Org/10.1016/j.chemphyslip.2008.04.004.
44. Kahn, J. N., Hsu, M. J., Racine, F., Giacobbe, R., & Motyl, M. (2006). Caspofungin susceptibility in Aspergillus and non-Aspergillus molds: inhibition of glucan synthase and reduction of beta-D-1,3 glucan levels in culture. Antimicrobial agents and chemotherapy, 50(6), 2214-2216. https://doi.org/10.1128/AAC.01610-05.
45. Kalvodova, L., Sampaio, J. L., Cordo, S., Ejsing, C. S., Shevchenko, A., & Simons, K. (2009). The lipidomes of vesicular stomatitis virus, semliki forest virus, and the host plasma membrane analyzed by quantitative shotgun mass spectrometry. Journal of virology, 83(16), 7996-8003. https://doi.org/10.1128/JVI.00635-09.
46. Kang, B. R., Park, J. S., & Jung, W. J. (2021). Antiviral activity by lecithin-induced fengycin lipopeptides as a potent key substrate against Cucumber mosaic virus. Microbial pathogenesis, 155, 104910. https://doi.Org/10.1016/j.micpath.2021.104910.
47. Kanj, S. S., & Kanafani, Z. A. (2011). Current concepts in antimicrobial therapy against resistant gram-negative organisms: extended-spectrum beta-lactamase-producing Enterobacteriaceae, carbapenem-resistant Enterobacteriaceae, and multidrug-resistant Pseudomonas aeruginosa. Mayo Clinic proceedings, 86(3), 250-259. https://doi.org/10.4065/mcp.2010.0674.
48. Kanlayavattanakul, M., & Lourith, N. (2010). Lipopeptides in cosmetics. International journal of cosmetic science, 32(1), 1-8. https://doi.Org/10.1111/j.1468-2494.2009.00543.x.
49. Khan, M. S., Dosoky, N. S., & Williams, J. D. (2013). Engineering lipid bilayer membranes for protein studies. International journal of molecular sciences, 14(11), 21561-21597. https://doi.org/10.3390/ijms141121561.
50. Kielian M. (2014). Mechanisms of Virus Membrane Fusion Proteins. Annual review of virology, 1(1), 171-189. https://doi.org/10.1146/annurev-virology-031413-085521.
51. Klebe, G., & Schlitzer, M. (2011). M2-hihibitoren und Neuraminidase-Inhibitoren [M2 inhibitors and neuraminidase inhibitors]. Pharmazie in unserer Zeit, 40(2), 144-150. https://doi.org/10.1002/pauz.201100410.
52. Korenblum, E., de Araujo, L. V., Guimaraes, C. R., de Souza, L. M., Sassaki, G., Abreu, F., Nitschke, M., Lins, U., Freire, D. M., Barreto-Bergter, E., & Seldin, L. (2012). Purification and characterization of a surfactin-like molecule produced by Bacillus sp.H20-1 and its antagonistic effect against sulfate reducing bacteria. BMC microbiology, 12, 252. https://doi.org/10.1186/1471 -2180-12-252.
53. Kracht, M., Rokos, H., Ozel, M., Kowall, M., Pauli, G., & Vater, J. (1999). Antiviral and hemolytic activities of surfactin isoforms and their methyl ester derivatives. The Journal of antibiotics, 52(7), 613 619. https://doi.org/10.7164/antibiotics.52.613.
54. Lai, A. L., & Freed, J. H. (2021). SARS-CoV-2 Fusion Peptide has a Greater Membrane Perturbating Effect than SARS-CoV with Highly Specific Dependence on Ca2. Journal of molecular biology, 433(10), 166946. https://doi.Org/10.1016/j.jmb.2021.166946.
55. Lai, A. L., Millet, J. K., Daniel, S., Freed, J. H., & Whittaker, G. R. (2017). The SARS-CoV Fusion Peptide Forms an Extended Bipartite Fusion Platform that Perturbs Membrane Order in a Calcium-Dependent Manner. Journal of molecular biology, 429(24), 3875 3892. https://doi.Org/10.1016/j.jmb.2017.10.017.
56. Lentz B. R. (2007). PEG as a tool to gain insight into membrane fusion. European biophysics journal: EBJ, 36(4-5), 315-326. https://doi.org/10.1007/s00249-006-0097-z.
57. Leventis, R.,  Fuller, N., Rand, R. P., & Silvius, J. R. (1986). Divalent cation induced fusion and lipid lateral segregation in phosphatidylcholine-phosphatidic acid vesicles. Biochemistry, 25(22), 6978-6987. https://doi.org/10.1021/bi00370a600.
Fuller, N., Rand, R. P., & Silvius, J. R. (1986). Divalent cation induced fusion and lipid lateral segregation in phosphatidylcholine-phosphatidic acid vesicles. Biochemistry, 25(22), 6978-6987. https://doi.org/10.1021/bi00370a600.
58. Liao, G., Shi, Т., & Xie, J. (2012). Regulation mechanisms underlying the biosynthesis of daptomycin and related lipopeptides. Journal of cellular biochemistry, 113(3), 735-741. https://doi.org/10.1002/jcb.23414.
59. Lu, J. W., Chen, Y. C, Huang, С. K., Lin, К. С, & Ho, Y. J. (2021). Synergistic in-vitro antiviral effects of combination treatment using anidulafungin and T-1105 against Zika virus infection. Antiviral research, 195, 105188. https://doi.Org/10.1016/j.antiviral.2021.105188.
60. Maget-Dana, R., & Peypoux, F. (1994). Iturins, a special class of pore-forming lipopeptides: biological and physicochemical properties. Toxicology, 87(1-3), 151-174. https://doi.org/10.1016/0300-483x(94)90159-7.
61. Makovitzki, A., Viterbo, A., Brotman, Y., Chet, I., & Shai, Y. (2007). Inhibition of fungal and bacterial plant pathogens in vitro and in planta with ultrashort cationic lipopeptides. Applied and environmental microbiology, 73(20), 6629-6636.
62. Mares, J., Kumaran, S., Gobbo, M., & Zerbe, O. (2009). Interactions of lipopolysaccharide and polymyxin studied by NMR spectroscopy. The Journal of biological chemistry, 284(17), 11498-11506. https://doi.org/10.1074/jbc.M806587200.
63. Marrink, S. J., Corradi, V., Souza, P., Ingolfsson, H. L, Tieleman, D. P., & Sansom, M. (2019). Computational Modeling of Realistic Cell Membranes. Chemical reviews, 119(9), 6184-6226. https://doi.org/10.1021/acs.chemrev.8b00460.
64. Matyus, E.,  Fidy, J., & Tieleman, D. P. (2008). Structure and dynamics of the antifungal molecules Syringotoxin-B and Syringopeptin-25A from molecular dynamics simulation. European biophysics journal: EBJ, 37(4), 495-502. https://doi.org/10.1007/s00249-007-0242-3
Fidy, J., & Tieleman, D. P. (2008). Structure and dynamics of the antifungal molecules Syringotoxin-B and Syringopeptin-25A from molecular dynamics simulation. European biophysics journal: EBJ, 37(4), 495-502. https://doi.org/10.1007/s00249-007-0242-3
65. Meena, K. R., & Kanwar, S. S. (2015). Lipopeptides as the antifungal and antibacterial agents: applications in food safety and therapeutics. BioMed research international, 2015, 473050. https://doi.org/10.1155/2015/473050.
66. Miao, V.,  M. F., Brian, P., Brost, R., Penn, J., Whiting, A., Martin, S., Ford, R., Parr, I., Bouchard, M., Silva, C. J., Wrigley, S. K., & Baltz, R. H. (2005). Daptomycin biosynthesis in Streptomyces roseosporus: cloning and analysis of the gene cluster and revision of peptide stereochemistry. Microbiology (Reading, England), 151 (Pt 5), 1507-1523. https://doi.org/10.1099/mic0.27757-0.
M. F., Brian, P., Brost, R., Penn, J., Whiting, A., Martin, S., Ford, R., Parr, I., Bouchard, M., Silva, C. J., Wrigley, S. K., & Baltz, R. H. (2005). Daptomycin biosynthesis in Streptomyces roseosporus: cloning and analysis of the gene cluster and revision of peptide stereochemistry. Microbiology (Reading, England), 151 (Pt 5), 1507-1523. https://doi.org/10.1099/mic0.27757-0.
67. Mikamo, H., Sato, Y., & Tamaya, T. (2000). In vitro antifungal activity of FK463, a new water-soluble echinocandin-like lipopeptide. The Journal of antimicrobial chemotherapy, 46(3), 485-487. https://doi.org/10.1093/jac/46.3.485.
68. Montal, M., Mueller, P. Formation of bimolecular membranes from lipid monolayers and a study of their electrical properties. Proceedings of the National Academy of Sciences USA, 1972, 69(12), 3561-3566. https://doi: 10.1073/pnas.69.12.3561.
69. Moyne, A. L., Shelby, R., Cleveland, Т. E., & Tuzun, S. (2001). Bacillomycin D: an iturin with antifungal activity against Aspergillus flavus. Journal of applied microbiology, 90(4), 622-629. https://doi.Org/10.1046/j.1365-2672.2001.01290.x.
70. Munusamy, S., Conde, R., Bertrand, В., & Munoz-Garay, C. (2020). Biophysical approaches for exploring lipopeptide-lipid interactions. Biochimie, 170, 173-202. https://doi.org/10.1016/j.biochi.2020.01.009.
71. Murdoch, D., & Plosker, G. L. (2004). Anidulafungin. Drugs, 64(19), 2249-2260. https://doi.org/10.2165/00003495-200464190-00011.
72. Nasir, M. N., & Besson, F. (2012). Interactions of the antifungal mycosubtilin with ergosterol-containing interfacial monolayers. Biochimica et biophysica acta, 1818(5), 1302 1308. https://doi.Org/10.1016/j.bbamem.2012.01.020.
73. Nasir, M. N., Laurent, P., Flore, C, Lins, L., Ongena, M., & Deleu, M. (2013). Analysis of calcium-induced effects on the conformation of fengycin. Spectrochimica acta. Part A, Molecular and biomolecular spectroscopy, 110, 450-457. https://doi.Org/10.1016/j.saa.2013.03.063.
74. Park, W. В., Kwon, N. J., Choi, S. J., Kang, С. K., Choe, P. G, Kim, J. Y., Yun, J., Lee, G. W., Seong, M. W., Kim, N. J., Seo, J. S., & Oh, M. D. (2020). Virus Isolation from the First Patient with SARS-CoV-2 in Korea. Journal of Korean medical science, 35(7), e84. https://doi.org/10.3346/jkms.2020.35.e84.
75. Pathak, К. V., Keharia, FL, Gupta, K., Thakur, S. S., & Balaram, P. (2012). Lipopeptides from the banyan endophyte, Bacillus subtilis Kl: mass spectrometric characterization of a library of fengycins. Journal of the American Society for Mass Spectrometry, 23(10), 1716-1728. https://doi.org/10.1007/sl3361-012-0437-4.
76. Pattnaik, G. P., & Chakraborty, H. (2018). Coronin 1 derived tryptophan-aspartic acid containing peptides inhibit membrane fusion. Chemistry and physics of lipids, 217, 35^12. https://doi.Org/10.1016/j.chemphyslip.2018.10.005.
77. Pfaller, M. A., Boyken, L., Hollis, R. J., Messer, S. A., Tendolkar, S., & Diekema, D. J. (2005). In vitro activities of anidulafungin against more than 2,500 clinical isolates of Candida spp., including 315 isolates resistant to fluconazole. Journal of clinical microbiology, 43(11), 5425-5427. https://doi.org/10.1128/JCM.43.ll.5425-5427.2005.
78. Pfaller, M. A., Boyken, L., Hollis, R. J., Messer, S. A., Tendolkar, S., & Diekema, D. J. (2006). In vitro susceptibilities of Candida spp.to caspofungin: four years of global surveillance. Journal of clinical microbiology, 44(3), 760 763. https://doi.Org/10.1128/JCM.44.3.760-763.2006.
79. Pizzorno, A., Abed, Y., & Boivin, G. (2011). Influenza drug resistance. Seminars in respiratory and critical care medicine, 32(4), 409-422. https://doi.org/10.1055/s-0031-1283281.
80. Raaijmakers, J. M., De Bruijn, I., Nybroe, O., & Ongena, M. (2010). Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS microbiology reviews, 34(6), 1037-1062. https://doi.Org/10.llll/j.1574-6976.2010.00221.x.
81. Regine, M. D., Ptak, M., Peypoux, F., & Michel, G. (1985). Pore-forming properties of iturin A; a lipopeptide antibiotic. Biochim. Biophys. Acta, 815, 405-409.
82. Rey, F. A., & Lok, S. M. (2018). Common Features of Enveloped Viruses and Implications for Immunogen Design for Next-Generation Vaccines. Cell, 172(6), 1319 1334. https://doi.Org/10.1016/j.cell.2018.02.054.
83. Risselada, H. J., Bubnis, G., &  (2014). Expansion of the fusion stalk and its implication for biological membrane fusion. Proceedings of the National Academy of Sciences of the United States of America, 111(30), 11043-11048. https://doi.org/10.1073/pnas. 1323221111.
(2014). Expansion of the fusion stalk and its implication for biological membrane fusion. Proceedings of the National Academy of Sciences of the United States of America, 111(30), 11043-11048. https://doi.org/10.1073/pnas. 1323221111.
84. Riva, L., Yuan, S., Yin, X., Martin-Sancho, L., Matsunaga, N., Pache, L., Burgstaller-Muehlbacher, S., De Jesus, P. D., Teriete, P., Hull, M. V., Chang, M. W., Chan, J. F., Cao, J., Poon, V. K., Herbert, K. M., Cheng, K., Nguyen, Т. H., Rubanov, A., Pu, Y., Nguyen, C,... Chanda, S. K. (2020). Discovery of SARS-CoV-2 antiviral drugs through large-scale compound repurposing. Nature, 586(7827), 113-119. https://doi.org/10.1038/s41586-020-2577-l.
85. Romero, D., de Vicente, A., Rakotoaly, R. H., Dufour, S. E., Veening, J. W., Arrebola, E., Cazorla, F. M., Kuipers, O. P., Paquot, M., & Perez-Garcia, A. (2007). The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Molecular plant-microbe interactions: MPMI, 20(4), 430-440. https://doi.org/10.1094/MPMI-20-4-0430.
86. Schneider, Т., Muller, A., Miess, H., & Gross, H. (2014). Cyclic lipopeptides as antibacterial agents - potent antibiotic activity mediated by intriguing mode of actions. International journal of medical microbiology: IJMM, 304(1), 37-43. https://doi.Org/10.1016/j.ijmm.2013.08.009.
87. Serra, M. D., Fagiuoli, G., Nordera, P., Bernhart, L, Volpe, C. D., Di Giorgio, D.,... & Menestrina, G. (1999). The interaction of lipodepsipeptide toxins from Pseudomonas syringae pv. syringae with biological and model membranes: a comparison of syringotoxin, syringomycin, and two syringopeptins. Molecular plant-microbe interactions, 12(5), 391-400.
88. Seydlova, G., & Svobodova, J. (2011). Surfactin-novel solutions for global issues. Bio Eng Trends Re s Technol, 13, 305-330.
& Svobodova, J. (2011). Surfactin-novel solutions for global issues. Bio Eng Trends Re s Technol, 13, 305-330.
89. Shekunov, E. V., Efimova, S. S., Yudintceva, N. M., Muryleva, A. A., Zarubaev, V. V., Slita, A. V., & Ostroumova, O. S. (2021). Plant Alkaloids Inhibit Membrane Fusion Mediated by Calcium and Fragments of MERS-CoV and SARS-CoV/SARS-CoV-2 Fusion Peptides. Biomedicines, 9(10), 1434. https://doi.org/10.3390/biomedicines9101434.
90. Silverman, J. A., Perlmutter, N. G., & Shapiro, H. M. (2003). Correlation of daptomycin bactericidal activity and membrane depolarization in Staphylococcus aureus. Antimicrobial agents and chemotherapy, 47(8), 2538-2544. https://doi.Org/10.1128/AAC.47.8.2538-2544.2003.
91. Sorensen, K. N., Kim, K. H., & Takemoto, J. Y. (1996). hi vitro antifungal and fungicidal activities and erythrocyte toxicities of cyclic lipodepsinonapeptides produced by Pseudomonas syringae pv. syringae. Antimicrobial agents and chemotherapy, 40(12), 2710-2713.
92. St Vincent, M. R., Colpitts, С. C, Ustinov, A. V., Muqadas, M., Joyce, M. A., Barsby, N. L., Epand, R. F., Epand, R. M., Khramyshev, S. A., Valueva, O. A., Korshun, V. A., Tyrrell, D. L., & Schang, L. M. (2010). Rigid amphipathic fusion inhibitors, small molecule antiviral compounds against enveloped viruses. Proceedings of the National Academy of Sciences of the United States of America, 107(40), 17339-17344. https://doi.org/10.1073/pnas.1010026107.
93. Stevens, D. A., Ichinomiya, M., Koshi, Y., & Horiuchi, H. (2006). Escape of Candida from caspofungin inhibition at concentrations above the MIC (paradoxical effect) accomplished by increased cell wall chitin; evidence for beta-l,6-glucan synthesis inhibition by caspofungin. Antimicrobial agents and chemotherapy, 50(9), 3160-3161. https://doi.org/10.1128/AAC.00563-06.
94. Storm, D. R., Rosenthal, K. S., & Swanson, P. E. (1977). Polymyxin and related peptide antibiotics. Annual review of biochemistry, 46, 723 763. https://doi.org/10.1146/annurev.bi.46.070177.003.
95. Streit, J. M., Jones, R. N., & Sader, H. S. (2004). Daptomycin activity and spectrum: a worldwide sample of 6737 clinical Gram-positive organisms. The Journal of antimicrobial chemotherapy, 53(4), 669-674. https://doi.org/10.1093/jac/dkhl43.
96. Tang, Q., Bie, X., Lu, Z., Lv, F., Tao, Y., & Qu, X. (2014). Effects of fengycin from Bacillus subtilis ftnbJ on apoptosis and necrosis in Rhizopus stolonifer. Journal of microbiology (Seoul, Korea), 52(8), 675-680. https://doi.org/10.1007/sl2275-014-3605-3.
97. Tang, Т., Bidon, M., Jaimes, J. A., Whittaker, G. R., & Daniel, S. (2020). Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antiviral research, 178, 104792. https://doi.Org/10.1016/j.antiviral.2020.104792.
98. Teissier, E., Penin, F., & Pecheur, E. I. (2010). Targeting cell entry of enveloped viruses as an antiviral strategy. Molecules (Basel, Switzerland), 16(1), 221-250. https://doi.org/10.3390/moleculesl6010221
99. Velkov, Т., Roberts, K. D., Nation, R. L., Thompson, P. E., & Li, J. (2013). Pharmacology of polymyxins: new insights into an 'old' class of antibiotics. Future microbiology, 8(6), 711-724. https://doi.org/10.2217/fmb.13.39.
100. Vergoten, G., & Bailly, C. (2021). In silico analysis of echinocandins binding to the main proteases of coronaviruses PEDV (3CLpro) and SARS-CoV-2 (Mpro). hi silico pharmacology, 9(1), 41. https://doi.org/10.1007/s40203-021-00101-l.
101.  Badosa, E., Montesinos, E., Planas, M., & Feliu, L. (2016). Synthetic Cyclolipopeptides Selective against Microbial, Plant and Animal Cell Targets by Incorporation of D-Amino Acids or Histidine. PloS one, 11(3), e0151639. https://doi.org/10.1371/journal.pone.0151639
Badosa, E., Montesinos, E., Planas, M., & Feliu, L. (2016). Synthetic Cyclolipopeptides Selective against Microbial, Plant and Animal Cell Targets by Incorporation of D-Amino Acids or Histidine. PloS one, 11(3), e0151639. https://doi.org/10.1371/journal.pone.0151639
102. Vollenbroich, D., Ozel, M., Vater, J., Kamp, R. M., & Pauli, G. (1997). Mechanism of inactivation of enveloped viruses by the biosurfactant surfactin from Bacillus subtilis. Biologicals: journal of the International Association of Biological Standardization, 25(3), 289-297.
103. Wei, X., Decker, J. M., Liu, H., Zhang, Z., Arani, R. В., Kilby, J. M., Saag, M. S., Wu, X., Shaw, G. M., & Kappes, J. C. (2002). Emergence of resistant human immunodeficiency virus type 1 in patients receiving fusion inhibitor (T-20) monotherapy. Antimicrobial agents and chemotherapy, 46(6), 1896-1905. https://doi.Org/10.1128/AAC.46.6.1896-1905.2002.
104. Xia, В., Luo, M., Pang, L., Liu, X., & Yi, Y. (2021). Lipopeptides against COVID-19 RNA-dependent RNA polymerase using molecular docking. Biomedical journal, 44(6 Suppl 1), S15-S24. https://doi.Org/10.1016/j.bj.2021.11.010.
105. Yamaguchi, H., Hiratani, Т., Baba, M., & Osumi, M. (1985). Effect of aculeacin A, a wall-active antibiotic, on synthesis of the yeast cell wall. Microbiology and immunology, 29(7), 609 623. https://doi.Org/10.1111/j.1348-0421.1985.tb00865.x.
106. Yoshino, N., Endo, M., Kanno, H., Matsukawa, N., Tsutsumi, R., Takeshita, R., & Sato, S. (2013). Polymyxins as novel and safe mucosal adjuvants to induce humoral immune responses in mice. PLoS One, 8(4), e61643.
107. Yu, G. Y., Sinclair, J. В., Hartman, G. L., & Bertagnolli, B. L. (2002). Production of iturin A by Bacillus amyloliquefaciens suppressing Rhizoctonia solani. Soil Biology and Biochemistry, 34(7), 955-963.
108. Yuan, L., Zhang, S., Wang, Y., Li, Y., Wang, X., & Yang, Q. (2018). Surfactin Inhibits Membrane Fusion during Invasion of Epithelial Cells by Enveloped Viruses. Journal of virology, 92(21), e00809-18. https://doi.org/10.1128/JVI.00809-18.
109. Yuan, L., Zhang, S., Peng, J., Li, Y., Yang, Q. (2019). Synthetic surfactin analogues have improved anti-PEDV properties. PLoS One, 14(4), e0215227. https: //doi. org/10.1371/journal.pone. 0215227.
110. Zeriouh, H., Romero, D., Garcia-Gutierrez, L., Cazorla, F. M., de Vicente, A., & Perez -Garcia, A. (2011). The iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Molecular plant-microbe interactions: MPMI, 24(12), 1540-1552. https://doi.org/10.1094/MPMI-06-11-0162.
111. Zhang, Т., Muraih, J. K., MacCormick, В., Silverman, J., & Palmer, M. (2014). Daptomycin forms cation- and size-selective pores in model membranes. Biochimica et biophysica acta, 1838(10), 2425-2430. https://doi.Org/10.1016/j.bbamem.2014.05.014.
112. Zhao, H., Shao, D., Jiang, C., Shi, J., Li, Q., Huang, Q., Rajoka, M., Yang, H., & Jin, M. (2017). Biological activity of lipopeptides from Bacillus. Applied microbiology and biotechnology, 101(15), 5951-5960. https://doi.org/10.1007/s00253-017-8396-0.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ, ИМЕЮЩАЯ ВЫСОКУЮ КОНЦЕНТРАЦИЮ ИТУРИНОВЫХ ЛИПОПЕПТИДОВ | 2018 |
|
RU2801578C1 |
| ПОВЕРХНОСТНО-АКТИВНЫЕ ВЕЩЕСТВА Bacillus sp. И ВКЛЮЧАЮЩАЯ ИХ КОМПОЗИЦИЯ | 2012 |
|
RU2605625C2 |
| Способ получения суммарной фракции липопептидов бактерий Bacillus subtilis MG-8 ВКПМ В-12476 и его использование в качестве профилактического средства от болезней сельскохозяйственных птиц | 2021 |
|
RU2770481C1 |
| Однослойные липосомы, содержащие микафунгин | 2024 |
|
RU2835492C1 |
| Рекомбинантный штамм Sen-Sdelta(M) вируса Сендай, продуцирующий S белок SARS-CoV-2 и используемый для получения интраназальной вакцины против COVID-19 | 2024 |
|
RU2837536C1 |
| Штамм гибридных клеток животных Mus musculus 2E1B5 - продуцент моноклонального антитела к рецептор-связывающему домену белка S вируса SARS-CoV-2 | 2021 |
|
RU2771288C1 |
| Тест-система для обнаружения РНК вируса SARS-CoV-2 в биоматериале от животных, пищевых продуктах и объектах окружающей среды методом полимеразной цепной реакции в режиме реального времени | 2021 |
|
RU2756474C1 |
| ЦИКЛОДЕКАПЕПТИДНЫЙ АНТИБИОТИК ШИРОКОГО СПЕКТРА АНТАГОНИСТИЧЕСКОГО ДЕЙСТВИЯ ЛАТЕРОЦИДИН, ШТАММ БАКТЕРИЙ BREVIBACILLUS LATEROSPORUS, ОБЛАДАЮЩИЙ ШИРОКИМ СПЕКТРОМ АНТАГОНИСТИЧЕСКОГО ДЕЙСТВИЯ - ПРОДУЦЕНТ АНТИБИОТИКА ЛАТЕРОЦИДИНА | 2002 |
|
RU2229520C1 |
| Модифицированный антисмысловой олигонуклеотид против вируса SARS-CoV-2 | 2020 |
|
RU2750584C1 |
| Способ получения самособирающегося октамерного белкового комплекса на основе апоферритина | 2023 |
|
RU2834642C1 |
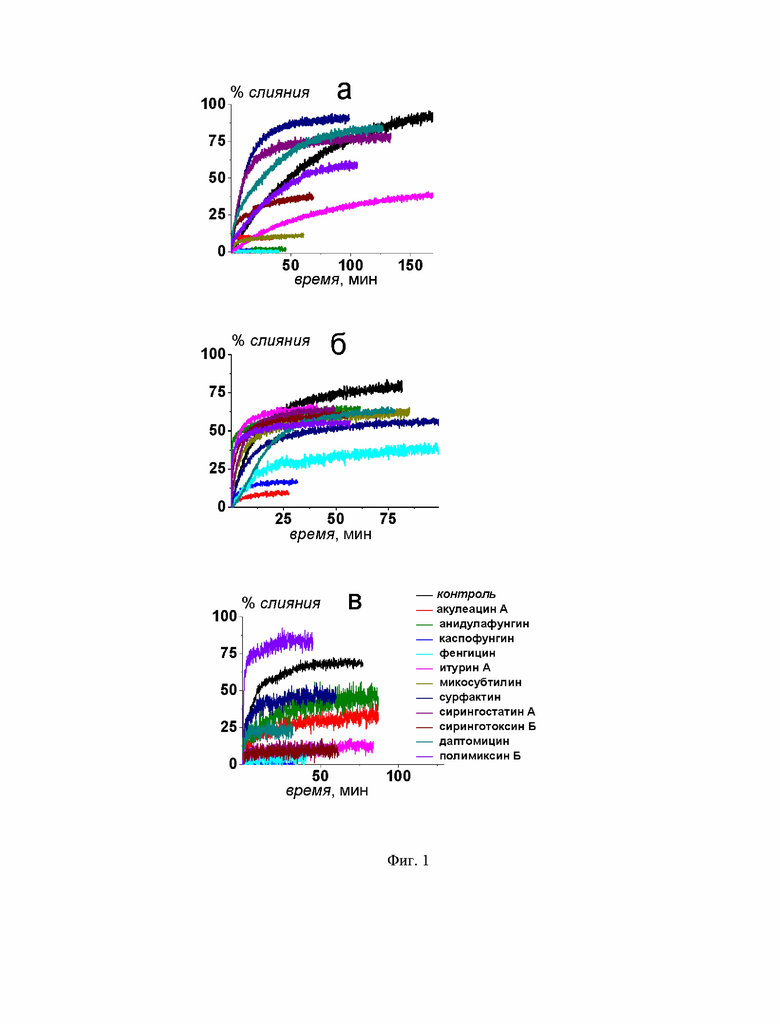
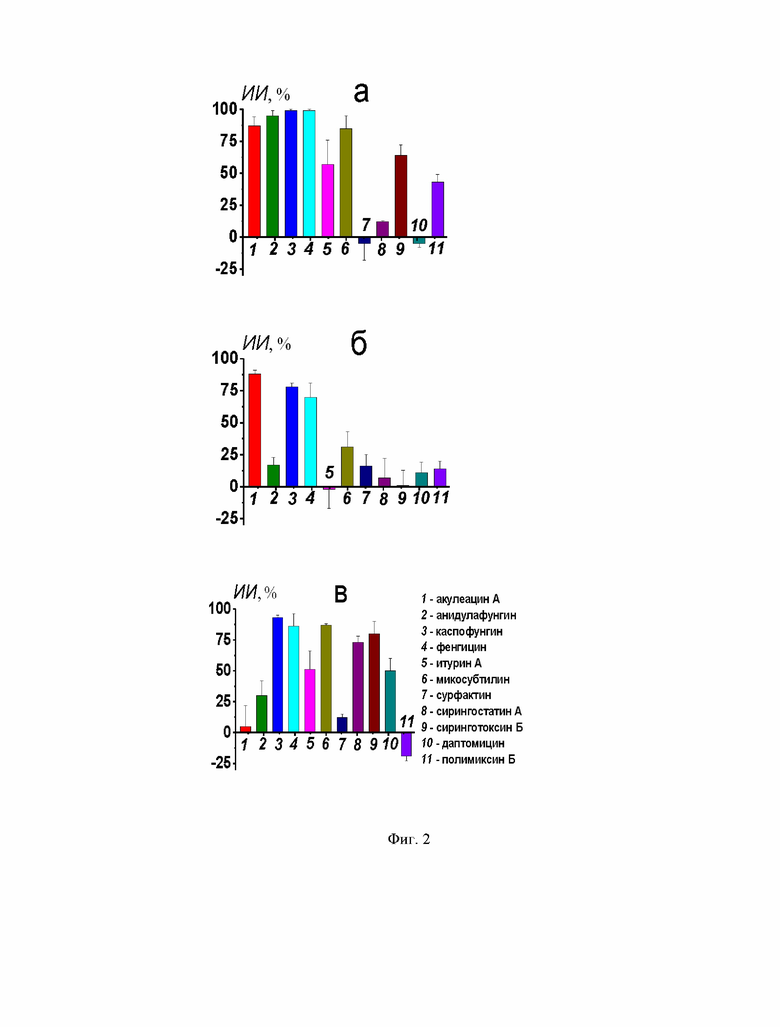
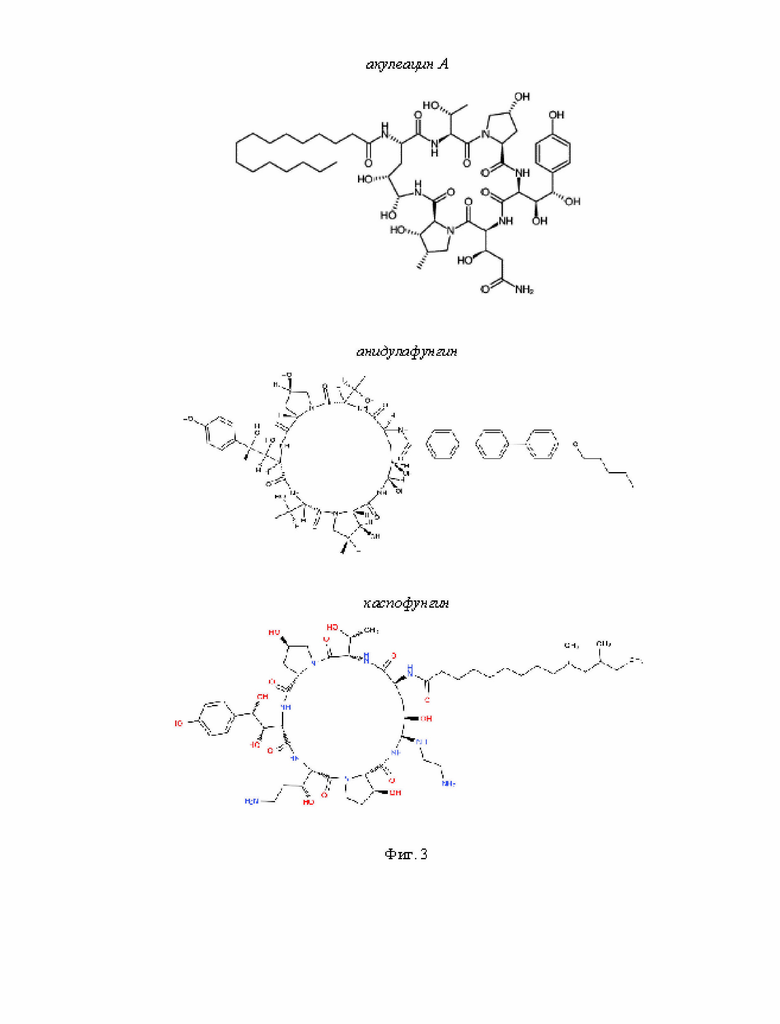
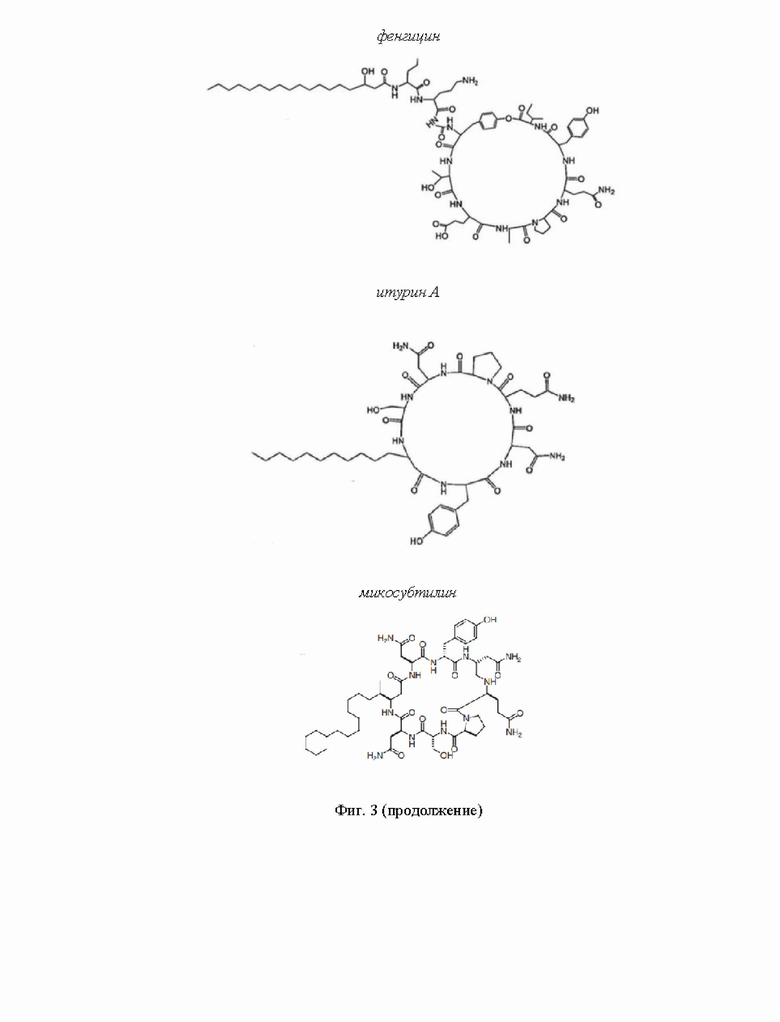
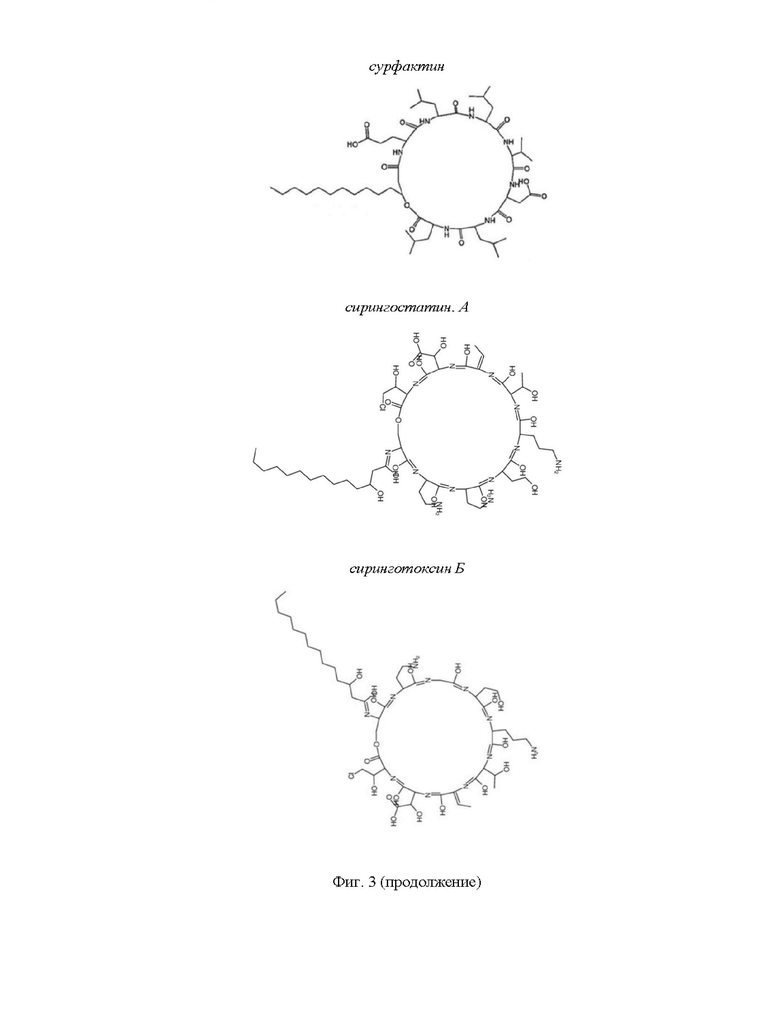
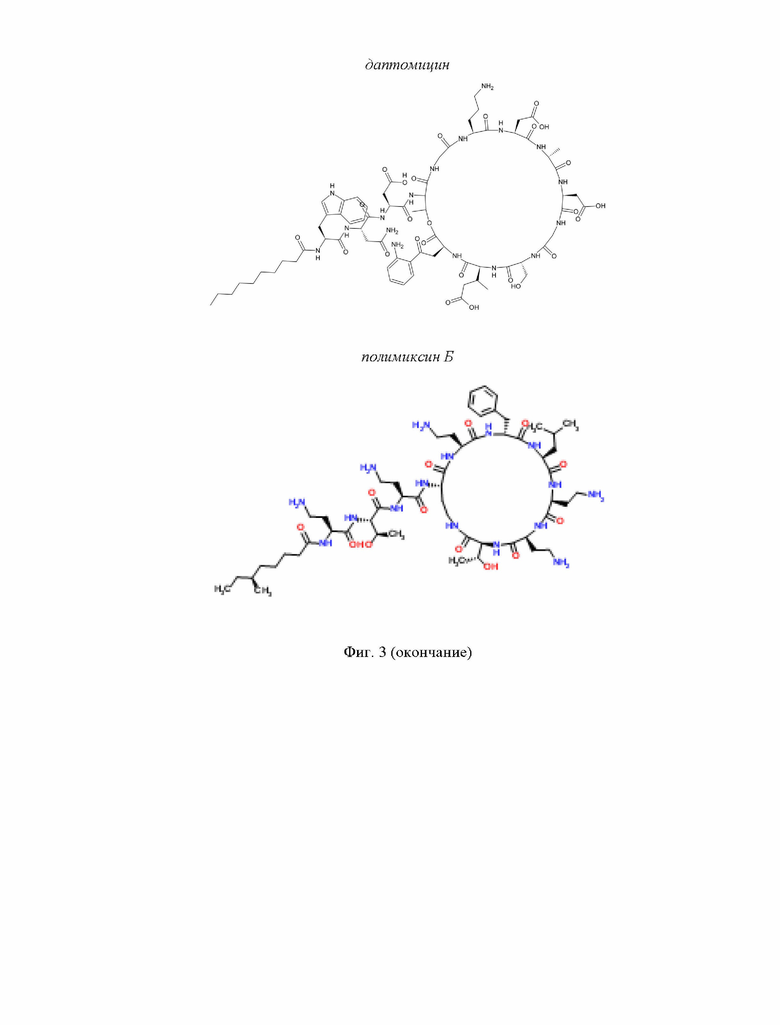
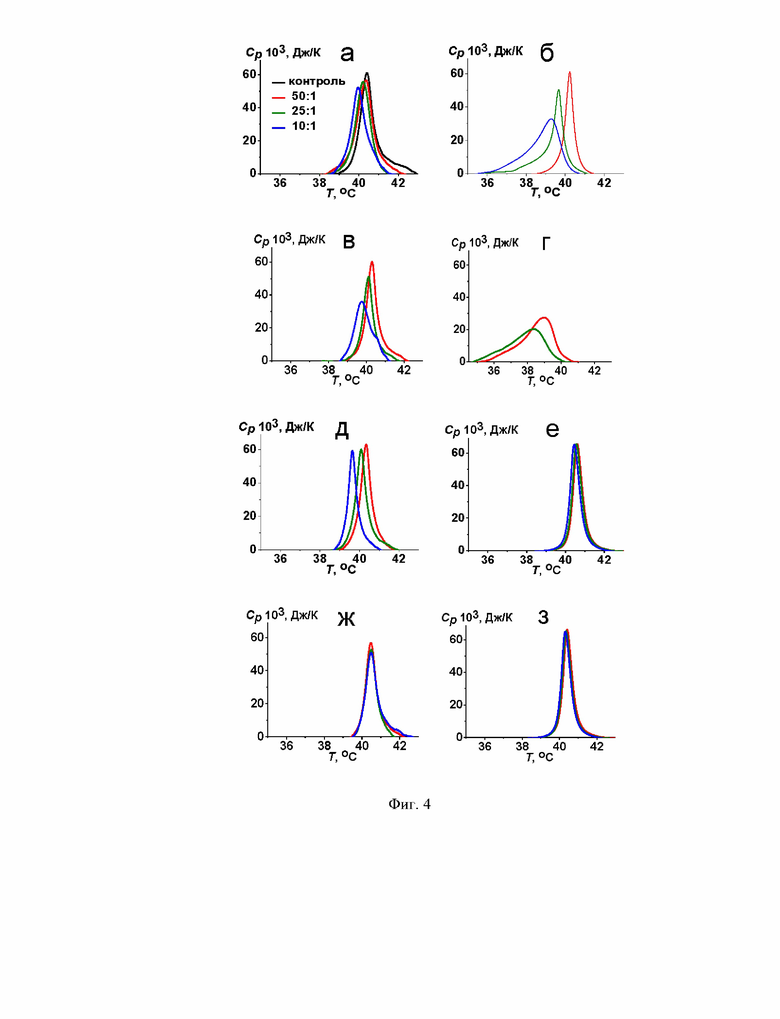
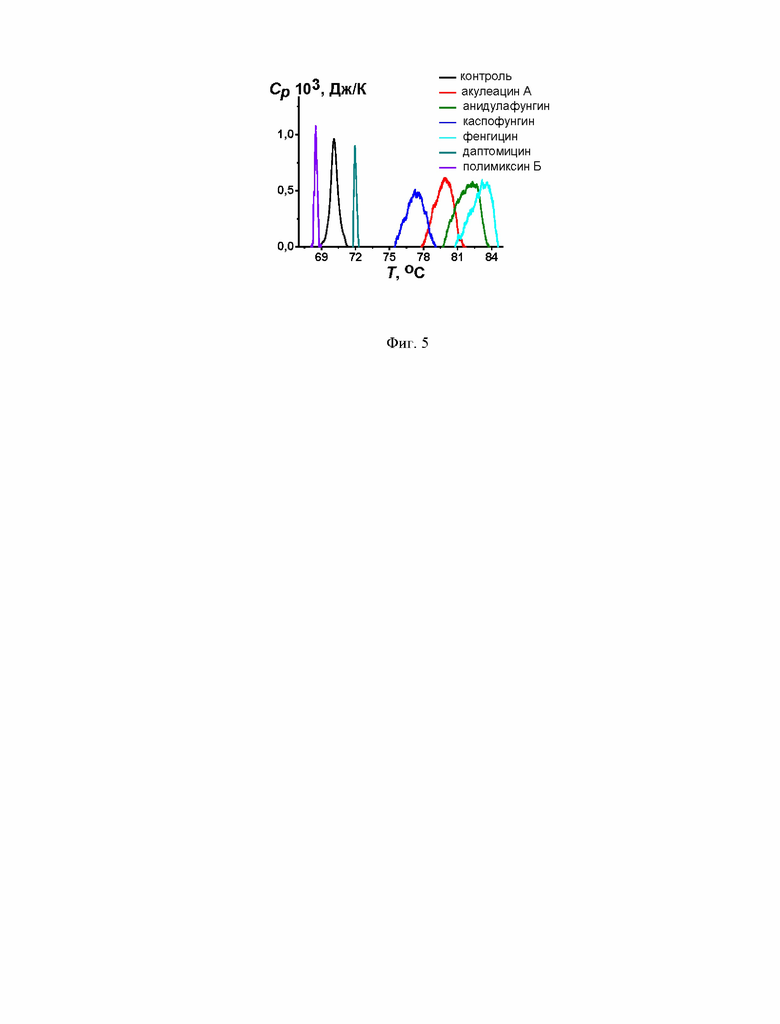
Изобретение относится к биотехнологии. Описано применение противомикробных циклических липопептидов в качестве ингибиторов слияния мембран, где липопептиды выбраны из группы, содержащей акулеацин А, анидулафунгин, каспофунгин, фенгицин, итурин А, микосубтилин и сиринготоксин Б. В одном из вариантов реализации описано применение липопептидов для ингибирования слияния вирусных частиц SARS-CoV-2 с клетками. Изобретение расширяет арсенал средств, ингибирующих слияние клеточных мембран. 1 з.п. ф-лы, 5 ил., 4 табл., 2 пр.
1. Применение липопептида, выбранного из группы, содержащей акулеацин А, анидулафунгин, каспофунгин, фенгицин, итурин А, микосубтилин, сиринготоксин Б, для ингибирования слияния мембран.
2. Применение по п.1, характеризующееся тем, что липопептид выбран из группы, содержащей акулеацин А, анидулафунгин, итурин А, микосубтилин, а ингибирование слияния мембран представляет собой ингибирование слияния вирусных частиц SARS-CoV-2 с клетками.
| EP 3613762 A1, 26.02.2020 | |||
| WO 2009092611 A1, 30.07.2009 | |||
| CN 111675752 A, 18.09.2020 | |||
| CN 108727475 B, 25.08.2020 | |||
| Валиахметов А | |||
| Я | |||
| Геометрия мембраны и функции белков // Биологические мембраны, 2008, Т | |||
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| Пуговица | 0 |
|
SU83A1 |
| Андрианов А | |||
| М | |||
| и др | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

