Изобретение относится к области микробной биотехнологии, более детально, к генной инженерии, в частности к лабораторному способу повышения эффективности биосинтеза бациллярной рибонуклеазы, которая может найти применение в научных исследованиях в области молекулярной биологии для очистки биопрепаратов от РНК, а в медицине - как противовирусное средство, при этом заявленный способ может стать основой для промышленного производства бациллярной рибонуклеазы, для чего требуется доработка технологии под требования заказчика и проведение экспериментов в промышленных условиях.
Рибонуклеазы - это ферменты, расщепляющие рибонуклеиновые кислоты (РНК), поэтому они находят применение в научных исследованиях в качестве реагента для удаления РНК. Кроме того, рибонуклеазы способны разрушать геном РНК-содержащих вирусов и используются в медицине как противовирусные средства. На рынке присутствует препарат «Рибонуклеаза» производства ООО «САМСОН-МЕД» (г. Санкт-Петербург, Россия), входящий в справочник лекарственных препаратов VIDAL и рекомендованный для лечения клещевого энцефалита. Препараты рибонуклеаз для научных исследований производят «ФЭНЧЕН ГРУПП» (FENGCHEN GROUP CO., LTD, Циндао, Китай), «Термо Фишер Сайентифик» (Thermo Fisher Scientific Inc., Уолтем, США), «Гуанчжоу Маген Байотекнолоджи» (Guangzhou Magen Biotechnology Co., Ltd, Гуандун, Китай). При этом в настоящее время все коммерческие препараты рибонуклеаз представляют собой ферменты животного происхождения, которые получают из поджелудочной железы быка. Однако в организме человека присутствует белок ингибитор рибонуклеаз животного происхождения RI, блокирующий их биологическую активность. Кроме того, использование органов животных в качестве источника получения препаратов рибонуклеаз трудозатратно и неэффективно. Бактериальные рибонуклеазы обладают такой же активностью, как и рибонуклеазы животных, но при этом не подвержены блокирующему действию белка ингибитора RI. Кроме того, получение рибонуклеаз из бактериальных клеток проще и дешевле. Бактерии рода Bacillus являются природными продуцентами рибонуклеаз, именуемыми бациллярными рибонуклеазами. Предлагаемое изобретение представляет собой способ повышения эффективности биосинтеза бациллярных рибонуклеаз и может быть использовано для их биотехнологического производства с целью дальнейшего применения в медицине и молекулярной биологии.
Далее заявителем представлены использованные в тексте термины и определения:
Бациллярные рибонуклеазы - рибонуклеазы бактерий рода Bacillus.
Бактериофаг (фаг) - вирус, поражающий бактерии.
Гомологичная рекомбинация - обмен нуклеотидными последовательностями между двумя сходными или идентичными хромосомами.
Барстар - внутриклеточный белок ингибитор рибонуклеаз бактерий рода Bacillus.
ИПТГ - изопропил-β-D-1-тиогалактопиранозид.
LB - питательная среда Луриа-Бертани.
мкл - микролитр.
п.о. - пара оснований.
Промотор - нуклеотидная последовательность, расположенная в начале гена и запускающая его транскрипцию.
Репрессор - белок, блокирующий транскрипцию гена.
Транскрипция - прочтение гена, перенос генетической информации с ДНК на РНК.
Культуральная жидкость - жидкая среда, получаемая при культивировании бактерий, содержащая остаточные питательные вещества и продукты метаболизма этих бактерий.
Ингибитор рибонуклеаз RI - белок, присутствующий в организме человека и блокирующий активность рибонуклеаз животного происхождения.
Из исследованного заявителем уровня техники известно, что бациллярные рибонуклеазы обладают противоопухолевыми и противовирусными свойствами [Surchenko Y., Dudkina E., Nadyrova A. I., Ulyanova V., Zelenikhin P. V., Ilinskaya, O. N. Cytotoxic potential of novel bacillary ribonucleases balnase and balifase (Цитотоксический потенциал новых бациллярных рибонуклеаз бальназы и балифазы) // BioNanoScience. - 2020. - V. 10. - P. 409-415.; Шах Махмуд Р., Гарифулина К.И., Ульянова В.В., Евтюгин В.Г., Миндубаева Л.Н., Хазиева Л.Р., Дудкина Е.В., Вершинина В.И., Колпаков А.И., Ильинская О.Н. Бактериофаги почвенных бацилл: новый поливалентный фаг Bacillus altitudinis // Молекулярная генетика, микробиология и вирусология. - 2017. - № 35, вып. 2. - С. 59-64]. В опухолевых и вирус-зараженных клетках рибонуклеазы разрушают молекулы РНК, что приводит к остановке синтеза белков, нарушению передачи сигналов, и как следствие приводит к их гибели.
Для получения бациллярных рибонуклеаз используют природные штаммы-продуценты, секретирующие (выделяющие) фермент в культуральную жидкость. Бациллярные рибонуклеазы, как конечный, целевой продукт, получают из культуральной жидкости путем их выделения и последующей очистки с использованием ионообменной хроматографии, например, - с использованием фосфоцеллюлозы Р11 [Dudkina E., Ulyanova V., Shah Mahmud R., Khodzhaeva V., Dao L., Vershinina V., Kolpakov A., Ilinskaya O. Three-step procedure for preparation of pure Bacillus altitudinis ribonuclease (Трехэтапная процедура получения чистой рибонуклеазы Bacillus altitudinis) // FEBS Open Bio. - 2016. - V. 6, I. 1. - P. 24-32]. Краткая сущность статьи заключается в том, что бациллярную рибонуклеазу получают из культуральной жидкости бактерий Bacillus altitudinis в три этапа: на первом этапе культуральную жидкость пропускают через положительно заряженный сорбент (ДЕАЕ-целлюлозу) для удаления примесных белков, на втором этапе рибонуклеазу, оставшуюся в культуральной жидкости, извлекают путем сорбции на отрицательно заряженном сорбенте (фосфоцеллюлозе Р11), а на третьем этапе проводят дополнительную очистку рибонуклеазы с использованием высоко-эффективной жидкостной хроматографии.
О выходе рибонуклеазы - количестве фермента, полученного в ходе очистки, судят по её удельной активности, которая рассчитывается как отношение активности фермента к его количеству. За единицу активности принимают количество фермента, которое в стандартных условиях катализирует превращение 1 мкмоль субстрата за 1 мин. Поэтому для увеличения выхода рибонуклеазы необходимо повысить ее удельную ферментативную активность в культуральной жидкости. Выход рибонуклеазы как конечного, целевого продукта зависит от нескольких факторов:
1) количества рибонуклеазы, наработанного бактериями в процессе биосинтеза;
2) эффективности секреции (выделения) рибонуклеазы из клеток;
3) качества очистки рибонуклеазы.
По мнению заявителей, наиболее существенным фактором, определяющим выход рибонуклеазы, является этап биосинтеза (наработки) рибонуклеазы бактериями. Природные штаммы-продуценты рибонуклеазы обладают низким уровнем ее биосинтеза, что приводит к низкому выходу фермента и делает нерентабельным промышленное производство препарата. Поэтому для промышленного производства рибонуклеазы актуальным является поиск вариантов повышения эффективности биосинтеза рибонуклеазы в клетках штамма-продуцента.
Из исследованного уровня техники заявителем выявлены аналоги предлагаемого изобретения. При описании аналогов в заявленном техническом решении заявителем использована оригинальная терминология, приведённая в описаниях аналогов.
Из исследованного заявителем уровня техники выявлено изобретение «Способ получения щелочной внеклеточной рибонуклеазы Bacillus intermedius 7р» по А.с. СССР SU 1138411. МПК C12N 9/22 (2000-01-01), приоритет от 18.05.1983. Сущность способа заключается в культивировании продуцента на среде, содержащей пептон, глюкозу и минеральные соли, отличается тем, что, с целью увеличения активности фермента, в культуральную жидкость вносят дактиномицин в количестве 0,075-0,125 мкг/мл через 6 - 7 часов (далее по тексту - ч) от начала культивирования в фазе замедления роста микробной популяции.
Недостатком известного способа является недостаточный (низкий) выход рибонуклеазы - целевого продукта, а также необходимость использования при культивировании штамма-продуцента антибиотика дактиномицина, обладающего токсичным эффектом в отношении клеток животных и растений, что снижает эффективность его практического применения по назначению.
Из исследованного заявителем уровня техники выявлено изобретение «Способ получения внеклеточной щелочной рибонуклеазы Bacillus intermedius 7р» по А.с. СССР SU 1010125, МПК C12N 9/22 (2000-01-01), C12N 1/20 (2000-01-01), приоритет от 23.12.1980. Сущность способа заключается в глубинном культивировании продуцента на питательной среде, содержащей источники углерода, азота, минеральные соли, биостимулятор, коагуляцию биомассы и кислых белков подкислением до рН 2,6 - 3,0, осаждение белка сульфатом аммония и фракционирование фермента на фосфоцеллюлозе, отличается тем, что, с цепью повышения активности рибонуклеазы в культуральной жидкости и выхода целевого продукта, в качестве биостимулятора используют дрожжевой экстракт или кислотный экстракт продукта «Витамин В12 кормовой» в количествах, соответственно равных 0,05 и 0,30 вес. % от среды.
Недостатком известного способа является недостаточный (низкий) выход рибонуклеазы - целевого продукта, что ограничивает область применения рибонуклеазы по назначению.
Из исследованного заявителем уровня техники выявлено изобретение «Рекомбинантная плазмида, обеспечивающая синтез барназы в клетках Escherichia coli, штамм Escherichia coli - продуцент барназы и способ получения барназы» по патенту России RU 2650871. МПК C12N 15/52 (2006-01-01), C12N 15/63 (2006-01-01), C12N 15/70 (2006-01-01), C12N 9/22 (2006-01-01), C12N 1/21 (2006-01-01), C12R 1/19 (2006-01-01), приоритет от 17.01.2007. Сущность известного изобретения в том, что рекомбинантная плазмида pET-STII-Ba, обеспечивающая синтез в клетках Escherichia coli барназы, имеющую аминокислотную последовательность SEQ ID №3, причем указанная плазмида получена путем клонирования рекомбинантного фрагмента ДНК с нуклеотидной последовательностью SEQ ID № 1 в плазмидный вектор pET39b по сайтам рестрикции NdeI / HindIII. Рекомбинантный штамм Escherichia coli BL21(DE3)/pET-STII-Ba, содержащий рекомбинантную плазмиду pET-STII-Ba по п.1 - продуцент барназы. Способ получения барназы, имеющей аминокислотную последовательность SEQ ID № 3, из биомассы штамма Escherichia coli BL21(DE3)/pET-STII-Ba по п.2, включающий выделение белка из периплазмы клеток, выращенных при пониженной температуре (плюс 25°С), и последующую его очистку при помощи хроматографии на фосфоцеллюлозе и катионообменнике.
Недостатком известного технического решения является необходимость использования для продукции рибонуклеазы гетерологичной (чужеродной) системы биосинтеза на основе бактерий Escherichia coli, при использовании которой (системы биосинтеза) существует ненулевая вероятность образования белка с измененной пространственной структурой, приводящей к нарушению функциональной активности целевого белка. Кроме того, повышенный синтез чужеродных белков в клетках Escherichia coli нередко приводит к их (чужеродных белков) внутриклеточному накоплению в агрегированной форме, что пагубно влияет на физиологические свойства целевого белка и требует проведения дополнительных этапов восстановления его (белка) пространственной структуры. Из-за наличия эндотоксина в клеточной стенке Escherichia coli требуется проведение дополнительных этапов очистки, направленных на удаление эндотоксина, что осложняет, т.е. делает не технологичным производство конечного продукта для его использования в промышленных и/или медицинских целях при использовании по назначению ввиду необходимости проведения дополнительных дорогостоящих этапов очистки.
Наиболее близким по сущности к заявляемому изобретению, выбранным заявителем в качестве прототипа, является изобретение «Стрептомицинустойчивый штамм Bacillus sp. ВКПМ В-9862 - продуцент внеклеточной щелочной рибонуклеазы» по патенту RU 2384619. Сущностью известного технического решения является штамм бактерий Bacillus sp. ВКПМ В-9862 - стрептомицинустойчивый продуцент внеклеточной щелочной рибонуклеазы.
Недостатком прототипа является то, что оно основано на модификации генома бактерий путем химического мутагенеза. Для этого бактерии-продуценты выращивают на среде с различными концентрациями антибиотика стрептомицина до приобретения ими (бактериями) устойчивости к антибиотику. При химическом мутагенезе в геноме случайным образом возникают и накапливаются мутации (ошибки), которые иногда приводят к повышению эффективности биосинтеза рибонуклеазы до 6 раз по сравнению с природным продуцентом, при этом, такое повышение биосинтеза рибонуклеазы происходит спонтанно, неуправляемо и непредсказуемо, что является существенным недостатком для управляемого промышленного производства препарата.
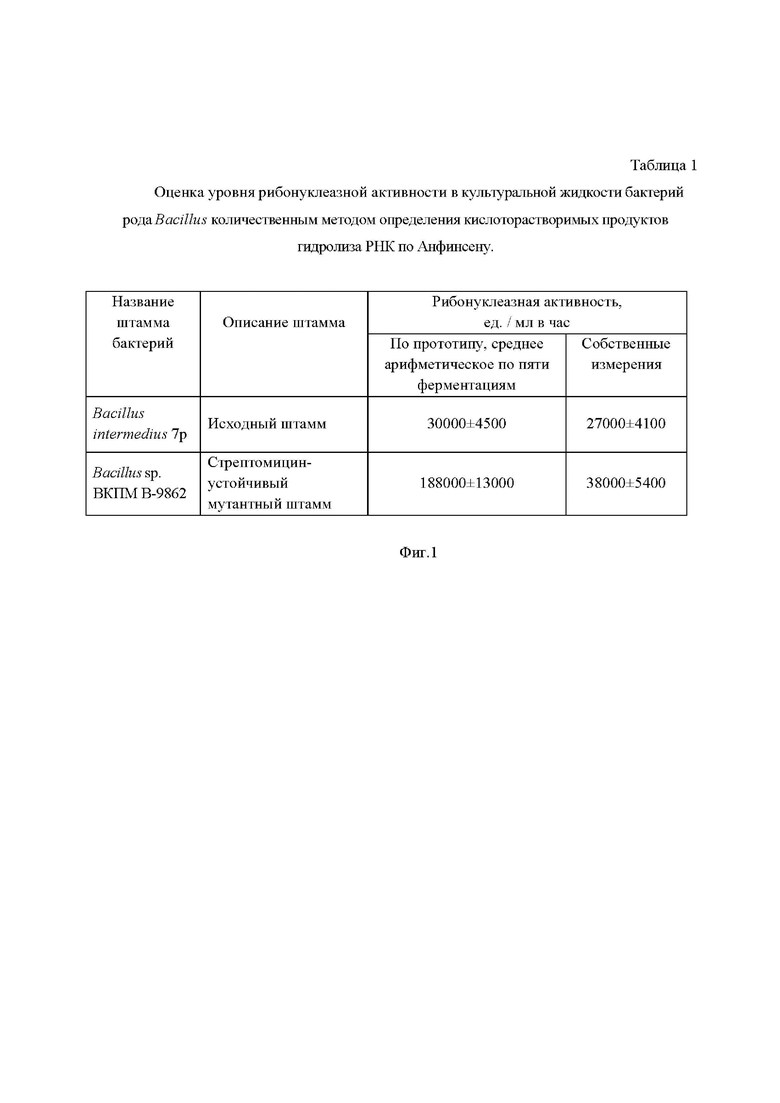
Заявителем выполнена оценка (см. Фиг. 1) рибонуклеазной активности стрептомицинустойчивого штамма Bacillus sp. ВКПМ В-9862 количественным методом определения кислоторастворимых продуктов гидролиза РНК по Анфинсену, как указано в прототипе.
На Фиг. 1 показан результат экспериментальной оценки рибонуклеазной активности прототипа - стрептомицинустойчивого штамма Bacillus sp. ВКПМ В-9862, где рибонуклеазная активность стрептомицинустойчивого штамма снизилась по сравнению с заявленной в прототипе активностью (188000 ед. / мл в час) и достигла значений, близких исходному штамму-продуценту Bacillus intermedius 7p (30000 ед. / мл в час). На дату представления заявочных материалов, уровень активности стрептомицинустойчивого штамма (38000 ед. / мл в час) превышает уровень активности природного продуцента (27000 ед. / мл в час) не в 6 раз (188000:30000 = 6,3), а только в 1,4 раза (38000:27000 = 1,4). Таким образом, заявителем экспериментально установлено, что в действительности уровень рибонуклеазной активности стрептомицинустойчивого штамма Bacillus sp. ВКПМ В-9862 существенно ниже по сравнению с заявленным в прототипе. По мнению заявителя, у стрептомицинустойчивого штамма с течением времени, вероятно, произошло восстановление генетических мутаций, накопленных в процессе химического мутагенеза, что привело к снижению эффективности биосинтеза рибонуклеазы до исходного уровня.
Кроме того, существенным недостатком известного технического решения по патенту RU 2384619 является то, что при использовании химического мутагенеза генетические мутации в геноме бактерий происходят неуправляемо, случайным образом, то есть имеют вероятностный характер. При вероятностном характере образующихся вследствие химического мутагенеза бактерий является существенным недостатком для контролируемого промышленного производства препарата. Кроме того, известное техническое решение может быть использовано применительно только к природному продуценту и не применимо для генно-инженерных штаммов с заданным генотипом, созданных для облегчения процесса выделения и очистки белков, так как будет неуправляемо и неконтролируемо изменять их геном и нарушать ранее созданный генотип.
Технической проблемой, решаемой изобретением и его техническим результатом является повышение эффективности биосинтеза бациллярной рибонуклеазы природными и/или генно-инженерными штаммами бактерий рода Bacillus за счет направленной модификации их (бактерий) геномов.
Сущностью заявленного технического решения является лабораторный способ биосинтеза бациллярной рибонуклеазы, заключающийся в использовании штаммов бактерий рода Bacillus с модифицированным геномом в качестве продуцента, характеризующийся тем, что модификацию проводят путем направленной инактивации гена spo0A, кодирующего белок-репрессор биосинтеза рибонуклеазы, заменяя ген spo0A в ходе гомологичной рекомбинации на ген устойчивости к антибиотику канамицину в геноме природного штамма Bacillus altitudinis B-388, выращивание модифицированного штамма проводят на низкофосфатной питательной среде. Лабораторный способ биосинтеза бациллярной рибонуклеазы, заключающийся в использовании в качестве продуцента штаммов бактерий рода Bacillus с модифицированным геномом, характеризующийся тем, что непосредственно в хромосому бактерий рода Bacillus, лишенных внеклеточных протеаз и не секретирующих собственных рибонуклеаз, вносят ген рибонуклеазы совместно с геном её внутриклеточного ингибитора – барстара, а затем в полученном генноинженерном штамме проводят модификацию путем направленной инактивации гена spo0A, кодирующего белок-репрессор биосинтеза рибонуклеазы, заменяя ген spo0A в ходе гомологичной рекомбинации на ген устойчивости к антибиотику канамицину, выращивание модифицированного штамма бактерий проводят на низкофосфатной питательной среде.
Заявленное техническое решение иллюстрируется Фиг.1 – Фиг.3.
На Фиг. 1 представлена таблица, в которой приведена оценка уровня рибонуклеазной активности в культуральной жидкости бактерий рода Bacillus количественным методом определения кислоторастворимых продуктов гидролиза РНК по Анфинсену.
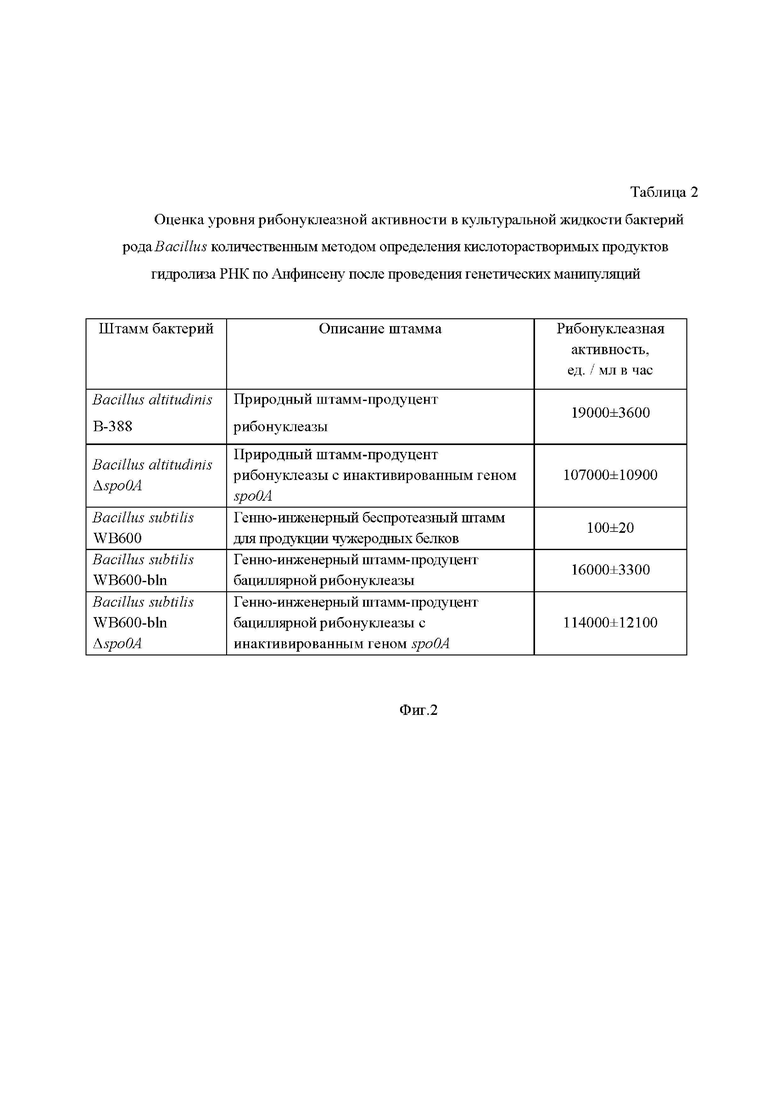
На Фиг. 2 представлена таблица, в которой приведена оценка уровня рибонуклеазной активности в культуральной жидкости бактерий рода Bacillus количественным методом определения кислоторастворимых продуктов гидролиза РНК по Анфинсену после проведения генетических манипуляций.
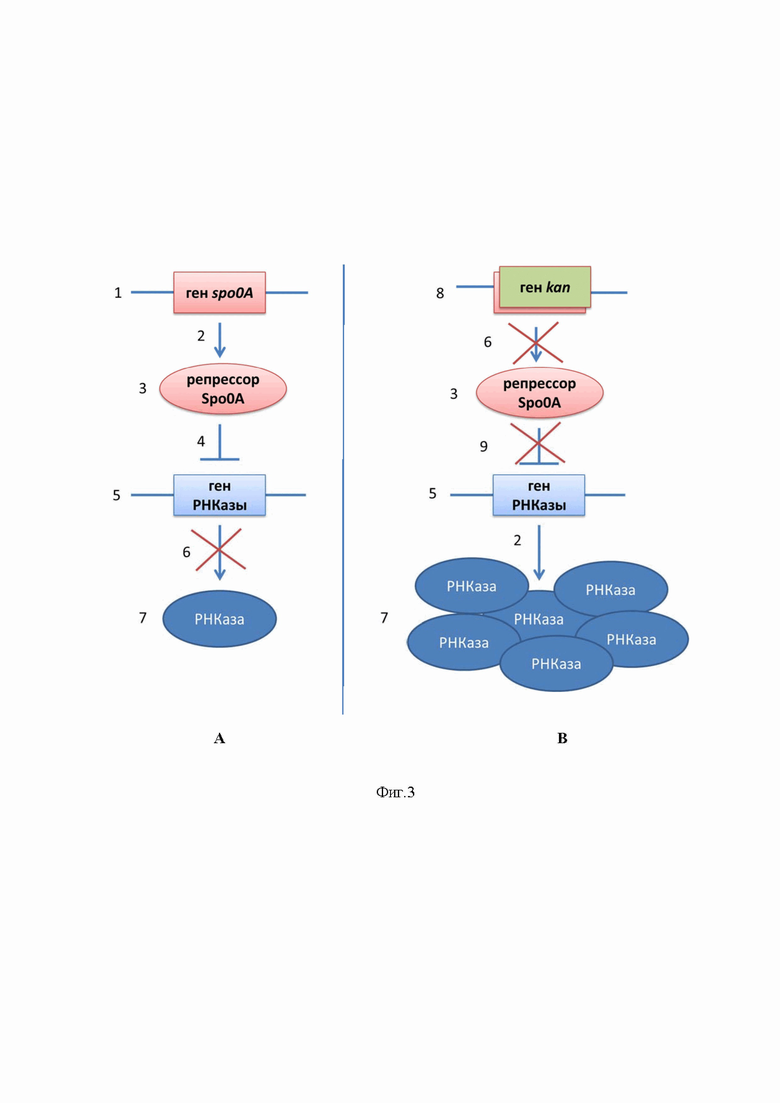
На Фиг. 3 представлена схема, иллюстрирующая способ повышения эффективности биосинтеза бациллярной рибонуклеазы путем инактивации гена spo0A, кодирующего белок-репрессор биосинтеза рибонуклеазы в геноме бактерий рода Bacillus, где позиции обозначают следующее:
А – схема биосинтеза бациллярной рибонуклеазы бактериями рода Bacillus в естественных условиях. В – схема биосинтеза бациллярной рибонуклеазы бактериями рода Bacillus после модификации их генома.
1 – ген spo0A в геноме бактерий рода Bacillus.
2 – образование белка с кодирующего его гена.
3 – белок-репрессор биосинтеза рибонуклеазы Spo0A.
4 – репрессия (блокировка) транскрипции гена.
5 – ген рибонуклеазы в геноме бактерий рода Bacillus.
6 – остановка образования белка с кодирующего его гена.
7 – бациллярная рибонуклеаза.
8 – ген устойчивости к антибиотику канамицину, замещающий ген spo0A в геноме бактерий рода Bacillus.
9 – остановка репрессии транскрипции гена.
В естественных условиях (А) в бактериях рода Bacillus с расположенного в геноме гена spo0A (1) образуется (2) белок-репрессор Spo0A (3), который блокирует (4) транскрипцию гена рибонуклеазы (5), в результате чего в клетках бактерий происходит остановка биосинтеза (6) бациллярной рибонуклеазы (7). В ходе модификации (В) генома бактерий рода Bacillus путем инактивации гена spo0A за счет его замены в ходе гомологичной рекомбинации на ген устойчивости к антибиотику канамицину (8) прекращается образование (6) белка-репрессора Spo0A (3), что снимает блокировку (9) транскрипции гена рибонуклеазы (5) и приводит к образованию (2) бациллярной рибонуклеазы (7) в повышенных количествах.
Далее заявителем приведено описание заявленного технического решения.
Выявленная проблема решается и заявленный технический результат достигается тем, что в геном природных и/или генно-инженерных штаммов бактерий рода Bacillus вносят направленные мутации, приводящие к инактивации гена spo0A, кодирующего белок-репрессор биосинтеза бациллярной рибонуклеазы, что приводит к увеличению выхода рибонуклеазы в 6 раз.
В варианте использования природных штаммов бактерий рода Bacillus в качестве продуцента бациллярной рибонуклеазы модификацию бактериального генома выполняют путем инактивации гена spo0A, заменяя его в ходе гомологичной рекомбинации на ген устойчивости к антибиотику канамицину.
В варианте использования в качестве продуцента бациллярной рибонуклеазы генно-инженерных штаммов бактерий рода Bacillus, не имеющих собственных рибонуклеаз, сначала в их геном вводят ген бациллярной рибонуклеазы совместно с геном её внутриклеточного ингибитора барстара, а затем в полученном штамме проводят инактивацию гена spo0A, кодирующего белок-репрессор биосинтеза бациллярной рибонуклеазы, заменяя его в ходе гомологичной рекомбинации на ген устойчивости к антибиотику канамицину.
Достижение заявленного технического результата оценивают, выращивая полученные модифицированные штаммы на низкофосфатной питательной среде и определяя уровень рибонуклеазной активности в культуральной жидкости.
По потребности модифицированные штаммы в необходимом количестве выращивают на питательной среде и используют в качестве продуцента для производства бациллярной рибонуклеазы. При этом следует обратить внимание на то, что в отличие от прототипа, где генетические мутации в результате химического мутагенеза в геноме бактерий происходят неуправляемо, случайным образом, заявленный способ характеризуется повышением эффективности биосинтеза бациллярной рибонуклеазы, основанным на направленной управляемой модификации генома природных и/или генно-инженерных штаммов бактерий рода Bacillus, что гарантированно обеспечивает увеличение выхода рибонуклеазы, так как заявленная модификация бактериального генома носит необратимый характер и введенные мутации не подлежат восстановлению, что устраняет присущий прототипу существенный недостаток, связанный с нестабильностью его генетической модификации, и гарантирует стабильное повышение уровня биосинтеза бациллярной рибонуклеазы. При чём возможность применения заявленного способа, как к природным, так и генно-инженерным штаммам бактерий рода Bacillus, созданным для облегчения выделения и очистки чужеродных белков, обеспечивает расширение спектра продуцентов для производства препарата бациллярной рибонуклеазы, в отличие от прототипа, обеспечивающего биосинтез бациллярной рибонуклеазы при использовании только природного штамма бактерий рода Bacillus.
Более подробно для достижения заявленного технического результата - повышения эффективности биосинтеза бациллярных рибонуклеаз, в природных штаммах бактерий рода Bacillus проводят направленную инактивацию гена spo0A, кодирующего белок-репрессор биосинтеза бациллярной рибонуклеазы. При этом, инактивацию гена spo0A выполняют известным способом, например, с помощью гомологичной рекомбинации [Kim J.H., Kim B.G. Application of LFH-PCR for the disruption of spoIIIE and spoIIIG of B. subtilis (Применение LFH-ПЦР для разрушения spoIIIE и spoIIIG у B. subtilis) // Biotechnol. Bioprocess Eng. - 2000. - V. 5. - P. 327-331], обеспечивающей замену гена spo0A на ген устойчивости к антибиотику канамицину, что приводит к инактивации гена spo0A.
Для повышения эффективности биосинтеза бациллярных рибонуклеаз в генно-инженерных штаммах бактерий рода Bacillus, не имеющих собственных рибонуклеаз, сначала в геном бактерий вводят ген рибонуклеазы совместно с геном её внутриклеточного ингибитора барстара для защиты клеток от токсического действия рибонуклеазы, а затем в полученном штамме проводят инактивацию гена spo0A, кодирующего белок-репрессор биосинтеза бациллярной рибонуклеазы, аналогичным (вышеописанным) способом.
Далее, в целях гарантированного получения результатов, заявителем проводится проверка успешности реализации заявленного способа повышения эффективности биосинтеза бациллярной рибонуклеазы, для чего полученные штаммы (с инактивированным геном spo0A) выращивают на питательной среде, например, на низкофосфатной питательной среде, содержащей 2,00 % пептона, 1,00 % глюкозы, 0,01 % CaCl2 × 2H2O, 0,03 % MgSO4 × 7H2O, 0,01 % MnSO4, 0,30 % NaCl, в течение 24 ч. Активность оценивают известным способом, например, количественным методом определения кислоторастворимых продуктов гидролиза РНК по Анфинсену, как указано в прототипе. Инактивация гена spo0A приводит к снятию репрессии с биосинтеза рибонуклеазы, при этом активность рибонуклеазы в культуральной жидкости возрастает до 6 раз (6-ти кратно) по сравнению с исходным продуцентом. Таким образом, бактерии рода Bacillus с подтверждённым повышенным уровнем биосинтеза рибонуклеазы направляют для использования по назначению.
Повышение удельной активности рибонуклеазы в культуральной жидкости доказывает повышение выхода целевого продукта - рибонуклеазы - в процессе её биосинтеза в клетках бактерий рода Bacillus, что и является доказательством достижения технического результата заявленного технического решения.
Заявленное техническое решение иллюстрируется примерами практического осуществления.
Пример 1. Лабораторный способ повышения эффективности биосинтеза бациллярной рибонуклеазы с использованием природного штамма бактерий Bacillus altitudinis B-388
Для повышения эффективности биосинтеза бациллярной рибонуклеазы в природном штамме бактерий рода Bacillus, используют, например, штамм Bacillus altitudinis B-388. Геном штамма Bacillus altitudinis B-388 модифицируют путем инактивации гена spo0A, кодирующего белок-репрессор биосинтеза бациллярной рибонуклеазы. Гена spo0A инактивируют путем его замены на ген kan, кодирующий канамицин нуклеотидилтрансферазу, обеспечивающую устойчивость бактерий к антибиотику канамицину. Инактивацию проводят известным способом [Niaudet B., Jannière L., Ehrlich S.D. Integration of linear, heterologous DNA molecules into the Bacillus subtilis chromosome: mechanism and use in induction of predictable rearrangements (Интеграция линейных гетерологичных молекул ДНК в хромосому Bacillus subtilis: механизм и использование для индукции предсказуемых перестроек) // J. Bacteriol. - 1985. - V. 163. - P. 111-120]. Для инактивации получают генетическую конструкцию, состоящую из трех фрагментов: 1) нуклеотидной последовательности, расположенной в геноме Bacillus altitudinis справа от гена spo0A; 2) гена устойчивости к антибиотику канамицину; 3) нуклеотидной последовательности, расположенной в геноме Bacillus altitudinis слева от гена spo0A. Фрагменты (части от целого) генетической конструкции получают с помощью полимеразной цепной реакции (ПЦР), используя специально сконструированные олигонуклеотиды. Олигонуклеотиды для амплификации трёх фрагментов конструируют известным способом [Nelson M.D., Fitch D.H. Overlap extension PCR: an efficient method for transgene construction (ПЦР с удлинением перекрытия: эффективный метод конструирования трансгена) // Methods Mol Biol. - 2011. - V. 772. - P. 459-470] таким образом, чтобы полученные в ходе ПЦР нуклеотидные последовательности имели комплементарные (соответствующие) друг другу участки. Генетическую конструкцию получают, например, следующим путем.
1) Для получения первого фрагмента нуклеотидную последовательность размером 1427 п.о., расположенную в геноме Bacillus altitudinis B-388 справа от гена spo0A, получают с помощью пары олигонуклеотидов:
spo_up_for 5'-ATATATCAGCATTCCAACAACTGTC-3'
и
spo_up_rev 5'-AAGCATCCTTCAATCCTCTCTTCTGAGTGTTCAATTCGAC-3'. Здесь и далее подчеркнута часть последовательности олигонуклеотидов, не соответствующая ДНК-матрице, но комплементарная другим частям создаваемой генетической конструкции.
Реакционная смесь содержит:
15,5 мкл деионизованной воды;
2,5 мкл 10-кратного TersusPlus-буфера («Евроген», Россия);
0,5 мкл 10 мМ смеси dNTPs («Евроген», Россия);
по 2,5 мкл каждого из олигонуклеотидов spo_up_for и spo_up_rev с концентрацией 10 мкМ;
1,0 мкл геномной ДНК с концентрацией 20 нг/мкл;
0,5 мкл термостабильной ДНК полимеразы Tersus («Евроген», Россия).
Условия проведения ПЦР: температура крышки плюс 105°C; предварительная денатурация +95°C, 180 сек (3,0 мин); денатурация +95°С, 60 сек (1,0 мин); отжиг олигонуклеотидов +64°С, 30 сек (0,5 мин); элонгация ДНК +72°С, 90 сек (1,5 мин); всего 30 циклов.
ПЦР продукты анализируют при помощи электрофореза в 1,5% агарозном геле и выявляют фрагмент ДНК размером около 1500 п.о., который очищают с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
2) Для получения второго фрагмента - гена устойчивости к антибиотику - используют олигонуклеотиды:
kan_for 5'-TTGAACACTCAGAAGAGGATTGAAGGATGCTTAGGAAGAC-3' и
kan_rev 5'-CATTTCTTTCATGCTTTAAATGGCCCGATTTAAGCACAC-3'.
В качестве ДНК-матрицы для синтеза гена kan используют плазмидную ДНК, например, pUB110.
Реакционная смесь содержит:
16,0 мкл деионизованной воды;
2,5 мкл 10-кратного TersusPlus-буфера («Евроген», Россия);
0,5 мкл 10 мМ смеси dNTPs («Евроген», Россия);
по 2,5 мкл каждого из олигонуклеотидов kan_for и kan_rev с концентрацией 10 мкМ;
0,5 мкл плазмидной ДНК pUB110 с концентрацией 10 нг/мкл;
0,5 мкл термостабильной ДНК полимеразы Tersus («Евроген», Россия).
Условия проведения ПЦР: температура крышки плюс 105°C; предварительная денатурация +95°C, 60 сек (1,0 мин); денатурация +95°С, 30 сек (0,5 мин); отжиг олигонуклеотидов +64°С, 30 сек (0,5 мин); элонгация ДНК +72°С, 60 сек (1,0 мин); всего 30 циклов.
ПЦР продукты анализируют при помощи электрофореза в 1,5% агарозном геле и выявляют фрагмент ДНК размером около 1000 п.о. (пар оснований), который очищают с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
3) Для получения третьего фрагмента нуклеотидную последовательность размером 1427 п.о., расположенную в геноме Bacillus altitudinis B-388 справа от гена spo0A, используют пары олигонуклеотидов:
spo_do_for 5'-CTTAAATCGGGCCATTTAAAGCATGAAAGAAATGAATAGGAC-3', и
spo_do_rev 5'-GAATACGTCTGCTGCTCCAC-3'.
Реакционная смесь содержит:
15,5 мкл деионизованной воды;
2,5 мкл 10-кратного TersusPlus-буфера («Евроген», Россия);
0,5 мкл 10 мМ смеси dNTPs («Евроген», Россия);
по 2,5 мкл каждого из олигонуклеотидов spo_do_for и spo_do_rev с концентрацией 10 мкМ;
1,0 мкл геномной ДНК с концентрацией 20 нг/мкл;
0,5 мкл термостабильной ДНК полимеразы Tersus («Евроген», Россия).
Условия проведения ПЦР: температура крышки плюс 105°C; предварительная денатурация +95°C, 180 сек (3,0 мин); денатурация +95°С, 60 сек (1,0 мин); отжиг олигонуклеотидов +64°С, 30 сек (0,5); элонгация ДНК +72°С, 90 сек (1,5 мин); всего 30 циклов.
ПЦР продукты анализируют при помощи электрофореза в 1,5% агарозном геле и выявляют фрагмент ДНК размером около 1500 п.о., который очищают с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
Затем три фрагмента объединяют с помощью ПЦР с расширением перекрытия в единую конструкцию с использованием олигонуклеотидов spo_up_for и spo_do_rev.
Реакционная смесь содержит:
14,0 мкл деионизованной воды;
2,5 мкл 10-кратного TersusPlus-буфера (Евроген, Россия);
0,5 мкл 10 мМ смеси dNTPs (Евроген, Россия);
2,5 мкл фрагмента 1 с концентрацией 20 нг/мкл;
1,8 мкл фрагмента 2 с концентрацией 20 нг/мкл;
2,2 мкл фрагмента 3 с концентрацией 20 нг/мкл;
0,5 мкл термостабильной ДНК полимеразы Tersus (Евроген, Россия).
Условия проведения реакции: температура крышки плюс 105°C; предварительная денатурация +95°C, 60 сек (1,0 мин); денатурация +95°С, 30 сек (0,5 мин); отжиг олигонуклеотидов +64°С, 30 сек (0,5 мин); элонгация ДНК +72°С, 120 сек (2,0 мин); всего 15 циклов.
По завершении реакции к реакционной смеси сразу добавляют по 0,5 мкл олигонуклеотидов spo_up_for и spo_do_rev с концентрацией 10 мкМ и снова проводят ПЦР: температура крышки плюс 105°C; предварительная денатурация +95°C, 60 сек (1,0 мин); денатурация +95°С, 30 сек (0,5 мин); отжиг олигонуклеотидов +64°С, 30 сек (0,5 мин); элонгация ДНК +72°С, 220 сек (3,7 мин); всего 30 циклов.
ПЦР продукт, объединяющий вышеуказанные три фрагмента, размером 3661 п.о. анализируют при помощи электрофореза в 1,5% агарозном геле, очищают из агарозного геля с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя, и используют для генетической трансформации (встраивания) бактерий Bacillus altitudinis.
Генетическую трансформацию бактерий проводят известным способом [Molecular biological methods for Bacillus (Молекулярно-биологические методы для Bacillus) / ed. C.R. Harwood and S.M. Cutting, John Wiley; Chicohester, 1990. - 581 p.]. В результате генетической трансформации линейный фрагмент ДНК проникает в клетки Bacillus altitudinis и происходит двойной кроссинговер (обмен) между хромосомной ДНК и гомологичными (сходными) областями линейного фрагмента ДНК, что приводит к замене гена spo0A на ген устойчивости к антибиотику канамицину. Клоны, в которых произошла успешная замена, отбирают по приобретению бактериями способности расти на среде с канамицином и утрате способности к образованию спор. Полученные клоны анализируют на наличие рибонуклеазной активности в культуральной жидкости бактерий по количеству кислоторастворимых продуктов гидролиза РНК и отбирают клоны с максимальной активностью, а значит, и с повышенным биосинтезом рибонуклеазы. Бактерии рода Bacillus с подтверждённым повышенным уровнем биосинтеза рибонуклеазы направляют для использования по назначению.
В Таблице 2 показан результат оценки рибонуклеазной активности в культуральной жидкости природного штамма бактерий рода Bacillus после проведения заявленных генетических манипуляций. Инактивация гена spo0A в природном штамме Bacillus altitudinis B-388 привела к увеличению уровня рибонуклеазной активности в 5,6 раз (107000:19000 = 5,6 ≈ 6). Таким образом, заявленный способ повышения эффективности биосинтеза рибонуклеазы позволяет увеличить выход рибонуклеазы в 5,6 раз (≈ в 6 раз) по сравнению с природным штаммом (Таблица 2).
Таблица 2 показывает и доказывает, что заявленный способ обеспечивает достижение заявленной цели технического решения - повышение эффективности биосинтеза и выхода целевого продукта при использовании природного штамма как продуцента рибонуклеазы.
Пример 2. Лабораторный способ повышения эффективности биосинтеза бациллярной рибонуклеазы с использованием генно-инженерного беспротеазного штамма бактерий Bacillus subtilis WB600
Для создания генно-инженерного продуцента бациллярной рибонуклеазы используют штамм бактерий рода Bacillus, не секретирующих собственных рибонуклеаз и обладающий заданными свойствами. Например, для продукции рибонуклеазы используют штамм Bacillus subtilis WB600, лишенный 6 внеклеточных протеаз (безпротеазный штамм), что предотвращает нежелательную деградацию рибонуклеазы в культуральной жидкости. Ген рибонуклеазы доставляют в клетки бактерий в составе генетической конструкции совместно с геном внутриклеточного ингибитора барстара для защиты от её (рибонуклеазы) токсичного действия. Для доставки генетической конструкции используют плазмиду, не способную к репликации (воспроизведению) внутри клеток бактерий рода Bacillus и обеспечивающую её (конструкции) интеграцию в участок бактериальной хромосомы, не существенный для роста и развития бактерий. Например, используют плазмиду pDG1662, которая способствует встраиванию генетической конструкции совместно с геном устойчивости к антибиотику хлорамфениколу для последующего отбора клонов в локус (участок) amyE, отвечающий за синтез α-амилазы. Для создания плазмиды pDG1662-bln, несущей генетическую конструкцию, полный ген рибонуклеазы амплифицируют с помощью ПЦР с геномной ДНК Bacillus altitudinis B-388, а полный ген барстара - с геномной ДНК Bacillus amyloliquefaciens H2. Для амплификации используют две пары олигонуклеотидов:
bln_bam_for 5'-CAGTCGGGATCCGTTTACGTCATCTAGAGACAGTAG-3',
bln_hind_rev 5'-GGAAGTCAAGCTTACAACCGTTTGAGGAGTACGTAC-3'
и
brs_hind_for 5'-GGAAGTCAAGCTTGTCCGGTCCAATCTGCAG-3',
brs_eco_rev 5'-GAGTGAGAATTCGGGTTTGTGTTTCCATATTGTTC-3',
содержащие, соответственно, участки распознавания для эндонуклеаз рестрикции BamHI, HindIII и HindIII, EcoRI, что позволяет лигировать (объединять) гены рибонуклеазы и барстара с плазмидой pDG1662, рестрицированной (разрезанной) по сайтам BamHI и EcoRI.
1) Ген рибонуклеазы получают с использованием геномной ДНК Bacillus altitudinis B-388 и олигонуклеотидов bln_bam_for и bln_hind_rev.
Реакционная смесь содержит:
31,0 мкл деионизованной воды;
5,0 мкл 10-кратного TersusPlus-буфера («Евроген», Россия);
1,0 мкл 10 мМ смеси dNTPs («Евроген», Россия);
по 5,0 мкл каждого из олигонуклеотидов bln_bam_for и bln_hind_rev с концентрацией 10 мкМ;
2,0 мкл геномной ДНК концентрацией 20 нг/мкл;
1,0 мкл термостабильной ДНК полимеразы Tersus («Евроген», Россия).
Условия проведения ПЦР: температура крышки плюс 105°C; предварительная денатурация +95°C, 180 сек (3,0 мин); денатурация +95°С, 60 сек (1,0 мин); отжиг олигонуклеотидов +68°С, 30 сек (0,5 мин); элонгация ДНК +72°С, 45 сек (0,75 мин); всего 30 циклов.
ПЦР продукты анализируют при помощи электрофореза в 2% агарозном геле и выявляют фрагмент ДНК размером около 750 п.о., который очищают с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
Проводят рестрикцию очищенного гена рибонуклеазы с использованием эндонуклеаз BamHI и HindIII. Реакционная смесь содержит:
2,0 мкл деионизованной воды;
5,0 мкл 10-кратного SE-буфера Rose («СибЭнзим», Россия);
40,0 мкл ПЦР-продукта с концентрацией 50 нг/мкл;
по 1,5 мкл эндонуклеаз рестрикции BamHI и HindIII («СибЭнзим», Россия).
Условия проведения реакции: инкубация в течение 3 ч при температуре +37°С.
Рестрицированный ПЦР-продукт очищают с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
2) Ген барстара получают с использованием геномной ДНК Bacillus amyloliquefaciens H2 и олигонуклеотидов brs_hind_for и brs_eco_rev. Реакционная смесь содержит:
15,5 мкл деионизованной воды;
2,5 мкл 10-кратного TersusPlus-буфера («Евроген», Россия);
0,5 мкл 10 мМ смеси dNTPs («Евроген», Россия);
по 2,5 мкл каждого из олигонуклеотидов brs_hind_for и brs_eco_rev с концентрацией 10 мкМ;
1,0 мкл геномной ДНК концентрацией 20 нг/мкл;
0,5 мкл термостабильной ДНК полимеразы Tersus («Евроген», Россия).
Условия проведения ПЦР: температура крышки плюс 105°C; предварительная денатурация +95°C, 180 сек (3,0 мин); денатурация +95°С, 60 сек (1,0 мин); отжиг олигонуклеотидов +58°С, 30 сек (0,5 мин); элонгация ДНК +72°С, 30 сек (0,5 мин); всего 30 циклов.
ПЦР продукты анализируют при помощи электрофореза в 2% агарозном геле и выявляют фрагмент ДНК размером около 450 п.о., который очищают с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
Проводят рестрикцию очищенного гена барстара с использованием эндонуклеаз HindIII и EcoRI.
Реакционная смесь содержит:
2,0 мкл деионизованной воды;
5,0 мкл 10-кратного SE-буфера W («СибЭнзим», Россия);
40,0 мкл ПЦР-продукта с концентрацией 50 нг/мкл;
по 1,5 мкл эндонуклеаз рестрикции HindIII и EcoRI («СибЭнзим», Россия).
Условия проведения реакции: инкубация в течение 3 ч при температуре плюс 37°С.
Рестрицированный ПЦР-продукт очищают с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
3) Плазмиду pDG1662 гидролизуют эндонуклеазами BamHI и EcoRI. Реакционная смесь содержит:
6,0 мкл 10-кратного SE-буфера W («СибЭнзим», Россия);
50,0 мкл плазмидной ДНК с концентрацией 150 нг/мкл;
по 2,0 мкл эндонуклеаз рестрикции HindIII и EcoRI («СибЭнзим», Россия).
Условия проведения реакции: инкубация в течение 5 ч при температуре +37°С.
Реакционную смесь анализируют при помощи электрофореза в 1% агарозном геле, очищают из агарозного геля фрагмент размером около 7000 п.о. с помощью набора Cleanup Mini («Евроген», Россия), руководствуясь инструкцией производителя.
Затем проводят лигирование трех рестрицированных фрагментов: гена бациллярной рибонуклеазы, гена барстара и плазмиды pDG1662 с получением плазмиды pDG1662-bln-brs. Реакционная смесь содержит:
1,4 мкл деионизованной воды;
1,0 мкл 10-кратного SE-буфера ДНК лигазы T4 («СибЭнзим», Россия);
5,0 мкл рестрицированной плазмидной ДНК с концентрацией 20 нг/мкл;
1,0 мкл рестрицированного продукта ПЦР-амплификации гена бациллярной рибонуклеазы с концентрацией 20 нг/мкл;
0,6 мкл рестрицированного продукта ПЦР-амплификации гена барстара с концентрацией 20 нг/мкл;
1,0 мкл Т4 ДНК лигазы («СибЭнзим», Россия).
Условия проведения реакции: инкубация в течение 16 ч при температуре +4°С.
Полученной лигазной смесью трансформируют компетентные клетки штамма Escherichia coli DH5α известным способом [Molecular cloning. A laboratory manual (Молекулярное клонирование. Лабораторное руководство) by T Maniatis, E F Fritsch and J Sambrook, Cold Spring Harbor Laboratory, New York. - 1982. - 545 p.]. Клоны выращивают на среде с ампициллином (100 мкг/мл) и используют для выделения плазмидной ДНК с помощью коммерческих наборов. Правильность сборки плазмиды pDG1662-bln-brs выявляют и подтверждают с помощью известных методов: ПЦР, рестрикционного анализа и пиросеквенирования (определения последовательности нуклеотидов в молекуле ДНК).
Полученной плазмидой pDG1662-bln-brs трансформируют компетентные клетки бактерий Bacillus subtilis WB600 известным способом [Molecular biological methods for Bacillus (Молекулярно-биологические методы для Bacillus) / ed. C.R. Harwood and S.M. Cutting, John Wiley; Chicohester, 1990. - 581 p.]. В ходе трансформации происходит интеграция части плазмиды pDG1662-bln-brs, содержащей гены рибонуклеазы и барстара и ген устойчивости к хлорамфениколу, в локус amyE хромосомы. Известным методом реплик отбирают клоны, которые способны расти на среде с хлорамфениколом и продуцировать внеклеточную рибонуклеазу, и не способны расти на среде со спектиномицином (за счет потери части плазмиды при рекомбинации) и образовывать внеклеточную амилазу (за счет интеграции в локус amyE). Внеклеточную рибонуклеазную активность определяют по кислоторастворимым продуктам гидролиза РНК, а внеклеточную амилазу - по продуктам гидролиза крахмала [Molecular biological methods for Bacillus (Молекулярно-биологические методы для Bacillus) / ed. C.R. Harwood and S.M. Cutting, John Wiley; Chicohester, 1990. - 581 p.]. В результате получают генно-инженерный штамм Bacillus subtilis WB600, в геном которого встроен ген рибонуклеазы с геном барстара.
Затем в полученном генно-инженерный штамме проводят инактивацию гена spo0A, кодирующего репрессор биосинтеза рибонуклеазы, способом, описанным в ПРИМЕРЕ 1. По уровню рибонуклеазной активности отбирают клоны с повышенным биосинтезом рибонуклеазы. Бактерии рода Bacillus с подтверждённым повышенным уровнем биосинтеза рибонуклеазы направляют для использования по назначению.
В Таблице 2 показан результат оценки рибонуклеазной активности в культуральной жидкости бактерий рода Bacillus после проведения заявленных генетических манипуляций. Генно-инженерный беспротеазный штамм Bacillus subtilis WB600, выбранный для продукции рибонуклеазы, обладает крайне низким исходным уровнем рибонуклеазной активности (100 ед./мл в час). При встраивании в его геном гена бациллярной рибонуклеазы рибонуклеазная активность увеличилась до 16000 ед./мл в час (штамм Bacillus subtilis WB600-bln). Инактивация гена spo0A в генно-инженерном штамме-продуценте рибонуклеазы (штамм Bacillus subtilis WB600-bln Δspo0A) привела к увеличению уровня рибонуклеазной активности в 7,1 раз по сравнению с генно-инженерным штаммом Bacillus subtilis WB600-bln (114000:16000 = 7,1) и в 6 раз (114000:19000 = 6,0) по сравнению с природным продуцентом Bacillus altitudinis B-388. Следовательно, предлагаемый способ повышения эффективности биосинтеза бациллярной рибонуклеазы позволяет увеличить выход рибонуклеазы в 6 раз (шестикратно) по сравнению с природным продуцентом Bacillus altitudinis.
Таблица 2 показывает и доказывает, что заявленный способ обеспечивает решение выявленной технической проблемы и достижение заявленного технического результата - повышение эффективности биосинтеза бациллярной рибонуклеазы природными и/или генно-инженерными штаммами бактерий рода Bacillus за счет направленной модификации их (бактерий) геномов и выхода целевого продукта при использовании генно-инженерного штамма как продуцента бациллярной рибонуклеазы.
По мнению заявителя, заявленное техническое решение удовлетворяет предъявляемому к изобретениям условию патентоспособности «новизна», так как при определении уровня техники не обнаружены технические решения и средства, совпадающие с заявленным по существенным признакам, включенным в формулу изобретения, включая характеристику назначения.
Заявленное техническое решение удовлетворяет предъявляемому к изобретениям условию патентоспособности «изобретательский уровень», поскольку из исследованного уровня техники не выявлены способы повышения эффективности биосинтеза бациллярной рибонуклеазы путем направленной модификации генома бактерий рода Bacillus, устраняющей репрессию биосинтеза рибонуклеазы, имеющие признаки, совпадающие с отличительными признаками заявленного изобретения, и не установлена известность влияния отличительных признаков на указанный технический результат.
Заявленное техническое решение удовлетворяет предъявляемому к изобретениям условию патентоспособности «промышленная применимость», так как может быть реализовано на любом специализированном предприятии с использованием стандартного оборудования, известных отечественных материалов и технологий. Заявленное техническое решение апробировано в условиях лаборатории, в результате чего подтверждено достижение заявленного технического результата.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ БАРНАЗЫ В КЛЕТКАХ ESCHERICHIA COLI, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ БАРНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ БАРНАЗЫ. | 2017 |
|
RU2650871C1 |
| СТРЕПТОМИЦИНУСТОЙЧИВЫЙ ШТАММ Bacillus sp. ВКПМ В-9862 - ПРОДУЦЕНТ ВНЕКЛЕТОЧНОЙ ЩЕЛОЧНОЙ РИБОНУКЛЕАЗЫ | 2008 |
|
RU2384619C1 |
| ШТАММ БАКТЕРИЙ Serratia species, ЯВЛЯЮЩИЙСЯ ПРОДУЦЕНТОМ ВНЕКЛЕТОЧНОЙ РИБОНУКЛЕАЗЫ И ДЕЗОКСИРИБОНУКЛЕАЗЫ, ОБЛАДАЮЩИХ ПРОТИВОВИРУСНОЙ АКТИВНОСТЬЮ | 2013 |
|
RU2528064C1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ПУРИНОВОГО НУКЛЕОЗИДА 5'-АМИНОИМИДАЗОЛ-4-КАРБОКСАМИДРИБОЗИДА (АИКАР) И ШТАММ БАКТЕРИЙ BACILLUS SUBTILIS - ПРОДУЦЕНТ АИКАР | 2008 |
|
RU2405833C2 |
| СПОСОБ ПРОДУКЦИИ ПУРИНОВЫХ НУКЛЕОЗИДОВ И НУКЛЕОТИДОВ МЕТОДОМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ Escherichia ИЛИ Bacillus | 2006 |
|
RU2365622C2 |
| Рекомбинантная плазмида pUSB2-AmQ, обеспечивающая синтез белка альфа-амилазы Bacillus amyloliquefaciens, и штамм Bacillus subtilis/pUSB2-AmQ - продуцент белка альфа-амилазы Bacillus amyloliquefaciens | 2020 |
|
RU2747627C1 |
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКОЙ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ydiB, СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА НИЗШИХ АЛКИЛОВ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИНА | 2007 |
|
RU2355759C1 |
| БАКТЕРИЯ Bacillus subtilis, ПРОДУЦИРУЮЩАЯ 5`-АМИНОИМИДАЗОЛ-4-КАРБОКСАМИДРИБОЗИД (АИКАР), И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА АИКАР ПУТЕМ КУЛЬТИВИРОВАНИЯ ТАКОЙ БАКТЕРИИ | 2013 |
|
RU2542387C1 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН pnp, - ПРОДУЦЕНТ L-ТРЕОНИНА, И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2005 |
|
RU2330883C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН hipA | 2005 |
|
RU2320718C2 |
Изобретения относятся к области биотехнологии, в частности к генной инженерии. Предложены лабораторные способы биосинтеза бациллярной рибонуклеазы. Способы предусматривают модификацию штаммов бактерий рода Bacillus путем направленной инактивации гена spo0A, заменяя его в ходе гомологичной рекомбинации на ген устойчивости к канамицину, и выращивание модифицированного штамма на низкофосфатной питательной среде. Изобретения позволяют увеличить выход рибонуклеазы более чем в 5,6 раз. 2 н.п. ф-лы, 3 ил., 2 пр.
1. Лабораторный способ биосинтеза бациллярной рибонуклеазы, заключающийся в использовании штаммов бактерий рода Bacillus с модифицированным геномом в качестве продуцента, отличающийся тем, что модификацию проводят путем направленной инактивации гена spo0A, кодирующего белок-репрессор биосинтеза рибонуклеазы, заменяя ген spo0A в ходе гомологичной рекомбинации на ген устойчивости к антибиотику канамицину в геноме природного штамма Bacillus altitudinis B-388, выращивание модифицированного штамма проводят на низкофосфатной питательной среде.
2. Лабораторный способ биосинтеза бациллярной рибонуклеазы, заключающийся в использовании в качестве продуцента штаммов бактерий рода Bacillus с модифицированным геномом, отличающийся тем, что непосредственно в хромосому бактерий рода Bacillus, лишенных внеклеточных протеаз и не секретирующих собственных рибонуклеаз, вносят ген рибонуклеазы совместно с геном её внутриклеточного ингибитора – барстара, а затем в полученном генноинженерном штамме проводят модификацию путем направленной инактивации гена spo0A, кодирующего белок-репрессор биосинтеза рибонуклеазы, заменяя ген spo0A в ходе гомологичной рекомбинации на ген устойчивости к антибиотику канамицину, выращивание модифицированного штамма бактерий проводят на низкофосфатной питательной среде.
| Ульянова В.В | |||
| и др | |||
| Влияние Spo0A и AbrB белков на экспрессию генов гуанилспецифичных рибонуклеаз Bacillus intermedius и Bacillus pumilus в рекомбинантных штаммах Bacillus subtilis | |||
| МИКРОБИОЛОГИЯ, 2007, т.76, No.5, с.1-6 | |||
| JP 2011010634 A, 20.01.2011 | |||
| СТРЕПТОМИЦИНУСТОЙЧИВЫЙ ШТАММ Bacillus sp. ВКПМ В-9862 - ПРОДУЦЕНТ ВНЕКЛЕТОЧНОЙ ЩЕЛОЧНОЙ РИБОНУКЛЕАЗЫ | 2008 |
|
RU2384619C1 |
| KIM J.H., KIM B.G | |||
| Application of LFH-PCR for the disruption of | |||

