Изобретение относится к генетической конструкции (плазмиде), обеспечивающей синтез рекомбинантного белка - альфа-амилазы Bacillus amyloliquefaciens, предназначен для использования в пищевой (хлебобулочной), спиртовой промышленности и производстве моющих средств, и рекомбинантному штамму Bacillus subtilis - продуценту белка альфа-амилазы, который может быть использован в биотехнологии и пищевой промышленности.
Альфа-амилаза – фермент, широко использующийся в промышленном масштабе с целью гидролиза крахмала для получения различных форм глюкозы и фруктозы. Альфа-амилаза является основным ферментом пищевой, спиртовой промышленности и при производстве моющих средств.
В хлебопекарной отрасли добавление Альфа-амилазы позволяет улучшить свойства теста. Альфа-амилазы катализируют расщепления внутренних 1,4-α-гликозидных связей в амилозе и амилопектине крахмала муки в более короткие декстрины, которые используются дрожжами в качестве питательных веществ в процессе брожения. Результатом является увеличение объема буханки, улучшение текстуры продукта и его вкусовых характеристик [1].
В пивоварении и спиртовой промышленности применение амилаз позволяет осуществлять более глубокий ферментативный гидролиз углеводов (в частности, крахмала), тем самым полнее сбродить сусло до основного продукта - этилового спирта. В пивоварении использование ферментных препаратов позволяет экономить до 165 г ячменя при производстве каждого декалитра пива. Применение амилазы при производстве спирта дает возможность полностью отказаться от зернового солода и одновременно увеличить выход спирта из сырья на 1,5% при снижении себестоимости декалитра спирта [2].
В производстве моющих веществ препараты альфа-амилазы используются как добавка для удаления углеводных загрязнений. Удаление крахмалосодержащих пятен с одежды и фарфора (при использовании посудомоечных машин) стало трудной задачей в домашнем хозяйстве. Даже при температуре выше 50°С крахмал практически нерастворим и его трудно удалить, и обычно встречаются остаточные крахмальные пятна. Решением этой проблемы может быть добавление препаратов амилазы к моющим средствам. Стирка белья и мытье посуды выполняются в довольно жестких, с точки зрения стабильности ферментов, условиях: высокая температура, щелочной рН и присутствие сильно окисляющихся соединений. Помимо способности противостоять этой среде, амилаза также должна быть совместима с поверхностно-активными веществами, хелатирующими кальций агентами и протеазами, присутствующими в моющем средстве.
Нативная (природная) альфа-амилаза имеет низкую активность и имеет недостаточное эффективное действие в моющих средствах, особенно если они используется на промышленных установках. Наиболее перспективными выглядят термоустойчивые амилолитические препараты. Применение таких ферментов позволяет увеличивать реакционную температуру (до 90°С), что позволяет минимизировать риск микробного загрязнения, уменьшает время реакции, что позволяет уменьшить расход энергии [3].
Привлекательной системой для получения препаратов альфа-амилазы является Bacillus subtilis. Эта система обладает преимуществами в качестве продуцента: общепризнанный безопасный статус (GRAS), легкие генетические манипуляции, использование данного организма позволяет получать рекомбинантный фермент в секретируемой форме, за счет этого снижаются расходы на выделение и очистку конечного продукта. Эффективность синтеза и секреции рекомбинантных белков в настоящее время может быть изменена в значительной степени за счет генно-инженерных манипуляций, конструирования экспрессионной кассеты, включающей последовательность промотера и сигнального пептида.
Известны патентные заявки в которых описываются разработки векторов для продуцента Bacillus subtilis с целью получения ферментов: альфа-амилазы, бета-лактамазы и альфа- бета- интерферона (US 5010000, С12Р, 21/00, C12N, 15/00, 1/20, 9/86, опубл. 23.04.1991) (US 5010015, С12Р, 21/00, C12N, 15/00, 1/20, 9/86, опубл. 23.04.1991). В патенте не делают акцент на термостабильность, активность, а так же на получение большого количества белка, здесь рассматривается Bacillus subtilis, как новый продуцент этих ферментов
Известны заявки на получение препарата альфа-амилазы в системе Bacillus amyloliquefaciens (RU 203689, C12N, 9/28, 15/32, 15/56,15/75, опубл. 30.11.1993) и в системе Bacillus subtilis (SU 1717633, A1, C12N, 15/18, опубл. 07.03.1992). Продуцент альфа-амилазы Bacillus amyloliquefaciens является устойчивым к фагам и позволяет получать высокий выход фермента. Альфа-амилаза полученная в системе Bacillus subtilis является конструкцией полученной из фрагментов генов альфа-амилазы Bacillus licheniformis и альфа-амилазы Bacillus amyloliquefaciens, это позволяет повысить термостабильность конечного продукта.
Известна заявка на изобретение препарата протеазы и способ его получения (WO 91/02792, A1, C12N, 9/54, 15/57, 15/75, C11D, 3/386, опубл. 07.03.1991 г.) [6] у конечного продукта заявлена повышенная активность, устойчивость при высоких температурах и рН=7-10, а так же увеличенный выход фермента для использования в промышленности. В качестве продуцента используется культура Bacillus subtilis. Заявленные показатели достигаются за счет слияний между геном щелочной протеазы АТСС 53926 В. licheniformis и геном щелочной протеазы В. lentus, что позволяет получить ген несущий регуляторную область кодирующую сигнальный пептид, последовательность кодирующую пропептид и зрелую протеазу.
В другом патенте (RU 2124559 С1, C12N, 15/76, 15/67, 15/66, С12Р, 21/02, опубл. 10.01.1999 г.) [5] в качестве продуцента альфа-амилазы предлагают использовать бактерии рода Streptomyces. Увеличение секреции фермента, предлагается получить за счет SAF-промотора (фактор активации вторичного метаболизма), он обладает плейотропным действием и более эффективен в экспрессии белка, чем другие известные промоторы. Разработка направлена только на повышение экспрессии белка, упуская из внимания улучшение свойств (например, термостабильность или повышение активности). В предлагаемом изобретении увеличение термостабильности и устойчивости к щелочным рН достигается за счет использования гена альфа-амилазы из Bacillus amyloliquefaciens, а повышение выхода фермента за счет использования синтетического промотера Bacillus subtilis.
Наиболее близким аналогом (прототипом) является заявка (RU 2661790, C2, C12N, 9/50, 15/75, 15/63, 15/62, 15/70, опубл. 19.07.2018 г.) [4] в которой предлагается экспрессионный вектор для получения различных вариантов белков, в том числе и альфа-амилазы. Предлагается несколько вариантов сигнальных пептидов для экспрессионной плазмиды, которые улучшают внеклеточную секрецию белка. Таким образом, авторы предлагают использовать в качестве продуцентов бактерии рода Bacillus или рода Actinomycetales. Так как в патенте предложена универсальная конструкция, эффективность этой конструкции для секреции амилазы может быть ограничена. Тогда как преимуществом изобретения является специализированная конструкция с «сильным» промотором и лидерной последовательностью амилазы, что обеспечивает эффективность секреции альфа-амилазы. Кроме того, предлагаемый вектор обладает высокой стабильностью при использовании в системе Bacillus.
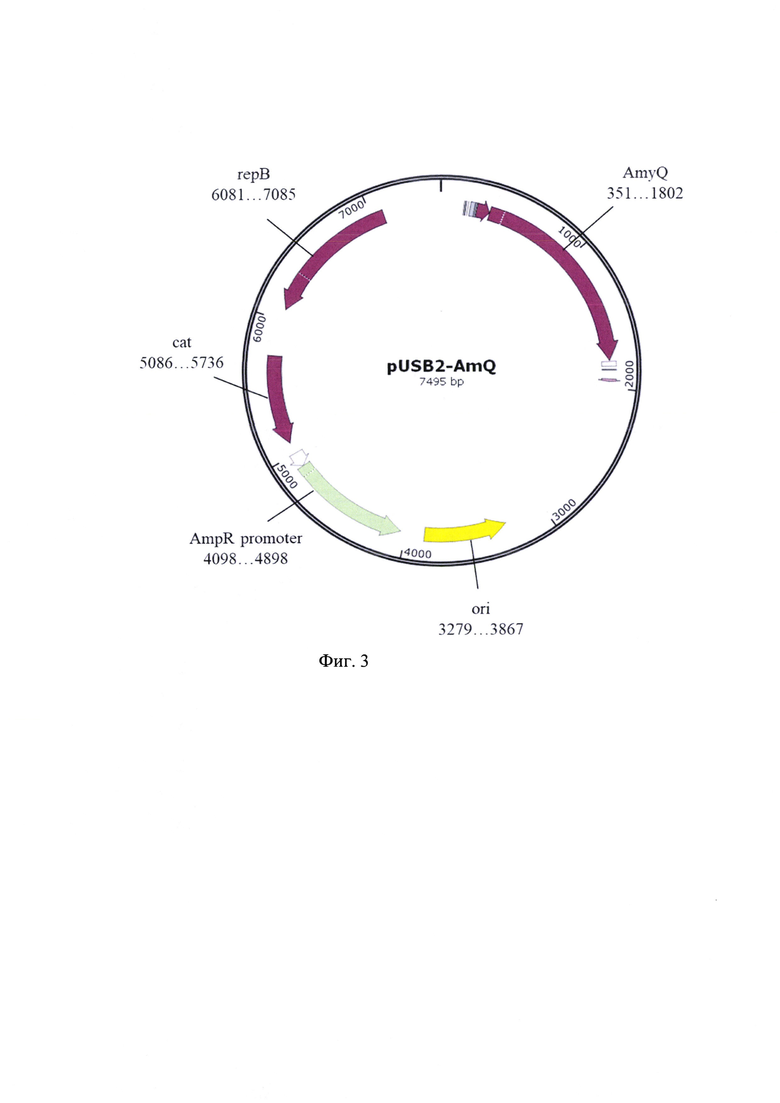
Указанный технический результат достигается конструированием рекомбинантной плазмиды pUSB2-AmQ, размером 7517 пар оснований (п.о.) характеризующейся физической и генетической картой, представленной на фиг. - 3, обеспечивающей синтез рекомбинантного белка альфа-амилазы Bacillus amyloliquefaciens 55 кДа в клетках Bacillus subtilis, содержащей рекомбинантный ген белка альфа-амилазы Bacillus amyloliquefaciens, имеющей нуклеотидную последовательность SEQ ID NO:l от нуклеотида в положении 351 п.о. до нуклеотида в положении 1802 п.о., кодирующей целевой белок альфа-амилазы Bacillus amyloliquefaciensc полной аминокислотной последовательностью SEQ ID NO: 2, встроенный по сайтам рестрикции AhlI и BseX3I в область полилинкера вектора pUSB2, содержащая сильный промотор и сигнальную последовательность, что обеспечивает эффективную наработку рекомбинантных белков и их экспорт в культуральную среду.
Указанный технический результат достигается также созданием рекомбинантного штамма Bacillus subtilis WB800N/pUSB2-AmQ - продуцента белка альфа-амилазы Bacillus amyloliquefaciens, полученного трансформацией рекомбинантной плазмидой pUSB2-AmQ по п. 2.
Заявляемый штамм Bacillus subtilis WB800N/pUSB2-AmQ - продуцент рекомбинатной альфа-амилазы, обеспечивающей катализ гидролиза крахмала и гликогена до олигосахаридов, и предназначенный для использования в детергентах, получают следующим образом.
Для работы была выбрана альфа-амилаза Bacillus amyloliquefaciens. Это выбор был продиктован рядом параметров, которые характерны для этой альфа-амилазы, в частности термостабильность и устойчивость в щелочных средах.
Нуклеотидная последовательность, кодирующую альфа-амилазу Bacillus amyloliquefaciens была взята из базы данных GenBank [https://www.ncbi.nlm.nih.gov/]. Для обеспечения высокого и стабильного выхода целевого продукта провели оптимизацию кодонного состава под систему экспрессии штамма Bacillus subtilis на сервере ThermoFisher [https://www.thermofisher.com/]. После оптимизации был проведен анализ различия частот использования кодонов, а так же негативных элементов с помощью серверов GenScript [https://www.genscript.com/tools/rare-codon-analysis] и EFMCalculator [http://barricklab.org/django/efm/]. Индекс Кодонной Адаптации (CAI) гена составил 0,93 (CAI=1,0 считается идеальным). Оптимизированная последовательность была синтезирована (SEQ.1).
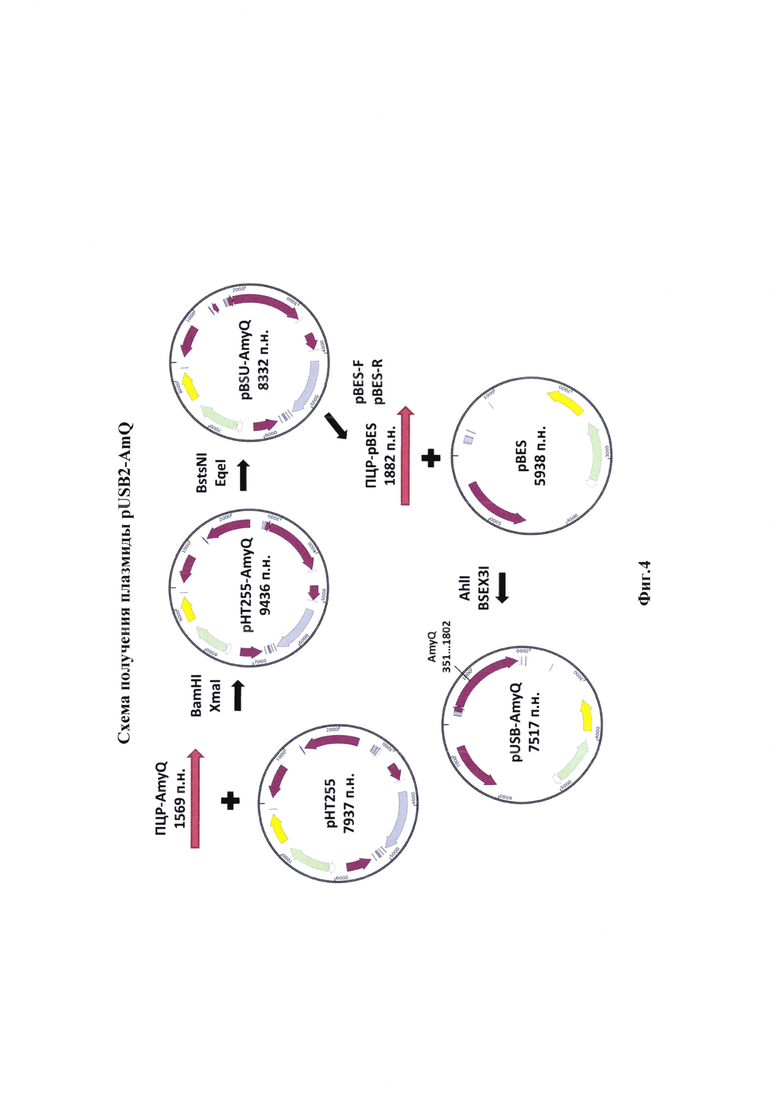
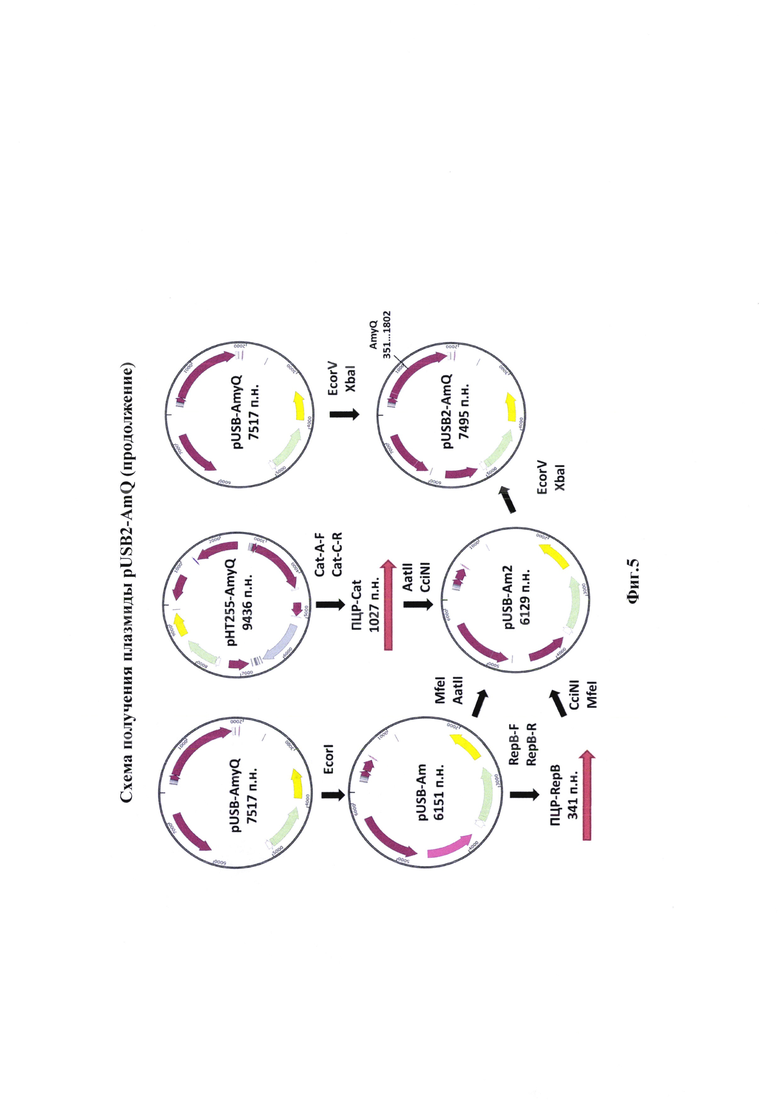
Для обеспечения синтеза был сконструирован вектор pUSB2 (фиг. 4, 5). Для его конструирования были проведены последовательные встройки и удаления нуклеотидных последовательностей кодирующих регуляторные факторы (фиг. 4, 5).
В качестве продуцента была выбрана культура Bacillus subtilis, штамм WB800N которая обладает преимуществами в качестве продуцента: общепризнанный безопасный статус (GRAS), легкие генетические манипуляции, так же использование данного организма позволит получать рекомбинантный фермент в секретируемой форме, за счет этого снижаются расходы на выделение и очистку конечного продукта.
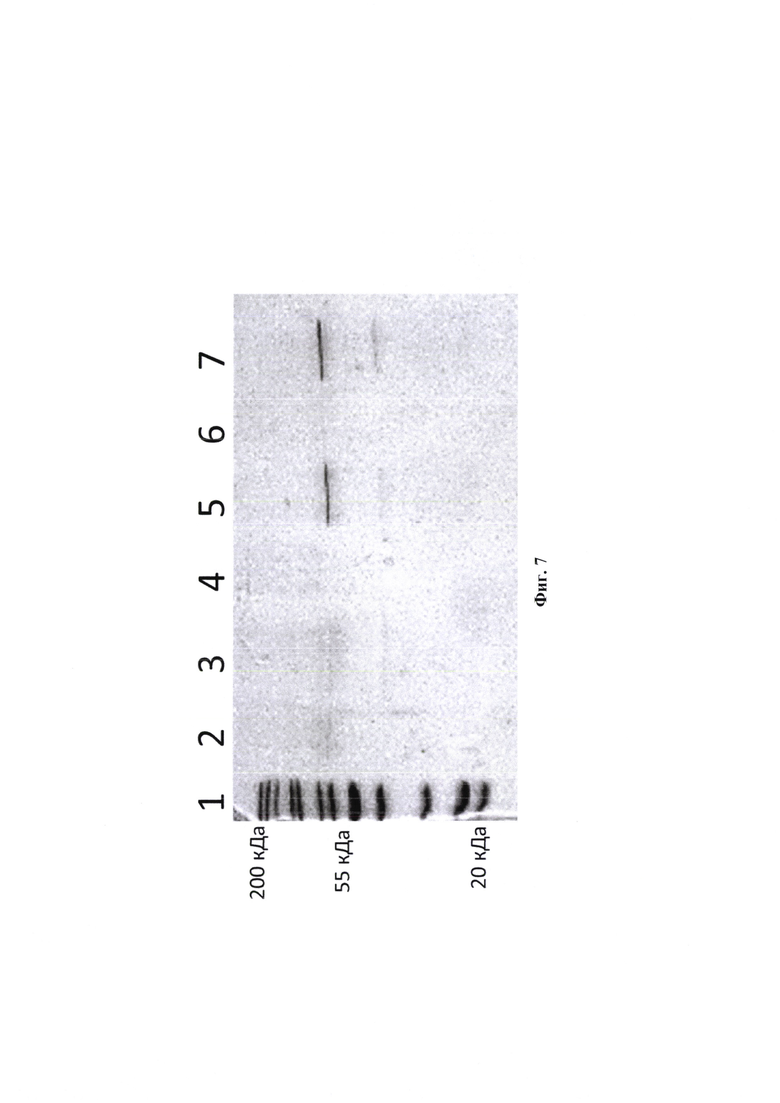
При культивировании, штамм Bacillus subtilis WB800N/pUSB2-AmQ, обеспечивает синтез рекомбинантной альфа-амилазы 90% от общего секретируемого белка Bacillus subtilis (фиг. 7).
Указанный технический результат достигается благодаря оптимизации кодоного состава, наличие сильного промотора, который включает оптимизированную последовательность -35, -10 и +1 и отсутствие последовательности кодирующую белок репрессор LacI. Наличие белка репрессора LacI в составе экспрессионной кассеты обеспечивает контроль синтеза мРНК, но, в случае постоянного синтеза при интеграции, он оказывает отрицательное влияние.
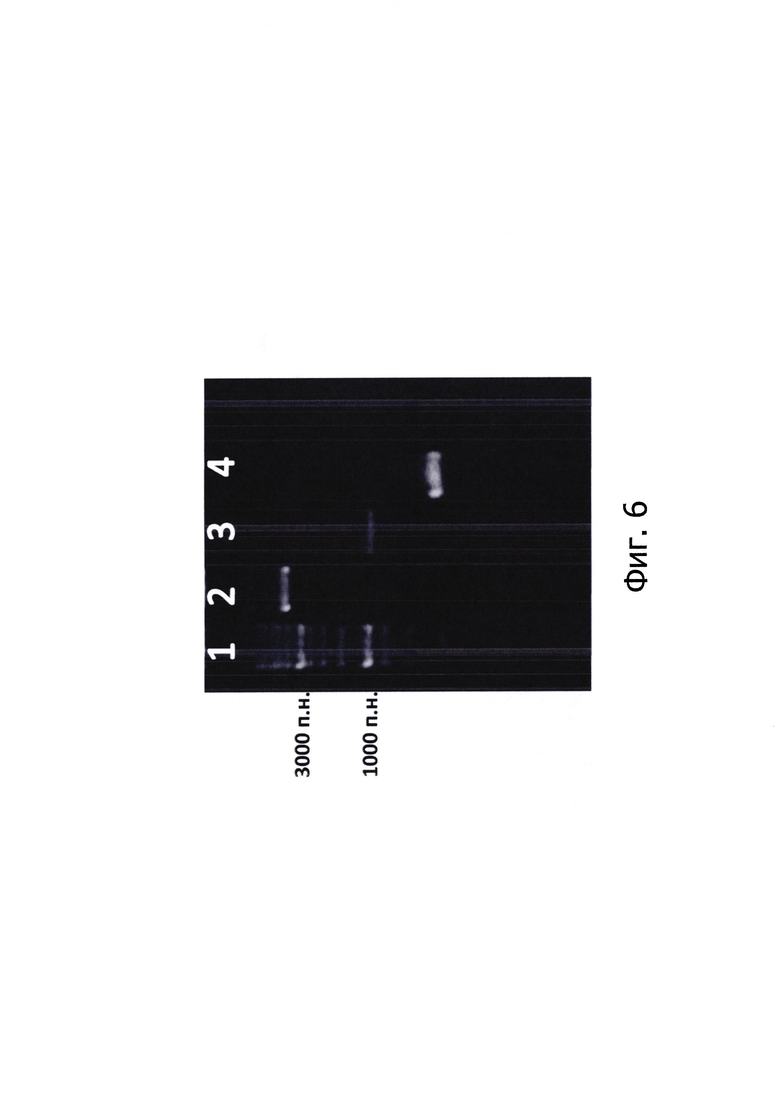
Изобретение иллюстрируется следующими графическими фигурами. На фиг. 1 приведена нуклеотидная последовательность гена рекомбинантного белка альфа-амилазы. На фиг. 2 представлена схема альфа-амилазы Bacillus amyloliquefaciens, показаны сигнальная последовательность и зрелый фермент. На фиг. 3 изображена физическая и генетическая карта рекомбинантной плазмиды pUSB2-AmQ, содержащая последовательность белка альфа-амилазы Bacillus amyloliquefaciens. На фиг. 4, 5 изображено последовательное клонирование для получения плазмиды pUSB2-AmQ. На фиг. 6 представлена фотография электрофоретического разделения препаратов ДНК 1 - маркер М12; 2 - рекомбинантная плазмида pUSB-AmyQ, 3 - ПЦР-продукт Cat (последовательность хлорамфеникол ацетилтрансферазы); 4 - ПЦР-продукт Rep (последовательность репликативного белка. На фиг. 7 представлена фотография электрофоретического разделения препарата белка альфа-амилазы Bacillus amyloliquefaciens: 1 - маркер молекулярной массы; 2 - культуральная жидкость исходной культуры Bacillus subtilis (без плазмид); 3 - культуральная жидкость Bacillus subtilis содержащая плазмиду рНТ255; 4 -культуральная жидкость Bacillus subtilis содержащая плазмиду рНТ255-AmyQ без добавления индуктора; 5 - культуральная жидкость Bacillus subtilis содержащая плазмиду pHT255-AmyQ после индукции; 6 - культуральная жидкость Bacillus subtilis содержащая плазмиду pUSB2-AmQ после 3х часов культивирования; 7 - культуральная жидкость Bacillus subtilis содержащая плазмиду pUSB2-AmQ после 20 часов культивирования.
Для лучшего понимания сущности предлагаемого изобретения ниже приведены примеры его осуществления. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам [7].
Пример 1. Синтез, клонирование и анализ фрагмента ДНК, включающего кодирующую последовательность гена альфа-амилазы.
Заявленный технический результат достигается за счет того, что заявляемая плазмида содержит следующие существенные для ее функционирования и экспрессии целевого белка структурные элементы: наличие сильного промотора, который включает оптимизированную последовательность -35, -10 и +1 и отсутствие последовательности кодирующую белок репрессор LacI. Наличие белка репрессора LacI в составе экспрессионной кассеты обеспечивает контроль синтеза мРНК, но, в случае постоянного синтеза при интеграции, он оказывает отрицательное влияние.
Оптимизированная нуклеотидная последовательность гена альфа-амилазы AmyQ была синтезирована на автоматическом синтезаторе ABI 3400 DNA/RNA Synthesizer. Синтезированный ген был клонирован в составе клонирующего вектора pGH («ДНК синтез», г. Москва). Синтезированную плазмидную ДНК pGHAmyQ использовали в качестве матрицы для амплификации последовательности гена AmyQ. С помощью праймеров AmyQ-F и AmyQ-R в состав последовательности были внесены сайты гидролиза (таблица 1). Для проведения амплификации была использована высокоточная Pfu-полимераза (ООО «СибЭнзим»).

ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 20 нг ДНК (плазмида pGHAmyQ), 10 пкМ каждого праймера (AmyQ-F, AmyQ-R), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ), 5 мкл10× буфера РпдДНК-полимеразы и 0.25 ед. Pfu ДНК-полимеразы. Реакцию осуществляли при следующих параметрах: денатурация и плавление 5 мин при 96°С, 30 циклов в которые входили денатурация и плавление 15 сек при 96°С, отжиг 20 с при 58°С и элонгация 2 мин при 72°С, продолжительность этапа элонгации программы рассчитывалась из расчета 1 минута на 1000 п.н. После завершения амплификации достройка концов ДНК осуществлялась при 72°С в течении 5 минут.
Готовый ПЦР-продукт и предварительно наработанную и очищенную плазмиду рНТ255, использованную для встройки обрабатывали эндонуклеазами рестрикции BamHI и XmaI («СибЭнзим», г. Новосибирск). Реакцию ферментативного гидролиза проводили в условиях рекомендованных производителем. В реакционной смеси на 25 мкл, содержащей 3 мкг ДНК добавляли 20 ед.а. фермента BamHI и фермента XmaI. Реакцию лигирования ДНК проводили с использованием ДНК-лигазы бактериофага Т4 («СибЭнзим», г. Новосибирск). Лигазной смесью трансформировали Е. coli штамм Stbl3.
Отбор клонов на наличие вставки проводили при помощи ПЦР с колонии. Параметры реакции подбирали согласно длине нуклеотидной вставки (таблица 1). Реакционная смесь 50 мкл содержала 10 пкМ каждого праймера (AmyQ-F, AmyQ-R), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ), 5 мкл 10×буфера Pfu ДНК-полимеразы, 0.25 ед. Pfu ДНК-полимеразы и одну трансформированную колонию с плазмидой pHT255-AmyQ. Реакцию осуществляли при следующих параметрах: денатурирация и плавление 5 мин при 96°С, 30 циклов в которые входили денатурация и плавление 15 сек при 96°С, отжиг 20 с при 58°С и элонгация 2 мин при 72°С, продолжительность подбирали из расчета 1 минута на 1000 п.н. После завершения амплификации достройка концов ДНК осуществлялась при 72°С в течение 5 минут. Разделение продуктов амплификации проводили в 1%-м агарозном геле с последующим окрашиванием бромистым этидием (0,5 мкг/мл)
Положительные колонии, вносили в 5 мл среды LB с ампициллином (50 мкг/мл) и растили в течение ночи при 37°С при 170 об/мин. Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNA minikit фирмы «Qiagen» согласно рекомендациям производителя. Структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). При проведении анализа использовался набор CEQ2000 Dye Terminator Cycle Sequencing Kit и 16-капилярный автоматический секвенатор ABI 3130x1. Для выравнивания нуклеотидных и соответствующих им аминокислотных последовательностей данных участков использовалась программа BioEdit.
Полученная рекомбинантная плазмида pHT255-AmyQ (фиг. 4) стала первым этапом работы. Она содержит сильный промотор, который включает оптимизированную последовательность -35, -10 и +1, а также операторные области необходимые для строгой регуляции синтеза мРНК встраиваемых генов.
Следующим этапом стало удаление последовательности, кодирующей белок репрессор LacI. Полученную плазмиду pHT255-AmyQ обрабатывали эндонуклеазами BstsNI и EqeI(«СибЭнзим», г. Новосибирск).
После обработки эндонуклеазами рестрикции полученную ДНК очищали при помощи разделение в 1% геле агарозы. Бенд соответствующий целевому продукту вектор рНТ255 без последовательности гена белка репрессора (общая длина 8332 п.н.) вырезали и очищали при помощи набора для выделения продуктов ПЦР из реакционных смесей (Евроген, Москва). Затем проводили реакцию лигирования. Продуктами лигазной реакции трансформировали клетки Е. coli штамм NebStable.
Структуру полученных плазмид подтверждали секвенированием, как описано выше.
Наличие белка репрессора LacI в составе экспрессионной кассеты обеспечивает контроль синтеза мРНК, но, в случае постоянного синтеза при интеграции, он оказывает отрицательное влияние. В результате была получена плазмида pBSU-AmyQ (фиг. 4).
Полученную плазмиду pBSU-AmyQ использовали в качестве матрицы для амплификации продукта несущий альфа-амилазу и «сильный» синтетический промотер. Для проведения ПЦР использовали праймеры, несущие сайты гидролиза AhlI и BseX3I (таблица 2).

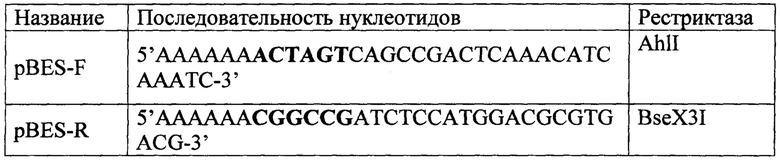
Условия ПЦР были следующими, смесь содержала 20 нгр ДНК плазмиды pBSU-AmyQ, 10 пкМ каждого праймера (AmyQ-F, AmyQ-R), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ), 5 мкл 10× буфера Pfu ДНК-полимеразы и 0.25 ед. Pfu ДНК-полимеразы. Реакцию осуществляли при следующих параметрах: денатурировали 3 мин при 95°С, амплифицировали в течении 30 циклов в которые входили денатурация 30 сек при 95°С, отжиг 30 с при 59°С и элонгация 3 мин 50 сек при 72°С, продолжительность подбиралась из расчета 1 минута на 1000 п. н. После завершения амплификации достройка концов ДНК осуществлялась при 72°С в течении 2 минут.
Готовый ПЦР-продукт и предварительно наработанную и очищенную акцепторную плазмиду pBE-S обрабатывали эндонуклеазами рестрикции AhlI и BseX3I («СибЭнзим», г. Новосибирск). Реакцию ферментативного гидролиза проводили в условиях, рекомендованных производителем. В реакционной смеси на 25 мкл 3 мкг ДНК обрабатывались 20 ед.а. фермента AhlI и фермента BseX3I. После обработки эндонуклеазами рестрикции полученную ДНК очищали при помощи разделение в 1% геле агарозы. Бенд соответствующий целевому продукту вырезали и очищали при помощи набора для выделения продуктов ПЦР из реакционных смесей (Евроген, Москва). Затем проводили реакцию лигирования. Продуктами лигазной реакции трансформировали клетки Е. coli штамм NebStable. Первичную проверку на наличие вставки проводили при помощи ПЦР с колонии. Разделение продуктов амплификации проводили в 1%-м агарозном геле с последующим окрашиванием бромистым этидием (0,5 мкг/мл).
Положительные колонии, вносили в 5 мл среды LB с ампициллином (50 мкг/мл) и растили в течение ночи при 37°С при 170 об/мин. Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNAminikit фирмы «Qiagen» согласно рекомендациям производителя. Структуру полученных плазмид подтверждали секвенированием, как было описано выше.
Для оптимизации структуры плазмидного вектора было решено изменить один из генов резистентности, вместо последовательности гена устойчивости к канамицину было решено включить ген устойчивости к хлормафениколу. Полученную плазмиду pUSB-AmyQ обрабатывали эндонуклеазой EcoRI («СибЭнзим», г. Новосибирск). Затем проводили реакцию лигирования. Продуктами лигазной реакции трансформировали клетки Е. coli штамм NebStable. Проверку на наличие вставки проводили при помощи рестрикционного анализа с эндонуклеазой VneI.
Полученная плазмиду pUSB-Am использовали в качестве матрицы для амплификации репликативного белка герВ. Для проведения ПЦР использовали праймеры, несущие сайты гидролиза CciNI и MfeI (таблица 3).

Условия ПЦР были следующими, смесь содержала 20 нгр ДНК плазмиды pUSB-Am, 10 пкМ каждого праймера (RepB-F, RepB-R), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ), 5 мкл 10× буфера Pfu ДНК-полимеразы и 0.25 ед. Pfu ДНК-полимеразы. Реакцию осуществляли при следующих параметрах: денатурировали 3 мин при 95°С, амплифицировали в течении 30 циклов в которые входили денатурация 30 сек при 95°С, отжиг 30 сек при 56°С и элонгация 50 сек при 72°С, продолжительность подбиралась из расчета 1 минута на 1000 п.н. После завершения амплификации достройка концов ДНК осуществлялась при 72°С в течении 1 мин 30 сек.
Последовательность устойчивости к хлорамфениколу (Cat) получали путем амплификации с матрицы, в качестве которой была использована плазмида pHT255-AmyQ. Для проведения ПЦР использовали праймеры, несущие сайты гидролиза AatII и CciNI (таблица 4).

Условия ПЦР были следующими, смесь содержала 20 нгр ДНК плазмиды pBSU-AmyQ, 10 пкМ каждого праймера (Cat-A-F, Cat-C-R), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ), 5 мкл 10× буфера Pfu ДНК-полимеразы и 0.25 ед. Pfu ДНК-полимеразы. Реакцию осуществляли при следующих параметрах: денатурировали 4 мин при 95°С, амплифицировали в течении 30 циклов в которые входили денатурация 30 сек при 95°С, отжиг 30 сек при 56°С и элонгация 2 мин 10 сек при 72°С, продолжительность подбиралась из расчета 1 минута на 1000 п.н. После завершения амплификации достройка концов ДНК осуществлялась при 72°С в течении 2 мин 30 сек.
Готовые ПЦР-продукты и предварительно наработанную и очищенную плазмиду pUSB-Am обрабатывали эндонуклеазами рестрикции AatII, CciNI и MfeI. («СибЭнзим», г. Новосибирск). Реакцию ферментативного гидролиза проводили в условиях, рекомендованных производителем. В реакционной смеси на 25 мкл 3 мкг ДНК обрабатывались 20 ед.а. фермента AatII, фермента CciNI и фермента MfeI.
После обработки эндонуклеазами рестрикции полученную ДНК очищали при помощи разделение в 1% геле агарозы. Бенд соответствующий целевому продукту вырезали и очищали при помощи набора для выделения продуктов ПЦР из реакционных смесей (Евроген, Москва). Затем проводили реакцию лигирования. Продуктами лигазной реакции трансформировали клетки Е. coli штамм NebStable. Первичную проверку на наличие вставки проводили при помощи ПЦР с колонии. Разделение продуктов амплификации проводили в 1%-м агарозном геле с последующим окрашиванием бромистым этидием (0,5 мкг/мл).
Положительные колонии, вносили в 5 мл среды LB с ампициллином (50 мкг/мл) и растили в течение ночи при 37°С при 170 об/мин. Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNAminikit фирмы «Qiagen» согласно рекомендациям производителя. Структуру полученных плазмид подтверждали секвенированием.
Последним этапом стала встройка гена альфа-амилазы в полученный оптимизированный вектор pUSB-Am2. Плазмида pUSB-Am2 и плазмида pUSB-AmyQ обрабатывались эндонуклеазами рестрикции EcoRV и Xba.
После обработки эндонуклеазами рестрикции полученную ДНК очищали при помощи разделение в 1% геле агарозы. Бенд соответствующий целевому продукту вырезали и очищали при помощи набора для выделения продуктов ПЦР из реакционных смесей (Евроген, Москва). Затем проводили реакцию лигирования. Продуктами лигазной реакции трансформировали клетки Е. coli штамм NebStable. Проверку на наличие вставки проводили при помощи рестрикционного анализа с эндонуклеазой BamHI.
Положительные колонии, вносили в 5 мл среды LB с ампициллином (50 мкг/мл) и растили в течение ночи при 37°С при 170 об/мин. Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNAminikit фирмы «Qiagen» согласно рекомендациям производителя. Структуру полученных плазмид подтверждали секвенированием.
В завершении была получена оригинальная плазмида pUSB2-AmQ (фиг. 3) содержащая сильный промотор, что обеспечивает эффективную наработку рекомбинантных белков и экспорт в культуральную среду.
Секвенирование плазмидной ДНК положительных клонов в районе встройки гена альфа-амилазы позволило отобрать клоны с отсутствием дефектов встраиваемых генов (вставки, делеции, замены), после чего из отобранных клонов была наработана и выделена плазмидная ДНК (фиг.).
Пример 2. Получение рекомбинантного штамма Bacillus subtilis.
Полученной рекомбинантной плазмидой pUSB2-AmQ была проведена трансформация методом электропарации клеток B.subtilis WB800N. Культуру клеток в количестве 50 мл подращивали до ODλ.600=0.2. о.е. Затем в культуру добавили 0,5 гр. трионина, 1 гр. глицил-глицина, 0,05 гр. триптофана и 15 мкл TWEEN 80 и инкубировали в течении часа при 30°С. После культуру клеток инкубировали 20 мин на льду. 50 мл культуры центрифугировали 10 мин 5000g при 4°С на центрифуге AvantiJ-30I. Осажденную биомассу Дважды промыли в 5 мл ЕР Buffer (0,5 М трегалоза, 0,5 М сорбитол, 0,5 М маннитол, 0,7 мМ MgCl, 0,5 мМ K2HPO4, 0,5 мМ KH2PO4, рН=7,4). После промывки ресуспендировали в 0,5 мл EPBuffer. 100 мкл суспензии клеток перенесли в 2 мл холодные кюветы для электропорации и добавили ДНК с расчетом 25 нг/мкл. Произвели электропорацию при 2,5 kV на электропораторе MicroPulser. К про электропорированным клеткам добавили питательную среду YTx2 с 0,5 М сорбитолом и 0,38 М маннитолом. Инкубировали 3 часа при 37°С. 50 и 200 мкл культуры высевали на чашки с агаризованной средой YTx2 содержащие 5 мкг/мл хлорамфеникола. Чашки помещали в термостат 30°С и инкубировали в течении ночи. На следующий день были идентифицированы отдельные колонии, содержащие плазмиду pUSB2-AmQ.
Пример 3. Наработка биомассы pUSB2-AmQ и выделение рекомбинантной альфа-амилазы
Полученные клоны B.subtilis штамма WB800N после электропорации рекомбинантной плазмидой pUSB2-AmQ наращивали в 5 мл среды YT×2 содержащей 5 мкг/мл хлорамфеникола в течение ночи. Полученную ночную культуру добавляли по 1,5 мл в колбы объемом 750 мл содержащих 150 мл среды YT×2 и растили до плотности ODλ600=0,6. После этого продолжали культивирование в течение 16 часов при температуре 30°С, 170 об/мин. Биомассу бактериальных клеток отделяли от культуральной среды центрифугированием (7000 об/мин), в течение 10 мин, 4°С.
Культуральную среду анализировали в 12% ПААГ на наличие целевого белка. Было показано, что целевой белок секретируется в культуральную среду (фиг. 7).
Пример 4 Определение активности рекомбинантной альфа-амилазы
Определение активности проводилось с помощью набора «Альфа-АМИЛАЗА-1-ОЛЬВЕКС» (ООО «ОЛЬВЕКС ДИАГНОСТИКУМ», Россия) предназначенного для количественного определения активности альфа-амилазы методом Каравея. Проведение анализа включает в себя приготовление образцов опытной и контрольной пробы. Для каждой пробы подготавливалось 0,5 мл рабочего раствора субстрата (РРС) включающий в себя 24 части буферного раствора (200 мМ фосфатный буфер с рН=6,90; 77 мМ NaCl; азид натрия 0,2 г/л) и 1 части концентрированного субстрата (крахмал картофельный 10 г/л; 100 мМ NaOH). РРС прогревали при температуре 37°С в течение 5 минут. К прогретому РРС для опытной пробы добавили 10 мкл исследуемого образца. РРС для опытной и контрольной пробы снова прогревали при 37°С в течение 5 минут. К прогретым пробам добавили 4 мл рабочего раствора соляной кислоты, приготовленный в соотношении 1:15 из дистиллированной воды и 1,6 М соляной кислоты. К контрольной пробе добавили 10 мкл исследуемого образца. Далее контрольную и опытную пробу смешивали с рабочим раствором йода, приготовленный в соотношении 7:1:2 из дистиллированной воды, концентрированного раствора йода (127 мМ KI, 35,06 мМ йод кристаллический) и 3,87 мМ раствора KF. Пробы тщательно перемешивали. У готовых проб измеряли оптическую плотность против дистиллированной воды при длине волны 630 нм в кювете с длиной оптического пути 1,0 см. Измерения проводились на спектрофотометре SmartSpecPlusBIORAD. Расчет активности (А, мг/с*л)) проводили по формуле:
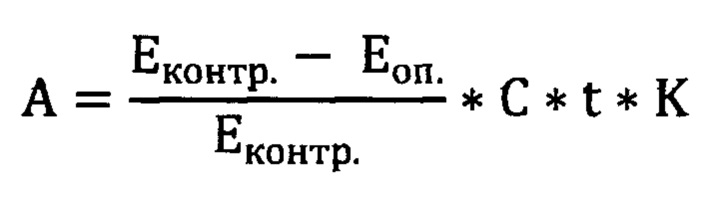
где:
Еоп. - оптическая плотность опытной пробы,
Еконтр. - оптическая плотность контрольной пробы,
С - коэффициент пересчета на 1 мг крахмала
t - коэффициент пересчета на 1 сек инкубации
К - коэффициент пересчета на 1 л биологической жидкости.
Аналитические характеристики:
Линейность - от 3,0 до 55,0 мг/с*л.
Чувствительность - не более 2,0 мг/(с*л).
Коэффициент вариации результатов - не более 10%.

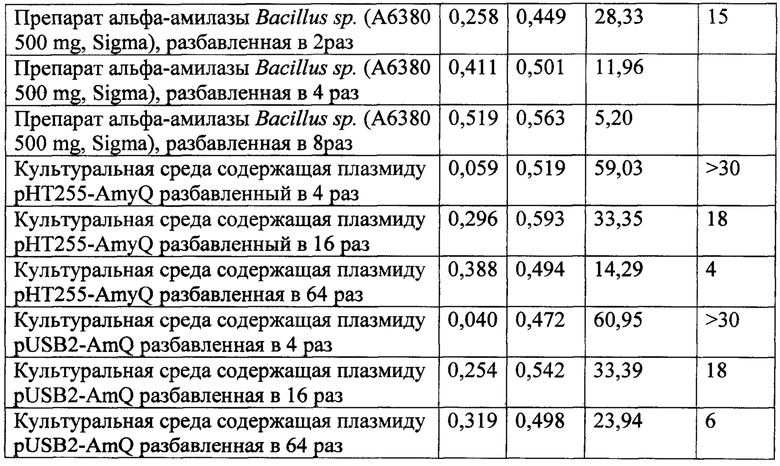
В культуральной среде Bacillus subtilis, не содержащей плазмиды, альфа-амилазной активности не обнаружено. Культуральная среда, содержащая плазмиду pHT255-AmyQ и культуральная среда содержащая плазмиду pUSB2-AmQ показали активность альфа-амилазы 18 ед/мл, в качестве сравнения использовали препарат альфа-амилазы Bacillus sp. (А6380 500 mg, Sigma).
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Никитина Е.В., Решетник О.А., Губайдуллин Р.А. Биотехнологические аспекты применения амилолитических ферментов в пищевой промышленности // Вестник Казанского технологического университета. 2013. Т. 16. №. 13.
2. Высоцкая Н.И., Голета М.В., Каравская Л.И. Применение амилолитических ферментов в процессе спиртового брожения // Научное сообщество студентов XXI столетия. Естественные науки: сб. ст. по мат. XL VIII междунар. студ. науч.-практ. конф. №1(47). URL: https://sibac.info/archive/nature/l(47).pdf (дата обращения: 14.02.2019)
3. Borchert Т.V. et al. Oxidation stable amylases for detergents //Progress in Biotechnology. Elsevier, 1995. Vol. 10. P. 175-179.
4. Заявка РФ 2018/2661790, C2, C12N, 9/50, 15/75, 15/63, 15/62, 15/70, опубл. 19.07.2018 г.
5. Заявка РФ 1999/2124559, CI, C12N, 15/76, 15/67, 15/66, С12Р, 21/02, опубл. 10.01.1999 г.
6. Заявка WO 1991/02792, A1, C12N, 9/54, 15/57, 15/75, C11D, 3/386, опубл. 07.03.1991 г.
7. Маниатис Т., Фрич Э, Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д. Гловера, Пер. с англ. Москва. Мир. 1988. 479 с.
8. Saiki R.K., Gelfand D.H., Stoffel S., Sharf S.J., Higuehi R., Horn G.T., Mullis K.B., Eriich H.A. Science, 1988. V. 239. №4839. P. 487-491
9. Sanger F., Nicklen S., Coulson A.R. Proc. Nat. Acad. Sci. USA. 1977. V. 74. P. 5463-5467.
Приложение
<110>АлтГУ
<120>Рекомбинантная плазмида pUSB2-AmQ, обеспечивающая синтез белка альфа-амилазы Bacillus amyloliquefaciens, и штамм Bacillus subtilis/pUSB2-AmQ – продуцент белка альфа-амилазы Bacillus amyloliquefaciens
<160> SEQ ID NO 2
<210> SEQ ID NO:1
<211> 1545
<212> DNA
<213> Artificial Sequence
<220>
<223>Нуклеотидная последовательность гена AmQ рекомбинантного белка альфа-амилазы Bacillus amyloliquefaciens.
<400> 1
ATGATCCAAAAACGCAAACGCACAGTTAGCTTTAGACTGGTTCTGATGTGCACACTGCTGTTTGTTTCACTGCCGATTACAAAAACATCAGCAGTTAATGGCACACTGATGCAGTATTTTGAATGGTATACACCGAATGATGGCCAACATTGGAAAAGACTGCAAAATGATGCAGAACATCTGAGCGATATTGGCATTACAGCAGTTTGGATTCCGCCTGCATATAAAGGCCTGTCACAATCAGATAATGGCTATGGACCGTATGATCTTTATGATCTGGGCGAATTTCAACAGAAAGGCACAGTCAGAACAAAATATGGCACAAAATCAGAACTGCAAGATGCAATTGGCTCACTGCATTCAAGAAATGTTCAAGTTTATGGCGACGTCGTCCTGAATCATAAAGCAGGCGCAGATGCAACAGAAGATGTTACAGCGGTTGAAGTTAATCCGGCAAATCGCAATCAAGAAACGAGCGAAGAATATCAGATTAAAGCGTGGACAGATTTTCGCTTTCCTGGCAGAGGCAATACATATTCAGACTTTAAATGGCATTGGTATCACTTTGATGGCGCAGATTGGGATGAAAGCAGAAAAATTTCACGCATCTTCAAATTTCGCGGAGAAGGCAAAGCATGGGATTGGGAAGTTTCATCAGAAAACGGCAACTATGACTATCTGATGTATGCGGATGTCGATTATGATCATCCTGATGTTGTTGCGGAAACAAAgAAATGGGGCATTTGGTATGCAAATGAACTGTCACTGGATGGCTTTAGAATTGATGCAGCGAAACACATCAAATTCTCATTTCTGAGAGATTGGGTTCAAGCAGTTAGACAAGCAACAGGCAAAGAGATGTTTACAGTTGCAGAATATTGGCAGAATAACGCAGGCAAACTGGAAAACTATCTGAACAAAACAAGCTTCAACCAGAGCGTTTTTGATGTTCCGCTGCATTTTAATCTGCAAGCAGCATCATCACAAGGCGGAGGCTATGATATGCGCAGACTGCTGGATGGCACAGTTGTTTCAAGACATCCGGAAAAAGCAGTCACATTTGTCGAAAACCATGATACACAACCGGGACAATCACTGGAATCAACAGTTCAAACATGGTTCAAACCGCTGGCATATGCATTTATTCTGACAAGAGAATCAGGCTATCCGCAAGTTTTcTACGGCGATATGTATGGAACAAAAGGCACGAGCCCGAAAGAAATTCCGTCACTGAAAGATAACATTGAGCCGATTCTTAAGGCACGCAAAGAATATGCGTATGGACCGCAACATGATTACATTGATCATCCGGACGTTATTGGCTGGACACGCGAAGGCGATTCATCAGCAGCAAAATCAGGCCTGGCAGCACTGATTACAGATGGCCCTGGCGGATCAAAAAGAATGTATGCAGGCCTGAAAAATGCTGGCGAAACATGGTATGATATTACAGGCAATAGAAGCGACACAGTCAAAATTGGATCAGATGGCTGGGGAGAATTTCATGTTAATGATGGCTCAGTCAGCATCTACGTCCAGAAATAA (1545 нуклеотида)
<210> SEQ ID NO:2
<211>514
<212> DNA
<213> Artificial Sequence
<220>
<223>Аминокислотная последовательность AmQ рекомбинантного белка альфа-амилазы Bacillus amyloliquefaciens.
<400> 2
MIQKRKRTVSFRLVLMCTLLFVSLPITKTSAVNGTLMQYFEWYTPNDGQHWKRLQNDAEHLSDIGITAVWIPPAYKGLSQSDNGYGPYDLYDLGEFQQKGTVRTKYGTKSELQDAIGSLHSRNVQVYGDVVLNHKAGADATEDVTAVEVNPANRNQETSEEYQIKAWTDFRFPGRGNTYSDFKWHWYHFDGADWDESRKISRIFKFRGEGKAWDWEVSSENGNYDYLMYADVDYDHPDVVAETKKWGIWYANELSLDGFRIDAAKHIKFSFLRDWVQAVRQATGKEMFTVAEYWQNNAGKLENYLNKTSFNQSVFDVPLHFNLQAASSQGGGYDMRRLLDGTVVSRHPEKAVTFVENHDTQPGQSLESTVQTWFKPLAYAFILTRESGYPQVFYGDMYGTKGTSPKEIPSLKDNIEPILKARKEYAYGPQHDYIDHPDVIGWTREGDSSAAKSGLAALITDGPGGSKRMYAGLKNAGETWYDITGNRSDTVKIGSDGWGEFHVNDGSVSIYVQK
| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантная плазмида для экспрессии альфа-амилазы из Bacillus sp. H3 и штамм дрожжей Saccharomyces cerevisiae - продуцент альфа-амилазы из Bacillus sp. H3, обладающий регулируемой мутабельностью | 2023 |
|
RU2833838C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBANP 464, КОДИРУЮЩАЯ МЕТАЛЛОПРОТЕИНАЗУ BACILLUS AMYLOLIQUEFACIENS A-50 И ШТАММ БАКТЕРИЙ BACILLUS AMYLOLIQUEFACIENS - ПРОДУЦЕНТ МЕТАЛЛОПРОТЕИНАЗЫ. | 1989 |
|
RU1679800C |
| ВАРИАНТЫ АЛЬФА-АМИЛАЗЫ | 2000 |
|
RU2231547C2 |
| ШТАММ Bacillus amyloliquefaciens - ПРОДУЦЕНТ АЛЬФА-АМИЛАЗЫ Bacillus amyloliquefaciens | 2010 |
|
RU2455352C1 |
| Фрагмент ДНК ALV 7, кодирующий синтез альфа-амилазы, способ его конструирования и штамм бактерий BacILLUS SUвтILIS - продуцент альфа-амилазы | 1990 |
|
SU1717633A1 |
| Штамм бактерий BacILLUS амYLоLIQUеFасIеNS-продуцент @ -амилазы BacILLUS LIснеNIFоRмIS | 1991 |
|
SU1788966A3 |
| Лабораторный способ биосинтеза бациллярной рибонуклеазы (варианты) | 2024 |
|
RU2835675C1 |
| УЛУЧШЕННАЯ ПРОДУКЦИЯ БЕЛКА В BACILLUS | 2008 |
|
RU2515112C2 |
| Рекомбинантная плазмида pBU-Sav, обеспечивающая синтез и секрецию белка щелочной протеазы в системе Bacillus subtilis | 2023 |
|
RU2816513C1 |
| ВАРИАНТЫ АЛЬФА-АМИЛАЗЫ С ИЗМЕНЕННЫМИ СВОЙСТВАМИ | 2008 |
|
RU2499044C2 |


Изобретение относится к биотехнологии. Заявлена плазмида pUSB2-AmQ. Размер плазмиды pUSB2-AmQ 7495 пар оснований. Плазмида pUSB2-AmQ содержит ряд структурных элементов. Синтетический промотор обеспечивает эффективную транскрипцию контролируемой мРНК. Рекомбинантный ген α-амилазы В. amyloliquefaciens кодирует целевой рекомбинантный белок. Последовательность bla кодирует белок бета-лактамазу. Последовательность bla является селективным маркером для трансформации штаммов Е. coli. Участок repB обеспечивает инициацию репликации в штаммах В. subtilis. Хлорамфениколацетилаза является селективным маркером для трансформации штаммов В. subtilis. Рекомбинантный штамм Bacillus subtilis WB800N/pUSB2-AmQ получен трансформацией исходного штамма плазмидой pUSB2-AmQ. Штамм обеспечивает продукцию α-амилазы. При культивировании штамм Bacillus subtilis WB800N/pUSB2-AmQ обеспечивает синтез рекомбинантной альфа-амилазы 90% от общего секретируемого белка Bacillus subtilis. 2 н.п. ф-лы, 7 ил., 5 табл., 4 пр.
1. Рекомбинантная плазмида pUSB2-AmQ размером 7495 пар оснований (п.о.), обеспечивающая синтез рекомбинантного белка α-амилазы Bacillus amyloliquefaciens размером 55 кДа в клетках Bacillus subtilis, характеризующаяся физической и генетической картой, представленной на фиг. 3, содержащая: синтетический промотор, обеспечивающий эффективную транскрипцию контролируемой мРНК; рекомбинантный ген α-амилазы Bacillus amyloliquefaciens с нуклеотидной последовательностью SEQ ID NO: 1, встроенный по сайтам рестрикции Ahll и BseX3I в область полилинкера вектора pUSB и кодирующий целевой рекомбинантный белок; последовательность bla, кодирующую белок бета-лактамазу, являющуюся селективным маркером для трансформации штаммов Е. coli; участок repB, обеспечивающий инициацию репликации в штаммах Bacillus subtilis; белок устойчивости к хлорамфениколу Cat (chloramphenicol acetylase).
2. Рекомбинантный штамм Bacillus subtilis WB800N/pUSB2-AmQ - продуцент рекомбинантного белка α-амилазы Bacillus amyloliquefaciens, полученный трансформацией рекомбинантной плазмидой pUSB2-AmQ по п. 1.
| ЭКСПРЕССИОННЫЕ ВЕКТОРЫ ДЛЯ УЛУЧШЕННОЙ СЕКРЕЦИИ БЕЛКА | 2012 |
|
RU2661790C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СПОСОБ ЭКСПРЕССИИ В КЛЕТКАХ STREPTOMYCES ГЕНА α -АМИЛАЗЫ | 1990 |
|
RU2124559C1 |
| SAIKI R.K | |||
| et al | |||
| Primer-Directed Enzymatic Amplification of DNA with a Thermostable DNA Polymerase | |||
| Science, 1988, 239, 487-491 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |

