ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к способам и композициям, которые являются полезными для лечения рака. В частности, настоящее изобретение относится к способам идентификации и направленного действия на неоантигены, присутствующие в опухоли.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Известно, что гетерогенность внутри опухоли (ITH) и мутационный ландшафт опухоли могут влиять на способность иммунной системы реагировать на рак.
Например, генетическая нестабильность играет важную роль в способности опухолевых клеток вырабатывать ускользающие мутанты, которые уклоняются от иммунной элиминации. Эта фундаментальная характеристика опухолевых клеток является основной причиной того, что многие многообещающие способы иммунотерапии, предназначенные для выработки мощного Т-клеточного иммунитета против специфических опухолевых антигенов, в конечном итоге терпят неудачу, и это создает существенную проблему для успешных стратегий разработки вакцин против рака. Таким образом, иммунотерапия, предназначенная для выработки антигенспецифического Т-клеточного иммунитета против опухолей, парадоксальна в том смысле, что специфический для опухоли иммунитет, который эффективно элиминирует опухоль, в то же время оказывает селективное давление, которое способствует возникновению ускользающих опухолевых мутантов, которые устойчивы к элиминации Т-клетками. Многочисленные сообщения указывают на то, что опухоли избегают иммунной элиминации в результате селективного роста опухолевых клеток, экспрессирующих случайные мутации, которые вызывают либо инициацию, либо сайленсинг генов посредством точечных мутаций, мутаций со сдвигом рамки считывания, геномных транслокаций, вставок или делеций.
Гетерогенность опухоли характеризуется тем фактом, что различные опухолевые клетки могут проявлять различные морфологические и фенотипические профили, включая различия в морфологии клеток, экспрессии генов, генетических и эпигенетических мутациях, метаболизме, подвижности, пролиферации и метастатическом потенциале. Это явление возникает как между опухолями (межопухолевая гетерогенность), так и внутри опухолей (внутриопухолевая гетерогенность или ITH). Гетерогенность раковых клеток создает серьезные проблемы при разработке эффективных стратегий лечения.
В качестве примера, гетерогенные опухоли могут проявлять различную чувствительность к цитотоксическим или направленным на опухоль лекарственным средствам в различных клональных популяциях. Это объясняется взаимодействиями клонов, которые могут ингибировать или изменять терапевтическую эффективность, что создает проблему для успешной терапии гетерогенных опухолей (и их гетерогенных метастазов).
Введение лекарственных средств в гетерогенные опухоли в редких случаях будет уничтожать все опухолевые клетки. Первоначальная гетерогенная опухолевая популяция может быть узким местом, так что выживут немногие клетки, устойчивые к лекарственному средству. Это позволяет резистентным опухолевым популяциям реплицироваться и повторно вызывать рост опухоли с помощью механизма эволюции ветвления. Возникающая в результате опухоль с вновь образованными популяциями также может быть гетерогенной и будет устойчива к исходной используемой лекарственной терапии. Возникающая в результате опухоль с вновь образованными популяциями может также возвратиться в более агрессивной форме.
Введение цитотоксических лекарственных средств часто приводит к первоначальному уменьшению опухоли. Это означает разрушение исходных неустойчивых субклональных популяций в гетерогенной опухоли с сохранением только устойчивых клонов. Эти устойчивые клоны теперь имеют селективное преимущество на фоне химиотерапии и могут реплицироваться с повторным заселением опухоли. Репликация, вероятно, возникает с помощью механизма эволюции ветвления, способствуя гетерогенности опухоли. Репопулированная опухоль может оказаться более агрессивной. Это объясняется избирательным преимуществом опухолевых клеток, устойчивых к лекарственным средствам, и дополнительными генетическими изменениями, которые происходят во время терапии и течения заболевания.
В патенте WO 2014/168874 описан способ создания персонализированной неопластической вакцины для индивидуума, у которого диагностировано наличие неоплазии, который включает идентификацию множества мутаций в неоплазии, анализ множества мутаций для идентификации подмножества, по меньшей мере, пяти неоантигенных мутаций, как предсказывается, кодирующих пептиды неоантигенов, и получение на основе идентифицированного подмножества персонализированной неопластической вакцины.
Таким образом, существует потребность в альтернативных методах лечения рака, в частности гетерогенных опухолей.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения определили, что стволовые мутации, то есть мутации, присутствующие, по существу, во всех опухолевых клетках в гетерогенной опухоли, могут быть идентифицированы посредством взятия образцов из множества областей опухоли или с помощью подходов к идентификации клональных мутаций в одиночных биопсиях. Например, фракция раковых клеток, описывающая фракцию раковых клеток, несущих мутацию, может быть определена для того, чтобы отличить неоантигены, которые могут присутствовать в каждой раковой клетке в опухоли (стволовые неоантигены) от неоантигенов, присутствующих только в субпопуляции опухолевых клеток (неоантигенов ветвления). Используемый в настоящем документе термин «стволовые мутации» является синонимом термина «клональные мутации». Они оба предназначены для определения мутаций, присутствующих практически во всех опухолевых клетках в гетерогенной опухоли. Используемый в настоящем документе термин «мутации ветвления» является синонимом термина «субклональные мутации». Они оба предназначены для определения мутаций, присутствующих в субпопуляции опухолевых клеток. Введение терапевтических Т-клеток, которые направленно действуют на стволовые неоантигены, а не на неоантигены ветвления, или введение вакцин, как описано в настоящем документе, позволяет выработать эффективный иммунный ответ против всей опухоли и, таким образом, снижает риск сохранения резистентных клеток, репопулирующих опухоль.
Таким образом, в первом аспекте настоящее изобретение относится к способу идентификации стволового неоантигена в опухоли от индивидуума, который включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию.
Во втором аспекте настоящее изобретение относится к способу идентификации стволового неоантигена в опухоли от индивидуума, который включает стадии:
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию.
Стволовая мутация может представлять собой мутацию с вариантом одного нуклеотида или со вставкой/делецией, или мутацию сайта сплайсинга, приводящую к изменению аминокислотной последовательности (кодирующую мутацию).
Мутации могут быть идентифицированы секвенированием Exome, RNA-seq, секвенированием целого генома и/или секвенированием панелей генов-мишеней. Подходящие способы известны в данной области техники. Описания секвенирования Exome и RNA-seq представлены в статьях Boa et al. (Cancer Informatics. 2014;13 (Suppl 2):67-82) и Ares et al. (Cold Sping Harb Protoc. Nov 3; 2014(11):1139-48); соответственно. Описания панелей секвенирования генов-мишеней можно найти, например, в статьях Kammermeier et al. (J Med Genet. 2014 Nov; 51(11):748-55) и Yap KL et al. (Clin Cancer Res. 2014, 20:6605). Смотри также Meyerson et al., Nat. Rev. Genetics, 2010 и Mardis, Annu Rev Anal Chem, 2013. Панели секвенирования генов-мишеней также являются коммерчески доступными (например, как описано Biocompare (http://www.biocompare.com/Editorial-Articles/161 194-Build-Your-Own-Gene-Panels-with-These-Custom-NGS-Targrting-Tools /).
Способ может включать стадию оценки профиля аллелей HLA индивидуума для определения будет ли пептид стволового неоантигена связываться с молекулой MHC индивидуума. Подходящие способы известны в данной области техники, например, в OptiType, Szolek et al., 2014.
В третьем аспекте настоящее изобретение относится к способу получения популяции Т-клеток, которые направлены на стволовой неоантиген в опухоли от индивидуума, включающему стадии:
i) идентификации Т-клетки из образца, выделенного от индивидуума, которая способна специфически узнавать пептид стволового неоантигена; и
ii) наращивания Т-клеток для получения популяции Т-клеток, которая направленно действует на стволовой неоантиген.
Специалист в данной области техники должен понимать, что в настоящем документе ссылки на Т-клетку, «узнающую» стволовой неоантиген или пептид стволового неоантигена, включают узнавание в форме пептид стволового неоантигена:комплекс MHC.
Изобретение также относится к способу идентификации Т-клетки, специфичной для стволового неоантигена, который включает следующие стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, который представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
iv) идентификации в образце от указанного индивидуума Т-клетки, способной специфически узнавать стволовой неоантиген как стволовой неоантиген, специфичный для Т-клетки.
Образец, в котором идентифицирована Т-клетка, может представлять собой образец крови, образец опухоли, образец лимфоузла, ассоциированного с опухолью, или образец участка метастазирования.
Изобретение также относится к способу получения популяции Т-клеток, которые направлены на стволовой неоантиген в опухоли, который включает стадии:
(а) идентификации стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
b) идентификации Т-клетки из образца, выделенного от индивидуума, которая способна специфически узнавать указанный стволовой неоантиген; и
c) наращивания Т-клеток для получения популяции Т-клеток, которая направлена на стволовой неоантиген.
В изобретении также предлагается популяция Т-клеток, которая получена или может быть получена способом, который включает стадии:
(а) идентификации стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) определения стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
b) идентификации Т-клетки из образца, выделенного от индивидуума, которая способна специфически узнавать указанный стволовой неоантиген; и
c) наращивания Т-клеток для получения популяции Т-клеток, которая направлена на стволовой неоантиген.
Таким образом, полученная популяция Т-клеток обогащена увеличенным количеством Т-клеток, которые направлены на стволовые неоантигены (например, по сравнению с образцом, выделенным от индивидуума).
Образец может представлять собой опухоль, кровь, ткань или мононуклеарные клетки периферической крови от индивидуума.
Стволовой неоантиген может быть образован стволовой мутацией, идентифицированной в соответствии со способом в соответствии с первым или вторым аспектом изобретения.
Популяция Т-клеток может включать CD8+ Т-клетки, CD4+ Т-клетки или CD8+ и CD4+ Т-клетки.
Способ может включать предоставление, по меньшей мере, первой и второй Т-клеток, где первая Т-клетка направлена на первый стволовой неоантиген, возникающий в результате первой стволовой мутации, а вторая Т-клетка направлена на второй стволовой неоантиген, возникающий в результате второй стволовой мутации.
В четвертом аспекте настоящее изобретение относится к композиции Т-клеток, которая включает Т-клетку, специфичную для стволового неоантигена, или популяцию Т-клеток, как описано в настоящем документе.
Стволовой неоантиген может быть идентифицирован способом в соответствии с первым или вторым аспектом настоящего изобретения.
Т-клетка, специфичная для стволового неоантигена, может представлять собой Т-клетку, как определено в третьем аспекте настоящего изобретения.
Т-клетка, специфичная для стволового неоантигена, может экспрессировать рецептор химерного антигена (CAR) или рецептор Т-клеток (TCR) или рецептор Т-клеток с повышенной аффинностью (TCR), который специфически связывает стволовой неоантиген или пептид стволового неоантигена (т. е. пептид, происходящий от стволового неоантигена), как далее обсуждается в настоящем документе ниже.
Способы создания TCRs и TCRs с повышенной аффинностью известны в данной области техники. TCRs с повышенной аффинностью представляют собой TCRs с повышенной аффинностью к комплексу пептид-MHC. Способы включают, например, выделение генов TCRs, которые кодируют TCRs из образцов пациента (например, из периферической крови пациента или TILs), и улучшения аффинности TCR к комплексу пептид-MHC посредством модификации последовательностей TCRs (например, путем мутагенеза in vitro и выбора TCRs с повышенной аффинностью (или со зрелой аффинностью)). Способы введения таких генов TCR в Т-клетки известны в данной области техники. Способы идентификации TCRs с оптимальной аффинностью, включающие иммунизацию антигеннегативных гуманизированных трансгенных мышей, которые имеют разнообразный репертуар TCRs человека (например, TCR/MHC гуманизированных мышей, таких как мыши ABabDII), антигеном, и выделение антигенспецифических TCRs от таких иммунизированных трансгенных мышей, также известны в данной области техники (смотри Obenaus M et al., Nat Biotechnol. 33(4):402-7, 2015).
В пятом аспекте настоящее изобретение относится к мультимеру MHC, включающему пептид стволового неоантигена, где стволовой неоантиген идентифицируют способом в соответствии с первым или вторым аспектом настоящего изобретения. Мультимеры MHC и способы их использования для выделения Т-клеток известны в данной области техники, например, как описано в статье Hadrup, Nature Methods 6:520-526 2009; и Andersen, Nature Protocol 7:891-902, 2012.
Мультимеры MHC, как описано в настоящем документе, могут быть использованы в способах по изобретению, например, в методах идентификации T-клеток NES. Мультимеры MHC можно использовать для идентификации, наращивания или обогащения Т-клеток NES с помощью способов, описанных в настоящем документе, например, способов получения T-клеток или популяции, или композиции T-клеток, как описано в настоящем документе.
В шестом аспекте настоящее изобретение относится к вакцине, включающей пептид стволового неоантигена из стволового неоантигена, идентифицированного с помощью способа в соответствии с первым или вторым аспектом настоящего изобретения. Как обсуждалось в настоящем документе, вакцина стволового неоантигена согласно изобретению может быть доставлена в виде вакцины дендритных клеток, пульсированных или нагруженных стволовым неоантигеном или генетически модифицированных (посредством переноса ДНК или РНК), для экспрессии одного, двух или более стволовых неоантигенов.
В седьмом аспекте настоящее изобретение относится к композиции Т-клеток в соответствии с четвертым аспектом настоящего изобретения для использования при лечении рака.
В восьмом аспекте настоящее изобретение относится к Т-клетке, определенной в третьем или четвертом аспекте настоящего изобретения, для использования при получении лекарственного средства для лечения рака.
В девятом аспекте настоящее изобретение относится к способу лечения рака у индивидуума, который включает введение индивидууму композиции Т-клеток в соответствии с четвертым аспектом настоящего изобретения.
Способ может включать следующие стадии:
(i) выделения Т-клетки, содержащей образец от индивидуума;
(ii) идентификации и наращивания популяции Т-клеток, которые направлены на стволовой неоантиген; и
(iii) введения клеток из (ii) индивидууму.
Способ может включать следующие стадии:
(i) выделения образца, содержащего Т-клетки;
(ii) конструирования Т-клетки для экспрессии CAR или TCR, который узнает указанный стволовой неоантиген, как описано в настоящем документе, для получения популяции T-клеток, которые направлены на стволовой неоантиген; и
(iii) введения клеток из (ii) индивидууму.
В одном аспекте способ также охватывает стадию идентификации стволового неоантигена, как описано в настоящем документе, то есть настоящее изобретение относится к способу лечения рака у индивидуума, где указанный способ включает:
(а) идентификацию стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, который представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
b) идентификацию Т-клетки из образца, выделенного от индивидуума, которая способна специфически узнавать указанный стволовой неоантиген;
c) наращивание Т-клеток для обеспечения популяции Т-клеток, которая направлена на стволовой неоантиген; и
d) введение указанной популяции T-клеток указанному индивидууму.
Способ может включать стадии:
(а) идентификации стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, который представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
(b) обеспечения образцом, содержащим Т-клетки;
(c) конструирования Т-клетки для экспрессии CAR или TCR, который узнает указанный стволовой неоантиген, для обеспечения популяции T-клеток, которые направлены на стволовой неоантиген; и
(d) введения указанной популяции Т-клеток индивидууму.
В одном аспекте Т-клетка сконструирована для экспрессии CAR или TCR с повышенной аффинностью, как описано в настоящем документе.
Изобретение также относится к способу лечения пациента, который болен раком, включающему:
(i) выявление пациента, который болен раком; и
(ii) введение указанному пациенту Т-клетки или популяции Т-клеток, как определено в настоящем документе.
Стволовой неоантиген, Т-клетка или популяция Т-клеток могут быть идентифицированы или получены в соответствии с аспектами изобретения, как описано в настоящем документе.
Образец может представлять собой образец опухоли, образец крови или образец ткани или образец мононуклеарных клеток периферической крови от индивидуума.
В десятом аспекте настоящее изобретение относится к способу лечения рака у индивидуума, который включает введение вакцины в соответствии с шестым аспектом настоящего изобретения индивидууму.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 - Способ разработки для прогнозирования и идентификации Т-клеток, реагирующих на неоантиген, в образцах NSCLC. Exome seq и RNAseq используют для определения неоэпитопов стволовых мутаций и мутаций ветвления. Прогнозируют связывание мутантных пептидов или пептидов дикого типа с HLA пациента, и выбирают пептиды с прогнозируемой высокой аффинностью (зеленый) к HLA (низкая IC50) и пептиды с высокой аффинностью в качестве мутантных и низкой в качестве пептидов дикого типа (синий) для создания флуоресцентных MHC-мультимеров для использования при идентификации Т-клеток NES в образцах опухоли.
Фигура 2 - Иллюстрация различия между стволовыми мутациями и мутациями ветвления в образце, выделенном из рака легких, рака мозга и нормальной ткани легких и мозга.
Фигура 3 - Иллюстрация способа разработки для идентификации стволовых мутаций в опухоли с целью идентификации Т-клеток, специфичных для неоантигена.
Фигура 4 - (A) Идентификация Т-клеток, специфичных для неоантигена, на ранней стадии развития рака легкого. Показаны спрогнозированные аффинности мутанта (ось Y) по сравнению с диким типом (ось X), полученные из данных секвенирования экзонов. Красные точки указывают на прогнозируемые пептиды с высоким баллом (с низкой аффинностью) для HLA пациента с формой WT и низким баллом (с высокой аффинностью) с мутантной формой. (B) Наращенные in vitro TILs окрашивали флуоресцентными тетрамерами, нагруженными спрогнозированными мутантными пептидами или контрольными пептидами цитомегаловируса (CMV), и анализировали с помощью проточной цитометрии. Т-клетки, реагирующие на CMV, выявляли с равной частотой (0,2-0,3%) в опухоли и нормальном легком.
Фигура 5 - (A) Мышей нагружали гетерогенной смесью опухолей (B16/B16-OVA), содержащих клональный (tyrpl) и субклональный (OVA) неоантиген, и опухолям давали расти без лечения, и мышей должны были забить между 20 и 30 днями после нагрузки опухолями. (B) Мыши, которых нагружали гетерогенной смесью опухолей (B16/B16-OVA), но лечили Tg-клетками TRP1 TCR, направленными на клональный неоантиген, были способны отторгать свои опухоли. (C) Мышей лечили Т-клетками OTII TCR Tg, направленными на субклональный неоантиген, ни одна из мышей не была способна отторгать свою опухоль. (D) Фигура демонстрирует способность Т-клеток OTII TCR Tg отторгать созданные опухоли, когда все клетки в опухоли экспрессируют неоантиген OVA. Каждая линия в каждом графике представляет собой независимую мышь. Для этих экспериментов использовали 6 мышей на группу. (E) Показаны все экспериментальные группы и средний размер опухолей в каждой группе.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Стволовой неоантиген
Настоящее изобретение относится к способу прогнозирования и идентификации в опухоли стволовых (клональных) неоантигенов и неоантигенов ветвления (субклональных).
«Неоантиген» представляет собой специфичный для опухоли антиген, который возникает вследствие мутации внутри раковой клетки. Таким образом, неоантиген не экспрессируется здоровыми клетками индивидуума. По существу одним преимуществом терапевтической направленности на стволовой неоантиген является более низкие уровни прогнозируемой токсичности, поскольку терапия не направлена на здоровые клетки.
Многие антигены, экспрессируемые раковыми клетками, являются собственными антигенами, которые избирательно экспрессируются или гиперэкспрессируются на раковых клетках. Клеточная иммунотерапия с трудом может быть направлена на эти собственные антигены, потому что они требуют преодоления как центральной толерантности (при которой аутореактивные Т-клетки удаляются в тимусе во время развития), так и периферической толерантности (при которой зрелые Т-клетки подавляются регуляторными механизмами).
Эти механизмы толерантности могут быть отменены путем направленного действия на неоантигены. В частности, немолчащие мутации, которые происходят в раковых клетках, могут приводить к экспрессии белков раковой клеткой, которые не экспрессируются здоровыми клетками. Эти измененные белки не распознаются иммунной системой как «собственные антигены».
Поскольку неоантигены не распознаются как «собственные антигены», Т-клетки, которые способны направленно действовать на неоантигены, не подвержены механизмам центральной и периферической толерантности в той же степени, что и Т-клетки, которые распознают собственные антигены.
Неоантиген, описанный в настоящем документе, может возникнуть в результате любой немолчащей мутации, которая изменяет белок, экспрессируемый раковой клеткой, по сравнению с немутированным белком, экспрессируемым здоровой клеткой дикого типа.
«Мутация» относится к разнице в нуклеотидной последовательности (например, ДНК или РНК) в опухолевой клетке по сравнению со здоровой клеткой одного и того же индивидуума. Различие в нуклеотидной последовательности может привести к экспрессии белка, который не экспрессируется здоровой клеткой одного и того же индивидуума.
Например, мутация может представлять собой вариацию одного нуклеотида (SNV), вариации множественных нуклеотидов, мутацию с делецией, мутацию со вставкой, транслокацию, миссенс-мутацию или мутацию сайта сплайсинга, приводящую к изменению аминокислотной последовательности (кодирующую мутацию). Известно, что в раковых клетках может происходить дупликация генома. Мутация, которая происходит до события дупликации генома, поэтому будет присутствовать в раковой клетке в удвоенном количестве относительно количества копии мутации, которая произошла после события дупликации. Если событие дупликации генома представляет собой стволовое событие, присутствующее в каждой клетке, неоантигены, возникающие до дупликации генома, будут представлять собой предпочтительную неоантигенную мишень по указанным причинам. В предпочтительном варианте осуществления стволовой неоантиген согласно изобретению представляет собой неоантиген, присутствующий в области генома, в которой редко происходит потеря количества копий.
В конкретных вариантах осуществления мутация, в результате которой продуцируется неоантиген, представляет собой SNV.
Разные области опухолей могут быть морфологически различными. Кроме того, мутационная гетерогенность может возникать внутри опухоли и может быть связана с различиями в прогнозе для опухоли и в потенциальной способности опухолевых клеток избегать иммунных методов лечения, направленных на мутации, которые отсутствуют во всех или в большинстве опухолевых клеток.
Авторы настоящего изобретения определили, что гетерогенность внутри опухоли может быть причиной вариации между неоантигенами, экспрессируемыми в разных областях опухоли и между различными клетками в опухоли. В частности, изобретатели установили, что в пределах опухоли определенные неоантигены экспрессируются во всех областях и по существу во всех клетках опухоли, тогда как другие неоантигены экспрессируются только в подмножестве областей опухоли и клеток.
Таким образом, «стволовой» или «клональный» неоантиген представляет собой неоантиген, который эффективно экспрессируется в пределах всей опухоли и кодируется по существу каждой опухолевой клеткой. «Неоантиген ветвления» или «субклональный неоантиген» представляет собой неоантиген, который экспрессируется в подмножестве или в доле клеток или областей в опухоли.
«Присутствует в пределах всей опухоли», «эффективно экспрессируется в пределах всей опухоли» и «кодируется по существу каждой опухолевой клеткой», может означать, что стволовой неоантиген экспрессируется во всех областях опухоли, из которых анализируются образцы.
Должно быть понятно, что определение того, что мутация «кодируется в пределах по существу каждой опухолевой клетки», относится к статистическому расчету и, следовательно, подвержено статистическому анализу и пороговым значениям.
Аналогичным образом, определение того, что стволовой неоантиген «эффективно экспрессируется в пределах всей опухоли» относится к статистическому расчету и, следовательно, подвержено статистическому анализу и пороговым значениям.
Эффективно экспрессируется, по существу, в каждой опухолевой клетке или, по существу, во всех опухолевых клетках означает, что мутация присутствует во всех опухолевых клетках, проанализированных в образце, как определено с использованием соответствующих статистических методов.
В качестве примера, фракция раковых клеток (CCF), описывающая долю раковых клеток, которые несут мутацию, может быть использована для определения, являются ли мутации стволовыми мутациями или мутациями ветвления. Например, фракция раковых клеток может быть определена путем интеграции частоты вариации аллелей с количеством копий и оценками чистоты, как описано Landau et al. (Cell. 2013 Feb 14;152(4):714-26). Определение CCF продемонстрировано в примерах, описанных в настоящем документе.
Вкратце, значения CCF рассчитывают для всех мутаций, идентифицированных во всех без исключения анализируемых областях опухоли. Если использована только одна область (т.е. только один образец), будет получен только один набор значений CCF. Это даст информацию о том, какие мутации присутствуют во всех опухолевых клетках в пределах этой области опухоли, и тем самым даст указание на то, является ли мутация стволовой или мутацией ветвления. Все субклональные мутации (т. CCF<1) в области опухоли определяют как мутации ветвления, в то время как клональные мутации с CCF=1 определяют как стволовые.
Как установлено, определение стволовой мутации подвержено статистическому анализу и пороговому значению. Таким образом, мутация может быть идентифицирована как стволовая, если она определена как имеющая 95% доверительный интервал CCF≥0,75, например 0,80, 0,85, 0,90, 0,95, 1,00 или >1,00. Напротив, мутация может быть идентифицирована как мутация ветвления, если она определена как имеющая 95% доверительный интервал CCF≤0,75, например 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35, 0,30, 0,25, 0,20, 0,15, 0,10, 0,05, 0,01 в любом анализируемом образце.
Должно быть понятно, что точность метода идентификации стволовых мутаций увеличивается путем идентификации клональных мутаций для более одного образца, выделенного из опухоли.
В одном варианте осуществления способы могут включать идентификацию множества, т.е. более одного клонального неоантигена.
В одном варианте осуществления количество клональных неоантигенов составляет 2-1000. Например, количество клональных неоантигенов может составлять 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950 или 1000, например, количество клональных неоантигенов может составлять от 2 до 100.
В предпочтительном варианте осуществления в способе может предлагаться множество или популяция, т.е. более одной Т-клетки, где множество Т-клеток включает Т-клетку, которая узнает клональный неоантиген, и Т-клетку, которая узнает отличный клональный неоантиген. Таким образом, способ обеспечивает множеством Т-клеток, которые узнают различные клональные неоантигены.
В предпочтительном варианте осуществления количество клональных неоантигенов, узнаваемых множеством Т-клеток, составляет 2-1000. Например, количество узнаваемых клональных неоантигенов может составлять 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950 или 1000, например, количество узнаваемых клональных неоантигенов может составлять от 2 до 100.
В одном аспекте множество Т-клеток узнает один и тот же стволовой неоантиген.
Образцы опухолей
Способ по первому аспекту настоящего изобретения включает стадию определения мутаций, присутствующих, по существу, во всех раковых клетках, выделенных из опухоли. Ссылки в настоящем документе на «по существу, во всех» предназначены для охвата большинства опухолевых клеток у индивидуума. Например, это может включать 60-100% клеток, например, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% опухолевых клеток у индивидуума.
Получение биопсийного материала и образцов из опухолей является общепринятой практикой в данной области техники и может быть выполнено в соответствии с любым подходящим методом, и такие методы должны быть известны специалисту в данной области техники.
Опухоль может представлять собой солидную опухоль или несолидную опухоль.
Способ по настоящему изобретению может включать, например, определение мутаций, присутствующих в раковых клетках из одной или более областей опухолей, выделенных из опухоли.
Например, способ может включать определение мутаций, присутствующих, по меньшей мере, в двух, по меньшей мере, в трех, по меньшей мере, в четырех, по меньшей мере, в пяти, по меньшей мере, в шести, по меньшей мере, в семи, по меньшей мере, в восьми, по меньшей мере, в девяти или, по меньшей мере, в десяти или более биопсий, полученных из опухоли. Способ также может быть использован для определения магистральных (стволовых) мутаций в одной биопсии.
Индивидуальные образцы опухоли могут быть выделены из разных областей, расположенных по всей опухоли в пределах первичного места локализации или между первичным местом локализации и метастазами или в пределах метастаза или между метастазами. Например, определение мутаций, присутствующих в опухолях, которые, как известно, проявляют морфологически разнородную гистологию в разных областях, может включать определение мутаций, присутствующих в ряде индивидуальных образцов, выделенных из морфологически разнородных областей.
Образец может представлять собой образцом крови. Например, образец крови может включать циркулирующую опухолевую ДНК, циркулирующие опухолевые клетки или экзосомы, включающие опухолевую ДНК.
Определение мутаций, присутствующих в образце опухоли, может быть выполнено, например, путем сравнения последовательностей ДНК и/или РНК, выделенных из образцов опухоли, и сравниваемых образцов здоровой ткани от одного и того же индивидуума, путем секвенирования Exome, секвенирования целого генома, секвенирования панели генов-мишеней и/или РНК-Seq. Описания секвенирования Exome и RNA-seq представлены в статьях Boa et al. (Cancer Informatics. 2014; 13(Suppl 2):67-82.) и Ares et al. (Cold Spring Harb Protoc. 2014 Nov 3;2014(1 1 ):1 139-48); соответственно.
Выравнивание последовательностей для идентификации различий нуклеотидов (например, SNVs) в ДНК и/или РНК из образца опухоли по сравнению с ДНК и/или РНК из неопухолевого образца может быть осуществлено с использованием способов, которые известны в данной области техники. Например, различия нуклеотидов по сравнению с эталонным образцом могут быть выявлены с использованием метода, описанного Koboldt et al. (Genome Res.; 2012; 22: 568-576). Эталонным образцом может служить последовательность ДНК и/или РНК зародышевой линии.
Аллели HLA
Т-клетки, которые специфически узнают неоантиген, обозначают в настоящем документе как специфические для неоантигена (NES) Т-клетки.
Антигены презентируются Т-клеткам в контексте пептидов, происходящих от антигена, связанных с молекулами главного комплекса гистосовместимости (МНС).
Таким образом, стволовой неоантиген может быть узнан NES Т-клеткой в виде пептида, происходящего от стволового неоантигена (называемого в настоящем документе «пептидом стволового неоантигена»), представляемого молекулой MHC.
Пептид стволового неоантигена представляет собой пептид, который происходит от участка полипептида, который включает мутацию, специфичную для раковых клеток. Фактически пептиды стволового неоантигена не должны происходить от полипептидов, кодируемых геномом здоровых клеток.
Белки класса I МНС образуют функциональный рецептор на большинстве клеток организма, имеющих ядро. В HLA есть 3 главных гена класса MHC: HLA-A, HLA-B, HLA-C и три минорных гена HLA-E, HLA-F и HLA-G. β2-микроглобулин связывается с главными и минорными субъединицами генов с получением гетеродимера.
Пептиды, которые связываются с молекулами МНС класса I, обычно составляют от 7 до 13, более обычно от 8 до 11 аминокислот в длину. Связывание пептида стабилизируется на двух его концах контактами между атомами в основной цепи пептида и инвариантными сайтами в пептидсвязывающей канавке всех молекул МНС класса I. На обоих концах канавки имеются инвариантные сайты, которые связывают амино- и карбоксильные концы пептида. Вариации длины пептида аккомодируются перегибанием в пептидной цепи часто у остатков пролина или глицина, что обеспечивает необходимую гибкость.
Существует 3 главных и 2 минорных белка MHC класса II, кодируемых HLA. Гены класса II объединяются с образованием гетеродимерных (αβ) белковых рецепторов, которые обычно экспрессируются на поверхности антигенпрезентирующих клеток.
Пептиды, которые связываются с молекулами МНС класса II, обычно составляют от 8 до 20 аминокислот в длину, более обычно от 10 до 17 аминокислот в длину и могут быть более длинными (например, до 40 аминокислот). Эти пептиды лежат вдоль пептидсвязывающей канавки MHC II в растянутой конформации, которая (в отличие от пептидсвязывающей канавки MHC класса I) открыта с обоих концов. Пептид удерживается на месте главным образом за счет контактов атомов основной цепи с консервативными остатками, которые выстилают пептидсвязывающую канавку.
Способы по настоящему изобретению могут включать стадию оценки аллелей HLA индивидуума для определения того, будет ли пептид стволового неоантигена связываться с молекулой MHC, экспрессируемой у индивидуума.
Профиль аллелей HLA индивидуума может быть определен способами, которые известны в данной области техники. Например, профиль HLA индивидуума может быть определен HLA-серотипированием и/или секвенированием генов HLA. HLA-фенотипирование с помощью ПЦР с одиночным аллель-специфическим праймером (SSP-PCR) является альтернативной стратегией для определения профиля HLA индивидуума.
В представленных примерах профиль HLA индивидуума определен путем секвенирования локуса HLA и обработки с использованием алгоритма прогнозирования Optitype для определения типа HLA каждого индивидуума (Szolek et al., Bioinformatics; 2014; 30(23):3310-3316).
Связывание пептида с конкретной молекулой MHC может быть предсказано с использованием способов, которые известны в данной области техники. Примеры способов предсказания связывания МНС включают способы, которые описаны Lundegaard et al. (Nucleic Acids Res. 2008:W509-12.2008 & Bioinformatics. 2008 Jun 1;24(11):1397-8) и Shen et al. (Proteome Sci. 2013 Nov 7; 11 (Suppl 1):S15).
Способы по настоящему изобретению могут включать определение пептида стволового неоантигена, который, как прогнозируется, связывается с молекулой MHC, экспрессируемой индивидуумом. В частности, способы могут включать стадию определения и выбора пептида стволового неоантигена, который, как предполагается, прочно связывается с молекулой МНС, экспрессируемой индивидуумом. Точное определение «прочного связывания» должно зависеть от метода, используемого для прогнозирования связывающего взаимодействия с MHC (смотри, например, Lundegaard et al. (как представлено выше)). Тем не менее, во всех случаях выбранный пептид стволового неоантигена должен быть предсказан как способный к связыванию и к презентации в контексте молекулы MHC, экспрессируемой индивидуумом.
Аффинность связывания с пептидом стволового неоантигена может быть ниже 500 нМ. Под «высокой аффинностью» может пониматься аффинность связывания от 0 до 50 нМ. В других вариантах осуществления пептид стволового неоантигена может связываться с молекулой MHC с промежуточной аффинностью от 50 до 150 нМ аффинности связывания или с низкой аффинностью от 150 до 500 нМ аффинности связывания.
В некоторых вариантах осуществления пептид стволового неоантигена, как предполагается, может связываться с молекулой MHC с высокой аффинностью, в то время как соответствующий пептид дикого типа (например, эквивалентный пептид, происходящий из того же участка соответствующего полипептида дикого типа), как предполагается, связывается с той же молекулой MHC с низкой аффинностью.
Популяция Т-клеток
Настоящее изобретение также относится к способу предоставления популяции Т-клеток, которые направлены на стволовой неоантиген из опухоли.
Популяция Т-клеток может включать CD8+ Т-клетки, CD4+ Т-клетки или CD8+ и CD4+ Т-клетки.
Вспомогательные Т-хелперные клетки (клетки ТН) помогают другим лейкоцитам в иммунных процессах, включая созревание В-клеток до плазматических клеток и В-клеток памяти, и активацию цитотоксических Т-клеток и макрофагов. TH-клетки экспрессируют CD4 на своей поверхности. TH-клетки становятся активированными, когда происходит презентация им пептидных антигенов молекулами MHC класса II на поверхности антигенпрезентирующих клеток (APCs). Эти клетки могут дифференцироваться в один из нескольких подтипов, включая TH1, TH2, TH3, TH17, Th9 или TFH, которые секретируют различные цитокины для облегчения различных типов иммунных ответов.
Цитотоксические Т-клетки (клетки ТС или CTLs) уничтожают инфицированные вирусом клетки и опухолевые клетки, а также участвуют в отторжении трансплантата. CTLs экспрессируют на своей поверхности CD8. Эти клетки узнают свои мишени путем связывания с антигеном, ассоциированным с МНС класса I, который присутствует на поверхности всех клеток, имеющих ядро. С помощью IL-10, аденозина и других молекул, секретируемых регуляторными Т-клетками, клетки CD8+ могут быть инактивированы, что предотвращает аутоиммунные заболевания.
Популяции Т-клеток, получаемые в соответствии с настоящим изобретением, могут быть обогащены Т-клетками, которые являются специфическими, т.е. направлены на стволовые неоантигены. Это означает, что популяция Т-клеток, которая получена в соответствии с настоящим изобретением, будет иметь увеличенное количество Т-клеток, которые направлены на один или более стволовых неоантигенов. Например, популяция Т-клеток по настоящему изобретению будет иметь увеличенное количество Т-клеток, которые направлены на стволовой неоантиген, по сравнению с Т-клетками в образце, выделенном от индивидуума. Говоря иначе, состав популяции Т-клеток будет отличаться от состава «природной» популяции Т-клеток (т.е. популяции, которая не подвергалась стадиям идентификации и наращивания, обсуждаемым в настоящем документе), тем, что процент или доля Т-клеток, которые направлены на стволовой неоантиген, будет увеличена.
Популяции Т-клеток, получаемые в соответствии с настоящим изобретением, могут быть обогащены Т-клетками, которые являются специфическими, т.е. направлены на стволовые неоантигены (т.е. клональные неоантигены, так как используемые в настоящем документе термины «стволовой» неоантиген и «клональный» неоантиген являются эквивалентными, и термины неоантиген «ветвления» и «субклональный» неоантиген являются эквивалентными), и могут иметь отношение Т-клеток, которые направлены на стволовые неоантигены, к Т-клеткам, которые направлены на неоантигены ветвления, которое будет выше в пользу Т-клеток, которые направлены на стволовые неоантигены, по сравнению с Т-клетками в образце, выделенном от индивидуума.
Это означает, что популяция Т-клеток, получаемая в соответствии с настоящим изобретением, будет иметь увеличенное количество Т-клеток, которые направлены на один или более стволовых неоантигенов. Например, популяция Т-клеток по настоящему изобретению будет иметь увеличенное количество Т-клеток, которые направлены на стволовой неоантиген, по сравнению с Т-клетками в образце, выделенном от индивидуума. Говоря иначе, состав популяции Т-клеток будет отличаться от состава «природной» популяции Т-клеток (т.е. популяции, которая не подвергалась стадиям идентификации и наращивания, обсуждаемым в настоящем документе), тем, что процент или доля Т-клеток, которые направлены на стволовой неоантиген, будет увеличена, а отношение Т-клеток в популяции, которые направлены на стволовые неоантигены, к Т-клеткам, которые направлены на неоантигены ветвления, будет выше в пользу Т-клеток, которые направлены на стволовые неоантигены.
Популяция Т-клеток согласно изобретению может содержать, по меньшей мере, приблизительно 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100% Т-клеток, которые направлены на стволовой неоантиген. Например, популяция Т-клеток может содержать приблизительно 0,2%-5%, 5%-10%, 10-20%, 20-30%, 30-40%, 40-50%, 50-70% или 70-100% Т-клеток, которые направлены на стволовой неоантиген. В одном аспекте популяция Т-клеток содержит, по меньшей мере, приблизительно 1, 2, 3, 4 или 5% Т-клеток, которые направлены на стволовой неоантиген, например, по меньшей мере, приблизительно 2% или, по меньшей мере, 2% Т-клеток, которые направлены на стволовой неоантиген.
Альтернативно полагается, что популяция Т-клеток может содержать не более приблизительно 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 99,1, 99,2, 99,3, 99,4, 99,5, 99,6, 99,7, 99,8% Т-клеток, которые не направлены на стволовой неоантиген. Например, популяция Т-клеток может содержать не более 95%-99,8%, 90%-95%, 80-90%, 70-80%, 60-70%, 50-60%, 30-50% или 0-30% Т-клеток, которые не направлены на стволовой неоантиген. В одном аспекте популяция Т-клеток содержит не более приблизительно 99, 98, 97, 96 или 95% Т-клеток, которые не направлены на стволовой неоантиген, например, не более приблизительно 98% или 95% Т-клеток, которые не направлены стволовой неоантиген.
Наращенная популяция Т-клеток, реагирующих на стволовой неоантиген, может иметь более высокую активность, чем ненаращенная популяция Т-клеток, например, при использовании пептида стволового неоантигена. Ссылка на «активность» может представлять собой ответ популяции Т-клеток на повторную стимуляцию пептидом стволового неоантигена, например, пептидом, соответствующим пептиду, используемому для наращивания, или смесью пептидов стволового неоантигена. Подходящие способы анализа ответа известны в данной области техники. Например, может быть измерена продукция цитокинов (например, может быть измерена продукция IL2 или IFNγ). Ссылка на «более высокую активность» включает, например, увеличение активности в 1-5, 5-10, 10-20, 20-50, 50-100, 100-500, 500-1000 раз. В одном аспекте активность может быть более чем в 1000 раз более высокой.
В предпочтительном варианте осуществления в изобретении предлагается множество или популяция, т.е. более одной Т-клетки, где множество Т-клеток включает Т-клетку, которая распознает клональный неоантиген, и Т-клетку, которая распознает отличный клональный неоантиген. Таким образом, в изобретении предлагается множество Т-клеток, которые распознают различные клональные неоантигены. Различные Т-клетки во множестве или в популяции могут альтернативно иметь разные TCRs, которые узнают один и тот же стволовой неоантиген.
В предпочтительном варианте осуществления количество клональных неоантигенов, узнаваемых множеством Т-клеток, составляет от 2 до 1000. Например, количество узнаваемых клональных неоантигенов может составлять 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950 или 1000, предпочтительно от 2 до 100. Может существовать множество Т-клеток с различными TCRs, но которые узнают один и тот же клональный неоантиген.
Популяция Т-клеток может полностью или в основном состоять из CD8+ Т-клеток или полностью или в основном состоять из смеси CD8+ Т-клеток и CD4+ Т-клеток, или полностью или в основном состоять из CD4+ Т-клеток.
В конкретных вариантах осуществления популяция Т-клеток создана из Т-клеток, выделенных от индивидуума с опухолью.
Например, популяция Т-клеток может быть создана из Т-клеток в образце, выделенном от индивидуума с опухолью. Образец может представлять собой образец опухоли, образец периферической крови или образец из других тканей индивидуума.
В конкретном варианте осуществления популяция Т-клеток создана из образца из опухоли, в которой идентифицирован стволовой неоантиген. Другими словами, популяцию Т-клеток выделяют из образца, происходящего из опухоли пациента, подлежащего лечению. Такие Т-клетки обозначаются в настоящем документе как «инфильтрирующие опухоль лимфоциты» (TILs).
Т-клетки могут быть выделены с использованием способов, которые хорошо известны в данной области техники. Например, Т-клетки могут быть очищены из суспензий отдельных клеток, полученных из образцов, на основе экспрессии CD3, CD4 или CD8. Т-клетки из образцов могут быть обогащены путем прохождения через градиент фиколл-пака.
Наращивание Т-клеток NES может быть выполнено с использованием способов, которые известны в данной области техники. Например, Т-клетки NES могут быть наращены в культуре ex vivo в условиях, которые, как известно, обеспечивают митогенные стимулы для Т-клеток. В качестве примера, Т-клетки NES можно культивировать с цитокинами, такими как IL-2 или с митогенными антителами, такими как анти-CD3 и/или CD28. Т-клетки NES также можно культивировать совместно с облученными антигенпрезентирующими клетками (APCs), такими как дендритные клетки, пульсированные пептидами, содержащими идентифицированные стволовые мутации, в качестве отдельных стимуляторов или в виде пулов стимулирующих стволовых неоантигенов или пептидов.
Наращивание Т-клеток NES может быть выполнено с использованием способов, которые известны в данной области техники, включая, например, использование искусственных антигенпрезентирующих клеток (aAPCs), например, которые обеспечивают дополнительными костимулирующими сигналами, и аутологичных РВМСs, которые представляют соответствующие пептиды. В качестве примера, аутологичные PBMCs могут быть пульсированы пептидами, содержащими стволовые мутации, как обсуждалось в настоящем документе, в виде единственных стимуляторов, или, альтернативно, в виде пулов стимулирующих пептидов стволового неоантигена.
Изобретение относится к способу получения композиции, включающей антигенпрезентирующую клетку и стволовой неоантиген или пептид стволового неоантигена. Стволовой неоантиген может быть идентифицирован в соответствии со способами по настоящему изобретению. В одном варианте осуществления указанный способ включает следующие стадии:
(а) идентификации стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
b) получения композиции, включающей указанный стволовой неоантиген или пептид стволового неоантигена и антигенпрезентирующую клетку.
Изобретение также относится к композиции, включающей антигенпрезентирующую клетку, например, дендритную клетку и стволовой неоантиген или пептид стволового неоантигена. Стволовой неоантиген может быть идентифицирован в соответствии со способами по изобретению, как описано в настоящем документе.
Композиция может быть получена в соответствии со способом, описанным в настоящем документе. Композицию можно также использовать в способах по изобретению, описанных в настоящем документе, например, в способах получения Т-клетки или популяции или композиции Т-клеток, как описано в настоящем документе.
Композиции, описанные в настоящем документе, могут представлять собой фармацевтическую композицию, которая дополнительно включает фармацевтически приемлемый носитель, разбавитель или наполнитель. Фармацевтическая композиция может необязательно включать один или более дополнительных фармацевтически активных полипептидов и/или соединений. Такая композиция может, например, находиться в форме, подходящей для внутривенной инфузии.
Изобретение также относится к способу обеспечения популяцией Т-клеток, направленных на стволовой неоантиген в опухоли, который включает стадии:
i) идентификации Т-клетки из образца, выделенного от индивидуума, которая способна специфически узнавать указанный стволовой неоантиген; и
ii) наращивания Т-клеток для обеспечения популяции Т-клеток, которые направлены на стволовой неоантиген, где наращивание Т-клеток обеспечивают путем совместного культивирования с антигенпрезентирующими клетками, которые представляют пептиды стволового неоантигена, происходящие от указанного стволового неоантигена.
Полученную популяцию T-клеток обогащают Т-клетками, которые направлены на стволовые неоантигены.
В одном аспекте антигенпрезентирующие клетки пульсированы или нагружены указанным пептидом.
Изобретение также относится к композиции Т-клеток, которая включает популяцию Т-клеток, специфичных для стволового неоантигена, где указанную популяцию Т-клеток, специфичных для стволового неоантигена, получают путем совместного культивирования Т-клеток с антигенпрезентирующими клетками, которые презентируют пептиды неоантигена.
В одном аспекте антигенпрезентирующая клетка представляет собой дендритную клетку. В одном аспекте антигенпрезентирующую клетку облучают.
В одном аспекте антигенпрезентирующая клетка представляет собой клетку, способную представлять соответствующий пептид, например, в правильном контексте HLA. Такая клетка может представлять собой аутологичную активированную PBMC, экспрессирующую аутологичную молекулу HLA, или неаутологичную клетку, экспрессирующую набор подобранных HLAs. В одном аспекте искусственную антигенпрезентирующую клетку облучают.
Т-клетки NES также могут быть обогащены путем исходной стимуляции TILs стволовыми неоантигенами в присутствии или в отсутствие экзогенных APCs с последующей поликлональной стимуляцией и экспансией с помощью цитокинов, таких как IL-2, или с помощью митогенных антител, таких как анти-CD3 и/или CD28. Такие способы известны в данной области техники. Например, смотри Forget et al. J Immunother. 2014 Nov-Dec; 37(9):448-60, Donia et al. Cytotherapy. 2014 Aug; 16(8):1117-20, Donia et al. Scand J Immunol. 2012 Feb; 75(2):157-67 и Ye et al. J Transl Med. 2011 Aug 9; 9:131.
Идентификация Т-клеток NES в смешанной исходной популяции Т-клеток может быть проведена с использованием способов, которые известны в данной области техники. Например, Т-клетки NES могут быть идентифицированы с использованием мультимеров MHC, включающих пептид стволового неоантигена, идентифицированный способом по настоящему изобретению.
Мультимеры MHC представляют собой олигомерные формы молекул MHC, предназначенные для идентификации и выделения Т-клеток с высокой аффинностью к специфическим антигенам среди большой группы неродственных Т-клеток. Мультимеры могут быть использованы для экспонирования MHC класса 1, MHC класса 2 или неклассических молекул (например, CD1d).
Наиболее часто используемые мультимеры MHC представляют собой тетрамеры. Их обычно получают путем биотинилирования растворимых мономеров МНС, которые обычно получают рекомбинантно в эукариотических или бактериальных клетках. Затем эти мономеры связываются с остовом, с таким как стрептавидин или авидин, с созданием четырехвалентной структуры. Эти остовы конъюгируют с флуорохромами для последующего выделения связанных Т-клеток, например, с помощью проточной цитометрии.
Изобретение относится к мультимеру MHC, включающему пептид стволового неоантигена. Стволовой неоантиген может быть идентифицирован с помощью способа согласно изобретению, как описано в настоящем документе.
В одном аспекте настоящее изобретение относится к способу получения мультимера MHC, который может быть использован в соответствии с изобретением, как описано в настоящем документе. Указанный способ включает стадии:
(а) идентификации стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
(b) получения пептида стволового неоантигена из указанного стволового неоантигена; и
(c) получения мультимера MHC, включающего указанный пептид стволового неоантигена.
Мультимеры MHC в соответствии с изобретением могут быть использованы в способах идентификации, выделения, наращивания или иного получения Т-клетки, популяции или композиции Т-клеток в соответствии с настоящим изобретением. Пептиды стволового неоантигена могут быть синтезированы с использованием способов, которые известны в данной области техники.
Термин «пептид» используют в нормальном смысле для обозначения ряда остатков, обычно L-аминокислот, связанных друг с другом, как правило, пептидными связями между α-аминогруппами и карбоксильными группами соседних аминокислот. Термин включает модифицированные пептиды и синтетические пептидные аналоги.
Пептид может быть получен с использованием химических методов (Peptide Chemistry, A practical Textbook. Mikos Bodansky, Springer-Verlag, Berlin.). Например, пептиды могут быть синтезированы твердофазными методами (Roberge JY et al (1995) Science 269: 202-204), отщеплены от смолы и очищены препаративной высокоэффективной жидкостной хроматографией (например, Creighton (1983) Proteins Structures And Molecular Principles, WH Freeman and Co, New York NY). Автоматический синтез может быть достигнут, например, с использованием пептидного синтезатора ABI 43 1 A (Perkin Elmer) в соответствии с инструкциями производителя.
Альтернативно пептид может быть получен рекомбинантными способами или путем расщепления полипептида, который представляет собой или включает неоантиген. Состав пептида может быть подтвержден аминокислотным анализом или секвенированием (например, с помощью процедуры деградации Эдмана).
Пептид стволового неоантигена может включать специфическую для раковой клетки мутацию/стволовую мутацию (например, немолчащую аминокислотную замену, кодируемую SNV) в любом положении остатка в пептиде. В качестве примера, пептид, который способен связываться с молекулой МНС класса I, обычно имеет длину от 7 до 13 аминокислот. Таким образом, аминокислотная замена может присутствовать в положении 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или 13 пептида, включающего тринадцать аминокислот.
В следующем аспекте могут быть использованы более длинные пептиды, например, из 27-31 аминокислот, и мутация может происходить в любом положении, например, в центре пептида, например, в положениях 13, 14, 15 или 16, она также может быть использована для стимуляции как CD4+, так и CD8+ клеток для узнавания клональных неоантигенов.
Настоящее изобретение дополнительно относится к мультимеру MHC, включающему пептид стволового неоантигена, как определено в настоящем документе.
Композиция Т-клеток
Настоящее изобретение дополнительно относится к композиции Т-клеток, которая включает Т-клетку, специфичную для стволового неоантигена.
Композиция Т-клеток может представлять собой фармацевтическую композицию, включающую множество специфических для неоантигена Т-клеток, как определено в настоящем документе. Фармацевтическая композиция может дополнительно включать фармацевтически приемлемый носитель, разбавитель или наполнитель. Фармацевтическая композиция может необязательно включать один или более дополнительных фармацевтически активных полипептидов и/или соединений. Такая композиция может, например, находиться в форме, подходящей для внутривенной инфузии.
В предпочтительном варианте осуществления настоящего изобретения индивидуумом, описанным в настоящем документе, является млекопитающее, предпочтительно человек, кошка, собака, лошадь, осел, овца, свинья, коза, корова, мышь, крыса, кролик или морская свинка, но наиболее предпочтительным индивидуумом является человек.
Способы по изобретению, описанные выше, могут быть использованы in vitro, ex vivo или in vivo, например, либо для обработки in situ, либо для обработки ex vivo с последующим введением обработанных клеток в организм.
В определенных аспектах в соответствии с настоящим изобретением, как описано в настоящем документе, Т-клетку или популяцию или композицию Т-клеток повторно вводят индивидууму, например, после выделения и наращивания Т-клеток, как описано в настоящем документе. Подходящие способы достижения этого должны быть известны специалисту в данной области техники. Например, способы получения, отбора и наращивания Т-клеток известны в данной области техники, смотри, например, статьи Dudley J Immunother. 2003; 26(4):332-342, и Rosenberg et al. 2011 Clin Cancer Res:17(13):4550-7. Способы повторного введения Т-клеток описаны, например, в статьях Dudley et al. Clin Cancer Res. 2010 Dec 15; 16(24): 6122-6131, 2011 и Rooney et al. Blood. 1998 Sep 1;92(5):1549-55.
Т-клетка, специфичная для стволового неоантигена, может представлять собой любую Т-клетку, которая способна узнавать стволовой неоантиген (т.е. Т-клетку NES).
Например, Т-клетка NES может представлять собой Т-клетку, полученную способом по настоящему изобретению.
Т-клетка NES может представлять собой сконструированную Т-клетку. Например, Т-клетка NES может экспрессировать химерный рецептор антигена (CAR) или рецептор Т-клеток (TCR), который специфически связывается со стволовым неоантигеном или пептидом стволового неоантигена (например, рецептор Т-клеток (TCR) с повышенной аффинностью, который специфически связывается со стволовым неоантигеном или пептидом стволового неоантигена).
CARs представляют собой белки, которые в своем обычном формате придают специфичность моноклонального антитела (mAb) эффекторной функции Т-клетки. Их обычной формой является форма трансмембранного домена типа 1 белка с аминоконцом, узнающим антиген, спейсером, трансмембранным доменом, причем весь трансмембранный домен связан с эндодоменом соединения, которое передает сигналы выживания и активации Т-клеток.
В наиболее распространенной форме этих молекул используются одноцепочечные вариабельные фрагменты (scFv), происходящие от моноклональных антител, для узнавания антигена-мишени. scFv соединяют через спейсер и трансмембранный домен с сигнальным эндодоменом. Такие молекулы приводят к активации Т-клетки в ответ на узнавание с помощью scFv ее мишени. Когда Т-клетки экспрессируют такой CAR, они узнают и уничтожают клетки-мишени, которые экспрессируют антиген-мишень. Разработано несколько CARs против антигенов, ассоциированных с опухолями, и методы адаптивного переноса с использованием таких Т-клеток, экспрессирующих CARs, в настоящее время проходят клинические испытания для лечения различных видов рака.
TCRs с повышенной аффинностью создают путем идентификации клона Т-клеток, из которого клонируют α и β цепи TCR с желаемой специфичностью мишени. Затем кандидатный TCR подвергают ПЦР-направленному мутагенезу в областях, определяющих комплементарность α и β цепей. Мутации в каждой области CDR подвергают скринингу для выбора мутантов с повышенной аффинностью по сравнению с нативным TCR. После завершения главные кандидаты клонируют в векторы для обеспечения функционального тестирования в Т-клетках, экспрессирующих TCR с повышенной аффинностью.
Т-клетки NES могут нести TCRs с высокой аффинностью, и, следовательно, повышение аффинности может не потребоваться. TCRs с высокой аффинностью могут быть выделены из T-клеток NES от индивидуума и могут не нуждаться в повышении аффинности.
Кандидатные клоны Т-клеток, способные связывать пептид стволового неоантигена, могут быть идентифицированы с использованием мультимеров MHC, включающих пептид стволового неоантигена, например, как описано в настоящем документе.
Идентифицированные TCRs и/или CARs, которые специфически направлены на пептид стволового неоантигена или на стволовой неоантиген, могут быть экспрессированы в аутологичных Т-клетках от индивидуума с использованием способов, которые известны в данной области техники, например, путем введения ДНК или РНК, кодирующей TCR или CAR, одним из многих способов, включая трансдукцию вирусным вектором, трансфекцию ДНК или РНК.
Аутологичные Т-клетки могут быть из образца, выделенного от индивидуума, как описано в настоящем документе.
Изобретение охватывает Т-клетку, как описано в настоящем документе, например, сконструированную Т-клетку.
Вакцина
Настоящее изобретение относится к вакцине, включающей стволовой неоантиген или пептид стволового неоантигена, как определено в настоящем документе. Например, стволовой неоантиген или пептид стволового неоантигена может быть идентифицирован с помощью способа по настоящему изобретению.
В одном аспекте настоящего изобретения вакцина может включать более одного отличного стволового неоантигена или пептида стволового неоантигена, например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 различных пептидов. Стволовой неоантиген может также находиться в форме белка.
В одном варианте осуществления вакцина может включать полипептид, который включает описанный в настоящем документе стволовой неоантиген. В одном варианте осуществления изобретения вакцина может включать более одного отличного полипептида, каждый из которых включает стволовой неоантиген, например 2, 3, 4, 5, 6, 7, 8, 9 или 10 различных полипептидов.
Вакцина может представлять собой фармацевтическую композицию, которая дополнительно включает фармацевтически приемлемый носитель, разбавитель или наполнитель. Фармацевтическая композиция может необязательно включать один или более дополнительных фармацевтически активных полипептидов и/или соединений. Такая композиция может, например, быть в форме, подходящей для внутривенной инфузии. Смотри, например, Butterfield, BMJ. 2015 22;350 для обсуждения вакцин против рака.
В частности, вакцина может дополнительно включать адъювант. Примеры адъювантов включают, но не ограничены этим, соли алюминия, масляные эмульсии и бактериальные компоненты (например, ЛПС и липосомы).
Подходящие дозы пептидов в вакцине могут быть определены специалистом в данной области техники. Доза может зависеть от пептида, который должен использоваться. Для использования пептида in vivo может быть использована доза in vivo 0,1-4000 мкг, например, 0,1-2000 мкг, 0,1-1000 мкг или 0,1-500 мкг, например, 0,1-100 мкг.
Вакцина в соответствии с изобретением, как обсуждалось в настоящем документе, может приводить к выработке иммунного ответа у индивидуума. «Иммунный ответ», который может быть выработан, может представлять собой гуморальный и/или опосредованный клетками иммунитет, например, стимуляцию продукции антител или стимуляцию цитотоксических или киллерных клеток, которые могут узнавать и разрушать (или иным образом удалять) клетки, экспрессирующие на своей поверхности антигены, соответствующие антигенам в вакцине. Термин «стимуляция иммунного ответа», таким образом, включает все типы иммунных ответов и механизмы их стимуляции и охватывает стимулирующие CTLs, которые входят в предпочтительный аспект изобретения. Предпочтительным стимулируемым иммунным ответом являются цитотоксические CD8+ Т-клетки и хелперные CD4+ Т-клетки. Величина иммунного ответа может быть оценена с помощью маркеров иммунного ответа, например, секретируемых молекул, таких как IL-2 или IFNγ, или генерации антигенспецифических Т-клеток.
Кроме того, вакцина против стволового неоантигена может быть доставлена в форме клетки, такой как антигенпрезентирующая клетка, например, в форме вакцины дендритных клеток. Антигенпрезентирующая клетка, такая как дендритная клетка, может быть пульсирована или нагружена стволовым неоантигеном или пептидом стволового неоантигена, или генетически модифицирована (посредством переноса ДНК или РНК) для экспрессии одного, двух или более стволовых неоантигенов или пептидов стволовых неоантигенов (смотри, например, Butterfield 2015 выше; Palucka 2013, выше), например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 стволовых неоантигенов или пептидов стволовых антигенов. Способы получения вакцин дендритных клеток известны в данной области техники.
Подходящие вакцины также могут быть в форме вакцин ДНК или РНК, относящихся к стволовым неоантигенам или пептидам стволовых неоантигенов, как описано в настоящем документе. Например, ДНК или РНК, кодирующая один или более стволовых неоантигенов, или пептидов или белков, полученных из них, может быть использована в качестве вакцины, например, путем прямой инъекции индивидууму. Например, можно использовать ДНК или РНК, кодирующую 2, 3, 4, 5, 6, 7, 8, 9 или 10 стволовых неоантигенов или пептидов или белков, происходящих от них. Один или более стволовых неоантигенов или пептидов стволовых неоантигенов могут быть доставлены с помощью бактериального или вирусного вектора, содержащего последовательности ДНК или РНК, которые кодируют один или более стволовых неоантигенов или пептидов стволовых неоантигенов.
Вакцины, как описано в настоящем документе, могут вводиться любым подходящим способом. Например, можно использовать любой подходящий механизм доставки, известный в данной области техники. Вакцина может включать использование векторной системы доставки, или векторная система доставки может быть не востребована. Векторы могут быть вирусными или бактериальными. В качестве системы доставки можно использовать липосомы. Можно использовать также вакцины Listeria или электропорацию.
Таким образом, настоящее изобретение дополнительно относится к клетке, экспрессирующей стволовой неоантиген или пептид стволового неоантигена на своей поверхности (или внутриклеточно), или к популяции таких клеток, где клетку или популяцию можно получить (или получают) способами, определенными в настоящем документе. В предпочтительном варианте осуществления клетка представляет собой антигенпрезентирующую клетку, такую как дендритная клетка.
Для введения клеток in vivo, как описано в настоящем документе, может быть использован любой способ введения популяции клеток, который является общим или стандартным в данной области техники, например, с помощью инъекции или инфузии соответствующим путем. В одном аспекте вводят от 1×104 до 1×108 клеток на кг массы испытуемого (например, от 1,4×104 до 2,8×106 на кг массы человека). В одном аспекте вводят приблизительно или не более 107 клеток на кг массы индивидуума. Так, например, у человека дозу 0,1-20×107 клеток на кг массы индивидуума можно вводить как единицу дозы, т.е. на дозу, например, в качестве дозы Т-клеток или дозы вакцинации. В одном аспекте вводят от 1×104 до 1×105 клеток, от 1×105 до 1×106 клеток, от 1×106 до 1×107 клеток или от 1×107 до 1×108 клеток, на кг массы индивидуума. При применении вакцинации можно использовать 1-20×106 клеток на дозу. Доза может быть повторена в более поздние сроки, если это необходимо.
Вакцина согласно изобретению может быть использована для лечения рака.
Изобретение также относится к способу лечения рака у индивидуума, включающему введение вакцины, как описано в настоящем документе, указанному индивидууму. Способ может дополнительно включать стадию идентификации индивидуума, у которого обнаружен рак.
В следующем аспекте изобретение относится к способу получения вакцины, включающей пептид стволового неоантигена или стволовой неоантиген, причем указанный способ включает стадии:
(а) идентификации стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
(b) получения пептида стволового неоантигена или стволового неоантигена из указанного стволового неоантигена; и
(c) получения вакцины с указанным пептидом стволового неоантигена или с белком стволового неоантигена.
В предпочтительном аспекте изобретения получение вакцины включает получение вакцины дендритных клеток, где указанная дендритная клетка представляет стволовой неоантиген или пептид стволового неоантигена.
Белок стволового неоантигена можно также использовать в вакцинах и способах, относящихся к вакцинации в соответствии с изобретением.
В следующем аспекте изобретение относится к способу получения вакцины, включающей молекулу ДНК или РНК, кодирующую пептид стволового неоантигена или стволовой неоантиген, причем указанный способ включает стадии:
(а) идентификации стволового неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию;
или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации стволовой мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации стволового неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает стволовую мутацию; и
(b) получения молекулы ДНК или РНК, кодирующей пептид стволового неоантигена или стволовой неоантиген; и
(c) получения вакцины с указанной молекулой ДНК или РНК.
Вакцина может быть доставлена подходящими способами, как описано в настоящем документе выше.
В одном аспекте вакцинация является терапевтической вакцинацией. В этом аспекте вакцину вводят индивидууму, у которого обнаружен рак, для лечения рака.
В следующем аспекте вакцинация представляет собой профилактическую вакцинацию. В этом аспекте вакцину вводят индивидууму, который может подвергаться риску развития рака.
В одном аспекте вакцину вводят индивидууму, у которого ранее был рак, и у которого существует риск рецидива рака.
Вакцина также может быть представлена в форме ДНК или РНК, кодирующей один или более пептидов или белков стволового неоантигена, и доставляемой дополнительными способами, включая, но, не ограничиваясь этим, вирусные векторы, антигенпрезентирующие клетки и электропорацию.
РАК
Т-клетки, которые специфически направлены на стволовой неоантиген, могут быть использованы в методах лечения рака.
«Лечить» относится к терапевтическому использованию композиции Т-клеток в соответствии с настоящим изобретением. В настоящем документе композиция Т-клеток может быть введена индивидууму, имеющему существующее заболевание или состояние, чтобы уменьшить, снизить или улучшить, по меньшей мере, один симптом, связанный с заболеванием, и/или замедлить, уменьшить или блокировать прогрессию заболевания.
Рак может представлять собой, например, рак мочевого пузыря, желудка, пищевода, рак молочной железы, колоректальный рак, рак шейки матки, рак яичников, рак эндометрия, рак почек (почечноклеточный), рак легких (мелкоклеточный, немелкоклеточный и мезотелиому), рак мозга (например, глиомы, астроцитомы, глиобластомы), меланому, лимфому, типы рака тонкой кишки (двенадцатиперстной и тощей), лейкемию, рак поджелудочной железы, гепатобилиарные опухоли, типы рака половых клеток, рак предстательной железы, типы рака головы и шеи, рак щитовидной железы и саркомы. В предпочтительном аспекте изобретения рак представляет собой рак легкого, предпочтительно немелкоклеточный рак легкого. В другом аспекте изобретения рак представляет собой меланому.
Лечение с использованием композиций и способов по настоящему изобретению также может охватывать направленность на циркулирующие опухолевые клетки и/или метастазы, происходящие из опухоли.
Лечение композицией Т-клеток по настоящему изобретению, направленное на один или более стволовых неоантигенов, может помочь предотвратить развитие резистентных к терапии опухолевых клеток, что часто возникает при стандартных подходах.
Способы и применение для лечения рака в соответствии с настоящим изобретением могут быть выполнены в сочетании с дополнительной терапией рака. В частности, композиции Т-клеток в соответствии с настоящим изобретением можно вводить в сочетании с лечением блокадой контрольных точек, костимулирующими антителами, химиотерапией и/или лучевой терапией, направленной терапией или лечением моноклональными антителами.
Ингибиторы контрольных точек включают, но не ограничиваются этим, например, ингибиторы PD-1, ингибиторы PD-L1, ингибиторы Lag-3, ингибиторы Tim-3, ингибиторы TIGIT, ингибиторы BTLA и ингибиторы CTLA-4. Костимулирующие антитела доставляют позитивные сигналы через иммунорегуляторные рецепторы, включая, но, не ограничиваясь этим, ICOS, CD137, CD27 OX-40 и GITR. В предпочтительном варианте осуществления ингибитор контрольной точки представляет собой ингибитор CTLA-4.
Химиотерапевтический агент при использовании в настоящем документе относится к агенту, который разрушает клетку, то есть к агенту, который снижает жизнеспособность клетки. Химиотерапевтический агент может представлять собой цитотоксическое лекарственное средство. Предлагаемый химиотерапевтический агент включает без ограничения алкилирующие агенты, антрациклины, эпотилоны, производные нитрозомочевины, этиленимины/метилмеламин, алкилсульфонаты, алкилирующие агенты, антиметаболиты, аналоги пиримидина, эпиподофилотоксины, ферменты, такие как L-аспарагиназа; модификаторы биологического ответа, такие как IFNα, IL-2, G-CSF и GM-CSF; координационные комплексы платины, такие как цисплатин, оксалиплатин и карбоплатин, антрацендионы, замещенную мочевину, такую как гидроксимочевина, производные метилгидразина, включая N-метилгидразин (MIH) и прокарбазин, супрессорные агенты коры надпочечников, такие как митотан (о,ρ'-DDD) и аминоглутетимид; гормоны и антагонисты, включая антагонисты адренокортикостероидов, такие как преднизон и его эквиваленты, дексаметазон и аминоглютетимид; прогестин, такой как гидроксипрогестерона капроат, медроксипрогестерона ацетат и мегестрола ацетат; эстроген, такой как диэтилстильбэстрол и эквиваленты этинилэстрадиола; антиэстроген, такой как тамоксифен; андрогены, включая пропионат тестостерона и флуоксиместерон/эквиваленты; антиандрогены, такие как флутамид, аналоги гонадотропин-рилизинг гормонов и лейпролид; и нестероидные антиандрогены, такие как флутамид.
«В сочетании» может относиться к введению дополнительных терапевтических агентов до, одновременно или после введения композиции Т-клеток в соответствии с настоящим изобретением.
В дополнение или в качестве альтернативы сочетания с ингибированием контрольных точек композиция Т-клеток по настоящему изобретению может быть также генетически модифицирована для придания им устойчивости к иммунным контрольным точкам с использованием методов редактирования генома, включая, но, не ограничиваясь этим, TALEN и Crispr/Cas. Такие методы известны в данной области техники, смотри, например, патент US20140120622. Методы редактирования генома могут быть использованы для предотвращения экспрессии иммунных контрольных точек, экспрессируемых Т-клетками, включая, но, не ограничиваясь этим, PD-1, Lag-3, Tim-3, TIGIT, BTLA CTLA-4 и их сочетания. Т-клетка, как обсуждалось в настоящем документе, может быть модифицирована любым из этих методов.
Т-клетку в соответствии с настоящим изобретением можно также генетически модифицировать для экспрессии молекул, повышающих хоминг в опухоли, и/или для доставки медиаторов воспаления в микроокружение опухоли, включая без ограничения цитокины, растворимые иммуномодулирующие рецепторы и/или лиганды.
Изобретение будет далее описано с помощью примеров, которые предназначены для помощи специалисту в данной области техники в осуществлении изобретения и никоим образом не предназначены для ограничения объема изобретения.
ПРИМЕРЫ
Пример 1 - Идентификация стволовых неоантигенов в опухолях немелкоклеточного рака легкого
Образцы опухоли из опухоли немелкоклеточного рака легкого (NSCLC) подвергали глубокому анализу последовательности экзонов для определения степени внутриопухолевой гетерогенности (ITH), мутационной нагрузки в каждой области опухоли и дифференциации мутаций, присутствующих во всех опухолевых клетках, от мутаций, присутствующих только в субпопуляции. Параллельно суспензии одиночных клеток, полученные из тех же областей опухоли, обрабатывали, аликвотировали и хранили для последующего анализа и экспансии in vitro.
Идентификация вариантов одиночных нуклеотидов по данным секвенирования экзома
Секвенирование экзома проводили на образцах из множества областей, выделенных из опухолей NSCLC. Считывания исходных спаренных концов (100 п.о.) в формате FastQ, сгенерированные контуром lllumina, выравнивали с полной сборкой генома hg19 (включая неизвестные контиги), полученной из пакета GATK 2.8, с использованием bwa mem (bwa-0.7.7) (Li and Durbin; 2009; Bioinformatics; 25(14):1754-60). Инструменты Picard v1.107 затем применяли для очистки, сортировки и объединения файлов из одной и той же области пациента и для удаления повторных считываний (http://broadinstitute.github.io/picard). Показатели контроля качества получали с использованием сочетания инструментов Picard (1.107), GATK (2.8.1) и FastQC (0.10.1) (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/).
SAMtools mpileup (0.1.16) (Li et al.; Bioinformatics; 2009; 25(16); 2078-2079) использовали для определения нереференсных положений в образцах опухоли и зародышевой линии. Основания с баллами Phred <20 или считывания с качеством картирования <20 игнорировали. Вычисление BAQ отключали, и коэффициент качества усеченного картирования устанавливали на 50.
Соматические варианты одиночных нуклеотидов (SNVs) между опухолью и соответствующей зародышевой линией определяли с использованием VarScan2 somatic (v2.3.6) (Koboldt et al.; Genome Res. 2012, 22: 568-576), используя выход SAMtools mpileup. Использовали параметры по умолчанию за исключением минимального покрытия для образца зародышевой линии, установленного на 10, минимальная частота варианта составляла 0,01 и чистота опухоли 0,5. Для извлечения соматических вариантов затем использовали VarScan2 processSomatic.
Полученные вызовы SNV отфильтровывали на ложно положительные срабатывания с использованием связанного с Varscan2's скрипта fpfilter.pl, выполнив первый запуск данных через bam-readcount (0.5.1). Кроме того, применяли дополнительную фильтрацию, с помощью которой варианты принимали только в том случае, если они присутствуют в≥5 считываниях и с частотой вариантного аллеля (VAF) ≥5%, по меньшей мере, в одной области опухоли при VAF зародышевой линии≤1%. Если был найден вариант, отвечающий этим критериям в одной области, то порог VAF снижали до≥1% для обнаружения вариантов с низкой частотой в других областях опухоли.
Анализ количества копий
Анализируемый экзом образца SNP и данные о количестве копий из пары опухоль-нормальная ткань генерировали с использованием VarScan2 (v2.3.6). Varscan2 для анализа числа копий запускали с использованием параметров по умолчанию, за исключением минимального покрытия (8) и отношения данных. Отношение данных рассчитывали на основе каждого образца, как описано в статье (Koboldt et al.; Genome Res.; 2012; 22: 568-576). Выходной сигнал от Varscan затем обрабатывали с использованием пакета Sequenza R 2.1.1 для предоставления сегментированных данных числа копий, и оценивали насыщенность клетками и плоидность для всех образцов на основе данных последовательности экзома. Использовали следующие настройки: breaks.method='full', gamma=40, kmin=5, gamma.pcf=200, kmin.pcf=200.
Анализ РНК-seq
Считывания исходных спаренных концов обрезали и выравнивали относительно эталонного генома и транскриптома человека с использованием Tophat (версия 1.3.3) (Trapnell et al.; 2009; Bioinformatics; 25(9):1105-11). Затем величины экспрессии вычисляли как фрагменты на килобазу экзона на миллион картируемых фрагментов (FPKM) с использованием Cufflinks (Trapnell et al.; 2010; Nat Biotech; 28(5); 511-5) с применением нормирования верхних квартилей, коррекции смещения фрагментов и многократной коррекции.
Идентификация стволовых SNV
Набор идентифицированных SNV классифицировали как стволовые варианты или варианты ветвления на основе оценок их фракций раковых клеток (CCF) во всех секвенированных областях опухоли. Вкратце, CCF, описывающую долю раковых клеток, несущих мутацию, рассчитывают путем суммирования числа копий и чистоту оценивают с использованием частот вариантных аллелей.
Для каждого варианта ожидаемую частоту вариантных аллелей (VAF), учитывая CCF, рассчитывали следующим образом:
VAF(CCF)=p*CCF/CPNnorm(1-p)+p*CPNmut
где CPNmut соответствует количеству локальных копий опухоли, p представляет собой чистоту опухоли, и CPNnorm представляет собой количество локальных копий спаренного нормального образца.
Для данной мутации с альтернативными считываниями «a» и глубиной «N» вероятность данной CCF оценивали с использованием биномиального распределения
P(CCF)= бином(a|N, VAF(CCF)).
Значения CCF затем вычисляли по равномерной сетке 100 значений CCF (0,01, 1) и затем нормировали для получения апостериорного распределения.
Любой SNV, присутствующий в клонах (95% доверительный интервал CCF≥0,75) во всех областях опухоли, рассматривали как стволовой. Напротив, любой SNV, присутствующий только в подмножестве областей опухоли или с 95% доверительным интервалом CCF≤0,75 в любой области, рассматривали как вариант ветвления (смотри фигуру 2 для обобщения).
Пример 2 - Прогнозирования типов HLA
Для каждого индивидуума файлы FASTQ секвенирования полного генома зародышевой линии, сопоставляли картированием с эталонным файлом FASTA, содержащим последовательности для известных аллелей HLA. Картирование выполняли с использованием Razers3 (Weese et al.; Bioinformatics; 2012; 28(20): 2592-2599) с порогом процента идентичности 90, с максимумом одного попадания и диапазоном расстояний 0. После картирования сгенерированные файлы SAM преобразовывали в FASTQ и использовали в качестве входных данных для алгоритма прогнозирования Optitype (Szolek et al.; Bioinformatics; 2014; 30(23):3310-3316) с параметрами по умолчанию. Optitype генерирует предсказанный 4-значный тип HLA разрешения для каждого пациента, который сохраняли для использования при прогнозировании связывания HLA.
Прогнозирование связывания HLA
Кодирующие мутации, запускаемые во множестве областей полного экзома опухоли, использовали для генерации всех возможных 9-11-членных мутантных пептидов из неоантигена, захватывая мутированную аминокислоту в каждом положении n-члена.
Таким образом, для данной мутации SNV получено в целом 446 пептидов. Кроме того, получены также соответствующие пептиды дикого типа.
В качестве входных данных для NetMHC (Lundegaard et al.; 2008; Nucleic Acids Res; 36:W509-12) использовали файл fasta, содержащий все последовательности пептидов мутантного и дикого типа и спрогнозированный 4-значный тип HLA, который прогнозирует аффинность связывания каждого пептида со специфическими аллелями HLA пациента. Пептиды, которые, как предполагается, связываются при <500 нМ, классифицировали как предполагаемые слабые неоантигены, тогда как пептиды, которые связываются при <50 нМ, классифицировали как предполагаемые сильные неоантигены. Кроме того, для каждого пептида рассчитывали дельта-балл, отражающий различие в связывании между мутантными пептидами и пептидами дикого типа (смотри фигуру 3).
Пример 3 - Идентификация предполагаемых стволовых неоантигенов
Все предполагаемые неоантигены классифицировали как стволовые неоантигены или как неоантигены ветвления на основе фракции их раковых клеток в секвенированных областях опухоли (как описано в примере 1). Связывающие пептиды, которые происходят от мутации, обнаруженной в каждой секвенированной области опухоли, идентифицировали как потенциальные стволовые неоантигены (как описано в примере 2).
Фильтрация предполагаемых стволовых неоантигенов с использованием данных RNAseq
Все предполагаемые стволовые неоантигены дополнительно фильтровали с использованием данных RNAseq. В частности, среднюю длину транскрипта использовали для преобразования из рассчитанного FPKM в TPM (транскрипты на миллион) и идентификации предполагаемого стволового неоантигена как неоантигена, который экспрессируется с медианой более 10 TPM.
Пример 4. Получение и наращивание инфильтрирующих опухоль лимфоцитов (TILs) из образцов NSCLC
Опухоли измельчали в стерильных условиях с последующим ферментативным гидролизом (RPMI-1640 с либеразой TL (Roche) и ДНКазой I (Roche) высокой чистоты для исследования) при 37°C в течение 30 минут перед механическим разделением тканей с использованием gentleMACS (Miltenyi Biotech). Полученные суспензии отдельных клеток обогащали лейкоцитами путем прохождения через градиент фиколл-пака. Живые клетки подсчитывали и замораживали при -80°С перед переносом в жидкий азот. TILs наращивали с использованием протокола быстрого наращивания (REP) в колбах T25, содержащих среду EX-VIVO, дополненную 10% сывороткой человека, растворимым анти-CD3 (OKT3), 6000 МЕ/мл рекомбинантным (rhlL-2) человека и 2×107 облученными РВМС, объединенными от 3 аллогенных здоровых доноров. Как только экспансия была очевидна, каждые три дня добавляли свежие среды, содержащие rhlL-2 3000 МЕ/мл. После 2 недель наращивания TILs подсчитывали, подвергали фенотипированию и замораживали при -80°C перед использованием в соответствующих анализах или для долгосрочного хранения в жидком азоте.
Идентификация Т-клеток, специфичных для неоантигена (NES), с использованием растворимых мультимеров пептид/HLA
Синтезировали наиболее ранжированные последовательности пептидов неоантигенов (n=240) и использовали для получения флуоресцентномеченных, полученных индивидуально мультимеров MHC для идентификации Т-клеток NES в наращенных инфильтрировавших опухоль лимфоцитах (TILs). Наращенные in vitro TILs окрашивали флуоресцентными, полученными индивидуально мультимерами MHC, нагруженными спрогнозированными мутантными пептидами или контрольными пептидами цитомегаловируса (CMV), и анализировали с помощью многоцветной проточной цитометрии (смотри фигуру 4).
Пример 5 - Киллерная активность T-клеток NES ex vivo и in vivo
Линии аутологичных опухолевых клеток и наращенные Т-клетки NES использовали для демонстрации способности этих Т-клеток уничтожать опухолевые мишени in vitro и in vivo.
Идентифицировали линии опухолевых клеток, несущие стволовые мутации, и аутологичные наращенные Т-клетки NES. Фиксированное количество опухолевых клеток высевали на 96-луночные планшеты с варьирующим количеством Т-клеток NES. Киллерную активность Т-клеток NES in vitro оценивали в разные моменты времени (через 4-16 часов после стандартных анализов с помощью проточной цитометрии). Для анализов in vivo иммунодефицитных мышей (NSG) подкожно нагружали линиями опухолевых клеток и после приживления либо оставляли без лечения, либо они получали в./в. инфузию 1-5×106 аутологичных T-клеток NES, наращенных in vitro.
Рост опухоли в группах с лечением и без него измеряли с течением времени (каждые 3 дня).
Пример 6 - Т-клетки, направленные на клональный неоантиген, способствуют отторжению сформировавшихся гетерогенных опухолей, в то время как Т-клетки, направленные на субклональный неоантиген, не могут этого сделать
Для сравнения противоопухолевой активности in vivo Т-клеток, направленных на клональные неоантигены, по сравнению с клетками, направленными на субклональные неоантигены, изобретатели использовали мышиную модель меланомы (линия B16) и трансгенные Т-клетки с рецептором Т-клеток (TCR Tg), узнающие клональный неоантиген (trp1) или субклональный неоантиген (OTII).
Т-клетки, специфичные для неоантигенов: CD4+ Trp1 TCR Tg-клетки узнают пептид, происходящий от Tyrpl, антигена, присутствующего во всех клетках меланомы B16. Trp1 TCR Tg-клетки генерируются у мышей с отсутствием Tyrpl, поэтому они узнают Tyrpl как неоантиген.
CD4+ OTII TCR Tg-клетки узнают пептид, происходящий от овальбумина (OVA), модельного неоантигена, который может быть искусственно введен генной инженерией в линии опухолевых клеток B16.
Для моделирования клональных и субклональных неоантигенов изобретатели использовали клетки меланомы B16 мыши (экспрессирующие Tyrpl) и клетки B16-OVA, также экспрессирующие Tyrpl, но дополнительно трансдуцированные для экспрессии OVA. Путем смешивания этих двух клеточных линий изобретатели создали модель, в которой Tyrpl представляет собой клональный неоантиген (экспрессируемый всеми опухолевыми клетками) и OVA - субклональный неоантиген (экспрессируемый только некоторыми опухолевыми клетками).
Вкратце мышам B6 в день 0 вводили смесь 12,5×104 B16 и 12,5×104 клеток B16-OVA. На 8-й день после инокуляции опухоли и когда опухоли становились пальпируемыми, мышей подвергали сублетальному облучению (5 Гр), и они получали внутривенную (в./в.) инфузию 30×104 клеток CD4 OT-II (реактивных в отношении субклонального неоантигена) или 6×104 CD4 клеток Trp-1 (реактивных в отношении клонального неоантигена) и 0,2 мг антител против CTLA-4 внутрибрюшинно (в./б.). Мыши получали две дополнительные дозы антител против CTLA-4 на 11 и 14 день (0,1 мг). В качестве контроля изобретатели использовали группу мышей, нагруженных опухолью, но оставленных без лечения.
Способность клеток OTII вызывать отторжение опухолей B16-OVA демонстрировали в дополнительной группе мышей, инокулированных только 25×104 клетками B16-OVA (в этом случае OVA экспрессируется всеми клетками B16-OVA в опухолевой массе, следовательно, представляет собой клональный неоантиген). На 8-й день мышей подвергали сублетальному облучению (5 Гр), и они получали 30×104 клеток CD4 OT-II в./в. и 0,2 мг антитела против CTLA-4 в./б. Мыши получали две дополнительные дозы антител против CTLA-4 на 11 и 14 день (0,1 мг).
Результаты
У контрольной группы мышей, нагруженных гетерогенной смесью опухолей (B16/B16-OVA), содержащей и клональный (tyrpl), и субклональный (OVA) неоантигены, и оставленных без лечения, опухоли росли, и их необходимо было забить между 20 и 30 днями после нагрузки опухолью (фиг. 5А). Поразительно, что все мыши, нагруженные смесью опухолей B16/B16-OVA, но которых лечили TRP1 TCR Tg-клетками, направленными на клональный неоантиген, были в состоянии отторгать свои опухоли (фиг. 5B).
Напротив, когда мышей лечили OTII TCR Tg-T-клетками, направленными на субклональный неоантиген, ни одна из мышей не обладала способность отторгать свою опухоль (фиг. 1C). В этой группе наблюдалась небольшая задержка прогрессии опухоли, предполагающая потенциальный контроль клетками B16-OVA, экспрессирующими субклональный неоантиген, с последующей прогрессией из-за невозможности отторгания опухолевых клеток, не экспрессирующих OVA (субклональный неоантиген) (фиг. 5C). Наконец (фиг. 5D) демонстрирует способность OTII TCR Tg-T-клеток отторгать прижившиеся опухоли, когда все клетки в опухоли экспрессируют неоантиген OVA. Каждая линия каждого графика представляет собой независимую мышь. Для этих экспериментов использовали 6 мышей на группу. На (фиг. 5Е) показаны все экспериментальные группы и средний размер опухолей в каждой группе.
Данные демонстрируют превосходную способность Т-клеток, направленных на клональный неоантиген, отторгать прижившиеся опухоли по сравнению с Т-клетками, направленными только на субклональные неоантигены, не экспрессируемые всеми опухолевыми клетками в пределах гетерогенной опухолевой массы.
Все документы, упомянутые в настоящем описании, включены в него в качестве ссылки в полном объеме с обращением особого внимания на предмет, для которого они указаны. Различные модификации и варианты описанных способов и систем изобретения будут очевидны специалистам в данной области техники без отхода от объема и сущности изобретения. Хотя изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что заявляемое изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. Действительно, различные модификации описанных способов осуществления изобретения, которые очевидны для специалистов в области молекулярной биологии, клеточной иммунологии или смежных областей, предназначены для включения в объем последующей формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| МЕТОД | 2016 |
|
RU2739036C2 |
| Выбор раковых мутаций для создания персонализированной противораковой вакцины | 2019 |
|
RU2809620C2 |
| СПОСОБЫ ПРОГНОЗИРОВАНИЯ ПРИМЕНИМОСТИ БЕЛКОВ ИЛИ БЕЛКОВЫХ ФРАГМЕНТОВ ДЛЯ ИММУНОТЕРАПИИ | 2017 |
|
RU2782336C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ПРИМЕНЕНИЕМ АКТИВИРОВАННЫХ T-КЛЕТОК | 2016 |
|
RU2729362C2 |
| ИДЕНТИФИКАЦИЯ, ПРОИЗВОДСТВО И ПРИМЕНЕНИЕ НЕОАНТИГЕНОВ | 2016 |
|
RU2729116C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ПРОИЗВОДСТВА Т-КЛЕТОК | 2018 |
|
RU2793344C2 |
| Способы и композиции для лечения опухолей | 2017 |
|
RU2741786C2 |
| СПОСОБЫ ПРОГНОЗИРОВАНИЯ ПРИМЕНИМОСТИ СПЕЦИФИЧНЫХ ДЛЯ ЗАБОЛЕВАНИЯ АМИНОКИСЛОТНЫХ МОДИФИКАЦИЙ ДЛЯ ИММУНОТЕРАПИИ | 2018 |
|
RU2799341C2 |
| Способ отбора неоэпитопов | 2019 |
|
RU2826184C2 |
| СОСТАВЫ ВАКЦИН ПРОТИВ НЕОПЛАЗИИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2016 |
|
RU2753246C2 |
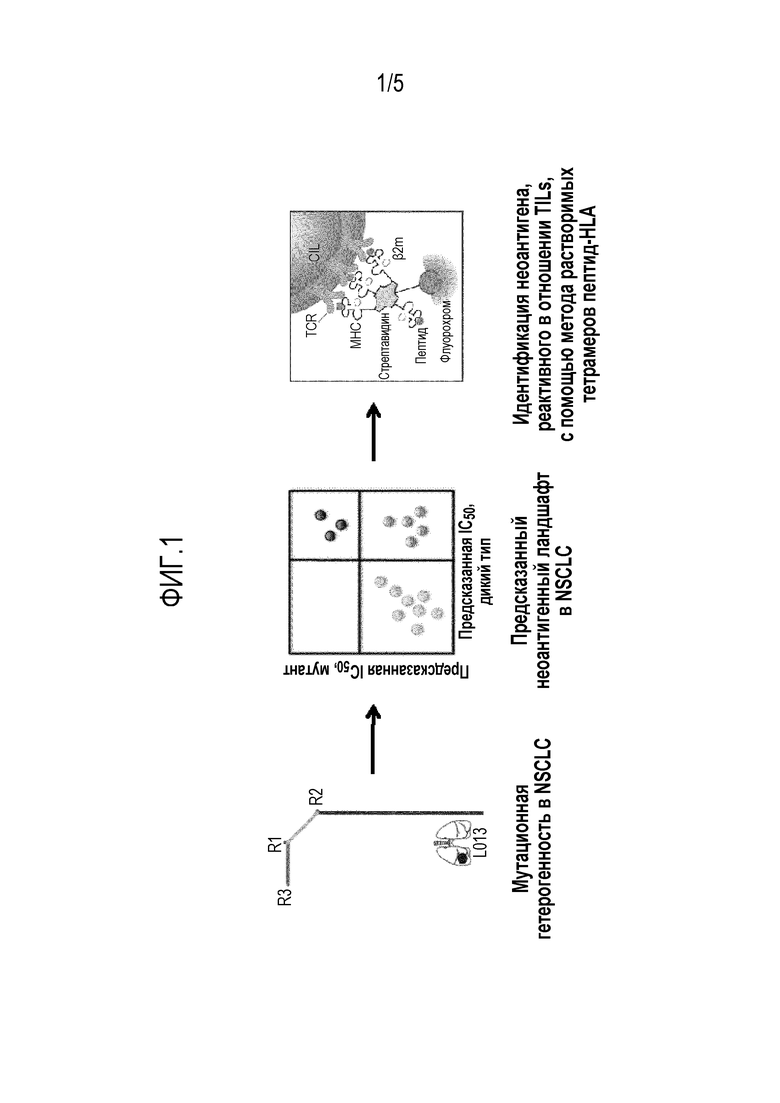
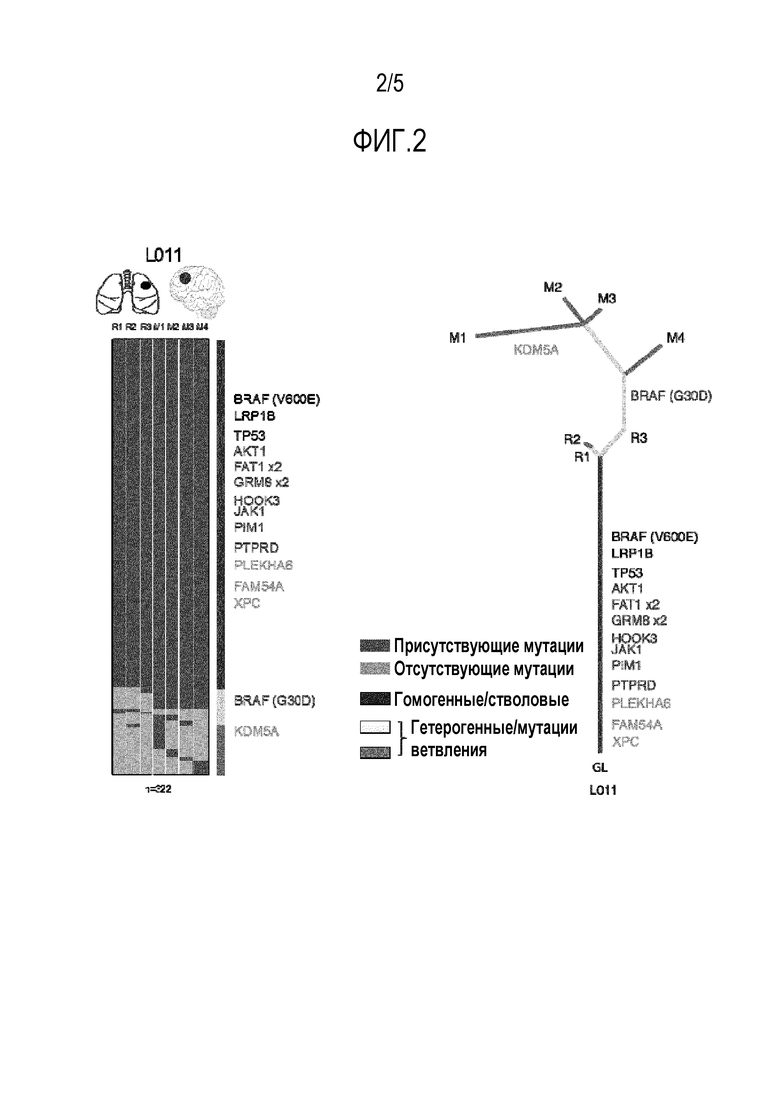
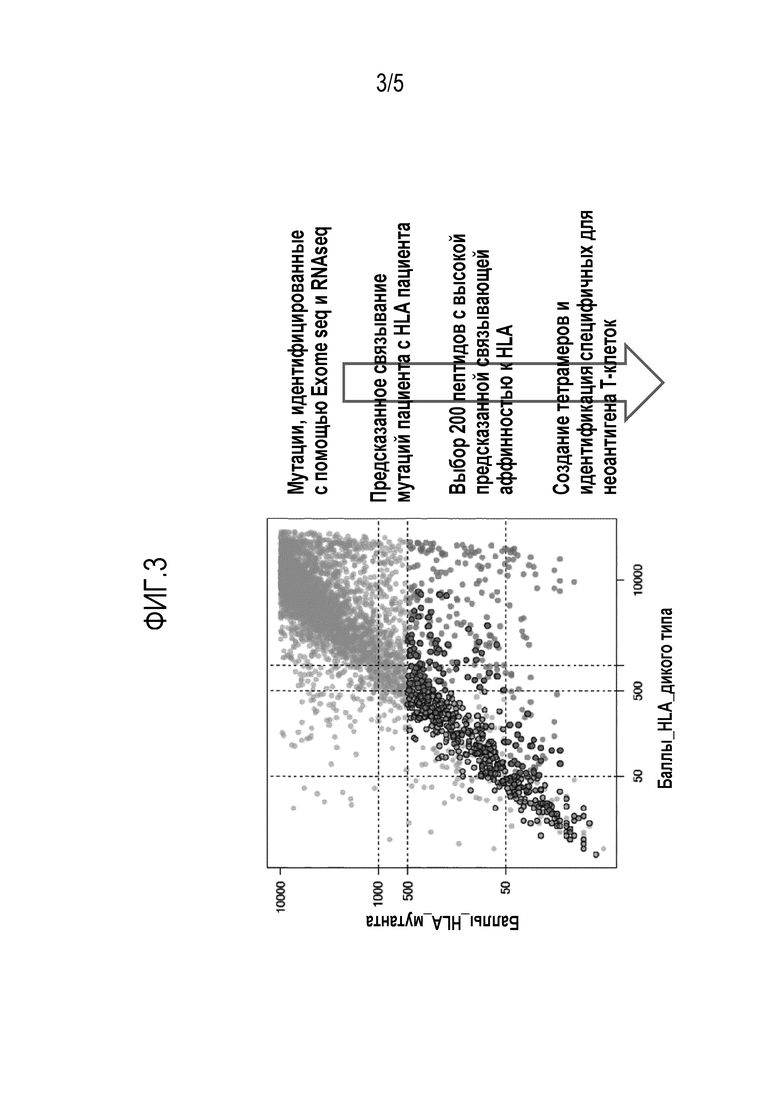
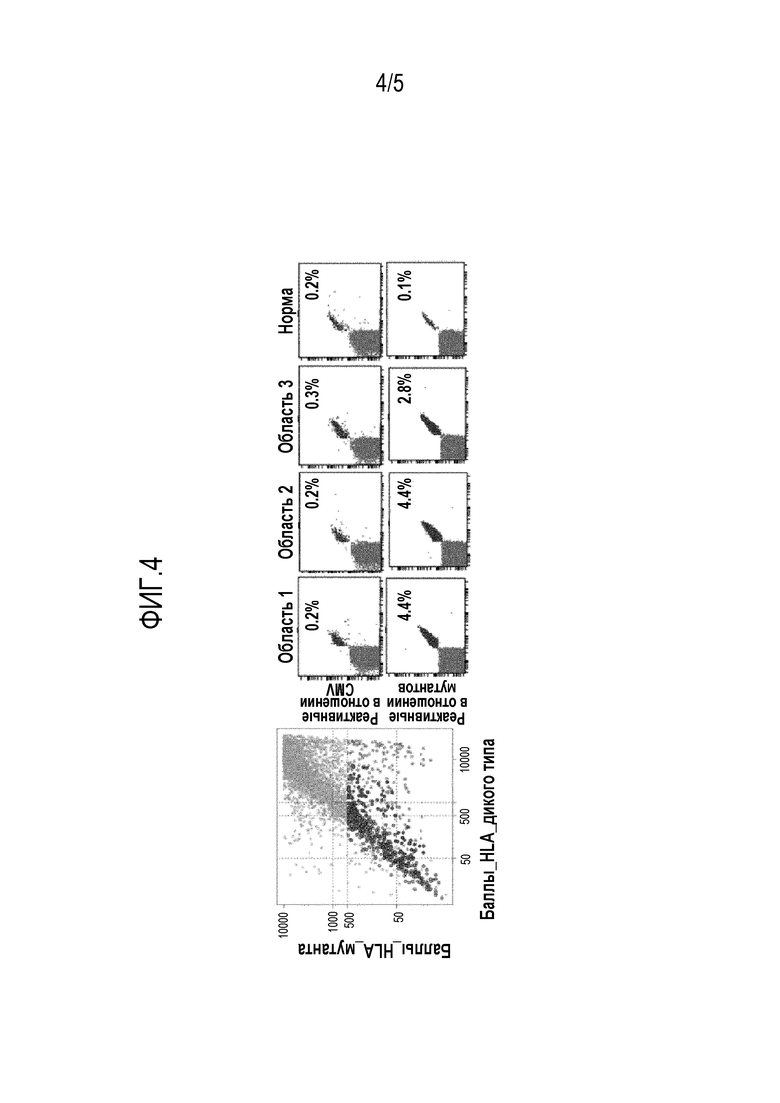
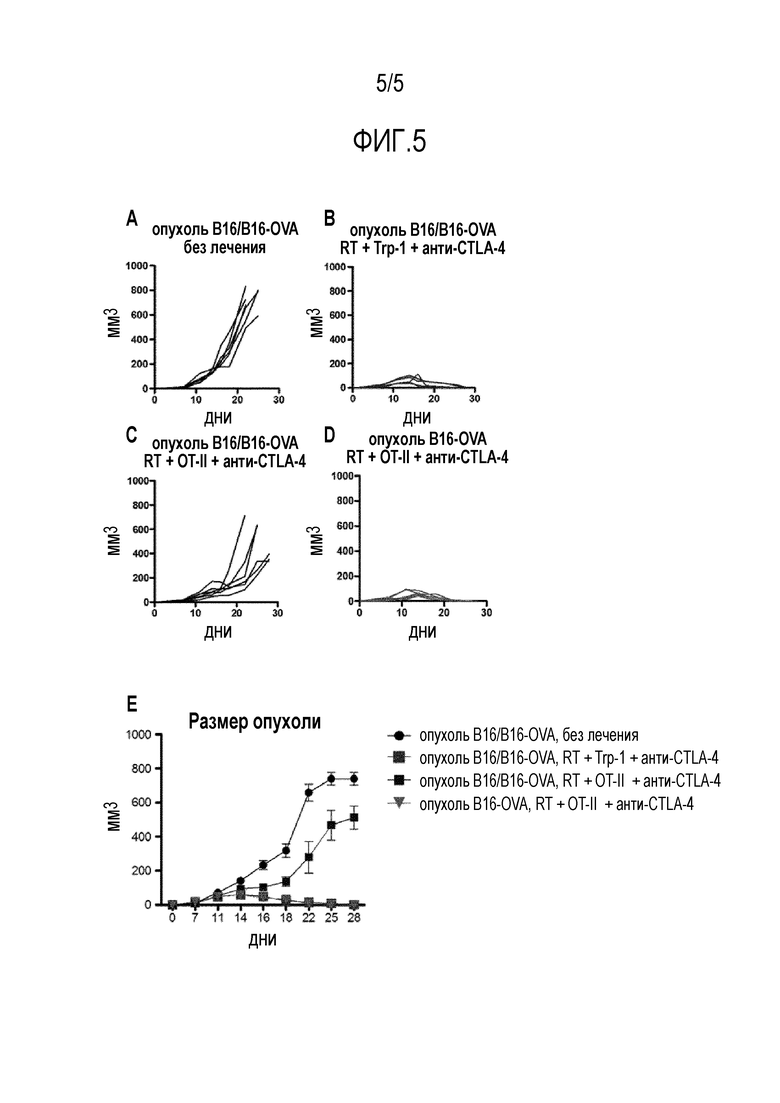
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая композицию Т-клеток для применения в лечении рака у индивидуума, способ лечения рака у индивидуума (варианты). Композиция Т-клеток включает селективно наращенные Т-клетки, которые направлены на клональные неоантигены, характерные для рака индивидуума, где клональный неоантиген имеет 95% доверительный интервал фракции раковых клеток (CCF), больший или равный 0,75. В одном из вариантов реализации рак представляет собой немелкоклеточный рак легкого (NSCLC) или меланому. Изобретение расширяет арсенал средств для лечения рака. 3 н. и 20 з.п. ф-лы, 5 ил., 6 пр.
1. Композиция Т-клеток для применения в лечении рака у индивидуума, которая включает селективно наращенные Т-клетки, которые направлены на клональные неоантигены, характерные для рака индивидуума, где клональный неоантиген имеет 95% доверительный интервал фракции раковых клеток (CCF), больший или равный 0,75.
2. Композиция Т-клеток по п. 1, которая обогащена Т-клетками, которые специфичны в отношении клональных неоантигенов.
3. Композиция Т-клеток по пп. 1 и 2, где наращенная популяция клональных Т-клеток, реагирующих на стволовой неоантиген, имеет более высокую активность, чем ненаращенная популяция Т-клеток, как измерено с помощью ответа указанной популяции Т-клеток на повторную стимуляцию пептидом клонального неоантигена.
4. Композиция Т-клеток по п. 3, где указанную активность измеряют посредством измерения продукции цитокинов, и где более высокая активность представляет собой увеличение активности в 5-10 или более раз.
5. Композиция Т-клеток по п. 1, где указанные Т-клетки были селективно наращены способом, который включает стадии:
i) предоставления образца, включающего популяцию Т-клеток, которая способна специфически узнавать указанный неоантиген; и
ii) совместного культивирования указанной популяции Т-клеток с композицией, включающей указанный неоантиген или пептид неоантигена, и с антигенпрезентирующей клеткой;
где указанный неоантиген был идентифицирован в образце опухоли от указанного индивидуума как представляющий собой клональный неоантиген.
6. Композиция Т-клеток по п. 5, где указанные Т-клетки избирательно наращивают с использованием множества клональных неоантигенов или пептидов клональных неоантигенов, где каждый из указанных пептидов включает отличную стволовую мутацию.
7. Композиция Т-клеток по п. 6, где указанное множество пептидов включает от 2 до 100 пептидов.
8. Композиция Т-клеток по любому из пп. 5-7, где указанная антигенпрезентирующая клетка нагружена или пульсирована пептидом, происходящим из указанного клонального неоантигена.
9. Композиция Т-клеток по любому из пп. 5-8, где указанная антигенпрезентирующая клетка представляет собой дендритную клетку.
10. Композиция Т-клеток по любому из пп. 5-9, где указанную популяцию Т-клеток культивируют совместно с IL-2 или антителами к CD3 и/или CD28.
11. Композиция Т-клеток по любому из пп. 5-10, где указанную популяцию Т-клеток выделяют из образца опухоли пациента, подлежащего лечению.
12. Композиция Т-клеток по п. 11, где указанная популяция Т-клеток содержит инфильтрирующие опухоль лимфоциты (TIL).
13. Композиция Т-клеток по любому из пп. 5-12, где указанная популяция Т-клеток содержит включает CD8+ Т-клетки, CD4+ Т-клетки или CD8+ и CD4+ T-клетки.
14. Композиция Т-клеток по любому из пп. 5-13, где обеспечивают по меньшей мере первую и вторую Т-клетки, где указанная первая Т-клетка направлена на первый клональный неоантиген, созданный первой клональной мутацией, а указанная вторая Т-клетка направлена на второй клональный неоантиген, созданный второй клональной мутацией.
15. Композиция Т-клеток по любому из пп. 5-14, где указанную наращенную популяцию Т-клеток обогащают Т-клетками, которые направлены на клональные неоантигены, по сравнению с начальным образцом.
16. Композиция Т-клеток по любому предшествующему пункту, где указанная композиция Т-клеток используется в сочетании с ингибитором контрольных точек.
17. Композиция Т-клеток по любому предшествующему пункту, где рак представляет собой немелкоклеточный рак легкого (NSCLC) или меланому.
18. Способ лечения рака у индивидуума, который включает введение композиции Т-клеток по любому предшествующему пункту индивидууму.
19. Способ по п. 18, который включает следующие стадии:
(i) выделение образца, содержащего Т-клетки, от индивидуума;
(ii) идентификация и наращивание популяции Т-клеток, которые направлены на клональный неоантиген; и
(iii) введение клеток из (ii) индивидууму.
20. Способ лечения рака у индивидуума, где указанный способ включает:
(а) идентификацию клонального неоантигена в опухоли от индивидуума, которая включает стадии:
i) определения мутаций, присутствующих в образце, выделенном из опухоли; и
ii) идентификации клональной мутации, которая представляет собой мутацию, присутствующую, по существу, во всех опухолевых клетках; и
iii) идентификации клонального неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает клональную мутацию;
где клональный неоантиген имеет 95% доверительный интервал фракции раковых клеток (CCF), больший или равный 0,75; или
i) определения мутаций, присутствующих во множестве образцов, выделенных из опухоли;
ii) идентификации клональной мутации, которая представляет собой мутацию, присутствующую во всех образцах;
iii) идентификации клонального неоантигена, который представляет собой антиген, кодируемый последовательностью, которая включает клональную мутацию;
где клональный неоантиген имеет 95% доверительный интервал фракции раковых клеток (CCF), больший или равный 0,75; и
b) идентификацию Т-клетки из образца, выделенного от индивидуума, которая способна специфически узнавать указанный клональный неоантиген;
c) наращивание Т-клеток для получения популяции Т-клеток, которые направлены на клональный неоантиген; и
d) введение указанной популяции Т-клеток указанному индивидууму.
21. Способ по любому из пп. 19, 20, в котором образец представляет собой образец опухоли, крови или ткани от индивидуума.
22. Способ по любому из пп. 18-21, в котором популяцию Т-клеток вводят в сочетании с ингибитором контрольных точек.
23. Способ по любому из пп. 18-21, где рак представляет собой немелкоклеточный рак легкого (NSCLC) или меланому.
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Т-КЛЕТОЧНАЯ ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ РАССЕЯННОГО СКЛЕРОЗА | 1998 |
|
RU2209633C2 |

