Область техники, к которой относится изобретение
Изобретение относится к области медицины и молекулярной биологии, в частности, к использованию генноинженерных биотехнологий в получении препаратов для лечения и экстренной профилактики COVID-19 путем нейтрализации компонентов вируса SARS- CoV- 2, отвечающих за его взаимодействие с клетками организма человека и репликацию в них при условии пролонгированного системного действия в тест-системах in vivo.
Уровень техники
К августу 2022 года инфекция, вызванная коронавирусом 2 и проявляющаяся тяжелым острым респираторным синдромом (SARS-CoV-2), служила причиной более 6,4 миллиона смертей по всему миру[1].Более того, проблемой стала длительность заболевания или пост-инфекционный синдром COVID-19 (post-acute COVID-19) — это состояние, характеризующееся различными симптомами, включая усталость, одышку, когнитивные нарушения и психические расстройства, которые сохраняются через четыре недели после появления острых симптомов[2]. Существует также такое понятие как постковидный синдром – симптомо-комплекс, развившийся во время или после заболевания, вызванного новой коронавирусной инфекцией, продолжающийся более 12 недель и не являющийся следствием альтернативного диагноза [3].Исследования показали, что 10–55 % пациентов с COVID-19, в том числе с легкой или бессимптомной формой, страдают от подобных изнурительных симптомов[4]. В настоящее время установлено также, что вакцинация против COVID-19 до заражения SARS-CoV-2 связана с более низким риском длительного заболевания COVID, однако существует довольно значительная когорта людей, прошедших вакцинацию уже после заражения, у которых длительное течение этой инфекции не предупреждалосьвакцинацией и не сопровождалось изменениями в ее симптоматике [5]. По состоянию на 10 февраля 2023 г. во всем мире было введено более 6 миллионов доз вакцины против COVID-19, и, по оценкам, 200 миллионов человек живут с продолжающимися симптомами этого заболевания [6]. В соответствии с современными взглядами на этиопатогенетические механизмы развития COVID-19 названные проявления не связаны с длительностью пребывания коронавируса в организме человека и тяжестью вызываемых им поражений, но, несомненно, обусловлены определенными системными эффектами, запускаемыми этим возбудителем [7, 8].
Отмеченные особенности делают актуальными исследования по получению препаратов для лечения и экстренной профилактики COVID-19, которые были бы направлены на значительное подавление инвазивной и репликативной активности SARS- CoV-2 и снижали бы эффективность его длительного системного воздействия на чувствительный организм.
На современном этапе изучаются два различных подхода к перепрофилированию традиционных лекарств и разработке новых терапевтических препаратов: предотвращение проникновения вируса в клетки хозяина и подавление различных этапов репликации вируса внутри клеток [7].
Так, известно, что коронавирусы SARS-CoV-2 нацелены на рецепторы ангиотензинпревращающего фермента 2 (АСЕ2), сверхэкспрессированные в клетках тканей легких и желудочно-кишечного тракта в организме человека. Рецептор-связывающий домен (RBD) белка-шипа (S) на поверхности коронавируса связывается с ACE2 на плазматической мембране инфицированных клеток, инициируя рецептор-опосредованный эндоцитоз.В настоящее время изучаются три стратегии, позволяющие заблокировать это взаимодействие и устранить эту инфекцию [8].Во-первых, можно вводить большое количество растворимых миметиков RBD для взаимодействия с рецепторами ACE2, предотвращая связывание вируса и его проникновение в клетки.Во-вторых, для выполнения аналогичной задачи можно вводить антитела или фрагменты одноцепочечных антител (scFv) против рецепторов ACE2.Третья стратегия нацелена непосредственно на RBD коронавируса, используя, в первую очередь, препараты антител или их фрагментов [7].Кроме того, предлагаются лекарственные средства, которые могут блокировать одинили несколько этапов репликации вируса внутри клетки: предотвращать эндоцитоз, ингибировать созревание эндосом и высвобождение вирусного генома, а также репликацию РНК, транскрипцию и трансляцию вирусных белков, чаще всего, путем блокады соответствующих ферментных систем [9-16].
Что касается нейтрализующих RBD агентов, то они включают в себя, помимо моноклональных и поликлональных антител [17, 18, 19], пептиды [20, 21, 22], реконвалесцентную плазму [23, 24] и др.
Несмотря на обширные исследования по разработке различных противовирусных препаратов, особенно, способных ингибировать связывание вируса SARS-CoV-2 с рецепторами ACE2, некоторые аспекты этих подходов потенциально могут ограничивать их эффективность [7].Во-первых, нативные миметики, антитела и их фрагменты обладают относительно низкой стабильностью в кровотоке и могут снижать свою специфическую активность при транспортировке к первичным мишеням (рецепторам ACE2 в клетках легких или RBD на поверхности вируса) [25].Кроме того, антитела представляют собой довольно большие молекулы, что препятствует их проникновению в ткани, а также доступу к стерически экранированным эпитопам [26].Другие ограничения включают возможное связывание лекарств с участками рецепторов ACE2 или RBD, которые не конкурируют с вирусом или рецептором и не препятствуют проникновению вируса [27].
Учитывая эти особенности моноклональных антител, многие исследователи, работающие в этом направлении, сосредоточились на получении однодоменных антител и других рекомбинантных белков с меньшей молекулярной массой, о чем свидетельствуют многочисленные отечественные и зарубежные патенты, например: патенты РФ 2763001 от 24.12.2021 г. [28], 2802825 от 29.08.2022 г. [29], 2794141 от 11.04.2023 г. [30], 2798508 от 23.06.2023 г. [31], а также патенты CША US2023331824 - 2023-10-19 [32], US2023374116 - 2023-11-23 [33], Израиля IL305301 - 2023-10-01 [33], Канады CA3160388 - 2023-11-25 [35], Китая CN114072422 - 2021-12-09 [36], Корейской Республики KR1020220064331 – 2022-05-19 [37], Японии US2023348573-2023-11-02 [38].
В последнее время гораздо большее внимание привлекают пептиды.
Пептиды отличаются малой молекулярной массой, высокой специфичностью и аффинитетом, протеолитической стабильностью [20]. Их относительная простота и экономичность в производстве еще больше повышают их привлекательность. В то же время, несмотря на то, что вируснейтрализующие пептиды (VNPs) обладают многими преимуществами, им недостает эффекторных функций и авидности, присущих иммуноглобулинам [39, 40]. Это наводит на мысль о потенциальном усилении их возможностей, помимо нейтрализации, за счет конъюгации с активными фрагментами. Кроме того, по сравнению с иммуноглобулинами, применение пептидных препаратов часто сталкивается с проблемами из-за более короткого времени циркуляции и ограничений в способах введения [20, 41].
Существует множество способов получения специфичных к RBDSARS-CoV-2 вируснейтрализующих пептидов, отраженных в современной патентной базе, в том числе: CN112513076 – 2022-03-31 [42], JP2022060186 – 2022-04-07 [43], CN113072623 – 2022-01-11 [44], MX2022012545 - 2022-11-07 [45], US2023322865 - 2023-10-12 [46], US2023365659-2023-11-16 [47].
Так, например, американскими исследователями была создана панель коротких минипротеинов, LCB, состоящих из 56 аминокислот, которые с высоким сродством (особенно LCB1) связывали рецептор-связывающий домен (RBD) S белка SARS-CoV-2 и проявляли мощный вируснейтрализующий эффект на модели трансгенных мышей, экспрессирующих человеческий ACE2 (hACE2) [48]. Для распространения вируснейтрализующего действия этих минипептидов на продолжающееся до настоящего времени появление новых вариантов вируса SARS-CoV-2 потребовались разнообразные модификации молекулярной структуры LCB1 c учетом механизмов устойчивости коронавирусов к данному пептиду [49, 50].В способы получения таких модифицированных препаратов входят, в частности: получение укороченных пептидных вариантов LCB1, устойчивых к протеолитической деградации [51]; конъюгация целевого пептида с липопептидами, усиливающими его связывание с вирусными и клеточными мембранами [52]; нативное химическое лигирование с использованием метода конденсации пептидных фрагментов в водных растворах для преодоления ограничения длины молекулы пептида [53]; использование комплекса пептидов с разными сайтами взаимодействия с RBD и другими компонентами вируса [54].
Последнее направление в последние годы развивается особенно активно, а в качестве одного из компонентов комбинированных препаратов рассматриваются, в частности, микробные рибонуклеазы (РНКазы). РНКазы — это ферменты, расщепляющие РНК, способные проникать в клетки млекопитающих, проявляя цитотоксичность и создавая препятствия для размножения вирусов, как путем разрушения их РНК, так и путем разрушения клеток, необходимых для их репликации. Это приводит к весьма разнообразным биологическим последствиям, используемым в разработке ряда стратегий противораковой и противовирусной терапии. Наиболее известными с этой точки зрения микробными РНКазами являются барназа, продуцируемая бактериями вида Bacillusamyloliquefaciens, и биназа, образуемая бактериями вида Bacilluspumilis [55].
Барназа представляет собой внеклеточную рибонуклеазу (РНКазу), секретируемую во внеклеточное пространство в виде одноцепочечного белка из 110 аминокислот.Барназа известна своей высокой стабильностью, благодаря отсутствию дисульфидных связей, посттрансляционных модификаций, двухвалентных катионов или других непептидных компонентов, необходимых для ее функции. У бактерий вида Bacillusamyloliquefaciens вырабатывается еще и барстар – специфический внутриклеточный ингибитор барназы.Барстар прочно связывается с барназой и тем самым ингибирует ее внутриклеточную ферментативную активность, защищая клетки от повреждающего действия этой РНКазы [56]. Комплекс барназы и барстара довольно стабилен и распадается только в присутствии сильно диссоциирующих агентов, таких как 5 М гуанидин HCl или 0,1% додецилсульфат натрия. Было показано, что барстар покрывает активный центр барназы, защищая тем самым бактериальную клетку от рибонуклеазы, которая может случайно синтезироваться на свободных рибосомах в цитоплазме [57]. При этом N- и C-концы как барназы, так и барстара, расположены далеко от границы их взаимодействия [58], поэтому оба белка могут быть слиты с другими функциональными модулями без потери функции или сродства барназы и барстара.Этот механизм контроля повреждающего действия барназы успешно используется в настоящее время, например, при разработке противораковых средств [59, 60].
Целью данного изобретения служила разработка способа получения химерного полипептида c расширенным спектром анти-SARS-CoV-2 активности вследствие наличия в его составе как RBD-узнающего пептида LCB1, так и барназы, и пролонгирования противовирусного эффекта и придания ему системного характера in vivo путем интеграции в структуру этого химерного полипептида альбуминсвязывающего домена.
Прототипом данного изобретения может служить патент РФ 2778942 от 29.08.2022 [61], который касается создания рекомбинантной плазмиды, штамма-продуцента и способа получения рекомбинантного белка с условным названием Таргевир.
Таргевир – химерный бифункциональный белок, состоящий из целевого полипептида LCB1 или LCB3 и высокоактивной бактериальной рибонуклеазы барназы, соединенных гибким пептидным линкером. Этот рекомбинантный белок обладает способностью специфически связываться с вирионами SARS-CoV-2, имеющими на своей поверхности S-белок, и блокировать их связывание с рецептором ACE-2, представленным на поверхности клеток, чувствительных к заражению вирусом. При интернализации в клетки вместе с вирусом Таргевир обладает статическим воздействием за счет гидролиза вирусной РНК и последующего прекращения биосинтеза белка, необходимого для синтеза новых вирионов. При взаимодействии PAB с природным ингибитором барназы – рекомбинантным белком барстар– цитотоксический эффект Таргевир моментально отменяется, что может быть использовано в случае гиперчувствительной реакции организма на РНКазную активность или передозировки лекарственного средства. Сочетание таких свойств у PAB выгодно отличает его от всех известных и применяемых в клинической практике аналогов.
В то же время Таргевир не лишен определенных недостатков. Его относительно небольшая молекулярная масса (21,7 кДа) ограничивает пути введения этого химерного белка и продолжительность его действия в организме. Отказ от интраназального введения в пользу парентерального применения позволил бы перейти к нейтрализации SARS-CoV-2 не только в месте входных ворот, но и на системном уровне. Однако такой подход поставил бы еще одну задачу – задержку целевого белка в кровотоке, что, в свою очередь, потребовало бы увеличения периода полураспада препарата в организме.
Большое количество исследований продемонстрировали, что способность белков к связи с альбумином обладает потенциалом более длительного периода полураспада терапевтических белков [62].Известно, что альбумин имеет чрезвычайно длительный период полувыведения из кровообращения, составляющий у человека 19 дней, вследствие размера, превышающего порог почечной фильтрации, а также благодаря рН-зависимому связыванию с неонатальным Fc-рецептором (FcRn), который обеспечивает механизм для избавления альбумина от лизосомального пути деградации [63].В результате связь с альбумином может быть использована для продления периода полувыведения лекарств. Важно отметить, что сайт связывания FcRn на альбумине расположен в домене III, который не перекрывается и не мешает его связыванию с комплементарными белками [64].
Для технического решения проблемы увеличения периода полураспада целевого комплекса пептидов в организме была избрана стратегия нековалентного присоединения химерного белка, включающего комплекс пептида LCB1 и барназы (Ва) к альбумину, используя альбуминсвязывающий домен (ABD). Этот домен, состоящий из 46 а.о., первоначально был получен из стрептококкового белка G и впоследствии усилен с помощью белковой инженерии для достижения исключительного фемтомолярного сродства к сывороточному альбумину человека (HSA) [65].
В процессе поэтапного выполнения патентных исследований осуществлялись:
Настоящее изобретение решает техническую проблему получения рекомбинантного белка PAB, имеющего последовательность SEQ ID NO:2, на основе адресного полипептида LCB1 с последовательностью SEQIDNO: 3[48], альбумин-связывающего домена (ABD) с последовательностью SEQIDNO: 5 и барназы (Ba) с последовательностью SEQ ID NO:7, соединенных между собой гибкими пептидными линкерами с последовательностями SEQ ID NO:4 и SEQ ID NO:6, способного блокировать связывание с клетками и/или препятствовать размножению вируса SARS-CoV-2 и имеющего сильно увеличенное время циркуляции в кровотоке в сравнении с Таргевир [61].
Поставленная техническая проблема достигается за счет:
1) рекомбинантного белка PAB, имеющего последовательность SEQ ID NO: 2, состоящего из адресного полипептида LCB1 с последовательностью SEQ ID NO: 3, ABDc последовательностью SEQ ID NO:5 и барназы с последовательностью SEQ ID NO: 7, соединенных гибкими пептидными линкерами с последовательностями SEQ ID NO: 4 и SEQ ID NO: 6, обладающий способностью эффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2 и характеризующийся длительным временем циркуляции в кровотоке, а также его применения в качестве вирус-нейтрализующего средства против SARS-CoV-2.
2) рекомбинантной плазмиды pET39b-LCB1-ABD-Ba, содержащей ген PAB и ген барстара в составе плазмидного вектора экспрессии pET39b.
3) Штамма E. Coli BL21(DE3)/pET39b-LCB1-ABD-Ba, получаемого путем трансформации штамма BL21(DE3) плазмидой pET39b-LCB1-ABD-Ba и отбора клона трансформанта с наиболее высоким уровнем синтеза PAB.
4) Способа получения из биомассы штаммов Escherichia coli BL21(DE3)/pET39b-LCB1-ABD-Ba рекомбинантного белка PAB.
В результате решения заявленной технической проблемы получают следующий технический результат:
1) высокий уровень индуцируемого синтеза (40% от суммарного клеточного белка) и стабильную продукциюPAB, который обеспечивается штаммом E. coli BL21(DE3)/pET39b-LCB1-ABD-Ba;
2) высокую специфическую антивирусную активность PAB;
3) пролонгирование времени циркуляции и противовирусного эффекта РАВ в кровотоке в сравнении с аналогом Targevir1.
Раскрытие изобретения
Получен рекомбинантный белок PAB, имеющий последовательность SEQ ID NO: 2, состоящий из адресного полипептида LCB1 с последовательностью SEQ ID NO: 3, ABDc последовательностью SEQ ID NO:5 и барназы с последовательностью SEQ ID NO: 7, соединенных гибкими пептидными линкерами с последовательностями SEQ ID NO: 4 и SEQ ID NO: 6, который обладает способностью эффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2 и имеет продолжительное время циркуляции в кровотоке.
Показано применение рекомбинантного белка PAB, имеющего последовательность SEQ ID NO:2, в качестве вируснейтрализующего средства в отношении SARS-CoV-2.Показано пролонгирование времени циркуляции и противовирусного эффекта РАВ в кровотоке в сравнении с аналогом Targevir1.
Разработан способ получения PAB, – химерного белка, состоящего из адресного полипептида LCB1 [48], специфичного к рецептор-связывающему домену (RBD) S-белка SARS-CoV-2, ABD [67] и высокоактивной бактериальной рибонуклеазы барназы, соединенных между собой гибкими пептидными линкерами. PAB обладает способностью специфически связываться с вирионами SARS-CoV-2, имеющими на своей поверхности S-белок, и блокировать их связывание с рецептором ACE-2, представленным на поверхности клеток, чувствительных к заражению вирусом. При интернализации в клетки вместе с вирусом обладает вирус статическим действием за счет гидролиза вирусной РНК барназой и последующего прекращения биосинтеза белка, необходимого для синтеза новых вирионов. Вместе с тем наличие в составе PAB альбумин-связывающего домена позволило существенно продлить его время циркуляции белка в кровотоке, пролонгируя противовирусный эффект и придавая ему системный характер in vivo. Сочетание таких свойств PAB выгодно отличает его от всех известных и применяемых в клинической практике аналогов.
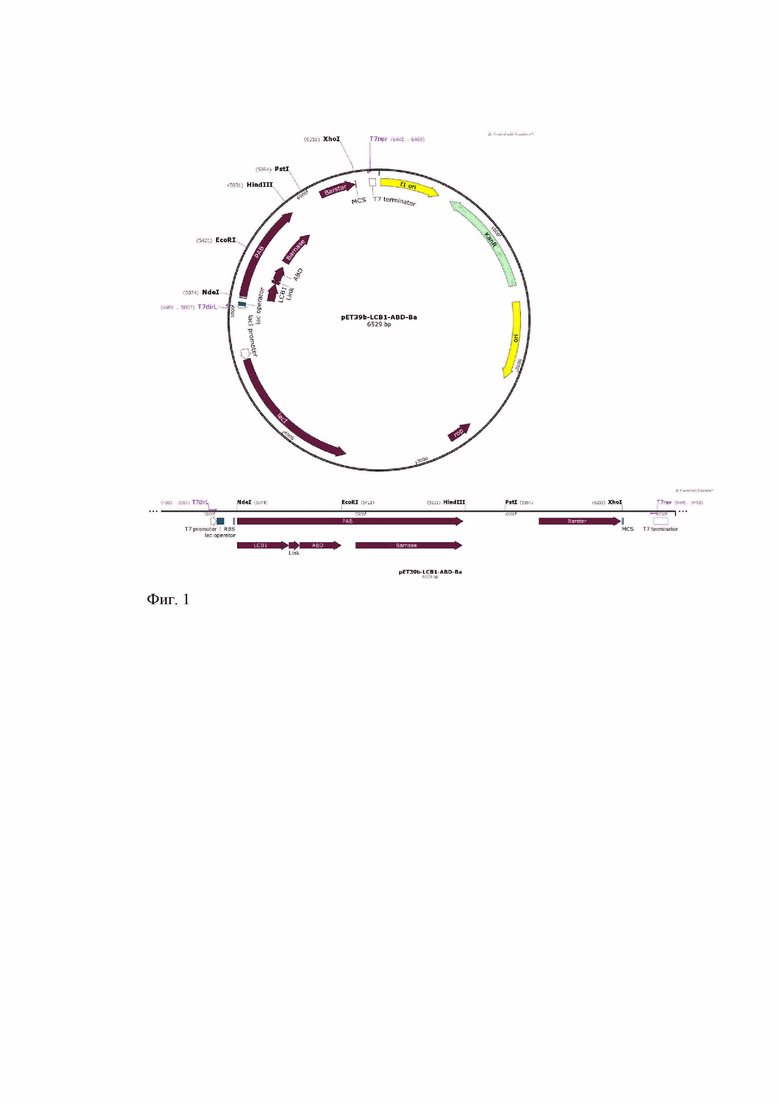
Экспрессирующую рекомбинантную плазмиду pET39b-LCB1-ABD-Ba получают путем клонирования последовательности, кодирующей PAB и барстар, в вектор pET39b (pET System Manual, 10th Edition Rev.B 0403, Novagen Inc. (2003)) по сайтам рестрикции NdeI/XhoI (фиг. 1). Для преодоления чрезвычайной токсичности рибонуклеазы барназы для бактериальной клетки ген целевого белка ставят под контроль индуцируемого промотора фага Т7, кроме того, в конструкцию включают ген ингибитора барназы, барстара, под собственным промотором. В процессе культивирования ингибитор барстар, синтезирующийся в бактериальной клетке одновременно с PAB, полностью ингибирует ее РНКазную активность, что позволяет проводить наработку клеток штамма-продуцента, но требует последующей очистки PAB от ингибитора барстара.
Путем трансформации клеток штамма Escherichia coli BL21(DE3) [68] сконструированной плазмидой pET39b-LCB1-ABD-Ba или pET39b-LCB3-Ba, отбора и культивирования клонов трансформантов с высоким уровнем синтеза гибридного белка получают рекомбинантный штамм Escherichia coli BL21(DE3)/pET39b-LCB1-ABD-Ba– продуцент PAB. Синтез PAB в полученном рекомбинантном штамме осуществляется при культивировании на обычных селективных средах с добавлением индуктора изопропил-D-тиогалактозида (ИПТГ) или лактозы.
В способе получения рекомбинантного PAB биомассу рекомбинантного штамма E. coli-продуцента PAB разрушают ультразвуком в лизирующем буфере, содержащем Tris-HCl и хлорид натрия. Целевой белок, изначально снабженный олигогистидиновой последовательностью SEQID: 8, выделяют при помощи металлохелатной аффинной хроматографии. В бактериальных клетках PAB образует чрезвычайно устойчивый комплекс со своим ингибитором барстаром (KD 10-13 M-1) и в стандартной процедуре аффинной хроматографии на Ni2+-сефарозе выделяется совместно с ним в виде прочного комплекса, поэтому основной проблемой при очистке является отделение ингибитора барстара. Очистку PAB от барстара осуществляют в денатурирующих для комплекса PAB-барстар условиях с использованием 6 М гуанидингидрохлорида и 0.5 М NaCl. Затем PAB дочищают при помощи анионообменной хроматографии.
Таким образом, настоящее изобретение включает 5 объектов:
Первый объект –рекомбинантный белок PAB, имеющий последовательность SEQ ID NO: 2, который обладает активностью к рецептор-связывающему домену S-белка SARS-CoV-2 и характеризующийся длительным временем циркуляции в кровотоке.
Второй объект – применение указанных белков в качестве вируснейтрализующего средства в отношении SARS-CoV-2
Третий объект– рекомбинантная плазмида pET39b-LCB1-ABD-Ba, обеспечивающая синтез PAB в клетках Escherichiacoli и состоящая из фрагмента ДНК с последовательностью SEQ ID NO: 1и фрагмента плазмиды pET39b.
Четвертый объект – рекомбинантный штамм Escherichia coli BL21 (DE3)/pET39b-LCB1-ABD-Ba.
Пятый объект–способ получения рекомбинантного белка PAB с использованием штамма EscherichiacoliBL21 (DE3)/pET39b-LCB1-ABD-Ba с высокой степенью очистки.
Изобретение иллюстрируется следующими фигурами:
Фиг. 1 – Физическая и генетическая карты вектора pET39b-LCB1-ABD-Ba. Обозначено положение гена PAB, других элементов вектора, уникальных сайтов рестрикции.
Фиг. 2 –Гель-электрофорез PAB после осаждения сульфатом аммония. Стрелкой обозначено положение PAB. Дорожка PAB –образец PAB, дорожка МW – маркермолекулярных масс.
Фиг. 3 – Результаты металлохелатной аффинной хроматографии на HisTrapHP, 5 мл. Гель-электрофорез фракций 16-20, собранных в ходе элюции PAB с колонки, дорожка МW– маркермолекулярных масс, стрелкой обозначено положение PAB.
Фиг. 4– Результаты финишной анионообменной хроматографии на HiTrapQHP, 5 мл. Гель-электрофорез фракций 7-9, собранных в ходе элюции PAB с колонки, дорожка МW – маркермолекулярных масс, стрелкой обозначено положение PAB.
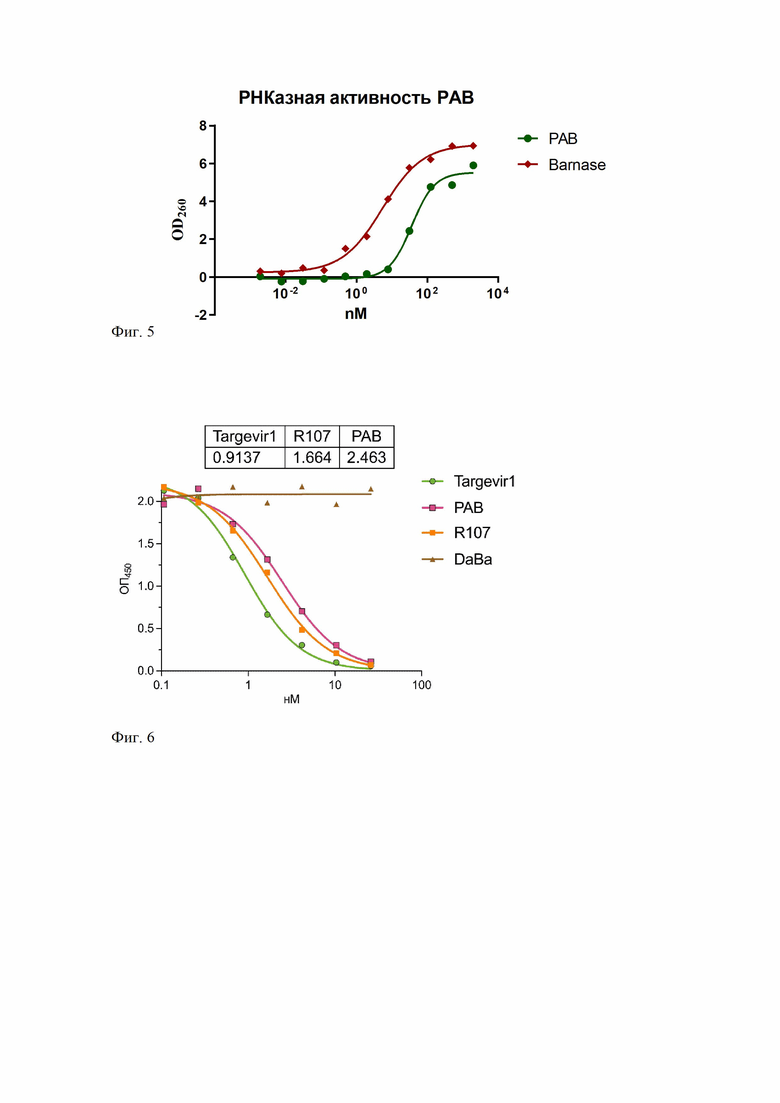
Фиг. 5 – Определение РНКазной активности PAB в сравнении с барназой дикого типа методом кислотно-растворимого осадка.
Фиг. 6 – Изучение конкурентного связывания PAB и RBD с рекомбинантным АСЕ - 2 методом ИФА
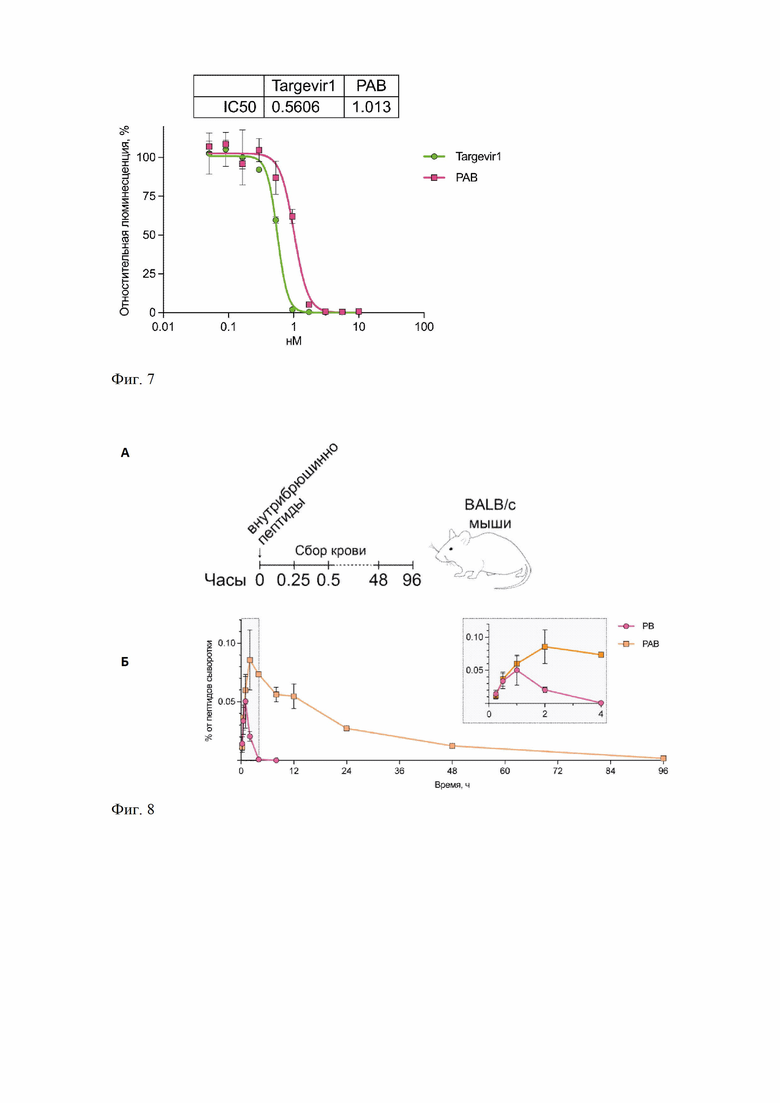
Фиг. 7 – Зависимость интенсивности люминисценции клеток линии ACE2-HEK293T, экспрессирующих на поверхность рецептор ACE-2 человека, от концентрации исследуемых белков в псевдовирусной системе.
Фиг. 8 –Определение времени циркуляции РАВ в крови мышей
А. Схема эксперимента по исследованию фармакокинетики препаратов Targevir1 и PAB.
Б. График содержания химерных пептидов в крови мышей после внутрибрюшинного введения. На вертикальной оси (оси Y) графика отражены значения площадей пиков, соответствующих полипептидам в кровяной сыворотке, приведенные к площади сигнала остальных белков.
Осуществление изобретения.
Пример 1. Получение рекомбинантной плазмиды pET39b-LCB1-ABD-Ba.
Плазмиду pET39b-LCB1-ABD-Ba получают путем клонирования фрагмента ДНК с последовательностью SEQ ID NO: 1 в вектор pET39b (pET System Manual,10th Edition Rev.B 0403, Novagen Inc. (2003)) по сайтам рестрикции NdeI/XhoI (Фиг. 1).
Пример 2. Получение рекомбинантного штамма – продуцента PAB.
Рекомбинантной плазмидой pET39b-LCB1-ABD-Ba трансформируют штамм E. coliBL21(DE3) [F-, ompT, hsdSB (rB-, mB-), dcm, gal, λ(DE3)][68]. Выбор данного штамма в качестве реципиента для продукции PAB обусловлен тем, что он синтезирует РНК-полимеразу фага Т7, а также обладает сниженной протеазной активностью, что способствует повышению выхода целевых белков.
Поскольку первичные трансформанты DE3 – реципиентов могу заметно отличаться по уровню экспрессии целевого белка [69] для селекции наиболее активного продуцента PAB и параметров индукции синтеза PAB отдельные клоны трансформантов выращивают в условиях «автоиндукции» [70] в течение 24-48 часов при 25°С с периодическим отбором аликвот суспензии клеток. Для получения грубого экстракта осадок клеток из 100-200 мклкультуры суспендируют в 100 мкл лизирующего буфера для нанесения на ДСН-ПААГ, прогревают 3 мин при 85°С, клеточный дебрис удаляют центрифугированием (12 000 об/мин, 5 мин) и 5 мкл полученного экстракта анализируют путем электрофореза в 15% денатурирующем полиакриламидном геле.
Появление отчетливой полосы размером порядка 27,8кДадля PAB 3 в образцах анализируемых штаммов при ее отсутствии в контрольном реципиентном штамме BL21(DE3) принимают за доказательство способности штамма синтезировать PAB.
Клон трансформанта, отличающийся наибольшим выходом PAB, обозначают BL21(DE3)/pET39b-LCB1-ABD-Ba.
Выход PAB в полученных нами штаммах E. Coli BL21(DE3)/pET39b-LCB1-ABD-Ba и E. coliBL21(DE3)/pET39b-LCB3-Ba составляет 40% от суммарного белка клетки.
Клетки полученных рекомбинантных штаммов Escherichia coli BL21(DE3)/pET39b-LCB1-ABD-Ba характеризуются следующими признаками.
Морфологические признаки: Клетки имеют продолговатую палочковидную форму, при делении не почкуются.
Культуральные признаки: Клетки хорошо растут на обычно используемых питательных средах. Время генерации около 30 мин в жидкой LB-среде. На 2-2,5% питательном агаре "Difco" образуются круглые, гладкие, желтоватые колонии с ровными краями. При выращивании на жидких LB- и YT-средах образуется интенсивная ровная мутность.
Физиолого-биохимические признаки: Оптимальная температура культивирования – от 25 до 37°C, оптимум рН - 7,6. Источником азота служат органические соединения (в виде триптона, дрожжевого экстракта).
Уровень синтеза PAB в сконструированных штаммах составляет около 40мг/л при титре культуры 1×109кл/мл, что следует из данных определения PAB в образцах биомассы штамма-продуцента с помощью электрофореза в 15% ДСН-ПААГ.
Пример 3. Получение биомассы рекомбинантного штамма – продуцента PAB.
Клетки E. Coli BL21(DE3) трансформируют соответствующей плазмидой pET39b-LCB1-ABD-Ba или pET39b-LCB3-Ba по методу [71].Клетки из глицеринового стока, хранящегося в морозильнике, рассевают на поверхности чашки с питательной агаризованной средой. Инкубируют ночь при 37ºС. Собирают около 20-30 колоний стеклянной палочкой и суспендируют в 100 мкл 0.1 М СaCl2, содержащем 1 мг PEG 8000 (pH 8.0). Инкубируют во льду в течение 5 мин. Добавляют плазмидную ДНК в количестве 5 нг и продолжают инкубацию во льду еще в течение 8 мин. Подвергают клетки тепловому шоку при 42ºС в течение 30 сек и затем возвращают в лед на 2 мин. Добавляют 900 мкл среды YT (8 г бакто-триптона, 5 г дрожжевого экстракта, 5 г NaCl на 1 л воды) и высевают 100 мкл суспензии на чашки с агаризованной средой и канамицином (25 мг/л).
Для получения посевного материала собирают стеклянной палочкой около 50 колоний с чашек Петри, засеянных трансформантами, и суспендируют в 200 мкл среды YT. Суспензию клеток переносят в 200 мл жидкой автоиндукционной питательной среды ZYM-5052 [70], содержащей канамицин (50 мг/л), и культивируют при температуре 37°С при интенсивной аэрации (скорость качалки 180 об/мин) в течение суток.
Выход биомассы обычно составляет от 10.0 до 15.0 г/л культуральной среды.
Пример 4. Выделение и очистка рекомбинантного белка PAB
Биомассу ресуспендируют в 20 мл раствора 200 мM Трис-HCl, pH 8.0 и добавляют лизоцим до конечной концентрации 60 мкг/мл. Суспензию разбавляют в 2 раза дистиллированной водой, добавляют PMSF до конечной концентрации 100 мкМ и инкубируют при комнатной температуре в течение 30 мин, периодически перемешивая. Суспензию переносят в ледяную баню и обрабатывают ультразвуком при помощи SonicsVibra-Cell VCX 130, добиваясь полного лизиса клеток (общее время обработки 5 раз по 2 мин, ‒ циклы по 10 сек с 10 секундным интервалом, мощность 80W). Клеточный лизат центрифугируют в течение 30 мин при 18500 gи декантируют надосадочную жидкость, содержащую растворимую фракцию целевого белка. К осветленному лизату при постоянном перемешивании добавляют кристаллический сернокислый аммоний до 60% от насыщения. Суспензию оставляют на ночь в холодильнике. Типичный результат электрофоретического анализа PAB после осаждения сульфатом аммония представлен в Фиг. 2.
Суспензию разливают по стаканам и центрифугируют в течение 30 мин при 18500 g. Надосадочную жидкость сливают, а осадок растворяют в растворе I (20 мМ NaPi, pH 7.5, 500 мМ NaCl, 30 мМ имидазол) и фильтруют через мембрану с размером пор 0.22 мкм (MilliPore).
Раствор, содержащий целевой белок, наносят на колонку HisTrap HP, 5мл, уравновешенную 10-кратным объемом раствора I, со скоростью 5 мл/мин, затем колонку промывают от примесных белков 10 объемами раствора I с той же скоростью. Для денатурации комплекса PAB: барстар, иммобилизованного на колонке, носитель промывают 25 объёмами раствора I(без имидазола) с 6 М гуанидин гидрохлоридом со скоростью 1 мл/мин. PAB, освобожденный от ингибитора барстар, ренатурируют непосредственно на колонкелинейным ниспадающим градиентом гуанидин гидрохлорида (60 объемов колонки) от 6 М до 0 М при скорости 1 мл/мин.
После ренатурации PAB, иммобилизованного на колонке, ее промывают 10 объёмами раствора II(20 мM NaPi, pH 7.5, 100 мM NaCl) со скоростью 5 мл/мин. PAB элюируют с колонки раствором II с 250 мМ имидазолом с той же скоростью. Типичные результаты анализа фракций, собранных в ходе элюции PAB с металлохелатного аффинногосорбента, а также профиль хроматографической элюции, представлены в Фиг. 3.
Полученный после аффинной хроматографии элюат разбавляют раствором III (20 мМ NaPi, pH 7.5) в 5 раз и наносят со скоростью 5 мл/мин на колонку HiTrapQHP, 5 мл, уравновешенную 10-кратным объемом раствора III. Колонку промывают 10 объёмами раствора III, а затем элюируют PAB со скоростью 5 мл/мин линейным градиентом NaCl от 0 до 500 мМ в растворе III. Объем градиента составляет 20 объемов колонки. Фракции, содержащие PAB, объединяют и стерильно фильтруют через мембрану с диаметром пор 0,22 микрона. Концентрацию белка определяют методом УФ-спектроскопии, используя молярный коэффициент экстинкции ε280 = 36840M-1cм-1. Типичные результаты анализа фракций, собранных в ходе элюции PAB с анионообменного сорбента, а также профиль хроматографической элюции, представлены в Фиг. 4.
Пример 5. Проверка ферментативной (РНКазной) активности PAB.
Сохранение функций барназы как токсического агента в составе PAB проверяют путем определения ферментативной РНКазной активности. Для этого используют модифицированный сравнительный метод кислоторастворимого осадка. В качестве субстрата используют дрожжевую РНК.
Исследуемую пробу белка растворяют в концентрации 1,25 мкМ в буферном растворе 0,125М Трис-HCl, pH 8.5 и затем получают серию последовательных 5-кратных разведений образца в том же буфере. В качестве отрицательного контрольного образца используют буферный раствор 0,125М Трис-HCl, pH 8.5. В качестве положительных контрольных образцов используют образцы барназы дикого типа с известной молярной концентрацией. К 40 мкл каждого образца добавляют 160 мкл раствора дрожжевой РНК (в концентрации 2 г/л) в 0,125М Трис-HCl, pH 8.5 на льду. Реакционную смесь инкубируют15 минут при +37°С. Реакцию останавливают добавлением 200 мкл 6% хлорной кислоты и инкубируют смесь 15 мин при 0°С. После чего центрифугируют при 16 000g в течение 10 минут. Супернатанты разбавляют в 10 раз и измеряют оптическую плотность при 260 нм (OD260) относительно контрольного образца. Большим значениям оптической плотности соответствует большая рибонуклеазная активность. 1 ед. акт. рибонуклеазы в исследуемой пробе соответствует приращению OD260 на 0,05 единиц.
Величина РНКазной активности PAB обычно колеблется в диапазоне от4.8 × 106 до 11,3 × 106ед. акт./нмоль, что составляет от 20 до 70% рибонуклеазной активности рекомбинантной барназы из E. coli(Фиг. 5).
Пример 6. Изучение связывания RBD с АСЕ-2 в присутствии PAB методом конкурентного ИФА
Исследования связывания рекомбинантного RBD с рекомбинантным внеклеточным доменом ангеотензинпревращающего фермента-2 человека (АСЕ-2) в присутствии PAB или Targevir1 проводят методом конкурентной ИФА как описано в [73]. При этом используют следующие контроли:
(1) моноклональное антитело мыши R107 (кат. R107, HyTest), специфичное к RBD-домену, в качестве положительного контроля. Это антитело, как было заявлено производителем, блокирует связывание RBD с ACE-2;
(2) рекомбинантный белок Darpin9-29-Barnase (DaBa) [74] в качестве отрицательного контроля. Этот контроль был выбран в силу схожести его структуры с PAB.
В лунки 96-луночных планшетов NuncMaxiSorp (Nunc, США) вносят по 100 мкл раствора рекомбинантного белка RBD в концентрации 1 мкг/мл в фосфатно-солевом буфере (PBS) и инкубируют в течение 16 ч при 4°С без перемешивания. По окончании лунки промывают 5 раз промывочным раствором (PBS, 0.1% Tween 20), вносят по 200 мкл блокирующего раствора (фосфатно-солевой буфер (PBS), 0.1% Tween 20, 3% БСА), и инкубируют в течение 1 ч при 37°С без перемешивания. По окончании лунки промывают 5 раз промывочным раствором (PBS, 0.1% Tween 20), вносят по 100 мкл блокирующего раствора, содержащего один из исследуемых белков (LB1, DaBa и R107) в концентрациях 20, 8, 3.2, 1.3, 0.5, 0.2, 0.08 и 0 нМ для каждого из исследуемых белков и инкубируют в термостатируемом шейкере при 700 об/мин в течение 30 мин при 37°С. По окончании лунки промывают 5 раз промывочным раствором (PBS, 0.1% Tween 20) и вносят по 100 мкл блокирующего раствора, содержащего рекомбинантный внеклеточный домен ангеотензинпревращающего фермента-2 человека, содержащего 3xFLAG эпитоп, в концентрации 0.2 мкг/мл. Планшет инкубируют в термостатируемом шейкере при 700 об/мин в течение 30 мин при 37°С.По окончании инкубации лунки промывают 5 раз промывочным раствором (PBS, 0.1% Tween20) и вносят по 100 мкл блокирующего раствора, содержащего моноклональное антитело мыши, специфичное к 3xFLAG эпитопу, коньюгированного с пероксидазой хрена (SigmaAldrich, США, кат. # A8592), в разведении 1:10 000. Планшет инкубируют в термостатируемом шейкере при 700 об/мин в течение 30 мин при 37°С. По окончании инкубации лунки промывают 5 раз промывочным раствором (PBS, 0.1% Tween 20), вносят по 100 мкл раствора субстрата ТМБ и инкубируют в темноте 10 мин. Реакцию останавливают внесением в лунки планшета по 100 мкл 10% раствора фосфорной кислоты и измеряют абсорбцию при 450 нм на планшетном спектрофотометре. Строят кривую зависимости величин ОП450 от концентрации исследуемых веществ. Обработку данных и расчет значений полумаксимальной эффективной концентрации (EC50) проводили с помощью программного обеспечения GraphPadPrism 8.
PAB и Targevir1 блокируют связывание внеклеточного домена ангеотензинпревращающего фермента-2 человека (АСЕ2) с RBD, иммобилизованном на стенках лунок планшета, концентрационно-зависимым способом, как и коммерческое моноклональное антитело мыши R107 (кат. R107, HyTest), специфичное к RBD-домену. (Фиг. 6). В тех же условиях Darpin9.29-Barnase (DaBa), выступавший в качестве отрицательного контроля, не влияет на связывание RBDcACE-2.
Пример 7. Изучение вируснейтрализующего действия PAB с использованием псевдовирусной системы
Вируснейтрализующее действие PAB изучали в, так называемой, псевдовирусной системе. Для этого была создана линия клеток HEK293T-ACE-2, несущая на своей поверхности ACE-2, а также получены рекомбинантные лентивирусные частицы, псевдотипированные S-белком SARS-CoV-2 и содержащие в качестве репортерного гена ген люциферазы светлячка [72]. Детекция вируснейтрализующей активности происходит за счет измерения люминесценции клетками, несущими на своей поверхности АСЕ2, при добавлении к ним псевдовирусных частиц, прединкубированных с испытуемыми веществами. При этом чем больше вируснейтрализующая активность испытуемого вещества, тем меньше уровень детектируемой люминесценции.
Линию клеток HEK293T-ACE-2, гиперэкспрессирующую на поверхности рецептор ACE-2 человека, получали путем трансдукции HEK293T псевдовирусными частицами с мембранным гликопротеином VSV-g (гликопротеин G вируса везикулярного стоматита), содержащими в геноме гены рецептора ACE-2 человека и зеленого флуоресцентного белка GFP. После трех последовательных обогащений трансдуцированных клеток, чередующихся пассажированием, методом проточной цитофлуориметрии были отобраны клетки, стабильно экспрессирующие GFP. Присутствие ACE2 на поверхности пула клеток подтверждали проточной цитофлуориметрией. Клетки инкубировали с рекомбинантным RBD-Fc с дальнейшей визуализацией поликлональными антителами козы, кроссреактивными с Fc-доменом человека, коньюгированными с флуоресцентной меткой DL650, испускание которой детектируется в канале APC. Согласно полученным с помощью проточной цитофлуориметрии данным 96% клеток несут на своей поверхности АСЕ2. Полученные клетки ACE2-HEK293T используют далее в тестах.
Получение рекомбинантных лентивирусных частиц, псевдотипированных S-белком SARS-CoV-2 и содержащих в качестве репортерного гена ген люциферазы светлячка, проводили как описано в работе [72]. Псевдовирус способен проникать в клетки HEK293T-ACE-2, несущие на своей поверхности АСЕ2, где на рекомбинантном РНК-геноме синтезируется ДНК провируса, содержащая маркерный ген люциферазы светлячка, встраивание которого в геном клетки-мишени придает ей способность люминесцировать.
В тесте на вируснейтрализующую активность в качестве контроля выступал Targevir1, для которого ранее [31] показана эффективная вируснейтрализующая активность в псевдовирусной системе.
Тест на нейтрализующую активность проводили следующим образом. Клетки ACE2-HEK293T рассевали в лунки 96-луночной планшета в количестве 5000 клеток на 100 мкл среды DMEM с 10% FBS (Gibco) и инкубировали в течение 24 часов при 37°C, 5% CO2. Лентивирусы, псевдотипированные S-белком SARS-CoV-2 вносили по 50 мкл в лунки 96-луночной плашки, добавляли к ним тестируемые вещества (PAB и PB) в концентрациях 23, 2.3, 0.23, 0.023, 0,0023, 0,00023 нМ и инкубировали при 37°C, 5% CO2 в течение 1часа. Из лунок 96-луночной плашки с клетками ACE2-HEK293T удаляли по 50 мкл культуральной среды, добавляли прединкубированные смеси псевдовирусных частиц с указанными концентрациями для каждого из тестируемых веществ и инкубировали при 37°C, 5% CO2. Через 24 часа проводили смену среды, и через 48 культуральную среду удаляли, клетки лизировали с помощью GLO lysisbuffer (кат. E2661, Promega), смешивали с субстратом Bright-Glo™ LuciferaseAssay System (кат. E2620, Promega), измеряли уровень люминесценции и строили график зависимости уровня люминисценции в зависимости от концентрации тестируемого вещества. Эксперимент проводили в трех повторностях для каждого из исследуемых веществ. Обработку данных и расчет значений полумаксимальной ингибирующей концентрации (IC50) проводили с помощью программного обеспечения GraphPadPrism 8.
PAB, а также Targevir1, характеризующийся отсутствием в полипептидной последовательности альбуминсвязывающего домена (ABD), одинаково эффективно нейтрализуют вирус (Фиг. 7) в псевдовирусной системе.
Пример 8. Изучение связывания РАВ с человеческим альбумином при помощи метода плазмонного резонанса.
Изучение связывания PAB с сывороточными альбуминами человека, быка и мыши (ЧСА, БСА и МСА соответственно) проводят с использованием оптических биосенсоров Biacore X100 и Biacore 8K (GE Heathcare, США) и стандартных сенсорных чипов CM5, покрытых карбоксиметилированным декстраном (Cytiva, США) при температуре 25°C. В качестве рабочего буфера использовали PBS-буфер (10 мМ Na2HPO4, 1,8 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, pH 7,4) («Cytiva», США). В качестве контролей выступали белковые компоненты, из которых построен PAB, а именно, PA и АБД, способные связываться с альбумином.
Альбумины (ЧСА, БСА и МСА) и тример S-белка иммобилизуют на поверхности чипа CM5. Иммобилизацию осуществляют с помощью карбодиимидной реакции. Поверхность чипа активируют 7-минутным введением смеси 0,2 М 1-этил-3-(3-диметиламинопропил)карбодиимид и 0,05 М N-гидроксисукцинимид (Cytiva, США) 1:1. Иммобилизованные белки растворяют в натрий-ацетатном буфере (10 мМ CH3COONa) при pH 4,5 (альбумины) или 5,0 (S-белок). Концентрация белка в буфере для иммобилизации составляла 50 мкг/мл. Время контакта раствора белка с поверхностью чипа в процессе иммобилизации составляло 7 минут при скорости потока 5 мкл/мин. Средний конечный уровень иммобилизации составил 6000 ± 2000 RU для альбуминов и 17 000 ± 3 000 RU для S-белка соответственно.
Для коррекции эффектов неспецифического связывания анализируемых веществ с поверхностью чипа использовался эталонный канал без иммобилизованного белка. Для регистрации межмолекулярных взаимодействий растворы PAB, PA и АБД в буфере PBS вводили в течение 3 минут со скоростью потока 5 мкл/мин, диссоциацию регистрировали в течение 5–15 минут. Концентрации растворов находились в диапазоне 25–400 нМ (для взаимодействия с S-белком) или 100–16000 нМ (для взаимодействия с альбуминами). При регистрации взаимодействия химер с альбуминами проводили две инъекции глицинового буфера (рН 2,5) для регенерации поверхности чипа (в течение 30 с, скорость потока 30 мкл/мин).
Полученные сенсорограммы обрабатывались в программном обеспечении BIAevaluation 4.1.1 (с использованием модели взаимодействия 1:1 с массообменом или модели стационарного сродства для многоциклового анализа и модели кинетического титрования для одноциклового анализа) или в программном обеспечении Biacore Insight, версия 3.0.11.15423 (с использованием модели взаимодействия 1:1 по умолчанию с моделью массообмена или стационарной модели сродства для многоциклового анализа и модели кинетического титрования для одноциклового анализа). Описание методов связывания 1:1 и кинетического титрования (модель одного цикла) дано в [Karlsson R. etal. - Анализ серии кинетических титрований с использованием аффинных биосенсоров. Аналитическая биохимия 349:136-147; (2006) https://doi.org/10.1016/j.ab.2005.09.034]. Полученные значения равновесных констант диссоциации комплекса KD приведены в табл. 1. Как можно видеть из представленных данных аффинность связывания АБД с альбуминами максимальна для ЧСА, снижается в случае МСА и значительно снижается в случае БСА. В случае введения в N-концевую часть АБД последовательности пептида LCB1 (РА) аффинность связывания АБД с ЧСА также снижается. Дальнейшее снижение аффинности связывания АБД с ЧСА наблюдается в случае одновременного введения в N-концевую часть АБД последовательности пептида LCB1 и в С-концевую часть последовательности барназы (PAB). Тем не менее значение константы диссоциации комплекса АБД с ЧСА KD(1.17±0.13) *10-9 М остается достаточно низким, лежащим в наномолярном диапазоне, что с сравнимо аффинностью антител, применяемых для терапии в настоящее время и что обеспечивает достаточную прочность связывания комплекса ЧСА:PAB и, соответственно может обеспечить пролонгированное время содержания PAB в кровотоке.
Табл. 1 Значения равновесных констант диссоциации комплекса KD. KD – равновесная константа диссоциации комплекса, рассчитанная как = kd/ka, ka – константа скорости ассоциации комплекса, kd – константа скорости диссоциации комплекса
Пример 9. Определение времени циркуляции РАВ в крови мышей.
Время полужизни PAB определяли in vivo в крови мышей линии BALB/c. При этом в качестве контроля выступал белок Targevir 1, аналогичный по структуре PAB, но не имеющий в своем составе ABD [30].
Мышам вводили внутрибрюшинно эквимолярные дозы двух разных белков PAB (в дозе 400 мг/кг) и Targevir1 (в дозе 300 мг/кг). Через 15 мин, 30 мин, 12 ч и далее каждые 12 часов вплоть до 96 часов проводили отбор 10 мкл крови из ретроорбитального синуса (Фиг. 8А). В качестве контроля проводили отбор крови указанным способом у интактных мышей линии BALB/c. Измерение уровней исследуемых белков в крови мышей проводили методом нано-жидкостной хроматографии, сопряженным с электрораспылительным ионизационным тандемным масс-спектрометрическим анализом (nano-LC-ESI-MS/MS).
Результаты исследования представлены на фигуре 8Б. На вертикальной оси (оси Y) графика отражены значения площадей пиков, соответствующих полипептидам в кровяной сыворотке, приведенные к площади сигнала остальных сывороточных белков. Зафиксировано, более чем 24-кратное увеличение (96 ч против 4 ч) продолжительности полужизни полипептида PAB, имеющего в своем составе ABD, чем у контрольного белка Targevir 1 (Фиг. 8Б).
Литература
1. World Health Organization. WHO Coronavirus (COVID-19) Dashboard. WHO Coronavirus (COVID-19) Dashboard 2022. https://covid19.who.int/ (accessed March 10, 2022).
2. Crook H., Raza S., Nowell J., Young M., Edison P. Long covid-mechanisms, risk factors, and management. BMJ. 2021; 374: n1648.
3. Асфандиярова Н.С. Постковидный синдром. Клиническая медицина. 2021; 99 (7-8): 429-435.
4. van Kessel S.A.M., Olde Hartman T.C., Lucassen P.L.B.J., van Jaarsveld C.H.M. Post-acute and long-COVID-19 symptoms in patients with mild diseases: a systematic review. Fam Pract. 2022; 39 (1): 159–167.
5. Watanabe A., Iwagami M., Yasuhara J., Takagi H., Kuno T. Protective effect of COVID-19 vaccination against long COVID syndrome: A systematic review and meta-analysis. Vaccine. 2023; 41 (11): 1783-1790.
6. O’Keefe L.C. Post-Acute COVID 19 Syndrome, ‘Long COVID’. Workplace Health Saf. 2023; 71 (5): 263.
7. Marzoog B.A. Coagulopathy and brain injury pathogenesis in post-Covid-19 syndrome. Cardiovasc Hematol Agents Med Chem. 2022; 20 (3): 178-188.
8. Wesselingh R. Prevalence, pathogenesis and spectrum of neurological symptoms in COVID-19 and post-COVID-19 syndrome: a narrative review. Med J Aust. 2023; 219 (5): 230-236.
9. Majumder J., Minko T. Recent developments on therapeutic and diagnostic approaches for COVID-19. AAPS J. 2021; 23 (1): 14.
10. Kruse R. Therapeutic strategies in an outbreak scenario to treat the novel coronavirus originating in Wuhan, China [version 2; peer review: 2 approved]. F1000Research. 2020; 9: 72.
11. Eurosurveillance editorial team note from the editors: World Health Organization declares novel coronavirus (2019-nCoV) sixth public health emergency of international concern. Euro Surveil. 2020; 25 (5): 200131e.
12. Cucinotta D., Vanelli M. WHO declares COVID-19 a pandemic. Acta Biomed. 2020; 91(1): 157–160.
13. Chinese Clinical Trial Registry. A randomized controlled trial for the efficacy and safety of baloxavirmarboxil, favipiravir tablets in novel coronavirus pneumonia (COVID-19) patients who are still positive on virus detection under the current antiviral therapy. 2020. https://www.chictr.org.cn/showprojen.aspx?proj=49013. Accessed 11/23/2020.
14. Beigel J.H., Tomashek K.M., Dodd L.E., Mehta A.K., Zingman B.S., Kalil A.C., et al. Remdesivir for the treatment of COVID-19 - preliminary report. N Engl J Med. 2020; 383 (19): 1813-1826.
15. Gao J., Tian Z., Yang X. Breakthrough: chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Biosci Trends. 2020; 14 (1): 72–73.
16. Gautret P., Lagier J.C., Parola P., Hoang V.T., Meddeb L., Mailhe M., et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: results of an open-label non-randomized clinical trial. Int J Antimicrob Agents. 2020; 56 (1): 105949.
17. Tong S., Su Y., Yu Y., Wu C., Chen J., Wang S., et al. Ribavirin therapy for severe COVID-19: a retrospective cohort study. Int J Antimicrob Agents. 2020; 56: 106114.
18. Vankadari N. Arbidol: a potential antiviral drug for the treatment of SARS-CoV-2 by blocking trimerization of the spike glycoprotein. Int J Antimicrob Agents. 2020; 56 (2): 105998.
19. Guo Y., Huang L., Zhang G., Yao Y., Zhou H., Shen S., et al. A SARS-CoV-2 neutralizing antibody with extensive Spike binding coverage and modified for optimal therapeutic outcomes. Nat Commun. 2021; 12 (1): 2623.
20. Baum A., Ajithdoss D., Copin R., Zhou A., Lanza K., Negron N., et al. REGN-COV2 antibodies prevent and treat SARS-CoV-2 infection in rhesus macaques and hamsters. Science. 2020; 370 (6520): 1110-1115.
21. Chen Y., Zhao X., Zhou H., Zhu H., Jiang S., Wang P. Broadly neutralizing antibodies to SARS-CoV-2 and other human coronaviruses. Nat Rev Immunol. 2023; 23 (3): 189-199.
22. Quagliata M., Papini A.M., Rovero P. Chemically modified antiviral peptides against SARS-CoV-2. J Pept Sci. 2023: e3541.
23. Li T., Kan Q., Ge J., Wan Z., Yuan M., Huang Y., et al. A novel linear and broadly neutralizing peptide in the SARS-CoV-2 S2 protein for universal vaccine development. Cell Mol Immunol.2021; 18 (11): 2563-2565.
24. Karoyan P., Vieillard V., Gómez-Morales L., Odile E., Guihot A., Luyt C.-E., et al.Human ACE2 peptide-mimics block SARS-CoV-2 pulmonary cells infection. Commun Biol. 2021; 4 (1): 197.
25. Aboul-Fotouh S., Mahmoud A.N., Elnahas E.M., Habib M.Z., Abdelraouf S.M. What are the current anti-COVID-19 drugs? From traditional to smart molecular mechanisms, Virol J. 2023; 20 (1): 241.
26. Senefeld J.W., Franchini M., Mengoli C., Cruciani M., Zani M., Gorman E.K., et al.COVID-19 convalescent plasma for the treatment of immunocompromised patients: A systematic review and meta-analysis. JAMA Netw Open; 2023; 6 (1): e2250647.
27. Correia I.R. Stability of IgG isotypes in serum. mAbs. 2010; 2 (3): 221–232.
28. Weißenborn L., Richel E., Hüseman H., Welzer J., Beck S., Schäfer S., et al. Smaller, stronger, more stable: peptide variants of a SARS-CoV-2 neutralizing miniprotein. Int J Mol Sci. 2022; 23 (11): 6309.
29. Rogers T.F., Zhao F., Huang D., Beutler N., Burns A., He W.T., et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science. 2020; 369 (6506): 956–963.
30. Патент РФ 2763001. Однодоменное антитело и его модификации, специфически связывающиеся с RBDS белка вируса SARS - CoV - 2 , и способ их применения для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS - CoV – 2. Авторы: Фаворская И.А., Щебляков Д.В., Есмагамбетов И.Б., Деркаев А.А., Алексеева Т.А., Рябова Е.И. и др. Патентообладатель: ФГБОУ Национальный исследовательский центр эпидемиологии и микробиологии имени почетного академика Н.Ф. Гамалеи» Минздрава России. Опубл.24.12.2021 г. Бюл. № 36.
31. Патент РФ 2802825. Рекомбинантный белок, связывающийся с RBDS-белка SARS - CoV – 2. Авторы: Алексеева Л.Г., Бобик Т.В., Габибов А.Г., Деев С.М., Коновалова Е.В., Лукьянова Т.И. и др. Патентообладатель: ФГБУН Институт биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова Российской академии наук (ИБХ РАН). Опубл. 29.08.2022 г. Бюл. № 25.
32. Патент РФ 2794141. Однодоменные наноантитела против шиповидного белка вируса SARS-CoV-2. Авторы: Чумаков С.П., Жучков В.А. Патентообладатель: Чумаков С.П. Опубл. 11.04.2023 г. Бюл. № 11.
33. Патент РФ 2798508. Плазмидная генетическая конструкция pET21_Ab_CoV-2_1.3, обеспечивающая экспрессию рекомбинантного белка AB_COV-2_1.3 в прокариотической системе E. coli, и рекомбинантный белок AB_COV-2_1.3, обладающий свойствами однодоменного наноантитела против SARS - CoV – 2. Авторы: Арипов В.С., Волкова Н.В., Меркульева Ю.А., Несмеянова В.С., Исаева А.А., Щербаков Д.Н. и др. Патентообладатель: ФБУН ГНЦ ВБ "Вектор" Роспотребнадзора. Опубл. 23.06.2023 г. Бюл. № 18.
34. Patent US2023331824. Single-domain antibodies that bind SARS-COV-2. Inventors:Chait B.T., Rout M.P., Aitchison J., Mast F.D., Olivier J.P., Fenyo D. Applicants: UnivRockefeller [US]; Univ New York [US]; Seattle Childrens Hospital D/B/A SeattleChildrens Res Institute [US]. Publ. 2023-10-19.
35. Patent US2023374116. Single domain antibodies that target SARS-COV-2. Inventors: Huo J., Owens R., Naismith J. Applicants: The Rosalind [GB]; The Chancellor Masters Franklin Inst. [GB]; Univ Oxford Innovation LTD and Scholars of the Univ of Oxford. Publ. 2023-11-23.
36. Patent IL305301. Single domain antibodies that neutralize sars-cov-2. Applicants: US Health [US]. Publ. 2023-10-01.
37. Patent CA3160388. Anti-SARS-COV-2 spike receptor binding domain (RBD)single domain antibodies and methods of use thereof. Inventors: Dormeshkin D., Shapira M. Applicants: Dormeshkin D., Shapira M. Publ. 2023-11-25.
38. Patent CN111647077. SARS-COV-2 spike protein binding molecule and application thereof. Inventors: Zhang J. Applicants: Shenzhen Immunotherapy Biotech CO., LTD. [CN]. Publ. 09.12.2021.
39. Patent KR1020220064331. Single domain antibody against SARS-COV-2, and use thereof. Inventors: Seong S.Y., Kim J.H., Lee S.B., Jeong H.H., Yang H.Y., Kim B.J., et al. Applicants: Shaperon Inc. [KR], Huvetbio Inc. [KR], Seoul National Univercity R&DB Foundation [KR]. Publ. 2022-05-19.
40. Patent JPWO2022065445. SARS-COV-2 neutralizing antibody or fragment thereof. Inventors: Takeshita M., Takeuchi T., Suzuki K., Saya H., Takahashi Y., Moriyama S., et al. Applicants: Univ Keio [JP]; Japan as represented by Director General of Nat Institute of infectious diseases [JP]; Riken [JP]; Univ Shiga Medical Science [JP]; Univ Tokyo [JP]. Publ. 2023-11-02.
41. Vidarsson G., Dekkers G., Rispens T. IgG subclasses and allotypes: from structure to effector functions. Front Immunol. 2014; 5: 520.
42. Oostindie S.C., Lazar G.A., Schuurman J., Parren P.W.H.I. Avidity in antibody effector functions and biotherapeutic drug design. Nat Rev Drug Discov. 2022; 21 (10): 715-735.
43. Araste F., Abnous K., Hashemi M., Taghdisi S.M., Ramezani M., Alibolandi M.Peptide-based targeted therapeutics: Focus on cancer treatment. J Control Release. 2018; 292: 141-162.
44. Patent CN112513076. SARS-COV-2 spike protein binding molecule and use thereof. Inventors: Zhang J. Applicants: Shenzhen Immunotherapy Biotech CO., LTD. [CN]. Publ. 2022-03-31.
45. Patent JP2022060186. SARS-COV-2-binding peptide. Inventors: Masaki H., Kumachi S., Yonehara R., Matsumura Y., Kawano H., Tojo T., et al. Applicants: EpsilonMolecular Engineering, Inc. [JP], THE Kitasato Institute [JP], Kao Corpiration [JP]. Publ. 2022-04-07.
46. Patent CN113072623. Preparation method for interfering peptide targeting SARS-COV-2Nproteinandapplicationthereof.Inventors:ZhouF.,DaiT.,WangS., Zhang L. Applicants: Soochow University [CN]. Publ. 2022-11-03.
47. Patent MX2022012545. Peptides and commbinations of peptides for use in immunotherapy against an infection by SARS-COV-2 (COVID-19). Inventors: Nelde A., Walz J., Rammensee H.-G., Bilich T. Applicants: Univ TuebingenMedizinischeFakultaet [DE]. Publ. 2022-11-07.
48. Patent US2023322865. Peptides and combinations of peptides for use in immunotherapy against an infection by Sars-COV-2 (COVID-19). Inventors: Walz J., Nelde A., Rammensee H.-G., Bilich T. Applicants: P Univ TuebingenMedizinischeFakultaet [DE]. Publ. 2023-10-12.
49. Patent US2023365659. SARS-COV-2-binding peptide. Inventors: Masaki H.,Kumachi S., Yonehara R., Matsumsra Y., Kawano H., Tojo T., et al. Applicants: Epsilon Molecular Eng Inc [JP]; The Kitasato Inst [JP]; Kao Corp [JP]. Publ. 2023-11-16.
50. Case J.B., Chen R.E., Cao L., Ying B., Winkler E.S., Johnson M., et al. Ultrapotent miniproteins targeting the SARS-CoV-2 receptor-binding domain protect against infection and disease. Cell Host Microbe. 2021; 29 (7): 1151-1161.e5.
51. Wu T., Zhu Y., Liu N., Hu Y., Chong H., He Y. Resistance profile and mechanism of severe acute respiratory syndrome coronavirus-2 variants to LCB1 inhibitor targeting the spike receptor-binding motif. Front Microbiol. 2022; 13: 1022006.
52. Jawad B., Adhikari P., Cheng K., Podgornik R., Ching W.-Y. Computational design of miniproteins as SARS-CoV-2 therapeutic inhibitors. Int J Mol Sci. 2022; 23 (2): 838.
53. Chattaraj R., Kim C.Y., Lee D., Hammer D.A. Recombinant protein micelles to block transduction by SARS-CoV-2 pseudovirus. ACS Nano. 2022; 16 (10): 17466-17477.
54. Zhu Y., Li M., Liu N., Wu T., Han X., Zhao G., et al. Development of highly effective LCB1-based lipopeptides targeting the spike receptor-binding motif of SARS-CoV-2. Antiviral Res. 2023; 211: 105541.
55. Chen K., Yu F.-Q., Zhang Y.-N., Fang G.-M. Total chemical synthesis of a SARS-CoV-2 miniprotein inhibitor LCB1. Methods Mol Biol. 2022; 2530: 19-31.
56. Prasad R., Ajith H., Chandrakumaran N.K., Khangar P.D., Mohan A., Nelson-Sathi S. In silico study identifies peptide inhibitors that negate the effect of non-synonymous mutations in major drug targets of SARS-CoV-2 variants. J Biomol Struct Dyn. 2023; 41 (19): 9551-9561.
57. Ulyanova V., Vershinina V., Ilinskaya O. Barnase and binase: twins with distinct fates. FEBS J. 2011; 278 (19): 3633-3643.
58. Hartley R.W. Barnase and barstar: Two small proteins to fold and fit together. Trends Biochem. Sci. 1989; 14: 450–454.
59. Hartley R.W. Barnase-barstar interaction. In: Methods in Enzymology. Academic Press Inc.; New York, NY, USA. 2001; 341: 599–611.
60. Guillet V., Lapthorn A., Hartley R., Mauguen Y. Recognition between a bacterial ribonuclease, barnase, and its natural inhibitor, barstar. Structure. 1993; 1: 165–176.
61. Shilova O., Kotelnikova P., Proshkina G., Shramova E., Deyev S. Barnase-barstar pair: Contemporary application in cancer research and nanotechnology. Molecules. 2021; 26 (22): 6785.
62. Edelweiss Е., Balandin Т.G., Ivanova J.L., Lutsenko G.V., Leonova O.G., Popenko V.I. Barnase as a new therapeutic agent triggering apoptosis in human cancer cells. PLoS ONE. 2008; 3 (6): e2434.
63. Патент РФ 2778942. Рекомбинантный белок, связывающийся с RBDS-белка SARS-CoV-2. Авторы: Алексеева Л.Г., Бобик Т.В., Габибов А.Г., Деев С.М., Коновалова Е.В., Лукьянова Т.И. и др. Патентообладатель: ФГБУН Институт биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова Российской академии наук (ИБХ РАН). Опубл. 29.08.2022 г. Бюл. № 25.
64. Kontermann R.E. Strategies for extended serum half-life of protein therapeutics. CurrOpinBiotechnol. 2011; 22: 868–876.
65. Sleep D., Cameron J., Evans L.R. Albumin as a versatile platform for drug half-life extension. BiochimBiophys Acta. 2013; 1830 (12): 5526-5534.
66. Nilvebrant J., Hober S. The albumin-binding domain as a scaffold for protein engineering. Comput Struct Biotechnol J. 2013; 6: e201303009.
67. Jonsson A., Dogan J., Herne N., Abrahmsen L., Nygren P.-A. Engineering of a femtomolar affinity binding protein to human serum albumin. Protein EngDes Sel. 2008; 21:515–527.
68. Studier F.W., Moffatt B.A. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol. 1986;189(1):113-130.
69. Vethanayagam J.G., Flower A.M. Decreased gene expression from T7 promoters may be due to impaired production of active T7 RNA polymerase. Microb Cell Fact. 2005;4(1):3.
70. Studier F.W. Protein production by auto-induction in high density shaking cultures. Protein Expr Purif. 2005;41(1):207-234.
71. Kurien B.T., Scofield R.H. Polyethylene glycol-mediated bacterial colony transformation. Biotechniques. 1995;18(6):1023-1026.
72. Kruglova N., Siniavin A., Gushchin V., Mazurov D. Different neutralization sensitivity of SARS-cov-2 cell-to-cell and cell-free modes of infection to convalescent sera. Viruses. 2021; 13(6): 1133.
73. Kostin N.N., Bobik T.V., Skryabin G.A., Simonova M.A., Knorre V.D., Abrikosova V.A., Mokrushina Y.A., Smirnov I.V., Aleshenko N.L., Kruglova N.A., Mazurov D.V., Nikitin A.E., Gabibov A.G., An ELISA Platform for the quantitativeqnalysis of SARS-CoV-2 RBD-neutralizing antibodies as a nalternativetomonitoring of the virus-neutralizingactivity. Acta Naturae. 2022; 14: 109–119.
74. Патент РФ 2610179. Способ получения рекомбинантного противоопухолевого токсина на основе белков барназа-барстари адресного полипептида дарпина с эффектом моментальной отмены цитотоксического действия. Авторы: Деев С.М., Шульга А.А., Стремовский О.А., Прошкина Г.М., Лебеденко Е.К. Патентообладатель: ФГБУН Институт биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова Российской академии наук (ИБХ РАН). Опубл. 29.08.2022 г. Бюл. № 25.
--->
Перечень последовательностей
Sequence Listing Information:
DTD Version: V1_3
File Name: PAB1.xml
Software Name: WIPO Sequence
Software Version: 2.3.0
Production Date: 2023-12-17
General Information:
Current application / IP Office: RU
Current application / Application number: 10
Current application / Filing date: 2023-12-17
Current application / Applicant file reference: PAB
Earliest priority application / IP Office: RU
Earliest priority application / Application number: 1
Earliest priority application / Filing date: 2023-12-17
Applicant name: ИБХ РАН
Applicant name / Language: ru
Applicant name / Name Latin: IBCh RAS
Inventiontitle: ПОЛИПЕПТИД С АНТИ-SARS-CoV-2 АКТИВНОСТЬЮ И
ПРОЛОНГИРОВАННЫМ СИСТЕМНЫМ ДЕЙСТВИЕМ INVIVO( ru )
Sequence Total Quantity: 9
Sequences:
Sequence Number (ID): 1
Length: 1284
Molecule Type: DNA
Features Location/Qualifiers:
- source, 1..1284
>mol_type, other DNA
> organism, Escherichia coli
- CDS, 4..750
>protein_id, 2
> translation,
MDKEWILQKIYEIMRLLDELGHAEASMRVSDLIYEFMKKGDERLLEEAERLLEEVERGSSSGSSSGSSSL
AEAKVLANRELDKYGVSDFYKRLINKAKTVEGVEALKLHILAALPEFPKPSTPPGSSGGAPAQVINTFDG
VADYLQTYHKLPDNYITKSEAQALGWVASKGNLADVAPGKSIGGDIFSNREGKLPGKSGRTWREADINYT
SGFRNSDRILYSSDWLIYKTTDHYQTFTKIRGAPHHHHH
- CDS, 4..174
>protein_id, 3
> translation,
MDKEWILQKIYEIMRLLDELGHAEASMRVSDLIYEFMKKGDERLLEEAERLLEEVER
- CDS, 175..210
>protein_id, 4
> translation, GSSSGSSSGSSS
- CDS, 211..348
>protein_id, 5
> translation, LAEAKVLANRELDKYGVSDFYKRLINKAKTVEGVEALKLHILAALP
- CDS, 349..396
>protein_id, 6
> translation, EFPKPSTPPGSSGGAP
- CDS, 397..726
>protein_id, 7
> translation,
AQVINTFDGVADYLQTYHKLPDNYITKSEAQALGWVASKGNLADVAPGKSIGGDIFSNREGKLPGKSGRT
WREADINYTSGFRNSDRILYSSDWLIYKTTDHYQTFTKIR
- CDS, 736..750
>protein_id, 8
> translation, HHHHH
- CDS, 1004..1276
>protein_id, 9
> translation,
MKKAVINGEQIRSISDLHQTLKKELALPEYYGENLDALWDCLTGWVEYPLVLEWRQFEQSKQLTENGAES
VLQVFREAKAEGCDITIILS
Residues:
catatggataaagaatggattctgcaaaaaatttacgagatcatgcgtctgctggacgaa 60
ctgggccacgcggaggcttccatgcgtgtgagcgatctgatctatgagttcatgaagaaa 120
ggtgacgaacgtctgctggaagaagcagaacgcctgctggaagaagttgaacgcggttct 180
tcttccggctcctcctctggtagcagcagcctggcagaagctaaagtgctggcaaaccgt 240
gaactggacaagtatggtgtatctgatttctacaaacgcctgattaacaaagcgaaaacc 300
gttgaaggcgttgaggctctgaaactgcacatcctggcggccctgccggaattccccaaa 360
ccttctactccacccgggagcagtggtggcgcgcctgcacaggttatcaacacgtttgac 420
ggggttgcggattatcttcagacatatcataagctacctgataattacattacaaaatca 480
gaagcacaagccctcggctgggtggcatcaaaagggaaccttgcagacgtcgctccgggg 540
aaaagcatcggcggagacatcttctcaaacagggaaggcaaactcccgggcaaaagcgga 600
cgaacatggcgtgaagcggatattaactatacatcaggcttcagaaattcagaccggatt 660
ctttactcaagcgactggctgatttacaaaacaacggaccattatcagacctttacaaaa 720
atcagaggcgcgccgcaccatcatcaccattgacgtctaagcttataacgaaaaaaacgg 780
cttcctgcggaggccgtttttttcagctttacataaagtgtgtaataaatttttcttcaa 840
actctgatcggtcaatttcactttccggctctagagtccggtccaatctgcagccgtccg 900
agacaggaggacatcgtccagctgaaaccggggcagaatccggccatttctgaagagaaa 960
aatggtaaactgatagaataaaatcataagaaaggagccgcacatgaaaaaagcagtcat 1020
taacggggaacaaatcagaagtatcagcgacctccaccagacattgaaaaaggagcttgc 1080
ccttccggaatactacggtgaaaacctggacgctttatgggattgtctgaccggatgggt 1140
ggagtacccgctcgttttggaatggaggcagtttgaacaaagcaagcagctgactgaaaa 1200
tggcgccgagagtgtgcttcaggttttccgtgaagcgaaagcggaaggctgcgacatcac 1260
catcatactttcttaatactcgag
1284
Sequence Number (ID): 2
Length: 249
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..249
>mol_type, protein
> organism, Escherichia coli
Residues:
MDKEWILQKI YEIMRLLDEL GHAEASMRVS DLIYEFMKKG DERLLEEAER
LLEEVERGSS 60
SGSSSGSSSL AEAKVLANRE LDKYGVSDFY KRLINKAKTV EGVEALKLHI
LAALPEFPKP 120
STPPGSSGGA PAQVINTFDG VADYLQTYHK LPDNYITKSE AQALGWVASK
GNLADVAPGK 180
SIGGDIFSNR EGKLPGKSGR TWREADINYT SGFRNSDRIL YSSDWLIYKT
TDHYQTFTKI 240
RGAPHHHHH
249
Sequence Number (ID): 3
Length: 57
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..57
>mol_type, protein
> organism, Escherichia coli
Residues:
MDKEWILQKI YEIMRLLDEL GHAEASMRVS DLIYEFMKKG DERLLEEAER LLEEVER
57
Sequence Number (ID): 4
Length: 12
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..12
>mol_type, protein
> organism, Escherichia coli
Residues:
GSSSGSSSGS SS
12
Sequence Number (ID): 5
Length: 46
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..46
>mol_type, protein
> organism, Escherichia coli
Residues:
LAEAKVLANR ELDKYGVSDF YKRLINKAKT VEGVEALKLH ILAALP
46
Sequence Number (ID): 6
Length: 16
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..16
>mol_type, protein
> organism, Escherichia coli
Residues:
EFPKPSTPPG SSGGAP
16
Sequence Number (ID): 7
Length: 110
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..110
>mol_type, protein
> organism, Escherichia coli
Residues:
AQVINTFDGV ADYLQTYHKL PDNYITKSEA QALGWVASKG NLADVAPGKS
IGGDIFSNRE 60
GKLPGKSGRT WREADINYTS GFRNSDRILY SSDWLIYKTT DHYQTFTKIR 110
Sequence Number (ID): 8
Length: 5
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..5
>mol_type, protein
> organism, Escherichia coli
Residues:
HHHHH
5
Sequence Number (ID): 9
Length: 90
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..90
>mol_type, protein
> organism, Escherichia coli
Residues:
MKKAVINGEQ IRSISDLHQT LKKELALPEY YGENLDALWD CLTGWVEYPL
VLEWRQFEQS 60
KQLTENGAESVLQVFREAKAEGCDITIILS
90
END
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный белок, связывающийся с RBD S-белка SARS-CoV-2 | 2022 |
|
RU2778942C1 |
| ИММУНОЛИПОСОМА, СВЯЗЫВАЮЩАЯСЯ С RBD S-БЕЛКА SARS-COV-2 | 2023 |
|
RU2827165C1 |
| Рекомбинантный RBD S-белок коронавируса SARS-CoV-2 и способ его получения | 2023 |
|
RU2833839C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ БАРНАЗЫ В КЛЕТКАХ ESCHERICHIA COLI, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ БАРНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ БАРНАЗЫ. | 2017 |
|
RU2650871C1 |
| Широко нейтрализующее антитело против SARS-CoV-2 | 2022 |
|
RU2810476C1 |
| Плазмидная генетическая конструкция pET21a_SKP, обеспечивающая экспрессию в прокариотической системе E.coli рекомбинантного белка SKP, и рекомбинантный белок SKP, обладающий широконейтрализующими свойствами однодоменного наноантитела против SARS-CoV-2 | 2024 |
|
RU2839376C1 |
| СПОСОБ ИСПОЛЬЗОВАНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ SARS-COV-2 В СОСТАВЕ ТЕСТ-СИСТЕМЫ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА С ОПРЕДЕЛЕНИЕМ УРОВНЕЙ АНТИТЕЛ КЛАССОВ IgM, IgG, IgA В СЫВОРОТКЕ/ПЛАЗМЕ КРОВИ БОЛЬНЫХ COVID-19 | 2020 |
|
RU2730897C1 |
| Плазмидная генетическая конструкция pET21_Ab_CoV-2_1.3, обеспечивающая экспрессию рекомбинантного белка AB_COV-2_1.3 в прокариотической системе E. coli, и рекомбинантный белок AB_COV-2_1.3, обладающий свойствами однодоменного наноантитела против SARS-CoV-2 | 2022 |
|
RU2798508C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ НЕЙТРАЛИЗУЮЩИХ АНТИТЕЛ К SARS-CoV-2 В СЫВОРОТКЕ ИЛИ ПЛАЗМЕ КРОВИ ЛЮДЕЙ, ПЕРЕНЕСШИХ COVID-19 ИЛИ ПРИВИТЫХ ВАКЦИНАМИ ДЛЯ ПРОФИЛАКТИКИ НОВОЙ КОРОНАВИРУСНОЙ ИНФЕКЦИИ COVID-19, С ИСПОЛЬЗОВАНИЕМ НАБОРА РЕАГЕНТОВ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА, СОДЕРЖАЩЕГО РЕКОМБИНАНТНЫЙ РЕЦЕПТОР-СВЯЗЫВАЮЩИЙ ДОМЕН (RBD) ПОВЕРХНОСТНОГО ГЛИКОПРОТЕИНА S КОРОНАВИРУСА SARS-COV-2 И РЕКОМБИНАНТНЫЙ ЧЕЛОВЕЧЕСКИЙ РЕЦЕПТОР АСЕ2 | 2021 |
|
RU2784655C1 |
| Искусственная генетическая конструкция для гетерологической экспрессии рецептор-связывающего домена S-белка в слитной полипептидной цепи с нуклеокапсидным белком | 2022 |
|
RU2801597C1 |

Изобретение относится к области белковой инженерии, биотехнологии и медицине. Описан рекомбинантный белок, представленный на SEQ ID NO: 2, обладающий способностью эффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2. Представлена рекомбинантная плазмида pET39b-LCB1-ABD-Ba, обеспечивающая синтез в клетках Escherichia coli рекомбинантного белка, полученная клонированием фрагмента ДНК, представленной на SEQ ID NO: 1, в плазмидный вектор pET39b по сайтам рестрикции NdeI/XhoI. Раскрыт рекомбинантный штамм Escherichia coli BL21(DE3)/pET39b-LCB1-ABD-Ba, содержащий рекомбинантную плазмиду pET39b-LCB1-ABD-Ba по п. 2 – продуцент рекомбинантного белка. Описан способ получения рекомбинантного белка из биомассы рекомбинантного штамма Escherichia coli BL21(DE3)/pET39b-LCB1-ABD-Ba, при котором осуществляют разрушение клеток дезинтеграцией, удаляют клеточный дебрис, выделяют белок при помощи двухстадийной хроматографии, включающей стадию белковой денатурации/ренатурации на металлохелатном аффинном сорбенте и очистку при помощи анионообменной хроматографии. Изобретение может найти применение в биотехнологии и медицине при создании лекарств против КОВИД-19. Изобретение позволяет получать высокий уровень индуцируемого синтеза - 40% от суммарного клеточного белка и стабильную продукцию, высокую специфическую антивирусную активность, пролонгирование времени циркуляции и противовирусного эффекта в кровотоке в сравнении с аналогом Targevir1. 4 н.п. ф-лы, 8 ил., 1 табл., 9 пр.
1. Рекомбинантный белок, представленный на SEQ ID NO: 2, обладающий способностью эффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2.
2. Рекомбинантная плазмида pET39b-LCB1-ABD-Ba, обеспечивающая синтез в клетках Escherichia coli рекомбинантного белка по п.1, полученная клонированием фрагмента ДНК, представленной на SEQ ID NO: 1, в плазмидный вектор pET39b по сайтам рестрикции NdeI/XhoI.
3. Рекомбинантный штамм Escherichia coli BL21(DE3)/pET39b-LCB1-ABD-Ba, содержащий рекомбинантную плазмиду pET39b-LCB1-ABD-Ba по п. 2 – продуцент рекомбинантного белка по п.1.
4. Способ получения рекомбинантного белка по п. 1 из биомассы рекомбинантного штамма Escherichia coli BL21(DE3)/pET39b-LCB1-ABD-Ba по п. 3, при котором осуществляют разрушение клеток дезинтеграцией, удаляют клеточный дебрис, выделяют белок по п.1 при помощи двухстадийной хроматографии, включающей стадию белковой денатурации/ренатурации на металлохелатном аффинном сорбенте, и очистку при помощи анионообменной хроматографии.

