Изобретение относится к области медицины, вирусологии, микробиологии, молекулярной генетики и биотехнологии и может быть использовано в медицинской промышленности при производстве живых вакцин для профилактики и лечения инфекций, вызванных коронавирусом. Коронавирусы (Coronaviridae) — семейство вирусов, включающее на январь 2020 года 40 видов РНК-содержащих вирусов, объединённых в четыре подсемейства, которые поражают человека и животных. Название связано со строением вируса, который имеет шиповидные отростки. Они напоминают солнечную корону и содержат рецептор-связывающие белки на поверхности, которые связывают трансмембранные рецепторы клеток.
Известно, что ученые из Китая расшифровали и опубликовали 12 января 2020 г. в международной базе данных геном нового коронавируса [1- Guo YR, Cao QD, Hong ZS, Tan YY, Chen SD, Jin HJ, Tan KS, Wang DY, Yan Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak - an update on the status. Mil Med Res. 2020 Mar 13;7(1):11. doi: 10.1186/s40779-020-00240-0. PMID: 32169119; PMCID: PMC7068984.] и что в настоящее время во всем мире учеными ведутся интенсивные исследования по разработке вакцины от коронавируса SARS-CoV-2 и их лабораторные и тестовые испытании (краткая информация о коронавирусе человека представлена в Приложении к описанию изобретения).
Известен пробиотический штамм Enterococcus faecium L3 ND-79 ВНИИСХМ, предназначенный для изготовления лечебно-профилактических средств и продуктов питания лечебно-профилактического назначения, обладающий устойчивостью к широкому спектру антибиотиков, более выраженным антагонизмом к патогенным бактериям, высокой жизнеспособностью и высоким уровнем продукции витаминов группы "В" и фолиевой кислоты; известный штамм положен в основу заявляемого изобретения [2 – патент РФ №2220199 на штамм энтерококков Enterococcus faecium l-3 для изготовления лечебно-профилактических средств (патентообладатели и авторы Алехина Г.Г. и Суворов А.Н.)].
Известна созданная на основе этого штамма [2] живая вакцина для профилактики вагинальной инфекции, вызванной Streptococcus agalactiae (СГВ). На основе штамма пробиотика Еnterococcus faecium L3 была сконструирована первая генно-инженерная живая вакцина со встроенным в хромосому участком гена белка патогенного СГВ. [3-Т.В.Гупалова, Г.Ф.Леонтьева, Е.И.Ермоленко, К.Б.Грабоская, Т.А. Крамская, А.Н.Цапиева, И.В.Королева, А.Н.Суворов. Создание и опыт применения живой вакцины на основе штамма пробиотика Enterococcus Faecium L3 для профилактики вагинальной инфекции, вызванной Streptococcus agalactiae, Медицинский академический журнал, 2013, 13(2), С. 64-70].
Известен способ, основанный на введении bac гена патогенных СГВ в хромосомную ДНК в области кодирования белков пилей пробиотического штамма Enterococcus faecium L3 с последующей экспрессией белка Bac, кодируемого этим геном, в пилях. Пили энтерококков выступают за пределы бактериальных клеток и способны проникать сквозь капсулу, которая экранирует большинство бактериальных белков-антигенов. Пили состоят из белковых мономеров, способных к агрегации. За счет этого процесса может увеличиваться доза чужеродного антигена, встроенного в белок пилей. Последнее должно способствовать увеличению титров антител, специфичных к встраиваемому в структуру поверхностного белка энтерококка полипептидного фрагмента штамма патогенного СГВ.
Известный способ введения гена патогенных стрептококков в пили пробиотического штамма Enterococcus faecium L3 интересен тем, что рекомбинантный белок экспрессируется не в цитоплазме, а на поверхности бактерии [4- патент РФ №2640250 «Способ введения генов патогенных стрептококков в хромосомную ДНК пробиотического штамма Еnterococcus faecium l3 для экспрессии в пилях», МПК:A61K39/092; авторы- Суворов А.Н. и др.].
Известно, что штамм Еnterococcus faecium L3 обладает выраженной антагонистической активностью в отношении грамположительных и грамотрицательных бактерий, способностью восстанавливать микробиоценоз кишечника на фоне дисбиотических состояний [5 - Yermolenko E., Suvorov A., Chernush A., et al. International Congress Series, 1289: 363-366 (2006)], а также оказывать иммуномодулирующее действие на организм хозяина [6 - TarasovaE., YermolenkoE. , DonetsV., SundukovaZ. , BochkarevaA. , BorsсhevI. , SuvorovaM. , Ilyasov. I., Simanenkov V., Suvorov A. The influence of probiotic enetrococci on the microbiota and cytokines expression in rats with dysbiosis induced by antibiotics// Beneficial Microbes. -2010- Т. 1. -№ 3.- С. 265-270].
Осуществлено физическое картирование штамма Еnterococcus faecium L3 [7 - Suvorov A., Simanenkov V., Gromova L. et al. Prebiotics and probiotics potencial for human health, 104-112, (2011)]. В экспериментах на здоровых самках мышей показано, что интравагинальное введение высоких доз штамма Еnterococcus faecium L3 не только не оказывает токсического действия на организм, но не влияет на состояние слизистой оболочки влагалища [8 - Суворов А., Алехина Г.Г., Пигаревский П.В. и др. Гастробюллетень, 4: 29-31, (2001)] и способствует экспрессии IL-10 клетками слизистой оболочки влагалища крыс с экспериментальным вагинитом, вызванном S.agalactiae и Staphylococcus aureus [6 - TarasovaE., YermolenkoE. , DonetsV., SundukovaZ. , BochkarevaA. , BorsсhevI. , SuvorovaM. , IlyasovI., SimanenkovV., SuvorovA. The influence of probiotic enetrococci on the microbiota and cytokines expression in rats with dysbiosis induced by antibiotics// Beneficial Microbes. -2010.- Т. 1. -№ 3.- С. 265-270].
В штамме Еnterococcus faecium L3, как и у других грамположительных бактерий, имеются пили, которые представляют собой фимбрии длиной 0,3-3 μm и диаметром 2-10nm [9 - Telford JL, Barocchi MA, Margarit I, Rappuoli R, Grandi G. Pili in gram-positive pathogens. Nat Rev Microbiol. -2006- T.4, №7-С. 509-519. doi: 10.1038/nrmicro1443. PMID: 16778837]. Это длинные белок-подобные полимеры, тянущиеся на поверхности бактерий, представляют собой субъединицы белка пилина, соединенные ковалентной связью. Они играют большую роль в адгезии колонизации хозяина. Пили являются высокоиммуногенными структурами, которые находятся под селективным давлением иммунных реакций хозяина [10 - Danne C, Dramsi S. Pili of gram-positive bacteria: roles in host colonization. Res Microbiol. 2012 Nov-Dec;163(9-10):645-58. doi: 10.1016/j.resmic.2012.10.012. Epub 2012 Oct 29. PMID: 23116627]. Принцип модификации пилей энтерококков вакцинными антигенами является приоритетным подходом при создании эффективных живых вакцин благодаря экспозиции целевого антигена на поверхности энтерококка.
Известен способ создания живой вакцины за счет введения генов патогенных стрептококков в хромосомную ДНК пробиотического штамма Enterococcus faecium L3 для экспрессии в пилях, и живая вакцины на основе модифицированного штамма пробиотиков Еnterococcus faecium L3 для профилактики инфекции, вызванной Streptococcus pneumonie [11 - Суворов А.Н., Гупалова Т.В., Кулешевич Е.В., Леонтьева Г.Ф., Крамская Т.А, Патент РФ№ 2701733], который принят в качестве прототипа.
Недостатком известного способа [11]является сложность конструирования такой вакцины за счет большого количества стадий необходимых для создания суммарного фрагмента ДНК, состоящего из двух отдельных фрагментов гена пробиотика Enterococcus faecium L3 и химерного гена Streptococcus pneumonie-pspf.
Указанный недостаток устраняется в заявленном способе создания живой вакцины за счет замены химерного гена Streptococcus pneumoniae-pspf на фрагмент гена covid-19 методом клонирования, что сокращает получение вакцины в отличие от прототипа, а также позволяет создавать любые другие вакцинные кандидаты на этой платформе. Существенно, что созданный вакцинный кандидат в отличие от вакцины-прототипа, предназначен для обеспечения специфической профилактики заболеваний, вызванных вирусом SARS-Cov-2, а не Streptococcus pneumoniae.
Техническим результатом изобретения является значительное упрощение способа получения живой вакцины против коронавирусной инфекции.
Указанный технический результат достигается тем, что в способе создания живой вакцины против коронавирусной инфекции COVID-19 на основе пробиотического штамма Enterococcus faecium L3 (по патенту РФ №2220199), модифицированного в результате электропорации культуры энтерококков штамма Enterococcus faecium L3 рекомбинантной плазмидной ДНК pentF-covid-19, содержащей последовательность SEQ ID No:1 в соответствии с заявленным изобретением, для создания SEQ ID No:1 в плазмиде pentF-pspf (по патенту РФ №2701733), участок pspf был заменен на фрагмент гена шиповидного белка SARS-CoV-2, при этом ДНК pentF-covid-19 кодирует аминокислотную последовательность SEQ ID No:2, которая выполняет функцию шиповидного белка вируса SARS-CoV-2, способного осуществлять стимуляцию гуморального и клеточного иммунитета в отношении вируса SARS-CoV-2 .При этом задача генетической части работы состояла в осуществлении интеграции участка ДНК вируса в структуру гена поверхностного белка пробиотика без нарушения открытой рамки считывания и повреждения участков кодирования компонентов, участвующих в процессинге поверхностного белка PilF.
Кроме того, указанный технический результат достигается тем, что живая вакцина Enterococcus faecium L3-pentF-covid-19, в которой штамм Enterococcus faecium экспрессирует ген слитого белка, способного индуцировать образование антител, причем образующиеся специфические антитела классов А и G в отношении инфекции, в соответствии с заявленным изобретением, в плазмидную ДНК вводят фрагмент гена, кодирующего шиповидный белок вируса SARS-CoV-2, способного осуществлять стимуляцию гуморального и клеточного иммунитета в отношении вируса SARS-CoV-2, т.е созданием живой вакцины против коронавируса SARS-CoV-2, обладающей выраженным иммуногенным эффектом.
В известном способе, использованном в качестве прототипа[11], на основе штамма пробиотика Еnterococcus faecium L3 была сконструирована генно-инженерная живая вакцина со встроенным в хромосому, в области, кодирующей ген пилей, участком химерного гена pspf, сконструированного на основе генов патогенности Streptococcus pneumoniae. Штамм Еnterococcus faecium L3-PSPF+ был получен в результате трансформации плазмидой pentF-pspf штамма Enterococcus faecium L3.
Интраназальное введение вакцины Еnterococcus faecium L3-PSPF+ стимулировало развитие специфического системного и местного иммунного ответа. На модели интраназальной летальной пневмококковой инфекции у мышей линии СВА было показано, что трехкратная интраназальная вакцинация созданной живой пробиотической вакциной повышала защиту от летальной пневмококковой инфекции.
При реализации заявленного способа создания живой вакцины на основе пробиотического штамма Еnterococcus faecium L3 был использован синтезированный фрагмент ДНК шиповидного белка коронавируса, который был смоделирован авторами патента, а затем синтезирован компанией Евроген. При этом в плазмиде pentF-pspf участок гена pspf был заменен на фрагмент гена шиповидного белка.
В итоге реализации этого способа достигается решение конечной технической задачи заявленного изобретения, заключающееся в создании живой вакцины против коронавируса SARS-CoV-2, обладающей выраженными иммуногенными свойствам, на основе биологически активного штамма Еnterococcus faecium L3 за счет замещения в структуре участка его гена пилей на участок гена корановируса SARS-CoV-2. Создание такой вакцины против инфекции, вызываемой корановирусом, основано на введении участка ДНК, кодирующего антигенный фрагмент иммуногенных эпитопов вируса SARS-CoV-2, в структуру гена пилей пробиотика.
Реализация указанной технической задачи поясняется конкретными примерами введения гена covid-19 коронавируса в хромосомную ДНК в области, кодирующей ген пилей, пробиотического штамма Еnterococcus faecium L3 для экспрессии в пилях иммуногенного участка шиповидного белка SARS-CoV-2:
а) получение слитого гена entF-covid-19 и его клонирование
б) выявление бактериальных клонов, содержащих ген слитого белка COVID-19+ в хромосоме
в) пероральная вакцинация мышей живой пробиотической вакциной
г) определение COVID19+-специфических антител в крови и вагинальных смывах.
д) выявление клеточного звена специфического иммунного ответа на пероральную вакцинацию живой противовирусной вакциной.
Технической сущностью предлагаемого изобретения является создание живой вакцины против коронавируса SARS-CoV-2, обладающей выраженными иммуногенными свойствами.
Пероральное введение вакцины Еnterococcus faecium L3 с геном вируса SARS-CoV-2 стимулировало развитие специфического системного и местного иммунного ответа. На модели интраназальной летальной коронавирусной инфекции у мышей было показано, что трехкратная пероральная вакцинация созданной живой пробиотической вакциной повышала защиту от летальной коронавирусной инфекции.
Конструирование живой вакцины на основе биологически активного пробиотического штамма Еnterococcus faecium L3 за счет включения в его пили антигена SARS-CoV-2 позволило объединить в одном препарате эффективность полезных свойств пробиотика и специфического антигенного стимула.
Для получения слитого гена entF-covid-19 и его клонирования были сконструированы ДНК праймеры, представленные в таблице «Олигонуклеотидные праймеры», в которой в графе 1 приведены 8 названий. праймеров; в графе 2 приведена их ориентация по отношению к положительной цепочке ДНК.; а в графе 3 приведены нуклеотидные последовательности, которые были сконструированы в соответствии с направлением цепочки ДНК последовательность от 5' к 3'.
Олигонуклеотидные праймеры
Таблица
Нуклеотидные последовательности в графе 3 с названиями К1 и К2 (в графе 1), с участками, выделенными подчеркиванием, указывают на соответствующие им сайты гидролиза рестрикционными эндонуклеазами.
Фрагмент гена covid-19 был синтезирован с последующим клонированием в векторную плазмидную ДНК pAL2-T, фирмой Евроген. Для удобства последующего клонирования сайты рестрикции для NdEI и EcoRI были вставлены во фрагмент гена covid-19 при его синтезе.
Рекомбинантная плазмидная ДНК pentF-pspf, представляющая собой плазмидную ДНК, полученную в результате вставки суммарного фрагмента ДНК, состоящего из двух отдельных фрагментов гена пробиотика Enterococcus faecium L3 и фрагмента гена pspf, описанная в прототипе [11-патент РФ 2701733], была использована для заявляемой конструкции. Из плазмидной ДНК pentF-pspf была удалена вставка гена пневмококка pspf и заменена на вставку гена covid-19. Для этого плазмидную ДНК pentF-psp гидролизовали ферментами NdEI и EcoRI, сайты для которых ограничивают последовательность гена pspf (Фиг.1). Такие же сайты рестрикции были заложены в синтезированный фрагмент гена covid-19. Полученный верхний фрагмент после гидролиза был использован для дальнейшего клонирования.
Плазмидная ДНК pAL2-T, содержащая фрагмент гена covid-19, была гидролизована ферментами NdEI и EcoRI. Клонирование рестрицированного фрагмента, кодирующего COVID-19, осуществлялся использованием верхнего фрагмента после гидролиза плазмиды pentF-pspf. Продукты рестрикции разделялись с помощью электрофореза в 1% агарозном геле. ДНК рестрикты выделялись из агарозы с помощью набора «QIAquick Gel Extraction Kit» (Qiagen, USA), которые затем лигировали и трансформировали в гетерологичную систему E.coli DH5α. Среда для отбора содержала 500 мкг/мл эритромицина. В результате трансформации был получен один клон. Из этого клона была выделена плазмидная ДНК. Наличие вставки подтверждалось гидролизом плазмиды ферментами NdEI и EcoRI (Фиг.2).
Продукты ПЦР плазмидной ДНК, полученные с праймерами K1 и K2,и SeqF и K2 были просеквенированы. Результаты сиквенса показали правильность последовательности плазмидной ДНК, которая содержала нужные фрагмнты ДНК энтерококка и ДНК коронавируса (перечень последовательностей SEQIDNo:1).
В результате клонирования были получена плазмида pentF-covid-19 спрогнозируемой вставкой и геном устойчивости к эритромицину.
В результате электропорации энтерококков созданной интегративной плазмидой было получено 14 трансформантов. Они были проверены в реакции ПЦР с праймерами K1 и K2. Для этого из трансформантов выделяли ДНК, используя набор ДНК-экспресс (Литех, Россия). Положительный ответ дали 13трансформантов, все кроме 12. (Фиг3, верхний ряд)
Для определения интеграции плазмидной ДНК pentF-covid-19 в хромосомную ДНК энтерококка была проведена амплификация ДНК, выделенных из 13 клонов с праймерами B1 и K2. Интеграция плазмидной ДНК pentF-covid-19 в хромосомную ДНК энтерококка была обнаружена только у двух трансформантов (Фиг3, дорожки 22 и 23,нижний ряд). Секвенирование ДНК, выделенной из одного из положительных клонов, обозначенного, как COVID 19+,было проведено с праймерами, соответствующими последовательности гена covid-19 (праймер K2) и последовательности хромосомной ДНК энтерококков (праймер B1) для подтверждения интеграции плазмидной ДНК pentF-covid-19 в хромосомную ДНК энтерококка (перечень последовательностей SEQIDNo:1).
На Фиг.4 показан результат после проведения ПЦР с ДНК клона COVID 19+ с разными парами праймеров. На Фиг. 4 приведены результаты ПЦР с хромосомной ДНК энтерококка и плазмидной ДНК pentF-covid-19.
Клон COVID 19+, экспрессирующий ген COVID-19 белка, был выбран в качестве вакцинного препарата для дальнейшего исследования.
Ниже приведены конкретные примеры, подтверждающие экспериментально проведение стадий получения живой вакцины против коронавирусной инфекции
Пример 1.
Синтез фрагмента ДНК коронавируса и его клонирование в плазмидный вектор pAL2-T
Синтез фрагмента ДНК коронавируса и его клонирование в плазмидный вектор pAL2-T было проведено фирмой Евроген. Для удобства последующего клонирования сайты рестрикции для NdEI и EcoRI были вставлены во фрагмент гена covid-19 при его синтезе.
Пример 2.
Получение слитого гена entF-covid-19 и его клонирование.
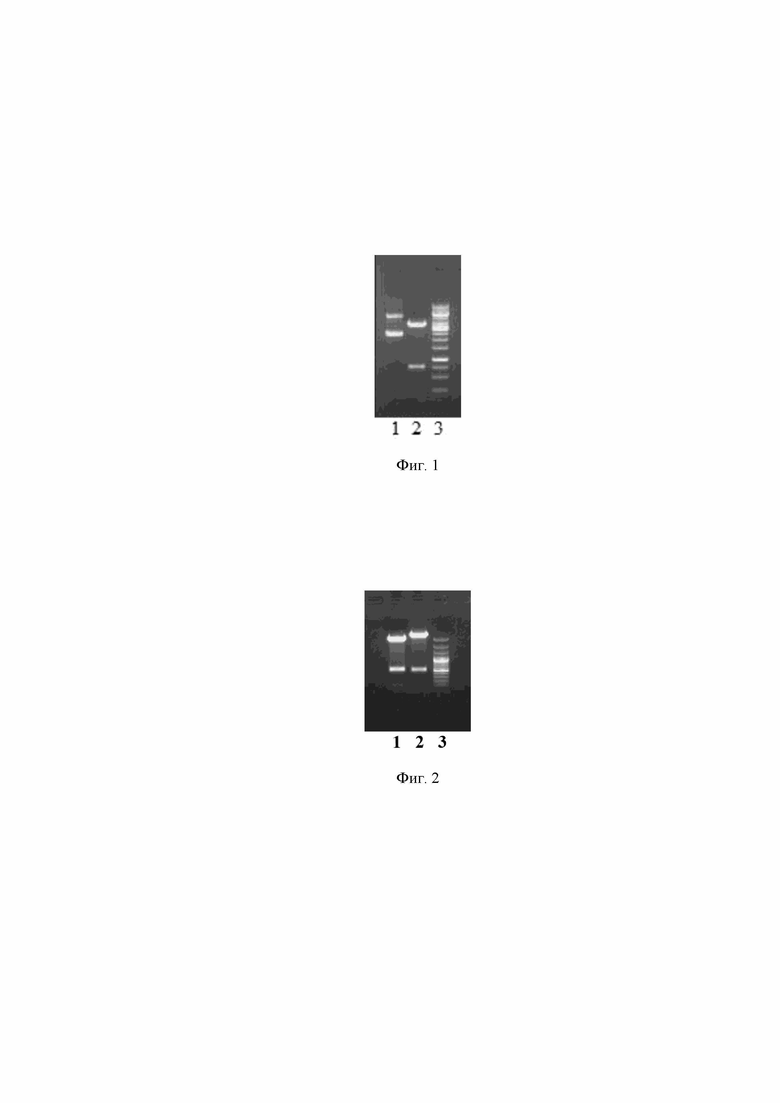
Рекомбинантная плазмидная ДНК pentF-pspf, представляющая собой плазмидную ДНК, полученную в результате вставки суммарного фрагмента ДНК, состоящего из двух отдельных фрагментов гена пробиотика Enterococcus faecium L3 и фрагмента гена pspf, описанная в прототипе [11- патент РФ 2701733], была использована для создания слитого гена entF-covid-19. Для этого из плазмидной ДНК pentF-psp f была вырезана вставка гена пневмококка pspf, проведен гидролиз плазмидной ДНК pentF-pspf ферментами NdEI и EcoRI (Фиг.1). Полученный верхний фрагмент после гидролиза был использован для дальнейшего клонирования.
На Фиг. 1 представлена электрофореграмма рестрицированной плазмидной ДНК pentF-pspf ферментами NdEI и EcoRI., где:
1 - плазмида pentF-pspf - исходная
2 - плазмида pentF-pspf рестрицированная
3 - 100 п.н. ДНК - маркер (сверху вниз: 3000, 2000, 1000, 900, 800, 700, 600, 500, 400, 300, 200 и 100 нуклеотидных пар).
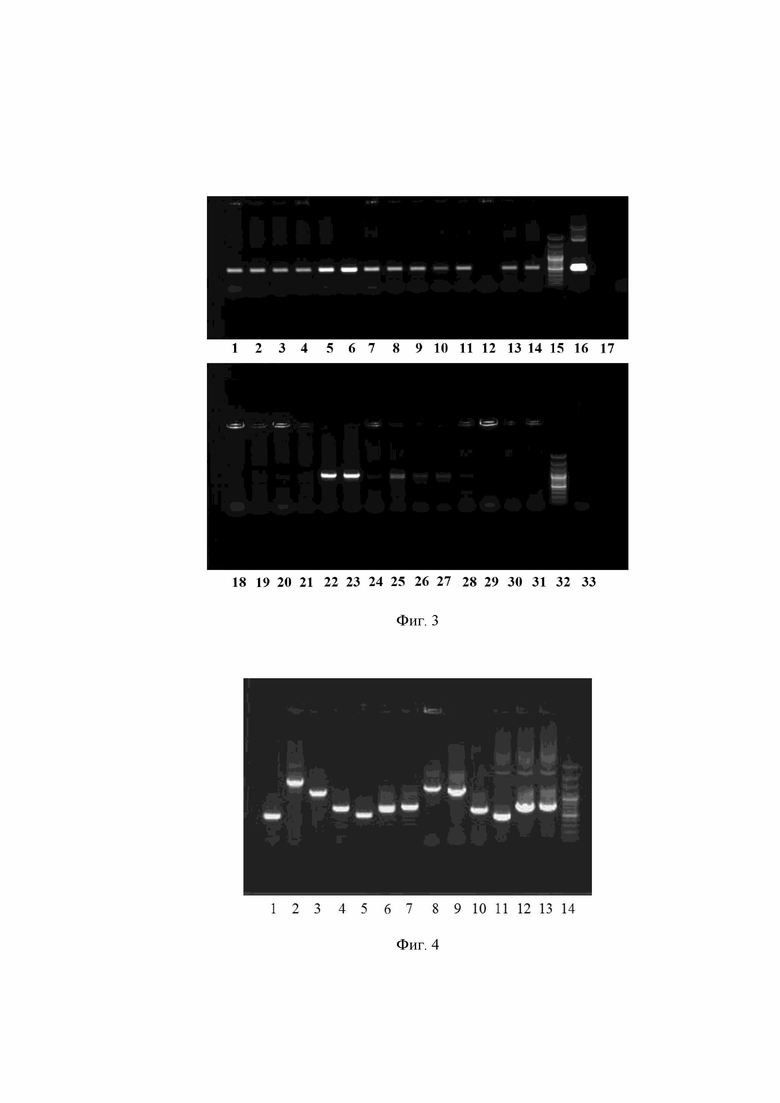
Плазмидная ДНК pAL2-T, содержащая фрагмент гена covid-19, была гидролизована ферментами NdEI и EcoRI (Фиг.2). Клонирование рестрицированного фрагмента гена, кодирующего COVID-19, осуществлялось с использованием верхнего фрагмента после гидролиза плазмиды pentF-pspf. Продукты гидролиза рестрикионными эндонуклеазами были разделены с помощью электрофореза в 1% агарозном геле. ДНК гидролизаты были выделены из агарозы с помощью набора «QIAquick Gel Extraction Kit» (Qiagen, USA), лигированы и трансформированы в гетерологичную систему E.coli DH5α. Среда для отбора клонов E.coli с плазмидой содержала 500 мкг/мл эритромицина. В результате трансформации был получен один клон. Из этого клона была выделена плазмидная ДНК. Наличие вставки подтверждалось гидролизом плазмиды ферментами NdEI и EcoRI (Фиг.2).
На Фиг. 2 представлена электрофореграмма рестрицированных пазмид NdEI и EcoRI, где:
1 - плазмидная ДНК pAL2-T, содержащая фрагмент гена covid-19
2 - плазмидная ДНК pentF-covid-19
3 - 100 п.н. ДНК - маркер (сверху вниз: 3000, 2000, 1000, 900, 800, 700, 600, 500, 400, 300, 200 и 100 нуклеотидных пар).
Пример 3.
Трансформация энтерококков методом электропорации.
Для электропорации энтерококков культуру Еnterococcus faecium L3 сеяли в 3 мл бульона Tood-Hewitt (THB) («HiMedia», Индия) и выращивали в течение ночи при 37°С, затем пересевали в 50 мл бульона THB 1 мл ночной культуры и выращивали ее до оптической плотности при длине волны 650 нм 0.3. После этого культуру помещали в лед и затем отмывали трижды в 20 мл 10% глицерола при температуре 4°С, полученный осадок суспендировали в 0.5 мл стерильного раствора глицерола, переосаждали и конечный осадок ресуспендировали в 0.3 мл того же раствора, разлили по 50 мкл в пробирки и проводили электропорацию в кювете с расстоянием между электродами 1 мм при напряжении 2100 В. Во льду к клеткам добавили ДНК (полученную интегративную плазмиду pentF-covid-19 плазмиду, 300 нг). Оптимальная продолжительность импульса составила 4-5 миллисекунд. После проведения разряда тока в кювету добавляли 1 мл THB, инкубировали 1 час и высевали на чашки с селективной средой, которая содержала 10 мкг/мл эритромицина. Затем ожидали появления трансформантов через 24 часа.
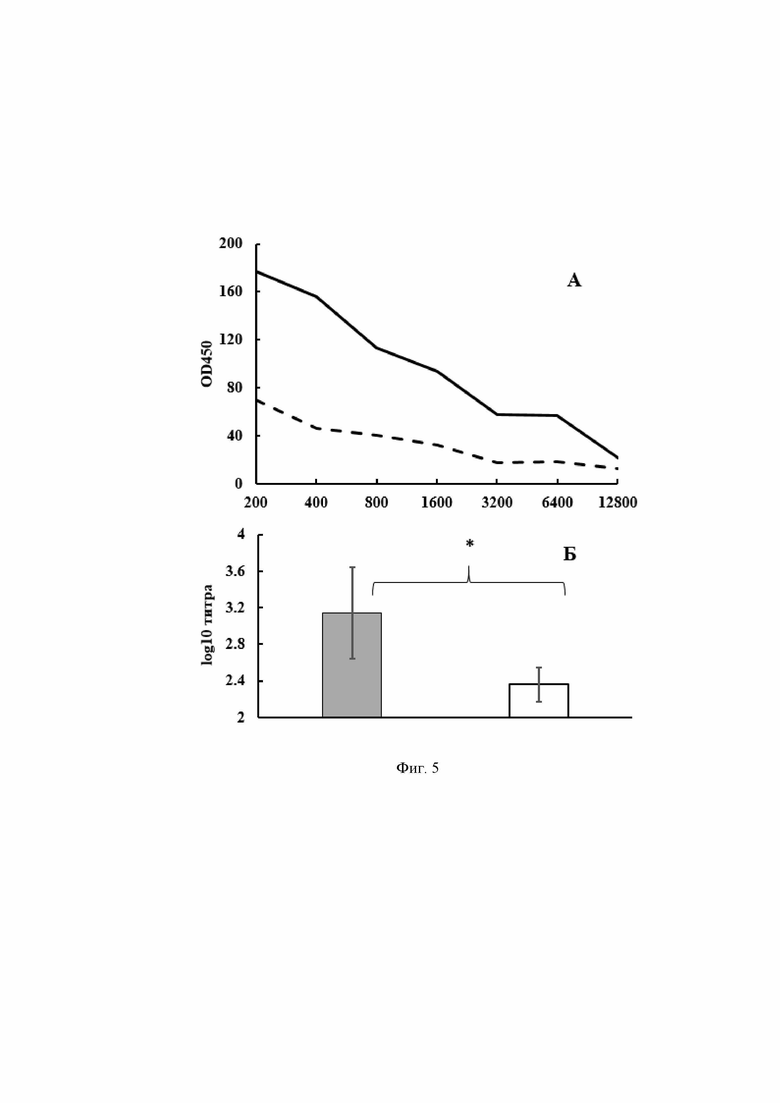
В результате электропорации энтерококков, созданной интегративной плазмидой, было получено 14 клонов трансформантов. Они были проверены в реакции ПЦР с праймерами K1 и K2. Для этого из трансформантов выделяли ДНК, используя набор ДНК-экспресс (Литех, Россия). Положительный ответ дали 13трансформантов (Фиг. 3, верхний ряд).
На Фиг. 3 в верхнем ряду представлена электрофореграмма амплифицированных ДНК-фрагментов с праймерами К1 и К2.
1 – 14 – продукты ПЦР ДНК из полученных 14 клонов.
15 - 100 п.н. ДНК - маркер (сверху вниз: 3000, 2000, 1000, 900, 800, 700, 600, 500, 400, 300, 200 и 100 нуклеотидных пар).
16 – продукт ПЦР плазмидной ДНК pentF-covid-19
17 - продукт ПЦР без добавления ДНК.
Была проведена амплификация ДНК, выделенных из 13 клонов для определения интеграции плазмидной ДНК pentF- covid-19 в хромосомную ДНК энтерококка с праймерами B1 и K2. Интеграция плазмидной ДНК pentF- covid-19 в хромосомную ДНК энтерококка была обнаружена только у двух трансформантов (Фиг. 3, нижний ряд).
На. Фиг. 3 в нижнем ряду представлена электрофореграмма амплифицированных ДНК-фрагментов с праймерами В1 и К2.
18 – 31 – продукты ПЦР ДНК из полученных 14 клонов.
32 - 100 п.н. ДНК - маркер (сверху вниз: 3000, 2000, 1000, 900, 800, 700, 600, 500, 400, 300, 200 и 100 нуклеотидных пар).
33 - продукт ПЦР без добавления ДНК.
Секвенирование ДНК, выделенной из одного из положительных клонов, обозначенного, как COVID 19+,было проведено с ДНК праймерами, соответствующими последовательности гена covid-19 (праймер K2) и последовательности хромосомной ДНК энтерококков (праймер B1) для подтверждения интеграции плазмидной ДНК pentF- covid-19 в хромосомную ДНК энтерококка (перечень последовательностей SEQ ID No: 1).
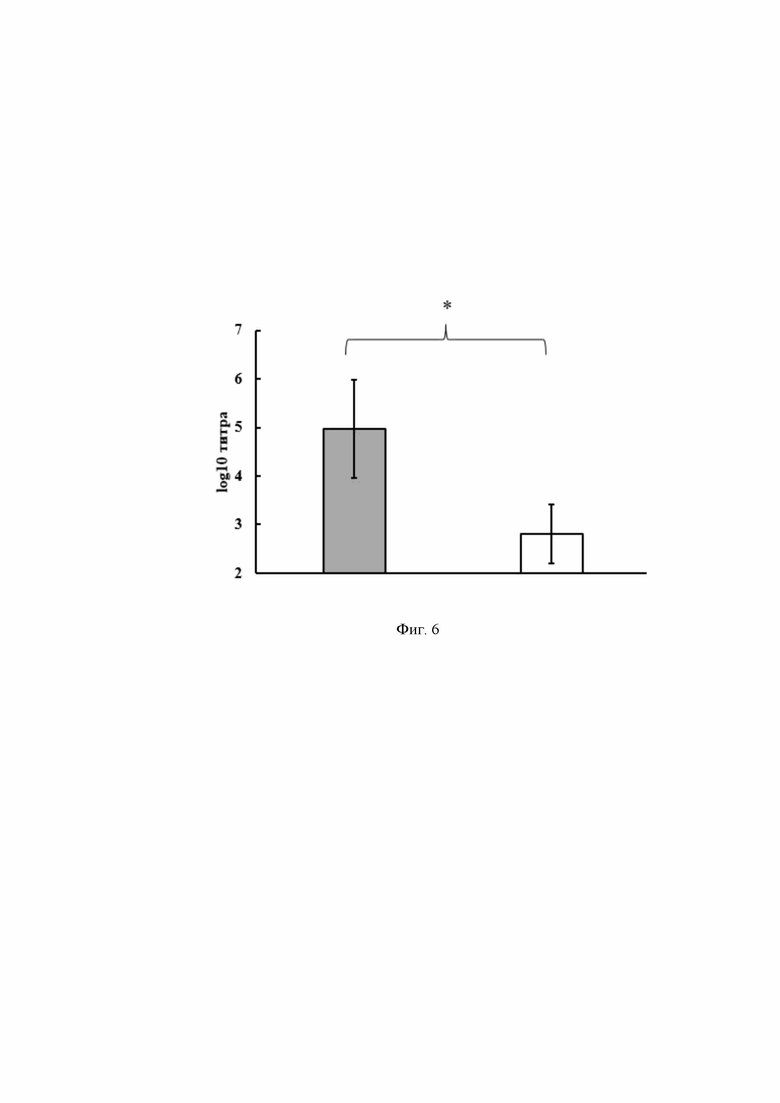
На Фиг. 4 показан результат, полученный после проведения ПЦР с использованием ДНК клона COVID 19+ с разными парами праймеров. На Фиг. 4 приведены результаты ПЦР с хромосомной ДНК энтерококка и плазмидной ДНК pentF- covid-19.
На Фиг. 4 представлена электрофореграмма амплифицированных ДНК-фрагментов с разными парами праймеров, где:
1 – ДНКклона COVID 19+ с праймерами K1 и K2
2 - ДНКклона COVID 19+ с праймерами А1 и D2
3 – ДНКклона COVID 19+ с праймерами B1и K2
4 – ДНКклона COVID 19+ с праймерами K1 и E2
5 – ДНКклона COVID 19+ с праймерами E1 и K2
6 – ДНКклона COVID 19+ с праймерами E1 и E2
7 – ДНКклона COVID 19+ с праймерами seq F и K2
8 – ДНКклона COVID 19+ с праймерами B1 и E2
9 – ДНК L3 с праймерами B1 и E2
10 – ДНК L3 с праймерами E1 и E2
11 – ДНК pentF- covid-19 с праймерами K1 и K2
12 – ДНК pentF- covid-19 с праймерами seqF и K2
13 – ДНК pentF- covid-19 с праймерами E1 и E2
14 - 100 п.н. ДНК - маркер (сверху вниз: 3000, 2000, 1000, 900, 800, 700, 600, 500, 400, 300, 200 и 100 нуклеотидных пар).
Клон COVID 19+, экспрессирующий ген COVID-19белка, выбран в качестве вакцинного препарата для дальнейшего исследования.
Пример 4.
Пероральная вакцинация мышей живой пробиотической вакциной.
Мышам, линии Balb/c (n=10), самкам в возрасте 9-10 недель с помощью адаптера вводили перорально E.faecium L3 (n=10) и COVID 19+ (n=10). Курс вакцинации состоял из трех циклов иммунизации с интервалом в две недели. Каждый цикл включал трехкратное ежедневное введение 0,5 мл взвеси бактерий в дозе 109 КОЕ/мышь. Для вакцинации исходный и модифицированный варианты E.faecium культивировали в среде BL в течение 18 часов при 37°С в аэробных условиях. E.faecium, модифицированный участком гена коронавируса, культивировали в присутствии эритромицина в концентрации 5 мкг/мл. Суспензию клеток отмывали дважды в ЗФФР и вводили мышам.
Пример 5.
Определение COVID-19+ - специфических антител в крови мышей и анализ специфических цитотоксических лимфоцитов (ЦТЛ).
Через две недели после окончания второго этапа иммунизации, то есть через шесть недель от начала иммунизации, у мышей забирали кровь для оценки уровня специфического гуморального иммунного ответа и получали селезенки для анализа специфических ЦТЛ.
Уровень специфических IgG и IgA определяли общепринятым методом ИФА. На дно 96-луночного планшета сорбировали рекомбинантный белок S (аналог вирусной вставки в энтерококк) в концентрации 0,5 мкг/мл в течение ночи при 4°С. Последовательные 4-кратные разведения сыворотки вносили в лунки в объеме 100 мкл и инкубировали 1 час при 37°С. Избыток антител отмывали ЗФФР с 0,5% Tween 20. Специфические антитела выявляли с помощью вторичных антител, конъюгированных с пероксидазой хрена. Для идентификации антител класса A использовали козий анти мышь –IgA-HRP конъюгат (Sigma), антител класса G - козий анти мышь–IgG-HRP конъюгат (Sigma). Раствор конъюгата вносили в объеме 100 мкл и инкубировали 1 час при 37°С. Планшеты отмывали четыре раза в ЗФФР с 0,5% Tween 20, связанный конъюгат выявляли в цветной реакции с ТМБ, интенсивность окраски оценивали при длине волны 450 нм.
Анализ сывороток показал, что в крови вакцинированных животных присутствовали антитела класса G, специфичные в отношении вирусного белка S, аналогичного вставке в энтерококк. Титры антител колебались в пределах 1: 400- 1:25600.
На Фиг. 5 представлено содержание иммуноглобулинов класса G в сыворотке мышей через 6 недель после начала вакцинации.
А – средние кривые титрования
На графике 1 - представлена кривая титрования сыворотки, полученной от мышей, получавших COVID 19+
На графике 2 представлена кривая титрования сыворотки, полученной от мышей, получавших штамм E.faecium L3.
Б – средние титры специфического IgG
Столбец 1 – титр специфического IgG из сыворотки мышей, получавших COVID 19+
Столбец 2 - титр специфического IgG из сыворотки мышей, получавших E.faecium L3
* - отличия между группами достоверны р≤0,05
Определение уровня специфических антител класса А позволило установить, что вакцинация приводит к появлению в сыворотке крови мышей специфических IgA в высоких титрах, значения колеблются в пределах 1: 10000- 1: 500000, что существенно превышает показатели для специфических IgG.
На Фиг. 6 представлено содержание иммуноглобулинов класса A в сыворотке мышей через 6 недель после начала вакцинации, где в столбце 1 представлены средние титры специфического IgA из сыворотки мышей, получавших COVID 19+; в столбце 2 средний титр специфического IgA из сыворотки мышей, получавших E.faecium L3
* - отличия между группами достоверны р≤0,05.
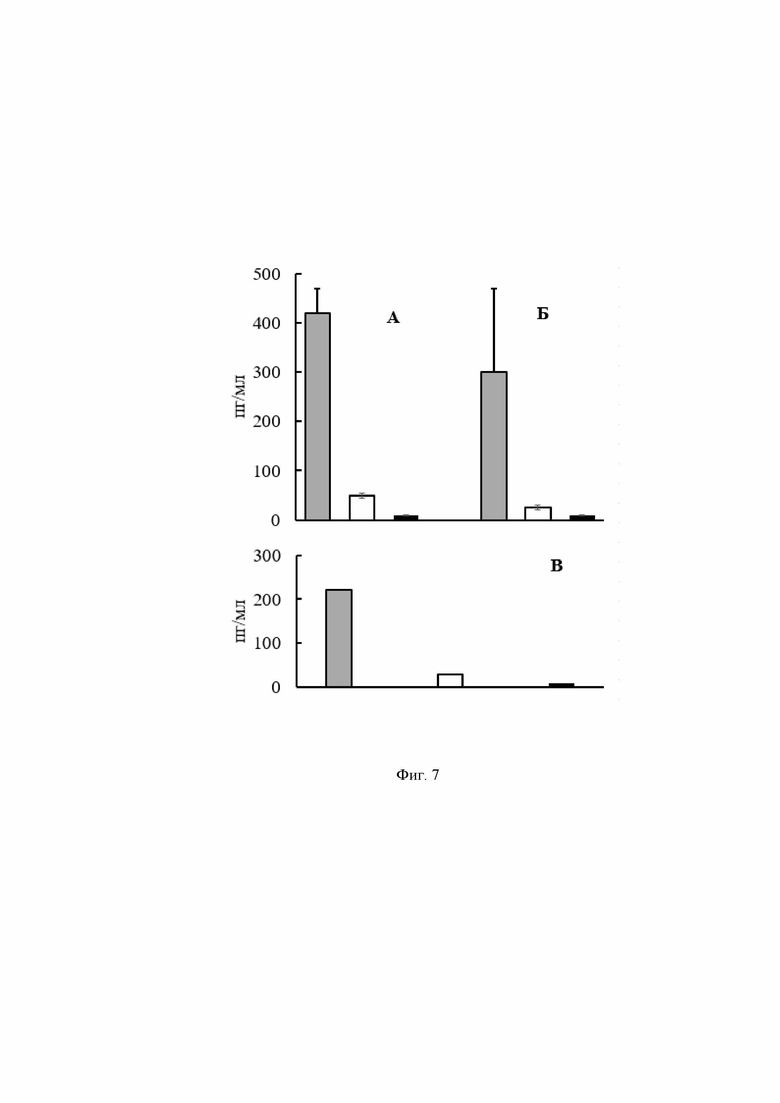
Анализ клеточного звена специфического иммунного ответа на пероральную вакцинацию живой противовирусной вакциной проводили по оценке интенсивности индукции интерферона гамма после контакта суспензии клеток селезенки с S белком SARS-Cov-2.
Суспензии отдельных клеток селезенки иммунизированных мышей готовили путем протирания селезенок через сито BD Falcon (BD Biosciences, Бедфорд, США). Клетки отмывали средой RPMI-1640 (2000 об/мин, 5 мин) и ресуспендировали в полной среде RPMI-1640, содержащей 10 % фетальной телячьей сыворотки. Подсчет числа клеток и их жизнеспособность проводили, используя метод с 0.4% трипановым синим. Пул клеток от трех мышей, рассевали в 24-луночные планшеты в посевной дозе 105 клеток/мл в трех повторах. После этого клетки стимулировали рекомбинантным пептидом COVID 19+ в дозе 1-0,5-0,1 мкг/мл. После 6 дней культивирования при 37°С в атмосфере CO2 супернатанты собирали и содержание интерферона (IFN) -гамма определяли с помощью ИФА набора (Invitrogen, Waltham, Massachusetts, USA) в соответствии с инструкциями производителя.
На Фиг. 7 представлена концентрация внеклеточного ИФН-γ в супернатанте культуры клеток.
А - стимуляция рекомбинантным пептидом COVID 19+ в дозе 1 мкг/мл.
Столбец 1 – концентрация внеклеточного ИФН-γ в супернатанте культуры клеток мышей, получавших COVID 19+
Столбец 2 – концентрация внеклеточного ИФН-γ в супернатанте культуры клеток мышей, получавших L3
Столбец 3 - концентрация внеклеточного ИФН-γ в супернатанте культуры клеток интактных мышей
Б - стимуляция рекомбинантным пептидом COVID 19+ в дозе 0,1мкг/мл
Столбец 1 – концентрация внеклеточного ИФН-γ в супернатанте культуры клеток мышей, получавших COVID 19+
Столбец 2 – концентрация внеклеточного ИФН-γ в супернатанте культуры клеток мышей, получавших L3
Столбец 3 - концентрация внеклеточного ИФН-γ в супернатанте культуры клеток интактных мышей
В - концентрация внеклеточного ИФН-γ в супернатанте культуры клеток в отсутствие стимуляции рекомбинантным пептидом COVID 19+
Столбец 1 – концентрация внеклеточного ИФН-γ в супернатанте культуры клеток мышей, получавших COVID 19+
Столбец 2 – концентрация внеклеточного ИФН-γ в супернатанте культуры клеток мышей, получавших L3
Столбец 3 - концентрация внеклеточного ИФН-γ в супернатанте культуры клеток интактных мышей.
Полученные данные указывают на то, что на 6 день после внесения специфического антигена в суспензию спленоцитов мышей, вакцинированных живой пробиотической вакциной, в среде накапливается интерферон гамма в количестве, которое достоверно превышает аналогичный показатель для контрольных проб. Последнее свидетельствует о том, что курс пероральной вакцинации живой пробиотической вакциной стимулирует цитотоксические лимфоциты Th1, которые координируют специфические клеточные реакции, в частности, активность, специфичную в отношении S белка вируса.
Как показывают результаты экспериментов, заявленное изобретение позволяет создать эффективную живую вакцину с профилактическим эффектом в отношении инфекции COVID-19.
Список используемых источников информации
1. Guo YR, Cao QD, Hong ZS, Tan YY, Chen SD, Jin HJ, Tan KS, Wang DY, Yan Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak - an update on the status. Mil Med Res. 2020 Mar 13;7(1):11. doi: 10.1186/s40779-020-00240-0. PMID: 32169119; PMCID: PMC7068984.
2. Патент РФ №2220199 «Штамм энтерококков enterococcus faeciuml-3 для изготовления лечебно-профилактических средств и продуктов питания лечебно-профилактического назначения»; МПК: A23C 9/12, A61K 35/74 (патентообладатели и авторы: Алехина Г.Г. и Суворов А.Н.).
3. Гупалова Т.В. и др. «Создание и опыт применения живой вакцины на основе штамма пробиотика Enterococcus faecium L3 для профилактики вагинальной инфекции, вызванной Streptococcus agalactiae». Медицинский академический журнал, 2013.
4. Патент РФ №2640250 «Способ введения генов патогенных стрептококков в хромосомную днк пробиотического штамма Еnterococcus faecium l3 для экспрессии в пилях», МПК: A61K39/092; (авторы Суворов А.Н. и др.).
5. Yermolenko E., Suvorov A., Chernush A., et al. International Congress Series, 1289: 363-366; 2006.
6. Tarasova E., Yermolenko E., Donets V. et al. Beneficial Microbs, 1: 265-270, (2010).
7. Suvorov A., Simanenkov V., Gromova L. et al. Prebiotics and probiotics potencial for human health, 104-112, (2011).
8. Суворов А., Алехина Г.Г., Пигаревский П.В. и др. Гастробюллетень, 4: 29-31, (2001).
9. Telford JL, Barocchi MA, Margarit I et al. Nature Reviews Microbiology 4: 509-519, (2006).
10. Camille Danne, Shaynoor Dramsi. Research in Microbiology, 163: 645-658, (2012).
11. Патент РФ № 2701733 (Авторы – Суворов А.Н., Гупалова Т.В., Кулешевич Е.В., Леонтьева Г.Ф., Крамская Т.А.) - прототип.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>
<120> Способ создания живой вакцины против коронавирусной инфекции COVID-19
на основе пробиотического штамма Enterococcus faecium L3 и живая вакцина
Enterococcus faecium L3-pentF-covid-19
<140>
<141>
<150>
<151>
<160> 1
<170>
<210> 1
<211> 1599
<212> Нуклеотидная последовательность
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность гена covid-19 коронавируса COVID-19,
экспрессированная в системе Escherichia coli
<400> 1
Atgagagcagctggtattgagttgaatgatacatttctatctatttacagtttaaatgga 60
cagtatcagcaacgtgtgtcttggtataatgacaataatgaatctgtcggtgaacgtaat 120
attgatatgagagaatttgttgggtatgaaaaaatgggtagcttaccttattttgtcaca 180
acagatacagcatgtgcagaatacaaagctcctgcgttatcaacaaacaatttaacttca 240
aaagtagtgggaggacgtgcagaaaaggcttatagctcgaatgatcatttcaccgatgtt 300
gtaggagctgatacttatcacagaagtggtgtaacgtatacgcttcaaggcgcttcccca 360
acattcatgattggcgcaaatacgaatagtatgatgtttagctttgatactgcattgcta 420
tggacaccacaaccatcgaagcctacaaaagaagtgtttaacaaagctaatactgaagag 480
gcagcacacaatattgacaaaaaagtgattccacaaggatcagatgtttactatcatatt 540
catcaaaagtttgatgcattaacagtcaacacaatgaacaaatacaaatcatttaaaatc 600
actgatacctttgacagcaaaaattttgatatggtatcggatgggaaaaactatgatggc 660
gcattgcatatgggtttccaacccactaatggtgttggttaccaaccatacagagtagta 720
gtactttcttttgaacttctacatgcaccagcaactgtttgtggacctaaaaagtctact 780
aatttggttaaaaacaaatgtgtcaatttcaacttcaatggtttaacaggcacaggtgtt 840
cttactgagtctaacaaaaagtttctgcctttccaacaatttggcagagacattgctgac 900
actactgatgctgtccgtgatccacagacacttgagattcttgacattacaccatgttct 960
tttggtggtgtcagtgttataacaccaggaacaaatacttctaaccaggttgctgttctt 1020
tatcaggatgttaactgcacagaagtccctgttgctattcatgcagatcaacttactcct 1080
acttggcgtgtttattctacaggttctaatgtttttcaaacacgtgcaggctgtttaata 1140
ggggctgaacatgtcaacaacgaattctacgaaaaaaacctgaaagaatacaccgacaag 1200
ctggataaactcgagaaaggttctgcgcgagtgatagatatgagtacaggaaaagatatt 1260
acttcagaaggtacactaacctatgatagcaatttaagaacgctgaaatgggaagcttcc 1320
gctgatttcttaagtaaaaatcttttagatggacgagaaattcaactgatatttacagct 1380
aaaactccactacagtcagaaaaaaatattgataaccaagccgtagcagcagtagagaat 1440
gtttctaataaaacgaatgttgtaacaattggtgttgatcctaacttaccacaagtcatt 1500
gttcctagaacaggttctacacatttagtaacaattttagtggttagcttagtattactt 1560
gtgttagctactcttagttatgtggctctgaagttcaat 1599
<210> 2
<211> 533
<212> Аминокислотная последовательность
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 2
MetArgAlaAlaGlyIleGluLeuAsnAspThrPheLeuSerIleTyr
5 10 15
SerLeuAsnGlyGlnTyrGlnGlnArgValSerTrpTyrAsnAspAsn
20 25 30
AsnGluSerValGlyGluArgAsnIleAspMetArgGluPheValGly
35 40 45
TyrGluLysMetGlySerLeuProTyrPheValThrThrAspThrAla
50 55 60
CysAlaGluTyrLysAlaProAlaLeuSerThrAsnAsnLeuThrSer
65 70 75 80
LysValValGlyGlyArgAlaGluLysAlaTyrSerSerAsnAspHis
85 90 95
PheThrAspValValGlyAlaAspThrTyrHisArgSerGlyValThr
100 105 110
TyrThrLeuGlnGlyAlaSerProThrPheMetIleGlyAlaAsnThr
115 120 125
AsnSerMetMetPheSerPheAspThrAlaLeuLeuTrpThrProGln
130 135 140
ProSerLysProThrLysGluValPheAsnLysAlaAsnThrGluGlu
145 150 155 160
AlaAlaHisAsnIleAspLysLysValIleProGlnGlySerAspVal
165 170 175
TyrTyrHisIleHisGlnLysPheAspAlaLeuThrValAsnThrMet
180 185 190
AsnLysTyrLysSerPheLysIleThrAspThrPheAspSerLysAsn
195 200 205
PheAspMetValSerAspGlyLysAsnTyrAspGlyAlaLeuHisMet
210 215 220
GlyPheGlnProThrAsnGlyValGlyTyrGlnProTyrArgValVal
225 230 235 240
ValLeuSerPheGluLeuLeuHisAlaProAlaThrValCysGlyPro
245 250 255
LysLysSerThrAsnLeuValLysAsnLysCysValAsnPheAsnPhe
260 265 270
AsnGlyLeuThrGlyThrGlyValLeuThrGluSerAsnLysLysPhe
275 280 285
LeuProPheGlnGlnPheGlyArgAspIleAlaAspThrThrAspAla
290 295 300
ValArgAspProGlnThrLeuGluIleLeuAspIleThrProCysSer
305 310 315 320
PheGlyGlyValSerValIleThrProGlyThrAsnThrSerAsnGln
325 330 335
ValAlaValLeuTyrGlnAspValAsnCysThrGluValProValAla
340 345 350
IleHisAlaAspGlnLeuThrProThrTrpArgValTyrSerThrGly
355 360 365
SerAsnValPheGlnThrArgAlaGlyCysLeuIleGlyAlaGluHis
370 375 380
ValAsnAsnGluPheTyrGluLysAsnLeuLysGluTyrThrAspLys
385 390 395 400
LeuAspLysLeuGluLysGlySerAlaArgValIleAspMetSerThr
405 410 415
GlyLysAspIleThrSerGluGlyThrLeuThrTyrAspSerAsnLeu
420 425 430
ArgThrLeuLysTrpGluAlaSerAlaAspPheLeuSerLysAsnLeu
435 440 445
LeuAspGlyArgGluIleGlnLeuIlePheThrAlaLysThrProLeu
450 455 460
GlnSerGluLysAsnIleAspAsnGlnAlaValAlaAlaValGluAsn
465 470 475 480
ValSerAsnLysThrAsnValValThrIleGlyValAspProAsnLeu
485 490 495
ProGlnValIleValProArgThrGlySerThrHisLeuValThrIle
500 505 510
LeuValValSerLeuValLeuLeuValLeuAlaThrLeuSerTyrVal
515 520 525
AlaLeuLysPheAsn
530 533
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ создания рекомбинантного штамма энтерококка L3-SARSN1 на основе биологически активного штамма Enterococcus faecium L3 | 2022 |
|
RU2820058C1 |
| Способ создания рекомбинантного штамма энтерококка L3-SARS"XBB.1.5" на основе биологически активного штамма Enterococcus faecium L3 | 2023 |
|
RU2840971C1 |
| Способ создания живого штамма энтерококка L3-SARS на основе биологически активного штамма Е. faecium L3 | 2021 |
|
RU2782529C1 |
| Живая вакцина на основе штамма пробиотиков ENTEROCOCCUS FAECIUM L3 для профилактики инфекции, вызванной STREPTOCOCCUS PNEUMONIE | 2018 |
|
RU2701733C1 |
| СПОСОБ ВВЕДЕНИЯ ГЕНОВ ПАТОГЕННЫХ СТРЕПТОКОККОВ В ХРОМОСОМНУЮ ДНК ПРОБИОТИЧЕСКОГО ШТАММА ENTEROCOCCUS FAECIUM L3 ДЛЯ ЭКСПРЕССИИ В ПИЛЯХ | 2015 |
|
RU2640250C2 |
| Рекомбинантная ДНК, обеспечивающая получение рекомбинантного белка Cov1, обладающего иммуногенными свойствами в отношении вируса SARS-CoV-2 | 2021 |
|
RU2776484C1 |
| Вакцинная композиция против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743595C1 |
| Вакцина против COVID-19 на основе PIV5 | 2021 |
|
RU2838536C1 |
| Штамм энтерококков Enterococcus faecium L-3 для лечения и профилактики бактериальных и вирусных инфекций с использованием изготовленных на его основе лечебно-профилактических средств и продуктов питания и способ культивирования штамма энтерококков Enterococcus faecium L-3 | 2022 |
|
RU2800359C1 |
| Пептидные иммуногены, используемые в качестве компонентов вакцинной композиции против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743594C1 |
Изобретение относится к области медицины, вирусологии, микробиологии, молекулярной генетики и биотехнологии. Представлена живая вакцина Enterococcus faecium L3-pentF-covid-19, содержащая клон энтерококков COVID 19+ с введенным в его геном участком ДНК SEQ ID No: 1, обеспечивающий экспрессию белка, имеющего аминокислотную последовательность SEQID No: 2, стимулирующая гуморальный и клеточный иммунитет в отношении вируса SARS-CoV-2. Также представлен способ получения указанной вакцины на основе пробиотического штамма Enterococcus faecium L3, модифицированного в результате электропорации культуры энтерококков Enterococcus faecium L3 рекомбинантной плазмидной ДНК pentF-covid-19, SEQ ID No: 1, отличающийся тем, что в плазмиде pentF-pspf, участок pspf заменен на фрагмент гена шиповидного белка SARS-CoV-2, при этом ДНК pentF-covid-19 кодирует аминокислотную последовательность SEQ ID No: 2, которая выполняет функцию шиповидного белка вируса SARS-CoV-2, способного осуществлять стимуляцию гуморального и клеточного иммунитета в отношении вируса SARS-CoV-2. Пероральное введение вакцины Еnterococcus faecium pentF-covid-19 стимулирует развитие специфического системного и местного иммунного ответа, что проявляется выработкой специфических иммуноглобулинов классов G и А, а также повышенной продукцией интерферона гамма у вакцинированных животных. 2 н.п. ф-лы, 7 ил., 1 табл., 5 пр.
1. Способ создания живой вакцины против коронавирусной инфекции COVID-19 на основе пробиотического штамма Enterococcus faecium L3, модифицированного в результате электропорации культуры энтерококков Enterococcus faecium L3 рекомбинантной плазмидной ДНК pentF-covid-19, SEQ ID No: 1, отличающийся тем, что в плазмиде pentF-pspf участок pspf заменен на фрагмент гена шиповидного белка SARS-CoV-2, при этом ДНК pentF-covid-19 кодирует аминокислотную последовательность SEQ ID No: 2, которая выполняет функцию шиповидного белка вируса SARS-CoV-2, способного осуществлять стимуляцию гуморального и клеточного иммунитета в отношении вируса SARS-CoV-2.
2. Живая вакцина Enterococcus faecium L3-pentF-covid-19, полученная способом по п. 1 и содержащая клон энтерококков COVID 19+ с введенным в его геном участком ДНК SEQ ID No: 1, обеспечивающий экспрессию белка, имеющего аминокислотную последовательность SEQ ID No: 2, способна стимулировать гуморальный и клеточный иммунитет в отношении вируса SARS-CoV-2.

