ДАННЫЕ О ПРОДОЛЖАЮЩЕЙ ЗАЯВКЕ
Данная заявка испрашивает приоритет по предварительной заявке на патент США под №63/080862, поданной 21 сентября 2020 года, и предварительной заявке на патент США под серийным №63/217361, поданной 1 июля 2021 года, каждая из которых включена в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Данная заявка содержит перечень последовательностей, поданный в электронной форме посредством EFS Web в Ведомство по патентам и товарным знакам США в виде текстового файла с кодировкой ASCII под названием "0235-000294WO01_ST25", имеющего размер 38 килобайтов и созданного 15 сентября 2021 года. Информация, содержащаяся в перечне последовательностей, включена в данный документ посредством ссылки.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
SARS-CoV-2 представляет собой новый коронавирус, который был впервые идентифицирован в Ухане, Китай, в декабре 2019 года в качестве причины пневмонии и впоследствии распространился по всему миру, вызвав пандемию COVID-19. Вирусом было инфицировано более чем 221 миллион человек во всем мире, по состоянию на 8 сентября 2021 г. он стал причиной более чем 4574000 случаев смерти и может продолжать распространяться в отсутствие коллективного иммунитета (см. во всемирной сети по адресу who.int/emergencies/diseases/novel-coronavirus-2019). Социальное дистанцирование, использование средств индивидуальной защиты и повсеместное тестирование с отслеживанием контактов, карантинные процедуры и ограниченные запасы единственной вакцины, одобренной FDA (Управлением по контролю качества пищевых продуктов и лекарственных средств), и вакцин, одобренных FDA в соответствии с разрешением на экстренное применение (EUA), в настоящее время являются единственными мерами, доступными для ограничения распространения вируса. Срочно необходимы вакцины, которые предупреждают смертность и снижают передачу инфекции.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение предусматривает вектор экспрессии на основе вируса, содержащий геном вируса парагриппа 5 (PIV5), содержащий гетерологичную нуклеотидную последовательность, обеспечивающую экспрессию гетерологичного полипептида, где гетерологичный полипептид предусматривает шиповидный (S) белок коронавируса.
В некоторых аспектах вектора экспрессии на основе вируса S-белок коронавируса предусматривает S-белок коронавируса из коронавируса 2 тяжелого острого респираторного синдрома (SARS-CoV-2).
В некоторых аспектах вектора экспрессии на основе вируса цитоплазматический хвост S-белка коронавируса был заменен на цитоплазматический хвост белка слияния (F) PIV5.
В некоторых аспектах вектора экспрессии на основе вируса S-белок коронавируса включает S-белок коронавируса из коронавируса 2 тяжелого острого респираторного синдрома (SARS-CoV-2), и где цитоплазматический хвост S-белка коронавируса был заменен на цитоплазматический хвост белка слияния (F) PIV5.
В некоторых аспектах вектора экспрессии на основе вируса гетерологичный полипептид предусматривает шиповидный (S) белок коронавируса, который предусматривает наличие мутаций по аминокислотному остатку W886 и/или F888. В некоторых аспектах аминокислотная замена по аминокислотному остатку W886 предусматривает замену триптофана (W) на аргинин (R) и/или аминокислотная замена по аминокислотному остатку W888 предусматривает замену фенилаланина (F) на аргинин (R).
В некоторых аспектах вектора экспрессии на основе вируса по настоящему изобретению гетерологичная нуклеотидная последовательность встроена между геном низкомолекулярного гидрофобного белка (SH) и геном гемагглютинин-нейраминидазы (HN) в геноме PIV5.
В некоторых аспектах вектора экспрессии на основе вируса по настоящему изобретению гетерологичная нуклеотидная последовательность заменяет нуклеотидную последовательность гена SH.
В некоторых аспектах вектора экспрессии на основе вируса по настоящему изобретению гетерологичная нуклеотидная последовательность встроена между геном гемагглютинин нейраминидазы (HN) и геном высокомолекулярного белка РНК полимеразы (L) в геноме PIV5.
В некоторых аспектах вектора экспрессии на основе вируса по настоящему изобретению гетерологичная нуклеотидная последовательность встроена скорее ближе к лидерной последовательности, чем между геном гемагглютинин нейраминидазы (HN) и геном высокомолекулярного белка РНК полимеразы (L) в геноме PIV5; встроена выше по ходу транскрипции от гена нуклеокапсидного белка (NP) в геноме PIV5; встроена непосредственно ниже по ходу транскрипции от лидерной последовательности в геноме PIV5; встроена между геном белка слияния (F) и геном SH в геноме PIV5; встроена между геном VP и геном матриксного (М) белка в геноме PIV5; встроена между геном М и геном F в геноме PIV5; встроена между геном нуклеокапсидного (NP) белка и геном V/P в геноме PIV5; встроена между лидерной последовательностью и геном нуклеокапсидного белка (NP) в геноме PIV5; встроена туда, где часть гена F или HN PIV5 была заменена на гетерологичную нуклеотидную последовательность; встроена в пределах нуклеотидной последовательности гена SH, встроена в пределах нуклеотидной последовательности гена NP, встроена в пределах нуклеотидной последовательности гена V/P, встроена в пределах нуклеотидной последовательности гена М, встроена в пределах нуклеотидной последовательности гена F, встроена в пределах нуклеотидной последовательности гена HN, и/или встроена в пределах нуклеотидной последовательности гена L.
В некоторых аспектах вектора экспрессии на основе вируса по настоящему изобретению геном PIV5 дополнительно предусматривает наличие одной или нескольких мутаций. В некоторых аспектах одна или несколько мутаций включают мутацию гена V/P, мутацию общего N-конца белков V и Р, мутацию остатков 26, 32, 33, 50, 102 и/или 157 общего N-конца белков V и Р, мутацию, обеспечивающую отсутствие С-конца белка V, мутацию, обеспечивающую отсутствие низкомолекулярного гидрофобного (SH) белка, мутацию белка слияния (F), мутацию фосфопротеина (Р), мутацию высокомолекулярного белка РНК полимеразы (L), мутацию, обеспечивающую встраивание остатков из вируса парагриппа собак, мутацию, индуцирующую апоптоз, или их комбинацию. В некоторых аспектах одна или несколько мутаций включают PIV5VΔC, PIV5ΔSH, PIV5-P-S308G или их комбинацию.
Настоящее изобретение предусматривает вирусную частицу, содержащую вектор экспрессии на основе вируса, как описано в данном документе.
Настоящее изобретение предусматривает композицию на основе вектора экспрессии на основе вируса, описанного в данном документе, или вирусной частицы, как описано в данном документе.
Настоящее изобретение предусматривает способ обеспечения экспрессии гетерологичного шиловидного (S) гликопротеина коронавируса в клетке, при этом способ включает приведение клетки в контакт с вектором экспрессии на основе вируса, вирусной частицей или композицией, как описано в данном документе.
Настоящее изобретение предусматривает способ индуцирования иммунного ответа у субъекта на шиловидный (S) гликопротеин коронавируса, при этом способ включает введение субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции, как описано в данном документе. В некоторых аспектах иммунный ответ включает гуморальный иммунный ответ и/или клеточный иммунный ответ. В некоторых аспектах вектор экспрессии на основе вируса, вирусную частицу или композицию вводят интраназально, внутримышечно, местно или перорально.
Настоящее изобретение предусматривает способ вакцинирования субъекта против коронавирусного заболевания 2019 г. (COVID-19), при этом способ включает введение субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции, как описано в данном документе. В некоторых аспектах вектор экспрессии на основе вируса, вирусную частицу или композицию вводят интраназально, внутримышечно, местно или перорально.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На ФИГ. 1 показана схема клонирования плазмиды для pDA27. На ФИГ. 2 показана конструкция CVX-UGA1, полученная из pDA27.
На ФИГ. 3 показана карта плазмиды, демонстрирующая фрагменты секвенирования.
На ФИГ. 4 показан процесс конструирования вакцины CVX-UGA1 с использованием плазмиды рАВ76.
На ФИГ. 5 показана общая схема "спасения" вируса и очистки с использованием бляшкообразования.
На ФИГ. 6 показаны изображения, полученные с помощью иммунофлуоресцентного анализа клеток MDBK, инфицированных посредством CVX-UGA1 и окрашенных в отношении S SARS-CoV-2 с помощью моноклонального антитела к шиповидному белку SI SARS-CoV-2.
На ФИГ. 7 показана карта плазмиды pDA16.
На ФИГ. 8 показана карта плазмиды nCOV19-S-human-Genscript.
На ФИГ. 9 показаны карта и последовательность плазмиды pDA27 (CVX-UGA1) (SEQ ID NO: 1).
На ФИГ. 10 показана карта плазмиды pCAGGS-NP.
На ФИГ. 11 показана карта плазмиды pCAGGS-Р.
На ФИГ. 12 показана карта плазмиды pCAGGS-L.
На ФИГ. 13 показана карта плазмиды pBH437-pCAGGS-T7.
На ФИГ. 14 показана карта плазмиды рВН276.
На ФИГ. 15 показана карта плазмиды рВН161.
На ФИГ. 16 показана карта плазмиды pCH10.
На ФИГ. 17 показана карта плазмиды рАВ76.
На ФИГ. 18А ФИГ. 18F показано получение CVXGA1 и анализ CVXGA1 в клетках культур тканей и мышах. На ФИГ. 18А показано схематическое изображение CVXGA1. PIV5 содержит семь генов и кодирует 8 белков: NP, V, Р, М, F, SH, HN и L. Лидерная и трейлерная последовательности важны для синтеза вирусной РНК. S SARS-CoV-2 содержит сигнальный пептид (SP), рецепторсвязывающий домен (RBD), пептид слияния (FP), транс мембранный домен (ТМ) и цитоплазматический хвост (СТ). CVXGA1 содержит S, СТ которого был заменен на таковой из F PIV5 и который был встроен между SH и HN PIV5. На ФИГ. 18 В показано выявление экспрессии S. Для выявления экспрессии SARS-CoV-2-S были проведены иммунофлуоресцентные анализы. Клетки MDBK подвергали имитационному инфицированию или инфицировали при значении множественности заражения (MOI), составляющем 1, посредством PIV5 или CVXGA1. Через два дня после инфицирования клетки фиксировали, пермеабилизировали и окрашивали антителами, специфичными в отношении PIV5-V/P (антитела к вектору) и S SARS-CoV-2. Для выявления экспрессии с использованием иммуноблоттинга клетки Vero инфицировали посредством PIV5, CVXGA1 при 1 MOI и лизировали через 24 часа после инфицирования (hpi). Лизаты разделяли на геле SDS-PAGE и подвергали иммуноблоттингу с использованием антитела к S SARS-CoV-2 и антитела к PIV5-V/P. На ФИГ. 18С показано образование синцития с помощью CVXGA1. Клетки Vero инфицировали посредством PIV5 или CVXGA1 при значении MOI, составляющем 0,1. Через 48 HPI клетки фотографировали. На ФИГ. 18D показаны титры антител к S у мышей BALB/c, иммунизированных с помощью CVXGA1. Мышей BALB/c (n = 5 на группу) иммунизировали через интраназальный (IN) путь с помощью CVXGA1 в дозе 104, 105 и 106 бляшкообразующих единиц (БОЕ). Через 28 дней после иммунизации (dpi) у мышей брали кровь и измеряли титры антител к S с использованием ELISA. На ФИГ. 18Е показаны титры антител к рецепторсвязывающему домену (RBD) у мышей BALB/c, иммунизированных с помощью CVXGA1. Титры RBD для образцов на ФИГ. 18D определяли с использованием ELISA. На ФИГ. 18F показаны титры нейтрализации у мышей BALB/c, иммунизированных с помощью CVXGA1. Титры нейтрализации для образцов на ФИГ. 18D определяли, как описано в разделе "Материалы и способы" из примера 2.
На ФИГ. 19А - ФИГ. 19D показано, что интраназальная вакцинация с помощью CVXGA1 обеспечивает уменьшение клинических и патологических проявлений заболевания у мышей BALB/c, сенсибилизированных посредством Ad-hACE2, после инфицирования с помощью SARS-CoV-2. На ФИГ. 19А показано схематическое изображение экспериментального протокола исследования вакцины CVXGA1 на мышах BALB/c. Группы обработки (Gp) включают следующие: Gp 1 иммунизация с помощью DMEM посредством внутримышечного (IM) пути; Gp 2 - иммунизация с помощью CVXGA1 посредством интраназального (IN) пути; Gp 3 - инактивированный УФ-излучением SARS-CoV-2, который вводили посредством IM пути с квасцовым адъювантом. Затем всех животных сенсибилизировали к инфекции SARS-CoV-2 посредством Ad-hACE2 и через 5 дней инокулировали с помощью 1×105 БОЕ SARS CoV-2. Указаны другие измерения и число животных на группу. Дни после иммунизации обозначены как dpi. На ФИГ. 19В показаны результаты, полученные в модели инфекции SARS-CoV-2 на мышах. Мышей BALB/c, трансдуцированных с помощью Ad5 hACE2, инфицировали интраназально посредством 1×105 БОЕ SARS-CoV-2 в 50 мкл DMEM. Изменения веса у мышей в возрасте 6-8 недель отслеживали ежедневно (DMEM: n=6, CVXGA1: n=14, SARS-CoV-2, инактивированный УФ-излучением: n=6 мышей на группу). Мыши, иммунизированные с помощью CVXGA1, продемонстрировали значительно меньшую потерю веса. *Р-значение < 0,05; **Р значение < 0,005; критерий Манна Уитни. На ФИГ. 19С показано, что вакцина CVXGA1 ослабляет тяжесть легочного проявления заболевания. Результаты гистопатологического оценивания легочного проявления заболевания. Мононуклеарные клеточные инфильтраты присутствовали во всех группах, в то время как гранулоцитарная инфильтрация увеличивалась у мышей, иммунизированных IM посредством инактивированного УФ-излучением SARS-CoV-2 с последующим введением второй бустерной дозы, n=4. На ФИГ. 19D показано, что вакцина CVXGA1 ослабляет легочное проявление заболевания. Через пять дней после инфицирования с помощью SARS-CoV-2 легкие собирали, фиксировали в цинковом формалине и заливали в парафин. Представлено репрезентативное окрашивание легких с помощью гематоксилина и эозина. Эозинофильные инфильтраты были отмечены в группе с инактивированным УФ излучением SARS-CoV-2 в день 5 после инфицирования с помощью SARS-CoV-2 (указаны стрелками на нижней панели и вставке). Количественные данные по эозинофильным инфильтратам представлены на графике справа. Масштабный отрезок = 255 и 51 мкм, верхняя и нижняя панели соответственно.
На ФИГ. 20А - ФИГ. 20F представлена иммуногенность и эффективность CVXGA1 у хорьков. На ФИГ. 20А показано схематическое изображение иммунизации хорьков и заражения вирусом. Группы из шести хорьков иммунизировали с помощью CVXGA1 или подвергали имитационной иммунизации (PBS) в дозе 106 БОЕ интраназально. После иммунизации осуществляли назальные смывы для определения того, выделяется ли вакцина в среду, и еженедельно осуществляли сбор крови. Хорьков заражали через 39 дней после заражения (dpc) посредством 4×105 БОЕ SARS-CoV-2 интраназально. Назальные смывы и ректальные мазки выполняли через 1, 3, 5 и 7 dpc. Половину (n=6) хорьков гуманно усыпляли через четыре дня после инфицирования. Исследование завершали через 7 dpc. На ФИГ. 20 В показаны титры IgG к RBD в сыворотке крови у хорьков после IN иммунизации. Титры IgG к RBD в сыворотке крови оценивали посредством ELISA. Показаны титры через 28 dpi. На ФИГ. 20С показаны титры нейтрализации после иммунизации. Титры нейтрализации антител к SARS-CoV-2 через 28 dpi определяли с использованием псевдочастицы VSV-CoV-SARS2-S. +RHS (положительная восстановленная сыворотка крови человека) использовалась в качестве положительного контроля, а отдельно среды использовались в качестве отрицательного контроля. На ФИГ. 20D показана вирусная РНК в назальных смывах после заражения. Количество вирусных РНК в назальных смывах через 1, 3, 5 и 7 dpc определяли с использованием qRT-PCR, как описано в разделе "Материалы и способы". На ФИГ. 20Е показана вирусная РНК в легком после заражения. Количество вирусных РНК в легких через 4 и 7 dpc определяли с использованием qRT PCR. На ФИГ. 20F показана вирусная РНК в трахее после заражения. Количество вирусных РНК в трахее через 4 и 7 dpc определяли с использованием qRT-PCR.
На ФИГ. 21А - ФИГ. 21Е представлена иммуногенность и эффективность CVXGA1 у кошек. На ФИГ. 21А представлено схематическое изображение иммунизации и заражения. Кошек в группе из 4 иммунизировали с помощью PBS, CVXGA1 в дозе 106 БОЕ посредством интраназального или подкожного (subQ) пути. В группе subQ кошек повторно иммунизировали посредством такой же дозы через 21 dpi. Через 42 дня после первоначальной иммунизации кошек заражали с помощью 4×105 БОЕ SARS-CoV-2 посредством внутривенного пути. Назальные смывы и ректальные мазки собирали через день до окончания эксперимента через 14 dpc. На ФИГ. 21В показан титр антител к S у кошек после иммунизации. Титры IgG к S в сыворотке крови определяли посредством ELISA. Показаны титры в сыворотке крови через 28 dpi. На ФИГ. 21С показан титр антител к RBD у кошек после иммунизации. Титры антител к RBD в сыворотке крови определяли с использованием ELISA. Были показаны титры в сыворотке крови через 28 dpi. На ФИГ. 21D показан титр IgA к S в назальных смывах после иммунизации. Титры IgA к S в назальных смывах определяли с использованием ELISA. Были показаны титры через 28 dpi. На ФИГ. 21Е показаны титры вируса в назальных смывах после заражения. Инфекционные частицы SARS-CoV-2, полученные в назальных смывах кошек, количественно определяли в виде фокусобразующих единиц (FFU) и представляли в виде logio FFU/мл после заражения вирусом.
На ФИГ. 22А - ФИГ. 22С представлена иммуногенность CVCGA1 у мышей С57В16. На ФИГ. 22А представлено схематическое изображение иммунизации. Мышей С57В16 (n=5 на группу) иммунизировали интраназальным (IN) путем посредством PBS, PIV5 в дозе 106 бляшкообразующих единиц (БОЕ) или CVXGA1 в дозе 6×103 БОЕ. Через 28 дней после иммунизации мышей умерщвляли. Собирали терминальную кровь и спленоциты. На ФИГ. 22 В показаны титры антител к S у мышей. Титры антител к S измеряли с использованием ELISA. На ФИГ. 22С показан клеточный иммунный ответ. Клеточный иммунный ответ определяли с использованием Elispot. Спленоциты стимулировали с помощью библиотеки пептидов S-белка SARS-CoV-2. Результаты были представлены в виде числа клеток, секретирующих IFN-γ, на 106 спленоцитов. Планки погрешностей представляют стандартную ошибку среднего. * Значимость (Р<0,05) при сравнении группы введения PIV5 и группы введения вакцины.
На ФИГ. 23А - ФИГ 23С представлена защита мышей, сенсибилизированных в отношении Ad-hACE2, с помощью CVXGA1. На ФИГ. 23А представлено схематическое изображение экспериментального протокола исследования вакцины на основе PIV5 против SARS-CoV-2 у мышей BALB/c. Группы обработки включают следующие: группа (Gp) 1 - иммунизация с помощью DMEM посредством IN пути; Gp 2 - иммунизация с помощью CVXGA1 посредством 5×103 БОЕ IN; Gp 3 - иммунизация с помощью CVXGA1 посредством 5×104 БОЕ IN. Затем всех животных сенсибилизировали к инфекции SARS-CoV-2 посредством Ad-hACE2 и через 5 дней инокулировали с помощью 1×105 БОЕ SARS-CoV-2. Указаны другие измерения и число животных на группу. Дни после иммунизации обозначены как dpi. На ФИГ. 23В показаны результаты, полученные у мышей, иммунизированных с помощью CVXGA1 с последующим инфицированием с помощью SARS-CoV-2. Мышей BALB/c, трансдуцированных с помощью Ad5-hACE2, инфицировали интраназально посредством 1×105 БОЕ SARS-CoV-2 в 50 мкл DMEM. Мониторинг изменения веса у мышей в возрасте от 6 до 8 недель осуществляли ежедневно (n=5 мышей на группу). Мыши, иммунизированные посредством 5×104 БОЕ CVXGA1, продемонстрировали меньшую потерю веса, чем мыши, получавшие 5×103 БОЕ (*Р-значение < 0,05; **Р значение < 0,005; критерий Манна Уитни). На ФИГ. 23С показано, что интраназальная иммунизация с помощью CVXGA1 снижает титры SARS-CoV-2 в легочной ткани. Для получения кинетики вируса легкие собирали и гомогенизировали через 1 dpi, а вирус титровали посредством анализа бляшкообразования. Титры выражали в виде БОЕ на легочную ткань (n=3 мыши на группу). Мыши, иммунизированные посредством 5×104 БОЕ CVXGA1, продемонстрировали большее снижение титров вируса в легочной ткани (*Р-значение < 0,05; **Р значение < 0,005; критерий Манна Уитни).
На ФИГ. 24А - ФИГ. 24В представлена иммуногенность инактивированного SARS-CoV-2 у мышей. На ФИГ. 24А показаны уровни антител к S у мышей после иммунизации в режиме "прайм-буст" посредством инактивированного SARS-CoV-2. Уровни антител к S в сыворотке крови мышей, иммунизированных с помощью инактивированного SARS-CoV-2, как описано на ФИГ. 19А, определяли с использованием ELISA. На ФИГ. 24В показаны уровни антител к RBD у мышей после иммунизации в режиме "прайм-буст" посредством инактивированного SARS-CoV-2. Уровни антител к RBD в сыворотке крови мышей, иммунизированных с помощью инактивированного SARS-CoV-2, как описано на ФИГ. 19А, определяли с использованием ELISA.
На ФИГ. 25А - ФИГ. 25Е представлена иммуногенность и эффективность CVXGA1 у хорьков. На ФИГ. 25А показан титр IgG к S в сыворотке крови после иммунизации. Уровни антител к S в сыворотке крови через 7, 28 dpi и 7 dpc определяли с использованием ELISA и наносили на график. На ФИГ. 25В показаны титры IgA к S в назальных смывах. Уровни IgA в назальных смывах, собранных через 28 dpi, измеряли с использованием ELISA. На ФИГ. 25 показано выявление вируса и вирусной РНК в назальном смыве через 5 dpc. Титры SARS-CoV-2 в назальных смывах через 5 dpc определяли с использованием иммунофлуоресцентного анализа после анализа бляшкообразования и отображали в виде FFU (фокусобразующей единицы флуоресценции) на миллилитр. Результаты qRT-PCR для назальных смывов через 5 dpc были такими же, как на ФИГ. 20D. На ФИГ. 25D показано выявление антигена SARS-CoV-2 в легких через 4 dpc. Срезы легких из хорьков, взятые через 4 дня после заражения, фиксировали в формалине и обрабатывали для иммуногистохимии в отношении антигена N SARS-CoV2. На ФИГ. 25Е представлено сравнение титров антител к S до и после заражения. Уровни антител к S у хорьков через 28 dpi (пунктирная линия) и 7 dpc (сплошная линия) определяли с использованием ELISA.
На ФИГ. 26А - ФИГ. 26D представлена иммуногенность и эффективность CVXGA1 у кошек. На ФИГ. 26А показаны титры нейтрализующих антител. Сыворотку крови, собранную через 28 dpi, анализировали с использованием псевдочастицы VSV CoV-SARS2-S. +RHS (положительная восстановленная сыворотка крови человека) использовалась в качестве положительного контроля. На ФИГ. 26В показано выявление вирусной РНК в назальных смывах. Количество вирусных РНК в назальных смывах через 1, 3, 5, 7, 9, 11, 13 и 14 dpc определяли с использованием qRT PCR. На ФИГ. 26С представлено сравнение титров антител к S до и после заражения. Уровни антител к S у хорьков через 41 dpi и 14 dpc определяли с использованием ELISA. На ФИГ. 26D представлено сравнение титров антител к N до и после заражения. Уровни антител к N у хорьков в 41 dpi и 14 dpc определяли с использованием ELISA.
На ФИГ. 27 показаны титры нейтрализации у мышей BALB/c, иммунизированных с помощью CVXGA1. Титры нейтрализации образцов определяли, как описано в разделе "Материалы и способы". Планки погрешностей представляют стандартную ошибку среднего, и Р значения были рассчитаны посредством однофакторного ANOVA. *Р значение < 0,05, ***Р-значение < 0,001.
На ФИГ. 28А - ФИГ. 28С показаны результаты интраназальной вакцинации с помощью CVXGA1, защищающей трансгенных мышей K18-hACE2 от инфекции SARS-CoV-2. Мыши получали DMEM интраназально; иммунизацию с помощью CVXGA1 посредством интраназального (106 БОЕ, IN) пути; инактивированный УФ излучением SARS-CoV-2 (UV-SARS2), введенный посредством IM пути с квасцовым адъювантом с последующей бустерной дозой через 21 dpi; PIV5 дикого типа (106 БОЕ) посредством IN пути. Через пять недель после иммунизации мышей K18-hACE2 интраназально инфицировали посредством 4×104 БОЕ SARS-CoV-2 в 50 мкл DMEM. Планки погрешностей представляют стандартную ошибку среднего. На ФИГ. 28А показана потеря веса и на ФИГ. 26В показаны результаты в виде выживания трансгенных мышей K18-hACE2. Изменения веса у мышей в возрасте 6-8 недель отслеживали ежедневно (DMEM: n=9, CVXGA1: n=8, инактивированный УФ излучением SARS-CoV-2: n=10 мышей на группу, PIV5: n=10). Мыши, иммунизированные с помощью CVXGA1, не теряли в весе и выживали. Все мыши, обработанные с помощью DMEM и PIV5, погибали. Одна мышь, обработанная инактивированным УФ излучением SARS-CoV-2, выжила. Дни после инфицирования с помощью SARS-CoV-2 обозначены в виде дней p.i. На ФИГ. 28А и ФИГ. 28 В показано, что вакцина CVXGA1 снижает титр вируса в легочной ткани и предупреждает его распространение в головной мозг. Через пять дней после инфицирования с помощью SARS-CoV-2 из групп в А и В были получены ткани легкого (n=4) и головного мозга (n=4) и определены их титры. Подробности см. в разделе "Материалы и способы". Предел выявления (LOD): планки погрешностей представляют стандартную ошибку среднего. **Р значение < 0,01; однофакторный ANOVA с апостериорным критерием Даннетта.
На ФИГ. 29А - ФИГ. 29D показан гистопатологический анализ инфекции SARS-CoV-2 у трансгенных мышей К18 hACE2, иммунизированных с помощью CVXGA1. На ФИГ. 29А ФИГ. 29С показана гистология фиксированных легочных тканей через пять дней после инфицирования с помощью SARS-CoV-2. Репрезентативные изображения мышей, инфицированных с помощью SARS-CoV-2, которые получали DMEM, CVXGA1, UV-SARS2 или PIV5. На ФИГ. 29А показано репрезентативное распределение вирусного антигена (белок N, коричневый краситель) в тканях легких из указанных групп (n=4 на группу). Можно заметить, что в группе CVXGA1 в результате иммуноокрашивания получены редкие очаги, тогда как в других группах получено диффузное иммуноокрашивание. Масштабный отрезок = 220 мкм. На ФИГ. 29В показаны ткани, окрашенные гематоксилином и эозином (n=4 на группу). Эозинофильные инфильтраты отмечались только в группе UV-SARS2 (стрелки на вставке, масштабный отрезок = 45 мкм). На ФИГ. 29С показаны ткани, окрашенные гематоксилином и эозином (n=4 на группу). Интерстициальное легочное проявление заболевания было ослаблено в случае CVXGA1. Масштабный отрезок = 90 мкм. На ФИГ. 29D показано гистопатологическое оценивание легочных тканей (подробности см. в разделе "Способы", n=4 на группу). Ткани из всех 4 групп поддавали балльной оценке посредством ординальной шкалы на наличие эозинофильных инфильтратов, периваскулярных инфильтратов и интерстициального легочного проявления заболевания. Планки погрешностей представляют стандартную ошибку среднего. *Р<0,05,**Р<0,01, однофакторный ANOVA с апостериорным критерием Даннетта.
На ФИГ. 30А – ФИГ. 30С показана иммуногенность CVXGA1 у хорьков. На ФИГ. 30А показано схематическое изображение иммунизации хорьков и заражения вирусом. Группы из шести хорьков иммунизировали с помощью CVXGA1 или подвергали имитационной иммунизации (PBS) в дозе 106 БОЕ интраназально. Хорьков заражали через 39 дней после иммунизации (dpi) посредством 4×105 БОЕ SARS-CoV-2 интраназально. Назальные смывы и ректальные мазки выполняли через 1, 3, 5 и 7 дней после заражения (dpc). Половину (n=6) хорьков гуманно усыпляли через четыре дня после заражения. Исследование завершали через 7 dpc. На ФИГ. 30В показано IgG к RBD в сыворотке крови у хорьков после иммунизации. Титры IgG к RBD в сыворотке крови оценивали посредством ELISA. Показаны титры через 28 dpi. Планки погрешностей представляют стандартную ошибку среднего. На ФИГ. 30С показаны титры нейтрализации после иммунизации. Титры нейтрализации антител к SARS-CoV-2 через 28 dpi определяли с использованием вируса VSV-CoV-SARS2-S. +RHS (положительная восстановленная сыворотка крови человека) использовалась в качестве положительного контроля, а отдельно среды использовались в качестве отрицательного контроля. Планки погрешностей представляют стандартную ошибку среднего, и Р значения были рассчитаны посредством однофакторного ANOVA. ****Р значение < 0,0001.
На ФИГ. 31А - ФИГ. 31С показана эффективность CVXGA1 у хорьков. На ФИГ. 31А показано выявление вирусной РНК в 1, 3, 5 и 7 назальных смывах после заражения посредством qRT-PCR. Вирусную РНК количественно определяли в виде количества копий генома на мл и представляли в виде logio геномов/мл, как описано в разделе "Материалы и способы". Каждый символ представляет отдельное животное (6 PBS и 6 CVXGA1). Животные, получавшие PBS, обозначены черными символами, а животные, вакцинированные с помощью CVXGA1, обозначены красными символами. 1 БОЕ/ПЦР реакция (гхп) обозначено для дифференциации между инфекционными вирусными частицами и неинфекционными остатками генома. Предел выявления (LOD) указан самой нижней пунктирной линией. На ФИГ. 31В показано выявление вирусной РНК в тканях легких, собранных через 4, 7 dpc, посредством qRT PCR. Вирусную РНК определяли количественно и представляли в виде копий генома на реакцию (rxn). На ФИГ. 31С показано выявление вирусной РНК в тканях трахеи, собранных через 4, 7 dpc, посредством qRT-PCR. Вирусную РНК определяли количественно и представляли в виде копий генома на реакцию (rxn).
На ФИГ. 32А 32В показана эффективность блокады передачи путем иммунизации с помощью CVXGA1 у хорьков. На ФИГ. 32А показано схематическое изображение иммунизации хорьков, заражения вирусом и передачи. Группы из шести хорьков иммунизировали с помощью CVXGA1, пустого вектора на основе вируса или подвергали имитационной иммунизации (PBS) в дозе 106 БОЕ интраназально. Хорьков заражали через 42 dpi посредством 4×105 БОЕ SARS-CoV-2 интраназально. Назальные смывы собирали через 1, 3, 5, 7, 9 и 11 dpc. Половину хорьков гуманно усыпляли через четыре дня после заражения. Исследование завершали через 11 dpc. Через 2 dpc одного зараженного хорька располагали совместно с одним не подвергнутым обработке хорьком, носовые смывы собирали у не подвергнутых обработке хорьков через 1, 3, 5, 7 и 9 дней после расположения совместно (3, 5, 7, 9 и 11 dpc соответственно). На ФИГ. 32В показано выявление SARS-CoV-2 через 1, 3, 5, 7, 9 и 11 dpc в назальных смывах. Живой вирус выявляли с использованием анализа определения фокусобразующих единиц (FFA, FFU/мл), как описано в разделе "Материалы и способы". Каждый символ представляет отдельное животное. Предел выявления (LOD) указан самой нижней пунктирной линией.
На ФИГ. 33А ФИГ. 33С показана иммуногенность CVXGA1 у мышей C57BL/6. На ФИГ. 33А показано схематическое изображение иммунизации. Мышей C57BL/6 (n=5 на группу) иммунизировали посредством интраназального (IN) пути посредством PBS, PIV5 в дозе 106 бляшкообразующих единиц (БОЕ) или CVXGA1 в дозе 6×103 БОЕ. Через 28 дней после иммунизации мышей умерщвляли. Собирали терминальную кровь и спленоциты. На ФИГ. 33В показаны титры антител к S у мышей. Титры антител к S измеряли с использованием ELISA. Планки погрешностей показывают стандартную ошибку среднего. На ФИГ. 33С показан клеточный иммунный ответ. Клеточный иммунный ответ определяли с использованием анализа Elispot. Спленоциты стимулировали с помощью библиотеки пептидов S-белка SARS-CoV-2. Результаты были представлены в виде числа клеток, секретирующих IFN-γ, на 106 спленоцитов. Планки погрешностей показывают стандартную ошибку среднего. * Значимость (Р<0,05) при сравнении группы введения PIV5 и группы введения вакцины. Дисперсионный анализ (ANOVA) с последующим применением критерия множественных сравнений Даннета.
На ФИГ. 34А - ФИГ. 34Е показана интраназальная вакцинация с помощью CVXGA1, защищающая трансгенных мышей К18-hACE2 от летальной инфекции SARS-CoV-2. Мыши получали иммунизацию с помощью DMEM посредством внутримышечного (IM) пути; иммунизацию с помощью CVXGA1 посредством интраназального (IN) пути; инактивированный УФ-излучением SARS-CoV-2 (UV-SARS2), введенный посредством IM пути с квасцовым адъювантом с последующей второй бустерной дозой через 14 dpi. Через четыре недели после иммунизации мышей K18-hACE2 интраназально инфицировали посредством 106 БОЕ SARS-CoV-2 в 50 мкл DMEM. На ФИГ. 34А показана потеря веса, и на ФИГ. 34В показаны результаты в виде выживания трансгенных мышей K18-hACE2. Изменения веса у мышей в возрасте 6-8 недель отслеживали ежедневно (DMEM: n=5, CVXGA1: n=6, инактивированный УФ-излучением SARS-CoV-2: n=5 мышей на группу). Хотя все мыши характеризовались потерей веса, выживали только мыши, иммунизированные с помощью CVXGA1. Дни после инфицирования с помощью SARS-CoV-2 обозначены в виде dpi. *Р-значение < 0,05; **Р-значение < 0,005; двухфакторный ANOVA. На ФИГ. 34С и 34D представлен анализ, который показывает, что вакцина CVXGA1 снижает титр вируса в легочной ткани и предупреждает его распространение в головной мозг. Через пять дней после инфицирования с помощью SARS-CoV-2 были получены ткани легкого (n=4) и головного мозга (n=4) и определены титры вируса в них. (N/D: не поддается выявлению) *Р-значение < 0,05; **Р-значение < 0,005; однофакторный ANOVA. На ФИГ. 34Е показаны гистология и оценивание легочных тканей через пять дней после заражения с помощью SARS-CoV-2. Левые панели: репрезентативная гистология групп DMEM, CVXGA1 или UV SARS2. Эозинофильные инфильтраты (стрелки на вставке) отмечались в группе UV-SARS2. Количественные данные по эозинофильным инфильтратам представлены на графике справа. **** Р < 0,0001, при сравнении UV-SARS2 с DMEM или CVXGA1, однофакторный ANOVA. Масштабный отрезок = 387 и 78 мкм (верхняя и нижняя панели соответственно).
На ФИГ. 35А ФИГ. 35D показаны иммуногенность и эффективность CVXGA1 у хорьков. На ФИГ. 35А показана репликация CVXGA1 после иммунизации в верхних дыхательных путях хорька. Назальные смывы хорьков собирали через 3, 7 и 14 дней после иммунизации. Анализы бляшкообразования проводили в клетках Vero. CVXGA1 в назальных смывах количественно определяли в виде бляшкообразующих единиц на мл (БОЕ/мл). На ФИГ. 35 В показан титр IgG к S в сыворотке крови после иммунизации. Уровни антител к S в сыворотке крови через 7, 14, 21, 28 dpi и 7 dpc определяли с использованием ELISA. Точки данных представляют средние значения +/-SD полученных в двух повторностях значений для отдельных животных (n=6 dpi; n=3 dpc). **** Р<0,0001, двухфакторный дисперсионный анализ с критерием множественных сравнений Сидака. На ФИГ. 35С показаны титры IgA к S в назальных смывах. Уровни IgA в назальных смывах, собранных через 28 dpi, измеряли с использованием ELISA. Точки данных представляют объединенные образцы (n=6) в трех повторностях +/- стандартное отклонение. На ФИГ. 35D показано сравнение ответов в виде IgG к S в сыворотке крови до и после заражения с помощью SARS-CoV-2. Уровни IgG к S у хорьков через 28 dpi (пунктирная линия) и 7 dpc (сплошная линия) определяли с использованием ELISA. Точки данных представляют средние значения +/-SD полученных в двух повторностях значений для отдельных животных (n=6 dpi; n=3 dpc). **** р<0,0001, двухфакторный дисперсионный анализ с критерием множественных сравнений Сидака.
На ФИГ. 36А - ФИГ. 36С показано помещение ABSL3 для инфицирования хорька с помощью SARS-CoV-2. На ФИГ. 36А показана схема помещения для эксперимента, описанного на ФИГ. 31. Исследования с заражением выполняли в помещениях BSL3 Ag. Помещения оснащены системой подачи воздуха с однократной очисткой на НЕРА-фильтре и выпуска с двукратной очисткой на НЕРА фильтре и в них поддерживается отрицательное давление по сравнению с внешними "чистыми" коридорами. Отдельно вентилируемые клетки (IVC), выполненные с использованием твердых доньев для клеток и пластиковой собственно клетки (Tecniplast) были укомплектованы системой подачи и выпуска воздуха с очисткой на НЕРА фильтре. На ФИГ. 36В показано репрезентативное изображение стандартной клетки для хорьков от Allentown, Inc. Каждый стандартный стеллаж для хорьков состоит из шести отдельных клеток, выполненных с использованием пластиковых вставок для клеток с перфорированными нижними элементами и съемными дверцами из нержавеющей стали, пластиковыми поддонами для экскрементов, J-образными кормушками и автоматической системой подачи воды с поилками типа Lixit.
На ФИГ. 36С показаны расположения групп внутри стеллажей клеток. Не подвергнутых обработке хорьков помещали в IVC перед размещением совместно с зараженными хорьками в обычных (открытых) клетках. Отдельные клетки помечали с указанием группы в отношении обработки вакциной и идентификацией животного, где буквой "С" обозначали животных, не подвергнутых обработке путем контакта. Gp3 представляет собой кандидатную вакцину, которая не действовала столь же эффективно, как CVXGA1: у этих хорьков заражение вирусом в носовых полостях наблюдалось через 3 dpc.
На ФИГ. 37А ФИГ. 37В показаны уровни вирусной РНК у хорьков, иммунизированных с помощью CVXGA1, и их прямые контакты. На ФИГ. 37А показаны уровни РНК в назальных смывах. Выявление вирусной РНК в назальных смывах, собранных через 1, 3, 5, 7 и 9 dpc, посредством qRT-PCR. Вирусную РНК количественно определяли с использованием qRT PCR. Каждая точка представляет отдельное животное, 1 БОЕ/ПЦР реакция (гхп) обозначено для дифференциации между инфекционными вирусными частицами и неинфекционными остатками генома. Предел выявления (LOD) указан самой нижней пунктирной линией. На ФИГ. 37В показаны уровни РНК в легких зараженных хорьков. Образцы легких собирали в дни 4 и 10 dpc. Вирусную РНК количественно определяли посредством qRT PCR и БОЕ/мл рассчитывали по стандартной кривой. Каждая точка представляет отдельное животное, и 1 БОЕ/мл обозначено для дифференциации между инфекционными вирусными частицами и неинфекционными остатками генома. Предел выявления (LOD) указан самой нижней пунктирной линией.
На ФИГ. 38 показано схематическое изображение конструкции вектора CVX-UGA2.
ПОДРОБНОЕ ОПИСАНИЕ
С использованием настоящего изобретения были созданы конструкции на основе вируса парагриппа типа 5 (PIV5), экспрессирующие шиловидный (S) белок оболочки SARS-CoV-2, для использования в качестве вакцин против COVID. Эти конструкции демонстрируют эффективность в качестве вакцин, при этом интраназальная иммунизация посредством однократной дозы индуцирует установление стерильного иммунитета у хорьков и кошек.
Коронавирусное заболевание 2019 г. (COVID 19) является новым инфекционным заболеванием, которое в настоящее время распространяется по всему миру. Оно вызывается новым коронавирусом - коронавирусом 2 тяжелого острого респираторного синдрома (SARS-CoV-2) (Zhu et al. N EnglJ Med 382, 727-733 (2020)). SARS-CoV-2 был впервые идентифицирован в Ухане, Китай, в декабре 2019 года и впоследствии распространился по всему миру, вызвав пандемию COVID-19. Вирусом было инфицировано более чем 221 миллион человек во всем мире, по состоянию на 8 сентября 2021 г. он стал причиной более чем 4574000 случаев смерти и может продолжать распространяться в отсутствие коллективного иммунитета (см. во всемирной сети по адресу who.int/emergencies/diseases/novel-coronavirus-2019). Социальное дистанцирование и повсеместное тестирование с отслеживанием контактов и карантинными процедурами в настоящее время являются единственными мерами, доступными для ограничения распространения вируса. Срочно необходима вакцина для защиты от SARS-CoV-2, чтобы предупредить дальнейшие случаи смертельного исхода и уменьшить передачу.
SARS-CoV-2 представляет собой вирус с одноцепочечной РНК, заключенной в оболочку, принадлежащий к семейству р коронавирусов (Lu et al. Lancet 395, 565-574 (2020)). Подход с метагеномным секвенированием следующего поколения на основе РНК применялся для определения характеристик его полного генома, длина которого составляет 29881 нуклеотид (nt) (последовательность под номером доступа в GenBank MN908947), кодирующего 9860 аминокислот (Chen et al. EmergMicrobes Infect 9, 313-319 (2020)). Полногеномные секвенированные геномы, доступные в GenBank, включают изолят WHU01 2019-nCoV (номер доступа в GenBank MN988668) и NC_045512 для SARS CoV-2, оба изолята из Уханя, Китай, и по меньшей мере семь дополнительных последовательностей (MN938384.1, MN975262.1, MN985325.1, MN988713.1, MN994467.1, MN994468.1 и MN997409.1), которые являются идентичными на > 99,9%.
Вирус парагриппа 5 (PIV5), РНК вирус с отрицательной цепью, является представителем рода Rubulavirus из семейства Paramyxoviridae, которое включает многие важные патогены человека и животных, такие как вирус эпидемического паротита, вирус парагриппа человека типа 2 и типа 4, вирус болезни Ньюкасла, вирус Сендай, HPIV3, вирус кори, вирус чумы собак, вирус чумы крупного рогатого скота и респираторно-синцитиальный вирус. PIV5 был ранее известен как вирус 5 обезьянообразных (SV5). Хотя PIV5 является вирусом, который инфицирует многих животных и людей, никакие известные симптомы или заболевания у людей не ассоциировались с PIV5. В отличие от большинства парамиксовирусов, PIV5 инфицирует нормальные клетки с небольшим цитопатическим эффектом. Являясь геномом вируса с отрицательной цепью геном PIV5 очень стабилен. Поскольку PIV5 не имеет фазы ДНК в своем жизненном цикле и реплицируется исключительно в цитоплазме, то PIV5 не может интегрироваться в геном хозяина. Следовательно, использование PIV5 в качестве вектора позволяет избежать возможных непредвиденных последствий генетических модификаций ДНК клеток хозяев. PIV5 может расти до высоких титров в клетках, включая клетки Vero, которые были одобрены ВОЗ и FDA для получения вакцин. Таким образом, PIV5 имеет много преимуществ в качестве вакцинного вектора.
Вакцинный вектор на основе PIV5 по настоящему изобретению может быть на основе любого из множества штаммов дикого типа, мутантных или рекомбинантных (rPIV5) штаммов. Штаммы дикого типа включают без ограничения штаммы PIV5 W3A, WR (номер согласно АТСС® VR-288™), штамм вируса парагриппа собак 78-238 (номер согласно АТСС VR-1573) (Evermann etal. Arch Virol 68, 165-172 (1981); Evermann et al. J Am Vet Med Assoc 177, 1132-1134 (1980)), штамм вируса парагриппа собак D008 (номер согласно АТСС VR-399) (Binn et al. Proc Soc Exp Biol Med 126, 140-145 (1967)), MIL, DEN, LN, MEL, криптовирус, CPI+, CPI-, H221, 78524, Tl и SER. См., например, (Baumgartner et al. Intervirology 27, 218-223 (1987); Chatziandreou et A.J Gen Virol 85, 3007-3016 (2004); Choppin Virology 23, 224-233 (1964)). Дополнительно можно использовать штаммы PIV5, используемые в коммерческих вакцинах против питомникового кашля, таких как, например, вакцины BI, FD, Merck и Merial.
Вакцинный вектор на основе PIV5 по настоящему изобретению может быть сконструирован с использованием любого из множества способов, включая без ограничения систему реверсивной генетики, более подробно описанную в Не et al. (Virology; 237(2):249-60, 1997). PIV5 кодирует восемь вирусных белков. Нуклеокапсидный белок (NP), фосфопротеин (Р) и высокомолекулярный белок РНК-полимеразы (L) важны для транскрипции и репликации вирусного РНК генома. Белок V играет важные роли в вирусном патогенезе, а также в синтезе вирусной РНК. Белок слияния (F), представляющий собой гликопротеин, опосредует как слияние клетки с клеткой, так и слияние вируса с клеткой рН-независимым образом, что необходимо для проникновения вируса в клетки. Были определены структуры белка F и идентифицированы критически важные аминокислотные остатки для эффективного слияния. Гликопротеин гемагглютинин нейраминидаза (HN) также участвует в проникновении вируса в клетки хозяева и высвобождении вируса из них. Матриксный (М) белок играет важную роль в сборке и почковании вируса. Гидрофобный (SH) белок представляет собой гидрофобный интегральный мембранный белок из 44 остатков и в мембранах ориентирован своим N-концом к цитоплазме. В отношении обзоров по молекулярной биологии парамиксовирусов см., например, (Lamb Fields Virology: Sixth Edition 1, 957-995 (2013); Whelan et al. Biology of Negative Strand RNA Viruses: The Power of Reverse Genetics, 61-119 (2004)).
Что касается вакцинных векторов на основе PIV5 по настоящему изобретению, то гетерологичная нуклеотидная последовательность, кодирующая шиловидный (S) белок коронавируса, включая без ограничения S-белок SARS-CoV-2, встроена в геном PIV5. Проникновение коронавируса в клетки-хозяева опосредуется транс мембранным гликопротеином S (Tortorici et al. Adv Virus Res 105, 93-116 (2019)). Поскольку гликопротеин S коронавируса экспонируется на поверхности и опосредует проникновение в клетки-хозяева, то он является основной мишенью для нейтрализующих антител при инфекции и находится в центре внимания при разработке терапевтических средств и вакцин. Шиловидный S-белок SARS-CoV-2 состоит из двух субъединиц - S1 и S2. Субъединица S1 содержит рецепторсвязывающий домен, который распознает рецептор хозяина, представляющий собой ангиотензинпревращающий фермент 2, и связывается с ним, в то время как субъединица S2 опосредует слияние вируса с клеточной мембраной, образуя пучок из шести спиралей посредством домена с двумя гептадными повторами (Huang et al. ActaPharmacologica Sinica 41, 1141-1149 (2020)).
Общая длина S SARS-CoV-2 составляет 1273 аминокислоты (aa) и состоит из сигнального пептида (аминокислоты 1-13), расположенного HaN-конце, субъединицы S1 (14-685 остатков) и субъединицы S2 (686-1273 остатка); последние две области отвечают за связывание с рецептором и слияние с мембраной соответственно. В субъединице S1 имеется N концевой домен (14 305 остатков) и рецепторсвязывающий домен (RBD, 319 541 остаток); пептид слияния (FP) (788-806 остатков), гептапептидный повтор последовательности 1 (HR1) (912-984 остатка), HR2 (1163-1213 остатков), домен ТМ (1213 1237 остатков) и цитоплазматический домен (1237 1273 остатка) составляют субъединицу S2 (Xia et al. CellMolImmunol 17, 765-767 (2020)).
В некоторых вакцинных векторах на основе PIV5 по настоящему изобретению гетерологичная нуклеотидная последовательность, кодирующая шиповидный (S) белок коронавируса, включая без ограничения S-белок SARS-CoV-2, была модифицирован таким образом, что цитоплазматический хвост белка S коронавируса был заменен на цитоплазматический хвост белка слияния (F) PIV5. Пример такой конструкции на основе PIV5 включает конструкцию на основе PIV5 CVX-GA1, также обозначаемую в данном документе как CVXGA1, CVX-UGA1, pDA27 или DA27. Карта плазмиды CVX-GA1 показана на ФИГ. 2 и 9, при этом последовательность конструкции включена на ФИГ. 9.
В некоторых вакцинных векторах на основе PIV5 по настоящему изобретению гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, была модифицирована таким образом, что S-белок характеризуется наличием аминокислотной замены по аминокислотному остатку W886 и/или F888. В некоторых аспектах аминокислотная замена по аминокислотному остатку W886 предусматривает замену триптофана (W) на аргинин (R), и/или аминокислотная замена по аминокислотному остатку W888 предусматривает замену фенилаланина (F) на аргинин (R).
В некоторых вакцинных векторах на основе PIV5 по настоящему изобретению гетерологичная нуклеотидная последовательность, кодирующая шиповидный (S) белок коронавируса, включая без ограничения S-белок SARS-CoV-2, предусматривает наличие как модификации, такой, что цитоплазматический хвост S-белка коронавируса заменяется на цитоплазматический хвост белка слияния (F) PIV5, так и аминокислотной замены по аминокислотному остатку W886 и/или F888. В некоторых аспектах аминокислотная замена по аминокислотному остатку W886 предусматривает замену триптофана (W) на аргинин (R), и/или аминокислотная замена по аминокислотному остатку W888 предусматривает замену фенилаланина (F) на аргинин (R). Пример такой конструкции на основе PIV5 включает конструкцию на основе PIV5 CVX-GA2, также обозначаемую в данном документе как CVXGA2 или CVX-UGA2. Карта плазмиды CVX-GA1 показана на ФИГ. 38.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена в любое из множества местоположений в геноме PIV5.
В некоторых предпочтительных вариантах осуществления гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена между геном низкомолекулярного гидрофобного белка (SH) и геном гемагглютинин нейраминидазы (HN) в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена между геном гемагглютинин-нейраминидазы (HN) и геном высокомолекулярного белка РНК полимеразы (L) в геноме PIV5. В некоторых вариантах осуществления гетерологичная нуклеотидная последовательность не встроена в местоположение между геном гемагглютинин-нейраминидазы (HN) и геном высокомолекулярного белка РНК-полимеразы (L) в геноме PIV5. В некоторых вариантах осуществления гетерологичная нуклеотидная последовательность встроена в местоположение, отличное от местоположения между геном гемагглютинин нейраминидазы (HN) и геном высокомолекулярного белка РНК-полимеразы (L) в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена между геном нуклеокапсидного белка (NP) и геном V/P в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена между геном М и геном F в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена между геном F и геном SH в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена между геном VP и геном матриксного белка (М) в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена между лидерной последовательностью и геном нуклеокапсидного белка (NP) в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена непосредственно ниже по ходу транскрипции от лидерной последовательности в геноме PIV5.
Гетерологичная нуклеотидная последовательность, кодирующая S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, может быть встроена с заменой всего или части гена PIV5 в геноме PIV5. Например, гетерологичная нуклеотидная последовательность может заменять ген F, HN или SH в геноме PIV5. Гетерологичная нуклеотидная последовательность может быть встроена в пределах гена PIV5, что приводит к экспрессии химерного полипептида. Например, гетерологичная нуклеотидная последовательность может быть встроена в пределах нуклеотидной последовательности гена SH, в пределах нуклеотидной последовательности гена NP, в пределах нуклеотидной последовательности гена V/P, в пределах нуклеотидной последовательности гена М, в пределах нуклеотидной последовательности гена F, в пределах нуклеотидной последовательности гена HN и/или в пределах нуклеотидной последовательности гена L в геноме PIV5.
Вакцина на основе вируса PIV5 по настоящему изобретению может также характеризоваться наличием мутации, изменения или делеции, касающихся одного или нескольких из этих восьми белков, в геноме PIV5. Например, вектор экспрессии на основе вируса PIV5 может предусматривать наличие одной или нескольких мутаций, включая без ограничения любую из тех, которые описаны в данном документе. В некоторых аспектах комбинация из двух или более (двух, трех, четырех, пяти, шести, семи или больше) мутаций может быть предпочтительной и может демонстрировать повышенную активность.
Мутация включает без ограничения мутацию гена V/P, мутацию общего N конца белков V и Р, мутацию остатков 26, 32, 33, 50, 102 и/или 157 общего N-конца белков V и Р, мутацию, обеспечивающую отсутствие С-конца белка V, мутацию, обеспечивающую отсутствие низкомолекулярного гидрофобного (SH) белка, мутацию белка слияния (F), мутацию фосфопротеина (Р), мутацию высокомолекулярного белка РНК-полимер азы (L), мутацию, обеспечивающую встраивание остатков из вируса парагриппа собак, и/или мутацию, обеспечивающую усиление образования синцития.
Мутация может включать без ограничения rPIV5-V/P-CPI-, rPIV5-CPI-, rPIV5-CPI+, rPIVSV ΔC, rPIV-Rev, rPIVS RL, rPIVS-P-S157A, rPIV5-P-S308A, rPIVS-L-A1981D и rPIV5-F S443P, rPIVS-MDA7, rPIV5-ΔSH-CPI, rPIV5 ΔSH-Rev и их комбинации.
PIV5 может инфицировать клетки по продуктивному типу с небольшим цитопатическим эффектом (СРЕ) во многих типах клеток. В некоторых типах клеток инфицирование с помощью PIV5 вызывает образование синцитиев, т.е. слияние многих клеток вместе, что приводит к гибели клеток. Мутация может предусматривать наличие одной или нескольких мутаций, которые способствуют образованию синцитиев (см., например, (Paterson et al. Virology 270, 17-30 (2000))).
Белок V PIV5 играет критически важную роль в блокировании апоптоза, индуцированного вирусом. Рекомбинантный PIV5, лишенный консервативного, богатого на цистеин С конца (rPIV5V ΔС) белка V, индуцирует апоптоз в различных клетках посредством внутреннего апоптотического пути, который вероятно инициируется стрессовым воздействием на эндоплазматический ретикулум (ER) (Sun et al. Journal of virology 78, 5068-5078 (2004)). Мутантный рекомбинантный PIV5 с мутациями на N-конце продуктов гена V/P, таких как rPIV5-CPI-, также индуцирует апоптоз (Wansley et al. J Virol 76, 10109-10121 (2002)). Мутация включает без ограничения rPIVS ΔSH, rPIV5-CPI, rPIVSVΔC и их комбинации.
Также в настоящее изобретение включены вирионы и инфекционные вирусные частицы, которые содержат геном PIV5, содержащий гетерологичную нуклеотидную последовательность, кодирующую S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2.
Также в настоящее изобретение включены композиции, содержащие одну или несколько конструкций на основе вируса или вир ионов PIV5, как описано в данном документе. Такая композиция может содержать фармацевтически приемлемый носитель. Как используется, термин "фармацевтически приемлемый носитель" относится к одному или нескольким совместимым твердым или жидким наполнителям, разбавителям или инкапсулирующим веществам, подходящим для введения человеку или другому позвоночному животному. Такой носитель может являться апирогенным. Настоящее изобретение также включает способы получения и применения векторов на основе вируса и композиций, описанных в данном документе.
Композиции по настоящему изобретению могут быть составлены в фармацевтические препараты в различных формах, адаптированных к выбранному пути введения. Специалисту в данной области техники будет понятно, что композиция будет варьироваться в зависимости от способа введения и единицы дозирования.
Средства по настоящему изобретению можно вводить различными путями, включая без ограничения внутривенную, местную, пероральную, интраназальную, подкожную, внутрибрюшинную, внутримышечную и внутриопухолевую доставку. В некоторых аспектах средства по настоящему изобретению могут быть составлены для контролируемого или замедленного высвобождения. Одно преимущество интраназальной иммунизации заключается в способности индуцировать иммунный ответ слизистой оболочки.
Также в настоящее изобретение включены способы получения и применения векторов экспрессии на основе вируса PIV5, включая без ограничения любые из описанных в данном документе.
Например, настоящее изобретение включает способы обеспечения экспрессии S-белка коронавируса, включая без ограничения S-белок SARS-CoV-2, в клетке путем приведения клетки в контакт с вектором экспрессии на основе вируса PIV5, вирусной частицей или композицией, как описано в данном документе, или инфицирования клетки таковыми.
Настоящее изобретение предусматривает способы индуцирования у субъекта иммунного ответа на S-белок коронавируса, включая без ограничения S-белок SARS-CoV-2, посредством введения субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции, как описано в данном документе. Иммунный ответ может включать гуморальный иммунный ответ и/или клеточный иммунный ответ. Иммунный ответ может усиливать врожденный и/или адаптивный иммунный ответ.
Настоящее изобретение предусматривает способы экспрессии гетерологичного S-белка коронавируса, включая без ограничения S-белок SARS-CoV-2, у субъекта посредством введения субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции, как описано в данном документе.
Настоящее изобретение предусматривает способы вакцинирования субъекта посредством введения субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции, как описано в данном документе.
В способах по настоящему изобретению можно применять любой из множества способов введения. Например, введение может быть внутривенным, местным, пероральным, интраназальным, подкожным, внутрибрюшинным, внутримышечным, внутриопухолевым, in ovo, материнским и т.п. В некоторых аспектах введение осуществляется на поверхность слизистой оболочки. Вакцину можно вводить посредством методик массового введения, как, например, путем помещения вакцины в питьевую воду или путем распыления в окружающую среду животных. При ведении путем инъекции иммуногенную композицию или вакцину можно вводить парентерально. Парентеральное введение включает, например, введение путем внутривенной, подкожной, внутримышечной или внутрибрюшинной инъекции.
Средство по настоящему изобретению можно вводить сразу, в виде однократной дозы, или можно вводить в виде нескольких доз, вводимых через определенные интервалы времени. Например, средства по настоящему изобретению можно вводить повторно, например по меньшей мере 2, 3, 4, 5, 6, 7, 8 или больше раз. Понятно, что точная дозировка и продолжительность лечения зависят от заболевания, в отношении которого осуществляют лечение, и могут быть определены опытным путем с применением известных протоколов тестирования или путем экстраполяции данных из тестов in vivo или in vitro. Следует отметить, что концентрации и значения дозировки также могут варьироваться в зависимости от тяжести состояния, подлежащего облегчению. Кроме того, следует понимать, что для любого конкретного субъекта конкретные схемы введения доз должны корректироваться с течением времени в соответствии с индивидуальными потребностями и профессиональным мнением лица, вводящего композиции или контролирующего их введение, и что любые диапазоны концентраций, указанные в данном документе, являются только иллюстративными и не предназначены для ограничения объема или практического применения заявленных композиций и способов.
В некоторых терапевтических вариантах осуществления "эффективное количество" средства представляет собой количество, которое приводит к снижению по меньшей мере одного параметра патологического состояния. Таким образом, например, в некоторых аспектах настоящего изобретения эффективное количество представляет собой количество, эффективное для достижения снижения на по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20% или по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90% или по меньшей мере приблизительно 95% по сравнению с ожидаемым снижением параметра у индивидуума, не получавшего лечение с помощью средства.
В некоторых аспектах в настоящем изобретении могут применяться любые конструкции и способы на основе PIV5, описанные в WO 2013/112690 и WO 2013/112720 (которые таким образом включены в данный документ посредством ссылки в их полном объеме).
Как используется в данном документе, термин "субъект" означает организм, включая например млекопитающее. Млекопитающее включает без ограничения человека, примата, отличного от человека, и других позвоночных, отличных от человека. Субъект может являться "индивидуумом", "пациентом" или "хозяином". Позвоночные, отличные от человека, включают сельскохозяйственных животных (например, корову, лошадь, козу и свинью), домашнее животное или животное-компаньона, таких как без ограничения собака или кошка, и лабораторных животных. Субъекты, отличные от человека, также включают приматов, отличных от человека, а также грызунов, таких как без ограничения крыса или мышь. Субъекты, отличные от человека, также включают без ограничения домашнюю птицу, лошадей, коров, свиней, коз, собак, кошек, морских свинок, хомяков, норок и кроликов.
Как используется в данном документе, термин "in vitro" относится к культуре клеток, a "in vivo" относится к телу субъекта. Как используется в данном документе, термин "выделенный" относится к материалу, который был либо извлечен из его естественной среды (например, естественной среды, если он встречается в природе), либо получен с использованием рекомбинантных методик, либо синтезирован химическим или ферментативным путем и таким образом изменен "рукой человека" относительно его естественного состояния.
Иллюстративные варианты осуществления настоящего изобретения включают без ограничения следующее.
1. Вектор экспрессии на основе вируса, содержащий геном вируса парагриппа 5 (PIV5), который содержит гетерологичную нуклеотидную последовательность, обеспечивающую экспрессию гетерологичного полипептида, где гетерологичный полипептид предусматривает шиповидный (S) белок коронавируса.
2. Вектор экспрессии на основе вируса согласно варианту осуществления 1, где S-белок коронавируса предусматривает S-белок коронавируса из коронавируса 2 тяжелого острого респираторного синдрома (SARS-CoV-2).
3. Вектор экспрессии на основе вируса согласно варианту осуществления 1, где цитоплазматический хвост S-белка коронавируса заменен на цитоплазматический хвост белка слияния (F) PIV5.
4. Вектор экспрессии на основе вируса согласно варианту осуществления 1, где S-белок коронавируса предусматривает S-белок коронавируса из коронавируса 2 тяжелого острого респираторного синдрома (SARS-CoV-2), и где цитоплазматический хвост S-белка коронавируса заменен на цитоплазматический хвост белка слияния (F) PIV5.
5. Вектор экспрессии на основе вируса по любому из вариантов осуществления 1-4, где гетерологичный полипептид предусматривает шиповидный (S) белок коронавируса, который предусматривает наличие мутаций по аминокислотному остатку W886 и/или F888.
6. Вектор экспрессии на основе вируса согласно варианту осуществления 5, где аминокислотная замена по аминокислотному остатку W886 предусматривает замену триптофана (W) на аргинин (R), и/или аминокислотная замена по аминокислотному остатку W888 предусматривает замену фенилаланина (F) на аргинин (R).
7. Вектор экспрессии на основе вируса согласно любому из вариантов осуществления 1-6, где гетерологичная нуклеотидная последовательность встроена между геном низкомолекулярного гидрофобного белка (SH) и геном гемагглютинин нейраминидазы (HN) в геноме PIV5.
8. Вектор экспрессии на основе вируса согласно любому из вариантов осуществления 16, где гетерологичная нуклеотидная последовательность заменяет нуклеотидную последовательность гена SH.
9. Вектор экспрессии на основе вируса согласно любому из вариантов осуществления 16, где гетерологичная нуклеотидная последовательность встроена между геном гемагглютинин нейраминидазы (HN) и геном высокомолекулярного белка РНК полимеразы (L) в геноме PIV5.
10. Вектор экспрессии на основе вируса согласно любому из вариантов осуществления 16, где гетерологичная нуклеотидная последовательность встроена скорее ближе к лидерной последовательности, чем между геном гемагглютинин нейраминидазы (HN) и геном высокомолекулярного белка РНК-полимеразы (L) в геноме PIV5; встроена выше по ходу транскрипции от гена нуклеокапсидного белка (NP) в геноме PIV5; встроена непосредственно ниже по ходу транскрипции от лидерной последовательности в геноме PIV5; встроена между геном белка слияния (F) и геном SH в геноме PIV5; встроена между геном VP и геном матриксного (М) белка в геноме PIV5; встроена между геном М и геном F в геноме PIV5; встроена между геном нуклеокапсидного (NP) белка и геном V/P в геноме PIV5; встроена между лидерной последовательностью и геном нуклеокапсидного белка (NP) в геноме PIV5; встроена туда, где часть гена F или HN PIV5 была заменена на гетерологичную нуклеотидную последовательность; встроена в пределах нуклеотидной последовательности гена SH, встроена в пределах нуклеотидной последовательности гена NP, встроена в пределах нуклеотидной последовательности гена V/P, встроена в пределах нуклеотидной последовательности гена М, встроена в пределах нуклеотидной последовательности гена F, встроена в пределах нуклеотидной последовательности гена HN, и/или встроена в пределах нуклеотидной последовательности гена L.
11. Вектор экспрессии на основе вируса согласно любому из вариантов осуществления 110, где геном PIV5 дополнительно предусматривает наличие одной или нескольких мутаций.
12. Вектор экспрессии на основе вируса согласно варианту осуществления 11, где одна или несколько мутаций предусматривают мутацию гена V/P, мутацию общего N-конца белков V и Р, мутацию остатков 26, 32, 33, 50, 102 и/или 157 общего N конца белков V и Р, мутацию, обеспечивающую отсутствие С конца белка V, мутацию, обеспечивающую отсутствие низкомолекулярного гидрофобного (SH) белка, мутацию белка слияния (F), мутацию фосфопротеина (Р), мутацию высокомолекулярного белка РНК полимеразы (L), мутацию, обеспечивающую встраивание остатков из вируса парагриппа собак, мутацию, индуцирующую апоптоз, или их комбинацию.
13. Вектор экспрессии на основе вируса согласно вариантам осуществления 11 или 12, где одна или несколько мутаций предусматривают PIV5VΔC, PIV5ΔSH, PIV5-Р-S308G или их комбинацию.
14. Вирусная частица, содержащая вектор экспрессии на основе вируса согласно любому из вариантов осуществления 113.
15. Композиция, содержащая вектор экспрессии на основе вируса согласно любому из вариантов осуществления 1-13 и/или вирусную частицу согласно пункту 14.
16. Способ обеспечения экспрессии гетерологичного шиповидного (S) гликопротеина коронавируса в клетке, при этом способ включает приведение клетки в контакт с вектором экспрессии на основе вируса, вирусной частицей или композицией согласно любому из вариантов осуществления 1-15.
17. Способ индуцирования иммунного ответа у субъекта на шиловидный (S) гликопротеин коронавируса, при этом способ включает введение субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции согласно любому из вариантов осуществления 1-15.
18. Способ согласно варианту осуществления 17, где иммунный ответ предусматривает гуморальный иммунный ответ и/или клеточный иммунный ответ.
19. Способ вакцинирования субъекта против коронавирусного заболевания 2019 г. (COVID-19), при этом способ включает введение субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции согласно любому из вариантов осуществления 1-15.
20. Способ согласно любому из вариантов осуществления 17 19, где вектор экспрессии на основе вируса, вирусную частицу или композицию вводят интраназально, внутримышечно, местно или перорально.
Термин "и/или" означает один или все из перечисленных элементов или комбинацию любых двух или более из перечисленных элементов.
Слова "предпочтительный" и "предпочтительно" относятся к вариантам осуществления настоящего изобретения, которые могут предоставлять определенные преимущества в определенных обстоятельствах. Однако другие варианты осуществления также могут быть предпочтительными в тех же или других обстоятельствах. Кроме того, перечисление одного или нескольких предпочтительных вариантов осуществления не означает, что другие варианты не являются применимыми, и не предполагает исключения других вариантов осуществления из объема настоящего изобретения.
Термины "содержит" и его варианты не имеют ограничивающего значения в случаях, когда эти термины встречаются в описании изобретения и формуле изобретения.
Если не указано иное, то формы существительного в единственном числе и выражение "по меньшей мере один" используются взаимозаменяемо и означают один или более чем один.
Если не указано иное, то все числа, отражающие количества компонентов, молекулярные массы и т.д., которые используются в описании и формуле изобретения, следует понимать во всех случаях как модифицированные термином "приблизительно". Соответственно, если в иных случаях не указано противоположное, то числовые параметры, изложенные в описании и формуле изобретения, являются приблизительными значениями, которые могут варьироваться в зависимости от требуемых свойств, к достижению которых стремятся в настоящем изобретении. По крайней мере, и не в качестве попытки ограничить основные принципы эквивалентов объемом формулы изобретения, каждый числовой параметр следует интерпретировать по меньшей мере с учетом количества описанных значимых цифр и путем применения стандартных методик округления.
Несмотря на то, что числовые диапазоны и параметры, представляющие широкий объем настоящего изобретения, являются приблизительными значениями, числовые значения, изложенные в конкретных примерах, представлены настолько точно, насколько это возможно. Все числовые значения, тем не менее, по своему существу включают диапазон, неизбежно обусловленный стандартным отклонением, присутствующим в соответствующих им тестовых измерениях.
Для любого способа, раскрытого в данном документе, который включает отдельные стадии, эти стадии можно осуществлять в любом удобном порядке. И, при необходимости, две или больше стадий в любой комбинации можно осуществлять одновременно.
В описании представлены иллюстративные варианты осуществления. В нескольких местах в настоящей заявке представлено руководство в виде списка примеров, каковые примеры можно применять в различных комбинациях. В каждом случае изложенный список служит только в качестве типичной группы и не должен толковаться как исключающий список.
Все заголовки приведены для удобства читателя и не должны использоваться для ограничения значения текста, который следует после заголовка, если это не указано.
Настоящее изобретение проиллюстрировано следующими примерами. Следует понимать, что конкретные примеры, материалы, количества и процедуры следует толковать в широком смысле в соответствии с объемом и сущностью настоящего изобретения, изложенными в данном документе.
ПРИМЕРЫ
Пример 1
Конструирование CVX-UGA1
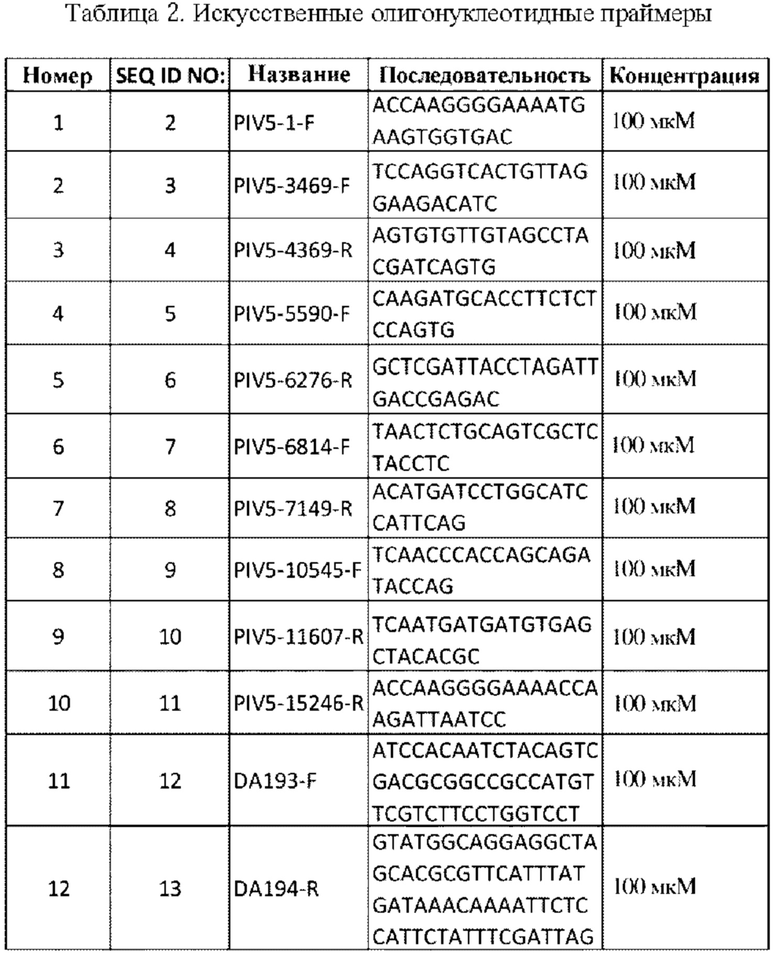
CVX-UGA1 получали из плазмиды pDA27 (pDA27). Плазмида pDA27 содержит ген SARS-CoV-2S из плазмиды nC0V19S human Genscript (многокопийная плазмида из GenScript) с заменой цитоплазматического хвоста на цитоплазматический хвост F PIV5, который был встроен в каркас плазмиды pDA16 (малокопийная плазмида) между генами SH и HN. Последовательность гена S была основана на MN908947 и кодон-оптимизирована для экспрессии у человека. Плазмида pDA16 содержит полный геном вируса PIV5 и ген устойчивости к хлорамфениколу. Последовательность малокопийной плазмиды pDA16 была полностью верифицированной.
Процесс получения pDA27 показан на ФИГ. 1. Ген SARS-CoV-2-S с заменой цитоплазматического хвоста на F хвост PIV5 получали посредством ПЦР из плазмиды nCOV19-S-human-Genscript. Праймеры DA193 F и DA194 R, используемые для реакции ПЦР, имеют области, перекрывающиеся с плазмидой pDA16, а праймеры DA194-R содержат последовательность F-хвоста PIV5. В качестве каркаса использовали плазмида pDA16. Плазмиду расщепляли с помощью рестрикционных ферментов NotI и Mlul для удаления предыдущего гена, встроенного между генами SH и HN PIV5. Плазмида pDA27 была сконструирована посредством сборки методом Гибсона. Продукт ПЦР, представляющий собой SARS-CoV-2-S-Ftail, объединяли с расщепленной pDA16 и мастер-миксом для сборки методом Гибсона и инкубировали при температуре 50°С в течение 1 часа.
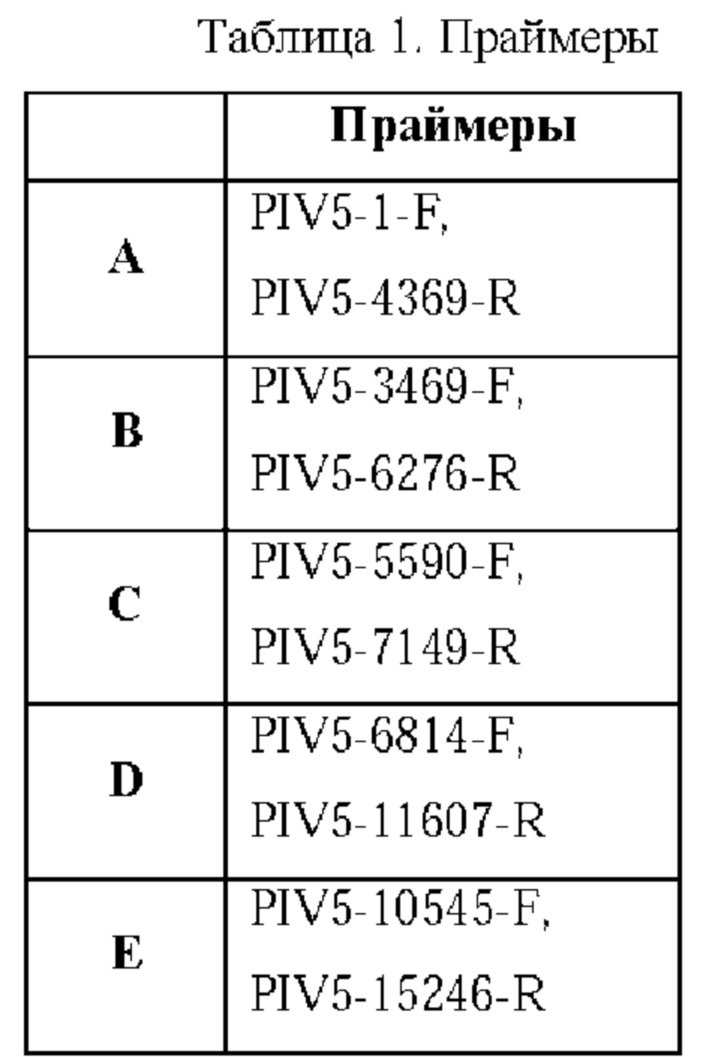
На ФИГ. 2 показано, где S из SARS-CoV-2 был встроен в геном PIV5 в pDA27 (CVX-UGA1). Затем новой плазмидой pDA27 трансформировали компетентные клетки ТОР10, полученные от Thermo Fisher Scientific, и высевали на чашки с агаром LB, содержащим хлорамфеникол. Одну колонию размножали в среде LB, содержащей хлорамфеникол, и плазмиду очищали и секвенировали посредством ПЦР амплификации 5 фрагментов, показанных на ФИГ. 3, с использованием праймеров, представленных в таблице 1 и таблице 2.
Плазмиду отправляли на глубокое секвенирование, где получали полную последовательность плазмиды, а не только части с геномом PIV5(SH-HN)-CoV-2-S-Ftail, и последовательность верифицировали. После подтверждения наличия правильной последовательности осуществляли "спасение" вируса. Данный процесс показан на ФИГ. 5.
Альтернативным способом конструирования вакцины CVX-UGA1 является конструирование плазмиды рАВ76. Эта плазмида содержит точно такой же геном PIV5(SH-HN)-SARS-CoV-2-S, как pDA27, но с многокопийным каркасом (pUC19), содержащим ген устойчивости к ампициллину. Процесс для рАВ76 показан на ФИГ. 4.
Плазмиду рСН10 использовали для получения многокопийного каркаса pUC19. Плазмиду расщепляли рестрикционными ферментами RsrII и AatII.
Плазмиду pDA27 (малокопийную) использовали для получения PIV5(SH-HN)-SARS-CoV-2-S-Ftail. Плазмиду расщепляли с плазмидой pDA27 рестрикционными ферментами RsrII и Aatll.
Продукты расщепления (многокопийный каркас pUC19 и PIV5(SH-HN)-SARS-CoV-2-S-Ftail) лигировали в соответствии с протоколом быстрого лигирования с получением конечной плазмиды.
Любой из процессов, показанных на ФИГ. 1 или ФИГ. 2, можно использовать для получения конечной плазмиды, используемой для получения посевного материала CVX-UGA1. Геномы PIV5(SH-HN)-SARS-CoV-2-S-Ftail в pDA27 и рАВ76 являются идентичными. Единственное различие состоит в том, что pDA27 представляет собой малокопийную плазмиду (устойчивую к хлорамфениколу), а рАВ76 представляет собой многокопийную плазмиду (устойчивую к ампициллину).
Для "спасения" рекомбинантного вектора на основе вируса клетки ВНК 21 (полученные из АТСС) трансфицировали плазмидой pDA27 вместе с плазмидами, кодирующими белки NP, Р, L PIV5 и РНК-полимеразу Т7, что обеспечивало возможность "спасения" рекомбинантного вируса из надосадочной жидкости. Плазмидами для "спасения" являются CAGGS-NP, pCAGGS-P, CAGGS-L и рВН437-Т7. Полная последовательность плазмид для "спасения" была подтверждена перед использованием для "спасения" вирусов. Средами, используемыми для "спасения" вирусов, были среды DMEM (получено от Gibco) с ТРВ (получено от BD), FBS (получено от HyClone) и PenStrep (получено от Lonza).
После семи дней инкубации получали 5 мл надосадочной жидкости, содержащей вектор на основе вируса, которую смешивали с 10Х SPG и хранили при температуре -80°С. Аликвоты этого замороженного маточного раствора подвергали серийному разбавлению для проведения анализа бляшкообразования на клетках Vero (полученных от АТСС) в 6 луночных планшетах с целью получения 6 лунок с одной бляшкой. Наконечник пипетки на 1000 мкл использовали для прокола одной бляшки и ресуспендирования в средах DMEM, содержащих FBS и PenStrep. Затем ресуспендированную бляшку использовали для инфицирования не подвергнутых обработке клеток Vero в чашке диаметром 6 см. Через 7 дней надосадочную жидкость (5 мл) из чашки диаметром 6 см, содержащую продукт одного раунда очистки с использованием бляшкообразования, смешивали с 10Х SPG и хранили при температуре -80°С. Часть надосадочной жидкости (140 мкл) использовали для экстракции РНК и RT-PCR для верификации последовательности. RT-PCR осуществляли с теми же праймерами, что и представленные в таблице 1.
Для "спасения" CVX-UGA1 с получением маточного посевного материала бессывороточные клетки 293Т (полученные из АТСС) трансфицировали плазмидой DA27 или АВ76 вместе с плазмидами, кодирующими белки NP, Р, L PIV5 и РНК-полимеразу Т7, что обеспечивало возможность "спасения" рекомбинантного вируса из надосадочной жидкости. На следующий день трансфицированные бессывороточные клетки 293Т обрабатывают трипсином и совместно культивируют с клетками Vero-SF (бессывороточными) в чашке диаметром 10 см, а затем инкубируют в течение семи дней. После семи дней инкубации получали 5 мл надосадочной жидкости, содержащей вектор на основе вируса, которую смешивали с 10Х SPG и хранили при температуре -80°С. Аликвоты этого замороженного маточного раствора подвергали серийному разбавлению для проведения анализа бляшкообразования на клетках Vero-SF в 6 луночных планшетах с получением 6 лунок с одной бляшкой. Наконечник пипетки на 1000 мкл используется для прокола одной бляшки и ресуспендирования в средах VP-SFM. Затем ресуспендированные бляшки используются для инфицирования не подвергнутых обработке клеток Vero-SF в чашке диаметром 6 см. Через 7 дней надосадочную жидкость (5 мл) из чашки диаметром 6 см, содержащую продукт одного раунда очистки с использованием бляшкообразования, смешивали с 10Х SPG и хранили при температуре 80°С. Аликвоты из начального цикла очистки с использованием бляшкообразования используются в качестве инокулята, представляющего собой посевной материал, для изготовления вакцины в виде предшествующего посевного материала для получения маточного посевного материала в соответствующей требованиям GMP системе с использованием проверенных полученных в соответствии с требованиями GMP банков клеток Vero-SF в бессывороточных средах (среды VP-SFM).
"Спасение" вируса CVX-UGA1 может осуществляться с помощью либо pDA27, либо рАВ76, поскольку они имеют точный геном PIV5(SH-HN)-SARS-CoV-2-S-Ftail. Единственное различие состоит в том, что pDA27 является малокопийной плазмидой, а рАВ76 является многокопийной плазмидой.
Плазмиды, используемые в будущем "спасении" для получения CVX-UGA1, будут получать с использованием продуктов, не содержащих компонентов животного происхождения. Цель состоит в том, чтобы получить CVX-UGA1 без продуктов животного происхождения, за исключением клеток (бессывороточных 293Т и бессывороточных Vero, и без продуктов животного происхождения).
Продуцирование SARS-CoV-2-3-белка конструкцией в составе посевного материала подтверждалось посредством иммунофлуоресцентного анализа. Это показано на ФИГ. 6. Первичное антитело: моноклональное антитела кролика к шиловидному белку S1 SARS-CoV-2 от Sino Biological, и вторичное антитело: антитело козы к IgG кролика (H+L), меченое Су3, от KPL.
Выявление S SARS-CoV-2 в CVXUGA1 инфицированных клетках MDBK посредством иммунофлуоресцентного анализа. S SARS-CoV-2 был выявлен с использованием моноклонального антитела к шиповидному белку S1 SARS-CoV-2, а затем антитела козы к IgG кролика, меченого Су3, в качестве вторичного. Инфицированные клетки также метили антителом к PIV5-P/V (РК) в качестве контроля, а затем вторичным антителом, представляющим собой антитело козы к IgG мыши, H&L (FITC). DAPI (4,6-диамидино-2-фенилиндол; синий) использовался в качестве ядерного красителя. Для будущих экспериментов вместо клеток MDBK будут использоваться клетки Vero-SF.
Карта плазмиды pDA16 показана на ФИГ. 7. Эта плазмида содержит геном PIV5 плюс ген PCDV1. Она является малокопийной плазмидой с геном устойчивости к хлор амфе николу.
Карта плазмиды nCOV19-S-human-Genscript показана на ФИГ. 8. Это многокопийная плазмида и она содержит ген устойчивости к ампициллину. Она была получена от GenScript.
Карта и последовательность плазмиды pDA27 (CVX-UGA1) показаны на ФИГ. 9. Эта плазмида представляет собой малокопийную плазмиду и она содержит ген устойчивости к хлорамфениколу.
Карта плазмиды pCAGGS-NP показана на ФИГ. 10. Эта плазмида представляет собой многокопийную плазмиду и она содержит ген устойчивости к ампициллину. Последовательность ДНК NP PIV5 из плазмиды рВН276 субклонировали в плазмиду для экспрессии pCAGGS. Более подробно см. в работе Schmitt et al., 2002, J Virol 76, 3952 3964,doi:10.1128/jvi.76.8.3952-3964.2002.
Карта плазмиды pCAGGS P показана на ФИГ. 11. Эта плазмида представляет собой многокопийную плазмиду и она содержит ген устойчивости к ампициллину. Последовательность ДНК Р PIV5 из плазмиды рВН276 субклонировали в плазмиду для экспрессии pCAGGS. Более подробно см. в работе Waning et al., 2002, J Virol; 76:9284-9297,doi:10.1128/jvi.76.18.9284-9297.2002.
Карта плазмиды pCAGGS L показана на ФИГ. 12. Эта плазмида представляет собой многокопийную плазмиду и она содержит ген устойчивости к ампициллину. Последовательность ДНК L PIV5 из плазмиды рВН276 субклонировали в плазмиду для экспрессии pCAGGS. Более подробно см. в работе Waning et al., 2002, J Virol; 76:9284-9297,doi:10.1128/jvi.76.18.9284-9297.2002.
Карта плазмиды pBH437-pCAGGS-T7 показана на ФИГ. 13. Эта плазмида представляет собой многокопийную плазмиду и она содержит ген устойчивости к ампициллину. Последовательность ДНК полимеразы Т7 из плазмиды рВН161 субклонировали в плазмиду для экспрессии pCAGGS.
Карта плазмиды рВН276 показана на ФИГ. 14. Эта плазмида содержит геном PIV5. Она является многокопийной плазмидой с геном устойчивости к ампициллину. Более подробно см. в работе Не et al., 1997, Virology; 237:249-260, doi:10.1006/viro.l997.8801.
Карта плазмиды рВН161 показана на ФИГ. 15. Эта плазмида представляет собой многокопийную плазмиду и содержит ген устойчивости к ампициллину. Более подробно см. в работе Не et al., 1997, Protein Expression and Purification; 9:142-151, doi:10.1006/prep. 1996.0663.
Карта плазмиды рСН10 показана на ФИГ. 16. Эта плазмида содержит геном PIV5. Она является многокопийной плазмидой с геном устойчивости к ампициллину.
Карта плазмиды рАВ76 показана на ФИГ. 17. Эта плазмида представляет собой многокопийную плазмиду и она содержит ген устойчивости к ампициллину.
Протоколы
Процедура субкультивирования для Vero-SF
Объемы, используемые в данном протоколе, предназначены для колбы объемом 75 см2.
Удаляют и отбрасывают культуральную среду с использованием пастеровской пипетки и вакуумного насоса.
В течение короткого времени промывают клеточный слой с помощью 10 мл фосфатно солевого буфера Дульбекко (DPBS) без Ca++/Mg++.
Добавляют в колбу 5 мл TrypLE. Инкубируют при температуре 37°С до отделения клеток. Осуществляют визуальный контроль клеточного монослоя с помощью инвертированного микроскопа, чтобы убедиться в полном отделении клеток от поверхности колбы.
Во избежание образования скоплений в ходе ожидания отделения клеток не осуществляют перемешивания клеток путем постукивания по колбе или ее встряхивания. Клетки, которые трудно отделить, могут быть помещены в условия с температурой 37°С для облегчения диспергирования.
Добавляют в колбу 10 мл предварительно нагретой среды VP-SFM и аспирируют клетки, осторожно пипетируя с помощью серологической пипетки объемом 10 мл. Наклоняют колбу во всех направлениях, чтобы тщательно промыть колбу.
Переносят клеточную суспензию в коническую пробирку объемом 15 мл и откручивают при 125 g в течение 10 минут.
Удаляют надосадочную жидкость и ресуспендируют клетки в свежей VP-SFM. Добавляют соответствующие аликвоты клеточной суспензии в новые сосуды для культивирования.
Инкубируют культуры при температуре 37°С.
Лучше всего клетки растут после первого субкультивирования.
Коэффициент субкультивирования: разбавление 1:4.
Обновление среды: два раза в неделю.
Процедура субкультивирования для 293Т-SF
Объемы, используемые в данном протоколе, предназначены для колбы объемом 75 см2.
Удаляют и отбрасывают культуральную среду с использованием пастеровской пипетки и вакуумного насоса.
В течение короткого времени промывают клеточный слой с помощью 10 мл фосфатно солевого буфера Дульбекко (DPBS) без Ca++/Mg++.
Добавляют в колбу 10 мл предварительно нагретой среды для HEK293 BalanCD и ресуспендируют клетки, осторожно пипетируя с помощью серологической пипетки объемом 10 мл.
Переносят клеточную суспензию в коническую пробирку объемом 15 мл и откручивают при 125 g в течение 10 минут.
Отбрасывают надосадочную жидкость и ресуспендируют клетки в свежей среде для HEK293 BalanCD. Добавляют соответствующие аликвоты клеточной суспензии в новые сосуды для культивирования.
Инкубируют культуры при температуре 37°С.
Лучше всего клетки растут после первого субкультивирования.
Коэффициент субкультивирования: разбавление 1:4.
Обновление среды: два раза в неделю.
Процедура субкультивирования клеток ВНК 21 и Vero
Объемы, используемые в данном протоколе, предназначены для колбы объемом 75 см2.
Удаляют и отбрасывают культуральную среду с использованием пастеровской пипетки и вакуумного насоса (IV-18).
В течение короткого времени промывают клеточный слой с помощью 10 мл фосфатно-солевого буфера Дульбекко (DPBS) без Ca++/Mg++.
Добавляют в колбу 5 мл трипсина/EDTA. Наклоняют колбу, чтобы покрыть поверхность, и удаляют излишек.
Инкубируют при температуре 37°С до отделения клеток. Осуществляют визуальный контроль клеточного монослоя с помощью инвертированного микроскопа, чтобы убедиться в полном отделении клеток от поверхности колбы.
Во избежание образования скоплений в ходе ожидания отделения клеток не осуществляют перемешивания клеток путем постукивания по колбе или ее встряхивания. Клетки, которые трудно отделить, могут быть помещены в условия с температурой 37°С для облегчения диспергирования.
Добавляют в колбу 20 мл предварительно нагретой соответствующей культуральной среды и ресуспендируют клетки с использованием серологической пипетки объемом 10 мл. Наклоняют колбу во всех направлениях, чтобы тщательно промыть колбу. Добавляют соответствующие аликвоты клеточной суспензии в новые сосуды для культивирования.
Инкубируют культуры при температуре 37°С.
Культуральной средой для ВНК является среда DMEM+TPB+FBS+PenStrep, а для Vero - среда DMEM+FBS+PenStrep.
"Спасение" PIV5 (трансфекция)
За 1 день до этого:
высевают клетки ВНК-21 в чашке для культур тканей диаметром 60 мм при разбавлении 1:8 (конфлюентность 75% на следующий день).
В данный день:
смешивают плазмидную ДНК, pCAGGs NP, pCAGGs Р, pCAGGs-L и pBH437 (pCAGGS-T7) в микроцентрифужной пробирке объемом 1,6 мл в боксе биологической безопасности.
Между добавлениями осуществляют перемешивание путем пипетирования.
Добавляют буфер JetPRIME и реагент для трансфекции JetPRIME.
Инкубируют при комнатной температуре в течение 15 минут.
Добавляют по каплям смесь для трансфекции к клеткам.
Осуществляют мониторинг клеток в течение следующих 7 дней в отношении образования синцитиев с использованием EVOS.
Сразу после обнаружения синцитиев начинают собирать 1 мл среды в криопробирку и смешивают с 10Х SPG.
Хранят при температуре -80°С.
Совместное культивирование для "спасения" PIV5 (трансфекция).
За 1 день до этого:
высевают клетки 293Т SF в 6 луночной чашке для культур тканей при разбавлении 1:6 (конфлюентность приблизительно 75%).
Высевают клетки Vero-SF в чашке диаметром 10 мм для культур тканей при разбавлении 1:20 (конфлюентность приблизительно 50%).
В данный день:
смешивают плазмидную ДНК, pCAGGs NP, pCAGGs Р, pCAGGs-L и pBH437 (pCAGGs-T7) в микроцентрифужной пробирке объемом 1,6 мл, содержащей бессывороточную Opti-MEM, в боксе биологической безопасности.
Между добавлениями осуществляют перемешивание путем пипетирования.
Добавляют реагент Р3000.
В другую микроцентрифужную пробирку объемом 1,6 мл, содержащую бессывороточную Opti-MEM, добавляют реагент для трансфекции Lipofectamine3000.
Инкубируют при комнатной температуре в течение 20 минут.
Добавляют по каплям смесь для трансфекции к клеткам. На следующий день:
293T-SF трипсинизируют, хорошо ресуспендируют в 1 мл VP-SFM, перемещают в отдельную коническую пробирку объемом 15 мл.
Трипсинизируют 10 см Vero-SF, ресуспендируют в 3 мл VP-SFM, перемещают в коническую пробирку объемом 15 мл.
Добавляют 6 мл VP-SFM в коническую пробирку объемом 15 мл.
Хорошо перемешивают и добавляют полные 10 мл из конической пробирки объемом 15 мл в новую чашку диаметром 10 см.
Осуществляют мониторинг клеток в течение следующих 7 дней в отношении образования синцитиев с использованием EVOS.
Сразу после обнаружения синцитиев начинают собирать 1 мл среды в криопробирку и смешивают с 10Х SPG.
Хранят при температуре -80°С.
Очистка вируса с использованием бляшкообразования
Берут 1 мл замороженных аликвот для "спасения" и проводят анализ бляшкообразования.
Через 7 дней протыкают отдельную бляшку с использованием наконечника пипетки объемом 1000 мкл и добавляют в чашку диаметром 6 см с клетками Vero-SF, высеянными с обеспечением 90% конфлюентности за день до этого, следуя протоколу в приложении IV-1. Используют среду VP-SFM.
В день 7 отбирают образец в виде 140 мкл среды и используют его для выделения
РНК с использованием мини-набора для выделения вирусной РНК QIAamp Viral RNA Mini Kit.
Собирают остаток среды (4,8 мл) в криопробирку и смешивают с 10Х SPG. Хранят при температуре -80°С.
Анализ бляшкообразования для PIV5
Все стадии выполняют в боксе биологической безопасности.
За 1 день до этого:
высевают клетки Vero-SF в 6 луночный планшет для культивирования в разбавлении 1:3 (конфлюентность 95% на следующий день).
В данный день:
размораживают вирус PIV5 при температуре 37°С в инкубаторе и хранят на льду после оттаивания.
Пока вирус оттаивает, готовят 10-кратные разбавления в 96-луночном планшете, начиная с неразбавленного образца и заканчивая разбавлением 10-5 с использованием многоканальной пипетки, и помечают 6-луночные планшеты названием вируса, номером пассажа, датой сбора и коэффициентом разбавления следующим образом.
Добавляют 200 мкл неразбавленного образца в первую лунку.
Добавляют 180 мкл инфекционной среды в лунки 2-6.
Берут 20 мкл из первой лунки, переносят во вторую лунку и тщательно перемешивают с использованием функции пипетирования + смешивания в многоканальной пипетке. Отбрасывают наконечники.
Повторяют предыдущую стадию еще 5 раз.
Путем аспирации отделяют среды от клеток с использованием пастеровской пипетки и вакуумного насоса. Добавляют 900 мкл среды VP-SFM в каждую лунку 6-луночного планшета для культивирования.
Инфицируют 6-луночный планшет для культивирования с помощью 100 мкл разбавления вируса на лунку. Используют один и тот же наконечник, переходя от наименее к наиболее концентрированному раствору, и инкубируют в течение 1 часа.
Аспирируют среды через 1 час после инфицирования.
Добавляют 4 мл покровного слоя в каждую лунку. Покровный слой: для 1 6-луночного планшета для культивирования смешивают 13 мл 2% агара LMP и 13 мл 2Х сред VP-SFM.
Инкубируют при температуре 37°С в течение 7 дней.
Выращивание вируса PIV5.
За 1 день до этого:
высевают клетки Vero-SF в колбу с площадью роста клеток 175 см2 в разбавлении 1:3 (конфлюентность 95% на следующий день).
В данный день:
получают инокулят. Вирус при MOI = 0,01 в 20 мл сред VP-SFM.
Путем аспирации отделяют поддерживающие среды от клеток с использованием пастеровской пипетки и вакуумного насоса.
Инфицируют клетки Vero-SF в течение 2 часов при температуре 37°С.
Взбалтывают планшет каждые 15 минут.
Аспирируют инокулят и добавляют 40 мл сред VP-SFM.
Инкубируют при температуре 37°С в течение 7 дней. День 7:
собранные среды помещают в коническую пробирку объемом 50 мл и центрифугируют при 1500 об./мин. в течение 10 минут при температуре 4°С для удаления клеточного дебриса.
Переносят 38 мл надосадочной жидкости в новую коническую пробирку объемом 50 мл.
Добавляют 3,8 мл 10Х SPG и получают аликвоты по 1 мл в криопробирках. Обозначьте пробирки следующим образом:
название вируса, № бляшки;
пассаж клеток;
дата, инициалы.
Хранят аликвоты вируса при температуре -80°С.
Пример 2
Интраназальная иммунизация посредством однократной дозы вакцины COVID-19 на основе вируса парагриппа 5 создает стерильный иммунитет в носовых полостях хорьков и кошек.
SARS-CoV-2 представляет собой новый бета коронавирус и является причиной пандемии COVID-19. Срочно необходима вакцина для защиты от инфекции SARS-CoV-2, чтобы уменьшить распространение и ограничить дальнейшую смертность. Многочисленные кандидатные вакцины оценивают для определения эффективных средств борьбы с пандемией. Верхние дыхательные пути являются первоначальным участком инфекции SARS-CoV-2 и для многих инфицированных индивидуумов остаются основным участком репликации вируса. Высокие вирусные нагрузки и выделение в среду из этой области могут начаться за несколько дней до появления симптомов и могут продолжаться в течение нескольких дней после начала заболевания. Осуществление контроля инфекции в этих участках имеет решающее значение для борьбы с пандемией. На сегодняшний день ни одна вакцина не обеспечила стерильного иммунитета в верхних дыхательных путях, например в носовой полости крупных животных моделей. Данный пример оптимизировал кандидатную вакцину в мышиных моделях и впоследствии демонстрирует, что интраназальная иммунизация однократной дозой вируса парагриппа 5 (PIV5), экспрессирующего S-белок SARS-CoV-2, индуцирует стерильный иммунитету хорьков и кошек. Эта стратегия вакцинации через слизистые оболочки ингибировала репликацию SARS-CoV-2 в верхних дыхательных путях, предупреждая тем самым распространение инфекции в нижние дыхательные пути. Что наиболее важно, кандидатная вакцина, которая индуцирует стерильный иммунитет в верхних дыхательных путях, вероятно ограничит передачу, снижая тем самым количество инфекций SARS-CoV-2 в популяциях.
Синоназальный эпителий верхних дыхательных путей является начальным участком инфекции SARS-CoV-2 и для многих индивидуумов остается основным участком репликации вируса. Высокие уровни репликации вируса наблюдаются в верхних дыхательных путях (Wolfel et al. Nature 581, 465-469 (2020)), выделение из носоглотки в среду может продолжаться в течение нескольких дней после первоначального проявления (Li et al. Journal of Medical Virology 92, 2286-2287 (2020)). Прогрессирующие проявления пневмонии в нижних дыхательных путях, острый респираторный дистресс-синдром (ARDS) и дыхательная недостаточность вносят большой вклад в заболеваемость и смертность вследствие COVID-19, и появляется все больше свидетельств поражения других систем органов, помимо легких. Поскольку верхние и нижние дыхательные пути являются критически важными участками патогенеза COVID-19, то прекращение репликации вируса в верхних дыхательных путях до его распространения в легкие является важной целью защитной вакцины. Ведущие вакцины на сегодняшний день обеспечивали развитие устойчивых ответов в виде выработки антител у иммунизированных животных и в клинических испытаниях с участием людей, а также снижали репликацию SARS-CoV-2 в нижних дыхательных путях в моделях на приматах, отличных от человека (Corbett et al. N Engl J Med 383, 1544-1555 (2020); Gao et al. Science 369, 77 81 (2020); Mercado et A. Nature, (2020); van Doremalen et al. bioRxiv, (2020); Wang et al. Cell 182, 713-721 e719 (2020)). Однако сложно было добиться полной защиты верхних дыхательных путей, что важно для предупреждения передачи SARS-CoV-2. В настоящее время нет сообщений о том, могут ли эти кандидатные вакцины блокировать передачу.
Вирус парагриппа типа 5 (PIV5) представляет собой РНК, содержащий вирус с отрицательной цепью из семейства Paramyxoviridae, который оценивался в качестве вакцинного вектора для гриппа, респираторно синцитиального вируса (RSV), бешенства и различных других патогенов (Chen et al. J Virol 87, 2986-2993 (2013); Li et al. mBio 11, (2020); Mooney et al. J Virol 87, 363-371 (2013); Phan et al. Vaccine 32, 3050-3057 (2014); Phan et al. J Virol 91, (2017)). В животных моделях PIV5 является безопасным и не ассоциирован с каким либо заболеванием, за исключением питомникового кашля у собак (Cornwell etal. Vet Rec 98, 301-302 (1976)). Интраназально вводимые вакцины против питомникового кашля, содержащие живой PIV5, используются в течение более четырех десятилетий с превосходными показателями безопасности. Собаки, иммунизированные вакцинами против питомникового кашля, могут выделять PIV5 в среду в течение 5 дней, и это было безопасным для людей при тесном контакте с иммунизированными животными (Kontor et al. American journal of veterinary research 42, 1694-1698 (1981); Chen et al. PLoS One 7, e50144 (2012)). PIV5 особенно хорошо подходит в качестве вакцинного вектора для респираторных заболеваний, поскольку при интраназальном введении он вызывает локальные защитные ответы в виде IgA в дыхательных путях, а также системные врожденные и адаптивные иммунные ответы (Wang et al. J Virol 91, (2017)). Недавно однократная интраназальная доза вакцины MERS на основе PIV5 индуцировала нейтрализующие антитела и клеточные иммунные ответы и обеспечивала 100% защиту против заражения летальной дозой MERS-CoV в мышиной модели MERS (Li et al. mBio 11, (2020)). Данный пример предусматривает многообещающую кандидатную вакцину для защиты против SARS-CoV-2.
С использованием стратегии, аналогичной той, которая использовалась при разработке вакцины против MERS CoV на основе PIV5, был создан PIV5, обеспечивающий экспрессию шиловидного (S) белка SARS-CoV-2 (названный CVXGA1) (ФИГ. 18А), и при этом экспрессия его гликопротеина S подтвердилась посредством иммунофлуоресценции и вестерн-блоттинга (ФИГ. 18В). Кроме полноразмерного S в клетках, инфицированных CVXGA1, также был выявлен продукт расщепления, S1, что указывает на то, что S подвергался процессингу (ФИГ. 18В). S способствует слиянию клетки с клеткой и вируса с клеткой для облегчения проникновения вируса, а иммунизация посредством S создает защитный иммунитет против SARS-CoV-2. Считается, что нативная (до слияния) конформация S индуцирует требуемый иммунитет для оптимальной защиты. Чтобы проверить, является ли S, экспрессируемый CVXGA1, функциональным, клетки Vero, которые экспрессируют рецептор АСЕ2, инфицировали CVXGA1. Образование синцитиев (слияние клетки с клеткой) наблюдалось только в клетках, инфицированных CVXGA1, что указывает на то, что S-белок, экспрессируемый CVXGA1, является функциональным (ФИГ. 18С). Для исследования иммуногенности CVXGA1 иммунизированных мышей интраназально иммунизировали различными дозами CVXGA1. Дозозависимое повышение содержания антител к S было выявлено у мышей BALB/c после однократного интраназального введения (ФИГ. 18D). Аналогично, антитела, распознающие рецепторсвязывающий домен (RBD) S, выявлялись дозозависимым образом у мышей BALB/c, иммунизированных с помощью CVXGA1 (ФИГ. 18Е). Кроме того, нейтрализующие антитела к SARS-CoV-2 были выявлены у мышей BALB/c, иммунизированных дозозависимым образом (ФИГ. 18F). Значительное образование антител и клеточные ответы наблюдались у мышей C57BL/6 после однократной интраназальной (IN) иммунизации дозой, составляющей всего лишь 6×103 бляшкообразующих единиц (БОЕ) (ФИГ. 22). Эти результаты свидетельствуют о том, что иммунизация однократной интраназальной дозой CVXGA1 обеспечивала развитие устойчивого иммунного ответа у мышей.
Для определения эффективности CVXGA1 использовали три взаимодополняющие животные модели. У мышей BALB/c отсутствует функциональный рецептор АСЕ2 для SARS-CoV-2 и поэтому они устойчивы к инфекции. Введение человеческого АСЕ2 путем трансдукции аденовирусного вектора (Ad5-hACE2) обеспечивало сенсибилизацию мышей к инфекции, обусловленной SARS-CoV-2, что приводило к временной легочной инфекции. Эту мышиную модель использовали для оценки эффективности CVXGA1. Интраназальная вакцинация с помощью CVXGA1 защищала мышей от потери веса (ФИГ. 23), а также снижала титры в легочной ткани и защищала ткани от гистопатологических изменений. Отсутствие полной защиты у мышей может быть связано с более низкими дозами (наиболее высокая на уровне 5×104 БОЕ на мышь), используемыми в этих экспериментах. В качестве альтернативы возможно, что трансдукция Ad5-hACE2 может являться фактором, способствующим отсутствию стерильного иммунитета в этой модели. В то время как иммунизация посредством инактивированного УФ-излучением SARS-CoV-2 приводила к выработке более высоких титров антител к S, нейтрализующих антител выявлено не было (ФИГ. 24), а иммунизация инактивированным SARS-CoV-2 не защищала мышей от заражения с помощью SARS-CoV-2 (ФИГ. 19В). Хотя CVXGA1 обеспечивал выработку более низких титров антител к S, чем инактивированный SARS-CoV-2, он защищал мышей лучше, чем инактивированный SARS-CoV-2, что позволяет предположить, что клеточный иммунитет, создаваемый CVXGA1, вероятно сыграл роль в данном исходе. Это согласуется с отсутствием стерильного иммунитета в легких мышей, иммунизированных с помощью CVXGA1, сенсибилизированных посредством Ad5-ACE2 и зараженных SARS-CoV-2.
Ранее на мышах сравнивали вакцину MERS на основе PIV5 с инактивированной вакциной MERS-CoV, и было обнаружено, что иммунизация инактивированной вакциной MERS-CoV вызывает иммунный ответ по типу гиперчувствительности, о чем свидетельствует миграция эозинофилов в легкие после заражения вирусом. Подобно этому наблюдению, иммунизация посредством инактивированного УФ излучением SARS-CoV-2 была ассоциирована с эозинофильными инфильтратами, в то время как у мышей, иммунизированных с помощью CVXGA1, такие инфильтраты отсутствовали после заражения, что свидетельствует о том, что CVXGA1 не вызывал ответа по типу гипер чувствительно ста (ФИГ. 19С, D). Напротив, все группы обработки характеризовались одинаковыми степенями инфильтрации мононуклеарными клетками (ФИГ. 19С).
Хорьки являются широко используемой моделью респираторных инфекций человека, они восприимчивы к инфекции SARS-CoV-2 и могут передавать вирус другим животным при непосредственном контакте и воздушно-капельным путем (Kim et al. Cell Host Microbe, (2020); Shi et al. Science, (2020); Richard et al. Nat Commun 11, 3496 (2020)). Для исследования эффективности CVXGA1 у хорьков животных интраназально иммунизировали с помощью PBS или CVXGA1 (ФИГ. 20А). Иммунизация с помощью CVXGA1 обеспечивала развитие устойчивых гуморальных ответов, о чем свидетельствуют высокие титры IgG к S (ФИГ. 25А), IgG к RBD (ФИГ. 20 В) и нейтрализующих антител на ФИГ. 20С. Низкие уровни IgA к S были выявлены в назальных смывах (ФИГ. 25В). При интраназальном заражении с помощью SARS-CoV-2 вирусная РНК не выявлялась в назальных смывах хорьков, иммунизированных с помощью CVXGA1 (ФИГ. 20D). Напротив, вирусная РНК была выявлена у всех хорьков, подвергнутых имитационной иммунизации (ФИГ. 20D и таблицу 3). Чтобы сравнить выявление вирусной РНК с анализом инфекционных фокусобразующих единиц, инфекционный вирус в назальных смывах через 5 дней после заражения (dpc) определяли с использованием анализа инфекционных фокусобразующих единиц.
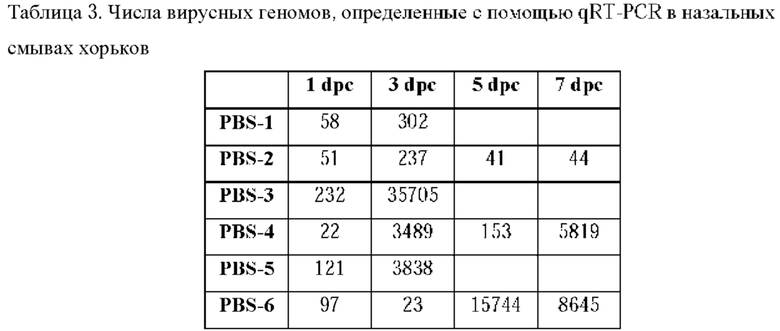
Выявление вирусного РНК генома посредством qRT PCR было сопоставимо с анализом инфекционных фокусобразующих единиц (ФИГ. 25С). N-белок SARS-CoV-2 был выявлен в дыхательных путях и паренхиме хорьков, иммунизированных с помощью PBS и инфицированных SARS-CoV-2, через 4 dpc (ФИГ. 25D). Вирусная РНК также была выявлена в трахее и легких хорьков, подвергнутых имитационной иммунизации, но не в трахее и легких хорьков, иммунизированных с помощью CVXGA1 (ФИГ. 20Е и 20F). Для оценки уровня воздействия SARS-CoV-2 у хорьков уровни антител к S-антител определяли количественно во время окончания эксперимента (7 dpc). Уровни антител к S в группе, иммунизированной с помощью CVXGA1, увеличились после заражения (ФИГ. 25Е), что свидетельствует об анамнестическом характере ответа и подверганию воздействию антигена S. Эти результаты свидетельствуют о том, что иммунизация с помощью CVXGA1 индуцировала стерильный иммунитет после заражения с помощью SARS-CoV-2, что является наиболее важным, в носовой полости, - критически важном участке для репликации и передачи SARS-CoV-2.
Кошки от природы восприимчивы к инфекции SARS-CoV-2 и в лабораторных условиях могут передавать вирус.Чтобы определить, блокирует ли иммунизация с помощью CVXGA1 инфекцию верхних дыхательных путей, кошек иммунизировали с помощью CVXGA1 интраназально (IN) и подкожно (subQ) без вспомогательного вещества (ФИГ. 21А). Для усиления иммунных ответов группе, иммунизированной посредством subQ, осуществляли бустерную иммунизацию (ФИГ. 21А). Как subQ (в режиме "прайм-буст"), так и IN иммунизация обеспечивали развитие аналогичных ответо] в виде выработки антител (ФИГ. 21В, и 21С, и ФИГ. 26А). Следует отметить, что IN иммунизация обеспечивала выработку поддающегося выявлению IgA к S в назальных смывах (ФИГ. 21D). После заражения все кошки в группе с имитацией иммунизации (PBS) характеризовались высокими титрами SARS-CoV-2 в назальных смывах (в среднем свыше 6,27 × 104 FFU/мл) через 1 день после заражения. Особенно интересно то, что в группе IN иммунизации у двух из четырех кошек (50%) вирус не выявлялся, а две из четырех характеризовались 230-кратным снижением титров SARS-CoV-2 (в среднем 2,7×102 FFU/мл). Интересно, что в то время как subQ иммунизация обеспечивала титры сывороточных антител, подобные таковым при IN иммунизации, она не защищала от инфекции носовых полостей через 1 dpc (6,16×104FFU/мл). Тем не менее, у кошек, иммунизированных subQ, инфекция проходила быстрее, чем у кошек, подвергнутых имитационной иммунизации: к 5 dpc у 3 из 4 кошек вирус не был выявлен, в то время как все животные, которых подвергали имитационной иммунизации, оставались инфицированными (ФИГ. 21Е). К 7 dpc в назальных смывах у всех кошек вирус выявлен не был. Эти результаты были подтверждены с использованием qRT-PCR для вирусной РНК (ФИГ. 26 В). Примечательным и согласующимся с предыдущими сообщениями является то, что хотя живой вирус не был выявлен в назальных смывах через 7 dpc, вирусные геномы выявляли до 11 dpc в группе PBS.
Для определения подверженности животного воздействию репликации SARS-CoV-2 в конце эксперимента (14 dpc) измеряли уровни антител к N. Антитела к N были выявлены у всех кошек через 14 dpc; кошки, иммунизированные посредством PBS и subQ, характеризовались более высокими уровнями антител к N, чем животные, иммунизированные IN (ФИГ. 26С). В соответствии с уровнем воздействия SARS-CoV-2, IN иммунизированные животные характеризовались повышением титров антител к S после заражения (ФИГ. 26D). Вывод о том, что subQ иммунизация обеспечивала развитие ответов в виде выработки антител к S, сравнимых с таковыми при IN иммунизации, но не предупреждала инфекцию в носовых полостях, согласуется с наблюдениями того, что ответы в виде высокого содержания антител в сыворотке крови не предупреждают инфекцию носовых полостей. При оценке иммунных ответов отдельных животных не наблюдалось корреляции между титрами сывороточных антител и защитой носовых полостей, что еще раз подтверждает то, что титры антител к S не являются прогностическим фактором защиты с ино назально го тракта (таблица 4).
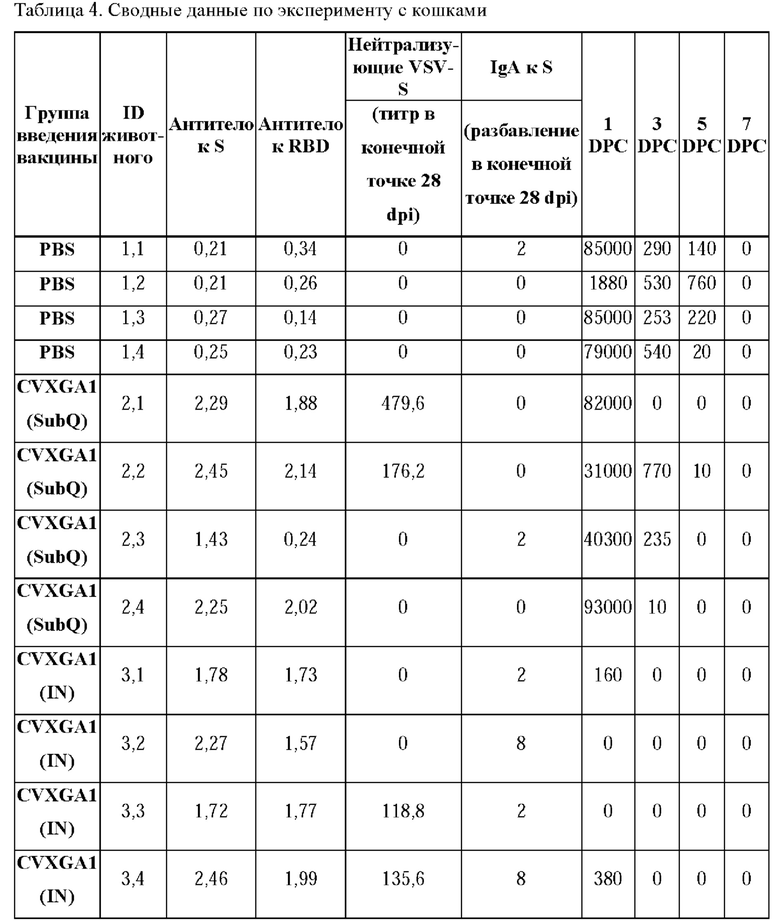
Эта работа демонстрирует, что интраназальная вакцина на основе PIV5 против COVID-19 может создавать стерильный иммунитет у животных. Дальнейшая разработка этой кандидатной вакцины может обеспечить вакцину против COVID-19, которая могла бы блокировать передачу SARS-CoV-2. Недорогостоящее производство вакцин против питомникового кашля существовало на протяжении десятилетий. Безопасная, эффективная и недорогая вакцина на основе вектора PIV5 против COVID-19 может способствовать борьбе с пандемией COVID-19.
МАТЕРИАЛЫ И СПОСОБЫ
Вирус и клетки
Плазмида CVXGA1, кодирующая полноразмерный геном PIV5 с SARS-CoV-2-S, цитоплазматический хвост которого был заменен на хвост F из PIV5, и который был встроен между SH и HN PIV5. "Спасение" вируса осуществляли, как описано ранее (Li et al., 2013, J Virol; 87:354-362, doi:10.1128/JVI.02321-12). Вкратце, плазмидой CVXGA1, a также четырьмя хелперными плазмидами, pPIVS-NP, pPIV5 P, pPIV5 L и плазмидой pT7 полимер азы, кодирующими белки NP, Р, L и РНК полимеразу Т7 соответственно, совместно трансфицировали клетки BHK21 при конфлюентности 90% в плашках диаметром 6 см с липофектамином 3000 (Invitrogen). Вирус, выпущенный в среды, амплифицировали в клетках Vero. На восстановление вируса указывает образование синцитиев в клетках Vero. Затем вирус очищали посредством отбора бляшки в виде отдельной бляшки из клеток Vero. Полноразмерные геномы клонированного методом бляшкообразования одиночного клона вируса CVXGA1 секвенировали, как описано (Li et al., 2013, J Virol; 87:354-362, doi: 10.1128/JVI.02321-12). Вирусы выращивали в клетках Vero в течение 5-7 дней с использованием DMEM, содержащей 2% FBS. Среды собирали и осаждали при 3000 об./мин. для удаления клеточного дебриса с использованием настольной центрифуги Sorvall в течение 10 мин. Вирусную надосадочную жидкость дополняли 10% сахарозо-фосфатно-глутаматным (SPG) буфером, быстро замораживали в жидком азоте и хранили при температуре 80°С немедленно после сбора.
Надосадочные жидкости вируса SARS-CoV-2 или гомогената ткани подвергали серийному разбавлению в DMEM. Инокулировали 12-луночные планшеты с клетками VeroE6 при температуре 37°С в 5% СО2 в течение 1 ч. и осторожно встряхивали каждые 15 мин. После удаления инокулята планшеты покрывали 1,2% агарозой, содержащей 4% FBS. Через три дня покровные слои удаляли и бляшки визуализировали с использованием окрашивания 0,1% кристаллическим фиолетовым. Вирусные титры количественно определяли в виде БОЕ/мл ткани. SARS-CoV-2 был инактивирован посредством воздействия УФ-излучения в течение 1 ч. При использовании мощности 4,016 мкВт/см2.
Все клеточные линии инкубировали при температуре 37°С в 5% СО2. Клетки Vero выращивали в среде Игла, модифицированной по Дульбекко (DMEM), с добавлением 5% эмбриональной бычьей сыворотки (FBS), 100 МЕ/мл пенициллина и 100 мкг/мл стрептомицина. Клетки ВНК21 выращивали в DMEM, содержащей 10% триптозофосфатного бульона (ТРВ), 5% эмбриональной бычьей сыворотки (FBS), 100 МЕ/мл пенициллина и 100 мкг/мл стрептомицина.
Иммунофлуоресценция и иммуноблоттинг
Иммунофлуоресцентный анализ экспрессии SARS-CoV-2-S осуществляли в клетках MDBK в 24 луночных планшетах, которые были инфицированы CVXGA1 или PIV5, при MOI 1. Через 2 дня после инфицирования (dpi) клетки промывали с помощью PBS, а затем фиксировали в 2% формальдегиде. Клетки пермеабилизировали в 0,1% растворе PBS-сапонин и инкубировали в течение 1 hpi с антителом к SARS-CoV-2-S (Sino Biological, номер по каталогу 40150 R007) и антителом к PIV5-V/P при разбавлении 1:200, а затем в клетки добавляли вторичные антитела в виде меченого флуоресцеинизотиоцианатом (FITC) антитела козы к IgG кролика (KPL, номер по каталогу 02-15-16) и меченого Су3 антитела козы к IgG мыши (KPL, номер по каталогу 072-01-18-06). Клетки инкубировали в течение 30 мин., а также исследовали и фотографировали с использованием флуоресцентного микроскопа (Advanced Microscopy Group).
Иммуноблоттинг осуществляли на клетках Vero в 12-луночных планшетах, которые были инфицированы PIV5 или CVXGA1 при MOI 1. Через 24 hpi для лизиса клеток использовали буфер Лэммли для образцов (Bio Rad, номер по каталогу 1610737) с 5% р меркаптоэтанола. Лизаты анализировали на геле SDS-PAGE и подвергали иммуноблотингу с антителом к SARS-CoV-2-S (Sigma, номер по каталогу ZHU1076) и антителом к PIV5-V/P.
Очистка S и RBD S
Была синтезирована плазмида (Twist Bioscience), кодирующая кДНК для эктодомена шиловидного белка SARS-CoV-2, стабилизированного в конформации до слияния (Wrapp et al. 2020, Science; 367:1260-1263, doi:10.1126/science.abb2507), и клонирована в вектор pTwist CMV Hygro. Плазмида, кодирующая домен связывания рецептора мономерного шиловидного белка, была получена от BEI Resources (NR-52309). Плазмиды размножали путем трансформации клеток Escherichia coli DH5a с помощью 100 мкг/мл ампициллина (Thermo Fisher Scientific), используемого для селекции. Плазмиды очищали с использованием макси-набора для плазмид EZNA (Omega Biotek) в соответствии с протоколом производителя. На каждый литр смеси для трансфекции смешивали 1 мг плазмидной ДНК с 4 мг полиэтиленимина с молекулярной массой 25000 (PEI; PolySciences Inc.) в 66 мл среды для культивирования клеток Opti-MEM (Gibco). Через 30 мин. смесь ДНК-PEI добавляли к клеткам HEK293F (1 миллион клеток/мл) в среде Freestyle 293 (Gibco). Через 5-7 дней культуры центрифугировали для осаждения клеток и надосадочные жидкости фильтровали через стерильный фильтр с размером пор 0,45 мкм. Рекомбинантные белки очищали из отфильтрованных надосадочных жидкостей культур с использованием колонок HisTrap Excel (GE Healthcare Life Sciences). Каждую колонку хранили в 20% этаноле и промывали 5 объемами колонки (CV) промывочного буфера (20 мМ Tris, рН 7,5, 500 мМ NaCl и 20 мМ имидазола) перед загрузкой образцов в колонку. После нанесения образца колонки промывали с помощью 10 CV промывочного буфера. Белки элюировали из колонки с помощью 6 CV элюирующего буфера (20 мМ Tris, рН 7,5, 500 мМ NaCl и 250 мМ имидазола). Белки концентрировали и буфер заменяли на фосфатно солевой буфер (PBS) с использованием центрифужных фильтров Amicon Ultra-15 с отсечкой 30 кДа (Millipore Sigma).
ELISA
Для количественного определения гуморального ответа на антитела к SARS-CoV-2-S после примирующей и бустерной вакцинации сыворотку крови мыши анализировали посредством ELISA. 96-луночные микротитровальные планшеты IMMULON® 2НВ покрывали посредством 100 мкл очищенного SARS-CoV-2-S при концентрации 1 мкг/мл. Сыворотку крови подвергали серийному двукратному разбавлению и инкубировали в планшетах в течение 2 ч. Вторичное антитело, представляющее собой антитело козы к IgG мыши, меченое пероксидазой хрена (Southern Biotech, Бирмингем, Алабама), разбавляли 1:2500 и инкубировали в лунках в течение 1 ч. Планшеты проявляли с использованием субстрата пероксидазы KPL SureBlue Reserve ТМВ Microwell Peroxidase Substrate (SeraCare Life Sciences, Inc., Милфорд, Массачусетс, США), и значения OD450 регистрировали с помощью спектрофотометра для микропланшетов BioTek Epoch (BioTek, Винуски, Вермонт). Конечные точки содержания антител рассчитывали в виде log10 наиболее высокого разбавления в сыворотке крови, при котором OD450 превышала более чем на два стандартных отклонения среднюю OD450 сыворотки крови, не подвергнутой обработке.
Для выявления IgG хорька 96 луночные микропланшеты для ELISA со средним связыванием (Greiner Bio-One 655001) покрывали посредством 20 мкг либо полноразмерного шиповидного белка SARS-Cov-2, либо рецептор-связывающего домена SARS-Cov-2 (RBD) (Amanaetal. 2020, NatMed; 26:1033-1036, doi:10.1038/s41591 020 0913 15) в стерильном 1x фосфатно солевом буфере (Corning, 21-040-CV) на ночь при температуре 4°С. Планшеты промывали 3х с помощью 300 мкл 0,05% PBS-T с использованием автоматической мойки для планшетов (BioTek 405 TS Washer). Все промывки выполнялись с использованием одной и той же методики. Лунки блокировали посредством 200 мкл блокирующего буфера (0,5% бычьего сывороточного альбумина + 3% обезжиренного сухого молока в 0,05% PBS-T), инкубировали 2 часа при комнатной температуре, затем промывали. Сыворотку крови, инактивированную нагреванием, разбавляли в блокирующем буфере, добавляли по 100 мкл в соответствующие лунки и инкубировали в течение 2 часов при комнатной температуре с последующей стадией промывки. Добавляли по 100 мкл на лунку антитела козы к IgG хорька, конъюгированного с HRP (Bethyl Laboratories, А140-108Р), разбавленного 1:5000-1:10000 в блокирующем буфере, и инкубировали в течение 1 часа при комнатной температуре. Планшеты промывали и осушивали постукиванием для удаления остатков растворов. Таблетки SIGMAFAST™ OPD (дигидрохлорид о-фенилендиамина) (Sigma-Aldrich, Р9187) для выявления пероксидазной активности получали в 20 мл DI Н2О и добавляли по 100 мкл на лунку. После 8 минутного периода проявления добавляли 50 мкл 1 н. H2SO4 для остановки реакции. Планшеты немедленно считывали при поглощении 490 нм (BioTek, прибор Cytation7). Фоновый сигнал рассчитывали по средним значениям поглощения, полученным в лунках, покрытых белком для захвата, в которые вводили антитело козы к IgG хорька, конъюгированное с HRP.
Для выявления IgG кошек 96 луночные микропланшеты для ELISA покрывали и блокировали так же, как и для анализа IgG хорька. Сыворотку крови, инактивированную нагреванием, разбавляли в блокирующем буфере, добавляли по 100 мкл в соответствующие лунки и инкубировали в течение 2 часов при комнатной температуре. Планшеты промывали и добавляли по 100 мкл на лунку антитела козы к IgG кошки, конъюгированного с HRP (Bethyl Laboratories, А20-120Р), разбавленного 1:10000 в блокирующем буфере, и инкубировали в течение 1 часа при комнатной температуре. Планшеты промывали и осушивали постукиванием для удаления остатков растворов. Планшеты проявляли и считывали так же, как для и ELISA IgG хорька. Фоновый сигнал рассчитывали по средним значениям поглощения, полученным в лунках, покрытых белком для захвата, в которые вводили антитело козы к IgG кошки, конъюгированное с HRP.
Для выявления IgA кошек микропланшеты для ELISA покрывали, блокировали и промывали так же, как и для выявления IgG хорька. Двукратные серийные разбавления каждого образца назального смыва (собранного в PBS) осуществляли в блокирующем буфере и добавляли по 100 мкл в соответствующие лунки. Разбавления назальных смывов инкубировали 2 часа при комнатной температуре, промывали и добавляли по 100 мкл на лунку антитела козы к IgA кошки, конъюгированного с HRP (Bethyl Laboratories, А20-101Р), разбавленного 1:10000 в блокирующем буфере, и инкубировали 1 час при комнатной температуре. Планшеты промывали и осушивали постукиванием для удаления остатков растворов. Планшеты проявляли и считывали так же, как для и ELISA IgG хорька. Фоновый сигнал рассчитывали по средним значениям поглощения, полученным из покрытых шиловидным белком контрольных лунок только с антителом козы к IgA кошки, конъюгированным с HRP.
Для выявления IgA хорька микропланшеты для ELISA покрывали, блокировали и промывали так же, как и для выявления IgG хорька. Двукратные серийные разбавления объединенного в пул инактивированного нагреванием образца назального смыва (собранного в PBS) осуществляли в блокирующем буфере и добавляли по 100 мкл в соответствующие лунки. Разбавления назальных смывов инкубировали 2 часа при комнатной температуре, промывали и добавляли по 100 мкл на лунку антитела козы к IgA хорька, конъюгированного с АР (Rockland, 618 105 006), разбавленного 1:2000 в блокирующем буфере, и инкубировали 1 час при комнатная температура. Планшеты промывали и осушивали постукиванием для удаления остатков растворов. Таблетки SIGMAFAST™ п-нитрофенилфосфата (Sigma-Aldrich, N1891) для выявления активности щелочной фосфатазы получали в 5 мл DI Н2О и добавляли по 200 мкл на лунку. Планшеты заворачивали в фольгу для защиты от света и инкубировали 30 45 минут при комнатной температуре. После 8 минутного периода проявления добавляли 50 мкл 1 н. H2SO4 для остановки реакции. Планшеты немедленно считывали при поглощении 405 нм (BioTek, прибор Cytation7). Фоновый сигнал рассчитывали по средним значениям поглощения, полученным в лунках, покрытых белком для захвата, в которые вводили антитело козы к IgA хорька, конъюгированное с АР.
ELISPOT-анализ интерферона-γ (IFN-γ)
Для анализа клеточных иммунных ответов использовали набор для выявления IFN-γ мыши BD™ ELISPOT Mouse IFN-γ (BD Biosciences, Сан-Хосе, Калифорния, США). Планшеты BD™ ELISPOT покрывали очищенным антителом к IFN-γ мыши за 24 часа до проведения анализа. Селезенки мышей (n=5 на группу) собирали через 28 дней после иммунизации и помещали в конические пробирки объемом 15 мл, содержащие 5 мл HBSS. Спленоциты получали путем продавливания селезенки через клеточное сито с размером ячеек 70 мкм, инкубирования их с буфером для лизиса АСК, промывания их с помощью HBSS и ресуспендирования в полной опухолевой среде (СТМ) до концентрации 5×106 клеток/мл. Раствор антител для захвата удаляли из планшетов, а затем планшеты 5-6 раз промывали с помощью PBS. Затем планшеты блокировали с помощью СТМ в течение 90 минут.Блокирующий раствор отбрасывали и в лунки добавляли по 0,1 мкг пептидов S-белка SARS-CoV-2, покрывающих весь белок, в 50 мкл СТМ. В планшеты добавляли 50 мкл спленоцитов (2,5×105 клеток/лунка) и инкубировали при температуре 37°С, 5% СО2, в течение 48 часов. Пятна подвергали иммуноокрашиванию в соответствии с инструкцией по эксплуатации набора BD™ ELISPOT и подсчитывали с использованием анализатора IMMUNOSPOT® (Cellular Technology Limited, CTL). Результаты были представлены в виде числа клеток, секретирующих IFN-γ, на 106 спленоцитов.
Иммунизация и инфицирование мышей с помощью SARS-CoV-2
Иммунизация. В данном исследовании использовали самок мышей (Envigo) в возрасте от шести до восьми недель. Мышей анестезировали посредством внутрибрюшинной инъекции 250 мкл 2,2,2-трибромэтанола в трет-амиловом спирте (авертине) и интраназально инокулировали посредством 50 мкл PBS или 104, 105, 106 БОЕ CVXGA1. Через 28 дней после иммунизации мышей подвергали эвтаназии, собирали сыворотку крови посредством кардиоторакального кровотечения и собирали селезенки. Мышей содержали и иммунизировали в помещениях с повышенным уровнем биобезопасности 2 в изоляторах, снабженных НЕРА-фильтром. Все эксперименты проводились в соответствии с протоколами, одобренными Институциональным комитетом по уходу за животными и их использованию в Университете Джорджии.
106 БОЕ инактивированного УФ-излучением SARS-CoV-2 смешивали 1:1 (об./об.) с квасцовым адъювантом (Thermo, номер по каталогу 77161) в объеме 200 мкл и вводили мышам внутримышечно.
Сенсибилизация мышей с помощью Ad5-АСЕ2. Вектор Ad5TiACE2 был описан ранее (Jia et al., 2005, J Virol; 79:14614-14621, doi:10.1128/JVI.79.23.14614-14621.2005 и Sun et al. 2020, Cell, doi: 10.1016/j.cell.2020.06.010). Конструирование вирусного вектора осуществляют с использованием системы VVC RAPAD™ System (Anderson et al., 2000, Gene Ther; 7:1034-1038, doi: 10.1038/sj.gt.3301197). Мышей анестезировали смесью кетамин/ксилазин (87,5 мг/кг кетамина/ 12,5 мг/кг ксилазина) и интраназально трансдуцировали посредством 2 5×108 БОЕ Ad5-hACE2 в 75 мкл DMEM.
Инфицирование мышей. Мышей слегка анестезировали смесью кетамин/ксилазин и интраназально инокулировали указанным количеством SARS-CoV-2 в общем объеме 50 мкл DMEM. Ежедневно контролировали вес и состояние здоровья животных. Все эксперименты с SARS-CoV-2 проводились в лаборатории уровня биобезопасности 3 (BSL3). Через пять дней после трансдукции мышей анестезировали смесью кетамин/ксилазин (87,5 мг/кг кетамина/12,5 мг/кг ксилазина) и инфицировали интраназально посредством 105 БОЕ SARS-CoV-2 (изолят USA-WA1/2020 № BEI NR-52281) в 50 мкл DMEM. Мониторинг и взвешивание мышей осуществляли ежедневно.
Иммунизация и инфицирование хорьков с помощью SARS-CoV-2
Двенадцать самцов и самок (700-2000 г) черных хорьков в возрасте от восьми до девяти месяцев получали от Triple F Farms и акклиматизировали в вивариях UGA. Перед прибытием хорьков подвергали стерилизации и удалению пахучих желез. После периода акклиматизации продолжительностью не менее семи дней хорьков анестезировали для подкожного размещения тепловых транспондеров между лопатками. Имплантируемые программируемые температурные транспондеры (Bio Medic Data Systems, США) надежно прикрепляются к ткани в месте имплантации и дают возможность считывать температуру в диапазоне 90-110 градусов по Фаренгейту. Одновременно собирали цельную кровь для серологического анализа на исходном уровне и брали мазок кала для анализа выделения вируса в среду.
Все экспериментальные процедуры проводились в биологических вивариях Университета Джорджии. Животных содержали парами со свободным доступом к пище и воде. Все эксперименты были одобрены IACUC Колледжа ветеринарной медицины Университета Джорджии. Все процедуры, включая сбор носовых смывов и крови, выполнялись под кратковременной газовой анестезией изофлураном.
Для инфекционного заражения с помощью SARS-CoV-2 животных перемещали в помещение Исследовательского центра охраны здоровья животных Университета Джорджии, обеспечивающее уровень биологической безопасности животных 3 (ABSL3). Все эксперименты были одобрены IACUC Колледжа ветеринарной медицины Университета Джорджии.
Группы из шести хорьков вакцинировали интраназально (IN) посредством 5×106 БОЕ CVXGA1 в 1,0 мл стерильного фосфатно солевого буфера (PBS), распределяя по 500 мкл на ноздрю. После вакцинации животных подвергали ежедневному мониторингу на наличие клинических признаков, включая выделения из носа, чихание, диарею, вялость, увеличение частоты дыхания и дыхательного усилия (заложенность), цианоз, неврологические изменения и ответ на внешние раздражители. Температуру тела отслеживали с использованием имплантированных датчиков температуры. Назальные смывы и фекальные мазки собирали через 3, 7, 14, 21 и 28 дней после иммунизации (dpi) для оценки выделения вакцины в среду. Цельную кровь получали еженедельно после иммунизации. Мононуклеарные клетки периферической крови (РВМС) выделяли и криоконсервировали для дальнейшего анализа. Для инфекционного заражения с помощью SARS-CoV-2 4×105 БОЕ доставляли посредством интраназального пути. Опять таки, физические наблюдения, взвешивания и измерения температуры проводили ежедневно.
Подгруппу из шести животных гуманно усыпляли и вскрывали в день 4 после заражения, а оставшихся животных (n=6) гуманно усыпляли в день 7 после заражения. Все вскрытия проводили под руководством ветеринарного патологоанатома, получившего профессиональную сертификацию. Образцы тканей (трахея и легкое) собирали для определения патологии и вирусной нагрузки. Осуществляли конечный сбор крови для выделения сыворотки крови и РВМС.
Иммунизация и инфицирование кошек с помощью SARS-CoV-2
Двенадцать полувзрослых (в возрасте от семи до девяти месяцев) здоровых самцов домашних короткошерстных кошек (DSH) получали от MBR Wavery LLC. После периода акклиматизации самцы кошек были хирургически кастрированы сертифицированным ветеринарным хирургом. Всех стерилизованных самцов содержали в одиночестве до полного восстановления после хирургического вмешательства, что определял сертифицированный ветеринарный врач для лабораторных животных. Перед иммунизацией кошек анестезировали с использованием инъецируемых анестетиков для подкожного размещения имплантируемых программируемых температурных транспондеров IPTT 300 (Bio Medic Data Systems, США) между лопатками. Одновременно собирали образцы цельной крови и кала. Отдельную сыворотку крови проверяли на наличие антител, представляющих собой IgG, реагирующие с белком SARS-CoV-2, посредством ELISA. Фекальные мазки оценивали посредством qRT PCR на наличие кишечного коронавируса кошек.
Все процедуры с кошками, проведенные до инфекционного заражения, проводились в биологическом виварии Университета Джорджии. Животных содержали группами или парами в зависимости от пола и темперамента. Кошек кормили коммерчески доступным сухим кормом и им обеспечивали свободный доступ к воде.
Группы кошек (n=4/группа) вакцинировали либо интраназально (IN), либо подкожно (SQ) посредством 1×106 БОЕ CVXGA1, суспендированных в стерильном фосфатно-солевом буфере (PBS). Интраназальную вакцинацию осуществляли с использованием объема 0,5 мл, распределяемого по 250 мкл на ноздрю. Подкожную доставку осуществляли с использованием общего объема, составляющего 1,0 мл. Подкожно вакцинированных животных подвергали бустерной иммунизации через двадцать один день после первичной инъекции. После вакцинации животных подвергали ежедневному мониторингу на наличие клинических признаков, включая выделения из носа, чихание, диарею, вялость, увеличение частоты дыхания и дыхательного усилия (заложенность), цианоз, неврологическое изменение и ответ на внешние раздражители.
Температуру тела отслеживали с использованием имплантированных датчиков температуры. Назальные смывы и фекальные мазки собирали через 1, 3, 7, 14, 21 и 28 дней после вакцинации (dpv) для оценки выделения вакцины в среду. Цельную кровь получали еженедельно после иммунизации. Мононуклеарные клетки периферической крови (РВМС) выделяли и криоконсервировали для дальнейшего анализа. Для инфекционного заражения SARS-CoV-2 доставляли посредством интраназального пути в дозе 4×105 БОЕ в объеме 1,0 мл. Всех животных (n=12) гуманно усыпляли через четырнадцать дней после заражения. Вскрытия проводили под руководством сертифицированного ветеринарного патологоанатома. В дополнение к тканям осуществляли конечный сбор крови для выделения сыворотки крови и РВМС.
Гистология и иммуногистохимия
Животных анестезировали и транскардиально перфузировали посредством PBS. Ткани легких собирали, фиксировали (10% нейтральный забуференный формалин), обезвоживали с помощью серии ванн со спиртом и ксилолом, заливали парафином, делали срезы (~4 мкм) и окрашивали красителями, представляющими собой гематоксилин и эозин (НЕ). Ткани исследовались сертифицированным патологоанатом по схеме с маскировкой данных и в соответствии с принципами воспроизводимых оценок тканей (Meyerholz and Beck, 2018, Invest; 98:844-855, doi:10.1038/s41374-018-0057-0). Периваскулярную эозинофильную инфильтрацию оценивали, как описано ранее (Fuentes et al., 2015, J Virol; 89:8193 8205, doi: 10.1128/JVI.00133 15). Вкратце, периваскулярные области с клеточной инфильтрацией выбирались случайным образом (n=20/легкое) патологоанатомом, для которого данные были замаскированы, а количество эозинофилов подсчитывалось и усреднялось для окончательной балльной оценки для каждого легкого. Легкие поддавали балльной оценке на наличие мононуклеарных инфильтратов, при этом балл 0 представлял значения параметров в пределах нормы, 1 представлял наличие небольших агрегатов в перибронхиальном и периваскулярном участках, 2 представлял наличие периваскулярных агрегатов и агрегатов, расположенных вокруг дыхательных путей, заполняющих периваскулярное пространство, и 3 представлял балл 2 плюс расширяющиеся прослойки инфильтратов в перегородки и очаги консолидации в областях легкого соответственно. Легкие поддавали балльной оценке на наличие гранулоцитарных инфильтратов, при этом баллы являлись следующими: 0 - параметры в пределах нормы; 1 - разрозненные PMN, секвестрированные в перегородках; 2 - балл 1 плюс одиночные PMN, проникающие в воздушные прослойки; 3 балл 2 плюс мелкие агрегаты в сосудах и в воздушных прослойках соответственно.
Нейтрализация SARS-COV2-S
VSV, псевдотипированный SARS-CoV-2 (VSV-S), описанный ранее (Nie et al., 2020, EmergMicrobes Infect; 9:680-686, oi:10.1080/22221751.2020.1743767), использовался для анализа нейтрализации. Частицы VSV-S титровали посредством TCID50 в 96-луночном планшете в клетках Vero для определения оптимального числа частиц для нейтрализации. Их разбавляли в 2% FBS, 1% P/S в средах DMEM. Титр определяли с использованием аналитической системы для выявления с помощью люциферазы Firefly Bio-Glo Luciferase Assay System от Promega. На каждом планшете использовался контроль частиц, который состоял из разбавленных частиц и стерильного PBS (бессывороточного) в соотношении 1:1. На каждом планшете также использовался отрицательный контроль, состоящий из стерильного PBS и 2% FBS, 1% P/S в средах DMEM для учета фонового уровня.
Сыворотку крови инактивировали нагреванием при 56°С в течение 45 минут перед анализом нейтрализации. Затем его подвергали серийному разбавлению в стерильном PBS с использованием разбавлений 1:2, начиная с 1:50 (кошки) или 1:100 (хорьки), в 96 луночном необработанном планшете, не содержащем клеток. Образец восстановленной сыворотки крови человека (+RHS) использовали в качестве положительного контроля в каждом анализе и последовательно разбавляли 1:2, начиная с 1:100. Образцы и контроли получали в четырех повторностях.
Добавляли 50 мкл разбавленных частиц VSV-S в 96 луночный планшет, содержащий 50 мкл разбавленной сыворотке крови, и инкубировали в течение 1 часа при 5% СО2 при температуре 37°С. Через 1 ч. среду аспирировали из белого 96-луночного планшета, содержащего клетки Vero при конфлюентности 90-100%, и к клеткам добавляли смесь частиц и сыворотки крови (100 мкл). Планшет инкубировали при 5% СО2 при температуре 37°С в течение 18-24 ч. На следующий день образцы уравновешивали при комнатной температуре в течение 10-20 минут и в каждую лунку добавляли по 50 мкл реагента Bio-Glo. Планшеты немедленно считывали с применением протокола Bio-Glo на люминометре GloMax от Promega.
Для нейтрализации посредством SARS-CoV-2 образцы в сыворотке крови инактивировали нагреванием при 56°С в течение 30 минут перед анализом нейтрализации. Затем их подвергали серийному разбавлению в стерильной DMEM и смешивали с равным объемом DMEM, содержащим ~20 БОЕ SARS-CoV-2. После инкубации при температуре 37°С в течение 1 ч. аликвоты добавляли в клетки Vero Е6 в 12 луночных планшетах и инкубировали при температуре 37°С в 5% СО2 в течение 1 ч. После удаления инокулята чашки покрывали 1,2% агарозой, содержащей 4% FBS. После дальнейшей инкубации при температуре 37°С в 5% СО2 в течение 2 дней покровные слои удаляли и бляшки визуализировали путем окрашивания 0,1% кристаллическим фиолетовым.
qPCR выделения SARS-CoV-2 в среду из назальной полости
Построение стандартной кривой. Для размножения вируса, используемого для построения стандартной кривой, клетки Vero Е6 инфицировали при MOI примерно 0,001 посредством SARS-CoV-2 USA-WA01/2020 (BEI Resources, № по каталогу NR52281), пассаж 1, и инкубировали при температуре 37°С + 5% СО2. Через 48 часов после инфицирования вирус собирали и титровали посредством анализа бляшкообразования в клетках Vero Е6 с авицел содержащим покровным слоем, состоящим из 1,2% авицела + 0,5Х DMEM + 1% FBS + 0,5Х антибиотик/антимикотик. Вирус в концентрации 1,1×106 БОЕ/мл или 1,44×107 БОЕ/мл для образцов из хорька и кошки соответственно смешивали 1:1 с реагентом TRIzol (ThermoFisher Scientific) для инактивации и сохранения генетического материала. С использованием набора Quick RNA Viral Kit (Zymo Research) РНК экстрагировали из 600 мкл и элюировали в 50 мкл TRIzol. РНК подвергали серийному разбавлению 1:10. Использовали по 10 мкл каждого разбавления вируса в 40 мкл реакционной смеси для qPCR с 10 мкл мастер-микса TaqPath 1 -Step RT-qPCR Master Mix (ThermoFisher Scientific) и 3 мкл смеси праймер/зонд nCov N1 из набора EUA CDC SARS 2 (Integrated DNA Technologies). Все реакции qPCR анализировали с использованием Agilent Мх3000Р.
Назальный смыв хорька для qPCR. Полученный образец назального смыва объемом 500 мкл смешивали 1:1 с реагентом DNA/RNA Shield (Zymo Research). С использованием набора Quick RNA Viral Kit (Zymo Research) РНК экстрагировали из 600 мкл и элюировали в 15 мкл реагента DNA/RNA Shield. Использовали по 2 нг каждого образца назального смыва в 40 мкл реакционной смеси для qPCR с 10 мкл мастер микса TaqPath 1 Step RT qPCR Master Mix (ThermoFisher Scientific) и 3 мкл праймера/зонда nCov N1 из набора EUA CDC SARS-2 (Integrated DNA Technologies). Количество копий генома на реакцию qPCR рассчитывали с использованием стандартной кривой. Соотношение РНК, используемой в реакции qPCR, рассчитывали путем деления 2 нг на общее количество нг РНК на образец. Для расчета количества копий генома для каждого образца количество копий генома на реакцию qPCR умножали на соотношение РНК, используемой в реакции, и при этом количество копий генома на мл вычисляли путем обратных вычислений с использованием описанных объемов разбавления и экстракции. Образцы со значениями СТ свыше 37 считались ПЦР отрицательными. Для каждого эксперимента с qPCR предельное значение СТ, составляющее 1 FFU/реакция (rxn), устанавливали с использованием стандартной кривой, а образцам со значением СТ ниже 37, но выше порогового значения 1 FFU/rxn, присваивали значение копий генома/мл 0,5 для цели построения графика. Линии отсечки для 1 FFU/мл и ПЦР положительных результатов были нанесены на уровне у=1 и у=0,1 соответственно.
qPCR назальных смывов кошек
Полученный образец назального смыва объемом 500 мкл смешивали 1:1 с реагентом DNA/RNA Shield (Zymo Research). С использованием набора Quick RNA Viral Kit (Zymo Research) РНК экстрагировали из 600 мкл и элюировали в 15 мкл реагента DNA/RNA Shield. Использовали по 1 мкл каждого образца назального смыва в 40 мкл реакционной смеси для qPCR с 10 мкл мастер микса TaqPath 1 Step RT qPCR Master Mix (ThermoFisher Scientific) и 3 мкл смеси праймер/зонд nCov N1 из набора EUA CDC SARS 2 (Integrated DNA Technologies).
Количество копий генома на реакцию qPCR рассчитывали с использованием стандартной кривой. Количество копий генома на мл рассчитывали путем обратных вычислений с использованием описанных объемов разбавления и экстракции. Образцы со значениями СТ свыше 37 считались ПЦР-отрицательными. Для каждого эксперимента с qPCR предельное значение СТ, составляющее 1 FFU/rxn, устанавливали с использованием стандартной кривой, а образцам со значением СТ ниже 37, но выше порогового значения 1 FFU/rxn, присваивали значение копий генома/мл 0,5 для цели построения графика. Линии отсечки для 1 FFU/мл и ПЦР-положительных результатов были нанесены на уровне у=1 и у=0,1 соответственно.
Анализ для определения количества фокусобразующих единиц
Количественную оценку инфекционного SARS-Cov-2 выполняли посредством фокусобразующих единиц (FFU) в клетках Vero Е6. Вкратце, 96 луночные планшеты для культивирования клеток засевали из расчета 3,2×105 клеток/лунка и инкубировали в течение ночи. Конфлюэнтные монослои инкубировали с 10-кратными серийными разбавлениями инокулята в культуральных средах DMEM, предусматривающих среды с 2% FBS и 1х смеси антибиотик/антимикотик. Питательные среды из планшетов для культивирования, 50 мкл инокулята инкубировали в течение 1 часа, 37°С, 5% СО2 при определенной влажности. Покровные среды, 0,8% метилцеллюлозу в культуральных средах, наносили в количестве 150 мкл/лунка и планшеты возвращали в условия с температурой 37°С, 5% СО2 при определенной влажности на 20-24 часа. Для колориметрического проявления фокусобразующих единиц (FFU) содержащий метилцеллюлозу покровный слой декантировали и планшеты трижды промывали с помощью lx PBS. Фиксирующий раствор (80% метанола, 20% ацетона) добавляли в объеме 100 мкл и обеспечивали инкубирование в течение минимум 10 минут при комнатной температуре перед погружением в фиксирующий раствор и удалением из помещений BSL3. Первичное антитело (конъюгированное с HRP, 1СО2, 1,4 мкг/мл) разбавляли до 1:1000 в блокирующем буфере (0,1% Tween 20, 5% NFDM, 5% BSA) при 75 мкл/лунка и инкубировали при комнатной температуре в течение 45 минут. Далее первичный раствор декантировали и планшеты дважды промывали с помощью 0,1% PBS-Т. Дополнительную стадию промывки осуществляли с помощью проявляющего раствора ТМВ с dH2 0,75 мкл/лунка и инкубировали при комнатной температуре в течение 1 часа. Осуществляли заключительную стадию промывки с помощью dH2O, планшеты высушивали на воздухе и изображения получали посредством прибора Cytation7 от BioTek для ручного количественного определения FFU.
Пример 3
Защита мышей К18 hACE2 и хорьков от заражения с помощью SARS-CoV-2 посредством осуществляемой через слизистые оболочки иммунизации однократной дозой вакцины против COVID-19 на основе вируса парагриппа 5
В данном примере был создан рекомбинантный PIV5, содержащий полноразмерный шиповидный белок SARS-CoV-2 с цитоплазматическим хвостом, замененным на хвост F-белка PIV5 (CVXGA1), и протестирована его эффективность в качестве вакцины у мышей и хорьков.
Создание и анализ PIV5, обеспечивающего экспрессию S-белка SARS-CoV-2 (CVXGA1)
С использованием стратегии, аналогичной той, которая использовалась при разработке вакцины против MERS-CoV на основе PIV5, был создан PIV5, обеспечивающий экспрессию шиповидного (S) белка SARS-CoV-2 (названный CVXGA1) (ФИГ. 18А). Определение последовательности вирусного генома посредством секвенирования с помощью RT PCR, а также подтвержденная экспрессия гликопротеина S в клетках, инфицированных CVXGA1, с использованием иммунофлуоресценции и вестерн-блоттинга (ФИГ. 18В). Кроме полноразмерного S в клетках, инфицированных CVXGA1, также был выявлен продукт расщепления, S1, что указывает на то, что S подвергался процессингу (ФИГ. 18В). S-белок опосредует слияние клетки с клеткой и вируса с клеткой для обеспечения проникновения вируса, а иммунизация посредством S создает защитный иммунитет против SARS-CoV-2 (Corbett et al. N Engl J Med, (2020)). Считается, что нативная (предусматривающая белки в конформации до слияния) конформация S индуцирует требуемый иммунитет для оптимальной защиты (Corbett et al. N Engl J Med, (2020)). Чтобы проверить, является ли S-белок, экспрессируемый посредством вектора PIV5, функциональным, клетки Vero, которые экспрессируют рецептор АСЕ2, инфицировали CVXGA1. Образование синцитиев (слияние клетки с клеткой) наблюдалось только в клетках, инфицированных CVXGA1, что указывает на функциональную экспрессию S-белка (ФИГ. 18С).
CVXGA1 обеспечивал развитие гуморальных и клеточных иммунных ответов у мышей.
Для исследования антигенности CVXGA1 мышей интраназально иммунизировали рядом инокулятов CVXGA1. Дозозависимое повышение содержания антител к S было выявлено у мышей BALB/c после однократного интраназально го введения (ФИГ. 18D). Аналогично, антитела, распознающие рецепторсвязывающий домен (RBD) S, выявлялись дозозависимым образом у мышей BALB/c, иммунизированных с помощью CVXGA1 (ФИГ. 18Е). Кроме того, нейтрализующие антитела к SARS-CoV-2 были выявлены у мышей BALB/c, иммунизированных дозозависимым образом (ФИГ. 27). Значительное образование антител и клеточные ответы наблюдались у мышей C57BL/6 после однократной интраназальной (IN) иммунизации дозой, составляющей всего лишь 6×103 бляшкообразующих единиц (БОЕ) CVXGA1 (ФИГ. 33). Эти результаты свидетельствуют о том, что иммунизация однократной интраназальной дозой CVXGA1 обеспечивала развитие устойчивого иммунного ответа у мышей.
CVXGA1 защищал мышей с человеческим рецептором АСЕ2 от летального заражения с помощью SARS-CoV2.
Для определения эффективности CVXGA1 использовали две взаимодополняющие животные модели: модель тяжелого заболевания с использованием трансгенных мышей и модель инфекции верхних дыхательных путей с использованием хорьков. У мышей отсутствует функциональный рецептор АСЕ2 для SARS-CoV-2 и вследствие этого они являются устойчивыми к инфекции. Для исследования эффективности вакцины в очень строгой модели использовали трансгенных мышей с человеческим АСЕ2 (hACE2). Недавно было продемонстрировано, что у мышей, экспрессирующих hACE2 под контролем промотора цитокератина 18 (K18), первоначально разработанного для исследования инфекции SARS-CoV (McCray et al. J Virol 81, 813-821 (2007)), развивается тяжелое заболевание легких в ответ на инфицирование с помощью SARS-CoV-2 (Jiang et al. Cell 182, 50-58 e58 (2020); Zheng et al. Nature, (2020)). Инокуляция мышей K18 hACE2 посредством 105 БОЕ SARS-CoV-2 приводит к 100% смертности, заболеванию легких с признаками диффузного повреждения альвеол и разной степенью распространения в ЦНС. Согласно оценкам летальная доза 50% (LDso) должна составлять 104 БОЕ (Jiang et al. Cell 182, 50-58 e58 (2020); Zheng et al. Nature, (2020)). Мышей K18-hACE2 иммунизировали интраназально однократной дозой CVXGA1 (106 БОЕ), а через 4 недели заражали посредством 4×104 БОЕ SARS-CoV-2. Другая группа мышей К18-АСЕ2 была иммунизирована внутримышечно с помощью инактивированного УФ излучением SARS CoV-2 с последующим осуществлением бустерной иммунизации через 2 недели. Невакцинированные мыши получали внутримышечно DMEM. Вторую контрольную группу иммунизировали интраназально однократной дозой вектора PIV5 интраназально (106 БОЕ). В ответ на заражение с помощью SARS-CoV-2 контрольная группа DMEM потеряла вес и погибла от инфекции через 7 дней после заражения (dpc) (ФИГ. 28А, 28В). Мыши, иммунизированные с помощью инактивированного УФ излучением SARS-CoV-2 или PIV5, потеряли столько же веса, сколько и в группе DMEM. Все мыши, иммунизированные PIV5, умерли до 7 dpc, а также умерли 4 из 6 мышей, иммунизированных с помощью инактивированного УФ излучением SARS-CoV-2 (ФИГ. 28А, 28 В). Мыши, иммунизированные с помощью CVXGA1, не потеряли в весе и все они выжили (ФИГ. 28А, 28В).
Через 5 dpc титры вируса в легочной ткани мышей, иммунизированных с помощью DMEM, инактивированного УФ излучением SARS-CoV-2 или PIV5, были сходными (ФИГ. 28С), и при этом все три группы продемонстрировали признаки инфекции головного мозга (ФИГ. 28D). В отличие от этого, у мышей, иммунизированных с помощью CVXGA1, не было выявляемого SARS-CoV-2 в легочной ткани, что свидетельствует о снижении титра вируса на ~5 log (ФИГ. 28С). Примечательно, что у 4 из 4 мышей, иммунизированных с помощью CVXGA1, вирус в ткани головного мозга не был выявлен (ФИГ. 28D) после заражения с помощью SARS-CoV-2.
Через пять дней после заражения с помощью SARS-CoV-2 вирусный антиген (N белок) был локализован в тканях легких. Контрольные животные, обработанные с помощью DMEM, характеризовались диффузным распространением окрашивания антигена в дыхательных путях и паренхиматозных эпителиальных клетках легких (ФИГ. 29А). Напротив, у мышей, иммунизированных с помощью CVXGA1, обнаруживались только необычные небольшие очаги положительных в отношении N-белка клеток, предположительно указывающих на первоначальные участки инфекции, которые не прогрессировали (ФИГ. 29А). В тканях легких из групп обработки с помощью как инактивированного УФ-излучением SARS-CoV-2, так и PIV5, наблюдались диффузно расположенные антиген-положительные клетки (ФИГ. 29А).
Срезы легочной ткани животных, инфицированных SARS-CoV-2, исследовали и поддавали балльной оценке на наличие периваскулярных эозинофильных инфильтратов. Через 5 дней после инфицирования у мышей, иммунизированных с помощью инактивированного УФ-излучением SARS-CoV-2, отчетливо наблюдалась миграция эозинофилов, которая отсутствовала в других группах (ФИГ. 29В, вставка со стрелками). Ткани легких также поддавали балльной оценке на наличие периваскулярных инфильтратов, состоящих из воспалительных клеток. Периваскулярные легочные инфильтраты, в случае если они состоят в основном из лимфоидных клеток, могут являться благоприятным признаком наличия предшествующего антигенного воздействия и эффективной вакцинации. Как показано на ФИГ. 29 В, по сравнению с контрольной группой DMEM, у мышей, иммунизированных с помощью CVXGA1, наблюдалось значительное увеличение периваскулярных инфильтратов, состоящих из лимфоидных клеток.
Наличие интерстициального заболевания часто является признаком тяжелой вирусной пневмонии. Ткани легких исследовали и поддавали балльной оценке на наличие интерстициального заболевания (Н-балл), определяемого по наличию инфильтрации альвеолярных перегородок, распространения в воздушные прослойки и связанных с ними ателектаза и отека. По сравнению с другими группами обработки, у мышей, иммунизированных с помощью CVXGA1, наблюдалось наименьшее количество признаков интерстициального заболевания через 5 дней после заражения с помощью SARS-CoV-2 (ФИГ. 29С). Гистопатологические балльные оценки эозинофильных инфильтратов, периваскулярных инфильтратов и тяжести интерстициального заболевания легких представлены на ФИГ. 29D. Дополнительно была исследована эффективность иммунизации с помощью CVXGA1 в отношении высокой дозы заражения (106 БОЕ SARS-CoV-2 на мышь), и при этом было обнаружено, что иммунизация с помощью CVXGA1 защищала 100% мышей от этого летального заражения (ФИГ. 34).
CVXGA1 защищал хорьков от инфекции SARS-CoV-2
Хорьки являются широко используемой моделью респираторных инфекций человека, они восприимчивы к инфекции SARS-CoV-2 и могут передавать вирус другим животным при непосредственном контакте и воздушно-капельным путем (Kim et al. Cell Host Microbe, (2020); Shi et al. Science, (2020); Richard et al. Nat Commun 11, 3496 (2020)). Для тестирования эффективности CVXGA1 у хорьков животных интраназально иммунизировали с помощью PBS или CVXGA1 (ФИГ. 30А). CVXGA1 реплицировался в носовых полостях хорьков в дни 3 и 7 после иммунизации с пиковым титром в назальных смывах, составляющим более 104 БОЕ/мл, и при этом CVXGA1 исчезал через 14 дней после иммунизации (ФИГ. 35А). Иммунизация с помощью CVXGA1 обеспечивала развитие устойчивых гуморальных ответов, о чем свидетельствуют высокие титры IgG к S (ФИГ. 35В), IgG к RBD (ФИГ. 30В) и нейтрализующих антител на ФИГ. 30С. Низкие уровни IgA к S были выявлены в назальных смывах (ФИГ. 35С). При интраназальном заражении с помощью SARS-CoV-2 геномная РНК вируса не выявлялась в назальных смывах хорьков, иммунизированных с помощью CVXGA1 (ФИГ. 31 А). Напротив, с течением времени вирусная РНК была выявлена у всех хорьков, подвергнутых имитационной иммунизации, (ФИГ. 31 А). Вирусная РНК также была выявлена в трахее и легких хорьков, подвергнутых имитационной иммунизации, но не в трахее и легких хорьков, иммунизированных с помощью CVXGA1 (ФИГ. 31 В, С). Для оценки уровня воздействия SARS-CoV-2 у хорьков уровни антител к S определяли количественно во время окончания эксперимента (7 dpc). Уровни IgG к S в группе, иммунизированной с помощью CVXGA1, увеличились после заражения (ФИГ. 35D), что свидетельствует об анамнестическом характере ответа и подверганию воздействию антигена S. Эти результаты свидетельствуют о том, что иммунизация с помощью CVXGA1 заметно снижает репликацию SARS-CoV-2 в верхних дыхательных путях - участке, имеющем решающее значение для возникновения SARS-CoV-2.
Для определения того, может ли иммунизация с помощью CVXGA1 блокировать передачу, хорьков иммунизировали однократной дозой CVXGA1, как и прежде. Контрольных животных иммунизировали с помощью PBS или пустого вирусного вектора PIV5 (ФИГ. 32А). Через 42 дня после IN иммунизации хорьков заражали SARS-CoV-2 и назальные смывы собирали через 1, 3, 5, 7, 9 и 11 dpc (ФИГ. 32А). Не подвергнутых обработке хорьков содержали вместе с зараженными хорьками при соотношении не подвергнутых обработке и инфицированных хорьков, составляющем 1:1, начиная с 2 dpc (2 хорька на клетку). В соответствии с правилами защиты животных все хорьки содержались в открытых клетках в помещении BSL3 (ФИГ. 36). Все хорьки в группах с PBS и пустым вектором PIV5 заразились, на что указывало выявление живого SARS-CoV-2 в назальных выделениях с пиком через 3 dpc (ФИГ. 32 В). Все не подвергнутые обработке хорьки, содержащиеся вместе с группами хорьков, инфицированных с помощью PBS и пустого вектора PIV5, заразились SARS-CoV-2 с пиковыми титрами, выявленными через примерно 5 дней после совместного содержания. Титры у подвергнутых воздействию не подвергнутых обработке животных достигали таких же уровней, как и у непосредственно инфицированных животных, что соответствовало 100% передаче (ФИГ. 32В). Как и раньше (ФИГ. 32В), вирус не был выявлен в носовых полостях хорьков, иммунизированных с помощью CVXGA1, после заражения. Вирус не был выявлен у не подвергнутых обработке хорьков, содержащихся совместно с CVXGA1 иммунизированными и зараженными SARS-CoV-2 хорьками в течение первых 5 дней после совместного содержания (ФИГ. 32В), что позволяет предположить, что непосредственный контакт с CVXGA1 иммунизированными и зараженными хорьками не приводил к передаче SARS-CoV-2 хорькам, ранее не подвергнутым обработке. Через 7 дней после расположения совместно у хорьков, расположенных совместно с животными, иммунизированными с помощью CVXGA1, развилась инфекция, вероятно попавшая из окружающей среды, которая включала инфицированных хорьков в соседних клетках (ФИГ. 36).
Эта работа демонстрирует, что мукозальная вакцина против COVID-19 на основе PIV5 может предупреждать смертельное заболевание в мышиной модели, и заметно снижать репликацию вируса в верхних дыхательных путях, и ингибировать передачу у хорьков. Хотя ни одна животная модель не воспроизводит полностью все признаки тяжелой инфекции SARS CoV-2 у людей, у мышей K18-hACE2 поддерживается устойчивая репликация вируса и развивается дозозависимое тяжелое заболевание, однако на сегодняшний день передача в этой модели не исследовалась (Jiang et al. Cell 182, 50-58 e58 (2020); Zheng et al. Nature, (2020)). Согласно оценке, у мышей K18-hACE2 летальная доза 50% (LD50) должна составлять 104 БОЕ (Zheng et al. Nature, (2020); Winkler et al. Nat Immunol 21, 1327-1335 (2020)). Иммунизация в режиме "прайм-буст" мышей K18-hACE2 с помощью инактивированного УФ-излучением SARS-CoV-2 не обеспечила защиты 100% мышей от летального заражения. Следует отметить, что в случае использования при заражении примерно 100 LD50 (106 БОЕ) мыши, иммунизированные инактивированный вирусом, теряли больше веса, чем контрольная группа с PBS (ФИГ. 34). Подобно предыдущим наблюдениям с вакциной на основе инактивированного УФ излучением MERS-CoV, иммунизация с помощью инактивированного УФ-излучением SARS-CoV-2 с последующим заражением с помощью 106 БОЕ SARS-CoV-2 запускала миграцию эозинофилов, что согласуется с наличием реакции по типу гиперчувствительности (ФИГ. 34Е). Мыши К18 hACE2, получавшие сублетальную инокуляцию с помощью SARS-CoV-2, выживали после последующего заражения летальной дозой, но вирус выявлялся в легочной ткани выживших мышей через 5 dpc Qiang et al. Cell 182, 50-58 e58 (2020)). Bee мыши, иммунизированные посредством однократной дозы CVXGA1, выживали при заражении дозой, составляющей примерно 100 LD50 (106 БОЕ) без выявляемого вируса в головном мозге и, что наиболее важно, у 75% мышей не был выявлен вирус в легочной ткани через 5 дней после заражения, что демонстрирует мощную защитную эффективность CVXGA1.
Создание нативной конфигурации вирусного гликопротеина в качестве антигена требуется для максимизации защитных иммунных ответов и может представлять сложную задачу. Инактивация вируса часто приводит к нежелательным изменениям в конформации антигена. Для получения оптимальной структуры очищенных вирусных гликопротеинов часто вводят мутации (Wrapp et al. Science 367, 1260-1263 (2020)). Ранее было продемонстрировано, что PIV5 является превосходным вектором для представления F-белка RSV. Клетки, экспрессирующие F, после инфицирования рекомбинантным PIV5, содержащим F, обеспечивают сохранение такой же конформации, как и у нативного F белка в RSV инфицированных клетках (Wang et al. J Virol 91, (2017)), что демонстрирует то, что вектор на основе живого вируса PIV5 может соответствующим образом экспрессировать нативные вирусные гликопротеины. Экспрессия полноразмерного S-белка SARS-CoV-2 в клетках, инфицированных CVXGA1, вызывала образование синцитиев (ФИГ. 18С), что согласуется с проявлением нативного, компетентного в отношении слияния S-белка. PIV5 проникает в клетки хозяева посредством F и HN-опосредованного слияния мембран: HN связывается с остатками сиаловой кислоты на белках клеточной поверхности, a F способствует слиянию мембран. Поскольку остатки сиаловой кислоты встречаются повсеместно, то известно, что PIV5 инфицирует практически все клетки млекопитающих. Таким образом, экспрессия функционального S SARS-CoV-2 не увеличивает клеточный тропизм CVXGA1, поскольку CVXGA1 экспрессирует S наряду с F и HN (ФИГ. 18А). Кроме того, ускоренное образование синцитиев, которому способствует CVXGA1 (PIV5 также вызывает образование синцитиев в клетках Vero Е6, хотя и с меньшей скоростью), вряд ли вызовет повреждение, поскольку синцитии чаще наблюдаются в клетках, выращиваемых в монослойной культуре, и редко встречаются в первичных эпителиальных клетках, которые обычно хорошо дифференцированы и поляризованы.
Хорьки очень восприимчивы к инфекции SARS-CoV-2 и легко передают вирус при непосредственном контакте и воздушно-капельным путем (Kim et al. Cell Host Microbe, (2020); Shi et al. Science, (2020); Richard et al. Nat Commun 11, 3496 (2020)). Непосредственный контакт является более эффективным путем передачи, чем опосредованный (воздушно-капельный) путь. В опубликованной работе одного инфицированного хорька содержали вместе с одним или двумя не подвергнутыми обработке животными для исследования передачи инфекции при непосредственном контакте (Kim et al. Cell Host Microbe, (2020); Richard et al. Nat Commun 11, 3496 (2020)). Для тестирования вакцины в устойчивой модели передачи одного не подвергнутого обработке хорька содержали вместе с одним инфицированным хорьком (ФИГ. 32А и ФИГ. 36). Иммунизация посредством однократной IN дозы CVXGA1 полностью защищала от инфекции SARS-CoV-2: вирусная РНК не была выявлена в носовых полостях иммунизированных животных (ФИГ. 31 А). Важно отметить, что иммунизация однократной дозой CVXGA1 ингибировала непосредственную передачу SARS-CoV-2 (ФИГ. 32). Поскольку хорьки содержались в открытых клетках, то животные, содержащиеся совместно с хорьками, иммунизированными с помощью CVXGA1, заражались через 7 и 9 дней после расположения совместно, вероятно посредством передачи через окружающую среду из других клеток. Также возможно, что поздние инфекции не подвергнутых обработке хорьков, содержащихся вместе с хорьками, иммунизированными с помощью CVXGA1, являлись результатом передачи хорьками, иммунизированными с помощью CVXGA1 и зараженными SARS-CoV-2, таким образом, CVXGA1 просто задерживал передачу SARS-CoV-2.
Однако вирус не был выявлен в назальных смывах или легких хорьков, иммунизированных с помощью CVXGA1 (ФИГ. 37), что предполагает то, что вероятным источником вируса для совместно содержащихся хорьков были инфицированные хорьки, содержавшиеся в соседних открытых клетках. Отсутствие вирусной РНК у хорьков, иммунизированных с помощью CVXGA1, после контрольного заражения подтверждает, что позднее инфицирование хорьков, содержащихся совместно с хорьками, иммунизированными с помощью CVXGA1, произошло от других инфицированных хорьков (ФИГ. 37). Было два возможных пути заражения не подвергнутых обработке хорьков, содержащихся вместе с хорьками, иммунизированными с помощью CVXGA1/зараженными SARS-CoV-2, в более поздние моменты времени. Один заключается в том, что хорьки заразились воздушно капельным путем передачи от инфицированных хорьков в соседних клетках, поскольку они содержались в открытых клетках (ФИГ. 36). Другой возможностью было заражение в ходе осуществления действий, связанных с уходом, и посредством оборудования: процедура, предписанная IACUC, требует замены клеток для животных каждые 7 дней. Возможно что хорьки, содержащиеся вместе, получали инфекцию в результате непосредственного контакта с зараженным оборудованием во время такой замены клеток. Наблюдалось несоответствие между уровнем содержания вирусной РНК и живого вируса в группе PBS (ФИГ. 32В и ФИГ. 37): титры живых вирусов достигали пика в день 3 и являлись не поддающимися выявлению в день 7, в то время как вирусная РНК впервые достигала пика в день 3 и характеризовалась высоким уровнем в день 7. Этот отсроченный клиренс вирусной РНК может отражать методологические различия, при которых qRT-PCR может выявлять остаточную вирусную РНК. Интраназальная иммунизация с помощью CVXGA1 обеспечивала развитие устойчивых ответов в виде выработки сывороточных антител у мышей и хорьков. Важно отметить, что IN иммунизация с помощью CVXGA1 индуцировала образование IgA слизистых оболочек дыхательных путей у хорьков, что согласуется с предыдущим отчетом о том, что PIV5 является действенным вектором для разработки вакцины против респираторных инфекций, включая вирус гриппа и респираторно-синцитиальный вирус (RSV) (Wang et al. J Virol 91, (2017); Mooney et al. J Virol 91, (2017)). IgA слизистых оболочек также был выявлен у обезьян, иммунизированных интраназально посредством PIV5, обеспечивающего экспрессию F RSV (Wanget al. J Virol 91, (2017)). Было высказано предположение, что в качестве живого вирусного вектора CVXGA1 может обеспечивали развитие устойчивых клеточных иммунных ответов и, как видно на ФИГ. 33, у мышей были выявлены клеточные иммунные ответы. Вакцины против питомникового кашля производились недорого и безопасно использовались в течение многих десятилетий, а производственные мощности и системы доставки вакцин для PIV5 хорошо зарекомендовали себя. Дальнейшая разработка вакцины на основе вектора PIV5 может помочь в блокировании передачи SARS-CoV-2 и контроле пандемии COVID-19.
МАТЕРИАЛЫ И СПОСОБЫ
Получение вируса и клеток, способ иммуноблоттинга, очистка S и RBD из S, протокол ELISA и протокол анализа для определения количества фокусобразующих единиц аналогичны тем, которые описаны в примере 2.
Иммунофлуоресценция
Иммунофлуоресцентное определение локализации экспрессии белка SARS-CoV-2 S осуществляли в 24-луночных планшетах в клетках MDBK, инфицированных с помощью CVXGA1 или PIV5, при MOI 1. Через 2 дня после инфицирования клетки промывали фосфатно солевым буфером (PBS), а затем фиксировали в 2% формальдегиде. Клетки пермеабилизировали в 0,1% растворе сапонина в PBS и инкубировали в течение 1 ч. с антителом к S SARS-CoV-2 (Sino Biological, №по каталогу 40150-R007) и антителом к PIV5-V/P при разбавлении 1:200, а затем к клеткам добавляли вторичные антитела в виде меченого флуоресцеинизотиоцианатом (FITC) антитела козы к IgG кролика (KPL, № по каталогу 02-15-16) и меченого Су3 антитела козы к IgG мыши (KPL, №по каталогу 072-01-18-06) с 2 каплями/мл 4',6-диамидино-2-фенилиндола (DAPI) (реагент NucBlue Live Cell Stain ReadyProbes, Life Technologies Corporation, Юджин, Орегон, США). Клетки инкубировали в течение 30 мин., а также исследовали и фотографировали с использованием конфокального флуоресцентного микроскопа Nikon Eclipse Ti.
Иммунизация и инфицирование мышей с помощью SARS-CoV-2
В этих исследованиях использовали самок мышей (Envigo) в возрасте от шести до восьми недель или мышей K18 hACE2 (B6.Cg.Tg(K18 hACE2)2Prlmn/J, Jackson Laboratory). Мышей анестезировали посредством внутрибрюшинной инъекции 250 мкл 2,2,2-трибромэтанола в трет-амиловом спирте (авертине) и интраназально инокулировали посредством 50 мкл PBS (или DMEM) или 104, 105, 106 БОЕ CVXGA1 или 106 БОЕ вектора PIV5. Через двадцать восемь дней после иммунизации мышей подвергали эвтаназии, собирали сыворотку крови посредством кардиоторакального кровотечения и собирали селезенки. Мышей содержали и иммунизировали в помещениях с повышенным уровнем биобезопасности 2 в изоляторах, снабженных НЕРА-фильтром. Все эксперименты проводились в соответствии с протоколами, одобренными Институциональными комитетами по уходу за животными и их использованию в Университете Джорджии и Университете Айовы.
106 БОЕ инактивированного УФ-излучением SARS-CoV-2 смешивали 1:1 (об./об.) с квасцовым адъювантом (Thermo, номер по каталогу 77161) в объеме 200 мкл и вводили мышам внутримышечно.
Инфицирование мышей. Мышей слегка анестезировали смесью кетамин/ксилазин и интраназально инокулировали указанным количеством SARS-CoV-2 в общем объеме 50 мкл DMEM. Ежедневно контролировали вес и состояние здоровья животных. Все эксперименты с SARS-CoV-2 проводились в лаборатории уровня биобезопасности 3 (BSL3).
Иммунизация и инфицирование хорьков с помощью SARS-CoV-2
Двенадцать самцов и самок (700-2000 г) черных хорьков в возрасте от восьми до девяти месяцев получали от Triple F Farms и акклиматизировали в вивариях UGA. Перед прибытием хорьков подвергали стерилизации и удалению пахучих желез. После периода акклиматизации продолжительностью не менее семи дней хорьков анестезировали для подкожного размещения тепловых транспондеров между лопатками. Имплантируемые программируемые температурных транспондеры (Bio Medic Data Systems, США) были надежно закреплены к ткани в участке имплантации и давали возможность считывать температуру в диапазоне 90-110°F. Одновременно собирали цельную кровь для получения данных серологии на исходном уровне и брали мазок кала для анализа выделения вируса в среду.
Процедуры перед заражением проводились в биологических вивариях Университета Джорджии. Животных содержали парами со свободным доступом к пище и воде. После вакцинации процедуры, включая получение назальных смывов и сборы крови, проводили на анестезированных животных с использованием смеси кетамин/ксилазин (15-20 мг/кг/1-2 мг/кг ксилазина), доставляемой IM. Хорьков подвергали ежедневному мониторингу на наличие клинических признаков. Значения веса и температуры регистрировали, как минимум, во время каждой процедуры под седацией.
Заражение вирусом хорьков
Для заражения с помощью SARS-CoV-2 животных перемещали в помещение Исследовательского центра охраны здоровья животных Университета Джорджии, обеспечивающее уровень биологической безопасности животных 3 (ABSL3). Процедуры, выполняемые после заражения вирусом, проводили под кратковременной газовой анестезией изофлураном.
Группы из шести хорьков вакцинировали интраназально (IN) посредством 1×106 БОЕ CVXGA1 в 1,0 мл стерильного PBS, распределяя по 500 мкл на ноздрю. После вакцинации животных подвергали ежедневному мониторингу на наличие клинических признаков, включая выделения из носа, чихание, диарею, вялость, увеличение частоты дыхания и дыхательного усилия (заложенность), цианоз, неврологические изменения и измененные ответы на внешние раздражители. Температуру тела отслеживали с использованием имплантированных датчиков температуры. Назальные смывы и фекальные мазки собирали через 3, 7, 14, 21 и 28 дней после иммунизации (dpi) для оценки выделения вакцины в среду. Цельную кровь получали еженедельно после иммунизации. Мононуклеарные клетки периферической крови (РВМС) выделяли и криоконсервировали для дальнейшего анализа. Для инфекционного заражения с помощью SARS-CoV-2 1×106 БОЕ доставляли посредством интраназального пути. Опять-таки, физические наблюдения, взвешивания и измерения температуры выполнялись ежедневно. Подгруппу из шести животных гуманно усыпляли и вскрывали в день 4 после заражения, а оставшихся животных (n=6) гуманно усыпляли в день 7 после заражения. Все вскрытия проводили под руководством ветеринарного патологоанатома, получившего профессиональную сертификацию. Образцы тканей (трахея и легкое) собирали для определения патологии и вирусной нагрузки. Осуществляли конечный сбор крови для выделения сыворотки крови и РВМС. Для исследования передачи через 2 дня после заражения (dpc) двух зараженных хорьков размещали совместно с одним не подвергнутым обработке хорьком.
Совместное содержание хорьков
Через два дня после заражения с помощью SARS-CoV-2 не подвергнутого обработке хорька размещали вместе с одним инфицированным хорьком. Второго зараженного хорька перемещали в изолированную клетку до вскрытия через 4 дня после заражения.
Все эксперименты были одобрены IACUC Колледжа ветеринарной медицины Университета Джорджии.
Гистология и иммуногистохимия
Мышей анестезировали и транскардиально перфузировали посредством PBS. Ткани легких собирали, фиксировали в 10% нейтральном забуференном формалине, обезвоживали с помощью серии ванн со спиртом и ксилолом, заливали парафином, делали срезы ~4 мкм и окрашивали красителями, представляющими собой гематоксилин и эозин (НЕ). Ткани исследовались сертифицированным патологоанатом по схеме с маскировкой данных и в соответствии с принципами воспроизводимых балльных оценок тканей (Meyerholz et al. Lab Invest 98, 844-855 (2018)). Периваскулярную эозинофильную инфильтрацию оценивали, как описано ранее (Li et al. mBio 11, (2020)). Вкратце, периваскулярные области с клеточной инфильтрацией выбирались случайным образом (n=20/легкое), а количество эозинофилов подсчитывалось и усреднялось для окончательной балльной оценки для каждого легкого. Периваскулярные лимфоидные агрегаты поддавали балльной оценке посредством ординальной шкалы: 0 - отсутствуют, 1
немногочисленные одиночные клетки, 2 от умеренно мелких до средних агрегатов, или 3 устойчивые агрегаты, образующие периферические периваскулярные манжетки со сдавливанием прилегающей паренхимы. Интерстициальное заболевание поддавали балльной оценке посредством ординальной шкалы с использованием модифицированного Н балла: 0 отсутствует, 1 мелкие рассеянные клетки в перегородках, 2 умеренно инфильтрует перегородки и распространяется в просвет, или 3 от умеренных до выраженных инфильтратов в перегородках и просвете с ассоциированными уплотнением/ателектазом и/или отеком. Для каждого уровня регистрировали % пораженных легких. Окончательный модифицированный Н балл для каждого образца легкого рассчитывали следующим образом: % пораженных × балл для каждого уровня, суммируется, а затем делится на 100 с получением балла от 0 до 3.
Иммуногистохимический анализ выполняли, как описано ранее (Zheng et al. Nature 589, 603-607 (2021)). Вкратце, за первичным антителом к N-белку SARS-CoV-2 (разбавление 1:20000 × 60 мин., 40143-R019, SinoBiological) следовали Rabbit Envision (Dako) и диаминобензидин (DAB, Dako) в качестве хромогена с гематоксилином в качестве контрастного красителя. Оценку иммуноокрашивания посредством ординальной шкалы проводили на основе распределения: 0 - отсутствует, 1 - от 0-25%, 2 - 26-50%, 3 - 51-75% и 4 ->75% легочных полей в срезе ткани.
Нейтрализация S SARS-CoV-2
Для анализа нейтрализации использовали описанный ранее VSV, псевдотипированный с помощью SARS-CoV-2 (VSV-S) (Havranek et al. Viruses 12, (2020)). Частицы VSV-S титровали посредством TCIDso в 96-луночном планшете в клетках Vero для определения оптимального числа частиц для нейтрализации. Их разбавляли в 2% FBS, 1% P/S в средах DMEM. Титр определяли с использованием аналитической системы для выявления с помощью люциферазы Firefly Bio Glo Luciferase Assay System от Promega. Ha каждом планшете использовался контроль частиц, который состоял из разбавленных частиц и стерильного PBS (бессывороточного) в соотношении 1:1. На каждом планшете также использовался отрицательный контроль, состоящий из стерильного PBS и 2% FBS, 1% P/S в средах DMEM для учета фонового уровня.
Сыворотку крови инактивировали нагреванием при 56°С в течение 45 минут перед анализом нейтрализации. Затем его подвергали серийному разбавлению в стерильном PBS с использованием разбавлений 1:2, начиная с 1:100 (хорьки) в 96 луночном необработанном планшете, не содержащем клеток. Образец восстановленной сыворотки крови человека (+RHS) использовали в качестве положительного контроля в каждом анализе и последовательно разбавляли 1:2, начиная с 1:100. Образцы и контроли получали в четырех повторностях.
Добавляли 50 мкл разбавленных частиц VSV-S в 96-луночный планшет, содержащий 50 мкл разбавленной сыворотки крови, и инкубировали в течение 1 часа при 5% СО2 при температуре 37°С. Через 1 час среды аспирировали из белого 96 луночного планшета, содержащего клетки Vero при конфлюентности 90-100%, и к клеткам добавляли смесь частиц и сыворотки крови (100 мкл). Планшет инкубировали при 5% СО2 при температуре 37°С в течение 18 24 часов. На следующий день среды аспирировали и в каждую лунку добавляли по 50 мкл реагента Nano-glo. Планшеты встряхивали, а затем немедленно считывали с применением протокола для люциферазы Renilla на люминометре GloMax от Promega.
Для нейтрализации в случае SARS-CoV-2 образцы сыворотки крови инактивировали нагреванием при 56°С в течение 30 минут перед анализом нейтрализации. Затем их серийно разбавляли в стерильной DMEM и смешивали с равным объемом DMEM, содержащей -20 БОЕ SARS-CoV-2. После инкубации при температуре 37°С в течение 1 часа аликвоты добавляли в клетки Vero Е6 в 12 луночных планшетах и инкубировали при температуре 37°С в 5% СО2 в течение 1 ч. После удаления инокулята планшеты покрывали 1,2% агарозой, содержащей 4% FBS. После дальнейшей инкубации при температуре 37°С в 5% СО2 в течение 2 дней покровные слои удаляли и бляшки визуализировали путем окрашивания 0,1% кристаллическим фиолетовым.
qPCR выделения SARS-CoV-2 в среду из назальной полости
Для получения стандартной кривой для РНК-генома SARS-CoV-2 1,1×106 БОЕ/мл или 1,44×107 БОЕ/мл для стандартов хорька и кошки соответственно инактивировали путем разбавления 1:1 (об./об.) реагентом TRIzol (ThermoFisher Scientific) в соответствии с протоколами инактивации. РНК экстрагировали из 600 мкл разбавленного вируса с использованием набора Direct-zol RNA Miniprep Plus (Zymo Research) и элюировали в 50 мкл воды, не содержащей ДНК/РНК, в соответствии с протоколом. Чистоту РНК (А260/А280) и концентрацию оценивали с использованием спектрофотометра/флуорометра DeNovix DS-11 FX+ (DeNovix). РНК хранили при температуре -80С. Для получения стандартной кривой РНК разбавляли в 10 раз и 10 мкл каждого разбавления РНК использовали в 40 мкл реакционной смеси для qPCR с 10 мкл мастер-микса TaqPath 1 -Step RT qPCR Master Mix (ThermoFisher Scientific) и 3 мкл праймера/зонда nCov N1 из набора EUA CDC SARS-2 (Integrated DNA Technologies). Для образцов назальных смывов 500 мкл восстановленного образца назального смыва или ректального мазка инактивировали путем разбавления 1:1 с помощью 2х реагента DNA/RNA Shield (Zymo Research) и хранили при -20°С до экстракции. РНК экстрагировали из 600 мкл разбавленного образца с использованием набора Quick RNA Viral Kit (Zymo Research) и элюировали в 15 мкл воды, не содержащей ДНК/РНК, в соответствии с протоколом. Чистоту РНК (А260/А280) и концентрацию оценивали с использованием спектрофотометра/флуорометра DeNovix DS-11 FX+. РНК хранили при температуре -80°С. Все реакции qPCR анализировали с использованием Agilent Мх3000Р.
qPCR назального смыва хорька. qPCR осуществляли с использованием мастер-микса TaqPath 1 Step RT qPCR Master Mix (ThermoFisher Scientific) на системе для qPCR Мх3000Р от Agilent. Реакционная смесь каждого назального смыва состояла из 10 мкл TaqPath, 3 мкл смеси праймер/зонд, 17 мкл воды и 10 мкл образца, разбавленного до 2 нг, с конечным объемом реакционной смеси 40 мкл на лунку. Праймеры, специфические в отношении гена N nCoV, были следующими:
прямой, 5'-GACCCCAAAATCAGCGAAAT-3' (SEQ ID NO: 14);
обратный, 5'-TCTGGTTACTGCCAGTTGAATCTG - 3' (SEQ ID NO: 15) и
зонд (5' - (FAM)ACCCCGCATTACGTTTGGTGGACC(BHQ1) - 3' (SEQ ID NO: 16), и их приобретали в Integrated DNA Technologies в виде части набора RUO CDC SARS 2. Термический профиль состоял из 1 цикла продолжительностью 2 минуты при 25°С, 1 цикла продолжительностью 15 минут при 50°С, 1 цикла продолжительностью 2 минуты при 95С и 50 циклов по 3 секунды при 95°С, затем 30 секунд при 55°С. qPCR анализы для каждого планшета включали стандарты РНК (10 кратные разбавления по 10 мкл на реакцию в двух повторностях) без матричного контроля, без контроля полимеразы и с использованием образа с добавкой вирусной РНК в качестве положительного контроля. Концентрацию, представленную в БОЕ/мл, в каждом образце определяли с использованием исходной концентрации, представленной в БОЕ/мл, маточного раствора вируса, который использовали для получения стандартной кривой для РНК. Соотношение РНК, используемое в реакции qPCR, рассчитывали путем деления 2 нг на общую концентрацию РНК на образец. Полученное из стандартной кривой значение в БОЕ/мл умножали на соотношение РНК, используемое в реакции, и общее содержание, представленное в БОЕ/мл, в образце рассчитывали путем обратных вычислений с использованием описанных объемов разбавления и экстракции. Образцы со значениями Ct более 37 считались ПЦР-отрицательными. Для каждого планшета для qPCR значение Ct, составляющее 1 БОЕ/ПЦР-реакция (rxn), было определено на основе стандартной кривой и установлено как у=0, предел выявления был установлен на уровне концентрации на стандартной кривой при значении Ct 37, и для сравнения была включена доза заражения. Все образцы со значениями в диапазоне от 1 БОЕ/rxn до предела выявления считаются ПЦР-положительными, но содержащими неинфекционный геномный материал.
qPCR тканей хорька
Образцы тканей инактивировали с помощью 1х реагента DNA/RNA Shield в соотношении 1 мл Shield на 100 мг ткани. Ткань гомогенизировали с использованием шарикового гомогенизатора (TissueLyser II, Qiagen) при частоте 30 в течение 1,5 минуты и гомогенат хранили при 20°С до экстракции. Процесс экстракции был таким же, как и для назальных смывов. Ткани хорька анализировали с применением вышеуказанного протокола со следующими исключениями. Каждая реакционная смесь состояла из 6,66 мкл TaqPath, 2 мкл смеси праймер/зонд, 20,34 мкл воды и 1 мкл образца с конечным объемом реакционной смеси 30 мкл на лунку. HPRT использовали в качестве гена домашнего хозяйства, необходимого чтобы удостовериться в присутствии РНК ткани при отсутствии вирусной РНК.
Специфические в отношении гена HPRT праймеры (прямой, 5' -CACTGGGAAAACAATGCAGA - 3' (SEQ ID NO: 17) и обратный, 5' -ACAAAGTCAGGTTTATAGCCAACA - 3' (SEQ ID NO: 18)) были синтезированы компанией Integrated DNA Technologies. Специфический в отношении гена зонд TaqMan MGB 5' - NED-TGCTGGTGAAgAGGACCCCTCG-MGBNFQ - 3' (SEQ ID N0:19) был синтезирован компанией Applied Biosystems. Флуоресценцию HPRT считывали с использованием спектров поглощения и испускания HEX. Концентрацию, представленную в БОЕ/мл, в каждом образце определяли с использованием исходной концентрации, представленной в БОЕ/мл, маточного раствора вируса, который использовали для получения стандартной кривой для РНК. Общее содержание, представленное в БОЕ/мл, в образце рассчитывали путем обратных вычислений с использованием объемов разбавления и экстракции, описанных для БОЕ/мл гомогената. Образцы со значениями Ct более 37 считались ПЦР-отрицательными. Для каждого планшета для qPCR значение Ct, составляющее 1 БОЕ/мл, было определено на основе стандартной кривой и установлено как у=0, предел выявления был установлен на уровне концентрации на стандартной кривой при значении Ct 37. Все образцы со значениями в диапазоне от 1 БОЕ/мл до предела выявления считаются ПЦР-положительными, но содержащими неинфекционный геномный материал. С целью графического представления значениям СТ ниже 37, но выше порогового значения 1 БОЕ/rxn было присвоено значение количества копий генома/мл, равное 0,5.
Пример 4
Мутации по аминокислотному остатку W886 и/или F888
Следуя процедурам, подробно описанным в предыдущих примерах, была получена конструкция, в которой S-белок коронавируса дополнительно содержит мутации по аминокислотному остатку W886 и/или F888. Схематическое изображение этой конструкции показано на ФИГ. 38. Например, аминокислотная замена по аминокислотному остатку W886 может представлять собой замену триптофана (W) на аргинин (R), и/или аминокислотная замена по аминокислотному остатку W888 может представлять собой замену фенилаланина (F) на аргинин (R).
Полное раскрытие всех патентов, заявок на патенты и публикаций, а также материалов, доступных в электронном виде (включающих, например, нуклеотидные последовательности, предоставленные, например, в GenBank и RefSeq, и аминокислотные последовательности, предоставленные, например, в SwissProt, PIR, PRF, PDB, и продукты трансляции аннотированных кодирующих участков в GenBank и RefSeq), цитируемых в данном документе, включено посредством ссылки. В случае, если существует какое-либо несоответствие между раскрытием настоящей заявки и раскрытием (раскрытиями) любого из документов, включенных в данный документ посредством ссылки, раскрытие настоящей заявки имеет преимущественную силу. Вышеизложенные подробное описание и примеры были представлены исключительно для четкости понимания. На основании этого не должны истолковываться никакие излишние ограничения. Настоящее изобретение не ограничивается приведенными и описанными подробными деталями, а что касается вариантов, очевидных для специалиста в данной области, то они будут включены в настоящее изобретение, определенное формулой изобретения.
Текст перечня последовательностей в свободном формате
SEQ ID NO: 1 Последовательность плазмиды pDA27 (CVX-UGA1)
SEQ ID NO: 2-19 Искусственные олигонуклеотидные праймеры
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> University of Georgia Research Foundation
Исследовательский Фонд Университета Джорджии Инк.
Хэ Бяо
<120> ВАКЦИНА ПРОТИВ COVID-19 НА ОСНОВЕ PIV5
<130> 0235.000294WO01
<150> 63/080,862
<151> 2020-09-21
<150> 63/217,361
<151> 2021-07-01
<160> 19
<170> PatentIn версия 3.5
<210> 1
<211> 25935
<212> ДНК
<213> Искусственная
<220>
<223> Плазмида pDA27 (CVX-UGA1)
<400> 1
gatctgacgt cgaattctaa tacgactcac tatagggacc agggggaaaa cgaagtggtg 60
actcaaatca tcgaagaccc tcgagattac ataggtccgg aacctatggc cttcgtgacc 120
gacctcgagt cagagtagtt caataaggac ctatcaagtt tgggcaattt ttcgtccccg 180
acacaaaaat gtcatccgtg cttaaagcat atgagcgatt cacactcact caagaactgc 240
aagatcagag tgaggaaggt acaatcccac ctacaacact aaaaccggta atcagggtat 300
ttatactaac ctctaataac ccagagctaa gatcccggct tcttctattc tgcctacgga 360
ttgttctcag taatggtgca agggattccc atcgctttgg agcattactt acaatgtttt 420
cgctaccatc agccacaatg ctcaatcatg tcaaattagc tgaccagtca ccagaagctg 480
atatcgaaag ggtagagatc gatggctttg aggagggatc attccgctta atccccaatg 540
ctcgttcagg tatgagccgt ggagagatca atgcctatgc tgcacttgca gaagatctac 600
ctgacacact aaaccatgca acaccttttg ttgattccga agtcgaggga actgcatggg 660
atgagattga gactttctta gatatgtgtt acagtgtcct aatgcaggca tggatagtga 720
cttgcaagtg catgactgcg ccagaccaac ctgctgcttc tattgagaaa cgcctgcaaa 780
aatatcgtca gcaaggcagg atcaacccga gatatctcct gcaaccggag gctcgaagaa 840
taatccagaa tgtaatccga aagggaatgg tggtcagaca tttcctcacc tttgaactgc 900
agcttgcccg agcacaaagc cttgtatcaa ataggtatta tgctatggta ggggatgttg 960
gaaagtatat agagaattgt ggaatgggag gcttcttttt gacactaaaa tatgcattag 1020
gaaccagatg gcccacactt gctttagctg cattttcagg agagctaaca aagctaaagt 1080
ccctcatggc attataccag acccttggtg agcaggcccg atatttggcc ctattggagt 1140
caccacattt gatggatttt gctgcagcaa actacccact gctatatagc tatgctatgg 1200
gaataggcta tgtgttagat gtcaacatga ggaactacgc tttctccaga tcatacatga 1260
ataagacata tttccaattg ggaatggaaa ctgcaagaaa acaacagggt gcagttgaca 1320
tgaggatggc agaagatctc ggtctaactc aagccgaacg caccgagatg gcaaatacac 1380
ttgccaaatt gaccacagca aatcgagggg cagacaccag gggaggagtc aacccgttct 1440
catctatcac tgggacaact caggtgcccg ctgcagcaac aggtgacaca ttcgagagtt 1500
acatggcagc ggatcgactg aggcagagat atgctgatgc aggcacccac gatgatgaga 1560
tgccaccatt ggaagaggag gaagaggacg acacatctgc aggtccacgc actggactaa 1620
ctcttgaaca agtggccttg gacatccaga acgcagcagt tggagctccc atccatacag 1680
atgacctgaa tgccgcactg ggtgatcttg acatctagac aattcagatc ccaatcttaa 1740
atcgacacac ctaattgacc agttagatgg aactacagtg gattccataa ggttcctgcc 1800
taccatcggc ttttaagaaa aaaataggcc cggacgggtt agcaacaagc gactgccgat 1860
gccaataaca caatccacaa tctacaatgg atcccactga tctgagcttc tccccagatg 1920
agatcaataa gctcatagag acaggcctga atactgtgga gtattttact tcccaacaag 1980
tcacaggaac atcctctctt ggaaagaata caataccacc aggggtcaca ggactactaa 2040
ccaatgctgc agaggcaaag atccaagagt caaccaacca tcagaagggt tcagttggtg 2100
ggggcgcaaa accaaagaaa ccgcgaccaa aaattgccat tgtgccagca gatgacaaaa 2160
cggtgcccgg aaagccgatc ccaaaccctc tattaggtct ggactccacc ccgagcaccc 2220
aaactgtgct tgatctaagt gggaaaacat taccatcagg atcctataag ggggttaaac 2280
ttgcgaaatt tggaaaagaa aatctgatga cacggttcat cgaggaaccc agagagaatc 2340
ctatcgcaac caattccccc atcgatttta agaggggcag ggataccggc gggttccata 2400
gaagggagta ctcaatcgga tgggtgggag atgaagtcaa ggtcactgag tggtgcaatc 2460
catcctgttc tccaatcacc gctgcagcaa ggcgatttga atgcacttgt caccagtgtc 2520
cagtcacttg ctctgaatgt gaacgagata cttaatacag tgagaaattt ggactctcgg 2580
atgaatcaac tggagacaaa agtagatcgc attctctcat ctcagtctct aatccagacc 2640
atcaagaatg acatagttgg acttaaagca gggatggcta ctttagaagg aatgattaca 2700
actgtgaaaa tcatggaccc gggagttccc agtaatgtta ctgtggaaga tgtacgcaag 2760
aaactaagta accatgctgt tgttgtgcca gaatcattca atgatagttt cttgactcaa 2820
tctgaagatg taatttcact tgatgagttg gctcgaccaa ctgcaacaag tgttaagaag 2880
attgtcagga aggttcctcc tcagaaggat ctgactggat tgaagatcac actagagcaa 2940
ttggcaaagg attgcatcag caaaccgaag atgagggaag agtatctcct caaaatcaac 3000
caggcttcca gtgaggctca gctaattgac ctcaagaaag caatcatccg cagtgcaatt 3060
tgatcaagaa acacccaatt acactacact ggtatgacac tgtactaacc ctgagggttt 3120
tagaaaaaac gattaacgat aaataagccc gaacactaca caccacccga ggcagccatg 3180
ccatccatca gcatccccgc agaccccacc aatccacgtc aatcaataaa agcgttccca 3240
attgtgatca accgtgatgg gggtgagaaa ggtcgcttgg ttaaacaact acgcacaacc 3300
tacttgaatg acctagatac tcatgagcca ctggtgacat tcgtaaatac ttatggattc 3360
atctacgaac aggatcgggg aaataccatt gtcggagagg atcaacatgg gaagaaaaga 3420
gaggctgtga ctgctgcaat ggttaccctt ggatgtgggc ctaatctacc atcattaggg 3480
aatgtcctgg gacaactgag tgaattccag gtcattgtta ggaagacatc cagcaaagcg 3540
gaagagatgg tctttgaaat tgttaagtat ccgagaatat ttcggggtca tacattaatc 3600
cagaaaggac tagtctgtgt ctccgcagaa aaatttgtta agtcaccagg gaaagtacaa 3660
tctggaatgg actatctctt cattccgaca tttctgtcag tgacttactg tccagctgca 3720
atcaaatttc aggtacctgg ccccatgttg aaaatgagat caagatacac tcagagctta 3780
caacttgaac taatgataag aatcctgtgt aagcccgatt cgccacttat gaaggtccat 3840
atccctgaca aggaaggaag aggatgtctt gtatcagtat ggttgcatgt atgcaatatc 3900
ttcaaatcag gaaacaagaa tggcagtgag tggcaggaat actggatgag aaagtgtgct 3960
aacatgcaac ttgaagtgtc gattgcagat atgtggggac caactatcat aattcatgcc 4020
agaggtcaca ttcccaaaag tgctaagttg ttttttggaa agggtggatg gagctgccat 4080
ccacttcacg aagttgttcc aagtgtcact aaaacactat ggtccgtggg ctgtgagatt 4140
acaaaggcga aggcaataat acaagagagt agcatctctc ttctcgtgga gactactgac 4200
atcataagtc caaaagtcaa aatttcatct aagcatcgcc gctttgggaa atcaaattgg 4260
ggtctgttca agaaaaccaa atcactgccc aacctgacgg agctggaatg actgaccccc 4320
aatcgagact acaccacctc aaactatagg tgggtggtac ctcagtgatt aatctcgtaa 4380
gcactgatcg taggctacaa cacactaata ttatccagat tagagagctt aattagctct 4440
gtattaataa taacactact attccaataa ctggaatcac cagcttgatt tatctccaaa 4500
atgattcaaa gaaaacaaat catattaaga ctatcctaag cacgaaccca tatcgtcctt 4560
caaatcatgg gtactataat tcaatttctg gtggtctcct gtctattggc aggagcaggc 4620
agtcttgatc cagcagccct catgcaaatc ggtgtcattc caacaaatgt ccggcaactt 4680
atgtattata ctgaggcctc atcagcattc attgttgtga agttaatgcc tacaattgac 4740
tcgccgatta gtggatgtaa tataacatca atttcaagct ataatgcaac agtgacaaaa 4800
ctcctacagc cgatcggtga gaatttggag acgattagga accagttgat tccaactcgg 4860
aggagacgcc ggtttgcagg ggtggtgatt ggattagctg cattaggagt agctactgcc 4920
gcacaggtca ctgccgcagt agcactagta aaggcaaatg aaaatgctgc ggctatactc 4980
aatctcaaaa atgcaatcca aaaaacaaat gcagcagttg cagatgtggt ccaggccaca 5040
caatcactag gaacagcagt tcaagcagtt caagatcaca taaacagtgt ggtaagtcca 5100
gcaattacag cagccaattg taaagcccaa gatgctatca ttggctcaat cctcaatctc 5160
tatttgaccg agttgacaac tatcttccac aatcaaatta caaaccctgc attgagtcct 5220
attacaattc aagctttaag gatcctacta gggagtacct tgccgactgt ggtcgaaaaa 5280
tctttcaata cccagataag tgcggctgag cttctctcat cagggttatt gacaggccag 5340
attgtgggat tagatttgac ctatatgcag atggtcataa aaattgagct gccaacttta 5400
actgtacaac ctgcaaccca gatcatagat ctggccacca tttctgcatt cattaacaat 5460
caagaagtca tggcccaatt accaacacgt gttattgtga ctggcagctt gatccaagcc 5520
tatcccgcat cgcaatgcac tattacaccc aacactgtgt actgtaggta taatgatgcc 5580
caagtactct cagatgatac gatggcttgc ctccaaggta acttgacaag atgcaccttc 5640
tctccagtgg ttgggagctt tctcactcga ttcgtgctgt tcgatggaat agtttatgca 5700
aattgcaggt cgatgttgtg caagtgcatg cagcctgctg ctgtgatcct acagccgagt 5760
tcatcccctg taactgtcat tgacatgtac aaatgtgtga gtctgcagct tgacaatctc 5820
agattcacca tcactcaatt ggccaatgta acctacaata gcaccatcaa gcttgaaaca 5880
tcccagatct tgcctattga tccgttggat atatcccaga atctagctgc ggtgaataag 5940
agtctaagtg atgcactaca acacttagca caaagtgaca catacctttc tgcaatcaca 6000
tcagctacga ctacaagtgt attatccata atagcaatct gtcttggatc gttaggttta 6060
atattaataa tcttgctcag tgtagttgtg tggaagttat tgaccattgt cgctgctaat 6120
cgaaatagaa tggagaattt tgtttatcat aattcagcat tccaccactc acgatctgat 6180
ctcagtgaga aaaatcaacc tgcaactctt ggaacaagat aagacagtca tccattagta 6240
atttttaaga aaaaaacgat aggaccgaaa ctagtattga aagaaccgtc tcggtcaatc 6300
taggtaatcg agctgctacc gtctcggaaa gctcaaatca tgctgcctga tccggaagat 6360
ccggaaagca aaaaagctac aaggagaaca ggaaacctaa ttatctgctt cctattcatc 6420
ttctttccgt ttgtaaactt cattgttcca actctaagac acttgctgtc ctaacacctg 6480
ctataggcta tccactgcat catctctttt taagaaaaaa ataggcccgg acgggttagc 6540
aacaagcgac tgccggtgcc aacaacacaa tccacaatct acagtcgacg cggccgccat 6600
gttcgtcttc ctggtcctgc tgcctctggt ctcctcacag tgcgtcaatc tgacaactcg 6660
gactcagctg ccacctgctt atactaatag cttcaccaga ggcgtgtact atcctgacaa 6720
ggtgtttaga agctccgtgc tgcactctac acaggatctg tttctgccat tctttagcaa 6780
cgtgacctgg ttccacgcca tccacgtgag cggcaccaat ggcacaaagc ggttcgacaa 6840
tcccgtgctg ccttttaacg atggcgtgta cttcgcctct accgagaaga gcaacatcat 6900
cagaggctgg atctttggca ccacactgga ctccaagaca cagtctctgc tgatcgtgaa 6960
caatgccacc aacgtggtca tcaaggtgtg cgagttccag ttttgtaatg atcccttcct 7020
gggcgtgtac tatcacaaga acaataagag ctggatggag tccgagttta gagtgtattc 7080
tagcgccaac aactgcacat ttgagtacgt gagccagcct ttcctgatgg acctggaggg 7140
caagcagggc aatttcaaga acctgaggga gttcgtgttt aagaatatcg acggctactt 7200
caaaatctac tctaagcaca cccccatcaa cctggtgcgc gacctgcctc agggcttcag 7260
cgccctggag cccctggtgg atctgcctat cggcatcaac atcacccggt ttcagacact 7320
gctggccctg cacagaagct acctgacacc cggcgactcc tctagcggat ggaccgccgg 7380
cgctgccgcc tactatgtgg gctacctcca gccccggacc ttcctgctga agtacaacga 7440
gaatggcacc atcacagacg cagtggattg cgccctggac cccctgagcg agacaaagtg 7500
tacactgaag tcctttaccg tggagaaggg catctatcag acatccaatt tcagggtgca 7560
gccaaccgag tctatcgtgc gctttcctaa tatcacaaac ctgtgcccat ttggcgaggt 7620
gttcaacgca acccgcttcg ccagcgtgta cgcctggaat aggaagcgga tcagcaactg 7680
cgtggccgac tatagcgtgc tgtacaactc cgcctctttc agcaccttta agtgctatgg 7740
cgtgtccccc acaaagctga atgacctgtg ctttaccaac gtctacgccg attctttcgt 7800
gatcaggggc gacgaggtgc gccagatcgc ccccggccag acaggcaaga tcgcagacta 7860
caattataag ctgccagacg atttcaccgg ctgcgtgatc gcctggaaca gcaacaatct 7920
ggattccaaa gtgggcggca actacaatta tctgtaccgg ctgtttagaa agagcaatct 7980
gaagcccttc gagagggaca tctctacaga aatctaccag gccggcagca ccccttgcaa 8040
tggcgtggag ggctttaact gttatttccc actccagtcc tacggcttcc agcccacaaa 8100
cggcgtgggc tatcagcctt accgcgtggt ggtgctgagc tttgagctgc tgcacgcccc 8160
agcaacagtg tgcggcccca agaagtccac caatctggtg aagaacaagt gcgtgaactt 8220
caacttcaac ggcctgaccg gcacaggcgt gctgaccgag tccaacaaga agttcctgcc 8280
atttcagcag ttcggcaggg acatcgcaga taccacagac gccgtgcgcg acccacagac 8340
cctggagatc ctggacatca caccctgctc tttcggcggc gtgagcgtga tcacacccgg 8400
caccaataca agcaaccagg tggccgtgct gtatcaggac gtgaattgta ccgaggtgcc 8460
cgtggctatc cacgccgatc agctgacccc aacatggcgg gtgtacagca ccggctccaa 8520
cgtcttccag acaagagccg gatgcctgat cggagcagag cacgtgaaca attcctatga 8580
gtgcgacatc ccaatcggcg ccggcatctg tgcctcttac cagacccaga caaactctcc 8640
cagaagagcc cggagcgtgg cctcccagtc tatcatcgcc tataccatgt ccctgggcgc 8700
cgagaacagc gtggcctact ctaacaatag catcgccatc ccaaccaact tcacaatctc 8760
tgtgaccaca gagatcctgc ccgtgtccat gaccaagaca tctgtggact gcacaatgta 8820
tatctgtggc gattctaccg agtgcagcaa cctgctgctc cagtacggca gcttttgtac 8880
ccagctgaat agagccctga caggcatcgc cgtggagcag gataagaaca cacaggaggt 8940
gttcgcccag gtgaagcaaa tctacaagac cccccctatc aaggactttg gcggcttcaa 9000
tttttcccag atcctgcctg atccatccaa gccttctaag cggagcttta tcgaggacct 9060
gctgttcaac aaggtgaccc tggccgatgc cggcttcatc aagcagtatg gcgattgcct 9120
gggcgacatc gcagccaggg acctgatctg cgcccagaag tttaatggcc tgaccgtgct 9180
gccacccctg ctgacagatg agatgatcgc acagtacaca agcgccctgc tggccggcac 9240
catcacatcc ggatggacct tcggcgcagg agccgccctc cagatcccct ttgccatgca 9300
gatggcctat aggttcaacg gcatcggcgt gacccagaat gtgctgtacg agaaccagaa 9360
gctgatcgcc aatcagttta actccgccat cggcaagatc caggacagcc tgtcctctac 9420
agccagcgcc ctgggcaagc tccaggatgt ggtgaatcag aacgcccagg ccctgaatac 9480
cctggtgaag cagctgagca gcaacttcgg cgccatctct agcgtgctga atgacatcct 9540
gagccggctg gacaaggtgg aggcagaggt gcagatcgac cggctgatca ccggccggct 9600
ccagagcctc cagacctatg tgacacagca gctgatcagg gccgccgaga tcagggccag 9660
cgccaatctg gcagcaacca agatgtccga gtgcgtgctg ggccagtcta agagagtgga 9720
cttttgtggc aagggctatc acctgatgtc cttccctcag tctgccccac acggcgtggt 9780
gtttctgcac gtgacctacg tgcccgccca ggagaagaac ttcaccacag cccctgccat 9840
ctgccacgat ggcaaggccc actttccaag ggagggcgtg ttcgtgtcca acggcaccca 9900
ctggtttgtg acacagcgca atttctacga gccccagatc atcaccacag acaacacctt 9960
cgtgagcggc aactgtgacg tggtcatcgg catcgtgaac aataccgtgt atgatccact 10020
ccagcccgag ctggacagct ttaaggagga gctggataag tatttcaaga atcacacctc 10080
ccctgacgtg gatctgggcg acatcagcgg catcaatgcc tccgtggtga acatccagaa 10140
ggagatcgac cgcctgaacg aggtggctaa gaatctgaac gagagcctga tcgacctcca 10200
ggagctgggc aagtatgagc agtacatcaa gtggccctgg tacatctggc tgggcttcat 10260
cgccggcctg atcgccatcg tgatggtgac catcatgctg tgctgtatga catcctgctg 10320
ttcttgcctg aagggctgct gtagctgtgg ctcctgctgt aagttattga ccattgtcgt 10380
tgctaatcga aatagaatgg agaattttgt ttatcataaa tgaacgcgtg ctagcctcct 10440
gccatacttc ctactcacat catatctatt ttaaagaaaa aataggcccg aacactaatc 10500
gtgccggcag tgccactgca cacacaacac tacacataca atacactaca atggttgcag 10560
aagatgcccc tgttaggggc acttgccgag tattatttcg aacaacaact ttaatttttc 10620
tatgcacact attagcatta agcatctcta tcctttatga gagtttaata acccaaaagc 10680
aaatcatgag ccaagcaggc tcaactggat ctaattctag attaggaagt atcactgatc 10740
ttcttaataa tattctctct gtcgcaaatc agattatata taactctgca gtcgctctac 10800
ctctacaatt ggacactctt gaatcaacac tccttacagc cattaagtct cttcaaacaa 10860
gtgacaagct agaacagaac tgctcgtggg gtgctgcact gattaatgat aatagataca 10920
ttaatggcat caatcagttc tatttctcaa ttgctgaggg tcgcaagctg acacttggcc 10980
cacttcttaa tatacctagt ttcattccaa ctgccacgac accagagggc tgcaccagga 11040
tcccatcatt ctcgctcacc aagacacact ggtgttatac acacaatgtt atcctgaatg 11100
gatgccagga tcatgtatcc tcaaatcaat ttgtttccat gggaatcatt gaacccactt 11160
ctgccgggtt tccatccttt cgaaccctaa agactctata tctcagcgat ggggtcaatc 11220
gtaagagctg ctctatcagt acagttccgg ggggttgtat gatgtactgt ttcgtctcta 11280
ctcaaccaga gagggatgac tacttttcta ccgctcctcc agaacaacga attattataa 11340
tgtactataa tgatacaatc gtggagcgca taattaatcc acccggggta ctagatgtat 11400
gggcaacatt gaacccagga acaggaagcg gggtatatta tttaggttgg gtactctttc 11460
caatatatgg cggcgtgatt aaagatacga gtttatggaa taatcaagca aataaatact 11520
ttatccccca gatggttgct gctctctgct cacaaaacca ggcaactcaa gtccaaaatg 11580
ctaagtcatc atactatagc agctggtttg gcaatcgaat gattcagtct gggatcctgg 11640
catgtcctct tcaacaggat ctaaccaatg agtgtttagt tctgcccttt tctaatgatc 11700
aggtgcttat gggtgctgaa gggagattat acatgtatgg tgactcggtg tattactacc 11760
aaagaagcaa tagttggtgg cctatgacca tgctgtataa ggtaaccata acattcacta 11820
atggtcagcc atctgctata tcagctcaga atgtgcccac acagcaggtc cctagacctg 11880
ggacaggaag ctgctctgca acaaatagat gtcccggttt ttgcttgaaa ggagtgtatg 11940
ctgatgcctg gttactgacc aacccttcgt ctaccagtac atttggatca gaagcaacct 12000
tcactggttc ttatctcaac gcagcaactc agcgtatcaa tccgacgatg tatatcgcga 12060
acaacacaca gatcataagc tcacagcaat ttggatcaag cggtcaagaa gcagcatatg 12120
gccacacaac ttgttttagg gacacaggct ctgttatggt atactgtatc tatattattg 12180
aattgtcctc atctctctta ggacaatttc agattgtccc atttatccgt caggtgacac 12240
tatcctaaag gcagaagcct ccaggtctga cccagccaat caaagcatta taccagacca 12300
tggaatgcat accaaacatt attgacacta atgacacaca aaattggttt taagaaaaac 12360
caagagaaca ataggccaga atggctgggt ctcgggagat attactccct gaagtccatc 12420
tcaattcacc aattgtaaag cataagctat actattacat tctacttgga aacctcccaa 12480
atgagatcga cattgacgat ttaggtccat tacataatca aaattggaat caaatagcac 12540
atgaagagtc taacttagcc caacgcttgg taaatgtaag aaattttcta attacccaca 12600
tccctgatct tagaaagggc cattggcaag agtatgtcaa tgtaatactg tggccgcgaa 12660
ttcttccctt gatcccggat tttaaaatca atgaccaatt gcctctactc aaaaattggg 12720
acaagttagt taaagaatca tgttcagtaa tcaatgcggg tacttcccag tgcattcaga 12780
atctcagcta tggactgaca ggtcgtggga acctctttac acgatcacgt gaactctctg 12840
gtgaccgcag ggatattgat cttaagacgg ttgtggcagc atggcatgac tcagactgga 12900
aaagaataag tgatttttgg attatgatca aattccagat gagacaatta attgttaggc 12960
aaacagatca taatgatcct gatttaatca cgtatatcga aaatagagaa ggcataatca 13020
tcataacccc tgaactggta gcattattta acactgagaa tcatacacta acatacatga 13080
cctttgaaat tgtactgatg gtttcagata tgtacgaagg tcgtcacaac attttatcac 13140
tatgcacagt tagcacttac ctgaatcctc tgaagaaaag aataacatat ttattgagcc 13200
ttgtagataa cttagctttt cagataggtg atgctgtata taacataatt gctttgctag 13260
aatcctttgt atatgcacag ttgcaaatgt cagatcccat cccagaactc agaggacaat 13320
tccatgcatt cgtatgttct gagattcttg atgcactaag gggaactaat agtttcaccc 13380
aggatgaatc aagaactgtg acaaccaatt tgatatcccc attccaagat ctgaccccag 13440
atcttacggc tgaattgctc tgtataatga ggctttgggg acaccccatg ctcaccgcca 13500
gtcaagctgc gggaaaggta cgcgagtcca tgtgtgctgg aaaagtatta gactttccca 13560
ccattatgaa aacactagcc tttttccata ctattctgat caatggatac aggaggaagc 13620
atcatggagt atggccaccc ttaaacttac cgggtaatgc ttcaaagggt ctcacggaac 13680
ttatgaatga caatactgag ataagctatg aattcacact taagcattgg aaggaaatct 13740
ctcttataaa attcaagaaa tgttttgatg cagacgcagg tgaggaactc agtatattta 13800
tgaaagataa agcaattagt gccccaaaac aagattggat gagtgtgttt agaagaagcc 13860
taatcaaaca gcgccatcag catcatcagg tccccctacc aaatccattc aatcgacggc 13920
tattgctaaa ctttctcgga gatgacaaat tcgacccgaa tgtggagcta cagtatgtaa 13980
catcaggtga gtatctacat gatgacacgt tttgtgcatc atattcacta aaagagaagg 14040
aaattaaacc tgatggtcga atttttgcaa agttgactaa gagaatgaga tcatgtcaag 14100
ttatagcaga atctctttta gcgaaccatg ctgggaagtt aatgaaagag aatggtgttg 14160
tgatgaatca gctatcatta acaaaatcac tattaacaat gagtcagatt ggaataatat 14220
ccgagaaagc tagaaaatcg actcgagata acataaatca acctggtttc cagaatatcc 14280
agagaaataa atcacatcac tccaagcaag tcaatcagcg agatccaagt gatgactttg 14340
aattggcagc atctttttta actactgatc tcaaaaaata ttgtttacaa tggaggtacc 14400
agacaattat cccatttgct caatcattaa acagaatgta tggttatcct catctctttg 14460
agtggattca cttacggcta atgcgtagta cactttacgt gggggatccc ttcaacccac 14520
cagcagatac cagtcaattt gatctagata aagtaattaa tggagatatc ttcattgtat 14580
cacccagagg tggaattgaa gggctatgtc aaaaggcttg gacaatgata tctatctctg 14640
tgataattct atctgccaca gagtctggca cacgagtaat gagtatggtg cagggagata 14700
atcaagcaat tgctgtcacc acacgagtac caaggagcct gccgactctt gagaaaaaga 14760
ctattgcttt tagatcttgt aatctattct ttgagaggtt aaaatgtaat aattttggat 14820
taggtcacca tttgaaagaa caagagacta tcattagttc tcacttcttt gtttatagca 14880
agagaatatt ctatcagggg aggattctaa cgcaagcctt aaaaaatgct agtaagctct 14940
gcttgacagc tgatgtccta ggagaatgta cccaatcatc atgttctaat cttgcaacta 15000
ctgtcatgag gttaactgag aatggtgttg aaaaagatat ctgtttctac ttgaatatct 15060
atatgaccat caaacagctc tcctatgata tcatcttccc tcaagtgtca attcctggag 15120
atcagatcac attagaatac ataaataatc cacacctggt atcacgattg gctcttctgc 15180
catcccagct aggaggtcta aactacctgt catgcagtag gctgttcaat cgaaacatag 15240
gcgacccggt ggtttccgca gttgcagatc ttaagagatt aattaaatca ggatgtatgg 15300
attactggat cctttataac ttattaggga gaaaaccggg aaacggctca tgggctactt 15360
tagcagctga cccgtactca atcaatatag agtatcaata ccccccaact acagctctta 15420
agaggcacac ccaacaagct ctgatggaac tcagtacgaa tccaatgtta cgtggcatat 15480
tctctgacaa tgcacaggca gaagaaaata atcttgctag atttctcctg gatagggagg 15540
tgatctttcc gcgtgtagct cacatcatca ttgagcaaac cagtgtcggg aggagaaaac 15600
agattcaagg atatttggat tcaactagat cgataatgag taaatcacta gaaattaagc 15660
ccttgtccaa taggaagctt aatgaaatac tggattacaa catcaattac ctagcttaca 15720
atttggcatt actcaagaat gctattgaac ctccgactta tttgaaagca atgactcttg 15780
aaacatgtag catcgacatt gcaaggagcc tccggaagct ctcctgggcc ccactcttgg 15840
gtgggagaaa tcttgaagga ttagagacgc cagatcccat tgaaattact gcaggagcat 15900
taattgttgg atcgggctac tgtgaacagt gtgctgcagg agacaatcga ttcacatggt 15960
ttttcttgcc atctggtatc gagataggag gggatccccg tgataatcct cctatccgtg 16020
taccgtacat tggctccagg actgatgaga ggagggtagc ctcaatggca tacatcaggg 16080
gtgcctcgag tagcctaaaa gcagttctta gactggcggg agtgtacatc tgggcattcg 16140
gagatactct ggagaattgg atagatgcac tggatttgtc tcacactaga gttaacatca 16200
cacttgaaca gctgcaatcc ctcaccccac ttccaacctc tgccaatcta acccatcggt 16260
tggatgatgg cacaactacc ctaaagttta ctcctgcgag ctcttacacc ttttcaagtt 16320
tcactcatat atcaaatgat gagcaatacc tgacaattaa tgacaaaact gcagattcaa 16380
atataatcta ccaacagtta atgatcactg gactcggaat tttagaaaca tggaataatc 16440
ccccaatcaa tagaacattc gaagaatcta ccctacattt gcacactggt gcatcatgtt 16500
gtgtccgacc tgtggactcc tgcatcatct cagaagcatt aacagtcaag ccacatatta 16560
cagtaccgta cagcaataaa tttgtatttg atgaagaccc gctatctgaa tatgagactg 16620
caaaactgga atcgttatca tttcaagccc aattaggcaa cattgatgct gtagatatga 16680
caggtaaatt aacattattg tcccaattca ctgcaaggca gattattaat gcaatcactg 16740
gactcgatga gtctgtctct cttactaatg atgccattgt tgcatcagac tatgtctcca 16800
attggattag tgaatgcatg tataccaaat tagatgaatt atttatgtat tgtgggtggg 16860
aactactatt ggaactatcc tatcaaatgt attatctgag ggtagttggg tggagtaata 16920
tagtggatta ttcttacatg atcttgagaa gaatcccggg tgcagcatta aacaatctgg 16980
catctacatt aagtcatcca aaacttttcc gacgagctat caacctagat atagttgccc 17040
ccttaaatgc tcctcatttt gcatctctgg actacatcaa gatgagtgtg gatgcaatac 17100
tctggggctg taaaagagtc atcaatgtgc tctccaatgg aggggactta gaattagttg 17160
tgacatctga agatagcctt attctcagtg accgatccat gaatctcatt gcaaggaaat 17220
taactttatt atcactgatt caccataatg gtttggaact accaaagatt aaggggttct 17280
ctcctgatga gaagtgtttc gctttgacag aatttttgag gaaagtggtg aactcagggt 17340
tgagttcaat agagaaccta tcaaatttta tgtacaatgt ggagaaccca cggcttgcag 17400
cattcgccag caacaattac tacctgacca gaaaattatt gaattcaata cgagatactg 17460
agtcgggtca agtagcagtc acctcatatt atgaatcatt agaatatatt gatagtctta 17520
agctaacccc acatgtgcct ggtacctcat gcattgagga tgatagtcta tgtacaaatg 17580
attacataat ctggatcata gagtctaatg caaacttgga gaagtatcca attccaaata 17640
gccctgagga tgattccaat ttccataact ttaagttgaa tgctccatcg caccatacct 17700
tacgcccatt agggttgtca tcaactgctt ggtataaggg tataagctgt tgcaggtacc 17760
ttgagcgatt aaagctacca caaggtgatc atttatatat tgcagaaggt agtggtgcca 17820
gtatgacaat catagaatac ctattcccag gaagaaagat atattacaat tctttattta 17880
gtagtggtga caatccccca caaagaaatt atgcaccaat gcctactcag ttcattgaga 17940
gtgtcccata caagctctgg caagcacaca cagatcaata tcccgagatt tttgaggact 18000
tcatccctct atggaacgga aatgccgcca tgactgacat aggaatgaca gcttgtgtag 18060
aattcatcat caatcgagtc ggcccaagga cttgcagttt agtacatgta gatttggaat 18120
caagtgcaag cttaaatcaa caatgcctgt caaagccgat aattaatgct atcatcactg 18180
ctacaactgt tttgtgccct catggggtgc ttattctgaa atatagttgg ttgccattta 18240
ctagatttag tactttgatc actttcttat ggtgctactt tgagagaatc actgttctta 18300
ggagcacata ttctggtcca gctaatcatg aggtttattt aatttgtatc cttgccaaca 18360
actttgcatt ccagactgtc tcgcaggcaa caggaatggc gatgacttta accgatcaag 18420
ggtttacttt gatatcacct gaaagaataa atcagtattg ggatggtcac ttgaagcaag 18480
aacgtatcgt agcagaagca attgataagg tggttctagg agaagatgct ctattcaatt 18540
cgagtgataa tgaattaatt ctcaaatgtg gagggacacc aaatgcacgg aatcttatcg 18600
atatcgagcc agtcgcaact ttcatagaat ttgaacaact gatctgcaca atgttgacaa 18660
cccacttgaa ggaaataatt gatataacaa ggtctggaac ccaggattat gaaagtttat 18720
tactcactcc ttacaattta ggtcttcttg gtaaaatcag tacgatagtg agattattaa 18780
cagaaaggat tctaaatcat actatcagga attggttgat cctcccacct tcgctccgga 18840
tgatcgtgaa gcaggacttg gaattcggca tattcaggat tacttccatc ctcaattctg 18900
atcggttcct gaagctttct ccaaatagga aatacttgat tacacaatta actgcaggct 18960
acattaggaa attgattgag ggggattgta atatcgatct aaccagacct atccaaaaac 19020
aaatctggaa agcattaggt tgtgtagtct attgtcacga tccagtagat caaagggaat 19080
caacagagtt tattgatata aatattaatg aagaaataga ccgcgggatc gatggcgagg 19140
aaatctaaat atatcaagaa tcagaattag tttaagaaaa aagaagagga ttaatcttgg 19200
ttttcccctt ggtgggtcgg catggcatct ccacctcctc gcggtccgac ctgggcatcc 19260
gaaggaggac gcacgtccac tcggatggct aagggagcgg ccggggatcc ggctgctaac 19320
aaagcccgaa aggaagctga gttggctgct gccaccgctg agcaataact agcataaccc 19380
cttggggcct ctaaacgggt cttgaggggt tttttgctga aaggaggaac tatatccgga 19440
tcatgtgagc aaaaggccag caaaaggcca ggaaccgtaa aaaggccgcg ttgctggcgt 19500
ttttccatag gctccgcccc cctgacgagc aaagcttctc gaccaattct catgtttgac 19560
agcttatcat cgaatttctg ccattcatcc gcttattatc acttattcag gcgtagcaac 19620
caggcgttta agggcaccaa taactgcctt aaaaaaatta cgccccgccc tgccactcat 19680
cgcagtactg ttgtaattca ttaagcattc tgccgacatg gaagccatca caaacggcat 19740
gatgaacctg aatcgccagc ggcatcagca ccttgtcgcc ttgcgtataa tatttgccca 19800
tggtgaaaac gggggcgaag aagttgtcca tattggccac gtttaaatca aaactggtga 19860
aactcaccca gggattggct gagacgaaaa acatattctc aataaaccct ttagggaaat 19920
aggccaggtt ttcaccgtaa cacgccacat cttgcgaata tatgtgtaga aactgccgga 19980
aatcgtcgtg gtattcactc cagagcgatg aaaacgtttc agtttgctca tggaaaacgg 20040
tgtaacaagg gtgaacacta tcccatatca ccagctcacc gtctttcatt gccatacgga 20100
attccggatg agcattcatc aggcgggcaa gaatgtgaat aaaggccgga taaaacttgt 20160
gcttattttt ctttacggtc tttaaaaagg ccgtaatatc cagctgaacg gtctggttat 20220
aggtacattg agcaactgac tgaaatgcct caaaatgttc tttacgatgc cattgggata 20280
tatcaacggt ggtatatcca gtgatttttt tctccatttt agcttcctta gctcctgaaa 20340
atctcgataa ctcaaaaaat acgcccggta gtgatcttat ttcattatgg tgaaagttgg 20400
aacctcttac gtgccgatca acgtctcatt ttcgccaaaa gttggcccag ggcttcccgg 20460
tatcaacagg gacaccagga tttatttatt ctgcgaagtg atcttccgtc acaggtattt 20520
attcgcgata agctcatgga gcggcgtaac cgtcgcacag gaaggacaga gaaagcgcgg 20580
atctgggaag tgacggacag aacggtcagg acctggattg gggaggcggt tgccgccgct 20640
gctgctgacg gtgtgacgtt ctctgttccg gtcacaccac atacgttccg ccattcctat 20700
gcgatgcaca tgctgtatgc cggtataccg ctgaaagttc tgcaaagcct gatgggacat 20760
aagtccatca gttcaacgga agtctacacg aaggtttttg cgctggatgt ggctgcccgg 20820
caccgggtgc agtttgcgat gccggagtct gatgcggttg cgatgctgaa acaattatcc 20880
tgagaataaa tgccttggcc tttatatgga aatgtggaac tgagtggata tgctgttttt 20940
gtctgttaaa cagagaagct ggctgttatc cactgagaag cgaacgaaac agtcgggaaa 21000
atctcccatt atcgtagaga tccgcattat taatctcagg agcctgtgta gcgtttatag 21060
gaagtagtgt tctgtcatga tgcctgcaag cggtaacgaa aacgatttga atatgccttc 21120
aggaacaata gaaatcttcg tgcggtgtta cgttgaagtg gagcggatta tgtcagcaat 21180
ggacagaaca acctaatgaa cacagaacca tgatgtggtc tgtcctttta cagccagtag 21240
tgctcgccgc agtcgagcga cagggcgaag ccctcgagtg agcgaggaag caccagggaa 21300
cagcacttat atattctgct tacacacgat gcctgaaaaa acttcccttg gggttatcca 21360
cttatccacg gggatatttt tataattatt ttttttatag tttttagatc ttctttttta 21420
gagcgccttg taggccttta tccatgctgg ttctagagaa ggtgttgtga caaattgccc 21480
tttcagtgtg acaaatcacc ctcaaatgac agtcctgtct gtgacaaatt gcccttaacc 21540
ctgtgacaaa ttgccctcag aagaagctgt tttttcacaa agttatccct gcttattgac 21600
tcttttttat ttagtgtgac aatctaaaaa cttgtcacac ttcacatgga tctgtcatgg 21660
cggaaacagc ggttatcaat cacaagaaac gtaaaaatag cccgcgaatc gtccagtcaa 21720
acgacctcac tgaggcggca tatagtctct cccgggatca aaaacgtatg ctgtatctgt 21780
tcgttgacca gatcagaaaa tctgatggca ccctacagga acatgacggt atctgcgaga 21840
tccatgttgc taaatatgct gaaatattcg gattgacctc tgcggaagcc agtaaggata 21900
tacggcaggc attgaagagt ttcgcgggga aggaagtggt tttttatcgc cctgaagagg 21960
atgccggcga tgaaaaaggc tatgaatctt ttccttggtt tatcaaacgt gcgcacagtc 22020
catccagagg gctttacagt gtacatatca acccatatct cattcccttc tttatcgggt 22080
tacagaaccg gtttacgcag tttcggctta gtgaaacaaa agaaatcacc aatccgtatg 22140
ccatgcgttt atacgaatcc ctgtgtcagt atcgtaagcc ggatggctca ggcatcgtct 22200
ctctgaaaat cgactggatc atagagcgtt accagctgcc tcaaagttac cagcgtatgc 22260
ctgacttccg ccgccgcttc ctgcaggtct gtgttaatga gatcaacagc agaactccaa 22320
tgcgcctctc atacattgag aaaaagaaag gccgccagac gactcatatc gtattttcct 22380
tccgcgatat cacttccatg acgacaggat agtctgaggg ttatctgtca cagatttgag 22440
ggtggttcgt cacatttgtt ctgacctact gagggtaatt tgtcacagtt ttgctgtttc 22500
cttcagcctg catggatttt ctcatacttt ttgaactgta atttttaagg aagccaaatt 22560
tgagggcagt ttgtcacagt tgatttcctt ctctttccct tcgtcatgtg acctgatatc 22620
gggggttagt tcgtcatcat tgatgagggt tgattatcac agtttattac tctgaattgg 22680
ctatccgcgt gtgtacctct acctggagtt tttcccacgg tggatatttc ttcttgcgct 22740
gagcgtaaga gctatctgac agaacagttc ttctttgctt cctcgccagt tcgctcgcta 22800
tgctcggtta cacggctgcg gcgagcgcta gtgataataa gtgactgagg tatgtgctct 22860
tcttatctcc ttttgtagtg ttgctcttat tttaaacaac tttgcggttt tttgatgact 22920
ttgcgatttt gttgttgctt tgcagtaaat tgcaagattt aataaaaaaa cgcaaagcaa 22980
tgattaaagg atgttcagaa tgaaactcat ggaaacactt aaccagtgca taaacgctgg 23040
tcatgaaatg acgaaggcta tcgccattgc acagtttaat gatgacagcc cggaagcgag 23100
gaaaataacc cggcgctgga gaataggtga agcagcggat ttagttgggg tttcttctca 23160
ggctatcaga gatgccgaga aagcagggcg actaccgcac ccggatatgg aaattcgagg 23220
acgggttgag caacgtgttg gttatacaat tgaacaaatt aatcatatgc gtgatgtgtt 23280
tggtacgcga ttgcgacgtg ctgaagacgt atttccaccg gtgatcgggg ttgctgccca 23340
taaaggtggc gtttacaaaa cctcagtttc tgttcatctt gctcaggatc tggctctgaa 23400
ggggctacgt gttttgctcg tggaaggtaa cgacccccag ggaacagcct caatgtatca 23460
cggatgggta ccagatcttc atattcatgc agaagacact ctcctgcctt tctatcttgg 23520
ggaaaaggac gatgtcactt atgcaataaa gcccacttgc tggccggggc ttgacattat 23580
tccttcctgt ctggctctgc accgtattga aactgagtta atgggcaaat ttgatgaagg 23640
taaactgccc accgatccac acctgatgct ccgactggcc attgaaactg ttgctcatga 23700
ctatgatgtc atagttattg acagcgcgcc taacctgggt atcggcacga ttaatgtcgt 23760
atgtgctgct gatgtgctga ttgttcccac gcctgctgag ttgtttgact acacctccgc 23820
actgcagttt ttcgatatgc ttcgtgatct gctcaagaac gttgatctta aagggttcga 23880
gcctgatgta cgtattttgc ttaccaaata cagcaatagt aatggctctc agtccccgtg 23940
gatggaggag caaattcggg atgcctgggg aagcatggtt ctaaaaaatg ttgtacgtga 24000
aacggatgaa gttggtaaag gtcagatccg gatgagaact gtttttgaac aggccattga 24060
tcaacgctct tcaactggtg cctggagaaa tgctctttct atttgggaac ctgtctgcaa 24120
tgaaattttc gatcgtctga ttaaaccacg ctgggagatt agataatgaa gcgtgcgcct 24180
gttattccaa aacatacgct caatactcaa ccggttgaag atacttcgtt atcgacacca 24240
gctgccccga tggtggattc gttaattgcg cgcgtaggag taatggctcg cggtaatgcc 24300
attactttgc ctgtatgtgg tcgggatgtg aagtttactc ttgaagtgct ccggggtgat 24360
agtgttgaga agacctctcg ggtatggtca ggtaatgaac gtgaccagga gctgcttact 24420
gaggacgcac tggatgatct catcccttct tttctactga ctggtcaaca gacaccggcg 24480
ttcggtcgaa gagtatctgg tgtcatagaa attgccgatg ggagtcgccg tcgtaaagct 24540
gctgcactta ccgaaagtga ttatcgtgtt ctggttggcg agctggatga tgagcagatg 24600
gctgcattat ccagattggg taacgattat cgcccaacaa gtgcttatga acgtggtcag 24660
cgttatgcaa gccgattgca gaatgaattt gctggaaata tttctgcgct ggctgatgcg 24720
gaaaatattt cacgtaagat tattacccgc tgtatcaaca ccgccaaatt gcctaaatca 24780
gttgttgctc ttttttctca ccccggtgaa ctatctgccc ggtcaggtga tgcacttcaa 24840
aaagccttta cagataaaga ggaattactt aagcagcagg catctaacct tcatgagcag 24900
aaaaaagctg gggtgatatt tgaagctgaa gaagttatca ctcttttaac ttctgtgctt 24960
aaaacgtcat ctgcatcaag aactagttta agctcacgac atcagtttgc tcctggagcg 25020
acagtattgt ataagggcga taaaatggtg cttaacctgg acaggtctcg tgttccaact 25080
gagtgtatag agaaaattga ggccattctt aaggaacttg aaaagccagc accctgatgc 25140
gaccacgttt tagtctacgt ttatctgtct ttacttaatg tcctttgtta caggccagaa 25200
agcataactg gcctgaatat tctctctggg cccactgttc cacttgtatc gtcggtctga 25260
taatcagact gggaccacgg tcccactcgt atcgtcggtc tgattattag tctgggacca 25320
cggtcccact cgtatcgtcg gtctgattat tagtctggga ccacggtccc actcgtatcg 25380
tcggtctgat aatcagactg ggaccacggt cccactcgta tcgtcggtct gattattagt 25440
ctgggaccat ggtcccactc gtatcgtcgg tctgattatt agtctgggac cacggtccca 25500
ctcgtatcgt cggtctgatt attagtctgg aaccacggtc ccactcgtat cgtcggtctg 25560
attattagtc tgggaccacg gtcccactcg tatcgtcggt ctgattatta gtctgggacc 25620
acgatcccac tcgtgttgtc ggtctgatta tcggtctggg accacggtcc cacttgtatt 25680
gtcgatcaga ctatcagcgt gagactacga ttccatcaat gcctgtcaag ggcaagtatt 25740
gacatgtcgt cgtaacctgt agaacggagt aacctcggtg tgcggttgta tgcctgctgt 25800
ggattgctgc tgtgtcctgc ttatccacaa cattttgcgc acggttatgt ggacaaaata 25860
cctggttacc caggccgtgc cggcacgtta accgggctgc atccgatgca agtgtgtcgc 25920
tgtcgagttt aaacg 25935
<210> 2
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 2
accaagggga aaatgaagtg gtgac 25
<210> 3
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 3
tccaggtcac tgttaggaag acatc 25
<210> 4
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 4
agtgtgttgt agcctacgat cagtg 25
<210> 5
<211> 23
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 5
caagatgcac cttctctcca gtg 23
<210> 6
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 6
gctcgattac ctagattgac cgagac 26
<210> 7
<211> 23
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 7
taactctgca gtcgctctac ctc 23
<210> 8
<211> 23
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 8
acatgatcct ggcatccatt cag 23
<210> 9
<211> 22
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 9
tcaacccacc agcagatacc ag 22
<210> 10
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 10
tcaatgatga tgtgagctac acgc 24
<210> 11
<211> 26
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 11
accaagggga aaaccaagat taatcc 26
<210> 12
<211> 49
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 12
atccacaatc tacagtcgac gcggccgcca tgttcgtctt cctggtcct 49
<210> 13
<211> 104
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 13
gtatggcagg aggctagcac gcgttcattt atgataaaca aaattctcca ttctatttcg 60
attagcaacg acaatggtca ataacttaca gcaggagcca cagc 104
<210> 14
<211> 20
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 14
gaccccaaaa tcagcgaaat 20
<210> 15
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 15
tctggttact gccagttgaa tctg 24
<210> 16
<211> 29
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 16
tttccccgca ttacgtttgg tggaccggg 29
<210> 17
<211> 20
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 17
cactgggaaa acaatgcaga 20
<210> 18
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 18
acaaagtcag gtttatagcc aaca 24
<210> 19
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 19
ttttgctggt gaagaggacc cctcg 25
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Вакцина против гриппа типа А, гриппа типа B и COVID-19 | 2021 |
|
RU2751485C1 |
| Искусственный ген, кодирующий бицистронную структуру, образованную последовательностями рецептор-связующего домена гликопротеина S коронавируса SARS-CoV-2, P2A-пептида и гликопротеина G VSV, рекомбинантная плазмида pStem-rVSV-Stbl_RBD_SC2, обеспечивающая экспрессию искусственного гена и рекомбинантный штамм вируса везикулярного стоматита rVSV-Stbl_RBD_SC2, используемого для создания вакцины против коронавируса SARS-CoV-2 | 2020 |
|
RU2733831C1 |
| РЕКОМБИНАНТНЫЙ ВИРУС ГРИППА, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ПРОФИЛАКТИКИ COVID-19 И ГРИППА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2802058C1 |
| Рекомбинантный вакцинный штамм для живой интраназальной вакцины, обеспечивающей сочетанную профилактику гриппозной и коронавирусной инфекций | 2022 |
|
RU2782531C1 |
| Рекомбинантный штамм Sen-Sdelta(M) вируса Сендай, продуцирующий S белок SARS-CoV-2 и используемый для получения интраназальной вакцины против COVID-19 | 2024 |
|
RU2837536C1 |
| Искусственный ген Stbl_RBD_TrM_SC2, кодирующий бицистронную структуру, образованную последовательностями рецепторсвязывающего домена гликопротеина S коронавируса SARS-CoV-2, трансмембранного региона, P2A-пептида и гликопротеина G VSV, рекомбинантная плазмида pStem-rVSV-Stbl_RBD_TrM_SC2, обеспечивающая экспрессию искусственного гена, и рекомбинантный штамм вируса везикулярного стоматита rVSV-Stbl_RBD_TrM_SC2, используемый для создания вакцины против коронавируса SARS-CoV-2 | 2020 |
|
RU2733832C1 |
| Пептидные иммуногены и вакцинная композиция против коронавирусной инфекции COVID-19 с использованием пептидных иммуногенов | 2020 |
|
RU2743593C1 |
| Средство для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2 в жидкой форме (варианты) | 2021 |
|
RU2743963C1 |
| Вакцинная композиция против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743595C1 |
| Пептидные иммуногены, используемые в качестве компонентов вакцинной композиции против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743594C1 |










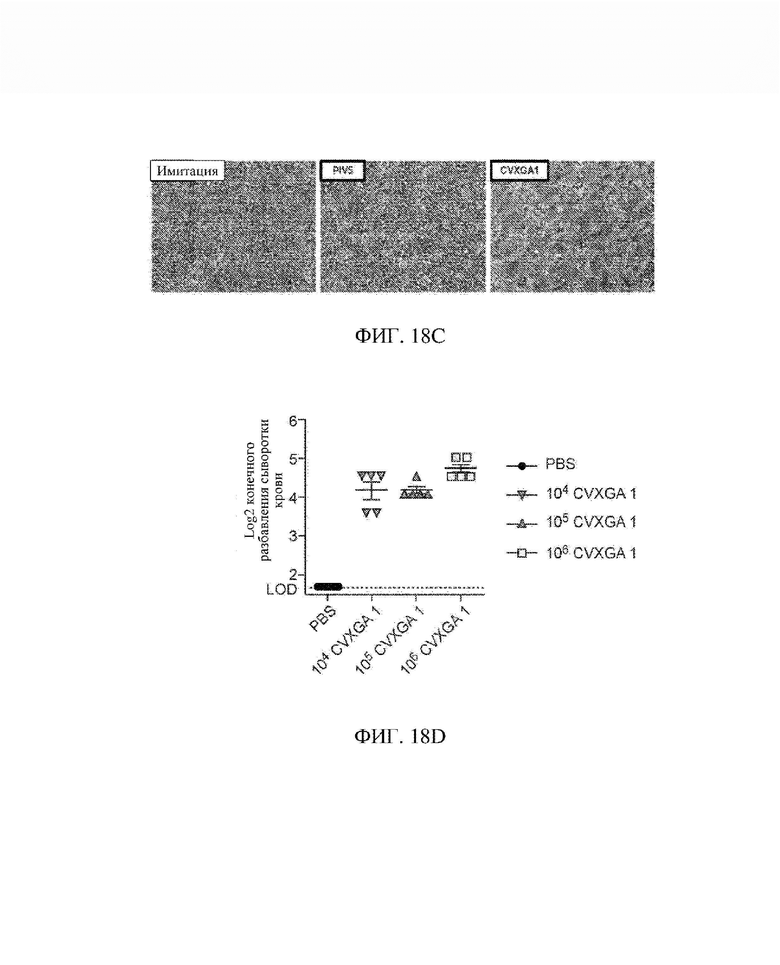
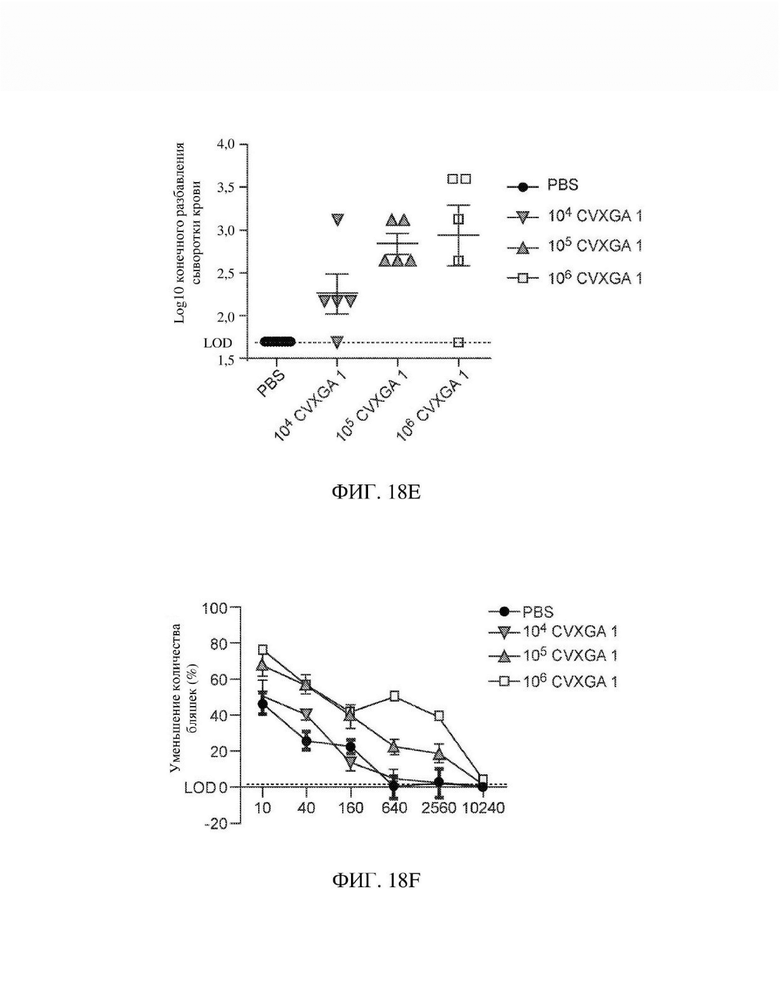
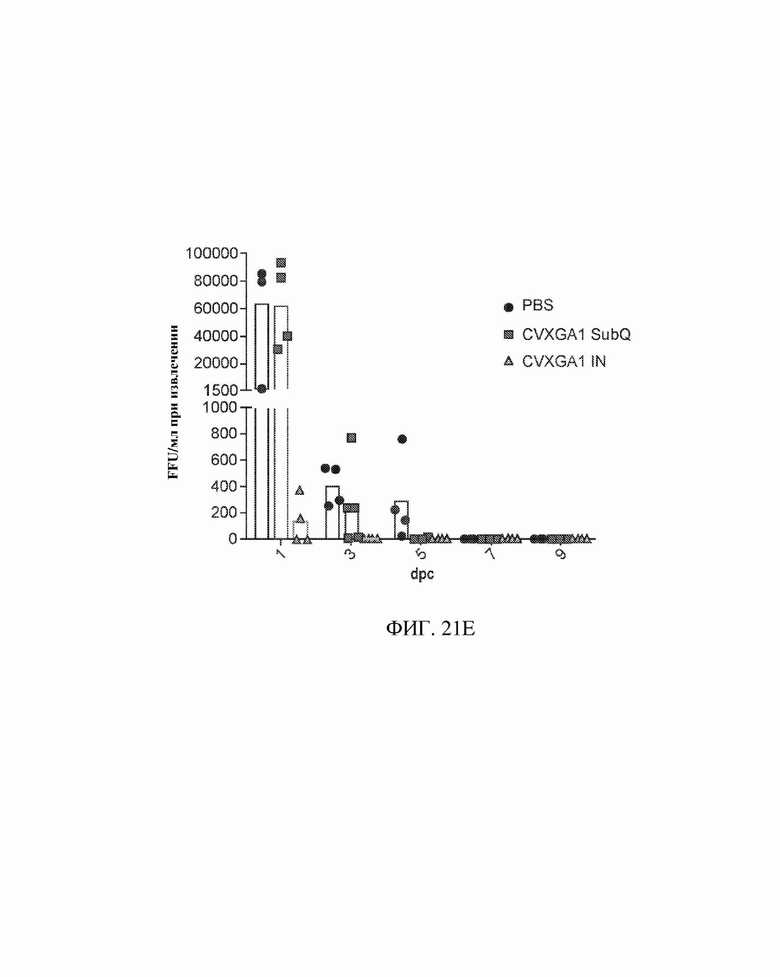
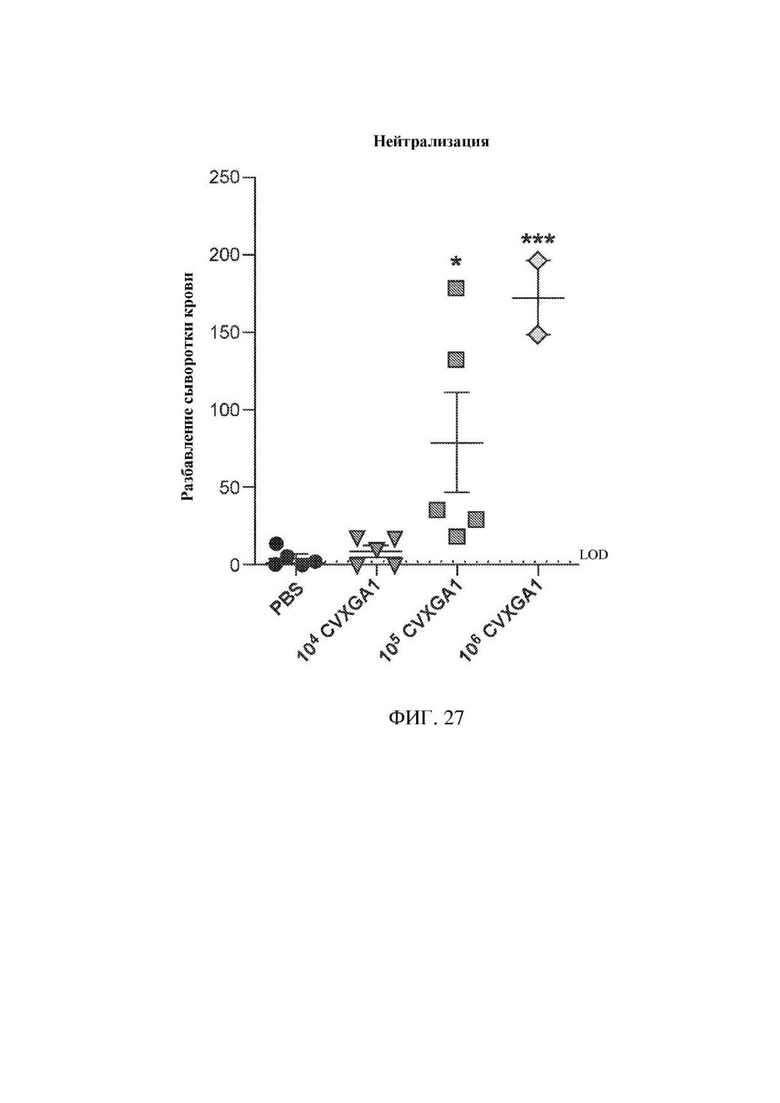
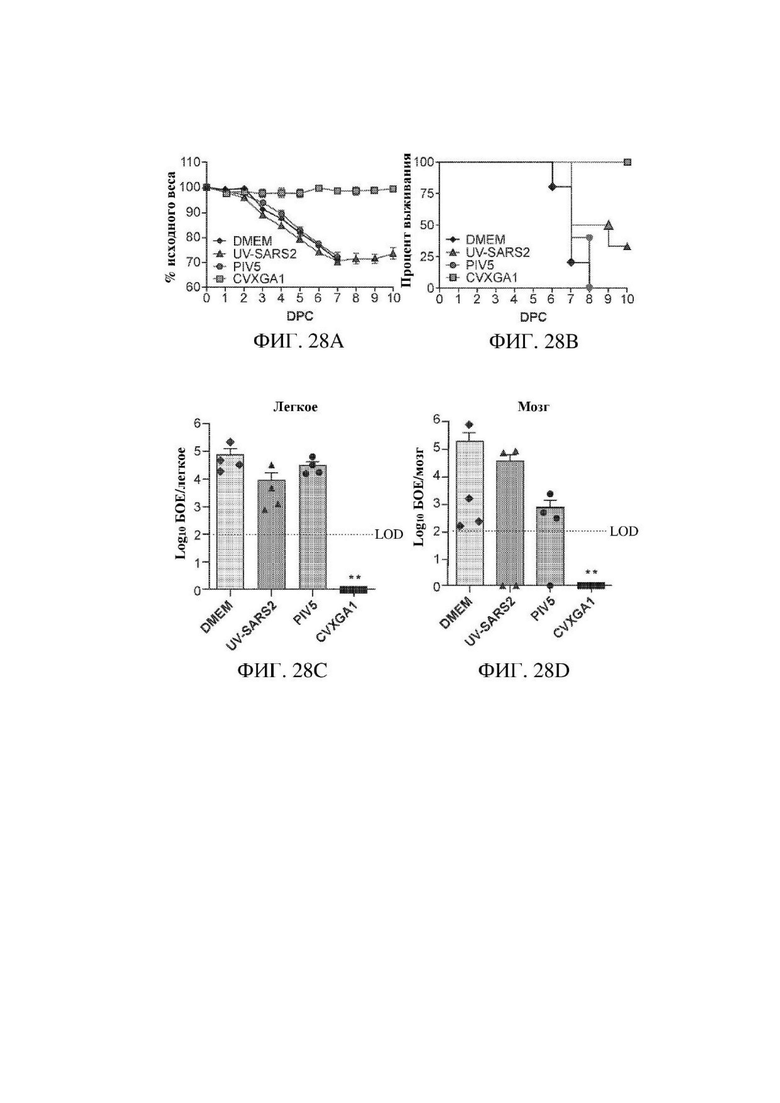
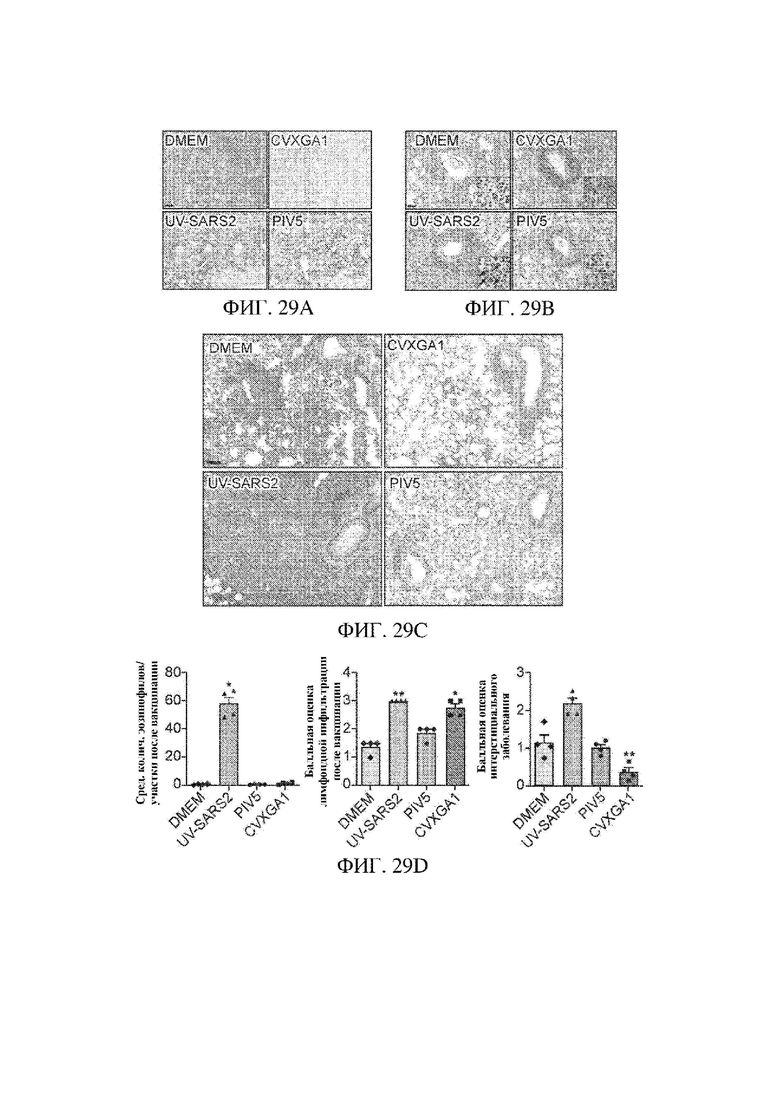
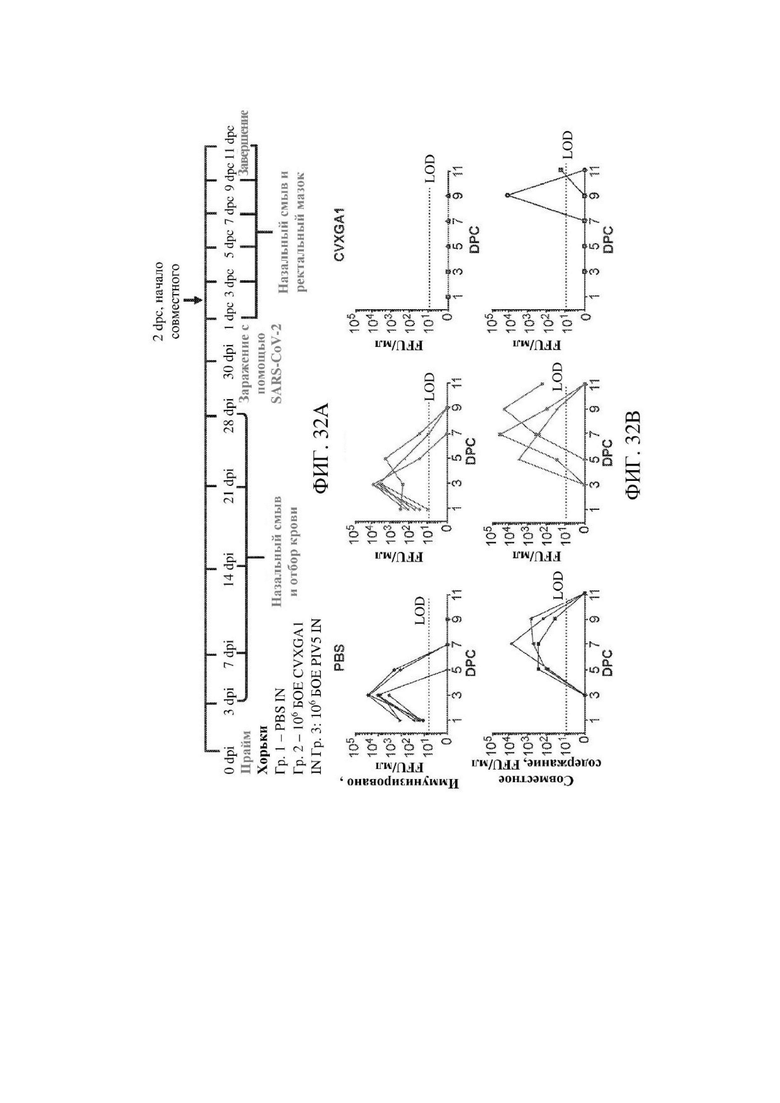
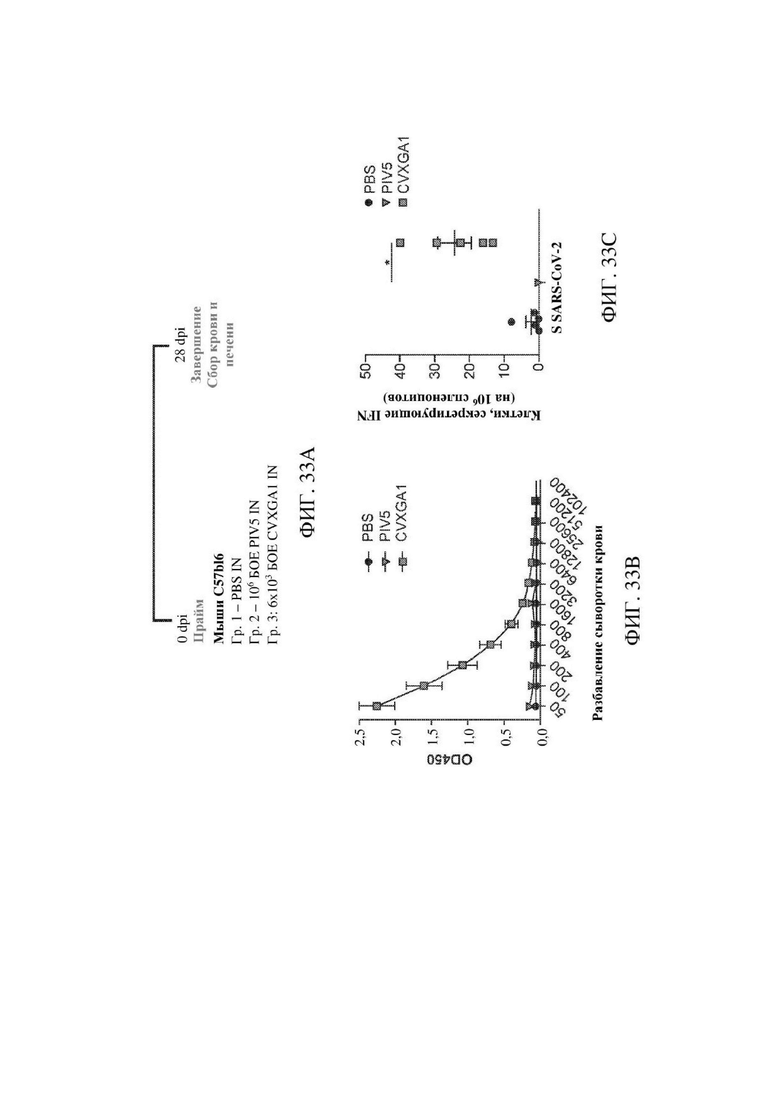
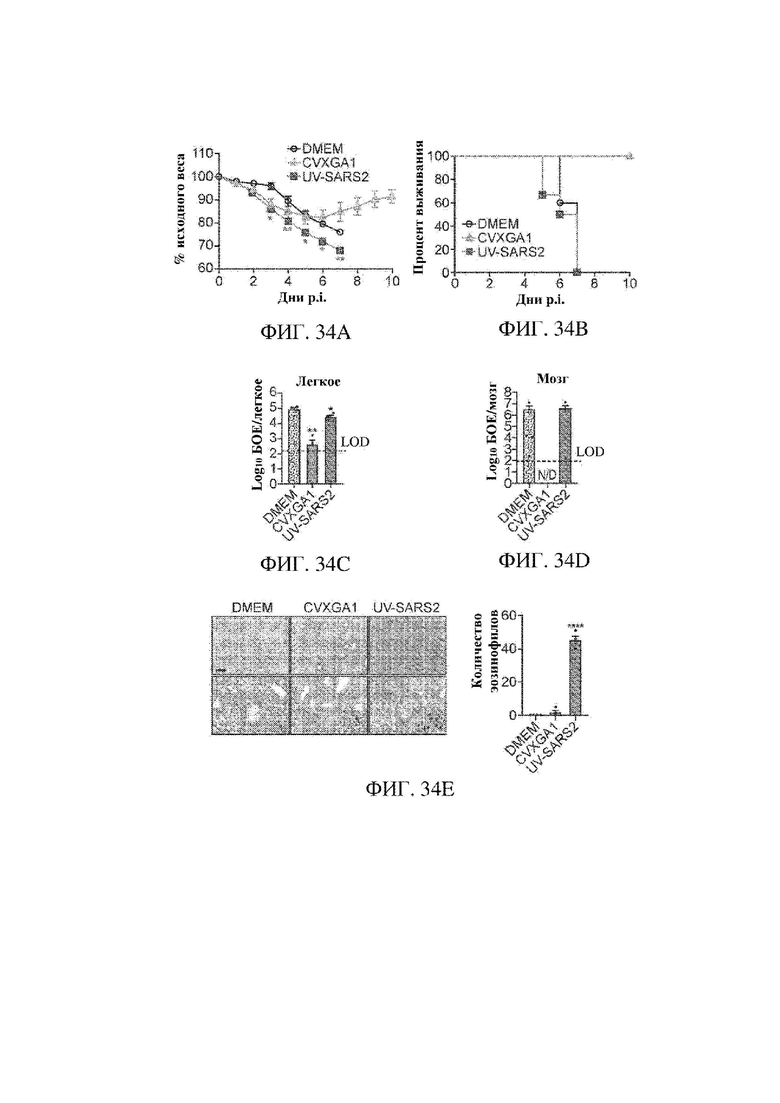
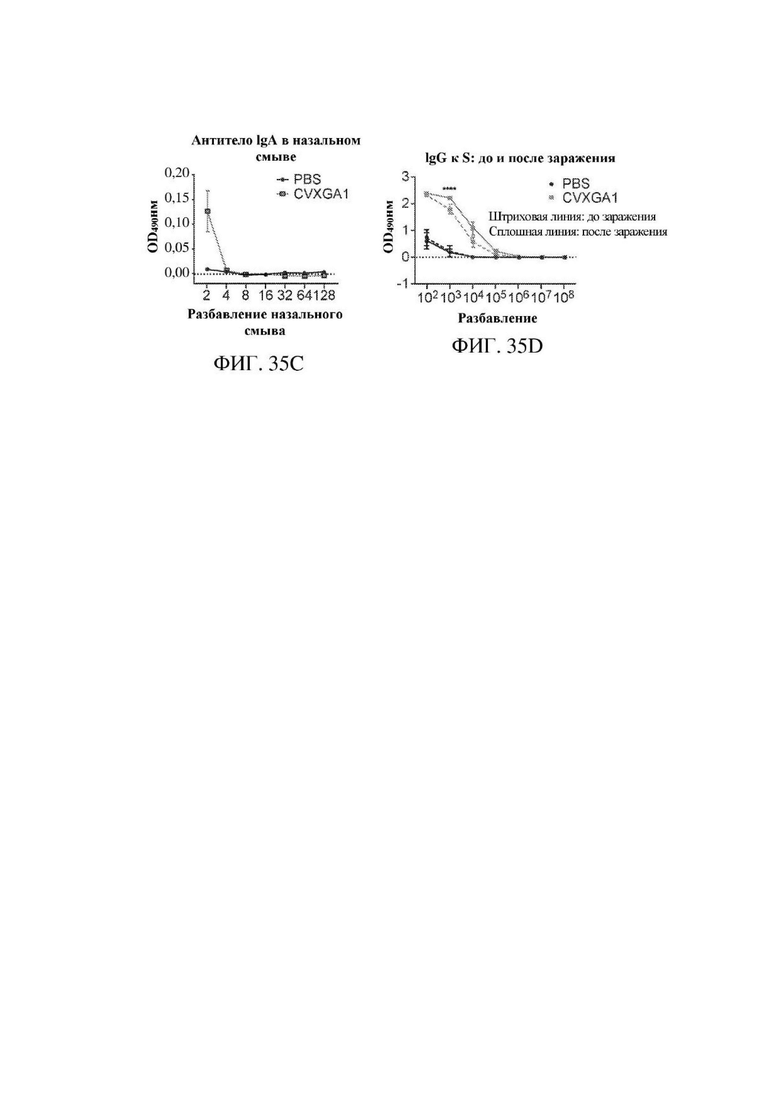
Изобретение относится к биотехнологии, вирусологии и медицине. Предложен вектор экспрессии на основе вируса, содержащий геном вируса парагриппа 5 (PIV5) Вектор содержит гетерологичную нуклеотидную последовательность, обеспечивающую экспрессию S-белка коронавируса из коронавируса 2 тяжелого острого респираторного синдрома (SARS-CoV-2). Гетерологичная нуклеотидная последовательность встроена между геном низкомолекулярного гидрофобного белка (SH) и геном гемагглютинин-нейраминидазы (HN) в геноме PIV5, и цитоплазматический хвост S-белка коронавируса был заменен на цитоплазматический хвост белка слияния (F) PIV5. Предложены: вирусная частица для экспрессии гетерологичного шиповидного (S) гликопротеина коронавируса, содержащая вектор экспрессии; композиция для экспрессии гетерологичного шиповидного (S) гликопротеина коронавируса, содержащая вектор экспрессии и/или вирусную частицу; способ обеспечения экспрессии гетерологичного шиповидного (S) гликопротеина коронавируса в клетке; способ индуцирования иммунного ответа у субъекта на шиповидный (S) гликопротеин коронавируса; способ вакцинирования субъекта против коронавирусного заболевания 2019 г. (COVID-19), при этом способ включает введение субъекту вектора экспрессии на основе вируса, вирусной частицы или композиции. Изобретение предусматривает конструкции на основе вируса парагриппа типа 5 (PIV5), обеспечивающие экспрессию шиповидного (S) белка оболочки SARS-CoV-2, для использования в качестве безопасных, стабильных и эффективных вакцин против COVID-19. 6 н. и 4 з.п. ф-лы, 90 ил., 4 табл., 4 пр.
1. Вектор экспрессии на основе вируса, содержащий геном вируса парагриппа 5 (PIV5), который содержит гетерологичную нуклеотидную последовательность, обеспечивающую экспрессию гетерологичного полипептида, где гетерологичный полипептид предусматривает шиповидный (S) белок коронавируса; где S-белок коронавируса предусматривает S-белок коронавируса из коронавируса 2 тяжелого острого респираторного синдрома (SARS-CoV-2), где гетерологичная нуклеотидная последовательность встроена между геном низкомолекулярного гидрофобного белка (SH) и геном гемагглютинин-нейраминидазы (HN) в геноме PIV5, и где цитоплазматический хвост S-белка коронавируса был заменен на цитоплазматический хвост белка слияния (F) PIV5.
2. Вектор экспрессии на основе вируса по п. 1, где шиповидный (S) белок коронавируса предусматривает наличие мутаций по аминокислотному остатку W886 и/или F888, предпочтительно где аминокислотная замена по аминокислотному остатку W886 предусматривает замену триптофана (W) на аргинин (R), и/или аминокислотная замена по аминокислотному остатку W888 предусматривает замену фенилаланина (F) на аргинин (R).
3. Вектор экспрессии на основе вируса по любому из пп. 1 или 2, где геном PIV5 дополнительно предусматривает наличие одной или нескольких мутаций, предпочтительно где одна или несколько мутаций предусматривают мутацию гена V/P, мутацию общего N-конца белков V и P, мутацию остатков 26, 32, 33, 50, 102 и/или 157 общего N-конца белков V и P, мутацию, обеспечивающую отсутствие С-конца белка V, мутацию, обеспечивающую отсутствие низкомолекулярного гидрофобного (SH) белка, мутацию белка слияния (F), мутацию фосфопротеина (P), мутацию высокомолекулярного белка РНК-полимеразы (L), мутацию, обеспечивающую встраивание остатков из вируса парагриппа собак, мутацию, индуцирующую апоптоз, или их комбинацию, более предпочтительно где одна или несколько мутаций предусматривают PIV5VΔC, PIV5ΔSH, PIV5-P-S308G или их комбинацию.
4. Вирусная частица для экспрессии гетерологичного шиповидного (S) гликопротеина коронавируса, содержащая вектор экспрессии на основе вируса по любому из пп. 1-3.
5. Композиция для экспрессии гетерологичного шиповидного (S) гликопротеина коронавируса, содержащая вектор экспрессии на основе вируса по любому из пп. 1-3 и/или вирусную частицу по п. 4.
6. Способ обеспечения экспрессии гетерологичного шиповидного (S) гликопротеина коронавируса в клетке, при этом способ включает приведение клетки в контакт с вектором экспрессии на основе вируса по любому из пп. 1-3, вирусной частицей по п. 4 или композицией по п. 5.
7. Способ индуцирования иммунного ответа у субъекта на шиповидный (S) гликопротеин коронавируса, при этом способ включает введение субъекту вектора экспрессии на основе вируса по любому из пп. 1-3, вирусной частицы по п. 4 или композиции по п. 5.
8. Способ по п. 7, где иммунный ответ предусматривает гуморальный иммунный ответ и/или клеточный иммунный ответ.
9. Способ вакцинирования субъекта против коронавирусного заболевания 2019 г. (COVID-19), при этом способ включает введение субъекту вектора экспрессии на основе вируса по любому из пп. 1-3, вирусной частицы по п. 4 или композиции по п. 5.
10. Способ по любому из пп. 7-9, где вектор экспрессии на основе вируса, вирусную частицу или композицию вводят интраназально, внутримышечно, местно или перорально.
| Kun Li et al., Single-Dose, Intranasal Immunization with Recombinant Parainfluenza Virus 5 Expressing Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Spike Protein Protects Mice from Fatal MERS-CoV Infection, mBio, March/April 2020, v | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Alexandra C | |||
| WALLS et al., Tectonic conformational changes of a coronavirus spike | |||

