Изобретение относится в целом к области биотехнологии, конкретно-к кодон-оптимизированным нуклеотидными последовательностям генов ERVW-1 и MYMK.
Сконструирована экспрессионная кассета, содержащая кодон-оптимизированные последовательности генов ERVW-1 и MYMK, разделенные нуклеотидной последовательностью IRES или нуклеотидной последовательностью, определяющей аминокислотную последовательность P2A, E2A, F2A либо T2A пептидов, с целью экспрессии белков Syncytin-1. Генетическая конструкция (экспрессионная кассета) используется в составе плазмидного вектора. Изобретение позволяет ускорить процесс слияния миогенных клеток, что повысит эффективность генно-клеточной терапии наследственных миодистрофий.
Область техники
Изобретение относится к области генной инженерии, более точно - к кодон-оптимизированным нуклеотидным последовательностям генов MYMK и ERVW-1, разделенных последовательностью 2А пептидов или IRES, к генетическим конструкциям, содержащим кодон оптимизированные нуклеотидные последовательности генов MYMK и ERVW-1.
Изобретение может быть использовано для модификации мышечных клеток в условиях in vitro с целью повышения способности вышеуказанных клеток к слиянию. Более детально заявленное техническое решение относится к генетическим (плазмидным) конструкциям, предназначенным для генетической модификации клеток в целях генно-клеточной терапии, более точно – для усиления слияния мышечных клеток в программе генно-клеточной терапии мышечных дистрофий.
Далее, в целях исключения неоднозначного понимания текста, заявителем приведены использованные в заявочном материале термины и их расшифровка:
кДНК - комплементарная дезоксирибонуклеиновая кислота;
мРНК - матричная рибонуклеиновая кислота;
Саморасщепляющиеся 2А пептиды - класс пептидов длиной 18–22 аминокислот, которые располагаются между двумя целевыми пептидами и обеспечивают их расщепление друг от друга во время синтеза белка путем рибосомного скиппинга;
E2A - саморасщепляющийся 2А пептид из equine rhinitis virus, применяющийся для соединения двух генов в одну генетическую кассету;
F2A - саморасщепляющийся 2А пептид из foot-and-mouth disease virus, применяющийся для соединения двух генов в одну генетическую кассету;
IRES - внутренний сайт входа рибосомы, который обеспечивает трансляцию, независимую от 5'-конца РНК и применяется для соединения двух генов в одну генетическую кассету;
P2A - саморасщепляющийся 2А пептид из porcine teschovirus-1, применяющийся для соединения двух генов в одну генетическую кассету;
T2A - саморасщепляющийся 2А пептид из thosea asigna virus, применяющийся для соединения двух генов в одну генетическую кассету.
П.н. - пара нуклеотидов, основная структурная единица ДНК
Экспрессионная кассета - это тип генетической конструкции, которая включает набор последовательностей ДНК, необходимых для транскрипции и трансляции определенного гена. Она обычно состоит из следующих элементов: 1. Кодирующая последовательность; 2. Промотор; 3. Регуляторные элементы последовательности; 4. Сигналы окончания транскрипции.
Прогрессирующие мышечные дистрофии представляют собой группу из приблизительно 50 генетически обусловленных заболеваний, причиной которых являются мутации в генах, кодирующих различные белки мышечной ткани. Заболевания проявляются постепенной потерей мышечной ткани, что приводит к нарастающей слабости, инвалидности, и, зачастую, к фатальным осложнениям и смерти. Наиболее распространенной является мышечная дистрофия Дюшенна, встречающаяся со средней частотой 1 случай на 3500 новорожденных мальчиков.
Миодистрофии обусловлены мутациями в различных генах, которые могут являться белками внеклеточного матрикса (коллаген IV типа, мерозин), белками базальной мембраны и сарколеммы (интегрины альфа7 и альфа9), сарколеммы (дистрофин, дисферлин, саркогликаны), ферментами саркоплазмы (гликозилтрансферазы), ферментами эндоплазматической сети (селенопротеин N1), ферментами кариолеммы (эмерин, ламины А/С), белками митохондрий (холинкиназа). Мутации, приводящие к нарушению нормальной последовательности генов, вызывают нарушение синтеза белков, что приводит к полному либо частичному выпадению функций, выполняемых этой молекулой. Дисфункциональное мышечное волокно подвергается некрозу, что сопровождается воспалением - естественным ответом организма на некроз, и репаративной регенерацией. В конечном итоге указанные процессы приводят к атрофии и фиброзу скелетной мышечной ткани. В случаях экспрессии белков в других тканях помимо мышечной, процессы мышечной атрофии сопровождаются иными клиническими проявлениями - кардиомиопатией, энцефалопатией, и др.
В клинической практике используется несколько подходов, предназначенных для лечения мышечных дистрофий. К ним относятся:
- применение глюкокортикоидов;
- метаболическая терапия;
- немедикаментозные методы лечения;
- экзон-скиппинг.
Глюкокортикоидные препараты преднизолон и дефлазакорт традиционно являются препаратами первой линии в терапии мышечных дистрофий [Клинический протокол диагностики и лечения: прогрессирующая мышечная дистрофия Дюшенна/Беккера. Казахстан, 2016]. Положительное влияние стероидов на мышечную ткань связано с их способностью активировать сигнальный путь NFAT, что помогает снизить скорость дегенерации мышечных волокон. Также стероиды подавляют воспалительный ответ иммунной системы, возникающий как реакция на некроз мышечного волокна. Согласно альтернативной гипотезе, стероиды способны усиливать пролиферацию миобластов, и тем самым ускорять регенерацию мышечных волокон. [Angelini C. The role of corticosteroids in muscular dystrophy: a critical appraisal. Muscle Nerve. 2007 Oct;36(4):424-35]. Однако, несмотря на оказываемый положительный эффект, глюкокортикоиды не останавливают процесс дегенерации мышечных волокон, и никаким образом не влияют на причины заболевания.
Также применяется метаболическая терапия, направленная на улучшение обменных процессов в скелетных мышцах, миокарде и печени, а именно-угнетает образование кетокислот, снижает выработку лактата и частично устраняет побочные эффекты глюкокортикоидов. Примерами препаратов являются витамины группы В, витамин D3, левокарнитин и препараты кальция. Однако, современные клинические исследования демонстрируют неэффективность подобного подхода [Cedillo R. E. E. et al. L-carnitine supplementation in duchenne muscular dystrophy steroid-naïve patients: A pilot study. August 2013Current Topics in Nutraceutical Research 11(3):97-102].
К немедикаментозным методам лечения относился физическая реабилитация, лечебная физкультура, использование ортопедических приспособлений, таких как ортезы типа «лодыжка-стопа» или «голень-лодыжка-стопа», и другие методы, направленные на предотвращение формирования мышечных контрактур. При сформировавшихся контрактурах применяется оперативное лечение [Клинический протокол диагностики и лечения: прогрессирующая мышечная дистрофия Дюшенна/Беккера. Казахстан, 2016].
Пропуск экзонов осуществляется путем синтетических последовательностей нуклеотидов, комплементарным целевой последовательности нуклеотидов в пре-мРНК (антисмысловые нуклеотиды). При связывании антисмыслового нуклеотида с мутантной последовательностью в пре-мРНК, в ходе сплайсинга эта последовательность не включается в зрелую мРНК, что приводит к синтезу укороченного, но функционально полноценного белка, способного частично выполнять свои функции. Коммерчески доступны препараты на основе фосфородиамидат-морфолиноолигомерных антисмысловых нуклеотидов, такие как голодирсен, этеплирсен, касимерсен и вилтоларсен, однако к моменту подачи заявки клинические испытания вышеописанных препаратов не завершены, достоверные доказательства эффективности отсутствуют. [Center for drug evaluation and research application number: 211970Orig1s000 https://www.accessdata.fda.gov/drugsatfda_docs/nda/2019/211970Orig1s000OtherActionLtrs.pdf дата доступа 01.06.2021].
Таким образом, имеющиеся в настоящий момент методы лечения прогрессирующих мышечных дистрофий являются симптоматическими либо патогенетическими, либо направленными на лечение поздних осложнений.
Возможными методами, способными восстановить эндогенный синтез полноценного белка, являются:
-генная терапия (генный трансфер: вирусные векторы, невирусные конструкции: плазмиды, транспозоны, искусственные хромосомы);
-коррекция генома;
-генно-клеточная терапия [Деев Р. В. и др. Генно-клеточная терапия наследственных заболеваний мышечной системы: современное состояние вопроса. Гены & Клетки: Том IX, №4, 2014 год, стр.: 6-33].
Генная терапия является терапевтическим подходом в основе которого лежит метод доставки в организм больного человека генетической последовательности, которая приведет к выработке функционального продукта гена в количестве, достаточном для компенсации нарушенной функции. К методикам генной терапии относится доставка гена при помощи невирусных методов (плазмиды, транспозоны, искусственные хромосомы), либо использование вирусного вектора. Однако подобный подход сопряжен с высокой вирусной нагрузкой, существующим иммунитетом к вирусным капсидам, а также недостаточной емкостью вирусных конструкций, неспособных вместить последовательности необходимых генов.
Коррекция генома подразумевает коррекцию поврежденной последовательности ДНК у пациента. Существует несколько потенциально возможных подходов: нуклеазы на основе цинковых пальцев (ZFN), эффекторные нуклеазы, сходные с факторами транскрипции (TALEN), и система CRISPR-Cas9. Применение описанных методов пока не показало достаточного для функциональной коррекции уровня синтеза белка, кроме этого, точность связывания подобных систем с ДНК рецепиента остается под вопросом [Kosicki, M., Tomberg, K. & Bradley, A. Repair of double-strand breaks induced by CRISPR–Cas9 leads to large deletions and complex rearrangements. Nat Biotechnol 36, 765–771 (2018). https://doi.org/10.1038/nbt.4192].
Плазмидные векторы в свою очередь не производят изменений в ДНК реципиента, поскольку находятся в цитоплазме в виде эписомы. Поскольку в физиологических условиях ДНК за редкими исключениями не обладает иммуногенностью [Su KY, Pisetsky DS. The role of extracellular DNA in autoimmunity in SLE. Scand J Immunol. 2009 Sep;70(3):175-83. doi], возможно системное использование высоких доз плазмидного вектора. Также, в связи с отсутствием инициации вторичного иммунного ответа, один и тот же плазмидный вектор может быть применен многократно. Однако, время жизни плазмидного вектора в клетке ограничена ферментативной активностью внутриклеточных ДНКаз, неизбежно разрушающих экзогенную последовательность ДНК. Указанная особенность может потребовать регулярной доставки вектора в организм, либо может быть использована для транзиторной экспрессии трансгена.
Методика применения искусственных хромосом заключается в создании носителя генетической информации, обладающего теломерами и центромерой, и несущего целевую генетическую информацию. Таким образом, при попадании в клетку искусственная хромосома будет способна к самовоспроизведению. Создание подобных конструкций весьма ресурсоёмкий процесс, по сравнению с другими методами. Кроме этого, искусственные хромосомы не способны самостоятельно проникать в клетку, поэтому их клиническое применение целесообразно только в составе генно-клеточной терапии.
Генно-клеточная терапия заключается в доставке в организм пациента клеток, содержащих генетический материал без патогенных мутаций. Для этого производят изоляцию аутологичных клеток, которые могут являться миобластами, мезоангиобластами, другой миогенной клеточной линией, либо служить источником индуцированных плюрипотентных стволовых клеток. В дальнейшем клеточная линия подвергается последовательно экспансии - увеличению численности клеток, и генной модификации тем или иным методом, после чего клетка приобретает способность к синтезу полноценного полнофункционального трансгена. В качестве генной модификации может использоваться коррекция генома, искусственные хромосомы, вирусные интегрирующиеся векторы, либо другие способы, в зависимости от характера мутации. Затем модифицированные клетки вводятся обратно в организм пациента, где сливаются с мышечными волокнами. Таким образом, клетки являются вектором для доставки нуклеотидной последовательности в организм пациента [Деев Р. В. и др. Генно-клеточная терапия наследственных заболеваний мышечной системы: современное состояние вопроса. Гены & Клетки: Том IX, №4, 2014 год, стр.: 6-33].
Фактором, ограничивающим эффективность такой терапии, является низкая эффективность слияния: значительная часть миобластов не проникает в периферические ткани из сосудистого русла. Также, среди клеток, доставленных в мышцы посредством местной инъекции, около 50% миобластов выходят из клеточного цикла и не способны слиться с мышечным волокном. При системной доставке клеток (внутривенно либо внутриартериально) до 90 % клеток погибают, до 5 % выходят из клеточного цикла и лишь 5% претерпевают слияние мышечным волокном [Beauchamp J.R., Morgan J.E., Pagel C.N., Partridge T.A. Dynamics of myoblast transplantation reveal a discrete minority of precursors with stem cell-like properties as the myogenic source. J. Cell Biol. 1999; 144: 1113-22.] Использование клеток, способных к миграции через сосудистую стенку, таких как мезоангиобласты, позволяет снизить уровень клеточной гибели в системном кровотоке, однако, процент слияния всё равно остается низким [Sampaolesi M., Torrente Y., Innocenzi A. et al. Cell therapy of alpha-sarcoglycan null dystrophic mice through intra-arterial delivery of mesoangioblasts. Science 2003; 301: 487-92]. Возможным решением вышеописанных проблем является искусственное усиление процессов слияния клеток.
Fusion-феноменом или клеточным слиянием называется процесс объединения двух или более клеток в одну более крупную многоядерную структуру - симпласт. Этот процесс в ходе нормального и патологического гистогенеза может быть обнаружен у многих организмов, включая человека. Слияние является одним из ключевых этапов цито- и гистогенеза при формировании поперечнополосатой скелетной мышечной ткани. При этом ключевыми молекулами, регулирующим слияние цитоплазматических мембран, являются мембранные белки Syncitin-1 и Myomaker, кодируемые соответственно генами ERVW-1 и MYMK [Буев Д. О. и др. Fusion-феномен в нормальном гистогенезе и при патологии: часть 1. Гены & Клетки: Том XIII, №2, 2018 год, стр.: 13-20].
Заявителем проведен анализ существующего уровня техники по патентной и научной информации в области усиления слияния миобластов и других клеток.
Из исследованного уровня техники выявлено изобретение, описанное в заявке на патент WO 2014/210448 A1. Сущностью изобретения является клетка, трансформированная экзогенной нуклеиновой кислотой, кодирующей полипептид Myomaker под контролем промотора, активного в данной клетке. Клетка может являться человеческой клеткой, немиогенной клеткой, такой, как фибробласт, клетка костного мозга или клетка крови. Экзогенная нуклеиновая кислота может быть под контролем конститутивного либо индуцибельного промотора. Экзогенная нуклеиновая кислота может быть встроена в хромосому указанной клетки, либо находится в цитоплазме в форме эписомы. Клетка может экспрессировать детектируемый маркер. Клетка может быть трансформирована другим целевым геном, помимо Myomaker. Также метод подготовки партнера для слияния немиогенной клетки, включающий перенос в немиогенную клетку нуклеиновой кислоты, кодирующей белок Myomaker под контролем промотора, активного в указанной клетке. Клетка может быть стабильно трансформирована либо транзиторно трансфицирована. Метод может включать перенос в указанную клетку нуклеиновой кислоты, кодирующей детектируемый маркер или достаточной для того, чтобы являться маркером. Экзогенная нуклеиновая кислота может быть под контролем конститутивного промотора или индуцированного промотора. Клетка может быть человеческой. Клетка может являться фибробластом, клеткой костного мозга или клеткой крови. Экзогенная нуклеиновая кислота может кодировать маркер селекции. Клетка может быть трансформирована другим геном, помимо Myomaker.
Также метод слияния немиогенной клетки с мышцей, включающий: (а) предоставление немиогенной клетке способности экспрессировать экзогенный белок Myomaker и (б) контакт указанной немиогенной клетки с мышцей, где указанная немиогенная клетка будет сливаться с указанной мышцей. Немиогенная клетка может являться человеческой клеткой, и/или быть фибробластом, клеткой костного мозга, или клеткой крови. Шаг (б) может быть выполнен in vivo или in vitro. Немиогенная клетка может экспресировать маркер детекции и маркер селекции. Мышечная клетка может являться изолированной мышечной клеткой или находиться в интактной мышечной ткани. Мышечная клетка может являться миобластом. Также метод доставки целевого гена в мышечную клетку, включающий (а) предоставление немышечной клетке способности экспрессировать Myomaker, в то время как указанная немиогенная клетка экспрессирует целевой ген, и (б) контакт вышеописанной немиогенной клетки с мышцей, где указанная немиогенная клетка сливается с мышечной клеткой и доставляет целевой ген в вышеуказанную мышечную клетку. Немиогенная клетка может являться человеческой клеткой и/или может являться фибробластом, клеткой костного мозга или клеткой крови. Шаг (б) может быть выполнен in vivo или in vitro. Немиогенная клетка может экспрессировать маркер детекции и/или маркер селекции. Мышечная клетка может являться изолированной мышечной клеткой или мышечной клеткой в интактной мышечной ткани. Мышечная клетка может являться миобластом. Мышечная клетка может иметь патологический фенотип, а целевой ген может корректировать данный генотип. Патологический фенотип может быть пониженной экспрессией или полным отсутствием нормального продукта гена, или экспрессией дефектного продукта гена. Патологический фенотип может быть ассоцииорован с врожденной миопатией, саркопенией, боковым амиотрофическим склерозом, мышечной дистрофией, болезнью Помпе или рабдомиосаркомой. Немиогенная клетка может быть доставлена в мышечную ткань субъекта при помощи внутримышечной инъекции. Доставку возможно повторить как минимум один раз. Указанный субъект может получать вторичную терапию. Немиогенная клетка может быть доставлена в мышцу ex vivo и впоследствии имплантирована в мышечную ткань субъекта.
Таким образом, в изобретении в целом описывается методика генно-клеточной терапии, при которой в качестве вектора используются немиогенные клетки, экспрессирующие увеличенное количество белка Myomaker. Недостатком данного технического решения является тот факт, что миобласты обладают специфическими мембранными и цитоплазматическими белками, участвующими в процессе слияния, в то время как у немиогенных клеток, предлагаемых авторами вышеописанного технического решения, подобных механизмов не имеется. Кроме того, слияние немиогенных клеток с мышечными волокнами является нефизиологическим процессом. Помимо этого, авторы изобретения используют лишь один из мембранных белков, влияющих на процесс слияния миобластов.
Из исследованного уровня техники выявлено изобретение, описанное в патенте US9506033B2 «Compositions and methods for inducing myoblast differentiation and myotube formation» Сущностью изобретения является метод индукции дифференцировки миобластов в миоцит у млекопитающего, включающий а) выбор млекопитающего, имеющего мышечного заболевание, и б) доставка выбранному млекопитающего олигонуклеотида, который может являться ингибирующей РНК, антисмысловым нуклеотидом, или рибозимом, который снижает уровень экспрессии мРНК Map4k4 миобласте млекопитающего, в количестве, достаточном для индукции дифференцировки миобласта в миоцит у выбранного млекопитающего, где олигонуклеотид модифицирован основным фрагментом, углеводным фрагментом, или фосфатной основой; метод, где выбранное млекопитающее является человеком; метод, где мышечное заболевание является мышечной атрофией, мышечной слабостью, миопатией, синдромом хронической усталости, фибромиалгией, мышечной дистрофией, спинальной мышечной атрофией, дистальной мышечной дистрофией, дерматомизитом, полимиозитом, ревматической полимиалгией, разрывом мышцы или растяжением мышцы; метод, при котором доставка олигонуклеотида приводит к лечению мышечного заболевания у выбранного млекопитающего; метод, при котором доставка вызывает снижение мРНК Map4k4 в миобласте млекопитающего, и усиливает экспрессию одного из следующих белков в миобласте: Myf5, креатинкиназа, кальсеквестрин-1, пептидиларгининдезаминаза, CD24a. Также метод индукции миобласта или миоцита в организме млекопитающего, имеющего мыщечное заболевание, с целью формирования миотубы, включающий а) выбор млекопитающего, имеющего мышечное заболевание, и б) доставка выбранному млекопитающему олигонуклеотида, являющегося ингибирующей РНК, антисмысловым нуклеотидом или рибозимом, снижающим уровень экспрессии мРНК Map4k4 в миобласте или миоците, в количестве, достаточном для индукции двух или более миоцитов в теле млекопитающего, приводящей к формированию миотиуб, где олигонуклеотид модифицирован основным фрагментом, углеводным фрагментом, или фосфатной основой; метод, где выбранное млекопитающее является человеком, метод, где выбранное млекопитающее имеет мышечное заболевание, метод, где мышечное заболевание является мышечной атрофией, мышечной слабостью, миопатией, синдромом хронической усталости, фибромиалгией, мышечной дистрофией, спинальной мышечной атрофией, дистальной мышечной дистрофией, дерматомизитом, полимиозитом, ревматической полимиалгией, разрывом мышцы или растяжением мышцы; метод, где выбранным млекопитающим является человек; метод, при котором доставка олигонуклеотида выбранному млекопитающему приводит к лечению заболевания; метод, где олигонуклеотидом является ингибирующая РНК, малая ингибирующая РНК.
Таким образом, в изобретении описана технология индукции дифференцировки миобластов и миоцитов в миотубы при помощи олигонуклеотидов, снижающих уровень Map4k4. Однако, данная методика активирует дифференцировку миобластов в миотубы, и никаким образом не влияет на процесс слияния клеток.
Из исследованного уровня техники выявлено изобретение, описанное в патенте US20190382732A1 «Compositions and methods relating to myomixer-promoted muscle cell fusion». Сущностью изобретения является клетка, трансформированная экзогенной нуклеиновой кислотой, кодирующей полипептид Myomixer под контролем промотора, активного в данной клетке. Клетка может являться клеткой человека, немиогенной клеткой, фибробластом, клеткой костного мозга или клеткой крови. Экзогенная нуклеиновая кислота может быть под контролем конститутивного или индуцибельного промотора. Клетка в дальнейшем может быть трансформирована экзогенной нуклеиновой кислотой, кодирующей Myomaker под контролем промотора, активного в данной клетке. Экзогенная нуклеиновая кислота может быть встроена в хромосому либо находиться в клетке в виде эписомы. Клетка может экспрессировать маркер детекции и/или маркер селекции. Клетка может быть трансформирована другим геном, помимо Myomixer, таким как терапевтический ген или маркерный ген.
Также метод подготовки партнера немиогенной клетки, включающий перенос в немиогенную клетку нуклеиновой кислоты, кодирующей протеин Myomixer под контролем промотора, активного в указанной клетке. Клетка может быть стабильно трансформирована, или транзиторно трансфицирована. Метод может также включать перенос в клетку нуклеиновой кислоты, кодирующей маркер детекции. Экзогенная нуклеиновая кислота может находиться под контролем конститутивного или индуцибельного промотора. Клетка может быть в последующем трансформирована нуклеиновой кислотой, кодирующей Myomaker под контролем промотора, активного в данной клетке. Клетка может быть клеткой человека, фибробластом, клеткой костного мозга или клеткой крови. Экзогенная нуклеиновая кислота может кодировать маркер селекции. Клетка может быть трансформирована другим геном, таким как терапевтический или маркерный ген.
Также метод слияния немиогенной клетки с миогенной, включающий в себя а) предоставление немиогенной клетке возможности экспрессировать Myomixer и Myomaker, и б) контакт указанной немиогенной клетки с мышцей, после чего немиогенная клетка сольется с указанной мышечной клеткой. Немиогенная клетка может быть человеческой клеткой, клеткой костного мозга или клеткой крови. Шаг б может быть выполнен in vivo или in vitro. Немиогенная клетка может экспрессировать маркер детекции или маркер селекции. Немиогенная клетка может быть трансформирована экзогенной кислотой, кодирующей полипептид Myomaker под контролем промотора, активного в указанной клетке. Миогенная клетка может быть изолированной, может находится в интаткной мышечной ткани и/или являться миобластом.
Также метод доставки целевого гена в мышечную клетку, включающий а) предоставдение немиогенной клетки способности экспрессировать протеины Myomaker или Myomixer, в то время немиогенная клетка экспрессирует целевой ген, и б) контакт указанной немиогенной клетки с миогенной, где указанная немиогенная клетка, экспрессирующая белки Myomaker и Myomixer сливается с указанной миогенной и доставляет указанный целевой ген в указанную миогенную клетку. Немиогенной клеткой может являться клетка человека, фибробласт, клетка костного мозга или клетка крови. Шаг б может быть выполнен in vivo и in vitro. Немиогенная клетка может экспрессировать маркер детекции или маркер селекции. Немиогенная клетка может быть впоследствии трансформирована нуклеиновой кислотой, кодирующей полипептид Myomaker под контролем промотора, активного в указанной клетке. Миогенная клетка может являться изолированной мышечной клеткой, либо может находиться в интактной мышечной ткани. Мышечная клетка может демонстрировать патологический фенотип, и указанный ген будет корректировать этот фенотип, в случае если фенотипом является сниженная экспрессия или отсутствие нормального продукта гена, или экспрессия дефектного продукта гена. Патологический фенотип может быть ассоциирован с врожденной миопатией, саркопенией, боковым амиотрофическим склерозом, мышечной дистрофией, болезнью Помпе или рабдомиосаркомой. Немиогенная клетка может быть доставлена в поврежденную мышцу при помощи внутримышечной инъекции, и доставка может быть повторена как минимум однократно, в то время как субъект может получать вторичную терапию. Немиогенная клетка может быть доставлена в указанную мышцу ex vivo с последующей имплантацией субъекту.
Также экспрессионная кассета, содержащая полипептид Myomixer под контролем промотора, активного в эукариотической клетке. Экзогенная нуклеиновая кислота может находится под контролем индуцибельного или конститутивного промотора. Экспрессионная кассета может кодировать маркер детекции и/или маркер селекции. Экспрессионная кассета может содержать другой ген, помимо Myomaker или Myomixer. Экспрессионная кассета может быть встроена в вектор репликации, такой как вирусный (к примеру, ретровирусный, аденовирусный, векторн на основе аденоассоциированного вируса) или невирусный вектор (к примеру, липосома или наночастица).
Также метод коррекции генетического дефекта в клетке, включающий а) получение немиогенной клетки субъекта, б) введение в указанную немиогенную клетку одной или нескольких экспрессионных кассет, экспрессирующих экзогенные Myomixer и Myomaker и один или несколько терапевтических генов, и в) контакт указанной немиогенной клетки с мышечной клеткой, имеющей генетический дефект, после чего указанная немиогенная сливается с указанной мышцей и доставляет целевой ген в мышцу, тем самым корректируя генетический дефект. Немиогенная клетка может являться клеткой человека, фибробластом, клеткой костного мозга или клеткой крови. Шаги б и в могут быть выполнены in vivo или in vitro, или шаг б может быть выполнен in vitro, в то время как шаг в-in vivo. Немиогенная клетка может экспрессировать маркер селекции или детекции. Терапевтический ген может являться Cas9 вкупе как минимум с одной sgРНК. Миогенная клетка может являться изолированной клеткой, либо находиться в интактной мышце. Генетический дефект может быть ассоциирован мышечной дистрофией Дюшенна, наследственной миопатией, болезнью Помпе, боковым амиотрофическим склерозом. Немиогенная клетка может быть доставлена в поврежденную мышцу при помощи внутримышечной инъекции. Доставка может быть повторена как минимум однократно. Субъект может получать вторичную терапию. Немиогенная клетка может контактировать с указанной мышечной клеткой ex vivo, и впоследствии импплантирована субъекту. Мышечная клетка может находиться в интактной мышце. Мышечная клетка может быть миобластом. Экспрессионная кассета может содержать конститутивный или индуцибельный промотор. Экспрессионная кассета может кодировать маркер селекции или маркер детекции. Экспрессионная кассета может содержаться в реплицируемом векторе, таком как вирусный вектор (например, ретровирусный, аденовирусный, или вектор на основе аденоассоциированного вируса) или невирусном векторе, таком как липосома или наночастица.
Таким образом, в изобретении описана методика модификации немиогенных клеток полипептидами Myomaker и Myomixer. Однако, авторы не указывают возможности модификации миогенных линий клеток, которые будут являться более физиологичными векторами, по сравнению с другими линиями клеток.
Таким образом, были выявлены аналоги только по назначению. Аналогов заявленных кодон-оптимизированных нуклеотидных последовательностей генов MYMK и ERVW-1, разделенных последовательностью 2А (или аналогичных) пептидов, заявителем не выявлено.
Целью и техническим результатом заявленного технического решения является разработка кодон-оптимизированных нуклеотидных последовательностей генов, кодирующих белки Myomaker и Syncytin-1 (MYMK и ERVW-1 соответственно), а также мультицистронных кассет, созданных на основе описанных выше генов, предназначенных для экспрессии белков Syncitin-1 и Myomaker в количестве, достаточном для увеличения интенсивности процессов слияния миобластов с мышечными волокнами.
Сущностью заявленного технического решения является генетическая конструкция (экспрессионная кассета), содержащая кодон-оптимизированные нуклеотидные последовательности генов ERVW-1 и MYMK, разделенные нуклеотидной последовательностью IRES, или кодирующей P2А, E2А, F2А или T2А пептид, используемая для экспрессии белков Syncytin-1 и Myomaker в миогенной клетке, и предназначенная для усиления способности миогенных клеток к слиянию. Также использование плазмиды, кодирующей ген ERVW-1 или использование плазмиды, кодирующей ген MYMK, для экспрессии белков Syncytin-1 или Myomaker в миогенной клетке, с целью усиления способности миогенных клеток к слиянию.
Список иллюстраций:
Фиг. 1. Генетическая карта конструкции MYMK-P2A-ERVW-1.
Фиг. 2. Фазово-контрастные изображения миотуб, сформированных в интактной клеточной культуре С2С12, индекс слияния 28,1 %, (слева), фазово-контрастное изображение миотуб, сформированных в клеточной С2С12 после трансфекции плазмидой MYMK-P2A-ERVW-1, индекс слияния 41,7 % (справа).
Заявленное изобретение осуществляется в 4 этапа в нижеприведенной последовательности, а именно:
1. Этап-оптимизация кодонного состава нуклеотидных последовательностей генов MYMK и ERVW-1;
2. Этап: создание донорной плазмиды (вектор);
3. Этап: клонирование заявленной последовательности MYMK -P2A- ERVW-1 из донорной плазмиды в экспрессионную векторную плазмиду.
4. Этап: анализ функциональной активности кодон-оптимизированных последовательностей генов MYMK и ERVW-1 на клеточных культурах in vitro.
Далее заявителем приведено подробное описание этапов осуществления заявленного технического решения:
1 этап-оптимизация кодонного состава нуклеотидных последовательностей генов MYMK и ERVW-1.
Проводят кодонную оптимизацию нуклеотидных последовательностей генов. Для кодонной оптимизации генов ERVW-1 и MYMK используют различные алгоритмы, такие как OptimumGene (GeneScript). Алгоритм OptimumGene учитывает содержание CpG-нуклеотидов, смещение кодонов, вторичную структуру мРНК, тандемные повторы, преждевременные сайты полиаденилирования, дополнительные сайты связывания с рибосомой, сайты рестрикции. В качестве матрицы для кодонной оптимизации используют нуклеотидные последовательности мРНК генов MYMK (GeneBank Gene ID: 389827, 666 пар нуклеотидов (п.н.)) и ERVW-1 (GeneBank Gene ID: 30816, 1617 п.н.). Дикий тип нуклеотидных последовательностей кодирующей части генов MYMK и ERVW-1 содержит тандем редких кодонов, которые могут остановить трансляцию или снизить ее эффективность. При оптимизации кодонного состава дикого типа гена MYMK улучшается индекс адаптации кодонов CAI (англ. Codon Adaptation Index) с 0,82 до 0,94. При оптимизации кодонного состава дикого типа гена и ERVW-1 улучшается индекс адаптации кодонов CAI с 0,67 до 0,94. Для увеличения стабильности мРНК оптимизируют GC-состав путем удаления протяженных участков с высоким содержанием GC-пар. Кроме того, в процессе оптимизации удаляют потенциальные цис-действующие сайты.
Заявителем установлено, что в результате кодонной оптимизации аминокислотные последовательности генов MYMK и ERVW-1 не изменяются и составляют 221 и 538 аминокислотных остатков, соответственно.
Далее проводят синтез de novo нуклеотидных последовательностей кДНК генов MYMK (SEQ ID NO: 1) и ERVW-1 (SEQ ID NO: 2), разделенных общеизвестными нуклеотидными последовательностями саморасщепляющегося пептида, из ряда 2А, P2A, E2A, F2A, T2A или нуклеотидной последовательностью IRES.
2 этап: создание донорной плазмиды (вектор)
Создают донорную плазмиду, например pUC57-MYMK-P2A-ERVW-1, содержащую нуклеотидные последовательности генов MYMK и ERVW-1, разделенные нуклеотидной последовательностью Р2А пептида, путем субклонирования нуклеотидных последовательности генов MYMK и ERVW-1, разделенных нуклеотидной последовательностью Р2А пептида в плазмидный вектор pUC57 по сайтам рестрикции XbaI/BamHI. Правильность сборки генетической конструкции pUC57-MYMK-P2A-ERVW-1 подтверждается рестрикционным анализом и секвенированием нуклеотидных последовательностей генов MYMK и ERVW-1.
3 этап: клонирование заявленной последовательности MYMK-P2A-ERVW-1 из донорной плазмиды в экспрессионную векторную плазмиду
Проводят субклонирование генов MYMK и ERVW-1, разделенных нуклеотидной последовательностью Р2А пептида из плазмиды pUC57-MYMK-P2A-ERVW-1 в экспрессионный плазмидный вектор по сайтам рестрикции HindIII/NotI. Правильность сборки результирующей экспрессионной генетической конструкции pMYMK-P2A-ERVW-1 (Фиг. 1.) подтверждается рестрикционным анализом и секвенированием нуклеотидных последовательностей генов MYMK и ERVW-1.
4 этап: анализ функциональной активности кодон-оптимизированных последовательностей генов MYMK и ERVW-1 на клеточных культурах in vitro.
Проводят трансфекцию клеточной линии C2C12 генетической конструкцией pMYMK-P2A-ERVW-1 при помощи трансфекционного агента Lipofectamine 3000 с использованием стандартного протокола трансфекции. Культивируют траснфицированные клетки на низкосывороточной среде, например DMEM HG+2% FBS. На 8 сутки после трансфекции проводят подсчет клеток, многоядерных структур, число ядер в многоядерных структурах. Вычисляют индекс слияния по формуле Fi=(Nm-Qm)/(Nt-1)×100% , где Fi-индекс слияния, Nm-число ядер в многоядерных структурах, Qm - количество многоядерных структур, Nt - общее число ядер в поле зрения. Экспрессия генов MYMK и ERVW-1 подтверждается иммуноцитохимическим анализом с использованием первичных антител к белкам Myomaker и Syncytin-1.
В эксперименте с использованием клеточной линии C2C12 трансфекция плазмидой MYMK-P2A-ERVW-1 привела к экспрессии обоих целевых белков, что было подтверждено Вестерн-блот анализом. При обработке клеточных лизатов специфическими антителами к Myomaker и Syncytin-1 в образцах трансфицированных клеток наблюдались две отчетливо выраженные полосы:
Полоса, соответствующая белку Myomaker, имела молекулярную массу примерно 25 кДа, что соответствует расчетной массе белка из 221 аминокислотного остатка.
Полоса, соответствующая белку Syncytin-1, была обнаружена в диапазоне 60–70 кДа, что согласуется с его молекулярной массой (538 аминокислот с учетом возможных посттрансляционных модификаций).
Эти результаты свидетельствуют о корректном функционировании саморасщепляющегося пептида P2A, обеспечивающего независимую и стехиометричную экспрессию Myomaker и Syncytin-1, и подтверждают эффективность данной генетической конструкции для усиления процессов слияния миогенных клеток.
Таким образом, из вышеизложенного можно сделать общий вывод, что заявителем достигнуты поставленные цели и заявленный технический результат, а именно разработаны:
- кодон-оптимизированные последовательности генов, кодирующие белки Syncytin-1 и Myomaker (ERVW-1 и MYMK соответственно)
- мультицистронные кассеты MYMK-P2A-ERVW-1, MYMK-E2A-ERVW-1, MYMK-F2A-ERVW-1, MYMK-T2A-ERVW-1 и MYMK-IRES-ERVW-1, созданные на основе вышеуказанных последовательностей, и предназначенные для усиления слияния миогенных клеток.
Экспериментально доказано, что использование заявленного технического решения позволяет увеличить индекс слияния клеток.
Заявленное техническое решение удовлетворяет критерию «новизна», предъявляемому к изобретениям, так как из исследованного уровня техники не выявлены источники, в которых описаны признаки, совпадающие по исполняемой ими функции и форме выполнения этих признаков, перечисленные в формуле изобретения, включая характеристику назначения.
Заявленное техническое решение удовлетворяет критерию «изобретательский уровень», предъявляемому к изобретениям, поскольку из исследованного уровня техники заявителем не выявлены технические решения, характеризующиеся использованием кодон-оптимизированных последовательностей генов ERVW-1 и MYMK соответственно, а также с использованием генетических конструкций, содержащие вышеописанные последовательности (SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3) для использования с целью усиления слияния миогенных клеток. Кроме того, заявленное техническое решение, по мнению заявителя, не является очевидным для специалиста, так как обеспечивает реализацию задачи по преодолению фундаментальной проблемы генно-клеточной терапии заболеваний мышечной ткани.
Заявленное техническое решение удовлетворяет критерию «промышленная применимость», предъявляемому к изобретениям, так как оно может быть использовано в промышленных масштабах для создания препаратов, предназначенных для комплексной генно-клеточной терапии заболеваний мышечной ткани.
--->
<?xml version="1.0" encoding="ISO-8859-1"?>
<!DOCTYPE ST26SequenceListing SYSTEM "ST26SequenceListing_V1_3.dtd"
PUBLIC "-//WIPO//DTD Sequence Listing 1.3//EN">
<ST26SequenceListing productionDate="2025-02-25"
softwareVersion="2.2.0" softwareName="WIPO Sequence"
fileName="Генетическая конструкция и способ экспрессии ERVW-1 и
MYMK.xml" dtdVersion="V1_3">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText/>
<FilingDate/>
</ApplicationIdentification>
<ApplicantFileReference>Генетическая конструкция и способ экспрессии
REVW-1 и MYMK</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное бюджетное
научное учреждение «Российский научный центр хирургии имени академика
Б.В. Петровского» </ApplicantName>
<ApplicantNameLatin>Federal State Budgetary Scientific Institution
"Russian Scientific Center of Surgery named after Academician B.V.
Petrovsky"</ApplicantNameLatin>
<InventionTitle languageCode="ru">Генетическая конструкция и способ
экспрессии ERVW-1 и MYMK</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>666</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..666</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>unassigned DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>unidentified</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgggcaccctggtggcaaagctgctgctgccaacactgagctccctggcct
ttctgccaaccgtgagcatcgcagcaaagcggagatttcacatggaggccatggtgtacctgttcacact
gttctttgtggccctgcaccacgcatgcaacggaccaggactgagcgtgctgtgctttatgcggcacgac
atcctggagtacttcagcgtgtatggcaccgccctgtctatgtgggtgagcctgatggccctggccgact
tcgatgagcccaagaggtccacctttgtgatgttcggcgtgctgacaatcgccgtgaggatctatcacga
ccgctggggctacggcgtgtattctggccctatcggcacagccatcctgatcatcgccgccaagtggctg
cagaagatgaaggagaagaagggcctgtaccctgataagtccgtgtatacacagcagatcggaccaggac
tgtgctttggcgccctggccctgatgctgcggttctttttcgaggactgggattacacctatgtgcactc
cttttaccactgtgccctggccatgtctttcgtgctgctgctgcccaaagtgaataagaaggcaggaagc
ccaggaacccctgcaaagctggattgctccacactgtgctgtgcctgcgtgtga</INSDSeq_sequen
ce>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>1617</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1617</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>unassigned DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>unidentified</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atggccctgccttaccacatcttcctgtttacagtgctgctgcccagcttca
ccctgacagcaccacctccatgcaggtgtatgaccagctcctctccctaccaggagtttctgtggcggat
gcagagacccggcaacatcgacgccccttcctataggtctctgagcaagggcaccccaacattcaccgcc
cacacacacatgccccgcaattgctaccacagcgccaccctgtgcatgcacgccaacacacactattgga
ccggcaagatgatcaatccatcttgcccaggaggactgggagtgaccgtgtgctggacctactttacaca
gaccggaatgagcgacggaggaggagtgcaggatcaggcacgggagaagcacgtgaaggaagtgatctct
cagctgacaagagtgcacggcaccagctccccttataagggcctggatctgagcaagctgcacgagaccc
tgcggacacacaccagactggtgagcctgttcaacaccacactgacaggcctgcacgaggtgtccgccca
gaacccaaccaattgctggatctgtctgcctctgaacttccgcccatacgtgagcatccccgtgcctgag
cagtggaacaatttttccaccgagatcaataccacatctgtgctggtgggccccctggtgagcaacctgg
agatcacacacacctccaatctgacctgcgtgaagttttctaacaccacatataccacaaatagccagtg
catccggtgggtgacaccccctacccagatcgtgtgcctgcctagcggcatcttcttcgtgtgcggcacc
tccgcctacagatgtctgaacggctctagcgagtctatgtgcttcctgagcttcctggtgccacccatga
caatctataccgagcaggacctgtacagctatgtgatctccaagcccaggaataagcgcgtgcctatcct
gccattcgtgatcggagcaggcgtgctgggcgccctgggaacaggcatcggcggcatcaccacatccacc
cagttttactataagctgtctcaggagctgaacggcgacatggagagggtggccgattctctggtgaccc
tgcaggaccagctgaacagcctggccgccgtggtgctgcagaatcggagagccctggatctgctgacagc
agagaggggaggaacctgcctgttcctgggcgaggagtgctgttactatgtgaaccagtccggcatcgtg
acagagaaggtgaaggagatcagggacaggatccagaggagggcagaggagctgaggaataccggaccat
ggggcctgctgtctcagtggatgccatggatcctgcccttcctgggacctctggcagccatcatcctgct
gctgctgttcggcccctgtatctttaacctgctggtgaatttcgtgagctctagaatcgaggccgtgaag
ctgcagatggagcccaagatgcagtccaagacaaagatctaccggagacctctggataggccagcatccc
ctcgctctgacgtgaacgatatcaagggaacccctccagaggagatctccgccgcacagccactgctgag
acccaattctgccggcagctcctga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>4990</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..4990</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>unassigned DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>unidentified</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tccgctagaagctgcatgtctaagctagacccttcagattaaaaataactga
ggtaagggcctgggtaggggaggtggtgtgagacgctcctgtctctcctctatctgcccatcggcccttt
ggggaggaggaatgtgcccaaggactaaaaaaaggccatggagccagaggggcgagggcaacagaccttt
catgggcaaaccttggggccctgctgtctagcatgccccactacgggtctaggctgcccatgtaaggagg
caaggcctggggacacccgagatgcctggttataattaacccagacatgtggctgcccccccccccccaa
cacctgctgcctctaaaaataaccctgtccctggtggatcccctgcatgcgaagatcttcgaacaaggct
gtgggggactgagggcaggctgtaacaggcttgggggccagggcttatacgtgcctgggactcccaaagt
attactgttccatgttcccggcgaagggccagctgtcccccgccagctagactcagcacttagtttagga
accagtgagcaagtcagcccttggggcagcccatacaaggccatggggctgggcaagctgcacgcctggg
tccggggtgggcacggtgcccgggcaacgagctgaaagctcatctgctctcaggggcccctccctgggga
cagcccctcctggctagtcacaccctgtaggctcctctatataacccaggggcacaggggctgccctcat
tctaccaccacctccacagcacgagctagacgtaagtatcaaggttacaagacaggtttaaggagaccaa
tagaaactgggcttgtcgagacagagaagactcttgcgtttctgataggcacctattggtcttactgaca
tccactttgcctttctctccacagccaagctggctagcgtttaaacttaagcttgccaccatgggcaccc
tggtggcaaagctgctgctgccaacactgagctccctggcctttctgccaaccgtgagcatcgcagcaaa
gcggagatttcacatggaggccatggtgtacctgttcacactgttctttgtggccctgcaccacgcatgc
aacggaccaggactgagcgtgctgtgctttatgcggcacgacatcctggagtacttcagcgtgtatggca
ccgccctgtctatgtgggtgagcctgatggccctggccgacttcgatgagcccaagaggtccacctttgt
gatgttcggcgtgctgacaatcgccgtgaggatctatcacgaccgctggggctacggcgtgtattctggc
cctatcggcacagccatcctgatcatcgccgccaagtggctgcagaagatgaaggagaagaagggcctgt
accctgataagtccgtgtatacacagcagatcggaccaggactgtgctttggcgccctggccctgatgct
gcggttctttttcgaggactgggattacacctatgtgcactccttttaccactgtgccctggccatgtct
ttcgtgctgctgctgcccaaagtgaataagaaggcaggaagcccaggaacccctgcaaagctggattgct
ccacactgtgctgtgcctgcgtgggaagcggagctactaacttcagcctgctgaagcaggctggagacgt
ggaggagaaccctggacctatggccctgccttaccacatcttcctgtttacagtgctgctgcccagcttc
accctgacagcaccacctccatgcaggtgtatgaccagctcctctccctaccaggagtttctgtggcgga
tgcagagacccggcaacatcgacgccccttcctataggtctctgagcaagggcaccccaacattcaccgc
ccacacacacatgccccgcaattgctaccacagcgccaccctgtgcatgcacgccaacacacactattgg
accggcaagatgatcaatccatcttgcccaggaggactgggagtgaccgtgtgctggacctactttacac
agaccggaatgagcgacggaggaggagtgcaggatcaggcacgggagaagcacgtgaaggaagtgatctc
tcagctgacaagagtgcacggcaccagctccccttataagggcctggatctgagcaagctgcacgagacc
ctgcggacacacaccagactggtgagcctgttcaacaccacactgacaggcctgcacgaggtgtccgccc
agaacccaaccaattgctggatctgtctgcctctgaacttccgcccatacgtgagcatccccgtgcctga
gcagtggaacaatttttccaccgagatcaataccacatctgtgctggtgggccccctggtgagcaacctg
gagatcacacacacctccaatctgacctgcgtgaagttttctaacaccacatataccacaaatagccagt
gcatccggtgggtgacaccccctacccagatcgtgtgcctgcctagcggcatcttcttcgtgtgcggcac
ctccgcctacagatgtctgaacggctctagcgagtctatgtgcttcctgagcttcctggtgccacccatg
acaatctataccgagcaggacctgtacagctatgtgatctccaagcccaggaataagcgcgtgcctatcc
tgccattcgtgatcggagcaggcgtgctgggcgccctgggaacaggcatcggcggcatcaccacatccac
ccagttttactataagctgtctcaggagctgaacggcgacatggagagggtggccgattctctggtgacc
ctgcaggaccagctgaacagcctggccgccgtggtgctgcagaatcggagagccctggatctgctgacag
cagagaggggaggaacctgcctgttcctgggcgaggagtgctgttactatgtgaaccagtccggcatcgt
gacagagaaggtgaaggagatcagggacaggatccagaggagggcagaggagctgaggaataccggacca
tggggcctgctgtctcagtggatgccatggatcctgcccttcctgggacctctggcagccatcatcctgc
tgctgctgttcggcccctgtatctttaacctgctggtgaatttcgtgagctctagaatcgaggccgtgaa
gctgcagatggagcccaagatgcagtccaagacaaagatctaccggagacctctggataggccagcatcc
cctcgctctgacgtgaacgatatcaagggaacccctccagaggagatctccgccgcacagccactgctga
gacccaattctgccggcagctcctgagcggccgctcgagtctagagggcccgtttaaacccgctaacttg
tttattgcagcttataatggttacaaataaagcaatagcatcacaaatttcacaaataaagcattttttt
cactgcattctagttgtggtttgtccaaactcatcaatgtatcttaaggcggtgcgcggaacccctattt
gtttatttttctaaatacattcaaatatgtatccgctcatgagacaataaccctgataaatgcttcaata
atattgaaaaaggaagagtcgatggatcgtttcgcatgattgaacaagatggattgcacgcaggttctcc
ggccgcttgggtggagaggctattcggctatgactgggcacaacagacaatcggctgctctgatgccgcc
gtgttccggctgtcagcgcaggggcgcccggttctttttgtcaagaccgacctgtccggtgccctgaatg
aactgcaagacgaggcagcgcggctatcgtggctggccacgacgggcgttccttgcgcagctgtgctcga
cgttgtcactgaagcgggaagggactggctgctattgggcgaagtgccggggcaggatctcctgtcatct
caccttgctcctgccgagaaagtatccatcatggctgatgcaatgcggcggctgcatacgcttgatccgg
ctacctgcccattcgaccaccaagcgaaacatcgcatcgagcgagcacgtactcggatggaagccggtct
tgtcgatcaggatgatctggacgaagagcatcaggggctcgcgccagccgaactgttcgccaggctcaag
gcgagcatgcccgacggcgaggatctcgtcgtgacccatggcgatgcctgcttgccgaatatcatggtgg
aaaatggccgcttttctggattcatcgactgtggccggctgggtgtggcggaccgctatcaggacatagc
gttggctacccgtgatattgctgaagagcttggcggcgaatgggctgaccgcttcctcgtgctttacggt
atcgccgctcccgattcgcagcgcatcgccttctatcgccttcttgacgagttcttctgagcgggactct
ggggttttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcg
gtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcaga
taccaaatactgtccttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctac
atacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggttg
gactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacagccca
gcttggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcc
cgaagggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagctt
ccagggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttt
tgtgatgctcgtcaggggggcggagcctatggaaaaac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОДОН-ОПТИМИЗИРОВАННАЯ КДНК, КОДИРУЮЩАЯ ДИСФЕРЛИН ЧЕЛОВЕКА, ГЕННО-ИНЖЕНЕРНАЯ КОНСТРУКЦИЯ, РЕКОМБИНАНТНЫЙ АДЕНОВИРУС И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ДИСФЕРЛИНОПАТИЙ | 2012 |
|
RU2527073C2 |
| СИНТЕТИЧЕСКАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ), ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ КЛЕТОК МЛЕКОПИТАЮЩЕГО, КЛЕТКА МЛЕКОПИТАЮЩЕГО ХОЗЯИНА И СПОСОБ ЭКСПРЕССИИ ЧЕЛОВЕЧЕСКОГО БЕЛКА АНТИГЕНА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА-2 (HER2/neu) ИЛИ ЕГО УКОРОЧЕННОЙ ФОРМЫ | 2004 |
|
RU2397249C2 |
| НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ С ДЕФИЦИТОМ ДПНРНК | 2014 |
|
RU2721855C2 |
| СПОСОБ БЛОКИРОВАНИЯ АКТИВНОСТИ ACTIVIN С ПОМОЩЬЮ НОГГИН2 | 2005 |
|
RU2354662C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ НАРУШЕНИЙ И ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С RDH12 | 2017 |
|
RU2764920C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДОСТАВКИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА | 1994 |
|
RU2174845C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок SMN1, и ее применение | 2020 |
|
RU2742837C1 |
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2711147C2 |
| ПРОДУКЦИЯ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ФУКОЗИЛИРОВАНИЕМ | 2008 |
|
RU2479629C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДИФИКАЦИИ ЦЕЛЕВОГО ЛОКУСА | 2015 |
|
RU2704283C2 |






Изобретение относится к области биотехнологии. Описана генетическая конструкция pMYMK-P2A-ERVW-1, представленная на фиг. 1, для раздельной экспрессии генов MYMK SEQ ID NO: 1 и ERVW-1 SEQ ID NO: 2 и усиления слияния мышечных клеток. Техническим результатом заявленного технического решения является разработка кодон-оптимизированных нуклеотидных последовательностей генов, кодирующих белки Myomaker и Syncytin-1 (MYMK и ERVW-1 соответственно), а также мультицистронных кассет, созданных на основе описанных выше генов, предназначенных для экспрессии белков Syncitin-1 и Myomaker в количестве, достаточном для увеличения интенсивности процессов слияния миобластов с мышечными волокнами. 2 ил., 1 пр.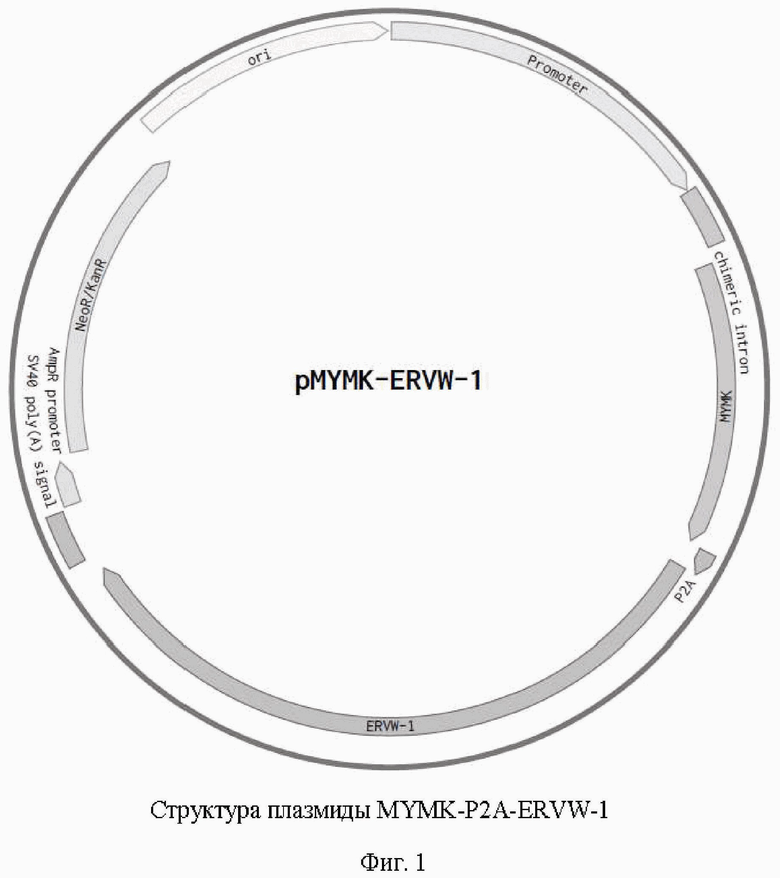
Генетическая конструкция pMYMK-P2A-ERVW-1, представленная на фиг. 1, для раздельной экспрессии генов MYMK SEQ ID NO: 1 и ERVW-1 SEQ ID NO: 2 и усиления слияния мышечных клеток.
| ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В КЛЕТКАХ СНО РЕКОМБИНАНТНОГО ФОЛЛИКУЛОСТИМУЛИРУЮЩЕГО ГОРМОНА (ФСГ) ЧЕЛОВЕКА, ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В КЛЕТКАХ СНО БЕТА-СУБЪЕДИНИЦЫ РЕКОМБИНАНТНОГО ФСГ ЧЕЛОВЕКА, КЛЕТКА СНО - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ФСГ ЧЕЛОВЕКА И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО ГОРМОНА | 2013 |
|
RU2560596C2 |
| CN 102242143 A 16.11.2011 | |||
| Camacho A | |||
| et al | |||
| Carey-Fineman-Ziter syndrome: a MYMK-related myopathy mimicking brainstem dysgenesis | |||
| Journal of Neuromuscular Diseases, 2020, V.7(3), P | |||
| Переставная шейка для вала | 1921 |
|
SU309A1 |

