Изобретение относится к микробиологической промышленности, а именно к способам получения из микроорганизмов ферментов, катализирующих синтез веществ, которые широко применяются в исследованиях по биохимии, молекулярной биологии и генетической инженерии. К таким ферментам принадлежат S-аденозилметионинсинтетаза (АТФ: 1-метионин S-аденозилтрансфераза, ЕС 2.5.1.6), катализирующая реакцию
1-метионин + АТФ 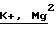 S-аденозил-1-метионин+ ФФ4 + Ф4˙S-аденозилметионин (S AM) является активированным донором метильных групп и играет ключевую роль в метилтрансферазных реакциях, среди которых следует отметить энзиматическое метилирование белков и нуклеиновых кислот, метилирование низкомолекулярных веществ (фосфолипиды, стероиды), образование циклопропановых жирных кислот. Кроме того, SАМ является предшественником важных клеточных компонентов (спермидин и спермин). Участвуя в энзиматическом метилировании ДНК и белков, SАМ играет весьма существенную роль в процессах модификации-рестрикции ДНК и хемотаксисе.
S-аденозил-1-метионин+ ФФ4 + Ф4˙S-аденозилметионин (S AM) является активированным донором метильных групп и играет ключевую роль в метилтрансферазных реакциях, среди которых следует отметить энзиматическое метилирование белков и нуклеиновых кислот, метилирование низкомолекулярных веществ (фосфолипиды, стероиды), образование циклопропановых жирных кислот. Кроме того, SАМ является предшественником важных клеточных компонентов (спермидин и спермин). Участвуя в энзиматическом метилировании ДНК и белков, SАМ играет весьма существенную роль в процессах модификации-рестрикции ДНК и хемотаксисе.
В настоящее время SАМ широко используют в молекулярной биологии и генетической инженерии для избирательного включения меченых радиоактивными изотопами метильных групп в ДНК и защиты ДНК от соответствующих рестрикционных эндонуклеаз. Меченые по метильной группе радиоактивные препараты SАМ также широко используются для определения активности различных метилтрансфераз.
Цель изобретения - повышение содержания целевого продукта в биомассе.
Способ заключается в следующем.
В состав минеральной питательной среды с метанолом для штамма Methylophilus methanolovorus BKM B-1337 Д используют концентрации минеральных солей Mg2+ и Fe2+, 0,100-0,350 г/л MgSO4 ˙7 H2О и 0,007-0,030 г/л FeSO4 ˙7 Н2О. Эти количества способствуют изменению консистенции биомассы из желеобразной в плотную, а также значительному снижению потерь клеток в процессе отмыва бактерий от культуральной жидкости, при сохранении высокой активности SАМ-синтетазы.
Величина лаг-фазы роста бактерий и конечный выход биомассы коррелируют с концентрацией метанола в среде (0,3-1,0% ).
Концентрация метанола в минеральной среде 0,5% является оптимальной для получения биомассы штамма-продуцента. Более низкие концентрации этого источника углерода и энергии дают меньший выход биомассы. Увеличение концентрации метанола в среде удлиняет лаг-фазу роста клеток. В этом случае полученное за 18 ч количество биомассы будет ниже.
Культивируют штамм-продуцент Methylophilus methanolovorus ВКМ В-1447 Д. Выбор данного штамма обусловлен его способностью использовать дешевый непищевой субстрат - метанол, отсутствием потребности в ростовых факторах (витамины, дрожжевой экстракт, аминокислоты), высокой активностью SAМ-синтетазы в бесклеточных экстрактах, составляющей 5-10 ед/мг белка. Продуцент был найден в результате скрининга среди дрожжей (представители родов Sacharomyces, Pichia, Hansenula, Candida, Torulopsis), а также среди факультативных и облигатных метилотрофных бактерий (представители родов Pseudomonas, Methylomonas, Methylophilus, Methylobacillus).
Штамм М. methaholovorus БКМ В-1447 Д выделен из природных источников, отселекционирован в процессе длительного пассирования на жидких средах с метанолом и рассевов на агаризованную синтетическую среду с метанолом. Штамм депонирован во Всесоюзной коллекции микроорганизмов при Институте биохимии и физиологии микроорганизмов АН СССР под номером ВКМ В-1447 Д.
Морфологические признаки. Клетки одиночные или в парах, палочковидной формы (0,4-0,8˙ 1,0 - 4,5 мкм), подвижные благодаря наличию полярного жгутика. Пигмент, спор, капсул и слизи не образуют.
Культуральные признаки. Колонии на синтетической агаризованной среде с метанолом на 3 сутки: округлые, бесцветные, край ровный, до 1 мм в диаметре, гладкие, выпуклые, блестящие структура однородная, консистенция вязкая. На мясо-пептонных средах не растет. На скошенном агаре с метанолом растет в виде гладкого стелющегося полупрозрачного штриха. Желатину не разжижает. Окраска по Граму - отрицательная.
Физиологические признаки. Облигатный метилитроф, использует метанол в качестве источника углерода и энергии. Неспособен расти на других одноуглеродных (метан, формальдегид, метиламин, диметиламин, триметиламин, формамид, диметилформамид) и полиуглеродных субстратах (спирты, сахара, органические кислоты, аминокислоты). Нейтрофил, хорошо растет при значениях рН, близких к нейтральному. Температурный оптимум 30-37оС. Нитраты восстанавливает до нитритов. Сероводород и индол не образует. Тесты на каталазу и оксидазу положительные. Полисахариды в среде не накапливает. Молярное содержание ГЦ в ДНК 52 мол. % . Источниками азота могут служить соли аммония и нитраты. В дополнительных факторах роста не нуждается.
Биохимическая характеристика. Штамм обладает дегидрогеназами метанола, формальдегида и формиата, проявляющими активность с феназинметосульфатом как акцептором электронов. Кроме того имеет НАД-зависимую формальдегиддегидрогеназу, не требующую восстановленного глутатиона. Отсутствуют специфические ферменты серинового цикла оксипируватредуктаза, серии-глиоксилатаминотроансфераза) и рибулозодифосфатного цикла (фосфорибулокиназа, рибуолозодифосфаткарбоксилаза). Углерод метанола ассимилируется в основном на уровне окисленности формальдегида гексулофосфатным путем (обладает высокоактивной 3-гексулозофосфатсинтазой). Имеет высокие активности НАД(Ф)-зависимых дегидрогеназ глюкозо-6-фосфата и 6-фосфоглюконата, осуществляет циклическое окисление формальдегида до СО2 в диссимиляционной ветви гексулозофосфатного цикла. Характеризуется наличием альдолазы 2-кето-3-дезокси-6-фосфоглюконтата, но не имеет 6-фосфофруктокиназы, фруктозо-1,6-дифосфатальдолазы, пируваткиназы, фруктозо-1,6-дифосфатазы, седогептулозо-1,7-дифосфотазы, ФЕП-карбоксикиназы, ФЕП-синтетазы, пируватфосфаткиназы, изюцитратлиазы и мелатсинтазы. Цитратный цикл разомкнут на уровне α -кетоглутаратдегидрогеназы. Фиксация СО2 катализируется карбоксилазами пирувата и фосфоенолпирувата. Первичная ассимиляция аммония осуществляется глутаматдегидрогеназой.
Активность SАМ-синтетазы определяют в клетках М. methanolovorus ВКМ В-1447Д, выращенных на минеральной среде, содержащей метанол. Клетки отделяют от среды центрифугированием (5000 g, 10 мин, 0-2оС). Бесклеточный экстракт получают путем экструзионной дезинтеграции клеток на прессе типа Хьюза при -55оС и последующего центрифугирования гомогената. Полученный экстракт используют в качестве источника SАМ-синтазы. За единицу активности SАМ-синтезаты принимают такое количество фермента, которое способно синтезировать 1 нмоль SАМ за 1 мин.
П р и м е р 1. Methylophilus methanovorus БКМ В-1447Д выращивают в колбах объемом 750 мл, заполненных 200 мл среды следующего состава, г/л: KH2PO4 2,0; NaCl 0,5; (NH4)2 SO4 2,0; MgSO4 0,175; FeSO4-7 H2О 0,01, метанол 0,5; дистиллированная вода, рН среды доводят до 7,2 10% -ным раствором NaOH перед стерилизацией. Метанол добавляют в среду после стерилизации. Бактерии культивируют при 30оС на качалке (140 об/мин). После 18 ч выращивания клетки отделяют от культуральной жидкости центрифугированием (5000 g, 10 мин, 0-2оС), промывают 50 мМ трис-НСl-буфера, рН 8,0 и снова центрифугируют в том же режиме. Клетки 1 г суспендируют в буферном растворе (10 мл), содержащем 10 мМ трис-НСl, рН 8,2, 10 мМ MgClz, 1 мМ меркаптоэтанола, 20 мкг/мл фенилметилсульфонилфторида, глицерин до конечной концентрации 15% и разрушают на прессе типа Хьюза с рабочим давлением 3000 кг/см2 при температуре около -50оС. Гомогенат центрифугируют (5000 g, 1 ч, 0оС), осадок отбрасывают и в супернатанте определяют активность SАМ-синтетазы. Реакцию проводят в реакционной смеси (100 мкл), содержащей 100 мМ трис-НСl-буфера, рН 8,2, 100 мМ KCl, 20 мМ MgCl2, 3 мМ АТФ и 1,3 мМ L-(метил-14С) метионин (уд. активность 160 мкюри-мМ), бесклеточный экстракт (150-300 мкг белка). Реакцию проводят в течение 10 мин и останавливают добавлением 50 мМ ЭДТА (35 мкл на 15 мкл реакционной смеси). Радиоактивный АМ отделяют от реакционной смеси на ионообменной бумаге "Ватман" Р-81 в Н+-форме. Для перевода в Н+-форму бумагу предварительно выдерживают 30 мин в 4% -ной уксусной кислоте, далее отмывают дистиллированной водой и высушивают на воздухе. Пробу с реакционной смесью (20 мкл) наносят на обработанную бумагу "Ватман" Р-81 (3х3) см, подсушивают и отмывают от непрореагировавшего метионина дистиллированной водой (1 л на 4 пробы). Фильтры высушивают под инфракрасной лампой, помещают в сцинтилляционную смесь, содержащую 4 г ППО и 0,5 г ПОПОП в 1 л толуола и просчитывают на жидкостном спектрометре SL-30 "интертехник". Активность SАМ-синтетазы 10 нмоль/мин/мг белка.
П р и м е р 2. Способ осуществляют, как описано в примере 1, используя для выращивания продуцента среду, содержащую, г/л MgSO4 ˙7 Н2О 0,1 и FeSО4 ˙7 Н2О 0,007 (содержание других компонентов среды не изменяют). Активность SАМ-синтетазы 6 ед. /мг белка.
Уменьшение концентрации Fe2+ и Mg2+ в среде культивирования приводит к получению желеобразной биомассы в результате выделения клетками М. methanolovirus полисахарида. В такой биомассе затруднено определение активности SАМ-синтетазы. Кроме того, за счет увеличения потерь биомассы в процессе отмыва бактерий от культуральной жидкости уменьшается количество клеток, используемых для выделения и определения активности SАМ-синтетазы.
П р и м е р 3. Способ осуществляют, как описано в примере 1, используя для выращивания продуцента среду, содержащую, г/л: MgSO4 ˙7 Н2О 0,350 и FeSO4 ˙7 Н2О 0,030. Активность SАМ-синтетазы 7,3 ед. /мг белка.
Увеличение концентрации Mg2+ и Fe2+ в среде культивирования вызывает удлинение лаг-фазы роста клеток и снижение выхода клеточной биомассы в 1,3-1,7 раза.
Таким образом, содержание АМ-синтетазы в биомассе предлагаемого продуцента больше, чем в биомассе известных продуцентов:
Продуцент Содержание
SАМ-синтетазы
в биомассе,
ед. /мг белка
Пекарские дрожжи 0,3-0,6 Escherichia coli и 5,8 M. methanolovorus БКМ В-1447 Д 6,0-10,0
Кроме того, для выращивания предлагаемого продуцента требуется намного более дешевая среда, содержащая только метанол в качестве источника углерода и энергии, как витаминов, сахаров и каких-либо биологических добавок. (56) S. H. Mudd, G. L. Cantoni "Activation of methionine for transmethylation", I. Biol. Chem. , 1958, 231, 481-491.
P. K. Chiang, G. L. Cantoni, "Activatiom of methionine for transmethylation, " I. Biol. Chem, 1977, 252, 4506-4513.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИЙ METHYLOBACTERIUM SPECIES, РАЗЛАГАЮЩИЙ МЕТАНОЛ И/ИЛИ ФОРМАЛЬДЕГИД | 1995 |
|
RU2077580C1 |
| Способ получения глутаматдегидрогеназы | 1982 |
|
SU1017730A1 |
| Способ хранения микроорганизмов | 1989 |
|
SU1671682A1 |
| ШТАММ БАКТЕРИЙ METHYLOBACILLUS METHANOLIVORANS GSA - ПРОДУЦЕНТ КОРМОВОГО БЕЛКА | 2023 |
|
RU2808127C1 |
| Консорциум штаммов бактерий РSеNDомоNаS Sp. и МетнYLовасILLUS метнаNоLоVоRUS, разлагающий метилацетат | 1989 |
|
SU1687608A1 |
| Штамм облигатно-метилотрофной бактерии @ @ ВСБ-792 | 1980 |
|
SU989867A1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pACYC-LANS(KM), ШТАММ Escherichia coli BL21(DE3), ТРАНСФОРМИРОВАННЫЙ РЕКОМБИНАНТНОЙ ДНК pACYC-LANS(KM), И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ L-АСПАРАГИНАЗЫ Erwinia carotovora | 2010 |
|
RU2441916C1 |
| ПРОМЫШЛЕННЫЙ СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ФЕРМЕНТА ПЕНИЦИЛЛИН G АЦИЛАЗЫ ESCHERICHIA COLI | 2020 |
|
RU2729410C1 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |
СПОСОБ ПОЛУЧЕНИЯ S - АДЕНОЗИЛМЕТИОНИНСИНТЕТАЗЫ, включающий культивирование штамма - продуцента на питательной среде, содержащей источник углерода и энергии, а также минеральные соли, с последующим выделением бесклеточного экстракта, отличающийся тем, что, с целью повышения содержания целевого продукта в биомассе, в качестве штамма - продуцента используют штамм Methylophilus methanolovorus BKM B-1447Д, а культивирование ведут на питательной среде, содержащей в качестве источника углерода и энергии метанол, а в качестве минеральных солей - хлористый натрий, однозамещенный фосфорно-кислый калий, серно-кислый аммоний, сернокислый магний и сернокислое железо, при следующем соотношении компонентов, г/л:
Метанол 3 - 10
Хлористый натрий 0,3 - 0,7
Однозамещенный фосфорно-кислый калий 1,7 - 2,3
Сернокислый аммоний 1,7 - 2,3
Сернокислый магний 0,100 - 0,350
Сернокислое железо 0,007 - 0,030
